
CHLAMYDIATRACHOMATISに対する免疫化
【課題】Chlamydia trachomatisの公開されたゲノム配列からの、免疫化およびワクチン接種のための有用な抗原の提供。
【解決手段】ワクチン作製および開発のため、および/または診断目的のための、特定なC.trachomatisタンパク質配列。当該タンパク質のアミノ酸配列に対して50%以上の配列同一性を有するアミノ酸配列を含むタンパク質、または、特定なC.trachomatisタンパク質のフラグメントを含むタンパク質である、免疫原性組成物。
【解決手段】ワクチン作製および開発のため、および/または診断目的のための、特定なC.trachomatisタンパク質配列。当該タンパク質のアミノ酸配列に対して50%以上の配列同一性を有するアミノ酸配列を含むタンパク質、または、特定なC.trachomatisタンパク質のフラグメントを含むタンパク質である、免疫原性組成物。
【発明の詳細な説明】
【技術分野】
【0001】
本明細書中で引用される全ての文献は、その全体が参考として援用される。
【0002】
(技術分野)
本発明は、クラミジア感染、特に、Chlamydia trachmatisによる感染に対する免疫化の分野に存在する。
【背景技術】
【0003】
(背景技術)
Chlamydiaeは、地域性の性感染症および種々の他の疾患症候群に寄与する真核生物細胞の偏性細胞内寄生生物である。これらは、排他的な真正細菌の分科を占め、他のいずれの既知の生物に対しても密接な関係を有さず、これらは、単一のファミリー(Chlamydiaceae)を含むこれら独自の目(Chlamydiales)に分類され、次いで単一の属(Chlamydia、Chlamydophilaともいわれる)を含む。Chlamydiaeは、これらの固有の生活環が特に特徴的であり、この生活環において、細菌は、2つの形態学的に別々の形態:細胞外感染性形態(基本小体、EB)と細胞内非感染性形態(網状体(reticulate bodies)、RB)との間で交替する。この生活環は、RBのEBへの再編成で完了し、EBは、引き続き、分裂した宿主細胞がさらなる細胞を感染する準備をさせておく。
【0004】
4つのクラミジア種(C.trachomatis、C.pneumoniae、C.pecorumおよびC.psittaci)が現在知られており[例えば、参考文献1,2](非特許文献1、2)、ゲノム配列は、公表されている[参考文献3〜9](非特許文献3〜6、特許文献1〜3)。
【0005】
C.trachomatisのヒトの血清改変体(「血清型亜型」)は、2つのバイオ改変体(「biovar」)に分けられる。血清型亜型A〜Kは、主に眼の組織(A〜C)または泌尿生殖器管(D〜K)において上皮性感染を誘発する。血清型亜型L1、L2およびL3は、浸潤性の性病性リンパ肉芽腫(LGV)の因子である。
【0006】
クラミジア感染自体が疾患を生じるが、幾人かの患者においては、症候群の重篤度は、実際には、異常な宿主免疫応答に起因していると考えられる。感染を排除することの失敗は持続的な免疫刺激を生じ、そして宿主を助けることよりも、これは、重篤な結果(不妊および失明を含む)を伴う慢性感染を生じる[10](非特許文献7)。さらに、天然のクラミジア感染により与えられる防御は、通常、不完全で、一過性で、そして株特異的である。
【0007】
疾患の深刻な性質に起因して、適切なワクチンを提供することが所望される。これらは、(a)クラミジア感染に対する、またはクラミジア誘導性疾患に対する免疫のために(予防ワクチン接種)、あるいは(b)確立された慢性クラミジア感染の根絶のために(治療ワクチン接種)有用であり得る。しかし、細胞内寄生生物であることから、この細菌は、一般に抗体媒介免疫応答を逃れ得る。
【0008】
種々の抗原性タンパク質が、C.trachomatisについて記載されており、そして特に細胞表面が、詳細な検索の標的となっている[例えば、1,11](非特許文献1、8)。これらとしては、例えば、pgp3[12、13、14](非特許文献9、特許文献4、5)、MOMP[15](非特許文献10)、Hsp60(GroEL)[16](非特許文献11)およびHsp70(DnaK様)[17](非特許文献12)が挙げられる。しかし、これらの全てが有効なワクチンであると証明されたわけではなく、そして、ワクチン開発における使用に適切な抗原および免疫原を提供するため、天然の感染の間に免疫応答を惹起するC.trachomatis抗原を同定することが本発明の目的である。C.trachomatisの診断(例えば、免疫診断)に有用な抗原を同定することは、さらなる目的である。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】International patent application WO99/27105
【特許文献2】International patent application WO00/27994
【特許文献3】International patent application WO99/28475
【特許文献4】EP-A-0499681
【特許文献5】International patent application WO95/28487
【非特許文献】
【0010】
【非特許文献1】Raulston (1995) Mol Microbiol 15:607-616
【非特許文献2】Everett (2000) Vet Microbiol 75: 109-126
【非特許文献3】Kalman et al. (1999) Nature Genetics 21:385-389
【非特許文献4】Read et al. (2000) Nucleic Acids Res 28:1397-1406
【非特許文献5】Shirai et al. (2000) Nucleic Acids Res 28:2311-2314
【非特許文献6】Stephens et al. (1998) Science 282:754-759
【非特許文献7】Ward (1995) Apmis. 103:769-96.
【非特許文献8】Moulder (1991) Microbiol Rev 55(1):143-190.
【非特許文献9】Comanducci et al. (1994) Infect Immun 62(12):5491-5497.
【非特許文献10】Murdin et al. (1993) Infect Immun 61:4406-4414
【非特許文献11】Cerrone et al. (1991) Infect Immun 59(1):79-90.
【非特許文献12】Raulston et al. (1993) J. Biol. Chem. 268:23139-23147.
【発明の概要】
【課題を解決するための手段】
【0011】
参考文献18は、種々のC.pneumoniae由来のタンパク質を開示する。これらは、免疫反応性、免疫受容可能および/または基本小体に存在するとして、実験的に検証された。このタンパク質のこれらの性質は、ゲノム配列情報から推論できなかった。参考文献18は、これらのタンパク質が、C.pneumoniaeが主要な病巣であるChlamydia細菌に起因する感染症の処置または予防に使用され得ることを開示する。C.pneumoniaeタンパク質はまた、種間交差反応性に起因する、他の種のChlamydiaによる感染症を処置または予防するために使用され得る。
【0012】
全ゲノムの比較[3、4、5]によって示されるように、C.pneumoniaeはC.trachomatisと密接に関連する。
【0013】
本発明は、参考文献18に開示されるC.pneumoniaeタンパク質に対応するC.tracomatisタンパク質(配列番号1〜261の奇数番号)に関する。これらのタンパク質は、Chlamidia細菌(特にC.trachomatisにおいて)に起因する感染症の処置および予防において使用され得る。特に好ましいタンパク質は、以前に「仮定タンパク質」として注釈されたもの(本明細書中の表Iを参照のこと)か、または細胞質位置を有すると考えられたものである。
(C.trachomatisタンパク質)
本発明は、アミノ酸配列(配列番号1〜261の奇数番号)のうちの1つ以上を含むタンパク質を提供する。
【0014】
本発明はまた、1つ以上の配列番号1〜261の奇数番号のアミノ酸配列と少なくともx%配列同一性を共有する配列を含むタンパク質を提供する。特定の配列に依存して、xは、好ましくは、50%以上(例えば、60%、70%、80%、90%、95%、99%以上)である。これらは、変異体および対立遺伝子改変体を含む。代表的には、2つのタンパク質間の50%以上の同一性は、機能的同等性の指標であると考えられる。タンパク質間の同一性は、好ましくは、パラメーターのギャップオープンペナルティー=12およびギャップ伸長ペナルティー=1を用いるアフィンギャップ検索を使用して、MPSRCHプログラム(Oxford Molecular)において実行されるように、Smith−Waterman相同性検索アルゴリズムによって決定される。
【0015】
本発明は、配列番号1〜261の奇数番号のアミノ酸配列のフラグメントを含むタンパク質をさらに提供する。これらのフラグメントは、配列から少なくともn連続するアミノ酸を含むはずであり、そして特定の配列に依存して、nは、7以上(例えば、8、10、12、14、16、18、20、30、40、50、75、100、150、200以上)である。好ましくは、これらのフラグメントは、配列から1つ以上のエピトープを含む。他の好ましいフラグメントは、シグナルペプチドを省略する。
【0016】
本発明のタンパク質は、種々の手段(例えば、化学合成(少なくとも一部)、プロテアーゼを用いるより長いポリペプチドの消化、RNAからの翻訳、細胞培養物(例えば、組換え発現物、C.trachomatis培養物)からの精製など)によって調製され得る。E.coliにおける異種発現は、好ましい調製用経路である。
【0017】
本発明のタンパク質は種々の形態(例えば、ネイティブ、融合、グリコシル化、非グリコシル化、脂溶化(lipidate)など)を取り得る。
【0018】
本発明のタンパク質は、好ましくは、実質的に純粋な形態(すなわち、他のC.trachomatisまたは宿主細胞タンパク質を実質的に含まない)で調製される。
【0019】
本発明のタンパク質は、固体支持体に結合され得る。これらは検出可能な標識(例えば、放射活性標識もしくは蛍光標識、またはビオチン標識)を含み得る。
【0020】
本発明のタンパク質は、好ましくはChlamydialタンパク質である。
【0021】
(C.trachomatis核酸)
本発明は、配列番号2〜262の偶数番号のヌクレオチド配列を含むタンパク質を提供する。
【0022】
本発明はまた、配列番号2〜262の偶数番号のヌクレオチド配列の1つ以上と少なくともx%配列同一性を共有する配列を含む核酸を提供する。特定の配列に依存して、xは、好ましくは、50%以上(例えば、60%、70%、80%、90%、95%、99%以上)である。
【0023】
さらに、本発明は、偶数のヌクレオチド配列である配列番号2〜262を含む核酸にハイブリダイズし得る核酸を提供する。ハイブリダイゼーション反応は、異なる「ストリンジェンシー」の条件下で行われ得る。ハイブリダイゼーション反応のストリンジェンシーを増す条件は、広く知られており、当該分野で公開されている。関連する条件の例としては、(ストリンジェンシーを増大させるために)以下が挙げられる:25℃、37℃、50℃、55℃および68℃のインキュベーション温度;10×SSC、6×SSC、1×SSC、0.1×SSCの緩衝液濃度および他の緩衝液系を使用するそれらの等価物;0%、25%、50%および75%のホルムアミド濃度;5分〜24時間のインキュベーション時間;1回、2回、またはそれ以上の洗浄工程;1分、2分または15分の洗浄インキュベーション時間;ならびに6×SSC、1×SSC、0.1×SSCの洗浄溶液濃度または脱イオン水。いくつかの実施形態において、本発明の単離された核酸は、低いストリンジェンシー条件下で選択的にハイブリダイズする;他の実施形態において、これは、中間のストリンジェンシー条件下で選択的にハイブリダイズする;他の実施形態において、これは、高いストリンジェンシー条件下で選択的にハイブリダイズする。低いストリンジェンシーハイブリダイゼーション条件の例示的なセットは、50℃および10×SSCである。中間のストリンジェンシーハイブリダイゼーション条件の例示的なセットは、55℃、1×SSCである。高いストリンジェンシーハイブリダイゼーション条件の例示的なセットは、68℃および0.1×SSCである。
【0024】
偶数のヌクレオチド配列である配列番号2〜262のフラグメントを含む核酸もまた、提供される。これらは、C.trachomatis配列に由来する少なくともn個の連続した核酸ヌクレオチドを含むべきであり、そして特定の配列に依存して、nは、7以上(例えば、10、12、14、15、18、20、25、30、35、40、50、75、100、200、300以上)である。
【0025】
さらなる局面に従って、本発明は、本発明のタンパク質およびタンパク質フラグメントをコードする核酸を提供する。
【0026】
本発明が上記の配列に相補的な配列を含む核酸を(例えば、アンチセンスまたはプローブ化目的のために)提供する。
【0027】
本発明の核酸は、もちろん、多数の方法で(例えば、化学合成によって(少なくとも一部)、より長いポリヌクレオチドを制限酵素を使用して消化することによって、ゲノムライブラリーまたはcDNAライブラリーから、生物体自体からなど)調製され得る。
【0028】
本発明の核酸は、種々の形態(例えば、一本鎖、二本鎖、直鎖状、環状、ベクター、プライマー、プローブなど)を採り得る。
【0029】
本発明の核酸は、固体支持体(例えば、ビーズ、プレート、フィルター、フィルム、スライドガラス、樹脂など)に結合され得る。本発明の核酸は、検出可能な標識(例えば、放射活性標識または蛍光標識、あるいはビオチン標識)を含み得る。これは、核酸検出技術(例えば、その核酸が、プライマーであるか、またはPCR、LCR、TMA、NASBA、bDNAなどのような技術において使用するためのプローブとしてである場合)において使用される場合に特に有用である。
【0030】
本発明の核酸は、好ましくは、クラミジア核酸である。
【0031】
用語「核酸」は、DNA、RNA、DNA/RNAハイブリッド、およびDNAまたはRNAのアナログ(例えば、改変された骨格もしくは塩基を含むもの)、そしてまたペプチド核酸(PNA)などを含む。
【0032】
本発明の核酸は、単離され得、そして実質的な純度で得られ得、一般的には、インタクトな染色体以外として得られ得る。通常ポリヌクレオチドは、実質的に他の天然に存在する核酸配列なしに得られ、一般には、少なくとも約50%(重量で)純粋、通常は、少なくとも約90%純粋である。
【0033】
核酸は、例えば、以下のために使用され得る:ポリペプチドを生成するため;生物学的サンプル中の核酸の検出のためのプローブとして;ポリペプチドのさらなるコピーを生成するため;リボザイムまたはアンチセンスオリゴヌクレオチドを生成するため;および一本鎖DNAプローブとしてまたは三本鎖形成オリゴヌクレオチドとしてなど。
【0034】
本発明は、本発明のヌクレオチド配列を含むベクター(例えば、クローニングベクターまたは発現ベクター)およびそれで形質転換された宿主細胞を提供する。
【0035】
(組成物)
さらなる局面に従って、本発明は、本発明に従うタンパク質および/または核酸を含む組成物を提供する。これらの組成物は、好ましくは、免疫原性組成物(例えば、ワクチン)であり、免疫およびワクチン接種の目的に適している。本発明のワクチンは、予防的または治療的であり得、そして代表的に、(a)クラミジア付着、(b)クラミジア侵入、および/または(c)宿主細胞内での首尾良い複製を阻害し得る抗体を誘導し得る抗原を含む。これらのワクチンは、好ましくは、宿主からのクラミジアクリアランスに必要である任意の細胞媒介性T細胞応答を誘導する。
【0036】
本発明はまた、医薬としての(例えば、ワクチンとしての)使用のための本発明に従う核酸およびタンパク質を提供する。
【0037】
本発明はまた、Chlamydiaに起因する感染を処置または予防するための医薬(例えば、ワクチンまたは免疫原性組成物)の製造における本発明に従う核酸またはタンパク質の使用を提供する。Chlamydiaは、一般的にC.trachomatisであるが、種間交差反応に起因して、これはまた、C.pneumoniae、C.pecorum、またはC.psittaciであり得る。予防のために、その医薬は、好ましくは、ChlamydiaのEB形態に対して特異的な免疫応答を誘発する;処置のためには、その医薬は、好ましくは、ChlamydiaのRB形態に対して特異的な免疫応答を誘発する。
【0038】
本発明はまた、Chlamydia trachomatis基本小体を中和するための医薬(例えば、ワクチンまたは免疫原性組成物)の製造において、本発明に従う核酸またえはタンパク質の使用を提供する。
【0039】
本発明はまた、患者(例えば、ヒト)を処置(例えば、免疫化)する方法を提供し、この方法は、本発明に従う治療的に有効な量の核酸またはタンパク質を患者に投与する工程を包含する。
【0040】
本発明はまた、患者における免疫応答を惹起する方法を提供し、この方法は、本発明に従う免疫学的に有効な量の核酸またはタンパク質を患者に投与する工程を包含する。この免疫応答は、患者において抗体を惹起し、そして/または細胞性免疫応答(例えば、CTL応答)を惹起することを包含する。この免疫応答は、EBタンパク質またはRBタンパク質に特異的であり得るか、または宿主細胞質中で発現されるタンパク質に対して特異的であり得る。抗体応答は。好ましくはEBに対して特異的であるのに対して、細胞免疫応答は、好ましくは、細胞質タンパク質に対して、好ましくやRBタンパク質に対して特異的である。
【0041】
本発明はまた、本発明のタンパク質を認識する抗体を惹起する方法を提供し、この方法は、Chlamydiaの基本小体または網様体を患者に投与する工程を包含する。この抗体は、好ましくは、EBに特異的である。
【0042】
本発明はまた、C.trachomatis感染を中和するための方法を提供し、この方法は、本発明のタンパク質、核酸または抗体を患者に投与する工程を包含する。この方法は、好ましくは、EB感染性を中和する。
【0043】
本発明はまた、生物学的サンプルにおけるChlamydia EBまたはRBを検出するための方法を提供し、この方法は、このサンプルと、本発明の抗体とを接触させる工程を包含する。このサンプルは、血液サンプル、別の体液、または組織サンプルであり得る。この方法は、クラミジア感染を診断するために使用され得る。
【0044】
本発明の免疫原性組成物はまた、以下の抗原のうちの1つ以上を含み得る:
−Helicobacter pylori由来のタンパク質抗原(例えば、VacA、CagA、NAP、HopX、HopY{例えば、WO98/04702})および/またはウレアーゼ。
【0045】
−N.meningitidis血清群B由来のタンパク質抗原(例えば、WO99/24578、WO99/36544、WO99/57280、WO00/22430、Tettelinら(2000)Science 287:1809−1815、Pizzaら(2000)Science 287:1816−1820およびWO96/29412)であり、タンパク質「287」および誘導体は、特に好ましい。
【0046】
−N.meningitidis血清群B由来の外膜ベシクル(OMV)調製物(例えば、WO01/52885;Bjuneら(1991)Lancet 338(8775):1093−1096;Fukasawaら(1999)Vaccine 17:2951−2958;Rosenqvistら(1998)Dev.Biol.Stand.92:323−333などにおいて開示される)。
【0047】
−N.meningitidis血清群A、C、W135および/またはY由来のサッカリド抗原(例えば、Costantinoら(1992)Vaccine 10:691−698において開示された、血清群C由来のオリゴサッカリド{Costantinoら(1999)Vaccine 17:1251−1263もまた参照のこと})。
【0048】
Streptococcus pneumoniae由来のサッカリド抗原{例えば、Watson(2000)Pediatr Infect Dis J 19:331−332;Rubin(2000)Pediatr Clin North Am 47:269−285,v;Jedrzejas(2001)Microbiol Mol Biol Rev 65:187−207}。
−B型肝炎ウイルス由来の抗原(例えば、表面抗原および/またはコア抗原){例えば、Gerlichら(1990)Vaccine 補遺8:S63〜68および79〜80}。
−C型肝炎ウイルス由来の抗原{例えば、Hsuら(1999)Clin Liver Dis 3:901〜915}。
−Bordetella pertussis由来の抗原(例えば、B.pertussis由来の百日咳ホロトキシン(holotoxin)(PT)および線維状赤血球凝集素(FHA)、必要に応じてペルタクチンおよび/または凝集原2および凝集原3とも組み合わされる{Gustafssonら(1996)N.Engl.J.Med.334:349−355;Rappuoliら(1991)TIBTECH 9:232−238}。
−ジフテリア抗原(例えば、ジフテリアトキソイド{例えば、Vaccines(1988)の第3章PlotkinおよびMortimer編 ISBN 0−7216−1946−0}例えば、CRM197変異体{例えば、Del Guidiceら(1998)Molecular Aspects of Medicine 19:1−70}。
−破傷風抗原(例えば、破傷風トキソイド{例えば、PlotkinおよびMortimerの第4章}。
−Haemophilus influenzae B由来のサッカリド抗原。
−N.gonorrhoeae由来の抗原{例えば、WO99/24578、WO99/36544、WO99/57280}。
−Chlamydia pneumoniae由来の抗原{例えば、PCT/IB01/01445;Kalmanら(1999)Nature Genetics 21:385−389;Readら(2000)Nucleic Acids Res 28:1397−406;Shiraiら(2000)J.Infect.Dis.181(補遺3):S524−S527;WO99/27105;WO00/27994;WO00/37494}。
−Chlamydia trachomatis由来の抗原{例えば、WO99/28475}。
−Porphyromonas gingivalis由来の抗原{例えば、Rossら(2001)Vaccine 19:4135−4142}。
−ポリオ抗原{例えば、Sutterら(2000)Pediatr Clin North Am 47:287−308;ZimmermanおよびSpann(1999)Am Fain Physician 59:113−118,125−126}(例えば、IPVまたはOPV)。
−狂犬病抗原{例えば、Dreesen(1997)Vaccine 補遺15:S2−6}(例えば、凍結乾燥されて非活性化されたウイルス{例えば、MMWR Morb Mortal Wkly Rep 1998 Jan 16;47(1):12,19;RabAvertTM}。
−麻疹抗原、耳下腺炎抗原および/または風疹抗原{例えば、PlotkinおよびMortimerの9、10および11章}。
−インフルエンザ抗原{例えば、PlotkinおよびMortimerの19章}。(例えば、赤血球凝集素および/またはノイラミニダーゼ表面蛋白質)。
−Moraxella catarrhalis由来の抗原{例えば、McMichael(2000) Vaccine 19 補遺1:S101−107}。
−Staphylococcus aureus由来の抗原{例えば、Kurodaら(2001)Lancet 357(9264):1225−1240;1218−1219頁もまた参照のこと}。
−Streptococcus agalactiae由来の抗原{例えば、WO02/34771を参照のこと}。
−Streptococcus pyogeraes由来の抗原{例えば、WO02/34771を参照のこと}。
【0049】
サッカリド抗原または糖鎖抗原が含まれる場合、免疫原性を高めるために、その抗原は、好ましくは、キャリアタンパク質に結合される{例えば、Ramsayら(2001)Lancet 357(9251):195−196;Lindberg(1999)Vaccine 17 補遺2:S28−36;Conjugate Vaccines(Cruseら編)ISBN 3805549326,特に10巻:48−114等}。好ましいキャリアタンパク質は、細菌毒素または細菌トキソイドである(例えば、ジフテリアトキソイドまたは破傷風トキソイド)。CRM97ジフテリアトキソイドが、特に好ましい。他の適切なキャリアタンパク質としては、N.meningitidis外膜タンパク質{例えば、EP−0372501}、合成ペプチド{例えば、EP−0378881,EP−0427347}、熱ショックタンパク質{例えば、WO93/17712}、百日咳タンパク質{例えば、WO98/58668;EP−0471177}、H.influenzae由来のタンパク質D{例えば、WO00/56360}、C.difficile由来の毒素Aまたは毒素B{例えば、WO00/61761}などが挙げられる。任意の適切な結合反応が、必要に応じて任意の適切なリンカーと共に使用され得る。
【0050】
毒素タンパク質抗原は、必要に応じて、解毒され得る(例えば、百日咳毒素の解毒は、化学的手段および/または遺伝的手段による)。
【0051】
ジフテリア抗原が組成物に含まれる場合、破傷風抗原および百日咳抗原をも含むことが好ましい。同様に、破傷風抗原が含まれる場合、ジフテリア抗原および百日咳抗原をも含むことが好ましい。同様に、百日咳抗原が含まれる場合、ジフテリア抗原および破傷風抗原をも含むことが好ましい。
【0052】
抗原は、好ましくはアルミニウム塩に吸着される。
【0053】
組成物中の抗原は、代表的には、各々少なくとも1μg/mlの濃度で存在する。一般的に、任意の所定の抗原の濃度は、その抗原に対する免疫応答を誘発するのに十分である。
【0054】
本発明はまた、本発明の2つ以上のタンパク質を含む組成物を提供する。
【0055】
(プロセス)
本発明は、本発明のタンパク質を産生するプロセスを提供し、このプロセスは、タンパク質発現を誘導する条件下で、本発明に従う宿主細胞を培養する工程を包含する。
【0056】
本発明は、本発明のタンパク質または核酸を生成するプロセスを提供し、ここで、このタンパク質または核酸は、化学的手段を使用して一部または全体的に合成される。
【0057】
本発明は、サンプル中のC.trachomatisを検出するためのプロセスを提供し、ここで、このサンプルは、本発明のタンパク質に結合する抗体と接触される。
【0058】
本発明を実行するために(例えば、免疫化について開示される配列を利用するために)使用され得る標準的な技術および手順の要約は、以下の通りである。この要約は、本発明を制限するのではなく、むしろ使用され得る実施例を示すが、必要とされない。
【0059】
(概要)
本発明の実施は、他に示されなければ、当該分野の技術の範囲内である、分子生物学、微生物学、組換えDNA、および免疫学の従来の技術を使用する。このような技術は以下の文献で十分説明されている(例えば、Sambrook Molecular Cloning;A Laboratory Manual 第2版(1989)および第3版(2001);DNA Cloning,Volumes I and ii(D.N Glover編 1985);Oligonucleotide Synthesis(M.J.Gait編 1984);Nucleic Acid Hybridization(B.D.HamesおよびS.J.Higgins編 1984);Transcription and Translation(B.D.HamesおよびS.J.Higgins編 1984);Animal Cell Culture(R.I.Freshney編 1986);Immobilized Cells and Enzymes(IRL Press,1986);B.Perbal,A Practial Guide to Molecular Cloning(1984);the Methods in Enzymology series(Academic Press,Inc.),特に154巻および155巻;Gene Transfer Vectors for Mammalian Cells(J.H.MillerおよびM.P.Calos編 1987,Cold Spring Harbor Laboratory);MayerおよびWalker編(1987),Immunochemical Methods in Cell and Molecular Biology(Academic Press,London);Scopes,(1987)Protein Purification:Principles and Practice,第2版(Springer−Verlag,N.Y.)、およびHandbook of Experimental Immunology,Volumes I−IV(D.M.WeirおよびC.C.Blackwell編 1986)。
【0060】
ヌクレオチドおよびアミノ酸についての標準的な略語が、本明細書において使用される。
【図面の簡単な説明】
【0061】
【図1A】図1Aは、実施例1のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図1B】図1Bは、実施例1のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図1C】図1Cは、実施例1のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図2A】図2Aは、実施例2のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図2B】図2Bは、実施例2のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図2C】図2Cは、実施例2のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図3A】図3Aは、実施例3のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図3B】図3Bは、実施例3のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図3C】図3Cは、実施例3のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図3D】図3Dは、実施例3のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4A】図4Aは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4B】図4Bは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4C】図4Cは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4D】図4Dは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4E】図4Eは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5A】図5Aは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5B】図5Bは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5C】図5Cは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5D】図5Dは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5E】図5Eは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図6A】図6Aは、実施例6のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図6B】図6Bは、実施例6のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図6C】図6Cは、実施例6のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図7A】図7Aは、実施例7のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図7B】図7Bは、実施例7のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図8A】図8Aは、実施例8のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図8B】図8Bは、実施例8のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図8C】図8Cは、実施例8のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図9A】図9Aは、実施例9のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図9B】図9Bは、実施例9のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図9C】図9Cは、実施例9のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図9D】図9Dは、実施例9のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図10A】図10Aは、実施例10のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図10B】図10Bは、実施例10のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図10C】図10Cは、実施例10のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図11A】図11Aは、実施例11のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図11B】図11Bは、実施例11のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図11C】図11Cは、実施例11のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12A】図12Aは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12B】図12Bは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12C】図12Cは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12D】図12Dは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12E】図12Eは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図13A】図13Aは、実施例13のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図13B】図13Bは、実施例13のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図13C】図13Cは、実施例13のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図14A】図14Aは、実施例14のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図14B】図14Bは、実施例14のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図14C】図14Cは、実施例14のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図15A】図15Aは、実施例15のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図15B】図15Bは、実施例15のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図15C】図15Cは、実施例15のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図16A】図16Aは、実施例16のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図16B】図16Bは、実施例16のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図16C】図16Cは、実施例16のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図17】図17は、実施例17のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図18A】図18Aは、実施例18のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図18B】図18Bは、実施例18のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図19】図19は、実施例19のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図20A】図20Aは、実施例20のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図20B】図20Bは、実施例20のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図21】図21は、実施例21のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図22】図22は、実施例22のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図23A】図23Aは、実施例23のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図23B】図23Bは、実施例23のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図23C】図23Cは、実施例23のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図24A】図24Aは、実施例24のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図24B】図24Bは、実施例24のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図24C】図24Cは、実施例24のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図24D】図24Dは、実施例24のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図25A】図25Aは、実施例25のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図25B】図25Bは、実施例25のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図25C】図25Cは、実施例25のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図26A】図26Aは、実施例26のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図26B】図26Bは、実施例26のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27A】図27Aは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27B】図27Bは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27C】図27Cは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27D】図27Dは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27E】図27Eは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図28A】図28Aは、実施例28のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図28B】図28Bは、実施例28のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図29A】図29Aは、実施例29のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図29B】図29Bは、実施例29のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図30】図30は、実施例30のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図31】図31は、実施例31のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図32】図32は、実施例32のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図33A】図33Aは、実施例33のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図33B】図33Bは、実施例33のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図34A】図34Aは、実施例34のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図34B】図34Bは、実施例34のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図35A】図35Aは、実施例35のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図35B】図35Bは、実施例35のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図36】図36は、実施例36のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図37A】図37Aは、実施例37のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図37B】図37Bは、実施例37のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図37C】図37Cは、実施例37のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図38A】図38Aは、実施例38のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図38B】図38Bは、実施例38のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図39】図39は、実施例39のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図40】図40は、実施例40のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図41】図41は、実施例41のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図42】図42は、実施例42のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図43】図43は、実施例43のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図44A】図44Aは、実施例44のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図44B】図44Bは、実施例44のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【発明を実施するための形態】
【0062】
(定義)
Xを含む組成物は、組成物中の全X+Yの少なくとも85重量%がXであるとき、Yを「実質的に含まない」。好ましくは、Xは、組成物中の全X+Yの少なくとも約90重量%を、さらに好ましくは少なくとも約95重量%または99重量%さえを含む。
【0063】
用語「含む(comprising)」は、「含む(including)」および「からなる」を意味する。例えば、Xを「含む」組成物は、もっぱらXからなり得るか、またはX+YのようなXに付加したものも含み得る。
【0064】
用語「異種」とは、天然では共に見られない2つの生物学的成分をいう。この成分は、宿主細胞、遺伝子、または調節領域(例えば、プロモーター)であり得る。異種成分は天然では共に見られないが、遺伝子に対して異種のプロモーターがその遺伝子に作動可能に連結されるときのように、それらは共に機能し得る。別の例は、クラミジア配列がマウス宿主細胞に対して異種であることである。さらなる例は、天然においてみられない配置で単一タンパク質に組み立てられる同じタンパク質または異なるタンパク質由来の2つのエピトープである。
【0065】
「複製起点」とは、発現ベクターのような、ポリヌクレオチドの複製を開始または調節するポリヌクレオチド配列である。複製起点は、細胞内でのポリヌクレオチド複製の自律性ユニットとして振る舞い、それ自体の制御下で複製し得る。複製起点は、ベクターが特定の宿主細胞において複製するために必要であり得る。特定の複製起点を有せば、発現ベクターは、細胞内の適切なタンパク質の存在下で高いコピー数で再生され得る。起点の例は、酵母において有効な自律性複製配列;およびCOS−7細胞で有効であるウイルスT抗原である。
【0066】
「変異体」配列は、ネイティブの配列または開示された配列とは異なるが配列同一性を有するDNA配列、RNA配列またはアミノ酸配列として規定される。特定の配列に依存して、ネイティブの配列または開示された配列と変異体配列との間の配列同一性の程度は、好ましくは50%より大きい(例えば、上記のSmith−Watermanアルゴリズムを使用して算出して60%、70%、80%、90%、95%、99%またはそれ以上)。本明細書中で使用される場合、本明細書で核酸分子配列が提供される核酸分子、または領域の「対立遺伝子改変体」は、別のもしくは2番目の単離体のゲノム中の同じ遺伝子座で本質的に生じる、核酸分子、または領域および、例えば、変異または組換えにより生ずる天然変化に起因して、類似するが、しかし同一でない核酸配列を有する核酸分子、または領域である。コード領域の対立遺伝子改変体は、代表的には、比較される遺伝子によってコードされるタンパク質の活性と類似した活性を有するタンパク質をコードする。対立遺伝子改変体はまた、遺伝子の5’または3’非翻訳領域(例えば、調節制御領域)での変化を含み得る(例えば、米国特許第5,753,235号を参照のこと)。
【0067】
(発現系)
クラミジアヌクレオチド配列は、種々の異なる発現系;例えば、哺乳動物細胞、バキュロウイルス、植物、細菌、および酵母と共に使用される発現系において発現され得る。
【0068】
(i.哺乳動物系)
哺乳動物発現系は当該分野において公知である。哺乳動物プロモーターは、哺乳動物RNAポリメラーゼを結合し、コード配列(例えば、構造遺伝子)のmRNAへの下流(3’)転写を開始し得る任意のDNA配列である。プロモーターは、転写開始領域(これはコード配列の5’末端の近位に通常位置する)およびTATAボックス(転写開始部位の25〜30塩基対(bp)上流に通常位置する)を有する。TATAボックスは、その正しい部位においてRNAポリメラーゼIIにRNA合成を開始させるよう指示すると考えられている。哺乳動物プロモーターはまた、TATAボックスの100〜200bp上流以内に通常位置する上流プロモーターエレメントを含む。上流プロモーターエレメントは、転写が開始され、そしていずれかの方向において作用し得る速度を決定する{Sambrookら(1989)「Expression of Cloned Genes in Mammalian Cells」 Molecular Cloning:A Laboratory Manual、第2版}。
【0069】
哺乳動物ウイルス遺伝子は、しばしば高度に発現され、そして広い宿主範囲を有する;従って、哺乳動物ウイルス遺伝子をコードする配列は、特に有用なプロモーター配列を提供する。例としては、SV40初期プロモーター、マウス乳癌ウイルスLTRプロモーター、アデノウイルス主要後期プロモーター(Ad MLP)、および単純疱疹ウイルスプロモーターが挙げられる。さらに、マウスのメタロチオネイン遺伝子のような非ウイルス性遺伝子に由来する配列もまた、有用なプロモーター配列を提供する。発現は、構成性であるかまたは調節される(誘導可能)かのいずれかであり得る、プロモーターに依存して、ホルモン応答性細胞においてグルココルチコイドで誘導され得る。
【0070】
上記に記載されるプロモーターエレメントと組み合わされるエンハンサーエレメント(エンハンサー)の存在は、通常発現レベルを増大させる。エンハンサーは、相同性プロモーターまたは異種プロモーターに連結されるとき、転写を1000倍まで刺激し得る調節DNA配列であり、合成は、通常のRNA開始部位で開始する。エンハンサーはまた、それらが、通常方向もしくは反対(flipped)方向のいずれかで転写開始部位より上流または下流に、もしくはプロモーターから1000ヌクレオチドを超える距離に位置するとき、活性である(Maniatisら(1987) Science 236:1237;Albertsら(1989)Molecular Biology of the Cell,第2版)。ウイルス由来のエンハンサーエレメントは、それらが通常、より広い宿主範囲を有するので、特に有用であり得る。例としては、SV40初期遺伝子エンハンサー(Dijkemaら(1985)EMBO J.4:761)およびラウス肉腫ウイルスの長末端反復(LTR)に由来するエンハンサー/プロモーター(Gormanら(1982)PNAS USA 79:6777)およびヒトサイトメガウイルスに由来するエンハンサー/プロモーター(Boshartら(1985)Cell 41:521)を包含する。さらに、いくつかのエンハンサーは調節可能であり、そしてホルモンまたは金属イオンのような誘導因子の存在下のみで活性になる(Sassone−CorsiおよびBorelli(1986)Trends Genet.2:215;Maniatisら(1987)Science 236:1237)。
【0071】
DNA分子は、哺乳動物細胞において細胞内で発現され得る。プロモーター配列は、組換えタンパク質のN末端における最初のアミノ酸が常にATG開始コドンによりコードされるメチオニンである場合、DNA分子と直接連結され得る。所望される場合、N末端は、臭化シアンとのインビトロでのインキュベーションによりタンパク質から切断され得る。
【0072】
あるいは、外来タンパク質もまた、哺乳動物細胞において外来タンパク質の分泌を提供するリーダー配列フラグメントで構成される融合タンパク質をコードするキメラDNA分子を作製することにより、細胞から成長培地へ分泌され得る。好ましくは、インビボまたはインビトロのいずれかにおいて切断され得る、リーダーフラグメントおよび外来遺伝子との間にコードされるプロセシング部位が存在する。リーダー配列フラグメントは通常、細胞からのタンパク質の分泌を指示する疎水性アミノ酸で構成される単一のペプチドをコードする。アデノウイルス3部分からなるリーダーは、哺乳動物細胞における外来タンパク質の分泌を提供するリーダー配列の例である。
【0073】
通常、哺乳動物細胞によって認識される転写終結配列およびポリアデニル化配列は、翻訳終止コドンの3’側に存在する調節領域であり、従って、プロモーターエレメントと共に、コード配列に隣接する。成熟mRNAの3’末端は、部位特異的転写後切断およびポリアデニル化により形成される(Birnstielら(1985)Cell 41:349;ProudfootおよびWhitelaw(1988)「Termination and 3’end processing of eukaryotic RNA.」Transcription and splicing(B.D.HamesおよびD.M.Glover編);Proudfoot(1989)Trends Biochem.Sci.14:105)。これらの配列は、mRNAの転写を指示し、そのmRNAは、そのDNAにコードされるポリペプチドに翻訳され得る。転写終結/ポリアデニル化シグナルの例としては、SV40由来のシグナルが挙げられる(Sambrookら(1989)「Expression of cloned genes in cultured mammalian cells.」Molecular Cloning:A Laboratory Manual)。
【0074】
通常、プロモーター、ポリアデニル化シグナル、および転写終結配列を含む上記の構成要素は、発現構築物内に共に導入される。エンハンサー、機能的なスプライスドナー部位および受容部位を有するイントロン、ならびにリーダー配列もまた、所望される場合、発現構築物内に含まれ得る。発現構築物はしばしば、哺乳動物細胞または細菌のような宿主内で安定的に維持し得る染色体外エレメント(例えば、プラスミド)のような、レプリコン内で維持される。哺乳動物の複製系としては、複製にトランス作用性の因子を必要とする、動物ウイルス由来の複製系が挙げられる。例えば、SV40(Gluzman(1981)Cell 23:175)、あるいはポリオーマウイルスのようなパポバウイルスの複製系を含むプラスミドは、適切なウイルスのT抗原の存在下で極めて高いコピー数で複製する。哺乳動物レプリコンのさらなる例としては、ウシパピローマウイルスおよびエプスタイン−バーウイルス由来のレプリコンが挙げられる。さらに、このレプリコンは、二つの複製系を有し得、従って、例えば、発現用の哺乳動物細胞内で、ならびにクローニングおよび増幅用の原核生物の宿主内で、そのレプリコンを維持することが可能である。このような哺乳動物−細菌シャトルベクターの例としては、pMT2(Kaufmanら(1989)Mol.Cell.Biol.9:946)およびpHEBO(Shimizuら(1986)Mol.Cell.Biol.6:1074)が挙げられる。
【0075】
使用される形質転換の手順は、形質転換される宿主に依存する。異種のポリヌクレオチドの哺乳動物細胞への導入方法は、当該分野で公知であり、その方法としては、デキストラン媒介トランスフェクション、リン酸カルシウム沈殿、ポリブレン媒介トランスフェクション、プロトプラスト融合、エレクトロポレーション、リポソーム内でのポリヌクレオチドの封入、およびDNAの核内への直接微量注入が挙げられる。
【0076】
発現用宿主として利用可能な哺乳動物細胞株は、当該分野で公知であり、そのような細胞株としては、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、乳仔ハムスター腎臓(BHK)細胞、サル腎臓細胞(COS)、ヒト肝細胞癌細胞(例えば、Hep G2)および多くの他の細胞株を含むが、これらに限定しない、American Type Culture Collection(ATCC)から入手可能な多くの不死化細胞株が含まれる。
【0077】
(ii バキュロウイルス系)
タンパク質をコードしているポリヌクレオチドはまた、適切な昆虫の発現ベクター内に挿入され得、そしてそのベクター内で、制御エレメントに作動可能に連結される。ベクターの構築には、当該分野で公知の技術を使用する。一般に、その発現系の構成要素として、以下のものが挙げられる:バキュロウイルスゲノムのフラグメント、および発現させる異種遺伝子の挿入用の簡便な制限部位の両方を含む転移ベクター(通常は細菌プラスミド;転移ベクター内のバキュロウイルスに特異的なフラグメントに相同性のある配列を有する野生型バキュロウイルス(これは、バキュロウイルスゲノム内への異種遺伝子の相同組換えを可能にする);ならびに適切な昆虫宿主細胞および成長培地。
【0078】
転移ベクターにタンパク質をコードするDNA配列を挿入した後、そのベクターおよび野生型ウイルスゲノムを、昆虫宿主細胞にトランスフェクトし、そこでベクターとウイルスゲノムを組換え可能にする。パッケージングされた組換えウイルスは発現され、そして組換えプラークが同定されそして精製される。バキュロウイルス/昆虫細胞発現系に対する材料および方法は、特に、Invitrogen、San Diego CAからキット形態(「MaxBac」キット)で市販される。これらの技術は、一般に当業者に公知であり、そしてSummersならびにSmith,Texas Agricultural Experiment Station Bulletin No.1555(1987)(以下、「Summers and Smith」)に十分に記載されている。
【0079】
タンパク質をコードするDNA配列をバキュロウイルスゲノムに挿入するの前に、プロモーター配列、リーダー配列(所望される場合は)、目的のコード配列、および転写終結配列を含む上記の構成要素を、通常、中間転移構築物(転移ベクター)に構築される。この構築物は、単一の遺伝子および作動可能に連結された調節エレメント;操作可能に連結された調節エレメントのセットを各々が所有する複数の遺伝子;あるいは同じ調節エレメントのセットにより調節される複数の遺伝子を含み得る。中間転移構築物は、しばしば、細菌のような宿主内で安定的に維持し得る染色体外エレメント(例えば、プラスミド)のような、レプリコン内で維持される。レプリコンは、複製系を有しており、従って、クローニングおよび増幅に適切な宿主内で維持されることを可能にする。
【0080】
現在、外来遺伝子のAcNPVへの導入のために最も一般に使用される転移ベクターは、pAc373である。当業者に公知の多くの他のベクターもまた設計され、これらのものとして、例えば、pVL985(ポリへドリンの開始コドンをATGからATTに変更し、そしてそのATTから32塩基対下流に、BamHIクローニング部位を導入する;LuckowおよびSummers,Virology(1989)17:31を参照のこと)が挙げられる。
【0081】
そのプラスミドも通常、ポリへドリンポリアデニル化シグナル(Millerら(1988)Ann.Rev.Microbiol.,42:177)、および原核生物のアンピシリン耐性(amp)遺伝子、および大腸菌においての選抜ならびに増殖のための複製起点を含む。
【0082】
バキュロウイルス転移ベクターは通常、バキュロウイルスプロモーターを含む。バキュロウイルスプロモーターは、バキュロウイルスRNAポリメラーゼに結合し、そしてコード配列(例えば、構造遺伝子)のmRNAへの下流(5’から3’)方向の転写を開始し得る任意のDNA配列である。プロモーターは、通常、コード配列の5’末端に近接して存在する転写開始領域を有する。この転写開始領域は通常、RNAポリメラーゼ結合部位および転写開始部位を含む。バキュロウイルス転移ベクターはまた、エンハンサーと呼ばれる第二のドメインを有し得、存在する場合は、通常、構造遺伝子に対し遠位にある。発現は調節され得るか、あるいは構成的であり得る。
【0083】
ウイルスの感染周期の後期で大量に転写される構造遺伝子は、特に有用なプロモーター配列を提供する。例としては、ウイルス多面体タンパク質をコードする遺伝子(Friesenら(1986)「The Regulation of Baculovirus Gene Expression,」:The Molecular Biology of Baculoviruses(Walter Doerfler編);EPO公開番号127839および155476;ならびにp10タンパク質をコードする遺伝子(Vlakら,(1988),J.Gen.Virol.69:765)由来の配列が挙げられる。
【0084】
適切なシグナル配列をコードするDNAは、分泌昆虫タンパク質、あるいはバキュロウイルスポリへドリン遺伝子(Carbonellら,(1998)Gene,73:409)のような、分泌バキュロウイルスタンパク質についての遺伝子から由来し得る。あるいは、哺乳動物細胞の翻訳後修飾(例えば、シグナルペプチド切断、タンパク質分解性切断、およびリン酸化)のシグナルは、昆虫細胞に認識されると思われ、そして分泌および核蓄積に必要なシグナルはまた、無脊椎動物細胞と脊椎動物細胞との間で保存されると思われるので、ヒトインターフェロンα(Maedaら,(1985),Nature 315:592);ヒトガストリン放出ペプチド(Lebacq−Verheydenら,(1988),Molec.Cell.Biol.8:3129);ヒトIL−2(Smithら,(1985)Proc.Nat’l Acad.Sci.USA、82:8404);マウスIL−3(Miyajimaら,(1987)Gene 58:273);およびヒトグルコセレブロシダーゼ(Martinら,(1988)DNA、7:99)をコードする遺伝子由来のような、非昆虫起源のリーダーも、昆虫での分泌を与えるために使用され得る。
【0085】
組換えポリペプチドあるいは組換えポリタンパク質は、細胞内に発現され得、あるいは適切な調節配列と共に発現される場合、分泌され得る。非融合の外来タンパク質の優れた細胞内発現には通常、ATG開始シグナルに先行する適切な翻訳開始シグナルを含む短いリーダー配列を理想的には有する異種遺伝子が必要である。所望であれば、N末端のメチオニンは、臭化シアンとのインビトロインキュベーションにより、成熟タンパク質から切断され得る。
【0086】
あるいは、自然に分泌されない組換えポリタンパク質あるいは組換えタンパク質は、昆虫において外来タンパク質の分泌を与えるリーダー配列フラグメントからなる、融合タンパク質をコードするキメラDNA分子を作成することにより、昆虫細胞から分泌され得る。リーダー配列フラグメントは通常、タンパク質の小胞体内への輸送を指示する疎水性アミノ酸からなるシグナルペプチドをコードしている。
【0087】
タンパク質の前駆体である発現産物をコードするDNA配列および/または遺伝子の挿入後、昆虫細胞宿主に、転移ベクターの異種DNAおよび野生型バキュロウイルスのゲノムDNAを同時形質転換(通常は、同時トランスフェクションによって)する。構築物のプロモーターおよび転写終結配列は、通常バキュロウイルスゲノムの2〜5kbの区域を含む。バキュロウイルスウイルスの望ましい部位に異種DNAを導入する方法は、当該分野で公知である(SummersおよびSmith、上記;Juら(1987);Smithら,Mol.Cell.Biol.(1983)3:2156;ならびにLuckowおよびSummers(1989)を参照のこと)。例えば、その挿入物は、相同二重交差組換え(homologous double crossover recombination)により、ポリへドリン遺伝子のような遺伝子内にあり得る;挿入物はまた、所望のバキュロウイルス遺伝子に設計された制限酵素部位内にあり得る。Millerら(1989)、Bioessays 4;91。発現ベクター内のポリへドリン遺伝子の代わりにクローン化される場合のDNA配列は、ポリへドリン特異的配列により5’および3’の両側で隣接され、そしてポリへドリンプロモーターの下流に位置される。
【0088】
新規に形成されたバキュロウイルス発現ベクターは続いて、感染性の組換えバキュロウイルス内にパッケージされる。相同組換えは、低い頻度で起こる(約1%〜約5%の間);従って、同時トランスフェクション後に産生されたウイルスの大半は、依然野生型ウイルスである。従って、方法には、組換えウイルスの同定が必要となる。その発現系の利点は、組換えウイルスを区別させ得る可視的スクリーニングである。天然のウイルスにより産生されるポリへドリンタンパク質は、ウイルス感染後の後期で、その感染された細胞の核内で非常に高いレベルで産生される。蓄積されたポリへドリンタンパク質は、閉塞体を形成し、またそれは包理された粒子を含む。これらの閉塞体は、最大15μmの大きさで、高度に屈折し、明るく輝く外見を与え、容易に光学顕微鏡下で可視化される。組換えウイルスに感染された細胞は、閉塞体を欠く。組換えウイルスと野生型ウイルスを区別するために、トランスフェクションの上清を、当業者に公知の技術により昆虫細胞の単層にプラーク形成させた。すなわち、プラークを、光学顕微鏡下で閉塞体の存在(野性型ウイルスを示す)または非存在(組換えウイルスを示す)によりスクリーニングする。「Current Protocols in Microbiology」2巻(Ausubelら編)16.8(増補10、1990);SummersおよびSmith、上記;Millerら(1989)。
【0089】
組換えバキュロウイルス発現ベクターは、いくつかの昆虫細胞への感染用に開発された。例えば、組換えバキュロウイルスは、特に以下に示すもののために開発された:Aedes aegypti、Autographa californica、Bombyx mori、Drosophila melanogaster、Spodoptera frugiperda、およびTrichoplusia ni(WO89/046699;Carbonellら,(1985)J.Virol.56:153;Wright(1986)Nature 321:718;Smithら,(1983)Mol.Cell.Biol.3:2156;およびFraserら,(1989)In Vitro Cell.Dev.Biol.25:225を一般に参照のこと)。
【0090】
細胞および細胞培養培地は、バキュロウイルス/発現系における異種ポリペプチドの直接発現および融合発現の両方のために市販される;細胞培養技術は、一般に当業者に公知である。例えば、SummersおよびSmith(上記)を参照のこと。
【0091】
次いで、改変した昆虫細胞を、適切な栄養培地で増殖し、その改変した昆虫宿主内に存在するプラスミドの安定的維持を可能にする。発現産物の遺伝子が、誘導性の制御下にある場合、宿主は高密度で増殖され得、そして発現は誘導され得る。あるいは、発現が構成的である場合、その産物は培地中に連続的に発現され、そして栄養培地を連続的に循環し、目的産物を取り出し、そして枯渇した栄養を補給する必要がある。その産物は、クロマトグラフィー(例えば、HPLC、アフィニティークロマトグラフィー、イオン交換クロマトグラフィーなど);電気泳動;密度勾配遠心;溶媒抽出などのような技術により精製され得る。適切には、その産物をさらに精製し、必要ならば、培地中に分泌されたか、または昆虫細胞の溶解物から生じた任意の昆虫タンパク質を実質的に除去し、宿主細片(例えば、タンパク質、脂質および多糖類)を少なくとも実質的に含まない産物を提供する。
【0092】
タンパク質の発現を得るために、形質転換体に由来する組換え宿主細胞は、組換えタンパク質をコードする配列の発現を可能にする条件下でインキュベートされる。これらの条件は、選択された宿主細胞に依存して変動する。しかし、その条件は、当該分野で公知の条件に基づいて、当業者に容易に確かめられる。
【0093】
(iii 植物系)
当該分野で公知の多くの植物細胞培養および全植物遺伝子発現系が存在する。例示的な植物細胞遺伝子発現系としては、米国特許第5,693,506号;米国特許第5,659,122号;米国特許第5,608,143号のような特許に記載されるものが挙げられる。植物細胞培養における遺伝子発現の別の例は、Zenk,Phytochemistry 30:3861−3863(1991)に記載された。植物タンパク質のシグナルペプチドの記載は、上記の参考文献に加え、以下に示すものの中においても確認される;Vaulcombeら,Mol.Gen.Genet.209:33−40(1987);Chandlerら,Plant Molecular Biology 3:407−418(1984);Rogers,J.Biol.Chem.260:3731−3738(1985);Rothsteinら,Gene 55:353−356(1987);Whittierら,Nucleic Acids Research 15:2515−2535(1987);Wirselら,Molecular Microbiology 3:3−14(1989);Yuら,Gene 122:247−253(1992)。植物ホルモン(ジベレリン酸およびジベレリン酸により誘導される分泌酵素)による植物遺伝子発現の調節の記載は、R.L.JonesおよびJ.MacMillin,Gibberellins:Advanced Plant Physiology,Malcolm B.Wilkins編 1984 Pitman Publishing Limited,London,21−52頁の中に確認され得る。他の代謝調節性遺伝子が記載される参考文献は以下である;Sheen,Plant Cell,2:1027−1038(1990);Maasら,EMBO J.9:3447−3452(1990);BenkelおよびHickey、Proc.Natl.Acad.Sci.84:1337−1339(1987)。
【0094】
代表的に、当該分野で公知の技術を使用して、所望のポリヌクレオチド配列は、植物内で操作するために設計された遺伝子調節エレメントを含む発現カセットの中に挿入される。その発現カセットは、植物宿主内での発現に適切な発現カセットの上流および下流にコンパニオン配列を有する所望の発現ベクターの中に挿入される。そのコンパニオン配列は、プラスミドまたはウイルス起源のものであり、そしてそのベクターが、細菌のような本来のクローニング宿主から、所望の植物宿主へDNAを移動させるために必要とされる特徴をベクターに提供する。基本的な細菌/植物ベクター構築物は、好ましくは、広い宿主範囲の原核生物の複製起点;原核生物の選択マーカー;および、アグロバクテリウムの形質転換については、アグロバクテリウム媒介転位のためのT DNA配列を植物染色体に提供する。異種遺伝子が容易に検出に従順しない場合は、好ましくは、その構築物はまた、植物細胞が形質転換されたかどうかを決定するために適した選択マーカー遺伝子を有する。適切なマーカーの一般的な総説は、例えば、イネ科のメンバーについては、WilminkおよびDons,1993,Plant Mol.Biol.Reptr,11(2):165−185に見られる。
【0095】
植物ゲノムへの異種配列の組み込みを可能にするのに適した配列もまた、推奨される。これらは、植物ゲノム内へ異種発現カセットのランダム挿入を可能にする相同組換えのためのトランスポゾン配列など、およびTi配列を含み得る。適切な原核生物選択マーカーとしては、アンピシリンまたはテトラサイクリンのような抗生物質に対する耐性が挙げられる。別の機能をコードしている他のDNA配列もまた、そのベクターの中に存在し得、当該分野で公知である。
【0096】
本発明の核酸分子は、目的のタンパク質の発現用の発現カセットに含まれ得る。2つ以上も可能であるが、通常はただ一つの発現カセットである。組換え発現カセットは、異種タンパクのコード配列に加え、以下のエレメントを含む;プロモーター領域、植物5’非翻訳配列、構造遺伝子がそれを備えているかどうかに依存して、開始コドン、ならびに転写および翻訳終結配列。そのカセットの5’および3’末端の独特な制限酵素部位は、既存のベクター内への容易な挿入を可能にする。
【0097】
異種のコード配列は、本発明に関係する任意のタンパク質のための配列であり得る。目的のタンパクをコードする配列は、そのタンパク質の適切なプロセッシングおよび輸送を可能にするシグナルペプチドをコードし、そして通常、本発明の所望のタンパク質の膜への結合を生じ得る任意の配列を欠いている。翻訳開始領域は、大部分は、発芽中に発現および輸送される遺伝子のためのものであるから、輸送を提供するシグナルペプチドを使用することにより、それはまた、目的のタンパク質の輸送を提供し得る。この方法で、目的のタンパク質は、それらが発現される細胞から輸送され、そして効率的に回収され得る。代表的には、種子における分泌は、種子の胚乳内へ、アリューロン層あるいは胚盤上皮層を通過する。タンパク質がそれが産生された細胞から分泌される必要がない場合、このことは組換えタンパク質の分離および精製を容易にする。
【0098】
所望の遺伝子産物の最終的な発現が、真核生物におけるものであるので、クローン化した遺伝子の任意の部分が、イントロンのような、宿主のスプライセオソーム(splicosome)機構により、プロセッシングされる配列を含むか否かを決定することが望ましい。そのような場合、「イントロン」領域の部位特異的変異誘発は、仮性イントロンコードとして遺伝的情報の一部を欠失することを防ぐために実施され得る。ReedおよびManiatis、Cell 41:95−105(1985)。
【0099】
ベクターは、組換えDNAを機械的に転移するためにマイクロピペットを用いて植物細胞内に直接的に微量注入され得る(Crossway、Mol.Gen.Genet,202:179−185、1985)。遺伝子物質はまた、ポリエチレングリコールを用いて植物細胞内に転移され得る(Krensら、Nature,296、72−74、1982)。核酸部分の導入の別の方法は、小さいビーズまたは微粒子のいずれかのマトリックスの内部に、あるいは表面に核酸を有する小さな微粒子による高速バリスティック(ballistic)穿通法である(Kleinら,Nature,327,70−73,1987ならびにKnudsenおよびMuller,1991,Planta,185:330−336は、大麦胚乳の粒子の照射(bombardment)によりトランスジェニック大麦を作製することを示している)。さらに別の導入方法は、他の物体、いずれかのミニ細胞、細胞、リソソームあるいは他の易融な脂肪表面体とのプロトプラストの融合である(Fraleyら、Proc.Natl.Acad.Sci.USA,79,1859−1863,1982)。
【0100】
ベクターはまた、エレクトロポレーションにより植物細胞内に導入され得る(Frommら、Proc.Natl.Acad.Sci.USA 82:5824,1985)。この技術において、植物プロトプラストは、遺伝子構築物を含むプラスミドの存在中でエレクトロポレート(電気穿孔)される。高い電界の強さの電気インパルス(衝撃)により、生体膜を可逆的に通過できるようにし、プラスミドの導入を可能にする。エレクトロポレートされた植物プロトプラストは細胞壁を再形成し、分裂し、植物カルス形成する。
【0101】
プロトプラストが単離され得、そして培養されて完全な再生植物を与え得る全ての植物は、本発明により形質転換され得、これによって移入した遺伝子を保持する完全な植物が再生される。実際に、全ての植物は、さとうきび、甜菜、綿、果実および他の樹木、マメ科植物および野菜の全ての主要な種を含むがそれらに限定されない培養細胞あるいは組織から再生され得る。いくつかの適応した植物としては、例えば、以下の属由来の種が挙げられる;Fragaria,Lotus,Medicago,Onobrychis,Trifolium,Trigonella,Vigna,Citrus,Linum,Geranium,Manihot,Daucus,Arabidopsis,Brassica,Raphanus,Sinapis,Atropa,Capsicum,Datura,Hyoscyamus,Lycopersion,Nicotiana,Solanum,Petunia,Digitalis,Majorana,Cichorium,Helianthus,Lactuca,Bromus,Asparagus,Antirrhinum,Hererocallis,Nemesia,Pelargonium,Panicum,Pennisetum,Ranunculus,Senecio,Salpiglossis,Cucumis,Browaalia,Glycine,Lolium,Zea,Triticum,Sorghum,およびDatura。
【0102】
再生のための手法は、植物の種によって変化するが、しかし一般に異種遺伝子のコピーを含む形質転換されたプロトプラストの懸濁液が最初に提供される。カルス組織は形成され、そしてシュート(苗条)がカルスから誘導され得、続いて発根される。あるいは、胚形成がプロトプラスト懸濁液から誘導され得る。これらの胚は、天然の胚として発芽し、植物を形成する。培養培地は、一般に種々のアミノ酸、ならびにオーキシンおよびサイトカイニンのようなホルモンを含有する。またグルタミン酸およびプロリンを培地に添加することは、特にコーン(穀草)およびアルファルファのような種にとって有用である。シュートおよび根は通常、同時に発生する。効果的な再生は培地、遺伝子型、および培養遍歴に依存する。これらの3つの変数が制御される場合は、再生は十分に再現性がありそして繰り返し可能である。
【0103】
いくつかの植物細胞培養系において、本発明の所望のタンパク質は排出され、あるいはこのタンパク質は植物全体から抽出され得る。本発明の所望のタンパク質が培地内に分泌される場合、これは回収され得る。あるいは、胚および胚のない不完全な種子または他の植物組織は、機械的に破壊され細胞間および組織間の任意の分泌されたタンパク質を放出し得る。その混合物は緩衝液に懸濁され、可溶タンパクを回収し得る。次いで、慣用的なタンパク質分離および精製方法は組換えタンパク質を精製するために使用される。時間、温度、pH、酸素、および容量のパラメーターは、慣用的な方法によって、異種タンパク質の発現および回収を最適化するように調整される。
【0104】
(iv.細菌系)
細菌の発現技術は、当該分野で公知である。細菌のプロモーターは、細菌のRNAポリメラーゼに結合し得、そしてコード配列(例えば、構造遺伝子)の下流方向(3’方向)へのmRNAへの転写を開始し得る任意のDNA配列である。プロモーターは、通常、コード配列の5’末端に近接して存在する転写開始領域を有する。この転写開始領域は、通常、RNAポリメラーゼ結合部位および転写開始部位を含む。細菌のプロモーターはまた、オペレーターと呼ばれる第二のドメインを有し、RNA合成が始まる近接のRNAポリメラーゼ結合部位と重複している。オペレーターは、遺伝子リプレッサータンパク質が、オペレーターに結合し、そのため特定の遺伝子の転写を抑制し得るような、負の調節された(誘導性の)転写を可能にする。構成的発現は、オペレーターのような負の調節エレメントの非存在下で起こり得る。さらに、正の調節は、遺伝子アクチベーター(活性化因子)タンパク質結合配列により達成され得、その配列が存在する場合は通常、RNAポリメラーゼ結合配列の(5’)側に近接している。遺伝子アクチベータータンパク質の例としては、異化(カタボライト)活性化タンパク質(CAP)があり、それはEscherichia coli(E.Coli)におけるlacオペロンの転写の開始を助ける(Raibaudら(1984)Annu.Rev.Genet.18:173)。それゆえ、調節される発現は、正または負のいずれかであり得、従って転写を増強するかまたは低下し得るかのいずれかである。
【0105】
代謝経路の酵素をコードする配列は、特に有用なプロモーター配列を提供する。例としては、ガラクトース、ラクトース(lac)(Changら(1997)Nature 198:1056)、およびマルトースのような糖代謝の酵素由来のプロモーター配列が挙げられる。さらなる例としては、トリプトファン(trp)(Goeddelら(1980)Nuc.Acids Res.8:4057;Yelvertonら(1981)Nucl.Acids Res.9:731;米国特許第4,738,921号;EP−A−0036776号およびEP−A−0121775)のような生合成酵素由来のプロモーター配列が挙げられる。g−ラクタマーゼ(g−laotamase)(bla)プロモーター系(Weissmann(1981)「The cloning of Interferon and other mistakes.」Interferon 3(I.Gresser編))、バクテリオファージλPL(Shimatakeら(1981)Nature 292:128)およびT5(米国特許第4,689,406号)プロモーター系もまた有用なプロモーター配列を提供する。
【0106】
さらに、天然に存在しない合成プロモーターもまた、細菌のプロモーターとして機能する。例えば、ある細菌(バクテリア)あるいはバクテリオファージプロモーターの転写活性化配列は、別のバクテリアあるいはバクテリオファージプロモーターのオペロン配列と結合し得、合成ハイブリッドプロモーターを形成する。(米国特許第4,551,433号)。例えば、tacプロモーターは、lacリプレッサーにより調節されるtrpプロモーター配列およびlacオペロン配列の両方から構成される、ハイブリッドtrp−lacプロモーターである(Amannら(1983)Gene 25:167;de Boerら(1983)Proc.Natl.Acad.Sci.80:21)。さらに、ある細菌のプロモーターは、細菌のRNAポリメラーゼに結合し、そして転写を開始させる能力を有する非細菌起源の天然に存在するプロモーターを含み得る。非細菌起源の天然に存在するプロモーターはまた、原核生物内でいくらかの遺伝子の高いレベルでの発現を生じるために、適合性のあるRNAポリメラーゼと結合され得る。バクテリオファージT7 RNAポリメラーゼ/プロモーター系は、連結したプロモーター系の例である(Studierら(1986)J.Mol.Biol.189:113;Taborら(1985)Proc Natl.Acad.Sci.82:1074)。さらに、ハイブリッドプロモーターはまた、バクテリオファージプロモーターおよびE.coliオペレーター領域から構成され得る(EPO−A−0267851)。
【0107】
機能性のプロモーター配列に加え、効果的なリボソーム結合部位もまた、原核生物における外来遺伝子の発現に有用である。E.coliにおいて、リボソーム結合部位は、シャイン−ダルガノ(SD)配列と呼ばれ、そして開始コドン(ATG)および開始コドンの3〜11ヌクレオチド上流に位置する長さ3〜9ヌクレオチドの配列を含む(Shineら(1975)Nature 254:34)。SD配列は、SD配列とE.coliの16S rRNAの3’側との間の塩基対形成によりmRNAのリボソームへの結合を促進すると考えられている(Steitzら(1979)「Genetic Signals and nucleotide sequences in messenger RNA」Biological Regulation and Development:Gene Expression(R.F.Goldbergerら編))。弱いリボソーム結合部位を有する真核遺伝子および原核遺伝子の発現のためには(Sambrookら(1989)「Expression of cloned genes in Escherichia coli」Molecular Cloning:A Laboratory Manual)。
【0108】
DNA分子は、細胞内で発現され得る。プロモーター配列は、直接的にそのDNA分子と連結され得、この場合、N末端の最初のアミノ酸は、常に、ATG開始コドンによりコードされるメチオニンである。所望される場合、N末端のメチオニンは、臭化シアンとのインビトロインキュベーション、あるいは細菌のメチオニンN末端ペプチダーゼとのインビボまたはインビトロインキュベーションのいずれかによりタンパク質から切断され得る(EPO−A−0219237)。
【0109】
融合タンパク質は、直接的発現に代わるものを提供する。通常、内在性の細菌のタンパク質、あるいは他の安定なタンパク質のN末端部分をコードするDNA配列は、異種のコード配列の5’末端と融合される。発現の際に、この構築物は、2つのアミノ酸配列の融合を提供する。例えば、バクテリオファージλ細胞遺伝子は、外来遺伝子の5’末端と連結し得、そして細菌において発現され得る。生じた融合タンパク質は、好ましくは、外来遺伝子由来のバクテリオファージタンパク質を切断するための切断酵素(第Xa因子)用の部位を保持する(Nagaiら(1984)Nature 309:810)。融合タンパク質はまた、lacZ(Jiaら(1987)Gene 60:197)、trpE(Allenら(1987)J.Biotechnol.5:93;Makoffら(1989)J.Gen.Microbiol.135:11)、およびChey(EP−A−0324647)遺伝子由来の配列を用いて作製され得る。2つのアミノ酸配列の接合部でのDNA配列は、切断部位をコードしてもよいし、あるいはコードしなくてもよい。別の例としては、ユビキチン融合タンパク質である。そのような融合タンパク質は、好ましくは、外来タンパク質からユビキチンを切断するための切断酵素(例えば、ユビキチン特異的切断プロテアーゼ)用の部位を保持するユビキチン領域と共に作製され得る。この方法を通して、天然外来タンパク質は分離され得る(Millerら(1989)Bio/Technology 7:698)。
【0110】
あるいは、外来タンパク質はまた、細菌における外来タンパク質の分泌を提供するシグナルペプチド配列フラグメントから構成される、融合タンパク質をコードするキメラDNA分子を作製することによって、細胞から分泌され得る(米国特許第4,336,336号)。シグナル配列フラグメントは、通常、細胞からのタンパク質の分泌を指向する、疎水性アミノ酸から構成されるシグナルペプチドをコードする。このタンパク質は、増殖培地(グラム陽性細菌)、または細胞の内膜と外膜との間に位置する細胞周辺腔(グラム陰性細菌)のいずれかへ分泌される。好ましくは、インビボまたはインビトロのいずれかで切断され得る、このシグナルペプチドフラグメントと外来遺伝子との間にコードされるプロセシング部位がある。
【0111】
適切なシグナル配列をコードするDNAは、E.coli外膜タンパク質遺伝子(ompA)(Masuiら(1983)、Experimetal Manipulation of Gene Expression;Ghrayebら(1984)EMBO J.3:2437)およびE.coliアルカリホスファターゼシグナル配列(phoA)(Okaら(1985)Proc.Natl.Acad.Sci.82:7212)のような、分泌性細菌タンパク質に関する遺伝子由来であり得る。さらなる例として、種々のBacillus株由来のα−アミラーゼ遺伝子のシグナル配列は、B.subtilis由来の異種タンパク質を分泌するために使用され得る(Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582;EP−A−0244042)。
【0112】
通常、細菌によって認識される転写終結配列は、翻訳終止コドンの3’側に位置する調節領域であり、そして従って、プロモーターとともに、コード配列に隣接する。これらの配列は、そのDNAによってコードされるポリペプチドへと翻訳され得るmRNAの転写を指向する。転写終結配列は、しばしば、転写の終結を補助するステムループ構造を形成し得る、約50ヌクレオチドのDNA配列を含む。例は、強力なプロモーターを有する遺伝子(例えば、E.coliのtrp遺伝子および他の生合成遺伝子)由来の転写終結配列を含む。
【0113】
通常、上記の成分(プロモーター、シグナル配列(もし所望ならば)、目的のコード配列、および転写終結配列を含む)は、組み立てられて発現構築物となる。発現構築物は、しばしば、宿主(例えば細菌)における安定な保持が可能である染色体外エレメント(例えばプラスミド)のような、レプリコンに保持される。このレプリコンは複製系を有し、従って、このことによって、発現またはクローニングおよび増幅のいずれかのために、レプリコンが原核生物宿主において保持されることを可能になる。さらに、レプリコンは、高コピー数プラスミドまたは低コピー数プラスミドのいずれかであり得る。高コピー数プラスミドは、一般に約5〜約200、そして通常約10〜約150の範囲のコピー数を有する。高コピー数プラスミドを含む宿主は、好ましくは少なくとも約10個、そしてより好ましくは少なくとも約20個のプラスミドを含む。高コピー数ベクターまたは低コピー数ベクターのいずれかが選択され得、それは、宿主に対するベクターおよび外来タンパク質の効果に依存する。
【0114】
あるいは、発現構築物は、組み込みベクターを用いて、細菌のゲノムへ組み込まれ得る。組み込みベクターは、通常、ベクターが組み込むのを可能にする、細菌の染色体と相同な少なくとも1つの配列を含む。組み込みは、ベクターにおける相同なDNAと細菌の染色体との間の組換えから生じるようである。例えば、種々のBacillus株からのDNAによって構築される組み込みベクターは、Bacillus染色体に組み込まれる(EP−A−0 127 328)。組み込みベクターはまた、バクテリオファージまたはトランスポゾン配列から構成され得る。
【0115】
通常、染色体外発現構築物および組み込み発現構築物は、選択マーカーを含んで、形質転換された細菌株の選択を可能にし得る。選択マーカーは、細菌宿主において発現され得、そして細菌が薬物(例えば、アンピシリン、クロラムフェニコール、エリスロマイシン、カナマイシン(ネオマイシン)、およびテトラサイクリン)に耐性になるようにする遺伝子を含み得る(Daviesら(1978)Annu.Rev.Microbiol.32:469)。選択マーカーはまた、ヒスチジン、トリプトファン、およびロイシンの生合成経路における生合成遺伝子のような、生合成遺伝子を含み得る。
【0116】
あるいは、上記の成分のいくつかは、形質転換ベクターにおいて組み立てられ得る。形質転換ベクターは、通常、上記のように、レプリコンにおいて保持されるか、または組み込みベクターへと開発されるかのいずれかの選択マーカー(market)から構成され得る。
【0117】
発現ベクターまたは形質転換ベクターは、染色体外レプリコンまたは組み込みベクターのいずれも、多くの細菌への形質転換のために開発されてきた。例えば、発現ベクターは、とりわけ、以下の細菌のために開発されてきた:Bacillus subtilis(Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582;EP−A−0 036 259およびEP−A−0 063 953;WO 84/04541)、Escherichia coli(Shimatakeら(1981)Nature 292:128;Amannら(1985)Gene 40:183;Studierら(1986)J.Mol.Biol.189:113;EP−A−0 036 776、EP−A−0 136 829およびEP−A−0 136 907)、Streptococcus cremoris(Powellら(1988)Appl.Environ.Microbiol.54:655);Streptococcus lividans(Powellら(1988)Appl.Environ.Microbiol.54:655)、Streptomyces lividans(米国特許第4,745,056号)。
【0118】
外来DNAを細菌宿主へ導入する方法は、当該分野において周知であり、そして通常、CaCl2または他の薬剤(例えば、2価の陽イオンおよびDMSO)のいずれかで処理された細菌の形質転換を含む。DNAはまた、エレクトロポレーションによって、細菌細胞へ導入され得る。形質転換の手順は、通常、形質転換される細菌の種によって変化する。例えば以下を参照のこと:(Massonら(1989)FEMS Microbiol.Lett.60:273;Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582;EP−A−0 036 259およびEP−A−063 953;WO 84/04541、Bacillus)、(Millerら(1988)Proc.Natl.Acad.Sci.85:856;Wangら(1990)J.Bacteriol.172:949、Campylobacter)、(Cohenら(1973)Proc.Natl.Acad.Sci.69:2110;Dowerら(1988)Nucleic Acids Res.16:6127;Kushner(1978)「ColE1由来のプラスミドによるEscherichia coliの形質転換のための改良された方法」Genetic Engineering:Proceedings of the International Symposium on Genetic Engineering(H.W.BoyerおよびS.Nicosia編);Mandelら(1970)J.Mol.Biol.53:159;Taketo(1988)Biochim.Biophys.Acta 949:318;Escherichia)、(Chassyら(1987)FEMS Microbiol.Lett.44:173 Lactobacillus);(Fiedlerら(1988)Anal.Biochem 170:38、Pseudomonas);(Augustinら(1990)FEMS Microbiol.Lett.66:203、Staphylococcus)、(Baranyら(1980)J.Bacteriol.144:698;Harlander(1987)「エレクトロポレーションによるStreptococcus lactisの形質転換」Streptococcal Genetics(J.FerrettiおよびR.Curtiss III編);Perryら(1981)Infect.Immun.32:1295;Powellら(1988)Appl.Environ.Microbiol.54:655;Somkutiら(1987)Proc.4th Evr.Cong.Biotechnology 1:412、Streptococcus)。
【0119】
(v.酵母発現)
酵母発現系もまた、当業者に公知である。酵母プロモーターは、酵母RNAポリメラーゼに結合可能であり、そしてコード配列(例えば、構造遺伝子)からmRNAへの下流の(3’側の)転写を開始し得る、任意のDNA配列である。プロモーターは、通常、コード配列の5’末端の近位に位置する転写開始領域を有する。この転写開始領域は、通常、RNAポリメラーゼ結合部位(「TATAボックス」)および転写開始部位を含む。酵母プロモーターはまた、上流アクチベーター配列(UAS)と呼ばれる第2のドメインを有し得、これは、もし存在するならば、通常、構造遺伝子とは遠位である。このUASは、調節された(誘導できた)発現を可能にする。構成的発現は、UASの非存在下で生じる。調節される発現は、正または負のいずれかであり得、それによって転写を増加させるかまたは減少させるかのいずれかであり得る。
【0120】
酵母は、活性な代謝経路を有する発酵性生物であり、従って、代謝経路における酵素をコードする配列は、特に有用なプロモーター配列を提供する。例としては、以下が挙げられる:アルコールデヒドロゲナーゼ(ADH)(EP−A−0 284 044)、エノラーゼ、グルコキナーゼ、グルコース−6−リン酸イソメラーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(GAPまたはGAPDH)、ヘキソキナーゼ、ホスホフルクトキナーゼ、3−ホスホグリセリン酸ムターゼ、およびピルビン酸キナーゼ(PyK)(EPO−A−0 329 203)。酵母PHO5遺伝子はまた、酸性ホスファターゼをコードし、有用なプロモーター配列を提供する(Myanoharaら(1983)Proc.Natl.Acad.Sci.USA 80:1)。
【0121】
さらに、天然には生じない合成プロモーターもまた、酵母のプロモーターとして機能する。例えば、ある1つの酵母プロモーターのUAS配列は、別の酵母プロモーターの転写活性化領域と連結され得、合成ハイブリッドプロモーターを生成し得る。このようなハイブリッドプロモーターの例は、GAP転写活性化領域と連結されるADH調節配列(米国特許第4,876,197号および同第4,880,734号)を含む。ハイブリッドプロモーターの他の例は、ADH2、GAL4、GAL10、またはPHO5遺伝子のいずれかの調節配列からなり、GAPまたはPyK(EP−A−0 164 556)のような解糖酵素遺伝子の転写活性化領域に結合されているプロモーターを含む。さらに、酵母プロモーターは、酵母RNAポリメラーゼと結合し、そして転写を開始する能力を有する、天然に生じる非酵母起源のプロモーターを含み得る。このようなプロモーターの例としては、とりわけ、以下が挙げられる:(Cohenら(1980)Proc.Natl.Acad.Sci.USA 77:1078;Henikoffら(1981)Nature 283:835;Hollenbergら(1981)Curr.Topics Microbiol.Immunol.96:119;Hollenbergら(1979)「酵母Saccharomyces cerevisiaeにおける細菌の抗生物質耐性遺伝子の発現」、Plasmids of Medical,Environmental and Commercial Importance(K.N.TimmisおよびA.Puhler編);Mercerau−Puigalonら(1980)Gene 11:163;Panthierら(1980)Curr.Genet.2:109;)。
【0122】
DNA分子は、酵母において、細胞内で発現され得る。プロモーター配列は、DNA分子と直接連結され得、その場合、組換えタンパク質のN末端にある最初のアミノ酸は常にATG開始コドンによってコードされているメチオニンである。もし所望ならば、N末端のメチオニンは、臭化シアンとのインビトロインキュベーションによって、タンパク質から切断され得る。
【0123】
融合タンパク質は、酵母発現系について、ならびに哺乳動物、バキュロウイルス、および細菌の発現系において、代替物を提供する。通常、内因性酵母タンパク質、または他の安定なタンパク質のN末端部分をコードするDNA配列は、異種コード配列の5’末端に融合される。発現の際に、この構築物は、2つのアミノ酸配列の融合物を提供する。例えば、酵母またはヒトのスーパーオキシドジスムターゼ(SOD)遺伝子は、外来遺伝子の5’末端に連結され、そして酵母において発現し得る。2つのアミノ酸配列の連結部にあるDNA配列は、切断部位をコードしてもよいし、コードしなくてもよい。例えば、EP−A−0 196 056を参照のこと。別の例はユビキチン融合タンパク質である。このような融合タンパク質は、プロセシング酵素(例えば、ユビキチン特異的プロセシングプロテアーゼが)外来タンパク質からユビキチンを切断するための部位を好ましくは保持する、ユビキチン領域を伴って作製される。従って、この方法により、ネイティブ外来タンパク質は、単離され得る(例えば、WO88/024066)。
【0124】
あるいは、外来タンパク質はまた、酵母における外来タンパク質の分泌を提供する、リーダー配列フラグメントから構成される融合タンパク質をコードする、キメラDNA分子を作製することによって、細胞から増殖培地へ分泌され得る。好ましくは、インビボまたはインビトロのいずれかで切断され得る、リーダーフラグメントと外来遺伝子との間にコードされるプロセシング部位が存在する。リーダー配列フラグメントは、細胞からのタンパク質の分泌を指向する、疎水性アミノ酸から構成されるシグナルペプチドを、通常コードする。
【0125】
適切なシグナル配列をコードするDNAは、分泌性酵母タンパク質に関する遺伝子由来であり得、その遺伝子は例えば、インベルターゼ遺伝子(EP−A−0012873;JPO62,096,086)およびA因子遺伝子(米国特許第4,588,684号)である。あるいは、インターフェロンリーダーのような、酵母における分泌もまた提供する、非酵母起源のリーダーが存在する(EP−A−0060057)。
【0126】
好ましいクラスの分泌リーダーは、酵母α因子遺伝子のフラグメントを使用するリーダーであり、これは「プレ」シグナル配列、および「プロ」領域の両方を含む。用いられ得るα因子フラグメントの型は、完全長の、プレ−プロα因子リーダー(約83アミノ酸残基)および短縮されたα因子リーダー(通常約25〜約50アミノ酸残基)を含む(米国特許第4,546,083号および同第4,870,008号;EP−A−0 324 274)。分泌を提供するα因子リーダーフラグメントを使用するさらなるリーダーは、第1の酵母のプレ配列を有するが、第2の酵母α因子からのプロ領域を有しないで作製される、ハイブリッドα因子リーダーを含む(例えば、WO89/02463を参照のこと)。
【0127】
通常、酵母に認識される転写終結配列は、翻訳終止コドンの3’側に位置する調節領域であり、従って、プロモーターと共にコード配列に隣接する。これらの配列は、そのDNAにコードされるポリペプチドへと翻訳され得る、mRNAの転写を指向する。転写終結配列および他の酵母に認識される終結配列の例は、例えば、解糖酵素をコードする転写終結配列である。
【0128】
通常、上記の成分(プロモーター、リーダー(もし所望ならば)、目的のコード配列、および転写終結配列を含む)は、組み立てられて発現構築物になる。発現構築物は、宿主(例えば、酵母または細菌)において安定に保持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコンにおいてしばしば保持される。このレプリコンは、2つの複製系を有し得、従って、例えば、発現のために酵母において、ならびにクローニングおよび増幅のために原核生物宿主において保持されることを可能にする。このような酵母−細菌シャトルベクターの例としては、以下が挙げられる:YEp24(Botsteinら(1979)Gene 8:17〜24)、pCl/1(Brakeら(1984)Proc.Natl.Acad.Sci.USA 81:4642〜4646)、およびYRp17(Stinchcombら(1982)J.Mol.Biol.158:157)。さらに、レプリコンは、高コピー数プラスミドまたは低コピー数プラスミドのいずれかであり得る。高コピー数プラスミドは、一般に約5〜約200、そして通常約10〜約150の範囲のコピー数を有する。高コピー数プラスミドを含む宿主は、好ましくは少なくとも約10、そしてより好ましくは少なくとも約20を有する。高コピー数ベクターまたは低コピー数ベクターのいずれかが選択され得、それは、宿主に対するベクターおよび外来タンパク質の効果に依存する。例えば、Brakeら、前出を参照のこと。
【0129】
あるいは、発現構築物は、組み込みベクターを用いて、酵母のゲノムへ組み込まれ得る。組み込みベクターは、通常、ベクターが組み込むのを可能にする、酵母の染色体と相同な少なくとも1つの配列を含み、そして好ましくは、発現構築物に隣接する2つの相同配列を含む。組み込みは、ベクターにおける相同なDNAと酵母の染色体との間の組換えから生じるようである(Orr−Weaverら(1983)Methods in Enzymol.101:228〜245)。組み込みベクターは、そのベクター中に含有するために適切な相同配列を選択することによって、酵母における特定の遺伝子座を指向され得る。Orr−Weaverら、前出を参照のこと。1つ以上の発現構築物が組み込まれ得、産生される組換えタンパク質のレベルにおそらく影響を与え得る(Rineら(1983)Proc.Natl.Acad.Sci.USA 80:6750)。ベクターに含まれる染色体配列は、ベクターにおける単一セグメント(ベクター全体の組み込みを生じる)、または染色体における隣接セグメントに相同でかつベクターにおける発現構築物に隣接する2つのセグメント(発現構築物のみの安定した組み込みを生じ得る)のいずれかとして生じ得る。
【0130】
通常、染色体外発現構築物および組み込み発現構築物は、選択マーカーを含み得、形質転換された酵母株の選択を可能にする。選択マーカーは、酵母宿主において発現され得る生合成遺伝子(例えば、ADE2、HIS4、LEU2、TRP1、およびALG7)、ならびにG418耐性遺伝子(それぞれ、酵母細胞がツニカマイシンおよびG418に耐性になるようにする)を含み得る。さらに、適切な選択マーカーはまた、金属のような毒性化合物の存在下において増殖する能力を、酵母に提供し得る。例えば、CUP1の存在は、酵母が、銅イオンの存在下において増殖することを可能にする(Buttら(1987)Microbiol.Rev.51:351)。
【0131】
あるいは、上記成分のうちのいくつかは、組み立てられて形質転換ベクターになり得る。形質転換ベクターは、通常、上記のように、レプリコンにおいて保持される選択マーカーから構成されるか、または組み込みベクターへと構築されるかのいずれかである。
【0132】
発現ベクターおよび形質転換ベクターは、染色体外レプリコンまたは組み込みベクターのいずれかであり、多くの酵母への形質転換のために開発されてきた。例えば、発現ベクターは、とりわけ、以下の酵母のために開発されてきた:Candida albicans(Kurtzら(1986)Mol.Cell.Biol.6:142)、Candida maltosa(Kunzeら(1985)J.Basic Microbiol.25:141)。Hansenula polymorpha(Gleesonら(1986)J.Gen.Microbiol.132:3459;Roggenkampら(1986)Mol.Gen.Genet.202:302)、Kluyveromyces fragilis(Dasら(1984)J.Bacteriol.158:1165)、Kluyveromyces lactis(De Louvencourtら(1983)J.Bacteriol.154:737;Van den Bergら(1990)Bio/Technology 8:135)、Pichia guillerimondii(Kunzeら(1985)J.Basic Microbiol.25:141)、Pichia pastoris(Creggら(1985)Mol.Cell.Biol.5:3376;米国特許第4,837,148号および同第4,929,555号)、Saccharomyces cerevisiae(Hinnenら(1978)Proc.Natl.Acad.Sci.USA 75:1929;Itoら(1983)J.Bacteriol.153:163)、Schizosaccharomyces pombe(BeachおよびNurse(1981)Nature 300:706)、およびYarrowia lipolytica(Davidowら(1985)Curr.Genet.10:380471;Gaillardinら(1985)Curr.Genet.10:49)。
【0133】
外来DNAを酵母宿主へ導入する方法は、当該分野において周知であり、そして通常、スフェロプラストの、またはアルカリ陽イオンで処理されたインタクトな酵母細胞のいずれかの形質転換を含む。形質転換の手順は、通常、形質転換される酵母の種によって変化する。例えば以下を参照のこと:(Kurtzら(1986)Mol.Cell.Biol.6:142;Kunzeら(1985)J.Basic Microbiol.25:141;Candida);(Gleesonら(1986)J.Gen.Microbiol.132:3459;Roggenkampら(1986)Mol.Gen.Genet.202:302;Hansenula);(Dasら(1984)J.Bacteriol.158:1165;De Louvencourtら(1983)J.Bacteriol.154:1165;Van den Bergら(1990)Bio/Technology 8:135;Kluyveromyces);(Creggら(1985)Mol.Cell.Biol.5:3376;Kunzeら(1985)J.Basic Microbiol.25:141;米国特許第4,837,148号および同第4,929,555号;Pichia);(Hinnenら(1978)Proc.Natl.Acad.Sci.USA 75:1929;Itoら(1983)J.Bacteriol.153:163 Saccharomyces);(BeachおよびNurse(1981)Nature 300:706;Schizosaccharomyces);(Davidowら(1985)Curr.Genet.10:39;Gaillardinら(1985)Curr.Genet.10:49;Yarrowia)。
【0134】
(薬学的組成物)
薬学的組成物は、本発明のポリペプチドおよび/または核酸を含み得る。この薬学的組成物は、治療上有効な量の、本願発明のポリペプチド、抗体、またはポリヌクレオチドのいずれかを含む。
【0135】
本明細書において使用される用語「治療上有効な量」とは、所望の疾患または状態を処置、改善、または予防するための治療薬剤の量、または、検出可能な治療効果または予防効果を示すための治療薬剤の量をいう。この効果は、例えば、キメラマーカーまたは抗原レベルによって検出され得る。治療効果はまた、体温低下のような、身体の症状における減少を含む。被験体に関する正確な有効量は、被験体の大きさおよび健康、状態の性質および程度、および投与のために選択される治療または治療の組合せに依存する。従って、あらかじめ正確な有効量を特定することは有用ではない。しかし、所定の情況のための有効量は、慣用的な実験によって決定され得、そして臨床医の判断内である。
【0136】
本発明の目的のために、有効な用量は、DNA構築物が投与される個体において、約0.01mg/kg〜50mg/kgまたは0.05mg/kg〜約10mg/kgのDNA構築物である。
【0137】
薬学的組成物はまた、薬学的に受容可能なキャリアを含み得る。用語「薬学的に受容可能なキャリア」とは、抗体またはポリペプチド、遺伝子、および他の治療薬剤のような、治療薬剤の投与のためのキャリアをいう。この用語は、この組成物を与えられる個体に有害な抗体の産生をそれ自体誘導せず、そして過度の毒性を伴わずに投与され得る、任意の薬学的キャリアをいう。適切なキャリアは、タンパク質、多糖、ポリ乳酸、ポリグリコール酸、ポリマーアミノ酸、アミノ酸コポリマー、および不活性ウイルス粒子のように、大きく、遅く代謝される高分子であり得る。このようなキャリアは、当業者に周知である。
【0138】
薬学的に受容可能な塩が、その中で使用され得る。例えば、塩酸塩、臭化水素酸塩、リン酸塩、硫酸塩などのような鉱酸塩;および酢酸塩、プロピオン酸塩、マロン酸塩、安息香酸塩などのような有機酸の塩である。薬学的に受容可能な賦形剤の徹底的な議論は、Remington’s Pharmaceutical Sciences(Mack Pub.Co.、N.J.1991)にて利用可能である。
【0139】
治療組成物における薬学的に受容可能なキャリアは、水、生理的食塩水、グリセロールおよびエタノールのような液体を含み得る。さらに、湿潤剤または乳化剤、pH緩衝物質などのような補助物質が、このようなビヒクルに存在し得る。代表的には、治療組成物は、液体溶液または懸濁液のいずれかの、注射可能物質として調製される;注射前に液体ビヒクルに溶解または懸濁するのに適切な固体形態もまた、調製され得る。リポソームは、薬学的に受容可能なキャリアの定義中に含まれる。
【0140】
(送達方法)
一旦処方されると、本発明の組成物は、その被験体へ直接投与され得る。処置される被験体は、動物であり得;特に、ヒト被験体が処置され得る。
【0141】
その組成物の直接送達は、一般的に、皮下に、腹腔内に、静脈内に、または筋肉内のいずれかでの注入によって達成されるか、あるいは、組織の間質空間へ送達される。この組成物はまた、病巣へ投与され得る。他の投与様式には、経口投与、および肺投与、坐剤、および経皮(transdermal)適用または経皮(transcutaneous)適用(例えば、WO98/20734を参照のこと)、針、および遺伝子銃またはハイポスプレー(hypospray)が含まれる。投薬処置は、単回用量スケジュール、または多数回用量スケジュールであり得る。
【0142】
(ワクチン)
本発明に従うワクチンは、予防的(すなわち、感染を予防するため)または治療(すなわち、感染後の疾患を処置するため)のいずれかであり得る。
【0143】
このようなワクチンは、免疫抗原、免疫原、ポリペプチド、タンパク質または核酸を、通常「薬学的に受容可能なキャリア」とともに含み、このキャリア自体は、その組成物を受ける個体に有害である抗体の産生を誘発しない任意のキャリアを含む。適切なキャリアは、代表的に、大きく、ゆっくり代謝される高分子(例えば、タンパク質、ポリサッカリド、ポリ乳酸、ポリグリコール酸、ポリマーアミノ酸、アミノ酸コポリマー、脂質凝集物(例えば、油小滴またはリポソーム)、および不活性ウイルス粒子である。このようなキャリアは、当業者に周知である。さらに、これらのキャリアは免疫刺激剤(「アジュバント」)として機能し得る。さらに、この抗原または免疫原は、細菌毒素(例えば、ジフテリア、破傷風、コレラ、H.pyloriなどの病原因子からの毒素)と結合体化され得る。
【0144】
この組成物の効力を増強するために好ましいアジュバントとしては、(1)アルミニウム塩(「明礬」)(例えば、水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムなど)、(2)o/wエマルジョン処方物(他の特定の免疫刺激剤(例えば、ムラミルペプチド(以下を参照のこと)または細菌細胞壁成分)を伴うか伴わない)が挙げられるが、それらに限定されず、例えば、以下:(a)5%スクアレン、0.5% Tween 80、および0.5% Span85(必要に応じて、種々の量のMTP−PE(以下を参照のこと)を含有するが、必要ではない)を含み、モデル110Y微小流体化器(Microfluidics、Newton、MA)のような微小流体化器を用いてミクロン未満の粒子へと処方されたMF59TM(WO90/14837;Vaccine design:the subunit and adjuvant approachの第10章、PowellおよびNewman編、Plenum Press 1995);(b)ミクロン未満のエマルジョンへと微小流体化されたか、またはボルテックスして、より大きな粒子径エマルジョンを生成したかのいずれかである、10%スクアレン、0.4%Tween80、5%プルロニックブロックポリマーL121、およびthr−MDP(以下を参照のこと)を含有するSAF、ならびに(c)2%スクアレン、0.2%Tween80、およびモノホスホリルリピドA(MPL)、トレハロースジミコレート(TDM)、および細胞壁骨格(CWS)好ましくはMPLおよびCWS(DetoxTM)からなる群由来の1つ以上の細菌細胞壁成分を含むRibiTMアジュバント系(RAS)、(Ribi Immunochem、Hamilton、MT);(3)サポニンアジュバント(例えば、StimulonTM)(Cambridge Bioscience、Worcester、MA)を使用し得るか、またはそれから粒子(例えば、ISCOM(免疫刺激性複合体)を生成し得る;(4)完全フロイントアジュバント(CFA)および不完全フロイントアジュバント(IFA);(5)サイトカイン(例えば、インターロイキン(例えば、IL−1、IL−2、IL−4、IL−5、IL−6、IL−7、IL−12など)、インターフェロン(例えば、γインターフェロン)、マクロファージコロニー刺激因子(M−CSF)、腫瘍壊死因子(TNF)など;および(6)その組成物の効力を強化するための免疫刺激因子として作用する他の物質である。ミョウバンおよびMF59TMが好ましい。
【0145】
上記で言及したように、ムラミルペプチドとしては、N−アセチル−ムラミル−L−スレオニル−D−イソグルタミン(thr−MDP)、N−アセチル−ノルムラミル−L−アラニル−D−イソグルタミン(ノル−MDP)、N−アセチルムラミル−L−アラニル−D−イソグルタミニル−L−アラニン−2−(1’−2’−ジパルミトイル−sn−グリセロ−3−ヒドロキシホスホリルオキシ)−エチルアミン(MTP−PE)などが挙げられるがそれらに限定されない。
【0146】
免疫原性組成物(例えば、免疫抗原/免疫原/ポリペプチド/タンパク質/核酸、薬学的に受容可能なキャリア、およびアジュバント)は、代表的に、希釈剤(例えば、水、生理食塩水、グリセロール、エタノールなど)を含有する。さらに、補助物質(例えば、湿潤剤または乳化剤、pH緩衝物質など)は、このようなビヒクルにおいて存在し得る。
【0147】
代表的に、免疫原性組成物は、液体溶液または懸濁物として、注射剤として調製され;注射前に液体ビヒクル中の溶液または懸濁物として適切な固体形態もまた調製され得る。この調製物はまた、薬学的に受容可能なキャリアの下で、上記に記載のように、アジュバント効果の強化のために乳化され得るかまたはリポソーム中にカプセル化され得る。
【0148】
ワクチンとして使用される免疫原性組成物は、免疫学的有効量の抗原性または免疫原性のポリペプチド、および任意の他の上記の成分を必要に応じて含む。「免疫学的有効量」とは、その量の個体への投与が、単回用量であれ、一連の(用量の)一部としてであれ、処置または予防に有効であることを意味する。この量は、処置される個体の健康および身体状態、処置される個体の分類学上の群(例えば、非ヒト霊長類、霊長類など)、個体の免疫系が抗体を合成する能力、所望される保護の程度、そのワクチンの処方物、処置する医師の医療的状況の評価、および他の関連する因子に依存して変動する。その量は、比較的広い範囲に入り、この量が慣用的な試行を通して決定され得ることが予想される。
【0149】
免疫学的組成物は、従来のように、非経口的(例えば、皮下、筋肉内または経皮(transudermally)/経皮(transucutaneously)のいずれかでの注射による)((例えば、WO98/20734)に投与される)。他の投与様式に適切なさらなる処方物は、経口処方物および肺処方物、坐剤、および経皮適用を含む。投薬処置は、単回用量スケジュールまたは多数回用量スケジュールであり得る。ワクチンは、他の免疫調節剤とともに投与され得る。
【0150】
タンパク質ベースのワクチンの代替として、DNAワクチンを使用し得る(例えば、RobinsonおよびTorres(1997)Seminars in Immunology 9:271−283;Donnellyら(1997)Annu Rev Immunol 15:617−648;本明細書中後半部を参照のこと)。
【0151】
(遺伝子送達ビヒクル)
本発明の治療剤のコード配列を含む、哺乳動物における発現のためにその哺乳動物へ送達される構築物の送達のための遺伝子治療ビヒクルは、局所または全身的のいずれかで投与され得る。これらの構築物は、ウイルスベクターアプローチまたは非ウイルスベクターアプローチを、インビボまたはエキソビボの様式で利用し得る。このようなコード配列の発現は、内因性哺乳動物プロモーターまたは外因性プロモーターを用いて誘導され得る。このコード配列のインビボでの発現は、構成性または調節性のいずれかであり得る。
【0152】
本発明は、意図された核酸配列を発現し得る遺伝子送達ビヒクルを含む。この遺伝子送達ビヒクルは、好ましくは、ウイルスベクター、およびより好ましくはレトロウイルスベクター、アデノウイルスベクター、アデノ随伴ウイルス(AAV)ベクター、ヘルペスウイルスベクターまたはαウイルスベクターである。このウイルスベクターはまた、アストロウイルスベクター、コロナウイルスベクター、オルトミクソウイルスベクター、パポバウイルスベクター、パラミクソウイルスベクター、パルボウイルスベクター、ピコルナウイルスベクター、ポックスウイルスベクター、またはトガウイルスベクターであり得る。一般的には、Jolly(1994)Cancer Gene Therapy 1:51−64;Kimura(1994)Human Gene Therapy 5:845−852;Connelly(1995)Human Gene Therapy 6:185−193;およびKaplitt(1994)Nature Genetics 6:148−153を参照のこと。
【0153】
レトロウイルスベクターは、当該分野で周知であり、そして本発明者らは、任意のレトロウイルス遺伝子治療ベクターが本発明において使用可能であることを意図する。これには、B型、C型およびD型のレトロウイルス、異種栄養性ウイルス(例えば、NZB−X1、NZB−X2およびNZB9−1(O’Neill(1985)J.Virol.53:160を参照のこと)多栄養性レトロウイルス(例えば、MCFおよびMCF−MLV(Kelly(1983)J.Virol.45:291を参照のこと)、スプマウイルスおよびレンチウイルスが含まれる。RNA Tumor Viruses、第2版、Cold Spring Harobor Laboratory、1985を参照のこと。
【0154】
レトロウイルス遺伝子治療ベクターの部分は、異なるレトロウイルスに由来し得る。例えば、レトロウイルスLTRは、マウス肉腫ウイルスに由来し得、tRNA結合部位はラウス肉腫ウイルスに由来し得、パッケージングシグナルはマウス白血病ウイルスに由来し得、そして第二の鎖合成の起源はトリ白血病ウイルスに由来し得る。
【0155】
これらの組換えレトロウイルスベクターを使用して、適切なパッケージング細胞株へそれらを導入することによって形質導入適合性レトロウイルスベクター粒子を生成し得る(米国特許第5,591,624号を参照のこと)。レトロウイルスベクターは、レトロウイルス粒子へのキメラインテグラーゼ酵素の組込みによって宿主細胞DNAへの部位特異的組込みについて構築され得る(WO96/37626号を参照のこと)。この組換えウイルスベクターは複製欠損組換えウイルスであることが好ましい。
【0156】
上記に記載のレトロウイルスベクターを伴う使用について適切なパッケージング細胞株は、当該分野で周知であり、容易に調製され(WO95/30763号およびWO92/05266号を参照のこと)、そしてこれを使用して、組換えベクター粒子の生産のためのプロデューサー細胞株(これは、ベクター細胞株または「VCL」とも称される)を作製し得る。好ましくは、このパッケージング細胞株は、ヒトの親細胞(例えば、HT1080細胞)またはミンク親細胞株から作製され、これは、ヒト血清における不活化を除去する。
【0157】
レトロウイルス遺伝子治療ベクターの構築のために好ましいレトロウイルスは、トリ白血病ウイルス、ウシ白血病ウイルス、マウス白血病ウイルス、ミンク細胞フォーカス形成ウイルス、マウス肉腫ウイルス、細網内皮症ウイルス、およびラウス肉腫ウイルスを含む。特に好ましいマウス白血病ウイルスは、4070Aおよび1504A(HartleyおよびRowe(1976)J. Virol.19:19−25)、Abelson(ATCC番号VR−999)、Friend(ATCC番号VR−245)、Graffi、Gross(ATCC番号VR−590)、Kirsten、Harvey肉腫ウイルスおよびRauscher(ATCC番号VR−998)およびモロニーマウス白血病ウイルス(ATCC番号VR−190)を含む。このようなレトロウイルスベクターは、寄託機関または収集機関(例えば、アメリカンタイプカルチャーコレクション(「ATCC」)、Rockville、Maryland)から入手し得るか、または一般に利用可能な技術を用いて公知の供給源から単離され得る。
【0158】
本発明において使用可能な例示的な公知のレトロウイルス遺伝子治療ベクターは、特許出願GB2200651、EP0415731、EP0345242、EP0334301、WO89/02468;WO89/05349、WO89/09271、WO90/02806、WO90/07936、WO94/03622、WO93/25698、WO93/25234、WO93/11230、WO93/10218、WO91/02805、WO91/02825、WO95/07994、米国特許第5,219,740号、同4,405,712号、同4,861,719号、同4,980,289号、同4,777,127号、同5,591,624号に記載されるものを含む。Vile(1993)Cancer Res 53:3860−3864;Vile(1993)Cancer Res.53:962−967;Ram(1993)Cancer Res 53(1993)83−88;Takamiya(1992)J Neurosci Res 33:493−503;Baba(1993)J Neurosurg 79:729−735;Mann(1983)Cell 33:153;Cane(1984)Proc Natl Acad Sci 81;6349;およびMiller(1990)Human Gene Therapy 1もまた参照のこと。
【0159】
ヒトアデノウイルス遺伝子治療ベクターもまた当該分野で公知であり、そして本発明において使用可能である。例えば、Berkner(1988)Biotechniques 6:616およびRosenfeld(1991)Science 252:431;ならびにWO93/07283、WO93/06223、およびWO93/07282を参照のこと。本発明において使用可能な例示的な公知のアデノウイルス遺伝子治療ベクターは、上記に参照される文書およびWO94/12649、WO93/03769、WO93/19191、WO94/28938、WO95/11984、WO95/00655、WO95/27071、WO95/29993、WO95/34671、WO96/05320、WO94/08026、WO94/11506、WO93/06223、WO94/24299、WO95/14102、WO95/24297、WO95/02697、WO94/28152、WO94/24299、WO95/09241,WO95/25807、WO95/05835、WO94/18922およびWO95/09654において記載されるものを含む。あるいは、Curiel(1992)Hum.Gene Ther.3:147−154に記載されるような殺傷したアデノウイルスに連結したDNAの投与が使用され得る。本発明の遺伝子送達ビヒクルはまた、アデノウイルス随伴ウイルス(AAV)ベクターを含む。本発明における使用のためのこのようなベクターの主要なおよび好ましい例は、Srivastava WO93/09239に開示されるAAV−2ベースのベクターである。最も好ましいAAVベクターは、2つのAAV逆方向末端反復を含む。ここで、ネイティブD配列は、ヌクレオチドの置換によって改変され、その結果、少なくとも5つのネイティブなヌクレオチドおよび18までのネイティブヌクレオチド、好ましくは少なくとも10のネイティブヌクレオチドから18までのネイティブヌクレオチド、最も好ましくは10のネイティブヌクレオチドが維持され、そしてD配列の残りのヌクレオチドが欠失またはネイティブでないヌクレオチドで置換されている。AAV逆方向末端反復のネイティブなD配列は、各AAV逆方向末端反復において(すなわち、各末端に1つの配列が存在する)20の連続するヌクレオチドの配列であって、これは、HP形成に関与しない。ネイティブでない置換ヌクレオチドは、同じ位置でのネイティブなD配列に見出されるヌクレオチド以外の任意のヌクレオチドであり得る。他の使用可能な例示的なAAVベクターは、、pWP−19、pWN−1であり、これらは両方ともNahreini(1993)Gene 124:257−262に開示される。このようなAAVベクターの別の例は、psub201(Samulski(1987)J.Virol.61:3096を参照のこと)である。別の例示的なAAVベクターは、Double−D ITRベクターである。Double−D ITRベクターの構築は、米国特許第5,478,745号に開示される。なお他のベクターは、Carter 米国特許第4,797,368号およびMuzyczka 米国特許第5,139,941号、Chartejee 米国特許第5,474,935号ならびにKotin WO94/288157に開示されるものである。本発明において使用可能なAAVベクターのなおさらなる例は、SSV9AFABTKneoであり、これは、AFPエンハンサーおよびアルブミンプロモーターを含み、そして肝臓において優性に発現を指向する。その構造および構築は、Su(1996)Human Gene Therapy 7:463−470に開示される。さらなるAAV遺伝子治療ベクターは、米国特許第5,354,678号、同5,173,414号、同5,139,941号、および同5,252,479号に記載される。
【0160】
本発明の遺伝子治療ベクターはまた、ヘルペスウイルスベクターを含む。主要なおよび好ましい例は、チミジンキナーゼポリペプチドをコードする配列を含む単純ヘルペスウイルスベクター(例えば、米国特許第5,288,641号、およびEP0176170(Roizman)に開示されるもの)である。さらなる例示的な単純ヘルペスウイルスベクターは、WO95/04139(Wistar)に開示されるHFEM/ICP6−lacZ、Geller(1988)Science 241:1667−1669ならびにWO90/09441およびWO92/07945に記載されるpHSVlac、Fink(1992)Human Gene Therapy 3:11−19に記載されるHSV Us3::pgC−lacZ、ならびにEP0453242(Breakefieled)に記載されるHSV 7134、2RH 105およびGAL4、ならびにATCCに受託番号ATCC VR−977およびATCC VR−260として寄託されたものを含む。
【0161】
意図されるのはまた、本発明において使用され得るαウイルス遺伝子治療ベクターである。好ましいαウイルスベクターは、シンドビスウイルスベクターである。トガウイルス、セムリキ森林ウイルス(ATCC VR−67;ATCC VR−1247)、Middlebergウイルス(ATCC VR−370)、ロスリバーウイルス(ATCC VR−373;ATCC VR−1246)、ベネズエラウマ脳脊髄炎ウイルス(ATCC VR−923;ATCC VR−1250;ATCC VR−1249;ATCC VR−532)および米国特許第5,0091,309号、同5,217,879号、およびWO92/10578に記載されるもの。より詳細には、米国特許出願第08/405,627号(1995年3月15日出願)、WO94/21792号、WO92/10578号、WO95/07994号、米国特許第5,091,309号、および米国特許第5,217,879号に記載されるそれらのαウイルスベクターが使用可能である。このようなαウイルスは、ATCC、Rockville、Marylandのような寄託機関または収集機関から入手し得るか、または一般的に利用可能な技術を用いて公知の供給源から単離され得る。好ましくは、細胞傷害性が減少したαウイルスベクターを使用する(米国特許仮出願08/679640号を参照のこと)。
【0162】
DNAベクター系(例えば、真核細胞層状発現系)もまた、本発明の核酸の発現について有用である。真核生物層状発現系の詳細な説明についてはWO95/07994を参照のこと。好ましくは、本発明の真核細胞層状発現系はαウイルスベクターに由来し、そして最も好ましくはシンドビスウイルスベクターに由来する。
【0163】
本発明における使用に適切な他のウイルスベクターは、ポリオウイルス(例えば、ATCC VR−58およびEvans、Nature339(1989)385およびSabin(1973)J.Biol.Standardization 1:115に記載されるもの;ライノウイルス、例えば、ATCC VR−1110およびArnold(1990)J Cell Biochem L401に記載されるもの;ポックスウイルス(例えば、カナリアポックスルウイルスまたはワクシニアウイルス(例えば、ATCC VR−111およびATCC VR−2010ならびにFisher−Hoch(1989)Proc Natl Acad Sci 86:317;Flexner(1989)Ann NY Acad Sci 569:86、Flexner(1990)Vaccine 8:17;米国特許第4,603,112号および同4,769,330号ならびにWO89/01973号に記載されるもの));SV40ウイルス(例えば、ATCC VR−305およびMulligan(1979)Nature 277:108およびMadzak(1992)J Gen Virol 73:1533に記載されるもの);インフルエンザウイルス(例えば、ATCC VR−797および米国特許第5,166,057号およびEnami(1990)Proc Natl Acad Sci .87:3802−3805;EnamiおよびPalese(1991)J Virol 65:2711−2713およびLuytjes(1989)Cell 59:110(McMichael(1983)NEJ Med 309:13ならびにYap(1978)Nature 273:238およびNature(1979)277:108もまた参照のこと)に記載されるような逆遺伝子技術を使用して作製した組換えインフルエンザウイルス);EP−0386882およびBuchschachler(1992)J.Virol.66:2731に記載されるようなヒト免疫不全ウイルス;麻疹ウイルス(例えば、ATCC VR−67およびVR−1247ならびにEP−0440219に記載されるもの);アウラウイルス(例えば、ATCC VR−368);ベバルウイルス(例えば、ATCC VR−600およびATCC VR−1240);カバス(Cabassou)ウイルス(例えば、ATCC VR−922);チクングンヤウイルス(例えば、ATCC VR−64およびATCC VR−1241);フォートモーガン(Fort Morgan)ウイルス(例えば、ATCC VR−924);ゲタウイルス(例えば、ATCC VR−369およびATCC VR−1243);キジラガハ(Kyzylagach)ウイルス(例えば、ATCC VR−927);マヤロウイルス(例えば、ATCC VR−66);ムカンボウイルス(例えば、ATCC VR−580およびATCC VR−1244);ヌヅムウイルス(例えば、ATCC VR−371);ピクスナウイルス(例えば、ATCC VR−372およびATCC VR−1245);トナテ(Tonate)ウイルス(例えば、ATCC VR−925);トリニティウイルス(例えば、ATCC VR−469);ユナウイルス(例えば、ATCC VR−374);ワタロアウイルス(例えば、ATCC VR−926);Y−62−33ウイルス(例えば、ATCC VR−375);オニオンウイルス、東部ウマ脳脊髄炎ウイルス(例えば、ATCC VR−65およびATCC VR−1242);西部ウマ脳脊髄炎ウイルス(例えば、ATCC VR−70、ATCC VR−1251、ATCC VR−622およびATCC VR−1252);ならびにコロナウイルス(例えば、ATCC VR−740)およびHamre(1966)Proc Soc Exp Biol Med 121:190に記載のものに由来するウイルスベクターを含む。
【0164】
本発明の組成物の細胞への送達は、上記に言及したウイルスベクターに限定されない。他の送達方法および媒体が使用され得る(例えば、核酸発現ベクター、殺傷したアデノウイルスに連結したかまたは連結してないポリカチオン性縮合DNA単独(例えば、米国特許出願番号08/366,787(1994年12月30日出願)およびCuriel(1992)Hum Gene Ther 3:147−154を参照のこと)、リガンド連結DNA(例えば、Wu(1989)J Biol Chem 264:16985−16987を参照のこと)、真核生物細胞送達ビヒクル細胞(例えば、米国特許出願番号08/240,030(1994年5月9日出願)および米国特許出願番号08/404,796を参照のこと)、光重合化ヒドロゲル物質の沈着、手動の遺伝子送達粒子銃(米国特許第5,149,655号に記載されるような)、米国特許第5,206,152およびWO92/11033に記載されるような電離放射線、核酸電荷中和または細胞膜との融合)。さらなるアプローチは、Philip(1994)Mol Cell Biol 14:2411−2418およびWoffendin(1994)Proc Natl Acad Sci 91:1581−1585に記載される。
【0165】
粒子媒介遺伝子送達が使用され得る(例えば、米国特許出願60/023,867号を参照のこと)。手短には、配列を、高レベル発現のための従来の制御配列を含む従来のベクターに挿入し得、次いで細胞標的化リガンド(例えば、アシアロオロソムコイド(WuおよびWu(1987)J.Biol.Chem.262:4429−4432に記載されるような)、Hucked(1990)Biochem Pharmacol 40:253−263に記載されるようなインスリン、Plank(1992)Bioconjugate Chem 3:533−539に記載されるようなガラクトース、ラクトースまたはトランスフェリン)に連結された合成遺伝子送達分子(例えば、ポリリジン、プロタミンおよびアルブミンのような重合DNA結合カチオン)とともにインキュベートされ得る。
【0166】
裸のDNAもまた使用され得る。例示的な裸のDNA導入方法は、WO90/11092および米国特許第5,580,859号に記載される。取り込み効率は、生体分解性のラテックスビーズを用いて改良され得る。DNAコートラテックスビーズは、ビーズによるエンドサイトーシス開始の後に効率よく細胞へと輸送される。この方法は、ビーズを処理して疎水性を高め、それによってエンドソームの破壊および細胞質へのDNAの放出を容易にすることによってさらに改良され得る。
【0167】
遺伝子送達ビヒクルとして作用し得るリポソームは、米国特許第5,422,120号、WO95/13796、WO94/23697、WO91/14445、およびEP524,968に記載される。米国特許出願60/023,867に記載されるように、非ウイルス性送達において、ポリペプチドをコードする核酸配列は、高レベル発現のための従来の制御配列を含む従来のベクターへと挿入され得、次いで細胞標的化リガンド(例えば、アシアロオロソムコイド、インスリン、ガラクトース、ラクトースまたはトランスフェリン)に連結された、重合性DNA結合カチオン(例えば、ポリリジン、プロタミン、およびアルブミン)のような合成遺伝子伝達分子とともにインキュベートされ得る。他の送達系は、種々の組織特異的または普遍的作用性のプロモーターの制御下に遺伝子を含むDNAをカプセル化するためのリポソームの使用を含む。さらに、使用に適切な非ウイルス送達は、機械的送達系(例えば、Woffendinら(1994)Proc.Natl.Acad.Sci.USA 91(24):11581−11585に記載されるアプローチを含む。さらに、コード配列およびそのようなものの発現産物は、光重合化ヒドロゲル物質の沈着を介して送達され得る。コード配列の送達について使用され得る、遺伝子送達のための他の従来の方法は、例えば、手動の遺伝子送達粒子銃(米国特許第5,149,655号に記載されるような);移入された遺伝子を活性化するための電離放射線の使用(米国特許第5,206,152号およびWO92/11033に記載されるような)を含む。
【0168】
例示的なリポソームおよびポリカチオン性遺伝子送達ビヒクルは、米国特許第5,422,120号および同4,762,915号;WO95/13796;WO94/23697;およびWO91/14445;EP−0524968;およびStryer、Biochemistry、236−240頁(1975)、W.H.Freeman、San Francisco;Szoka(1980)Biochem Biophys Acta 600:1;Bayer(1979) Biochem Biophys Acta 550:464;Rivnay(1987)Meth Enzymol 149:119;Wang(1987)Proc Natl Acad Sci 84:7851;Plant(1989)Anal Biochem 176:420に記載されるものである。
【0169】
ポリヌクレオチド組成物は、治療有効量(この用語は上記に定義されるとおりである)の遺伝子治療ビヒクルを含み得る。本発明の目的のために、有効用量は、投与される個体において、約0.01mg/kg〜50mg/kgまたは0.05mg/kg〜約10mg/kgのDNA構築物である。
【0170】
(送達方法)
一旦処方されると、本発明のポリヌクレオチド組成物は、(1)被験体に直接;(2)エキソビボで被験体由来の細胞に送達されて;または(3)組換えタンパク質の発現のためにインビトロで、投与され得る。処置される被験体は、哺乳動物または鳥類であり得る。ヒト被験体もまた処置され得る。
【0171】
この組成物の直接送達は、皮下、腹腔内、静脈内、または筋肉内の注射によって、または組織の間質空間への送達のいずれかによって一般的に達成される。この組成物はまた、病巣へ投与され得る。他の投与様式は、経口投与または肺投与、坐剤、および経皮(transdermal)または経皮(transcutaneous)適用(例えば、WO98/20734を参照のこと)、針、および遺伝子銃またはハイポスプレーを含む。投薬治療は、単回用量スケジュールまたは多数回用量スケジュールであり得る。
【0172】
エキソビボ送達および被験体への形質転換細胞の再移植のための方法は、当該分野で公知であり、そして例えばWO93/14778に記載されている。エキソビボ適用に有用である細胞の例は、例えば、幹細胞、特に、造血細胞、リンパ細胞、マクロファージ、樹状細胞または腫瘍細胞を含む。
【0173】
一般的に、エキソビボ適用およびインビトロ適用の両方のための核酸の送達は、以下の手順:例えば、デキストラン媒介トランスフェクション、リン酸カルシウム沈降、ポリブレン媒介トランスフェクション、原形質融合、エレクトロポレーション、ポリヌクレオチドのリポソーム内へのカプセル化、およびDNAの核への直接の微量注入(これらはすべて当該分野で周知である)で達成され得る。
【0174】
(ポリヌクレオチドおよびポリペプチドの薬学的組成物)
上記に記載の薬学的に受容可能なキャリアおよび塩に加えて、以下のさらなる薬剤がポリヌクレオチド組成物および/またはポリペプチド組成物とともに使用され得る。
【0175】
(A.ポリペプチド)
1つの例は、限定することなく以下を包含する:アシアロオロソムコイド(ASOR);トランスフェリン;アシアロ糖タンパク質;抗体;抗体フラグメント;フェリチン;インターロイキン;インターフェロン;顆粒球マクロファージコロニー刺激因子(GM−CSF)、顆粒球コロニー刺激因子(G−CSF)、マクロファージコロニー刺激因子(M−CSF)、幹細胞因子およびエリスロポエチン。ウイルス抗原(例えば、エンベロープタンパク質)もまた、使用され得る。また、他の侵襲性生物由来のタンパク質(例えば、RIIとして知られるPlasmodium falciparumの環境スポロゾイト(circumsporozoite)タンパク質由来の17アミノ酸ペプチド)。
【0176】
(B.ホルモン、ビタミンなど)
包含され得る他の群は、例えば、ホルモン、ステロイド、アンドロゲン、エストロゲン、甲状腺ホルモン、またはビタミン、葉酸である。
【0177】
(C.ポリアルキレン、ポリサッカリドなど)
また、ポリアルキレングリコールが、所望のポリヌクレオチド/ポリペプチドとともに含有され得る。好ましい実施態様において、ポリアルキレングリコールは、ポリエチレングリコールである。さらに、モノサッカリド、ジサッカリド、またはポリサッカリドが含有され得る。この局面の好ましい実施態様において、このポリサッカリドは、デキストランまたはDEAE−デキストランである。また、キトサンおよびポリ(乳酸−コ−グリコリド)。
【0178】
(D.脂質およびリポソーム)
所望のポリヌクレオチド/ポリペプチドはまた、被験体またはそれに由来する細胞への送達の前に、脂質中にカプセル化され得るか、またはリポソーム中にパッケージングされ得る。
【0179】
脂質カプセル化は、一般的に、核酸に安定に結合し得るか、または核酸を捕捉もしくは維持し得るリポソームを用いて達成される。縮合ポリヌクレオチドの脂質調製物に対する比は、変動し得るが、一般的に約1:1(mgDNA:マイクロモル脂質)であるか、またはより多くの脂質である。核酸の送達のためのキャリアとしてのリポソーム使用の概説については、HugおよびSleight(1991)Biochim.Biophys.Acta.1097:1−17;Straubinger(1983)Meth.Enzymol.101:512−527を参照のこと。
【0180】
本発明における使用のためのリポソーム調製物は、カチオン性(正に荷電した)、アニオン性(負に荷電した)および中性の調製物を包含する。カチオン性リポソームは、機能的な形態で、プラスミドDNA(Felgner(1987)Proc.Natl.Acad.Sci.USA 84:7413−7416);mRNA(Malone(1989)Proc.Natl.Acad.Sci.USA 86:6077−6081;および精製した転写因子(Debs(1990)J.Biol.Chem.265:10189−10192)の細胞内送達を媒介することが示されている。
【0181】
カチオン性リポソームは容易に入手可能である。例えば、N{1−2,3−ジオレイルオキシ)プロピル}−N,N,N−トリエチルアンモニウム(DOTMA)リポソームは、GIBCO BRL、Grand Island、NYからの商標リポフェクチン(Lipofectin)の下で入手可能である(Fegner前出もまた参照のこと)。他の市販されているリポソームとしては、トランスフェクテース(transfectace)(DDAB/DOPE)およびDOTAP/DOPE(Boerhinger)が挙げられる。他のカチオン性リポソームは、当該分野で周知の技法を使用して、容易に利用可能な物質から調製され得る。例えば、DOTAP(1,2−ビス(オレイルオキシ)−3−(トリメチルアンモニオ)プロパン)リポソームの合成の記載について、Szoka(1978)Proc.Natl.Acad.Sci.USA 75:4194−4198;WO90/11092を参照のこと。
【0182】
同様に、アニオン性リポソームおよび中性リポソームは、例えば、Avanti Polar Lipids(Birmingham,AL)から容易に入手可能であるか、または容易に入手可能な物質を使用してたやすく調製され得る。このような物質には、とりわけ、ホスファチジルコリン、コレステロール、ホスファチジルエタノールアミン、ジオレオイルホスファチジルコリン(DOPC)、ジオレオイルホスファチジルグリセロール(DOPG)、ジオレオイルホスファチジルエタノールアミン(DOPE)が含まれる。これらの物質はまた、適切な比率でDOTMAおよびDOTAPの出発物質と混合され得る。これらの物質を使用してリポソームを作製する方法は、当該分野で周知である。
【0183】
このリポソームは、多重膜のベシクル(MLV)、小さな単一膜ベシクル(SUV)、または大きな単一膜のベシクル(LUV)を含み得る。種々のリポソーム−核酸複合体は、当該分野で公知の方法を使用して調製される。例えば、Straubinger(1983)Meth.Immunol.101:512−527;Szoka(1978)Proc.Natl.Acad.Sci.USA 75:4194−4198;Papahadjopoulos(1975)Biochim.Biophys.Acta 394:483;Wilson(1979)Cell 17:77);DeamerおよびBangham(1976)Biochim.Biophys.Acta 443:629;Ostro(1977)Biochem.Biophys.Res.Commun.76:836;Fraley(1979)Proc.Natl.Acad.Sci.USA 76:3348);EnochおよびStrittmatter(1979)Proc.Natl.Acad.Sci.USA 76:145;Fraley(1980)J.Biol.Chem.(1980)255:10431;SzokaおよびPapahadjopoulos(1978)Proc.Natl.Acad.Sci.USA 75:145;ならびにSchaefer−Ridder(1982)Science 215:166を参照のこと。
【0184】
(E.リポタンパク質)
さらに、リポタンパク質が、送達されるポリヌクレオチド/ポリペプチドとともに含まれ得る。利用されるリポタンパク質の例としては、キロミクロン、HDL、IDL、LDL、およびVLDLが挙げられる。これらのタンパク質の変異体、フラグメント、または融合物もまた、使用され得る。また、天然に存在するリポタンパク質の改変体(例えば、アセチル化されたLDL)が使用され得る。これらのリポタンパク質は、リポタンパク質レセプターを発現する細胞へ、ポリヌクレオチドの送達を標的化し得る。好ましくは、リポタンパク質が、送達されるポリヌクレオチドとともに含まれる場合、他の標的化リガンドはその組成物中には含まれない。
【0185】
天然に存在するリポタンパク質は、脂質部分およびタンパク質部分を含む。このタンパク質部分は、アポタンパク質として知られる。現在では、アポタンパク質A、B、C、D、およびEが単離および同定されている。少なくともこれらの2つはいくつかのタンパク質を含み、ローマ数字、AI、AII、AIV;CI、CII、CIIIによって命名されている。
【0186】
1つのリポタンパク質は、1を超えるアポタンパク質を含み得る。例えば、天然に存在するキロミクロンはA、B、C、およびEからなり、そして時間が経てばこれらのリポタンパク質はAを欠失し、そしてCおよびEアポタンパク質を獲得する。VLDLは、A、B、C、およびEアポタンパク質を含み、LDLはアポタンパク質Bを含み;そしてHDLはアポタンパク質A、C、およびEを含む。
【0187】
これらのアポタンパク質のアミノ酸は公知であり、そして例えば、Breslow(1985)Annu Rev.Biochem 54:699;Law(1986)Adv.Exp.Med.Biol.151:162;Chen(1986)J Biol Chem 261:12918;Kane(1980)Proc Natl Acad Sci USA 77:2465;Utermann(1984)Hum Genet 65:232に記載されている。
【0188】
リポタンパク質は、トリグリセリド、コレステロール(遊離およびエステル)、およびリン脂質を含む、種々の脂質を含む。この脂質の組成は、天然に存在するリポタンパク質において変化する。例えば、キロミクロンは主としてトリグリセリドを含む。天然に存在するリポタンパク質の脂質含有物のより詳細な記載は、例えば、Meth.Enzymol.128(1986)に見いだされ得る。この脂質の組成は、レセプター結合活性についてアポタンパク質の立体構造において補助するために選択される。脂質組成はまた、ポリヌクレオチド結合分子との疎水性相互作用および会合を容易にするように選択され得る。
【0189】
天然に存在するリポタンパク質は、例えば、血清から超遠心分離によって単離され得る。そのような方法は、Meth.Enzymol.(前出);Pitas(1980)J.Biochem.253:5454−5460およびMahey(1979)J Clin.Invest 64:743−750に記載される。リポタンパク質はまた、インビトロまたは所望の宿主細胞中のアポタンパク質遺伝子の発現による組換え方法によって産生され得る。例えば、Atkinson(1986)Annu Rev Biophys Chem 15:403およびRadding(1958)Biochim Biophys Acta 30:443を参照のこと。リポタンパク質はまた、Biomedical Techniologies,Inc.,Stoughton,Massachusetts,USAのような商業的な供給者から購入され得る。さらなるリポタンパク質の記載は、Zuckermannら、PCT/US97/14465に見い出され得る。
【0190】
(F.ポリカチオン性薬剤)
ポリカチオン性薬剤は、送達される所望のポリヌクレオチド/ポリペプチドを有する組成物中に、リポタンパク質を伴って、またはリポタンパク質を伴わずに含まれ得る。
【0191】
ポリカチオン性薬剤は、代表的には、生理的に適切なpHにおいて正味の正電荷を示し、そして所望の位置への送達を容易にするための核酸の電荷を中和し得る。これらの薬剤は、インビトロ、エキソビボ、およびインビボ適用のいずれもを有する。ポリカチオン性薬剤は、生きている被験体に、筋肉内、皮下などのいずれかで核酸を送達するために使用され得る。
【0192】
以下は、ポリカチオン性薬剤としての有用なポリペプチドの例である:ポリリジン、ポリアルギニン、ポリオルニチン、およびプロタミン。他の例は、ヒストン、プロタミン、ヒト血清アルブミン、DNA結合タンパク質、非ヒストン染色体タンパク質、DNAウイルス由来のコートタンパク質(例えば、X174)を含む。転写因子もまた、DNAに結合するドメインを含み、従って核酸縮合薬剤として有用であり得る。手短に言えば、転写因子(例えば、C/CEBP、c−jun、c−fos、AP−1、AP−2、AP−3、CPF、Prot−1、Sp−1、Oct−1、Oct−2、CREP、およびTFIID)は、DNA配列に結合する塩基性ドメインを含む。
【0193】
有機ポリカチオン性薬剤は、スペルミン、スペルミジン、およびプトレシン(purtrescine)を含む。
【0194】
ポリカチオン性薬剤の大きさおよびその物理的特性は、上記の表から外挿されて、他のポリペプチドポリカチオン性薬剤が構築され得るか、または合成ポリカチオン性薬剤が産生され得る。
【0195】
有用な合成ポリカチオン性薬剤は、例えば、DEAE−デキストラン、ポリブレンを含む。LipofectinTM、およびlipofectAMINETMは、ポリヌクレオチド/ポリペプチドと組み合わせた場合にポリカチオン性複合体を形成するモノマーである。
【0196】
(核酸ハイブリダイゼーション)
「ハイブリダイゼーション」とは、水素結合による2つの核酸配列の互いの会合をいう。代表的には、1つの配列は、固体支持体に固定され、そして他方は溶液中で遊離している。次いで、2つの配列は、水素結合に好ましい条件下で互いに接触される。この結合に影響を与える因子は以下を含む:溶媒のタイプおよび容量;反応温度;ハイブリダイゼーションの時間;撹拌;液体相の配列の固体支持体への非特異的な付着をブロックする薬剤(Denhardt’s試薬またはBLOTTO);配列の濃度;配列の会合の速度を増大させる化合物(硫酸デキストランまたはポリエチレングリコール)の使用;およびハイブリダイゼーション後の洗浄条件のストリンジェンシー。Sambrookら(前出)第2巻、第9章、9.47〜9.57頁を参照のこと。
【0197】
「ストリンジェンシー」とは、異なる配列よりも、非常に類似する配列の会合に好ましいハイブリダイゼーション反応における条件をいう。例えば、研究中のハイブリッドの計算されたTmより約120〜200℃低い温度および塩濃度の組み合わせが選択されるべきである。温度および塩条件はしばしば、フィルターに固定したゲノムDNAのサンプルが目的の配列にハイブリダイズし、次いで異なるストリンジェンシーの条件下で洗浄される、予備的な実験において経験的に決定され得る。Sambrookら、9.50頁を参照のこと。
【0198】
例えば、サザンブロットを行う場合、考慮する変数は、(1)ブロットされるDNAの複雑さ、および(2)プローブと検出される配列との間の相同性である。研究されるフラグメント全量は、プラスミドまたはファージ消化物については0.1〜1μg、高度に複雑な真核生物ゲノム中の単一コピーの遺伝子については10−9〜10−8gまで、10倍変化し得る。より低い複雑さのポリヌクレオチドについては、実質的により短いブロッティング、ハイブリダイゼーション、および曝露回数、より少量の出発ポリヌクレオチド、およびより低い非活性のプローブが使用され得る。例えば、単一コピーの酵母遺伝子は、1μgの酵母DNAで開始し、2時間ブロットし、そして4〜8時間108cpm/μgのプローブを用いてハイブリダイズして、わずか1時間の曝露時間を用いて検出され得る。単一コピーの哺乳動物遺伝子について、保存性のアプローチは、10μgのDNAで開始し、一晩ブロットし、そして108cpm/μgより多いプローブを用いて10%硫酸デキストランの存在下で一晩ハイブリダイズし、約24時間露光時間を生じる。
【0199】
いくつかの因子が、プローブと目的のフラグメントとの間のDNA−DNAハイブリッドの融解温度(Tm)、ならびに、結果として、ハイブリダイゼーションおよび洗浄についての適切な条件に影響を与え得る。多くの場合において、そのプローブはフラグメントに対して100%相同なわけではない。他の共通して直面する変数には、長さ、ハイブリダイズする配列の全G+C含量、ならびにイオン強度およびハイブリダイゼーション緩衝液のホルムアミド含量が含まれる。これらのすべての因子の影響は、一つの式によって近似され得る:
Tm=81+16.6(log10Ci)+0.4{(G+C)%}−0.6(ホルムアミド%)−600/n−1.5(ミスマッチ%)。
ここでCiは、塩濃度(一価イオン)であり、そしてnは塩基対内のハイブリッドの長さである(MeinkothおよびWahl(1984)Anal.Biochem.138:267/284からわずかに改変した)。
【0200】
ハイブリダイゼーション実験の設計において、核酸ハイブリダイゼーションに影響を与えるいくつかの因子が簡便に変更され得る。ハイブリダイゼーションおよび洗浄の温度ならびに洗浄時の塩濃度を調整するのが最も単純である。ハイブリダイゼーション温度(すなわち、ストリンジェンシー)が上昇するにつれて、非相同的な鎖の間で起こるハイブリダイゼーションは起こりにくくなるようであり、結果として、バックグラウンドが減少する。放射標識したプローブが固定化されたフラグメントと完全に相同ではない場合(遺伝子ファミリーおよび種間のハイブリダイゼーション実験における場合で頻繁であるように)、ハイブリダイゼーション温度は低下されなければならず、そしてバックグラウンドが増大する。洗浄の温度は、類似の様式で、ハイブリダイズするバンドの強度、およびバックグラウンドの程度に影響を与える。洗浄のストリンジェンシーはまた、塩濃度の減少とともに増大する。
【0201】
一般的に、50%ホルミアミドの存在下で都合よいハイブリダイゼーション温度は、標的フラグメントに95%〜100%相同であるプローブについて42℃、90%〜95%相同性では37℃、85%〜90%相同性については32℃である。より低い相同性については、上記の式を用いて、適切にホルムアミド含量が低くされ、そして温度が調整されるべきである。プローブと標的フラグメントとの間の相同性が未知である場合、最も単純なアプローチは、ともにストリンジェントではないハイブリダイゼーション条件および洗浄条件で開始することである。オートラジオグラフィー後に非特異的バンドまたは高いバックグラウンドが観察される場合、フィルターは高ストリンジェンシーで洗浄され得、そして再び露光され得る。露光のために必要な時間がこのアプローチを非実用的にする場合、いくつかのハイブリダイゼーションおよび/または洗浄ストリンジェンシーが並行して試験されるべきである。
【0202】
(核酸プローブアッセイ)
本発明に従う核酸プローブを利用する、PCR、分枝DNAプローブアッセイ、またはブロッティング技術のような方法は、cDNAまたはmRNAの存在を決定し得る。プローブは、検出されるに十分に安定な、二重鎖または二本鎖複合体を形成し得る場合に、本発明の配列に「ハイブリダイズする」といわれる。
【0203】
核酸プローブは、本発明のクラミジアの(Chlamydial)ヌクレオチド配列(センス鎖およびアンチセンス鎖の両方を含む)にハイブリダイズする。多くの異なるヌクレオチド配列がアミノ酸配列をコードするが、ネイティブなクラミジアの配列は、細胞に存在する実際の配列であるので、好ましい。mRNAは、コード配列を表し、従ってプローブはコード配列に相補的であるべきであり、一本鎖cDNAはmRNAに相補的であり、従ってcDNAプローブは非コード配列に相補的であるべきである。
【0204】
プローブ配列はクラミジアの配列(またはその相補物)に同一である必要はない。核酸プローブが標的ヌクレオチドと検出され得る二重鎖を形成し得る場合、配列および長さのいくらかの変動は、アッセイの感度の増加をもたらし得る。また、核酸プローブは、形成された二重鎖を安定化するためにさらなるヌクレオチドを含み得る。さらなるクラミジアの配列もまた、形成された二重鎖を検出するための標識としての一助となり得る。例えば、非相補的ヌクレオチド配列が、そのプローブの5’末端に付着され得、ここでそのプローブ配列の残りはクラミジアの配列に相補的である。あるいは、プローブ配列が、クラミジアの配列とハイブリダイズし、そしてそれによって検出され得る二重鎖を形成するためにクラミジアの配列との十分な相補性を有する場合、非相補的塩基またはより長い配列は、プローブ中に分散され得る。
【0205】
プローブの正確な長さおよび配列は、ハイブリダイゼーション条件(例えば、温度、塩条件など)に依存する。例えば、診断的適用については、分析物の配列の複雑さに依存して、核酸プローブは、代表的には、少なくとも10〜20ヌクレオチド、好ましくは15〜25、そしてより好ましくは少なくとも30ヌクレオチド以上を含むが、これよりも短くてもよい。短いプライマーは、一般的には、鋳型との安定なハイブリッド複合体を形成するのにより低い温度を必要とする。
【0206】
プローブは、合成的手順(例えば、Matteucciらのトリエステル法、(J.Am.Chem.Soc.(1981)103:3185))、またはUrdeaら(Proc.Natl.Acad.Sci.USA(1983)80:7461)に従って、または市販の自動オリゴヌクレオチド合成機を使用して、産生され得る。
【0207】
プローブの化学的性質は、優先度に従って選択され得る。特定の適用については、DNAまたはRNAが適切である。他の適用については、改変(例えば、ホスホロチオエートまたはメチルホスホネートのようなバックボーンの改変)が組み込まれ得、インビボの半減期を増大させるために使用され得、RNA親和性を変化させ、ヌクレアーゼ耐性などを増大させるなどを行い(例えば、AgrawalおよびIyer(1995)Curr Opin Biotechnol 6:12−19;Agrawal(1996)TIBTECH 14:376−387);ペプチド核酸のようなアナログもまた使用され得る(例えば、Corey(1997)TIBTECH15 224−229;Buchardtら(1993)TIBTECH 11:384−386を参照のこと)。
【0208】
あるいは、ポリメラーゼ連鎖反応(PCR)は、少量の標的核酸を検出する別の周知の手段である。そのアッセイは、Mullisら、(Meth.Enzymol.(1987)155:335−350);米国特許第4,683,195号および同第4,683,202号に記載されている。2つの「プライマー」ヌクレオチドは、標的核酸とハイブリダイズし、そして反応を開始するために使用される。このプライマーは、増殖標的(またはその相補物)の配列にハイブリダイズしない、二重鎖の安定性を補助するための、または、例えば、首尾よい制限部位を組み込むための配列を含む。代表的には、このような配列は、所望のクラミジアの配列に隣接する。
【0209】
熱安定性のポリメラーゼは、もともとの標的核酸を鋳型として使用して、プライマーから標的核酸のコピーを作製する。標的核酸の閾値量がポリメラーゼによって産生された後、それらはより従来的な方法(例えば、サザンブロット)によって検出され得る。サザンブロット法を使用する場合、標識されたプローブは、クラミジアの配列(またはその相補物)にハイブリダイズする。
【0210】
また、mRNAまたはcDNAは、Sambrookら(前出)に記載される、従来的なブロッティング技術によって検出され得る。mRNA、またはmRNAからポリメラーゼ酵素を使用して生成されたcDNAは、ゲル電気泳動を使用して精製および分離され得る。次いで、ゲル上のこの核酸は、ニトロセルロースのような固体支持体にブロットされる。この固体支持体は、標識されたプローブに曝露され、次いですべてのハイブリダイズしていないプローブを洗浄して除去する。次に、標識プローブを含む二重鎖を検出する。代表的には、そのプローブは、放射活性部分で標識される。
【0211】
ウエスタンブロットのために、2つのサンプルを各タンパク質について試験した。対の左側のレーンは、前免疫血清で染色した膜ストリップを使用し、一方、右側のレーンは、免疫血清で染色した膜ストリップを使用した。図1〜5、35B、37B、38Bおよび39のウエスタンブロットにおいて、マーカーは、66kDa、45kDa、30kDa、20.1kDaおよび14.4kDaにある。図6〜16、20B、23C、24D、27E、38A、40、41、42および43のウエスタンブロットにおいて、マーカーは、172.6kDa、111.4kDa、79.6kDa、61.3kDa、49.0kDa、36.4kDa、24.7kDa、19.2kDaおよび13.1kDaにある。
【0212】
図1〜5のウエスタンブロットにおいて、レーン2および3は、GST融合コントロール抗原に対して惹起されたコントロール血清を示す。図1〜5のウエスタンブロットにおいて、レーン4および5は、Hisタグ化コントロール抗原に対して惹起されたコントロール血清を含む。
【0213】
低分子量マーカーは、精製ゲルのレーン1で泳動される。
【0214】
(発明を実施するための様式)
表Iは、参考文献18由来のC.pneumoniaeタンパク質の名称、このタンパク質についてのGenBank登録番号および表題、本発明の対応するC.trachomatisタンパク質についてのGenBank登録番号および表題、ならびにこれらのC.trachomatisタンパク質についての配列番号(配列番号1〜262、奇数はアミノ酸配列であり、そして偶数はヌクレオチド配列である)を与える。これらは、対応するC.pneumoniaeタンパク質について参考文献18に記載されたのと同じ方法で、発現および使用され得る。C.trachomatisタンパク質は、診断目的および免疫原性目的のために有用である。これらの特性は、配列のみからでは明らかでない。
【0215】
種々の試験を使用して、本発明のタンパク質のインビボでの免疫原性を評価し得る。例えば、これらのタンパク質は、組換え的に発現され得、そしてイムノブロットによって患者の血清をスクリーニングするために使用され得る。タンパク質と患者の血清との間のポジティブ反応は、この患者が、問題のタンパク質に対する免疫応答を以前に生じていたことを示す(すなわち、このタンパク質は、免疫原である)。この方法はまた、免疫優性タンパク質を同定するためにも使用され得る。
【0216】
この組換えタンパク質はまた、例えば、マウスにおいて、抗体を調製するために簡便に使用され得る。これらは、(例えば、インタクトなクラミジアに対するウエスタンブロットにおいてこれらの抗体を使用することによって)タンパク質がC.trachomatisの細胞表面上に位置することを直接確認するために使用され得る。標識された抗体(例えば、FACSのための蛍光標識)は、インタクトな細菌と共にインキュベートされ得、そしてこの細菌表面上の標識の存在が、タンパク質の位置を確認する。FACSの図は、アッセイにおいて使用されるChlamydia調製物の散乱プロファイル、組換え抗原に対する抗体が、Chlamydia細胞に結合する場合に得られるピークシフト(白抜き領域=コントロールサンプル;塗りつぶした領域=抗体と反応したサンプル)、定量的Kolmogorov−Smirnov(K−S)統計分析、およびFACS分析ソフトウェアの出力を示す。
【実施例】
【0217】
(実施例1)
CT242(配列番号57および配列番号58)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図1A;レーン4および5、クロマトグラフィー画分1および2、予測された分子量42.4kDa)として、およびHisタグ化融合タンパク質(図1B;レーン2〜4、クロマトグラフィー画分1、2および3、予測された分子量16.4kDa)としての両方で精製された。
【0218】
この組換えタンパク質を、マウスを免疫化するために使用し、このマウスの血清を、ウエスタンブロットで使用した(図1C:Hisタグ化:レーン12および13;GST融合:レーン20および21)。レーン12は、Hisタグ化CT242についての、前免疫血清で染色した膜ストリップを示し、一方、レーン13は、Hisタグ化CT242についての、免疫血清で染色した膜ストリップを示す。レーン20は、GST融合CT242についての前免疫血清で染色した膜ストリップを示し、レーン21は、GST融合CT242についての免疫血清で染色した膜ストリップを示す。
【0219】
これらの実験は、CT242が表面曝露されておりかつ免疫アクセス可能(immunoaccessible)なタンパク質であること、およびこのCT242が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0220】
(実施例2)
CT045(配列番号71および配列番号72)は、E.coli中で発現された。この組換え産物は、Hisタグ化融合タンパク質(図2A;レーン4〜6、クロマトグラフィー画分1、2および3、予測された分子量55.8kDa)として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図2B:レーン8および9)およびFACS分析(図2C、K−S値16.81)のために使用した。
【0221】
これらの実験は、CT045が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT045が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0222】
(実施例3)
CT381(配列番号105および配列番号106)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図3A;レーン2および3、クロマトグラフィー画分1および2、予測された分子量52.7kDa)およびHisタグ化融合タンパク質(図3A;レーン7〜9、クロマトグラフィー画分1、2および3、予測された分子量26.7kDa)として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図3B:Hisタグ化:レーン6および7;GST融合:レーン16および17)およびFACS分析(図3C、GSTタグ化、K−S値35.98;図3D:Hisタグ化、K−S値32.54)のために使用した。
【0223】
これらの実験は、CT381が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT381が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0224】
(実施例4)
CT396(配列番号107および配列番号108)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図4A;レーン6および7、クロマトグラフィー画分1および2、予測された分子量99.5kDa)およびHisタグ化融合タンパク質(図4B;レーン5〜7、クロマトグラフィー画分1、2および3、予測された分子量73.5kDa)として精製された。この組換えHisタグ化タンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図4C:レーン14および15)に使用した。この組換えHisタグ化タンパク質および組換えGST融合タンパク質をまた、FACS分析(図4D:Hisタグ化、K−S値34.50;図4E:GST融合、K−S値32.76)のために使用した。
【0225】
これらの実験は、CT396が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT396が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0226】
(実施例5)
CT398(配列番号111および配列番号112)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図5A;レーン8および9、クロマトグラフィー画分1および2、予測された分子量54.8kDa)およびHisタグ化融合タンパク質(図5B;レーン8〜10、クロマトグラフィー画分1、2および3、予測された分子量28.8kDa)として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図5C:Hisタグ化:レーン10および11;GST融合:レーン18および19)およびFACS分析(図5D:GST融合、K−S値31.24;図5E:Hisタグ化、K−S値26.10)のために使用した。
【0227】
これらの実験は、CT398が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT398が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0228】
(実施例6)
CT089(配列番号61および配列番号62)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図6C;レーン2、クロマトグラフィー画分1、予測された分子量70.8kDa)およびHisタグ化融合タンパク質(図6C;レーン3、4および5、クロマトグラフィー画分1、2および3、予測された分子量44.8kDa)として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図6A:GST融合:レーン14および15;Hisタグ化:レーン16および17)およびFACS分析(図6B:Hisタグ化、K−S値26.59)のために使用した。
【0229】
これらの実験は、CT089が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT089が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0230】
(実施例7)
CT443(配列番号125および配列番号126)は、E.coli中で発現された。この組換え産物は、Hisタグ化融合タンパク質として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図7A:レーン10および11)およびFACS分析(図7B:K−S値21.28)のために使用した。
【0231】
これらの実験は、CT443が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT443が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0232】
(実施例8)
CT541(配列番号149および配列番号150)を、E.coliにおいて発現させた。この組換え産物を、GST融合タンパク質として精製した(図8C:レーン2および3、クロマトグラフィー画分1および2、予測分子量51.6kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図8A:レーン6および7)ならびにFACS分析(図8B:K−S値9.94)のために使用した。
【0233】
これらの実験は、CT541が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0234】
(実施例9)
CT547(配列番号151および配列番号152)を、E.coliにおいて発現させた。この組換え産物を、GST融合タンパク質(図9D:レーン4および5、クロマトグラフィー画分1および2、予測分子量58.3kDa)およびHisタグ化融合タンパク質の両方として精製した。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図9A:Hisタグ化:レーン20および21)ならびにFACS分析(図9B:GST融合:K−S値14.60および15.57;図9C:Hisタグ化:K−S値28.21)のために使用した。
【0235】
これらの実験は、CT547が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0236】
(実施例10)
CT587(配列番号189および配列番号190)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図10C:レーン5、6および7、クロマトグラフィー画分1、2および3、予測分子量47.5kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図10A:レーン12および13)ならびにFACS分析(図10B:Hisタグ化:K−S値20.85)のために使用した。
【0237】
これらの実験は、CT587が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0238】
(実施例11)
CT266(配列番号77および配列番号78)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図11C:レーン11および12、クロマトグラフィー画分1および2、予測分子量44.1kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図11A:レーン4および5)ならびにFACS分析(図11B:K−S値21.29)のために使用した。
【0239】
これらの実験は、CT266が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0240】
(実施例12)
CT444(配列番号127および配列番号128)を、E.coliにおいて発現させた。この組換え産物を、GST融合タンパク質(図12B:レーン2および3、クロマトグラフィー画分1および2、予測分子量87.3kDa)およびHisタグ化融合タンパク質(図12C:レーン3および4、クロマトグラフィー画分2および3、予測分子量9.0kDa)として精製した。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図12A:レーン16および17)ならびにFACS分析(図12D:GST融合:K−S値14.98;図12E:Hisタグ化:K−S値13.28)のために使用した。
【0241】
これらの実験は、CT444が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0242】
(実施例13)
CT559(配列番号199および配列番号200)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化タンパク質として精製した(図13C:レーン2、3および4、クロマトグラフィー画分1、2および3、予測分子量34.9kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図13A:レーン6および7)ならびにFACS分析(図13B:K−S値23.21)のために使用した。
【0243】
これらの実験は、CT559が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0244】
(実施例14)
CT681(配列番号155および配列番号156)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図14C:レーン5および6、クロマトグラフィー画分1および2、予測分子量41.8kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図14A:レーン10および11)ならびにFACS分析(図14B:K−S値34.66)のために使用した。
【0245】
これらの実験は、CT681が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0246】
(実施例15)
CT713(配列番号201および配列番号202)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図15B:レーン4、5および6、クロマトグラフィー画分1、2および3、予測分子量35.4kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図15A:レーン12および13)ならびにFACS分析(図15C:K−S値25.82)のために使用した。
【0247】
これらの実験は、CT713が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0248】
(実施例16)
CT823(配列番号229および配列番号230)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図16C:レーン7、8および9、クロマトグラフィー画分1、2および3、予測分子量53.9kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図16A:レーン14および15)ならびにFACS分析(図16B:K−S値26.62)のために使用した。
【0249】
これらの実験は、CT823が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0250】
(実施例17)
CT114(配列番号243および配列番号244)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図17:レーン6および7、クロマトグラフィー画分1および2、予測分子量48.5kDa)。
【0251】
(実施例18)
CT198(配列番号43および配列番号44)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図18A:レーン6、クロマトグラフィー画分1、予測分子量56.3kDa)。このHisタグ化組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、FACS分析のために使用した(図18B)。
【0252】
これらの実験は、CT198がEB異種集団のほんの一部において存在することを示す(クラミジア調製物が通常そうであるように)。これが存在する場合、これは、表面に露出された免疫アクセス可能なタンパク質である。これらの特性は、配列単独からは明らかではない。
【0253】
(実施例19)
CT241(配列番号55および配列番号56)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図19:レーン4、クロマトグラフィー画分3、予測分子量85.3kDa)。
【0254】
(実施例20)
CT350(配列番号27および配列番号28)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図20A:レーン2、3および4、クロマトグラフィー画分1、2および3、予測分子量61.3kDa)およびGST融合タンパク質(図20A:レーン7、8および9、クロマトグラフィー画分1、2および3、予測分子量87.3kDa)の両方として精製した。これらの組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図20B:Hisタグ化:レーン4および5;GSTタグ化:レーン8および9)のために使用した。
【0255】
(実施例21)
CT351(配列番号25および配列番号26)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図21:レーン2および3、クロマトグラフィー画分1および2、推定分子量 76.1kDa)として精製した。
【0256】
(実施例22)
CT391(配列番号251および配列番号252)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図22:レーン8および9、クロマトグラフィー画分1および2、推定分子量 32.6kDa)として精製した。
【0257】
(実施例23)
CT077(配列番号65および配列番号66)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図23:レーン2および3、クロマトグラフィー画分1および2、推定分子量 59.7kDa)、およびHisタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図23C:レーン6および7)、およびFACS分析(図23B、Hisタグ化:K−S値 9.17)のために使用した。
【0258】
これらの実験は、CT077が、表面露出された免疫接近可能なタンパク質であること、およびこのCT077が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0259】
(実施例24)
CT181(配列番号245および配列番号246)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図24A:レーン4、クロマトグラフィー画分1、推定分子量 50.1kDa)、およびHisタグ化融合タンパク質(図24B:レーン2、3および4、クロマトグラフィー画分1、2および3,推定分子量 32.0kDa)の両方として精製した。このGSTタグ化組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図24D:レーン4および5(矢印で示される))、およびFACS分析(図24C、K−S値 7.62)のために使用した。
【0260】
これらの実験は、CT181が、表面露出された免疫接近可能なタンパク質であること、およびこのCT181が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0261】
(実施例25)
CT589(配列番号185および配列番号186)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図25A:レーン4および5、クロマトグラフィー画分1および2、推定分子量 89.4kDa)、およびHisタグ化融合タンパク質(図25B:レーン2および3、クロマトグラフィー画分1および2、推定分子量 63.4kDa)の両方として精製した。このHisタグ化組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、FACS分析(図25C)のために使用した。
【0262】
これらの実験は、CT589が、EB異種集団の一部のみにしか存在しないことを示す(クラミジア調製物が通常そうであるように)このCT589が存在する場合、このCT589は、表面露出された免疫接近可能なタンパク質である。これらの特性は、配列のみからは示されない。
【0263】
(実施例26)
CT597(配列番号179および配列番号180)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図26A:レーン5および6、クロマトグラフィー画分1および2、推定分子量 36.0kDa)、およびHisタグ化融合タンパク質(図26B:レーン2、3および4、クロマトグラフィー画分1、2および3、推定分子量 10.3kDa)の両方として精製した。
【0264】
(実施例27)
CT623(配列番号163および配列番号164)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図27A:レーン3および4、クロマトグラフィー画分1および2、推定分子量 71.8kDa)、およびHisタグ化融合タンパク質(図27B:レーン2、3および4、クロマトグラフィー画分1、2および3、推定分子量 45.8kDa)の両方として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図27E:GSTタグ化、レーン4(矢印で示される);Hisタグ化、レーン13(矢印で示される))、およびFACS分析(図27C:GSTタグ化:K−S値 15.89;図27D:Hisタグ化:K−S値 20.27)のために使用した。
【0265】
これらの実験は、CT623が、表面露出された免疫接近可能なタンパク質であること、およびこのCT623が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0266】
(実施例28)
CT700(配列番号261および配列番号262)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図28A:レーン5、6および7、クロマトグラフィー画分1、2および3、推定分子量 73.7kDa)として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、FACS分析(図28B:K−S値 8.72)のために使用した。
【0267】
これらの実験は、CT700が、表面露出された免疫接近可能なタンパク質であること、およびこのCT700が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0268】
(実施例29)
CT761(配列番号217および配列番号218)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図29A:レーン6および7、クロマトグラフィー画分1および2、推定分子量 63.9kDa)として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、FACS分析(図29B:K−S値 11.45)のために使用した。
【0269】
これらの実験は、CT761が、表面露出された免疫接近可能なタンパク質であること、およびこのCT761が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0270】
(実施例30)
CT415(配列番号117および配列番号118)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図30:レーン3および4、クロマトグラフィー画分1および2、推定分子量 55.4kDa)として精製した。
【0271】
(実施例31)
CT454(配列番号253および配列番号254)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図31:レーン2および3、クロマトグラフィー画分1および2、推定分子量 56.2kDa)として精製した。
【0272】
(実施例32)
CT467(配列番号129および配列番号130)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図32:レーン3および4、クロマトグラフィー画分1および2、推定分子量 65.6kDa)として精製した。
【0273】
(実施例33)
CT551(配列番号257および配列番号258)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図33A:レーン5、6および7、クロマトグラフィー画分1、2および3、推定分子量 34.1kDa)、およびGSTタグ化融合タンパク質(図33B:レーン4および5、クロマトグラフィー画分1および2、推定分子量 60.1kDa)の両方として精製した。
【0274】
(実施例34)
CT567(配列番号195および配列番号196)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図34A:レーン8および9、クロマトグラフィー画分1および2、推定分子量 44.0kDa)、およびHisタグ化融合タンパク質(図34B:レーン7および8、クロマトグラフィー画分1および2、推定分子量 18.3kDa)の両方として精製した。
【0275】
(実施例35)
CT569(配列番号193および配列番号194)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図35A:レーン2、3および4、クロマトグラフィー画分1、2および3、推定分子量 11.2kDa)として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図35B:レーン8および9、矢印で示される)で使用した。
【0276】
(実施例36)
CT647(配列番号169および配列番号170)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図36:レーン6および7、クロマトグラフィー画分1および2、推定分子量 45.7kDa)として精製した。
【0277】
(実施例37)
CT600(配列番号173および配列番号174)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図37A:レーン5、6および7、クロマトグラフィー画分1、2および3、推定分子量 19.5kDa)として精製した。この組換え産物を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図37B:レーン10および11、矢印で示される)、およびFACS分析(図37C、K−S値 10.46)のために使用した。
【0278】
これらの実験は、CT600が、表面露出された免疫接近可能なタンパク質であること、およびこのCT600が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0279】
(実施例38)
CT279(配列番号247および配列番号248)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質、Hisタグ化融合タンパク質として精製した。これらの組換えHisタグ化タンパク質および組換えGSTタグ化タンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図38A:Hisタグ化:レーン5(矢印で示される);図38B:GSTタグ化:レーン12および13(矢印で示される))において使用した。
【0280】
(実施例39)
CT560(配列番号259および配列番号260)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した。この組換えHisタグ化タンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図39:レーン6および7(矢印で示される))において使用した。
【0281】
(実施例40)
CT389(配列番号249および配列番号250)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図40:レーン16および17(矢印で示される))において使用した。
【0282】
(実施例41)
CT456(配列番号255および配列番号256)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図41:レーン2および3(矢印で示される))において使用した。
【0283】
(実施例42)
CT622(配列番号161および配列番号162)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図42:レーン9(矢印で示される))において使用した。
【0284】
(実施例43)
CT759(配列番号213および配列番号214)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図43:レーン8および9(矢印で示される))において使用した。
【0285】
(実施例44)
インビトロ中和アッセイは、培養物中の真核細胞に対するC.trachomatis 感染性を阻害するために、本発明の種々の組換えタンパク質で免疫されたマウスから得られた血清の能力を示し、このアッセイを、LLCMK2(アカゲザル腎臓内皮)細胞を使用して実施した。マウスポリクローナル血清の連続4倍希釈を、SP(スクロース−リン酸)緩衝液中に調製した。全EBに対するマウス抗血清を、ポジティブコントロールとして使用し、前免疫血清およびSP緩衝液のみを、ネガティブコントロールとして使用した。C.trachomatis(血液型亜型D)から精製されたEBを、SP緩衝液中に希釈し、3×105IFU/mlを含むようにし、この懸濁液の10μlを、最終容量100μlで各血清希釈液に添加した。抗体EB相互作用が、37℃で30分間進行するようにした。次いで、各サンプルからの100μlの反応混合液を、96ウェルマイクロタイタープレート中のPBS洗浄LLCMK2細胞単層の上に添加し、805×gで37℃で1時間遠心分離した。全ての血清およびコントロールを、2連のサンプルで試験した。過剰の接種物の除去後、細胞を、PBSで一度リンスし、20%FCSおよび1μgシクロヘキシミドを補充した200μlのDMEM培地を補充し、そして37℃で48時間インキュベートした。細胞をメタノールで固定し、進行性細胞内クラミジア感染によって生成された代表的な細胞質封入物を、抗クラミジアフルオレセイン結合モノクローナル抗体(Meridian Diagnostics)で染色した。宿主細胞に対する十分な希釈およびEB比において、観察された封入物の数を、宿主細胞感染を最初に首尾よく確立し得た生存クラミジアの数に等しいと考えられる(これらは、Inclusion Forming Units,IFUと名づけられる)。フルオレセイン標識封入物を、40×の倍率で1ウェル当たり4つの顕微鏡視野において計数した。抗体相互作用に起因する感染性の阻害を、SPコントロール(緩衝液のみ)に比較した場合、平均IFUのパーセント減少として計算した。共通の実施に従って、血清を、これらが50%以上の感染性の減少を引き起こし得る場合、「中和」として標識し、全スクリーニングアッセイ(例えば、宿主細胞の交換、またはケラミジア単離物、または感染性接種物の調製における環境条件のバリエーション)の複雑性を考慮して、より低い程度のEB感染性を阻害し得る血清はまた、さらなる研究のためのワクチン候補として考慮されるべきである。図44Aは、中和ポジティブ血清から得られた結果の例を示し、一方、図44Bは、中和ネガティブ血清から得られた結果の例を示す。
【0286】
インビトロ中和アッセイを、実施例1〜10、13〜22、24〜26および29〜37において言及される組換えタンパク質で免疫されたマウスから得られた血清を使用して実施した。結果を、表IIに示す。これらの結果は、CT045、CT242、CT381.CT396、CT398、CT467、CT547、CT587およびCT681が、C.trachomatisによる感染を予防するワクチンに対する、特に良好な候補の全てであることを示す。これらの特性は、配列単独からは示されない。
【0287】
さらなる実験において、C.trachomatisに対して惹起された血清を、交差中和活性について、C.pneumoniae EBに対して試験した。手順は、上記されるとおりであったが、C.pneumoniaeから精製されたEBを、SP緩衝液中に希釈し、3×106IFU/mlを含むようにし、この懸濁物の10μlを、最終容量100μlで各血清希釈物に添加した。CT242およびCT467を使用して得られた血清は、C.pneumoniae EBを交差中和し得た。
【0288】
本発明が例示のためのみに記載され、その改変がなされ得、一方で、本発明の意図および範囲内のままであることは、理解される。
【0289】
【数1】

【0290】
【表1】
【0291】
【表2】
【0292】
【表3】
【0293】
【表4】
【0294】
【表5】
【技術分野】
【0001】
本明細書中で引用される全ての文献は、その全体が参考として援用される。
【0002】
(技術分野)
本発明は、クラミジア感染、特に、Chlamydia trachmatisによる感染に対する免疫化の分野に存在する。
【背景技術】
【0003】
(背景技術)
Chlamydiaeは、地域性の性感染症および種々の他の疾患症候群に寄与する真核生物細胞の偏性細胞内寄生生物である。これらは、排他的な真正細菌の分科を占め、他のいずれの既知の生物に対しても密接な関係を有さず、これらは、単一のファミリー(Chlamydiaceae)を含むこれら独自の目(Chlamydiales)に分類され、次いで単一の属(Chlamydia、Chlamydophilaともいわれる)を含む。Chlamydiaeは、これらの固有の生活環が特に特徴的であり、この生活環において、細菌は、2つの形態学的に別々の形態:細胞外感染性形態(基本小体、EB)と細胞内非感染性形態(網状体(reticulate bodies)、RB)との間で交替する。この生活環は、RBのEBへの再編成で完了し、EBは、引き続き、分裂した宿主細胞がさらなる細胞を感染する準備をさせておく。
【0004】
4つのクラミジア種(C.trachomatis、C.pneumoniae、C.pecorumおよびC.psittaci)が現在知られており[例えば、参考文献1,2](非特許文献1、2)、ゲノム配列は、公表されている[参考文献3〜9](非特許文献3〜6、特許文献1〜3)。
【0005】
C.trachomatisのヒトの血清改変体(「血清型亜型」)は、2つのバイオ改変体(「biovar」)に分けられる。血清型亜型A〜Kは、主に眼の組織(A〜C)または泌尿生殖器管(D〜K)において上皮性感染を誘発する。血清型亜型L1、L2およびL3は、浸潤性の性病性リンパ肉芽腫(LGV)の因子である。
【0006】
クラミジア感染自体が疾患を生じるが、幾人かの患者においては、症候群の重篤度は、実際には、異常な宿主免疫応答に起因していると考えられる。感染を排除することの失敗は持続的な免疫刺激を生じ、そして宿主を助けることよりも、これは、重篤な結果(不妊および失明を含む)を伴う慢性感染を生じる[10](非特許文献7)。さらに、天然のクラミジア感染により与えられる防御は、通常、不完全で、一過性で、そして株特異的である。
【0007】
疾患の深刻な性質に起因して、適切なワクチンを提供することが所望される。これらは、(a)クラミジア感染に対する、またはクラミジア誘導性疾患に対する免疫のために(予防ワクチン接種)、あるいは(b)確立された慢性クラミジア感染の根絶のために(治療ワクチン接種)有用であり得る。しかし、細胞内寄生生物であることから、この細菌は、一般に抗体媒介免疫応答を逃れ得る。
【0008】
種々の抗原性タンパク質が、C.trachomatisについて記載されており、そして特に細胞表面が、詳細な検索の標的となっている[例えば、1,11](非特許文献1、8)。これらとしては、例えば、pgp3[12、13、14](非特許文献9、特許文献4、5)、MOMP[15](非特許文献10)、Hsp60(GroEL)[16](非特許文献11)およびHsp70(DnaK様)[17](非特許文献12)が挙げられる。しかし、これらの全てが有効なワクチンであると証明されたわけではなく、そして、ワクチン開発における使用に適切な抗原および免疫原を提供するため、天然の感染の間に免疫応答を惹起するC.trachomatis抗原を同定することが本発明の目的である。C.trachomatisの診断(例えば、免疫診断)に有用な抗原を同定することは、さらなる目的である。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】International patent application WO99/27105
【特許文献2】International patent application WO00/27994
【特許文献3】International patent application WO99/28475
【特許文献4】EP-A-0499681
【特許文献5】International patent application WO95/28487
【非特許文献】
【0010】
【非特許文献1】Raulston (1995) Mol Microbiol 15:607-616
【非特許文献2】Everett (2000) Vet Microbiol 75: 109-126
【非特許文献3】Kalman et al. (1999) Nature Genetics 21:385-389
【非特許文献4】Read et al. (2000) Nucleic Acids Res 28:1397-1406
【非特許文献5】Shirai et al. (2000) Nucleic Acids Res 28:2311-2314
【非特許文献6】Stephens et al. (1998) Science 282:754-759
【非特許文献7】Ward (1995) Apmis. 103:769-96.
【非特許文献8】Moulder (1991) Microbiol Rev 55(1):143-190.
【非特許文献9】Comanducci et al. (1994) Infect Immun 62(12):5491-5497.
【非特許文献10】Murdin et al. (1993) Infect Immun 61:4406-4414
【非特許文献11】Cerrone et al. (1991) Infect Immun 59(1):79-90.
【非特許文献12】Raulston et al. (1993) J. Biol. Chem. 268:23139-23147.
【発明の概要】
【課題を解決するための手段】
【0011】
参考文献18は、種々のC.pneumoniae由来のタンパク質を開示する。これらは、免疫反応性、免疫受容可能および/または基本小体に存在するとして、実験的に検証された。このタンパク質のこれらの性質は、ゲノム配列情報から推論できなかった。参考文献18は、これらのタンパク質が、C.pneumoniaeが主要な病巣であるChlamydia細菌に起因する感染症の処置または予防に使用され得ることを開示する。C.pneumoniaeタンパク質はまた、種間交差反応性に起因する、他の種のChlamydiaによる感染症を処置または予防するために使用され得る。
【0012】
全ゲノムの比較[3、4、5]によって示されるように、C.pneumoniaeはC.trachomatisと密接に関連する。
【0013】
本発明は、参考文献18に開示されるC.pneumoniaeタンパク質に対応するC.tracomatisタンパク質(配列番号1〜261の奇数番号)に関する。これらのタンパク質は、Chlamidia細菌(特にC.trachomatisにおいて)に起因する感染症の処置および予防において使用され得る。特に好ましいタンパク質は、以前に「仮定タンパク質」として注釈されたもの(本明細書中の表Iを参照のこと)か、または細胞質位置を有すると考えられたものである。
(C.trachomatisタンパク質)
本発明は、アミノ酸配列(配列番号1〜261の奇数番号)のうちの1つ以上を含むタンパク質を提供する。
【0014】
本発明はまた、1つ以上の配列番号1〜261の奇数番号のアミノ酸配列と少なくともx%配列同一性を共有する配列を含むタンパク質を提供する。特定の配列に依存して、xは、好ましくは、50%以上(例えば、60%、70%、80%、90%、95%、99%以上)である。これらは、変異体および対立遺伝子改変体を含む。代表的には、2つのタンパク質間の50%以上の同一性は、機能的同等性の指標であると考えられる。タンパク質間の同一性は、好ましくは、パラメーターのギャップオープンペナルティー=12およびギャップ伸長ペナルティー=1を用いるアフィンギャップ検索を使用して、MPSRCHプログラム(Oxford Molecular)において実行されるように、Smith−Waterman相同性検索アルゴリズムによって決定される。
【0015】
本発明は、配列番号1〜261の奇数番号のアミノ酸配列のフラグメントを含むタンパク質をさらに提供する。これらのフラグメントは、配列から少なくともn連続するアミノ酸を含むはずであり、そして特定の配列に依存して、nは、7以上(例えば、8、10、12、14、16、18、20、30、40、50、75、100、150、200以上)である。好ましくは、これらのフラグメントは、配列から1つ以上のエピトープを含む。他の好ましいフラグメントは、シグナルペプチドを省略する。
【0016】
本発明のタンパク質は、種々の手段(例えば、化学合成(少なくとも一部)、プロテアーゼを用いるより長いポリペプチドの消化、RNAからの翻訳、細胞培養物(例えば、組換え発現物、C.trachomatis培養物)からの精製など)によって調製され得る。E.coliにおける異種発現は、好ましい調製用経路である。
【0017】
本発明のタンパク質は種々の形態(例えば、ネイティブ、融合、グリコシル化、非グリコシル化、脂溶化(lipidate)など)を取り得る。
【0018】
本発明のタンパク質は、好ましくは、実質的に純粋な形態(すなわち、他のC.trachomatisまたは宿主細胞タンパク質を実質的に含まない)で調製される。
【0019】
本発明のタンパク質は、固体支持体に結合され得る。これらは検出可能な標識(例えば、放射活性標識もしくは蛍光標識、またはビオチン標識)を含み得る。
【0020】
本発明のタンパク質は、好ましくはChlamydialタンパク質である。
【0021】
(C.trachomatis核酸)
本発明は、配列番号2〜262の偶数番号のヌクレオチド配列を含むタンパク質を提供する。
【0022】
本発明はまた、配列番号2〜262の偶数番号のヌクレオチド配列の1つ以上と少なくともx%配列同一性を共有する配列を含む核酸を提供する。特定の配列に依存して、xは、好ましくは、50%以上(例えば、60%、70%、80%、90%、95%、99%以上)である。
【0023】
さらに、本発明は、偶数のヌクレオチド配列である配列番号2〜262を含む核酸にハイブリダイズし得る核酸を提供する。ハイブリダイゼーション反応は、異なる「ストリンジェンシー」の条件下で行われ得る。ハイブリダイゼーション反応のストリンジェンシーを増す条件は、広く知られており、当該分野で公開されている。関連する条件の例としては、(ストリンジェンシーを増大させるために)以下が挙げられる:25℃、37℃、50℃、55℃および68℃のインキュベーション温度;10×SSC、6×SSC、1×SSC、0.1×SSCの緩衝液濃度および他の緩衝液系を使用するそれらの等価物;0%、25%、50%および75%のホルムアミド濃度;5分〜24時間のインキュベーション時間;1回、2回、またはそれ以上の洗浄工程;1分、2分または15分の洗浄インキュベーション時間;ならびに6×SSC、1×SSC、0.1×SSCの洗浄溶液濃度または脱イオン水。いくつかの実施形態において、本発明の単離された核酸は、低いストリンジェンシー条件下で選択的にハイブリダイズする;他の実施形態において、これは、中間のストリンジェンシー条件下で選択的にハイブリダイズする;他の実施形態において、これは、高いストリンジェンシー条件下で選択的にハイブリダイズする。低いストリンジェンシーハイブリダイゼーション条件の例示的なセットは、50℃および10×SSCである。中間のストリンジェンシーハイブリダイゼーション条件の例示的なセットは、55℃、1×SSCである。高いストリンジェンシーハイブリダイゼーション条件の例示的なセットは、68℃および0.1×SSCである。
【0024】
偶数のヌクレオチド配列である配列番号2〜262のフラグメントを含む核酸もまた、提供される。これらは、C.trachomatis配列に由来する少なくともn個の連続した核酸ヌクレオチドを含むべきであり、そして特定の配列に依存して、nは、7以上(例えば、10、12、14、15、18、20、25、30、35、40、50、75、100、200、300以上)である。
【0025】
さらなる局面に従って、本発明は、本発明のタンパク質およびタンパク質フラグメントをコードする核酸を提供する。
【0026】
本発明が上記の配列に相補的な配列を含む核酸を(例えば、アンチセンスまたはプローブ化目的のために)提供する。
【0027】
本発明の核酸は、もちろん、多数の方法で(例えば、化学合成によって(少なくとも一部)、より長いポリヌクレオチドを制限酵素を使用して消化することによって、ゲノムライブラリーまたはcDNAライブラリーから、生物体自体からなど)調製され得る。
【0028】
本発明の核酸は、種々の形態(例えば、一本鎖、二本鎖、直鎖状、環状、ベクター、プライマー、プローブなど)を採り得る。
【0029】
本発明の核酸は、固体支持体(例えば、ビーズ、プレート、フィルター、フィルム、スライドガラス、樹脂など)に結合され得る。本発明の核酸は、検出可能な標識(例えば、放射活性標識または蛍光標識、あるいはビオチン標識)を含み得る。これは、核酸検出技術(例えば、その核酸が、プライマーであるか、またはPCR、LCR、TMA、NASBA、bDNAなどのような技術において使用するためのプローブとしてである場合)において使用される場合に特に有用である。
【0030】
本発明の核酸は、好ましくは、クラミジア核酸である。
【0031】
用語「核酸」は、DNA、RNA、DNA/RNAハイブリッド、およびDNAまたはRNAのアナログ(例えば、改変された骨格もしくは塩基を含むもの)、そしてまたペプチド核酸(PNA)などを含む。
【0032】
本発明の核酸は、単離され得、そして実質的な純度で得られ得、一般的には、インタクトな染色体以外として得られ得る。通常ポリヌクレオチドは、実質的に他の天然に存在する核酸配列なしに得られ、一般には、少なくとも約50%(重量で)純粋、通常は、少なくとも約90%純粋である。
【0033】
核酸は、例えば、以下のために使用され得る:ポリペプチドを生成するため;生物学的サンプル中の核酸の検出のためのプローブとして;ポリペプチドのさらなるコピーを生成するため;リボザイムまたはアンチセンスオリゴヌクレオチドを生成するため;および一本鎖DNAプローブとしてまたは三本鎖形成オリゴヌクレオチドとしてなど。
【0034】
本発明は、本発明のヌクレオチド配列を含むベクター(例えば、クローニングベクターまたは発現ベクター)およびそれで形質転換された宿主細胞を提供する。
【0035】
(組成物)
さらなる局面に従って、本発明は、本発明に従うタンパク質および/または核酸を含む組成物を提供する。これらの組成物は、好ましくは、免疫原性組成物(例えば、ワクチン)であり、免疫およびワクチン接種の目的に適している。本発明のワクチンは、予防的または治療的であり得、そして代表的に、(a)クラミジア付着、(b)クラミジア侵入、および/または(c)宿主細胞内での首尾良い複製を阻害し得る抗体を誘導し得る抗原を含む。これらのワクチンは、好ましくは、宿主からのクラミジアクリアランスに必要である任意の細胞媒介性T細胞応答を誘導する。
【0036】
本発明はまた、医薬としての(例えば、ワクチンとしての)使用のための本発明に従う核酸およびタンパク質を提供する。
【0037】
本発明はまた、Chlamydiaに起因する感染を処置または予防するための医薬(例えば、ワクチンまたは免疫原性組成物)の製造における本発明に従う核酸またはタンパク質の使用を提供する。Chlamydiaは、一般的にC.trachomatisであるが、種間交差反応に起因して、これはまた、C.pneumoniae、C.pecorum、またはC.psittaciであり得る。予防のために、その医薬は、好ましくは、ChlamydiaのEB形態に対して特異的な免疫応答を誘発する;処置のためには、その医薬は、好ましくは、ChlamydiaのRB形態に対して特異的な免疫応答を誘発する。
【0038】
本発明はまた、Chlamydia trachomatis基本小体を中和するための医薬(例えば、ワクチンまたは免疫原性組成物)の製造において、本発明に従う核酸またえはタンパク質の使用を提供する。
【0039】
本発明はまた、患者(例えば、ヒト)を処置(例えば、免疫化)する方法を提供し、この方法は、本発明に従う治療的に有効な量の核酸またはタンパク質を患者に投与する工程を包含する。
【0040】
本発明はまた、患者における免疫応答を惹起する方法を提供し、この方法は、本発明に従う免疫学的に有効な量の核酸またはタンパク質を患者に投与する工程を包含する。この免疫応答は、患者において抗体を惹起し、そして/または細胞性免疫応答(例えば、CTL応答)を惹起することを包含する。この免疫応答は、EBタンパク質またはRBタンパク質に特異的であり得るか、または宿主細胞質中で発現されるタンパク質に対して特異的であり得る。抗体応答は。好ましくはEBに対して特異的であるのに対して、細胞免疫応答は、好ましくは、細胞質タンパク質に対して、好ましくやRBタンパク質に対して特異的である。
【0041】
本発明はまた、本発明のタンパク質を認識する抗体を惹起する方法を提供し、この方法は、Chlamydiaの基本小体または網様体を患者に投与する工程を包含する。この抗体は、好ましくは、EBに特異的である。
【0042】
本発明はまた、C.trachomatis感染を中和するための方法を提供し、この方法は、本発明のタンパク質、核酸または抗体を患者に投与する工程を包含する。この方法は、好ましくは、EB感染性を中和する。
【0043】
本発明はまた、生物学的サンプルにおけるChlamydia EBまたはRBを検出するための方法を提供し、この方法は、このサンプルと、本発明の抗体とを接触させる工程を包含する。このサンプルは、血液サンプル、別の体液、または組織サンプルであり得る。この方法は、クラミジア感染を診断するために使用され得る。
【0044】
本発明の免疫原性組成物はまた、以下の抗原のうちの1つ以上を含み得る:
−Helicobacter pylori由来のタンパク質抗原(例えば、VacA、CagA、NAP、HopX、HopY{例えば、WO98/04702})および/またはウレアーゼ。
【0045】
−N.meningitidis血清群B由来のタンパク質抗原(例えば、WO99/24578、WO99/36544、WO99/57280、WO00/22430、Tettelinら(2000)Science 287:1809−1815、Pizzaら(2000)Science 287:1816−1820およびWO96/29412)であり、タンパク質「287」および誘導体は、特に好ましい。
【0046】
−N.meningitidis血清群B由来の外膜ベシクル(OMV)調製物(例えば、WO01/52885;Bjuneら(1991)Lancet 338(8775):1093−1096;Fukasawaら(1999)Vaccine 17:2951−2958;Rosenqvistら(1998)Dev.Biol.Stand.92:323−333などにおいて開示される)。
【0047】
−N.meningitidis血清群A、C、W135および/またはY由来のサッカリド抗原(例えば、Costantinoら(1992)Vaccine 10:691−698において開示された、血清群C由来のオリゴサッカリド{Costantinoら(1999)Vaccine 17:1251−1263もまた参照のこと})。
【0048】
Streptococcus pneumoniae由来のサッカリド抗原{例えば、Watson(2000)Pediatr Infect Dis J 19:331−332;Rubin(2000)Pediatr Clin North Am 47:269−285,v;Jedrzejas(2001)Microbiol Mol Biol Rev 65:187−207}。
−B型肝炎ウイルス由来の抗原(例えば、表面抗原および/またはコア抗原){例えば、Gerlichら(1990)Vaccine 補遺8:S63〜68および79〜80}。
−C型肝炎ウイルス由来の抗原{例えば、Hsuら(1999)Clin Liver Dis 3:901〜915}。
−Bordetella pertussis由来の抗原(例えば、B.pertussis由来の百日咳ホロトキシン(holotoxin)(PT)および線維状赤血球凝集素(FHA)、必要に応じてペルタクチンおよび/または凝集原2および凝集原3とも組み合わされる{Gustafssonら(1996)N.Engl.J.Med.334:349−355;Rappuoliら(1991)TIBTECH 9:232−238}。
−ジフテリア抗原(例えば、ジフテリアトキソイド{例えば、Vaccines(1988)の第3章PlotkinおよびMortimer編 ISBN 0−7216−1946−0}例えば、CRM197変異体{例えば、Del Guidiceら(1998)Molecular Aspects of Medicine 19:1−70}。
−破傷風抗原(例えば、破傷風トキソイド{例えば、PlotkinおよびMortimerの第4章}。
−Haemophilus influenzae B由来のサッカリド抗原。
−N.gonorrhoeae由来の抗原{例えば、WO99/24578、WO99/36544、WO99/57280}。
−Chlamydia pneumoniae由来の抗原{例えば、PCT/IB01/01445;Kalmanら(1999)Nature Genetics 21:385−389;Readら(2000)Nucleic Acids Res 28:1397−406;Shiraiら(2000)J.Infect.Dis.181(補遺3):S524−S527;WO99/27105;WO00/27994;WO00/37494}。
−Chlamydia trachomatis由来の抗原{例えば、WO99/28475}。
−Porphyromonas gingivalis由来の抗原{例えば、Rossら(2001)Vaccine 19:4135−4142}。
−ポリオ抗原{例えば、Sutterら(2000)Pediatr Clin North Am 47:287−308;ZimmermanおよびSpann(1999)Am Fain Physician 59:113−118,125−126}(例えば、IPVまたはOPV)。
−狂犬病抗原{例えば、Dreesen(1997)Vaccine 補遺15:S2−6}(例えば、凍結乾燥されて非活性化されたウイルス{例えば、MMWR Morb Mortal Wkly Rep 1998 Jan 16;47(1):12,19;RabAvertTM}。
−麻疹抗原、耳下腺炎抗原および/または風疹抗原{例えば、PlotkinおよびMortimerの9、10および11章}。
−インフルエンザ抗原{例えば、PlotkinおよびMortimerの19章}。(例えば、赤血球凝集素および/またはノイラミニダーゼ表面蛋白質)。
−Moraxella catarrhalis由来の抗原{例えば、McMichael(2000) Vaccine 19 補遺1:S101−107}。
−Staphylococcus aureus由来の抗原{例えば、Kurodaら(2001)Lancet 357(9264):1225−1240;1218−1219頁もまた参照のこと}。
−Streptococcus agalactiae由来の抗原{例えば、WO02/34771を参照のこと}。
−Streptococcus pyogeraes由来の抗原{例えば、WO02/34771を参照のこと}。
【0049】
サッカリド抗原または糖鎖抗原が含まれる場合、免疫原性を高めるために、その抗原は、好ましくは、キャリアタンパク質に結合される{例えば、Ramsayら(2001)Lancet 357(9251):195−196;Lindberg(1999)Vaccine 17 補遺2:S28−36;Conjugate Vaccines(Cruseら編)ISBN 3805549326,特に10巻:48−114等}。好ましいキャリアタンパク質は、細菌毒素または細菌トキソイドである(例えば、ジフテリアトキソイドまたは破傷風トキソイド)。CRM97ジフテリアトキソイドが、特に好ましい。他の適切なキャリアタンパク質としては、N.meningitidis外膜タンパク質{例えば、EP−0372501}、合成ペプチド{例えば、EP−0378881,EP−0427347}、熱ショックタンパク質{例えば、WO93/17712}、百日咳タンパク質{例えば、WO98/58668;EP−0471177}、H.influenzae由来のタンパク質D{例えば、WO00/56360}、C.difficile由来の毒素Aまたは毒素B{例えば、WO00/61761}などが挙げられる。任意の適切な結合反応が、必要に応じて任意の適切なリンカーと共に使用され得る。
【0050】
毒素タンパク質抗原は、必要に応じて、解毒され得る(例えば、百日咳毒素の解毒は、化学的手段および/または遺伝的手段による)。
【0051】
ジフテリア抗原が組成物に含まれる場合、破傷風抗原および百日咳抗原をも含むことが好ましい。同様に、破傷風抗原が含まれる場合、ジフテリア抗原および百日咳抗原をも含むことが好ましい。同様に、百日咳抗原が含まれる場合、ジフテリア抗原および破傷風抗原をも含むことが好ましい。
【0052】
抗原は、好ましくはアルミニウム塩に吸着される。
【0053】
組成物中の抗原は、代表的には、各々少なくとも1μg/mlの濃度で存在する。一般的に、任意の所定の抗原の濃度は、その抗原に対する免疫応答を誘発するのに十分である。
【0054】
本発明はまた、本発明の2つ以上のタンパク質を含む組成物を提供する。
【0055】
(プロセス)
本発明は、本発明のタンパク質を産生するプロセスを提供し、このプロセスは、タンパク質発現を誘導する条件下で、本発明に従う宿主細胞を培養する工程を包含する。
【0056】
本発明は、本発明のタンパク質または核酸を生成するプロセスを提供し、ここで、このタンパク質または核酸は、化学的手段を使用して一部または全体的に合成される。
【0057】
本発明は、サンプル中のC.trachomatisを検出するためのプロセスを提供し、ここで、このサンプルは、本発明のタンパク質に結合する抗体と接触される。
【0058】
本発明を実行するために(例えば、免疫化について開示される配列を利用するために)使用され得る標準的な技術および手順の要約は、以下の通りである。この要約は、本発明を制限するのではなく、むしろ使用され得る実施例を示すが、必要とされない。
【0059】
(概要)
本発明の実施は、他に示されなければ、当該分野の技術の範囲内である、分子生物学、微生物学、組換えDNA、および免疫学の従来の技術を使用する。このような技術は以下の文献で十分説明されている(例えば、Sambrook Molecular Cloning;A Laboratory Manual 第2版(1989)および第3版(2001);DNA Cloning,Volumes I and ii(D.N Glover編 1985);Oligonucleotide Synthesis(M.J.Gait編 1984);Nucleic Acid Hybridization(B.D.HamesおよびS.J.Higgins編 1984);Transcription and Translation(B.D.HamesおよびS.J.Higgins編 1984);Animal Cell Culture(R.I.Freshney編 1986);Immobilized Cells and Enzymes(IRL Press,1986);B.Perbal,A Practial Guide to Molecular Cloning(1984);the Methods in Enzymology series(Academic Press,Inc.),特に154巻および155巻;Gene Transfer Vectors for Mammalian Cells(J.H.MillerおよびM.P.Calos編 1987,Cold Spring Harbor Laboratory);MayerおよびWalker編(1987),Immunochemical Methods in Cell and Molecular Biology(Academic Press,London);Scopes,(1987)Protein Purification:Principles and Practice,第2版(Springer−Verlag,N.Y.)、およびHandbook of Experimental Immunology,Volumes I−IV(D.M.WeirおよびC.C.Blackwell編 1986)。
【0060】
ヌクレオチドおよびアミノ酸についての標準的な略語が、本明細書において使用される。
【図面の簡単な説明】
【0061】
【図1A】図1Aは、実施例1のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図1B】図1Bは、実施例1のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図1C】図1Cは、実施例1のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図2A】図2Aは、実施例2のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図2B】図2Bは、実施例2のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図2C】図2Cは、実施例2のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図3A】図3Aは、実施例3のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図3B】図3Bは、実施例3のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図3C】図3Cは、実施例3のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図3D】図3Dは、実施例3のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4A】図4Aは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4B】図4Bは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4C】図4Cは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4D】図4Dは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図4E】図4Eは、実施例4のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5A】図5Aは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5B】図5Bは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5C】図5Cは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5D】図5Dは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図5E】図5Eは、実施例5のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図6A】図6Aは、実施例6のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図6B】図6Bは、実施例6のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図6C】図6Cは、実施例6のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図7A】図7Aは、実施例7のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図7B】図7Bは、実施例7のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図8A】図8Aは、実施例8のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図8B】図8Bは、実施例8のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図8C】図8Cは、実施例8のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図9A】図9Aは、実施例9のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図9B】図9Bは、実施例9のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図9C】図9Cは、実施例9のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図9D】図9Dは、実施例9のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図10A】図10Aは、実施例10のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図10B】図10Bは、実施例10のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図10C】図10Cは、実施例10のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図11A】図11Aは、実施例11のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図11B】図11Bは、実施例11のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図11C】図11Cは、実施例11のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12A】図12Aは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12B】図12Bは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12C】図12Cは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12D】図12Dは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図12E】図12Eは、実施例12のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図13A】図13Aは、実施例13のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図13B】図13Bは、実施例13のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図13C】図13Cは、実施例13のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図14A】図14Aは、実施例14のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図14B】図14Bは、実施例14のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図14C】図14Cは、実施例14のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図15A】図15Aは、実施例15のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図15B】図15Bは、実施例15のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図15C】図15Cは、実施例15のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図16A】図16Aは、実施例16のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図16B】図16Bは、実施例16のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図16C】図16Cは、実施例16のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図17】図17は、実施例17のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図18A】図18Aは、実施例18のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図18B】図18Bは、実施例18のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図19】図19は、実施例19のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図20A】図20Aは、実施例20のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図20B】図20Bは、実施例20のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図21】図21は、実施例21のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図22】図22は、実施例22のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図23A】図23Aは、実施例23のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図23B】図23Bは、実施例23のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図23C】図23Cは、実施例23のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図24A】図24Aは、実施例24のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図24B】図24Bは、実施例24のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図24C】図24Cは、実施例24のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図24D】図24Dは、実施例24のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図25A】図25Aは、実施例25のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図25B】図25Bは、実施例25のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図25C】図25Cは、実施例25のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図26A】図26Aは、実施例26のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図26B】図26Bは、実施例26のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27A】図27Aは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27B】図27Bは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27C】図27Cは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27D】図27Dは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図27E】図27Eは、実施例27のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図28A】図28Aは、実施例28のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図28B】図28Bは、実施例28のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図29A】図29Aは、実施例29のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図29B】図29Bは、実施例29のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図30】図30は、実施例30のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図31】図31は、実施例31のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図32】図32は、実施例32のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図33A】図33Aは、実施例33のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図33B】図33Bは、実施例33のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図34A】図34Aは、実施例34のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図34B】図34Bは、実施例34のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図35A】図35Aは、実施例35のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図35B】図35Bは、実施例35のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図36】図36は、実施例36のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図37A】図37Aは、実施例37のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図37B】図37Bは、実施例37のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図37C】図37Cは、実施例37のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図38A】図38Aは、実施例38のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図38B】図38Bは、実施例38のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図39】図39は、実施例39のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図40】図40は、実施例40のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図41】図41は、実施例41のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図42】図42は、実施例42のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図43】図43は、実施例43のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図44A】図44Aは、実施例44のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【図44B】図44Bは、実施例44のデータを示す。図はゲルの図であり、レーン1とは、その図の左側のレーンである。
【発明を実施するための形態】
【0062】
(定義)
Xを含む組成物は、組成物中の全X+Yの少なくとも85重量%がXであるとき、Yを「実質的に含まない」。好ましくは、Xは、組成物中の全X+Yの少なくとも約90重量%を、さらに好ましくは少なくとも約95重量%または99重量%さえを含む。
【0063】
用語「含む(comprising)」は、「含む(including)」および「からなる」を意味する。例えば、Xを「含む」組成物は、もっぱらXからなり得るか、またはX+YのようなXに付加したものも含み得る。
【0064】
用語「異種」とは、天然では共に見られない2つの生物学的成分をいう。この成分は、宿主細胞、遺伝子、または調節領域(例えば、プロモーター)であり得る。異種成分は天然では共に見られないが、遺伝子に対して異種のプロモーターがその遺伝子に作動可能に連結されるときのように、それらは共に機能し得る。別の例は、クラミジア配列がマウス宿主細胞に対して異種であることである。さらなる例は、天然においてみられない配置で単一タンパク質に組み立てられる同じタンパク質または異なるタンパク質由来の2つのエピトープである。
【0065】
「複製起点」とは、発現ベクターのような、ポリヌクレオチドの複製を開始または調節するポリヌクレオチド配列である。複製起点は、細胞内でのポリヌクレオチド複製の自律性ユニットとして振る舞い、それ自体の制御下で複製し得る。複製起点は、ベクターが特定の宿主細胞において複製するために必要であり得る。特定の複製起点を有せば、発現ベクターは、細胞内の適切なタンパク質の存在下で高いコピー数で再生され得る。起点の例は、酵母において有効な自律性複製配列;およびCOS−7細胞で有効であるウイルスT抗原である。
【0066】
「変異体」配列は、ネイティブの配列または開示された配列とは異なるが配列同一性を有するDNA配列、RNA配列またはアミノ酸配列として規定される。特定の配列に依存して、ネイティブの配列または開示された配列と変異体配列との間の配列同一性の程度は、好ましくは50%より大きい(例えば、上記のSmith−Watermanアルゴリズムを使用して算出して60%、70%、80%、90%、95%、99%またはそれ以上)。本明細書中で使用される場合、本明細書で核酸分子配列が提供される核酸分子、または領域の「対立遺伝子改変体」は、別のもしくは2番目の単離体のゲノム中の同じ遺伝子座で本質的に生じる、核酸分子、または領域および、例えば、変異または組換えにより生ずる天然変化に起因して、類似するが、しかし同一でない核酸配列を有する核酸分子、または領域である。コード領域の対立遺伝子改変体は、代表的には、比較される遺伝子によってコードされるタンパク質の活性と類似した活性を有するタンパク質をコードする。対立遺伝子改変体はまた、遺伝子の5’または3’非翻訳領域(例えば、調節制御領域)での変化を含み得る(例えば、米国特許第5,753,235号を参照のこと)。
【0067】
(発現系)
クラミジアヌクレオチド配列は、種々の異なる発現系;例えば、哺乳動物細胞、バキュロウイルス、植物、細菌、および酵母と共に使用される発現系において発現され得る。
【0068】
(i.哺乳動物系)
哺乳動物発現系は当該分野において公知である。哺乳動物プロモーターは、哺乳動物RNAポリメラーゼを結合し、コード配列(例えば、構造遺伝子)のmRNAへの下流(3’)転写を開始し得る任意のDNA配列である。プロモーターは、転写開始領域(これはコード配列の5’末端の近位に通常位置する)およびTATAボックス(転写開始部位の25〜30塩基対(bp)上流に通常位置する)を有する。TATAボックスは、その正しい部位においてRNAポリメラーゼIIにRNA合成を開始させるよう指示すると考えられている。哺乳動物プロモーターはまた、TATAボックスの100〜200bp上流以内に通常位置する上流プロモーターエレメントを含む。上流プロモーターエレメントは、転写が開始され、そしていずれかの方向において作用し得る速度を決定する{Sambrookら(1989)「Expression of Cloned Genes in Mammalian Cells」 Molecular Cloning:A Laboratory Manual、第2版}。
【0069】
哺乳動物ウイルス遺伝子は、しばしば高度に発現され、そして広い宿主範囲を有する;従って、哺乳動物ウイルス遺伝子をコードする配列は、特に有用なプロモーター配列を提供する。例としては、SV40初期プロモーター、マウス乳癌ウイルスLTRプロモーター、アデノウイルス主要後期プロモーター(Ad MLP)、および単純疱疹ウイルスプロモーターが挙げられる。さらに、マウスのメタロチオネイン遺伝子のような非ウイルス性遺伝子に由来する配列もまた、有用なプロモーター配列を提供する。発現は、構成性であるかまたは調節される(誘導可能)かのいずれかであり得る、プロモーターに依存して、ホルモン応答性細胞においてグルココルチコイドで誘導され得る。
【0070】
上記に記載されるプロモーターエレメントと組み合わされるエンハンサーエレメント(エンハンサー)の存在は、通常発現レベルを増大させる。エンハンサーは、相同性プロモーターまたは異種プロモーターに連結されるとき、転写を1000倍まで刺激し得る調節DNA配列であり、合成は、通常のRNA開始部位で開始する。エンハンサーはまた、それらが、通常方向もしくは反対(flipped)方向のいずれかで転写開始部位より上流または下流に、もしくはプロモーターから1000ヌクレオチドを超える距離に位置するとき、活性である(Maniatisら(1987) Science 236:1237;Albertsら(1989)Molecular Biology of the Cell,第2版)。ウイルス由来のエンハンサーエレメントは、それらが通常、より広い宿主範囲を有するので、特に有用であり得る。例としては、SV40初期遺伝子エンハンサー(Dijkemaら(1985)EMBO J.4:761)およびラウス肉腫ウイルスの長末端反復(LTR)に由来するエンハンサー/プロモーター(Gormanら(1982)PNAS USA 79:6777)およびヒトサイトメガウイルスに由来するエンハンサー/プロモーター(Boshartら(1985)Cell 41:521)を包含する。さらに、いくつかのエンハンサーは調節可能であり、そしてホルモンまたは金属イオンのような誘導因子の存在下のみで活性になる(Sassone−CorsiおよびBorelli(1986)Trends Genet.2:215;Maniatisら(1987)Science 236:1237)。
【0071】
DNA分子は、哺乳動物細胞において細胞内で発現され得る。プロモーター配列は、組換えタンパク質のN末端における最初のアミノ酸が常にATG開始コドンによりコードされるメチオニンである場合、DNA分子と直接連結され得る。所望される場合、N末端は、臭化シアンとのインビトロでのインキュベーションによりタンパク質から切断され得る。
【0072】
あるいは、外来タンパク質もまた、哺乳動物細胞において外来タンパク質の分泌を提供するリーダー配列フラグメントで構成される融合タンパク質をコードするキメラDNA分子を作製することにより、細胞から成長培地へ分泌され得る。好ましくは、インビボまたはインビトロのいずれかにおいて切断され得る、リーダーフラグメントおよび外来遺伝子との間にコードされるプロセシング部位が存在する。リーダー配列フラグメントは通常、細胞からのタンパク質の分泌を指示する疎水性アミノ酸で構成される単一のペプチドをコードする。アデノウイルス3部分からなるリーダーは、哺乳動物細胞における外来タンパク質の分泌を提供するリーダー配列の例である。
【0073】
通常、哺乳動物細胞によって認識される転写終結配列およびポリアデニル化配列は、翻訳終止コドンの3’側に存在する調節領域であり、従って、プロモーターエレメントと共に、コード配列に隣接する。成熟mRNAの3’末端は、部位特異的転写後切断およびポリアデニル化により形成される(Birnstielら(1985)Cell 41:349;ProudfootおよびWhitelaw(1988)「Termination and 3’end processing of eukaryotic RNA.」Transcription and splicing(B.D.HamesおよびD.M.Glover編);Proudfoot(1989)Trends Biochem.Sci.14:105)。これらの配列は、mRNAの転写を指示し、そのmRNAは、そのDNAにコードされるポリペプチドに翻訳され得る。転写終結/ポリアデニル化シグナルの例としては、SV40由来のシグナルが挙げられる(Sambrookら(1989)「Expression of cloned genes in cultured mammalian cells.」Molecular Cloning:A Laboratory Manual)。
【0074】
通常、プロモーター、ポリアデニル化シグナル、および転写終結配列を含む上記の構成要素は、発現構築物内に共に導入される。エンハンサー、機能的なスプライスドナー部位および受容部位を有するイントロン、ならびにリーダー配列もまた、所望される場合、発現構築物内に含まれ得る。発現構築物はしばしば、哺乳動物細胞または細菌のような宿主内で安定的に維持し得る染色体外エレメント(例えば、プラスミド)のような、レプリコン内で維持される。哺乳動物の複製系としては、複製にトランス作用性の因子を必要とする、動物ウイルス由来の複製系が挙げられる。例えば、SV40(Gluzman(1981)Cell 23:175)、あるいはポリオーマウイルスのようなパポバウイルスの複製系を含むプラスミドは、適切なウイルスのT抗原の存在下で極めて高いコピー数で複製する。哺乳動物レプリコンのさらなる例としては、ウシパピローマウイルスおよびエプスタイン−バーウイルス由来のレプリコンが挙げられる。さらに、このレプリコンは、二つの複製系を有し得、従って、例えば、発現用の哺乳動物細胞内で、ならびにクローニングおよび増幅用の原核生物の宿主内で、そのレプリコンを維持することが可能である。このような哺乳動物−細菌シャトルベクターの例としては、pMT2(Kaufmanら(1989)Mol.Cell.Biol.9:946)およびpHEBO(Shimizuら(1986)Mol.Cell.Biol.6:1074)が挙げられる。
【0075】
使用される形質転換の手順は、形質転換される宿主に依存する。異種のポリヌクレオチドの哺乳動物細胞への導入方法は、当該分野で公知であり、その方法としては、デキストラン媒介トランスフェクション、リン酸カルシウム沈殿、ポリブレン媒介トランスフェクション、プロトプラスト融合、エレクトロポレーション、リポソーム内でのポリヌクレオチドの封入、およびDNAの核内への直接微量注入が挙げられる。
【0076】
発現用宿主として利用可能な哺乳動物細胞株は、当該分野で公知であり、そのような細胞株としては、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、乳仔ハムスター腎臓(BHK)細胞、サル腎臓細胞(COS)、ヒト肝細胞癌細胞(例えば、Hep G2)および多くの他の細胞株を含むが、これらに限定しない、American Type Culture Collection(ATCC)から入手可能な多くの不死化細胞株が含まれる。
【0077】
(ii バキュロウイルス系)
タンパク質をコードしているポリヌクレオチドはまた、適切な昆虫の発現ベクター内に挿入され得、そしてそのベクター内で、制御エレメントに作動可能に連結される。ベクターの構築には、当該分野で公知の技術を使用する。一般に、その発現系の構成要素として、以下のものが挙げられる:バキュロウイルスゲノムのフラグメント、および発現させる異種遺伝子の挿入用の簡便な制限部位の両方を含む転移ベクター(通常は細菌プラスミド;転移ベクター内のバキュロウイルスに特異的なフラグメントに相同性のある配列を有する野生型バキュロウイルス(これは、バキュロウイルスゲノム内への異種遺伝子の相同組換えを可能にする);ならびに適切な昆虫宿主細胞および成長培地。
【0078】
転移ベクターにタンパク質をコードするDNA配列を挿入した後、そのベクターおよび野生型ウイルスゲノムを、昆虫宿主細胞にトランスフェクトし、そこでベクターとウイルスゲノムを組換え可能にする。パッケージングされた組換えウイルスは発現され、そして組換えプラークが同定されそして精製される。バキュロウイルス/昆虫細胞発現系に対する材料および方法は、特に、Invitrogen、San Diego CAからキット形態(「MaxBac」キット)で市販される。これらの技術は、一般に当業者に公知であり、そしてSummersならびにSmith,Texas Agricultural Experiment Station Bulletin No.1555(1987)(以下、「Summers and Smith」)に十分に記載されている。
【0079】
タンパク質をコードするDNA配列をバキュロウイルスゲノムに挿入するの前に、プロモーター配列、リーダー配列(所望される場合は)、目的のコード配列、および転写終結配列を含む上記の構成要素を、通常、中間転移構築物(転移ベクター)に構築される。この構築物は、単一の遺伝子および作動可能に連結された調節エレメント;操作可能に連結された調節エレメントのセットを各々が所有する複数の遺伝子;あるいは同じ調節エレメントのセットにより調節される複数の遺伝子を含み得る。中間転移構築物は、しばしば、細菌のような宿主内で安定的に維持し得る染色体外エレメント(例えば、プラスミド)のような、レプリコン内で維持される。レプリコンは、複製系を有しており、従って、クローニングおよび増幅に適切な宿主内で維持されることを可能にする。
【0080】
現在、外来遺伝子のAcNPVへの導入のために最も一般に使用される転移ベクターは、pAc373である。当業者に公知の多くの他のベクターもまた設計され、これらのものとして、例えば、pVL985(ポリへドリンの開始コドンをATGからATTに変更し、そしてそのATTから32塩基対下流に、BamHIクローニング部位を導入する;LuckowおよびSummers,Virology(1989)17:31を参照のこと)が挙げられる。
【0081】
そのプラスミドも通常、ポリへドリンポリアデニル化シグナル(Millerら(1988)Ann.Rev.Microbiol.,42:177)、および原核生物のアンピシリン耐性(amp)遺伝子、および大腸菌においての選抜ならびに増殖のための複製起点を含む。
【0082】
バキュロウイルス転移ベクターは通常、バキュロウイルスプロモーターを含む。バキュロウイルスプロモーターは、バキュロウイルスRNAポリメラーゼに結合し、そしてコード配列(例えば、構造遺伝子)のmRNAへの下流(5’から3’)方向の転写を開始し得る任意のDNA配列である。プロモーターは、通常、コード配列の5’末端に近接して存在する転写開始領域を有する。この転写開始領域は通常、RNAポリメラーゼ結合部位および転写開始部位を含む。バキュロウイルス転移ベクターはまた、エンハンサーと呼ばれる第二のドメインを有し得、存在する場合は、通常、構造遺伝子に対し遠位にある。発現は調節され得るか、あるいは構成的であり得る。
【0083】
ウイルスの感染周期の後期で大量に転写される構造遺伝子は、特に有用なプロモーター配列を提供する。例としては、ウイルス多面体タンパク質をコードする遺伝子(Friesenら(1986)「The Regulation of Baculovirus Gene Expression,」:The Molecular Biology of Baculoviruses(Walter Doerfler編);EPO公開番号127839および155476;ならびにp10タンパク質をコードする遺伝子(Vlakら,(1988),J.Gen.Virol.69:765)由来の配列が挙げられる。
【0084】
適切なシグナル配列をコードするDNAは、分泌昆虫タンパク質、あるいはバキュロウイルスポリへドリン遺伝子(Carbonellら,(1998)Gene,73:409)のような、分泌バキュロウイルスタンパク質についての遺伝子から由来し得る。あるいは、哺乳動物細胞の翻訳後修飾(例えば、シグナルペプチド切断、タンパク質分解性切断、およびリン酸化)のシグナルは、昆虫細胞に認識されると思われ、そして分泌および核蓄積に必要なシグナルはまた、無脊椎動物細胞と脊椎動物細胞との間で保存されると思われるので、ヒトインターフェロンα(Maedaら,(1985),Nature 315:592);ヒトガストリン放出ペプチド(Lebacq−Verheydenら,(1988),Molec.Cell.Biol.8:3129);ヒトIL−2(Smithら,(1985)Proc.Nat’l Acad.Sci.USA、82:8404);マウスIL−3(Miyajimaら,(1987)Gene 58:273);およびヒトグルコセレブロシダーゼ(Martinら,(1988)DNA、7:99)をコードする遺伝子由来のような、非昆虫起源のリーダーも、昆虫での分泌を与えるために使用され得る。
【0085】
組換えポリペプチドあるいは組換えポリタンパク質は、細胞内に発現され得、あるいは適切な調節配列と共に発現される場合、分泌され得る。非融合の外来タンパク質の優れた細胞内発現には通常、ATG開始シグナルに先行する適切な翻訳開始シグナルを含む短いリーダー配列を理想的には有する異種遺伝子が必要である。所望であれば、N末端のメチオニンは、臭化シアンとのインビトロインキュベーションにより、成熟タンパク質から切断され得る。
【0086】
あるいは、自然に分泌されない組換えポリタンパク質あるいは組換えタンパク質は、昆虫において外来タンパク質の分泌を与えるリーダー配列フラグメントからなる、融合タンパク質をコードするキメラDNA分子を作成することにより、昆虫細胞から分泌され得る。リーダー配列フラグメントは通常、タンパク質の小胞体内への輸送を指示する疎水性アミノ酸からなるシグナルペプチドをコードしている。
【0087】
タンパク質の前駆体である発現産物をコードするDNA配列および/または遺伝子の挿入後、昆虫細胞宿主に、転移ベクターの異種DNAおよび野生型バキュロウイルスのゲノムDNAを同時形質転換(通常は、同時トランスフェクションによって)する。構築物のプロモーターおよび転写終結配列は、通常バキュロウイルスゲノムの2〜5kbの区域を含む。バキュロウイルスウイルスの望ましい部位に異種DNAを導入する方法は、当該分野で公知である(SummersおよびSmith、上記;Juら(1987);Smithら,Mol.Cell.Biol.(1983)3:2156;ならびにLuckowおよびSummers(1989)を参照のこと)。例えば、その挿入物は、相同二重交差組換え(homologous double crossover recombination)により、ポリへドリン遺伝子のような遺伝子内にあり得る;挿入物はまた、所望のバキュロウイルス遺伝子に設計された制限酵素部位内にあり得る。Millerら(1989)、Bioessays 4;91。発現ベクター内のポリへドリン遺伝子の代わりにクローン化される場合のDNA配列は、ポリへドリン特異的配列により5’および3’の両側で隣接され、そしてポリへドリンプロモーターの下流に位置される。
【0088】
新規に形成されたバキュロウイルス発現ベクターは続いて、感染性の組換えバキュロウイルス内にパッケージされる。相同組換えは、低い頻度で起こる(約1%〜約5%の間);従って、同時トランスフェクション後に産生されたウイルスの大半は、依然野生型ウイルスである。従って、方法には、組換えウイルスの同定が必要となる。その発現系の利点は、組換えウイルスを区別させ得る可視的スクリーニングである。天然のウイルスにより産生されるポリへドリンタンパク質は、ウイルス感染後の後期で、その感染された細胞の核内で非常に高いレベルで産生される。蓄積されたポリへドリンタンパク質は、閉塞体を形成し、またそれは包理された粒子を含む。これらの閉塞体は、最大15μmの大きさで、高度に屈折し、明るく輝く外見を与え、容易に光学顕微鏡下で可視化される。組換えウイルスに感染された細胞は、閉塞体を欠く。組換えウイルスと野生型ウイルスを区別するために、トランスフェクションの上清を、当業者に公知の技術により昆虫細胞の単層にプラーク形成させた。すなわち、プラークを、光学顕微鏡下で閉塞体の存在(野性型ウイルスを示す)または非存在(組換えウイルスを示す)によりスクリーニングする。「Current Protocols in Microbiology」2巻(Ausubelら編)16.8(増補10、1990);SummersおよびSmith、上記;Millerら(1989)。
【0089】
組換えバキュロウイルス発現ベクターは、いくつかの昆虫細胞への感染用に開発された。例えば、組換えバキュロウイルスは、特に以下に示すもののために開発された:Aedes aegypti、Autographa californica、Bombyx mori、Drosophila melanogaster、Spodoptera frugiperda、およびTrichoplusia ni(WO89/046699;Carbonellら,(1985)J.Virol.56:153;Wright(1986)Nature 321:718;Smithら,(1983)Mol.Cell.Biol.3:2156;およびFraserら,(1989)In Vitro Cell.Dev.Biol.25:225を一般に参照のこと)。
【0090】
細胞および細胞培養培地は、バキュロウイルス/発現系における異種ポリペプチドの直接発現および融合発現の両方のために市販される;細胞培養技術は、一般に当業者に公知である。例えば、SummersおよびSmith(上記)を参照のこと。
【0091】
次いで、改変した昆虫細胞を、適切な栄養培地で増殖し、その改変した昆虫宿主内に存在するプラスミドの安定的維持を可能にする。発現産物の遺伝子が、誘導性の制御下にある場合、宿主は高密度で増殖され得、そして発現は誘導され得る。あるいは、発現が構成的である場合、その産物は培地中に連続的に発現され、そして栄養培地を連続的に循環し、目的産物を取り出し、そして枯渇した栄養を補給する必要がある。その産物は、クロマトグラフィー(例えば、HPLC、アフィニティークロマトグラフィー、イオン交換クロマトグラフィーなど);電気泳動;密度勾配遠心;溶媒抽出などのような技術により精製され得る。適切には、その産物をさらに精製し、必要ならば、培地中に分泌されたか、または昆虫細胞の溶解物から生じた任意の昆虫タンパク質を実質的に除去し、宿主細片(例えば、タンパク質、脂質および多糖類)を少なくとも実質的に含まない産物を提供する。
【0092】
タンパク質の発現を得るために、形質転換体に由来する組換え宿主細胞は、組換えタンパク質をコードする配列の発現を可能にする条件下でインキュベートされる。これらの条件は、選択された宿主細胞に依存して変動する。しかし、その条件は、当該分野で公知の条件に基づいて、当業者に容易に確かめられる。
【0093】
(iii 植物系)
当該分野で公知の多くの植物細胞培養および全植物遺伝子発現系が存在する。例示的な植物細胞遺伝子発現系としては、米国特許第5,693,506号;米国特許第5,659,122号;米国特許第5,608,143号のような特許に記載されるものが挙げられる。植物細胞培養における遺伝子発現の別の例は、Zenk,Phytochemistry 30:3861−3863(1991)に記載された。植物タンパク質のシグナルペプチドの記載は、上記の参考文献に加え、以下に示すものの中においても確認される;Vaulcombeら,Mol.Gen.Genet.209:33−40(1987);Chandlerら,Plant Molecular Biology 3:407−418(1984);Rogers,J.Biol.Chem.260:3731−3738(1985);Rothsteinら,Gene 55:353−356(1987);Whittierら,Nucleic Acids Research 15:2515−2535(1987);Wirselら,Molecular Microbiology 3:3−14(1989);Yuら,Gene 122:247−253(1992)。植物ホルモン(ジベレリン酸およびジベレリン酸により誘導される分泌酵素)による植物遺伝子発現の調節の記載は、R.L.JonesおよびJ.MacMillin,Gibberellins:Advanced Plant Physiology,Malcolm B.Wilkins編 1984 Pitman Publishing Limited,London,21−52頁の中に確認され得る。他の代謝調節性遺伝子が記載される参考文献は以下である;Sheen,Plant Cell,2:1027−1038(1990);Maasら,EMBO J.9:3447−3452(1990);BenkelおよびHickey、Proc.Natl.Acad.Sci.84:1337−1339(1987)。
【0094】
代表的に、当該分野で公知の技術を使用して、所望のポリヌクレオチド配列は、植物内で操作するために設計された遺伝子調節エレメントを含む発現カセットの中に挿入される。その発現カセットは、植物宿主内での発現に適切な発現カセットの上流および下流にコンパニオン配列を有する所望の発現ベクターの中に挿入される。そのコンパニオン配列は、プラスミドまたはウイルス起源のものであり、そしてそのベクターが、細菌のような本来のクローニング宿主から、所望の植物宿主へDNAを移動させるために必要とされる特徴をベクターに提供する。基本的な細菌/植物ベクター構築物は、好ましくは、広い宿主範囲の原核生物の複製起点;原核生物の選択マーカー;および、アグロバクテリウムの形質転換については、アグロバクテリウム媒介転位のためのT DNA配列を植物染色体に提供する。異種遺伝子が容易に検出に従順しない場合は、好ましくは、その構築物はまた、植物細胞が形質転換されたかどうかを決定するために適した選択マーカー遺伝子を有する。適切なマーカーの一般的な総説は、例えば、イネ科のメンバーについては、WilminkおよびDons,1993,Plant Mol.Biol.Reptr,11(2):165−185に見られる。
【0095】
植物ゲノムへの異種配列の組み込みを可能にするのに適した配列もまた、推奨される。これらは、植物ゲノム内へ異種発現カセットのランダム挿入を可能にする相同組換えのためのトランスポゾン配列など、およびTi配列を含み得る。適切な原核生物選択マーカーとしては、アンピシリンまたはテトラサイクリンのような抗生物質に対する耐性が挙げられる。別の機能をコードしている他のDNA配列もまた、そのベクターの中に存在し得、当該分野で公知である。
【0096】
本発明の核酸分子は、目的のタンパク質の発現用の発現カセットに含まれ得る。2つ以上も可能であるが、通常はただ一つの発現カセットである。組換え発現カセットは、異種タンパクのコード配列に加え、以下のエレメントを含む;プロモーター領域、植物5’非翻訳配列、構造遺伝子がそれを備えているかどうかに依存して、開始コドン、ならびに転写および翻訳終結配列。そのカセットの5’および3’末端の独特な制限酵素部位は、既存のベクター内への容易な挿入を可能にする。
【0097】
異種のコード配列は、本発明に関係する任意のタンパク質のための配列であり得る。目的のタンパクをコードする配列は、そのタンパク質の適切なプロセッシングおよび輸送を可能にするシグナルペプチドをコードし、そして通常、本発明の所望のタンパク質の膜への結合を生じ得る任意の配列を欠いている。翻訳開始領域は、大部分は、発芽中に発現および輸送される遺伝子のためのものであるから、輸送を提供するシグナルペプチドを使用することにより、それはまた、目的のタンパク質の輸送を提供し得る。この方法で、目的のタンパク質は、それらが発現される細胞から輸送され、そして効率的に回収され得る。代表的には、種子における分泌は、種子の胚乳内へ、アリューロン層あるいは胚盤上皮層を通過する。タンパク質がそれが産生された細胞から分泌される必要がない場合、このことは組換えタンパク質の分離および精製を容易にする。
【0098】
所望の遺伝子産物の最終的な発現が、真核生物におけるものであるので、クローン化した遺伝子の任意の部分が、イントロンのような、宿主のスプライセオソーム(splicosome)機構により、プロセッシングされる配列を含むか否かを決定することが望ましい。そのような場合、「イントロン」領域の部位特異的変異誘発は、仮性イントロンコードとして遺伝的情報の一部を欠失することを防ぐために実施され得る。ReedおよびManiatis、Cell 41:95−105(1985)。
【0099】
ベクターは、組換えDNAを機械的に転移するためにマイクロピペットを用いて植物細胞内に直接的に微量注入され得る(Crossway、Mol.Gen.Genet,202:179−185、1985)。遺伝子物質はまた、ポリエチレングリコールを用いて植物細胞内に転移され得る(Krensら、Nature,296、72−74、1982)。核酸部分の導入の別の方法は、小さいビーズまたは微粒子のいずれかのマトリックスの内部に、あるいは表面に核酸を有する小さな微粒子による高速バリスティック(ballistic)穿通法である(Kleinら,Nature,327,70−73,1987ならびにKnudsenおよびMuller,1991,Planta,185:330−336は、大麦胚乳の粒子の照射(bombardment)によりトランスジェニック大麦を作製することを示している)。さらに別の導入方法は、他の物体、いずれかのミニ細胞、細胞、リソソームあるいは他の易融な脂肪表面体とのプロトプラストの融合である(Fraleyら、Proc.Natl.Acad.Sci.USA,79,1859−1863,1982)。
【0100】
ベクターはまた、エレクトロポレーションにより植物細胞内に導入され得る(Frommら、Proc.Natl.Acad.Sci.USA 82:5824,1985)。この技術において、植物プロトプラストは、遺伝子構築物を含むプラスミドの存在中でエレクトロポレート(電気穿孔)される。高い電界の強さの電気インパルス(衝撃)により、生体膜を可逆的に通過できるようにし、プラスミドの導入を可能にする。エレクトロポレートされた植物プロトプラストは細胞壁を再形成し、分裂し、植物カルス形成する。
【0101】
プロトプラストが単離され得、そして培養されて完全な再生植物を与え得る全ての植物は、本発明により形質転換され得、これによって移入した遺伝子を保持する完全な植物が再生される。実際に、全ての植物は、さとうきび、甜菜、綿、果実および他の樹木、マメ科植物および野菜の全ての主要な種を含むがそれらに限定されない培養細胞あるいは組織から再生され得る。いくつかの適応した植物としては、例えば、以下の属由来の種が挙げられる;Fragaria,Lotus,Medicago,Onobrychis,Trifolium,Trigonella,Vigna,Citrus,Linum,Geranium,Manihot,Daucus,Arabidopsis,Brassica,Raphanus,Sinapis,Atropa,Capsicum,Datura,Hyoscyamus,Lycopersion,Nicotiana,Solanum,Petunia,Digitalis,Majorana,Cichorium,Helianthus,Lactuca,Bromus,Asparagus,Antirrhinum,Hererocallis,Nemesia,Pelargonium,Panicum,Pennisetum,Ranunculus,Senecio,Salpiglossis,Cucumis,Browaalia,Glycine,Lolium,Zea,Triticum,Sorghum,およびDatura。
【0102】
再生のための手法は、植物の種によって変化するが、しかし一般に異種遺伝子のコピーを含む形質転換されたプロトプラストの懸濁液が最初に提供される。カルス組織は形成され、そしてシュート(苗条)がカルスから誘導され得、続いて発根される。あるいは、胚形成がプロトプラスト懸濁液から誘導され得る。これらの胚は、天然の胚として発芽し、植物を形成する。培養培地は、一般に種々のアミノ酸、ならびにオーキシンおよびサイトカイニンのようなホルモンを含有する。またグルタミン酸およびプロリンを培地に添加することは、特にコーン(穀草)およびアルファルファのような種にとって有用である。シュートおよび根は通常、同時に発生する。効果的な再生は培地、遺伝子型、および培養遍歴に依存する。これらの3つの変数が制御される場合は、再生は十分に再現性がありそして繰り返し可能である。
【0103】
いくつかの植物細胞培養系において、本発明の所望のタンパク質は排出され、あるいはこのタンパク質は植物全体から抽出され得る。本発明の所望のタンパク質が培地内に分泌される場合、これは回収され得る。あるいは、胚および胚のない不完全な種子または他の植物組織は、機械的に破壊され細胞間および組織間の任意の分泌されたタンパク質を放出し得る。その混合物は緩衝液に懸濁され、可溶タンパクを回収し得る。次いで、慣用的なタンパク質分離および精製方法は組換えタンパク質を精製するために使用される。時間、温度、pH、酸素、および容量のパラメーターは、慣用的な方法によって、異種タンパク質の発現および回収を最適化するように調整される。
【0104】
(iv.細菌系)
細菌の発現技術は、当該分野で公知である。細菌のプロモーターは、細菌のRNAポリメラーゼに結合し得、そしてコード配列(例えば、構造遺伝子)の下流方向(3’方向)へのmRNAへの転写を開始し得る任意のDNA配列である。プロモーターは、通常、コード配列の5’末端に近接して存在する転写開始領域を有する。この転写開始領域は、通常、RNAポリメラーゼ結合部位および転写開始部位を含む。細菌のプロモーターはまた、オペレーターと呼ばれる第二のドメインを有し、RNA合成が始まる近接のRNAポリメラーゼ結合部位と重複している。オペレーターは、遺伝子リプレッサータンパク質が、オペレーターに結合し、そのため特定の遺伝子の転写を抑制し得るような、負の調節された(誘導性の)転写を可能にする。構成的発現は、オペレーターのような負の調節エレメントの非存在下で起こり得る。さらに、正の調節は、遺伝子アクチベーター(活性化因子)タンパク質結合配列により達成され得、その配列が存在する場合は通常、RNAポリメラーゼ結合配列の(5’)側に近接している。遺伝子アクチベータータンパク質の例としては、異化(カタボライト)活性化タンパク質(CAP)があり、それはEscherichia coli(E.Coli)におけるlacオペロンの転写の開始を助ける(Raibaudら(1984)Annu.Rev.Genet.18:173)。それゆえ、調節される発現は、正または負のいずれかであり得、従って転写を増強するかまたは低下し得るかのいずれかである。
【0105】
代謝経路の酵素をコードする配列は、特に有用なプロモーター配列を提供する。例としては、ガラクトース、ラクトース(lac)(Changら(1997)Nature 198:1056)、およびマルトースのような糖代謝の酵素由来のプロモーター配列が挙げられる。さらなる例としては、トリプトファン(trp)(Goeddelら(1980)Nuc.Acids Res.8:4057;Yelvertonら(1981)Nucl.Acids Res.9:731;米国特許第4,738,921号;EP−A−0036776号およびEP−A−0121775)のような生合成酵素由来のプロモーター配列が挙げられる。g−ラクタマーゼ(g−laotamase)(bla)プロモーター系(Weissmann(1981)「The cloning of Interferon and other mistakes.」Interferon 3(I.Gresser編))、バクテリオファージλPL(Shimatakeら(1981)Nature 292:128)およびT5(米国特許第4,689,406号)プロモーター系もまた有用なプロモーター配列を提供する。
【0106】
さらに、天然に存在しない合成プロモーターもまた、細菌のプロモーターとして機能する。例えば、ある細菌(バクテリア)あるいはバクテリオファージプロモーターの転写活性化配列は、別のバクテリアあるいはバクテリオファージプロモーターのオペロン配列と結合し得、合成ハイブリッドプロモーターを形成する。(米国特許第4,551,433号)。例えば、tacプロモーターは、lacリプレッサーにより調節されるtrpプロモーター配列およびlacオペロン配列の両方から構成される、ハイブリッドtrp−lacプロモーターである(Amannら(1983)Gene 25:167;de Boerら(1983)Proc.Natl.Acad.Sci.80:21)。さらに、ある細菌のプロモーターは、細菌のRNAポリメラーゼに結合し、そして転写を開始させる能力を有する非細菌起源の天然に存在するプロモーターを含み得る。非細菌起源の天然に存在するプロモーターはまた、原核生物内でいくらかの遺伝子の高いレベルでの発現を生じるために、適合性のあるRNAポリメラーゼと結合され得る。バクテリオファージT7 RNAポリメラーゼ/プロモーター系は、連結したプロモーター系の例である(Studierら(1986)J.Mol.Biol.189:113;Taborら(1985)Proc Natl.Acad.Sci.82:1074)。さらに、ハイブリッドプロモーターはまた、バクテリオファージプロモーターおよびE.coliオペレーター領域から構成され得る(EPO−A−0267851)。
【0107】
機能性のプロモーター配列に加え、効果的なリボソーム結合部位もまた、原核生物における外来遺伝子の発現に有用である。E.coliにおいて、リボソーム結合部位は、シャイン−ダルガノ(SD)配列と呼ばれ、そして開始コドン(ATG)および開始コドンの3〜11ヌクレオチド上流に位置する長さ3〜9ヌクレオチドの配列を含む(Shineら(1975)Nature 254:34)。SD配列は、SD配列とE.coliの16S rRNAの3’側との間の塩基対形成によりmRNAのリボソームへの結合を促進すると考えられている(Steitzら(1979)「Genetic Signals and nucleotide sequences in messenger RNA」Biological Regulation and Development:Gene Expression(R.F.Goldbergerら編))。弱いリボソーム結合部位を有する真核遺伝子および原核遺伝子の発現のためには(Sambrookら(1989)「Expression of cloned genes in Escherichia coli」Molecular Cloning:A Laboratory Manual)。
【0108】
DNA分子は、細胞内で発現され得る。プロモーター配列は、直接的にそのDNA分子と連結され得、この場合、N末端の最初のアミノ酸は、常に、ATG開始コドンによりコードされるメチオニンである。所望される場合、N末端のメチオニンは、臭化シアンとのインビトロインキュベーション、あるいは細菌のメチオニンN末端ペプチダーゼとのインビボまたはインビトロインキュベーションのいずれかによりタンパク質から切断され得る(EPO−A−0219237)。
【0109】
融合タンパク質は、直接的発現に代わるものを提供する。通常、内在性の細菌のタンパク質、あるいは他の安定なタンパク質のN末端部分をコードするDNA配列は、異種のコード配列の5’末端と融合される。発現の際に、この構築物は、2つのアミノ酸配列の融合を提供する。例えば、バクテリオファージλ細胞遺伝子は、外来遺伝子の5’末端と連結し得、そして細菌において発現され得る。生じた融合タンパク質は、好ましくは、外来遺伝子由来のバクテリオファージタンパク質を切断するための切断酵素(第Xa因子)用の部位を保持する(Nagaiら(1984)Nature 309:810)。融合タンパク質はまた、lacZ(Jiaら(1987)Gene 60:197)、trpE(Allenら(1987)J.Biotechnol.5:93;Makoffら(1989)J.Gen.Microbiol.135:11)、およびChey(EP−A−0324647)遺伝子由来の配列を用いて作製され得る。2つのアミノ酸配列の接合部でのDNA配列は、切断部位をコードしてもよいし、あるいはコードしなくてもよい。別の例としては、ユビキチン融合タンパク質である。そのような融合タンパク質は、好ましくは、外来タンパク質からユビキチンを切断するための切断酵素(例えば、ユビキチン特異的切断プロテアーゼ)用の部位を保持するユビキチン領域と共に作製され得る。この方法を通して、天然外来タンパク質は分離され得る(Millerら(1989)Bio/Technology 7:698)。
【0110】
あるいは、外来タンパク質はまた、細菌における外来タンパク質の分泌を提供するシグナルペプチド配列フラグメントから構成される、融合タンパク質をコードするキメラDNA分子を作製することによって、細胞から分泌され得る(米国特許第4,336,336号)。シグナル配列フラグメントは、通常、細胞からのタンパク質の分泌を指向する、疎水性アミノ酸から構成されるシグナルペプチドをコードする。このタンパク質は、増殖培地(グラム陽性細菌)、または細胞の内膜と外膜との間に位置する細胞周辺腔(グラム陰性細菌)のいずれかへ分泌される。好ましくは、インビボまたはインビトロのいずれかで切断され得る、このシグナルペプチドフラグメントと外来遺伝子との間にコードされるプロセシング部位がある。
【0111】
適切なシグナル配列をコードするDNAは、E.coli外膜タンパク質遺伝子(ompA)(Masuiら(1983)、Experimetal Manipulation of Gene Expression;Ghrayebら(1984)EMBO J.3:2437)およびE.coliアルカリホスファターゼシグナル配列(phoA)(Okaら(1985)Proc.Natl.Acad.Sci.82:7212)のような、分泌性細菌タンパク質に関する遺伝子由来であり得る。さらなる例として、種々のBacillus株由来のα−アミラーゼ遺伝子のシグナル配列は、B.subtilis由来の異種タンパク質を分泌するために使用され得る(Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582;EP−A−0244042)。
【0112】
通常、細菌によって認識される転写終結配列は、翻訳終止コドンの3’側に位置する調節領域であり、そして従って、プロモーターとともに、コード配列に隣接する。これらの配列は、そのDNAによってコードされるポリペプチドへと翻訳され得るmRNAの転写を指向する。転写終結配列は、しばしば、転写の終結を補助するステムループ構造を形成し得る、約50ヌクレオチドのDNA配列を含む。例は、強力なプロモーターを有する遺伝子(例えば、E.coliのtrp遺伝子および他の生合成遺伝子)由来の転写終結配列を含む。
【0113】
通常、上記の成分(プロモーター、シグナル配列(もし所望ならば)、目的のコード配列、および転写終結配列を含む)は、組み立てられて発現構築物となる。発現構築物は、しばしば、宿主(例えば細菌)における安定な保持が可能である染色体外エレメント(例えばプラスミド)のような、レプリコンに保持される。このレプリコンは複製系を有し、従って、このことによって、発現またはクローニングおよび増幅のいずれかのために、レプリコンが原核生物宿主において保持されることを可能になる。さらに、レプリコンは、高コピー数プラスミドまたは低コピー数プラスミドのいずれかであり得る。高コピー数プラスミドは、一般に約5〜約200、そして通常約10〜約150の範囲のコピー数を有する。高コピー数プラスミドを含む宿主は、好ましくは少なくとも約10個、そしてより好ましくは少なくとも約20個のプラスミドを含む。高コピー数ベクターまたは低コピー数ベクターのいずれかが選択され得、それは、宿主に対するベクターおよび外来タンパク質の効果に依存する。
【0114】
あるいは、発現構築物は、組み込みベクターを用いて、細菌のゲノムへ組み込まれ得る。組み込みベクターは、通常、ベクターが組み込むのを可能にする、細菌の染色体と相同な少なくとも1つの配列を含む。組み込みは、ベクターにおける相同なDNAと細菌の染色体との間の組換えから生じるようである。例えば、種々のBacillus株からのDNAによって構築される組み込みベクターは、Bacillus染色体に組み込まれる(EP−A−0 127 328)。組み込みベクターはまた、バクテリオファージまたはトランスポゾン配列から構成され得る。
【0115】
通常、染色体外発現構築物および組み込み発現構築物は、選択マーカーを含んで、形質転換された細菌株の選択を可能にし得る。選択マーカーは、細菌宿主において発現され得、そして細菌が薬物(例えば、アンピシリン、クロラムフェニコール、エリスロマイシン、カナマイシン(ネオマイシン)、およびテトラサイクリン)に耐性になるようにする遺伝子を含み得る(Daviesら(1978)Annu.Rev.Microbiol.32:469)。選択マーカーはまた、ヒスチジン、トリプトファン、およびロイシンの生合成経路における生合成遺伝子のような、生合成遺伝子を含み得る。
【0116】
あるいは、上記の成分のいくつかは、形質転換ベクターにおいて組み立てられ得る。形質転換ベクターは、通常、上記のように、レプリコンにおいて保持されるか、または組み込みベクターへと開発されるかのいずれかの選択マーカー(market)から構成され得る。
【0117】
発現ベクターまたは形質転換ベクターは、染色体外レプリコンまたは組み込みベクターのいずれも、多くの細菌への形質転換のために開発されてきた。例えば、発現ベクターは、とりわけ、以下の細菌のために開発されてきた:Bacillus subtilis(Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582;EP−A−0 036 259およびEP−A−0 063 953;WO 84/04541)、Escherichia coli(Shimatakeら(1981)Nature 292:128;Amannら(1985)Gene 40:183;Studierら(1986)J.Mol.Biol.189:113;EP−A−0 036 776、EP−A−0 136 829およびEP−A−0 136 907)、Streptococcus cremoris(Powellら(1988)Appl.Environ.Microbiol.54:655);Streptococcus lividans(Powellら(1988)Appl.Environ.Microbiol.54:655)、Streptomyces lividans(米国特許第4,745,056号)。
【0118】
外来DNAを細菌宿主へ導入する方法は、当該分野において周知であり、そして通常、CaCl2または他の薬剤(例えば、2価の陽イオンおよびDMSO)のいずれかで処理された細菌の形質転換を含む。DNAはまた、エレクトロポレーションによって、細菌細胞へ導入され得る。形質転換の手順は、通常、形質転換される細菌の種によって変化する。例えば以下を参照のこと:(Massonら(1989)FEMS Microbiol.Lett.60:273;Palvaら(1982)Proc.Natl.Acad.Sci.USA 79:5582;EP−A−0 036 259およびEP−A−063 953;WO 84/04541、Bacillus)、(Millerら(1988)Proc.Natl.Acad.Sci.85:856;Wangら(1990)J.Bacteriol.172:949、Campylobacter)、(Cohenら(1973)Proc.Natl.Acad.Sci.69:2110;Dowerら(1988)Nucleic Acids Res.16:6127;Kushner(1978)「ColE1由来のプラスミドによるEscherichia coliの形質転換のための改良された方法」Genetic Engineering:Proceedings of the International Symposium on Genetic Engineering(H.W.BoyerおよびS.Nicosia編);Mandelら(1970)J.Mol.Biol.53:159;Taketo(1988)Biochim.Biophys.Acta 949:318;Escherichia)、(Chassyら(1987)FEMS Microbiol.Lett.44:173 Lactobacillus);(Fiedlerら(1988)Anal.Biochem 170:38、Pseudomonas);(Augustinら(1990)FEMS Microbiol.Lett.66:203、Staphylococcus)、(Baranyら(1980)J.Bacteriol.144:698;Harlander(1987)「エレクトロポレーションによるStreptococcus lactisの形質転換」Streptococcal Genetics(J.FerrettiおよびR.Curtiss III編);Perryら(1981)Infect.Immun.32:1295;Powellら(1988)Appl.Environ.Microbiol.54:655;Somkutiら(1987)Proc.4th Evr.Cong.Biotechnology 1:412、Streptococcus)。
【0119】
(v.酵母発現)
酵母発現系もまた、当業者に公知である。酵母プロモーターは、酵母RNAポリメラーゼに結合可能であり、そしてコード配列(例えば、構造遺伝子)からmRNAへの下流の(3’側の)転写を開始し得る、任意のDNA配列である。プロモーターは、通常、コード配列の5’末端の近位に位置する転写開始領域を有する。この転写開始領域は、通常、RNAポリメラーゼ結合部位(「TATAボックス」)および転写開始部位を含む。酵母プロモーターはまた、上流アクチベーター配列(UAS)と呼ばれる第2のドメインを有し得、これは、もし存在するならば、通常、構造遺伝子とは遠位である。このUASは、調節された(誘導できた)発現を可能にする。構成的発現は、UASの非存在下で生じる。調節される発現は、正または負のいずれかであり得、それによって転写を増加させるかまたは減少させるかのいずれかであり得る。
【0120】
酵母は、活性な代謝経路を有する発酵性生物であり、従って、代謝経路における酵素をコードする配列は、特に有用なプロモーター配列を提供する。例としては、以下が挙げられる:アルコールデヒドロゲナーゼ(ADH)(EP−A−0 284 044)、エノラーゼ、グルコキナーゼ、グルコース−6−リン酸イソメラーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(GAPまたはGAPDH)、ヘキソキナーゼ、ホスホフルクトキナーゼ、3−ホスホグリセリン酸ムターゼ、およびピルビン酸キナーゼ(PyK)(EPO−A−0 329 203)。酵母PHO5遺伝子はまた、酸性ホスファターゼをコードし、有用なプロモーター配列を提供する(Myanoharaら(1983)Proc.Natl.Acad.Sci.USA 80:1)。
【0121】
さらに、天然には生じない合成プロモーターもまた、酵母のプロモーターとして機能する。例えば、ある1つの酵母プロモーターのUAS配列は、別の酵母プロモーターの転写活性化領域と連結され得、合成ハイブリッドプロモーターを生成し得る。このようなハイブリッドプロモーターの例は、GAP転写活性化領域と連結されるADH調節配列(米国特許第4,876,197号および同第4,880,734号)を含む。ハイブリッドプロモーターの他の例は、ADH2、GAL4、GAL10、またはPHO5遺伝子のいずれかの調節配列からなり、GAPまたはPyK(EP−A−0 164 556)のような解糖酵素遺伝子の転写活性化領域に結合されているプロモーターを含む。さらに、酵母プロモーターは、酵母RNAポリメラーゼと結合し、そして転写を開始する能力を有する、天然に生じる非酵母起源のプロモーターを含み得る。このようなプロモーターの例としては、とりわけ、以下が挙げられる:(Cohenら(1980)Proc.Natl.Acad.Sci.USA 77:1078;Henikoffら(1981)Nature 283:835;Hollenbergら(1981)Curr.Topics Microbiol.Immunol.96:119;Hollenbergら(1979)「酵母Saccharomyces cerevisiaeにおける細菌の抗生物質耐性遺伝子の発現」、Plasmids of Medical,Environmental and Commercial Importance(K.N.TimmisおよびA.Puhler編);Mercerau−Puigalonら(1980)Gene 11:163;Panthierら(1980)Curr.Genet.2:109;)。
【0122】
DNA分子は、酵母において、細胞内で発現され得る。プロモーター配列は、DNA分子と直接連結され得、その場合、組換えタンパク質のN末端にある最初のアミノ酸は常にATG開始コドンによってコードされているメチオニンである。もし所望ならば、N末端のメチオニンは、臭化シアンとのインビトロインキュベーションによって、タンパク質から切断され得る。
【0123】
融合タンパク質は、酵母発現系について、ならびに哺乳動物、バキュロウイルス、および細菌の発現系において、代替物を提供する。通常、内因性酵母タンパク質、または他の安定なタンパク質のN末端部分をコードするDNA配列は、異種コード配列の5’末端に融合される。発現の際に、この構築物は、2つのアミノ酸配列の融合物を提供する。例えば、酵母またはヒトのスーパーオキシドジスムターゼ(SOD)遺伝子は、外来遺伝子の5’末端に連結され、そして酵母において発現し得る。2つのアミノ酸配列の連結部にあるDNA配列は、切断部位をコードしてもよいし、コードしなくてもよい。例えば、EP−A−0 196 056を参照のこと。別の例はユビキチン融合タンパク質である。このような融合タンパク質は、プロセシング酵素(例えば、ユビキチン特異的プロセシングプロテアーゼが)外来タンパク質からユビキチンを切断するための部位を好ましくは保持する、ユビキチン領域を伴って作製される。従って、この方法により、ネイティブ外来タンパク質は、単離され得る(例えば、WO88/024066)。
【0124】
あるいは、外来タンパク質はまた、酵母における外来タンパク質の分泌を提供する、リーダー配列フラグメントから構成される融合タンパク質をコードする、キメラDNA分子を作製することによって、細胞から増殖培地へ分泌され得る。好ましくは、インビボまたはインビトロのいずれかで切断され得る、リーダーフラグメントと外来遺伝子との間にコードされるプロセシング部位が存在する。リーダー配列フラグメントは、細胞からのタンパク質の分泌を指向する、疎水性アミノ酸から構成されるシグナルペプチドを、通常コードする。
【0125】
適切なシグナル配列をコードするDNAは、分泌性酵母タンパク質に関する遺伝子由来であり得、その遺伝子は例えば、インベルターゼ遺伝子(EP−A−0012873;JPO62,096,086)およびA因子遺伝子(米国特許第4,588,684号)である。あるいは、インターフェロンリーダーのような、酵母における分泌もまた提供する、非酵母起源のリーダーが存在する(EP−A−0060057)。
【0126】
好ましいクラスの分泌リーダーは、酵母α因子遺伝子のフラグメントを使用するリーダーであり、これは「プレ」シグナル配列、および「プロ」領域の両方を含む。用いられ得るα因子フラグメントの型は、完全長の、プレ−プロα因子リーダー(約83アミノ酸残基)および短縮されたα因子リーダー(通常約25〜約50アミノ酸残基)を含む(米国特許第4,546,083号および同第4,870,008号;EP−A−0 324 274)。分泌を提供するα因子リーダーフラグメントを使用するさらなるリーダーは、第1の酵母のプレ配列を有するが、第2の酵母α因子からのプロ領域を有しないで作製される、ハイブリッドα因子リーダーを含む(例えば、WO89/02463を参照のこと)。
【0127】
通常、酵母に認識される転写終結配列は、翻訳終止コドンの3’側に位置する調節領域であり、従って、プロモーターと共にコード配列に隣接する。これらの配列は、そのDNAにコードされるポリペプチドへと翻訳され得る、mRNAの転写を指向する。転写終結配列および他の酵母に認識される終結配列の例は、例えば、解糖酵素をコードする転写終結配列である。
【0128】
通常、上記の成分(プロモーター、リーダー(もし所望ならば)、目的のコード配列、および転写終結配列を含む)は、組み立てられて発現構築物になる。発現構築物は、宿主(例えば、酵母または細菌)において安定に保持され得る染色体外エレメント(例えば、プラスミド)のような、レプリコンにおいてしばしば保持される。このレプリコンは、2つの複製系を有し得、従って、例えば、発現のために酵母において、ならびにクローニングおよび増幅のために原核生物宿主において保持されることを可能にする。このような酵母−細菌シャトルベクターの例としては、以下が挙げられる:YEp24(Botsteinら(1979)Gene 8:17〜24)、pCl/1(Brakeら(1984)Proc.Natl.Acad.Sci.USA 81:4642〜4646)、およびYRp17(Stinchcombら(1982)J.Mol.Biol.158:157)。さらに、レプリコンは、高コピー数プラスミドまたは低コピー数プラスミドのいずれかであり得る。高コピー数プラスミドは、一般に約5〜約200、そして通常約10〜約150の範囲のコピー数を有する。高コピー数プラスミドを含む宿主は、好ましくは少なくとも約10、そしてより好ましくは少なくとも約20を有する。高コピー数ベクターまたは低コピー数ベクターのいずれかが選択され得、それは、宿主に対するベクターおよび外来タンパク質の効果に依存する。例えば、Brakeら、前出を参照のこと。
【0129】
あるいは、発現構築物は、組み込みベクターを用いて、酵母のゲノムへ組み込まれ得る。組み込みベクターは、通常、ベクターが組み込むのを可能にする、酵母の染色体と相同な少なくとも1つの配列を含み、そして好ましくは、発現構築物に隣接する2つの相同配列を含む。組み込みは、ベクターにおける相同なDNAと酵母の染色体との間の組換えから生じるようである(Orr−Weaverら(1983)Methods in Enzymol.101:228〜245)。組み込みベクターは、そのベクター中に含有するために適切な相同配列を選択することによって、酵母における特定の遺伝子座を指向され得る。Orr−Weaverら、前出を参照のこと。1つ以上の発現構築物が組み込まれ得、産生される組換えタンパク質のレベルにおそらく影響を与え得る(Rineら(1983)Proc.Natl.Acad.Sci.USA 80:6750)。ベクターに含まれる染色体配列は、ベクターにおける単一セグメント(ベクター全体の組み込みを生じる)、または染色体における隣接セグメントに相同でかつベクターにおける発現構築物に隣接する2つのセグメント(発現構築物のみの安定した組み込みを生じ得る)のいずれかとして生じ得る。
【0130】
通常、染色体外発現構築物および組み込み発現構築物は、選択マーカーを含み得、形質転換された酵母株の選択を可能にする。選択マーカーは、酵母宿主において発現され得る生合成遺伝子(例えば、ADE2、HIS4、LEU2、TRP1、およびALG7)、ならびにG418耐性遺伝子(それぞれ、酵母細胞がツニカマイシンおよびG418に耐性になるようにする)を含み得る。さらに、適切な選択マーカーはまた、金属のような毒性化合物の存在下において増殖する能力を、酵母に提供し得る。例えば、CUP1の存在は、酵母が、銅イオンの存在下において増殖することを可能にする(Buttら(1987)Microbiol.Rev.51:351)。
【0131】
あるいは、上記成分のうちのいくつかは、組み立てられて形質転換ベクターになり得る。形質転換ベクターは、通常、上記のように、レプリコンにおいて保持される選択マーカーから構成されるか、または組み込みベクターへと構築されるかのいずれかである。
【0132】
発現ベクターおよび形質転換ベクターは、染色体外レプリコンまたは組み込みベクターのいずれかであり、多くの酵母への形質転換のために開発されてきた。例えば、発現ベクターは、とりわけ、以下の酵母のために開発されてきた:Candida albicans(Kurtzら(1986)Mol.Cell.Biol.6:142)、Candida maltosa(Kunzeら(1985)J.Basic Microbiol.25:141)。Hansenula polymorpha(Gleesonら(1986)J.Gen.Microbiol.132:3459;Roggenkampら(1986)Mol.Gen.Genet.202:302)、Kluyveromyces fragilis(Dasら(1984)J.Bacteriol.158:1165)、Kluyveromyces lactis(De Louvencourtら(1983)J.Bacteriol.154:737;Van den Bergら(1990)Bio/Technology 8:135)、Pichia guillerimondii(Kunzeら(1985)J.Basic Microbiol.25:141)、Pichia pastoris(Creggら(1985)Mol.Cell.Biol.5:3376;米国特許第4,837,148号および同第4,929,555号)、Saccharomyces cerevisiae(Hinnenら(1978)Proc.Natl.Acad.Sci.USA 75:1929;Itoら(1983)J.Bacteriol.153:163)、Schizosaccharomyces pombe(BeachおよびNurse(1981)Nature 300:706)、およびYarrowia lipolytica(Davidowら(1985)Curr.Genet.10:380471;Gaillardinら(1985)Curr.Genet.10:49)。
【0133】
外来DNAを酵母宿主へ導入する方法は、当該分野において周知であり、そして通常、スフェロプラストの、またはアルカリ陽イオンで処理されたインタクトな酵母細胞のいずれかの形質転換を含む。形質転換の手順は、通常、形質転換される酵母の種によって変化する。例えば以下を参照のこと:(Kurtzら(1986)Mol.Cell.Biol.6:142;Kunzeら(1985)J.Basic Microbiol.25:141;Candida);(Gleesonら(1986)J.Gen.Microbiol.132:3459;Roggenkampら(1986)Mol.Gen.Genet.202:302;Hansenula);(Dasら(1984)J.Bacteriol.158:1165;De Louvencourtら(1983)J.Bacteriol.154:1165;Van den Bergら(1990)Bio/Technology 8:135;Kluyveromyces);(Creggら(1985)Mol.Cell.Biol.5:3376;Kunzeら(1985)J.Basic Microbiol.25:141;米国特許第4,837,148号および同第4,929,555号;Pichia);(Hinnenら(1978)Proc.Natl.Acad.Sci.USA 75:1929;Itoら(1983)J.Bacteriol.153:163 Saccharomyces);(BeachおよびNurse(1981)Nature 300:706;Schizosaccharomyces);(Davidowら(1985)Curr.Genet.10:39;Gaillardinら(1985)Curr.Genet.10:49;Yarrowia)。
【0134】
(薬学的組成物)
薬学的組成物は、本発明のポリペプチドおよび/または核酸を含み得る。この薬学的組成物は、治療上有効な量の、本願発明のポリペプチド、抗体、またはポリヌクレオチドのいずれかを含む。
【0135】
本明細書において使用される用語「治療上有効な量」とは、所望の疾患または状態を処置、改善、または予防するための治療薬剤の量、または、検出可能な治療効果または予防効果を示すための治療薬剤の量をいう。この効果は、例えば、キメラマーカーまたは抗原レベルによって検出され得る。治療効果はまた、体温低下のような、身体の症状における減少を含む。被験体に関する正確な有効量は、被験体の大きさおよび健康、状態の性質および程度、および投与のために選択される治療または治療の組合せに依存する。従って、あらかじめ正確な有効量を特定することは有用ではない。しかし、所定の情況のための有効量は、慣用的な実験によって決定され得、そして臨床医の判断内である。
【0136】
本発明の目的のために、有効な用量は、DNA構築物が投与される個体において、約0.01mg/kg〜50mg/kgまたは0.05mg/kg〜約10mg/kgのDNA構築物である。
【0137】
薬学的組成物はまた、薬学的に受容可能なキャリアを含み得る。用語「薬学的に受容可能なキャリア」とは、抗体またはポリペプチド、遺伝子、および他の治療薬剤のような、治療薬剤の投与のためのキャリアをいう。この用語は、この組成物を与えられる個体に有害な抗体の産生をそれ自体誘導せず、そして過度の毒性を伴わずに投与され得る、任意の薬学的キャリアをいう。適切なキャリアは、タンパク質、多糖、ポリ乳酸、ポリグリコール酸、ポリマーアミノ酸、アミノ酸コポリマー、および不活性ウイルス粒子のように、大きく、遅く代謝される高分子であり得る。このようなキャリアは、当業者に周知である。
【0138】
薬学的に受容可能な塩が、その中で使用され得る。例えば、塩酸塩、臭化水素酸塩、リン酸塩、硫酸塩などのような鉱酸塩;および酢酸塩、プロピオン酸塩、マロン酸塩、安息香酸塩などのような有機酸の塩である。薬学的に受容可能な賦形剤の徹底的な議論は、Remington’s Pharmaceutical Sciences(Mack Pub.Co.、N.J.1991)にて利用可能である。
【0139】
治療組成物における薬学的に受容可能なキャリアは、水、生理的食塩水、グリセロールおよびエタノールのような液体を含み得る。さらに、湿潤剤または乳化剤、pH緩衝物質などのような補助物質が、このようなビヒクルに存在し得る。代表的には、治療組成物は、液体溶液または懸濁液のいずれかの、注射可能物質として調製される;注射前に液体ビヒクルに溶解または懸濁するのに適切な固体形態もまた、調製され得る。リポソームは、薬学的に受容可能なキャリアの定義中に含まれる。
【0140】
(送達方法)
一旦処方されると、本発明の組成物は、その被験体へ直接投与され得る。処置される被験体は、動物であり得;特に、ヒト被験体が処置され得る。
【0141】
その組成物の直接送達は、一般的に、皮下に、腹腔内に、静脈内に、または筋肉内のいずれかでの注入によって達成されるか、あるいは、組織の間質空間へ送達される。この組成物はまた、病巣へ投与され得る。他の投与様式には、経口投与、および肺投与、坐剤、および経皮(transdermal)適用または経皮(transcutaneous)適用(例えば、WO98/20734を参照のこと)、針、および遺伝子銃またはハイポスプレー(hypospray)が含まれる。投薬処置は、単回用量スケジュール、または多数回用量スケジュールであり得る。
【0142】
(ワクチン)
本発明に従うワクチンは、予防的(すなわち、感染を予防するため)または治療(すなわち、感染後の疾患を処置するため)のいずれかであり得る。
【0143】
このようなワクチンは、免疫抗原、免疫原、ポリペプチド、タンパク質または核酸を、通常「薬学的に受容可能なキャリア」とともに含み、このキャリア自体は、その組成物を受ける個体に有害である抗体の産生を誘発しない任意のキャリアを含む。適切なキャリアは、代表的に、大きく、ゆっくり代謝される高分子(例えば、タンパク質、ポリサッカリド、ポリ乳酸、ポリグリコール酸、ポリマーアミノ酸、アミノ酸コポリマー、脂質凝集物(例えば、油小滴またはリポソーム)、および不活性ウイルス粒子である。このようなキャリアは、当業者に周知である。さらに、これらのキャリアは免疫刺激剤(「アジュバント」)として機能し得る。さらに、この抗原または免疫原は、細菌毒素(例えば、ジフテリア、破傷風、コレラ、H.pyloriなどの病原因子からの毒素)と結合体化され得る。
【0144】
この組成物の効力を増強するために好ましいアジュバントとしては、(1)アルミニウム塩(「明礬」)(例えば、水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムなど)、(2)o/wエマルジョン処方物(他の特定の免疫刺激剤(例えば、ムラミルペプチド(以下を参照のこと)または細菌細胞壁成分)を伴うか伴わない)が挙げられるが、それらに限定されず、例えば、以下:(a)5%スクアレン、0.5% Tween 80、および0.5% Span85(必要に応じて、種々の量のMTP−PE(以下を参照のこと)を含有するが、必要ではない)を含み、モデル110Y微小流体化器(Microfluidics、Newton、MA)のような微小流体化器を用いてミクロン未満の粒子へと処方されたMF59TM(WO90/14837;Vaccine design:the subunit and adjuvant approachの第10章、PowellおよびNewman編、Plenum Press 1995);(b)ミクロン未満のエマルジョンへと微小流体化されたか、またはボルテックスして、より大きな粒子径エマルジョンを生成したかのいずれかである、10%スクアレン、0.4%Tween80、5%プルロニックブロックポリマーL121、およびthr−MDP(以下を参照のこと)を含有するSAF、ならびに(c)2%スクアレン、0.2%Tween80、およびモノホスホリルリピドA(MPL)、トレハロースジミコレート(TDM)、および細胞壁骨格(CWS)好ましくはMPLおよびCWS(DetoxTM)からなる群由来の1つ以上の細菌細胞壁成分を含むRibiTMアジュバント系(RAS)、(Ribi Immunochem、Hamilton、MT);(3)サポニンアジュバント(例えば、StimulonTM)(Cambridge Bioscience、Worcester、MA)を使用し得るか、またはそれから粒子(例えば、ISCOM(免疫刺激性複合体)を生成し得る;(4)完全フロイントアジュバント(CFA)および不完全フロイントアジュバント(IFA);(5)サイトカイン(例えば、インターロイキン(例えば、IL−1、IL−2、IL−4、IL−5、IL−6、IL−7、IL−12など)、インターフェロン(例えば、γインターフェロン)、マクロファージコロニー刺激因子(M−CSF)、腫瘍壊死因子(TNF)など;および(6)その組成物の効力を強化するための免疫刺激因子として作用する他の物質である。ミョウバンおよびMF59TMが好ましい。
【0145】
上記で言及したように、ムラミルペプチドとしては、N−アセチル−ムラミル−L−スレオニル−D−イソグルタミン(thr−MDP)、N−アセチル−ノルムラミル−L−アラニル−D−イソグルタミン(ノル−MDP)、N−アセチルムラミル−L−アラニル−D−イソグルタミニル−L−アラニン−2−(1’−2’−ジパルミトイル−sn−グリセロ−3−ヒドロキシホスホリルオキシ)−エチルアミン(MTP−PE)などが挙げられるがそれらに限定されない。
【0146】
免疫原性組成物(例えば、免疫抗原/免疫原/ポリペプチド/タンパク質/核酸、薬学的に受容可能なキャリア、およびアジュバント)は、代表的に、希釈剤(例えば、水、生理食塩水、グリセロール、エタノールなど)を含有する。さらに、補助物質(例えば、湿潤剤または乳化剤、pH緩衝物質など)は、このようなビヒクルにおいて存在し得る。
【0147】
代表的に、免疫原性組成物は、液体溶液または懸濁物として、注射剤として調製され;注射前に液体ビヒクル中の溶液または懸濁物として適切な固体形態もまた調製され得る。この調製物はまた、薬学的に受容可能なキャリアの下で、上記に記載のように、アジュバント効果の強化のために乳化され得るかまたはリポソーム中にカプセル化され得る。
【0148】
ワクチンとして使用される免疫原性組成物は、免疫学的有効量の抗原性または免疫原性のポリペプチド、および任意の他の上記の成分を必要に応じて含む。「免疫学的有効量」とは、その量の個体への投与が、単回用量であれ、一連の(用量の)一部としてであれ、処置または予防に有効であることを意味する。この量は、処置される個体の健康および身体状態、処置される個体の分類学上の群(例えば、非ヒト霊長類、霊長類など)、個体の免疫系が抗体を合成する能力、所望される保護の程度、そのワクチンの処方物、処置する医師の医療的状況の評価、および他の関連する因子に依存して変動する。その量は、比較的広い範囲に入り、この量が慣用的な試行を通して決定され得ることが予想される。
【0149】
免疫学的組成物は、従来のように、非経口的(例えば、皮下、筋肉内または経皮(transudermally)/経皮(transucutaneously)のいずれかでの注射による)((例えば、WO98/20734)に投与される)。他の投与様式に適切なさらなる処方物は、経口処方物および肺処方物、坐剤、および経皮適用を含む。投薬処置は、単回用量スケジュールまたは多数回用量スケジュールであり得る。ワクチンは、他の免疫調節剤とともに投与され得る。
【0150】
タンパク質ベースのワクチンの代替として、DNAワクチンを使用し得る(例えば、RobinsonおよびTorres(1997)Seminars in Immunology 9:271−283;Donnellyら(1997)Annu Rev Immunol 15:617−648;本明細書中後半部を参照のこと)。
【0151】
(遺伝子送達ビヒクル)
本発明の治療剤のコード配列を含む、哺乳動物における発現のためにその哺乳動物へ送達される構築物の送達のための遺伝子治療ビヒクルは、局所または全身的のいずれかで投与され得る。これらの構築物は、ウイルスベクターアプローチまたは非ウイルスベクターアプローチを、インビボまたはエキソビボの様式で利用し得る。このようなコード配列の発現は、内因性哺乳動物プロモーターまたは外因性プロモーターを用いて誘導され得る。このコード配列のインビボでの発現は、構成性または調節性のいずれかであり得る。
【0152】
本発明は、意図された核酸配列を発現し得る遺伝子送達ビヒクルを含む。この遺伝子送達ビヒクルは、好ましくは、ウイルスベクター、およびより好ましくはレトロウイルスベクター、アデノウイルスベクター、アデノ随伴ウイルス(AAV)ベクター、ヘルペスウイルスベクターまたはαウイルスベクターである。このウイルスベクターはまた、アストロウイルスベクター、コロナウイルスベクター、オルトミクソウイルスベクター、パポバウイルスベクター、パラミクソウイルスベクター、パルボウイルスベクター、ピコルナウイルスベクター、ポックスウイルスベクター、またはトガウイルスベクターであり得る。一般的には、Jolly(1994)Cancer Gene Therapy 1:51−64;Kimura(1994)Human Gene Therapy 5:845−852;Connelly(1995)Human Gene Therapy 6:185−193;およびKaplitt(1994)Nature Genetics 6:148−153を参照のこと。
【0153】
レトロウイルスベクターは、当該分野で周知であり、そして本発明者らは、任意のレトロウイルス遺伝子治療ベクターが本発明において使用可能であることを意図する。これには、B型、C型およびD型のレトロウイルス、異種栄養性ウイルス(例えば、NZB−X1、NZB−X2およびNZB9−1(O’Neill(1985)J.Virol.53:160を参照のこと)多栄養性レトロウイルス(例えば、MCFおよびMCF−MLV(Kelly(1983)J.Virol.45:291を参照のこと)、スプマウイルスおよびレンチウイルスが含まれる。RNA Tumor Viruses、第2版、Cold Spring Harobor Laboratory、1985を参照のこと。
【0154】
レトロウイルス遺伝子治療ベクターの部分は、異なるレトロウイルスに由来し得る。例えば、レトロウイルスLTRは、マウス肉腫ウイルスに由来し得、tRNA結合部位はラウス肉腫ウイルスに由来し得、パッケージングシグナルはマウス白血病ウイルスに由来し得、そして第二の鎖合成の起源はトリ白血病ウイルスに由来し得る。
【0155】
これらの組換えレトロウイルスベクターを使用して、適切なパッケージング細胞株へそれらを導入することによって形質導入適合性レトロウイルスベクター粒子を生成し得る(米国特許第5,591,624号を参照のこと)。レトロウイルスベクターは、レトロウイルス粒子へのキメラインテグラーゼ酵素の組込みによって宿主細胞DNAへの部位特異的組込みについて構築され得る(WO96/37626号を参照のこと)。この組換えウイルスベクターは複製欠損組換えウイルスであることが好ましい。
【0156】
上記に記載のレトロウイルスベクターを伴う使用について適切なパッケージング細胞株は、当該分野で周知であり、容易に調製され(WO95/30763号およびWO92/05266号を参照のこと)、そしてこれを使用して、組換えベクター粒子の生産のためのプロデューサー細胞株(これは、ベクター細胞株または「VCL」とも称される)を作製し得る。好ましくは、このパッケージング細胞株は、ヒトの親細胞(例えば、HT1080細胞)またはミンク親細胞株から作製され、これは、ヒト血清における不活化を除去する。
【0157】
レトロウイルス遺伝子治療ベクターの構築のために好ましいレトロウイルスは、トリ白血病ウイルス、ウシ白血病ウイルス、マウス白血病ウイルス、ミンク細胞フォーカス形成ウイルス、マウス肉腫ウイルス、細網内皮症ウイルス、およびラウス肉腫ウイルスを含む。特に好ましいマウス白血病ウイルスは、4070Aおよび1504A(HartleyおよびRowe(1976)J. Virol.19:19−25)、Abelson(ATCC番号VR−999)、Friend(ATCC番号VR−245)、Graffi、Gross(ATCC番号VR−590)、Kirsten、Harvey肉腫ウイルスおよびRauscher(ATCC番号VR−998)およびモロニーマウス白血病ウイルス(ATCC番号VR−190)を含む。このようなレトロウイルスベクターは、寄託機関または収集機関(例えば、アメリカンタイプカルチャーコレクション(「ATCC」)、Rockville、Maryland)から入手し得るか、または一般に利用可能な技術を用いて公知の供給源から単離され得る。
【0158】
本発明において使用可能な例示的な公知のレトロウイルス遺伝子治療ベクターは、特許出願GB2200651、EP0415731、EP0345242、EP0334301、WO89/02468;WO89/05349、WO89/09271、WO90/02806、WO90/07936、WO94/03622、WO93/25698、WO93/25234、WO93/11230、WO93/10218、WO91/02805、WO91/02825、WO95/07994、米国特許第5,219,740号、同4,405,712号、同4,861,719号、同4,980,289号、同4,777,127号、同5,591,624号に記載されるものを含む。Vile(1993)Cancer Res 53:3860−3864;Vile(1993)Cancer Res.53:962−967;Ram(1993)Cancer Res 53(1993)83−88;Takamiya(1992)J Neurosci Res 33:493−503;Baba(1993)J Neurosurg 79:729−735;Mann(1983)Cell 33:153;Cane(1984)Proc Natl Acad Sci 81;6349;およびMiller(1990)Human Gene Therapy 1もまた参照のこと。
【0159】
ヒトアデノウイルス遺伝子治療ベクターもまた当該分野で公知であり、そして本発明において使用可能である。例えば、Berkner(1988)Biotechniques 6:616およびRosenfeld(1991)Science 252:431;ならびにWO93/07283、WO93/06223、およびWO93/07282を参照のこと。本発明において使用可能な例示的な公知のアデノウイルス遺伝子治療ベクターは、上記に参照される文書およびWO94/12649、WO93/03769、WO93/19191、WO94/28938、WO95/11984、WO95/00655、WO95/27071、WO95/29993、WO95/34671、WO96/05320、WO94/08026、WO94/11506、WO93/06223、WO94/24299、WO95/14102、WO95/24297、WO95/02697、WO94/28152、WO94/24299、WO95/09241,WO95/25807、WO95/05835、WO94/18922およびWO95/09654において記載されるものを含む。あるいは、Curiel(1992)Hum.Gene Ther.3:147−154に記載されるような殺傷したアデノウイルスに連結したDNAの投与が使用され得る。本発明の遺伝子送達ビヒクルはまた、アデノウイルス随伴ウイルス(AAV)ベクターを含む。本発明における使用のためのこのようなベクターの主要なおよび好ましい例は、Srivastava WO93/09239に開示されるAAV−2ベースのベクターである。最も好ましいAAVベクターは、2つのAAV逆方向末端反復を含む。ここで、ネイティブD配列は、ヌクレオチドの置換によって改変され、その結果、少なくとも5つのネイティブなヌクレオチドおよび18までのネイティブヌクレオチド、好ましくは少なくとも10のネイティブヌクレオチドから18までのネイティブヌクレオチド、最も好ましくは10のネイティブヌクレオチドが維持され、そしてD配列の残りのヌクレオチドが欠失またはネイティブでないヌクレオチドで置換されている。AAV逆方向末端反復のネイティブなD配列は、各AAV逆方向末端反復において(すなわち、各末端に1つの配列が存在する)20の連続するヌクレオチドの配列であって、これは、HP形成に関与しない。ネイティブでない置換ヌクレオチドは、同じ位置でのネイティブなD配列に見出されるヌクレオチド以外の任意のヌクレオチドであり得る。他の使用可能な例示的なAAVベクターは、、pWP−19、pWN−1であり、これらは両方ともNahreini(1993)Gene 124:257−262に開示される。このようなAAVベクターの別の例は、psub201(Samulski(1987)J.Virol.61:3096を参照のこと)である。別の例示的なAAVベクターは、Double−D ITRベクターである。Double−D ITRベクターの構築は、米国特許第5,478,745号に開示される。なお他のベクターは、Carter 米国特許第4,797,368号およびMuzyczka 米国特許第5,139,941号、Chartejee 米国特許第5,474,935号ならびにKotin WO94/288157に開示されるものである。本発明において使用可能なAAVベクターのなおさらなる例は、SSV9AFABTKneoであり、これは、AFPエンハンサーおよびアルブミンプロモーターを含み、そして肝臓において優性に発現を指向する。その構造および構築は、Su(1996)Human Gene Therapy 7:463−470に開示される。さらなるAAV遺伝子治療ベクターは、米国特許第5,354,678号、同5,173,414号、同5,139,941号、および同5,252,479号に記載される。
【0160】
本発明の遺伝子治療ベクターはまた、ヘルペスウイルスベクターを含む。主要なおよび好ましい例は、チミジンキナーゼポリペプチドをコードする配列を含む単純ヘルペスウイルスベクター(例えば、米国特許第5,288,641号、およびEP0176170(Roizman)に開示されるもの)である。さらなる例示的な単純ヘルペスウイルスベクターは、WO95/04139(Wistar)に開示されるHFEM/ICP6−lacZ、Geller(1988)Science 241:1667−1669ならびにWO90/09441およびWO92/07945に記載されるpHSVlac、Fink(1992)Human Gene Therapy 3:11−19に記載されるHSV Us3::pgC−lacZ、ならびにEP0453242(Breakefieled)に記載されるHSV 7134、2RH 105およびGAL4、ならびにATCCに受託番号ATCC VR−977およびATCC VR−260として寄託されたものを含む。
【0161】
意図されるのはまた、本発明において使用され得るαウイルス遺伝子治療ベクターである。好ましいαウイルスベクターは、シンドビスウイルスベクターである。トガウイルス、セムリキ森林ウイルス(ATCC VR−67;ATCC VR−1247)、Middlebergウイルス(ATCC VR−370)、ロスリバーウイルス(ATCC VR−373;ATCC VR−1246)、ベネズエラウマ脳脊髄炎ウイルス(ATCC VR−923;ATCC VR−1250;ATCC VR−1249;ATCC VR−532)および米国特許第5,0091,309号、同5,217,879号、およびWO92/10578に記載されるもの。より詳細には、米国特許出願第08/405,627号(1995年3月15日出願)、WO94/21792号、WO92/10578号、WO95/07994号、米国特許第5,091,309号、および米国特許第5,217,879号に記載されるそれらのαウイルスベクターが使用可能である。このようなαウイルスは、ATCC、Rockville、Marylandのような寄託機関または収集機関から入手し得るか、または一般的に利用可能な技術を用いて公知の供給源から単離され得る。好ましくは、細胞傷害性が減少したαウイルスベクターを使用する(米国特許仮出願08/679640号を参照のこと)。
【0162】
DNAベクター系(例えば、真核細胞層状発現系)もまた、本発明の核酸の発現について有用である。真核生物層状発現系の詳細な説明についてはWO95/07994を参照のこと。好ましくは、本発明の真核細胞層状発現系はαウイルスベクターに由来し、そして最も好ましくはシンドビスウイルスベクターに由来する。
【0163】
本発明における使用に適切な他のウイルスベクターは、ポリオウイルス(例えば、ATCC VR−58およびEvans、Nature339(1989)385およびSabin(1973)J.Biol.Standardization 1:115に記載されるもの;ライノウイルス、例えば、ATCC VR−1110およびArnold(1990)J Cell Biochem L401に記載されるもの;ポックスウイルス(例えば、カナリアポックスルウイルスまたはワクシニアウイルス(例えば、ATCC VR−111およびATCC VR−2010ならびにFisher−Hoch(1989)Proc Natl Acad Sci 86:317;Flexner(1989)Ann NY Acad Sci 569:86、Flexner(1990)Vaccine 8:17;米国特許第4,603,112号および同4,769,330号ならびにWO89/01973号に記載されるもの));SV40ウイルス(例えば、ATCC VR−305およびMulligan(1979)Nature 277:108およびMadzak(1992)J Gen Virol 73:1533に記載されるもの);インフルエンザウイルス(例えば、ATCC VR−797および米国特許第5,166,057号およびEnami(1990)Proc Natl Acad Sci .87:3802−3805;EnamiおよびPalese(1991)J Virol 65:2711−2713およびLuytjes(1989)Cell 59:110(McMichael(1983)NEJ Med 309:13ならびにYap(1978)Nature 273:238およびNature(1979)277:108もまた参照のこと)に記載されるような逆遺伝子技術を使用して作製した組換えインフルエンザウイルス);EP−0386882およびBuchschachler(1992)J.Virol.66:2731に記載されるようなヒト免疫不全ウイルス;麻疹ウイルス(例えば、ATCC VR−67およびVR−1247ならびにEP−0440219に記載されるもの);アウラウイルス(例えば、ATCC VR−368);ベバルウイルス(例えば、ATCC VR−600およびATCC VR−1240);カバス(Cabassou)ウイルス(例えば、ATCC VR−922);チクングンヤウイルス(例えば、ATCC VR−64およびATCC VR−1241);フォートモーガン(Fort Morgan)ウイルス(例えば、ATCC VR−924);ゲタウイルス(例えば、ATCC VR−369およびATCC VR−1243);キジラガハ(Kyzylagach)ウイルス(例えば、ATCC VR−927);マヤロウイルス(例えば、ATCC VR−66);ムカンボウイルス(例えば、ATCC VR−580およびATCC VR−1244);ヌヅムウイルス(例えば、ATCC VR−371);ピクスナウイルス(例えば、ATCC VR−372およびATCC VR−1245);トナテ(Tonate)ウイルス(例えば、ATCC VR−925);トリニティウイルス(例えば、ATCC VR−469);ユナウイルス(例えば、ATCC VR−374);ワタロアウイルス(例えば、ATCC VR−926);Y−62−33ウイルス(例えば、ATCC VR−375);オニオンウイルス、東部ウマ脳脊髄炎ウイルス(例えば、ATCC VR−65およびATCC VR−1242);西部ウマ脳脊髄炎ウイルス(例えば、ATCC VR−70、ATCC VR−1251、ATCC VR−622およびATCC VR−1252);ならびにコロナウイルス(例えば、ATCC VR−740)およびHamre(1966)Proc Soc Exp Biol Med 121:190に記載のものに由来するウイルスベクターを含む。
【0164】
本発明の組成物の細胞への送達は、上記に言及したウイルスベクターに限定されない。他の送達方法および媒体が使用され得る(例えば、核酸発現ベクター、殺傷したアデノウイルスに連結したかまたは連結してないポリカチオン性縮合DNA単独(例えば、米国特許出願番号08/366,787(1994年12月30日出願)およびCuriel(1992)Hum Gene Ther 3:147−154を参照のこと)、リガンド連結DNA(例えば、Wu(1989)J Biol Chem 264:16985−16987を参照のこと)、真核生物細胞送達ビヒクル細胞(例えば、米国特許出願番号08/240,030(1994年5月9日出願)および米国特許出願番号08/404,796を参照のこと)、光重合化ヒドロゲル物質の沈着、手動の遺伝子送達粒子銃(米国特許第5,149,655号に記載されるような)、米国特許第5,206,152およびWO92/11033に記載されるような電離放射線、核酸電荷中和または細胞膜との融合)。さらなるアプローチは、Philip(1994)Mol Cell Biol 14:2411−2418およびWoffendin(1994)Proc Natl Acad Sci 91:1581−1585に記載される。
【0165】
粒子媒介遺伝子送達が使用され得る(例えば、米国特許出願60/023,867号を参照のこと)。手短には、配列を、高レベル発現のための従来の制御配列を含む従来のベクターに挿入し得、次いで細胞標的化リガンド(例えば、アシアロオロソムコイド(WuおよびWu(1987)J.Biol.Chem.262:4429−4432に記載されるような)、Hucked(1990)Biochem Pharmacol 40:253−263に記載されるようなインスリン、Plank(1992)Bioconjugate Chem 3:533−539に記載されるようなガラクトース、ラクトースまたはトランスフェリン)に連結された合成遺伝子送達分子(例えば、ポリリジン、プロタミンおよびアルブミンのような重合DNA結合カチオン)とともにインキュベートされ得る。
【0166】
裸のDNAもまた使用され得る。例示的な裸のDNA導入方法は、WO90/11092および米国特許第5,580,859号に記載される。取り込み効率は、生体分解性のラテックスビーズを用いて改良され得る。DNAコートラテックスビーズは、ビーズによるエンドサイトーシス開始の後に効率よく細胞へと輸送される。この方法は、ビーズを処理して疎水性を高め、それによってエンドソームの破壊および細胞質へのDNAの放出を容易にすることによってさらに改良され得る。
【0167】
遺伝子送達ビヒクルとして作用し得るリポソームは、米国特許第5,422,120号、WO95/13796、WO94/23697、WO91/14445、およびEP524,968に記載される。米国特許出願60/023,867に記載されるように、非ウイルス性送達において、ポリペプチドをコードする核酸配列は、高レベル発現のための従来の制御配列を含む従来のベクターへと挿入され得、次いで細胞標的化リガンド(例えば、アシアロオロソムコイド、インスリン、ガラクトース、ラクトースまたはトランスフェリン)に連結された、重合性DNA結合カチオン(例えば、ポリリジン、プロタミン、およびアルブミン)のような合成遺伝子伝達分子とともにインキュベートされ得る。他の送達系は、種々の組織特異的または普遍的作用性のプロモーターの制御下に遺伝子を含むDNAをカプセル化するためのリポソームの使用を含む。さらに、使用に適切な非ウイルス送達は、機械的送達系(例えば、Woffendinら(1994)Proc.Natl.Acad.Sci.USA 91(24):11581−11585に記載されるアプローチを含む。さらに、コード配列およびそのようなものの発現産物は、光重合化ヒドロゲル物質の沈着を介して送達され得る。コード配列の送達について使用され得る、遺伝子送達のための他の従来の方法は、例えば、手動の遺伝子送達粒子銃(米国特許第5,149,655号に記載されるような);移入された遺伝子を活性化するための電離放射線の使用(米国特許第5,206,152号およびWO92/11033に記載されるような)を含む。
【0168】
例示的なリポソームおよびポリカチオン性遺伝子送達ビヒクルは、米国特許第5,422,120号および同4,762,915号;WO95/13796;WO94/23697;およびWO91/14445;EP−0524968;およびStryer、Biochemistry、236−240頁(1975)、W.H.Freeman、San Francisco;Szoka(1980)Biochem Biophys Acta 600:1;Bayer(1979) Biochem Biophys Acta 550:464;Rivnay(1987)Meth Enzymol 149:119;Wang(1987)Proc Natl Acad Sci 84:7851;Plant(1989)Anal Biochem 176:420に記載されるものである。
【0169】
ポリヌクレオチド組成物は、治療有効量(この用語は上記に定義されるとおりである)の遺伝子治療ビヒクルを含み得る。本発明の目的のために、有効用量は、投与される個体において、約0.01mg/kg〜50mg/kgまたは0.05mg/kg〜約10mg/kgのDNA構築物である。
【0170】
(送達方法)
一旦処方されると、本発明のポリヌクレオチド組成物は、(1)被験体に直接;(2)エキソビボで被験体由来の細胞に送達されて;または(3)組換えタンパク質の発現のためにインビトロで、投与され得る。処置される被験体は、哺乳動物または鳥類であり得る。ヒト被験体もまた処置され得る。
【0171】
この組成物の直接送達は、皮下、腹腔内、静脈内、または筋肉内の注射によって、または組織の間質空間への送達のいずれかによって一般的に達成される。この組成物はまた、病巣へ投与され得る。他の投与様式は、経口投与または肺投与、坐剤、および経皮(transdermal)または経皮(transcutaneous)適用(例えば、WO98/20734を参照のこと)、針、および遺伝子銃またはハイポスプレーを含む。投薬治療は、単回用量スケジュールまたは多数回用量スケジュールであり得る。
【0172】
エキソビボ送達および被験体への形質転換細胞の再移植のための方法は、当該分野で公知であり、そして例えばWO93/14778に記載されている。エキソビボ適用に有用である細胞の例は、例えば、幹細胞、特に、造血細胞、リンパ細胞、マクロファージ、樹状細胞または腫瘍細胞を含む。
【0173】
一般的に、エキソビボ適用およびインビトロ適用の両方のための核酸の送達は、以下の手順:例えば、デキストラン媒介トランスフェクション、リン酸カルシウム沈降、ポリブレン媒介トランスフェクション、原形質融合、エレクトロポレーション、ポリヌクレオチドのリポソーム内へのカプセル化、およびDNAの核への直接の微量注入(これらはすべて当該分野で周知である)で達成され得る。
【0174】
(ポリヌクレオチドおよびポリペプチドの薬学的組成物)
上記に記載の薬学的に受容可能なキャリアおよび塩に加えて、以下のさらなる薬剤がポリヌクレオチド組成物および/またはポリペプチド組成物とともに使用され得る。
【0175】
(A.ポリペプチド)
1つの例は、限定することなく以下を包含する:アシアロオロソムコイド(ASOR);トランスフェリン;アシアロ糖タンパク質;抗体;抗体フラグメント;フェリチン;インターロイキン;インターフェロン;顆粒球マクロファージコロニー刺激因子(GM−CSF)、顆粒球コロニー刺激因子(G−CSF)、マクロファージコロニー刺激因子(M−CSF)、幹細胞因子およびエリスロポエチン。ウイルス抗原(例えば、エンベロープタンパク質)もまた、使用され得る。また、他の侵襲性生物由来のタンパク質(例えば、RIIとして知られるPlasmodium falciparumの環境スポロゾイト(circumsporozoite)タンパク質由来の17アミノ酸ペプチド)。
【0176】
(B.ホルモン、ビタミンなど)
包含され得る他の群は、例えば、ホルモン、ステロイド、アンドロゲン、エストロゲン、甲状腺ホルモン、またはビタミン、葉酸である。
【0177】
(C.ポリアルキレン、ポリサッカリドなど)
また、ポリアルキレングリコールが、所望のポリヌクレオチド/ポリペプチドとともに含有され得る。好ましい実施態様において、ポリアルキレングリコールは、ポリエチレングリコールである。さらに、モノサッカリド、ジサッカリド、またはポリサッカリドが含有され得る。この局面の好ましい実施態様において、このポリサッカリドは、デキストランまたはDEAE−デキストランである。また、キトサンおよびポリ(乳酸−コ−グリコリド)。
【0178】
(D.脂質およびリポソーム)
所望のポリヌクレオチド/ポリペプチドはまた、被験体またはそれに由来する細胞への送達の前に、脂質中にカプセル化され得るか、またはリポソーム中にパッケージングされ得る。
【0179】
脂質カプセル化は、一般的に、核酸に安定に結合し得るか、または核酸を捕捉もしくは維持し得るリポソームを用いて達成される。縮合ポリヌクレオチドの脂質調製物に対する比は、変動し得るが、一般的に約1:1(mgDNA:マイクロモル脂質)であるか、またはより多くの脂質である。核酸の送達のためのキャリアとしてのリポソーム使用の概説については、HugおよびSleight(1991)Biochim.Biophys.Acta.1097:1−17;Straubinger(1983)Meth.Enzymol.101:512−527を参照のこと。
【0180】
本発明における使用のためのリポソーム調製物は、カチオン性(正に荷電した)、アニオン性(負に荷電した)および中性の調製物を包含する。カチオン性リポソームは、機能的な形態で、プラスミドDNA(Felgner(1987)Proc.Natl.Acad.Sci.USA 84:7413−7416);mRNA(Malone(1989)Proc.Natl.Acad.Sci.USA 86:6077−6081;および精製した転写因子(Debs(1990)J.Biol.Chem.265:10189−10192)の細胞内送達を媒介することが示されている。
【0181】
カチオン性リポソームは容易に入手可能である。例えば、N{1−2,3−ジオレイルオキシ)プロピル}−N,N,N−トリエチルアンモニウム(DOTMA)リポソームは、GIBCO BRL、Grand Island、NYからの商標リポフェクチン(Lipofectin)の下で入手可能である(Fegner前出もまた参照のこと)。他の市販されているリポソームとしては、トランスフェクテース(transfectace)(DDAB/DOPE)およびDOTAP/DOPE(Boerhinger)が挙げられる。他のカチオン性リポソームは、当該分野で周知の技法を使用して、容易に利用可能な物質から調製され得る。例えば、DOTAP(1,2−ビス(オレイルオキシ)−3−(トリメチルアンモニオ)プロパン)リポソームの合成の記載について、Szoka(1978)Proc.Natl.Acad.Sci.USA 75:4194−4198;WO90/11092を参照のこと。
【0182】
同様に、アニオン性リポソームおよび中性リポソームは、例えば、Avanti Polar Lipids(Birmingham,AL)から容易に入手可能であるか、または容易に入手可能な物質を使用してたやすく調製され得る。このような物質には、とりわけ、ホスファチジルコリン、コレステロール、ホスファチジルエタノールアミン、ジオレオイルホスファチジルコリン(DOPC)、ジオレオイルホスファチジルグリセロール(DOPG)、ジオレオイルホスファチジルエタノールアミン(DOPE)が含まれる。これらの物質はまた、適切な比率でDOTMAおよびDOTAPの出発物質と混合され得る。これらの物質を使用してリポソームを作製する方法は、当該分野で周知である。
【0183】
このリポソームは、多重膜のベシクル(MLV)、小さな単一膜ベシクル(SUV)、または大きな単一膜のベシクル(LUV)を含み得る。種々のリポソーム−核酸複合体は、当該分野で公知の方法を使用して調製される。例えば、Straubinger(1983)Meth.Immunol.101:512−527;Szoka(1978)Proc.Natl.Acad.Sci.USA 75:4194−4198;Papahadjopoulos(1975)Biochim.Biophys.Acta 394:483;Wilson(1979)Cell 17:77);DeamerおよびBangham(1976)Biochim.Biophys.Acta 443:629;Ostro(1977)Biochem.Biophys.Res.Commun.76:836;Fraley(1979)Proc.Natl.Acad.Sci.USA 76:3348);EnochおよびStrittmatter(1979)Proc.Natl.Acad.Sci.USA 76:145;Fraley(1980)J.Biol.Chem.(1980)255:10431;SzokaおよびPapahadjopoulos(1978)Proc.Natl.Acad.Sci.USA 75:145;ならびにSchaefer−Ridder(1982)Science 215:166を参照のこと。
【0184】
(E.リポタンパク質)
さらに、リポタンパク質が、送達されるポリヌクレオチド/ポリペプチドとともに含まれ得る。利用されるリポタンパク質の例としては、キロミクロン、HDL、IDL、LDL、およびVLDLが挙げられる。これらのタンパク質の変異体、フラグメント、または融合物もまた、使用され得る。また、天然に存在するリポタンパク質の改変体(例えば、アセチル化されたLDL)が使用され得る。これらのリポタンパク質は、リポタンパク質レセプターを発現する細胞へ、ポリヌクレオチドの送達を標的化し得る。好ましくは、リポタンパク質が、送達されるポリヌクレオチドとともに含まれる場合、他の標的化リガンドはその組成物中には含まれない。
【0185】
天然に存在するリポタンパク質は、脂質部分およびタンパク質部分を含む。このタンパク質部分は、アポタンパク質として知られる。現在では、アポタンパク質A、B、C、D、およびEが単離および同定されている。少なくともこれらの2つはいくつかのタンパク質を含み、ローマ数字、AI、AII、AIV;CI、CII、CIIIによって命名されている。
【0186】
1つのリポタンパク質は、1を超えるアポタンパク質を含み得る。例えば、天然に存在するキロミクロンはA、B、C、およびEからなり、そして時間が経てばこれらのリポタンパク質はAを欠失し、そしてCおよびEアポタンパク質を獲得する。VLDLは、A、B、C、およびEアポタンパク質を含み、LDLはアポタンパク質Bを含み;そしてHDLはアポタンパク質A、C、およびEを含む。
【0187】
これらのアポタンパク質のアミノ酸は公知であり、そして例えば、Breslow(1985)Annu Rev.Biochem 54:699;Law(1986)Adv.Exp.Med.Biol.151:162;Chen(1986)J Biol Chem 261:12918;Kane(1980)Proc Natl Acad Sci USA 77:2465;Utermann(1984)Hum Genet 65:232に記載されている。
【0188】
リポタンパク質は、トリグリセリド、コレステロール(遊離およびエステル)、およびリン脂質を含む、種々の脂質を含む。この脂質の組成は、天然に存在するリポタンパク質において変化する。例えば、キロミクロンは主としてトリグリセリドを含む。天然に存在するリポタンパク質の脂質含有物のより詳細な記載は、例えば、Meth.Enzymol.128(1986)に見いだされ得る。この脂質の組成は、レセプター結合活性についてアポタンパク質の立体構造において補助するために選択される。脂質組成はまた、ポリヌクレオチド結合分子との疎水性相互作用および会合を容易にするように選択され得る。
【0189】
天然に存在するリポタンパク質は、例えば、血清から超遠心分離によって単離され得る。そのような方法は、Meth.Enzymol.(前出);Pitas(1980)J.Biochem.253:5454−5460およびMahey(1979)J Clin.Invest 64:743−750に記載される。リポタンパク質はまた、インビトロまたは所望の宿主細胞中のアポタンパク質遺伝子の発現による組換え方法によって産生され得る。例えば、Atkinson(1986)Annu Rev Biophys Chem 15:403およびRadding(1958)Biochim Biophys Acta 30:443を参照のこと。リポタンパク質はまた、Biomedical Techniologies,Inc.,Stoughton,Massachusetts,USAのような商業的な供給者から購入され得る。さらなるリポタンパク質の記載は、Zuckermannら、PCT/US97/14465に見い出され得る。
【0190】
(F.ポリカチオン性薬剤)
ポリカチオン性薬剤は、送達される所望のポリヌクレオチド/ポリペプチドを有する組成物中に、リポタンパク質を伴って、またはリポタンパク質を伴わずに含まれ得る。
【0191】
ポリカチオン性薬剤は、代表的には、生理的に適切なpHにおいて正味の正電荷を示し、そして所望の位置への送達を容易にするための核酸の電荷を中和し得る。これらの薬剤は、インビトロ、エキソビボ、およびインビボ適用のいずれもを有する。ポリカチオン性薬剤は、生きている被験体に、筋肉内、皮下などのいずれかで核酸を送達するために使用され得る。
【0192】
以下は、ポリカチオン性薬剤としての有用なポリペプチドの例である:ポリリジン、ポリアルギニン、ポリオルニチン、およびプロタミン。他の例は、ヒストン、プロタミン、ヒト血清アルブミン、DNA結合タンパク質、非ヒストン染色体タンパク質、DNAウイルス由来のコートタンパク質(例えば、X174)を含む。転写因子もまた、DNAに結合するドメインを含み、従って核酸縮合薬剤として有用であり得る。手短に言えば、転写因子(例えば、C/CEBP、c−jun、c−fos、AP−1、AP−2、AP−3、CPF、Prot−1、Sp−1、Oct−1、Oct−2、CREP、およびTFIID)は、DNA配列に結合する塩基性ドメインを含む。
【0193】
有機ポリカチオン性薬剤は、スペルミン、スペルミジン、およびプトレシン(purtrescine)を含む。
【0194】
ポリカチオン性薬剤の大きさおよびその物理的特性は、上記の表から外挿されて、他のポリペプチドポリカチオン性薬剤が構築され得るか、または合成ポリカチオン性薬剤が産生され得る。
【0195】
有用な合成ポリカチオン性薬剤は、例えば、DEAE−デキストラン、ポリブレンを含む。LipofectinTM、およびlipofectAMINETMは、ポリヌクレオチド/ポリペプチドと組み合わせた場合にポリカチオン性複合体を形成するモノマーである。
【0196】
(核酸ハイブリダイゼーション)
「ハイブリダイゼーション」とは、水素結合による2つの核酸配列の互いの会合をいう。代表的には、1つの配列は、固体支持体に固定され、そして他方は溶液中で遊離している。次いで、2つの配列は、水素結合に好ましい条件下で互いに接触される。この結合に影響を与える因子は以下を含む:溶媒のタイプおよび容量;反応温度;ハイブリダイゼーションの時間;撹拌;液体相の配列の固体支持体への非特異的な付着をブロックする薬剤(Denhardt’s試薬またはBLOTTO);配列の濃度;配列の会合の速度を増大させる化合物(硫酸デキストランまたはポリエチレングリコール)の使用;およびハイブリダイゼーション後の洗浄条件のストリンジェンシー。Sambrookら(前出)第2巻、第9章、9.47〜9.57頁を参照のこと。
【0197】
「ストリンジェンシー」とは、異なる配列よりも、非常に類似する配列の会合に好ましいハイブリダイゼーション反応における条件をいう。例えば、研究中のハイブリッドの計算されたTmより約120〜200℃低い温度および塩濃度の組み合わせが選択されるべきである。温度および塩条件はしばしば、フィルターに固定したゲノムDNAのサンプルが目的の配列にハイブリダイズし、次いで異なるストリンジェンシーの条件下で洗浄される、予備的な実験において経験的に決定され得る。Sambrookら、9.50頁を参照のこと。
【0198】
例えば、サザンブロットを行う場合、考慮する変数は、(1)ブロットされるDNAの複雑さ、および(2)プローブと検出される配列との間の相同性である。研究されるフラグメント全量は、プラスミドまたはファージ消化物については0.1〜1μg、高度に複雑な真核生物ゲノム中の単一コピーの遺伝子については10−9〜10−8gまで、10倍変化し得る。より低い複雑さのポリヌクレオチドについては、実質的により短いブロッティング、ハイブリダイゼーション、および曝露回数、より少量の出発ポリヌクレオチド、およびより低い非活性のプローブが使用され得る。例えば、単一コピーの酵母遺伝子は、1μgの酵母DNAで開始し、2時間ブロットし、そして4〜8時間108cpm/μgのプローブを用いてハイブリダイズして、わずか1時間の曝露時間を用いて検出され得る。単一コピーの哺乳動物遺伝子について、保存性のアプローチは、10μgのDNAで開始し、一晩ブロットし、そして108cpm/μgより多いプローブを用いて10%硫酸デキストランの存在下で一晩ハイブリダイズし、約24時間露光時間を生じる。
【0199】
いくつかの因子が、プローブと目的のフラグメントとの間のDNA−DNAハイブリッドの融解温度(Tm)、ならびに、結果として、ハイブリダイゼーションおよび洗浄についての適切な条件に影響を与え得る。多くの場合において、そのプローブはフラグメントに対して100%相同なわけではない。他の共通して直面する変数には、長さ、ハイブリダイズする配列の全G+C含量、ならびにイオン強度およびハイブリダイゼーション緩衝液のホルムアミド含量が含まれる。これらのすべての因子の影響は、一つの式によって近似され得る:
Tm=81+16.6(log10Ci)+0.4{(G+C)%}−0.6(ホルムアミド%)−600/n−1.5(ミスマッチ%)。
ここでCiは、塩濃度(一価イオン)であり、そしてnは塩基対内のハイブリッドの長さである(MeinkothおよびWahl(1984)Anal.Biochem.138:267/284からわずかに改変した)。
【0200】
ハイブリダイゼーション実験の設計において、核酸ハイブリダイゼーションに影響を与えるいくつかの因子が簡便に変更され得る。ハイブリダイゼーションおよび洗浄の温度ならびに洗浄時の塩濃度を調整するのが最も単純である。ハイブリダイゼーション温度(すなわち、ストリンジェンシー)が上昇するにつれて、非相同的な鎖の間で起こるハイブリダイゼーションは起こりにくくなるようであり、結果として、バックグラウンドが減少する。放射標識したプローブが固定化されたフラグメントと完全に相同ではない場合(遺伝子ファミリーおよび種間のハイブリダイゼーション実験における場合で頻繁であるように)、ハイブリダイゼーション温度は低下されなければならず、そしてバックグラウンドが増大する。洗浄の温度は、類似の様式で、ハイブリダイズするバンドの強度、およびバックグラウンドの程度に影響を与える。洗浄のストリンジェンシーはまた、塩濃度の減少とともに増大する。
【0201】
一般的に、50%ホルミアミドの存在下で都合よいハイブリダイゼーション温度は、標的フラグメントに95%〜100%相同であるプローブについて42℃、90%〜95%相同性では37℃、85%〜90%相同性については32℃である。より低い相同性については、上記の式を用いて、適切にホルムアミド含量が低くされ、そして温度が調整されるべきである。プローブと標的フラグメントとの間の相同性が未知である場合、最も単純なアプローチは、ともにストリンジェントではないハイブリダイゼーション条件および洗浄条件で開始することである。オートラジオグラフィー後に非特異的バンドまたは高いバックグラウンドが観察される場合、フィルターは高ストリンジェンシーで洗浄され得、そして再び露光され得る。露光のために必要な時間がこのアプローチを非実用的にする場合、いくつかのハイブリダイゼーションおよび/または洗浄ストリンジェンシーが並行して試験されるべきである。
【0202】
(核酸プローブアッセイ)
本発明に従う核酸プローブを利用する、PCR、分枝DNAプローブアッセイ、またはブロッティング技術のような方法は、cDNAまたはmRNAの存在を決定し得る。プローブは、検出されるに十分に安定な、二重鎖または二本鎖複合体を形成し得る場合に、本発明の配列に「ハイブリダイズする」といわれる。
【0203】
核酸プローブは、本発明のクラミジアの(Chlamydial)ヌクレオチド配列(センス鎖およびアンチセンス鎖の両方を含む)にハイブリダイズする。多くの異なるヌクレオチド配列がアミノ酸配列をコードするが、ネイティブなクラミジアの配列は、細胞に存在する実際の配列であるので、好ましい。mRNAは、コード配列を表し、従ってプローブはコード配列に相補的であるべきであり、一本鎖cDNAはmRNAに相補的であり、従ってcDNAプローブは非コード配列に相補的であるべきである。
【0204】
プローブ配列はクラミジアの配列(またはその相補物)に同一である必要はない。核酸プローブが標的ヌクレオチドと検出され得る二重鎖を形成し得る場合、配列および長さのいくらかの変動は、アッセイの感度の増加をもたらし得る。また、核酸プローブは、形成された二重鎖を安定化するためにさらなるヌクレオチドを含み得る。さらなるクラミジアの配列もまた、形成された二重鎖を検出するための標識としての一助となり得る。例えば、非相補的ヌクレオチド配列が、そのプローブの5’末端に付着され得、ここでそのプローブ配列の残りはクラミジアの配列に相補的である。あるいは、プローブ配列が、クラミジアの配列とハイブリダイズし、そしてそれによって検出され得る二重鎖を形成するためにクラミジアの配列との十分な相補性を有する場合、非相補的塩基またはより長い配列は、プローブ中に分散され得る。
【0205】
プローブの正確な長さおよび配列は、ハイブリダイゼーション条件(例えば、温度、塩条件など)に依存する。例えば、診断的適用については、分析物の配列の複雑さに依存して、核酸プローブは、代表的には、少なくとも10〜20ヌクレオチド、好ましくは15〜25、そしてより好ましくは少なくとも30ヌクレオチド以上を含むが、これよりも短くてもよい。短いプライマーは、一般的には、鋳型との安定なハイブリッド複合体を形成するのにより低い温度を必要とする。
【0206】
プローブは、合成的手順(例えば、Matteucciらのトリエステル法、(J.Am.Chem.Soc.(1981)103:3185))、またはUrdeaら(Proc.Natl.Acad.Sci.USA(1983)80:7461)に従って、または市販の自動オリゴヌクレオチド合成機を使用して、産生され得る。
【0207】
プローブの化学的性質は、優先度に従って選択され得る。特定の適用については、DNAまたはRNAが適切である。他の適用については、改変(例えば、ホスホロチオエートまたはメチルホスホネートのようなバックボーンの改変)が組み込まれ得、インビボの半減期を増大させるために使用され得、RNA親和性を変化させ、ヌクレアーゼ耐性などを増大させるなどを行い(例えば、AgrawalおよびIyer(1995)Curr Opin Biotechnol 6:12−19;Agrawal(1996)TIBTECH 14:376−387);ペプチド核酸のようなアナログもまた使用され得る(例えば、Corey(1997)TIBTECH15 224−229;Buchardtら(1993)TIBTECH 11:384−386を参照のこと)。
【0208】
あるいは、ポリメラーゼ連鎖反応(PCR)は、少量の標的核酸を検出する別の周知の手段である。そのアッセイは、Mullisら、(Meth.Enzymol.(1987)155:335−350);米国特許第4,683,195号および同第4,683,202号に記載されている。2つの「プライマー」ヌクレオチドは、標的核酸とハイブリダイズし、そして反応を開始するために使用される。このプライマーは、増殖標的(またはその相補物)の配列にハイブリダイズしない、二重鎖の安定性を補助するための、または、例えば、首尾よい制限部位を組み込むための配列を含む。代表的には、このような配列は、所望のクラミジアの配列に隣接する。
【0209】
熱安定性のポリメラーゼは、もともとの標的核酸を鋳型として使用して、プライマーから標的核酸のコピーを作製する。標的核酸の閾値量がポリメラーゼによって産生された後、それらはより従来的な方法(例えば、サザンブロット)によって検出され得る。サザンブロット法を使用する場合、標識されたプローブは、クラミジアの配列(またはその相補物)にハイブリダイズする。
【0210】
また、mRNAまたはcDNAは、Sambrookら(前出)に記載される、従来的なブロッティング技術によって検出され得る。mRNA、またはmRNAからポリメラーゼ酵素を使用して生成されたcDNAは、ゲル電気泳動を使用して精製および分離され得る。次いで、ゲル上のこの核酸は、ニトロセルロースのような固体支持体にブロットされる。この固体支持体は、標識されたプローブに曝露され、次いですべてのハイブリダイズしていないプローブを洗浄して除去する。次に、標識プローブを含む二重鎖を検出する。代表的には、そのプローブは、放射活性部分で標識される。
【0211】
ウエスタンブロットのために、2つのサンプルを各タンパク質について試験した。対の左側のレーンは、前免疫血清で染色した膜ストリップを使用し、一方、右側のレーンは、免疫血清で染色した膜ストリップを使用した。図1〜5、35B、37B、38Bおよび39のウエスタンブロットにおいて、マーカーは、66kDa、45kDa、30kDa、20.1kDaおよび14.4kDaにある。図6〜16、20B、23C、24D、27E、38A、40、41、42および43のウエスタンブロットにおいて、マーカーは、172.6kDa、111.4kDa、79.6kDa、61.3kDa、49.0kDa、36.4kDa、24.7kDa、19.2kDaおよび13.1kDaにある。
【0212】
図1〜5のウエスタンブロットにおいて、レーン2および3は、GST融合コントロール抗原に対して惹起されたコントロール血清を示す。図1〜5のウエスタンブロットにおいて、レーン4および5は、Hisタグ化コントロール抗原に対して惹起されたコントロール血清を含む。
【0213】
低分子量マーカーは、精製ゲルのレーン1で泳動される。
【0214】
(発明を実施するための様式)
表Iは、参考文献18由来のC.pneumoniaeタンパク質の名称、このタンパク質についてのGenBank登録番号および表題、本発明の対応するC.trachomatisタンパク質についてのGenBank登録番号および表題、ならびにこれらのC.trachomatisタンパク質についての配列番号(配列番号1〜262、奇数はアミノ酸配列であり、そして偶数はヌクレオチド配列である)を与える。これらは、対応するC.pneumoniaeタンパク質について参考文献18に記載されたのと同じ方法で、発現および使用され得る。C.trachomatisタンパク質は、診断目的および免疫原性目的のために有用である。これらの特性は、配列のみからでは明らかでない。
【0215】
種々の試験を使用して、本発明のタンパク質のインビボでの免疫原性を評価し得る。例えば、これらのタンパク質は、組換え的に発現され得、そしてイムノブロットによって患者の血清をスクリーニングするために使用され得る。タンパク質と患者の血清との間のポジティブ反応は、この患者が、問題のタンパク質に対する免疫応答を以前に生じていたことを示す(すなわち、このタンパク質は、免疫原である)。この方法はまた、免疫優性タンパク質を同定するためにも使用され得る。
【0216】
この組換えタンパク質はまた、例えば、マウスにおいて、抗体を調製するために簡便に使用され得る。これらは、(例えば、インタクトなクラミジアに対するウエスタンブロットにおいてこれらの抗体を使用することによって)タンパク質がC.trachomatisの細胞表面上に位置することを直接確認するために使用され得る。標識された抗体(例えば、FACSのための蛍光標識)は、インタクトな細菌と共にインキュベートされ得、そしてこの細菌表面上の標識の存在が、タンパク質の位置を確認する。FACSの図は、アッセイにおいて使用されるChlamydia調製物の散乱プロファイル、組換え抗原に対する抗体が、Chlamydia細胞に結合する場合に得られるピークシフト(白抜き領域=コントロールサンプル;塗りつぶした領域=抗体と反応したサンプル)、定量的Kolmogorov−Smirnov(K−S)統計分析、およびFACS分析ソフトウェアの出力を示す。
【実施例】
【0217】
(実施例1)
CT242(配列番号57および配列番号58)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図1A;レーン4および5、クロマトグラフィー画分1および2、予測された分子量42.4kDa)として、およびHisタグ化融合タンパク質(図1B;レーン2〜4、クロマトグラフィー画分1、2および3、予測された分子量16.4kDa)としての両方で精製された。
【0218】
この組換えタンパク質を、マウスを免疫化するために使用し、このマウスの血清を、ウエスタンブロットで使用した(図1C:Hisタグ化:レーン12および13;GST融合:レーン20および21)。レーン12は、Hisタグ化CT242についての、前免疫血清で染色した膜ストリップを示し、一方、レーン13は、Hisタグ化CT242についての、免疫血清で染色した膜ストリップを示す。レーン20は、GST融合CT242についての前免疫血清で染色した膜ストリップを示し、レーン21は、GST融合CT242についての免疫血清で染色した膜ストリップを示す。
【0219】
これらの実験は、CT242が表面曝露されておりかつ免疫アクセス可能(immunoaccessible)なタンパク質であること、およびこのCT242が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0220】
(実施例2)
CT045(配列番号71および配列番号72)は、E.coli中で発現された。この組換え産物は、Hisタグ化融合タンパク質(図2A;レーン4〜6、クロマトグラフィー画分1、2および3、予測された分子量55.8kDa)として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図2B:レーン8および9)およびFACS分析(図2C、K−S値16.81)のために使用した。
【0221】
これらの実験は、CT045が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT045が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0222】
(実施例3)
CT381(配列番号105および配列番号106)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図3A;レーン2および3、クロマトグラフィー画分1および2、予測された分子量52.7kDa)およびHisタグ化融合タンパク質(図3A;レーン7〜9、クロマトグラフィー画分1、2および3、予測された分子量26.7kDa)として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図3B:Hisタグ化:レーン6および7;GST融合:レーン16および17)およびFACS分析(図3C、GSTタグ化、K−S値35.98;図3D:Hisタグ化、K−S値32.54)のために使用した。
【0223】
これらの実験は、CT381が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT381が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0224】
(実施例4)
CT396(配列番号107および配列番号108)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図4A;レーン6および7、クロマトグラフィー画分1および2、予測された分子量99.5kDa)およびHisタグ化融合タンパク質(図4B;レーン5〜7、クロマトグラフィー画分1、2および3、予測された分子量73.5kDa)として精製された。この組換えHisタグ化タンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図4C:レーン14および15)に使用した。この組換えHisタグ化タンパク質および組換えGST融合タンパク質をまた、FACS分析(図4D:Hisタグ化、K−S値34.50;図4E:GST融合、K−S値32.76)のために使用した。
【0225】
これらの実験は、CT396が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT396が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0226】
(実施例5)
CT398(配列番号111および配列番号112)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図5A;レーン8および9、クロマトグラフィー画分1および2、予測された分子量54.8kDa)およびHisタグ化融合タンパク質(図5B;レーン8〜10、クロマトグラフィー画分1、2および3、予測された分子量28.8kDa)として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図5C:Hisタグ化:レーン10および11;GST融合:レーン18および19)およびFACS分析(図5D:GST融合、K−S値31.24;図5E:Hisタグ化、K−S値26.10)のために使用した。
【0227】
これらの実験は、CT398が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT398が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0228】
(実施例6)
CT089(配列番号61および配列番号62)は、E.coli中で発現された。この組換え産物は、GST融合タンパク質(図6C;レーン2、クロマトグラフィー画分1、予測された分子量70.8kDa)およびHisタグ化融合タンパク質(図6C;レーン3、4および5、クロマトグラフィー画分1、2および3、予測された分子量44.8kDa)として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図6A:GST融合:レーン14および15;Hisタグ化:レーン16および17)およびFACS分析(図6B:Hisタグ化、K−S値26.59)のために使用した。
【0229】
これらの実験は、CT089が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT089が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0230】
(実施例7)
CT443(配列番号125および配列番号126)は、E.coli中で発現された。この組換え産物は、Hisタグ化融合タンパク質として精製された。この組換えタンパク質を、マウスを免疫化するために使用し、この血清を、ウエスタンブロット(図7A:レーン10および11)およびFACS分析(図7B:K−S値21.28)のために使用した。
【0231】
これらの実験は、CT443が表面曝露されておりかつ免疫アクセス可能なタンパク質であること、およびこのCT443が有用な免疫原であることを示す。これらの特性は、配列単独では明らかでない。
【0232】
(実施例8)
CT541(配列番号149および配列番号150)を、E.coliにおいて発現させた。この組換え産物を、GST融合タンパク質として精製した(図8C:レーン2および3、クロマトグラフィー画分1および2、予測分子量51.6kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図8A:レーン6および7)ならびにFACS分析(図8B:K−S値9.94)のために使用した。
【0233】
これらの実験は、CT541が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0234】
(実施例9)
CT547(配列番号151および配列番号152)を、E.coliにおいて発現させた。この組換え産物を、GST融合タンパク質(図9D:レーン4および5、クロマトグラフィー画分1および2、予測分子量58.3kDa)およびHisタグ化融合タンパク質の両方として精製した。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図9A:Hisタグ化:レーン20および21)ならびにFACS分析(図9B:GST融合:K−S値14.60および15.57;図9C:Hisタグ化:K−S値28.21)のために使用した。
【0235】
これらの実験は、CT547が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0236】
(実施例10)
CT587(配列番号189および配列番号190)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図10C:レーン5、6および7、クロマトグラフィー画分1、2および3、予測分子量47.5kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図10A:レーン12および13)ならびにFACS分析(図10B:Hisタグ化:K−S値20.85)のために使用した。
【0237】
これらの実験は、CT587が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0238】
(実施例11)
CT266(配列番号77および配列番号78)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図11C:レーン11および12、クロマトグラフィー画分1および2、予測分子量44.1kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図11A:レーン4および5)ならびにFACS分析(図11B:K−S値21.29)のために使用した。
【0239】
これらの実験は、CT266が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0240】
(実施例12)
CT444(配列番号127および配列番号128)を、E.coliにおいて発現させた。この組換え産物を、GST融合タンパク質(図12B:レーン2および3、クロマトグラフィー画分1および2、予測分子量87.3kDa)およびHisタグ化融合タンパク質(図12C:レーン3および4、クロマトグラフィー画分2および3、予測分子量9.0kDa)として精製した。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図12A:レーン16および17)ならびにFACS分析(図12D:GST融合:K−S値14.98;図12E:Hisタグ化:K−S値13.28)のために使用した。
【0241】
これらの実験は、CT444が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0242】
(実施例13)
CT559(配列番号199および配列番号200)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化タンパク質として精製した(図13C:レーン2、3および4、クロマトグラフィー画分1、2および3、予測分子量34.9kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図13A:レーン6および7)ならびにFACS分析(図13B:K−S値23.21)のために使用した。
【0243】
これらの実験は、CT559が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0244】
(実施例14)
CT681(配列番号155および配列番号156)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図14C:レーン5および6、クロマトグラフィー画分1および2、予測分子量41.8kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図14A:レーン10および11)ならびにFACS分析(図14B:K−S値34.66)のために使用した。
【0245】
これらの実験は、CT681が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0246】
(実施例15)
CT713(配列番号201および配列番号202)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図15B:レーン4、5および6、クロマトグラフィー画分1、2および3、予測分子量35.4kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図15A:レーン12および13)ならびにFACS分析(図15C:K−S値25.82)のために使用した。
【0247】
これらの実験は、CT713が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0248】
(実施例16)
CT823(配列番号229および配列番号230)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図16C:レーン7、8および9、クロマトグラフィー画分1、2および3、予測分子量53.9kDa)。この組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図16A:レーン14および15)ならびにFACS分析(図16B:K−S値26.62)のために使用した。
【0249】
これらの実験は、CT823が、表面に露出された免疫アクセス可能なタンパク質であり、そして有用な免疫原であることを示す。これらの特性は、配列単独からは明らかではない。
【0250】
(実施例17)
CT114(配列番号243および配列番号244)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図17:レーン6および7、クロマトグラフィー画分1および2、予測分子量48.5kDa)。
【0251】
(実施例18)
CT198(配列番号43および配列番号44)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図18A:レーン6、クロマトグラフィー画分1、予測分子量56.3kDa)。このHisタグ化組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、FACS分析のために使用した(図18B)。
【0252】
これらの実験は、CT198がEB異種集団のほんの一部において存在することを示す(クラミジア調製物が通常そうであるように)。これが存在する場合、これは、表面に露出された免疫アクセス可能なタンパク質である。これらの特性は、配列単独からは明らかではない。
【0253】
(実施例19)
CT241(配列番号55および配列番号56)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した(図19:レーン4、クロマトグラフィー画分3、予測分子量85.3kDa)。
【0254】
(実施例20)
CT350(配列番号27および配列番号28)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図20A:レーン2、3および4、クロマトグラフィー画分1、2および3、予測分子量61.3kDa)およびGST融合タンパク質(図20A:レーン7、8および9、クロマトグラフィー画分1、2および3、予測分子量87.3kDa)の両方として精製した。これらの組換えタンパク質を使用してマウスを免疫し、このマウスの血清を、ウェスタンブロット(図20B:Hisタグ化:レーン4および5;GSTタグ化:レーン8および9)のために使用した。
【0255】
(実施例21)
CT351(配列番号25および配列番号26)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図21:レーン2および3、クロマトグラフィー画分1および2、推定分子量 76.1kDa)として精製した。
【0256】
(実施例22)
CT391(配列番号251および配列番号252)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図22:レーン8および9、クロマトグラフィー画分1および2、推定分子量 32.6kDa)として精製した。
【0257】
(実施例23)
CT077(配列番号65および配列番号66)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図23:レーン2および3、クロマトグラフィー画分1および2、推定分子量 59.7kDa)、およびHisタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図23C:レーン6および7)、およびFACS分析(図23B、Hisタグ化:K−S値 9.17)のために使用した。
【0258】
これらの実験は、CT077が、表面露出された免疫接近可能なタンパク質であること、およびこのCT077が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0259】
(実施例24)
CT181(配列番号245および配列番号246)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図24A:レーン4、クロマトグラフィー画分1、推定分子量 50.1kDa)、およびHisタグ化融合タンパク質(図24B:レーン2、3および4、クロマトグラフィー画分1、2および3,推定分子量 32.0kDa)の両方として精製した。このGSTタグ化組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図24D:レーン4および5(矢印で示される))、およびFACS分析(図24C、K−S値 7.62)のために使用した。
【0260】
これらの実験は、CT181が、表面露出された免疫接近可能なタンパク質であること、およびこのCT181が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0261】
(実施例25)
CT589(配列番号185および配列番号186)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図25A:レーン4および5、クロマトグラフィー画分1および2、推定分子量 89.4kDa)、およびHisタグ化融合タンパク質(図25B:レーン2および3、クロマトグラフィー画分1および2、推定分子量 63.4kDa)の両方として精製した。このHisタグ化組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、FACS分析(図25C)のために使用した。
【0262】
これらの実験は、CT589が、EB異種集団の一部のみにしか存在しないことを示す(クラミジア調製物が通常そうであるように)このCT589が存在する場合、このCT589は、表面露出された免疫接近可能なタンパク質である。これらの特性は、配列のみからは示されない。
【0263】
(実施例26)
CT597(配列番号179および配列番号180)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図26A:レーン5および6、クロマトグラフィー画分1および2、推定分子量 36.0kDa)、およびHisタグ化融合タンパク質(図26B:レーン2、3および4、クロマトグラフィー画分1、2および3、推定分子量 10.3kDa)の両方として精製した。
【0264】
(実施例27)
CT623(配列番号163および配列番号164)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図27A:レーン3および4、クロマトグラフィー画分1および2、推定分子量 71.8kDa)、およびHisタグ化融合タンパク質(図27B:レーン2、3および4、クロマトグラフィー画分1、2および3、推定分子量 45.8kDa)の両方として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図27E:GSTタグ化、レーン4(矢印で示される);Hisタグ化、レーン13(矢印で示される))、およびFACS分析(図27C:GSTタグ化:K−S値 15.89;図27D:Hisタグ化:K−S値 20.27)のために使用した。
【0265】
これらの実験は、CT623が、表面露出された免疫接近可能なタンパク質であること、およびこのCT623が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0266】
(実施例28)
CT700(配列番号261および配列番号262)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図28A:レーン5、6および7、クロマトグラフィー画分1、2および3、推定分子量 73.7kDa)として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、FACS分析(図28B:K−S値 8.72)のために使用した。
【0267】
これらの実験は、CT700が、表面露出された免疫接近可能なタンパク質であること、およびこのCT700が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0268】
(実施例29)
CT761(配列番号217および配列番号218)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図29A:レーン6および7、クロマトグラフィー画分1および2、推定分子量 63.9kDa)として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、FACS分析(図29B:K−S値 11.45)のために使用した。
【0269】
これらの実験は、CT761が、表面露出された免疫接近可能なタンパク質であること、およびこのCT761が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0270】
(実施例30)
CT415(配列番号117および配列番号118)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図30:レーン3および4、クロマトグラフィー画分1および2、推定分子量 55.4kDa)として精製した。
【0271】
(実施例31)
CT454(配列番号253および配列番号254)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図31:レーン2および3、クロマトグラフィー画分1および2、推定分子量 56.2kDa)として精製した。
【0272】
(実施例32)
CT467(配列番号129および配列番号130)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図32:レーン3および4、クロマトグラフィー画分1および2、推定分子量 65.6kDa)として精製した。
【0273】
(実施例33)
CT551(配列番号257および配列番号258)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図33A:レーン5、6および7、クロマトグラフィー画分1、2および3、推定分子量 34.1kDa)、およびGSTタグ化融合タンパク質(図33B:レーン4および5、クロマトグラフィー画分1および2、推定分子量 60.1kDa)の両方として精製した。
【0274】
(実施例34)
CT567(配列番号195および配列番号196)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図34A:レーン8および9、クロマトグラフィー画分1および2、推定分子量 44.0kDa)、およびHisタグ化融合タンパク質(図34B:レーン7および8、クロマトグラフィー画分1および2、推定分子量 18.3kDa)の両方として精製した。
【0275】
(実施例35)
CT569(配列番号193および配列番号194)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図35A:レーン2、3および4、クロマトグラフィー画分1、2および3、推定分子量 11.2kDa)として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図35B:レーン8および9、矢印で示される)で使用した。
【0276】
(実施例36)
CT647(配列番号169および配列番号170)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質(図36:レーン6および7、クロマトグラフィー画分1および2、推定分子量 45.7kDa)として精製した。
【0277】
(実施例37)
CT600(配列番号173および配列番号174)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質(図37A:レーン5、6および7、クロマトグラフィー画分1、2および3、推定分子量 19.5kDa)として精製した。この組換え産物を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図37B:レーン10および11、矢印で示される)、およびFACS分析(図37C、K−S値 10.46)のために使用した。
【0278】
これらの実験は、CT600が、表面露出された免疫接近可能なタンパク質であること、およびこのCT600が有用な免疫原であることを示す。これらの特性は、配列のみからは示されない。
【0279】
(実施例38)
CT279(配列番号247および配列番号248)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質、Hisタグ化融合タンパク質として精製した。これらの組換えHisタグ化タンパク質および組換えGSTタグ化タンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図38A:Hisタグ化:レーン5(矢印で示される);図38B:GSTタグ化:レーン12および13(矢印で示される))において使用した。
【0280】
(実施例39)
CT560(配列番号259および配列番号260)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した。この組換えHisタグ化タンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図39:レーン6および7(矢印で示される))において使用した。
【0281】
(実施例40)
CT389(配列番号249および配列番号250)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図40:レーン16および17(矢印で示される))において使用した。
【0282】
(実施例41)
CT456(配列番号255および配列番号256)を、E.coliにおいて発現させた。この組換え産物を、GSTタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図41:レーン2および3(矢印で示される))において使用した。
【0283】
(実施例42)
CT622(配列番号161および配列番号162)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図42:レーン9(矢印で示される))において使用した。
【0284】
(実施例43)
CT759(配列番号213および配列番号214)を、E.coliにおいて発現させた。この組換え産物を、Hisタグ化融合タンパク質として精製した。この組換えタンパク質を使用して、マウスを免疫し、このマウスの血清を、ウエスタンブロット(図43:レーン8および9(矢印で示される))において使用した。
【0285】
(実施例44)
インビトロ中和アッセイは、培養物中の真核細胞に対するC.trachomatis 感染性を阻害するために、本発明の種々の組換えタンパク質で免疫されたマウスから得られた血清の能力を示し、このアッセイを、LLCMK2(アカゲザル腎臓内皮)細胞を使用して実施した。マウスポリクローナル血清の連続4倍希釈を、SP(スクロース−リン酸)緩衝液中に調製した。全EBに対するマウス抗血清を、ポジティブコントロールとして使用し、前免疫血清およびSP緩衝液のみを、ネガティブコントロールとして使用した。C.trachomatis(血液型亜型D)から精製されたEBを、SP緩衝液中に希釈し、3×105IFU/mlを含むようにし、この懸濁液の10μlを、最終容量100μlで各血清希釈液に添加した。抗体EB相互作用が、37℃で30分間進行するようにした。次いで、各サンプルからの100μlの反応混合液を、96ウェルマイクロタイタープレート中のPBS洗浄LLCMK2細胞単層の上に添加し、805×gで37℃で1時間遠心分離した。全ての血清およびコントロールを、2連のサンプルで試験した。過剰の接種物の除去後、細胞を、PBSで一度リンスし、20%FCSおよび1μgシクロヘキシミドを補充した200μlのDMEM培地を補充し、そして37℃で48時間インキュベートした。細胞をメタノールで固定し、進行性細胞内クラミジア感染によって生成された代表的な細胞質封入物を、抗クラミジアフルオレセイン結合モノクローナル抗体(Meridian Diagnostics)で染色した。宿主細胞に対する十分な希釈およびEB比において、観察された封入物の数を、宿主細胞感染を最初に首尾よく確立し得た生存クラミジアの数に等しいと考えられる(これらは、Inclusion Forming Units,IFUと名づけられる)。フルオレセイン標識封入物を、40×の倍率で1ウェル当たり4つの顕微鏡視野において計数した。抗体相互作用に起因する感染性の阻害を、SPコントロール(緩衝液のみ)に比較した場合、平均IFUのパーセント減少として計算した。共通の実施に従って、血清を、これらが50%以上の感染性の減少を引き起こし得る場合、「中和」として標識し、全スクリーニングアッセイ(例えば、宿主細胞の交換、またはケラミジア単離物、または感染性接種物の調製における環境条件のバリエーション)の複雑性を考慮して、より低い程度のEB感染性を阻害し得る血清はまた、さらなる研究のためのワクチン候補として考慮されるべきである。図44Aは、中和ポジティブ血清から得られた結果の例を示し、一方、図44Bは、中和ネガティブ血清から得られた結果の例を示す。
【0286】
インビトロ中和アッセイを、実施例1〜10、13〜22、24〜26および29〜37において言及される組換えタンパク質で免疫されたマウスから得られた血清を使用して実施した。結果を、表IIに示す。これらの結果は、CT045、CT242、CT381.CT396、CT398、CT467、CT547、CT587およびCT681が、C.trachomatisによる感染を予防するワクチンに対する、特に良好な候補の全てであることを示す。これらの特性は、配列単独からは示されない。
【0287】
さらなる実験において、C.trachomatisに対して惹起された血清を、交差中和活性について、C.pneumoniae EBに対して試験した。手順は、上記されるとおりであったが、C.pneumoniaeから精製されたEBを、SP緩衝液中に希釈し、3×106IFU/mlを含むようにし、この懸濁物の10μlを、最終容量100μlで各血清希釈物に添加した。CT242およびCT467を使用して得られた血清は、C.pneumoniae EBを交差中和し得た。
【0288】
本発明が例示のためのみに記載され、その改変がなされ得、一方で、本発明の意図および範囲内のままであることは、理解される。
【0289】
【数1】
【0290】
【表1】
【0291】
【表2】
【0292】
【表3】
【0293】
【表4】
【0294】
【表5】
【特許請求の範囲】
【請求項1】
Chlamydia trachomatisに起因する感染の処置または予防のための医薬の製造におけるタンパク質の使用であって、該タンパク質が、(a)配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号17、配列番号19、配列番号21、配列番号23、配列番号25、配列番号27、配列番号29、配列番号31、配列番号33、配列番号35、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号53、配列番号55、配列番号57、配列番号59、配列番号61、配列番号63、配列番号65、配列番号67、配列番号69、配列番号71、配列番号73、配列番号75、配列番号77、配列番号79、配列番号81、配列番号83、配列番号85、配列番号87、配列番号89、配列番号91、配列番号93、配列番号95、配列番号97、配列番号99、配列番号101、配列番号103、配列番号105、配列番号107、配列番号109、配列番号111、配列番号113、配列番号115、配列番号117、配列番号119、配列番号121、配列番号123、配列番号125、配列番号127、配列番号129、配列番号131、配列番号133、配列番号135、配列番号137、配列番号139、配列番号141、配列番号143、配列番号145、配列番号147、配列番号149、配列番号151、配列番号153、配列番号155、配列番号157、配列番号159、配列番号161、配列番号163、配列番号165、配列番号167、配列番号169、配列番号171、配列番号173、配列番号175、配列番号177、配列番号179、配列番号181、配列番号183、配列番号185、配列番号187、配列番号189、配列番号191、配列番号193、配列番号
195、配列番号197、配列番号199、配列番号201、配列番号203、配列番号205、配列番号207、配列番号209、配列番号211、配列番号213、配列番号215、配列番号217、配列番号219、配列番号221、配列番号223、配列番号225、配列番号227、配列番号229、配列番号231、配列番号233、配列番号235、配列番号237、配列番号239、配列番号241、配列番号243、配列番号245、配列番号247、配列番号249、配列番号251、配列番号253、配列番号255、配列番号257、配列番号259および配列番号261からなる群より選択されるアミノ酸配列を含むタンパク質、(b)(a)のアミノ酸配列に対して50%以上の配列同一性を有するアミノ酸配列を含むタンパク質、または(c)(a)のアミノ酸配列のフラグメントを含むタンパク質である、使用。
【請求項2】
Chlamydia trachomatisに起因する感染の処置または予防のための医薬の製造における核酸の使用であって、該核酸が、(a)配列番号2、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号18、配列番号20、配列番号22、配列番号24、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号56、配列番号58、配列番号60、配列番号62、配列番号64、配列番号66、配列番号68、配列番号70、配列番号72、配列番号74、配列番号76、配列番号78、配列番号80、配列番号82、配列番号84、配列番号86、配列番号88、配列番号90、配列番号92、配列番号94、配列番号96、配列番号98、配列番号100、配列番号102、配列番号104、配列番号106、配列番号108、配列番号110、配列番号112、配列番号114、配列番号116、配列番号118、配列番号120、配列番号122、配列番号124、配列番号126、配列番号128、配列番号130、配列番号132、配列番号134、配列番号136、配列番号138、配列番号140、配列番号142、配列番号144、配列番号146、配列番号148、配列番号150、配列番号152、配列番号154、配列番号156、配列番号158、配列番号160、配列番号162、配列番号164、配列番号166、配列番号168、配列番号170、配列番号172、配列番号174、配列番号176、配列番号178、配列番号180、配列番号182、配列番号184、配列番号186、配列番号188、配列番号190、配列番号192、配列番号194、配列番号196、配列番号198、配列番号200、配列番号202、配列番号204、配列番号206、配列番号208、配列番号210、配列番号212、配列番号214、配列番号216、配列番号218、配列番号220、配列番号222、配列番号224、配列番号226、配列番号228、配列番号230、配列番号232、配列番号234、配列番号236、配列番号238、配列番号240、配列番号242、配列番号244、配列番号246、配列番号248、配列番号250、配列番号252、配列番号254、配列番号256、配列番号258、配列番号260および配列番号262からなる群より選択されるヌクレオチド配列を含む核酸である、使用。
【請求項3】
および242、(b)(a)のヌクレオチド配列に対して50%以上の配列同一性を有するヌクレオチド配列を含む核酸、または(c)(a)ヌクレオチド配列のフラグメントを含む核酸。
【請求項4】
請求項1または2に記載の使用であって、感染が、Chlamydia基本小体に対して特異的な免疫応答を誘発する医薬によって処置または予防される、使用。
【請求項5】
Chlamydia trachomatis基本小体を中和するための医薬の製造における、請求項1に記載のタンパク質または請求項2に記載の核酸の使用。
【請求項6】
免疫原性組成物であって、タンパク質およびアジュバントを含み、該タンパク質が、(a)配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号17、配列番号19、配列番号21、配列番号23、配列番号25、配列番号27、配列番号29、配列番号31、配列番号33、配列番号35、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号53、配列番号55、配列番号57、配列番号59、配列番号61、配列番号63、配列番号65、配列番号67、配列番号69、配列番号71、配列番号73、配列番号75、配列番号77、配列番号79、配列番号81、配列番号83、配列番号85、配列番号87、配列番号89、配列番号91、配列番号93、配列番号95、配列番号97、配列番号99、配列番号101、配列番号103、配列番号105、配列番号107、配列番号109、配列番号111、配列番号113、配列番号115、配列番号117、配列番号119、配列番号121、配列番号123、配列番号125、配列番号127、配列番号129、配列番号131、配列番号133、配列番号135、配列番号137、配列番号139、配列番号141、配列番号143、配列番号145、配列番号147、配列番号149、配列番号151、配列番号153、配列番号155、配列番号157、配列番号159、配列番号161、配列番号163、配列番号165、配列番号167、配列番号169、配列番号171、配列番号173、配列番号175、配列番号177、配列番号179、配列番号181、配列番号
183、配列番号185、配列番号187、配列番号189、配列番号191、配列番号193、配列番号195、配列番号197、配列番号199、配列番号201、配列番号203、配列番号205、配列番号207、配列番号209、配列番号211、配列番号213、配列番号215、配列番号217、配列番号219、配列番号221、配列番号223、配列番号225、配列番号227、配列番号229、配列番号231、配列番号233、配列番号235、配列番号237、配列番号239、配列番号241、配列番号243、配列番号245、配列番号247、配列番号249、配列番号251、配列番号253、配列番号255、配列番号257、配列番号259および配列番号261からなる群より選択されるアミノ酸配列を含むタンパク質、(b)(a)のアミノ酸配列に対して50%以上の配列同一性を有するアミノ酸配列を含むタンパク質、または(c)(a)のアミノ酸配列のフラグメントを含むタンパク質である、免疫原性組成物。
【請求項7】
免疫原性組成物であって、核酸およびアジュバントを含み、該核酸が、(a)配列番号2、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号18、配列番号20、配列番号22、配列番号24、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号56、配列番号58、配列番号60、配列番号62、配列番号64、配列番号66、配列番号68、配列番号70、配列番号72、配列番号74、配列番号76、配列番号78、配列番号80、配列番号82、配列番号84、配列番号86、配列番号88、配列番号90、配列番号92、配列番号94、配列番号96、配列番号98、配列番号100、配列番号102、配列番号104、配列番号106、配列番号108、配列番号110、配列番号112、配列番号114、配列番号116、配列番号118、配列番号120、配列番号122、配列番号124、配列番号126、配列番号128、配列番号130、配列番号132、配列番号134、配列番号136、配列番号138、配列番号140、配列番号142、配列番号144、配列番号146、配列番号148、配列番号150、配列番号152、配列番号154、配列番号156、配列番号158、配列番号160、配列番号162、配列番号164、配列番号166、配列番号168、配列番号170、配列番号172、配列番号174、配列番号176、配列番号178、配列番号180、配列番号182、配列番号184、配列番号186、配列番号188、配列番号190、配列番号192、配列番号194、配列番号196、配列番号198、配列番号200、配列番号202、配列番号204、配列番号206、配列番号208、配列番号210、配列番号212、配列番号214、配列番号216、配列番号218、配列番号220、配列番号222、配列番号224、配列番号226、配列番号228、配列番号230、配列番号232、配列番号234、配列番号236、配列番号238、配列番号240、配列番号242、配列番号244、配列番号246、配列番号248、配列番号250、配列番号252、配列番号254、配列番号256、配列番号258、配列番号260および配列番号262からなる群より選択されるヌクレオチド配列を含む核酸、(b)(a)のヌクレオチド配列に対して50%以上の配列同一性を有するヌクレオチド配列を含む核酸、または(c)(a)ヌクレオチド配列のフラグメントを含む核酸である、免疫原性組成物。
【請求項8】
医薬品としての使用のための、請求項5または請求項6に記載の組成物。
【請求項9】
患者におけるC.trachomatis感染性を中和する方法であって、請求項5または請求項6に記載の組成物、あるいは請求項1に記載のタンパク質を認識する抗体を、前記患者に投与する工程を包含する、方法。
【請求項10】
Chlamydia trachomatisに対して患者を免疫する方法であって、請求項5または請求項6に記載の組成物を該患者に投与する工程を包含する、方法。
【請求項11】
Chlamydia trachomatis基本小体に特異的な抗体を惹起する方法であって、請求項5または請求項6に記載の組成物を該患者に投与する工程を包含する、方法。
【請求項12】
請求項1に記載のタンパク質を認識する抗体を惹起する方法であって、Chlamydia trachomatis基本小体を、該患者に投与する工程を包含する、方法。
【請求項13】
生物学的サンプルにおけるChlamydia trachomatis基本小体を検出するための方法であって、該サンプルを、請求項1に記載のタンパク質を認識する抗体と接触させる工程を包含する、方法。
【請求項1】
Chlamydia trachomatisに起因する感染の処置または予防のための医薬の製造におけるタンパク質の使用であって、該タンパク質が、(a)配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号17、配列番号19、配列番号21、配列番号23、配列番号25、配列番号27、配列番号29、配列番号31、配列番号33、配列番号35、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号53、配列番号55、配列番号57、配列番号59、配列番号61、配列番号63、配列番号65、配列番号67、配列番号69、配列番号71、配列番号73、配列番号75、配列番号77、配列番号79、配列番号81、配列番号83、配列番号85、配列番号87、配列番号89、配列番号91、配列番号93、配列番号95、配列番号97、配列番号99、配列番号101、配列番号103、配列番号105、配列番号107、配列番号109、配列番号111、配列番号113、配列番号115、配列番号117、配列番号119、配列番号121、配列番号123、配列番号125、配列番号127、配列番号129、配列番号131、配列番号133、配列番号135、配列番号137、配列番号139、配列番号141、配列番号143、配列番号145、配列番号147、配列番号149、配列番号151、配列番号153、配列番号155、配列番号157、配列番号159、配列番号161、配列番号163、配列番号165、配列番号167、配列番号169、配列番号171、配列番号173、配列番号175、配列番号177、配列番号179、配列番号181、配列番号183、配列番号185、配列番号187、配列番号189、配列番号191、配列番号193、配列番号
195、配列番号197、配列番号199、配列番号201、配列番号203、配列番号205、配列番号207、配列番号209、配列番号211、配列番号213、配列番号215、配列番号217、配列番号219、配列番号221、配列番号223、配列番号225、配列番号227、配列番号229、配列番号231、配列番号233、配列番号235、配列番号237、配列番号239、配列番号241、配列番号243、配列番号245、配列番号247、配列番号249、配列番号251、配列番号253、配列番号255、配列番号257、配列番号259および配列番号261からなる群より選択されるアミノ酸配列を含むタンパク質、(b)(a)のアミノ酸配列に対して50%以上の配列同一性を有するアミノ酸配列を含むタンパク質、または(c)(a)のアミノ酸配列のフラグメントを含むタンパク質である、使用。
【請求項2】
Chlamydia trachomatisに起因する感染の処置または予防のための医薬の製造における核酸の使用であって、該核酸が、(a)配列番号2、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号18、配列番号20、配列番号22、配列番号24、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号56、配列番号58、配列番号60、配列番号62、配列番号64、配列番号66、配列番号68、配列番号70、配列番号72、配列番号74、配列番号76、配列番号78、配列番号80、配列番号82、配列番号84、配列番号86、配列番号88、配列番号90、配列番号92、配列番号94、配列番号96、配列番号98、配列番号100、配列番号102、配列番号104、配列番号106、配列番号108、配列番号110、配列番号112、配列番号114、配列番号116、配列番号118、配列番号120、配列番号122、配列番号124、配列番号126、配列番号128、配列番号130、配列番号132、配列番号134、配列番号136、配列番号138、配列番号140、配列番号142、配列番号144、配列番号146、配列番号148、配列番号150、配列番号152、配列番号154、配列番号156、配列番号158、配列番号160、配列番号162、配列番号164、配列番号166、配列番号168、配列番号170、配列番号172、配列番号174、配列番号176、配列番号178、配列番号180、配列番号182、配列番号184、配列番号186、配列番号188、配列番号190、配列番号192、配列番号194、配列番号196、配列番号198、配列番号200、配列番号202、配列番号204、配列番号206、配列番号208、配列番号210、配列番号212、配列番号214、配列番号216、配列番号218、配列番号220、配列番号222、配列番号224、配列番号226、配列番号228、配列番号230、配列番号232、配列番号234、配列番号236、配列番号238、配列番号240、配列番号242、配列番号244、配列番号246、配列番号248、配列番号250、配列番号252、配列番号254、配列番号256、配列番号258、配列番号260および配列番号262からなる群より選択されるヌクレオチド配列を含む核酸である、使用。
【請求項3】
および242、(b)(a)のヌクレオチド配列に対して50%以上の配列同一性を有するヌクレオチド配列を含む核酸、または(c)(a)ヌクレオチド配列のフラグメントを含む核酸。
【請求項4】
請求項1または2に記載の使用であって、感染が、Chlamydia基本小体に対して特異的な免疫応答を誘発する医薬によって処置または予防される、使用。
【請求項5】
Chlamydia trachomatis基本小体を中和するための医薬の製造における、請求項1に記載のタンパク質または請求項2に記載の核酸の使用。
【請求項6】
免疫原性組成物であって、タンパク質およびアジュバントを含み、該タンパク質が、(a)配列番号1、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号17、配列番号19、配列番号21、配列番号23、配列番号25、配列番号27、配列番号29、配列番号31、配列番号33、配列番号35、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号53、配列番号55、配列番号57、配列番号59、配列番号61、配列番号63、配列番号65、配列番号67、配列番号69、配列番号71、配列番号73、配列番号75、配列番号77、配列番号79、配列番号81、配列番号83、配列番号85、配列番号87、配列番号89、配列番号91、配列番号93、配列番号95、配列番号97、配列番号99、配列番号101、配列番号103、配列番号105、配列番号107、配列番号109、配列番号111、配列番号113、配列番号115、配列番号117、配列番号119、配列番号121、配列番号123、配列番号125、配列番号127、配列番号129、配列番号131、配列番号133、配列番号135、配列番号137、配列番号139、配列番号141、配列番号143、配列番号145、配列番号147、配列番号149、配列番号151、配列番号153、配列番号155、配列番号157、配列番号159、配列番号161、配列番号163、配列番号165、配列番号167、配列番号169、配列番号171、配列番号173、配列番号175、配列番号177、配列番号179、配列番号181、配列番号
183、配列番号185、配列番号187、配列番号189、配列番号191、配列番号193、配列番号195、配列番号197、配列番号199、配列番号201、配列番号203、配列番号205、配列番号207、配列番号209、配列番号211、配列番号213、配列番号215、配列番号217、配列番号219、配列番号221、配列番号223、配列番号225、配列番号227、配列番号229、配列番号231、配列番号233、配列番号235、配列番号237、配列番号239、配列番号241、配列番号243、配列番号245、配列番号247、配列番号249、配列番号251、配列番号253、配列番号255、配列番号257、配列番号259および配列番号261からなる群より選択されるアミノ酸配列を含むタンパク質、(b)(a)のアミノ酸配列に対して50%以上の配列同一性を有するアミノ酸配列を含むタンパク質、または(c)(a)のアミノ酸配列のフラグメントを含むタンパク質である、免疫原性組成物。
【請求項7】
免疫原性組成物であって、核酸およびアジュバントを含み、該核酸が、(a)配列番号2、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号18、配列番号20、配列番号22、配列番号24、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号56、配列番号58、配列番号60、配列番号62、配列番号64、配列番号66、配列番号68、配列番号70、配列番号72、配列番号74、配列番号76、配列番号78、配列番号80、配列番号82、配列番号84、配列番号86、配列番号88、配列番号90、配列番号92、配列番号94、配列番号96、配列番号98、配列番号100、配列番号102、配列番号104、配列番号106、配列番号108、配列番号110、配列番号112、配列番号114、配列番号116、配列番号118、配列番号120、配列番号122、配列番号124、配列番号126、配列番号128、配列番号130、配列番号132、配列番号134、配列番号136、配列番号138、配列番号140、配列番号142、配列番号144、配列番号146、配列番号148、配列番号150、配列番号152、配列番号154、配列番号156、配列番号158、配列番号160、配列番号162、配列番号164、配列番号166、配列番号168、配列番号170、配列番号172、配列番号174、配列番号176、配列番号178、配列番号180、配列番号182、配列番号184、配列番号186、配列番号188、配列番号190、配列番号192、配列番号194、配列番号196、配列番号198、配列番号200、配列番号202、配列番号204、配列番号206、配列番号208、配列番号210、配列番号212、配列番号214、配列番号216、配列番号218、配列番号220、配列番号222、配列番号224、配列番号226、配列番号228、配列番号230、配列番号232、配列番号234、配列番号236、配列番号238、配列番号240、配列番号242、配列番号244、配列番号246、配列番号248、配列番号250、配列番号252、配列番号254、配列番号256、配列番号258、配列番号260および配列番号262からなる群より選択されるヌクレオチド配列を含む核酸、(b)(a)のヌクレオチド配列に対して50%以上の配列同一性を有するヌクレオチド配列を含む核酸、または(c)(a)ヌクレオチド配列のフラグメントを含む核酸である、免疫原性組成物。
【請求項8】
医薬品としての使用のための、請求項5または請求項6に記載の組成物。
【請求項9】
患者におけるC.trachomatis感染性を中和する方法であって、請求項5または請求項6に記載の組成物、あるいは請求項1に記載のタンパク質を認識する抗体を、前記患者に投与する工程を包含する、方法。
【請求項10】
Chlamydia trachomatisに対して患者を免疫する方法であって、請求項5または請求項6に記載の組成物を該患者に投与する工程を包含する、方法。
【請求項11】
Chlamydia trachomatis基本小体に特異的な抗体を惹起する方法であって、請求項5または請求項6に記載の組成物を該患者に投与する工程を包含する、方法。
【請求項12】
請求項1に記載のタンパク質を認識する抗体を惹起する方法であって、Chlamydia trachomatis基本小体を、該患者に投与する工程を包含する、方法。
【請求項13】
生物学的サンプルにおけるChlamydia trachomatis基本小体を検出するための方法であって、該サンプルを、請求項1に記載のタンパク質を認識する抗体と接触させる工程を包含する、方法。
【図1A】

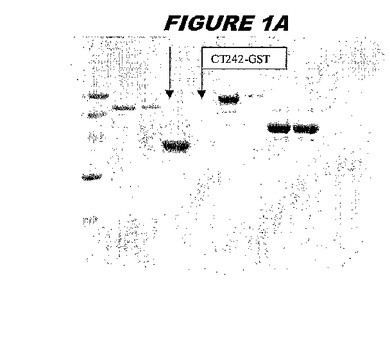
【図1B】
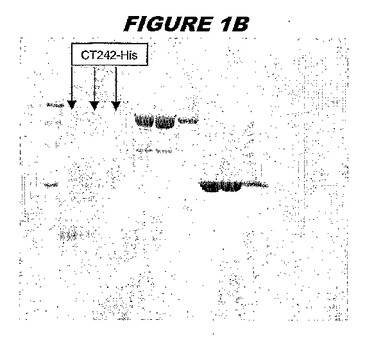
【図1C】
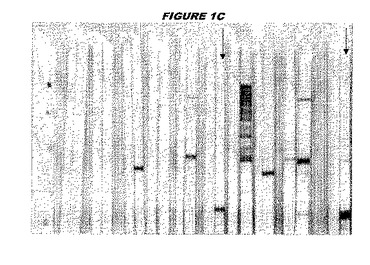
【図2A】
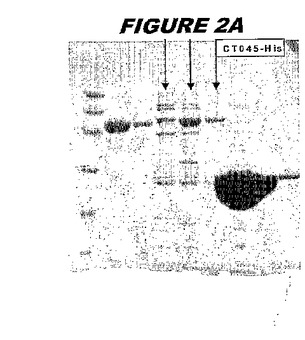
【図2B】
【図2C】
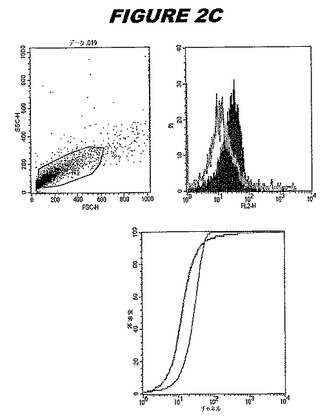
【図3A】
【図3B】
【図3C】
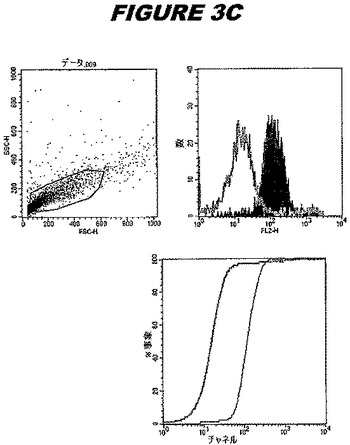
【図3D】
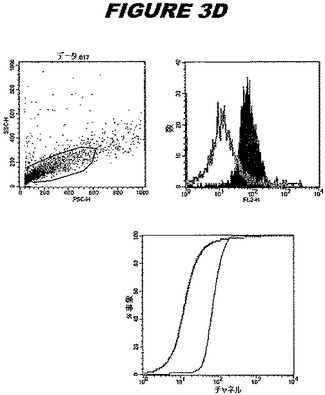
【図4A】
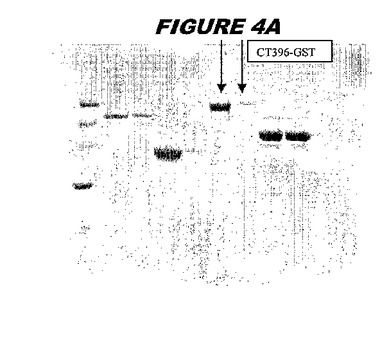
【図4B】
【図4C】
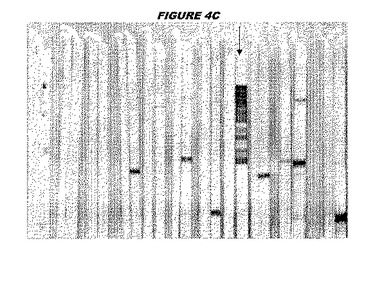
【図4D】
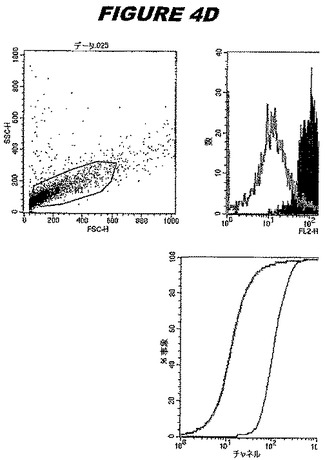
【図4E】
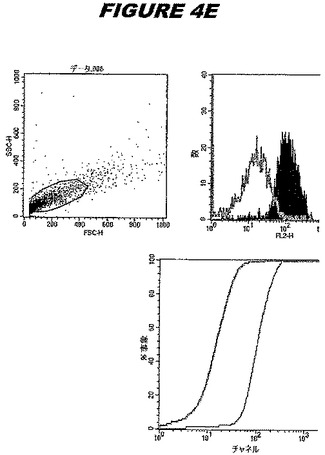
【図5A】

【図5B】
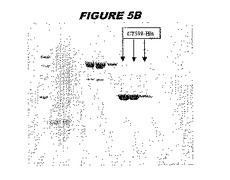
【図5C】
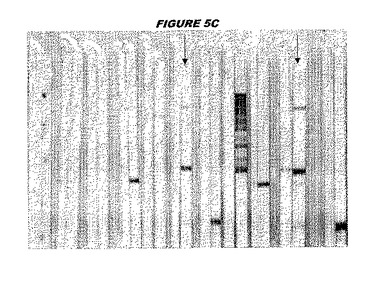
【図5D】
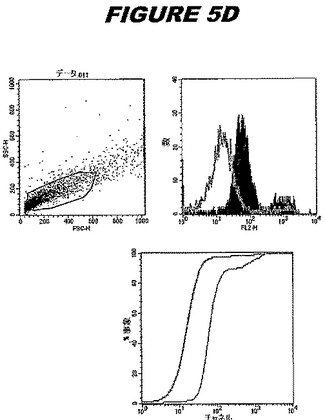
【図5E】
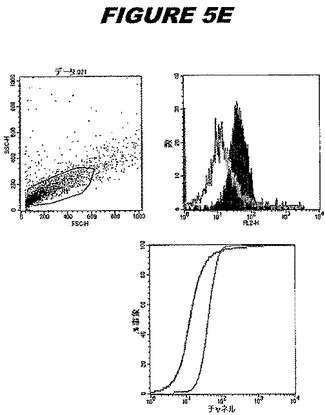
【図6A】
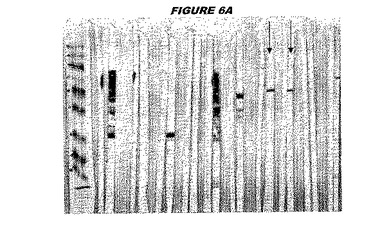
【図6B】
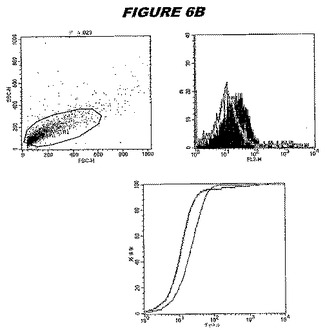
【図6C】
【図7A】
【図7B】
【図8A】
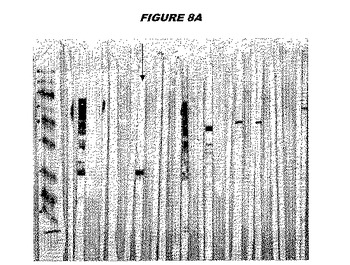
【図8B】
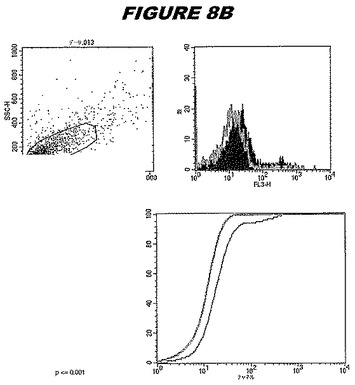
【図8C】
【図9A】
【図9B】
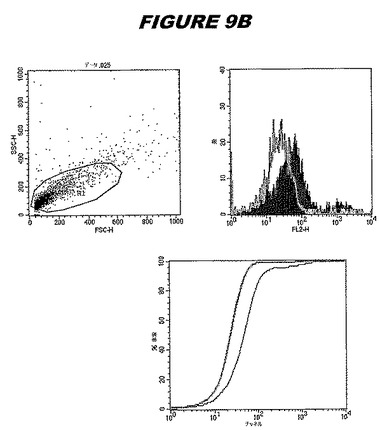
【図9C】
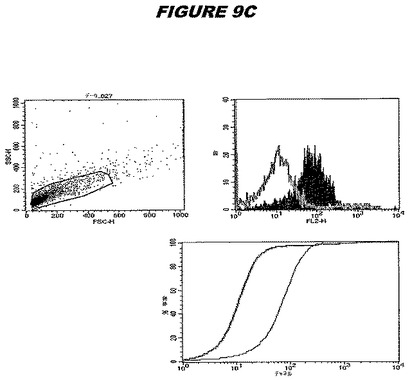
【図9D】
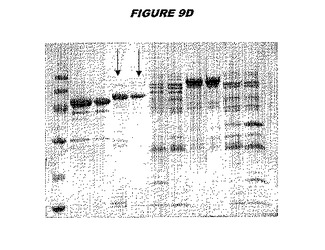
【図10A】
【図10B】
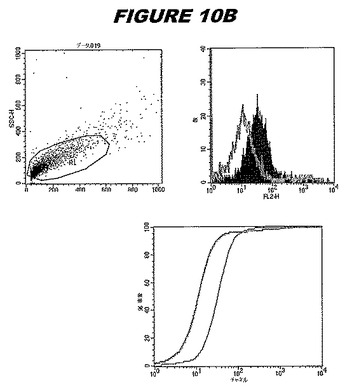
【図10C】
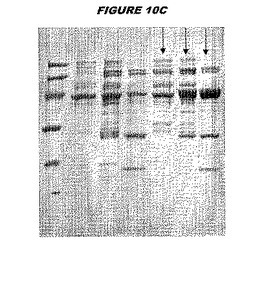
【図11A】
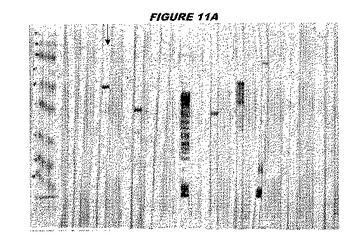
【図11B】
【図11C】
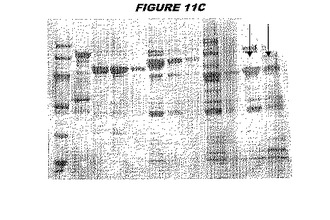
【図12A】
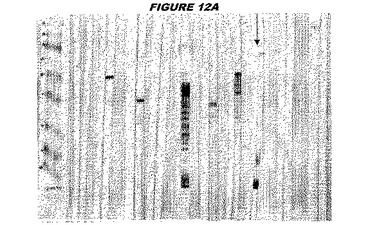
【図12B】
【図12C】
【図12D】
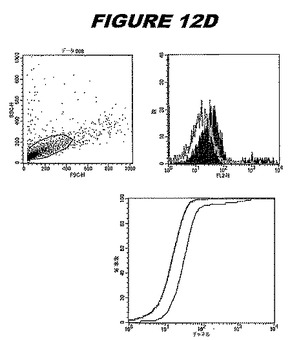
【図12E】
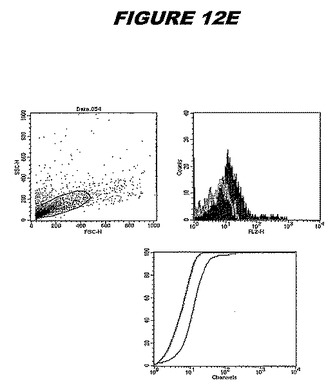
【図13A】
【図13B】
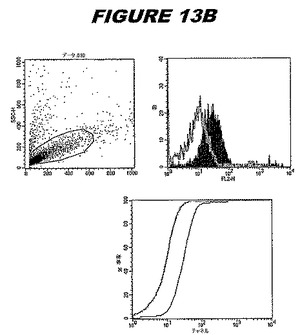
【図13C】
【図14A】
【図14B】
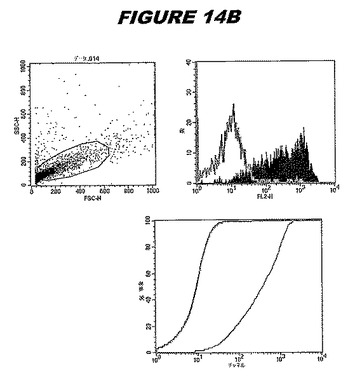
【図14C】
【図15A】
【図15B】
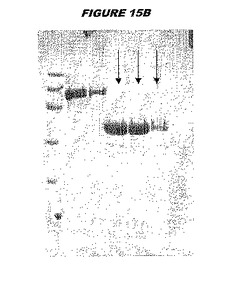
【図15C】
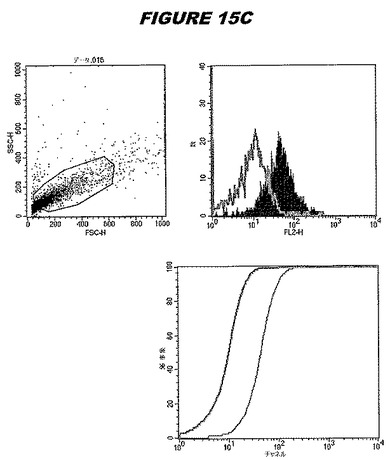
【図16A】
【図16B】
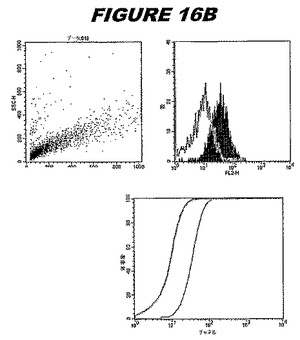
【図16C】
【図17】
【図18A】
【図18B】
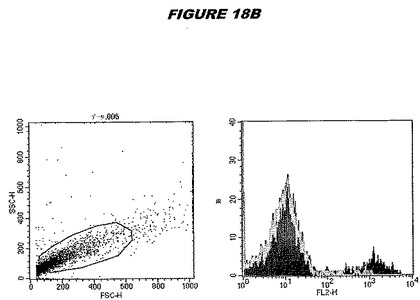
【図19】

【図20A】
【図20B】

【図21】
【図22】

【図23A】

【図23B】

【図23C】

【図24A】

【図24B】
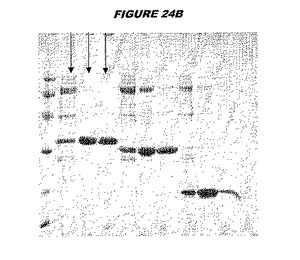
【図24C】
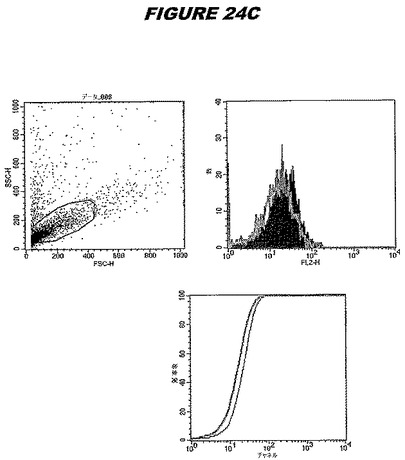
【図24D】

【図25A】
【図25B】
【図25C】
【図26A】

【図26B】
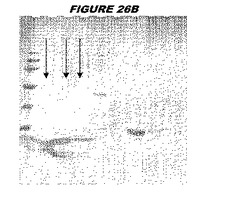
【図27A】
【図27B】

【図27C】
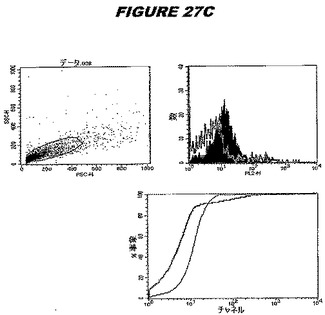
【図27D】
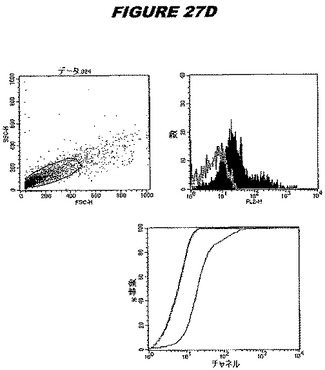
【図27E】

【図28A】
【図28B】
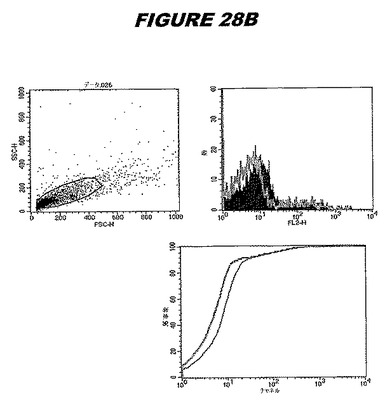
【図29A】
【図29B】
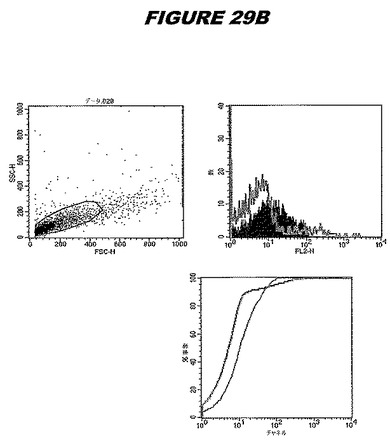
【図30】
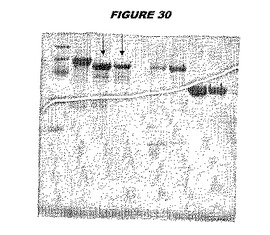
【図31】
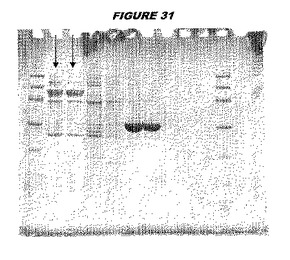
【図32】
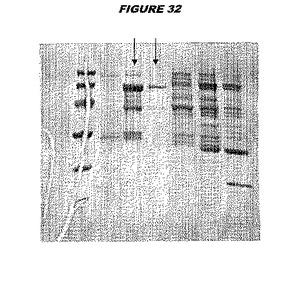
【図33A】
【図33B】
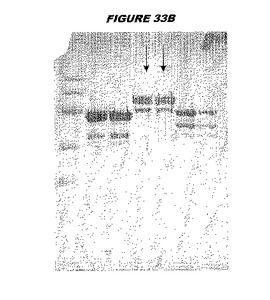
【図34A】
【図34B】

【図35A】
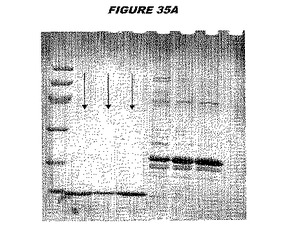
【図35B】

【図36】
【図37A】
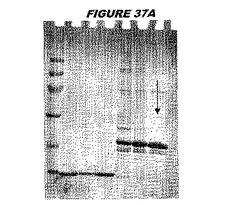
【図37B】
【図37C】
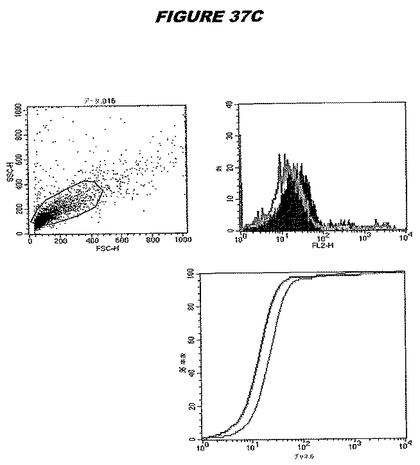
【図38A】
【図38B】
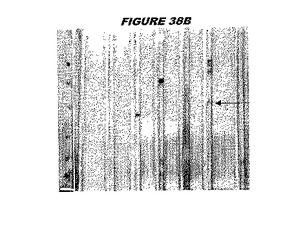
【図39】
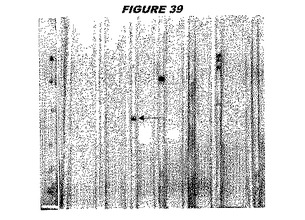
【図40】

【図41】

【図42】
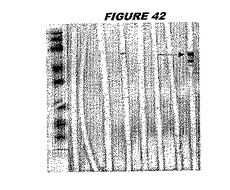
【図43】

【図44A】
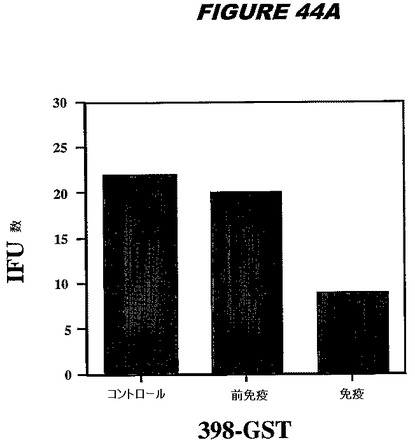
【図44B】
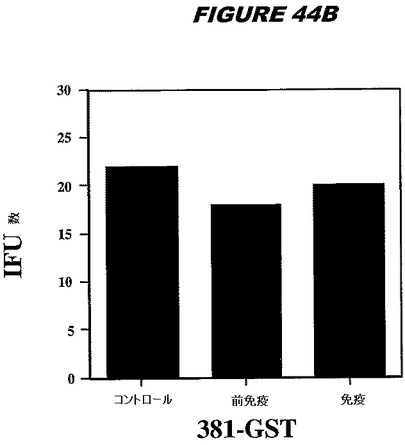
【図1B】
【図1C】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図8A】
【図8B】
【図8C】
【図9A】
【図9B】
【図9C】
【図9D】
【図10A】
【図10B】
【図10C】
【図11A】
【図11B】
【図11C】
【図12A】
【図12B】
【図12C】
【図12D】
【図12E】
【図13A】
【図13B】
【図13C】
【図14A】
【図14B】
【図14C】
【図15A】
【図15B】
【図15C】
【図16A】
【図16B】
【図16C】
【図17】
【図18A】
【図18B】
【図19】
【図20A】
【図20B】
【図21】
【図22】
【図23A】
【図23B】
【図23C】
【図24A】
【図24B】
【図24C】
【図24D】
【図25A】
【図25B】
【図25C】
【図26A】
【図26B】
【図27A】
【図27B】
【図27C】
【図27D】
【図27E】
【図28A】
【図28B】
【図29A】
【図29B】
【図30】
【図31】
【図32】
【図33A】
【図33B】
【図34A】
【図34B】
【図35A】
【図35B】
【図36】
【図37A】
【図37B】
【図37C】
【図38A】
【図38B】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44A】
【図44B】
【公開番号】特開2011−250797(P2011−250797A)
【公開日】平成23年12月15日(2011.12.15)
【国際特許分類】
【出願番号】特願2011−170941(P2011−170941)
【出願日】平成23年8月4日(2011.8.4)
【分割の表示】特願2008−236179(P2008−236179)の分割
【原出願日】平成14年12月12日(2002.12.12)
【出願人】(592243793)ノバルティス ヴァクシンズ アンド ダイアグノスティクス エスアールエル (107)
【Fターム(参考)】
【公開日】平成23年12月15日(2011.12.15)
【国際特許分類】
【出願日】平成23年8月4日(2011.8.4)
【分割の表示】特願2008−236179(P2008−236179)の分割
【原出願日】平成14年12月12日(2002.12.12)
【出願人】(592243793)ノバルティス ヴァクシンズ アンド ダイアグノスティクス エスアールエル (107)
【Fターム(参考)】
[ Back to top ]