
CRIg及びC3b:CRIg複合体の結晶構造
本発明は、マクロファージ特異的レセプターCRIg(以前はSTIgMAと呼ばれていたもの)と、補体C3のC3b及びC3cサブユニットとのその複合体(C3b:CRIg及びC3c:CRIg複合体)の結晶構造の決定に関する。本発明は更にCRIgアゴニスト及びアンタゴニストを含むCRIgに構造的及び/又は機能的に関連する分子をスクリーニングし同定するためのCRIg又はC3b:CRIg複合体の結晶構造の使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、マクロファージ特異的レセプターCRIg(以前はSTIgMAと呼ばれていたもの)と、補体C3のC3b及びC3cサブユニットとのその複合体(C3b:CRIg及びC3c:CRIg複合体)の結晶構造の決定に関する。本発明は更にCRIgアゴニスト及びアンタゴニストを含むCRIgに構造的及び/又は機能的に関連する分子をスクリーニングし同定するためのCRIg又はC3b:CRIg複合体の結晶構造の使用に関する。
【背景技術】
【0002】
補体系は、通常は不活性なプロ酵素形態で存在している一連の血清糖タンパク質から構成される複雑な酵素カスケードである。C3は、補体の古典的、レクチン及び代替経路の3つの主要な経路の収束点を表すので、補体活性化において重要な役割を果たしている。C3は20種を超えるタンパク質と(M. J. Walport, N Engl J Med 344, 1058 (2001)及び(M. J. Walport, N Engl J Med 344, 1140 (2001))タンパク分解酵素C3転換酵素(J. Janssen等, Nature 437, 505 (2005))によるその切断後にのみさらされる多数の別個の結合部位を介して相互作用する。最初のタンパク質切断段階はアナフィラトキシンとも呼ばれる小さい(9kDa)C3aペプチドと大きな(177kDa)C3b断片を生成する。C3からC3a及びC3bへの切断は、分子を粒子表面に共有的に結合させることができるC3b中の埋設チオ-エステル結合を露出させる立体構造変化を誘発する。「オプソニン化」と言われるこの過程は、粒子を標的にしてマクロファージ特異的レセプターに結合させ、ついで系から粒子を除去する。
【0003】
最近、CRIgがC3b/iC3bオプソニン化粒子に対するレセプターとして同定された(K. Y. Helmy等, Cell 124, 915 (2006))。CRIgはC3b、iC3b、並びにC3cに結合するが、不活性な前駆体C3との複合体を形成することはできない。
【0004】
オプソニンとしてのその機能に加えて、C3bは補体のC3及びC5転換酵素のサブユニットである。C3bはセリンプロテアーゼ因子Bに結合する(W. Vogt, G. Schmidt, B. Von Buttlar, L. Dieminger, Immunology 34, 29 (1978);Z. Fishelson, M. K. Pangburn, H. J. Muller-Eberhard, J Biol Chem 258, 741 1 (1983))。この複合体は、D因子の補充とD因子による活性化後に、代替経路(AP)の活性なC3転換酵素であるC3bBb複合体を形成する。C3転換酵素への第二C3b分子の付加は、実験的動物モデルにおいて炎症を媒介し、ヒト自己免疫及び炎症性疾患を媒介する際に重要な役割を果たすことが証明されている(M. J. Walport, K. A. Davies, B. J. Morley, M. Botto, Ann N YAcadSci 815, 267 (1997))補体の代替経路のC5転換酵素である(C3b)2Bbの形成を生じる。代替経路転換酵素のC3bサブユニットは各基質C3又はC5に対してドッキング部位となり、転換酵素の触媒活性に必要とされる(N. Rawal, M. K. Pangburn, J Immunol 164, 1379 (2000))。補体の代替経路は、代替、古典及びマンノース結合レクチン経路の3つの経路の何れかを通して開始される補体活性化を増幅させる。その結果、代替経路の調節因子である補体H因子の欠損が炎症の増幅を招く(N. Rougier等, J Am Soc Nephrol 9, 2318 (1998);M. A. Abrera-Abeleda等 J Med Genet (Nov 18, 2005))。よって、代替経路補体転換酵素によるC3及びC5の切断がオプソニン化、転換酵素の生成及び炎症を生じさせる。
【0005】
転換酵素の中心的成分として、C3bは数多くの補体調節因子の標的となる。これらには補体レセプター1(CR1)、メンブレンコファクタープロテイン(MCP)、崩壊促進因子(DAF)、補体関連レセプターy(Crry)、H因子及びI因子(D. Hourcade, V. M. Holers, J. P. Atkinson, Adv Immunol 45, 381 (1989))がある。I因子の結合はタンパク質分解を介してC3bの不活性化を生じてC3b及びC3f(2kDa)を形成し、チオ-エステルドメイン(TED)を含み標的に結合したままであるC3dg(40kDa)と、C3c(135kDa)を最終的に生成する。C3及びC3cの結晶構造に関する最近の報告は、C3が活性化される機序の我々の理解を顕著に向上させた(上掲のB. J. Janssen等, 2005)。C3cはC3と比較して著しい構造変化を示し、複数のタンパク質分解切断段階の結果を反映している。これらの構造研究はC3活性化及び調節の立体構造依存的機序を示唆しているが、活性化種C3bの立体構造に対する回答を提供するものではない。
【発明の概要】
【0006】
本開示は、病原体の排除に必要なマクロファージ補体レセプターCRIgの結晶構造を提供する。また、CRIgとの複合体でのC3b及びC3cの結晶構造が提供される。天然細胞性レセプター及び補体活性化の調節物質との複合体での転換酵素サブユニットのこの第一構造が補体活性化及び調節の機序の我々の理解を改善する。本発明はまたCRIgアゴニスト及びアンタゴニストを含むCRIgに機能的に関連した分子を同定するために使用することができる、CRIgのC3b結合領域、及びそのような結合に関与するC3b内のドメイン及び配列についての情報を提供する。
【0007】
一態様では、本発明は、天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体により形成された結晶に関する。
【0008】
一実施態様では、該結晶は、格子定数(セル定数)a=30.3Å、b=50.8Å、c=62.0Å、及びP212121の空間群をおよそ有している。
【0009】
他の実施態様では、天然配列CRIgポリペプチドは、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される。
【0010】
更に他の実施態様では、CRIgの機能的断片は天然配列CRIgの細胞外ドメイン(ECD)配列である。
【0011】
更なる実施態様では、結晶は約1から40Åまでの分解能、又は少なくとも約5Å、又は少なくとも4Å又は少なくとも3Åの分解能で原子座標を決定するためにX線を回折する。
【0012】
また更なる実施態様では、本発明はアペンディクス1に示された構造座標を有するCRIg結晶に関する。
【0013】
他の態様では、本発明は上述のような結晶を含んでなる組成物に関する。
【0014】
また他の態様では、本発明は、天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体と、補体因子C3bとの複合体の結晶形態に関する。
【0015】
一実施態様では、複合体の結晶形態は、格子定数a=97.6Å、b=255.7Å、c=180.3Å及びC2221の空間群をおよそ有している。
【0016】
他の実施態様では、複合体の結晶形態は、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドの一つを含む。
【0017】
更なる実施態様では、機能的CRIg断片は天然配列CRIgの細胞外ドメイン(ECD)配列である。
【0018】
また更なる実施態様では、複合体の結晶形態は、約1から40Åまでの分解能、又は少なくとも約5Å、又は少なくとも4Å又は少なくとも3Åの分解能で原子座標を決定するためにX線を回折する。
【0019】
更に他の実施態様では、CRIg:C3b複合体の結晶形態はアペンディクス2に示された構造座標を有する。
【0020】
更なる態様では、本発明は、天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体と、補体因子C3cとの複合体の結晶形態に関する。
【0021】
特定の実施態様では、CRIg:C3b複合体の結晶形態は、格子定数a=382.8Å、b=65.0Å、c=147.2Å、β=102.7、及びC2の空間群をおよそ有している。
【0022】
他の実施態様では、CRIg:C3C複合体中に存在する天然配列CRIgポリペプチドは、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される。
【0023】
また他の実施態様では、複合体中に存在するCRIg機能的断片は天然配列CRIgの細胞外ドメイン(ECD)配列である。
【0024】
更なる実施態様では、CRIg:C3c複合体の結晶形態は、約1から40Åまでの分解能、又は少なくとも約5Å、又は少なくとも4Å又は少なくとも3Åの分解能で原子座標を決定するためにX線を回折する。
【0025】
特定の実施態様では、CRIg:C3c複合体はアペンディクス3に示された構造座標を有する。
【0026】
更なる態様では、本発明は、配列番号2のCRIgポリペプチド又はその保存的置換のC3b結合部位の少なくとも一部を含んでなる分子又は分子複合体であって、結合部位が、8、14−15、41−42、44、45−47、50、52、54−61、62、64、85−87、89、95、99、105、107−110、及び111からなる群から選択される少なくとも一つのアミノ酸残基を含み、結合部位がアペンディクス2に記載の構造座標によって表されるアミノ酸の原子を表す点から4.7Å未満の距離を有する点の集合によって定まる分子又は分子複合体に関する。
【0027】
他の態様では、本発明は、点の少なくとも一部が、C3bに対するCRIg結合部位を定める少なくともコアアミノ酸の骨格原子の位置を表すアペンディクス2の構造座標から誘導されている点の三次元立体構造に関する。点の三次元立体構造は、限定するものではないが、C3bに対するCRIg結合部位が空間群対称C2221を有する結晶を形成する、ホログラフィー像、立体ダイアグラム、モデル、又はコンピュータ・ディスプレイ画像として示されうる。
【0028】
また他の態様では、本発明は、機械可読データでコードされたデータストレージ材を含む機械可読データストレージ媒体であって、該データを使用するための指示書を用いてプログラムされた機械が、C3b結合部位の少なくとも一部を含んでなる少なくとも一つの分子又は分子複合体の三次元グラフィック図を表示し、点の集合によって定まった結合部位が、アペンディクス2に記載された構造座標によって表されるアミノ酸の原子を表す点から約4.7Å未満の距離を有している媒体に関する。
【0029】
更なる態様では、本発明は、CRIgのC3b結合部位を模倣する化合物を設計することを含むCRIgアゴニストを同定する方法に関する。
【0030】
本発明は更に(a)計算又は実験手段を用いて化学物質とCRIgポリペプチド又はC3b:CRIg複合体の三次元構造の間のフィッティング操作を実施し;(b)工程(a)で得られたデータを解析して化学物質と天然CRIg又はC3b:CRIg複合体の間の結合の特徴を決定することにより同定された化学物質に関する。
【0031】
特定の実施態様では、上記の化学物質はCRIgとC3bの間の結合をインビボ又はインビトロで妨害する。
【0032】
本発明は更に天然配列CRIg分子又はその保存的アミノ酸置換変異体のC3b結合領域の少なくとも一部を含む分子又は分子複合体に関する。
【図面の簡単な説明】
【0033】
本特許又は出願ファイルはカラーで作製した少なくとも一図面を含んでいる。カラー図面の本特許又は特許出願のコピーは請求し必要な料金を支払えば特許庁から提供されるものである。
【図1】C3及びC3b:CRIg及びC3c:CRIg複合体のドメイン構造及び全体構造。(A)C3のドメイン組織、β鎖を緑色、α鎖を青緑色(ANAドメイン)、オレンジ色(CUB)、青色(TED)、及び紫色で示す。赤色の球は、チオ-エステルの位置を示している。(B)天然C3(左側)、CRIgとの複合体としてのC3b(中央)、及びCRIgとの複合体としてのC3cのリボン図。図1Aのカラースキームを使用し、CRIgの表面を黄色で示す。C3とC3bを比較する場合CUB及びTEDドメインの大きな動きに留意のこと。活性化後、TEDはC3bの残りと比較して移動し回転し、Cys988は80Åを越えて移動する。(C)図1Bの概略図;病原体に対するC3bの潜在的な共有結合がC3B:CRIg複合体において示されている。
【0034】
【図2】CRIgはC3b及びC3cのβ鎖キーリングの中心に結合する。(A)C3c及びCRIgの複合体。C3cの表面を白色にしたα鎖の残基と共に示し、β鎖のMG1、MG2、MG3、MG4、MG6、及びLNKドメインはそれぞれ白色、オレンジ色、赤色、淡緑色、青色及び暗緑色にした。CRIgはリボン図として示し、全てのβ鎖及びN末端及びC末端を標識した。(B)図2AからY軸の回りに180度回転させたCRIgの表面図。C3c/C3bに対して4.5A未満の距離である全原子は、図2Aにおいてそれらが接触している残基と同様に着色した。(C)CRIg野生型及びカラーのコードされたCRIg変異体の相対的親和性。親和性は、プレート上の被覆C3bへの結合についてCRIg-LFHとの様々なCRIgタンパク質の競合によって決定した。CRIg-LFH結合は抗FLAG抗体を使用して検出した。(D)図2Cに示した結合実験に使用したCRIg変異を示すCRIgの表面。変異された残基は、図2Cにおいてそれらが表す結合曲線に従って着色した。
【0035】
【図3】CRIgは補体の代替(AP)経路を阻害するが、古典(CP)経路を阻害しない。(A)CRIgはAPを介したC5b-9沈着を阻害するが、CP又はMBL経路のものは阻害しない。補体経路はLPS(AP)又はIgM(CP)で被覆されたプレート上で再構成され、C1q欠損(AP)又はfB欠損(CP)血清と共にインキュベートした。補体活性化は、C5b-9への結合HRPOコンジュゲート抗体の検出について色素源によって決定した。APアッセイにおけるCRIgのIC50値は1.49±0.19μM(n=3)である。(B)CRIgは液相C3転換酵素活性を阻害する。C3、fB及びfDを、増大する濃度のhuCRIg(L)-ECDの存在下でベロナールバッファー中でインキュベートした。反応をEDTAで停止させ、切断産物をクーマシー染色ゲル(左パネル)又はELISA(右パネル)で可視化した。CRIg-ECDに対するIC50値は0.39±0.14μM(n=3)である。(C)CRIgはザイモサン粒子の表面上に集められたC5転換酵素を阻害する。ザイモサン粒子はfB及びfDの存在下でC3bでオプソニン化され、C5転換酵素を生成する。固定濃度のC5を、増加する濃度のCRIg wt又はCRIg変異体C(CRIg(C))の存在下で転換酵素と混合した。転換酵素活性は、溶血アッセイを使用して液相中に生産されたC5b6を測定することによって決定した。IC50値:CRIg(Wt)1.63±0.62μM(n=3)、CRIg(E)6.52±3.21μM(n=3)。
【0036】
【図4】全身的に投与されたCRIg-Fcは抗体誘発関節炎に伴う局所的及び全身的補体活性化を阻害する。(A)コラーゲンでの二次免疫から7日後の、既に確認された疾患のmuCRIg-Fcで治療されたマウスにおける炎症の減少(組織学的スコア)及び骨変形の減少(関節皮質骨体積JCBVの増加)。64日目に中足骨関節を常套的なH&E組織検査、又はマイクロコンピュータ断層撮影(マイクロCT)のために準備した。(B)抗コラーゲンII型抗体での受動免疫後3日(左側パネル)又は5日後(右側パネル)に開始する12mg/kgのmuCRIg-Fcの3回の毎週の投与を受けたマウスにおける臨床スコアの減少。(C)CRIg-Fcでの処置は、関節の軟骨表面上でのIgG2aではなくC3断片の蓄積を阻害する。組み合わさった画像では、赤色がIgG2a抗コラーゲン抗体の存在を示し、緑色が軟骨表面上の補体C3産物の存在を示している。(D)既に確認された疾患のCRIg-Fcで治療されたマウスの関節中におけるC3へのCRIg-Fcの結合(緑色)。尺度バー=20um。(E)上側パネル:受動免疫前の日に開始したCRIg-Fc処置マウスにおける関節中でのC3a生産の阻害。下側パネル:CRIg-Fcで処置されたマウスの血清中におけるC5aレベルの有意な減少。統計的解析(スチューデントt検定):* p<0.01、** p<0.001、** p<0.0001。
【0037】
【図5】CRIgは、転換酵素の非触媒サブユニットへのC5の結合を阻害することによりC5転換酵素を阻害する。(A)CRIgはC5のC3b2への結合をブロックするがC3bC4bへの結合はブロックしない。C3b、C3b2及びC3bC4bをポリクローナル抗C3抗体で捕捉し、C5と増加濃度のCRIg(wt)又は変異体CRIg(C)との混合物と共にインキュベートした。転換酵素サブユニットに結合したC5を、ポリクローナル抗C5抗体及びHRPOコンジュゲート二次抗体で検出した。C5結合のCRIg阻害のIC50は0.88±0.22μMであった。(B)CRIgは精製C5転換酵素のVmaxを減少させる。固定濃度のCRIgを、増加濃度のC5の存在下でザイモサン結合C5転換酵素と混合した。ミカエリスメンテン式に基づいて計算したKm及びVmax値は、CRIg(wt)の不在下で0.99及び4.01であり、0.8uMのRIg(wt)の存在下で1.70及び2.16であった。
【0038】
【図6】C3及びC3b間のドメイン配置の差。示されているのはCUB、TED及びMG8-ドメインのリボン図である。(A)C3及びC3b(共にオレンジ色)のCUBドメインに基づく重ね合わせ。この重ね合わせは、C3b中のCUBドメインがC3のMG8ドメインに関して移動する一方(淡紫色で示す)、TEDドメインがC3活性化時に大きな回転を受けることを示している。(C)TEDドメインに基づく重ね合わせ。C3bは暗色の、C3は明るい青色で示す。C3b構造において完全に開放しているC3中の非常にコンパクトなドメイン配置に留意のこと。Cys988の[位置は赤色の球で示す。
【0039】
【図7】C3b中のTEDとArg80との間の相互作用。該図はプログラムpymol(DeLano, M.C.及びCao, Y., Neuroimaging Clin. N. Am. 12(1):21-34 (2002))で計算した静電ポテンシャルに従って着色したTEDの表面図を示している。赤色及び青色は負及び正に荷電したパッチをそれぞれ示している。R80の側鎖原子は球で表している。Arg80はTEDの表面の高度に負に荷電した溝に近接していることに留意のこと。
【0040】
【図8】異なった種からのCRIgの配列アラインメント。C3bと接触している残基は図2Aに従って着色した。残基番号はシグナル配列後のヒトCRIgを意味する。二次的構造要素は上の配列アラインメントに矢印(ストランド)及びボックス(ヘリックス)として示す。
【0041】
【図9】CRIgはC3b及びC3cと同様の親和性を有している。親和性は固定濃度のCRIg-LFHとの増加濃度のC3b又はC3cの競合によって決定した。CRIg-LFH結合は、HRPOにコンジュゲートした抗FLAG抗体を使用して検出し、TNB反応産物の吸光度を415nmの波長で分光測定により測定した。
【0042】
【図10】CRIg結合部位の近傍におけるC3及びC3b間の局所的差。ここに示したC3及びC3bの重ね合わせはMG6ドメインのCα原子に基づく。C3の一部は白色で示し、α'NTドメインは青色で示す。C3b:CRIg複合体は緑色(C3bのβ鎖)、紫色(C3bのα鎖)で示し、α'NTドメインは青色で示す。CRIgは黄色で示す。C3活性化時に起こる大きな立体構造変化は赤色の矢印で示す。α'NTドメインの大きな動きに留意のこと。MG6に関するMG3の回転と、螺旋セグメント(577:590)の動きはC3b及びC3cへのCRIgの結合に必要とされている。
【0043】
【図11】CRIgはヒト及びマウス血清中における代替経路を選択的に阻害する。(A)CRIgはC1q欠損血清中においてウサギ赤血球の溶血を阻止するが(代替経路、左側パネル)、fB欠損血清中におけるIgMオプソニン化ヒツジ赤血球の溶血は阻止しない(古典経路、右側パネル)。E-IgMとErを、異なった濃度のCRIg-Fc又はCRIg-ECD融合タンパク質を含む5%血清中でインキュベートした。氷冷EDTAを加えることによって溶血を停止させ、415nmにて分光計で上清中のO.Dを決定することによって溶血量を測定した。IC50値:CRIg-Fc 127±49nM(n=6)、CRIg-ECD 640±123nM。(B)CRIgは古典経路によって開示される補体の代替経路を阻害する。古典経路の補体成分をSRBC上に集めてEAC34(AP、左側パネル)又はEAC1234(CP、右側パネル)を生成する。溶血は(A)に記載されるようにして決定した。(C)マウスCRIg-Fcはマウス血清中の代替経路溶血を阻害する。Erを、増加濃度のマウスCRIg-Fcを含む30%マウス血清と共にインキュベートした。溶血は(A)に記載されるようにして決定した。(D)CRIgはマンノース結合レクチン経路を阻害しない。MBL経路活性化によって誘導されるC5b−9沈着を、415nmの波長で分光測定により測定した。
【0044】
【図12】CRIgはコラーゲン誘発関節炎を阻害する。(A)ウシコラーゲンを用いて0日目と23日目に2回マウスを免役することによって関節炎を誘発させた。疾患が既に確認されたときに24日目(予防的)又は45日目(矢印)に開始して毎週3回muCRIg-Fcでマウスを処置した。臨床スコアは足首の厚みを反映させたものである。関節炎の誘発後70日目に、マウスを安楽死させ、関節皮質骨体積(JCBV)を測定し(B)、H&E染色切片について炎症の徴候を評価した(C)。(D)CRIg-Fcで処置したマウスの関節におけるIL-6(左側パネル)及びIL-1b(右側パネル)の減少。関節炎の誘発後、表示した時点でマウスを安楽死させ、関節を解剖し、サイトカイン測定のために加工した。LPS処置(最初の時点)によるサイトカインレベルの増加はmuCRIg-Fc処置には影響を受けない。統計:* p<0.01、** p<0.001、スチューデントt検定。
【0045】
【図13】(A)CRIgはH因子のコファクター活性に影響しないし、内因的なコファクター活性を有してもいない。左側パネル:固定濃度のC3bを4μMのCRIg(wt)又はCRIg(C)の存在下で増加濃度のH因子と混合した。コファクター活性を、iC3bからC3bを区別するためにクーマシー染色SDSゲル上の反応産物を可視化することによってモニターした。右側パネル:固定濃度のC3b及びfIをfH又は増加濃度のCRIgと混合した。(B)CRIgはC3bに結合するB因子を阻害しない。固定濃度のB因子を増加濃度のCRIgと混合した。結合したB因子の量を、fBに対するポリクローナル抗体及び二次HRPOコンジュゲートabを使用して決定した。(C)CRIgはC3転換酵素を崩壊させない。C3転換酵素は、C3b、B因子及びD因子をインキュベートすることによってマキシソーププレート上に形成された。ついで、そのプレートを増加濃度のCRIg又はH因子と共にインキュベートした。残りの結合したB因子は、抗B因子抗体及びHRPOコンジュゲート二次抗体で検出した。示される結果は3回の独立した実験を代表するものである。
【0046】
【図14】CRIg ECDは、C3bへの結合についてCRIg-LFHと競合できるがC5はできない。CRIg-LFHと増加濃度のCRIg-ECD又はC5の混合物を、C3bを被覆したマキシソルブプレートに加えた。残っているCRIg-LFH結合を抗flag抗体を用いて検出した。
【発明を実施するための形態】
【0047】
好ましい実施態様の詳細な説明
定義
特に他の定義をしない限り、ここで使用される技術的及び科学的用語は、この発明が属する分野の当業者が通常理解するところと同じ意味を持つ。例えば、Singleton等, Dictionary of Microbiology and Molecular Biology 2版, J. Wiley & Sons (New York, NY 1994); Sambrook等, Molecular Cloning, A Laboratory Manual, Cold Springs Harbor Press (Cold Springs Harbor, NY 1989 )を参照のこと。本発明のために、次の用語を下記のように定義する。
【0048】
「CRIg」、「PRO362」、「JAM4」及び「STIgMA」は、互いに交換可能に使用され、天然配列及び変異体CRIgポリペプチドを意味する。
【0049】
「天然配列」CRIgは、その調製態様にかかわらず、天然由来のCRIgと同一のアミノ酸配列を有するポリペプチドである。しかして、天然配列CRIgは、天然から単離されうるか、又は組換え及び/又は合成手段によって製造されうる。「天然配列CRIg」なる用語は、CRIgの天然に生じる切断又は分泌型(例えば細胞外ドメイン配列)、天然に生じる変異体型(例えば選択的スプライシング型)及びCRIgの天然に生じる対立遺伝子変異体を特に包含する。天然配列CRIgポリペプチドとその断片は、配列番号1の321アミノ酸長のヒトCRIgポリペプチドで、N末端シグナル配列を含むもの又は含まないもの、1位の開始メチオニンを含むもの又は含まないもの、配列番号1のおよそ277から307のアミノ酸位置の膜貫通ドメインの何れか又は全てを含むもの又は含まないものを特に包含する。天然配列CRIgポリペプチドとその断片は、配列番号2の完全長399アミノ酸長ヒトCRIgポリペプチド(huCRIg、又はhuCRIg-long)で、N末端シグナル配列を含むもの又は含まないもの、1位の開始メチオニンを含むもの又は含まないもの、配列番号2のおよそ277から307のアミノ酸位置の膜貫通ドメインの何れか又は全てを含むもの又は含まないものを特に包含する。また更なる実施態様では、天然配列CRIgポリペプチドとその断片は、ヒトCRIgポリペプチドの305アミノ酸の短い形態(huCRIg-short、配列番号3)で、N末端シグナル配列を含むもの又は含まないもの、1位の開始メチオニンを含むもの又は含まないもの、配列番号3のおよそ183から213のアミノ酸位置の膜貫通ドメインの何れか又は全てを含むもの又は含まないものである。異なった実施態様では、天然配列CRIgポリペプチド又はその断片は、配列番号4の280アミノ酸長の完全長マウスCRIgポリペプチド(muCRIg)で、N末端シグナル配列を含むもの又は含まないもの、1位の開始メチオニンを含むもの又は含まないもの、配列番号4のおよそ181から211のアミノ酸位置の膜貫通ドメインの何れか又は全てを含むもの又は含まないものである。天然配列CRIgポリペプチド又はその断片は、配列番号5のラットCRIgポリペプチドで、N末端シグナル配列を含むもの又は含まないもの、開始メチオニンを含むもの又は含まないもの、及び膜貫通ドメインの何れか又は全てを含むもの又は含まないものでありうる。該用語は更に配列番号6のウシCRIgポリペプチド及び配列番号7のサルCRIgポリペプチドで、それぞれN末端シグナル配列を含むもの又は含まないもの、開始メチオニンを含むもの又は含まないもの、及び各配列中に膜貫通ドメインの何れか又は全てを含むもの又は含まないものを特に含む。
【0050】
「CRIg変異体」とは、天然配列CRIgポリペプチドと少なくとも約80%のアミノ酸配列同一性を有する以下に定義する活性なCRIgポリペプチドを意味する。特定の実施態様では、CRIg変異体は天然配列の成熟完全長CRIgポリペプチドの配列と少なくとも約80%のアミノ酸配列相同性を有している。かかるCRIgポリペプチドには、例えば、天然配列のN又はC末端において、一又は複数のアミノ酸残基が挿入され、置換され、及び/又は欠失されたCRIgポリペプチドが含まれる。他の変異体は天然ポリペプチド配列の膜貫通領域内に一又は複数のアミノ酸が挿入され、置換され、及び/又は欠失されたものである。
【0051】
通常、CRIg変異体は、配列番号1−7の何れかのものからの成熟アミノ酸配列と少なくとも約80%アミノ酸配列同一性、又は少なくとも約85%アミノ酸配列同一性、又は少なくとも約90%アミノ酸配列同一性、又は少なくとも約95%アミノ酸配列同一性、又は少なくとも約98%アミノ酸配列同一性、又は少なくとも約99%アミノ酸配列同一性を有する。好ましくは、最も高い度合いの配列同一性は細胞外ドメイン(ECD)内(配列番号1又は2のアミノ酸1又は約21からX、ここで、Xは271から281の任意のアミノ酸残基;又は配列番号3のアミノ酸1又は約21からX、ここで、Xは178から186の任意のアミノ酸残基、あるいは配列番号4のアミノ酸1又は約21からX、ここでXは176から184の任意のアミノ酸残基である)で生じる。
【0052】
CRIg(PRO362)「細胞外ドメイン」又は「ECD」は、各全長分子の膜貫通及び細胞質ドメインが本質的にないCRIgポリペプチドの形態を意味する。通常、CRIg ECDはこのような膜貫通及び/又は細胞質ドメインを1%未満、好ましくはこのようなドメインを0.5%未満有している。上で検討したように、場合によっては、CRIg ECDは、配列番号1、2、3、又は4のアミノ酸残基1又は約21からXを含み、ここで、Xは配列番号1又は2の約271から281の任意のアミノ酸残基、配列番号3の約178から186の任意のアミノ酸、及び配列番号4の約176から184の任意のアミノ酸を含む。
【0053】
CRIg配列に関する「パーセント(%)アミノ酸配列同一性」とは、配列を整列させ、最大のパーセント配列同一性を得るために必要ならば間隙を導入し、如何なる保存的置換も配列同一性の一部と考えないとした後の、CRIg配列のアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセントとして定義される。パーセントアミノ酸配列同一性を決定する目的のためのアラインメントは、当業者の技量の範囲にある様々な方法、例えばBLAST、BLAST-2、ALIGN、又はMegalign(DNASTAR)ソフトウエアのような公に入手可能なコンピュータソフトウエアを使用することにより達成可能である。当業者であれば、比較される配列の完全長に対して最大のアラインメントを達成するために必要な任意のアルゴリズムを含む、アラインメントを測定するための適切なパラメータを決定することができる。別段の記載がなされない限り、アラインメントは、選択されたソフトウェアのデフォルトパラメータを使用して実施される。ついで、酸配列同一性は、より長い配列に対して計算される。つまり、より短い配列が、より長い配列の一部と100%の配列同一性を示すとしても、全体の配列同一性は100%未満になるであろう。
【0054】
CRIgコード配列に対する「パーセント(%)核酸配列同一性」は、配列を整列させ、最大のパーセント配列同一性を得るために必要ならば間隙を導入した後、CRIgコード配列のヌクレオチドと同一である候補配列中のヌクレオチドのパーセントとして定義される。パーセント核酸配列同一性を決定する目的のためのアラインメントは、当業者の知る範囲にある種々の方法、例えばBLAST、BLAST-2、ALIGN又はMegalign(DNASTAR)ソフトウエアのような公に入手可能なコンピュータソフトウエアを使用することにより達成可能である。当業者であれば、比較される配列の完全長に対して最大のアラインメントを達成するために必要な任意のアルゴリズムを含む、アラインメントを測定するための適切なパラメータを決定することができる。ついで、酸配列同一性は、より長い配列に対して計算される。つまり、より短い配列が、より長い配列の一部と100%の配列同一性を示すとしても、全体の配列同一性は100%未満になるであろう。
【0055】
「単離された」核酸分子は、同定され、核酸の天然供給源に通常付随している少なくとも一つの汚染核酸分子から分離された核酸分子である。単離された核酸分子は、天然に見出される形態あるいは設定以外のものである。ゆえに、単離された核酸分子は、天然の細胞中に存在する核酸分子とは区別される。しかし、単離された核酸分子は、例えば、核酸分子が天然の細胞のものとは異なった染色体位置にあるコード化ポリペプチドを通常発現する細胞に含まれる核酸分子を含む。
【0056】
「単離された」CRIgポリペプチドコード核酸分子は、同定され、CRIgコード核酸の天然供給源に通常付随している少なくとも一つの汚染核酸分子から分離された核酸分子である。単離されたCRIgポリペプチドコード核酸分子は、天然に見出される形態あるいは設定以外のものである。ゆえに、単離されたCRIgポリペプチドコード核酸分子は、天然の細胞中に存在する核酸分子とは区別される。しかし、単離されたCRIgコード核酸分子は、例えば、核酸分子が天然の細胞のものとは異なった染色体位置にあるCRIgを通常発現する細胞に含まれるCRIgコード核酸分子を含む。
【0057】
「補体関連疾患」なる用語は、ここでは最も広い意味で使用され、例えば補体欠損症のような病因が補体系の活性化の異常性を含む全ての疾患及び病理学的症状を含む。該用語はC3転換酵素の阻害から恩恵を受ける疾患及び病理学的症状を含む。該用語は、代替補体経路の選択的阻害を含む阻害から恩恵を受ける疾患及び病理学的症状を更に含む。補体関連疾患には、限定するものではないが、炎症性疾患及び自己免疫疾患、例えばリウマチ様関節炎(RA)、急性呼吸促迫症候群(ARDS)、虚血及び再灌流後の遠隔組織傷害、心肺バイパス手術中の補体活性化、皮膚筋炎、天疱瘡、ループス腎炎及びその結果の糸球体腎炎及び脈管炎、心肺バイパス、心臓麻痺誘発性冠血管内皮機能不全、II型膜性増殖性糸球体腎炎、IgA腎症、急性腎不全、クリオグロブリン血症、抗リン脂質症候群、黄斑変性疾患及び他の補体関連の眼の症状、例えば加齢性黄斑変性症(AMD)、脈絡膜血管新生(CNV)、ブドウ膜炎、糖尿病及び他の虚血関連網膜症、眼内炎、及び他の眼内血管新生疾患、例えば糖尿病性黄斑浮腫、病的近視、フォン・ヒッペル・リンドウ病、眼のヒストプラスマ症、網膜中心静脈閉塞症(CRVO)、角膜血管新生、網膜血管新生、並びに異物移植、超急性拒絶反応、血液透析、慢性閉塞肺促迫症候群(COPD)、喘息、及び誤嚥性肺炎が含まれる。
【0058】
「補体関連の眼の症状」なる用語は最も広い意味で使用され、古典及び代替経路、特に補体の代替経路を含む病因が補体に関連する全ての眼の症状を含む。補体関連の眼の症状には、限定するものではないが、黄斑変性疾患、例えばドライ及びウェット(非滲出性及び滲出性)形態を含む加齢性黄斑変性症(AMD)、脈絡膜血管新生(CNV)、ブドウ膜炎、糖尿病及び他の虚血関連網膜症、及び他の眼内血管新生疾患、例えば糖尿病性黄斑浮腫、病的近視、フォン・ヒッペル・リンドウ病、眼のヒストプラスマ症、網膜中心静脈閉塞症(CRVO)、角膜血管新生、及び網膜血管新生が含まれる。補体関連の眼の症状の好ましい群には、非滲出性(ウェット)及び滲出性(ドライ又は萎縮性)AMDを含む加齢性黄斑変性症(AMD)、脈絡膜血管新生(CNV)、糖尿病性網膜症(DR)、及び眼内炎が含まれる。
【0059】
「炎症性疾患」及び「炎症性障害」なる用語は、ここでは置き換え可能に使用され、哺乳動物の免疫系の成分が哺乳動物の罹患率に寄与する免疫反応を生じ、媒介し又は寄与等する疾患又は障害を意味する。また含まれるものは、炎症反応の減少が疾患の進行に寛解効果を有する疾患である。この用語に含まれるものは、自己免疫疾患を含む免疫媒介炎症疾患である。
【0060】
「T細胞媒介」疾患という用語は、T細胞が直接的又は間接的に哺乳動物の病的状態を媒介するか又は寄与等する疾患を意味する。T細胞媒介疾患は細胞媒介効果、リンホカイン媒介効果等に、また例えばT細胞により分泌されるリンホカインによりB細胞が刺激されるならばB細胞に関連した効果にさえも関連している。
【0061】
その幾つかがT細胞媒介性である免疫関連及び炎症疾患の例には、限定するものではないが、炎症性腸疾患(IBD)、全身性紅斑性狼瘡、リウマチ様関節炎、若年性慢性関節炎、脊椎関節症、全身性硬化症(強皮症)、特発性炎症ミオパシー(皮膚筋炎、多発性筋炎)、シェーグレン症候群、全身性血管炎、サルコイドーシス、自己免疫性溶血性貧血(免疫性汎血球減少症、発作性夜間血色素尿症)、自己免疫性血小板減少(特発性血小板減少性紫斑病、免疫媒介血小板減少)、甲状腺炎(グレーブス疾患、ハシモト甲状腺炎、若年性リンパ球性甲状腺炎、萎縮性甲状腺炎)、真性糖尿病、免疫介在性腎疾患(糸球体腎炎、尿細管間質性腎炎)、中枢及び末梢神経系の脱髄疾患、例えば多発性硬化症、多発性神経障害、肝胆汁性疾患、例えば感染性肝炎(A型、B型、C型、D型、E型肝炎及び他の非肝親和性(nonhepatotropic)ウィルス)、自己免疫慢性活性肝炎、原発性胆汁性肝硬変症、肉芽腫性肝炎、及び硬化性胆管炎、炎症性及び線維性肺疾患(例えば嚢胞性線維症)、グルテン過敏性腸疾患、ウィップル病、水疱性皮膚疾患、多形性紅斑及び接触性皮膚炎を含む自己免疫又は免疫媒介皮膚疾患、乾癬、肺のアレルギー性疾患、例えば好球性肺炎、特発性肺線維症及び過敏性肺炎、拒絶反応及び移植片対宿主疾患を含む移植関連疾患が含まれる。アルツハイマー病、及びアテローム性動脈硬化症が含まれる。
【0062】
ここで使用される「腫瘍」は、悪性であろうと良性であろうとあらゆる新生物細胞成長及び増殖、及びあらゆる前癌状態の細胞及び組織を意味する。
【0063】
「癌」及び「癌性」という用語は、典型的には調節されない細胞増殖を特徴とする、哺乳動物における生理学的状態を指すか記述する。癌の例には、これらに限定されるものではないが、癌腫、リンパ腫、芽細胞腫、肉腫、及び白血病が含まれる。このような癌のより特定の例には、乳癌、前立腺癌、大腸癌、扁平細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸管癌、卵巣癌、肝臓癌、膀胱癌、肝癌、結腸直腸癌、子宮内膜癌、唾液腺癌、腎臓癌、肝癌、外陰癌、甲状腺癌、肝臓癌及び様々なタイプの頭頸部癌が含まれる。
【0064】
「治療」とは、疾患の病理の進展阻止又は変更を行なう本発明により実施される介入である。従って、「治療」は治療的処置及び予防的又は保護的手段の両方を指す。治療が必要なものは、既に疾患に罹患しているもの並びに疾患が防止されるべきものを含む。免疫関連疾患の治療では、治療薬は直接的に免疫反応の成分の反応の大きさを変え、又は他の治療剤、例えば抗生物質、抗真菌剤、抗炎症剤、化学療法剤等による治療に対する疾患の感受性を高めることができる。
【0065】
補体関連疾患のような疾患の「病理」は、患者の良好な健康を危険にさらす全ての現象を含む。これには、限定されるものではないが、異常又は制御不能な細胞増殖(好中球、好酸球、単球性、リンパ性細胞)、抗体産生、自己抗体産生、補体産生、正常に機能している隣接細胞の妨害、サイトカイン又は他の分泌産物の異常なレベルでの放出、あらゆる炎症性又は免疫性反応の抑制又は悪化、炎症性細胞(好中球、好酸球、単球性、リンパ性)の組織空間への浸潤等が含まれる。
【0066】
ここで使用される「哺乳動物」なる用語は、ヒト、家庭及び農業用動物、動物園、スポーツ、又はペット動物、例えばウマ、ブタ、ウシ、イヌ、ネコ及びケナガイタチなどを含む哺乳類に分類される任意の動物を意味する。本発明の好ましい実施態様では、哺乳動物はヒトである。
【0067】
一又は複数の更なる治療薬と「組み合わせた」投与とは、同時(同時期)及び任意の順序での連続した投与を含む。
【0068】
「サイトカイン」という用語は、一つの細胞集団から放出されるタンパク質であって、他の細胞に対して細胞間メディエータとして作用するものの包括的な用語である。このようなサイトカインの例としては、リンフォカイン、モノカイン、及び伝統的なポリペプチドホルモンを挙げることができる。サイトカインには、成長ホルモン、例えばヒト成長ホルモン、N-メチオニルヒト成長ホルモン、及びウシ成長ホルモン、副甲状腺ホルモン、チロキシン、インスリン、プロインスリン、リラクシン、プロリラクシン、卵胞刺激ホルモン(FSH)のような糖タンパク質ホルモン、副甲状腺刺激ホルモン(TSH)、及び黄体形成ホルモン(LH)、肝臓成長因子、線維芽細胞増殖因子、プロラクチン、胎盤ラクトゲン、腫瘍壊死因子-α及び-β、ミュラー阻害物質、マウス性腺刺激ホルモン関連ペプチド、インヒビン、アクチビン、血管内皮増殖因子、インテグリン、トロンボポエチン(TPO)、神経成長因子、例えばNGF-β血小板増殖因子、トランスフォーミング増殖因子(TGF)、例えばTGF-α及びTGF-β、インスリン様増殖因子I及びII、エリスロポイエチン(EPO)、骨誘導因子、インターフェロンα、β、γのようなインターフェロン;マクロファージCSF(M-CSF)のようなコロニー刺激因子(CSF)、顆粒球マクロファージCSF(GM-CSF)、及び顆粒球CSF(G-CSF);IL-1、IL-1α、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-8、IL-9、IL-11、IL-12等のインターロイキン(IL)、腫瘍壊死因子、例えばTNF-α又はTNF-β、及びLIF及びキットリガンド(KL)を含む他のポリペプチド因子が含まれる。ここで使用される場合、サイトカインなる用語は天然源由来あるいは組換え細胞培養由来のタンパク質及び天然配列サイトカインの生物的に活性な等価物を含む。
【0069】
「治療的有効量」は、標的の疾患又は症状、例えば補体関連疾患又は症状、又は癌の状態、例えば病理において測定可能な改善を達成するために必要とされる活性なCRIg、CRIgアゴニスト及びアンタゴニストの量である。
【0070】
「コントロール配列」という用語は、特定の宿主生物において作用可能に結合したコード配列を発現するために必要なDNA配列を指す。例えば原核生物に好適なコントロール配列は、プロモーター、場合によってはオペレータ配列、及びリボソーム結合部位を含む。真核生物の細胞は、プロモーター、ポリアデニル化シグナル、及びエンハンサーを利用することが知られている。
【0071】
核酸は、他の核酸配列と機能的な関係にあるときに「作用可能に結合」している。例えば、プレ配列又は分泌リーダーのDNAは、ポリペプチドの分泌に参画するプレタンパク質として発現されているならば、そのポリペプチドのDNAに作用可能に結合している;プロモーター又はエンハンサーは、配列の転写に影響を及ぼすならば、コード配列に作用可能に結合している;又はリボソーム結合部位は、もしそれが翻訳を容易にするような位置にあるなら、コード配列と作用可能に結合している。一般的に、「作用可能に結合している」とは、結合したDNA配列が近接しており、分泌リーダーの場合には近接していて読みフェーズにあることを意味する。しかしながら、エンハンサーは必ずしも近接している必要はない。結合は簡便な制限部位でのライゲーションにより達成される。そのような部位が存在しない場合は、合成オリゴヌクレオチドアダプター又はリンカーが常套的な手法に従って使用される。
【0072】
ハイブリダイゼーション反応の「ストリンジェンシー」は、当業者によって容易に決定され、一般的にプローブ長、洗浄温度、及び塩濃度に依存する経験的な計算である。一般に、プローブが長くなると適切なアニーリングのための温度が高くなり、プローブが短くなると温度は低くなる。ハイブリダイゼーションは、一般的に、相補的鎖がその融点に近いがそれより低い環境に存在する場合における変性DNAの再アニールする能力に依存する。プローブとハイブリダイゼーション可能な配列との間の所望の相同性の程度が高くなると、使用できる相対温度が高くなる。その結果、より高い相対温度は、反応条件をよりストリンジェントにするが、低い温度はストリンジェンシーを低下させる。ハイブリダイゼーション反応のストリンジェンシーの更なる詳細及び説明は、Ausubel等, Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995)を参照のこと。
【0073】
ここで定義される「ストリンジェントな条件」又は「高度のストリンジェント条件」は、(1)洗浄のために低イオン強度及び高温度を用いるもの、例えば、50℃において0.015Mの塩化ナトリウム/0.0015Mのクエン酸ナトリウム/0.1%のドデシル硫酸ナトリウムを用いるもの;(2)ハイブリダイゼーション中にホルムアミド等の変性剤を用いるもの、例えば、42℃において50%(v/v)ホルムアミド、0.1%ウシ血清アルブミン/0.1%フィコール/0.1%のポリビニルピロリドン/50mMのpH6.5のリン酸ナトリウムバッファー、及び750mMの塩化ナトリウム、75mMのクエン酸ナトリウムを用いるもの;又は(3)42℃において50%ホルムアミド、5×SSC(0.75MのNaCl、0.075Mのクエン酸ナトリウム)、50mMのリン酸ナトリウム(pH6.8)、0.1%のピロリン酸ナトリウム、5×デンハード液、超音波処理サケ精子DNA(50μg/ml)、0.1%SDS、及び10%のデキストラン硫酸を用い、42℃において0.2×SSC(塩化ナトリウム/クエン酸ナトリウム)中で、55℃において50%ホルムアミド中で洗浄した後、55℃においてEDTAを含む0.1×SSCからなる高ストリンジェント洗浄を用いるものによって同定できる。
【0074】
「中程度のストリンジェント条件」は、Sambrook等, Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989に記載されているように特定され、上記のものよりストリンジェンシーが低い洗浄液及びハイブリダイゼーション条件(例えば、温度、イオン強度及び%SDS)の使用を含む。中程度のストリンジェント条件は、20%ホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハード液、10%デキストラン硫酸、及び20mg/mLの変性剪断サケ精子DNAを含む溶液中の37℃での終夜のインキュベーション後に、37〜50℃にて1×SSC中でフィルター洗浄を行うという条件である。プローブ長などの因子に必要に応じて適合させるには、どのようにして温度、イオン強度等を調節するかは当業者であれば分かるであろう。
【0075】
「エピトープタグ」なる用語は、ここで用いられるときは、「タグポリペプチド」に融合した本発明のポリペプチドを含んでなるキメラポリペプチドを指す。タグポリペプチドは、その抗体が産生され得るエピトープを提供するに十分な数の残基を有しているが、その長さは融合するポリペプチドの活性を阻害しないよう充分に短い。また、タグポリペプチドは、好ましくは、抗体が他のエピトープと実質的に交差反応をしないようにかなり独特である。適切なタグポリペプチドは、一般に、少なくとも6のアミノ酸残基、通常は約8〜約50のアミノ酸残基(好ましくは約10〜約20の残基)を有する。
【0076】
本発明のCRIgポリペプチドの変異体に関し、「活性な」又は「活性」は、本発明の天然又は天然に生じるポリペプチドの生物学的及び/又は免疫学的活性を保持するポリペプチドの形態を意味する。好ましい生物学的活性は、C3bに結合し、及び/又は補体又は補体活性化に影響を及ぼし、特に代替の補体経路及び/又はC3転換酵素を阻害する能力である。3転換酵素の阻害は、例えばコラーゲン-又は抗体-誘発関節炎の間における正常な血清中でのC3代謝回転の阻害、又は関節炎の間接におけるC3沈着の阻害を測定することにより測定することができる。
【0077】
CRIgの生物学的活性を模倣し、ここに開示されたスクリーニングアッセイによって同定することができる抗体、ポリペプチド又は他の分子(例えば有機又は無機小分子、ペプチド等々)に関する「生物学的活性」とは、C3bに結合し、及び/又は補体又は補体活性化に影響を及ぼし、特に代替の補体経路及び/又はC3転換酵素を阻害するそのような分子の能力を指す。
【0078】
CRIg「アゴニスト」なる用語は最も広い意味で使用され、天然配列CRIgポリペプチドの(上述の)定性的生物学的活性を模倣する任意の分子を含む。
【0079】
CRIg「アンタゴニスト」なる用語は最も広い意味で使用され、天然配列CRIgポリペプチドのような天然ポリペプチドの定性的生物学的活性を部分的に又は完全にブロックし、阻害し、又は中和する任意の分子を含む。
【0080】
適切なアゴニスト又はアンタゴニスト分子には、アゴニスト又はアンタゴニスト抗体又は抗体断片、本発明の天然ポリペプチドの断片、融合体又はアミノ酸配列変異体、ペプチド、小有機分子を含む小分子等々が特に含まれる。
【0081】
「小分子」はここでは約600以下、好ましくは約1000ダルトン以下の分子量を有するものと定義されている。
【0082】
「抗体」という用語は最も広い意味において使用され、限定するものではないが、単一の抗CRIgモノクローナル抗体(アゴニスト、アンタゴニスト、及び中和抗体を含む)、及び多エピトープ特異性を持つ抗CRIg抗体組成物を包含する。ここで使用される「モノクローナル抗体」という用語は、実質的に均一な抗体の集団、すなわち、構成する個々の抗体が、少量存在しうる自然に生じる可能性のある突然変異を除いて同一である集団から得られる抗体を意味する。
【0083】
「抗体」(Abs)及び「免疫グロブリン」(Igs)は同じ構造的特徴を有する糖タンパク質である。抗体は特定の抗原に対して結合特異性を示すが、免疫グロブリンは抗体と抗原特異性を欠く抗体様分子の双方を含む。後者の種類のポリペプチドは例えばリンパ系によって低レベルで、ミエローマによって増大したレベルで生産される。「抗体」なる用語は最も広い意味で使用され、限定するものではないが、無傷のモノクローナル抗体、ポリクローナル抗体、少なくとも2つの無傷抗体から形成された多重特異的抗体(例えば二重特異的抗体)、及びそれらが所望の生物学的活性を示す限り抗体断片を特に包含する。
【0084】
「天然抗体」及び「天然免疫グロブリン」は、通常、2つの同一の軽(L)鎖及び2つの同一の重(H)鎖からなる、約150000ダルトンのヘテロ四量体糖タンパク質である。各軽鎖は一つの共有ジスルフィド結合により重鎖に結合しており、ジスルフィド結合の数は、異なった免疫グロブリンアイソタイプの重鎖間で変わる。また各重鎖と軽鎖は、規則的に離間した鎖内ジスルフィド結合を有している。各重鎖は、多くの定常ドメインが続く可変ドメイン(VH)を一端に有する。各軽鎖は、一端に可変ドメイン(VL)を、他端に定常ドメインを有し;軽鎖の定常ドメインは重鎖の第一定常ドメインと整列し、軽鎖の可変ドメインは重鎖の可変ドメインと整列している。特定のアミノ酸残基が、軽鎖及び重鎖可変ドメイン間の界面を形成すると考えられている。
【0085】
「可変」という用語は、可変ドメインのある部分が、抗体間で配列が広範囲に異なっており、その特定の抗原に対する各特定の抗体の結合性と特異性に使用されていることを意味する。しかしながら、可変性は抗体の可変ドメインにわたって一様には分布していない。それは、軽鎖及び重鎖の可変ドメインの両方の相補性決定領域(CDR)又は高頻度可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存された部分はフレームワーク領域(FR)と呼ばれる。天然の重鎖及び軽鎖の可変ドメインは、βシート構造を結合し、ある場合にはその一部を形成するループを形成する、3つのCDRにより連結されたβシート配置を主にとる4つのFR領域を含んでいる。各鎖のCDRは、FR領域によって近接して互いに保持され、他の鎖のCDRと共に、抗体の抗原結合部位の形成に寄与している(Kabat等, NIH Publ. No.91-3242, Vol. I, pp. 647-669 (1991)を参照)。定常ドメインは、抗体の抗原への結合に直接関与しているものではないが、様々なエフェクター機能、例えば抗体依存性細胞障害活性への抗体の関与を示す。
【0086】
「抗体断片」は、無傷の抗体の一部、好ましくは無傷の抗体の抗原結合又は可変領域を含む。抗体断片の例は、Fab、Fab’、F(ab’)2、及びFv断片;ダイアボディ(diabodies);線形抗体(Zapata等, Protein Eng. 8(10): 1057-1062 [1995]);一本鎖抗体分子;及び抗体断片から形成された多重特異性抗体を含む。
【0087】
抗体のパパイン消化は、「Fab」断片と呼ばれる二つの同一の抗原結合断片を生成し、その各々は単一の抗原結合部位、及び残基「Fc」断片を持つ。該名称「Fc」は容易に結晶化する能力を反映している。ペプシン処理は、二つの抗原結合部位を持ち、なお抗原と交差結合可能なF(ab’)2断片を生じる。
【0088】
「Fv」は、完全な抗原認識及び結合部位を含む最小の抗体断片である。この領域は、強固に非共有結合した1本の重鎖と1本の軽鎖の可変領域の二量体からなる。この構造において各可変ドメインの三つのCDRが相互作用してVH−VL二量体の表面に抗原結合部位を形成する。集合的には、6つのCDRが抗体に対する抗原結合特異性を与える。しかしながら、単一の可変ドメイン(又は抗原に特異的な三つのCDRのみを含んでなるFvの半分)でさえ、結合部位全体よりは低い親和性であるが、抗原を認識し結合する能力を有している。
【0089】
またFab断片は、軽鎖の定常ドメイン及び重鎖の第一定常ドメイン(CH1)も含む。Fab’断片は、抗体ヒンジ領域からの一又は複数のシステインを含む重鎖CH1ドメインのカルボキシ末端に数残基が付加されていることでFab断片と相違する。Fab’-SHは、定常ドメインのシステイン残基が遊離のチオール基を持つFab’に対するここでの表記である。F(ab’)2抗体断片は、最初は間にヒンジシステインを有しているFab’断片の対として生成された。抗体断片の他の化学的結合もまた知られている。
【0090】
任意の脊椎動物種からの抗体(免疫グロブリン)の「軽鎖」は、それらの定常ドメインのアミノ酸配列に基づいて、カッパ(κ)及びラムダ(λ)と呼ばれる二つの明らかに異なる型の一方に分類できる。
【0091】
それらの重鎖の定常ドメインのアミノ酸配列によって、免疫グロブリンは異なるクラスに分類できる。免疫グロブリンの五つの主要なクラス:IgA、IgD、IgE、IgG及びIgMがあり、それらの幾つかは更にサブクラス(アイソタイプ)、例えばIgG1、IgG2、IgG3、IgG4、IgA及びIgA2に分類される。免疫グロブリンの異なったクラスに対応する重鎖定常ドメインはそれぞれγ、μ、δ、α、及びεと呼ばれる。異なったクラスの免疫グロブリンのサブユニット構造及び三次元立体構造はよく知られている。
【0092】
ここで使用される「モノクローナル抗体」という用語は、実質的に均一な抗体の集団、すなわち、構成する個々の抗体が、少量存在しうる自然に生じる可能性のある突然変異を除いて同一である集団から得られる抗体を称する。モノクローナル抗体は高度に特異的であり、一つの抗原部位に対応する。更に、異なる決定基(エピトープ)に対応する異なる抗体を典型的に含む通常の(ポリクローナル)抗体とは異なり、各モノクローナル抗体は抗原の単一の決定基に向く。特異性に加えて、他の免疫グロブリンによって汚染されていないハイブリドーマ培養によって合成されることで、モノクローナル抗体は有利である。「モノクローナル」との形容は、実質的に均一な抗体集団から得られたという抗体の性質を示し、特定の方法で抗体を生産しなければならないことを意味するものではない。例えば、本発明において使用されるモノクローナル抗体は、Kohler等によって最初にNature 256:495[1975]に記載されたハイブリドーマ法によって作ることができるか、あるいは組換えDNA法(例えば米国特許第4816567号を参照)によって作ることができる。「モノクローナル抗体」は例えば Clackson等, Nature 352:624-628 (1991) 及び Marks等, J.Mol.Biol. 222:581-597 (1991)に記載された技術を用いてファージ抗体ライブラリーから単離してもよい。また、ファージミッド及びファージベクターの利用による抗体の調製が記載されている米国特許第5750373号、第5571698号、第5403484号及び第5223409号を参照のこと。
【0093】
ここで、モノクローナル抗体は、特に重鎖及び/又は軽鎖の一部が特定の種から誘導されたか又は特定の抗体クラス又はサブクラスに属する抗体の対応する配列と同一又は相同である一方、鎖の残りの部分は他の種から誘導されたか又は特定の抗体クラス又はサブクラスに属する抗体の対応する配列と同一又は相同である「キメラ」抗体(免疫グロブリン)、並びにそれらが所望の生物学的活性を示す限りそれらの抗体の断片を含む(米国特許第4816567号;Morrison等,Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984))。
【0094】
非ヒト(例えばマウス)抗体の「ヒト化」形とは、キメラ免疫グロブリン、免疫グロブリン鎖又はその断片(例えばFv、Fab、Fab’、F(ab’)2あるいは抗体の他の抗原結合サブ配列)であって、非ヒト免疫グロブリンに由来する最小配列を含むものである。大抵は、ヒト化抗体はレシピエントの相補性決定領域(CDR)の残基が、マウス、ラット又はウサギのような所望の特異性、親和性及び能力を有する非ヒト種(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(レシピエント抗体)である。ある場合には、ヒト免疫グロブリンのFvフレームワーク領域(FR)残基はまた対応する非ヒト残基によって置換されうる。更に、ヒト化抗体は、レシピエント抗体にも、移入されたCDRもしくはフレームワーク配列にも見出されない残基を含んでいてもよい。これらの変更は、抗体の性能を更に洗練させ、最大化するために行なわれる。一般に、ヒト化抗体は、全てあるいはほとんど全てのCDR領域が非ヒト免疫グロブリンのものに対応し、全てあるいはほとんど全てのFR領域がヒト免疫グロブリン配列のものである、少なくとも一つ、典型的には二つの可変ドメインの実質的に全てを含む。ヒト化抗体は、最適には免疫グロブリン定常領域(Fc)、典型的にはヒトの免疫グロブリンの定常領域の少なくとも一部を含んでなる。さらなる詳細については、Jones等, Nature, 321:522-525 (1986);Riechmann等, Nature, 332:323-329 [1988];及びPresta, Curr. Op Struct. Biol., 2:593-596 (1992)を参照のこと。ヒト化抗体は、抗体の抗原結合領域が、マカクザルを対象の抗原で免疫化することにより生成された抗体に由来している「霊長類化」抗体を含む。旧世界ザル由来の残基を含む抗体もまた本発明において可能である。例えば米国特許第5658570号、第5693780号、第5681722号、第5750105号及び第5756096号を参照のこと。
【0095】
「一本鎖Fv」又は「sFv」抗体断片は、抗体のVH及びVLドメインを含み、これらのドメインは単一のポリペプチド鎖に存在する。好ましくは、Fvポリペプチドは、sFvが抗原結合にとって望ましい構造の形成を可能にするVH及びVLドメイン間のポリペプチドリンカーを更に含む。sFvの概説については、Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg及びMoore編, Springer-Verlag, New York, pp. 269-315 (1994)を参照のこと。
【0096】
「ダイアボディ」という用語は、二つの抗原結合部位を持つ小抗体断片を指し、その断片は同じポリペプチド鎖(VH-VL)内で軽鎖可変ドメイン(VL)に結合した重鎖可変ドメイン(VH)を含む。同じ鎖の二つのドメイン間に対形成するには短すぎるリンカーを用いることにより、ドメインは強制的に他の鎖の相補的ドメインと対形成して二つの抗原結合部位をつくりだす。ダイアボディは、例えば、EP404097;WO93/11161;及びHollinger等, Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)に更に詳しく記載されている。
【0097】
抗体のような「単離された」ポリペプチドとは、その自然環境の成分から同定され分離され及び/又は回収されたものを意味する。その自然環境の汚染成分とは、その抗体の診断又は治療への使用を典型的には妨害する物質であり、酵素、ホルモン、及び他のタンパク質様又は非タンパク質様溶質を含み得る。好ましい実施形態では、抗体を含むポリペプチドは、(1)ローリー(Lowry)法による測定で化合物の95重量%よりも大きな割合まで、最も好ましくは99重量%よりも大きな割合まで、(2)スピニングカップシークエネーターを使用することにより、少なくとも15残基のN末端あるいは内部アミノ酸配列を得るのに充分な程度まで、あるいは(3)クーマシーブルー又は好ましくは銀染色を用いた還元又は非還元条件下でのSDS-PAGEによる均一性まで精製される。単離された化合物、例えば抗体又は他のポリペプチドには、化合物の自然環境の少なくとも一つの成分が存在しないため、組換え細胞内のインサイツの化合物が含まれる。しかしながら、通常は、単離された化合物は少なくとも一つの精製工程により調製される。
【0098】
「標識」なる語は、ここで用いられる場合、「標識した」化合物が生成されるように、化合物、例えば抗体又はポリペプチドに直接又は間接的に抱合させた検出可能な化合物又は組成物を意味する。標識は、それ自身検出可能でもよく(例えば、放射性標識又は蛍光標識)、又は酵素標識の場合、検出可能な基質化合物又は組成物の化学変換を触媒しうる。
【0099】
「固相」とは、本発明の化合物が付着することのできる非水性マトリクスを意味する。ここで包含される固相の例は、部分的又は全体的に、ガラス(例えば、孔制御ガラス)、多糖類(例えばアガロース)、ポリアクリルアミド、ポリスチレン、ポリビニルアルコール及びシリコーンから形成されたものを含む。ある実施形態では、内容に応じて、固相はアッセイプレートのウェルを含み得;その他では精製カラム(例えばアフィニティークロマトグラフィーカラム)とすることもできる。また、この用語は、米国特許第4275149号に記載されたような、別個の粒子の不連続な固相も包含する。
【0100】
「リポソーム」は、哺乳動物への薬物(ここに開示する抗ErbB2抗体及び場合によっては化学療法剤)の輸送に有用である種々の型の脂質、リン脂質及び/又は界面活性剤からなる小型の小胞である。リポソームの成分は、通常は生体膜の脂質配列に類似する二層形態に配列させる。
【0101】
ここで用いられる場合、「イムノアドヘシン」なる用語は、異種タンパク質(「アドヘシン」)の結合特異性と免疫グロブリン定常ドメインのエフェクター機能とを併せ持つ抗体様分子を指す。構造的には、イムノアドヘシンは、所望の結合特異性を持ち、抗体の抗原認識及び結合部位以外である(つまり「異種の」)アミノ酸配列と、免疫グロブリン定常ドメイン配列との融合物を含む。イムノアドヘシン分子のアドへシン部分は、典型的には少なくともレセプター又はリガンドの結合部位を含む隣接アミノ酸配列である。イムノアドヘシンの免疫グロブリン定常ドメイン配列は、IgG-1、IgG-2、IgG-3又はIgG-4サブタイプ、IgA(IgA-1及びIgA-2を含む)、IgE、IgD又はIgMなどの任意の免疫グロブリンから得ることができる。
【0102】
「慢性」投与とは、延長された期間の間、所望の効果を維持するように、急性態様とは異なる連続態様で薬剤を投与することを意味する。
【0103】
「間欠」投与とは、中断がなく連続的になされるのではなく、むしろ周期的な性質の治療法である。
【0104】
一又は複数の更なる治療剤「との併用での」投与は同時(同時期)と任意の順序の連続投与を含む。
【0105】
「患者」は脊椎動物、好ましくは哺乳動物、より好ましくはヒトである。
【0106】
「哺乳動物」なる用語は、ここでは限定するものではないが、ヒト、牧畜、家畜、及び動物園の動物、スポーツ、又はペット動物、例えばヒツジ、イヌ、ウマ、ネコ、ウシ等々を含む哺乳動物として分類される任意の動物を指すために使用される。
【0107】
「有効量」とは有益な又は所望される治療的(予防を含む)結果を得るのに十分な量である。有効量は一又は複数回の投与で投与できる。
【0108】
(発明を実施するための形態)
本発明の実施には、別段の記載がない限り、当業者の技量の範囲にある(組換え技術を含む)分子生物学、微生物学、細胞生物学、生化学及び免疫学の一般的な技術を用いる。かかる技術は例えば、“Molecular Cloning: A Laboratory Manual”, 2版 (Sambrook等, 1989);“Oligonucleotide Synthesis” (M.J. Gait編, 1984);“Animal Cell Culture” (R.I. Freshney編, 1987);“Methods in Enzymology” (Academic Press, Inc.);“Handbook of Experimental Immunology”, 4版 (D.M. Weir & C.C. Blackwell編, Blackwell Science Inc., 1987);“Gene Transfer Vectors for Mammalian Cells” (J.M. Miller & M.P. Calos編, 1987);“Current Protocols in Molecular Biology” (F.M. Ausubel等編, 1987);“PCR: The Polymerase Chain Reaction”, (Mullis等編, 1994);及び“Current Protocols in Immunology” (J.E. Coligan等編, 1991)のような文献に十分に説明されている。
【0109】
CRIg及びC3b:CRIh及びC3c複合体の結晶構造
CRIg分子を含むポリペプチドは一次アミノ酸配列及びポリペプチドを囲む環境によって決定される3次元構造を有している。この3次元構造はポリペプチドの活性、安定性、結合親和性、結合特異性、及び他の生化学的特質を樹立する。よって、タンパク質の3次元構造の知識はその生物学的活性を模倣し、阻害し又は改善する薬剤を設計する際の大きな指針を提供する。
【0110】
ポリペプチドの3次元構造は多くの方法で決定することができる。最も精確な方法の多くはx線結晶解析(Van Holde, Physical Biochemistry (Prentice Hall: N.J., 1971), pp. 221-239)を用いる。この技術はx線又は他の形態の放射線を回折する結晶格子の能力に依存している。巨大分子の3次元構造を決定するために適した回折実験は典型的には高品質の結晶を必要とする。不幸にも、かかる結晶はIGF-1並びに興味のある多くの他のタンパク質には利用できなかった。結晶は例えばM−CSF(EP668914B1)、CD40リガンド(WO97/00895)、及びBC2Fab断片(WO99/01476)に対して記載されている。
【0111】
結晶性タンパク質及びポリペプチドを調製するための様々な方法が当該分野で知られている(McPherson等., “Preparation and Analysis of Protein Crystals,” McPherson (Robert E. Krieger Publishing Company, Malabar, FL, 1989);Weber, Advances in Protein Chemistry, 41: 1-36 (1991);米国特許第4672108号及び第4833233号)。ポリペプチドを結晶化させるための複数のアプローチ法が存在しているが、特に結晶がx線回折研究に適したものでなければならない場合には、合理的な成功の可能性をもたらす単一セットの条件は存在しない。構造に関する精確な情報をもたらす十分なサイズと分解能の結晶を得るには顕著な努力が必要とされる。例えば、十分な純度のタンパク質がひとたび得られれば、x線回折と続く構造解明に有用なサイズと透明度になるまで結晶化されなければならない。更に、標的タンパク質のアミノ酸配列は知られていても、この配列情報からタンパク質の結晶構造を正確に予測することはできない。また配列情報から、タンパク質とCRIgやC3bのようなそれが相互作用する結合対との間の構造的、コンフォメーション的及び化学的相互作用の理解ができるようになるものでもない。よって、結晶構造はドラッグデザイン及び発見の分野で豊富な価値ある情報をもたらしうるが、CRIgのようなある種の生物学的に関連した化合物の結晶は当業者が直ぐに利用できるものではない。CRIgの高品質の回折結晶は、その生物学的役割を更に理解し、限定するものではないがアゴニスト及びアンタゴニスト抗体を含むCRIg変異体、アゴニスト及び淡タゴニストを設計するために重要になるその3次元構造の決定に役立つ。
【0112】
構造情報の提供に加えて、結晶性ポリペプチドは他の利点をもたらす。例えば、結晶化方法自体はポリペプチドを更に精製し、均一性の古典的条件の一つを満たす。実際、結晶化は、HPLC、透析、一般的なカラムクロマトグラフィー等々の他の精製方法によっては除去されない不純物を除去する無比の精製品質をもたらすことがよくある。更に、結晶性ポリペプチドはしばしば周囲温度で安定であり、プロテアーゼの汚染や溶液保存に伴う他の分解を受けない。結晶性ポリペプチドはまた医薬製剤としても有用でありうる。最後に、一般に結晶化技術には、他の安定化法(例えば凍結乾燥)に付随する変性のような問題が概してない。
【0113】
本発明はCRIg単独とC3b及びC3cそれぞれとの複合体の結晶構造を提供する。C3b:CRIg複合体の構造は、C3活性化時に生じ補体レセプターCRIgの結合部位を定める立体構造変化を完全に明らかにする。また、代替経路転換酵素のタンパク質分解活性に対するこの相互作用の結果を調べる。
【0114】
すなわち、本発明は、およそ次の寸法:a=30.3Å、b=50.8Å、c=62.0Åを有するCRIg結晶を提供する。該結晶は、P212121の対称、又は空間群を有している。CRIg結晶の構造座標はアペンディクス1に提供する。
【0115】
本発明は更におよそ次の寸法:a=97.6Å、b=255.7Å、c=180.3Å及びC2221の空間群を有しているC3b:CRIg複合体の結晶構造を提供する。CRIg結晶の構造座標はアペンディクス2に提供し、そのリボン構造は図1Bに示す。
【0116】
本発明はまたおよそ次の寸法:a=382.8Å、b=65.0Å、c=147.2Å、β=102.7、及びC2の空間群を有しているC3c:CRIg複合体の結晶構造を提供する。C3c:CRIg結晶の構造座標はアペンディクス3に提供し、そのリボン構造は図1Cに示す。
【0117】
残基1〜645からなるC3bは、5つのマクログロブリン様ドメイン(MG1-MG5)、及び第6MGドメインのN末端半分、リンカー領域(LNK)(図1A及び1B)に折り畳まれる。免疫グロブリントポロジーMGドメインは「キーリング」様形式に配置され、約10Å幅で30Å長の中央溝の回りを円で囲んでいる。C3bのα鎖は残基729:1641からなり、C3と比較してC3a又はANAドメインを欠いている。α'NTドメインと呼ばれるC3bα鎖のN末端残基(729:745)は、伸展したコンフォメーションを採用しており、第6MGドメインの第2の半分を形成する残基につながっている。これに続いて、α鎖は間にCUBドメインとチオ-エステルドメイン(TED)が挿入された2つの更なるMGドメイン(MG7及びMG8)を含んでいる。最後に、α鎖のC末端の170残基はいわゆる「アンカー領域」とC345Cドメインを形成している。
【0118】
実施例1に記載されるように、C3bのコンフォメーションは活性化の際に劇的な変化を受ける。TEDはCUB及びNG8ドメインから離れる方向に回転し、完全に異なったコンフォメーションでTED及びCUBドメインを連結する残基を残す。このコンフォメーション変化の一部として、C3bの988位のCys残基がC3構造中のその最初の保護されたネストから露出した位置まで移動する。
【0119】
実施例1に記載された結晶化研究から、CRIgに対する結合部位がC3bのCUB及びTEDドメインとは反対側にあり、CRIgをTEDドメインから約40Åのところに位置させていることが明らかになった(図1A及び1Bを参照)。
【0120】
CRIgとC3c又はC3bの間の結合界面は大きく、不連続で、全体で約2670Å2の溶媒露出面積を埋めている(図2A)。CRIgはC3bと接触し、A’、G、G、C、C’、C’’シートからの残基が、ストランドC’とC”の近傍の残基及びループによって形成された相互作用の大部分を有する。C’とC”を連結するヘアピンループは、C3bのキーリング形状のβ鎖の中心に形成された裂け目中に突出している。界面のC3b側では、ドメインMG3、MG4、MG5、MG6、LNK及びMG6が全てCRIg結合に寄与し、埋まった界面のそれぞれ約30%と40%に対してMG3とMG6が原因となっている(図2Bを参照)。MG3ドメインはC3b(及びC3c)へのCRIgの結合のゲーティングに重要な役割を担っていると考えられる。また、C3分子の残りと比較してMG3の比較的小さいがなお有意な15°の回転が、CRIg結合部位を構成する他のMGドメインのリングに関したMG3の相対的再配向の原因である。従って、LNK領域の螺旋部分の動きと連係して、この回転がCRIgをC3b及びC3cに結合させる。これらの中程度の変化がないと、C3はCRIgにはラッチされないであろう。
【0121】
これらの構造-機能の知見はCRIgの数個のアミノ酸置換変異体を試験することによって確認した。NG6及びMG3結合ドメインにおける変異はC3bへのCRIgの結合を効果的に抑止することが確認された。これに対して、反対側のCRIg表面残基の変異は野生型CRIgの活性に効果はなかった。
【0122】
CRIgアゴニスト及びアンタゴニストのデザイン、調製及びスクリーニング
本発明は、例えば補体関連疾患又は障害、例えば免疫複合体及び自己免疫疾患、及び補体介在炎症組織損傷を含む様々な炎症症状の治療に有用性があるCRIgアゴニストを同定するスクリーニングアッセイを含む。補体関連疾患の病理は様々であり、長期間又は短期間の補体活性化、カスケード全体、カスケードの一つだけ(例えば古典又は代替経路)、カスケードの幾つかの成分だけの活性化等々を含むかもしれない。ある疾患では、補体断片の補体生物学的活性が組織損傷及び疾患を生じさせる。従って、補体の阻害剤は高い治療的潜在性を有している。代替経路の選択的阻害剤は、古典経路による血液からの病原体及び他の生物の除去が手を付けられていないままであるので、特に有用であろう。
【0123】
CRIgポリペプチドのアゴニストは天然配列CRIgポリペプチドの定性的生物学的活性を模倣する。好ましくは、生物学的活性はC3bに結合し、及び/又は補体又は補体活性化に影響を及ぼし、特に代替補体経路及び/又はC3転換酵素を阻害する能力である。アゴニストには、例えばイムノアドヘシン、ペプチド模倣薬、天然CRIgの定性的生物学的活性を模倣する非ペプチド小有機分子並びに抗体断片を含むアゴニスト抗体が含まれる。
【0124】
イムノアドヘシンは、異種タンパク質(「アドヘシン」)の結合特異性と免疫グロブリン定常ドメインのエフェクター機能とを兼ね備えた抗体様分子である。構造的には、イムノアドヘシンは、抗体の抗原認識及び結合部位以外(すなわち「異種性の」もの)である所望の結合特異性を有するアミノ酸配列と、免疫グロブリン定常ドメイン配列の融合体を含んでなる。イムノアドヘシン分子のアドヘシン部分は典型的にはレセプター又はリガンドの結合部位を少なくとも含んでなる連続するアミノ酸配列である。イムノアドヘシンの免疫グロブリン定常ドメイン配列は、任意の免疫グロブリン、例えばIgG-1、IgG-2、IgG-3、又はIgG-4サブタイプ、IgA(IgA-1及びIgA-2を含む)、IgE、IgD又はIgMから得られうる。
【0125】
ペプチド模倣薬は、例えば、該化合物がここに記載のCRIg生物学的活性を保持しているならば非天然発生アミノ酸を含むペプチドを含む。同様に、ペプチド模倣薬及びアナログは、本発明のCRIgポリペプチドの重要な構造的エレメントの構造を模倣しCRIg生物学的活性を保持している非アミノ酸化学構造を含みうる。「ペプチド」なる用語は、約50アミノ酸残基未満、及び好ましくは40アミノ酸残基未満の制約された(すなわち、例えば、βターン又はβプリーツシートを先導するアミノ酸の存在、又は例えば、ジスルフィド結合Cys残基の存在によって環状化する構造のある要素を有する)あるいは非制約(例えば直鎖状)アミノ酸配列で、その二量体又はその中間のような多量体を包含するものを指すようにここでは使用される。約40アミノ酸残基未満のペプチドでは、約10及び約30アミノ酸残基の間のペプチド、特に約20アミノ酸残基のペプチドが好ましい。しかしながら、本開示を読めば、当業者ならば、ペプチドを識別するのは、特定のペプチドの長さではなく、そのC3bへ結合しC3転換酵素、特に代替補体経路のC3転換酵素を阻害する能力であることを認めるであろう。
【0126】
CRIgアゴニストのスクリーニングと同定は、CRIg及びC3b:CRIg複合体の結晶構造の開示と、CRIgとC3b間の結合界面の同定によって非常に容易になる。この情報によりCRIgのC3b結合部位を模倣する化合物の設計が可能になる。
【0127】
また、CRIgアゴニストは、(a)計算又は実験手段を用いて化学物質とCRIgポリペプチド又はC3b:CRIg複合体の三次元構造の間のフィッティング操作を実施し;(b)工程(a)で得られたデータを解析して化学物質と天然CRIg又はC3b:CRIg複合体の間の結合の特徴を決定することにより同定することができる。この情報に基づいて、アゴニスト候補を合成することができ、そのアゴニスト特性をCRIg活性の生物学的アッセイにおいて実証することができる。
【0128】
特定の実施態様では、アゴニストは、天然配列CRIg分子又はその保存的アミノ酸置換変異体のC3b結合領域の少なくとも一部を含む化学物質であろう。
【0129】
ペプチド模倣薬は固相ペプチド合成法を用いて簡便に調製できる(Merrifield, J. Am. Chem. Soc., 85: 2149(1964);Houghten, Proc. Natl. Acad. Sci. USA 82: 5132 (1985))。固相合成法は、保護されたアミノ酸を不活性固形担体に結合させることにより推定ペプチドのカルボキシ末端において開始される。不活性な固形担体は、開始アミノ酸のC末端のアンカーとして働くことのできる任意の高分子であり得る。典型的には、高分子担体は、架橋高分子樹脂(例えば、ポリアミド又はポリスチレン樹脂)である。一実施態様では、C末端アミノ酸はポリスチレン樹脂に連結されてベンジルエステルを形成する。高分子支持体は、ペプチド合成の阻害アミノ酸のα-アミノ基を脱保護するのに使用される条件下でペプチドアンカー結合が安定であるように選択される。塩基不安定α保護基が使用されるならば、ペプチドと固形担体の間に酸不安定結合を使用するのが望ましい。例えば、酸不安定エーテル樹脂は、塩基不安定Fmocアミノ酸ペプチド合成に効果的である。あるいは、ペプチドアンカー結合及び差次的に酸分解に移行するα-保護基が使用できる。例えば、フェニルアセトアミドメチル(Pam)樹脂のようなアミノメチル樹脂は、Boc-アミノ酸ペプチド合成に関連して良好に働く。開始アミノ酸が不活性固形担体に連結された後、開始アミノ酸のαアミノ酸保護基を例えば塩化メチレン中のトリフルオロ酢酸(TFA)で除去し、例えばトリエチルアミン(TEA)で中和する。開始アミノ酸のαアミノ基の脱保護に続いて、合成における次のα-アミノ及び側鎖保護したアミノ酸を添加する。次に残りのα-アミノ酸と、必要ならば側鎖保護されたアミノ酸を縮合により所定の順序で続けて結合させ、固形担体に結合した中間体化合物を得る。あるいは、ペプチド断片を伸長している固相ペプチド鎖に付加する前に、幾つかのアミ酸を互いに結合させて所望のペプチド断片を形成してもよい。
【0130】
2つのアミノ酸、又はアミノ酸とペプチド、又はペプチドとペプチドの間の縮合反応は、通常の縮合方法、例えばアジド法、混合酸無水物法、DCC(N,N'-ジシクロヘキシルカルボジイミド)又はDIC(N,N'-ジイソプロピルカルボジイミド)法、活性エステル法、p-ニトロフェニルエステル法、BOP(ベンゾトリアゾール-1-イル-オキシ-トリス[ジメチルアミノ]ホスホニウムヘキサフルオロホスフェート)法、N-ヒドロキシコハク酸イミドエステル法など、及びウッドワード試薬K法等に従って実施される。
【0131】
ペプチドの化学合成に共通なのは、アミノ酸の任意の反応性側鎖基を適切な保護基で保護することである。最終的には、これらの保護基は、所望のポリペプチド鎖が連続的に組み立てられた後に除去される。更に共通なのは、アミノ酸又はペプチド断片上のαアミノ基を保護するが、アミノ酸又はペプチド断片のC末端カルボキシル基は成長固形相ポリペプチド鎖の遊離N末端アミノ基と反応し、続いてαアミノ酸基を選択的に除去し、次のアミノ酸又はペプチド断片を固相ペプチド鎖に付加させる。従って、ペプチド合成において、ペプチド鎖の所望の配列に位置するアミノ酸残基の各々を含み、その個々の残基が側鎖保護基を担持する中間体化合物が生成されるのが一般的である。これらの保護基は、実質的に同時に除去でき、固相から取り外した後に所望のポリペプチド生成物が生成される。
【0132】
α-及びε-アミノ酸側鎖を、ベンジルオキシカルボニル(Zと略)、イソニコチニルオキシカルボニル(iNOC)、o-クロロベンジルオキシカルボニル[Z(2Cl)]、p-ニトロベンジルオキシカルボニル[Z(NO2)]、p-メトキシベンジルオキシカルボニル[Z(OMe)]、t-ブトキシカルボニル(Boc)、t-アミルオキシカルボニル(Aoc)、イソボロニルオキシカルボニル、アダマンチルオキシカルボニル、2-(4-ビフェニル)-2-プロピル-オキシカルボニル(Bpoc)、9-フルオレニルメトキシカルボニル(Fmoc)、メチルスルホンエトキシカルボニル(Msc)、トリフルオロアセチル、フタリル、ホルミル、2-ニトロフェニルスルフェニル(NPS)、ジフェニルホスフィノチオイル(Ppt)、及びジメチロホスフィノチオイル(Mpt)基などで保護できる。
【0133】
カルボキシ官能基の保護基としては、ベンジルエステル(OBzl)、シクロヘキシルエステル(Chx)、4-ニトロベンジルエステル(ONb)、t-ブチルエステル(Obut)、4-ピリジルメチルエステル(OPic)等が例示される。アミノ及びカルボキシル基以外の官能基を有するアルギニン、システイン、及びセリンなどの特定のアミノ酸は、適当な保護基で保護するのが望ましいことが多い。例えば、アルギニンのグアニジノ基は、ニトロ、p-トルエンスルホニル、ベンジルオキシカルボニル、アダマンチルオキシカルボニル、p-メトキシベンゼンスルホニル、4-メトキシ-2,6-ジメチルベンゼンスルホニル(Nds)、1,3,5-トリメチルフェニルスルホニル(Mts)等で保護される。システインのチオール基は、p-メトキシベンジル、トリチルなどで保護されうる。
【0134】
上述の多くの阻害アミノ酸は、Novabiochem(San Diego, Calif.)、Bachem CA(Terrence, Calif.)又はPeninsula Labs(Belmont, Calif.)のような市販元から得ることができる。
【0135】
所望のアミノ酸配列が完成した後、ペプチドは固形担体から切断され、回収され、精製されうる。ペプチドは、ペプチド固相結合を崩壊することのできる試薬により固形担体から取り除かれ、場合によっては、ペプチドの阻害側鎖官能基を脱保護する。一実施態様では、ペプチドは、残っている側鎖保護基も取り除く液体フッ化水素酸(HF)での酸分解により固相から取り外す。好ましくは、ポリペプチド中の残基のアルキル化(例えば、メチオニン、システイン、及びチロシン残基のアルキル化)を回避するために、酸分解反応混合物はチオ-クレゾール及びクレゾール捕捉剤を含む。HF切断後、樹脂はエーテルで洗浄され、遊離ペプチドが酢酸溶液の連続洗浄で固相から抽出される。組み合わされた洗浄液は凍結乾燥され、ペプチドが精製される。
【0136】
天然配列CRIgポリペプチドのアンタゴニストは過剰な補体活性化及び亢進された補体介在宿主防御メカニズムから恩恵を受ける症状の治療に有用性が見出される。CRIgに対する抗体のようなCRIgアンタゴニストは、腫瘍(癌)治療における免疫アジュバント治療に使用することができる。T細胞がヒト腫瘍特異的抗原を認識することは現在は十分に確立されている。遺伝子のMAGE、BAGE及びGAGEファミリーによりコードされる腫瘍抗原の一グループは全ての成人の正常組織においてサイレントであるが、腫瘍、例えばメラノーマ、肺腫瘍、頭頸部腫瘍、膀胱癌においては有意な量で発現される。DeSmet, C.等(1996) Proc. Natl. Acad. Sci.93:7149。T細胞の同時刺激により、インビトロ及びインビボの双方において、腫瘍の退行及び抗腫瘍反応が誘導されることが示されている。Melero. I.等, Nature Medicine (1997) 3:682;Kwon. E.D.等, Proc. Natl. Acad. Sci.USA (1997) 94:8099;Lynch, D.H.等, Nature Medicine (1997) 3:625;Finn, O.J.及びLotze. M.T., J. Immunol. (1998) 21:114。本発明によって設計され同定されたCRIgアンタゴニストはアジュバントとして単独で又は増殖調節剤、細胞毒性薬又は化学療法剤と共に投与することができ、T細胞増殖/活性化及び腫瘍抗原に対する抗腫瘍反応を刺激する。増殖調節剤、細胞毒性薬又は化学療法剤は既知の投与方法を使用して一般的な量で投与することができる。本発明のCRIgアンタゴニストによる免疫刺激活性により、増殖調節剤、細胞毒性薬又は化学療法剤の量を減らすことができ、よって、潜在的に患者に対する毒性を低下させることができる。
【0137】
ある種のマクロファージは腫瘍根絶に関わっているけれども、多くの固形腫瘍は腫瘍増殖を支援するマクロファージを含んでいることが知られている(Bingle等, J Pathol 196:254-265 (2002);Mantovani等, Trends Immunol 23:549-555 (2002))。これらのマクロファージはその表面にCRIgを含みうる。CRIgの補体活性化を阻害する能力をブロックする抗体を用いて腫瘍細胞上の補体を活性化させ、補体介在溶解により腫瘍を根絶するのを補助することができよう。このアプローチ法はCRIg陽性マクロファージを含む腫瘍に特に有用であることが予想される。
【0138】
アンタゴニストの好ましい群には天然CRIgに特異的に結合しその生物学的活性を阻害する抗体が含まれる。
【0139】
抗体(アゴニストとアンタゴニストの双方)の例には、ポリクローナル、モノクローナル、ヒト化、二重特異的及びヘテロコンジュゲート抗体、及び抗体断片が含まれる。
【0140】
本発明のポリペプチドを認識してそれに結合し、又はそれに対するアンタゴニストとして作用する抗体は、あるいはモノクローナル抗体であってもよい。モノクローナル抗体は、Kohler及びMilstein, Nature, 256:495 (1975)に記載されているようなハイブリドーマ法を使用することで調製することができる。ハイブリドーマ法では、マウス、ハムスター又は他の適切な宿主動物を典型的には免疫剤により免疫化することで、免疫剤に特異的に結合する抗体を生成するかあるいは生成可能なリンパ球を誘発する。あるいは、リンパ球をインビトロで免疫化することもできる。
【0141】
免疫剤は、典型的には本発明のCRIgポリペプチド、その抗原性断片又は融合タンパク質を含む。一般に、ヒト由来の細胞が望まれる場合には末梢血リンパ球(「PBL」)が使用され、あるいは非ヒト哺乳動物源が望まれている場合は、脾臓細胞又はリンパ節細胞が使用される。ついで、ポリエチレングリコール等の適当な融合剤を用いてリンパ球を不死化株化細胞と融合させ、ハイブリドーマ細胞を形成する[Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103]。不死化株化細胞は、通常は、形質転換した哺乳動物細胞、特に齧歯動物、ウシ、及びヒト由来の骨髄腫細胞である。通常、ラット又はマウスの骨髄腫株化細胞が使用される。ハイブリドーマ細胞は、好ましくは、未融合の不死化細胞の生存又は増殖を阻害する一又は複数の物質を含有する適切な培地で培養される。例えば、親細胞が、酵素のヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRT又はHPRT)を欠いていると、ハイブリドーマの培地は、典型的には、ヒポキサチン、アミノプテリン及びチミジンを含み(「HAT培地」)、この物質がHGPRT欠乏性細胞の増殖を阻止する。
【0142】
好ましい不死化株化細胞は、効率的に融合し、選択された抗体生成細胞による安定した高レベルの抗体発現を支援し、HAT培地のような培地に対して感受性であるものである。より好ましい不死化株化細胞はマウス骨髄腫株であり、これは例えばカリフォルニア州サンディエゴのSalk Institute Cell Distribution Centerやバージニア州マナッサスのアメリカン・タイプ・カルチャー・コレクションより入手可能である。ヒトモノクローナル抗体を生成するためのヒト骨髄腫及びマウス-ヒト異種骨髄腫株化細胞も開示されている[Kozbor, J. Immunol., 133:3001 (1984);Brodeur等, Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp.51-63]。
【0143】
ついでハイブリドーマ細胞が培養される培養培地を、本発明のポリペプチドに対する又は本発明のポリペプチドと同様の活性を有するモノクローナル抗体の存在について検定することができる。好ましくは、ハイブリドーマ細胞によって生成されたモノクローナル抗体の結合特異性は免疫沈降又はラジオイムノアッセイ(RIA)や酵素結合免疫測定法(ELISA)等のインビトロ結合検定法によって測定する。このような技術及びアッセイは当該分野において知られている。モノクローナル抗体の結合親和性は、例えば、Munson及びPollard, Anal. Biochem., 107:220 (1980)によるスキャッチャード解析法によって測定することができる。
【0144】
所望のハイブリドーマ細胞が同定された後、クローンを限界希釈法によりサブクローニングし、標準的な方法で増殖させることができる[上掲のGoding]。この目的のための適当な培地には、例えば、ダルベッコの改変イーグル培地及びRPMI-1640倍地が含まれる。あるいは、ハイブリドーマ細胞は哺乳動物においてインビボで腹水として増殖させることもできる。
【0145】
サブクローンによって分泌されたモノクローナル抗体は、例えばプロテインA−セファロース法、ヒドロキシルアパタイトクロマトグラフィー法、ゲル電気泳動法、透析法又はアフィニティークロマトグラフィー等の一般的な免疫グロブリン精製法によって培養培地又は腹水液から単離又は精製されうる。
【0146】
また、モノクローナル抗体は、組換えDNA法、例えば米国特許第4816567号に記載された方法により作成することができる。本発明のモノクローナル抗体をコードするDNAは、常套的な方法を用いて(例えば、マウス抗体の重鎖及び軽鎖をコードする遺伝子に特異的に結合可能なオリゴヌクレオチドプローブを使用して)、容易に単離し配列決定することができる。本発明のハイブリドーマ細胞はそのようなDNAの好ましい供給源となる。ひとたび単離されたら、DNAは発現ベクター内に配することができ、これが宿主細胞、例えばサルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、あるいは免疫グロブリンタンパク質をさもなければ生成しない骨髄腫細胞内に形質移入され、組換え宿主細胞内でモノクローナル抗体の合成をすることができる。また、DNAは、例えば相同マウス配列に換えてヒト重鎖及び軽鎖定常ドメインのコード配列を置換することにより[上掲の米国特許第4816567号;Morrison等]、又は免疫グロブリンコード配列に非免疫グロブリンポリペプチドのコード配列の一部又は全部を共有結合することにより修飾することができる。このような非免疫グロブリンポリペプチドは、本発明の抗体の定常ドメインに置換でき、あるいは本発明の抗体の一つの抗原結合部位の可変ドメインに置換でき、キメラ性二価抗体を生成する。
【0147】
抗体は好ましくは一価抗体である。一価抗体の調製方法は当該分野においてよく知られている。例えば、一つの方法は免疫グロブリン軽鎖と修飾重鎖の組換え発現を含む。重鎖は一般的に、重鎖の架橋を防止するようにFc領域の任意の点で切断される。あるいは、関連するシステイン残基を他のアミノ酸残基で置換するか欠失させて架橋を防止する。
【0148】
一価抗体の調製には、またインビトロ法が適している。抗体の消化による、その断片、特にFab断片の生成は、当該分野において知られている慣用的技術を使用して達成することができる。
【0149】
本発明の抗体は、更にヒト化抗体又はヒト抗体を含みうる。非ヒト(例えばマウス)抗体のヒト化型とは、キメラ免疫グロブリン、免疫グロブリン鎖又はその断片(例えばFv、Fab、Fab’、F(ab’)2あるいは抗体の他の抗原結合サブ配列)であって、非ヒト免疫グロブリンに由来する最小配列を含むものである。ヒト化抗体はレシピエントの相補性決定領域(CDR)の残基が、マウス、ラット又はウサギのような所望の特異性、親和性及び能力を有する非ヒト種(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(レシピエント抗体)を含む。幾つかの例では、ヒト免疫グロブリンのFvフレームワーク残基は、対応する非ヒト残基によって置換されている。また、ヒト化抗体は、レシピエント抗体にも、移入されたCDRもしくはフレームワーク配列にも見出されない残基を含んでいてもよい。一般に、ヒト化抗体は、全てあるいは殆ど全てのCDR領域が非ヒト免疫グロブリンのものに対応し、全てあるいは殆ど全てのFR領域がヒト免疫グロブリンコンセンサス配列のものである、少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含む。ヒト化抗体は、最適には免疫グロブリン定常領域(Fc)、典型的にはヒトの免疫グロブリンの定常領域の少なくとも一部を含んでなる[Jones等, Nature, 321:522-525 (1986);Riechmann等, Nature, 332:323-329 (1988);及びPresta, Curr. Op Struct. Biol., 2:593-596 (1992)]。
【0150】
非ヒト抗体をヒト化する方法はこの分野でよく知られている。一般的に、ヒト化抗体には非ヒト由来の一つ又は複数のアミノ酸残基が導入される。これら非ヒトアミノ酸残基は、しばしば、典型的には「移入」可変ドメインから得られる「移入」残基と称される。ヒト化は基本的にウィンター(Winter)及び共同研究者[Jones等, Nature, 321:522-525 (1986);Riechmann等, Nature, 332:323-327 (1988);Verhoeyen等, Science, 239:1534-1536 (1988)]の方法に従って、齧歯類CDR又はCDR配列をヒト抗体の対応する配列の代わりに用いることにより実施される。よって、このような「ヒト化」抗体は、無傷のヒト可変ドメインより実質的に少ない分が非ヒト種由来の対応する配列で置換されたキメラ抗体(米国特許第4816567号)である。実際には、ヒト化抗体は典型的には幾つかのCDR残基及び場合によっては幾つかのFR残基が齧歯類抗体の類似する部位からの残基によって置換されたヒト抗体である。
【0151】
また、ヒト抗体は、ファージ表示ライブラリー[Hoogenboom及びWinter, J. Mol. Biol., 227:381(1991);Marks等, J. Mol. Biol., 222:581 (1991)]を含むこの分野で知られた種々の方法を用いて作成することもできる。また、Cole等及びBoerner等の方法も、ヒトモノクローナル抗体の調製に利用することができる[Cole等, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985)及びBoerner等, J. Immunol., 147(1):86-95 (1991)]。同様に、ヒト抗体はヒト免疫グロブリン座位をトランスジェニック動物、例えば内在性免疫グロブリン遺伝子は部分的又は完全に不活性化されたマウスに導入することにより産生することができる。投与の際に、遺伝子再配列、組立、及び抗体レパートリーを含むあらゆる観点においてヒトに見られるものに非常に類似しているヒト抗体の生産が観察される。このアプローチは、例えば米国特許第5545807号;同第5545806号;同第5569825号;同第5625126号;同第5633425号;同第5661016号、及び次の科学文献:Marks等, Bio/Technology 10, 779-783 (1992);Lonberg等, Nature 368 856-859 (1994);Morrison, Nature 368, 812-13 (1994);Fishwild等, Nature Biotechnology 14, 845-51 (1996);Neuberger, Nature Biotechnology 14, 826 (1996);Lonberg及びHuszar, Intern. Rev. Immunol. 13 65-93 (1995)に記載されている。
【0152】
二重特異性抗体は、少なくとも2つの異なる抗原に対して結合特異性を有するモノクローナル抗体、好ましくはヒトもしくはヒト化抗体である。本発明の場合、結合特異性の一方は本発明のポリペプチドに対するものであり、他方は任意の他の抗原、好ましくは細胞表面タンパク質又はレセプター又はレセプターサブユニットに対してであり得る。
【0153】
二重特異性抗体を作製する方法は当該分野において知られている。伝統的には、二重特異性抗体の組換え生産は、二つの重鎖が異なる特異性を持つ二つの免疫グロブリン重鎖/軽鎖対の同時発現に基づく(Milstein及びCuello, Nature, 305:537-539 [1983])。免疫グロブリンの重鎖と軽鎖を無作為に取り揃えるため、これらハイブリドーマ(クアドローマ)は10種の異なる抗体分子の潜在的混合物を生成し、その内の一種のみが正しい二重特異性構造を有する。正しい分子の精製は、アフィニティークロマトグラフィー工程によって通常達成される。同様の手順が1993年5月13日公開の国際公開第93/08829号、及びTraunecker等, EMBO J.,10:3655-3656 (1991)に開示されている。
【0154】
所望の結合特異性(抗体-抗原結合部位)を有する抗体可変ドメインを免疫グロブリン定常ドメイン配列に融合できる。融合は、好ましくは少なくともヒンジ部、CH2及びCH3領域の一部を含む免疫グロブリン重鎖定常ドメインとのものである。少なくとも一つの融合には軽鎖結合に必要な部位を含む第一の重鎖定常領域(CH1)が存在することが望ましい。免疫グロブリン重鎖融合をコードするDNA、及び望むのであれば免疫グロブリン軽鎖を、別々の発現ベクターに挿入し、適当な宿主生物に同時形質移入する。二重特異性抗体を作成するための更なる詳細については、例えば、Suresh等, Methods in Enzymology, 121:210 (1986)を参照されたい。
【0155】
ヘテロコンジュゲート抗体は、2つの共有結合した抗体からなる。このような抗体は、例えば、免疫系細胞を不要な細胞に対してターゲティングさせるため(米国特許第4676980号)及びHIV感染の治療のために(国際公開第91/00360;国際公開第92/200373;欧州特許第03089号)提案されている。この抗体は、架橋剤に関連したものを含む合成タンパク化学における既知の方法を使用して、インビトロで調製することができると考えられる。例えば、ジスルフィド交換反応を使用するか又はチオエーテル結合を形成することによって、免疫毒素を作成することができる。この目的に対して好適な試薬の例には、イミノチオレート及びメチル-4-メルカプトブチルイミダート、及び例えば米国特許第4676980号に開示されたものが含まれる。
【0156】
本発明の抗体をエフェクター機能について改変し、例えば免疫関連疾患の治療における抗体の効力を向上させることは望ましい。例えば、システイン残基をFc領域に導入し、それにより、この領域に鎖間ジスルフィド結合を形成するようにしてもよい。そのようにして生成された同種二量体抗体は、向上したインターナリゼーション能力及び/又は増加した補体媒介細胞殺傷及び抗体依存細胞性細胞障害性(ADCC)を有する可能性がある。Caron等, J. Exp. Med. 176: 1191-1195 (1992)及びShopes, B. J. Immunol. 148: 2918-2922 (1992)参照。また、向上した抗腫瘍活性を持つ同種二量体抗体は、Wolff等, Cancer Research 53: 2560-2565 (1993)に記載されている異種二官能性架橋を用いて調製することができる。あるいは、抗体は、2つのFc領域を有するように加工して、それにより補体溶解及びADCC能力を向上させることもできる。Stevenson等, Anti-Cancer Drug Design 3: 219-230 (1989)参照。
【0157】
また、本発明は、化学療法剤、毒素(例えば、細菌、真菌、植物又は動物由来の酵素活性毒素、又はその断片)などの細胞傷害性剤、あるいは放射性同位体(つまり、放射性コンジュゲート)と抱合している抗体を含む免疫複合体にも関する。
【0158】
このような免疫複合体の生成に有用な化学療法剤は上述の通りである。用いることのできる酵素活性毒素及びその断片には、ジフテリアA鎖、ジフテリア毒素の非結合活性断片、(緑膿菌からの)外毒素A鎖、リシンA鎖、アブリンA鎖、モデクシン(modeccin)A鎖、アルファ-サルシン、アレウリテス・フォーディ(Aleurites fordii)タンパク質、ジアンチン(dianthin)タンパク質、フィトラカ・アメリカーナ(Phytolaca americana)タンパク質(PAPI、PAPII、及びPAP-S)、モモルディカ・チャランチア(momordica charantia)インヒビター、クルシン(curcin)、クロチン(crotin)、サパオナリア・オフィシナリス(sapaonaria officinalis)インヒビター、ゲロニン(gelonin)、ミトゲリン(mitogellin)、レストリクトシン(restrictocin)、フェノマイシン(phenomycin)、エノマイシン(enomycin)及びトリコテセン(tricothecene)が含まれる。放射性コンジュゲート抗体の生成には、様々な放射性ヌクレオチドが利用可能である。例としては、212Bi、131I、131In、90Y及び186Reが含まれる。
【0159】
抗体及び細胞傷害性薬の複合体は、種々の二官能性タンパク質カップリング剤、例えば、N-スクシンイミジル-3-(2-ピリジルジチオール)プロピオナート(SPDP)、イミノチオラン(IT)、イミドエステルの二官能性誘導体(ジメチルアジピミデートHCL等)、活性エステル(ジスクシンイミジルスベレート等)、アルデヒド(グルタルアルデヒド等)、ビス-アジド化合物(ビス(p-アジドベンゾイル)ヘキサンジアミン等)、ビス-ジアゾニウム誘導体(ビス-(p-ジアゾニウムベンゾイル)-エチレンジアミン等)、ジイソシアネート(トリエン2,6-ジイソシアネート等)、及びビス-活性フッ素化合物(1,5-ジフルオロ-2,4-ジニトロベンゼン等)を用いて作製できる。例えば、リシン免疫毒素は、Vitetta等, Science 238: 1098 (1987)に記載されているように調製することができる。カーボン-14標識1-イソチオシアナトベンジル-3-メチルジエチレントリアミン五酢酸(MX-DTPA)は、放射性ヌクレオチドの抗体への抱合のためのキレート剤の例である。国際公開94/11026参照。
【0160】
他の実施態様では、組織の予備標的化で使用するために、抗体は「レセプター」(ストレプトアビジン等)に抱合されてもよく、抗体-レセプター複合体は患者に投与され、ついで清澄化剤を用いて未結合複合体を循環から除去し、次に細胞傷害性薬(例えば、放射性ヌクレオチド等)に抱合された「リガンド」(アビジン等)を投与する。
【0161】
CRIg及びそのアゴニスト及びアンタゴニストは、天然CRIg分子の生物学的活性を模倣し又は阻害するかどうかを決定するために様々なインビトロ及びインビボアッセイにおいて試験することができる。
【0162】
補体活性化、特に代替補体経路を阻害するその能力の結果として、CRIgポリペプチドは、補体関連疾患及び病理状態の予防及び/又は治療において有用性が見出される。このような疾患及び症状には、限定するものではないが、補体関連の炎症性及び自己免疫疾患が含まれる。
【0163】
補体関連疾患の特定の例には、限定するものではないが、リウマチ様関節炎(RA)、急性呼吸促迫症候群(ARDS)、虚血及び再灌流後の遠隔組織傷害、心肺バイパス手術中の補体活性化、皮膚筋炎、天疱瘡、ループス腎炎及びその結果の糸球体腎炎及び脈管炎、心肺バイパス、心臓麻痺誘発性冠血管内皮機能不全、II型膜性増殖性糸球体腎炎、IgA腎症、急性腎不全、クリオグロブリン血症、抗リン脂質症候群、黄斑変性疾患及び他の補体関連の眼の症状、非滲出性及び滲出性AMDを含む加齢性黄斑変性症(AMD)、糖尿病性網膜症(DR)、眼内炎、ブドウ膜炎、同種移植、異物移植、超急性拒絶反応、血液透析、慢性閉塞肺促迫症候群(COPD)、喘息、遺伝性血管浮腫、発作性夜間血色素尿症、アルツハイマー病、アテローム性動脈硬化症、誤嚥性肺炎、蕁麻疹、例えば慢性特発性蕁麻疹、溶血性尿毒症症候群、子宮内膜症、心原性ショック、虚血再かん流傷害、多発性硬化症(MS)が含まれる。
【0164】
AMDは、60歳以上の個人における不可逆的な視力障害の主たる原因である黄斑の加齢性変性症である。非滲出性(ドライ)及び滲出性(ウェット)AMDの2つのタイプのAMDが存在している。ドライ又は非滲出型は中心網膜(黄斑)並びにRPE上の沈積物(ドルーゼ)の根底にある網膜色素上皮(RPE)内の萎縮及び異常肥大変化を含む。非滲出型ARMDの患者は、脈絡膜新生血管膜(CNVM)と呼ばれる異常な血管が網膜、漏出液及び血液の下で発達し、最終的には網膜内及び網膜下で失明に至る円板状瘢痕を生じるARMDのウェット又は滲出型まで進行しうる。通常は滲出型ARMDの前躯体である非滲出型ARMDはより一般的である。非滲出型ARMDの提示は様々である;ハードドルーゼ、ソフトドルーゼ、RPE地図状萎縮、及び色素凝集が存在しうる。補体成分はAMDの早期にRPE上に付着し、ドルーゼの主要構成成分である。補体H因子(CFH)多形性がAMDの起因リスクの50%を占めることが最近報告されている(Klein等, Science 308:385-9 (2005))。
【0165】
CRIgは、カテゴリー3及びカテゴリー4AMDを含む高リスクのAMDの治療において特に有用である。カテゴリー3AMDは、双方の眼に進行AMDがなく、少なくとも一つの眼が20/32又はそれ以上の視力、少なくとも一つの大きなドルーゼ(例えば125μm)、広範な(ドルーゼ面積で測定)中間ドルーゼ、又は黄斑の中心を含まない地図状萎縮(GA)又はこれらの任意の組合せを有することによって特徴付けられる。カテゴリー4高リスクAMDは、20/32又はそれ以上の視力によって特徴付けられ、インデックス眼において進行AMD(黄斑の中心又は脈絡膜血管新生の特徴を含むGA)はない。他眼は進行AMD、又はAMD黄斑症に帰す20/32未満の視力によって特徴付けられている。典型的には、未治療ならば、高リスクAMDは、カテゴリー1又は2(高リスクではない)AMDの進行速度よりも約10−30倍高い速度で脈絡膜血管新生(CNV)に速やかに進行する。従って、CRIgはCNVとAMDの予防と治療に有用性が見出される。
【0166】
補体関連疾患の例としての炎症性症状のより広範なリストには、例えば炎症性腸疾患(IBD)、全身性エリテマトーデス、リウマチ様関節炎、若年性慢性関節炎、脊椎関節症、全身性硬化症(強皮症)、特発性炎症ミオパシー(皮膚筋炎、多発性筋炎)、シェーグレン症候群、全身性血管炎、サルコイドーシス、自己免疫性溶血性貧血(免疫再生不良性貧血、発作性夜間血色素尿)、自己免疫性血小板減少(特発性血小板減少性紫斑病、免疫仲介血小板減少)、甲状腺炎(グレーブス疾患、ハシモト甲状腺炎、若年性リンパ球性甲状腺炎、萎縮性甲状腺炎)、真性糖尿病、免疫仲介腎疾患(糸球体腎炎、尿細管間質性腎炎)、中枢及び末梢神経系の脱髄疾患、例えば多発性硬化症、特発性脱随性多発神経障害、又はギラン-バレー症候群、及び慢性炎症脱随性多発神経障害、肝胆道疾患、例えば感染性肝炎(A型、B型、C型、D型、E型肝炎及び他の非肝親和性(nonhepatotropic)ウイルス)、自己免疫慢性活動性肝炎、原発性胆汁性肝硬変症、肉芽腫性肝炎、及び硬化性胆管炎、炎症性腸疾患(潰瘍性大腸炎;クローン病)、グルテン過敏性腸疾患、及びウィップル病、水疱性皮膚疾患、多形性紅斑及び接触性皮膚炎を含む自己免疫又は免疫媒介皮膚疾患、乾癬、アレルギー性疾患、例えば喘息、アレルギー性鼻炎、アトピー性皮膚炎、食物過敏症及び蕁麻疹、肺の免疫疾患、例えば好球性肺炎、特発性肺線維症及び過敏性肺炎、拒絶反応及び移植片対宿主疾患を含む移植関連疾患が含まれる。
【0167】
全身性エリテマトーデスでは、疾患の中心的な媒介物は自己タンパク質/組織に対する自己反応性抗体の産生と、続いての免疫媒介炎症の発生である。抗体は直接的又は間接的に組織傷害を媒介する。Tリンパ球は組織損傷に直接的には関与していることは示されていないが、Tリンパ球は自己反応性抗体の発生に必要である。よって、疾患の発生はTリンパ球に依存している。腎臓、肺、筋骨格、皮膚粘膜、眼、中枢神経系、心臓血管系、胃腸管、骨髄及び血液を含む複数の器官及び系が臨床的に冒される。
【0168】
リウマチ様関節炎(RA)は、主に複数の関節の滑膜に関連する慢性全身性自己免疫炎症疾患であり、結果として関節軟骨に傷害が生じる。病原はTリンパ球依存性であり、リウマチ因子、自己IgGに対する自己抗体の生成に付随し、結果として滑液及び血液において高レベルに達する免疫複合体を生成する。関節中のこれらの複合体は、滑膜中へのリンパ球及び単球の顕著な浸潤と、続いての顕著な滑膜変化を誘発し;多数の好中球の添加により同様の細胞で浸潤されるならば、関節空間/液でもしかりである。冒されている組織は、多くの場合対称的なパターンで、主に関節である。しかしながら、二つの主な形態の関節外疾患もまた生じる。一形態は進行中の進行性関節疾患及び肺線維症の典型的病巣、血管炎、及び皮膚潰瘍を伴う関節外障害の発生である。関節外疾患の第二の形態はいわゆるフェルティー症候群であり、これは、RA疾患過程の末期、時には関節疾患が鎮静した後に生じ、好中球減少、血小板減少及び脾肥大の存在に関与する。これには、梗塞、皮膚潰瘍及び壊疽の形成を伴う複数の器官において血管炎が付随する。多くの場合、患者には、発病している関節上にある皮下組織にリウマチ様小結節が発達し;その小結節は、末期には混合炎症細胞浸潤に包囲された壊死性中心を有する。RAにおいて生じる可能性のある他の徴候には:心外膜炎、胸膜炎、冠動脈炎、肺線維症を伴う間質性肺炎、乾性角結膜炎、及びリウマチ様小結節が含まれる。
【0169】
若年性慢性関節炎は、多くの場合16歳未満で発症する慢性特発性炎症疾患である。その表現型はRAと幾つかの類似点があり;リウマチ因子が陽性である患者の中には若年性リウマチ様関節炎に分類されるものもいる。この疾患は三つの主要なカテゴリー:小関節(pauarticular)、多関節及び全身性に細分類される。関節炎は重度で典型的には破壊的であり、関節強直症及び遅延成長に至る。他の徴候には慢性前部ブドウ膜炎及び全身性アミロイド症が含まれる。
【0170】
脊椎関節症は、幾つかの共通した臨床的特徴とHLA-B27遺伝子産物の発現との共通の関連性を持つ疾患のグループである。該疾患には:強直性脊椎炎、ライター症候群(反応性関節炎)、炎症性大腸疾患に関連した関節炎、乾癬に関連した脊椎炎、若年発生脊椎関節症及び未分化脊椎関節症が含まれる。顕著な特徴には、脊椎炎を伴うか伴わない仙腸関節炎;炎症非対称性関節炎;HLA-B27(クラスI MHCのHLA-B座位にある血清学的に定義された対立遺伝子)との関連;眼の炎症、及び他のリウマチ疾患に関連した自己抗体の不在が含まれる。疾患の誘導に対する鍵として最も関わっている細胞はCD8+Tリンパ球で、クラスI MHC分子により提示される抗原を標的としている細胞である。CD8+T細胞は、MHCクラスI分子により発現された外来ペプチドであるかのように、クラスI MHC対立遺伝子HLA-B27に対して反応する。HLA-B27のエピトープが細菌性又は他の微生物の抗原性エピトープを模倣し、よってCD8+細胞の反応を誘発すると仮定されている。
【0171】
全身性硬化症(強皮症)は病因がよく知られていない。疾患の顕著な特徴は皮膚の硬結であり;これは活性な炎症プロセスにより誘発されると思われる。強皮症は局部的又は全身性であり:血管病巣が共通しており、微小血管系における内皮細胞傷害が全身性硬化症の発達における初期の重要な事象であり;血管傷害は免疫媒介されうる。免疫学的基準は、皮膚病巣における単核細胞浸潤の存在と、多くの患者において抗細胞核抗体の存在によって導かれる。多くの場合、ICAM-1が皮膚病巣の線維芽細胞の細胞表面でアップレギュレーションされ、これらの細胞とのT細胞の相互作用が疾患の病因においてある役割を担っていることが示唆される。関連する他の器官には:胃腸管:結果的に異常なぜん動/運動性となる平滑筋萎縮症及び線維症:腎臓:小弓形及び小葉間動脈に影響を及ぼし、結果として腎皮質の血流が低下し、タンパク尿、高窒素血尿及び高血圧になる同心性内皮下内膜増殖:骨格筋:萎縮、間質性線維症:炎症:肺:間質性肺炎及び間質性線維症:及び心臓:収縮バンド壊死、瘢痕/線維症が含まれる。
【0172】
皮膚筋炎、多発性筋炎及び他のものを含む特発性炎症ミオパシーは病因がよく知られていない慢性筋肉炎症疾患であり、筋肉の弱化に至る。筋肉損傷/炎症は多くの場合対照的で進行性である。自己抗体が多くの形態と関連している。これらの筋炎特異的自己抗体は、タンパク質合成に関与する成分、タンパク質及びRNAに対して産生されその機能を阻害する。
【0173】
シェーグレン症候群は、免疫媒介炎症と、涙腺及び唾液腺の続く機能破壊によるものである。この疾患は炎症結合組織疾患に関連するか又はそれに伴う場合がある。この疾患は、双方とも小RNA-タンパク質複合体であるRo及びLa抗原に対する自己抗体産生に関連している。病巣は乾性角結膜炎、口内乾燥症で、胆汁性硬変、末梢又は感覚ニューロパシー、及び明白な紫斑病を含む他の徴候又は関連を伴うものに至る。
【0174】
全身性血管炎症は一次病巣が炎症で、続いて血管にダメージを受け、結果として冒された脈管により供給される組織に虚血/壊死/変性が生じ、幾つかのケースでは最終的な末端器官機能障害になるといった疾患である。また、血管炎(vasculitides)は他の免疫炎症媒介疾患、例えばリウマチ様関節炎、全身性硬化症等、特に免疫複合体の生成に関連した疾患等の続発症として又は二次病変として生じる場合がある。原発性全身性血管炎症グループの疾患には:全身壊死性血管炎:多動脈炎結節(polyarteritis nodosa)、アレルギー性脈管炎及び肉芽腫症、多脈管炎:ヴェゲナー肉芽腫症;リンパ腫様肉芽腫症:及び巨細胞動脈炎が含まれる。その他の血管炎には:粘膜皮膚リンパ節症候群(MLNS又は川崎病)、隔離されたCNS血管炎、ベーチェット(Behet's)病、閉塞性血栓性血管炎(バージャー病)及び皮膚壊死性細静脈炎(venulitis)が含まれる。列挙した血管炎のほとんどの種類の発病メカニズムは、主に脈管壁に免疫グロブリン複合体が付着し、続いてADCC、補体活性又は双方を介して炎症反応が誘発されることによると考えられている。
【0175】
サルコイドーシスは、体内のほとんど全ての組織中における類上皮細胞肉芽腫の存在により特徴づけられる病因がよく知られていない病状であり;肺の関与が最も一般的である。病因は疾患部位に活性マクロファージ及びリンパ球が残留していることに関連しており、続いてこれらの細胞型より放出される局部的又は全身的活性産物の放出の結果として慢性続発症が生じる。
【0176】
自己免疫性溶血性貧血、免疫再生不良性貧血、及び発作性夜間血色素尿を含む自己免疫溶性血性貧血は、赤血球(幾つかの場合においては血小板もまた含む他の血液細胞)表面で発現した抗原と反応する抗体が産出される結果によるものであり、補体媒介溶解及び/又はADCC/Fc-レセプター-媒介メカニズムを介して、その抗体被覆細胞の除去に反映される。
【0177】
他の臨床的環境における血小板減少性紫斑病及び免疫仲介血小板減少を含む自己免疫性血小板減少では、血小板破壊/除去が、抗体又は補体が血小板に接合し、続いて補体溶解、ADCC又はFc-レセプター-媒介メカニズムにより除去される結果として生じる。
【0178】
グレーブス疾患、ハシモト甲状腺炎、若年性リンパ球性甲状腺炎、及び萎縮性甲状腺炎を含む甲状腺炎は、甲状腺内に存在し多くの場合甲状腺に特異的なタンパク質と反応する抗体の産生を伴う、甲状腺抗原に対する自己免疫反応の結果によるものである。自然のモデル:ラット(BUF及びBBラット)及びチキン(肥満チキン種);誘導性モデル:サイログロブリン、甲状腺ミクロソーム抗原(甲状腺ペルオキシダーゼ)のいずれかでの動物の免疫化を含む実験用モデルが存在する。
【0179】
I型真性糖尿病又はインスリン依存性糖尿病は膵臓ランゲルハンス島β細胞の自己免疫破壊であり;この破壊は自己抗体及び自己反応性T細胞により媒介される。また、インスリン又はインスリンレセプターに対する抗体は、インスリン-非反応性の表現型をつくりだすことができる。
【0180】
糸球体腎炎及び尿細管間質性腎炎を含む免疫仲介腎疾患は、腎抗原に対する自己反応性抗体又はT細胞が産生される結果として直接的に、又は他の非腎抗原に対して反応性である、腎臓中の抗体及び/又は免疫複合体の沈着の結果として間接的に、腎組織に抗体又はT細胞媒介傷害が生じることによるものである。よって、免疫複合体の生成をもたらす他の免疫媒介疾患により、間接的続発症として免疫媒介腎疾患も誘発しうる。直接的及び間接的免疫メカニズムの双方により、結果として、器官機能が損なわれ、幾つかの場合では腎臓機能不全に進行する、病巣発達を腎組織に生じさせ/誘発する炎症反応が生じる。体液及び細胞免疫メカニズムの双方が障害の発病に関与しうる。
【0181】
多発性硬化症;特発性脱随性多発神経障害又はギラン-バレー症候群;及び慢性炎症脱随性多発神経障害を含む中枢及び末梢神経系の脱髄疾患は、自己免疫に原因を有し、オリゴデンドロサイト又はミエリンに直接的に引き起こされる損傷の結果として神経脱髄が生じると考えられている。MSにおいては、疾患の誘発及び進行がTリンパ球に依存することを示唆する証拠がある。多発性硬化症は、Tリンパ球依存性であり、再発性弛緩経路又は慢性進行経路のいずれかを有する脱髄疾患である。病因はよく知られていないが、ウイルス感染、遺伝的素因、環境及び自己免疫性の全てが寄与している。病巣は優勢なT細胞媒介小膠細胞の湿潤と、浸潤しているマイクロファージを含み;CD4+Tリンパ球は病巣における優勢な細胞型である。オリゴデンドロサイトの細胞死と続く脱髄のメカニズムはよく知られていないが、Tリンパ球により推進されていると思われる。
【0182】
好球性肺炎;特発性肺線維症及び過敏性肺炎を含む炎症及び線維症の肺疾患には、調節されない免疫炎症反応が関連している。その反応の阻害は治療的に有益であろう。
【0183】
水疱性皮膚疾患、多形性紅斑及び接触性皮膚炎を含む自己免疫又は免疫媒介皮膚疾患は自己抗体により媒介され、その発病はTリンパ球依存性である。
【0184】
乾癬はTリンパ球媒介炎症疾患である。病巣にはTリンパ球、マクロファージ及び抗原プロセシング細胞及びある種の好中球の浸潤が含まれる。喘息;アレルギー性鼻炎;アトピー性皮膚炎;食物過敏症及び蕁麻疹等を含むアレルギー性疾患はTリンパ球依存性である。これらの疾患はTリンパ球誘発性炎症、IgE媒介炎症、又は双方の組合せにより主に媒介される。
【0185】
拒絶反応及び移植片対宿主疾患(GVHD)を含む移植関連疾患はTリンパ球依存性であり;Tリンパ球の機能を阻害することで改善される。
【0186】
CRIgの上に列挙した治療的用途に鑑みて、CRIgの潜在的アゴニストは様々な細胞ベースアッセイ及び補体関連、炎症及び/又は自己免疫疾患又は障害の動物モデルで評価することができる。
【0187】
よって、例えば、関節炎の予防及び/又は治療における効力はコラーゲン誘発関節炎モデル(Terato等 Brit. J. Rheum. 35:828-838 (1966))において評価することができる。潜在的な関節炎予防/治療薬は、Terato等, J. Immunol. 148:2103-8 (1992);Terato等, Autoimmunity 22:137-47 (1995)に記載されたように、4種のモノクローナル抗体のカクテルの静脈内注射により誘発される抗体媒介関節炎モデルにおいてスクリーニングできる。またここの実施例4を参照のこと。関節炎の予防及び/又は治療のための候補は、例えばTNF-αトランスジェニックマウス(Taconic)のようなトランスジェニック動物モデルで研究することがまたできる。これらの動物は、ヒトリウマチ様関節炎の病因に結びつけられているサイトカインであるヒト腫瘍壊死因子(TNF-α)を発現する。これらのマウスにおけるTNF-αの発現は、前肢及び後肢の重症の慢性関節炎を生じ、炎症性関節炎の簡単なマウスモデルを提供する。
【0188】
最近、乾癬の動物モデルがまた開発された。よって、Asebia(ab)、鱗状皮膚(fsn)及び慢性増殖性皮膚炎(cpd)は乾癬様皮膚変化を持つ自然マウス突然変異である。例えばインターフェロン-γ、インターロイキン-1α、ケラチノサイト増殖因子、トランスフォーミング増殖因子-α、インターフェロン-6、血管内皮増殖因子、又は骨形成タンパク質6のようなサイトカインの皮膚性過剰発現があるトランスジェニックマウスをまた使用して、インビボで乾癬を研究し、感染治療のための治療薬を同定することができる。乾癬様病巣はまたPL/J系統に戻し交配したβ2-インテグリン低形質マウス及びβ1-インテグリントランスジェニックマウス、CD4+/CD45RBhiTリンパ球で再構成されたscid/scidマウス並びにHLA-B27/hβ2mトランスジェニックラットにおいて記載されている。免疫不全マウスに移植されたヒト皮膚を使用する異種移植モデルもまた知られている。よって、本発明の化合物は、マウスが乾癬に類似した組織病理学的皮膚病巣を示すSchon, M.P.等, Nat. Med. (1997) 3:183により記載されているscid/scidマウスモデルにおいて試験することができる。他の適切なモデルは、Nickoloff, B.J.等, Am. J. Path. (1995) 146:580に記載されているようにして調製されるヒトskin/scidマウスキメラである。更なる詳細については、例えばSchon, M.P., J Invest Dermatology 112:405-410 (1999)を参照のこと。
【0189】
組換え(トランスジェニック)動物モデルは、対象の遺伝子のコード部分を、トランスジェニック動物作製のための標準的技術を用いて、対象動物のゲノム中に導入することにより操作できる。トランスジェニック操作の標的となりうる動物は、限定されないが、マウス、ラット、ウサギ、モルモット、ヒツジ、ヤギ、ブタ、及び非-ヒト霊長類、例えばヒヒ、チンパンジー及び他のサルを含む。このような動物に導入遺伝子を導入するための当該分野で知られた技術は、前核マイクロインジェクション(Hoppe及びWanger, 米国特許第4873191号);胚系列へのレトロウイルス媒介遺伝子転移(例えば、Van der Putten等, Proc. Natl. Acad. Sci. USA 82, 6148-615 [1985]);胚性幹細胞での遺伝子ターゲティング(Thompson等, Cell 56, 313-321 [1989]);胚のエレクトロポレーション(Lo, Mol. Cel. Biol. 3, 1803-1814 [1983]);精子媒介遺伝子導入(Lavitrano等, Cell 57, 717-73 [1989])を含む。概説としては、例えば、米国特許第4736866号を参照のこと。
【0190】
本発明の目的のために、トランスジェニック動物は、その細胞の一部にのみ導入遺伝子を有するもの(「モザイク動物」)を含む。導入遺伝子は、単一の導入遺伝子として、又はコンカテマー、例えば頭部と頭部又は頭部と尾部の直列型として組み込まれうる。特定の細胞型への導入遺伝子の選択的導入も、例えばLasko等, Proc. Natl. Acad. Sci. USA 89, 6232-636 (1992)の技術に従えば、可能である。
【0191】
トランスジェニック動物における導入遺伝子の発現は、標準的技術によってモニターできる。例えば、導入遺伝子の組込みの確認にはサザンブロット分析又はPCR増幅が用いられる。ついで、mRNA発現のレベルは、インサイツハイブリダイゼーション、ノーザンブロット分析、PCR、又は免疫組織化学などの技術を用いて分析できる。
【0192】
例えば免疫細胞の特定の細胞への浸潤を確定するための組織学的検査によって、免疫疾患病理の徴候について該動物を更に調べることができる。補体及び古典的及び代替経路を含む補体活性化又はT細胞増殖に対する効果の度合いを決定するためにCRIg又は候補アゴニストでトランスジェニック動物を治療する遮断実験もまた行うことができる。これらの実験では、本発明のポリペプチドと結合する遮断抗体が動物へ投与され、興味ある生物学的効果がモニターされる。
【0193】
あるいは、「ノックアウト」動物は、CRIgポリペプチドをコードする内在性の遺伝子と、動物の胚性細胞へ導入された同ポリペプチドをコードする改変ゲノムDNAとの間の相同組換えの結果として、CRIgをコードする欠陥又は改変遺伝子を有するものとして構築することができる。例えば、CRIgをコードするcDNAは、確立されている技術によって、CRIgをコードするゲノムDNAのクローニングに利用できる。CRIgをコードするゲノムDNAの一部は、その他の遺伝子、例えば組込みをモニターするために利用できる選択可能マーカーをコードする遺伝子によって置換え又は除去できる。典型的には、ベクターには数キロベースの未変化フランキングDNA(5'と3'末端の両方)が含まれる[例えば、相同組換えベクターの説明についてはThomas及びCapecchi, Cell, 51:503(1987)を参照]。ベクターを胚性幹細胞株へ(例えば電気穿孔法等によって)導入し、導入されたDNAが内在性DNAと相同的に組換えられた細胞を選択する[例えば、Li等, Cell, 69:915 (1992)参照]。選択された細胞は次に動物(例えばマウス又はラット)の胚盤胞内に注入され、集合キメラを形成する[例えば、Bradley, Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson編 (IRL, Oxford, 1987), pp. 113-152参照]。その後、キメラ性胚を適切な偽妊娠の雌性乳母に移植し、出産させて「ノックアウト」動物をつくり出す。その胚細胞に相同組換えされたDNAを有する子孫は標準的な技術により同定でき、それらを使用して動物の全細胞が相同組換えされたDNAを含む動物を飼育することができる。ノックアウト動物は、例えば、CRIgポリペプチドが不在であることによるある種の病理的状態及びその病理的状態の発達に対する防御能力によって特徴付けることができる。
【0194】
よって、潜在的CRIgの生物学的活性は、以下の実施例7に記載されたように、マウスCRIgノックアウトマウスにおいて更に研究することができる。
【0195】
抗原誘発性気道反応亢進、好酸球増加症及び炎症が卵白アルブミンで動物を感作させ、エアゾールにより送達される同じタンパク質に動物を暴露することで誘発される喘息モデルが記載されている。幾つかの動物モデル(モルモット、ラット、非ヒト霊長類)では、エアゾール抗原への暴露時にヒトのアトピー性喘息に類似した徴候を示す。マウスモデルはヒト喘息の特徴の多くを有している。喘息治療における活性と有効性についてCRIg及びCRIgアゴニストを試験するのに適した手順は、Wolyniec, W.W.等, Am. J. Respir. Cell Mol. Biol. (1998) 18:777とそこに引用されている文献に記載されている。
【0196】
接触性過敏症は、細胞媒介免疫機能の単純なインビボアッセイである。この手順において、遅発型過敏反応を生じさせる外因性ハプテンに表皮細胞を暴露し、反応を測定して定量する。接触過敏症は最初の感作段階に顕在化段階が続く。顕在化段階は表皮細胞がそれらが過去に接触したことのある抗原に遭遇したときに生じる。腫れと炎症が生じ、ヒトアレルギー性接触皮膚炎の優れたモデルが作製される。適切な手順は、Current Protocols in Immunology, J.E. Coligan, A.M.Kruisbeek, D.H.Marglies, E. M.Shevach, 及びW.Strober編, John Wiley & Sons, Inc, unit4.2に詳細に記載されている。また、Grabbe, S.及びSchwarz, T. Immun. Today 19(1):37-44(1998)も参照のこと。
【0197】
移植片対宿主疾患は、免疫適格細胞が免疫抑制され又は耐性のある患者に移植された場合に生じる。ドナー細胞は宿主抗原を認識しそれに反応する。反応は生命に危険性のある重度の炎症から、下痢や体重の減少等の軽度の場合まで多様である。移植片対宿主疾患モデルはMHC抗原及び少量の移植抗原に対するT細胞反応性を評価する手段を提供する。適切な手順は上記のCurrent Protocols in Immunology, unit4.3.に詳細に記載されている。
【0198】
皮膚同種移植片拒絶のための動物モデルは、抗ウイルス及び腫瘍免疫におけるそれらの役割の指標であり尺度であるT細胞がインビボで組織の破壊を媒介する能力を試験する手段である。最も一般的で容認されているモデルではマウスの尾-皮膚移植片が使用される。繰り返し実験により、皮膚同種移植片拒絶がT細胞、ヘルパーT細胞及びキラー-エフェクターT細胞により媒介されるが、抗体では媒介されないことが分かった。Auchincloss, H. Jr.及びSachs, D.H., Fundamental Immunology, 2nd ed., W.E.Paul ed., Raven Press, NY, 1989, 889-992。適切な手順は上掲のCurrent Protocols in Immunology, unit4.4.に詳細に記載されている。CRIg及びCRIgアゴニストの試験に使用可能な他の移植拒絶モデルは、Tanabe, M.等, Transplantation (1994) 58:23及びTinubu, S.A.等, J. Immunol. (1994) 4330-4338により記載されている同種心臓移植片モデルである。
【0199】
遅発型過敏症の動物モデルは、細胞媒介免疫機能のアッセイをまた提供する。遅発型過敏反応は、抗原投与後の経過した時間まで、ピークに達しない炎症により特徴付けられるT細胞媒介インビボ免疫反応である。これらの反応はまた組織特異的自己免疫疾患、例えば多発性硬化症(MS)及び実験的自己免疫脳脊髄炎(EAE、MS用のモデル)で生じる。適切な手順は上掲のCurrent Protocols in Immunology, unit4.5.に詳細に記載されている。
【0200】
EAEは、T細胞及び単核細胞の炎症と、続いての中枢神経系における軸索の脱髄により特徴付けられるT細胞媒介自己免疫疾患である。EAEは、一般的にヒトにおけるMSの関連動物モデルであると考えられている。Bolton. C., Multiple Sclerosis (1995)1:143。急性及び再発寛解型モデルの双方が開発されている。CRIg及びそのアゴニスト及びアンタゴニストは上掲のCurrent Protocols in Immunology, unit15.1及び15.2.に記載されているプロトコールを使用して、免疫媒介脱髄疾患に対するT細胞刺激又は阻害活性を試験することができる。また、Duncan. I.D.等, Molec. Med. Today (1997) 554-561に記載されているようにして、オリゴデンドロサイト又はシュワン細胞が中枢神経系に移植されたミエリン疾患モデルも参照されたい。
【0201】
年齢関連性黄斑変性(AMD)の動物モデルは、Ccl-2又はCcr-2遺伝子に無発現変異を有するマウスからなる。これらのマウスは網膜色素上皮(RPE)の中のリポフスチン及びその下のドルーゼの蓄積、光受容体萎縮及び脈絡膜血管新生(CNV)を含むAMDの主要な特徴を発症する。これらの特徴は6ヶ月の年齢を越えて発達する。CRIg及びCRIgアゴニストはドルーゼの形成、光受容体及び脈絡膜血管新生について試験することができる。
【0202】
心筋虚血再灌流のモデルはマウス又はラットで実施することができる。動物を気管切開し、小動物人工呼吸器で酸素供給する。ポリエチレンカテーテルを平均動脈圧の測定のために内頸動脈と外頸静脈内に配する。心筋虚血再灌流は、冠動脈左前下行枝(LAD)を6-O縫合によって結紮することによって開始される。虚血は血管を完全に閉塞するためにLADの回りに可逆的な結紮糸を締めることによってつくり出される。結紮糸は30分後に除去され、心臓は4時間灌流される。CRIg及びCRIgアゴニストは、心臓梗塞サイズ、心臓クレアチンキナーゼ活性、ミエロペルオキシダーゼ活性及び抗C3抗体を使用する免疫組織化学の測定によってその効力を試験することができる。
【0203】
糖尿病性網膜症のモデルはストレプトゾトシンでのマウス又はラットの治療を含む。CRIg及びCRIgアゴニストは、 細静脈拡張、網膜内微小血管異常、及び網膜及び硝子体腔の血管新生に対するその効果について試験することができる。
【0204】
膜性増殖性糸球体腎炎のモデルは次のようにして樹立することができる:雌マウスをCFA中の0.5mgコントロールウサギIgGを用いて腹腔内免疫する(−7日目)。7日後に(0日目)、1mgのウサギ抗マウス糸球体基底膜(GBM)抗体を尾静脈から静脈内注射する。血清中の抗ウサギIgG抗体の上昇をELISAによって測定する。24時間の尿試料を代謝ケージ中のマウスから集め、マウスの腎臓機能を、血中尿素窒素に加えて尿中タンパク質を測定することによって評価する。
【0205】
薬学的組成物
本発明に従って同定されたCRIgアゴニスト及びアンタゴニストは、炎症性疾患の治療のために、薬学的組成物の形態で投与することができる。
【0206】
治療製剤は、所望される度合いの純度を持つ活性分子を、凍結乾燥製剤又は水溶液の形態で、最適な製薬上許容される担体、賦形剤又は安定化剤と混合することにより調製されて保存される(Remington's Pharmaceutical Science 16版, Osol, A.編 [1980])。許容される担体、賦形剤、又は安定化剤は、用いられる用量及び濃度で受容者に非毒性であり、リン酸塩、クエン酸塩、及び他の有機酸などの緩衝液;アスコルビン酸及びメチオニンを含む酸化防止剤;保存料(塩化オクタデシルジメチルベンジルアンモニウム;塩化ヘキサメトニウム;塩化ベンザルコニウム;塩化ベンゼトニウム;フェノール;ブチル又はベンジルアルコール;メチル又はプロピルパラベン等のアルキルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3-ペンタノール;及びm-クレゾールなど);低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチン、又は免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、又はリジン等のアミノ酸;グルコース、マンノース、又はデキストリンを含む単糖類、二糖類、及び他の炭水化物、EDTA等のキレート剤、スクロース、マンニトール、トレハロース又はソルビトールなどの糖;ナトリウムなどの塩形成対イオン;金属錯体(例えば、Zn-タンパク質錯体)及び/又はトゥイーン(TWEENTM)、プルロニクス(PLURONICSTM)、及びポリエチレングリコール(PEG)等の非イオン性界面活性剤を含む。
【0207】
リポフェクション又はリポソームを使用して、細胞中にポリペプチド、抗体、又は抗体断片を送達することができる。抗体断片が使用される場合、標的タンパク質の結合ドメインに特異的に結合する最小の断片が好ましい。例えば、抗体の可変領域配列に基づいて、標的タンパク質配列に結合する能力を保持するペプチド分子を設計することができる。このようなペプチドは化学的に合成し、及び/又は組換えDNA技術(Marasco等, Proc. Natl. Acad. Sci. USA 90:7889-7893 [1993])により生産することができる。
【0208】
ここでの製剤は、治療すべき特定の徴候に必要な場合に一つ以上の活性化合物、好ましくは互いに悪影響を及ぼさない相補的活性を持つものも含んでいてもよい。あるいは、又はそれに加えて、組成物は、細胞毒性薬、サイトカイン又は増殖阻害剤を含んでもよい。このような分子は、適切には、意図する目的に有効な量の組み合わせで存在する。
【0209】
また、活性分子は、例えばコアセルベーション技術により又は界面重合により調製されたマイクロカプセル、例えば、各々ヒドロキシメチルセルロース又はゼラチン-マイクロカプセル及びポリ(メタクリル酸メチル)マイクロカプセル中、コロイド状薬物送達系(例えば、リポソーム、アルブミンミクロスフィア、マイクロエマルション、ナノ粒子及びナノカプセル)中、又はマイクロエマルション中に包括されていてもよい。このような技術は、Remington's Pharmaceutical Science 16版, Osol, A.編(1980)に開示されている。
【0210】
インビボ投与に使用される製剤は無菌でなければならない。これは、滅菌濾過膜を通した濾過により容易に達成される。
【0211】
徐放性製剤を調製することもできる。徐放性製剤の好適な例は、抗体を含む固体疎水性ポリマーの半透性マトリクスを含み、このマトリクスは成形品、例えばフィルム、又はマイクロカプセルの形態である。除放性マトリクスの例は、ポリエステル、ヒドロゲル(例えば、ポリ(2-ヒドロキシエチル-メタクリレート)又はポリ(ビニルアルコール))、ポリラクチド(米国特許第3773919号)、L-グルタミン酸とγ-エチル-L-グルタマートのコポリマー、非分解性エチレン-酢酸ビニル、LUPRON DEPOTTM(乳酸-グリコール酸コポリマーと酢酸リュープロリドからなる注射可能なミクロスフィア)等の分解性乳酸-グリコール酸コポリマー、ポリ-(D)-3-ヒドロキシブチル酸を含む。エチレン-酢酸ビニル及び乳酸-グリコール酸などのポリマーは分子を100日に亘って放出することができるが、ある種のヒドロゲルはより短時間でタンパク質を放出する。カプセル化された抗体が身体内に長時間残ると、それらは37℃の水分に露出されることにより変性又は凝集し、その結果、生物学的活性の低下及び起こりうる免疫原性の変化をもたらす。合理的な方法は、含まれる機構に依存する安定化について工夫することができる。例えば、凝集機構がチオ-ジスルフィド交換を通した分子間S-S結合形成であると発見された場合、安定化はスルフヒドリル残基の修飾、酸性溶液からの凍結乾燥、水分含有量の制御、適切な添加剤の付加、及び特異的ポリマーマトリクス組成物の開発によって達成されうる。
【0212】
次の実施例は例証のためにのみ提供するものであり、本発明の範囲を限定するものでは決してない。
【0213】
本明細書において引用された全ての特許及び文献はその全体を出典明示によりここに援用される。
【実施例】
【0214】
実施例1
CRIg及びC3b:CRIg複合体の結晶構造の決定
材料と方法
a.成熟ヒトCRIgタンパク質の生産と精製
配列番号2(配列番号8)の成熟ヒトCRIgの残基1〜119をコードするDNA断片をpET28b発現ベクター(Novagen)のNdeI/BamHI部位中にクローニングし、トロンビン切断部位が続くN末端Hisタグを有する融合体をつくった。Ni-アフィニティークロマトグラフィーを使用した精製後に、融合タンパク質をトロンビンで消化させ、サイズ排除クロマトグラフィーを使用して更に精製した。最終のタンパク質原液は、10mMのHepes、50mMのNaCl、pH7.2中に20mg/mlのタンパク質濃度を有していた。セレノメチオニン標識タンパク質を、標準的なプロトコルを使用して得た。
【0215】
精製されたCRIgを5倍モル過剰でC3c又はC3bと混合し、氷上で30分間インキュベートした。その試料を、サイズ排除クロマトグラフィーを使用して精製し、25mMのHepes、pH7.2、50mMのNaCl中で10−20mg/mlまで濃縮した。
【0216】
b.結晶化
全ての結晶は懸滴蒸気拡散法を使用して19℃で成長させた。
【0217】
CRIgを、30%のPEG4000、0.1Mの酢酸ナトリウム、及び0.2Mの酢酸アンモニウムを含むリザーバの等容量でタンパク質溶液を等しく混合することによって結晶化させた。結晶は3日後に形成された。12%のPEG20000、0.1MのMES,pH6.5を含むリザーバ溶液と1:1の比でタンパク質溶液(20mg/ml)を混合することによってC3c:CRIg複合体の結晶を得た。12%のPEG20000、0.1MのMES,pH6.5を含むリザーバ溶液の等容量とタンパク質溶液(10mg/ml)を混合することによってC3b:CRIg複合体の結晶を得た。結晶は3日後に形成された。
【0218】
c.データ収集、構造解明及び緻密化
データ収集のために、20%グリセロールを添加した溶液含有リザーバ中に結晶を短時間浸漬した後、液体窒素でフラッシュ凍結させた。全てのデータはALSビームライン5.0.2.で集め、HKL2000を使用して処理した(Otwinowski, Z.及びMinor, W., Methods Enzymol. 276:307-326 (1997))。未複合体化CRIgの結晶は1.2Åの分解能で回折し、a=30.2、b=50.7及びc=61.9Aの格子パラメータの空間群P212121に属する。構造は複数の異常な分散系及びプログラムauto-SHARP(G. Bricogne, C. Vonrhein, C. Flensburg, M. Schiltz, W. Paciorek, Acta Crystallogr D Biol Crystallogr 59, 2023 (2003))を用いて解明した。 Refmac(G. N. Murshudov, A. A. Vagin, E. J. Dodson, Acta Crystallogr D Biol Crystallogr 53, 240 (1997))を用いる緻密化とプログラムO(T. A. Jones, J. Y. Zou, S. W. Cowan, Kjeldgaard, Acta Crystallogr A 47 ( Pt 2), 110 (1991))を用いたマニュアルでの調節によって、14.9%のRcrystと17.7%のRfreeのモデルを得た。C3cの結晶は3.1Åに回折し、a=382Å、b=65.0Å、c=147.2Å及びβ=102.7°の格子パラメータの空間群C2に属し、非対称ユニットに二つの複合体を含んでいる。構造はAMoRe(Navazza J., Acta Crystallogr. A 50:157-163 (1994))及び未結合CRIg及びC3Cモデル(pdbコード2A74)を使用して解明した。非結晶学的対称制約を適用する緻密化後の最終のRcryst及びRfreeはそれぞれ23.7%及び29.7%であった。C3b:CRIg複合体の結晶は4.1Å分解能に回折し、a=97.6Å、b=255.7Å、c=180.3Åの格子パラメータの空間群C22221に属する。C3c:CRIg複合体を使用する分子置換後、CUBドメイン(pdbコード2A73)とTEDドメイン(1C3D)を電子密度マップ中にマニュアルでドッキングさせた。剛性体緻密化後、TED及びCUBドメインを連結する領域並びに2つのグリコシル化部位の明確な密度は解釈可能な密度を有しており、電子密度中に嵌り込むことができた。最終モデルは、C345CドメインのないCRIg及びC3bの複合体を含む;後者のドメインは非常に弱い密度を有しており、図1に対して0.0に設定された占有率でモデルに含められた。最終Rcyst及びRfreeはそれぞれ25.2%及び33.3%であった。
【0219】
結果
a.構造の決定
CRIgはC3b及びiC3bと複合体を形成するが、親分子C3に結合することができないことが最近確定された(K. Y. Helmy等, Cell 124, 915 (2006))。CRIgの選択性に対する構造的基礎をより良好に理解するために、その未結合状態におけるCRIgの結晶構造を高分解能で決定し、またC3b及びC3cとの複合体についても決定した(それぞれ4.1Å及び3.2Å)。未結合CRIgの構造を、3波長異常分散(MAD)フェージングを使用して解明した(Hendrickson, W.A.等, Proteins 4:77-88 (1988))。つづいて、複合体の構造を、サーチモデルとして、CRIg及びC3(2A73)、C3c(2A74)、並びにC3d(1C3D)の構造の様々なドメイン及び断片の座標を使用して分子置換によって解明した。C3c:CRIg構造は非対称ユニットに2分子を含み、非結晶学的対称制約を使用する緻密化後に、最終のR及びRfreeはそれぞれ23.6%及び29.5%であった。C3c:CRIg複合体の限られた分解能のため広範な位置的緻密化は可能ではない。しかしながら、様々なリンカーのモデル化及び個々のドメインの剛性体緻密化後、最終モデルのR及びRfreeはそれぞれ24.8%及び33.1%であり、構造が限られた分解能を考えると良い品質であることを示している。
【0220】
b. C3bの全体構造
C3bの個々のドメインの構造は最近報告されたC3構造(上掲のJanssen等;D. Hourcade, V. M. Holers, J. P. Atkinson, Adv Immunol 45, 381 (1989))と同様であるが、これらのドメインの配置はC3と比較するとかなり異なっている。簡単に述べると、残基1〜645からなるC3bのβ鎖は、5つのマクログロブリン様ドメイン(MG1-MG5)、第6MGドメインのN末端半分、リンカー領域(LNK)(図1A、B)に折り畳まれる。免疫グロブリントポロジーMGドメインは「キーリング」様形式に配置され、約10Å幅で30Å長の中央溝の回りを円で囲んでいる。C3bのα鎖は残基729:1641からなり、C3と比較してC3a又はANAドメインを欠いている。α'NTドメインと呼ばれるC3bα鎖のN末端残基(729:745)は、伸展したコンフォメーションを採用し、第6MGドメインの第2の半分を形成する残基につながっている。これに続いて、α鎖は間にCUBドメインとチオ-エステルドメイン(TED)が挿入された2つの更なるMGドメイン(MG7及びMG8)を含んでいる。最後に、α鎖のC末端の170残基はいわゆる「アンカー領域」とC345Cドメインを形成している。
【0221】
c. C3活性化時のコンフォメーション変化
C3、C3b、及びC3cの構造の比較は、大きなコンフォメーション変化、つまりC3のC3bとC3aへの開裂が活性化後直ぐに生じることを示している(図1A、B)。C3中のMG3及びMG8ドメイン間に割り込んでいるANAドメインの放出により(上掲のF. Fredslund, J等)MG3とMG8が回転させられる。これらのコンフォメーション変化によりMG7ドメインが更に回転し、C3をC3cと比較した場合、上掲のJannsen等に記載されたα'NTドメインの興味深い動きが生じる。C3cの場合と同様に、α'NTはC3におけるよりもC3bにおいて更に溶媒露出性であり、補体活性化のレセプター及び調節因子、CR1、CFH及びB因子のための多くの潜在的結合部位を示す(J. D. Lambris等, J Immunol 156, 4821 (1996);A. Taniguchi-Sidle, D. E. Isenman, J Immunol 153, 5285 (1994);A. E. Oran, D. E. Isenman, J Biol Chem 274, 5120 (1999))。しかしながら、C3活性化によって誘導される最も大きな動きはCUB及びTEDモジュールに関する。C3中のTED及びMG8に対して密接に充填されているCUBはMG8に対して約25Å動き(図1B及び図6)、C3b構造中においてMG2と緩い相互作用のみを形成する。C3においてTEDは、残基Cys988及びGln991の間に形成されたチオ-エステル結合を溶媒から保護するMG8ドメイン及びCUBに対して堅固に埋もれている。活性化の際、TEDはCUB及びMG8ドメインから離れる方向に回転しTED及びCUBドメインを連結する残基を完全に異なったコンフォメーションで残す;これらの変化の過程において、Cys988は、C3構造中のその最初の保護された巣からC3構造中のその顕著に露出した位置まで80Åを越えて移動する。
【0222】
C3は炎症疾患の発生率に影響を及ぼす2つの一般的なアロタイプで存在している(J. E. Finn等, Nephrol Dial Transplant 9, 1564 (1994))。これらの変異体は単一のアミノ酸が異なる。更に一般的なアロタイプのC3Fは、80位に大きい正荷電のアルギニンを有する一方、C3Sは小さい非荷電のグリシンを有している。残基R80と回りのループはC3又はC3cの構造では相互作用を形成せず(上掲のJanssen等)、R80と相互作用する他のタンパク質はこれまで同定されていない。C3bにおいて、このループはMG1とTEDドメイン間の多くの接触の原因であり、R80はTEDの表面上の負に荷電した溝に近接して位置している(図7)。R80及びTEDドメイン間の相互作用がC3S及びC3Fアロタイプ間の観察された差異の原因であるかどうかを確認するためには更なる実験が必要である。
【0223】
C3d及びC3におけるαヘリックストロイドTEDモジュールは多くの有意な差を示している(上掲のJanssen等)。最も重要な点は、−粒子表面へのTEDドメインの共有結合に必要とされる−残基His1104及びGlu1106が互いに離れ、またCys988から離れていることである。しかしながら、C3dにおいては、これらの残基は非常に近接しており、これは共有結合反応のための重要な必要条件である。ここに提示されたC3b:CRIg複合体の分解能は個々のアミノ酸側鎖の正確な位置の検討を可能にするものではない一方、C3b中のTEDのコンフォメーションがC3d構造に非常に類似していることは我々の構造から明らかである。C3とC3b間の著しいコンフォメーションシフトにより、結合反応に重要なTED残基が溶媒露出性になり、C3b分子の遠位端に槍の穂先のように位置せしめられる。
【0224】
d. CRIgはC3b及びC3cのβ鎖に結合する
配列解析によって予測されるように(K. Langnaese等, Biochim Biophys Acta 1492, 522 (2000))、CRIgのN末端ドメインは、位相幾何学的には、免疫グロブリン様ドメインのIgVファミリーに属するが、プロテインデータバンクに現在寄託されている他のIgドメインと僅か約20%の配列同一性を共有しているだけである。この折り畳みファミリーの全てのメンバーと同様に、CRIgドメインは2つのβシートによって形成されている。これらのシートの一方はストランドA’、G、F、C、C’及びC’’によって構成され、他方はストランドB、E、及びDによって構成される(図2A及び図8)。該ドメインは、ストランドB及びFを内部で連結する2つのシステイン残基間に形成された正準なIg様ジスルフィド結合によって更に安定化されている。
【0225】
CRIgは同一の形式でC3bとC3cに係合し、両方のCRIg複合体中に存在する全ての接触ドメインの配置は非常に類似している。これに沿って、C3bとC3cに対するCRIgの親和性は比較できる(図9)。C3b:CRIg及びC3c:CRIg構造の重ね合わせは、β鎖の642の共通のCa位置に対しては0.6Å未満で、C3cとC3bのα鎖の479Ca原子に対しては0.5Å未満のRMSDになる。
【0226】
これまでに同定された全ての他の補体レセプターとは異なり(J. D. Lambris等, J Immunol 156, 4821 (1996);A. Taniguchi-Sidle, D. E. Isenman, J Immunol 153, 5285 (1994);A. E. Oran, D. E. Isenman, J Biol Chem 274, 5120 (1999))、CRIgは主にC3bのβ鎖に結合する。その未結合状態及び互いの複合体でのC3c及びCRIg構造の比較から、結合を亢進させるために2分子の何れにおいても有意なコンフォメーションの適応が必要とはされないことが明らかになった。CRIgへの未結合(上掲のJanssen等)及び結合双方のC3c構造における963の共通のCa位置は0.8ÅのRMSDで重ね合わさり、最も大きな動きは可動性領域で生じ、ただ一つの小さなループの移動がMG6ドメインで生じる(結果を示さず)。CRIgに対する結合部位は、CUB及びTEDドメインとはC3bの反対側にあり、TEDドメインから離れる方向におよそ40Å離れてCRIgを位置させている。
【0227】
CRIgとC3c又はC3bの間の結合界面は大きく、不連続で、全体で約2670Å2の溶媒露出面積を埋めている(図2A)。CRIgはC3bと接触し、A’、G、G、C、C’、C’’シートからの残基が、ストランドC’とC”の近傍の残基及びループによって形成された相互作用の大部分を有する。C’とC”を連結するヘアピンループは、C3bのキーリング形状のβ鎖の中心に形成された裂け目中に突出している。界面のC3b側では、ドメインMG3、MG4、MG5、MG6、LNK及びMG6が全てCRIg結合に寄与し、埋まった界面のそれぞれ約30%と40%に対してMG3とMG6が原因となっている(図2B)。C3及びC3b構造の比較は、C3b及びC3cへのCRIgの結合のゲーティングにおけるMG3ドメインの重要な役割を示唆している。C3活性化時に生じる大規模なドメイン移動とは別に、機能的な重要性を有することがある数種のより小さいドメインシフトがある。これらの間のキーは、比較的僅かであるが、C3分子の残りと比較してなお有意な15°のMG3の回転である。この回転は、CRIg結合部位を構成する他のMGドメインのリングに関したMG3の相対的再配向の原因であり、LNK領域の螺旋部分の動きと連係して(図9)、CRIgをC3b及びC3cに結合させる;これらの中程度の変化がないと、C3はCRIgにはラッチされないであろう。
【0228】
重要な結合パラメータを次の表1にまとめる。

【0229】
実施例2
CRIg変異体
構造複合体によって明らかになった結合界面の重要性を機能的に検証するために、次のCRIg変異体をデザインし、製造し、C3bに結合するそれらの能力を試験した:次の表2において、番号はシグナル配列切断部位の開始位置を示す。
表2
変異体 アミノ酸置換
A E85A M86A
B H57A Q59A
C V107S D109R I111E
D L34S K36D L38H
E P91A T93A D95A
コントロール H21A T23A D25A
【0230】
変異体を、QuikChange部位特異的突然変異誘発キット(Stratagene)を使用して作製し、プライマーは製造者の指示に従って設計した。二重又は三重変異体を用いた結合実験の結果は結晶学的に定めたタンパク質-タンパク質接触と密接に一致している(図2B)。MG6及びMG3結合ドメイン(E85A M86A及びH57A Q59A)における変異がCRIgのC3bへの結合を最も効果的に抑止し(IC50値はwtCRIgと比較して>10000及び100倍高かった)、CRIg結合におけるこれらのドメインの相互作用の重要性を示している。これに対して、結合界面とは反対側に位置するCRIg表面残基における変異は、野生型分子と同様の結合活性を示し、これらの変異が分子の全体の折り畳みを変えないことを確認した。また、円二色性分光法を更に使用して、全ての変異タンパク質が正しく折り畳まれたことを確認した(結果は示さず)。
【0231】
実施例3
C3bへのCRIgの結合は補体の代替経路を阻害する
AP及び古典/MBL経路双方のC5転換酵素がC3bサブユニットを含んでいるので(Pangburn及びMuller-Eberhard, Biochem J., 235:723 (1986))、我々は、CRIgがそのC3bへの結合を介して補体活性化の何れかの経路を調節できるかどうかを問うた。CRIgは代替経路を介する補体活性化を阻止したが、古典又はMBL経路はしなかった(図3a)。この阻害活性はC3b結合活性を欠いたCRIg変異体では失われており、CRIg-C3b結合が転換酵素活性を乱す直接の原因であることを確認した。CRIgによるAPの選択的阻害は、代替及び古典/MBL経路を区別する細胞ベースアッセイで更に確認した。IC50は、CRIgがIgGのFc部分に融合された場合に2倍まで減少した(図S6A)。CRIgは代替経路の選択的インヒビターであるので、CRIgは古典経路によって開始される代替経路溶血を阻害することができる(図S6B)。代替経路阻害は、マウスCRIgにおいて保持されており(図S3C)、CRIgの補体阻害活性は系統的に保存されていることを確認した。
【0232】
CRIgが転換酵素を阻害する分子メカニズムを更に決定するために、C3及びC5転換酵素のC3bサブユニットとのCRIgの相互作用を精製した成分を使用して研究した。CRIgは、減少したC3a生産によって示されるように液相C3転換酵素を阻害することができた(図3B)。CRIgはまたザイモサン粒子の表面に集まったC5転換酵素の酵素活性を減少させた(図3C)。様々なCRIg変異体の阻害活性はC3bサブユニットへのその親和性と相関しており、C3bへのCRIgの結合がC5転換酵素の活性を直接調節することを示している。これは、C3bβ鎖へのCRIgの結合が代替経路の転換酵素によってC3とC5の双方の切断を妨害することを示している。
【0233】
実施例4
CRIgは関節炎のマウスモデルにおいて補体活性化を阻害する
材料と方法
a.マウスにおける関節炎の誘発
コラーゲン誘発関節炎を、他に記載されているようにして(K. H. Barck等, Arthritis Rheum 50, 3377 (2004))実施した。マウスに、21日目に開始して4mg/kgのCRIg-Ig融合タンパク質を、又は一次免疫後36日目に開始して12mg/kgの融合タンパク質を皮下的に注射した。CRIg-Ig融合タンパク質では、マウスCRIg細胞外ドメインをIgG1分子のマウスFc部分に融合させている。抗体誘発関節炎は関節炎誘発性混合物を使用してTeraoと同僚の方法(K. Terato等, J Exp Med 162, 637 (1985))によって誘発させた。簡単に言えば、マウスに尾静脈から2mgの抗CIIAb(2mg/500μl/体)を静脈内注射し、3日後に25μgのLPS(25μg/100μl/体)を腹腔内注射した。骨体積の測定と組織検査を、他に記載されているようにして(上掲のBarck等)実施した。
【0234】
b.関節炎の後足におけるサイトカイン濃度の測定
屠殺したマウスの後足蹠を毛の生え際で切断し、液体N2で凍結させた。ポリトロンホモジナイザー(KINEMATICA)を使用して25ml当たり1完全プロテアーゼインヒビター錠剤(Roche)を補填した氷冷RIPA可溶化バッファー中で足蹠をホモジナイズさせた。均質化に使用したPBSの体積を、バッファー1ミリリットル当たり75mgの組織に調節した。ホモジネートを1870×gで15分間遠心分離し、上清を13230×gで5分間遠心分離した。その上清をELISA分析にかけた。マウスIL-1β及びIL-6(BD DuoSet)及びmC3a(Bachem)のELISAキットを、各ELISAキットのプロトコルに従って使用した。C5aは、製造者のプロトコル(BD)に従ってC5a-desargのネオ-エピトープに対する特異的抗体のセットを使用してELISAによって測定した。上清中の全タンパク質濃度はBCAキット(Pierce)を使用して測定した。サイトカインとケモカインの濃度は、タンパク質1ミリグラム当たりのピコグラムで表した。
【0235】
c.ELISA解析
C3a及びC5aのELISAは、BDからの捕捉及び検出抗体を使用して、又はマウスELISAキットを使用して確立した。Weilisa全補体スクリーン:プロトコルはキットによって提供された指示書に従った。CPの潜在的なCRIg阻害を評価するために、B因子枯渇血清を使用した。APのCRIg阻害を決定するために、C2枯渇血清を使用した。
【0236】
d.関節炎の後足におけるサイトカイン濃度の測定
屠殺したマウスの後足蹠を毛の生え際で切断し、液体N2で凍結させた。ポリトロンホモジナイザー(KINEMATICA)を使用して25ml当たり1完全プロテアーゼインヒビター錠剤(Roche)を補填した氷冷RIPA可溶化バッファー中で足蹠をホモジナイズさせた。均質化に使用したPBSの体積を、バッファー1ミリリットル当たり75mgの組織に調節した。ホモジネートを1870×gで15分間遠心分離し、上清を13230×gで5分間遠心分離した。その上清をELISA分析にかけた。マウスIL-1β及びIL-6(BD DuoSet)及びmC3a(Bachem)のELISAキットを、各ELISAキットのプロトコルに従って使用した。C5aは、製造者のプロトコル(BD)に従ってC5a-desargのネオ-エピトープに対する特異的抗体のセットを使用してELISAによって測定した。上清中の全タンパク質濃度はBCAキット(Pierce)を使用して測定した。サイトカインとケモカインの濃度は、タンパク質1ミリグラム当たりのピコグラムで表した。
【0237】
結果
補体活性化の可溶型調節因子は炎症に至る補体活性化を阻害する強力な治療的ツールであることが証明された(H. F. Weisman等, Science 249, 146 (1990);B. P. Morgan, C. L. Harris, Mol Immunol 40, 159 (2003))。CRIgの治療上の潜在性を、コラーゲン自己抗体及び補体活性化が疾患の原因に寄与するリウマチ様関節炎の二つのマウスモデルで試験した(A. Aggarwal等, Rheumatology (Oxford) 39, 189 (2000);S. Solomon等, Arthritis Res Ther 7, 129 (2005);K. Terato等, J Immunol 148, 2103 (Apr 1, 1992))。インビボでのその薬理学的半減期を延長させるために、muCRIgの細胞外ドメインをマウスIgG1のFc部分に融合させた。CRIgは、関節炎の臨床的徴候の前又は後で与えられたとき、関節膨張、組織学的スコア及び骨量減少を有意に減少させた(図4A及びB、S12)。その転換酵素活性の阻害に沿って、CRIgは、軟骨表面上のC3沈着の減少によって示されるように(図4C)、関節において補体活性化を有意に減少させた。既に確認された疾患を持つマウスでは、CRIgは関節の裏にあるC3と共に同時局在化され(図4D)、それが転換酵素に結合し、転換酵素を局所的に阻害することができることを示している。また、C3aの局所レベル及びC5aの全身レベルは、コントロール-Fc処置マウスにおけるよりもCRIg-Fc処置において有意に低くなっていた(図4E)。これらの結果は、CRIgが、局所的及び全身性補体活性化を阻害することによって疾患のエフェクター相において関節炎を阻害することを示している。CRIgの保護活性は、Fcレセプター結合を消失させるCRIg-FcのFc部分の変異が、wtFcタンパク質に融合したCRIgと等価な活性を有していたので、Fcレセプター機能と独立であった(結果は示さず)。これらの結果に基づくと、CRIgの単一IgV様ドメインは補体介在疾患において代替経路を阻害する有望なタンパク質治療剤となりうる。
【0238】
実施例5
CRIgは基質-酵素結合の阻害によりAP転換酵素を阻害する
材料と方法
a.崩壊促進アッセイ
代替経路DAAに対するマイクロタイタープレートアッセイを過去に記載されているようにして(M. Krych-Goldberg等, J Biol Chem 274, 31160 (1999))実施した。マイクロタイタープレートを、リン酸緩衝生理食塩水中4μg/mlのC3bで一晩被覆した。プレートを、1%のウシ血清アルブミンを含むリン酸緩衝生理食塩水で37℃にて2時間ブロックし、71mMのNaCl、0.05%のTween20及び4%のBSAを含むベロナールバッファー中の40ngのB因子、10ngのD因子、及び0.8mMのNiCl2と共に室温で2時間インキュベートした(バッファーの濃度と組成はプロトコルでチェック)。ついで、ウェルを、0.05%のTween20を含むPBS(PBST)中のH因子又はCRIg-HisECDと共に室温で15分間インキュベートして、プレート上のC3bからBbを解離させた。ついで、Bbを、1:5000の希釈のPBST中ヤギ抗ヒトB因子ポリクローナル抗体(Kent)と1:3000の希釈のPBST中西洋わさびペルオキシダーゼ結合ロバ抗ヤギ抗体(Caltag)と共に連続1時間インキュベートして検出した(希釈と二次抗体をプロトコルでチェック)。色はO-フェニレンジアミンで発色させた。このアッセイにおいて、H因子は予想された通りに崩壊促進活性のメディエーターとして挙動した。
【0239】
b.C3b-C4b及びC3b-C3b二量体調製及び結合/競合アッセイ
C3bC4bヘテロ二量体を次の変更をして(S. Meri, M.K. Pangburn, Eur. J. Immunol. 20:2555 (1990))本質的に他に記載されているようにして(S. Meri, M. K. Pangburn, Eur J Immunol 20, 2555 (1990))作製した。C3b-C4bヘテロ二量体とC3bホモ二量体へのCRIgの結合を決定するために、マキシソープ96ウェルマイクロタイタープレートを、PBS中5ug/mlのヤギ抗C3ポリクローナル抗体(ICN)で4℃にて一晩被覆し、PBSTで3回洗浄した。ついで、プレートを室温で2時間、250μlのPBS/1%BSAでブロックし、滴定量のhCRIg-L LFHを、1%BSAを含むPBST中で1時間加えた。プレートを再びPBSTで3回洗浄し、CRIg結合を、PBST/1%BSA中の1:20000希釈のHRPにコンジュゲートしたマウス抗FLAG M2抗体(SIGMA)で検出した。ついで、プレートをPBSTで3回洗浄し、100ulのTMB基質溶液(KPL)で展開させ、50ulの2N H2SO4で停止させた。吸光度を450nmで読み取った。C5がこの形態においてC3b-C4bヘテロ二量体及びC3bホモ二量体の双方に結合することがまた決定された。CRIgがC3b-C4b又はC3b-C3bへのC5の結合をブロックすることができるかどうかを決定するために、補体二量体を捕捉し上述のようにブロックした。20mMのTRIS pH7.5、20mMのMgCl2、20mMのCaCl2、150mMのNaCl、0.05%のTween20及び1%のBSAを含むバッファーに希釈して、400nMのC5を滴定濃度のhCRIg-S HisECD又はC3bに結合しないこの融合体の変異型と前もって混合し、室温で1時間、捕捉した二量体に加えた。ついで、ウェルをPBSTで3回洗浄し、C5を、1:5000のPBST希釈のマウス抗ヒトC5(Quidel)と1:3000のヤギF'ab抗マウス抗体(Caltag)の段階的添加によって検出した。該プレートを上のようにして発色させ、2つのウェルのA415を平均化し単一の点と考えた。
【0240】
結果
補体活性化の全ての既知の調節因子は、C3のI因子介在性切断のコファクターとして作用し、又はC3bへの結合についてBbの競合体であり、よって転換酵素活性を除去する(D. Hourcade等, Adv Immunol 45, 381 (1989);T. Seya, J. P. Atkinson, Biochem J 264, 581 (1989);X. Sun等, Proc Natl Acad Sci U S A 96, 628 (1999))。これに対して、CRIgは崩壊活性もコファクター活性も示さない。また、CRIgは、H因子、B因子又はプロパージンのC3bへの結合と干渉しない(図S13A−C、結果は示さず)。CRIgがAP補体活性化を阻害するメカニズムを更に探究するために、我々はCRIgがC5転換酵素の触媒性BbサブユニットとよりもむしろC3bの機能と干渉するかどうかを決定した。代替経路転換酵素の非触媒性C3bサブユニットは基質C3及びC5の結合の原因である。ついで、転換酵素のBb因子サブユニットはプロテアーゼとして作用することができ、結合した基質を切断する(W. Vogt等, Immunology 34, 29 (1978))。その結果、転換酵素の活性はC3bに対する基質の親和性に直接的に比例する(N. Rawal, M. K. Pangburn, J Immunol 164, 1379 (2000))。CRIgはAP転換酵素のC3b2サブユニットへのC5の結合を防止するが、APに対するその選択性と一致して、C5転換酵素(図5A)はCP転換酵素のC3bC4bサブユニットに対して効果は有していない。C5転換酵素の活性に対するCRIgの結合の重要性を、精製されたC5転換酵素で更に決定した(N. Rawal, M. K. Pangburn, J Biol Chem 273, 16828 (1998))。CRIgの存在下ではs、酵素のVmaxは〜50%減少し(図5B)、これは、CRIgが基質濃度とは独立にC3bへのC5の最大結合能を阻害することを示している。これは、CRIgによるC3bへのC5の結合の立体的又はアロステリックな阻害の指標であり、C3bへの結合に対するC5とのCRIgの直接の競合ではない。これは、C5がC3bへの結合についてCRIgと競合できない結合アッセイからの結果によって更に裏付けられている(図14)。よって、C3bのキーリング様β鎖構造の中心に結合したCRIgは転換酵素へのC5の結合をブロックすることができ、よって酵素の活性を阻害する。これは、C5-転換酵素のその基質との相互作用に対するC3bのβ鎖の重要性を示している(N. Rawal, M. K. Pangburn, J Biol Chem 273, 16828 (1998))。
【技術分野】
【0001】
本発明は、マクロファージ特異的レセプターCRIg(以前はSTIgMAと呼ばれていたもの)と、補体C3のC3b及びC3cサブユニットとのその複合体(C3b:CRIg及びC3c:CRIg複合体)の結晶構造の決定に関する。本発明は更にCRIgアゴニスト及びアンタゴニストを含むCRIgに構造的及び/又は機能的に関連する分子をスクリーニングし同定するためのCRIg又はC3b:CRIg複合体の結晶構造の使用に関する。
【背景技術】
【0002】
補体系は、通常は不活性なプロ酵素形態で存在している一連の血清糖タンパク質から構成される複雑な酵素カスケードである。C3は、補体の古典的、レクチン及び代替経路の3つの主要な経路の収束点を表すので、補体活性化において重要な役割を果たしている。C3は20種を超えるタンパク質と(M. J. Walport, N Engl J Med 344, 1058 (2001)及び(M. J. Walport, N Engl J Med 344, 1140 (2001))タンパク分解酵素C3転換酵素(J. Janssen等, Nature 437, 505 (2005))によるその切断後にのみさらされる多数の別個の結合部位を介して相互作用する。最初のタンパク質切断段階はアナフィラトキシンとも呼ばれる小さい(9kDa)C3aペプチドと大きな(177kDa)C3b断片を生成する。C3からC3a及びC3bへの切断は、分子を粒子表面に共有的に結合させることができるC3b中の埋設チオ-エステル結合を露出させる立体構造変化を誘発する。「オプソニン化」と言われるこの過程は、粒子を標的にしてマクロファージ特異的レセプターに結合させ、ついで系から粒子を除去する。
【0003】
最近、CRIgがC3b/iC3bオプソニン化粒子に対するレセプターとして同定された(K. Y. Helmy等, Cell 124, 915 (2006))。CRIgはC3b、iC3b、並びにC3cに結合するが、不活性な前駆体C3との複合体を形成することはできない。
【0004】
オプソニンとしてのその機能に加えて、C3bは補体のC3及びC5転換酵素のサブユニットである。C3bはセリンプロテアーゼ因子Bに結合する(W. Vogt, G. Schmidt, B. Von Buttlar, L. Dieminger, Immunology 34, 29 (1978);Z. Fishelson, M. K. Pangburn, H. J. Muller-Eberhard, J Biol Chem 258, 741 1 (1983))。この複合体は、D因子の補充とD因子による活性化後に、代替経路(AP)の活性なC3転換酵素であるC3bBb複合体を形成する。C3転換酵素への第二C3b分子の付加は、実験的動物モデルにおいて炎症を媒介し、ヒト自己免疫及び炎症性疾患を媒介する際に重要な役割を果たすことが証明されている(M. J. Walport, K. A. Davies, B. J. Morley, M. Botto, Ann N YAcadSci 815, 267 (1997))補体の代替経路のC5転換酵素である(C3b)2Bbの形成を生じる。代替経路転換酵素のC3bサブユニットは各基質C3又はC5に対してドッキング部位となり、転換酵素の触媒活性に必要とされる(N. Rawal, M. K. Pangburn, J Immunol 164, 1379 (2000))。補体の代替経路は、代替、古典及びマンノース結合レクチン経路の3つの経路の何れかを通して開始される補体活性化を増幅させる。その結果、代替経路の調節因子である補体H因子の欠損が炎症の増幅を招く(N. Rougier等, J Am Soc Nephrol 9, 2318 (1998);M. A. Abrera-Abeleda等 J Med Genet (Nov 18, 2005))。よって、代替経路補体転換酵素によるC3及びC5の切断がオプソニン化、転換酵素の生成及び炎症を生じさせる。
【0005】
転換酵素の中心的成分として、C3bは数多くの補体調節因子の標的となる。これらには補体レセプター1(CR1)、メンブレンコファクタープロテイン(MCP)、崩壊促進因子(DAF)、補体関連レセプターy(Crry)、H因子及びI因子(D. Hourcade, V. M. Holers, J. P. Atkinson, Adv Immunol 45, 381 (1989))がある。I因子の結合はタンパク質分解を介してC3bの不活性化を生じてC3b及びC3f(2kDa)を形成し、チオ-エステルドメイン(TED)を含み標的に結合したままであるC3dg(40kDa)と、C3c(135kDa)を最終的に生成する。C3及びC3cの結晶構造に関する最近の報告は、C3が活性化される機序の我々の理解を顕著に向上させた(上掲のB. J. Janssen等, 2005)。C3cはC3と比較して著しい構造変化を示し、複数のタンパク質分解切断段階の結果を反映している。これらの構造研究はC3活性化及び調節の立体構造依存的機序を示唆しているが、活性化種C3bの立体構造に対する回答を提供するものではない。
【発明の概要】
【0006】
本開示は、病原体の排除に必要なマクロファージ補体レセプターCRIgの結晶構造を提供する。また、CRIgとの複合体でのC3b及びC3cの結晶構造が提供される。天然細胞性レセプター及び補体活性化の調節物質との複合体での転換酵素サブユニットのこの第一構造が補体活性化及び調節の機序の我々の理解を改善する。本発明はまたCRIgアゴニスト及びアンタゴニストを含むCRIgに機能的に関連した分子を同定するために使用することができる、CRIgのC3b結合領域、及びそのような結合に関与するC3b内のドメイン及び配列についての情報を提供する。
【0007】
一態様では、本発明は、天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体により形成された結晶に関する。
【0008】
一実施態様では、該結晶は、格子定数(セル定数)a=30.3Å、b=50.8Å、c=62.0Å、及びP212121の空間群をおよそ有している。
【0009】
他の実施態様では、天然配列CRIgポリペプチドは、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される。
【0010】
更に他の実施態様では、CRIgの機能的断片は天然配列CRIgの細胞外ドメイン(ECD)配列である。
【0011】
更なる実施態様では、結晶は約1から40Åまでの分解能、又は少なくとも約5Å、又は少なくとも4Å又は少なくとも3Åの分解能で原子座標を決定するためにX線を回折する。
【0012】
また更なる実施態様では、本発明はアペンディクス1に示された構造座標を有するCRIg結晶に関する。
【0013】
他の態様では、本発明は上述のような結晶を含んでなる組成物に関する。
【0014】
また他の態様では、本発明は、天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体と、補体因子C3bとの複合体の結晶形態に関する。
【0015】
一実施態様では、複合体の結晶形態は、格子定数a=97.6Å、b=255.7Å、c=180.3Å及びC2221の空間群をおよそ有している。
【0016】
他の実施態様では、複合体の結晶形態は、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドの一つを含む。
【0017】
更なる実施態様では、機能的CRIg断片は天然配列CRIgの細胞外ドメイン(ECD)配列である。
【0018】
また更なる実施態様では、複合体の結晶形態は、約1から40Åまでの分解能、又は少なくとも約5Å、又は少なくとも4Å又は少なくとも3Åの分解能で原子座標を決定するためにX線を回折する。
【0019】
更に他の実施態様では、CRIg:C3b複合体の結晶形態はアペンディクス2に示された構造座標を有する。
【0020】
更なる態様では、本発明は、天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体と、補体因子C3cとの複合体の結晶形態に関する。
【0021】
特定の実施態様では、CRIg:C3b複合体の結晶形態は、格子定数a=382.8Å、b=65.0Å、c=147.2Å、β=102.7、及びC2の空間群をおよそ有している。
【0022】
他の実施態様では、CRIg:C3C複合体中に存在する天然配列CRIgポリペプチドは、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される。
【0023】
また他の実施態様では、複合体中に存在するCRIg機能的断片は天然配列CRIgの細胞外ドメイン(ECD)配列である。
【0024】
更なる実施態様では、CRIg:C3c複合体の結晶形態は、約1から40Åまでの分解能、又は少なくとも約5Å、又は少なくとも4Å又は少なくとも3Åの分解能で原子座標を決定するためにX線を回折する。
【0025】
特定の実施態様では、CRIg:C3c複合体はアペンディクス3に示された構造座標を有する。
【0026】
更なる態様では、本発明は、配列番号2のCRIgポリペプチド又はその保存的置換のC3b結合部位の少なくとも一部を含んでなる分子又は分子複合体であって、結合部位が、8、14−15、41−42、44、45−47、50、52、54−61、62、64、85−87、89、95、99、105、107−110、及び111からなる群から選択される少なくとも一つのアミノ酸残基を含み、結合部位がアペンディクス2に記載の構造座標によって表されるアミノ酸の原子を表す点から4.7Å未満の距離を有する点の集合によって定まる分子又は分子複合体に関する。
【0027】
他の態様では、本発明は、点の少なくとも一部が、C3bに対するCRIg結合部位を定める少なくともコアアミノ酸の骨格原子の位置を表すアペンディクス2の構造座標から誘導されている点の三次元立体構造に関する。点の三次元立体構造は、限定するものではないが、C3bに対するCRIg結合部位が空間群対称C2221を有する結晶を形成する、ホログラフィー像、立体ダイアグラム、モデル、又はコンピュータ・ディスプレイ画像として示されうる。
【0028】
また他の態様では、本発明は、機械可読データでコードされたデータストレージ材を含む機械可読データストレージ媒体であって、該データを使用するための指示書を用いてプログラムされた機械が、C3b結合部位の少なくとも一部を含んでなる少なくとも一つの分子又は分子複合体の三次元グラフィック図を表示し、点の集合によって定まった結合部位が、アペンディクス2に記載された構造座標によって表されるアミノ酸の原子を表す点から約4.7Å未満の距離を有している媒体に関する。
【0029】
更なる態様では、本発明は、CRIgのC3b結合部位を模倣する化合物を設計することを含むCRIgアゴニストを同定する方法に関する。
【0030】
本発明は更に(a)計算又は実験手段を用いて化学物質とCRIgポリペプチド又はC3b:CRIg複合体の三次元構造の間のフィッティング操作を実施し;(b)工程(a)で得られたデータを解析して化学物質と天然CRIg又はC3b:CRIg複合体の間の結合の特徴を決定することにより同定された化学物質に関する。
【0031】
特定の実施態様では、上記の化学物質はCRIgとC3bの間の結合をインビボ又はインビトロで妨害する。
【0032】
本発明は更に天然配列CRIg分子又はその保存的アミノ酸置換変異体のC3b結合領域の少なくとも一部を含む分子又は分子複合体に関する。
【図面の簡単な説明】
【0033】
本特許又は出願ファイルはカラーで作製した少なくとも一図面を含んでいる。カラー図面の本特許又は特許出願のコピーは請求し必要な料金を支払えば特許庁から提供されるものである。
【図1】C3及びC3b:CRIg及びC3c:CRIg複合体のドメイン構造及び全体構造。(A)C3のドメイン組織、β鎖を緑色、α鎖を青緑色(ANAドメイン)、オレンジ色(CUB)、青色(TED)、及び紫色で示す。赤色の球は、チオ-エステルの位置を示している。(B)天然C3(左側)、CRIgとの複合体としてのC3b(中央)、及びCRIgとの複合体としてのC3cのリボン図。図1Aのカラースキームを使用し、CRIgの表面を黄色で示す。C3とC3bを比較する場合CUB及びTEDドメインの大きな動きに留意のこと。活性化後、TEDはC3bの残りと比較して移動し回転し、Cys988は80Åを越えて移動する。(C)図1Bの概略図;病原体に対するC3bの潜在的な共有結合がC3B:CRIg複合体において示されている。
【0034】
【図2】CRIgはC3b及びC3cのβ鎖キーリングの中心に結合する。(A)C3c及びCRIgの複合体。C3cの表面を白色にしたα鎖の残基と共に示し、β鎖のMG1、MG2、MG3、MG4、MG6、及びLNKドメインはそれぞれ白色、オレンジ色、赤色、淡緑色、青色及び暗緑色にした。CRIgはリボン図として示し、全てのβ鎖及びN末端及びC末端を標識した。(B)図2AからY軸の回りに180度回転させたCRIgの表面図。C3c/C3bに対して4.5A未満の距離である全原子は、図2Aにおいてそれらが接触している残基と同様に着色した。(C)CRIg野生型及びカラーのコードされたCRIg変異体の相対的親和性。親和性は、プレート上の被覆C3bへの結合についてCRIg-LFHとの様々なCRIgタンパク質の競合によって決定した。CRIg-LFH結合は抗FLAG抗体を使用して検出した。(D)図2Cに示した結合実験に使用したCRIg変異を示すCRIgの表面。変異された残基は、図2Cにおいてそれらが表す結合曲線に従って着色した。
【0035】
【図3】CRIgは補体の代替(AP)経路を阻害するが、古典(CP)経路を阻害しない。(A)CRIgはAPを介したC5b-9沈着を阻害するが、CP又はMBL経路のものは阻害しない。補体経路はLPS(AP)又はIgM(CP)で被覆されたプレート上で再構成され、C1q欠損(AP)又はfB欠損(CP)血清と共にインキュベートした。補体活性化は、C5b-9への結合HRPOコンジュゲート抗体の検出について色素源によって決定した。APアッセイにおけるCRIgのIC50値は1.49±0.19μM(n=3)である。(B)CRIgは液相C3転換酵素活性を阻害する。C3、fB及びfDを、増大する濃度のhuCRIg(L)-ECDの存在下でベロナールバッファー中でインキュベートした。反応をEDTAで停止させ、切断産物をクーマシー染色ゲル(左パネル)又はELISA(右パネル)で可視化した。CRIg-ECDに対するIC50値は0.39±0.14μM(n=3)である。(C)CRIgはザイモサン粒子の表面上に集められたC5転換酵素を阻害する。ザイモサン粒子はfB及びfDの存在下でC3bでオプソニン化され、C5転換酵素を生成する。固定濃度のC5を、増加する濃度のCRIg wt又はCRIg変異体C(CRIg(C))の存在下で転換酵素と混合した。転換酵素活性は、溶血アッセイを使用して液相中に生産されたC5b6を測定することによって決定した。IC50値:CRIg(Wt)1.63±0.62μM(n=3)、CRIg(E)6.52±3.21μM(n=3)。
【0036】
【図4】全身的に投与されたCRIg-Fcは抗体誘発関節炎に伴う局所的及び全身的補体活性化を阻害する。(A)コラーゲンでの二次免疫から7日後の、既に確認された疾患のmuCRIg-Fcで治療されたマウスにおける炎症の減少(組織学的スコア)及び骨変形の減少(関節皮質骨体積JCBVの増加)。64日目に中足骨関節を常套的なH&E組織検査、又はマイクロコンピュータ断層撮影(マイクロCT)のために準備した。(B)抗コラーゲンII型抗体での受動免疫後3日(左側パネル)又は5日後(右側パネル)に開始する12mg/kgのmuCRIg-Fcの3回の毎週の投与を受けたマウスにおける臨床スコアの減少。(C)CRIg-Fcでの処置は、関節の軟骨表面上でのIgG2aではなくC3断片の蓄積を阻害する。組み合わさった画像では、赤色がIgG2a抗コラーゲン抗体の存在を示し、緑色が軟骨表面上の補体C3産物の存在を示している。(D)既に確認された疾患のCRIg-Fcで治療されたマウスの関節中におけるC3へのCRIg-Fcの結合(緑色)。尺度バー=20um。(E)上側パネル:受動免疫前の日に開始したCRIg-Fc処置マウスにおける関節中でのC3a生産の阻害。下側パネル:CRIg-Fcで処置されたマウスの血清中におけるC5aレベルの有意な減少。統計的解析(スチューデントt検定):* p<0.01、** p<0.001、** p<0.0001。
【0037】
【図5】CRIgは、転換酵素の非触媒サブユニットへのC5の結合を阻害することによりC5転換酵素を阻害する。(A)CRIgはC5のC3b2への結合をブロックするがC3bC4bへの結合はブロックしない。C3b、C3b2及びC3bC4bをポリクローナル抗C3抗体で捕捉し、C5と増加濃度のCRIg(wt)又は変異体CRIg(C)との混合物と共にインキュベートした。転換酵素サブユニットに結合したC5を、ポリクローナル抗C5抗体及びHRPOコンジュゲート二次抗体で検出した。C5結合のCRIg阻害のIC50は0.88±0.22μMであった。(B)CRIgは精製C5転換酵素のVmaxを減少させる。固定濃度のCRIgを、増加濃度のC5の存在下でザイモサン結合C5転換酵素と混合した。ミカエリスメンテン式に基づいて計算したKm及びVmax値は、CRIg(wt)の不在下で0.99及び4.01であり、0.8uMのRIg(wt)の存在下で1.70及び2.16であった。
【0038】
【図6】C3及びC3b間のドメイン配置の差。示されているのはCUB、TED及びMG8-ドメインのリボン図である。(A)C3及びC3b(共にオレンジ色)のCUBドメインに基づく重ね合わせ。この重ね合わせは、C3b中のCUBドメインがC3のMG8ドメインに関して移動する一方(淡紫色で示す)、TEDドメインがC3活性化時に大きな回転を受けることを示している。(C)TEDドメインに基づく重ね合わせ。C3bは暗色の、C3は明るい青色で示す。C3b構造において完全に開放しているC3中の非常にコンパクトなドメイン配置に留意のこと。Cys988の[位置は赤色の球で示す。
【0039】
【図7】C3b中のTEDとArg80との間の相互作用。該図はプログラムpymol(DeLano, M.C.及びCao, Y., Neuroimaging Clin. N. Am. 12(1):21-34 (2002))で計算した静電ポテンシャルに従って着色したTEDの表面図を示している。赤色及び青色は負及び正に荷電したパッチをそれぞれ示している。R80の側鎖原子は球で表している。Arg80はTEDの表面の高度に負に荷電した溝に近接していることに留意のこと。
【0040】
【図8】異なった種からのCRIgの配列アラインメント。C3bと接触している残基は図2Aに従って着色した。残基番号はシグナル配列後のヒトCRIgを意味する。二次的構造要素は上の配列アラインメントに矢印(ストランド)及びボックス(ヘリックス)として示す。
【0041】
【図9】CRIgはC3b及びC3cと同様の親和性を有している。親和性は固定濃度のCRIg-LFHとの増加濃度のC3b又はC3cの競合によって決定した。CRIg-LFH結合は、HRPOにコンジュゲートした抗FLAG抗体を使用して検出し、TNB反応産物の吸光度を415nmの波長で分光測定により測定した。
【0042】
【図10】CRIg結合部位の近傍におけるC3及びC3b間の局所的差。ここに示したC3及びC3bの重ね合わせはMG6ドメインのCα原子に基づく。C3の一部は白色で示し、α'NTドメインは青色で示す。C3b:CRIg複合体は緑色(C3bのβ鎖)、紫色(C3bのα鎖)で示し、α'NTドメインは青色で示す。CRIgは黄色で示す。C3活性化時に起こる大きな立体構造変化は赤色の矢印で示す。α'NTドメインの大きな動きに留意のこと。MG6に関するMG3の回転と、螺旋セグメント(577:590)の動きはC3b及びC3cへのCRIgの結合に必要とされている。
【0043】
【図11】CRIgはヒト及びマウス血清中における代替経路を選択的に阻害する。(A)CRIgはC1q欠損血清中においてウサギ赤血球の溶血を阻止するが(代替経路、左側パネル)、fB欠損血清中におけるIgMオプソニン化ヒツジ赤血球の溶血は阻止しない(古典経路、右側パネル)。E-IgMとErを、異なった濃度のCRIg-Fc又はCRIg-ECD融合タンパク質を含む5%血清中でインキュベートした。氷冷EDTAを加えることによって溶血を停止させ、415nmにて分光計で上清中のO.Dを決定することによって溶血量を測定した。IC50値:CRIg-Fc 127±49nM(n=6)、CRIg-ECD 640±123nM。(B)CRIgは古典経路によって開示される補体の代替経路を阻害する。古典経路の補体成分をSRBC上に集めてEAC34(AP、左側パネル)又はEAC1234(CP、右側パネル)を生成する。溶血は(A)に記載されるようにして決定した。(C)マウスCRIg-Fcはマウス血清中の代替経路溶血を阻害する。Erを、増加濃度のマウスCRIg-Fcを含む30%マウス血清と共にインキュベートした。溶血は(A)に記載されるようにして決定した。(D)CRIgはマンノース結合レクチン経路を阻害しない。MBL経路活性化によって誘導されるC5b−9沈着を、415nmの波長で分光測定により測定した。
【0044】
【図12】CRIgはコラーゲン誘発関節炎を阻害する。(A)ウシコラーゲンを用いて0日目と23日目に2回マウスを免役することによって関節炎を誘発させた。疾患が既に確認されたときに24日目(予防的)又は45日目(矢印)に開始して毎週3回muCRIg-Fcでマウスを処置した。臨床スコアは足首の厚みを反映させたものである。関節炎の誘発後70日目に、マウスを安楽死させ、関節皮質骨体積(JCBV)を測定し(B)、H&E染色切片について炎症の徴候を評価した(C)。(D)CRIg-Fcで処置したマウスの関節におけるIL-6(左側パネル)及びIL-1b(右側パネル)の減少。関節炎の誘発後、表示した時点でマウスを安楽死させ、関節を解剖し、サイトカイン測定のために加工した。LPS処置(最初の時点)によるサイトカインレベルの増加はmuCRIg-Fc処置には影響を受けない。統計:* p<0.01、** p<0.001、スチューデントt検定。
【0045】
【図13】(A)CRIgはH因子のコファクター活性に影響しないし、内因的なコファクター活性を有してもいない。左側パネル:固定濃度のC3bを4μMのCRIg(wt)又はCRIg(C)の存在下で増加濃度のH因子と混合した。コファクター活性を、iC3bからC3bを区別するためにクーマシー染色SDSゲル上の反応産物を可視化することによってモニターした。右側パネル:固定濃度のC3b及びfIをfH又は増加濃度のCRIgと混合した。(B)CRIgはC3bに結合するB因子を阻害しない。固定濃度のB因子を増加濃度のCRIgと混合した。結合したB因子の量を、fBに対するポリクローナル抗体及び二次HRPOコンジュゲートabを使用して決定した。(C)CRIgはC3転換酵素を崩壊させない。C3転換酵素は、C3b、B因子及びD因子をインキュベートすることによってマキシソーププレート上に形成された。ついで、そのプレートを増加濃度のCRIg又はH因子と共にインキュベートした。残りの結合したB因子は、抗B因子抗体及びHRPOコンジュゲート二次抗体で検出した。示される結果は3回の独立した実験を代表するものである。
【0046】
【図14】CRIg ECDは、C3bへの結合についてCRIg-LFHと競合できるがC5はできない。CRIg-LFHと増加濃度のCRIg-ECD又はC5の混合物を、C3bを被覆したマキシソルブプレートに加えた。残っているCRIg-LFH結合を抗flag抗体を用いて検出した。
【発明を実施するための形態】
【0047】
好ましい実施態様の詳細な説明
定義
特に他の定義をしない限り、ここで使用される技術的及び科学的用語は、この発明が属する分野の当業者が通常理解するところと同じ意味を持つ。例えば、Singleton等, Dictionary of Microbiology and Molecular Biology 2版, J. Wiley & Sons (New York, NY 1994); Sambrook等, Molecular Cloning, A Laboratory Manual, Cold Springs Harbor Press (Cold Springs Harbor, NY 1989 )を参照のこと。本発明のために、次の用語を下記のように定義する。
【0048】
「CRIg」、「PRO362」、「JAM4」及び「STIgMA」は、互いに交換可能に使用され、天然配列及び変異体CRIgポリペプチドを意味する。
【0049】
「天然配列」CRIgは、その調製態様にかかわらず、天然由来のCRIgと同一のアミノ酸配列を有するポリペプチドである。しかして、天然配列CRIgは、天然から単離されうるか、又は組換え及び/又は合成手段によって製造されうる。「天然配列CRIg」なる用語は、CRIgの天然に生じる切断又は分泌型(例えば細胞外ドメイン配列)、天然に生じる変異体型(例えば選択的スプライシング型)及びCRIgの天然に生じる対立遺伝子変異体を特に包含する。天然配列CRIgポリペプチドとその断片は、配列番号1の321アミノ酸長のヒトCRIgポリペプチドで、N末端シグナル配列を含むもの又は含まないもの、1位の開始メチオニンを含むもの又は含まないもの、配列番号1のおよそ277から307のアミノ酸位置の膜貫通ドメインの何れか又は全てを含むもの又は含まないものを特に包含する。天然配列CRIgポリペプチドとその断片は、配列番号2の完全長399アミノ酸長ヒトCRIgポリペプチド(huCRIg、又はhuCRIg-long)で、N末端シグナル配列を含むもの又は含まないもの、1位の開始メチオニンを含むもの又は含まないもの、配列番号2のおよそ277から307のアミノ酸位置の膜貫通ドメインの何れか又は全てを含むもの又は含まないものを特に包含する。また更なる実施態様では、天然配列CRIgポリペプチドとその断片は、ヒトCRIgポリペプチドの305アミノ酸の短い形態(huCRIg-short、配列番号3)で、N末端シグナル配列を含むもの又は含まないもの、1位の開始メチオニンを含むもの又は含まないもの、配列番号3のおよそ183から213のアミノ酸位置の膜貫通ドメインの何れか又は全てを含むもの又は含まないものである。異なった実施態様では、天然配列CRIgポリペプチド又はその断片は、配列番号4の280アミノ酸長の完全長マウスCRIgポリペプチド(muCRIg)で、N末端シグナル配列を含むもの又は含まないもの、1位の開始メチオニンを含むもの又は含まないもの、配列番号4のおよそ181から211のアミノ酸位置の膜貫通ドメインの何れか又は全てを含むもの又は含まないものである。天然配列CRIgポリペプチド又はその断片は、配列番号5のラットCRIgポリペプチドで、N末端シグナル配列を含むもの又は含まないもの、開始メチオニンを含むもの又は含まないもの、及び膜貫通ドメインの何れか又は全てを含むもの又は含まないものでありうる。該用語は更に配列番号6のウシCRIgポリペプチド及び配列番号7のサルCRIgポリペプチドで、それぞれN末端シグナル配列を含むもの又は含まないもの、開始メチオニンを含むもの又は含まないもの、及び各配列中に膜貫通ドメインの何れか又は全てを含むもの又は含まないものを特に含む。
【0050】
「CRIg変異体」とは、天然配列CRIgポリペプチドと少なくとも約80%のアミノ酸配列同一性を有する以下に定義する活性なCRIgポリペプチドを意味する。特定の実施態様では、CRIg変異体は天然配列の成熟完全長CRIgポリペプチドの配列と少なくとも約80%のアミノ酸配列相同性を有している。かかるCRIgポリペプチドには、例えば、天然配列のN又はC末端において、一又は複数のアミノ酸残基が挿入され、置換され、及び/又は欠失されたCRIgポリペプチドが含まれる。他の変異体は天然ポリペプチド配列の膜貫通領域内に一又は複数のアミノ酸が挿入され、置換され、及び/又は欠失されたものである。
【0051】
通常、CRIg変異体は、配列番号1−7の何れかのものからの成熟アミノ酸配列と少なくとも約80%アミノ酸配列同一性、又は少なくとも約85%アミノ酸配列同一性、又は少なくとも約90%アミノ酸配列同一性、又は少なくとも約95%アミノ酸配列同一性、又は少なくとも約98%アミノ酸配列同一性、又は少なくとも約99%アミノ酸配列同一性を有する。好ましくは、最も高い度合いの配列同一性は細胞外ドメイン(ECD)内(配列番号1又は2のアミノ酸1又は約21からX、ここで、Xは271から281の任意のアミノ酸残基;又は配列番号3のアミノ酸1又は約21からX、ここで、Xは178から186の任意のアミノ酸残基、あるいは配列番号4のアミノ酸1又は約21からX、ここでXは176から184の任意のアミノ酸残基である)で生じる。
【0052】
CRIg(PRO362)「細胞外ドメイン」又は「ECD」は、各全長分子の膜貫通及び細胞質ドメインが本質的にないCRIgポリペプチドの形態を意味する。通常、CRIg ECDはこのような膜貫通及び/又は細胞質ドメインを1%未満、好ましくはこのようなドメインを0.5%未満有している。上で検討したように、場合によっては、CRIg ECDは、配列番号1、2、3、又は4のアミノ酸残基1又は約21からXを含み、ここで、Xは配列番号1又は2の約271から281の任意のアミノ酸残基、配列番号3の約178から186の任意のアミノ酸、及び配列番号4の約176から184の任意のアミノ酸を含む。
【0053】
CRIg配列に関する「パーセント(%)アミノ酸配列同一性」とは、配列を整列させ、最大のパーセント配列同一性を得るために必要ならば間隙を導入し、如何なる保存的置換も配列同一性の一部と考えないとした後の、CRIg配列のアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセントとして定義される。パーセントアミノ酸配列同一性を決定する目的のためのアラインメントは、当業者の技量の範囲にある様々な方法、例えばBLAST、BLAST-2、ALIGN、又はMegalign(DNASTAR)ソフトウエアのような公に入手可能なコンピュータソフトウエアを使用することにより達成可能である。当業者であれば、比較される配列の完全長に対して最大のアラインメントを達成するために必要な任意のアルゴリズムを含む、アラインメントを測定するための適切なパラメータを決定することができる。別段の記載がなされない限り、アラインメントは、選択されたソフトウェアのデフォルトパラメータを使用して実施される。ついで、酸配列同一性は、より長い配列に対して計算される。つまり、より短い配列が、より長い配列の一部と100%の配列同一性を示すとしても、全体の配列同一性は100%未満になるであろう。
【0054】
CRIgコード配列に対する「パーセント(%)核酸配列同一性」は、配列を整列させ、最大のパーセント配列同一性を得るために必要ならば間隙を導入した後、CRIgコード配列のヌクレオチドと同一である候補配列中のヌクレオチドのパーセントとして定義される。パーセント核酸配列同一性を決定する目的のためのアラインメントは、当業者の知る範囲にある種々の方法、例えばBLAST、BLAST-2、ALIGN又はMegalign(DNASTAR)ソフトウエアのような公に入手可能なコンピュータソフトウエアを使用することにより達成可能である。当業者であれば、比較される配列の完全長に対して最大のアラインメントを達成するために必要な任意のアルゴリズムを含む、アラインメントを測定するための適切なパラメータを決定することができる。ついで、酸配列同一性は、より長い配列に対して計算される。つまり、より短い配列が、より長い配列の一部と100%の配列同一性を示すとしても、全体の配列同一性は100%未満になるであろう。
【0055】
「単離された」核酸分子は、同定され、核酸の天然供給源に通常付随している少なくとも一つの汚染核酸分子から分離された核酸分子である。単離された核酸分子は、天然に見出される形態あるいは設定以外のものである。ゆえに、単離された核酸分子は、天然の細胞中に存在する核酸分子とは区別される。しかし、単離された核酸分子は、例えば、核酸分子が天然の細胞のものとは異なった染色体位置にあるコード化ポリペプチドを通常発現する細胞に含まれる核酸分子を含む。
【0056】
「単離された」CRIgポリペプチドコード核酸分子は、同定され、CRIgコード核酸の天然供給源に通常付随している少なくとも一つの汚染核酸分子から分離された核酸分子である。単離されたCRIgポリペプチドコード核酸分子は、天然に見出される形態あるいは設定以外のものである。ゆえに、単離されたCRIgポリペプチドコード核酸分子は、天然の細胞中に存在する核酸分子とは区別される。しかし、単離されたCRIgコード核酸分子は、例えば、核酸分子が天然の細胞のものとは異なった染色体位置にあるCRIgを通常発現する細胞に含まれるCRIgコード核酸分子を含む。
【0057】
「補体関連疾患」なる用語は、ここでは最も広い意味で使用され、例えば補体欠損症のような病因が補体系の活性化の異常性を含む全ての疾患及び病理学的症状を含む。該用語はC3転換酵素の阻害から恩恵を受ける疾患及び病理学的症状を含む。該用語は、代替補体経路の選択的阻害を含む阻害から恩恵を受ける疾患及び病理学的症状を更に含む。補体関連疾患には、限定するものではないが、炎症性疾患及び自己免疫疾患、例えばリウマチ様関節炎(RA)、急性呼吸促迫症候群(ARDS)、虚血及び再灌流後の遠隔組織傷害、心肺バイパス手術中の補体活性化、皮膚筋炎、天疱瘡、ループス腎炎及びその結果の糸球体腎炎及び脈管炎、心肺バイパス、心臓麻痺誘発性冠血管内皮機能不全、II型膜性増殖性糸球体腎炎、IgA腎症、急性腎不全、クリオグロブリン血症、抗リン脂質症候群、黄斑変性疾患及び他の補体関連の眼の症状、例えば加齢性黄斑変性症(AMD)、脈絡膜血管新生(CNV)、ブドウ膜炎、糖尿病及び他の虚血関連網膜症、眼内炎、及び他の眼内血管新生疾患、例えば糖尿病性黄斑浮腫、病的近視、フォン・ヒッペル・リンドウ病、眼のヒストプラスマ症、網膜中心静脈閉塞症(CRVO)、角膜血管新生、網膜血管新生、並びに異物移植、超急性拒絶反応、血液透析、慢性閉塞肺促迫症候群(COPD)、喘息、及び誤嚥性肺炎が含まれる。
【0058】
「補体関連の眼の症状」なる用語は最も広い意味で使用され、古典及び代替経路、特に補体の代替経路を含む病因が補体に関連する全ての眼の症状を含む。補体関連の眼の症状には、限定するものではないが、黄斑変性疾患、例えばドライ及びウェット(非滲出性及び滲出性)形態を含む加齢性黄斑変性症(AMD)、脈絡膜血管新生(CNV)、ブドウ膜炎、糖尿病及び他の虚血関連網膜症、及び他の眼内血管新生疾患、例えば糖尿病性黄斑浮腫、病的近視、フォン・ヒッペル・リンドウ病、眼のヒストプラスマ症、網膜中心静脈閉塞症(CRVO)、角膜血管新生、及び網膜血管新生が含まれる。補体関連の眼の症状の好ましい群には、非滲出性(ウェット)及び滲出性(ドライ又は萎縮性)AMDを含む加齢性黄斑変性症(AMD)、脈絡膜血管新生(CNV)、糖尿病性網膜症(DR)、及び眼内炎が含まれる。
【0059】
「炎症性疾患」及び「炎症性障害」なる用語は、ここでは置き換え可能に使用され、哺乳動物の免疫系の成分が哺乳動物の罹患率に寄与する免疫反応を生じ、媒介し又は寄与等する疾患又は障害を意味する。また含まれるものは、炎症反応の減少が疾患の進行に寛解効果を有する疾患である。この用語に含まれるものは、自己免疫疾患を含む免疫媒介炎症疾患である。
【0060】
「T細胞媒介」疾患という用語は、T細胞が直接的又は間接的に哺乳動物の病的状態を媒介するか又は寄与等する疾患を意味する。T細胞媒介疾患は細胞媒介効果、リンホカイン媒介効果等に、また例えばT細胞により分泌されるリンホカインによりB細胞が刺激されるならばB細胞に関連した効果にさえも関連している。
【0061】
その幾つかがT細胞媒介性である免疫関連及び炎症疾患の例には、限定するものではないが、炎症性腸疾患(IBD)、全身性紅斑性狼瘡、リウマチ様関節炎、若年性慢性関節炎、脊椎関節症、全身性硬化症(強皮症)、特発性炎症ミオパシー(皮膚筋炎、多発性筋炎)、シェーグレン症候群、全身性血管炎、サルコイドーシス、自己免疫性溶血性貧血(免疫性汎血球減少症、発作性夜間血色素尿症)、自己免疫性血小板減少(特発性血小板減少性紫斑病、免疫媒介血小板減少)、甲状腺炎(グレーブス疾患、ハシモト甲状腺炎、若年性リンパ球性甲状腺炎、萎縮性甲状腺炎)、真性糖尿病、免疫介在性腎疾患(糸球体腎炎、尿細管間質性腎炎)、中枢及び末梢神経系の脱髄疾患、例えば多発性硬化症、多発性神経障害、肝胆汁性疾患、例えば感染性肝炎(A型、B型、C型、D型、E型肝炎及び他の非肝親和性(nonhepatotropic)ウィルス)、自己免疫慢性活性肝炎、原発性胆汁性肝硬変症、肉芽腫性肝炎、及び硬化性胆管炎、炎症性及び線維性肺疾患(例えば嚢胞性線維症)、グルテン過敏性腸疾患、ウィップル病、水疱性皮膚疾患、多形性紅斑及び接触性皮膚炎を含む自己免疫又は免疫媒介皮膚疾患、乾癬、肺のアレルギー性疾患、例えば好球性肺炎、特発性肺線維症及び過敏性肺炎、拒絶反応及び移植片対宿主疾患を含む移植関連疾患が含まれる。アルツハイマー病、及びアテローム性動脈硬化症が含まれる。
【0062】
ここで使用される「腫瘍」は、悪性であろうと良性であろうとあらゆる新生物細胞成長及び増殖、及びあらゆる前癌状態の細胞及び組織を意味する。
【0063】
「癌」及び「癌性」という用語は、典型的には調節されない細胞増殖を特徴とする、哺乳動物における生理学的状態を指すか記述する。癌の例には、これらに限定されるものではないが、癌腫、リンパ腫、芽細胞腫、肉腫、及び白血病が含まれる。このような癌のより特定の例には、乳癌、前立腺癌、大腸癌、扁平細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸管癌、卵巣癌、肝臓癌、膀胱癌、肝癌、結腸直腸癌、子宮内膜癌、唾液腺癌、腎臓癌、肝癌、外陰癌、甲状腺癌、肝臓癌及び様々なタイプの頭頸部癌が含まれる。
【0064】
「治療」とは、疾患の病理の進展阻止又は変更を行なう本発明により実施される介入である。従って、「治療」は治療的処置及び予防的又は保護的手段の両方を指す。治療が必要なものは、既に疾患に罹患しているもの並びに疾患が防止されるべきものを含む。免疫関連疾患の治療では、治療薬は直接的に免疫反応の成分の反応の大きさを変え、又は他の治療剤、例えば抗生物質、抗真菌剤、抗炎症剤、化学療法剤等による治療に対する疾患の感受性を高めることができる。
【0065】
補体関連疾患のような疾患の「病理」は、患者の良好な健康を危険にさらす全ての現象を含む。これには、限定されるものではないが、異常又は制御不能な細胞増殖(好中球、好酸球、単球性、リンパ性細胞)、抗体産生、自己抗体産生、補体産生、正常に機能している隣接細胞の妨害、サイトカイン又は他の分泌産物の異常なレベルでの放出、あらゆる炎症性又は免疫性反応の抑制又は悪化、炎症性細胞(好中球、好酸球、単球性、リンパ性)の組織空間への浸潤等が含まれる。
【0066】
ここで使用される「哺乳動物」なる用語は、ヒト、家庭及び農業用動物、動物園、スポーツ、又はペット動物、例えばウマ、ブタ、ウシ、イヌ、ネコ及びケナガイタチなどを含む哺乳類に分類される任意の動物を意味する。本発明の好ましい実施態様では、哺乳動物はヒトである。
【0067】
一又は複数の更なる治療薬と「組み合わせた」投与とは、同時(同時期)及び任意の順序での連続した投与を含む。
【0068】
「サイトカイン」という用語は、一つの細胞集団から放出されるタンパク質であって、他の細胞に対して細胞間メディエータとして作用するものの包括的な用語である。このようなサイトカインの例としては、リンフォカイン、モノカイン、及び伝統的なポリペプチドホルモンを挙げることができる。サイトカインには、成長ホルモン、例えばヒト成長ホルモン、N-メチオニルヒト成長ホルモン、及びウシ成長ホルモン、副甲状腺ホルモン、チロキシン、インスリン、プロインスリン、リラクシン、プロリラクシン、卵胞刺激ホルモン(FSH)のような糖タンパク質ホルモン、副甲状腺刺激ホルモン(TSH)、及び黄体形成ホルモン(LH)、肝臓成長因子、線維芽細胞増殖因子、プロラクチン、胎盤ラクトゲン、腫瘍壊死因子-α及び-β、ミュラー阻害物質、マウス性腺刺激ホルモン関連ペプチド、インヒビン、アクチビン、血管内皮増殖因子、インテグリン、トロンボポエチン(TPO)、神経成長因子、例えばNGF-β血小板増殖因子、トランスフォーミング増殖因子(TGF)、例えばTGF-α及びTGF-β、インスリン様増殖因子I及びII、エリスロポイエチン(EPO)、骨誘導因子、インターフェロンα、β、γのようなインターフェロン;マクロファージCSF(M-CSF)のようなコロニー刺激因子(CSF)、顆粒球マクロファージCSF(GM-CSF)、及び顆粒球CSF(G-CSF);IL-1、IL-1α、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-8、IL-9、IL-11、IL-12等のインターロイキン(IL)、腫瘍壊死因子、例えばTNF-α又はTNF-β、及びLIF及びキットリガンド(KL)を含む他のポリペプチド因子が含まれる。ここで使用される場合、サイトカインなる用語は天然源由来あるいは組換え細胞培養由来のタンパク質及び天然配列サイトカインの生物的に活性な等価物を含む。
【0069】
「治療的有効量」は、標的の疾患又は症状、例えば補体関連疾患又は症状、又は癌の状態、例えば病理において測定可能な改善を達成するために必要とされる活性なCRIg、CRIgアゴニスト及びアンタゴニストの量である。
【0070】
「コントロール配列」という用語は、特定の宿主生物において作用可能に結合したコード配列を発現するために必要なDNA配列を指す。例えば原核生物に好適なコントロール配列は、プロモーター、場合によってはオペレータ配列、及びリボソーム結合部位を含む。真核生物の細胞は、プロモーター、ポリアデニル化シグナル、及びエンハンサーを利用することが知られている。
【0071】
核酸は、他の核酸配列と機能的な関係にあるときに「作用可能に結合」している。例えば、プレ配列又は分泌リーダーのDNAは、ポリペプチドの分泌に参画するプレタンパク質として発現されているならば、そのポリペプチドのDNAに作用可能に結合している;プロモーター又はエンハンサーは、配列の転写に影響を及ぼすならば、コード配列に作用可能に結合している;又はリボソーム結合部位は、もしそれが翻訳を容易にするような位置にあるなら、コード配列と作用可能に結合している。一般的に、「作用可能に結合している」とは、結合したDNA配列が近接しており、分泌リーダーの場合には近接していて読みフェーズにあることを意味する。しかしながら、エンハンサーは必ずしも近接している必要はない。結合は簡便な制限部位でのライゲーションにより達成される。そのような部位が存在しない場合は、合成オリゴヌクレオチドアダプター又はリンカーが常套的な手法に従って使用される。
【0072】
ハイブリダイゼーション反応の「ストリンジェンシー」は、当業者によって容易に決定され、一般的にプローブ長、洗浄温度、及び塩濃度に依存する経験的な計算である。一般に、プローブが長くなると適切なアニーリングのための温度が高くなり、プローブが短くなると温度は低くなる。ハイブリダイゼーションは、一般的に、相補的鎖がその融点に近いがそれより低い環境に存在する場合における変性DNAの再アニールする能力に依存する。プローブとハイブリダイゼーション可能な配列との間の所望の相同性の程度が高くなると、使用できる相対温度が高くなる。その結果、より高い相対温度は、反応条件をよりストリンジェントにするが、低い温度はストリンジェンシーを低下させる。ハイブリダイゼーション反応のストリンジェンシーの更なる詳細及び説明は、Ausubel等, Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995)を参照のこと。
【0073】
ここで定義される「ストリンジェントな条件」又は「高度のストリンジェント条件」は、(1)洗浄のために低イオン強度及び高温度を用いるもの、例えば、50℃において0.015Mの塩化ナトリウム/0.0015Mのクエン酸ナトリウム/0.1%のドデシル硫酸ナトリウムを用いるもの;(2)ハイブリダイゼーション中にホルムアミド等の変性剤を用いるもの、例えば、42℃において50%(v/v)ホルムアミド、0.1%ウシ血清アルブミン/0.1%フィコール/0.1%のポリビニルピロリドン/50mMのpH6.5のリン酸ナトリウムバッファー、及び750mMの塩化ナトリウム、75mMのクエン酸ナトリウムを用いるもの;又は(3)42℃において50%ホルムアミド、5×SSC(0.75MのNaCl、0.075Mのクエン酸ナトリウム)、50mMのリン酸ナトリウム(pH6.8)、0.1%のピロリン酸ナトリウム、5×デンハード液、超音波処理サケ精子DNA(50μg/ml)、0.1%SDS、及び10%のデキストラン硫酸を用い、42℃において0.2×SSC(塩化ナトリウム/クエン酸ナトリウム)中で、55℃において50%ホルムアミド中で洗浄した後、55℃においてEDTAを含む0.1×SSCからなる高ストリンジェント洗浄を用いるものによって同定できる。
【0074】
「中程度のストリンジェント条件」は、Sambrook等, Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989に記載されているように特定され、上記のものよりストリンジェンシーが低い洗浄液及びハイブリダイゼーション条件(例えば、温度、イオン強度及び%SDS)の使用を含む。中程度のストリンジェント条件は、20%ホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハード液、10%デキストラン硫酸、及び20mg/mLの変性剪断サケ精子DNAを含む溶液中の37℃での終夜のインキュベーション後に、37〜50℃にて1×SSC中でフィルター洗浄を行うという条件である。プローブ長などの因子に必要に応じて適合させるには、どのようにして温度、イオン強度等を調節するかは当業者であれば分かるであろう。
【0075】
「エピトープタグ」なる用語は、ここで用いられるときは、「タグポリペプチド」に融合した本発明のポリペプチドを含んでなるキメラポリペプチドを指す。タグポリペプチドは、その抗体が産生され得るエピトープを提供するに十分な数の残基を有しているが、その長さは融合するポリペプチドの活性を阻害しないよう充分に短い。また、タグポリペプチドは、好ましくは、抗体が他のエピトープと実質的に交差反応をしないようにかなり独特である。適切なタグポリペプチドは、一般に、少なくとも6のアミノ酸残基、通常は約8〜約50のアミノ酸残基(好ましくは約10〜約20の残基)を有する。
【0076】
本発明のCRIgポリペプチドの変異体に関し、「活性な」又は「活性」は、本発明の天然又は天然に生じるポリペプチドの生物学的及び/又は免疫学的活性を保持するポリペプチドの形態を意味する。好ましい生物学的活性は、C3bに結合し、及び/又は補体又は補体活性化に影響を及ぼし、特に代替の補体経路及び/又はC3転換酵素を阻害する能力である。3転換酵素の阻害は、例えばコラーゲン-又は抗体-誘発関節炎の間における正常な血清中でのC3代謝回転の阻害、又は関節炎の間接におけるC3沈着の阻害を測定することにより測定することができる。
【0077】
CRIgの生物学的活性を模倣し、ここに開示されたスクリーニングアッセイによって同定することができる抗体、ポリペプチド又は他の分子(例えば有機又は無機小分子、ペプチド等々)に関する「生物学的活性」とは、C3bに結合し、及び/又は補体又は補体活性化に影響を及ぼし、特に代替の補体経路及び/又はC3転換酵素を阻害するそのような分子の能力を指す。
【0078】
CRIg「アゴニスト」なる用語は最も広い意味で使用され、天然配列CRIgポリペプチドの(上述の)定性的生物学的活性を模倣する任意の分子を含む。
【0079】
CRIg「アンタゴニスト」なる用語は最も広い意味で使用され、天然配列CRIgポリペプチドのような天然ポリペプチドの定性的生物学的活性を部分的に又は完全にブロックし、阻害し、又は中和する任意の分子を含む。
【0080】
適切なアゴニスト又はアンタゴニスト分子には、アゴニスト又はアンタゴニスト抗体又は抗体断片、本発明の天然ポリペプチドの断片、融合体又はアミノ酸配列変異体、ペプチド、小有機分子を含む小分子等々が特に含まれる。
【0081】
「小分子」はここでは約600以下、好ましくは約1000ダルトン以下の分子量を有するものと定義されている。
【0082】
「抗体」という用語は最も広い意味において使用され、限定するものではないが、単一の抗CRIgモノクローナル抗体(アゴニスト、アンタゴニスト、及び中和抗体を含む)、及び多エピトープ特異性を持つ抗CRIg抗体組成物を包含する。ここで使用される「モノクローナル抗体」という用語は、実質的に均一な抗体の集団、すなわち、構成する個々の抗体が、少量存在しうる自然に生じる可能性のある突然変異を除いて同一である集団から得られる抗体を意味する。
【0083】
「抗体」(Abs)及び「免疫グロブリン」(Igs)は同じ構造的特徴を有する糖タンパク質である。抗体は特定の抗原に対して結合特異性を示すが、免疫グロブリンは抗体と抗原特異性を欠く抗体様分子の双方を含む。後者の種類のポリペプチドは例えばリンパ系によって低レベルで、ミエローマによって増大したレベルで生産される。「抗体」なる用語は最も広い意味で使用され、限定するものではないが、無傷のモノクローナル抗体、ポリクローナル抗体、少なくとも2つの無傷抗体から形成された多重特異的抗体(例えば二重特異的抗体)、及びそれらが所望の生物学的活性を示す限り抗体断片を特に包含する。
【0084】
「天然抗体」及び「天然免疫グロブリン」は、通常、2つの同一の軽(L)鎖及び2つの同一の重(H)鎖からなる、約150000ダルトンのヘテロ四量体糖タンパク質である。各軽鎖は一つの共有ジスルフィド結合により重鎖に結合しており、ジスルフィド結合の数は、異なった免疫グロブリンアイソタイプの重鎖間で変わる。また各重鎖と軽鎖は、規則的に離間した鎖内ジスルフィド結合を有している。各重鎖は、多くの定常ドメインが続く可変ドメイン(VH)を一端に有する。各軽鎖は、一端に可変ドメイン(VL)を、他端に定常ドメインを有し;軽鎖の定常ドメインは重鎖の第一定常ドメインと整列し、軽鎖の可変ドメインは重鎖の可変ドメインと整列している。特定のアミノ酸残基が、軽鎖及び重鎖可変ドメイン間の界面を形成すると考えられている。
【0085】
「可変」という用語は、可変ドメインのある部分が、抗体間で配列が広範囲に異なっており、その特定の抗原に対する各特定の抗体の結合性と特異性に使用されていることを意味する。しかしながら、可変性は抗体の可変ドメインにわたって一様には分布していない。それは、軽鎖及び重鎖の可変ドメインの両方の相補性決定領域(CDR)又は高頻度可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存された部分はフレームワーク領域(FR)と呼ばれる。天然の重鎖及び軽鎖の可変ドメインは、βシート構造を結合し、ある場合にはその一部を形成するループを形成する、3つのCDRにより連結されたβシート配置を主にとる4つのFR領域を含んでいる。各鎖のCDRは、FR領域によって近接して互いに保持され、他の鎖のCDRと共に、抗体の抗原結合部位の形成に寄与している(Kabat等, NIH Publ. No.91-3242, Vol. I, pp. 647-669 (1991)を参照)。定常ドメインは、抗体の抗原への結合に直接関与しているものではないが、様々なエフェクター機能、例えば抗体依存性細胞障害活性への抗体の関与を示す。
【0086】
「抗体断片」は、無傷の抗体の一部、好ましくは無傷の抗体の抗原結合又は可変領域を含む。抗体断片の例は、Fab、Fab’、F(ab’)2、及びFv断片;ダイアボディ(diabodies);線形抗体(Zapata等, Protein Eng. 8(10): 1057-1062 [1995]);一本鎖抗体分子;及び抗体断片から形成された多重特異性抗体を含む。
【0087】
抗体のパパイン消化は、「Fab」断片と呼ばれる二つの同一の抗原結合断片を生成し、その各々は単一の抗原結合部位、及び残基「Fc」断片を持つ。該名称「Fc」は容易に結晶化する能力を反映している。ペプシン処理は、二つの抗原結合部位を持ち、なお抗原と交差結合可能なF(ab’)2断片を生じる。
【0088】
「Fv」は、完全な抗原認識及び結合部位を含む最小の抗体断片である。この領域は、強固に非共有結合した1本の重鎖と1本の軽鎖の可変領域の二量体からなる。この構造において各可変ドメインの三つのCDRが相互作用してVH−VL二量体の表面に抗原結合部位を形成する。集合的には、6つのCDRが抗体に対する抗原結合特異性を与える。しかしながら、単一の可変ドメイン(又は抗原に特異的な三つのCDRのみを含んでなるFvの半分)でさえ、結合部位全体よりは低い親和性であるが、抗原を認識し結合する能力を有している。
【0089】
またFab断片は、軽鎖の定常ドメイン及び重鎖の第一定常ドメイン(CH1)も含む。Fab’断片は、抗体ヒンジ領域からの一又は複数のシステインを含む重鎖CH1ドメインのカルボキシ末端に数残基が付加されていることでFab断片と相違する。Fab’-SHは、定常ドメインのシステイン残基が遊離のチオール基を持つFab’に対するここでの表記である。F(ab’)2抗体断片は、最初は間にヒンジシステインを有しているFab’断片の対として生成された。抗体断片の他の化学的結合もまた知られている。
【0090】
任意の脊椎動物種からの抗体(免疫グロブリン)の「軽鎖」は、それらの定常ドメインのアミノ酸配列に基づいて、カッパ(κ)及びラムダ(λ)と呼ばれる二つの明らかに異なる型の一方に分類できる。
【0091】
それらの重鎖の定常ドメインのアミノ酸配列によって、免疫グロブリンは異なるクラスに分類できる。免疫グロブリンの五つの主要なクラス:IgA、IgD、IgE、IgG及びIgMがあり、それらの幾つかは更にサブクラス(アイソタイプ)、例えばIgG1、IgG2、IgG3、IgG4、IgA及びIgA2に分類される。免疫グロブリンの異なったクラスに対応する重鎖定常ドメインはそれぞれγ、μ、δ、α、及びεと呼ばれる。異なったクラスの免疫グロブリンのサブユニット構造及び三次元立体構造はよく知られている。
【0092】
ここで使用される「モノクローナル抗体」という用語は、実質的に均一な抗体の集団、すなわち、構成する個々の抗体が、少量存在しうる自然に生じる可能性のある突然変異を除いて同一である集団から得られる抗体を称する。モノクローナル抗体は高度に特異的であり、一つの抗原部位に対応する。更に、異なる決定基(エピトープ)に対応する異なる抗体を典型的に含む通常の(ポリクローナル)抗体とは異なり、各モノクローナル抗体は抗原の単一の決定基に向く。特異性に加えて、他の免疫グロブリンによって汚染されていないハイブリドーマ培養によって合成されることで、モノクローナル抗体は有利である。「モノクローナル」との形容は、実質的に均一な抗体集団から得られたという抗体の性質を示し、特定の方法で抗体を生産しなければならないことを意味するものではない。例えば、本発明において使用されるモノクローナル抗体は、Kohler等によって最初にNature 256:495[1975]に記載されたハイブリドーマ法によって作ることができるか、あるいは組換えDNA法(例えば米国特許第4816567号を参照)によって作ることができる。「モノクローナル抗体」は例えば Clackson等, Nature 352:624-628 (1991) 及び Marks等, J.Mol.Biol. 222:581-597 (1991)に記載された技術を用いてファージ抗体ライブラリーから単離してもよい。また、ファージミッド及びファージベクターの利用による抗体の調製が記載されている米国特許第5750373号、第5571698号、第5403484号及び第5223409号を参照のこと。
【0093】
ここで、モノクローナル抗体は、特に重鎖及び/又は軽鎖の一部が特定の種から誘導されたか又は特定の抗体クラス又はサブクラスに属する抗体の対応する配列と同一又は相同である一方、鎖の残りの部分は他の種から誘導されたか又は特定の抗体クラス又はサブクラスに属する抗体の対応する配列と同一又は相同である「キメラ」抗体(免疫グロブリン)、並びにそれらが所望の生物学的活性を示す限りそれらの抗体の断片を含む(米国特許第4816567号;Morrison等,Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984))。
【0094】
非ヒト(例えばマウス)抗体の「ヒト化」形とは、キメラ免疫グロブリン、免疫グロブリン鎖又はその断片(例えばFv、Fab、Fab’、F(ab’)2あるいは抗体の他の抗原結合サブ配列)であって、非ヒト免疫グロブリンに由来する最小配列を含むものである。大抵は、ヒト化抗体はレシピエントの相補性決定領域(CDR)の残基が、マウス、ラット又はウサギのような所望の特異性、親和性及び能力を有する非ヒト種(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(レシピエント抗体)である。ある場合には、ヒト免疫グロブリンのFvフレームワーク領域(FR)残基はまた対応する非ヒト残基によって置換されうる。更に、ヒト化抗体は、レシピエント抗体にも、移入されたCDRもしくはフレームワーク配列にも見出されない残基を含んでいてもよい。これらの変更は、抗体の性能を更に洗練させ、最大化するために行なわれる。一般に、ヒト化抗体は、全てあるいはほとんど全てのCDR領域が非ヒト免疫グロブリンのものに対応し、全てあるいはほとんど全てのFR領域がヒト免疫グロブリン配列のものである、少なくとも一つ、典型的には二つの可変ドメインの実質的に全てを含む。ヒト化抗体は、最適には免疫グロブリン定常領域(Fc)、典型的にはヒトの免疫グロブリンの定常領域の少なくとも一部を含んでなる。さらなる詳細については、Jones等, Nature, 321:522-525 (1986);Riechmann等, Nature, 332:323-329 [1988];及びPresta, Curr. Op Struct. Biol., 2:593-596 (1992)を参照のこと。ヒト化抗体は、抗体の抗原結合領域が、マカクザルを対象の抗原で免疫化することにより生成された抗体に由来している「霊長類化」抗体を含む。旧世界ザル由来の残基を含む抗体もまた本発明において可能である。例えば米国特許第5658570号、第5693780号、第5681722号、第5750105号及び第5756096号を参照のこと。
【0095】
「一本鎖Fv」又は「sFv」抗体断片は、抗体のVH及びVLドメインを含み、これらのドメインは単一のポリペプチド鎖に存在する。好ましくは、Fvポリペプチドは、sFvが抗原結合にとって望ましい構造の形成を可能にするVH及びVLドメイン間のポリペプチドリンカーを更に含む。sFvの概説については、Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg及びMoore編, Springer-Verlag, New York, pp. 269-315 (1994)を参照のこと。
【0096】
「ダイアボディ」という用語は、二つの抗原結合部位を持つ小抗体断片を指し、その断片は同じポリペプチド鎖(VH-VL)内で軽鎖可変ドメイン(VL)に結合した重鎖可変ドメイン(VH)を含む。同じ鎖の二つのドメイン間に対形成するには短すぎるリンカーを用いることにより、ドメインは強制的に他の鎖の相補的ドメインと対形成して二つの抗原結合部位をつくりだす。ダイアボディは、例えば、EP404097;WO93/11161;及びHollinger等, Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)に更に詳しく記載されている。
【0097】
抗体のような「単離された」ポリペプチドとは、その自然環境の成分から同定され分離され及び/又は回収されたものを意味する。その自然環境の汚染成分とは、その抗体の診断又は治療への使用を典型的には妨害する物質であり、酵素、ホルモン、及び他のタンパク質様又は非タンパク質様溶質を含み得る。好ましい実施形態では、抗体を含むポリペプチドは、(1)ローリー(Lowry)法による測定で化合物の95重量%よりも大きな割合まで、最も好ましくは99重量%よりも大きな割合まで、(2)スピニングカップシークエネーターを使用することにより、少なくとも15残基のN末端あるいは内部アミノ酸配列を得るのに充分な程度まで、あるいは(3)クーマシーブルー又は好ましくは銀染色を用いた還元又は非還元条件下でのSDS-PAGEによる均一性まで精製される。単離された化合物、例えば抗体又は他のポリペプチドには、化合物の自然環境の少なくとも一つの成分が存在しないため、組換え細胞内のインサイツの化合物が含まれる。しかしながら、通常は、単離された化合物は少なくとも一つの精製工程により調製される。
【0098】
「標識」なる語は、ここで用いられる場合、「標識した」化合物が生成されるように、化合物、例えば抗体又はポリペプチドに直接又は間接的に抱合させた検出可能な化合物又は組成物を意味する。標識は、それ自身検出可能でもよく(例えば、放射性標識又は蛍光標識)、又は酵素標識の場合、検出可能な基質化合物又は組成物の化学変換を触媒しうる。
【0099】
「固相」とは、本発明の化合物が付着することのできる非水性マトリクスを意味する。ここで包含される固相の例は、部分的又は全体的に、ガラス(例えば、孔制御ガラス)、多糖類(例えばアガロース)、ポリアクリルアミド、ポリスチレン、ポリビニルアルコール及びシリコーンから形成されたものを含む。ある実施形態では、内容に応じて、固相はアッセイプレートのウェルを含み得;その他では精製カラム(例えばアフィニティークロマトグラフィーカラム)とすることもできる。また、この用語は、米国特許第4275149号に記載されたような、別個の粒子の不連続な固相も包含する。
【0100】
「リポソーム」は、哺乳動物への薬物(ここに開示する抗ErbB2抗体及び場合によっては化学療法剤)の輸送に有用である種々の型の脂質、リン脂質及び/又は界面活性剤からなる小型の小胞である。リポソームの成分は、通常は生体膜の脂質配列に類似する二層形態に配列させる。
【0101】
ここで用いられる場合、「イムノアドヘシン」なる用語は、異種タンパク質(「アドヘシン」)の結合特異性と免疫グロブリン定常ドメインのエフェクター機能とを併せ持つ抗体様分子を指す。構造的には、イムノアドヘシンは、所望の結合特異性を持ち、抗体の抗原認識及び結合部位以外である(つまり「異種の」)アミノ酸配列と、免疫グロブリン定常ドメイン配列との融合物を含む。イムノアドヘシン分子のアドへシン部分は、典型的には少なくともレセプター又はリガンドの結合部位を含む隣接アミノ酸配列である。イムノアドヘシンの免疫グロブリン定常ドメイン配列は、IgG-1、IgG-2、IgG-3又はIgG-4サブタイプ、IgA(IgA-1及びIgA-2を含む)、IgE、IgD又はIgMなどの任意の免疫グロブリンから得ることができる。
【0102】
「慢性」投与とは、延長された期間の間、所望の効果を維持するように、急性態様とは異なる連続態様で薬剤を投与することを意味する。
【0103】
「間欠」投与とは、中断がなく連続的になされるのではなく、むしろ周期的な性質の治療法である。
【0104】
一又は複数の更なる治療剤「との併用での」投与は同時(同時期)と任意の順序の連続投与を含む。
【0105】
「患者」は脊椎動物、好ましくは哺乳動物、より好ましくはヒトである。
【0106】
「哺乳動物」なる用語は、ここでは限定するものではないが、ヒト、牧畜、家畜、及び動物園の動物、スポーツ、又はペット動物、例えばヒツジ、イヌ、ウマ、ネコ、ウシ等々を含む哺乳動物として分類される任意の動物を指すために使用される。
【0107】
「有効量」とは有益な又は所望される治療的(予防を含む)結果を得るのに十分な量である。有効量は一又は複数回の投与で投与できる。
【0108】
(発明を実施するための形態)
本発明の実施には、別段の記載がない限り、当業者の技量の範囲にある(組換え技術を含む)分子生物学、微生物学、細胞生物学、生化学及び免疫学の一般的な技術を用いる。かかる技術は例えば、“Molecular Cloning: A Laboratory Manual”, 2版 (Sambrook等, 1989);“Oligonucleotide Synthesis” (M.J. Gait編, 1984);“Animal Cell Culture” (R.I. Freshney編, 1987);“Methods in Enzymology” (Academic Press, Inc.);“Handbook of Experimental Immunology”, 4版 (D.M. Weir & C.C. Blackwell編, Blackwell Science Inc., 1987);“Gene Transfer Vectors for Mammalian Cells” (J.M. Miller & M.P. Calos編, 1987);“Current Protocols in Molecular Biology” (F.M. Ausubel等編, 1987);“PCR: The Polymerase Chain Reaction”, (Mullis等編, 1994);及び“Current Protocols in Immunology” (J.E. Coligan等編, 1991)のような文献に十分に説明されている。
【0109】
CRIg及びC3b:CRIh及びC3c複合体の結晶構造
CRIg分子を含むポリペプチドは一次アミノ酸配列及びポリペプチドを囲む環境によって決定される3次元構造を有している。この3次元構造はポリペプチドの活性、安定性、結合親和性、結合特異性、及び他の生化学的特質を樹立する。よって、タンパク質の3次元構造の知識はその生物学的活性を模倣し、阻害し又は改善する薬剤を設計する際の大きな指針を提供する。
【0110】
ポリペプチドの3次元構造は多くの方法で決定することができる。最も精確な方法の多くはx線結晶解析(Van Holde, Physical Biochemistry (Prentice Hall: N.J., 1971), pp. 221-239)を用いる。この技術はx線又は他の形態の放射線を回折する結晶格子の能力に依存している。巨大分子の3次元構造を決定するために適した回折実験は典型的には高品質の結晶を必要とする。不幸にも、かかる結晶はIGF-1並びに興味のある多くの他のタンパク質には利用できなかった。結晶は例えばM−CSF(EP668914B1)、CD40リガンド(WO97/00895)、及びBC2Fab断片(WO99/01476)に対して記載されている。
【0111】
結晶性タンパク質及びポリペプチドを調製するための様々な方法が当該分野で知られている(McPherson等., “Preparation and Analysis of Protein Crystals,” McPherson (Robert E. Krieger Publishing Company, Malabar, FL, 1989);Weber, Advances in Protein Chemistry, 41: 1-36 (1991);米国特許第4672108号及び第4833233号)。ポリペプチドを結晶化させるための複数のアプローチ法が存在しているが、特に結晶がx線回折研究に適したものでなければならない場合には、合理的な成功の可能性をもたらす単一セットの条件は存在しない。構造に関する精確な情報をもたらす十分なサイズと分解能の結晶を得るには顕著な努力が必要とされる。例えば、十分な純度のタンパク質がひとたび得られれば、x線回折と続く構造解明に有用なサイズと透明度になるまで結晶化されなければならない。更に、標的タンパク質のアミノ酸配列は知られていても、この配列情報からタンパク質の結晶構造を正確に予測することはできない。また配列情報から、タンパク質とCRIgやC3bのようなそれが相互作用する結合対との間の構造的、コンフォメーション的及び化学的相互作用の理解ができるようになるものでもない。よって、結晶構造はドラッグデザイン及び発見の分野で豊富な価値ある情報をもたらしうるが、CRIgのようなある種の生物学的に関連した化合物の結晶は当業者が直ぐに利用できるものではない。CRIgの高品質の回折結晶は、その生物学的役割を更に理解し、限定するものではないがアゴニスト及びアンタゴニスト抗体を含むCRIg変異体、アゴニスト及び淡タゴニストを設計するために重要になるその3次元構造の決定に役立つ。
【0112】
構造情報の提供に加えて、結晶性ポリペプチドは他の利点をもたらす。例えば、結晶化方法自体はポリペプチドを更に精製し、均一性の古典的条件の一つを満たす。実際、結晶化は、HPLC、透析、一般的なカラムクロマトグラフィー等々の他の精製方法によっては除去されない不純物を除去する無比の精製品質をもたらすことがよくある。更に、結晶性ポリペプチドはしばしば周囲温度で安定であり、プロテアーゼの汚染や溶液保存に伴う他の分解を受けない。結晶性ポリペプチドはまた医薬製剤としても有用でありうる。最後に、一般に結晶化技術には、他の安定化法(例えば凍結乾燥)に付随する変性のような問題が概してない。
【0113】
本発明はCRIg単独とC3b及びC3cそれぞれとの複合体の結晶構造を提供する。C3b:CRIg複合体の構造は、C3活性化時に生じ補体レセプターCRIgの結合部位を定める立体構造変化を完全に明らかにする。また、代替経路転換酵素のタンパク質分解活性に対するこの相互作用の結果を調べる。
【0114】
すなわち、本発明は、およそ次の寸法:a=30.3Å、b=50.8Å、c=62.0Åを有するCRIg結晶を提供する。該結晶は、P212121の対称、又は空間群を有している。CRIg結晶の構造座標はアペンディクス1に提供する。
【0115】
本発明は更におよそ次の寸法:a=97.6Å、b=255.7Å、c=180.3Å及びC2221の空間群を有しているC3b:CRIg複合体の結晶構造を提供する。CRIg結晶の構造座標はアペンディクス2に提供し、そのリボン構造は図1Bに示す。
【0116】
本発明はまたおよそ次の寸法:a=382.8Å、b=65.0Å、c=147.2Å、β=102.7、及びC2の空間群を有しているC3c:CRIg複合体の結晶構造を提供する。C3c:CRIg結晶の構造座標はアペンディクス3に提供し、そのリボン構造は図1Cに示す。
【0117】
残基1〜645からなるC3bは、5つのマクログロブリン様ドメイン(MG1-MG5)、及び第6MGドメインのN末端半分、リンカー領域(LNK)(図1A及び1B)に折り畳まれる。免疫グロブリントポロジーMGドメインは「キーリング」様形式に配置され、約10Å幅で30Å長の中央溝の回りを円で囲んでいる。C3bのα鎖は残基729:1641からなり、C3と比較してC3a又はANAドメインを欠いている。α'NTドメインと呼ばれるC3bα鎖のN末端残基(729:745)は、伸展したコンフォメーションを採用しており、第6MGドメインの第2の半分を形成する残基につながっている。これに続いて、α鎖は間にCUBドメインとチオ-エステルドメイン(TED)が挿入された2つの更なるMGドメイン(MG7及びMG8)を含んでいる。最後に、α鎖のC末端の170残基はいわゆる「アンカー領域」とC345Cドメインを形成している。
【0118】
実施例1に記載されるように、C3bのコンフォメーションは活性化の際に劇的な変化を受ける。TEDはCUB及びNG8ドメインから離れる方向に回転し、完全に異なったコンフォメーションでTED及びCUBドメインを連結する残基を残す。このコンフォメーション変化の一部として、C3bの988位のCys残基がC3構造中のその最初の保護されたネストから露出した位置まで移動する。
【0119】
実施例1に記載された結晶化研究から、CRIgに対する結合部位がC3bのCUB及びTEDドメインとは反対側にあり、CRIgをTEDドメインから約40Åのところに位置させていることが明らかになった(図1A及び1Bを参照)。
【0120】
CRIgとC3c又はC3bの間の結合界面は大きく、不連続で、全体で約2670Å2の溶媒露出面積を埋めている(図2A)。CRIgはC3bと接触し、A’、G、G、C、C’、C’’シートからの残基が、ストランドC’とC”の近傍の残基及びループによって形成された相互作用の大部分を有する。C’とC”を連結するヘアピンループは、C3bのキーリング形状のβ鎖の中心に形成された裂け目中に突出している。界面のC3b側では、ドメインMG3、MG4、MG5、MG6、LNK及びMG6が全てCRIg結合に寄与し、埋まった界面のそれぞれ約30%と40%に対してMG3とMG6が原因となっている(図2Bを参照)。MG3ドメインはC3b(及びC3c)へのCRIgの結合のゲーティングに重要な役割を担っていると考えられる。また、C3分子の残りと比較してMG3の比較的小さいがなお有意な15°の回転が、CRIg結合部位を構成する他のMGドメインのリングに関したMG3の相対的再配向の原因である。従って、LNK領域の螺旋部分の動きと連係して、この回転がCRIgをC3b及びC3cに結合させる。これらの中程度の変化がないと、C3はCRIgにはラッチされないであろう。
【0121】
これらの構造-機能の知見はCRIgの数個のアミノ酸置換変異体を試験することによって確認した。NG6及びMG3結合ドメインにおける変異はC3bへのCRIgの結合を効果的に抑止することが確認された。これに対して、反対側のCRIg表面残基の変異は野生型CRIgの活性に効果はなかった。
【0122】
CRIgアゴニスト及びアンタゴニストのデザイン、調製及びスクリーニング
本発明は、例えば補体関連疾患又は障害、例えば免疫複合体及び自己免疫疾患、及び補体介在炎症組織損傷を含む様々な炎症症状の治療に有用性があるCRIgアゴニストを同定するスクリーニングアッセイを含む。補体関連疾患の病理は様々であり、長期間又は短期間の補体活性化、カスケード全体、カスケードの一つだけ(例えば古典又は代替経路)、カスケードの幾つかの成分だけの活性化等々を含むかもしれない。ある疾患では、補体断片の補体生物学的活性が組織損傷及び疾患を生じさせる。従って、補体の阻害剤は高い治療的潜在性を有している。代替経路の選択的阻害剤は、古典経路による血液からの病原体及び他の生物の除去が手を付けられていないままであるので、特に有用であろう。
【0123】
CRIgポリペプチドのアゴニストは天然配列CRIgポリペプチドの定性的生物学的活性を模倣する。好ましくは、生物学的活性はC3bに結合し、及び/又は補体又は補体活性化に影響を及ぼし、特に代替補体経路及び/又はC3転換酵素を阻害する能力である。アゴニストには、例えばイムノアドヘシン、ペプチド模倣薬、天然CRIgの定性的生物学的活性を模倣する非ペプチド小有機分子並びに抗体断片を含むアゴニスト抗体が含まれる。
【0124】
イムノアドヘシンは、異種タンパク質(「アドヘシン」)の結合特異性と免疫グロブリン定常ドメインのエフェクター機能とを兼ね備えた抗体様分子である。構造的には、イムノアドヘシンは、抗体の抗原認識及び結合部位以外(すなわち「異種性の」もの)である所望の結合特異性を有するアミノ酸配列と、免疫グロブリン定常ドメイン配列の融合体を含んでなる。イムノアドヘシン分子のアドヘシン部分は典型的にはレセプター又はリガンドの結合部位を少なくとも含んでなる連続するアミノ酸配列である。イムノアドヘシンの免疫グロブリン定常ドメイン配列は、任意の免疫グロブリン、例えばIgG-1、IgG-2、IgG-3、又はIgG-4サブタイプ、IgA(IgA-1及びIgA-2を含む)、IgE、IgD又はIgMから得られうる。
【0125】
ペプチド模倣薬は、例えば、該化合物がここに記載のCRIg生物学的活性を保持しているならば非天然発生アミノ酸を含むペプチドを含む。同様に、ペプチド模倣薬及びアナログは、本発明のCRIgポリペプチドの重要な構造的エレメントの構造を模倣しCRIg生物学的活性を保持している非アミノ酸化学構造を含みうる。「ペプチド」なる用語は、約50アミノ酸残基未満、及び好ましくは40アミノ酸残基未満の制約された(すなわち、例えば、βターン又はβプリーツシートを先導するアミノ酸の存在、又は例えば、ジスルフィド結合Cys残基の存在によって環状化する構造のある要素を有する)あるいは非制約(例えば直鎖状)アミノ酸配列で、その二量体又はその中間のような多量体を包含するものを指すようにここでは使用される。約40アミノ酸残基未満のペプチドでは、約10及び約30アミノ酸残基の間のペプチド、特に約20アミノ酸残基のペプチドが好ましい。しかしながら、本開示を読めば、当業者ならば、ペプチドを識別するのは、特定のペプチドの長さではなく、そのC3bへ結合しC3転換酵素、特に代替補体経路のC3転換酵素を阻害する能力であることを認めるであろう。
【0126】
CRIgアゴニストのスクリーニングと同定は、CRIg及びC3b:CRIg複合体の結晶構造の開示と、CRIgとC3b間の結合界面の同定によって非常に容易になる。この情報によりCRIgのC3b結合部位を模倣する化合物の設計が可能になる。
【0127】
また、CRIgアゴニストは、(a)計算又は実験手段を用いて化学物質とCRIgポリペプチド又はC3b:CRIg複合体の三次元構造の間のフィッティング操作を実施し;(b)工程(a)で得られたデータを解析して化学物質と天然CRIg又はC3b:CRIg複合体の間の結合の特徴を決定することにより同定することができる。この情報に基づいて、アゴニスト候補を合成することができ、そのアゴニスト特性をCRIg活性の生物学的アッセイにおいて実証することができる。
【0128】
特定の実施態様では、アゴニストは、天然配列CRIg分子又はその保存的アミノ酸置換変異体のC3b結合領域の少なくとも一部を含む化学物質であろう。
【0129】
ペプチド模倣薬は固相ペプチド合成法を用いて簡便に調製できる(Merrifield, J. Am. Chem. Soc., 85: 2149(1964);Houghten, Proc. Natl. Acad. Sci. USA 82: 5132 (1985))。固相合成法は、保護されたアミノ酸を不活性固形担体に結合させることにより推定ペプチドのカルボキシ末端において開始される。不活性な固形担体は、開始アミノ酸のC末端のアンカーとして働くことのできる任意の高分子であり得る。典型的には、高分子担体は、架橋高分子樹脂(例えば、ポリアミド又はポリスチレン樹脂)である。一実施態様では、C末端アミノ酸はポリスチレン樹脂に連結されてベンジルエステルを形成する。高分子支持体は、ペプチド合成の阻害アミノ酸のα-アミノ基を脱保護するのに使用される条件下でペプチドアンカー結合が安定であるように選択される。塩基不安定α保護基が使用されるならば、ペプチドと固形担体の間に酸不安定結合を使用するのが望ましい。例えば、酸不安定エーテル樹脂は、塩基不安定Fmocアミノ酸ペプチド合成に効果的である。あるいは、ペプチドアンカー結合及び差次的に酸分解に移行するα-保護基が使用できる。例えば、フェニルアセトアミドメチル(Pam)樹脂のようなアミノメチル樹脂は、Boc-アミノ酸ペプチド合成に関連して良好に働く。開始アミノ酸が不活性固形担体に連結された後、開始アミノ酸のαアミノ酸保護基を例えば塩化メチレン中のトリフルオロ酢酸(TFA)で除去し、例えばトリエチルアミン(TEA)で中和する。開始アミノ酸のαアミノ基の脱保護に続いて、合成における次のα-アミノ及び側鎖保護したアミノ酸を添加する。次に残りのα-アミノ酸と、必要ならば側鎖保護されたアミノ酸を縮合により所定の順序で続けて結合させ、固形担体に結合した中間体化合物を得る。あるいは、ペプチド断片を伸長している固相ペプチド鎖に付加する前に、幾つかのアミ酸を互いに結合させて所望のペプチド断片を形成してもよい。
【0130】
2つのアミノ酸、又はアミノ酸とペプチド、又はペプチドとペプチドの間の縮合反応は、通常の縮合方法、例えばアジド法、混合酸無水物法、DCC(N,N'-ジシクロヘキシルカルボジイミド)又はDIC(N,N'-ジイソプロピルカルボジイミド)法、活性エステル法、p-ニトロフェニルエステル法、BOP(ベンゾトリアゾール-1-イル-オキシ-トリス[ジメチルアミノ]ホスホニウムヘキサフルオロホスフェート)法、N-ヒドロキシコハク酸イミドエステル法など、及びウッドワード試薬K法等に従って実施される。
【0131】
ペプチドの化学合成に共通なのは、アミノ酸の任意の反応性側鎖基を適切な保護基で保護することである。最終的には、これらの保護基は、所望のポリペプチド鎖が連続的に組み立てられた後に除去される。更に共通なのは、アミノ酸又はペプチド断片上のαアミノ基を保護するが、アミノ酸又はペプチド断片のC末端カルボキシル基は成長固形相ポリペプチド鎖の遊離N末端アミノ基と反応し、続いてαアミノ酸基を選択的に除去し、次のアミノ酸又はペプチド断片を固相ペプチド鎖に付加させる。従って、ペプチド合成において、ペプチド鎖の所望の配列に位置するアミノ酸残基の各々を含み、その個々の残基が側鎖保護基を担持する中間体化合物が生成されるのが一般的である。これらの保護基は、実質的に同時に除去でき、固相から取り外した後に所望のポリペプチド生成物が生成される。
【0132】
α-及びε-アミノ酸側鎖を、ベンジルオキシカルボニル(Zと略)、イソニコチニルオキシカルボニル(iNOC)、o-クロロベンジルオキシカルボニル[Z(2Cl)]、p-ニトロベンジルオキシカルボニル[Z(NO2)]、p-メトキシベンジルオキシカルボニル[Z(OMe)]、t-ブトキシカルボニル(Boc)、t-アミルオキシカルボニル(Aoc)、イソボロニルオキシカルボニル、アダマンチルオキシカルボニル、2-(4-ビフェニル)-2-プロピル-オキシカルボニル(Bpoc)、9-フルオレニルメトキシカルボニル(Fmoc)、メチルスルホンエトキシカルボニル(Msc)、トリフルオロアセチル、フタリル、ホルミル、2-ニトロフェニルスルフェニル(NPS)、ジフェニルホスフィノチオイル(Ppt)、及びジメチロホスフィノチオイル(Mpt)基などで保護できる。
【0133】
カルボキシ官能基の保護基としては、ベンジルエステル(OBzl)、シクロヘキシルエステル(Chx)、4-ニトロベンジルエステル(ONb)、t-ブチルエステル(Obut)、4-ピリジルメチルエステル(OPic)等が例示される。アミノ及びカルボキシル基以外の官能基を有するアルギニン、システイン、及びセリンなどの特定のアミノ酸は、適当な保護基で保護するのが望ましいことが多い。例えば、アルギニンのグアニジノ基は、ニトロ、p-トルエンスルホニル、ベンジルオキシカルボニル、アダマンチルオキシカルボニル、p-メトキシベンゼンスルホニル、4-メトキシ-2,6-ジメチルベンゼンスルホニル(Nds)、1,3,5-トリメチルフェニルスルホニル(Mts)等で保護される。システインのチオール基は、p-メトキシベンジル、トリチルなどで保護されうる。
【0134】
上述の多くの阻害アミノ酸は、Novabiochem(San Diego, Calif.)、Bachem CA(Terrence, Calif.)又はPeninsula Labs(Belmont, Calif.)のような市販元から得ることができる。
【0135】
所望のアミノ酸配列が完成した後、ペプチドは固形担体から切断され、回収され、精製されうる。ペプチドは、ペプチド固相結合を崩壊することのできる試薬により固形担体から取り除かれ、場合によっては、ペプチドの阻害側鎖官能基を脱保護する。一実施態様では、ペプチドは、残っている側鎖保護基も取り除く液体フッ化水素酸(HF)での酸分解により固相から取り外す。好ましくは、ポリペプチド中の残基のアルキル化(例えば、メチオニン、システイン、及びチロシン残基のアルキル化)を回避するために、酸分解反応混合物はチオ-クレゾール及びクレゾール捕捉剤を含む。HF切断後、樹脂はエーテルで洗浄され、遊離ペプチドが酢酸溶液の連続洗浄で固相から抽出される。組み合わされた洗浄液は凍結乾燥され、ペプチドが精製される。
【0136】
天然配列CRIgポリペプチドのアンタゴニストは過剰な補体活性化及び亢進された補体介在宿主防御メカニズムから恩恵を受ける症状の治療に有用性が見出される。CRIgに対する抗体のようなCRIgアンタゴニストは、腫瘍(癌)治療における免疫アジュバント治療に使用することができる。T細胞がヒト腫瘍特異的抗原を認識することは現在は十分に確立されている。遺伝子のMAGE、BAGE及びGAGEファミリーによりコードされる腫瘍抗原の一グループは全ての成人の正常組織においてサイレントであるが、腫瘍、例えばメラノーマ、肺腫瘍、頭頸部腫瘍、膀胱癌においては有意な量で発現される。DeSmet, C.等(1996) Proc. Natl. Acad. Sci.93:7149。T細胞の同時刺激により、インビトロ及びインビボの双方において、腫瘍の退行及び抗腫瘍反応が誘導されることが示されている。Melero. I.等, Nature Medicine (1997) 3:682;Kwon. E.D.等, Proc. Natl. Acad. Sci.USA (1997) 94:8099;Lynch, D.H.等, Nature Medicine (1997) 3:625;Finn, O.J.及びLotze. M.T., J. Immunol. (1998) 21:114。本発明によって設計され同定されたCRIgアンタゴニストはアジュバントとして単独で又は増殖調節剤、細胞毒性薬又は化学療法剤と共に投与することができ、T細胞増殖/活性化及び腫瘍抗原に対する抗腫瘍反応を刺激する。増殖調節剤、細胞毒性薬又は化学療法剤は既知の投与方法を使用して一般的な量で投与することができる。本発明のCRIgアンタゴニストによる免疫刺激活性により、増殖調節剤、細胞毒性薬又は化学療法剤の量を減らすことができ、よって、潜在的に患者に対する毒性を低下させることができる。
【0137】
ある種のマクロファージは腫瘍根絶に関わっているけれども、多くの固形腫瘍は腫瘍増殖を支援するマクロファージを含んでいることが知られている(Bingle等, J Pathol 196:254-265 (2002);Mantovani等, Trends Immunol 23:549-555 (2002))。これらのマクロファージはその表面にCRIgを含みうる。CRIgの補体活性化を阻害する能力をブロックする抗体を用いて腫瘍細胞上の補体を活性化させ、補体介在溶解により腫瘍を根絶するのを補助することができよう。このアプローチ法はCRIg陽性マクロファージを含む腫瘍に特に有用であることが予想される。
【0138】
アンタゴニストの好ましい群には天然CRIgに特異的に結合しその生物学的活性を阻害する抗体が含まれる。
【0139】
抗体(アゴニストとアンタゴニストの双方)の例には、ポリクローナル、モノクローナル、ヒト化、二重特異的及びヘテロコンジュゲート抗体、及び抗体断片が含まれる。
【0140】
本発明のポリペプチドを認識してそれに結合し、又はそれに対するアンタゴニストとして作用する抗体は、あるいはモノクローナル抗体であってもよい。モノクローナル抗体は、Kohler及びMilstein, Nature, 256:495 (1975)に記載されているようなハイブリドーマ法を使用することで調製することができる。ハイブリドーマ法では、マウス、ハムスター又は他の適切な宿主動物を典型的には免疫剤により免疫化することで、免疫剤に特異的に結合する抗体を生成するかあるいは生成可能なリンパ球を誘発する。あるいは、リンパ球をインビトロで免疫化することもできる。
【0141】
免疫剤は、典型的には本発明のCRIgポリペプチド、その抗原性断片又は融合タンパク質を含む。一般に、ヒト由来の細胞が望まれる場合には末梢血リンパ球(「PBL」)が使用され、あるいは非ヒト哺乳動物源が望まれている場合は、脾臓細胞又はリンパ節細胞が使用される。ついで、ポリエチレングリコール等の適当な融合剤を用いてリンパ球を不死化株化細胞と融合させ、ハイブリドーマ細胞を形成する[Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103]。不死化株化細胞は、通常は、形質転換した哺乳動物細胞、特に齧歯動物、ウシ、及びヒト由来の骨髄腫細胞である。通常、ラット又はマウスの骨髄腫株化細胞が使用される。ハイブリドーマ細胞は、好ましくは、未融合の不死化細胞の生存又は増殖を阻害する一又は複数の物質を含有する適切な培地で培養される。例えば、親細胞が、酵素のヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRT又はHPRT)を欠いていると、ハイブリドーマの培地は、典型的には、ヒポキサチン、アミノプテリン及びチミジンを含み(「HAT培地」)、この物質がHGPRT欠乏性細胞の増殖を阻止する。
【0142】
好ましい不死化株化細胞は、効率的に融合し、選択された抗体生成細胞による安定した高レベルの抗体発現を支援し、HAT培地のような培地に対して感受性であるものである。より好ましい不死化株化細胞はマウス骨髄腫株であり、これは例えばカリフォルニア州サンディエゴのSalk Institute Cell Distribution Centerやバージニア州マナッサスのアメリカン・タイプ・カルチャー・コレクションより入手可能である。ヒトモノクローナル抗体を生成するためのヒト骨髄腫及びマウス-ヒト異種骨髄腫株化細胞も開示されている[Kozbor, J. Immunol., 133:3001 (1984);Brodeur等, Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp.51-63]。
【0143】
ついでハイブリドーマ細胞が培養される培養培地を、本発明のポリペプチドに対する又は本発明のポリペプチドと同様の活性を有するモノクローナル抗体の存在について検定することができる。好ましくは、ハイブリドーマ細胞によって生成されたモノクローナル抗体の結合特異性は免疫沈降又はラジオイムノアッセイ(RIA)や酵素結合免疫測定法(ELISA)等のインビトロ結合検定法によって測定する。このような技術及びアッセイは当該分野において知られている。モノクローナル抗体の結合親和性は、例えば、Munson及びPollard, Anal. Biochem., 107:220 (1980)によるスキャッチャード解析法によって測定することができる。
【0144】
所望のハイブリドーマ細胞が同定された後、クローンを限界希釈法によりサブクローニングし、標準的な方法で増殖させることができる[上掲のGoding]。この目的のための適当な培地には、例えば、ダルベッコの改変イーグル培地及びRPMI-1640倍地が含まれる。あるいは、ハイブリドーマ細胞は哺乳動物においてインビボで腹水として増殖させることもできる。
【0145】
サブクローンによって分泌されたモノクローナル抗体は、例えばプロテインA−セファロース法、ヒドロキシルアパタイトクロマトグラフィー法、ゲル電気泳動法、透析法又はアフィニティークロマトグラフィー等の一般的な免疫グロブリン精製法によって培養培地又は腹水液から単離又は精製されうる。
【0146】
また、モノクローナル抗体は、組換えDNA法、例えば米国特許第4816567号に記載された方法により作成することができる。本発明のモノクローナル抗体をコードするDNAは、常套的な方法を用いて(例えば、マウス抗体の重鎖及び軽鎖をコードする遺伝子に特異的に結合可能なオリゴヌクレオチドプローブを使用して)、容易に単離し配列決定することができる。本発明のハイブリドーマ細胞はそのようなDNAの好ましい供給源となる。ひとたび単離されたら、DNAは発現ベクター内に配することができ、これが宿主細胞、例えばサルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、あるいは免疫グロブリンタンパク質をさもなければ生成しない骨髄腫細胞内に形質移入され、組換え宿主細胞内でモノクローナル抗体の合成をすることができる。また、DNAは、例えば相同マウス配列に換えてヒト重鎖及び軽鎖定常ドメインのコード配列を置換することにより[上掲の米国特許第4816567号;Morrison等]、又は免疫グロブリンコード配列に非免疫グロブリンポリペプチドのコード配列の一部又は全部を共有結合することにより修飾することができる。このような非免疫グロブリンポリペプチドは、本発明の抗体の定常ドメインに置換でき、あるいは本発明の抗体の一つの抗原結合部位の可変ドメインに置換でき、キメラ性二価抗体を生成する。
【0147】
抗体は好ましくは一価抗体である。一価抗体の調製方法は当該分野においてよく知られている。例えば、一つの方法は免疫グロブリン軽鎖と修飾重鎖の組換え発現を含む。重鎖は一般的に、重鎖の架橋を防止するようにFc領域の任意の点で切断される。あるいは、関連するシステイン残基を他のアミノ酸残基で置換するか欠失させて架橋を防止する。
【0148】
一価抗体の調製には、またインビトロ法が適している。抗体の消化による、その断片、特にFab断片の生成は、当該分野において知られている慣用的技術を使用して達成することができる。
【0149】
本発明の抗体は、更にヒト化抗体又はヒト抗体を含みうる。非ヒト(例えばマウス)抗体のヒト化型とは、キメラ免疫グロブリン、免疫グロブリン鎖又はその断片(例えばFv、Fab、Fab’、F(ab’)2あるいは抗体の他の抗原結合サブ配列)であって、非ヒト免疫グロブリンに由来する最小配列を含むものである。ヒト化抗体はレシピエントの相補性決定領域(CDR)の残基が、マウス、ラット又はウサギのような所望の特異性、親和性及び能力を有する非ヒト種(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(レシピエント抗体)を含む。幾つかの例では、ヒト免疫グロブリンのFvフレームワーク残基は、対応する非ヒト残基によって置換されている。また、ヒト化抗体は、レシピエント抗体にも、移入されたCDRもしくはフレームワーク配列にも見出されない残基を含んでいてもよい。一般に、ヒト化抗体は、全てあるいは殆ど全てのCDR領域が非ヒト免疫グロブリンのものに対応し、全てあるいは殆ど全てのFR領域がヒト免疫グロブリンコンセンサス配列のものである、少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含む。ヒト化抗体は、最適には免疫グロブリン定常領域(Fc)、典型的にはヒトの免疫グロブリンの定常領域の少なくとも一部を含んでなる[Jones等, Nature, 321:522-525 (1986);Riechmann等, Nature, 332:323-329 (1988);及びPresta, Curr. Op Struct. Biol., 2:593-596 (1992)]。
【0150】
非ヒト抗体をヒト化する方法はこの分野でよく知られている。一般的に、ヒト化抗体には非ヒト由来の一つ又は複数のアミノ酸残基が導入される。これら非ヒトアミノ酸残基は、しばしば、典型的には「移入」可変ドメインから得られる「移入」残基と称される。ヒト化は基本的にウィンター(Winter)及び共同研究者[Jones等, Nature, 321:522-525 (1986);Riechmann等, Nature, 332:323-327 (1988);Verhoeyen等, Science, 239:1534-1536 (1988)]の方法に従って、齧歯類CDR又はCDR配列をヒト抗体の対応する配列の代わりに用いることにより実施される。よって、このような「ヒト化」抗体は、無傷のヒト可変ドメインより実質的に少ない分が非ヒト種由来の対応する配列で置換されたキメラ抗体(米国特許第4816567号)である。実際には、ヒト化抗体は典型的には幾つかのCDR残基及び場合によっては幾つかのFR残基が齧歯類抗体の類似する部位からの残基によって置換されたヒト抗体である。
【0151】
また、ヒト抗体は、ファージ表示ライブラリー[Hoogenboom及びWinter, J. Mol. Biol., 227:381(1991);Marks等, J. Mol. Biol., 222:581 (1991)]を含むこの分野で知られた種々の方法を用いて作成することもできる。また、Cole等及びBoerner等の方法も、ヒトモノクローナル抗体の調製に利用することができる[Cole等, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985)及びBoerner等, J. Immunol., 147(1):86-95 (1991)]。同様に、ヒト抗体はヒト免疫グロブリン座位をトランスジェニック動物、例えば内在性免疫グロブリン遺伝子は部分的又は完全に不活性化されたマウスに導入することにより産生することができる。投与の際に、遺伝子再配列、組立、及び抗体レパートリーを含むあらゆる観点においてヒトに見られるものに非常に類似しているヒト抗体の生産が観察される。このアプローチは、例えば米国特許第5545807号;同第5545806号;同第5569825号;同第5625126号;同第5633425号;同第5661016号、及び次の科学文献:Marks等, Bio/Technology 10, 779-783 (1992);Lonberg等, Nature 368 856-859 (1994);Morrison, Nature 368, 812-13 (1994);Fishwild等, Nature Biotechnology 14, 845-51 (1996);Neuberger, Nature Biotechnology 14, 826 (1996);Lonberg及びHuszar, Intern. Rev. Immunol. 13 65-93 (1995)に記載されている。
【0152】
二重特異性抗体は、少なくとも2つの異なる抗原に対して結合特異性を有するモノクローナル抗体、好ましくはヒトもしくはヒト化抗体である。本発明の場合、結合特異性の一方は本発明のポリペプチドに対するものであり、他方は任意の他の抗原、好ましくは細胞表面タンパク質又はレセプター又はレセプターサブユニットに対してであり得る。
【0153】
二重特異性抗体を作製する方法は当該分野において知られている。伝統的には、二重特異性抗体の組換え生産は、二つの重鎖が異なる特異性を持つ二つの免疫グロブリン重鎖/軽鎖対の同時発現に基づく(Milstein及びCuello, Nature, 305:537-539 [1983])。免疫グロブリンの重鎖と軽鎖を無作為に取り揃えるため、これらハイブリドーマ(クアドローマ)は10種の異なる抗体分子の潜在的混合物を生成し、その内の一種のみが正しい二重特異性構造を有する。正しい分子の精製は、アフィニティークロマトグラフィー工程によって通常達成される。同様の手順が1993年5月13日公開の国際公開第93/08829号、及びTraunecker等, EMBO J.,10:3655-3656 (1991)に開示されている。
【0154】
所望の結合特異性(抗体-抗原結合部位)を有する抗体可変ドメインを免疫グロブリン定常ドメイン配列に融合できる。融合は、好ましくは少なくともヒンジ部、CH2及びCH3領域の一部を含む免疫グロブリン重鎖定常ドメインとのものである。少なくとも一つの融合には軽鎖結合に必要な部位を含む第一の重鎖定常領域(CH1)が存在することが望ましい。免疫グロブリン重鎖融合をコードするDNA、及び望むのであれば免疫グロブリン軽鎖を、別々の発現ベクターに挿入し、適当な宿主生物に同時形質移入する。二重特異性抗体を作成するための更なる詳細については、例えば、Suresh等, Methods in Enzymology, 121:210 (1986)を参照されたい。
【0155】
ヘテロコンジュゲート抗体は、2つの共有結合した抗体からなる。このような抗体は、例えば、免疫系細胞を不要な細胞に対してターゲティングさせるため(米国特許第4676980号)及びHIV感染の治療のために(国際公開第91/00360;国際公開第92/200373;欧州特許第03089号)提案されている。この抗体は、架橋剤に関連したものを含む合成タンパク化学における既知の方法を使用して、インビトロで調製することができると考えられる。例えば、ジスルフィド交換反応を使用するか又はチオエーテル結合を形成することによって、免疫毒素を作成することができる。この目的に対して好適な試薬の例には、イミノチオレート及びメチル-4-メルカプトブチルイミダート、及び例えば米国特許第4676980号に開示されたものが含まれる。
【0156】
本発明の抗体をエフェクター機能について改変し、例えば免疫関連疾患の治療における抗体の効力を向上させることは望ましい。例えば、システイン残基をFc領域に導入し、それにより、この領域に鎖間ジスルフィド結合を形成するようにしてもよい。そのようにして生成された同種二量体抗体は、向上したインターナリゼーション能力及び/又は増加した補体媒介細胞殺傷及び抗体依存細胞性細胞障害性(ADCC)を有する可能性がある。Caron等, J. Exp. Med. 176: 1191-1195 (1992)及びShopes, B. J. Immunol. 148: 2918-2922 (1992)参照。また、向上した抗腫瘍活性を持つ同種二量体抗体は、Wolff等, Cancer Research 53: 2560-2565 (1993)に記載されている異種二官能性架橋を用いて調製することができる。あるいは、抗体は、2つのFc領域を有するように加工して、それにより補体溶解及びADCC能力を向上させることもできる。Stevenson等, Anti-Cancer Drug Design 3: 219-230 (1989)参照。
【0157】
また、本発明は、化学療法剤、毒素(例えば、細菌、真菌、植物又は動物由来の酵素活性毒素、又はその断片)などの細胞傷害性剤、あるいは放射性同位体(つまり、放射性コンジュゲート)と抱合している抗体を含む免疫複合体にも関する。
【0158】
このような免疫複合体の生成に有用な化学療法剤は上述の通りである。用いることのできる酵素活性毒素及びその断片には、ジフテリアA鎖、ジフテリア毒素の非結合活性断片、(緑膿菌からの)外毒素A鎖、リシンA鎖、アブリンA鎖、モデクシン(modeccin)A鎖、アルファ-サルシン、アレウリテス・フォーディ(Aleurites fordii)タンパク質、ジアンチン(dianthin)タンパク質、フィトラカ・アメリカーナ(Phytolaca americana)タンパク質(PAPI、PAPII、及びPAP-S)、モモルディカ・チャランチア(momordica charantia)インヒビター、クルシン(curcin)、クロチン(crotin)、サパオナリア・オフィシナリス(sapaonaria officinalis)インヒビター、ゲロニン(gelonin)、ミトゲリン(mitogellin)、レストリクトシン(restrictocin)、フェノマイシン(phenomycin)、エノマイシン(enomycin)及びトリコテセン(tricothecene)が含まれる。放射性コンジュゲート抗体の生成には、様々な放射性ヌクレオチドが利用可能である。例としては、212Bi、131I、131In、90Y及び186Reが含まれる。
【0159】
抗体及び細胞傷害性薬の複合体は、種々の二官能性タンパク質カップリング剤、例えば、N-スクシンイミジル-3-(2-ピリジルジチオール)プロピオナート(SPDP)、イミノチオラン(IT)、イミドエステルの二官能性誘導体(ジメチルアジピミデートHCL等)、活性エステル(ジスクシンイミジルスベレート等)、アルデヒド(グルタルアルデヒド等)、ビス-アジド化合物(ビス(p-アジドベンゾイル)ヘキサンジアミン等)、ビス-ジアゾニウム誘導体(ビス-(p-ジアゾニウムベンゾイル)-エチレンジアミン等)、ジイソシアネート(トリエン2,6-ジイソシアネート等)、及びビス-活性フッ素化合物(1,5-ジフルオロ-2,4-ジニトロベンゼン等)を用いて作製できる。例えば、リシン免疫毒素は、Vitetta等, Science 238: 1098 (1987)に記載されているように調製することができる。カーボン-14標識1-イソチオシアナトベンジル-3-メチルジエチレントリアミン五酢酸(MX-DTPA)は、放射性ヌクレオチドの抗体への抱合のためのキレート剤の例である。国際公開94/11026参照。
【0160】
他の実施態様では、組織の予備標的化で使用するために、抗体は「レセプター」(ストレプトアビジン等)に抱合されてもよく、抗体-レセプター複合体は患者に投与され、ついで清澄化剤を用いて未結合複合体を循環から除去し、次に細胞傷害性薬(例えば、放射性ヌクレオチド等)に抱合された「リガンド」(アビジン等)を投与する。
【0161】
CRIg及びそのアゴニスト及びアンタゴニストは、天然CRIg分子の生物学的活性を模倣し又は阻害するかどうかを決定するために様々なインビトロ及びインビボアッセイにおいて試験することができる。
【0162】
補体活性化、特に代替補体経路を阻害するその能力の結果として、CRIgポリペプチドは、補体関連疾患及び病理状態の予防及び/又は治療において有用性が見出される。このような疾患及び症状には、限定するものではないが、補体関連の炎症性及び自己免疫疾患が含まれる。
【0163】
補体関連疾患の特定の例には、限定するものではないが、リウマチ様関節炎(RA)、急性呼吸促迫症候群(ARDS)、虚血及び再灌流後の遠隔組織傷害、心肺バイパス手術中の補体活性化、皮膚筋炎、天疱瘡、ループス腎炎及びその結果の糸球体腎炎及び脈管炎、心肺バイパス、心臓麻痺誘発性冠血管内皮機能不全、II型膜性増殖性糸球体腎炎、IgA腎症、急性腎不全、クリオグロブリン血症、抗リン脂質症候群、黄斑変性疾患及び他の補体関連の眼の症状、非滲出性及び滲出性AMDを含む加齢性黄斑変性症(AMD)、糖尿病性網膜症(DR)、眼内炎、ブドウ膜炎、同種移植、異物移植、超急性拒絶反応、血液透析、慢性閉塞肺促迫症候群(COPD)、喘息、遺伝性血管浮腫、発作性夜間血色素尿症、アルツハイマー病、アテローム性動脈硬化症、誤嚥性肺炎、蕁麻疹、例えば慢性特発性蕁麻疹、溶血性尿毒症症候群、子宮内膜症、心原性ショック、虚血再かん流傷害、多発性硬化症(MS)が含まれる。
【0164】
AMDは、60歳以上の個人における不可逆的な視力障害の主たる原因である黄斑の加齢性変性症である。非滲出性(ドライ)及び滲出性(ウェット)AMDの2つのタイプのAMDが存在している。ドライ又は非滲出型は中心網膜(黄斑)並びにRPE上の沈積物(ドルーゼ)の根底にある網膜色素上皮(RPE)内の萎縮及び異常肥大変化を含む。非滲出型ARMDの患者は、脈絡膜新生血管膜(CNVM)と呼ばれる異常な血管が網膜、漏出液及び血液の下で発達し、最終的には網膜内及び網膜下で失明に至る円板状瘢痕を生じるARMDのウェット又は滲出型まで進行しうる。通常は滲出型ARMDの前躯体である非滲出型ARMDはより一般的である。非滲出型ARMDの提示は様々である;ハードドルーゼ、ソフトドルーゼ、RPE地図状萎縮、及び色素凝集が存在しうる。補体成分はAMDの早期にRPE上に付着し、ドルーゼの主要構成成分である。補体H因子(CFH)多形性がAMDの起因リスクの50%を占めることが最近報告されている(Klein等, Science 308:385-9 (2005))。
【0165】
CRIgは、カテゴリー3及びカテゴリー4AMDを含む高リスクのAMDの治療において特に有用である。カテゴリー3AMDは、双方の眼に進行AMDがなく、少なくとも一つの眼が20/32又はそれ以上の視力、少なくとも一つの大きなドルーゼ(例えば125μm)、広範な(ドルーゼ面積で測定)中間ドルーゼ、又は黄斑の中心を含まない地図状萎縮(GA)又はこれらの任意の組合せを有することによって特徴付けられる。カテゴリー4高リスクAMDは、20/32又はそれ以上の視力によって特徴付けられ、インデックス眼において進行AMD(黄斑の中心又は脈絡膜血管新生の特徴を含むGA)はない。他眼は進行AMD、又はAMD黄斑症に帰す20/32未満の視力によって特徴付けられている。典型的には、未治療ならば、高リスクAMDは、カテゴリー1又は2(高リスクではない)AMDの進行速度よりも約10−30倍高い速度で脈絡膜血管新生(CNV)に速やかに進行する。従って、CRIgはCNVとAMDの予防と治療に有用性が見出される。
【0166】
補体関連疾患の例としての炎症性症状のより広範なリストには、例えば炎症性腸疾患(IBD)、全身性エリテマトーデス、リウマチ様関節炎、若年性慢性関節炎、脊椎関節症、全身性硬化症(強皮症)、特発性炎症ミオパシー(皮膚筋炎、多発性筋炎)、シェーグレン症候群、全身性血管炎、サルコイドーシス、自己免疫性溶血性貧血(免疫再生不良性貧血、発作性夜間血色素尿)、自己免疫性血小板減少(特発性血小板減少性紫斑病、免疫仲介血小板減少)、甲状腺炎(グレーブス疾患、ハシモト甲状腺炎、若年性リンパ球性甲状腺炎、萎縮性甲状腺炎)、真性糖尿病、免疫仲介腎疾患(糸球体腎炎、尿細管間質性腎炎)、中枢及び末梢神経系の脱髄疾患、例えば多発性硬化症、特発性脱随性多発神経障害、又はギラン-バレー症候群、及び慢性炎症脱随性多発神経障害、肝胆道疾患、例えば感染性肝炎(A型、B型、C型、D型、E型肝炎及び他の非肝親和性(nonhepatotropic)ウイルス)、自己免疫慢性活動性肝炎、原発性胆汁性肝硬変症、肉芽腫性肝炎、及び硬化性胆管炎、炎症性腸疾患(潰瘍性大腸炎;クローン病)、グルテン過敏性腸疾患、及びウィップル病、水疱性皮膚疾患、多形性紅斑及び接触性皮膚炎を含む自己免疫又は免疫媒介皮膚疾患、乾癬、アレルギー性疾患、例えば喘息、アレルギー性鼻炎、アトピー性皮膚炎、食物過敏症及び蕁麻疹、肺の免疫疾患、例えば好球性肺炎、特発性肺線維症及び過敏性肺炎、拒絶反応及び移植片対宿主疾患を含む移植関連疾患が含まれる。
【0167】
全身性エリテマトーデスでは、疾患の中心的な媒介物は自己タンパク質/組織に対する自己反応性抗体の産生と、続いての免疫媒介炎症の発生である。抗体は直接的又は間接的に組織傷害を媒介する。Tリンパ球は組織損傷に直接的には関与していることは示されていないが、Tリンパ球は自己反応性抗体の発生に必要である。よって、疾患の発生はTリンパ球に依存している。腎臓、肺、筋骨格、皮膚粘膜、眼、中枢神経系、心臓血管系、胃腸管、骨髄及び血液を含む複数の器官及び系が臨床的に冒される。
【0168】
リウマチ様関節炎(RA)は、主に複数の関節の滑膜に関連する慢性全身性自己免疫炎症疾患であり、結果として関節軟骨に傷害が生じる。病原はTリンパ球依存性であり、リウマチ因子、自己IgGに対する自己抗体の生成に付随し、結果として滑液及び血液において高レベルに達する免疫複合体を生成する。関節中のこれらの複合体は、滑膜中へのリンパ球及び単球の顕著な浸潤と、続いての顕著な滑膜変化を誘発し;多数の好中球の添加により同様の細胞で浸潤されるならば、関節空間/液でもしかりである。冒されている組織は、多くの場合対称的なパターンで、主に関節である。しかしながら、二つの主な形態の関節外疾患もまた生じる。一形態は進行中の進行性関節疾患及び肺線維症の典型的病巣、血管炎、及び皮膚潰瘍を伴う関節外障害の発生である。関節外疾患の第二の形態はいわゆるフェルティー症候群であり、これは、RA疾患過程の末期、時には関節疾患が鎮静した後に生じ、好中球減少、血小板減少及び脾肥大の存在に関与する。これには、梗塞、皮膚潰瘍及び壊疽の形成を伴う複数の器官において血管炎が付随する。多くの場合、患者には、発病している関節上にある皮下組織にリウマチ様小結節が発達し;その小結節は、末期には混合炎症細胞浸潤に包囲された壊死性中心を有する。RAにおいて生じる可能性のある他の徴候には:心外膜炎、胸膜炎、冠動脈炎、肺線維症を伴う間質性肺炎、乾性角結膜炎、及びリウマチ様小結節が含まれる。
【0169】
若年性慢性関節炎は、多くの場合16歳未満で発症する慢性特発性炎症疾患である。その表現型はRAと幾つかの類似点があり;リウマチ因子が陽性である患者の中には若年性リウマチ様関節炎に分類されるものもいる。この疾患は三つの主要なカテゴリー:小関節(pauarticular)、多関節及び全身性に細分類される。関節炎は重度で典型的には破壊的であり、関節強直症及び遅延成長に至る。他の徴候には慢性前部ブドウ膜炎及び全身性アミロイド症が含まれる。
【0170】
脊椎関節症は、幾つかの共通した臨床的特徴とHLA-B27遺伝子産物の発現との共通の関連性を持つ疾患のグループである。該疾患には:強直性脊椎炎、ライター症候群(反応性関節炎)、炎症性大腸疾患に関連した関節炎、乾癬に関連した脊椎炎、若年発生脊椎関節症及び未分化脊椎関節症が含まれる。顕著な特徴には、脊椎炎を伴うか伴わない仙腸関節炎;炎症非対称性関節炎;HLA-B27(クラスI MHCのHLA-B座位にある血清学的に定義された対立遺伝子)との関連;眼の炎症、及び他のリウマチ疾患に関連した自己抗体の不在が含まれる。疾患の誘導に対する鍵として最も関わっている細胞はCD8+Tリンパ球で、クラスI MHC分子により提示される抗原を標的としている細胞である。CD8+T細胞は、MHCクラスI分子により発現された外来ペプチドであるかのように、クラスI MHC対立遺伝子HLA-B27に対して反応する。HLA-B27のエピトープが細菌性又は他の微生物の抗原性エピトープを模倣し、よってCD8+細胞の反応を誘発すると仮定されている。
【0171】
全身性硬化症(強皮症)は病因がよく知られていない。疾患の顕著な特徴は皮膚の硬結であり;これは活性な炎症プロセスにより誘発されると思われる。強皮症は局部的又は全身性であり:血管病巣が共通しており、微小血管系における内皮細胞傷害が全身性硬化症の発達における初期の重要な事象であり;血管傷害は免疫媒介されうる。免疫学的基準は、皮膚病巣における単核細胞浸潤の存在と、多くの患者において抗細胞核抗体の存在によって導かれる。多くの場合、ICAM-1が皮膚病巣の線維芽細胞の細胞表面でアップレギュレーションされ、これらの細胞とのT細胞の相互作用が疾患の病因においてある役割を担っていることが示唆される。関連する他の器官には:胃腸管:結果的に異常なぜん動/運動性となる平滑筋萎縮症及び線維症:腎臓:小弓形及び小葉間動脈に影響を及ぼし、結果として腎皮質の血流が低下し、タンパク尿、高窒素血尿及び高血圧になる同心性内皮下内膜増殖:骨格筋:萎縮、間質性線維症:炎症:肺:間質性肺炎及び間質性線維症:及び心臓:収縮バンド壊死、瘢痕/線維症が含まれる。
【0172】
皮膚筋炎、多発性筋炎及び他のものを含む特発性炎症ミオパシーは病因がよく知られていない慢性筋肉炎症疾患であり、筋肉の弱化に至る。筋肉損傷/炎症は多くの場合対照的で進行性である。自己抗体が多くの形態と関連している。これらの筋炎特異的自己抗体は、タンパク質合成に関与する成分、タンパク質及びRNAに対して産生されその機能を阻害する。
【0173】
シェーグレン症候群は、免疫媒介炎症と、涙腺及び唾液腺の続く機能破壊によるものである。この疾患は炎症結合組織疾患に関連するか又はそれに伴う場合がある。この疾患は、双方とも小RNA-タンパク質複合体であるRo及びLa抗原に対する自己抗体産生に関連している。病巣は乾性角結膜炎、口内乾燥症で、胆汁性硬変、末梢又は感覚ニューロパシー、及び明白な紫斑病を含む他の徴候又は関連を伴うものに至る。
【0174】
全身性血管炎症は一次病巣が炎症で、続いて血管にダメージを受け、結果として冒された脈管により供給される組織に虚血/壊死/変性が生じ、幾つかのケースでは最終的な末端器官機能障害になるといった疾患である。また、血管炎(vasculitides)は他の免疫炎症媒介疾患、例えばリウマチ様関節炎、全身性硬化症等、特に免疫複合体の生成に関連した疾患等の続発症として又は二次病変として生じる場合がある。原発性全身性血管炎症グループの疾患には:全身壊死性血管炎:多動脈炎結節(polyarteritis nodosa)、アレルギー性脈管炎及び肉芽腫症、多脈管炎:ヴェゲナー肉芽腫症;リンパ腫様肉芽腫症:及び巨細胞動脈炎が含まれる。その他の血管炎には:粘膜皮膚リンパ節症候群(MLNS又は川崎病)、隔離されたCNS血管炎、ベーチェット(Behet's)病、閉塞性血栓性血管炎(バージャー病)及び皮膚壊死性細静脈炎(venulitis)が含まれる。列挙した血管炎のほとんどの種類の発病メカニズムは、主に脈管壁に免疫グロブリン複合体が付着し、続いてADCC、補体活性又は双方を介して炎症反応が誘発されることによると考えられている。
【0175】
サルコイドーシスは、体内のほとんど全ての組織中における類上皮細胞肉芽腫の存在により特徴づけられる病因がよく知られていない病状であり;肺の関与が最も一般的である。病因は疾患部位に活性マクロファージ及びリンパ球が残留していることに関連しており、続いてこれらの細胞型より放出される局部的又は全身的活性産物の放出の結果として慢性続発症が生じる。
【0176】
自己免疫性溶血性貧血、免疫再生不良性貧血、及び発作性夜間血色素尿を含む自己免疫溶性血性貧血は、赤血球(幾つかの場合においては血小板もまた含む他の血液細胞)表面で発現した抗原と反応する抗体が産出される結果によるものであり、補体媒介溶解及び/又はADCC/Fc-レセプター-媒介メカニズムを介して、その抗体被覆細胞の除去に反映される。
【0177】
他の臨床的環境における血小板減少性紫斑病及び免疫仲介血小板減少を含む自己免疫性血小板減少では、血小板破壊/除去が、抗体又は補体が血小板に接合し、続いて補体溶解、ADCC又はFc-レセプター-媒介メカニズムにより除去される結果として生じる。
【0178】
グレーブス疾患、ハシモト甲状腺炎、若年性リンパ球性甲状腺炎、及び萎縮性甲状腺炎を含む甲状腺炎は、甲状腺内に存在し多くの場合甲状腺に特異的なタンパク質と反応する抗体の産生を伴う、甲状腺抗原に対する自己免疫反応の結果によるものである。自然のモデル:ラット(BUF及びBBラット)及びチキン(肥満チキン種);誘導性モデル:サイログロブリン、甲状腺ミクロソーム抗原(甲状腺ペルオキシダーゼ)のいずれかでの動物の免疫化を含む実験用モデルが存在する。
【0179】
I型真性糖尿病又はインスリン依存性糖尿病は膵臓ランゲルハンス島β細胞の自己免疫破壊であり;この破壊は自己抗体及び自己反応性T細胞により媒介される。また、インスリン又はインスリンレセプターに対する抗体は、インスリン-非反応性の表現型をつくりだすことができる。
【0180】
糸球体腎炎及び尿細管間質性腎炎を含む免疫仲介腎疾患は、腎抗原に対する自己反応性抗体又はT細胞が産生される結果として直接的に、又は他の非腎抗原に対して反応性である、腎臓中の抗体及び/又は免疫複合体の沈着の結果として間接的に、腎組織に抗体又はT細胞媒介傷害が生じることによるものである。よって、免疫複合体の生成をもたらす他の免疫媒介疾患により、間接的続発症として免疫媒介腎疾患も誘発しうる。直接的及び間接的免疫メカニズムの双方により、結果として、器官機能が損なわれ、幾つかの場合では腎臓機能不全に進行する、病巣発達を腎組織に生じさせ/誘発する炎症反応が生じる。体液及び細胞免疫メカニズムの双方が障害の発病に関与しうる。
【0181】
多発性硬化症;特発性脱随性多発神経障害又はギラン-バレー症候群;及び慢性炎症脱随性多発神経障害を含む中枢及び末梢神経系の脱髄疾患は、自己免疫に原因を有し、オリゴデンドロサイト又はミエリンに直接的に引き起こされる損傷の結果として神経脱髄が生じると考えられている。MSにおいては、疾患の誘発及び進行がTリンパ球に依存することを示唆する証拠がある。多発性硬化症は、Tリンパ球依存性であり、再発性弛緩経路又は慢性進行経路のいずれかを有する脱髄疾患である。病因はよく知られていないが、ウイルス感染、遺伝的素因、環境及び自己免疫性の全てが寄与している。病巣は優勢なT細胞媒介小膠細胞の湿潤と、浸潤しているマイクロファージを含み;CD4+Tリンパ球は病巣における優勢な細胞型である。オリゴデンドロサイトの細胞死と続く脱髄のメカニズムはよく知られていないが、Tリンパ球により推進されていると思われる。
【0182】
好球性肺炎;特発性肺線維症及び過敏性肺炎を含む炎症及び線維症の肺疾患には、調節されない免疫炎症反応が関連している。その反応の阻害は治療的に有益であろう。
【0183】
水疱性皮膚疾患、多形性紅斑及び接触性皮膚炎を含む自己免疫又は免疫媒介皮膚疾患は自己抗体により媒介され、その発病はTリンパ球依存性である。
【0184】
乾癬はTリンパ球媒介炎症疾患である。病巣にはTリンパ球、マクロファージ及び抗原プロセシング細胞及びある種の好中球の浸潤が含まれる。喘息;アレルギー性鼻炎;アトピー性皮膚炎;食物過敏症及び蕁麻疹等を含むアレルギー性疾患はTリンパ球依存性である。これらの疾患はTリンパ球誘発性炎症、IgE媒介炎症、又は双方の組合せにより主に媒介される。
【0185】
拒絶反応及び移植片対宿主疾患(GVHD)を含む移植関連疾患はTリンパ球依存性であり;Tリンパ球の機能を阻害することで改善される。
【0186】
CRIgの上に列挙した治療的用途に鑑みて、CRIgの潜在的アゴニストは様々な細胞ベースアッセイ及び補体関連、炎症及び/又は自己免疫疾患又は障害の動物モデルで評価することができる。
【0187】
よって、例えば、関節炎の予防及び/又は治療における効力はコラーゲン誘発関節炎モデル(Terato等 Brit. J. Rheum. 35:828-838 (1966))において評価することができる。潜在的な関節炎予防/治療薬は、Terato等, J. Immunol. 148:2103-8 (1992);Terato等, Autoimmunity 22:137-47 (1995)に記載されたように、4種のモノクローナル抗体のカクテルの静脈内注射により誘発される抗体媒介関節炎モデルにおいてスクリーニングできる。またここの実施例4を参照のこと。関節炎の予防及び/又は治療のための候補は、例えばTNF-αトランスジェニックマウス(Taconic)のようなトランスジェニック動物モデルで研究することがまたできる。これらの動物は、ヒトリウマチ様関節炎の病因に結びつけられているサイトカインであるヒト腫瘍壊死因子(TNF-α)を発現する。これらのマウスにおけるTNF-αの発現は、前肢及び後肢の重症の慢性関節炎を生じ、炎症性関節炎の簡単なマウスモデルを提供する。
【0188】
最近、乾癬の動物モデルがまた開発された。よって、Asebia(ab)、鱗状皮膚(fsn)及び慢性増殖性皮膚炎(cpd)は乾癬様皮膚変化を持つ自然マウス突然変異である。例えばインターフェロン-γ、インターロイキン-1α、ケラチノサイト増殖因子、トランスフォーミング増殖因子-α、インターフェロン-6、血管内皮増殖因子、又は骨形成タンパク質6のようなサイトカインの皮膚性過剰発現があるトランスジェニックマウスをまた使用して、インビボで乾癬を研究し、感染治療のための治療薬を同定することができる。乾癬様病巣はまたPL/J系統に戻し交配したβ2-インテグリン低形質マウス及びβ1-インテグリントランスジェニックマウス、CD4+/CD45RBhiTリンパ球で再構成されたscid/scidマウス並びにHLA-B27/hβ2mトランスジェニックラットにおいて記載されている。免疫不全マウスに移植されたヒト皮膚を使用する異種移植モデルもまた知られている。よって、本発明の化合物は、マウスが乾癬に類似した組織病理学的皮膚病巣を示すSchon, M.P.等, Nat. Med. (1997) 3:183により記載されているscid/scidマウスモデルにおいて試験することができる。他の適切なモデルは、Nickoloff, B.J.等, Am. J. Path. (1995) 146:580に記載されているようにして調製されるヒトskin/scidマウスキメラである。更なる詳細については、例えばSchon, M.P., J Invest Dermatology 112:405-410 (1999)を参照のこと。
【0189】
組換え(トランスジェニック)動物モデルは、対象の遺伝子のコード部分を、トランスジェニック動物作製のための標準的技術を用いて、対象動物のゲノム中に導入することにより操作できる。トランスジェニック操作の標的となりうる動物は、限定されないが、マウス、ラット、ウサギ、モルモット、ヒツジ、ヤギ、ブタ、及び非-ヒト霊長類、例えばヒヒ、チンパンジー及び他のサルを含む。このような動物に導入遺伝子を導入するための当該分野で知られた技術は、前核マイクロインジェクション(Hoppe及びWanger, 米国特許第4873191号);胚系列へのレトロウイルス媒介遺伝子転移(例えば、Van der Putten等, Proc. Natl. Acad. Sci. USA 82, 6148-615 [1985]);胚性幹細胞での遺伝子ターゲティング(Thompson等, Cell 56, 313-321 [1989]);胚のエレクトロポレーション(Lo, Mol. Cel. Biol. 3, 1803-1814 [1983]);精子媒介遺伝子導入(Lavitrano等, Cell 57, 717-73 [1989])を含む。概説としては、例えば、米国特許第4736866号を参照のこと。
【0190】
本発明の目的のために、トランスジェニック動物は、その細胞の一部にのみ導入遺伝子を有するもの(「モザイク動物」)を含む。導入遺伝子は、単一の導入遺伝子として、又はコンカテマー、例えば頭部と頭部又は頭部と尾部の直列型として組み込まれうる。特定の細胞型への導入遺伝子の選択的導入も、例えばLasko等, Proc. Natl. Acad. Sci. USA 89, 6232-636 (1992)の技術に従えば、可能である。
【0191】
トランスジェニック動物における導入遺伝子の発現は、標準的技術によってモニターできる。例えば、導入遺伝子の組込みの確認にはサザンブロット分析又はPCR増幅が用いられる。ついで、mRNA発現のレベルは、インサイツハイブリダイゼーション、ノーザンブロット分析、PCR、又は免疫組織化学などの技術を用いて分析できる。
【0192】
例えば免疫細胞の特定の細胞への浸潤を確定するための組織学的検査によって、免疫疾患病理の徴候について該動物を更に調べることができる。補体及び古典的及び代替経路を含む補体活性化又はT細胞増殖に対する効果の度合いを決定するためにCRIg又は候補アゴニストでトランスジェニック動物を治療する遮断実験もまた行うことができる。これらの実験では、本発明のポリペプチドと結合する遮断抗体が動物へ投与され、興味ある生物学的効果がモニターされる。
【0193】
あるいは、「ノックアウト」動物は、CRIgポリペプチドをコードする内在性の遺伝子と、動物の胚性細胞へ導入された同ポリペプチドをコードする改変ゲノムDNAとの間の相同組換えの結果として、CRIgをコードする欠陥又は改変遺伝子を有するものとして構築することができる。例えば、CRIgをコードするcDNAは、確立されている技術によって、CRIgをコードするゲノムDNAのクローニングに利用できる。CRIgをコードするゲノムDNAの一部は、その他の遺伝子、例えば組込みをモニターするために利用できる選択可能マーカーをコードする遺伝子によって置換え又は除去できる。典型的には、ベクターには数キロベースの未変化フランキングDNA(5'と3'末端の両方)が含まれる[例えば、相同組換えベクターの説明についてはThomas及びCapecchi, Cell, 51:503(1987)を参照]。ベクターを胚性幹細胞株へ(例えば電気穿孔法等によって)導入し、導入されたDNAが内在性DNAと相同的に組換えられた細胞を選択する[例えば、Li等, Cell, 69:915 (1992)参照]。選択された細胞は次に動物(例えばマウス又はラット)の胚盤胞内に注入され、集合キメラを形成する[例えば、Bradley, Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson編 (IRL, Oxford, 1987), pp. 113-152参照]。その後、キメラ性胚を適切な偽妊娠の雌性乳母に移植し、出産させて「ノックアウト」動物をつくり出す。その胚細胞に相同組換えされたDNAを有する子孫は標準的な技術により同定でき、それらを使用して動物の全細胞が相同組換えされたDNAを含む動物を飼育することができる。ノックアウト動物は、例えば、CRIgポリペプチドが不在であることによるある種の病理的状態及びその病理的状態の発達に対する防御能力によって特徴付けることができる。
【0194】
よって、潜在的CRIgの生物学的活性は、以下の実施例7に記載されたように、マウスCRIgノックアウトマウスにおいて更に研究することができる。
【0195】
抗原誘発性気道反応亢進、好酸球増加症及び炎症が卵白アルブミンで動物を感作させ、エアゾールにより送達される同じタンパク質に動物を暴露することで誘発される喘息モデルが記載されている。幾つかの動物モデル(モルモット、ラット、非ヒト霊長類)では、エアゾール抗原への暴露時にヒトのアトピー性喘息に類似した徴候を示す。マウスモデルはヒト喘息の特徴の多くを有している。喘息治療における活性と有効性についてCRIg及びCRIgアゴニストを試験するのに適した手順は、Wolyniec, W.W.等, Am. J. Respir. Cell Mol. Biol. (1998) 18:777とそこに引用されている文献に記載されている。
【0196】
接触性過敏症は、細胞媒介免疫機能の単純なインビボアッセイである。この手順において、遅発型過敏反応を生じさせる外因性ハプテンに表皮細胞を暴露し、反応を測定して定量する。接触過敏症は最初の感作段階に顕在化段階が続く。顕在化段階は表皮細胞がそれらが過去に接触したことのある抗原に遭遇したときに生じる。腫れと炎症が生じ、ヒトアレルギー性接触皮膚炎の優れたモデルが作製される。適切な手順は、Current Protocols in Immunology, J.E. Coligan, A.M.Kruisbeek, D.H.Marglies, E. M.Shevach, 及びW.Strober編, John Wiley & Sons, Inc, unit4.2に詳細に記載されている。また、Grabbe, S.及びSchwarz, T. Immun. Today 19(1):37-44(1998)も参照のこと。
【0197】
移植片対宿主疾患は、免疫適格細胞が免疫抑制され又は耐性のある患者に移植された場合に生じる。ドナー細胞は宿主抗原を認識しそれに反応する。反応は生命に危険性のある重度の炎症から、下痢や体重の減少等の軽度の場合まで多様である。移植片対宿主疾患モデルはMHC抗原及び少量の移植抗原に対するT細胞反応性を評価する手段を提供する。適切な手順は上記のCurrent Protocols in Immunology, unit4.3.に詳細に記載されている。
【0198】
皮膚同種移植片拒絶のための動物モデルは、抗ウイルス及び腫瘍免疫におけるそれらの役割の指標であり尺度であるT細胞がインビボで組織の破壊を媒介する能力を試験する手段である。最も一般的で容認されているモデルではマウスの尾-皮膚移植片が使用される。繰り返し実験により、皮膚同種移植片拒絶がT細胞、ヘルパーT細胞及びキラー-エフェクターT細胞により媒介されるが、抗体では媒介されないことが分かった。Auchincloss, H. Jr.及びSachs, D.H., Fundamental Immunology, 2nd ed., W.E.Paul ed., Raven Press, NY, 1989, 889-992。適切な手順は上掲のCurrent Protocols in Immunology, unit4.4.に詳細に記載されている。CRIg及びCRIgアゴニストの試験に使用可能な他の移植拒絶モデルは、Tanabe, M.等, Transplantation (1994) 58:23及びTinubu, S.A.等, J. Immunol. (1994) 4330-4338により記載されている同種心臓移植片モデルである。
【0199】
遅発型過敏症の動物モデルは、細胞媒介免疫機能のアッセイをまた提供する。遅発型過敏反応は、抗原投与後の経過した時間まで、ピークに達しない炎症により特徴付けられるT細胞媒介インビボ免疫反応である。これらの反応はまた組織特異的自己免疫疾患、例えば多発性硬化症(MS)及び実験的自己免疫脳脊髄炎(EAE、MS用のモデル)で生じる。適切な手順は上掲のCurrent Protocols in Immunology, unit4.5.に詳細に記載されている。
【0200】
EAEは、T細胞及び単核細胞の炎症と、続いての中枢神経系における軸索の脱髄により特徴付けられるT細胞媒介自己免疫疾患である。EAEは、一般的にヒトにおけるMSの関連動物モデルであると考えられている。Bolton. C., Multiple Sclerosis (1995)1:143。急性及び再発寛解型モデルの双方が開発されている。CRIg及びそのアゴニスト及びアンタゴニストは上掲のCurrent Protocols in Immunology, unit15.1及び15.2.に記載されているプロトコールを使用して、免疫媒介脱髄疾患に対するT細胞刺激又は阻害活性を試験することができる。また、Duncan. I.D.等, Molec. Med. Today (1997) 554-561に記載されているようにして、オリゴデンドロサイト又はシュワン細胞が中枢神経系に移植されたミエリン疾患モデルも参照されたい。
【0201】
年齢関連性黄斑変性(AMD)の動物モデルは、Ccl-2又はCcr-2遺伝子に無発現変異を有するマウスからなる。これらのマウスは網膜色素上皮(RPE)の中のリポフスチン及びその下のドルーゼの蓄積、光受容体萎縮及び脈絡膜血管新生(CNV)を含むAMDの主要な特徴を発症する。これらの特徴は6ヶ月の年齢を越えて発達する。CRIg及びCRIgアゴニストはドルーゼの形成、光受容体及び脈絡膜血管新生について試験することができる。
【0202】
心筋虚血再灌流のモデルはマウス又はラットで実施することができる。動物を気管切開し、小動物人工呼吸器で酸素供給する。ポリエチレンカテーテルを平均動脈圧の測定のために内頸動脈と外頸静脈内に配する。心筋虚血再灌流は、冠動脈左前下行枝(LAD)を6-O縫合によって結紮することによって開始される。虚血は血管を完全に閉塞するためにLADの回りに可逆的な結紮糸を締めることによってつくり出される。結紮糸は30分後に除去され、心臓は4時間灌流される。CRIg及びCRIgアゴニストは、心臓梗塞サイズ、心臓クレアチンキナーゼ活性、ミエロペルオキシダーゼ活性及び抗C3抗体を使用する免疫組織化学の測定によってその効力を試験することができる。
【0203】
糖尿病性網膜症のモデルはストレプトゾトシンでのマウス又はラットの治療を含む。CRIg及びCRIgアゴニストは、 細静脈拡張、網膜内微小血管異常、及び網膜及び硝子体腔の血管新生に対するその効果について試験することができる。
【0204】
膜性増殖性糸球体腎炎のモデルは次のようにして樹立することができる:雌マウスをCFA中の0.5mgコントロールウサギIgGを用いて腹腔内免疫する(−7日目)。7日後に(0日目)、1mgのウサギ抗マウス糸球体基底膜(GBM)抗体を尾静脈から静脈内注射する。血清中の抗ウサギIgG抗体の上昇をELISAによって測定する。24時間の尿試料を代謝ケージ中のマウスから集め、マウスの腎臓機能を、血中尿素窒素に加えて尿中タンパク質を測定することによって評価する。
【0205】
薬学的組成物
本発明に従って同定されたCRIgアゴニスト及びアンタゴニストは、炎症性疾患の治療のために、薬学的組成物の形態で投与することができる。
【0206】
治療製剤は、所望される度合いの純度を持つ活性分子を、凍結乾燥製剤又は水溶液の形態で、最適な製薬上許容される担体、賦形剤又は安定化剤と混合することにより調製されて保存される(Remington's Pharmaceutical Science 16版, Osol, A.編 [1980])。許容される担体、賦形剤、又は安定化剤は、用いられる用量及び濃度で受容者に非毒性であり、リン酸塩、クエン酸塩、及び他の有機酸などの緩衝液;アスコルビン酸及びメチオニンを含む酸化防止剤;保存料(塩化オクタデシルジメチルベンジルアンモニウム;塩化ヘキサメトニウム;塩化ベンザルコニウム;塩化ベンゼトニウム;フェノール;ブチル又はベンジルアルコール;メチル又はプロピルパラベン等のアルキルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3-ペンタノール;及びm-クレゾールなど);低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチン、又は免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、又はリジン等のアミノ酸;グルコース、マンノース、又はデキストリンを含む単糖類、二糖類、及び他の炭水化物、EDTA等のキレート剤、スクロース、マンニトール、トレハロース又はソルビトールなどの糖;ナトリウムなどの塩形成対イオン;金属錯体(例えば、Zn-タンパク質錯体)及び/又はトゥイーン(TWEENTM)、プルロニクス(PLURONICSTM)、及びポリエチレングリコール(PEG)等の非イオン性界面活性剤を含む。
【0207】
リポフェクション又はリポソームを使用して、細胞中にポリペプチド、抗体、又は抗体断片を送達することができる。抗体断片が使用される場合、標的タンパク質の結合ドメインに特異的に結合する最小の断片が好ましい。例えば、抗体の可変領域配列に基づいて、標的タンパク質配列に結合する能力を保持するペプチド分子を設計することができる。このようなペプチドは化学的に合成し、及び/又は組換えDNA技術(Marasco等, Proc. Natl. Acad. Sci. USA 90:7889-7893 [1993])により生産することができる。
【0208】
ここでの製剤は、治療すべき特定の徴候に必要な場合に一つ以上の活性化合物、好ましくは互いに悪影響を及ぼさない相補的活性を持つものも含んでいてもよい。あるいは、又はそれに加えて、組成物は、細胞毒性薬、サイトカイン又は増殖阻害剤を含んでもよい。このような分子は、適切には、意図する目的に有効な量の組み合わせで存在する。
【0209】
また、活性分子は、例えばコアセルベーション技術により又は界面重合により調製されたマイクロカプセル、例えば、各々ヒドロキシメチルセルロース又はゼラチン-マイクロカプセル及びポリ(メタクリル酸メチル)マイクロカプセル中、コロイド状薬物送達系(例えば、リポソーム、アルブミンミクロスフィア、マイクロエマルション、ナノ粒子及びナノカプセル)中、又はマイクロエマルション中に包括されていてもよい。このような技術は、Remington's Pharmaceutical Science 16版, Osol, A.編(1980)に開示されている。
【0210】
インビボ投与に使用される製剤は無菌でなければならない。これは、滅菌濾過膜を通した濾過により容易に達成される。
【0211】
徐放性製剤を調製することもできる。徐放性製剤の好適な例は、抗体を含む固体疎水性ポリマーの半透性マトリクスを含み、このマトリクスは成形品、例えばフィルム、又はマイクロカプセルの形態である。除放性マトリクスの例は、ポリエステル、ヒドロゲル(例えば、ポリ(2-ヒドロキシエチル-メタクリレート)又はポリ(ビニルアルコール))、ポリラクチド(米国特許第3773919号)、L-グルタミン酸とγ-エチル-L-グルタマートのコポリマー、非分解性エチレン-酢酸ビニル、LUPRON DEPOTTM(乳酸-グリコール酸コポリマーと酢酸リュープロリドからなる注射可能なミクロスフィア)等の分解性乳酸-グリコール酸コポリマー、ポリ-(D)-3-ヒドロキシブチル酸を含む。エチレン-酢酸ビニル及び乳酸-グリコール酸などのポリマーは分子を100日に亘って放出することができるが、ある種のヒドロゲルはより短時間でタンパク質を放出する。カプセル化された抗体が身体内に長時間残ると、それらは37℃の水分に露出されることにより変性又は凝集し、その結果、生物学的活性の低下及び起こりうる免疫原性の変化をもたらす。合理的な方法は、含まれる機構に依存する安定化について工夫することができる。例えば、凝集機構がチオ-ジスルフィド交換を通した分子間S-S結合形成であると発見された場合、安定化はスルフヒドリル残基の修飾、酸性溶液からの凍結乾燥、水分含有量の制御、適切な添加剤の付加、及び特異的ポリマーマトリクス組成物の開発によって達成されうる。
【0212】
次の実施例は例証のためにのみ提供するものであり、本発明の範囲を限定するものでは決してない。
【0213】
本明細書において引用された全ての特許及び文献はその全体を出典明示によりここに援用される。
【実施例】
【0214】
実施例1
CRIg及びC3b:CRIg複合体の結晶構造の決定
材料と方法
a.成熟ヒトCRIgタンパク質の生産と精製
配列番号2(配列番号8)の成熟ヒトCRIgの残基1〜119をコードするDNA断片をpET28b発現ベクター(Novagen)のNdeI/BamHI部位中にクローニングし、トロンビン切断部位が続くN末端Hisタグを有する融合体をつくった。Ni-アフィニティークロマトグラフィーを使用した精製後に、融合タンパク質をトロンビンで消化させ、サイズ排除クロマトグラフィーを使用して更に精製した。最終のタンパク質原液は、10mMのHepes、50mMのNaCl、pH7.2中に20mg/mlのタンパク質濃度を有していた。セレノメチオニン標識タンパク質を、標準的なプロトコルを使用して得た。
【0215】
精製されたCRIgを5倍モル過剰でC3c又はC3bと混合し、氷上で30分間インキュベートした。その試料を、サイズ排除クロマトグラフィーを使用して精製し、25mMのHepes、pH7.2、50mMのNaCl中で10−20mg/mlまで濃縮した。
【0216】
b.結晶化
全ての結晶は懸滴蒸気拡散法を使用して19℃で成長させた。
【0217】
CRIgを、30%のPEG4000、0.1Mの酢酸ナトリウム、及び0.2Mの酢酸アンモニウムを含むリザーバの等容量でタンパク質溶液を等しく混合することによって結晶化させた。結晶は3日後に形成された。12%のPEG20000、0.1MのMES,pH6.5を含むリザーバ溶液と1:1の比でタンパク質溶液(20mg/ml)を混合することによってC3c:CRIg複合体の結晶を得た。12%のPEG20000、0.1MのMES,pH6.5を含むリザーバ溶液の等容量とタンパク質溶液(10mg/ml)を混合することによってC3b:CRIg複合体の結晶を得た。結晶は3日後に形成された。
【0218】
c.データ収集、構造解明及び緻密化
データ収集のために、20%グリセロールを添加した溶液含有リザーバ中に結晶を短時間浸漬した後、液体窒素でフラッシュ凍結させた。全てのデータはALSビームライン5.0.2.で集め、HKL2000を使用して処理した(Otwinowski, Z.及びMinor, W., Methods Enzymol. 276:307-326 (1997))。未複合体化CRIgの結晶は1.2Åの分解能で回折し、a=30.2、b=50.7及びc=61.9Aの格子パラメータの空間群P212121に属する。構造は複数の異常な分散系及びプログラムauto-SHARP(G. Bricogne, C. Vonrhein, C. Flensburg, M. Schiltz, W. Paciorek, Acta Crystallogr D Biol Crystallogr 59, 2023 (2003))を用いて解明した。 Refmac(G. N. Murshudov, A. A. Vagin, E. J. Dodson, Acta Crystallogr D Biol Crystallogr 53, 240 (1997))を用いる緻密化とプログラムO(T. A. Jones, J. Y. Zou, S. W. Cowan, Kjeldgaard, Acta Crystallogr A 47 ( Pt 2), 110 (1991))を用いたマニュアルでの調節によって、14.9%のRcrystと17.7%のRfreeのモデルを得た。C3cの結晶は3.1Åに回折し、a=382Å、b=65.0Å、c=147.2Å及びβ=102.7°の格子パラメータの空間群C2に属し、非対称ユニットに二つの複合体を含んでいる。構造はAMoRe(Navazza J., Acta Crystallogr. A 50:157-163 (1994))及び未結合CRIg及びC3Cモデル(pdbコード2A74)を使用して解明した。非結晶学的対称制約を適用する緻密化後の最終のRcryst及びRfreeはそれぞれ23.7%及び29.7%であった。C3b:CRIg複合体の結晶は4.1Å分解能に回折し、a=97.6Å、b=255.7Å、c=180.3Åの格子パラメータの空間群C22221に属する。C3c:CRIg複合体を使用する分子置換後、CUBドメイン(pdbコード2A73)とTEDドメイン(1C3D)を電子密度マップ中にマニュアルでドッキングさせた。剛性体緻密化後、TED及びCUBドメインを連結する領域並びに2つのグリコシル化部位の明確な密度は解釈可能な密度を有しており、電子密度中に嵌り込むことができた。最終モデルは、C345CドメインのないCRIg及びC3bの複合体を含む;後者のドメインは非常に弱い密度を有しており、図1に対して0.0に設定された占有率でモデルに含められた。最終Rcyst及びRfreeはそれぞれ25.2%及び33.3%であった。
【0219】
結果
a.構造の決定
CRIgはC3b及びiC3bと複合体を形成するが、親分子C3に結合することができないことが最近確定された(K. Y. Helmy等, Cell 124, 915 (2006))。CRIgの選択性に対する構造的基礎をより良好に理解するために、その未結合状態におけるCRIgの結晶構造を高分解能で決定し、またC3b及びC3cとの複合体についても決定した(それぞれ4.1Å及び3.2Å)。未結合CRIgの構造を、3波長異常分散(MAD)フェージングを使用して解明した(Hendrickson, W.A.等, Proteins 4:77-88 (1988))。つづいて、複合体の構造を、サーチモデルとして、CRIg及びC3(2A73)、C3c(2A74)、並びにC3d(1C3D)の構造の様々なドメイン及び断片の座標を使用して分子置換によって解明した。C3c:CRIg構造は非対称ユニットに2分子を含み、非結晶学的対称制約を使用する緻密化後に、最終のR及びRfreeはそれぞれ23.6%及び29.5%であった。C3c:CRIg複合体の限られた分解能のため広範な位置的緻密化は可能ではない。しかしながら、様々なリンカーのモデル化及び個々のドメインの剛性体緻密化後、最終モデルのR及びRfreeはそれぞれ24.8%及び33.1%であり、構造が限られた分解能を考えると良い品質であることを示している。
【0220】
b. C3bの全体構造
C3bの個々のドメインの構造は最近報告されたC3構造(上掲のJanssen等;D. Hourcade, V. M. Holers, J. P. Atkinson, Adv Immunol 45, 381 (1989))と同様であるが、これらのドメインの配置はC3と比較するとかなり異なっている。簡単に述べると、残基1〜645からなるC3bのβ鎖は、5つのマクログロブリン様ドメイン(MG1-MG5)、第6MGドメインのN末端半分、リンカー領域(LNK)(図1A、B)に折り畳まれる。免疫グロブリントポロジーMGドメインは「キーリング」様形式に配置され、約10Å幅で30Å長の中央溝の回りを円で囲んでいる。C3bのα鎖は残基729:1641からなり、C3と比較してC3a又はANAドメインを欠いている。α'NTドメインと呼ばれるC3bα鎖のN末端残基(729:745)は、伸展したコンフォメーションを採用し、第6MGドメインの第2の半分を形成する残基につながっている。これに続いて、α鎖は間にCUBドメインとチオ-エステルドメイン(TED)が挿入された2つの更なるMGドメイン(MG7及びMG8)を含んでいる。最後に、α鎖のC末端の170残基はいわゆる「アンカー領域」とC345Cドメインを形成している。
【0221】
c. C3活性化時のコンフォメーション変化
C3、C3b、及びC3cの構造の比較は、大きなコンフォメーション変化、つまりC3のC3bとC3aへの開裂が活性化後直ぐに生じることを示している(図1A、B)。C3中のMG3及びMG8ドメイン間に割り込んでいるANAドメインの放出により(上掲のF. Fredslund, J等)MG3とMG8が回転させられる。これらのコンフォメーション変化によりMG7ドメインが更に回転し、C3をC3cと比較した場合、上掲のJannsen等に記載されたα'NTドメインの興味深い動きが生じる。C3cの場合と同様に、α'NTはC3におけるよりもC3bにおいて更に溶媒露出性であり、補体活性化のレセプター及び調節因子、CR1、CFH及びB因子のための多くの潜在的結合部位を示す(J. D. Lambris等, J Immunol 156, 4821 (1996);A. Taniguchi-Sidle, D. E. Isenman, J Immunol 153, 5285 (1994);A. E. Oran, D. E. Isenman, J Biol Chem 274, 5120 (1999))。しかしながら、C3活性化によって誘導される最も大きな動きはCUB及びTEDモジュールに関する。C3中のTED及びMG8に対して密接に充填されているCUBはMG8に対して約25Å動き(図1B及び図6)、C3b構造中においてMG2と緩い相互作用のみを形成する。C3においてTEDは、残基Cys988及びGln991の間に形成されたチオ-エステル結合を溶媒から保護するMG8ドメイン及びCUBに対して堅固に埋もれている。活性化の際、TEDはCUB及びMG8ドメインから離れる方向に回転しTED及びCUBドメインを連結する残基を完全に異なったコンフォメーションで残す;これらの変化の過程において、Cys988は、C3構造中のその最初の保護された巣からC3構造中のその顕著に露出した位置まで80Åを越えて移動する。
【0222】
C3は炎症疾患の発生率に影響を及ぼす2つの一般的なアロタイプで存在している(J. E. Finn等, Nephrol Dial Transplant 9, 1564 (1994))。これらの変異体は単一のアミノ酸が異なる。更に一般的なアロタイプのC3Fは、80位に大きい正荷電のアルギニンを有する一方、C3Sは小さい非荷電のグリシンを有している。残基R80と回りのループはC3又はC3cの構造では相互作用を形成せず(上掲のJanssen等)、R80と相互作用する他のタンパク質はこれまで同定されていない。C3bにおいて、このループはMG1とTEDドメイン間の多くの接触の原因であり、R80はTEDの表面上の負に荷電した溝に近接して位置している(図7)。R80及びTEDドメイン間の相互作用がC3S及びC3Fアロタイプ間の観察された差異の原因であるかどうかを確認するためには更なる実験が必要である。
【0223】
C3d及びC3におけるαヘリックストロイドTEDモジュールは多くの有意な差を示している(上掲のJanssen等)。最も重要な点は、−粒子表面へのTEDドメインの共有結合に必要とされる−残基His1104及びGlu1106が互いに離れ、またCys988から離れていることである。しかしながら、C3dにおいては、これらの残基は非常に近接しており、これは共有結合反応のための重要な必要条件である。ここに提示されたC3b:CRIg複合体の分解能は個々のアミノ酸側鎖の正確な位置の検討を可能にするものではない一方、C3b中のTEDのコンフォメーションがC3d構造に非常に類似していることは我々の構造から明らかである。C3とC3b間の著しいコンフォメーションシフトにより、結合反応に重要なTED残基が溶媒露出性になり、C3b分子の遠位端に槍の穂先のように位置せしめられる。
【0224】
d. CRIgはC3b及びC3cのβ鎖に結合する
配列解析によって予測されるように(K. Langnaese等, Biochim Biophys Acta 1492, 522 (2000))、CRIgのN末端ドメインは、位相幾何学的には、免疫グロブリン様ドメインのIgVファミリーに属するが、プロテインデータバンクに現在寄託されている他のIgドメインと僅か約20%の配列同一性を共有しているだけである。この折り畳みファミリーの全てのメンバーと同様に、CRIgドメインは2つのβシートによって形成されている。これらのシートの一方はストランドA’、G、F、C、C’及びC’’によって構成され、他方はストランドB、E、及びDによって構成される(図2A及び図8)。該ドメインは、ストランドB及びFを内部で連結する2つのシステイン残基間に形成された正準なIg様ジスルフィド結合によって更に安定化されている。
【0225】
CRIgは同一の形式でC3bとC3cに係合し、両方のCRIg複合体中に存在する全ての接触ドメインの配置は非常に類似している。これに沿って、C3bとC3cに対するCRIgの親和性は比較できる(図9)。C3b:CRIg及びC3c:CRIg構造の重ね合わせは、β鎖の642の共通のCa位置に対しては0.6Å未満で、C3cとC3bのα鎖の479Ca原子に対しては0.5Å未満のRMSDになる。
【0226】
これまでに同定された全ての他の補体レセプターとは異なり(J. D. Lambris等, J Immunol 156, 4821 (1996);A. Taniguchi-Sidle, D. E. Isenman, J Immunol 153, 5285 (1994);A. E. Oran, D. E. Isenman, J Biol Chem 274, 5120 (1999))、CRIgは主にC3bのβ鎖に結合する。その未結合状態及び互いの複合体でのC3c及びCRIg構造の比較から、結合を亢進させるために2分子の何れにおいても有意なコンフォメーションの適応が必要とはされないことが明らかになった。CRIgへの未結合(上掲のJanssen等)及び結合双方のC3c構造における963の共通のCa位置は0.8ÅのRMSDで重ね合わさり、最も大きな動きは可動性領域で生じ、ただ一つの小さなループの移動がMG6ドメインで生じる(結果を示さず)。CRIgに対する結合部位は、CUB及びTEDドメインとはC3bの反対側にあり、TEDドメインから離れる方向におよそ40Å離れてCRIgを位置させている。
【0227】
CRIgとC3c又はC3bの間の結合界面は大きく、不連続で、全体で約2670Å2の溶媒露出面積を埋めている(図2A)。CRIgはC3bと接触し、A’、G、G、C、C’、C’’シートからの残基が、ストランドC’とC”の近傍の残基及びループによって形成された相互作用の大部分を有する。C’とC”を連結するヘアピンループは、C3bのキーリング形状のβ鎖の中心に形成された裂け目中に突出している。界面のC3b側では、ドメインMG3、MG4、MG5、MG6、LNK及びMG6が全てCRIg結合に寄与し、埋まった界面のそれぞれ約30%と40%に対してMG3とMG6が原因となっている(図2B)。C3及びC3b構造の比較は、C3b及びC3cへのCRIgの結合のゲーティングにおけるMG3ドメインの重要な役割を示唆している。C3活性化時に生じる大規模なドメイン移動とは別に、機能的な重要性を有することがある数種のより小さいドメインシフトがある。これらの間のキーは、比較的僅かであるが、C3分子の残りと比較してなお有意な15°のMG3の回転である。この回転は、CRIg結合部位を構成する他のMGドメインのリングに関したMG3の相対的再配向の原因であり、LNK領域の螺旋部分の動きと連係して(図9)、CRIgをC3b及びC3cに結合させる;これらの中程度の変化がないと、C3はCRIgにはラッチされないであろう。
【0228】
重要な結合パラメータを次の表1にまとめる。
【0229】
実施例2
CRIg変異体
構造複合体によって明らかになった結合界面の重要性を機能的に検証するために、次のCRIg変異体をデザインし、製造し、C3bに結合するそれらの能力を試験した:次の表2において、番号はシグナル配列切断部位の開始位置を示す。
表2
変異体 アミノ酸置換
A E85A M86A
B H57A Q59A
C V107S D109R I111E
D L34S K36D L38H
E P91A T93A D95A
コントロール H21A T23A D25A
【0230】
変異体を、QuikChange部位特異的突然変異誘発キット(Stratagene)を使用して作製し、プライマーは製造者の指示に従って設計した。二重又は三重変異体を用いた結合実験の結果は結晶学的に定めたタンパク質-タンパク質接触と密接に一致している(図2B)。MG6及びMG3結合ドメイン(E85A M86A及びH57A Q59A)における変異がCRIgのC3bへの結合を最も効果的に抑止し(IC50値はwtCRIgと比較して>10000及び100倍高かった)、CRIg結合におけるこれらのドメインの相互作用の重要性を示している。これに対して、結合界面とは反対側に位置するCRIg表面残基における変異は、野生型分子と同様の結合活性を示し、これらの変異が分子の全体の折り畳みを変えないことを確認した。また、円二色性分光法を更に使用して、全ての変異タンパク質が正しく折り畳まれたことを確認した(結果は示さず)。
【0231】
実施例3
C3bへのCRIgの結合は補体の代替経路を阻害する
AP及び古典/MBL経路双方のC5転換酵素がC3bサブユニットを含んでいるので(Pangburn及びMuller-Eberhard, Biochem J., 235:723 (1986))、我々は、CRIgがそのC3bへの結合を介して補体活性化の何れかの経路を調節できるかどうかを問うた。CRIgは代替経路を介する補体活性化を阻止したが、古典又はMBL経路はしなかった(図3a)。この阻害活性はC3b結合活性を欠いたCRIg変異体では失われており、CRIg-C3b結合が転換酵素活性を乱す直接の原因であることを確認した。CRIgによるAPの選択的阻害は、代替及び古典/MBL経路を区別する細胞ベースアッセイで更に確認した。IC50は、CRIgがIgGのFc部分に融合された場合に2倍まで減少した(図S6A)。CRIgは代替経路の選択的インヒビターであるので、CRIgは古典経路によって開始される代替経路溶血を阻害することができる(図S6B)。代替経路阻害は、マウスCRIgにおいて保持されており(図S3C)、CRIgの補体阻害活性は系統的に保存されていることを確認した。
【0232】
CRIgが転換酵素を阻害する分子メカニズムを更に決定するために、C3及びC5転換酵素のC3bサブユニットとのCRIgの相互作用を精製した成分を使用して研究した。CRIgは、減少したC3a生産によって示されるように液相C3転換酵素を阻害することができた(図3B)。CRIgはまたザイモサン粒子の表面に集まったC5転換酵素の酵素活性を減少させた(図3C)。様々なCRIg変異体の阻害活性はC3bサブユニットへのその親和性と相関しており、C3bへのCRIgの結合がC5転換酵素の活性を直接調節することを示している。これは、C3bβ鎖へのCRIgの結合が代替経路の転換酵素によってC3とC5の双方の切断を妨害することを示している。
【0233】
実施例4
CRIgは関節炎のマウスモデルにおいて補体活性化を阻害する
材料と方法
a.マウスにおける関節炎の誘発
コラーゲン誘発関節炎を、他に記載されているようにして(K. H. Barck等, Arthritis Rheum 50, 3377 (2004))実施した。マウスに、21日目に開始して4mg/kgのCRIg-Ig融合タンパク質を、又は一次免疫後36日目に開始して12mg/kgの融合タンパク質を皮下的に注射した。CRIg-Ig融合タンパク質では、マウスCRIg細胞外ドメインをIgG1分子のマウスFc部分に融合させている。抗体誘発関節炎は関節炎誘発性混合物を使用してTeraoと同僚の方法(K. Terato等, J Exp Med 162, 637 (1985))によって誘発させた。簡単に言えば、マウスに尾静脈から2mgの抗CIIAb(2mg/500μl/体)を静脈内注射し、3日後に25μgのLPS(25μg/100μl/体)を腹腔内注射した。骨体積の測定と組織検査を、他に記載されているようにして(上掲のBarck等)実施した。
【0234】
b.関節炎の後足におけるサイトカイン濃度の測定
屠殺したマウスの後足蹠を毛の生え際で切断し、液体N2で凍結させた。ポリトロンホモジナイザー(KINEMATICA)を使用して25ml当たり1完全プロテアーゼインヒビター錠剤(Roche)を補填した氷冷RIPA可溶化バッファー中で足蹠をホモジナイズさせた。均質化に使用したPBSの体積を、バッファー1ミリリットル当たり75mgの組織に調節した。ホモジネートを1870×gで15分間遠心分離し、上清を13230×gで5分間遠心分離した。その上清をELISA分析にかけた。マウスIL-1β及びIL-6(BD DuoSet)及びmC3a(Bachem)のELISAキットを、各ELISAキットのプロトコルに従って使用した。C5aは、製造者のプロトコル(BD)に従ってC5a-desargのネオ-エピトープに対する特異的抗体のセットを使用してELISAによって測定した。上清中の全タンパク質濃度はBCAキット(Pierce)を使用して測定した。サイトカインとケモカインの濃度は、タンパク質1ミリグラム当たりのピコグラムで表した。
【0235】
c.ELISA解析
C3a及びC5aのELISAは、BDからの捕捉及び検出抗体を使用して、又はマウスELISAキットを使用して確立した。Weilisa全補体スクリーン:プロトコルはキットによって提供された指示書に従った。CPの潜在的なCRIg阻害を評価するために、B因子枯渇血清を使用した。APのCRIg阻害を決定するために、C2枯渇血清を使用した。
【0236】
d.関節炎の後足におけるサイトカイン濃度の測定
屠殺したマウスの後足蹠を毛の生え際で切断し、液体N2で凍結させた。ポリトロンホモジナイザー(KINEMATICA)を使用して25ml当たり1完全プロテアーゼインヒビター錠剤(Roche)を補填した氷冷RIPA可溶化バッファー中で足蹠をホモジナイズさせた。均質化に使用したPBSの体積を、バッファー1ミリリットル当たり75mgの組織に調節した。ホモジネートを1870×gで15分間遠心分離し、上清を13230×gで5分間遠心分離した。その上清をELISA分析にかけた。マウスIL-1β及びIL-6(BD DuoSet)及びmC3a(Bachem)のELISAキットを、各ELISAキットのプロトコルに従って使用した。C5aは、製造者のプロトコル(BD)に従ってC5a-desargのネオ-エピトープに対する特異的抗体のセットを使用してELISAによって測定した。上清中の全タンパク質濃度はBCAキット(Pierce)を使用して測定した。サイトカインとケモカインの濃度は、タンパク質1ミリグラム当たりのピコグラムで表した。
【0237】
結果
補体活性化の可溶型調節因子は炎症に至る補体活性化を阻害する強力な治療的ツールであることが証明された(H. F. Weisman等, Science 249, 146 (1990);B. P. Morgan, C. L. Harris, Mol Immunol 40, 159 (2003))。CRIgの治療上の潜在性を、コラーゲン自己抗体及び補体活性化が疾患の原因に寄与するリウマチ様関節炎の二つのマウスモデルで試験した(A. Aggarwal等, Rheumatology (Oxford) 39, 189 (2000);S. Solomon等, Arthritis Res Ther 7, 129 (2005);K. Terato等, J Immunol 148, 2103 (Apr 1, 1992))。インビボでのその薬理学的半減期を延長させるために、muCRIgの細胞外ドメインをマウスIgG1のFc部分に融合させた。CRIgは、関節炎の臨床的徴候の前又は後で与えられたとき、関節膨張、組織学的スコア及び骨量減少を有意に減少させた(図4A及びB、S12)。その転換酵素活性の阻害に沿って、CRIgは、軟骨表面上のC3沈着の減少によって示されるように(図4C)、関節において補体活性化を有意に減少させた。既に確認された疾患を持つマウスでは、CRIgは関節の裏にあるC3と共に同時局在化され(図4D)、それが転換酵素に結合し、転換酵素を局所的に阻害することができることを示している。また、C3aの局所レベル及びC5aの全身レベルは、コントロール-Fc処置マウスにおけるよりもCRIg-Fc処置において有意に低くなっていた(図4E)。これらの結果は、CRIgが、局所的及び全身性補体活性化を阻害することによって疾患のエフェクター相において関節炎を阻害することを示している。CRIgの保護活性は、Fcレセプター結合を消失させるCRIg-FcのFc部分の変異が、wtFcタンパク質に融合したCRIgと等価な活性を有していたので、Fcレセプター機能と独立であった(結果は示さず)。これらの結果に基づくと、CRIgの単一IgV様ドメインは補体介在疾患において代替経路を阻害する有望なタンパク質治療剤となりうる。
【0238】
実施例5
CRIgは基質-酵素結合の阻害によりAP転換酵素を阻害する
材料と方法
a.崩壊促進アッセイ
代替経路DAAに対するマイクロタイタープレートアッセイを過去に記載されているようにして(M. Krych-Goldberg等, J Biol Chem 274, 31160 (1999))実施した。マイクロタイタープレートを、リン酸緩衝生理食塩水中4μg/mlのC3bで一晩被覆した。プレートを、1%のウシ血清アルブミンを含むリン酸緩衝生理食塩水で37℃にて2時間ブロックし、71mMのNaCl、0.05%のTween20及び4%のBSAを含むベロナールバッファー中の40ngのB因子、10ngのD因子、及び0.8mMのNiCl2と共に室温で2時間インキュベートした(バッファーの濃度と組成はプロトコルでチェック)。ついで、ウェルを、0.05%のTween20を含むPBS(PBST)中のH因子又はCRIg-HisECDと共に室温で15分間インキュベートして、プレート上のC3bからBbを解離させた。ついで、Bbを、1:5000の希釈のPBST中ヤギ抗ヒトB因子ポリクローナル抗体(Kent)と1:3000の希釈のPBST中西洋わさびペルオキシダーゼ結合ロバ抗ヤギ抗体(Caltag)と共に連続1時間インキュベートして検出した(希釈と二次抗体をプロトコルでチェック)。色はO-フェニレンジアミンで発色させた。このアッセイにおいて、H因子は予想された通りに崩壊促進活性のメディエーターとして挙動した。
【0239】
b.C3b-C4b及びC3b-C3b二量体調製及び結合/競合アッセイ
C3bC4bヘテロ二量体を次の変更をして(S. Meri, M.K. Pangburn, Eur. J. Immunol. 20:2555 (1990))本質的に他に記載されているようにして(S. Meri, M. K. Pangburn, Eur J Immunol 20, 2555 (1990))作製した。C3b-C4bヘテロ二量体とC3bホモ二量体へのCRIgの結合を決定するために、マキシソープ96ウェルマイクロタイタープレートを、PBS中5ug/mlのヤギ抗C3ポリクローナル抗体(ICN)で4℃にて一晩被覆し、PBSTで3回洗浄した。ついで、プレートを室温で2時間、250μlのPBS/1%BSAでブロックし、滴定量のhCRIg-L LFHを、1%BSAを含むPBST中で1時間加えた。プレートを再びPBSTで3回洗浄し、CRIg結合を、PBST/1%BSA中の1:20000希釈のHRPにコンジュゲートしたマウス抗FLAG M2抗体(SIGMA)で検出した。ついで、プレートをPBSTで3回洗浄し、100ulのTMB基質溶液(KPL)で展開させ、50ulの2N H2SO4で停止させた。吸光度を450nmで読み取った。C5がこの形態においてC3b-C4bヘテロ二量体及びC3bホモ二量体の双方に結合することがまた決定された。CRIgがC3b-C4b又はC3b-C3bへのC5の結合をブロックすることができるかどうかを決定するために、補体二量体を捕捉し上述のようにブロックした。20mMのTRIS pH7.5、20mMのMgCl2、20mMのCaCl2、150mMのNaCl、0.05%のTween20及び1%のBSAを含むバッファーに希釈して、400nMのC5を滴定濃度のhCRIg-S HisECD又はC3bに結合しないこの融合体の変異型と前もって混合し、室温で1時間、捕捉した二量体に加えた。ついで、ウェルをPBSTで3回洗浄し、C5を、1:5000のPBST希釈のマウス抗ヒトC5(Quidel)と1:3000のヤギF'ab抗マウス抗体(Caltag)の段階的添加によって検出した。該プレートを上のようにして発色させ、2つのウェルのA415を平均化し単一の点と考えた。
【0240】
結果
補体活性化の全ての既知の調節因子は、C3のI因子介在性切断のコファクターとして作用し、又はC3bへの結合についてBbの競合体であり、よって転換酵素活性を除去する(D. Hourcade等, Adv Immunol 45, 381 (1989);T. Seya, J. P. Atkinson, Biochem J 264, 581 (1989);X. Sun等, Proc Natl Acad Sci U S A 96, 628 (1999))。これに対して、CRIgは崩壊活性もコファクター活性も示さない。また、CRIgは、H因子、B因子又はプロパージンのC3bへの結合と干渉しない(図S13A−C、結果は示さず)。CRIgがAP補体活性化を阻害するメカニズムを更に探究するために、我々はCRIgがC5転換酵素の触媒性BbサブユニットとよりもむしろC3bの機能と干渉するかどうかを決定した。代替経路転換酵素の非触媒性C3bサブユニットは基質C3及びC5の結合の原因である。ついで、転換酵素のBb因子サブユニットはプロテアーゼとして作用することができ、結合した基質を切断する(W. Vogt等, Immunology 34, 29 (1978))。その結果、転換酵素の活性はC3bに対する基質の親和性に直接的に比例する(N. Rawal, M. K. Pangburn, J Immunol 164, 1379 (2000))。CRIgはAP転換酵素のC3b2サブユニットへのC5の結合を防止するが、APに対するその選択性と一致して、C5転換酵素(図5A)はCP転換酵素のC3bC4bサブユニットに対して効果は有していない。C5転換酵素の活性に対するCRIgの結合の重要性を、精製されたC5転換酵素で更に決定した(N. Rawal, M. K. Pangburn, J Biol Chem 273, 16828 (1998))。CRIgの存在下ではs、酵素のVmaxは〜50%減少し(図5B)、これは、CRIgが基質濃度とは独立にC3bへのC5の最大結合能を阻害することを示している。これは、CRIgによるC3bへのC5の結合の立体的又はアロステリックな阻害の指標であり、C3bへの結合に対するC5とのCRIgの直接の競合ではない。これは、C5がC3bへの結合についてCRIgと競合できない結合アッセイからの結果によって更に裏付けられている(図14)。よって、C3bのキーリング様β鎖構造の中心に結合したCRIgは転換酵素へのC5の結合をブロックすることができ、よって酵素の活性を阻害する。これは、C5-転換酵素のその基質との相互作用に対するC3bのβ鎖の重要性を示している(N. Rawal, M. K. Pangburn, J Biol Chem 273, 16828 (1998))。
【特許請求の範囲】
【請求項1】
天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体により形成された結晶。
【請求項2】
格子定数a=30.3Å、b=50.8Å、c=62.0Å及びP212121の空間群をおよそ有している請求項1に記載の結晶。
【請求項3】
上記天然配列CRIgポリペプチドが、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される請求項1に記載の結晶。
【請求項4】
上記機能的断片が天然配列CRIgの細胞外ドメイン(ECD)配列である請求項1に記載の結晶。
【請求項5】
結晶が約1から40Åまでの分解能で原子座標を決定するためにX線を回折する請求項1に記載の結晶。
【請求項6】
結晶が少なくとも約5Åの分解能で原子座標を決定するためにX線を回折する請求項1に記載の結晶。
【請求項7】
上記分解能が少なくとも4Åである請求項6に記載の結晶。
【請求項8】
上記分解能が少なくとも3Åである請求項7に記載の結晶。
【請求項9】
アペンディクス1に示された構造座標を有するCRIg結晶。
【請求項10】
請求項1から9の何れか一項に記載の結晶を含んでなる組成物。
【請求項11】
天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体と、補体因子C3bとの複合体の結晶形態。
【請求項12】
格子定数a=97.6Å、b=255.7Å、c=180.3Å及びC2221の空間群をおよそ有している請求項11に記載の結晶形態。
【請求項13】
上記天然配列CRIgポリペプチドが、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される請求項11に記載の結晶形態。
【請求項14】
上記機能的断片が天然配列CRIgの細胞外ドメイン(ECD)配列である請求項11に記載の結晶形態。
【請求項15】
約1から40Åまでの分解能で原子座標を決定するためにX線を回折する請求項11に記載の結晶形態。
【請求項16】
少なくとも約5Åの分解能で原子座標を決定するためにX線を回折する請求項11に記載の結晶形態。
【請求項17】
上記分解能が少なくとも4Åである請求項16に記載の結晶。
【請求項18】
上記分解能が少なくとも3Åである請求項17に記載の結晶。
【請求項19】
アペンディクス2に示された構造座標を有する請求項11に記載の結晶形態。
【請求項20】
天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体と、補体因子C3cとの複合体の結晶形態。
【請求項21】
格子定数a=382.8Å、b=65.0Å、c=147.2Å、β=102.7、及びC2の空間群をおよそ有している請求項20に記載の結晶形態。
【請求項22】
上記天然配列CRIgポリペプチドが、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される請求項20に記載の結晶形態。
【請求項23】
上記機能的断片が天然配列CRIgの細胞外ドメイン(ECD)配列である請求項20に記載の結晶形態。
【請求項24】
約1から40Åまでの分解能で原子座標を決定するためにX線を回折する請求項20に記載の結晶形態。
【請求項25】
少なくとも約5Åの分解能で原子座標を決定するためにX線を回折する請求項20に記載の結晶形態。
【請求項26】
上記分解能が少なくとも4Åである請求項25に記載の結晶。
【請求項27】
上記分解能が少なくとも3Åである請求項26に記載の結晶。
【請求項28】
アペンディクス3に示された構造座標を有する請求項20に記載の結晶形態。
【請求項29】
配列番号2のCRIgポリペプチド又はその保存的置換のC3b結合部位の少なくとも一部を含んでなる分子又は分子複合体であって、結合部位が、8、14−15、41−42、44、45−47、50、52、54−61、62、64、85−87、89、95、99、105、107−110、及び111からなる群から選択される少なくとも一つのアミノ酸残基を含み、結合部位がアペンディクス2に記載の構造座標によって表されるアミノ酸の原子を表す点から4.7Å未満の距離を有する点の集合によって定まる分子又は分子複合体。
【請求項30】
上記結合部位が上記アミノ酸残基の実質的に全てを含む請求項29に記載の分子又は分子複合体。
【請求項31】
点の少なくとも一部が、C3bに対するCRIg結合部位を定める少なくともコアアミノ酸の骨格原子の位置を表すアペンディクス2の構造座標から誘導されている点の三次元立体構造。
【請求項32】
C3bに対するCRIg結合部位が空間群対称C2221を有する結晶を形成する、ホログラフィー像、立体ダイアグラム、モデル、又はコンピュータ・ディスプレイ画像として示された請求項31に記載の点の三次元立体構造。
【請求項33】
機械可読データでコードされたデータストレージ材を含む機械可読データストレージ媒体であって、該データを使用するための指示書を用いてプログラムされた機械が、C3b結合部位の少なくとも一部を含んでなる少なくとも一つの分子又は分子複合体の三次元グラフィック図を表示し、点の集合によって定まった結合部位が、アペンディクス2に記載された構造座標によって表されるアミノ酸の原子を表す点から約4.7Å未満の距離を有している媒体。
【請求項34】
CRIgのC3b結合部位を模倣する化合物を設計することを含むCRIgアゴニストを同定する方法。
【請求項35】
(a)計算又は実験手段を用いて化学物質とCRIgポリペプチド又はC3b:CRIg複合体の三次元構造の間のフィッティング操作を実施し;(b)工程(a)で得られたデータを解析して化学物質と天然CRIg又はC3b:CRIg複合体の間の結合の特徴を決定することにより同定された化学物質。
【請求項36】
CRIgとC3bの間の結合をインビボ又はインビトロで妨害する請求項35に記載の化学物質。
【請求項37】
天然配列CRIg分子又はその保存的アミノ酸置換変異体のC3b結合領域の少なくとも一部を含む分子又は分子複合体。
【請求項1】
天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体により形成された結晶。
【請求項2】
格子定数a=30.3Å、b=50.8Å、c=62.0Å及びP212121の空間群をおよそ有している請求項1に記載の結晶。
【請求項3】
上記天然配列CRIgポリペプチドが、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される請求項1に記載の結晶。
【請求項4】
上記機能的断片が天然配列CRIgの細胞外ドメイン(ECD)配列である請求項1に記載の結晶。
【請求項5】
結晶が約1から40Åまでの分解能で原子座標を決定するためにX線を回折する請求項1に記載の結晶。
【請求項6】
結晶が少なくとも約5Åの分解能で原子座標を決定するためにX線を回折する請求項1に記載の結晶。
【請求項7】
上記分解能が少なくとも4Åである請求項6に記載の結晶。
【請求項8】
上記分解能が少なくとも3Åである請求項7に記載の結晶。
【請求項9】
アペンディクス1に示された構造座標を有するCRIg結晶。
【請求項10】
請求項1から9の何れか一項に記載の結晶を含んでなる組成物。
【請求項11】
天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体と、補体因子C3bとの複合体の結晶形態。
【請求項12】
格子定数a=97.6Å、b=255.7Å、c=180.3Å及びC2221の空間群をおよそ有している請求項11に記載の結晶形態。
【請求項13】
上記天然配列CRIgポリペプチドが、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される請求項11に記載の結晶形態。
【請求項14】
上記機能的断片が天然配列CRIgの細胞外ドメイン(ECD)配列である請求項11に記載の結晶形態。
【請求項15】
約1から40Åまでの分解能で原子座標を決定するためにX線を回折する請求項11に記載の結晶形態。
【請求項16】
少なくとも約5Åの分解能で原子座標を決定するためにX線を回折する請求項11に記載の結晶形態。
【請求項17】
上記分解能が少なくとも4Åである請求項16に記載の結晶。
【請求項18】
上記分解能が少なくとも3Åである請求項17に記載の結晶。
【請求項19】
アペンディクス2に示された構造座標を有する請求項11に記載の結晶形態。
【請求項20】
天然配列CRIgポリペプチド又はその機能的断片又は保存的アミノ酸置換変異体と、補体因子C3cとの複合体の結晶形態。
【請求項21】
格子定数a=382.8Å、b=65.0Å、c=147.2Å、β=102.7、及びC2の空間群をおよそ有している請求項20に記載の結晶形態。
【請求項22】
上記天然配列CRIgポリペプチドが、配列番号1、2及び3のヒトCRIgポリペプチド;配列番号4のマウスCRIgポリペプチド;配列番号5のラットCRIgポリペプチド;配列番号6のウシCRIgポリペプチド;及び配列番号7のサルCRIgポリペプチドからなる群から選択される請求項20に記載の結晶形態。
【請求項23】
上記機能的断片が天然配列CRIgの細胞外ドメイン(ECD)配列である請求項20に記載の結晶形態。
【請求項24】
約1から40Åまでの分解能で原子座標を決定するためにX線を回折する請求項20に記載の結晶形態。
【請求項25】
少なくとも約5Åの分解能で原子座標を決定するためにX線を回折する請求項20に記載の結晶形態。
【請求項26】
上記分解能が少なくとも4Åである請求項25に記載の結晶。
【請求項27】
上記分解能が少なくとも3Åである請求項26に記載の結晶。
【請求項28】
アペンディクス3に示された構造座標を有する請求項20に記載の結晶形態。
【請求項29】
配列番号2のCRIgポリペプチド又はその保存的置換のC3b結合部位の少なくとも一部を含んでなる分子又は分子複合体であって、結合部位が、8、14−15、41−42、44、45−47、50、52、54−61、62、64、85−87、89、95、99、105、107−110、及び111からなる群から選択される少なくとも一つのアミノ酸残基を含み、結合部位がアペンディクス2に記載の構造座標によって表されるアミノ酸の原子を表す点から4.7Å未満の距離を有する点の集合によって定まる分子又は分子複合体。
【請求項30】
上記結合部位が上記アミノ酸残基の実質的に全てを含む請求項29に記載の分子又は分子複合体。
【請求項31】
点の少なくとも一部が、C3bに対するCRIg結合部位を定める少なくともコアアミノ酸の骨格原子の位置を表すアペンディクス2の構造座標から誘導されている点の三次元立体構造。
【請求項32】
C3bに対するCRIg結合部位が空間群対称C2221を有する結晶を形成する、ホログラフィー像、立体ダイアグラム、モデル、又はコンピュータ・ディスプレイ画像として示された請求項31に記載の点の三次元立体構造。
【請求項33】
機械可読データでコードされたデータストレージ材を含む機械可読データストレージ媒体であって、該データを使用するための指示書を用いてプログラムされた機械が、C3b結合部位の少なくとも一部を含んでなる少なくとも一つの分子又は分子複合体の三次元グラフィック図を表示し、点の集合によって定まった結合部位が、アペンディクス2に記載された構造座標によって表されるアミノ酸の原子を表す点から約4.7Å未満の距離を有している媒体。
【請求項34】
CRIgのC3b結合部位を模倣する化合物を設計することを含むCRIgアゴニストを同定する方法。
【請求項35】
(a)計算又は実験手段を用いて化学物質とCRIgポリペプチド又はC3b:CRIg複合体の三次元構造の間のフィッティング操作を実施し;(b)工程(a)で得られたデータを解析して化学物質と天然CRIg又はC3b:CRIg複合体の間の結合の特徴を決定することにより同定された化学物質。
【請求項36】
CRIgとC3bの間の結合をインビボ又はインビトロで妨害する請求項35に記載の化学物質。
【請求項37】
天然配列CRIg分子又はその保存的アミノ酸置換変異体のC3b結合領域の少なくとも一部を含む分子又は分子複合体。
【図1】

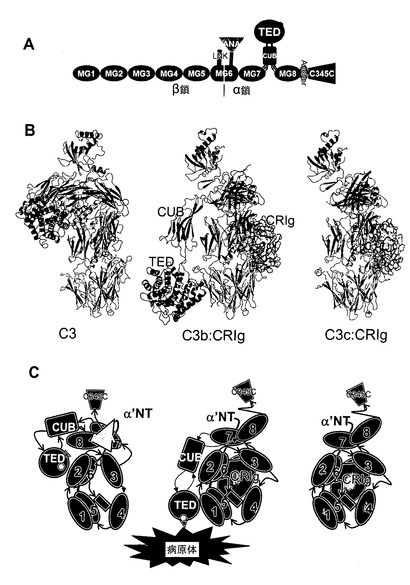
【図2】
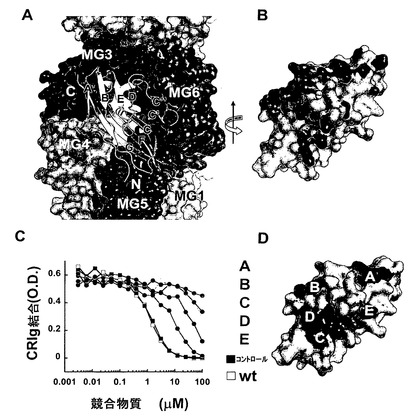
【図3】
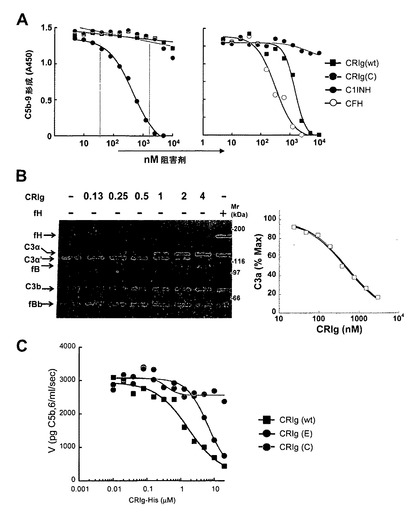
【図4】
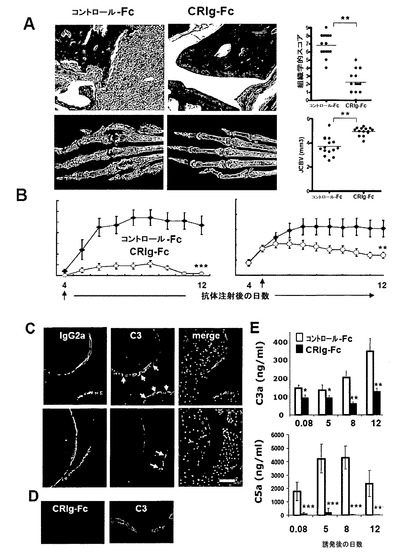
【図5】
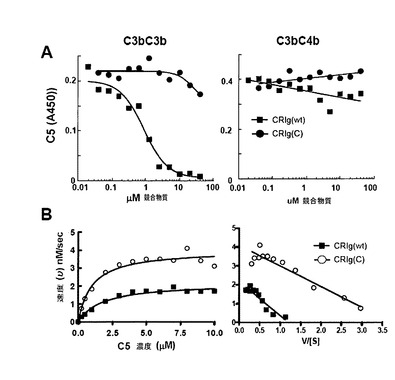
【図6】
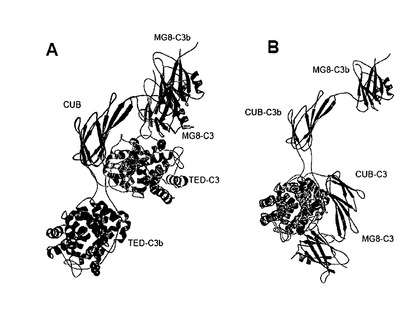
【図7】

【図8】
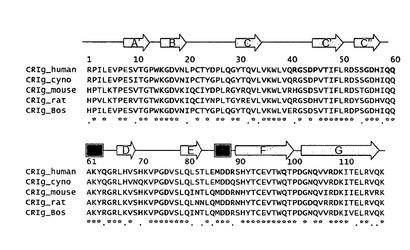
【図9】
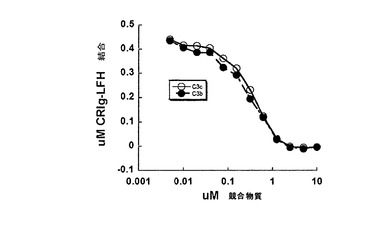
【図10】
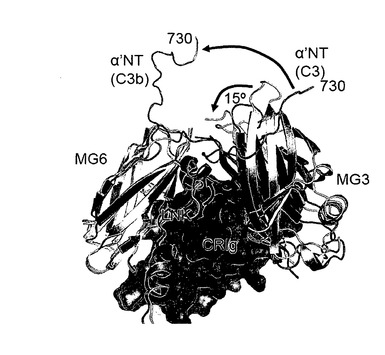
【図11】
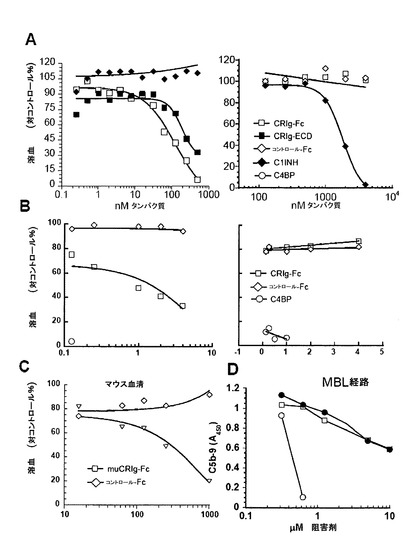
【図12】
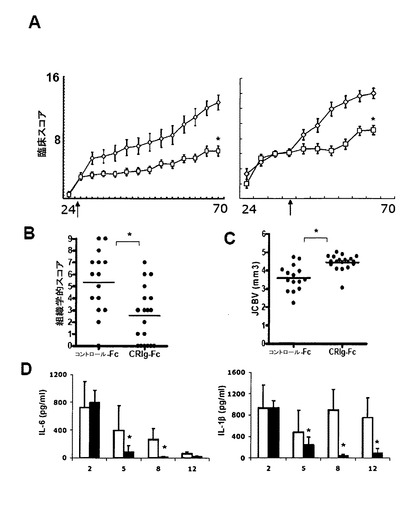
【図13】
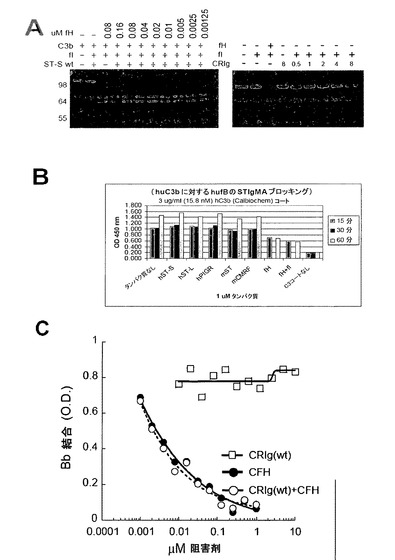
【図14】
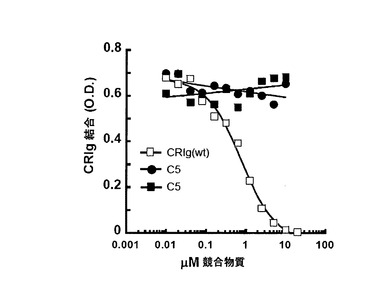
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公表番号】特表2010−505743(P2010−505743A)
【公表日】平成22年2月25日(2010.2.25)
【国際特許分類】
【出願番号】特願2009−513268(P2009−513268)
【出願日】平成19年5月30日(2007.5.30)
【国際出願番号】PCT/US2007/012816
【国際公開番号】WO2008/036135
【国際公開日】平成20年3月27日(2008.3.27)
【出願人】(509012625)ジェネンテック インコーポレイテッド (357)
【Fターム(参考)】
【公表日】平成22年2月25日(2010.2.25)
【国際特許分類】
【出願日】平成19年5月30日(2007.5.30)
【国際出願番号】PCT/US2007/012816
【国際公開番号】WO2008/036135
【国際公開日】平成20年3月27日(2008.3.27)
【出願人】(509012625)ジェネンテック インコーポレイテッド (357)
【Fターム(参考)】
[ Back to top ]