
CA6抗原特異的細胞傷害性コンジュゲート、および該コンジュゲートを用いる方法
【課題】性細胞表面上に発現される分子/受容体を認識する抗体を含む細胞結合性剤、および癌性細胞の表面上に発現される分子/受容体を特異的にターゲティングする細胞傷害性剤を含む、新規細胞傷害性コンジュゲートを提供する。
【解決手段】CA6グリコトープを認識し、これに結合する、モノクローナル抗体、およびそのエピトープ結合性断片。ネズミ抗CA6モノクローナル抗体DS6(DS6抗体)、及び前記DS6抗体のヒト化型または表面再構成(resurfaced)型、ならびにそのエピトープ結合性断片。
【解決手段】CA6グリコトープを認識し、これに結合する、モノクローナル抗体、およびそのエピトープ結合性断片。ネズミ抗CA6モノクローナル抗体DS6(DS6抗体)、及び前記DS6抗体のヒト化型または表面再構成(resurfaced)型、ならびにそのエピトープ結合性断片。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
[01]本発明は、ネズミ抗CA6グリコトープ・モノクローナル抗体、およびそのヒト化型またはその表面再構成(resurfaced)型に関する。本発明はまた、抗CA6グリコトープ・モノクローナル抗体のエピトープ結合性断片に、ならびに抗CA6グリコトープ・モノクローナル抗体のヒト化型または表面再構成型のエピトープ結合性断片にも関する。
【0002】
[02]本発明は、細胞結合性剤および細胞傷害性剤を含む細胞傷害性コンジュゲート、該コンジュゲートを含む療法組成物、細胞増殖の阻害および疾患の治療において該コンジュゲートを用いるための方法、ならびに該細胞傷害性コンジュゲートを含むキットにさらに関する。特に、細胞結合性剤は、CA6グリコトープを認識し、そしてこれに結合する、モノクローナル抗体またはそのエピトープ結合性断片、あるいはそのヒト化型または表面再構成型である。
【背景技術】
【0003】
発明の背景
[03]周囲の非癌性細胞および組織に害を及ぼすことなく、ターゲット癌細胞を特異的に破壊する、抗癌療法剤を開発する数多くの試みがなされてきている。こうした療法剤は、ヒト患者における癌の治療を非常に改善する潜在能力を有する。
【0004】
[04]1つの有望なアプローチは、モノクローナル抗体などの細胞結合性剤と、細胞傷害性薬剤を連結することであった(Selaら, Immunoconjugates中 189−216(C. Vogel監修 1987); Ghoseら, Targeted Drugs中 1−22(E. Goldberg監修 1983); Dienerら, Antibody mediated delivery systems中 1−23(J. Rodwell監修 1988); Pieterszら, Antibody mediated delivery systems中 25−53(J. Rodwell監修 1988); Bumolら, Antibody mediated delivery systems中 55−79(J. Rodwell監修 1988)。細胞結合性剤の選択次第で、こうした細胞の表面上に発現される分子の発現プロフィールに基づいて、癌性細胞の特定の種類のみを認識し、そしてこうした種類のみに結合するように、これらの細胞傷害性コンジュゲートを設計することも可能である。
【0005】
[05]多様なネズミ・モノクローナル抗体に連結された、メトトレキセート、ダウノルビシン、ドキソルビシン、ビンクリスチン、ビンブラスチン、メルファラン、マイトマイシンC、およびクロラムブシルなどの細胞傷害性薬剤が、こうした細胞傷害性コンジュゲートで用いられてきている。いくつかの場合、血清アルブミン(Garnettら, 46 Cancer Res. 2407−2412(1986); Ohkawaら 23 Cancer Immunol. Immunother. 81−86(1986); Endoら, 47 Cancer Res. 1076−1080(1980))、デキストラン(Hurwitzら, 2 Appl. Biochem. 25−35(1980); Manabiら, 34 Biochem. Pharmacol. 289−291(1985); Dillmanら, 46 Cancer Res. 4886−4891(1986); Shovalら, 85 Proc. Nat
l. Acad. Sci. 8276−8280(1988))、またはポリグルタミン酸(Tsukadaら, 73 J. Natl. Canc. Inst. 721−729(1984); Katoら 27 J. Med. Chem. 1602−1607(1984); Tsukadaら, 52 Br. J. Cancer 111−116(1985))などの中間キャリアー分子を通じて、薬剤分子が抗体分子に連結された。
【0006】
[06]ある程度有望であることが示されている1つの特異的コンジュゲートの例として、結腸直腸腫瘍および膵臓腫瘍上に発現される抗原であるCanAgに対して向けられるC242抗体、ならびにメイタンシン誘導体DM1のコンジュゲートがある(Liuら, Proc Natl Acad Sci USA, 93: 8618−8623(1996))。このコンジュゲートのin vitro評価によって、細胞表面上に発現されるCanAgに向かうその結合親和性が高く、見かけのKd値が3x10−11Mであり、そしてCanAg陽性細胞に対する細胞傷害能が高く、IC50が6x10−11Mであることが示された。この細胞傷害性は、過剰な非コンジュゲート化抗体によって遮断されたため、そして抗原陰性細胞が該コンジュゲートに対して100倍以上感受性が低かったため、該細胞傷害性は抗原依存性である。それぞれのターゲット細胞に向かう親和性が高く、そしてかつ抗原選択的細胞傷害性が高い、抗体−DM1コンジュゲートの他の例には、ヒトCD56に対する抗体のヒト化型、huN901;ヒトCD33に対する抗体のヒト化型、huMy9−6;CanAg Muc1エピトープに対する抗体のヒト化型、huC242;PSMAに対する脱免疫化(deimmunized)抗体、huJ591;Her2/neuに対するヒト化抗体、トラスツズマブ;およびCD44v6に対するヒト化抗体、ビバツズマブが含まれる。
【0007】
[07]癌患者を治療するのに用いる方法を、続けて改善する際に、特定の種類の癌性細胞を特異的に認識するさらなる細胞傷害性コンジュゲートの開発が重要であろう。
【発明の開示】
【発明が解決しようとする課題】
【0008】
[08]この目的に向けて、本発明は、癌性細胞表面上に発現される分子/受容体を認識し、そしてこれに結合する抗体の開発に、そして抗体などの細胞結合性剤、および癌性細胞の表面上に発現される分子/受容体を特異的にターゲティングする細胞傷害性剤を含む、新規細胞傷害性コンジュゲートの開発に関する。
【0009】
[09]より具体的には、本発明は、癌性細胞によって発現されるMuc1ムチン受容体上の新規CA6シアログリコトープの性質決定に、そしてMuc1ムチンの新規CA6シアログリコトープを認識し、そして細胞傷害性剤の背景で、CA6グリコトープを発現している細胞の増殖を阻害するために使用可能である抗体、好ましくはヒト化抗体の提供に関する。
【課題を解決するための手段】
【0010】
発明の概要
[10]本発明には、Muc1ムチン受容体の新規CA6シアログリコトープを特異的に認識し、そして結合する、抗体またはそのエピトープ結合性断片が含まれる。別の態様において、本発明には、Muc1ムチン受容体の新規CA6シアログリコトープ(「CA6グリコトープ」)を認識する、ヒト化抗体またはそのエピトープ結合性断片が含まれる。
【0011】
[11]好ましい態様において、本発明には、ネズミ抗CA6モノクローナル抗体DS6(「DS6抗体」)、および当該抗体またはそのエピトープ結合性断片の、表面に曝露
される残基が、既知のヒト抗体表面により密接に似るように、軽鎖および重鎖両方において置換されている、DS6抗体の表面再構成型またはヒト化型が含まれる。本発明のヒト化抗体およびそのエピトープ結合性断片は、投与されるヒト被験体において、完全ネズミ型よりも、はるかにより免疫原性でない(または完全に非免疫原性である)点で、改善された特性を有する。したがって、本発明のヒト化DS6抗体およびそのエピトープ結合性断片は、ヒトに免疫原性でない一方、Muc1ムチン受容体上の新規シアログリコトープ、すなわちCA6グリコトープを特異的に認識する。ヒト化抗体およびそのエピトープ結合性断片を、メイタンシノイドなどの薬剤にコンジュゲート化させて、Muc1 CA6シアログリコトープに薬剤をターゲティングすることによって、抗原を発現している細胞に向かう特異的細胞傷害性を有するプロドラッグを形成してもよい。したがって、こうした抗体および小さい非常に毒性である薬剤(例えばメイタンシノイド、タキサン、およびCC−1065類似体)を含む細胞傷害性コンジュゲートを、乳房腫瘍および卵巣腫瘍などの腫瘍治療のための療法剤として用いてもよい。
【0012】
[12]本発明のDS6抗体のヒト化型は、軽鎖および重鎖可変領域両方のそれぞれのアミノ酸配列、軽鎖および重鎖可変領域の遺伝子のDNA配列、CDRの同定、その表面アミノ酸の同定、ならびに組換え型におけるその発現のための手段の開示に関して、本明細書において完全に性質決定されている。
【0013】
[13]1つの態様において、配列番号1〜3によって示されるアミノ酸配列:
【0014】
【化1】

【0015】
を有するCDRを含む重鎖を有し、そして配列番号4〜6によって示されるアミノ酸配列:
【0016】
【化2】
【0017】
を有するCDRを含む軽鎖を有する、ヒト化DS6抗体またはそのエピトープ結合性断片を提供する。
[14]配列番号7または配列番号8によって示されるアミノ酸配列:
【0018】
【化3】
【0019】
と、少なくとも90%の配列同一性を共有するアミノ酸配列を有する軽鎖可変領域を有する、ヒト化DS6抗体およびそのエピトープ結合性断片もまた提供する。
[15]同様に、配列番号9、配列番号10、または配列番号11によって示されるアミノ酸配列:
【0020】
【化4】
【0021】
と、少なくとも90%の配列同一性を共有するアミノ酸配列を有する重鎖可変領域を有する、ヒト化DS6抗体およびエピトープ結合性断片を提供する。
[16]別の態様において、配列番号8に対応するアミノ酸配列
【0022】
【化5】
【0023】
を有するヒト化または表面再構成軽鎖可変領域を有する、ヒト化DS6抗体およびそのエピトープ結合性断片を提供する。
[17]同様に、それぞれ、配列番号10または配列番号11に対応するアミノ酸配列:
【0024】
【化6】
【0025】
を有するヒト化または表面再構成重鎖可変領域を有する、ヒト化DS6抗体およびそのエピトープ結合性断片を提供する。
[18]本発明のヒト化DS6抗体およびそのエピトープ結合性断片にはまた、ヒト残基への変更が必要な、CDRの5オングストローム以内に見られるネズミ表面フレームワーク残基に相当する、表1の星印の残基によって定義される、1以上の位での軽鎖および/または重鎖アミノ酸残基中の置換をも含むことができる。例えば、ネズミ配列(配列番号7)中の最初のアミノ酸残基Qは、抗体をヒト化するため、E(配列番号8)によって置換されている。しかし、この残基はCDRに近接しているため、抗体親和性を維持するために、ネズミ残基Qへの復帰突然変異が必要でありうる。
表1
【0026】
【表1】
【0027】
[19]3つの最も相同なヒト可変領域表面残基と並列させて、muDS6可変領域表面残基を示す表2に、これをさらに示す。表1のアミノ酸残基は、表2の下線のアミノ酸残基に対応する。
表2
【0028】
【表2】
【0029】
[20]本発明は、(1)CA6グリコトープを認識し、そして該グリコトープに結合する細胞結合性剤、および(2)細胞傷害性剤を含む、細胞傷害性コンジュゲートをさらに提供する。本発明の細胞傷害性コンジュゲートが有効な殺傷剤を形成するように、細胞傷害性コンジュゲートにおいて、細胞結合性剤はCA6グリコトープに対して高い親和性を有し、そして細胞傷害性剤はCA6グリコトープを発現している細胞に対して高い度合いの細胞傷害性を有する。
【0030】
[21]好ましい態様において、細胞結合性剤は、抗CA6抗体またはそのエピトープ結合性断片、より好ましくはヒト化抗CA6抗体またはそのエピトープ結合性断片であり、ここで細胞傷害性剤が、直接、あるいは切断可能または切断不能リンカーを介して、抗体またはそのエピトープ結合性断片に共有結合している。より好ましい態様において、細胞結合性剤は、ヒト化DS6抗体またはそのエピトープ結合性断片であり、そして細胞傷害性剤は、タキソール、メイタンシノイド、CC−1065またはCC−1065類似体である。
【0031】
[22]本発明の好ましい態様において、細胞結合性剤はヒト化抗CA6抗体であり、そして細胞傷害性剤はメイタンシノイドまたはタキサンなどの細胞傷害性薬剤である。
[23]より好ましくは、細胞結合性剤はヒト化抗CA6抗体DS6であり、そして細胞傷害性剤はDM1またはDM4などのメイタンシン化合物である。
【0032】
[24]本発明にはまた、CA6グリコトープを発現している細胞の増殖を阻害するための方法も含まれる。好ましい態様において、CA6グリコトープを発現している細胞の増殖を阻害するための方法は、in vivoで起こり、そして細胞の死を生じるが、in vitroおよびex vivo適用もまた含まれる。
【0033】
[25]本発明はまた、細胞傷害性コンジュゲート、および薬学的に許容しうるキャリアーまたは賦形剤を含む療法組成物も提供する。
[26]本発明には、療法組成物を用いて、癌を有する被験体を治療する方法がさらに含まれる。好ましい態様において、細胞傷害性コンジュゲートは、抗CA6抗体および細胞傷害性剤を含む。より好ましい態様において、細胞傷害性コンジュゲートは、ヒト化DS6抗体−DM1コンジュゲート、ヒト化DS6抗体−DM4またはヒト化DS6抗体−タキサン・コンジュゲートを含み、そして該コンジュゲートは、薬学的に許容しうるキャリアーまたは賦形剤と一緒に投与される。
【0034】
[27]本発明にはまた、抗CA6抗体−細胞傷害性剤コンジュゲートおよび使用のための説明書を含むキットも含まれる。好ましい態様において、抗CA6抗体はヒト化DS6抗体であり、細胞傷害性剤はDM1またはDM4などのメイタンシン化合物、あるいは
タキサンであり、そして説明書は、癌を有する被験体の治療において、コンジュゲートを使用するためのものである。キットにはまた、コンジュゲートが凍結乾燥状態または濃縮型であるならば希釈剤などの、薬学的に許容しうる配合物の調製に必要な構成要素、および配合物の投与に必要な構成要素も含まれていてもよい。
【0035】
[28]本発明にはまた、CA6グリコトープに特異的に結合し、そして該グリコトープを認識する抗体の誘導体も含まれる。好ましい態様において、抗体誘導体は、CA6グリコトープに結合する抗体の表面を再構成するかまたは該抗体をヒト化することによって調製され、ここで該誘導体は宿主に対して減少した免疫原性を有する。
【0036】
[29]本発明は、研究または診断適用において使用するため、さらに標識されているヒト化抗体またはその断片をさらに提供する。好ましい態様において、標識は、放射標識、蛍光体、発色団、造影剤または金属イオンである。
【0037】
[30]前記の標識ヒト化抗体またはそのエピトープ結合性断片を、癌を有すると推測される被験体に投与し、そして被験体体内の標識分布を測定するかまたは監視する、診断法もまた、提供する。
【0038】
[31]本発明はまた、本発明のヒト化抗体コンジュゲートを、単独で、あるいは他の細胞傷害性剤または療法剤と組み合わせて投与することによって、癌を有する被験体を治療するための方法もまた提供する。癌は、CA6が発現されている、例えば乳癌、結腸癌、卵巣癌、子宮内膜癌、骨肉腫、子宮頸癌、前立腺癌、肺癌、滑膜癌、膵臓癌、肉腫または癌腫、あるいはCA6グリコトープが主に発現されている、まだ決定されていない他の癌の1以上であってもよい。
【0039】
[32]別に言及しない限り、本明細書に引用するすべての参考文献および特許は、本明細書に援用される。
【0040】
発明の詳細な説明
[63]本発明は、他の特徴の中でも、抗CA6モノクローナル抗体、抗CA6ヒト化抗体、および抗CA6抗体の断片を提供する。本発明の抗体および抗体断片の各々は、細胞表面上のCA6グリコトープを特異的に認識し、そして該グリコトープに結合するよう設計される。CA6は、多くのヒト腫瘍:漿液性卵巣癌の95%、子宮内膜様卵巣癌の50%、子宮頸部新生物の50%、子宮内膜新生物の69%、外陰新生物の80%、乳癌の60%、膵臓腫瘍の67%、および尿路上皮腫瘍の48%によって発現されることが知られるが、正常ヒト組織によっては滅多に発現されない。
【0041】
[64]Kearseら, Int. J. Cancer 88(6):866−872(2000)による報告は、ハイブリドーマ上清を用いてCA6エピトープを性質決定した際、CA6エピトープが見られるタンパク質を、CA6エピトープを含有するN連結炭水化物を有する80kDaタンパク質と誤認した。その後、本発明者らは、精製DS6を用いて、250kDaの非ジスルフィド連結糖タンパク質より大きい、O連結炭水化物上にCA6エピトープが見られることを立証した。さらに、該糖タンパク質は、ムチン、Muc1と同定された。異なるMuc1アレルは、可変数タンデム反復(VNTR)ドメインにおいて、多様な数のタンデム反復を有するため、細胞はしばしば、異なるサイズの2つの別個のMuc1タンパク質を発現する(Taylor−Papadimitriou, Biochim. Biophys. Acta 1455(2−3):301−13(1999)。VNTRドメイン中の反復数が異なり、さらにグリコシル化も異なるため、Muc1の分子量は細胞株間で多様である。
【0042】
[65]過ヨウ素酸に対するCA免疫反応性の感受性は、CA6が炭水化物エピトープ「グリコトープ」であることを示す。コレラ菌(Vibrio cholerae)由来のノイラミニダーゼでの処理に対する、CA6免疫反応性のさらなる感受性は、CA6エピトープがシアル酸依存性グリコトープであり、したがって、「シアログリコトープ」であることを示す。
【0043】
[66]CA6の性質決定の詳細を実施例2に見出しうる(以下を参照されたい)。CA6に関するさらなる詳細は、WO 02/16401; Wennerbergら, Am. J. Pathol. 143(4):1050−1054(1993); Smithら, Human Antibodies 9:61−65(1999); Kearseら, Int. J. Cancer 88(6):866−872(2000); Smithら, Int. J. Gynecol. Pathol. 20(3):260−6(2001);およびSmithら, Appl. Immunohistochem. Mol. Morphol. 10(2):152−8(2002)に見出されうる。
【0044】
[67]本発明にはまた、2つの主な構成要素を含む細胞傷害性コンジュゲートも含まれる。第一の構成要素は、CA6グリコトープを認識し、そして該グリコトープに結合する、細胞結合性剤である。細胞傷害性コンジュゲートが、意図する細胞のみを認識し、そして該細胞に結合するように、細胞結合性剤は、高い度合いの特異性で、Muc1上のCA6シアログリコトープを認識すべきである。高い度合いの特異性は、コンジュゲートが、非特異的結合から生じる副作用をほとんど伴わずに、ターゲティングされた方式で作用することを可能にするであろう。
【0045】
[68]別の態様において、コンジュゲートの細胞傷害性薬剤部分が細胞に対して作用するのを可能にするのに十分な期間、そして/または細胞によってコンジュゲートが内部移行されるのを可能にするのに十分な期間、コンジュゲートがターゲット細胞と接触するように、本発明の細胞結合性剤はまた、CA6グリコエピトープを高い度合いの親和性でも認識するであろう。
【0046】
[69]好ましい態様において、細胞傷害性コンジュゲートは、細胞結合性剤として、抗CA6抗体、より好ましくはネズミDS6抗CA6モノクローナル抗体を含む。より好ましい態様において、細胞傷害性コンジュゲートは、ヒト化DS6抗体またはそのエピトープ結合性断片を含む。DS6抗体は、高い度合いの特異性でCA6を認識することが可能であり、そして細胞傷害性剤を、ターゲティング方式で、癌細胞などの異常な細胞または組織に向ける。
【0047】
[70]本発明の細胞傷害性コンジュゲートの第二の構成要素は、細胞傷害性剤である。好ましい態様において、細胞傷害性剤は、タキソール、DM1またはDM4などのメイタンシノイド、CC−1065またはCC−1065類似体である。好ましい態様において、本発明の細胞結合性剤は、直接、あるいは切断可能または切断不能リンカーを介して、細胞傷害性剤に共有結合している。
【0048】
[71]細胞結合性剤、細胞傷害性剤、およびリンカーを、以下により詳細に論じる。
細胞結合性剤
[72]療法剤としての本発明の化合物の有効性は、適切な細胞結合性剤の注意深い選択次第である。細胞結合性剤は、現在知られるかまたは知られるようになる、いかなる種類のものであってもよく、そしてこうした剤には、ペプチドおよび非ペプチドが含まれる。細胞結合性剤は、特異的または非特異的方式のいずれかで、細胞に結合可能な、いかなる化合物であってもよい。一般的に、これらは、抗体(特にモノクローナル抗体)、リン
ホカイン、ホルモン、増殖因子、ビタミン、栄養輸送分子(トランスフェリンなど)、あるいは他の任意の細胞結合性分子または物質であってもよい。
【0049】
[73]使用可能な細胞結合性剤のより具体的な例には:
(a)ポリクローナル抗体;
(b)モノクローナル抗体;
(c)Fab、Fab’、およびF(ab’)2、Fvなどの抗体断片(Parham, J. Immunol. 131:2895−2902(1983); Springら J. Immunol. 113:470−478(1974); Nisonoffら, Arch. Biochem. Biophys. 89:230−244(1960));
(d)インターフェロン(例えばアルファ、ベータ、ガンマ);
(e)IL−2、IL−3、IL−4、IL−6などのリンホカイン;
(f)インスリン、TRH(甲状腺刺激ホルモン放出ホルモン)、MSH(メラニン形成細胞刺激ホルモン)、アンドロゲンおよびエストロゲンなどのステロイドホルモンなどのホルモン;
(g)EGF、TGF−アルファ、FGF、VEGF、G−CSF、M−CSFおよびGM−CSFなどの増殖因子およびコロニー刺激因子(Burgess, Immunology Today 5:155−158(1984));
(h)トランスフェリン(O’Keefeら J. Biol. Chem. 260:932−937(1985));ならびに
(i)葉酸などのビタミン
が含まれる。
【0050】
抗体
[74]適切な細胞結合性剤の選択は、ターゲティングしようとする特定の細胞集団に応じた選択の問題であるが、一般的に、適切なものが入手可能であるか、または調製可能であるならば、抗体、より好ましくはモノクローナル抗体が好ましい。
【0051】
[75]モノクローナル抗体技術によって、特異的モノクローナル抗体の形で、非常に特異的な細胞結合性剤の産生が可能になる。当該技術分野に特によく知られるのは、マウス、ラット、ハムスター、または任意の他の哺乳動物を、損なわれていない(intact)ターゲット細胞、ターゲット細胞から単離された抗原、完全ウイルス、弱毒化完全ウイルス、およびウイルス・コートタンパク質などのウイルスタンパク質などの、関心対象の抗原で免疫することによって産生される、モノクローナル抗体を生成するための技術である。感作されたヒト細胞もまた使用可能である。モノクローナル抗体を生成する別の方法は、scFv(一本鎖可変領域)、特にヒトscFvの、ファージライブラリーの使用である(例えば、Griffithsら、米国特許第5,885,793号および第5,969,108号; McCaffertyら、WO 92/01047; Limingら、WO 99/06587を参照されたい)。
【0052】
[76]典型的な抗体は、ジスルフィド結合によって連結される2つの同一の重鎖および2つの同一の軽鎖で構成される。可変領域は、抗体の中で配列が異なり、そして抗原に対する特定の抗体各々の結合および特異性において協調する、抗体重鎖および軽鎖の部分である。可変性は、通常、抗体可変領域全体に均一に分布しているのではない。典型的には、軽鎖および重鎖可変領域両方における、相補性決定領域(CDR)または超可変領域と呼ばれる可変領域の3つのセグメント内に集中している。可変領域のより高く保存される部分は、フレームワーク領域と呼ばれる。重鎖および軽鎖の可変領域は4つのフレームワーク領域を含み、これらは大部分、ベータ−シート立体配置を採用し、各フレームワーク領域は3つのCDRによって連結され、CDRは、ベータ−シート構造を連結するルー
プを形成し、そしていくつかの場合、ベータ−シート構造の一部を形成する。各鎖中のCDRは、フレームワーク領域によって非常に近接して保持され、そして他の鎖に由来するCDRとともに、抗体の抗原結合部位の形成に寄与する(E. A. Kabatら Sequences of Proteins of Immunological Interest, 第5版, 1991, NIH)。
【0053】
[77]定常領域は、重鎖の一部である。抗体の抗原への結合に直接関与してはいないが、抗体依存性細胞傷害性における抗体の関与など、多様なエフェクター機能を示す。
[78]本発明で使用するのに適したモノクローナル抗体には、ネズミDS6モノクローナル抗体(米国特許第6,596,503号; ATCC寄託番号PTA−4449)が含まれる。
【0054】
ヒト化または表面再構成DS抗体
[79]好ましくは、本発明の細胞結合性剤として、ヒト化抗CA6抗体を用いる。こうしたヒト化抗体の好ましい態様は、ヒト化DS6抗体またはそのエピトープ結合性断片である。
【0055】
[80]ヒト化の目的は、抗体の完全な抗原結合親和性および特異性を維持しつつ、ヒトに導入するために、ネズミ抗体などの異種抗体の免疫原性を減少させることである。
[81]表面再構成およびCDR移植などのいくつかの技術を用いて、ヒト化抗体を産生してもよい。本明細書において、表面再構成技術は、分子モデリング、統計分析および突然変異誘発の組み合わせを用いて、抗体可変領域の非CDR表面を改変し、ターゲット宿主の既知の抗体の表面に似せる。
【0056】
[82]抗体の表面再構成のための戦略および方法、ならびに異なる宿主内で抗体の免疫原性を減少させるための他の方法が、その全体が本明細書に援用される、米国特許第5,639,641号(Pedersenら)に開示される。簡潔には、好ましい方法において、(1)抗体重鎖および軽鎖可変領域のプールの位置アライメントを生成して、重鎖および軽鎖可変領域フレームワーク表面曝露位置のセットを生じ、ここで、すべての可変領域のアライメント位置は、少なくとも約98%同一である;(2)重鎖および軽鎖可変領域フレームワーク表面曝露アミノ酸残基のセットを、げっ歯類抗体(またはその断片)に関して定義し;(3)げっ歯類表面曝露アミノ酸残基セットに、最も密接に同一である、重鎖および軽鎖可変領域フレームワーク表面曝露アミノ酸残基のセットを同定し;(4)げっ歯類抗体の相補性決定領域のいずれかの残基のいずれかの原子から5Å以内にあるアミノ酸残基を除いて、工程(2)で定義した、重鎖および軽鎖可変領域フレームワーク表面曝露アミノ酸残基のセットを、工程(3)で同定した、重鎖および軽鎖可変領域フレームワーク表面曝露アミノ酸残基のセットで置換し;そして(5)結合特異性を有するヒト化げっ歯類抗体を産生する。
【0057】
[83]CDR移植(EP 0 239 400; WO 91/09967;米国特許第5,530,101号;および第5,585,089号)、表面加工(veneering)または表面再構成(EP 0 592 106; EP 0 519 596;
Padlan E. A., 1991, Molecular Immunology 28(4/5):489−498; Studnicka G. M.ら, 1994, Protein Engineering 7(6):805−814; Roguska M.A.ら, 1994, PNAS 91:969−973)、および鎖シャフリング(米国特許第5,565,332号)を含む、多様な他の技術を用いて、抗体をヒト化してもよい。ファージディスプレイ法を含む、当該技術分野に知られる多様な方法によって、ヒト抗体を作製してもよい。米国特許第4,444,887号、第4,716,111号、第5,545,806号、および第5,814,318号;ならびに国際
特許出願公報WO 98/46645、WO 98/50433、WO 98/24893、WO 98/16654、WO 96/34096、WO 96/33735、およびWO 91/10741(前記文献は、その全体が本明細書に援用される)もまた参照されたい。
【0058】
[84]好ましい態様において、本発明は、Muc1ムチン上の新規シアログリコトープ(CA6グリコトープ)を認識するヒト化抗体またはその断片を提供する。別の態様において、ヒト化抗体またはそのエピトープ結合性断片は、CA6グリコトープを発現している細胞の増殖を阻害するさらなる能力を有する。
【0059】
[85]より好ましい態様において、DS6抗体の表面再構成型またはヒト化型を提供し、ここで、抗体またはその断片の表面曝露残基は、既知のヒト抗体表面により密接に似るように、軽鎖および重鎖両方で置換される。本発明のヒト化DS6抗体またはそのエピトープ結合性断片は、改善された特性を有する。例えば、ヒト化DS6抗体またはそのエピトープ結合性断片は、Muc1ムチン上の新規シアログリコトープ(CA6グリコトープ)を特異的に認識する。より好ましくは、ヒト化DS6抗体またはそのエピトープ結合性断片は、CD6グリコトープを発現している細胞の増殖を阻害するさらなる能力を有する。ヒト化抗体またはそのエピトープ結合性断片を、メイタンシノイドなどの薬剤にコンジュゲート化して、薬剤を新規Muc1シアログリコトープ、CA6にターゲティングすることによって、抗原発現細胞に向けて特異的細胞傷害性を有するプロドラッグを形成してもよい。こうした抗体、および小さく非常に毒性である薬剤(例えばメイタンシノイド、タキサン、およびCC−1065類似体)を含む細胞傷害性コンジュゲートを、乳房腫瘍および卵巣腫瘍などの腫瘍の治療のための療法剤として用いてもよい。
【0060】
[86]DS6抗体のヒト化型はまた、軽鎖および重鎖可変領域両方のそれぞれのアミノ酸配列、軽鎖および重鎖可変領域の遺伝子のDNA配列、CDRの同定、その表面アミノ酸の同定、ならびに組換え型におけるその発現のための手段の開示に関しても、本明細書において完全に性質決定されている。
【0061】
[87]1つの態様において、配列番号1〜3によって示されるアミノ酸配列:
【0062】
【化7】
【0063】
を有するCDRを含む重鎖を有する、ヒト化抗体またはそのエピトープ結合性断片を提供する。
[88]AbMモデリングソフトウェアによって重鎖CDRを決定すると、重鎖CDRは配列番号20〜22:
【0064】
【化8】
【0065】
によって示される。
[89]同じ態様において、ヒト化抗体またはそのエピトープ結合性断片は、配列番号4〜6によって示されるアミノ酸配列:
【0066】
【化9】
【0067】
を有するCDRを含む、軽鎖を有する。
[90]配列番号7または8によって示されるアミノ酸配列:
【0068】
【化10】
【0069】
と、少なくとも90%の配列同一性を共有するアミノ酸配列を有する軽鎖可変領域を有する、ヒト化抗体およびそのエピトープ結合性断片もまた提供する。
[91]同様に、配列番号9、配列番号10、または配列番号11によって示されるアミノ酸配列:
【0070】
【化11】
【0071】
と、少なくとも90%の配列同一性を共有するアミノ酸配列を有する重鎖可変領域を有する、ヒト化抗体およびそのエピトープ結合性断片を提供する。
[92]別の態様において、配列番号8に対応するアミノ酸配列
【0072】
【化12】
【0073】
を有するヒト化または表面再構成軽鎖可変領域を有する、ヒト化抗体およびそのエピトープ結合性断片を提供する。
[93]同様に、配列番号10または配列番号11に対応するアミノ酸配列:
【0074】
【化13】
【0075】
を有するヒト化または表面再構成重鎖可変領域を有する、ヒト化抗体およびそのエピトープ結合性断片を提供する。
[94]本発明のヒト化抗体およびそのエピトープ結合性断片にはまた、muDS6の結合親和性および特異性を維持するため、CDRに近接するヒト表面アミノ酸残基が、表1のアステリスクで印を付けた残基によって定義される1以上の位(Kabatの番号付け)で、対応するmuDS6表面残基によって置換されている、軽鎖および/または重鎖可変領域型も含まれてもよい。
表1
【0076】
【表3】
【0077】
[95]DS6抗体軽鎖および重鎖、ならびにそのヒト化型の一次アミノ酸配列およびDNA配列を本明細書に開示する。しかし、本発明の範囲は、これらの配列を含む抗体および断片に限定されない。その代わり、Muc1受容体上のユニークな腫瘍特異的グリコ
トープとしてのCA6に特異的に結合するすべての抗体および断片が、本発明に含まれる。好ましくは、CA6に特異的に結合する抗体および断片はまた、受容体の生物学的活性もアンタゴナイズする。より好ましくは、こうした抗体は、さらに、アゴニスト活性を実質的に回避する。したがって、本発明の抗体および抗体断片は、骨格、CDR、および/または軽鎖および重鎖のアミノ酸配列が、DS6抗体またはそのヒト化誘導体と異なり、そしてなお、本発明の範囲内に属することも可能である。
【0078】
[96]モデリングによって、DS6抗体のCDRが同定され、そしてその分子構造が予測されている。ここでも、CDRはエピトープ認識に重要であるが、本発明の抗体および断片には本質的でない。したがって、例えば本発明の抗体の親和性成熟によって産生される、改善された特性を有する抗体および断片を提供する。
【0079】
[97]DS6が由来しているようである、マウス軽鎖IgVκ ap4生殖系列遺伝子および重鎖IgVh J558.41生殖系列遺伝子を、DS6抗体の配列と整列させて、図11に示す。この比較によって、CDR中のいくつかを含めて、DS6抗体中のありうる体細胞突然変異が同定される。
【0080】
[98]DS6抗体の重鎖および軽鎖可変領域の配列、ならびにDS6抗体のCDRの配列は、先には知られておらず、そしてこれらを図9Aおよび9Bに示す。こうした情報を用いて、DS6抗体のヒト化型を生じてもよい。
【0081】
抗体断片
[99]本発明の抗体には、上に論じる全長抗体ならびにエピトープ結合性断片の両方が含まれる。本明細書において、「抗体断片」には、全長抗体によって認識されるエピトープに結合する能力を保持する抗体のいかなる部分も含まれ、これらは一般的に「エピトープ結合性断片」と称される。抗体断片の例には、限定されるわけではないが、Fab、Fab’およびF(ab’)2、Fd、一本鎖Fv(scFv)、一本鎖抗体、ジスルフィド連結Fv(dsFv)、ならびにVLまたはVH領域のいずれかを含む断片が含まれる。一本鎖抗体を含む、エピトープ結合性断片は、単独で、あるいは以下:ヒンジ領域、CH1、CH2、およびCH3ドメインのすべてまたは一部と組み合わせて、可変領域(単数または複数)を含んでもよい。
【0082】
[100]こうした断片は、1つまたは両方のFab断片またはF(ab’)2断片を含有してもよい。好ましくは、抗体断片は、完全抗体の6つのCDRすべてを含有するが、こうした領域すべてより少ない、例えば、3つ、4つまたは5つのCDRを含有する断片もまた、機能する。さらに、断片は、以下の免疫グロブリンクラス:IgG、IgM、IgA、IgD、またはIgE、およびそのサブクラスの任意の1つのメンバーであっても、またはそれらを組み合わせてもよい。
【0083】
[101]パパイン(Fab断片)またはペプシン(F(ab’)2断片)などの酵素を用いたタンパク質分解切断によって、FabおよびF(ab’)2断片を産生してもよい。
【0084】
[102]一本鎖FV(scFv)断片は、抗体軽鎖可変領域(VL)の少なくとも1つの断片に連結された抗体重鎖可変領域(VH)の少なくとも1つの断片を含有する、エピトープ結合性断片である。リンカーは、一本鎖抗体断片が由来する完全抗体のターゲット分子結合特異性を維持するために、連結された場合、(VL)および(VH)領域の適切な三次元フォールディングを確実にするように選択された、短い、柔軟なペプチドであってもよい。(VL)または(VH)配列のカルボキシル末端は、リンカーによって、相補(VL)または(VH)配列のアミノ酸末端に共有結合されてもよい。
【0085】
[103]本発明の一本鎖抗体断片は、本明細書に記載する完全抗体の可変または相補性決定領域(CDR)の少なくとも1つを有するが、こうした抗体の定常ドメインのいくつかまたはすべてを欠く、アミノ酸配列を含有する。これらの定常ドメインは、抗原結合には必要でないが、完全抗体の構造の主要な部分を構成する。したがって、一本鎖抗体断片は、定常ドメインの一部またはすべてを含有する抗体の使用と関連する問題のいくつかを克服しうる。例えば、一本鎖抗体断片は、生物学的分子および重鎖定常領域の間の望ましくない相互作用、または他の望ましくない生物学的活性がない傾向がある。さらに、一本鎖抗体断片は、完全抗体よりかなり小さく、そしてしたがって、完全抗体よりも高い毛細管透過性を有し、一本鎖抗体断片が、より効率的にターゲット抗原結合部位に局在し、そして結合することを可能にしうる。また、原核細胞において、比較的大規模に抗体断片を産生し、したがってその産生を容易にすることも可能である。さらに、一本鎖抗体断片が比較的小さいサイズであることから、これらは完全抗体よりも、レシピエントにおいて、免疫応答を誘発する可能性がより低くなる。
【0086】
[104]分子クローニング、抗体ファージディスプレイライブラリーまたは当業者に周知の類似の技術によって、一本鎖抗体断片を生成してもよい。真核細胞、または細菌を含む原核細胞において、これらのタンパク質を産生してもよい。また、当該技術分野に知られる多様なファージディスプレイ法を用いて、本発明のエピトープ結合性断片を生成してもよい。ファージディスプレイ法において、抗体ドメインをコードするポリヌクレオチド配列を所持するファージ粒子の表面上に、機能する抗体ドメインがディスプレイされる。特に、こうしたファージを利用して、レパートリーまたはコンビナトリアル抗体ライブラリー(例えばヒトまたはネズミ)から発現されるエピトープ結合性ドメインをディスプレイしてもよい。抗原を用いて、例えば固体表面またはビーズに結合したかまたは捕捉された標識抗原を用いて、関心対象の抗原に結合するエピトープ結合性ドメインを発現するファージを選択するかまたは同定してもよい。これらの方法で用いるファージは、ファージ遺伝子IIIまたは遺伝子VIIIタンパク質のいずれかに組換え的に融合された、Fab、Fvまたはジスルフィド安定化Fv抗体ドメインを含むファージから発現される、fdおよびM13結合ドメインを含む、典型的には繊維状ファージである。
【0087】
[105]本発明のエピトープ結合性断片を作製するために使用可能なファージディスプレイ法の例には、各々、その全体が本明細書に援用される、Brinkmanら, 1995, J. Immunol. Methods 182:41−50; Amesら, 1995, J. Immunol. Methods 184:177−186; Kettleboroughら, 1994, Eur. J. Immunol.
24:952−958; Persicら, 1997, Gene 187:9−18; Burtonら, 1994, Advances in Immunology
57:191−280; PCT出願第PCT/GB91/01134号; PCT公報WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401;ならびに米国特許第5,698,426号;第5,223,409号;第5,403,484号;第5,580,717号;第5,427,908号;第5,750,753号;第5,821,047号;第5,571,698号;第5,427,908号;第5,516,637号;第5,780,225号;第5,658,727号;第5,733,743号および第5,969,108号に開示されるものが含まれる。
【0088】
[106]ファージ選択後、断片をコードするファージ領域を単離し、そしてこれを用いて、例えば以下に詳細に記載するような組換えDNA技術を用い、哺乳動物細胞、昆虫細胞、植物細胞、酵母、および細菌を含む、選択した宿主における発現を通じて、エピト
ープ結合性断片を生成してもよい。例えば、PCT公報WO 92/22324; Mullinaxら, 1992, BioTechniques 12(6):864−869; Sawaiら, 1995, AJRI 34:26−34;ならびにBetterら, 1988, Science 240:1041−1043に開示されるものなど、当該技術分野に知られる方法を用いて、Fab、Fab’およびF(ab’)2断片を組換え的に産生する技術もまた使用してもよく;前記参考文献は、その全体が本明細書に援用される。一本鎖Fvおよび抗体を産生するために使用可能な技術の例には、米国特許第4,946,778号および第5,258,498号; Hustonら, 1991, Methods in Enzymology 203:46−88; Shuら, 1993, PNAS 90:7995−7999; Skerraら, 1988, Science 240:1038−1040に記載されるものが含まれる。
【0089】
機能的同等物
[107]本発明の範囲内にやはり含まれるのは、抗CA6抗体およびヒト化抗CA6抗体の機能的同等物である。用語「機能的同等物」には、例えば、相同配列を持つ抗体、キメラ抗体、人工的抗体および修飾抗体が含まれ、各機能同等物は、CA6に結合する能力によって定義される。当業者は、「抗体断片」と称される分子群および「機能的同等物」と称される群には重複があることを理解するであろう。機能的同等物を産生する方法は、例えば、本明細書にそれぞれの全体が援用される、PCT出願WO 93/21319、欧州特許出願第239,400号; PCT出願WO 89/09622;欧州特許出願338,745;および欧州特許出願EP 332,424に開示される。
【0090】
[108]相同配列を持つ抗体は、本発明の抗CA6抗体およびヒト化抗CA6抗体のアミノ酸配列と配列相同性を有するアミノ酸配列を持つ抗体である。好ましくは、相同性は、本発明の抗CA6抗体およびヒト化抗CA6抗体の可変領域のアミノ酸配列との相同性である。「配列相同性」は、本明細書において、アミノ酸配列に適用された際、例えば、PearsonおよびLipman, Proc. Natl. Acad. Sci. USA 85, 2444−2448(1988)にしたがったFASTA検索法によって決定した際、少なくとも約90%、91%、92%、93%、または94%の配列相同性、そしてより好ましくは、少なくとも約95%、96%、97%、98%、または99%の配列相同性を持つ配列と定義される。
【0091】
[109]本明細書において、キメラ抗体は、抗体の異なる部分が異なる動物種に由来するものである。例えば、ヒト免疫グロブリン定常領域と対形成した、ネズミ・モノクローナル抗体由来の可変領域を有する抗体がある。キメラ抗体を産生するための方法が当該技術分野に知られる。例えば、その全体が本明細書に援用される、Morrison, 1985, Science 229:1202; Oiら, 1986, BioTechniques 4:214; Gilliesら, 1989, J. Immunol. Methods 125:191−202;米国特許第5,807,715号;第4,816,567号;および第4,816,397号を参照されたい。
【0092】
[110]キメラ抗体のヒト化型は、ヒト・フレームワークドメイン内で、例えばマウス抗体の相補性決定領域を代用することによって、作製される。例えばPCT公報第WO92/22653号を参照されたい。ヒト化キメラ抗体は、好ましくは、対応するヒト抗体領域に実質的にまたはもっぱら由来する、相補性決定領域(CDR)以外の定常領域および可変領域、ならびにヒト以外の哺乳動物に実質的にまたはもっぱら由来するCDRを有する。
【0093】
[111]人工的な抗体には、各々、抗原結合能を有する、scFv断片、ディアボディ、トリアボディ、テトラボディおよびmruが含まれる(Winter, G.および
Milstein, C, 1991, Nature 349: 293−299; Hudson, P.J., 1999, Current Opinion in Immunology 11:548−557による概説を参照されたい)。一本鎖Fv断片(scFv)において、抗体のVHおよびVLドメインは、柔軟なペプチドによって連結される。典型的には、このリンカーペプチドは、長さ約15アミノ酸残基である。リンカーがはるかに小さいならば、例えば5アミノ酸であるならば、二価scFv二量体であるディアボディが形成される。リンカーが3アミノ酸残基未満に減少する場合、トリアボディおよびテトラボディと呼ばれる三量体および四量体構造が形成される。抗体の最少の結合単位は、別個に使用可能であるのに十分な特異的認識および結合を有するCDR、典型的には重鎖のCDR2である。こうした断片は、分子認識単位またはmruと呼ばれる。いくつかのこうしたmruは、短いリンカーペプチドと一緒に連結されて、したがって単一のmruより高いアビディティーを持つ人工的な結合タンパク質を形成してもよい。
【0094】
[112]本出願の機能的同等物にはまた、修飾抗体、例えば抗体への任意の種類の分子の共有結合によって修飾された抗体も含まれる。例えば、修飾抗体には、例えば、グリコシル化、アセチル化、PEG化、リン酸化、アミド化、既知の保護基/ブロッキング基による誘導体化、タンパク質分解切断、細胞リガンドまたは他のタンパク質への連結等によって修飾されている抗体が含まれる。こうした共有結合は、抗体が抗イディオタイプ応答を生じるのを妨げない。限定されるわけではないが、特異的化学切断、アセチル化、ホルミル化、ツニカマイシンの代謝合成等を含む、既知の技術によって、これらの修飾を行ってもよい。さらに、修飾抗体は、1以上の非古典的アミノ酸を含有してもよい。
【0095】
[113]異なるフレームワーク内の異なる鎖上の異なるCDRを交換することによって、機能的同等物を生じてもよい。したがって、例えば、IgG1〜4、IgM、IgA1〜2、IgD、IgE抗体タイプおよびアイソタイプを産生可能な、異なる重鎖の置換によって、所定のCDRセットに関して、抗体の異なるクラスにすることも可能である。同様に、完全に合成であるフレームワーク内に、所定のCDRセットを包埋することによって、本発明の範囲内の人工的抗体を産生してもよい。
【0096】
[114]当該技術分野に知られる非常に多様な方法を用いて、CDRの特定のセットに隣接する可変および/または定常領域配列内の突然変異、欠失および/または挿入によって、機能的同等物を容易に産生可能である。
【0097】
[115]本発明の抗体断片および機能的同等物は、DS6抗体に比較した際、CA6に検出可能な度合いで結合する分子を含む。検出可能な度合いの結合には、CA6に対するネズミDS6抗体の結合能の少なくとも10〜100%、好ましくは少なくとも50%、60%または70%、より好ましくは少なくとも75%、80%、85%、90%、95%または99%の範囲のすべての値が含まれる。
【0098】
改善された抗体
[116]CDRは、エピトープ認識および抗体結合に、最も重要である。しかし、抗体がその同族(cognate)エピトープを認識し、そして結合する能力に干渉することなく、CDRを含む残基に変化を作製してもよい。例えば、エピトープ認識に影響を及ぼさず、さらにエピトープに対する抗体の結合親和性を増加させる変化を作製してもよい。
【0099】
[117]したがって、本発明の範囲内にやはり含まれるのは、好ましくは増加した親和性で、やはりCA6を特異的に認識し、そしてこれに結合する、ネズミ抗体およびヒト化抗体の両方の改善型である。
【0100】
[118]いくつかの研究によって、一次抗体配列の知識に基づいて、結合および発現レベルなどの特性に対して、抗体の配列の多様な位で、1以上のアミノ酸変化を導入する影響が調べられてきている(Yang, W. P.ら, 1995, J. Mol.
Biol., 254, 392−403; Rader, C.ら, 1998, Proc. Natl. Acad. Sci. USA, 95, 8910−8915; Vaughan, T. J.ら, 1998, Nature Biotechnology, 16, 535−539)。
【0101】
[119]これらの研究において、オリゴヌクレオチドが仲介する部位特異的突然変異誘発、カセット突然変異誘発、変異性(error−prone)PCR、DNAシャフリング、または大腸菌(E. coli)の突然変異誘発株などの方法を用いて、CDR1、CDR2、CDR3、またはフレームワーク領域において、重鎖および軽鎖遺伝子の配列を変化させることによって、一次抗体の同等物が生成されてきている(Vaughan, T. J.ら, 1998, Nature Biotechnology, 16, 535−539; Adey, N. B.ら, 1996, 第16章, pp. 277−291, “Phage Display of Peptides and Proteins”中, Kay, B. K.ら監修, Academic Press)。一次抗体配列を変化させるこれらの方法は、二次抗体の改善された親和性を生じている(Gram, H.ら, 1992, Proc. Natl. Acad. Sci. USA, 89, 3576−3580; Boder, E. T.ら, 2000, Proc. Natl. Acad. Sci. USA, 97, 10701−10705; Davies, J.およびRiechmann, L., 1996, Immunotechnolgy, 2, 169−179; Thompson, J.ら, 1996, J. Mol. Biol., 256, 77−88; Short, M. K.ら, 2002, J. Biol. Chem., 277, 16365−16370; Furukawa, K.ら, 2001, J.
Biol. Chem., 276, 27622−27628)。
【0102】
[120]抗体の1以上のアミノ酸残基を変化させる類似の指令戦略によって、本発明で記載する抗体配列を用いて、CA6に対する改善された親和性を含む、改善された機能を持つ抗CA6抗体を開発してもよい。
【0103】
[121]改善された抗体にはまた、動物免疫、ハイブリドーマ形成、および特定の性質を持つ抗体に関する選択の標準的技術によって調製される、改善された性質を有する抗体が含まれる。
【0104】
細胞傷害性剤
[122]本発明の細胞傷害性コンジュゲートで用いる細胞傷害性剤は、細胞の死を生じるか、細胞の死を誘導するか、または何らかの方式で細胞生存度を減少させる、いかなる化合物であってもよい。好ましい細胞傷害性剤には、例えば、以下に定義される、メイタンシノイドおよびメイタンシノイド類似体、タキソイド、CC−1065およびCC−1065類似体、ドラスタチンおよびドラスタチン類似体が含まれる。これらの細胞傷害性剤を、本明細書に開示するような抗体、抗体断片、機能的同等物、改善された抗体およびその類似体にコンジュゲート化する。
【0105】
[123]in vitro法によって、細胞傷害性コンジュゲートを調製してもよい。抗体に薬剤またはプロドラッグを連結するため、連結基を用いる。適切な連結基が当該技術分野に周知であり、そしてこうした連結基には、ジスルフィド基、チオエーテル基、酸不安定性基、光解離基、ペプチダーゼ不安定性基およびエステラーゼ不安定性基が含まれる。好ましい連結基はジスルフィド基およびチオエーテル基である。例えば、ジスルフ
ィド交換反応を用いて、あるいは抗体および薬剤またはプロドラッグ間にチオエーテル結合を形成することによって、コンジュゲートを構築してもよい。
【0106】
メイタンシノイド
[124]細胞傷害性コンジュゲートを形成するため、本発明で使用可能な細胞傷害性剤の中に、メイタンシノイドおよびメイタンシノイド類似体がある。適切なメイタンシノイドの例には、メイタンシノールおよびメイタンシノール類似体が含まれる。メイタンシノイドは、微小管形成を阻害し、そして哺乳動物細胞に対して非常に毒性である薬剤である。
【0107】
[125]適切なメイタンシノール類似体の例には、修飾芳香族環を有するものおよび他の位で修飾を有するものが含まれる。こうした適切なメイタンシノイドが、米国特許第4,424,219号;第4,256,746号;第4,294,757号;第4,307,016号;第4,313,946号;第4,315,929号;第4,331,598号;第4,361,650号;第4,362,663号;第4,364,866号;第4,450,254号;第4,322,348号;第4,371,533号;第6,333,410号;第5,475,092号;第5,585,499号;および第5,846,545号に開示される。
【0108】
[126]修飾芳香族環を有するメイタンシノールの適切な類似体の特定の例には:
[127](1)C−19−デクロロ(米国特許第4,256,746号)(アンサマイトシンP2のLAH還元によって調製);
[128](2)C−20−ヒドロキシ(またはC−20−デメチル)+/−C−19−デクロロ(米国特許第4,361,650号および第4,307,016号)(ストレプトミセス属(Streptomyces)またはアクチノミセス属(Actinomyces)を用いた脱メチル化、あるいはLAHを用いた脱クロロ化によって調製);および
[129](3)C−20−デメトキシ、C−20−アシルオキシ(−OCOR)、+/−デクロロ(米国特許第4,294,757号)(塩化アシルを用いたアシル化によって調製)
が含まれる。
【0109】
[130]他の位の修飾を有するメイタンシノールの適切な類似体の特定の例には:
[131](1)C−9−SH(米国特許第4,424,219号)(H2SまたはP2S5とメイタンシノールの反応によって調製);
[132](2)C−14−アルコキシメチル(デメトキシ/CH2OR)(米国特許第4,331,598号);
[133](3)C−14−ヒドロキシメチルまたはアシルオキシメチル(CH2OHまたはCH2OAc)(米国特許第4,450,254号)(ノカルジア属(Nocardia)から調製);
[134](4)C−15−ヒドロキシ/アシルオキシ(米国特許第4,364,866号)(ストレプトミセス属によるメイタンシノールの変換によって調製);
[135](5)C−15−メトキシ(米国特許第4,313,946号および第4,315,929号)(トレウィア・ヌディフロラ(Trewia nudiflora)から単離);
[136](6)C−18−N−デメチル(米国特許第4,362,663号および第4,322,348号)(ストレプトミセス属によるメイタンシノールの脱メチル化によって調製);および
[137](7)4,5−デオキシ(米国特許第4,371,533号)(メイタンシノールの三塩化チタン/LAH還元によって調製)
が含まれる。
【0110】
[138]好ましい態様において、本発明の細胞傷害性コンジュゲートは、細胞傷害性剤として、以前、N2’−デアセチル−N2’−(3−メルカプト−1−オキソプロピル)−メイタンシンと称された、チオール含有メイタンシノイド(DM1)を利用する。DM1は、以下の構造式(I):
【0111】
【化14】
【0112】
によって示される。
[139]別の好ましい態様において、本発明の細胞傷害性コンジュゲートは、細胞傷害性剤として、チオール含有メイタンシノイド、N2’−デアセチル−N2’−(4−メチル−4−メルカプト−1−オキソペンチル)−メイタンシンを利用する。DM4は、以下の構造式(II):
【0113】
【化15】
【0114】
によって示される。
[140]本発明のさらなる態様において、イオウ原子を所持する炭素原子上に、モノまたはジアルキル置換を所持する、チオールおよびジスルフィド含有メイタンシノイドを含む、他のメイタンシンを用いてもよい。これらには、C−3、C−14ヒドロキシメチル、C−15ヒドロキシ、またはC−20デスメチルに、立体障害を受けた(hindered)スルフヒドリル基を所持するアシル基を含むアシル化アミノ酸側鎖を有するメイタンシノイドが含まれ、ここで、チオール官能性を所持するアシル基の炭素原子は、1つまたは2つの置換基を有し、前記置換基は、CH3、C2H5、1〜10の炭素原子を有する直鎖もしくは分枝鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する環状アルキルもしくはアルケニル、フェニル、置換フェニル、または複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、置換基の1つはHであってもよく、そ
してアシル基は、カルボニル官能性およびイオウ原子間に少なくとも3つの炭素原子の長さの直鎖を有する。
【0115】
[141]こうしたさらなるメイタンシンには、式(III)によって示される化合物:
【0116】
【化16】
【0117】
式中:
Y’は
(CR7CR8)l(CR9=CR10)pC=CqAr(CR5CR6)mDu(CR11=CR12)r(C=C)sBt(CR3CR4)nCR1R2SZ
を示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
A、B、Dは、3〜10炭素原子を有するシクロアルキルもしくはシクロアルケニル、単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
R3、R4、R5、R6、R7、R8、R9、R11、およびR12は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、m、n、o、p、q、r、s、およびtは、各々独立に、0または1〜5の整数であり、但し、どの時点においても、l、m、n、o、p、q、r、s、およびtの少なくとも2つはゼロではなく;そして
Zは、H、SRまたは−CORであり、式中、Rは、1〜10炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルである
が含まれる。
【0118】
[142]式(III)の好ましい態様には:
R1がHであり、R2がメチルであり、そしてZがHである
R1およびR2がメチルであり、そしてZがHである
R1がHであり、R2がメチルであり、そしてZが−SCH3である
R1およびR2がメチルであり、そしてZが−SCH3である
式(III)の化合物が含まれる。
【0119】
[143]こうしたさらなるメイタンシンにはまた、式(IV−L)、(IV−D)、または(IV−D,L):
【0120】
【化17】
【0121】
式中:
Yは、(CR7CR8)l(CR5CR6)m(CR3CR4)nCR1R2SZを示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニル、または複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
R3、R4、R5、R6、R7およびR8は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニル、または複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、mおよびnは、各々独立に、1〜5の整数であり、そしてさらに、nはゼロであってもよく;
Zは、H、SRまたは−CORであり、式中、Rは、1〜10炭素原子を有する直鎖もしくは分枝鎖アルキルもしくはアルケニル、3〜10炭素原子を有する環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;そして
Mayは、C−3、C−14ヒドロキシメチル、C−15ヒドロキシまたはC−20デスメチルに側鎖を所持するメイタンシノイドを示す
によって示される化合物も含まれる。
【0122】
[144]式(IV−L)、(IV−D)および(IV−D,L)の好ましい態様には:
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり、lおよびmが各々1であり、nがゼロであり、そしてZがHである
R1およびR2がメチルであり、R5、R6、R7、R8が各々Hであり、lおよびmが1であり、nがゼロであり、そしてZがHである
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり、lおよびmが各々1であり、nがゼロであり、そしてZが−SCH3である
R1およびR2がメチルであり、R5、R6、R7、R8が各々Hであり、lおよびmが1であり、nがゼロであり、そしてZが−SCH3である
式(IV−L)、(IV−D)および(IV−D,L)の化合物が含まれる。
【0123】
[145]好ましくは、細胞傷害性剤は、式(IV−L)によって示される。
[146]こうしたさらなるメイタンシンにはまた、式(V):
【0124】
【化18】
【0125】
式中:
Yは、(CR7CR8)l(CR5CR6)m(CR3CR4)nCR1R2SZを示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
R3、R4、R5、R6、R7およびR8は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニル、または複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、mおよびnは、各々独立に、1〜5の整数であり、そしてさらに、nはゼロであってもよく;そして
Zは、H、SRまたは−CORであり、式中、Rは、1〜10炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルである
によって示される化合物も含まれる。
【0126】
[147]式(V)の好ましい態様には:
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり;lおよびmが各々1であり;nがゼロであり;そしてZがHである
R1およびR2がメチルであり;R5、R6、R7、R8が各々Hであり、lおよびmが1であり;nがゼロであり;そしてZがHである
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり、lおよびmが各々1であり;nがゼロであり;そしてZが−SCH3である
R1およびR2がメチルであり、R5、R6、R7、R8が各々Hであり、lおよびmが1であり、nがゼロであり、そしてZが−SCH3である
式(V)の化合物が含まれる。
【0127】
[148]こうしたさらなるメイタンシンにはさらに、式(VI−L)、(VI−D)、または(VI−D,L):
【0128】
【化19】
【0129】
式中:
Y2は、(CR7CR8)l(CR5CR6)m(CR3CR4)nCR1R2SZ2を示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
R3、R4、R5、R6、R7およびR8は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖、環状アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、mおよびnは、各々独立に、1〜5の整数であり、そしてさらに、nはゼロであってもよく;
Z2は、SRまたはCORであり、式中、Rは、1〜10炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;そして
Mayはメイタンシノイドである
によって示される化合物が含まれる。
【0130】
[149]こうしたさらなるメイタンシンにはまた、式(VII)によって示される化合物:
【0131】
【化20】
【0132】
式中:
Y2’は
(CR7CR8)l(CR9=CR10)p(C=C)qAr(CR5CR6)mDu(CR11=CR12)r(C=C)sBt(CR3CR4)nCR1R2SZ2
を示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖もしくは分枝鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
A、B、およびDは、各々独立に、3〜10炭素原子を有するシクロアルキルまたはシクロアルケニル、単純または置換アリール、または複素環芳香族またはヘテロシクロアルキルラジカルであり;
R3、R4、R5、R6、R7、R8、R9、R11、およびR12は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、m、n、o、p、q、r、s、およびtは、各々独立に、0または1〜5の整数であり、但し、どの時点においても、l、m、n、o、p、q、r、s、およびtの少なくとも2つはゼロではなく;そして
Z2は、SRまたは−CORであり、式中、Rは、1〜10炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルである
も含まれる。
【0133】
[150]式(VII)の好ましい態様には:
R1がHであり、そしてR2がメチルである
式(VII)の化合物が含まれる。
【0134】
[151]上述のメイタンシノイドを、抗CA6抗体DS6、あるいはその相同体または断片にコンジュゲート化してもよく、ここで、抗体は、メイタンシノイドのC−3、C−14ヒドロキシメチル、C−15ヒドロキシまたはC−20デスメチルに見られるアシ
ル化アミノ酸側鎖のアシル基上に存在するチオールまたはジスルフィド官能性を用いて、メイタンシノイドに連結され、そしてアシル化アミノ酸側鎖のアシル基が、1つまたは2つの置換基を有する炭素原子に位置する、チオールまたはジスルフィド官能性を有し、前記置換基は、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、置換基の1つはHであってもよく、そしてアシル基は、カルボニル官能性およびイオウ原子間に少なくとも3つの炭素原子の長さの直鎖を有する。
【0135】
[152]本発明の好ましいコンジュゲートは、式(VIII):
【0136】
【化21】
【0137】
式中:
Y1’は
(CR7CR8)l(CR9=CR10)p(C=C)qAr(CR5CR6)mDu(CR11=CR12)r(C=C)sBt(CR3CR4)nCR1R2S−
を示し、
式中:
A、B、およびDは、各々独立に、3〜10炭素原子を有するシクロアルキルもしくはシクロアルケニル、単純もしくは置換アリール、または複素環芳香族またはヘテロシクロアルキルラジカルであり;
R3、R4、R5、R6、R7、R8、R9、R11、およびR12は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;そして
l、m、n、o、p、q、r、s、およびtは、各々独立に、0または1〜5の整数であり、但し、どの時点においても、l、m、n、o、p、q、r、s、およびtの少なくとも2つはゼロではない
のメイタンシノイドにコンジュゲート化された、抗抗CA6抗体DS6、あるいはその相同体または断片を含むものである。
【0138】
[153]好ましくは、R1はHであり、そしてR2はメチルであるか、またはR1およびR2はメチルである。
[154]本発明のさらにより好ましいコンジュゲートは、式(IX−L)、(IX−D)、または(IX−D,L):
【0139】
【化22】
【0140】
式中:
Y1は、(CR7CR8)l(CR5CR6)m(CR3CR4)nCR1R2S−を示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニル、複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
R3、R4、R5、R6、R7およびR8は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、mおよびnは、各々独立に、1〜5の整数であり、そしてさらに、nはゼロであってもよく;そして
Mayは、C−3、C−14ヒドロキシメチル、C−15ヒドロキシまたはC−20デスメチルで側鎖を所持するメイタンシノールを示す
のメイタンシノイドにコンジュゲート化された、抗CA6抗体DS6、あるいはその相同体または断片を含むものである。
【0141】
[155]式(IX−L)、(IX−D)および(IX−D,L)の好ましい態様には:
R1がHであり、そしてR2がメチルであるか、またはR1およびR2がメチルである、
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり;lおよびmが各々1であり;nが0である、
R1およびR2がメチルであり;R5、R6、R7、およびR8が各々Hであり;lおよびmが1であり;nが0である
式(IX−L)、(IX−D)および(IX−D,L)の化合物が含まれる。
【0142】
[156]好ましくは、細胞傷害性剤は、式(IX−L)によって示される。
[157]本発明のさらに好ましいコンジュゲートは、式(X):
【0143】
【化23】
【0144】
式中、置換基は、上記式(IX)に関して定義されるとおりである
のメイタンシノイドにコンジュゲート化された、抗CA6抗体DS6、あるいはその相同体または断片を含むものである。
【0145】
[158]特に好ましいのは、R1がHであり、R2がメチルであり、R5、R6、R7およびR8が各々Hであり、lおよびmが各々1であり、そしてnが0である、上記化合物のいずれかである。
【0146】
[159]さらに特に好ましいのは、R1およびR2がメチルであり、R5、R6、R7およびR8が、各々、Hであり、lおよびmが1であり、そしてnが0である、上記化合物のいずれかである。
【0147】
[160]さらに、L−アミノアシル立体異性体が好ましい。
[161]係属中の米国特許出願第10/849,136号、2004年5月20日出願に解説される各メイタンシノイドもまた、本発明の細胞傷害性コンジュゲートに使用可能である。米国特許出願第10/849,136号の全開示が、本明細書に援用される。
【0148】
ジスルフィド含有連結基
[162]DS6抗体などの細胞結合性剤にメイタンシノイドを連結するため、メイタンシノイドは連結基を含む。連結部分は、完全活性メイタンシノイドの放出を可能にする化学結合を特定の部位に含有する。適切な化学結合は、当該技術分野に周知であり、そしてこれには、ジスルフィド結合、酸不安定性結合、光解離結合、ペプチダーゼ不安定性結合およびエステラーゼ不安定性結合が含まれる。好ましいのはジスルフィド結合である。
【0149】
[163]連結部分はまた、反応性化学基も含む。好ましい態様において、反応性化学基を、ジスルフィド結合連結部分を介して、メイタンシノイドに共有結合させてもよい。
[164]特に好ましい反応性化学基は、N−スクシンイミジルエステルおよびN−スルホスクシンイミジルエステルである。
【0150】
[165]反応性化学基を含有する連結部分を含む、特に好ましいメイタンシノイドは、連結部分がジスルフィド結合を含有し、そして化学反応基がN−スクシンイミジルまたはN−スルホスクシンイミジルエステルを含む、メイタンシノールおよびその類似体のC−3エステルである。
【0151】
[166]メイタンシノイド上の多くの位が、連結部分を化学的に連結する位として働きうる。例えば、ヒドロキシル基を有するC−3位、ヒドロキシメチルで修飾されたC−14位、ヒドロキシで修飾されたC−15位、およびヒドロキシ基を有するC−20位がすべて、有用であると期待される。しかし、C−3位が好ましく、そしてメイタンシノールのC−3位が特に好ましい。
【0152】
[167]連結部分を有するメイタンシノールのエステルの合成を、ジスルフィド結合含有連結部分の観点で記載する一方、当業者は、他の化学結合(上述のようなもの)を持つ連結部分もまた、本発明で使用可能であり、他のメイタンシノイドも使用可能であることを理解するであろう。他の化学結合の特定の例には、酸不安定性結合、光解離結合、ペプチダーゼ不安定性結合およびエステラーゼ不安定性結合が含まれる。本明細書に援用される、米国特許第5,208,020号の開示は、こうした結合を所持するメイタンシノイドの産生を解説する。
【0153】
[168]反応性基を所持するジスルフィド部分を有するメイタンシノイドおよびメイタンシノイド誘導体の合成が、各々、本明細書に援用される、米国特許第6,441,163号および第6,333,410号、ならびに米国出願第10/161,651号に記載される。
【0154】
[169]DM1などの反応性基含有メイタンシノイドを、DS6抗体などの抗体と反応させて、細胞傷害性コンジュゲートを産生する。HPLCによって、またはゲルろ過によって、これらのコンジュゲートを精製してもよい。
【0155】
[170]こうした抗体メイタンシノイド・コンジュゲートを産生するためのいくつかの優れたスキームが、各々、その全体が本明細書に援用される、米国特許第6,333,410号、ならびに米国出願第09/867,598号、第10/161,651号および第10/024,290号に提供される。
【0156】
[171]一般的に、水性緩衝液中の抗体溶液を、反応性基を所持するジスルフィド部分を有する、モル過剰のメイタンシノイドとインキュベーションしてもよい。過剰なアミン(エタノールアミン、タウリンなど)の添加によって、反応混合物の反応を停止してもよい。次いで、ゲルろ過によって、メイタンシノイド−抗体コンジュゲートを精製してもよい。
【0157】
[172]252nmおよび280nmの吸光度の比を、分光光度的に測定することによって、抗体分子あたりに結合するメイタンシノイド分子の数を決定することができる。平均1〜10のメイタンシノイド分子/抗体分子が好ましい。
【0158】
[173]メイタンシノイド薬剤と抗体のコンジュゲートを、in vitroで、多様な望ましくない細胞株の増殖を抑制する能力に関して、評価することができる。例えば、ヒト類表皮癌細胞株A−431、ヒト小細胞肺癌細胞株SW2、ヒト乳房腫瘍細胞株SKBR3およびバーキットリンパ腫細胞株Namalwaなどの細胞株は、これらの化合物の細胞傷害性の評価のため、容易に使用可能である。評価しようとする細胞を化合物に24時間曝露して、そして既知の方法による直接アッセイにおいて、細胞の生存率を測定することができる。次いで、アッセイ結果からIC50値を計算することができる。
【0159】
PEG含有連結基
[174]メイタンシノイドをまた、米国出願第10/024,290号に示されるように、PEG連結基を用いて、細胞結合性剤に連結してもよい。これらのPEG連結基は
、水中および非水性溶媒中の両方で、可溶性であり、そしてこれを用いて、1以上の細胞傷害性剤を、細胞結合性剤に連結してもよい。典型的なPEG連結基には、一端の官能性スルフヒドリル基またはジスルフィド基、およびもう一端の活性エステルを通じて、リンカーの反対の端で、細胞傷害性剤および細胞結合性剤に結合する、ヘテロ二官能性PEGリンカーが含まれる。
【0160】
[175]PEG連結基を用いた、細胞傷害性コンジュゲートの合成の一般的な例として、特定の詳細に関して、米国出願第10/024,290号に再び言及する。合成は、反応性PEG部分を所持する1以上の細胞傷害性剤と細胞結合性剤の反応で始まり、この反応が、細胞結合性剤のアミノ酸残基による、各反応性PEG部分の末端活性エステルの置換を生じ、PEG連結基を通じて細胞結合性剤に共有結合した1以上の細胞傷害性剤を含む、細胞傷害性コンジュゲートを生じる。
【0161】
タキサン
[176]本発明記載の細胞傷害性コンジュゲートで用いる細胞傷害性剤はまた、タキサンまたはその誘導体であってもよい。
【0162】
[177]タキサンは、細胞傷害性天然産物であるパクリタキセル(タキソール)、および半合成誘導体であるドセタキセル(タキソテール)を含む化合物ファミリーであり、これらの2つの化合物は、癌の治療に広く用いられている。タキサンは、チューブリンの脱分極を阻害し、細胞死を生じる、紡錘体毒物である。ドセタキセルおよびパクリタキセルは、癌治療に有用な剤であるが、正常細胞に対して非特異的に毒性であるため、その抗腫瘍活性は限定される。さらに、パクリタキセルおよびドセタキセルのような化合物は、それ自体、細胞結合性剤のコンジュゲートにおいて使用するには、十分に強力ではない。
【0163】
[178]細胞傷害性コンジュゲートの調製に使用するのに好ましいタキサンは、式(XI)のタキサン:
【0164】
【化24】
【0165】
である。
[179]抗体などの細胞結合性剤にタキサンをコンジュゲート化するための方法とともに、本発明の細胞傷害性コンジュゲートにおいて使用可能なタキサンを合成するための方法が、米国特許第5,416,064号、第5,475,092号、第6,340,701号、第6,372,738号および第6,436,931号、ならびに米国出願第10/024,290号、第10/144,042号、第10/207,814号、第10/210,112号および第10/369,563号に詳細に記載される。
【0166】
CC−1065類似体
[180]本発明記載の細胞傷害性コンジュゲートで使用する細胞傷害性剤はまた、CC−1065またはその誘導体であってもよい。
【0167】
[181]CC−1065は、ストレプトミセス・ゼレンシス(Streptomyces zelensis)の培養ブロスから単離される、強力な抗腫瘍抗生物質である。CC−1065は、ドキソルビシン、メトトレキセートおよびビンクリスチンなどの、一般的に用いられる抗癌薬剤よりも、in vitroで、約1000倍強力である(B.K. Bhuyanら, Cancer Res., 42, 3532−3537(1982))。CC−1065およびその類似体は、米国特許第6,372,738号、第6,340,701号、第5,846,545号および第5,585,499号に開示される。
【0168】
[182]CC−1065の細胞傷害能は、そのアルキル化活性、およびそのDNA結合またはDNA挿入活性と相関付けられてきている。これらの2つの活性は、分子の別個の部分に属する。したがって、アルキル化活性は、シクロプロパピロロインドール(CPI)・サブユニットに含有され、そしてDNA結合活性は、2つのピロロインドール・サブユニット中に属する。
【0169】
[183]CC−1065は、細胞傷害性剤として特定の魅力的な特徴を有するが、療法的使用においては限界を有する。マウスにCC−1065を投与すると、遅延型肝毒性が引き起こされ、12.5μg/kgの単回静脈内用量後、第50日での死亡を導く{V. L. Reynoldsら, J. Antibiotics, XXIX, 319−334(1986)}。このことから、遅延型毒性を引き起こさない類似体を開発する努力に拍車がかかり、そしてCC−1065をモデルとするより単純な類似体の合成が記載されてきている{M.A. Warpehoskiら, J. Med. Chem., 31, 590−603(1988)}。
【0170】
[184]別の一連の類似体において、CPI部分がシクロプロパベンズインドール(CBI)部分によって置換された{D.L. Bogerら, J. Org. Chem., 55, 5823−5833, (1990), D.L. Bogerら, BioOrg. Med. Chem. Lett., 1, 115−120(1991)}。これらの化合物は、マウスにおいて、遅延型毒性を引き起こすことなく、親薬剤の高いin vitro強度を維持する。CC−1065同様、これらの化合物は、共有結合性の方式でDNAの副溝に結合して、細胞死を生じる、アルキル化剤である。しかし、最も有望な類似体、アドゼレシンおよびカルゼレシンの臨床的評価は、期待はずれの結果を導いた{B.F. Fosterら, Investigational New Drugs, 13, 321−326(1996); I. Wolffら, Clin. Cancer Res., 2,1717−1723(1996)}。これらの薬剤は、全身毒性が高いため、劣った療法効果を示す。
【0171】
[185]腫瘍部位へのターゲティング送達を通じて、in vivo分布を変化させ、ターゲティングされない組織に対するより低い毒性、そしてしたがって、より低い全身毒性を生じることによって、CC−1065類似体の療法的有効性を非常に改善することも可能である。この目的を達成するため、腫瘍細胞を特異的にターゲティングする、細胞結合性剤とCC−1065の類似体および誘導体のコンジュゲートが記載されてきている{米国特許第5,475,092号;第5,585,499号;第5,846,545号}。これらのコンジュゲートは、典型的には、in vitroで高いターゲット特異的細胞傷害性を示し、そしてマウスにおけるヒト腫瘍異種移植片モデルにおいて、ひときわ優れた抗腫瘍活性を示す{R.V.J. Chariら, Cancer Res., 55, 4079−4084(1995)}。
【0172】
[186]抗体などの細胞結合性剤に類似体をコンジュゲート化するための方法とともに、本発明の細胞傷害性コンジュゲートで使用可能なCC−1065類似体を合成するための方法が、米国特許第5,475,092号、第5,846,545号、第5,585,499号、第6,534,660号および第6,586,618号に、そして米国出願第10/116,053号および第10/265,452号に、詳細に記載される。
【0173】
他の薬剤
[187]メトトレキセート、ダウノルビシン、ドキソルビシン、ビンクリスチン、ビンブラスチン、メルファラン、マイトマイシンC、クロラムブシル、カリケアマイシン、チューブリシン(tubulysin)およびチューブリシン類似体、デュオカルマイシンおよびデュオカルマイシン類似体、ドラスタチンおよびドラスタチン類似体などの薬剤もまた、本発明のコンジュゲートの調製に適している。また、血清アルブミンなどの仲介キャリアー分子を通じて、抗体分子に薬剤分子を連結させてもよい。米国第09/740991号に記載されるように、ドキソルビシンおよびダウノルビシン化合物もまた、有用な細胞傷害性剤でありうる。
【0174】
療法組成物
[188]本発明はまた:
(a)1以上の細胞傷害性コンジュゲートの有効量、および
(b)薬学的に許容しうるキャリアー
を含む療法組成物も提供する。
【0175】
[189]同様に、本発明は、選択した細胞集団の増殖を阻害するための方法であって、ターゲット細胞、またはターゲット細胞を含有する組織を、有効量の細胞傷害性コンジュゲート、または細胞傷害性コンジュゲートを含む療法剤と、単独で、あるいは他の細胞傷害性剤または療法剤と組み合わせて、接触させることを含む、前記方法を提供する。
【0176】
[190]本発明はまた、本発明の療法組成物を用いて、癌を有する被験体を治療するための方法も含む。
[191]先に記載される方法によって、細胞傷害性コンジュゲートを、in vitro強度および特異性に関して評価することができる(例えば、R.V.J. Chariら, Cancer Res. 55:4079−4084(1995)を参照されたい)。やはり先に記載される方法によって、マウスにおけるヒト腫瘍異種移植モデルにおいて、抗腫瘍活性を評価することができる(例えば、Liuら, Proc. Natl. Acad. Sci. 93:8618−8623(1996)を参照されたい)。
【0177】
[192]適切な薬学的に許容しうるキャリアーが周知であり、そして臨床状況が保証するように、一般の当業者によって決定可能である。本明細書において、キャリアーには希釈剤および賦形剤が含まれる。
【0178】
[193]適切なキャリアー、希釈剤および/または賦形剤の例には:(1)約1mg/ml〜25mg/mlのヒト血清アルブミンを含有するかまたは含有しない、ダルベッコのリン酸緩衝生理食塩水、pH〜7.4、(2)0.9%生理食塩水(0.9%w/v塩化ナトリウム(NaCl))、および(3)5%(w/v)デキストロースが含まれ;そしてまた、こうしたキャリアーは、トリプタミンなどの酸化防止剤、およびTween20などの安定化剤を含有してもよい。
【0179】
[194]選択した細胞集団の増殖を阻害するための方法を、in vitro、in
vivo、またはex vivoで実施することができる。本明細書において、増殖の
阻害は、短期間であれ、または長期間であれ、細胞の増殖の遅延、細胞生存度の減少、細胞死を引き起こすこと、細胞溶解、および細胞死の誘導を意味する。
【0180】
[195]in vitro使用の例には、疾患細胞または悪性細胞を殺すための、同じ患者への移植前の自己骨髄の治療;適格性T細胞を殺し、そして移植片対宿主病(GVHD)を防止するための、移植前の骨髄の治療;ターゲット抗原を発現していない望ましい変異体以外のすべての細胞を殺すか;または望ましくない抗原を発現する変異体を殺すための、細胞培養の治療が含まれる。
【0181】
[196]非臨床的in vitro使用の条件は、一般の当業者によって容易に決定される。
[197]臨床的ex vivo使用の例は、癌治療において、または自己免疫疾患治療において、自己移植前に、骨髄から腫瘍細胞またはリンパ球を取り除くこと、あるいは移植片対宿主病(GVHD)を防止するため、移植前に、自己または同種骨髄または組織からT細胞または他のリンパ球を取り除くことである。治療を以下のように行ってもよい。骨髄を患者または他の個体から採取し、そして次いで、本発明の細胞傷害性剤を添加した、血清を含有する培地中でインキュベーションする。濃度は、約10μM〜1pMの範囲であり、約37℃で約30分間〜約48時間、インキュベーションする。濃度およびインキュベーション時間、すなわち用量の正確な条件は、一般の当業者によって容易に決定される。インキュベーション後、血清を含有する培地で、骨髄細胞を洗浄し、そして既知の方法にしたがって、i.v.注入によって、患者に戻す。患者が骨髄採取時および処置細胞の再注入の間に切除化学療法または全身照射過程などの他の治療を受ける状況下では、標準的医学装置を用いて、液体窒素中で処置骨髄細胞を凍結保存してもよい。
【0182】
[198]臨床的in vivo使用のため、本発明の細胞傷害性コンジュゲートを溶液として供給して、無菌性および内毒素レベルに関して試験する。細胞傷害性コンジュゲート投与の適切なプロトコルの例は、以下のとおりである。コンジュゲートを毎週、4週間、i.v.ボーラスとして投与する。ボーラス用量は、生理食塩水50〜100ml中で投与され、これに5〜10mlのヒト血清アルブミンを添加してもよい。投薬量は、i.v.投与あたり10μg〜100mgである(1日あたり、100ng〜1mg/kgの範囲)。より好ましくは、投薬量は、50μg〜30mgの範囲であろう。最も好ましくは、投薬量は、1mg〜20mgの範囲であろう。治療4週後、患者に、毎週、治療を投与し続けてもよい。臨床状況が保証するように、投与経路、賦形剤、希釈剤、投薬量、時間等に関して、特定の臨床プロトコルを、一般の当業者が決定してもよい。
【0183】
[199]選択した細胞集団を殺すin vivo法またはex vivo法にしたがって治療可能な医学的条件の例には、CA6が発現されている、例えば肺、乳房、結腸、前立腺、腎臓、膵臓、卵巣、子宮頸部およびリンパ器官の癌、骨肉腫、滑膜癌、肉腫または癌腫、ならびにCA6グリコトープが主に発現されている、まだ決定されていない他の癌を含む、いかなる種類の悪性腫瘍も含まれ;全身性狼瘡(systemic lupus)、関節リウマチ、および多発性硬化症などの自己免疫疾患;腎移植拒絶、肝移植拒絶、肺移植拒絶、心臓移植拒絶、および骨髄移植拒絶などの移植片拒絶;移植片対宿主病;mV感染、HIV感染、AIDS等のウイルス感染;ならびにランブルべん毛虫症、アメーバ症、住血吸虫症などの寄生虫感染、および一般の当業者によって決定されるような他のものが含まれる。
【0184】
キット
[200]本発明にはまた、例えば記載する細胞傷害性コンジュゲート、および特定の細胞種を殺すための細胞傷害性コンジュゲートの使用のための説明書を含むキットも含まれる。使用説明書には、in vitro、in vivoまたはex vivoで細胞
傷害性コンジュゲートを使用するための指示が含まれてもよい。
【0185】
[201]典型的には、キットは、細胞傷害性コンジュゲートを含有する区画を有するであろう。細胞傷害性コンジュゲートは、凍結乾燥型、液体型、またはキットに含まれるよう受け入れ可能な他の型であってもよい。キットはまた、キット中の使用説明書に記載される方法を実施するのに必要なさらなる要素、例えば凍結乾燥粉末を再構成するための滅菌溶液、患者に投与する前に、細胞傷害性コンジュゲートと組み合わせるためのさらなる剤、および患者にコンジュゲートを投与するのに役立つツールも含有してもよい。
【0186】
さらなる態様
[202]本発明は、研究または診断適用で使用するためにさらに標識されている、モノクローナル抗体、ヒト化抗体およびそのエピトープ結合性断片をさらに提供する。好ましい態様において、標識は、放射標識、蛍光体、発色団、造影剤または金属イオンである。
【0187】
[203]前記の標識ヒト化抗体またはそのエピトープ結合性断片を、癌を有すると推測される被験体に投与し、そして被験体体内の標識分布を測定するかまたは監視する、診断法もまた、提供する。
【実施例】
【0188】
実施例
[204]本発明の広い範囲は、以下の実施例を参照すると最適に理解され、これらは本発明を特定の態様に限定することを意図しない。
【0189】
実施例1:フローサイトメトリー結合アッセイによる、抗原陽性および陰性細胞株の同定
[205]フローサイトメトリー分析を用いて、DS6エピトープ、CA6を細胞表面に局在させた。OVCAR5(Kearseら, Int. J. Cancer 88(6):866−872(2000))、OVCAR8およびIGROV1細胞(M. Seiden, Massachusetts General Hospital)を除いて、ヒト細胞株をアメリカン・タイプ・カルチャー・コレクション(ATCC)から得た。本明細書において、以後、培地と称する、4mM L−グルタミン、50U/mlペニシリン、50μg/mlストレプトマイシン(Cambrex Bio Science、メイン州ロックランド)および10%v/vウシ胎児血清(Atlas Biologicals、コロラド州フォートコリンズ)を補ったRPMI1640中で、すべての細胞を増殖させた。細胞を37℃、5%CO2加湿インキュベーター中で維持した。
【0190】
[206]96ウェルプレート中、FACS緩衝液(2%ヤギ血清、RPMI)中で調製した連続希釈濃度のDS6抗体と、細胞(1〜2x10−5細胞/ウェル)を氷上で3〜4時間インキュベーションした。卓上遠心分離装置中、1500rpm、4℃で5分間、細胞を回転させて落とした。培地を取り除いた後、ウェルに150μlのFACS緩衝液を再度満たした。次いで、洗浄工程を反復した。FITC標識ヤギ抗マウスIgG(Jackson Immunoresearch)を、FACS緩衝液に対して1:100に希釈し、そして氷上で1時間、細胞とインキュベーションした。プレートをホイルで覆い、シグナルの光退色を防いだ。2回の洗浄後、細胞を1%ホルムアルデヒドで固定し、そしてフローサイトメーター上で分析した。
【0191】
[207]腫瘍免疫組織化学から予測されるように、CA6エピトープは、主に、卵巣、乳房、子宮頸部、および膵臓起源の細胞株で見られる(表3)。しかし、他の腫瘍種のいくつかの細胞株が、限定されたCA6発現を示した。DS6抗体は、135.6pMの見かけのKDで結合する(PC−3細胞において、表3)。抗原陽性細胞株における結合
曲線(図1)の最大平均蛍光(表3)は、相対的抗原密度を示唆する。
表3
【0192】
【表4】
【0193】
*平均最大相対的平均蛍光
【0194】
実施例2:DS6エピトープの性質決定
[208]タンパク質分解(プロナーゼおよびプロテイナーゼK)および/または糖分解(ノイラミニダーゼおよび過ヨウ素酸)処理で消化した、CA6陽性細胞溶解物(Caov−3)のドットブロットを免疫ブロッティングすることによって、DS6抗原、CA6の特性を分析した。陽性対照には、多様なエピトープ種を認識する他の抗体を、抗原陽性細胞株の溶解物に対して試験した(Caov−3およびCM1;Colo205およびC242;SKMEL28およびR24)。CM1は、Muc−1の可変数タンデム反復ドメイン(VNTR)のタンパク質エピトープを認識する抗体であり、そしてしたがって、タンパク質エピトープに関する対照を提供する。C242は、Muc−1上の新規結腸直腸癌特異的シアル酸依存性グリコトープ(CanAg)に結合し、これはタンパク質上のグリコトープに関する対照を提供する。R24は、黒色腫に特異的なGD3ガングリオシドに結合し、そしてしたがって、非タンパク質骨格上のグリコトープに関する対照を提供する。
【0195】
[209]Caov−3、Colo205、およびSKMEL28細胞を15cm組織培養プレート中にプレーティングした。培地(30ml/プレート)を溶解前日に新しくした。修飾RIPA緩衝液(50mM Tris−HCl pH7.6、150mM NaCl、5mM EDTA、1%NP40、0.25%デオキシコール酸ナトリウム)、プロテアーゼ阻害剤(PMSF、ペプスタチンA、ロイペプチン、およびアプロチニン)、およびPBSを氷上であらかじめ冷却した。培地をプレートから吸引した後、10mlの冷却PBSで細胞を2回洗浄した。続くすべての工程を、氷上および/または4℃の低
温室で行った。PBSの最後の洗浄液を吸引した後、溶解緩衝液(最終濃度1mM PMSF、1μMペプスタチンA、10μg/mlロイペプチン、および2μg/mlアプロチニンまで、プロテアーゼ阻害剤を新鮮に添加した、RIPA緩衝液)1〜2ml中で細胞を溶解した。細胞リフターを用いて、プレートから溶解物をこそげ落とし、そして18G針で懸濁物を上下に(5〜10回)ピペッティングすることによって、すりつぶした(triturated)。溶解物を10分間回転させ、そして次いで、最大出力で10分間微量遠心分離装置中で遠心分離した(13K rpm)。ペレットを廃棄し、そして次いで、ブラッドフォード・タンパク質アッセイキット(Biorad)を用いて上清をアッセイした。
【0196】
[210]乾いた0.2μmニトロセルロース膜上に、溶解物(2μl)を直接ピペッティングした。スポットをおよそ30分間風乾させた。膜を、各々、単一のスポットを含有するピースに分けた。プロナーゼ(1mg/ml酵素、50mM Tris pH7.5、5mM CaCl2)、プロテイナーゼK(1mg/ml酵素、50mM Tris
pH7.5、5mM CaCl2)、ノイラミニダーゼ(20mU/ml酵素、50mM酢酸ナトリウムpH5、5mM CaCl2、100μg/ml BSA)または過ヨウ素酸(20mM、0.5M酢酸ナトリウムpH5)の存在下で、スポットを37℃で1時間インキュベーションした。Roche(酵素)およびVWR(過ヨウ素酸)から試薬を購入した。T−TBS洗浄緩衝液(0.1%Tween20、1xTBS)中で膜を洗浄し(5分間)、ブロッキング緩衝液(3%BSA、T−TBS)中で、室温で2時間ブロッキングし、そしてブロッキング緩衝液中、2μg/mlの一次抗体(すなわちDS6、CM1、C242、R24)で一晩インキュベーションした。T−TBS中で5分間、膜を3回洗浄し、そして次いで、HRPコンジュゲート化ヤギ抗マウス(またはヒト)IgG二次抗体(Jackson Immunoresearch;ブロッキング緩衝液中、1:2000希釈)と、室温で1時間インキュベーションした。免疫ブロットを3回洗浄し、そしてECL系(Amersham)を用いて現像した。
【0197】
[211]消化した対照溶解物の免疫ブロット(図2)は、タンパク質分解処理によってCM1シグナルが破壊される一方、糖分解消化のシグナルは、タンパク質エピトープを認識する抗体に関して予測されるように、影響を受けなかったことを示した。C242シグナルは、タンパク質上に見られるグリコトープを認識する抗体に関して予測されるように、タンパク質分解処理または糖分解処理のいずれによっても破壊された。タンパク質分解処理によって影響を受けなかったR24シグナルは、ガングリオシドを認識する抗体に関して予測されるように、ノイラミニダーゼ処理または過ヨウ素酸処理で消滅した。消化したCaov−3溶解物のDS6免疫ブロットは、タンパク質分解化合物および糖分解化合物のいずれかでの処理に際して、シグナル喪失を示した。したがって、C242同様、DS6は、タンパク性コア上の炭水化物エピトープに結合する。さらに、DS6免疫ブロット中のシグナルは、ノイラミニダーゼ処理に感受性であった。したがって、CanAg同様、CA6は、シアル酸依存性グリコトープである。
【0198】
[212]CA6が炭水化物性であることを確認するため、Caov−3溶解物をPVDF膜上にスポッティングし、そして化学的脱グリコシル化剤、トリフルオロメタンスルホン酸(TFMSA)で、窒素下、周囲温度で5分間、処理した。ブロットをT−TBSで洗浄し、そしてCM1またはDS6のいずれかで免疫ブロッティングした(図3)。DS6シグナルは、酸処理で破壊され、CA6がグリコトープであるさらなる証拠が提供された。TFSMA処理に際してCM1シグナルが増進することから、酸処理がフィルター上のタンパク質に影響を及ぼさないことが示され、そして糖分解処理によって、CM1が認識するタンパク質エピトープが暴露されることが示唆される。
【0199】
[213]CA6が属する炭水化物の構造をさらに解明するため、ドットブロットをN
−グリカナーゼ、O−グリカナーゼ、および/またはシアリダーゼ(図4)で消化した。Caov−3細胞溶解物(100μg、30μl)を、SDSおよびβ−メルカプトエタノールを含有する変性緩衝液(Glyko)2.5μlと100℃で5分間インキュベーションした。次いで、変性溶解物を1μlのN−グリカナーゼ、O−グリカナーゼ、および/またはシアリダーゼA(Glyko)を用いて、37℃で1時間消化した。次いで、消化した溶解物をニトロセルロース上にスポッティングし(2μl)、そして上述のように免疫ブロッティングした。
【0200】
[214]N−グリカナーゼは、DS6免疫ブロットシグナルに対して明らかな影響がなかった。しかし、シアリダーゼで消化した試料はまったくシグナルを生じなかった。O−グリカナーゼは、シアリダーゼでの前処理なしには、シアル化されたO連結炭水化物を消化不能であるため、O−グリカナーゼ単独でプロセシングした試料のDS6シグナルは、影響を受けないであろう。対照的に、N−グリカナーゼは、活性のため、いかなるグリコシド酵素での前処理も必要としない。N−グリカナーゼ処理がDS6シグナルに影響を及ぼさないという事実は、CA6エピトープが、シアル化O連結炭水化物鎖上に存在する可能性が最も高いことを示唆する。
【0201】
実施例3:CA6エピトープが見出される抗原の解明
[215]CA6シアログリコトープが見出される抗原を同定するため、SDS−PAGEおよびウェスタンブロッティングによって、DS6免疫沈降物を分析した。細胞溶解物上清(1ml/試料;3〜5mgタンパク質)をプロテインGビーズ(30μl)であらかじめきれいにし、回転させながら4℃で1〜2時間、1mlのRIPA緩衝液で平衡化させた。続くすべての工程を、氷上および/または4℃の低温室で行った。あらかじめきれいにしたビーズを、微量遠心分離装置中、簡単に(2〜3秒)回転させて落とした。あらかじめきれいにした上清を、新しい試験管に移し、そして回転させながら、2μgのDS6と一晩インキュベーションした。新鮮な、平衡化プロテインGビーズ(30μl)を溶解物に添加し、そして回転させながら、1時間インキュベーションした。ビーズ−溶解物懸濁物を、微量遠心分離装置中、簡単に回転させて落とし、そして場合によって、免疫沈降後の溶解物の試料を採取した。1mlのRIPA緩衝液で、ビーズを5〜10回洗浄した。
【0202】
[216]次いで、免疫沈降したDS6試料を、30μlのノイラミニダーゼ(20mUノイラミニダーゼ(Roche)、50mM酢酸ナトリウムpH5、5mM CaCl2、100μg/ml BSA)または30μlの過ヨウ素酸(20mM過ヨウ素酸(VWR)、0.5M酢酸ナトリウムpH5)で、37℃で1時間消化した。次いで、これらを、2x試料装填緩衝液(β−メルカプトエタノールを含有する)30μl中に再懸濁した。ビーズを5分間煮沸し、そして装填緩衝液上清を、4〜12%または4〜20%Tris−グリシン・ゲル(Invitrogen)上に装填した。Laemmli電気泳動緩衝液中、125Vで1.5時間、ゲルを泳動した。Mini Trans−blotトランスファー装置(Biorad)を用いて、0.2μmニトロセルロース膜(Invitrogen)上に、ゲル試料を20mAで一晩トランスファーした。実施例2に上述するように、DS6で膜を免疫ブロッティングした。
【0203】
[217]あるいは、免疫沈降したビーズをまず変性させ、そして次いで、N−グリカナーゼ、O−グリカナーゼおよび/またはシアリダーゼA(Glyko)で酵素的に消化した。27μlのインキュベーション緩衝液および2μlの変性溶液(Glyko)中にビーズを再懸濁し、そして100℃で5分間インキュベーションした。室温に冷却した後、界面活性剤溶液(2μl)を添加し、そして試料を1μlのN−グリカナーゼ、O−グリカナーゼ、および/またはシアリダーゼAと、37℃で4時間インキュベーションした。5x試料装填緩衝液(7μl)を添加した後、試料を5分間煮沸した。試料をSDS−
PAGEに供し、そして上述のように、免疫ブロッティングした。
【0204】
[218]DS6は、抗原陽性細胞溶解物中に見られうる、>250kDaのタンパク質バンドを免疫沈降する(図5A、BおよびC)。ある細胞株(すなわちT−47D)では、ダブレットが観察される。>250kDaバンドは、ノイラミニダーゼまたは過ヨウ素酸で処理したCaov−3免疫沈降物では消失し(図5AおよびB)、CA6エピトープが、>250kDaバンド上に属することが示唆される。>250kDaバンドはまた、免疫沈降物のN−グリカナーゼ処理には感受性ではないことも示され、このことは、CA6がO連結炭水化物上に属することと一致する(図5F)。250kDaバンドがCA6抗原であることをさらに支持するのは、DS6が、DS6抗原陰性細胞から、こうしたバンドを免疫沈降させないという事実である(図5DおよびE)。
【0205】
[219]いくつかの系列の証拠が、CA6抗原がMuc1であることを示唆した。分子量が高く、そしてO連結炭水化物特異的糖分解酵素に感受性であるため、CA6抗原がムチンである可能性が高いようである。ムチン過剰発現は、腫瘍、特に乳房および卵巣の腫瘍でよく特徴付けられており、これは、DS6の主な腫瘍反応性と一致する。さらに、CA6は、CanAg(Muc1上のシアログリコトープ)同様、過塩素酸沈殿に感受性ではなく、CA6抗原が、非常にO−グリコシル化されていることが示唆される。いくつかのDS6発現細胞株において、DS6が、>250kDaのダブレットを免疫沈降させた観察から、CA6がMuc1であることが示唆された。ヒトにおけるMuc1の特質は、タンデム反復数が異なる2つの別個のMuc1アレルの存在であり、これは異なる分子量の2つのMuc1タンパク質の発現を生じる。
【0206】
[220]CA6がMuc1上に見られるかどうかを試験するため、Caov−3溶解物からのDS6免疫沈降物を、SDS−PAGEに供し、そしてDS6、またはMuc1VNTR抗体、CM1のいずれかで免疫ブロッティングした。図6Aに見られうるように、CM1は、DS6によって免疫沈降される>250kDaバンドと強く反応する。図6Bでは、HeLa細胞溶解物からのDS6およびCM1免疫沈降物は、DS6またはCM1のいずれかで免疫ブロッティングした際、同じ>250kDaダブレットを示す。これらの結果によって、CA6エピトープは実際、Muc−1タンパク質上に位置することが示される。HeLa(およびT−47D)細胞に見られるDS6ダブレットは、Muc−1発現が、異なる数のタンデム反復を有する別個のアレルによって指示されるという事実によって説明可能である。
【0207】
[221]CM1およびDS6は、同じMuc−1タンパク質に結合するが、これらは別個のエピトープである。トリフルオロメタンスルホン酸(TFMSA)によるCaov−3溶解物ドットブロットの化学的脱グリコシル化によって、DS6シグナルが消滅した(図3)。しかし、この同じ処理は、CM1シグナルを増進させた。脱グリコシル化が、CM1抗体の隠れたエピトープを暴露した可能性もある。さらに、DS6およびCM1のフローサイトメトリー結合結果の比較(表4)によって、CA6エピトープが、Muc1を発現するすべての細胞上には存在しないことが示される。高レベルのMuc1 CanAgシアログリコトープを発現することが知られる細胞株であるColo205上に、CA6エピトープが発現されないことに注目すると興味深い(表3)。
表4
【0208】
【表5】
【0209】
*MMF=最大平均相対的蛍光
【0210】
実施例4:脱落(shed)CA6エピトープの定量的分析
[222]CA6エピトープは、多くの癌患者において、血流内に脱落することが知られる分子であるMuc1上に属するため、こうしたレベルがDS6抗体療法を禁ずるほどであるかどうかを決定するために、定量的アプローチを行った。抗原に循環抗体が結合すると、血液からの免疫複合体の迅速なクリアランスにつながると考えられる。投与した抗体用量のかなりの部分が循環から迅速に除去されるならば、腫瘍に到達する量は、減少する可能性が高く、抗体療法剤の抗腫瘍活性の減少を生じる。抗体を非常に強力な細胞傷害性化合物にコンジュゲート化すると、コンジュゲートの迅速なクリアランスが、非特異的な毒性を潜在的に増加させうる。したがって、DS6−DM1などの抗体−小薬剤コンジュゲートの場合、高レベルの脱落抗原は、抗腫瘍効果を減少させ、そしてかつ用量を制限する毒性を増加させると予期されうる。
【0211】
[223]抗体療法剤の最近の臨床試験は、薬物動態に対する脱落抗原濃度の影響に関する情報を生じた。例えば、her2/neuを発現する転移性乳癌治療に使用する抗体であるトラスツズマブ(ハーセプチン)を用いた臨床試験において、トラスツズマブ・クリアランスの薬物動態は、脱落Her2/neuレベルが500ng/ml未満である場合、改変されないことが示された(Pegramら, J. Clin. Oncol.
16(8):2659−71(1998))。脱落Her2/neuの分子量を110,000ダルトンと仮定すると、4.5nM未満の脱落Her2/neuのモル濃度は、薬物動態にほとんど影響を及ぼさないようである。
【0212】
[224]別の例において、カンツズマブ・マータンシン(cantuzumab mertansine)(huC242−DMl)を用いた臨床試験によって、処置前脱落CanAg(C242エピトープ)レベルおよび抗体クリアランスの薬物動態に相関がないことが示された(Tolcherら, J. Clin. Oncol. 21(2):211−22(2003))。CanAgエピトープは、DS6によって認識されるCA6エピトープ同様、Muc1上のユニークな腫瘍特異的O連結シアログリコトープである。しかし、CanAgエピトープは不均一性であるため、モルで定量化するのが困難である。一般集団において、Muc1アレルは、可変数タンデム反復(VNTR)ドメイン中のタンデム反復数に応じて、長さが多様である。O連結グリコシル化のためのいくつかの部位が、各タンデム反復中に存在する。CanAg発現の複雑さに加えて、生得的なグリコシルトランスフェラーゼ活性には、細胞間変動がある。したがって、単一の患者においてであってさえ、Muc1分子あたり、広い範囲のCanAgエピトープがありうる。さらに、Muc1分子あたりのCanAgエピトープの比は、患者集団に渡って異なるであろう。このため、CanAgエピトープを含む脱落Muc1が、C242に捕捉され、
そしてビオチン化C242/ストレプトアビジンHRP系によって検出される、サンドイッチELISAによって、血清試料中の脱落CanAgを測定する。Muc1のモル濃度によるのではなく、血清1mlあたりのエピトープ数に比例する標準化単位で、脱落CanAgを定量化する。同様に、脱落CA6エピトープの定量化に関して、類似の状況が生じる。対照的に、トラスツズマブに関しては、脱落her2/neu分子あたりにただ1つのエピトープしかなく、脱落抗原の定量化は非常に単純化される。
【0213】
[225]トラスツズマブおよびカンツズマブ・マータンシンを用いた臨床試験で見られるものに、CA6脱落エピトープレベルを関連させるため、Muc1上のシアログリコトープなどの複雑な脱落エピトープのモル濃度を得るための方法を開発した。まず、DS6に関する単純サンドイッチELISAアッセイを確立した。代表的なアッセイを図7Aに示す。DS6を用いて、CA6エピトープを有するMuc1を捕捉した。各Muc1分子は複数のCA6エピトープを有するため、ビオチン化DS6を、トレーサー抗体としてもまた用いた。基質としてABTSを用い、ストレプトアビジン−HRPによって、捕捉されたCA6に結合したビオチン化DS6を検出した。卵巣癌患者血清から、または乳癌患者において脱落Muc1を監視するために用いられる商業的に入手可能なMuc1試験キット(CA15−3)に由来する標準から、CA6エピトープを捕捉した。DS6単位/mlを、恣意的に、CA15−3標準単位/mlに等しく設定した。
【0214】
[226]図7Bにおいて、CA15−3標準を用いる、DS6サンドイッチELISAの結果を示す。生じる曲線は、CA15−3アッセイにおいて、CA15−3標準で得られるものと非常に類似である。DS6単位/mlを、CA6のモル濃度に変換するため、シグナルをDS6のピコグラムに変換する、ビオチン化DS6に関する標準曲線が必要である。CA6エピトープおよびビオチン化DS6抗体間は1対1の化学量論であり、そしてビオチン化DS6の分子量は160,000ダルトンであると仮定して、添加した試料体積あたりに捕捉されるCA6のモル数を計算してもよい。
【0215】
[227]図8AおよびBにおいて、ビオチン化DS6に関する標準曲線を生じる、2つの代替手段を示す。図8Aにおいて、ヤギ抗マウスIgGポリクローナル抗体を用いて、ビオチン化DS6を捕捉し、これを次に、図7に示すサンドイッチELISAアッセイで用いるものと同一の方式で検出する。図8Bに示す方法において、ビオチン化DS6を、ELISAプレート上に直接プレーティングし、そして図8Aにおけるように検出する。図8Cにおいて見られるように、各方法によって生成されるビオチン化DS6標準曲線は、十分一致している。
【0216】
[228]表5において、多様な脱落抗原に関する卵巣癌患者血清試料の分析を示す。一般的に、CA125 ELISAを用いて、脱落CA125単位/mlを測定することによって、卵巣癌患者の治療を監視する。血清試料を用いて、CA125状態を提供する。一般的に、CA15−3 ELISAを用いて、DS6によって認識されるものと異なるエピトープを認識する捕捉および検出抗体を用い、脱落Muc1の単位/mlを測定することによって、乳癌患者の治療を監視する。表5において、卵巣癌患者血清試料において、CA15−3を測定する。
表5
【0217】
【表6】
【0218】
1商業的ELISAキットによって測定
2商業的CA15−3標準によって測定(1 CA15−3 U=1 DS6 U)
3ヤギ抗マウスIgGおよびビオチン−DS6標準曲線
4ビオチン−DS6標準曲線
【0219】
[229]表5に報告するCA15−3値に関しては、CanAg Dagnosticsの商業的に入手可能なCA15−3酵素免疫アッセイキットを用いた。DS6単位/mlに関しては、DS6サンドイッチELISAにおいて、CA15−3標準(CanAg DiagnosticsのCA15−3酵素免疫アッセイキット由来)を用いて標準曲線を生成した。DS6単位/mlを、恣意的に、CA15−3単位/mlに等しく設定した。最後の2つの列では、図8Cに示すビオチン化DS6標準曲線を用いて、ピコモル(pM)脱落CA6を計算した。
【0220】
[230]CanAgレベルの定量的分析に関して、CanAg血清レベルは、処置前にカンツズマブ・マータンシン臨床試験に参加した患者に関して報告されたものであった(Tolcherら, J. Clin. Oncol. 21(2):211−22(2003))。DS6に関して記載されるものと類似のELISAアッセイを用いて、CanAg標準を用い、CanAg標準曲線を作成した。C242を用いて、CanAg標準を捕捉した。ビオチン化C242トレーサーを用いて、その後、基質としてABTSを用い、ストレプトアビジン−HRPで発色させて、捕捉されたCanAgの検出を達成した。ビオチン化DS6に関して行ったように、ビオチン化C242標準曲線を構築して、単位/mlを循環CanAgエピトープのモル濃度に変換することを可能にした。表6において、カンツズマブ・マータンシン臨床試験患者由来のCanAgレベルを、循環CanAgの対応する計算モル濃度と一緒に報告する。
表6
【0221】
【表7】
【0222】
1サンドイッチELISAによって測定した、循環CanAgの処置前レベル
2ヤギ抗マウスIgGおよびビオチン−C242標準曲線
3ビオチン−C242標準曲線
【0223】
[231]卵巣癌患者における脱落CA6のpMレベルと、CanAg陽性癌患者における脱落CanAgに関して計算したものの比較によって、一般的に、脱落CA6レベルは、脱落CanAgレベルと類似であることが示される。さらに、16の卵巣癌患者血清試料のうち2つのみが、4.5nMより高いCA6レベルを潜在的に有し(シグナルが標準曲線の範囲から外れている、血清試料5および9)、このレベルは、これより高い場合、Her2/neu陽性乳癌患者での臨床試験において、改変されたハーセプチン薬物動態が観察されたレベルであった。4.5nMを超えるCanAgレベルは、37の臨床試験患者のうち3人でのみ見られた。この臨床試験において、脱落CanAgレベルおよびカンツズマブ・マータンシンのより迅速なクリアランスには相関はなかった。しかし、最高のCanAgレベル(31240U/ml)の患者は、注入後8時間のみ試料採取された。これらの結果によって、Muc1の特定のエピトープ、例えばCA6およびCanAgは、癌患者において脱落するものの、抗体療法治療を禁じるレベルまでは脱落しないことが示される。
【0224】
実施例6:ネズミDS6抗体可変領域のクローニング
[232]DS6などのネズミ・モノクローナル抗体は、ヒト免疫系によって異質(foreign)と認識されるため、臨床設定において、有用性が限定されている。患者は、ヒト抗マウス抗体(HAMA)を迅速に発展させ、ネズミ抗体の迅速なクリアランスを生じる。このため、ネズミDS6(muDS6)の可変領域の表面を再構成して、ヒト化DS6(huDS6)抗体を生じた。
【0225】
[233]RT−PCRによって、ネズミDS6抗体可変領域をクローニングした。Qiagen RNeasyミニプレップキットを用いて、DS6ハイブリドーマ細胞の集密T175フラスコから総RNAを精製した。UV分光光度測定によってRNA濃度を測定し、そしてGibco Superscript IIキットおよびランダム六量体プライマーを用いて、4〜5μg総RNAでRT反応を行った。
【0226】
[234]Wang Zら, J Immunol Methods. Jan 13;233(l−2):167−77(2000)に記載されるものに基き、縮重プライマーを用いて、PCR反応を行った。RT反応混合物を縮重PCR反応に直接用いた。3’軽鎖プライマー、HindKL、
(TATAGAGCTCAAGCTTGGATGGTGGGAAGATGGATACAGTTGGTGC)(配列番号25)
および3’重鎖プライマー、BamIgG1、
(GGAGGATCCATAGACAGATGGGGGTGTCGTTTTGGC)(配列番号26)
を用い、そして5’端に関しては、PCRプライマーは、軽鎖に関して、Sac1MK
(GGGAGCTCGAYATTGTGMTSACMCARWCTMCA)(配列番号27)
そして重鎖に関して、EcoR1MH1
(CTTCCGGAATTCSARGTNMAGCTGSAGSAGTC)(配列番号28)およびEcoR1MH2(CTTCCGGAATTCSARGTNMAGCTGSAGSAGTCWGG)(配列番号29)の等量混合物であった(混合塩基:H=A+T+C、S=G+C、Y=C+T、K=G+T、M=A+C、R=A+G、W=A+T、V=A+C+G、N=A+T+G+C)。
【0227】
[235]10%DMSOを補った以外、PCR反応は標準的であった(50μl反応混合物は、最終濃度1x反応緩衝液(ROCHE)、各2mMのdNTP、各1mMのプライマー、2μl RT反応、5μl DMSO、および0.5μl Taq(ROCHE)を含有した)。Wang Zら(J Immunol Methods. Jan 13;233(1−2):167−77(2000))から適応させたプログラム:1)94℃、3分間;2)94℃、15秒間;3)45℃、1分間;4)72℃、2分間;5)工程2に戻って29周期;6)72℃、10分間の最終伸長工程で終了を用いて、MJ
researchサーモサイクラー上で、PCR反応を行った。PCRプライマーによって生成されたPCR産物を、制限酵素を用いて、pBluescript II SK+(Stratagene)内にクローニングした。Seqwright配列決定サービスによって、重鎖および軽鎖クローンを配列決定した。
【0228】
[236]5’端cDNA配列を確認するため、さらなるPCRおよびクローニングを行った。縮重PCRクローンから決定したDS6軽鎖および重鎖cDNA配列をNCBIのBlast検索ウェブサイト内に打ち込み、そして提示されたシグナル配列を持つネズミ抗体配列を記録した。関連DNA配列の間で保存されたストレッチを用いて、これらのシグナルペプチドからPCRプライマーを設計した。EcoRI制限部位をリーダー配列
プライマーに付加し(表7)、そしてこれらを、上述のようなRT−PCR反応に用いた。
【0229】
【表8】
【0230】
[237]いくつかの個々の軽鎖および重鎖クローンを配列決定して同定し、そしてポリメラーゼが生成する可能性がある配列エラーを回避した。軽鎖および重鎖RT−PCRクローンの両方に関して、単一の配列のみが得られた。これらの配列は、シグナル配列内まで拡大してネズミDS6軽鎖および重鎖配列を増幅可能なプライマーを設計するのに十分であった。これらの追加PCR反応由来の続くクローンによって、元来の縮重プライマーによって改変されている可変領域の5’端配列が確認された。多様なcDNAクローンからの累積結果によって、図9に示す最終ネズミDS6軽鎖および重鎖配列が提供された。KabatおよびAbM定義を用いて、3つの軽鎖および重鎖CDRが同定された(図9および10)。NCBI IgBlastデータベースの検索によって、muDS6抗体軽鎖可変領域が、ネズミIgVκ ap4生殖系列遺伝子に由来する可能性が最も高く、一方、重鎖可変領域が、ネズミIgVh J558.41生殖系列遺伝子に由来する可能性が最も高いことが示される(図11)。
【0231】
実施例7:DS6抗体の可変領域表面残基の決定
[238]Pedersenら(1994)およびRoguskaら(1996)によって記載される抗体表面再構成技術は、ネズミ抗体可変配列の表面残基を予測することから始まる。表面残基は、その総表面積の少なくとも30%が水分子に対してアクセス可能であるアミノ酸と定義される。muDS6に関する表面残基を発見するために解明された構造がないため、本発明者らは、127の抗体構造ファイルセット中、最も相同な配列を持つ10の抗体を整列させた(図12)。これらの整列配列に関して、各Kabat位の溶媒アクセス可能性を平均した(図13AおよびB)。
【0232】
[239]いずれかの側に隣接する2つの同一残基を含有する抗体サブセットを比較することによって、25%〜35%の間の平均アクセス可能性を持つ表面位を、第二周期の分析に供した(図13AおよびB)。第二周期の分析後、muDS6重鎖に関して、21の予測される表面残基は、23に増加し、30%より高い表面アクセス可能性を持つと予測される残基のリストに、Tyr3およびLys23が加わった。本発明者らの表面再構成抗体の大部分では、重鎖CDR1のKabat定義を用いたが、DS6に関しては、計算中、不注意にAbM定義を用いたため、重鎖残基T28は、Kabat定義でフレームワーク表面残基と定義されうるようには、定義されなかった。Ala80の予測される表面アクセス可能性が、第二周期分析において、30.5%から27.8%に減少したため、軽鎖表面位の数は、16から15に減少した。総合すると、muDS6重鎖および軽鎖可変配列は、38の予測される表面アクセス可能フレームワーク残基を有する。
【0233】
実施例8:ヒト抗体選択
[240]ネズミDS6可変領域の表面位を、Kabatデータベース中のヒト抗体配列中の対応する位に比較した(Johnson G, Wu TT. Nucleic Acids Res. Jan 1;29(1):205−6(2001))。抗体データベース管理ソフトウェアSR(Searle、1998)を用いて、天然重鎖および軽鎖ヒト抗体対から表面残基を抽出し、そして整列させた。CDRの5Å以内にある位に特に考慮を払い、最も同一の表面残基を持つヒト抗体可変領域表面を選択して、ネズミDS6抗体可変領域表面残基を置換した。
【0234】
実施例9:キメラおよびヒト化抗体の発現ベクター
[241]軽鎖および重鎖対形成配列を、単一哺乳動物発現ベクター内にクローニングした。ヒト可変配列のPCRプライマーは、制限部位を生成し、pBluescriptIIクローニングベクターにヒトシグナル配列を付加することを可能にした。次いで、軽鎖または重鎖に関して、それぞれ、EcoRIおよびBsiWIまたはHidIIIおよびApaIを持つ哺乳動物発現プラスミド内に、可変配列をクローニングしてもよい(図14)。軽鎖可変配列を、インフレームでヒトIgカッパ定常領域上にクローニングし、そして重鎖可変配列を、ヒトIgガンマ1定常領域配列内にクローニングした。最終発現プラスミドにおいて、ヒトCMVプロモーターが、軽鎖および重鎖両方のcDNA配列の発現を駆動する。
【0235】
実施例10:DS6活性に負に影響を及ぼしうる残基の同定
[242]今日まで、ヒト化の大部分では、潜在的に問題がある残基として、CDRに近接する残基を同定するため、対象抗体の分子モデルが構築されてきた。作業を開始する表面再構成された抗体の数が増加しつつあるため、歴史的な経験は、問題を予測するのに、モデルを構築するのと少なくとも同程度に有効であり、DS6に関しては、分子モデルを構築しなかった。その代わり、ネズミDS6表面残基を、先に表面再構成された抗体と比較し、そして抗体の結合能に影響を及ぼすリスクが低い残基から高い残基を同定した。
【0236】
[243]入手可能な解明された抗体構造および先のヒト化に由来する分子モデルの両方において、類似の残基セットが、CDRの5Å以内にあると繰り返し同定された。このデータを用いて、表1は、CDRに近接する可能性があり、そしておそらく5Å以内である、ネズミDS6残基を提供する。これらの位の多くが、先のヒト化でもまた変更されているが、これまでに結合活性の喪失を生じたのは、重鎖74位のみであった。ネズミ抗体の結合活性を保存するため、huC242およびhuB4の両方において、この位でネズミ残基を保持した。一方、この同じ位が、ヒト化6.2G5C6において、対応するヒト残基に変更されたが、活性の喪失は伴わなかった(6.2G5C6は、しばしば、単に抗C6と称される抗IGF1−R抗体である)。表1中のいかなる残基もヒト化抗体において問題を提示しうるが、重鎖残基P73が、この位での先の経験から、特に懸念される。
【0237】
実施例11:最も相同なヒト表面の選択
[244]SRソフトウェアを用いて、Kabat抗体配列データベースから、muDS6の表面を再構成するための候補ヒト抗体表面を引き出した。このソフトウェアは、抗体データベースに対して、特定の残基位のみを検索するインターフェースを提供する。天然の対を保持するため、軽鎖および重鎖両方の表面残基を一緒に比較した。配列同一性のランク順に、Kabatデータベース由来の最も相同なヒト表面を整列させた。SR Kabatデータベースソフトウェアによって並列した際の上位3つの表面を表2に示す。次いで、表面を比較して、表1に同定する残基に対して最も少ない変化しか必要としないのはどのヒト表面かを同定した。抗Rh(D)抗体、28E4(Boucherら、1997)は、最少数の表面残基変化しか必要とせず(総数11)、そしてこれらの残基のう
ち3つのみが、潜在的な問題の残基のリストに含まれた。28E4抗体は、最も相同なヒト表面を提供するため、これが、muDS6の表面を再構成する最適な候補である。
【0238】
実施例12:ヒト化DS6抗体のためのDNA配列の構築
[245]PCR突然変異誘発を用いて、DS6のための11の表面残基変化を作製した。ネズミDS6可変領域cDNAクローンに対して、PCR突然変異誘発を行って、表面を再構成したヒトDS6遺伝子を構築した。以下の表8に示すように、ヒト化プライマーセットを設計して、表面再構成DS6に必要なアミノ酸変化を作製した。
表8
【0239】
【表9】
【0240】
[246]10%DMSOを補った以外、PCR反応は標準的であった(50μl反応混合物は、最終濃度1x反応緩衝液(ROCHE)、各2mMのdNTP、各1mMのプライマー、100ngのテンプレート、5μl DMSO、および0.5μl Taq(ROCHE)を含有した)。以下のプログラム:1)94℃、1分間;2)94℃、15秒間;3)55℃、1分間;4)72℃、1分間;5)工程2に戻って29周期;6)72℃、4分間の最終伸長工程で終了を用いて、MJ Researchサーモサイクラー上で、PCR反応を行った。PCR産物を、対応する制限酵素で消化して、そしてpBluescriptクローニングベクター内にクローニングした。クローンを配列決定して、アミノ酸変化を確認した。
【0241】
[247]重鎖残基P73を変化させると、過去に問題が引き起こされたため、重鎖の2つの型、ヒト28E4 T73を持つもの、およびネズミP73を保持するものを構築した。ヒト化DS6のどちらの型でも、他の10の表面残基をネズミからヒト28E4残基に変化させた(表2)。通常の命名法に忠実にしたがい、11のヒト表面残基すべてを有するため、最もヒトであるのは、1.0型である。さらなる型が必要とされる場合のために、ネズミP73を保持する重鎖型を1.2と名付け、したがって、1.1型は、最大数のネズミ残基を含有する型のために取っておく。図15AおよびBにおいて、2つのヒト化型のアミノ酸配列を、ネズミDS6アミノ酸配列と整列させて示す。一過性および安定トランスフェクションのため、ヒト化DS6抗体遺伝子の両方を、抗体発現プラスミド(図14)内にクローニングした。ヒト化型1.01および1.21軽鎖可変領域のcDNAおよびアミノ酸配列は、同じであり、そしてこれらを図16に示す。ヒト化型1.01および1.21の重鎖cDNAおよびアミノ酸配列を図17AおよびBに示す。
【0242】
実施例13:CHO細胞におけるhuDS6の発現および精製、ならびに親和性測定
[248]ヒト化DS6型が、muDS6の結合親和性を保持するかどうかを決定するため、抗体を発現し、そして精製することが必要であった。ヒト定常領域およびネズミ可変領域を有するDS6のキメラ型(chDS6)で、あるいはhuDS6v1.01またはhuDS6v1.21で、CHO細胞を安定トランスフェクションした。
【0243】
[249]CHODG44細胞(4.32x106細胞/プレート)を、非選択培地(4mM L−グルタミン、50U/mlペニシリン、50μg/mlストレプトマイシン、および10%v/v FBSを補った、アルファMEM+ヌクレオチド(Gibco))中、15cmプレートに植え付け、そして37℃、5%CO2加湿インキュベーター中に入れた。翌日、Polyfect Transfectionに関してQiagenが推奨するプロトコルの修飾型を用いて、chDS6発現プラスミドで細胞をトランスフェクションした。非選択培地を細胞から吸引した。7mlのあらかじめ温めた(37℃)PBSでプレートを洗浄し、そして20mlの非選択培地を再補充した。プラスミドDNA(11μg)を800μlのハイブリドーマSFM(Gibco)内に希釈した。次いで、70μlのPolyfect(Qiagen)をDNA/SFM混合物に添加した。次いで、Polyfect混合物を数秒間穏やかにボルテックスし、そして周囲温度で10分間インキュベーションした。非選択培地(2.7ml)を混合物に添加した。この最終混合物を、プレーティングした細胞と24時間インキュベーションした。
【0244】
[250]トランスフェクション混合物/培地をプレートから取り除き、そして次いで、細胞をトリプシン処理し、そして計数した。次いで、多様な密度(1800、600、200、および67細胞/ウェル)で、96ウェルプレート(250μl/ウェル)中、細胞を選択培地(4mM L−グルタミン、50U/mlペニシリン、50μg/mlストレプトマイシン、10%v/v FBS、1.25mg/ml G418を補った、アルファMEM−ヌクレオチド(Gibco))中でプレーティングした。必要であれば培地を補って、細胞を2〜3週間インキュベーションした。定量的ELISAを用いて、抗体産生レベルに関してウェルをスクリーニングした。Immulon 2HB 96ウェルプレートを、ヤギ抗ヒトIgG F(ab)2抗体(Jackson Immunoresearch;100μlの50mM炭酸ナトリウム緩衝液pH9.6中、1μg/ウェル)でコーティングし、そして振盪しながら、周囲温度で1.5時間インキュベーションした。すべての続く工程を、周囲温度で行った。ウェルをT−TBS(0.1%Tween−20、TBS)で2回洗浄し、そして200μlのブロッキング緩衝液(1%BSA、T−TBS)で1時間ブロッキングした。ウェルをT−TBSで2回洗浄した。別個のプレート中で、抗体標準、EM164(100ng/ml)および培養上清を、ブロッキング緩衝液中で連続希釈した(1:2または1:3)。これらの希釈物(100μl)を、ELISAプレートに移し、そして1時間インキュベーションした。ウェルをT−TBSで3回洗浄し、そしてブロッキング緩衝液中で1:3000希釈した100μlのヤギ抗ヒトIgG Fc−AP(Jackson ImmunoResearch)と45分間インキュベーションした。T−TBSで5回洗浄した後、100μlのPNPP発色試薬(10mg/ml PNPP(p−ニトロフェニルリン酸、二ナトリウム塩;Pierce)、0.1MジエタノールアミンpH10.3緩衝液)を用いて、ウェルを25分間、発色させた。ELISAプレート読取装置中、405nmでの吸光度を測定した。標準曲線の直線部分での(培養上清の)吸光度読取り値を用いて、抗体レベルを決定した。
【0245】
[251]次いで、ELISAによって同定された最大産生クローンをサブクローニングし、拡大し、そして凍結細胞ストックを調製した。
[252]huDS6v1.01およびhuDS6v1.21の発現に関しては、DG44 CHO細胞(Lawrence Chasin博士、ニューヨーク州コロンビア大学)を、リボヌクレオチドおよびデオキシリボヌクレオチドを含むアルファMEM(Gibcoカタログ番号12571、ニューヨーク州グランドアイランド)中で培養した。培
地に10%ウシ胎児血清(HyCloneカタログ番号SH30071.03、ユタ州ローガン)、1%ゲンタマイシン(Mediatechカタログ番号30−005−CR、バージニア州ハーンドーン)、および2mM L−グルタミン(L−glut)(BioWhittakerカタログ番号17−605E、メリーランド州ウォーカーズビル)を補った。この配合物をCHO完全培地と名付けた。
【0246】
[253]DG44 CHO細胞(5x106)を、50μgのhuDS6プラスミドDNAでトランスフェクションした。トランスフェクション前に、トリプシン(Gibcoカタログ番号15090−046、ニューヨーク州グランドアイランド)でフラスコから細胞を取り除き、そしてリボヌクレオシドおよびデオキシリボヌクレオシドを欠く、非補充アルファMEM(Gibcoカタログ番号12561、ニューヨーク州グランドアイランド)で2回洗浄した。これを洗浄培地と名付けた。0.4cmギャップの電極キュベット(BioRadカタログ番号1652088、ペンシルバニア州ハーキュルス)中、細胞をプラスミドDNAと混合した。これらを氷上に2分間置き、そして次いで、BioRadエレクトロポレーション装置中、1,000μFおよび260ボルトでパルス処理した。エレクトロポレーション後、氷上で2分間、細胞をインキュベーションした。次いで、CHO完全培地中、細胞を5つの24ウェルプレート(Costarカタログ番号3524)中にプレーティングし、そして5%CO2の37℃インキュベーター中で維持した。48時間後、ウェルから培地を取り除いた。洗浄培地でウェルを1回リンスし、そして1%ゲンタマイシン、2mM L−glut、10%透析ウシ胎児血清(Gibcoカタログ番号26400−044、ニューヨーク州グランドアイランド)、および1.25mg/mlジェネティシン(G418)(Gibcoカタログ番号11811、ニューヨーク州グランドアイランド)を補った、リボヌクレオシドおよびデオキシリボヌクレオシドを含まないアルファMEM(Gibcoカタログ番号12561、ニューヨーク州グランドアイランド)を供給した。この完全配合物を、選択培地と名付けた。選択培地中でおよそ2週間、クローンをインキュベーションし、この時点で、定量的ELISAによって、抗体産生に関してスクリーニングした。次いで、最高産生クローンをサブクローニングし、拡大し、そして凍結細胞ストックを調製した。
【0247】
[254]精製に十分な量の抗体を産生するため、Ultra Low IgG FBS(Gibco)を補った選択培地30mlを含む15cmプレート(〜1x106細胞/プレート)上に、細胞を拡大し、そして1週間インキュベーションした。250mlの円錐管内に培養上清を収集し、卓上遠心分離装置中で回転して落とし(2000rpm、5分間、4℃)、そして次いで、0.2μmフィルター装置を通じて滅菌ろ過した。
【0248】
[255]DS6を精製するため、NaOHのペレットをろ過培養上清に添加して、最終pH8.0にした。HiTrap rプロテインAカラム(Amersham)を、20〜50カラム体積の結合緩衝液で平衡化した。蠕動ポンプを用いて、上清をカラム上に装填した。次いで、カラムを50カラム体積の結合緩衝液で洗浄した。溶出緩衝液(100mM酢酸、50mM NaCl、pH3)を用いて、結合した抗体を、カラムから、フラクションコレクター中にセッティングした試験管内に溶出させた。中和緩衝液(2M K2HPO4、pH10.0)を用いて、溶出した抗体を中和し、そしてPBS中で一晩透析した。0.2μmシリンジフィルターを通じて、透析抗体をろ過した。280nmでの吸光度を測定して、最終タンパク質濃度を決定した。
【0249】
[256]精製huIgGの親和性を、フローサイトメトリーによって、muDS6と比較した。最初の実験セットでは、CA6発現細胞株、KBへの直接結合を測定した。図18に示すように、muDS6、chDS6、huDS5v1.01およびhuDS6v1.21は、非常に類似の親和性を示し、見かけのKdが、それぞれ、0.82nM、0.69nM、0.82および0.85nMであり、表面再構成がCDRを妨害しなかった
ことが示唆された。huDS6型が、muDS6の親和性を保持することを確認するため、競合的結合実験を行った。この形式の利点は、ネズミおよびヒト抗体の両方に関して、同じ検出系;すなわち、ビオチン−muDS6/ストレプトアビジン−DTAFを使用することである。muDS6、chDS6、huDS6v1.01およびhuDS6v1.21がビオチン−DS6と競合する能力を比較する、競合結合アッセイの結果を図19に示す。見かけのEC50は、muDS6、chDS6、huDS6v1.01、およびhuDS6v1.21に関して、それぞれ、1.9nM、1.7nM、3.0および1.9nMである。これらの結果は、ヒト化DS6を産生するためのmuDS6の表面再構成が、結合親和性の減少をほとんど引き起こさないことを示す。
【0250】
実施例14:muDS6−DM1細胞傷害性コンジュゲートの調製
[257]8倍モル過剰のN−スクシンイミジル−4−(2−ピリジルジチオ)ペンタノエート(SPP)を用いて、muDS6抗体(8mg/ml)を修飾して、ジチオピリジル基を導入した。95%v/v緩衝液A(50mM KPi、50mM NaCl、2mM EDTA、pH6.5)および5%v/v DMA中で、室温で2時間、反応を行った。NAPまたはSephadex G25カラム(緩衝液A中で平衡化)を通じて、わずかに膨らんだ(turgid)反応混合物をゲルろ過した。280nmで抗体の吸光度を、そして280および343nmで、DTTで放出される2−メルカプトピリジン(Spy)を測定することによって、修飾の度合いを決定した。次いで、SPyよりも1.7倍モル過剰のN2’−デアセチル−N2’−(3−メルカプト−1−オキソプロピル)−メイタンシン(L−DM1)を用いて、修飾muDS6を2.5mg Ab/mlでコンジュゲート化した。DMA(3%v/v)を含む緩衝液A(97%v/v)中で反応を行った。反応を室温で一晩、〜20時間、インキュベーションした。不透明な反応混合物を遠心分離し(1162xg、10分間)、そして次いで、緩衝液B(1xPBS pH6.5)中で平衡化したNAP−25またはS300(Tandem 3、3x26/10脱塩カラム、G25媒体)カラムを通じて上清をゲルろ過した。ペレットを廃棄した。0.22μm Millex−GVフィルターを用いて、コンジュゲートを滅菌ろ過し、そしてSlide−A−Lyzerを用いて、緩衝液B中で透析した。ろ過物質の252nmおよび280nm両方の吸光度を測定することによって、muDS6の分子あたりに連結されたDM1分子の数を決定した。DM1/Ab比は、4.36であることが見出され、そしてコンジュゲート化MUDS6の工程収率は55%であった。コンジュゲート化抗体濃度は1.32mg/mlであった。サイズ排除クロマトグラフィー(SEC)によって、精製コンジュゲートを生化学的に性質決定して、そして92%単量体であることを見出した。精製コンジュゲート中のDM1の分析によって、99%が抗体に共有結合していることが示された。図20において、muDS6−DM1コンジュゲートおよび非修飾muDS6のCaov−3細胞へのフローサイトメトリー結合によって、muDS6のコンジュゲート化が親和性のわずかな喪失しか生じないことが示される。
【0251】
実施例15:muDS6−DM1のin vitro細胞傷害性
[258]裸の抗体として、muDS6は、細胞培養において、増殖または成長阻害活性を示さなかった(図21)。しかし、ヤギ抗マウスIgG重鎖および軽鎖に対するDM1コンジュゲートの存在下で、muDS6を細胞とインキュベーションすると、muDS6は、このコンジュゲートを細胞にターゲティングし、そして送達するのに非常に有効であり、間接的な細胞傷害性を生じた(図21)。裸のmuDS6の生得的な活性をさらに試験するため、muDS6を用いた、補体依存性細胞傷害性(CDC)アッセイを行った。200μlのRHBP培地(RPMI−1640、0.1%BSA、20mM HEPES(pH7.2〜7.4)、100U/mlペニシリンおよび100μg/mlストレプトマイシン)中、5%ヒトまたはウサギ血清および多様な希釈のmuDS6の存在下で、HPACおよびZR−75−1細胞(25000細胞/ウェル)を96ウェルプレートにプレーティングした。細胞を37℃で2時間インキュベーションした。次いで、Ala
mar Blue(最終濃度10%)試薬(Biosource)を上清に添加した。蛍光を測定する前に、5〜24時間、細胞をインキュベーションした。ネズミDS6は、補体依存性細胞傷害性(CDC)アッセイにおいて、影響を持たなかった(図22)。これによって、muDS6の療法的適用が、毒性エフェクター分子のコンジュゲート化を必要とするであろうことが示唆される。
【0252】
[259]多様なDS6陽性細胞株において、2つの異なるアッセイ形式を用いて、メイタンシノイド・コンジュゲート化muDS6抗体の細胞傷害性を調べた。培地中で希釈した2mlのコンジュゲート中、6ウェルプレート上に細胞(1000〜2500細胞/ウェル)をプレーティングする、コロニー形成アッセイを行った。一般的に、3x10−11M〜3x10−9Mの間のいくつかの濃度で、細胞を連続してコンジュゲートに曝露し、そして37℃、6%CO2加湿チャンバー中で、5〜9日間、インキュベーションした。PBSでウェルを洗浄し、そして1%w/vクリスタルバイオレット/10%v/vホルムアルデヒド/PBS溶液で、コロニーを染色した。次いで、再蒸留水を用いて、未結合染色剤を徹底的にウェルから洗い落とし、そしてプレートを乾燥させた。Leica
StereoZoom4解剖顕微鏡を用いて、コロニーを計数した。
【0253】
[260]コロニーの数/プレーティングした細胞数として、プレーティング効率(PE)を計算した。処理細胞のPE/非処理細胞のPEとして、生存率を計算した。細胞の生存率対コンジュゲートのモル濃度をグラフで示すことによって、IC50濃度を決定した。コロニー形成アッセイにおいて(図23)、muDS6−DM1は、Caov−3細胞を殺すのに有効であり、概算されるIC50は800pMであった。抗原陰性細胞、A375は、試験したmuDS6−DM1の最高の濃度である、3x10−9Mの濃度のコンジュゲートによって、わずかにしか影響を受けず、コンジュゲートの細胞殺傷活性が、抗原発現細胞に特異的に向けられることを立証した。しかし、メイタンシンに対する見かけの感受性にもかかわらず、多くの他のDS6陽性細胞株は、免疫コンジュゲートに対して、特に感受性ではなかった。すべての子宮頸部細胞株(HeLa、KB、およびWISH)は、コンジュゲートに対して感受性であり、一方、卵巣および乳房細胞株の選ばれた数のみが、何らかの細胞傷害性効果を示した。膵臓細胞株は、いずれも、影響を受けないようであった。
【0254】
[261]MTTアッセイにおいて、1000〜5000細胞/ウェルの密度で、細胞を96ウェルプレートに植え付けた。200μlの培地中、裸のmuDS6またはmuDS6−DM1免疫コンジュゲートのいずれかの連続希釈とともに、細胞をプレーティングした。試料を3つ組で実験した。次いで、細胞および抗体/コンジュゲート混合物を2〜7日間インキュベーションし、この時点で、MTT([3(4,5−ジメチルチアゾール−2−イル)−2,5−ジフェニルテトラゾリウムブロミド])アッセイによって、細胞生存度を評価した。MTT(50μg/ウェル)を培養上清に添加して、そして37℃で3〜4時間インキュベーションさせた。培地を取り除き、そしてMTTホルマザンをDMSO(175μl/ウェル)中に可溶化した。540〜545nmの吸光度を測定した。MTT細胞生死判別アッセイにおいて(図24C)、免疫コンジュゲートは、Caov−3細胞を有効に殺すことが可能であり、概算されるIC50は1.61nMであった。まったく影響を及ぼさなかった裸の抗体に比較して、最高濃度のコンジュゲートを含むウェルは、生存細胞をまったく含有しなかった(図21および24)。
【0255】
[262]他の細胞株に対するMTTアッセイの結果は、わずかに異なった(図24A、B、およびD〜I)。多くの場合、ある程度の細胞傷害性が見られるが、コンジュゲートは、全細胞集団を完全に殺すことは不可能であった(WISH細胞を除く)。BT−20、OVCAR5、およびHPAC細胞が特に抵抗性であった:最高コンジュゲート濃度(32nM)のウェルで、50%を超える細胞がなお生存可能であった。
【0256】
実施例16:in vivoコンジュゲート抗腫瘍活性
[263]muDS6−DM1コンジュゲートのin vivo活性を立証するため、SCIDマウスにおいて、ヒト腫瘍異種移植片を確立した。ヒト子宮頸癌細胞株KBの皮下モデルを開発した。KB細胞をin vitroで増殖させ、収集し、そして各マウスの右肩下に、100μlの血清不含培地中、5x106細胞を注射し、そして平均腫瘍体積144±125mm3になるまで6日間増殖させ、この時点で薬剤処置を開始した。マウスに、PBS、150μg/kg DM1のコンジュゲート、または225μg/kg
DM1のコンジュゲート(群あたり2匹のマウス)のいずれかを静脈内に毎日5日間投与した。処置中、毒性応答を毎日監視した。研究の間中、腫瘍体積(図25A)および対応する体重(図25B)を監視した。
【0257】
[264]PBS対照で処置したKB腫瘍は、約4日間の倍増時間で、迅速に増殖した。対照的に、コンジュゲートで処置したマウスの両群は、225μg/kgおよび150μg/kg用量群で、それぞれ、処置開始の14日後および18日後に、完全腫瘍退行を示す。150μg/kg用量では、腫瘍遅延はおよそ70日であった。225μg/kgでの処置は、第120日に研究を終了した際、腫瘍再発の証拠がまったくなかったように、治癒を生じた。図25Bに見られるように、150μg/kg群のマウスは、体重喪失をまったく示さず、用量がよく許容されることを示した。より高い用量では、マウスは、体重の一時的な3%の減少しか経験しない。5日間の処置経過中、マウスは毒性の可視徴候を示さなかった。総合すると、この研究は、muDS6−DM1処置が、非毒性用量で、KB異種移植片腫瘍のマウスを治癒させうることを立証する。
【0258】
[265]一団の皮下異種移植片モデルに対して、muDS6−DM1活性をさらに試験した(図26を参照されたい)。異種移植片を作製するのに用いた腫瘍細胞株は、さまざまなin vitroメイタンシン感度およびCA6エピトープ密度を示した(以下の表9)。OVCAR5細胞およびTOV−21Gは、卵巣腫瘍細胞株であり;HPACは膵臓腫瘍細胞株であり;HeLaは子宮頸部腫瘍細胞株である。OVCAR5およびTOV−21G細胞は、低い表面CA6発現を有し;HeLa細胞は、中間レベルの表面CA6発現を有し;HPAC細胞は、高いCA6表面発現密度を有する。TOV−21GおよびHPAC細胞はメイタンシン感受性であり;OVCAR5およびHeLa細胞はメイタンシン感受性が2〜7倍低かった。
表9
【0259】
【表10】
【0260】
*平均最大相対的平均蛍光
【0261】
[266]4つの細胞株をin vitroで増殖させ、収集し、そして100μlの血清不含培地中の1x107細胞を、各マウスの右肩下(モデルあたり6匹のマウス)に注射し、そしてOVCAR5の試験群および対照群では、それぞれ57.6±6.7および90.2±13.4mm3、HPACの試験群および対照群では、それぞれ147.1±29.6および176.2±18.9mm3、HeLaの試験群および対照群では、それぞれ194.3±37.2および201.7±71.7mm3、そしてTOV−21Gの試験群および対照群では、それぞれ96.6±22.8および155.6±13.4mm3の平均腫瘍体積になるまで、6日間増殖させ、この時点で薬剤処置を開始した。各モデルに関して、3匹の対照マウスをPBSの2回の毎週用量で処置し、そして3匹の試験マウスを静脈内でのコンジュゲート(600μg/kg DM1)の2回の毎週用量で処置した。処置中、毒性応答を毎日監視し、そして研究の間中、腫瘍体積および体重を監視した。多様なモデルのコンジュゲート有効性を、図26A、C、E、およびGにグラフで示し、そして対応する体重を図26B、D、F、およびHにプロットする。OVCAR5、TOV−21G、およびHPAC細胞株は、各モデルに関するPBS対照で見られうるように、攻撃的な腫瘍を形成する。HeLaモデルは、指数関数的な増殖を開始するまでに、約3週間の遅延期間を有した。すべてのモデルにおいて、DS6−DM1コンジュゲート処置は、すべてのマウスにおいて、完全腫瘍退行を生じた。TOV−21G、HPAC、およびHeLaモデルに関して、マウスは、第61日に、腫瘍不含のままである。OVCAR5モデルにおいて、腫瘍接種後、約45日で、腫瘍が再発した。したがって、このモデルにおいて、muDS6−DM1処置は、およそ34日間の腫瘍増殖遅延を生じる。OVCAR5細胞がよりメイタンシン感受性でなく、そして低いCA6エピトープ発現を有するため、増殖遅延は有意である。CA6エピトープ密度がより高いか、またはモデルがより高いメイタンシン感受性を有するモデルにおいては、腫瘍退行はより強固である。2回の用量しか投与されないことに注目することが重要である。体重喪失が観察されないため、本研究で用いる投薬スケジュールは、マウスにとっては明らかに毒性でない。治癒は、さらなるまたはより高いコンジュゲート用量で達成可能である可能性がある。
【0262】
[267]ヒト卵巣癌は、大部分、腹膜の疾患である。OVCAR5細胞は、SCIDマウスにおいて、腹腔内(IP)モデルとして攻撃的に増殖して、ヒト疾患と類似の方式で、腫瘍節を形成し、そして腹水を産生する。IPモデルにおいて活性を立証するため、
muDS6−DM1を用いて、OVCAR5 IP腫瘍を所持するマウスを処置した(図27)。OVCAR5細胞をin vitroで増殖させ、採取し、そして100μlの血清不含培地中、1x107細胞を腹腔内注射した。腫瘍を6日間増殖させ、この時点で処置を開始した。600μg/kg DM1の用量で、PBSまたはDS6−DM1コンジュゲートのいずれかで、マウスを2週間、毎週処置し、そして腹腔疾患から生じる体重喪失に関して監視した。第28日までに、マウスのPBS群は、20%より多く体重を失い、そして安楽死させられた。20%を超える体重を喪失した後、第45日に、処置群を屠殺した。この研究によって、OVCAR5細胞がメイタンシンにより感受性でなく、そして細胞あたりより少ないCA6エピトープを有するという事実にもかかわらず、muDS6−DM1は、攻撃的OVCAR5 IPモデルにおいて、腫瘍増殖を遅延させることが可能であることが立証される。用いた投薬スケジュールは、毒性の可視徴候を誘発しなかったため、さらなる用量およびより高い用量を用いて、さらなる腫瘍増殖遅延または治癒を達成することも可能である可能性が高い。
【0263】
実施例17:DS6−SPP−MM1−202タキソイド細胞傷害性コンジュゲートの合成および性質決定
[268]N−スルホスクシンイミジル4−ニトロ−2−ピリジル−ペンタノエート(SSNPP)リンカーを用いて、muDS6を修飾した。90%緩衝液A、10%DMA中の50mgのmuDS6 Abに、DMA中の10当量のSSNPPを添加した。Abの最終濃度は、8mg/mlであった。反応を室温で4時間攪拌し、次いでG25クロマトグラフィーによって精製した。280nm(抗体)および325(リンカー)の吸光度を用いて、抗体修飾の度合いを分光光度的に測定し、そして3.82リンカー/抗体を有することが見出された。抗体の回収は、43.3mgであり、87%収率を生じた。muDS6−ニトロSPPを、タキソイドMM1−202(1812P.16)とコンジュゲート化した。90%緩衝液A、10%DM1中、42mgスケールで、コンジュゲート化を行った。約20時間の期間に渡って、0.43当量/リンカー(各アリコット)の4アリコットでタキソイドを添加した。この時点までに、反応は、著しく曇った。G25精製後、約64%の収率で回収された、生じたコンジュゲートは、約4.3タキソイド/Abを有し、そして約1当量の未反応リンカーが残った。未反応リンカーの反応を停止するため、1当量のシステイン/未反応リンカーを、一晩攪拌しながら、コンジュゲートに添加した。システインを添加した際、明確な黄色がかった色合いが顕著であり、これはチオピリジンの放出を示す。次いで、反応溶液を緩衝液B/0.01%Tween20中で透析し、その後、緩衝液Bのみでさらに数日に渡って透析した。最終コンジュゲートは、2.86薬剤/抗体を有した。抗体回収は14.7mgであり、全体で35%の収率を生じた。SECによってコンジュゲートをさらに生化学的に性質決定し、そして89%単量体、10.5%二量体および0.5%のより高い分子量の凝集体を有することが見出された。
【0264】
[269]HeLa細胞に対する、muDS6−SPP−MM1−202タキソイド対muDS6抗体の結合を比較するフローサイトメトリー分析の結果を、図28に示す。結果によって、muDS6がタキサンにコンジュゲート化された際、結合活性を保持することが示される。
【0265】
実施例18:ヒト化DS6コンジュゲートのin vitroおよびin vivo活性
[270]huDS6v1.01−SPDB−DM4コンジュゲートを構築した。このコンジュゲートは、コンジュゲートのリンカー/メイタンシン薬剤部分の構造がジスルフィド結合周囲で異なることを除いて、実施例14に記載するmuDS6−SPP−DM1コンジュゲートと類似であり;muDS6−SPP−DM1コンジュゲートは、リンカーの抗体側でジスルフィド炭素上に1つのメチル基立体障害を有し、一方、SPDB−DM4コンジュゲートは、リンカーのメイタンシン側でジスルフィド炭素上に2つのメチル基立体障害を有する。
【0266】
[271]8倍モル過剰のN−スクシンイミジル−4−(2−ピリジルジチオ)ブタノエート(SPDB)を用いて、huDS6v1.01抗体(8mg/ml)を修飾して、ジチオピリジル基を導入した。95%v/v緩衝液A(50mM KPi、50mM NaCl、2mM EDTA、pH6.5)および5%v/vエタノール中で、室温で1.5時間、反応を行った。15mlのSephadex G25カラム(緩衝液A中で平衡化)を通じて、反応混合物をゲルろ過した。280nmで抗体の吸光度を、そして280および343nmで、DTTで放出される2−メルカプトピリジン(Spy)を測定することによって、修飾の度合いを決定した。次いで、SPyよりも1.7倍モル過剰のN2’−デアセチル−N2’−(4−メチル−4−メルカプト−1−オキソペンチル)−メイタンシン(L−DM4)を用いて、修飾DS6を1.8mg Ab/mlでコンジュゲート化した。DMA(3%v/v)を含む緩衝液A(97%v/v)中で反応を行った。反応を室温で一晩、〜20時間、インキュベーションした。muDS6のコンジュゲート化と対照的に、反応混合物は透明であり、そして直ちに、クエン酸緩衝液(20mMクエン酸、135mM NaCl、pH5.5)中で平衡化されたNAP 15ml G25カラムを通じたゲルろ過を経た。0.22μm Millex−GVフィルターを用いて、コンジュゲートを滅菌ろ過した。ろ過物質の252nmおよび280nm両方の吸光度を測定することによって、DS6の分子あたりに連結したDM4分子の数を決定した。DM4/Ab比は、3.2であることが見出され、そしてコンジュゲート化DS6の工程収率は69%であった。コンジュゲート化抗体濃度は1.51mg/mlであった。サイズ排除クロマトグラフィー(SEC)によって、精製コンジュゲートを生化学的に性質決定して、そして92.5%単量体であることを見出した。精製コンジュゲート中のDM4の分析によって、>99%が抗体に共有結合していることが示された。
【0267】
[272]図29Aにおいて、huDS6v1.01−DM4コンジュゲートおよび非修飾DS6のKB細胞へのフローサイトメトリー結合によって、huDS6v1.01のコンジュゲート化が親和性の喪失を本質的に生じないことを示す。CaOv−3細胞ではなく、KB細胞を用いたことを除いて、図20に関して記載したのと本質的に同様に、結合を行った。本質的に図24Gに記載するように、huDS6v1.01のin vitro細胞傷害性を試験した。huDS6v1.01はWISH細胞を4.4x10−10MのIC50で殺し、一方、非コンジュゲート化huDS6v1.01は細胞傷害性活性をまったく示さなかった。
【0268】
[273]huDS6v1.01−DM4のin vivo活性を、HPAC膵臓モデルに対して試験した。HPAC細胞を第0日に接種し、そして免疫コンジュゲート処置を第13日に投与した。腫瘍体積が1000cm3を越えた時点で、PBS対照動物を安楽死させた。コンジュゲートを200μg/kgまたは600μg/kg DM4のいずれかの用量で投与し、これはそれぞれ、15mg/kgおよび45mg/kgの抗体濃度に対応した。研究の経過中、マウスの腫瘍体積(図30A)および体重(図30B)を監視した。huDS6v1.01−DM4は、200μg/kg DM4で強力な抗腫瘍活性を示し、すべてのマウスは、完全な腫瘍退行を達成した。HPAC異種移植片上では発現されない抗原を認識する対照B4−DM4コンジュゲートは、200μg/kgで、本質的に活性を持たなかった。マウスの体重喪失が欠如している(図30B)ことから、200μg/kgコンジュゲートでの処置は、最大許容用量未満であることが示される。この結果によって、DS6のヒト化型は、メイタンシノイド薬剤のターゲティング送達を仲介し、強力な抗腫瘍活性を生じることが可能であることが立証される。
【図面の簡単な説明】
【0269】
【図1】[33]図1は、DS6抗体が、選択した癌細胞株の表面に結合する能力を決定するために行った研究の結果を示す。DS6一次抗体およびFITCコンジュゲート化抗マウスIgG(H+L)二次抗体とインキュベーションした細胞株の蛍光を、フローサイトメトリーによって測定した。DS6抗体はCaov−3(図1A)およびT−47D(図1B)細胞に結合し、見かけのKdは、それぞれ、1.848nMおよび2.586nMであった。抗原陰性細胞株、SK−OV−3(図1C)およびColo205(図1D)は、抗原特異的結合をまったく示さなかった。
【図2】[34]図2は、エピトープ発現のドットブロット分析の結果を示す。Caov−3(図2Aおよび図2B)、SKMEL28(図2C)、およびColo205(図2D)細胞溶解物を、ニトロセルロース膜上に個々にスポッティングし、そして次いで、プロナーゼ、プロテイナーゼK、ノイラミニダーゼまたは過ヨウ素酸と、個々にインキュベーションした。次いで、膜をDS6抗体(図2A)、CM1抗体(図2B)、R24抗体(図2C)、またはC242抗体(図2D)で免疫ブロッティングした。
【図3】[35]図3は、DS6抗原発現のドットブロット分析の結果を示す。Caov−3細胞溶解物を、PVDF膜上に個々にスポッティングし、そして次いで、トリフルオロメタンスルホン酸(TFMSA)の存在下でインキュベーションした。次いで、膜を、CM1抗体(1および2)またはDS6(3および4)で免疫ブロッティングした。
【図4】[36]図4は、DS6抗原のグリコトープ分析の結果を示す。N−グリカナーゼ(「N−gly」)、O−グリカナーゼ(「O−gly」)、および/またはシアリダーゼ(「S」)であらかじめ処理したCaov−3溶解物を、ニトロセルロース上にスポッティングし、そして次いで、DS−6抗体またはCM1抗体(Muc−1 VNTR)で免疫ブロッティングした。
【図5−1】[37]図5は、DS6抗原のウェスタンブロット分析の結果を示す。細胞溶解物を、DS6抗体で免疫沈降(「IP」)し、そして免疫ブロッティングした。抗原は、抗原陽性Caov−3(図5Aおよび図5B)およびT47D(図5C)細胞で観察される>250kDaタンパク質バンドに対応する。抗原陰性SK−OV−3(図5D)およびColo205(図5E)細胞株は、このバンドを示さない。免疫沈降後、Caov−3細胞溶解物のプロテインGビーズを、(図5A)ノイラミニダーゼ(「N」)または(図5B)過ヨウ素酸(「PA」)とインキュベーションした。抗体(「α」)、IP前(「Lys」)およびIP後フロースルー(「FT」)溶解物対照を同じゲル上で泳動した。Caov−3免疫沈降物を、N−グリカナーゼ(「N−gly」)、O−グリカナーゼ(「O−gly」)、および/またはシアリダーゼ(「S」)ともまたインキュベーションし(図5Fを参照されたい)、この場合、ブロットを代わりに、ビオチン化DS6およびストレプトアビジン−HRPで探査した(probed)。
【図5−2】[37]図5は、DS6抗原のウェスタンブロット分析の結果を示す。細胞溶解物を、DS6抗体で免疫沈降(「IP」)し、そして免疫ブロッティングした。抗原は、抗原陽性Caov−3(図5Aおよび図5B)およびT47D(図5C)細胞で観察される>250kDaタンパク質バンドに対応する。抗原陰性SK−OV−3(図5D)およびColo205(図5E)細胞株は、このバンドを示さない。免疫沈降後、Caov−3細胞溶解物のプロテインGビーズを、(図5A)ノイラミニダーゼ(「N」)または(図5B)過ヨウ素酸(「PA」)とインキュベーションした。抗体(「α」)、IP前(「Lys」)およびIP後フロースルー(「FT」)溶解物対照を同じゲル上で泳動した。Caov−3免疫沈降物を、N−グリカナーゼ(「N−gly」)、O−グリカナーゼ(「O−gly」)、および/またはシアリダーゼ(「S」)ともまたインキュベーションし(図5Fを参照されたい)、この場合、ブロットを代わりに、ビオチン化DS6およびストレプトアビジン−HRPで探査した(probed)。
【図6】[38]図6は、Caov−3(図6A)およびHeLa(図6B)細胞溶解物に対する、DS6抗体およびCM1抗体の免疫沈降および/または免疫ブロットの結果を示す。CM1およびDS6ウェスタンブロットシグナルの重複は、DS6抗原がMuc1タンパク質上にあることを示す。HeLa溶解物において、Muc1ダブレットは、タンデム反復数が異なる別個のアレルによって指示されるMuc1発現から生じる。
【図7】[39]図7は、DS6抗体サンドイッチELISA設計(図7A)および標準曲線(図7B)を示す。商業的に入手可能なCA15−3標準の既知の濃度を用いて、標準曲線を生じた(1 CA15−3単位=1 DS6単位)。
【図8】[40]図8は、定量的ELISA標準曲線を示す。プレーティングされたヤギ抗マウスIgGによって捕捉される(図8A)か、またはELISAプレート上に直接結合する(図8B)か、いずれかの、既知の濃度のビオチン−DS6を用いて、検出抗体(ストレプトアビジン−HRP/ビオチン−DS6)シグナルの標準曲線(図8C)を決定した。
【図9】[41]図9は、ネズミDS6抗体の軽鎖(図9A)および重鎖(図9B)可変領域のcDNAおよびアミノ酸配列を示す。各配列中の3つのCDRを下線で示す(Kabat定義)。
【図10】[42]図10は、Kabat定義によって決定されるネズミDS6抗体の軽鎖(図10A)および重鎖(図10B)CDRを示す。AbMモデリングソフトウェアは、重鎖CDRのわずかに異なる定義を生じる(図10C)。
【図11】[43]図11は、IgVκap4(配列番号23)およびIgVh J588.41(配列番号24)遺伝子の生殖系列配列と並列させた、ネズミDS6抗体の軽鎖(「muDS6LC」)(配列番号7の残基1〜95)および重鎖(「muDS6HC」)(配列番号9の残基1〜98)アミノ酸配列を示す。灰色は配列相違を示す。
【図12】[44]図12は、Brookhavenデータベースにおいて、解明された構造ファイルを有する、ネズミDS6(muDS6)軽鎖(「muDS6LC」)および重鎖(「muDS6HC」)配列に最も相同な10の軽鎖および重鎖抗体配列を示す。最も相同であるものから最も相同でないものの順に、配列を並列させる。
【図13−1】[45]図13は、ネズミDS6抗体軽鎖可変領域のどのフレームワーク残基が表面にアクセス可能であるかを予測する、表面アクセス可能性データおよび計算を示す。25〜35%の平均表面アクセス可能性を持つ位に印を付け(*??*)、そしてこれを第二周期の分析に供した。DS6抗体軽鎖可変領域(図13A)および重鎖可変領域(図13B)。〓
【図13−2】[45]図13は、ネズミDS6抗体軽鎖可変領域のどのフレームワーク残基が表面にアクセス可能であるかを予測する、表面アクセス可能性データおよび計算を示す。25〜35%の平均表面アクセス可能性を持つ位に印を付け(*??*)、そしてこれを第二周期の分析に供した。DS6抗体軽鎖可変領域(図13A)および重鎖可変領域(図13B)。
【図14】[46]図14は、prDS6 v1−0哺乳動物発現プラスミドマップを示す。このプラスミドを用いて、組換えキメラおよびヒト化DS6抗体を構築し、そして発現させた。
【図15】[47]図15は、ネズミ(「muDS6」)およびヒト化(「huDS6」)(1.01および1.21)DS6抗体軽鎖(図15A)および重鎖(図15B)可変ドメインのアミノ酸配列を示す。
【図16】[48]図16は、ヒト化DS6抗体(「huDS6」)(1.01および1.21)の軽鎖可変領域のcDNAおよびアミノ酸配列を示す。
【図17】[49]図17は、ヒト化DS6抗体(「huDS6」)1.01(図17A)および1.21(図17B)の重鎖可変領域のcDNAおよびアミノ酸配列を示す。
【図18】[50]図18は、KB細胞に対して行ったアッセイからの、ネズミDS6(muDS6)、キメラDS6(chDS6)、およびヒトDS6 1.01型(huDS6 v1.01)およびhuDS6 1.21型(huDS6 v1.21)のフローサイトメトリー結合曲線を示す。ネズミ、キメラ、およびヒトv1.01およびv1.21 DS6抗体のアビディティー(muDS6=0.82nM、chDS6=0.69nM、huDS6v1.01=0.82nMおよびhuDS6v1.21=0.85nM)は同程度であり、表面再構成がアビディティーを減少させなかったことを示す。
【図19】[51]図19は、muDS6、chDS6、huDS6 v1.01およびhuDS6 v1.21抗体とビオチン化muDS6の競合結合アッセイの結果を示す。多様な濃度の裸のmuDS6、chDS6、huDS6v1.01およびhuDS6v1.21を、2nMのビオチン−muDS6およびストレプトアビジン−DTAF二次抗体と合わせた。すべての抗体のIC50(muDS6=1.9nM、chDS6=1.7nM、huDS6v1.01=3.0nM、およびhuDS6v1.21=1.9nM)は類似であり、これは、ヒト化がアビディティーを減少させなかったことを示す。
【図20】[52]図20は、DS6抗体−DM1コンジュゲートに対する非コンジュゲート化DS6抗体の結合親和性決定の結果を示す。この結果は、DM1コンジュゲート化が、抗体の結合親和性に不都合に影響を及ぼさないことを示した。DS6抗体−DM1コンジュゲートの見かけのKd(3.902nM)(「DS6−DM1」)は、裸の抗体(2.020nM)(「DS6」)よりわずかに高かった。
【図21】[53]図21は、抗マウスIgG(H+L)DM1コンジュゲート(2°Ab−DM1)の存在下または非存在下で、DS6抗体を用いた、間接的細胞生死判別アッセイの結果を示す。抗原陽性Caov−3細胞は、二次コンジュゲートの存在下(「DS6+2°Ab−DM1」)でのみ、DS6抗体依存性方式で殺された(IC50=424.9pM)。
【図22】[54]図22は、muDS6抗体の補体依存性細胞傷害性(CDC)アッセイの結果を示す。結果は、HPAC(図22A)およびZR−75−1(図22B)細胞に対して、CDCが仲介するDS6抗体の効果がないことを示す。
【図23−1】[55]図23は、未結合(free)のメイタンシンに対する、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。コロニー形成アッセイにおいて、DS6抗原陽性卵巣癌(図23A)、乳癌(図23B)、子宮頸癌(図23C)、および膵臓癌(図23D)細胞株を、DS6抗体−DM1コンジュゲートに対する連続曝露の細胞傷害性に関して試験した(左のパネル)。未結合メイタンシンへの72時間の曝露によって、メイタンシン感受性に関して、これらの細胞株を同様に試験した(右のパネル)。試験した卵巣癌細胞株は、OVCAR5、TOV−21G、Caov−4およびCaov−3であった。試験した乳癌細胞株は、T47D、BT−20およびBT−483であった。試験した子宮頸癌細胞株は、KB、HeLaおよびWISHであった。試験した膵臓癌細胞株は、HPAC、Hs766TおよびHPAF−IIであった。
【図23−2】[55]図23は、未結合(free)のメイタンシンに対する、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。コロニー形成アッセイにおいて、DS6抗原陽性卵巣癌(図23A)、乳癌(図23B)、子宮頸癌(図23C)、および膵臓癌(図23D)細胞株を、DS6抗体−DM1コンジュゲートに対する連続曝露の細胞傷害性に関して試験した(左のパネル)。未結合メイタンシンへの72時間の曝露によって、メイタンシン感受性に関して、これらの細胞株を同様に試験した(右のパネル)。試験した卵巣癌細胞株は、OVCAR5、TOV−21G、Caov−4およびCaov−3であった。試験した乳癌細胞株は、T47D、BT−20およびBT−483であった。試験した子宮頸癌細胞株は、KB、HeLaおよびWISHであった。試験した膵臓癌細胞株は、HPAC、Hs766TおよびHPAF−IIであった。
【図23−3】[55]図23は、未結合(free)のメイタンシンに対する、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。コロニー形成アッセイにおいて、DS6抗原陽性卵巣癌(図23A)、乳癌(図23B)、子宮頸癌(図23C)、および膵臓癌(図23D)細胞株を、DS6抗体−DM1コンジュゲートに対する連続曝露の細胞傷害性に関して試験した(左のパネル)。未結合メイタンシンへの72時間の曝露によって、メイタンシン感受性に関して、これらの細胞株を同様に試験した(右のパネル)。試験した卵巣癌細胞株は、OVCAR5、TOV−21G、Caov−4およびCaov−3であった。試験した乳癌細胞株は、T47D、BT−20およびBT−483であった。試験した子宮頸癌細胞株は、KB、HeLaおよびWISHであった。試験した膵臓癌細胞株は、HPAC、Hs766TおよびHPAF−IIであった。
【図23−4】[55]図23は、未結合(free)のメイタンシンに対する、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。コロニー形成アッセイにおいて、DS6抗原陽性卵巣癌(図23A)、乳癌(図23B)、子宮頸癌(図23C)、および膵臓癌(図23D)細胞株を、DS6抗体−DM1コンジュゲートに対する連続曝露の細胞傷害性に関して試験した(左のパネル)。未結合メイタンシンへの72時間の曝露によって、メイタンシン感受性に関して、これらの細胞株を同様に試験した(右のパネル)。試験した卵巣癌細胞株は、OVCAR5、TOV−21G、Caov−4およびCaov−3であった。試験した乳癌細胞株は、T47D、BT−20およびBT−483であった。試験した子宮頸癌細胞株は、KB、HeLaおよびWISHであった。試験した膵臓癌細胞株は、HPAC、Hs766TおよびHPAF−IIであった。
【図24】[56]図24は、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。MTT細胞生死判別アッセイにおいて、ヒト卵巣癌(図24A、図24Bおよび図24C)、乳癌(図24Dおよび図24E)、子宮頸癌(図24Fおよび図24G)、および膵臓癌(図24Hおよび図24I)細胞は、DS6抗体−DM1コンジュゲート依存性方式で、殺された。裸のDS6は、これらの細胞の増殖に不都合に影響を及ぼさず、これは、細胞傷害性には、DM1コンジュゲート化が必要であることを示した。
【図25】[57]図25Aは、確立された皮下KB腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートのin vivo抗腫瘍有効性研究の結果を示す。腫瘍細胞を、第0日に接種し、そして最初の処置を第6日に与えた。免疫コンジュゲート処置を、総数5用量で毎日続けた。腫瘍体積が1500mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを150または225μg/kg DM1の用量で投与し、これはそれぞれ、5.7および8.5mg/kgの抗体濃度に対応した。研究経過中、マウスの体重(図25B)を監視した。
【図26−1】[58]図26は、確立された皮下腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートの抗腫瘍有効性研究の結果を示す。第0日に、OVCAR5(図26Aおよび図26B)、TOV−21G(図26Cおよび図26D)、HPAC(図26Eおよび図26F)、およびHeLa(図26Gおよび図26H)細胞を接種し、そして免疫コンジュゲート処置を、第6日および第13日に与えた。腫瘍体積が1000mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを600μg/kg DM1の用量で投与し、これは、27.7mg/kgの抗体濃度に対応した。研究経過中、マウスの腫瘍体積(図26A、図26C、図26E、および図26G)および体重(図26B、図26D、図26F、および図26H)を監視した。
【図26−2】[58]図26は、確立された皮下腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートの抗腫瘍有効性研究の結果を示す。第0日に、OVCAR5(図26Aおよび図26B)、TOV−21G(図26Cおよび図26D)、HPAC(図26Eおよび図26F)、およびHeLa(図26Gおよび図26H)細胞を接種し、そして免疫コンジュゲート処置を、第6日および第13日に与えた。腫瘍体積が1000mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを600μg/kg DM1の用量で投与し、これは、27.7mg/kgの抗体濃度に対応した。研究経過中、マウスの腫瘍体積(図26A、図26C、図26E、および図26G)および体重(図26B、図26D、図26F、および図26H)を監視した。
【図26−3】[58]図26は、確立された皮下腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートの抗腫瘍有効性研究の結果を示す。第0日に、OVCAR5(図26Aおよび図26B)、TOV−21G(図26Cおよび図26D)、HPAC(図26Eおよび図26F)、およびHeLa(図26Gおよび図26H)細胞を接種し、そして免疫コンジュゲート処置を、第6日および第13日に与えた。腫瘍体積が1000mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを600μg/kg DM1の用量で投与し、これは、27.7mg/kgの抗体濃度に対応した。研究経過中、マウスの腫瘍体積(図26A、図26C、図26E、および図26G)および体重(図26B、図26D、図26F、および図26H)を監視した。
【図26−4】[58]図26は、確立された皮下腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートの抗腫瘍有効性研究の結果を示す。第0日に、OVCAR5(図26Aおよび図26B)、TOV−21G(図26Cおよび図26D)、HPAC(図26Eおよび図26F)、およびHeLa(図26Gおよび図26H)細胞を接種し、そして免疫コンジュゲート処置を、第6日および第13日に与えた。腫瘍体積が1000mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを600μg/kg DM1の用量で投与し、これは、27.7mg/kgの抗体濃度に対応した。研究経過中、マウスの腫瘍体積(図26A、図26C、図26E、および図26G)および体重(図26B、図26D、図26F、および図26H)を監視した。
【図27】[59]図27は、腹腔内OVCAR5腫瘍に対する、muDS6抗体−DM1コンジュゲートのin vivo有効性研究の結果を示す。第0日に、腫瘍細胞を腹腔内注射し、そして第6日および第13日に、免疫コンジュゲート処置を与えた。体重喪失が20%を超えた時点で、動物を安楽死させた。
【図28】[60]図28は、HeLa細胞に対する、裸のおよびタキサン−コンジュゲート化DS6抗体の結合親和性の研究由来のフローサイトメトリー結合曲線を示す。タキサン(MM1−202)−コンジュゲート化は、抗体の結合親和性に不都合に影響を及ぼさない。DS6−MM1−202コンジュゲートの見かけのKd(1.24nM)は、裸のDS6抗体(620pM)よりもわずかに大きかった。
【図29】[61]図29は、ヒト化DS6 1.01型抗体コンジュゲートのin vitro結合および強度を示す。DM4とhuDS6v1.01のコンジュゲート化は、KB細胞に対するhuDS6v1.01のアビディティーにほとんど影響を及ぼさない(図29A)。huDS6v1.01−DM4は、DS6を発現しているWISH細胞に向かって、強力なin vitro細胞傷害性を示し、IC50は0.44nMであった(図29B)。
【図30】[62]図30は、HPAC膵臓癌モデルにおける、huDS6v1.01−DM4コンジュゲートを用いたin vivo有効性研究の結果を示す。huDS6v1.01−DM4は、強力な抗腫瘍活性を示し、一方、HPACモデル中にターゲットが発現されていないB4−DM4対照コンジュゲートは、本質的にまったく活性を持たなかった(図30A)。200μg/kgの投与用量は、体重喪失の欠如によって示されるように、動物に対して毒性ではなかった(図30B)。
【技術分野】
【0001】
発明の分野
[01]本発明は、ネズミ抗CA6グリコトープ・モノクローナル抗体、およびそのヒト化型またはその表面再構成(resurfaced)型に関する。本発明はまた、抗CA6グリコトープ・モノクローナル抗体のエピトープ結合性断片に、ならびに抗CA6グリコトープ・モノクローナル抗体のヒト化型または表面再構成型のエピトープ結合性断片にも関する。
【0002】
[02]本発明は、細胞結合性剤および細胞傷害性剤を含む細胞傷害性コンジュゲート、該コンジュゲートを含む療法組成物、細胞増殖の阻害および疾患の治療において該コンジュゲートを用いるための方法、ならびに該細胞傷害性コンジュゲートを含むキットにさらに関する。特に、細胞結合性剤は、CA6グリコトープを認識し、そしてこれに結合する、モノクローナル抗体またはそのエピトープ結合性断片、あるいはそのヒト化型または表面再構成型である。
【背景技術】
【0003】
発明の背景
[03]周囲の非癌性細胞および組織に害を及ぼすことなく、ターゲット癌細胞を特異的に破壊する、抗癌療法剤を開発する数多くの試みがなされてきている。こうした療法剤は、ヒト患者における癌の治療を非常に改善する潜在能力を有する。
【0004】
[04]1つの有望なアプローチは、モノクローナル抗体などの細胞結合性剤と、細胞傷害性薬剤を連結することであった(Selaら, Immunoconjugates中 189−216(C. Vogel監修 1987); Ghoseら, Targeted Drugs中 1−22(E. Goldberg監修 1983); Dienerら, Antibody mediated delivery systems中 1−23(J. Rodwell監修 1988); Pieterszら, Antibody mediated delivery systems中 25−53(J. Rodwell監修 1988); Bumolら, Antibody mediated delivery systems中 55−79(J. Rodwell監修 1988)。細胞結合性剤の選択次第で、こうした細胞の表面上に発現される分子の発現プロフィールに基づいて、癌性細胞の特定の種類のみを認識し、そしてこうした種類のみに結合するように、これらの細胞傷害性コンジュゲートを設計することも可能である。
【0005】
[05]多様なネズミ・モノクローナル抗体に連結された、メトトレキセート、ダウノルビシン、ドキソルビシン、ビンクリスチン、ビンブラスチン、メルファラン、マイトマイシンC、およびクロラムブシルなどの細胞傷害性薬剤が、こうした細胞傷害性コンジュゲートで用いられてきている。いくつかの場合、血清アルブミン(Garnettら, 46 Cancer Res. 2407−2412(1986); Ohkawaら 23 Cancer Immunol. Immunother. 81−86(1986); Endoら, 47 Cancer Res. 1076−1080(1980))、デキストラン(Hurwitzら, 2 Appl. Biochem. 25−35(1980); Manabiら, 34 Biochem. Pharmacol. 289−291(1985); Dillmanら, 46 Cancer Res. 4886−4891(1986); Shovalら, 85 Proc. Nat
l. Acad. Sci. 8276−8280(1988))、またはポリグルタミン酸(Tsukadaら, 73 J. Natl. Canc. Inst. 721−729(1984); Katoら 27 J. Med. Chem. 1602−1607(1984); Tsukadaら, 52 Br. J. Cancer 111−116(1985))などの中間キャリアー分子を通じて、薬剤分子が抗体分子に連結された。
【0006】
[06]ある程度有望であることが示されている1つの特異的コンジュゲートの例として、結腸直腸腫瘍および膵臓腫瘍上に発現される抗原であるCanAgに対して向けられるC242抗体、ならびにメイタンシン誘導体DM1のコンジュゲートがある(Liuら, Proc Natl Acad Sci USA, 93: 8618−8623(1996))。このコンジュゲートのin vitro評価によって、細胞表面上に発現されるCanAgに向かうその結合親和性が高く、見かけのKd値が3x10−11Mであり、そしてCanAg陽性細胞に対する細胞傷害能が高く、IC50が6x10−11Mであることが示された。この細胞傷害性は、過剰な非コンジュゲート化抗体によって遮断されたため、そして抗原陰性細胞が該コンジュゲートに対して100倍以上感受性が低かったため、該細胞傷害性は抗原依存性である。それぞれのターゲット細胞に向かう親和性が高く、そしてかつ抗原選択的細胞傷害性が高い、抗体−DM1コンジュゲートの他の例には、ヒトCD56に対する抗体のヒト化型、huN901;ヒトCD33に対する抗体のヒト化型、huMy9−6;CanAg Muc1エピトープに対する抗体のヒト化型、huC242;PSMAに対する脱免疫化(deimmunized)抗体、huJ591;Her2/neuに対するヒト化抗体、トラスツズマブ;およびCD44v6に対するヒト化抗体、ビバツズマブが含まれる。
【0007】
[07]癌患者を治療するのに用いる方法を、続けて改善する際に、特定の種類の癌性細胞を特異的に認識するさらなる細胞傷害性コンジュゲートの開発が重要であろう。
【発明の開示】
【発明が解決しようとする課題】
【0008】
[08]この目的に向けて、本発明は、癌性細胞表面上に発現される分子/受容体を認識し、そしてこれに結合する抗体の開発に、そして抗体などの細胞結合性剤、および癌性細胞の表面上に発現される分子/受容体を特異的にターゲティングする細胞傷害性剤を含む、新規細胞傷害性コンジュゲートの開発に関する。
【0009】
[09]より具体的には、本発明は、癌性細胞によって発現されるMuc1ムチン受容体上の新規CA6シアログリコトープの性質決定に、そしてMuc1ムチンの新規CA6シアログリコトープを認識し、そして細胞傷害性剤の背景で、CA6グリコトープを発現している細胞の増殖を阻害するために使用可能である抗体、好ましくはヒト化抗体の提供に関する。
【課題を解決するための手段】
【0010】
発明の概要
[10]本発明には、Muc1ムチン受容体の新規CA6シアログリコトープを特異的に認識し、そして結合する、抗体またはそのエピトープ結合性断片が含まれる。別の態様において、本発明には、Muc1ムチン受容体の新規CA6シアログリコトープ(「CA6グリコトープ」)を認識する、ヒト化抗体またはそのエピトープ結合性断片が含まれる。
【0011】
[11]好ましい態様において、本発明には、ネズミ抗CA6モノクローナル抗体DS6(「DS6抗体」)、および当該抗体またはそのエピトープ結合性断片の、表面に曝露
される残基が、既知のヒト抗体表面により密接に似るように、軽鎖および重鎖両方において置換されている、DS6抗体の表面再構成型またはヒト化型が含まれる。本発明のヒト化抗体およびそのエピトープ結合性断片は、投与されるヒト被験体において、完全ネズミ型よりも、はるかにより免疫原性でない(または完全に非免疫原性である)点で、改善された特性を有する。したがって、本発明のヒト化DS6抗体およびそのエピトープ結合性断片は、ヒトに免疫原性でない一方、Muc1ムチン受容体上の新規シアログリコトープ、すなわちCA6グリコトープを特異的に認識する。ヒト化抗体およびそのエピトープ結合性断片を、メイタンシノイドなどの薬剤にコンジュゲート化させて、Muc1 CA6シアログリコトープに薬剤をターゲティングすることによって、抗原を発現している細胞に向かう特異的細胞傷害性を有するプロドラッグを形成してもよい。したがって、こうした抗体および小さい非常に毒性である薬剤(例えばメイタンシノイド、タキサン、およびCC−1065類似体)を含む細胞傷害性コンジュゲートを、乳房腫瘍および卵巣腫瘍などの腫瘍治療のための療法剤として用いてもよい。
【0012】
[12]本発明のDS6抗体のヒト化型は、軽鎖および重鎖可変領域両方のそれぞれのアミノ酸配列、軽鎖および重鎖可変領域の遺伝子のDNA配列、CDRの同定、その表面アミノ酸の同定、ならびに組換え型におけるその発現のための手段の開示に関して、本明細書において完全に性質決定されている。
【0013】
[13]1つの態様において、配列番号1〜3によって示されるアミノ酸配列:
【0014】
【化1】
【0015】
を有するCDRを含む重鎖を有し、そして配列番号4〜6によって示されるアミノ酸配列:
【0016】
【化2】
【0017】
を有するCDRを含む軽鎖を有する、ヒト化DS6抗体またはそのエピトープ結合性断片を提供する。
[14]配列番号7または配列番号8によって示されるアミノ酸配列:
【0018】
【化3】
【0019】
と、少なくとも90%の配列同一性を共有するアミノ酸配列を有する軽鎖可変領域を有する、ヒト化DS6抗体およびそのエピトープ結合性断片もまた提供する。
[15]同様に、配列番号9、配列番号10、または配列番号11によって示されるアミノ酸配列:
【0020】
【化4】
【0021】
と、少なくとも90%の配列同一性を共有するアミノ酸配列を有する重鎖可変領域を有する、ヒト化DS6抗体およびエピトープ結合性断片を提供する。
[16]別の態様において、配列番号8に対応するアミノ酸配列
【0022】
【化5】
【0023】
を有するヒト化または表面再構成軽鎖可変領域を有する、ヒト化DS6抗体およびそのエピトープ結合性断片を提供する。
[17]同様に、それぞれ、配列番号10または配列番号11に対応するアミノ酸配列:
【0024】
【化6】
【0025】
を有するヒト化または表面再構成重鎖可変領域を有する、ヒト化DS6抗体およびそのエピトープ結合性断片を提供する。
[18]本発明のヒト化DS6抗体およびそのエピトープ結合性断片にはまた、ヒト残基への変更が必要な、CDRの5オングストローム以内に見られるネズミ表面フレームワーク残基に相当する、表1の星印の残基によって定義される、1以上の位での軽鎖および/または重鎖アミノ酸残基中の置換をも含むことができる。例えば、ネズミ配列(配列番号7)中の最初のアミノ酸残基Qは、抗体をヒト化するため、E(配列番号8)によって置換されている。しかし、この残基はCDRに近接しているため、抗体親和性を維持するために、ネズミ残基Qへの復帰突然変異が必要でありうる。
表1
【0026】
【表1】
【0027】
[19]3つの最も相同なヒト可変領域表面残基と並列させて、muDS6可変領域表面残基を示す表2に、これをさらに示す。表1のアミノ酸残基は、表2の下線のアミノ酸残基に対応する。
表2
【0028】
【表2】
【0029】
[20]本発明は、(1)CA6グリコトープを認識し、そして該グリコトープに結合する細胞結合性剤、および(2)細胞傷害性剤を含む、細胞傷害性コンジュゲートをさらに提供する。本発明の細胞傷害性コンジュゲートが有効な殺傷剤を形成するように、細胞傷害性コンジュゲートにおいて、細胞結合性剤はCA6グリコトープに対して高い親和性を有し、そして細胞傷害性剤はCA6グリコトープを発現している細胞に対して高い度合いの細胞傷害性を有する。
【0030】
[21]好ましい態様において、細胞結合性剤は、抗CA6抗体またはそのエピトープ結合性断片、より好ましくはヒト化抗CA6抗体またはそのエピトープ結合性断片であり、ここで細胞傷害性剤が、直接、あるいは切断可能または切断不能リンカーを介して、抗体またはそのエピトープ結合性断片に共有結合している。より好ましい態様において、細胞結合性剤は、ヒト化DS6抗体またはそのエピトープ結合性断片であり、そして細胞傷害性剤は、タキソール、メイタンシノイド、CC−1065またはCC−1065類似体である。
【0031】
[22]本発明の好ましい態様において、細胞結合性剤はヒト化抗CA6抗体であり、そして細胞傷害性剤はメイタンシノイドまたはタキサンなどの細胞傷害性薬剤である。
[23]より好ましくは、細胞結合性剤はヒト化抗CA6抗体DS6であり、そして細胞傷害性剤はDM1またはDM4などのメイタンシン化合物である。
【0032】
[24]本発明にはまた、CA6グリコトープを発現している細胞の増殖を阻害するための方法も含まれる。好ましい態様において、CA6グリコトープを発現している細胞の増殖を阻害するための方法は、in vivoで起こり、そして細胞の死を生じるが、in vitroおよびex vivo適用もまた含まれる。
【0033】
[25]本発明はまた、細胞傷害性コンジュゲート、および薬学的に許容しうるキャリアーまたは賦形剤を含む療法組成物も提供する。
[26]本発明には、療法組成物を用いて、癌を有する被験体を治療する方法がさらに含まれる。好ましい態様において、細胞傷害性コンジュゲートは、抗CA6抗体および細胞傷害性剤を含む。より好ましい態様において、細胞傷害性コンジュゲートは、ヒト化DS6抗体−DM1コンジュゲート、ヒト化DS6抗体−DM4またはヒト化DS6抗体−タキサン・コンジュゲートを含み、そして該コンジュゲートは、薬学的に許容しうるキャリアーまたは賦形剤と一緒に投与される。
【0034】
[27]本発明にはまた、抗CA6抗体−細胞傷害性剤コンジュゲートおよび使用のための説明書を含むキットも含まれる。好ましい態様において、抗CA6抗体はヒト化DS6抗体であり、細胞傷害性剤はDM1またはDM4などのメイタンシン化合物、あるいは
タキサンであり、そして説明書は、癌を有する被験体の治療において、コンジュゲートを使用するためのものである。キットにはまた、コンジュゲートが凍結乾燥状態または濃縮型であるならば希釈剤などの、薬学的に許容しうる配合物の調製に必要な構成要素、および配合物の投与に必要な構成要素も含まれていてもよい。
【0035】
[28]本発明にはまた、CA6グリコトープに特異的に結合し、そして該グリコトープを認識する抗体の誘導体も含まれる。好ましい態様において、抗体誘導体は、CA6グリコトープに結合する抗体の表面を再構成するかまたは該抗体をヒト化することによって調製され、ここで該誘導体は宿主に対して減少した免疫原性を有する。
【0036】
[29]本発明は、研究または診断適用において使用するため、さらに標識されているヒト化抗体またはその断片をさらに提供する。好ましい態様において、標識は、放射標識、蛍光体、発色団、造影剤または金属イオンである。
【0037】
[30]前記の標識ヒト化抗体またはそのエピトープ結合性断片を、癌を有すると推測される被験体に投与し、そして被験体体内の標識分布を測定するかまたは監視する、診断法もまた、提供する。
【0038】
[31]本発明はまた、本発明のヒト化抗体コンジュゲートを、単独で、あるいは他の細胞傷害性剤または療法剤と組み合わせて投与することによって、癌を有する被験体を治療するための方法もまた提供する。癌は、CA6が発現されている、例えば乳癌、結腸癌、卵巣癌、子宮内膜癌、骨肉腫、子宮頸癌、前立腺癌、肺癌、滑膜癌、膵臓癌、肉腫または癌腫、あるいはCA6グリコトープが主に発現されている、まだ決定されていない他の癌の1以上であってもよい。
【0039】
[32]別に言及しない限り、本明細書に引用するすべての参考文献および特許は、本明細書に援用される。
【0040】
発明の詳細な説明
[63]本発明は、他の特徴の中でも、抗CA6モノクローナル抗体、抗CA6ヒト化抗体、および抗CA6抗体の断片を提供する。本発明の抗体および抗体断片の各々は、細胞表面上のCA6グリコトープを特異的に認識し、そして該グリコトープに結合するよう設計される。CA6は、多くのヒト腫瘍:漿液性卵巣癌の95%、子宮内膜様卵巣癌の50%、子宮頸部新生物の50%、子宮内膜新生物の69%、外陰新生物の80%、乳癌の60%、膵臓腫瘍の67%、および尿路上皮腫瘍の48%によって発現されることが知られるが、正常ヒト組織によっては滅多に発現されない。
【0041】
[64]Kearseら, Int. J. Cancer 88(6):866−872(2000)による報告は、ハイブリドーマ上清を用いてCA6エピトープを性質決定した際、CA6エピトープが見られるタンパク質を、CA6エピトープを含有するN連結炭水化物を有する80kDaタンパク質と誤認した。その後、本発明者らは、精製DS6を用いて、250kDaの非ジスルフィド連結糖タンパク質より大きい、O連結炭水化物上にCA6エピトープが見られることを立証した。さらに、該糖タンパク質は、ムチン、Muc1と同定された。異なるMuc1アレルは、可変数タンデム反復(VNTR)ドメインにおいて、多様な数のタンデム反復を有するため、細胞はしばしば、異なるサイズの2つの別個のMuc1タンパク質を発現する(Taylor−Papadimitriou, Biochim. Biophys. Acta 1455(2−3):301−13(1999)。VNTRドメイン中の反復数が異なり、さらにグリコシル化も異なるため、Muc1の分子量は細胞株間で多様である。
【0042】
[65]過ヨウ素酸に対するCA免疫反応性の感受性は、CA6が炭水化物エピトープ「グリコトープ」であることを示す。コレラ菌(Vibrio cholerae)由来のノイラミニダーゼでの処理に対する、CA6免疫反応性のさらなる感受性は、CA6エピトープがシアル酸依存性グリコトープであり、したがって、「シアログリコトープ」であることを示す。
【0043】
[66]CA6の性質決定の詳細を実施例2に見出しうる(以下を参照されたい)。CA6に関するさらなる詳細は、WO 02/16401; Wennerbergら, Am. J. Pathol. 143(4):1050−1054(1993); Smithら, Human Antibodies 9:61−65(1999); Kearseら, Int. J. Cancer 88(6):866−872(2000); Smithら, Int. J. Gynecol. Pathol. 20(3):260−6(2001);およびSmithら, Appl. Immunohistochem. Mol. Morphol. 10(2):152−8(2002)に見出されうる。
【0044】
[67]本発明にはまた、2つの主な構成要素を含む細胞傷害性コンジュゲートも含まれる。第一の構成要素は、CA6グリコトープを認識し、そして該グリコトープに結合する、細胞結合性剤である。細胞傷害性コンジュゲートが、意図する細胞のみを認識し、そして該細胞に結合するように、細胞結合性剤は、高い度合いの特異性で、Muc1上のCA6シアログリコトープを認識すべきである。高い度合いの特異性は、コンジュゲートが、非特異的結合から生じる副作用をほとんど伴わずに、ターゲティングされた方式で作用することを可能にするであろう。
【0045】
[68]別の態様において、コンジュゲートの細胞傷害性薬剤部分が細胞に対して作用するのを可能にするのに十分な期間、そして/または細胞によってコンジュゲートが内部移行されるのを可能にするのに十分な期間、コンジュゲートがターゲット細胞と接触するように、本発明の細胞結合性剤はまた、CA6グリコエピトープを高い度合いの親和性でも認識するであろう。
【0046】
[69]好ましい態様において、細胞傷害性コンジュゲートは、細胞結合性剤として、抗CA6抗体、より好ましくはネズミDS6抗CA6モノクローナル抗体を含む。より好ましい態様において、細胞傷害性コンジュゲートは、ヒト化DS6抗体またはそのエピトープ結合性断片を含む。DS6抗体は、高い度合いの特異性でCA6を認識することが可能であり、そして細胞傷害性剤を、ターゲティング方式で、癌細胞などの異常な細胞または組織に向ける。
【0047】
[70]本発明の細胞傷害性コンジュゲートの第二の構成要素は、細胞傷害性剤である。好ましい態様において、細胞傷害性剤は、タキソール、DM1またはDM4などのメイタンシノイド、CC−1065またはCC−1065類似体である。好ましい態様において、本発明の細胞結合性剤は、直接、あるいは切断可能または切断不能リンカーを介して、細胞傷害性剤に共有結合している。
【0048】
[71]細胞結合性剤、細胞傷害性剤、およびリンカーを、以下により詳細に論じる。
細胞結合性剤
[72]療法剤としての本発明の化合物の有効性は、適切な細胞結合性剤の注意深い選択次第である。細胞結合性剤は、現在知られるかまたは知られるようになる、いかなる種類のものであってもよく、そしてこうした剤には、ペプチドおよび非ペプチドが含まれる。細胞結合性剤は、特異的または非特異的方式のいずれかで、細胞に結合可能な、いかなる化合物であってもよい。一般的に、これらは、抗体(特にモノクローナル抗体)、リン
ホカイン、ホルモン、増殖因子、ビタミン、栄養輸送分子(トランスフェリンなど)、あるいは他の任意の細胞結合性分子または物質であってもよい。
【0049】
[73]使用可能な細胞結合性剤のより具体的な例には:
(a)ポリクローナル抗体;
(b)モノクローナル抗体;
(c)Fab、Fab’、およびF(ab’)2、Fvなどの抗体断片(Parham, J. Immunol. 131:2895−2902(1983); Springら J. Immunol. 113:470−478(1974); Nisonoffら, Arch. Biochem. Biophys. 89:230−244(1960));
(d)インターフェロン(例えばアルファ、ベータ、ガンマ);
(e)IL−2、IL−3、IL−4、IL−6などのリンホカイン;
(f)インスリン、TRH(甲状腺刺激ホルモン放出ホルモン)、MSH(メラニン形成細胞刺激ホルモン)、アンドロゲンおよびエストロゲンなどのステロイドホルモンなどのホルモン;
(g)EGF、TGF−アルファ、FGF、VEGF、G−CSF、M−CSFおよびGM−CSFなどの増殖因子およびコロニー刺激因子(Burgess, Immunology Today 5:155−158(1984));
(h)トランスフェリン(O’Keefeら J. Biol. Chem. 260:932−937(1985));ならびに
(i)葉酸などのビタミン
が含まれる。
【0050】
抗体
[74]適切な細胞結合性剤の選択は、ターゲティングしようとする特定の細胞集団に応じた選択の問題であるが、一般的に、適切なものが入手可能であるか、または調製可能であるならば、抗体、より好ましくはモノクローナル抗体が好ましい。
【0051】
[75]モノクローナル抗体技術によって、特異的モノクローナル抗体の形で、非常に特異的な細胞結合性剤の産生が可能になる。当該技術分野に特によく知られるのは、マウス、ラット、ハムスター、または任意の他の哺乳動物を、損なわれていない(intact)ターゲット細胞、ターゲット細胞から単離された抗原、完全ウイルス、弱毒化完全ウイルス、およびウイルス・コートタンパク質などのウイルスタンパク質などの、関心対象の抗原で免疫することによって産生される、モノクローナル抗体を生成するための技術である。感作されたヒト細胞もまた使用可能である。モノクローナル抗体を生成する別の方法は、scFv(一本鎖可変領域)、特にヒトscFvの、ファージライブラリーの使用である(例えば、Griffithsら、米国特許第5,885,793号および第5,969,108号; McCaffertyら、WO 92/01047; Limingら、WO 99/06587を参照されたい)。
【0052】
[76]典型的な抗体は、ジスルフィド結合によって連結される2つの同一の重鎖および2つの同一の軽鎖で構成される。可変領域は、抗体の中で配列が異なり、そして抗原に対する特定の抗体各々の結合および特異性において協調する、抗体重鎖および軽鎖の部分である。可変性は、通常、抗体可変領域全体に均一に分布しているのではない。典型的には、軽鎖および重鎖可変領域両方における、相補性決定領域(CDR)または超可変領域と呼ばれる可変領域の3つのセグメント内に集中している。可変領域のより高く保存される部分は、フレームワーク領域と呼ばれる。重鎖および軽鎖の可変領域は4つのフレームワーク領域を含み、これらは大部分、ベータ−シート立体配置を採用し、各フレームワーク領域は3つのCDRによって連結され、CDRは、ベータ−シート構造を連結するルー
プを形成し、そしていくつかの場合、ベータ−シート構造の一部を形成する。各鎖中のCDRは、フレームワーク領域によって非常に近接して保持され、そして他の鎖に由来するCDRとともに、抗体の抗原結合部位の形成に寄与する(E. A. Kabatら Sequences of Proteins of Immunological Interest, 第5版, 1991, NIH)。
【0053】
[77]定常領域は、重鎖の一部である。抗体の抗原への結合に直接関与してはいないが、抗体依存性細胞傷害性における抗体の関与など、多様なエフェクター機能を示す。
[78]本発明で使用するのに適したモノクローナル抗体には、ネズミDS6モノクローナル抗体(米国特許第6,596,503号; ATCC寄託番号PTA−4449)が含まれる。
【0054】
ヒト化または表面再構成DS抗体
[79]好ましくは、本発明の細胞結合性剤として、ヒト化抗CA6抗体を用いる。こうしたヒト化抗体の好ましい態様は、ヒト化DS6抗体またはそのエピトープ結合性断片である。
【0055】
[80]ヒト化の目的は、抗体の完全な抗原結合親和性および特異性を維持しつつ、ヒトに導入するために、ネズミ抗体などの異種抗体の免疫原性を減少させることである。
[81]表面再構成およびCDR移植などのいくつかの技術を用いて、ヒト化抗体を産生してもよい。本明細書において、表面再構成技術は、分子モデリング、統計分析および突然変異誘発の組み合わせを用いて、抗体可変領域の非CDR表面を改変し、ターゲット宿主の既知の抗体の表面に似せる。
【0056】
[82]抗体の表面再構成のための戦略および方法、ならびに異なる宿主内で抗体の免疫原性を減少させるための他の方法が、その全体が本明細書に援用される、米国特許第5,639,641号(Pedersenら)に開示される。簡潔には、好ましい方法において、(1)抗体重鎖および軽鎖可変領域のプールの位置アライメントを生成して、重鎖および軽鎖可変領域フレームワーク表面曝露位置のセットを生じ、ここで、すべての可変領域のアライメント位置は、少なくとも約98%同一である;(2)重鎖および軽鎖可変領域フレームワーク表面曝露アミノ酸残基のセットを、げっ歯類抗体(またはその断片)に関して定義し;(3)げっ歯類表面曝露アミノ酸残基セットに、最も密接に同一である、重鎖および軽鎖可変領域フレームワーク表面曝露アミノ酸残基のセットを同定し;(4)げっ歯類抗体の相補性決定領域のいずれかの残基のいずれかの原子から5Å以内にあるアミノ酸残基を除いて、工程(2)で定義した、重鎖および軽鎖可変領域フレームワーク表面曝露アミノ酸残基のセットを、工程(3)で同定した、重鎖および軽鎖可変領域フレームワーク表面曝露アミノ酸残基のセットで置換し;そして(5)結合特異性を有するヒト化げっ歯類抗体を産生する。
【0057】
[83]CDR移植(EP 0 239 400; WO 91/09967;米国特許第5,530,101号;および第5,585,089号)、表面加工(veneering)または表面再構成(EP 0 592 106; EP 0 519 596;
Padlan E. A., 1991, Molecular Immunology 28(4/5):489−498; Studnicka G. M.ら, 1994, Protein Engineering 7(6):805−814; Roguska M.A.ら, 1994, PNAS 91:969−973)、および鎖シャフリング(米国特許第5,565,332号)を含む、多様な他の技術を用いて、抗体をヒト化してもよい。ファージディスプレイ法を含む、当該技術分野に知られる多様な方法によって、ヒト抗体を作製してもよい。米国特許第4,444,887号、第4,716,111号、第5,545,806号、および第5,814,318号;ならびに国際
特許出願公報WO 98/46645、WO 98/50433、WO 98/24893、WO 98/16654、WO 96/34096、WO 96/33735、およびWO 91/10741(前記文献は、その全体が本明細書に援用される)もまた参照されたい。
【0058】
[84]好ましい態様において、本発明は、Muc1ムチン上の新規シアログリコトープ(CA6グリコトープ)を認識するヒト化抗体またはその断片を提供する。別の態様において、ヒト化抗体またはそのエピトープ結合性断片は、CA6グリコトープを発現している細胞の増殖を阻害するさらなる能力を有する。
【0059】
[85]より好ましい態様において、DS6抗体の表面再構成型またはヒト化型を提供し、ここで、抗体またはその断片の表面曝露残基は、既知のヒト抗体表面により密接に似るように、軽鎖および重鎖両方で置換される。本発明のヒト化DS6抗体またはそのエピトープ結合性断片は、改善された特性を有する。例えば、ヒト化DS6抗体またはそのエピトープ結合性断片は、Muc1ムチン上の新規シアログリコトープ(CA6グリコトープ)を特異的に認識する。より好ましくは、ヒト化DS6抗体またはそのエピトープ結合性断片は、CD6グリコトープを発現している細胞の増殖を阻害するさらなる能力を有する。ヒト化抗体またはそのエピトープ結合性断片を、メイタンシノイドなどの薬剤にコンジュゲート化して、薬剤を新規Muc1シアログリコトープ、CA6にターゲティングすることによって、抗原発現細胞に向けて特異的細胞傷害性を有するプロドラッグを形成してもよい。こうした抗体、および小さく非常に毒性である薬剤(例えばメイタンシノイド、タキサン、およびCC−1065類似体)を含む細胞傷害性コンジュゲートを、乳房腫瘍および卵巣腫瘍などの腫瘍の治療のための療法剤として用いてもよい。
【0060】
[86]DS6抗体のヒト化型はまた、軽鎖および重鎖可変領域両方のそれぞれのアミノ酸配列、軽鎖および重鎖可変領域の遺伝子のDNA配列、CDRの同定、その表面アミノ酸の同定、ならびに組換え型におけるその発現のための手段の開示に関しても、本明細書において完全に性質決定されている。
【0061】
[87]1つの態様において、配列番号1〜3によって示されるアミノ酸配列:
【0062】
【化7】
【0063】
を有するCDRを含む重鎖を有する、ヒト化抗体またはそのエピトープ結合性断片を提供する。
[88]AbMモデリングソフトウェアによって重鎖CDRを決定すると、重鎖CDRは配列番号20〜22:
【0064】
【化8】
【0065】
によって示される。
[89]同じ態様において、ヒト化抗体またはそのエピトープ結合性断片は、配列番号4〜6によって示されるアミノ酸配列:
【0066】
【化9】
【0067】
を有するCDRを含む、軽鎖を有する。
[90]配列番号7または8によって示されるアミノ酸配列:
【0068】
【化10】
【0069】
と、少なくとも90%の配列同一性を共有するアミノ酸配列を有する軽鎖可変領域を有する、ヒト化抗体およびそのエピトープ結合性断片もまた提供する。
[91]同様に、配列番号9、配列番号10、または配列番号11によって示されるアミノ酸配列:
【0070】
【化11】
【0071】
と、少なくとも90%の配列同一性を共有するアミノ酸配列を有する重鎖可変領域を有する、ヒト化抗体およびそのエピトープ結合性断片を提供する。
[92]別の態様において、配列番号8に対応するアミノ酸配列
【0072】
【化12】
【0073】
を有するヒト化または表面再構成軽鎖可変領域を有する、ヒト化抗体およびそのエピトープ結合性断片を提供する。
[93]同様に、配列番号10または配列番号11に対応するアミノ酸配列:
【0074】
【化13】
【0075】
を有するヒト化または表面再構成重鎖可変領域を有する、ヒト化抗体およびそのエピトープ結合性断片を提供する。
[94]本発明のヒト化抗体およびそのエピトープ結合性断片にはまた、muDS6の結合親和性および特異性を維持するため、CDRに近接するヒト表面アミノ酸残基が、表1のアステリスクで印を付けた残基によって定義される1以上の位(Kabatの番号付け)で、対応するmuDS6表面残基によって置換されている、軽鎖および/または重鎖可変領域型も含まれてもよい。
表1
【0076】
【表3】
【0077】
[95]DS6抗体軽鎖および重鎖、ならびにそのヒト化型の一次アミノ酸配列およびDNA配列を本明細書に開示する。しかし、本発明の範囲は、これらの配列を含む抗体および断片に限定されない。その代わり、Muc1受容体上のユニークな腫瘍特異的グリコ
トープとしてのCA6に特異的に結合するすべての抗体および断片が、本発明に含まれる。好ましくは、CA6に特異的に結合する抗体および断片はまた、受容体の生物学的活性もアンタゴナイズする。より好ましくは、こうした抗体は、さらに、アゴニスト活性を実質的に回避する。したがって、本発明の抗体および抗体断片は、骨格、CDR、および/または軽鎖および重鎖のアミノ酸配列が、DS6抗体またはそのヒト化誘導体と異なり、そしてなお、本発明の範囲内に属することも可能である。
【0078】
[96]モデリングによって、DS6抗体のCDRが同定され、そしてその分子構造が予測されている。ここでも、CDRはエピトープ認識に重要であるが、本発明の抗体および断片には本質的でない。したがって、例えば本発明の抗体の親和性成熟によって産生される、改善された特性を有する抗体および断片を提供する。
【0079】
[97]DS6が由来しているようである、マウス軽鎖IgVκ ap4生殖系列遺伝子および重鎖IgVh J558.41生殖系列遺伝子を、DS6抗体の配列と整列させて、図11に示す。この比較によって、CDR中のいくつかを含めて、DS6抗体中のありうる体細胞突然変異が同定される。
【0080】
[98]DS6抗体の重鎖および軽鎖可変領域の配列、ならびにDS6抗体のCDRの配列は、先には知られておらず、そしてこれらを図9Aおよび9Bに示す。こうした情報を用いて、DS6抗体のヒト化型を生じてもよい。
【0081】
抗体断片
[99]本発明の抗体には、上に論じる全長抗体ならびにエピトープ結合性断片の両方が含まれる。本明細書において、「抗体断片」には、全長抗体によって認識されるエピトープに結合する能力を保持する抗体のいかなる部分も含まれ、これらは一般的に「エピトープ結合性断片」と称される。抗体断片の例には、限定されるわけではないが、Fab、Fab’およびF(ab’)2、Fd、一本鎖Fv(scFv)、一本鎖抗体、ジスルフィド連結Fv(dsFv)、ならびにVLまたはVH領域のいずれかを含む断片が含まれる。一本鎖抗体を含む、エピトープ結合性断片は、単独で、あるいは以下:ヒンジ領域、CH1、CH2、およびCH3ドメインのすべてまたは一部と組み合わせて、可変領域(単数または複数)を含んでもよい。
【0082】
[100]こうした断片は、1つまたは両方のFab断片またはF(ab’)2断片を含有してもよい。好ましくは、抗体断片は、完全抗体の6つのCDRすべてを含有するが、こうした領域すべてより少ない、例えば、3つ、4つまたは5つのCDRを含有する断片もまた、機能する。さらに、断片は、以下の免疫グロブリンクラス:IgG、IgM、IgA、IgD、またはIgE、およびそのサブクラスの任意の1つのメンバーであっても、またはそれらを組み合わせてもよい。
【0083】
[101]パパイン(Fab断片)またはペプシン(F(ab’)2断片)などの酵素を用いたタンパク質分解切断によって、FabおよびF(ab’)2断片を産生してもよい。
【0084】
[102]一本鎖FV(scFv)断片は、抗体軽鎖可変領域(VL)の少なくとも1つの断片に連結された抗体重鎖可変領域(VH)の少なくとも1つの断片を含有する、エピトープ結合性断片である。リンカーは、一本鎖抗体断片が由来する完全抗体のターゲット分子結合特異性を維持するために、連結された場合、(VL)および(VH)領域の適切な三次元フォールディングを確実にするように選択された、短い、柔軟なペプチドであってもよい。(VL)または(VH)配列のカルボキシル末端は、リンカーによって、相補(VL)または(VH)配列のアミノ酸末端に共有結合されてもよい。
【0085】
[103]本発明の一本鎖抗体断片は、本明細書に記載する完全抗体の可変または相補性決定領域(CDR)の少なくとも1つを有するが、こうした抗体の定常ドメインのいくつかまたはすべてを欠く、アミノ酸配列を含有する。これらの定常ドメインは、抗原結合には必要でないが、完全抗体の構造の主要な部分を構成する。したがって、一本鎖抗体断片は、定常ドメインの一部またはすべてを含有する抗体の使用と関連する問題のいくつかを克服しうる。例えば、一本鎖抗体断片は、生物学的分子および重鎖定常領域の間の望ましくない相互作用、または他の望ましくない生物学的活性がない傾向がある。さらに、一本鎖抗体断片は、完全抗体よりかなり小さく、そしてしたがって、完全抗体よりも高い毛細管透過性を有し、一本鎖抗体断片が、より効率的にターゲット抗原結合部位に局在し、そして結合することを可能にしうる。また、原核細胞において、比較的大規模に抗体断片を産生し、したがってその産生を容易にすることも可能である。さらに、一本鎖抗体断片が比較的小さいサイズであることから、これらは完全抗体よりも、レシピエントにおいて、免疫応答を誘発する可能性がより低くなる。
【0086】
[104]分子クローニング、抗体ファージディスプレイライブラリーまたは当業者に周知の類似の技術によって、一本鎖抗体断片を生成してもよい。真核細胞、または細菌を含む原核細胞において、これらのタンパク質を産生してもよい。また、当該技術分野に知られる多様なファージディスプレイ法を用いて、本発明のエピトープ結合性断片を生成してもよい。ファージディスプレイ法において、抗体ドメインをコードするポリヌクレオチド配列を所持するファージ粒子の表面上に、機能する抗体ドメインがディスプレイされる。特に、こうしたファージを利用して、レパートリーまたはコンビナトリアル抗体ライブラリー(例えばヒトまたはネズミ)から発現されるエピトープ結合性ドメインをディスプレイしてもよい。抗原を用いて、例えば固体表面またはビーズに結合したかまたは捕捉された標識抗原を用いて、関心対象の抗原に結合するエピトープ結合性ドメインを発現するファージを選択するかまたは同定してもよい。これらの方法で用いるファージは、ファージ遺伝子IIIまたは遺伝子VIIIタンパク質のいずれかに組換え的に融合された、Fab、Fvまたはジスルフィド安定化Fv抗体ドメインを含むファージから発現される、fdおよびM13結合ドメインを含む、典型的には繊維状ファージである。
【0087】
[105]本発明のエピトープ結合性断片を作製するために使用可能なファージディスプレイ法の例には、各々、その全体が本明細書に援用される、Brinkmanら, 1995, J. Immunol. Methods 182:41−50; Amesら, 1995, J. Immunol. Methods 184:177−186; Kettleboroughら, 1994, Eur. J. Immunol.
24:952−958; Persicら, 1997, Gene 187:9−18; Burtonら, 1994, Advances in Immunology
57:191−280; PCT出願第PCT/GB91/01134号; PCT公報WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401;ならびに米国特許第5,698,426号;第5,223,409号;第5,403,484号;第5,580,717号;第5,427,908号;第5,750,753号;第5,821,047号;第5,571,698号;第5,427,908号;第5,516,637号;第5,780,225号;第5,658,727号;第5,733,743号および第5,969,108号に開示されるものが含まれる。
【0088】
[106]ファージ選択後、断片をコードするファージ領域を単離し、そしてこれを用いて、例えば以下に詳細に記載するような組換えDNA技術を用い、哺乳動物細胞、昆虫細胞、植物細胞、酵母、および細菌を含む、選択した宿主における発現を通じて、エピト
ープ結合性断片を生成してもよい。例えば、PCT公報WO 92/22324; Mullinaxら, 1992, BioTechniques 12(6):864−869; Sawaiら, 1995, AJRI 34:26−34;ならびにBetterら, 1988, Science 240:1041−1043に開示されるものなど、当該技術分野に知られる方法を用いて、Fab、Fab’およびF(ab’)2断片を組換え的に産生する技術もまた使用してもよく;前記参考文献は、その全体が本明細書に援用される。一本鎖Fvおよび抗体を産生するために使用可能な技術の例には、米国特許第4,946,778号および第5,258,498号; Hustonら, 1991, Methods in Enzymology 203:46−88; Shuら, 1993, PNAS 90:7995−7999; Skerraら, 1988, Science 240:1038−1040に記載されるものが含まれる。
【0089】
機能的同等物
[107]本発明の範囲内にやはり含まれるのは、抗CA6抗体およびヒト化抗CA6抗体の機能的同等物である。用語「機能的同等物」には、例えば、相同配列を持つ抗体、キメラ抗体、人工的抗体および修飾抗体が含まれ、各機能同等物は、CA6に結合する能力によって定義される。当業者は、「抗体断片」と称される分子群および「機能的同等物」と称される群には重複があることを理解するであろう。機能的同等物を産生する方法は、例えば、本明細書にそれぞれの全体が援用される、PCT出願WO 93/21319、欧州特許出願第239,400号; PCT出願WO 89/09622;欧州特許出願338,745;および欧州特許出願EP 332,424に開示される。
【0090】
[108]相同配列を持つ抗体は、本発明の抗CA6抗体およびヒト化抗CA6抗体のアミノ酸配列と配列相同性を有するアミノ酸配列を持つ抗体である。好ましくは、相同性は、本発明の抗CA6抗体およびヒト化抗CA6抗体の可変領域のアミノ酸配列との相同性である。「配列相同性」は、本明細書において、アミノ酸配列に適用された際、例えば、PearsonおよびLipman, Proc. Natl. Acad. Sci. USA 85, 2444−2448(1988)にしたがったFASTA検索法によって決定した際、少なくとも約90%、91%、92%、93%、または94%の配列相同性、そしてより好ましくは、少なくとも約95%、96%、97%、98%、または99%の配列相同性を持つ配列と定義される。
【0091】
[109]本明細書において、キメラ抗体は、抗体の異なる部分が異なる動物種に由来するものである。例えば、ヒト免疫グロブリン定常領域と対形成した、ネズミ・モノクローナル抗体由来の可変領域を有する抗体がある。キメラ抗体を産生するための方法が当該技術分野に知られる。例えば、その全体が本明細書に援用される、Morrison, 1985, Science 229:1202; Oiら, 1986, BioTechniques 4:214; Gilliesら, 1989, J. Immunol. Methods 125:191−202;米国特許第5,807,715号;第4,816,567号;および第4,816,397号を参照されたい。
【0092】
[110]キメラ抗体のヒト化型は、ヒト・フレームワークドメイン内で、例えばマウス抗体の相補性決定領域を代用することによって、作製される。例えばPCT公報第WO92/22653号を参照されたい。ヒト化キメラ抗体は、好ましくは、対応するヒト抗体領域に実質的にまたはもっぱら由来する、相補性決定領域(CDR)以外の定常領域および可変領域、ならびにヒト以外の哺乳動物に実質的にまたはもっぱら由来するCDRを有する。
【0093】
[111]人工的な抗体には、各々、抗原結合能を有する、scFv断片、ディアボディ、トリアボディ、テトラボディおよびmruが含まれる(Winter, G.および
Milstein, C, 1991, Nature 349: 293−299; Hudson, P.J., 1999, Current Opinion in Immunology 11:548−557による概説を参照されたい)。一本鎖Fv断片(scFv)において、抗体のVHおよびVLドメインは、柔軟なペプチドによって連結される。典型的には、このリンカーペプチドは、長さ約15アミノ酸残基である。リンカーがはるかに小さいならば、例えば5アミノ酸であるならば、二価scFv二量体であるディアボディが形成される。リンカーが3アミノ酸残基未満に減少する場合、トリアボディおよびテトラボディと呼ばれる三量体および四量体構造が形成される。抗体の最少の結合単位は、別個に使用可能であるのに十分な特異的認識および結合を有するCDR、典型的には重鎖のCDR2である。こうした断片は、分子認識単位またはmruと呼ばれる。いくつかのこうしたmruは、短いリンカーペプチドと一緒に連結されて、したがって単一のmruより高いアビディティーを持つ人工的な結合タンパク質を形成してもよい。
【0094】
[112]本出願の機能的同等物にはまた、修飾抗体、例えば抗体への任意の種類の分子の共有結合によって修飾された抗体も含まれる。例えば、修飾抗体には、例えば、グリコシル化、アセチル化、PEG化、リン酸化、アミド化、既知の保護基/ブロッキング基による誘導体化、タンパク質分解切断、細胞リガンドまたは他のタンパク質への連結等によって修飾されている抗体が含まれる。こうした共有結合は、抗体が抗イディオタイプ応答を生じるのを妨げない。限定されるわけではないが、特異的化学切断、アセチル化、ホルミル化、ツニカマイシンの代謝合成等を含む、既知の技術によって、これらの修飾を行ってもよい。さらに、修飾抗体は、1以上の非古典的アミノ酸を含有してもよい。
【0095】
[113]異なるフレームワーク内の異なる鎖上の異なるCDRを交換することによって、機能的同等物を生じてもよい。したがって、例えば、IgG1〜4、IgM、IgA1〜2、IgD、IgE抗体タイプおよびアイソタイプを産生可能な、異なる重鎖の置換によって、所定のCDRセットに関して、抗体の異なるクラスにすることも可能である。同様に、完全に合成であるフレームワーク内に、所定のCDRセットを包埋することによって、本発明の範囲内の人工的抗体を産生してもよい。
【0096】
[114]当該技術分野に知られる非常に多様な方法を用いて、CDRの特定のセットに隣接する可変および/または定常領域配列内の突然変異、欠失および/または挿入によって、機能的同等物を容易に産生可能である。
【0097】
[115]本発明の抗体断片および機能的同等物は、DS6抗体に比較した際、CA6に検出可能な度合いで結合する分子を含む。検出可能な度合いの結合には、CA6に対するネズミDS6抗体の結合能の少なくとも10〜100%、好ましくは少なくとも50%、60%または70%、より好ましくは少なくとも75%、80%、85%、90%、95%または99%の範囲のすべての値が含まれる。
【0098】
改善された抗体
[116]CDRは、エピトープ認識および抗体結合に、最も重要である。しかし、抗体がその同族(cognate)エピトープを認識し、そして結合する能力に干渉することなく、CDRを含む残基に変化を作製してもよい。例えば、エピトープ認識に影響を及ぼさず、さらにエピトープに対する抗体の結合親和性を増加させる変化を作製してもよい。
【0099】
[117]したがって、本発明の範囲内にやはり含まれるのは、好ましくは増加した親和性で、やはりCA6を特異的に認識し、そしてこれに結合する、ネズミ抗体およびヒト化抗体の両方の改善型である。
【0100】
[118]いくつかの研究によって、一次抗体配列の知識に基づいて、結合および発現レベルなどの特性に対して、抗体の配列の多様な位で、1以上のアミノ酸変化を導入する影響が調べられてきている(Yang, W. P.ら, 1995, J. Mol.
Biol., 254, 392−403; Rader, C.ら, 1998, Proc. Natl. Acad. Sci. USA, 95, 8910−8915; Vaughan, T. J.ら, 1998, Nature Biotechnology, 16, 535−539)。
【0101】
[119]これらの研究において、オリゴヌクレオチドが仲介する部位特異的突然変異誘発、カセット突然変異誘発、変異性(error−prone)PCR、DNAシャフリング、または大腸菌(E. coli)の突然変異誘発株などの方法を用いて、CDR1、CDR2、CDR3、またはフレームワーク領域において、重鎖および軽鎖遺伝子の配列を変化させることによって、一次抗体の同等物が生成されてきている(Vaughan, T. J.ら, 1998, Nature Biotechnology, 16, 535−539; Adey, N. B.ら, 1996, 第16章, pp. 277−291, “Phage Display of Peptides and Proteins”中, Kay, B. K.ら監修, Academic Press)。一次抗体配列を変化させるこれらの方法は、二次抗体の改善された親和性を生じている(Gram, H.ら, 1992, Proc. Natl. Acad. Sci. USA, 89, 3576−3580; Boder, E. T.ら, 2000, Proc. Natl. Acad. Sci. USA, 97, 10701−10705; Davies, J.およびRiechmann, L., 1996, Immunotechnolgy, 2, 169−179; Thompson, J.ら, 1996, J. Mol. Biol., 256, 77−88; Short, M. K.ら, 2002, J. Biol. Chem., 277, 16365−16370; Furukawa, K.ら, 2001, J.
Biol. Chem., 276, 27622−27628)。
【0102】
[120]抗体の1以上のアミノ酸残基を変化させる類似の指令戦略によって、本発明で記載する抗体配列を用いて、CA6に対する改善された親和性を含む、改善された機能を持つ抗CA6抗体を開発してもよい。
【0103】
[121]改善された抗体にはまた、動物免疫、ハイブリドーマ形成、および特定の性質を持つ抗体に関する選択の標準的技術によって調製される、改善された性質を有する抗体が含まれる。
【0104】
細胞傷害性剤
[122]本発明の細胞傷害性コンジュゲートで用いる細胞傷害性剤は、細胞の死を生じるか、細胞の死を誘導するか、または何らかの方式で細胞生存度を減少させる、いかなる化合物であってもよい。好ましい細胞傷害性剤には、例えば、以下に定義される、メイタンシノイドおよびメイタンシノイド類似体、タキソイド、CC−1065およびCC−1065類似体、ドラスタチンおよびドラスタチン類似体が含まれる。これらの細胞傷害性剤を、本明細書に開示するような抗体、抗体断片、機能的同等物、改善された抗体およびその類似体にコンジュゲート化する。
【0105】
[123]in vitro法によって、細胞傷害性コンジュゲートを調製してもよい。抗体に薬剤またはプロドラッグを連結するため、連結基を用いる。適切な連結基が当該技術分野に周知であり、そしてこうした連結基には、ジスルフィド基、チオエーテル基、酸不安定性基、光解離基、ペプチダーゼ不安定性基およびエステラーゼ不安定性基が含まれる。好ましい連結基はジスルフィド基およびチオエーテル基である。例えば、ジスルフ
ィド交換反応を用いて、あるいは抗体および薬剤またはプロドラッグ間にチオエーテル結合を形成することによって、コンジュゲートを構築してもよい。
【0106】
メイタンシノイド
[124]細胞傷害性コンジュゲートを形成するため、本発明で使用可能な細胞傷害性剤の中に、メイタンシノイドおよびメイタンシノイド類似体がある。適切なメイタンシノイドの例には、メイタンシノールおよびメイタンシノール類似体が含まれる。メイタンシノイドは、微小管形成を阻害し、そして哺乳動物細胞に対して非常に毒性である薬剤である。
【0107】
[125]適切なメイタンシノール類似体の例には、修飾芳香族環を有するものおよび他の位で修飾を有するものが含まれる。こうした適切なメイタンシノイドが、米国特許第4,424,219号;第4,256,746号;第4,294,757号;第4,307,016号;第4,313,946号;第4,315,929号;第4,331,598号;第4,361,650号;第4,362,663号;第4,364,866号;第4,450,254号;第4,322,348号;第4,371,533号;第6,333,410号;第5,475,092号;第5,585,499号;および第5,846,545号に開示される。
【0108】
[126]修飾芳香族環を有するメイタンシノールの適切な類似体の特定の例には:
[127](1)C−19−デクロロ(米国特許第4,256,746号)(アンサマイトシンP2のLAH還元によって調製);
[128](2)C−20−ヒドロキシ(またはC−20−デメチル)+/−C−19−デクロロ(米国特許第4,361,650号および第4,307,016号)(ストレプトミセス属(Streptomyces)またはアクチノミセス属(Actinomyces)を用いた脱メチル化、あるいはLAHを用いた脱クロロ化によって調製);および
[129](3)C−20−デメトキシ、C−20−アシルオキシ(−OCOR)、+/−デクロロ(米国特許第4,294,757号)(塩化アシルを用いたアシル化によって調製)
が含まれる。
【0109】
[130]他の位の修飾を有するメイタンシノールの適切な類似体の特定の例には:
[131](1)C−9−SH(米国特許第4,424,219号)(H2SまたはP2S5とメイタンシノールの反応によって調製);
[132](2)C−14−アルコキシメチル(デメトキシ/CH2OR)(米国特許第4,331,598号);
[133](3)C−14−ヒドロキシメチルまたはアシルオキシメチル(CH2OHまたはCH2OAc)(米国特許第4,450,254号)(ノカルジア属(Nocardia)から調製);
[134](4)C−15−ヒドロキシ/アシルオキシ(米国特許第4,364,866号)(ストレプトミセス属によるメイタンシノールの変換によって調製);
[135](5)C−15−メトキシ(米国特許第4,313,946号および第4,315,929号)(トレウィア・ヌディフロラ(Trewia nudiflora)から単離);
[136](6)C−18−N−デメチル(米国特許第4,362,663号および第4,322,348号)(ストレプトミセス属によるメイタンシノールの脱メチル化によって調製);および
[137](7)4,5−デオキシ(米国特許第4,371,533号)(メイタンシノールの三塩化チタン/LAH還元によって調製)
が含まれる。
【0110】
[138]好ましい態様において、本発明の細胞傷害性コンジュゲートは、細胞傷害性剤として、以前、N2’−デアセチル−N2’−(3−メルカプト−1−オキソプロピル)−メイタンシンと称された、チオール含有メイタンシノイド(DM1)を利用する。DM1は、以下の構造式(I):
【0111】
【化14】
【0112】
によって示される。
[139]別の好ましい態様において、本発明の細胞傷害性コンジュゲートは、細胞傷害性剤として、チオール含有メイタンシノイド、N2’−デアセチル−N2’−(4−メチル−4−メルカプト−1−オキソペンチル)−メイタンシンを利用する。DM4は、以下の構造式(II):
【0113】
【化15】
【0114】
によって示される。
[140]本発明のさらなる態様において、イオウ原子を所持する炭素原子上に、モノまたはジアルキル置換を所持する、チオールおよびジスルフィド含有メイタンシノイドを含む、他のメイタンシンを用いてもよい。これらには、C−3、C−14ヒドロキシメチル、C−15ヒドロキシ、またはC−20デスメチルに、立体障害を受けた(hindered)スルフヒドリル基を所持するアシル基を含むアシル化アミノ酸側鎖を有するメイタンシノイドが含まれ、ここで、チオール官能性を所持するアシル基の炭素原子は、1つまたは2つの置換基を有し、前記置換基は、CH3、C2H5、1〜10の炭素原子を有する直鎖もしくは分枝鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する環状アルキルもしくはアルケニル、フェニル、置換フェニル、または複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、置換基の1つはHであってもよく、そ
してアシル基は、カルボニル官能性およびイオウ原子間に少なくとも3つの炭素原子の長さの直鎖を有する。
【0115】
[141]こうしたさらなるメイタンシンには、式(III)によって示される化合物:
【0116】
【化16】
【0117】
式中:
Y’は
(CR7CR8)l(CR9=CR10)pC=CqAr(CR5CR6)mDu(CR11=CR12)r(C=C)sBt(CR3CR4)nCR1R2SZ
を示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
A、B、Dは、3〜10炭素原子を有するシクロアルキルもしくはシクロアルケニル、単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
R3、R4、R5、R6、R7、R8、R9、R11、およびR12は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、m、n、o、p、q、r、s、およびtは、各々独立に、0または1〜5の整数であり、但し、どの時点においても、l、m、n、o、p、q、r、s、およびtの少なくとも2つはゼロではなく;そして
Zは、H、SRまたは−CORであり、式中、Rは、1〜10炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルである
が含まれる。
【0118】
[142]式(III)の好ましい態様には:
R1がHであり、R2がメチルであり、そしてZがHである
R1およびR2がメチルであり、そしてZがHである
R1がHであり、R2がメチルであり、そしてZが−SCH3である
R1およびR2がメチルであり、そしてZが−SCH3である
式(III)の化合物が含まれる。
【0119】
[143]こうしたさらなるメイタンシンにはまた、式(IV−L)、(IV−D)、または(IV−D,L):
【0120】
【化17】
【0121】
式中:
Yは、(CR7CR8)l(CR5CR6)m(CR3CR4)nCR1R2SZを示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニル、または複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
R3、R4、R5、R6、R7およびR8は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニル、または複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、mおよびnは、各々独立に、1〜5の整数であり、そしてさらに、nはゼロであってもよく;
Zは、H、SRまたは−CORであり、式中、Rは、1〜10炭素原子を有する直鎖もしくは分枝鎖アルキルもしくはアルケニル、3〜10炭素原子を有する環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;そして
Mayは、C−3、C−14ヒドロキシメチル、C−15ヒドロキシまたはC−20デスメチルに側鎖を所持するメイタンシノイドを示す
によって示される化合物も含まれる。
【0122】
[144]式(IV−L)、(IV−D)および(IV−D,L)の好ましい態様には:
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり、lおよびmが各々1であり、nがゼロであり、そしてZがHである
R1およびR2がメチルであり、R5、R6、R7、R8が各々Hであり、lおよびmが1であり、nがゼロであり、そしてZがHである
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり、lおよびmが各々1であり、nがゼロであり、そしてZが−SCH3である
R1およびR2がメチルであり、R5、R6、R7、R8が各々Hであり、lおよびmが1であり、nがゼロであり、そしてZが−SCH3である
式(IV−L)、(IV−D)および(IV−D,L)の化合物が含まれる。
【0123】
[145]好ましくは、細胞傷害性剤は、式(IV−L)によって示される。
[146]こうしたさらなるメイタンシンにはまた、式(V):
【0124】
【化18】
【0125】
式中:
Yは、(CR7CR8)l(CR5CR6)m(CR3CR4)nCR1R2SZを示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
R3、R4、R5、R6、R7およびR8は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニル、または複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、mおよびnは、各々独立に、1〜5の整数であり、そしてさらに、nはゼロであってもよく;そして
Zは、H、SRまたは−CORであり、式中、Rは、1〜10炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルである
によって示される化合物も含まれる。
【0126】
[147]式(V)の好ましい態様には:
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり;lおよびmが各々1であり;nがゼロであり;そしてZがHである
R1およびR2がメチルであり;R5、R6、R7、R8が各々Hであり、lおよびmが1であり;nがゼロであり;そしてZがHである
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり、lおよびmが各々1であり;nがゼロであり;そしてZが−SCH3である
R1およびR2がメチルであり、R5、R6、R7、R8が各々Hであり、lおよびmが1であり、nがゼロであり、そしてZが−SCH3である
式(V)の化合物が含まれる。
【0127】
[148]こうしたさらなるメイタンシンにはさらに、式(VI−L)、(VI−D)、または(VI−D,L):
【0128】
【化19】
【0129】
式中:
Y2は、(CR7CR8)l(CR5CR6)m(CR3CR4)nCR1R2SZ2を示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
R3、R4、R5、R6、R7およびR8は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖、環状アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、mおよびnは、各々独立に、1〜5の整数であり、そしてさらに、nはゼロであってもよく;
Z2は、SRまたはCORであり、式中、Rは、1〜10炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;そして
Mayはメイタンシノイドである
によって示される化合物が含まれる。
【0130】
[149]こうしたさらなるメイタンシンにはまた、式(VII)によって示される化合物:
【0131】
【化20】
【0132】
式中:
Y2’は
(CR7CR8)l(CR9=CR10)p(C=C)qAr(CR5CR6)mDu(CR11=CR12)r(C=C)sBt(CR3CR4)nCR1R2SZ2
を示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖もしくは分枝鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
A、B、およびDは、各々独立に、3〜10炭素原子を有するシクロアルキルまたはシクロアルケニル、単純または置換アリール、または複素環芳香族またはヘテロシクロアルキルラジカルであり;
R3、R4、R5、R6、R7、R8、R9、R11、およびR12は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、m、n、o、p、q、r、s、およびtは、各々独立に、0または1〜5の整数であり、但し、どの時点においても、l、m、n、o、p、q、r、s、およびtの少なくとも2つはゼロではなく;そして
Z2は、SRまたは−CORであり、式中、Rは、1〜10炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、あるいは単純もしくは置換アリールまたは複素環芳香族またはヘテロシクロアルキルラジカルである
も含まれる。
【0133】
[150]式(VII)の好ましい態様には:
R1がHであり、そしてR2がメチルである
式(VII)の化合物が含まれる。
【0134】
[151]上述のメイタンシノイドを、抗CA6抗体DS6、あるいはその相同体または断片にコンジュゲート化してもよく、ここで、抗体は、メイタンシノイドのC−3、C−14ヒドロキシメチル、C−15ヒドロキシまたはC−20デスメチルに見られるアシ
ル化アミノ酸側鎖のアシル基上に存在するチオールまたはジスルフィド官能性を用いて、メイタンシノイドに連結され、そしてアシル化アミノ酸側鎖のアシル基が、1つまたは2つの置換基を有する炭素原子に位置する、チオールまたはジスルフィド官能性を有し、前記置換基は、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、置換基の1つはHであってもよく、そしてアシル基は、カルボニル官能性およびイオウ原子間に少なくとも3つの炭素原子の長さの直鎖を有する。
【0135】
[152]本発明の好ましいコンジュゲートは、式(VIII):
【0136】
【化21】
【0137】
式中:
Y1’は
(CR7CR8)l(CR9=CR10)p(C=C)qAr(CR5CR6)mDu(CR11=CR12)r(C=C)sBt(CR3CR4)nCR1R2S−
を示し、
式中:
A、B、およびDは、各々独立に、3〜10炭素原子を有するシクロアルキルもしくはシクロアルケニル、単純もしくは置換アリール、または複素環芳香族またはヘテロシクロアルキルラジカルであり;
R3、R4、R5、R6、R7、R8、R9、R11、およびR12は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;そして
l、m、n、o、p、q、r、s、およびtは、各々独立に、0または1〜5の整数であり、但し、どの時点においても、l、m、n、o、p、q、r、s、およびtの少なくとも2つはゼロではない
のメイタンシノイドにコンジュゲート化された、抗抗CA6抗体DS6、あるいはその相同体または断片を含むものである。
【0138】
[153]好ましくは、R1はHであり、そしてR2はメチルであるか、またはR1およびR2はメチルである。
[154]本発明のさらにより好ましいコンジュゲートは、式(IX−L)、(IX−D)、または(IX−D,L):
【0139】
【化22】
【0140】
式中:
Y1は、(CR7CR8)l(CR5CR6)m(CR3CR4)nCR1R2S−を示し、
式中:
R1およびR2は、各々独立に、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニル、複素環芳香族またはヘテロシクロアルキルラジカルであり、そしてさらに、R2はHであってもよく;
R3、R4、R5、R6、R7およびR8は、各々独立に、H、CH3、C2H5、1〜10の炭素原子を有する直鎖アルキルもしくはアルケニル、3〜10の炭素原子を有する分枝鎖もしくは環状アルキルもしくはアルケニル、フェニル、置換フェニルまたは複素環芳香族またはヘテロシクロアルキルラジカルであり;
l、mおよびnは、各々独立に、1〜5の整数であり、そしてさらに、nはゼロであってもよく;そして
Mayは、C−3、C−14ヒドロキシメチル、C−15ヒドロキシまたはC−20デスメチルで側鎖を所持するメイタンシノールを示す
のメイタンシノイドにコンジュゲート化された、抗CA6抗体DS6、あるいはその相同体または断片を含むものである。
【0141】
[155]式(IX−L)、(IX−D)および(IX−D,L)の好ましい態様には:
R1がHであり、そしてR2がメチルであるか、またはR1およびR2がメチルである、
R1がHであり、R2がメチルであり、R5、R6、R7、およびR8が各々Hであり;lおよびmが各々1であり;nが0である、
R1およびR2がメチルであり;R5、R6、R7、およびR8が各々Hであり;lおよびmが1であり;nが0である
式(IX−L)、(IX−D)および(IX−D,L)の化合物が含まれる。
【0142】
[156]好ましくは、細胞傷害性剤は、式(IX−L)によって示される。
[157]本発明のさらに好ましいコンジュゲートは、式(X):
【0143】
【化23】
【0144】
式中、置換基は、上記式(IX)に関して定義されるとおりである
のメイタンシノイドにコンジュゲート化された、抗CA6抗体DS6、あるいはその相同体または断片を含むものである。
【0145】
[158]特に好ましいのは、R1がHであり、R2がメチルであり、R5、R6、R7およびR8が各々Hであり、lおよびmが各々1であり、そしてnが0である、上記化合物のいずれかである。
【0146】
[159]さらに特に好ましいのは、R1およびR2がメチルであり、R5、R6、R7およびR8が、各々、Hであり、lおよびmが1であり、そしてnが0である、上記化合物のいずれかである。
【0147】
[160]さらに、L−アミノアシル立体異性体が好ましい。
[161]係属中の米国特許出願第10/849,136号、2004年5月20日出願に解説される各メイタンシノイドもまた、本発明の細胞傷害性コンジュゲートに使用可能である。米国特許出願第10/849,136号の全開示が、本明細書に援用される。
【0148】
ジスルフィド含有連結基
[162]DS6抗体などの細胞結合性剤にメイタンシノイドを連結するため、メイタンシノイドは連結基を含む。連結部分は、完全活性メイタンシノイドの放出を可能にする化学結合を特定の部位に含有する。適切な化学結合は、当該技術分野に周知であり、そしてこれには、ジスルフィド結合、酸不安定性結合、光解離結合、ペプチダーゼ不安定性結合およびエステラーゼ不安定性結合が含まれる。好ましいのはジスルフィド結合である。
【0149】
[163]連結部分はまた、反応性化学基も含む。好ましい態様において、反応性化学基を、ジスルフィド結合連結部分を介して、メイタンシノイドに共有結合させてもよい。
[164]特に好ましい反応性化学基は、N−スクシンイミジルエステルおよびN−スルホスクシンイミジルエステルである。
【0150】
[165]反応性化学基を含有する連結部分を含む、特に好ましいメイタンシノイドは、連結部分がジスルフィド結合を含有し、そして化学反応基がN−スクシンイミジルまたはN−スルホスクシンイミジルエステルを含む、メイタンシノールおよびその類似体のC−3エステルである。
【0151】
[166]メイタンシノイド上の多くの位が、連結部分を化学的に連結する位として働きうる。例えば、ヒドロキシル基を有するC−3位、ヒドロキシメチルで修飾されたC−14位、ヒドロキシで修飾されたC−15位、およびヒドロキシ基を有するC−20位がすべて、有用であると期待される。しかし、C−3位が好ましく、そしてメイタンシノールのC−3位が特に好ましい。
【0152】
[167]連結部分を有するメイタンシノールのエステルの合成を、ジスルフィド結合含有連結部分の観点で記載する一方、当業者は、他の化学結合(上述のようなもの)を持つ連結部分もまた、本発明で使用可能であり、他のメイタンシノイドも使用可能であることを理解するであろう。他の化学結合の特定の例には、酸不安定性結合、光解離結合、ペプチダーゼ不安定性結合およびエステラーゼ不安定性結合が含まれる。本明細書に援用される、米国特許第5,208,020号の開示は、こうした結合を所持するメイタンシノイドの産生を解説する。
【0153】
[168]反応性基を所持するジスルフィド部分を有するメイタンシノイドおよびメイタンシノイド誘導体の合成が、各々、本明細書に援用される、米国特許第6,441,163号および第6,333,410号、ならびに米国出願第10/161,651号に記載される。
【0154】
[169]DM1などの反応性基含有メイタンシノイドを、DS6抗体などの抗体と反応させて、細胞傷害性コンジュゲートを産生する。HPLCによって、またはゲルろ過によって、これらのコンジュゲートを精製してもよい。
【0155】
[170]こうした抗体メイタンシノイド・コンジュゲートを産生するためのいくつかの優れたスキームが、各々、その全体が本明細書に援用される、米国特許第6,333,410号、ならびに米国出願第09/867,598号、第10/161,651号および第10/024,290号に提供される。
【0156】
[171]一般的に、水性緩衝液中の抗体溶液を、反応性基を所持するジスルフィド部分を有する、モル過剰のメイタンシノイドとインキュベーションしてもよい。過剰なアミン(エタノールアミン、タウリンなど)の添加によって、反応混合物の反応を停止してもよい。次いで、ゲルろ過によって、メイタンシノイド−抗体コンジュゲートを精製してもよい。
【0157】
[172]252nmおよび280nmの吸光度の比を、分光光度的に測定することによって、抗体分子あたりに結合するメイタンシノイド分子の数を決定することができる。平均1〜10のメイタンシノイド分子/抗体分子が好ましい。
【0158】
[173]メイタンシノイド薬剤と抗体のコンジュゲートを、in vitroで、多様な望ましくない細胞株の増殖を抑制する能力に関して、評価することができる。例えば、ヒト類表皮癌細胞株A−431、ヒト小細胞肺癌細胞株SW2、ヒト乳房腫瘍細胞株SKBR3およびバーキットリンパ腫細胞株Namalwaなどの細胞株は、これらの化合物の細胞傷害性の評価のため、容易に使用可能である。評価しようとする細胞を化合物に24時間曝露して、そして既知の方法による直接アッセイにおいて、細胞の生存率を測定することができる。次いで、アッセイ結果からIC50値を計算することができる。
【0159】
PEG含有連結基
[174]メイタンシノイドをまた、米国出願第10/024,290号に示されるように、PEG連結基を用いて、細胞結合性剤に連結してもよい。これらのPEG連結基は
、水中および非水性溶媒中の両方で、可溶性であり、そしてこれを用いて、1以上の細胞傷害性剤を、細胞結合性剤に連結してもよい。典型的なPEG連結基には、一端の官能性スルフヒドリル基またはジスルフィド基、およびもう一端の活性エステルを通じて、リンカーの反対の端で、細胞傷害性剤および細胞結合性剤に結合する、ヘテロ二官能性PEGリンカーが含まれる。
【0160】
[175]PEG連結基を用いた、細胞傷害性コンジュゲートの合成の一般的な例として、特定の詳細に関して、米国出願第10/024,290号に再び言及する。合成は、反応性PEG部分を所持する1以上の細胞傷害性剤と細胞結合性剤の反応で始まり、この反応が、細胞結合性剤のアミノ酸残基による、各反応性PEG部分の末端活性エステルの置換を生じ、PEG連結基を通じて細胞結合性剤に共有結合した1以上の細胞傷害性剤を含む、細胞傷害性コンジュゲートを生じる。
【0161】
タキサン
[176]本発明記載の細胞傷害性コンジュゲートで用いる細胞傷害性剤はまた、タキサンまたはその誘導体であってもよい。
【0162】
[177]タキサンは、細胞傷害性天然産物であるパクリタキセル(タキソール)、および半合成誘導体であるドセタキセル(タキソテール)を含む化合物ファミリーであり、これらの2つの化合物は、癌の治療に広く用いられている。タキサンは、チューブリンの脱分極を阻害し、細胞死を生じる、紡錘体毒物である。ドセタキセルおよびパクリタキセルは、癌治療に有用な剤であるが、正常細胞に対して非特異的に毒性であるため、その抗腫瘍活性は限定される。さらに、パクリタキセルおよびドセタキセルのような化合物は、それ自体、細胞結合性剤のコンジュゲートにおいて使用するには、十分に強力ではない。
【0163】
[178]細胞傷害性コンジュゲートの調製に使用するのに好ましいタキサンは、式(XI)のタキサン:
【0164】
【化24】
【0165】
である。
[179]抗体などの細胞結合性剤にタキサンをコンジュゲート化するための方法とともに、本発明の細胞傷害性コンジュゲートにおいて使用可能なタキサンを合成するための方法が、米国特許第5,416,064号、第5,475,092号、第6,340,701号、第6,372,738号および第6,436,931号、ならびに米国出願第10/024,290号、第10/144,042号、第10/207,814号、第10/210,112号および第10/369,563号に詳細に記載される。
【0166】
CC−1065類似体
[180]本発明記載の細胞傷害性コンジュゲートで使用する細胞傷害性剤はまた、CC−1065またはその誘導体であってもよい。
【0167】
[181]CC−1065は、ストレプトミセス・ゼレンシス(Streptomyces zelensis)の培養ブロスから単離される、強力な抗腫瘍抗生物質である。CC−1065は、ドキソルビシン、メトトレキセートおよびビンクリスチンなどの、一般的に用いられる抗癌薬剤よりも、in vitroで、約1000倍強力である(B.K. Bhuyanら, Cancer Res., 42, 3532−3537(1982))。CC−1065およびその類似体は、米国特許第6,372,738号、第6,340,701号、第5,846,545号および第5,585,499号に開示される。
【0168】
[182]CC−1065の細胞傷害能は、そのアルキル化活性、およびそのDNA結合またはDNA挿入活性と相関付けられてきている。これらの2つの活性は、分子の別個の部分に属する。したがって、アルキル化活性は、シクロプロパピロロインドール(CPI)・サブユニットに含有され、そしてDNA結合活性は、2つのピロロインドール・サブユニット中に属する。
【0169】
[183]CC−1065は、細胞傷害性剤として特定の魅力的な特徴を有するが、療法的使用においては限界を有する。マウスにCC−1065を投与すると、遅延型肝毒性が引き起こされ、12.5μg/kgの単回静脈内用量後、第50日での死亡を導く{V. L. Reynoldsら, J. Antibiotics, XXIX, 319−334(1986)}。このことから、遅延型毒性を引き起こさない類似体を開発する努力に拍車がかかり、そしてCC−1065をモデルとするより単純な類似体の合成が記載されてきている{M.A. Warpehoskiら, J. Med. Chem., 31, 590−603(1988)}。
【0170】
[184]別の一連の類似体において、CPI部分がシクロプロパベンズインドール(CBI)部分によって置換された{D.L. Bogerら, J. Org. Chem., 55, 5823−5833, (1990), D.L. Bogerら, BioOrg. Med. Chem. Lett., 1, 115−120(1991)}。これらの化合物は、マウスにおいて、遅延型毒性を引き起こすことなく、親薬剤の高いin vitro強度を維持する。CC−1065同様、これらの化合物は、共有結合性の方式でDNAの副溝に結合して、細胞死を生じる、アルキル化剤である。しかし、最も有望な類似体、アドゼレシンおよびカルゼレシンの臨床的評価は、期待はずれの結果を導いた{B.F. Fosterら, Investigational New Drugs, 13, 321−326(1996); I. Wolffら, Clin. Cancer Res., 2,1717−1723(1996)}。これらの薬剤は、全身毒性が高いため、劣った療法効果を示す。
【0171】
[185]腫瘍部位へのターゲティング送達を通じて、in vivo分布を変化させ、ターゲティングされない組織に対するより低い毒性、そしてしたがって、より低い全身毒性を生じることによって、CC−1065類似体の療法的有効性を非常に改善することも可能である。この目的を達成するため、腫瘍細胞を特異的にターゲティングする、細胞結合性剤とCC−1065の類似体および誘導体のコンジュゲートが記載されてきている{米国特許第5,475,092号;第5,585,499号;第5,846,545号}。これらのコンジュゲートは、典型的には、in vitroで高いターゲット特異的細胞傷害性を示し、そしてマウスにおけるヒト腫瘍異種移植片モデルにおいて、ひときわ優れた抗腫瘍活性を示す{R.V.J. Chariら, Cancer Res., 55, 4079−4084(1995)}。
【0172】
[186]抗体などの細胞結合性剤に類似体をコンジュゲート化するための方法とともに、本発明の細胞傷害性コンジュゲートで使用可能なCC−1065類似体を合成するための方法が、米国特許第5,475,092号、第5,846,545号、第5,585,499号、第6,534,660号および第6,586,618号に、そして米国出願第10/116,053号および第10/265,452号に、詳細に記載される。
【0173】
他の薬剤
[187]メトトレキセート、ダウノルビシン、ドキソルビシン、ビンクリスチン、ビンブラスチン、メルファラン、マイトマイシンC、クロラムブシル、カリケアマイシン、チューブリシン(tubulysin)およびチューブリシン類似体、デュオカルマイシンおよびデュオカルマイシン類似体、ドラスタチンおよびドラスタチン類似体などの薬剤もまた、本発明のコンジュゲートの調製に適している。また、血清アルブミンなどの仲介キャリアー分子を通じて、抗体分子に薬剤分子を連結させてもよい。米国第09/740991号に記載されるように、ドキソルビシンおよびダウノルビシン化合物もまた、有用な細胞傷害性剤でありうる。
【0174】
療法組成物
[188]本発明はまた:
(a)1以上の細胞傷害性コンジュゲートの有効量、および
(b)薬学的に許容しうるキャリアー
を含む療法組成物も提供する。
【0175】
[189]同様に、本発明は、選択した細胞集団の増殖を阻害するための方法であって、ターゲット細胞、またはターゲット細胞を含有する組織を、有効量の細胞傷害性コンジュゲート、または細胞傷害性コンジュゲートを含む療法剤と、単独で、あるいは他の細胞傷害性剤または療法剤と組み合わせて、接触させることを含む、前記方法を提供する。
【0176】
[190]本発明はまた、本発明の療法組成物を用いて、癌を有する被験体を治療するための方法も含む。
[191]先に記載される方法によって、細胞傷害性コンジュゲートを、in vitro強度および特異性に関して評価することができる(例えば、R.V.J. Chariら, Cancer Res. 55:4079−4084(1995)を参照されたい)。やはり先に記載される方法によって、マウスにおけるヒト腫瘍異種移植モデルにおいて、抗腫瘍活性を評価することができる(例えば、Liuら, Proc. Natl. Acad. Sci. 93:8618−8623(1996)を参照されたい)。
【0177】
[192]適切な薬学的に許容しうるキャリアーが周知であり、そして臨床状況が保証するように、一般の当業者によって決定可能である。本明細書において、キャリアーには希釈剤および賦形剤が含まれる。
【0178】
[193]適切なキャリアー、希釈剤および/または賦形剤の例には:(1)約1mg/ml〜25mg/mlのヒト血清アルブミンを含有するかまたは含有しない、ダルベッコのリン酸緩衝生理食塩水、pH〜7.4、(2)0.9%生理食塩水(0.9%w/v塩化ナトリウム(NaCl))、および(3)5%(w/v)デキストロースが含まれ;そしてまた、こうしたキャリアーは、トリプタミンなどの酸化防止剤、およびTween20などの安定化剤を含有してもよい。
【0179】
[194]選択した細胞集団の増殖を阻害するための方法を、in vitro、in
vivo、またはex vivoで実施することができる。本明細書において、増殖の
阻害は、短期間であれ、または長期間であれ、細胞の増殖の遅延、細胞生存度の減少、細胞死を引き起こすこと、細胞溶解、および細胞死の誘導を意味する。
【0180】
[195]in vitro使用の例には、疾患細胞または悪性細胞を殺すための、同じ患者への移植前の自己骨髄の治療;適格性T細胞を殺し、そして移植片対宿主病(GVHD)を防止するための、移植前の骨髄の治療;ターゲット抗原を発現していない望ましい変異体以外のすべての細胞を殺すか;または望ましくない抗原を発現する変異体を殺すための、細胞培養の治療が含まれる。
【0181】
[196]非臨床的in vitro使用の条件は、一般の当業者によって容易に決定される。
[197]臨床的ex vivo使用の例は、癌治療において、または自己免疫疾患治療において、自己移植前に、骨髄から腫瘍細胞またはリンパ球を取り除くこと、あるいは移植片対宿主病(GVHD)を防止するため、移植前に、自己または同種骨髄または組織からT細胞または他のリンパ球を取り除くことである。治療を以下のように行ってもよい。骨髄を患者または他の個体から採取し、そして次いで、本発明の細胞傷害性剤を添加した、血清を含有する培地中でインキュベーションする。濃度は、約10μM〜1pMの範囲であり、約37℃で約30分間〜約48時間、インキュベーションする。濃度およびインキュベーション時間、すなわち用量の正確な条件は、一般の当業者によって容易に決定される。インキュベーション後、血清を含有する培地で、骨髄細胞を洗浄し、そして既知の方法にしたがって、i.v.注入によって、患者に戻す。患者が骨髄採取時および処置細胞の再注入の間に切除化学療法または全身照射過程などの他の治療を受ける状況下では、標準的医学装置を用いて、液体窒素中で処置骨髄細胞を凍結保存してもよい。
【0182】
[198]臨床的in vivo使用のため、本発明の細胞傷害性コンジュゲートを溶液として供給して、無菌性および内毒素レベルに関して試験する。細胞傷害性コンジュゲート投与の適切なプロトコルの例は、以下のとおりである。コンジュゲートを毎週、4週間、i.v.ボーラスとして投与する。ボーラス用量は、生理食塩水50〜100ml中で投与され、これに5〜10mlのヒト血清アルブミンを添加してもよい。投薬量は、i.v.投与あたり10μg〜100mgである(1日あたり、100ng〜1mg/kgの範囲)。より好ましくは、投薬量は、50μg〜30mgの範囲であろう。最も好ましくは、投薬量は、1mg〜20mgの範囲であろう。治療4週後、患者に、毎週、治療を投与し続けてもよい。臨床状況が保証するように、投与経路、賦形剤、希釈剤、投薬量、時間等に関して、特定の臨床プロトコルを、一般の当業者が決定してもよい。
【0183】
[199]選択した細胞集団を殺すin vivo法またはex vivo法にしたがって治療可能な医学的条件の例には、CA6が発現されている、例えば肺、乳房、結腸、前立腺、腎臓、膵臓、卵巣、子宮頸部およびリンパ器官の癌、骨肉腫、滑膜癌、肉腫または癌腫、ならびにCA6グリコトープが主に発現されている、まだ決定されていない他の癌を含む、いかなる種類の悪性腫瘍も含まれ;全身性狼瘡(systemic lupus)、関節リウマチ、および多発性硬化症などの自己免疫疾患;腎移植拒絶、肝移植拒絶、肺移植拒絶、心臓移植拒絶、および骨髄移植拒絶などの移植片拒絶;移植片対宿主病;mV感染、HIV感染、AIDS等のウイルス感染;ならびにランブルべん毛虫症、アメーバ症、住血吸虫症などの寄生虫感染、および一般の当業者によって決定されるような他のものが含まれる。
【0184】
キット
[200]本発明にはまた、例えば記載する細胞傷害性コンジュゲート、および特定の細胞種を殺すための細胞傷害性コンジュゲートの使用のための説明書を含むキットも含まれる。使用説明書には、in vitro、in vivoまたはex vivoで細胞
傷害性コンジュゲートを使用するための指示が含まれてもよい。
【0185】
[201]典型的には、キットは、細胞傷害性コンジュゲートを含有する区画を有するであろう。細胞傷害性コンジュゲートは、凍結乾燥型、液体型、またはキットに含まれるよう受け入れ可能な他の型であってもよい。キットはまた、キット中の使用説明書に記載される方法を実施するのに必要なさらなる要素、例えば凍結乾燥粉末を再構成するための滅菌溶液、患者に投与する前に、細胞傷害性コンジュゲートと組み合わせるためのさらなる剤、および患者にコンジュゲートを投与するのに役立つツールも含有してもよい。
【0186】
さらなる態様
[202]本発明は、研究または診断適用で使用するためにさらに標識されている、モノクローナル抗体、ヒト化抗体およびそのエピトープ結合性断片をさらに提供する。好ましい態様において、標識は、放射標識、蛍光体、発色団、造影剤または金属イオンである。
【0187】
[203]前記の標識ヒト化抗体またはそのエピトープ結合性断片を、癌を有すると推測される被験体に投与し、そして被験体体内の標識分布を測定するかまたは監視する、診断法もまた、提供する。
【実施例】
【0188】
実施例
[204]本発明の広い範囲は、以下の実施例を参照すると最適に理解され、これらは本発明を特定の態様に限定することを意図しない。
【0189】
実施例1:フローサイトメトリー結合アッセイによる、抗原陽性および陰性細胞株の同定
[205]フローサイトメトリー分析を用いて、DS6エピトープ、CA6を細胞表面に局在させた。OVCAR5(Kearseら, Int. J. Cancer 88(6):866−872(2000))、OVCAR8およびIGROV1細胞(M. Seiden, Massachusetts General Hospital)を除いて、ヒト細胞株をアメリカン・タイプ・カルチャー・コレクション(ATCC)から得た。本明細書において、以後、培地と称する、4mM L−グルタミン、50U/mlペニシリン、50μg/mlストレプトマイシン(Cambrex Bio Science、メイン州ロックランド)および10%v/vウシ胎児血清(Atlas Biologicals、コロラド州フォートコリンズ)を補ったRPMI1640中で、すべての細胞を増殖させた。細胞を37℃、5%CO2加湿インキュベーター中で維持した。
【0190】
[206]96ウェルプレート中、FACS緩衝液(2%ヤギ血清、RPMI)中で調製した連続希釈濃度のDS6抗体と、細胞(1〜2x10−5細胞/ウェル)を氷上で3〜4時間インキュベーションした。卓上遠心分離装置中、1500rpm、4℃で5分間、細胞を回転させて落とした。培地を取り除いた後、ウェルに150μlのFACS緩衝液を再度満たした。次いで、洗浄工程を反復した。FITC標識ヤギ抗マウスIgG(Jackson Immunoresearch)を、FACS緩衝液に対して1:100に希釈し、そして氷上で1時間、細胞とインキュベーションした。プレートをホイルで覆い、シグナルの光退色を防いだ。2回の洗浄後、細胞を1%ホルムアルデヒドで固定し、そしてフローサイトメーター上で分析した。
【0191】
[207]腫瘍免疫組織化学から予測されるように、CA6エピトープは、主に、卵巣、乳房、子宮頸部、および膵臓起源の細胞株で見られる(表3)。しかし、他の腫瘍種のいくつかの細胞株が、限定されたCA6発現を示した。DS6抗体は、135.6pMの見かけのKDで結合する(PC−3細胞において、表3)。抗原陽性細胞株における結合
曲線(図1)の最大平均蛍光(表3)は、相対的抗原密度を示唆する。
表3
【0192】
【表4】
【0193】
*平均最大相対的平均蛍光
【0194】
実施例2:DS6エピトープの性質決定
[208]タンパク質分解(プロナーゼおよびプロテイナーゼK)および/または糖分解(ノイラミニダーゼおよび過ヨウ素酸)処理で消化した、CA6陽性細胞溶解物(Caov−3)のドットブロットを免疫ブロッティングすることによって、DS6抗原、CA6の特性を分析した。陽性対照には、多様なエピトープ種を認識する他の抗体を、抗原陽性細胞株の溶解物に対して試験した(Caov−3およびCM1;Colo205およびC242;SKMEL28およびR24)。CM1は、Muc−1の可変数タンデム反復ドメイン(VNTR)のタンパク質エピトープを認識する抗体であり、そしてしたがって、タンパク質エピトープに関する対照を提供する。C242は、Muc−1上の新規結腸直腸癌特異的シアル酸依存性グリコトープ(CanAg)に結合し、これはタンパク質上のグリコトープに関する対照を提供する。R24は、黒色腫に特異的なGD3ガングリオシドに結合し、そしてしたがって、非タンパク質骨格上のグリコトープに関する対照を提供する。
【0195】
[209]Caov−3、Colo205、およびSKMEL28細胞を15cm組織培養プレート中にプレーティングした。培地(30ml/プレート)を溶解前日に新しくした。修飾RIPA緩衝液(50mM Tris−HCl pH7.6、150mM NaCl、5mM EDTA、1%NP40、0.25%デオキシコール酸ナトリウム)、プロテアーゼ阻害剤(PMSF、ペプスタチンA、ロイペプチン、およびアプロチニン)、およびPBSを氷上であらかじめ冷却した。培地をプレートから吸引した後、10mlの冷却PBSで細胞を2回洗浄した。続くすべての工程を、氷上および/または4℃の低
温室で行った。PBSの最後の洗浄液を吸引した後、溶解緩衝液(最終濃度1mM PMSF、1μMペプスタチンA、10μg/mlロイペプチン、および2μg/mlアプロチニンまで、プロテアーゼ阻害剤を新鮮に添加した、RIPA緩衝液)1〜2ml中で細胞を溶解した。細胞リフターを用いて、プレートから溶解物をこそげ落とし、そして18G針で懸濁物を上下に(5〜10回)ピペッティングすることによって、すりつぶした(triturated)。溶解物を10分間回転させ、そして次いで、最大出力で10分間微量遠心分離装置中で遠心分離した(13K rpm)。ペレットを廃棄し、そして次いで、ブラッドフォード・タンパク質アッセイキット(Biorad)を用いて上清をアッセイした。
【0196】
[210]乾いた0.2μmニトロセルロース膜上に、溶解物(2μl)を直接ピペッティングした。スポットをおよそ30分間風乾させた。膜を、各々、単一のスポットを含有するピースに分けた。プロナーゼ(1mg/ml酵素、50mM Tris pH7.5、5mM CaCl2)、プロテイナーゼK(1mg/ml酵素、50mM Tris
pH7.5、5mM CaCl2)、ノイラミニダーゼ(20mU/ml酵素、50mM酢酸ナトリウムpH5、5mM CaCl2、100μg/ml BSA)または過ヨウ素酸(20mM、0.5M酢酸ナトリウムpH5)の存在下で、スポットを37℃で1時間インキュベーションした。Roche(酵素)およびVWR(過ヨウ素酸)から試薬を購入した。T−TBS洗浄緩衝液(0.1%Tween20、1xTBS)中で膜を洗浄し(5分間)、ブロッキング緩衝液(3%BSA、T−TBS)中で、室温で2時間ブロッキングし、そしてブロッキング緩衝液中、2μg/mlの一次抗体(すなわちDS6、CM1、C242、R24)で一晩インキュベーションした。T−TBS中で5分間、膜を3回洗浄し、そして次いで、HRPコンジュゲート化ヤギ抗マウス(またはヒト)IgG二次抗体(Jackson Immunoresearch;ブロッキング緩衝液中、1:2000希釈)と、室温で1時間インキュベーションした。免疫ブロットを3回洗浄し、そしてECL系(Amersham)を用いて現像した。
【0197】
[211]消化した対照溶解物の免疫ブロット(図2)は、タンパク質分解処理によってCM1シグナルが破壊される一方、糖分解消化のシグナルは、タンパク質エピトープを認識する抗体に関して予測されるように、影響を受けなかったことを示した。C242シグナルは、タンパク質上に見られるグリコトープを認識する抗体に関して予測されるように、タンパク質分解処理または糖分解処理のいずれによっても破壊された。タンパク質分解処理によって影響を受けなかったR24シグナルは、ガングリオシドを認識する抗体に関して予測されるように、ノイラミニダーゼ処理または過ヨウ素酸処理で消滅した。消化したCaov−3溶解物のDS6免疫ブロットは、タンパク質分解化合物および糖分解化合物のいずれかでの処理に際して、シグナル喪失を示した。したがって、C242同様、DS6は、タンパク性コア上の炭水化物エピトープに結合する。さらに、DS6免疫ブロット中のシグナルは、ノイラミニダーゼ処理に感受性であった。したがって、CanAg同様、CA6は、シアル酸依存性グリコトープである。
【0198】
[212]CA6が炭水化物性であることを確認するため、Caov−3溶解物をPVDF膜上にスポッティングし、そして化学的脱グリコシル化剤、トリフルオロメタンスルホン酸(TFMSA)で、窒素下、周囲温度で5分間、処理した。ブロットをT−TBSで洗浄し、そしてCM1またはDS6のいずれかで免疫ブロッティングした(図3)。DS6シグナルは、酸処理で破壊され、CA6がグリコトープであるさらなる証拠が提供された。TFSMA処理に際してCM1シグナルが増進することから、酸処理がフィルター上のタンパク質に影響を及ぼさないことが示され、そして糖分解処理によって、CM1が認識するタンパク質エピトープが暴露されることが示唆される。
【0199】
[213]CA6が属する炭水化物の構造をさらに解明するため、ドットブロットをN
−グリカナーゼ、O−グリカナーゼ、および/またはシアリダーゼ(図4)で消化した。Caov−3細胞溶解物(100μg、30μl)を、SDSおよびβ−メルカプトエタノールを含有する変性緩衝液(Glyko)2.5μlと100℃で5分間インキュベーションした。次いで、変性溶解物を1μlのN−グリカナーゼ、O−グリカナーゼ、および/またはシアリダーゼA(Glyko)を用いて、37℃で1時間消化した。次いで、消化した溶解物をニトロセルロース上にスポッティングし(2μl)、そして上述のように免疫ブロッティングした。
【0200】
[214]N−グリカナーゼは、DS6免疫ブロットシグナルに対して明らかな影響がなかった。しかし、シアリダーゼで消化した試料はまったくシグナルを生じなかった。O−グリカナーゼは、シアリダーゼでの前処理なしには、シアル化されたO連結炭水化物を消化不能であるため、O−グリカナーゼ単独でプロセシングした試料のDS6シグナルは、影響を受けないであろう。対照的に、N−グリカナーゼは、活性のため、いかなるグリコシド酵素での前処理も必要としない。N−グリカナーゼ処理がDS6シグナルに影響を及ぼさないという事実は、CA6エピトープが、シアル化O連結炭水化物鎖上に存在する可能性が最も高いことを示唆する。
【0201】
実施例3:CA6エピトープが見出される抗原の解明
[215]CA6シアログリコトープが見出される抗原を同定するため、SDS−PAGEおよびウェスタンブロッティングによって、DS6免疫沈降物を分析した。細胞溶解物上清(1ml/試料;3〜5mgタンパク質)をプロテインGビーズ(30μl)であらかじめきれいにし、回転させながら4℃で1〜2時間、1mlのRIPA緩衝液で平衡化させた。続くすべての工程を、氷上および/または4℃の低温室で行った。あらかじめきれいにしたビーズを、微量遠心分離装置中、簡単に(2〜3秒)回転させて落とした。あらかじめきれいにした上清を、新しい試験管に移し、そして回転させながら、2μgのDS6と一晩インキュベーションした。新鮮な、平衡化プロテインGビーズ(30μl)を溶解物に添加し、そして回転させながら、1時間インキュベーションした。ビーズ−溶解物懸濁物を、微量遠心分離装置中、簡単に回転させて落とし、そして場合によって、免疫沈降後の溶解物の試料を採取した。1mlのRIPA緩衝液で、ビーズを5〜10回洗浄した。
【0202】
[216]次いで、免疫沈降したDS6試料を、30μlのノイラミニダーゼ(20mUノイラミニダーゼ(Roche)、50mM酢酸ナトリウムpH5、5mM CaCl2、100μg/ml BSA)または30μlの過ヨウ素酸(20mM過ヨウ素酸(VWR)、0.5M酢酸ナトリウムpH5)で、37℃で1時間消化した。次いで、これらを、2x試料装填緩衝液(β−メルカプトエタノールを含有する)30μl中に再懸濁した。ビーズを5分間煮沸し、そして装填緩衝液上清を、4〜12%または4〜20%Tris−グリシン・ゲル(Invitrogen)上に装填した。Laemmli電気泳動緩衝液中、125Vで1.5時間、ゲルを泳動した。Mini Trans−blotトランスファー装置(Biorad)を用いて、0.2μmニトロセルロース膜(Invitrogen)上に、ゲル試料を20mAで一晩トランスファーした。実施例2に上述するように、DS6で膜を免疫ブロッティングした。
【0203】
[217]あるいは、免疫沈降したビーズをまず変性させ、そして次いで、N−グリカナーゼ、O−グリカナーゼおよび/またはシアリダーゼA(Glyko)で酵素的に消化した。27μlのインキュベーション緩衝液および2μlの変性溶液(Glyko)中にビーズを再懸濁し、そして100℃で5分間インキュベーションした。室温に冷却した後、界面活性剤溶液(2μl)を添加し、そして試料を1μlのN−グリカナーゼ、O−グリカナーゼ、および/またはシアリダーゼAと、37℃で4時間インキュベーションした。5x試料装填緩衝液(7μl)を添加した後、試料を5分間煮沸した。試料をSDS−
PAGEに供し、そして上述のように、免疫ブロッティングした。
【0204】
[218]DS6は、抗原陽性細胞溶解物中に見られうる、>250kDaのタンパク質バンドを免疫沈降する(図5A、BおよびC)。ある細胞株(すなわちT−47D)では、ダブレットが観察される。>250kDaバンドは、ノイラミニダーゼまたは過ヨウ素酸で処理したCaov−3免疫沈降物では消失し(図5AおよびB)、CA6エピトープが、>250kDaバンド上に属することが示唆される。>250kDaバンドはまた、免疫沈降物のN−グリカナーゼ処理には感受性ではないことも示され、このことは、CA6がO連結炭水化物上に属することと一致する(図5F)。250kDaバンドがCA6抗原であることをさらに支持するのは、DS6が、DS6抗原陰性細胞から、こうしたバンドを免疫沈降させないという事実である(図5DおよびE)。
【0205】
[219]いくつかの系列の証拠が、CA6抗原がMuc1であることを示唆した。分子量が高く、そしてO連結炭水化物特異的糖分解酵素に感受性であるため、CA6抗原がムチンである可能性が高いようである。ムチン過剰発現は、腫瘍、特に乳房および卵巣の腫瘍でよく特徴付けられており、これは、DS6の主な腫瘍反応性と一致する。さらに、CA6は、CanAg(Muc1上のシアログリコトープ)同様、過塩素酸沈殿に感受性ではなく、CA6抗原が、非常にO−グリコシル化されていることが示唆される。いくつかのDS6発現細胞株において、DS6が、>250kDaのダブレットを免疫沈降させた観察から、CA6がMuc1であることが示唆された。ヒトにおけるMuc1の特質は、タンデム反復数が異なる2つの別個のMuc1アレルの存在であり、これは異なる分子量の2つのMuc1タンパク質の発現を生じる。
【0206】
[220]CA6がMuc1上に見られるかどうかを試験するため、Caov−3溶解物からのDS6免疫沈降物を、SDS−PAGEに供し、そしてDS6、またはMuc1VNTR抗体、CM1のいずれかで免疫ブロッティングした。図6Aに見られうるように、CM1は、DS6によって免疫沈降される>250kDaバンドと強く反応する。図6Bでは、HeLa細胞溶解物からのDS6およびCM1免疫沈降物は、DS6またはCM1のいずれかで免疫ブロッティングした際、同じ>250kDaダブレットを示す。これらの結果によって、CA6エピトープは実際、Muc−1タンパク質上に位置することが示される。HeLa(およびT−47D)細胞に見られるDS6ダブレットは、Muc−1発現が、異なる数のタンデム反復を有する別個のアレルによって指示されるという事実によって説明可能である。
【0207】
[221]CM1およびDS6は、同じMuc−1タンパク質に結合するが、これらは別個のエピトープである。トリフルオロメタンスルホン酸(TFMSA)によるCaov−3溶解物ドットブロットの化学的脱グリコシル化によって、DS6シグナルが消滅した(図3)。しかし、この同じ処理は、CM1シグナルを増進させた。脱グリコシル化が、CM1抗体の隠れたエピトープを暴露した可能性もある。さらに、DS6およびCM1のフローサイトメトリー結合結果の比較(表4)によって、CA6エピトープが、Muc1を発現するすべての細胞上には存在しないことが示される。高レベルのMuc1 CanAgシアログリコトープを発現することが知られる細胞株であるColo205上に、CA6エピトープが発現されないことに注目すると興味深い(表3)。
表4
【0208】
【表5】
【0209】
*MMF=最大平均相対的蛍光
【0210】
実施例4:脱落(shed)CA6エピトープの定量的分析
[222]CA6エピトープは、多くの癌患者において、血流内に脱落することが知られる分子であるMuc1上に属するため、こうしたレベルがDS6抗体療法を禁ずるほどであるかどうかを決定するために、定量的アプローチを行った。抗原に循環抗体が結合すると、血液からの免疫複合体の迅速なクリアランスにつながると考えられる。投与した抗体用量のかなりの部分が循環から迅速に除去されるならば、腫瘍に到達する量は、減少する可能性が高く、抗体療法剤の抗腫瘍活性の減少を生じる。抗体を非常に強力な細胞傷害性化合物にコンジュゲート化すると、コンジュゲートの迅速なクリアランスが、非特異的な毒性を潜在的に増加させうる。したがって、DS6−DM1などの抗体−小薬剤コンジュゲートの場合、高レベルの脱落抗原は、抗腫瘍効果を減少させ、そしてかつ用量を制限する毒性を増加させると予期されうる。
【0211】
[223]抗体療法剤の最近の臨床試験は、薬物動態に対する脱落抗原濃度の影響に関する情報を生じた。例えば、her2/neuを発現する転移性乳癌治療に使用する抗体であるトラスツズマブ(ハーセプチン)を用いた臨床試験において、トラスツズマブ・クリアランスの薬物動態は、脱落Her2/neuレベルが500ng/ml未満である場合、改変されないことが示された(Pegramら, J. Clin. Oncol.
16(8):2659−71(1998))。脱落Her2/neuの分子量を110,000ダルトンと仮定すると、4.5nM未満の脱落Her2/neuのモル濃度は、薬物動態にほとんど影響を及ぼさないようである。
【0212】
[224]別の例において、カンツズマブ・マータンシン(cantuzumab mertansine)(huC242−DMl)を用いた臨床試験によって、処置前脱落CanAg(C242エピトープ)レベルおよび抗体クリアランスの薬物動態に相関がないことが示された(Tolcherら, J. Clin. Oncol. 21(2):211−22(2003))。CanAgエピトープは、DS6によって認識されるCA6エピトープ同様、Muc1上のユニークな腫瘍特異的O連結シアログリコトープである。しかし、CanAgエピトープは不均一性であるため、モルで定量化するのが困難である。一般集団において、Muc1アレルは、可変数タンデム反復(VNTR)ドメイン中のタンデム反復数に応じて、長さが多様である。O連結グリコシル化のためのいくつかの部位が、各タンデム反復中に存在する。CanAg発現の複雑さに加えて、生得的なグリコシルトランスフェラーゼ活性には、細胞間変動がある。したがって、単一の患者においてであってさえ、Muc1分子あたり、広い範囲のCanAgエピトープがありうる。さらに、Muc1分子あたりのCanAgエピトープの比は、患者集団に渡って異なるであろう。このため、CanAgエピトープを含む脱落Muc1が、C242に捕捉され、
そしてビオチン化C242/ストレプトアビジンHRP系によって検出される、サンドイッチELISAによって、血清試料中の脱落CanAgを測定する。Muc1のモル濃度によるのではなく、血清1mlあたりのエピトープ数に比例する標準化単位で、脱落CanAgを定量化する。同様に、脱落CA6エピトープの定量化に関して、類似の状況が生じる。対照的に、トラスツズマブに関しては、脱落her2/neu分子あたりにただ1つのエピトープしかなく、脱落抗原の定量化は非常に単純化される。
【0213】
[225]トラスツズマブおよびカンツズマブ・マータンシンを用いた臨床試験で見られるものに、CA6脱落エピトープレベルを関連させるため、Muc1上のシアログリコトープなどの複雑な脱落エピトープのモル濃度を得るための方法を開発した。まず、DS6に関する単純サンドイッチELISAアッセイを確立した。代表的なアッセイを図7Aに示す。DS6を用いて、CA6エピトープを有するMuc1を捕捉した。各Muc1分子は複数のCA6エピトープを有するため、ビオチン化DS6を、トレーサー抗体としてもまた用いた。基質としてABTSを用い、ストレプトアビジン−HRPによって、捕捉されたCA6に結合したビオチン化DS6を検出した。卵巣癌患者血清から、または乳癌患者において脱落Muc1を監視するために用いられる商業的に入手可能なMuc1試験キット(CA15−3)に由来する標準から、CA6エピトープを捕捉した。DS6単位/mlを、恣意的に、CA15−3標準単位/mlに等しく設定した。
【0214】
[226]図7Bにおいて、CA15−3標準を用いる、DS6サンドイッチELISAの結果を示す。生じる曲線は、CA15−3アッセイにおいて、CA15−3標準で得られるものと非常に類似である。DS6単位/mlを、CA6のモル濃度に変換するため、シグナルをDS6のピコグラムに変換する、ビオチン化DS6に関する標準曲線が必要である。CA6エピトープおよびビオチン化DS6抗体間は1対1の化学量論であり、そしてビオチン化DS6の分子量は160,000ダルトンであると仮定して、添加した試料体積あたりに捕捉されるCA6のモル数を計算してもよい。
【0215】
[227]図8AおよびBにおいて、ビオチン化DS6に関する標準曲線を生じる、2つの代替手段を示す。図8Aにおいて、ヤギ抗マウスIgGポリクローナル抗体を用いて、ビオチン化DS6を捕捉し、これを次に、図7に示すサンドイッチELISAアッセイで用いるものと同一の方式で検出する。図8Bに示す方法において、ビオチン化DS6を、ELISAプレート上に直接プレーティングし、そして図8Aにおけるように検出する。図8Cにおいて見られるように、各方法によって生成されるビオチン化DS6標準曲線は、十分一致している。
【0216】
[228]表5において、多様な脱落抗原に関する卵巣癌患者血清試料の分析を示す。一般的に、CA125 ELISAを用いて、脱落CA125単位/mlを測定することによって、卵巣癌患者の治療を監視する。血清試料を用いて、CA125状態を提供する。一般的に、CA15−3 ELISAを用いて、DS6によって認識されるものと異なるエピトープを認識する捕捉および検出抗体を用い、脱落Muc1の単位/mlを測定することによって、乳癌患者の治療を監視する。表5において、卵巣癌患者血清試料において、CA15−3を測定する。
表5
【0217】
【表6】
【0218】
1商業的ELISAキットによって測定
2商業的CA15−3標準によって測定(1 CA15−3 U=1 DS6 U)
3ヤギ抗マウスIgGおよびビオチン−DS6標準曲線
4ビオチン−DS6標準曲線
【0219】
[229]表5に報告するCA15−3値に関しては、CanAg Dagnosticsの商業的に入手可能なCA15−3酵素免疫アッセイキットを用いた。DS6単位/mlに関しては、DS6サンドイッチELISAにおいて、CA15−3標準(CanAg DiagnosticsのCA15−3酵素免疫アッセイキット由来)を用いて標準曲線を生成した。DS6単位/mlを、恣意的に、CA15−3単位/mlに等しく設定した。最後の2つの列では、図8Cに示すビオチン化DS6標準曲線を用いて、ピコモル(pM)脱落CA6を計算した。
【0220】
[230]CanAgレベルの定量的分析に関して、CanAg血清レベルは、処置前にカンツズマブ・マータンシン臨床試験に参加した患者に関して報告されたものであった(Tolcherら, J. Clin. Oncol. 21(2):211−22(2003))。DS6に関して記載されるものと類似のELISAアッセイを用いて、CanAg標準を用い、CanAg標準曲線を作成した。C242を用いて、CanAg標準を捕捉した。ビオチン化C242トレーサーを用いて、その後、基質としてABTSを用い、ストレプトアビジン−HRPで発色させて、捕捉されたCanAgの検出を達成した。ビオチン化DS6に関して行ったように、ビオチン化C242標準曲線を構築して、単位/mlを循環CanAgエピトープのモル濃度に変換することを可能にした。表6において、カンツズマブ・マータンシン臨床試験患者由来のCanAgレベルを、循環CanAgの対応する計算モル濃度と一緒に報告する。
表6
【0221】
【表7】
【0222】
1サンドイッチELISAによって測定した、循環CanAgの処置前レベル
2ヤギ抗マウスIgGおよびビオチン−C242標準曲線
3ビオチン−C242標準曲線
【0223】
[231]卵巣癌患者における脱落CA6のpMレベルと、CanAg陽性癌患者における脱落CanAgに関して計算したものの比較によって、一般的に、脱落CA6レベルは、脱落CanAgレベルと類似であることが示される。さらに、16の卵巣癌患者血清試料のうち2つのみが、4.5nMより高いCA6レベルを潜在的に有し(シグナルが標準曲線の範囲から外れている、血清試料5および9)、このレベルは、これより高い場合、Her2/neu陽性乳癌患者での臨床試験において、改変されたハーセプチン薬物動態が観察されたレベルであった。4.5nMを超えるCanAgレベルは、37の臨床試験患者のうち3人でのみ見られた。この臨床試験において、脱落CanAgレベルおよびカンツズマブ・マータンシンのより迅速なクリアランスには相関はなかった。しかし、最高のCanAgレベル(31240U/ml)の患者は、注入後8時間のみ試料採取された。これらの結果によって、Muc1の特定のエピトープ、例えばCA6およびCanAgは、癌患者において脱落するものの、抗体療法治療を禁じるレベルまでは脱落しないことが示される。
【0224】
実施例6:ネズミDS6抗体可変領域のクローニング
[232]DS6などのネズミ・モノクローナル抗体は、ヒト免疫系によって異質(foreign)と認識されるため、臨床設定において、有用性が限定されている。患者は、ヒト抗マウス抗体(HAMA)を迅速に発展させ、ネズミ抗体の迅速なクリアランスを生じる。このため、ネズミDS6(muDS6)の可変領域の表面を再構成して、ヒト化DS6(huDS6)抗体を生じた。
【0225】
[233]RT−PCRによって、ネズミDS6抗体可変領域をクローニングした。Qiagen RNeasyミニプレップキットを用いて、DS6ハイブリドーマ細胞の集密T175フラスコから総RNAを精製した。UV分光光度測定によってRNA濃度を測定し、そしてGibco Superscript IIキットおよびランダム六量体プライマーを用いて、4〜5μg総RNAでRT反応を行った。
【0226】
[234]Wang Zら, J Immunol Methods. Jan 13;233(l−2):167−77(2000)に記載されるものに基き、縮重プライマーを用いて、PCR反応を行った。RT反応混合物を縮重PCR反応に直接用いた。3’軽鎖プライマー、HindKL、
(TATAGAGCTCAAGCTTGGATGGTGGGAAGATGGATACAGTTGGTGC)(配列番号25)
および3’重鎖プライマー、BamIgG1、
(GGAGGATCCATAGACAGATGGGGGTGTCGTTTTGGC)(配列番号26)
を用い、そして5’端に関しては、PCRプライマーは、軽鎖に関して、Sac1MK
(GGGAGCTCGAYATTGTGMTSACMCARWCTMCA)(配列番号27)
そして重鎖に関して、EcoR1MH1
(CTTCCGGAATTCSARGTNMAGCTGSAGSAGTC)(配列番号28)およびEcoR1MH2(CTTCCGGAATTCSARGTNMAGCTGSAGSAGTCWGG)(配列番号29)の等量混合物であった(混合塩基:H=A+T+C、S=G+C、Y=C+T、K=G+T、M=A+C、R=A+G、W=A+T、V=A+C+G、N=A+T+G+C)。
【0227】
[235]10%DMSOを補った以外、PCR反応は標準的であった(50μl反応混合物は、最終濃度1x反応緩衝液(ROCHE)、各2mMのdNTP、各1mMのプライマー、2μl RT反応、5μl DMSO、および0.5μl Taq(ROCHE)を含有した)。Wang Zら(J Immunol Methods. Jan 13;233(1−2):167−77(2000))から適応させたプログラム:1)94℃、3分間;2)94℃、15秒間;3)45℃、1分間;4)72℃、2分間;5)工程2に戻って29周期;6)72℃、10分間の最終伸長工程で終了を用いて、MJ
researchサーモサイクラー上で、PCR反応を行った。PCRプライマーによって生成されたPCR産物を、制限酵素を用いて、pBluescript II SK+(Stratagene)内にクローニングした。Seqwright配列決定サービスによって、重鎖および軽鎖クローンを配列決定した。
【0228】
[236]5’端cDNA配列を確認するため、さらなるPCRおよびクローニングを行った。縮重PCRクローンから決定したDS6軽鎖および重鎖cDNA配列をNCBIのBlast検索ウェブサイト内に打ち込み、そして提示されたシグナル配列を持つネズミ抗体配列を記録した。関連DNA配列の間で保存されたストレッチを用いて、これらのシグナルペプチドからPCRプライマーを設計した。EcoRI制限部位をリーダー配列
プライマーに付加し(表7)、そしてこれらを、上述のようなRT−PCR反応に用いた。
【0229】
【表8】
【0230】
[237]いくつかの個々の軽鎖および重鎖クローンを配列決定して同定し、そしてポリメラーゼが生成する可能性がある配列エラーを回避した。軽鎖および重鎖RT−PCRクローンの両方に関して、単一の配列のみが得られた。これらの配列は、シグナル配列内まで拡大してネズミDS6軽鎖および重鎖配列を増幅可能なプライマーを設計するのに十分であった。これらの追加PCR反応由来の続くクローンによって、元来の縮重プライマーによって改変されている可変領域の5’端配列が確認された。多様なcDNAクローンからの累積結果によって、図9に示す最終ネズミDS6軽鎖および重鎖配列が提供された。KabatおよびAbM定義を用いて、3つの軽鎖および重鎖CDRが同定された(図9および10)。NCBI IgBlastデータベースの検索によって、muDS6抗体軽鎖可変領域が、ネズミIgVκ ap4生殖系列遺伝子に由来する可能性が最も高く、一方、重鎖可変領域が、ネズミIgVh J558.41生殖系列遺伝子に由来する可能性が最も高いことが示される(図11)。
【0231】
実施例7:DS6抗体の可変領域表面残基の決定
[238]Pedersenら(1994)およびRoguskaら(1996)によって記載される抗体表面再構成技術は、ネズミ抗体可変配列の表面残基を予測することから始まる。表面残基は、その総表面積の少なくとも30%が水分子に対してアクセス可能であるアミノ酸と定義される。muDS6に関する表面残基を発見するために解明された構造がないため、本発明者らは、127の抗体構造ファイルセット中、最も相同な配列を持つ10の抗体を整列させた(図12)。これらの整列配列に関して、各Kabat位の溶媒アクセス可能性を平均した(図13AおよびB)。
【0232】
[239]いずれかの側に隣接する2つの同一残基を含有する抗体サブセットを比較することによって、25%〜35%の間の平均アクセス可能性を持つ表面位を、第二周期の分析に供した(図13AおよびB)。第二周期の分析後、muDS6重鎖に関して、21の予測される表面残基は、23に増加し、30%より高い表面アクセス可能性を持つと予測される残基のリストに、Tyr3およびLys23が加わった。本発明者らの表面再構成抗体の大部分では、重鎖CDR1のKabat定義を用いたが、DS6に関しては、計算中、不注意にAbM定義を用いたため、重鎖残基T28は、Kabat定義でフレームワーク表面残基と定義されうるようには、定義されなかった。Ala80の予測される表面アクセス可能性が、第二周期分析において、30.5%から27.8%に減少したため、軽鎖表面位の数は、16から15に減少した。総合すると、muDS6重鎖および軽鎖可変配列は、38の予測される表面アクセス可能フレームワーク残基を有する。
【0233】
実施例8:ヒト抗体選択
[240]ネズミDS6可変領域の表面位を、Kabatデータベース中のヒト抗体配列中の対応する位に比較した(Johnson G, Wu TT. Nucleic Acids Res. Jan 1;29(1):205−6(2001))。抗体データベース管理ソフトウェアSR(Searle、1998)を用いて、天然重鎖および軽鎖ヒト抗体対から表面残基を抽出し、そして整列させた。CDRの5Å以内にある位に特に考慮を払い、最も同一の表面残基を持つヒト抗体可変領域表面を選択して、ネズミDS6抗体可変領域表面残基を置換した。
【0234】
実施例9:キメラおよびヒト化抗体の発現ベクター
[241]軽鎖および重鎖対形成配列を、単一哺乳動物発現ベクター内にクローニングした。ヒト可変配列のPCRプライマーは、制限部位を生成し、pBluescriptIIクローニングベクターにヒトシグナル配列を付加することを可能にした。次いで、軽鎖または重鎖に関して、それぞれ、EcoRIおよびBsiWIまたはHidIIIおよびApaIを持つ哺乳動物発現プラスミド内に、可変配列をクローニングしてもよい(図14)。軽鎖可変配列を、インフレームでヒトIgカッパ定常領域上にクローニングし、そして重鎖可変配列を、ヒトIgガンマ1定常領域配列内にクローニングした。最終発現プラスミドにおいて、ヒトCMVプロモーターが、軽鎖および重鎖両方のcDNA配列の発現を駆動する。
【0235】
実施例10:DS6活性に負に影響を及ぼしうる残基の同定
[242]今日まで、ヒト化の大部分では、潜在的に問題がある残基として、CDRに近接する残基を同定するため、対象抗体の分子モデルが構築されてきた。作業を開始する表面再構成された抗体の数が増加しつつあるため、歴史的な経験は、問題を予測するのに、モデルを構築するのと少なくとも同程度に有効であり、DS6に関しては、分子モデルを構築しなかった。その代わり、ネズミDS6表面残基を、先に表面再構成された抗体と比較し、そして抗体の結合能に影響を及ぼすリスクが低い残基から高い残基を同定した。
【0236】
[243]入手可能な解明された抗体構造および先のヒト化に由来する分子モデルの両方において、類似の残基セットが、CDRの5Å以内にあると繰り返し同定された。このデータを用いて、表1は、CDRに近接する可能性があり、そしておそらく5Å以内である、ネズミDS6残基を提供する。これらの位の多くが、先のヒト化でもまた変更されているが、これまでに結合活性の喪失を生じたのは、重鎖74位のみであった。ネズミ抗体の結合活性を保存するため、huC242およびhuB4の両方において、この位でネズミ残基を保持した。一方、この同じ位が、ヒト化6.2G5C6において、対応するヒト残基に変更されたが、活性の喪失は伴わなかった(6.2G5C6は、しばしば、単に抗C6と称される抗IGF1−R抗体である)。表1中のいかなる残基もヒト化抗体において問題を提示しうるが、重鎖残基P73が、この位での先の経験から、特に懸念される。
【0237】
実施例11:最も相同なヒト表面の選択
[244]SRソフトウェアを用いて、Kabat抗体配列データベースから、muDS6の表面を再構成するための候補ヒト抗体表面を引き出した。このソフトウェアは、抗体データベースに対して、特定の残基位のみを検索するインターフェースを提供する。天然の対を保持するため、軽鎖および重鎖両方の表面残基を一緒に比較した。配列同一性のランク順に、Kabatデータベース由来の最も相同なヒト表面を整列させた。SR Kabatデータベースソフトウェアによって並列した際の上位3つの表面を表2に示す。次いで、表面を比較して、表1に同定する残基に対して最も少ない変化しか必要としないのはどのヒト表面かを同定した。抗Rh(D)抗体、28E4(Boucherら、1997)は、最少数の表面残基変化しか必要とせず(総数11)、そしてこれらの残基のう
ち3つのみが、潜在的な問題の残基のリストに含まれた。28E4抗体は、最も相同なヒト表面を提供するため、これが、muDS6の表面を再構成する最適な候補である。
【0238】
実施例12:ヒト化DS6抗体のためのDNA配列の構築
[245]PCR突然変異誘発を用いて、DS6のための11の表面残基変化を作製した。ネズミDS6可変領域cDNAクローンに対して、PCR突然変異誘発を行って、表面を再構成したヒトDS6遺伝子を構築した。以下の表8に示すように、ヒト化プライマーセットを設計して、表面再構成DS6に必要なアミノ酸変化を作製した。
表8
【0239】
【表9】
【0240】
[246]10%DMSOを補った以外、PCR反応は標準的であった(50μl反応混合物は、最終濃度1x反応緩衝液(ROCHE)、各2mMのdNTP、各1mMのプライマー、100ngのテンプレート、5μl DMSO、および0.5μl Taq(ROCHE)を含有した)。以下のプログラム:1)94℃、1分間;2)94℃、15秒間;3)55℃、1分間;4)72℃、1分間;5)工程2に戻って29周期;6)72℃、4分間の最終伸長工程で終了を用いて、MJ Researchサーモサイクラー上で、PCR反応を行った。PCR産物を、対応する制限酵素で消化して、そしてpBluescriptクローニングベクター内にクローニングした。クローンを配列決定して、アミノ酸変化を確認した。
【0241】
[247]重鎖残基P73を変化させると、過去に問題が引き起こされたため、重鎖の2つの型、ヒト28E4 T73を持つもの、およびネズミP73を保持するものを構築した。ヒト化DS6のどちらの型でも、他の10の表面残基をネズミからヒト28E4残基に変化させた(表2)。通常の命名法に忠実にしたがい、11のヒト表面残基すべてを有するため、最もヒトであるのは、1.0型である。さらなる型が必要とされる場合のために、ネズミP73を保持する重鎖型を1.2と名付け、したがって、1.1型は、最大数のネズミ残基を含有する型のために取っておく。図15AおよびBにおいて、2つのヒト化型のアミノ酸配列を、ネズミDS6アミノ酸配列と整列させて示す。一過性および安定トランスフェクションのため、ヒト化DS6抗体遺伝子の両方を、抗体発現プラスミド(図14)内にクローニングした。ヒト化型1.01および1.21軽鎖可変領域のcDNAおよびアミノ酸配列は、同じであり、そしてこれらを図16に示す。ヒト化型1.01および1.21の重鎖cDNAおよびアミノ酸配列を図17AおよびBに示す。
【0242】
実施例13:CHO細胞におけるhuDS6の発現および精製、ならびに親和性測定
[248]ヒト化DS6型が、muDS6の結合親和性を保持するかどうかを決定するため、抗体を発現し、そして精製することが必要であった。ヒト定常領域およびネズミ可変領域を有するDS6のキメラ型(chDS6)で、あるいはhuDS6v1.01またはhuDS6v1.21で、CHO細胞を安定トランスフェクションした。
【0243】
[249]CHODG44細胞(4.32x106細胞/プレート)を、非選択培地(4mM L−グルタミン、50U/mlペニシリン、50μg/mlストレプトマイシン、および10%v/v FBSを補った、アルファMEM+ヌクレオチド(Gibco))中、15cmプレートに植え付け、そして37℃、5%CO2加湿インキュベーター中に入れた。翌日、Polyfect Transfectionに関してQiagenが推奨するプロトコルの修飾型を用いて、chDS6発現プラスミドで細胞をトランスフェクションした。非選択培地を細胞から吸引した。7mlのあらかじめ温めた(37℃)PBSでプレートを洗浄し、そして20mlの非選択培地を再補充した。プラスミドDNA(11μg)を800μlのハイブリドーマSFM(Gibco)内に希釈した。次いで、70μlのPolyfect(Qiagen)をDNA/SFM混合物に添加した。次いで、Polyfect混合物を数秒間穏やかにボルテックスし、そして周囲温度で10分間インキュベーションした。非選択培地(2.7ml)を混合物に添加した。この最終混合物を、プレーティングした細胞と24時間インキュベーションした。
【0244】
[250]トランスフェクション混合物/培地をプレートから取り除き、そして次いで、細胞をトリプシン処理し、そして計数した。次いで、多様な密度(1800、600、200、および67細胞/ウェル)で、96ウェルプレート(250μl/ウェル)中、細胞を選択培地(4mM L−グルタミン、50U/mlペニシリン、50μg/mlストレプトマイシン、10%v/v FBS、1.25mg/ml G418を補った、アルファMEM−ヌクレオチド(Gibco))中でプレーティングした。必要であれば培地を補って、細胞を2〜3週間インキュベーションした。定量的ELISAを用いて、抗体産生レベルに関してウェルをスクリーニングした。Immulon 2HB 96ウェルプレートを、ヤギ抗ヒトIgG F(ab)2抗体(Jackson Immunoresearch;100μlの50mM炭酸ナトリウム緩衝液pH9.6中、1μg/ウェル)でコーティングし、そして振盪しながら、周囲温度で1.5時間インキュベーションした。すべての続く工程を、周囲温度で行った。ウェルをT−TBS(0.1%Tween−20、TBS)で2回洗浄し、そして200μlのブロッキング緩衝液(1%BSA、T−TBS)で1時間ブロッキングした。ウェルをT−TBSで2回洗浄した。別個のプレート中で、抗体標準、EM164(100ng/ml)および培養上清を、ブロッキング緩衝液中で連続希釈した(1:2または1:3)。これらの希釈物(100μl)を、ELISAプレートに移し、そして1時間インキュベーションした。ウェルをT−TBSで3回洗浄し、そしてブロッキング緩衝液中で1:3000希釈した100μlのヤギ抗ヒトIgG Fc−AP(Jackson ImmunoResearch)と45分間インキュベーションした。T−TBSで5回洗浄した後、100μlのPNPP発色試薬(10mg/ml PNPP(p−ニトロフェニルリン酸、二ナトリウム塩;Pierce)、0.1MジエタノールアミンpH10.3緩衝液)を用いて、ウェルを25分間、発色させた。ELISAプレート読取装置中、405nmでの吸光度を測定した。標準曲線の直線部分での(培養上清の)吸光度読取り値を用いて、抗体レベルを決定した。
【0245】
[251]次いで、ELISAによって同定された最大産生クローンをサブクローニングし、拡大し、そして凍結細胞ストックを調製した。
[252]huDS6v1.01およびhuDS6v1.21の発現に関しては、DG44 CHO細胞(Lawrence Chasin博士、ニューヨーク州コロンビア大学)を、リボヌクレオチドおよびデオキシリボヌクレオチドを含むアルファMEM(Gibcoカタログ番号12571、ニューヨーク州グランドアイランド)中で培養した。培
地に10%ウシ胎児血清(HyCloneカタログ番号SH30071.03、ユタ州ローガン)、1%ゲンタマイシン(Mediatechカタログ番号30−005−CR、バージニア州ハーンドーン)、および2mM L−グルタミン(L−glut)(BioWhittakerカタログ番号17−605E、メリーランド州ウォーカーズビル)を補った。この配合物をCHO完全培地と名付けた。
【0246】
[253]DG44 CHO細胞(5x106)を、50μgのhuDS6プラスミドDNAでトランスフェクションした。トランスフェクション前に、トリプシン(Gibcoカタログ番号15090−046、ニューヨーク州グランドアイランド)でフラスコから細胞を取り除き、そしてリボヌクレオシドおよびデオキシリボヌクレオシドを欠く、非補充アルファMEM(Gibcoカタログ番号12561、ニューヨーク州グランドアイランド)で2回洗浄した。これを洗浄培地と名付けた。0.4cmギャップの電極キュベット(BioRadカタログ番号1652088、ペンシルバニア州ハーキュルス)中、細胞をプラスミドDNAと混合した。これらを氷上に2分間置き、そして次いで、BioRadエレクトロポレーション装置中、1,000μFおよび260ボルトでパルス処理した。エレクトロポレーション後、氷上で2分間、細胞をインキュベーションした。次いで、CHO完全培地中、細胞を5つの24ウェルプレート(Costarカタログ番号3524)中にプレーティングし、そして5%CO2の37℃インキュベーター中で維持した。48時間後、ウェルから培地を取り除いた。洗浄培地でウェルを1回リンスし、そして1%ゲンタマイシン、2mM L−glut、10%透析ウシ胎児血清(Gibcoカタログ番号26400−044、ニューヨーク州グランドアイランド)、および1.25mg/mlジェネティシン(G418)(Gibcoカタログ番号11811、ニューヨーク州グランドアイランド)を補った、リボヌクレオシドおよびデオキシリボヌクレオシドを含まないアルファMEM(Gibcoカタログ番号12561、ニューヨーク州グランドアイランド)を供給した。この完全配合物を、選択培地と名付けた。選択培地中でおよそ2週間、クローンをインキュベーションし、この時点で、定量的ELISAによって、抗体産生に関してスクリーニングした。次いで、最高産生クローンをサブクローニングし、拡大し、そして凍結細胞ストックを調製した。
【0247】
[254]精製に十分な量の抗体を産生するため、Ultra Low IgG FBS(Gibco)を補った選択培地30mlを含む15cmプレート(〜1x106細胞/プレート)上に、細胞を拡大し、そして1週間インキュベーションした。250mlの円錐管内に培養上清を収集し、卓上遠心分離装置中で回転して落とし(2000rpm、5分間、4℃)、そして次いで、0.2μmフィルター装置を通じて滅菌ろ過した。
【0248】
[255]DS6を精製するため、NaOHのペレットをろ過培養上清に添加して、最終pH8.0にした。HiTrap rプロテインAカラム(Amersham)を、20〜50カラム体積の結合緩衝液で平衡化した。蠕動ポンプを用いて、上清をカラム上に装填した。次いで、カラムを50カラム体積の結合緩衝液で洗浄した。溶出緩衝液(100mM酢酸、50mM NaCl、pH3)を用いて、結合した抗体を、カラムから、フラクションコレクター中にセッティングした試験管内に溶出させた。中和緩衝液(2M K2HPO4、pH10.0)を用いて、溶出した抗体を中和し、そしてPBS中で一晩透析した。0.2μmシリンジフィルターを通じて、透析抗体をろ過した。280nmでの吸光度を測定して、最終タンパク質濃度を決定した。
【0249】
[256]精製huIgGの親和性を、フローサイトメトリーによって、muDS6と比較した。最初の実験セットでは、CA6発現細胞株、KBへの直接結合を測定した。図18に示すように、muDS6、chDS6、huDS5v1.01およびhuDS6v1.21は、非常に類似の親和性を示し、見かけのKdが、それぞれ、0.82nM、0.69nM、0.82および0.85nMであり、表面再構成がCDRを妨害しなかった
ことが示唆された。huDS6型が、muDS6の親和性を保持することを確認するため、競合的結合実験を行った。この形式の利点は、ネズミおよびヒト抗体の両方に関して、同じ検出系;すなわち、ビオチン−muDS6/ストレプトアビジン−DTAFを使用することである。muDS6、chDS6、huDS6v1.01およびhuDS6v1.21がビオチン−DS6と競合する能力を比較する、競合結合アッセイの結果を図19に示す。見かけのEC50は、muDS6、chDS6、huDS6v1.01、およびhuDS6v1.21に関して、それぞれ、1.9nM、1.7nM、3.0および1.9nMである。これらの結果は、ヒト化DS6を産生するためのmuDS6の表面再構成が、結合親和性の減少をほとんど引き起こさないことを示す。
【0250】
実施例14:muDS6−DM1細胞傷害性コンジュゲートの調製
[257]8倍モル過剰のN−スクシンイミジル−4−(2−ピリジルジチオ)ペンタノエート(SPP)を用いて、muDS6抗体(8mg/ml)を修飾して、ジチオピリジル基を導入した。95%v/v緩衝液A(50mM KPi、50mM NaCl、2mM EDTA、pH6.5)および5%v/v DMA中で、室温で2時間、反応を行った。NAPまたはSephadex G25カラム(緩衝液A中で平衡化)を通じて、わずかに膨らんだ(turgid)反応混合物をゲルろ過した。280nmで抗体の吸光度を、そして280および343nmで、DTTで放出される2−メルカプトピリジン(Spy)を測定することによって、修飾の度合いを決定した。次いで、SPyよりも1.7倍モル過剰のN2’−デアセチル−N2’−(3−メルカプト−1−オキソプロピル)−メイタンシン(L−DM1)を用いて、修飾muDS6を2.5mg Ab/mlでコンジュゲート化した。DMA(3%v/v)を含む緩衝液A(97%v/v)中で反応を行った。反応を室温で一晩、〜20時間、インキュベーションした。不透明な反応混合物を遠心分離し(1162xg、10分間)、そして次いで、緩衝液B(1xPBS pH6.5)中で平衡化したNAP−25またはS300(Tandem 3、3x26/10脱塩カラム、G25媒体)カラムを通じて上清をゲルろ過した。ペレットを廃棄した。0.22μm Millex−GVフィルターを用いて、コンジュゲートを滅菌ろ過し、そしてSlide−A−Lyzerを用いて、緩衝液B中で透析した。ろ過物質の252nmおよび280nm両方の吸光度を測定することによって、muDS6の分子あたりに連結されたDM1分子の数を決定した。DM1/Ab比は、4.36であることが見出され、そしてコンジュゲート化MUDS6の工程収率は55%であった。コンジュゲート化抗体濃度は1.32mg/mlであった。サイズ排除クロマトグラフィー(SEC)によって、精製コンジュゲートを生化学的に性質決定して、そして92%単量体であることを見出した。精製コンジュゲート中のDM1の分析によって、99%が抗体に共有結合していることが示された。図20において、muDS6−DM1コンジュゲートおよび非修飾muDS6のCaov−3細胞へのフローサイトメトリー結合によって、muDS6のコンジュゲート化が親和性のわずかな喪失しか生じないことが示される。
【0251】
実施例15:muDS6−DM1のin vitro細胞傷害性
[258]裸の抗体として、muDS6は、細胞培養において、増殖または成長阻害活性を示さなかった(図21)。しかし、ヤギ抗マウスIgG重鎖および軽鎖に対するDM1コンジュゲートの存在下で、muDS6を細胞とインキュベーションすると、muDS6は、このコンジュゲートを細胞にターゲティングし、そして送達するのに非常に有効であり、間接的な細胞傷害性を生じた(図21)。裸のmuDS6の生得的な活性をさらに試験するため、muDS6を用いた、補体依存性細胞傷害性(CDC)アッセイを行った。200μlのRHBP培地(RPMI−1640、0.1%BSA、20mM HEPES(pH7.2〜7.4)、100U/mlペニシリンおよび100μg/mlストレプトマイシン)中、5%ヒトまたはウサギ血清および多様な希釈のmuDS6の存在下で、HPACおよびZR−75−1細胞(25000細胞/ウェル)を96ウェルプレートにプレーティングした。細胞を37℃で2時間インキュベーションした。次いで、Ala
mar Blue(最終濃度10%)試薬(Biosource)を上清に添加した。蛍光を測定する前に、5〜24時間、細胞をインキュベーションした。ネズミDS6は、補体依存性細胞傷害性(CDC)アッセイにおいて、影響を持たなかった(図22)。これによって、muDS6の療法的適用が、毒性エフェクター分子のコンジュゲート化を必要とするであろうことが示唆される。
【0252】
[259]多様なDS6陽性細胞株において、2つの異なるアッセイ形式を用いて、メイタンシノイド・コンジュゲート化muDS6抗体の細胞傷害性を調べた。培地中で希釈した2mlのコンジュゲート中、6ウェルプレート上に細胞(1000〜2500細胞/ウェル)をプレーティングする、コロニー形成アッセイを行った。一般的に、3x10−11M〜3x10−9Mの間のいくつかの濃度で、細胞を連続してコンジュゲートに曝露し、そして37℃、6%CO2加湿チャンバー中で、5〜9日間、インキュベーションした。PBSでウェルを洗浄し、そして1%w/vクリスタルバイオレット/10%v/vホルムアルデヒド/PBS溶液で、コロニーを染色した。次いで、再蒸留水を用いて、未結合染色剤を徹底的にウェルから洗い落とし、そしてプレートを乾燥させた。Leica
StereoZoom4解剖顕微鏡を用いて、コロニーを計数した。
【0253】
[260]コロニーの数/プレーティングした細胞数として、プレーティング効率(PE)を計算した。処理細胞のPE/非処理細胞のPEとして、生存率を計算した。細胞の生存率対コンジュゲートのモル濃度をグラフで示すことによって、IC50濃度を決定した。コロニー形成アッセイにおいて(図23)、muDS6−DM1は、Caov−3細胞を殺すのに有効であり、概算されるIC50は800pMであった。抗原陰性細胞、A375は、試験したmuDS6−DM1の最高の濃度である、3x10−9Mの濃度のコンジュゲートによって、わずかにしか影響を受けず、コンジュゲートの細胞殺傷活性が、抗原発現細胞に特異的に向けられることを立証した。しかし、メイタンシンに対する見かけの感受性にもかかわらず、多くの他のDS6陽性細胞株は、免疫コンジュゲートに対して、特に感受性ではなかった。すべての子宮頸部細胞株(HeLa、KB、およびWISH)は、コンジュゲートに対して感受性であり、一方、卵巣および乳房細胞株の選ばれた数のみが、何らかの細胞傷害性効果を示した。膵臓細胞株は、いずれも、影響を受けないようであった。
【0254】
[261]MTTアッセイにおいて、1000〜5000細胞/ウェルの密度で、細胞を96ウェルプレートに植え付けた。200μlの培地中、裸のmuDS6またはmuDS6−DM1免疫コンジュゲートのいずれかの連続希釈とともに、細胞をプレーティングした。試料を3つ組で実験した。次いで、細胞および抗体/コンジュゲート混合物を2〜7日間インキュベーションし、この時点で、MTT([3(4,5−ジメチルチアゾール−2−イル)−2,5−ジフェニルテトラゾリウムブロミド])アッセイによって、細胞生存度を評価した。MTT(50μg/ウェル)を培養上清に添加して、そして37℃で3〜4時間インキュベーションさせた。培地を取り除き、そしてMTTホルマザンをDMSO(175μl/ウェル)中に可溶化した。540〜545nmの吸光度を測定した。MTT細胞生死判別アッセイにおいて(図24C)、免疫コンジュゲートは、Caov−3細胞を有効に殺すことが可能であり、概算されるIC50は1.61nMであった。まったく影響を及ぼさなかった裸の抗体に比較して、最高濃度のコンジュゲートを含むウェルは、生存細胞をまったく含有しなかった(図21および24)。
【0255】
[262]他の細胞株に対するMTTアッセイの結果は、わずかに異なった(図24A、B、およびD〜I)。多くの場合、ある程度の細胞傷害性が見られるが、コンジュゲートは、全細胞集団を完全に殺すことは不可能であった(WISH細胞を除く)。BT−20、OVCAR5、およびHPAC細胞が特に抵抗性であった:最高コンジュゲート濃度(32nM)のウェルで、50%を超える細胞がなお生存可能であった。
【0256】
実施例16:in vivoコンジュゲート抗腫瘍活性
[263]muDS6−DM1コンジュゲートのin vivo活性を立証するため、SCIDマウスにおいて、ヒト腫瘍異種移植片を確立した。ヒト子宮頸癌細胞株KBの皮下モデルを開発した。KB細胞をin vitroで増殖させ、収集し、そして各マウスの右肩下に、100μlの血清不含培地中、5x106細胞を注射し、そして平均腫瘍体積144±125mm3になるまで6日間増殖させ、この時点で薬剤処置を開始した。マウスに、PBS、150μg/kg DM1のコンジュゲート、または225μg/kg
DM1のコンジュゲート(群あたり2匹のマウス)のいずれかを静脈内に毎日5日間投与した。処置中、毒性応答を毎日監視した。研究の間中、腫瘍体積(図25A)および対応する体重(図25B)を監視した。
【0257】
[264]PBS対照で処置したKB腫瘍は、約4日間の倍増時間で、迅速に増殖した。対照的に、コンジュゲートで処置したマウスの両群は、225μg/kgおよび150μg/kg用量群で、それぞれ、処置開始の14日後および18日後に、完全腫瘍退行を示す。150μg/kg用量では、腫瘍遅延はおよそ70日であった。225μg/kgでの処置は、第120日に研究を終了した際、腫瘍再発の証拠がまったくなかったように、治癒を生じた。図25Bに見られるように、150μg/kg群のマウスは、体重喪失をまったく示さず、用量がよく許容されることを示した。より高い用量では、マウスは、体重の一時的な3%の減少しか経験しない。5日間の処置経過中、マウスは毒性の可視徴候を示さなかった。総合すると、この研究は、muDS6−DM1処置が、非毒性用量で、KB異種移植片腫瘍のマウスを治癒させうることを立証する。
【0258】
[265]一団の皮下異種移植片モデルに対して、muDS6−DM1活性をさらに試験した(図26を参照されたい)。異種移植片を作製するのに用いた腫瘍細胞株は、さまざまなin vitroメイタンシン感度およびCA6エピトープ密度を示した(以下の表9)。OVCAR5細胞およびTOV−21Gは、卵巣腫瘍細胞株であり;HPACは膵臓腫瘍細胞株であり;HeLaは子宮頸部腫瘍細胞株である。OVCAR5およびTOV−21G細胞は、低い表面CA6発現を有し;HeLa細胞は、中間レベルの表面CA6発現を有し;HPAC細胞は、高いCA6表面発現密度を有する。TOV−21GおよびHPAC細胞はメイタンシン感受性であり;OVCAR5およびHeLa細胞はメイタンシン感受性が2〜7倍低かった。
表9
【0259】
【表10】
【0260】
*平均最大相対的平均蛍光
【0261】
[266]4つの細胞株をin vitroで増殖させ、収集し、そして100μlの血清不含培地中の1x107細胞を、各マウスの右肩下(モデルあたり6匹のマウス)に注射し、そしてOVCAR5の試験群および対照群では、それぞれ57.6±6.7および90.2±13.4mm3、HPACの試験群および対照群では、それぞれ147.1±29.6および176.2±18.9mm3、HeLaの試験群および対照群では、それぞれ194.3±37.2および201.7±71.7mm3、そしてTOV−21Gの試験群および対照群では、それぞれ96.6±22.8および155.6±13.4mm3の平均腫瘍体積になるまで、6日間増殖させ、この時点で薬剤処置を開始した。各モデルに関して、3匹の対照マウスをPBSの2回の毎週用量で処置し、そして3匹の試験マウスを静脈内でのコンジュゲート(600μg/kg DM1)の2回の毎週用量で処置した。処置中、毒性応答を毎日監視し、そして研究の間中、腫瘍体積および体重を監視した。多様なモデルのコンジュゲート有効性を、図26A、C、E、およびGにグラフで示し、そして対応する体重を図26B、D、F、およびHにプロットする。OVCAR5、TOV−21G、およびHPAC細胞株は、各モデルに関するPBS対照で見られうるように、攻撃的な腫瘍を形成する。HeLaモデルは、指数関数的な増殖を開始するまでに、約3週間の遅延期間を有した。すべてのモデルにおいて、DS6−DM1コンジュゲート処置は、すべてのマウスにおいて、完全腫瘍退行を生じた。TOV−21G、HPAC、およびHeLaモデルに関して、マウスは、第61日に、腫瘍不含のままである。OVCAR5モデルにおいて、腫瘍接種後、約45日で、腫瘍が再発した。したがって、このモデルにおいて、muDS6−DM1処置は、およそ34日間の腫瘍増殖遅延を生じる。OVCAR5細胞がよりメイタンシン感受性でなく、そして低いCA6エピトープ発現を有するため、増殖遅延は有意である。CA6エピトープ密度がより高いか、またはモデルがより高いメイタンシン感受性を有するモデルにおいては、腫瘍退行はより強固である。2回の用量しか投与されないことに注目することが重要である。体重喪失が観察されないため、本研究で用いる投薬スケジュールは、マウスにとっては明らかに毒性でない。治癒は、さらなるまたはより高いコンジュゲート用量で達成可能である可能性がある。
【0262】
[267]ヒト卵巣癌は、大部分、腹膜の疾患である。OVCAR5細胞は、SCIDマウスにおいて、腹腔内(IP)モデルとして攻撃的に増殖して、ヒト疾患と類似の方式で、腫瘍節を形成し、そして腹水を産生する。IPモデルにおいて活性を立証するため、
muDS6−DM1を用いて、OVCAR5 IP腫瘍を所持するマウスを処置した(図27)。OVCAR5細胞をin vitroで増殖させ、採取し、そして100μlの血清不含培地中、1x107細胞を腹腔内注射した。腫瘍を6日間増殖させ、この時点で処置を開始した。600μg/kg DM1の用量で、PBSまたはDS6−DM1コンジュゲートのいずれかで、マウスを2週間、毎週処置し、そして腹腔疾患から生じる体重喪失に関して監視した。第28日までに、マウスのPBS群は、20%より多く体重を失い、そして安楽死させられた。20%を超える体重を喪失した後、第45日に、処置群を屠殺した。この研究によって、OVCAR5細胞がメイタンシンにより感受性でなく、そして細胞あたりより少ないCA6エピトープを有するという事実にもかかわらず、muDS6−DM1は、攻撃的OVCAR5 IPモデルにおいて、腫瘍増殖を遅延させることが可能であることが立証される。用いた投薬スケジュールは、毒性の可視徴候を誘発しなかったため、さらなる用量およびより高い用量を用いて、さらなる腫瘍増殖遅延または治癒を達成することも可能である可能性が高い。
【0263】
実施例17:DS6−SPP−MM1−202タキソイド細胞傷害性コンジュゲートの合成および性質決定
[268]N−スルホスクシンイミジル4−ニトロ−2−ピリジル−ペンタノエート(SSNPP)リンカーを用いて、muDS6を修飾した。90%緩衝液A、10%DMA中の50mgのmuDS6 Abに、DMA中の10当量のSSNPPを添加した。Abの最終濃度は、8mg/mlであった。反応を室温で4時間攪拌し、次いでG25クロマトグラフィーによって精製した。280nm(抗体)および325(リンカー)の吸光度を用いて、抗体修飾の度合いを分光光度的に測定し、そして3.82リンカー/抗体を有することが見出された。抗体の回収は、43.3mgであり、87%収率を生じた。muDS6−ニトロSPPを、タキソイドMM1−202(1812P.16)とコンジュゲート化した。90%緩衝液A、10%DM1中、42mgスケールで、コンジュゲート化を行った。約20時間の期間に渡って、0.43当量/リンカー(各アリコット)の4アリコットでタキソイドを添加した。この時点までに、反応は、著しく曇った。G25精製後、約64%の収率で回収された、生じたコンジュゲートは、約4.3タキソイド/Abを有し、そして約1当量の未反応リンカーが残った。未反応リンカーの反応を停止するため、1当量のシステイン/未反応リンカーを、一晩攪拌しながら、コンジュゲートに添加した。システインを添加した際、明確な黄色がかった色合いが顕著であり、これはチオピリジンの放出を示す。次いで、反応溶液を緩衝液B/0.01%Tween20中で透析し、その後、緩衝液Bのみでさらに数日に渡って透析した。最終コンジュゲートは、2.86薬剤/抗体を有した。抗体回収は14.7mgであり、全体で35%の収率を生じた。SECによってコンジュゲートをさらに生化学的に性質決定し、そして89%単量体、10.5%二量体および0.5%のより高い分子量の凝集体を有することが見出された。
【0264】
[269]HeLa細胞に対する、muDS6−SPP−MM1−202タキソイド対muDS6抗体の結合を比較するフローサイトメトリー分析の結果を、図28に示す。結果によって、muDS6がタキサンにコンジュゲート化された際、結合活性を保持することが示される。
【0265】
実施例18:ヒト化DS6コンジュゲートのin vitroおよびin vivo活性
[270]huDS6v1.01−SPDB−DM4コンジュゲートを構築した。このコンジュゲートは、コンジュゲートのリンカー/メイタンシン薬剤部分の構造がジスルフィド結合周囲で異なることを除いて、実施例14に記載するmuDS6−SPP−DM1コンジュゲートと類似であり;muDS6−SPP−DM1コンジュゲートは、リンカーの抗体側でジスルフィド炭素上に1つのメチル基立体障害を有し、一方、SPDB−DM4コンジュゲートは、リンカーのメイタンシン側でジスルフィド炭素上に2つのメチル基立体障害を有する。
【0266】
[271]8倍モル過剰のN−スクシンイミジル−4−(2−ピリジルジチオ)ブタノエート(SPDB)を用いて、huDS6v1.01抗体(8mg/ml)を修飾して、ジチオピリジル基を導入した。95%v/v緩衝液A(50mM KPi、50mM NaCl、2mM EDTA、pH6.5)および5%v/vエタノール中で、室温で1.5時間、反応を行った。15mlのSephadex G25カラム(緩衝液A中で平衡化)を通じて、反応混合物をゲルろ過した。280nmで抗体の吸光度を、そして280および343nmで、DTTで放出される2−メルカプトピリジン(Spy)を測定することによって、修飾の度合いを決定した。次いで、SPyよりも1.7倍モル過剰のN2’−デアセチル−N2’−(4−メチル−4−メルカプト−1−オキソペンチル)−メイタンシン(L−DM4)を用いて、修飾DS6を1.8mg Ab/mlでコンジュゲート化した。DMA(3%v/v)を含む緩衝液A(97%v/v)中で反応を行った。反応を室温で一晩、〜20時間、インキュベーションした。muDS6のコンジュゲート化と対照的に、反応混合物は透明であり、そして直ちに、クエン酸緩衝液(20mMクエン酸、135mM NaCl、pH5.5)中で平衡化されたNAP 15ml G25カラムを通じたゲルろ過を経た。0.22μm Millex−GVフィルターを用いて、コンジュゲートを滅菌ろ過した。ろ過物質の252nmおよび280nm両方の吸光度を測定することによって、DS6の分子あたりに連結したDM4分子の数を決定した。DM4/Ab比は、3.2であることが見出され、そしてコンジュゲート化DS6の工程収率は69%であった。コンジュゲート化抗体濃度は1.51mg/mlであった。サイズ排除クロマトグラフィー(SEC)によって、精製コンジュゲートを生化学的に性質決定して、そして92.5%単量体であることを見出した。精製コンジュゲート中のDM4の分析によって、>99%が抗体に共有結合していることが示された。
【0267】
[272]図29Aにおいて、huDS6v1.01−DM4コンジュゲートおよび非修飾DS6のKB細胞へのフローサイトメトリー結合によって、huDS6v1.01のコンジュゲート化が親和性の喪失を本質的に生じないことを示す。CaOv−3細胞ではなく、KB細胞を用いたことを除いて、図20に関して記載したのと本質的に同様に、結合を行った。本質的に図24Gに記載するように、huDS6v1.01のin vitro細胞傷害性を試験した。huDS6v1.01はWISH細胞を4.4x10−10MのIC50で殺し、一方、非コンジュゲート化huDS6v1.01は細胞傷害性活性をまったく示さなかった。
【0268】
[273]huDS6v1.01−DM4のin vivo活性を、HPAC膵臓モデルに対して試験した。HPAC細胞を第0日に接種し、そして免疫コンジュゲート処置を第13日に投与した。腫瘍体積が1000cm3を越えた時点で、PBS対照動物を安楽死させた。コンジュゲートを200μg/kgまたは600μg/kg DM4のいずれかの用量で投与し、これはそれぞれ、15mg/kgおよび45mg/kgの抗体濃度に対応した。研究の経過中、マウスの腫瘍体積(図30A)および体重(図30B)を監視した。huDS6v1.01−DM4は、200μg/kg DM4で強力な抗腫瘍活性を示し、すべてのマウスは、完全な腫瘍退行を達成した。HPAC異種移植片上では発現されない抗原を認識する対照B4−DM4コンジュゲートは、200μg/kgで、本質的に活性を持たなかった。マウスの体重喪失が欠如している(図30B)ことから、200μg/kgコンジュゲートでの処置は、最大許容用量未満であることが示される。この結果によって、DS6のヒト化型は、メイタンシノイド薬剤のターゲティング送達を仲介し、強力な抗腫瘍活性を生じることが可能であることが立証される。
【図面の簡単な説明】
【0269】
【図1】[33]図1は、DS6抗体が、選択した癌細胞株の表面に結合する能力を決定するために行った研究の結果を示す。DS6一次抗体およびFITCコンジュゲート化抗マウスIgG(H+L)二次抗体とインキュベーションした細胞株の蛍光を、フローサイトメトリーによって測定した。DS6抗体はCaov−3(図1A)およびT−47D(図1B)細胞に結合し、見かけのKdは、それぞれ、1.848nMおよび2.586nMであった。抗原陰性細胞株、SK−OV−3(図1C)およびColo205(図1D)は、抗原特異的結合をまったく示さなかった。
【図2】[34]図2は、エピトープ発現のドットブロット分析の結果を示す。Caov−3(図2Aおよび図2B)、SKMEL28(図2C)、およびColo205(図2D)細胞溶解物を、ニトロセルロース膜上に個々にスポッティングし、そして次いで、プロナーゼ、プロテイナーゼK、ノイラミニダーゼまたは過ヨウ素酸と、個々にインキュベーションした。次いで、膜をDS6抗体(図2A)、CM1抗体(図2B)、R24抗体(図2C)、またはC242抗体(図2D)で免疫ブロッティングした。
【図3】[35]図3は、DS6抗原発現のドットブロット分析の結果を示す。Caov−3細胞溶解物を、PVDF膜上に個々にスポッティングし、そして次いで、トリフルオロメタンスルホン酸(TFMSA)の存在下でインキュベーションした。次いで、膜を、CM1抗体(1および2)またはDS6(3および4)で免疫ブロッティングした。
【図4】[36]図4は、DS6抗原のグリコトープ分析の結果を示す。N−グリカナーゼ(「N−gly」)、O−グリカナーゼ(「O−gly」)、および/またはシアリダーゼ(「S」)であらかじめ処理したCaov−3溶解物を、ニトロセルロース上にスポッティングし、そして次いで、DS−6抗体またはCM1抗体(Muc−1 VNTR)で免疫ブロッティングした。
【図5−1】[37]図5は、DS6抗原のウェスタンブロット分析の結果を示す。細胞溶解物を、DS6抗体で免疫沈降(「IP」)し、そして免疫ブロッティングした。抗原は、抗原陽性Caov−3(図5Aおよび図5B)およびT47D(図5C)細胞で観察される>250kDaタンパク質バンドに対応する。抗原陰性SK−OV−3(図5D)およびColo205(図5E)細胞株は、このバンドを示さない。免疫沈降後、Caov−3細胞溶解物のプロテインGビーズを、(図5A)ノイラミニダーゼ(「N」)または(図5B)過ヨウ素酸(「PA」)とインキュベーションした。抗体(「α」)、IP前(「Lys」)およびIP後フロースルー(「FT」)溶解物対照を同じゲル上で泳動した。Caov−3免疫沈降物を、N−グリカナーゼ(「N−gly」)、O−グリカナーゼ(「O−gly」)、および/またはシアリダーゼ(「S」)ともまたインキュベーションし(図5Fを参照されたい)、この場合、ブロットを代わりに、ビオチン化DS6およびストレプトアビジン−HRPで探査した(probed)。
【図5−2】[37]図5は、DS6抗原のウェスタンブロット分析の結果を示す。細胞溶解物を、DS6抗体で免疫沈降(「IP」)し、そして免疫ブロッティングした。抗原は、抗原陽性Caov−3(図5Aおよび図5B)およびT47D(図5C)細胞で観察される>250kDaタンパク質バンドに対応する。抗原陰性SK−OV−3(図5D)およびColo205(図5E)細胞株は、このバンドを示さない。免疫沈降後、Caov−3細胞溶解物のプロテインGビーズを、(図5A)ノイラミニダーゼ(「N」)または(図5B)過ヨウ素酸(「PA」)とインキュベーションした。抗体(「α」)、IP前(「Lys」)およびIP後フロースルー(「FT」)溶解物対照を同じゲル上で泳動した。Caov−3免疫沈降物を、N−グリカナーゼ(「N−gly」)、O−グリカナーゼ(「O−gly」)、および/またはシアリダーゼ(「S」)ともまたインキュベーションし(図5Fを参照されたい)、この場合、ブロットを代わりに、ビオチン化DS6およびストレプトアビジン−HRPで探査した(probed)。
【図6】[38]図6は、Caov−3(図6A)およびHeLa(図6B)細胞溶解物に対する、DS6抗体およびCM1抗体の免疫沈降および/または免疫ブロットの結果を示す。CM1およびDS6ウェスタンブロットシグナルの重複は、DS6抗原がMuc1タンパク質上にあることを示す。HeLa溶解物において、Muc1ダブレットは、タンデム反復数が異なる別個のアレルによって指示されるMuc1発現から生じる。
【図7】[39]図7は、DS6抗体サンドイッチELISA設計(図7A)および標準曲線(図7B)を示す。商業的に入手可能なCA15−3標準の既知の濃度を用いて、標準曲線を生じた(1 CA15−3単位=1 DS6単位)。
【図8】[40]図8は、定量的ELISA標準曲線を示す。プレーティングされたヤギ抗マウスIgGによって捕捉される(図8A)か、またはELISAプレート上に直接結合する(図8B)か、いずれかの、既知の濃度のビオチン−DS6を用いて、検出抗体(ストレプトアビジン−HRP/ビオチン−DS6)シグナルの標準曲線(図8C)を決定した。
【図9】[41]図9は、ネズミDS6抗体の軽鎖(図9A)および重鎖(図9B)可変領域のcDNAおよびアミノ酸配列を示す。各配列中の3つのCDRを下線で示す(Kabat定義)。
【図10】[42]図10は、Kabat定義によって決定されるネズミDS6抗体の軽鎖(図10A)および重鎖(図10B)CDRを示す。AbMモデリングソフトウェアは、重鎖CDRのわずかに異なる定義を生じる(図10C)。
【図11】[43]図11は、IgVκap4(配列番号23)およびIgVh J588.41(配列番号24)遺伝子の生殖系列配列と並列させた、ネズミDS6抗体の軽鎖(「muDS6LC」)(配列番号7の残基1〜95)および重鎖(「muDS6HC」)(配列番号9の残基1〜98)アミノ酸配列を示す。灰色は配列相違を示す。
【図12】[44]図12は、Brookhavenデータベースにおいて、解明された構造ファイルを有する、ネズミDS6(muDS6)軽鎖(「muDS6LC」)および重鎖(「muDS6HC」)配列に最も相同な10の軽鎖および重鎖抗体配列を示す。最も相同であるものから最も相同でないものの順に、配列を並列させる。
【図13−1】[45]図13は、ネズミDS6抗体軽鎖可変領域のどのフレームワーク残基が表面にアクセス可能であるかを予測する、表面アクセス可能性データおよび計算を示す。25〜35%の平均表面アクセス可能性を持つ位に印を付け(*??*)、そしてこれを第二周期の分析に供した。DS6抗体軽鎖可変領域(図13A)および重鎖可変領域(図13B)。〓
【図13−2】[45]図13は、ネズミDS6抗体軽鎖可変領域のどのフレームワーク残基が表面にアクセス可能であるかを予測する、表面アクセス可能性データおよび計算を示す。25〜35%の平均表面アクセス可能性を持つ位に印を付け(*??*)、そしてこれを第二周期の分析に供した。DS6抗体軽鎖可変領域(図13A)および重鎖可変領域(図13B)。
【図14】[46]図14は、prDS6 v1−0哺乳動物発現プラスミドマップを示す。このプラスミドを用いて、組換えキメラおよびヒト化DS6抗体を構築し、そして発現させた。
【図15】[47]図15は、ネズミ(「muDS6」)およびヒト化(「huDS6」)(1.01および1.21)DS6抗体軽鎖(図15A)および重鎖(図15B)可変ドメインのアミノ酸配列を示す。
【図16】[48]図16は、ヒト化DS6抗体(「huDS6」)(1.01および1.21)の軽鎖可変領域のcDNAおよびアミノ酸配列を示す。
【図17】[49]図17は、ヒト化DS6抗体(「huDS6」)1.01(図17A)および1.21(図17B)の重鎖可変領域のcDNAおよびアミノ酸配列を示す。
【図18】[50]図18は、KB細胞に対して行ったアッセイからの、ネズミDS6(muDS6)、キメラDS6(chDS6)、およびヒトDS6 1.01型(huDS6 v1.01)およびhuDS6 1.21型(huDS6 v1.21)のフローサイトメトリー結合曲線を示す。ネズミ、キメラ、およびヒトv1.01およびv1.21 DS6抗体のアビディティー(muDS6=0.82nM、chDS6=0.69nM、huDS6v1.01=0.82nMおよびhuDS6v1.21=0.85nM)は同程度であり、表面再構成がアビディティーを減少させなかったことを示す。
【図19】[51]図19は、muDS6、chDS6、huDS6 v1.01およびhuDS6 v1.21抗体とビオチン化muDS6の競合結合アッセイの結果を示す。多様な濃度の裸のmuDS6、chDS6、huDS6v1.01およびhuDS6v1.21を、2nMのビオチン−muDS6およびストレプトアビジン−DTAF二次抗体と合わせた。すべての抗体のIC50(muDS6=1.9nM、chDS6=1.7nM、huDS6v1.01=3.0nM、およびhuDS6v1.21=1.9nM)は類似であり、これは、ヒト化がアビディティーを減少させなかったことを示す。
【図20】[52]図20は、DS6抗体−DM1コンジュゲートに対する非コンジュゲート化DS6抗体の結合親和性決定の結果を示す。この結果は、DM1コンジュゲート化が、抗体の結合親和性に不都合に影響を及ぼさないことを示した。DS6抗体−DM1コンジュゲートの見かけのKd(3.902nM)(「DS6−DM1」)は、裸の抗体(2.020nM)(「DS6」)よりわずかに高かった。
【図21】[53]図21は、抗マウスIgG(H+L)DM1コンジュゲート(2°Ab−DM1)の存在下または非存在下で、DS6抗体を用いた、間接的細胞生死判別アッセイの結果を示す。抗原陽性Caov−3細胞は、二次コンジュゲートの存在下(「DS6+2°Ab−DM1」)でのみ、DS6抗体依存性方式で殺された(IC50=424.9pM)。
【図22】[54]図22は、muDS6抗体の補体依存性細胞傷害性(CDC)アッセイの結果を示す。結果は、HPAC(図22A)およびZR−75−1(図22B)細胞に対して、CDCが仲介するDS6抗体の効果がないことを示す。
【図23−1】[55]図23は、未結合(free)のメイタンシンに対する、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。コロニー形成アッセイにおいて、DS6抗原陽性卵巣癌(図23A)、乳癌(図23B)、子宮頸癌(図23C)、および膵臓癌(図23D)細胞株を、DS6抗体−DM1コンジュゲートに対する連続曝露の細胞傷害性に関して試験した(左のパネル)。未結合メイタンシンへの72時間の曝露によって、メイタンシン感受性に関して、これらの細胞株を同様に試験した(右のパネル)。試験した卵巣癌細胞株は、OVCAR5、TOV−21G、Caov−4およびCaov−3であった。試験した乳癌細胞株は、T47D、BT−20およびBT−483であった。試験した子宮頸癌細胞株は、KB、HeLaおよびWISHであった。試験した膵臓癌細胞株は、HPAC、Hs766TおよびHPAF−IIであった。
【図23−2】[55]図23は、未結合(free)のメイタンシンに対する、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。コロニー形成アッセイにおいて、DS6抗原陽性卵巣癌(図23A)、乳癌(図23B)、子宮頸癌(図23C)、および膵臓癌(図23D)細胞株を、DS6抗体−DM1コンジュゲートに対する連続曝露の細胞傷害性に関して試験した(左のパネル)。未結合メイタンシンへの72時間の曝露によって、メイタンシン感受性に関して、これらの細胞株を同様に試験した(右のパネル)。試験した卵巣癌細胞株は、OVCAR5、TOV−21G、Caov−4およびCaov−3であった。試験した乳癌細胞株は、T47D、BT−20およびBT−483であった。試験した子宮頸癌細胞株は、KB、HeLaおよびWISHであった。試験した膵臓癌細胞株は、HPAC、Hs766TおよびHPAF−IIであった。
【図23−3】[55]図23は、未結合(free)のメイタンシンに対する、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。コロニー形成アッセイにおいて、DS6抗原陽性卵巣癌(図23A)、乳癌(図23B)、子宮頸癌(図23C)、および膵臓癌(図23D)細胞株を、DS6抗体−DM1コンジュゲートに対する連続曝露の細胞傷害性に関して試験した(左のパネル)。未結合メイタンシンへの72時間の曝露によって、メイタンシン感受性に関して、これらの細胞株を同様に試験した(右のパネル)。試験した卵巣癌細胞株は、OVCAR5、TOV−21G、Caov−4およびCaov−3であった。試験した乳癌細胞株は、T47D、BT−20およびBT−483であった。試験した子宮頸癌細胞株は、KB、HeLaおよびWISHであった。試験した膵臓癌細胞株は、HPAC、Hs766TおよびHPAF−IIであった。
【図23−4】[55]図23は、未結合(free)のメイタンシンに対する、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。コロニー形成アッセイにおいて、DS6抗原陽性卵巣癌(図23A)、乳癌(図23B)、子宮頸癌(図23C)、および膵臓癌(図23D)細胞株を、DS6抗体−DM1コンジュゲートに対する連続曝露の細胞傷害性に関して試験した(左のパネル)。未結合メイタンシンへの72時間の曝露によって、メイタンシン感受性に関して、これらの細胞株を同様に試験した(右のパネル)。試験した卵巣癌細胞株は、OVCAR5、TOV−21G、Caov−4およびCaov−3であった。試験した乳癌細胞株は、T47D、BT−20およびBT−483であった。試験した子宮頸癌細胞株は、KB、HeLaおよびWISHであった。試験した膵臓癌細胞株は、HPAC、Hs766TおよびHPAF−IIであった。
【図24】[56]図24は、DS6抗体−DM1コンジュゲートのin vitro細胞傷害性アッセイの結果を示す。MTT細胞生死判別アッセイにおいて、ヒト卵巣癌(図24A、図24Bおよび図24C)、乳癌(図24Dおよび図24E)、子宮頸癌(図24Fおよび図24G)、および膵臓癌(図24Hおよび図24I)細胞は、DS6抗体−DM1コンジュゲート依存性方式で、殺された。裸のDS6は、これらの細胞の増殖に不都合に影響を及ぼさず、これは、細胞傷害性には、DM1コンジュゲート化が必要であることを示した。
【図25】[57]図25Aは、確立された皮下KB腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートのin vivo抗腫瘍有効性研究の結果を示す。腫瘍細胞を、第0日に接種し、そして最初の処置を第6日に与えた。免疫コンジュゲート処置を、総数5用量で毎日続けた。腫瘍体積が1500mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを150または225μg/kg DM1の用量で投与し、これはそれぞれ、5.7および8.5mg/kgの抗体濃度に対応した。研究経過中、マウスの体重(図25B)を監視した。
【図26−1】[58]図26は、確立された皮下腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートの抗腫瘍有効性研究の結果を示す。第0日に、OVCAR5(図26Aおよび図26B)、TOV−21G(図26Cおよび図26D)、HPAC(図26Eおよび図26F)、およびHeLa(図26Gおよび図26H)細胞を接種し、そして免疫コンジュゲート処置を、第6日および第13日に与えた。腫瘍体積が1000mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを600μg/kg DM1の用量で投与し、これは、27.7mg/kgの抗体濃度に対応した。研究経過中、マウスの腫瘍体積(図26A、図26C、図26E、および図26G)および体重(図26B、図26D、図26F、および図26H)を監視した。
【図26−2】[58]図26は、確立された皮下腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートの抗腫瘍有効性研究の結果を示す。第0日に、OVCAR5(図26Aおよび図26B)、TOV−21G(図26Cおよび図26D)、HPAC(図26Eおよび図26F)、およびHeLa(図26Gおよび図26H)細胞を接種し、そして免疫コンジュゲート処置を、第6日および第13日に与えた。腫瘍体積が1000mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを600μg/kg DM1の用量で投与し、これは、27.7mg/kgの抗体濃度に対応した。研究経過中、マウスの腫瘍体積(図26A、図26C、図26E、および図26G)および体重(図26B、図26D、図26F、および図26H)を監視した。
【図26−3】[58]図26は、確立された皮下腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートの抗腫瘍有効性研究の結果を示す。第0日に、OVCAR5(図26Aおよび図26B)、TOV−21G(図26Cおよび図26D)、HPAC(図26Eおよび図26F)、およびHeLa(図26Gおよび図26H)細胞を接種し、そして免疫コンジュゲート処置を、第6日および第13日に与えた。腫瘍体積が1000mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを600μg/kg DM1の用量で投与し、これは、27.7mg/kgの抗体濃度に対応した。研究経過中、マウスの腫瘍体積(図26A、図26C、図26E、および図26G)および体重(図26B、図26D、図26F、および図26H)を監視した。
【図26−4】[58]図26は、確立された皮下腫瘍異種移植片に対する、DS6抗体−DM1コンジュゲートの抗腫瘍有効性研究の結果を示す。第0日に、OVCAR5(図26Aおよび図26B)、TOV−21G(図26Cおよび図26D)、HPAC(図26Eおよび図26F)、およびHeLa(図26Gおよび図26H)細胞を接種し、そして免疫コンジュゲート処置を、第6日および第13日に与えた。腫瘍体積が1000mm3を超えた時点で、PBS対照動物を安楽死させた。コンジュゲートを600μg/kg DM1の用量で投与し、これは、27.7mg/kgの抗体濃度に対応した。研究経過中、マウスの腫瘍体積(図26A、図26C、図26E、および図26G)および体重(図26B、図26D、図26F、および図26H)を監視した。
【図27】[59]図27は、腹腔内OVCAR5腫瘍に対する、muDS6抗体−DM1コンジュゲートのin vivo有効性研究の結果を示す。第0日に、腫瘍細胞を腹腔内注射し、そして第6日および第13日に、免疫コンジュゲート処置を与えた。体重喪失が20%を超えた時点で、動物を安楽死させた。
【図28】[60]図28は、HeLa細胞に対する、裸のおよびタキサン−コンジュゲート化DS6抗体の結合親和性の研究由来のフローサイトメトリー結合曲線を示す。タキサン(MM1−202)−コンジュゲート化は、抗体の結合親和性に不都合に影響を及ぼさない。DS6−MM1−202コンジュゲートの見かけのKd(1.24nM)は、裸のDS6抗体(620pM)よりもわずかに大きかった。
【図29】[61]図29は、ヒト化DS6 1.01型抗体コンジュゲートのin vitro結合および強度を示す。DM4とhuDS6v1.01のコンジュゲート化は、KB細胞に対するhuDS6v1.01のアビディティーにほとんど影響を及ぼさない(図29A)。huDS6v1.01−DM4は、DS6を発現しているWISH細胞に向かって、強力なin vitro細胞傷害性を示し、IC50は0.44nMであった(図29B)。
【図30】[62]図30は、HPAC膵臓癌モデルにおける、huDS6v1.01−DM4コンジュゲートを用いたin vivo有効性研究の結果を示す。huDS6v1.01−DM4は、強力な抗腫瘍活性を示し、一方、HPACモデル中にターゲットが発現されていないB4−DM4対照コンジュゲートは、本質的にまったく活性を持たなかった(図30A)。200μg/kgの投与用量は、体重喪失の欠如によって示されるように、動物に対して毒性ではなかった(図30B)。
【特許請求の範囲】
【請求項1】
少なくとも1つの重鎖可変領域またはその断片および少なくとも1つの軽鎖可変領域またはその断片を含む、抗体またはそのエピトープ結合性断片であって、前記重鎖可変領域またはその断片が、配列番号9、配列番号10、および配列番号11からなる群より選択されるアミノ酸配列:
【化1】
に対し、少なくとも90%の配列同一性を有する、前記抗体またはそのエピトープ結合性断片。
【請求項2】
前記重鎖可変領域またはその断片が、配列番号9、配列番号10、または配列番号11のアミノ酸配列に対し、少なくとも95%の配列同一性を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項3】
前記重鎖可変領域またはその断片が、配列番号9、配列番号10、または配列番号11のアミノ酸配列を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項4】
前記軽鎖可変領域またはその断片が、配列番号7または配列番号8によって示されるアミノ酸配列:
【化2】
に対し、少なくとも90%の配列同一性を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項5】
前記軽鎖可変領域またはその断片が、配列番号7または配列番号8のアミノ酸配列に対
し、少なくとも95%の配列同一性を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項6】
前記軽鎖可変領域またはその断片が、配列番号7または配列番号8のアミノ酸配列を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項7】
前記重鎖可変領域またはその断片が、配列番号9、配列番号10、または配列番号11のアミノ酸配列を有し、そして前記軽鎖可変領域またはその断片が、配列番号7または配列番号8のアミノ酸配列を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項8】
請求項1記載の抗体またはそのエピトープ結合性断片をコードするポリヌクレオチド。
【請求項9】
請求項4記載の抗体またはそのエピトープ結合性断片をコードするポリヌクレオチド。
【請求項10】
請求項7記載の抗体またはそのエピトープ結合性断片をコードするポリヌクレオチド。
【請求項11】
請求項1記載の抗体またはそのエピトープ結合性断片の重鎖をコードするポリヌクレオチド。
【請求項12】
請求項8のポリヌクレオチドを含む発現ベクター。
【請求項13】
請求項9のポリヌクレオチドを含む発現ベクター。
【請求項14】
請求項10のポリヌクレオチドを含む発現ベクター。
【請求項15】
請求項12の発現ベクターを含む宿主細胞。
【請求項16】
請求項13の発現ベクターを含む宿主細胞。
【請求項17】
請求項14の発現ベクターを含む宿主細胞。
【請求項18】
抗体またはそのエピトープ結合性断片を調製する方法であって、請求項15の宿主細胞を、前記抗体またはそのエピトープ結合性断片の発現を促進する条件下で培養し、そして該細胞培養から前記ポリペプチドを回収する
ここで、前記抗体またはそのエピトープ結合性断片が、少なくとも1つの重鎖可変領域またはその断片および少なくとも1つの軽鎖可変領域またはその断片を含み、前記重鎖可変領域またはその断片が、配列番号9、配列番号10、および配列番号11からなる群より選択されるアミノ酸配列:
【化3】
に対し、少なくとも90%の配列同一性を有する
工程を含む、前記方法。
【請求項19】
抗体またはそのエピトープ結合性断片を調製する方法であって、請求項16の宿主細胞を、前記抗体またはそのエピトープ結合性断片の発現を促進する条件下で培養し、そして該細胞培養から前記ポリペプチドを回収する
ここで、前記重鎖可変領域またはその断片が、配列番号9、配列番号10、および配列番号11からなる群より選択されるアミノ酸配列:
【化4】
に対し、少なくとも90%の配列同一性を有し、そして
前記軽鎖可変領域またはその断片が、配列番号7または配列番号8によって示されるアミノ酸配列:
【化5】
に対し、少なくとも90%の配列同一性を有する
工程を含む、前記方法。
【請求項20】
抗体またはそのエピトープ結合性断片を調製する方法であって、請求項17の宿主細胞を、前記抗体またはそのエピトープ結合性断片の発現を促進する条件下で培養し、そして該細胞培養から前記ポリペプチドを回収する
ここで、前記重鎖可変領域またはその断片が、配列番号9、配列番号10、または配列番号11のアミノ酸配列を有し、そして前記軽鎖可変領域またはその断片が、配列番号7または配列番号8のアミノ酸配列を有する
工程を含む、前記方法。
【請求項1】
少なくとも1つの重鎖可変領域またはその断片および少なくとも1つの軽鎖可変領域またはその断片を含む、抗体またはそのエピトープ結合性断片であって、前記重鎖可変領域またはその断片が、配列番号9、配列番号10、および配列番号11からなる群より選択されるアミノ酸配列:
【化1】
に対し、少なくとも90%の配列同一性を有する、前記抗体またはそのエピトープ結合性断片。
【請求項2】
前記重鎖可変領域またはその断片が、配列番号9、配列番号10、または配列番号11のアミノ酸配列に対し、少なくとも95%の配列同一性を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項3】
前記重鎖可変領域またはその断片が、配列番号9、配列番号10、または配列番号11のアミノ酸配列を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項4】
前記軽鎖可変領域またはその断片が、配列番号7または配列番号8によって示されるアミノ酸配列:
【化2】
に対し、少なくとも90%の配列同一性を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項5】
前記軽鎖可変領域またはその断片が、配列番号7または配列番号8のアミノ酸配列に対
し、少なくとも95%の配列同一性を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項6】
前記軽鎖可変領域またはその断片が、配列番号7または配列番号8のアミノ酸配列を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項7】
前記重鎖可変領域またはその断片が、配列番号9、配列番号10、または配列番号11のアミノ酸配列を有し、そして前記軽鎖可変領域またはその断片が、配列番号7または配列番号8のアミノ酸配列を有する、請求項1の抗体またはそのエピトープ結合性断片。
【請求項8】
請求項1記載の抗体またはそのエピトープ結合性断片をコードするポリヌクレオチド。
【請求項9】
請求項4記載の抗体またはそのエピトープ結合性断片をコードするポリヌクレオチド。
【請求項10】
請求項7記載の抗体またはそのエピトープ結合性断片をコードするポリヌクレオチド。
【請求項11】
請求項1記載の抗体またはそのエピトープ結合性断片の重鎖をコードするポリヌクレオチド。
【請求項12】
請求項8のポリヌクレオチドを含む発現ベクター。
【請求項13】
請求項9のポリヌクレオチドを含む発現ベクター。
【請求項14】
請求項10のポリヌクレオチドを含む発現ベクター。
【請求項15】
請求項12の発現ベクターを含む宿主細胞。
【請求項16】
請求項13の発現ベクターを含む宿主細胞。
【請求項17】
請求項14の発現ベクターを含む宿主細胞。
【請求項18】
抗体またはそのエピトープ結合性断片を調製する方法であって、請求項15の宿主細胞を、前記抗体またはそのエピトープ結合性断片の発現を促進する条件下で培養し、そして該細胞培養から前記ポリペプチドを回収する
ここで、前記抗体またはそのエピトープ結合性断片が、少なくとも1つの重鎖可変領域またはその断片および少なくとも1つの軽鎖可変領域またはその断片を含み、前記重鎖可変領域またはその断片が、配列番号9、配列番号10、および配列番号11からなる群より選択されるアミノ酸配列:
【化3】
に対し、少なくとも90%の配列同一性を有する
工程を含む、前記方法。
【請求項19】
抗体またはそのエピトープ結合性断片を調製する方法であって、請求項16の宿主細胞を、前記抗体またはそのエピトープ結合性断片の発現を促進する条件下で培養し、そして該細胞培養から前記ポリペプチドを回収する
ここで、前記重鎖可変領域またはその断片が、配列番号9、配列番号10、および配列番号11からなる群より選択されるアミノ酸配列:
【化4】
に対し、少なくとも90%の配列同一性を有し、そして
前記軽鎖可変領域またはその断片が、配列番号7または配列番号8によって示されるアミノ酸配列:
【化5】
に対し、少なくとも90%の配列同一性を有する
工程を含む、前記方法。
【請求項20】
抗体またはそのエピトープ結合性断片を調製する方法であって、請求項17の宿主細胞を、前記抗体またはそのエピトープ結合性断片の発現を促進する条件下で培養し、そして該細胞培養から前記ポリペプチドを回収する
ここで、前記重鎖可変領域またはその断片が、配列番号9、配列番号10、または配列番号11のアミノ酸配列を有し、そして前記軽鎖可変領域またはその断片が、配列番号7または配列番号8のアミノ酸配列を有する
工程を含む、前記方法。
【図1】

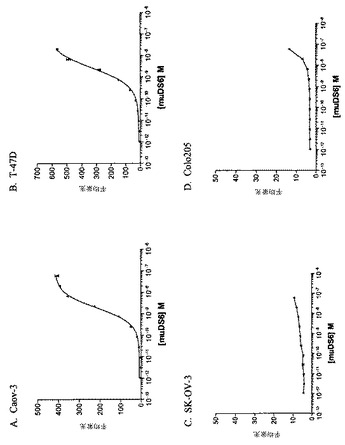
【図2】
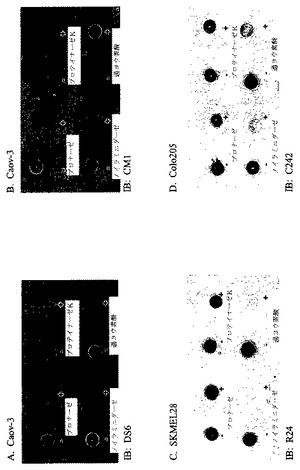
【図4】

【図5−1】
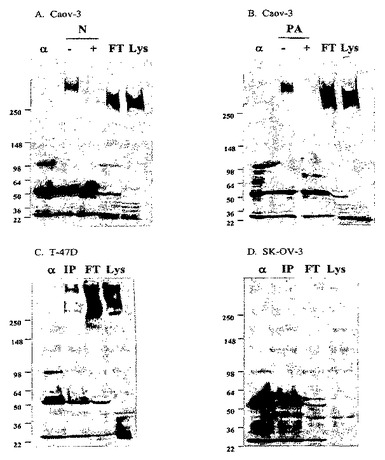
【図5−2】
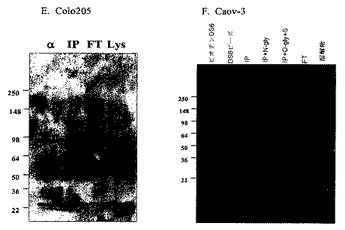
【図6】
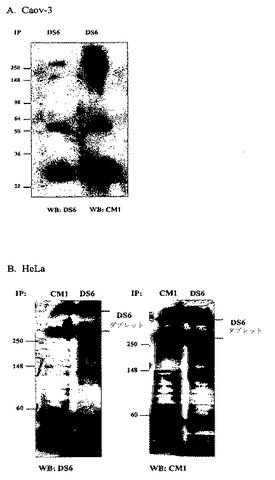
【図7】
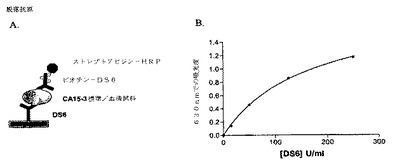
【図8】
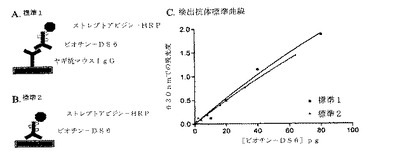
【図9】

【図10】

【図11】

【図12】

【図13−1】
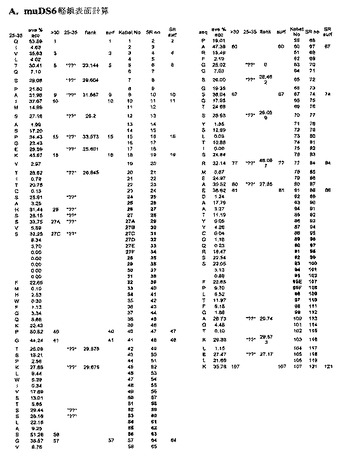
【図13−2】
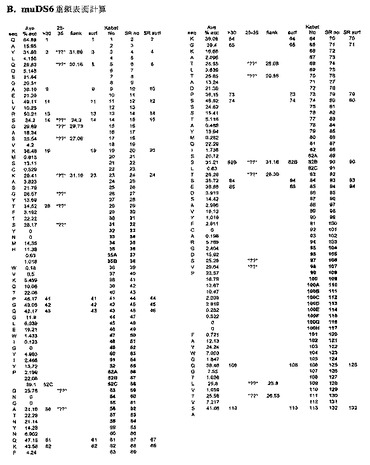
【図14】
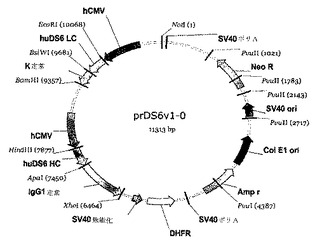
【図15】

【図16】

【図17】

【図18】
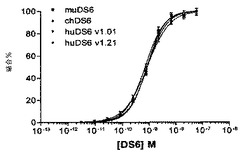
【図19】
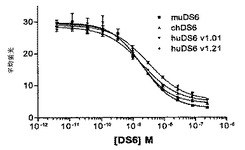
【図20】
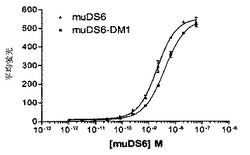
【図21】
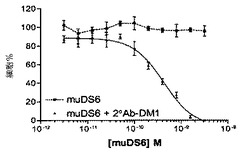
【図22】
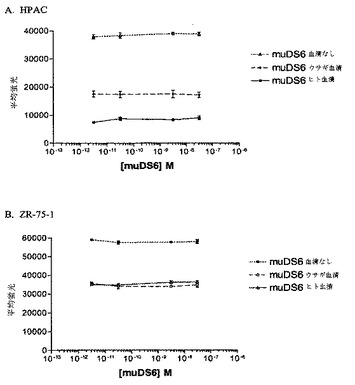
【図23−1】
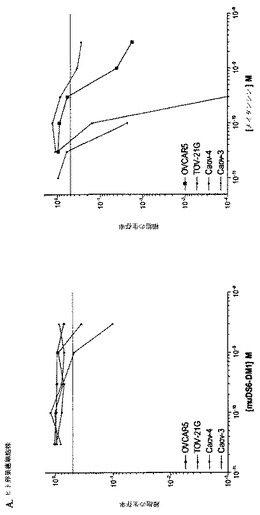
【図23−2】
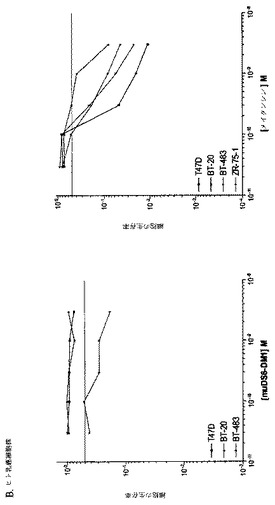
【図23−3】
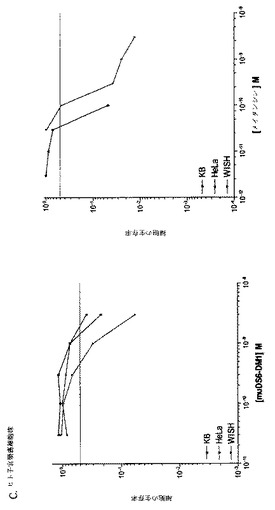
【図23−4】
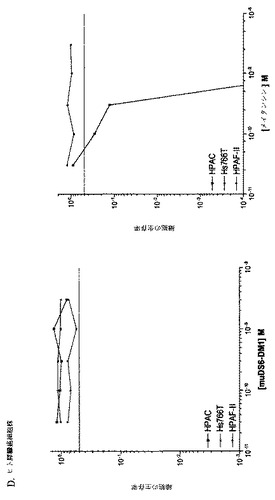
【図24】
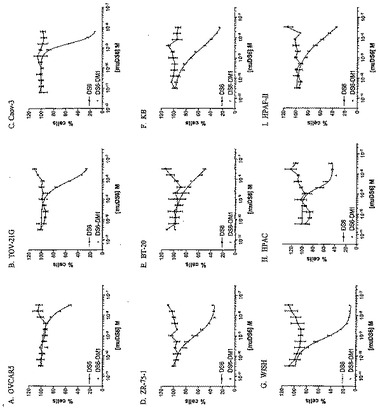
【図25】
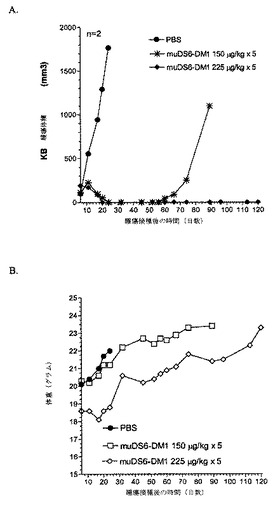
【図26−1】
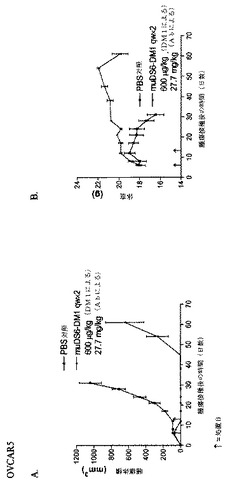
【図26−2】
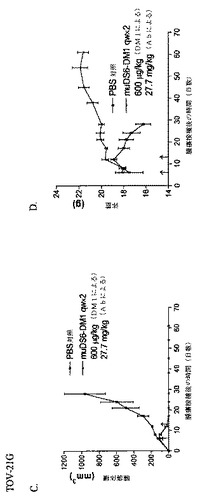
【図26−3】
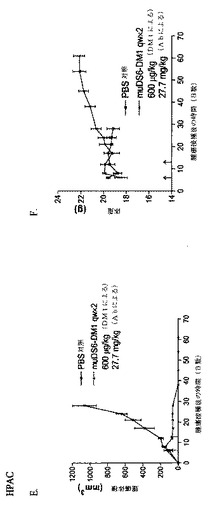
【図26−4】
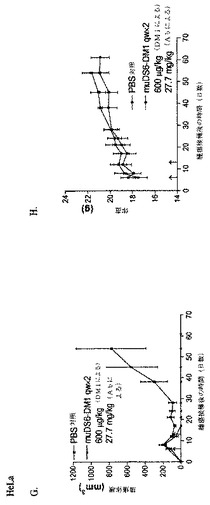
【図27】
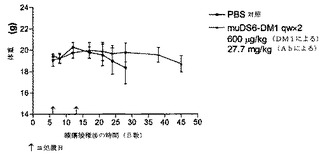
【図28】
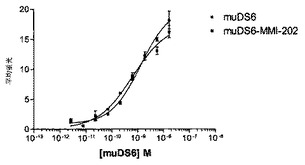
【図29】
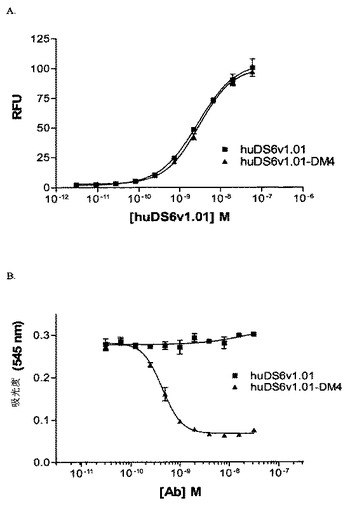
【図30】
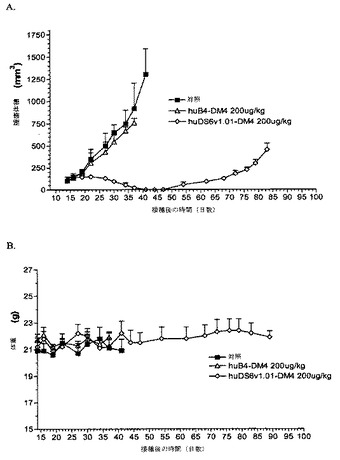
【図3】
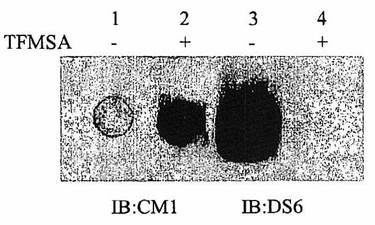
【図2】
【図4】
【図5−1】
【図5−2】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13−1】
【図13−2】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23−1】
【図23−2】
【図23−3】
【図23−4】
【図24】
【図25】
【図26−1】
【図26−2】
【図26−3】
【図26−4】
【図27】
【図28】
【図29】
【図30】
【図3】
【公開番号】特開2012−161321(P2012−161321A)
【公開日】平成24年8月30日(2012.8.30)
【国際特許分類】
【出願番号】特願2012−80686(P2012−80686)
【出願日】平成24年3月30日(2012.3.30)
【分割の表示】特願2008−527882(P2008−527882)の分割
【原出願日】平成17年8月22日(2005.8.22)
【出願人】(504039155)イミュノジェン・インコーポレーテッド (36)
【Fターム(参考)】
【公開日】平成24年8月30日(2012.8.30)
【国際特許分類】
【出願日】平成24年3月30日(2012.3.30)
【分割の表示】特願2008−527882(P2008−527882)の分割
【原出願日】平成17年8月22日(2005.8.22)
【出願人】(504039155)イミュノジェン・インコーポレーテッド (36)
【Fターム(参考)】
[ Back to top ]