
C4bpコアタンパク質及び単量体抗原を含む生成物及びその使用
本発明は、C4bpコアタンパク質と単量体抗原を含み、好ましくは融合タンパク質の形の生成物を提供する。単量体抗原はマラリア及びインフルエンザ抗原を含む。C4bpコアタンパク質により、前記単量体抗原を多量体の複合体に組立てられたもの又はそれらの混合物が提供される。この複合体はワクチンとして有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アジュバントと抗原を含む融合タンパク質などの高分子の集合体(assembly)に関し、この集合体は、抗原単独と比較した場合に、抗原に対する高まった免疫反応を誘導する。
【背景技術】
【0002】
アジュバントは抗原に対する免疫反応を高め、従ってワクチンに有用である。しかしながら、ヒトの使用に承認されているアジュバントは限られた数しかなく、動物での研究からもっと強いアジュバントが知られており、ヒトにおける使用に安全なより強い免疫学的アジュバントの必要性が明らかに存在する。最近の総説については、「Advances in vaccine adjuvants」(Nature Biotechnology, 1999, Volume 17, p1075-1081)及び「Recent advances in the discovery and delivery of vaccine adjuvants」(Nature Reviews in Drug Discovery, 2003, Volume 2, p727-735)を参照されたい。
【0003】
補体系は、外来抗原に対する免疫系の応答に重要な一連の血清タンパク質から成る。補体系は、その初期の成分が切断され、その生成物が単独で又は他のタンパク質と一緒に、別の補体タンパク質を活性化してタンパク質分解カスケードを生じ、活性化される。補体系の活性化は、様々な反応、例えば血管透過性の上昇、食細胞の走化性、炎症細胞の活性化、異物のオプソニン化、細胞の直接死及び組織損傷を引き起こす。補体系の活性化は抗原-抗体複合体により誘導されるか(古典経路)、又は通常のゆっくりとした活性化が、細菌及びウイルス等の侵入生物の細胞壁の存在下で増幅される(代替経路)。補体系は、古典経路及び代替経路の両方の中心のタンパク質であるC3が関与している特定の経路を介して、細胞免疫系と相互作用する。C3のタンパク質分解活性化により、大きいフラグメント(C3b)が生じ、侵入生物又は外来細胞の細胞表面タンパク質などの外部の求核物質と共有結合的に反応することができる化学的に反応性の内在性チオールエステル結合が露出する。その結果、潜在的な抗原がC3bで「標識(tag)」され、更にタンパク質分解を受けてiC3b及びC3d,gになる間そのタンパク質に付着したままになる。後者のフラグメントは、それぞれ、補体レセプターCR3及びCR2に対するリガンドである(CR2はCD21ともいう)。従って、C3bで抗原が標識されると、これらのレセプターを持っている免疫系の細胞のための標的メカニズムが生じうる。
【0004】
このようなターゲット化が免疫反応の増強に重要であるということは、マウスにおいて循環するC3を枯渇させ、次に抗原(ヒツジ赤血球)に暴露した実験により、初めて示された。C3を除去すると、前記抗原に対する抗体反応が減少した(M.B. Pepys, J. Exp. Med., 140, 126-145, 1974)。C3の役割は、C3、又はC3bを生ずる補体カスケードの上流成分であるC2及びC4の遺伝子が欠損している動物の研究により確認された(J.M. Ahearn及びD.T. Fearon, Adv. Immunol., 46, 183-219, 1989)。最近、モデル抗原とマウスC3dフラグメント配列の2つより多くのコピーを直線状に結合させると、修飾されていない抗原対照と比較して、マウスの抗体反応が非常に大きく上昇(1000〜10000倍)したことが示された(P.W. Dempseyら, Science, 271, 348-350, 1996; WO96/17625, PCT/GB95/02851)。この上昇は、ヒトに使用するには毒性が強すぎるフロイントの完全アジュバントなどの従来のアジュバントを使用することなく行うことができた。この顕著な効果のメカニズムは、多価のC3d構築物とB細胞上のCR2が高い親和性で結合し、次に、CR2と、別のB細胞膜タンパク質CD19及び膜結合イムノグロブリンが同時連結し、B細胞の核にシグナルを発する、と実証された。
【0005】
しかしながら、C3dの3つのコピーを含む均質な組換えタンパク質を大量に製造するのは難しいことが分かっている。主な問題点は、
i) (3つの)繰り返し配列を含む構築物の遺伝的不安定性、及び
ii) 大腸菌(Escherichia coli)の中に形成された封入体由来の組換えタンパク質の折りたたみ(又は可溶化及び再折りたたみ)
である。
【0006】
C3d遺伝子の繰り返しコピーを含む構築物の遺伝的不安定性を最小限にするために採られた1つの方法が、国際公開WO 99/35260及びWO 01/77324に開示されている。これらの出願に開示されている技術は、C3dの繰り返し配列をコードするDNAの同一でない配列を使用するものである。
【0007】
国際公開WO 91/11461では、補体4結合タンパク質(C4bp)を用いる多量体化系が開示されている。ヒトC4b-結合タンパク質(C4BP)は、7つの同一のα鎖と1つのβ鎖でできたクモ状構造を有する高分子量(570kDa)の血漿糖タンパク質である。C4bpのα鎖は、分子が多量体に組み立てられる機能を担っているC末端コア領域を有する。標準的なモデルによれば、C4bpモノマーの位置+498のシステインが、別のモノマーの位置+510のシステインとジスルフィド結合を形成する。7つのα鎖のみを含むより小さい形状のものもヒトの血漿中に発見されている。この血漿糖タンパク質の本来の機能は、補体の活性化の古典経路を抑制することである。
【0008】
C4bpのα鎖のほとんどは、補体コントロールタンパク質(complement control protein, CCP)の繰り返しとして知られる長さ約60アミノ酸の8つの直列に並んだドメインから構成される。国際公開WO91/11461は、C4bpの全部もしくは一部及び対象となる生物学的タンパク質を含む融合タンパク質を作るのに、C4bpタンパク質の多量体化する能力が使えることを提案している。国際公開WO91/11461に開示された融合タンパク質では、1つより多くのCCP繰り返し(SCRとしても知られている)を含めることが好ましい。
【0009】
国際公開WO91/11461は、融合タンパク質をワクチンとして使用できることを示唆している。B型肝炎e抗原のフラグメントと融合した少なくとも1つのC4bp SCR領域を含むいくつかの特定のタンパク質が作られた。使用されたe抗原のフラグメントは、多量体構造を形成できるコア抗原フラグメントである。
【0010】
Libyh M. T.ら(1997, Blood, 90, 3978-3983)は、多量体化を妨げずに、(C末端の57アミノ酸のみを残して)すべてのCCPを除去することができることを示した。このC4bpのC末端領域を、本明細書においてはC4bpコアという。
【0011】
また、ヒト293細胞系統で発現された自己集合性の(self-assembling)多量体の可溶性CD4-C4bp融合タンパク質も、Shinyaら(1999, Biomed & Pharmacother, Vol. 53: 471)によって示されている。
【0012】
また、C4bpの使用は、Oudinら(2000, Journal of Immunology, Vol. 164:1505)に開示されている。
【0013】
Christiansenら(2000, Journal of Virology, Vol. 74:4672)は、CD46-C4bp融合タンパク質の治療用途について議論している。
【0014】
国際公開WO2004/020639は、異種のポリペプチドと融合していてもよいC4bpα鎖のC末端コアタンパク質の足場(scaffold)を含む組換え融合タンパク質を原核生物の宿主で得る方法を提供し、前記組換え融合タンパク質は、原核生物の宿主細胞内で可溶性の形で多量体を形成することができる。
【発明の開示】
【課題を解決するための手段】
【0015】
本発明は、特定のクラスのC4bp融合タンパク質に関する新規な知見に基づく。前述の先行技術の幾つかは、C4bp部分は本質的に不活性担体であることに基づき、対象の治療用タンパク質を運搬するのにC4bp融合タンパク質を使用することを提案している。
【0016】
前述の国際公開WO91/11461は、多量体形成性抗原を含む融合タンパク質を例示している。
【0017】
一方、本発明は、多量体を自然に形成する抗原はC4bpコアが多量体に組み立てられるのを事実上妨害するので、このような抗原はC4bpコアとの融合に望ましくないという認識に基づく。更に、驚くべきことに、C4bpコアと単量体抗原の融合は、抗原に対する強い免疫反応を引き起こすことが示された。後述の実施例において、単量体抗原を含む融合タンパク質は、抗原をフロイントのアジュバントと共に注射したときの低い力価の非抑制的な抗体応答と比較して、より高い力価の抑制的な抗体応答を引き起こしたことを示す。
【0018】
従って、本発明は、単量体抗原をC4bpコアタンパク質と組合わせて複合体にすることにより、その抗原の免疫原性を増加させる方法を提供する。好ましい方法において、単量体抗原をC4bpコアタンパク質と共有結合させる。極めて好ましい方法において、単量体抗原をC4bpコアタンパク質と遺伝学的に融合させる。
【0019】
また、本発明は、抗原に対する高い力価の抗体を誘導する方法、及び受動免疫による感染性で悪性の疾患の予防及び/又は治療における、前記方法の使用により製造した高力価の抗血清の使用を提供する。好ましい方法において、抗原に対する高力価の抗体を、過免疫の血清又は血漿のイムノグロブリン分画を単離することにより部分的に精製し、極めて好ましい方法において、過免疫の血清又は血漿のイムノグロブリン分画を、その抗血清が感染性または悪性の疾患を予防又は治療するために使用される同じ種の個体から単離する。
【0020】
従って、本発明は、
C4bpコアタンパク質、及び
単量体抗原
を含む生成物を提供する。
【0021】
第一及び第二成分は、融合タンパク質の形であってもよい。或いは、これらは、第一成分のアミノ酸側鎖を介して、又は特異的に第二成分の化学的結合を可能にするように第一成分に付加されたアミノ酸の側鎖を介して、化学的に結合してもよい。
【0022】
第一及び第二成分は、堅固に、しかし非共有結合で結合してもよい。例えば、第一成分のアミノ酸の側鎖は、付加的なビオチン基を持つように修飾でき、このビオチンを使用してストレプトアビジン(ストレプトアビジンは第二成分)と結合させることができるし、又はストレプトアビジンと融合した抗原をこのビオチンを介して第一成分と結合させることもできる。別の可能性において、ビオチン化した抗原とビオチン化した第一成分を、ストレプトアビジンを添加して、生じた複合体を精製することにより、共にしっかりと、しかし非共有結合的に保持することができる。
【0023】
疑いを回避するために、「第一」及び「第二」成分の意味は、2つの成分の生成物における特定の直線的な順序を意味しないし、表さない。2つの成分はどんな順序で結合してもよい。
【0024】
従って、両方の成分がポリペプチドであり、生成物が融合タンパク質として作られる場合、2つの成分のN末端からC末端への順序はどんな順序でもよい。
【0025】
更に、本発明は、前記第一及び第二成分の融合タンパク質をコードする核酸を提供する。また本発明は、前記核酸を含むベクター及びそのベクターを有する宿主細胞を提供する。
【0026】
別の実施形態において、本発明は、
C4bpコアタンパク質、及び
ポリペプチドの単量体抗原、
を含む生成物を製造する方法であって、融合タンパク質の形態で2つの成分をコードする核酸を発現させる工程、及び前記生成物を回収する工程を含む方法を提供する。
【0027】
別の実施形態において、本発明は、
C4bpコアタンパク質;
非ポリペプチドの単量体抗原、
を含む生成物を製造する方法であって、C4bpコアタンパク質をコードする核酸を発現させる工程、前記コアタンパク質を前記抗原に結合させる工程、及び前記生成物を回収する工程を含む方法を提供する。
【0028】
生成物を製造する方法は、真核細胞又は原核細胞で行うことができる。
【0029】
また本発明は、本発明による生成物の有効量を患者に投与することを含む抗原に対する免疫反応を誘導する方法を提供する。
【0030】
また本発明は、人体又は動物体の治療方法、特に免疫反応を誘導する方法のための本発明の生成物の使用を提供する。
【0031】
更に本発明は、製薬上許容できる担体又は希釈剤と組合わせた本発明の生成物を含む医薬組成物を提供する。
【0032】
更に本発明は、病原体に対する受動免疫で使用するための感染防御免疫血清を調製する方法を提供し、前記方法は、動物に本発明の生成物をワクチン接種する工程、該動物から抗血清を回収する工程を含む。次に抗血清を、ヒト患者の受動免疫の方法に使用することができる。ヒト患者は病原体に感染した患者又はリスクのある患者である。
【発明を実施するための最良の形態】
【0033】
C4bpα鎖のコアタンパク質
本明細書では、C4bpα鎖のコアタンパク質を「C4bpコアタンパク質」若しくは「コアタンパク質」、又は「C4bp足場(scaffold)」とする。この用語は互換的に使用される。このタンパク質は哺乳動物のC4bpコアタンパク質であっても、又は多量体を形成でき、アジュバントとして作用できる(C4bpコアタンパク質の)フラグメントであっても、あるいは多量体を形成でき、アジュバントとして作用できる(C4bpコアタンパク質の)合成若しくはキメラ変異体であってもよい。
【0034】
本発明において、C4bpコアタンパク質又はコアタンパク質を含むC4bpα鎖のフラグメントはアジュバントとして働き、本明細書で更に詳細に説明される。配列番号1のヒトC4bpコアタンパク質は、多量体を形成することが当該技術分野で知られている、全長C4bpタンパク質配列のアミノ酸+493〜+549に対応する。
【0035】
更に本発明は、抗原の免疫原性を増強するためのC4bpコアの誘導体の使用を含む。このような誘導体は、誘導体の突然変異体を含み、本明細書で説明されるアジュバントの性質の維持を条件として、アミノ酸の欠失、付加、特にシステイン残基の付加、又は置換、C4bpファミリーの異なるメンバーの一部分の融合により形成されたハイブリッド又はキメラ分子、及び/又は環状の並べ替えられた(circular permutated)タンパク質足場を含む。
【0036】
また本発明は、複数の種由来のC4bpコア配列のアライメントに基づく人工の共通(コンセンサス)C4bp配列も使用できる。非常に多くあるであろう、このクラスのキメラ分子の一例を以下に挙げる(配列番号20、図1)。
【0037】
従って、アジュバントは、C4bpコア及び場合によってはコアと融合させた1つ以上のSCRでよい。
【0038】
特に好ましい実施形態において、本発明の生成物のC4bp成分は、C4bpα鎖のコアタンパク質、即ち、C4bp SCR配列に結合していない、本明細書で定義したコアタンパク質である。このような形態において、C4bpコアは、望ましくは、配列番号1の残基1〜57又はそのホモログの対応する残基、或いは配列番号1の少なくとも47アミノ酸のフラグメント若しくはそのホモログから成る。
【0039】
更に、本発明の生成物のC4bpコアは、可撓性のリンカーなどのN末端又はC末端伸長部分を含んでもよい。一般的に、このようなリンカーは、長さが1〜20の少数のアミノ酸、例えば長さ2〜10アミノ酸である。このようなリンカーの1つは、(Glym-Ser)nリンカーであり、ここで、mとnは、それぞれ独立して1〜4である。これらは、当該技術分野で、互いにタンパク質ドメインを付着させるのに使用される。従って第一成分をこのようなリンカーで第二成分と連結することができる。
【0040】
第一成分がC4bpコアである場合、第一成分は生成物のC末端にあることが好ましい。
【0041】
多くの哺乳動物C4bpタンパク質の配列は、当該技術分野で利用可能である。これらには、ヒトC4bpコアタンパク質(配列番号1)が含まれる。当該技術分野で利用可能な、ヒトC4bpコアタンパク質のホモログがたくさんある。2つのタイプのホモログがある:オルソログとパラログである。オルソログは、異なる生物のホモログ遺伝子と定義され、即ち、そのホモログ遺伝子は、その遺伝子を発生させた種分化現象が起こった(coincident)共通の祖先を持つ。パラログは、遺伝子、染色体又はゲノム重複に由来する同一生物のホモログ遺伝子として定義され、即ち、その遺伝子の共通の祖先は、最後の種分化現象後に発生した。
【0042】
例えば、GenBank、生のゲノムトレース及びEST(発現遺伝子配列断片)データベースの検索は、チンパンジー、アカゲザル、ウサギ、ラット、イヌ、ウマ、マウス、モルモット、ブタ及びウシを含む種の、哺乳動物C4bpコア配列番号1のホモログタンパク質を示唆する。配列番号1のC4bpのパラログ及びオルソログは、図1のアライメントに含まれている。
【0043】
全部で配列番号1〜19のアライメントを図1に示した。全部の19の配列は、C末端では変化がより大きいが、高いレベルの類似性を有することを示す。BLASTなどの市販されている検索プログラムを使ってDNA又はタンパク質配列のデータベースを検索することによって別のC4bpコアタンパク質を同定することができる。
【0044】
所望の哺乳動物源由来のC4bpタンパク質がデータベースで利用できない場合、当該技術分野で十分に確立されている日常的なクローニング方法を使って得ることができる。基本的に、このような技術は、入手可能なC4bpコアタンパク質の1つをコードする核酸をプローブとして使い、対象の他の種由来のC4bpコアタンパク質の配列を回収し決定する工程を含む。広範多様な技術がこのために利用でき、例えば、ゲノムDNA又はmRNAの適切な供給源(例えば胚又は活発に分裂している分化細胞又は腫瘍細胞)を使用する遺伝子のPCR増幅及びクローニング、又は、哺乳動物由来のcDNAライブラリー(例えば上述した供給源の1つに由来するcDNAライブラリー)を得る工程、中〜高ストリンジェントな条件(例えば0.03M塩化ナトリウム、0.03Mクエン酸ナトリウム、約50℃〜約60℃)下で公知のC4bp核酸で前記ライブラリーを探索する工程、及びその哺乳動物のC4bpタンパク質の全部又は一部をコードするcDNAを回収する工程を含む方法である。部分的なcDNAが得られる場合、全長コード配列をプライマー伸長技術により決定することができる。
【0045】
多量体を形成できるC4bpコアタンパク質のフラグメントは、少なくとも47アミノ酸、好ましくは少なくとも50アミノ酸を含んでよい。このフラグメントの、多量体を形成する能力は、本発明に従って、原核生物の宿主細胞でフラグメントを発現させ、全57アミノ酸C4bpコアの多量体化を生じる条件下でC4bpフラグメントを回収し、フラグメントが更に多量体を形成するかどうかを決定することにより、テストできる。望ましくは、C4bpコアのフラグメントは、配列番号1の少なくとも残基6〜52又はそのホモログの対応する残基を含む。
【0046】
また、同様に多量体を形成する能力を保持しているC4bpコアの変異体及び多量体を形成できるフラグメントも(フラグメントについては上述したように決定できる)、使用することができる。好ましくは、変異体は、野生型哺乳動物のC4bpコア又はその多量体形成性フラグメントに対して、少なくとも70%、より好ましくは少なくとも80%、更に好ましくは少なくとも90%、例えば少なくとも95%又は最も好ましくは少なくとも98%の配列同一性を有する。
【0047】
ある態様において、C4bpコアは、配列番号1の位置12に現れるグリシン、位置28に現れるアラニン、位置29、34、36及び41に現れるロイシン、及び位置32に現れるチロシン及び位置33に現れるリジン、及び好ましくは位置6と18に現れる2つのシステイン残基を含むコアであろう。望ましくは、変異体はこれらの残基の間に相対的な間隔を保持するであろう。
【0048】
上記に特定した同一性の程度は、配列番号1〜20のうちの1つ又はそれらの多量体形成性フラグメントに対するものである。
【0049】
最も好ましくは、特定した同一性の程度は、配列番号1又はその多量体形成性フラグメントに対するものである。
【0050】
配列の同一性の程度は、Accelrys (以前はGenetics Computer Group, Madison, WI)から入手可能な、当該技術分野で広く使用されるアルゴリズムの「ウイスコンシンパッケージ(Wisconsin package)」の一部であるアルゴリズムGAPにより決定することができる。GAPはNeedlemanとWunschのアルゴリズムを使用し、マッチする数を最大にし、ギャップの数を最小にするように2つの完全な配列を整列させる。GAPは、同じような長さの、短く非常に密接な配列のアライメントに有用であるので、配列が前述の同一性レベルに当たる場合に決定するのに適切である。GAPはデフォルトパラメーターで使用できる。
【0051】
哺乳動物のC4bpコアタンパク質の合成変異体は、C-又はN-終端に、1つ以上のアミノ酸置換、欠失、挿入又は付加を有する変異体を含む。置換が特に考えられる。置換は保存性の置換を含む。保存性の置換の例として、しばしばデイホフ(Dayhoff)グループと呼ばれる類似のアミノ酸のグループに留意した置換が挙げられる。これらを以下に挙げる。
【0052】
グループ1 D, E, N, Q
グループ2 I, L, V, M
グループ3 F, Y, W
グループ4 K, R, H
グループ5 S, P, T, A, G
グループ6 C
従って、作成し、多量体を形成する能力およびアジュバントとして作用する能力をテストすることができる、C4bpコアタンパク質のフラグメントと変異体の例として、以下の表1に示す配列番号37〜44が挙げられる。
【表1】

【0053】
A=配列番号、B=配列、C=%同一性、同じ長さの配列番号1のフラグメントを参照することにより計算した。
【0054】
配列を欠失させる場合、N-又はC-末端の切断は別として、欠失は、近接又は非近接でありうる、1つのみ、2つ又は3つの欠失に限定されることが好ましいであろう。
【0055】
コアタンパク質配列に挿入する場合、又はN-又はC-末端を伸長する場合、挿入及び伸長は、コアタンパク質の大きさが、野生型配列の長さを20アミノ酸より多く超えないような、好ましくは15アミノ酸以下、より好ましくは10アミノ酸以下になるような数に限定されることが好ましいであろう。従って、配列番号1の場合、コアタンパク質は、挿入又は伸長により改変される場合、77アミノ酸以下の長さが望ましいであろう。
【0056】
抗原
抗原は、抗体又はT細胞受容体により認識される能力を持つ分子である。しかしながら、すべての抗原が免疫原というわけではない。免疫原は免疫反応を引き出す物質である。ある態様において、本発明は免疫原でない抗原を免疫原にすることができ、弱い免疫原である抗原をより強い免疫原にすることができる。
【0057】
本発明の重要な特徴は、C4bpコアと遺伝学的に融合させることにより抗原が作られる場合、単量体抗原が非常に好ましい、ということである。なぜならば単量体抗原は、C4bpコアタンパク質が組み立てられてオリゴマーになり、その結果機能的な形になるのを妨げないからである。
【0058】
しかしながら、それとは別の態様において、抗原がC4bpコアタンパク質と化学的に又は非共有結合的に結合する場合、その抗原は単量体でなくてよい。
【0059】
従って、単量体抗原は以下の2つの主要グループに分類できる。
【0060】
1) 天然の状態で多量体(即ち、二量体又はそれより高次の多量体)である親タンパク質のフラグメント又は変異体である抗原だが、親タンパク質がこのような多量体を形成する条件下では、抗原そのものは多量体を形成しない、抗原。
【0061】
2) 天然の状態でモノマーである、抗原。
【0062】
両方のタイプの抗原の例については、本明細書で以下に更に議論する。
【0063】
単量体抗原は、DNAの一本鎖断片上にコードされ、このDNAがC4bpコアタンパク質をコードするDNAと融合されて、その後タンパク質に翻訳される場合、抗原は、抗原上の唯一の点を介して単一のC4bpコアタンパク質鎖と結合されるという点で共通している。このような抗原の簡単な例は、ニワトリ卵白のリゾチームであろう。全長リゾチームオープンリーディングフレームをコードするcDNAは、生じた融合タンパク質のC4bp部分の組み立てが妨げられないようにして、C4bpコアオープンリーティングフレームと融合することができる。
【0064】
生合成の後、C4bpコアと融合した単一のポリペプチド鎖を加工することができ、例えばタンパク質分解酵素により、ポリペプチド鎖内に新しいN-及びC-終末を作ることができる。タンパク質分解による切断により生じた2つ以上の鎖が、例えばジスルフィド結合を介して、互いに付着したままである場合、C4bp融合タンパク質は、プロセシングの終わりの時点でタンパク質と付着しており、通常は単量体とみなされないであろう。しかしながら、本発明の目的において、このタイプのタンパク質は、単一のオープンリーディングフレーム内で単一の融合タンパク質としてコードされるので、単量体だと考えられる。このタイプの例としては、生合成の後に加工されて、ジスルフィド結合により結合されたA及びBと呼ばれる2つの鎖になるプロインシュリンであろう。プロインシュリンのCペプチドといわれるフラグメントは、前駆体融合タンパク質のタンパク質分解酵素による加工の後に除去される。
【0065】
単量体抗原は、天然の状態で必ずしも単量体でないタンパク質に由来してもよい。すなわち、自然界においてポリマー状態で見出される多くの抗原は、例えばタンパク質工学技術により単量体になるように改変することができる。3つの例がある。このような抗原の一例はインフルエンザウイルス赤血球凝集素タンパク質に由来するものである。この抗原は、天然の状態で複合体化した三量体構造を形成することがよく知られている(Wilsonら、Nature 289, 366-373, 1981)。しかしながら、分子の三量体化を引き起こすコイルドコイルを除去することにより、単量体のフラグメントを得ることが可能である。具体的な例が、Jeon及びArnonの研究(Viral Immunology 15, 165-176, 2002)により提供される。この著者らは赤血球凝集素の球状領域のみを包囲するフラグメントを得るために、赤血球凝集素の残基96〜261のみを使用した。
【0066】
別の例は、マラリア原虫メロゾイト表面タンパク質1(MSP1)である。この大きい(約200kDa)タンパク質は、マラリア感染の血液段階を担うメロゾイトの表面を飾っている。通常、C末端のGPIアンカー(GPIはグリコシルホスファチジルイノシトール)を介してメロゾイトの表面に固定されている。このGPIアンカーの前には、一続きの疎水性アミノ酸配列部分(stretch)がある。このアンカーのため、全長MSP1、又はMSP1.19と呼ばれるC末端フラグメント(メロゾイトが赤血球に侵入するときでも膜と結合したままである)のいずれもが、事実上、単量体の状態で発見されていない。同じことが、単一の疎水性膜貫通領域を有する多くの膜タンパク質に当てはまる。本発明は、これらの疎水性の部分を欠失させることにより最も良好に行われる。MSP1.19タンパク質とC4bpコアタンパク質の融合を説明する以下の実施例を参照されたい。
【0067】
従って、本発明の1つの好ましい態様において、本発明の生成物は、C4bpコアタンパク質と融合したマラリア原虫MSP1単量体抗原性フラグメントの融合体である。マラリア原虫MSP1抗原性フラグメントは、約50〜約200アミノ酸、好ましくは約50〜約150アミノ酸を含む。抗原性フラグメントは、どのマラリア原虫種由来でもよく、例えばPlasmodium falciparum、Plasmodium vivax、Plasmodium ovale、Plasmodium malariae(これらのすべてはヒトに病気を生じさせることができる)又はPlasmodium yoeliiである。
【0068】
欠失は、普通はオリゴマー性のタンパク質を単量体にする最も簡単な方法であるが、幾つかの場合においては、1つ以上のアミノ酸を変異することで十分であろう。この一例は、Cpn10タンパク質であり、主要なイソ型のC4bpコアと同様に天然の状態では七量体のタンパク質である。Cpn10の一個のアミノ酸の突然範囲により、Cpn10が、C4bpコアタンパク質との融合に適した、単量体の突然変異体に変換される(Guidryら、BMC Biochemistry 4, 14-26, 2003)。このタンパク質を単量体化するための別の方法は、N-末端又はC-末端のアミノ酸を欠失させ(Llorcaら、Biochem. Biophysica Acta 1337, 47-56, 1997; Seale及びHorowitz, J. Biol. Chem. 270, 30268-30270, 1995)、これにより、サブユニット間の相互作用の原因である領域を欠失させる方法である。
【0069】
一般的に、組み立てられてオリゴマー構造(例えばウイルスのカプシドタンパク質)になる強い傾向を有し、その結果、融合されるC4bpコアタンパク質の集合体を破壊するタンパク質に関して、タンパク質-タンパク質相互作用を生じさせる領域を欠失させる原理又は境界面の残基を突然変異させる原理を、単量体のタンパク質を得るのに適用できる。
【0070】
抗原は、2つのカテゴリーに分類することができ、両方とも本発明に用いるのに適する。第一のカテゴリーは、外来性抗原であり、感染性の生物に見られるすべての分子を含む。細菌の免疫原、寄生虫の免疫原及びウイルスの免疫原は、ワクチンとして有用な多量体の又はヘテロ多量体のC4bp融合タンパク質を作るのに、ポリペプチド部分として有用である。
【0071】
これらの免疫原の細菌源として、細菌性肺炎、髄膜炎、コレラ、ジフテリア、百日咳、破傷風、結核及びハンセン氏病の原因である細菌源が挙げられる。
【0072】
寄生虫源として、トリパノゾーマ及びリーシュマニア種と同様に、Plasmodiumなどのマラリア原虫が挙げられる。
【0073】
ウイルス源として、ポックスウイルス、例えば痘瘡ウイルス、牛痘ウイルス及びオルフウイルス;ヘルペスウイルス、例えば単純ヘルペスウイルスタイプ1及び2、B-ウイルス、水疱瘡ウイルス、サイトメガロウイルス、及びエプスタイン・バーウイルス;アデノウイルス、例えばマストアデノウイルス;パポバウイルス、例えばHPV16等のパピローマウイルス、BK及びJCウイルス等のポリオーマウイルス;パルボウイルス、例えばアデノ随伴ウイルス;レオウイルス、例えばレオウイルス1、2、及び3;オルビウイルス、例えばコロラドダニ熱;ロタウイルス、例えばヒトロタウイルス;αウイルス、例えば東部脳炎ウイルス及びベネズエラ脳炎ウイルス;ルビウイルス、例えば風疹;フラビウイルス、例えば黄熱病ウイルス、デング熱ウイルス、日本脳炎ウイルス、ダニ媒介性脳炎ウイルス及びC型肝炎ウイルス;コロナウイルス、例えばヒトコロナウイルス;パラミクソウイルス、例えばパラインフルエンザ1、2、3及び4及びおたふくかぜ;はしかウイルス(morbillivirus)、例えば麻疹ウイルス(measles virus);肺炎ウイルス、例えば呼吸器合胞体ウイルス;ベシキュロウイルス(vesiculovirus)、例えば水疱性口内炎ウイルス;リッサウイルス、例えば狂犬病ウイルス;オルトミクソウイルス、例えばA型及びB型インフルエンザ;ブンヤウイルス、例えばラクロスウイルス;フレボウイルス、例えばリフトバレー熱ウイルス;ナイロウイルス(nairovirus)、例えばコンゴ出血熱ウイルス(Congo hemorrhagic fever virus);ヘパドナウイルス科、例えばB型肝炎;アレナウイルス、例えば1cmウイルス(1cm virus)、ラッサ熱ウイルス(Lasso virus)及びフニンウイルス(Junin virus);レトロウイルス、例えば、HTLV I、HTLV II、HIV-1及びHIV-2;エンテロウイルス、例えばポリオウイルス1、- 2及び3、コクサッキーウイルス、エコーウイルス、ヒトエンテロウイルス、A型肝炎ウイルス、E型肝炎ウイルス及びノーウォークウイルス;ライノウイルス、例えばヒトライノウイルス;及びフィロウイルス科、例えばマールブルグ(病)ウイルス及びエボラウイルスが挙げられる。
【0074】
これらの細菌、ウイルス及び寄生虫源に由来する抗原は、ワクチンとして有用な多量体のタンパク質の製造に使用することができる。多量体は、異なる抗原を有するモノマーの混合物を含んでもよい。
【0075】
これらの細菌、ウイルス及び寄生虫源に由来する抗原は、通常宿主に存在せず、宿主ゲノムにコードされていないので、外来性抗原として考えられる。対照的に、内在性抗原は通常宿主に存在するか、もしくは宿主ゲノムにコードされているか、又はその両方である。内在性抗原に対する免疫反応を起こす能力は、その抗原を持つ腫瘍を治療したり、又は腫瘍の成長因子を中和するのに有用である。内在性抗原の第一タイプの例は、HER2であり、ハーセプチンと呼ばれるモノクローナル抗体の標的である。内在性抗原の第二の(成長因子の)タイプの例は、前立腺の幾つかの悪性腫瘍に栄養作用のある、ゴナドトロピン放出ホルモン(GnRHと呼ばれる)である。
【0076】
本発明を使って作られる免疫原は、研究又は治療目的に使用できる。例えば、研究の適用例として、ゲノム配列データで予測される遺伝子産物に対する抗血清を作ることが挙げられる。この要望は、細菌などの原核生物並びに真菌及び哺乳動物を含む真核生物の遺伝子産物に当てはまる。抗原は、ワクチンの技術分野における通常のあらゆるサイズ、即ち短いペプチドから非常に大きいタンパク質のサイズが可能である。
【0077】
非ポリペプチド免疫原は、例えば、炭水化物又は核酸でよい。ナイセリア(Neisseria)種又はストレプトコッカス・ニューモニア(Streptococcus pneumoniae)種の多糖外被は、本発明の目的に使用できる炭水化物の例である。
【0078】
非ポリペプチド免疫原が本発明の生成物の一部である場合、その免疫原は、日常的な合成方法を使って、生成物の第一成分と共有結合させることができる。一般的に、免疫原は、第一成分を含むC4bpコアタンパク質のN-又はC-末端のどちらかと結合させるか、又はアミノ酸側鎖基(例えばリジンのε-アミノ基又はシステインのチオール基)又はそれらの組合わせと結合させることができる。融合タンパク質当たり1つより多くの免疫原を加えてもよい。結合を促進するため、例えばN-又はC-終端として、システイン残基をC4bpコアタンパク質に加えることができる。
【0079】
本発明は、免疫反応の発生において多くの利点がある。例えば、多量体の使用により、多くの抗原を同時に免疫系に対して提示できる。このことは、単一の生物又は多くの異なる生物に存在し得る1つより多くのエピトープに対する免疫反応を引き起こすことができる多価ワクチンの調製を可能にする。
【0080】
その結果、更なる態様において、単量体抗原は、2種類の異なる生物に由来するか又は同じ生物の2つの異なるタンパク質に由来する、2つの異なるエピトープを含む合成抗原であってもよい。後者の例は、スポロゾイト抗原配列、例えば、MSP1配列と結合したスポロゾイト周囲タンパク質由来の2つ以上のNANP繰り返し配列の融合体である。後者の第二の例は、単量体のインフルエンザ赤血球凝集素フラグメントと融合したNeirynckら(Nature Medicine 5, 1157-1163, 1999)により開示されたM2e配列の融合体である。
【0081】
従って、本発明により形成されたワクチンは、1種類より多くの疾患に対する同時ワクチン接種に使用することができ、または所定の病原体の複数のエピトープを同時に標的にするのに使用できる。エピトープは単一のモノマーユニット内又はヘテロ多量体を提供するために組合わされる異なるモノマーユニット上に存在しうる。
【0082】
特にC4bpコア融合タンパク質は、免疫化の状況において有用であり、コアタンパク質は免疫化される者の血清又は血漿に通常存在するからであり、またコアタンパク質はそれ自身に対する免疫反応を惹起しないからである。C4bpタンパク質は多くの哺乳動物種で知られており、標準的な遺伝子クローニング技術を使って当業者により、哺乳動物種における適切なホモログを見出すことができる。
【0083】
核酸
本発明の生成物は、タンパク質をコードする核酸構築物を使って、原核生物又は真核生物の宿主細胞で融合タンパク質を発現させることにより作ることができる。抗原がポリペプチドの場合、核酸配列からの融合タンパク質の発現を、本発明の生成物を作るのに使用できる。
【0084】
従って、本発明は、本発明の生成物をコードする核酸構築物(一般的にはDNA又はRNA)を提供する。
【0085】
一般的に、構築物は複製可能なベクターの形であり、このベクター内ではタンパク質をコードする配列が、所望の宿主細胞でのタンパク質の発現に適切なプロモーターと有効に結合している。
【0086】
ベクターは、複製開始点及び任意にプロモーターの制御因子を含んでいることができる。ベクターは、1つ以上の選択マーカー遺伝子を含むことができる。当該技術分野で公知の広範多様な原核細胞又は真核細胞の発現ベクターがあり、本発明は、当業者の個々の選択により、あらゆるベクターを利用することができる。
【0087】
広範多様な原核生物の宿主細胞を本発明の方法に使用することができる。これらの宿主として、大腸菌類、シュードモナス属、桿菌属、乳酸桿菌属、好熱菌(Thermophilus)、サルモネラ属、腸内細菌科又は放線菌類の菌系統が挙げられる。例えば、エシェリキア属の大腸菌(E. coli)が本発明の方法に使用される場合、使用するこの細菌の好ましい菌系統として、国際公開WO98/02559で開示され供されるC41(DE3)、C43(DE3)又はCO214(DE3)を含むBL21(DE3)の派生系統(derivative)が挙げられよう。
【0088】
更に好ましくは、プロファージDE3を欠くこれらの系統の派生系統は、プロモーターがT7プロモーターではない場合に使用できる。
【0089】
原核生物のベクターとして、細菌プラスミドベクター、例えば大腸菌由来のプラスミド(ColEI、pCR1、pBR322、pMB9及びその派生プラスミドを含む)、広い宿主範囲のプラスミド、例えばRP4、;ファージDNA、例えばλファージの多数の派生ファージ、例えばNM989、及び他のDNAファージ、例えばM13及び繊維状一本鎖のDNAファージが挙げられる。これらのベクター及び他のベクターは、場合によってプロモーターと連結した本発明の核酸を導入するために、標準的な組換えDNA方法論を使って操作することができる。
【0090】
プロモーターは誘導プロモーター(inducible promoter)でよい。適切なプロモーターとして、T7プロモーター、tacプロモーター、trpプロモーター、λプロモーターPL又はPR及び当業者に周知の他のプロモーターが挙げられる。
【0091】
また、広範多様な真核生物の宿主細胞も使用でき、例えば酵母、昆虫及び哺乳動物の細胞が挙げられる。哺乳動物の細胞として、CHO及びマウス細胞、アフリカミドリザルの細胞、例えばCOS-1、及びヒト細胞が挙げられる。
【0092】
タンパク質の発現に適切な、多くの真核生物のベクターが知られている。これらのベクターは、真核生物の細胞ゲノムに染色体性(chromosomally)に組み込まれるように設計してもよいし、又は過剰染色体性(extrachromosomally)に維持されるように設計してもよいし、又は真核細胞に一過性(transiently)にしか維持されないように設計してもよい。核酸を、例えば、CMVプロモーターを含む強力なウイルスプロモーター、及びSV40 T-抗原プロモーター又はレトロウイルスLTRなどの適切なプロモーターと、場合により結合させてもよい。
【0093】
本発明の生成物を得るために、本発明のベクターを運ぶ宿主細胞は、タンパク質の発現に適切な条件下で培養することができ、そのタンパク質を培養培地の細胞から回収することができる。
【0094】
細胞培養
本発明による融合タンパク質をコードするプラスミドは、従来の形質転換技術を使って宿主細胞に導入し、その細胞を融合タンパク質の生成を促進する条件下で培養することができる。誘導プロモーターを使う場合、細胞は誘導物質の不存在下で最初は培養し、次に、タンパク質の回収率を最大にするために、誘導物質を一度添加して高い濃度で生育させることができる。
【0095】
細胞培養条件は、当該技術分野で広く知られており、そのような公知の手順に従って用いればよい。
【0096】
国際公開WO91/11461は、原核生物の宿主細胞をC4bpベースのタンパク質の生成に使用できることを示唆しているが、そのような生成の実験による実証はされていない。
【0097】
最近、原核生物の発現系で生成されたC4bpコアと融合したタンパク質は、その機能的活性を保持していることが分かった。これは国際公開WO2004/020639に開示されており、その内容は参照によって本明細書に組み込まれる。このような方法は、本発明の融合タンパク質の生成に使用できる。
【0098】
培養物からのタンパク質の回収
細胞を生育してタンパク質の生成が可能になると、タンパク質を細胞から回収することができる。本発明者らは、驚くべきことに、タンパク質が可溶性のままであることを見出したので、細胞を通常どおり遠心沈殿して、超音波処理で細胞を溶解でき、例えば、そのタンパク質画分の可溶性を維持し、更に速いスピード(例えば1時間15000rpm)で遠心分離した後の上清にこの画分を残すことができる。
【0099】
上清のタンパク質画分の融合タンパク質は、標準的なタンパク質クロマトグラフィー技術の適切な組み合わせにより、更に精製できる。本発明者らは、イオン交換クロマトグラフィーに続いてゲルろ過クロマトグラフィーを使用した。他のグロマトグラフィー技術、例えばアフィニティークロマトグラフィーも使用できる。
【0100】
ある実施形態において、本発明者らは、溶解物の遠心分離又は他の精製工程のいずれかの後に上清サンプルを加熱することにより、タンパク質の回収を助けるであろうことを見出した。サンプルは約10〜30分間、約70〜80℃に加熱できる。
【0101】
意図するタンパク質の使用に応じて、タンパク質を、追加の精製工程、例えば透析、又は濃縮工程、例えば凍結乾燥してもよい。
【0102】
組成物及びその使用
本発明による生成物を、医薬組成物の形に調製することができる。生成物を、1種類以上の製薬上許容できる担体又は希釈剤と一緒に存在させることができる。組成物を、意図する使用及び生成物の投与ルートに従って調製できる。従って、本発明は、本発明の生成物を多量体の形で、1種類以上の製薬上許容できる担体又は希釈剤を一緒に含む組成物を提供し、また、ヒト又は動物患者の治療又は予防のための免疫療法における、そのような組成物の使用を提供する。
【0103】
製薬上許容できる担体又は希釈剤として、経口、直腸、経鼻、局所的(頬側及び舌下を含む)、膣又は非経口的(皮下、筋肉内、静脈内、皮内、くも膜下腔内及び硬膜外を含む)投与に適切な製剤で使用されるものが挙げられる。これらの製剤を、便利なように投薬単位剤形にすることができ、製薬業界で周知の方法により調製できる。
【0104】
液体状の医薬的に投与可能な組成物は、例えば水、生理的食塩水に溶性のブドウ糖、グリセロール、エタノール等の担体に、任意の医薬品アジュバントと本発明の融合タンパク質を、例えば溶解、分散などにより調製し、溶液又は懸濁液の形にすることができる。所望であれば、投与される組成物をpH緩衝化剤等の物質で補助してもよい。このような投薬剤形を調製する実際の方法は当業者に公知であり又は明白であろう。例えば、Remington’s Pharmaceutical Sciences, Mack Publishing Company, Easton, Pennsylvania, 第19版, 1995を参照されたい。
【0105】
本発明による組成物は、1種類以上のアジュバント、例えば水酸化アルミニウム又はリン酸カルシウムなどの無機塩、又はIL-12若しくはGM-CSFなどのサイトカインを追加で含めてもよい。適切なアジュバントの詳しいリストは、Singh及びO’Hagan, Nature Biotechnology, 17, 1075-1081, 1999の表1であり、この記載は参照によって本明細書に組みこまれる。
【0106】
本発明による生成物は、望ましくは組成物又は製剤の形で、ヒト又は動物患者にこれらの生成物又は組成物を投与することにより、本明細書に開示した治療方法に使用することができる。治療を受ける患者の症状を軽減する有効量は、治療を受ける患者とその状態を考慮して医師により決定されるであろう。0.25〜95%の範囲の有効成分を含み、残りが無毒性担体から作られた剤形又は組成物を調製できる。
【0107】
非経口投与は、一般的に、皮下注射、筋肉注射又は静脈内注射のいずれかの注射により特徴付けられる。注射可能物質は、溶液又は懸濁液として、注射前に液体で溶液又は懸濁液にするのに適した固形として、あるいは乳濁液として、従来の形状に調製できる。適切な賦形剤として、例えば水、生理食塩水、ブドウ糖、グリセロール、エタノール等がある。つい最近考案された非経口投与の方法は、投与量の一定レベルを維持するような、緩徐放出性(slow-release)系又は持効性(sustained-release)系の埋込み(implantation)を用いる。米国特許第3,710,795号を参照されたい。
【0108】
生成物の投与量は、抗原の性質に依存するであろうし、従来のワクチン製剤の抗原の投与の現行手法に従って決定すればよい。
【0109】
受動免疫法
更なる態様において、本発明は、本発明の生成物を宿主患者にワクチン接種することにより得られた抗体を含む免疫血清で患者を受動免疫する方法を提供する。宿主患者はヒト又は非ヒト哺乳動物でよい。従って、更なる態様において、本発明は、このような方法により得られた免疫血清、及び人体又は動物体の治療方法におけるそのような免疫血清の使用を提供する。
【0110】
DNAワクチン
別の態様において、本発明は、人体又は動物体の治療に使用するための、本発明の組換え融合タンパク質生成物をコードする核酸配列を含む真核生物の発現ベクターを提供する。
【0111】
このような治療は、免疫反応を上昇させる目的で抗原をコードする核酸配列を導入することにより、その治療効果を上げるであろう。核酸の運搬は、プラスミドベクター(「裸(naked)」の又は処方される形)又は組換え発現ベクターを使って行うことができる。DNAワクチン接種の概説については、Ada G.及びRamshaw I, in Expert Opinion in Emerging Drugs 8, 27-35, 2003を参照されたい。
【0112】
遺伝子運搬に利用される様々なウイルスベクターとして、アデノウイルス、ヘルペスウイルス、ワクシニアウイルス又はレトロウイルス等のRNAウイルスが挙げられる。レトロウイルスベクターは、マウス又は鳥類のレトロウイルスの誘導株でもよい。一本鎖の外来遺伝子を導入できるレトロウイルスベクターの例として、モロニーマウス白血病ウイルス(MoMuLV)、ハーベイマウス肉腫ウイルス(HaMuSV)、マウス乳癌ウイルス(MuMTV)、及びラウス肉腫ウイルス(RSV)が挙げられるが、これらに限定されない。患者がヒトの場合、ギボンザル(gibbon ape)白血病ウイルス(GaLV)などのベクターを利用することができる。
【0113】
ベクターは、転写の制御配列、特にRNA合成の開始を誘導するのに十分なプロモーター領域を含むであろう。適切な真核生物のプロモーターとして、マウスメタロチオネインI遺伝子のプロモーター(Hamerら, 1982, J. Molec. Appl. Genet. 1: 273);ヘルペスウイルスのTKプロモーター(McKnight, 1982, Cell 31: 355);SV40初期プロモーター(Benoistら, 1981, Nature 290: 304);ラウス肉腫ウイルスプロモーター(Gormanら, 1982, Proc. Natl. Acad. Sci. USA 79: 6777);及びサイトメガロウイルスプロモーター(Foeckingら, 1980, Gene 45: 101)が挙げられる。
【0114】
患者への本発明のこのような態様のベクターの投与、即ちプラスミドベクターとして又はウイルスベクターの一部としての投与は、多くの異なるルートにより影響されうる。プラスミドDNAは、直接的又は間接的に運搬するために、「裸」でもよいし、陽イオン性及び中性脂質(リポソーム)で処方してもよいし、マイクロカプセル化してもよい。また、DNA配列は、直接的又は間接的に運搬するために使用できるウイルス(例えばアデノウイルス、レトロウイルス、ヘルペスウイルス、ポックスウイルス)ベクター内に含めてもよい。運搬ルートは、経口、筋肉内、皮内(Sato, Y.ら, 1996, Science 273: 352-354)、静脈内、動脈内、くも膜下腔内、肝臓内、吸入、膣内点滴注入(Bagarazziら, 1997, J Med. Primatol. 26:27)、直腸内、腫瘍内又は腹腔内が挙げられるが、これらに限定されない。
【0115】
従って、本発明は、治療用ポリペプチドを発現させ、治療効果を有するように、即ち抗原に対する免疫反応を誘導するように、幾つかの細胞をDNAベクターで形質移入可能にするのに有用な医薬組成物として本明細書で開示されるベクターを含む。本発明による医薬組成物は、本発明による構築物を、溶剤、担体、送達系、賦形剤及び添加物又は補助剤を使って、患者に投与するのに適切な形にすることにより、調製される。頻繁に使用される溶剤として滅菌水及び食塩水(緩衝化したもの又はしないもの)が挙げられる。ある担体として、バイオリスティカルに(biolistically)(即ち、気体の圧力下で)運搬される金の粒子が挙げられる。他に頻繁に使用される担体又は運搬システムとして、陽イオン性リポソーム、渦巻型(cochleate)又はマイクロカプセルが挙げられ、これらを溶液として提供してもよいし、運搬カプセル内に封入し又は食べ物に組み込んでもよい。
【0116】
遺伝子運搬ベクターの投与における別の製剤として、リポソームが挙げられる。リポソームのカプセル化により、ポリヌクレオチド及び発現ベクターの投与のための別の製剤が提供される。リポソームは、水溶性のコンパートメントを取り囲む1種類以上の脂質二重層から成る微細小胞である。一般的には、Bakker-Woudenbergら, 1993, Eur. J. Clin. Microbiol. Infect. Dis. 12 (Suppl. 1): S61、及びKim, 1993, Drugs 46: 618を参照されたい。リポソームは組成の点で細胞膜と似ており、その結果、リポソームを安全に投与できるし、生分解性である。調製方法に応じて、リポソームは単層にも多層にもでき、0.02μMから10μM以上の範囲で直径の大きさを変えることができる。例えば、Machyら, 1987, LIPOSOMES IN CELL BIOLOGY AND PHARMACOLOGY (John Libbey)、及び Ostroら, 1989, American J. Hosp. Phann. 46: 1576を参照されたい。
【0117】
発現ベクターは、標準的な技術を使ってリポソーム内にカプセル化できる。異なるリポソームの組成の種類と合成方法は当業者に公知である。例えば、米国特許US-A-4,844,904、US-A-5,000,959、US-A-4,863,740、US-A-5,589,466、US-A-5,580,859及びUS-A-4,975,282を参照されたい。これらはすべて参照によって本明細書に組み込まれる。
【0118】
一般的に、投与されるリポソームカプセル化ベクターの投与量は、患者の年齢、体重、身長、性別、一般的な病状及びこれまでの病歴などの因子に応じて変えられるであろう。特定の剤形における投与量の範囲は適切な動物モデルを使って決定することができる。
【0119】
本発明を以下の実施例により説明する。
【実施例1】
【0120】
熱帯熱マラリア原虫MSP1.19-ウサギC4bp融合タンパク質
本実施例は、(熱帯熱マラリア原虫MSP1のアミノ酸1567-1661を含む)単量体抗原とウサギコアC4bpタンパク質の融合を説明する。AVD174と名づけた融合タンパク質を、細菌株C41(DE3)で発現し、精製した。融合タンパク質のみを、アジュバンドを添加せずにウサギを免疫するのに使用した。
【0121】
クローニング
MSP1タンパク質の残基1567-1661をコードする合成294bp DNAフラグメントを、NdeI及びBamHIで消化し、あらかじめNdeI及びBamHIで消化したpAVD181に連結した。これにより、T7後期プロモーターの下流のウサギC4bpα鎖のC末端57残基と融合した95アミノ酸のMSP1.19タンパク質フラグメントをコードするオープンリーディングフレームを作製した。pAVD174と名づけた構築物をDNA配列決定で調べた。
【0122】
AVD174融合タンパク質をコードする核酸配列は、以下の通りである。
【0123】
atgttaaacatttcccagcaccagtgcgttaagaaacagtgcccgcagaa
ctctggttgtttccgtcatctggacgagcgtgaagagtgcaaatgtctgc
tgaactacaaacaggaaggtgataaatgtgttgagaacccaaacccgacc
tgtaacgaaaacaacggcggttgtgacgctgatgctaaatgcaccgagga
agacagcggttctaacggtaagaaaatcacctgcgagtgtactaaaccgg
actcctacccgctgttcgacggtatcttttgctccGGATCCGAGGTCCCG
GAAGGCTGTGAGCAGGTGCAAGCGGGTCGCCGTCTCATGCAGTGTCTCGC
AGACCCATACGAAGTGAAAATGGCCCTGGAGGTCTACAAGCTGTCTCTGG
AGATTGAACTCCTGGAACTGCAGCGCGATAAGGCACGTAAAAGCTCTGTG
CTGCGCCAGCTGTAA (配列番号21)
当該構築物によりコードされる融合タンパク質AVD174のアミノ酸配列は、以下の通りである。
【0124】
MLNISQHQCV KKQCPQNSGC FRHLDEREEC KCLLNYKQEG DKCVENPNPT CNENNGGCDA DAKCTEEDSG SNGKKITCEC TKPDSYPLFD GIFCSGSEVP EGCEQVQAGR RLMQCLADPY EVKMALEVYK LSLEIELLEL QRDKARKSSV LRQL (配列番号22)
配列番号22の残基1-95は、熱帯熱マラリア原虫MSP1(単量体抗原)の残基1567-1661に対応し、配列番号22の残基98-154は、ウサギC4bpコアタンパク質の57残基に対応する。GSリンカー配列は、2つの成分の間に現れる。
【0125】
このタンパク質は、推定分子量17,319ダルトン、理論pIは5.05である。
【0126】
発現
熱帯熱マラリア原虫-ウサギC4bpコアタンパク質をコードするプラスミドpAVD174を、大腸菌株C41(DE3)で発現した。形質転換した細胞を、37℃でOD600が約0.6になるまでLB培地で生育させ、次に最終濃度0.5mMになるようにIPTGで誘導し、培養物を37℃で更に3時間生育させ、この時点で細胞を遠心分離によって回収した。
【0127】
AVD174融合タンパク質の精製
タンパク質AVD174を1LのC41(DE3)細胞から精製した。細胞を20mM MES pH6.5及び5mM EDTAを含むバッファーで超音波処理で溶解した後、可溶性画分にすべての融合タンパク質が検出された。遠心分離後の上清をHitrapSカラムに載せた。
【0128】
陽イオン性カラム(HiTrap S)
カラムを20mM MES pH 6.5、20mM EDTAバッファー(バッファーA)で平衡化した。タンパク質を、バッファーAからバッファーB(バッファーAプラス0.5M NaCl)へ、10カラム量のグラジエントで溶出した。AVD174を濃度約200mM NaClで溶出した。
【0129】
AVD174を含むHiTrapS画分を、Millipore concentrator (カットオフ30 K)を使って濃縮し、次にゲルろ過カラムに載せた。
【0130】
ゲルろ過カラム(Superdex 200 26/60 prep grade)
Superdex 200 26/60カラムを20mM Trisバッファー pH8, 150mM NaClで平衡化し、HiTrapS画分から得られた濃縮したAVD174タンパク質を載せた。
【0131】
タンパク質は2つのピークに溶出された。正確に折りたたまれ集合したタンパク質は、156mlのところで溶出されたのに対し、120mlのところで溶出しているもっと早い小さいピークのものは、正確に集合しても折りたたまれてもいない。
【0132】
生物物理学的性質決定
ジスルフィド結合(マウスコアタンパク質を除くすべてに存在する。図1参照)を含むC4bp融合タンパク質のオリゴマー状態は、還元剤β-メルカプトエタノール(BME)の存在下及び不存在下で、SDS-PAGEゲル上のタンパク質の挙動を比較することによって簡単に調べることができる。AVD177タンパク質は、BMEの不存在下で見かけの大きさが約140kDaであり、一方BME存在下では還元されて、見かけの大きさがちょうど20kDaの辺りで流れる。
【0133】
質量分析
AVD174タンパク質を、BMEによる還元及びN-エチルマレイミド(NEM)によるアルキル化の後、エレクトロスプレー質量分析法により試験を行った。結果は、分子量が19,072Daであると測定されたタンパク質に14個のNEM分子(各125Da)が付加したことを示した。
【0134】
エンドトキシンレベル
精製されたタンパク質のエンドトキシンのレベルは、BiowhittakerのLAL(カブトガニ血球抽出成分(limulus amoebocyte lysate))テストキットを使い、タンパク質1mg当たり21EUであることが測定された。
【実施例2】
【0135】
熱帯熱マラリア原虫MSP1.19-ヒトC4bp融合タンパク質
本実施例は、単量体抗原(熱帯熱マラリア原虫MSP1のアミノ酸1567-1661を含む)とヒトC4bpコアタンパク質の融合を示す。融合タンパク質を、細菌株C41(DE3)で発現させ精製した。アジュバンドを添加することなくヒトを免疫するのに、この融合タンパク質のみを使用することができる。
【0136】
クローニング
MSP1タンパク質の残基1567-1661をコードする合成294bpDNAフラグメントをNdeI及びBamHIで消化して、あらかじめNdeI及びBamHIで消化したpHVD181に連結した。これにより、T7後期プロモーターの下流のヒトC4bpα鎖のC末端57残基に融合した95アミノ酸MSP1.19タンパク質フラグメントをコードするオープンリーディングフレームを作製した。この構築物をpAVD177とし、DNA配列決定により調べた。
【0137】
AVD177融合タンパク質をコードする核酸配列は以下の通りである。
【0138】
atgttaaacatttcccagcaccagtgcgttaagaaacagtgcccgcagaa
ctctggttgtttccgtcatctggacgagcgtgaagagtgcaaatgtctgc
tgaactacaaacaggaaggtgataaatgtgttgagaacccaaacccgacc
tgtaacgaaaacaacggcggttgtgacgctgatgctaaatgcaccgagga
agacagcggttctaacggtaagaaaatcacctgcgagtgtactaaaccgg
actcctacccgctgttcgacggtatcttttgctccGGATCCgagaccccc
gaaggctgtgaacaagtgctcacaggcaaaagactcatgcagtgtctccc
aaacccagaggatgtgaaaatggccctggaggtatataagctgtctctgg
aaattgaacaactggaactacagagagacagcgcaagacaatccactttg
gataaagaactataa (配列番号23)
この構築物によりコードされる融合タンパク質AVD177のアミノ酸配列は以下の通りである。
【0139】
MLNISQHQCV KKQCPQNSGC FRHLDEREEC KCLLNYKQEG DKCVENPNPT CNENNGGCDA DAKCTEEDSG SNGKKITCEC TKPDSYPLFD GIFCSGSETP EGCEQVLTGK RLMQCLPNPE DVKMALEVYK LSLEIEQLEL QRDSARQSTL DKEL (配列番号24)
配列番号24の残基1-95は、熱帯熱マラリア原虫MSP1(単量体抗原)の残基1567-1661に対応し、配列番号24の残基98-154は、ヒトC4bpコアタンパク質の57残基に対応する。GSリンカー配列は、これらの2つの成分の間に現れる。
【0140】
タンパク質は、推定分子量17,261ダルトン、理論pIは4.72である。
【0141】
発現
タンパク質の発現を図2に示す。3つの異なる細菌株、即ちC41(DE3)、BL21(DE3)及びC43(DE3)で、非誘導(U)、37℃又は30℃で誘導後のAVD177タンパク質の発現を示す、SDS-PAGEゲルを提示する。ゲルのレーンは以下の通りである。
【0142】
レーン1:分子量マーカー(降順: 66, 60, 46, 36, 28, 20, 14, 12, 6 kDa)
レーン2:C41(DE3) 誘導前
レーン3:C41(DE3) 37℃で誘導後3時間;
レーン4:C41(DE3) 30℃で誘導後3時間;
レーン5:BL21(DE3) 誘導前
レーン6:BL21(DE3) 37℃で誘導後3時間;
レーン7:BL21(DE3) 30℃で誘導後3時間;
レーン8:C43(DE3) 誘導前
レーン9:C43(DE3) 37℃で誘導後3時間;
レーン10:C43(DE3) 30℃で誘導後3時間。
【0143】
図に示すように、C41(DE3)と細菌株C43(DE3)で良好な発現が得られた。その一方、テストされた条件下では、細菌株BL21(DE3)で発現は見られなかった。図2を参照のこと。培養物を、光学密度(OD600)が約0.6になるまで30℃と37℃のLB培地で生育させ、次に、最終濃度0.5mMになるようにIPTGの添加により発現を誘導した。
【0144】
AVD177融合タンパク質の精製
タンパク質AVD177を、37℃で誘導後、3時間生育した1LのC41(DE3)細胞から精製した。細菌ペレットを溶解した後、可溶性画分にすべての融合タンパク質を検出した。細胞を、20mM MES pH6.5及び5mM EDTAを含むバッファーで超音波処理により溶解した。遠心分離後上清をMonoSカラムに載せた。
【0145】
陽イオンカラム(Mono S HR 10/10)
カラムを20 mM MES pH 6.5, 20mM EDTAバッファー(バッファーA)で平衡化した。タンパク質をバッファーAから、バッファーAに0.5M NaClを加えたバッファーBへ、10カラム量のグラジエントで溶出した。AVD177を濃度約200mM NaClで溶出した。
【0146】
AVD177を含むMonoS画分をMillipore concentrator(カットオフ30 K)を使って濃縮し、次にゲルろ過カラムに載せた。
【0147】
ゲルろ過カラム(Superdex 200 26/60 prep grade)
Superdex 200 26/60カラムを20mM TrisバッファーpH8、150mM NaClで平衡化し、MonoS画分から得られた濃縮したAVD177タンパク質を載せた。
【0148】
タンパク質が2つのピークに溶出された。正確に折りたたまれ集合したタンパク質は150mlのところで溶出され、一方、115mlのところで溶出されたもっと早い小さいピークは、正確に集合しても折りたたまれてもいない。
【0149】
生物物理学的性質決定
ジスルフィド結合(マウスコアタンパク質以外のすべてに存在する:図1参照)を含むC4bp融合タンパク質のオリゴマー状態を、還元剤β-メルカプトエタノール(BME)の存在下及び不存在下においてSDS-PAGEゲルのタンパク質の挙動を比較することにより簡単に検査できる。結果を図3に示す。図3において、SDS-PAGEゲルは、分子量マーカー(降順:66, 60, 46, 36, 28, 20, 14, 12, 6 kDa)で分けられた、還元条件下(左+BME)及び非還元条件下(右-BME)でのAVD177タンパク質の挙動を示す。図3から分かるように、AVD177タンパク質は見かけの大きさがBME不存在下で約140kDaであり、一方BEM存在下では還元されて、見かけの大きさがちょうど20kDa辺りで流れる。
【0150】
質量分析
AVD177タンパク質を、BMEで還元し、N-エチルマレイミド(NEM)でアルキル化した後、エレクトロスプレー質量分析法でテストした。結果は、分子量が19,015Daと測定されたタンパク質に14個のNEM分子(各125Da)が付加したことを示した。
【0151】
エンドトキシンレベル
精製タンパク質のエンドトキシンレベルをBiowhittakerのLAL(カブトガニ血球抽出成分)テストキットを使って測定し、タンパク質1mg当たり38EUであった。
【実施例3】
【0152】
突然変異熱帯熱マラリア原虫MSP1.19-ウサギC4bp融合タンパク質
一例として、第二の熱帯熱マラリア原虫MSP1.19-ウサギC4bpタンパク質を本実施例で説明する。該タンパク質は、主に、単量体抗原遺伝子においてpAVD174及びpAVD177に対して別々のコドンの利用を有する点で異なり、また、3つのアミノ酸変化を含む(Uthaipibullら, J Mol Biol. 307, 1381-1394, 2001に記載されている)。この融合タンパク質をAVD178とした。
【0153】
AVD178融合タンパク質をコードする核酸配列は、以下の通りである。
【0154】
atgctgaatatttcccagcaccagtgcgtaaagaaacagtgtcctcagaa
ctctggttgcttccgccatctggacgaacgcgaatattgcaaatgccgtc
tgaactacaaacaggaaggtgacaagtgcgttctgaacccgaacccaact
tgtaacgagaacaacggtggctgcgatgctgatgctaaatgcactgaaga
agacagcggttctaacggcaaaaaaatcacctgcgagtgcaccaaaccgg
acagctatccgctgttcgacggcattttttgttctggatccGAGGTCCCG
GAAGGCTGTGAGCAGGTGCAAGCGGGTCGCCGTCTCATGCAGTGTCTCGC
AGACCCATACGAAGTGAAAATGGCCCTGGAGGTCTACAAGCTGTCTCTGG
AGATTGAACTCCTGGAACTGCAGCGCGATAAGGCACGTAAAAGCTCTGTG
CTGCGCCAGCTGTAA (配列番号25)
この構築物によりコードされる融合タンパク質AVD178のアミノ酸配列は、以下の通りである。
【0155】
MLNISQHQCVKKQCPQNSGCFRHLDEREYCKCRLNYKQEGDKCVLNPNPTCNENNGGCDADAKCTEEDSGSNGKKITCECTKPDSYPLFDGIFCSGSEVPEGCEQVQAGRRLMQCLADPYEVKMALEVYKLSLEIELLELQRDKARKSSVLRQL (配列番号26)
3つの突然変異アミノ酸は太字及び下線部である。
【0156】
配列番号25の残基1-95は、3つの突然変異を有する熱帯熱マラリア原虫MSP1(単量体抗原)の残基1567-1661に対応し、配列番号24の残基98-154は、ウサギC4bpコアタンパク質の57残基に対応する。GSリンカー配列はこれら2つの成分の間に現れる。
【0157】
AVD178の発現、精製及び性質決定
基本的に、AVD174及びAVD177タンパク質について記載したように行った。HiTrapSカラムからの溶出は約200mMであった。ゲルろ過では、オリゴマータンパク質は159mlの量のところで溶出された。質量分析では、単量体あたり14 NEM残基で、分子量は19,133Daであった。エンドトキシンレベルは、タンパク質1mg当たり58EUと測定された。
【実施例4】
【0158】
マウスマラリア原虫(Plasmodium yoelii)MSP1.19-マウスC4bp融合タンパク質
クローニング
AVD108タンパク質を以下のようにして調製した:マウスマラリア原虫(Plasmodium yoelii)由来のMSP1.19と、MSP1.33の一部をコードする合成DNAフラグメントを、マウスC4bpα鎖のC末端54アミノ酸から上流のNdeI-BamHIフラグメントとしてクローニングした。
【0159】
融合タンパク質AVD108をコードする核酸配列は、以下の通りである。
【0160】
ATGAGATCTCACATTGCCTCTATTGCTTTGAACAACTTGAACAAGTCTGG
TTTGGTAGGAGAAGGTGAGTCTAAGAAGATTTTGGCTAAGATGCTGAACA
TGGACGGTATGGACTTGTTGGGTGTTGACCCTAAGCATGTTTGTGTTGAC
ACTAGAGACATTCCTAAGAACGCTGGATGTTTCAGAGACGACAACGGTAC
TGAAGAGTGGAGATGTTTGTTGGGTTACAAGAAGGGTGAGGGTAACACCT
GCGTTGAGAACAACAACCCTACTTGCGACATCAACAACGGTGGATGTGAC
CCAACCGCCTCTTGTCAAAACGCTGAATCTACCGAAAACTCCAAGAAGAT
TATTTGCACCTGTAAGGAACCAACCCCTAACGCCTACTACGAGGGTGTTT
TCTGTTCTTCTTCCGGATCCGAGGCCTCTGAAGACCTTAAGCCTGCGCTT
ACAGGCAACAAGACCATGCAGTATGTGCCAAATTCACACGATGTGAAAAT
GGCTCTGGAGATCTACAAGCTGACTCTGGAGGTTGAACTACTACAGCTCC
AGATACAAAAGGAGAAACACACTGAAGCACACTAA (配列番号27)
タンパク質AVD108のアミノ酸配列は、以下の通りである。
【0161】
MRSHIASIAL NNLNKSGLVG EGESKKILAK MLNMDGMDLL GVDPKHVCVD TRDIPKNAGC FRDDNGTEEW RCLLGYKKGE GNTCVENNNP TCDINNGGCD PTASCQNAES TENSKKIICT CKEPTPNAYY EGVFCSSSGS EASEDLKPAL TGNKTMQYVP NSHDVKMALE IYKLTLEVEL LQLQIQKEKH TEAH (配列番号28)
配列番号28の残基3-138はマウスマラリア原虫MSP1(単量体抗原)の残基1619-1753に対応し、配列番号28の残基141-194は、ウサギC4bpコアタンパク質の54残基に対応する。GSリンカー配列はこれらの2つの成分の間に現れ、短い制限酵素認識部位をコードする配列は第一成分に先行する。
【0162】
AVD108の発現
タンパク質AVD108を大腸菌株C41(DE3)で発現させた。3Lの培養物を、LB培地で37℃、光学密度(OD600)約0.6まで生育させ、次に最終濃度0.7mMになるようにIPTGを添加することにより発現を誘導した。誘導4時間後、細胞を遠心分離により回収した。細胞をバッファーA(50mM Tris pH9, 5mM EDTA)で溶解し、残骸を遠心分離で除去した。
【0163】
AVD108の精製及び性質決定
タンパク質AVD108を、4つのカラムクロマトグラフィー工程を使って精製した。第一の陰イオン交換クロマトグラフィー工程では、DEAE HR16/10カラムを使用した。タンパク質をバッファーAで載せて、バッファーAに1M NaClを加えたバッファーBのグラジエントで溶出した。AVD108は180-300mM NaCl間の幅の広いピークで溶出された。
【0164】
第2の疎水性相互作用クロマトグラフィーの工程では、DEAEカラムから得られたAVD108を含むプールした画分をMacro-Prep Phenyl Sepharoseカラムに載せ、塩が1M〜0M NaClに減少するグラジエントで溶出した。最後の2つのクロマトグラフィーの工程では、AVD108タンパク質をSuperdex S200 26/60カラムのゲルろ過により精製した。最初に、タンパク質を最終濃度8Mになるように尿素を添加して変性させ、4℃で一晩、インキュベーションした。モノマーはこのカラムから203mlの量のところで溶出された。バッファーH(20mM Tris pH7.5, 150mM NaCl)で、4℃で一晩、透析により再生した後、同じカラムに載せる前に、今度は、バッファーHで平衡化した。今度は、オリゴマーが164mlの量のところで溶出された。質量分析により、質量は21,257Daと測定された。BiowhittakerのLALテストキットによるエンドトキシンレベルは、タンパク質1mg当たり4EUであった。
【実施例5】
【0165】
熱帯熱マラリア原虫MSP1.19-ウサギC4bp融合タンパク質を使用する免疫化
免疫化
実施例1で述べたように調製したAVD174タンパク質を、3羽のニュージーランドホワイト(NZW)ウサギに免疫するのに使用した。免疫化のスケジュールは以下の通りであった:各ウサギは2週間のインターバルで4回の注射(換言すれば、0、14、28及び42日目)を受けた。各注射は、皮下にし、公知のアジュバントを添加せず、緩衝化した等張生理食塩水に345μg(又は20nmol)のタンパク質を含有した。
【0166】
並行して、3羽のNZWウサギをフロイントアジュバントと212μg(又は20nmol)のAVD172タンパク質で免疫した。AVD172はAVD174と同一であるが、ウサギC4bpからC末端57アミノ酸を欠いている。それは以下のアミノ酸配列である。
【0167】
MLNISQHQCV KKQCPQNSGC FRHLDEREEC KCLLNYKQEG DKCVENPNPT CNENNGGCDA DAKCTEEDSG SNGKKITCEC TKPDSYPLFD GIFCS (配列番号29)
免疫化スケジュールは以下の通りであった:各ウサギは2週間のインターバルで4回の注射(換言すれば、0、14、28及び42日目)を受けた。各ウサギにとって最初の注射は、完全フロイントアジュバントで、皮内に投与され、一方、その後の3回の注射は、不完全フロイントアジュバントで与えられ、皮下に投与された。
【0168】
抗体価
熱帯熱マラリア原虫メロゾイトの表面上のMSP1に対する、最後の注射後1週間(即ち63日目)の抗体価を、熱帯熱マラリア原虫に感染した赤血球のアセトン固定化スメアの間接免疫蛍光(P. yoeliiについて、Lingら、Vaccine 15, 1562-1567, 1997に記載されている)で測定した。
【0169】
AVD174で免疫化したウサギの2つの最も高い力価は、1/81,920であった。フロイントアジュバントのAVD172タンパク質で免疫したウサギでは、2つの最も高い力価は1/20,480であった。
【0170】
これは、単量体抗原とC4bpコアの融合により、フロイントアジュバントの同じ単量体抗原を投与するよりも、高い抗体価を誘導することができることを立証した。
【0171】
生成された抗体の性質決定
高力価の抗体は、感染を予防又は治療するのに十分ではないだろうし、抗原による免疫化で生成された抗体の特異性は、決定的に重要であることが知られている。Guevara Patinoら(J.Exp.Med. 186, 1689-1699, 1997)は、MSP1.19に対するブロッキング抗体又は抑制抗体をアッセイする方法を詳細に述べている。これらの方法は、抑制抗体が存在するか(MSP1.42をMSP1.33及びMSP1.19にプロセシングするのをブロックし、赤血球への侵入を防ぐので有用である)を見るのに使用された。
【0172】
抑制抗体は、AVD174タンパク質によって誘導された抗体中に見出されただけである。AVD172タンパク質で免疫したウサギの抗血清中には何も見つからなかった。一方、マラリアはヒトに自然感染でき、有害であるが、フロイントアジュバントのAVD172は、ブロッキング抗体を誘導することができる(Guevara Patinoら, op. cit.参照)。
【実施例6】
【0173】
マウスマラリア原虫MSP1.19-マウスC4bp融合タンパク質を使用する免疫化
マウスの免疫化
実施例5で述べたように調製したAVD108タンパク質を、6匹のBALB/cマウスに免疫するのに使用した。アジュバントは使用せず、前記タンパク質を緩衝化された等張生理食塩水に存在させた。40μg(1.9nmol)のタンパク質を1回の注射に使用した。各マウスに皮下で4週間インターバルで3回(換言すれば、0、28及び56日目に)注射した。
【0174】
並行して、6匹のBALB/cマウスを、23μg(1.9nmol)のAVD183タンパク質で免疫した。AVD183タンパク質はAVD108と同じであるが、マウスC4bpのC末端の54アミノ酸が欠けている(即ちP.yoeliiのMSP1.19タンパク質単独である)。23μgの該タンパク質(AVD108に使用したのと同じ、緩衝化した等張生理食塩水に存在する)を、1回の注射で使用した。各マウスに、皮下に3回、0、28及び56日目に注射した。
【0175】
抗体価
マウスマラリア原虫のメロゾイトの表面にあるMSP1に対する、最後の注射後2週間(即ち70日目)の抗体価を、マウスマラリア原虫に感染した赤血球のアセトン固定化スメアの間接免疫蛍光(Lingら Vaccine 15, 1562-1567, 1997に記載されている)によって測定した。
【0176】
結果は、AVD108タンパク質で免疫した6匹のマウスのうち5匹が、1/40,960の抗体価を有し、6匹目のマウスは1/10,240の力価であった。一方、MSP1に対する抗体は、1/80の希釈で、AVD183タンパク質で免疫したマウスのどれにも検出されなかった。このことは、単量体のMSP1.19抗原とC4bpコアの融合により、得られた抗体の力価が500倍まで上昇できたことを立証した。
【0177】
寄生虫の暴露
前述のように免疫した6匹のマウスのグループ両方を、致死量5000のマウスマラリア原虫感染赤血球に暴露した。このアッセイも、Lingら(op. cit.)により説明されている。AVD183タンパク質で免疫した6匹のマウスは、寄生虫の暴露7日間以内に全部死んだ。他方で、AVD108で免疫した6匹のマウスのうち5匹は生き残り、それらの血液中に(顕微鏡で検査される薄層の血液スメアのギムザ染色で評価したところ)寄生虫はいなかった。70日目の力価が1/10,240であった6匹目のマウスは、暴露後19日で死んだ。このマウスの赤血球の70%以上に、19日目のギムザ染色で感染が見られた。
【0178】
一方、この暴露実験は、公知のアジュバンドの不存在下でC4bpコアタンパク質のみと融合した単量体抗原で免疫化すると、他の場合には致命的なマラリア原虫の感染から保護することができることを立証した。これは、現在のところ、公知のアジュバンドを伴わない単一のタンパク質だけを使ってマラリア原虫の感染に対する成功したワクチン接種の最初の実例を表すと信じられる。
【実施例7】
【0179】
インフルエンザ血球凝集素-C4bp融合タンパク質
本実施例は、単量体抗原(インフルエンザAウイルスのHA1血球凝集素タンパク質の残基91-261を含む)と、ヒト、ウサギ及びマウスのコアC4bpタンパク質の融合を説明する。通常、全長HA1は、ビリオンの細胞表面で三量体に集合し、そのため、このペプチドフラグメントのみを使うことにより、三量体を単量体抗原に有効に変化させる。これらの融合タンパク質をAVD272〜AVD274とし、細菌株C41(DE3)で発現し、精製する。これらの融合タンパク質のみを、アジュバントを添加することなくマウスとウサギを免疫するのに使用する。
【0180】
AVD272では、HA1フラグメント(Jeon及びArnon, Viral Immunology, 15, 165-176, 2002に記載されている)をマウスC4bp足場と融合する。
【0181】
AVD272のアミノ酸配列は以下の通りである。
【0182】
kafsncypyd vpdyaslrsl vassgtlefi tegftwtgvt qnggsnackr gpgsgffsrl nwltksgsty pvlnvtmpnn dnfdklyiwg ihhpstnqeq tslyvqasgr vtvstrrsqq tiipnigsrp wvrglssris iywtivkpgd vlvinsngnl iaprgyfkmr GSEASEDLKP ALTGNKTMQY VPNSHDVKMA LEIYKLTLEV ELLQLQIQKE KHTEAH (配列番号30)
AVD273では、HA1フラグメントをウサギC4bp足場と融合する。
【0183】
AVD273のアミノ酸配列は以下の通りである。
【0184】
kafsncypyd vpdyaslrsl vassgtlefi tegftwtgvt qnggsnackr gpgsgffsrl nwltksgsty pvlnvtmpnn dnfdklyiwg ihhpstnqeq tslyvqasgr vtvstrrsqq tiipnigsrp wvrglssris iywtivkpgd vlvinsngnl iaprgyfkmr GSEVPEGCEQ VQAGRRLMQC LADPYEVKMA LEVYKLSLEI ELLELQRDKA RKSSVLRQL (配列番号31)
AVD274では、HA1フラグメントをヒトC4bp足場と融合した。
【0185】
AVD274のアミノ酸配列は以下の通りである。
【0186】
kafsncypyd vpdyaslrsl vassgtlefi tegftwtgvt qnggsnackr gpgsgffsrl nwltksgsty pvlnvtmpnn dnfdklyiwg ihhpstnqeq tslyvqasgr vtvstrrsqq tiipnigsrp wvrglssris iywtivkpgd vlvinsngnl iaprgyfkmr GSETPEGCEQ VLTGKRLMQC LPNPEDVKMA LEVYKLSLEI EQLELQRDSA RQSTLDKEL (配列番号32)
マウスを、皮下で3回、2週間のインターバルで(即ち0、14及び28日目に)アジュバントを添加しないAVD272タンパク質(2nmol)で免疫し、かなりの抗体価(原則的に、Jeonら, op.cit.により説明されているように測定)を示し、また、マウスに適応したA/PR/8/34株の5 LD50の投与量での致死的な暴露に対して十分に保護することを示す。
【実施例8】
【0187】
インフルエンザM2ペプチド-C4bp融合タンパク質
本実施例は、単量体抗原(インフルエンザA型ウイルスのM2タンパク質の残基2-24、即ち細胞外部分を含む)とヒト、ウサギ及びマウスのコアC4bpタンパク質の融合を説明する。通常、全長M2はビリオン及び感染細胞で四量体に集合し、そのためこのペプチドフラグメントのみを使うことにより、四量体を単量体抗原に有効に変化させる。融合タンパク質をAVD275〜AVD278とし、細菌株C41(DE3)で発現し、精製した。これらの融合タンパク質のみを、アジュバンドを添加せずにマウスとウサギに免疫するのに使用した。
【0188】
AVD275では、細胞外M2ペプチド(Neirynckら、Nature Medicine 5, 1157- 1163, 1999に記載されている)をマウスC4bp足場と融合した。
【0189】
AVD275のアミノ酸配列は以下の通りである。
【0190】
SLLTEVETPI RNEWGCRCND SSDGSEASED LKPALTGNKT MQYVPNSHDV KMALEIYKLT LEVELLQLQI QKEKHTEAH (配列番号33)
AVD276では、細胞外M2ペプチドをウサギC4bp足場と融合した。
【0191】
AVD276のアミノ酸配列は以下の通りである。
【0192】
SLLTEVETPI RNEWGCRCND SSDGSEVPEG CEQVQAGRRL MQCLADPYEV KMALEVYKLS LEIELLELQR DKARKSSVLR QL (配列番号34)
AVD277では、細胞外M2ペプチドをヒトC4bp足場と融合した。
【0193】
AVD277のアミノ酸配列は以下の通りである。
【0194】
SLLTEVETPI RNEWGCRCND SSDGSETPEG CEQVLTGKRL MQCLPNPEDV KMALEVYKLS LEIEQLELQR DSARQSTLDK EL (配列番号35)
AVD278では、両方のシステインがセリン残基で置換された細胞外M2ペプチドの変異体を、ヒトC4bp足場と融合した。
【0195】
AVD278のアミノ酸配列は以下の通りである。
【0196】
SLLTEVETPI RNEWGSRSND SSDGSETPEG CEQVLTGKRL MQCLPNPEDV KMALEVYKLS LEIEQLELQR DSARQSTLDK EL (配列番号36)
マウスを2週間のインターバルで3回(即ち0、14及び28日目)、アジュバントを添加せずにAVD275タンパク質(2nmol)を皮下で免疫したことは、かなりの抗体価(原則として、Neirynckら, op.cit.により説明されているように測定)を示し、また、マウスに適応したA/PR/8/34株を5 LD50の投与量での致死的な暴露に対して十分に保護することを示す。
【図面の簡単な説明】
【0197】
【図1】C4bpコアタンパク質のアライメントを示す。
【図2】大腸菌のタンパク質の発現の結果を示す。
【図3】還元及び非還元条件下でゲルに流した、本発明のタンパク質の比較を示す。
【技術分野】
【0001】
本発明は、アジュバントと抗原を含む融合タンパク質などの高分子の集合体(assembly)に関し、この集合体は、抗原単独と比較した場合に、抗原に対する高まった免疫反応を誘導する。
【背景技術】
【0002】
アジュバントは抗原に対する免疫反応を高め、従ってワクチンに有用である。しかしながら、ヒトの使用に承認されているアジュバントは限られた数しかなく、動物での研究からもっと強いアジュバントが知られており、ヒトにおける使用に安全なより強い免疫学的アジュバントの必要性が明らかに存在する。最近の総説については、「Advances in vaccine adjuvants」(Nature Biotechnology, 1999, Volume 17, p1075-1081)及び「Recent advances in the discovery and delivery of vaccine adjuvants」(Nature Reviews in Drug Discovery, 2003, Volume 2, p727-735)を参照されたい。
【0003】
補体系は、外来抗原に対する免疫系の応答に重要な一連の血清タンパク質から成る。補体系は、その初期の成分が切断され、その生成物が単独で又は他のタンパク質と一緒に、別の補体タンパク質を活性化してタンパク質分解カスケードを生じ、活性化される。補体系の活性化は、様々な反応、例えば血管透過性の上昇、食細胞の走化性、炎症細胞の活性化、異物のオプソニン化、細胞の直接死及び組織損傷を引き起こす。補体系の活性化は抗原-抗体複合体により誘導されるか(古典経路)、又は通常のゆっくりとした活性化が、細菌及びウイルス等の侵入生物の細胞壁の存在下で増幅される(代替経路)。補体系は、古典経路及び代替経路の両方の中心のタンパク質であるC3が関与している特定の経路を介して、細胞免疫系と相互作用する。C3のタンパク質分解活性化により、大きいフラグメント(C3b)が生じ、侵入生物又は外来細胞の細胞表面タンパク質などの外部の求核物質と共有結合的に反応することができる化学的に反応性の内在性チオールエステル結合が露出する。その結果、潜在的な抗原がC3bで「標識(tag)」され、更にタンパク質分解を受けてiC3b及びC3d,gになる間そのタンパク質に付着したままになる。後者のフラグメントは、それぞれ、補体レセプターCR3及びCR2に対するリガンドである(CR2はCD21ともいう)。従って、C3bで抗原が標識されると、これらのレセプターを持っている免疫系の細胞のための標的メカニズムが生じうる。
【0004】
このようなターゲット化が免疫反応の増強に重要であるということは、マウスにおいて循環するC3を枯渇させ、次に抗原(ヒツジ赤血球)に暴露した実験により、初めて示された。C3を除去すると、前記抗原に対する抗体反応が減少した(M.B. Pepys, J. Exp. Med., 140, 126-145, 1974)。C3の役割は、C3、又はC3bを生ずる補体カスケードの上流成分であるC2及びC4の遺伝子が欠損している動物の研究により確認された(J.M. Ahearn及びD.T. Fearon, Adv. Immunol., 46, 183-219, 1989)。最近、モデル抗原とマウスC3dフラグメント配列の2つより多くのコピーを直線状に結合させると、修飾されていない抗原対照と比較して、マウスの抗体反応が非常に大きく上昇(1000〜10000倍)したことが示された(P.W. Dempseyら, Science, 271, 348-350, 1996; WO96/17625, PCT/GB95/02851)。この上昇は、ヒトに使用するには毒性が強すぎるフロイントの完全アジュバントなどの従来のアジュバントを使用することなく行うことができた。この顕著な効果のメカニズムは、多価のC3d構築物とB細胞上のCR2が高い親和性で結合し、次に、CR2と、別のB細胞膜タンパク質CD19及び膜結合イムノグロブリンが同時連結し、B細胞の核にシグナルを発する、と実証された。
【0005】
しかしながら、C3dの3つのコピーを含む均質な組換えタンパク質を大量に製造するのは難しいことが分かっている。主な問題点は、
i) (3つの)繰り返し配列を含む構築物の遺伝的不安定性、及び
ii) 大腸菌(Escherichia coli)の中に形成された封入体由来の組換えタンパク質の折りたたみ(又は可溶化及び再折りたたみ)
である。
【0006】
C3d遺伝子の繰り返しコピーを含む構築物の遺伝的不安定性を最小限にするために採られた1つの方法が、国際公開WO 99/35260及びWO 01/77324に開示されている。これらの出願に開示されている技術は、C3dの繰り返し配列をコードするDNAの同一でない配列を使用するものである。
【0007】
国際公開WO 91/11461では、補体4結合タンパク質(C4bp)を用いる多量体化系が開示されている。ヒトC4b-結合タンパク質(C4BP)は、7つの同一のα鎖と1つのβ鎖でできたクモ状構造を有する高分子量(570kDa)の血漿糖タンパク質である。C4bpのα鎖は、分子が多量体に組み立てられる機能を担っているC末端コア領域を有する。標準的なモデルによれば、C4bpモノマーの位置+498のシステインが、別のモノマーの位置+510のシステインとジスルフィド結合を形成する。7つのα鎖のみを含むより小さい形状のものもヒトの血漿中に発見されている。この血漿糖タンパク質の本来の機能は、補体の活性化の古典経路を抑制することである。
【0008】
C4bpのα鎖のほとんどは、補体コントロールタンパク質(complement control protein, CCP)の繰り返しとして知られる長さ約60アミノ酸の8つの直列に並んだドメインから構成される。国際公開WO91/11461は、C4bpの全部もしくは一部及び対象となる生物学的タンパク質を含む融合タンパク質を作るのに、C4bpタンパク質の多量体化する能力が使えることを提案している。国際公開WO91/11461に開示された融合タンパク質では、1つより多くのCCP繰り返し(SCRとしても知られている)を含めることが好ましい。
【0009】
国際公開WO91/11461は、融合タンパク質をワクチンとして使用できることを示唆している。B型肝炎e抗原のフラグメントと融合した少なくとも1つのC4bp SCR領域を含むいくつかの特定のタンパク質が作られた。使用されたe抗原のフラグメントは、多量体構造を形成できるコア抗原フラグメントである。
【0010】
Libyh M. T.ら(1997, Blood, 90, 3978-3983)は、多量体化を妨げずに、(C末端の57アミノ酸のみを残して)すべてのCCPを除去することができることを示した。このC4bpのC末端領域を、本明細書においてはC4bpコアという。
【0011】
また、ヒト293細胞系統で発現された自己集合性の(self-assembling)多量体の可溶性CD4-C4bp融合タンパク質も、Shinyaら(1999, Biomed & Pharmacother, Vol. 53: 471)によって示されている。
【0012】
また、C4bpの使用は、Oudinら(2000, Journal of Immunology, Vol. 164:1505)に開示されている。
【0013】
Christiansenら(2000, Journal of Virology, Vol. 74:4672)は、CD46-C4bp融合タンパク質の治療用途について議論している。
【0014】
国際公開WO2004/020639は、異種のポリペプチドと融合していてもよいC4bpα鎖のC末端コアタンパク質の足場(scaffold)を含む組換え融合タンパク質を原核生物の宿主で得る方法を提供し、前記組換え融合タンパク質は、原核生物の宿主細胞内で可溶性の形で多量体を形成することができる。
【発明の開示】
【課題を解決するための手段】
【0015】
本発明は、特定のクラスのC4bp融合タンパク質に関する新規な知見に基づく。前述の先行技術の幾つかは、C4bp部分は本質的に不活性担体であることに基づき、対象の治療用タンパク質を運搬するのにC4bp融合タンパク質を使用することを提案している。
【0016】
前述の国際公開WO91/11461は、多量体形成性抗原を含む融合タンパク質を例示している。
【0017】
一方、本発明は、多量体を自然に形成する抗原はC4bpコアが多量体に組み立てられるのを事実上妨害するので、このような抗原はC4bpコアとの融合に望ましくないという認識に基づく。更に、驚くべきことに、C4bpコアと単量体抗原の融合は、抗原に対する強い免疫反応を引き起こすことが示された。後述の実施例において、単量体抗原を含む融合タンパク質は、抗原をフロイントのアジュバントと共に注射したときの低い力価の非抑制的な抗体応答と比較して、より高い力価の抑制的な抗体応答を引き起こしたことを示す。
【0018】
従って、本発明は、単量体抗原をC4bpコアタンパク質と組合わせて複合体にすることにより、その抗原の免疫原性を増加させる方法を提供する。好ましい方法において、単量体抗原をC4bpコアタンパク質と共有結合させる。極めて好ましい方法において、単量体抗原をC4bpコアタンパク質と遺伝学的に融合させる。
【0019】
また、本発明は、抗原に対する高い力価の抗体を誘導する方法、及び受動免疫による感染性で悪性の疾患の予防及び/又は治療における、前記方法の使用により製造した高力価の抗血清の使用を提供する。好ましい方法において、抗原に対する高力価の抗体を、過免疫の血清又は血漿のイムノグロブリン分画を単離することにより部分的に精製し、極めて好ましい方法において、過免疫の血清又は血漿のイムノグロブリン分画を、その抗血清が感染性または悪性の疾患を予防又は治療するために使用される同じ種の個体から単離する。
【0020】
従って、本発明は、
C4bpコアタンパク質、及び
単量体抗原
を含む生成物を提供する。
【0021】
第一及び第二成分は、融合タンパク質の形であってもよい。或いは、これらは、第一成分のアミノ酸側鎖を介して、又は特異的に第二成分の化学的結合を可能にするように第一成分に付加されたアミノ酸の側鎖を介して、化学的に結合してもよい。
【0022】
第一及び第二成分は、堅固に、しかし非共有結合で結合してもよい。例えば、第一成分のアミノ酸の側鎖は、付加的なビオチン基を持つように修飾でき、このビオチンを使用してストレプトアビジン(ストレプトアビジンは第二成分)と結合させることができるし、又はストレプトアビジンと融合した抗原をこのビオチンを介して第一成分と結合させることもできる。別の可能性において、ビオチン化した抗原とビオチン化した第一成分を、ストレプトアビジンを添加して、生じた複合体を精製することにより、共にしっかりと、しかし非共有結合的に保持することができる。
【0023】
疑いを回避するために、「第一」及び「第二」成分の意味は、2つの成分の生成物における特定の直線的な順序を意味しないし、表さない。2つの成分はどんな順序で結合してもよい。
【0024】
従って、両方の成分がポリペプチドであり、生成物が融合タンパク質として作られる場合、2つの成分のN末端からC末端への順序はどんな順序でもよい。
【0025】
更に、本発明は、前記第一及び第二成分の融合タンパク質をコードする核酸を提供する。また本発明は、前記核酸を含むベクター及びそのベクターを有する宿主細胞を提供する。
【0026】
別の実施形態において、本発明は、
C4bpコアタンパク質、及び
ポリペプチドの単量体抗原、
を含む生成物を製造する方法であって、融合タンパク質の形態で2つの成分をコードする核酸を発現させる工程、及び前記生成物を回収する工程を含む方法を提供する。
【0027】
別の実施形態において、本発明は、
C4bpコアタンパク質;
非ポリペプチドの単量体抗原、
を含む生成物を製造する方法であって、C4bpコアタンパク質をコードする核酸を発現させる工程、前記コアタンパク質を前記抗原に結合させる工程、及び前記生成物を回収する工程を含む方法を提供する。
【0028】
生成物を製造する方法は、真核細胞又は原核細胞で行うことができる。
【0029】
また本発明は、本発明による生成物の有効量を患者に投与することを含む抗原に対する免疫反応を誘導する方法を提供する。
【0030】
また本発明は、人体又は動物体の治療方法、特に免疫反応を誘導する方法のための本発明の生成物の使用を提供する。
【0031】
更に本発明は、製薬上許容できる担体又は希釈剤と組合わせた本発明の生成物を含む医薬組成物を提供する。
【0032】
更に本発明は、病原体に対する受動免疫で使用するための感染防御免疫血清を調製する方法を提供し、前記方法は、動物に本発明の生成物をワクチン接種する工程、該動物から抗血清を回収する工程を含む。次に抗血清を、ヒト患者の受動免疫の方法に使用することができる。ヒト患者は病原体に感染した患者又はリスクのある患者である。
【発明を実施するための最良の形態】
【0033】
C4bpα鎖のコアタンパク質
本明細書では、C4bpα鎖のコアタンパク質を「C4bpコアタンパク質」若しくは「コアタンパク質」、又は「C4bp足場(scaffold)」とする。この用語は互換的に使用される。このタンパク質は哺乳動物のC4bpコアタンパク質であっても、又は多量体を形成でき、アジュバントとして作用できる(C4bpコアタンパク質の)フラグメントであっても、あるいは多量体を形成でき、アジュバントとして作用できる(C4bpコアタンパク質の)合成若しくはキメラ変異体であってもよい。
【0034】
本発明において、C4bpコアタンパク質又はコアタンパク質を含むC4bpα鎖のフラグメントはアジュバントとして働き、本明細書で更に詳細に説明される。配列番号1のヒトC4bpコアタンパク質は、多量体を形成することが当該技術分野で知られている、全長C4bpタンパク質配列のアミノ酸+493〜+549に対応する。
【0035】
更に本発明は、抗原の免疫原性を増強するためのC4bpコアの誘導体の使用を含む。このような誘導体は、誘導体の突然変異体を含み、本明細書で説明されるアジュバントの性質の維持を条件として、アミノ酸の欠失、付加、特にシステイン残基の付加、又は置換、C4bpファミリーの異なるメンバーの一部分の融合により形成されたハイブリッド又はキメラ分子、及び/又は環状の並べ替えられた(circular permutated)タンパク質足場を含む。
【0036】
また本発明は、複数の種由来のC4bpコア配列のアライメントに基づく人工の共通(コンセンサス)C4bp配列も使用できる。非常に多くあるであろう、このクラスのキメラ分子の一例を以下に挙げる(配列番号20、図1)。
【0037】
従って、アジュバントは、C4bpコア及び場合によってはコアと融合させた1つ以上のSCRでよい。
【0038】
特に好ましい実施形態において、本発明の生成物のC4bp成分は、C4bpα鎖のコアタンパク質、即ち、C4bp SCR配列に結合していない、本明細書で定義したコアタンパク質である。このような形態において、C4bpコアは、望ましくは、配列番号1の残基1〜57又はそのホモログの対応する残基、或いは配列番号1の少なくとも47アミノ酸のフラグメント若しくはそのホモログから成る。
【0039】
更に、本発明の生成物のC4bpコアは、可撓性のリンカーなどのN末端又はC末端伸長部分を含んでもよい。一般的に、このようなリンカーは、長さが1〜20の少数のアミノ酸、例えば長さ2〜10アミノ酸である。このようなリンカーの1つは、(Glym-Ser)nリンカーであり、ここで、mとnは、それぞれ独立して1〜4である。これらは、当該技術分野で、互いにタンパク質ドメインを付着させるのに使用される。従って第一成分をこのようなリンカーで第二成分と連結することができる。
【0040】
第一成分がC4bpコアである場合、第一成分は生成物のC末端にあることが好ましい。
【0041】
多くの哺乳動物C4bpタンパク質の配列は、当該技術分野で利用可能である。これらには、ヒトC4bpコアタンパク質(配列番号1)が含まれる。当該技術分野で利用可能な、ヒトC4bpコアタンパク質のホモログがたくさんある。2つのタイプのホモログがある:オルソログとパラログである。オルソログは、異なる生物のホモログ遺伝子と定義され、即ち、そのホモログ遺伝子は、その遺伝子を発生させた種分化現象が起こった(coincident)共通の祖先を持つ。パラログは、遺伝子、染色体又はゲノム重複に由来する同一生物のホモログ遺伝子として定義され、即ち、その遺伝子の共通の祖先は、最後の種分化現象後に発生した。
【0042】
例えば、GenBank、生のゲノムトレース及びEST(発現遺伝子配列断片)データベースの検索は、チンパンジー、アカゲザル、ウサギ、ラット、イヌ、ウマ、マウス、モルモット、ブタ及びウシを含む種の、哺乳動物C4bpコア配列番号1のホモログタンパク質を示唆する。配列番号1のC4bpのパラログ及びオルソログは、図1のアライメントに含まれている。
【0043】
全部で配列番号1〜19のアライメントを図1に示した。全部の19の配列は、C末端では変化がより大きいが、高いレベルの類似性を有することを示す。BLASTなどの市販されている検索プログラムを使ってDNA又はタンパク質配列のデータベースを検索することによって別のC4bpコアタンパク質を同定することができる。
【0044】
所望の哺乳動物源由来のC4bpタンパク質がデータベースで利用できない場合、当該技術分野で十分に確立されている日常的なクローニング方法を使って得ることができる。基本的に、このような技術は、入手可能なC4bpコアタンパク質の1つをコードする核酸をプローブとして使い、対象の他の種由来のC4bpコアタンパク質の配列を回収し決定する工程を含む。広範多様な技術がこのために利用でき、例えば、ゲノムDNA又はmRNAの適切な供給源(例えば胚又は活発に分裂している分化細胞又は腫瘍細胞)を使用する遺伝子のPCR増幅及びクローニング、又は、哺乳動物由来のcDNAライブラリー(例えば上述した供給源の1つに由来するcDNAライブラリー)を得る工程、中〜高ストリンジェントな条件(例えば0.03M塩化ナトリウム、0.03Mクエン酸ナトリウム、約50℃〜約60℃)下で公知のC4bp核酸で前記ライブラリーを探索する工程、及びその哺乳動物のC4bpタンパク質の全部又は一部をコードするcDNAを回収する工程を含む方法である。部分的なcDNAが得られる場合、全長コード配列をプライマー伸長技術により決定することができる。
【0045】
多量体を形成できるC4bpコアタンパク質のフラグメントは、少なくとも47アミノ酸、好ましくは少なくとも50アミノ酸を含んでよい。このフラグメントの、多量体を形成する能力は、本発明に従って、原核生物の宿主細胞でフラグメントを発現させ、全57アミノ酸C4bpコアの多量体化を生じる条件下でC4bpフラグメントを回収し、フラグメントが更に多量体を形成するかどうかを決定することにより、テストできる。望ましくは、C4bpコアのフラグメントは、配列番号1の少なくとも残基6〜52又はそのホモログの対応する残基を含む。
【0046】
また、同様に多量体を形成する能力を保持しているC4bpコアの変異体及び多量体を形成できるフラグメントも(フラグメントについては上述したように決定できる)、使用することができる。好ましくは、変異体は、野生型哺乳動物のC4bpコア又はその多量体形成性フラグメントに対して、少なくとも70%、より好ましくは少なくとも80%、更に好ましくは少なくとも90%、例えば少なくとも95%又は最も好ましくは少なくとも98%の配列同一性を有する。
【0047】
ある態様において、C4bpコアは、配列番号1の位置12に現れるグリシン、位置28に現れるアラニン、位置29、34、36及び41に現れるロイシン、及び位置32に現れるチロシン及び位置33に現れるリジン、及び好ましくは位置6と18に現れる2つのシステイン残基を含むコアであろう。望ましくは、変異体はこれらの残基の間に相対的な間隔を保持するであろう。
【0048】
上記に特定した同一性の程度は、配列番号1〜20のうちの1つ又はそれらの多量体形成性フラグメントに対するものである。
【0049】
最も好ましくは、特定した同一性の程度は、配列番号1又はその多量体形成性フラグメントに対するものである。
【0050】
配列の同一性の程度は、Accelrys (以前はGenetics Computer Group, Madison, WI)から入手可能な、当該技術分野で広く使用されるアルゴリズムの「ウイスコンシンパッケージ(Wisconsin package)」の一部であるアルゴリズムGAPにより決定することができる。GAPはNeedlemanとWunschのアルゴリズムを使用し、マッチする数を最大にし、ギャップの数を最小にするように2つの完全な配列を整列させる。GAPは、同じような長さの、短く非常に密接な配列のアライメントに有用であるので、配列が前述の同一性レベルに当たる場合に決定するのに適切である。GAPはデフォルトパラメーターで使用できる。
【0051】
哺乳動物のC4bpコアタンパク質の合成変異体は、C-又はN-終端に、1つ以上のアミノ酸置換、欠失、挿入又は付加を有する変異体を含む。置換が特に考えられる。置換は保存性の置換を含む。保存性の置換の例として、しばしばデイホフ(Dayhoff)グループと呼ばれる類似のアミノ酸のグループに留意した置換が挙げられる。これらを以下に挙げる。
【0052】
グループ1 D, E, N, Q
グループ2 I, L, V, M
グループ3 F, Y, W
グループ4 K, R, H
グループ5 S, P, T, A, G
グループ6 C
従って、作成し、多量体を形成する能力およびアジュバントとして作用する能力をテストすることができる、C4bpコアタンパク質のフラグメントと変異体の例として、以下の表1に示す配列番号37〜44が挙げられる。
【表1】
【0053】
A=配列番号、B=配列、C=%同一性、同じ長さの配列番号1のフラグメントを参照することにより計算した。
【0054】
配列を欠失させる場合、N-又はC-末端の切断は別として、欠失は、近接又は非近接でありうる、1つのみ、2つ又は3つの欠失に限定されることが好ましいであろう。
【0055】
コアタンパク質配列に挿入する場合、又はN-又はC-末端を伸長する場合、挿入及び伸長は、コアタンパク質の大きさが、野生型配列の長さを20アミノ酸より多く超えないような、好ましくは15アミノ酸以下、より好ましくは10アミノ酸以下になるような数に限定されることが好ましいであろう。従って、配列番号1の場合、コアタンパク質は、挿入又は伸長により改変される場合、77アミノ酸以下の長さが望ましいであろう。
【0056】
抗原
抗原は、抗体又はT細胞受容体により認識される能力を持つ分子である。しかしながら、すべての抗原が免疫原というわけではない。免疫原は免疫反応を引き出す物質である。ある態様において、本発明は免疫原でない抗原を免疫原にすることができ、弱い免疫原である抗原をより強い免疫原にすることができる。
【0057】
本発明の重要な特徴は、C4bpコアと遺伝学的に融合させることにより抗原が作られる場合、単量体抗原が非常に好ましい、ということである。なぜならば単量体抗原は、C4bpコアタンパク質が組み立てられてオリゴマーになり、その結果機能的な形になるのを妨げないからである。
【0058】
しかしながら、それとは別の態様において、抗原がC4bpコアタンパク質と化学的に又は非共有結合的に結合する場合、その抗原は単量体でなくてよい。
【0059】
従って、単量体抗原は以下の2つの主要グループに分類できる。
【0060】
1) 天然の状態で多量体(即ち、二量体又はそれより高次の多量体)である親タンパク質のフラグメント又は変異体である抗原だが、親タンパク質がこのような多量体を形成する条件下では、抗原そのものは多量体を形成しない、抗原。
【0061】
2) 天然の状態でモノマーである、抗原。
【0062】
両方のタイプの抗原の例については、本明細書で以下に更に議論する。
【0063】
単量体抗原は、DNAの一本鎖断片上にコードされ、このDNAがC4bpコアタンパク質をコードするDNAと融合されて、その後タンパク質に翻訳される場合、抗原は、抗原上の唯一の点を介して単一のC4bpコアタンパク質鎖と結合されるという点で共通している。このような抗原の簡単な例は、ニワトリ卵白のリゾチームであろう。全長リゾチームオープンリーディングフレームをコードするcDNAは、生じた融合タンパク質のC4bp部分の組み立てが妨げられないようにして、C4bpコアオープンリーティングフレームと融合することができる。
【0064】
生合成の後、C4bpコアと融合した単一のポリペプチド鎖を加工することができ、例えばタンパク質分解酵素により、ポリペプチド鎖内に新しいN-及びC-終末を作ることができる。タンパク質分解による切断により生じた2つ以上の鎖が、例えばジスルフィド結合を介して、互いに付着したままである場合、C4bp融合タンパク質は、プロセシングの終わりの時点でタンパク質と付着しており、通常は単量体とみなされないであろう。しかしながら、本発明の目的において、このタイプのタンパク質は、単一のオープンリーディングフレーム内で単一の融合タンパク質としてコードされるので、単量体だと考えられる。このタイプの例としては、生合成の後に加工されて、ジスルフィド結合により結合されたA及びBと呼ばれる2つの鎖になるプロインシュリンであろう。プロインシュリンのCペプチドといわれるフラグメントは、前駆体融合タンパク質のタンパク質分解酵素による加工の後に除去される。
【0065】
単量体抗原は、天然の状態で必ずしも単量体でないタンパク質に由来してもよい。すなわち、自然界においてポリマー状態で見出される多くの抗原は、例えばタンパク質工学技術により単量体になるように改変することができる。3つの例がある。このような抗原の一例はインフルエンザウイルス赤血球凝集素タンパク質に由来するものである。この抗原は、天然の状態で複合体化した三量体構造を形成することがよく知られている(Wilsonら、Nature 289, 366-373, 1981)。しかしながら、分子の三量体化を引き起こすコイルドコイルを除去することにより、単量体のフラグメントを得ることが可能である。具体的な例が、Jeon及びArnonの研究(Viral Immunology 15, 165-176, 2002)により提供される。この著者らは赤血球凝集素の球状領域のみを包囲するフラグメントを得るために、赤血球凝集素の残基96〜261のみを使用した。
【0066】
別の例は、マラリア原虫メロゾイト表面タンパク質1(MSP1)である。この大きい(約200kDa)タンパク質は、マラリア感染の血液段階を担うメロゾイトの表面を飾っている。通常、C末端のGPIアンカー(GPIはグリコシルホスファチジルイノシトール)を介してメロゾイトの表面に固定されている。このGPIアンカーの前には、一続きの疎水性アミノ酸配列部分(stretch)がある。このアンカーのため、全長MSP1、又はMSP1.19と呼ばれるC末端フラグメント(メロゾイトが赤血球に侵入するときでも膜と結合したままである)のいずれもが、事実上、単量体の状態で発見されていない。同じことが、単一の疎水性膜貫通領域を有する多くの膜タンパク質に当てはまる。本発明は、これらの疎水性の部分を欠失させることにより最も良好に行われる。MSP1.19タンパク質とC4bpコアタンパク質の融合を説明する以下の実施例を参照されたい。
【0067】
従って、本発明の1つの好ましい態様において、本発明の生成物は、C4bpコアタンパク質と融合したマラリア原虫MSP1単量体抗原性フラグメントの融合体である。マラリア原虫MSP1抗原性フラグメントは、約50〜約200アミノ酸、好ましくは約50〜約150アミノ酸を含む。抗原性フラグメントは、どのマラリア原虫種由来でもよく、例えばPlasmodium falciparum、Plasmodium vivax、Plasmodium ovale、Plasmodium malariae(これらのすべてはヒトに病気を生じさせることができる)又はPlasmodium yoeliiである。
【0068】
欠失は、普通はオリゴマー性のタンパク質を単量体にする最も簡単な方法であるが、幾つかの場合においては、1つ以上のアミノ酸を変異することで十分であろう。この一例は、Cpn10タンパク質であり、主要なイソ型のC4bpコアと同様に天然の状態では七量体のタンパク質である。Cpn10の一個のアミノ酸の突然範囲により、Cpn10が、C4bpコアタンパク質との融合に適した、単量体の突然変異体に変換される(Guidryら、BMC Biochemistry 4, 14-26, 2003)。このタンパク質を単量体化するための別の方法は、N-末端又はC-末端のアミノ酸を欠失させ(Llorcaら、Biochem. Biophysica Acta 1337, 47-56, 1997; Seale及びHorowitz, J. Biol. Chem. 270, 30268-30270, 1995)、これにより、サブユニット間の相互作用の原因である領域を欠失させる方法である。
【0069】
一般的に、組み立てられてオリゴマー構造(例えばウイルスのカプシドタンパク質)になる強い傾向を有し、その結果、融合されるC4bpコアタンパク質の集合体を破壊するタンパク質に関して、タンパク質-タンパク質相互作用を生じさせる領域を欠失させる原理又は境界面の残基を突然変異させる原理を、単量体のタンパク質を得るのに適用できる。
【0070】
抗原は、2つのカテゴリーに分類することができ、両方とも本発明に用いるのに適する。第一のカテゴリーは、外来性抗原であり、感染性の生物に見られるすべての分子を含む。細菌の免疫原、寄生虫の免疫原及びウイルスの免疫原は、ワクチンとして有用な多量体の又はヘテロ多量体のC4bp融合タンパク質を作るのに、ポリペプチド部分として有用である。
【0071】
これらの免疫原の細菌源として、細菌性肺炎、髄膜炎、コレラ、ジフテリア、百日咳、破傷風、結核及びハンセン氏病の原因である細菌源が挙げられる。
【0072】
寄生虫源として、トリパノゾーマ及びリーシュマニア種と同様に、Plasmodiumなどのマラリア原虫が挙げられる。
【0073】
ウイルス源として、ポックスウイルス、例えば痘瘡ウイルス、牛痘ウイルス及びオルフウイルス;ヘルペスウイルス、例えば単純ヘルペスウイルスタイプ1及び2、B-ウイルス、水疱瘡ウイルス、サイトメガロウイルス、及びエプスタイン・バーウイルス;アデノウイルス、例えばマストアデノウイルス;パポバウイルス、例えばHPV16等のパピローマウイルス、BK及びJCウイルス等のポリオーマウイルス;パルボウイルス、例えばアデノ随伴ウイルス;レオウイルス、例えばレオウイルス1、2、及び3;オルビウイルス、例えばコロラドダニ熱;ロタウイルス、例えばヒトロタウイルス;αウイルス、例えば東部脳炎ウイルス及びベネズエラ脳炎ウイルス;ルビウイルス、例えば風疹;フラビウイルス、例えば黄熱病ウイルス、デング熱ウイルス、日本脳炎ウイルス、ダニ媒介性脳炎ウイルス及びC型肝炎ウイルス;コロナウイルス、例えばヒトコロナウイルス;パラミクソウイルス、例えばパラインフルエンザ1、2、3及び4及びおたふくかぜ;はしかウイルス(morbillivirus)、例えば麻疹ウイルス(measles virus);肺炎ウイルス、例えば呼吸器合胞体ウイルス;ベシキュロウイルス(vesiculovirus)、例えば水疱性口内炎ウイルス;リッサウイルス、例えば狂犬病ウイルス;オルトミクソウイルス、例えばA型及びB型インフルエンザ;ブンヤウイルス、例えばラクロスウイルス;フレボウイルス、例えばリフトバレー熱ウイルス;ナイロウイルス(nairovirus)、例えばコンゴ出血熱ウイルス(Congo hemorrhagic fever virus);ヘパドナウイルス科、例えばB型肝炎;アレナウイルス、例えば1cmウイルス(1cm virus)、ラッサ熱ウイルス(Lasso virus)及びフニンウイルス(Junin virus);レトロウイルス、例えば、HTLV I、HTLV II、HIV-1及びHIV-2;エンテロウイルス、例えばポリオウイルス1、- 2及び3、コクサッキーウイルス、エコーウイルス、ヒトエンテロウイルス、A型肝炎ウイルス、E型肝炎ウイルス及びノーウォークウイルス;ライノウイルス、例えばヒトライノウイルス;及びフィロウイルス科、例えばマールブルグ(病)ウイルス及びエボラウイルスが挙げられる。
【0074】
これらの細菌、ウイルス及び寄生虫源に由来する抗原は、ワクチンとして有用な多量体のタンパク質の製造に使用することができる。多量体は、異なる抗原を有するモノマーの混合物を含んでもよい。
【0075】
これらの細菌、ウイルス及び寄生虫源に由来する抗原は、通常宿主に存在せず、宿主ゲノムにコードされていないので、外来性抗原として考えられる。対照的に、内在性抗原は通常宿主に存在するか、もしくは宿主ゲノムにコードされているか、又はその両方である。内在性抗原に対する免疫反応を起こす能力は、その抗原を持つ腫瘍を治療したり、又は腫瘍の成長因子を中和するのに有用である。内在性抗原の第一タイプの例は、HER2であり、ハーセプチンと呼ばれるモノクローナル抗体の標的である。内在性抗原の第二の(成長因子の)タイプの例は、前立腺の幾つかの悪性腫瘍に栄養作用のある、ゴナドトロピン放出ホルモン(GnRHと呼ばれる)である。
【0076】
本発明を使って作られる免疫原は、研究又は治療目的に使用できる。例えば、研究の適用例として、ゲノム配列データで予測される遺伝子産物に対する抗血清を作ることが挙げられる。この要望は、細菌などの原核生物並びに真菌及び哺乳動物を含む真核生物の遺伝子産物に当てはまる。抗原は、ワクチンの技術分野における通常のあらゆるサイズ、即ち短いペプチドから非常に大きいタンパク質のサイズが可能である。
【0077】
非ポリペプチド免疫原は、例えば、炭水化物又は核酸でよい。ナイセリア(Neisseria)種又はストレプトコッカス・ニューモニア(Streptococcus pneumoniae)種の多糖外被は、本発明の目的に使用できる炭水化物の例である。
【0078】
非ポリペプチド免疫原が本発明の生成物の一部である場合、その免疫原は、日常的な合成方法を使って、生成物の第一成分と共有結合させることができる。一般的に、免疫原は、第一成分を含むC4bpコアタンパク質のN-又はC-末端のどちらかと結合させるか、又はアミノ酸側鎖基(例えばリジンのε-アミノ基又はシステインのチオール基)又はそれらの組合わせと結合させることができる。融合タンパク質当たり1つより多くの免疫原を加えてもよい。結合を促進するため、例えばN-又はC-終端として、システイン残基をC4bpコアタンパク質に加えることができる。
【0079】
本発明は、免疫反応の発生において多くの利点がある。例えば、多量体の使用により、多くの抗原を同時に免疫系に対して提示できる。このことは、単一の生物又は多くの異なる生物に存在し得る1つより多くのエピトープに対する免疫反応を引き起こすことができる多価ワクチンの調製を可能にする。
【0080】
その結果、更なる態様において、単量体抗原は、2種類の異なる生物に由来するか又は同じ生物の2つの異なるタンパク質に由来する、2つの異なるエピトープを含む合成抗原であってもよい。後者の例は、スポロゾイト抗原配列、例えば、MSP1配列と結合したスポロゾイト周囲タンパク質由来の2つ以上のNANP繰り返し配列の融合体である。後者の第二の例は、単量体のインフルエンザ赤血球凝集素フラグメントと融合したNeirynckら(Nature Medicine 5, 1157-1163, 1999)により開示されたM2e配列の融合体である。
【0081】
従って、本発明により形成されたワクチンは、1種類より多くの疾患に対する同時ワクチン接種に使用することができ、または所定の病原体の複数のエピトープを同時に標的にするのに使用できる。エピトープは単一のモノマーユニット内又はヘテロ多量体を提供するために組合わされる異なるモノマーユニット上に存在しうる。
【0082】
特にC4bpコア融合タンパク質は、免疫化の状況において有用であり、コアタンパク質は免疫化される者の血清又は血漿に通常存在するからであり、またコアタンパク質はそれ自身に対する免疫反応を惹起しないからである。C4bpタンパク質は多くの哺乳動物種で知られており、標準的な遺伝子クローニング技術を使って当業者により、哺乳動物種における適切なホモログを見出すことができる。
【0083】
核酸
本発明の生成物は、タンパク質をコードする核酸構築物を使って、原核生物又は真核生物の宿主細胞で融合タンパク質を発現させることにより作ることができる。抗原がポリペプチドの場合、核酸配列からの融合タンパク質の発現を、本発明の生成物を作るのに使用できる。
【0084】
従って、本発明は、本発明の生成物をコードする核酸構築物(一般的にはDNA又はRNA)を提供する。
【0085】
一般的に、構築物は複製可能なベクターの形であり、このベクター内ではタンパク質をコードする配列が、所望の宿主細胞でのタンパク質の発現に適切なプロモーターと有効に結合している。
【0086】
ベクターは、複製開始点及び任意にプロモーターの制御因子を含んでいることができる。ベクターは、1つ以上の選択マーカー遺伝子を含むことができる。当該技術分野で公知の広範多様な原核細胞又は真核細胞の発現ベクターがあり、本発明は、当業者の個々の選択により、あらゆるベクターを利用することができる。
【0087】
広範多様な原核生物の宿主細胞を本発明の方法に使用することができる。これらの宿主として、大腸菌類、シュードモナス属、桿菌属、乳酸桿菌属、好熱菌(Thermophilus)、サルモネラ属、腸内細菌科又は放線菌類の菌系統が挙げられる。例えば、エシェリキア属の大腸菌(E. coli)が本発明の方法に使用される場合、使用するこの細菌の好ましい菌系統として、国際公開WO98/02559で開示され供されるC41(DE3)、C43(DE3)又はCO214(DE3)を含むBL21(DE3)の派生系統(derivative)が挙げられよう。
【0088】
更に好ましくは、プロファージDE3を欠くこれらの系統の派生系統は、プロモーターがT7プロモーターではない場合に使用できる。
【0089】
原核生物のベクターとして、細菌プラスミドベクター、例えば大腸菌由来のプラスミド(ColEI、pCR1、pBR322、pMB9及びその派生プラスミドを含む)、広い宿主範囲のプラスミド、例えばRP4、;ファージDNA、例えばλファージの多数の派生ファージ、例えばNM989、及び他のDNAファージ、例えばM13及び繊維状一本鎖のDNAファージが挙げられる。これらのベクター及び他のベクターは、場合によってプロモーターと連結した本発明の核酸を導入するために、標準的な組換えDNA方法論を使って操作することができる。
【0090】
プロモーターは誘導プロモーター(inducible promoter)でよい。適切なプロモーターとして、T7プロモーター、tacプロモーター、trpプロモーター、λプロモーターPL又はPR及び当業者に周知の他のプロモーターが挙げられる。
【0091】
また、広範多様な真核生物の宿主細胞も使用でき、例えば酵母、昆虫及び哺乳動物の細胞が挙げられる。哺乳動物の細胞として、CHO及びマウス細胞、アフリカミドリザルの細胞、例えばCOS-1、及びヒト細胞が挙げられる。
【0092】
タンパク質の発現に適切な、多くの真核生物のベクターが知られている。これらのベクターは、真核生物の細胞ゲノムに染色体性(chromosomally)に組み込まれるように設計してもよいし、又は過剰染色体性(extrachromosomally)に維持されるように設計してもよいし、又は真核細胞に一過性(transiently)にしか維持されないように設計してもよい。核酸を、例えば、CMVプロモーターを含む強力なウイルスプロモーター、及びSV40 T-抗原プロモーター又はレトロウイルスLTRなどの適切なプロモーターと、場合により結合させてもよい。
【0093】
本発明の生成物を得るために、本発明のベクターを運ぶ宿主細胞は、タンパク質の発現に適切な条件下で培養することができ、そのタンパク質を培養培地の細胞から回収することができる。
【0094】
細胞培養
本発明による融合タンパク質をコードするプラスミドは、従来の形質転換技術を使って宿主細胞に導入し、その細胞を融合タンパク質の生成を促進する条件下で培養することができる。誘導プロモーターを使う場合、細胞は誘導物質の不存在下で最初は培養し、次に、タンパク質の回収率を最大にするために、誘導物質を一度添加して高い濃度で生育させることができる。
【0095】
細胞培養条件は、当該技術分野で広く知られており、そのような公知の手順に従って用いればよい。
【0096】
国際公開WO91/11461は、原核生物の宿主細胞をC4bpベースのタンパク質の生成に使用できることを示唆しているが、そのような生成の実験による実証はされていない。
【0097】
最近、原核生物の発現系で生成されたC4bpコアと融合したタンパク質は、その機能的活性を保持していることが分かった。これは国際公開WO2004/020639に開示されており、その内容は参照によって本明細書に組み込まれる。このような方法は、本発明の融合タンパク質の生成に使用できる。
【0098】
培養物からのタンパク質の回収
細胞を生育してタンパク質の生成が可能になると、タンパク質を細胞から回収することができる。本発明者らは、驚くべきことに、タンパク質が可溶性のままであることを見出したので、細胞を通常どおり遠心沈殿して、超音波処理で細胞を溶解でき、例えば、そのタンパク質画分の可溶性を維持し、更に速いスピード(例えば1時間15000rpm)で遠心分離した後の上清にこの画分を残すことができる。
【0099】
上清のタンパク質画分の融合タンパク質は、標準的なタンパク質クロマトグラフィー技術の適切な組み合わせにより、更に精製できる。本発明者らは、イオン交換クロマトグラフィーに続いてゲルろ過クロマトグラフィーを使用した。他のグロマトグラフィー技術、例えばアフィニティークロマトグラフィーも使用できる。
【0100】
ある実施形態において、本発明者らは、溶解物の遠心分離又は他の精製工程のいずれかの後に上清サンプルを加熱することにより、タンパク質の回収を助けるであろうことを見出した。サンプルは約10〜30分間、約70〜80℃に加熱できる。
【0101】
意図するタンパク質の使用に応じて、タンパク質を、追加の精製工程、例えば透析、又は濃縮工程、例えば凍結乾燥してもよい。
【0102】
組成物及びその使用
本発明による生成物を、医薬組成物の形に調製することができる。生成物を、1種類以上の製薬上許容できる担体又は希釈剤と一緒に存在させることができる。組成物を、意図する使用及び生成物の投与ルートに従って調製できる。従って、本発明は、本発明の生成物を多量体の形で、1種類以上の製薬上許容できる担体又は希釈剤を一緒に含む組成物を提供し、また、ヒト又は動物患者の治療又は予防のための免疫療法における、そのような組成物の使用を提供する。
【0103】
製薬上許容できる担体又は希釈剤として、経口、直腸、経鼻、局所的(頬側及び舌下を含む)、膣又は非経口的(皮下、筋肉内、静脈内、皮内、くも膜下腔内及び硬膜外を含む)投与に適切な製剤で使用されるものが挙げられる。これらの製剤を、便利なように投薬単位剤形にすることができ、製薬業界で周知の方法により調製できる。
【0104】
液体状の医薬的に投与可能な組成物は、例えば水、生理的食塩水に溶性のブドウ糖、グリセロール、エタノール等の担体に、任意の医薬品アジュバントと本発明の融合タンパク質を、例えば溶解、分散などにより調製し、溶液又は懸濁液の形にすることができる。所望であれば、投与される組成物をpH緩衝化剤等の物質で補助してもよい。このような投薬剤形を調製する実際の方法は当業者に公知であり又は明白であろう。例えば、Remington’s Pharmaceutical Sciences, Mack Publishing Company, Easton, Pennsylvania, 第19版, 1995を参照されたい。
【0105】
本発明による組成物は、1種類以上のアジュバント、例えば水酸化アルミニウム又はリン酸カルシウムなどの無機塩、又はIL-12若しくはGM-CSFなどのサイトカインを追加で含めてもよい。適切なアジュバントの詳しいリストは、Singh及びO’Hagan, Nature Biotechnology, 17, 1075-1081, 1999の表1であり、この記載は参照によって本明細書に組みこまれる。
【0106】
本発明による生成物は、望ましくは組成物又は製剤の形で、ヒト又は動物患者にこれらの生成物又は組成物を投与することにより、本明細書に開示した治療方法に使用することができる。治療を受ける患者の症状を軽減する有効量は、治療を受ける患者とその状態を考慮して医師により決定されるであろう。0.25〜95%の範囲の有効成分を含み、残りが無毒性担体から作られた剤形又は組成物を調製できる。
【0107】
非経口投与は、一般的に、皮下注射、筋肉注射又は静脈内注射のいずれかの注射により特徴付けられる。注射可能物質は、溶液又は懸濁液として、注射前に液体で溶液又は懸濁液にするのに適した固形として、あるいは乳濁液として、従来の形状に調製できる。適切な賦形剤として、例えば水、生理食塩水、ブドウ糖、グリセロール、エタノール等がある。つい最近考案された非経口投与の方法は、投与量の一定レベルを維持するような、緩徐放出性(slow-release)系又は持効性(sustained-release)系の埋込み(implantation)を用いる。米国特許第3,710,795号を参照されたい。
【0108】
生成物の投与量は、抗原の性質に依存するであろうし、従来のワクチン製剤の抗原の投与の現行手法に従って決定すればよい。
【0109】
受動免疫法
更なる態様において、本発明は、本発明の生成物を宿主患者にワクチン接種することにより得られた抗体を含む免疫血清で患者を受動免疫する方法を提供する。宿主患者はヒト又は非ヒト哺乳動物でよい。従って、更なる態様において、本発明は、このような方法により得られた免疫血清、及び人体又は動物体の治療方法におけるそのような免疫血清の使用を提供する。
【0110】
DNAワクチン
別の態様において、本発明は、人体又は動物体の治療に使用するための、本発明の組換え融合タンパク質生成物をコードする核酸配列を含む真核生物の発現ベクターを提供する。
【0111】
このような治療は、免疫反応を上昇させる目的で抗原をコードする核酸配列を導入することにより、その治療効果を上げるであろう。核酸の運搬は、プラスミドベクター(「裸(naked)」の又は処方される形)又は組換え発現ベクターを使って行うことができる。DNAワクチン接種の概説については、Ada G.及びRamshaw I, in Expert Opinion in Emerging Drugs 8, 27-35, 2003を参照されたい。
【0112】
遺伝子運搬に利用される様々なウイルスベクターとして、アデノウイルス、ヘルペスウイルス、ワクシニアウイルス又はレトロウイルス等のRNAウイルスが挙げられる。レトロウイルスベクターは、マウス又は鳥類のレトロウイルスの誘導株でもよい。一本鎖の外来遺伝子を導入できるレトロウイルスベクターの例として、モロニーマウス白血病ウイルス(MoMuLV)、ハーベイマウス肉腫ウイルス(HaMuSV)、マウス乳癌ウイルス(MuMTV)、及びラウス肉腫ウイルス(RSV)が挙げられるが、これらに限定されない。患者がヒトの場合、ギボンザル(gibbon ape)白血病ウイルス(GaLV)などのベクターを利用することができる。
【0113】
ベクターは、転写の制御配列、特にRNA合成の開始を誘導するのに十分なプロモーター領域を含むであろう。適切な真核生物のプロモーターとして、マウスメタロチオネインI遺伝子のプロモーター(Hamerら, 1982, J. Molec. Appl. Genet. 1: 273);ヘルペスウイルスのTKプロモーター(McKnight, 1982, Cell 31: 355);SV40初期プロモーター(Benoistら, 1981, Nature 290: 304);ラウス肉腫ウイルスプロモーター(Gormanら, 1982, Proc. Natl. Acad. Sci. USA 79: 6777);及びサイトメガロウイルスプロモーター(Foeckingら, 1980, Gene 45: 101)が挙げられる。
【0114】
患者への本発明のこのような態様のベクターの投与、即ちプラスミドベクターとして又はウイルスベクターの一部としての投与は、多くの異なるルートにより影響されうる。プラスミドDNAは、直接的又は間接的に運搬するために、「裸」でもよいし、陽イオン性及び中性脂質(リポソーム)で処方してもよいし、マイクロカプセル化してもよい。また、DNA配列は、直接的又は間接的に運搬するために使用できるウイルス(例えばアデノウイルス、レトロウイルス、ヘルペスウイルス、ポックスウイルス)ベクター内に含めてもよい。運搬ルートは、経口、筋肉内、皮内(Sato, Y.ら, 1996, Science 273: 352-354)、静脈内、動脈内、くも膜下腔内、肝臓内、吸入、膣内点滴注入(Bagarazziら, 1997, J Med. Primatol. 26:27)、直腸内、腫瘍内又は腹腔内が挙げられるが、これらに限定されない。
【0115】
従って、本発明は、治療用ポリペプチドを発現させ、治療効果を有するように、即ち抗原に対する免疫反応を誘導するように、幾つかの細胞をDNAベクターで形質移入可能にするのに有用な医薬組成物として本明細書で開示されるベクターを含む。本発明による医薬組成物は、本発明による構築物を、溶剤、担体、送達系、賦形剤及び添加物又は補助剤を使って、患者に投与するのに適切な形にすることにより、調製される。頻繁に使用される溶剤として滅菌水及び食塩水(緩衝化したもの又はしないもの)が挙げられる。ある担体として、バイオリスティカルに(biolistically)(即ち、気体の圧力下で)運搬される金の粒子が挙げられる。他に頻繁に使用される担体又は運搬システムとして、陽イオン性リポソーム、渦巻型(cochleate)又はマイクロカプセルが挙げられ、これらを溶液として提供してもよいし、運搬カプセル内に封入し又は食べ物に組み込んでもよい。
【0116】
遺伝子運搬ベクターの投与における別の製剤として、リポソームが挙げられる。リポソームのカプセル化により、ポリヌクレオチド及び発現ベクターの投与のための別の製剤が提供される。リポソームは、水溶性のコンパートメントを取り囲む1種類以上の脂質二重層から成る微細小胞である。一般的には、Bakker-Woudenbergら, 1993, Eur. J. Clin. Microbiol. Infect. Dis. 12 (Suppl. 1): S61、及びKim, 1993, Drugs 46: 618を参照されたい。リポソームは組成の点で細胞膜と似ており、その結果、リポソームを安全に投与できるし、生分解性である。調製方法に応じて、リポソームは単層にも多層にもでき、0.02μMから10μM以上の範囲で直径の大きさを変えることができる。例えば、Machyら, 1987, LIPOSOMES IN CELL BIOLOGY AND PHARMACOLOGY (John Libbey)、及び Ostroら, 1989, American J. Hosp. Phann. 46: 1576を参照されたい。
【0117】
発現ベクターは、標準的な技術を使ってリポソーム内にカプセル化できる。異なるリポソームの組成の種類と合成方法は当業者に公知である。例えば、米国特許US-A-4,844,904、US-A-5,000,959、US-A-4,863,740、US-A-5,589,466、US-A-5,580,859及びUS-A-4,975,282を参照されたい。これらはすべて参照によって本明細書に組み込まれる。
【0118】
一般的に、投与されるリポソームカプセル化ベクターの投与量は、患者の年齢、体重、身長、性別、一般的な病状及びこれまでの病歴などの因子に応じて変えられるであろう。特定の剤形における投与量の範囲は適切な動物モデルを使って決定することができる。
【0119】
本発明を以下の実施例により説明する。
【実施例1】
【0120】
熱帯熱マラリア原虫MSP1.19-ウサギC4bp融合タンパク質
本実施例は、(熱帯熱マラリア原虫MSP1のアミノ酸1567-1661を含む)単量体抗原とウサギコアC4bpタンパク質の融合を説明する。AVD174と名づけた融合タンパク質を、細菌株C41(DE3)で発現し、精製した。融合タンパク質のみを、アジュバンドを添加せずにウサギを免疫するのに使用した。
【0121】
クローニング
MSP1タンパク質の残基1567-1661をコードする合成294bp DNAフラグメントを、NdeI及びBamHIで消化し、あらかじめNdeI及びBamHIで消化したpAVD181に連結した。これにより、T7後期プロモーターの下流のウサギC4bpα鎖のC末端57残基と融合した95アミノ酸のMSP1.19タンパク質フラグメントをコードするオープンリーディングフレームを作製した。pAVD174と名づけた構築物をDNA配列決定で調べた。
【0122】
AVD174融合タンパク質をコードする核酸配列は、以下の通りである。
【0123】
atgttaaacatttcccagcaccagtgcgttaagaaacagtgcccgcagaa
ctctggttgtttccgtcatctggacgagcgtgaagagtgcaaatgtctgc
tgaactacaaacaggaaggtgataaatgtgttgagaacccaaacccgacc
tgtaacgaaaacaacggcggttgtgacgctgatgctaaatgcaccgagga
agacagcggttctaacggtaagaaaatcacctgcgagtgtactaaaccgg
actcctacccgctgttcgacggtatcttttgctccGGATCCGAGGTCCCG
GAAGGCTGTGAGCAGGTGCAAGCGGGTCGCCGTCTCATGCAGTGTCTCGC
AGACCCATACGAAGTGAAAATGGCCCTGGAGGTCTACAAGCTGTCTCTGG
AGATTGAACTCCTGGAACTGCAGCGCGATAAGGCACGTAAAAGCTCTGTG
CTGCGCCAGCTGTAA (配列番号21)
当該構築物によりコードされる融合タンパク質AVD174のアミノ酸配列は、以下の通りである。
【0124】
MLNISQHQCV KKQCPQNSGC FRHLDEREEC KCLLNYKQEG DKCVENPNPT CNENNGGCDA DAKCTEEDSG SNGKKITCEC TKPDSYPLFD GIFCSGSEVP EGCEQVQAGR RLMQCLADPY EVKMALEVYK LSLEIELLEL QRDKARKSSV LRQL (配列番号22)
配列番号22の残基1-95は、熱帯熱マラリア原虫MSP1(単量体抗原)の残基1567-1661に対応し、配列番号22の残基98-154は、ウサギC4bpコアタンパク質の57残基に対応する。GSリンカー配列は、2つの成分の間に現れる。
【0125】
このタンパク質は、推定分子量17,319ダルトン、理論pIは5.05である。
【0126】
発現
熱帯熱マラリア原虫-ウサギC4bpコアタンパク質をコードするプラスミドpAVD174を、大腸菌株C41(DE3)で発現した。形質転換した細胞を、37℃でOD600が約0.6になるまでLB培地で生育させ、次に最終濃度0.5mMになるようにIPTGで誘導し、培養物を37℃で更に3時間生育させ、この時点で細胞を遠心分離によって回収した。
【0127】
AVD174融合タンパク質の精製
タンパク質AVD174を1LのC41(DE3)細胞から精製した。細胞を20mM MES pH6.5及び5mM EDTAを含むバッファーで超音波処理で溶解した後、可溶性画分にすべての融合タンパク質が検出された。遠心分離後の上清をHitrapSカラムに載せた。
【0128】
陽イオン性カラム(HiTrap S)
カラムを20mM MES pH 6.5、20mM EDTAバッファー(バッファーA)で平衡化した。タンパク質を、バッファーAからバッファーB(バッファーAプラス0.5M NaCl)へ、10カラム量のグラジエントで溶出した。AVD174を濃度約200mM NaClで溶出した。
【0129】
AVD174を含むHiTrapS画分を、Millipore concentrator (カットオフ30 K)を使って濃縮し、次にゲルろ過カラムに載せた。
【0130】
ゲルろ過カラム(Superdex 200 26/60 prep grade)
Superdex 200 26/60カラムを20mM Trisバッファー pH8, 150mM NaClで平衡化し、HiTrapS画分から得られた濃縮したAVD174タンパク質を載せた。
【0131】
タンパク質は2つのピークに溶出された。正確に折りたたまれ集合したタンパク質は、156mlのところで溶出されたのに対し、120mlのところで溶出しているもっと早い小さいピークのものは、正確に集合しても折りたたまれてもいない。
【0132】
生物物理学的性質決定
ジスルフィド結合(マウスコアタンパク質を除くすべてに存在する。図1参照)を含むC4bp融合タンパク質のオリゴマー状態は、還元剤β-メルカプトエタノール(BME)の存在下及び不存在下で、SDS-PAGEゲル上のタンパク質の挙動を比較することによって簡単に調べることができる。AVD177タンパク質は、BMEの不存在下で見かけの大きさが約140kDaであり、一方BME存在下では還元されて、見かけの大きさがちょうど20kDaの辺りで流れる。
【0133】
質量分析
AVD174タンパク質を、BMEによる還元及びN-エチルマレイミド(NEM)によるアルキル化の後、エレクトロスプレー質量分析法により試験を行った。結果は、分子量が19,072Daであると測定されたタンパク質に14個のNEM分子(各125Da)が付加したことを示した。
【0134】
エンドトキシンレベル
精製されたタンパク質のエンドトキシンのレベルは、BiowhittakerのLAL(カブトガニ血球抽出成分(limulus amoebocyte lysate))テストキットを使い、タンパク質1mg当たり21EUであることが測定された。
【実施例2】
【0135】
熱帯熱マラリア原虫MSP1.19-ヒトC4bp融合タンパク質
本実施例は、単量体抗原(熱帯熱マラリア原虫MSP1のアミノ酸1567-1661を含む)とヒトC4bpコアタンパク質の融合を示す。融合タンパク質を、細菌株C41(DE3)で発現させ精製した。アジュバンドを添加することなくヒトを免疫するのに、この融合タンパク質のみを使用することができる。
【0136】
クローニング
MSP1タンパク質の残基1567-1661をコードする合成294bpDNAフラグメントをNdeI及びBamHIで消化して、あらかじめNdeI及びBamHIで消化したpHVD181に連結した。これにより、T7後期プロモーターの下流のヒトC4bpα鎖のC末端57残基に融合した95アミノ酸MSP1.19タンパク質フラグメントをコードするオープンリーディングフレームを作製した。この構築物をpAVD177とし、DNA配列決定により調べた。
【0137】
AVD177融合タンパク質をコードする核酸配列は以下の通りである。
【0138】
atgttaaacatttcccagcaccagtgcgttaagaaacagtgcccgcagaa
ctctggttgtttccgtcatctggacgagcgtgaagagtgcaaatgtctgc
tgaactacaaacaggaaggtgataaatgtgttgagaacccaaacccgacc
tgtaacgaaaacaacggcggttgtgacgctgatgctaaatgcaccgagga
agacagcggttctaacggtaagaaaatcacctgcgagtgtactaaaccgg
actcctacccgctgttcgacggtatcttttgctccGGATCCgagaccccc
gaaggctgtgaacaagtgctcacaggcaaaagactcatgcagtgtctccc
aaacccagaggatgtgaaaatggccctggaggtatataagctgtctctgg
aaattgaacaactggaactacagagagacagcgcaagacaatccactttg
gataaagaactataa (配列番号23)
この構築物によりコードされる融合タンパク質AVD177のアミノ酸配列は以下の通りである。
【0139】
MLNISQHQCV KKQCPQNSGC FRHLDEREEC KCLLNYKQEG DKCVENPNPT CNENNGGCDA DAKCTEEDSG SNGKKITCEC TKPDSYPLFD GIFCSGSETP EGCEQVLTGK RLMQCLPNPE DVKMALEVYK LSLEIEQLEL QRDSARQSTL DKEL (配列番号24)
配列番号24の残基1-95は、熱帯熱マラリア原虫MSP1(単量体抗原)の残基1567-1661に対応し、配列番号24の残基98-154は、ヒトC4bpコアタンパク質の57残基に対応する。GSリンカー配列は、これらの2つの成分の間に現れる。
【0140】
タンパク質は、推定分子量17,261ダルトン、理論pIは4.72である。
【0141】
発現
タンパク質の発現を図2に示す。3つの異なる細菌株、即ちC41(DE3)、BL21(DE3)及びC43(DE3)で、非誘導(U)、37℃又は30℃で誘導後のAVD177タンパク質の発現を示す、SDS-PAGEゲルを提示する。ゲルのレーンは以下の通りである。
【0142】
レーン1:分子量マーカー(降順: 66, 60, 46, 36, 28, 20, 14, 12, 6 kDa)
レーン2:C41(DE3) 誘導前
レーン3:C41(DE3) 37℃で誘導後3時間;
レーン4:C41(DE3) 30℃で誘導後3時間;
レーン5:BL21(DE3) 誘導前
レーン6:BL21(DE3) 37℃で誘導後3時間;
レーン7:BL21(DE3) 30℃で誘導後3時間;
レーン8:C43(DE3) 誘導前
レーン9:C43(DE3) 37℃で誘導後3時間;
レーン10:C43(DE3) 30℃で誘導後3時間。
【0143】
図に示すように、C41(DE3)と細菌株C43(DE3)で良好な発現が得られた。その一方、テストされた条件下では、細菌株BL21(DE3)で発現は見られなかった。図2を参照のこと。培養物を、光学密度(OD600)が約0.6になるまで30℃と37℃のLB培地で生育させ、次に、最終濃度0.5mMになるようにIPTGの添加により発現を誘導した。
【0144】
AVD177融合タンパク質の精製
タンパク質AVD177を、37℃で誘導後、3時間生育した1LのC41(DE3)細胞から精製した。細菌ペレットを溶解した後、可溶性画分にすべての融合タンパク質を検出した。細胞を、20mM MES pH6.5及び5mM EDTAを含むバッファーで超音波処理により溶解した。遠心分離後上清をMonoSカラムに載せた。
【0145】
陽イオンカラム(Mono S HR 10/10)
カラムを20 mM MES pH 6.5, 20mM EDTAバッファー(バッファーA)で平衡化した。タンパク質をバッファーAから、バッファーAに0.5M NaClを加えたバッファーBへ、10カラム量のグラジエントで溶出した。AVD177を濃度約200mM NaClで溶出した。
【0146】
AVD177を含むMonoS画分をMillipore concentrator(カットオフ30 K)を使って濃縮し、次にゲルろ過カラムに載せた。
【0147】
ゲルろ過カラム(Superdex 200 26/60 prep grade)
Superdex 200 26/60カラムを20mM TrisバッファーpH8、150mM NaClで平衡化し、MonoS画分から得られた濃縮したAVD177タンパク質を載せた。
【0148】
タンパク質が2つのピークに溶出された。正確に折りたたまれ集合したタンパク質は150mlのところで溶出され、一方、115mlのところで溶出されたもっと早い小さいピークは、正確に集合しても折りたたまれてもいない。
【0149】
生物物理学的性質決定
ジスルフィド結合(マウスコアタンパク質以外のすべてに存在する:図1参照)を含むC4bp融合タンパク質のオリゴマー状態を、還元剤β-メルカプトエタノール(BME)の存在下及び不存在下においてSDS-PAGEゲルのタンパク質の挙動を比較することにより簡単に検査できる。結果を図3に示す。図3において、SDS-PAGEゲルは、分子量マーカー(降順:66, 60, 46, 36, 28, 20, 14, 12, 6 kDa)で分けられた、還元条件下(左+BME)及び非還元条件下(右-BME)でのAVD177タンパク質の挙動を示す。図3から分かるように、AVD177タンパク質は見かけの大きさがBME不存在下で約140kDaであり、一方BEM存在下では還元されて、見かけの大きさがちょうど20kDa辺りで流れる。
【0150】
質量分析
AVD177タンパク質を、BMEで還元し、N-エチルマレイミド(NEM)でアルキル化した後、エレクトロスプレー質量分析法でテストした。結果は、分子量が19,015Daと測定されたタンパク質に14個のNEM分子(各125Da)が付加したことを示した。
【0151】
エンドトキシンレベル
精製タンパク質のエンドトキシンレベルをBiowhittakerのLAL(カブトガニ血球抽出成分)テストキットを使って測定し、タンパク質1mg当たり38EUであった。
【実施例3】
【0152】
突然変異熱帯熱マラリア原虫MSP1.19-ウサギC4bp融合タンパク質
一例として、第二の熱帯熱マラリア原虫MSP1.19-ウサギC4bpタンパク質を本実施例で説明する。該タンパク質は、主に、単量体抗原遺伝子においてpAVD174及びpAVD177に対して別々のコドンの利用を有する点で異なり、また、3つのアミノ酸変化を含む(Uthaipibullら, J Mol Biol. 307, 1381-1394, 2001に記載されている)。この融合タンパク質をAVD178とした。
【0153】
AVD178融合タンパク質をコードする核酸配列は、以下の通りである。
【0154】
atgctgaatatttcccagcaccagtgcgtaaagaaacagtgtcctcagaa
ctctggttgcttccgccatctggacgaacgcgaatattgcaaatgccgtc
tgaactacaaacaggaaggtgacaagtgcgttctgaacccgaacccaact
tgtaacgagaacaacggtggctgcgatgctgatgctaaatgcactgaaga
agacagcggttctaacggcaaaaaaatcacctgcgagtgcaccaaaccgg
acagctatccgctgttcgacggcattttttgttctggatccGAGGTCCCG
GAAGGCTGTGAGCAGGTGCAAGCGGGTCGCCGTCTCATGCAGTGTCTCGC
AGACCCATACGAAGTGAAAATGGCCCTGGAGGTCTACAAGCTGTCTCTGG
AGATTGAACTCCTGGAACTGCAGCGCGATAAGGCACGTAAAAGCTCTGTG
CTGCGCCAGCTGTAA (配列番号25)
この構築物によりコードされる融合タンパク質AVD178のアミノ酸配列は、以下の通りである。
【0155】
MLNISQHQCVKKQCPQNSGCFRHLDEREYCKCRLNYKQEGDKCVLNPNPTCNENNGGCDADAKCTEEDSGSNGKKITCECTKPDSYPLFDGIFCSGSEVPEGCEQVQAGRRLMQCLADPYEVKMALEVYKLSLEIELLELQRDKARKSSVLRQL (配列番号26)
3つの突然変異アミノ酸は太字及び下線部である。
【0156】
配列番号25の残基1-95は、3つの突然変異を有する熱帯熱マラリア原虫MSP1(単量体抗原)の残基1567-1661に対応し、配列番号24の残基98-154は、ウサギC4bpコアタンパク質の57残基に対応する。GSリンカー配列はこれら2つの成分の間に現れる。
【0157】
AVD178の発現、精製及び性質決定
基本的に、AVD174及びAVD177タンパク質について記載したように行った。HiTrapSカラムからの溶出は約200mMであった。ゲルろ過では、オリゴマータンパク質は159mlの量のところで溶出された。質量分析では、単量体あたり14 NEM残基で、分子量は19,133Daであった。エンドトキシンレベルは、タンパク質1mg当たり58EUと測定された。
【実施例4】
【0158】
マウスマラリア原虫(Plasmodium yoelii)MSP1.19-マウスC4bp融合タンパク質
クローニング
AVD108タンパク質を以下のようにして調製した:マウスマラリア原虫(Plasmodium yoelii)由来のMSP1.19と、MSP1.33の一部をコードする合成DNAフラグメントを、マウスC4bpα鎖のC末端54アミノ酸から上流のNdeI-BamHIフラグメントとしてクローニングした。
【0159】
融合タンパク質AVD108をコードする核酸配列は、以下の通りである。
【0160】
ATGAGATCTCACATTGCCTCTATTGCTTTGAACAACTTGAACAAGTCTGG
TTTGGTAGGAGAAGGTGAGTCTAAGAAGATTTTGGCTAAGATGCTGAACA
TGGACGGTATGGACTTGTTGGGTGTTGACCCTAAGCATGTTTGTGTTGAC
ACTAGAGACATTCCTAAGAACGCTGGATGTTTCAGAGACGACAACGGTAC
TGAAGAGTGGAGATGTTTGTTGGGTTACAAGAAGGGTGAGGGTAACACCT
GCGTTGAGAACAACAACCCTACTTGCGACATCAACAACGGTGGATGTGAC
CCAACCGCCTCTTGTCAAAACGCTGAATCTACCGAAAACTCCAAGAAGAT
TATTTGCACCTGTAAGGAACCAACCCCTAACGCCTACTACGAGGGTGTTT
TCTGTTCTTCTTCCGGATCCGAGGCCTCTGAAGACCTTAAGCCTGCGCTT
ACAGGCAACAAGACCATGCAGTATGTGCCAAATTCACACGATGTGAAAAT
GGCTCTGGAGATCTACAAGCTGACTCTGGAGGTTGAACTACTACAGCTCC
AGATACAAAAGGAGAAACACACTGAAGCACACTAA (配列番号27)
タンパク質AVD108のアミノ酸配列は、以下の通りである。
【0161】
MRSHIASIAL NNLNKSGLVG EGESKKILAK MLNMDGMDLL GVDPKHVCVD TRDIPKNAGC FRDDNGTEEW RCLLGYKKGE GNTCVENNNP TCDINNGGCD PTASCQNAES TENSKKIICT CKEPTPNAYY EGVFCSSSGS EASEDLKPAL TGNKTMQYVP NSHDVKMALE IYKLTLEVEL LQLQIQKEKH TEAH (配列番号28)
配列番号28の残基3-138はマウスマラリア原虫MSP1(単量体抗原)の残基1619-1753に対応し、配列番号28の残基141-194は、ウサギC4bpコアタンパク質の54残基に対応する。GSリンカー配列はこれらの2つの成分の間に現れ、短い制限酵素認識部位をコードする配列は第一成分に先行する。
【0162】
AVD108の発現
タンパク質AVD108を大腸菌株C41(DE3)で発現させた。3Lの培養物を、LB培地で37℃、光学密度(OD600)約0.6まで生育させ、次に最終濃度0.7mMになるようにIPTGを添加することにより発現を誘導した。誘導4時間後、細胞を遠心分離により回収した。細胞をバッファーA(50mM Tris pH9, 5mM EDTA)で溶解し、残骸を遠心分離で除去した。
【0163】
AVD108の精製及び性質決定
タンパク質AVD108を、4つのカラムクロマトグラフィー工程を使って精製した。第一の陰イオン交換クロマトグラフィー工程では、DEAE HR16/10カラムを使用した。タンパク質をバッファーAで載せて、バッファーAに1M NaClを加えたバッファーBのグラジエントで溶出した。AVD108は180-300mM NaCl間の幅の広いピークで溶出された。
【0164】
第2の疎水性相互作用クロマトグラフィーの工程では、DEAEカラムから得られたAVD108を含むプールした画分をMacro-Prep Phenyl Sepharoseカラムに載せ、塩が1M〜0M NaClに減少するグラジエントで溶出した。最後の2つのクロマトグラフィーの工程では、AVD108タンパク質をSuperdex S200 26/60カラムのゲルろ過により精製した。最初に、タンパク質を最終濃度8Mになるように尿素を添加して変性させ、4℃で一晩、インキュベーションした。モノマーはこのカラムから203mlの量のところで溶出された。バッファーH(20mM Tris pH7.5, 150mM NaCl)で、4℃で一晩、透析により再生した後、同じカラムに載せる前に、今度は、バッファーHで平衡化した。今度は、オリゴマーが164mlの量のところで溶出された。質量分析により、質量は21,257Daと測定された。BiowhittakerのLALテストキットによるエンドトキシンレベルは、タンパク質1mg当たり4EUであった。
【実施例5】
【0165】
熱帯熱マラリア原虫MSP1.19-ウサギC4bp融合タンパク質を使用する免疫化
免疫化
実施例1で述べたように調製したAVD174タンパク質を、3羽のニュージーランドホワイト(NZW)ウサギに免疫するのに使用した。免疫化のスケジュールは以下の通りであった:各ウサギは2週間のインターバルで4回の注射(換言すれば、0、14、28及び42日目)を受けた。各注射は、皮下にし、公知のアジュバントを添加せず、緩衝化した等張生理食塩水に345μg(又は20nmol)のタンパク質を含有した。
【0166】
並行して、3羽のNZWウサギをフロイントアジュバントと212μg(又は20nmol)のAVD172タンパク質で免疫した。AVD172はAVD174と同一であるが、ウサギC4bpからC末端57アミノ酸を欠いている。それは以下のアミノ酸配列である。
【0167】
MLNISQHQCV KKQCPQNSGC FRHLDEREEC KCLLNYKQEG DKCVENPNPT CNENNGGCDA DAKCTEEDSG SNGKKITCEC TKPDSYPLFD GIFCS (配列番号29)
免疫化スケジュールは以下の通りであった:各ウサギは2週間のインターバルで4回の注射(換言すれば、0、14、28及び42日目)を受けた。各ウサギにとって最初の注射は、完全フロイントアジュバントで、皮内に投与され、一方、その後の3回の注射は、不完全フロイントアジュバントで与えられ、皮下に投与された。
【0168】
抗体価
熱帯熱マラリア原虫メロゾイトの表面上のMSP1に対する、最後の注射後1週間(即ち63日目)の抗体価を、熱帯熱マラリア原虫に感染した赤血球のアセトン固定化スメアの間接免疫蛍光(P. yoeliiについて、Lingら、Vaccine 15, 1562-1567, 1997に記載されている)で測定した。
【0169】
AVD174で免疫化したウサギの2つの最も高い力価は、1/81,920であった。フロイントアジュバントのAVD172タンパク質で免疫したウサギでは、2つの最も高い力価は1/20,480であった。
【0170】
これは、単量体抗原とC4bpコアの融合により、フロイントアジュバントの同じ単量体抗原を投与するよりも、高い抗体価を誘導することができることを立証した。
【0171】
生成された抗体の性質決定
高力価の抗体は、感染を予防又は治療するのに十分ではないだろうし、抗原による免疫化で生成された抗体の特異性は、決定的に重要であることが知られている。Guevara Patinoら(J.Exp.Med. 186, 1689-1699, 1997)は、MSP1.19に対するブロッキング抗体又は抑制抗体をアッセイする方法を詳細に述べている。これらの方法は、抑制抗体が存在するか(MSP1.42をMSP1.33及びMSP1.19にプロセシングするのをブロックし、赤血球への侵入を防ぐので有用である)を見るのに使用された。
【0172】
抑制抗体は、AVD174タンパク質によって誘導された抗体中に見出されただけである。AVD172タンパク質で免疫したウサギの抗血清中には何も見つからなかった。一方、マラリアはヒトに自然感染でき、有害であるが、フロイントアジュバントのAVD172は、ブロッキング抗体を誘導することができる(Guevara Patinoら, op. cit.参照)。
【実施例6】
【0173】
マウスマラリア原虫MSP1.19-マウスC4bp融合タンパク質を使用する免疫化
マウスの免疫化
実施例5で述べたように調製したAVD108タンパク質を、6匹のBALB/cマウスに免疫するのに使用した。アジュバントは使用せず、前記タンパク質を緩衝化された等張生理食塩水に存在させた。40μg(1.9nmol)のタンパク質を1回の注射に使用した。各マウスに皮下で4週間インターバルで3回(換言すれば、0、28及び56日目に)注射した。
【0174】
並行して、6匹のBALB/cマウスを、23μg(1.9nmol)のAVD183タンパク質で免疫した。AVD183タンパク質はAVD108と同じであるが、マウスC4bpのC末端の54アミノ酸が欠けている(即ちP.yoeliiのMSP1.19タンパク質単独である)。23μgの該タンパク質(AVD108に使用したのと同じ、緩衝化した等張生理食塩水に存在する)を、1回の注射で使用した。各マウスに、皮下に3回、0、28及び56日目に注射した。
【0175】
抗体価
マウスマラリア原虫のメロゾイトの表面にあるMSP1に対する、最後の注射後2週間(即ち70日目)の抗体価を、マウスマラリア原虫に感染した赤血球のアセトン固定化スメアの間接免疫蛍光(Lingら Vaccine 15, 1562-1567, 1997に記載されている)によって測定した。
【0176】
結果は、AVD108タンパク質で免疫した6匹のマウスのうち5匹が、1/40,960の抗体価を有し、6匹目のマウスは1/10,240の力価であった。一方、MSP1に対する抗体は、1/80の希釈で、AVD183タンパク質で免疫したマウスのどれにも検出されなかった。このことは、単量体のMSP1.19抗原とC4bpコアの融合により、得られた抗体の力価が500倍まで上昇できたことを立証した。
【0177】
寄生虫の暴露
前述のように免疫した6匹のマウスのグループ両方を、致死量5000のマウスマラリア原虫感染赤血球に暴露した。このアッセイも、Lingら(op. cit.)により説明されている。AVD183タンパク質で免疫した6匹のマウスは、寄生虫の暴露7日間以内に全部死んだ。他方で、AVD108で免疫した6匹のマウスのうち5匹は生き残り、それらの血液中に(顕微鏡で検査される薄層の血液スメアのギムザ染色で評価したところ)寄生虫はいなかった。70日目の力価が1/10,240であった6匹目のマウスは、暴露後19日で死んだ。このマウスの赤血球の70%以上に、19日目のギムザ染色で感染が見られた。
【0178】
一方、この暴露実験は、公知のアジュバンドの不存在下でC4bpコアタンパク質のみと融合した単量体抗原で免疫化すると、他の場合には致命的なマラリア原虫の感染から保護することができることを立証した。これは、現在のところ、公知のアジュバンドを伴わない単一のタンパク質だけを使ってマラリア原虫の感染に対する成功したワクチン接種の最初の実例を表すと信じられる。
【実施例7】
【0179】
インフルエンザ血球凝集素-C4bp融合タンパク質
本実施例は、単量体抗原(インフルエンザAウイルスのHA1血球凝集素タンパク質の残基91-261を含む)と、ヒト、ウサギ及びマウスのコアC4bpタンパク質の融合を説明する。通常、全長HA1は、ビリオンの細胞表面で三量体に集合し、そのため、このペプチドフラグメントのみを使うことにより、三量体を単量体抗原に有効に変化させる。これらの融合タンパク質をAVD272〜AVD274とし、細菌株C41(DE3)で発現し、精製する。これらの融合タンパク質のみを、アジュバントを添加することなくマウスとウサギを免疫するのに使用する。
【0180】
AVD272では、HA1フラグメント(Jeon及びArnon, Viral Immunology, 15, 165-176, 2002に記載されている)をマウスC4bp足場と融合する。
【0181】
AVD272のアミノ酸配列は以下の通りである。
【0182】
kafsncypyd vpdyaslrsl vassgtlefi tegftwtgvt qnggsnackr gpgsgffsrl nwltksgsty pvlnvtmpnn dnfdklyiwg ihhpstnqeq tslyvqasgr vtvstrrsqq tiipnigsrp wvrglssris iywtivkpgd vlvinsngnl iaprgyfkmr GSEASEDLKP ALTGNKTMQY VPNSHDVKMA LEIYKLTLEV ELLQLQIQKE KHTEAH (配列番号30)
AVD273では、HA1フラグメントをウサギC4bp足場と融合する。
【0183】
AVD273のアミノ酸配列は以下の通りである。
【0184】
kafsncypyd vpdyaslrsl vassgtlefi tegftwtgvt qnggsnackr gpgsgffsrl nwltksgsty pvlnvtmpnn dnfdklyiwg ihhpstnqeq tslyvqasgr vtvstrrsqq tiipnigsrp wvrglssris iywtivkpgd vlvinsngnl iaprgyfkmr GSEVPEGCEQ VQAGRRLMQC LADPYEVKMA LEVYKLSLEI ELLELQRDKA RKSSVLRQL (配列番号31)
AVD274では、HA1フラグメントをヒトC4bp足場と融合した。
【0185】
AVD274のアミノ酸配列は以下の通りである。
【0186】
kafsncypyd vpdyaslrsl vassgtlefi tegftwtgvt qnggsnackr gpgsgffsrl nwltksgsty pvlnvtmpnn dnfdklyiwg ihhpstnqeq tslyvqasgr vtvstrrsqq tiipnigsrp wvrglssris iywtivkpgd vlvinsngnl iaprgyfkmr GSETPEGCEQ VLTGKRLMQC LPNPEDVKMA LEVYKLSLEI EQLELQRDSA RQSTLDKEL (配列番号32)
マウスを、皮下で3回、2週間のインターバルで(即ち0、14及び28日目に)アジュバントを添加しないAVD272タンパク質(2nmol)で免疫し、かなりの抗体価(原則的に、Jeonら, op.cit.により説明されているように測定)を示し、また、マウスに適応したA/PR/8/34株の5 LD50の投与量での致死的な暴露に対して十分に保護することを示す。
【実施例8】
【0187】
インフルエンザM2ペプチド-C4bp融合タンパク質
本実施例は、単量体抗原(インフルエンザA型ウイルスのM2タンパク質の残基2-24、即ち細胞外部分を含む)とヒト、ウサギ及びマウスのコアC4bpタンパク質の融合を説明する。通常、全長M2はビリオン及び感染細胞で四量体に集合し、そのためこのペプチドフラグメントのみを使うことにより、四量体を単量体抗原に有効に変化させる。融合タンパク質をAVD275〜AVD278とし、細菌株C41(DE3)で発現し、精製した。これらの融合タンパク質のみを、アジュバンドを添加せずにマウスとウサギに免疫するのに使用した。
【0188】
AVD275では、細胞外M2ペプチド(Neirynckら、Nature Medicine 5, 1157- 1163, 1999に記載されている)をマウスC4bp足場と融合した。
【0189】
AVD275のアミノ酸配列は以下の通りである。
【0190】
SLLTEVETPI RNEWGCRCND SSDGSEASED LKPALTGNKT MQYVPNSHDV KMALEIYKLT LEVELLQLQI QKEKHTEAH (配列番号33)
AVD276では、細胞外M2ペプチドをウサギC4bp足場と融合した。
【0191】
AVD276のアミノ酸配列は以下の通りである。
【0192】
SLLTEVETPI RNEWGCRCND SSDGSEVPEG CEQVQAGRRL MQCLADPYEV KMALEVYKLS LEIELLELQR DKARKSSVLR QL (配列番号34)
AVD277では、細胞外M2ペプチドをヒトC4bp足場と融合した。
【0193】
AVD277のアミノ酸配列は以下の通りである。
【0194】
SLLTEVETPI RNEWGCRCND SSDGSETPEG CEQVLTGKRL MQCLPNPEDV KMALEVYKLS LEIEQLELQR DSARQSTLDK EL (配列番号35)
AVD278では、両方のシステインがセリン残基で置換された細胞外M2ペプチドの変異体を、ヒトC4bp足場と融合した。
【0195】
AVD278のアミノ酸配列は以下の通りである。
【0196】
SLLTEVETPI RNEWGSRSND SSDGSETPEG CEQVLTGKRL MQCLPNPEDV KMALEVYKLS LEIEQLELQR DSARQSTLDK EL (配列番号36)
マウスを2週間のインターバルで3回(即ち0、14及び28日目)、アジュバントを添加せずにAVD275タンパク質(2nmol)を皮下で免疫したことは、かなりの抗体価(原則として、Neirynckら, op.cit.により説明されているように測定)を示し、また、マウスに適応したA/PR/8/34株を5 LD50の投与量での致死的な暴露に対して十分に保護することを示す。
【図面の簡単な説明】
【0197】
【図1】C4bpコアタンパク質のアライメントを示す。
【図2】大腸菌のタンパク質の発現の結果を示す。
【図3】還元及び非還元条件下でゲルに流した、本発明のタンパク質の比較を示す。
【特許請求の範囲】
【請求項1】
C4bpコアタンパク質と単量体抗原を含む生成物。
【請求項2】
前記C4bpコアが、配列番号1の残基1-57若しくはそのホモログの対応する残基、又は配列番号1の少なくとも47のアミノ酸のフラグメント若しくはそのホモログから成る、請求項1に記載の生成物。
【請求項3】
前記ホモログが、配列番号2〜20のいずれか1つである、請求項2に記載の生成物。
【請求項4】
前記ホモログが、配列番号1〜20のいずれか1つに対して少なくとも70%のアミノ酸同一性を有する変異体である、請求項2又は3に記載の生成物。
【請求項5】
前記単量体抗原が、前記C4bpコアタンパク質のN-又はC-末端と融合している、請求項1〜4のいずれか1項に記載の生成物。
【請求項6】
前記融合が、可撓性のリンカーを介する、請求項5に記載の生成物。
【請求項7】
前記単量体抗原が、マラリア原虫メロゾイト表面タンパク質1の単量体抗原性フラグメントである、請求項1〜6のいずれか1項に記載の生成物。
【請求項8】
前記単量体抗原が、インフルエンザウイルス赤血球凝集素タンパク質又はインフルエンザM2eペプチドの単量体抗原性フラグメントである、請求項1〜6のいずれか1項に記載の生成物。
【請求項9】
製薬上許容できる希釈剤、担体又はアジュバントと一緒に請求項1〜8のいずれか1項に記載の生成物を含む、組成物。
【請求項10】
人体又は動物体を治療する方法に使用するための、請求項1〜8のいずれか1項に記載の生成物又は請求項9に記載の組成物。
【請求項11】
請求項7に記載の生成物の有効量を個体に投与することを含む、マラリアの免疫療法。
【請求項12】
前記個体がマラリア原虫に感染している、請求項11に記載の方法。
【請求項13】
予防ワクチン接種における請求項11に記載の方法。
【請求項14】
マラリアの治療又は予防のための、請求項7に記載の生成物。
【請求項15】
マラリアの治療用又は予防用医薬の製造のための、請求項7に記載の生成物の使用。
【請求項16】
請求項7に記載の生成物を非ヒト哺乳動物に導入する工程、及び該哺乳動物から免疫血清を回収する工程を含む、マラリア原虫(Plasmodium parasite)に対する抗体を製造する方法。
【請求項17】
請求項1〜5のいずれか1項に記載の生成物を宿主対象にワクチン接種して得られた抗体を含む免疫血清を、患者に投与することを含む、患者の病気に対する受動免疫方法。
【請求項18】
請求項16に記載の方法に従って製造された免疫血清の有効量を、ヒトに投与することを含む、ヒト患者のマラリアの受動免疫療法。
【請求項19】
ヒト患者のマラリアの免疫療法に使用するための、請求項16に記載の方法により得られた免疫血清。
【請求項20】
請求項8に記載の生成物の有効量を個体に投与することを含む、インフルエンザの免疫療法。
【請求項21】
前記個体がインフルエンザウイルスに感染している、請求項20に記載の方法。
【請求項22】
予防ワクチン接種における請求項20に記載の方法。
【請求項23】
インフルエンザの治療又は予防のための、請求項8に記載の生成物。
【請求項24】
インフルエンザの治療用又は予防用医薬の製造のための、請求項8に記載の生成物の使用。
【請求項25】
請求項8に記載の生成物を非ヒト哺乳動物に導入する工程、及び該哺乳動物から免疫血清を回収する工程を含む、インフルエンザに対する抗体を製造する方法。
【請求項26】
請求項25に記載の方法に従って製造された免疫血清の有効量を、ヒトに投与することを含む、ヒト患者のインフルエンザの受動免疫療法。
【請求項27】
ヒト患者のインフルエンザの免疫療法に使用するための、請求項25に記載の方法により得られた免疫血清。
【請求項28】
C4bpコアタンパク質と、
非ポリペプチド単量体抗原、
を含む生成物の製造方法であって、
前記一方の成分をコードする核酸を発現する工程、前記他方の成分と前記融合タンパク質を結合させる工程、及び前記生成物を回収する工程を含む、前記方法。
【請求項29】
C4bpコアタンパク質と、
ポリペプチドの単量体抗原、
の融合物を含む生成物を製造する方法であって、
該融合物をコードする核酸を発現させる工程及び前記生成物を回収する工程を含む、前記方法。
【請求項30】
前記核酸を原核生物の宿主細胞で発現させる、請求項28又は29に記載の方法。
【請求項31】
前記融合タンパク質を多量体の形で回収する、請求項30に記載の方法。
【請求項32】
抗原とC4bpコアタンパク質を結合させる工程を含む、単量体抗原の免疫原性を増強させる方法。
【請求項33】
宿主細胞内で機能的なプロモーターと有効に結合している、
C4bpコアタンパク質と、
ポリペプチドの単量体抗原、
の融合タンパク質をコードする核酸配列を含む発現ベクター。
【請求項34】
前記C4bpコアタンパク質が、請求項2〜5のいずれか1項に記載のタンパク質である、請求項33に記載の発現ベクター。
【請求項35】
前記単量体抗原が、請求項7又は8に記載の抗原である、請求項33又は34に記載の発現ベクター。
【請求項36】
請求項33〜35のいずれか1項に記載の発現ベクターで形質転換された細菌の宿主細胞。
【請求項37】
請求項33〜35のいずれか1項に記載のベクターで形質転換された真核生物の宿主細胞。
【請求項38】
人体又は動物体の治療方法における、請求項33〜35のいずれか1項に記載の発現ベクターの使用。
【請求項1】
C4bpコアタンパク質と単量体抗原を含む生成物。
【請求項2】
前記C4bpコアが、配列番号1の残基1-57若しくはそのホモログの対応する残基、又は配列番号1の少なくとも47のアミノ酸のフラグメント若しくはそのホモログから成る、請求項1に記載の生成物。
【請求項3】
前記ホモログが、配列番号2〜20のいずれか1つである、請求項2に記載の生成物。
【請求項4】
前記ホモログが、配列番号1〜20のいずれか1つに対して少なくとも70%のアミノ酸同一性を有する変異体である、請求項2又は3に記載の生成物。
【請求項5】
前記単量体抗原が、前記C4bpコアタンパク質のN-又はC-末端と融合している、請求項1〜4のいずれか1項に記載の生成物。
【請求項6】
前記融合が、可撓性のリンカーを介する、請求項5に記載の生成物。
【請求項7】
前記単量体抗原が、マラリア原虫メロゾイト表面タンパク質1の単量体抗原性フラグメントである、請求項1〜6のいずれか1項に記載の生成物。
【請求項8】
前記単量体抗原が、インフルエンザウイルス赤血球凝集素タンパク質又はインフルエンザM2eペプチドの単量体抗原性フラグメントである、請求項1〜6のいずれか1項に記載の生成物。
【請求項9】
製薬上許容できる希釈剤、担体又はアジュバントと一緒に請求項1〜8のいずれか1項に記載の生成物を含む、組成物。
【請求項10】
人体又は動物体を治療する方法に使用するための、請求項1〜8のいずれか1項に記載の生成物又は請求項9に記載の組成物。
【請求項11】
請求項7に記載の生成物の有効量を個体に投与することを含む、マラリアの免疫療法。
【請求項12】
前記個体がマラリア原虫に感染している、請求項11に記載の方法。
【請求項13】
予防ワクチン接種における請求項11に記載の方法。
【請求項14】
マラリアの治療又は予防のための、請求項7に記載の生成物。
【請求項15】
マラリアの治療用又は予防用医薬の製造のための、請求項7に記載の生成物の使用。
【請求項16】
請求項7に記載の生成物を非ヒト哺乳動物に導入する工程、及び該哺乳動物から免疫血清を回収する工程を含む、マラリア原虫(Plasmodium parasite)に対する抗体を製造する方法。
【請求項17】
請求項1〜5のいずれか1項に記載の生成物を宿主対象にワクチン接種して得られた抗体を含む免疫血清を、患者に投与することを含む、患者の病気に対する受動免疫方法。
【請求項18】
請求項16に記載の方法に従って製造された免疫血清の有効量を、ヒトに投与することを含む、ヒト患者のマラリアの受動免疫療法。
【請求項19】
ヒト患者のマラリアの免疫療法に使用するための、請求項16に記載の方法により得られた免疫血清。
【請求項20】
請求項8に記載の生成物の有効量を個体に投与することを含む、インフルエンザの免疫療法。
【請求項21】
前記個体がインフルエンザウイルスに感染している、請求項20に記載の方法。
【請求項22】
予防ワクチン接種における請求項20に記載の方法。
【請求項23】
インフルエンザの治療又は予防のための、請求項8に記載の生成物。
【請求項24】
インフルエンザの治療用又は予防用医薬の製造のための、請求項8に記載の生成物の使用。
【請求項25】
請求項8に記載の生成物を非ヒト哺乳動物に導入する工程、及び該哺乳動物から免疫血清を回収する工程を含む、インフルエンザに対する抗体を製造する方法。
【請求項26】
請求項25に記載の方法に従って製造された免疫血清の有効量を、ヒトに投与することを含む、ヒト患者のインフルエンザの受動免疫療法。
【請求項27】
ヒト患者のインフルエンザの免疫療法に使用するための、請求項25に記載の方法により得られた免疫血清。
【請求項28】
C4bpコアタンパク質と、
非ポリペプチド単量体抗原、
を含む生成物の製造方法であって、
前記一方の成分をコードする核酸を発現する工程、前記他方の成分と前記融合タンパク質を結合させる工程、及び前記生成物を回収する工程を含む、前記方法。
【請求項29】
C4bpコアタンパク質と、
ポリペプチドの単量体抗原、
の融合物を含む生成物を製造する方法であって、
該融合物をコードする核酸を発現させる工程及び前記生成物を回収する工程を含む、前記方法。
【請求項30】
前記核酸を原核生物の宿主細胞で発現させる、請求項28又は29に記載の方法。
【請求項31】
前記融合タンパク質を多量体の形で回収する、請求項30に記載の方法。
【請求項32】
抗原とC4bpコアタンパク質を結合させる工程を含む、単量体抗原の免疫原性を増強させる方法。
【請求項33】
宿主細胞内で機能的なプロモーターと有効に結合している、
C4bpコアタンパク質と、
ポリペプチドの単量体抗原、
の融合タンパク質をコードする核酸配列を含む発現ベクター。
【請求項34】
前記C4bpコアタンパク質が、請求項2〜5のいずれか1項に記載のタンパク質である、請求項33に記載の発現ベクター。
【請求項35】
前記単量体抗原が、請求項7又は8に記載の抗原である、請求項33又は34に記載の発現ベクター。
【請求項36】
請求項33〜35のいずれか1項に記載の発現ベクターで形質転換された細菌の宿主細胞。
【請求項37】
請求項33〜35のいずれか1項に記載のベクターで形質転換された真核生物の宿主細胞。
【請求項38】
人体又は動物体の治療方法における、請求項33〜35のいずれか1項に記載の発現ベクターの使用。
【図1】


【図2】
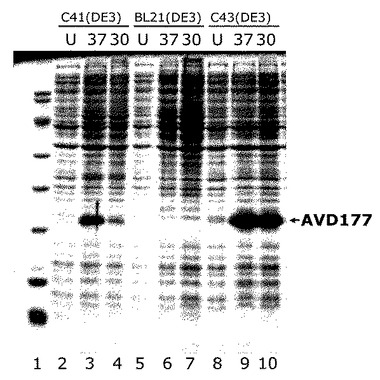
【図3】
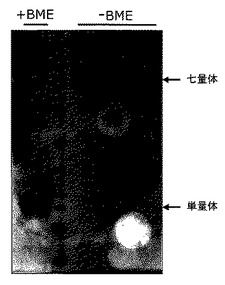
【図2】
【図3】
【公表番号】特表2007−528210(P2007−528210A)
【公表日】平成19年10月11日(2007.10.11)
【国際特許分類】
【出願番号】特願2006−523080(P2006−523080)
【出願日】平成16年8月12日(2004.8.12)
【国際出願番号】PCT/IB2004/002717
【国際公開番号】WO2005/014654
【国際公開日】平成17年2月17日(2005.2.17)
【出願人】(505051792)
【Fターム(参考)】
【公表日】平成19年10月11日(2007.10.11)
【国際特許分類】
【出願日】平成16年8月12日(2004.8.12)
【国際出願番号】PCT/IB2004/002717
【国際公開番号】WO2005/014654
【国際公開日】平成17年2月17日(2005.2.17)
【出願人】(505051792)
【Fターム(参考)】
[ Back to top ]