
D−アミノ酸からのL−アミノ酸の製造方法
本発明は、開始生物に対して、D−アミノ酸オキシダーゼ、アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物、及び前記微生物の使用下でのD−アミノ酸からのL−アミノ酸の製造方法に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、開始生物に対して、D−アミノ酸オキシダーゼ、L−アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物、及び前記微生物の使用下でのD−アミノ酸からのL−アミノ酸の製造方法に関する。
【0002】
多種の天然アミノ酸が今日ではエナンチオマー純粋な形で、遺伝子的に最適化された細菌を用いた発酵により製造される(de Graaf AA, Eggeling L, Sahm H. 2001. Metabolic engineering for L-lysine production by Corynebacterium glutamicum. Adv Biochem Eng Biotechnol 73:9-29; Sahm H, Eggeling L, Eikmanns B, Kramer R. 1996. Construction of L-lysine-, L-threonine-, and L-isoleucine-overproducing strains of Corynebacterium glutamicum. Ann N Y Acad Sci 782:25-39)。
【0003】
もっともこの方法により、全てのタンパク質性アミノ酸が製造されるわけではなく、また非常に少ない非天然アミノ酸及びD−アミノ酸のみが製造される。エナンチオマー純粋なアミノ酸(このアミノ酸のうちいくらかが一年につき数トンの規模において使用される)の化学的合成は非常に高価であるので、酵素による方法の系列が開発された。この方法は、アシラーゼ、アミダーゼ、エステラーゼ、ヒタンドイナーゼ、アミノ酸オキシダーゼ及びプロテアーゼの助けによる動力学的なラセミ体分割から、リアーゼ、アミノトランスフェラーゼ及びデヒドロゲナーゼを用いたエナンチオ選択的な合成にまで達する(Schmid A. et al. (2002) "The use of enzymes in the chemical industry in Europe" Curr Opin Biotechnol 13(4):359-66)。
【0004】
エナンチオ選択的な合成の他に、ダイナミックに動力学的なラセミ体分割は、このラセミ体分割の際この不所望のエナンチオマーはin situでラセミ化されるが、特に有効である。ダイナミックに動力学的なラセミ体分割の際のように、D−アミノ酸オキシダーゼ又はL−アミノ酸オキシダーゼと、この生じたイミノ酸の再度のアミノ酸への非選択的な化学的還元との組み合わせによっても、原則的に100%の収率が達成されてよい。しかしながらこの還元剤、例えばNaBH4は、少なくとも25当量の過剰量で使用されることが望ましく、これはこの変形を非常に高価にする(Enright et al., "Stereoinversion of beta- and gamma- substituted alpha -amino acids using a chemo-enzymatic oxidation-reduction procedure", Chemical Communications (2003), (20), 2636-2637)。
【0005】
アミノ酸デヒドロゲナーゼによるα−ケト酸のアミン化は一般的に公知である。無論このエダクトは、例えば相応するラセミ体のアミノ酸よりも数倍高価である。
【0006】
アミノ酸オキシダーゼとアミノ酸デヒドロゲナーゼとのカップリングにより、この相応するケト酸をしかしながら、in situでアミノ酸から産生してよい。この両方の酵素が、反対のエナンチオ選択性を有する場合には、D−アミノ酸は完全にL−アミノ酸に、又はL−アミノ酸は完全にD−アミノ酸に変換されてよい。ラセミ体から出発する場合には、これらからエナンチオマー純粋な化合物が製造されてよい。
【0007】
補基質NADHはこの際、酵素を用いて再生されることが望ましく、というのは前記補基質は化学量論量で用いるにはあまりに高価であるからである。特に適しているのは、このための酵素、例えばギ酸デヒドロゲナーゼ及びリンゴ酸デヒドロゲナーゼ(脱カルボキシル性)であり、前記酵素は、これらの基質から二酸化炭素を放出し、従ってこの反応を不可逆にする。(Hanson, R. L. et al. "Enzymatic synthesis of L-6-hydroxynorleucine", Bioorganic & Medicinal Chemistry (1999), 7(10), 2247-2252及びNakajima, N. et al., "Enzymatic conversion of racemic methionine to the L-enantiomer", Journal of the Chemical Society, Chemical Communications (1990), (13), 947-8)。
【0008】
【表1】
【0009】
迅速かつ完全な変換のために、無細胞系においてはしかしながら、付加的にカタラーゼが添加されていることが望ましく、というのは前記アミノ酸オキシダーゼにより触媒される酸化的な一部工程では、過酸化水素が生じ、これはケト酸の脱カルボキシル及び前記酵素の不活化を生じるからである(Trost, E.-M.; Fischer, L., Minimization of by-product formation during D-amino acid oxidase catalyzed racemate resolution of D/L-amino acids, Journal of Molecular Catalysis B: Enzymatic (2002), 19-20 189-195)。ラセミ体のアミノ酸のエナンチオマー純粋なアミノ酸への変換は、この系により>99%ee及び>95%の収率が可能である。
【0010】
この方法はしかしながら高価であり、というのは四つの異なる酵素が別々に製造されかつ単離されなくてはならないからである。
【0011】
本発明の課題は、D−アミノ酸のL−アミノ酸への変換に必要な酵素の高価な単離及び調整を回避する方法の提供である。
【0012】
本発明の主題は有利には、開始生物、例えば野生型に対して、D−アミノ酸オキシダーゼ、アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物の静止細胞である。オキシダーゼ及びデヒドロゲナーゼはこの際、上述の酵素の変換すべき基質及び基質スペクトルを考慮して組み合わせられる。この際前記開始生物は、この上述の酵素を天然には含有するものではない。
【0013】
補基質NADHのための再生酵素として、例えばギ酸デヒドロゲナーゼ、リンゴ酸デヒドロゲナーゼ(DE10240603)又はアルコールデヒドロゲナーゼを使用してよい。
【0014】
これらの酵素をコードするポリヌクレオチドの由来は一般的に、前記の組み換え微生物の属又は種に制限されない。
【0015】
宿主生物の形質転換のためのこの遺伝子は、この由来を考慮することなしに選択されてよい。
【0016】
前記遺伝子の由来は、微生物、真菌又は酵母、特に微生物、特にD−AAOのためのアルスロバクター・プロトホルミエ(Arthrobacter protophormiae)又はトリゴノプシス・バリアビリス(Trigonopsis variablilis)、LeuDHのためのバシラス・セレウスであってよい。
【0017】
宿主生物として、有利には、安定な発現系が存在する微生物、例えばバシラス、様々な酵母、スタフィロコッカス又はストレプトマイセス、特に大腸菌が用いられる。
【0018】
アミノ酸デヒドロゲナーゼとして、特に適するのは、バシラス種からのL−ロイシンデヒドロゲナーゼ(EP0792933)、ロドコッカス種からのグルタミン酸デヒドロゲナーゼ、L−フェニルアラニンデヒドロゲナーゼ、サーモアクチノマイセス・インテルメジウス(Thermoactinomyces intermedius)又はバシラス株からのL−アラニンデヒドロゲナーゼである。
【0019】
これらは、還元によりアミン化するケト酸に依存して選択される。
【0020】
D−アミノ酸オキシダーゼ又はD−アミノ酸オキシダーゼをコードするポリヌクレオチドは、本発明により特に、酵母、ロトトルラ・グラシリス(Rhototorula gracilis)(US6,187,574)から、トリゴノプシス・バリアビリス(Long-Liu Lin et al., Enzyme and Microbial Technol. 27 (2000), 482-491)、カンジダ種、真菌、ニューロスポラ・クラッサ、ベルリカリウム・ルテオラルボ(Verlicullium luteoralbo)及び様々なフザリウム種に、及びD−アミノ酸オキシダーゼはアルスロバクター・プロトホルミエ(EP1375649)に由来する。
【0021】
様々なD−AAOsの基質に関する概要は、Gabler et al.(Enzyme Microbiol. Technol. 27(8): 605-611 (2000))に見出される。EP0897006A1では、属する遺伝子を有するロドスポリジウム(Rhodosporidium)からのD−AAOが記載されている。変換すべき基質に依存して、前記D−アミノ酸オキシダーゼ及びアミノ酸デヒドロゲナーゼをコードする遺伝子が選択され、そして前述の宿主株は前記遺伝子により形質転換される。
【0022】
従って、例えばDL−メチオニン又はDL−ロイシンの脱ラセミ化(Entracemisierung)のためには、有利にはアルスロバクター・プロトホルミエ(EP1375649A)からのD−アミノ酸オキシダーゼ及びバシラス・セレウスからのロイシンデヒドロゲナーゼをコードする遺伝子が大腸菌に過剰発現される。
【0023】
以下の表は、本発明により使用してよい例示的な遺伝子及びこれに属する酵素を有する:
表1:D−アミノ酸オキシダーゼ
受託番号
gi32140775 アルスロバクター・プロトホルミエ D−アミノ酸オキシダーゼ
gi1616634 トリゴノプシス・バリアビリス D−アミノ酸オキシダーゼ
gi27806895 ウシ D−アスパラギン酸オキシダーゼ
EP0897006 ロドスポリジウム D−アミノ酸オキシダーゼ
US6,187,574 ロトトルラ・グラシリス D−アミノ酸オキシダーゼ
表2:L−アミノ酸デヒドロゲナーゼ
受託番号
gi6741938 バシラス・セレウス L−ロイシンデヒドロゲナーゼ
gi625925 ロドコッカス種 L−フェニルアラニンデヒドロゲナーゼ
−−−−ホモ・サピエンス L−リシンデヒドロゲナーゼ
gi16080244 バシラス・ズブチリス L−アラニンデヒドロゲナーゼ
gi118533 ウシ グルタミン酸デヒドロゲナーゼ。
【0024】
この含有されるDNA配列は、次いで公知のアルゴリズム又は配列分析プログラムを用いて、例えばStaden(Nucleic Acids Research 14, 217-232(1986))、Marck(Nucleic Acids Research 16, 1829-1836 (1988))によるプログラム、又はButler(Methods of Biochemical Analysis 39, 74-97 (1998))によるGCG−プログラムにより調査されてよい。
【0025】
ポリメラーゼ連鎖反応(PCR)を用いたDNA配列の増幅のための指示を、当業者は特に、Gaitによるマニュアル:Oligonucleotide synthesis: A Practical Approach (IRL Press, Oxford, UK, 1984)及びNewton及びGrahamによる:PCR(Spektrum Akademischer Verlag, Heidelberg, Deutschland, 1994)に見出す。
【0026】
本発明のその他の対象は、前記の選択した宿主株において一般的に自発的に複製可能なベクターであって、相互に和合性でありかつ、本発明により必要とされる酵素をコードする少なくとも1つの遺伝子を有するベクターの調整である。
【0027】
ベクターDNAは、真核細胞又は原核細胞中に、公知の形質転換技術により導入されてよい。
【0028】
二つの酵素、特に、例えばリンゴ酸デヒドロゲナーゼ及びアミノ酸デヒドロゲナーゼ又はリンゴ酸デヒドロゲナーゼ及びD−アミノ酸オキシダーゼをコードするヌクレオチド配列を含有するベクターが有利である。
【0029】
有利には、ベクター上でアミノ酸デヒドロゲナーゼ及びD−アミノ酸オキシダーゼをコードするヌクレオチド配列の組み合わせでもある。
【0030】
系中でなお欠けている酵素に対するヌクレオチド配列を、次いで更なるベクターに見出す。
【0031】
一般的に、良好に発現可能である遺伝子を低コピー数を有するベクター中に、より弱い発現能力を有する遺伝子をより高いコピー数及び/又は強力なプロモーターを有するベクター上でクローニングするように行われる。この宿主細胞は、前記ベクターにより、前記宿主細胞が開始生物と比較して、D−アミノ酸からL−アミノ酸への変換のために必要な3つ又は場合により4つの酵素をコードするヌクレオチド配列のそれぞれ少なくとも1つの付加的なコピーを含有するように形質転換される。
【0032】
この処置により、上述のヌクレオチド配列を過剰発現することが成功する。
【0033】
過剰発現の達成のために、前記の相応する遺伝子のコピー数を増加させるか、又は前記遺伝子は、この構造遺伝子の上流に存在するこのプロモーター領域又は調節領域又はこのリボソーム結合部位で変異されてよい。同様に、前記構造遺伝子の上流に構成される発現カセットが作用する。誘導可能なプロモーターにより、この発現を、発酵によるアミノ酸産生の過程において上昇させることが付加的に可能である。m−RNAの寿命の延長のための処置により、同様に前記発現が改善される。更に、この酵素タンパク質の崩壊の防止により、同様に前記酵素活性は増強される。前記遺伝子又は遺伝子構築物は、様々なコピー数を有するプラスミド中に存在するか又は染色体に組み込まれそして増幅されてよい。又は、更に、この遺伝子の過剰発現は、この培地組成及び培養操作の変更により達成されてよい。
【0034】
このための指示を、当業者は特に、Martin et al.(Bio/Technology 5, 137-146 (1987))、Guerrero et al.(Gene 138, 35-41 (1994))、Tsuchiya及びMorinaga(Bio/Technology 6, 428-430 (1988))、Eikmanns et al.(Gene 102, 93-98 (1991))で、ヨーロッパ特許公報0472869で、US特許4,601,893で、Schwarzer及びPuehler(Bio/Technology 9, 84-87 (1991)で、Reinscheid et al.(Applied and Environmental Microbiology 60, 126-132 (1994))で、LaBarre et al.(Journal of Bacteriology 175, 1001-1007 (1993))で、特許出願WO96/15246で、Malumbres et al.(Gene 134, 15 - 24 (1993))で、日本国公開公報JP−A−10−229891で、Jensen及びHammer(Biotechnology and Bioengineering 58, 191-195 (1998))で、Makrides(Microbiological Reviews 60:512-538 (1996))で、及び遺伝子学及び分子生物学の公知の教本で見出す。
【0035】
前記過剰発現は、この相応する酵素の、細胞内活性又は濃度の上昇を生じる。
【0036】
この上昇は、この形質転換体に基づく生物(開始生物)中の酵素の濃度又は活性と比較して、一般的に少なくとも10〜500%、特に50〜500%、又は100〜500%、最高で1000〜2000%までにある。
【0037】
本発明の対象は、エナンチオ選択的な酵素による合成経路の使用下でのD−アミノ酸からL−アミノ酸の製造において、形質転換すべき開始生物に比較して、D−アミノ酸オキシダーゼ、L−アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物を、D−アミノ酸を有する溶液と反応させ、この生じるL−アミノ酸を単離することを特徴とする、D−アミノ酸からL−アミノ酸を製造するための方法でもある。
【0038】
有利には、補基質を再生する酵素としてのリンゴ酸酵素の過剰発現であり、この際、この変換すべきD−アミノ酸を含有する、ホールセル触媒(Ganzzellkatalysator)を含有する緩衝水溶液中に、同時にL−リンゴ酸塩又はL−リンゴ酸を、このD−アミノ酸の変換すべき量において、少なくとも等モル量、有利には1.5〜6倍モルにおいて見出す。
【0039】
場合により、カタラーゼも過酸化物分解酵素として過剰発現させる。適しているのは、様々な生物からのカタラーゼ、例えば大腸菌(カタラーゼ HPII(ヒドロキシペルオキシダーゼII)受託番号:gi115722)からの酵素である。
【0040】
前記D−アミノ酸の変換は有利には、静止細胞を用いて行われる。静止細胞とは、生存可能であるが、所定の条件下では増加しない細胞が理解される。
【0041】
アミノ酸とは、この関連において、天然及び非天然に存在するα−アミノ酸が理解され、これは例えば、Beyer-Walter, Lehrbuch der organischen Chemie, S. Hirzel Verlag Stuttgart,第22版,1991, p.822参照、に記載されている。
【0042】
有利には、以下のグループから選択されたD−アミノ酸及びL−アミノ酸の混合物、これらのラセミ体、又はこの純粋なD−エナンチオマーが使用される:リシン、アルギニン、フェニルアラニン、バリン、オルニチン、ロイシン、ヒスチジン、ノルロイシン、チロシン、アラニン、グルタミン酸、及びセファロスポリン、特にメチオニン。
【0043】
特定の酵素が、この様々なD−アミノ酸の変換のために特に適していることが示される(参照、Gabler et al., 2000)。従って、アルスロバクター・プロトホルミエからのD−AAOは、特に塩基性及び疎水性のアミノ酸の変換に適する。
【0044】
前記の適した酵素及びヌクレオチド配列は、一般的には公知技術から公知である。
【0045】
本発明による方法では、単独の酵素の単離が回避される。更なる利点は、本発明により使用された細胞(ホールセル触媒)は、この反応後容易に分離されてよいことから生じる。
【0046】
単離された酵素を用いた、D−メチオニンからL−メチオニンへの変換の際には、公知技術によれば、これに対してカタラーゼを添加しなくてはならない(Nakajima et al., 1990)。
【0047】
本発明により使用される微生物は同様に、本発明の対象であり、かつ連続的に又は非連続的に回分法(バッチ培養)又は流加回分法(流加法)又は反復した流加回分法(反復流加法)において培養されてよい。公知の培養方法に関する概要は、Chmiel(Bioprozesstechnik 1. Einfuehrung in die Bioverfahrenstechnik (Gustav Fischer Verlag, Stuttgart, 1991))による教本又はStorhasによる教本(Bioreaktoren und periphere Einrichtungen (Vieweg Verlag, Braunschweig/Wiesbaden, 1994))に記載されている。
【0048】
使用すべき培地は適したように、前記のそれぞれの株の要求を満足しなくてはならない。様々な微生物の培地の記載は、American Society for Bacteriologyの教本「Manual of Methods for General Bacteriology」(Washington D.C., USA, 1981)に含まれている。
【0049】
炭素供給源として、糖及び炭水化物、例えばグルコース、サッカロース、ラクトース、フルクトース、マルトース、糖蜜、デンプン及びセルロース、油脂、例えば大豆油、ヒマワリ油、ピーナッツ油、ヤシ脂、脂肪酸、例えばパルミチン酸、ステアリン酸及びリノール酸、アルコール、例えばグリセリン及びエタノール、及び有機酸、例えば酢酸が使用されてよい。これらの物質は単独で又は混合物として使用されてよい。
【0050】
窒素供給源として、有機窒素含有化合物、例えばペプトン、酵母エキス、肉エキス、麦芽エキス、コーンスティープリカー、大豆粉、及び尿素又は無機化合物、例えば硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム、炭酸アンモニウム、及び硝酸アンモニウムが使用されてよい。前記窒素供給源は、単独で又は混合物として使用されてよい。
【0051】
リン供給源として、リン酸、リン酸水素カリウム又はリン酸水素二カリウム又はこの相応するナトリウム含有塩が使用されてよい。前記培地は更に、金属塩、例えば硫酸マグネシウム又は硫酸鉄を含有することが必要であり、これらは成長に必要である。最後に、必須栄養成長物質、例えばアミノ酸及びビタミンを上述の物質に加えて使用してよい。上述の使用物質は、1回のバッチの形で培養物に添加するか、適した方法で、この培養の間に供給されてよい。
【0052】
前記培養物のpH調節のために、塩基性化合物、例えば水酸化ナトリウム、水酸化カリウム、アンモニア、又はアンモニア水、又は酸性化合物、例えばリン酸、又は硫酸を適した方法で使用する。泡の発生の調節のために、消泡剤、例えば脂肪酸ポリグリコールエステルが使用されてよい。プラスミドの安定性の維持のために前記溶媒に、適した選択的に作用する物質、例えば抗生物質を添加してよい。好気的条件を維持するために、酸素又は酸素含有混合物、例えば空気を前記培養物に導入する。前記培養物の温度は通常は、20〜45℃、有利には25〜40℃である。この培養は、対数増殖期を経過している限りは継続される。この目標は通常は、10〜20時間の間に達成される。これに引き続き、この細胞を有利には、回収し、洗浄し、緩衝液中に懸濁液としてpH値6〜9、特に6.8〜7.9で取り込む。この細胞濃度は、1〜6%、特に1.5〜4%(湿分質量/v)に達する。前記細胞は一般的に、物理的又は化学的方法により、例えばトルエンで、例えばWilms et al., J. Biotechnol., 86巻 (2001), 19-30が記載しているように、この変換すべきD−アミノ酸がこの細胞壁を透過し、かつL−アミノ酸が流出するように透過処理されている。
【0053】
この懸濁液は次いで、D−アミノ酸及びL−リンゴ酸塩又はL−リンゴ酸を含有する溶液と混合される。この変換は、10〜40℃、特に25〜36℃で、pH値6.8〜8.9、有利には7.5〜8.5で行われる。前記変換は70〜100℃への加熱により停止された。
【0054】
実施例1
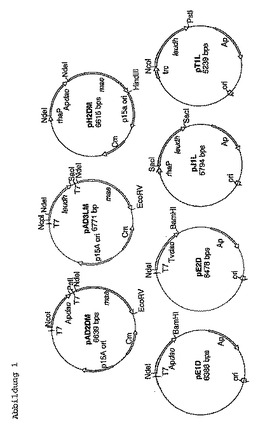
プラスミドの構築
前記の反応原理を確認するために、プラスミドの系列を構築し、この系列は以下の酵素の遺伝子を様々な組み合わせで有する:リンゴ酸酵素(MAE)、ロイシン−デヒドロゲナーゼ(LeuDH)、アロスロバクター・プロトホルミエからのD−AAO(ApD−AAO)及びトリゴノプシス・バリアビリスからのD−AAO(TvD−AAO)。このために、表3に表示したプライマー及びプラスミドを使用した。
【0055】
ApD−AAO(Apdao; gi:32140775)、TvD−AAO(Tvdao; gi:1616634)及びリンゴ酸酵素(mae; gi:1787752)の遺伝子の増幅のために、アルスロバクター・プロトホルミエ、トリゴノプシス・バリアビリス及び大腸菌K12からのゲノムDNAをテンプレートとして使用した。ロイシンデヒドロゲナーゼ(leudh; gi:6741938)の遺伝子を、プラスミドpT1L(=pLeuB,(Ansorge MB, Kula MR. (2000), Investigating expression systems for the stable large-scale production of recombinant L-leucine-dehydrogenase from Bacillus cereus in Escherichia coli. Appl Microbiol Biotechnol 53:668-73))と共に、テンプレートとして増幅した。
【0056】
適したプライマー(表3に表示したプライマー)により、PCRを用いて、クローニングのための制限酵素切断部位を導入した。フォワードプライマーDAAOTvforNdeIを用いて、Tvdao−遺伝子中のイントロンを除去した。
【0057】
pH1M中のApdao−遺伝子のクローニングの際に、シャイン−ダルガルノ配列及び付加的な制限酵素切断部位(EcoRV、PstI及びNotIのため)を、PCRを用いて、リバースプライマーApNdeIrev(表1)を使用することにより導入した。
【0058】
表3
プラスミド及びプライマー
【表2】
【0059】
実施例2
発現株の構築
プラスミドの適した組み合わせにより、表4に表示した組み換え大腸菌株を構築し、前記株はアミノ酸オキシダーゼ、ロイシンデヒドロゲナーゼ及びリンゴ酸酵素のためのそれぞれの遺伝子を含有する。
出発株として使用:
NovagenのBL21(DE3)
及びStratgeneのJM109。
【0060】
表4
大腸菌株
【表3】
【0061】
この株の名称(1〜8)は、以下の実施例においてこの表に相応して維持する。
【0062】
実施例3
細胞内酵素活性の検証
表4に相応する株を、標準的な条件下で、ルリア−ベルタニ(LB)培地中でpH7.5で培養した。形質転換したプラスミドに依存して、前記培地に100μg ml-1のアンピシリン及び/又は34μg ml-1のクロラムフェニコールを添加した。この酵素発現のために、前記の組み換え大腸菌株を、好気性条件下で20mlの培地を有する100mlの振盪フラスコ中で培養させた。この細胞を37℃で、ロータリー振盪機で、200rpmでインキュベーションし、かつ約0.5のOD550で、100μMのIPTG及び/又は0.2%のラムノースにより誘導した。このインキュベーション後、前記株を次いで30℃で更にインキュベーションした。
【0063】
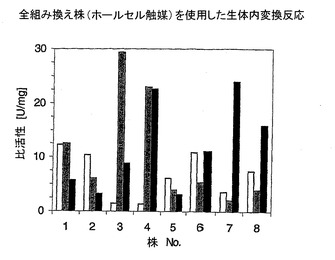
図2には、測定した酵素活性を提示した。
【0064】
白:D−AAO;灰色:MAE;黒:LeuDH。
【0065】
株6は、ラムノースプロモーターの作用を示した。
【0066】
実施例4
全組み換え株による生体内変換反応
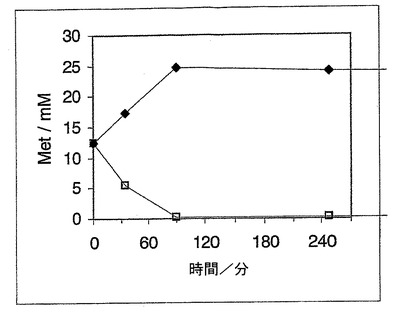
A)D,L−メチオニン
脱ラセミ化のための例として、メチオニンのラセミ体を基質として使用し、プラスミドpAD3LM(ロイシンデヒドロゲナーゼ及びリンゴ酸酵素のための遺伝子を有する)及びpE1D(アルスロバクター・プロトホルミエ−D−アミノ酸オキシダーゼのための遺伝子を有する)の両方を含有する全組み換え大腸菌細胞株1(表2に相応)により変換した。HPLCを用いて、このオキシダーゼにより触媒したD−Metの分解及びこの還元によるアミン化により触媒したL−Metの合成を追跡した。詳しくは、以下を使用した:
25mMのD、L−Met、100mMのL−リンゴ酸塩、0.7MのNH4Cl、50mMのトリス、10mMのMgCl2、終pH8.0。前記細胞を、50mMのTEA/HCl緩衝液、pH7.6中に懸濁し、10μl/mlのトルエンと混合し、この懸濁液を30分間30℃で撹拌し、次いで変換のために使用した。この変換における細胞濃度は、1mlの終容量中で3.3%(湿分質量/v)を有した。前記変換は30℃で、1000rpmでの振盪下で、サーモミキサー5436(Eppendorf社)中で行われた。試料を、5分間の95℃での加熱により停止させ、この透明な上清をHPLCを用いて分析した。
【0067】
D−Met及びL−Metの濃度決定のために、前記試料を希釈後誘導体化した。ここに、pH10.4の100mMのナトリウム−ホウ酸緩衝液中の260mMのイソブチリル−L−システイン及び170mMのo−フタルジアルデヒドの溶液20μlを添加した。HPLC分離条件は、刊行されたように励行した(Krieg L, et al. (2002) Screening for amidases: isolation and characterization of a novel D-amidase from Variovorax paradoxus. Adv Synth Catal 344(9):965-73)。
【0068】
D−Metの分解及びL−Metの形成を、図3に概要して示した。L−Metのエナンチオマー純粋な生成物を、終濃度25mMで得た。
【0069】
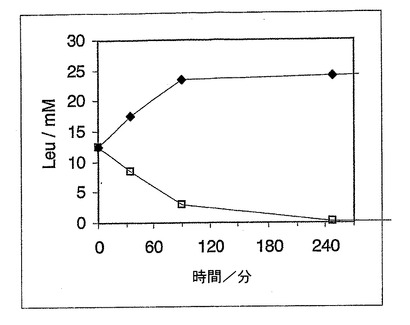
B)D,L−ロイシン
A部で記載したのと同様にして、25mMのD,L−ロイシンの溶液を、前記したのと同一の微生物で変換させた。この変換自体及びこのHPLC−分析の詳細は、前述した実施例に相応する。
【0070】
D−Leuの分解及びL−Leuの形成を、図4に概要して示した。L−Leuのエナンチオマー純粋な生成物を、終濃度25mMで得た。
【図面の簡単な説明】
【0071】
【図1】図1は、使用したプラスミドを示す図である。
【図2】図2は、測定した酵素活性を示す図である。
【図3】図3は、D−Metの分解及びL−Metの形成を示す図である。
【図4】図4は、D−Leuの分解及びL−Leuの形成を示す図である。
【技術分野】
【0001】
本発明は、開始生物に対して、D−アミノ酸オキシダーゼ、L−アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物、及び前記微生物の使用下でのD−アミノ酸からのL−アミノ酸の製造方法に関する。
【0002】
多種の天然アミノ酸が今日ではエナンチオマー純粋な形で、遺伝子的に最適化された細菌を用いた発酵により製造される(de Graaf AA, Eggeling L, Sahm H. 2001. Metabolic engineering for L-lysine production by Corynebacterium glutamicum. Adv Biochem Eng Biotechnol 73:9-29; Sahm H, Eggeling L, Eikmanns B, Kramer R. 1996. Construction of L-lysine-, L-threonine-, and L-isoleucine-overproducing strains of Corynebacterium glutamicum. Ann N Y Acad Sci 782:25-39)。
【0003】
もっともこの方法により、全てのタンパク質性アミノ酸が製造されるわけではなく、また非常に少ない非天然アミノ酸及びD−アミノ酸のみが製造される。エナンチオマー純粋なアミノ酸(このアミノ酸のうちいくらかが一年につき数トンの規模において使用される)の化学的合成は非常に高価であるので、酵素による方法の系列が開発された。この方法は、アシラーゼ、アミダーゼ、エステラーゼ、ヒタンドイナーゼ、アミノ酸オキシダーゼ及びプロテアーゼの助けによる動力学的なラセミ体分割から、リアーゼ、アミノトランスフェラーゼ及びデヒドロゲナーゼを用いたエナンチオ選択的な合成にまで達する(Schmid A. et al. (2002) "The use of enzymes in the chemical industry in Europe" Curr Opin Biotechnol 13(4):359-66)。
【0004】
エナンチオ選択的な合成の他に、ダイナミックに動力学的なラセミ体分割は、このラセミ体分割の際この不所望のエナンチオマーはin situでラセミ化されるが、特に有効である。ダイナミックに動力学的なラセミ体分割の際のように、D−アミノ酸オキシダーゼ又はL−アミノ酸オキシダーゼと、この生じたイミノ酸の再度のアミノ酸への非選択的な化学的還元との組み合わせによっても、原則的に100%の収率が達成されてよい。しかしながらこの還元剤、例えばNaBH4は、少なくとも25当量の過剰量で使用されることが望ましく、これはこの変形を非常に高価にする(Enright et al., "Stereoinversion of beta- and gamma- substituted alpha -amino acids using a chemo-enzymatic oxidation-reduction procedure", Chemical Communications (2003), (20), 2636-2637)。
【0005】
アミノ酸デヒドロゲナーゼによるα−ケト酸のアミン化は一般的に公知である。無論このエダクトは、例えば相応するラセミ体のアミノ酸よりも数倍高価である。
【0006】
アミノ酸オキシダーゼとアミノ酸デヒドロゲナーゼとのカップリングにより、この相応するケト酸をしかしながら、in situでアミノ酸から産生してよい。この両方の酵素が、反対のエナンチオ選択性を有する場合には、D−アミノ酸は完全にL−アミノ酸に、又はL−アミノ酸は完全にD−アミノ酸に変換されてよい。ラセミ体から出発する場合には、これらからエナンチオマー純粋な化合物が製造されてよい。
【0007】
補基質NADHはこの際、酵素を用いて再生されることが望ましく、というのは前記補基質は化学量論量で用いるにはあまりに高価であるからである。特に適しているのは、このための酵素、例えばギ酸デヒドロゲナーゼ及びリンゴ酸デヒドロゲナーゼ(脱カルボキシル性)であり、前記酵素は、これらの基質から二酸化炭素を放出し、従ってこの反応を不可逆にする。(Hanson, R. L. et al. "Enzymatic synthesis of L-6-hydroxynorleucine", Bioorganic & Medicinal Chemistry (1999), 7(10), 2247-2252及びNakajima, N. et al., "Enzymatic conversion of racemic methionine to the L-enantiomer", Journal of the Chemical Society, Chemical Communications (1990), (13), 947-8)。
【0008】
【表1】
【0009】
迅速かつ完全な変換のために、無細胞系においてはしかしながら、付加的にカタラーゼが添加されていることが望ましく、というのは前記アミノ酸オキシダーゼにより触媒される酸化的な一部工程では、過酸化水素が生じ、これはケト酸の脱カルボキシル及び前記酵素の不活化を生じるからである(Trost, E.-M.; Fischer, L., Minimization of by-product formation during D-amino acid oxidase catalyzed racemate resolution of D/L-amino acids, Journal of Molecular Catalysis B: Enzymatic (2002), 19-20 189-195)。ラセミ体のアミノ酸のエナンチオマー純粋なアミノ酸への変換は、この系により>99%ee及び>95%の収率が可能である。
【0010】
この方法はしかしながら高価であり、というのは四つの異なる酵素が別々に製造されかつ単離されなくてはならないからである。
【0011】
本発明の課題は、D−アミノ酸のL−アミノ酸への変換に必要な酵素の高価な単離及び調整を回避する方法の提供である。
【0012】
本発明の主題は有利には、開始生物、例えば野生型に対して、D−アミノ酸オキシダーゼ、アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物の静止細胞である。オキシダーゼ及びデヒドロゲナーゼはこの際、上述の酵素の変換すべき基質及び基質スペクトルを考慮して組み合わせられる。この際前記開始生物は、この上述の酵素を天然には含有するものではない。
【0013】
補基質NADHのための再生酵素として、例えばギ酸デヒドロゲナーゼ、リンゴ酸デヒドロゲナーゼ(DE10240603)又はアルコールデヒドロゲナーゼを使用してよい。
【0014】
これらの酵素をコードするポリヌクレオチドの由来は一般的に、前記の組み換え微生物の属又は種に制限されない。
【0015】
宿主生物の形質転換のためのこの遺伝子は、この由来を考慮することなしに選択されてよい。
【0016】
前記遺伝子の由来は、微生物、真菌又は酵母、特に微生物、特にD−AAOのためのアルスロバクター・プロトホルミエ(Arthrobacter protophormiae)又はトリゴノプシス・バリアビリス(Trigonopsis variablilis)、LeuDHのためのバシラス・セレウスであってよい。
【0017】
宿主生物として、有利には、安定な発現系が存在する微生物、例えばバシラス、様々な酵母、スタフィロコッカス又はストレプトマイセス、特に大腸菌が用いられる。
【0018】
アミノ酸デヒドロゲナーゼとして、特に適するのは、バシラス種からのL−ロイシンデヒドロゲナーゼ(EP0792933)、ロドコッカス種からのグルタミン酸デヒドロゲナーゼ、L−フェニルアラニンデヒドロゲナーゼ、サーモアクチノマイセス・インテルメジウス(Thermoactinomyces intermedius)又はバシラス株からのL−アラニンデヒドロゲナーゼである。
【0019】
これらは、還元によりアミン化するケト酸に依存して選択される。
【0020】
D−アミノ酸オキシダーゼ又はD−アミノ酸オキシダーゼをコードするポリヌクレオチドは、本発明により特に、酵母、ロトトルラ・グラシリス(Rhototorula gracilis)(US6,187,574)から、トリゴノプシス・バリアビリス(Long-Liu Lin et al., Enzyme and Microbial Technol. 27 (2000), 482-491)、カンジダ種、真菌、ニューロスポラ・クラッサ、ベルリカリウム・ルテオラルボ(Verlicullium luteoralbo)及び様々なフザリウム種に、及びD−アミノ酸オキシダーゼはアルスロバクター・プロトホルミエ(EP1375649)に由来する。
【0021】
様々なD−AAOsの基質に関する概要は、Gabler et al.(Enzyme Microbiol. Technol. 27(8): 605-611 (2000))に見出される。EP0897006A1では、属する遺伝子を有するロドスポリジウム(Rhodosporidium)からのD−AAOが記載されている。変換すべき基質に依存して、前記D−アミノ酸オキシダーゼ及びアミノ酸デヒドロゲナーゼをコードする遺伝子が選択され、そして前述の宿主株は前記遺伝子により形質転換される。
【0022】
従って、例えばDL−メチオニン又はDL−ロイシンの脱ラセミ化(Entracemisierung)のためには、有利にはアルスロバクター・プロトホルミエ(EP1375649A)からのD−アミノ酸オキシダーゼ及びバシラス・セレウスからのロイシンデヒドロゲナーゼをコードする遺伝子が大腸菌に過剰発現される。
【0023】
以下の表は、本発明により使用してよい例示的な遺伝子及びこれに属する酵素を有する:
表1:D−アミノ酸オキシダーゼ
受託番号
gi32140775 アルスロバクター・プロトホルミエ D−アミノ酸オキシダーゼ
gi1616634 トリゴノプシス・バリアビリス D−アミノ酸オキシダーゼ
gi27806895 ウシ D−アスパラギン酸オキシダーゼ
EP0897006 ロドスポリジウム D−アミノ酸オキシダーゼ
US6,187,574 ロトトルラ・グラシリス D−アミノ酸オキシダーゼ
表2:L−アミノ酸デヒドロゲナーゼ
受託番号
gi6741938 バシラス・セレウス L−ロイシンデヒドロゲナーゼ
gi625925 ロドコッカス種 L−フェニルアラニンデヒドロゲナーゼ
−−−−ホモ・サピエンス L−リシンデヒドロゲナーゼ
gi16080244 バシラス・ズブチリス L−アラニンデヒドロゲナーゼ
gi118533 ウシ グルタミン酸デヒドロゲナーゼ。
【0024】
この含有されるDNA配列は、次いで公知のアルゴリズム又は配列分析プログラムを用いて、例えばStaden(Nucleic Acids Research 14, 217-232(1986))、Marck(Nucleic Acids Research 16, 1829-1836 (1988))によるプログラム、又はButler(Methods of Biochemical Analysis 39, 74-97 (1998))によるGCG−プログラムにより調査されてよい。
【0025】
ポリメラーゼ連鎖反応(PCR)を用いたDNA配列の増幅のための指示を、当業者は特に、Gaitによるマニュアル:Oligonucleotide synthesis: A Practical Approach (IRL Press, Oxford, UK, 1984)及びNewton及びGrahamによる:PCR(Spektrum Akademischer Verlag, Heidelberg, Deutschland, 1994)に見出す。
【0026】
本発明のその他の対象は、前記の選択した宿主株において一般的に自発的に複製可能なベクターであって、相互に和合性でありかつ、本発明により必要とされる酵素をコードする少なくとも1つの遺伝子を有するベクターの調整である。
【0027】
ベクターDNAは、真核細胞又は原核細胞中に、公知の形質転換技術により導入されてよい。
【0028】
二つの酵素、特に、例えばリンゴ酸デヒドロゲナーゼ及びアミノ酸デヒドロゲナーゼ又はリンゴ酸デヒドロゲナーゼ及びD−アミノ酸オキシダーゼをコードするヌクレオチド配列を含有するベクターが有利である。
【0029】
有利には、ベクター上でアミノ酸デヒドロゲナーゼ及びD−アミノ酸オキシダーゼをコードするヌクレオチド配列の組み合わせでもある。
【0030】
系中でなお欠けている酵素に対するヌクレオチド配列を、次いで更なるベクターに見出す。
【0031】
一般的に、良好に発現可能である遺伝子を低コピー数を有するベクター中に、より弱い発現能力を有する遺伝子をより高いコピー数及び/又は強力なプロモーターを有するベクター上でクローニングするように行われる。この宿主細胞は、前記ベクターにより、前記宿主細胞が開始生物と比較して、D−アミノ酸からL−アミノ酸への変換のために必要な3つ又は場合により4つの酵素をコードするヌクレオチド配列のそれぞれ少なくとも1つの付加的なコピーを含有するように形質転換される。
【0032】
この処置により、上述のヌクレオチド配列を過剰発現することが成功する。
【0033】
過剰発現の達成のために、前記の相応する遺伝子のコピー数を増加させるか、又は前記遺伝子は、この構造遺伝子の上流に存在するこのプロモーター領域又は調節領域又はこのリボソーム結合部位で変異されてよい。同様に、前記構造遺伝子の上流に構成される発現カセットが作用する。誘導可能なプロモーターにより、この発現を、発酵によるアミノ酸産生の過程において上昇させることが付加的に可能である。m−RNAの寿命の延長のための処置により、同様に前記発現が改善される。更に、この酵素タンパク質の崩壊の防止により、同様に前記酵素活性は増強される。前記遺伝子又は遺伝子構築物は、様々なコピー数を有するプラスミド中に存在するか又は染色体に組み込まれそして増幅されてよい。又は、更に、この遺伝子の過剰発現は、この培地組成及び培養操作の変更により達成されてよい。
【0034】
このための指示を、当業者は特に、Martin et al.(Bio/Technology 5, 137-146 (1987))、Guerrero et al.(Gene 138, 35-41 (1994))、Tsuchiya及びMorinaga(Bio/Technology 6, 428-430 (1988))、Eikmanns et al.(Gene 102, 93-98 (1991))で、ヨーロッパ特許公報0472869で、US特許4,601,893で、Schwarzer及びPuehler(Bio/Technology 9, 84-87 (1991)で、Reinscheid et al.(Applied and Environmental Microbiology 60, 126-132 (1994))で、LaBarre et al.(Journal of Bacteriology 175, 1001-1007 (1993))で、特許出願WO96/15246で、Malumbres et al.(Gene 134, 15 - 24 (1993))で、日本国公開公報JP−A−10−229891で、Jensen及びHammer(Biotechnology and Bioengineering 58, 191-195 (1998))で、Makrides(Microbiological Reviews 60:512-538 (1996))で、及び遺伝子学及び分子生物学の公知の教本で見出す。
【0035】
前記過剰発現は、この相応する酵素の、細胞内活性又は濃度の上昇を生じる。
【0036】
この上昇は、この形質転換体に基づく生物(開始生物)中の酵素の濃度又は活性と比較して、一般的に少なくとも10〜500%、特に50〜500%、又は100〜500%、最高で1000〜2000%までにある。
【0037】
本発明の対象は、エナンチオ選択的な酵素による合成経路の使用下でのD−アミノ酸からL−アミノ酸の製造において、形質転換すべき開始生物に比較して、D−アミノ酸オキシダーゼ、L−アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物を、D−アミノ酸を有する溶液と反応させ、この生じるL−アミノ酸を単離することを特徴とする、D−アミノ酸からL−アミノ酸を製造するための方法でもある。
【0038】
有利には、補基質を再生する酵素としてのリンゴ酸酵素の過剰発現であり、この際、この変換すべきD−アミノ酸を含有する、ホールセル触媒(Ganzzellkatalysator)を含有する緩衝水溶液中に、同時にL−リンゴ酸塩又はL−リンゴ酸を、このD−アミノ酸の変換すべき量において、少なくとも等モル量、有利には1.5〜6倍モルにおいて見出す。
【0039】
場合により、カタラーゼも過酸化物分解酵素として過剰発現させる。適しているのは、様々な生物からのカタラーゼ、例えば大腸菌(カタラーゼ HPII(ヒドロキシペルオキシダーゼII)受託番号:gi115722)からの酵素である。
【0040】
前記D−アミノ酸の変換は有利には、静止細胞を用いて行われる。静止細胞とは、生存可能であるが、所定の条件下では増加しない細胞が理解される。
【0041】
アミノ酸とは、この関連において、天然及び非天然に存在するα−アミノ酸が理解され、これは例えば、Beyer-Walter, Lehrbuch der organischen Chemie, S. Hirzel Verlag Stuttgart,第22版,1991, p.822参照、に記載されている。
【0042】
有利には、以下のグループから選択されたD−アミノ酸及びL−アミノ酸の混合物、これらのラセミ体、又はこの純粋なD−エナンチオマーが使用される:リシン、アルギニン、フェニルアラニン、バリン、オルニチン、ロイシン、ヒスチジン、ノルロイシン、チロシン、アラニン、グルタミン酸、及びセファロスポリン、特にメチオニン。
【0043】
特定の酵素が、この様々なD−アミノ酸の変換のために特に適していることが示される(参照、Gabler et al., 2000)。従って、アルスロバクター・プロトホルミエからのD−AAOは、特に塩基性及び疎水性のアミノ酸の変換に適する。
【0044】
前記の適した酵素及びヌクレオチド配列は、一般的には公知技術から公知である。
【0045】
本発明による方法では、単独の酵素の単離が回避される。更なる利点は、本発明により使用された細胞(ホールセル触媒)は、この反応後容易に分離されてよいことから生じる。
【0046】
単離された酵素を用いた、D−メチオニンからL−メチオニンへの変換の際には、公知技術によれば、これに対してカタラーゼを添加しなくてはならない(Nakajima et al., 1990)。
【0047】
本発明により使用される微生物は同様に、本発明の対象であり、かつ連続的に又は非連続的に回分法(バッチ培養)又は流加回分法(流加法)又は反復した流加回分法(反復流加法)において培養されてよい。公知の培養方法に関する概要は、Chmiel(Bioprozesstechnik 1. Einfuehrung in die Bioverfahrenstechnik (Gustav Fischer Verlag, Stuttgart, 1991))による教本又はStorhasによる教本(Bioreaktoren und periphere Einrichtungen (Vieweg Verlag, Braunschweig/Wiesbaden, 1994))に記載されている。
【0048】
使用すべき培地は適したように、前記のそれぞれの株の要求を満足しなくてはならない。様々な微生物の培地の記載は、American Society for Bacteriologyの教本「Manual of Methods for General Bacteriology」(Washington D.C., USA, 1981)に含まれている。
【0049】
炭素供給源として、糖及び炭水化物、例えばグルコース、サッカロース、ラクトース、フルクトース、マルトース、糖蜜、デンプン及びセルロース、油脂、例えば大豆油、ヒマワリ油、ピーナッツ油、ヤシ脂、脂肪酸、例えばパルミチン酸、ステアリン酸及びリノール酸、アルコール、例えばグリセリン及びエタノール、及び有機酸、例えば酢酸が使用されてよい。これらの物質は単独で又は混合物として使用されてよい。
【0050】
窒素供給源として、有機窒素含有化合物、例えばペプトン、酵母エキス、肉エキス、麦芽エキス、コーンスティープリカー、大豆粉、及び尿素又は無機化合物、例えば硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム、炭酸アンモニウム、及び硝酸アンモニウムが使用されてよい。前記窒素供給源は、単独で又は混合物として使用されてよい。
【0051】
リン供給源として、リン酸、リン酸水素カリウム又はリン酸水素二カリウム又はこの相応するナトリウム含有塩が使用されてよい。前記培地は更に、金属塩、例えば硫酸マグネシウム又は硫酸鉄を含有することが必要であり、これらは成長に必要である。最後に、必須栄養成長物質、例えばアミノ酸及びビタミンを上述の物質に加えて使用してよい。上述の使用物質は、1回のバッチの形で培養物に添加するか、適した方法で、この培養の間に供給されてよい。
【0052】
前記培養物のpH調節のために、塩基性化合物、例えば水酸化ナトリウム、水酸化カリウム、アンモニア、又はアンモニア水、又は酸性化合物、例えばリン酸、又は硫酸を適した方法で使用する。泡の発生の調節のために、消泡剤、例えば脂肪酸ポリグリコールエステルが使用されてよい。プラスミドの安定性の維持のために前記溶媒に、適した選択的に作用する物質、例えば抗生物質を添加してよい。好気的条件を維持するために、酸素又は酸素含有混合物、例えば空気を前記培養物に導入する。前記培養物の温度は通常は、20〜45℃、有利には25〜40℃である。この培養は、対数増殖期を経過している限りは継続される。この目標は通常は、10〜20時間の間に達成される。これに引き続き、この細胞を有利には、回収し、洗浄し、緩衝液中に懸濁液としてpH値6〜9、特に6.8〜7.9で取り込む。この細胞濃度は、1〜6%、特に1.5〜4%(湿分質量/v)に達する。前記細胞は一般的に、物理的又は化学的方法により、例えばトルエンで、例えばWilms et al., J. Biotechnol., 86巻 (2001), 19-30が記載しているように、この変換すべきD−アミノ酸がこの細胞壁を透過し、かつL−アミノ酸が流出するように透過処理されている。
【0053】
この懸濁液は次いで、D−アミノ酸及びL−リンゴ酸塩又はL−リンゴ酸を含有する溶液と混合される。この変換は、10〜40℃、特に25〜36℃で、pH値6.8〜8.9、有利には7.5〜8.5で行われる。前記変換は70〜100℃への加熱により停止された。
【0054】
実施例1
プラスミドの構築
前記の反応原理を確認するために、プラスミドの系列を構築し、この系列は以下の酵素の遺伝子を様々な組み合わせで有する:リンゴ酸酵素(MAE)、ロイシン−デヒドロゲナーゼ(LeuDH)、アロスロバクター・プロトホルミエからのD−AAO(ApD−AAO)及びトリゴノプシス・バリアビリスからのD−AAO(TvD−AAO)。このために、表3に表示したプライマー及びプラスミドを使用した。
【0055】
ApD−AAO(Apdao; gi:32140775)、TvD−AAO(Tvdao; gi:1616634)及びリンゴ酸酵素(mae; gi:1787752)の遺伝子の増幅のために、アルスロバクター・プロトホルミエ、トリゴノプシス・バリアビリス及び大腸菌K12からのゲノムDNAをテンプレートとして使用した。ロイシンデヒドロゲナーゼ(leudh; gi:6741938)の遺伝子を、プラスミドpT1L(=pLeuB,(Ansorge MB, Kula MR. (2000), Investigating expression systems for the stable large-scale production of recombinant L-leucine-dehydrogenase from Bacillus cereus in Escherichia coli. Appl Microbiol Biotechnol 53:668-73))と共に、テンプレートとして増幅した。
【0056】
適したプライマー(表3に表示したプライマー)により、PCRを用いて、クローニングのための制限酵素切断部位を導入した。フォワードプライマーDAAOTvforNdeIを用いて、Tvdao−遺伝子中のイントロンを除去した。
【0057】
pH1M中のApdao−遺伝子のクローニングの際に、シャイン−ダルガルノ配列及び付加的な制限酵素切断部位(EcoRV、PstI及びNotIのため)を、PCRを用いて、リバースプライマーApNdeIrev(表1)を使用することにより導入した。
【0058】
表3
プラスミド及びプライマー
【表2】
【0059】
実施例2
発現株の構築
プラスミドの適した組み合わせにより、表4に表示した組み換え大腸菌株を構築し、前記株はアミノ酸オキシダーゼ、ロイシンデヒドロゲナーゼ及びリンゴ酸酵素のためのそれぞれの遺伝子を含有する。
出発株として使用:
NovagenのBL21(DE3)
及びStratgeneのJM109。
【0060】
表4
大腸菌株
【表3】
【0061】
この株の名称(1〜8)は、以下の実施例においてこの表に相応して維持する。
【0062】
実施例3
細胞内酵素活性の検証
表4に相応する株を、標準的な条件下で、ルリア−ベルタニ(LB)培地中でpH7.5で培養した。形質転換したプラスミドに依存して、前記培地に100μg ml-1のアンピシリン及び/又は34μg ml-1のクロラムフェニコールを添加した。この酵素発現のために、前記の組み換え大腸菌株を、好気性条件下で20mlの培地を有する100mlの振盪フラスコ中で培養させた。この細胞を37℃で、ロータリー振盪機で、200rpmでインキュベーションし、かつ約0.5のOD550で、100μMのIPTG及び/又は0.2%のラムノースにより誘導した。このインキュベーション後、前記株を次いで30℃で更にインキュベーションした。
【0063】
図2には、測定した酵素活性を提示した。
【0064】
白:D−AAO;灰色:MAE;黒:LeuDH。
【0065】
株6は、ラムノースプロモーターの作用を示した。
【0066】
実施例4
全組み換え株による生体内変換反応
A)D,L−メチオニン
脱ラセミ化のための例として、メチオニンのラセミ体を基質として使用し、プラスミドpAD3LM(ロイシンデヒドロゲナーゼ及びリンゴ酸酵素のための遺伝子を有する)及びpE1D(アルスロバクター・プロトホルミエ−D−アミノ酸オキシダーゼのための遺伝子を有する)の両方を含有する全組み換え大腸菌細胞株1(表2に相応)により変換した。HPLCを用いて、このオキシダーゼにより触媒したD−Metの分解及びこの還元によるアミン化により触媒したL−Metの合成を追跡した。詳しくは、以下を使用した:
25mMのD、L−Met、100mMのL−リンゴ酸塩、0.7MのNH4Cl、50mMのトリス、10mMのMgCl2、終pH8.0。前記細胞を、50mMのTEA/HCl緩衝液、pH7.6中に懸濁し、10μl/mlのトルエンと混合し、この懸濁液を30分間30℃で撹拌し、次いで変換のために使用した。この変換における細胞濃度は、1mlの終容量中で3.3%(湿分質量/v)を有した。前記変換は30℃で、1000rpmでの振盪下で、サーモミキサー5436(Eppendorf社)中で行われた。試料を、5分間の95℃での加熱により停止させ、この透明な上清をHPLCを用いて分析した。
【0067】
D−Met及びL−Metの濃度決定のために、前記試料を希釈後誘導体化した。ここに、pH10.4の100mMのナトリウム−ホウ酸緩衝液中の260mMのイソブチリル−L−システイン及び170mMのo−フタルジアルデヒドの溶液20μlを添加した。HPLC分離条件は、刊行されたように励行した(Krieg L, et al. (2002) Screening for amidases: isolation and characterization of a novel D-amidase from Variovorax paradoxus. Adv Synth Catal 344(9):965-73)。
【0068】
D−Metの分解及びL−Metの形成を、図3に概要して示した。L−Metのエナンチオマー純粋な生成物を、終濃度25mMで得た。
【0069】
B)D,L−ロイシン
A部で記載したのと同様にして、25mMのD,L−ロイシンの溶液を、前記したのと同一の微生物で変換させた。この変換自体及びこのHPLC−分析の詳細は、前述した実施例に相応する。
【0070】
D−Leuの分解及びL−Leuの形成を、図4に概要して示した。L−Leuのエナンチオマー純粋な生成物を、終濃度25mMで得た。
【図面の簡単な説明】
【0071】
【図1】図1は、使用したプラスミドを示す図である。
【図2】図2は、測定した酵素活性を示す図である。
【図3】図3は、D−Metの分解及びL−Metの形成を示す図である。
【図4】図4は、D−Leuの分解及びL−Leuの形成を示す図である。
【特許請求の範囲】
【請求項1】
開始生物に対して、D−アミノ酸オキシダーゼ、L−アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する、組み換え微生物。
【請求項2】
静止細胞からなる、請求項1記載の微生物。
【請求項3】
ギ酸デヒドロゲナーゼの増加した濃度又は活性を有する、請求項1又は2記載の微生物。
【請求項4】
リンゴ酸デヒドロゲナーゼの増加した濃度又は活性を有する、請求項1又は2記載の微生物。
【請求項5】
アルコールデヒドロゲナーゼの増加した濃度又は活性を有する、請求項1又は2記載の微生物。
【請求項6】
L−ロイシンデヒドロゲナーゼの増加した濃度又は活性を有する、請求項1から5までのいずれか1項記載の微生物。
【請求項7】
フェニルアラニンデヒドロゲナーゼ、アラニンデヒドロゲナーゼ、グルタミン酸デヒドロゲナーゼのグループから選択されたL−アミノ酸デヒドロゲナーゼの増加した濃度又は活性を有する、請求項1から5までのいずれか1項記載の微生物。
【請求項8】
前述の酵素をコードする複数の遺伝子が、他の属の微生物、真菌又は酵母に由来する、請求項1から4までのいずれか1項記載の微生物。
【請求項9】
D−アミノ酸オキシダーゼ、L−アミノ酸デヒドロゲナーゼ、補酵素NADHを再生する酵素及び場合によりカタラーゼの細胞内活性が、前記酵素をコードするヌクレオチド配列のコピー数を増加させるか、又は強力なプロモーターを使用するか、又は両者を組み合わせることにより開始生物に対して増加する、請求項1から8までのいずれか1項記載の微生物。
【請求項10】
D−アミノ酸オキシダーゼ、アミノ酸デヒドロゲナーゼ又は補基質NADHを再生する酵素をコードするヌクレオチド配列の1つ又は複数を有する、ベクター。
【請求項11】
D−アミノ酸デヒドロゲナーゼ及びリンゴ酸デヒドロゲナーゼをコードするヌクレオチド配列を含有する、請求項10記載のベクター。
【請求項12】
アミノ酸デヒドロゲナーゼ及びリンゴ酸デヒドロゲナーゼをコードするヌクレオチド配列を含有する、請求項10記載のベクター。
【請求項13】
アミノ酸デヒドロゲナーゼ及びD−アミノ酸オキシダーゼをコードするヌクレオチド配列を含有する、請求項10記載のベクター。
【請求項14】
開始生物に比較して、上述の酵素をコードするヌクレオチド配列のそれぞれ少なくとも1つの付加的なコピーを有するように、請求項10から13までのいずれか1項記載のベクターを用いて形質変換された微生物。
【請求項15】
エナンチオ選択的な酵素による合成経路の使用下でD−アミノ酸からL−アミノ酸を製造するための方法であって、開始生物(例えば野生型)に対して、アミノ酸オキシダーゼ、アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物を、D−アミノ酸を有する溶液と反応させ、この生じるL−アミノ酸を単離することを特徴とする、D−アミノ酸からL−アミノ酸を製造するための方法。
【請求項16】
請求項1から9及び14のいずれか1項記載の微生物を使用することを特徴とする、請求項15記載の方法。
【請求項17】
補基質NADHを再生する酵素として、リンゴ酸デヒドロゲナーゼを選択し、前記のD−アミノ酸を含有する溶液に、L−リンゴ酸塩又はL−リンゴ酸を添加することを特徴とする、請求項15又は16記載の方法。
【請求項18】
前記微生物の静止細胞を使用することを特徴とする、請求項15から17までのいずれか1項記載の方法。
【請求項19】
使用した細胞の細胞壁が、化学的又は物理的処置により、前記溶液からのD−アミノ酸の取り込みのために透過性であることを特徴とする、請求項15から18までのいずれか1項記載の方法。
【請求項20】
リシン、アルギニン、フェニルアラニン、バリン、オルニチン、ロイシン、ヒスチジン、ノルロイシン、チロシン、アラニン、グルタミン酸、セファロスポリン、特にメチオニンのグループから選択されたD−アミノ酸又はラセミ体のDL−アミノ酸を使用することを特徴とする、請求項15から19までのいずれか1項記載の方法。
【請求項1】
開始生物に対して、D−アミノ酸オキシダーゼ、L−アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する、組み換え微生物。
【請求項2】
静止細胞からなる、請求項1記載の微生物。
【請求項3】
ギ酸デヒドロゲナーゼの増加した濃度又は活性を有する、請求項1又は2記載の微生物。
【請求項4】
リンゴ酸デヒドロゲナーゼの増加した濃度又は活性を有する、請求項1又は2記載の微生物。
【請求項5】
アルコールデヒドロゲナーゼの増加した濃度又は活性を有する、請求項1又は2記載の微生物。
【請求項6】
L−ロイシンデヒドロゲナーゼの増加した濃度又は活性を有する、請求項1から5までのいずれか1項記載の微生物。
【請求項7】
フェニルアラニンデヒドロゲナーゼ、アラニンデヒドロゲナーゼ、グルタミン酸デヒドロゲナーゼのグループから選択されたL−アミノ酸デヒドロゲナーゼの増加した濃度又は活性を有する、請求項1から5までのいずれか1項記載の微生物。
【請求項8】
前述の酵素をコードする複数の遺伝子が、他の属の微生物、真菌又は酵母に由来する、請求項1から4までのいずれか1項記載の微生物。
【請求項9】
D−アミノ酸オキシダーゼ、L−アミノ酸デヒドロゲナーゼ、補酵素NADHを再生する酵素及び場合によりカタラーゼの細胞内活性が、前記酵素をコードするヌクレオチド配列のコピー数を増加させるか、又は強力なプロモーターを使用するか、又は両者を組み合わせることにより開始生物に対して増加する、請求項1から8までのいずれか1項記載の微生物。
【請求項10】
D−アミノ酸オキシダーゼ、アミノ酸デヒドロゲナーゼ又は補基質NADHを再生する酵素をコードするヌクレオチド配列の1つ又は複数を有する、ベクター。
【請求項11】
D−アミノ酸デヒドロゲナーゼ及びリンゴ酸デヒドロゲナーゼをコードするヌクレオチド配列を含有する、請求項10記載のベクター。
【請求項12】
アミノ酸デヒドロゲナーゼ及びリンゴ酸デヒドロゲナーゼをコードするヌクレオチド配列を含有する、請求項10記載のベクター。
【請求項13】
アミノ酸デヒドロゲナーゼ及びD−アミノ酸オキシダーゼをコードするヌクレオチド配列を含有する、請求項10記載のベクター。
【請求項14】
開始生物に比較して、上述の酵素をコードするヌクレオチド配列のそれぞれ少なくとも1つの付加的なコピーを有するように、請求項10から13までのいずれか1項記載のベクターを用いて形質変換された微生物。
【請求項15】
エナンチオ選択的な酵素による合成経路の使用下でD−アミノ酸からL−アミノ酸を製造するための方法であって、開始生物(例えば野生型)に対して、アミノ酸オキシダーゼ、アミノ酸デヒドロゲナーゼ、補基質NADHを再生する酵素及び場合によりカタラーゼの増加した濃度又は活性を有する組み換え微生物を、D−アミノ酸を有する溶液と反応させ、この生じるL−アミノ酸を単離することを特徴とする、D−アミノ酸からL−アミノ酸を製造するための方法。
【請求項16】
請求項1から9及び14のいずれか1項記載の微生物を使用することを特徴とする、請求項15記載の方法。
【請求項17】
補基質NADHを再生する酵素として、リンゴ酸デヒドロゲナーゼを選択し、前記のD−アミノ酸を含有する溶液に、L−リンゴ酸塩又はL−リンゴ酸を添加することを特徴とする、請求項15又は16記載の方法。
【請求項18】
前記微生物の静止細胞を使用することを特徴とする、請求項15から17までのいずれか1項記載の方法。
【請求項19】
使用した細胞の細胞壁が、化学的又は物理的処置により、前記溶液からのD−アミノ酸の取り込みのために透過性であることを特徴とする、請求項15から18までのいずれか1項記載の方法。
【請求項20】
リシン、アルギニン、フェニルアラニン、バリン、オルニチン、ロイシン、ヒスチジン、ノルロイシン、チロシン、アラニン、グルタミン酸、セファロスポリン、特にメチオニンのグループから選択されたD−アミノ酸又はラセミ体のDL−アミノ酸を使用することを特徴とする、請求項15から19までのいずれか1項記載の方法。
【図1】

【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公表番号】特表2007−522810(P2007−522810A)
【公表日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願番号】特願2006−553471(P2006−553471)
【出願日】平成17年1月27日(2005.1.27)
【国際出願番号】PCT/EP2005/000768
【国際公開番号】WO2005/090590
【国際公開日】平成17年9月29日(2005.9.29)
【出願人】(501073862)デグサ ゲーエムベーハー (837)
【氏名又は名称原語表記】Degussa GmbH
【住所又は居所原語表記】Bennigsenplatz 1, D−40474 Duesseldorf, Germany
【Fターム(参考)】
【公表日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願日】平成17年1月27日(2005.1.27)
【国際出願番号】PCT/EP2005/000768
【国際公開番号】WO2005/090590
【国際公開日】平成17年9月29日(2005.9.29)
【出願人】(501073862)デグサ ゲーエムベーハー (837)
【氏名又は名称原語表記】Degussa GmbH
【住所又は居所原語表記】Bennigsenplatz 1, D−40474 Duesseldorf, Germany
【Fターム(参考)】
[ Back to top ]