
EBウィルスにより誘発される腫瘍に対する融合ポリペプチドとコリシンIA変異体
本発明は、EBウィルスに対する抗体または模倣抗体と、E1、Ia、Ib、A、B、Nおよびそれらの変異体から選択される、イオンチャネルを形成するコリシンとを含む、EBウィルスにより誘発される腫瘍に対する融合ポリペプチド、を提供する。本発明は、G11A、H22G、A26G、V31L、およびH40Dの変異を含むコリシンIa変異体も提供する。また、本発明は、この融合ポリペプチドの、遺伝子、ベクター、調製方法および使用も提供し、また、この変異体の、遺伝子および使用も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗腫瘍剤の分野に関し、より具体的には、EBウィルスが原因である腫瘍に対する新規ポリペプチド、ならびに、その使用および生産方法、に関する。
【背景技術】
【0002】
抗生物質研究の領域においては、研究は、同種の異種の株間の相互殺傷機構をまねた新しい抗生物質の開発へと向けられてきた。自然界には、細菌の細胞膜上に直接的にイオンチャネルを形成することにより細胞を殺傷する細菌毒素が数多く存在する。そうした毒素のモデル例が、大腸菌により分泌される細菌毒素であるコリシンである。コリシンIaは、1952年にJacobにより発見され、それ以降、何世代にもわたる努力を経て、Qiu et al.(Major transmembrane movement associated with colicin Ia channel gating. J. Gen. Physiology, 107:313‐328 (1996))は、人工の脂質二重膜中に形成されたイオンチャネルが開いている又は閉じられているときの、コリシンIaの膜貫通型立体構造をついに明らかにした。これは、分子レベルでの新しい抗生物質の設計および製造のための基礎をもたらすものである。その後、コリシンポリペプチドと、白色連鎖球菌またはブドウ球菌の誘引物質などのシグナルペプチドとの結合により作製された、コリシンを目的の細菌の細胞膜へと向かわせ、形成された膜貫通型イオンチャネルを通っての細胞含有物の漏出により細胞を殺傷するポリペプチド分子が存在している。
【0003】
悪性腫瘍は人間の健康に大きな脅威を与えるものである。毎年、世界中で七百万人が悪性腫瘍により亡くなっており、その六分の一は中国におけるものである。現在、悪性腫瘍は、我が国における死亡原因の第二位となっている。悪性腫瘍の病因、病原および臨床症状が明確に解明されてはいないため、予防および治療は効果的ではない。抗腫瘍剤は腫瘍の治療において重要である。それらにより一部の腫瘍に対しての治療効果は達成されるが、例えば不十分な腫瘍選択性、免疫学的抑制、有害反応、薬剤耐性等の、いくつかの欠点が残っている。
【0004】
EBウィルスが原因であるバーキットリンパ腫、ホジキンリンパ腫および上咽頭癌の細胞の表面は、EBウィルスの特異的表面抗原を有する。したがって、EBウィルス表面抗原は、そうした腫瘍細胞の特異的マーカーとみなすことができる。EBウィルスが原因である腫瘍に対する薬剤について、中国特許第ZL200410081446.8号の発明では、コリシンと、EBウィルス表面抗原を認識する抗体模倣物との結合により形成される抗腫瘍ポリペプチドが開示されている。この抗腫瘍ポリペプチドは、体内の、EBウィルスが原因であるがん細胞を、特異的に殺傷することができるものであり、正常細胞に対して害がなく、その殺傷能力は他の抗腫瘍剤の数倍であり、腫瘍選択性、薬剤耐性、がん細胞が殺傷されるときの正常組織の障害などの問題を克服する。Xiao‐Qing Qiu et al.(Xiao‐Qing Qiu et al., 2007, Small antibody mimetics comprising two complementarity‐determining regions and a framework region for tumor targeting, Nature Biotechnology 25, 921‐929, 1 August 2007)は、一連の抗体模倣物とコリシンとにより構築される抗腫瘍ポリペプチドの殺傷効果を比較し、VHCDR1‐VHFR2‐VLCDR3およびVLCDR1‐VHFR2‐VHCDR3の抗体模倣物とコリシンとにより構築される抗腫瘍ポリペプチドが、優れた殺傷能力を有することを発見している。この研究は、EBウィルスが原因である腫瘍に対するポリペプチドの調製のための、より多くの候補抗体模倣物をもたらす。
【0005】
しかしながら、上述の抗腫瘍ポリペプチドについては、コリシンの疎水性末端が、過敏性を有し得るいくつかのアミノ酸残基を有しているため、コリシンのポリペプチドを含む薬品は、in vivoでの異常な免疫応答をより容易に引き起こす可能性がある。多くのがん患者の代謝機構は、がん細胞からの妨害に起因して異常になっており、彼らはポリペプチドの薬に対するアレルギー応答を起こしやすく、それ故、こうした薬によって治療することができない、と報告されている。したがって、より安全な、かつ、より多くの患者に適した抗がん薬を得るためには、コリシンポリペプチドを改良する必要がある。
【発明の概要】
【発明が解決しようとする課題】
【0006】
上述の先行技術の欠点に基づいて、本発明は、EBウィルスが原因である腫瘍に対する新規ポリペプチド、ならびに、その使用および生産方法を提供し、ひいては、高い殺傷能力、および、高い特異性を有し、かつ、アレルギーの可能性の低い、EBウィルスが原因である腫瘍の治療のための薬品を提供する。
【0007】
イオンチャネルを形成することのできるコリシンの変異ポリペプチドと、抗EBウィルス抗体ポリペプチドまたは抗EBウィルス抗体模倣物ポリペプチドとの機能可能な連結により形成される、EBウィルスが原因である腫瘍に対する新規ポリペプチドであって、前記イオンチャネルを形成することのできるコリシンの変異ポリペプチドが、野生型コリシンE1、Ia、Ib、A、B、Nまたはそれらの水溶性チャネルドメインのペプチド鎖の、G11A、H22G、A26G、V31L、およびH40Dのアミノ酸残基の変異により得られるものであり、前記抗EBウィルス抗体のポリペプチドのアミノ酸配列が、ATCC HB‐168のハイブリドーマにより分泌されるモノクローナル抗体のポリペプチドと同じである、新規ポリペプチド。
【0008】
前記抗体模倣物のポリペプチドが、抗EBウィルス抗体の、重鎖のCDR1領域と、重鎖のCDR1‐CDR2連結ペプチドセグメントと、軽鎖のCDR3との結合ペプチドである。
【0009】
前記イオンチャネルを形成することのできるコリシンの変異ポリペプチドが、野生型コリシンIaの変異により得られるものである。
【0010】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドが、配列番号29に示されているアミノ酸配列を有する。
【0011】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドをコードする遺伝子。
【0012】
配列番号30に示されているヌクレオチド配列を有する、前記遺伝子。
【0013】
前記遺伝子を含む組換えプラスミド。
【0014】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドの生産方法であって、前記組換えプラスミドを、発現のために発現系に形質転換する工程、および、発現したポリペプチドを単離する工程、を含む、生産方法。
【0015】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドの、EBウィルスが原因である腫瘍の治療および予防のための医薬の生産における使用。
【0016】
そのアミノ酸配列が、配列番号24に示されているものである、コリシンIaの変異ポリペプチド。
【0017】
コリシンIaの変異ポリペプチドをコードする遺伝子。
【0018】
前記遺伝子を、そのペプチドを誘導する遺伝子と機能可能に連結し、発現ベクターにクローニングし、次いで、前記発現ベクターを発現系に形質転換し、その発現したポリペプチドを単離する、ペプチド医薬の生産における、前記遺伝子の使用。
【課題を解決するための手段】
【0019】
本発明は、イオンチャネルを形成することのできるコリシンの変異ポリペプチドと、抗EBウィルス抗体のポリペプチドまたは抗EBウィルス抗体模倣物のポリペプチドとにより形成される、EBウィルスが原因である腫瘍に対する新規ポリペプチド、を提供する。野生型コリシンポリペプチド分子中には、過敏性を有し得るいくつかのアミノ酸残基が存在するため、イオンチャネル構築物を形成することのできるコリシンのポリペプチド分子において、本発明は、アレルギー応答を容易に引き起こし得る疎水性の領域のアミノ酸残基を選択的に変異させている。例えば、本発明の好ましい一実施形態においては、コリシンIaのポリペプチドの変異の部位は以下である:G11A、H22G、A26G、V31LおよびH40D。コリシンIaのポリペプチドまたは変異体Iaのポリペプチドの注射によりそれぞれ免疫したマウスにおいては、その実験データは、変異体Iaのポリペプチドを注射したマウスによりもたらされる血清力価は前者よりも数桁低い、すなわち、免疫応答のレベルがより低いことを示しており、これは、この変異ポリペプチドが、細胞膜にイオンチャネルを形成するという機能を保持していながらも、アレルギーの可能性を低減することを実証するものである。実験は、本発明の組換えポリペプチドの殺傷能力は影響を受けないことを示し、これは、変異体アミノ酸残基が、コリシンのイオンチャネルを形成する機能に影響を与えないことを意味している。本発明により提供される、EBウィルスが原因である腫瘍に対する新規ポリペプチドにおいては、抗EBウィルス抗体のポリペプチドまたは抗EBウィルス抗体模倣物のポリペプチドがEBウィルスによる腫瘍細胞の表面抗原を認識することを通じて、コリシンの変異ポリペプチドは標的細胞の膜にターゲティングされ、コリシンの変異ポリペプチド膜貫通型イオンチャネルドメインの疎水性の領域が腫瘍細胞の細胞膜へと挿入され、イオンチャネルを形成し、よって、腫瘍細胞が細胞内容物の漏出によって死ぬ。抗EBウィルス抗体のポリペプチドのアミノ酸配列は、ATCC HB‐168のハイブリドーマにより分泌される抗体のポリペプチドのアミノ酸配列を完全に参照する。
【0020】
本発明の一実施形態においては、上述の抗EBウィルス抗体模倣物のポリペプチドと、コリシンの変異ポリペプチドのカルボキシル末端との機能的連結により得られる、本発明の低分子量の抗腫瘍ポリペプチドが好ましい。つまり、そのような低分子量の模倣ポリペプチドは、抗EBウィルス抗体のポリペプチドの、VHCDR1領域、VLCDR3領域、VHCDR1‐VHCDR2の連結ペプチドセグメントと、軽鎖のVLCDR3の結合により得られる、VHCDR1‐VHFR2‐VLCDR3のペプチド鎖を含む。抗体模倣物の新規抗腫瘍ペプチド1のアミノ酸配列を、配列番号25に示す。この抗体模倣物は、30個未満のアミノ酸しか含んでおらず、アミノ酸150個の天然の抗体よりもずっと低い分子量を有する。これは、抗腫瘍ポリペプチドの分子量を大幅に減少させて、本発明の抗腫瘍ポリペプチドの組織浸透能に貢献するものでありながらも、抗原認識の要件を満たす。
【0021】
本発明のもう1つの目的は、本発明の抗腫瘍ポリペプチドをコードする遺伝子配列を提供することである。本発明の抗腫瘍ポリペプチドの遺伝子は、コリシンの変異ポリペプチドをコードする遺伝子と、抗EBウィルス抗体のポリペプチドまたはその抗体模倣物のポリペプチドをコードする遺伝子との機能可能な連結により形成され、ここで、コリシンのポリペプチド、および、抗EBウィルス抗体の遺伝子配列は、当該分野において公知であり、コリシンの変異ポリペプチドの遺伝子は、コリシンポリペプチドの遺伝子の対応するコドンにおける、以下の点変異により得られるものである:G11A、H22G、A26G、V31LおよびH40D。遺伝コードの縮重の結果として、当業者は、アミノ酸配列を変えることなく、本発明の抗腫瘍ポリペプチドをコードするヌクレオチド配列を調整してもよい。
【0022】
本発明の組換えプラスミドは、野生型コリシンの遺伝子が組み込まれた元のベクターが、二本鎖ヌクレオチドに部位特異的に変異導入され、変異標的部位に変異コドンにより挿入され、それにより、コリシン変異ポリペプチドの遺伝子を含む変異体ベクターを得る、ということを意味するものである。同じプロセスの部位特異的突然変異誘発により、抗EBウィルス抗体の抗体模倣物をコードする遺伝子を、前記コリシン変異ポリペプチドの遺伝子のカルボキシル末端へと挿入し、本発明の組換えプラスミドを得る。当該元のベクターpSELECTTM‐1は、プロメガ社から購入したものであり、コリシンIaおよび免疫タンパク質の遺伝子を有する。部位特異的突然変異誘発のプロセスは、Strategene社からのキットの指示書に従ったものである。本発明は、コリシンの変異ポリペプチドを調製するべく、いくつかの部位特異的突然変異誘発を行なうが、ここでは、5つのコドンを部位特異的に変異させる。よって、5対のプライマー配列が設計されている(配列番号1〜10)。本発明の実施例においては、6対のプライマー配列が、抗体模倣物の遺伝子用に設計されている(配列番号11〜22)。
【0023】
本発明は、上記で得られる組換えベクターを、改変細菌 大腸菌BL21(DE3)中へと形質転換すること、陽性クローンを選抜すること、陽性クローンにより発現した当該タンパク質を単離および精製すること、それにより、本発明のEBウィルスが原因である腫瘍に対する新規ポリペプチドを得ること、を含む、本発明の抗腫瘍ポリペプチドの生産のための方法も提供する。
【0024】
本発明により提供される、EBウィルスが原因である腫瘍に対する新規ポリペプチドは、EBウィルスが原因である腫瘍の治療および予防のための医薬の生産において用いることができる。本発明において得られる新規抗生物質のポリペプチドを、薬剤的に許容できる担体もしくは賦形剤または他の所望により選択される成分中へと加えることにより、臨床上適切な医薬組成物を作製することができる。
【0025】
本発明は、コリシンIaの変異ポリペプチドのアミノ酸配列および遺伝子配列も提供する。この変異ポリペプチドは、本発明において用いることができ、他の標的化ポリペプチドを用いる抗体ポリペプチドの構築においても用いることができる。本発明中の実施例3の実験データは、当該変異ポリペプチドを含むペプチド医薬の免疫原性が低いこと、および、他の標的化ポリペプチドを用いる当該変異ポリペプチドにより形成される抗体ポリペプチドが殺菌能を有するとのこと、を証明するものである。当該生産方法は、当該分野における常用の実験プロセスである。
【0026】
本発明により提供される新規抗腫瘍ポリペプチドは、特許第ZL200410081446.8号に開示されている抗腫瘍ポリペプチドの利点、すなわち、高い特異性のターゲティング、正常細胞に対する安全性、および、薬剤耐性を生じさせにくい、といった利点を有する。それと同時に、本発明の抗腫瘍ポリペプチドは、アレルギー応答を引き起こしやすいアミノ酸残基に変異を受けたものであり、そうした変異ポリペプチドを含む当該抗腫瘍ポリペプチドの免疫原性は低減される。すなわち、アレルギー反応の可能性が低下する。そうしたポリペプチドの医薬の、使用安全性、および、腫瘍を殺傷するという効果は改善される。これは、また、コリシンポリペプチドを含む他の医薬の改良のための良い例でもあり得る。
【図面の簡単な説明】
【0027】
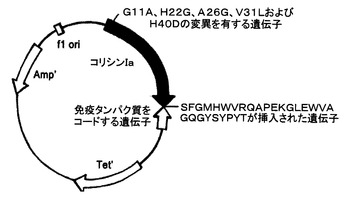
【図1】抗体模倣物VHCDR1‐VHFR2‐VLCDR3のポリペプチドの遺伝子と、コリシンIaの変異ポリペプチドの遺伝子とを含む、組換えプラスミドpCHCEB11の構造の略図である。
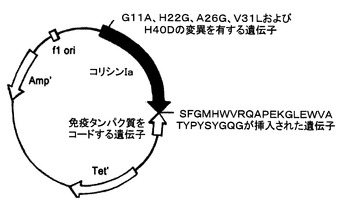
【図2】抗体模倣物VHCDR1‐VHFR2‐(Rev)VLCDR3のポリペプチドの遺伝子と、コリシンIaの変異ポリペプチドの遺伝子とを含む、組換えプラスミドpCHCEB22の構造の略図である。
【図3】コリシンIaの変異ポリペプチドの感作効果の実験1を示す図である。 (A)致死量のMRSA(ATCC BAA42)を腹腔内注射した昆明(Kunming)マウスを、(1)コントロールグループ、(2)アンピシリングループ、(3)黄色ブドウ球菌に対するポリペプチド(ZL01128836.1)のグループ、(4)黄色ブドウ球菌に対するポリペプチド1のグループ、へとランダムにグループ分けする。 (B)14日後、昆明マウスの新たな一群を、コントロールグループおよびアンピシリングループへとグループ分けする。黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループ由来の生き残ったマウスを、黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループへとグループ分けし、当該実験を繰り返す。 (C)41日後、昆明マウスの新たな一群を、(1)コントロールグループ、(2)レボフロキサシンのグループ、(3)セフトリアキソンナトリウムのグループ、へとグループ分けし、また、(4)黄色ブドウ球菌に対するポリペプチドのグループおよび(5)黄色ブドウ球菌に対するポリペプチド1のグループ由来の生き残ったマウスは、緑膿菌に対するポリペプチド2のグループおよび緑膿菌に対するポリペプチド1のグループへとグループ分けする。
【図4】コリシンIaの変異ポリペプチドの低感作効果の実験2を示す図である。 (A)黄色ブドウ球菌に対するポリペプチド/緑膿菌に対するポリペプチド2のグループの血清、1:50,000の力価; (B)黄色ブドウ球菌に対するポリペプチド1/緑膿菌に対するポリペプチド1のグループの血清、1:50,000の力価。 (1)第1週、(2)第2週、(3)第7週の血清、(4)ネガティブコントロール。
【図5】EBウィルスが原因であるバーキットリンパ腫に対する、新規抗腫瘍ポリペプチドのin vitroにおける殺傷効果の比較を示す図である。 (A)コントロールグループ、(B)新規抗腫瘍ポリペプチド1処理グループ、(C)新規抗腫瘍ポリペプチド2処理グループ。
【図6】EBウィルスが原因であるバーキットリンパ腫の細胞および他の腫瘍細胞に対する、新規抗腫瘍ポリペプチドのin vitroにおける殺傷効果を示す図である。 (A)バーキットリンパ腫のEBV陽性細胞、 (B)バーキットリンパ腫のEBV陰性細胞、 (C)AIDS患者由来の悪性リンパ肉腫のEBV陽性細胞。 (1)コントロールグループ、(2)新規抗腫瘍ポリペプチド1処理グループ。
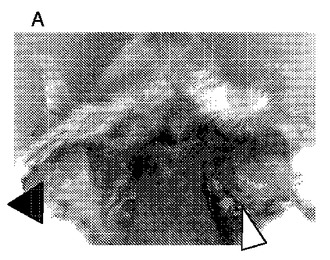
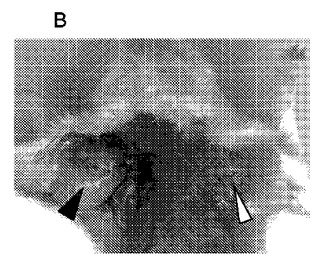
【図7】EBウィルスが原因であるバーキットリンパ腫の細胞を移植したヌードマウス由来の成長した固形腫瘍に対する新規抗腫瘍ポリペプチドの殺傷効果を示す図である。 (A)コントロールグループ、 (B)新規抗腫瘍ポリペプチド1治療グループ、に由来するすべてのSCID免疫不全マウスの、両方の脇の下の側面部中に、バーキットリンパ腫の細胞を接種する。左矢印はEBV陰性リンパ肉腫、右矢印はEBV陽性リンパ肉腫である。
【図8】EBウィルスが原因であるバーキットリンパ腫の細胞を移植したヌードマウス由来の成長した固形腫瘍に対する新規抗腫瘍ポリペプチドの殺傷効果を示す図である。 (A)コントロールマウスのEBV陰性リンパ肉腫の切片、(B)コントロールマウスのEBV陽性リンパ肉腫の切片、(C)新規抗腫瘍ポリペプチド1治療マウスのEBV陰性リンパ肉腫の切片、(D)新規抗腫瘍ポリペプチド1治療マウスのEBV陽性リンパ肉腫の切片。
【発明を実施するための形態】
【0028】
以下、本発明の好ましい実施形態を記述することにより、および、添付の図面を参照して、本発明を説明する。
【0029】
本発明において使用される元のベクターpSELECTTM‐1は、プロメガ社から購入したものである。
【0030】
大腸菌BL21(DE3)という改変細菌は、ノバジェン社から購入したものである。
【実施例】
【0031】
(実施例1)
変異体コリシンIaをコードする遺伝子を含む組換えプラスミドの構築。
元のベクターは、プラスミドpSELECTTM‐1(8.3kb)(プロメガ社から購入)であり、これは、コリシンIaおよび免疫タンパク質の遺伝子を有する。変異体アミノ酸をコードする配列番号1〜10に示されているオリゴヌクレオチドプライマーの配列を、それぞれ、二本鎖オリゴヌクレオチド部位特異的突然変異誘発技術(QuickChangeTMキット、Strategene社)により、野生型コリシンIaの遺伝子へと機能可能に連結し、コリシンIaの変異ポリペプチドをコードする配列番号23に示されている遺伝子、および、変異体プラスミド、を得る。その後、この変異体プラスミドに、配列番号26または配列番号28の遺伝子を、コリシンIaの変異ポリペプチドの遺伝子のI626のコドンの後に挿入し、EBウィルスが原因である腫瘍に対する新規ポリペプチドのための、2種類の組換えプラスミドpCHCEB11(図1に示されている)およびpCHCEB22(図2に示されている)を得る。組換えプラスミド中の、EBウィルスに対する抗体をコードする遺伝子の調製のために設計された6対のオリゴヌクレオチドプライマーの配列が配列番号11〜22に示されている。前記組換えプラスミドを、大腸菌BL21(DE3)(ノバジェン社から購入)という改変細菌中へとトランスフェクションし、ポリペプチドを調製する。得られるポリペプチドは、配列表の、配列番号25(以下、「新規抗腫瘍ポリペプチド1」という)および配列番号27(以下、「新規抗腫瘍ポリペプチド2」という)に示されたものである。
【0032】
二本鎖オリゴヌクレオチド部位特異的突然変異誘発のプロセスは、Strategene QuickChang部位特異的突然変異誘発キット(カタログ番号200518)に従ったものである。
1. 部位特異的突然変異誘発のための反応物の調製:
5μlの10×緩衝液
2μl(10ng)の、コリシンIaの野生型のポリペプチドの遺伝子および免疫タンパク質の遺伝子を有する元のプラスミドpSELECTTM‐1。
1.25μl(125ng)の、設計された5’‐3’オリゴヌクレオチドプライマー
1.25μl(125ng)の、設計された3’‐5’オリゴヌクレオチドプライマー
1μlのdNTP
再蒸留水50μl
1μl pfu
(プラスミド、プライマーおよび再蒸留水以外は、前記キットにより提供されるものである)
2. PCR増幅、増幅条件:95℃で35秒間の変性、53℃で70秒間のアニール、および68℃で17分間の伸長、を25サイクル;
3. 1μlのエンドヌクレアーゼDpn1を加えて、親DNA鎖を消化(37℃、1時間)し、1μlの反応物と50μlのXL1‐Blueコンピテント細胞とを、共に、氷上で30分間インキュベートし、42℃での45秒間のヒートショックの後に、氷中で2分間インキュベートする;
4. 0.5mlの培養培地NZYを加え、37℃および220rpmにて1時間振盪する。50〜100μlの反応物をまく(1%寒天および50μg/mlアンピシリンを加えたLB培地、37℃で一晩);
5. 18時間後にコロニーをピックアップする。プラスミドを抽出し、配列決定を行い、変異が成功していることを確認する;
6. 複数の部位における変異によって最終的に得られた50ngの組換えプラスミドを、40μlの大腸菌BL‐21(DE3)コンピテント細胞と共に氷上で5分間インキュベートし、42℃で30秒間ヒートショックを与え、氷中で2分間インキュベートする。ノバジェン社からの160μlの培養培地SOCを加え、37℃および220rpmにて1時間振盪した後にまく(1%寒天および50μg/mlアンピシリンを加えたLB培地、37℃で一晩)。
7. 増幅のためにシングルコロニーをピックアップし、8〜16リットルFB培地、250rpm、30℃で4〜5時間とし、42℃でヒートショックを与え、250rpmにて30分間、37℃にて2時間。細菌を、6000gおよび4℃での20分間の遠心分離により沈殿させる。4℃の50mMホウ酸緩衝液(2mM EDTA+2mM DTT)および50〜80mlの細菌懸濁液は、250mlの0.2M PMSFを加え、超音波処理により処理する(4℃、400W、2分)。細菌破砕物(debris)を高速遠心分離する(4℃、75,000g、90分)。その上清には、500万単位の硫酸ストレプトマイシンを加え、DNAを沈殿させる。15000gおよび4℃での10分間の遠心分離による沈殿の後、その上清を、4℃にて、50mMホウ酸緩衝液中、分子量15,000の透析バッグ中で一晩透析する。再び15000gおよび4℃での10分間の遠心分離による沈殿の後、その上清を、CMイオン交換カラムにロードする。このカラムを、0.1〜0.3M NaCl+50mMホウ酸緩衝液の勾配を用いて溶出し、組換え抗腫瘍ポリペプチドを得る。
【0033】
部位特異的突然変異誘発のために設計されたプライマーの配列は以下の通りである:
【0034】
配列番号1、コリシンの遺伝子におけるG11Aの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
cgt att aca aat ccc GCA gca gaa tcg ctg ggg
配列番号2、コリシンの遺伝子におけるG11Aの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
ccc cag cga ttc tgc TGC ggg att tgt aat acg
配列番号3、コリシンの遺伝子におけるH22Gの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
gat tca gat ggc GGT aaa tta tgg gtg
配列番号4、コリシンの遺伝子におけるH22Gの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
cac cca taa ttt ACC gcc atc tga atc
配列番号5、コリシンの遺伝子におけるA26Gの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
gaaa ttatgGGTgt tgatatttat
配列番号6、コリシンの遺伝子におけるA26Gの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
ataaatatacaacACCcataatttc
配列番号7、コリシンの遺伝子におけるV31Lの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
gt tgatatttat CTC aaccctc cacgtgtc
配列番号8、コリシンの遺伝子におけるV31Lの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
gacacgtggagggttGAGataaatatcaac
配列番号9、コリシンの遺伝子におけるH40Dの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
cgtgtcga tgtctttGATggtaccccgc ctgcat
配列番号10、コリシンの遺伝子におけるH40Dの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
atgcaggcggggtaccATCaaagacatcgacacg
【0035】
配列番号11、組換えプラスミドpCHCEB11中のVHCDR1の遺伝子のためのプライマー5’‐3’:
gcg aat aag ttc tgg ggt att TCC TTC GGT ATG CAT TGG GTG CGTCAGtaa ata aaa tat aag aca ggc
配列番号12、組換えプラスミドpCHCEB11中のVHCDR1の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta CTG ACG CAC CCA ATG CAT ACC GAA GGA aat acc cca gaa ctt att cgc
配列番号13、組換えプラスミドpCHCEB11中のVHFR2の遺伝子のためのプライマー5’‐3’:
ggt atg cat tgg gtg cgt cag GCC CCC GAG AAA GGT CTG GAG TGG GTG GCC taa ata aaa tat aag aca ggc
配列番号14、組換えプラスミドpCHCEB11中のVHFR2の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta GGC CAC CCA CTC CAG ACCT TTT CTC GGG GGC ctg acg cac cca atg cat acc
配列番号15、組換えプラスミドpCHCEB11中の(Rev)VlCDR3の遺伝子のためのプライマー5’‐3’:
aaa ggt ctg gag tgg gtg gcc ACC TAC CCC TAC TCC TAC GGT CAG GGT taa ata aaa tat aag aca ggc
配列番号16、組換えプラスミドpCHCEB11中の(Rev)VLCDR3の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta ACC CTG ACC GTA GGA GTA GGG GGT ggc cac cca ctc cag acc ttt
配列番号17、組換えプラスミドpCHCEB22中のVHCDR1の遺伝子のためのプライマー5’‐3’:
gcg aat aag ttc tgg ggt att TCC TTC GGT ATG CAT TGG GTG CGT CAG taa ata aaa tat aag aca ggc
配列番号18、組換えプラスミドpCHCEB22中のVHCDR1の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta CTG ACG CAC CCA ATG CAT ACC GAA GGA aat acc cca gaa ctt att cgc
配列番号19、組換えプラスミドpCHCEB22中のVHFR2の遺伝子のためのプライマー5’‐3’:
ggt atg cat tgg gtg cgt cag GCC CCC GAG AAA GGT CTG GAG TGG GTG GCC taa ataaaa tat aag aca ggc
配列番号20、組換えプラスミドpCHCEB22中のVHFR2の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta GGC CAC CCA CTC CAG ACCT TTT CTC GGG GGC ctg acg cac cca atg cat acc
配列番号21、組換えプラスミドpCHCEB22中のVLCDR3の遺伝子のためのプライマー5’‐3’:
aaa ggt ctg gag tgg gtg gcc GGT CAG GGT TAC TCC TAC CCC TAC ACC taa ata aaa tat aag aca ggc
配列番号22、組換えプラスミドpCHCEB22中のVLCDR3の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta GGT GTA GGG GTA GGA GTA ACC CTG ACC ggc cac cca ctc cag acc ttt
【0036】
(実施例2)
組換えプラスミドpCHCEB11およびpCHCEB22から調製した新規抗腫瘍ポリペプチドの免疫効果の観察。
実施例1において得られた組換えプラスミドpCHCEB11およびpCHCEB22から調製した新規抗腫瘍ポリペプチド1および新規抗腫瘍ポリペプチド2、ならびに、発明者が所有する、先の発明(ZL200410081446.8)からの抗腫瘍ポリペプチド1および抗腫瘍ポリペプチド2、により、マウスを免疫する。上述の各タンパク質をアジュバントと混合する。プライミング投与量およびブースト投与量は、各マウスにつき50μg(0.5ml)の1回の腹腔内注射、2週間の間隔をあけて合計5回の注射、である。血清力価は、間接ELISA法により決定する。本発明により調製した新規抗腫瘍ポリペプチド1および2により免疫したマウスの力価は、10−3〜10−4の範囲であり、一方、抗腫瘍ポリペプチド1および抗腫瘍ポリペプチド2により免疫したマウスの力価は、10−4〜10−5の範囲である。
本発明の新規抗腫瘍ポリペプチドにより誘発される過敏性反応の可能性は、野生型コリシンIaを含む抗腫瘍ポリペプチドにより誘発される過敏性反応の可能性よりも1桁〜2桁低い、とのことがわかる。
【0037】
(実施例3)
新規抗腫瘍ポリペプチドを形成するコリシンIaの変異ポリペプチドの低感作効果の実験。
実施例1の、コリシンIaの変異ポリペプチドの変異体プラスミド(水溶性チャネルドメインのペプチド鎖において、G11A、H22G、A26G、V31L、およびH40Dのアミノ酸残基に変異を受けている)を、変異ポリペプチドのN末端またはC末端にて、黄色ブドウ球菌のフェロモンAgrD1(YSTCDFIM)に機能可能に連結し、2種類の抗菌ポリペプチドを得る。当該変異体コリシンIaのカルボキシル末端におけるAgrD1の連結により得られるポリペプチドを、黄色ブドウ球菌に対するポリペプチド1と名付け、当該変異体コリシンIaのアミノ末端におけるAgrD1の連結により得られるポリペプチドを、緑膿菌に対するポリペプチド1と名付ける。野生型コリシンIaについてのプラスミドを、アミノ末端にて、黄色ブドウ球菌のフェロモンAgrD1(YSTCDFIM)に連結し、緑膿菌に対するポリペプチド2を得る。
【0038】
実験1:昆明マウスの一群に、致死量のMRSA(ATCC BAA42)を腹腔内注射し、(1)コントロールグループ、(2)アンピシリングループ、(3)黄色ブドウ球菌に対するポリペプチドのグループ、(4)黄色ブドウ球菌に対するポリペプチド1のグループ、へとランダムにグループ分けする。各グループは、10匹のマウスからなる。
処理方法:
致死量のMRSA(ATCC BAA42)の腹腔内注射の1時間後に:
コントロールグループ:0.5mlの0.3M NaCl+50mMホウ酸緩衝液を、尾静脈を介して1回注射する;
アンピシリンのグループ:2.5mg/kgのアンピシリンを、尾静脈を介して1回注射する;
黄色ブドウ球菌に対するポリペプチドのグループ:6mg/kgの、本発明者が所有する、黄色ブドウ球菌に対するポリペプチド(ZL01128836.1)を、尾静脈を介して1回注射する;
黄色ブドウ球菌に対するポリペプチド1のグループ:6mg/kgの、黄色ブドウ球菌に対するポリペプチド1を、尾静脈を介して1回を注射する;
結果:コントロールグループおよびアンピシリンのグループのマウスは、すべて、2日以内に死亡。黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループの85%のマウスは生き残る。
【0039】
実験2:実験1の14日後に、昆明マウスの新たな一群を、コントロールグループおよびアンピシリンのグループへとグループ分けする。黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループ由来の生き残ったマウスを、黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループへとグループ分けし、上記の実験を繰り返す。コントロールグループおよびアンピシリンのグループのマウスは、すべて、2日以内に死亡。黄色ブドウ球菌に対するポリペプチドのグループの75%のマウスは生き残り、黄色ブドウ球菌に対するポリペプチド1のグループの90%のマウスは生き残る。
【0040】
実験3:実験1の41日後に、昆明マウスの新たな一群を、コントロールグループ、レボフロキサシンのグループ、およびセフトリアキソンナトリウムのグループへとグループ分けする。黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループ由来の生き残ったマウスは、緑膿菌に対するポリペプチド2のグループおよび緑膿菌に対するポリペプチド1のグループへとグループ分けする。
マウスに、致死量の多剤耐性緑膿菌(四川大学華西医院実験医学科からの臨床分離株13578)を腹腔内注射する。1時間の後、
コントロールグループに、0.5mlの0.3M NaCl+50mMホウ酸緩衝液を、尾静脈を介して1回注射する;
レボフロキサシンのグループに、5mg/kgのレボフロキサシンを、尾静脈を介して1回注射する;
セフトリアキソンナトリウムのグループに、30mg/kgのセフトリアキソンナトリウムを、尾静脈を介して1回注射する;
緑膿菌に対するポリペプチド2のグループに、8mg/kgの、緑膿菌に対するポリペプチド2を、尾静脈を介して1回注射する;
緑膿菌に対するポリペプチド1のグループに、8mg/kgの、緑膿菌に対するポリペプチド2を、尾静脈を介して1回注射する。
コントロールグループおよびレボフロキサシンのグループのマウスは、すべて、1日以内に死亡。セフトリアキソンナトリウムのグループの25%のマウスは生き残る。緑膿菌に対するポリペプチド2のグループの60%のマウスは生き残る。緑膿菌に対するポリペプチド1のグループのマウスは、すべて、生き残る。宿主の抗体が、変異ポリペプチドの殺傷効果を、野生型ポリペプチドの殺傷効果よりも低く妨害をすることが示されている。
図3を参照のこと。
【0041】
この実験の第1週、第2週および第7週において、黄色ブドウ球菌に対するポリペプチドのグループ/緑膿菌に対するポリペプチド2のグループ、および、黄色ブドウ球菌に対するポリペプチド1のグループ/緑膿菌に対するポリペプチド1のグループ、に由来する、生き残ったマウスの血清について、血液中の抗体を検出するために、間接ELISA法により分析する。酵素標識プレートのウェルを、野生型コリシンIaおよびコリシンIaの変異ポリペプチドで、100ng/ウェルでコーティングする。一次抗体は、黄色ブドウ球菌に対するポリペプチドのグループ/緑膿菌に対するポリペプチド2のグループ、および、黄色ブドウ球菌に対するポリペプチド1のグループ/緑膿菌に対するポリペプチド1のグループ、に由来する、生き残ったマウスの血清である。二次抗体は、ヤギ抗マウス標識抗体である。一次抗体のネガティブコントロールは、5%ミルク‐PBSである。1:50,000の力価の結果は、以下の通りである(図4参照):
【0042】
【表1】

【0043】
本発明により調製されたコリシンIaの変異ポリペプチドにより誘発される宿主の過敏感反応の可能性は、野生型コリシンIaにより誘発される宿主の過敏感反応の可能性よりも低いことが示されている。
【0044】
(実施例4)
EBウィルスが原因であるバーキットリンパ腫に対する新規抗腫瘍ポリペプチドのin vitroにおける殺傷効果。
EBV陽性細胞株およびEBV陰性細胞株は、米国のATCCからの、標準的な細胞株である。
細胞培養:起こしたRaji細胞の懸濁液0.1mlを、ゆっくりと、培養ディッシュ中の3mlの1640液体培地(10%血清を加えたもの)へと加え(希釈率、1:30)、混合し、CO2を含有する37℃インキュベーター中で培養する。当該EBV陽性細胞株は、ATCC CCL‐86(世界中の研究室において使用されている標準的なバーキットリンパ腫細胞。Raji細胞。12歳のアフリカの少年から1963年に単離されたもの)である。
この試験細胞を、3つのグループにグループ分けする。
グループ1は、ブランクのグループであり、抗腫瘍ポリペプチドを含有しない保存液(10mMPB+0.2M NaClリン酸緩衝液(pH7.4))が加えられたものである。
グループ2は、200μg/mlの新規抗腫瘍ポリペプチド1(プラスミドpCHCEB11、保存液、10mMPB+0.2M NaClリン酸緩衝液、pH7.4)が加えられたものである。
グループ3は、200μg/mlの新規抗腫瘍ポリペプチド2(プラスミドpCHCEB22、保存液、10mMPB+0.2M NaClリン酸緩衝液、pH7.4)が加えられたものである。
24時間の培養後、培養ディッシュに、上述の処理剤を加える。これらの処理剤の添加の72時間後、この培養ディッシュに、20μlの100μMolヨウ化プロピジウム(PI)を加え、10分後、この培養ディッシュを顕微鏡下で観察する。その結果が示すのは、ブランクのグループの細胞はよく成長し、新規抗腫瘍ポリペプチド1のグループの細胞の大部分はPIにより赤く染色されるということであり、この抗腫瘍ポリペプチドによって、細胞膜が破壊されることを示しており、これは腫瘍細胞の死を導く。死細胞の数を比較すると、2種類の新規抗腫瘍ポリペプチドのうちで、新規抗腫瘍ポリペプチド2の効果はあまり良くない。図5を参照のこと。
【0045】
(実施例5)
EBウィルスが原因であるバーキットリンパ腫の細胞および他の腫瘍細胞に対する新規抗腫瘍ポリペプチドのin vitroにおける殺傷効果についての多重蛍光染色の観察。
細胞培養の条件は実施例2と同じである。本実験では、次の3種類の細胞株を使用する:EBウィルス陽性細胞株:ATCC CCL‐86(Raji細胞。バーキットリンパ腫細胞);EBウィルスおよびカポジ肉腫ウィルスが陽性の、46歳のAIDS男性患者由来の悪性リンパ肉腫細胞の株である、ATCC CRL‐2230;EBウィルス陽性細胞株:ATCC CRL‐1648(CA‐46。アメリカのバーキットリンパ腫患者の腹水から単離された細胞)。
各株は、2つの試験グループにグループ分けする。グループ1は、新規抗腫瘍ポリペプチドを含有しない保存液(10mMPB+0.2M NaClリン酸緩衝液(pH7.4))が加えられたものである。グループ2は、200μg/mlの新規抗腫瘍ポリペプチド1(プラスミドpCHCEB11)が加えられたものであり、保存液は10mMPB+0.2M NaClリン酸緩衝液、pH7.4である。
24時間の培養後、培養ディッシュに、上述のグループの処理剤を加える。これらの処理剤の添加の72時間後、この培養ディッシュに、2種類の蛍光色素、すなわち、20μlの50μMol FITCと、20μlの50uMol ローダミン123とを加え、10分後、この培養ディッシュを顕微鏡オリンパスIX‐71下で観察する。
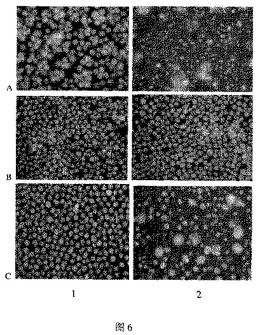
その結果、EBV陰性腫瘍細胞の株は、新規抗腫瘍ポリペプチド1の処理後、よく成長すること、及び、EBV陽性腫瘍細胞の株は、いずれの株も、大部分の細胞がミトコンドリアおよび核を消失し、膨張し、ネクローシスを起こし、それらのうちのほとんどが死んでいることを示す。実施例4のPI染色実験と比較すると、明らかに、多重蛍光染色実験からの結果は、EBウィルス陽性腫瘍細胞に対する新規抗腫瘍ポリペプチド1の強力な殺傷効果を、より明確に示しているようである。図6を参照のこと。
EBV陰性腫瘍細胞はよく成長し、当該新規抗腫瘍ポリペプチドは、細胞膜中にEBウィルスの表面抗原を有しない細胞を攻撃しないことを意味する。本発明の新規抗腫瘍ポリペプチドは理想的なターゲティング特異性と安全性とを有している、とのことが示唆される。
【0046】
(実施例6)
EBウィルスが原因であるバーキットリンパ腫の細胞を移植したヌードマウスの体内で成長した固形腫瘍に対する新規抗腫瘍ポリペプチドの殺傷効果。
SCID免疫不全マウスは、中国科学院上海実験動物センターから購入したものである。マウスは、標準飼育要件に従って飼育する。水、敷き藁および飼料は、すべて、高温またはUV光により滅菌する。マウスは、相対的無菌条件下で、1週間飼育し、異常性が全くみとめられない場合に、本接種実験に使用する。
対数期にあるRaji(ATCC CCL‐86)および1648(ATCC CRL‐1648)の細胞の懸濁液を、50mlの遠心管中に集め、4℃で遠心する。次いで、その上清を捨てる。細胞を、1640液体培養培地(仔ウシ血清を加えたもの)に再懸濁して、1.0×107細胞/mlにする。0.1mlのRajiの細胞懸濁液をマウスの左腋窩、また、0.1mlの1648(ATCC CRL‐1648)の細胞懸濁液をマウスの右腋窩に、皮下注射する。
注射の3〜4日後、腫瘍は成長して約2×2mmとなる。腫瘍を有するマウスを、以下のグループに分ける:
(グループA)コントロールグループとしての、抗腫瘍ポリペプチドを含有しない保存液(10mM PBS+0.2M NaClリン酸緩衝液(pH7.4));
(グループB)処理グループとしての、新規抗腫瘍ポリペプチド1(プラスミドpCHCEB11)、300μg/マウス/日(25gとして計算)で20日間連続。
各グループの10匹のマウスに、1日2回、20日間連続で、0.5ml皮下注射する。マウスの挙動は、毎日観察および記録する。腫瘍の大きさについては、2日ごとに測定し、写真撮影する。
その結果(図7参照)は、新規抗腫瘍ポリペプチドのグループBの腫瘍の成長が有意に阻害され、7匹のマウスの腫瘍は消失し、残りの3匹のマウスの腫瘍は、コントロールグループのそれよりも明らかに小さいことを示す。新規抗腫瘍ポリペプチドは、リンパ肉腫のEBV陽性細胞により引き起こされるマウス体内の固形腫瘍の成長を阻害するのに効果的である。しかし、当該新規抗腫瘍ポリペプチドは、リンパ肉腫のEBV陰性細胞により引き起こされるマウス体内の固形腫瘍の成長の阻害には効果がない。
【0047】
(実施例7)
腫瘍消失のin vivo実験の病理学的観察
腫瘍の組織病理学的観察:実施例6の実験の終わりにマウスを屠殺する。腫瘍を抽出し、10%ホルマリン中で固定する。パラフィンスライスを、HE染色し、常用の光学顕微鏡法に基づき観察する。
顕微鏡下で観察すると、コントロールグループのマウス由来の固形腫瘍は活発に増殖しており、;当該新規抗腫瘍ポリペプチドのグループのマウス由来のEBV陽性固形腫瘍の細胞は著しく縮小している。切片中のこれら細胞塊のほとんどは壊死した腫瘍細胞であり、多量の、腫瘍周辺のリンパ球浸潤が観察される。この組織病理学的結果は、20日間の処理の間に、新規抗腫瘍ポリペプチドが、固形腫瘍中の腫瘍細胞をほぼすべて殺傷したことを示唆している(図8、Dを参照のこと)。
【技術分野】
【0001】
本発明は、抗腫瘍剤の分野に関し、より具体的には、EBウィルスが原因である腫瘍に対する新規ポリペプチド、ならびに、その使用および生産方法、に関する。
【背景技術】
【0002】
抗生物質研究の領域においては、研究は、同種の異種の株間の相互殺傷機構をまねた新しい抗生物質の開発へと向けられてきた。自然界には、細菌の細胞膜上に直接的にイオンチャネルを形成することにより細胞を殺傷する細菌毒素が数多く存在する。そうした毒素のモデル例が、大腸菌により分泌される細菌毒素であるコリシンである。コリシンIaは、1952年にJacobにより発見され、それ以降、何世代にもわたる努力を経て、Qiu et al.(Major transmembrane movement associated with colicin Ia channel gating. J. Gen. Physiology, 107:313‐328 (1996))は、人工の脂質二重膜中に形成されたイオンチャネルが開いている又は閉じられているときの、コリシンIaの膜貫通型立体構造をついに明らかにした。これは、分子レベルでの新しい抗生物質の設計および製造のための基礎をもたらすものである。その後、コリシンポリペプチドと、白色連鎖球菌またはブドウ球菌の誘引物質などのシグナルペプチドとの結合により作製された、コリシンを目的の細菌の細胞膜へと向かわせ、形成された膜貫通型イオンチャネルを通っての細胞含有物の漏出により細胞を殺傷するポリペプチド分子が存在している。
【0003】
悪性腫瘍は人間の健康に大きな脅威を与えるものである。毎年、世界中で七百万人が悪性腫瘍により亡くなっており、その六分の一は中国におけるものである。現在、悪性腫瘍は、我が国における死亡原因の第二位となっている。悪性腫瘍の病因、病原および臨床症状が明確に解明されてはいないため、予防および治療は効果的ではない。抗腫瘍剤は腫瘍の治療において重要である。それらにより一部の腫瘍に対しての治療効果は達成されるが、例えば不十分な腫瘍選択性、免疫学的抑制、有害反応、薬剤耐性等の、いくつかの欠点が残っている。
【0004】
EBウィルスが原因であるバーキットリンパ腫、ホジキンリンパ腫および上咽頭癌の細胞の表面は、EBウィルスの特異的表面抗原を有する。したがって、EBウィルス表面抗原は、そうした腫瘍細胞の特異的マーカーとみなすことができる。EBウィルスが原因である腫瘍に対する薬剤について、中国特許第ZL200410081446.8号の発明では、コリシンと、EBウィルス表面抗原を認識する抗体模倣物との結合により形成される抗腫瘍ポリペプチドが開示されている。この抗腫瘍ポリペプチドは、体内の、EBウィルスが原因であるがん細胞を、特異的に殺傷することができるものであり、正常細胞に対して害がなく、その殺傷能力は他の抗腫瘍剤の数倍であり、腫瘍選択性、薬剤耐性、がん細胞が殺傷されるときの正常組織の障害などの問題を克服する。Xiao‐Qing Qiu et al.(Xiao‐Qing Qiu et al., 2007, Small antibody mimetics comprising two complementarity‐determining regions and a framework region for tumor targeting, Nature Biotechnology 25, 921‐929, 1 August 2007)は、一連の抗体模倣物とコリシンとにより構築される抗腫瘍ポリペプチドの殺傷効果を比較し、VHCDR1‐VHFR2‐VLCDR3およびVLCDR1‐VHFR2‐VHCDR3の抗体模倣物とコリシンとにより構築される抗腫瘍ポリペプチドが、優れた殺傷能力を有することを発見している。この研究は、EBウィルスが原因である腫瘍に対するポリペプチドの調製のための、より多くの候補抗体模倣物をもたらす。
【0005】
しかしながら、上述の抗腫瘍ポリペプチドについては、コリシンの疎水性末端が、過敏性を有し得るいくつかのアミノ酸残基を有しているため、コリシンのポリペプチドを含む薬品は、in vivoでの異常な免疫応答をより容易に引き起こす可能性がある。多くのがん患者の代謝機構は、がん細胞からの妨害に起因して異常になっており、彼らはポリペプチドの薬に対するアレルギー応答を起こしやすく、それ故、こうした薬によって治療することができない、と報告されている。したがって、より安全な、かつ、より多くの患者に適した抗がん薬を得るためには、コリシンポリペプチドを改良する必要がある。
【発明の概要】
【発明が解決しようとする課題】
【0006】
上述の先行技術の欠点に基づいて、本発明は、EBウィルスが原因である腫瘍に対する新規ポリペプチド、ならびに、その使用および生産方法を提供し、ひいては、高い殺傷能力、および、高い特異性を有し、かつ、アレルギーの可能性の低い、EBウィルスが原因である腫瘍の治療のための薬品を提供する。
【0007】
イオンチャネルを形成することのできるコリシンの変異ポリペプチドと、抗EBウィルス抗体ポリペプチドまたは抗EBウィルス抗体模倣物ポリペプチドとの機能可能な連結により形成される、EBウィルスが原因である腫瘍に対する新規ポリペプチドであって、前記イオンチャネルを形成することのできるコリシンの変異ポリペプチドが、野生型コリシンE1、Ia、Ib、A、B、Nまたはそれらの水溶性チャネルドメインのペプチド鎖の、G11A、H22G、A26G、V31L、およびH40Dのアミノ酸残基の変異により得られるものであり、前記抗EBウィルス抗体のポリペプチドのアミノ酸配列が、ATCC HB‐168のハイブリドーマにより分泌されるモノクローナル抗体のポリペプチドと同じである、新規ポリペプチド。
【0008】
前記抗体模倣物のポリペプチドが、抗EBウィルス抗体の、重鎖のCDR1領域と、重鎖のCDR1‐CDR2連結ペプチドセグメントと、軽鎖のCDR3との結合ペプチドである。
【0009】
前記イオンチャネルを形成することのできるコリシンの変異ポリペプチドが、野生型コリシンIaの変異により得られるものである。
【0010】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドが、配列番号29に示されているアミノ酸配列を有する。
【0011】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドをコードする遺伝子。
【0012】
配列番号30に示されているヌクレオチド配列を有する、前記遺伝子。
【0013】
前記遺伝子を含む組換えプラスミド。
【0014】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドの生産方法であって、前記組換えプラスミドを、発現のために発現系に形質転換する工程、および、発現したポリペプチドを単離する工程、を含む、生産方法。
【0015】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドの、EBウィルスが原因である腫瘍の治療および予防のための医薬の生産における使用。
【0016】
そのアミノ酸配列が、配列番号24に示されているものである、コリシンIaの変異ポリペプチド。
【0017】
コリシンIaの変異ポリペプチドをコードする遺伝子。
【0018】
前記遺伝子を、そのペプチドを誘導する遺伝子と機能可能に連結し、発現ベクターにクローニングし、次いで、前記発現ベクターを発現系に形質転換し、その発現したポリペプチドを単離する、ペプチド医薬の生産における、前記遺伝子の使用。
【課題を解決するための手段】
【0019】
本発明は、イオンチャネルを形成することのできるコリシンの変異ポリペプチドと、抗EBウィルス抗体のポリペプチドまたは抗EBウィルス抗体模倣物のポリペプチドとにより形成される、EBウィルスが原因である腫瘍に対する新規ポリペプチド、を提供する。野生型コリシンポリペプチド分子中には、過敏性を有し得るいくつかのアミノ酸残基が存在するため、イオンチャネル構築物を形成することのできるコリシンのポリペプチド分子において、本発明は、アレルギー応答を容易に引き起こし得る疎水性の領域のアミノ酸残基を選択的に変異させている。例えば、本発明の好ましい一実施形態においては、コリシンIaのポリペプチドの変異の部位は以下である:G11A、H22G、A26G、V31LおよびH40D。コリシンIaのポリペプチドまたは変異体Iaのポリペプチドの注射によりそれぞれ免疫したマウスにおいては、その実験データは、変異体Iaのポリペプチドを注射したマウスによりもたらされる血清力価は前者よりも数桁低い、すなわち、免疫応答のレベルがより低いことを示しており、これは、この変異ポリペプチドが、細胞膜にイオンチャネルを形成するという機能を保持していながらも、アレルギーの可能性を低減することを実証するものである。実験は、本発明の組換えポリペプチドの殺傷能力は影響を受けないことを示し、これは、変異体アミノ酸残基が、コリシンのイオンチャネルを形成する機能に影響を与えないことを意味している。本発明により提供される、EBウィルスが原因である腫瘍に対する新規ポリペプチドにおいては、抗EBウィルス抗体のポリペプチドまたは抗EBウィルス抗体模倣物のポリペプチドがEBウィルスによる腫瘍細胞の表面抗原を認識することを通じて、コリシンの変異ポリペプチドは標的細胞の膜にターゲティングされ、コリシンの変異ポリペプチド膜貫通型イオンチャネルドメインの疎水性の領域が腫瘍細胞の細胞膜へと挿入され、イオンチャネルを形成し、よって、腫瘍細胞が細胞内容物の漏出によって死ぬ。抗EBウィルス抗体のポリペプチドのアミノ酸配列は、ATCC HB‐168のハイブリドーマにより分泌される抗体のポリペプチドのアミノ酸配列を完全に参照する。
【0020】
本発明の一実施形態においては、上述の抗EBウィルス抗体模倣物のポリペプチドと、コリシンの変異ポリペプチドのカルボキシル末端との機能的連結により得られる、本発明の低分子量の抗腫瘍ポリペプチドが好ましい。つまり、そのような低分子量の模倣ポリペプチドは、抗EBウィルス抗体のポリペプチドの、VHCDR1領域、VLCDR3領域、VHCDR1‐VHCDR2の連結ペプチドセグメントと、軽鎖のVLCDR3の結合により得られる、VHCDR1‐VHFR2‐VLCDR3のペプチド鎖を含む。抗体模倣物の新規抗腫瘍ペプチド1のアミノ酸配列を、配列番号25に示す。この抗体模倣物は、30個未満のアミノ酸しか含んでおらず、アミノ酸150個の天然の抗体よりもずっと低い分子量を有する。これは、抗腫瘍ポリペプチドの分子量を大幅に減少させて、本発明の抗腫瘍ポリペプチドの組織浸透能に貢献するものでありながらも、抗原認識の要件を満たす。
【0021】
本発明のもう1つの目的は、本発明の抗腫瘍ポリペプチドをコードする遺伝子配列を提供することである。本発明の抗腫瘍ポリペプチドの遺伝子は、コリシンの変異ポリペプチドをコードする遺伝子と、抗EBウィルス抗体のポリペプチドまたはその抗体模倣物のポリペプチドをコードする遺伝子との機能可能な連結により形成され、ここで、コリシンのポリペプチド、および、抗EBウィルス抗体の遺伝子配列は、当該分野において公知であり、コリシンの変異ポリペプチドの遺伝子は、コリシンポリペプチドの遺伝子の対応するコドンにおける、以下の点変異により得られるものである:G11A、H22G、A26G、V31LおよびH40D。遺伝コードの縮重の結果として、当業者は、アミノ酸配列を変えることなく、本発明の抗腫瘍ポリペプチドをコードするヌクレオチド配列を調整してもよい。
【0022】
本発明の組換えプラスミドは、野生型コリシンの遺伝子が組み込まれた元のベクターが、二本鎖ヌクレオチドに部位特異的に変異導入され、変異標的部位に変異コドンにより挿入され、それにより、コリシン変異ポリペプチドの遺伝子を含む変異体ベクターを得る、ということを意味するものである。同じプロセスの部位特異的突然変異誘発により、抗EBウィルス抗体の抗体模倣物をコードする遺伝子を、前記コリシン変異ポリペプチドの遺伝子のカルボキシル末端へと挿入し、本発明の組換えプラスミドを得る。当該元のベクターpSELECTTM‐1は、プロメガ社から購入したものであり、コリシンIaおよび免疫タンパク質の遺伝子を有する。部位特異的突然変異誘発のプロセスは、Strategene社からのキットの指示書に従ったものである。本発明は、コリシンの変異ポリペプチドを調製するべく、いくつかの部位特異的突然変異誘発を行なうが、ここでは、5つのコドンを部位特異的に変異させる。よって、5対のプライマー配列が設計されている(配列番号1〜10)。本発明の実施例においては、6対のプライマー配列が、抗体模倣物の遺伝子用に設計されている(配列番号11〜22)。
【0023】
本発明は、上記で得られる組換えベクターを、改変細菌 大腸菌BL21(DE3)中へと形質転換すること、陽性クローンを選抜すること、陽性クローンにより発現した当該タンパク質を単離および精製すること、それにより、本発明のEBウィルスが原因である腫瘍に対する新規ポリペプチドを得ること、を含む、本発明の抗腫瘍ポリペプチドの生産のための方法も提供する。
【0024】
本発明により提供される、EBウィルスが原因である腫瘍に対する新規ポリペプチドは、EBウィルスが原因である腫瘍の治療および予防のための医薬の生産において用いることができる。本発明において得られる新規抗生物質のポリペプチドを、薬剤的に許容できる担体もしくは賦形剤または他の所望により選択される成分中へと加えることにより、臨床上適切な医薬組成物を作製することができる。
【0025】
本発明は、コリシンIaの変異ポリペプチドのアミノ酸配列および遺伝子配列も提供する。この変異ポリペプチドは、本発明において用いることができ、他の標的化ポリペプチドを用いる抗体ポリペプチドの構築においても用いることができる。本発明中の実施例3の実験データは、当該変異ポリペプチドを含むペプチド医薬の免疫原性が低いこと、および、他の標的化ポリペプチドを用いる当該変異ポリペプチドにより形成される抗体ポリペプチドが殺菌能を有するとのこと、を証明するものである。当該生産方法は、当該分野における常用の実験プロセスである。
【0026】
本発明により提供される新規抗腫瘍ポリペプチドは、特許第ZL200410081446.8号に開示されている抗腫瘍ポリペプチドの利点、すなわち、高い特異性のターゲティング、正常細胞に対する安全性、および、薬剤耐性を生じさせにくい、といった利点を有する。それと同時に、本発明の抗腫瘍ポリペプチドは、アレルギー応答を引き起こしやすいアミノ酸残基に変異を受けたものであり、そうした変異ポリペプチドを含む当該抗腫瘍ポリペプチドの免疫原性は低減される。すなわち、アレルギー反応の可能性が低下する。そうしたポリペプチドの医薬の、使用安全性、および、腫瘍を殺傷するという効果は改善される。これは、また、コリシンポリペプチドを含む他の医薬の改良のための良い例でもあり得る。
【図面の簡単な説明】
【0027】
【図1】抗体模倣物VHCDR1‐VHFR2‐VLCDR3のポリペプチドの遺伝子と、コリシンIaの変異ポリペプチドの遺伝子とを含む、組換えプラスミドpCHCEB11の構造の略図である。
【図2】抗体模倣物VHCDR1‐VHFR2‐(Rev)VLCDR3のポリペプチドの遺伝子と、コリシンIaの変異ポリペプチドの遺伝子とを含む、組換えプラスミドpCHCEB22の構造の略図である。
【図3】コリシンIaの変異ポリペプチドの感作効果の実験1を示す図である。 (A)致死量のMRSA(ATCC BAA42)を腹腔内注射した昆明(Kunming)マウスを、(1)コントロールグループ、(2)アンピシリングループ、(3)黄色ブドウ球菌に対するポリペプチド(ZL01128836.1)のグループ、(4)黄色ブドウ球菌に対するポリペプチド1のグループ、へとランダムにグループ分けする。 (B)14日後、昆明マウスの新たな一群を、コントロールグループおよびアンピシリングループへとグループ分けする。黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループ由来の生き残ったマウスを、黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループへとグループ分けし、当該実験を繰り返す。 (C)41日後、昆明マウスの新たな一群を、(1)コントロールグループ、(2)レボフロキサシンのグループ、(3)セフトリアキソンナトリウムのグループ、へとグループ分けし、また、(4)黄色ブドウ球菌に対するポリペプチドのグループおよび(5)黄色ブドウ球菌に対するポリペプチド1のグループ由来の生き残ったマウスは、緑膿菌に対するポリペプチド2のグループおよび緑膿菌に対するポリペプチド1のグループへとグループ分けする。
【図4】コリシンIaの変異ポリペプチドの低感作効果の実験2を示す図である。 (A)黄色ブドウ球菌に対するポリペプチド/緑膿菌に対するポリペプチド2のグループの血清、1:50,000の力価; (B)黄色ブドウ球菌に対するポリペプチド1/緑膿菌に対するポリペプチド1のグループの血清、1:50,000の力価。 (1)第1週、(2)第2週、(3)第7週の血清、(4)ネガティブコントロール。
【図5】EBウィルスが原因であるバーキットリンパ腫に対する、新規抗腫瘍ポリペプチドのin vitroにおける殺傷効果の比較を示す図である。 (A)コントロールグループ、(B)新規抗腫瘍ポリペプチド1処理グループ、(C)新規抗腫瘍ポリペプチド2処理グループ。
【図6】EBウィルスが原因であるバーキットリンパ腫の細胞および他の腫瘍細胞に対する、新規抗腫瘍ポリペプチドのin vitroにおける殺傷効果を示す図である。 (A)バーキットリンパ腫のEBV陽性細胞、 (B)バーキットリンパ腫のEBV陰性細胞、 (C)AIDS患者由来の悪性リンパ肉腫のEBV陽性細胞。 (1)コントロールグループ、(2)新規抗腫瘍ポリペプチド1処理グループ。
【図7】EBウィルスが原因であるバーキットリンパ腫の細胞を移植したヌードマウス由来の成長した固形腫瘍に対する新規抗腫瘍ポリペプチドの殺傷効果を示す図である。 (A)コントロールグループ、 (B)新規抗腫瘍ポリペプチド1治療グループ、に由来するすべてのSCID免疫不全マウスの、両方の脇の下の側面部中に、バーキットリンパ腫の細胞を接種する。左矢印はEBV陰性リンパ肉腫、右矢印はEBV陽性リンパ肉腫である。
【図8】EBウィルスが原因であるバーキットリンパ腫の細胞を移植したヌードマウス由来の成長した固形腫瘍に対する新規抗腫瘍ポリペプチドの殺傷効果を示す図である。 (A)コントロールマウスのEBV陰性リンパ肉腫の切片、(B)コントロールマウスのEBV陽性リンパ肉腫の切片、(C)新規抗腫瘍ポリペプチド1治療マウスのEBV陰性リンパ肉腫の切片、(D)新規抗腫瘍ポリペプチド1治療マウスのEBV陽性リンパ肉腫の切片。
【発明を実施するための形態】
【0028】
以下、本発明の好ましい実施形態を記述することにより、および、添付の図面を参照して、本発明を説明する。
【0029】
本発明において使用される元のベクターpSELECTTM‐1は、プロメガ社から購入したものである。
【0030】
大腸菌BL21(DE3)という改変細菌は、ノバジェン社から購入したものである。
【実施例】
【0031】
(実施例1)
変異体コリシンIaをコードする遺伝子を含む組換えプラスミドの構築。
元のベクターは、プラスミドpSELECTTM‐1(8.3kb)(プロメガ社から購入)であり、これは、コリシンIaおよび免疫タンパク質の遺伝子を有する。変異体アミノ酸をコードする配列番号1〜10に示されているオリゴヌクレオチドプライマーの配列を、それぞれ、二本鎖オリゴヌクレオチド部位特異的突然変異誘発技術(QuickChangeTMキット、Strategene社)により、野生型コリシンIaの遺伝子へと機能可能に連結し、コリシンIaの変異ポリペプチドをコードする配列番号23に示されている遺伝子、および、変異体プラスミド、を得る。その後、この変異体プラスミドに、配列番号26または配列番号28の遺伝子を、コリシンIaの変異ポリペプチドの遺伝子のI626のコドンの後に挿入し、EBウィルスが原因である腫瘍に対する新規ポリペプチドのための、2種類の組換えプラスミドpCHCEB11(図1に示されている)およびpCHCEB22(図2に示されている)を得る。組換えプラスミド中の、EBウィルスに対する抗体をコードする遺伝子の調製のために設計された6対のオリゴヌクレオチドプライマーの配列が配列番号11〜22に示されている。前記組換えプラスミドを、大腸菌BL21(DE3)(ノバジェン社から購入)という改変細菌中へとトランスフェクションし、ポリペプチドを調製する。得られるポリペプチドは、配列表の、配列番号25(以下、「新規抗腫瘍ポリペプチド1」という)および配列番号27(以下、「新規抗腫瘍ポリペプチド2」という)に示されたものである。
【0032】
二本鎖オリゴヌクレオチド部位特異的突然変異誘発のプロセスは、Strategene QuickChang部位特異的突然変異誘発キット(カタログ番号200518)に従ったものである。
1. 部位特異的突然変異誘発のための反応物の調製:
5μlの10×緩衝液
2μl(10ng)の、コリシンIaの野生型のポリペプチドの遺伝子および免疫タンパク質の遺伝子を有する元のプラスミドpSELECTTM‐1。
1.25μl(125ng)の、設計された5’‐3’オリゴヌクレオチドプライマー
1.25μl(125ng)の、設計された3’‐5’オリゴヌクレオチドプライマー
1μlのdNTP
再蒸留水50μl
1μl pfu
(プラスミド、プライマーおよび再蒸留水以外は、前記キットにより提供されるものである)
2. PCR増幅、増幅条件:95℃で35秒間の変性、53℃で70秒間のアニール、および68℃で17分間の伸長、を25サイクル;
3. 1μlのエンドヌクレアーゼDpn1を加えて、親DNA鎖を消化(37℃、1時間)し、1μlの反応物と50μlのXL1‐Blueコンピテント細胞とを、共に、氷上で30分間インキュベートし、42℃での45秒間のヒートショックの後に、氷中で2分間インキュベートする;
4. 0.5mlの培養培地NZYを加え、37℃および220rpmにて1時間振盪する。50〜100μlの反応物をまく(1%寒天および50μg/mlアンピシリンを加えたLB培地、37℃で一晩);
5. 18時間後にコロニーをピックアップする。プラスミドを抽出し、配列決定を行い、変異が成功していることを確認する;
6. 複数の部位における変異によって最終的に得られた50ngの組換えプラスミドを、40μlの大腸菌BL‐21(DE3)コンピテント細胞と共に氷上で5分間インキュベートし、42℃で30秒間ヒートショックを与え、氷中で2分間インキュベートする。ノバジェン社からの160μlの培養培地SOCを加え、37℃および220rpmにて1時間振盪した後にまく(1%寒天および50μg/mlアンピシリンを加えたLB培地、37℃で一晩)。
7. 増幅のためにシングルコロニーをピックアップし、8〜16リットルFB培地、250rpm、30℃で4〜5時間とし、42℃でヒートショックを与え、250rpmにて30分間、37℃にて2時間。細菌を、6000gおよび4℃での20分間の遠心分離により沈殿させる。4℃の50mMホウ酸緩衝液(2mM EDTA+2mM DTT)および50〜80mlの細菌懸濁液は、250mlの0.2M PMSFを加え、超音波処理により処理する(4℃、400W、2分)。細菌破砕物(debris)を高速遠心分離する(4℃、75,000g、90分)。その上清には、500万単位の硫酸ストレプトマイシンを加え、DNAを沈殿させる。15000gおよび4℃での10分間の遠心分離による沈殿の後、その上清を、4℃にて、50mMホウ酸緩衝液中、分子量15,000の透析バッグ中で一晩透析する。再び15000gおよび4℃での10分間の遠心分離による沈殿の後、その上清を、CMイオン交換カラムにロードする。このカラムを、0.1〜0.3M NaCl+50mMホウ酸緩衝液の勾配を用いて溶出し、組換え抗腫瘍ポリペプチドを得る。
【0033】
部位特異的突然変異誘発のために設計されたプライマーの配列は以下の通りである:
【0034】
配列番号1、コリシンの遺伝子におけるG11Aの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
cgt att aca aat ccc GCA gca gaa tcg ctg ggg
配列番号2、コリシンの遺伝子におけるG11Aの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
ccc cag cga ttc tgc TGC ggg att tgt aat acg
配列番号3、コリシンの遺伝子におけるH22Gの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
gat tca gat ggc GGT aaa tta tgg gtg
配列番号4、コリシンの遺伝子におけるH22Gの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
cac cca taa ttt ACC gcc atc tga atc
配列番号5、コリシンの遺伝子におけるA26Gの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
gaaa ttatgGGTgt tgatatttat
配列番号6、コリシンの遺伝子におけるA26Gの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
ataaatatacaacACCcataatttc
配列番号7、コリシンの遺伝子におけるV31Lの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
gt tgatatttat CTC aaccctc cacgtgtc
配列番号8、コリシンの遺伝子におけるV31Lの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
gacacgtggagggttGAGataaatatcaac
配列番号9、コリシンの遺伝子におけるH40Dの変異のために設計されたオリゴヌクレオチドプライマー5’‐3’:
cgtgtcga tgtctttGATggtaccccgc ctgcat
配列番号10、コリシンの遺伝子におけるH40Dの変異のために設計されたオリゴヌクレオチドプライマー3’‐5’:
atgcaggcggggtaccATCaaagacatcgacacg
【0035】
配列番号11、組換えプラスミドpCHCEB11中のVHCDR1の遺伝子のためのプライマー5’‐3’:
gcg aat aag ttc tgg ggt att TCC TTC GGT ATG CAT TGG GTG CGTCAGtaa ata aaa tat aag aca ggc
配列番号12、組換えプラスミドpCHCEB11中のVHCDR1の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta CTG ACG CAC CCA ATG CAT ACC GAA GGA aat acc cca gaa ctt att cgc
配列番号13、組換えプラスミドpCHCEB11中のVHFR2の遺伝子のためのプライマー5’‐3’:
ggt atg cat tgg gtg cgt cag GCC CCC GAG AAA GGT CTG GAG TGG GTG GCC taa ata aaa tat aag aca ggc
配列番号14、組換えプラスミドpCHCEB11中のVHFR2の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta GGC CAC CCA CTC CAG ACCT TTT CTC GGG GGC ctg acg cac cca atg cat acc
配列番号15、組換えプラスミドpCHCEB11中の(Rev)VlCDR3の遺伝子のためのプライマー5’‐3’:
aaa ggt ctg gag tgg gtg gcc ACC TAC CCC TAC TCC TAC GGT CAG GGT taa ata aaa tat aag aca ggc
配列番号16、組換えプラスミドpCHCEB11中の(Rev)VLCDR3の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta ACC CTG ACC GTA GGA GTA GGG GGT ggc cac cca ctc cag acc ttt
配列番号17、組換えプラスミドpCHCEB22中のVHCDR1の遺伝子のためのプライマー5’‐3’:
gcg aat aag ttc tgg ggt att TCC TTC GGT ATG CAT TGG GTG CGT CAG taa ata aaa tat aag aca ggc
配列番号18、組換えプラスミドpCHCEB22中のVHCDR1の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta CTG ACG CAC CCA ATG CAT ACC GAA GGA aat acc cca gaa ctt att cgc
配列番号19、組換えプラスミドpCHCEB22中のVHFR2の遺伝子のためのプライマー5’‐3’:
ggt atg cat tgg gtg cgt cag GCC CCC GAG AAA GGT CTG GAG TGG GTG GCC taa ataaaa tat aag aca ggc
配列番号20、組換えプラスミドpCHCEB22中のVHFR2の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta GGC CAC CCA CTC CAG ACCT TTT CTC GGG GGC ctg acg cac cca atg cat acc
配列番号21、組換えプラスミドpCHCEB22中のVLCDR3の遺伝子のためのプライマー5’‐3’:
aaa ggt ctg gag tgg gtg gcc GGT CAG GGT TAC TCC TAC CCC TAC ACC taa ata aaa tat aag aca ggc
配列番号22、組換えプラスミドpCHCEB22中のVLCDR3の遺伝子のためのプライマー3’‐5’:
gcc tgt ctt ata ttt tat tta GGT GTA GGG GTA GGA GTA ACC CTG ACC ggc cac cca ctc cag acc ttt
【0036】
(実施例2)
組換えプラスミドpCHCEB11およびpCHCEB22から調製した新規抗腫瘍ポリペプチドの免疫効果の観察。
実施例1において得られた組換えプラスミドpCHCEB11およびpCHCEB22から調製した新規抗腫瘍ポリペプチド1および新規抗腫瘍ポリペプチド2、ならびに、発明者が所有する、先の発明(ZL200410081446.8)からの抗腫瘍ポリペプチド1および抗腫瘍ポリペプチド2、により、マウスを免疫する。上述の各タンパク質をアジュバントと混合する。プライミング投与量およびブースト投与量は、各マウスにつき50μg(0.5ml)の1回の腹腔内注射、2週間の間隔をあけて合計5回の注射、である。血清力価は、間接ELISA法により決定する。本発明により調製した新規抗腫瘍ポリペプチド1および2により免疫したマウスの力価は、10−3〜10−4の範囲であり、一方、抗腫瘍ポリペプチド1および抗腫瘍ポリペプチド2により免疫したマウスの力価は、10−4〜10−5の範囲である。
本発明の新規抗腫瘍ポリペプチドにより誘発される過敏性反応の可能性は、野生型コリシンIaを含む抗腫瘍ポリペプチドにより誘発される過敏性反応の可能性よりも1桁〜2桁低い、とのことがわかる。
【0037】
(実施例3)
新規抗腫瘍ポリペプチドを形成するコリシンIaの変異ポリペプチドの低感作効果の実験。
実施例1の、コリシンIaの変異ポリペプチドの変異体プラスミド(水溶性チャネルドメインのペプチド鎖において、G11A、H22G、A26G、V31L、およびH40Dのアミノ酸残基に変異を受けている)を、変異ポリペプチドのN末端またはC末端にて、黄色ブドウ球菌のフェロモンAgrD1(YSTCDFIM)に機能可能に連結し、2種類の抗菌ポリペプチドを得る。当該変異体コリシンIaのカルボキシル末端におけるAgrD1の連結により得られるポリペプチドを、黄色ブドウ球菌に対するポリペプチド1と名付け、当該変異体コリシンIaのアミノ末端におけるAgrD1の連結により得られるポリペプチドを、緑膿菌に対するポリペプチド1と名付ける。野生型コリシンIaについてのプラスミドを、アミノ末端にて、黄色ブドウ球菌のフェロモンAgrD1(YSTCDFIM)に連結し、緑膿菌に対するポリペプチド2を得る。
【0038】
実験1:昆明マウスの一群に、致死量のMRSA(ATCC BAA42)を腹腔内注射し、(1)コントロールグループ、(2)アンピシリングループ、(3)黄色ブドウ球菌に対するポリペプチドのグループ、(4)黄色ブドウ球菌に対するポリペプチド1のグループ、へとランダムにグループ分けする。各グループは、10匹のマウスからなる。
処理方法:
致死量のMRSA(ATCC BAA42)の腹腔内注射の1時間後に:
コントロールグループ:0.5mlの0.3M NaCl+50mMホウ酸緩衝液を、尾静脈を介して1回注射する;
アンピシリンのグループ:2.5mg/kgのアンピシリンを、尾静脈を介して1回注射する;
黄色ブドウ球菌に対するポリペプチドのグループ:6mg/kgの、本発明者が所有する、黄色ブドウ球菌に対するポリペプチド(ZL01128836.1)を、尾静脈を介して1回注射する;
黄色ブドウ球菌に対するポリペプチド1のグループ:6mg/kgの、黄色ブドウ球菌に対するポリペプチド1を、尾静脈を介して1回を注射する;
結果:コントロールグループおよびアンピシリンのグループのマウスは、すべて、2日以内に死亡。黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループの85%のマウスは生き残る。
【0039】
実験2:実験1の14日後に、昆明マウスの新たな一群を、コントロールグループおよびアンピシリンのグループへとグループ分けする。黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループ由来の生き残ったマウスを、黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループへとグループ分けし、上記の実験を繰り返す。コントロールグループおよびアンピシリンのグループのマウスは、すべて、2日以内に死亡。黄色ブドウ球菌に対するポリペプチドのグループの75%のマウスは生き残り、黄色ブドウ球菌に対するポリペプチド1のグループの90%のマウスは生き残る。
【0040】
実験3:実験1の41日後に、昆明マウスの新たな一群を、コントロールグループ、レボフロキサシンのグループ、およびセフトリアキソンナトリウムのグループへとグループ分けする。黄色ブドウ球菌に対するポリペプチドのグループおよび黄色ブドウ球菌に対するポリペプチド1のグループ由来の生き残ったマウスは、緑膿菌に対するポリペプチド2のグループおよび緑膿菌に対するポリペプチド1のグループへとグループ分けする。
マウスに、致死量の多剤耐性緑膿菌(四川大学華西医院実験医学科からの臨床分離株13578)を腹腔内注射する。1時間の後、
コントロールグループに、0.5mlの0.3M NaCl+50mMホウ酸緩衝液を、尾静脈を介して1回注射する;
レボフロキサシンのグループに、5mg/kgのレボフロキサシンを、尾静脈を介して1回注射する;
セフトリアキソンナトリウムのグループに、30mg/kgのセフトリアキソンナトリウムを、尾静脈を介して1回注射する;
緑膿菌に対するポリペプチド2のグループに、8mg/kgの、緑膿菌に対するポリペプチド2を、尾静脈を介して1回注射する;
緑膿菌に対するポリペプチド1のグループに、8mg/kgの、緑膿菌に対するポリペプチド2を、尾静脈を介して1回注射する。
コントロールグループおよびレボフロキサシンのグループのマウスは、すべて、1日以内に死亡。セフトリアキソンナトリウムのグループの25%のマウスは生き残る。緑膿菌に対するポリペプチド2のグループの60%のマウスは生き残る。緑膿菌に対するポリペプチド1のグループのマウスは、すべて、生き残る。宿主の抗体が、変異ポリペプチドの殺傷効果を、野生型ポリペプチドの殺傷効果よりも低く妨害をすることが示されている。
図3を参照のこと。
【0041】
この実験の第1週、第2週および第7週において、黄色ブドウ球菌に対するポリペプチドのグループ/緑膿菌に対するポリペプチド2のグループ、および、黄色ブドウ球菌に対するポリペプチド1のグループ/緑膿菌に対するポリペプチド1のグループ、に由来する、生き残ったマウスの血清について、血液中の抗体を検出するために、間接ELISA法により分析する。酵素標識プレートのウェルを、野生型コリシンIaおよびコリシンIaの変異ポリペプチドで、100ng/ウェルでコーティングする。一次抗体は、黄色ブドウ球菌に対するポリペプチドのグループ/緑膿菌に対するポリペプチド2のグループ、および、黄色ブドウ球菌に対するポリペプチド1のグループ/緑膿菌に対するポリペプチド1のグループ、に由来する、生き残ったマウスの血清である。二次抗体は、ヤギ抗マウス標識抗体である。一次抗体のネガティブコントロールは、5%ミルク‐PBSである。1:50,000の力価の結果は、以下の通りである(図4参照):
【0042】
【表1】
【0043】
本発明により調製されたコリシンIaの変異ポリペプチドにより誘発される宿主の過敏感反応の可能性は、野生型コリシンIaにより誘発される宿主の過敏感反応の可能性よりも低いことが示されている。
【0044】
(実施例4)
EBウィルスが原因であるバーキットリンパ腫に対する新規抗腫瘍ポリペプチドのin vitroにおける殺傷効果。
EBV陽性細胞株およびEBV陰性細胞株は、米国のATCCからの、標準的な細胞株である。
細胞培養:起こしたRaji細胞の懸濁液0.1mlを、ゆっくりと、培養ディッシュ中の3mlの1640液体培地(10%血清を加えたもの)へと加え(希釈率、1:30)、混合し、CO2を含有する37℃インキュベーター中で培養する。当該EBV陽性細胞株は、ATCC CCL‐86(世界中の研究室において使用されている標準的なバーキットリンパ腫細胞。Raji細胞。12歳のアフリカの少年から1963年に単離されたもの)である。
この試験細胞を、3つのグループにグループ分けする。
グループ1は、ブランクのグループであり、抗腫瘍ポリペプチドを含有しない保存液(10mMPB+0.2M NaClリン酸緩衝液(pH7.4))が加えられたものである。
グループ2は、200μg/mlの新規抗腫瘍ポリペプチド1(プラスミドpCHCEB11、保存液、10mMPB+0.2M NaClリン酸緩衝液、pH7.4)が加えられたものである。
グループ3は、200μg/mlの新規抗腫瘍ポリペプチド2(プラスミドpCHCEB22、保存液、10mMPB+0.2M NaClリン酸緩衝液、pH7.4)が加えられたものである。
24時間の培養後、培養ディッシュに、上述の処理剤を加える。これらの処理剤の添加の72時間後、この培養ディッシュに、20μlの100μMolヨウ化プロピジウム(PI)を加え、10分後、この培養ディッシュを顕微鏡下で観察する。その結果が示すのは、ブランクのグループの細胞はよく成長し、新規抗腫瘍ポリペプチド1のグループの細胞の大部分はPIにより赤く染色されるということであり、この抗腫瘍ポリペプチドによって、細胞膜が破壊されることを示しており、これは腫瘍細胞の死を導く。死細胞の数を比較すると、2種類の新規抗腫瘍ポリペプチドのうちで、新規抗腫瘍ポリペプチド2の効果はあまり良くない。図5を参照のこと。
【0045】
(実施例5)
EBウィルスが原因であるバーキットリンパ腫の細胞および他の腫瘍細胞に対する新規抗腫瘍ポリペプチドのin vitroにおける殺傷効果についての多重蛍光染色の観察。
細胞培養の条件は実施例2と同じである。本実験では、次の3種類の細胞株を使用する:EBウィルス陽性細胞株:ATCC CCL‐86(Raji細胞。バーキットリンパ腫細胞);EBウィルスおよびカポジ肉腫ウィルスが陽性の、46歳のAIDS男性患者由来の悪性リンパ肉腫細胞の株である、ATCC CRL‐2230;EBウィルス陽性細胞株:ATCC CRL‐1648(CA‐46。アメリカのバーキットリンパ腫患者の腹水から単離された細胞)。
各株は、2つの試験グループにグループ分けする。グループ1は、新規抗腫瘍ポリペプチドを含有しない保存液(10mMPB+0.2M NaClリン酸緩衝液(pH7.4))が加えられたものである。グループ2は、200μg/mlの新規抗腫瘍ポリペプチド1(プラスミドpCHCEB11)が加えられたものであり、保存液は10mMPB+0.2M NaClリン酸緩衝液、pH7.4である。
24時間の培養後、培養ディッシュに、上述のグループの処理剤を加える。これらの処理剤の添加の72時間後、この培養ディッシュに、2種類の蛍光色素、すなわち、20μlの50μMol FITCと、20μlの50uMol ローダミン123とを加え、10分後、この培養ディッシュを顕微鏡オリンパスIX‐71下で観察する。
その結果、EBV陰性腫瘍細胞の株は、新規抗腫瘍ポリペプチド1の処理後、よく成長すること、及び、EBV陽性腫瘍細胞の株は、いずれの株も、大部分の細胞がミトコンドリアおよび核を消失し、膨張し、ネクローシスを起こし、それらのうちのほとんどが死んでいることを示す。実施例4のPI染色実験と比較すると、明らかに、多重蛍光染色実験からの結果は、EBウィルス陽性腫瘍細胞に対する新規抗腫瘍ポリペプチド1の強力な殺傷効果を、より明確に示しているようである。図6を参照のこと。
EBV陰性腫瘍細胞はよく成長し、当該新規抗腫瘍ポリペプチドは、細胞膜中にEBウィルスの表面抗原を有しない細胞を攻撃しないことを意味する。本発明の新規抗腫瘍ポリペプチドは理想的なターゲティング特異性と安全性とを有している、とのことが示唆される。
【0046】
(実施例6)
EBウィルスが原因であるバーキットリンパ腫の細胞を移植したヌードマウスの体内で成長した固形腫瘍に対する新規抗腫瘍ポリペプチドの殺傷効果。
SCID免疫不全マウスは、中国科学院上海実験動物センターから購入したものである。マウスは、標準飼育要件に従って飼育する。水、敷き藁および飼料は、すべて、高温またはUV光により滅菌する。マウスは、相対的無菌条件下で、1週間飼育し、異常性が全くみとめられない場合に、本接種実験に使用する。
対数期にあるRaji(ATCC CCL‐86)および1648(ATCC CRL‐1648)の細胞の懸濁液を、50mlの遠心管中に集め、4℃で遠心する。次いで、その上清を捨てる。細胞を、1640液体培養培地(仔ウシ血清を加えたもの)に再懸濁して、1.0×107細胞/mlにする。0.1mlのRajiの細胞懸濁液をマウスの左腋窩、また、0.1mlの1648(ATCC CRL‐1648)の細胞懸濁液をマウスの右腋窩に、皮下注射する。
注射の3〜4日後、腫瘍は成長して約2×2mmとなる。腫瘍を有するマウスを、以下のグループに分ける:
(グループA)コントロールグループとしての、抗腫瘍ポリペプチドを含有しない保存液(10mM PBS+0.2M NaClリン酸緩衝液(pH7.4));
(グループB)処理グループとしての、新規抗腫瘍ポリペプチド1(プラスミドpCHCEB11)、300μg/マウス/日(25gとして計算)で20日間連続。
各グループの10匹のマウスに、1日2回、20日間連続で、0.5ml皮下注射する。マウスの挙動は、毎日観察および記録する。腫瘍の大きさについては、2日ごとに測定し、写真撮影する。
その結果(図7参照)は、新規抗腫瘍ポリペプチドのグループBの腫瘍の成長が有意に阻害され、7匹のマウスの腫瘍は消失し、残りの3匹のマウスの腫瘍は、コントロールグループのそれよりも明らかに小さいことを示す。新規抗腫瘍ポリペプチドは、リンパ肉腫のEBV陽性細胞により引き起こされるマウス体内の固形腫瘍の成長を阻害するのに効果的である。しかし、当該新規抗腫瘍ポリペプチドは、リンパ肉腫のEBV陰性細胞により引き起こされるマウス体内の固形腫瘍の成長の阻害には効果がない。
【0047】
(実施例7)
腫瘍消失のin vivo実験の病理学的観察
腫瘍の組織病理学的観察:実施例6の実験の終わりにマウスを屠殺する。腫瘍を抽出し、10%ホルマリン中で固定する。パラフィンスライスを、HE染色し、常用の光学顕微鏡法に基づき観察する。
顕微鏡下で観察すると、コントロールグループのマウス由来の固形腫瘍は活発に増殖しており、;当該新規抗腫瘍ポリペプチドのグループのマウス由来のEBV陽性固形腫瘍の細胞は著しく縮小している。切片中のこれら細胞塊のほとんどは壊死した腫瘍細胞であり、多量の、腫瘍周辺のリンパ球浸潤が観察される。この組織病理学的結果は、20日間の処理の間に、新規抗腫瘍ポリペプチドが、固形腫瘍中の腫瘍細胞をほぼすべて殺傷したことを示唆している(図8、Dを参照のこと)。
【特許請求の範囲】
【請求項1】
イオンチャネルを形成することのできるコリシンの変異ポリペプチドと、抗EBウィルス抗体のポリペプチドまたは抗EBウィルス抗体模倣物のポリペプチドとの機能可能な連結により形成される、EBウィルスが原因である腫瘍に対する新規ポリペプチドであって、前記イオンチャネルを形成することのできるコリシンの変異ポリペプチドが、野生型コリシンE1、Ia、Ib、A、B、Nまたはそれらの水溶性チャネルドメインのペプチド鎖の、G11A、H22G、A26G、V31L、およびH40Dのアミノ酸残基の変異により得られるものであり、前記抗EBウィルス抗体のポリペプチドのアミノ酸配列が、ATCC HB‐168のハイブリドーマにより分泌されるモノクローナル抗体のポリペプチドと同じである、新規ポリペプチド。
【請求項2】
前記抗体模倣物のポリペプチドが、抗EBウィルス抗体の、重鎖のCDR1領域と、重鎖のCDR1‐CDR2連結ペプチドセグメントと、軽鎖のCDR3との結合ペプチドである、請求項1に記載の、EBウィルスが原因である腫瘍に対する新規ポリペプチド。
【請求項3】
前記イオンチャネルを形成することのできるコリシンの変異ポリペプチドが、野生型コリシンIaの変異により得られるものである、請求項2に記載の、EBウィルスが原因である腫瘍に対する新規ポリペプチド。
【請求項4】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドが、配列番号29に示されるアミノ酸配列を有する、請求項3に記載の、EBウィルスが原因である腫瘍に対する新規ポリペプチド。
【請求項5】
請求項1〜4のいずれか1項に記載のEBウィルスが原因である腫瘍に対する新規ポリペプチドをコードする遺伝子。
【請求項6】
配列番号30に示されているヌクレオチド配列を有する、請求項5に記載の遺伝子。
【請求項7】
請求項5に記載の遺伝子を含む組換えプラスミド。
【請求項8】
請求項1〜4のいずれか1項に記載のEBウィルスが原因である腫瘍に対する新規ポリペプチドの生産方法であって、請求項7に記載の組換えプラスミドを、発現のために発現系へと形質転換する工程、および、発現したポリペプチドを単離する工程、を含む生産方法。
【請求項9】
請求項1〜4のいずれか1項に記載のEBウィルスが原因である腫瘍に対する新規ポリペプチドの、EBウィルスが原因である腫瘍の治療および予防のための医薬の生産における使用。
【請求項10】
アミノ酸配列が、配列番号24に示されているものである、コリシンIaの変異ポリペプチド。
【請求項11】
請求項10に記載のコリシンIaの変異ポリペプチドをコードする遺伝子。
【請求項12】
請求項11に記載の遺伝子の使用であって、前記遺伝子を、前記ペプチドを誘導する遺伝子と機能可能に連結し、発現ベクターにクローニングし、次いで、前記発現ベクターを、発現系に形質転換し、発現したポリペプチドを単離する、ペプチド医薬の生産における、前記遺伝子の使用。
【請求項1】
イオンチャネルを形成することのできるコリシンの変異ポリペプチドと、抗EBウィルス抗体のポリペプチドまたは抗EBウィルス抗体模倣物のポリペプチドとの機能可能な連結により形成される、EBウィルスが原因である腫瘍に対する新規ポリペプチドであって、前記イオンチャネルを形成することのできるコリシンの変異ポリペプチドが、野生型コリシンE1、Ia、Ib、A、B、Nまたはそれらの水溶性チャネルドメインのペプチド鎖の、G11A、H22G、A26G、V31L、およびH40Dのアミノ酸残基の変異により得られるものであり、前記抗EBウィルス抗体のポリペプチドのアミノ酸配列が、ATCC HB‐168のハイブリドーマにより分泌されるモノクローナル抗体のポリペプチドと同じである、新規ポリペプチド。
【請求項2】
前記抗体模倣物のポリペプチドが、抗EBウィルス抗体の、重鎖のCDR1領域と、重鎖のCDR1‐CDR2連結ペプチドセグメントと、軽鎖のCDR3との結合ペプチドである、請求項1に記載の、EBウィルスが原因である腫瘍に対する新規ポリペプチド。
【請求項3】
前記イオンチャネルを形成することのできるコリシンの変異ポリペプチドが、野生型コリシンIaの変異により得られるものである、請求項2に記載の、EBウィルスが原因である腫瘍に対する新規ポリペプチド。
【請求項4】
前記EBウィルスが原因である腫瘍に対する新規ポリペプチドが、配列番号29に示されるアミノ酸配列を有する、請求項3に記載の、EBウィルスが原因である腫瘍に対する新規ポリペプチド。
【請求項5】
請求項1〜4のいずれか1項に記載のEBウィルスが原因である腫瘍に対する新規ポリペプチドをコードする遺伝子。
【請求項6】
配列番号30に示されているヌクレオチド配列を有する、請求項5に記載の遺伝子。
【請求項7】
請求項5に記載の遺伝子を含む組換えプラスミド。
【請求項8】
請求項1〜4のいずれか1項に記載のEBウィルスが原因である腫瘍に対する新規ポリペプチドの生産方法であって、請求項7に記載の組換えプラスミドを、発現のために発現系へと形質転換する工程、および、発現したポリペプチドを単離する工程、を含む生産方法。
【請求項9】
請求項1〜4のいずれか1項に記載のEBウィルスが原因である腫瘍に対する新規ポリペプチドの、EBウィルスが原因である腫瘍の治療および予防のための医薬の生産における使用。
【請求項10】
アミノ酸配列が、配列番号24に示されているものである、コリシンIaの変異ポリペプチド。
【請求項11】
請求項10に記載のコリシンIaの変異ポリペプチドをコードする遺伝子。
【請求項12】
請求項11に記載の遺伝子の使用であって、前記遺伝子を、前記ペプチドを誘導する遺伝子と機能可能に連結し、発現ベクターにクローニングし、次いで、前記発現ベクターを、発現系に形質転換し、発現したポリペプチドを単離する、ペプチド医薬の生産における、前記遺伝子の使用。
【図1】

【図2】
【図3】

【図4】

【図5A】

【図5B】

【図5C】

【図6】
【図7A】
【図7B】
【図8A】
【図8B】

【図8C】

【図8D】

【図2】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【図6】
【図7A】
【図7B】
【図8A】
【図8B】
【図8C】
【図8D】
【公表番号】特表2013−514060(P2013−514060A)
【公表日】平成25年4月25日(2013.4.25)
【国際特許分類】
【出願番号】特願2012−543448(P2012−543448)
【出願日】平成22年2月26日(2010.2.26)
【国際出願番号】PCT/CN2010/070762
【国際公開番号】WO2011/072501
【国際公開日】平成23年6月23日(2011.6.23)
【出願人】(512159786)プロテイン デザイン ラブ リミテッド (1)
【Fターム(参考)】
【公表日】平成25年4月25日(2013.4.25)
【国際特許分類】
【出願日】平成22年2月26日(2010.2.26)
【国際出願番号】PCT/CN2010/070762
【国際公開番号】WO2011/072501
【国際公開日】平成23年6月23日(2011.6.23)
【出願人】(512159786)プロテイン デザイン ラブ リミテッド (1)
【Fターム(参考)】
[ Back to top ]