
D−グルコースの存在下でペントース糖を代謝する酵母菌株を同定・使用するための材料及び方法
本明細書において開示されているものは、酵母の大部分の菌株がD−グルコース以外の糖において増殖することを阻害する量の2−デオキシ−グルコース及び/又はD−グルコースの存在下で、D−グルコース以外の糖において増殖することができる酵母の変種、特にサッカロミセス・セレビシエ(Saccharomyces cerevisiae)の変種を作り出す及び/又は単離する材料及び方法である。そのような変種を単離するのに使用できる選択培地は、D−キシロース、L−グルタミン及び2−デオキシ−グルコースのようなペントース糖を含む。幾つかの菌株のGrr1及びRed遺伝子における突然変異も、2−デオキシ−グルコースの存在下でペントースのD−キシロースを含む糖において増殖することができる変種を産生する。
【発明の詳細な説明】
【技術分野】
【0001】
(優先権の主張)
本出願は、2009年2月25日に出願した米国仮出願第61/155,255号(この全ては参照として本明細書に組み込まれる)の利益を主張する。
【0002】
本発明は、相当なレベルのD−グルコースの存在下でD−グルコース以外の糖において増殖することができるサッカロミセス属(Saccharomyces)の菌株、並びに、その開発及び使用方法に関する。
【背景技術】
【0003】
サッカロミセス属の多様な種は、産業的に増殖される微生物のうち最も重要なものである。パンを発酵するため、ビール及びワインを生産するため、並びに食用香料及び微量栄養素の供給源として長い間使用されてきているが、これらの生物は、現在、糖原料からエタノールへの変換を促進することによって、燃料の生産において中心的な役割を果たしている。代謝的に複雑な生物である酵母は、好気的にも、また少なくとも数世代にわたって嫌気的も増殖することができる。商業的な製パン製造業を支援するために使用される酵母の生産のような商業的に増殖される場合、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)のような酵母を、通気発酵タンクにおいて増殖することができるこれらの条件下での酵母の増殖を制御して、酵母バイオマスの生産を増加することができる。これを達成することができる1つの方法は、D−グルコースのような糖の添加及び好気的に増殖することを促すための酵母への酸素移動の速さについて計画することである。サッカロミセス属の多様な菌株も、エタノールの生産を最大限にするように設計された条件下で増殖することができる。多くの場合、目的が糖からエタノールへの変換を最大限にすることである場合、発酵容器中の酸素レベルは、目的が嫌気性増殖に好ましいように酵母バイオマス生産を最大限にすることである場合の容器において使用される酸素のレベルに対して低減されうる。
【0004】
サッカロミセス属の大部分の菌株は、D−グルコースにおいて優先的に増殖するが、多くの菌株は、他の天然に生じるヘキソースにおいて、さらには幾つかの二糖類においても増殖することが知られている。異なる糖において、異なる酸素レベルの存在下で増殖するサッカロミセス属の異なる種の能力は、酵母が燃料産業で植物バイオマスからエタノールへの変換において現在果たしている中心的な役割を含む、商業的な有用性の多くを説明している。
【0005】
酵母によるエタノールの生産において一番良く知られている経路の1つは、エタノールへの6−炭素糖(ヘキソース)、特にD−グルコースの発酵である(図1)。エタノールの生産に広く使用されている供給原料の1つは多糖類のデンプンである。デンプンは、D−グルコースを含む単独ポリマーである。現在、米国において少なくともトウモロコシ由来のデンプンは、サッカロミセス・セレビシエによるエタノール生産にとって好ましい供給原料である。トウモロコシは、栄養素の高い作物であり、現在、トウモロコシの穀粒のみが、酵母を使用するエタノール発酵にとってデンプン/D−グルコースの適した供給源である。酵母に基づいたエタノール生産のための糖の別の供給源は、サトウキビである。サトウキビは、天然に発酵性糖が高く、酵母を使用するエタノールの生産にとって好ましい基質である。しかし、トウモロコシは、米国においてサトウキビよりも広範囲に栽培されている。気候のおかげで、このままの状態が続く可能性が高い。いずれにしても、トウモロコシに基づいたエタノール生産の持続可能性が疑問視されてきており、サトウキビは米国において実現可能な選択肢ではないので、バイオ燃料産業は、トウモロコシ及びサトウギビ以外の発酵性供給原料の他の供給源を探している。
【0006】
1つの大いにもてはやされている供給原料は、セルロースであり、トウモロコシよりも持続可能性があり、サトウキビよりも容易に入手可能であると考えられている。発酵性糖を生産するように加工されたセルロースは、恐らくエタノール生産の未来にとって最適な炭素供給源でありうる。酵母バイオマスを増加するため又はデンプン若しくはセルロースのような原料からエタノールを生産するために酵母を増殖させることは、バイオポリマーセルロースを、酵母により容易に発酵されうるD−グルコース、マルトース、三糖類及び四糖類のような糖単位に分解する前発酵加工工程を必要とする。
【0007】
供給源にかかわらず、6炭素糖、特にD−グルコースは、酵母に基づいた発酵にとって主なエネルギー供給源である。特徴決定されているサッカロミセス属の大部分の種は、D−グルコースにおいて優先的に増殖する。サッカロミセス属の多くの実験室由来菌株を含むこれらの菌株の多くは、D−グルコース以外のヘキソース、並びに二糖類及び三糖類において増殖することができる。しかし、D−グルコースにおいて優先的に増殖するサッカロミセス属は、ほぼ全ての産業的に重要な菌株を含むこの酵母の大部分の変種が異化代謝産物抑制を示すほど強力であり、すなわち、菌株は、供給原料において検出可能なレベルのD−グルコースがある限り、D−グルコース以外の糖を発酵しない。
【0008】
サッカロミセス属の検査した全てのものが、D−グルコースの存在下でD−グルコース以外の糖において活発に増殖し、糖からエタノールを生産することに無力であることは、D−グルコースを含む発酵性糖の混合物を含む任意の供給原料からの酵母バイオマス及び/又はエタノールの生産にとって残念なことである。例えば、D−グルコースはセルロース性バイオマスの発酵性成分への分解により遊離され、発酵性糖のミックスにおけるD−グルコースの存在は、他の糖のエタノールへの変換を著しく遅くする(図1)。
【0009】
セルロースからエタノールを生産する現在の技術的な障害にもかかわらず、2007年エネルギー独立性及び安全保障法(2007 Energy Independence and Security Act)(EISA2007)は、米国が、輸入石油に取って代わるために、セルロース性エタノールを生産する技術を急速に開発することを義務づける。したがって、産業的なサッカロミセス属の新規菌株の必要性及び有意な量のD−グルコースの存在下でもD−グルコースのみ以外の糖をバイオマス又はエタノールに容易に変換するこれらの産業的菌株を作り出す方法の必要性が存在する。本発明の幾つかの態様は、これらの必要性に対処している。
【発明の概要】
【発明が解決しようとする課題】
【0010】
そこで、本発明は、相当なレベルのD−グルコースの存在下でD−グルコース以外の糖において増殖することができるサッカロミセス属の菌株、並びに、その開発及び使用方法を提供する。
【課題を解決するための手段】
【0011】
本発明の幾つかの態様は、酵母を単離する方法であって、増殖培地を準備する工程(ここで培地は2−デオキシ−グルコース、キシロース及びグルタミンを含み、キシロースが唯一の炭素供給源である);培地に少なくとも1つの酵母菌株を接種する工程;培地から少なくとも1つの酵母細胞を単離する工程(ここで前記酵母細胞は、約0.1重量%の2−デオキシ−グルコースの存在下で唯一の炭素供給源としてのD−キシロースにおいて増殖する)を含む方法を含む。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。幾つかの実施態様では、培地に増殖する菌株は、約14日後にのみ培地において検出可能な増殖を示す。さらに別の実施態様において、検出可能な増殖の出現は、接種後、約21日後においてのみ生じる。
【0012】
幾つかの実施態様において、酵母菌株を選択するのに使用した培地は、約2.0重量%のキシロース及び約0.5重量%のグルタミンを含むが、これらの試薬の任意の濃度が増殖培地に添加されうる特定の菌株の増殖を支援するために十分である。幾つかの実施態様において、単離された酵母菌株は、サッカロミセス・セレビシエの一倍体(又は、半数体、haploid)、二倍体又は2を超える倍数性を有する菌株である。
【0013】
幾つかの実施態様において、培地から単離された酵母菌株は、少なくとも0.1重量%の2−デオキシ−グルコースの存在下でも少なくとも1つのペントース糖を代謝する。幾つかの実施態様において、供給原料(feed stock)は、少なくとも約0.03重量%の2−デオキシ−グルコースを含む。幾つかの実施態様において、菌株は、少なくとも0.1重量%の2−デオキシ−グルコースの存在下でD−グルコース以外の少なくとも1つのヘキソース糖を代謝する。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。幾つかの実施態様において、菌株は自然発生突然変異体である。さらに別の実施態様において、菌株は、突然変異を引き起こすことが知られている方法で酵母を処理することによるオープンリーディングフレームの標的化破壊のような、特定の事象によって作り出すことができる。突然変異誘発の速度を加速する方法及び試薬には、電離放射(ionizing radiation)、UV線、並びに、挿入剤(intercalating agent)、アルキル化剤(alkylating agent)、DNA付加物(DNA adduct)などのようなDNA構造に影響を与える試薬への暴露が含まれるが、これらに限定されない。
【0014】
本発明のさらに他の実施態様は、少なくとも0.1重量%の2−デオキシ−グルコースの存在下で唯一の炭素供給源の少なくとも1つのペントース糖において増殖するサッカロミセス・セレビシエの菌株を含む、酵母の変種を含む。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。幾つかの実施態様において、変種は、一倍体、二倍体であるか又は2を超える倍数性数を有する。幾つかの実施態様において、サッカロミセス・セレビシエの変種菌株は、JH015、CDXR2、Fermentis Ethanol Red reg1Δ及びFermentis Ethanol Red grr1−/−(GX1)などからなる群より選択される。
【0015】
さらに別の実施態様は、糖供給源を発酵する方法であって、サッカロミセス・セレビシエの少なくとも1つの菌株を準備する工程(ここでサッカロミセス・セレビシエの少なくとも1つの菌株は、少なくとも0.1重量%の2−デオキシ−グルコースの存在下で少なくとも1つのペントースにおいて増殖する);少なくとも1つの糖を含む供給原料を供給する工程;及び前記酵母菌株を供給原料において増殖する工程を含む方法である。幾つかの実施態様において、供給原料は、任意の追加の糖供給源の不在下で酵母菌株の増殖を支援するのに十分な量のD−グルコースを含む。さらに他の実施態様において、供給原料は、発酵性ペントース糖を含む。なお他の実施態様において、供給原料は、少なくとも約0.1重量%の2−デオキシ−グルコースを含む。幾つかの実施態様において、供給原料は、少なくとも約0.03重量%の2−デオキシ−グルコースを含む。他の実施態様において、供給原料は、D−グルコース以外の発酵性ヘキソース糖を含むが、別の実施態様では、供給原料は更にD−グルコースを含む。
【0016】
さらに別の態様は、酵母の突然変異菌株を作り出す方法であって、酵母の菌株、例えばGrr1及びReg1からなる群より選択される少なくとも1つの遺伝子と適格性があるサッカロミセス・セレビシエの一倍体、二倍体又はより高い倍数性の菌株を準備する工程;Grrl及びReg1の両方の活性を欠失して、突然変異菌株を作り出す工程;菌株を試験して、0.035重量%の2−デオキシ−グルコースの存在下でペントース糖において増殖するかを決定する工程を含む方法を含む。
【0017】
追加的な実施態様は、酵母菌株を選択又は同定する方法であって、YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271からなる群より選択される少なくとも1つのオープンリーディングフレームに突然変異を含むサッカロミセス・セレビシエの一倍体又はより高い変数性の菌株を準備する工程(ここで、オープンリーディングフレームは機能性遺伝子をコードし、オープンリーディングフレームにおける突然変異はオープンリーディングフレームにおいてコードされた遺伝子の活性を破壊する);サッカロミセス・セレビシエの前記菌株を、唯一の炭素供給源としてのキシロース及び0.1重量%の2−デオキシ−グルコースを含む培地において増殖させる工程;並びに培地に増殖したサッカロミセス・セレビシエの菌株を単離する工程を含む方法を含む。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。
【0018】
さらに他の実施態様は、サッカロミセス・セレビシエ酵母菌株の一倍体又はより高い変数性の菌株を単離する方法であって、YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wからなる群より選択されるオープンリーディングフレームによりコードされる少なくとも1つの遺伝子の機能性コピーを含む菌株を準備する工程;YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wからなる群より選択される少なくとも1つのオープンリーディングフレームに突然変異を導入して、サッカロミセス・セレビシエの突然変異体を産生する工程;選択培地においてサッカロミセス・セレビシエの突然変異体を増殖する工程(ここで選択培地は、唯一の炭素供給源としてのキシロース及び0.1重量%の2−デオキシ−グルコースを含む);並びに培地に増殖する突然変異体を単離する工程を含む方法を含む。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。
【0019】
本発明の幾つかの実施態様は、例えば2−デオキシ−グルコースの存在下及び代理に(by proxy)D−グルコースの存在でセルロース性バイオマスにおいて見出される幾つかのペントース糖を含むD−グルコース以外の糖において増殖する酵母菌株、例えばサッカロミセス属の産業用又は実験室用菌株を選択する方法を含む。他の実施態様は、2−デオキシ−グルコースの存在下でD−グルコース以外の糖に増殖するサッカロミセス属の産業用菌株を使用して、相当量のD−グルコースの存在下で追加の酵母バイオマス及び/又はD−グルコース以外の糖からエタノールのような発酵の最終産物若しくは副産物を生産することを含む。
【0020】
セルロース性エタノールを生産する優れた性能。セルロース性バイオマスは複数の糖を有し、最も重要ものはD−グルコース及びD−キシロースである。しかし、酵母代謝生理学は、多くの場合、他の多くの糖を除外したD−グルコースの調節的使用である異化代謝産物抑制(catabolite repression)を受ける。このことはD−キシロース又はD−キシルロースでは実証されていないが、本出願者たちは、ここで、D−キシロース及びD−キシルロースの利用も異化代謝産物抑制を受けることを示す。したがって、野生型酵母菌株はこれらのペントース糖の存在下でD−グルコースを優先的に代謝する。更に、本出願者たちは、ここで、サッカロミセス属の産業用菌株がヘキソース及びペントースの両方において異化代謝産物抑制を受けることを示す。これは、大部分の酵素菌株を使用するセルロース性エタノールにとって重要な技術上の障壁である。この複数糖発酵の障壁を克服するために、D−キシロースに対する異化代謝産物抑制を排除する必要がある。一つの実施態様は、以下の遺伝子GRR1、REG1及びHXK2の少なくとも1つを除去することによって異化代謝産物抑制を排除することを含む。本発明の幾つかの実施態様は、2−デオキシ−グルコースの存在下でD−グルコース以外の糖、特にペントースにおいて増殖する産業用酵母菌株、例えばサッカロミセス属を選択することを含む。幾つかの実施態様において、これは、相当レベルの2−デオキシ−D−グルコースの存在下でD−グルコール以外の糖供給源において増殖する産業用サッカロミセス属の変種を選択することを含む方法である。幾つかの実施態様において、これらの菌株は、以下の遺伝子GRR1、REG1及びHXK2の少なくとも1つを欠いているか又は少なくとも有効レベル形態(effective level form)を発現しない。さらに他の実施態様において、これらの遺伝子は、適切なレベルの活性タンパク質を産生しないように突然変異している場合がある。
【0021】
他の幾つかの実施態様は、相当量のD−グルコースの存在下でのD−グルコース以外の糖からのエタノールのような発酵の最終生成若しくは副生成を生産し、及び/又は、追加の酵母バイオマスを生産するため、2−デオキシ−グルコースの存在下でD−グルコース以外の糖において増殖するサッカロミセス属の産業用菌株を使用することを含む。
【0022】
幾つかの実施態様は、D−グルコースを含む混合糖を含む供給原料を発酵する方法であって、異なる代謝要件により複数の酵母菌株を使用して、バイオマス又はエタノールのような代謝産物のいずれかを効率的に生産する工程を含む方法を含む。幾つかの実施態様において、発酵は、酵母菌株の少なくとも2つの異なる菌株を使用して実施され、ここで少なくとも1つの菌株はD−グルコースにおいて優先的に増殖し、検出可能なレベルのD−グルコースの存在下で異化代謝産物抑制さえも示すことができるが、一方、少なくとも1つの他の菌株は、検出可能なレベルのD−グルコースの存在下であっても、活性化され(de-repressed)、D−グルコース以外の糖を発酵してエタノール及び/又はバイオマスにすることができる。幾つかの実施態様において、この方法は、D−グルコース及びD−キシロースのようなペントース糖の効率的な同時発酵を可能にする系をもたらすことができる。
【0023】
サッカロミセス・セレビシエ菌株CEN.PKgrr1Δの変種又はサッカロミセス・セレビシエ菌株Ethanol Red GX1の変種を含むサッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株であり、ここで、変種株の単一細胞分離株は固体培地において増殖し、その培地はD−キシロース及び2−デオキシ−グルコースを含み、培地の主要な炭素供給源は、糖D−キシロースであり、変種菌株の増殖は、培地において2日以内に強力なコロニーを生じる。
【0024】
さらに他の実施態様は、例えばサッカロミセス・セレビシエ菌株CEN.PKgrr1Δの変種又はサッカロミセス・セレビシエ菌株Ethanol Red GX1の変種を含むサッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株を含み、ここで、変種株の単一細胞分離株は、マルトース及び2−デオキシ−グルコースを含む固体培地において増殖し、培地の主要な炭素供給源は、マルトースが含まれるが、これに限定されない糖を含有するD−グルコースであることができ、変種菌株の増殖は、培地において2日以内に強力なコロニーを生じる。さらに他の実施態様は、サッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株を含み、ここで培地は、約2%のD−キシロース及び約0.1%の2−デオキシ−グルコースを含む。
【0025】
さらに他の実施態様は、約2%のマルトース、又は、糖を含有する別のD−グルコース及び約0.1%の2−デオキシ−グルコースを含む培地において増殖する変種を含む。
【0026】
さらに他の実施態様は、サッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株を含み、ここで変種菌株は、一倍体実験室用菌株CEN.PK誘導体CDXR1、CEN.PK(113−7D)、CDXR2及びCDXR3、並びに二倍体産業用酵母菌株Ethanol Red、GX1及びRX4からなる群より選択される。幾つかの実施態様において、開示されている方法を使用して選択された異化代謝産物抑制抵抗性サッカロミセス・セレビシエ菌株は、固体培地において増殖し、ここで培地は、主要な炭素供給源としてD−グルコースを含む。
【0027】
幾つかの実施態様は、サッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株を含み、ここで変種菌株は、2%のD−ガラクトース及び約0.03%の2−デオキシ−グルコースを含む固体培地において増殖し、D−ガラクトースは、培地における主要な炭素供給源である。
【0028】
さらに別の実施態様は、D−グルコース以外の糖において増殖する幾つかの産業用菌株を含むサッカロミセス・セレビシエの菌株を生産する方法であって、サッカロミセス・セレビシエの第1変種菌株を準備する工程(ここで前記変種は、2−デオキシ−D−グルコールの存在下でD−グルコース以外の糖、特にペントースにおいて増殖する);及びグルコース以外の糖を代謝する代謝経路から単離された少なくとも1つの遺伝子を第1変種菌株において過剰発現させて、第2変種菌株を形成する工程を含む方法を含む。
【0029】
追加的な実施態様は、D−グルコースの存在下でペントースにおいて増殖するサッカロミセス・セレビシエの変種菌株を生産する方法を含む。幾つかの実施態様において、第1変種菌株は、例えば、GX1、RX4、CDXR1、CEN.PK(113−7D)、CDXR2及びCDXR3からなる群より選択され、ここで菌株を、D−キシロース代謝経路から単離された少なくとも1つの遺伝子を発現するように適合させることができる。
【0030】
さらに他の実施態様は、D−グルコース以外の糖供給源においてサッカロミセス・セレビシエの菌株を増殖させる方法であって、サッカロミセス・セレビシエの第1菌株を準備する工程(ここで前記菌株は、2−デオキシ−D−グルコールの存在下でD−グルコース以外の糖供給源、特にペントースにおいて増殖する);グルコース以外の糖、特にペントースを代謝する代謝経路から単離された少なくとも1つの遺伝子を第1菌株において過剰発現させて、第2菌株を形成する工程;及び前記第菌株を培地において増殖する工程(ここで培地における主要な炭素供給源は、グルコース以外の糖、特にペントースである)を含む方法を含む。
【0031】
幾つかの実施態様において、第1菌株は、CDXR1、CEN.PK(113−7D)、CDXR2及びCDXR3からなる群より選択されるサッカロミセス・セレビシエ菌株CEN.PKの変種であり、少なくとも1つの過剰発現遺伝子は、D−キシロース代謝経路からのものである。
【0032】
さらに他の実施態様において、第1菌株は、GX1及びRX4からなる群より選択される産業用サッカロミセス・セレビシエ菌株Ethanol Redの変種であり、少なくとも1つの過剰発現遺伝子は、D−キシロース代謝経路からのものである。
【0033】
さらに追加的な実施態様は、サッカロミセス・セレビシエにおいて異化代謝産物抑制に関与するオープンリーディングフレームを同定する方法であって、D−グルコース又はD−グルコース及びD−キシロース又はD−キシロースの存在下でグルコース抑制を受けるサッカロミセス・セレビシエ菌株の第1変種を増殖する工程;D−グルコース、D−グルコース及びD−キシロース又はD−キシロースのいずれかの存在下でD−グルコース以外の主な炭素供給源において増殖された、グルコース抑制に非感受性のサッカロミセス・セレビシエ菌株の第2変種を蕃殖させる工程;並びに第1変種と第2変種のプロテオームを比較して、第1変種と第2変種のプロテオームの差を同定する工程を含む方法を含む。
【0034】
幾つかの実施態様において、D−キシロースは、マルトース又はマルトトリオースなどであるが、マルトース又はマルトトリオースに限定されない別の二次的な糖(secondary sugar)に代えられている。
【0035】
幾つかの実施形態において、第1及び第2変種のプロテオームの差が、以下のオープンリーディングフレーム:YLR063w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wの少なくとも1つの産物における差を示す。
【発明の効果】
【0036】
本発明によれば、サッカロミセス属の新規菌株は、有意な量のD−グルコースの存在下でもD−グルコースのみ以外の糖をバイオマス又はエタノールに容易に変換することができる。
【図面の簡単な説明】
【0037】
【図1】トウモロコシに基づいたデンプン及び植物バイオマスの両方をエタノールに変換する酵母の経路を表す概略図である。
【図2】エタノールに変換されるD−グルコース又はD−キシロースの経路を示す概略図である。出芽酵母細胞の写真である。
【図3】D−キシロースにおけるサッカロマイセス属の幾つかの菌株の増殖を示す植物である。
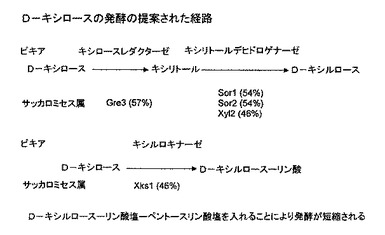
【図4】ビール酵母によるD−キシロースの発酵における提案される経路である。
【図5】YP+2%D−キシロースにおいて増殖しているCEN.PKの突然変異酵母コロニーの増殖を示すプレートの写真である。
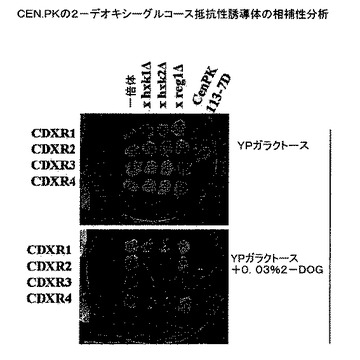
【図6】CEN.PKの2−デオキシ−グルコース抵抗性誘導体の相補性分析の結果を示す植物の写真である。
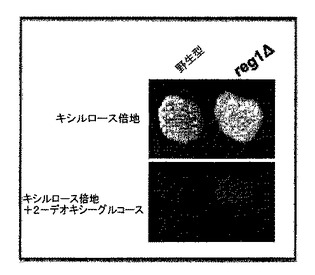
【図7】2−デオキシ−グルコースの存在下でD−キシルロース、D−キシロース混合物において増殖する酵母により示される植物の写真である。
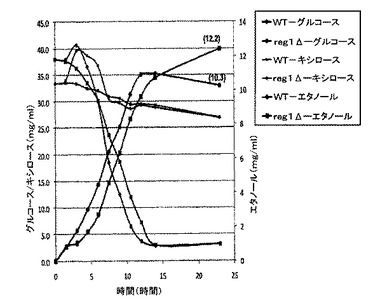
【図8】産業用酵母の多様な菌株によるトウモロコシ穂軸加水分解産物からエタノールへの発酵のグラフ図である。
【図9】PCR分析がGRR1遺伝子の欠失の同定を検査するために実施されることを示すセルである。
【図10】REG1遺伝子の欠失を検査するために実施されるPCR分析を示すセルである。
【図11】GX1によるマルトース及びD−グルコースの同時発酵を示すグラフである。
【発明を実施するための形態】
【0038】
新規技術の原理の理解を促進する目的において、ここで好ましい実施態様に対して参照を行い、それらを説明するために特定の言葉を使用する。それでも、それによって新規技術の範囲を制限することは意図されておらず、新規技術の原理に対するそのような変更、修正及び更なる適用は、新規技術が関連する当業者において通常起こることとして考慮されることが、理解されるべきである。
【0039】
本明細書で使用されるとき、特に指定のない限り、用語「約(about)」は、プラス又はマイナス20パーセントを意味し、例えば約1.0は、0.8〜1.2の範囲を包含する。
【0040】
本明細書で使用されるとき、特に指定のない限り、用語「検出可能な増殖(detectable growth)」は、増殖の証拠が人の裸眼による目視検査によって明らかになるまでを含む増殖を意味する。
【0041】
特に参照のない限り、遺伝子は、Demerec, M., Adelberg, E. A., Clark, A.J. & Hartman, P. E. in "A proposal for a uniform nomenclature in bacterial genetics". J. Gen. Microbiol 50, 1-14 (1968)により提案された命名法を使用して参照される。
【0042】
酵母サッカロミセス・セレビシエは、糖からエタノールへの発酵のために何世紀も前に栽培化(domesticated)された(図1及び2を参照すること)。酵母の栽培化は、D−グルコースをエタノールに変換するのに非常に効率的な産業用酵母菌株の世代をもたらした。有意には、これらの産業用酵母菌株は、一般に、大部分の微生物よりもはるかにエタノール耐性がある。この方法は、酵母増殖及び/又はエタノール生産のための糖が天然に最も豊富にあるヘキソースのうちの幾つか、特にD−グルコース、D−フルクトース及びD−マンノースであるときに、非常に効率的である。
【0043】
セルロースからエタノールへの産業用変換を実現可能な営利事業にするため、セルロースをグルコースのような発酵性糖に変換する方法を合理化することに大きな努力が注がれている。酵母に基づいたエタノール生産のためにセルロースから発酵性糖への変換を最適化することは重要であるが、植物材料に存在するD−グルコース以外の糖、特にD−キシロースからのエタノールの生産を合理化することも同様に重要である。植物材料において最も豊富な化合物は、グルコースポリマーセルロースであるが、有意な量の植物バイオマスが、キシランとして知られている糖ポリマーとして存在する(図1;例えばWarren 1996を参照すること)。事実、多くのセルロース供給源では、キシランは、ポリマーのバイオマスの20%超を占める。キシランそれ自体は、ペントース糖のD−キシロースの鎖から構成される。大部分のS.セレビシエの産業用及び実験室用菌株のような酵母による発酵のために、セルロースのようなキシランを最初にモノマーのD−キシロースに変換しなければならない。既に複雑な方法は、大部分の商業的に生産されたS.セレビシエの菌株がD−キシロースのようなペントース糖を効率的に発酵せず、特に供給原料中にD−グルコースが存在すると効率的に発酵しないので、発酵環境におけるD−グルコースの存在により更に複雑になる。
【0044】
S.セレビシエはD−キシロースを発酵することができないと広く考えられており、実際、酵母S.セレビシエは炭素供給源としてD−キシロースを利用しないことが、1970年代に報告された(Barnett 1976)。文献によると、酵母ピキア・スチピチス(Pichia stipitis)は、D−キシロースを発酵する能力を有さない。この情報に基づいて、幾つかの研究所は、S.セレビシエにおけるD−キシロースの利用に必要なピキア遺伝子を発現することにより、D−キシロースを発酵することができる酵母菌株の生成を試みた(例えば、Kotter and Ciriacy, 1993; Ho et al., 1998; Jin et al., 2003を参照すること)。改善されたD−キシロースの利用がそれぞれの場合において報告されたが、D−グルコースの存在下でのD−キシロース発酵の効率は著しく変動した。この観察を説明することができる幾つかの要因には、実験増殖及び前増殖の条件、並びにD−キシロース代謝経路の異種発現のレベルにおける差が含まれる。
【0045】
サッカロミセス・セレビシエについてのDr. Ho(Purdue U.)及びグラム(−)及びグラム(+)細菌の両方についてのDr. Ingram(U. of Florida)の研究は、D−キシロース(ピキア・スチピチス)の代謝及びエタノールの(多様な微生物からの)生産それぞれにおける他の生物の代謝経路内における外来性遺伝子の組み換え発現に集中してきた。
【0046】
あらゆる理論又は仮説に束縛されることなく、説明としてであって、限定ではなく、この観察には幾つかの見落とされた又は少なくとも正しく評価されていない理由がありうる。例えば、報告された結果と一致する1つの説明は、これらの実験に使用したD−キシロースの正確な組成における変動である。化学供給会社から購入した糖で実際に「純粋」であるものはほとんどない。純粋であるとして市販されている大部分の糖は、実際には約99又は98%の純度しかない。典型的には、糖における主な汚染物質は、極めて豊富にある糖D−グルコースである。酵母代謝の研究にとって残念なことに、0.1%の低さのD−グルコースのD−グルコース量は、例えばD−ガラクトースのようなD−グルコース以外の糖の利用に影響を及ぼすことが知られている。したがって、少なくとも幾つかの発表された研究において使用された純度98〜99%のD−キシロースの少なくとも一部は、実際はD−グルコースで汚染されていた可能性が高い。所定の酵母菌株がD−キシロースにおいて増殖することができるかを決定するように設計された研究において微量のD−グルコースによる汚染でも、これらの研究において観察された結果を歪めた可能性が十分にある。
【0047】
D−グルコースによる大部分の糖の汚染は、歴史的な問題である。例えば、D−ガラクトースを使用して酵母を確実に同定する能力は、0.01%未満のD−グルコースを含むD−ガラクトースの商業的導入(Sigma-Aldhch)の後でのみ生じた。残念なことに、D−グルコースで汚染されていないD−キシロースは、容易に入手することができない。したがって、幾つかの酵母菌株がD−キシロールにおいて増殖する能力を欠いているという報告は誤りである場合があり、このことは、文献における幾つかの矛盾した報告を説明する場合がある。
【0048】
キシロース発酵に関する文献において報告された不一致についてのさらに別の説明は、これらの研究に使用された多様な実験室用及び産業用酵母菌株の多様な遺伝子型組成に起因しうる。大部分の実験室用酵母菌株は、小数の祖先由来であったが(Mortimer and Johnston, 1986)、これらの子孫は、経時的に広く異なる増殖特性を発達させてきた(Winston et al., 1995; van Dijken et al., 2000)。事実、最近の論文は、S.セレビシエの少なくとも1つの菌株が、遺伝子修飾することなくD−キシロースにおいて増殖することができると、本出願者たちに示唆する場合がある(Sedlak and Ho, 2004; Figure 3; panel marked wild-type; Toivari et al. 2004を参照すること)。菌株MC996Aは、ピキアD−キシロース利用遺伝子を付加することなく、市販等級のD−キシロースにおいて増殖すると思われる。この菌株は、非常に強力な遺伝的背景を有することが示されている菌株のCEN.PKファミリーの誘導体である。これらの菌株の明らかに豊富な遺伝的多様性は、これらが広範囲の糖を発酵することを可能にする(van Dijkens ef al., 2000)。
【0049】
セルロース性バイオマスは複数の糖を含み、最も重要ものはD−グルコース及びD−キシロースである。しかし、酵母代謝生理学は、多くの場合、他の多くの糖を除外したD−グルコースの調節的使用である異化代謝産物抑制を受ける。このことはD−キシロース又はD−キシルロースでは実証されていないが、本明細書において実証されていることは、D−キシロース及びD−キシルロースの利用も異化代謝産物抑制下であることである。したがって、野生型酵母菌株はグルコースを優先的に代謝する。これは、大部分の酵素菌株を使用するセルロース性エタノールにとって重要な技術上の障壁である。この複数糖発酵の障壁を克服するために、D−キシロースに対する異化代謝産物抑制を排除する必要がある。
【0050】
D−グルコースを利用する能力を欠いている酵母菌株は、例えばヘキソキナーゼ遺伝子HXK1及びHXK2を欠いている菌株を作り出すことによって生成することができる。HXK2及びGRR1に突然変異を持つ酵母の菌株も本出願者たちのREG1突然変異体と同じように挙動することも、見出されている(データ示されず)。したがって、強力な野生型菌株を分析するか又はD−キシロース経路のピキア遺伝子を過剰発現する菌株を分析するかにかかわらず、D−グルコースの存在は、D−キシロースの利用を妨げる場合がある。大部分の酵母菌株はD−キシロースの分解に必要な遺伝子に高度に関連する遺伝子を有する。
【0051】
現在では、D−キシロースを発酵するS.セレビシエの存在を引用する報告でも、発酵が非常に非効率であることを認めている。例えば、Sedlak及びHo(2004)は、非常に少量のD−キシロースが、全てでなければ大部分のD−グルコースが利用される前に、彼らがD−キシロースを代謝できると考えるCEN.PK酵母菌株により消費されることを報告している。現在の文献は、D−キシロースのようなペントースにおいて増殖することができる酵母変種を同定する強力なスクリーニングを欠いていることを、明白に説明している。D−キシロースの発酵経路がS.セレビシエに存在すると仮定すると、いったん強力なスクリーニングが作り出されると、これらの経路を研究、改善及び増加することがかなり容易になる。
【0052】
ここで図3、パネルAを参照する。本明細書に開示されている結果は、キシロースにおいて増殖することができるCEN.PKの自然発生突然変異体をスクリーニングし、単離することが可能であることを示している。この結果は、D−キシロース発酵に必要なそれぞれの酵素に近接の相同体がサッカロミセスに存在することを示す(図4)、サッカロミセスゲノムの全配列の分析と一致する。
【0053】
図3を参照する。D−キシロースにおけるS288c及びCEN.PK誘導体の増殖。図3、パネルAでは、細胞をYP+2%D−キシロース培地にレプリカ平板培養し、30℃で4日間インキュベートした。図3、パネルBでは、細胞をYP+2%D−キシロース/0.1%2−デオキシグルコースにレプリカ平板培養し、30℃で10日間インキュベートした。パネルBの矢印は、2−デオキシ−グルコースの存在下で増殖するCEN.PKの自然発生突然変異体から生じたコロニーを指している。
【0054】
酵母細胞は、D−グルコース、D−フルクトース及びD−マンノースを発酵するのに極めて効率的であり、更に酵母は、他の多くの炭素供給源を除外してこれらの糖を発酵し、この現象は異化代謝産物抑制として知られている(Gancedo 1998において概説されている)。事実、研究されたS.セレビシエの実質的に全ての菌株は、今までのところ、D−グルコース、D−フルクトース及び/又はD−マンノースだけにエネルギーを、これらの糖が完全に又はほぼ完全に環境から排除されるまで依存している。例えば、わずかに微量のD−グルコース(<1%)がD−ガラクトースを汚染する場合、酵母は、D−グルコースの存在がなくなるまでD−ガラクトースを発酵しない。S.セレビシエによるD−キシロースの利用に関する文献における矛盾する報告は、異化代謝産物抑制を引き起こすD−グルコースによる増殖培地の汚染に起因する場合がある。
【0055】
幾つかの遺伝子における突然変異は、マルトース、スクロース及びD−ガラクトースに基づいた発酵の文脈において、実験室用菌株で異化代謝産物抑制を緩和したことを示した。異化代謝産物抑制に関与したと考えられる遺伝子には、GRR1、REG1およびHXK2が含まれる(Gancedo, 1998において概説されている)。特定の実験室用菌株内のこれらの遺伝子いずれかの損失は、幾つかの実験室用菌株の、D−ガラクトース又はスクロースのような他のヘキソースを伴ったD−グルコースの同時発酵を可能にする(Bailey and Woodward, 1984)。
【0056】
D−グルコースの存在下で幾つかの二次的な炭素供給源を利用する実験室用酵母由来の酵母細胞の能力についての1つの試験は、二次的な炭素供給源及び少量の2−デオキシ−グルコースの存在下で増殖する細胞の能力を測定することである。2−デオキシ−グルコースは、特定の実験室用菌株において、D−ガラクトース、マルトース及びスクロースのようなヘキソース糖に対してグルコース抑制を示すことが報告されているD−グルコースの非代謝性誘導体である。この現象は、D−ガラクトースによって実証されている(Bailey et al., 1982; Bailey and Woodward, 1984)。異化代謝産物抑制能力もあり、D−ガラクトース、マルトース又はスクロースを発酵することもできる、2−デオキシ−グルコースに暴露された酵母細胞は、2−デオキシ−グルコースの存在下で代替的な炭素供給源を利用することができないと考えられ、これらの菌株は、2−デオキシ−グルコースを代謝することができない。これらの条件は、細胞死をもたらすと報告されているが、2−デオキシ−グルコースに暴露された酵母細胞の正確な死亡原因は、依然として不明である(Raiser et al. 2008)。
【0057】
グルコース抑制は、それによりほとんどの炭素供給源の利用の前に、D−グルコースを培地から涸渇させなければならない、酵母における現象を説明する。グルコース抑制の十分に研究された調節物質は、代替的な炭素供給源の利用に関与する遺伝子の転写抑制因子として作用すると考えられているMig1転写因子であるが、MIG1の損失は、細胞を2−デオキシ−グルコースに対して抵抗性にしないことも報告されている(Schuller, 2003)。上記に記述されたように、GRR1、REG1又はHXK2の損失は、細胞を2−デオキシ−グルコースに対して抵抗性にする(Gancedo 1998)。Reg1は、PP1複合体をMig1に運ぶことによりMig1を調節すると考えられているPP1タンパク質ホスファターゼサブユニットであるが、他のタンパク質がグルコース抑制を達成するのに必要とする正確な機構はあまり明確ではない。マイクロアレイ分析も、Mig1がGrr1及びHxk2により調節されるプロセスのサブユニットのみに影響を及ぼすことを示す(Westergaard et al. 2006)。本明細書において実施及び報告されている実験結果のプロテオミクス分析は、異化代謝産物抑制の調節に対する大型の複製後成分が認識されていないことを示唆している。
【0058】
1に記述されているスクリーニングにより単離されたCEN.PKにおける自然発生突然変異は、REG1座位において不活性を示す(図6)。更に、REG1又は損失したときに同様の突然変異遺伝子型を有することが知られている他の2つの遺伝子、GRR1及びHXK2の標的化破壊も、この培地において増殖する能力を得た。REG1、GRR1又はHXK2の標的化破壊は、PCR仲介遺伝子破壊によって作り出される場合がある。これらの実験は、特定の遺伝子座位においてDNAの5’及び3’セグメントと同一であるプライマーを設計することによって実施することができるアミノグリコシド系抗生物質のノウルセオトリシンに対する抵抗性を付与する、ストレプトミセス・ノウルセオトリシイ(Streptomyces nourseothricii)のnat1遺伝子を使用して、上記に提示された遺伝子のいずれかを、酢酸リチウム形質転換法を使用する酵母の形質転換により欠失することができる。一倍体菌株において、単一対立遺伝子の破壊は、L−グルタミン、2%D−キシロース及び0.1%2−デオキシ−グルコースを補充したYP培地における増殖を可能にするのに十分である。しかし、異なる酵母菌株は、異なる濃度の2−デオキシ−グルコースに対して異なる感受性がある。
【0059】
大部分の産業用酵母菌株は二倍体である。したがって、これらの菌株における形質転換は、単一対立遺伝子の欠失を確実にするのみである。驚くべきことに、二倍体又は任意の更に高い倍数性の酵母の同じ遺伝子座位における他のコピーの再現性欠失は、2−デオキシ−グルコースと、マルトース、D−ガラクトース、スクロース、D−キシロース又はD−キシルロースのような糖が含まれるが、これらに限定されない主要炭素供給源として機能する二次的な糖とを含有する培地において異型接合体菌株を平板培養することにより達成できることが、本出願者たちにより見出された。
【0060】
ここで図3及び5を参照する。この現象もD−キシロースの文脈において実証された。図3に示されているように、S288c grr1Δ菌株、野生型CEN.PK菌株及びCEN.PK grr1Δ菌株を、2%D−キシロースを含有する培地で平板培養した場合、増殖を3つの菌株全てにおいて検出することができた。しかし、CEN.PK grr1Δ菌株のみが、2%D−キシロース及び0.1%2−デオキシ−グルコースを含有する培地において増殖を示す(図3)。経時的に、CEN.PK菌株の細胞の斑内では、単離されたコロニーが10〜15日後に出現し始めた。
【0061】
約21日後、これらのコロニーは、物理的に操作するのに十分な大きさになった。これらの単離されたコロニーは、2−デオキシ−グルコースの存在下でD−キシロースにおいて増殖する能力を得たCEN.PKの親菌株の自然発生突然変異体である(下記を参照すること)。この現象は、S288c grr1Δ誘導体では観察されない。これらの結果は、CEN.PK細胞と対照的に、GRR1遺伝子を欠失しても、S288c細胞は、2−デオキシ−グルコースの存在下でD−キシロースにおいて増殖できないことを示している。2−デオキシ−グルコース抵抗性/D−キシロース利用突然変異体がS288cにおいて発展する能力がないことは、S288cにより見られる少量の増殖がD−キシロース内の汚染量のグルコースの利用に起因しうることを示唆している。
【0062】
本明細書に開示されているスクリーニングにより単離された及び2−デオキシ−グルコースの存在下でD−キシロースにおいて増殖したサッカロミセス・セレビシエの2つの代表的な二倍体菌株CDXR2及びJH015は、2009年2月25日にアメリカン・タイプ・カルチャー・コレクションに寄託され、それぞれ受入番号PTA−9849及びPTA−9850が指定された。生物学的材料の寄託についてのブタペスト条約の規定において、これらの菌株は、本発明に対して特許が発行されると公に利用可能になる。
【0063】
酵母細胞代謝が利用を好ましい糖から好ましくない糖(例えば、D−ガラクトース及び恐らくはD−キシロース)へ変換するには、数時間かかる場合がある。この遅れは、D−キシロース分解に必要な酵素を過剰発現するように操作された酵母細胞においても生じる。糖の混合物を利用する産業用及び実験用の両方の酵母菌株において、D−キシロースの代謝は、混合物がD−グルコースをほぼ完全に欠くまで非常に非効率的であると思われる。
【0064】
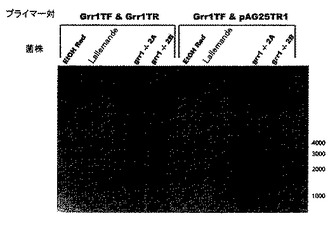
ここで図9を参照する。2−デオキシ−グルコースの存在下でキシロースにおいて増殖する能力により選択された二倍体産業用酵母菌株を、PCRを使用して分析した。3つのGrr1Tf及びGrr1TFプライマー対は、GRR1が無傷である場合、3810bpのPCR産物を産生するはずであり、GRR1が破壊されている場合、PCR産物は1400bpのはずである。Grr1TF&pAG25TR1プライマー対は、GRR1が無傷である場合、PCR産物を産生するはずはなく、GRR1がnat1遺伝子に代わっている場合は、約1200bpのPCR産物のはずである。PCR分析は、Fermentis Ethanol Redの誘導体であるgrr1−/−2A及びgrr1−/−2B菌株において、両方のGRR1遺伝子がnat1遺伝子に代わっていることを証明する。RC4がgrr1::NAT1を有する一倍体S288C誘導体であることに留意すること。以前の2つのPCR反応(10/18/04及び12/10/09)は、このPCR反応におけるgrr1−/−菌株について観察されたものと同じ産物を生じた。
【0065】
ここで図10を参照する。二倍体産業用菌株の同様の分析を実施して、Reg1遺伝子における変化を探した。Reg1 testA&Reg1 test IDプライマー対は、REG1が無傷である場合、約3600bpnのPCT産物を産生するはずであり、破壊されている場合、PCT産物は約1800bpのはずである。Reg1 testA&pAG25TR1プライマー対は、REG1が無傷である場合、PCR産物を産生するはずではなく、REG1がnat1遺伝子に代わっている場合、約1500bpのPCR産物のはずである。上記のPCRは、Fermentis Ethanol Redの誘導体であるreg1−/−1A及びreg1−/−1B菌株において、両方のREG1遺伝子がnat1遺伝子に代わっていることを証明する。grr1−/−2Aのようにnat1遺伝子を持つ菌株であっても、REG1 testA及びpAG25TR41プライマーによりPCR産物は作製されないことも、証明されている。
【0066】
D−グルコースの抑制効果の強度が、D−キシロースにより(Jin et al. 2004)並びにD−グルコースの異なる濃度により(Yin et al. 2003)調節される転写物の選択された群のレベルを含む表1に示されている。野生型細胞と比較した、2%D−グルコースにおいて増殖している間のGRR1及びHXK2の細胞突然変異体におけるこれらの転写物の調節も、含まれている(Westergaard et al. 2006)。表1から分かるように、一定レベルの低いD−グルコース(0.01%)であっても、異化代謝産物抑制を引き起こす。例えば、豊富なFBP1転写物は.01%〜1%D−グルコースで比較的一定のままであるが、2%D−キシロース又はGRR1を欠いている細胞における増殖により高く誘導されている。異化代謝産物抑制の排除は、混合糖供給原料をプロセスしてD−グルコース混合糖供給原料のレベルを、異化代謝産物抑制を誘導するレベルより下に低減することが可能である場合でも有用であるはずであり、それはプロセスされた混合糖原料からD−グルコースを実質的に涸渇させることは、時間がかかり、高価であるからである。
【0067】
【表1】

【0068】
本明細書に開示されているように、GRR1を欠いている細胞のプロテオミクス分析が実施され、このデータの分析は、少なくとも以下の2つの点について説明する。第1には、転写レベルは、必ずしもプロテオームの正確な状況を提供しているとは限らない(表1)。例えば、遺伝子HXT1及びHXT7の転写物はそれぞれ減少又は増加するが、対応するタンパク質は全く異なって挙動し、Hxt1タンパク質レベルは、野生型菌株をgrr1Δ菌株と比較すると変わらないままであるが、Hxt7タンパク質は、grr1Δでは減少している。これらの結果は、mRNAマイクロアレイデータをプロテオミクスデータの不在下で解釈する場合に、注意を払わなければならないことを示している。Hxt7がD−キシロースの重要な輸送体であると提案されているので、このことは本明細書において非常に重要である。
【0069】
第2には、このプロテオミクスデータは、Gre3タンパク質がgrr1Δ突然変異菌株において16×を超えて上昇していることを示す。GRE3は、D−キシロース分解の第1工程を実施するピキアキシロースレダクターゼの相同体をコードする。更に、Gre3における増加は、GRE3遺伝子の転写誘導の不在下で生じ、本出願者たちの実験においてタンパク質の直接的な検出によってのみ見ることができる。したがって、GRR1の不在は、更に細胞をD−キシロースの利用に向かわせる。これらの結果は、GRR1、HXK2及び最も可能性が高いものでREG1の損失は、完全に重複しているのでなければ非常の類似した反応をもたらしうることを示す。
【0070】
マイクロアレイ分析は、不偏様式で網羅的転写変化をモニターする方法を提供する。マイクロアレイ実験における本質的な前提は、mRNAレベルの差がタンパク質レベルの差に反映することである。しかし、全てのタンパク質がタンパク質活性を変化させることはない又は均一のレベルは転写制御における変化を必要とする。例えば、翻訳後仲介事象は、マイクロアレイ分析では「不可視」である顕著なプロテオミクス変化をもたらすことができる。幸いにも、マイクロアレイ分析とプロテオミクス分析の組み合わせを使用して、転写性を、豊富な特定のタンパク質における変化の原因である後転写調節事象と区別することができる。
【0071】
検出された2580個のタンパク質のうちの約1,200個のタンパク質についての定量データをまとめ(ここにデータを示す)、結果は、この方法にとって典型的である(de Godoy et al. 2006)。これらのプロテオミクスデータを、上記に記載された菌株及び培地条件(GRR1突然変異体における)を使用するマイクロアレイ分析と、また、野生型細胞をgrr1突然変異体及び他の異化代謝産物抑制突然変異体と比較した公表されたマイクロアレイ分析(Kodama et al., 2002; Kaniak et al., 2004; Westergaard et al., 2004)と組み合わせた。これらのデータセットの分析は、GRR1により影響を受けた調節の3つの特有のパターンを明らかにした(ここにデータを示す)。第1には、そのmRNAの豊富さがタンパク質産物の豊富さと相関する遺伝子が同定された。第2には、産物の豊富さの差がmRNA変化の不在下で生じる遺伝子が見出された。そのmRNAの豊富さが、産物の豊富さにおける同時変化なしに変化する遺伝子も同定された。これらの結果は、正確な条件下でこれらの技術を使用して、多様な活性代謝経路のプロテオームをプロテオミクス的に研究できることを示す。
【0072】
【表2】
【0073】
[実験]
1.S.セレビシエの異化代謝産物活性化一倍体自然発生突然変異体のスクリーニング
以下の実験を実施して、2−デオキシ−グルコースにおける酵母細胞の増殖を試験した。野生型酵母菌株を、2重量%のキシロース及び0.1重量%の2−デオキシ−グルコースを含む固体培地YPで平板培養した。しかし、酵母細胞の増殖は見られなかった。
【0074】
次に、0.5重量%のL−グルタミンを、そうでなければ同一の培地に加え、増殖を、接種の数日後に見出した。しかし、図3(パネルB)で見られるように、細胞はこの培地において容易に増殖しなかった。3週間の接種の後でのみ、他の酵母菌株由来の細胞ではなくCEN.PK誘導体が、予想外に、2−デオキシ−グルコースの存在下でキシロースにおいて増殖する能力を得た。これらの自然発生突然変異体は、最終的に、単一細胞から生じたと思われるコロニーを形成した。これらのコロニーは、自然発生突然変異誘発により誘導されたが、伝統的な化学的突然変異誘発要因又は照射がこのプロセスを増強することができる。
【0075】
2.一倍体突然変異体
4つの2−デオキシ−グルコース耐性/D−キシロース利用CEN.PKコロニーからの細胞を単離し、更なる研究のために取っておいた。ここで図5を参照すると、これらの突然変異CEN.PK菌株は、D−グルコース及びD−キシロースにおいて増殖し、D−キシロース+2−デオキシ−グルコース(DOG)培地に移されたとき、2日以内に強力な斑を生じる。
【0076】
3.相補性分析
単離された菌株である2−デオキシ−グルコース抵抗性CEN.PK誘導体を、酵母ノックアウトコレクション(Open Biosystems)からの特定の菌株と接合させた。4つの2−デオキシ−グルコース抵抗性突然変異菌株も、全て2%ガラクトース及び.03%2−デオキシ−グルコースを含有する培地において増殖した。したがって、S288c由来細胞がD−キシロースにおいて増殖しなかったので、相補性試験を、D−ガラクトース及び2−デオキシ−グルコースを含有する培地において実施した。分析は、2−デオキシ−グルコース表現型CDXR2及びCDXR4が、REG1における突然変異により補完されないことを実証し、CDXR2及びCDXR4がREG1の突然変異対立遺伝子を含有することを示した。これらの通常ではない表現型の原因であるこれらの菌株において、依然として未同定の変化も存在しうる。
【0077】
4.活性化一倍体産業用酵母菌株を得るためのスクリーニング及び標的化手法
一倍体実験室用菌株に使用したものと同様の手法を使用して、2−デオキシ−グルコースの存在下でD−キシロース及びD−キシルロースの混合物において強力に増殖する能力を自然発生的に得た修飾型の産業用二倍体菌株Fermentis Ethanol Redをスクリーニングし、単離することが可能であるはずである。
【0078】
Fermentis Ethanol Redのような幾つかの産業用一倍体菌株は、異株性であり、標準的な実験室業務の胞子形成及び四分子切開により一倍体として作製及び維持することができる。産業用二倍体菌株を一倍体に変換することは、グルコース抑制を付与する遺伝子の単一コピーしか破壊する必要がないので、2−デオキシグルコースの存在下でD−キシロースの混合物において強力に増殖するのに必要な特性を得る菌株の能力を増加する。
【0079】
活性化産業用二倍体菌株を、標的化遺伝子破壊、続く遺伝子の第2コピーの損失を選択することにより得ることもできる。例えば、菌株GX1(grr1−/−)は、GRR1遺伝子の2つのゲノムコピーのうちの1つを、ノウルセオトリシンN−セチルトランスフェラーゼ遺伝子(NAT1)遺伝子に代えることによって作り出すことができ、このことは、Fermentis Ethanol Red菌株においてアミノグリコシドノウルセオトリシンに対する抵抗性を付与する。2−デオキシ−グルコースと、マルトース、D−ガラクトース、スクロース、D−キシロース又はD−キシルロースのような主要な炭素供給源として機能する二次的な糖とを含有する培地において異型接合体菌株を平板培養することによって、GRR1遺伝子の両方のコピーを欠いている菌株を得た。
【0080】
ここで図7を参照する。産業用二倍体、野生型(Fermentis Ethanol Red)及びreg1Δ(Fermentis Ethanol Red誘導体)をYPDプレートにおいて2日間増殖させ、次にYP+0.4%キシルロース/3.6%キシロース(上側パネル)又はYP+0.4%キシルロース/3.6%キシロース+0.1%2−デオキシ−グルコースにおいて30℃で3日間レプリカ平板培養した。
【0081】
5.トウモロコシ穂軸加水分解産物における突然変異産業用二倍体菌株の増殖
グルコース及びキシロースの混合物において増殖する突然変異産業用二倍体の能力を試験した。産業用二倍体菌株、野生型(Fermentis Ethanol Red)及びreg1Δ(Fermentis Ethanol Red誘導体)菌株を、50mlのトウモロコシ穂軸加水分解産物に、出発密度の1×107細胞/mlで接種した。培養を30℃で振とうし、試料を指定された時間で取り出した。グルコース、キシロース及びエタノールの濃度を屈折率により分析した。ここで図8を参照する。突然変異菌株も、D−グルコース及びキシロースの両方を含むトウモロコシ穂軸由来のセルロース性混合物において、相当な率の糖をエタノールに変換する(約11%)。
【0082】
6.グルコース及びマルトースの両方を含む混合糖供給源における突然変異産業用二倍体酵母菌株の増殖
産業用二倍体菌株、野生型(Fermentis Ethanol Red)及びgrr1−/−(GX1)(Fermentis Ethanol Red誘導体)を、10g/Lの酵母抽出物、20g/Lのペプトン、50g/Lのグルコース及び50g/Lのマルトースを含有する50mlの培地に、出発密度の1×107細胞/mlで接種した。培養を30℃で振とうし、試料を指定された時間で取り出した。グルコース、キシロース及びエタノールの濃度を屈折率により分析した。図11に示されているように、この菌株も、両方の糖が同時に提供されたとき、D−グルコースよりも素速くマルトースを利用する。
【0083】
7.GRR1及びREG1破壊のPCR仲介高次構造
GRR1遺伝子のPCR分析に使用する条件は以下である。
PCRミックス RC4プログラム
5ul 10×Thermo Pol Rxn Buffer 94C−1分間
39ul 水 94C−50秒間|
1ul dNTP(各10mM、初期[]) 56C−1分間|30×
1.5ul 各プライマー(10mM、初期[]) 72C−3分間|
1ul ゲノムDNA
1ul Taq(NEB)
【0084】
以下のDNAプライマーをGRR1の分析に使用した。
配列番号1−Grr1TfF=5′GAAGCCCAAAAATTAAGGCATTGCA
配列番号2−Grr1TR=5′TTTGAAACTGTGTATAGAATGTTTCGC
配列番号3−pAG25Tr1=5′ATTACTTTCTGCGCACTTAACTTCG
これらの反応の結果を図9に示されているゲルにより示す。
【0085】
8.REG1破壊のPCR分析
REG1遺伝子のPCR分析に使用する条件は以下である。
PCRミックス RC4プログラム
5ul 10×Thermo Pol Rxn Buffer 94C−1分間
39ul 水 94C−50秒間|
1ul dNTP(各10mM、初期[]) 56C−1分間|30×
1.5ul 各プライマー(10mM、初期[]) 72C−3分間|
1ul ゲノムDNA
1ul Taq(NEB)
【0086】
以下のDNAプライマーをReg1の分析に使用した。
配列番号4−Reg1 testA=5′AGAATATACCATATAGGAGACGCGA
配列番号5−Reg1 testD=5′TACGACTATGGAAGCTCAAGAAGTT
配列番号6−pAG2tTr1=5′ATTACTTTCTGCGCACTTAACTTCG
これらの反応の結果を図10に示されているゲルにより示す。
【0087】
9.炭素代謝に関与していると考えられる酵母遺伝子の比較
遺伝子及びタンパク質発現の協調測定を、S288c由来菌株のgrr1Δ菌株で実施した。この菌株を、実験4に記載されたものと同じ方法を使用して作成した。ここで表1を参照する。主要な炭素代謝遺伝の選択されたリストの範囲内であっても、D−キシロースにおいて増殖した細胞とD−グルコースにおいて増殖したが、2つ主要な代謝抑制遺伝子であるHXK2又はGRR1を欠いている細胞の転写反応における大きな類似性を見ることができる。
【0088】
10.2−デオキシ−グルコースに抵抗性がある多様な単一遺伝子欠失菌株の同定
生酵母欠失突然変異体の一群をスクリーニングして、2−デオキシ−グルコースに対して抵抗性のあるものを同定した。この実験は、2%D−ガラクトース及び.03%2−デオキシ−グルコースを含有するYP培地においてOpen Biosystem酵母遺伝子欠失コレクションの細胞を増殖させ、30℃で3日接種した後の増殖を観察することによって実施した。D−ガラクトースを使用し、それはこの菌株は本出願者たちが試験したあらゆる条件下でD−キシロースにおいて増殖しないからである。この分析は、D−ガラクトースの存在下でD−キシロースの発酵をもたらしうる突然変異体のリストの拡大を本出願者たちにとって可能にする。結果を表2に表す。
【0089】
新規技術が図及び前記記載によって詳細に説明及び記載されてきたが、図及び前記記載は、例示的であり、限定的ではないことが考慮されるべきであり、好ましい実施態様のみが示され、記載されていること、並びに新規技術の精神の範囲内にある全ての変更及び修正が保護されることが望ましいことが理解される。同様に、新規技術が、特定の例、理論的議論、説明及び例示を使用して説明されたが、これらの説明及び付随の考察は、決して技術を制限するものとして解釈されるべきではない。本出願において参照されている全ての特許、特許出願、本文に対する参考文献、科学的な条約、公報などは、その全体が参照として本明細書に組み込まれる。
【0090】
[参考文献]
Bailey, R. B., Benitez, T., and Woodard, A. 1982. Saccharomyces cerevisiae mutants resistant to catabolite repression: Use in cheese whey hydrolysate fermentation. Appl. Environ. Microbiol. 44: 6310639.
Bailey, R. B. and Woodward, A. 1984. Isolation and characterization of a pleiotropic glucose repression resistant mutant of Saccharomyces cerevisiae. MoI. Gen. Genet. 793: 507-512.
Barnett, J.A. 1976. The utilization of sugars by yeast. Adv. Carbohydr. Chem. Biochem. 32: 126-228.
De Godoy, L. M. F., Olsen, J.V., de Souza, G.A., Li, G., Mortensen, P., and Mann, M. 2006. Status of complete proteome analysis by mass spectrometry: SILAC labeled yeast as a model system. Genome Biol. 7:R50.1-R50.15.
Gancedo, J. M. 1998. Yeast catabolite repression. Microbiol. MoI. Biol. Rev. 62: 334-361.
Gavin, A.-C, Aloy, P., Grandi, P., Krause, R., Boesche, M., et al. 2006. Proteome survey reveals modularity of the yeast cell machinery. Nature 440: 631-636.
Goldstein, A.I. and McCusker, J. H. 1999. Three new dominant resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 15: 1541-1553.
Ho, N.W.Y., Chen, Z., and Brainard, A.P. 1998. Genetically engineered Saccharomyces yeast capable of effective co-fermentation of glucose and xylose. Applied Env. Microbiol 64: 1852-1859.
Jin, Y.-S., Laplaza, J. M., and Jeffries, T.W. 2004. Saccharomyces cerevisiae engineered for xylose metabolism exhibits a respiratory response. Applied Env. Microbiol. 70: 6815-6825.
Kaniak, A., Xue, Z., Macool, D., Kim, J. -H., and Johnston, M. 2004. Regulatory network connecting two glucose signal transduction pathways in Saccharomyces cerevisiae. Euk. Cell 3: 221-231.
Keller, A., Nesvizhskii, A.I., Kolker, E., and Aebersold, R. 2003. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 74: 5383-5392.
Kodama, Y., Omura, F., Takahashi, K., Shirahige, K., and Ashikah, T. 2002. Genomewide expression analysis of genes affected by amino acid sensor Ssyi p in Saccharomyces cerevisiae. Curr. Genet. 41: 63-72.
Kotter, P. and Ciriacy, M. 1993. Xylose fermentation by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 38: 776-783.
Li, X., Zhang, H., Ranish, J.A., and Aebersold, R. 2003. Automated statistical analysis of protein abundance ratios from data generated by stable-isotope dilution and tandem mass spectrometry. Anal. Chem. 75: 6648-6657.
Lynd, L. R., Weimer, P.J., van ZyI, W.H., and Pretorius, I. S. 2002. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. MoI. Biol. Rev 66: 506-577.
Mathias, N., Johnson, S. L., Winey, M., Adams, A.E.M., Goetsch, L., Pringle, J. R., Byers, B., and Goebl, MG. 1996. Cdc53p acts in concert with Cdc4p and Cdc34p to control the G1 -to-S-phase transition and identifies a conserved family of proteins. MoI. Cell. Biol. 16: 6634-6643.
Mortimer, R. K. and Johnston, J. R. 1986. Genealogy of principal strains of the yeast genetic stock center. Genetics 113: 35-43.
Nesvizhskii, A.I., Keller, A., Kolker, E., and Aebersold, R. 2003. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 75: 4646-4658.
Neigeborn, L. and Carlson, M. Mutations causing constitutive invertase synthesis in yeast: Genetic interactions with snf mutations. Genetics 115: 247-253.
Ong, S. E., Blagoev, B., Kratchmarova, I., Kristensen, D. B., Steen, H., Pandey, A., Mann, M. 2002. Stable isotope labeling by amino acids in cell culture, SILAC, as a simple and accurate approach to expression proteomics. MoI. Cell. Proteomics 1: 376-386.
Phaff, H.J., Starmer, W.T., and Kurtzman, CP. 1999. Pichia lachancei sp. Nov., associate3d with several Hawaiian plant species. Int. J. Syst. Bacteriol. 49: 1295-1299.
Raiser, M., Wamelink, M. M., Struys, E.A., Joppich, C, Krobitsch, S., Jakobs, C, and Lehrach, H. 2008. A catabolic block does not sufficiently explain how 2-deoxy-glucose inhibits cell growth. Proc. Natl. Acad. Sci. USA 105: 17807-1781 1.
Schuller, H. -J. 2003. Transcriptional control of nonfermentative metabolism in the yeast Saccharomyces cerevisiae. Curr. Genet. 43: 139-160.
Schulte, F., Wieczorke, R., Hollenberg, C. P., and Boles, E. The HTR1 gene is a dominant negative mutant allele of MTH '1 and blocks Snf3- and Rgt2-dependent glucose signaling in yeast. J. Bac. 182: 540-542.
Sedlak, M. and Ho, N.W.Y. 2004. Characterization of the effectiveness of hexose transporters for transporting xylose during glucose and xylose co-fermentation by a recombinant Saccharomyces yeast. Yeast 21: 671-684.
Toivari, M. H., Salusjarvi, L., Ruohonen, L., and Penttila, M. 2004. Endogenous xylose pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 70: 3681-3686.
Tu, J. and Carlson, M. 1994. The GLC7 type 1 protein phosphatase is required for glucose repression in Saccharomyces cerevisiae. MoI. Cell. Biol. 14: 6789-6796.
Van Dijken, J. P., Bauer, J., Brambilla, L., Duboc, P. et al. 2000. An interlaboratory comparison of physiological and genetic properties of four Saccharomyces cerevisiae strains. Enz. Microb. Tech. 26: 706-714.
Van ZyI, C, Prior, B.A., Kilian, S. G., and Brandt, E.V. 1993. Role of D-ribose as a co-metabolite in D-xylose metabolism by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 59: 1487-1494.
Warren, R.A.J. 1996. Microbial hydrolysis of polysaccharides. Annu Rev. Microbiol. 50: 183-212.
Washburn, M. P., Wolters, D., Yates, J. R., III. 2001 . Large-scale analysis of the yeast proteome by multidimensional protein identification technology. Nat. Biotech. 19: 242-247.
Westergaard, S. L., Bro, C, Olsson, L., and Nielsen, J. 2004. Elucidation of the role of GrM p in glucose sensing by Saccharomyces cerevisiae through genome-wide transcriptional analysis. FEMS Yeast Res. 5: 193-204.
Westergaard, S. L., Oliveira, A.P., Bro, C, Olsson, L., and Nielsen, J. 2006. A systems biology approach to study glucose repression in the yeast Saccharomyces cerevisiae. Biotechnol. Bioeng. 1: 134-145.
Winston, F., Dollard, C, AND Ricupero-Hovasse, S. L. 1995. Construction of a set of convenient Saccharomyces cerevisiae strains that are isogenic to S288C. Yeast 1 1 : 53-55.
Yin, Z., Wilson, S., Hauser, N. C, Tournu, H., Hoheisel, J. D., and Brown, A.J. P. 2003. Glucose triggers different global responses in yeast, depending on the strength of the signal, and transiently stabilizes hbosomal protein mRNAs. MoI. Microbiol. 48: 713-724.
【技術分野】
【0001】
(優先権の主張)
本出願は、2009年2月25日に出願した米国仮出願第61/155,255号(この全ては参照として本明細書に組み込まれる)の利益を主張する。
【0002】
本発明は、相当なレベルのD−グルコースの存在下でD−グルコース以外の糖において増殖することができるサッカロミセス属(Saccharomyces)の菌株、並びに、その開発及び使用方法に関する。
【背景技術】
【0003】
サッカロミセス属の多様な種は、産業的に増殖される微生物のうち最も重要なものである。パンを発酵するため、ビール及びワインを生産するため、並びに食用香料及び微量栄養素の供給源として長い間使用されてきているが、これらの生物は、現在、糖原料からエタノールへの変換を促進することによって、燃料の生産において中心的な役割を果たしている。代謝的に複雑な生物である酵母は、好気的にも、また少なくとも数世代にわたって嫌気的も増殖することができる。商業的な製パン製造業を支援するために使用される酵母の生産のような商業的に増殖される場合、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)のような酵母を、通気発酵タンクにおいて増殖することができるこれらの条件下での酵母の増殖を制御して、酵母バイオマスの生産を増加することができる。これを達成することができる1つの方法は、D−グルコースのような糖の添加及び好気的に増殖することを促すための酵母への酸素移動の速さについて計画することである。サッカロミセス属の多様な菌株も、エタノールの生産を最大限にするように設計された条件下で増殖することができる。多くの場合、目的が糖からエタノールへの変換を最大限にすることである場合、発酵容器中の酸素レベルは、目的が嫌気性増殖に好ましいように酵母バイオマス生産を最大限にすることである場合の容器において使用される酸素のレベルに対して低減されうる。
【0004】
サッカロミセス属の大部分の菌株は、D−グルコースにおいて優先的に増殖するが、多くの菌株は、他の天然に生じるヘキソースにおいて、さらには幾つかの二糖類においても増殖することが知られている。異なる糖において、異なる酸素レベルの存在下で増殖するサッカロミセス属の異なる種の能力は、酵母が燃料産業で植物バイオマスからエタノールへの変換において現在果たしている中心的な役割を含む、商業的な有用性の多くを説明している。
【0005】
酵母によるエタノールの生産において一番良く知られている経路の1つは、エタノールへの6−炭素糖(ヘキソース)、特にD−グルコースの発酵である(図1)。エタノールの生産に広く使用されている供給原料の1つは多糖類のデンプンである。デンプンは、D−グルコースを含む単独ポリマーである。現在、米国において少なくともトウモロコシ由来のデンプンは、サッカロミセス・セレビシエによるエタノール生産にとって好ましい供給原料である。トウモロコシは、栄養素の高い作物であり、現在、トウモロコシの穀粒のみが、酵母を使用するエタノール発酵にとってデンプン/D−グルコースの適した供給源である。酵母に基づいたエタノール生産のための糖の別の供給源は、サトウキビである。サトウキビは、天然に発酵性糖が高く、酵母を使用するエタノールの生産にとって好ましい基質である。しかし、トウモロコシは、米国においてサトウキビよりも広範囲に栽培されている。気候のおかげで、このままの状態が続く可能性が高い。いずれにしても、トウモロコシに基づいたエタノール生産の持続可能性が疑問視されてきており、サトウキビは米国において実現可能な選択肢ではないので、バイオ燃料産業は、トウモロコシ及びサトウギビ以外の発酵性供給原料の他の供給源を探している。
【0006】
1つの大いにもてはやされている供給原料は、セルロースであり、トウモロコシよりも持続可能性があり、サトウキビよりも容易に入手可能であると考えられている。発酵性糖を生産するように加工されたセルロースは、恐らくエタノール生産の未来にとって最適な炭素供給源でありうる。酵母バイオマスを増加するため又はデンプン若しくはセルロースのような原料からエタノールを生産するために酵母を増殖させることは、バイオポリマーセルロースを、酵母により容易に発酵されうるD−グルコース、マルトース、三糖類及び四糖類のような糖単位に分解する前発酵加工工程を必要とする。
【0007】
供給源にかかわらず、6炭素糖、特にD−グルコースは、酵母に基づいた発酵にとって主なエネルギー供給源である。特徴決定されているサッカロミセス属の大部分の種は、D−グルコースにおいて優先的に増殖する。サッカロミセス属の多くの実験室由来菌株を含むこれらの菌株の多くは、D−グルコース以外のヘキソース、並びに二糖類及び三糖類において増殖することができる。しかし、D−グルコースにおいて優先的に増殖するサッカロミセス属は、ほぼ全ての産業的に重要な菌株を含むこの酵母の大部分の変種が異化代謝産物抑制を示すほど強力であり、すなわち、菌株は、供給原料において検出可能なレベルのD−グルコースがある限り、D−グルコース以外の糖を発酵しない。
【0008】
サッカロミセス属の検査した全てのものが、D−グルコースの存在下でD−グルコース以外の糖において活発に増殖し、糖からエタノールを生産することに無力であることは、D−グルコースを含む発酵性糖の混合物を含む任意の供給原料からの酵母バイオマス及び/又はエタノールの生産にとって残念なことである。例えば、D−グルコースはセルロース性バイオマスの発酵性成分への分解により遊離され、発酵性糖のミックスにおけるD−グルコースの存在は、他の糖のエタノールへの変換を著しく遅くする(図1)。
【0009】
セルロースからエタノールを生産する現在の技術的な障害にもかかわらず、2007年エネルギー独立性及び安全保障法(2007 Energy Independence and Security Act)(EISA2007)は、米国が、輸入石油に取って代わるために、セルロース性エタノールを生産する技術を急速に開発することを義務づける。したがって、産業的なサッカロミセス属の新規菌株の必要性及び有意な量のD−グルコースの存在下でもD−グルコースのみ以外の糖をバイオマス又はエタノールに容易に変換するこれらの産業的菌株を作り出す方法の必要性が存在する。本発明の幾つかの態様は、これらの必要性に対処している。
【発明の概要】
【発明が解決しようとする課題】
【0010】
そこで、本発明は、相当なレベルのD−グルコースの存在下でD−グルコース以外の糖において増殖することができるサッカロミセス属の菌株、並びに、その開発及び使用方法を提供する。
【課題を解決するための手段】
【0011】
本発明の幾つかの態様は、酵母を単離する方法であって、増殖培地を準備する工程(ここで培地は2−デオキシ−グルコース、キシロース及びグルタミンを含み、キシロースが唯一の炭素供給源である);培地に少なくとも1つの酵母菌株を接種する工程;培地から少なくとも1つの酵母細胞を単離する工程(ここで前記酵母細胞は、約0.1重量%の2−デオキシ−グルコースの存在下で唯一の炭素供給源としてのD−キシロースにおいて増殖する)を含む方法を含む。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。幾つかの実施態様では、培地に増殖する菌株は、約14日後にのみ培地において検出可能な増殖を示す。さらに別の実施態様において、検出可能な増殖の出現は、接種後、約21日後においてのみ生じる。
【0012】
幾つかの実施態様において、酵母菌株を選択するのに使用した培地は、約2.0重量%のキシロース及び約0.5重量%のグルタミンを含むが、これらの試薬の任意の濃度が増殖培地に添加されうる特定の菌株の増殖を支援するために十分である。幾つかの実施態様において、単離された酵母菌株は、サッカロミセス・セレビシエの一倍体(又は、半数体、haploid)、二倍体又は2を超える倍数性を有する菌株である。
【0013】
幾つかの実施態様において、培地から単離された酵母菌株は、少なくとも0.1重量%の2−デオキシ−グルコースの存在下でも少なくとも1つのペントース糖を代謝する。幾つかの実施態様において、供給原料(feed stock)は、少なくとも約0.03重量%の2−デオキシ−グルコースを含む。幾つかの実施態様において、菌株は、少なくとも0.1重量%の2−デオキシ−グルコースの存在下でD−グルコース以外の少なくとも1つのヘキソース糖を代謝する。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。幾つかの実施態様において、菌株は自然発生突然変異体である。さらに別の実施態様において、菌株は、突然変異を引き起こすことが知られている方法で酵母を処理することによるオープンリーディングフレームの標的化破壊のような、特定の事象によって作り出すことができる。突然変異誘発の速度を加速する方法及び試薬には、電離放射(ionizing radiation)、UV線、並びに、挿入剤(intercalating agent)、アルキル化剤(alkylating agent)、DNA付加物(DNA adduct)などのようなDNA構造に影響を与える試薬への暴露が含まれるが、これらに限定されない。
【0014】
本発明のさらに他の実施態様は、少なくとも0.1重量%の2−デオキシ−グルコースの存在下で唯一の炭素供給源の少なくとも1つのペントース糖において増殖するサッカロミセス・セレビシエの菌株を含む、酵母の変種を含む。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。幾つかの実施態様において、変種は、一倍体、二倍体であるか又は2を超える倍数性数を有する。幾つかの実施態様において、サッカロミセス・セレビシエの変種菌株は、JH015、CDXR2、Fermentis Ethanol Red reg1Δ及びFermentis Ethanol Red grr1−/−(GX1)などからなる群より選択される。
【0015】
さらに別の実施態様は、糖供給源を発酵する方法であって、サッカロミセス・セレビシエの少なくとも1つの菌株を準備する工程(ここでサッカロミセス・セレビシエの少なくとも1つの菌株は、少なくとも0.1重量%の2−デオキシ−グルコースの存在下で少なくとも1つのペントースにおいて増殖する);少なくとも1つの糖を含む供給原料を供給する工程;及び前記酵母菌株を供給原料において増殖する工程を含む方法である。幾つかの実施態様において、供給原料は、任意の追加の糖供給源の不在下で酵母菌株の増殖を支援するのに十分な量のD−グルコースを含む。さらに他の実施態様において、供給原料は、発酵性ペントース糖を含む。なお他の実施態様において、供給原料は、少なくとも約0.1重量%の2−デオキシ−グルコースを含む。幾つかの実施態様において、供給原料は、少なくとも約0.03重量%の2−デオキシ−グルコースを含む。他の実施態様において、供給原料は、D−グルコース以外の発酵性ヘキソース糖を含むが、別の実施態様では、供給原料は更にD−グルコースを含む。
【0016】
さらに別の態様は、酵母の突然変異菌株を作り出す方法であって、酵母の菌株、例えばGrr1及びReg1からなる群より選択される少なくとも1つの遺伝子と適格性があるサッカロミセス・セレビシエの一倍体、二倍体又はより高い倍数性の菌株を準備する工程;Grrl及びReg1の両方の活性を欠失して、突然変異菌株を作り出す工程;菌株を試験して、0.035重量%の2−デオキシ−グルコースの存在下でペントース糖において増殖するかを決定する工程を含む方法を含む。
【0017】
追加的な実施態様は、酵母菌株を選択又は同定する方法であって、YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271からなる群より選択される少なくとも1つのオープンリーディングフレームに突然変異を含むサッカロミセス・セレビシエの一倍体又はより高い変数性の菌株を準備する工程(ここで、オープンリーディングフレームは機能性遺伝子をコードし、オープンリーディングフレームにおける突然変異はオープンリーディングフレームにおいてコードされた遺伝子の活性を破壊する);サッカロミセス・セレビシエの前記菌株を、唯一の炭素供給源としてのキシロース及び0.1重量%の2−デオキシ−グルコースを含む培地において増殖させる工程;並びに培地に増殖したサッカロミセス・セレビシエの菌株を単離する工程を含む方法を含む。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。
【0018】
さらに他の実施態様は、サッカロミセス・セレビシエ酵母菌株の一倍体又はより高い変数性の菌株を単離する方法であって、YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wからなる群より選択されるオープンリーディングフレームによりコードされる少なくとも1つの遺伝子の機能性コピーを含む菌株を準備する工程;YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wからなる群より選択される少なくとも1つのオープンリーディングフレームに突然変異を導入して、サッカロミセス・セレビシエの突然変異体を産生する工程;選択培地においてサッカロミセス・セレビシエの突然変異体を増殖する工程(ここで選択培地は、唯一の炭素供給源としてのキシロース及び0.1重量%の2−デオキシ−グルコースを含む);並びに培地に増殖する突然変異体を単離する工程を含む方法を含む。幾つかの実施態様において、増殖培地は、約0.03重量%の2−デオキシ−グルコースを含む。
【0019】
本発明の幾つかの実施態様は、例えば2−デオキシ−グルコースの存在下及び代理に(by proxy)D−グルコースの存在でセルロース性バイオマスにおいて見出される幾つかのペントース糖を含むD−グルコース以外の糖において増殖する酵母菌株、例えばサッカロミセス属の産業用又は実験室用菌株を選択する方法を含む。他の実施態様は、2−デオキシ−グルコースの存在下でD−グルコース以外の糖に増殖するサッカロミセス属の産業用菌株を使用して、相当量のD−グルコースの存在下で追加の酵母バイオマス及び/又はD−グルコース以外の糖からエタノールのような発酵の最終産物若しくは副産物を生産することを含む。
【0020】
セルロース性エタノールを生産する優れた性能。セルロース性バイオマスは複数の糖を有し、最も重要ものはD−グルコース及びD−キシロースである。しかし、酵母代謝生理学は、多くの場合、他の多くの糖を除外したD−グルコースの調節的使用である異化代謝産物抑制(catabolite repression)を受ける。このことはD−キシロース又はD−キシルロースでは実証されていないが、本出願者たちは、ここで、D−キシロース及びD−キシルロースの利用も異化代謝産物抑制を受けることを示す。したがって、野生型酵母菌株はこれらのペントース糖の存在下でD−グルコースを優先的に代謝する。更に、本出願者たちは、ここで、サッカロミセス属の産業用菌株がヘキソース及びペントースの両方において異化代謝産物抑制を受けることを示す。これは、大部分の酵素菌株を使用するセルロース性エタノールにとって重要な技術上の障壁である。この複数糖発酵の障壁を克服するために、D−キシロースに対する異化代謝産物抑制を排除する必要がある。一つの実施態様は、以下の遺伝子GRR1、REG1及びHXK2の少なくとも1つを除去することによって異化代謝産物抑制を排除することを含む。本発明の幾つかの実施態様は、2−デオキシ−グルコースの存在下でD−グルコース以外の糖、特にペントースにおいて増殖する産業用酵母菌株、例えばサッカロミセス属を選択することを含む。幾つかの実施態様において、これは、相当レベルの2−デオキシ−D−グルコースの存在下でD−グルコール以外の糖供給源において増殖する産業用サッカロミセス属の変種を選択することを含む方法である。幾つかの実施態様において、これらの菌株は、以下の遺伝子GRR1、REG1及びHXK2の少なくとも1つを欠いているか又は少なくとも有効レベル形態(effective level form)を発現しない。さらに他の実施態様において、これらの遺伝子は、適切なレベルの活性タンパク質を産生しないように突然変異している場合がある。
【0021】
他の幾つかの実施態様は、相当量のD−グルコースの存在下でのD−グルコース以外の糖からのエタノールのような発酵の最終生成若しくは副生成を生産し、及び/又は、追加の酵母バイオマスを生産するため、2−デオキシ−グルコースの存在下でD−グルコース以外の糖において増殖するサッカロミセス属の産業用菌株を使用することを含む。
【0022】
幾つかの実施態様は、D−グルコースを含む混合糖を含む供給原料を発酵する方法であって、異なる代謝要件により複数の酵母菌株を使用して、バイオマス又はエタノールのような代謝産物のいずれかを効率的に生産する工程を含む方法を含む。幾つかの実施態様において、発酵は、酵母菌株の少なくとも2つの異なる菌株を使用して実施され、ここで少なくとも1つの菌株はD−グルコースにおいて優先的に増殖し、検出可能なレベルのD−グルコースの存在下で異化代謝産物抑制さえも示すことができるが、一方、少なくとも1つの他の菌株は、検出可能なレベルのD−グルコースの存在下であっても、活性化され(de-repressed)、D−グルコース以外の糖を発酵してエタノール及び/又はバイオマスにすることができる。幾つかの実施態様において、この方法は、D−グルコース及びD−キシロースのようなペントース糖の効率的な同時発酵を可能にする系をもたらすことができる。
【0023】
サッカロミセス・セレビシエ菌株CEN.PKgrr1Δの変種又はサッカロミセス・セレビシエ菌株Ethanol Red GX1の変種を含むサッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株であり、ここで、変種株の単一細胞分離株は固体培地において増殖し、その培地はD−キシロース及び2−デオキシ−グルコースを含み、培地の主要な炭素供給源は、糖D−キシロースであり、変種菌株の増殖は、培地において2日以内に強力なコロニーを生じる。
【0024】
さらに他の実施態様は、例えばサッカロミセス・セレビシエ菌株CEN.PKgrr1Δの変種又はサッカロミセス・セレビシエ菌株Ethanol Red GX1の変種を含むサッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株を含み、ここで、変種株の単一細胞分離株は、マルトース及び2−デオキシ−グルコースを含む固体培地において増殖し、培地の主要な炭素供給源は、マルトースが含まれるが、これに限定されない糖を含有するD−グルコースであることができ、変種菌株の増殖は、培地において2日以内に強力なコロニーを生じる。さらに他の実施態様は、サッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株を含み、ここで培地は、約2%のD−キシロース及び約0.1%の2−デオキシ−グルコースを含む。
【0025】
さらに他の実施態様は、約2%のマルトース、又は、糖を含有する別のD−グルコース及び約0.1%の2−デオキシ−グルコースを含む培地において増殖する変種を含む。
【0026】
さらに他の実施態様は、サッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株を含み、ここで変種菌株は、一倍体実験室用菌株CEN.PK誘導体CDXR1、CEN.PK(113−7D)、CDXR2及びCDXR3、並びに二倍体産業用酵母菌株Ethanol Red、GX1及びRX4からなる群より選択される。幾つかの実施態様において、開示されている方法を使用して選択された異化代謝産物抑制抵抗性サッカロミセス・セレビシエ菌株は、固体培地において増殖し、ここで培地は、主要な炭素供給源としてD−グルコースを含む。
【0027】
幾つかの実施態様は、サッカロミセス・セレビシエの異化代謝産物抑制抵抗性菌株を含み、ここで変種菌株は、2%のD−ガラクトース及び約0.03%の2−デオキシ−グルコースを含む固体培地において増殖し、D−ガラクトースは、培地における主要な炭素供給源である。
【0028】
さらに別の実施態様は、D−グルコース以外の糖において増殖する幾つかの産業用菌株を含むサッカロミセス・セレビシエの菌株を生産する方法であって、サッカロミセス・セレビシエの第1変種菌株を準備する工程(ここで前記変種は、2−デオキシ−D−グルコールの存在下でD−グルコース以外の糖、特にペントースにおいて増殖する);及びグルコース以外の糖を代謝する代謝経路から単離された少なくとも1つの遺伝子を第1変種菌株において過剰発現させて、第2変種菌株を形成する工程を含む方法を含む。
【0029】
追加的な実施態様は、D−グルコースの存在下でペントースにおいて増殖するサッカロミセス・セレビシエの変種菌株を生産する方法を含む。幾つかの実施態様において、第1変種菌株は、例えば、GX1、RX4、CDXR1、CEN.PK(113−7D)、CDXR2及びCDXR3からなる群より選択され、ここで菌株を、D−キシロース代謝経路から単離された少なくとも1つの遺伝子を発現するように適合させることができる。
【0030】
さらに他の実施態様は、D−グルコース以外の糖供給源においてサッカロミセス・セレビシエの菌株を増殖させる方法であって、サッカロミセス・セレビシエの第1菌株を準備する工程(ここで前記菌株は、2−デオキシ−D−グルコールの存在下でD−グルコース以外の糖供給源、特にペントースにおいて増殖する);グルコース以外の糖、特にペントースを代謝する代謝経路から単離された少なくとも1つの遺伝子を第1菌株において過剰発現させて、第2菌株を形成する工程;及び前記第菌株を培地において増殖する工程(ここで培地における主要な炭素供給源は、グルコース以外の糖、特にペントースである)を含む方法を含む。
【0031】
幾つかの実施態様において、第1菌株は、CDXR1、CEN.PK(113−7D)、CDXR2及びCDXR3からなる群より選択されるサッカロミセス・セレビシエ菌株CEN.PKの変種であり、少なくとも1つの過剰発現遺伝子は、D−キシロース代謝経路からのものである。
【0032】
さらに他の実施態様において、第1菌株は、GX1及びRX4からなる群より選択される産業用サッカロミセス・セレビシエ菌株Ethanol Redの変種であり、少なくとも1つの過剰発現遺伝子は、D−キシロース代謝経路からのものである。
【0033】
さらに追加的な実施態様は、サッカロミセス・セレビシエにおいて異化代謝産物抑制に関与するオープンリーディングフレームを同定する方法であって、D−グルコース又はD−グルコース及びD−キシロース又はD−キシロースの存在下でグルコース抑制を受けるサッカロミセス・セレビシエ菌株の第1変種を増殖する工程;D−グルコース、D−グルコース及びD−キシロース又はD−キシロースのいずれかの存在下でD−グルコース以外の主な炭素供給源において増殖された、グルコース抑制に非感受性のサッカロミセス・セレビシエ菌株の第2変種を蕃殖させる工程;並びに第1変種と第2変種のプロテオームを比較して、第1変種と第2変種のプロテオームの差を同定する工程を含む方法を含む。
【0034】
幾つかの実施態様において、D−キシロースは、マルトース又はマルトトリオースなどであるが、マルトース又はマルトトリオースに限定されない別の二次的な糖(secondary sugar)に代えられている。
【0035】
幾つかの実施形態において、第1及び第2変種のプロテオームの差が、以下のオープンリーディングフレーム:YLR063w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wの少なくとも1つの産物における差を示す。
【発明の効果】
【0036】
本発明によれば、サッカロミセス属の新規菌株は、有意な量のD−グルコースの存在下でもD−グルコースのみ以外の糖をバイオマス又はエタノールに容易に変換することができる。
【図面の簡単な説明】
【0037】
【図1】トウモロコシに基づいたデンプン及び植物バイオマスの両方をエタノールに変換する酵母の経路を表す概略図である。
【図2】エタノールに変換されるD−グルコース又はD−キシロースの経路を示す概略図である。出芽酵母細胞の写真である。
【図3】D−キシロースにおけるサッカロマイセス属の幾つかの菌株の増殖を示す植物である。
【図4】ビール酵母によるD−キシロースの発酵における提案される経路である。
【図5】YP+2%D−キシロースにおいて増殖しているCEN.PKの突然変異酵母コロニーの増殖を示すプレートの写真である。
【図6】CEN.PKの2−デオキシ−グルコース抵抗性誘導体の相補性分析の結果を示す植物の写真である。
【図7】2−デオキシ−グルコースの存在下でD−キシルロース、D−キシロース混合物において増殖する酵母により示される植物の写真である。
【図8】産業用酵母の多様な菌株によるトウモロコシ穂軸加水分解産物からエタノールへの発酵のグラフ図である。
【図9】PCR分析がGRR1遺伝子の欠失の同定を検査するために実施されることを示すセルである。
【図10】REG1遺伝子の欠失を検査するために実施されるPCR分析を示すセルである。
【図11】GX1によるマルトース及びD−グルコースの同時発酵を示すグラフである。
【発明を実施するための形態】
【0038】
新規技術の原理の理解を促進する目的において、ここで好ましい実施態様に対して参照を行い、それらを説明するために特定の言葉を使用する。それでも、それによって新規技術の範囲を制限することは意図されておらず、新規技術の原理に対するそのような変更、修正及び更なる適用は、新規技術が関連する当業者において通常起こることとして考慮されることが、理解されるべきである。
【0039】
本明細書で使用されるとき、特に指定のない限り、用語「約(about)」は、プラス又はマイナス20パーセントを意味し、例えば約1.0は、0.8〜1.2の範囲を包含する。
【0040】
本明細書で使用されるとき、特に指定のない限り、用語「検出可能な増殖(detectable growth)」は、増殖の証拠が人の裸眼による目視検査によって明らかになるまでを含む増殖を意味する。
【0041】
特に参照のない限り、遺伝子は、Demerec, M., Adelberg, E. A., Clark, A.J. & Hartman, P. E. in "A proposal for a uniform nomenclature in bacterial genetics". J. Gen. Microbiol 50, 1-14 (1968)により提案された命名法を使用して参照される。
【0042】
酵母サッカロミセス・セレビシエは、糖からエタノールへの発酵のために何世紀も前に栽培化(domesticated)された(図1及び2を参照すること)。酵母の栽培化は、D−グルコースをエタノールに変換するのに非常に効率的な産業用酵母菌株の世代をもたらした。有意には、これらの産業用酵母菌株は、一般に、大部分の微生物よりもはるかにエタノール耐性がある。この方法は、酵母増殖及び/又はエタノール生産のための糖が天然に最も豊富にあるヘキソースのうちの幾つか、特にD−グルコース、D−フルクトース及びD−マンノースであるときに、非常に効率的である。
【0043】
セルロースからエタノールへの産業用変換を実現可能な営利事業にするため、セルロースをグルコースのような発酵性糖に変換する方法を合理化することに大きな努力が注がれている。酵母に基づいたエタノール生産のためにセルロースから発酵性糖への変換を最適化することは重要であるが、植物材料に存在するD−グルコース以外の糖、特にD−キシロースからのエタノールの生産を合理化することも同様に重要である。植物材料において最も豊富な化合物は、グルコースポリマーセルロースであるが、有意な量の植物バイオマスが、キシランとして知られている糖ポリマーとして存在する(図1;例えばWarren 1996を参照すること)。事実、多くのセルロース供給源では、キシランは、ポリマーのバイオマスの20%超を占める。キシランそれ自体は、ペントース糖のD−キシロースの鎖から構成される。大部分のS.セレビシエの産業用及び実験室用菌株のような酵母による発酵のために、セルロースのようなキシランを最初にモノマーのD−キシロースに変換しなければならない。既に複雑な方法は、大部分の商業的に生産されたS.セレビシエの菌株がD−キシロースのようなペントース糖を効率的に発酵せず、特に供給原料中にD−グルコースが存在すると効率的に発酵しないので、発酵環境におけるD−グルコースの存在により更に複雑になる。
【0044】
S.セレビシエはD−キシロースを発酵することができないと広く考えられており、実際、酵母S.セレビシエは炭素供給源としてD−キシロースを利用しないことが、1970年代に報告された(Barnett 1976)。文献によると、酵母ピキア・スチピチス(Pichia stipitis)は、D−キシロースを発酵する能力を有さない。この情報に基づいて、幾つかの研究所は、S.セレビシエにおけるD−キシロースの利用に必要なピキア遺伝子を発現することにより、D−キシロースを発酵することができる酵母菌株の生成を試みた(例えば、Kotter and Ciriacy, 1993; Ho et al., 1998; Jin et al., 2003を参照すること)。改善されたD−キシロースの利用がそれぞれの場合において報告されたが、D−グルコースの存在下でのD−キシロース発酵の効率は著しく変動した。この観察を説明することができる幾つかの要因には、実験増殖及び前増殖の条件、並びにD−キシロース代謝経路の異種発現のレベルにおける差が含まれる。
【0045】
サッカロミセス・セレビシエについてのDr. Ho(Purdue U.)及びグラム(−)及びグラム(+)細菌の両方についてのDr. Ingram(U. of Florida)の研究は、D−キシロース(ピキア・スチピチス)の代謝及びエタノールの(多様な微生物からの)生産それぞれにおける他の生物の代謝経路内における外来性遺伝子の組み換え発現に集中してきた。
【0046】
あらゆる理論又は仮説に束縛されることなく、説明としてであって、限定ではなく、この観察には幾つかの見落とされた又は少なくとも正しく評価されていない理由がありうる。例えば、報告された結果と一致する1つの説明は、これらの実験に使用したD−キシロースの正確な組成における変動である。化学供給会社から購入した糖で実際に「純粋」であるものはほとんどない。純粋であるとして市販されている大部分の糖は、実際には約99又は98%の純度しかない。典型的には、糖における主な汚染物質は、極めて豊富にある糖D−グルコースである。酵母代謝の研究にとって残念なことに、0.1%の低さのD−グルコースのD−グルコース量は、例えばD−ガラクトースのようなD−グルコース以外の糖の利用に影響を及ぼすことが知られている。したがって、少なくとも幾つかの発表された研究において使用された純度98〜99%のD−キシロースの少なくとも一部は、実際はD−グルコースで汚染されていた可能性が高い。所定の酵母菌株がD−キシロースにおいて増殖することができるかを決定するように設計された研究において微量のD−グルコースによる汚染でも、これらの研究において観察された結果を歪めた可能性が十分にある。
【0047】
D−グルコースによる大部分の糖の汚染は、歴史的な問題である。例えば、D−ガラクトースを使用して酵母を確実に同定する能力は、0.01%未満のD−グルコースを含むD−ガラクトースの商業的導入(Sigma-Aldhch)の後でのみ生じた。残念なことに、D−グルコースで汚染されていないD−キシロースは、容易に入手することができない。したがって、幾つかの酵母菌株がD−キシロールにおいて増殖する能力を欠いているという報告は誤りである場合があり、このことは、文献における幾つかの矛盾した報告を説明する場合がある。
【0048】
キシロース発酵に関する文献において報告された不一致についてのさらに別の説明は、これらの研究に使用された多様な実験室用及び産業用酵母菌株の多様な遺伝子型組成に起因しうる。大部分の実験室用酵母菌株は、小数の祖先由来であったが(Mortimer and Johnston, 1986)、これらの子孫は、経時的に広く異なる増殖特性を発達させてきた(Winston et al., 1995; van Dijken et al., 2000)。事実、最近の論文は、S.セレビシエの少なくとも1つの菌株が、遺伝子修飾することなくD−キシロースにおいて増殖することができると、本出願者たちに示唆する場合がある(Sedlak and Ho, 2004; Figure 3; panel marked wild-type; Toivari et al. 2004を参照すること)。菌株MC996Aは、ピキアD−キシロース利用遺伝子を付加することなく、市販等級のD−キシロースにおいて増殖すると思われる。この菌株は、非常に強力な遺伝的背景を有することが示されている菌株のCEN.PKファミリーの誘導体である。これらの菌株の明らかに豊富な遺伝的多様性は、これらが広範囲の糖を発酵することを可能にする(van Dijkens ef al., 2000)。
【0049】
セルロース性バイオマスは複数の糖を含み、最も重要ものはD−グルコース及びD−キシロースである。しかし、酵母代謝生理学は、多くの場合、他の多くの糖を除外したD−グルコースの調節的使用である異化代謝産物抑制を受ける。このことはD−キシロース又はD−キシルロースでは実証されていないが、本明細書において実証されていることは、D−キシロース及びD−キシルロースの利用も異化代謝産物抑制下であることである。したがって、野生型酵母菌株はグルコースを優先的に代謝する。これは、大部分の酵素菌株を使用するセルロース性エタノールにとって重要な技術上の障壁である。この複数糖発酵の障壁を克服するために、D−キシロースに対する異化代謝産物抑制を排除する必要がある。
【0050】
D−グルコースを利用する能力を欠いている酵母菌株は、例えばヘキソキナーゼ遺伝子HXK1及びHXK2を欠いている菌株を作り出すことによって生成することができる。HXK2及びGRR1に突然変異を持つ酵母の菌株も本出願者たちのREG1突然変異体と同じように挙動することも、見出されている(データ示されず)。したがって、強力な野生型菌株を分析するか又はD−キシロース経路のピキア遺伝子を過剰発現する菌株を分析するかにかかわらず、D−グルコースの存在は、D−キシロースの利用を妨げる場合がある。大部分の酵母菌株はD−キシロースの分解に必要な遺伝子に高度に関連する遺伝子を有する。
【0051】
現在では、D−キシロースを発酵するS.セレビシエの存在を引用する報告でも、発酵が非常に非効率であることを認めている。例えば、Sedlak及びHo(2004)は、非常に少量のD−キシロースが、全てでなければ大部分のD−グルコースが利用される前に、彼らがD−キシロースを代謝できると考えるCEN.PK酵母菌株により消費されることを報告している。現在の文献は、D−キシロースのようなペントースにおいて増殖することができる酵母変種を同定する強力なスクリーニングを欠いていることを、明白に説明している。D−キシロースの発酵経路がS.セレビシエに存在すると仮定すると、いったん強力なスクリーニングが作り出されると、これらの経路を研究、改善及び増加することがかなり容易になる。
【0052】
ここで図3、パネルAを参照する。本明細書に開示されている結果は、キシロースにおいて増殖することができるCEN.PKの自然発生突然変異体をスクリーニングし、単離することが可能であることを示している。この結果は、D−キシロース発酵に必要なそれぞれの酵素に近接の相同体がサッカロミセスに存在することを示す(図4)、サッカロミセスゲノムの全配列の分析と一致する。
【0053】
図3を参照する。D−キシロースにおけるS288c及びCEN.PK誘導体の増殖。図3、パネルAでは、細胞をYP+2%D−キシロース培地にレプリカ平板培養し、30℃で4日間インキュベートした。図3、パネルBでは、細胞をYP+2%D−キシロース/0.1%2−デオキシグルコースにレプリカ平板培養し、30℃で10日間インキュベートした。パネルBの矢印は、2−デオキシ−グルコースの存在下で増殖するCEN.PKの自然発生突然変異体から生じたコロニーを指している。
【0054】
酵母細胞は、D−グルコース、D−フルクトース及びD−マンノースを発酵するのに極めて効率的であり、更に酵母は、他の多くの炭素供給源を除外してこれらの糖を発酵し、この現象は異化代謝産物抑制として知られている(Gancedo 1998において概説されている)。事実、研究されたS.セレビシエの実質的に全ての菌株は、今までのところ、D−グルコース、D−フルクトース及び/又はD−マンノースだけにエネルギーを、これらの糖が完全に又はほぼ完全に環境から排除されるまで依存している。例えば、わずかに微量のD−グルコース(<1%)がD−ガラクトースを汚染する場合、酵母は、D−グルコースの存在がなくなるまでD−ガラクトースを発酵しない。S.セレビシエによるD−キシロースの利用に関する文献における矛盾する報告は、異化代謝産物抑制を引き起こすD−グルコースによる増殖培地の汚染に起因する場合がある。
【0055】
幾つかの遺伝子における突然変異は、マルトース、スクロース及びD−ガラクトースに基づいた発酵の文脈において、実験室用菌株で異化代謝産物抑制を緩和したことを示した。異化代謝産物抑制に関与したと考えられる遺伝子には、GRR1、REG1およびHXK2が含まれる(Gancedo, 1998において概説されている)。特定の実験室用菌株内のこれらの遺伝子いずれかの損失は、幾つかの実験室用菌株の、D−ガラクトース又はスクロースのような他のヘキソースを伴ったD−グルコースの同時発酵を可能にする(Bailey and Woodward, 1984)。
【0056】
D−グルコースの存在下で幾つかの二次的な炭素供給源を利用する実験室用酵母由来の酵母細胞の能力についての1つの試験は、二次的な炭素供給源及び少量の2−デオキシ−グルコースの存在下で増殖する細胞の能力を測定することである。2−デオキシ−グルコースは、特定の実験室用菌株において、D−ガラクトース、マルトース及びスクロースのようなヘキソース糖に対してグルコース抑制を示すことが報告されているD−グルコースの非代謝性誘導体である。この現象は、D−ガラクトースによって実証されている(Bailey et al., 1982; Bailey and Woodward, 1984)。異化代謝産物抑制能力もあり、D−ガラクトース、マルトース又はスクロースを発酵することもできる、2−デオキシ−グルコースに暴露された酵母細胞は、2−デオキシ−グルコースの存在下で代替的な炭素供給源を利用することができないと考えられ、これらの菌株は、2−デオキシ−グルコースを代謝することができない。これらの条件は、細胞死をもたらすと報告されているが、2−デオキシ−グルコースに暴露された酵母細胞の正確な死亡原因は、依然として不明である(Raiser et al. 2008)。
【0057】
グルコース抑制は、それによりほとんどの炭素供給源の利用の前に、D−グルコースを培地から涸渇させなければならない、酵母における現象を説明する。グルコース抑制の十分に研究された調節物質は、代替的な炭素供給源の利用に関与する遺伝子の転写抑制因子として作用すると考えられているMig1転写因子であるが、MIG1の損失は、細胞を2−デオキシ−グルコースに対して抵抗性にしないことも報告されている(Schuller, 2003)。上記に記述されたように、GRR1、REG1又はHXK2の損失は、細胞を2−デオキシ−グルコースに対して抵抗性にする(Gancedo 1998)。Reg1は、PP1複合体をMig1に運ぶことによりMig1を調節すると考えられているPP1タンパク質ホスファターゼサブユニットであるが、他のタンパク質がグルコース抑制を達成するのに必要とする正確な機構はあまり明確ではない。マイクロアレイ分析も、Mig1がGrr1及びHxk2により調節されるプロセスのサブユニットのみに影響を及ぼすことを示す(Westergaard et al. 2006)。本明細書において実施及び報告されている実験結果のプロテオミクス分析は、異化代謝産物抑制の調節に対する大型の複製後成分が認識されていないことを示唆している。
【0058】
1に記述されているスクリーニングにより単離されたCEN.PKにおける自然発生突然変異は、REG1座位において不活性を示す(図6)。更に、REG1又は損失したときに同様の突然変異遺伝子型を有することが知られている他の2つの遺伝子、GRR1及びHXK2の標的化破壊も、この培地において増殖する能力を得た。REG1、GRR1又はHXK2の標的化破壊は、PCR仲介遺伝子破壊によって作り出される場合がある。これらの実験は、特定の遺伝子座位においてDNAの5’及び3’セグメントと同一であるプライマーを設計することによって実施することができるアミノグリコシド系抗生物質のノウルセオトリシンに対する抵抗性を付与する、ストレプトミセス・ノウルセオトリシイ(Streptomyces nourseothricii)のnat1遺伝子を使用して、上記に提示された遺伝子のいずれかを、酢酸リチウム形質転換法を使用する酵母の形質転換により欠失することができる。一倍体菌株において、単一対立遺伝子の破壊は、L−グルタミン、2%D−キシロース及び0.1%2−デオキシ−グルコースを補充したYP培地における増殖を可能にするのに十分である。しかし、異なる酵母菌株は、異なる濃度の2−デオキシ−グルコースに対して異なる感受性がある。
【0059】
大部分の産業用酵母菌株は二倍体である。したがって、これらの菌株における形質転換は、単一対立遺伝子の欠失を確実にするのみである。驚くべきことに、二倍体又は任意の更に高い倍数性の酵母の同じ遺伝子座位における他のコピーの再現性欠失は、2−デオキシ−グルコースと、マルトース、D−ガラクトース、スクロース、D−キシロース又はD−キシルロースのような糖が含まれるが、これらに限定されない主要炭素供給源として機能する二次的な糖とを含有する培地において異型接合体菌株を平板培養することにより達成できることが、本出願者たちにより見出された。
【0060】
ここで図3及び5を参照する。この現象もD−キシロースの文脈において実証された。図3に示されているように、S288c grr1Δ菌株、野生型CEN.PK菌株及びCEN.PK grr1Δ菌株を、2%D−キシロースを含有する培地で平板培養した場合、増殖を3つの菌株全てにおいて検出することができた。しかし、CEN.PK grr1Δ菌株のみが、2%D−キシロース及び0.1%2−デオキシ−グルコースを含有する培地において増殖を示す(図3)。経時的に、CEN.PK菌株の細胞の斑内では、単離されたコロニーが10〜15日後に出現し始めた。
【0061】
約21日後、これらのコロニーは、物理的に操作するのに十分な大きさになった。これらの単離されたコロニーは、2−デオキシ−グルコースの存在下でD−キシロースにおいて増殖する能力を得たCEN.PKの親菌株の自然発生突然変異体である(下記を参照すること)。この現象は、S288c grr1Δ誘導体では観察されない。これらの結果は、CEN.PK細胞と対照的に、GRR1遺伝子を欠失しても、S288c細胞は、2−デオキシ−グルコースの存在下でD−キシロースにおいて増殖できないことを示している。2−デオキシ−グルコース抵抗性/D−キシロース利用突然変異体がS288cにおいて発展する能力がないことは、S288cにより見られる少量の増殖がD−キシロース内の汚染量のグルコースの利用に起因しうることを示唆している。
【0062】
本明細書に開示されているスクリーニングにより単離された及び2−デオキシ−グルコースの存在下でD−キシロースにおいて増殖したサッカロミセス・セレビシエの2つの代表的な二倍体菌株CDXR2及びJH015は、2009年2月25日にアメリカン・タイプ・カルチャー・コレクションに寄託され、それぞれ受入番号PTA−9849及びPTA−9850が指定された。生物学的材料の寄託についてのブタペスト条約の規定において、これらの菌株は、本発明に対して特許が発行されると公に利用可能になる。
【0063】
酵母細胞代謝が利用を好ましい糖から好ましくない糖(例えば、D−ガラクトース及び恐らくはD−キシロース)へ変換するには、数時間かかる場合がある。この遅れは、D−キシロース分解に必要な酵素を過剰発現するように操作された酵母細胞においても生じる。糖の混合物を利用する産業用及び実験用の両方の酵母菌株において、D−キシロースの代謝は、混合物がD−グルコースをほぼ完全に欠くまで非常に非効率的であると思われる。
【0064】
ここで図9を参照する。2−デオキシ−グルコースの存在下でキシロースにおいて増殖する能力により選択された二倍体産業用酵母菌株を、PCRを使用して分析した。3つのGrr1Tf及びGrr1TFプライマー対は、GRR1が無傷である場合、3810bpのPCR産物を産生するはずであり、GRR1が破壊されている場合、PCR産物は1400bpのはずである。Grr1TF&pAG25TR1プライマー対は、GRR1が無傷である場合、PCR産物を産生するはずはなく、GRR1がnat1遺伝子に代わっている場合は、約1200bpのPCR産物のはずである。PCR分析は、Fermentis Ethanol Redの誘導体であるgrr1−/−2A及びgrr1−/−2B菌株において、両方のGRR1遺伝子がnat1遺伝子に代わっていることを証明する。RC4がgrr1::NAT1を有する一倍体S288C誘導体であることに留意すること。以前の2つのPCR反応(10/18/04及び12/10/09)は、このPCR反応におけるgrr1−/−菌株について観察されたものと同じ産物を生じた。
【0065】
ここで図10を参照する。二倍体産業用菌株の同様の分析を実施して、Reg1遺伝子における変化を探した。Reg1 testA&Reg1 test IDプライマー対は、REG1が無傷である場合、約3600bpnのPCT産物を産生するはずであり、破壊されている場合、PCT産物は約1800bpのはずである。Reg1 testA&pAG25TR1プライマー対は、REG1が無傷である場合、PCR産物を産生するはずではなく、REG1がnat1遺伝子に代わっている場合、約1500bpのPCR産物のはずである。上記のPCRは、Fermentis Ethanol Redの誘導体であるreg1−/−1A及びreg1−/−1B菌株において、両方のREG1遺伝子がnat1遺伝子に代わっていることを証明する。grr1−/−2Aのようにnat1遺伝子を持つ菌株であっても、REG1 testA及びpAG25TR41プライマーによりPCR産物は作製されないことも、証明されている。
【0066】
D−グルコースの抑制効果の強度が、D−キシロースにより(Jin et al. 2004)並びにD−グルコースの異なる濃度により(Yin et al. 2003)調節される転写物の選択された群のレベルを含む表1に示されている。野生型細胞と比較した、2%D−グルコースにおいて増殖している間のGRR1及びHXK2の細胞突然変異体におけるこれらの転写物の調節も、含まれている(Westergaard et al. 2006)。表1から分かるように、一定レベルの低いD−グルコース(0.01%)であっても、異化代謝産物抑制を引き起こす。例えば、豊富なFBP1転写物は.01%〜1%D−グルコースで比較的一定のままであるが、2%D−キシロース又はGRR1を欠いている細胞における増殖により高く誘導されている。異化代謝産物抑制の排除は、混合糖供給原料をプロセスしてD−グルコース混合糖供給原料のレベルを、異化代謝産物抑制を誘導するレベルより下に低減することが可能である場合でも有用であるはずであり、それはプロセスされた混合糖原料からD−グルコースを実質的に涸渇させることは、時間がかかり、高価であるからである。
【0067】
【表1】
【0068】
本明細書に開示されているように、GRR1を欠いている細胞のプロテオミクス分析が実施され、このデータの分析は、少なくとも以下の2つの点について説明する。第1には、転写レベルは、必ずしもプロテオームの正確な状況を提供しているとは限らない(表1)。例えば、遺伝子HXT1及びHXT7の転写物はそれぞれ減少又は増加するが、対応するタンパク質は全く異なって挙動し、Hxt1タンパク質レベルは、野生型菌株をgrr1Δ菌株と比較すると変わらないままであるが、Hxt7タンパク質は、grr1Δでは減少している。これらの結果は、mRNAマイクロアレイデータをプロテオミクスデータの不在下で解釈する場合に、注意を払わなければならないことを示している。Hxt7がD−キシロースの重要な輸送体であると提案されているので、このことは本明細書において非常に重要である。
【0069】
第2には、このプロテオミクスデータは、Gre3タンパク質がgrr1Δ突然変異菌株において16×を超えて上昇していることを示す。GRE3は、D−キシロース分解の第1工程を実施するピキアキシロースレダクターゼの相同体をコードする。更に、Gre3における増加は、GRE3遺伝子の転写誘導の不在下で生じ、本出願者たちの実験においてタンパク質の直接的な検出によってのみ見ることができる。したがって、GRR1の不在は、更に細胞をD−キシロースの利用に向かわせる。これらの結果は、GRR1、HXK2及び最も可能性が高いものでREG1の損失は、完全に重複しているのでなければ非常の類似した反応をもたらしうることを示す。
【0070】
マイクロアレイ分析は、不偏様式で網羅的転写変化をモニターする方法を提供する。マイクロアレイ実験における本質的な前提は、mRNAレベルの差がタンパク質レベルの差に反映することである。しかし、全てのタンパク質がタンパク質活性を変化させることはない又は均一のレベルは転写制御における変化を必要とする。例えば、翻訳後仲介事象は、マイクロアレイ分析では「不可視」である顕著なプロテオミクス変化をもたらすことができる。幸いにも、マイクロアレイ分析とプロテオミクス分析の組み合わせを使用して、転写性を、豊富な特定のタンパク質における変化の原因である後転写調節事象と区別することができる。
【0071】
検出された2580個のタンパク質のうちの約1,200個のタンパク質についての定量データをまとめ(ここにデータを示す)、結果は、この方法にとって典型的である(de Godoy et al. 2006)。これらのプロテオミクスデータを、上記に記載された菌株及び培地条件(GRR1突然変異体における)を使用するマイクロアレイ分析と、また、野生型細胞をgrr1突然変異体及び他の異化代謝産物抑制突然変異体と比較した公表されたマイクロアレイ分析(Kodama et al., 2002; Kaniak et al., 2004; Westergaard et al., 2004)と組み合わせた。これらのデータセットの分析は、GRR1により影響を受けた調節の3つの特有のパターンを明らかにした(ここにデータを示す)。第1には、そのmRNAの豊富さがタンパク質産物の豊富さと相関する遺伝子が同定された。第2には、産物の豊富さの差がmRNA変化の不在下で生じる遺伝子が見出された。そのmRNAの豊富さが、産物の豊富さにおける同時変化なしに変化する遺伝子も同定された。これらの結果は、正確な条件下でこれらの技術を使用して、多様な活性代謝経路のプロテオームをプロテオミクス的に研究できることを示す。
【0072】
【表2】
【0073】
[実験]
1.S.セレビシエの異化代謝産物活性化一倍体自然発生突然変異体のスクリーニング
以下の実験を実施して、2−デオキシ−グルコースにおける酵母細胞の増殖を試験した。野生型酵母菌株を、2重量%のキシロース及び0.1重量%の2−デオキシ−グルコースを含む固体培地YPで平板培養した。しかし、酵母細胞の増殖は見られなかった。
【0074】
次に、0.5重量%のL−グルタミンを、そうでなければ同一の培地に加え、増殖を、接種の数日後に見出した。しかし、図3(パネルB)で見られるように、細胞はこの培地において容易に増殖しなかった。3週間の接種の後でのみ、他の酵母菌株由来の細胞ではなくCEN.PK誘導体が、予想外に、2−デオキシ−グルコースの存在下でキシロースにおいて増殖する能力を得た。これらの自然発生突然変異体は、最終的に、単一細胞から生じたと思われるコロニーを形成した。これらのコロニーは、自然発生突然変異誘発により誘導されたが、伝統的な化学的突然変異誘発要因又は照射がこのプロセスを増強することができる。
【0075】
2.一倍体突然変異体
4つの2−デオキシ−グルコース耐性/D−キシロース利用CEN.PKコロニーからの細胞を単離し、更なる研究のために取っておいた。ここで図5を参照すると、これらの突然変異CEN.PK菌株は、D−グルコース及びD−キシロースにおいて増殖し、D−キシロース+2−デオキシ−グルコース(DOG)培地に移されたとき、2日以内に強力な斑を生じる。
【0076】
3.相補性分析
単離された菌株である2−デオキシ−グルコース抵抗性CEN.PK誘導体を、酵母ノックアウトコレクション(Open Biosystems)からの特定の菌株と接合させた。4つの2−デオキシ−グルコース抵抗性突然変異菌株も、全て2%ガラクトース及び.03%2−デオキシ−グルコースを含有する培地において増殖した。したがって、S288c由来細胞がD−キシロースにおいて増殖しなかったので、相補性試験を、D−ガラクトース及び2−デオキシ−グルコースを含有する培地において実施した。分析は、2−デオキシ−グルコース表現型CDXR2及びCDXR4が、REG1における突然変異により補完されないことを実証し、CDXR2及びCDXR4がREG1の突然変異対立遺伝子を含有することを示した。これらの通常ではない表現型の原因であるこれらの菌株において、依然として未同定の変化も存在しうる。
【0077】
4.活性化一倍体産業用酵母菌株を得るためのスクリーニング及び標的化手法
一倍体実験室用菌株に使用したものと同様の手法を使用して、2−デオキシ−グルコースの存在下でD−キシロース及びD−キシルロースの混合物において強力に増殖する能力を自然発生的に得た修飾型の産業用二倍体菌株Fermentis Ethanol Redをスクリーニングし、単離することが可能であるはずである。
【0078】
Fermentis Ethanol Redのような幾つかの産業用一倍体菌株は、異株性であり、標準的な実験室業務の胞子形成及び四分子切開により一倍体として作製及び維持することができる。産業用二倍体菌株を一倍体に変換することは、グルコース抑制を付与する遺伝子の単一コピーしか破壊する必要がないので、2−デオキシグルコースの存在下でD−キシロースの混合物において強力に増殖するのに必要な特性を得る菌株の能力を増加する。
【0079】
活性化産業用二倍体菌株を、標的化遺伝子破壊、続く遺伝子の第2コピーの損失を選択することにより得ることもできる。例えば、菌株GX1(grr1−/−)は、GRR1遺伝子の2つのゲノムコピーのうちの1つを、ノウルセオトリシンN−セチルトランスフェラーゼ遺伝子(NAT1)遺伝子に代えることによって作り出すことができ、このことは、Fermentis Ethanol Red菌株においてアミノグリコシドノウルセオトリシンに対する抵抗性を付与する。2−デオキシ−グルコースと、マルトース、D−ガラクトース、スクロース、D−キシロース又はD−キシルロースのような主要な炭素供給源として機能する二次的な糖とを含有する培地において異型接合体菌株を平板培養することによって、GRR1遺伝子の両方のコピーを欠いている菌株を得た。
【0080】
ここで図7を参照する。産業用二倍体、野生型(Fermentis Ethanol Red)及びreg1Δ(Fermentis Ethanol Red誘導体)をYPDプレートにおいて2日間増殖させ、次にYP+0.4%キシルロース/3.6%キシロース(上側パネル)又はYP+0.4%キシルロース/3.6%キシロース+0.1%2−デオキシ−グルコースにおいて30℃で3日間レプリカ平板培養した。
【0081】
5.トウモロコシ穂軸加水分解産物における突然変異産業用二倍体菌株の増殖
グルコース及びキシロースの混合物において増殖する突然変異産業用二倍体の能力を試験した。産業用二倍体菌株、野生型(Fermentis Ethanol Red)及びreg1Δ(Fermentis Ethanol Red誘導体)菌株を、50mlのトウモロコシ穂軸加水分解産物に、出発密度の1×107細胞/mlで接種した。培養を30℃で振とうし、試料を指定された時間で取り出した。グルコース、キシロース及びエタノールの濃度を屈折率により分析した。ここで図8を参照する。突然変異菌株も、D−グルコース及びキシロースの両方を含むトウモロコシ穂軸由来のセルロース性混合物において、相当な率の糖をエタノールに変換する(約11%)。
【0082】
6.グルコース及びマルトースの両方を含む混合糖供給源における突然変異産業用二倍体酵母菌株の増殖
産業用二倍体菌株、野生型(Fermentis Ethanol Red)及びgrr1−/−(GX1)(Fermentis Ethanol Red誘導体)を、10g/Lの酵母抽出物、20g/Lのペプトン、50g/Lのグルコース及び50g/Lのマルトースを含有する50mlの培地に、出発密度の1×107細胞/mlで接種した。培養を30℃で振とうし、試料を指定された時間で取り出した。グルコース、キシロース及びエタノールの濃度を屈折率により分析した。図11に示されているように、この菌株も、両方の糖が同時に提供されたとき、D−グルコースよりも素速くマルトースを利用する。
【0083】
7.GRR1及びREG1破壊のPCR仲介高次構造
GRR1遺伝子のPCR分析に使用する条件は以下である。
PCRミックス RC4プログラム
5ul 10×Thermo Pol Rxn Buffer 94C−1分間
39ul 水 94C−50秒間|
1ul dNTP(各10mM、初期[]) 56C−1分間|30×
1.5ul 各プライマー(10mM、初期[]) 72C−3分間|
1ul ゲノムDNA
1ul Taq(NEB)
【0084】
以下のDNAプライマーをGRR1の分析に使用した。
配列番号1−Grr1TfF=5′GAAGCCCAAAAATTAAGGCATTGCA
配列番号2−Grr1TR=5′TTTGAAACTGTGTATAGAATGTTTCGC
配列番号3−pAG25Tr1=5′ATTACTTTCTGCGCACTTAACTTCG
これらの反応の結果を図9に示されているゲルにより示す。
【0085】
8.REG1破壊のPCR分析
REG1遺伝子のPCR分析に使用する条件は以下である。
PCRミックス RC4プログラム
5ul 10×Thermo Pol Rxn Buffer 94C−1分間
39ul 水 94C−50秒間|
1ul dNTP(各10mM、初期[]) 56C−1分間|30×
1.5ul 各プライマー(10mM、初期[]) 72C−3分間|
1ul ゲノムDNA
1ul Taq(NEB)
【0086】
以下のDNAプライマーをReg1の分析に使用した。
配列番号4−Reg1 testA=5′AGAATATACCATATAGGAGACGCGA
配列番号5−Reg1 testD=5′TACGACTATGGAAGCTCAAGAAGTT
配列番号6−pAG2tTr1=5′ATTACTTTCTGCGCACTTAACTTCG
これらの反応の結果を図10に示されているゲルにより示す。
【0087】
9.炭素代謝に関与していると考えられる酵母遺伝子の比較
遺伝子及びタンパク質発現の協調測定を、S288c由来菌株のgrr1Δ菌株で実施した。この菌株を、実験4に記載されたものと同じ方法を使用して作成した。ここで表1を参照する。主要な炭素代謝遺伝の選択されたリストの範囲内であっても、D−キシロースにおいて増殖した細胞とD−グルコースにおいて増殖したが、2つ主要な代謝抑制遺伝子であるHXK2又はGRR1を欠いている細胞の転写反応における大きな類似性を見ることができる。
【0088】
10.2−デオキシ−グルコースに抵抗性がある多様な単一遺伝子欠失菌株の同定
生酵母欠失突然変異体の一群をスクリーニングして、2−デオキシ−グルコースに対して抵抗性のあるものを同定した。この実験は、2%D−ガラクトース及び.03%2−デオキシ−グルコースを含有するYP培地においてOpen Biosystem酵母遺伝子欠失コレクションの細胞を増殖させ、30℃で3日接種した後の増殖を観察することによって実施した。D−ガラクトースを使用し、それはこの菌株は本出願者たちが試験したあらゆる条件下でD−キシロースにおいて増殖しないからである。この分析は、D−ガラクトースの存在下でD−キシロースの発酵をもたらしうる突然変異体のリストの拡大を本出願者たちにとって可能にする。結果を表2に表す。
【0089】
新規技術が図及び前記記載によって詳細に説明及び記載されてきたが、図及び前記記載は、例示的であり、限定的ではないことが考慮されるべきであり、好ましい実施態様のみが示され、記載されていること、並びに新規技術の精神の範囲内にある全ての変更及び修正が保護されることが望ましいことが理解される。同様に、新規技術が、特定の例、理論的議論、説明及び例示を使用して説明されたが、これらの説明及び付随の考察は、決して技術を制限するものとして解釈されるべきではない。本出願において参照されている全ての特許、特許出願、本文に対する参考文献、科学的な条約、公報などは、その全体が参照として本明細書に組み込まれる。
【0090】
[参考文献]
Bailey, R. B., Benitez, T., and Woodard, A. 1982. Saccharomyces cerevisiae mutants resistant to catabolite repression: Use in cheese whey hydrolysate fermentation. Appl. Environ. Microbiol. 44: 6310639.
Bailey, R. B. and Woodward, A. 1984. Isolation and characterization of a pleiotropic glucose repression resistant mutant of Saccharomyces cerevisiae. MoI. Gen. Genet. 793: 507-512.
Barnett, J.A. 1976. The utilization of sugars by yeast. Adv. Carbohydr. Chem. Biochem. 32: 126-228.
De Godoy, L. M. F., Olsen, J.V., de Souza, G.A., Li, G., Mortensen, P., and Mann, M. 2006. Status of complete proteome analysis by mass spectrometry: SILAC labeled yeast as a model system. Genome Biol. 7:R50.1-R50.15.
Gancedo, J. M. 1998. Yeast catabolite repression. Microbiol. MoI. Biol. Rev. 62: 334-361.
Gavin, A.-C, Aloy, P., Grandi, P., Krause, R., Boesche, M., et al. 2006. Proteome survey reveals modularity of the yeast cell machinery. Nature 440: 631-636.
Goldstein, A.I. and McCusker, J. H. 1999. Three new dominant resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 15: 1541-1553.
Ho, N.W.Y., Chen, Z., and Brainard, A.P. 1998. Genetically engineered Saccharomyces yeast capable of effective co-fermentation of glucose and xylose. Applied Env. Microbiol 64: 1852-1859.
Jin, Y.-S., Laplaza, J. M., and Jeffries, T.W. 2004. Saccharomyces cerevisiae engineered for xylose metabolism exhibits a respiratory response. Applied Env. Microbiol. 70: 6815-6825.
Kaniak, A., Xue, Z., Macool, D., Kim, J. -H., and Johnston, M. 2004. Regulatory network connecting two glucose signal transduction pathways in Saccharomyces cerevisiae. Euk. Cell 3: 221-231.
Keller, A., Nesvizhskii, A.I., Kolker, E., and Aebersold, R. 2003. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 74: 5383-5392.
Kodama, Y., Omura, F., Takahashi, K., Shirahige, K., and Ashikah, T. 2002. Genomewide expression analysis of genes affected by amino acid sensor Ssyi p in Saccharomyces cerevisiae. Curr. Genet. 41: 63-72.
Kotter, P. and Ciriacy, M. 1993. Xylose fermentation by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 38: 776-783.
Li, X., Zhang, H., Ranish, J.A., and Aebersold, R. 2003. Automated statistical analysis of protein abundance ratios from data generated by stable-isotope dilution and tandem mass spectrometry. Anal. Chem. 75: 6648-6657.
Lynd, L. R., Weimer, P.J., van ZyI, W.H., and Pretorius, I. S. 2002. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. MoI. Biol. Rev 66: 506-577.
Mathias, N., Johnson, S. L., Winey, M., Adams, A.E.M., Goetsch, L., Pringle, J. R., Byers, B., and Goebl, MG. 1996. Cdc53p acts in concert with Cdc4p and Cdc34p to control the G1 -to-S-phase transition and identifies a conserved family of proteins. MoI. Cell. Biol. 16: 6634-6643.
Mortimer, R. K. and Johnston, J. R. 1986. Genealogy of principal strains of the yeast genetic stock center. Genetics 113: 35-43.
Nesvizhskii, A.I., Keller, A., Kolker, E., and Aebersold, R. 2003. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 75: 4646-4658.
Neigeborn, L. and Carlson, M. Mutations causing constitutive invertase synthesis in yeast: Genetic interactions with snf mutations. Genetics 115: 247-253.
Ong, S. E., Blagoev, B., Kratchmarova, I., Kristensen, D. B., Steen, H., Pandey, A., Mann, M. 2002. Stable isotope labeling by amino acids in cell culture, SILAC, as a simple and accurate approach to expression proteomics. MoI. Cell. Proteomics 1: 376-386.
Phaff, H.J., Starmer, W.T., and Kurtzman, CP. 1999. Pichia lachancei sp. Nov., associate3d with several Hawaiian plant species. Int. J. Syst. Bacteriol. 49: 1295-1299.
Raiser, M., Wamelink, M. M., Struys, E.A., Joppich, C, Krobitsch, S., Jakobs, C, and Lehrach, H. 2008. A catabolic block does not sufficiently explain how 2-deoxy-glucose inhibits cell growth. Proc. Natl. Acad. Sci. USA 105: 17807-1781 1.
Schuller, H. -J. 2003. Transcriptional control of nonfermentative metabolism in the yeast Saccharomyces cerevisiae. Curr. Genet. 43: 139-160.
Schulte, F., Wieczorke, R., Hollenberg, C. P., and Boles, E. The HTR1 gene is a dominant negative mutant allele of MTH '1 and blocks Snf3- and Rgt2-dependent glucose signaling in yeast. J. Bac. 182: 540-542.
Sedlak, M. and Ho, N.W.Y. 2004. Characterization of the effectiveness of hexose transporters for transporting xylose during glucose and xylose co-fermentation by a recombinant Saccharomyces yeast. Yeast 21: 671-684.
Toivari, M. H., Salusjarvi, L., Ruohonen, L., and Penttila, M. 2004. Endogenous xylose pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 70: 3681-3686.
Tu, J. and Carlson, M. 1994. The GLC7 type 1 protein phosphatase is required for glucose repression in Saccharomyces cerevisiae. MoI. Cell. Biol. 14: 6789-6796.
Van Dijken, J. P., Bauer, J., Brambilla, L., Duboc, P. et al. 2000. An interlaboratory comparison of physiological and genetic properties of four Saccharomyces cerevisiae strains. Enz. Microb. Tech. 26: 706-714.
Van ZyI, C, Prior, B.A., Kilian, S. G., and Brandt, E.V. 1993. Role of D-ribose as a co-metabolite in D-xylose metabolism by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 59: 1487-1494.
Warren, R.A.J. 1996. Microbial hydrolysis of polysaccharides. Annu Rev. Microbiol. 50: 183-212.
Washburn, M. P., Wolters, D., Yates, J. R., III. 2001 . Large-scale analysis of the yeast proteome by multidimensional protein identification technology. Nat. Biotech. 19: 242-247.
Westergaard, S. L., Bro, C, Olsson, L., and Nielsen, J. 2004. Elucidation of the role of GrM p in glucose sensing by Saccharomyces cerevisiae through genome-wide transcriptional analysis. FEMS Yeast Res. 5: 193-204.
Westergaard, S. L., Oliveira, A.P., Bro, C, Olsson, L., and Nielsen, J. 2006. A systems biology approach to study glucose repression in the yeast Saccharomyces cerevisiae. Biotechnol. Bioeng. 1: 134-145.
Winston, F., Dollard, C, AND Ricupero-Hovasse, S. L. 1995. Construction of a set of convenient Saccharomyces cerevisiae strains that are isogenic to S288C. Yeast 1 1 : 53-55.
Yin, Z., Wilson, S., Hauser, N. C, Tournu, H., Hoheisel, J. D., and Brown, A.J. P. 2003. Glucose triggers different global responses in yeast, depending on the strength of the signal, and transiently stabilizes hbosomal protein mRNAs. MoI. Microbiol. 48: 713-724.
【特許請求の範囲】
【請求項1】
増殖培地を準備する工程と、
培地に、少なくとも1つの酵母菌株を接種する工程と、
少なくとも1つの酵母細胞を培地から単離する工程と、を含む、酵母を単離する方法であって、
前記培地は、2−デオキシ−グルコース、キシロース及びグルタミンを含み、かつ、キシロースが唯一の炭素供給源であり、そして、
前記酵母細胞は、少なくとも約0.03重量%の2−デオキシ−グルコースの存在下で唯一の炭素供給源としてのD−キシロースにおいて増殖する、方法。
【請求項2】
菌株が約14日後にのみ培地において検出可能な増殖を示す、請求項1記載の方法。
【請求項3】
菌株が約21日後にのみ培地において検出可能な増殖を示す、請求項1記載の方法。
【請求項4】
培地が約2.0重量%のキシロース及び0.5重量%のグルタミンを含む、請求項1記載の方法。
【請求項5】
酵母がサッカロミセス属(Saccharomyces)の種である、請求項1記載の方法。
【請求項6】
酵母がサッカロミセス・セレビシエ(Saccharomyces cerevisiae)の菌株である、請求項6記載の方法。
【請求項7】
サッカロミセス・セレビシエの菌株が一倍体である、請求項4記載の方法。
【請求項8】
サッカロミセス・セレビシエの菌株が二倍体である、請求項6記載の方法。
【請求項9】
サッカロミセス・セレビシエの菌株が2を超える倍数性数を有する、請求項6記載の方法。
【請求項10】
培地から単離された酵母菌株が、少なくとも0.03重量%の2−デオキシ−グルコースの存在下で少なくとも1つのペントース糖を代謝する、請求項1記載の方法。
【請求項11】
培地から単離された酵母菌株が、少なくとも約0.03重量%の2−デオキシ−グルコースの存在下でD−グルコース以外の少なくとも1つのヘキソース糖を代謝する、請求項1記載の方法。
【請求項12】
培地から単離された酵母が自然発生突然変異体である、請求項1記載の方法。
【請求項13】
酵母を少なくとも1つの突然変異原に暴露する工程を更に含む、請求項1記載の方法。
【請求項14】
突然変異原が、電離放射線、UV線及び挿入剤、アルキル化剤、DNA付加物などのようなDNA構造に影響を与える試薬からなる群より選択される、請求項13記載の方法。
【請求項15】
少なくとも0.03重量%の2−デオキシ−グルコースの存在下で唯一の炭素供給源としての少なくとも1つのペントース糖において増殖するサッカロミセス・セレビシエの菌株を含む変種酵母。
【請求項16】
サッカロミセス・セレビシエの菌株が一倍体である、請求項15記載の変種酵母。
【請求項17】
サッカロミセス・セレビシエの菌株が二倍体である、請求項15記載の変種酵母。
【請求項18】
サッカロミセス・セレビシエの菌株が、JH015、CDXR2、Fermentis Ethanol Red reg1Δ及びFermentis Ethanol Red grr1−/−(GX1)からなる群より選択される、請求項15記載の変種酵母。
【請求項19】
サッカロミセス・セレビシエの少なくとも1つの菌株を準備する工程と、
少なくとも1つの糖を含む供給原料を供給する工程と、
前記酵母菌株を供給原料において増殖する工程と、
を含む、糖供給源を発酵する方法であって、
前記サッカロミセス・セレビシエの少なくとも1つの菌株は、少なくとも0.03重量%の2−デオキシ−グルコースの存在下で少なくとも1つのペントース糖において増殖する、方法。
【請求項20】
供給原料が、任意の追加の糖供給源の不在下で酵母菌株の増殖を支援するのに十分な量のD−グルコースを含む、請求項19記載の方法。
【請求項21】
供給原料が、前記酵母菌株により発酵されうるペントース糖を含む、請求項19記載の方法。
【請求項22】
供給原料が、前記酵母菌株及び少なくとも0.1重量%の2−デオキシ−グルコースにより発酵されうるペントース糖を含む、請求項19記載の方法。
【請求項23】
供給原料が、D−グルコース以外の発酵性ヘキソース糖を含む、請求項19記載の方法。
【請求項24】
供給原料がD−グルコースを更に含む、請求項23記載の方法。
【請求項25】
Grr1及びReg1からなる群より選択される少なくとも1つの遺伝子に適格性がある酵母菌株を準備する工程と、
Grr1及びReg1の両方の活性を欠失して、突然変異菌株を作り出す工程と、
菌株を試験して、少なくとも約0.03重量%の2−デオキシ−グルコースの存在下でペントース糖において増殖するかを決定する工程と、
を含む、酵母の突然変異菌株を作り出す方法。
【請求項26】
突然変異菌株が一倍体である、請求項25記載の方法。
【請求項27】
突然変異菌株が二倍体である、請求項25記載の方法。
【請求項28】
突然変異菌株が2を超える倍数性数を有する、請求項25記載の方法。
【請求項29】
YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271からなる群より選択される少なくとも1つのオープンリーディングフレームにおいて突然変異を含むサッカロミセス・セレビシエの菌株を準備する工程と、
サッカロミセス・セレビシエの前記菌株を、唯一の炭素供給源としてのキシロース及び少なくとも約0.03重量%の2−デオキシ−グルコースを含む培地において増殖させる工程と、
培地において増殖したサッカロミセス・セレビシエの菌株を単離する工程と、
を含む、酵母菌株を選択又は同定する方法であって、
前記オープンリーディングフレームは、機能性遺伝子をコードし、そして、前記オープンリーディングフレームにおける突然変異は、前記オープンリーディングフレームにおいてコードされた遺伝子の活性を破壊する、方法。
【請求項30】
YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wからなる群のオープンリーディングフレームによりコードされる少なくとも1つの遺伝子の機能性コピーを含む菌株を準備する工程と、
YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wからなる群より選択される少なくとも1つのオープンリーディングフレームに突然変異を導入して、サッカロミセス・セレビシエの突然変異体を産生する工程と、
サッカロミセス・セレビシエの突然変異体を選択培地において増殖させる工程と、
前記培地において増殖した突然変異体を単離する工程と、
を含む、酵母菌株を単離する方法であって、
前記培地は、唯一の炭素供給源としてのキシロース及び少なくとも約0.03重量%の2−デオキシ−グルコースを含む、方法。
【請求項1】
増殖培地を準備する工程と、
培地に、少なくとも1つの酵母菌株を接種する工程と、
少なくとも1つの酵母細胞を培地から単離する工程と、を含む、酵母を単離する方法であって、
前記培地は、2−デオキシ−グルコース、キシロース及びグルタミンを含み、かつ、キシロースが唯一の炭素供給源であり、そして、
前記酵母細胞は、少なくとも約0.03重量%の2−デオキシ−グルコースの存在下で唯一の炭素供給源としてのD−キシロースにおいて増殖する、方法。
【請求項2】
菌株が約14日後にのみ培地において検出可能な増殖を示す、請求項1記載の方法。
【請求項3】
菌株が約21日後にのみ培地において検出可能な増殖を示す、請求項1記載の方法。
【請求項4】
培地が約2.0重量%のキシロース及び0.5重量%のグルタミンを含む、請求項1記載の方法。
【請求項5】
酵母がサッカロミセス属(Saccharomyces)の種である、請求項1記載の方法。
【請求項6】
酵母がサッカロミセス・セレビシエ(Saccharomyces cerevisiae)の菌株である、請求項6記載の方法。
【請求項7】
サッカロミセス・セレビシエの菌株が一倍体である、請求項4記載の方法。
【請求項8】
サッカロミセス・セレビシエの菌株が二倍体である、請求項6記載の方法。
【請求項9】
サッカロミセス・セレビシエの菌株が2を超える倍数性数を有する、請求項6記載の方法。
【請求項10】
培地から単離された酵母菌株が、少なくとも0.03重量%の2−デオキシ−グルコースの存在下で少なくとも1つのペントース糖を代謝する、請求項1記載の方法。
【請求項11】
培地から単離された酵母菌株が、少なくとも約0.03重量%の2−デオキシ−グルコースの存在下でD−グルコース以外の少なくとも1つのヘキソース糖を代謝する、請求項1記載の方法。
【請求項12】
培地から単離された酵母が自然発生突然変異体である、請求項1記載の方法。
【請求項13】
酵母を少なくとも1つの突然変異原に暴露する工程を更に含む、請求項1記載の方法。
【請求項14】
突然変異原が、電離放射線、UV線及び挿入剤、アルキル化剤、DNA付加物などのようなDNA構造に影響を与える試薬からなる群より選択される、請求項13記載の方法。
【請求項15】
少なくとも0.03重量%の2−デオキシ−グルコースの存在下で唯一の炭素供給源としての少なくとも1つのペントース糖において増殖するサッカロミセス・セレビシエの菌株を含む変種酵母。
【請求項16】
サッカロミセス・セレビシエの菌株が一倍体である、請求項15記載の変種酵母。
【請求項17】
サッカロミセス・セレビシエの菌株が二倍体である、請求項15記載の変種酵母。
【請求項18】
サッカロミセス・セレビシエの菌株が、JH015、CDXR2、Fermentis Ethanol Red reg1Δ及びFermentis Ethanol Red grr1−/−(GX1)からなる群より選択される、請求項15記載の変種酵母。
【請求項19】
サッカロミセス・セレビシエの少なくとも1つの菌株を準備する工程と、
少なくとも1つの糖を含む供給原料を供給する工程と、
前記酵母菌株を供給原料において増殖する工程と、
を含む、糖供給源を発酵する方法であって、
前記サッカロミセス・セレビシエの少なくとも1つの菌株は、少なくとも0.03重量%の2−デオキシ−グルコースの存在下で少なくとも1つのペントース糖において増殖する、方法。
【請求項20】
供給原料が、任意の追加の糖供給源の不在下で酵母菌株の増殖を支援するのに十分な量のD−グルコースを含む、請求項19記載の方法。
【請求項21】
供給原料が、前記酵母菌株により発酵されうるペントース糖を含む、請求項19記載の方法。
【請求項22】
供給原料が、前記酵母菌株及び少なくとも0.1重量%の2−デオキシ−グルコースにより発酵されうるペントース糖を含む、請求項19記載の方法。
【請求項23】
供給原料が、D−グルコース以外の発酵性ヘキソース糖を含む、請求項19記載の方法。
【請求項24】
供給原料がD−グルコースを更に含む、請求項23記載の方法。
【請求項25】
Grr1及びReg1からなる群より選択される少なくとも1つの遺伝子に適格性がある酵母菌株を準備する工程と、
Grr1及びReg1の両方の活性を欠失して、突然変異菌株を作り出す工程と、
菌株を試験して、少なくとも約0.03重量%の2−デオキシ−グルコースの存在下でペントース糖において増殖するかを決定する工程と、
を含む、酵母の突然変異菌株を作り出す方法。
【請求項26】
突然変異菌株が一倍体である、請求項25記載の方法。
【請求項27】
突然変異菌株が二倍体である、請求項25記載の方法。
【請求項28】
突然変異菌株が2を超える倍数性数を有する、請求項25記載の方法。
【請求項29】
YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271からなる群より選択される少なくとも1つのオープンリーディングフレームにおいて突然変異を含むサッカロミセス・セレビシエの菌株を準備する工程と、
サッカロミセス・セレビシエの前記菌株を、唯一の炭素供給源としてのキシロース及び少なくとも約0.03重量%の2−デオキシ−グルコースを含む培地において増殖させる工程と、
培地において増殖したサッカロミセス・セレビシエの菌株を単離する工程と、
を含む、酵母菌株を選択又は同定する方法であって、
前記オープンリーディングフレームは、機能性遺伝子をコードし、そして、前記オープンリーディングフレームにおける突然変異は、前記オープンリーディングフレームにおいてコードされた遺伝子の活性を破壊する、方法。
【請求項30】
YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wからなる群のオープンリーディングフレームによりコードされる少なくとも1つの遺伝子の機能性コピーを含む菌株を準備する工程と、
YLRO63w、YMR167w、YPL176c、YPL123c、YPL121c、YBR242w、YBR422w、YHR012w、YHR103w、YHR154w、YCL048w、YLR133w、YOR138c、YOR177c、YDR269c、YIL064w、YOL101c、YML124C、YMR116C、YDR028c、YDR074c、YDL088c及びYGR271wからなる群より選択される少なくとも1つのオープンリーディングフレームに突然変異を導入して、サッカロミセス・セレビシエの突然変異体を産生する工程と、
サッカロミセス・セレビシエの突然変異体を選択培地において増殖させる工程と、
前記培地において増殖した突然変異体を単離する工程と、
を含む、酵母菌株を単離する方法であって、
前記培地は、唯一の炭素供給源としてのキシロース及び少なくとも約0.03重量%の2−デオキシ−グルコースを含む、方法。
【図1】


【図2】

【図3A】

【図3B】

【図4】
【図5】

【図6】
【図7】
【図8】
【図9】
【図10】

【図11】

【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2012−518437(P2012−518437A)
【公表日】平成24年8月16日(2012.8.16)
【国際特許分類】
【出願番号】特願2011−552159(P2011−552159)
【出願日】平成22年2月25日(2010.2.25)
【国際出願番号】PCT/US2010/025448
【国際公開番号】WO2010/099343
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(506265705)インディアナ ユニバーシティ リサーチ アンド テクノロジー コーポレイション (4)
【出願人】(312000985)
【出願人】(312000996)
【出願人】(312001007)
【出願人】(312001018)
【Fターム(参考)】
【公表日】平成24年8月16日(2012.8.16)
【国際特許分類】
【出願日】平成22年2月25日(2010.2.25)
【国際出願番号】PCT/US2010/025448
【国際公開番号】WO2010/099343
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(506265705)インディアナ ユニバーシティ リサーチ アンド テクノロジー コーポレイション (4)
【出願人】(312000985)
【出願人】(312000996)
【出願人】(312001007)
【出願人】(312001018)
【Fターム(参考)】
[ Back to top ]