
D−アミノアシラーゼ及びそれを用いたD−アミノ酸の製造方法
【課題】D−フェニルアラニン又はD−メチオニンの生成能に優れたD−アミノアシラーゼを提供すること。更に、当該酵素を利用したD−フェニルアラニン又はD−メチオニンの製造方法を提供する。
【解決手段】N-アシル-D-アミノ酸に作用してD−アミノ酸を生じる作用を有するD−アミノアシラーゼであって、特定のアミノ配列を含むD−アミノアシラーゼ、並びに、前記D-アミノアシラーゼを、N-アシル-D−フェニルアラニンに作用させ、生成するD−アミノ酸を回収する工程を含むD−フェニルアラニンの製造方法、及び、前記D−アミノアシラーゼを、N-アシル-D−メチオニンに作用させ、生成するD−メチオニンを回収する工程を含むD−メチオニンの製造方法。
【解決手段】N-アシル-D-アミノ酸に作用してD−アミノ酸を生じる作用を有するD−アミノアシラーゼであって、特定のアミノ配列を含むD−アミノアシラーゼ、並びに、前記D-アミノアシラーゼを、N-アシル-D−フェニルアラニンに作用させ、生成するD−アミノ酸を回収する工程を含むD−フェニルアラニンの製造方法、及び、前記D−アミノアシラーゼを、N-アシル-D−メチオニンに作用させ、生成するD−メチオニンを回収する工程を含むD−メチオニンの製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、D−フェニルアラニン又はD−メチオニンの生成能に優れたD−アミノアシラーゼに関する。
【0002】
更に、本発明は、当該D−アミノアシラーゼを用いたD−フェニルアラニン又はD−メチオニンの製造方法に関する。
【背景技術】
【0003】
近年、D−アミノ酸が医薬品等の原料として有効であることが明らかになり、光学的に純度の高いD−アミノ酸を安価に製造することが産業上重要な課題となっている。
【0004】
D−アミノ酸を製造する方法として、一般的には、化学合成したラセミ体を分割する方法が用いられており、特に副生成物や多量の廃溶媒を発生させない酵素法が注目されている。
【0005】
酵素法による製造方法としては、例えば、N−アセチル−D,L−アミノ酸にD−アミノアシラーゼを作用させ、D−アミノ酸を特異的に得る方法が知られている。
【0006】
また、D−アミノアシラーゼとして、ストレプトミセス・オリバゼウス(Streptomyces olivaceus)S62株(特許文献1参照)やアルカリゲネス・キシロースオキシダンス・サブスピーシーズ・キシロースオキシダンス(Alcaligenes xylosoxydanssubsp. xylosoxydans)A-6株(特許文献2参照)由来のD−アミノアシラーゼが報告されている。
【0007】
しかしながら、これらのD−アミノアシラーゼは、N−アセチル−D,L−アミノ酸の種類により反応特性が大きく異なり、D−アミノ酸の種類によって工業的生産が困難な場合があった。
【0008】
また、野生型D−アミノアシラーゼの変異体を用いて、D−アミノ酸を合成する方法も知られている。例えば、本発明者らは、N−アセチル−D−トリプトファンに対する触媒能が向上したD−アミノアシラーゼや、N−アセチル−D−アラニンに対する触媒能が向上したD−アミノアシラーゼを報告している(非特許文献1及び2参照)。
【0009】
しかしながら、D−フェニルアラニン又はD−メチオニンに対し、十分な触媒特性を有するD−アミノアシラーゼはこれまで知られていなかった。
【特許文献1】特開昭53−59092号公報
【特許文献2】特開平2−234677号公報
【非特許文献1】第14回日本生物工学会九州支部長崎大会講演要旨集(2007年)
【非特許文献2】日本農芸化学会2008年度大会講演要旨集(2008年3月5日発行)
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、D−フェニルアラニン又はD−メチオニンの生成能に優れたD−アミノアシラーゼ、更に、当該酵素を利用したD−フェニルアラニン又はD−メチオニンの製造方法を提供することを主な課題とする。
【課題を解決するための手段】
【0011】
本発明者は、上記課題を達成するために鋭意検討を重ねた結果、特定の配列を有する酵素が優れた作用を有することを見出し、更に鋭意検討を重ねて本発明を完成するに至った。
【0012】
即ち、本発明は、以下のD−アミノアシラーゼ及びそれを用いたD−アミノ酸の製造方法を提供する。
【0013】
項1:N−アシル-D-アミノ酸に作用してD-アミノ酸を生じる作用を有するD−アミノアシラーゼであって配列表の配列番号1に記載のアミノ配列を含むD−アミノアシラーゼ。
【0014】
項2:配列表の配列番号2記載のアミノ酸配列における297位のバリンがアラニンに置換された配列からなる項1に記載のD−アミノアシラーゼ。
【0015】
項3:項1又は2に記載のD−アミノアシラーゼをコードする遺伝子。
【0016】
項4:項3に記載の遺伝子を含有する組換えベクター。
【0017】
項5:項4に記載の組換えベクターで形質転換された形質転換体。
【0018】
項6:項1又は2に記載のD−アミノアシラーゼを、N-アシル-D−フェニルアラニンに作用させ、生成するD−フェニルアラニンを回収する工程を含むD−フェニルアラニンの製造方法。
【0019】
項7:項1又は2に記載のD−アミノアシラーゼを、N-アシル-D−メチオニンに作用させ、生成するD−メチオニンを回収する工程を含むD−メチオニンの製造方法。
【0020】
以下、本発明について、詳細に説明する。
【0021】
1.D−アミノアシラーゼ
本発明のD−アミノアシラーゼは、配列表の配列番号1(QDVSLDAYPYVAGSTMLKQDRALLAGRT)に記載のアミノ配列を含む。
【0022】
D−アミノアシラーゼは、N-アシル-D−アミノ酸に作用してD-アミノ酸を生成する反応を触媒する活性を有する酵素である。
【0023】
N−アシル-D−アミノ酸に作用してD-アミノ酸を生成する反応を触媒する活性は、公知の方法に従って測定することができ、例えば、以下のように測定することができる。
【0024】
酵素液、N-アシル-D-アミノ酸含有溶液、及び緩衝液を含む反応系で、30℃、10分間反応を行い、反応停止液の添加により反応を停止した後、生成したD-アミノ酸を、TNBS法(Biosci. Biotech.Biochem., 58, 24(1994))によって定量する。酵素活性は、30℃において1分間に1 μmolのD−アミノ酸の生成を触媒する酵素量を1単位(unit)として表す。
【0025】
より具体的には、実施例に記載の方法に従って、上記触媒活性を測定することができる。
【0026】
本発明のD−アミノアシラーゼは、特に、N-アシル-D−フェニルアラニンに作用してD−フェニルアラニンを生成する作用に優れる。またN−アシル-D-メチオニンに作用してD−メチオニンを生成する作用に優れる。
【0027】
特に、本発明のD−アミノアシラーゼは、N−アシル−D−フェニルアラニンからD−フェニルアラニンを生成、又は、N−アシル−D−メチオニンからD−メチオニンを生成する反応の触媒効率(kcat/Km)に優れ、野生型に対し、2倍以上の触媒効率を有する。
【0028】
本発明のD−アミノアシラーゼの一例として、配列表の配列番号2記載のアミノ酸配列において、297位のバリンがアラニンに置換されている配列からなるD−アミノアシラーゼ(以下、V297A変異酵素ともいう)を挙げることができる。
【0029】
配列表の配列番号2記載のアミノ酸配列で表される野生型D−アミノアシラーゼは、アルカリゲネス・キシロースオキシダンス・サブスピーシーズ・キシロースオキシダンスA-6(Alcaligenes xylosoxydans subsp. xylosoxydans A-6、以下、アルカリゲネスA-6ともいう)株が元来生産する酵素である。従って、本発明のV297A変異酵素は、アルカリゲネスA-6由来D−アミノアシラーゼの変異体とも換言できる。
【0030】
V297A変異酵素は、下記の理化学的性質を有する。
(1)分子量:(SDS-PAGEにより測定した分子量)約52,000
(2)等電点:約5.2
(3)至適pH:約7.0、安定pH:pH7〜8で最も安定
(4)至適温度:50℃
(5)熱安定性:40℃,10分の処理でほぼ安定
(6)基質特異性:N−アシル−D−フェニルアラニン及びN−アシル−D−メチオニンに対する加水分解作用に優れる
(7)金属イオンの作用:亜鉛イオン(Zn2+)を必須補因子として含有する。EDTAの添加により活性が50%程度低下する。
【0031】
また、本発明のD−アミノアシラーゼとして、V297A変異酵素において、配列表の配列番号1(QDVSLDAYPYVAGSTMLKQDRALLAGRT)に記載のアミノ配列部分を除く部分における1もしくは数個のアミノ酸が置換、欠失、付加及び/又は挿入された酵素であって、上記触媒特性を備えたものが挙げられる。
【0032】
また、本発明のD−アミノアシラーゼとして、Alcaligenes faecalis (アルカリゲネス フェカリス)DA1由来アミノアシラーゼの変異体であって、配列表の配列番号1(QDVSLDAYPYVAGSTMLKQDRALLAGRT)に記載の配列を有するものや、Variovorax paradoxus (バリオボラックス パラドクス)Iso1由来アミノアシラーゼの変異体であって、配列表の配列番号1(QDVSLDAYPYVAGSTMLKQDRALLAGRT)に記載の配列を有するものが挙げられる。
【0033】
本発明の酵素は、配列表の配列番号1に記載の配列を有するという特徴によって、基質のメチオニン又はフェニルアラニンが酵素の活性中心のポケット内に納まりやすくなっていると考えられ、このことが活性向上に寄与していると考えられる。
【0034】
2.製造方法
本発明のD-アミノアシラーゼは、公知の方法に従って、製造することができる。
【0035】
例えば、配列表の配列番号2に記載のアミノ酸配列をコードする遺伝子において、前記アミノ酸配列の297番目のバリン残基をコードする塩基配列を、アラニン残基をコードする塩基配列に置換した遺伝子を用いて、製造することができる。
【0036】
配列表の配列番号2に記載のアミノ酸配列をコードする遺伝子としては、配列表の配列番号3に記載の塩基配列を含む遺伝子、換言すると、アルカリゲネスA-6株由来 D-アミノアシラーゼ遺伝子を挙げることができる。
【0037】
配列表の配列番号3に記載の塩基配列からなる遺伝子において、297番目のバリン残基をコードする部分を、アラニン残基をコードする配列に置換する方法としては、例えば、Kunkelの方法 (T. A. Kunkel, Proc. Natl. Acad. Sci. USA, 82, 488 (1985))、ダブルプライマー法 (P. Carter, H. Bedouelle, G. Winter, Nucleic Acid Res., 13, 4431 (1985)) 、チオヌクレオチドを用いる方法 (J. W. Taylor, J. Ott, F. Eckstern, Nucleic Acid Res., 13, 8764 (1985))、カセット変異導入法(J. A. Wells, et al., Gene, 34, 315 (1985))等の方法を採ることができる。また、市販の遺伝子変異用キットを用いることも可能である。
【0038】
具体的には、D-アミノアシラーゼ遺伝子を含むプラスミドを鋳型とし、置換したい塩基を含む一対の変異プライマーと、DNAポリメラーゼを用いて、伸張反応を繰り返し、更に制限酵素を用いて鋳型を消化することにより、部位特異的変異された遺伝子を含むプラスミドを得て、目的とする変異が導入された遺伝子を得ることができる。更に具体的には、実施例に記載の方法を挙げることができる。
【0039】
上記のように変異を導入したD-アミノアシラーゼ遺伝子は、ベクターに導入して、組換えベクターを構築し、当該ベクターを用いて宿主の形質転換を行って、形質転換体を得る。
【0040】
ベクターの種類は、かかる変異導入遺伝子を発現可能なものであれば限定されず、例えば、ファージ、プラスミド、コスミド等を挙げることができる。具体的には、大腸菌由来のプラスミドを挙げることができる。
【0041】
組換えベクターの構築方法も、公知の方法に従って、行うことができる。ベクターには、遺伝子を効率良く発現させるために、プロモーター、選択マーカー、及び複製起点等を適宜含めることもできる。
【0042】
また、形質転換も、公知の方法に従って行うことができる。ベクターを導入する宿主は、選択したベクターの種類に応じて適宜決定することができる。例えば、大腸菌由来のベクターであれば、通常、宿主として大腸菌を用いる。
【0043】
得られた形質転換体を培地に培養し、かかる培養物からD-アミノアシラーゼ活性を有する酵素を回収することにより、本発明のD-アミノアシラーゼを取得することができる。
【0044】
培地の種類や培養温度及び培養時間は、宿主や発現ベクターの種類に応じて適宜設定される。大腸菌の場合は、通常、培養温度は20〜37℃、好ましくは30〜37℃程度であり、培養時間は、14〜24時間、好ましくは 16〜18時間程度である。
【0045】
また培養により得られる培養物から、酵素を回収する方法も、常法に従って行うことができる。
【0046】
例えば、宿主が菌体外にD-アミノアシラーゼを分泌する場合は、培養物を遠心分離し、得られた培養上清を粗酵素溶液として用いることができる。
【0047】
また、菌体外に分泌されない場合には、超音波破砕などによって菌体を破砕し、遠心分離を行って、粗酵素溶液を得ることができる。
【0048】
粗酵素溶液から、D-アミノアシラーゼを単離精製する方法も、常法に従って行うことができる。
【0049】
具体的には、硫酸アンモニウムを用いた塩析、電気泳動、アフィニティークロマトグラフィー、透析法、疎水クロマトグラフィー、又はこれらの組合せ等を用いることができる。
【0050】
このように、本発明のD-アミノアシラーゼは、D-アミノアシラーゼ遺伝子に変異を導入した遺伝子を構築することによって取得することができる。具体的には、実施例に記載されるように、アルカリゲネスA-6株由来 D-アミノアシラーゼ遺伝子に変異を導入した遺伝子を構築することによって取得することができる。また、アルカリゲネスA-6株由来D-アミノアシラーゼ遺伝子以外の遺伝子を用いる場合も、同様に実施することができる。
【0051】
また、例えば、配列番号2に記載のアミノ酸配列において、297番目のバリンがアラニンに置換された配列を、無細胞タンパク質合成法等を利用して合成することにより、取得することもできる。
【0052】
3.D−アミノ酸の製造方法
本発明のD−アミノアシラーゼを、N−アシル−D−フェニルアラニン又はそれを含む基質に接触させることにより、D−フェニルアラニンを効率よく製造することができる。
【0053】
また、本発明のD−アミノアシラーゼを、N−アシル−D−メチオニン又はそれを含む基質に接触させることにより、D−メチオニンを効率よく製造することができる。
【0054】
基質は、D−アミノアシラーゼが、N−アシル−D−フェニルアラニン又はN−アシル−D−メチオニンに作用可能であれば、D体とL体の混合物であってもよく、D体単独からなるものであってもよい。
【0055】
即ち、本発明のD-アミノアシラーゼを、N−アシル−DL−フェニルアラニンに接触させて、D−フェニルアラニンを製造してもよい。また、本発明のD−アミノアシラーゼをN−アシル−DL−メチオニンに接触させて、D−メチオニンを製造してもよい。本発明のD−アミノアシラーゼはD体に特異的に作用することから、基質をD体とL体に分離することなく、D−フェニルアラニン及びD−メチオニンを製造することができる。
【0056】
DL体におけるD体とL体との混合比率も特に限定されず、D体:L体の割合が10:90〜90:10程度のものであってもよく、50:50のラセミ体を用いてもよい。
【0057】
基質の濃度も特に制限されないが、通常0.1〜1M程度、好ましくは0.5〜1M、より好ましくは0.75〜1M程度である。
【0058】
また、基質は反応開始時に一括して添加することも可能であり、反応液中に分割して又は連続して添加することも可能である。
【0059】
D−アミノアシラーゼは、精製酵素の他、粗精製酵素の形態で用いてもよい。またD−アミノアシラーゼ産生能を有する形質転換体、又はその処理物として用いてもよい。すなわち、該D-アミノアシラーゼ産生能を有する形質転換体、又はその処理物を直接N-アシル-DL-アミノ酸に作用させて、D-アミノ酸を製造することも可能である。形質転換体の処理物としては、凍結融解処理、超音波処理、加圧処理、浸透圧差処理、磨砕処理などの物理的処理、リゾチームなどの細胞壁溶解酵素処理のような生化学的処理、もしくは界面活性剤、トルエン、キシレン、又はアセトンなどの有機溶媒との接触処理などの化学的処理等を行った形質転換体を挙げられる。
【0060】
また、酵素及び形質転換体は、安定化を目的として、固定化することもできる、例えば、イオン交換樹脂への結合によるポリアクリルアミドゲル法、含硫多糖ゲル法(カラギーナンゲル法)、アルギン酸ゲル法、寒天ゲル法などの公知の方法により固定化することができる。
【0061】
D-アミノアシラーゼの濃度も特に限定されず、用途ならびに使用時の条件によって0.1〜100 U/ml程度の範囲において適宜設定し得るが、通常10〜50U/mL、好ましくは20〜50U/mL、より好ましくは20〜30U/mL程度用いられる。
【0062】
反応温度は本発明の酵素がその活性を発現でき、上記反応のできる温度であれば特に制限されないが、通常20〜55℃、好ましくは30〜50℃、より好ましくは30〜40℃程度である。
【0063】
反応pHも、本発明の酵素がその活性を発現でき、反応できるpHであれば特に制限されないが、通常はpH6〜9、好ましくはpH7〜9、より好ましくは7〜8程度である。
【0064】
反応に用いる媒体としては、水或いは各種緩衝液からなる水性媒体を用いることができる。緩衝液としては、リン酸、トリス、クエン酸、酢酸、ホウ酸、グリシン、HEPES等から選択される1種又は2種以上を水に含有させた液を挙げることができる。
【0065】
また、反応は攪拌下でも行うことができ、静置下で行うこともできる。
【0066】
また、金属イオンの存在により酵素活性が阻害される場合には、反応液にEDTAなどのキレート剤を添加することができる。
【0067】
反応により生じたD-フェニルアラニン又はD−メチオニンは、公知の精製分離方法に従って回収することができる。精製分離方法としては、例えば、濃縮、等電点沈殿などによる直接結晶法や、イオン交換樹脂処理、膜分離などが挙げられる。
【0068】
反応後に残存するN-アシル-L-アミノ酸は、ラセミ化によってN-アシル-DL-アミノ酸とし、再利用することができる。N−アシル−L−アミノ酸をラセミ化する方法は公知であり、例えば、ラセマーゼを用いた酵素的方法が挙げられる。
【0069】
また、上記D−フェニルアラニン及びD−メチオニンの製造方法の逆反応を利用して、N−アシル−D−フェニルアラニン又はN−アシル−D−メチオニンを製造することもできる。
【0070】
例えば、上記本発明のD−アミノアシラーゼを、DL−フェニルアラニンに接触させ、生成するN−アシル−D−フェニルアラニンを回収することにより、N−アシル−D−フェニルアラニンを製造することができる。また、上記本発明のD−アミノアシラーゼを、DL−メチオニンに接触させ、生成するN−アシル−D−メチオニンを回収することにより、N−アシル−D−メチオニンを製造することができる。
【0071】
更に、本発明には、D−アミノアシラーゼ及びD−アミノアシラーゼを用いた物質の製造方法に係る公知技術を必要に応じて付加し得るものである。
【発明の効果】
【0072】
本発明により、Dフェニルアラニン及びD−メチオニンを効率良く製造できる技術が提供される。
【0073】
本発明の酵素は、N−アシル−D−フェニルアラニンからD−フェニルアラニンを生成する反応を触媒する活性に優れ、野生型に対し、2倍以上の触媒効率(kcat/Km)を有する。
【0074】
また、本発明の酵素は、N−アシル−D−メチオニンからD−メチオニンを生成する反応を触媒する活性に優れ、野生型に対し、2倍以上の触媒効率(kcat/Km)を有する。
【0075】
このような特徴によって、本発明の酵素を用いれば、N−アシル−D−フェニルアラニン及びN−アシル−D−メチオニンを含む基質から、D−フェニルアラニン及びD−メチオニンを効率良く取得することが可能になる。
【0076】
これにより、医薬品原料等として有用なD−フェニルアラニン及びD−メチオニンを、酵素を用いて工業的に生産することが可能になる。
【発明を実施するための最良の形態】
【0077】
以下、本発明を実施例や比較例等を用いてより詳細に説明するが、本発明はこれらの実施例に限定されることはない。
【0078】
なお、本発明において、「%」は、特に異ならない限り、「w/v(%)」(重量/容量(%))を意味する。
【実施例1】
【0079】
1−1.プラスミドの調製
Alcaligenes A-6株から染色体DNAを調製し、制限酵素Sau3AIにて部分消化して得られるDNA断片のうち、2〜9kbpのサイズのものをアガロースゲル電気泳動により回収した。この断片と制限酵素BamHIと脱リン酸化酵素で処理したpUC18をT4DNAリガーゼにより結合させた。このリガーゼ処理したDNA溶液で大腸菌JM109の形質転換を行った。β-D-thiogalactopyranoside(IPTG)と5-bromo-4-chloro-4-indolyl-β-D-galactoside(X-gal)を含むLB培地に植菌し、Alcaligenes A-6株の染色体DNA断片を有する形質転換体を取得した。得られた形質転換体をIPTGを含むLB培地に植菌し、30℃で生育させたのち、レプリカ法により、ろ紙に菌体(コロニー)を写し取った。菌体が張り付いたろ紙を基質(N-acetyl-D-leucine)および界面活性剤(cetyltrimethylammoniumbromide)の入ったHEPES緩衝液に浸し30分間浸した後、乾燥させた。次に、D-アミノ酸オキシダーゼ、ペルオキシダーゼ、o-ジアニシジンを含む溶液をろ紙に噴霧することにより、コロニーの周囲が赤色に呈色したものをD-アミノアシラーゼ遺伝子を含む形質転換体として取得した(Biosci. Biotech. Biochem., 59, (11), 2115-2119, 1995)。
D-アミノアシラーゼ遺伝子の開始コドンの上流に大腸菌用のSD配列、さらに上流にEcoRI認識部位、またストップコドンの下流にHindIII認識部位を、部位特異的変異導入法により挿入し、当該遺伝子を含むDNA断片を、EcoRIとHindIIIで処理したベクターpKK223-3に挿入することにより、pKNSD2を構築した(Protein Expression and Purification 7, 395-399, 1996)。
Mini-MTM Kitを用いてプラスミドを精製し、培地5 mlに一白金耳接種し、振とう培養 (30℃, 12hr, 200rpm.)後、遠心分離 (10,000rpm 、1min) して、1mlのミリQ水で洗浄し、再度遠心後、250μlのMX 1バッファーを加え菌体を完全に溶かした。次に、250μlのMX2バッファーを加え、溶菌液が透明になるまで穏やかに懸濁した後、室温で1〜5分間静置した。さらに350μl のMX3バッファーを加え懸濁し、遠心分離(10,000rpm 、10min)を行い、コレクションチューブの上に設置したMini-MTMカラムに上清を移した。再度遠心分離 (10,000rpm 、1min)し、カラムを素通りした溶液を捨て、0.5mlのWFバッファーを加え、遠心後、 0.7mlのWSバッファーを加え遠心分離を行なった。カラムを素通りした溶液を捨て、残存エタノールを取り除くために再度遠心分離(10,000rpm 、2min)した後、新しいチューブにカラムを移し変え、カラムのメンブレンの中央に50μlの溶出バッファーを加えた。2分間静止し、遠心分離(10,000rpm 、2min)後の溶出液を回収し、精製プラスミド(pKNSD2)を得た。
【0080】
1−2.部位特異的変異用プライマーの設計
上記のように調製したプラスミドにおいて、Alcaligenes A-6株由来D-アミノアシラーゼにおいて、297位のバリンがアラニンに変異するようにDNA塩基配列を設計した混合オリゴヌクレオチドをForward primerとした。
【0081】
また、Forward primerと相補的にDNA塩基配列を設計した混合オリゴヌクレオチドをReverse primerとした。プライマーの作成はクラボウ株式会社に委託した。
V297A変異酵素作製用プライマーの配列を以下に示す
(下線は置換アミノ酸に対応する塩基配列)
Forward primer:5’-tccggccagcagcgcgcggtcctgcttgag-3’(配列番号4)
Reverse primer:5’-ctcaagcaggaccgcgcgctgctggccgga-3’(配列番号5)
【0082】
また、比較のため、Alcaligenes A-6株由来D-アミノアシラーゼにおいて、298位のロイシンがメチオニンに変異した酵素(以下、L298M変異酵素)、及び298位のロイシンをトリプトファンに変異させた酵素(以下、L298W変異酵素)の作製用プライマーも同様に設計した。
【0083】
1−3.変異体D-アミノアシラーゼ構造遺伝子の増幅
上記1−2で調製した部位特異的変異用プライマーを用い、変異D-アミノアシラーゼ構造遺伝子の増幅を、QuickChange Site-Directed Mutagenesis Kitを用いて行った。1−1で調製したプラスミドpKNSD2を鋳型とし、1−2で作製したプライマーを用いてPCR反応を行った。
【0084】
PCRの反応液は、DNA 溶液1μl、Forward primer(10 pmol/μl) 1μl、Reverse Primer (10 pmol/μl)1μl、dNTP(2.5 mM) 1μl、PfuTurbo DNA polymerase (2.5U /μl) 1μl、10×reaction buffer 5μl、Distilled water 4μ1の組成のものを用いた。
【0085】
また、PCRの反応過程は、95℃×30secを1サイクル、95℃×30sec、50℃×1min、68℃×1min/kb of plasngthmid lengthを16サイクルとした。
【0086】
得られたPCR産物に、1μlのDpn I endonuclease(10U/μl)を加え、37℃で1時間反応させた。更に、プラスミドの増幅を行うために、得られたプラスミドによる大腸菌XL1 Blueの形質転換を行った。液体窒素中に保存してあるコンピテントセル(大腸菌 XL1 Blue) 100 μlの入ったエッペンドルフチューブを取り出し、氷中にて融解させた。融解させたコンピテントセルに、得られたプラスミド溶液の10分の1容を添加後、氷中で30分間静置し、その後42 ℃で1分間ヒートショックを行ない、すばやく氷冷した。このコンピテントセルに培地を900μl加え、37 ℃で1時間加温した。この溶液を、LB寒天培地(100μg/mlのアンピシリンを含む)に植菌し、37℃で一晩静置培養した。
【0087】
得られた組換え体大腸菌を、100μg/mlのアンピシリンを含むLB培地 5ml に植菌し、振とう培養 (30℃, 12hr, 200rpm.)した。培養液 1.5mlをエッペンドルフチューブに分注し、遠心分離 (4℃, 15,000rpm, 1min.)を行ない、菌体を得た。菌体を懸濁後、氷上に5分間静置した。次に1%SDS含有アルカリ溶液 400μlを添加し、染色体DNAの切断、混入を防ぐために、ゆるやかに攪拌し、氷上に5分間静置した。次いで、溶液を中和し、染色体DNAを凝集した。その後、遠心分離(4℃, 15,000rpm, 10min.)によって、上清を得た。この上清に対し、フェノール/クロロホルム抽出を2回行ない、タンパク質を除去した。完全にタンパク質が除去された溶液に、0.6倍量のイソプロパノールを添加し、遠心分離し、沈殿を採取した。沈殿を70%の冷エタノールでリンスし、エバポレーターを用いて、吸引乾燥した。これに、TE buffer 溶液30μlを添加し、RNase(終濃度10 μg/ml )処理を37℃で30分行なった。次いで、アガロースゲル電気泳動法によりプラスミドの確認を行なった。
【0088】
また、得られたプラスミドをMini-MTMキットを用いて精製し、塩基配列解析を行った。塩基配列解析は、島津バイオテックに依頼し、DNA sequence analyzer DSQ-2000L systemを使用して行った。
【0089】
その結果、Alcaligenes xylosoxydans subsp. xylosoxydans A-6株の塩基配列(配列表の配列番号3に記載の配列)において297位のバリンをコードする塩基GTGがGCGとなっていることが確認された。
【0090】
また、他の変異酵素作製用プライマーについても、同様に変異を確認した。
【0091】
1−4.大腸菌JM109の形質転換
1−3で変異導入が確認されたプラスミドを用いて、大腸菌JM109の形質転換を行った。コンピテントセル(大腸菌 JM109) 100 μlの入ったエッペンドルフチューブを取り出し、氷中にて融解させた。融解させたコンピテントセルに、約1〜10 ng/μlのプラスミド溶液の10分の1量を添加後、氷中で30分間静置し、その後42 ℃で1分間ヒートショックを行ない、すばやく氷冷した。このコンピテントセルに培地を900μl加え、37 ℃で1時間加温した。この溶液を、LB寒天培地(100μg/mlのアンピシリンを含む)に植菌し、37℃で一晩静置培養した。
【0092】
1−5.D-アミノアシラーゼの発現確認
1−4で得た組換え体大腸菌JM109を、50μg/mlのアンピシリンを含むLB培地5ml に植菌し、培養 (37℃, 20hr, 200rpm.)を行なった。培養終了後、遠心分離(15,000rpm、1min×3、4 ℃)により集菌した。この菌体を、10 mMリン酸カリウム緩衝液 (pH 7.0)で洗浄後、同緩衝液に再懸濁し、菌懸濁液とした。菌懸濁液を超音波処理(日本精機製作所、US300型、4 ℃, 10sec×3, 350-400μA)後、再度、遠心分離(15,000rpm、5 min、4 ℃)を行ない上清と沈殿を分け、沈殿は同緩衝液に再懸濁した。次いで、上清画分および沈殿画分をSDS-ポリアクリルアミドゲル電気泳動に供し、変異D-アミノアシラーゼの発現を確認した。
【0093】
1−6.D-アミノアシラーゼの精製
1−4で得られた組換え大腸菌JM109を300ml培養後、遠心分離(8000g,15min)によって菌体を得た。得られた菌体を10 mMリン酸カリウム緩衝液 (pH7.0)で洗浄し、同緩衝液に懸濁した。4℃に保ちながら日本精機製作所、U5300型を用いて超音波破砕 (4℃、350-400μA、15sec/time×6time)した後、遠心分離(8,000 g,15 min)によって得た上清を、10 mMリン酸カリウム緩衝液(pH7.0)に対して透析した。本操作で得た溶液を粗酵素標品とした。
【0094】
10mMリン酸カリウム緩衝液(pH 7.0)で平衡化したDEAE-Toyopearl 650Mカラム(3×12cm)に、粗酵素を吸着させた。吸着タンパク質を、まず50mM NaClを含む同緩衝液を用いて洗浄後、100mM NaClを含む同緩衝液を用いて溶出させ、活性画分を集めた。
【0095】
0.01mM塩化亜鉛と0.01%メルカプトエタノール,10%グリセロール(洗浄,溶出に用いる緩衝液にも同様に含む)および20 %飽和の硫酸アンモニウムを含む10 mMリン酸カリウム緩衝液(pH 7.0)で、平衡化したButyl-Cellulofineカラム(1.5×8 cm)に、20 %飽和の硫酸アンモニウムを添加したDEAE-Toyopearl 650M活性画分を、吸着させた。吸着タンパク質を、15%飽和の硫酸アンモニウムを含む同緩衝液で洗浄後、10%飽和の硫酸アンモニウムを含む同緩衝液を用いて溶出させた。10%飽和の硫酸アンモニウムを含む緩衝液で溶出した活性画分を集め、10 mMリン酸カリウム緩衝液(pH7.0)に対して透析した。本操作で得た溶液を精製酵素標品とした。
【0096】
また、L298M及びL298W変異酵素についても、同様の操作を行って、精製酵素を得た。
【0097】
2.酵素活性測定
上記1−6で用いた酵素を用い、0.1〜0.3 U/ml程度の酵素溶液を調製した。
【0098】
また、N-アシル-D-メチオニン又はN-アシル-D-フェニルアラニンを基質とし、濃度100mMの基質溶液を調製した。
【0099】
上記酵素溶液140μl、基質溶液40μl、及び1M のHEPES(2-[4-(2-hydroxyethyl)-1-piperazinyl]-ethanesulfonic acid)バッファー溶液20μlを混合し、反応溶液とした。
【0100】
反応は30℃で10分間行い、0.25 N NaOHを100 μl添加することで停止させた。
【0101】
反応停止10分後、0.125 M NaB4O7 を100 μl加え、続いてアミノ基検出試薬である0.2 M TNBSを40 μl添加することで発色させた。10分後、0.1 M NaH2PO4と0.1 M Na2SO3を19.7:0.3の割合で含む溶液800 μlを添加した。この溶液の520 nmにおける吸光度を測定し、反応液中のD-アミノ酸を定量した。なお、対照として基質を添加する前にNaOHを加えたものを用いた。
【0102】
また、1分間に1 μmolのD-アミノ酸を生成する酵素量を1 Uとした。
【0103】
また、基質濃度を8〜300mMに調製し、Lineweaver-Burkの逆数プロットを用いてVmax、Kmを算出した。また、kcatは、Vmaxと本酵素の分子量をもとに算出した。更に本酵素の触媒効率をkcat/Kmの形で求めた。
【0104】
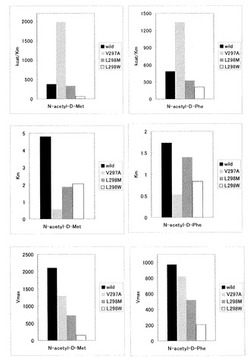
得られた結果を図1に示す。
【0105】
図1に示されるように、V297A変異酵素は、野生型に比べて触媒効率が格段に向上しており、他の変異酵素と比較しても格段に有利な効果を奏することがわかった。
【図面の簡単な説明】
【0106】
【図1】野生型酵素(Wild)、V297A変異酵素(V297A)、L298M変異酵素(L298M)及びL298W変異酵素(L298W)について、N-アシル-D−フェニルアラニン(N-acetyl-D-Phe)及びN-アシル-D−メチオニン(N-acetyl-D-Met)に対するKm、Vmax及びkcat/Kmを調べた結果を示す図面である。
【配列表フリーテキスト】
【0107】
配列番号1は、本発明の酵素が有するアミノ酸配列を示す。
配列番号2はAlcaligenes xylosoxydans subsp. xylosoxydans A-6由来D-aminoacylaseのアミノ酸配列を示す。
配列番号3は、Alcaligenes xylosoxydans subsp. xylosoxydans A-6由来D-aminoacylase遺伝子の塩基配列を示す。
配列番号4は、V297A変異酵素作製に用いたフォワードプライマーの配列を示す。
配列番号5は、V297A変異酵素作製に用いたリバースプライマーの配列を示す。
【技術分野】
【0001】
本発明は、D−フェニルアラニン又はD−メチオニンの生成能に優れたD−アミノアシラーゼに関する。
【0002】
更に、本発明は、当該D−アミノアシラーゼを用いたD−フェニルアラニン又はD−メチオニンの製造方法に関する。
【背景技術】
【0003】
近年、D−アミノ酸が医薬品等の原料として有効であることが明らかになり、光学的に純度の高いD−アミノ酸を安価に製造することが産業上重要な課題となっている。
【0004】
D−アミノ酸を製造する方法として、一般的には、化学合成したラセミ体を分割する方法が用いられており、特に副生成物や多量の廃溶媒を発生させない酵素法が注目されている。
【0005】
酵素法による製造方法としては、例えば、N−アセチル−D,L−アミノ酸にD−アミノアシラーゼを作用させ、D−アミノ酸を特異的に得る方法が知られている。
【0006】
また、D−アミノアシラーゼとして、ストレプトミセス・オリバゼウス(Streptomyces olivaceus)S62株(特許文献1参照)やアルカリゲネス・キシロースオキシダンス・サブスピーシーズ・キシロースオキシダンス(Alcaligenes xylosoxydanssubsp. xylosoxydans)A-6株(特許文献2参照)由来のD−アミノアシラーゼが報告されている。
【0007】
しかしながら、これらのD−アミノアシラーゼは、N−アセチル−D,L−アミノ酸の種類により反応特性が大きく異なり、D−アミノ酸の種類によって工業的生産が困難な場合があった。
【0008】
また、野生型D−アミノアシラーゼの変異体を用いて、D−アミノ酸を合成する方法も知られている。例えば、本発明者らは、N−アセチル−D−トリプトファンに対する触媒能が向上したD−アミノアシラーゼや、N−アセチル−D−アラニンに対する触媒能が向上したD−アミノアシラーゼを報告している(非特許文献1及び2参照)。
【0009】
しかしながら、D−フェニルアラニン又はD−メチオニンに対し、十分な触媒特性を有するD−アミノアシラーゼはこれまで知られていなかった。
【特許文献1】特開昭53−59092号公報
【特許文献2】特開平2−234677号公報
【非特許文献1】第14回日本生物工学会九州支部長崎大会講演要旨集(2007年)
【非特許文献2】日本農芸化学会2008年度大会講演要旨集(2008年3月5日発行)
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、D−フェニルアラニン又はD−メチオニンの生成能に優れたD−アミノアシラーゼ、更に、当該酵素を利用したD−フェニルアラニン又はD−メチオニンの製造方法を提供することを主な課題とする。
【課題を解決するための手段】
【0011】
本発明者は、上記課題を達成するために鋭意検討を重ねた結果、特定の配列を有する酵素が優れた作用を有することを見出し、更に鋭意検討を重ねて本発明を完成するに至った。
【0012】
即ち、本発明は、以下のD−アミノアシラーゼ及びそれを用いたD−アミノ酸の製造方法を提供する。
【0013】
項1:N−アシル-D-アミノ酸に作用してD-アミノ酸を生じる作用を有するD−アミノアシラーゼであって配列表の配列番号1に記載のアミノ配列を含むD−アミノアシラーゼ。
【0014】
項2:配列表の配列番号2記載のアミノ酸配列における297位のバリンがアラニンに置換された配列からなる項1に記載のD−アミノアシラーゼ。
【0015】
項3:項1又は2に記載のD−アミノアシラーゼをコードする遺伝子。
【0016】
項4:項3に記載の遺伝子を含有する組換えベクター。
【0017】
項5:項4に記載の組換えベクターで形質転換された形質転換体。
【0018】
項6:項1又は2に記載のD−アミノアシラーゼを、N-アシル-D−フェニルアラニンに作用させ、生成するD−フェニルアラニンを回収する工程を含むD−フェニルアラニンの製造方法。
【0019】
項7:項1又は2に記載のD−アミノアシラーゼを、N-アシル-D−メチオニンに作用させ、生成するD−メチオニンを回収する工程を含むD−メチオニンの製造方法。
【0020】
以下、本発明について、詳細に説明する。
【0021】
1.D−アミノアシラーゼ
本発明のD−アミノアシラーゼは、配列表の配列番号1(QDVSLDAYPYVAGSTMLKQDRALLAGRT)に記載のアミノ配列を含む。
【0022】
D−アミノアシラーゼは、N-アシル-D−アミノ酸に作用してD-アミノ酸を生成する反応を触媒する活性を有する酵素である。
【0023】
N−アシル-D−アミノ酸に作用してD-アミノ酸を生成する反応を触媒する活性は、公知の方法に従って測定することができ、例えば、以下のように測定することができる。
【0024】
酵素液、N-アシル-D-アミノ酸含有溶液、及び緩衝液を含む反応系で、30℃、10分間反応を行い、反応停止液の添加により反応を停止した後、生成したD-アミノ酸を、TNBS法(Biosci. Biotech.Biochem., 58, 24(1994))によって定量する。酵素活性は、30℃において1分間に1 μmolのD−アミノ酸の生成を触媒する酵素量を1単位(unit)として表す。
【0025】
より具体的には、実施例に記載の方法に従って、上記触媒活性を測定することができる。
【0026】
本発明のD−アミノアシラーゼは、特に、N-アシル-D−フェニルアラニンに作用してD−フェニルアラニンを生成する作用に優れる。またN−アシル-D-メチオニンに作用してD−メチオニンを生成する作用に優れる。
【0027】
特に、本発明のD−アミノアシラーゼは、N−アシル−D−フェニルアラニンからD−フェニルアラニンを生成、又は、N−アシル−D−メチオニンからD−メチオニンを生成する反応の触媒効率(kcat/Km)に優れ、野生型に対し、2倍以上の触媒効率を有する。
【0028】
本発明のD−アミノアシラーゼの一例として、配列表の配列番号2記載のアミノ酸配列において、297位のバリンがアラニンに置換されている配列からなるD−アミノアシラーゼ(以下、V297A変異酵素ともいう)を挙げることができる。
【0029】
配列表の配列番号2記載のアミノ酸配列で表される野生型D−アミノアシラーゼは、アルカリゲネス・キシロースオキシダンス・サブスピーシーズ・キシロースオキシダンスA-6(Alcaligenes xylosoxydans subsp. xylosoxydans A-6、以下、アルカリゲネスA-6ともいう)株が元来生産する酵素である。従って、本発明のV297A変異酵素は、アルカリゲネスA-6由来D−アミノアシラーゼの変異体とも換言できる。
【0030】
V297A変異酵素は、下記の理化学的性質を有する。
(1)分子量:(SDS-PAGEにより測定した分子量)約52,000
(2)等電点:約5.2
(3)至適pH:約7.0、安定pH:pH7〜8で最も安定
(4)至適温度:50℃
(5)熱安定性:40℃,10分の処理でほぼ安定
(6)基質特異性:N−アシル−D−フェニルアラニン及びN−アシル−D−メチオニンに対する加水分解作用に優れる
(7)金属イオンの作用:亜鉛イオン(Zn2+)を必須補因子として含有する。EDTAの添加により活性が50%程度低下する。
【0031】
また、本発明のD−アミノアシラーゼとして、V297A変異酵素において、配列表の配列番号1(QDVSLDAYPYVAGSTMLKQDRALLAGRT)に記載のアミノ配列部分を除く部分における1もしくは数個のアミノ酸が置換、欠失、付加及び/又は挿入された酵素であって、上記触媒特性を備えたものが挙げられる。
【0032】
また、本発明のD−アミノアシラーゼとして、Alcaligenes faecalis (アルカリゲネス フェカリス)DA1由来アミノアシラーゼの変異体であって、配列表の配列番号1(QDVSLDAYPYVAGSTMLKQDRALLAGRT)に記載の配列を有するものや、Variovorax paradoxus (バリオボラックス パラドクス)Iso1由来アミノアシラーゼの変異体であって、配列表の配列番号1(QDVSLDAYPYVAGSTMLKQDRALLAGRT)に記載の配列を有するものが挙げられる。
【0033】
本発明の酵素は、配列表の配列番号1に記載の配列を有するという特徴によって、基質のメチオニン又はフェニルアラニンが酵素の活性中心のポケット内に納まりやすくなっていると考えられ、このことが活性向上に寄与していると考えられる。
【0034】
2.製造方法
本発明のD-アミノアシラーゼは、公知の方法に従って、製造することができる。
【0035】
例えば、配列表の配列番号2に記載のアミノ酸配列をコードする遺伝子において、前記アミノ酸配列の297番目のバリン残基をコードする塩基配列を、アラニン残基をコードする塩基配列に置換した遺伝子を用いて、製造することができる。
【0036】
配列表の配列番号2に記載のアミノ酸配列をコードする遺伝子としては、配列表の配列番号3に記載の塩基配列を含む遺伝子、換言すると、アルカリゲネスA-6株由来 D-アミノアシラーゼ遺伝子を挙げることができる。
【0037】
配列表の配列番号3に記載の塩基配列からなる遺伝子において、297番目のバリン残基をコードする部分を、アラニン残基をコードする配列に置換する方法としては、例えば、Kunkelの方法 (T. A. Kunkel, Proc. Natl. Acad. Sci. USA, 82, 488 (1985))、ダブルプライマー法 (P. Carter, H. Bedouelle, G. Winter, Nucleic Acid Res., 13, 4431 (1985)) 、チオヌクレオチドを用いる方法 (J. W. Taylor, J. Ott, F. Eckstern, Nucleic Acid Res., 13, 8764 (1985))、カセット変異導入法(J. A. Wells, et al., Gene, 34, 315 (1985))等の方法を採ることができる。また、市販の遺伝子変異用キットを用いることも可能である。
【0038】
具体的には、D-アミノアシラーゼ遺伝子を含むプラスミドを鋳型とし、置換したい塩基を含む一対の変異プライマーと、DNAポリメラーゼを用いて、伸張反応を繰り返し、更に制限酵素を用いて鋳型を消化することにより、部位特異的変異された遺伝子を含むプラスミドを得て、目的とする変異が導入された遺伝子を得ることができる。更に具体的には、実施例に記載の方法を挙げることができる。
【0039】
上記のように変異を導入したD-アミノアシラーゼ遺伝子は、ベクターに導入して、組換えベクターを構築し、当該ベクターを用いて宿主の形質転換を行って、形質転換体を得る。
【0040】
ベクターの種類は、かかる変異導入遺伝子を発現可能なものであれば限定されず、例えば、ファージ、プラスミド、コスミド等を挙げることができる。具体的には、大腸菌由来のプラスミドを挙げることができる。
【0041】
組換えベクターの構築方法も、公知の方法に従って、行うことができる。ベクターには、遺伝子を効率良く発現させるために、プロモーター、選択マーカー、及び複製起点等を適宜含めることもできる。
【0042】
また、形質転換も、公知の方法に従って行うことができる。ベクターを導入する宿主は、選択したベクターの種類に応じて適宜決定することができる。例えば、大腸菌由来のベクターであれば、通常、宿主として大腸菌を用いる。
【0043】
得られた形質転換体を培地に培養し、かかる培養物からD-アミノアシラーゼ活性を有する酵素を回収することにより、本発明のD-アミノアシラーゼを取得することができる。
【0044】
培地の種類や培養温度及び培養時間は、宿主や発現ベクターの種類に応じて適宜設定される。大腸菌の場合は、通常、培養温度は20〜37℃、好ましくは30〜37℃程度であり、培養時間は、14〜24時間、好ましくは 16〜18時間程度である。
【0045】
また培養により得られる培養物から、酵素を回収する方法も、常法に従って行うことができる。
【0046】
例えば、宿主が菌体外にD-アミノアシラーゼを分泌する場合は、培養物を遠心分離し、得られた培養上清を粗酵素溶液として用いることができる。
【0047】
また、菌体外に分泌されない場合には、超音波破砕などによって菌体を破砕し、遠心分離を行って、粗酵素溶液を得ることができる。
【0048】
粗酵素溶液から、D-アミノアシラーゼを単離精製する方法も、常法に従って行うことができる。
【0049】
具体的には、硫酸アンモニウムを用いた塩析、電気泳動、アフィニティークロマトグラフィー、透析法、疎水クロマトグラフィー、又はこれらの組合せ等を用いることができる。
【0050】
このように、本発明のD-アミノアシラーゼは、D-アミノアシラーゼ遺伝子に変異を導入した遺伝子を構築することによって取得することができる。具体的には、実施例に記載されるように、アルカリゲネスA-6株由来 D-アミノアシラーゼ遺伝子に変異を導入した遺伝子を構築することによって取得することができる。また、アルカリゲネスA-6株由来D-アミノアシラーゼ遺伝子以外の遺伝子を用いる場合も、同様に実施することができる。
【0051】
また、例えば、配列番号2に記載のアミノ酸配列において、297番目のバリンがアラニンに置換された配列を、無細胞タンパク質合成法等を利用して合成することにより、取得することもできる。
【0052】
3.D−アミノ酸の製造方法
本発明のD−アミノアシラーゼを、N−アシル−D−フェニルアラニン又はそれを含む基質に接触させることにより、D−フェニルアラニンを効率よく製造することができる。
【0053】
また、本発明のD−アミノアシラーゼを、N−アシル−D−メチオニン又はそれを含む基質に接触させることにより、D−メチオニンを効率よく製造することができる。
【0054】
基質は、D−アミノアシラーゼが、N−アシル−D−フェニルアラニン又はN−アシル−D−メチオニンに作用可能であれば、D体とL体の混合物であってもよく、D体単独からなるものであってもよい。
【0055】
即ち、本発明のD-アミノアシラーゼを、N−アシル−DL−フェニルアラニンに接触させて、D−フェニルアラニンを製造してもよい。また、本発明のD−アミノアシラーゼをN−アシル−DL−メチオニンに接触させて、D−メチオニンを製造してもよい。本発明のD−アミノアシラーゼはD体に特異的に作用することから、基質をD体とL体に分離することなく、D−フェニルアラニン及びD−メチオニンを製造することができる。
【0056】
DL体におけるD体とL体との混合比率も特に限定されず、D体:L体の割合が10:90〜90:10程度のものであってもよく、50:50のラセミ体を用いてもよい。
【0057】
基質の濃度も特に制限されないが、通常0.1〜1M程度、好ましくは0.5〜1M、より好ましくは0.75〜1M程度である。
【0058】
また、基質は反応開始時に一括して添加することも可能であり、反応液中に分割して又は連続して添加することも可能である。
【0059】
D−アミノアシラーゼは、精製酵素の他、粗精製酵素の形態で用いてもよい。またD−アミノアシラーゼ産生能を有する形質転換体、又はその処理物として用いてもよい。すなわち、該D-アミノアシラーゼ産生能を有する形質転換体、又はその処理物を直接N-アシル-DL-アミノ酸に作用させて、D-アミノ酸を製造することも可能である。形質転換体の処理物としては、凍結融解処理、超音波処理、加圧処理、浸透圧差処理、磨砕処理などの物理的処理、リゾチームなどの細胞壁溶解酵素処理のような生化学的処理、もしくは界面活性剤、トルエン、キシレン、又はアセトンなどの有機溶媒との接触処理などの化学的処理等を行った形質転換体を挙げられる。
【0060】
また、酵素及び形質転換体は、安定化を目的として、固定化することもできる、例えば、イオン交換樹脂への結合によるポリアクリルアミドゲル法、含硫多糖ゲル法(カラギーナンゲル法)、アルギン酸ゲル法、寒天ゲル法などの公知の方法により固定化することができる。
【0061】
D-アミノアシラーゼの濃度も特に限定されず、用途ならびに使用時の条件によって0.1〜100 U/ml程度の範囲において適宜設定し得るが、通常10〜50U/mL、好ましくは20〜50U/mL、より好ましくは20〜30U/mL程度用いられる。
【0062】
反応温度は本発明の酵素がその活性を発現でき、上記反応のできる温度であれば特に制限されないが、通常20〜55℃、好ましくは30〜50℃、より好ましくは30〜40℃程度である。
【0063】
反応pHも、本発明の酵素がその活性を発現でき、反応できるpHであれば特に制限されないが、通常はpH6〜9、好ましくはpH7〜9、より好ましくは7〜8程度である。
【0064】
反応に用いる媒体としては、水或いは各種緩衝液からなる水性媒体を用いることができる。緩衝液としては、リン酸、トリス、クエン酸、酢酸、ホウ酸、グリシン、HEPES等から選択される1種又は2種以上を水に含有させた液を挙げることができる。
【0065】
また、反応は攪拌下でも行うことができ、静置下で行うこともできる。
【0066】
また、金属イオンの存在により酵素活性が阻害される場合には、反応液にEDTAなどのキレート剤を添加することができる。
【0067】
反応により生じたD-フェニルアラニン又はD−メチオニンは、公知の精製分離方法に従って回収することができる。精製分離方法としては、例えば、濃縮、等電点沈殿などによる直接結晶法や、イオン交換樹脂処理、膜分離などが挙げられる。
【0068】
反応後に残存するN-アシル-L-アミノ酸は、ラセミ化によってN-アシル-DL-アミノ酸とし、再利用することができる。N−アシル−L−アミノ酸をラセミ化する方法は公知であり、例えば、ラセマーゼを用いた酵素的方法が挙げられる。
【0069】
また、上記D−フェニルアラニン及びD−メチオニンの製造方法の逆反応を利用して、N−アシル−D−フェニルアラニン又はN−アシル−D−メチオニンを製造することもできる。
【0070】
例えば、上記本発明のD−アミノアシラーゼを、DL−フェニルアラニンに接触させ、生成するN−アシル−D−フェニルアラニンを回収することにより、N−アシル−D−フェニルアラニンを製造することができる。また、上記本発明のD−アミノアシラーゼを、DL−メチオニンに接触させ、生成するN−アシル−D−メチオニンを回収することにより、N−アシル−D−メチオニンを製造することができる。
【0071】
更に、本発明には、D−アミノアシラーゼ及びD−アミノアシラーゼを用いた物質の製造方法に係る公知技術を必要に応じて付加し得るものである。
【発明の効果】
【0072】
本発明により、Dフェニルアラニン及びD−メチオニンを効率良く製造できる技術が提供される。
【0073】
本発明の酵素は、N−アシル−D−フェニルアラニンからD−フェニルアラニンを生成する反応を触媒する活性に優れ、野生型に対し、2倍以上の触媒効率(kcat/Km)を有する。
【0074】
また、本発明の酵素は、N−アシル−D−メチオニンからD−メチオニンを生成する反応を触媒する活性に優れ、野生型に対し、2倍以上の触媒効率(kcat/Km)を有する。
【0075】
このような特徴によって、本発明の酵素を用いれば、N−アシル−D−フェニルアラニン及びN−アシル−D−メチオニンを含む基質から、D−フェニルアラニン及びD−メチオニンを効率良く取得することが可能になる。
【0076】
これにより、医薬品原料等として有用なD−フェニルアラニン及びD−メチオニンを、酵素を用いて工業的に生産することが可能になる。
【発明を実施するための最良の形態】
【0077】
以下、本発明を実施例や比較例等を用いてより詳細に説明するが、本発明はこれらの実施例に限定されることはない。
【0078】
なお、本発明において、「%」は、特に異ならない限り、「w/v(%)」(重量/容量(%))を意味する。
【実施例1】
【0079】
1−1.プラスミドの調製
Alcaligenes A-6株から染色体DNAを調製し、制限酵素Sau3AIにて部分消化して得られるDNA断片のうち、2〜9kbpのサイズのものをアガロースゲル電気泳動により回収した。この断片と制限酵素BamHIと脱リン酸化酵素で処理したpUC18をT4DNAリガーゼにより結合させた。このリガーゼ処理したDNA溶液で大腸菌JM109の形質転換を行った。β-D-thiogalactopyranoside(IPTG)と5-bromo-4-chloro-4-indolyl-β-D-galactoside(X-gal)を含むLB培地に植菌し、Alcaligenes A-6株の染色体DNA断片を有する形質転換体を取得した。得られた形質転換体をIPTGを含むLB培地に植菌し、30℃で生育させたのち、レプリカ法により、ろ紙に菌体(コロニー)を写し取った。菌体が張り付いたろ紙を基質(N-acetyl-D-leucine)および界面活性剤(cetyltrimethylammoniumbromide)の入ったHEPES緩衝液に浸し30分間浸した後、乾燥させた。次に、D-アミノ酸オキシダーゼ、ペルオキシダーゼ、o-ジアニシジンを含む溶液をろ紙に噴霧することにより、コロニーの周囲が赤色に呈色したものをD-アミノアシラーゼ遺伝子を含む形質転換体として取得した(Biosci. Biotech. Biochem., 59, (11), 2115-2119, 1995)。
D-アミノアシラーゼ遺伝子の開始コドンの上流に大腸菌用のSD配列、さらに上流にEcoRI認識部位、またストップコドンの下流にHindIII認識部位を、部位特異的変異導入法により挿入し、当該遺伝子を含むDNA断片を、EcoRIとHindIIIで処理したベクターpKK223-3に挿入することにより、pKNSD2を構築した(Protein Expression and Purification 7, 395-399, 1996)。
Mini-MTM Kitを用いてプラスミドを精製し、培地5 mlに一白金耳接種し、振とう培養 (30℃, 12hr, 200rpm.)後、遠心分離 (10,000rpm 、1min) して、1mlのミリQ水で洗浄し、再度遠心後、250μlのMX 1バッファーを加え菌体を完全に溶かした。次に、250μlのMX2バッファーを加え、溶菌液が透明になるまで穏やかに懸濁した後、室温で1〜5分間静置した。さらに350μl のMX3バッファーを加え懸濁し、遠心分離(10,000rpm 、10min)を行い、コレクションチューブの上に設置したMini-MTMカラムに上清を移した。再度遠心分離 (10,000rpm 、1min)し、カラムを素通りした溶液を捨て、0.5mlのWFバッファーを加え、遠心後、 0.7mlのWSバッファーを加え遠心分離を行なった。カラムを素通りした溶液を捨て、残存エタノールを取り除くために再度遠心分離(10,000rpm 、2min)した後、新しいチューブにカラムを移し変え、カラムのメンブレンの中央に50μlの溶出バッファーを加えた。2分間静止し、遠心分離(10,000rpm 、2min)後の溶出液を回収し、精製プラスミド(pKNSD2)を得た。
【0080】
1−2.部位特異的変異用プライマーの設計
上記のように調製したプラスミドにおいて、Alcaligenes A-6株由来D-アミノアシラーゼにおいて、297位のバリンがアラニンに変異するようにDNA塩基配列を設計した混合オリゴヌクレオチドをForward primerとした。
【0081】
また、Forward primerと相補的にDNA塩基配列を設計した混合オリゴヌクレオチドをReverse primerとした。プライマーの作成はクラボウ株式会社に委託した。
V297A変異酵素作製用プライマーの配列を以下に示す
(下線は置換アミノ酸に対応する塩基配列)
Forward primer:5’-tccggccagcagcgcgcggtcctgcttgag-3’(配列番号4)
Reverse primer:5’-ctcaagcaggaccgcgcgctgctggccgga-3’(配列番号5)
【0082】
また、比較のため、Alcaligenes A-6株由来D-アミノアシラーゼにおいて、298位のロイシンがメチオニンに変異した酵素(以下、L298M変異酵素)、及び298位のロイシンをトリプトファンに変異させた酵素(以下、L298W変異酵素)の作製用プライマーも同様に設計した。
【0083】
1−3.変異体D-アミノアシラーゼ構造遺伝子の増幅
上記1−2で調製した部位特異的変異用プライマーを用い、変異D-アミノアシラーゼ構造遺伝子の増幅を、QuickChange Site-Directed Mutagenesis Kitを用いて行った。1−1で調製したプラスミドpKNSD2を鋳型とし、1−2で作製したプライマーを用いてPCR反応を行った。
【0084】
PCRの反応液は、DNA 溶液1μl、Forward primer(10 pmol/μl) 1μl、Reverse Primer (10 pmol/μl)1μl、dNTP(2.5 mM) 1μl、PfuTurbo DNA polymerase (2.5U /μl) 1μl、10×reaction buffer 5μl、Distilled water 4μ1の組成のものを用いた。
【0085】
また、PCRの反応過程は、95℃×30secを1サイクル、95℃×30sec、50℃×1min、68℃×1min/kb of plasngthmid lengthを16サイクルとした。
【0086】
得られたPCR産物に、1μlのDpn I endonuclease(10U/μl)を加え、37℃で1時間反応させた。更に、プラスミドの増幅を行うために、得られたプラスミドによる大腸菌XL1 Blueの形質転換を行った。液体窒素中に保存してあるコンピテントセル(大腸菌 XL1 Blue) 100 μlの入ったエッペンドルフチューブを取り出し、氷中にて融解させた。融解させたコンピテントセルに、得られたプラスミド溶液の10分の1容を添加後、氷中で30分間静置し、その後42 ℃で1分間ヒートショックを行ない、すばやく氷冷した。このコンピテントセルに培地を900μl加え、37 ℃で1時間加温した。この溶液を、LB寒天培地(100μg/mlのアンピシリンを含む)に植菌し、37℃で一晩静置培養した。
【0087】
得られた組換え体大腸菌を、100μg/mlのアンピシリンを含むLB培地 5ml に植菌し、振とう培養 (30℃, 12hr, 200rpm.)した。培養液 1.5mlをエッペンドルフチューブに分注し、遠心分離 (4℃, 15,000rpm, 1min.)を行ない、菌体を得た。菌体を懸濁後、氷上に5分間静置した。次に1%SDS含有アルカリ溶液 400μlを添加し、染色体DNAの切断、混入を防ぐために、ゆるやかに攪拌し、氷上に5分間静置した。次いで、溶液を中和し、染色体DNAを凝集した。その後、遠心分離(4℃, 15,000rpm, 10min.)によって、上清を得た。この上清に対し、フェノール/クロロホルム抽出を2回行ない、タンパク質を除去した。完全にタンパク質が除去された溶液に、0.6倍量のイソプロパノールを添加し、遠心分離し、沈殿を採取した。沈殿を70%の冷エタノールでリンスし、エバポレーターを用いて、吸引乾燥した。これに、TE buffer 溶液30μlを添加し、RNase(終濃度10 μg/ml )処理を37℃で30分行なった。次いで、アガロースゲル電気泳動法によりプラスミドの確認を行なった。
【0088】
また、得られたプラスミドをMini-MTMキットを用いて精製し、塩基配列解析を行った。塩基配列解析は、島津バイオテックに依頼し、DNA sequence analyzer DSQ-2000L systemを使用して行った。
【0089】
その結果、Alcaligenes xylosoxydans subsp. xylosoxydans A-6株の塩基配列(配列表の配列番号3に記載の配列)において297位のバリンをコードする塩基GTGがGCGとなっていることが確認された。
【0090】
また、他の変異酵素作製用プライマーについても、同様に変異を確認した。
【0091】
1−4.大腸菌JM109の形質転換
1−3で変異導入が確認されたプラスミドを用いて、大腸菌JM109の形質転換を行った。コンピテントセル(大腸菌 JM109) 100 μlの入ったエッペンドルフチューブを取り出し、氷中にて融解させた。融解させたコンピテントセルに、約1〜10 ng/μlのプラスミド溶液の10分の1量を添加後、氷中で30分間静置し、その後42 ℃で1分間ヒートショックを行ない、すばやく氷冷した。このコンピテントセルに培地を900μl加え、37 ℃で1時間加温した。この溶液を、LB寒天培地(100μg/mlのアンピシリンを含む)に植菌し、37℃で一晩静置培養した。
【0092】
1−5.D-アミノアシラーゼの発現確認
1−4で得た組換え体大腸菌JM109を、50μg/mlのアンピシリンを含むLB培地5ml に植菌し、培養 (37℃, 20hr, 200rpm.)を行なった。培養終了後、遠心分離(15,000rpm、1min×3、4 ℃)により集菌した。この菌体を、10 mMリン酸カリウム緩衝液 (pH 7.0)で洗浄後、同緩衝液に再懸濁し、菌懸濁液とした。菌懸濁液を超音波処理(日本精機製作所、US300型、4 ℃, 10sec×3, 350-400μA)後、再度、遠心分離(15,000rpm、5 min、4 ℃)を行ない上清と沈殿を分け、沈殿は同緩衝液に再懸濁した。次いで、上清画分および沈殿画分をSDS-ポリアクリルアミドゲル電気泳動に供し、変異D-アミノアシラーゼの発現を確認した。
【0093】
1−6.D-アミノアシラーゼの精製
1−4で得られた組換え大腸菌JM109を300ml培養後、遠心分離(8000g,15min)によって菌体を得た。得られた菌体を10 mMリン酸カリウム緩衝液 (pH7.0)で洗浄し、同緩衝液に懸濁した。4℃に保ちながら日本精機製作所、U5300型を用いて超音波破砕 (4℃、350-400μA、15sec/time×6time)した後、遠心分離(8,000 g,15 min)によって得た上清を、10 mMリン酸カリウム緩衝液(pH7.0)に対して透析した。本操作で得た溶液を粗酵素標品とした。
【0094】
10mMリン酸カリウム緩衝液(pH 7.0)で平衡化したDEAE-Toyopearl 650Mカラム(3×12cm)に、粗酵素を吸着させた。吸着タンパク質を、まず50mM NaClを含む同緩衝液を用いて洗浄後、100mM NaClを含む同緩衝液を用いて溶出させ、活性画分を集めた。
【0095】
0.01mM塩化亜鉛と0.01%メルカプトエタノール,10%グリセロール(洗浄,溶出に用いる緩衝液にも同様に含む)および20 %飽和の硫酸アンモニウムを含む10 mMリン酸カリウム緩衝液(pH 7.0)で、平衡化したButyl-Cellulofineカラム(1.5×8 cm)に、20 %飽和の硫酸アンモニウムを添加したDEAE-Toyopearl 650M活性画分を、吸着させた。吸着タンパク質を、15%飽和の硫酸アンモニウムを含む同緩衝液で洗浄後、10%飽和の硫酸アンモニウムを含む同緩衝液を用いて溶出させた。10%飽和の硫酸アンモニウムを含む緩衝液で溶出した活性画分を集め、10 mMリン酸カリウム緩衝液(pH7.0)に対して透析した。本操作で得た溶液を精製酵素標品とした。
【0096】
また、L298M及びL298W変異酵素についても、同様の操作を行って、精製酵素を得た。
【0097】
2.酵素活性測定
上記1−6で用いた酵素を用い、0.1〜0.3 U/ml程度の酵素溶液を調製した。
【0098】
また、N-アシル-D-メチオニン又はN-アシル-D-フェニルアラニンを基質とし、濃度100mMの基質溶液を調製した。
【0099】
上記酵素溶液140μl、基質溶液40μl、及び1M のHEPES(2-[4-(2-hydroxyethyl)-1-piperazinyl]-ethanesulfonic acid)バッファー溶液20μlを混合し、反応溶液とした。
【0100】
反応は30℃で10分間行い、0.25 N NaOHを100 μl添加することで停止させた。
【0101】
反応停止10分後、0.125 M NaB4O7 を100 μl加え、続いてアミノ基検出試薬である0.2 M TNBSを40 μl添加することで発色させた。10分後、0.1 M NaH2PO4と0.1 M Na2SO3を19.7:0.3の割合で含む溶液800 μlを添加した。この溶液の520 nmにおける吸光度を測定し、反応液中のD-アミノ酸を定量した。なお、対照として基質を添加する前にNaOHを加えたものを用いた。
【0102】
また、1分間に1 μmolのD-アミノ酸を生成する酵素量を1 Uとした。
【0103】
また、基質濃度を8〜300mMに調製し、Lineweaver-Burkの逆数プロットを用いてVmax、Kmを算出した。また、kcatは、Vmaxと本酵素の分子量をもとに算出した。更に本酵素の触媒効率をkcat/Kmの形で求めた。
【0104】
得られた結果を図1に示す。
【0105】
図1に示されるように、V297A変異酵素は、野生型に比べて触媒効率が格段に向上しており、他の変異酵素と比較しても格段に有利な効果を奏することがわかった。
【図面の簡単な説明】
【0106】
【図1】野生型酵素(Wild)、V297A変異酵素(V297A)、L298M変異酵素(L298M)及びL298W変異酵素(L298W)について、N-アシル-D−フェニルアラニン(N-acetyl-D-Phe)及びN-アシル-D−メチオニン(N-acetyl-D-Met)に対するKm、Vmax及びkcat/Kmを調べた結果を示す図面である。
【配列表フリーテキスト】
【0107】
配列番号1は、本発明の酵素が有するアミノ酸配列を示す。
配列番号2はAlcaligenes xylosoxydans subsp. xylosoxydans A-6由来D-aminoacylaseのアミノ酸配列を示す。
配列番号3は、Alcaligenes xylosoxydans subsp. xylosoxydans A-6由来D-aminoacylase遺伝子の塩基配列を示す。
配列番号4は、V297A変異酵素作製に用いたフォワードプライマーの配列を示す。
配列番号5は、V297A変異酵素作製に用いたリバースプライマーの配列を示す。
【特許請求の範囲】
【請求項1】
N-アシル-D-アミノ酸に作用してD-アミノ酸を生じる作用を有するD−アミノアシラーゼであって配列表の配列番号1に記載のアミノ配列を含むD−アミノアシラーゼ。
【請求項2】
配列表の配列番号2記載のアミノ酸配列における297位のバリンがアラニンに置換された配列からなる請求項1に記載のD−アミノアシラーゼ。
【請求項3】
請求項1又は2に記載のD−アミノアシラーゼをコードする遺伝子。
【請求項4】
請求項3に記載の遺伝子を含有する組換えベクター。
【請求項5】
請求項4に記載の組換えベクターで形質転換された形質転換体。
【請求項6】
請求項1又は2に記載のD−アミノアシラーゼを、N-アシル−D−フェニルアラニンに作用させ、生成するD−フェニルアラニンを回収する工程を含むD−フェニルアラニンの製造方法。
【請求項7】
請求項1又は2に記載のD−アミノアシラーゼを、N-アシル−D−メチオニンに作用させ、生成するD−メチオニンを回収する工程を含むD−メチオニンの製造方法。
【請求項1】
N-アシル-D-アミノ酸に作用してD-アミノ酸を生じる作用を有するD−アミノアシラーゼであって配列表の配列番号1に記載のアミノ配列を含むD−アミノアシラーゼ。
【請求項2】
配列表の配列番号2記載のアミノ酸配列における297位のバリンがアラニンに置換された配列からなる請求項1に記載のD−アミノアシラーゼ。
【請求項3】
請求項1又は2に記載のD−アミノアシラーゼをコードする遺伝子。
【請求項4】
請求項3に記載の遺伝子を含有する組換えベクター。
【請求項5】
請求項4に記載の組換えベクターで形質転換された形質転換体。
【請求項6】
請求項1又は2に記載のD−アミノアシラーゼを、N-アシル−D−フェニルアラニンに作用させ、生成するD−フェニルアラニンを回収する工程を含むD−フェニルアラニンの製造方法。
【請求項7】
請求項1又は2に記載のD−アミノアシラーゼを、N-アシル−D−メチオニンに作用させ、生成するD−メチオニンを回収する工程を含むD−メチオニンの製造方法。
【図1】


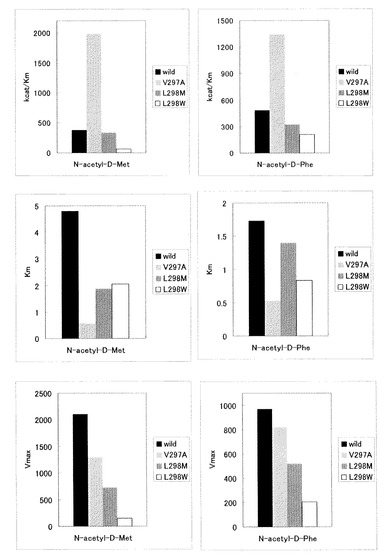
【公開番号】特開2010−154774(P2010−154774A)
【公開日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願番号】特願2008−333831(P2008−333831)
【出願日】平成20年12月26日(2008.12.26)
【出願人】(593006630)学校法人立命館 (359)
【Fターム(参考)】
【公開日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願日】平成20年12月26日(2008.12.26)
【出願人】(593006630)学校法人立命館 (359)
【Fターム(参考)】
[ Back to top ]