
CVDリスク評価のための生化学マーカー
プロテイナーゼによる動脈硬化性プラークの蛋白質、例えば、ルミカン、バーシカン、パールカン、デコリン、バイグリカン、III型コラーゲン、CRP、ApoE、又はエラスチンの切断によって形成されるネオエピトープを含むペプチド断片の定量のためのバイオアッセイの方法であって、尿又は血清のような試料を、ネオエピトープと反応する抗体と接触させるステップと、前記試料中の前記免疫学的結合パートナーのペプチド断片への結合レベルを定量するステップとを含む方法が開示される。アッセイは心血管疾患事象のリスクを予測する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アテローム性動脈硬化の進展及びプラーク不安定性から生じる心血管事象のリスクを示す生化学マーカーを含む、心血管疾患の診断目的及び疾患の進展の予後に有益な生化学マーカーの検出のためのアッセイに関する。
【背景技術】
【0002】
世界的に、心血管疾患(CVD)は罹病及び死亡の主因である。現在、診断と患者の異なるリスク群への分類及び低リスク患者の診断を可能にする効果的且つ非侵襲的診断法はない。診断及び予後ツールは、主に、単純なマーカー、例えば、年齢、喫煙並びに種々の脂質及びリポ蛋白質濃度の多変量解析から成る。
【0003】
CVDは複数の臨床症候群を包括し、それは主に狭心症、心筋梗塞(冠動脈血栓)及び脳卒中である。通常、これらの症候群はすべて複雑なアテローム性動脈硬化の続発症である。
【0004】
アテローム性動脈硬化は小児期に内膜肥厚から始まり、動脈内膜における脂肪線条に進行し、これらの病変はそれぞれI型、II型と特徴付けられる。脂肪線条はアテローム性動脈硬化の進展における最も初期の肉眼で視認可能な病変であり、すべての人種及び社会のすべてのヒトに生じる。非病原性状態では内皮細胞(EC)は白血球との付着性相互作用に抵抗性である。しかし、アテローム発生時の炎症性サイトカイン及び動脈壁における蓄積酸化リポ蛋白質の作用は、大動脈ECの表面上における接着分子、例えば、細胞間接着分子(ICAM)−1及び血管細胞接着分子(VCAM)−1の発現を引き起こす。これは、白血球の捕捉及び内皮表面を貫通して血管壁の内膜部分への移動を許容する。プラークの発達は、置換やアポトーシスを起こす平滑筋細胞(SMC)の増加を伴い、これはマトリックス代謝回転の増加をもたらす。コラーゲン合成の障害は線維性被膜の脆弱性及びより破裂しやすい動脈硬化性プラークをもたらし得るが、大部分の研究者らは、蛋白質分解酵素、例えば、マトリックスメタロプロテアーゼ(MMP)及び他のプロテアーゼの作用が、プラークの破裂のリスクに重大に寄与すると考えている(Clarkson and Kaplan 509−28)。
【0005】
プラークは2つの異なるタイプ、「不安定」及び「安定」プラークに分けられる。しかし、詳細な組織学的分析及び分子的理解のために、より詳細な分類が用いられる場合が多い。プラークの発達には3つの主要な段階、すなわち、初期、脂肪線条、及び、複雑性/進行性プラークがある(Stary H.C.)。
【0006】
動脈硬化性プラークは動脈内膜内で発達し、その組成及び構造により分類され得る。この分類では病変を8つのタイプに分類する(Stary H.C.)。
I.脂肪滴に満ち、脂肪滴によって拡大されたマクロファージ(マクロファージ泡沫細胞)が内膜に増加する。
II.マクロファージ泡沫細胞が内膜SMC内で脂肪滴とともにプロテオグリカン層の深部に蓄積する。泡沫細胞層は脂肪線条として視認可能である。II型病変では、単球がヒトアテロームにおいて過剰発現する単球走化性蛋白質(主にMCP−1)によって内膜を貫通する。病変の初期タイプ(I型及びII型)は幼児期に始まり得、必ずしもプラークの破裂には至らない。更に、アテローム性動脈硬化の発達はIII型病変の形成後に終了し得、プラークの形成は予測可能ではない(Stary H.C.)。
III.III型病変は脂肪線条(II型)とアテローム(IV型)との中間病変として判定される。これらの病変は細胞外脂質のプールを含み、これにより、内膜の筋弾性層の通常密接に隣接しているSMC間の空間を拡張する。物質のプールは通常ここに存在するプロテオグリカン及びコラーゲン線維に取って代わり得るが、これはアテローム発生のこの段階においてほとんど影響なく生じる。
IV.アテロームはアテローム性動脈硬化の最初の臨床徴候である。脂質の蓄積細胞外プールによる動脈内膜におけるSMCの移動及び内膜構造の崩壊が、IV型病変の顕著な特徴である。脂質コアの形成は、このSMC移動の最終結果である。脂質コアの形成は壁肥厚の増大の説明となる。脂質コアは深部内膜の大きな明瞭な輪郭をした領域であり、この動脈壁部分の通常の構造要素が高密度の泡沫細胞残遺物、遊離脂肪滴、コレステロール結晶及びカルシウム粒子に置換されている。通常、この領域に存在するSMCは、アテローム性動脈硬化のこの段階において減少し、又は完全に消失する。残遺SMCは広範に分散し、伸長細胞体及び、多くの場合、異常に厚い基底膜を生成している。この段階では脂質コアを覆う層の発達が始まる。この層はコラーゲン/プロテオグリカンに富む細胞間マトリックス、脂肪滴を有する又は有しないSMC、マクロファージ及び泡沫細胞から成る。
V.IV型病変に対する応答は、線維性「被膜」を形成する修復線維組織マトリックスの形成である。通常、これらの病変は脂質コア及び不規則に積み重なり合った修復組織の層から成る。血腫及び血栓形成のような事象がこれらのタイプの病変を更に複雑にし得る。致死的ではない場合、これらの病変合併症は病変に組み込まれ、コラーゲン及びプロテオグリカンから成る修復マトリックス組織の薄層に覆われる。細胞外マトリックス蛋白質であるコラーゲン及びプロテオグリカンの含量は被膜の形成時に動脈硬化性プラークにおいて増加する。
VI.内皮の異常、例えば、亀裂、びらん、潰瘍、血腫、血栓、出血は、組み合わされると、VI型病変と命名されるより複雑な病変型となり得る。
VII.病変は石灰化病変と称される場合が多く、病変の50%超がミネラルから成る。石灰化に加え、これらの病変は大量の修復線維結合組織を含む。ここに閉じ込められたSMCがアポトーシスを受けて分解すると、その石灰化小器官は石灰化の一部になる。
VIII.線維性病変が石灰性病変に続く。線維性病変は完全にコラーゲンから成り脂質を含み得ない(Stary H.C.)。
【0007】
心血管事象はプラークの破裂の結果である場合が多く、炎症及びプロテアーゼの放出が線維性被膜の肩領域を弱体化させ、プラークにおける脂肪質が壁在血栓を誘発する血液と接触することを許容する(Clarkson and Kaplan)。マトリックス生成の減少と併せたプロテアーゼ活性の増大による線維性被膜の菲薄化がプラーク不安定性の顕著な特徴であると考えられ、破裂のリスクを高める。プラークの不安定性及びその破裂リスクは臨床的に興味深い分野である。不安定プラーク(VP)の定義は標準化されていないが、安定プラークと比べて3つの組織学的特徴の存在を明言する一般的な見解の一致がある。
1)大きい脂質コア(総病変の40パーセント超)
2)薄い線維性被膜(65〜150マイクロメートル)
3)大量の急性炎症細胞
VPを定義するための主な基準には、活発な炎症(単球、マクロファージ及びT細胞の存在)、大きい脂質コアを有する薄い被膜、表在血小板凝集を伴う内皮の裸出、亀裂プラーク及び動脈の90%超の狭窄が含まれる。他の細かい基準には、表在石灰化結節、プラーク内出血、内皮機能障害及び外側の再構成が含まれる(Shin,Edelberg,and Hong)。
【0008】
プラーク合併症、不安定及び破裂は、医療処置及び/又は生活様式の変更によって抑制され得る。しかし、場合により、より侵襲的な方法、即ち、血管形成術又はバイパス手術が必要であり得る。
【0009】
現在、診断ツールは、依然として開発中である静止画像解析又はCVDリスクに関連する収縮期・拡張期血圧レベルのような低技術法に基づく。当該分野では高リスクにある患者をより良く同定し得る多変量解析の開発に多大な関心を向けてきた。そのようなモデルの1つがSCOREモデル(Systematic Coronary Risk Evaluationモデル)である。1994年に、そして2003年に改訂されたのであるが、欧州アテローム性動脈硬化学会(The European Atherosclerosis Society)、欧州心臓病学会(The European Society of Cardiology)及び欧州高血圧学会(The European Society of Hypertension)は、冠動脈心疾患の予防に関する一組の推奨を公表した。このガイドラインは、無症状の患者におけるCVDリスクを評価するために、即ち、無症状の高リスク患者の同定のために開発された複数の評価法に基づく。SCOREモデルは、性別、年齢、喫煙、収縮期血圧及び総コレステロール若しくはコレステロール/HDL比をリスク因子として一体化している(Graham et al.)。
【0010】
より詳細な診断を行うためにはSCOREモデルは十分ではなく、撮像法が用いられる。従って、撮像法は主に高リスク群の患者又は研究期間中に用いられる。
【0011】
<撮像法>
現在、冠動脈造影法(CAG)は狭窄の程度を判定するための判断基準となる撮像法である。CAGは二次元にて血管腔を撮像するが、血管腔のみに限定され、血管壁を撮像しないため、CAGは安定プラークを有する動脈と不安定プラークを有する動脈とを識別することができない。CAGは患者が手術(血管形成術又はバイパス手術)を必要とするかを判定するために用いられる場合が多い。血管の狭窄箇所が進行したプラークであるかを判定するためには、他の手法、即ち、血管内冠動脈超音波(IVUS)又は血管顕微法が必要である。
【0012】
IVUSはプラーク及び血管壁の二次元断面画像を提供し、血管壁及び形態及び石灰化の程度の特徴付けには良好であるが、病変における脂質を評価するには不向きな方法であると考えられている。しかし、IVUSは侵襲的であり、専門知識及び費用を必要とするため、その使用は広まっていない。血管顕微法はアテローム性動脈硬化を理解・同定するのに有用なもう1つの方法である。血管顕微法はプラーク表面の直接的可視化であり、プラークの色及び血栓を検出する能力を有する。しかし、血管顕微法は侵襲的で技術的に困難であり、これまでプラークの拡張の程度を検出することができていない。現在多くの関心を引いているもう1つの撮像法は磁気共鳴画像法(MRI)である。MRIは非侵襲的であり、脳卒中の高リスクにある頸動脈プラークを同定することができる。一方、小さいプラークサイズ及び冠動脈の位置のため、MRIは冠動脈を撮像するのに最適な手法ではない。他の撮像法、即ち、エラストグラフィ、サーモグラフィ及び光干渉断層撮影が開発中である(Schaar et al.)。
【0013】
前述の撮像法はすべて開発中であり、単独ではいずれも不安定プラークを同定することができないが、破裂前に分子事象及びプラーク代謝回転を理解するのに有用なツールである。現在、早期においてCVDを診断する唯一の機会は、問題となっている患者及びその患者の近親者の確立された冠動脈心疾患、末梢動脈疾患及び脳血管アテローム性動脈硬化症の一連のリスク因子を用いることである。
【0014】
<現在の生化学マーカー>
現在、アテローム性動脈硬化のリスク因子としていくつかの生化学マーカーが公知である。最近、血清中の生化学マーカー濃度の測定に多くの関心が向けられている。脂質、例えば、総コレステロール、低比重リポ蛋白コレステロール(LDL−C)及び高比重リポ蛋白コレステロール(HDL−C)並びに炎症マーカー、例えば、C反応性蛋白質(CRP)、インターロイキン6(IL−6)、インターロイキン18(IL−18)、腫瘍壊死因子アルファ(TNFα)、CD40、CD40リガンド(CD40L)など。
【0015】
リポ蛋白質の中で少なくとも2つの注目すべき進展があった。LDL粒子のサイズがアテローム性動脈硬化の進行を予測するように思われる。小さいLDL粒子の濃度上昇が大きい粒子の濃度上昇よりCVDリスクに関連する(Gardner,Fortmann,and Krauss)。
【0016】
HDL−Cのレベルはトリグリセリドに強く関連し、高レベルのトリグリセリドはCHDのより高いリスクと相関する。Jeppesen et al.(2003)によるコホート研究は、高TG/低HDL−CがIHD(虚血性心疾患)の最大のリスク因子であることを見出した(Jeppesen et al.)。
【0017】
脂質プロファイルはリスク因子の評価に重要であるが、プラーク代謝回転と関連する分子事象の理解及び測定を可能としない。多くの生化学マーカーがCVDのリスク因子として示唆されているが、疾患の具体的な産物ではない。これらにはCRP及び骨ナトリウム利尿ペプチド(BNP)が含まれる(表1参照)。表1はCVDの公知のマーカーの一部を概述している。
【0018】
【表1】

【0019】
従って、心血管事象のマーカーとして一連の異なる生化学マーカーが示唆されている。Wang et al(2006)は、表1に示される、Framingham研究に参加した3200人の患者における10個の異なる生化学マーカーを測定した。その結論は、10個の生化学マーカーの測定が標準的なリスク因子に加えて診断に適度に寄与するのみであるというものであった。10個の生化学マーカーの中でB型ナトリウム利尿ペプチドレベル、C反応性蛋白質レベル及び尿中アルブミン/クレアチニン比がマーカーと死亡/心血管事象との最大の相関を示した(Wang et al.)。
【0020】
<C反応性蛋白質>
C反応性蛋白質(CRP)は、異なる臨床状態、例えば、炎症、感染又は外傷に応答して肝臓によって産生される急性期血清蛋白質である(Gabay & Kushner 1999)。CRPの産生は罹患又は損傷組織から放出されるIL−6のようなサイトカインに誘発される。CRPの生理学的役割は未知であり、その炎症促進又は抗炎症作用に関する議論は進行中である。
【0021】
CRPがヒトにおけるCVDのリスク因子であるという証拠が蓄積している。Ridker et al.2002による研究では、急性心筋梗塞、脳卒中、冠血行再建又はCVDによる死亡の発生について8年間追跡した28,000人の健常女性から成る大集団において、CRPはLDLコレステロールより優れた将来の心血管事象の予測因子であることが示された。他の多くの研究もベースラインのCRPレベルが心血管事象の独立したリスク因子を構成することを報告している(Thompson et al.1995,Mendall et al.1996,Kuller et al.1996,Ridker et al.1997,Tracy et al.1997,Ridker et al.2000)。
【0022】
循環CRPはアテローム性動脈硬化プロセスにおいて生じる全般的な炎症を反映するだけであって、疾患の発病における能動的な構成成分ではないと推測されている。しかし、また、一連の証拠はCRPがアテローム発生に役割を有するという見解を支持している。第一に、CRPを生じさせる慢性感染症もCVDの増大リスクと関連する(Leinonen & Saikku 2002)。第二に、本発明者らは異なるレベルのアテローム性動脈硬化病変においてCRPを同定した(Reynolds & Vance 1987,Hatanaka et al.1995)。最後に、CRPはin vitroにてアテローム発生促進特性を有することが示されている:CRPは内皮細胞を活性化して接着分子を生成し得る(Pasceri et al.2000)。また、CRPは内皮細胞におけるeNOSの生成を減少させ(Venugopal et al.2002)、マクロファージによるLDLの取込みを増進し得る(Zwaka et al.2001)。
【0023】
<脳性ナトリウム利尿ペプチド>
脳性(B型)ナトリウム利尿ペプチド(BNP)は、心室における心筋細胞の過度の伸張に応答して心室によって分泌されるペプチドホルモンである。T−proBNP(不活性N末端断片)は活性ホルモン(BNP)とともにproBNPの切断と同時に血流に放出される。BNP及びNT−proBNPは心血管事象の潜在的生化学マーカーとして示唆されている(Wang et al.)。
【0024】
<ケモカイン>
ケモカインもCVDの潜在的マーカーである。ケモカインは炎症時に産生される低分子量サイトカインである。CVDに関連する1つの主要なケモカインは単球走化性蛋白質−1(MCP−1)である。MCP−1はアテローム性動脈硬化病変への単球の動員に初期の重要な役割を果たすように思われる。アテローム性動脈硬化のサルモデルを用いた研究では、MCP−1の血漿濃度はプラークサイズ及びプラーク合併症と高度に関連していた(Register et al.)。
【0025】
<コレステロールを含む脂質>
最近、血清中のコレステロール濃度の測定に多くの関心が向けられている(総コレステロール並びに低比重リポ蛋白コレステロール(LDL−C)及び高比重リポ蛋白コレステロール(HDL−C)の濃度)。リポ蛋白マーカーの中で少なくとも2つの注目すべき進展があった。第一に、LDL粒子サイズからアテローム性動脈硬化の進行の程度が予測されるように思われる。小LDL粒子の濃度上昇は、大粒子の濃度上昇よりCVDリスクに関連する(Gardner et al)。第二に、LDL粒子のオレイン酸コレステリル含量はCVDリスクの特に重要なマーカーになり得る。サルにおいて、オレイン酸コレステリルを有するリポ蛋白粒子コアの高濃度化は、より重度の冠動脈アテローム性硬化と強く且つ正に関連し(Rudel et al)、LDL及びHDLコレステロール濃度の寄与に相加的であった。実験動物におけるこれらの知見は、低比率のリノール酸コレステリル(逆に、高比率のオレイン酸コレステリル)を有する血漿リポ蛋白質が正常な対照と比べてCHD(冠動脈心疾患)の合併症を有する患者に典型であることを示した従前のヒト研究(Lawrie et al)によって更に支持される。
【0026】
HDL−Cのレベルはトリグリセリドと強く関連し、高レベルのトリグリセリドはCHDのより高いリスクと相関する。Jeppesen et al)によるコホート研究は、高TG/低HDL−CがIHD(虚血性心疾患)の最大のリスク因子であることを見出した。
【0027】
これらの脂質プロファイルはリスク因子の評価に重要であるが、プラーク代謝回転と関連する分子事象の理解及び測定を可能としない。多くの生化学マーカーがCVDのリスク因子として示唆されているが、これらは疾患の具体的な産物ではない。これらにはCRP及びApoEが含まれる。
【0028】
<リポ蛋白質>
CVDを予測するために最も一般的に用いられるバイオマーカーは、コレステロール濃度である(総コレステロール及びコレステロール/HDL比)。これらは他のリスク因子、例えば、血圧及びLDLレベルとともに用いられる。前述のSCOREモデルでは両方の因子が用いられる。LDLは血中にコレステロールを輸送し、酸化LDLの蓄積はアテローム性動脈硬化を促進し得るため、LDLレベルは重要である(Graham et al)。加えて、CHDとトリグリセリド(TG)レベルとの有意な関連が見出され、CHDの増大リスクはLDL−C及びHDL−Cレベルから独立して増大TGレベルと関連したが、コレステロールレベルはCVDの主要なリスク因子の1つであると見なされる(Jeppeson et al)。
【0029】
<APO−E>
アポリポ蛋白質Eはカイロミクロン、VLDL及びHDLに見出される。アポリポ蛋白質Eは主に肝臓で合成されるが、他の多くの臓器、例えば、脳、脾臓、腎臓でも合成される(Siest et al.1995)。ApoEは2つの受容体(LDL受容体及びApoE特異的カイロミクロンレムナント受容体)のリガンドとして作用することによってリポ蛋白代謝に重要な役割を果たす。ApoEとこれらの受容体との相互作用はコレステロールの代謝調節の基盤を付与する。apoE遺伝子座における遺伝子多型は3つの対立遺伝子(6つのapoE表現型を決定するε2,ε3及びε4)が大部分の集団に見出されることとなる。アイソフォームは112及び158位において1つのアミノ酸だけ互いに異なる。ApoE2は両方の残基においてシステインを有し、E4は両方の位置においてアルギニンを有する。ApoE3は112位においてシステインを、158位においてアルギニンを含む。対立遺伝子頻度は様々な集団において異なる。研究の中にはapoE多型とアテローム性動脈硬化との関連の可能性を評価したものがある。14の観察研究のメタアナリシスではε4対立遺伝子が男女の間で冠動脈疾患と関連することを示した(Wilson et al.1996)。更に、ε4対立遺伝子は頸動脈アテローム性硬化と関連している(Terry et al.1996,Cattin et al.1997,Haraki et al.2002)。
【0030】
ApoEは299アミノ酸長であり、リポ蛋白質、脂溶性ビタミン及びコレステロールをリンパ系へ、更には血液循環へ輸送する。ApoEは主に肝臓で合成される。現在、保存低比重リポ蛋白受容体遺伝子ファミリーに属するApoEの哺乳動物受容体は7つある。
【0031】
<更なる生化学マーカー>
微量アルブミン尿(Microalbuminurea)(アルブミン/クレアチニンレベル)も潜在的な独立したマーカーである。尿中アルブミン排泄率は腎臓の変化のマーカーであり、クレアチニンの微量の上昇と比べてアテローム性動脈硬化を示し得る(Wang et al.)。
【0032】
プロコラーゲンマーカーの中でIII型コラーゲン代謝回転速度のマーカー(PIIINP)は高血圧の予後マーカーとして研究され、心筋梗塞と関連している。Satta et al.は腹部大動脈瘤(AAA)と血中プロコラーゲン(PIIINP)の濃度との相関を検討した。彼らは、III型コラーゲン代謝回転がAAA患者において増加し、増進した合成、増進した分解又は両方の組合せによるものであり得ることを示した。同じ実験において、I型プロコラーゲン(PICP)のカルボキシ末端プロペプチドが測定され、動脈瘤嚢にI型コラーゲンの加速度的な合成はなかった。
【0033】
<プラークの蛋白質プロファイル>
ヒト動脈はより大きい若しくは弾性の動脈、中間若しくは筋性の動脈並びに小動脈に分けられ得る。血管壁は内弾性板及び外弾性板によって分離された内膜、中膜及び外膜から成る。内膜は結合組織、平滑筋細胞及び少数の分離マクロファージから成る。内膜の境界は内皮の管腔表面と内弾性板との間の層と定義され得る。
【0034】
動脈内膜は更に2つの層に分けられ得る。プロテオグリカン層と称される内層は豊富なプロテオグリカン、平滑筋細胞及びマクロファージから成る。下層、筋弾性層は豊富な平滑筋細胞及び弾性線維から成る。通常の状態では内膜の2つの層は光学顕微鏡によってどうにか視認可能であるが、内膜肥厚が生じると明確且つ著明である。中膜は動脈壁の筋部分であり、平滑筋細胞、エラスチン、コラーゲン原線維から成る。
【0035】
外膜、外層は高度に微小血管であり、コラーゲン、弾性線維、平滑筋細胞及びリンパ管を含む。
【0036】
ヒト動脈硬化性プラークは、線維状コラーゲン、エラスチン、プロテオグリカン及びSMCから成る線維性被膜に覆われた脂質に富むコアを特徴とする。プロテオグリカンヒアルロナンは、事象、例えば、脂質蓄積、血栓及び細胞増殖・移動を調節し、また、組織の物質特性に作用することによって病変発達に作用する可能性を有する細胞外マトリックスの主要な非線維状構成成分である(Wight 1995)。浸潤ApoE及びCRPも存在し、本発明者らは、アテローム性動脈硬化症の異なる段階において冠動脈の動脈硬化性プラークにおいて両方の局在を示した。??参照
【0037】
<ヒトにおけるApoE及びCRPの分布>
下記の表2はヒト体内のApoE及びCRPの分布を示す。
【0038】
【表2】
【0039】
下記の表3は、in vivo及び/又はin vitroにて示されるApoE及びCRPと蛋白質との公知の相互作用を示す。
【0040】
【表3】
【0041】
<ヒトにおけるコラーゲンの分布>
コラーゲンはヒトの身体に広範に分布し、即ち、ヒトの身体における蛋白質の質量の約30%はコラーゲンから成る。表4に主要なコラーゲンタイプがその主要な組織分布ととともに一覧されている。
【0042】
【表4】
【0043】
I型コラーゲンは最も豊富なコラーゲンであり、大部分の結合組織に見出される。I型コラーゲンは特に骨及び皮膚の構造に重要である。ヒトの身体におけるコラーゲンの主要含量は骨及び皮膚に分布し、主要なコラーゲン成分はI及びIII型コラーゲンである。III型コラーゲンは大動脈の主要構成成分であり、I型コラーゲンに富む組織にもより少ない量にて見出される。加えて、IV型コラーゲンは基底膜及び血管・神経周囲に見出される。V型コラーゲンの最も一般的な局在は、I及びIII型コラーゲンと関連して特徴的なコラーゲン原線維内である(Garrone et al)。
【0044】
一部のコラーゲンは限定された組織分布を有し、例えば、II型はほとんど軟骨に見出される(Mayne R.)。
【0045】
コラーゲン原線維は2つ以上のコラーゲンタイプから成る場合が多い。例えば、I型コラーゲン原線維は少量のIII,V及びXII型を含む場合が多く、一方、軟骨のII型コラーゲン原線維はIX及びXI型も含む。
【0046】
<動脈におけるコラーゲン>
動脈には6つのタイプのコラーゲンが見出され(I,III,IV,V,VI及びVIII型)、I及びIII型が最も豊富であり、コラーゲン含量の80〜90%である。I及びIII型は血管壁でも最も多い。I及びIII型は動脈壁の3つのすべての層内に異なる量にて共分布しているように思われ、I及びIII型コラーゲンの合成は内膜に位置している傾向にある(Mayne R)。
【0047】
<プラークにおけるコラーゲン及び他の構造蛋白質の代謝回転>
動脈硬化性プラークの発達の間、コラーゲンが線維性被膜に蓄積する(Stary H.C.)。Katsuda et al(1992)による研究では、I,III及びIV型コラーゲンがヒト大動脈組織における病変のすべての段階において肥厚内膜において見出された。VI型コラーゲンが内膜細胞の領域における基底膜に分布し、進展した病変では伸長SMC周囲にも検出された。I及びIII型の従前の研究はアテローム硬化性動脈壁における同等の分布のエビデンスを提供している(Shekhonin et al)。McCullagh et al(1980)によれば、III型が正常なヒト大動脈中膜において最も多いコラーゲンである(抽出可能なコラーゲンの約70%)。Eriksen et al(2006)による最近の研究は、狭窄の程度に依存するヒト大動脈弁における総コラーゲン含量の減少を見出した。狭窄の分子機構は、アテローム性動脈硬化に類似すると考えられている。健常な大動脈弁では、コラーゲン含量は主にI及びIII型である。狭窄の間、コラーゲンの総含量は減少し、それはおそらくI型コラーゲンの代謝回転の増大によって生じる。I型コラーゲンは総コラーゲンの約60〜70%を占め、一方、III型コラーゲンの割合は健常弁及び石灰化弁において30〜40%であった。
【0048】
V型コラーゲンも、進展したアテローム性動脈硬化病変において増加し、大動脈中膜及びプラークの内皮下領域において、細胞外マトリックス全体にわたって分布する(McCullagh et al)。
【0049】
動脈硬化性プラークに見出される主なコラーゲンタイプは、I及びIII型であるという見解の一致があるように思われ、それらが健常血管及びアテローム硬化性血管において同等に分布しているかは更に研究される必要がある。
【0050】
Katsuda et al(1992)による研究では、より進展した病変のアテロームの中心部にコラーゲンは検出されなかった。
【0051】
<エラスチン>
エラスチンは身体において最も安定な蛋白質の1つであり、その弾性及び弾力によって生じる大部分の結合組織に見出される。エラスチンは動脈壁の蛋白質含量を独占し、主要な細胞外マトリックス蛋白質である。
【0052】
エラスチンは弾性線維における主な構成成分であり、石灰化に関連する。血管石灰化は血管壁内の2つの異なる部位(内膜及び中膜)において生じる。内膜石灰化は主に壊死性コア内にてアテローム性動脈硬化に関連する。石灰化弾性線維はプラークが最も破裂しやすいプラーク肩部を構成し、弾性線維の石灰化がプラーク安定性に作用し得ることを示唆している(Bobryshev Y.V.)。アテローム性動脈硬化では、弾性線維の含量は脂質沈着とともに減少し、これはエラスチン分解酵素に対する感受性の亢進を生じさせる。これにより、コラーゲンとは対照的に、エラスチンの含量は、病変が発達するにつれて減少する。
【0053】
<ヒトにおけるエラスチンの分布>
表5はヒトの身体におけるエラスチンの分布を示す。
【0054】
【表5】
【0055】
表6はin vivo及び/又はin vitroにて示されるエラスチンと蛋白質との公知の相互作用を示す。
【0056】
【表6】
【0057】
<マトリックス構成成分としてのプロテオグリカン>
プロテオグリカン(PG)は、主に血管壁の細胞間マトリックスに局在する多糖蛋白質である(Salisbury and Wagner 1981)。PGは、結合領域を通じてコア蛋白質に共有結合したGAGと称される1つ以上の長い非分岐高ポリアニオン糖側鎖の存在を特徴とする高分子である。GAGの反復単位は、アミノ糖(N−アセチル−グルコサミン(GlcNAc)又はN−アセチル−ガラクトサミン(GaINAc))及びヘキスロン酸(グルクロン酸(glucouronic acid)(GlcA)又はイズロン酸(IdoA))から成る。反復単位における糖の一方又は両方は1つ以上の硫酸基を含む(Rodriguez−Lee 2007)。GAG鎖に加え、大部分のコア蛋白質はN−及び/又はO−結合オリゴ糖を保有する。
【0058】
<PGの分類及び命名>
PGは高分子の極めて異質の群である。単一タイプのコア蛋白質は結合GAG鎖の数及びタイプの点で異なり得る。鎖長及び鎖に沿った硫酸化残基の構成も異なる。
【0059】
反復二糖類単位の構造によってGAGの4つの主なクラスが識別される。コンドロイチン硫酸(CS)及びデルマタン硫酸(DS)、ヘパリン硫酸(HS)及びヘパリン、ヒアルロナン並びにケラチン硫酸(KS)。
【0060】
ヒアルロナンはGAGの最も単純なものである。他のすべてと対照的に、ヒアルロナンは硫酸化糖を含まない。その二糖類単位はすべて同一であり、その鎖長は膨大であり、ヒアルロナンはコア蛋白質に結合していない。
【0061】
KSは硫酸化ポリラクトサミン鎖である。KS−Iは元々角膜において説明され、コア蛋白質においてアスパラギン残基にN結合し、一方、KS−II又は軟骨KSはセリン又はスレオニン残基にO結合している(Funderburgh 2000)。
【0062】
PGはいくつかのパラメータによって分類され得る。
結合GAG鎖(CS/DS−又はHS含有PG)
細胞に関する組織分布(細胞外及び基底膜PG、細胞結合PG又は細胞内PG)
コア蛋白質相同性(ヒアレクタン(hyalectan)、小さなロイシンに富むPG(SLRP)
【0063】
コンドロイチン/デルマタン硫酸PG(バーシカン、アグリカン、ニューロカン及びブレビカン)はヒアルロナン結合プロテオグリカンのファミリーに属する。この遺伝子ファミリーは集合的にヒアレクタン(hyalectan)と称される。各ファミリーメンバーは特徴的な分布を有し、アグリカンは軟骨において顕著であり、ニューロカン及びブレビカンは中枢神経系において顕著であり、バーシカンは血管壁を含む様々な軟組織に存在する。バーシカンの遺伝子及び蛋白質構造はドメインテンプレートに従う。アミノ末端球状末端(G1)はGAGヒアルロナンに結合し、カルボキシ末端球状ドメイン(G3)は、2つの上皮増殖因子(EGF)ドメイン及び補体調節領域に隣接するC型レクチンから成る蛋白質のセレクチンファミリーに類似する。バーシカンコア蛋白質の中央領域は、バーシカンのCS結合領域を指定する2つの大きなエクソンにコードされる。エクソン7にコードされる領域はαGAGと称され、一方、エクソン8にコードされる領域はβGAGと称される。バーシカンの選択的スプライシングから4つのmRNA転写物が生じ、コア蛋白質の長さ及び結合GAGの数が異なるV0,V1,V2及びV3を生じさせる(Dours−Zimmermann and Zimmermann)。ヒトバーシカンにおける潜在的GAG結合部位の数は、V0では17〜23、V1では12〜15、V2では5〜8、V3では0である(Wight 617−23)。
【0064】
デコリン及びバイグリカンは、3つのクラス(I,II及びIII)及び異なるサブファミリーに分類される少なくとも9つのメンバーを含むSLRPファミリーのメンバーである。それらはすべて、強い蛋白質−蛋白質相互作用を成すロイシンに富むリピートを含む中央ドメインの強い存在を成すロイシンに富むリピートを含む中央ドメインの存在を特徴とする。デコリン及びバイグリカンはクラスIのメンバーであり、ファミリーの中で最も高い相同性を示し(約57%)、プロペプチドを有する唯一のSLRPである。プロペプチドは種にわたって高度に保存され、GAG鎖の合成に関与する最初の酵素であるキシロシルトランスフェラーゼの認識配列として機能し得る。
【0065】
バーシカン、デコリン及びバイグリカンは、哺乳動物動脈壁のマトリックスにおける主要なCS/DS PGである(Wight et al.1986)。バーシカンV0コア蛋白質のサイズは370kDaであり、36kDaのデコリン及び38kDaのバイグリカンよりほぼ10倍大きい。側鎖は広範なサイズを示すが、概して各々平均約40〜90kDaである。
【0066】
ヘパラン硫酸プロテオグリカン(HSPG)は、細胞結合及び細胞周囲PGの5つの異なるクラスに分類され、哺乳動物細胞表面、基底膜及びECMのHSの少なくとも95%を占める。細胞結合HSPGは膜内在性シンデカン及び固着グリピカンを含む。細胞周囲HSPGは主にパールカン、アグリンを含む。これらのPGは、インテグリンを介した細胞膜との密接な関連のために細胞周囲型と称される(Whitelock and Iozzo)。
【0067】
パールカンは、ほぼすべての基底膜並びに間葉系臓器及び結合組織に発現するモジュラーHSPGであり、脊椎及び無脊椎動物に見出される最大の単鎖ポリペプチドの1つである。パールカンの5つのモジュール及びそのHS側鎖は、例えば、線維芽細胞増殖因子2、血管内皮増殖因子(VEGF)、血小板由来増殖因子(PDGF)及び他のマトリックス蛋白質との多数の分子相互作用に関与する。ヒトパールカンのコア蛋白質は約470kDaであり、多くのO結合オリゴ糖及び4本のHS側鎖とともに800kDaを超える分子量に達し得る(Knox and Whitelock)。
【0068】
<プロテオグリカンの分布>
プロテオグリカン(PG)はヒトの身体のほぼあらゆる部位に分布する高分子である。PGの構造及びサイズは極めて異なる。すべてのPGの基本構造はコア蛋白質及び少なくとも1つであるが、多くの場合、多くの炭水化物鎖、すなわちグリコサミノグリカン(GAG)を含む。PGは細胞内、細胞表面及び細胞外マトリックスに見出され得る。PGの構造多様性は多くの生物学的機能を示唆するものであり、表7を参照されたい。
【0069】
【表7】
【0070】
<動脈におけるプロテオグリカン>
少なくとも5つのタイプのPGが動脈壁の細胞外マトリックスに存在する。ヒアルロナンと相互作用して大凝集体を形成するバーシカン;コラーゲン及びエラスチンのような原線維マトリックス構成成分と相互作用する、小さいロイシンに富むデコリン及びバイグリカン;基底層の構成成分であるヘパラン硫酸−パールカン並びにケラチン硫酸−ルミカン(Talusan et al.)。
【0071】
バーシカンはアテローム性動脈硬化病変に蓄積するいくつかのECM分子の1つである。多くの研究が、バーシカンが明らかにLDLに結合することができることを示すが、概して、バーシカンは壊死性コアの脂質に富む中央部には検出されない(Evanko et al.)。
【0072】
ルミカンはマクロファージに直接結合し、且つマクロファージの移動を増進することが示されている。従って、ルミカンは血管内膜におけるマクロファージの挙動に影響を及ぼすとともに、進展したアテローム性動脈硬化病変の特徴を示す壊死性コアの形成を刺激し得る(Funderburgh et al.1997)。
【0073】
バイグリカンは線維性被膜に見出される。バーシカン及びバイグリカンはLDLに対する親和性を有し、不溶性複合体を形成し、これはLDLの酸化を促進する。バイグリカンはリポ蛋白質を動脈壁に捕捉することによってアテローム性動脈硬化の発病に寄与し得る。動脈内膜のプロテオグリカン代謝の変化はアテローム性動脈硬化の初期病変を構成し、プロテオグリカンの蓄積はアテローム性動脈硬化の進行に基本的な役割を果たす(Kunz J.)。
【0074】
パールカンは、質量分析ベースの分析及び免疫組織化学によって内膜の細胞外マトリックスの中心成分の1つとしてヒト内膜過形成にて報告された。
【0075】
表8は、正常及びアテローム性動脈硬化血管におけるPGの免疫組織化学的染色でのいくつかのPGの分布を示す(Evanko et al)。
【0076】
【表8】
【0077】
<マトリックス再構築におけるプロテオグリカンの関与>
非ヒト霊長類におけるアテローム性動脈硬化の進行の研究は、特定のPGの蓄積が病変の重症度及び細胞・増殖因子の分布により異なることを示し、アテローム性動脈硬化の進行時に異なるPGが異なる役割を果たすことを示唆した。異なるレベルの特定のPGは、線維性マトリックス構成成分、例えば、エラスチン及びコラーゲンの構造配置の変化への寄与によって組織の物質特性に直接作用し得る。
【0078】
バーシカン及びヒアルロナンは、マトリックスにおける同様の局在を示し、アテローム発生におけるその2つの間の凝集体形成を示唆する。早期病変におけるバーシカン及びヒアルロナンの著明な増加は、早期アテローム性動脈硬化病変、例えば、SMC及び白血球の増殖・移動に役割を果たすことを示唆し得る。更に、バーシカン及びヒアルロナンはヒト再狭窄病変の主要なマトリックス構成成分であり、in vitroにて血管損傷後の新たな内膜肥厚に寄与することが示されている。また、早期アテローム発生における大量のバーシカンは、バーシカンのコンドロイチン硫酸鎖へのリポ蛋白質の結合により、細胞外マトリックスに脂質の捕捉を増大させやすくし得る。この見解は、移植動脈疾患におけるバーシカンとアポ蛋白質(a)及びアポリポ蛋白質Eとの共局在によって支持される(Evanko et al)。プラークからのバーシカンの喪失はマトリックス不安定をもたらし得る。
【0079】
このことは、血管損傷後に認められるバーシカン遺伝子のアップレギュレーションから更に明らかである。この場合、バーシカンは、アテローム性動脈硬化のすべての段階、すなわち、早期の発達プラークの内膜のみならず、進展した病変にわたり、脂質で満たされた壊死性コアの境界及びプラーク−血栓界面にても同定された(Wight and Merrilees 2005)。これらの所見は脂質の蓄積、炎症及び血栓にバーシカンが関与していると見なす。更に、バーシカンはECMのアセンブリ及び弾性線維原線維形成の調節に重要な役割を果たし、これは血管疾患時のECM再構築において基本的に重要である(Wight and Merrilees 2005)。
【0080】
動脈細胞生物学におけるバイグリカンの役割は不明瞭である。一部の免疫組織化学的研究は、ヒト再狭窄病変においてコラーゲンI及びIII染色とのバイグリカンの関連を示している(Evanko et al.)。
【0081】
マトリックス蛋白質としてのバイグリカンの重要性は、バイグリカン遺伝子のヌル変異に対してホモ接合性のBALB/cAマウスの作製によって更に述べられ、この場合、バイグリカン欠損雄マウスの50%が大動脈破裂の結果として寿命の最初の3カ月以内に突然死した。この所見は、大動脈壁の構造・機能的一体性にバイグリカンが不可欠であること、また、ヒトにおける大動脈解離・破裂の発病におけるバイグリカン遺伝子欠損の潜在的役割を示唆する(Heegaard et al.2007)。
【0082】
他の研究は、バイグリカンが霊長類動脈においてエラスチンと関連する主要なPGであることを示しており、これらの所見はヒト冠動脈疾患の所見に類似している(Evanko et al)。
【0083】
デコリンはコラーゲンに結合し、且つコラーゲン原線維形成を調節することが示されている(Brown and Vogel)(Danielson et al.)。
【0084】
<プロテアーゼのプロファイル>
プロテアーゼはペプチド結合を加水分解し、アテロームにおけるコラーゲン、プロテオグリカン及びエラスチンのような細胞外マトリックス蛋白質の分解に関与する。表9を参照されたい。動脈硬化性プラークには3つの主なタイプ(メタロプロテイナーゼ(即ち、MMP)、セリンプロテアーゼ及びシステインプロテアーゼ(即ち、カテプシン))が見出される。カテプシン及びMMPは、すべての細胞外マトリックス蛋白質の分解に関与する。プラーク安定性にはマトリックスが不可欠であるため、プロテアーゼによる線維性被膜からのその除去はプラーク破裂を引き起こし得る(Stary H.C)。
【0085】
表9に動脈硬化性プラークに見出される種々のプロテアーゼが一覧にされている。
【0086】
【表9】
【0087】
プラークにおけるMMP発現の主要源はマクロファージ及びSMC活性に関連する疑いがある。プラークにおけるマクロファージは豊富なMMP−1,−8,−9及び−13を含み、in situにてコラーゲン及びプロテオグリカン分解部位と共局在する(Kunz J.)。更に、自身のデータは動脈硬化性プラークにおけるMMP−8及びカテプシンKの局在を示唆する。
【0088】
<マトリックスメタロプロテアーゼ(MMP)>
MMPは、ECMの大部分の構成成分を分解することができるエンドペプチダーゼの大きな一群である。現在、25超のMMPが同定されている。メタロプロテアーゼは、金属原子、典型的には亜鉛を含む活性部位を特徴とし、チモーゲンとして分泌される。特定の組織阻害物質TIMPがMMPの活性を調節する。様々なMMPが動脈硬化性プラークに見出される。それらはほとんどの場合、SMC及びマクロファージにおいてプラーク肩部内にて線維性被膜に接するマクロファージにあり、線維性被膜内に同定されることは稀である(Kunz J.)。]
【0089】
MMPはその基質特異性によって異なる群に分類される。I,II,III及びV型コラーゲンのような線維状コラーゲン、更にプロテオグリカンも分解するコラゲナーゼ;プロテオグリカン、IV,V,VII型コラーゲン及びエラスチンを分解するゼラチナーゼ;プロテオグリカン及びエラスチンに対して活性を示すストロメライシン(Rouis M)。これらの3つのサブグループは動脈硬化性プラークにおけるマトリックス再構築に関して特に興味深い。
【0090】
<ゼラチナーゼ>
不溶性エラスチンは、ともにMMPのゼラチナーゼファミリーに属するMMP−2及び−9によって消化される。MMP−9は、動脈硬化性プラークのサイズ及び組成に作用する重要な役割を有する。不安定なヒト動脈硬化性プラーク及びプラークの脆弱領域では、MMP−9のより高い発現及び濃度が認められている。更に、MMP−9は、安定狭心症患者と比べて不安定狭心症患者においてより多く、冠動脈プラークにおいて細胞内に見出される(活発な合成を示す)。血中MMP−9濃度は、冠状動脈アテローム性動脈硬化と関連して上昇し、有害な心血管事象を予測する(Sundstrom and Vasan)。Kuzuya et al(2006)による最近の研究は、MMP−2が線維性被膜におけるSMCの蓄積に関与し、これにより、プラーク不安定性を誘発することを示す。
【0091】
<ストロメライシン>
MMP−3はストロメライシンプロテアーゼに属し、エラスチン及びプロテオグリカンを分解することができる。Yamada et al(2002)による研究は、MMP−3が女性における心筋梗塞の遺伝的リスクを予測する信頼できる手段であることが判明し得ることを示す。
【0092】
<コラゲナーゼ>
MMP−1,−8及び−13はすべて動脈硬化性プラークにおいて同定され、プロテオグリカン及びI,III型コラーゲンを分解する。
【0093】
MMP−1,−8及び−13はコラゲナーゼであり、コラーゲンを2つの断片に切断し、その断片はMMP−2,−3又は−9によって更に分解される。
【0094】
MMP−8は、ヒトアテロームには一般的に見出されないが動脈硬化性プラークにおいて同定されている好中球によって発現される。MMP−8は、I型コラーゲンを優先的に選択し(Herman et al)、MMP−1及び13より3倍大きいI型コラーゲンの分解活性を有するため、線維性被膜の分解に部分的に関与し得る。このことはTuru et al(2006)によって支持され、この研究では、血漿中のMMP−8含量は安定プラークを有する患者より不安定プラークを有する患者で著しく多い。
【0095】
MMP−13はバイグリカンに対する高特異性を有し、SLRPを切断することが報告されている。特定の切断部位(...G177/V178)におけるMMP−13によるバイグリカンの分解はMonfort et al.(2005)によって従前に示され、骨関節炎における軟骨分解の早期検出に重要な役割を果たすことが提案されている。
【0096】
<カテプシン>
ヒトシステインカテプシンは、カテプシンB,K,L及びSを含む11のメンバーから成り、主に細胞のエンドソーム/リソソームコンパートメント内に発現する。カテプシンはプロテオグリカン、コラーゲン及びエラスチンの加水分解を触媒することができる。
【0097】
腹部大動脈瘤(AAA)において、正常大動脈と比べて高レベルのカテプシンS,K及びLが見出された。正常ヒト血管SMCは免疫染色により検出可能なカテプシンKを含まないが、動脈硬化性プラーク内の細胞は明らかに陽性である。カテプシンKは、破裂しやすい領域、例えば、線維性被膜、プラーク肩部及びプラーク破裂の実際の部位に局在する(Chapman et al)。カテプシンSは動脈硬化性プラークにおいてエラスチン分解の増大領域と共局在することが見出され、アテローム性動脈硬化の減少がカテプシンS及びK欠損マウスにおいて認められる(Liu et al)。
【0098】
カテプシンL及びKは、いくつかのプロテオグリカン及びI,II型コラーゲンを分解し、カテプシンKは共有結合的に架橋された三重らせん内にて分解し、一方、カテプシンLは非らせんテロペプチド領域においてのみ切断する。カテプシンKは線維性被膜及びプラーク肩部に局在する。正常動脈におけるカテプシンKの発現は極めて低い。早期ヒトアテローム性動脈硬化病変は内膜及び中膜SMCにおけるカテプシンKの発現を示した。進展した動脈硬化性プラークにおいて、カテプシンKは主に線維性被膜のマクロファージ及びSMCに局在した(Lutgens et al)。カテプシンK蛋白質レベルは、正常動脈と比べてアテローム性動脈硬化病変において上昇したが、一方、カテプシンKのmRNAレベルは、アテローム性動脈硬化血管及び正常動脈において同様であった。更に、カテプシンKのmRNA及び蛋白質レベルは、早期アテローム性動脈硬化病変及び血栓を含む病変と比べて、進展しているが安定したヒト動脈硬化性プラークにおいて最大であることが示された(Chapman et al)。
【0099】
カテプシンSは、早期ヒトアテローム性動脈硬化病変及び脂肪線条において内膜及び中膜SMCにほんのわずかに発現する。進展したヒト動脈硬化性プラークにおいて、カテプシンSは線維性被膜のマクロファージ及びSMCに局在した。血管自体及びプラーク微小血管の管腔の内側を覆うECも、カテプシンSを発現した。更に、カテプシンSのmRNA及び蛋白質レベルは、正常動脈と比べてヒトアテロームにおいて増大した(Lutgens et al)。カテプシンSはプロテオグリカン、エラスチン及びコラーゲンを分解することができる(Liu et al)。
【発明の概要】
【0100】
現在、CVDリスクの決定は、アテローム性動脈硬化進行の後期(線維性プラーク破裂の有意なリスクがある時点)において行われている。早期及び後期におけるアテローム性動脈硬化又はCVDリスクに関する情報を提供する診断又は予後アッセイの必要性がある。Katsuda et al(1992)の知見は、進展病変からのコラーゲンの除去の酵素機構があることを示唆し、実際に動脈硬化におけるネオエピトープの主要な役割を示唆している。
【0101】
本発明は、プロテイナーゼによる動脈硬化性プラークの蛋白質の切断によって形成されるネオエピトープを含むペプチド断片の定量のためのバイオアッセイの方法を提供し、該方法は、該ペプチド断片を含む試料を、該ネオエピトープに対する特異的結合親和性を有する免疫学的結合パートナーと接触させるステップと、該試料中の該免疫学的結合パートナーのペプチド断片への結合レベルを定量するステップとを含む。
【0102】
該アッセイの結果は、特定の患者における動脈硬化性プラークの破裂のリスクの程度又は患者の動脈硬化性プラークの不安定状態を示す指標を生成し得る。
【0103】
閾値を越える該指標の値を有する患者は、プラーク撮像法(上述の撮像法を含む)による更なる検査又はアテローム性動脈硬化の処置のための薬剤の処方又はアテローム性動脈硬化の外科的処置を推奨され得、そのようなフォローアップ検査又は処置は本発明の方法の一部を成し得る。
【0104】
動脈硬化性プラークの蛋白質は、ルミカン、バーシカン、パールカン、デコリン、バイグリカン、III型コラーゲン、CRP、ApoE及びエラスチンを含む。I型コラーゲンは動脈硬化性プラークの蛋白質であるとは見なされない。動脈硬化性プラークに存在し、身体のどの部位よりも高い程度に動脈硬化性プラークにおいてプロテアーゼに晒される蛋白質が特に興味深い。
【0105】
該免疫学的結合パートナーは、C末端ネオエピトープ又はN末端ネオエピトープを含むペプチド断片に対する特異的結合親和性を有し得る。
【0106】
プロテオグリカンアッセイ
該ペプチド断片は、プロテオグリカンバーシカン(配列番号1)、ルミカン(配列番号2)、パールカン(配列番号3)、バイグリカン(配列番号4)及びデコリン(配列番号5)の断片でよく、それらはすべて正常及びアテローム性動脈硬化血管において同定される。プロテオグリカンは、エラスチン及びコラーゲンとともに、動脈硬化性プラーク及びプラーク被膜を構成する主な蛋白質の一部である。プロテオグリカンの含量は、アテローム性動脈硬化の進行時に異なり、その結果、プロテオグリカンの潜在的ネオエピトープは病期及び疾患進行の良好なマーカーになる。特に、バーシカン及びルミカンは多くの他の臓器には豊富ではないため、これにより、それらはより特異的な生化学マーカー候補になる。
【0107】
動脈硬化性プラークにおいて多くの異なるプロテアーゼが文献により報告されているように、複数の候補プロテアーゼがプラークにおけるプロテオグリカンの消化に関与し得る。おそらく、これは最終的にプラーク破裂に至る広範の複雑なプロセスの結果である。しかし、本発明者らの評価では、初期相は一定範囲のMMPから成り得、一方、後期はマトリックスのカテプシン分解により依存し得、病期に依存して異なるネオエピトーププロファイルとなる。本発明者らは、表4に一覧表示される酵素がルミカン、バーシカン、バイグリカン、パールカン及びデコリンを生じさせ、少なくとも以下の切断生成物を生じさせると判定した。
【0108】
【表10A】
【表10B】
【0109】
従って、本発明の方法では、好ましくは、該ペプチド断片は、上記部分配列のいずれの配列にもおける記号「*」を付された部位におけるプロテアーゼによるバーシカン、ルミカン、パールカン、デコリン又はバイグリカンの切断によって形成されるネオエピトープを含む。
【0110】
更に、本発明の方法では、好ましくは、該ペプチド断片は、バーシカン、ルミカン、デコリン、パールカン及びバイグリカンの以下の部分配列のいずれの配列にもおける部位における1つ(又はそれ以上)のプロテアーゼによるプロテオグリカン(バーシカン、ルミカン、パールカン、デコリン及びバイグリカン)の切断によって形成されるネオエピトープを含み、或いは免疫学的結合パートナーは以下の配列の1つと特異的に反応する。
【0111】
【表11】
【0112】
好ましくは、該免疫学的結合パートナーは、切断されていない状態のバーシカン、ルミカン、デコリン、パールカン及びバイグリカンと反応しない。好ましくは、該免疫学的結合パートナーは、生成断片のそれぞれのc末端及びN末端を過ぎて延長している場合、上記一覧の該配列と反応しない。
【0113】
免疫学的結合パートナーは、バーシカン、ルミカン、デコリン、パールカン及びバイグリカンタイプの切断によって形成されるC末端又はN末端ネオエピトープと特異的に反応する免疫学的結合パートナーでもよい。
【0114】
従って、好適な免疫学的結合パートナーは、ペプチドのN末端において表11における以下の配列のいずれかの又はペプチドのC末端において表12における以下の配列と特異的に反応し得る。
【0115】
【表12】
【0116】
【表13】
【0117】
同様にアッセイされ得るネオエピトープを決定する更なる切断部位は、プロテオグリカン又は他の動脈硬化性プラーク蛋白質を本明細書で述べられる任意の酵素と反応させ、そうして生成されるペプチドを単離して配列決定することによって同定され得る。
【0118】
<コラーゲンアッセイ>
該ペプチド断片はIII型コラーゲンの(配列番号153)、好ましくは、成熟III型コラーゲンの、即ち、III型コラーゲンプロペプチドの断片ではない断片でもよい。動脈硬化性プラークにおける主な蛋白質はI,III型コラーゲン及びエラスチンであり、一方、プロテオグリカンはプラークのマトリックスにわずかに寄与するのみである。動脈硬化性プラークに見出される3つの主要な蛋白質中、I及びIII型コラーゲンが優位であり、一方、エラスチンは動脈における蛋白質プロファイルを占めるが、プラークにおける主要な蛋白質成分ではない。I型コラーゲンはヒトの身体全体にわたって豊富であり、一方、III型はより限定された組織位置を有し、これにより、本発明者らの見解では生化学マーカーとしてより特異的な候補を構成する。
【0119】
動脈硬化性プラークにおける多くの異なるプロテアーゼが文献により報告されているように、複数の候補プロテアーゼがプラークにおけるコラーゲンの消化に寄与し得る。おそらく、これは、最終的にプラーク破裂に至る広範の複雑なプロセスの結果である。しかし、本発明者らの評価では、初期相は一定範囲のMMPから成り得、一方、後期はマトリックスのカテプシンK分解により依存し得、疾患レベルに依存して異なるネオエピトーププロファイルとなる。本発明者らは、以下の表に一覧表示される酵素が少なくとも以下の切断部位においてIII型コラーゲンを切断すると判定した。
【0120】
【表14A】
【表14B】
【表14C】
【表14D】
【表14E】
【0121】
従って、本発明の方法では、好ましくは、該ペプチド断片は、III型コラーゲンの上記部分配列のいずれの配列にもおける記号「*」を付された部位におけるプロテアーゼによるIII型コラーゲンの切断によって形成されるネオエピトープを含む。
【0122】
更に、本発明の方法では、好ましくは、該ペプチド断片は、*間におけるIII型コラーゲンの上記部分配列のいずれの配列にもおける部位における1つ(又はそれ以上)のプロテアーゼによるIII型コラーゲンの切断によって形成されるネオエピトープを含み、または免疫学的結合パートナーは上記表のエントリーにおける*間に伸長する配列と特異的に反応する。
【0123】
好ましくは、該免疫学的結合パートナーは、切断されていない状態のIII型コラーゲンと反応しない。好ましくは、該免疫学的結合パートナーは、それぞれの切断部位を過ぎて延長している場合、上記一覧の該配列と反応しない。
【0124】
免疫学的結合パートナーは、III型コラーゲンの切断によって形成されるC末端又はN末端ネオエピトープと特異的に反応する免疫学的結合パートナーでよい。
【0125】
従って、好適な免疫学的結合パートナーは、ペプチドのN末端において以下の配列(各配列の後に配列番号)又はペプチドのC末端において以下の配列と特異的に反応し得る。
【0126】
【表15】
【0127】
【表16】
【0128】
同様にアッセイされ得るネオエピトープを決定する更なる切断部位は、III型コラーゲン又は他の動脈硬化性プラーク蛋白質を本明細書で述べられる任意の酵素と反応させ、そうして生成されるペプチドを単離して配列決定することによって同定され得る。
【0129】
<CRP及びApoEアッセイ>
該ペプチド断片はCRP(配列番号658)又はApoE(配列番号659)の断片でもよい。好ましくは、ApoEでは、選択断片はApoEのすべての同定アイソタイプ、ε2,ε3及びε4において生じる。
【0130】
CRP及びApoEがヒトの身体全体にわたって豊富であるとしても、アテローム性動脈硬化組織におけるその局在により、それらは局所プロテアーゼの作用に晒される。これにより、これらの分子は生化学マーカーとして良好且つ特異的な候補である。
【0131】
動脈硬化性プラークにおける多くの異なるプロテアーゼが文献により報告されているように、複数の候補プロテアーゼがプラークにおけるCRP及びApoEの消化に寄与し得る。おそらく、これは、最終的にプラーク破裂に至る広範の複雑なプロセスの結果である。しかし、初期相は一定範囲のMMPから成り得、一方、後期はマトリックスのカテプシンK分解により依存し得、疾患レベルに依存して異なるネオエピトーププロファイルとなる。本発明者らは、純粋天然蛋白質の一定範囲のin vitro切断を通じて、以下の表に一覧表示される酵素が少なくとも以下の切断部位(*を付す)においてCRP及びApoEを切断すると判定した。
【0132】
【表17】
【0133】
従って、本発明の方法では、好ましくは、該ペプチド断片は、CRP及びApoEの以下の部分配列のいずれの配列にもおける記号「*」が付された部位におけるプロテアーゼによるCRP及びApoEの切断によって形成されるネオエピトープを含み、または免疫学的結合パートナーは以下の配列の1つにおける*間に画定される配列と特異的に反応する。
【0134】
【表18】
【0135】
従って、好適な免疫学的結合パートナーは、ペプチドのN末端において以下の配列又はペプチドのC末端において以下の配列と特異的に反応し得る。
【0136】
【表19】
【0137】
【表20】
【0138】
同様にアッセイされ得るネオエピトープを決定する更なる切断部位は、CRP及びApoEまたは他の動脈硬化性プラーク蛋白質を本明細書で述べられる任意の酵素と反応させ、そうして生成されるペプチドを単離して配列決定することによって同定され得る。
【0139】
<エラスチンアッセイ>
該ペプチドはエラスチン(配列番号735)の断片でもよい。エラスチンはヒトの身体全体にわたって豊富であるとしても、アテローム性動脈硬化組織におけるその局在はそれを局所プロテアーゼの作用に晒させ、そのためにこれらの分子は動脈硬化性プラーク代謝回転の生化学マーカーとして良好且つ特異的な候補である。
【0140】
動脈硬化性プラークにおける多くの異なるプロテアーゼが文献により報告されているように、複数の候補プロテアーゼがプラークにおけるエラスチンの消化に寄与し得る。おそらく、これは、最終的にプラーク破裂に至る広範の複雑なプロセスの結果である。しかし、初期相は一定範囲のMMPから成り得、一方、後期はマトリックスのカテプシンK分解により依存し得、疾患レベルに依存して異なるネオエピトーププロファイルとなる。本発明者らは、純粋天然蛋白質の一定範囲のin vitro切断を通じて、以下の表に一覧表示される酵素が少なくとも以下の切断部位(*を付す)においてエラスチンを切断すると判定した。
【0141】
【表21】
【0142】
従って、本発明の方法では、好ましくは、該ペプチド断片は、エラスチンの以下の部分配列のいずれの配列にもおける記号「*」が付された部位におけるプロテアーゼによるエラスチンの切断によって形成されるネオエピトープを含み、或いは免疫学的結合パートナーは以下の配列の1つにおける*間に画定される配列と特異的に反応する。
【0143】
【表22】
【0144】
従って、好適な免疫学的結合パートナーは、ペプチドのN末端において以下の配列又はペプチドのC末端において以下の配列と特異的に反応し得る。
【0145】
【表23】
【0146】
【表24】
【0147】
同様にアッセイされ得るネオエピトープを決定する更なる切断部位は、エラスチン又は他の動脈硬化性プラーク蛋白質を本明細書で述べられる任意の酵素と反応させ、そうして生成されるペプチドを単離して配列決定することによって同定され得る。
【0148】
上述の2つ以上のペプチドのアッセイは別個に行われ、それらの結果は組み合わせられ得、又は上述の2つ以上のペプチドはともに測定され得る。
【0149】
本発明によるアッセイの結果は、診断又は予後値の総合指数を形成するために1つ以上の他の測定バイオマーカーと組み合わせられ得る。
【0150】
本明細書で用いられる「免疫学的結合パートナー」という用語は、ポリクローナル及びモノクローナル抗体、更に抗体の特異的な結合断片、例えば、Fab又はF(ab’)2も含む。従って、該免疫学的結合パートナーはモノクローナル抗体又は特異的結合親和性を有するモノクローナル抗体の断片でよい。
【0151】
本明細書で用いられる「蛋白質」という用語は、リポ蛋白質及びプロテオグリカン並びに他の蛋白(または非蛋白)の天然複合体を含む。
【0152】
一般的に、本発明に従って不均一及び均一フォーマット、サンドイッチアッセイ、競合アッセイ、酵素結合アッセイ、放射免疫アッセイなどを含むあらゆる既知のイムノアッセイフォーマットが用いられ得る。従って、任意に、該方法は、該免疫学的結合パートナー及び競合物質が該試料の存在下にてインキュベートされ、競合物質が免疫学的結合パートナーと結合するために試料中のペプチド断片と競合する競合イムノアッセイとして行われる。
【0153】
該競合物質は、該ネオエピトープを示すためにネオエピトープが属する蛋白質の切断によって形成される合成ペプチド又は精製天然ペプチドでもよい。従って、ペプチドは、バーシカン、ルミカン、パールカン、デコリン、バイグリカン、III型コラーゲン、ApoE、CRP又はエラスチンに由来し得る。
【0154】
1つの好適な方法は、任意のこれらの蛋白質の断片のネオエピトープ若しくは動脈硬化性プラークに由来する他の蛋白質由来のペプチド断片上のネオエピトープに結合するモノクローナル抗体、又はその抗体結合断片を用いた競合イムノアッセイでよい。マイクロタイタープレートの固体表面上に被覆された、適切に選択された合成ペプチドは、モノクローナル抗体又は結合断片への結合について試料と競合し得る。或いは、モノクローナル抗体又は結合断片によって認識されるネオエピトープを保有する1つ以上のこれらの蛋白質由来の精製天然断片が、固体表面上に用いられ得る。更に別の代替例は、モノクローナル抗体又は結合断片を固体表面上に固定化し、次に、試料を、シグナル分子、例えば、西洋ワサビペルオキシダーゼ又はビオチンに適切に結合した合成ペプチドと共インキュベートすることである。試料は尿、血清、血液、血漿又は他の、例えば、動脈硬化性プラーク生検の試料でもよい。
【0155】
一部の好ましい方法では、試料は患者由来の試料であり、方法は、該ペプチド断片の該結合の定量レベルを、(a)比較できる健常患者及び/又は(b)アテローム性動脈硬化病態の特徴を示す数値と比較するステップと、任意に、高レベルの測定ペプチド(通常、高レベルの結合によって示される)をより重症度の該病態と関連付けるステップとを更に含む。
【0156】
本発明の一態様は、上述のネオエピトープを認識するモノクローナル抗体の開発に関する。これは、当該蛋白質分子のアミノ酸配列に由来する合成ペプチド(上記一覧表示された配列又はそこで終止する配列を含む)でマウスを免疫化し、選択マウス由来脾臓細胞を骨髄腫細胞に融合し、関連合成ペプチド上のネオエピトープへの結合についてモノクローナル抗体を試験することによって達成され得る。ネオエピトープに対する特異性は、合成ペプチドとの反応性及び(C末端ネオエピトープに対する)C末端延長形態の免疫ペプチド若しくは(N末端ネオエピトープに対する)N末端延長形態の免疫ペプチドとの反応性の欠如を必要とすることによって確実にされ得る。天然蛋白質に対する結合能の欠如を確定するために、ネオエピトープに対する抗体も評価され得る。或いは、ネオエピトープに対する特異性は、抗体の反応性が末端アミノ酸の1つに共有結合したビオチン又は他の官能基の存在が負に依存するよう求めることによって確実にされ得る。
【0157】
本発明は、上記に示される部分配列のいずれか1つの部分配列における末端部位におけるプロテアーゼによる該蛋白質の切断によって形成されるネオエピトープと特異的に免疫反応する免疫学的結合パートナーを含み、例えば、モノクローナル抗体又はその結合断片でもよい。
【0158】
本発明は、上記に示される部分配列のいずれか1つの部分配列における配列の末端部位における動脈硬化性プラーク蛋白質の切断によって形成されるC末端又はN末端ネオエピトープに対するモノクローナル抗体を産生する細胞株を含む。
【0159】
本発明は、上記に示されるこれらの蛋白質の部分配列のいずれか1つの部分配列における該蛋白質の切断によって形成されるC末端又はN末端ネオエピトープを含むペプチドを更に提供する。そのようなペプチドはイムノアッセイで用いるために、該ペプチドに対する免疫反応を生じさせるためにハプテンとして担体にコンジュゲートされ、又は固体表面に固定化され、又は検出可能なマーカーにコンジュゲートされ得る。
【0160】
本発明は、上記に示される部分配列のいずれか1つの部分配列における該蛋白質の切断によって形成されるC末端又はN末端ネオエピトープを含むペプチドをコードする単離核酸分子を更に含む。
【0161】
本発明は、発現シグナルと、上記に示される部分配列のいずれか1つの部分配列における該蛋白質の切断によって形成されるC末端又はN末端ネオエピトープを含むペプチドの発現をコードするコード配列とを含む核酸配列を含むベクターを更に含み、そのようなベクターで形質転換され、該ペプチドを発現する宿主細胞を更に含む。
【0162】
本発明の更に別の態様は、上述の方法を実施するために好都合に用いられ得るキットに関する。そのようなキットは、(1)合成ペプチドで被覆されたマイクロタイタープレートと、(2)該合成ペプチドと反応する本発明のモノクローナル抗体若しくは抗体結合断片と、(3)標識抗マウスIgG免疫グロブリンとを含み得る。或いは、そのようなキットは、(1)精製天然蛋白質断片で被覆されたマイクロタイタープレートと、(2)該蛋白質のいずれか1つの断片上のネオエピトープを認識し、該精製断片と反応するモノクローナル抗体と、(3)標識抗マウスIgG免疫グロブリンとを含み得る。或いは、そのようなキットは、(1)ストレプトアビジンで被覆されたマイクロタイタープレートと、(2)ビオチンに結合した合成ペプチドと、(3)該蛋白質断片上のネオエピトープを認識し、該合成ペプチドと反応するモノクローナル抗体と、(4)標識抗マウスIgG免疫グロブリンとを含み得る。更に別の代替例は、(1)ストレプトアビジンで被覆されたマイクロタイタープレートと、(2)ビオチンに結合した合成ペプチドと、(3)該蛋白質断片上のネオエピトープを認識し(該合成ペプチドと反応し)、西洋ワサビペルオキシダーゼにコンジュゲートしたモノクローナル抗体とを含むキットでよい。
【0163】
従って、本発明は、本明細書で述べられる免疫学的結合パートナー並びに該免疫学的結合パートナーに結合する競合物質、並びに、任意に、1つ以上の洗浄試薬、緩衝液、停止試薬、酵素標識、標識酵素のための基質、較正基準、抗マウス抗体及び該イムノアッセイを行うための使用説明書を含むイムノアッセイキットを含む。
【0164】
本明細書で述べられるアッセイは患者におけるアテローム性動脈硬化症の診断に有用である。加えて、検査は疾患の進行の評価及び治療に対する応答のモニタリングに有用である。本発明の免疫学的結合パートナーは、本明細書で述べられる動脈硬化性プラーク蛋白質の切断産物の存在又は位置を示すために免疫染色でも用いられ得る。
【0165】
本発明は、添付図面を参照して更に説明され、例示される。
【図面の簡単な説明】
【0166】
【図1】III型病変を含む大動脈試料に対してモノクローナルマウス抗体を用いたバイグリカン染色(それぞれ2,4,4及び10倍)を示す図である。
【図2】III型病変を含む大動脈試料に対してモノクローナルマウス抗体を用いたカテプシンK染色(それぞれ2,4,10及び10倍)を示す図である。
【図3】V型病変を含む大動脈試料に対してモノクローナルマウス抗体を用いたバイグリカン染色(それぞれ2,4,10及び10倍)を示す図である。
【図4】V型病変を含む大動脈試料に対してモノクローナルマウス抗体を用いたカテプシンK染色(それぞれ2,4,10及び10倍)を示す図である。
【図5】プロテアーゼ:MMP2、MMP3、MMP8、カテプシンK、カテプシンS、カテプシンB及びカテプシンLによって生じるバイグリカンの切断産物を示す図である。M=着色マーカー(Rainbow marker)、−enz=酵素消化なし、実施例2においてゲル上で泳動。
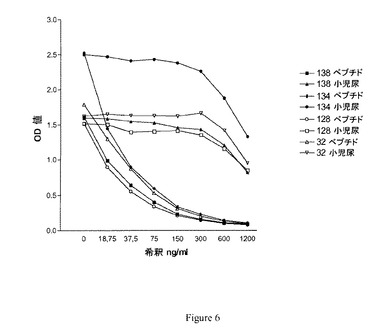
【図6】実施例4において得られた競合試験結果を示す図である。
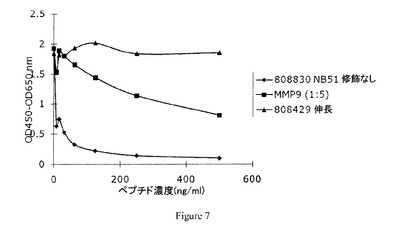
【図7】実施例4において得られた競合試験結果を示す図である。
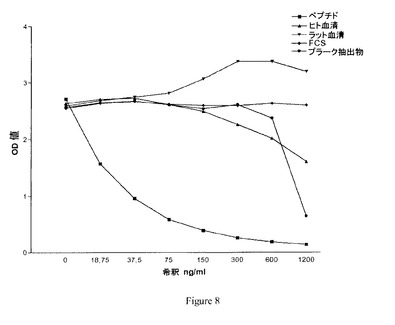
【図8】実施例4において得られた競合試験結果を示す図である。
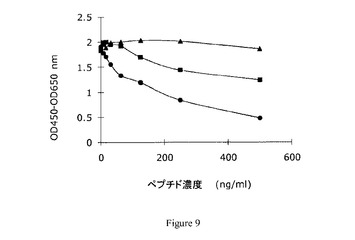
【図9】実施例6において得られた競合試験結果を示す図である。
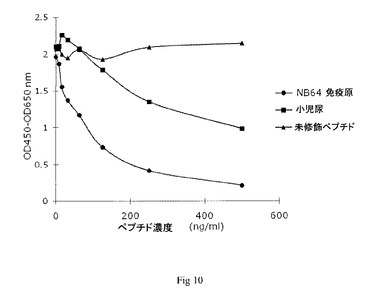
【図10】実施例7において得られた競合試験結果を示す図である。
【図11】実施例7において得られた競合試験結果を示す図である。
【実施例1】
【0167】
プロテオグリカン及びプロテアーゼの局在の分析のため、左冠動脈下行枝(LAD)由来のヒト動脈試料の免疫組織化学染色を行った。以下、カテプシンKプロテアーゼ及びバイグリカンの共局在が示される。
【0168】
図1及び2に見られる免疫組織化学染色はバイグリカン及びカテプシンKの共局在を明らかにした。これは、バイグリカンがカテプシンKの好ましい基質であることを示唆し得る。動脈硬化性プラークが形成された大動脈試料に対して同じ免疫組織化学染色を行い、この結果として正常な大動脈構造はマクロファージ泡沫細胞浸潤及び石灰化に置き換えられた。これらの免疫染色を図3及び4にまとめる。
【0169】
バイグリカン及びカテプシンKの免疫組織化学染色は進行性アテローム性動脈硬化病変に共局在することが示された。これらの結果はともにバイグリカンにおける特異的なカテプシンK切断部位の仮説を生じさせ、アテローム性動脈硬化病変におけるネオエピトープ生成の増加をもたらす。この仮説を試験するため、本発明者らはバイグリカンを異なるプロテアーゼで切断した。
【実施例2】
【0170】
分解断片の評価のためのバイグリカンの分解。ウシ関節軟骨由来のバイグリカン(B8041−Sigma−Aldrich社)をプロテアーゼ(MMP2、MMP3、MMP8、カテプシンK、カテプシンS、カテプシンB及びカテプシンL)によって切断した。上述のプロテアーゼの酵素切断によって生じたプロテオグリカンの断片を10%のNuPage(登録商標)Bis−Trisゲル上に分離し、その後、「Silver Express」−銀染色キット(Invitrogen社カタログ番号LC6100、ロット番号341099)によって銀染色した。蛋白質分解で得られたバイグリカンの分離及び銀染色の結果を図5に示す。
【実施例3】
【0171】
オボアルブミンにコンジュゲートしたIII型コラーゲン由来ペプチドでマウスを免疫化した。ビオチンにコンジュゲートしたスクリーニングペプチド配列との反応性について血清をスクリーニングした。モノクローナル抗体分泌クローンを作製し、スクリーニング配列を用いてスクリーニングした。III型コラーゲン由来の隣接配列で連続する標的ペプチドの伸長バージョン(除外ペプチド)との反応性の欠如及びナンセンスペプチドとの反応性の欠如についてクローンを調べた。
【0172】
標的配列、免疫原、スクリーニング配列及び除外配列は以下の通りであった。
【0173】
【表25】
【実施例4】
【0174】
<III型コラーゲンネオエピトープモノクローナル抗体のヒト尿との反応性>
免疫ペプチドを競合物質として用いて競合アッセイ形式にて実施例3からの選択モノクローナル抗体クローンのヒト尿との反応性を判定した。典型的な手順では、96ウェルストレプトアビジン被覆プレートを振盪しながら20℃でPBS−BTE中10ng/mLビオチンペプチドで30分間コートし、5倍洗浄緩衝液で洗浄した。20μlの希釈試料(尿又はペプチド溶液)を加えた。下記に詳述する100μLの未精製抗体溶液(細胞培養液由来の上清)を加えた。プレートを300rpmで振盪しながら20℃で1時間インキュベートし、次に、5倍洗浄緩衝液で洗浄した。100μLの二次抗体−POD(1:5000)を加え、300rpmで振盪しながら20℃で1時間インキュベートし、その後、5倍洗浄緩衝液で洗浄した。100μLのTMBを加え、300rpmで振盪しながら暗室で15分間インキュベートし、その後、停止液100μLを加えた。ELISAリーダー上で650nmを対照として450nmにてプレートを読み取った。従って、抗体に対するプレート上のペプチドと溶液中のペプチドとの間の競合が生じ、ペルオキシダーゼ発色反応によりプレート結合抗体を定量した。
【0175】
図6に4つの異なるクローンの結果が認められる。抗体が各々尿中の関連する配列を検出することがわかる。
【0176】
免疫ペプチドとMMP9によってin vitroにて切断された天然III型コラーゲンとの間の抗体結合の競合を試験するため、1つの選択クローンに対して更なる競合試験を行った。切断コラーゲン、ペプチドKNGETG及びその配列の伸長バージョンの結果を図7に示す。抗体が免疫ペプチド配列及び酵素切断コラーゲンに結合するが、伸長配列には結合しないことがわかる。
【0177】
図8に同じクローンに関する更なる競合試験が認められ、この場合、競合物質はそれぞれ、ペプチドKNGETG、ヒト血清、ラット血清、FCS(ウシ胎仔血清)及び動脈硬化性プラーク抽出物であった。抗体がペプチド、プラーク抽出物及びヒト血清と反応するが、ラット血清又はFCSとは反応しないことがわかる。
【実施例5】
【0178】
<デコリン、バイグリカン及びバーシカン配列に対する抗血清の産生>
抗血清を産生させ、実施例3のようにモノクローナル抗体を得たが、以下の免疫原、スクリーニング配列及び除外配列を用いた。
【0179】
【表26】
【実施例6】
【0180】
<デコリンネオエピトープモノクローナル抗体のヒト尿との反応性>
1つの抗デコリン未精製モノクローナル抗体(NB62)を用いて概して実施例5のように競合ELISAを行った。
【0181】
図9に結果を示す。これに対して抗体が産生され、選択されたペプチド配列及び尿に対する反応が認められるが、無関連ペプチド配列に対する反応は認められない。
【実施例7】
【0182】
<バーシカンネオエピトープモノクローナル抗体のヒト尿との反応性>
配列に対する2つの抗バーシカン未精製モノクローナル抗体(NB64)を用いて概して実施例5のように競合ELISAを行った。
【0183】
それぞれのクローンの結果を図10及び11に示す。それぞれの場合、これに対して抗体が産生され、選択されたペプチド配列及び尿に対する反応が認められるが、無関連ペプチド配列NB18に対する反応は認められない。
【0184】
本明細書では、特に明示されない限り、条件の1つのみが満たされることを要求する演算子「排他的or」と対照的に、「又は」という語は言及条件の一方又は両方が満たされる場合に真値を返す演算子の意味で用いられる。「含む(comprising)」という語は「〜から成る」を意味するというよりも「含む(including)」という意味で用いられる。上記に認められた従来の教示はすべて参照して本明細書に組み込まれる。
【0185】
[参考文献リスト]
【表27A】
【表27B】
【表27C】
【表27D】
【表27E】
【表27F】
【表27G】
【技術分野】
【0001】
本発明は、アテローム性動脈硬化の進展及びプラーク不安定性から生じる心血管事象のリスクを示す生化学マーカーを含む、心血管疾患の診断目的及び疾患の進展の予後に有益な生化学マーカーの検出のためのアッセイに関する。
【背景技術】
【0002】
世界的に、心血管疾患(CVD)は罹病及び死亡の主因である。現在、診断と患者の異なるリスク群への分類及び低リスク患者の診断を可能にする効果的且つ非侵襲的診断法はない。診断及び予後ツールは、主に、単純なマーカー、例えば、年齢、喫煙並びに種々の脂質及びリポ蛋白質濃度の多変量解析から成る。
【0003】
CVDは複数の臨床症候群を包括し、それは主に狭心症、心筋梗塞(冠動脈血栓)及び脳卒中である。通常、これらの症候群はすべて複雑なアテローム性動脈硬化の続発症である。
【0004】
アテローム性動脈硬化は小児期に内膜肥厚から始まり、動脈内膜における脂肪線条に進行し、これらの病変はそれぞれI型、II型と特徴付けられる。脂肪線条はアテローム性動脈硬化の進展における最も初期の肉眼で視認可能な病変であり、すべての人種及び社会のすべてのヒトに生じる。非病原性状態では内皮細胞(EC)は白血球との付着性相互作用に抵抗性である。しかし、アテローム発生時の炎症性サイトカイン及び動脈壁における蓄積酸化リポ蛋白質の作用は、大動脈ECの表面上における接着分子、例えば、細胞間接着分子(ICAM)−1及び血管細胞接着分子(VCAM)−1の発現を引き起こす。これは、白血球の捕捉及び内皮表面を貫通して血管壁の内膜部分への移動を許容する。プラークの発達は、置換やアポトーシスを起こす平滑筋細胞(SMC)の増加を伴い、これはマトリックス代謝回転の増加をもたらす。コラーゲン合成の障害は線維性被膜の脆弱性及びより破裂しやすい動脈硬化性プラークをもたらし得るが、大部分の研究者らは、蛋白質分解酵素、例えば、マトリックスメタロプロテアーゼ(MMP)及び他のプロテアーゼの作用が、プラークの破裂のリスクに重大に寄与すると考えている(Clarkson and Kaplan 509−28)。
【0005】
プラークは2つの異なるタイプ、「不安定」及び「安定」プラークに分けられる。しかし、詳細な組織学的分析及び分子的理解のために、より詳細な分類が用いられる場合が多い。プラークの発達には3つの主要な段階、すなわち、初期、脂肪線条、及び、複雑性/進行性プラークがある(Stary H.C.)。
【0006】
動脈硬化性プラークは動脈内膜内で発達し、その組成及び構造により分類され得る。この分類では病変を8つのタイプに分類する(Stary H.C.)。
I.脂肪滴に満ち、脂肪滴によって拡大されたマクロファージ(マクロファージ泡沫細胞)が内膜に増加する。
II.マクロファージ泡沫細胞が内膜SMC内で脂肪滴とともにプロテオグリカン層の深部に蓄積する。泡沫細胞層は脂肪線条として視認可能である。II型病変では、単球がヒトアテロームにおいて過剰発現する単球走化性蛋白質(主にMCP−1)によって内膜を貫通する。病変の初期タイプ(I型及びII型)は幼児期に始まり得、必ずしもプラークの破裂には至らない。更に、アテローム性動脈硬化の発達はIII型病変の形成後に終了し得、プラークの形成は予測可能ではない(Stary H.C.)。
III.III型病変は脂肪線条(II型)とアテローム(IV型)との中間病変として判定される。これらの病変は細胞外脂質のプールを含み、これにより、内膜の筋弾性層の通常密接に隣接しているSMC間の空間を拡張する。物質のプールは通常ここに存在するプロテオグリカン及びコラーゲン線維に取って代わり得るが、これはアテローム発生のこの段階においてほとんど影響なく生じる。
IV.アテロームはアテローム性動脈硬化の最初の臨床徴候である。脂質の蓄積細胞外プールによる動脈内膜におけるSMCの移動及び内膜構造の崩壊が、IV型病変の顕著な特徴である。脂質コアの形成は、このSMC移動の最終結果である。脂質コアの形成は壁肥厚の増大の説明となる。脂質コアは深部内膜の大きな明瞭な輪郭をした領域であり、この動脈壁部分の通常の構造要素が高密度の泡沫細胞残遺物、遊離脂肪滴、コレステロール結晶及びカルシウム粒子に置換されている。通常、この領域に存在するSMCは、アテローム性動脈硬化のこの段階において減少し、又は完全に消失する。残遺SMCは広範に分散し、伸長細胞体及び、多くの場合、異常に厚い基底膜を生成している。この段階では脂質コアを覆う層の発達が始まる。この層はコラーゲン/プロテオグリカンに富む細胞間マトリックス、脂肪滴を有する又は有しないSMC、マクロファージ及び泡沫細胞から成る。
V.IV型病変に対する応答は、線維性「被膜」を形成する修復線維組織マトリックスの形成である。通常、これらの病変は脂質コア及び不規則に積み重なり合った修復組織の層から成る。血腫及び血栓形成のような事象がこれらのタイプの病変を更に複雑にし得る。致死的ではない場合、これらの病変合併症は病変に組み込まれ、コラーゲン及びプロテオグリカンから成る修復マトリックス組織の薄層に覆われる。細胞外マトリックス蛋白質であるコラーゲン及びプロテオグリカンの含量は被膜の形成時に動脈硬化性プラークにおいて増加する。
VI.内皮の異常、例えば、亀裂、びらん、潰瘍、血腫、血栓、出血は、組み合わされると、VI型病変と命名されるより複雑な病変型となり得る。
VII.病変は石灰化病変と称される場合が多く、病変の50%超がミネラルから成る。石灰化に加え、これらの病変は大量の修復線維結合組織を含む。ここに閉じ込められたSMCがアポトーシスを受けて分解すると、その石灰化小器官は石灰化の一部になる。
VIII.線維性病変が石灰性病変に続く。線維性病変は完全にコラーゲンから成り脂質を含み得ない(Stary H.C.)。
【0007】
心血管事象はプラークの破裂の結果である場合が多く、炎症及びプロテアーゼの放出が線維性被膜の肩領域を弱体化させ、プラークにおける脂肪質が壁在血栓を誘発する血液と接触することを許容する(Clarkson and Kaplan)。マトリックス生成の減少と併せたプロテアーゼ活性の増大による線維性被膜の菲薄化がプラーク不安定性の顕著な特徴であると考えられ、破裂のリスクを高める。プラークの不安定性及びその破裂リスクは臨床的に興味深い分野である。不安定プラーク(VP)の定義は標準化されていないが、安定プラークと比べて3つの組織学的特徴の存在を明言する一般的な見解の一致がある。
1)大きい脂質コア(総病変の40パーセント超)
2)薄い線維性被膜(65〜150マイクロメートル)
3)大量の急性炎症細胞
VPを定義するための主な基準には、活発な炎症(単球、マクロファージ及びT細胞の存在)、大きい脂質コアを有する薄い被膜、表在血小板凝集を伴う内皮の裸出、亀裂プラーク及び動脈の90%超の狭窄が含まれる。他の細かい基準には、表在石灰化結節、プラーク内出血、内皮機能障害及び外側の再構成が含まれる(Shin,Edelberg,and Hong)。
【0008】
プラーク合併症、不安定及び破裂は、医療処置及び/又は生活様式の変更によって抑制され得る。しかし、場合により、より侵襲的な方法、即ち、血管形成術又はバイパス手術が必要であり得る。
【0009】
現在、診断ツールは、依然として開発中である静止画像解析又はCVDリスクに関連する収縮期・拡張期血圧レベルのような低技術法に基づく。当該分野では高リスクにある患者をより良く同定し得る多変量解析の開発に多大な関心を向けてきた。そのようなモデルの1つがSCOREモデル(Systematic Coronary Risk Evaluationモデル)である。1994年に、そして2003年に改訂されたのであるが、欧州アテローム性動脈硬化学会(The European Atherosclerosis Society)、欧州心臓病学会(The European Society of Cardiology)及び欧州高血圧学会(The European Society of Hypertension)は、冠動脈心疾患の予防に関する一組の推奨を公表した。このガイドラインは、無症状の患者におけるCVDリスクを評価するために、即ち、無症状の高リスク患者の同定のために開発された複数の評価法に基づく。SCOREモデルは、性別、年齢、喫煙、収縮期血圧及び総コレステロール若しくはコレステロール/HDL比をリスク因子として一体化している(Graham et al.)。
【0010】
より詳細な診断を行うためにはSCOREモデルは十分ではなく、撮像法が用いられる。従って、撮像法は主に高リスク群の患者又は研究期間中に用いられる。
【0011】
<撮像法>
現在、冠動脈造影法(CAG)は狭窄の程度を判定するための判断基準となる撮像法である。CAGは二次元にて血管腔を撮像するが、血管腔のみに限定され、血管壁を撮像しないため、CAGは安定プラークを有する動脈と不安定プラークを有する動脈とを識別することができない。CAGは患者が手術(血管形成術又はバイパス手術)を必要とするかを判定するために用いられる場合が多い。血管の狭窄箇所が進行したプラークであるかを判定するためには、他の手法、即ち、血管内冠動脈超音波(IVUS)又は血管顕微法が必要である。
【0012】
IVUSはプラーク及び血管壁の二次元断面画像を提供し、血管壁及び形態及び石灰化の程度の特徴付けには良好であるが、病変における脂質を評価するには不向きな方法であると考えられている。しかし、IVUSは侵襲的であり、専門知識及び費用を必要とするため、その使用は広まっていない。血管顕微法はアテローム性動脈硬化を理解・同定するのに有用なもう1つの方法である。血管顕微法はプラーク表面の直接的可視化であり、プラークの色及び血栓を検出する能力を有する。しかし、血管顕微法は侵襲的で技術的に困難であり、これまでプラークの拡張の程度を検出することができていない。現在多くの関心を引いているもう1つの撮像法は磁気共鳴画像法(MRI)である。MRIは非侵襲的であり、脳卒中の高リスクにある頸動脈プラークを同定することができる。一方、小さいプラークサイズ及び冠動脈の位置のため、MRIは冠動脈を撮像するのに最適な手法ではない。他の撮像法、即ち、エラストグラフィ、サーモグラフィ及び光干渉断層撮影が開発中である(Schaar et al.)。
【0013】
前述の撮像法はすべて開発中であり、単独ではいずれも不安定プラークを同定することができないが、破裂前に分子事象及びプラーク代謝回転を理解するのに有用なツールである。現在、早期においてCVDを診断する唯一の機会は、問題となっている患者及びその患者の近親者の確立された冠動脈心疾患、末梢動脈疾患及び脳血管アテローム性動脈硬化症の一連のリスク因子を用いることである。
【0014】
<現在の生化学マーカー>
現在、アテローム性動脈硬化のリスク因子としていくつかの生化学マーカーが公知である。最近、血清中の生化学マーカー濃度の測定に多くの関心が向けられている。脂質、例えば、総コレステロール、低比重リポ蛋白コレステロール(LDL−C)及び高比重リポ蛋白コレステロール(HDL−C)並びに炎症マーカー、例えば、C反応性蛋白質(CRP)、インターロイキン6(IL−6)、インターロイキン18(IL−18)、腫瘍壊死因子アルファ(TNFα)、CD40、CD40リガンド(CD40L)など。
【0015】
リポ蛋白質の中で少なくとも2つの注目すべき進展があった。LDL粒子のサイズがアテローム性動脈硬化の進行を予測するように思われる。小さいLDL粒子の濃度上昇が大きい粒子の濃度上昇よりCVDリスクに関連する(Gardner,Fortmann,and Krauss)。
【0016】
HDL−Cのレベルはトリグリセリドに強く関連し、高レベルのトリグリセリドはCHDのより高いリスクと相関する。Jeppesen et al.(2003)によるコホート研究は、高TG/低HDL−CがIHD(虚血性心疾患)の最大のリスク因子であることを見出した(Jeppesen et al.)。
【0017】
脂質プロファイルはリスク因子の評価に重要であるが、プラーク代謝回転と関連する分子事象の理解及び測定を可能としない。多くの生化学マーカーがCVDのリスク因子として示唆されているが、疾患の具体的な産物ではない。これらにはCRP及び骨ナトリウム利尿ペプチド(BNP)が含まれる(表1参照)。表1はCVDの公知のマーカーの一部を概述している。
【0018】
【表1】
【0019】
従って、心血管事象のマーカーとして一連の異なる生化学マーカーが示唆されている。Wang et al(2006)は、表1に示される、Framingham研究に参加した3200人の患者における10個の異なる生化学マーカーを測定した。その結論は、10個の生化学マーカーの測定が標準的なリスク因子に加えて診断に適度に寄与するのみであるというものであった。10個の生化学マーカーの中でB型ナトリウム利尿ペプチドレベル、C反応性蛋白質レベル及び尿中アルブミン/クレアチニン比がマーカーと死亡/心血管事象との最大の相関を示した(Wang et al.)。
【0020】
<C反応性蛋白質>
C反応性蛋白質(CRP)は、異なる臨床状態、例えば、炎症、感染又は外傷に応答して肝臓によって産生される急性期血清蛋白質である(Gabay & Kushner 1999)。CRPの産生は罹患又は損傷組織から放出されるIL−6のようなサイトカインに誘発される。CRPの生理学的役割は未知であり、その炎症促進又は抗炎症作用に関する議論は進行中である。
【0021】
CRPがヒトにおけるCVDのリスク因子であるという証拠が蓄積している。Ridker et al.2002による研究では、急性心筋梗塞、脳卒中、冠血行再建又はCVDによる死亡の発生について8年間追跡した28,000人の健常女性から成る大集団において、CRPはLDLコレステロールより優れた将来の心血管事象の予測因子であることが示された。他の多くの研究もベースラインのCRPレベルが心血管事象の独立したリスク因子を構成することを報告している(Thompson et al.1995,Mendall et al.1996,Kuller et al.1996,Ridker et al.1997,Tracy et al.1997,Ridker et al.2000)。
【0022】
循環CRPはアテローム性動脈硬化プロセスにおいて生じる全般的な炎症を反映するだけであって、疾患の発病における能動的な構成成分ではないと推測されている。しかし、また、一連の証拠はCRPがアテローム発生に役割を有するという見解を支持している。第一に、CRPを生じさせる慢性感染症もCVDの増大リスクと関連する(Leinonen & Saikku 2002)。第二に、本発明者らは異なるレベルのアテローム性動脈硬化病変においてCRPを同定した(Reynolds & Vance 1987,Hatanaka et al.1995)。最後に、CRPはin vitroにてアテローム発生促進特性を有することが示されている:CRPは内皮細胞を活性化して接着分子を生成し得る(Pasceri et al.2000)。また、CRPは内皮細胞におけるeNOSの生成を減少させ(Venugopal et al.2002)、マクロファージによるLDLの取込みを増進し得る(Zwaka et al.2001)。
【0023】
<脳性ナトリウム利尿ペプチド>
脳性(B型)ナトリウム利尿ペプチド(BNP)は、心室における心筋細胞の過度の伸張に応答して心室によって分泌されるペプチドホルモンである。T−proBNP(不活性N末端断片)は活性ホルモン(BNP)とともにproBNPの切断と同時に血流に放出される。BNP及びNT−proBNPは心血管事象の潜在的生化学マーカーとして示唆されている(Wang et al.)。
【0024】
<ケモカイン>
ケモカインもCVDの潜在的マーカーである。ケモカインは炎症時に産生される低分子量サイトカインである。CVDに関連する1つの主要なケモカインは単球走化性蛋白質−1(MCP−1)である。MCP−1はアテローム性動脈硬化病変への単球の動員に初期の重要な役割を果たすように思われる。アテローム性動脈硬化のサルモデルを用いた研究では、MCP−1の血漿濃度はプラークサイズ及びプラーク合併症と高度に関連していた(Register et al.)。
【0025】
<コレステロールを含む脂質>
最近、血清中のコレステロール濃度の測定に多くの関心が向けられている(総コレステロール並びに低比重リポ蛋白コレステロール(LDL−C)及び高比重リポ蛋白コレステロール(HDL−C)の濃度)。リポ蛋白マーカーの中で少なくとも2つの注目すべき進展があった。第一に、LDL粒子サイズからアテローム性動脈硬化の進行の程度が予測されるように思われる。小LDL粒子の濃度上昇は、大粒子の濃度上昇よりCVDリスクに関連する(Gardner et al)。第二に、LDL粒子のオレイン酸コレステリル含量はCVDリスクの特に重要なマーカーになり得る。サルにおいて、オレイン酸コレステリルを有するリポ蛋白粒子コアの高濃度化は、より重度の冠動脈アテローム性硬化と強く且つ正に関連し(Rudel et al)、LDL及びHDLコレステロール濃度の寄与に相加的であった。実験動物におけるこれらの知見は、低比率のリノール酸コレステリル(逆に、高比率のオレイン酸コレステリル)を有する血漿リポ蛋白質が正常な対照と比べてCHD(冠動脈心疾患)の合併症を有する患者に典型であることを示した従前のヒト研究(Lawrie et al)によって更に支持される。
【0026】
HDL−Cのレベルはトリグリセリドと強く関連し、高レベルのトリグリセリドはCHDのより高いリスクと相関する。Jeppesen et al)によるコホート研究は、高TG/低HDL−CがIHD(虚血性心疾患)の最大のリスク因子であることを見出した。
【0027】
これらの脂質プロファイルはリスク因子の評価に重要であるが、プラーク代謝回転と関連する分子事象の理解及び測定を可能としない。多くの生化学マーカーがCVDのリスク因子として示唆されているが、これらは疾患の具体的な産物ではない。これらにはCRP及びApoEが含まれる。
【0028】
<リポ蛋白質>
CVDを予測するために最も一般的に用いられるバイオマーカーは、コレステロール濃度である(総コレステロール及びコレステロール/HDL比)。これらは他のリスク因子、例えば、血圧及びLDLレベルとともに用いられる。前述のSCOREモデルでは両方の因子が用いられる。LDLは血中にコレステロールを輸送し、酸化LDLの蓄積はアテローム性動脈硬化を促進し得るため、LDLレベルは重要である(Graham et al)。加えて、CHDとトリグリセリド(TG)レベルとの有意な関連が見出され、CHDの増大リスクはLDL−C及びHDL−Cレベルから独立して増大TGレベルと関連したが、コレステロールレベルはCVDの主要なリスク因子の1つであると見なされる(Jeppeson et al)。
【0029】
<APO−E>
アポリポ蛋白質Eはカイロミクロン、VLDL及びHDLに見出される。アポリポ蛋白質Eは主に肝臓で合成されるが、他の多くの臓器、例えば、脳、脾臓、腎臓でも合成される(Siest et al.1995)。ApoEは2つの受容体(LDL受容体及びApoE特異的カイロミクロンレムナント受容体)のリガンドとして作用することによってリポ蛋白代謝に重要な役割を果たす。ApoEとこれらの受容体との相互作用はコレステロールの代謝調節の基盤を付与する。apoE遺伝子座における遺伝子多型は3つの対立遺伝子(6つのapoE表現型を決定するε2,ε3及びε4)が大部分の集団に見出されることとなる。アイソフォームは112及び158位において1つのアミノ酸だけ互いに異なる。ApoE2は両方の残基においてシステインを有し、E4は両方の位置においてアルギニンを有する。ApoE3は112位においてシステインを、158位においてアルギニンを含む。対立遺伝子頻度は様々な集団において異なる。研究の中にはapoE多型とアテローム性動脈硬化との関連の可能性を評価したものがある。14の観察研究のメタアナリシスではε4対立遺伝子が男女の間で冠動脈疾患と関連することを示した(Wilson et al.1996)。更に、ε4対立遺伝子は頸動脈アテローム性硬化と関連している(Terry et al.1996,Cattin et al.1997,Haraki et al.2002)。
【0030】
ApoEは299アミノ酸長であり、リポ蛋白質、脂溶性ビタミン及びコレステロールをリンパ系へ、更には血液循環へ輸送する。ApoEは主に肝臓で合成される。現在、保存低比重リポ蛋白受容体遺伝子ファミリーに属するApoEの哺乳動物受容体は7つある。
【0031】
<更なる生化学マーカー>
微量アルブミン尿(Microalbuminurea)(アルブミン/クレアチニンレベル)も潜在的な独立したマーカーである。尿中アルブミン排泄率は腎臓の変化のマーカーであり、クレアチニンの微量の上昇と比べてアテローム性動脈硬化を示し得る(Wang et al.)。
【0032】
プロコラーゲンマーカーの中でIII型コラーゲン代謝回転速度のマーカー(PIIINP)は高血圧の予後マーカーとして研究され、心筋梗塞と関連している。Satta et al.は腹部大動脈瘤(AAA)と血中プロコラーゲン(PIIINP)の濃度との相関を検討した。彼らは、III型コラーゲン代謝回転がAAA患者において増加し、増進した合成、増進した分解又は両方の組合せによるものであり得ることを示した。同じ実験において、I型プロコラーゲン(PICP)のカルボキシ末端プロペプチドが測定され、動脈瘤嚢にI型コラーゲンの加速度的な合成はなかった。
【0033】
<プラークの蛋白質プロファイル>
ヒト動脈はより大きい若しくは弾性の動脈、中間若しくは筋性の動脈並びに小動脈に分けられ得る。血管壁は内弾性板及び外弾性板によって分離された内膜、中膜及び外膜から成る。内膜は結合組織、平滑筋細胞及び少数の分離マクロファージから成る。内膜の境界は内皮の管腔表面と内弾性板との間の層と定義され得る。
【0034】
動脈内膜は更に2つの層に分けられ得る。プロテオグリカン層と称される内層は豊富なプロテオグリカン、平滑筋細胞及びマクロファージから成る。下層、筋弾性層は豊富な平滑筋細胞及び弾性線維から成る。通常の状態では内膜の2つの層は光学顕微鏡によってどうにか視認可能であるが、内膜肥厚が生じると明確且つ著明である。中膜は動脈壁の筋部分であり、平滑筋細胞、エラスチン、コラーゲン原線維から成る。
【0035】
外膜、外層は高度に微小血管であり、コラーゲン、弾性線維、平滑筋細胞及びリンパ管を含む。
【0036】
ヒト動脈硬化性プラークは、線維状コラーゲン、エラスチン、プロテオグリカン及びSMCから成る線維性被膜に覆われた脂質に富むコアを特徴とする。プロテオグリカンヒアルロナンは、事象、例えば、脂質蓄積、血栓及び細胞増殖・移動を調節し、また、組織の物質特性に作用することによって病変発達に作用する可能性を有する細胞外マトリックスの主要な非線維状構成成分である(Wight 1995)。浸潤ApoE及びCRPも存在し、本発明者らは、アテローム性動脈硬化症の異なる段階において冠動脈の動脈硬化性プラークにおいて両方の局在を示した。??参照
【0037】
<ヒトにおけるApoE及びCRPの分布>
下記の表2はヒト体内のApoE及びCRPの分布を示す。
【0038】
【表2】
【0039】
下記の表3は、in vivo及び/又はin vitroにて示されるApoE及びCRPと蛋白質との公知の相互作用を示す。
【0040】
【表3】
【0041】
<ヒトにおけるコラーゲンの分布>
コラーゲンはヒトの身体に広範に分布し、即ち、ヒトの身体における蛋白質の質量の約30%はコラーゲンから成る。表4に主要なコラーゲンタイプがその主要な組織分布ととともに一覧されている。
【0042】
【表4】
【0043】
I型コラーゲンは最も豊富なコラーゲンであり、大部分の結合組織に見出される。I型コラーゲンは特に骨及び皮膚の構造に重要である。ヒトの身体におけるコラーゲンの主要含量は骨及び皮膚に分布し、主要なコラーゲン成分はI及びIII型コラーゲンである。III型コラーゲンは大動脈の主要構成成分であり、I型コラーゲンに富む組織にもより少ない量にて見出される。加えて、IV型コラーゲンは基底膜及び血管・神経周囲に見出される。V型コラーゲンの最も一般的な局在は、I及びIII型コラーゲンと関連して特徴的なコラーゲン原線維内である(Garrone et al)。
【0044】
一部のコラーゲンは限定された組織分布を有し、例えば、II型はほとんど軟骨に見出される(Mayne R.)。
【0045】
コラーゲン原線維は2つ以上のコラーゲンタイプから成る場合が多い。例えば、I型コラーゲン原線維は少量のIII,V及びXII型を含む場合が多く、一方、軟骨のII型コラーゲン原線維はIX及びXI型も含む。
【0046】
<動脈におけるコラーゲン>
動脈には6つのタイプのコラーゲンが見出され(I,III,IV,V,VI及びVIII型)、I及びIII型が最も豊富であり、コラーゲン含量の80〜90%である。I及びIII型は血管壁でも最も多い。I及びIII型は動脈壁の3つのすべての層内に異なる量にて共分布しているように思われ、I及びIII型コラーゲンの合成は内膜に位置している傾向にある(Mayne R)。
【0047】
<プラークにおけるコラーゲン及び他の構造蛋白質の代謝回転>
動脈硬化性プラークの発達の間、コラーゲンが線維性被膜に蓄積する(Stary H.C.)。Katsuda et al(1992)による研究では、I,III及びIV型コラーゲンがヒト大動脈組織における病変のすべての段階において肥厚内膜において見出された。VI型コラーゲンが内膜細胞の領域における基底膜に分布し、進展した病変では伸長SMC周囲にも検出された。I及びIII型の従前の研究はアテローム硬化性動脈壁における同等の分布のエビデンスを提供している(Shekhonin et al)。McCullagh et al(1980)によれば、III型が正常なヒト大動脈中膜において最も多いコラーゲンである(抽出可能なコラーゲンの約70%)。Eriksen et al(2006)による最近の研究は、狭窄の程度に依存するヒト大動脈弁における総コラーゲン含量の減少を見出した。狭窄の分子機構は、アテローム性動脈硬化に類似すると考えられている。健常な大動脈弁では、コラーゲン含量は主にI及びIII型である。狭窄の間、コラーゲンの総含量は減少し、それはおそらくI型コラーゲンの代謝回転の増大によって生じる。I型コラーゲンは総コラーゲンの約60〜70%を占め、一方、III型コラーゲンの割合は健常弁及び石灰化弁において30〜40%であった。
【0048】
V型コラーゲンも、進展したアテローム性動脈硬化病変において増加し、大動脈中膜及びプラークの内皮下領域において、細胞外マトリックス全体にわたって分布する(McCullagh et al)。
【0049】
動脈硬化性プラークに見出される主なコラーゲンタイプは、I及びIII型であるという見解の一致があるように思われ、それらが健常血管及びアテローム硬化性血管において同等に分布しているかは更に研究される必要がある。
【0050】
Katsuda et al(1992)による研究では、より進展した病変のアテロームの中心部にコラーゲンは検出されなかった。
【0051】
<エラスチン>
エラスチンは身体において最も安定な蛋白質の1つであり、その弾性及び弾力によって生じる大部分の結合組織に見出される。エラスチンは動脈壁の蛋白質含量を独占し、主要な細胞外マトリックス蛋白質である。
【0052】
エラスチンは弾性線維における主な構成成分であり、石灰化に関連する。血管石灰化は血管壁内の2つの異なる部位(内膜及び中膜)において生じる。内膜石灰化は主に壊死性コア内にてアテローム性動脈硬化に関連する。石灰化弾性線維はプラークが最も破裂しやすいプラーク肩部を構成し、弾性線維の石灰化がプラーク安定性に作用し得ることを示唆している(Bobryshev Y.V.)。アテローム性動脈硬化では、弾性線維の含量は脂質沈着とともに減少し、これはエラスチン分解酵素に対する感受性の亢進を生じさせる。これにより、コラーゲンとは対照的に、エラスチンの含量は、病変が発達するにつれて減少する。
【0053】
<ヒトにおけるエラスチンの分布>
表5はヒトの身体におけるエラスチンの分布を示す。
【0054】
【表5】
【0055】
表6はin vivo及び/又はin vitroにて示されるエラスチンと蛋白質との公知の相互作用を示す。
【0056】
【表6】
【0057】
<マトリックス構成成分としてのプロテオグリカン>
プロテオグリカン(PG)は、主に血管壁の細胞間マトリックスに局在する多糖蛋白質である(Salisbury and Wagner 1981)。PGは、結合領域を通じてコア蛋白質に共有結合したGAGと称される1つ以上の長い非分岐高ポリアニオン糖側鎖の存在を特徴とする高分子である。GAGの反復単位は、アミノ糖(N−アセチル−グルコサミン(GlcNAc)又はN−アセチル−ガラクトサミン(GaINAc))及びヘキスロン酸(グルクロン酸(glucouronic acid)(GlcA)又はイズロン酸(IdoA))から成る。反復単位における糖の一方又は両方は1つ以上の硫酸基を含む(Rodriguez−Lee 2007)。GAG鎖に加え、大部分のコア蛋白質はN−及び/又はO−結合オリゴ糖を保有する。
【0058】
<PGの分類及び命名>
PGは高分子の極めて異質の群である。単一タイプのコア蛋白質は結合GAG鎖の数及びタイプの点で異なり得る。鎖長及び鎖に沿った硫酸化残基の構成も異なる。
【0059】
反復二糖類単位の構造によってGAGの4つの主なクラスが識別される。コンドロイチン硫酸(CS)及びデルマタン硫酸(DS)、ヘパリン硫酸(HS)及びヘパリン、ヒアルロナン並びにケラチン硫酸(KS)。
【0060】
ヒアルロナンはGAGの最も単純なものである。他のすべてと対照的に、ヒアルロナンは硫酸化糖を含まない。その二糖類単位はすべて同一であり、その鎖長は膨大であり、ヒアルロナンはコア蛋白質に結合していない。
【0061】
KSは硫酸化ポリラクトサミン鎖である。KS−Iは元々角膜において説明され、コア蛋白質においてアスパラギン残基にN結合し、一方、KS−II又は軟骨KSはセリン又はスレオニン残基にO結合している(Funderburgh 2000)。
【0062】
PGはいくつかのパラメータによって分類され得る。
結合GAG鎖(CS/DS−又はHS含有PG)
細胞に関する組織分布(細胞外及び基底膜PG、細胞結合PG又は細胞内PG)
コア蛋白質相同性(ヒアレクタン(hyalectan)、小さなロイシンに富むPG(SLRP)
【0063】
コンドロイチン/デルマタン硫酸PG(バーシカン、アグリカン、ニューロカン及びブレビカン)はヒアルロナン結合プロテオグリカンのファミリーに属する。この遺伝子ファミリーは集合的にヒアレクタン(hyalectan)と称される。各ファミリーメンバーは特徴的な分布を有し、アグリカンは軟骨において顕著であり、ニューロカン及びブレビカンは中枢神経系において顕著であり、バーシカンは血管壁を含む様々な軟組織に存在する。バーシカンの遺伝子及び蛋白質構造はドメインテンプレートに従う。アミノ末端球状末端(G1)はGAGヒアルロナンに結合し、カルボキシ末端球状ドメイン(G3)は、2つの上皮増殖因子(EGF)ドメイン及び補体調節領域に隣接するC型レクチンから成る蛋白質のセレクチンファミリーに類似する。バーシカンコア蛋白質の中央領域は、バーシカンのCS結合領域を指定する2つの大きなエクソンにコードされる。エクソン7にコードされる領域はαGAGと称され、一方、エクソン8にコードされる領域はβGAGと称される。バーシカンの選択的スプライシングから4つのmRNA転写物が生じ、コア蛋白質の長さ及び結合GAGの数が異なるV0,V1,V2及びV3を生じさせる(Dours−Zimmermann and Zimmermann)。ヒトバーシカンにおける潜在的GAG結合部位の数は、V0では17〜23、V1では12〜15、V2では5〜8、V3では0である(Wight 617−23)。
【0064】
デコリン及びバイグリカンは、3つのクラス(I,II及びIII)及び異なるサブファミリーに分類される少なくとも9つのメンバーを含むSLRPファミリーのメンバーである。それらはすべて、強い蛋白質−蛋白質相互作用を成すロイシンに富むリピートを含む中央ドメインの強い存在を成すロイシンに富むリピートを含む中央ドメインの存在を特徴とする。デコリン及びバイグリカンはクラスIのメンバーであり、ファミリーの中で最も高い相同性を示し(約57%)、プロペプチドを有する唯一のSLRPである。プロペプチドは種にわたって高度に保存され、GAG鎖の合成に関与する最初の酵素であるキシロシルトランスフェラーゼの認識配列として機能し得る。
【0065】
バーシカン、デコリン及びバイグリカンは、哺乳動物動脈壁のマトリックスにおける主要なCS/DS PGである(Wight et al.1986)。バーシカンV0コア蛋白質のサイズは370kDaであり、36kDaのデコリン及び38kDaのバイグリカンよりほぼ10倍大きい。側鎖は広範なサイズを示すが、概して各々平均約40〜90kDaである。
【0066】
ヘパラン硫酸プロテオグリカン(HSPG)は、細胞結合及び細胞周囲PGの5つの異なるクラスに分類され、哺乳動物細胞表面、基底膜及びECMのHSの少なくとも95%を占める。細胞結合HSPGは膜内在性シンデカン及び固着グリピカンを含む。細胞周囲HSPGは主にパールカン、アグリンを含む。これらのPGは、インテグリンを介した細胞膜との密接な関連のために細胞周囲型と称される(Whitelock and Iozzo)。
【0067】
パールカンは、ほぼすべての基底膜並びに間葉系臓器及び結合組織に発現するモジュラーHSPGであり、脊椎及び無脊椎動物に見出される最大の単鎖ポリペプチドの1つである。パールカンの5つのモジュール及びそのHS側鎖は、例えば、線維芽細胞増殖因子2、血管内皮増殖因子(VEGF)、血小板由来増殖因子(PDGF)及び他のマトリックス蛋白質との多数の分子相互作用に関与する。ヒトパールカンのコア蛋白質は約470kDaであり、多くのO結合オリゴ糖及び4本のHS側鎖とともに800kDaを超える分子量に達し得る(Knox and Whitelock)。
【0068】
<プロテオグリカンの分布>
プロテオグリカン(PG)はヒトの身体のほぼあらゆる部位に分布する高分子である。PGの構造及びサイズは極めて異なる。すべてのPGの基本構造はコア蛋白質及び少なくとも1つであるが、多くの場合、多くの炭水化物鎖、すなわちグリコサミノグリカン(GAG)を含む。PGは細胞内、細胞表面及び細胞外マトリックスに見出され得る。PGの構造多様性は多くの生物学的機能を示唆するものであり、表7を参照されたい。
【0069】
【表7】
【0070】
<動脈におけるプロテオグリカン>
少なくとも5つのタイプのPGが動脈壁の細胞外マトリックスに存在する。ヒアルロナンと相互作用して大凝集体を形成するバーシカン;コラーゲン及びエラスチンのような原線維マトリックス構成成分と相互作用する、小さいロイシンに富むデコリン及びバイグリカン;基底層の構成成分であるヘパラン硫酸−パールカン並びにケラチン硫酸−ルミカン(Talusan et al.)。
【0071】
バーシカンはアテローム性動脈硬化病変に蓄積するいくつかのECM分子の1つである。多くの研究が、バーシカンが明らかにLDLに結合することができることを示すが、概して、バーシカンは壊死性コアの脂質に富む中央部には検出されない(Evanko et al.)。
【0072】
ルミカンはマクロファージに直接結合し、且つマクロファージの移動を増進することが示されている。従って、ルミカンは血管内膜におけるマクロファージの挙動に影響を及ぼすとともに、進展したアテローム性動脈硬化病変の特徴を示す壊死性コアの形成を刺激し得る(Funderburgh et al.1997)。
【0073】
バイグリカンは線維性被膜に見出される。バーシカン及びバイグリカンはLDLに対する親和性を有し、不溶性複合体を形成し、これはLDLの酸化を促進する。バイグリカンはリポ蛋白質を動脈壁に捕捉することによってアテローム性動脈硬化の発病に寄与し得る。動脈内膜のプロテオグリカン代謝の変化はアテローム性動脈硬化の初期病変を構成し、プロテオグリカンの蓄積はアテローム性動脈硬化の進行に基本的な役割を果たす(Kunz J.)。
【0074】
パールカンは、質量分析ベースの分析及び免疫組織化学によって内膜の細胞外マトリックスの中心成分の1つとしてヒト内膜過形成にて報告された。
【0075】
表8は、正常及びアテローム性動脈硬化血管におけるPGの免疫組織化学的染色でのいくつかのPGの分布を示す(Evanko et al)。
【0076】
【表8】
【0077】
<マトリックス再構築におけるプロテオグリカンの関与>
非ヒト霊長類におけるアテローム性動脈硬化の進行の研究は、特定のPGの蓄積が病変の重症度及び細胞・増殖因子の分布により異なることを示し、アテローム性動脈硬化の進行時に異なるPGが異なる役割を果たすことを示唆した。異なるレベルの特定のPGは、線維性マトリックス構成成分、例えば、エラスチン及びコラーゲンの構造配置の変化への寄与によって組織の物質特性に直接作用し得る。
【0078】
バーシカン及びヒアルロナンは、マトリックスにおける同様の局在を示し、アテローム発生におけるその2つの間の凝集体形成を示唆する。早期病変におけるバーシカン及びヒアルロナンの著明な増加は、早期アテローム性動脈硬化病変、例えば、SMC及び白血球の増殖・移動に役割を果たすことを示唆し得る。更に、バーシカン及びヒアルロナンはヒト再狭窄病変の主要なマトリックス構成成分であり、in vitroにて血管損傷後の新たな内膜肥厚に寄与することが示されている。また、早期アテローム発生における大量のバーシカンは、バーシカンのコンドロイチン硫酸鎖へのリポ蛋白質の結合により、細胞外マトリックスに脂質の捕捉を増大させやすくし得る。この見解は、移植動脈疾患におけるバーシカンとアポ蛋白質(a)及びアポリポ蛋白質Eとの共局在によって支持される(Evanko et al)。プラークからのバーシカンの喪失はマトリックス不安定をもたらし得る。
【0079】
このことは、血管損傷後に認められるバーシカン遺伝子のアップレギュレーションから更に明らかである。この場合、バーシカンは、アテローム性動脈硬化のすべての段階、すなわち、早期の発達プラークの内膜のみならず、進展した病変にわたり、脂質で満たされた壊死性コアの境界及びプラーク−血栓界面にても同定された(Wight and Merrilees 2005)。これらの所見は脂質の蓄積、炎症及び血栓にバーシカンが関与していると見なす。更に、バーシカンはECMのアセンブリ及び弾性線維原線維形成の調節に重要な役割を果たし、これは血管疾患時のECM再構築において基本的に重要である(Wight and Merrilees 2005)。
【0080】
動脈細胞生物学におけるバイグリカンの役割は不明瞭である。一部の免疫組織化学的研究は、ヒト再狭窄病変においてコラーゲンI及びIII染色とのバイグリカンの関連を示している(Evanko et al.)。
【0081】
マトリックス蛋白質としてのバイグリカンの重要性は、バイグリカン遺伝子のヌル変異に対してホモ接合性のBALB/cAマウスの作製によって更に述べられ、この場合、バイグリカン欠損雄マウスの50%が大動脈破裂の結果として寿命の最初の3カ月以内に突然死した。この所見は、大動脈壁の構造・機能的一体性にバイグリカンが不可欠であること、また、ヒトにおける大動脈解離・破裂の発病におけるバイグリカン遺伝子欠損の潜在的役割を示唆する(Heegaard et al.2007)。
【0082】
他の研究は、バイグリカンが霊長類動脈においてエラスチンと関連する主要なPGであることを示しており、これらの所見はヒト冠動脈疾患の所見に類似している(Evanko et al)。
【0083】
デコリンはコラーゲンに結合し、且つコラーゲン原線維形成を調節することが示されている(Brown and Vogel)(Danielson et al.)。
【0084】
<プロテアーゼのプロファイル>
プロテアーゼはペプチド結合を加水分解し、アテロームにおけるコラーゲン、プロテオグリカン及びエラスチンのような細胞外マトリックス蛋白質の分解に関与する。表9を参照されたい。動脈硬化性プラークには3つの主なタイプ(メタロプロテイナーゼ(即ち、MMP)、セリンプロテアーゼ及びシステインプロテアーゼ(即ち、カテプシン))が見出される。カテプシン及びMMPは、すべての細胞外マトリックス蛋白質の分解に関与する。プラーク安定性にはマトリックスが不可欠であるため、プロテアーゼによる線維性被膜からのその除去はプラーク破裂を引き起こし得る(Stary H.C)。
【0085】
表9に動脈硬化性プラークに見出される種々のプロテアーゼが一覧にされている。
【0086】
【表9】
【0087】
プラークにおけるMMP発現の主要源はマクロファージ及びSMC活性に関連する疑いがある。プラークにおけるマクロファージは豊富なMMP−1,−8,−9及び−13を含み、in situにてコラーゲン及びプロテオグリカン分解部位と共局在する(Kunz J.)。更に、自身のデータは動脈硬化性プラークにおけるMMP−8及びカテプシンKの局在を示唆する。
【0088】
<マトリックスメタロプロテアーゼ(MMP)>
MMPは、ECMの大部分の構成成分を分解することができるエンドペプチダーゼの大きな一群である。現在、25超のMMPが同定されている。メタロプロテアーゼは、金属原子、典型的には亜鉛を含む活性部位を特徴とし、チモーゲンとして分泌される。特定の組織阻害物質TIMPがMMPの活性を調節する。様々なMMPが動脈硬化性プラークに見出される。それらはほとんどの場合、SMC及びマクロファージにおいてプラーク肩部内にて線維性被膜に接するマクロファージにあり、線維性被膜内に同定されることは稀である(Kunz J.)。]
【0089】
MMPはその基質特異性によって異なる群に分類される。I,II,III及びV型コラーゲンのような線維状コラーゲン、更にプロテオグリカンも分解するコラゲナーゼ;プロテオグリカン、IV,V,VII型コラーゲン及びエラスチンを分解するゼラチナーゼ;プロテオグリカン及びエラスチンに対して活性を示すストロメライシン(Rouis M)。これらの3つのサブグループは動脈硬化性プラークにおけるマトリックス再構築に関して特に興味深い。
【0090】
<ゼラチナーゼ>
不溶性エラスチンは、ともにMMPのゼラチナーゼファミリーに属するMMP−2及び−9によって消化される。MMP−9は、動脈硬化性プラークのサイズ及び組成に作用する重要な役割を有する。不安定なヒト動脈硬化性プラーク及びプラークの脆弱領域では、MMP−9のより高い発現及び濃度が認められている。更に、MMP−9は、安定狭心症患者と比べて不安定狭心症患者においてより多く、冠動脈プラークにおいて細胞内に見出される(活発な合成を示す)。血中MMP−9濃度は、冠状動脈アテローム性動脈硬化と関連して上昇し、有害な心血管事象を予測する(Sundstrom and Vasan)。Kuzuya et al(2006)による最近の研究は、MMP−2が線維性被膜におけるSMCの蓄積に関与し、これにより、プラーク不安定性を誘発することを示す。
【0091】
<ストロメライシン>
MMP−3はストロメライシンプロテアーゼに属し、エラスチン及びプロテオグリカンを分解することができる。Yamada et al(2002)による研究は、MMP−3が女性における心筋梗塞の遺伝的リスクを予測する信頼できる手段であることが判明し得ることを示す。
【0092】
<コラゲナーゼ>
MMP−1,−8及び−13はすべて動脈硬化性プラークにおいて同定され、プロテオグリカン及びI,III型コラーゲンを分解する。
【0093】
MMP−1,−8及び−13はコラゲナーゼであり、コラーゲンを2つの断片に切断し、その断片はMMP−2,−3又は−9によって更に分解される。
【0094】
MMP−8は、ヒトアテロームには一般的に見出されないが動脈硬化性プラークにおいて同定されている好中球によって発現される。MMP−8は、I型コラーゲンを優先的に選択し(Herman et al)、MMP−1及び13より3倍大きいI型コラーゲンの分解活性を有するため、線維性被膜の分解に部分的に関与し得る。このことはTuru et al(2006)によって支持され、この研究では、血漿中のMMP−8含量は安定プラークを有する患者より不安定プラークを有する患者で著しく多い。
【0095】
MMP−13はバイグリカンに対する高特異性を有し、SLRPを切断することが報告されている。特定の切断部位(...G177/V178)におけるMMP−13によるバイグリカンの分解はMonfort et al.(2005)によって従前に示され、骨関節炎における軟骨分解の早期検出に重要な役割を果たすことが提案されている。
【0096】
<カテプシン>
ヒトシステインカテプシンは、カテプシンB,K,L及びSを含む11のメンバーから成り、主に細胞のエンドソーム/リソソームコンパートメント内に発現する。カテプシンはプロテオグリカン、コラーゲン及びエラスチンの加水分解を触媒することができる。
【0097】
腹部大動脈瘤(AAA)において、正常大動脈と比べて高レベルのカテプシンS,K及びLが見出された。正常ヒト血管SMCは免疫染色により検出可能なカテプシンKを含まないが、動脈硬化性プラーク内の細胞は明らかに陽性である。カテプシンKは、破裂しやすい領域、例えば、線維性被膜、プラーク肩部及びプラーク破裂の実際の部位に局在する(Chapman et al)。カテプシンSは動脈硬化性プラークにおいてエラスチン分解の増大領域と共局在することが見出され、アテローム性動脈硬化の減少がカテプシンS及びK欠損マウスにおいて認められる(Liu et al)。
【0098】
カテプシンL及びKは、いくつかのプロテオグリカン及びI,II型コラーゲンを分解し、カテプシンKは共有結合的に架橋された三重らせん内にて分解し、一方、カテプシンLは非らせんテロペプチド領域においてのみ切断する。カテプシンKは線維性被膜及びプラーク肩部に局在する。正常動脈におけるカテプシンKの発現は極めて低い。早期ヒトアテローム性動脈硬化病変は内膜及び中膜SMCにおけるカテプシンKの発現を示した。進展した動脈硬化性プラークにおいて、カテプシンKは主に線維性被膜のマクロファージ及びSMCに局在した(Lutgens et al)。カテプシンK蛋白質レベルは、正常動脈と比べてアテローム性動脈硬化病変において上昇したが、一方、カテプシンKのmRNAレベルは、アテローム性動脈硬化血管及び正常動脈において同様であった。更に、カテプシンKのmRNA及び蛋白質レベルは、早期アテローム性動脈硬化病変及び血栓を含む病変と比べて、進展しているが安定したヒト動脈硬化性プラークにおいて最大であることが示された(Chapman et al)。
【0099】
カテプシンSは、早期ヒトアテローム性動脈硬化病変及び脂肪線条において内膜及び中膜SMCにほんのわずかに発現する。進展したヒト動脈硬化性プラークにおいて、カテプシンSは線維性被膜のマクロファージ及びSMCに局在した。血管自体及びプラーク微小血管の管腔の内側を覆うECも、カテプシンSを発現した。更に、カテプシンSのmRNA及び蛋白質レベルは、正常動脈と比べてヒトアテロームにおいて増大した(Lutgens et al)。カテプシンSはプロテオグリカン、エラスチン及びコラーゲンを分解することができる(Liu et al)。
【発明の概要】
【0100】
現在、CVDリスクの決定は、アテローム性動脈硬化進行の後期(線維性プラーク破裂の有意なリスクがある時点)において行われている。早期及び後期におけるアテローム性動脈硬化又はCVDリスクに関する情報を提供する診断又は予後アッセイの必要性がある。Katsuda et al(1992)の知見は、進展病変からのコラーゲンの除去の酵素機構があることを示唆し、実際に動脈硬化におけるネオエピトープの主要な役割を示唆している。
【0101】
本発明は、プロテイナーゼによる動脈硬化性プラークの蛋白質の切断によって形成されるネオエピトープを含むペプチド断片の定量のためのバイオアッセイの方法を提供し、該方法は、該ペプチド断片を含む試料を、該ネオエピトープに対する特異的結合親和性を有する免疫学的結合パートナーと接触させるステップと、該試料中の該免疫学的結合パートナーのペプチド断片への結合レベルを定量するステップとを含む。
【0102】
該アッセイの結果は、特定の患者における動脈硬化性プラークの破裂のリスクの程度又は患者の動脈硬化性プラークの不安定状態を示す指標を生成し得る。
【0103】
閾値を越える該指標の値を有する患者は、プラーク撮像法(上述の撮像法を含む)による更なる検査又はアテローム性動脈硬化の処置のための薬剤の処方又はアテローム性動脈硬化の外科的処置を推奨され得、そのようなフォローアップ検査又は処置は本発明の方法の一部を成し得る。
【0104】
動脈硬化性プラークの蛋白質は、ルミカン、バーシカン、パールカン、デコリン、バイグリカン、III型コラーゲン、CRP、ApoE及びエラスチンを含む。I型コラーゲンは動脈硬化性プラークの蛋白質であるとは見なされない。動脈硬化性プラークに存在し、身体のどの部位よりも高い程度に動脈硬化性プラークにおいてプロテアーゼに晒される蛋白質が特に興味深い。
【0105】
該免疫学的結合パートナーは、C末端ネオエピトープ又はN末端ネオエピトープを含むペプチド断片に対する特異的結合親和性を有し得る。
【0106】
プロテオグリカンアッセイ
該ペプチド断片は、プロテオグリカンバーシカン(配列番号1)、ルミカン(配列番号2)、パールカン(配列番号3)、バイグリカン(配列番号4)及びデコリン(配列番号5)の断片でよく、それらはすべて正常及びアテローム性動脈硬化血管において同定される。プロテオグリカンは、エラスチン及びコラーゲンとともに、動脈硬化性プラーク及びプラーク被膜を構成する主な蛋白質の一部である。プロテオグリカンの含量は、アテローム性動脈硬化の進行時に異なり、その結果、プロテオグリカンの潜在的ネオエピトープは病期及び疾患進行の良好なマーカーになる。特に、バーシカン及びルミカンは多くの他の臓器には豊富ではないため、これにより、それらはより特異的な生化学マーカー候補になる。
【0107】
動脈硬化性プラークにおいて多くの異なるプロテアーゼが文献により報告されているように、複数の候補プロテアーゼがプラークにおけるプロテオグリカンの消化に関与し得る。おそらく、これは最終的にプラーク破裂に至る広範の複雑なプロセスの結果である。しかし、本発明者らの評価では、初期相は一定範囲のMMPから成り得、一方、後期はマトリックスのカテプシン分解により依存し得、病期に依存して異なるネオエピトーププロファイルとなる。本発明者らは、表4に一覧表示される酵素がルミカン、バーシカン、バイグリカン、パールカン及びデコリンを生じさせ、少なくとも以下の切断生成物を生じさせると判定した。
【0108】
【表10A】
【表10B】
【0109】
従って、本発明の方法では、好ましくは、該ペプチド断片は、上記部分配列のいずれの配列にもおける記号「*」を付された部位におけるプロテアーゼによるバーシカン、ルミカン、パールカン、デコリン又はバイグリカンの切断によって形成されるネオエピトープを含む。
【0110】
更に、本発明の方法では、好ましくは、該ペプチド断片は、バーシカン、ルミカン、デコリン、パールカン及びバイグリカンの以下の部分配列のいずれの配列にもおける部位における1つ(又はそれ以上)のプロテアーゼによるプロテオグリカン(バーシカン、ルミカン、パールカン、デコリン及びバイグリカン)の切断によって形成されるネオエピトープを含み、或いは免疫学的結合パートナーは以下の配列の1つと特異的に反応する。
【0111】
【表11】
【0112】
好ましくは、該免疫学的結合パートナーは、切断されていない状態のバーシカン、ルミカン、デコリン、パールカン及びバイグリカンと反応しない。好ましくは、該免疫学的結合パートナーは、生成断片のそれぞれのc末端及びN末端を過ぎて延長している場合、上記一覧の該配列と反応しない。
【0113】
免疫学的結合パートナーは、バーシカン、ルミカン、デコリン、パールカン及びバイグリカンタイプの切断によって形成されるC末端又はN末端ネオエピトープと特異的に反応する免疫学的結合パートナーでもよい。
【0114】
従って、好適な免疫学的結合パートナーは、ペプチドのN末端において表11における以下の配列のいずれかの又はペプチドのC末端において表12における以下の配列と特異的に反応し得る。
【0115】
【表12】
【0116】
【表13】
【0117】
同様にアッセイされ得るネオエピトープを決定する更なる切断部位は、プロテオグリカン又は他の動脈硬化性プラーク蛋白質を本明細書で述べられる任意の酵素と反応させ、そうして生成されるペプチドを単離して配列決定することによって同定され得る。
【0118】
<コラーゲンアッセイ>
該ペプチド断片はIII型コラーゲンの(配列番号153)、好ましくは、成熟III型コラーゲンの、即ち、III型コラーゲンプロペプチドの断片ではない断片でもよい。動脈硬化性プラークにおける主な蛋白質はI,III型コラーゲン及びエラスチンであり、一方、プロテオグリカンはプラークのマトリックスにわずかに寄与するのみである。動脈硬化性プラークに見出される3つの主要な蛋白質中、I及びIII型コラーゲンが優位であり、一方、エラスチンは動脈における蛋白質プロファイルを占めるが、プラークにおける主要な蛋白質成分ではない。I型コラーゲンはヒトの身体全体にわたって豊富であり、一方、III型はより限定された組織位置を有し、これにより、本発明者らの見解では生化学マーカーとしてより特異的な候補を構成する。
【0119】
動脈硬化性プラークにおける多くの異なるプロテアーゼが文献により報告されているように、複数の候補プロテアーゼがプラークにおけるコラーゲンの消化に寄与し得る。おそらく、これは、最終的にプラーク破裂に至る広範の複雑なプロセスの結果である。しかし、本発明者らの評価では、初期相は一定範囲のMMPから成り得、一方、後期はマトリックスのカテプシンK分解により依存し得、疾患レベルに依存して異なるネオエピトーププロファイルとなる。本発明者らは、以下の表に一覧表示される酵素が少なくとも以下の切断部位においてIII型コラーゲンを切断すると判定した。
【0120】
【表14A】
【表14B】
【表14C】
【表14D】
【表14E】
【0121】
従って、本発明の方法では、好ましくは、該ペプチド断片は、III型コラーゲンの上記部分配列のいずれの配列にもおける記号「*」を付された部位におけるプロテアーゼによるIII型コラーゲンの切断によって形成されるネオエピトープを含む。
【0122】
更に、本発明の方法では、好ましくは、該ペプチド断片は、*間におけるIII型コラーゲンの上記部分配列のいずれの配列にもおける部位における1つ(又はそれ以上)のプロテアーゼによるIII型コラーゲンの切断によって形成されるネオエピトープを含み、または免疫学的結合パートナーは上記表のエントリーにおける*間に伸長する配列と特異的に反応する。
【0123】
好ましくは、該免疫学的結合パートナーは、切断されていない状態のIII型コラーゲンと反応しない。好ましくは、該免疫学的結合パートナーは、それぞれの切断部位を過ぎて延長している場合、上記一覧の該配列と反応しない。
【0124】
免疫学的結合パートナーは、III型コラーゲンの切断によって形成されるC末端又はN末端ネオエピトープと特異的に反応する免疫学的結合パートナーでよい。
【0125】
従って、好適な免疫学的結合パートナーは、ペプチドのN末端において以下の配列(各配列の後に配列番号)又はペプチドのC末端において以下の配列と特異的に反応し得る。
【0126】
【表15】
【0127】
【表16】
【0128】
同様にアッセイされ得るネオエピトープを決定する更なる切断部位は、III型コラーゲン又は他の動脈硬化性プラーク蛋白質を本明細書で述べられる任意の酵素と反応させ、そうして生成されるペプチドを単離して配列決定することによって同定され得る。
【0129】
<CRP及びApoEアッセイ>
該ペプチド断片はCRP(配列番号658)又はApoE(配列番号659)の断片でもよい。好ましくは、ApoEでは、選択断片はApoEのすべての同定アイソタイプ、ε2,ε3及びε4において生じる。
【0130】
CRP及びApoEがヒトの身体全体にわたって豊富であるとしても、アテローム性動脈硬化組織におけるその局在により、それらは局所プロテアーゼの作用に晒される。これにより、これらの分子は生化学マーカーとして良好且つ特異的な候補である。
【0131】
動脈硬化性プラークにおける多くの異なるプロテアーゼが文献により報告されているように、複数の候補プロテアーゼがプラークにおけるCRP及びApoEの消化に寄与し得る。おそらく、これは、最終的にプラーク破裂に至る広範の複雑なプロセスの結果である。しかし、初期相は一定範囲のMMPから成り得、一方、後期はマトリックスのカテプシンK分解により依存し得、疾患レベルに依存して異なるネオエピトーププロファイルとなる。本発明者らは、純粋天然蛋白質の一定範囲のin vitro切断を通じて、以下の表に一覧表示される酵素が少なくとも以下の切断部位(*を付す)においてCRP及びApoEを切断すると判定した。
【0132】
【表17】
【0133】
従って、本発明の方法では、好ましくは、該ペプチド断片は、CRP及びApoEの以下の部分配列のいずれの配列にもおける記号「*」が付された部位におけるプロテアーゼによるCRP及びApoEの切断によって形成されるネオエピトープを含み、または免疫学的結合パートナーは以下の配列の1つにおける*間に画定される配列と特異的に反応する。
【0134】
【表18】
【0135】
従って、好適な免疫学的結合パートナーは、ペプチドのN末端において以下の配列又はペプチドのC末端において以下の配列と特異的に反応し得る。
【0136】
【表19】
【0137】
【表20】
【0138】
同様にアッセイされ得るネオエピトープを決定する更なる切断部位は、CRP及びApoEまたは他の動脈硬化性プラーク蛋白質を本明細書で述べられる任意の酵素と反応させ、そうして生成されるペプチドを単離して配列決定することによって同定され得る。
【0139】
<エラスチンアッセイ>
該ペプチドはエラスチン(配列番号735)の断片でもよい。エラスチンはヒトの身体全体にわたって豊富であるとしても、アテローム性動脈硬化組織におけるその局在はそれを局所プロテアーゼの作用に晒させ、そのためにこれらの分子は動脈硬化性プラーク代謝回転の生化学マーカーとして良好且つ特異的な候補である。
【0140】
動脈硬化性プラークにおける多くの異なるプロテアーゼが文献により報告されているように、複数の候補プロテアーゼがプラークにおけるエラスチンの消化に寄与し得る。おそらく、これは、最終的にプラーク破裂に至る広範の複雑なプロセスの結果である。しかし、初期相は一定範囲のMMPから成り得、一方、後期はマトリックスのカテプシンK分解により依存し得、疾患レベルに依存して異なるネオエピトーププロファイルとなる。本発明者らは、純粋天然蛋白質の一定範囲のin vitro切断を通じて、以下の表に一覧表示される酵素が少なくとも以下の切断部位(*を付す)においてエラスチンを切断すると判定した。
【0141】
【表21】
【0142】
従って、本発明の方法では、好ましくは、該ペプチド断片は、エラスチンの以下の部分配列のいずれの配列にもおける記号「*」が付された部位におけるプロテアーゼによるエラスチンの切断によって形成されるネオエピトープを含み、或いは免疫学的結合パートナーは以下の配列の1つにおける*間に画定される配列と特異的に反応する。
【0143】
【表22】
【0144】
従って、好適な免疫学的結合パートナーは、ペプチドのN末端において以下の配列又はペプチドのC末端において以下の配列と特異的に反応し得る。
【0145】
【表23】
【0146】
【表24】
【0147】
同様にアッセイされ得るネオエピトープを決定する更なる切断部位は、エラスチン又は他の動脈硬化性プラーク蛋白質を本明細書で述べられる任意の酵素と反応させ、そうして生成されるペプチドを単離して配列決定することによって同定され得る。
【0148】
上述の2つ以上のペプチドのアッセイは別個に行われ、それらの結果は組み合わせられ得、又は上述の2つ以上のペプチドはともに測定され得る。
【0149】
本発明によるアッセイの結果は、診断又は予後値の総合指数を形成するために1つ以上の他の測定バイオマーカーと組み合わせられ得る。
【0150】
本明細書で用いられる「免疫学的結合パートナー」という用語は、ポリクローナル及びモノクローナル抗体、更に抗体の特異的な結合断片、例えば、Fab又はF(ab’)2も含む。従って、該免疫学的結合パートナーはモノクローナル抗体又は特異的結合親和性を有するモノクローナル抗体の断片でよい。
【0151】
本明細書で用いられる「蛋白質」という用語は、リポ蛋白質及びプロテオグリカン並びに他の蛋白(または非蛋白)の天然複合体を含む。
【0152】
一般的に、本発明に従って不均一及び均一フォーマット、サンドイッチアッセイ、競合アッセイ、酵素結合アッセイ、放射免疫アッセイなどを含むあらゆる既知のイムノアッセイフォーマットが用いられ得る。従って、任意に、該方法は、該免疫学的結合パートナー及び競合物質が該試料の存在下にてインキュベートされ、競合物質が免疫学的結合パートナーと結合するために試料中のペプチド断片と競合する競合イムノアッセイとして行われる。
【0153】
該競合物質は、該ネオエピトープを示すためにネオエピトープが属する蛋白質の切断によって形成される合成ペプチド又は精製天然ペプチドでもよい。従って、ペプチドは、バーシカン、ルミカン、パールカン、デコリン、バイグリカン、III型コラーゲン、ApoE、CRP又はエラスチンに由来し得る。
【0154】
1つの好適な方法は、任意のこれらの蛋白質の断片のネオエピトープ若しくは動脈硬化性プラークに由来する他の蛋白質由来のペプチド断片上のネオエピトープに結合するモノクローナル抗体、又はその抗体結合断片を用いた競合イムノアッセイでよい。マイクロタイタープレートの固体表面上に被覆された、適切に選択された合成ペプチドは、モノクローナル抗体又は結合断片への結合について試料と競合し得る。或いは、モノクローナル抗体又は結合断片によって認識されるネオエピトープを保有する1つ以上のこれらの蛋白質由来の精製天然断片が、固体表面上に用いられ得る。更に別の代替例は、モノクローナル抗体又は結合断片を固体表面上に固定化し、次に、試料を、シグナル分子、例えば、西洋ワサビペルオキシダーゼ又はビオチンに適切に結合した合成ペプチドと共インキュベートすることである。試料は尿、血清、血液、血漿又は他の、例えば、動脈硬化性プラーク生検の試料でもよい。
【0155】
一部の好ましい方法では、試料は患者由来の試料であり、方法は、該ペプチド断片の該結合の定量レベルを、(a)比較できる健常患者及び/又は(b)アテローム性動脈硬化病態の特徴を示す数値と比較するステップと、任意に、高レベルの測定ペプチド(通常、高レベルの結合によって示される)をより重症度の該病態と関連付けるステップとを更に含む。
【0156】
本発明の一態様は、上述のネオエピトープを認識するモノクローナル抗体の開発に関する。これは、当該蛋白質分子のアミノ酸配列に由来する合成ペプチド(上記一覧表示された配列又はそこで終止する配列を含む)でマウスを免疫化し、選択マウス由来脾臓細胞を骨髄腫細胞に融合し、関連合成ペプチド上のネオエピトープへの結合についてモノクローナル抗体を試験することによって達成され得る。ネオエピトープに対する特異性は、合成ペプチドとの反応性及び(C末端ネオエピトープに対する)C末端延長形態の免疫ペプチド若しくは(N末端ネオエピトープに対する)N末端延長形態の免疫ペプチドとの反応性の欠如を必要とすることによって確実にされ得る。天然蛋白質に対する結合能の欠如を確定するために、ネオエピトープに対する抗体も評価され得る。或いは、ネオエピトープに対する特異性は、抗体の反応性が末端アミノ酸の1つに共有結合したビオチン又は他の官能基の存在が負に依存するよう求めることによって確実にされ得る。
【0157】
本発明は、上記に示される部分配列のいずれか1つの部分配列における末端部位におけるプロテアーゼによる該蛋白質の切断によって形成されるネオエピトープと特異的に免疫反応する免疫学的結合パートナーを含み、例えば、モノクローナル抗体又はその結合断片でもよい。
【0158】
本発明は、上記に示される部分配列のいずれか1つの部分配列における配列の末端部位における動脈硬化性プラーク蛋白質の切断によって形成されるC末端又はN末端ネオエピトープに対するモノクローナル抗体を産生する細胞株を含む。
【0159】
本発明は、上記に示されるこれらの蛋白質の部分配列のいずれか1つの部分配列における該蛋白質の切断によって形成されるC末端又はN末端ネオエピトープを含むペプチドを更に提供する。そのようなペプチドはイムノアッセイで用いるために、該ペプチドに対する免疫反応を生じさせるためにハプテンとして担体にコンジュゲートされ、又は固体表面に固定化され、又は検出可能なマーカーにコンジュゲートされ得る。
【0160】
本発明は、上記に示される部分配列のいずれか1つの部分配列における該蛋白質の切断によって形成されるC末端又はN末端ネオエピトープを含むペプチドをコードする単離核酸分子を更に含む。
【0161】
本発明は、発現シグナルと、上記に示される部分配列のいずれか1つの部分配列における該蛋白質の切断によって形成されるC末端又はN末端ネオエピトープを含むペプチドの発現をコードするコード配列とを含む核酸配列を含むベクターを更に含み、そのようなベクターで形質転換され、該ペプチドを発現する宿主細胞を更に含む。
【0162】
本発明の更に別の態様は、上述の方法を実施するために好都合に用いられ得るキットに関する。そのようなキットは、(1)合成ペプチドで被覆されたマイクロタイタープレートと、(2)該合成ペプチドと反応する本発明のモノクローナル抗体若しくは抗体結合断片と、(3)標識抗マウスIgG免疫グロブリンとを含み得る。或いは、そのようなキットは、(1)精製天然蛋白質断片で被覆されたマイクロタイタープレートと、(2)該蛋白質のいずれか1つの断片上のネオエピトープを認識し、該精製断片と反応するモノクローナル抗体と、(3)標識抗マウスIgG免疫グロブリンとを含み得る。或いは、そのようなキットは、(1)ストレプトアビジンで被覆されたマイクロタイタープレートと、(2)ビオチンに結合した合成ペプチドと、(3)該蛋白質断片上のネオエピトープを認識し、該合成ペプチドと反応するモノクローナル抗体と、(4)標識抗マウスIgG免疫グロブリンとを含み得る。更に別の代替例は、(1)ストレプトアビジンで被覆されたマイクロタイタープレートと、(2)ビオチンに結合した合成ペプチドと、(3)該蛋白質断片上のネオエピトープを認識し(該合成ペプチドと反応し)、西洋ワサビペルオキシダーゼにコンジュゲートしたモノクローナル抗体とを含むキットでよい。
【0163】
従って、本発明は、本明細書で述べられる免疫学的結合パートナー並びに該免疫学的結合パートナーに結合する競合物質、並びに、任意に、1つ以上の洗浄試薬、緩衝液、停止試薬、酵素標識、標識酵素のための基質、較正基準、抗マウス抗体及び該イムノアッセイを行うための使用説明書を含むイムノアッセイキットを含む。
【0164】
本明細書で述べられるアッセイは患者におけるアテローム性動脈硬化症の診断に有用である。加えて、検査は疾患の進行の評価及び治療に対する応答のモニタリングに有用である。本発明の免疫学的結合パートナーは、本明細書で述べられる動脈硬化性プラーク蛋白質の切断産物の存在又は位置を示すために免疫染色でも用いられ得る。
【0165】
本発明は、添付図面を参照して更に説明され、例示される。
【図面の簡単な説明】
【0166】
【図1】III型病変を含む大動脈試料に対してモノクローナルマウス抗体を用いたバイグリカン染色(それぞれ2,4,4及び10倍)を示す図である。
【図2】III型病変を含む大動脈試料に対してモノクローナルマウス抗体を用いたカテプシンK染色(それぞれ2,4,10及び10倍)を示す図である。
【図3】V型病変を含む大動脈試料に対してモノクローナルマウス抗体を用いたバイグリカン染色(それぞれ2,4,10及び10倍)を示す図である。
【図4】V型病変を含む大動脈試料に対してモノクローナルマウス抗体を用いたカテプシンK染色(それぞれ2,4,10及び10倍)を示す図である。
【図5】プロテアーゼ:MMP2、MMP3、MMP8、カテプシンK、カテプシンS、カテプシンB及びカテプシンLによって生じるバイグリカンの切断産物を示す図である。M=着色マーカー(Rainbow marker)、−enz=酵素消化なし、実施例2においてゲル上で泳動。
【図6】実施例4において得られた競合試験結果を示す図である。
【図7】実施例4において得られた競合試験結果を示す図である。
【図8】実施例4において得られた競合試験結果を示す図である。
【図9】実施例6において得られた競合試験結果を示す図である。
【図10】実施例7において得られた競合試験結果を示す図である。
【図11】実施例7において得られた競合試験結果を示す図である。
【実施例1】
【0167】
プロテオグリカン及びプロテアーゼの局在の分析のため、左冠動脈下行枝(LAD)由来のヒト動脈試料の免疫組織化学染色を行った。以下、カテプシンKプロテアーゼ及びバイグリカンの共局在が示される。
【0168】
図1及び2に見られる免疫組織化学染色はバイグリカン及びカテプシンKの共局在を明らかにした。これは、バイグリカンがカテプシンKの好ましい基質であることを示唆し得る。動脈硬化性プラークが形成された大動脈試料に対して同じ免疫組織化学染色を行い、この結果として正常な大動脈構造はマクロファージ泡沫細胞浸潤及び石灰化に置き換えられた。これらの免疫染色を図3及び4にまとめる。
【0169】
バイグリカン及びカテプシンKの免疫組織化学染色は進行性アテローム性動脈硬化病変に共局在することが示された。これらの結果はともにバイグリカンにおける特異的なカテプシンK切断部位の仮説を生じさせ、アテローム性動脈硬化病変におけるネオエピトープ生成の増加をもたらす。この仮説を試験するため、本発明者らはバイグリカンを異なるプロテアーゼで切断した。
【実施例2】
【0170】
分解断片の評価のためのバイグリカンの分解。ウシ関節軟骨由来のバイグリカン(B8041−Sigma−Aldrich社)をプロテアーゼ(MMP2、MMP3、MMP8、カテプシンK、カテプシンS、カテプシンB及びカテプシンL)によって切断した。上述のプロテアーゼの酵素切断によって生じたプロテオグリカンの断片を10%のNuPage(登録商標)Bis−Trisゲル上に分離し、その後、「Silver Express」−銀染色キット(Invitrogen社カタログ番号LC6100、ロット番号341099)によって銀染色した。蛋白質分解で得られたバイグリカンの分離及び銀染色の結果を図5に示す。
【実施例3】
【0171】
オボアルブミンにコンジュゲートしたIII型コラーゲン由来ペプチドでマウスを免疫化した。ビオチンにコンジュゲートしたスクリーニングペプチド配列との反応性について血清をスクリーニングした。モノクローナル抗体分泌クローンを作製し、スクリーニング配列を用いてスクリーニングした。III型コラーゲン由来の隣接配列で連続する標的ペプチドの伸長バージョン(除外ペプチド)との反応性の欠如及びナンセンスペプチドとの反応性の欠如についてクローンを調べた。
【0172】
標的配列、免疫原、スクリーニング配列及び除外配列は以下の通りであった。
【0173】
【表25】
【実施例4】
【0174】
<III型コラーゲンネオエピトープモノクローナル抗体のヒト尿との反応性>
免疫ペプチドを競合物質として用いて競合アッセイ形式にて実施例3からの選択モノクローナル抗体クローンのヒト尿との反応性を判定した。典型的な手順では、96ウェルストレプトアビジン被覆プレートを振盪しながら20℃でPBS−BTE中10ng/mLビオチンペプチドで30分間コートし、5倍洗浄緩衝液で洗浄した。20μlの希釈試料(尿又はペプチド溶液)を加えた。下記に詳述する100μLの未精製抗体溶液(細胞培養液由来の上清)を加えた。プレートを300rpmで振盪しながら20℃で1時間インキュベートし、次に、5倍洗浄緩衝液で洗浄した。100μLの二次抗体−POD(1:5000)を加え、300rpmで振盪しながら20℃で1時間インキュベートし、その後、5倍洗浄緩衝液で洗浄した。100μLのTMBを加え、300rpmで振盪しながら暗室で15分間インキュベートし、その後、停止液100μLを加えた。ELISAリーダー上で650nmを対照として450nmにてプレートを読み取った。従って、抗体に対するプレート上のペプチドと溶液中のペプチドとの間の競合が生じ、ペルオキシダーゼ発色反応によりプレート結合抗体を定量した。
【0175】
図6に4つの異なるクローンの結果が認められる。抗体が各々尿中の関連する配列を検出することがわかる。
【0176】
免疫ペプチドとMMP9によってin vitroにて切断された天然III型コラーゲンとの間の抗体結合の競合を試験するため、1つの選択クローンに対して更なる競合試験を行った。切断コラーゲン、ペプチドKNGETG及びその配列の伸長バージョンの結果を図7に示す。抗体が免疫ペプチド配列及び酵素切断コラーゲンに結合するが、伸長配列には結合しないことがわかる。
【0177】
図8に同じクローンに関する更なる競合試験が認められ、この場合、競合物質はそれぞれ、ペプチドKNGETG、ヒト血清、ラット血清、FCS(ウシ胎仔血清)及び動脈硬化性プラーク抽出物であった。抗体がペプチド、プラーク抽出物及びヒト血清と反応するが、ラット血清又はFCSとは反応しないことがわかる。
【実施例5】
【0178】
<デコリン、バイグリカン及びバーシカン配列に対する抗血清の産生>
抗血清を産生させ、実施例3のようにモノクローナル抗体を得たが、以下の免疫原、スクリーニング配列及び除外配列を用いた。
【0179】
【表26】
【実施例6】
【0180】
<デコリンネオエピトープモノクローナル抗体のヒト尿との反応性>
1つの抗デコリン未精製モノクローナル抗体(NB62)を用いて概して実施例5のように競合ELISAを行った。
【0181】
図9に結果を示す。これに対して抗体が産生され、選択されたペプチド配列及び尿に対する反応が認められるが、無関連ペプチド配列に対する反応は認められない。
【実施例7】
【0182】
<バーシカンネオエピトープモノクローナル抗体のヒト尿との反応性>
配列に対する2つの抗バーシカン未精製モノクローナル抗体(NB64)を用いて概して実施例5のように競合ELISAを行った。
【0183】
それぞれのクローンの結果を図10及び11に示す。それぞれの場合、これに対して抗体が産生され、選択されたペプチド配列及び尿に対する反応が認められるが、無関連ペプチド配列NB18に対する反応は認められない。
【0184】
本明細書では、特に明示されない限り、条件の1つのみが満たされることを要求する演算子「排他的or」と対照的に、「又は」という語は言及条件の一方又は両方が満たされる場合に真値を返す演算子の意味で用いられる。「含む(comprising)」という語は「〜から成る」を意味するというよりも「含む(including)」という意味で用いられる。上記に認められた従来の教示はすべて参照して本明細書に組み込まれる。
【0185】
[参考文献リスト]
【表27A】
【表27B】
【表27C】
【表27D】
【表27E】
【表27F】
【表27G】
【特許請求の範囲】
【請求項1】
プロテイナーゼによる動脈硬化性プラークの蛋白質の切断によって形成されるネオエピトープを含むペプチド断片の定量のためのバイオアッセイの方法であって、前記ペプチド断片を含む試料を、前記ネオエピトープに対する特異的結合親和性を有する免疫学的結合パートナーと接触させるステップと、前記試料中の前記免疫学的結合パートナーのペプチド断片への結合レベルを定量するステップとを含む方法。
【請求項2】
前記免疫学的結合パートナーが、C末端ネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1に記載の方法。
【請求項3】
前記免疫学的結合パートナーが、N末端ネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1に記載の方法。
【請求項4】
前記免疫学的結合パートナーが、プロテアーゼによるCRPの以下の部分配列
【表1】
のいずれか1つの配列におけるCRPの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1〜3のいずれか一項に記載の方法。
【請求項5】
前記免疫学的結合パートナーが、プロテアーゼによるApoEの以下の部分配列
【表2】
のいずれか1つの配列におけるApoEの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1〜3のいずれか一項に記載の方法。
【請求項6】
前記免疫学的結合パートナーが、ペプチドのN末端において以下の配列
【表3】
のいずれかの配列に対する特異的結合親和性を有し、または、
ペプチドのC末端において以下の配列
【表4】
のいずれかの配列に対する特異的結合親和性を有する、請求項4または5に記載の方法。
【請求項7】
前記ペプチド断片が、ルミカン、バーシカン、パールカン、デコリン、バイグリカンの断片である、請求項1〜3のいずれか一項に記載の方法。
【請求項8】
前記免疫学的結合パートナーが、ルミカン、バーシカン、パールカン、デコリン及びバイグリカンの以下の部分配列
【表5】
のいずれか1つの配列における、プロテアーゼによるルミカン、バーシカン、パールカン、デコリン又はバイグリカンの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項7に記載の方法。
【請求項9】
前記免疫学的結合パートナーが、ペプチドのN末端において以下の配列
【表6】
のいずれかの配列に対する特異的結合親和性を有する、請求項8に記載の方法。
【請求項10】
前記免疫学的結合パートナーが、ペプチドのC末端において以下の配列
【表7】
のいずれかの配列に対する特異的結合親和性を有する、請求項8に記載の方法。
【請求項11】
前記免疫学的結合パートナーが、III型コラーゲンの以下の部分配列
【表8A】
【表8B】
【表8C】
【表8D】
のいずれか1つの配列における記号*を付された部位における、プロテアーゼによるIII型コラーゲンの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1〜3のいずれか一項に記載の方法。
【請求項12】
前記免疫学的結合パートナーが、ペプチドのN末端において以下の配列
【表9】
のいずれかの配列に対する特異的結合親和性を有し、または、
ペプチドのC末端において以下の配列
【表10】
のいずれかの配列に対する特異的結合親和性を有する、請求項11に記載の方法。
【請求項13】
前記免疫学的結合パートナーが、エラスチンの以下の部分配列
【表11】
のいずれか1つの配列における、プロテアーゼによるエラスチンの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1〜3のいずれか一項に記載の方法。
【請求項14】
前記免疫学的結合パートナーが、ペプチドのN末端において以下の配列
【表12】
のいずれかの配列に対する特異的結合親和性を有し、または、
ペプチドのC末端において以下の配列
【表13】
のいずれかの配列に対する特異的結合親和性を有する、請求項13に記載の方法。
【請求項15】
前記免疫学的結合パートナーが、ネオエピトープ配列を含む未切断の蛋白質には特異的に結合しない、請求項1〜14のいずれか一項に記載の方法。
【請求項16】
前記免疫学的結合パートナーが、特異的結合親和性を有するモノクローナル抗体またはモノクローナル抗体の断片である、請求項1〜15のいずれか一項に記載の方法。
【請求項17】
前記方法が、前記免疫学的結合パートナー及び競合物質が前記試料の存在下にてインキュベートされ、前記競合物質が試料中のペプチド断片と競合して前記免疫学的結合パートナーと結合する競合イムノアッセイとして行われる、請求項1〜16のいずれか一項に記載の方法。
【請求項18】
前記競合物質が合成ペプチドであるか、前記ネオエピトープが現れるように前記エピトープの由来する蛋白質が切断されることにより形成される精製天然ペプチドである、請求項17に記載の方法。
【請求項19】
前記試料が、尿、血清、血液又は血漿の試料である、請求項1から18のいずれか一項に記載の方法。
【請求項20】
前記試料が患者由来試料であり、前記方法が、前記ペプチド断片の結合の定量レベルを、(a)比較できる健常患者および/または(b)アテローム性動脈硬化病態の特徴を示す数値と比較するステップをさらに含む、請求項1〜19のいずれか一項に記載の方法。
【請求項21】
ルミカン、バーシカン、パールカン、デコリン、バイグリカン、III型コラーゲン、ApoE、CRP又はエラスチンのプロテイナーゼ切断によって形成される、C末端又はN末端ネオエピトープに対する免疫学的結合パートナー。
【請求項22】
請求項5〜14のいずれか一項に示される前記蛋白質の部分配列のいずれか1つの部分配列におけるプロテアーゼによる前記蛋白質の切断によって形成されるネオエピトープと特異的に免疫反応する、請求項21に記載の免疫学的結合パートナー。
【請求項23】
モノクローナル抗体又はその結合断片である、請求項21または22に記載の免疫学的結合パートナー。
【請求項24】
請求項23に記載のモノクローナル抗体を産生する細胞株。
【請求項25】
請求項5〜14のいずれか一項に示される前記蛋白質の部分配列のいずれか1つの部分配列におけるプロテアーゼによる前記蛋白質の切断によって形成される、C末端又はN末端ネオエピトープを含むペプチド。
【請求項26】
イムノアッセイで用いるために、前記ペプチドに対する免疫反応を生じさせるためにハプテンとして担体にコンジュゲートされ、または固体表面に固定化され、または検出可能なマーカーにコンジュゲートされた、請求項25に記載のペプチド。
【請求項27】
請求項5〜14のいずれか一項に示される前記蛋白質の部分配列のいずれか1つの部分配列におけるプロテアーゼによる前記蛋白質の切断によって形成される、C末端又はN末端ネオエピトープを含むペプチドをコードする単離核酸分子。
【請求項28】
発現シグナルと、請求項5〜14のいずれか一項に示される前記蛋白質の部分配列のいずれか1つの部分配列におけるプロテアーゼによる前記蛋白質の切断によって形成されるC末端又はN末端ネオエピトープを含むペプチドの発現をコードするコード配列とを含む、核酸配列を含むベクター。
【請求項29】
請求項28に記載のベクターで形質転換され、前記ペプチドを発現する宿主細胞。
【請求項30】
請求項21〜23のいずれか一項に記載の免疫学的結合パートナーと、前記免疫学的結合パートナーに結合する競合物質と、任意選択的に、1つ以上の洗浄試薬、緩衝液、停止試薬、酵素標識、標識酵素のための基質、較正基準、抗マウス抗体及びキットを用いてアッセイを行うための使用説明書とを含むイムノアッセイキット。
【請求項1】
プロテイナーゼによる動脈硬化性プラークの蛋白質の切断によって形成されるネオエピトープを含むペプチド断片の定量のためのバイオアッセイの方法であって、前記ペプチド断片を含む試料を、前記ネオエピトープに対する特異的結合親和性を有する免疫学的結合パートナーと接触させるステップと、前記試料中の前記免疫学的結合パートナーのペプチド断片への結合レベルを定量するステップとを含む方法。
【請求項2】
前記免疫学的結合パートナーが、C末端ネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1に記載の方法。
【請求項3】
前記免疫学的結合パートナーが、N末端ネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1に記載の方法。
【請求項4】
前記免疫学的結合パートナーが、プロテアーゼによるCRPの以下の部分配列
【表1】
のいずれか1つの配列におけるCRPの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1〜3のいずれか一項に記載の方法。
【請求項5】
前記免疫学的結合パートナーが、プロテアーゼによるApoEの以下の部分配列
【表2】
のいずれか1つの配列におけるApoEの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1〜3のいずれか一項に記載の方法。
【請求項6】
前記免疫学的結合パートナーが、ペプチドのN末端において以下の配列
【表3】
のいずれかの配列に対する特異的結合親和性を有し、または、
ペプチドのC末端において以下の配列
【表4】
のいずれかの配列に対する特異的結合親和性を有する、請求項4または5に記載の方法。
【請求項7】
前記ペプチド断片が、ルミカン、バーシカン、パールカン、デコリン、バイグリカンの断片である、請求項1〜3のいずれか一項に記載の方法。
【請求項8】
前記免疫学的結合パートナーが、ルミカン、バーシカン、パールカン、デコリン及びバイグリカンの以下の部分配列
【表5】
のいずれか1つの配列における、プロテアーゼによるルミカン、バーシカン、パールカン、デコリン又はバイグリカンの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項7に記載の方法。
【請求項9】
前記免疫学的結合パートナーが、ペプチドのN末端において以下の配列
【表6】
のいずれかの配列に対する特異的結合親和性を有する、請求項8に記載の方法。
【請求項10】
前記免疫学的結合パートナーが、ペプチドのC末端において以下の配列
【表7】
のいずれかの配列に対する特異的結合親和性を有する、請求項8に記載の方法。
【請求項11】
前記免疫学的結合パートナーが、III型コラーゲンの以下の部分配列
【表8A】
【表8B】
【表8C】
【表8D】
のいずれか1つの配列における記号*を付された部位における、プロテアーゼによるIII型コラーゲンの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1〜3のいずれか一項に記載の方法。
【請求項12】
前記免疫学的結合パートナーが、ペプチドのN末端において以下の配列
【表9】
のいずれかの配列に対する特異的結合親和性を有し、または、
ペプチドのC末端において以下の配列
【表10】
のいずれかの配列に対する特異的結合親和性を有する、請求項11に記載の方法。
【請求項13】
前記免疫学的結合パートナーが、エラスチンの以下の部分配列
【表11】
のいずれか1つの配列における、プロテアーゼによるエラスチンの切断によって形成されるネオエピトープを含むペプチド断片に対する特異的結合親和性を有する、請求項1〜3のいずれか一項に記載の方法。
【請求項14】
前記免疫学的結合パートナーが、ペプチドのN末端において以下の配列
【表12】
のいずれかの配列に対する特異的結合親和性を有し、または、
ペプチドのC末端において以下の配列
【表13】
のいずれかの配列に対する特異的結合親和性を有する、請求項13に記載の方法。
【請求項15】
前記免疫学的結合パートナーが、ネオエピトープ配列を含む未切断の蛋白質には特異的に結合しない、請求項1〜14のいずれか一項に記載の方法。
【請求項16】
前記免疫学的結合パートナーが、特異的結合親和性を有するモノクローナル抗体またはモノクローナル抗体の断片である、請求項1〜15のいずれか一項に記載の方法。
【請求項17】
前記方法が、前記免疫学的結合パートナー及び競合物質が前記試料の存在下にてインキュベートされ、前記競合物質が試料中のペプチド断片と競合して前記免疫学的結合パートナーと結合する競合イムノアッセイとして行われる、請求項1〜16のいずれか一項に記載の方法。
【請求項18】
前記競合物質が合成ペプチドであるか、前記ネオエピトープが現れるように前記エピトープの由来する蛋白質が切断されることにより形成される精製天然ペプチドである、請求項17に記載の方法。
【請求項19】
前記試料が、尿、血清、血液又は血漿の試料である、請求項1から18のいずれか一項に記載の方法。
【請求項20】
前記試料が患者由来試料であり、前記方法が、前記ペプチド断片の結合の定量レベルを、(a)比較できる健常患者および/または(b)アテローム性動脈硬化病態の特徴を示す数値と比較するステップをさらに含む、請求項1〜19のいずれか一項に記載の方法。
【請求項21】
ルミカン、バーシカン、パールカン、デコリン、バイグリカン、III型コラーゲン、ApoE、CRP又はエラスチンのプロテイナーゼ切断によって形成される、C末端又はN末端ネオエピトープに対する免疫学的結合パートナー。
【請求項22】
請求項5〜14のいずれか一項に示される前記蛋白質の部分配列のいずれか1つの部分配列におけるプロテアーゼによる前記蛋白質の切断によって形成されるネオエピトープと特異的に免疫反応する、請求項21に記載の免疫学的結合パートナー。
【請求項23】
モノクローナル抗体又はその結合断片である、請求項21または22に記載の免疫学的結合パートナー。
【請求項24】
請求項23に記載のモノクローナル抗体を産生する細胞株。
【請求項25】
請求項5〜14のいずれか一項に示される前記蛋白質の部分配列のいずれか1つの部分配列におけるプロテアーゼによる前記蛋白質の切断によって形成される、C末端又はN末端ネオエピトープを含むペプチド。
【請求項26】
イムノアッセイで用いるために、前記ペプチドに対する免疫反応を生じさせるためにハプテンとして担体にコンジュゲートされ、または固体表面に固定化され、または検出可能なマーカーにコンジュゲートされた、請求項25に記載のペプチド。
【請求項27】
請求項5〜14のいずれか一項に示される前記蛋白質の部分配列のいずれか1つの部分配列におけるプロテアーゼによる前記蛋白質の切断によって形成される、C末端又はN末端ネオエピトープを含むペプチドをコードする単離核酸分子。
【請求項28】
発現シグナルと、請求項5〜14のいずれか一項に示される前記蛋白質の部分配列のいずれか1つの部分配列におけるプロテアーゼによる前記蛋白質の切断によって形成されるC末端又はN末端ネオエピトープを含むペプチドの発現をコードするコード配列とを含む、核酸配列を含むベクター。
【請求項29】
請求項28に記載のベクターで形質転換され、前記ペプチドを発現する宿主細胞。
【請求項30】
請求項21〜23のいずれか一項に記載の免疫学的結合パートナーと、前記免疫学的結合パートナーに結合する競合物質と、任意選択的に、1つ以上の洗浄試薬、緩衝液、停止試薬、酵素標識、標識酵素のための基質、較正基準、抗マウス抗体及びキットを用いてアッセイを行うための使用説明書とを含むイムノアッセイキット。
【図6】

【図7】
【図8】
【図9】
【図10】
【図11】
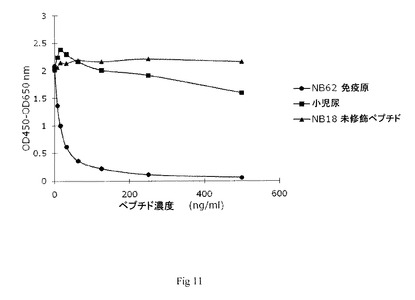
【図1】
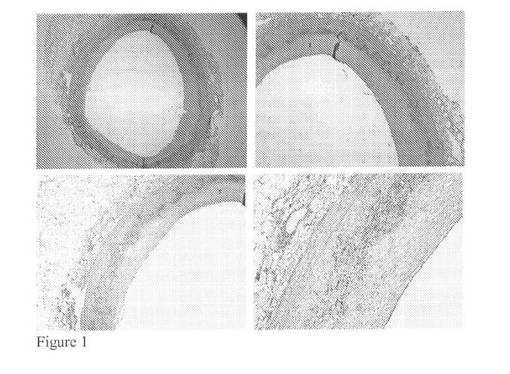
【図2】
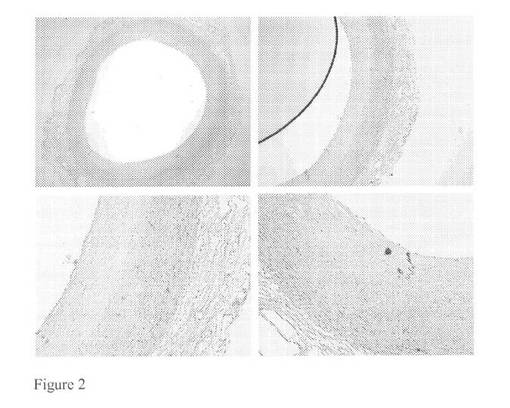
【図3】
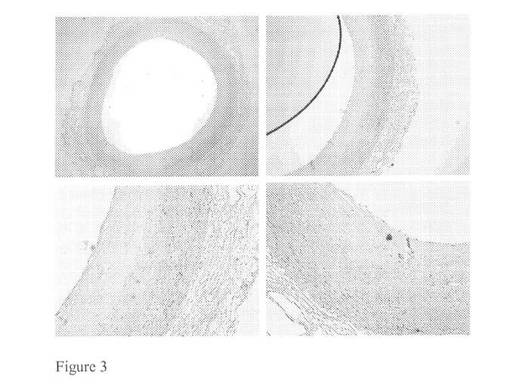
【図4】
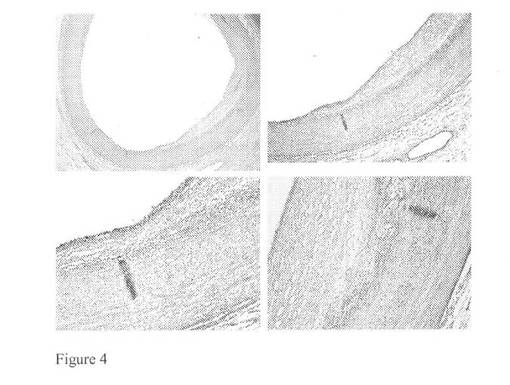
【図5】
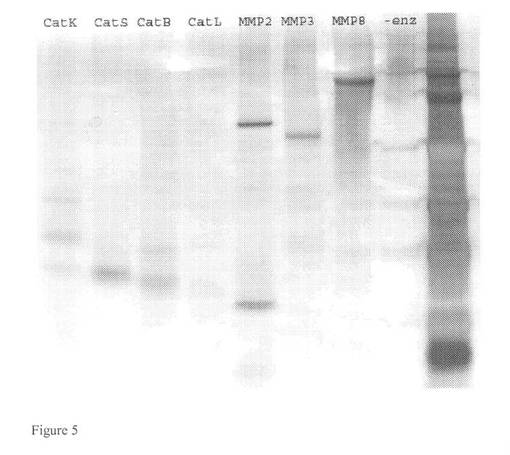
【図7】
【図8】
【図9】
【図10】
【図11】
【図1】
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2011−503564(P2011−503564A)
【公表日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願番号】特願2010−532571(P2010−532571)
【出願日】平成20年11月4日(2008.11.4)
【国際出願番号】PCT/EP2008/064946
【国際公開番号】WO2009/059972
【国際公開日】平成21年5月14日(2009.5.14)
【出願人】(503259129)ノルディック・ビオサイエンス・エー/エス (10)
【氏名又は名称原語表記】NORDIC BIOSCIENCE A/S
【Fターム(参考)】
【公表日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願日】平成20年11月4日(2008.11.4)
【国際出願番号】PCT/EP2008/064946
【国際公開番号】WO2009/059972
【国際公開日】平成21年5月14日(2009.5.14)
【出願人】(503259129)ノルディック・ビオサイエンス・エー/エス (10)
【氏名又は名称原語表記】NORDIC BIOSCIENCE A/S
【Fターム(参考)】
[ Back to top ]