
DNA配列決定方法
【課題】1本鎖フラグメントレベルでポリヌクレオチドの配列を決定するための改良された方法の提供。
【解決手段】ポリヌクレオチドの配列を決定するための方法であって、該方法は、ポリヌクレオチドと相互作用し、ポリヌクレオチドに沿って進行する酵素における立体配座の変化を検出することに貢献する。立体配座の変化に関する検出は、酵素に結合する蛍光団における変化を測定して行った。本発明の一つの態様によれば、ポリヌクレオチドの配列を決定するための方法は、酵素活性を誘導するのに十分な条件下で、標的ポリヌクレオチドを、該ポリヌクレオチドと相互作用し、かつポリヌクレオチドに沿って進行し得る酵素と、反応せしめ、酵素がポリヌクレオチドに沿って進行する時に、酵素における立体配座の変化を検出することを含む。
【解決手段】ポリヌクレオチドの配列を決定するための方法であって、該方法は、ポリヌクレオチドと相互作用し、ポリヌクレオチドに沿って進行する酵素における立体配座の変化を検出することに貢献する。立体配座の変化に関する検出は、酵素に結合する蛍光団における変化を測定して行った。本発明の一つの態様によれば、ポリヌクレオチドの配列を決定するための方法は、酵素活性を誘導するのに十分な条件下で、標的ポリヌクレオチドを、該ポリヌクレオチドと相互作用し、かつポリヌクレオチドに沿って進行し得る酵素と、反応せしめ、酵素がポリヌクレオチドに沿って進行する時に、酵素における立体配座の変化を検出することを含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明の分野
本発明は、ポリヌクレオチド配列決定に関する。
【0002】
本発明の技術分野
ポリヌクレオチドの配列を決定するための能力は、科学的に非常に重要である
。例えば、ヒトゲノムプロジェクトは30億塩基ものヒトゲノムにコードされた
DNA塩基の地図化および配列決定するための野望的な国際的努力である。完了
すると、得られる配列のデータベースは生物医学的研究に他に比肩できない有用
な道具となる。このプロジェクトを成功裡に完成さすのに主な障害となるのは、
配列決定の過程で使用される技術に関する。
【0003】
大規模なDNA配列決定のために一般的に用いられる主な方法は、連鎖終止反
応法である。この方法は、最初はサンガーおよびコールソン(Sanger et al., Pr
oc. Natl. Acad. Sci.USA, 1977; 74: 5463-5467)によって開発され、ポリメラ
ーゼ反応において未完成のポリヌクレオチド鎖に導入される4つのヌクレオシド
三リン酸のジデオキシ誘導体を使用することによる。導入に際し、該ジデオキシ
誘導体がポリメラーゼ反応を終了し、次いで、該生成物をゲル電気泳動によって
分離し、分析すると、特定のジデオキシ誘導体の鎖中に導入された部位が明らか
になる。
【0004】
この方法は、広く使用されており、また信頼できる結果を生むが、時間と人手
がかかり、費用が高くつく方法であると認識されている。
【0005】
別の配列決定方法がEP-A-0471732で提案されているが、これは分光
学的手段を使用して、ヌクレオチドが標的に相補的な未完成のポリヌクレオチド
鎖に導入されるのを検出する。該方法は鋳型およびプライマーとの固定化複合体
を基にしている。該複合体を別異のヌクレオチドのただ1つのみを含有するフロ
ーに暴露する。次いで、分光学的技術を用い、鋳型コピーのポリメラーゼ触媒に
よる増幅から生じる時間依存的シグナルを測定する。記載の分光学的技術は、エ
バネッセント波場内での分析物の変化を測定する表面プラスモン共鳴(SPR)分
光法および蛍光測定技術である。しかし、この方法には限界が認識されており、
SPR技術に関して最も深刻な問題は、コピー鎖の大きさが増すににつれて、シ
グナルの絶対サイズもエバネッセント波場から鎖の運動により増加し、増加量を
検出するのが困難となることである。該蛍光測定技術は、増加しつつある未完成
ポリヌクレオチド鎖上に導入された蛍光団からのバックグラウンドの干渉を増加
させる欠点をもっている。鎖が増すにつれ、バックグラウンド「ノイズ」が増加
し、各ヌクレオチド導入を検出に要する時間を増加する必要がある。これにより
、大きなポリヌクレオチドの配列決定について使用するのが非常に制限される。
【0006】
1本鎖フラグメントポリヌクレオチド配列決定アプローチは、WO-A-992
4797およびWO-A-9833939に概説され、両方法は1本鎖標識ヌクレ
オチド分子の蛍光検出を用いるものである。これらの1本鎖ヌクレオチドは、エ
クソヌクレアーゼ分子の作用によって、光トラップ(Jett, et al., J. Biomol.
Struc. Dyn, 1989;7:301-309)によるフロー中に保たれた該鋳型ポリヌクレオチ
ドから切断される。次いでこれらの切断されたヌクレオチド類はクウォーツ・フ
ロー・セル内で下方にフローし、レーザー励起に供され、次いで鋭敏な検出シス
テムで検出される。しかし、この方法の限界が認識されており、エクソヌクレア
ーゼ技術に関して最も重大な限界は、該標識ヌクレオチドがエクソヌクレアーゼ
酵素の進行能(プロセッシビリティ)に非常に影響するという事実である。該方法
の別の限界は、ポリヌクレオチドフラグメントを固定化するために使用されたビ
オチンビーズに該ヌクレオチドを「付着すること」、従ってヌクレオチドのフロ
ーがフェーズをはずれるようになること;最初の酵素の標識化過程の非効率およ
び長さの制限;および4つの異なる染料分子間の該励起「交差」の結果として非
常にエラー比が高くなることを包含する。
【0007】
そのため、好ましくは1本鎖フラグメントレベルでポリヌクレオチドの配列を
決定するための改良された方法、即ち該ポリヌクレオチドの配列決定に関するフ
ラグメントサイズと速度を顕著に増加させ、好ましくは自動化過程により行い、
現方法に関連する複雑性および費用を低下させる方法が必要である。
【発明の概要】
【課題を解決するための手段】
【0008】
本発明の要約
本発明は、標的ポリヌクレオチドの配列を、標的ポリヌクレオチドに結合し、
標的ポリヌクレオチドに沿って進行する酵素における立体配座の変化を測定する
ことによって、決定し得るということに基づいている。立体配座の変化が起こる
程度は、標的上のどの個々のヌクレオチドが該酵素と接触しているかによって異
なる。
【0009】
本発明の一つの態様によれば、ポリヌクレオチドの配列を決定するための方法
は、以下を包含する:
(i) 酵素活性を誘導するのに十分な条件下で、標的ポリヌクレオチドを、該
ポリヌクレオチドと相互作用し、かつポリヌクレオチドに沿って進行し得る酵素
と、反応せしめ、
(ii) 酵素がポリヌクレオチドに沿って進行する時に、酵素における立体配座
の変化を検出すること。
【0010】
好ましい実施態様において、酵素は相補鎖を伸長する過程で標的と相互作用す
るポリメラーゼ酵素である。酵素は、規定領域内に反応を局在化させるために固
体支持体上に固定化する。
【0011】
本発明の第2の実施態様によれば、酵素は最初に結合された検出し得る標識を
包含し、酵素が立体配座の変化を進行するにつれて、その特徴を変化させる。ま
た、酵素は、最初の標識と相互作用し得る2番目に結合した検出し得る標識を包
含する。ここで、相互作用の程度は酵素における立体配座の変化に依存する。通
常、最初の標識はエネルギー受容体であり、2番目の標識はエネルギー供与体で
あり、立体配座の変化を検出することは、2つの標識間のエネルギー遷移を測定
することにより行う。
【0012】
本発明のさらなる実施態様によれば、蛍光共鳴エネルギー遷移(FRET)を用
いて、標的ポリメラーゼと相互作用し、標的ポリメラーゼに沿って進行する酵素
における立体配座の変化を検出する。それによりポリヌクレオチドの配列を決定
する。蛍光共鳴エネルギー遷移は、酵素にそれぞれ結合しているFRET供与体
および受容体標識との間で行い得る。あるいは、標識の1つは酵素に結合し、も
う一方の標識はポリヌクレオチドに結合してもよい。
【0013】
さらなる実施態様によれば、ポリヌクレオチド配列を決定するために、標的ポ
リヌクレオチドと相互作用し、標的ポリヌクレオチドに沿って進行し得る検出可
能な標識酵素を使用する。ここで、該標識は、酵素がポリヌクレオチドに沿って
進行するにつれて、その検出し得る特性を変化させる。
【0014】
さらなる態様によれば、固体支持体は、標的ポリヌクレオチドと相互作用し、
標的ポリヌクレオチドに沿って進行し得る少なくとも1つの固定化酵素を包含す
る。酵素は1以上の検出し得る標識物によって標識されている。
【0015】
さらなる態様によれば、ポリヌクレオチドの配列を決定するためのシステムは
、上記のような固体支持体を包含し、標識を検出するための装置を包含する。
【0016】
本発明は、従来の配列決定技術を超えるいくつかの利点を提供する。ポリメラ
ーゼ酵素は、一旦ポリヌクレオチドの伸長ラウンドを始めると、鎖から脱落する
前までに数千のヌクレオチドを重合する。さらに、ある特定のポリメラーゼシス
テムは、鋳型分子を囲む「スライディング・クランプ」(例えば、ポリメラーゼ
III)を介して、または部分的に囲む分子フック(例えば、T7:チレオドキシン
複合体)を介して、鋳型ポリヌクレオチドにシステム自身を固着し、係留するこ
とができる。
【0017】
本発明は、1回の試行で、1秒あたり数百もの塩基対の速度で数十キロベース
(kb)またはそれ以上の配列決定をし得る。これは、DNAの1本鎖フラグメン
トに関する配列決定の結果である。DNAの1本鎖フラグメントの配列決定に関
する利点は、配列決定速度が利用された酵素システムによって決定され、間接的
総合的反応により決定されるのではないので相当速いことである。速い速度と同
じくらい重要なことは、DNAの大きなフラグメントを配列決定することができ
る能力である。これは、配列決定情報のメガベースセグメントを集めるのに必要
とされるサブクローニングの量および重複配列の数を顕著に低下させる。1本鎖
フラグメントアプローチのさらなる利点は、現在配列決定の努力を阻害するアク
リルアミドのような有害不用物の廃棄に関する問題を排除することである。
例えば、本願発明は以下の項目を提供する。
(項目1) 下記の過程:
(i) 酵素活性を誘導するのに十分な条件下で、標的ポリヌクレオチドを、該ポリヌクレオチドと相互作用し、かつポリヌクレオチドに沿って進行し得る酵素と、反応せしめ、
(ii) 酵素が該ポリヌクレオチドに沿って進行する時に、酵素における立体配座の変化を検出すること、
を含むポリヌクレオチドの配列を決定するための方法。
(項目2) 酵素がポリメラーゼ酵素である、項目1記載の方法。
(項目3) 酵素がヘリカーゼ酵素またはプライマーゼ酵素である、請求項1記載の方法。
(項目4) 酵素が固体支持上で固定化された、項目1〜3記載の方法。
(項目5) 固体支持上で固定化された複数の酵素を含む、項目4記載の方法。
(項目6) 酵素が、最初に結合した検出し得る標識を含み、酵素が立体配座の変化を進行させるにつれて、その特徴が変化する、項目1〜5記載の方法。
(項目7) 酵素が、最初の標識と相互作用し得る2番目に結合した検出し得る標識を包含し、相互作用の程度は酵素における立体配座の変化に依存する、項目6記載の方法。
(項目8) 2番目の検出し得る標識が、酵素と接触することになるヌクレオチドに結合している、項目6記載の方法。
(項目9) 最初の標識がエネルギー受容体であり、2番目の標識がエネルギー供与体であり、もしくは逆で、過程(ii)は2つの標識間のエネルギー遷移の変化を測定することにより行い得る、項目7または項目8記載の方法。
(項目10) 過程(ii)は共焦顕微鏡を用いて実施する、項目1〜9記載の方法。
(項目11) 過程(ii)は蛍光画像化によって実施する、項目10記載の方法。
(項目12) 過程(ii)は最初の標識の変化した特性に対する分極効果の結果を測定することによって実施する、項目10記載の方法。
(項目13) 過程(ii)は蛍光分極異方性によって実施する、項目10記載の方法。
(項目14) 標的ポリメラーゼと相互作用し、標的ポリメラーゼに沿って進行する酵素における立体配座の変化を検出して、それによってポリヌクレオチドの配列を決定するための蛍光共鳴エネルギー遷移(FRET)の使用。
(項目15) 酵素がポリメラーゼ酵素である、項目14記載の使用。
(項目16) 酵素が固体支持体上に固定化される、項目14または15記載の使用。
(項目17) ポリヌクレオチド配列を決定するために、標的ポリヌクレオチドと相互作用し、標的ポリヌクレオチドに沿って進行し得る検出可能な標識酵素の使用であって、酵素がポリヌクレオチドに沿って進行する時に、標識がその検出し得る特性を変化させる検出可能な標識酵素の使用。
(項目18) 標的ポリヌクレオチドと相互作用し、標的ポリヌクレオチドに沿って進行し得る少なくとも1つの固定酵素を包含し、酵素が1以上の検出し得る標識物によって標識される固体支持体。
(項目19) 酵素がポリメラーゼである、項目18記載の固体支持体。
(項目20) 標識が蛍光団である、項目18または19記載の固体支持体。
(項目21) 項目18〜21記載の固体支持体および標識を検出するための装置を含むポリヌクレオチドの配列を決定するためのシステム。
【図面の簡単な説明】
【0018】
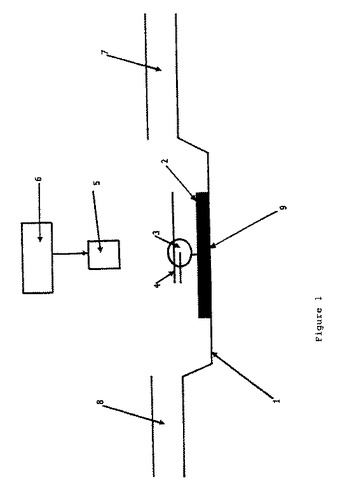
【図1】図1は、本発明で使用する共焦顕微鏡の配置のスキーム図である。
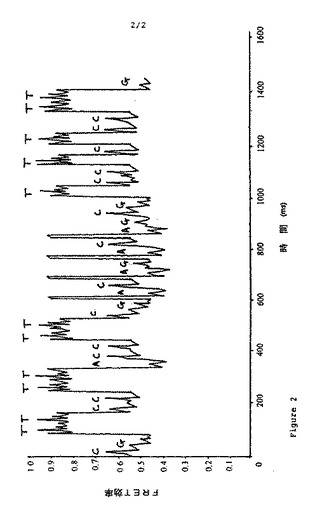
【図2】図2は、蛍光共鳴エネルギー遷移後にとったトレースを示すもので、各ピークは特定のヌクレオチドの検出を示す。
【発明を実施するための形態】
【0019】
本発明の説明
ポリヌクレオチドを配列決定するための本方法は、酵素と標的ポリヌクレオチ
ドとの間の立体配座の変化に関する分析を包含する。
【0020】
本明細書中で使用する「ポリヌクレオチド」なる用語は、広義に解釈されるべ
きであり、DNAおよびRNAならびに修飾DNAおよび修飾RNAを含み、さ
らに別のハイブリダイズする核酸様分子、例えばペプチドヌクレオチド核酸(P
NA)も含む。
【0021】
酵素は、ポリメラーゼ酵素であってもよく、立体配座の変化は、ポリメラーゼ
が標的ポリヌクレオチドに対し相補的な未完成鎖中にヌクレオチドを導入する場
合にもたらされる。立体配座の変化は、各々異なるヌクレオチド、A、T、Gま
たはCに対して異なり、そのため、該変化を測定することによりどのヌクレオチ
ドが組み込まれたかを同定するのである。
【0022】
あるいは、酵素は、ポリヌクレオチド、例えば、ヘリカーゼ酵素、プライマー
ゼおよびホロ酵素との相互作用に関与するものどれでもよい。酵素がポリヌクレ
オチドに沿って進行すると、その立体配座は、標的上のどのヌクレオチドに接触
するかに依存して変化する。
【0023】
酵素における立体配座の変化を検出する1つの方法は、適切なエネルギー供与
体標識と適切なエネルギー受容体標識との共鳴エネルギー遷移を測定することで
ある。一つの例において、供与体および受容体は酵素に各々結合し、標的ポリヌ
クレオチドとの相互作用によりもたらされる酵素における立体配座の変化が標識
の相対的位置を変える。位置付けにおける差異は、得られるエネルギー遷移を反
映しており、酵素と接触する特定のヌクレオチドの特徴となる。あるいは、1つ
の標識を酵素上に位置付け、もう一方の酵素を標的のヌクレオチド上または標的
に相補的な鎖上に導入されたヌクレオチド上に位置付けられ得る。
【0024】
蛍光共鳴エネルギー遷移(FRET)の使用は、本発明の好ましい実施態様であ
る。この技術は、2から8nmの距離を測定することができ、供与体蛍光団と受容
体蛍光団との距離依存性エネルギー遷移による。該技術は、優秀な静的共局在化
の能力を持つだけでなく、分子内および分子間FRETに対する2つの蛍光団の
距離または配向性における動的変化に対する情報も提供する。1つの供与体と1
つの受容体(一組のFRET)との間のエネルギー遷移についての最初の測定(Ha,
et al.,Proc. Natl. Acad. Sci. USA, 1996; 96:893)以来、該方法はリガンド
-受容体の共局在性(Schutz, et al., Biophys. J.,1998; 74:2223)を研究する
ため、触媒中の平衡タンパク質構造性変動および酵素−基質相互作用を調べるた
め(Ha, etal., 1999 上掲)、そして立体配座状態および溶液中の個々の拡散分
子の副固体数を同定するために使用されてきた。これらの変法の全ては、本発明
において適用し得るものと考えられる。
【0025】
本発明は、1つの標識のみを必要とする測定技術を使用しても実施し得る。1
分子レベルで酵素の局所環境における変化を測定することができる全てのシステ
ムは、本発明の許容される態様である。ポリヌクレオチドプロセシング酵素およ
び/またはその基質類と結合した1つの蛍光プローブの多様な特性は、本発明の
中で活用することができ、ヌクレオチドの合同事象に特異的である酵素システム
/分子環境中において、またはその非常に近接した環境において変動するものに
対するデータを提供する。そのような変動するものは、限定はしないが、分子相
互作用、酵素活性、反応速度論、立体配座動力学、運動の分子自由度、活性およ
び化学的静電的環境の変化を包含する。
【0026】
例えば、1つの蛍光団の吸収および放射遷移双極子を、分極化励起光を用いて
、または放出分極を分析することによって、または両方を用いて測定し得る。強
固に結合または回転拡散した係留標識の双極子配向性における一過性のバリエー
ションが、マクロ分子システムの角度運動またはサブユニットの1つについて報
告されている(Warshaw, et al., Proc. Natl. Acad. Sci.USA, 1998; 95:8034)
。これを本発明で適用し得る。
【0027】
本発明中で使用される標識は、当業者に既知である。好ましくは、標識は、Xu
e, et al.,Nature, 1995; 373:681に記載の蛍光標識である。または、蛍光酵素
、例えば緑色蛍光タンパク質 (Lu, et al., Science, 1998; 282:1877)を用い得
る。
【0028】
本発明の好ましい実施態様は、ポリヌクレオチドプロセシング酵素、例えばテ
トラメチルローダミン(TMR)に共有結合および部位特異的結合した小さな蛍光
分子の使用を包含する。蛍光標識を本発明で使用する場合、その検出は、励起波
長への反復暴露によって生じる光漂白に影響される。この問題を回避するための
1つの可能な方法は、多くの連鎖反応を実施するが、一度にただ少数のものにつ
いて蛍光シグナルを検出することである。この反復過程を用いて、シグナルの正
しい配列を測定し、ポリヌクレオチド配列を測定できる。例えば、固体支持体上
に複数の酵素を固定化し、それらを標的ポリヌクレオチドと接触させることによ
って、連続配列決定反応はほぼ同時に開始するはずである。蛍光の励起および検
出は、光漂白が明らかとなるまでのしばらくの間は、全体の反応の一部に局在す
ることがある。この時、励起および検出を反応の異なる部分に移し、配列決定を
継続することができる。全ての反応が同時性である場合、この正しい配列は、最
小の配列の再構築によって得られるはずである。
【0029】
標識は、共有結合または他の結合によって酵素と結合してもよい。多くのスト
ラテジーを、該標識を該酵素に結合するのに使用し得る。ストラテジーは、特定
のおよび直交染料標識化タンパク質のためのシステインおよびケトンハンドルを
導入する(Cornish,et al., Proc. Natl. Acad. Sci. USA, 1994; 91:2910)ため
に、部位指向性突然変異誘発および非天然アミノ酸突然変異誘発(Anthony-Cahil
, et al.,Trends Biochem. Sci., 1989;14:400)の使用を包含する。
【0030】
ポリヌクレオチドプロセシング酵素を標識するのに使用される別の予測される
実施態様は、当業者に既知の分子クローニング技術を介するプロセシング酵素(
例えば、ポリメラーゼ)との緑色蛍光タンパク質(GFP)の融合である(Pierce,
D.W. et al.,Nature, 1997; 388:338)。この技術は、立体配座の変化(Miyawaki
, et al.,Nature, 1997; 388:882)および局所pH変化(Llopis, et al., Proc.
Natl. Acad. Sci. USA, 1998; 95:6803)を測定するのに適用し得ることが分かっ
ている。
【0031】
固定化酵素における使用に適切な支持体は当業者に公知である。シリコン、ガ
ラスおよびセラミクッス材を用い得る。支持体は通常平らな表面である。酵素固
定は共有結合または別の手段によって実施し得る。例えば、共有結合リンカー分
子を使用して、適切に調製された酵素に結合する。結合方法は当業者に既知であ
る。
【0032】
固体支持体に固定化された酵素は1またはそれ以上であってよい。好ましい実
施態様において、複数の酵素を結合する。これは、多くの別々の反応をモニタリ
ングすることを可能にし、上述の光漂白問題を克服するのに有用である。
【0033】
種々の技術を用いて、酵素における立体配座の変化を測定することも可能であ
る。共鳴エネルギー遷移は、表面プラスモン共鳴(SPR)または蛍光表面プラス
モン共鳴技術によって測定し得る。
【0034】
しかし、「標識」との相互作用を介する放射変化を側定する別の技術またはエ
ネルギー変換器を考えてもよい。例えば、全内部反射率蛍光(TIRF)、漸先全
反射(ATR)、拡散全反射(FTR)、Brewster角度反射率、散乱全内部反射(S
TIR)、蛍光存在期間画像化顕微鏡および分光法(FLIMS)、蛍光分極異方
性(FPA)、蛍光分光法またはエバネッセント波楕円偏向法による分光法である
。
【0035】
本発明は、下記実施例で図面と併せて詳細に説明する。
【0036】
実施例
この実施例では、図1に示したような共焦蛍光配置を使用した。
図1に関して、その配置は、X、YおよびZ次元において高い解像度でスキャン
することができる走査テーブル(1)、プライマー−鋳型ポリヌクレオチド複合体
(4)およびヌクレオチド(9)緩衝液内の固定化されたポリメラーゼ分子(3)上に
導入する入口(8)を持つ微螢光測定フロー・セルシステムの一部であるクラスカ
バースリップ(2)、および排出物のための出口(7)からなる。供与体励起のため
のレーザー光源(6)からの入射光は、油浸漬対物レンズ(5)を介して伝達させる
。
【0037】
タンパク質接合
本実験において、テトラメチルローダミン(TMR、供与体)およびCy5(受
容体)は一組のFRETとして使用する。これは、それらの十分に分離した放出
波長 (>100nm)および大きなFoester半径を理由とする。
【0038】
NewEngland Biolabs (10 000 U/mlで与えられる)から入手したT7DNAポ
リメラーゼを用いた。T7(50μl)を、Vivaspin500(Vivaspin)中で、T7
DNA ポリメラーゼが入った貯蔵用緩衝液からDTTを除去するために、20
0mM 酢酸ナトリウム緩衝液(pH4)を4x 500μlに対して緩衝液交換した
。次いで、緩衝液を交換したT7 DNA ポリメラーゼ(50μl)を、酢酸ナト
リウム緩衝液(100μl,pH4)と飽和2-2-ジピリジル-ジスルフィド水性溶
液(50μl)に添加した。次いで、該反応を110分間放置し、343nmでの吸
収を記載した。最後に、該試料を前記のような200mMTris(pH8)に緩衝
液交換を行った(500μlで4回)。
【0039】
染料の結合を、ポリアクリルアミドゲル電気泳動を変性することで確認した。
Cy5スクシンイミジルエステル(分子プローブ)を、同じ標識化条件下でTMR
-T7DNAポリメラーゼに接合し、精製し、上記のように特性分析した。
【0040】
ポリメラーゼ固定化
ガラスカバースリップを、N-[(3-トリメトキシオキシシリル)プロピル] エ
チレンジアミン三酢酸塩によって修飾した。次いで、カバースリップを、修飾さ
れたガラス表面上に連続的に緩衝液が流れるようにしたフロー・セル配置中へ接
着した。 次いで、標識ポリメラーゼを緩衝液に添加し、タンパク質をガラス表
面上で固定するようにカバーガラス上に流した。
【0041】
次いで、タンパク質を、いずれかの一時でもレーザー励起体積下に1分子のみ
があるように、ガラス-水界面上に低密度に固定した。レーザー光(514 nm ア
ルゴンイオンレーザー、15μW、環状分極)を、スキャンステージにおける共焦
顕微鏡のエピ照明配置中に油浸漬対物レンズを用いて、0.4 μmスポットに集
束した。蛍光放出を同対物レンズによって集め、二色性ビーム分割(630nmで
のロングパス)によって2つに分割し、2つのAvalanchePhoto Diode (APD)計測
装置によって同時に検出した。
【0042】
585nmのバンドパスフィルターを供与体検出器の前に置き、650nmロン
グパスフィルターを受容体検出器の前に置いた。蛍光検出中スペクトル範囲を二
色性ビーム分割のカットオフ波長から十分除去するので、供与体および受容体シ
グナルの両方の検出効率に関する分極依存性は無視できる。高度に近いフィール
ド開口(NA)対物レンズによるこの分極混合は、大目にみてよいことが知られて
いる[(Ha etal, 上掲)]。
【0043】
供与体および受容体放射時間を得るために、受容体シグナルに対する探査条件
を、概説したように用い得る(Ha, et al., Appl. Phys. Lett., 1997; 70:782)
。この方法は、二重標識したタンパク質のスクリーニングに役立つ:受容体の直
接的な励起はせずに、FRETを受けるタンパク質のみが受容体シグナルを示し
得る。タンパク質をレーザースポット下でスクリーンし、位置し、配置すると、
供与体および受容体の時間トレース(5ms 集積化時間)を得た。捕捉時間は、光
漂白された標的タンパク質上で全て蛍光標識されるまで続いた。
【0044】
反応の開始
2種のオリゴヌクレオチドを、標準的なホスホラミダイト化学を用いて合成し
た。配列番号1として定義されるオリゴヌクレオチドは標的ポリヌクレオチドと
して用い、配列番号2として定義されるオリゴヌクレオチドはプライマーとして
用いた。2つのオリゴヌクレオチドをハイブダイズ条件下で反応し、標的−プラ
イマー複合体を形成した。
配列番号1
CAAGGAGAGGACGCTGTCTGTCGAAGGTAAGGAACGGACGAGAGAAGGGAGAG
配列番号2
CTCTCCCTTCTCTCGTC
【0045】
次いで、反応を、0.4mMの濃度で存在する4つのヌクレオチド(dGTP、
dCTP、dATPおよびdTTP)を有するフロー・セル中にプライム化DN
Aを注入して開始した。該フロー・セルを、25℃で修飾ペルレィエ(Peltier)
装置で維持した。
【0046】
また、酸素排除系[50μg/ml グルコースオキシダーゼ, 10 μg/mlカタラーゼ
, 18% (wt/wt)グルコース,1% (wt/vol) β-メルカプトエタノール]を用い、蛍
光存在時間(Funatsu, et al., Nature, 1995; 374:555-559)を長くした。
【0047】
FRETデータ分析
最初の試験は、点滅の起源、光漂白および三重項振幅を測定し(Ha, et al., C
hem. Phy., 1999; 247:107-118)、それら全ては、立体配座の変化の結果として
蛍光団間で変化する距離により、FRET効率の基本的な変化を干渉し得る。Ha
, et al., 1999(上掲)に記載のような供与体および受容体時間トレースからの
バックグラウンドシグナルの控除後、3重項振幅を除くために5点を平均した中
央値フィルタを適用した。次いで、供与体点滅結果による両検出器上の同時性ダ
ークカウントを示したデータ値は、時間トレースから無視した。
【0048】
受容体光漂白に対する供与体シグナル回復の量は、分子の量子収率およびその
全体的な検出効率に関する。
【0049】
次いで、エネルギー遷移効率時間トレースを得た。標的鎖配列番号1のポリメ
ライゼーション中の該FRET効率時間トレースを図2に示した。図2によると
、この配列は配列番号1の配列の相補物に対応する(右から左に読み、プライマ
ー配列とハイブリダイズする部分を除く)。
【符号の説明】
【0050】
(1)走査テーブル、(2)クラスカバースリップ、(3)ポリ
メラーゼ分子、(4)プライマー−鋳型ポリヌクレオチド複合体、(5)油浸漬対物
レンズ、(6)レーザー光源、(7)出口、(8)入口、(9)ヌクレオチド。
【化1】

【技術分野】
【0001】
本発明の分野
本発明は、ポリヌクレオチド配列決定に関する。
【0002】
本発明の技術分野
ポリヌクレオチドの配列を決定するための能力は、科学的に非常に重要である
。例えば、ヒトゲノムプロジェクトは30億塩基ものヒトゲノムにコードされた
DNA塩基の地図化および配列決定するための野望的な国際的努力である。完了
すると、得られる配列のデータベースは生物医学的研究に他に比肩できない有用
な道具となる。このプロジェクトを成功裡に完成さすのに主な障害となるのは、
配列決定の過程で使用される技術に関する。
【0003】
大規模なDNA配列決定のために一般的に用いられる主な方法は、連鎖終止反
応法である。この方法は、最初はサンガーおよびコールソン(Sanger et al., Pr
oc. Natl. Acad. Sci.USA, 1977; 74: 5463-5467)によって開発され、ポリメラ
ーゼ反応において未完成のポリヌクレオチド鎖に導入される4つのヌクレオシド
三リン酸のジデオキシ誘導体を使用することによる。導入に際し、該ジデオキシ
誘導体がポリメラーゼ反応を終了し、次いで、該生成物をゲル電気泳動によって
分離し、分析すると、特定のジデオキシ誘導体の鎖中に導入された部位が明らか
になる。
【0004】
この方法は、広く使用されており、また信頼できる結果を生むが、時間と人手
がかかり、費用が高くつく方法であると認識されている。
【0005】
別の配列決定方法がEP-A-0471732で提案されているが、これは分光
学的手段を使用して、ヌクレオチドが標的に相補的な未完成のポリヌクレオチド
鎖に導入されるのを検出する。該方法は鋳型およびプライマーとの固定化複合体
を基にしている。該複合体を別異のヌクレオチドのただ1つのみを含有するフロ
ーに暴露する。次いで、分光学的技術を用い、鋳型コピーのポリメラーゼ触媒に
よる増幅から生じる時間依存的シグナルを測定する。記載の分光学的技術は、エ
バネッセント波場内での分析物の変化を測定する表面プラスモン共鳴(SPR)分
光法および蛍光測定技術である。しかし、この方法には限界が認識されており、
SPR技術に関して最も深刻な問題は、コピー鎖の大きさが増すににつれて、シ
グナルの絶対サイズもエバネッセント波場から鎖の運動により増加し、増加量を
検出するのが困難となることである。該蛍光測定技術は、増加しつつある未完成
ポリヌクレオチド鎖上に導入された蛍光団からのバックグラウンドの干渉を増加
させる欠点をもっている。鎖が増すにつれ、バックグラウンド「ノイズ」が増加
し、各ヌクレオチド導入を検出に要する時間を増加する必要がある。これにより
、大きなポリヌクレオチドの配列決定について使用するのが非常に制限される。
【0006】
1本鎖フラグメントポリヌクレオチド配列決定アプローチは、WO-A-992
4797およびWO-A-9833939に概説され、両方法は1本鎖標識ヌクレ
オチド分子の蛍光検出を用いるものである。これらの1本鎖ヌクレオチドは、エ
クソヌクレアーゼ分子の作用によって、光トラップ(Jett, et al., J. Biomol.
Struc. Dyn, 1989;7:301-309)によるフロー中に保たれた該鋳型ポリヌクレオチ
ドから切断される。次いでこれらの切断されたヌクレオチド類はクウォーツ・フ
ロー・セル内で下方にフローし、レーザー励起に供され、次いで鋭敏な検出シス
テムで検出される。しかし、この方法の限界が認識されており、エクソヌクレア
ーゼ技術に関して最も重大な限界は、該標識ヌクレオチドがエクソヌクレアーゼ
酵素の進行能(プロセッシビリティ)に非常に影響するという事実である。該方法
の別の限界は、ポリヌクレオチドフラグメントを固定化するために使用されたビ
オチンビーズに該ヌクレオチドを「付着すること」、従ってヌクレオチドのフロ
ーがフェーズをはずれるようになること;最初の酵素の標識化過程の非効率およ
び長さの制限;および4つの異なる染料分子間の該励起「交差」の結果として非
常にエラー比が高くなることを包含する。
【0007】
そのため、好ましくは1本鎖フラグメントレベルでポリヌクレオチドの配列を
決定するための改良された方法、即ち該ポリヌクレオチドの配列決定に関するフ
ラグメントサイズと速度を顕著に増加させ、好ましくは自動化過程により行い、
現方法に関連する複雑性および費用を低下させる方法が必要である。
【発明の概要】
【課題を解決するための手段】
【0008】
本発明の要約
本発明は、標的ポリヌクレオチドの配列を、標的ポリヌクレオチドに結合し、
標的ポリヌクレオチドに沿って進行する酵素における立体配座の変化を測定する
ことによって、決定し得るということに基づいている。立体配座の変化が起こる
程度は、標的上のどの個々のヌクレオチドが該酵素と接触しているかによって異
なる。
【0009】
本発明の一つの態様によれば、ポリヌクレオチドの配列を決定するための方法
は、以下を包含する:
(i) 酵素活性を誘導するのに十分な条件下で、標的ポリヌクレオチドを、該
ポリヌクレオチドと相互作用し、かつポリヌクレオチドに沿って進行し得る酵素
と、反応せしめ、
(ii) 酵素がポリヌクレオチドに沿って進行する時に、酵素における立体配座
の変化を検出すること。
【0010】
好ましい実施態様において、酵素は相補鎖を伸長する過程で標的と相互作用す
るポリメラーゼ酵素である。酵素は、規定領域内に反応を局在化させるために固
体支持体上に固定化する。
【0011】
本発明の第2の実施態様によれば、酵素は最初に結合された検出し得る標識を
包含し、酵素が立体配座の変化を進行するにつれて、その特徴を変化させる。ま
た、酵素は、最初の標識と相互作用し得る2番目に結合した検出し得る標識を包
含する。ここで、相互作用の程度は酵素における立体配座の変化に依存する。通
常、最初の標識はエネルギー受容体であり、2番目の標識はエネルギー供与体で
あり、立体配座の変化を検出することは、2つの標識間のエネルギー遷移を測定
することにより行う。
【0012】
本発明のさらなる実施態様によれば、蛍光共鳴エネルギー遷移(FRET)を用
いて、標的ポリメラーゼと相互作用し、標的ポリメラーゼに沿って進行する酵素
における立体配座の変化を検出する。それによりポリヌクレオチドの配列を決定
する。蛍光共鳴エネルギー遷移は、酵素にそれぞれ結合しているFRET供与体
および受容体標識との間で行い得る。あるいは、標識の1つは酵素に結合し、も
う一方の標識はポリヌクレオチドに結合してもよい。
【0013】
さらなる実施態様によれば、ポリヌクレオチド配列を決定するために、標的ポ
リヌクレオチドと相互作用し、標的ポリヌクレオチドに沿って進行し得る検出可
能な標識酵素を使用する。ここで、該標識は、酵素がポリヌクレオチドに沿って
進行するにつれて、その検出し得る特性を変化させる。
【0014】
さらなる態様によれば、固体支持体は、標的ポリヌクレオチドと相互作用し、
標的ポリヌクレオチドに沿って進行し得る少なくとも1つの固定化酵素を包含す
る。酵素は1以上の検出し得る標識物によって標識されている。
【0015】
さらなる態様によれば、ポリヌクレオチドの配列を決定するためのシステムは
、上記のような固体支持体を包含し、標識を検出するための装置を包含する。
【0016】
本発明は、従来の配列決定技術を超えるいくつかの利点を提供する。ポリメラ
ーゼ酵素は、一旦ポリヌクレオチドの伸長ラウンドを始めると、鎖から脱落する
前までに数千のヌクレオチドを重合する。さらに、ある特定のポリメラーゼシス
テムは、鋳型分子を囲む「スライディング・クランプ」(例えば、ポリメラーゼ
III)を介して、または部分的に囲む分子フック(例えば、T7:チレオドキシン
複合体)を介して、鋳型ポリヌクレオチドにシステム自身を固着し、係留するこ
とができる。
【0017】
本発明は、1回の試行で、1秒あたり数百もの塩基対の速度で数十キロベース
(kb)またはそれ以上の配列決定をし得る。これは、DNAの1本鎖フラグメン
トに関する配列決定の結果である。DNAの1本鎖フラグメントの配列決定に関
する利点は、配列決定速度が利用された酵素システムによって決定され、間接的
総合的反応により決定されるのではないので相当速いことである。速い速度と同
じくらい重要なことは、DNAの大きなフラグメントを配列決定することができ
る能力である。これは、配列決定情報のメガベースセグメントを集めるのに必要
とされるサブクローニングの量および重複配列の数を顕著に低下させる。1本鎖
フラグメントアプローチのさらなる利点は、現在配列決定の努力を阻害するアク
リルアミドのような有害不用物の廃棄に関する問題を排除することである。
例えば、本願発明は以下の項目を提供する。
(項目1) 下記の過程:
(i) 酵素活性を誘導するのに十分な条件下で、標的ポリヌクレオチドを、該ポリヌクレオチドと相互作用し、かつポリヌクレオチドに沿って進行し得る酵素と、反応せしめ、
(ii) 酵素が該ポリヌクレオチドに沿って進行する時に、酵素における立体配座の変化を検出すること、
を含むポリヌクレオチドの配列を決定するための方法。
(項目2) 酵素がポリメラーゼ酵素である、項目1記載の方法。
(項目3) 酵素がヘリカーゼ酵素またはプライマーゼ酵素である、請求項1記載の方法。
(項目4) 酵素が固体支持上で固定化された、項目1〜3記載の方法。
(項目5) 固体支持上で固定化された複数の酵素を含む、項目4記載の方法。
(項目6) 酵素が、最初に結合した検出し得る標識を含み、酵素が立体配座の変化を進行させるにつれて、その特徴が変化する、項目1〜5記載の方法。
(項目7) 酵素が、最初の標識と相互作用し得る2番目に結合した検出し得る標識を包含し、相互作用の程度は酵素における立体配座の変化に依存する、項目6記載の方法。
(項目8) 2番目の検出し得る標識が、酵素と接触することになるヌクレオチドに結合している、項目6記載の方法。
(項目9) 最初の標識がエネルギー受容体であり、2番目の標識がエネルギー供与体であり、もしくは逆で、過程(ii)は2つの標識間のエネルギー遷移の変化を測定することにより行い得る、項目7または項目8記載の方法。
(項目10) 過程(ii)は共焦顕微鏡を用いて実施する、項目1〜9記載の方法。
(項目11) 過程(ii)は蛍光画像化によって実施する、項目10記載の方法。
(項目12) 過程(ii)は最初の標識の変化した特性に対する分極効果の結果を測定することによって実施する、項目10記載の方法。
(項目13) 過程(ii)は蛍光分極異方性によって実施する、項目10記載の方法。
(項目14) 標的ポリメラーゼと相互作用し、標的ポリメラーゼに沿って進行する酵素における立体配座の変化を検出して、それによってポリヌクレオチドの配列を決定するための蛍光共鳴エネルギー遷移(FRET)の使用。
(項目15) 酵素がポリメラーゼ酵素である、項目14記載の使用。
(項目16) 酵素が固体支持体上に固定化される、項目14または15記載の使用。
(項目17) ポリヌクレオチド配列を決定するために、標的ポリヌクレオチドと相互作用し、標的ポリヌクレオチドに沿って進行し得る検出可能な標識酵素の使用であって、酵素がポリヌクレオチドに沿って進行する時に、標識がその検出し得る特性を変化させる検出可能な標識酵素の使用。
(項目18) 標的ポリヌクレオチドと相互作用し、標的ポリヌクレオチドに沿って進行し得る少なくとも1つの固定酵素を包含し、酵素が1以上の検出し得る標識物によって標識される固体支持体。
(項目19) 酵素がポリメラーゼである、項目18記載の固体支持体。
(項目20) 標識が蛍光団である、項目18または19記載の固体支持体。
(項目21) 項目18〜21記載の固体支持体および標識を検出するための装置を含むポリヌクレオチドの配列を決定するためのシステム。
【図面の簡単な説明】
【0018】
【図1】図1は、本発明で使用する共焦顕微鏡の配置のスキーム図である。
【図2】図2は、蛍光共鳴エネルギー遷移後にとったトレースを示すもので、各ピークは特定のヌクレオチドの検出を示す。
【発明を実施するための形態】
【0019】
本発明の説明
ポリヌクレオチドを配列決定するための本方法は、酵素と標的ポリヌクレオチ
ドとの間の立体配座の変化に関する分析を包含する。
【0020】
本明細書中で使用する「ポリヌクレオチド」なる用語は、広義に解釈されるべ
きであり、DNAおよびRNAならびに修飾DNAおよび修飾RNAを含み、さ
らに別のハイブリダイズする核酸様分子、例えばペプチドヌクレオチド核酸(P
NA)も含む。
【0021】
酵素は、ポリメラーゼ酵素であってもよく、立体配座の変化は、ポリメラーゼ
が標的ポリヌクレオチドに対し相補的な未完成鎖中にヌクレオチドを導入する場
合にもたらされる。立体配座の変化は、各々異なるヌクレオチド、A、T、Gま
たはCに対して異なり、そのため、該変化を測定することによりどのヌクレオチ
ドが組み込まれたかを同定するのである。
【0022】
あるいは、酵素は、ポリヌクレオチド、例えば、ヘリカーゼ酵素、プライマー
ゼおよびホロ酵素との相互作用に関与するものどれでもよい。酵素がポリヌクレ
オチドに沿って進行すると、その立体配座は、標的上のどのヌクレオチドに接触
するかに依存して変化する。
【0023】
酵素における立体配座の変化を検出する1つの方法は、適切なエネルギー供与
体標識と適切なエネルギー受容体標識との共鳴エネルギー遷移を測定することで
ある。一つの例において、供与体および受容体は酵素に各々結合し、標的ポリヌ
クレオチドとの相互作用によりもたらされる酵素における立体配座の変化が標識
の相対的位置を変える。位置付けにおける差異は、得られるエネルギー遷移を反
映しており、酵素と接触する特定のヌクレオチドの特徴となる。あるいは、1つ
の標識を酵素上に位置付け、もう一方の酵素を標的のヌクレオチド上または標的
に相補的な鎖上に導入されたヌクレオチド上に位置付けられ得る。
【0024】
蛍光共鳴エネルギー遷移(FRET)の使用は、本発明の好ましい実施態様であ
る。この技術は、2から8nmの距離を測定することができ、供与体蛍光団と受容
体蛍光団との距離依存性エネルギー遷移による。該技術は、優秀な静的共局在化
の能力を持つだけでなく、分子内および分子間FRETに対する2つの蛍光団の
距離または配向性における動的変化に対する情報も提供する。1つの供与体と1
つの受容体(一組のFRET)との間のエネルギー遷移についての最初の測定(Ha,
et al.,Proc. Natl. Acad. Sci. USA, 1996; 96:893)以来、該方法はリガンド
-受容体の共局在性(Schutz, et al., Biophys. J.,1998; 74:2223)を研究する
ため、触媒中の平衡タンパク質構造性変動および酵素−基質相互作用を調べるた
め(Ha, etal., 1999 上掲)、そして立体配座状態および溶液中の個々の拡散分
子の副固体数を同定するために使用されてきた。これらの変法の全ては、本発明
において適用し得るものと考えられる。
【0025】
本発明は、1つの標識のみを必要とする測定技術を使用しても実施し得る。1
分子レベルで酵素の局所環境における変化を測定することができる全てのシステ
ムは、本発明の許容される態様である。ポリヌクレオチドプロセシング酵素およ
び/またはその基質類と結合した1つの蛍光プローブの多様な特性は、本発明の
中で活用することができ、ヌクレオチドの合同事象に特異的である酵素システム
/分子環境中において、またはその非常に近接した環境において変動するものに
対するデータを提供する。そのような変動するものは、限定はしないが、分子相
互作用、酵素活性、反応速度論、立体配座動力学、運動の分子自由度、活性およ
び化学的静電的環境の変化を包含する。
【0026】
例えば、1つの蛍光団の吸収および放射遷移双極子を、分極化励起光を用いて
、または放出分極を分析することによって、または両方を用いて測定し得る。強
固に結合または回転拡散した係留標識の双極子配向性における一過性のバリエー
ションが、マクロ分子システムの角度運動またはサブユニットの1つについて報
告されている(Warshaw, et al., Proc. Natl. Acad. Sci.USA, 1998; 95:8034)
。これを本発明で適用し得る。
【0027】
本発明中で使用される標識は、当業者に既知である。好ましくは、標識は、Xu
e, et al.,Nature, 1995; 373:681に記載の蛍光標識である。または、蛍光酵素
、例えば緑色蛍光タンパク質 (Lu, et al., Science, 1998; 282:1877)を用い得
る。
【0028】
本発明の好ましい実施態様は、ポリヌクレオチドプロセシング酵素、例えばテ
トラメチルローダミン(TMR)に共有結合および部位特異的結合した小さな蛍光
分子の使用を包含する。蛍光標識を本発明で使用する場合、その検出は、励起波
長への反復暴露によって生じる光漂白に影響される。この問題を回避するための
1つの可能な方法は、多くの連鎖反応を実施するが、一度にただ少数のものにつ
いて蛍光シグナルを検出することである。この反復過程を用いて、シグナルの正
しい配列を測定し、ポリヌクレオチド配列を測定できる。例えば、固体支持体上
に複数の酵素を固定化し、それらを標的ポリヌクレオチドと接触させることによ
って、連続配列決定反応はほぼ同時に開始するはずである。蛍光の励起および検
出は、光漂白が明らかとなるまでのしばらくの間は、全体の反応の一部に局在す
ることがある。この時、励起および検出を反応の異なる部分に移し、配列決定を
継続することができる。全ての反応が同時性である場合、この正しい配列は、最
小の配列の再構築によって得られるはずである。
【0029】
標識は、共有結合または他の結合によって酵素と結合してもよい。多くのスト
ラテジーを、該標識を該酵素に結合するのに使用し得る。ストラテジーは、特定
のおよび直交染料標識化タンパク質のためのシステインおよびケトンハンドルを
導入する(Cornish,et al., Proc. Natl. Acad. Sci. USA, 1994; 91:2910)ため
に、部位指向性突然変異誘発および非天然アミノ酸突然変異誘発(Anthony-Cahil
, et al.,Trends Biochem. Sci., 1989;14:400)の使用を包含する。
【0030】
ポリヌクレオチドプロセシング酵素を標識するのに使用される別の予測される
実施態様は、当業者に既知の分子クローニング技術を介するプロセシング酵素(
例えば、ポリメラーゼ)との緑色蛍光タンパク質(GFP)の融合である(Pierce,
D.W. et al.,Nature, 1997; 388:338)。この技術は、立体配座の変化(Miyawaki
, et al.,Nature, 1997; 388:882)および局所pH変化(Llopis, et al., Proc.
Natl. Acad. Sci. USA, 1998; 95:6803)を測定するのに適用し得ることが分かっ
ている。
【0031】
固定化酵素における使用に適切な支持体は当業者に公知である。シリコン、ガ
ラスおよびセラミクッス材を用い得る。支持体は通常平らな表面である。酵素固
定は共有結合または別の手段によって実施し得る。例えば、共有結合リンカー分
子を使用して、適切に調製された酵素に結合する。結合方法は当業者に既知であ
る。
【0032】
固体支持体に固定化された酵素は1またはそれ以上であってよい。好ましい実
施態様において、複数の酵素を結合する。これは、多くの別々の反応をモニタリ
ングすることを可能にし、上述の光漂白問題を克服するのに有用である。
【0033】
種々の技術を用いて、酵素における立体配座の変化を測定することも可能であ
る。共鳴エネルギー遷移は、表面プラスモン共鳴(SPR)または蛍光表面プラス
モン共鳴技術によって測定し得る。
【0034】
しかし、「標識」との相互作用を介する放射変化を側定する別の技術またはエ
ネルギー変換器を考えてもよい。例えば、全内部反射率蛍光(TIRF)、漸先全
反射(ATR)、拡散全反射(FTR)、Brewster角度反射率、散乱全内部反射(S
TIR)、蛍光存在期間画像化顕微鏡および分光法(FLIMS)、蛍光分極異方
性(FPA)、蛍光分光法またはエバネッセント波楕円偏向法による分光法である
。
【0035】
本発明は、下記実施例で図面と併せて詳細に説明する。
【0036】
実施例
この実施例では、図1に示したような共焦蛍光配置を使用した。
図1に関して、その配置は、X、YおよびZ次元において高い解像度でスキャン
することができる走査テーブル(1)、プライマー−鋳型ポリヌクレオチド複合体
(4)およびヌクレオチド(9)緩衝液内の固定化されたポリメラーゼ分子(3)上に
導入する入口(8)を持つ微螢光測定フロー・セルシステムの一部であるクラスカ
バースリップ(2)、および排出物のための出口(7)からなる。供与体励起のため
のレーザー光源(6)からの入射光は、油浸漬対物レンズ(5)を介して伝達させる
。
【0037】
タンパク質接合
本実験において、テトラメチルローダミン(TMR、供与体)およびCy5(受
容体)は一組のFRETとして使用する。これは、それらの十分に分離した放出
波長 (>100nm)および大きなFoester半径を理由とする。
【0038】
NewEngland Biolabs (10 000 U/mlで与えられる)から入手したT7DNAポ
リメラーゼを用いた。T7(50μl)を、Vivaspin500(Vivaspin)中で、T7
DNA ポリメラーゼが入った貯蔵用緩衝液からDTTを除去するために、20
0mM 酢酸ナトリウム緩衝液(pH4)を4x 500μlに対して緩衝液交換した
。次いで、緩衝液を交換したT7 DNA ポリメラーゼ(50μl)を、酢酸ナト
リウム緩衝液(100μl,pH4)と飽和2-2-ジピリジル-ジスルフィド水性溶
液(50μl)に添加した。次いで、該反応を110分間放置し、343nmでの吸
収を記載した。最後に、該試料を前記のような200mMTris(pH8)に緩衝
液交換を行った(500μlで4回)。
【0039】
染料の結合を、ポリアクリルアミドゲル電気泳動を変性することで確認した。
Cy5スクシンイミジルエステル(分子プローブ)を、同じ標識化条件下でTMR
-T7DNAポリメラーゼに接合し、精製し、上記のように特性分析した。
【0040】
ポリメラーゼ固定化
ガラスカバースリップを、N-[(3-トリメトキシオキシシリル)プロピル] エ
チレンジアミン三酢酸塩によって修飾した。次いで、カバースリップを、修飾さ
れたガラス表面上に連続的に緩衝液が流れるようにしたフロー・セル配置中へ接
着した。 次いで、標識ポリメラーゼを緩衝液に添加し、タンパク質をガラス表
面上で固定するようにカバーガラス上に流した。
【0041】
次いで、タンパク質を、いずれかの一時でもレーザー励起体積下に1分子のみ
があるように、ガラス-水界面上に低密度に固定した。レーザー光(514 nm ア
ルゴンイオンレーザー、15μW、環状分極)を、スキャンステージにおける共焦
顕微鏡のエピ照明配置中に油浸漬対物レンズを用いて、0.4 μmスポットに集
束した。蛍光放出を同対物レンズによって集め、二色性ビーム分割(630nmで
のロングパス)によって2つに分割し、2つのAvalanchePhoto Diode (APD)計測
装置によって同時に検出した。
【0042】
585nmのバンドパスフィルターを供与体検出器の前に置き、650nmロン
グパスフィルターを受容体検出器の前に置いた。蛍光検出中スペクトル範囲を二
色性ビーム分割のカットオフ波長から十分除去するので、供与体および受容体シ
グナルの両方の検出効率に関する分極依存性は無視できる。高度に近いフィール
ド開口(NA)対物レンズによるこの分極混合は、大目にみてよいことが知られて
いる[(Ha etal, 上掲)]。
【0043】
供与体および受容体放射時間を得るために、受容体シグナルに対する探査条件
を、概説したように用い得る(Ha, et al., Appl. Phys. Lett., 1997; 70:782)
。この方法は、二重標識したタンパク質のスクリーニングに役立つ:受容体の直
接的な励起はせずに、FRETを受けるタンパク質のみが受容体シグナルを示し
得る。タンパク質をレーザースポット下でスクリーンし、位置し、配置すると、
供与体および受容体の時間トレース(5ms 集積化時間)を得た。捕捉時間は、光
漂白された標的タンパク質上で全て蛍光標識されるまで続いた。
【0044】
反応の開始
2種のオリゴヌクレオチドを、標準的なホスホラミダイト化学を用いて合成し
た。配列番号1として定義されるオリゴヌクレオチドは標的ポリヌクレオチドと
して用い、配列番号2として定義されるオリゴヌクレオチドはプライマーとして
用いた。2つのオリゴヌクレオチドをハイブダイズ条件下で反応し、標的−プラ
イマー複合体を形成した。
配列番号1
CAAGGAGAGGACGCTGTCTGTCGAAGGTAAGGAACGGACGAGAGAAGGGAGAG
配列番号2
CTCTCCCTTCTCTCGTC
【0045】
次いで、反応を、0.4mMの濃度で存在する4つのヌクレオチド(dGTP、
dCTP、dATPおよびdTTP)を有するフロー・セル中にプライム化DN
Aを注入して開始した。該フロー・セルを、25℃で修飾ペルレィエ(Peltier)
装置で維持した。
【0046】
また、酸素排除系[50μg/ml グルコースオキシダーゼ, 10 μg/mlカタラーゼ
, 18% (wt/wt)グルコース,1% (wt/vol) β-メルカプトエタノール]を用い、蛍
光存在時間(Funatsu, et al., Nature, 1995; 374:555-559)を長くした。
【0047】
FRETデータ分析
最初の試験は、点滅の起源、光漂白および三重項振幅を測定し(Ha, et al., C
hem. Phy., 1999; 247:107-118)、それら全ては、立体配座の変化の結果として
蛍光団間で変化する距離により、FRET効率の基本的な変化を干渉し得る。Ha
, et al., 1999(上掲)に記載のような供与体および受容体時間トレースからの
バックグラウンドシグナルの控除後、3重項振幅を除くために5点を平均した中
央値フィルタを適用した。次いで、供与体点滅結果による両検出器上の同時性ダ
ークカウントを示したデータ値は、時間トレースから無視した。
【0048】
受容体光漂白に対する供与体シグナル回復の量は、分子の量子収率およびその
全体的な検出効率に関する。
【0049】
次いで、エネルギー遷移効率時間トレースを得た。標的鎖配列番号1のポリメ
ライゼーション中の該FRET効率時間トレースを図2に示した。図2によると
、この配列は配列番号1の配列の相補物に対応する(右から左に読み、プライマ
ー配列とハイブリダイズする部分を除く)。
【符号の説明】
【0050】
(1)走査テーブル、(2)クラスカバースリップ、(3)ポリ
メラーゼ分子、(4)プライマー−鋳型ポリヌクレオチド複合体、(5)油浸漬対物
レンズ、(6)レーザー光源、(7)出口、(8)入口、(9)ヌクレオチド。
【化1】
【特許請求の範囲】
【請求項1】
ポリヌクレオチドの配列を決定する方法であって、
(i)ポリメラーゼ酵素を標識されていない標的ポリヌクレオチドに接触させて複合体を形成する工程;
(ii)ポリメラーゼ活性を誘導するのに十分な条件下で、dATP、dTTP、dGTPおよびdCTPからなる群より選択される1つ以上の標識されていないヌクレオシド三リン酸に該複合体を接触させれう工程;および
(iii)該酵素が該標的ポリヌクレオチドに沿って進行する時にpHが変化することを検出することによって、該標的ポリヌクレオチドの配列を決定する工程、
を包含し、該pHの変化は、該標的ポリヌクレオチドに対して相補的な未完成のヌクレオチド鎖へのヌクレオシド三リン酸の取り込みから生じる、方法。
【請求項2】
前記酵素は、該酵素に対する緑色蛍光タンパク質の融合物を含む、請求項1に記載の方法。
【請求項3】
前記工程(ii)および(iii)は、同じ複合体を1回以上利用される、請求項1に記載の方法。
【請求項4】
前記未完成の鎖は、プライマー配列である、請求項1に記載の方法。
【請求項5】
前記検出工程は、フローセルで行われる、請求項1に記載の方法。
【請求項6】
前記酵素および/または前記複合体は、固体支持体上で固定される、請求項1に記載の方法。
【請求項7】
前記pHの変化における変化は、局所的なpHの変化である、請求項1に記載の方法。
【請求項1】
ポリヌクレオチドの配列を決定する方法であって、
(i)ポリメラーゼ酵素を標識されていない標的ポリヌクレオチドに接触させて複合体を形成する工程;
(ii)ポリメラーゼ活性を誘導するのに十分な条件下で、dATP、dTTP、dGTPおよびdCTPからなる群より選択される1つ以上の標識されていないヌクレオシド三リン酸に該複合体を接触させれう工程;および
(iii)該酵素が該標的ポリヌクレオチドに沿って進行する時にpHが変化することを検出することによって、該標的ポリヌクレオチドの配列を決定する工程、
を包含し、該pHの変化は、該標的ポリヌクレオチドに対して相補的な未完成のヌクレオチド鎖へのヌクレオシド三リン酸の取り込みから生じる、方法。
【請求項2】
前記酵素は、該酵素に対する緑色蛍光タンパク質の融合物を含む、請求項1に記載の方法。
【請求項3】
前記工程(ii)および(iii)は、同じ複合体を1回以上利用される、請求項1に記載の方法。
【請求項4】
前記未完成の鎖は、プライマー配列である、請求項1に記載の方法。
【請求項5】
前記検出工程は、フローセルで行われる、請求項1に記載の方法。
【請求項6】
前記酵素および/または前記複合体は、固体支持体上で固定される、請求項1に記載の方法。
【請求項7】
前記pHの変化における変化は、局所的なpHの変化である、請求項1に記載の方法。
【図1】

【図2】
【図2】
【公開番号】特開2012−196201(P2012−196201A)
【公開日】平成24年10月18日(2012.10.18)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−264043(P2011−264043)
【出願日】平成23年12月1日(2011.12.1)
【分割の表示】特願2001−528630(P2001−528630)の分割
【原出願日】平成12年10月6日(2000.10.6)
【出願人】(500169900)ジェン−プローブ・インコーポレーテッド (32)
【Fターム(参考)】
【公開日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願番号】特願2011−264043(P2011−264043)
【出願日】平成23年12月1日(2011.12.1)
【分割の表示】特願2001−528630(P2001−528630)の分割
【原出願日】平成12年10月6日(2000.10.6)
【出願人】(500169900)ジェン−プローブ・インコーポレーテッド (32)
【Fターム(参考)】
[ Back to top ]