
DNAセグメント及び多糖類の高生産方法
【課題】スフィンゴモナス菌株を操作してスフィンガンの大量生産菌を生産する方法、細菌がスフィンガンを増産する際に有用なDNAフラグメントの同定方法と利用方法、ならびに大量生産菌を提供する。
【解決手段】スフィンゴモナス菌株がスフィンガン多糖類を増産するために用いられるDNAセグメント又はフラグメントを、スフィンゴモナス菌株から単離し、その多数のコピーを他の菌株に挿入し、スフィンガン多糖類の大量生産菌を生産する。
【解決手段】スフィンゴモナス菌株がスフィンガン多糖類を増産するために用いられるDNAセグメント又はフラグメントを、スフィンゴモナス菌株から単離し、その多数のコピーを他の菌株に挿入し、スフィンガン多糖類の大量生産菌を生産する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、スフィンゴモナス属菌(Sphingomonas sp. )のスフィンガン(sphingan)多糖類の生合成生産に関与し、且つこのスフィンゴモナス属菌から単離されるDNA配列、及びそのフラグメントに関する。単離されたDNAフラグメントの多数のコピーをスフィンゴモナス属菌又は関連菌の同じ又は異なる菌株に挿入して、多糖類、好ましくはスフィンガンを増産することができる。このような操作により外因性DNAを含有する細菌の多糖類生産量は、同一の発酵条件下で、操作をしない細菌よりも有意に大きくなる。さらに、本発明はスフィンゴモナス属菌、その他の関連菌の菌株を操作して多糖類の大量生産菌とする方法、ならびにこのようにして操作された細菌に関する。さらにまた、スフィンゴモナス属菌がスフィンガン多糖類を増産するために有用なDNA配列の同定方法及び単離方法も説明する。
【背景技術】
【0002】
多くの微生物がエクソポリサッカライド類(exopolysaccharides)、あるいはEPSとしても知られている細胞外多糖類を生産する。エクソポリサッカライド類には、キサンタンガム(xanthan gum)及び「スフィンガン類」として知られている一群の多糖類が含まれるている。「スフィンガン類」はスフィンゴモナス属(genus Sphingomonas)のグラム陰性菌によって生産される。
【0003】
「スフィンガン類」は莢膜多糖類で、構造は互いに似ているが同一ではなく、スフィンゴモナス属細菌によって分泌される(Pollock, T.J.1993,J.Gen.Microbiol.,139:1939−1945)。スフィンガンはそれぞれ異なる側基を持っているが、主鎖のある場所では、L−ラムノースかL−マンノースのどちらかが存在する。L−マンノースそのものは天然では非常に稀である。ポリマーの水溶液には独特で有用な流動特性がある(Moorhouse,R.1987,”Structure/property relationships of a family of microbioal polysaccharides,”p189−206. In M.Yalpani(ed.),Industrial polysaccharides:genetic engineering,structure/property relation and applications.Elsevier Science Publishers B.V.,Amsterdamを参照されたい)。ポリマーの構造を変えることによって、どのようにして異なる流動特性が生じるのかまだ分かっていない。
【0004】
キサントモナス カンペストリス(Xanthomonas campestris)はグラム陰性菌で、エクソポリサッカライドの一つであるキサンタンガムを構成的に大量生産する(Jeanes, et al.,J. Appl. Polymer Sci.,5, 519〜526,1961)。キサンタンガムの生合成については、産業上の重要性からかなり詳細な研究がなされてきた。最近にいたり、新しい細菌性エクソポリサッカライドであるジェラン(gellan)がゲル化剤として開発された。ジェランは、S−88(Kang and Veeder, 米国特許第4, 535, 153号を参照)、ウエラン(welan)(Kang and Veeder, 米国特許第4, 342, 866号を参照)、NW11(Robison and Stipanovic, 米国特許第4,874, 044号を参照)、ラムザン(rhamsan)(Peik et al., 米国特許第4, 401, 760号を参照)、S−198(Peik etal., 米国特許第4, 529, 797号を参照)、S−657(Peiket al., 欧州特許出願第209277A1号を参照)、及びヘテロポリサッカライド−7(Kang and McNeely, 米国特許第4, 342, 866号を参照)を含む関連多糖類の一群に分類されている一つである。
【0005】
上記の引例ではスフィンガン多糖類組成に関する特許が数件含まれている。これらの特許はいずれも本発明の対象とはなんら関連性もない。
【0006】
【表1】

【0007】
スフィンガン多糖類の化学構造は、すべてある程度の関連性がある。スフィンガン各々の主鎖は、D−グルコース、D−グルクロン酸、L−マンノース及びL−ラムノースの四つの糖の関連性のある配列からなっている。スフィンガン群の多糖類は、ポリマーの主軸(主鎖)と側鎖を含む炭水化物によって互いに区別することができる。スフィンガンの構成糖には、アセチル基又はピルビル(pyruvyl)基が結合している場合もある。
【0008】
各種のスフィンガン類は、特殊ポリマーとして、あるいは繊維への応用、食品、化粧品、紙、ペンキ、セメントの添加剤、例えば粘度調整剤として、あるいは各種のコーティングへの応用において、さらには石油製品の接着剤や添加剤、特殊化学薬品として有用である。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明がなされるに至った研究の初期段階では、代表的なスフィンガン多糖類であるS−88の生合成第一工程が重要な課題であった。このスフィンガンは、スフィンゴモナス菌株S88によって生合成される。本研究以前では、スフィンガン以外の細菌性多糖類の生合成のすべてではないが、一部の製法において、リン酸イソプレニルを担体として使用されることが知られていた。例えば、X. カンペストリスが生合成するキサンタンガムの場合、キサンタンガムの主鎖にはグルコースしか含まれていないので、合成第一工程ではおそらくUDP−グルコースのグルコース−リン酸をC55−イソプレニルリン酸(IP)担体に転移させる必要がある。Ielpiら(FEBS Lett.,130, 253,1982及びJ. Bacteriol. ,175, 2490,1993)は、無細胞取り込み検定を行って、グルコースの次に第二のグルコース、その後、マンノース、グルクロン酸、そしてマンノースの順序でIP担体に連続添加して、キサンタンガムの繰り返し単位が構成されていることを確認した。まったく同様にして、エスチェリシア コリ(Escherichia coli、大腸菌)のコレニン酸のサブユニットの繰り返し単位も、まずグルコース−Pを担体IPに転移することにより構成されている(Johnson and Wilson, J. Bacteriol.,129, 225,1977)。対照的に、リゾビウム メリロチ(Rhizobium meliloti)によるスクシノグリカン(succinoglycan)多糖類の合成の場合には、まずガラクトース−PをIPに転移している(Tolmasky, et al.,J. Biol. Chem.,257, 6751,1982を参照)。しかしながら、イソプレニル担体は、デキストランあるいはレバン(levan)多糖類の合成には関与しないので、アルギン酸塩の合成におけるイソプレニル担体の役割は分かっていない。
【0010】
本発明の研究以前では、多糖類生合成の複合反応速度論(complex kinetics)から見て、担体の役割が重要であることについては分かっていなかった。さらに、スフィンゴモナス菌がスフィンガン多糖類を合成する全体を通して、イソプレニルリン酸担体がどんな役割を果たしているのかも分かっていなかった。
【0011】
同時にBacr でもありGum- でもある(バシトラシン耐性とキサンタンガム陰性)X. カンペストリス変異菌の特別クラスは、UDP−Glcのグルコース−PがIPに転移してGlc−PPIを形成するのに必要なgumD遺伝子中に認められることが、遺伝子相補性検定によって以前から証明されていた。(Pollock, et al.,1994, J. Bacteriol.,vol.176, pp. 6229〜6237, Vanderslice, et al.,”Genetic Engineering of Polysaccharidestructure in Xanthomonas campestris”, p. 145〜156, in V. Crescenzi, et al.,Biomedical and Biotechnological Advances in Industrial Polysaccharides, Gordonand Breach Science Publishers, New York and N. E. Harding and Y. N. Patel, 1993, Faseb Journal, Vol. 7, Number7)。後者の引例では、生産能がない変異菌がスフィンガンS−60の合成を復活できるDNAフラグメントを開示しているが、合成量が野生型菌株より大きいか、否かについては触れられていない。さらにまた、X. カンペストリスの野生型gumD遺伝子によって、スフィンゴモナス菌株のS88とNW11の相似菌であるBacr Sps- (スフィンガン多糖類陰性)変異菌がスフィンガン類の合成を復活できることが実験で証明された。さらに、Bacr Sps- スフィンゴモナス変異菌は、グルコース−PがIPに転移するのを阻害されるらしいと示唆された。
【0012】
本発明の目的は、スフィンゴモナス属菌から単離されて、多くの微生物、及び特に多くのスフィンゴモナス菌株が、スフィンガン多糖類を増産するために用いられるDNAセグメントを提供することにある。
【0013】
本発明の他の目的は、スフィンガン多糖類の生産量が操作されていない菌株よりも有意に大きい、微生物、及び特に多くのスフィンゴモナス菌株から得られる大量生産菌株を提供することにある。
【0014】
本発明のさらに他の目的は、スフィンガン多糖類大量生産菌である微生物の菌株、及び特にスフィンゴモナス属菌の菌株の生産方法を提供することにある。
【0015】
本発明のさらに他の目的は、DNAセグメントを単離してスフィンゴモナス菌株に挿入することにより、このようにして操作された微生物がスフィンガン多糖類大量生産菌となる方法を提供することにある。
【0016】
本発明のこれらの目的及び/又は他の目的は、次に述べる本発明の説明によって容易に理解することができる。
【課題を解決するための手段】
【0017】
本発明のDNA配列は、セグメントあるいはフラグメントのいずれも、スフィンガン生産能を持つ細菌、一般にはスフィンゴモナス菌株から単離する。このようにして得た遺伝子材料は、クローニングした後、多数のコピーをスフィンガン生産能を持つ、あるいは生産能をもたないスフィンゴモナス又は関連菌の変異体に組み入れる。このようなDNA配列は、スフィンガンを生産しない変異菌のスフィンガン生産を復活するのに有用であることが立証されている。さらに、予期していないことであったが、これらの変異菌はスフィンガンを生産できるばかりでなく、スフィンガン生産量が、本来スフィンガンを生産できる野生型菌株に期待されている生産量よりも有意に大きいことが見いだされた。
【0018】
さらにまた、DNAセグメントあるいはフラグメントをスフィンゴモナスの一菌株から単離し、多数のコピーをスフィンガン生産能を持つ、あるいは生産能を持たない同じスフィンゴモナス変異菌株又は異なる菌株に挿入すると、このようにして操作した細菌がスフィンガンの大量生産菌となることを発見できたことも、予期していないことであった。この発見は特に次の点で思いもよらないものであった。すなわち、DNAセグメント又はフラグメントを、例えば、スフィンゴモナスS60から単離して、スフィンゴモナスS88の野生型あるいは非ムコイド性変異菌に挿入すると、この操作によってS−88スフィンガンの大量生産菌ができ、しかも生産されるS−88は一般にS−60スフィンガンが混入していない。この相補性はスフィンゴモナスの各種菌株間でかなり広く適用したり(菌株間相補性)、またキサントモナス カンペストリスのキサンタンガム生産にも適用できる可能性がある(属間相補性)。
【0019】
さらにまた本発明者らは、スフィンガン生産能を持つスフィンゴモナス菌株からDNAセグメント又はフラグメントを単離して組み入れる操作によって、大量生産能を持つスフィンゴモナス菌株を生産する方法を発見した。スフィンガン生産能を持つ細菌から単離したDNAは、まずクローニングした後、スフィンガン生産能を持つスフィンゴモナス菌株、又はスフィンガン生産能を持つ菌株から派生した非ムコイド性変異体に挿入する。
【0020】
さらに本発明は、上記の単離したDNAセグメント又はフラグメントを挿入することにより得た、操作されているスフィンゴモナス菌を包含している。これらの操作されている細菌は、本発明により単離されたDNAセグメント又はフラグメントの多数のコピーを含有している。本発明により操作された細菌は、スフィンガンの大量生産菌である。
【0021】
本発明によれば、DNAフラグメントは、当業者が容易に利用できる技術により単離回収して、クローニングすることができる。その後、DNAの多数のコピーを、一般には染色体外又はプラスミドDNAとしてスフィンゴモナス属の細菌に挿入する。標的細菌に挿入した後、操作されている細菌により、同じ菌株の操作されていない、スフィンガン生産能を持つ細菌と同一の濃度で発酵させて、スフィンガン生産量を測定する。操作されていない、生産能を持つ菌株よりも、スフィンガン生産量が大きいものを大量生産菌として決定する。この方法を使えば、ほとんどすべてのスフィンガン生産能を持つスフィンゴモナス属菌から得られた、スフィンガン多糖類を増産できるDNA配列を容易に決定することができる。
【図面の簡単な説明】
【0022】
【図1】スフィンゴモナス菌株S88(ATCC寄託番号31554)の染色体DNAから単離した34kbpヌクレオチド単位のDNA配列を、制限酵素により消化した部位の概略図である。図1に示した多くのDNA配列をスフィンゴモナス菌に挿入して、これらのスフィンゴモナス菌がスフィンガンを増産する能力を検討した。つぎの数個の酵素の制限部位も、図1(さらに、図2、3、8、9及び10)に示す。B(BamHI)、Bg(Bg1II)、E(EcoRI)、H(HindIII)及びS(SalI)。図1に示すspsB領域は、タンパク質SpsBをコードするDNA配列に相当する。
【図2】スフィンゴモナス菌株S60(ATCC寄託番号31461)の染色体DNAから単離したDNA配列を(約28kbp単位)、制限酵素により消化した部位の概略図である。このDNA配列の制限部位をB、E及びH部位として図2に示す。sgeB領域は、タンパク質SgeBをコードするDNA配列に相当する。
【図3】スフィンゴモナス菌株NW11(ATCC寄託番号53272)の染色体DNAから単離したDNA配列(約33kbp単位)の制限酵素部位の概略図である。この配列の制限部位をE、H、B、Bg及びS部位として図3に示す。snwB領域は、タンパク質SnwBをコードするDNA配列に相当する。
【図4】本発明により生産する代表的な一スフィンガン多糖類の化学構造の概略図である。
【図5】本願の実施例6及び12で詳述するプラスミドpSEB24及びpSEB26の構成地図である。McsIは次の制限部位、即ちEcoRI、SmaI、BamHI、SAlI、PstI及びHindIIIを(上から時計廻りに)含んでいるマルチクローニング部位である。同様に、McsIIもHindIII、PstI、SalI、XbaI、BamHI、SmaI、SstI及びEcoRIを(上から時計廻りに)含んでいる。OriTは接合転移開始点で、OriVは広範囲宿主複製開始点、及びoriはpUC12と13から始まる複製開始点である。
【図6】スフィンゴモナス菌株S88(ATCC寄託番号31554)の染色体DNAから単離された34kbpヌクレオチド単位のDNAセグメントの制限酵素による切断部位の概略図である。次の数個の酵素の制限部位、即ちB(BamHI)、Bg(BglII)、E(EcoRI)、H(HindIII)及びS(SalI)が、図1に(さらには、図2及び図3でも)示されている。図6に(さらに、図1でも)示しているspsB領域は、タンパク質SpsBをコードするDNA配列に相当する。スフィンガン生合成に関与する他の「sps」遺伝子は、大文字G、S、R、Q、I、K、L、J、F、D、C及びEで示してある。rhsACBDはスフィンガン先駆物質dTDP−(L)ラムノースの合成に関与する遺伝子の地図上の位置を示す遺伝子の名称である。遺伝子32、26、31及び34は、まだ同定されていない翻訳の読み取り枠(ORF)であり、atrDB遺伝子は、スフィンガン合成に無関係の輸送機能をコードしている。「Sec」はスフィンガン類の分泌に必要な遺伝子を示し、「Trase」はスフィンガンの繰り返しサブユニットを組み立てるときに、ヌクレオチド−糖先駆物質からスフィンガンに糖を転移する酵素をコードしている遺伝子を示している。
【図7】スフィンゴモナス菌株S198(ATCC寄託番号31853)の染色体DNAから単離された(約42kbp単位の)DNAセグメントの制限酵素による切断部位の概略図である。図7では、このDNA配列の制限部位をH(HindIII)及びE(EcoRI)として表示してある。括弧内の間隔が詰って並んでいる部位の順序は、不明である。コスミドクローンc2、c3、c4、c5及びサブクローンL242の外側方向への広がりは、破線で示してある。四角で囲んでいる「B」領域は、S88遺伝子spsBの変異体を相補するタンパク質をコードしているDNA配列に相当する。
【図8】スフィンゴモナス菌株S7(ATCC寄託番号21423)の染色体DNAから単離された(約42kbp単位の)DNAセグメントの制限酵素による切断部位の概略図である。図8では、このDNA配列の制限部位をH(HindIII)、E(EcoRI)及びB(BamHI)として表示してある。コスミドクローンc1、c2、c3、及びc6の外側方向への広がりは、破線で示してある。クローンc2とc3は、図8の右側の領域を超えて伸びている。同様に、クローンc6は左側方向に伸びている。四角で囲んでいる「B」領域は、S88遺伝子spsBの変異体を相補するタンパク質をコードしているDNA配列に相当する。
【図9】スフィンガンS−88合成に用いられる遺伝子クラスターの概略図である。地図の下部には遺伝子名、境界、予定している機能、及び制限酵素による切断部位(BはBamHI、EはEcoRI、HはHindIIIである)でのヌクレオチドの位置を示している。図の上部に向って、自然Sps- 突然変異の名称と位置、Sps+ またはSps- の相補結果、及びプラスミドとS88染色体における特異的な挿入変異によるSps表現型を示している。mini−Tn10kanの挿入があるpZ167、pZ168、pZ180、pZ202及びpZ206は、プラスミドpRK311にクローニングされたc2セグメントにあり、2種の欠失菌株△Tn493か△Tn495のいずれかに導入された。同様にして、この他のプラスミドに対するトランスポゾン挿入はすべてc3セグメントに行い、欠失菌株である△Tn358と365に導入した。mini−Tn10kan挿入位置の精度は、配列決定された制限部位に対して±50bp、グラフの精度は±100bpである。 なお、外国書面出願時では、この図は、図10と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の上部を示してある。
【図10】スフィンガンS−88合成に用いられる遺伝子クラスターの概略図である。地図の上部には遺伝子名、境界、推定される機能、及び制限酵素による切断部位(BはBamHI、EはEcoRI、HはHindIIIである)でのヌクレオチドの位置を示している。クローニングされたフラグメント(c1△3、c2、c3、c4、c5及びc6)、サブクローニングされたセグメント(使用した制限酵素とkbp単位のおおよその長さに応じて名前を付けた)、及びS88染色体に生じた欠失(各線はDNAの存在を示す)は遺伝子地図の下に示している。 なお、外国書面出願時では、この図は、図9と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の下部を示してある。
【図11】SpsB及びグリコシル−IPトランスフェラーゼの推定アミノ酸配列のアラインメントである。右側の数は各配列の最右側のアミノ酸残基数である。各配列とも、ガラクトシル−IPトランスフェラーゼはSpsB配列の真上に示してあり、グリコシル−IPトランスフェラーゼはSpsB配列の下に示している。次の遺伝子産物は右側に示している。ExoYn、リゾビウムの一種(Rhizobium sp.)NGR234(Gray,et al.,1990,J.Bacteriol.,172:193)。CpsD、S.アガラクチエ(S.agalactiae)(Rubents,et al.,1993,Mol.Microbiol.,8:843)。RfbP、S.エンテリカ LT2(Jiang,et al.,1991,Mol.Microbiol.,5:695)。GumD、X.カンペストリス B1459S−4L(Capage,etal.,1987,International Patent WO/05938)。Pss4、R.レグミノサラム(R.leguminosarum)bv.vicae菌株VF39(GenBank寄託番号M93042)。Pss2、R.レグミノサラム bv.phaseoli(Borthakur,et al.,1988,Mol.Gen.Genet.,213:155)。記号について。|はSpsB及び上下各々に示すガラクトシル−IPトランスフェラーゼまたはグリコシル−IPトランスフェラーゼの同一アミノ酸を示す。:は次の関連したアミノ酸、即ちIFVWML、ST、QNED及びHKRのグループに基づいた保存的なアミノ酸置換を示している。アンダーラインをしてある配列は、疎水性アミノ酸約20個からなる連続セグメントである。
【図12】S.エンテリカ(Jiang,et al.,1991,Mol.Microbiol.,5:695)、及びX.カンペストリス(Koeplin,etal.,1993,J.Bacteriol.,175:7786−7792)のrhsA遺伝子産物と、dTDP−L−ラムノース生合成酵素のアラインメントを示す。記号は図11と同一である。 なお、外国書面出願時では、この図は、図13から図15と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の一部を示してある。
【図13】S.エンテリカ(Jiang,et al.,1991,Mol.Microbiol.,5:695)、及びX.カンペストリス(Koeplin,etal.,1993,J.Bacteriol.,175:7786−7792)のrhsB遺伝子産物と、dTDP−L−ラムノース生合成酵素のアラインメントを示す。記号は図11と同一である。 なお、外国書面出願時では、この図は、図12及び図14から図15と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の一部を示してある。
【図14】S.エンテリカ(Jiang,et al.,1991,Mol.Microbiol.,5:695)、及びX.カンペストリス(Koeplin,etal.,1993,J.Bacteriol.,175:7786−7792)のrhsC遺伝子産物と、dTDP−L−ラムノース生合成酵素のアラインメントを示す。記号は図11と同一である。 なお、外国書面出願時では、この図は、図12から図13及び図15と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の一部を示してある。
【図15】S.エンテリカ(Jiang,et al.,1991,Mol.Microbiol.,5:695)、及びX.カンペストリス(Koeplin,etal.,1993,J.Bacteriol.,175:7786−7792)のrhsD遺伝子産物と、dTDP−L−ラムノース生合成酵素のアラインメントを示す。記号は図11と同一である。 なお、外国書面出願時では、この図は、図12から図14と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の一部を示してある。
【発明を実施するための形態】
【0023】
次の用語は本発明の明細書全体を通して使用されるもので、その意味を次のように定義する。
【0024】
1. 「スフィンガン」なる語は、明細書全体を通して、スフィンゴモナス属細菌により分泌される、関連性はあるがお互いに別個であるエクソポリサッカライド群を意味する(Pollock, J. Gen. Microbiology,139:1939〜1945, 1993)。スフィンガン類の構造は、すべていくらかの関連性がある。スフィンガンおのおのの主鎖は、D−グルコース、D−グルクロン酸、L−マンノース、L−ラムノースの四つの糖の関連性のある配列からなっている。スフィンガン群の多糖類は、ポリマー主鎖と複数の側鎖を含む構成糖の配列によってお互いに区別することができる。スフィンガン多糖類はまた、その構成糖にアセチル基又はピルビル基が結合していてる場合もある(Mikolajczak, et al.,Appl. and Env. Microbiol.,60:402, 1994参照)。本発明のDNAセグメントとフラグメント及び一般方法を使用して生産した各種スフィンガンの化学構造の概略図は、おおむね図4に示してある。スフィンガン類の、ジェラン(S−60)、ウェラン(S−130)、ラムザン(S−194)、S−88、NW−11、S−198及びS−657の構造も、おおむね図4に示している。
【0025】
典型的には、スフィンガン多糖類群の各単体は、一般的に次の繰り返し化学構造によって表される。
【0026】
【化5】
【0027】
式中、Glcはグルコース、GlcAはグルクロン酸、Rhaはラムノース、Manはマンノースである。XはRhaでも、Manのいずれでもよい。Zは二番のGlc残基に結合している、α−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaのいずれかである。Wは1番のGlc残基に結合し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaのいずれかであって、下付き文字v及びyは0、0. 33、0. 5、0. 67又は1である。式中、ポリマーの還元末端は、主鎖のX残基の近くに位置している。本明細書で用いる「主鎖」なる語は、この構造からW鎖とZ鎖とを除いた、すなわちvとyが0に等しいときの残部を指す。ポリマーの「還元末端」は、糖単位が加えられるポリマーの末端である。
【0028】
スフィンガン多糖類群のうち、一部の単体はいろいろな位置でアセチル化されている。しかしながら、多糖類は常法により化学的に脱アシル化して、アシル基を除くことができる。例えば、ジェランにはウェランと同じ炭水化物の主鎖(すなわち、X=Rha)があるが、側鎖の糖が欠けており(すなわち、v=0及びy=0)、しかもグルコース残基1はグリセリン酸塩で完全に置換されている。ジェランのサブユニットの構造も、位置は分からないがどこかでアシル化されている。
【0029】
2. 「スフィンゴモナス」なる語は、明細書全体を通して、上記のエクソポリサッカライド類、すなわちスフィンガン類を生産するスフィンゴモナス属グラム陰性菌を意味する。本発明においては、スフィンゴモナス属グラム陰性菌の多くは、単離されたDNA配列源として、スフィンガン生産能を持つ他の菌株(好ましくは、スフィンゴモナス属のグラム陰性菌)に再挿入して、本発明のスフィンガン大量生産菌を生産するため、あるいは標的細菌として、これに外因性DNA配列を挿入してスフィンガン大量生産菌を生産するために用いることができる。
【0030】
グラム陰性菌のスフィンガン生産能を持つ科は、1993年にまずスフィンゴモナス属に分類されるものとして同定された。(Pollock, J. Gen.Microb. , 139, 1939,1993を参照)。しかしながら、各菌株をどの種に分類すべきかは、まだ正確には確立されていない。スフィンガン生産能を持つスフィンゴモナス菌株に最も近い種は、スフィンゴモナス ポーシモビリス(Sphingomonas paucimobilis)と考えられる。しかしながら、詳細な、決定的分類学分析結果を利用できる時まで、これらの菌株が上記の種に分類されるべきであると言うのは時期尚早である。スフィンゴモナス属のスフィンガン生産菌は当初数個の異なる属に分類されていたことを付言する。
【0031】
スフィンゴモナスのうち現在までに確認されている種としては、S. ポーシモビリス(S. paucimobilis)、S. パラポーシモビリス(S. parapaucimobilis)、S. アドヘイシバ(S. adhaesiva)、S. カプスラタ(S. capsulata)及びS. ヤノイクヤエ( S. yanoikuyae)が挙げられる(Yabuuchi, et al.,Microbiol. Immunol.,34, 99,1990を参照)。過去ではこれらスフィンゴモナスの種が、誤ってシュードモナス属(genus Pseudomonas)に分類されたこともあった。
【0032】
3. 「供与菌」及び「受容菌」なる語はおのおの、DNA配列を採取した細菌及びDNA配列を挿入又は組み入れる細菌を指すのに用いられる。
【0033】
4. 「菌株」又は「スフィンゴモナス菌株」なる語は、(化学構造に基づいて)特定のスフィンガンエクソポリサッカライドを生産するスフィンゴモナス属グラム陰性菌を指すのに用いる。スフィンガン生産能を持つスフィンゴモナス菌株を、簡単に、この菌株が生産するスフィンガン多糖類で呼ぶこととする。例えば、スフィンゴモナス菌株S88はスフィンガン多糖類S−88を生産し、スフィンゴモナス菌株S60はスフィンガン多糖類S−60(ジェラン)を生産するなどである。スフィンゴモナス菌株S88(ATCC No. 31554)、S60(ATCC No. 31461)、NW11(ATCC No. 53272)、S130(ATCC No. 31555)、S194(ATCC No. 31691)、S198(ATCC No. 31853)、S657(ATCC No. 53159)、及びS7(ATCC No. 21423)は、特に本発明に有用な代表的菌株である。
【0034】
5. 「大量生産菌」なる語は、明細書全体を通して、操作された細菌であって、同じ菌株又は異なる菌株のスフィンガン生産能を持つ細菌から単離したDNAセグメント又はフラグメントの多数のコピーを含有し、同じ菌株の操作されていない細菌、又は野生型細菌を同一又は実質的に同一の発酵条件で発酵して比較するとき、スフィンガン多糖類生産量が有意により大きい(重量/重量比で、少なくとも約5%以上)細菌を指すのに用いる。
【0035】
6. 「単離された」なる語は、微生物から採取して、少なくともある程度精製した、すなわち一以上の精製工程を通過したDNAを指すのに用いる。単離されたDNAは、実質的に純粋な形で用意しておくことが好ましい。すなわち、制限酵素によりフラグメントあるいはセグメントに切断したり、クローニングにより多数のコピーとしたり、又はプラスミドベクターその他の手段に挿入したり、又は細菌中に組み入れたりするDNAの性能に影響がない程度まで、汚染物質含有量を抑制する。
【0036】
7. 「DNA」又は「染色体DNA」なる語は、明細書全体を通して、スフィンゴモナスから単離されたDNAに関し、一般には微生物から単離する前に、スフィンゴモナス属菌の染色体又は内因性プラスミド中に見いだされるDNAを指すのに用いる。
【0037】
8. 「配列」なる語は、明細書全体を通して、ヌクレオチド単位又は制限酵素による切断部位のパターンによって同定されるDNAの特異的セグメントであって、一般的にはスフィンガン生産能を持つスフィンゴモナス属菌のDNAから制限酵素により単離し、このようにして得たDNA配列を細菌に挿入して大量生産菌を生産したり、あるいはさらに制限して前記の配列より小さいDNAの小部分、又はフラグメントを生産する配列を指すのに用いる。「部分」又は「フラグメント」なる語は、一般にDNAセグメントより小さいDNA配列を指すのに用いる。本発明に用いる好適なDNAセグメントは、グリコシルトランスフェラーゼ(グルコシル、ガラクトシル、ラムノシル及びグルクロノシルトランスフェラーゼ)、グリコシル−IPトランスフェラーゼ(グルコシル−IPトランスフェラーゼ、ガラクトシル−IPトランスフェラーゼ酵素などを含む)、ラムノースオペロン(あるラムノース含有スフィンガンへ挿入するためのラムノース前駆体の合成)、及び細菌の多糖類の分泌を含む様々なタンパク質をコードするセグメントである。
【0038】
9. 「挿入された」、「挿入する」、「組み入れられた」又は「組み入れる」なる語は、明細書全体を通して、スフィンガン生産能を持つスフィンゴモナス菌株の染色体DNA又はプラスミドDNAから単離したDNAセグメントを、スフィンガン生産能を持つ、同じあるいは異なる受容菌になるスフィンゴモナス菌株に転移する工程と結果を指す。結果として、転移されたDNAセグメントの少なくとも実質部分の多数のコピーを含有する大量生産菌株を得る。
【0039】
例を挙げると、まず単離されたDNAを当業者に周知の技術によりプラスミドベクター、例えば特にpRK311やpSEB24に導入し、クローニングした後、接合により受容菌となるスフィンゴモナス菌に転移する。受容菌となるスフィンゴモナス菌に挿入後、受容菌の細胞中で上記のDNAフラグメントを含むプラスミドベクターが複製されて、スフィンガン多糖類の大量生産に必要なDNAセグメントが数コピー(少なくとも2コピーで、通常は4〜20コピー)できる。プラスミドベクターの他に、バクテリオファージベクターや、トランスポゾンベクターを使ってもよい。
【0040】
多くのプラスミドベクターが、単離されたDNAセグメントやフラグメントを受容菌に挿入する際に好適である。上記で説明したpRK311やpSEB24の他に、特に、以下のプラスミドも有用である。不和合群P−1の広範囲宿主プラスミドであるRK2などや、そのプラスミドから派生したpRK290、pRK293、pRK404など(Ditta, et al.,Plasmid,Vol. 13, pp. 149〜153)、及びプラスミドの転移を可能にするプラスミドRP4のoriT遺伝子を含むその他の派生体であるpSUP101など(Simon, et al.,Bio/technology, November, 1983を参照)並びにプラスミドpLAFR1及びpLAFR3(Friedman, et al.,Gene, 18, 289, 1982)、及び不和合群Inc−Qの広範囲宿主プラスミドであるRSF1010や、そのプラスミドから派生したpMMB22及びpMMB66(Fuerste, et al.,Gene,48, 119, 1986)などである。
【0041】
プラスミドベクターを受容菌内に挿入する際に、接合を用いるのは一般に有効である。しかしながら、属が異なる細菌同志では、コンピテント細胞を精製DNAで形質転換させるのがより一般的である。
【0042】
スフィンゴモナスの場合、細菌にDNAフラグメントやプラスミドを導入するのに電気穿孔法も使われてきた(1992, Monteiro, et al.,J. of App. Bacteriol.,72, 423を参照)。この方法を使えば、単に単離されたDNAを細菌に添加し、電気穿孔で細胞膜を通して転移させるだけで、単離されたDNAセグメントやフラグメントの細胞コピーを2コピー以上受容菌となるスフィンゴモナス菌に組み入れることができる。
【0043】
前掲のMonteiroらは、DNAをスフィンゴモナスに導入する手段としての電気穿孔法を説明している。電気穿孔法の機能は、コンピテント細胞を化学的に処理して、例えば細胞を塩化カルシウムやルビジウム塩で処理した後に形質転換するのと同じである。形質転換させるDNAは標準法で精製するが、この場合プラスミドの形をとってもよいし、またとらなくてもよい。しかしながら、通常、二本鎖閉環型のDNAの場合、形質転換が最も効率的である。したがって、この場合、接合法でDNAをスフィンゴモナスに導入する必要もなければ、セグメントをクローニングしてからプラスミド、バクテリオファージあるいはトランスポゾンベクターに挿入する必要もない。しかしながら、単離されたDNAをまずプラスミドベクターに導入してから、単離DNAフラグメントを含有するプラスミドを細菌中に転移することが好ましい。
【0044】
受容菌となるスフィンゴモナス菌中で、プラスミドあるいはその他ベクター、例えばバクテリオファージ又はトランスポゾンベクターにDNAセグメントの担持を続ける必要はない。細菌DNA中にDNAセグメントのコピーをさらに導入して、細菌DNAを複製するのと同じメカニズムによりセグメントを世代毎に複製することは、常法として行われている。下記の実施例の部において、細菌DNAにDNAのコピーをさらに導入して、細菌DNAを複製するのと同じメカニズムによりセグメントを世代毎に複製する方法を詳述する二つの実施例を説明する。
【0045】
10. 「多数のコピー」なる語は、明細書全体を通して、少なくとも2コピー、好ましくは少なくとも4コピーをスフィンゴモナス細菌に組み入れる外因性DNA配列、フラグメント又はセグメント(前記のDNAの少なくとも実質部分)を指すのに用いられる。より好ましくは、スフィンゴモナス属菌に挿入するDNA配列、フラグメント又はセグメントのコピー数は、実際には約4から約20までの範囲である。ある例においては、DNA配列を1個のプラスミドベクターに組み入れて、接合によりスフィンゴモナス菌に転移し、受容菌中でプラスミドを複製して、2コピー以上のDNA配列、セグメント又はフラグメントを得ることもあることを付記する。
【0046】
11. 「生合成」なる語は、明細書全体を通して、スフィンゴモナス菌がスフィンガンを生物生産又は合成することを指すのに用いる。スフィンガン多糖類は、複数の細菌の酵素でコントロールされている一連の工程により炭水化物単位毎に合成される。
【0047】
12. 「操作されている」なる語は、明細書全体を通して、外因性DNA、好ましくはその多数のコピーが組み入れられている受容菌となるスフィンゴモナス菌を指すのに用いる。本発明の操作されている細菌とは、スフィンガン多糖類大量生産菌のことである。
【0048】
13. 「遺伝情報をコードする」なる語は、明細書全体を通して、特定の順序で並んでいるヌクレオチド単位の遺伝情報を内容とするDNA配列を指すのに用いる。DNA配列の遺伝情報(どんな長さでも)は、多数のコピーが組み入れられて、これにより得た操作されている細菌がスフィンガンを増産する場合、スフィンゴモナス菌によるスフィンガン合成に「有益又は不可欠」であると見なされる。「有益又は不可欠」なる語は、DNAがスフィンゴモナス細菌から単離され、遺伝情報をコードし、この遺伝情報の多数のコピーが組み入れられたスフィンゴモナス菌が、スフィンガン多糖類の大量生産菌に形質転換するDNAを指すのに用いる。本発明で使用する有益又は不可欠なDNAは、例えば、グルコシルIP−トランスフェラーゼ、ガラクトシルIP−トランスフェラーゼなど、グリコシルトランスフェラーゼの生合成のための遺伝子又はオペロン、及び、ラムノース、マンノース、グルコース、ガラクトースのような糖のシントンや、例えばdTDP−L−ラムノースなど、上記の糖の置換シントンの生合成のための遺伝子又はオペロン、及び、例えばポリメラーゼのようなスフィンガンを生産するための糖シントンの重合に関与する酵素の生合成のための遺伝子又はオペロン、及び、無処理の細胞構造から多糖類を分泌するための遺伝子又はオペロンを一以上含む。
【0049】
14. 「菌株間相補性」なる語は、第一のスフィンゴモナス菌株からDNA配列、セグメント又はフラグメントを単離して、異なる第二のスフィンゴモナス菌株に組み入れることを指すのに用いる。本発明の実施態様の一つとして、異なるスフィンゴモナス菌株からDNAフラグメントを取り、その多数のコピーを別のスフィンゴモナス菌株に組み入れると、スフィンガン多糖類の大量生産菌が生産できることを発見したことは、予期せざるところであった。本発明で有用なDNAフラグメントは、属間相補性(例えば、キサントモナス カンペストリス菌にキサンタンの増産をさせるなど)も示している。
【0050】
15. 「シントン」なる語は、本発明により細菌がスフィンガンを生合成する間に重合される糖又は糖単位を指すのに用いる。シントンは、スフィンガン多糖類の構成部分又は構成単位を含み、スフィンガン類の生合成に使用される糖成分、例えばアセチル化及びアシル化糖や関連先駆物質を含むグルコース、ガラクトース、ラムノース、マンノース、その他の糖シントン類などを含有する。
【0051】
16.「ラムノースオペロン」なる語は、本発明によるスフィンガン多糖類の生合成に利用される、ラムノース又はラムノースシントン類(dTDP−L−ラムノースなど)の生合成に関与する遺伝子やオペロンをコードするDNA配列を指すのに用いる。
【0052】
本発明は、スフィンガン生産能を持つ供与菌となるスフィンゴモナス菌からDNA配列を取って、多数のコピーを同じ菌株あるいは異なる菌株の受容菌となるスフィンゴモナス菌に組み入れると、この受容菌となるスフィンゴモナス菌がスフィンガン多糖類大量生産菌に形質転換するとの発見に関する。さらに、ある種のスフィンガン多糖類を生産する細菌からDNA配列を単離し、異なる菌株のスフィンゴモナス菌に多数のコピーを組み入れると、この異なる菌株から大量生産菌を生産することができるばかりでなく、この場合供与菌の特性を持つスフィンガン多糖類の混入が起こることがないことも発見されている。
【0053】
受容菌に組み入れられたDNA配列には、スフィンガン多糖類の生合成に有益又は不可欠の遺伝情報がコードされている。例えば、有益又は不可欠の遺伝情報は、細菌のスフィンガン生合成にいろいろな形で役割を果たしたり、あるいは関与する。外因性DNAは、例えば、律速酵素の過程に関与する酵素その他のタンパクの合成を発現したり、酵素、補因子、その他の生化学的成分の合成を誘導して多糖類を増産させたり、多糖類のサブユニットの連結を促進するポリメラーゼのような酵素の産生を増加させたり、あるいは一以上のリプレッサー遺伝子と結合したり、細菌から多糖類の分泌を促進させたり、通常、多糖類生合成の間は律速される工程の生産を阻害するリプレッサーの発現を押さえるなど、スフィンガン生合成に有益な効果をもたらすことができる。
【0054】
DNA配列は、当業者の標準的な技術や方法によりスフィンゴモナスの各種菌株から単離する。通常、細菌は(下記で詳細説明するように、約0. 5%より低い、好ましくは約0. 1%〜0. 2%のグルコース濃度を用いる標準発酵法により)培養され、高濃度の細菌を含有するブロスを作る。その後、細菌細胞を遠心分離し、DNA抽出のために再懸濁する。DNAは、まず混合液からタンパクを除いて、エタノール又はイソプロパノールにより高分子量のDNAを沈澱させることによって細菌から抽出する(Birnboim and Doly, Nucl. Acids Res.,7, 1513,1979を参照)。
【0055】
上記のように沈澱させたら、通常は単離されたDNAセグメントあるいはフラグメントのクローニングを行って、受容菌となるスフィンゴモナス菌挿入用のDNAを形成する。例として挙げると、上記した高分子量のDNA配列は、制限酵素(例えば、SalI酵素)により部分的に消化し、標準方法により電気泳動する(Loftus, et al.,BioTechniques, 12, 172,1992を参照)。電気泳動後、大きなDNAフラグメント(20kbp以上)をさらに精製する(抽出及び沈澱)。
【0056】
その後、細菌から単離したDNAフラグメントを、クローニングベクター(一般には、プラスミド)に直接挿入してDNAのクローニングを行うか、代わりに制限酵素によりさらに処理することにより、より小さなDNAフラグメントにして、クローニングベクターに挿入する。本発明のDNAクローニングは、当業者が標準としている一般的な技術と方法によって行う。本発明によれば多くの方法を用いてDNAセグメントのクローニングを行うことができ、したがって、本発明は、例えばプラスミドのクローニングベクターの使用に限定されない。例えば、DNAフラグメントをcharon 4A、EMBL3(Rodriguezand Denhardt, Vectors, Chapter 2, pg. 43, 1988, Butterworth Publishers, Boston)、又はP1(1990, Sternberg, Proc. Natl. Acad.Sci.,U.S.A.,87, 103〜107)などのバクテリオファージベクターに挿入してクローニングすることもできる。
【0057】
下記の実施例1及び12で詳述するように、まず、クローニングベクターに挿入するDNAフラグメントを用意する。本発明によれば、DNAセグメント又はフラグメント形成用として、数多くのクローニングベクターを使用することができる。しかしながら、本発明においては、DNAセグメント又はフラグメントのクローンを形成したプラスミドベクターをそのまま使用して、接合により外因性DNAを受容菌に挿入するのが有利であることが見いだされている。しかしながら、特にDNAを受容菌へ挿入する際に形質転換法を使用しようとする場合には、受容菌に接合するベクターとは異なる(プラスミドその他の)クローニングベクターを使用することもできる。
【0058】
クローニングベクターに挿入を終えたら、単離されたDNAを含有するベクターをバクテリオファージ中でパッケージングして、トランスフェクション法により細菌(一般に、大腸菌)に転移し、トランスフェクションされた細菌中で複製する。このようにして得たDNAクローンを含有する細菌細胞のコロニーをプールして、貯蔵するか、あるいは直接使用する。
【0059】
その後、DNAクローンのスクリーニングを行って、スフィンゴモナスによるスフィンガン生産に対するDNAフラグメントの相対的増産効果を決定する。スクリーニング法においては、DNAを適当なベクターに担持させた後、接合法(例えば、Ditta, et al.,Proc. Natl. Acad. Sci.,USA, 77, 7347,1980が説明している三元交配(tri−parental mating))により受容菌となるスフィンゴモナス菌株に挿入し、このようにして得たDNAの多数のコピーを含有する操作された細菌とそのスフィンガン生産を試験して、活性を決定する。
【0060】
スフィンガン生産に増産効果があると決定されたDNAセグメント又はフラグメントを、受容菌となるスフィンゴモナス菌株に転移して、前述のように転移されたDNAの少なくとも実質部分を少なくとも2コピー含有する大量生産菌株を生産する。
【0061】
本発明で使用する際に好ましいスクリーニング法を開発した。この方法では、DNAを生産能を持たない受容菌となるスフィンゴモナス菌株に挿入することにより、このDNA中にスフィンガン合成に有益又は不可欠である遺伝子が存在しているか、否かをスクリーニングするのである。すなわち、スフィンガン生産能を持つスフィンゴモナス菌株から、生産能を持たない変異菌(例えば、菌株S88のSps- Bacr )を派生させ、スクリーニング対象となるDNAの多数のコピーを含有するように操作する。グルコース1〜3%を含有する普通寒天平板培地でこのように操作した、生産能を持たない変異菌を増殖する。別に、生産能を持たないスフィンゴモナス変異菌を同一条件で増殖して、この変異菌と操作された細菌のコロニーの外観を比較する。このようにした後、一般的にスフィンゴモナス菌にスフィンガン合成を開始させるDNAの能力を、目視により測定する。
【0062】
スフィンガン生産活性を向上させるDNAセグメント又はフラグメントの能力は、培養皿上で容易に認識できる、スフィンガン生産能を持つ細菌とスフィンガン生産能を持たない変異菌の表現型の相違により判定される。例えば、スフィンガン生産能を持つスフィンゴモナス菌株はムコイドを産生するため、簡単な目視検査で容易に見分けることできるコロニーを形成することが多い(例えば、スフィンガン生産菌のコロニーが盛り上がって丸く、明るい輪で取り囲まれているのに対して、非生産菌のコロニーは平坦で、粗く、半透明である)。
【0063】
ある事例では、下記の実施例2で詳述するように、一つのスフィンゴモナス菌株において、スフィンガン生産能を持つ細菌とスフィンガン生産能を持たない細菌の表現型が容易にあるいは短時間で区別できない場合がある。このような場合には、生産性と非生産性の表現型の相違が目視検査で容易に識別できる細菌によって対象DNAの活性を判定するように、スクリーニング法を変更する。この本発明の実施態様では、本発明に有用なDNAフラグメントは菌株間でも、属間でも相補性を示す事実、及びこのDNAフラグメントの多数のコピーによって実際上すべてのスフィンゴモナス菌株がスフィンガンを増産することができる事実を利用するのである。
【0064】
本発明に有用なDNAセグメント又はフラグメントは、キサントモナス カンペストリスにおいても活性を示す。したがって、一以上のスフィンゴモナス菌株を使って容易にスクリーニングできなかったDNAフラグメントの多数のコピーを、キサンタン生産能を持たない、X. カンペストリス変異菌、例えば特にX59m31に組み入れて、目視検査によりキサンタン生産のスクリーニングを行うことができる。X59m31などの生産能を持たないX. カンペストリス変異菌は、バシトラシンにさらした後の生存菌を選び、このバシトラシン耐性変異菌にDNAフラグメントの重複コピーを組み入れ、YM寒天培養皿で培養し、形成されたコロニーの外観がムコイド性(生産菌)であるか、非ムコイド性(非生産菌)であるかを観察することにより、容易に得ることができる。(Pollock, et al.,1994 J. Bacteriol.,176, pp. 6229〜6237、及び米国特許第5, 338, 841号を参照)。スクリーニングされた細菌が多糖類(スフィンガン又はキサンタン)を増産するDNAは、菌株間あるいは属間相補性の証拠であって、異なるスフィンゴモナス菌株ばかりでなく、異なる属の細菌(キサントモナス)によるスフィンガン多糖類生産を増産させることができる。
【0065】
概して簡単に、スフィンガンの生産菌と非生産菌の表現型の相違を同定できる本発明の実施態様による簡便スクリーニング法を用いれば、通常の技術を有する当業者は容易に利用できるクローニング及び転移技術により、過度の又は理由のない実験を行うことなく、本発明で使用されるDNAセグメントやフラグメントを容易に得て、スフィンゴモナス菌によるスフィンガン多糖類を増産することができる。
【0066】
本発明の他の実施態様は、スフィンガン多糖類の増産に関する。スフィンガン多糖類を生産するためには、本発明の操作した細菌を周知の方法により適当な発酵条件下で培養する。操作されたスフィンゴモナス菌を培養する適当な培地又は発酵ブロスとしては、例えば、一般にグルコース、ラクトース、スクロース、マルトースやマルトデキストリンを含む炭水化物類などの炭素源、例えば無機アンモニウム、無機硝酸塩、有機アミノ酸類又は加水分解した酵母,大豆粉又はカゼインなどのタンパク性材料、蒸留業者によって提供される可溶性物質(distiller’s solubles)又はコーン スチープ リカー(cornsteep liquor)、無機塩類などの窒素源及びビタミン類を含有する水性培地が挙げられる。本発明では、非常に多種類の発酵培地を使ってスフィンガン類の生産を実施することができる。
【0067】
発酵ブロス中の炭水化物含量はいろいろあるが、通常は発酵培地に対して約1〜5重量%である。炭水化物類は発酵前あるいは発酵中のいずれかで全部を一どきに添加する。窒素用量は水性培地に対して約0. 01%〜約0. 4重量%の範囲である。炭素源と窒素源は1種のみの炭素源又は1種のみの窒素源を使用しても良いし、何種類かの炭素源又は窒素源を混合して使用してもよい。
【0068】
スフィンゴモナス菌の発酵に用いる無機塩類としては、ナトリウム、カリウム、アンモニウム、硝酸塩、カルシウム、リン酸塩、硫酸塩、塩化物、炭酸塩及びこれらに類似するイオン類を含有する塩類が挙げられる。マグネシウム、マンガン、コバルト、鉄、亜鉛、銅、モリブデン、ヨウ化物及びホウ酸塩などの微量金属も、含有すると有利である。また、ビオチン、葉酸塩、リポ酸塩、ナイアシンアミド、パントテン酸塩、ピリドキシン、リボフラビン、チアミン及びビタミンB12などのビタミン類、ならびにこれらの混合物もまた、有利に使用することができる。
【0069】
発酵温度は約25℃〜約35℃であるが、至適生産性を得るには約28℃〜約32℃の温度範囲がよい。接種源は、振盪フラスコ培養と小規模の浸漬攪伴発酵を含む、標準的な方法で容量をスケールアップして調製する。接種源の調製用培地は、生産用培地と同じものでも良いし、また当業者間で周知のいくつかの標準培地の一つであるルリア ブロス培地や、YM培地であってもよい。種培養では、炭水化物濃度を約1重量%より低くするとができる。所望の接種量を得るためには、種培養工程を一工程よりも多くすることができる。典型的な接種量は、最終発酵総量に対して約0. 5%〜約10%の範囲である。
【0070】
典型的には、発酵容器に内容物を攪伴する攪伴機を備える。容器に自動pH制御装置と自動発泡制御装置をつけてもよい。生産培地を容器に添加して、加熱によりその場で殺菌する。かわりに、添加前に、炭水化物や炭素源を別個に殺菌してもよい。予め増殖しておいた種培養を冷却してある培地(一般には、発酵温度の約28℃〜約32℃)に添加し、培養物を攪伴しながら約48〜約96時間発酵して、高粘度のブロスを調製する。標準法によりスフィンガン多糖類をアルコール、一般的にはイソプロパノールで沈澱させて、ブロスから回収する。
【0071】
本出願は、具体例として、数種の菌株、特にスフィンゴモナス菌株S88、S60、NW11、S198、S7、及びS194(それぞれ、寄託番号ATCC31554、ATCC31461、ATCC53272、ATCC31853、ATCC21423、ATCC31961としてアメリカン タイプ カルチャー コレクション(American Type Culture Collection)から利用することができる)から単離したDNAセグメントや、フラグメントを開示する。これらのDNAセグメントや、フラグメントを染色体外(プラスミド)DNAとして多数のコピーをスフィンゴモナス菌株中に組み入れることにより、これら菌株が生産するスフィンガンS−88、S−60及びNW−11を増産する際に有用であることが見いだされている。
【0072】
スフィンゴモナス菌株S88の場合、単離された染色体DNAセグメントの大きさは34kbpで、23から25の遺伝子を含む。図1のS88のクローン地図に示す通り、34kbp領域とは二つのクローン、c3とc2を合わせた広さである。この34kbpのDNA配列を複数のDNA配列に分けて、そのおのおのにcl△3、c2、c3、c4、c5、c6、H15. 6、B7. 1、B8.6、E5. 9、E1. 5、E2. 4、E4. 5、E6. 6、E12. 8など標識したものを、図1に示す。図6、図9及び図10は、単離されたS88染色体DNAをさらに詳細に示す。
【0073】
スフィンゴモナス菌株S60の場合、染色体DNAからc1、c2及びc3(図2を参照)を始めとする複数のDNA配列が単離された。配列c2をクローニングした後、pRK311ベクター中に置き、スフィンゴモナス菌株S88、S60及びNW11に挿入して、スフィンガン生産活性を評価した(実施例9に詳述してあるので、参照のこと)。
【0074】
スフィンゴモナス菌株S198、S7及びS194の場合、これらの菌株からもDNAセグメントを単離した(図7及び8を参照されたい)。これらの菌株からは、本願明細書で詳述する一般方法によりさらに多くのDNAセグメントを単離して、本発明により細菌のスフィンガン生産を増産するのに利用することができる。
【0075】
スフィンゴモナス菌株NW11の場合、DNAフラグメントcl、c2、c2. 1、c2. 2、c2Hd、c2H1、c2H2、c2H3、c2H10を単離した(図3参照)。配列c2. 2をクローニングした後、pRK311ベクター中に置いて、スフィンゴモナス菌株S88、S60及びNW11に挿入して、スフィンガン生産活性を評価した(実施例9に詳述してあるので、参照のこと)。
【0076】
スフィンゴモナス菌株S88、S60及びNW11から、上記で一般的に説明した方法により次のDNAセグメントを調製した。これらのDNAセグメントおのおのを(プラスミドベクター中の全長のDNAセグメントとして)、一以上のスフィンゴモナス菌株(野生型スフィンガン生産菌、又は野生型生産菌から派生した非ムコイド性変異菌)に挿入すると、それらの細菌はスフィンガン多糖類の大量生産菌に変わる。これらのDNAセグメントやフラグメントおのおのは、スフィンゴモナス菌株から単離されるDNAセグメント又はフラグメントから派生させ、前述したようにプラスミドベクター中に挿入する。DNAセグメント又はフラグメントの特徴は、制限酵素切断部位地図によって定まる(図1、2及び3)。
【0077】
菌株 S88
pRK311−S88cl△3
pRK311−S88c2
pRK311−S88c3
pRK311−S88c4
pRK311−S88c5
pSEB24−S88H15. 6
pSEB24−S88B8. 6
pSEB24−S88E4. 5
pSEB24−S88E6. 6
菌株 S60
pRK311−S60c2
菌株 NW11
pRK311−NW11c2. 2
【0078】
DNAクローンは、スフィンガン生産能を持つ野生型スフィンゴモナス菌、あるいはスフィンガン生産能を持たない変異菌に導入して、大量生産効果を実現することができる。例えば、DNAクローンの多数のコピーを、スフィンガン生産能を持つ野生型菌株や、スフィンガン生産能を持つ菌株から派生した非ムコイド性変異菌おのおのに導入する。このように操作した細菌は、同じ菌株由来のスフィンガン生産能を持つ野生型菌、あるいはスフィンガン生産能のない変異菌と比較すると、スフィンガンの大量生産菌である。
【0079】
DNAをスクリーニングして多数のコピーを、これらのスフィンゴモナス菌の菌株に導入したところ、予期しないところであったが、このようにして得た組換え菌のスフィンガン生産量は、一般的に野生型菌のスフィンガン生産水準よりも高かった。これは一般性を持つ現象であり、しかもこのスフィンガンの増産には菌株間の相補性が認められた。
【0080】
本発明においては、DNAクローンの多数のコピーを挿入して得る組換え菌は、野生型より高い水準のスフィンガン多糖類合成活性を持っている。また、本発明で有用なDNAセグメントは、菌株がスフィンガンを組み立てたり、生合成する初期工程で初めのグルコース残基をイソプレニルリン酸担体に結合するのに必要なタンパクをコードしているDNAフラグメントを始めとして、スフィンゴモナス菌株がスフィンガンを合成するのに有益あるいは不可欠な遺伝子を持っている。
【0081】
spsB遺伝子のDNA配列(配列番号1)と、SpsBタンパクの推定アミノ酸配列(配列番号2)も開示する。SpsBタンパク(又は、スフィンゴモナス菌株によっては、特にSgeB、SnwB、SneB、SssB、及びSrhBなどの相似タンパク)をコードしているDNAを含有するDNAフラグメントはすべて、多数のコピーを組み入れてスフィンゴモナス菌株を操作された菌とし、この操作された菌によってスフィンガンを増産することができる。
【0082】
同様に、S60スフィンゴモナスから単離されるsgeB遺伝子(SpsBに相似するSgeBタンパクをコードする)及びNW11スフィンゴモナスから単離されるsnwB遺伝子を含有するDNAセグメントやフラグメントについても同じことが言える。sgeB遺伝子(図2)は、S88染色体DNAのspsB遺伝子(及び、相当するsneB、sssB、srhB遺伝子、それらはそれぞれスフィンゴモナスのS198、S7及びS194株に由来し、SneB、SssB、SrhBタンパクをコードする)と(S88のspsB遺伝子に相当するフラグメントとのDNAハイブリッド形成に基づいて)相似するタンパクをコードしていると信じられている点で、お互いに相似している。図3に示すsnwB領域は、SnwBタンパクをコードするDNA配列に相当する。また、snwB遺伝子は、S88染色体DNAのspsB遺伝子及びS60染色体DNAのsgeB遺伝子と相似する。これらのDNAフラグメントは、上記で一般的に述べたようにプラスミド中に挿入して、スフィンゴモナスがスフィンガンを増産するための多数のコピーを形成する。
【0083】
スフィンゴモナスS88のspsB遺伝子はグルコシル−IPトランスフェラーゼをコードしていると信じられている。SpsBタンパクの推定アミノ酸配列は、異なる属の菌の推定グリコシル−IPトランスフェラーゼと、かなりの相同性があることが立証されている。この推定アミノ酸配列は一般にグリコシル−IPトランスフェラーゼをコードしていると信じられている他の遺伝子の配列と似ているが、この類似性がspsB遺伝子はグルコシル−IPトランスフェラーゼをコードするということの最も強い証拠となっている。実際、グルコシル−IPトランスフェラーゼのカルボキシル部分と、ガラクトシル−IPトランスフェラーゼのカルボキシル部分とには、かなりの相同性がある。アミノ末端領域にはこのように大きな相同性はないが、SpsBタンパクには膜貫通領域であると示唆されている疎水性の広がりが複数存在している点で、S. エンテリカ(S. enterica)のRfbPタンパクと似ている(1991, Jiang, etal.,Mol. Microbiol.,5, 695)。SpsBの疎水性領域は、35〜39の位置のアミノ酸(平均ハイドロパシー+2. 2)、68〜86の位置のアミノ酸(+1. 7)、105〜123の位置のアミノ酸(+2. 3)及び282〜303の位置のアミノ酸(+2. 9)を含んでいる。後者の疎水性領域の位置はこれらの関連遺伝子産物に共通していて、最大相同領域の隣に存在している。
【0084】
本発明の最良の実施態様において、DNAセグメントやフラグメントには、S88、S60、NW11、S130、S194、S198、S657及びS7を始めとするスフィンゴモナス菌の各種菌株の、グリコシル−IPトランスフェラーゼをコードするDNA配列が含有されている。これらのDNAセグメント及びフラグメントの多数のコピーを、受容菌となるスフィンゴモナス菌中で使用して、この受容菌がスフィンガンをより多く生産できる利得を実現することができる。グリコシルトランスフェラーゼ酵素をコードする遺伝子又はDNAフラグメントの組み込みに加えて、糖シントン又は糖前駆体(すなわち、スフィンガンポリサッカライドの構成部分を含み、スフィンガンの生合成に使われる糖成分、例えばグルコース、ガラクトース、ラムノース、マンノース、他の糖シントンや前駆体)をコードする遺伝子又はDNAフラグメント、又は、重合を促進したり、無処理の細胞構造からの多糖類分泌を促進する酵素や蛋白質をコードする遺伝子又はDNAフラグメントを好適に本発明で使うことができる。
【0085】
次の実施例により本発明を説明する。如何なる方法によっても実施例の説明を本発明の範囲を限定するものと誤解してはならない。
【実施例】
【0086】
次の実施例1〜21では、細菌の菌株、プラスミド及びバクテリオファージを表2に列記する。ルリア−ベルタニ培地及びYM培地を標準培地とする(Pollock,et al.,1994,J.Bacteriol.,176:6229−6237)。抗生物質(Sigma社)の用量は次の通りである。バシトラシン(Bac)は、特に本例では、73単位/mg又は0.01〜8mg/ml。リファンピシン(Rif)は、50μg/ml。ストレプトマイシン(Stm)は、50μg/ml。カナマイシン(Kan)は、25μg/ml。クロラムフェニコール(Cam)は、15μg/ml。テトラサイクリン(Tet)は、4〜12μg/ml。
【0087】
【表2】
【0088】
実施例1
スフィンゴモナスのDNAセグメントライブラリーの構成
スフィンゴモナス菌株から、スフィンガン類の合成に不可欠なDNAセグメントのクローンを形成した。次のようにして、DNAセグメントの完全なライブラリーを調製した。YM液体培地25ml中に菌株(この実施例ではS88)を置き、30℃で一晩振盪して、多量の細胞を含有する粘液性ブロスを作成した。
YM培地には、水1リットルあたり、バクト(Bacto)酵母エキス3g、バクト麦芽エキス3g、バクトペプトン(Difco)5g、及びD−グルコース( Difco)10gが含有されていた。アジ化ナトリウムを0. 01%まで加え、さらにスフィンガナーゼ(sphinganase)酵素(1994, Mikolajczak,et al.,Appl. Environ. Microbiol.,60,402)を37℃で8時間添加し、スフィンガンエクソポリサッカライドを消化して、粘度を部分的に低下させて細胞の増量を図った。細胞を遠心分離し、Birnboim and Doly,Nucl. Acids Res.,7,1513(1979)の方法によりDNAを抽出するために再懸濁した。ライセート(分離物)に等量のフェノール:CHC13:イソアミルアルコール(24:24:1)混液を加え、25℃で16時間緩やかに振とうし、その後、1容量のCHC13:イソアミルアルコール(24:1)混液を加え、25℃で3時間緩やかに振とうした。このようにして澄明としたライセートからタンパクを除いた。3Mの酢酸ナトリウム(pH5. 2)1/10容量を加え、高分子量のDNAを2容量のエタノールで沈澱させた後、乾燥して、さらにTE(10mMのトリス・HCl(pH8)、1mMのEDTA)0. 5ml中に再懸濁した。
【0089】
Loftus, Foster and Ross(BioTechniques, 12,172,1992)のコスミドクローニング計略により、S88DNAをSalI酵素により部分的に消化し、トリス・酢酸塩・EDTA緩衝液中、1%の低融点アガロースで電気泳動した。20kbpより大きなフラグメントをフェノール抽出とエタノール沈澱により精製した。このSalIで消化したS88DNAは、クレノウDNAポリメラーゼで処理して、dCMPとdTMPを付着端に添加し、70℃で20分間加熱した後、エタノールで沈澱させた。ベクタープラスミドpRK311(1985, Ditta, et al.,Plasmid,13, 149〜153)は、BamHI酵素で完全に消化し、65℃で15分間加熱して、フェノール抽出とエタノール沈澱により精製した。このようにしてBamHIで消化したpRK311DNAは、クレノウDNAポリメラーゼで処理して、dGMPとdAMPを添加して、上記のようにして精製した。T4DNAリガーゼによるライゲーション反応に含まれていたベクターと挿入フラグメントのモル量は等しかった。すべての制限酵素、クレノウDNAポリメラーゼ、及びT4DNAリガーゼはストラタジーン(Stratagene)社から入手し、このメーカーの反応条件を使用した。バクテリオファージ(ストラタジーン社のGigapackTMIIXL)中にパッケージングした後、ライゲーションした分子をトランスフェクションにより大腸菌DH5αTMに転移し、細胞を濃度4〜12μg/mlのテトラサイクリンを含有するLBプレート上に広げた。テトラサイクリン耐性コロニー1700個のうちの1つのライブラリー、及びテトラサイクリン耐性コロニー3400個のうちの1つのライブラリーのそれぞれを別個にプールして、凍結した。テトラサイクリン耐性コロニーは(試験した10個のうち10個が)内部にSalI制限酵素部位をもつ、長さ25〜30kbpの挿入断片を含有していた。
【0090】
同様にして、NW11、S60、S198、S194、及びS7を含む他の各種スフィンゴモナス菌株から、染色体DNAセグメントのライブラリーを作成した。この場合、クローニングしたDNAの供給源である細胞を、0. 5%w/v以下のグルコースを含む培地中で増殖させ、スフィンガナーゼ処理は省いた。
【0091】
実施例2
スフィンガンS−88生合成用DNAフラグメントの単離
プラスミド中にクローニングしたDNAのフラグメントのスクリーニングを行い、スフィンガン陰性変異菌がスフィンガン合成を復活するか、否かを観察することにより、スフィンガンS−88の合成に不可欠な遺伝子が存在するか、否かを判定した。かねて本発明者らは、バシトラシン500〜800μg/mlを含有するYM皿上で増殖できるスフィンゴモナスS88菌株の自然バシトラシン耐性変異菌のほとんどが、スフィンガン多糖類を生産できないことを見いだしていた(Pollock, et al.,1994, J. Bacteriol.,176, pp. 6229〜6237)。この所見に基づいてこの特別クラスの変異菌の簡便スクリーニング法を完成した。
【0092】
変異菌S88m260はこのSps- Bacr 群の代表的な変異菌である。S88m260その他のSps- Bacr 変異菌がエクソポリサッカライドを生産できなかった際のコロニーの外観は、野生型コロニーと比較して、平坦で、表面が粗く半透明であった。また、野生型のコロニーは光にかざして下から見ると狭い、光を屈折するハローで取り囲まれていた。これらの表現型の違いは、X. カンペストリスにおいて、野生型におびただしいムコイドがあるのに対して、対応するGum- 変異菌は平坦な外観を持っている違いほど明白ではなかった。コロニーの表現型は、培養物を液体YM培地で増殖し、エクソポリサッカライドをイソプロピルアルコールで沈澱し、乾燥計量して確認した。いくつかのSps- 変異菌はバシトラシンに感受性を持っていて、後にスフィンガン合成に不可欠であり、しかもバシトラシン耐性表現型に関する遺伝子とは区別されるべき遺伝子の特徴を備えていることが分かった。
【0093】
遺伝子ライブラリーからプラスミドDNAを取り、三元交配法により大腸菌からキサントモナスや、スフィンゴモナスに転移した(Ditta, et al.,Proc. Natl. Acad. Sci. USA,77, 7347, 1980)。Mob+ Tra- 組換えプラスミド(S88挿入断片を持つpRK311)を含有する供与菌の細胞、Mob+ Tra+ pRK2013プラスミドを含有するヘルパー細胞及びエクソポリサッカライド陰性の受容菌の細胞を比率5:2:10で混合した混液を、グルコースを入れない非選択的YMプレート中にスポットし、30℃で6〜16時間インキュベートした。次に、プレート中に、ヘルパー細胞と供与菌の細胞を排除するためのリファンピシン(50μg/ml)を入れ、かつ、組換えプラスミドを選択するために、pRK311用としてテトラサイクリン(4〜12μg/ml)、又はpSEB24用としてクラムフェニコール(20μg/ml)を入れた。これらのプレート上に上記の交配混液1白金耳を展開して、プラスミドを受容したキサントモナスの株を単離した。スフィンゴモナスは元来ストレプトマイシンに耐性を持っているので、スフィンゴモナスが受容菌のとき、供与菌及びヘルパー細胞に対してストレプトマイシンがスフィンゴモナスの選択に使われる。
【0094】
このようにして得たS88菌株のプラスミドを受容した株が、Sps+ (盛り上がって丸く不透明で、光にかざして下から見ると、明るい輪で取り囲まれている)あるいはSps- (平たく、粗く、半透明のコロニーで、輪がない)の、どちらであるか裸眼で判断して、その相補性を評価した。同様に、プラスミドを受容したキサントモナス菌から生育したGum+ のコロニーは、Gum- の受容菌の艶がなく、光らないコロニーと比べて、外観がムコイド性である。
【0095】
ライブラリー中のS88のクローンをS88の非ムコイド性変異菌中で直接同定しようとしたが、(ムコイドの表現型を明確に立証できなかったため)成功しなかった。そこでライブラリーを、一旦X. カンペストリスのgumD遺伝子の非ムコイド性変異体に転移してから、クローンを見いだす方法に切り替えた。これにより初期クローン「S88cl」を見いだすことができたが、これについては下記で詳しく説明する。
【0096】
S88遺伝子ライブラリーを大腸菌から、gumD遺伝子中にBacr Gum- が欠損しているX. カンペストリス菌株X59m31中に交配した(Pollock, et al.,J. Bacteriol.,176, pp. 6229〜6237, 1994; Thorneet al.,J. Bacteriol.,169,3593, 1987)。この属間交配では、いくつかのGum+ Tetr コロニーがYMプレート上で認められたが、このコロニーは約10-3〜10-4の頻度で出現すると考えられた。相補された変異体から、プラスミドをおのおの精製し、再び大腸菌に転移して制限酵素解析を行った。精製プラスミドをスフィンゴモナスS88m260に交配したところ、約10%の接合完了体(transconjugants)がSps+ となった。一つのプラスミド(pRK311−S88cl)を回収し、これを使用してさらに実験を続けた。プラスミドpRK311−S88clは、さらにいくつかのスフィンゴモナス菌株S88から別個に単離したBacr Sps- 変異菌、及びX. カンペストリスのBacr Gum- 変異菌とも相補した。属間交配ごとのプラスミドを受容した株が培地に分泌するエクソポリサッカライドを単離し、次に、薄層クロマトグラフィーを行い、酸水解物が受容菌の細胞の多糖類に存在している筈の糖残基を含有しているとの結果を得て、これらが多糖類であることを確認した。プラスミドpRK311−S88clは、スフィンゴモナスではスフィンガン合成を、X. カンペストリスではキサンタンガム合成をそれぞれ復活させた。これらの結果は、プラスミドpRK311−S88clは、Bacr 多糖類陰性変異菌が失っていたエクソポリサッカライド生合成機能をコードしていたこと、及びある属の遺伝子が他の属の失なわれていた機能を回復できたことを示している。セグメントS88clは、図1に示されるS88cl△3の最も右の末端に7. 5kbpのHindIIIフラグメントが付加されている点を除けば、S88cl△3と同一である。S88clからこの7. 5kbpセグメントを特異的に欠失させて、その派生物であるS88cl△3を産生した。
【0097】
上記で説明した、ガム生産(スフィンゴモナス属菌のスフィンガンガム、又はX. カンペストリスのキサンタンガム)の復活に有用なDNAフラグメントの測定方法には再現性があり、別個の実験では新しいクローンが単離された。スフィンゴモナスS88のSps- Bacs 変異菌76及び78を相補するセグメントを、クローンライブラリーから直接スクリーニングする際、S88clと部分的に重なり合っている三つの新しいクローンが形成された。これらのセグメントクローン3個は、おのおの長さが約23〜27kbであった。3個のセグメントのうち、二つは変異菌S88m260を相補した。制限酵素切断部位地図を図1及び図6に示す。
【0098】
上記の方法は、これらの菌株おのおののスフィンガンを増産するために有用な、スフィンゴモナス菌株S60、NW11、S198、S7、S194及びスフィンガン生産能を持つその他のスフィンガン菌株から単離したDNAセグメントの決定に用いられる。
【0099】
実施例3
spsB遺伝子のDNA配列とSpsBタンパクの推定アミノ酸配列
プラスミドpSEB24−S88E4. 5::Tn#72からサブクローニングした3300bpのフラグメントのうち、spsB領域の1950bpの二本鎖ヌクレオチド配列を得た。コード鎖の配列を配列表(配列番号1)に示す。1つの長い読み取り枠(open reading frame、ORF)があり、これをspsBと名付けた。コード領域はヌクレオチド361の位置のATGに始まり、1771の位置のTGA終止コドンまで続いていた。このORFはアミノ酸470個をコードし、ORFの先には推定リボソーム結合部位があった。標準三文字略記法による推定アミノ酸配列を配列表(配列番号2)に示す。
【0100】
実施例4
S88セグメントのクローンと、S88及びS60の染色体DNAとのDNA−DNAハイブリッド形成
プラスミドpRK311−S88cl△3のDNAクローンは、隣接するS88DNA配列から派生したものであることを立証するため、プラスミドS88cl△3DNAに標識を付け、スフィンゴモナス菌株S88、変異菌S88m260、及び野生型ジェラン生産菌であるS60から分離したDNA制限フラグメントとハイブリッド形成をおこなった。約1. 5、2. 4、4. 5、5. 9及び12kbpのEcoRIフラグメント中にハイブリッドを形成すると、野生型DNA及び変異菌S88DNA両方のDNAクローンの連続体と一致する。EcoRI部位の一つはベクターのマルチクローニング部位から始まっているので、重なり合っているクローンであるS88c2、S88c3、及びS88c4をEcoRIで消化すると、S88cl△3であることが証明されている最も左の6. 6kbpフラグメントは、実際には12. 8kbpとなる。S88DNAとジェランを生産するS60DNAとでハイブリッドを形成できることから、似ている遺伝子配列が存在しているが、遺伝子の組成が異なっている可能性があることが示唆された。これら二つのスフィンゴモナス菌株が分泌するエクソポリサッカライドの構造が似ているので、類似したトランスフェラーゼ遺伝子を持っていると考えられる。S88−S60の相同領域を図2に示す。別個のDNA相同性解析で、図3で示すように菌株NW11とS88の間の相同領域の位置を確定した。
【0101】
実施例5
菌株S60、NW11、S198、S7及びS194のスフィンガン生合成遺伝子クラスターのクローニング
スフィンゴモナスS60、NW11、S198、S7及びS194からDNAフラグメントを単離した。方法としては、菌株S88に関する上記実施例と同様にした。菌株S60、NW11、S198、S7及びS194のDNAフラグメントの制限部位地図は、それぞれ図2、図3、図9及び図10に示す。1種又は多種の制限酵素によるDNAの消化によって生じた制限酵素フラグメントの大きさを、2つの独立に単離されたクローンであるpRK311−S194c1及びpRK311−S194c2についてここで挙げる。この大きさは、そのフラグメントのアガロースゲル電気泳動の移動度を、大きさが既知のフラグメント、つまり、HindIIIで消化後のバクテリオファージラムダDNA、及びストラタジーンの「KbDNALadder」と比較して決定した。フラグメントの大きさは、上記の2つのクローンに含まれるDNA配列と一致する。
【0102】
スフィンゴモナスS194から単離したc1セグメントを含有するプラスミドpRK311−S194c1のフラグメントの大きさのデータを次に示す。pRK311−S194c1のフラメントの大きさは次の通りである。(EcoRI)>25、8.3、5.4及び1.5kbp。(EcoRI+HindIII)13.0、9.7、8.3、5.4、2.8及び1.5kbp。(HindIII)>25及び2.8kbp。(BamHI+HindIII)>25、5.8、3.9、2.0及び0.8kbp。(BamHI)>25、5.8及び4.7kbp。(BamHI+EcoRI+HindIII)13.0、8.3、5.8、5.4、3.9、2.0、1.5及び0.8kbp。(BamHI+EcoRI)15.0、8.3、5.8、5.4、4.7及び1.5kbp。pRK311ーS194c2のフラグメントサイズは次の通りである。(EcoRI)14.5、10.0、8.3、6.1、2.6及び1.35kbp。(EcoRI+HindIII)13、8.3、5.7、4.6、4.0、2.6、1.8、1.35、0.75、0.35.及び0.25kbp。(HindIII)>20、12.8、4.6、2.1、0.75及び0.25kbp。(BamHI+HindIII)>20、3.8、3.1、2.8、2.3、1.65、1.6、1.3、1.2、0.95、0.9、0.8及び0.25kbp。(BamHI)>20、5.8、3.1、2.8、2.6、2.3、2.0、1.6及び0.25kbp。(BamHI+EcoRI+HindIII)13、8.3、3.8、3.1、2.4、2.3、1.6、1.35、1.3、0.95、0.9、0.85、0.8、0.45、0.35及び0.3kbp。(BamHI+EcoRI)14.5、8.3、4.9、3.1、2.4、2.3、2.0、1.6、1.35、1.3、0.85、0.45及び0.2kbp。
【0103】
実施例6
プラスミドpSEB26の構成
特異的な複製機能と接合交配機能、並びに、スフィンゴモナス属に好適で、かつ、mini−Tn10kanと和合性がある薬剤耐性遺伝子を含有し、マルチクローニング部位を有する、プラスミドpSEB24とpSEB26(図5)を組み立てた。プラスミドpSEB24が広範囲宿主域を持つ反面、pSEB26は大腸菌では複製できるが、スフィンゴモナス属やキサントモナス属のどちらでも複製することができない。まず、スタフィロコッカス・アウレウスのプラスミドpC194(1982,Horinouchi and Weisblum,J.Bacteriol.,150,815)から取られた1031bpのHpaII−Sau3Aフラグメント上のCamr を、平滑末端に加工して、プラスミドpUC13(1982,Vieira and Messing,Gene,259)のXbaI平滑末端にライゲーションした。BamHI−SalIセグメント上のこのプラスミドからCamr カセットを取り除き、平滑末端にした後、pUC12中の独特なSspI部位と、二つあるPvuII部位のうちの最も近い部位(これも平滑末端)との間に挿入した。このようして、Ampr かつCamr であり、X−GalとIPTGとを添加すると青いコロニーを形成するプラスミドpSEB23を得て、pSEB24を構成するため、pSEB23から約2130bpのScaI−PvuIIフラグメントをとり、pMMB66EH(1986,Fuerste,et al.,Gene,48,119)のScaI−PvuII部分とライゲーションさせ、oriV(RSF1010からの広範囲宿主複製開始点)を保持させ、Ampr 遺伝子を再生した。プラスミドpNH−Kan/oriT(Hengen and Iyer,1992,Bio Techniques,13:57−62)のoriT配列を含む2700bpのHindIII−BamHIフラグメントは、3200bpのPvuIIで直鎖状にしたpSEB23プラスミドと、平滑末端同志で結合してpSEB26を形成した。BamHI−PvuIIのライゲーションにより再生したBamHI部位は、制限酵素処理に引き続き、付着末端にキャッピングを施し、再びライゲーションすることにより取り除かれた。
【0104】
実施例7
S88DNAフラグメントのコピーを菌株S88に導入したことによる多糖類S−88の増産
菌株S88から単離した図1に示すDNAセグメントの特異的な制限フラグメントを、DNAライゲーションによりマルチコピープラスミドベクター中に挿入した後、上記の実施例2で説明した三元交配により野生型菌株S88とS88の子である非ムコイド性変異菌に転移した。DNAクローンを変異菌に転移すると、多糖類合成が復活する。細菌を液体培地で培養した後、組換えプラスミド含有菌株とプラスミド遺伝子を付加しなかった菌株が生産したスフィンガン多糖類の累積量を測定した。じゃま板付きフラスコ(baffled flasks)中で振盪しながら30℃で24時間増殖した後、2容量のイソプロピルアルコールを加えて、エクソポリサッカライドを沈澱させた。各菌株ごとに2〜3回づつ独立に培養して、実験を行った。沈澱物をフィルターに集め、80℃で乾燥して、計量した。菌株毎の沈澱物の平均重量と標準偏差値を表3に示す。遺伝子のクローンのコピーを付加した組換え菌株のスフィンガンS−88生産量は、通常セットの生合成遺伝子しか持たなかった野生型菌株のそれよりも大きかった。
【0105】
【表3】
【0106】
制限酵素地図、spsB遺伝子のヌクレオチド配列及びこれを取り巻くDNAは、標準組換えDNA法によって約32kbの領域の新しいサブフラグメントを無数に形成するに充分な情報であることが、当業者にとり明白である(図6)。さらに、これらの新しいフラグメントを本明細書で説明した方法により試験して、スフィンガン多糖類を同様に増産するセグメントを同定できることも、当業者にとり明白である。表3から、このサブフラグメントが小断片であると(例えば、pSEB24−S88E12. 8)充分な生産刺激効果を示さないことが読み取れる。しかしながら、ほぼ全てのサブフラグメントは、増産を可能にしている。今日までの実験により、本発明者らは、pSEB24−S88B8. 6とpRK311−S88c3がスフィンガン生産に対して最大の刺激効果があると信じている。
【0107】
この増産法では、継続的に組換えプラスミドを選択するために、培養物に抗生物質を存在させておく必要はない。
【0108】
この増産法は、一以上のDNAフラグメントを染色体又は内因性の本来菌が持っていたプラスミドに挿入して、好ましくはこの染色体セグメントの単数又は複数の遺伝子をコードするDNAフラグメントを挿入することによって達成される。通常の技倆を持つ当業者であれば、内在の(resident)細菌染色体、内因性の本来菌が持っていたプラスミド、あるいは外因性プラスミドベクターに、スフィンガン生産能を持つ遺伝子を含有するDNAフラグメントを挿入することにより、スフィンガン生産能を持つ細菌にDNAフラグメントを導入することは容易である。本出願で立証されている試験結果では、このスフィンガン増産には、特定のプラスミドベクターの使用に対して依存性が認められないことは注目に値する。
【0109】
本発明の範囲には、上記で説明した遺伝子セグメント又は遺伝子群をコードするDNAフラグメントのどれかと、遺伝子の発現をコントロールすることで知られているDNA配列、本発明を限定することなく例を挙げると、大腸菌のlacプロモーターとを、融合させることも含まれている。
【0110】
実施例8
組換えS88菌株が生産したエクソポリサッカライドがスフィンガンS−88であることの同定
操作された菌株によって生産されたエクソポリサッカライドが受容菌の型と同一であることを確認するため、薄層クロマトグラフィーによって酸水解物中のモノサッカライド類を同定した。
【0111】
X. カンペストリスの細胞外キサンタンとスフィンゴモナスのスフィンガンS−88を、2〜3容量のイソプロピルアルコールにより沈澱させて、液体培地から分離し、80℃で乾燥して計量した。多糖類を、高性能液体クロマトグラフィー(HPLC)の水に5mg/mlの濃度で再懸濁した。0. 6ml超小型遠心分離管中で、無水トリフルオロ酢酸(88μl、Sigma ChemicalCo. から入手)を、75μlのHPLC用の水(Baker)及び225μlの多糖類(5mg/ml)と混合して、95℃で16時間インキュベートした。加水分解物を真空乾燥し、HPLCの水200μlに再懸濁して、新しい超小型遠心分離管に入れ、再び乾燥した後、HPLCの水45μlに懸濁した。サンプル(25μg/ml)は凍結して貯蔵した。糖標準液(D−グルコース、D−グルクロン酸、D−マンノース、L−マンノース、L−ラムノース及びL−フコース)を4mg/mlの濃度でHPLCの水に再懸濁した。コート済みで、チャンネルを形成した(channelled)シリカゲルクロマトグラフィープレート(Kieselgel 60 CF254、1020cm、E. Merck)を、一晩0. 3MのNaH2 PO4 中に浸漬した後、室温で30分間、さらに95℃で10分間乾燥した。サンプル1〜2μlをスポットし、室温で2. 5〜3時間アセトン40ml、ブタノール5ml及び脱イオン水5mlからなる溶媒混液にクロマトグラムをさらして、混液を上昇させた。プレートは65℃で3分間乾燥し、アセトン25ml、アニリン0. 5ml、ジフェニルアミン0. 5g及びリン酸3. 75mlからなる溶液に浸して染色した後、95℃で30分間乾燥した。X. カンペストリスがS88DNAフラグメントの受容菌である場合は、グルコース、マンノース及びグルクロン酸が存在した。スフィンゴモナス菌株S88がX. カンペストリスのgumD遺伝子の受容菌である場合は、ラムノース、グルコース、マンノース及びグルクロン酸がS−88エクソポリサッカライドとほぼ等量で存在した。
【0112】
実施例9
スフィンガンS−88、S−60、NW−11及びS−130の生産に対する、菌株S88、S60及びNW11から得たDNAフラグメントの刺激効果
上記の実施例5、6及び7で述べた一般法に従って菌株S88、S60及びNW11から得たDNAフラグメントを使用することにより、スフィンゴモナス菌株がスフィンガンS−88、ジェラン(S60)及びNW11を増産した。結果を次の表4に示す。
【0113】
【表4】
【0114】
一般法としては上記の実施例、特に実施例7で説明した技術や方法を用いて単離されたDNAフラグメントの多数のコピーを挿入することにより、特定のスフィンゴモナス菌株から単離されたDNAフラグメントを使って、異なるスフィンゴモナス菌株に(初めの菌株が生産するものとは異なる)スフィンガンを増産させようとするものである。
【0115】
本実験で確立された一般法においては、いずれかのスフィンゴモナス菌株のスフィンガン生産に不可欠な遺伝子材料を含有するDNAフラグメントを、適当なプラスミドベクターその他の手段に挿入する。このようにDNAを挿入したプラスミドベクターの多数のコピーを、DNAを単離したスフィゴモナスと同じ菌株あるいは異なる菌株のスフィンガン生産能を持つ菌又は非ムコイド性変異菌に導入する。この場合DNAが導入されたスフィンゴモナス菌株は、スフィンガン生産菌又はスフィンガン生産能を持つ菌のスフィンガン生産能を持たない変異菌のいずれでもよい。同一の発酵条件では、このように操作したスフィンゴモナス菌のスフィンガン生産量は一般に、操作しない、変異体ではないスフィンガン生産菌の生産量より大きい。
【0116】
次の実験により、DNAフラグメントの多数のコピーをスフィンゴモナス菌株その他の細菌に組み入れることは常法であることを証明する。
【0117】
実施例10
X. カンペストリスの染色体へのラクトース消費遺伝子(lactose−utilization gene)の挿入
制限酵素とDNAライゲーションを用いる標準クローニング法により、X. カンペストリス中で複製できないプラスミドに担持させた、X. カンペストリスの予めクローニングしたDNAセグメントに隣接して、トランスポゾンTn951から得られたラクトース消費遺伝子を挿入した(Thorne, et al.,J. Indust. Microbiol.,3, 321,1988)。その後、この組換えプラスミドを接合によりX. カンペストリス中に転移した。受容菌内では、ラクトース消費遺伝子の隣に位置するX. カンペストリスから得られた相同DNAが、通常の相同組換えによって細胞DNAと組換えられ、ラクトース消費遺伝子が細菌染色体と隣合わせに結合した。この結果、クローニングされたセグメントが細菌染色体に安定して挿入されることになった。このことは、上記の論文中で図を用いてより完全に説明されている。本発明以外でも、この方法によって遺伝子を細菌に挿入している例が数多く存在している。
【0118】
上記で説明したDNA組換えでは、事前に、プラスミドをベクターとして使用し、外因性DNAを細菌中に導入する必要がないことは注目に値する。DNAセグメントをバクテリオファージあるいはトランスポゾンに担持させると、部位特異的DNA組換えによって自然に細菌染色体に挿入されることが広く知られている。通常、バクテリオファージは特異的に1ないし2〜3個の位置で挿入するに過ぎないが、トランスポゾンは通常、多くの、実質的に任意の位置で挿入することができる。また、プラスミドなどのクローニングベクターと結合したDNAを細胞内に輸送する必要はない。DNAフラグメントは形質転換によって、細胞DNAと組換える能力を保持しながら細菌細胞内に入ることができることがよく知られている。
【0119】
実施例11
任意に選択された数個の異なる位置において、カナマイシン耐性をコードする遺伝子を、スフィンゴモナスS88菌DNAへ部位特異性挿入する
この実施例においては、クローニングされたDNAのセグメントをスフィンゴモナスの細胞DNAに挿入することは、容易に実施できるものであることを証明する。まず、S88E12. 8フラグメント(図1参照)を、プラスミドベクターpSEB26のマルチクローニング部位内のEcoRI部位にライゲーションした(図7)。プラスミドpSEB26は、図5で示すpSEB24のように、アンピシリン耐性遺伝子、クロラムフェニコール耐性遺伝子、マルチクローニング部位、Lacセグメント及びoriTを持っている(図5参照)。しかしながら、プラスミドpSEB24が広範囲宿主域のoriV配列を持つが、プラスミドpSEB26は狭い宿主域の複製開始点を持つということで、プラスミドpBR322とは異なる。pBR322の複製開始点は、大腸菌内におけるプラスミドの複製はできるが、スフィンゴモナス内では複製できない。したがって、プラスミドpSEB26内にクローニングされたDNA配列がスフィンゴモナス内で存続する唯一の方法は、プラスミドpSEB26が細胞あるいは培養物から消失する前に、プラスミドpSEB26を染色体や、内因性プラスミドなどの細菌DNAに組み込むことである。次に、このようにして得たpSEB26−S88E12. 8プラスミドを、常法によりトランスポゾンmini−Tn10kan(1991, Kleckner, et al.,204, pp. 139〜180)との突然変異生成にさらした。非抑制的な宿主HMS174における転位による突然変異誘発は、ラムダバクテリオファージNK1316によって担持されるTn10派生物103(mini−Tn10kan/Ptac−ATStransposase)による。その結果、カナマイシン耐性遺伝子がプラスミドに挿入された。また、S88E12. 8セグメント内のそれぞれ異なる位置にあるカナマイシン耐性遺伝子の数個の別個の挿入断片を単離した。次に、このようにして得た組換えプラスミドおのおのを、三元交配法によりスフィンゴモナスS88内に別個に転移して、カナマイシン耐性の子孫を選択した。事実上すべての場合、カナマイシン耐性遺伝子の両側に位置するS88DNA配列が受容細胞の染色体と組換えられ、カナマイシン耐性遺伝子が細菌染色体に組み込まれた。細菌染色体に組み込まれることにより、ベクタープラスミドがスフィンゴモナス内で複製することができないにも拘らず、カナマイシン遺伝子は存続することができた。
【0120】
この実施例は、外因性DNA配列(この場合、カナマイシン耐性遺伝子)を受容菌に導入して、導入遺伝子を細菌染色体に組み込むことができることを示している。組み込みの位置は、その遺伝子に隣接するDNA配列によって決定される。この実施例では、外因性遺伝子は菌株S88のE12. 8セグメント内の、相異なる部位に組み込まれた。相同組換えによる組み込みは、日常的に実施されている遺伝子操作法である。
【0121】
実施例12
DNAの配列決定の追加実験とその解析
図6に示すようにDNAの両鎖について、BamHI部位の1bpから28.804bpまでの間の配列を決定した。サンガー(Sanger)のジデオキシヌクレオチド鎖停止法(Sanger,et al.,1977,Proc.Nat.Acad.Sci.,74:5463−5467)が、エクソヌクレアーゼIIIとSIヌクレアーゼを用いてpBluescriptIIKS(+)上にクローニングされたDNAの配列決定に用いられた。内部シーケンシングプライマーも使用した。さらに、CoralSoftware社(San Diego)のSuperClone及びSuperSeeプログラム、並びにKyteand Doolittleの方法(Kyte and Doolittle,J.Mol.Biol.,157:105−132)により、膜貫通タンパク質領域の配列を解析した。また、「blastp」プログラム(Altschul,et al.,1990,J.Mol.Biol.,215:403−410)により、NCBIの膨大なデーターライブラリーから相同タンパク質セグメントを同定した。ナイロン膜とGeniusTM1キット(BoehringerMannheim)を用いる標準法により、DNAハイブリダイゼーションを行った。
【0122】
実施例13
スフィンガンS−88生合成に関与する遺伝子のクローニング
本実施例は一部を実施例2と同様にして行った。本発明者らは、かねてから、スフィンゴモナス菌株S88のスフィンガン多糖陰性(Sps- )変異株のほとんどは、バシトラシンを含有するYMプレートでも増殖できることを見いだしていた(Pollock,et al.,1994,J.Bacteriol.,176:6229−6237)。マッピング実験(後述)の結果、これらの変異株をすべてspsBと名付ける遺伝子に分類した。図9及び図10に列記した代表的なSpsB- 変異株(260、265及び102w)は、バシトラシン耐性(Bacr )でもあった。反対に、bac8菌株は、始めBacr として選択された代表的な菌株であったが、同時にSps- でもあることが示された。実施例2で述べたように、Sps- Bacr 変異株でスフィンガンS−88を形成しようとする試みは失敗に終り、代わりに野生型コロニーよりも平坦で、表面が粗く、半透明の外観を持つコロニーが得られ、いずれのコロニーも下から光にかざして見ると、光を屈折する狭いハローによって囲まれていた。アルコールに沈澱するエクソポリサッカライド類が出現しなかったので、Sps- 変異株はスフィンガン類を液体YM培地中に分泌できないものと判断した。Sps- Bacr 変異株の小さいフラクションは、第二の突然変異を持っていた。例えば、変異株102wのコロニーは黄色ではなく白色であった。変異株134ではspsBばかりでなく、rhsDでも変異があった。変異株54と302では、spsBのみでなくspsKもが欠損していた。図9の遺伝子地図の真上に示しているSps-変異には、自然発生のものと、紫外線又はエチルメタンスルホン酸にさらして変異させたものとがある。本実施例で実験したこの他の突然変異は、すべて(接頭字「Y」又は「B」を付けて、図9、10に示している)、トランポゾンmini−Tn101kanの無作意挿入により形成したものである。
【0123】
広範囲宿主コスミドpRK311に、スフィンゴモナス菌株S88から遺伝子ライブラリーを構築し、プールしておいたクローンを接合交配により大腸菌からS88m260に転移した。しかしながら、Sps+ のコロニーは、スクリーニングした103 〜104 のTetr プラスミドを受容した株に、またも、生じなかった。そこで、ライブラリーを予め単離してあったX.カンペストリスのBacr Gum- (gumD)変異株(Pollock,et al.,1994,J.Bacteriol.,176:6229−6237、及び、Thorne,et al.,1987,J.Bacteriol.,169:3593−3600)に転移した。gumD遺伝子は、キサンタンガムを組み立てる第一工程で、グルコース−6−リン酸をUDP−グルコースからイソプレニル−リン酸に転移するために必要である(Capage,et al.,October 1987,International PatentWO87/05938、及び、Ielpi,et al.,1993,J.Bacteriol.,175:2490−2500)。X.カンペストリスのgumD遺伝子を含有するDNAセグメントによって、スフィンゴモナスのBacr Sps- 変異株がスフィンガンS−88合成を回復できたことから、スフィンガンS−88の組み立ても、グルコースで始まり、スフィンゴモナスからの酵素は、X.カンペストリスのBacr Gum- 変異株を相補できるかもしれないと考えた(Pollock,et al.,1994,J.Bacteriol.,176:6229−6237)。属間交配からは、YMプレート上のTetr プラスミドを受容した株103 〜104 につき約1の頻度で、X.カンペストリスのGum+ コロニーが認められた。
【0124】
大腸菌を用いた形質転換によって、相補されたX.カンペストリス変異株からプラスミドを精製した後、スフィンゴモナスS88変異株260に交配した。プラスミドを受容した株のうち約5〜25%がSps+ となった。ベクター単独(pRK311)の場合の方が、より大きな組替えプラスミドの場合よりも、転移の頻度が100倍ないし1000倍高かった。ほとんどの組替えプラスミドはスフィンゴモナスと交配させると、大きな欠失が発生するが、pRK311−S88c1だけは、欠失を起こすことなくポリマー生産能を回復し、これを使用して実験を継続した。図1、図6、図9及び図10ではS88c1の21kbpからなる最左側部分をサブクローンc1△3として示している。プラスミドpRK311−S88c1によって、各々別個に単離した菌株S88のBacr Sps-変異株及びX.カンペストリスのBacr Gum- 変異株の数種が多糖の合成を回復した。属間交配の度に、プラスミドを受容した株が培養基中に分泌する多糖類を単離して、薄層クロマトグラフィーを実施し、酸水解物が受容細胞の多糖として期待されている中性糖を含有していることを確認した。プラスミドpRK311−S88c1を持つスフィンゴモナス変異株260が分泌するエキソポリサッカライドにはグルコース、マンノース及びラムノースが含有されていたが、プラスミドpRK311−S88c1を持つX.カンペストリス変異株m31からのものにはグルコースとマンノースしか含有されていなかった。さらに、加水分解条件下のため酸性糖の回収量は一貫して低かったが、多糖は各々グルクロン酸を含有していた。
【0125】
ライブラリーをスクリーニングしてSps- Bacr 変異株76及び78を相補するセグメント求め、図9及び図10の左側方向にクローニングした領域を広げていった。プラスミドを受容した株104 〜106 をスクリーニングして、部分的にS88c1セグメントと重なり合うさらに4個のクローン(S88c2、c3、c4及びc5)を得た。同様にして、Sps- 変異株43、71及び104を相補することにより、クローンc6を単離した。これら5個のクローニングされたセグメントは、各々長さが22〜28kbである。各セグメントの少なくとも1末端を図10に示す。さらに、クローンc2とサブクローンc1△3のどちらにも相補されない約15個からなる1群のSps- 変異株を同定した。これらの変異株の中で、c1△3よりも約8kbpだけ長く右側に広がっているc1全長クローンにより相補されたものは一つもなかった。また図3のc2よりも約18kbpだけ長く左側に広がっているc6によって相補されたものも一つとしてなかった。このように「同一連鎖群から外れた」(「unlinked」)1群の変異株は、スフィンガン合成に不可欠であるが、しかし図9及び図10に示す遺伝子クラスターとは直接に連続しない、さらに他の遺伝子の存在を示唆している。
【0126】
実施例14
機能的相補によるsps遺伝子のマッピング
Sps- 点変異株を接合の受容菌として用いる相補性検定により、spsG、spsK、spsF、spsD、spsC、spsE、spsB及びrhsD遺伝子の境界を決定した。結果を要約して図9に示す。サブクローニングされた小さいセグメント又はそれより大きいDNAセグメントのを持つ組替えプラスミドを、大腸菌の中でmini−Tn10kanによる挿入突然変異にさらし、菌株S88のSps- 変異株との交配によって転移した。これには交配可能な広範囲宿主プラスミドベクターであるpRK311及びpSEB24(図5)を使用した。担持しているプラスミドの耐薬性遺伝マーカーを受け取ったプラスミドを受容した株は、コロニーの外観によって、Sps+ 又はSps- と記録した。Bacr Sps- S88変異株は始めE4.5サブクローンとして位置づけ、その後、E4.5セグメントをmini−Tn10kanの無作意挿入変異にさらした。位置B231及びB230(図9及び図10)に挿入した時には、変異株260がスフィンガン合成を回復するのに影響しなかったが、B233、B239及びB238に挿入した時には相補が阻止された。変異株134は、セグメントB8.6によって相補されたが、E4.5又はE5.9のいずれによっても相補されなかった。変異株134では、spsB遺伝子と、さらに隣接のrhsD遺伝子の両方が欠損していた。B8.6セグメントをmini−Tn10kanによる突然変異にさらし、B441、B440、B438、B437及びB435に挿入することによる相補性のパターンを分析して、rhsD突然変異のより正確な位置を測定した。変異株54及び302も、spsKとspsB遺伝子が欠損している二重変異株であると考えられた。spsF変異株(図9の62、68及び94)は局在し、かつ、セグメントB12.6及びc1△3によって相補されず、またクローンc3とc5によってスフィンガンS−88の合成が回復されるため、隣接のspsDCEと分けた。E6.6フラグメントの挿入突然変異に続いて、連続しているspsD、spsC及びspsE遺伝子が相補された結果も併せて図9及び図10に示す。この相補結果から変異株76と78を含むグループと、変異株69、72、b104、3、9及び41を含むグループの二つのグループがあることが示唆された。その後、DNA配列の分析によって、後者のグループは連続している二つの別個の遺伝子、spsCとspsEとに分けられた。spsG突然変異(図9の11、43、71、81及び104)は、c6クローンとフラグメントB12.6のB4.5サブフラグメントとによって相補された(図9及び図10を参照されたい)。B12.6セグメントの挿入突然変異によって、プラスミドY652、Y635、Y636、Y653、Y640及びY641が産生された。このうち、Y652とY641のみがspsG変異株を相補することができた。
【0127】
実施例15
mini−Tn10kanを染色体とプラスミドに挿入したことによる表現型
S88DNAクローンのセグメントを交配可能な狭域宿主Camr プラスミドpSEB26(図9及び図10)にライゲーションし、大腸菌内でmini−Tn10kanによる挿入突然変異にさらした後、接合によって野生型(Sps+)スフィンゴモナス菌株S88に転移した。これらのプラスミドはスフィンゴモナス属では複製できないため、Kanr 遺伝子を維持するためには、細菌の染色体との組み替えが必要である。Kanr 及びCams である組み替え体はプラスミド配列を保持しないため、この群のみを選択した。これらのDNA置換の物理的構造の確認は行っていないが、常に同一のプラスミド、菌株及び選択方法を使用することによって、部位特異性な染色体欠失を生じさせ、制限地図作製とDNAハイブリダイゼーションによってこれらの二重組替えを確認した。Kanr Cams 染色体の組み替え体のコロニーは、Sps+ 又はSps- と判定した(挿入体には接頭字「c」を付けて図9に示している)。Sps- 表現型を示す変異株(cY776、cY757、cY771、cY770、cY676、cB589、cB583、cB580、cB579、cB300、cY726、cY725、cY676、cY673、cY721及びcY602)については、二重組替えが実際に発生したと信じることが、合理的である。しかしながら、相同領域の一個所にて組換えの行われたSps+ 組み替え体は、プラスミド全体が染色体に組み込まれたため、その染色体では、ある遺伝子は欠損し、ある遺伝子は正常のままである可能性がある。
【0128】
相同領域の二個所にて組替えを行う際の不確実性を排除するため、染色体に大きな部位特異的欠失をつくった後、mini−Tn10kanの単独挿入したS88c2の全DNAセグメント、又は、S88c3の全DNAセグメントのいずれかを持つ複製プラスミドを導入して、一部の遺伝子を不活性化させた。挿入位置とこれによるSps+ 又はSps- の表現型を、接頭字「p」を付けて図9に示す。染色体の変異戦略とプラスミドの変異戦略とはお互い整合し、必須(- )領域と非必須(+ )領域の両方が観察された。
【0129】
実施例16
DNA配列:G+C含量、希少コドンの使用及び翻訳開始配列
28、804bpのDNA配列について、両鎖を測定した(配列番号3を参照されたい)。このクラスター中の典型的なスフィンゴモナス遺伝子の平均プロフィル(及び標準偏差)を、G+C含量のゆがみ、希少コドンの頻度、及び「シャイン−ダルガノ(Shine−Dalgarno)」配列又は翻訳開始配列によって決定した。(表5)。各遺伝子とも第三コドンの位置でGあるいはCの頻度が高いことが特色となっていた。この研究の初期において、rhsACBDオペロンのコドン、及びspsB、D、C及びE遺伝子のコドン2500個を解析して、一群のスフィンゴモナス属のまれにしか使われないコドンを同定した。希少コドン各々が、この群に存在する比率は全体に対して0.2%以下で、群の構成はAGA、AGG、CGA、TGT、GGA、ATA、CTA、TTA、TTG、AAA、TTT、CCA、CCT、AGT、TCA、TCT、ACA及びACTを含んでいた。大腸菌においては、通常、16SrRNAの3’末端を相補する配列に連続する下流から翻訳が開始される(Shaine and Dalgarno,2974,Proc.Natl.,Acad.Sci.USA,71:134)。S.ポウシモビリスDSM1098の16SrRNAを相補する「シャイン−ダルガノ」の相同配列は、TAAGGAGGTGである(Moore,et al.,1993,Lett.Appli.Microbiol.,17:115−118)。
【0130】
このクラスター中の遺伝子が平均遺伝子プロフィルに合致し、かつ、変異してSps- 表現型が生じたならば、そのものこそ「sps」なる名称を与えることができる。しかしながら、本発明者らのタンパク質類似性サーチからは、spsG、I及びF遺伝子の機能について、有意の示唆を得ることができなかった。さらにまた、典型的な遺伝子プロフィルを満足する読み取り枠(ORF)が4個あったが、コンピューターデーターバンクではタンパク質配列について有意の類似性を発見することはできなかった。しかしながら、これらの推定遺伝子が変異しても、目に見えて多糖合成に変化をおこさなかったため、同定していないフレームとの意味から、「Urf」なる略記を付けた。4個のUrf配列(32、26、31及び34)があり、推定されるタンパク質のキロダルトン単位の大きさに応じて名称を付けた。
【0131】
【表5】
【0132】
実施例17
グリコシルーIPトランスフェラーゼをspsB遺伝子として同定
紫外線放射又は化学的変異原にさらした後に単離されたSps- 突然変異のほとんどは、spsB遺伝子上のものであった。グリコシル−IPトランスフェラーゼをコードすると信じられている他の遺伝子産物とSpsBの推定アミノ酸は非常に似ているので、このSpsBタンパク質はスフィンガンS−88を組み立てる第一工程を触媒すると信じられている。図11は、グリコシル−IPトランスフェラーゼとガラクトシル−IPトランスフェラーゼと推定されるアミノ酸配列のアラインメントである。これらのタンパク質のC末端半分は、お互いにかなり相同性がある。N末端領域ではこれほど大きな相同性はないが、SpsBタンパク質には膜貫通ドメインであると示唆されている数個の疎水性の領域があるので(図11のアンダーライン部分)、SpsBタンパク質はS.エンテリカのRfbPタンパク質(Jiang,et al.,1991,MolecularMicrobiology,5,695−713)と相似している。SpsBの疎水性ドメインはアミノ酸35〜59(+2.2平均ハイドロパシー)、68〜86(+1.7)、105〜123(+2.3)及び282〜303(+2.9)を含んでいる。また、SpsBタンパク質の最後の疎水性セグメントの位置は、これらの関連遺伝子産物に共通していて、相同性が最大である領域に続いているタンパク質の中央部分に所在している。
【0133】
DNA配列から推定されるドメインをコードするspsBを、相補性検定により確認した。E4.5セグメント中のmini−Tn10kanの挿入を変化させることより、Bacr Sps- 変異株の相補に干渉するかどうかを観察した。mini−Tn10kanの挿入部位と、相補の結果としての「+」又は「−」を図9のspsB遺伝子の上部に示す。三つの部位のmini−Tn10kanの挿入では、Sps- 変異株S88m260(B233、B239及びB238)にスフィンガン合成を回復することができなかった。その反面、B231とB230を含む数個の隣接する部位に挿入したところ失われていた機能を回復することができた。
【0134】
実施例18
スフィンゴモナスS88のラムノース生合成オペロン
rhsACBD遺伝子によってコードされているタンパク質の推定アミノ酸配列は、dTTP及びグルコース−1−リン酸から4工程でdTDP−L−ラムノースを合成する、S.エンテリカのグループBの酵素及びX.カンペストリスの酵素とよく似ている(図12から図15)。本発明者らは、グルコース−1−リン酸、rhsA遺伝子によってコードされているチミジリルトランスフェラーゼ(thymidylyltransferase)及びrhsB、C及びD遺伝子によってコードされている連続した触媒工程には、従来からの命名法を用いた。しかしながら、スフィンゴモナス属のオペロンは次の4点で独特である。第一に、遺伝子の順序ACBD→が、S.エンテリカ(BDAC→)や、X.カンペストリス(BACD→)のいずれとも異なる。第二に、シストロン間領域がほとんど存在していない。開始コドンと終止コドンとが重なり合っているか、間隔が狭く、rhsA−ATGA−rhsC−TGATCCATG−rhsB−TGATG−rhsDとなっている。第三に、rhsACBDの平均G+C含量が比較的高く(66%)、三番目のコドンの位置では特に高く(89%)、しかもオペロンを通じて一様である。第四に、このクラスター中の周囲の遺伝子でも、またスフィンゴモナス属において関係がない他の種の遺伝子でも同様にG+C含量が高い。
【0135】
当初rhsクラスターからは変異株(#134)一個のみが単離されたが、同時にspsB遺伝子でも第二の突然変異が生じたように見えた。Rhs- 突然変異は、致死性である可能性があると思われた。したがって、rhsクラスター内で単一の突然変異を特異的に操作することによって、スフィンガン合成が阻止されるかどうかを試験した。まず、rhs遺伝子の端から端までspsD遺伝子にわたって大きく染色体を欠失させたS88変異株を構築し(図10の△Tn365)、失われたDNAを持ち、特定部位にmini−Tn10kanを挿入したプラスミドを導入した。プラスミドの挿入位置がspsD、C、E、B又はrhsオペロンにある場合には、細胞はSps- のままであった。しかしながら、Urf32、26、31、34、atrD又はatrBのいずれかに挿入した場合は、欠失突然変異の相補に干渉せず、その細胞はSps+ となった。
【0136】
実施例19
スフィンゴモナスS88のグリコシルトランスフェラーゼ
spsQ、spsK及びspsLの3遺伝子がグリコシルトランスフェラーゼをコードしているように見受けられる。しかしながら、タンパク質がこれ以外のグルコシルトランスフェラーゼ及びラムノシルトランスフェラーゼ類に対して限られた局所的な相同性しか示さなかったため、グリコシルトランスフェラーゼ類の糖特異性は、配列解析のみで決定することはできなかった。他の研究者らも着目しているように、グリコシルトランスフェラーゼは、同一の結合における同一の糖に付着する、単一の細菌由来の酵素でも全く互いに異なる(Glucksman,et al.,1993,J.Bacteriol.,175:7045−7055)。推定spsQ遺伝子産物は、ラムノシルトランスフェラーゼをコードすると信じられている大腸菌K−12のgndに隣接するorf11(Stevenson,et al.,1994,J.Bacteriol., 174:4144−4156)に類似し、さらに、R.メリロチのExoO及びExoUグルコシルトランスフェラーゼ(Reuber and Walker,1993,Cell,74:269−280)とも類似していた。spsQ遺伝子はスフィンガンS−88合成に必須である。S88c2セグメントを有するプラスミド内のspsQ遺伝子に(図9のZ206)、mini−Tn10kanを挿入して、突然変異を起こしたプラスミドをspsGSRQI遺伝子の染色体を欠失しているS88細胞に導入した。受容細胞は生存可能であったが、多糖の合成は阻止された。推定spsL遺伝子産物は、ドッドマトリックス分析による比較では、spsQ産物と類似し、さらにS.エンテリカのラムノシルトランスフェラーゼ(RtbN)及びYersinia pseudotuberculosisの推定アベクオシルトランスフェラーゼ(abequosyl transferase)ともいくらか局所的に類似していた(Liu,et al.,1995.J.Bacteriol.,177:4084−4088)。spsK類似タンパク質の検索を行ったが、圧倒的多数が共通して推定UDP結合部位を含有するグリコシルトランスフェラーゼであるという僅かな類似性を認めたに過ぎなかった。これには、UDPが関与している可能性があるので、グルコシル又はグルクロノシルトランスフェラーゼであることが示唆されている。トランスポゾン挿入によらない変異株54と302同様、spsK遺伝子に特異的な挿入を行った変異株(図9のpY882)は、生存可能ではあったが、多糖を作ることはできなかった。
【0137】
実施例20
細菌による多糖の分泌
必須遺伝子産物の配列が類似していることから、各種細菌によって多糖類を分泌させる共通の機構が考えらる。この配列の比較(表6)では、spsD、spsC、spsE、spsJ、及びspsSの五つものsps遺伝子がスフィンガン類の分泌に関与しうることを示していた。R.メリロチの「エクソ」タンパク質など、かなりの情報が蓄積しているタンパク質の配列関係を要約して表6に示す。しかしながら、機能的に関係のあるタンパク質類は、この表が示しているものよりも大きい集団である。51、29及び22個のアミノ酸からなるSpsDタンパク質の各々異なる三つセグメントは、それぞれExoFと29%、31%及び36%の同一性があった。開始と終止コドンが重なり合い(TGATG)、しかも互いに連続するspsCとspsE遺伝子は、ExoP内の二つの異なるドメインに似たタンパク質をコードしている。類似するSpsC−ExoP配列には、最近、細菌性O−抗原の鎖長測定で示されたモチーフ(PX2 PX4 SPKXIIGXMXG)が含まれていた(Becker,et al.,1995,Mol.Microbiol.,16:l91−203)。92、30及び19個のアミノ酸からなるSpsCの三つのセグメントは、ExoPのN末端半分から取った同様な順序の配列に、それぞれ22%、30%及び42%同一であり、75及び98個のアミノ酸からなるSpsEの二つのセグメントは、ExoPのC末端半分に32%及び29%同一であった。37、20及び44個のアミノ酸からなるSpsSの三つのセグメントは、ExoTにそれぞれ38%、55%及び23%同一であった。推定SpsJタンパク質は、KpsT、BexA及びABC担体と推定ヌクレオチド結合ドメインを共有していて、お互いにある程度の類似性を持っていた。spsR遺伝子は、スフィンガン合成に必要ではないが、その遺伝子産物は、細菌性及び真菌性多糖リアーゼに僅かに類似していた。したがって、spsR遺伝子は、細胞あるいは基質の表面のいずれかからグルクロン酸含有スフィンガン類を放出したり、あるいはポリマーを炭素源として再使用することに重要であるかもしれない。
【0138】
図9及び図10に示すように、自然点突然変異や、spsD、spsC、spsE及びspsS遺伝子へのmini−Tn10kanの挿入では、生存可能であるが、培養上清中にスフィンガンS−88を蓄積しなかった。一方、R.メリロチの類似遺伝子の突然変異(Harding,et al.,1993,J.Gen.Microbiol.,139:447−457)、及びX.カンペストリスの類似遺伝子の突然変異は、致死性であった。spsJの染色体にmini−Tn10kanを挿入すると、Sps- となる。しかしながら、染色体が大きく欠失している変異菌株中のマルチコピープラスミドに保持されたspsJに、mini−Tn10kanを挿入すると、Sps+ か、Sps- のいづれかになった。
【0139】
【表6】
【0140】
各タンパク質の分泌の役割に対しての参考文献
ExoF,ExoP,ExoT(Becker et al.,1995,Mol.Microbiol.,16:191、Horinouchi andWeisblum,1982,J.Bacteriol.,150,815、及び、Reuber and Walker,1993,Cell,74:269)
GumB,GumC,GumJ(Glucksmann,et al.,1993,J.Bacteriol.,175:7033、Becker et al.,1995,Mol.Microbiol.,16:191、及び、Glucksmann,et al.,1993,J.Bacteriol.,175:7045)
KpsD and KpsT(Wunder,et al.,1994,J.Bacteriol.,176:4025、及び、Smith,et al.,1990,Mol.Microbiol.,4:1863)
BexD,BexC,and BexA(Kroll,et al.,1990.Mol.Microbiol.,4:1853)
【0141】
実施例21
溶菌性又は毒性タンパク質のABC輸送体
二つの連続した遺伝子であるatrBとatrDが、spsクラスター内に位置していた。この2つの遺伝子は、溶菌性又は毒素様タンパク質のABC輸送体や、輸送用補助タンパク質をコードしていると考えられる、これまでに、溶血素遺伝子(hlyA)が、現在ではスフィンゴモナス属として再分類されているシュードモナス ポーシモビリス(Pseudomonas paucimobilis)に同定された。本発明者らの菌株では、ヒツジ赤血球を含有する寒天プレート上で決定的な溶血現象を検出できなかったので、今回は「hly」なる略称を使用しなかった。atrB遺伝子のDNA配列から推定したアミノ酸の約48%は、百日咳菌(Bordetella pertussis)のシクロリシン(cyclolysin)ABC担体のアミノ酸と同一であった。atrB遺伝子産物は、それぞれ溶血素と白血球毒素を輸送する大腸菌のHlyBタンパク質及びパスツレラ ヘモリチカ(Pasteurella haemolytica)のLktBタンパク質と非常によく類似していた。また、atrBのC末端半分は、R.メリロチのNdvAタンパク質のC末端半分や、ヒト多剤耐性タンパク質Mdr1内の二つの繰り返しATP結合ドメインを含む、上記以外の多くのABC輸送体に類似していた。atrD遺伝子産物の配列は、大腸菌のHlyDタンパク質及びP.ヘモリチカのLktDタンパク質の配列と類似していた。他の属由来の関連輸送遺伝子とは違って、スフィンゴモナスatrB及びatrD遺伝子と隣接したり、あるいはspsクラスター内に存在する溶菌性又は毒性の類似遺伝子はなかった。
【0142】
結論
一つの属の細菌の多糖陰性変異株を第二の属のDNAで相互に遺伝的に相補することは、最初にキサントモナス属とリゾビウム属との間で立証された(Borthakur,et al.,1988,Mol.Gen.Genet.,213:155)。この初期の研究では、寒天プレート上で粘液性が回復したことが観察された。後年になって、X.カンペストリスのgumD遺伝子とスフィンゴモナス菌株S88のspsB遺伝子との間でお互に属間で相補がおこったことが報告され(Pollock,et al.,1994,J.Bacteriol.,176:6229)、本発明者らも組成分析によって、供与菌の相補遺伝子により、受容菌がエクソポリサッカライドの合成を回復したことを立証した。
【0143】
本願の実験では、spsB遺伝子がグルコシルIP−トランスフェラーゼをコードすることは、スフィンガン類生合成の重要な工程であることを証明している。したがって、本発明により受容菌に組み入れるDNAフラグメントは、グルコシルIP−トランスフェラーゼ酵素(グルコシル−トランスフェラーゼ、ガラクトシル−トランスフェラーゼ、又は関連IP−トランスフェラーゼのどれでもよい)をコードしている遺伝子を含有することが好ましい。例えば、スフィンガンS−88を組み立てる最初の工程は、グルコース−Pを担体IPに転移することである可能性が最も高いと信じられている。したがって、この生合成反応を促進する酵素をコードする遺伝子を、本発明によりDNAフラグメントに含有させることが有利である。
【0144】
本発明者らの研究により、スフィンゴモナスS88の大きなsps遺伝子クラスターには、dTDP−L−ラムノースの生合成をコードするより小さなオペロン(rhsACBD)のあることが証明された。rhsオペロンと他のラムノースオペロンの配列が類似していることは、スフィンゴモナス属がL−ラムノースの合成に同じ4つの酵素工程を有することを示唆すると共に、rhsオペロンがDNAフラグメント中において不可欠であることを立証している。
【0145】
侵襲してくる多くの病原菌がカプセルで保護されているように、各種のスフィンガンエクソポリサッカライド類も防御的性格として解釈することができる。また、エクソポリサッカライド類は、細胞を基質に付着させる役割を果たしているとも考えられる。スフィンゴモナス属のスフィンガン生産能を持つ他の菌株も、本願明細書で詳述したS88クラスターと同様に組織だった遺伝子クラスターがあるので、本願明細書の実施例で説明した結果から、通常の技術を持つ当業者が、すべてのスフィンゴモナス属細菌から実用に使用できるDNAフラグメントを得ることができる。
【0146】
スフィンガン生産能力を有する供与菌のスフィンゴモナス属細菌からスフィンガンの生合成に有用又は不可欠な遺伝情報をコードするDNAを単離して、受容菌の細菌に挿入することによって、スフィンガン生産能力がない細菌、又は通常の生産能力しか持たない細菌(受容菌)を、容易にスフィンガンガムの大量生産菌に作ることができることを本願明細書の開示により示した。大量生産菌は容易に得ることができるし、またその方法もスフィンガン生産能力を持っていさえすれば、スフィンゴモナス属のどの菌株にも一般適用することができる。
【0147】
本発明の実施態様の一部において、グリコシル−IPトランスフェラーゼをコードする(スフィンガン炭水化物を組み立てる第一工程)S88のspsBなどの遺伝子その他のDNAフラグメントを含有する供与菌から得たDNAフラグメントを、本発明で有利に使用している。本発明の他の実施態様、特にラムノースが多糖の形成に重要な糖シントン又は構築材料(例えば、スフィンガンの場合は、S−88、ジェランの場合は、S−60、及びウェランの場合は、S−130)である実施態様において、本発明の大量生産菌によるある種のスフィンガン多糖類の生産を最大に増産するために、ラムノースオペロン又は遺伝子(S−88のrhsABCD遺伝子など)を含めて使用することが好ましい。さらに、グルコシルトランスフェラーゼをコードする遺伝子又はその他のDNAフラグメント、例えばスフィンゴモナス菌株S88のspsQ、spsK及びspsL、あるいは形成される最終的な多糖を分泌のためにコードする遺伝子又はその他のDNAフラグメント(例えば、スフィンゴモナス属菌株S88のspsD、C、E、J及びS)を含めて有利に使用し、これらの酵素を最終的な多糖の生産に貢献させることができる。さらにまた、上記の酵素又は機能のすべてをコードするその他のDNAフラグメント又は遺伝子も、所望の多糖次第では有利に使用することができる。
【0148】
通常の技術を持つ当業者が、本明細書の開示に従って容易にDNAフラグメントを得ることができ、これらのフラグメントを受容菌に組み入れて、スフィンガン大量生産菌を作ることが可能であることに注目すべきである。これらの大量生産菌は公知の簡単な遺伝操作方法を使って容易に作ることができる。
【0149】
〔微生物の寄託〕
下記に記す上から6番目までの微生物は、特許手続上の微生物の寄託の国際承認に関するブダペスト条約に従い、メリーランド州20852 ロックビル、パークラウンドライブ 12301に所在する、アメリカン タイプ カルチャーコレクション(American type Culture Collection)に国際寄託されている。寄託物の利用可能性に対する制限は、寄託物についての特許の付与と同時に、取り消し不能条件を付してすべて撤廃される。下から3番目以降の微生物は、メリーランド州のロックビルのアメリカン タイプ カルチャー コレクションから公的に入手することができる。
【0150】
【表7】
【0151】
上記で説明した実施例及び実施態様は、例示によって本発明を説明することを目的とするものであって、如何なる方法によってもこれをもって本発明を制限するものと見なしてはならない。通常の技術を有する当業者によって、本発明で説明する事項及び対象に各種の修正また変更を行うことは可能であるが、これらはすべて本発明で予め考慮されていて、本発明の技術的範囲内に含まれている。
【技術分野】
【0001】
本発明は、スフィンゴモナス属菌(Sphingomonas sp. )のスフィンガン(sphingan)多糖類の生合成生産に関与し、且つこのスフィンゴモナス属菌から単離されるDNA配列、及びそのフラグメントに関する。単離されたDNAフラグメントの多数のコピーをスフィンゴモナス属菌又は関連菌の同じ又は異なる菌株に挿入して、多糖類、好ましくはスフィンガンを増産することができる。このような操作により外因性DNAを含有する細菌の多糖類生産量は、同一の発酵条件下で、操作をしない細菌よりも有意に大きくなる。さらに、本発明はスフィンゴモナス属菌、その他の関連菌の菌株を操作して多糖類の大量生産菌とする方法、ならびにこのようにして操作された細菌に関する。さらにまた、スフィンゴモナス属菌がスフィンガン多糖類を増産するために有用なDNA配列の同定方法及び単離方法も説明する。
【背景技術】
【0002】
多くの微生物がエクソポリサッカライド類(exopolysaccharides)、あるいはEPSとしても知られている細胞外多糖類を生産する。エクソポリサッカライド類には、キサンタンガム(xanthan gum)及び「スフィンガン類」として知られている一群の多糖類が含まれるている。「スフィンガン類」はスフィンゴモナス属(genus Sphingomonas)のグラム陰性菌によって生産される。
【0003】
「スフィンガン類」は莢膜多糖類で、構造は互いに似ているが同一ではなく、スフィンゴモナス属細菌によって分泌される(Pollock, T.J.1993,J.Gen.Microbiol.,139:1939−1945)。スフィンガンはそれぞれ異なる側基を持っているが、主鎖のある場所では、L−ラムノースかL−マンノースのどちらかが存在する。L−マンノースそのものは天然では非常に稀である。ポリマーの水溶液には独特で有用な流動特性がある(Moorhouse,R.1987,”Structure/property relationships of a family of microbioal polysaccharides,”p189−206. In M.Yalpani(ed.),Industrial polysaccharides:genetic engineering,structure/property relation and applications.Elsevier Science Publishers B.V.,Amsterdamを参照されたい)。ポリマーの構造を変えることによって、どのようにして異なる流動特性が生じるのかまだ分かっていない。
【0004】
キサントモナス カンペストリス(Xanthomonas campestris)はグラム陰性菌で、エクソポリサッカライドの一つであるキサンタンガムを構成的に大量生産する(Jeanes, et al.,J. Appl. Polymer Sci.,5, 519〜526,1961)。キサンタンガムの生合成については、産業上の重要性からかなり詳細な研究がなされてきた。最近にいたり、新しい細菌性エクソポリサッカライドであるジェラン(gellan)がゲル化剤として開発された。ジェランは、S−88(Kang and Veeder, 米国特許第4, 535, 153号を参照)、ウエラン(welan)(Kang and Veeder, 米国特許第4, 342, 866号を参照)、NW11(Robison and Stipanovic, 米国特許第4,874, 044号を参照)、ラムザン(rhamsan)(Peik et al., 米国特許第4, 401, 760号を参照)、S−198(Peik etal., 米国特許第4, 529, 797号を参照)、S−657(Peiket al., 欧州特許出願第209277A1号を参照)、及びヘテロポリサッカライド−7(Kang and McNeely, 米国特許第4, 342, 866号を参照)を含む関連多糖類の一群に分類されている一つである。
【0005】
上記の引例ではスフィンガン多糖類組成に関する特許が数件含まれている。これらの特許はいずれも本発明の対象とはなんら関連性もない。
【0006】
【表1】
【0007】
スフィンガン多糖類の化学構造は、すべてある程度の関連性がある。スフィンガン各々の主鎖は、D−グルコース、D−グルクロン酸、L−マンノース及びL−ラムノースの四つの糖の関連性のある配列からなっている。スフィンガン群の多糖類は、ポリマーの主軸(主鎖)と側鎖を含む炭水化物によって互いに区別することができる。スフィンガンの構成糖には、アセチル基又はピルビル(pyruvyl)基が結合している場合もある。
【0008】
各種のスフィンガン類は、特殊ポリマーとして、あるいは繊維への応用、食品、化粧品、紙、ペンキ、セメントの添加剤、例えば粘度調整剤として、あるいは各種のコーティングへの応用において、さらには石油製品の接着剤や添加剤、特殊化学薬品として有用である。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明がなされるに至った研究の初期段階では、代表的なスフィンガン多糖類であるS−88の生合成第一工程が重要な課題であった。このスフィンガンは、スフィンゴモナス菌株S88によって生合成される。本研究以前では、スフィンガン以外の細菌性多糖類の生合成のすべてではないが、一部の製法において、リン酸イソプレニルを担体として使用されることが知られていた。例えば、X. カンペストリスが生合成するキサンタンガムの場合、キサンタンガムの主鎖にはグルコースしか含まれていないので、合成第一工程ではおそらくUDP−グルコースのグルコース−リン酸をC55−イソプレニルリン酸(IP)担体に転移させる必要がある。Ielpiら(FEBS Lett.,130, 253,1982及びJ. Bacteriol. ,175, 2490,1993)は、無細胞取り込み検定を行って、グルコースの次に第二のグルコース、その後、マンノース、グルクロン酸、そしてマンノースの順序でIP担体に連続添加して、キサンタンガムの繰り返し単位が構成されていることを確認した。まったく同様にして、エスチェリシア コリ(Escherichia coli、大腸菌)のコレニン酸のサブユニットの繰り返し単位も、まずグルコース−Pを担体IPに転移することにより構成されている(Johnson and Wilson, J. Bacteriol.,129, 225,1977)。対照的に、リゾビウム メリロチ(Rhizobium meliloti)によるスクシノグリカン(succinoglycan)多糖類の合成の場合には、まずガラクトース−PをIPに転移している(Tolmasky, et al.,J. Biol. Chem.,257, 6751,1982を参照)。しかしながら、イソプレニル担体は、デキストランあるいはレバン(levan)多糖類の合成には関与しないので、アルギン酸塩の合成におけるイソプレニル担体の役割は分かっていない。
【0010】
本発明の研究以前では、多糖類生合成の複合反応速度論(complex kinetics)から見て、担体の役割が重要であることについては分かっていなかった。さらに、スフィンゴモナス菌がスフィンガン多糖類を合成する全体を通して、イソプレニルリン酸担体がどんな役割を果たしているのかも分かっていなかった。
【0011】
同時にBacr でもありGum- でもある(バシトラシン耐性とキサンタンガム陰性)X. カンペストリス変異菌の特別クラスは、UDP−Glcのグルコース−PがIPに転移してGlc−PPIを形成するのに必要なgumD遺伝子中に認められることが、遺伝子相補性検定によって以前から証明されていた。(Pollock, et al.,1994, J. Bacteriol.,vol.176, pp. 6229〜6237, Vanderslice, et al.,”Genetic Engineering of Polysaccharidestructure in Xanthomonas campestris”, p. 145〜156, in V. Crescenzi, et al.,Biomedical and Biotechnological Advances in Industrial Polysaccharides, Gordonand Breach Science Publishers, New York and N. E. Harding and Y. N. Patel, 1993, Faseb Journal, Vol. 7, Number7)。後者の引例では、生産能がない変異菌がスフィンガンS−60の合成を復活できるDNAフラグメントを開示しているが、合成量が野生型菌株より大きいか、否かについては触れられていない。さらにまた、X. カンペストリスの野生型gumD遺伝子によって、スフィンゴモナス菌株のS88とNW11の相似菌であるBacr Sps- (スフィンガン多糖類陰性)変異菌がスフィンガン類の合成を復活できることが実験で証明された。さらに、Bacr Sps- スフィンゴモナス変異菌は、グルコース−PがIPに転移するのを阻害されるらしいと示唆された。
【0012】
本発明の目的は、スフィンゴモナス属菌から単離されて、多くの微生物、及び特に多くのスフィンゴモナス菌株が、スフィンガン多糖類を増産するために用いられるDNAセグメントを提供することにある。
【0013】
本発明の他の目的は、スフィンガン多糖類の生産量が操作されていない菌株よりも有意に大きい、微生物、及び特に多くのスフィンゴモナス菌株から得られる大量生産菌株を提供することにある。
【0014】
本発明のさらに他の目的は、スフィンガン多糖類大量生産菌である微生物の菌株、及び特にスフィンゴモナス属菌の菌株の生産方法を提供することにある。
【0015】
本発明のさらに他の目的は、DNAセグメントを単離してスフィンゴモナス菌株に挿入することにより、このようにして操作された微生物がスフィンガン多糖類大量生産菌となる方法を提供することにある。
【0016】
本発明のこれらの目的及び/又は他の目的は、次に述べる本発明の説明によって容易に理解することができる。
【課題を解決するための手段】
【0017】
本発明のDNA配列は、セグメントあるいはフラグメントのいずれも、スフィンガン生産能を持つ細菌、一般にはスフィンゴモナス菌株から単離する。このようにして得た遺伝子材料は、クローニングした後、多数のコピーをスフィンガン生産能を持つ、あるいは生産能をもたないスフィンゴモナス又は関連菌の変異体に組み入れる。このようなDNA配列は、スフィンガンを生産しない変異菌のスフィンガン生産を復活するのに有用であることが立証されている。さらに、予期していないことであったが、これらの変異菌はスフィンガンを生産できるばかりでなく、スフィンガン生産量が、本来スフィンガンを生産できる野生型菌株に期待されている生産量よりも有意に大きいことが見いだされた。
【0018】
さらにまた、DNAセグメントあるいはフラグメントをスフィンゴモナスの一菌株から単離し、多数のコピーをスフィンガン生産能を持つ、あるいは生産能を持たない同じスフィンゴモナス変異菌株又は異なる菌株に挿入すると、このようにして操作した細菌がスフィンガンの大量生産菌となることを発見できたことも、予期していないことであった。この発見は特に次の点で思いもよらないものであった。すなわち、DNAセグメント又はフラグメントを、例えば、スフィンゴモナスS60から単離して、スフィンゴモナスS88の野生型あるいは非ムコイド性変異菌に挿入すると、この操作によってS−88スフィンガンの大量生産菌ができ、しかも生産されるS−88は一般にS−60スフィンガンが混入していない。この相補性はスフィンゴモナスの各種菌株間でかなり広く適用したり(菌株間相補性)、またキサントモナス カンペストリスのキサンタンガム生産にも適用できる可能性がある(属間相補性)。
【0019】
さらにまた本発明者らは、スフィンガン生産能を持つスフィンゴモナス菌株からDNAセグメント又はフラグメントを単離して組み入れる操作によって、大量生産能を持つスフィンゴモナス菌株を生産する方法を発見した。スフィンガン生産能を持つ細菌から単離したDNAは、まずクローニングした後、スフィンガン生産能を持つスフィンゴモナス菌株、又はスフィンガン生産能を持つ菌株から派生した非ムコイド性変異体に挿入する。
【0020】
さらに本発明は、上記の単離したDNAセグメント又はフラグメントを挿入することにより得た、操作されているスフィンゴモナス菌を包含している。これらの操作されている細菌は、本発明により単離されたDNAセグメント又はフラグメントの多数のコピーを含有している。本発明により操作された細菌は、スフィンガンの大量生産菌である。
【0021】
本発明によれば、DNAフラグメントは、当業者が容易に利用できる技術により単離回収して、クローニングすることができる。その後、DNAの多数のコピーを、一般には染色体外又はプラスミドDNAとしてスフィンゴモナス属の細菌に挿入する。標的細菌に挿入した後、操作されている細菌により、同じ菌株の操作されていない、スフィンガン生産能を持つ細菌と同一の濃度で発酵させて、スフィンガン生産量を測定する。操作されていない、生産能を持つ菌株よりも、スフィンガン生産量が大きいものを大量生産菌として決定する。この方法を使えば、ほとんどすべてのスフィンガン生産能を持つスフィンゴモナス属菌から得られた、スフィンガン多糖類を増産できるDNA配列を容易に決定することができる。
【図面の簡単な説明】
【0022】
【図1】スフィンゴモナス菌株S88(ATCC寄託番号31554)の染色体DNAから単離した34kbpヌクレオチド単位のDNA配列を、制限酵素により消化した部位の概略図である。図1に示した多くのDNA配列をスフィンゴモナス菌に挿入して、これらのスフィンゴモナス菌がスフィンガンを増産する能力を検討した。つぎの数個の酵素の制限部位も、図1(さらに、図2、3、8、9及び10)に示す。B(BamHI)、Bg(Bg1II)、E(EcoRI)、H(HindIII)及びS(SalI)。図1に示すspsB領域は、タンパク質SpsBをコードするDNA配列に相当する。
【図2】スフィンゴモナス菌株S60(ATCC寄託番号31461)の染色体DNAから単離したDNA配列を(約28kbp単位)、制限酵素により消化した部位の概略図である。このDNA配列の制限部位をB、E及びH部位として図2に示す。sgeB領域は、タンパク質SgeBをコードするDNA配列に相当する。
【図3】スフィンゴモナス菌株NW11(ATCC寄託番号53272)の染色体DNAから単離したDNA配列(約33kbp単位)の制限酵素部位の概略図である。この配列の制限部位をE、H、B、Bg及びS部位として図3に示す。snwB領域は、タンパク質SnwBをコードするDNA配列に相当する。
【図4】本発明により生産する代表的な一スフィンガン多糖類の化学構造の概略図である。
【図5】本願の実施例6及び12で詳述するプラスミドpSEB24及びpSEB26の構成地図である。McsIは次の制限部位、即ちEcoRI、SmaI、BamHI、SAlI、PstI及びHindIIIを(上から時計廻りに)含んでいるマルチクローニング部位である。同様に、McsIIもHindIII、PstI、SalI、XbaI、BamHI、SmaI、SstI及びEcoRIを(上から時計廻りに)含んでいる。OriTは接合転移開始点で、OriVは広範囲宿主複製開始点、及びoriはpUC12と13から始まる複製開始点である。
【図6】スフィンゴモナス菌株S88(ATCC寄託番号31554)の染色体DNAから単離された34kbpヌクレオチド単位のDNAセグメントの制限酵素による切断部位の概略図である。次の数個の酵素の制限部位、即ちB(BamHI)、Bg(BglII)、E(EcoRI)、H(HindIII)及びS(SalI)が、図1に(さらには、図2及び図3でも)示されている。図6に(さらに、図1でも)示しているspsB領域は、タンパク質SpsBをコードするDNA配列に相当する。スフィンガン生合成に関与する他の「sps」遺伝子は、大文字G、S、R、Q、I、K、L、J、F、D、C及びEで示してある。rhsACBDはスフィンガン先駆物質dTDP−(L)ラムノースの合成に関与する遺伝子の地図上の位置を示す遺伝子の名称である。遺伝子32、26、31及び34は、まだ同定されていない翻訳の読み取り枠(ORF)であり、atrDB遺伝子は、スフィンガン合成に無関係の輸送機能をコードしている。「Sec」はスフィンガン類の分泌に必要な遺伝子を示し、「Trase」はスフィンガンの繰り返しサブユニットを組み立てるときに、ヌクレオチド−糖先駆物質からスフィンガンに糖を転移する酵素をコードしている遺伝子を示している。
【図7】スフィンゴモナス菌株S198(ATCC寄託番号31853)の染色体DNAから単離された(約42kbp単位の)DNAセグメントの制限酵素による切断部位の概略図である。図7では、このDNA配列の制限部位をH(HindIII)及びE(EcoRI)として表示してある。括弧内の間隔が詰って並んでいる部位の順序は、不明である。コスミドクローンc2、c3、c4、c5及びサブクローンL242の外側方向への広がりは、破線で示してある。四角で囲んでいる「B」領域は、S88遺伝子spsBの変異体を相補するタンパク質をコードしているDNA配列に相当する。
【図8】スフィンゴモナス菌株S7(ATCC寄託番号21423)の染色体DNAから単離された(約42kbp単位の)DNAセグメントの制限酵素による切断部位の概略図である。図8では、このDNA配列の制限部位をH(HindIII)、E(EcoRI)及びB(BamHI)として表示してある。コスミドクローンc1、c2、c3、及びc6の外側方向への広がりは、破線で示してある。クローンc2とc3は、図8の右側の領域を超えて伸びている。同様に、クローンc6は左側方向に伸びている。四角で囲んでいる「B」領域は、S88遺伝子spsBの変異体を相補するタンパク質をコードしているDNA配列に相当する。
【図9】スフィンガンS−88合成に用いられる遺伝子クラスターの概略図である。地図の下部には遺伝子名、境界、予定している機能、及び制限酵素による切断部位(BはBamHI、EはEcoRI、HはHindIIIである)でのヌクレオチドの位置を示している。図の上部に向って、自然Sps- 突然変異の名称と位置、Sps+ またはSps- の相補結果、及びプラスミドとS88染色体における特異的な挿入変異によるSps表現型を示している。mini−Tn10kanの挿入があるpZ167、pZ168、pZ180、pZ202及びpZ206は、プラスミドpRK311にクローニングされたc2セグメントにあり、2種の欠失菌株△Tn493か△Tn495のいずれかに導入された。同様にして、この他のプラスミドに対するトランスポゾン挿入はすべてc3セグメントに行い、欠失菌株である△Tn358と365に導入した。mini−Tn10kan挿入位置の精度は、配列決定された制限部位に対して±50bp、グラフの精度は±100bpである。 なお、外国書面出願時では、この図は、図10と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の上部を示してある。
【図10】スフィンガンS−88合成に用いられる遺伝子クラスターの概略図である。地図の上部には遺伝子名、境界、推定される機能、及び制限酵素による切断部位(BはBamHI、EはEcoRI、HはHindIIIである)でのヌクレオチドの位置を示している。クローニングされたフラグメント(c1△3、c2、c3、c4、c5及びc6)、サブクローニングされたセグメント(使用した制限酵素とkbp単位のおおよその長さに応じて名前を付けた)、及びS88染色体に生じた欠失(各線はDNAの存在を示す)は遺伝子地図の下に示している。 なお、外国書面出願時では、この図は、図9と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の下部を示してある。
【図11】SpsB及びグリコシル−IPトランスフェラーゼの推定アミノ酸配列のアラインメントである。右側の数は各配列の最右側のアミノ酸残基数である。各配列とも、ガラクトシル−IPトランスフェラーゼはSpsB配列の真上に示してあり、グリコシル−IPトランスフェラーゼはSpsB配列の下に示している。次の遺伝子産物は右側に示している。ExoYn、リゾビウムの一種(Rhizobium sp.)NGR234(Gray,et al.,1990,J.Bacteriol.,172:193)。CpsD、S.アガラクチエ(S.agalactiae)(Rubents,et al.,1993,Mol.Microbiol.,8:843)。RfbP、S.エンテリカ LT2(Jiang,et al.,1991,Mol.Microbiol.,5:695)。GumD、X.カンペストリス B1459S−4L(Capage,etal.,1987,International Patent WO/05938)。Pss4、R.レグミノサラム(R.leguminosarum)bv.vicae菌株VF39(GenBank寄託番号M93042)。Pss2、R.レグミノサラム bv.phaseoli(Borthakur,et al.,1988,Mol.Gen.Genet.,213:155)。記号について。|はSpsB及び上下各々に示すガラクトシル−IPトランスフェラーゼまたはグリコシル−IPトランスフェラーゼの同一アミノ酸を示す。:は次の関連したアミノ酸、即ちIFVWML、ST、QNED及びHKRのグループに基づいた保存的なアミノ酸置換を示している。アンダーラインをしてある配列は、疎水性アミノ酸約20個からなる連続セグメントである。
【図12】S.エンテリカ(Jiang,et al.,1991,Mol.Microbiol.,5:695)、及びX.カンペストリス(Koeplin,etal.,1993,J.Bacteriol.,175:7786−7792)のrhsA遺伝子産物と、dTDP−L−ラムノース生合成酵素のアラインメントを示す。記号は図11と同一である。 なお、外国書面出願時では、この図は、図13から図15と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の一部を示してある。
【図13】S.エンテリカ(Jiang,et al.,1991,Mol.Microbiol.,5:695)、及びX.カンペストリス(Koeplin,etal.,1993,J.Bacteriol.,175:7786−7792)のrhsB遺伝子産物と、dTDP−L−ラムノース生合成酵素のアラインメントを示す。記号は図11と同一である。 なお、外国書面出願時では、この図は、図12及び図14から図15と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の一部を示してある。
【図14】S.エンテリカ(Jiang,et al.,1991,Mol.Microbiol.,5:695)、及びX.カンペストリス(Koeplin,etal.,1993,J.Bacteriol.,175:7786−7792)のrhsC遺伝子産物と、dTDP−L−ラムノース生合成酵素のアラインメントを示す。記号は図11と同一である。 なお、外国書面出願時では、この図は、図12から図13及び図15と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の一部を示してある。
【図15】S.エンテリカ(Jiang,et al.,1991,Mol.Microbiol.,5:695)、及びX.カンペストリス(Koeplin,etal.,1993,J.Bacteriol.,175:7786−7792)のrhsD遺伝子産物と、dTDP−L−ラムノース生合成酵素のアラインメントを示す。記号は図11と同一である。 なお、外国書面出願時では、この図は、図12から図14と合わせて一つの図であったが、大きさの関係上、日本語書面では、元の図の一部を示してある。
【発明を実施するための形態】
【0023】
次の用語は本発明の明細書全体を通して使用されるもので、その意味を次のように定義する。
【0024】
1. 「スフィンガン」なる語は、明細書全体を通して、スフィンゴモナス属細菌により分泌される、関連性はあるがお互いに別個であるエクソポリサッカライド群を意味する(Pollock, J. Gen. Microbiology,139:1939〜1945, 1993)。スフィンガン類の構造は、すべていくらかの関連性がある。スフィンガンおのおのの主鎖は、D−グルコース、D−グルクロン酸、L−マンノース、L−ラムノースの四つの糖の関連性のある配列からなっている。スフィンガン群の多糖類は、ポリマー主鎖と複数の側鎖を含む構成糖の配列によってお互いに区別することができる。スフィンガン多糖類はまた、その構成糖にアセチル基又はピルビル基が結合していてる場合もある(Mikolajczak, et al.,Appl. and Env. Microbiol.,60:402, 1994参照)。本発明のDNAセグメントとフラグメント及び一般方法を使用して生産した各種スフィンガンの化学構造の概略図は、おおむね図4に示してある。スフィンガン類の、ジェラン(S−60)、ウェラン(S−130)、ラムザン(S−194)、S−88、NW−11、S−198及びS−657の構造も、おおむね図4に示している。
【0025】
典型的には、スフィンガン多糖類群の各単体は、一般的に次の繰り返し化学構造によって表される。
【0026】
【化5】
【0027】
式中、Glcはグルコース、GlcAはグルクロン酸、Rhaはラムノース、Manはマンノースである。XはRhaでも、Manのいずれでもよい。Zは二番のGlc残基に結合している、α−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaのいずれかである。Wは1番のGlc残基に結合し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaのいずれかであって、下付き文字v及びyは0、0. 33、0. 5、0. 67又は1である。式中、ポリマーの還元末端は、主鎖のX残基の近くに位置している。本明細書で用いる「主鎖」なる語は、この構造からW鎖とZ鎖とを除いた、すなわちvとyが0に等しいときの残部を指す。ポリマーの「還元末端」は、糖単位が加えられるポリマーの末端である。
【0028】
スフィンガン多糖類群のうち、一部の単体はいろいろな位置でアセチル化されている。しかしながら、多糖類は常法により化学的に脱アシル化して、アシル基を除くことができる。例えば、ジェランにはウェランと同じ炭水化物の主鎖(すなわち、X=Rha)があるが、側鎖の糖が欠けており(すなわち、v=0及びy=0)、しかもグルコース残基1はグリセリン酸塩で完全に置換されている。ジェランのサブユニットの構造も、位置は分からないがどこかでアシル化されている。
【0029】
2. 「スフィンゴモナス」なる語は、明細書全体を通して、上記のエクソポリサッカライド類、すなわちスフィンガン類を生産するスフィンゴモナス属グラム陰性菌を意味する。本発明においては、スフィンゴモナス属グラム陰性菌の多くは、単離されたDNA配列源として、スフィンガン生産能を持つ他の菌株(好ましくは、スフィンゴモナス属のグラム陰性菌)に再挿入して、本発明のスフィンガン大量生産菌を生産するため、あるいは標的細菌として、これに外因性DNA配列を挿入してスフィンガン大量生産菌を生産するために用いることができる。
【0030】
グラム陰性菌のスフィンガン生産能を持つ科は、1993年にまずスフィンゴモナス属に分類されるものとして同定された。(Pollock, J. Gen.Microb. , 139, 1939,1993を参照)。しかしながら、各菌株をどの種に分類すべきかは、まだ正確には確立されていない。スフィンガン生産能を持つスフィンゴモナス菌株に最も近い種は、スフィンゴモナス ポーシモビリス(Sphingomonas paucimobilis)と考えられる。しかしながら、詳細な、決定的分類学分析結果を利用できる時まで、これらの菌株が上記の種に分類されるべきであると言うのは時期尚早である。スフィンゴモナス属のスフィンガン生産菌は当初数個の異なる属に分類されていたことを付言する。
【0031】
スフィンゴモナスのうち現在までに確認されている種としては、S. ポーシモビリス(S. paucimobilis)、S. パラポーシモビリス(S. parapaucimobilis)、S. アドヘイシバ(S. adhaesiva)、S. カプスラタ(S. capsulata)及びS. ヤノイクヤエ( S. yanoikuyae)が挙げられる(Yabuuchi, et al.,Microbiol. Immunol.,34, 99,1990を参照)。過去ではこれらスフィンゴモナスの種が、誤ってシュードモナス属(genus Pseudomonas)に分類されたこともあった。
【0032】
3. 「供与菌」及び「受容菌」なる語はおのおの、DNA配列を採取した細菌及びDNA配列を挿入又は組み入れる細菌を指すのに用いられる。
【0033】
4. 「菌株」又は「スフィンゴモナス菌株」なる語は、(化学構造に基づいて)特定のスフィンガンエクソポリサッカライドを生産するスフィンゴモナス属グラム陰性菌を指すのに用いる。スフィンガン生産能を持つスフィンゴモナス菌株を、簡単に、この菌株が生産するスフィンガン多糖類で呼ぶこととする。例えば、スフィンゴモナス菌株S88はスフィンガン多糖類S−88を生産し、スフィンゴモナス菌株S60はスフィンガン多糖類S−60(ジェラン)を生産するなどである。スフィンゴモナス菌株S88(ATCC No. 31554)、S60(ATCC No. 31461)、NW11(ATCC No. 53272)、S130(ATCC No. 31555)、S194(ATCC No. 31691)、S198(ATCC No. 31853)、S657(ATCC No. 53159)、及びS7(ATCC No. 21423)は、特に本発明に有用な代表的菌株である。
【0034】
5. 「大量生産菌」なる語は、明細書全体を通して、操作された細菌であって、同じ菌株又は異なる菌株のスフィンガン生産能を持つ細菌から単離したDNAセグメント又はフラグメントの多数のコピーを含有し、同じ菌株の操作されていない細菌、又は野生型細菌を同一又は実質的に同一の発酵条件で発酵して比較するとき、スフィンガン多糖類生産量が有意により大きい(重量/重量比で、少なくとも約5%以上)細菌を指すのに用いる。
【0035】
6. 「単離された」なる語は、微生物から採取して、少なくともある程度精製した、すなわち一以上の精製工程を通過したDNAを指すのに用いる。単離されたDNAは、実質的に純粋な形で用意しておくことが好ましい。すなわち、制限酵素によりフラグメントあるいはセグメントに切断したり、クローニングにより多数のコピーとしたり、又はプラスミドベクターその他の手段に挿入したり、又は細菌中に組み入れたりするDNAの性能に影響がない程度まで、汚染物質含有量を抑制する。
【0036】
7. 「DNA」又は「染色体DNA」なる語は、明細書全体を通して、スフィンゴモナスから単離されたDNAに関し、一般には微生物から単離する前に、スフィンゴモナス属菌の染色体又は内因性プラスミド中に見いだされるDNAを指すのに用いる。
【0037】
8. 「配列」なる語は、明細書全体を通して、ヌクレオチド単位又は制限酵素による切断部位のパターンによって同定されるDNAの特異的セグメントであって、一般的にはスフィンガン生産能を持つスフィンゴモナス属菌のDNAから制限酵素により単離し、このようにして得たDNA配列を細菌に挿入して大量生産菌を生産したり、あるいはさらに制限して前記の配列より小さいDNAの小部分、又はフラグメントを生産する配列を指すのに用いる。「部分」又は「フラグメント」なる語は、一般にDNAセグメントより小さいDNA配列を指すのに用いる。本発明に用いる好適なDNAセグメントは、グリコシルトランスフェラーゼ(グルコシル、ガラクトシル、ラムノシル及びグルクロノシルトランスフェラーゼ)、グリコシル−IPトランスフェラーゼ(グルコシル−IPトランスフェラーゼ、ガラクトシル−IPトランスフェラーゼ酵素などを含む)、ラムノースオペロン(あるラムノース含有スフィンガンへ挿入するためのラムノース前駆体の合成)、及び細菌の多糖類の分泌を含む様々なタンパク質をコードするセグメントである。
【0038】
9. 「挿入された」、「挿入する」、「組み入れられた」又は「組み入れる」なる語は、明細書全体を通して、スフィンガン生産能を持つスフィンゴモナス菌株の染色体DNA又はプラスミドDNAから単離したDNAセグメントを、スフィンガン生産能を持つ、同じあるいは異なる受容菌になるスフィンゴモナス菌株に転移する工程と結果を指す。結果として、転移されたDNAセグメントの少なくとも実質部分の多数のコピーを含有する大量生産菌株を得る。
【0039】
例を挙げると、まず単離されたDNAを当業者に周知の技術によりプラスミドベクター、例えば特にpRK311やpSEB24に導入し、クローニングした後、接合により受容菌となるスフィンゴモナス菌に転移する。受容菌となるスフィンゴモナス菌に挿入後、受容菌の細胞中で上記のDNAフラグメントを含むプラスミドベクターが複製されて、スフィンガン多糖類の大量生産に必要なDNAセグメントが数コピー(少なくとも2コピーで、通常は4〜20コピー)できる。プラスミドベクターの他に、バクテリオファージベクターや、トランスポゾンベクターを使ってもよい。
【0040】
多くのプラスミドベクターが、単離されたDNAセグメントやフラグメントを受容菌に挿入する際に好適である。上記で説明したpRK311やpSEB24の他に、特に、以下のプラスミドも有用である。不和合群P−1の広範囲宿主プラスミドであるRK2などや、そのプラスミドから派生したpRK290、pRK293、pRK404など(Ditta, et al.,Plasmid,Vol. 13, pp. 149〜153)、及びプラスミドの転移を可能にするプラスミドRP4のoriT遺伝子を含むその他の派生体であるpSUP101など(Simon, et al.,Bio/technology, November, 1983を参照)並びにプラスミドpLAFR1及びpLAFR3(Friedman, et al.,Gene, 18, 289, 1982)、及び不和合群Inc−Qの広範囲宿主プラスミドであるRSF1010や、そのプラスミドから派生したpMMB22及びpMMB66(Fuerste, et al.,Gene,48, 119, 1986)などである。
【0041】
プラスミドベクターを受容菌内に挿入する際に、接合を用いるのは一般に有効である。しかしながら、属が異なる細菌同志では、コンピテント細胞を精製DNAで形質転換させるのがより一般的である。
【0042】
スフィンゴモナスの場合、細菌にDNAフラグメントやプラスミドを導入するのに電気穿孔法も使われてきた(1992, Monteiro, et al.,J. of App. Bacteriol.,72, 423を参照)。この方法を使えば、単に単離されたDNAを細菌に添加し、電気穿孔で細胞膜を通して転移させるだけで、単離されたDNAセグメントやフラグメントの細胞コピーを2コピー以上受容菌となるスフィンゴモナス菌に組み入れることができる。
【0043】
前掲のMonteiroらは、DNAをスフィンゴモナスに導入する手段としての電気穿孔法を説明している。電気穿孔法の機能は、コンピテント細胞を化学的に処理して、例えば細胞を塩化カルシウムやルビジウム塩で処理した後に形質転換するのと同じである。形質転換させるDNAは標準法で精製するが、この場合プラスミドの形をとってもよいし、またとらなくてもよい。しかしながら、通常、二本鎖閉環型のDNAの場合、形質転換が最も効率的である。したがって、この場合、接合法でDNAをスフィンゴモナスに導入する必要もなければ、セグメントをクローニングしてからプラスミド、バクテリオファージあるいはトランスポゾンベクターに挿入する必要もない。しかしながら、単離されたDNAをまずプラスミドベクターに導入してから、単離DNAフラグメントを含有するプラスミドを細菌中に転移することが好ましい。
【0044】
受容菌となるスフィンゴモナス菌中で、プラスミドあるいはその他ベクター、例えばバクテリオファージ又はトランスポゾンベクターにDNAセグメントの担持を続ける必要はない。細菌DNA中にDNAセグメントのコピーをさらに導入して、細菌DNAを複製するのと同じメカニズムによりセグメントを世代毎に複製することは、常法として行われている。下記の実施例の部において、細菌DNAにDNAのコピーをさらに導入して、細菌DNAを複製するのと同じメカニズムによりセグメントを世代毎に複製する方法を詳述する二つの実施例を説明する。
【0045】
10. 「多数のコピー」なる語は、明細書全体を通して、少なくとも2コピー、好ましくは少なくとも4コピーをスフィンゴモナス細菌に組み入れる外因性DNA配列、フラグメント又はセグメント(前記のDNAの少なくとも実質部分)を指すのに用いられる。より好ましくは、スフィンゴモナス属菌に挿入するDNA配列、フラグメント又はセグメントのコピー数は、実際には約4から約20までの範囲である。ある例においては、DNA配列を1個のプラスミドベクターに組み入れて、接合によりスフィンゴモナス菌に転移し、受容菌中でプラスミドを複製して、2コピー以上のDNA配列、セグメント又はフラグメントを得ることもあることを付記する。
【0046】
11. 「生合成」なる語は、明細書全体を通して、スフィンゴモナス菌がスフィンガンを生物生産又は合成することを指すのに用いる。スフィンガン多糖類は、複数の細菌の酵素でコントロールされている一連の工程により炭水化物単位毎に合成される。
【0047】
12. 「操作されている」なる語は、明細書全体を通して、外因性DNA、好ましくはその多数のコピーが組み入れられている受容菌となるスフィンゴモナス菌を指すのに用いる。本発明の操作されている細菌とは、スフィンガン多糖類大量生産菌のことである。
【0048】
13. 「遺伝情報をコードする」なる語は、明細書全体を通して、特定の順序で並んでいるヌクレオチド単位の遺伝情報を内容とするDNA配列を指すのに用いる。DNA配列の遺伝情報(どんな長さでも)は、多数のコピーが組み入れられて、これにより得た操作されている細菌がスフィンガンを増産する場合、スフィンゴモナス菌によるスフィンガン合成に「有益又は不可欠」であると見なされる。「有益又は不可欠」なる語は、DNAがスフィンゴモナス細菌から単離され、遺伝情報をコードし、この遺伝情報の多数のコピーが組み入れられたスフィンゴモナス菌が、スフィンガン多糖類の大量生産菌に形質転換するDNAを指すのに用いる。本発明で使用する有益又は不可欠なDNAは、例えば、グルコシルIP−トランスフェラーゼ、ガラクトシルIP−トランスフェラーゼなど、グリコシルトランスフェラーゼの生合成のための遺伝子又はオペロン、及び、ラムノース、マンノース、グルコース、ガラクトースのような糖のシントンや、例えばdTDP−L−ラムノースなど、上記の糖の置換シントンの生合成のための遺伝子又はオペロン、及び、例えばポリメラーゼのようなスフィンガンを生産するための糖シントンの重合に関与する酵素の生合成のための遺伝子又はオペロン、及び、無処理の細胞構造から多糖類を分泌するための遺伝子又はオペロンを一以上含む。
【0049】
14. 「菌株間相補性」なる語は、第一のスフィンゴモナス菌株からDNA配列、セグメント又はフラグメントを単離して、異なる第二のスフィンゴモナス菌株に組み入れることを指すのに用いる。本発明の実施態様の一つとして、異なるスフィンゴモナス菌株からDNAフラグメントを取り、その多数のコピーを別のスフィンゴモナス菌株に組み入れると、スフィンガン多糖類の大量生産菌が生産できることを発見したことは、予期せざるところであった。本発明で有用なDNAフラグメントは、属間相補性(例えば、キサントモナス カンペストリス菌にキサンタンの増産をさせるなど)も示している。
【0050】
15. 「シントン」なる語は、本発明により細菌がスフィンガンを生合成する間に重合される糖又は糖単位を指すのに用いる。シントンは、スフィンガン多糖類の構成部分又は構成単位を含み、スフィンガン類の生合成に使用される糖成分、例えばアセチル化及びアシル化糖や関連先駆物質を含むグルコース、ガラクトース、ラムノース、マンノース、その他の糖シントン類などを含有する。
【0051】
16.「ラムノースオペロン」なる語は、本発明によるスフィンガン多糖類の生合成に利用される、ラムノース又はラムノースシントン類(dTDP−L−ラムノースなど)の生合成に関与する遺伝子やオペロンをコードするDNA配列を指すのに用いる。
【0052】
本発明は、スフィンガン生産能を持つ供与菌となるスフィンゴモナス菌からDNA配列を取って、多数のコピーを同じ菌株あるいは異なる菌株の受容菌となるスフィンゴモナス菌に組み入れると、この受容菌となるスフィンゴモナス菌がスフィンガン多糖類大量生産菌に形質転換するとの発見に関する。さらに、ある種のスフィンガン多糖類を生産する細菌からDNA配列を単離し、異なる菌株のスフィンゴモナス菌に多数のコピーを組み入れると、この異なる菌株から大量生産菌を生産することができるばかりでなく、この場合供与菌の特性を持つスフィンガン多糖類の混入が起こることがないことも発見されている。
【0053】
受容菌に組み入れられたDNA配列には、スフィンガン多糖類の生合成に有益又は不可欠の遺伝情報がコードされている。例えば、有益又は不可欠の遺伝情報は、細菌のスフィンガン生合成にいろいろな形で役割を果たしたり、あるいは関与する。外因性DNAは、例えば、律速酵素の過程に関与する酵素その他のタンパクの合成を発現したり、酵素、補因子、その他の生化学的成分の合成を誘導して多糖類を増産させたり、多糖類のサブユニットの連結を促進するポリメラーゼのような酵素の産生を増加させたり、あるいは一以上のリプレッサー遺伝子と結合したり、細菌から多糖類の分泌を促進させたり、通常、多糖類生合成の間は律速される工程の生産を阻害するリプレッサーの発現を押さえるなど、スフィンガン生合成に有益な効果をもたらすことができる。
【0054】
DNA配列は、当業者の標準的な技術や方法によりスフィンゴモナスの各種菌株から単離する。通常、細菌は(下記で詳細説明するように、約0. 5%より低い、好ましくは約0. 1%〜0. 2%のグルコース濃度を用いる標準発酵法により)培養され、高濃度の細菌を含有するブロスを作る。その後、細菌細胞を遠心分離し、DNA抽出のために再懸濁する。DNAは、まず混合液からタンパクを除いて、エタノール又はイソプロパノールにより高分子量のDNAを沈澱させることによって細菌から抽出する(Birnboim and Doly, Nucl. Acids Res.,7, 1513,1979を参照)。
【0055】
上記のように沈澱させたら、通常は単離されたDNAセグメントあるいはフラグメントのクローニングを行って、受容菌となるスフィンゴモナス菌挿入用のDNAを形成する。例として挙げると、上記した高分子量のDNA配列は、制限酵素(例えば、SalI酵素)により部分的に消化し、標準方法により電気泳動する(Loftus, et al.,BioTechniques, 12, 172,1992を参照)。電気泳動後、大きなDNAフラグメント(20kbp以上)をさらに精製する(抽出及び沈澱)。
【0056】
その後、細菌から単離したDNAフラグメントを、クローニングベクター(一般には、プラスミド)に直接挿入してDNAのクローニングを行うか、代わりに制限酵素によりさらに処理することにより、より小さなDNAフラグメントにして、クローニングベクターに挿入する。本発明のDNAクローニングは、当業者が標準としている一般的な技術と方法によって行う。本発明によれば多くの方法を用いてDNAセグメントのクローニングを行うことができ、したがって、本発明は、例えばプラスミドのクローニングベクターの使用に限定されない。例えば、DNAフラグメントをcharon 4A、EMBL3(Rodriguezand Denhardt, Vectors, Chapter 2, pg. 43, 1988, Butterworth Publishers, Boston)、又はP1(1990, Sternberg, Proc. Natl. Acad.Sci.,U.S.A.,87, 103〜107)などのバクテリオファージベクターに挿入してクローニングすることもできる。
【0057】
下記の実施例1及び12で詳述するように、まず、クローニングベクターに挿入するDNAフラグメントを用意する。本発明によれば、DNAセグメント又はフラグメント形成用として、数多くのクローニングベクターを使用することができる。しかしながら、本発明においては、DNAセグメント又はフラグメントのクローンを形成したプラスミドベクターをそのまま使用して、接合により外因性DNAを受容菌に挿入するのが有利であることが見いだされている。しかしながら、特にDNAを受容菌へ挿入する際に形質転換法を使用しようとする場合には、受容菌に接合するベクターとは異なる(プラスミドその他の)クローニングベクターを使用することもできる。
【0058】
クローニングベクターに挿入を終えたら、単離されたDNAを含有するベクターをバクテリオファージ中でパッケージングして、トランスフェクション法により細菌(一般に、大腸菌)に転移し、トランスフェクションされた細菌中で複製する。このようにして得たDNAクローンを含有する細菌細胞のコロニーをプールして、貯蔵するか、あるいは直接使用する。
【0059】
その後、DNAクローンのスクリーニングを行って、スフィンゴモナスによるスフィンガン生産に対するDNAフラグメントの相対的増産効果を決定する。スクリーニング法においては、DNAを適当なベクターに担持させた後、接合法(例えば、Ditta, et al.,Proc. Natl. Acad. Sci.,USA, 77, 7347,1980が説明している三元交配(tri−parental mating))により受容菌となるスフィンゴモナス菌株に挿入し、このようにして得たDNAの多数のコピーを含有する操作された細菌とそのスフィンガン生産を試験して、活性を決定する。
【0060】
スフィンガン生産に増産効果があると決定されたDNAセグメント又はフラグメントを、受容菌となるスフィンゴモナス菌株に転移して、前述のように転移されたDNAの少なくとも実質部分を少なくとも2コピー含有する大量生産菌株を生産する。
【0061】
本発明で使用する際に好ましいスクリーニング法を開発した。この方法では、DNAを生産能を持たない受容菌となるスフィンゴモナス菌株に挿入することにより、このDNA中にスフィンガン合成に有益又は不可欠である遺伝子が存在しているか、否かをスクリーニングするのである。すなわち、スフィンガン生産能を持つスフィンゴモナス菌株から、生産能を持たない変異菌(例えば、菌株S88のSps- Bacr )を派生させ、スクリーニング対象となるDNAの多数のコピーを含有するように操作する。グルコース1〜3%を含有する普通寒天平板培地でこのように操作した、生産能を持たない変異菌を増殖する。別に、生産能を持たないスフィンゴモナス変異菌を同一条件で増殖して、この変異菌と操作された細菌のコロニーの外観を比較する。このようにした後、一般的にスフィンゴモナス菌にスフィンガン合成を開始させるDNAの能力を、目視により測定する。
【0062】
スフィンガン生産活性を向上させるDNAセグメント又はフラグメントの能力は、培養皿上で容易に認識できる、スフィンガン生産能を持つ細菌とスフィンガン生産能を持たない変異菌の表現型の相違により判定される。例えば、スフィンガン生産能を持つスフィンゴモナス菌株はムコイドを産生するため、簡単な目視検査で容易に見分けることできるコロニーを形成することが多い(例えば、スフィンガン生産菌のコロニーが盛り上がって丸く、明るい輪で取り囲まれているのに対して、非生産菌のコロニーは平坦で、粗く、半透明である)。
【0063】
ある事例では、下記の実施例2で詳述するように、一つのスフィンゴモナス菌株において、スフィンガン生産能を持つ細菌とスフィンガン生産能を持たない細菌の表現型が容易にあるいは短時間で区別できない場合がある。このような場合には、生産性と非生産性の表現型の相違が目視検査で容易に識別できる細菌によって対象DNAの活性を判定するように、スクリーニング法を変更する。この本発明の実施態様では、本発明に有用なDNAフラグメントは菌株間でも、属間でも相補性を示す事実、及びこのDNAフラグメントの多数のコピーによって実際上すべてのスフィンゴモナス菌株がスフィンガンを増産することができる事実を利用するのである。
【0064】
本発明に有用なDNAセグメント又はフラグメントは、キサントモナス カンペストリスにおいても活性を示す。したがって、一以上のスフィンゴモナス菌株を使って容易にスクリーニングできなかったDNAフラグメントの多数のコピーを、キサンタン生産能を持たない、X. カンペストリス変異菌、例えば特にX59m31に組み入れて、目視検査によりキサンタン生産のスクリーニングを行うことができる。X59m31などの生産能を持たないX. カンペストリス変異菌は、バシトラシンにさらした後の生存菌を選び、このバシトラシン耐性変異菌にDNAフラグメントの重複コピーを組み入れ、YM寒天培養皿で培養し、形成されたコロニーの外観がムコイド性(生産菌)であるか、非ムコイド性(非生産菌)であるかを観察することにより、容易に得ることができる。(Pollock, et al.,1994 J. Bacteriol.,176, pp. 6229〜6237、及び米国特許第5, 338, 841号を参照)。スクリーニングされた細菌が多糖類(スフィンガン又はキサンタン)を増産するDNAは、菌株間あるいは属間相補性の証拠であって、異なるスフィンゴモナス菌株ばかりでなく、異なる属の細菌(キサントモナス)によるスフィンガン多糖類生産を増産させることができる。
【0065】
概して簡単に、スフィンガンの生産菌と非生産菌の表現型の相違を同定できる本発明の実施態様による簡便スクリーニング法を用いれば、通常の技術を有する当業者は容易に利用できるクローニング及び転移技術により、過度の又は理由のない実験を行うことなく、本発明で使用されるDNAセグメントやフラグメントを容易に得て、スフィンゴモナス菌によるスフィンガン多糖類を増産することができる。
【0066】
本発明の他の実施態様は、スフィンガン多糖類の増産に関する。スフィンガン多糖類を生産するためには、本発明の操作した細菌を周知の方法により適当な発酵条件下で培養する。操作されたスフィンゴモナス菌を培養する適当な培地又は発酵ブロスとしては、例えば、一般にグルコース、ラクトース、スクロース、マルトースやマルトデキストリンを含む炭水化物類などの炭素源、例えば無機アンモニウム、無機硝酸塩、有機アミノ酸類又は加水分解した酵母,大豆粉又はカゼインなどのタンパク性材料、蒸留業者によって提供される可溶性物質(distiller’s solubles)又はコーン スチープ リカー(cornsteep liquor)、無機塩類などの窒素源及びビタミン類を含有する水性培地が挙げられる。本発明では、非常に多種類の発酵培地を使ってスフィンガン類の生産を実施することができる。
【0067】
発酵ブロス中の炭水化物含量はいろいろあるが、通常は発酵培地に対して約1〜5重量%である。炭水化物類は発酵前あるいは発酵中のいずれかで全部を一どきに添加する。窒素用量は水性培地に対して約0. 01%〜約0. 4重量%の範囲である。炭素源と窒素源は1種のみの炭素源又は1種のみの窒素源を使用しても良いし、何種類かの炭素源又は窒素源を混合して使用してもよい。
【0068】
スフィンゴモナス菌の発酵に用いる無機塩類としては、ナトリウム、カリウム、アンモニウム、硝酸塩、カルシウム、リン酸塩、硫酸塩、塩化物、炭酸塩及びこれらに類似するイオン類を含有する塩類が挙げられる。マグネシウム、マンガン、コバルト、鉄、亜鉛、銅、モリブデン、ヨウ化物及びホウ酸塩などの微量金属も、含有すると有利である。また、ビオチン、葉酸塩、リポ酸塩、ナイアシンアミド、パントテン酸塩、ピリドキシン、リボフラビン、チアミン及びビタミンB12などのビタミン類、ならびにこれらの混合物もまた、有利に使用することができる。
【0069】
発酵温度は約25℃〜約35℃であるが、至適生産性を得るには約28℃〜約32℃の温度範囲がよい。接種源は、振盪フラスコ培養と小規模の浸漬攪伴発酵を含む、標準的な方法で容量をスケールアップして調製する。接種源の調製用培地は、生産用培地と同じものでも良いし、また当業者間で周知のいくつかの標準培地の一つであるルリア ブロス培地や、YM培地であってもよい。種培養では、炭水化物濃度を約1重量%より低くするとができる。所望の接種量を得るためには、種培養工程を一工程よりも多くすることができる。典型的な接種量は、最終発酵総量に対して約0. 5%〜約10%の範囲である。
【0070】
典型的には、発酵容器に内容物を攪伴する攪伴機を備える。容器に自動pH制御装置と自動発泡制御装置をつけてもよい。生産培地を容器に添加して、加熱によりその場で殺菌する。かわりに、添加前に、炭水化物や炭素源を別個に殺菌してもよい。予め増殖しておいた種培養を冷却してある培地(一般には、発酵温度の約28℃〜約32℃)に添加し、培養物を攪伴しながら約48〜約96時間発酵して、高粘度のブロスを調製する。標準法によりスフィンガン多糖類をアルコール、一般的にはイソプロパノールで沈澱させて、ブロスから回収する。
【0071】
本出願は、具体例として、数種の菌株、特にスフィンゴモナス菌株S88、S60、NW11、S198、S7、及びS194(それぞれ、寄託番号ATCC31554、ATCC31461、ATCC53272、ATCC31853、ATCC21423、ATCC31961としてアメリカン タイプ カルチャー コレクション(American Type Culture Collection)から利用することができる)から単離したDNAセグメントや、フラグメントを開示する。これらのDNAセグメントや、フラグメントを染色体外(プラスミド)DNAとして多数のコピーをスフィンゴモナス菌株中に組み入れることにより、これら菌株が生産するスフィンガンS−88、S−60及びNW−11を増産する際に有用であることが見いだされている。
【0072】
スフィンゴモナス菌株S88の場合、単離された染色体DNAセグメントの大きさは34kbpで、23から25の遺伝子を含む。図1のS88のクローン地図に示す通り、34kbp領域とは二つのクローン、c3とc2を合わせた広さである。この34kbpのDNA配列を複数のDNA配列に分けて、そのおのおのにcl△3、c2、c3、c4、c5、c6、H15. 6、B7. 1、B8.6、E5. 9、E1. 5、E2. 4、E4. 5、E6. 6、E12. 8など標識したものを、図1に示す。図6、図9及び図10は、単離されたS88染色体DNAをさらに詳細に示す。
【0073】
スフィンゴモナス菌株S60の場合、染色体DNAからc1、c2及びc3(図2を参照)を始めとする複数のDNA配列が単離された。配列c2をクローニングした後、pRK311ベクター中に置き、スフィンゴモナス菌株S88、S60及びNW11に挿入して、スフィンガン生産活性を評価した(実施例9に詳述してあるので、参照のこと)。
【0074】
スフィンゴモナス菌株S198、S7及びS194の場合、これらの菌株からもDNAセグメントを単離した(図7及び8を参照されたい)。これらの菌株からは、本願明細書で詳述する一般方法によりさらに多くのDNAセグメントを単離して、本発明により細菌のスフィンガン生産を増産するのに利用することができる。
【0075】
スフィンゴモナス菌株NW11の場合、DNAフラグメントcl、c2、c2. 1、c2. 2、c2Hd、c2H1、c2H2、c2H3、c2H10を単離した(図3参照)。配列c2. 2をクローニングした後、pRK311ベクター中に置いて、スフィンゴモナス菌株S88、S60及びNW11に挿入して、スフィンガン生産活性を評価した(実施例9に詳述してあるので、参照のこと)。
【0076】
スフィンゴモナス菌株S88、S60及びNW11から、上記で一般的に説明した方法により次のDNAセグメントを調製した。これらのDNAセグメントおのおのを(プラスミドベクター中の全長のDNAセグメントとして)、一以上のスフィンゴモナス菌株(野生型スフィンガン生産菌、又は野生型生産菌から派生した非ムコイド性変異菌)に挿入すると、それらの細菌はスフィンガン多糖類の大量生産菌に変わる。これらのDNAセグメントやフラグメントおのおのは、スフィンゴモナス菌株から単離されるDNAセグメント又はフラグメントから派生させ、前述したようにプラスミドベクター中に挿入する。DNAセグメント又はフラグメントの特徴は、制限酵素切断部位地図によって定まる(図1、2及び3)。
【0077】
菌株 S88
pRK311−S88cl△3
pRK311−S88c2
pRK311−S88c3
pRK311−S88c4
pRK311−S88c5
pSEB24−S88H15. 6
pSEB24−S88B8. 6
pSEB24−S88E4. 5
pSEB24−S88E6. 6
菌株 S60
pRK311−S60c2
菌株 NW11
pRK311−NW11c2. 2
【0078】
DNAクローンは、スフィンガン生産能を持つ野生型スフィンゴモナス菌、あるいはスフィンガン生産能を持たない変異菌に導入して、大量生産効果を実現することができる。例えば、DNAクローンの多数のコピーを、スフィンガン生産能を持つ野生型菌株や、スフィンガン生産能を持つ菌株から派生した非ムコイド性変異菌おのおのに導入する。このように操作した細菌は、同じ菌株由来のスフィンガン生産能を持つ野生型菌、あるいはスフィンガン生産能のない変異菌と比較すると、スフィンガンの大量生産菌である。
【0079】
DNAをスクリーニングして多数のコピーを、これらのスフィンゴモナス菌の菌株に導入したところ、予期しないところであったが、このようにして得た組換え菌のスフィンガン生産量は、一般的に野生型菌のスフィンガン生産水準よりも高かった。これは一般性を持つ現象であり、しかもこのスフィンガンの増産には菌株間の相補性が認められた。
【0080】
本発明においては、DNAクローンの多数のコピーを挿入して得る組換え菌は、野生型より高い水準のスフィンガン多糖類合成活性を持っている。また、本発明で有用なDNAセグメントは、菌株がスフィンガンを組み立てたり、生合成する初期工程で初めのグルコース残基をイソプレニルリン酸担体に結合するのに必要なタンパクをコードしているDNAフラグメントを始めとして、スフィンゴモナス菌株がスフィンガンを合成するのに有益あるいは不可欠な遺伝子を持っている。
【0081】
spsB遺伝子のDNA配列(配列番号1)と、SpsBタンパクの推定アミノ酸配列(配列番号2)も開示する。SpsBタンパク(又は、スフィンゴモナス菌株によっては、特にSgeB、SnwB、SneB、SssB、及びSrhBなどの相似タンパク)をコードしているDNAを含有するDNAフラグメントはすべて、多数のコピーを組み入れてスフィンゴモナス菌株を操作された菌とし、この操作された菌によってスフィンガンを増産することができる。
【0082】
同様に、S60スフィンゴモナスから単離されるsgeB遺伝子(SpsBに相似するSgeBタンパクをコードする)及びNW11スフィンゴモナスから単離されるsnwB遺伝子を含有するDNAセグメントやフラグメントについても同じことが言える。sgeB遺伝子(図2)は、S88染色体DNAのspsB遺伝子(及び、相当するsneB、sssB、srhB遺伝子、それらはそれぞれスフィンゴモナスのS198、S7及びS194株に由来し、SneB、SssB、SrhBタンパクをコードする)と(S88のspsB遺伝子に相当するフラグメントとのDNAハイブリッド形成に基づいて)相似するタンパクをコードしていると信じられている点で、お互いに相似している。図3に示すsnwB領域は、SnwBタンパクをコードするDNA配列に相当する。また、snwB遺伝子は、S88染色体DNAのspsB遺伝子及びS60染色体DNAのsgeB遺伝子と相似する。これらのDNAフラグメントは、上記で一般的に述べたようにプラスミド中に挿入して、スフィンゴモナスがスフィンガンを増産するための多数のコピーを形成する。
【0083】
スフィンゴモナスS88のspsB遺伝子はグルコシル−IPトランスフェラーゼをコードしていると信じられている。SpsBタンパクの推定アミノ酸配列は、異なる属の菌の推定グリコシル−IPトランスフェラーゼと、かなりの相同性があることが立証されている。この推定アミノ酸配列は一般にグリコシル−IPトランスフェラーゼをコードしていると信じられている他の遺伝子の配列と似ているが、この類似性がspsB遺伝子はグルコシル−IPトランスフェラーゼをコードするということの最も強い証拠となっている。実際、グルコシル−IPトランスフェラーゼのカルボキシル部分と、ガラクトシル−IPトランスフェラーゼのカルボキシル部分とには、かなりの相同性がある。アミノ末端領域にはこのように大きな相同性はないが、SpsBタンパクには膜貫通領域であると示唆されている疎水性の広がりが複数存在している点で、S. エンテリカ(S. enterica)のRfbPタンパクと似ている(1991, Jiang, etal.,Mol. Microbiol.,5, 695)。SpsBの疎水性領域は、35〜39の位置のアミノ酸(平均ハイドロパシー+2. 2)、68〜86の位置のアミノ酸(+1. 7)、105〜123の位置のアミノ酸(+2. 3)及び282〜303の位置のアミノ酸(+2. 9)を含んでいる。後者の疎水性領域の位置はこれらの関連遺伝子産物に共通していて、最大相同領域の隣に存在している。
【0084】
本発明の最良の実施態様において、DNAセグメントやフラグメントには、S88、S60、NW11、S130、S194、S198、S657及びS7を始めとするスフィンゴモナス菌の各種菌株の、グリコシル−IPトランスフェラーゼをコードするDNA配列が含有されている。これらのDNAセグメント及びフラグメントの多数のコピーを、受容菌となるスフィンゴモナス菌中で使用して、この受容菌がスフィンガンをより多く生産できる利得を実現することができる。グリコシルトランスフェラーゼ酵素をコードする遺伝子又はDNAフラグメントの組み込みに加えて、糖シントン又は糖前駆体(すなわち、スフィンガンポリサッカライドの構成部分を含み、スフィンガンの生合成に使われる糖成分、例えばグルコース、ガラクトース、ラムノース、マンノース、他の糖シントンや前駆体)をコードする遺伝子又はDNAフラグメント、又は、重合を促進したり、無処理の細胞構造からの多糖類分泌を促進する酵素や蛋白質をコードする遺伝子又はDNAフラグメントを好適に本発明で使うことができる。
【0085】
次の実施例により本発明を説明する。如何なる方法によっても実施例の説明を本発明の範囲を限定するものと誤解してはならない。
【実施例】
【0086】
次の実施例1〜21では、細菌の菌株、プラスミド及びバクテリオファージを表2に列記する。ルリア−ベルタニ培地及びYM培地を標準培地とする(Pollock,et al.,1994,J.Bacteriol.,176:6229−6237)。抗生物質(Sigma社)の用量は次の通りである。バシトラシン(Bac)は、特に本例では、73単位/mg又は0.01〜8mg/ml。リファンピシン(Rif)は、50μg/ml。ストレプトマイシン(Stm)は、50μg/ml。カナマイシン(Kan)は、25μg/ml。クロラムフェニコール(Cam)は、15μg/ml。テトラサイクリン(Tet)は、4〜12μg/ml。
【0087】
【表2】
【0088】
実施例1
スフィンゴモナスのDNAセグメントライブラリーの構成
スフィンゴモナス菌株から、スフィンガン類の合成に不可欠なDNAセグメントのクローンを形成した。次のようにして、DNAセグメントの完全なライブラリーを調製した。YM液体培地25ml中に菌株(この実施例ではS88)を置き、30℃で一晩振盪して、多量の細胞を含有する粘液性ブロスを作成した。
YM培地には、水1リットルあたり、バクト(Bacto)酵母エキス3g、バクト麦芽エキス3g、バクトペプトン(Difco)5g、及びD−グルコース( Difco)10gが含有されていた。アジ化ナトリウムを0. 01%まで加え、さらにスフィンガナーゼ(sphinganase)酵素(1994, Mikolajczak,et al.,Appl. Environ. Microbiol.,60,402)を37℃で8時間添加し、スフィンガンエクソポリサッカライドを消化して、粘度を部分的に低下させて細胞の増量を図った。細胞を遠心分離し、Birnboim and Doly,Nucl. Acids Res.,7,1513(1979)の方法によりDNAを抽出するために再懸濁した。ライセート(分離物)に等量のフェノール:CHC13:イソアミルアルコール(24:24:1)混液を加え、25℃で16時間緩やかに振とうし、その後、1容量のCHC13:イソアミルアルコール(24:1)混液を加え、25℃で3時間緩やかに振とうした。このようにして澄明としたライセートからタンパクを除いた。3Mの酢酸ナトリウム(pH5. 2)1/10容量を加え、高分子量のDNAを2容量のエタノールで沈澱させた後、乾燥して、さらにTE(10mMのトリス・HCl(pH8)、1mMのEDTA)0. 5ml中に再懸濁した。
【0089】
Loftus, Foster and Ross(BioTechniques, 12,172,1992)のコスミドクローニング計略により、S88DNAをSalI酵素により部分的に消化し、トリス・酢酸塩・EDTA緩衝液中、1%の低融点アガロースで電気泳動した。20kbpより大きなフラグメントをフェノール抽出とエタノール沈澱により精製した。このSalIで消化したS88DNAは、クレノウDNAポリメラーゼで処理して、dCMPとdTMPを付着端に添加し、70℃で20分間加熱した後、エタノールで沈澱させた。ベクタープラスミドpRK311(1985, Ditta, et al.,Plasmid,13, 149〜153)は、BamHI酵素で完全に消化し、65℃で15分間加熱して、フェノール抽出とエタノール沈澱により精製した。このようにしてBamHIで消化したpRK311DNAは、クレノウDNAポリメラーゼで処理して、dGMPとdAMPを添加して、上記のようにして精製した。T4DNAリガーゼによるライゲーション反応に含まれていたベクターと挿入フラグメントのモル量は等しかった。すべての制限酵素、クレノウDNAポリメラーゼ、及びT4DNAリガーゼはストラタジーン(Stratagene)社から入手し、このメーカーの反応条件を使用した。バクテリオファージ(ストラタジーン社のGigapackTMIIXL)中にパッケージングした後、ライゲーションした分子をトランスフェクションにより大腸菌DH5αTMに転移し、細胞を濃度4〜12μg/mlのテトラサイクリンを含有するLBプレート上に広げた。テトラサイクリン耐性コロニー1700個のうちの1つのライブラリー、及びテトラサイクリン耐性コロニー3400個のうちの1つのライブラリーのそれぞれを別個にプールして、凍結した。テトラサイクリン耐性コロニーは(試験した10個のうち10個が)内部にSalI制限酵素部位をもつ、長さ25〜30kbpの挿入断片を含有していた。
【0090】
同様にして、NW11、S60、S198、S194、及びS7を含む他の各種スフィンゴモナス菌株から、染色体DNAセグメントのライブラリーを作成した。この場合、クローニングしたDNAの供給源である細胞を、0. 5%w/v以下のグルコースを含む培地中で増殖させ、スフィンガナーゼ処理は省いた。
【0091】
実施例2
スフィンガンS−88生合成用DNAフラグメントの単離
プラスミド中にクローニングしたDNAのフラグメントのスクリーニングを行い、スフィンガン陰性変異菌がスフィンガン合成を復活するか、否かを観察することにより、スフィンガンS−88の合成に不可欠な遺伝子が存在するか、否かを判定した。かねて本発明者らは、バシトラシン500〜800μg/mlを含有するYM皿上で増殖できるスフィンゴモナスS88菌株の自然バシトラシン耐性変異菌のほとんどが、スフィンガン多糖類を生産できないことを見いだしていた(Pollock, et al.,1994, J. Bacteriol.,176, pp. 6229〜6237)。この所見に基づいてこの特別クラスの変異菌の簡便スクリーニング法を完成した。
【0092】
変異菌S88m260はこのSps- Bacr 群の代表的な変異菌である。S88m260その他のSps- Bacr 変異菌がエクソポリサッカライドを生産できなかった際のコロニーの外観は、野生型コロニーと比較して、平坦で、表面が粗く半透明であった。また、野生型のコロニーは光にかざして下から見ると狭い、光を屈折するハローで取り囲まれていた。これらの表現型の違いは、X. カンペストリスにおいて、野生型におびただしいムコイドがあるのに対して、対応するGum- 変異菌は平坦な外観を持っている違いほど明白ではなかった。コロニーの表現型は、培養物を液体YM培地で増殖し、エクソポリサッカライドをイソプロピルアルコールで沈澱し、乾燥計量して確認した。いくつかのSps- 変異菌はバシトラシンに感受性を持っていて、後にスフィンガン合成に不可欠であり、しかもバシトラシン耐性表現型に関する遺伝子とは区別されるべき遺伝子の特徴を備えていることが分かった。
【0093】
遺伝子ライブラリーからプラスミドDNAを取り、三元交配法により大腸菌からキサントモナスや、スフィンゴモナスに転移した(Ditta, et al.,Proc. Natl. Acad. Sci. USA,77, 7347, 1980)。Mob+ Tra- 組換えプラスミド(S88挿入断片を持つpRK311)を含有する供与菌の細胞、Mob+ Tra+ pRK2013プラスミドを含有するヘルパー細胞及びエクソポリサッカライド陰性の受容菌の細胞を比率5:2:10で混合した混液を、グルコースを入れない非選択的YMプレート中にスポットし、30℃で6〜16時間インキュベートした。次に、プレート中に、ヘルパー細胞と供与菌の細胞を排除するためのリファンピシン(50μg/ml)を入れ、かつ、組換えプラスミドを選択するために、pRK311用としてテトラサイクリン(4〜12μg/ml)、又はpSEB24用としてクラムフェニコール(20μg/ml)を入れた。これらのプレート上に上記の交配混液1白金耳を展開して、プラスミドを受容したキサントモナスの株を単離した。スフィンゴモナスは元来ストレプトマイシンに耐性を持っているので、スフィンゴモナスが受容菌のとき、供与菌及びヘルパー細胞に対してストレプトマイシンがスフィンゴモナスの選択に使われる。
【0094】
このようにして得たS88菌株のプラスミドを受容した株が、Sps+ (盛り上がって丸く不透明で、光にかざして下から見ると、明るい輪で取り囲まれている)あるいはSps- (平たく、粗く、半透明のコロニーで、輪がない)の、どちらであるか裸眼で判断して、その相補性を評価した。同様に、プラスミドを受容したキサントモナス菌から生育したGum+ のコロニーは、Gum- の受容菌の艶がなく、光らないコロニーと比べて、外観がムコイド性である。
【0095】
ライブラリー中のS88のクローンをS88の非ムコイド性変異菌中で直接同定しようとしたが、(ムコイドの表現型を明確に立証できなかったため)成功しなかった。そこでライブラリーを、一旦X. カンペストリスのgumD遺伝子の非ムコイド性変異体に転移してから、クローンを見いだす方法に切り替えた。これにより初期クローン「S88cl」を見いだすことができたが、これについては下記で詳しく説明する。
【0096】
S88遺伝子ライブラリーを大腸菌から、gumD遺伝子中にBacr Gum- が欠損しているX. カンペストリス菌株X59m31中に交配した(Pollock, et al.,J. Bacteriol.,176, pp. 6229〜6237, 1994; Thorneet al.,J. Bacteriol.,169,3593, 1987)。この属間交配では、いくつかのGum+ Tetr コロニーがYMプレート上で認められたが、このコロニーは約10-3〜10-4の頻度で出現すると考えられた。相補された変異体から、プラスミドをおのおの精製し、再び大腸菌に転移して制限酵素解析を行った。精製プラスミドをスフィンゴモナスS88m260に交配したところ、約10%の接合完了体(transconjugants)がSps+ となった。一つのプラスミド(pRK311−S88cl)を回収し、これを使用してさらに実験を続けた。プラスミドpRK311−S88clは、さらにいくつかのスフィンゴモナス菌株S88から別個に単離したBacr Sps- 変異菌、及びX. カンペストリスのBacr Gum- 変異菌とも相補した。属間交配ごとのプラスミドを受容した株が培地に分泌するエクソポリサッカライドを単離し、次に、薄層クロマトグラフィーを行い、酸水解物が受容菌の細胞の多糖類に存在している筈の糖残基を含有しているとの結果を得て、これらが多糖類であることを確認した。プラスミドpRK311−S88clは、スフィンゴモナスではスフィンガン合成を、X. カンペストリスではキサンタンガム合成をそれぞれ復活させた。これらの結果は、プラスミドpRK311−S88clは、Bacr 多糖類陰性変異菌が失っていたエクソポリサッカライド生合成機能をコードしていたこと、及びある属の遺伝子が他の属の失なわれていた機能を回復できたことを示している。セグメントS88clは、図1に示されるS88cl△3の最も右の末端に7. 5kbpのHindIIIフラグメントが付加されている点を除けば、S88cl△3と同一である。S88clからこの7. 5kbpセグメントを特異的に欠失させて、その派生物であるS88cl△3を産生した。
【0097】
上記で説明した、ガム生産(スフィンゴモナス属菌のスフィンガンガム、又はX. カンペストリスのキサンタンガム)の復活に有用なDNAフラグメントの測定方法には再現性があり、別個の実験では新しいクローンが単離された。スフィンゴモナスS88のSps- Bacs 変異菌76及び78を相補するセグメントを、クローンライブラリーから直接スクリーニングする際、S88clと部分的に重なり合っている三つの新しいクローンが形成された。これらのセグメントクローン3個は、おのおの長さが約23〜27kbであった。3個のセグメントのうち、二つは変異菌S88m260を相補した。制限酵素切断部位地図を図1及び図6に示す。
【0098】
上記の方法は、これらの菌株おのおののスフィンガンを増産するために有用な、スフィンゴモナス菌株S60、NW11、S198、S7、S194及びスフィンガン生産能を持つその他のスフィンガン菌株から単離したDNAセグメントの決定に用いられる。
【0099】
実施例3
spsB遺伝子のDNA配列とSpsBタンパクの推定アミノ酸配列
プラスミドpSEB24−S88E4. 5::Tn#72からサブクローニングした3300bpのフラグメントのうち、spsB領域の1950bpの二本鎖ヌクレオチド配列を得た。コード鎖の配列を配列表(配列番号1)に示す。1つの長い読み取り枠(open reading frame、ORF)があり、これをspsBと名付けた。コード領域はヌクレオチド361の位置のATGに始まり、1771の位置のTGA終止コドンまで続いていた。このORFはアミノ酸470個をコードし、ORFの先には推定リボソーム結合部位があった。標準三文字略記法による推定アミノ酸配列を配列表(配列番号2)に示す。
【0100】
実施例4
S88セグメントのクローンと、S88及びS60の染色体DNAとのDNA−DNAハイブリッド形成
プラスミドpRK311−S88cl△3のDNAクローンは、隣接するS88DNA配列から派生したものであることを立証するため、プラスミドS88cl△3DNAに標識を付け、スフィンゴモナス菌株S88、変異菌S88m260、及び野生型ジェラン生産菌であるS60から分離したDNA制限フラグメントとハイブリッド形成をおこなった。約1. 5、2. 4、4. 5、5. 9及び12kbpのEcoRIフラグメント中にハイブリッドを形成すると、野生型DNA及び変異菌S88DNA両方のDNAクローンの連続体と一致する。EcoRI部位の一つはベクターのマルチクローニング部位から始まっているので、重なり合っているクローンであるS88c2、S88c3、及びS88c4をEcoRIで消化すると、S88cl△3であることが証明されている最も左の6. 6kbpフラグメントは、実際には12. 8kbpとなる。S88DNAとジェランを生産するS60DNAとでハイブリッドを形成できることから、似ている遺伝子配列が存在しているが、遺伝子の組成が異なっている可能性があることが示唆された。これら二つのスフィンゴモナス菌株が分泌するエクソポリサッカライドの構造が似ているので、類似したトランスフェラーゼ遺伝子を持っていると考えられる。S88−S60の相同領域を図2に示す。別個のDNA相同性解析で、図3で示すように菌株NW11とS88の間の相同領域の位置を確定した。
【0101】
実施例5
菌株S60、NW11、S198、S7及びS194のスフィンガン生合成遺伝子クラスターのクローニング
スフィンゴモナスS60、NW11、S198、S7及びS194からDNAフラグメントを単離した。方法としては、菌株S88に関する上記実施例と同様にした。菌株S60、NW11、S198、S7及びS194のDNAフラグメントの制限部位地図は、それぞれ図2、図3、図9及び図10に示す。1種又は多種の制限酵素によるDNAの消化によって生じた制限酵素フラグメントの大きさを、2つの独立に単離されたクローンであるpRK311−S194c1及びpRK311−S194c2についてここで挙げる。この大きさは、そのフラグメントのアガロースゲル電気泳動の移動度を、大きさが既知のフラグメント、つまり、HindIIIで消化後のバクテリオファージラムダDNA、及びストラタジーンの「KbDNALadder」と比較して決定した。フラグメントの大きさは、上記の2つのクローンに含まれるDNA配列と一致する。
【0102】
スフィンゴモナスS194から単離したc1セグメントを含有するプラスミドpRK311−S194c1のフラグメントの大きさのデータを次に示す。pRK311−S194c1のフラメントの大きさは次の通りである。(EcoRI)>25、8.3、5.4及び1.5kbp。(EcoRI+HindIII)13.0、9.7、8.3、5.4、2.8及び1.5kbp。(HindIII)>25及び2.8kbp。(BamHI+HindIII)>25、5.8、3.9、2.0及び0.8kbp。(BamHI)>25、5.8及び4.7kbp。(BamHI+EcoRI+HindIII)13.0、8.3、5.8、5.4、3.9、2.0、1.5及び0.8kbp。(BamHI+EcoRI)15.0、8.3、5.8、5.4、4.7及び1.5kbp。pRK311ーS194c2のフラグメントサイズは次の通りである。(EcoRI)14.5、10.0、8.3、6.1、2.6及び1.35kbp。(EcoRI+HindIII)13、8.3、5.7、4.6、4.0、2.6、1.8、1.35、0.75、0.35.及び0.25kbp。(HindIII)>20、12.8、4.6、2.1、0.75及び0.25kbp。(BamHI+HindIII)>20、3.8、3.1、2.8、2.3、1.65、1.6、1.3、1.2、0.95、0.9、0.8及び0.25kbp。(BamHI)>20、5.8、3.1、2.8、2.6、2.3、2.0、1.6及び0.25kbp。(BamHI+EcoRI+HindIII)13、8.3、3.8、3.1、2.4、2.3、1.6、1.35、1.3、0.95、0.9、0.85、0.8、0.45、0.35及び0.3kbp。(BamHI+EcoRI)14.5、8.3、4.9、3.1、2.4、2.3、2.0、1.6、1.35、1.3、0.85、0.45及び0.2kbp。
【0103】
実施例6
プラスミドpSEB26の構成
特異的な複製機能と接合交配機能、並びに、スフィンゴモナス属に好適で、かつ、mini−Tn10kanと和合性がある薬剤耐性遺伝子を含有し、マルチクローニング部位を有する、プラスミドpSEB24とpSEB26(図5)を組み立てた。プラスミドpSEB24が広範囲宿主域を持つ反面、pSEB26は大腸菌では複製できるが、スフィンゴモナス属やキサントモナス属のどちらでも複製することができない。まず、スタフィロコッカス・アウレウスのプラスミドpC194(1982,Horinouchi and Weisblum,J.Bacteriol.,150,815)から取られた1031bpのHpaII−Sau3Aフラグメント上のCamr を、平滑末端に加工して、プラスミドpUC13(1982,Vieira and Messing,Gene,259)のXbaI平滑末端にライゲーションした。BamHI−SalIセグメント上のこのプラスミドからCamr カセットを取り除き、平滑末端にした後、pUC12中の独特なSspI部位と、二つあるPvuII部位のうちの最も近い部位(これも平滑末端)との間に挿入した。このようして、Ampr かつCamr であり、X−GalとIPTGとを添加すると青いコロニーを形成するプラスミドpSEB23を得て、pSEB24を構成するため、pSEB23から約2130bpのScaI−PvuIIフラグメントをとり、pMMB66EH(1986,Fuerste,et al.,Gene,48,119)のScaI−PvuII部分とライゲーションさせ、oriV(RSF1010からの広範囲宿主複製開始点)を保持させ、Ampr 遺伝子を再生した。プラスミドpNH−Kan/oriT(Hengen and Iyer,1992,Bio Techniques,13:57−62)のoriT配列を含む2700bpのHindIII−BamHIフラグメントは、3200bpのPvuIIで直鎖状にしたpSEB23プラスミドと、平滑末端同志で結合してpSEB26を形成した。BamHI−PvuIIのライゲーションにより再生したBamHI部位は、制限酵素処理に引き続き、付着末端にキャッピングを施し、再びライゲーションすることにより取り除かれた。
【0104】
実施例7
S88DNAフラグメントのコピーを菌株S88に導入したことによる多糖類S−88の増産
菌株S88から単離した図1に示すDNAセグメントの特異的な制限フラグメントを、DNAライゲーションによりマルチコピープラスミドベクター中に挿入した後、上記の実施例2で説明した三元交配により野生型菌株S88とS88の子である非ムコイド性変異菌に転移した。DNAクローンを変異菌に転移すると、多糖類合成が復活する。細菌を液体培地で培養した後、組換えプラスミド含有菌株とプラスミド遺伝子を付加しなかった菌株が生産したスフィンガン多糖類の累積量を測定した。じゃま板付きフラスコ(baffled flasks)中で振盪しながら30℃で24時間増殖した後、2容量のイソプロピルアルコールを加えて、エクソポリサッカライドを沈澱させた。各菌株ごとに2〜3回づつ独立に培養して、実験を行った。沈澱物をフィルターに集め、80℃で乾燥して、計量した。菌株毎の沈澱物の平均重量と標準偏差値を表3に示す。遺伝子のクローンのコピーを付加した組換え菌株のスフィンガンS−88生産量は、通常セットの生合成遺伝子しか持たなかった野生型菌株のそれよりも大きかった。
【0105】
【表3】
【0106】
制限酵素地図、spsB遺伝子のヌクレオチド配列及びこれを取り巻くDNAは、標準組換えDNA法によって約32kbの領域の新しいサブフラグメントを無数に形成するに充分な情報であることが、当業者にとり明白である(図6)。さらに、これらの新しいフラグメントを本明細書で説明した方法により試験して、スフィンガン多糖類を同様に増産するセグメントを同定できることも、当業者にとり明白である。表3から、このサブフラグメントが小断片であると(例えば、pSEB24−S88E12. 8)充分な生産刺激効果を示さないことが読み取れる。しかしながら、ほぼ全てのサブフラグメントは、増産を可能にしている。今日までの実験により、本発明者らは、pSEB24−S88B8. 6とpRK311−S88c3がスフィンガン生産に対して最大の刺激効果があると信じている。
【0107】
この増産法では、継続的に組換えプラスミドを選択するために、培養物に抗生物質を存在させておく必要はない。
【0108】
この増産法は、一以上のDNAフラグメントを染色体又は内因性の本来菌が持っていたプラスミドに挿入して、好ましくはこの染色体セグメントの単数又は複数の遺伝子をコードするDNAフラグメントを挿入することによって達成される。通常の技倆を持つ当業者であれば、内在の(resident)細菌染色体、内因性の本来菌が持っていたプラスミド、あるいは外因性プラスミドベクターに、スフィンガン生産能を持つ遺伝子を含有するDNAフラグメントを挿入することにより、スフィンガン生産能を持つ細菌にDNAフラグメントを導入することは容易である。本出願で立証されている試験結果では、このスフィンガン増産には、特定のプラスミドベクターの使用に対して依存性が認められないことは注目に値する。
【0109】
本発明の範囲には、上記で説明した遺伝子セグメント又は遺伝子群をコードするDNAフラグメントのどれかと、遺伝子の発現をコントロールすることで知られているDNA配列、本発明を限定することなく例を挙げると、大腸菌のlacプロモーターとを、融合させることも含まれている。
【0110】
実施例8
組換えS88菌株が生産したエクソポリサッカライドがスフィンガンS−88であることの同定
操作された菌株によって生産されたエクソポリサッカライドが受容菌の型と同一であることを確認するため、薄層クロマトグラフィーによって酸水解物中のモノサッカライド類を同定した。
【0111】
X. カンペストリスの細胞外キサンタンとスフィンゴモナスのスフィンガンS−88を、2〜3容量のイソプロピルアルコールにより沈澱させて、液体培地から分離し、80℃で乾燥して計量した。多糖類を、高性能液体クロマトグラフィー(HPLC)の水に5mg/mlの濃度で再懸濁した。0. 6ml超小型遠心分離管中で、無水トリフルオロ酢酸(88μl、Sigma ChemicalCo. から入手)を、75μlのHPLC用の水(Baker)及び225μlの多糖類(5mg/ml)と混合して、95℃で16時間インキュベートした。加水分解物を真空乾燥し、HPLCの水200μlに再懸濁して、新しい超小型遠心分離管に入れ、再び乾燥した後、HPLCの水45μlに懸濁した。サンプル(25μg/ml)は凍結して貯蔵した。糖標準液(D−グルコース、D−グルクロン酸、D−マンノース、L−マンノース、L−ラムノース及びL−フコース)を4mg/mlの濃度でHPLCの水に再懸濁した。コート済みで、チャンネルを形成した(channelled)シリカゲルクロマトグラフィープレート(Kieselgel 60 CF254、1020cm、E. Merck)を、一晩0. 3MのNaH2 PO4 中に浸漬した後、室温で30分間、さらに95℃で10分間乾燥した。サンプル1〜2μlをスポットし、室温で2. 5〜3時間アセトン40ml、ブタノール5ml及び脱イオン水5mlからなる溶媒混液にクロマトグラムをさらして、混液を上昇させた。プレートは65℃で3分間乾燥し、アセトン25ml、アニリン0. 5ml、ジフェニルアミン0. 5g及びリン酸3. 75mlからなる溶液に浸して染色した後、95℃で30分間乾燥した。X. カンペストリスがS88DNAフラグメントの受容菌である場合は、グルコース、マンノース及びグルクロン酸が存在した。スフィンゴモナス菌株S88がX. カンペストリスのgumD遺伝子の受容菌である場合は、ラムノース、グルコース、マンノース及びグルクロン酸がS−88エクソポリサッカライドとほぼ等量で存在した。
【0112】
実施例9
スフィンガンS−88、S−60、NW−11及びS−130の生産に対する、菌株S88、S60及びNW11から得たDNAフラグメントの刺激効果
上記の実施例5、6及び7で述べた一般法に従って菌株S88、S60及びNW11から得たDNAフラグメントを使用することにより、スフィンゴモナス菌株がスフィンガンS−88、ジェラン(S60)及びNW11を増産した。結果を次の表4に示す。
【0113】
【表4】
【0114】
一般法としては上記の実施例、特に実施例7で説明した技術や方法を用いて単離されたDNAフラグメントの多数のコピーを挿入することにより、特定のスフィンゴモナス菌株から単離されたDNAフラグメントを使って、異なるスフィンゴモナス菌株に(初めの菌株が生産するものとは異なる)スフィンガンを増産させようとするものである。
【0115】
本実験で確立された一般法においては、いずれかのスフィンゴモナス菌株のスフィンガン生産に不可欠な遺伝子材料を含有するDNAフラグメントを、適当なプラスミドベクターその他の手段に挿入する。このようにDNAを挿入したプラスミドベクターの多数のコピーを、DNAを単離したスフィゴモナスと同じ菌株あるいは異なる菌株のスフィンガン生産能を持つ菌又は非ムコイド性変異菌に導入する。この場合DNAが導入されたスフィンゴモナス菌株は、スフィンガン生産菌又はスフィンガン生産能を持つ菌のスフィンガン生産能を持たない変異菌のいずれでもよい。同一の発酵条件では、このように操作したスフィンゴモナス菌のスフィンガン生産量は一般に、操作しない、変異体ではないスフィンガン生産菌の生産量より大きい。
【0116】
次の実験により、DNAフラグメントの多数のコピーをスフィンゴモナス菌株その他の細菌に組み入れることは常法であることを証明する。
【0117】
実施例10
X. カンペストリスの染色体へのラクトース消費遺伝子(lactose−utilization gene)の挿入
制限酵素とDNAライゲーションを用いる標準クローニング法により、X. カンペストリス中で複製できないプラスミドに担持させた、X. カンペストリスの予めクローニングしたDNAセグメントに隣接して、トランスポゾンTn951から得られたラクトース消費遺伝子を挿入した(Thorne, et al.,J. Indust. Microbiol.,3, 321,1988)。その後、この組換えプラスミドを接合によりX. カンペストリス中に転移した。受容菌内では、ラクトース消費遺伝子の隣に位置するX. カンペストリスから得られた相同DNAが、通常の相同組換えによって細胞DNAと組換えられ、ラクトース消費遺伝子が細菌染色体と隣合わせに結合した。この結果、クローニングされたセグメントが細菌染色体に安定して挿入されることになった。このことは、上記の論文中で図を用いてより完全に説明されている。本発明以外でも、この方法によって遺伝子を細菌に挿入している例が数多く存在している。
【0118】
上記で説明したDNA組換えでは、事前に、プラスミドをベクターとして使用し、外因性DNAを細菌中に導入する必要がないことは注目に値する。DNAセグメントをバクテリオファージあるいはトランスポゾンに担持させると、部位特異的DNA組換えによって自然に細菌染色体に挿入されることが広く知られている。通常、バクテリオファージは特異的に1ないし2〜3個の位置で挿入するに過ぎないが、トランスポゾンは通常、多くの、実質的に任意の位置で挿入することができる。また、プラスミドなどのクローニングベクターと結合したDNAを細胞内に輸送する必要はない。DNAフラグメントは形質転換によって、細胞DNAと組換える能力を保持しながら細菌細胞内に入ることができることがよく知られている。
【0119】
実施例11
任意に選択された数個の異なる位置において、カナマイシン耐性をコードする遺伝子を、スフィンゴモナスS88菌DNAへ部位特異性挿入する
この実施例においては、クローニングされたDNAのセグメントをスフィンゴモナスの細胞DNAに挿入することは、容易に実施できるものであることを証明する。まず、S88E12. 8フラグメント(図1参照)を、プラスミドベクターpSEB26のマルチクローニング部位内のEcoRI部位にライゲーションした(図7)。プラスミドpSEB26は、図5で示すpSEB24のように、アンピシリン耐性遺伝子、クロラムフェニコール耐性遺伝子、マルチクローニング部位、Lacセグメント及びoriTを持っている(図5参照)。しかしながら、プラスミドpSEB24が広範囲宿主域のoriV配列を持つが、プラスミドpSEB26は狭い宿主域の複製開始点を持つということで、プラスミドpBR322とは異なる。pBR322の複製開始点は、大腸菌内におけるプラスミドの複製はできるが、スフィンゴモナス内では複製できない。したがって、プラスミドpSEB26内にクローニングされたDNA配列がスフィンゴモナス内で存続する唯一の方法は、プラスミドpSEB26が細胞あるいは培養物から消失する前に、プラスミドpSEB26を染色体や、内因性プラスミドなどの細菌DNAに組み込むことである。次に、このようにして得たpSEB26−S88E12. 8プラスミドを、常法によりトランスポゾンmini−Tn10kan(1991, Kleckner, et al.,204, pp. 139〜180)との突然変異生成にさらした。非抑制的な宿主HMS174における転位による突然変異誘発は、ラムダバクテリオファージNK1316によって担持されるTn10派生物103(mini−Tn10kan/Ptac−ATStransposase)による。その結果、カナマイシン耐性遺伝子がプラスミドに挿入された。また、S88E12. 8セグメント内のそれぞれ異なる位置にあるカナマイシン耐性遺伝子の数個の別個の挿入断片を単離した。次に、このようにして得た組換えプラスミドおのおのを、三元交配法によりスフィンゴモナスS88内に別個に転移して、カナマイシン耐性の子孫を選択した。事実上すべての場合、カナマイシン耐性遺伝子の両側に位置するS88DNA配列が受容細胞の染色体と組換えられ、カナマイシン耐性遺伝子が細菌染色体に組み込まれた。細菌染色体に組み込まれることにより、ベクタープラスミドがスフィンゴモナス内で複製することができないにも拘らず、カナマイシン遺伝子は存続することができた。
【0120】
この実施例は、外因性DNA配列(この場合、カナマイシン耐性遺伝子)を受容菌に導入して、導入遺伝子を細菌染色体に組み込むことができることを示している。組み込みの位置は、その遺伝子に隣接するDNA配列によって決定される。この実施例では、外因性遺伝子は菌株S88のE12. 8セグメント内の、相異なる部位に組み込まれた。相同組換えによる組み込みは、日常的に実施されている遺伝子操作法である。
【0121】
実施例12
DNAの配列決定の追加実験とその解析
図6に示すようにDNAの両鎖について、BamHI部位の1bpから28.804bpまでの間の配列を決定した。サンガー(Sanger)のジデオキシヌクレオチド鎖停止法(Sanger,et al.,1977,Proc.Nat.Acad.Sci.,74:5463−5467)が、エクソヌクレアーゼIIIとSIヌクレアーゼを用いてpBluescriptIIKS(+)上にクローニングされたDNAの配列決定に用いられた。内部シーケンシングプライマーも使用した。さらに、CoralSoftware社(San Diego)のSuperClone及びSuperSeeプログラム、並びにKyteand Doolittleの方法(Kyte and Doolittle,J.Mol.Biol.,157:105−132)により、膜貫通タンパク質領域の配列を解析した。また、「blastp」プログラム(Altschul,et al.,1990,J.Mol.Biol.,215:403−410)により、NCBIの膨大なデーターライブラリーから相同タンパク質セグメントを同定した。ナイロン膜とGeniusTM1キット(BoehringerMannheim)を用いる標準法により、DNAハイブリダイゼーションを行った。
【0122】
実施例13
スフィンガンS−88生合成に関与する遺伝子のクローニング
本実施例は一部を実施例2と同様にして行った。本発明者らは、かねてから、スフィンゴモナス菌株S88のスフィンガン多糖陰性(Sps- )変異株のほとんどは、バシトラシンを含有するYMプレートでも増殖できることを見いだしていた(Pollock,et al.,1994,J.Bacteriol.,176:6229−6237)。マッピング実験(後述)の結果、これらの変異株をすべてspsBと名付ける遺伝子に分類した。図9及び図10に列記した代表的なSpsB- 変異株(260、265及び102w)は、バシトラシン耐性(Bacr )でもあった。反対に、bac8菌株は、始めBacr として選択された代表的な菌株であったが、同時にSps- でもあることが示された。実施例2で述べたように、Sps- Bacr 変異株でスフィンガンS−88を形成しようとする試みは失敗に終り、代わりに野生型コロニーよりも平坦で、表面が粗く、半透明の外観を持つコロニーが得られ、いずれのコロニーも下から光にかざして見ると、光を屈折する狭いハローによって囲まれていた。アルコールに沈澱するエクソポリサッカライド類が出現しなかったので、Sps- 変異株はスフィンガン類を液体YM培地中に分泌できないものと判断した。Sps- Bacr 変異株の小さいフラクションは、第二の突然変異を持っていた。例えば、変異株102wのコロニーは黄色ではなく白色であった。変異株134ではspsBばかりでなく、rhsDでも変異があった。変異株54と302では、spsBのみでなくspsKもが欠損していた。図9の遺伝子地図の真上に示しているSps-変異には、自然発生のものと、紫外線又はエチルメタンスルホン酸にさらして変異させたものとがある。本実施例で実験したこの他の突然変異は、すべて(接頭字「Y」又は「B」を付けて、図9、10に示している)、トランポゾンmini−Tn101kanの無作意挿入により形成したものである。
【0123】
広範囲宿主コスミドpRK311に、スフィンゴモナス菌株S88から遺伝子ライブラリーを構築し、プールしておいたクローンを接合交配により大腸菌からS88m260に転移した。しかしながら、Sps+ のコロニーは、スクリーニングした103 〜104 のTetr プラスミドを受容した株に、またも、生じなかった。そこで、ライブラリーを予め単離してあったX.カンペストリスのBacr Gum- (gumD)変異株(Pollock,et al.,1994,J.Bacteriol.,176:6229−6237、及び、Thorne,et al.,1987,J.Bacteriol.,169:3593−3600)に転移した。gumD遺伝子は、キサンタンガムを組み立てる第一工程で、グルコース−6−リン酸をUDP−グルコースからイソプレニル−リン酸に転移するために必要である(Capage,et al.,October 1987,International PatentWO87/05938、及び、Ielpi,et al.,1993,J.Bacteriol.,175:2490−2500)。X.カンペストリスのgumD遺伝子を含有するDNAセグメントによって、スフィンゴモナスのBacr Sps- 変異株がスフィンガンS−88合成を回復できたことから、スフィンガンS−88の組み立ても、グルコースで始まり、スフィンゴモナスからの酵素は、X.カンペストリスのBacr Gum- 変異株を相補できるかもしれないと考えた(Pollock,et al.,1994,J.Bacteriol.,176:6229−6237)。属間交配からは、YMプレート上のTetr プラスミドを受容した株103 〜104 につき約1の頻度で、X.カンペストリスのGum+ コロニーが認められた。
【0124】
大腸菌を用いた形質転換によって、相補されたX.カンペストリス変異株からプラスミドを精製した後、スフィンゴモナスS88変異株260に交配した。プラスミドを受容した株のうち約5〜25%がSps+ となった。ベクター単独(pRK311)の場合の方が、より大きな組替えプラスミドの場合よりも、転移の頻度が100倍ないし1000倍高かった。ほとんどの組替えプラスミドはスフィンゴモナスと交配させると、大きな欠失が発生するが、pRK311−S88c1だけは、欠失を起こすことなくポリマー生産能を回復し、これを使用して実験を継続した。図1、図6、図9及び図10ではS88c1の21kbpからなる最左側部分をサブクローンc1△3として示している。プラスミドpRK311−S88c1によって、各々別個に単離した菌株S88のBacr Sps-変異株及びX.カンペストリスのBacr Gum- 変異株の数種が多糖の合成を回復した。属間交配の度に、プラスミドを受容した株が培養基中に分泌する多糖類を単離して、薄層クロマトグラフィーを実施し、酸水解物が受容細胞の多糖として期待されている中性糖を含有していることを確認した。プラスミドpRK311−S88c1を持つスフィンゴモナス変異株260が分泌するエキソポリサッカライドにはグルコース、マンノース及びラムノースが含有されていたが、プラスミドpRK311−S88c1を持つX.カンペストリス変異株m31からのものにはグルコースとマンノースしか含有されていなかった。さらに、加水分解条件下のため酸性糖の回収量は一貫して低かったが、多糖は各々グルクロン酸を含有していた。
【0125】
ライブラリーをスクリーニングしてSps- Bacr 変異株76及び78を相補するセグメント求め、図9及び図10の左側方向にクローニングした領域を広げていった。プラスミドを受容した株104 〜106 をスクリーニングして、部分的にS88c1セグメントと重なり合うさらに4個のクローン(S88c2、c3、c4及びc5)を得た。同様にして、Sps- 変異株43、71及び104を相補することにより、クローンc6を単離した。これら5個のクローニングされたセグメントは、各々長さが22〜28kbである。各セグメントの少なくとも1末端を図10に示す。さらに、クローンc2とサブクローンc1△3のどちらにも相補されない約15個からなる1群のSps- 変異株を同定した。これらの変異株の中で、c1△3よりも約8kbpだけ長く右側に広がっているc1全長クローンにより相補されたものは一つもなかった。また図3のc2よりも約18kbpだけ長く左側に広がっているc6によって相補されたものも一つとしてなかった。このように「同一連鎖群から外れた」(「unlinked」)1群の変異株は、スフィンガン合成に不可欠であるが、しかし図9及び図10に示す遺伝子クラスターとは直接に連続しない、さらに他の遺伝子の存在を示唆している。
【0126】
実施例14
機能的相補によるsps遺伝子のマッピング
Sps- 点変異株を接合の受容菌として用いる相補性検定により、spsG、spsK、spsF、spsD、spsC、spsE、spsB及びrhsD遺伝子の境界を決定した。結果を要約して図9に示す。サブクローニングされた小さいセグメント又はそれより大きいDNAセグメントのを持つ組替えプラスミドを、大腸菌の中でmini−Tn10kanによる挿入突然変異にさらし、菌株S88のSps- 変異株との交配によって転移した。これには交配可能な広範囲宿主プラスミドベクターであるpRK311及びpSEB24(図5)を使用した。担持しているプラスミドの耐薬性遺伝マーカーを受け取ったプラスミドを受容した株は、コロニーの外観によって、Sps+ 又はSps- と記録した。Bacr Sps- S88変異株は始めE4.5サブクローンとして位置づけ、その後、E4.5セグメントをmini−Tn10kanの無作意挿入変異にさらした。位置B231及びB230(図9及び図10)に挿入した時には、変異株260がスフィンガン合成を回復するのに影響しなかったが、B233、B239及びB238に挿入した時には相補が阻止された。変異株134は、セグメントB8.6によって相補されたが、E4.5又はE5.9のいずれによっても相補されなかった。変異株134では、spsB遺伝子と、さらに隣接のrhsD遺伝子の両方が欠損していた。B8.6セグメントをmini−Tn10kanによる突然変異にさらし、B441、B440、B438、B437及びB435に挿入することによる相補性のパターンを分析して、rhsD突然変異のより正確な位置を測定した。変異株54及び302も、spsKとspsB遺伝子が欠損している二重変異株であると考えられた。spsF変異株(図9の62、68及び94)は局在し、かつ、セグメントB12.6及びc1△3によって相補されず、またクローンc3とc5によってスフィンガンS−88の合成が回復されるため、隣接のspsDCEと分けた。E6.6フラグメントの挿入突然変異に続いて、連続しているspsD、spsC及びspsE遺伝子が相補された結果も併せて図9及び図10に示す。この相補結果から変異株76と78を含むグループと、変異株69、72、b104、3、9及び41を含むグループの二つのグループがあることが示唆された。その後、DNA配列の分析によって、後者のグループは連続している二つの別個の遺伝子、spsCとspsEとに分けられた。spsG突然変異(図9の11、43、71、81及び104)は、c6クローンとフラグメントB12.6のB4.5サブフラグメントとによって相補された(図9及び図10を参照されたい)。B12.6セグメントの挿入突然変異によって、プラスミドY652、Y635、Y636、Y653、Y640及びY641が産生された。このうち、Y652とY641のみがspsG変異株を相補することができた。
【0127】
実施例15
mini−Tn10kanを染色体とプラスミドに挿入したことによる表現型
S88DNAクローンのセグメントを交配可能な狭域宿主Camr プラスミドpSEB26(図9及び図10)にライゲーションし、大腸菌内でmini−Tn10kanによる挿入突然変異にさらした後、接合によって野生型(Sps+)スフィンゴモナス菌株S88に転移した。これらのプラスミドはスフィンゴモナス属では複製できないため、Kanr 遺伝子を維持するためには、細菌の染色体との組み替えが必要である。Kanr 及びCams である組み替え体はプラスミド配列を保持しないため、この群のみを選択した。これらのDNA置換の物理的構造の確認は行っていないが、常に同一のプラスミド、菌株及び選択方法を使用することによって、部位特異性な染色体欠失を生じさせ、制限地図作製とDNAハイブリダイゼーションによってこれらの二重組替えを確認した。Kanr Cams 染色体の組み替え体のコロニーは、Sps+ 又はSps- と判定した(挿入体には接頭字「c」を付けて図9に示している)。Sps- 表現型を示す変異株(cY776、cY757、cY771、cY770、cY676、cB589、cB583、cB580、cB579、cB300、cY726、cY725、cY676、cY673、cY721及びcY602)については、二重組替えが実際に発生したと信じることが、合理的である。しかしながら、相同領域の一個所にて組換えの行われたSps+ 組み替え体は、プラスミド全体が染色体に組み込まれたため、その染色体では、ある遺伝子は欠損し、ある遺伝子は正常のままである可能性がある。
【0128】
相同領域の二個所にて組替えを行う際の不確実性を排除するため、染色体に大きな部位特異的欠失をつくった後、mini−Tn10kanの単独挿入したS88c2の全DNAセグメント、又は、S88c3の全DNAセグメントのいずれかを持つ複製プラスミドを導入して、一部の遺伝子を不活性化させた。挿入位置とこれによるSps+ 又はSps- の表現型を、接頭字「p」を付けて図9に示す。染色体の変異戦略とプラスミドの変異戦略とはお互い整合し、必須(- )領域と非必須(+ )領域の両方が観察された。
【0129】
実施例16
DNA配列:G+C含量、希少コドンの使用及び翻訳開始配列
28、804bpのDNA配列について、両鎖を測定した(配列番号3を参照されたい)。このクラスター中の典型的なスフィンゴモナス遺伝子の平均プロフィル(及び標準偏差)を、G+C含量のゆがみ、希少コドンの頻度、及び「シャイン−ダルガノ(Shine−Dalgarno)」配列又は翻訳開始配列によって決定した。(表5)。各遺伝子とも第三コドンの位置でGあるいはCの頻度が高いことが特色となっていた。この研究の初期において、rhsACBDオペロンのコドン、及びspsB、D、C及びE遺伝子のコドン2500個を解析して、一群のスフィンゴモナス属のまれにしか使われないコドンを同定した。希少コドン各々が、この群に存在する比率は全体に対して0.2%以下で、群の構成はAGA、AGG、CGA、TGT、GGA、ATA、CTA、TTA、TTG、AAA、TTT、CCA、CCT、AGT、TCA、TCT、ACA及びACTを含んでいた。大腸菌においては、通常、16SrRNAの3’末端を相補する配列に連続する下流から翻訳が開始される(Shaine and Dalgarno,2974,Proc.Natl.,Acad.Sci.USA,71:134)。S.ポウシモビリスDSM1098の16SrRNAを相補する「シャイン−ダルガノ」の相同配列は、TAAGGAGGTGである(Moore,et al.,1993,Lett.Appli.Microbiol.,17:115−118)。
【0130】
このクラスター中の遺伝子が平均遺伝子プロフィルに合致し、かつ、変異してSps- 表現型が生じたならば、そのものこそ「sps」なる名称を与えることができる。しかしながら、本発明者らのタンパク質類似性サーチからは、spsG、I及びF遺伝子の機能について、有意の示唆を得ることができなかった。さらにまた、典型的な遺伝子プロフィルを満足する読み取り枠(ORF)が4個あったが、コンピューターデーターバンクではタンパク質配列について有意の類似性を発見することはできなかった。しかしながら、これらの推定遺伝子が変異しても、目に見えて多糖合成に変化をおこさなかったため、同定していないフレームとの意味から、「Urf」なる略記を付けた。4個のUrf配列(32、26、31及び34)があり、推定されるタンパク質のキロダルトン単位の大きさに応じて名称を付けた。
【0131】
【表5】
【0132】
実施例17
グリコシルーIPトランスフェラーゼをspsB遺伝子として同定
紫外線放射又は化学的変異原にさらした後に単離されたSps- 突然変異のほとんどは、spsB遺伝子上のものであった。グリコシル−IPトランスフェラーゼをコードすると信じられている他の遺伝子産物とSpsBの推定アミノ酸は非常に似ているので、このSpsBタンパク質はスフィンガンS−88を組み立てる第一工程を触媒すると信じられている。図11は、グリコシル−IPトランスフェラーゼとガラクトシル−IPトランスフェラーゼと推定されるアミノ酸配列のアラインメントである。これらのタンパク質のC末端半分は、お互いにかなり相同性がある。N末端領域ではこれほど大きな相同性はないが、SpsBタンパク質には膜貫通ドメインであると示唆されている数個の疎水性の領域があるので(図11のアンダーライン部分)、SpsBタンパク質はS.エンテリカのRfbPタンパク質(Jiang,et al.,1991,MolecularMicrobiology,5,695−713)と相似している。SpsBの疎水性ドメインはアミノ酸35〜59(+2.2平均ハイドロパシー)、68〜86(+1.7)、105〜123(+2.3)及び282〜303(+2.9)を含んでいる。また、SpsBタンパク質の最後の疎水性セグメントの位置は、これらの関連遺伝子産物に共通していて、相同性が最大である領域に続いているタンパク質の中央部分に所在している。
【0133】
DNA配列から推定されるドメインをコードするspsBを、相補性検定により確認した。E4.5セグメント中のmini−Tn10kanの挿入を変化させることより、Bacr Sps- 変異株の相補に干渉するかどうかを観察した。mini−Tn10kanの挿入部位と、相補の結果としての「+」又は「−」を図9のspsB遺伝子の上部に示す。三つの部位のmini−Tn10kanの挿入では、Sps- 変異株S88m260(B233、B239及びB238)にスフィンガン合成を回復することができなかった。その反面、B231とB230を含む数個の隣接する部位に挿入したところ失われていた機能を回復することができた。
【0134】
実施例18
スフィンゴモナスS88のラムノース生合成オペロン
rhsACBD遺伝子によってコードされているタンパク質の推定アミノ酸配列は、dTTP及びグルコース−1−リン酸から4工程でdTDP−L−ラムノースを合成する、S.エンテリカのグループBの酵素及びX.カンペストリスの酵素とよく似ている(図12から図15)。本発明者らは、グルコース−1−リン酸、rhsA遺伝子によってコードされているチミジリルトランスフェラーゼ(thymidylyltransferase)及びrhsB、C及びD遺伝子によってコードされている連続した触媒工程には、従来からの命名法を用いた。しかしながら、スフィンゴモナス属のオペロンは次の4点で独特である。第一に、遺伝子の順序ACBD→が、S.エンテリカ(BDAC→)や、X.カンペストリス(BACD→)のいずれとも異なる。第二に、シストロン間領域がほとんど存在していない。開始コドンと終止コドンとが重なり合っているか、間隔が狭く、rhsA−ATGA−rhsC−TGATCCATG−rhsB−TGATG−rhsDとなっている。第三に、rhsACBDの平均G+C含量が比較的高く(66%)、三番目のコドンの位置では特に高く(89%)、しかもオペロンを通じて一様である。第四に、このクラスター中の周囲の遺伝子でも、またスフィンゴモナス属において関係がない他の種の遺伝子でも同様にG+C含量が高い。
【0135】
当初rhsクラスターからは変異株(#134)一個のみが単離されたが、同時にspsB遺伝子でも第二の突然変異が生じたように見えた。Rhs- 突然変異は、致死性である可能性があると思われた。したがって、rhsクラスター内で単一の突然変異を特異的に操作することによって、スフィンガン合成が阻止されるかどうかを試験した。まず、rhs遺伝子の端から端までspsD遺伝子にわたって大きく染色体を欠失させたS88変異株を構築し(図10の△Tn365)、失われたDNAを持ち、特定部位にmini−Tn10kanを挿入したプラスミドを導入した。プラスミドの挿入位置がspsD、C、E、B又はrhsオペロンにある場合には、細胞はSps- のままであった。しかしながら、Urf32、26、31、34、atrD又はatrBのいずれかに挿入した場合は、欠失突然変異の相補に干渉せず、その細胞はSps+ となった。
【0136】
実施例19
スフィンゴモナスS88のグリコシルトランスフェラーゼ
spsQ、spsK及びspsLの3遺伝子がグリコシルトランスフェラーゼをコードしているように見受けられる。しかしながら、タンパク質がこれ以外のグルコシルトランスフェラーゼ及びラムノシルトランスフェラーゼ類に対して限られた局所的な相同性しか示さなかったため、グリコシルトランスフェラーゼ類の糖特異性は、配列解析のみで決定することはできなかった。他の研究者らも着目しているように、グリコシルトランスフェラーゼは、同一の結合における同一の糖に付着する、単一の細菌由来の酵素でも全く互いに異なる(Glucksman,et al.,1993,J.Bacteriol.,175:7045−7055)。推定spsQ遺伝子産物は、ラムノシルトランスフェラーゼをコードすると信じられている大腸菌K−12のgndに隣接するorf11(Stevenson,et al.,1994,J.Bacteriol., 174:4144−4156)に類似し、さらに、R.メリロチのExoO及びExoUグルコシルトランスフェラーゼ(Reuber and Walker,1993,Cell,74:269−280)とも類似していた。spsQ遺伝子はスフィンガンS−88合成に必須である。S88c2セグメントを有するプラスミド内のspsQ遺伝子に(図9のZ206)、mini−Tn10kanを挿入して、突然変異を起こしたプラスミドをspsGSRQI遺伝子の染色体を欠失しているS88細胞に導入した。受容細胞は生存可能であったが、多糖の合成は阻止された。推定spsL遺伝子産物は、ドッドマトリックス分析による比較では、spsQ産物と類似し、さらにS.エンテリカのラムノシルトランスフェラーゼ(RtbN)及びYersinia pseudotuberculosisの推定アベクオシルトランスフェラーゼ(abequosyl transferase)ともいくらか局所的に類似していた(Liu,et al.,1995.J.Bacteriol.,177:4084−4088)。spsK類似タンパク質の検索を行ったが、圧倒的多数が共通して推定UDP結合部位を含有するグリコシルトランスフェラーゼであるという僅かな類似性を認めたに過ぎなかった。これには、UDPが関与している可能性があるので、グルコシル又はグルクロノシルトランスフェラーゼであることが示唆されている。トランスポゾン挿入によらない変異株54と302同様、spsK遺伝子に特異的な挿入を行った変異株(図9のpY882)は、生存可能ではあったが、多糖を作ることはできなかった。
【0137】
実施例20
細菌による多糖の分泌
必須遺伝子産物の配列が類似していることから、各種細菌によって多糖類を分泌させる共通の機構が考えらる。この配列の比較(表6)では、spsD、spsC、spsE、spsJ、及びspsSの五つものsps遺伝子がスフィンガン類の分泌に関与しうることを示していた。R.メリロチの「エクソ」タンパク質など、かなりの情報が蓄積しているタンパク質の配列関係を要約して表6に示す。しかしながら、機能的に関係のあるタンパク質類は、この表が示しているものよりも大きい集団である。51、29及び22個のアミノ酸からなるSpsDタンパク質の各々異なる三つセグメントは、それぞれExoFと29%、31%及び36%の同一性があった。開始と終止コドンが重なり合い(TGATG)、しかも互いに連続するspsCとspsE遺伝子は、ExoP内の二つの異なるドメインに似たタンパク質をコードしている。類似するSpsC−ExoP配列には、最近、細菌性O−抗原の鎖長測定で示されたモチーフ(PX2 PX4 SPKXIIGXMXG)が含まれていた(Becker,et al.,1995,Mol.Microbiol.,16:l91−203)。92、30及び19個のアミノ酸からなるSpsCの三つのセグメントは、ExoPのN末端半分から取った同様な順序の配列に、それぞれ22%、30%及び42%同一であり、75及び98個のアミノ酸からなるSpsEの二つのセグメントは、ExoPのC末端半分に32%及び29%同一であった。37、20及び44個のアミノ酸からなるSpsSの三つのセグメントは、ExoTにそれぞれ38%、55%及び23%同一であった。推定SpsJタンパク質は、KpsT、BexA及びABC担体と推定ヌクレオチド結合ドメインを共有していて、お互いにある程度の類似性を持っていた。spsR遺伝子は、スフィンガン合成に必要ではないが、その遺伝子産物は、細菌性及び真菌性多糖リアーゼに僅かに類似していた。したがって、spsR遺伝子は、細胞あるいは基質の表面のいずれかからグルクロン酸含有スフィンガン類を放出したり、あるいはポリマーを炭素源として再使用することに重要であるかもしれない。
【0138】
図9及び図10に示すように、自然点突然変異や、spsD、spsC、spsE及びspsS遺伝子へのmini−Tn10kanの挿入では、生存可能であるが、培養上清中にスフィンガンS−88を蓄積しなかった。一方、R.メリロチの類似遺伝子の突然変異(Harding,et al.,1993,J.Gen.Microbiol.,139:447−457)、及びX.カンペストリスの類似遺伝子の突然変異は、致死性であった。spsJの染色体にmini−Tn10kanを挿入すると、Sps- となる。しかしながら、染色体が大きく欠失している変異菌株中のマルチコピープラスミドに保持されたspsJに、mini−Tn10kanを挿入すると、Sps+ か、Sps- のいづれかになった。
【0139】
【表6】
【0140】
各タンパク質の分泌の役割に対しての参考文献
ExoF,ExoP,ExoT(Becker et al.,1995,Mol.Microbiol.,16:191、Horinouchi andWeisblum,1982,J.Bacteriol.,150,815、及び、Reuber and Walker,1993,Cell,74:269)
GumB,GumC,GumJ(Glucksmann,et al.,1993,J.Bacteriol.,175:7033、Becker et al.,1995,Mol.Microbiol.,16:191、及び、Glucksmann,et al.,1993,J.Bacteriol.,175:7045)
KpsD and KpsT(Wunder,et al.,1994,J.Bacteriol.,176:4025、及び、Smith,et al.,1990,Mol.Microbiol.,4:1863)
BexD,BexC,and BexA(Kroll,et al.,1990.Mol.Microbiol.,4:1853)
【0141】
実施例21
溶菌性又は毒性タンパク質のABC輸送体
二つの連続した遺伝子であるatrBとatrDが、spsクラスター内に位置していた。この2つの遺伝子は、溶菌性又は毒素様タンパク質のABC輸送体や、輸送用補助タンパク質をコードしていると考えられる、これまでに、溶血素遺伝子(hlyA)が、現在ではスフィンゴモナス属として再分類されているシュードモナス ポーシモビリス(Pseudomonas paucimobilis)に同定された。本発明者らの菌株では、ヒツジ赤血球を含有する寒天プレート上で決定的な溶血現象を検出できなかったので、今回は「hly」なる略称を使用しなかった。atrB遺伝子のDNA配列から推定したアミノ酸の約48%は、百日咳菌(Bordetella pertussis)のシクロリシン(cyclolysin)ABC担体のアミノ酸と同一であった。atrB遺伝子産物は、それぞれ溶血素と白血球毒素を輸送する大腸菌のHlyBタンパク質及びパスツレラ ヘモリチカ(Pasteurella haemolytica)のLktBタンパク質と非常によく類似していた。また、atrBのC末端半分は、R.メリロチのNdvAタンパク質のC末端半分や、ヒト多剤耐性タンパク質Mdr1内の二つの繰り返しATP結合ドメインを含む、上記以外の多くのABC輸送体に類似していた。atrD遺伝子産物の配列は、大腸菌のHlyDタンパク質及びP.ヘモリチカのLktDタンパク質の配列と類似していた。他の属由来の関連輸送遺伝子とは違って、スフィンゴモナスatrB及びatrD遺伝子と隣接したり、あるいはspsクラスター内に存在する溶菌性又は毒性の類似遺伝子はなかった。
【0142】
結論
一つの属の細菌の多糖陰性変異株を第二の属のDNAで相互に遺伝的に相補することは、最初にキサントモナス属とリゾビウム属との間で立証された(Borthakur,et al.,1988,Mol.Gen.Genet.,213:155)。この初期の研究では、寒天プレート上で粘液性が回復したことが観察された。後年になって、X.カンペストリスのgumD遺伝子とスフィンゴモナス菌株S88のspsB遺伝子との間でお互に属間で相補がおこったことが報告され(Pollock,et al.,1994,J.Bacteriol.,176:6229)、本発明者らも組成分析によって、供与菌の相補遺伝子により、受容菌がエクソポリサッカライドの合成を回復したことを立証した。
【0143】
本願の実験では、spsB遺伝子がグルコシルIP−トランスフェラーゼをコードすることは、スフィンガン類生合成の重要な工程であることを証明している。したがって、本発明により受容菌に組み入れるDNAフラグメントは、グルコシルIP−トランスフェラーゼ酵素(グルコシル−トランスフェラーゼ、ガラクトシル−トランスフェラーゼ、又は関連IP−トランスフェラーゼのどれでもよい)をコードしている遺伝子を含有することが好ましい。例えば、スフィンガンS−88を組み立てる最初の工程は、グルコース−Pを担体IPに転移することである可能性が最も高いと信じられている。したがって、この生合成反応を促進する酵素をコードする遺伝子を、本発明によりDNAフラグメントに含有させることが有利である。
【0144】
本発明者らの研究により、スフィンゴモナスS88の大きなsps遺伝子クラスターには、dTDP−L−ラムノースの生合成をコードするより小さなオペロン(rhsACBD)のあることが証明された。rhsオペロンと他のラムノースオペロンの配列が類似していることは、スフィンゴモナス属がL−ラムノースの合成に同じ4つの酵素工程を有することを示唆すると共に、rhsオペロンがDNAフラグメント中において不可欠であることを立証している。
【0145】
侵襲してくる多くの病原菌がカプセルで保護されているように、各種のスフィンガンエクソポリサッカライド類も防御的性格として解釈することができる。また、エクソポリサッカライド類は、細胞を基質に付着させる役割を果たしているとも考えられる。スフィンゴモナス属のスフィンガン生産能を持つ他の菌株も、本願明細書で詳述したS88クラスターと同様に組織だった遺伝子クラスターがあるので、本願明細書の実施例で説明した結果から、通常の技術を持つ当業者が、すべてのスフィンゴモナス属細菌から実用に使用できるDNAフラグメントを得ることができる。
【0146】
スフィンガン生産能力を有する供与菌のスフィンゴモナス属細菌からスフィンガンの生合成に有用又は不可欠な遺伝情報をコードするDNAを単離して、受容菌の細菌に挿入することによって、スフィンガン生産能力がない細菌、又は通常の生産能力しか持たない細菌(受容菌)を、容易にスフィンガンガムの大量生産菌に作ることができることを本願明細書の開示により示した。大量生産菌は容易に得ることができるし、またその方法もスフィンガン生産能力を持っていさえすれば、スフィンゴモナス属のどの菌株にも一般適用することができる。
【0147】
本発明の実施態様の一部において、グリコシル−IPトランスフェラーゼをコードする(スフィンガン炭水化物を組み立てる第一工程)S88のspsBなどの遺伝子その他のDNAフラグメントを含有する供与菌から得たDNAフラグメントを、本発明で有利に使用している。本発明の他の実施態様、特にラムノースが多糖の形成に重要な糖シントン又は構築材料(例えば、スフィンガンの場合は、S−88、ジェランの場合は、S−60、及びウェランの場合は、S−130)である実施態様において、本発明の大量生産菌によるある種のスフィンガン多糖類の生産を最大に増産するために、ラムノースオペロン又は遺伝子(S−88のrhsABCD遺伝子など)を含めて使用することが好ましい。さらに、グルコシルトランスフェラーゼをコードする遺伝子又はその他のDNAフラグメント、例えばスフィンゴモナス菌株S88のspsQ、spsK及びspsL、あるいは形成される最終的な多糖を分泌のためにコードする遺伝子又はその他のDNAフラグメント(例えば、スフィンゴモナス属菌株S88のspsD、C、E、J及びS)を含めて有利に使用し、これらの酵素を最終的な多糖の生産に貢献させることができる。さらにまた、上記の酵素又は機能のすべてをコードするその他のDNAフラグメント又は遺伝子も、所望の多糖次第では有利に使用することができる。
【0148】
通常の技術を持つ当業者が、本明細書の開示に従って容易にDNAフラグメントを得ることができ、これらのフラグメントを受容菌に組み入れて、スフィンガン大量生産菌を作ることが可能であることに注目すべきである。これらの大量生産菌は公知の簡単な遺伝操作方法を使って容易に作ることができる。
【0149】
〔微生物の寄託〕
下記に記す上から6番目までの微生物は、特許手続上の微生物の寄託の国際承認に関するブダペスト条約に従い、メリーランド州20852 ロックビル、パークラウンドライブ 12301に所在する、アメリカン タイプ カルチャーコレクション(American type Culture Collection)に国際寄託されている。寄託物の利用可能性に対する制限は、寄託物についての特許の付与と同時に、取り消し不能条件を付してすべて撤廃される。下から3番目以降の微生物は、メリーランド州のロックビルのアメリカン タイプ カルチャー コレクションから公的に入手することができる。
【0150】
【表7】
【0151】
上記で説明した実施例及び実施態様は、例示によって本発明を説明することを目的とするものであって、如何なる方法によってもこれをもって本発明を制限するものと見なしてはならない。通常の技術を有する当業者によって、本発明で説明する事項及び対象に各種の修正また変更を行うことは可能であるが、これらはすべて本発明で予め考慮されていて、本発明の技術的範囲内に含まれている。
【特許請求の範囲】
【請求項1】
スフィンガンを生産するスフィンゴモナス属菌のDNAから単離されたDNA配列であって、受容菌となるスフィンゴモナス属菌に該DNA配列の多数コピーを組み入れたとき、該受容菌に関連するスフィンガン多糖類の大量生産菌を産生する、上記受容菌となるスフィンゴモナス属菌がスフィンガンを生合成する際に有益又は不可欠な遺伝情報をコードする単離されたDNA配列。
【請求項2】
ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス属の細菌株から単離された請求項1に記載のDNA配列。
【請求項3】
ATCC31554、ATCC31461、ATCC53272からなる群から選ばれたスフィンゴモナス属の細菌株から単離された請求項1に記載のDNA配列。
【請求項4】
スフィンゴモナス菌株ATCC31554から単離された請求項3に記載のDNA配列。
【請求項5】
S88cl△3、S88c2、及びS88c3から選ばれたセグメントの制限酵素地図を持つ請求項4に記載のDNA配列。
【請求項6】
該セグメントの一部分である請求項5に記載のDNA配列。
【請求項7】
スフィンゴモナス菌株ATCC31461から単離された請求項3に記載のDNA配列。
【請求項8】
S60c1、S60c2、S60c3からなる群から選ばれたセグメントの制限酵素地図を持つ請求項7に記載のDNA配列。
【請求項9】
該配列のフラグメントである請求項8に記載のDNA配列。
【請求項10】
スフィンゴモナス菌株ATCC53272から単離された請求項3に記載のDNA配列。
【請求項11】
NWc2.1及びNWc2.2からなる群から選ばれたセグメント制限酵素地図を持つ請求項1に記載のDNA配列。
【請求項12】
該セグメントのフラグメントである請求項11に記載のDNA配列。
【請求項13】
該スフィンガンが次の一般式で表される請求項1に記載のDNA配列。
【化1】
式中Glcはグルコースであり、GlcAはグルクロン酸であり、Rhaはラムノースであり、Manはマンノースであり、Xはラムノース又はマンノースである。ZはGlc残基2に付着し、かつ、Zはα−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaである。WはGlc残基1に付着し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaである。下付き文字v及びyは0、0. 33、0. 5、0. 67、又は、1である。ポリマーの還元末端は、主鎖のX残基の側であり、vとyが0のときにはW及びZは主鎖の上に存在しない。
【請求項14】
グリコシルトランスフェラーゼ酵素をコードする遺伝子を含有する請求項1に記載のDNA配列。
【請求項15】
該グリコシルトランスフェラーゼ酵素がグリコシルIP−トランスフェラーゼである請求項14に記載のDNA配列。
【請求項16】
ラムノースオペロン又はラムノース遺伝子を含有する請求項1に記載のDNA配列。
【請求項17】
供与菌となるスフィンガンを生成するスフィンゴモナス属菌のDNAからDNA配列を単離すること、該配列は、供与菌となるスフィンガン生産菌がスフィンガンを生合成する際に有益又は不可欠な遺伝情報をコードすること、受容菌に該配列を挿入すること、該受容菌は、該配列の多数のコピーを組み入れること、該供与菌に関連するスフィンガン多糖類の生産量が同一の発酵条件下で高まっていることを含むスフィンゴモナス属菌の菌株から派生した細菌を操作して、スフィンガン多糖類の大量生産菌とする方法。
【請求項18】
該DNA配列を挿入する該細菌が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、ATCC53272からなる群から選ばれたスフィンゴモナス菌株のメンバーである請求項17に記載の方法。
【請求項19】
該DNA配列を挿入する該細菌が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株のメンバーである請求項17に記載の方法。
【請求項20】
該DNA配列が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項17に記載の方法。
【請求項21】
該DNA配列が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項17に記載の方法。
【請求項22】
該DNA配列が、スフィンゴモナス菌株ATCC31554から単離された請求項18に記載の方法。
【請求項23】
該DNA配列が、S88cl△3、S88c2、及びS88c3から選ばれたセグメントの制限酵素地図を持つ請求項22に記載の方法。
【請求項24】
該DNA配列が、該セグメントのフラグメントである請求項23に記載の方法。
【請求項25】
該DNA配列が、スフィンゴモナス菌株ATCC31461から単離された請求項18に記載の方法。
【請求項26】
該DNA配列が、S60c1、S60c2、及びS60c3からなる群から選ばれたセグメントの制限酵素地図を持つ請求項25に記載の方法。
【請求項27】
該DNA配列が、該セグメントのフラグメントである請求項22に記載の方法。
【請求項28】
該DNA配列が、スフィンゴモナス菌株ATCC53272から単離された請求項18に記載の方法。
【請求項29】
該DNA配列が、セグメントNWc2.1又はNWc2.2の制限酵素地図を持つ請求項28に記載の方法。
【請求項30】
該DNA配列が、該セグメントのフラグメントである請求項29に記載の方法。
【請求項31】
該スフィンガンが次の一般式で表される請求項17に記載の方法。
【化2】
式中Glcはグルコースであり、GlcAはグルクロン酸であり、Rhaはラムノースであり、Manはマンノースであり、Xはラムノース又はマンノースである。ZはGlc残基2に付着し、かつ、Zはα−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaである。WはGlc残基1に付着し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaである。下付き文字v及びyは0、0. 33、0. 5、0. 67、又は、1である。ポリマーの還元末端は、主鎖のX残基の側であり、vとyが0のときにはW及びZは主鎖の上に存在しない。
【請求項32】
グリコシルトランスフェラーゼ酵素をコードする遺伝子を含有する請求項17に記載の方法。
【請求項33】
該グリコシルトランスフェラーゼ酵素がグリコシルIP−トランスフェラーゼである請求項32に記載の方法。
【請求項34】
ラムノースオペロン又はラムノース遺伝子を含有する請求項17に記載の方法。
【請求項35】
スフィンゴモナス属菌の菌株から派生する細菌であって、該細菌がスフィンガン多糖類の生合成に有益又は不可欠の遺伝情報をコードしているDNA配列の多数のコピーを含有し、該DNA配列がスフィンガン生産菌であってその供与菌となるスフィンゴモナス属菌の染色体DNA又はプラスミドDNAから単離され、受容菌に挿入されることにより該受容菌がスフィンガン多糖類の大量生産菌となる細菌。
【請求項36】
ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から派生する請求項35に記載の細菌。
【請求項37】
ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から派生する請求項35に記載の細菌。
【請求項38】
該DNA配列が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項35に記載の細菌。
【請求項39】
該菌株が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれる請求項35に記載の細菌。
【請求項40】
該菌株が、スフィンゴモナス菌株ATCC31554である請求項35に記載の細菌。
【請求項41】
該DNA配列が、S88cl△3、S88c2、及びS88c3から選ばれたセグメントの制限酵素地図を持つ請求項35に記載の細菌。
【請求項42】
該DNA配列が、該セグメントのフラグメントである請求項35に記載の細菌。
【請求項43】
該菌株が、ATCC31461である請求項35に記載の細菌。
【請求項44】
該DNA配列が、S60c1、S60c2、及びS60c3からなる群から選ばれたセグメントの制限酵素地図を持つ請求項35に記載の細菌。
【請求項45】
該DNA配列が、該セグメントのフラグメントである請求項35に記載の細菌。
【請求項46】
該菌株が、ATCC53272である請求項35に記載の細菌。
【請求項47】
該DNA配列が、セグメントNWc2.1又はNWc2.2の制限酵素地図を持つ請求項35に記載の細菌。
【請求項48】
該DNA配列が、S60c1、S60c2、及びS60c3からなる群から選ばれたセグメントの制限酵素地図を持つ請求項35に記載の細菌。
【請求項49】
該スフィンガンが次の一般式で表される請求項35に記載の細菌。
【化3】
式中Glcはグルコースであり、GlcAはグルクロン酸であり、Rhaはラムノースであり、Manはマンノースであり、Xはラムノース又はマンノースである。ZはGlc残基2に付着し、かつ、Zはα−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaである。WはGlc残基1に付着し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaである。下付き文字v及びyは0、0. 33、0. 5、0. 67、又は、1である。ポリマーの還元末端は、主鎖のX残基の側であり、vとyが0のときにはW及びZは主鎖の上に存在しない。
【請求項50】
グリコシルトランスフェラーゼ酵素をコードする遺伝子を含有する請求項35に記載の細菌。
【請求項51】
該グリコシルトランスフェラーゼ酵素がグリコシルIP−トランスフェラーゼである請求項50に記載の細菌。
【請求項52】
ラムノースオペロン又はラムノース遺伝子を含有する請求項35に記載の細菌。
【請求項53】
スフィンゴモナス属菌によってスフィンガンを増産する方法において、
1) スフィンガン生産菌であって供与菌となるスフィンゴモナス菌株から供与菌となるスフィンゴモナス属菌に、該スフィンガン生合成に有益又は不可欠の遺伝情報をコードしているDNA配列の多数のコピーを組み込み、
2) 工程1で単離した該DNA配列の多数のコピーを受容菌となるスフィンゴモナス菌に組み入れること、
3) 工程2から得られた細菌を発酵ブロス中で培養してスフィンガンを生産すること、
及び
4) 工程3からスフィンガンを単離すること
を含む方法。
【請求項54】
該DNA配列を挿入する該細菌が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株のメンバーである請求項53に記載の方法。
【請求項55】
該DNA配列を挿入する該細菌が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株のメンバーである請求項53に記載の方法。
【請求項56】
該DNA配列が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項53に記載の方法。
【請求項57】
該DNA配列が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項53に記載の方法。
【請求項58】
該DNA配列が、スフィンゴモナス菌株ATCC31554から単離された請求項53に記載の方法。
【請求項59】
該DNA配列が、S88cl△3、S88c2、及びS88c3から選ばれたセグメントの制限酵素地図を持つ請求項58に記載の方法。
【請求項60】
該DNA配列が、該セグメントのフラグメントである請求項59に記載の方法。
【請求項61】
該DNA配列が、スフィンゴモナス菌株ATCC31461から単離された請求項53に記載の方法。
【請求項62】
該DNA配列が、セグメントS60c1、S60c2、又はS60c3の制限酵素地図を持つ請求項61に記載の方法。
【請求項63】
該DNA配列が、該セグメントのフラグメントである請求項62に記載の方法。
【請求項64】
該DNA配列が、スフィンゴモナス菌株ATCC53272から単離された請求項53に記載の方法。
【請求項65】
該DNA配列が、セグメントNWc2.1又はNWc2.2の制限酵素地図を持つ請求項64に記載の方法。
【請求項66】
該DNA配列が、該セグメントのフラグメントである請求項65に記載の方法。
【請求項67】
該スフィンガンが次の一般式で表される請求項53に記載の方法。
【化4】
式中Glcはグルコースであり、GlcAはグルクロン酸であり、Rhaはラムノースであり、Manはマンノースであり、Xはラムノース又はマンノースである。ZはGlc残基2に付着し、かつ、Zはα−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaである。WはGlc残基1に付着し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaである。下付き文字v及びyは0、0. 33、0. 5、0. 67、又は、1である。ポリマーの還元末端は、主鎖のX残基の側であり、vとyが0のときにはW及びZは主鎖の上に存在しない。
【請求項68】
該DNA配列はグリコシルトランスフェラーゼ酵素をコードしている請求項53に記載の方法。
【請求項69】
該グリコシルトランスフェラーゼ酵素がグリコシルIP−トランスフェラーゼである請求項68に記載の方法。
【請求項70】
ラムノースオペロン又はラムノース遺伝子を含有する請求項53に記載の方法。
【請求項71】
該組み込みの工程1の前に、該DNA配列を単離する工程を、該方法がさらに含む請求項53に記載の方法。
【請求項72】
ATCC69735、ATCC69744、ATCC31853、ATCC21423、及びATCC31961からなる群から選ばれたスフィンゴモナス菌の微生物の生物学的に純粋な培養物。
【請求項73】
大腸菌であるATCC69732、ATCC69733、又はATCC69734の生物学的に純粋な培養物。
【請求項74】
配列番号3に示す配列と実質的に一致するDNA配列。
【請求項1】
スフィンガンを生産するスフィンゴモナス属菌のDNAから単離されたDNA配列であって、受容菌となるスフィンゴモナス属菌に該DNA配列の多数コピーを組み入れたとき、該受容菌に関連するスフィンガン多糖類の大量生産菌を産生する、上記受容菌となるスフィンゴモナス属菌がスフィンガンを生合成する際に有益又は不可欠な遺伝情報をコードする単離されたDNA配列。
【請求項2】
ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス属の細菌株から単離された請求項1に記載のDNA配列。
【請求項3】
ATCC31554、ATCC31461、ATCC53272からなる群から選ばれたスフィンゴモナス属の細菌株から単離された請求項1に記載のDNA配列。
【請求項4】
スフィンゴモナス菌株ATCC31554から単離された請求項3に記載のDNA配列。
【請求項5】
S88cl△3、S88c2、及びS88c3から選ばれたセグメントの制限酵素地図を持つ請求項4に記載のDNA配列。
【請求項6】
該セグメントの一部分である請求項5に記載のDNA配列。
【請求項7】
スフィンゴモナス菌株ATCC31461から単離された請求項3に記載のDNA配列。
【請求項8】
S60c1、S60c2、S60c3からなる群から選ばれたセグメントの制限酵素地図を持つ請求項7に記載のDNA配列。
【請求項9】
該配列のフラグメントである請求項8に記載のDNA配列。
【請求項10】
スフィンゴモナス菌株ATCC53272から単離された請求項3に記載のDNA配列。
【請求項11】
NWc2.1及びNWc2.2からなる群から選ばれたセグメント制限酵素地図を持つ請求項1に記載のDNA配列。
【請求項12】
該セグメントのフラグメントである請求項11に記載のDNA配列。
【請求項13】
該スフィンガンが次の一般式で表される請求項1に記載のDNA配列。
【化1】
式中Glcはグルコースであり、GlcAはグルクロン酸であり、Rhaはラムノースであり、Manはマンノースであり、Xはラムノース又はマンノースである。ZはGlc残基2に付着し、かつ、Zはα−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaである。WはGlc残基1に付着し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaである。下付き文字v及びyは0、0. 33、0. 5、0. 67、又は、1である。ポリマーの還元末端は、主鎖のX残基の側であり、vとyが0のときにはW及びZは主鎖の上に存在しない。
【請求項14】
グリコシルトランスフェラーゼ酵素をコードする遺伝子を含有する請求項1に記載のDNA配列。
【請求項15】
該グリコシルトランスフェラーゼ酵素がグリコシルIP−トランスフェラーゼである請求項14に記載のDNA配列。
【請求項16】
ラムノースオペロン又はラムノース遺伝子を含有する請求項1に記載のDNA配列。
【請求項17】
供与菌となるスフィンガンを生成するスフィンゴモナス属菌のDNAからDNA配列を単離すること、該配列は、供与菌となるスフィンガン生産菌がスフィンガンを生合成する際に有益又は不可欠な遺伝情報をコードすること、受容菌に該配列を挿入すること、該受容菌は、該配列の多数のコピーを組み入れること、該供与菌に関連するスフィンガン多糖類の生産量が同一の発酵条件下で高まっていることを含むスフィンゴモナス属菌の菌株から派生した細菌を操作して、スフィンガン多糖類の大量生産菌とする方法。
【請求項18】
該DNA配列を挿入する該細菌が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、ATCC53272からなる群から選ばれたスフィンゴモナス菌株のメンバーである請求項17に記載の方法。
【請求項19】
該DNA配列を挿入する該細菌が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株のメンバーである請求項17に記載の方法。
【請求項20】
該DNA配列が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項17に記載の方法。
【請求項21】
該DNA配列が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項17に記載の方法。
【請求項22】
該DNA配列が、スフィンゴモナス菌株ATCC31554から単離された請求項18に記載の方法。
【請求項23】
該DNA配列が、S88cl△3、S88c2、及びS88c3から選ばれたセグメントの制限酵素地図を持つ請求項22に記載の方法。
【請求項24】
該DNA配列が、該セグメントのフラグメントである請求項23に記載の方法。
【請求項25】
該DNA配列が、スフィンゴモナス菌株ATCC31461から単離された請求項18に記載の方法。
【請求項26】
該DNA配列が、S60c1、S60c2、及びS60c3からなる群から選ばれたセグメントの制限酵素地図を持つ請求項25に記載の方法。
【請求項27】
該DNA配列が、該セグメントのフラグメントである請求項22に記載の方法。
【請求項28】
該DNA配列が、スフィンゴモナス菌株ATCC53272から単離された請求項18に記載の方法。
【請求項29】
該DNA配列が、セグメントNWc2.1又はNWc2.2の制限酵素地図を持つ請求項28に記載の方法。
【請求項30】
該DNA配列が、該セグメントのフラグメントである請求項29に記載の方法。
【請求項31】
該スフィンガンが次の一般式で表される請求項17に記載の方法。
【化2】
式中Glcはグルコースであり、GlcAはグルクロン酸であり、Rhaはラムノースであり、Manはマンノースであり、Xはラムノース又はマンノースである。ZはGlc残基2に付着し、かつ、Zはα−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaである。WはGlc残基1に付着し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaである。下付き文字v及びyは0、0. 33、0. 5、0. 67、又は、1である。ポリマーの還元末端は、主鎖のX残基の側であり、vとyが0のときにはW及びZは主鎖の上に存在しない。
【請求項32】
グリコシルトランスフェラーゼ酵素をコードする遺伝子を含有する請求項17に記載の方法。
【請求項33】
該グリコシルトランスフェラーゼ酵素がグリコシルIP−トランスフェラーゼである請求項32に記載の方法。
【請求項34】
ラムノースオペロン又はラムノース遺伝子を含有する請求項17に記載の方法。
【請求項35】
スフィンゴモナス属菌の菌株から派生する細菌であって、該細菌がスフィンガン多糖類の生合成に有益又は不可欠の遺伝情報をコードしているDNA配列の多数のコピーを含有し、該DNA配列がスフィンガン生産菌であってその供与菌となるスフィンゴモナス属菌の染色体DNA又はプラスミドDNAから単離され、受容菌に挿入されることにより該受容菌がスフィンガン多糖類の大量生産菌となる細菌。
【請求項36】
ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から派生する請求項35に記載の細菌。
【請求項37】
ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から派生する請求項35に記載の細菌。
【請求項38】
該DNA配列が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項35に記載の細菌。
【請求項39】
該菌株が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれる請求項35に記載の細菌。
【請求項40】
該菌株が、スフィンゴモナス菌株ATCC31554である請求項35に記載の細菌。
【請求項41】
該DNA配列が、S88cl△3、S88c2、及びS88c3から選ばれたセグメントの制限酵素地図を持つ請求項35に記載の細菌。
【請求項42】
該DNA配列が、該セグメントのフラグメントである請求項35に記載の細菌。
【請求項43】
該菌株が、ATCC31461である請求項35に記載の細菌。
【請求項44】
該DNA配列が、S60c1、S60c2、及びS60c3からなる群から選ばれたセグメントの制限酵素地図を持つ請求項35に記載の細菌。
【請求項45】
該DNA配列が、該セグメントのフラグメントである請求項35に記載の細菌。
【請求項46】
該菌株が、ATCC53272である請求項35に記載の細菌。
【請求項47】
該DNA配列が、セグメントNWc2.1又はNWc2.2の制限酵素地図を持つ請求項35に記載の細菌。
【請求項48】
該DNA配列が、S60c1、S60c2、及びS60c3からなる群から選ばれたセグメントの制限酵素地図を持つ請求項35に記載の細菌。
【請求項49】
該スフィンガンが次の一般式で表される請求項35に記載の細菌。
【化3】
式中Glcはグルコースであり、GlcAはグルクロン酸であり、Rhaはラムノースであり、Manはマンノースであり、Xはラムノース又はマンノースである。ZはGlc残基2に付着し、かつ、Zはα−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaである。WはGlc残基1に付着し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaである。下付き文字v及びyは0、0. 33、0. 5、0. 67、又は、1である。ポリマーの還元末端は、主鎖のX残基の側であり、vとyが0のときにはW及びZは主鎖の上に存在しない。
【請求項50】
グリコシルトランスフェラーゼ酵素をコードする遺伝子を含有する請求項35に記載の細菌。
【請求項51】
該グリコシルトランスフェラーゼ酵素がグリコシルIP−トランスフェラーゼである請求項50に記載の細菌。
【請求項52】
ラムノースオペロン又はラムノース遺伝子を含有する請求項35に記載の細菌。
【請求項53】
スフィンゴモナス属菌によってスフィンガンを増産する方法において、
1) スフィンガン生産菌であって供与菌となるスフィンゴモナス菌株から供与菌となるスフィンゴモナス属菌に、該スフィンガン生合成に有益又は不可欠の遺伝情報をコードしているDNA配列の多数のコピーを組み込み、
2) 工程1で単離した該DNA配列の多数のコピーを受容菌となるスフィンゴモナス菌に組み入れること、
3) 工程2から得られた細菌を発酵ブロス中で培養してスフィンガンを生産すること、
及び
4) 工程3からスフィンガンを単離すること
を含む方法。
【請求項54】
該DNA配列を挿入する該細菌が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株のメンバーである請求項53に記載の方法。
【請求項55】
該DNA配列を挿入する該細菌が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株のメンバーである請求項53に記載の方法。
【請求項56】
該DNA配列が、ATCC31554、ATCC31461、ATCC31853、ATCC21423、ATCC31555、ATCC31961、ATCC53159、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項53に記載の方法。
【請求項57】
該DNA配列が、ATCC31554、ATCC31461、及びATCC53272からなる群から選ばれたスフィンゴモナス菌株から単離された請求項53に記載の方法。
【請求項58】
該DNA配列が、スフィンゴモナス菌株ATCC31554から単離された請求項53に記載の方法。
【請求項59】
該DNA配列が、S88cl△3、S88c2、及びS88c3から選ばれたセグメントの制限酵素地図を持つ請求項58に記載の方法。
【請求項60】
該DNA配列が、該セグメントのフラグメントである請求項59に記載の方法。
【請求項61】
該DNA配列が、スフィンゴモナス菌株ATCC31461から単離された請求項53に記載の方法。
【請求項62】
該DNA配列が、セグメントS60c1、S60c2、又はS60c3の制限酵素地図を持つ請求項61に記載の方法。
【請求項63】
該DNA配列が、該セグメントのフラグメントである請求項62に記載の方法。
【請求項64】
該DNA配列が、スフィンゴモナス菌株ATCC53272から単離された請求項53に記載の方法。
【請求項65】
該DNA配列が、セグメントNWc2.1又はNWc2.2の制限酵素地図を持つ請求項64に記載の方法。
【請求項66】
該DNA配列が、該セグメントのフラグメントである請求項65に記載の方法。
【請求項67】
該スフィンガンが次の一般式で表される請求項53に記載の方法。
【化4】
式中Glcはグルコースであり、GlcAはグルクロン酸であり、Rhaはラムノースであり、Manはマンノースであり、Xはラムノース又はマンノースである。ZはGlc残基2に付着し、かつ、Zはα−L−Rha−(1−−6)−α−L−Rha、α−L−Man、又はα−L−Rhaである。WはGlc残基1に付着し、かつ、Wはβ−D−Glc−(1−−6)−α−D−Glc、又はα−L−Rhaである。下付き文字v及びyは0、0. 33、0. 5、0. 67、又は、1である。ポリマーの還元末端は、主鎖のX残基の側であり、vとyが0のときにはW及びZは主鎖の上に存在しない。
【請求項68】
該DNA配列はグリコシルトランスフェラーゼ酵素をコードしている請求項53に記載の方法。
【請求項69】
該グリコシルトランスフェラーゼ酵素がグリコシルIP−トランスフェラーゼである請求項68に記載の方法。
【請求項70】
ラムノースオペロン又はラムノース遺伝子を含有する請求項53に記載の方法。
【請求項71】
該組み込みの工程1の前に、該DNA配列を単離する工程を、該方法がさらに含む請求項53に記載の方法。
【請求項72】
ATCC69735、ATCC69744、ATCC31853、ATCC21423、及びATCC31961からなる群から選ばれたスフィンゴモナス菌の微生物の生物学的に純粋な培養物。
【請求項73】
大腸菌であるATCC69732、ATCC69733、又はATCC69734の生物学的に純粋な培養物。
【請求項74】
配列番号3に示す配列と実質的に一致するDNA配列。
【図1】

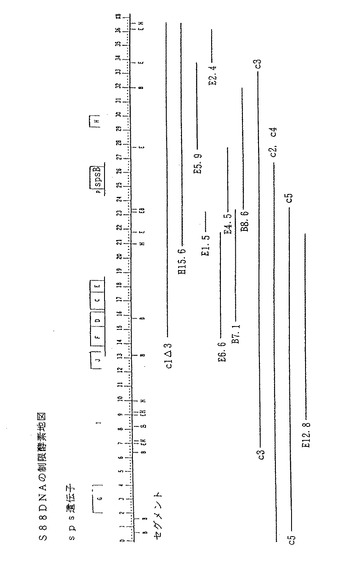
【図2】
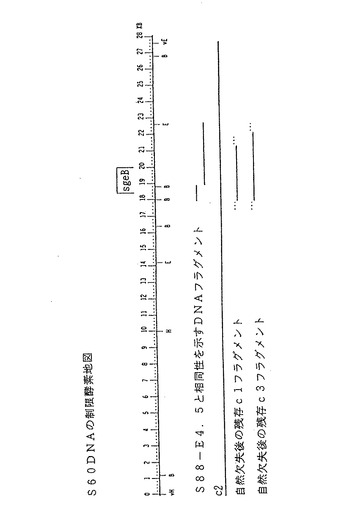
【図3】
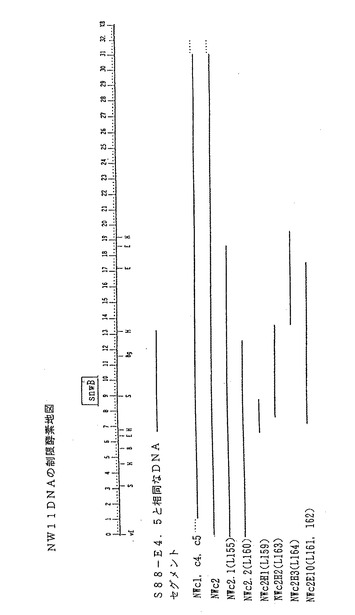
【図4】
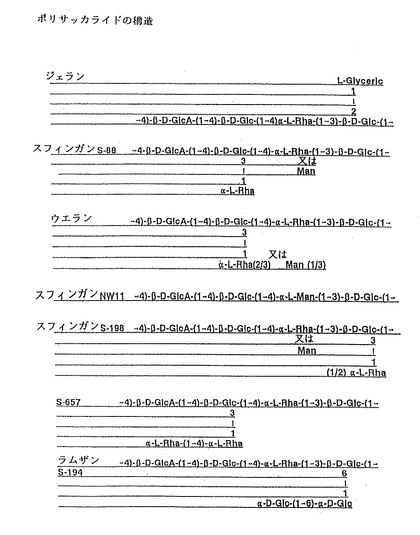
【図5】
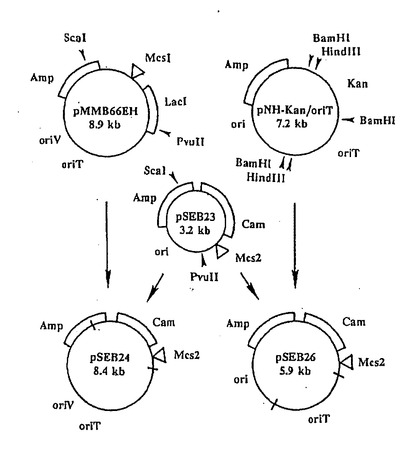
【図6】
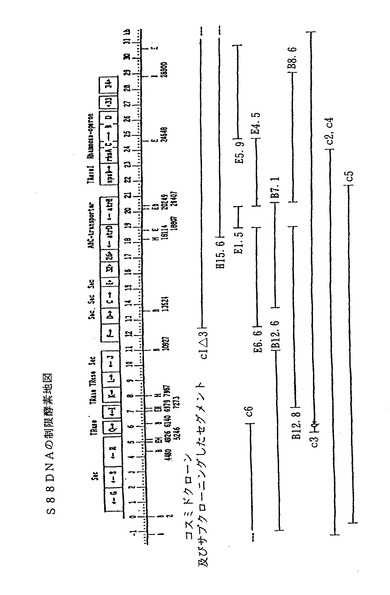
【図7】
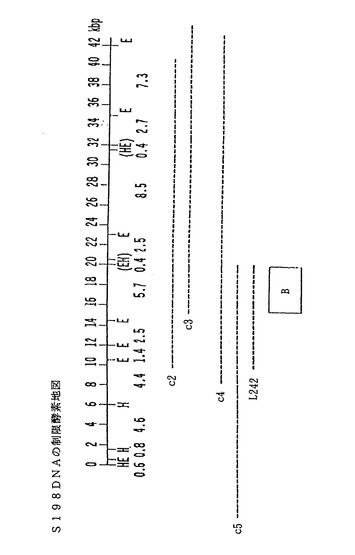
【図8】
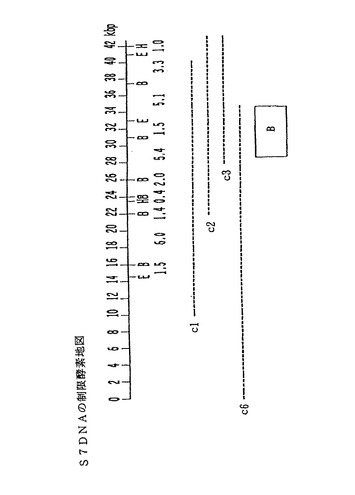
【図9】

【図10】

【図11】

【図12】
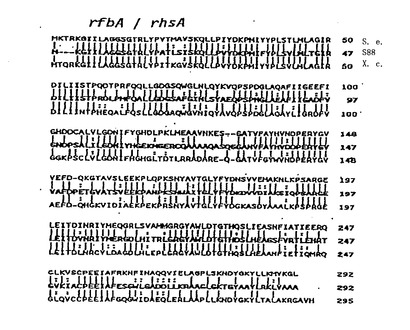
【図13】
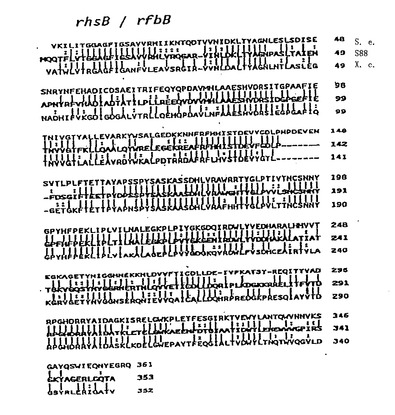
【図14】

【図15】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2011−155983(P2011−155983A)
【公開日】平成23年8月18日(2011.8.18)
【国際特許分類】
【出願番号】特願2011−62408(P2011−62408)
【出願日】平成23年3月22日(2011.3.22)
【分割の表示】特願平8−43977の分割
【原出願日】平成8年1月24日(1996.1.24)
【出願人】(506128352)シーピー・ケルコ・ユーエス・インコーポレーテッド (18)
【Fターム(参考)】
【公開日】平成23年8月18日(2011.8.18)
【国際特許分類】
【出願日】平成23年3月22日(2011.3.22)
【分割の表示】特願平8−43977の分割
【原出願日】平成8年1月24日(1996.1.24)
【出願人】(506128352)シーピー・ケルコ・ユーエス・インコーポレーテッド (18)
【Fターム(参考)】
[ Back to top ]