
FRETに基づく病原体の迅速診断
本発明は、微生物を検出するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、アミノ酸X1およびX2、またはX1、X2およびX3からなるジまたはトリペプチドに連結した一組の分子マーカーを含み、前記アミノ酸の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい、基質を含む。前記基質は、バチルス・アンスラシスの検出に使用することが好ましい。あるいは本発明は、微生物、より具体的にはP.エルギノーサを検出するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、グリシンアミノ酸からなるトリ、テトラまたはペンタペプチドに連結した一組の分子マーカーを含む基質を対象とする。本発明はさらに、本発明の基質を用いて微生物、具体的にはバチルス・アンスラシスおよびシュードモナス・エルギノーサを検出するための方法、およびこのような方法における基質の使用を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は診断法の分野に属し、より具体的には有害な化合物または生物の検出、より具体的には微生物、より具体的には炭疽菌(バチルス・アンスラシス(Bacillus anthracis))およびシュードモナス・エルギノーサ(Pseudomonas aeruginosa)の検出の分野に属する。
【背景技術】
【0002】
シュードモナス・エルギノーサは、肺管、尿路、火傷、創傷の感染症を引き起こす細菌性病原体であり、他の血液感染症も引き起こす。この菌は熱傷および外耳(外耳炎)の感染症の最も一般的な原因であり、医療機器(例えばカテーテル)で最も頻繁に見られる生着菌である。シュードモナスはまれな状況で市中感染性肺炎のほか人工呼吸器関連肺炎も引き起こすことができ、いくつかの研究において分離された最も一般的な媒介物の1つである。院内感染の10分の1はシュードモナスによるものである。
【0003】
臨床環境におけるP.エルギノーサの検出はなお主として、試料の細菌を選択プレート上で培養することにより、または菌体外多糖マトリックスなどの細菌の表面抗原への抗体を含む免疫学的検査に基づいて行われる。迅速な診断および検出は適切な治療を開始し、細菌の伝播を制御するのに重要である。
【0004】
炭疽は、胞子形成細菌のバチルス・アンスラシスにより引き起こされる人獣共通感染疾病である。B.アンスラシスの胞子は環境中で何年も生き残り、これは潜在的な感染源であることを表す。炭疽はヒトにおいて吸入性、胃腸性、皮膚性の3つの臨床形態で生じる。吸入性炭疽は、B.アンスラシス胞子の工業的処理または意図的な放出を通じたエアロゾル化によって生じる。胃腸性または中咽頭性の疾病形態は、加熱が不十分なまたは生の感染した肉の摂取によって生じる。皮膚性炭疽は自然獲得性の炭疽菌感染の最も一般的なタイプであり、通常、感染した動物由来の汚染された製品との皮膚接触後に生じる。歴史的に、皮膚性炭疽の致死率は抗生物質治療を伴う場合は<1%、抗生物質治療を伴わない場合は20%である(Brachman P.S.およびKaufmann A.F.、In: Evans A.S.、Brachman P.S.編、Bacterial infections of humans.、New York、Plenum Medical Book Company、1998:95〜111頁;Dixon T.C.他、N. Engl. J. Med.、1999;341:815〜26頁)。吸入性炭疽の致死率は、適切な抗生物質および支持療法を伴う場合でも高い。(Jernigan J.A.他、Emerg. Infect. Dis.、2001;7:933〜44頁)。アメリカで20世紀中に確認された吸入性炭疽の18症例において、全体の致死率は>75%であった。B.アンスラシスの胞子が郵便を通じて放出された2001年秋の生物テロ攻撃の後、吸入性炭疽患者の致死率は45%であった(11症例中5症例)(Jernigan D.B.他、Emerg. Infect. Dis.、2002;8:1019〜28頁)。炭疽の潜伏期間は通常<2週間だが、胞子の休眠と時間がかかる肺からの除去のため、吸入性炭疽の潜伏期間は数ヶ月間延長されうる。この遅延発症という現象は皮膚性または胃腸性の暴露では認められていない。皮膚損傷からの排出は潜在的に感染性であるが、ヒトからヒトへの伝染はほとんど報告されていない。吸入性炭疽のヒトからヒトへの伝染は記録されていない。
【0005】
B.アンスラシスは最も武器として用いられやすい生物剤の1つであるが、その理由は1)その胞子は高度に安定で、2)胞子は呼吸器経路を通じて感染可能で、3)結果として生じる吸入性疾病の致死率が高いことである。1979年の先のソビエト連邦における軍微生物学施設からの非意図的なB.アンスラシス胞子の放出では69人の死者を出す結果となった。(Meselson M.他、Science、1994;266:1202〜8頁)。B.アンスラシス胞子がアメリカの郵便システムを通じて2001年に分配された後の炭疽の流行は、テロの脅威としてのこの生物の危険性をさらに強調するものである。
【0006】
テロ攻撃の後、B.アンスラシス胞子への暴露が一次的および二次的なエアロゾルを通じて発生しうる。一次的なエアロゾルは、生物剤が散布装置を通じてまたは剤を含有する包装物の取扱を通じて(例えば郵便の機械的処理において)初期放出されることによって生じる、空気中での粒子の分散体である。二次的なエアロゾルは沈下した粒子の崩壊および再懸濁によって生じる。
【0007】
炭疽菌の存在を検出するシステムは数多く提案されている。染色などの古典的な技術は近年、PCR(例えばMakino,S.T.他、J.Clin.Microbiol. 31:547〜51頁、1993);US6,884,588;Qiagen's Real-Art(商標) B. anthracis PCR)およびイムノアッセイ(例えばSwiecki, M.K.他、J. Immunol. 176:6076〜84頁、2006;US6,828,110;Response Medical Corp.'s RAMP(商標) Anthrax Assay)などのより信頼できる分子診断法により置き換えられている。しかしながら、これらのシステムの多くはバチルス・アンスラシスの毒性株と無毒性株とを識別できない。炭疽菌の毒性は主としてpXO1プラスミドの存在により付与されるが、このプラスミド上には炭疽菌の有毒タンパク質遺伝子があり、これは致死因子(LF-タンパク質)と命名されるものである。
【0008】
比較的新規で、非常に頑強で高度に信頼できる、生物学的物質の可視化のための技術がFRET(蛍光共鳴エネルギー移動)技術である。このプロセスでは、エネルギー的に励起した蛍光体である「ドナー」からの光子が別の分子である「アクセプター」中の電子のエネルギー状態を励起状態のより高い振動準位に上昇させる。結果として、ドナー蛍光体のエネルギー準位は蛍光を発光することなく基底状態に戻る。アクセプターはこのように蛍光の消光物質として機能する。このメカニズムはドナー分子とアクセプター分子の間の距離により制限される。ドナー分子とアクセプター分子の間の典型的な有効距離は10から100Åの範囲にあり、これにより例えば核酸およびタンパク質の分子診断法において有用なものとなっている。
【0009】
FRET技術は、FRET成分が核酸、特にPCR産物から生じる核酸にカップリングすることを通じてバチルス・アンスラシスの検出に用いられている(Qi Y.他、Appl. Environm. Microbiol. 67:3720〜7頁、2001;Patra G.他、Annal. New York Acad. Sci. 969:106〜11頁、2002;Mathur, N.他、J. Sensors 2008、Art. ID #270475)。しかしながらPCRと組み合わせたFRETの使用は、アッセイを実施する技術者の高い技術レベルをなお要求する。最近FRET技術は、致死因子タンパク質(LF)のタンパク質分解特性を通じて、炭疽菌の検出に適用されている。このアッセイでは、標識された基質がLFにより切断可能な試料に添加された(Cummings, R.T.他、Proc. Natl. Acad. Sci. USA 99:6603〜6頁、2002)。しかしながらこのアッセイは、基質の切断特性が特異的でないため、偽陽性シグナルを増加させる。
【0010】
P.エルギノーサについては、DNAを基礎としたFRETに基づくシステムが記載されている(Mancini, n.他、2009、J. Clin. Microbiol.、doi:10.1128/JCM.00011-09)。しかしながら、このアッセイにおいては、十分なDNAを提供するためにDNA溶解ステップおよびPCRステップが行われなければならない。
【0011】
このようにB.アンスラシスおよびP.エルギノーサなどの病原性微生物を検出する改良されたシステムが依然として必要とされている。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】US6,884,588
【特許文献2】US6,828,110
【非特許文献】
【0013】
【非特許文献1】Brachman P.S.およびKaufmann A.F.、In: Evans A.S.、Brachman P.S.編、Bacterial infections of humans.、New York、Plenum Medical Book Company、1998:95〜111頁
【非特許文献2】Dixon T.C.他、N. Engl. J. Med.、1999; 341: 815〜26頁
【非特許文献3】Jernigan J.A.他、Emerg. Infect. Dis.、2001; 7: 933〜44頁
【非特許文献4】Jernigan D.B.他、Emerg. Infect. Dis.、2002;8:1019〜28頁
【非特許文献5】Meselson M.他、Science、1994; 266: 1202〜8頁
【非特許文献6】Makino, S.T.他、J. Clin. Microbiol. 31: 547〜51頁、1993
【非特許文献7】Swiecki, M.K.他、J. Immunol. 176: 6076〜84頁、2006
【非特許文献8】Qi Y.他、Appl. Environm. Microbiol. 67: 3720〜7頁、2001
【非特許文献9】Patra G.他、Annal. New York Acad. Sci. 969: 106〜11頁、2002
【非特許文献10】Mathur, N.他、J. Sensors 2008、Art. ID #270475
【非特許文献11】Cummings, R.T.他、Proc. Natl. Acad. Sci. USA 99: 6603〜6頁、2002
【非特許文献12】Mancini, n.他、2009、J. Clin. Microbiol.、doi: 10.1128/JCM. 00011-09
【非特許文献13】Marras, S.A.E.(Meth. MoI. Biol. 335:3〜16頁、2006)
【非特許文献14】Guarise, C.他、Proc. Natl. Acad. Sci. 103:3978〜82頁、2006
【発明の概要】
【発明が解決しようとする課題】
【0014】
本発明により解決しようとする課題は、病原性微生物、より具体的にはB.アンスラシスおよびP.エルギノーサのインサイチュの特異的な検出および診断を提供することである。
【課題を解決するための手段】
【0015】
本発明は、微生物、好ましくはB.アンスラシスまたはそれらの酵素を検出、診断または同定するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、アミノ酸X1およびX2、またはX1、X2およびX3からなるジまたはトリペプチドに連結した一組の分子マーカーを含み、前記アミノ酸の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい、基質を含む。かかる基質において好ましくは、D-アミノ酸は、中性アミノ酸、好ましくはDLeu、DVal、DIle、DGlyまたはDAla、より好ましくはDLeu、DValまたはDIleである。Lアミノ酸1つまたはLアミノ酸2つのうち1つがLeuである実施形態であるがさらに好ましい。前記一組の分子マーカーが、蛍光標識、好ましくはFITC(フルオレセイン-5-イソチオシアネート)および前記蛍光標識の消光物質、好ましくはDABCYL(4-((-4-(ジメチルアミノ)-フェニル)-アゾ)-安息香酸)を含む、本発明によるバチルス・アンスラシスを蛍光検出するための基質も好ましい。この場合、好ましくは、リンカーはアミノヘキソン酸(FITC用)およびリジン(DABCYL用)である。この場合、基質は、FITC-Ahx-X1-X2-X3-Lys-DABCYL(ここでX1、X2およびX3は上で定義されており、X3は存在しなくてもよく、Lysはリジンを表し、Ahxはアミノヘキソン酸を表す)であると定義されうる。より具体的には、本発明の基質は、次の表1からなる群から選択される。
【0016】
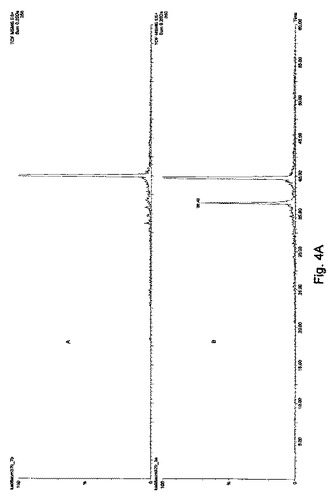
【表1】

【0017】
別の実施形態において、本発明による前記基質において、前記一組の分子マーカーは末端アセチル化システインを2つ含み、リンカーは存在しない。この場合、前記基質は、AcNH-Cys(S-Ac)-X1-X2-X3-Cys(S-Ac)-OH(ここでX1、X2およびX3は上で定義されており、X3は存在しなくてもよく、Cys(S-Ac)はチオールアセチル化システインを表し、AcNHはアセチル化アミノ末端を表す)であることが好ましい。より好ましくは、前記基質は次の表2からなる群から選択される。
【0018】
【表2】
【0019】
別の実施形態において、本発明は、試料中の微生物、特にバチルス・アンスラシスを検出するための方法であって、上で定義した基質を試料に添加する段階と、蛍光または色のシフトを検出する段階とを含む方法を含む。特定の実施形態において、蛍光シグナルは微生物細胞において細胞内に位置する。さらなる実施形態において、試料は、体液、粉末、水、食品、培地または任意の他の生物学的マトリックスから選択される。
【0020】
より具体的には、本発明は、試料中のバチルス・アンスラシスPXO1+を同定するための方法であって、上で定義した基質を試料に添加する段階と、蛍光、色のシフトもしくは特異的な質量スペクトルを検出するまたはキャピラリー電気泳動により検出を達成する段階とを含む方法に関する。
【0021】
本発明はさらにバチルス・アンスラシスを検出および診断するための、上で定義した基質の使用を含む。
【0022】
なお他の実施形態において、本発明は、微生物、より具体的にはP.エルギノーサを検出するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、グリシンアミノ酸からなるトリ、テトラまたはペンタペプチドに連結した一組の分子マーカーを含む基質を含む。より好ましくは、前記一組の分子マーカーは、蛍光標識、好ましくはFITC(フルオレセイン-5-イソチオシアネート)および前記蛍光標識の消光物質、好ましくはDABCYL(4-((-4-(ジメチルアミノ)-フェニル)-アゾ)-安息香酸)を含む。これらのマーカーに伴うリンカーは好ましくはアミノヘキソン酸(FITC用)およびリジン(DABCYL用)である。この点において、次の表3から選択される基質が最も好ましい。
【0023】
【表3】
【0024】
本発明はさらに、試料中の微生物、特にシュードモナス・エルギノーサを検出するための方法であって、上で定義した基質を試料に添加する段階と、蛍光または色のシフトを検出する段階とを含む方法を含む。本発明はまた、蛍光シグナルが微生物細胞において細胞内に位置する方法を提供する。好ましくは、これらの方法において試料は、体液、粉末、水、食品、培地または任意の他の生物学的マトリックスから選択される。さらに本発明は、シュードモナス・エルギノーサを検出および診断するための、上の定義による基質の使用に関する。
【発明の効果】
【0025】
第一に、本発明は、試料中の微生物、具体的には炭疽菌を特異的に検出する迅速で効率的な方法を提供する。さらに本発明は、PXO1プラスミドを保有する炭疽菌、すなわち毒性の炭疽菌を判別可能である。さらに、本検出方法は、炭疽菌を有する任意の試料において(前)分離の必要なく使用することができる。
【図面の簡単な説明】
【0026】
【図1A】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam1は(毒性)B.アンスラシス由来酵素により特異的に切断される。
【図1B】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam2は(毒性)B.アンスラシス由来酵素により特異的に切断される。弱いシグナルがB.セレウス(B. cereus)およびB.チューリンゲンシス(B. thuringiensis)で観察された。
【図1C】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。B.アンスラシス由来酵素はBikKam3を消化しない。その他、B.グロビギ(B. globigii)、C.ボツリヌム(C. botulinum)、P.エルギノーサ、V.コレラ(V. cholerae)およびE.ヘルビコラ(E. herbicola)由来の酵素は基質に対して活性を呈することから、酵素、より具体的にはこのアプローチによって幅広いスペクトルでの微生物検出が可能であることを示す(図3Cも参照)。
【図1D】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam4は(毒性)B.アンスラシス由来酵素により特異的に切断される。
【図1E】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam5は(毒性)B.アンスラシス由来酵素により特異的に切断される。いくらかの切断がV.コレラでも同様に観察された。
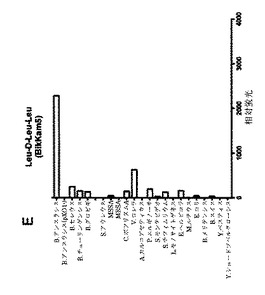
【図1F】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam6は(毒性)B.アンスラシス由来酵素により相対的に中程度であるものの、特異的に切断される。
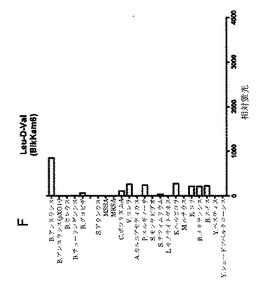
【図1G】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam7は(毒性)B.アンスラシス由来酵素により特異的に切断される。弱いシグナルがB.チューリンゲンシスで観察された。
【図1H】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam8を今回使用した微生物とともに用いた場合、有意な酵素活性を測定できなかった。
【図1I】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam9を今回使用した微生物とともに用いた場合、有意な酵素活性を測定できなかった。
【図1J】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。FITC-Ahx-Gly-Gly-Gly-Gly-Lys-DABCYLはP.エルギノーサ由来酵素により特異的に切断される。
【図2】BikKam1を切断する酵素はPXO1プラスミド上にあるが、致死因子ではないことを表す図である。A:基質BikKam1はいくつかの精製された酵素(致死因子、サーモリシン、コラゲナーゼ(クロストリジウム属菌由来)およびコラゲナーゼ(ビブリオ属菌由来))に添加された。全ての酵素についてシグナルは観察されず、これはLFではなくB.アンスラシス由来酵素に対するBikKam1の特異性を明確に示すものである。B:BikKam1はLFに添加された。追加されたLF阻害剤は基質の切断を有意に低下させず、これはLFではなくB.アンスラシス由来酵素に対するBikKam1の特異性を明確に示すものである。
【図3AB】図3Aは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam1は(毒性)B.アンスラシス由来酵素により特異的に切断される。いくらかの弱い活性がB.チューリンゲンシスでも同様に観察された。図3Bは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam2は(毒性)B.アンスラシス由来酵素およびB.チューリンゲンシス由来酵素により特異的に切断される。
【図3CD】図3Cは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam3は(毒性)B.アンスラシス由来酵素、B.チューリンゲンシス由来酵素およびV.コレラ由来酵素により特異的に切断される。いくらかの弱い活性がP.エルギノーサで観察された。これらのデータは、同じ基質がBHI中で培養された場合とヒト血清中で培養された場合とで異なった活性を呈する(図1Cと比較)という、本発明の基質/アプローチの異なった使用についての証拠を提供する。図3Dは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam4は(毒性)B.アンスラシス由来酵素およびB.チューリンゲンシス由来酵素により特異的に切断される。
【図3EF】図3Eは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam5は(毒性)B.アンスラシス由来酵素により特異的に切断される。いくらかの弱い活性がB.チューリンゲンシスで観察された。図3Fは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam6は(毒性)B.アンスラシス由来酵素により中程度に特異的に切断される。他の細菌でもいくらかの弱いバックグラウンドが60分時に観察された。例えば30分時などのより早い時点で測定しても、(毒性)B.アンスラシス以外の細菌ではバックグラウンドはほとんど見られなかった(非表示)。
【図3GH】図3Gは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam7は(毒性)B.アンスラシス由来酵素により、およびより低い程度でB.チューリンゲンシス由来酵素により、特異的に切断される。図3Hは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam8を今回使用した微生物とともに用いた場合、有意な酵素活性を測定できなかった。
【図3IJ】図3Iは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam9を今回使用した微生物とともに用いた場合、有意な酵素活性を測定できなかった。図3Jは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。FITC-Ahx-Gly-Gly-Gly-Gly-Lys-DABCYLはP.エルギノーサ由来酵素により特異的に切断される。
【図4A】[FITC]-Ahx-Leu-D-LeuのMH22+374.2生成イオン中の断片であるm/z132.1+348.1+390.1+503.2+616.2イオンの抽出されたイオンクロマトグラムである。上段パネルは非感染マウスの血清、下段パネルは感染マウスの血清である。
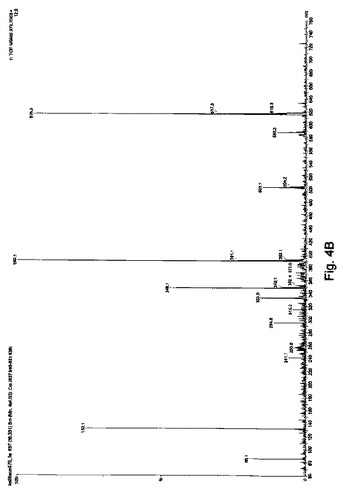
【図4B】[FITC]-Ahx-Leu-D-LeuのMH22+374.2生成イオンの質量スペクトルである。
【図5】B.セレウス細胞質における蛍光BikKam1断片検出の図である。詳細は本文参照。
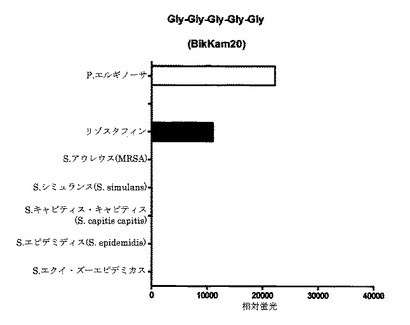
【図6A】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam20はP.エルギノーサ由来酵素およびリゾスタフィンにより特異的に切断される。
【図6B】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam21はP.エルギノーサ由来酵素およびリゾスタフィンにより特異的に切断される。BikKam20と比較して、P.エルギノーサでわずかな活性増加が観察される。
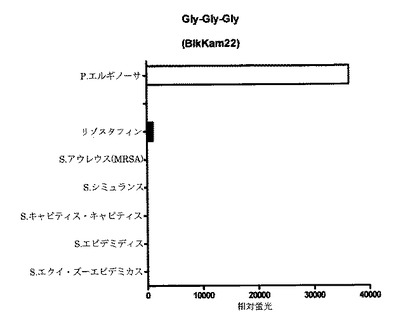
【図6C】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam22はP.エルギノーサ由来酵素により特異的に切断される。またリゾスタフィンによる多少の切断も検出することができた。BikKam20およびBikKam21と比較して、P.エルギノーサでわずかな活性増加が観察される。
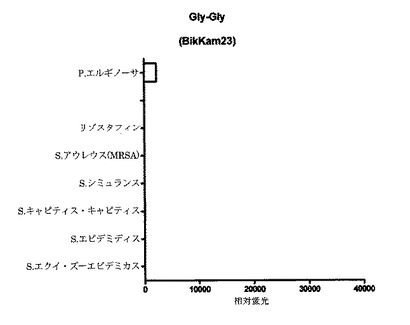
【図6D】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam23はP.エルギノーサ由来酵素により特異的に切断される。この基質の有効性はBikKam20〜BikKam22よりも低い。
【図6E】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam24を今回使用した微生物およびリゾスタフィンとともに用いた場合、有意な酵素活性を測定できなかった。
【発明を実施するための形態】
【0027】
本発明は、基質と細菌バチルス・アンスラシス中に存在するタンパク質分解酵素との間の相互作用に基づいて、炭疽菌を検出または診断するための方法を対象とする。検出を可能にするため、基質は、インタクトな基質とB.アンスラシスのプロテアーゼの特異的作用により切断された基質との差異を検出するのに使用される一組の分子マーカーを含む。
【0028】
微生物、より具体的にはB.アンスラシスを検出するための基質であって、リンカーを用いて、アミノ酸X1およびX2、またはX1、X2およびX3からなるジまたはトリペプチドに連結した一組の分子マーカーを含み、前記アミノ酸の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい、基質。
【0029】
基質の中央は、ロイシン、イソロイシン、バリン、グリシンまたはアラニン、より好ましくはロイシン、イソロイシンまたはバリンなどの中性アミノ酸であることが好ましいいずれか1つのD-アミノ酸を、別のD-またはL-アミノ酸、好ましくはL-ロイシンと、いずれかの方向(すなわちLeu-DLeuまたはDLeu-Leu)と組み合わせて構成される、2つまたは3つのアミノ酸の配列であり、このアミノ酸に一組の分子マーカーが、任意選択でリンカー分子を介して連結している。これらの配列は、これまでに知られていないB.アンスラシス酵素により非常に特異的に切断されるものと思われる。本明細書の実験部分において見られるように、この基質は近縁の細菌(B.チューリンゲンシスおよびB.セレウス)によりほとんど切断されず、B.グロビギ、B.スイス(B. suis)、E.コリ(E. coli)およびS.チフィムリウム(S. typhimurium)によって全く切断されない点で特異的である。
【0030】
またこれらの配列は、その内部における変化が特異的な特性の喪失を引き起こすという意味で特異的なように見える。本明細書の実験部分において見られるように、Leu-Leuを有する配列はB.アンスラシスによって切断されないが、いくつかの他の細菌(V.コレラ、P.エルギノーサ、E.ヘルビコラ、C.ボツリヌムおよびB.グロビギ)によって切断される。
【0031】
好ましいジ/トリペプチドは、ジペプチドLeu-DLeu(または逆のDLeu-Leu)である。この配列は最も性能の良いB.アンスラシスを検出するためのジ/トリペプチドであるだけでなく、炭疽菌のエクスビボ診断に用いられうるものと思われる。
【0032】
本発明のジペプチドに連結した分子マーカーは、インタクトな分子と切断された分子とを判別できる任意のマーカーのセットであってよい。簡単な検出方法は、例えば断片の質量がインタクトな分子の質量と容易に判別される質量分析であってもよい。かかるアッセイにおいて、2つの分子マーカーを持つこともまた必要ではなく、これは分子の一方にマーカーが存在すれば十分に検出できるためである。
【0033】
必要な場合、分子マーカーはリンカー分子またはリンカー部分を介してジまたはトリペプチドに連結されうる。
【0034】
好ましいのは切断されたペプチドの存在が直接的に目に見えるアッセイで、蛍光によるものなどがある。特に有利なのはFRET技術に基づくアッセイである。かかるアッセイにおいて、蛍光体は切断される予定の分子の一方に付着され、前記蛍光体の消光物質は分子の他方に付着されるが、インタクトな分子において消光物質はなお蛍光体を消光できる距離にある。通常FRETシステムにおいて、蛍光体と消光物質との間の距離は100Åを超えないものでありうる。蛍光体-消光物質ペアはいくつかの利用可能なペアから選択されてよく、例えばMarras, S.A.E.(Meth. Mol. Biol. 335:3〜16頁、2006)を参照すると、table 2、3および4にいくつかの蛍光体およびそれらの消光物質が記述されている。これら蛍光体および消光物質は全て本発明において用いられうる。好ましくは、FITC(フルオレセイン-5-イソチオシアネート)が蛍光体として、DABCYL(Dbc, 4-((-4-ジメチルアミノ)-フェニル)-アゾ)-安息香酸)が対応する消光物質として用いられる。
【0035】
次いで、蛍光体-消光物質ペアに隣接した基質は以下のように表現される。
マーカー1-(リンカー)-X1-X2-(リンカー)-マーカー2または
マーカー1-(リンカー)-X1-X2-X3-(リンカー)-マーカー2
ここでX1、X2またはX3の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい。
【0036】
蛍光体-消光物質ペアがFITCおよびDABCYLの場合、式は以下のように表される。
FITC-Ahx-X1-X2-(X3)-Lys-DABCYL
ここでX1、X2およびX3は上で定義されており、リンカー分子Ahxはアミノヘキソン酸、リンカー分子Lysはアミノ酸のリジンである。
【0037】
FRETアッセイにおいて、本発明による基質は試料に添加されて、蛍光は蛍光体がもはや消光物質により消光されないことを示すが、これは基質が切断されたことを意味し、B.アンスラシスが試料中に存在することを意味する。このように、これは簡単なアッセイであり、試料中の炭疽菌の存在の有無を直接視覚的に読み取ることによるものである。蛍光検出は(B.セレウスから分泌されたタンパク質分解酵素の存在に基づいて)培地中で行うことができるが、微生物自体の検出が実現可能であることも証明されている。我々の実験結果によると、本発明の基質タンパク質は細菌によって取り込まれて細胞内で切断されることで、細胞において蛍光を生成するものと思われる。このように本発明のアッセイは生きたB.セレウス細胞の蛍光染色にも用いられうる。
【0038】
あるいはプロテアーゼ活性の可視的検出は、システイン誘導体による金コロイドの色の変化に基づく新規アッセイを通じて行われうる(Guarise, C.他、Proc. Natl. Acad. Sci. 103:3978〜82頁、2006)。このアッセイにおいて、末端がアセチル化されたシステインを2つ持つペプチドがナノメーターサイズの金コロイドの凝集を妨げることで可視的な色のシフトを誘導する能力と、かかるアセチル化システイン末端を1つだけ有するペプチドの色の変化を誘導する能力の欠如との対比が用いられる。次いで、かかるアッセイのための本発明の基質は以下のように表される。
AcNH-Cys(S-Ac)-X1-X2-Cys(S-Ac)-OHまたは
AcNH-Cys(S-Ac)-X1-X2-X3-Cys(S-Ac)-OH
ここでX1、X2およびX3は上で定義されたものである。
【0039】
このアッセイにおいて、基質が試料に添加され、次いで試料および基質の全量が金ナノ粒子の懸濁液に添加される。金ナノ粒子の色が変わる場合、インタクトな基質はなお利用可能であり、これは基質が切断されていないことを意味し、B.アンスラシスが試料中に存在しないことを示す。色が変化しないことは切断された基質および炭疽菌の存在を示す。
【0040】
本発明の検出方法はB.アンスラシスの検出に適しているが、実施例中で数種の試料において示されるように、他の微生物も検出されうる。最も好ましくは、炭疽の疑いがある患者由来の試料が用いられる。かかる試料は血液、血清もしくは血漿試料であることができ、または例えば尿、唾液、リンパ液などの他の体液由来であってもよい。
【0041】
あるいは試料は土壌、水または空気などの環境資源から採取されうる。後者の場合、好ましくは空気を通して微生物を収集するフィルターが用いられる。言うまでもなく、あらゆる疑いのある粉末(郵送された封筒中の粉末など)、炭疽菌で汚染されうる食品または微生物が通常増殖される培地から試料を採取することも可能である。試料は採取後、本発明の基質と直接的に反応させることができ、蛍光または他の可視的なシグナルがB.アンスラシスが存在する試料において検出されうる。このようにして炭疽菌の存在の迅速診断がなされうる。
【0042】
さらにジペプチドの切断に関与するB.アンスラシス由来の酵素活性はPXO1プラスミド上に属すると思われる。このプラスミドは致死因子タンパク質などの炭疽菌の有毒遺伝子も抱えることから、本発明の検出方法はそれらの有害なB.アンスラシス菌株に対しても特異的である。それゆえに本発明の検出方法はB.アンスラシスPXO1+株を同定することもできる。
【0043】
B.アンスラシスの検出および診断のためのFRET基質を開発する中で、別の基質であるBikKam21がP.エルギノーサの検出において特異的であると思われた(図3J)。この基質の作用メカニズムをより理解するため、BikKam20からBikKam24(Table 1(表4))で表示されるBikKam21アナログが設計された。グリシン1つの追加または除去(BikKam20およびBikKam22)は、P.エルギノーサによる基質切断の増強をもたらした(図6A〜C)。一方、基質配列からもっとグリシンを除去した場合(BikKam23)、有意な活性の低下が観察された(図6D)。グリシンを1つだけ有するBikKam21アナログ(BikKam24)はP.エルギノーサ活性を呈さなかった(図6E)。
【0044】
P.エルギノーサ基質の切断はおそらくLasAによるものだが、これはP.エルギノーサの毒性において重要な酵素である。LasA以外に、BikKam20およびBikKam21のペプチド配列は細菌酵素であるリゾスタフィンによっても認識される。したがってBikKam21基質およびそのアナログの特異性がリゾスタフィン(Sigma)およびストレプトコッカス・エクイ・ズーエピデミカス(Streptococcus equi zooepidemicus)のようなリゾスタフィン産生細菌の培養上清を用いて調べられた。BikKam20およびBikKam21はリゾスタフィンにより切断された。基質を3つのグリシンに短くするとP.エルギノーサ活性の上昇およびリゾスタフィン活性の低下がもたらされた(図6C)。BikKam23はP.エルギノーサに対する活性がより低かったが、リゾスタフィンによって切断されなかった(図6D)。
【0045】
記述された基質のいくつかが精製されたリゾスタフィンによって切断された場合を除いて、リゾスタフィン産生細菌はいずれもBikKam基質を切断できなかった(図6)。結論として、FRET基質であるBikKam21からBikKam23はP.エルギノーサの検出および診断において用いられうるが、BikKam23はリゾスタフィンと相互作用しないことから最も好ましい。
【0046】
(実施例)
(実施例1)
材料と方法
細菌
Brain Heart Infusion(BHI)培地または70%ヒト血清(HuS)中、適切な増殖温度で細菌を一晩増殖させた。翌日、細菌を10分間10000rpmでの遠心分離によりペレット化した。上清を0.22uMフィルター(MilliPore)での濾過により滅菌した。質量分析の解析ならびにインビトロおよびエクスビボの結果の確認のため、B.アンスラシス感染を受けたマウスの血清をWaters Sep-Pak classic cartridge WAT051910 C18カラムで濾過した。
【0047】
試薬
FRETペプチドはPepScan(オランダ)により合成した。同一性および純度を質量分析および逆相HPLCにより確認した。炭疽菌致死因子(LF)はList Laboratoriesから購入した。サーモリシン、クロストリジウム由来コラゲナーゼおよびリゾスタフィンはSigmaから購入した。ビブリオ由来コラゲナーゼ/ディスパーゼはRocheから購入した。LF阻害剤12155はPyxis(オランダ)により合成した。9つの試験ペプチドを合成し、FITCおよびDABCYLと反応させて、以下の試薬を形成した。
【0048】
【表4】
【0049】
FRETアッセイ
アッセイ反応は、黒色ウェルで底部が透明の96ウェルプレート(Corning)中で行った。酵素アッセイのため、上述の試薬各々10uMを37℃でサーモリシン(1ug)、クロストリジウム由来コラゲナーゼ(10ug)、ビブリオ由来コラゲナーゼ/ディスパーゼ(5ug)またはLF(1ug)の存在下でインキュベートした。細菌上清中の酵素活性は、16uMのFRETペプチドを100uLの濾過された上清とともに37℃でインキュベートすることにより決定した。濾過されたBHI培地または70% HuSを対照として用いた。プレートは10分間隔でCytofluor 4000(Applied Biosystems)上で、485nmフィルターを用いた励起および530nmフィルターを用いた発光で読み取った。結果を図1、2および3に示す。
【0050】
質量分析
BikKam1のSepPak画分(Waters Sep-pak classic cartridge C18)を1)BHI中でB.アンスラシスを添加しておよび添加しないで、2)感染および対照マウスの血清とともにインキュベートしたものについて、[FITC]-Ahx-Leu-D-Leuおよび[dabcyl]-K(NH2)の存在を分析した。
【0051】
これらの試料はES-LC-MS/MSで以下の条件下で分析した。
【0052】
【表5】
【0053】
0.6ml/分の流速はLC-packingsスプリッターにより、およそ40μl/分に減速した。
カラム:PepMap C18;3μm;15cmx1mm サイズ:10 en 50μl
【0054】
[dabcyl]-K(NH2)の二価分子イオン(MH22+199.1)からコーン電圧20V、衝突エネルギー10〜11eVで生成したイオンのスペクトル(MS/MS)を記録した。[FITC]-Ahx-Leu-D-Leuの二価分子イオン(MH22+374.2)からのコーン電圧20〜25V、衝突エネルギー13eVでのスペクトルを記録した。
【0055】
アルゴンガス圧力は10-4mBarであった。
【0056】
図4の結果は非感染マウス(図4A)と比べてB.アンスラシス感染マウスにおいて追加ピークが観察されることを示しており、本実施例においては[FITC]-Ahx-Leu-D-Leuを表している。
【0057】
(実施例2)
材料と方法
B.セレウスおよびB.グロビギ細胞を5mlのBHI培地中で37℃で一晩増殖させた。翌日、細菌をBHI中で1:10に希釈して0.01MのBikKam1(PepScan、オランダ)存在下で増殖させた。増殖中、細胞試料を1時間間隔で採取した。細胞試料をPBSで2回洗浄し、20uLのPBSで再構成してガラススライド上にスポットした。陰性対照として、BikKam1を添加せずに16時間増殖させた細菌を用いた。蛍光顕微鏡検査はLeitz Orthoplan顕微鏡で1000倍の倍率を用いて行った。
【0058】
結果
2時間のインキュベーション後、蛍光のBikKam1断片をB.セレウスの細胞質中で検出できた(図5B)。4時間時では、蛍光のBikKam1断片は細胞質中に加えて、新たに分裂した側の細胞壁(側壁)中にも存在していた(図5C)。最終的に16時間のインキュベーション後、全てのBikKam1断片がB.セレウスの細胞質から側壁に移動した(図5D)。細菌B.グロビギはBikKam1基質を切断できないことから、陰性対照として用いた。予想された通り、BikKam1存在下で培養したB.グロビギは蛍光細菌にならなかった。また0時間時およびBikKam1を添加せずに増殖させた試料において、蛍光細菌を検出できなかった(図5A)。これらの結果は前述のFRETアッセイ、細菌酵素の検出に加えて、BikKam1基質を用いると細菌の可視化も可能であることを示す。
【技術分野】
【0001】
本発明は診断法の分野に属し、より具体的には有害な化合物または生物の検出、より具体的には微生物、より具体的には炭疽菌(バチルス・アンスラシス(Bacillus anthracis))およびシュードモナス・エルギノーサ(Pseudomonas aeruginosa)の検出の分野に属する。
【背景技術】
【0002】
シュードモナス・エルギノーサは、肺管、尿路、火傷、創傷の感染症を引き起こす細菌性病原体であり、他の血液感染症も引き起こす。この菌は熱傷および外耳(外耳炎)の感染症の最も一般的な原因であり、医療機器(例えばカテーテル)で最も頻繁に見られる生着菌である。シュードモナスはまれな状況で市中感染性肺炎のほか人工呼吸器関連肺炎も引き起こすことができ、いくつかの研究において分離された最も一般的な媒介物の1つである。院内感染の10分の1はシュードモナスによるものである。
【0003】
臨床環境におけるP.エルギノーサの検出はなお主として、試料の細菌を選択プレート上で培養することにより、または菌体外多糖マトリックスなどの細菌の表面抗原への抗体を含む免疫学的検査に基づいて行われる。迅速な診断および検出は適切な治療を開始し、細菌の伝播を制御するのに重要である。
【0004】
炭疽は、胞子形成細菌のバチルス・アンスラシスにより引き起こされる人獣共通感染疾病である。B.アンスラシスの胞子は環境中で何年も生き残り、これは潜在的な感染源であることを表す。炭疽はヒトにおいて吸入性、胃腸性、皮膚性の3つの臨床形態で生じる。吸入性炭疽は、B.アンスラシス胞子の工業的処理または意図的な放出を通じたエアロゾル化によって生じる。胃腸性または中咽頭性の疾病形態は、加熱が不十分なまたは生の感染した肉の摂取によって生じる。皮膚性炭疽は自然獲得性の炭疽菌感染の最も一般的なタイプであり、通常、感染した動物由来の汚染された製品との皮膚接触後に生じる。歴史的に、皮膚性炭疽の致死率は抗生物質治療を伴う場合は<1%、抗生物質治療を伴わない場合は20%である(Brachman P.S.およびKaufmann A.F.、In: Evans A.S.、Brachman P.S.編、Bacterial infections of humans.、New York、Plenum Medical Book Company、1998:95〜111頁;Dixon T.C.他、N. Engl. J. Med.、1999;341:815〜26頁)。吸入性炭疽の致死率は、適切な抗生物質および支持療法を伴う場合でも高い。(Jernigan J.A.他、Emerg. Infect. Dis.、2001;7:933〜44頁)。アメリカで20世紀中に確認された吸入性炭疽の18症例において、全体の致死率は>75%であった。B.アンスラシスの胞子が郵便を通じて放出された2001年秋の生物テロ攻撃の後、吸入性炭疽患者の致死率は45%であった(11症例中5症例)(Jernigan D.B.他、Emerg. Infect. Dis.、2002;8:1019〜28頁)。炭疽の潜伏期間は通常<2週間だが、胞子の休眠と時間がかかる肺からの除去のため、吸入性炭疽の潜伏期間は数ヶ月間延長されうる。この遅延発症という現象は皮膚性または胃腸性の暴露では認められていない。皮膚損傷からの排出は潜在的に感染性であるが、ヒトからヒトへの伝染はほとんど報告されていない。吸入性炭疽のヒトからヒトへの伝染は記録されていない。
【0005】
B.アンスラシスは最も武器として用いられやすい生物剤の1つであるが、その理由は1)その胞子は高度に安定で、2)胞子は呼吸器経路を通じて感染可能で、3)結果として生じる吸入性疾病の致死率が高いことである。1979年の先のソビエト連邦における軍微生物学施設からの非意図的なB.アンスラシス胞子の放出では69人の死者を出す結果となった。(Meselson M.他、Science、1994;266:1202〜8頁)。B.アンスラシス胞子がアメリカの郵便システムを通じて2001年に分配された後の炭疽の流行は、テロの脅威としてのこの生物の危険性をさらに強調するものである。
【0006】
テロ攻撃の後、B.アンスラシス胞子への暴露が一次的および二次的なエアロゾルを通じて発生しうる。一次的なエアロゾルは、生物剤が散布装置を通じてまたは剤を含有する包装物の取扱を通じて(例えば郵便の機械的処理において)初期放出されることによって生じる、空気中での粒子の分散体である。二次的なエアロゾルは沈下した粒子の崩壊および再懸濁によって生じる。
【0007】
炭疽菌の存在を検出するシステムは数多く提案されている。染色などの古典的な技術は近年、PCR(例えばMakino,S.T.他、J.Clin.Microbiol. 31:547〜51頁、1993);US6,884,588;Qiagen's Real-Art(商標) B. anthracis PCR)およびイムノアッセイ(例えばSwiecki, M.K.他、J. Immunol. 176:6076〜84頁、2006;US6,828,110;Response Medical Corp.'s RAMP(商標) Anthrax Assay)などのより信頼できる分子診断法により置き換えられている。しかしながら、これらのシステムの多くはバチルス・アンスラシスの毒性株と無毒性株とを識別できない。炭疽菌の毒性は主としてpXO1プラスミドの存在により付与されるが、このプラスミド上には炭疽菌の有毒タンパク質遺伝子があり、これは致死因子(LF-タンパク質)と命名されるものである。
【0008】
比較的新規で、非常に頑強で高度に信頼できる、生物学的物質の可視化のための技術がFRET(蛍光共鳴エネルギー移動)技術である。このプロセスでは、エネルギー的に励起した蛍光体である「ドナー」からの光子が別の分子である「アクセプター」中の電子のエネルギー状態を励起状態のより高い振動準位に上昇させる。結果として、ドナー蛍光体のエネルギー準位は蛍光を発光することなく基底状態に戻る。アクセプターはこのように蛍光の消光物質として機能する。このメカニズムはドナー分子とアクセプター分子の間の距離により制限される。ドナー分子とアクセプター分子の間の典型的な有効距離は10から100Åの範囲にあり、これにより例えば核酸およびタンパク質の分子診断法において有用なものとなっている。
【0009】
FRET技術は、FRET成分が核酸、特にPCR産物から生じる核酸にカップリングすることを通じてバチルス・アンスラシスの検出に用いられている(Qi Y.他、Appl. Environm. Microbiol. 67:3720〜7頁、2001;Patra G.他、Annal. New York Acad. Sci. 969:106〜11頁、2002;Mathur, N.他、J. Sensors 2008、Art. ID #270475)。しかしながらPCRと組み合わせたFRETの使用は、アッセイを実施する技術者の高い技術レベルをなお要求する。最近FRET技術は、致死因子タンパク質(LF)のタンパク質分解特性を通じて、炭疽菌の検出に適用されている。このアッセイでは、標識された基質がLFにより切断可能な試料に添加された(Cummings, R.T.他、Proc. Natl. Acad. Sci. USA 99:6603〜6頁、2002)。しかしながらこのアッセイは、基質の切断特性が特異的でないため、偽陽性シグナルを増加させる。
【0010】
P.エルギノーサについては、DNAを基礎としたFRETに基づくシステムが記載されている(Mancini, n.他、2009、J. Clin. Microbiol.、doi:10.1128/JCM.00011-09)。しかしながら、このアッセイにおいては、十分なDNAを提供するためにDNA溶解ステップおよびPCRステップが行われなければならない。
【0011】
このようにB.アンスラシスおよびP.エルギノーサなどの病原性微生物を検出する改良されたシステムが依然として必要とされている。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】US6,884,588
【特許文献2】US6,828,110
【非特許文献】
【0013】
【非特許文献1】Brachman P.S.およびKaufmann A.F.、In: Evans A.S.、Brachman P.S.編、Bacterial infections of humans.、New York、Plenum Medical Book Company、1998:95〜111頁
【非特許文献2】Dixon T.C.他、N. Engl. J. Med.、1999; 341: 815〜26頁
【非特許文献3】Jernigan J.A.他、Emerg. Infect. Dis.、2001; 7: 933〜44頁
【非特許文献4】Jernigan D.B.他、Emerg. Infect. Dis.、2002;8:1019〜28頁
【非特許文献5】Meselson M.他、Science、1994; 266: 1202〜8頁
【非特許文献6】Makino, S.T.他、J. Clin. Microbiol. 31: 547〜51頁、1993
【非特許文献7】Swiecki, M.K.他、J. Immunol. 176: 6076〜84頁、2006
【非特許文献8】Qi Y.他、Appl. Environm. Microbiol. 67: 3720〜7頁、2001
【非特許文献9】Patra G.他、Annal. New York Acad. Sci. 969: 106〜11頁、2002
【非特許文献10】Mathur, N.他、J. Sensors 2008、Art. ID #270475
【非特許文献11】Cummings, R.T.他、Proc. Natl. Acad. Sci. USA 99: 6603〜6頁、2002
【非特許文献12】Mancini, n.他、2009、J. Clin. Microbiol.、doi: 10.1128/JCM. 00011-09
【非特許文献13】Marras, S.A.E.(Meth. MoI. Biol. 335:3〜16頁、2006)
【非特許文献14】Guarise, C.他、Proc. Natl. Acad. Sci. 103:3978〜82頁、2006
【発明の概要】
【発明が解決しようとする課題】
【0014】
本発明により解決しようとする課題は、病原性微生物、より具体的にはB.アンスラシスおよびP.エルギノーサのインサイチュの特異的な検出および診断を提供することである。
【課題を解決するための手段】
【0015】
本発明は、微生物、好ましくはB.アンスラシスまたはそれらの酵素を検出、診断または同定するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、アミノ酸X1およびX2、またはX1、X2およびX3からなるジまたはトリペプチドに連結した一組の分子マーカーを含み、前記アミノ酸の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい、基質を含む。かかる基質において好ましくは、D-アミノ酸は、中性アミノ酸、好ましくはDLeu、DVal、DIle、DGlyまたはDAla、より好ましくはDLeu、DValまたはDIleである。Lアミノ酸1つまたはLアミノ酸2つのうち1つがLeuである実施形態であるがさらに好ましい。前記一組の分子マーカーが、蛍光標識、好ましくはFITC(フルオレセイン-5-イソチオシアネート)および前記蛍光標識の消光物質、好ましくはDABCYL(4-((-4-(ジメチルアミノ)-フェニル)-アゾ)-安息香酸)を含む、本発明によるバチルス・アンスラシスを蛍光検出するための基質も好ましい。この場合、好ましくは、リンカーはアミノヘキソン酸(FITC用)およびリジン(DABCYL用)である。この場合、基質は、FITC-Ahx-X1-X2-X3-Lys-DABCYL(ここでX1、X2およびX3は上で定義されており、X3は存在しなくてもよく、Lysはリジンを表し、Ahxはアミノヘキソン酸を表す)であると定義されうる。より具体的には、本発明の基質は、次の表1からなる群から選択される。
【0016】
【表1】
【0017】
別の実施形態において、本発明による前記基質において、前記一組の分子マーカーは末端アセチル化システインを2つ含み、リンカーは存在しない。この場合、前記基質は、AcNH-Cys(S-Ac)-X1-X2-X3-Cys(S-Ac)-OH(ここでX1、X2およびX3は上で定義されており、X3は存在しなくてもよく、Cys(S-Ac)はチオールアセチル化システインを表し、AcNHはアセチル化アミノ末端を表す)であることが好ましい。より好ましくは、前記基質は次の表2からなる群から選択される。
【0018】
【表2】
【0019】
別の実施形態において、本発明は、試料中の微生物、特にバチルス・アンスラシスを検出するための方法であって、上で定義した基質を試料に添加する段階と、蛍光または色のシフトを検出する段階とを含む方法を含む。特定の実施形態において、蛍光シグナルは微生物細胞において細胞内に位置する。さらなる実施形態において、試料は、体液、粉末、水、食品、培地または任意の他の生物学的マトリックスから選択される。
【0020】
より具体的には、本発明は、試料中のバチルス・アンスラシスPXO1+を同定するための方法であって、上で定義した基質を試料に添加する段階と、蛍光、色のシフトもしくは特異的な質量スペクトルを検出するまたはキャピラリー電気泳動により検出を達成する段階とを含む方法に関する。
【0021】
本発明はさらにバチルス・アンスラシスを検出および診断するための、上で定義した基質の使用を含む。
【0022】
なお他の実施形態において、本発明は、微生物、より具体的にはP.エルギノーサを検出するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、グリシンアミノ酸からなるトリ、テトラまたはペンタペプチドに連結した一組の分子マーカーを含む基質を含む。より好ましくは、前記一組の分子マーカーは、蛍光標識、好ましくはFITC(フルオレセイン-5-イソチオシアネート)および前記蛍光標識の消光物質、好ましくはDABCYL(4-((-4-(ジメチルアミノ)-フェニル)-アゾ)-安息香酸)を含む。これらのマーカーに伴うリンカーは好ましくはアミノヘキソン酸(FITC用)およびリジン(DABCYL用)である。この点において、次の表3から選択される基質が最も好ましい。
【0023】
【表3】
【0024】
本発明はさらに、試料中の微生物、特にシュードモナス・エルギノーサを検出するための方法であって、上で定義した基質を試料に添加する段階と、蛍光または色のシフトを検出する段階とを含む方法を含む。本発明はまた、蛍光シグナルが微生物細胞において細胞内に位置する方法を提供する。好ましくは、これらの方法において試料は、体液、粉末、水、食品、培地または任意の他の生物学的マトリックスから選択される。さらに本発明は、シュードモナス・エルギノーサを検出および診断するための、上の定義による基質の使用に関する。
【発明の効果】
【0025】
第一に、本発明は、試料中の微生物、具体的には炭疽菌を特異的に検出する迅速で効率的な方法を提供する。さらに本発明は、PXO1プラスミドを保有する炭疽菌、すなわち毒性の炭疽菌を判別可能である。さらに、本検出方法は、炭疽菌を有する任意の試料において(前)分離の必要なく使用することができる。
【図面の簡単な説明】
【0026】
【図1A】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam1は(毒性)B.アンスラシス由来酵素により特異的に切断される。
【図1B】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam2は(毒性)B.アンスラシス由来酵素により特異的に切断される。弱いシグナルがB.セレウス(B. cereus)およびB.チューリンゲンシス(B. thuringiensis)で観察された。
【図1C】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。B.アンスラシス由来酵素はBikKam3を消化しない。その他、B.グロビギ(B. globigii)、C.ボツリヌム(C. botulinum)、P.エルギノーサ、V.コレラ(V. cholerae)およびE.ヘルビコラ(E. herbicola)由来の酵素は基質に対して活性を呈することから、酵素、より具体的にはこのアプローチによって幅広いスペクトルでの微生物検出が可能であることを示す(図3Cも参照)。
【図1D】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam4は(毒性)B.アンスラシス由来酵素により特異的に切断される。
【図1E】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam5は(毒性)B.アンスラシス由来酵素により特異的に切断される。いくらかの切断がV.コレラでも同様に観察された。
【図1F】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam6は(毒性)B.アンスラシス由来酵素により相対的に中程度であるものの、特異的に切断される。
【図1G】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam7は(毒性)B.アンスラシス由来酵素により特異的に切断される。弱いシグナルがB.チューリンゲンシスで観察された。
【図1H】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam8を今回使用した微生物とともに用いた場合、有意な酵素活性を測定できなかった。
【図1I】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。BikKam9を今回使用した微生物とともに用いた場合、有意な酵素活性を測定できなかった。
【図1J】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明の基質のFRET蛍光シグナルの図である。FITC-Ahx-Gly-Gly-Gly-Gly-Lys-DABCYLはP.エルギノーサ由来酵素により特異的に切断される。
【図2】BikKam1を切断する酵素はPXO1プラスミド上にあるが、致死因子ではないことを表す図である。A:基質BikKam1はいくつかの精製された酵素(致死因子、サーモリシン、コラゲナーゼ(クロストリジウム属菌由来)およびコラゲナーゼ(ビブリオ属菌由来))に添加された。全ての酵素についてシグナルは観察されず、これはLFではなくB.アンスラシス由来酵素に対するBikKam1の特異性を明確に示すものである。B:BikKam1はLFに添加された。追加されたLF阻害剤は基質の切断を有意に低下させず、これはLFではなくB.アンスラシス由来酵素に対するBikKam1の特異性を明確に示すものである。
【図3AB】図3Aは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam1は(毒性)B.アンスラシス由来酵素により特異的に切断される。いくらかの弱い活性がB.チューリンゲンシスでも同様に観察された。図3Bは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam2は(毒性)B.アンスラシス由来酵素およびB.チューリンゲンシス由来酵素により特異的に切断される。
【図3CD】図3Cは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam3は(毒性)B.アンスラシス由来酵素、B.チューリンゲンシス由来酵素およびV.コレラ由来酵素により特異的に切断される。いくらかの弱い活性がP.エルギノーサで観察された。これらのデータは、同じ基質がBHI中で培養された場合とヒト血清中で培養された場合とで異なった活性を呈する(図1Cと比較)という、本発明の基質/アプローチの異なった使用についての証拠を提供する。図3Dは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam4は(毒性)B.アンスラシス由来酵素およびB.チューリンゲンシス由来酵素により特異的に切断される。
【図3EF】図3Eは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam5は(毒性)B.アンスラシス由来酵素により特異的に切断される。いくらかの弱い活性がB.チューリンゲンシスで観察された。図3Fは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam6は(毒性)B.アンスラシス由来酵素により中程度に特異的に切断される。他の細菌でもいくらかの弱いバックグラウンドが60分時に観察された。例えば30分時などのより早い時点で測定しても、(毒性)B.アンスラシス以外の細菌ではバックグラウンドはほとんど見られなかった(非表示)。
【図3GH】図3Gは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam7は(毒性)B.アンスラシス由来酵素により、およびより低い程度でB.チューリンゲンシス由来酵素により、特異的に切断される。図3Hは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam8を今回使用した微生物とともに用いた場合、有意な酵素活性を測定できなかった。
【図3IJ】図3Iは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。BikKam9を今回使用した微生物とともに用いた場合、有意な酵素活性を測定できなかった。図3Jは、ヒト血清中(エクスビボ)で増殖させた細菌の酵素とともに60分インキュベートした後の、基質のFRET蛍光シグナルの図である。FITC-Ahx-Gly-Gly-Gly-Gly-Lys-DABCYLはP.エルギノーサ由来酵素により特異的に切断される。
【図4A】[FITC]-Ahx-Leu-D-LeuのMH22+374.2生成イオン中の断片であるm/z132.1+348.1+390.1+503.2+616.2イオンの抽出されたイオンクロマトグラムである。上段パネルは非感染マウスの血清、下段パネルは感染マウスの血清である。
【図4B】[FITC]-Ahx-Leu-D-LeuのMH22+374.2生成イオンの質量スペクトルである。
【図5】B.セレウス細胞質における蛍光BikKam1断片検出の図である。詳細は本文参照。
【図6A】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam20はP.エルギノーサ由来酵素およびリゾスタフィンにより特異的に切断される。
【図6B】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam21はP.エルギノーサ由来酵素およびリゾスタフィンにより特異的に切断される。BikKam20と比較して、P.エルギノーサでわずかな活性増加が観察される。
【図6C】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam22はP.エルギノーサ由来酵素により特異的に切断される。またリゾスタフィンによる多少の切断も検出することができた。BikKam20およびBikKam21と比較して、P.エルギノーサでわずかな活性増加が観察される。
【図6D】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam23はP.エルギノーサ由来酵素により特異的に切断される。この基質の有効性はBikKam20〜BikKam22よりも低い。
【図6E】細菌増殖培地(BHI)中(インビトロ)で増殖させた細菌の酵素で処理してから60分後の、本発明のP.エルギノーサ基質のFRET蛍光シグナルの図である。BikKam24を今回使用した微生物およびリゾスタフィンとともに用いた場合、有意な酵素活性を測定できなかった。
【発明を実施するための形態】
【0027】
本発明は、基質と細菌バチルス・アンスラシス中に存在するタンパク質分解酵素との間の相互作用に基づいて、炭疽菌を検出または診断するための方法を対象とする。検出を可能にするため、基質は、インタクトな基質とB.アンスラシスのプロテアーゼの特異的作用により切断された基質との差異を検出するのに使用される一組の分子マーカーを含む。
【0028】
微生物、より具体的にはB.アンスラシスを検出するための基質であって、リンカーを用いて、アミノ酸X1およびX2、またはX1、X2およびX3からなるジまたはトリペプチドに連結した一組の分子マーカーを含み、前記アミノ酸の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい、基質。
【0029】
基質の中央は、ロイシン、イソロイシン、バリン、グリシンまたはアラニン、より好ましくはロイシン、イソロイシンまたはバリンなどの中性アミノ酸であることが好ましいいずれか1つのD-アミノ酸を、別のD-またはL-アミノ酸、好ましくはL-ロイシンと、いずれかの方向(すなわちLeu-DLeuまたはDLeu-Leu)と組み合わせて構成される、2つまたは3つのアミノ酸の配列であり、このアミノ酸に一組の分子マーカーが、任意選択でリンカー分子を介して連結している。これらの配列は、これまでに知られていないB.アンスラシス酵素により非常に特異的に切断されるものと思われる。本明細書の実験部分において見られるように、この基質は近縁の細菌(B.チューリンゲンシスおよびB.セレウス)によりほとんど切断されず、B.グロビギ、B.スイス(B. suis)、E.コリ(E. coli)およびS.チフィムリウム(S. typhimurium)によって全く切断されない点で特異的である。
【0030】
またこれらの配列は、その内部における変化が特異的な特性の喪失を引き起こすという意味で特異的なように見える。本明細書の実験部分において見られるように、Leu-Leuを有する配列はB.アンスラシスによって切断されないが、いくつかの他の細菌(V.コレラ、P.エルギノーサ、E.ヘルビコラ、C.ボツリヌムおよびB.グロビギ)によって切断される。
【0031】
好ましいジ/トリペプチドは、ジペプチドLeu-DLeu(または逆のDLeu-Leu)である。この配列は最も性能の良いB.アンスラシスを検出するためのジ/トリペプチドであるだけでなく、炭疽菌のエクスビボ診断に用いられうるものと思われる。
【0032】
本発明のジペプチドに連結した分子マーカーは、インタクトな分子と切断された分子とを判別できる任意のマーカーのセットであってよい。簡単な検出方法は、例えば断片の質量がインタクトな分子の質量と容易に判別される質量分析であってもよい。かかるアッセイにおいて、2つの分子マーカーを持つこともまた必要ではなく、これは分子の一方にマーカーが存在すれば十分に検出できるためである。
【0033】
必要な場合、分子マーカーはリンカー分子またはリンカー部分を介してジまたはトリペプチドに連結されうる。
【0034】
好ましいのは切断されたペプチドの存在が直接的に目に見えるアッセイで、蛍光によるものなどがある。特に有利なのはFRET技術に基づくアッセイである。かかるアッセイにおいて、蛍光体は切断される予定の分子の一方に付着され、前記蛍光体の消光物質は分子の他方に付着されるが、インタクトな分子において消光物質はなお蛍光体を消光できる距離にある。通常FRETシステムにおいて、蛍光体と消光物質との間の距離は100Åを超えないものでありうる。蛍光体-消光物質ペアはいくつかの利用可能なペアから選択されてよく、例えばMarras, S.A.E.(Meth. Mol. Biol. 335:3〜16頁、2006)を参照すると、table 2、3および4にいくつかの蛍光体およびそれらの消光物質が記述されている。これら蛍光体および消光物質は全て本発明において用いられうる。好ましくは、FITC(フルオレセイン-5-イソチオシアネート)が蛍光体として、DABCYL(Dbc, 4-((-4-ジメチルアミノ)-フェニル)-アゾ)-安息香酸)が対応する消光物質として用いられる。
【0035】
次いで、蛍光体-消光物質ペアに隣接した基質は以下のように表現される。
マーカー1-(リンカー)-X1-X2-(リンカー)-マーカー2または
マーカー1-(リンカー)-X1-X2-X3-(リンカー)-マーカー2
ここでX1、X2またはX3の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい。
【0036】
蛍光体-消光物質ペアがFITCおよびDABCYLの場合、式は以下のように表される。
FITC-Ahx-X1-X2-(X3)-Lys-DABCYL
ここでX1、X2およびX3は上で定義されており、リンカー分子Ahxはアミノヘキソン酸、リンカー分子Lysはアミノ酸のリジンである。
【0037】
FRETアッセイにおいて、本発明による基質は試料に添加されて、蛍光は蛍光体がもはや消光物質により消光されないことを示すが、これは基質が切断されたことを意味し、B.アンスラシスが試料中に存在することを意味する。このように、これは簡単なアッセイであり、試料中の炭疽菌の存在の有無を直接視覚的に読み取ることによるものである。蛍光検出は(B.セレウスから分泌されたタンパク質分解酵素の存在に基づいて)培地中で行うことができるが、微生物自体の検出が実現可能であることも証明されている。我々の実験結果によると、本発明の基質タンパク質は細菌によって取り込まれて細胞内で切断されることで、細胞において蛍光を生成するものと思われる。このように本発明のアッセイは生きたB.セレウス細胞の蛍光染色にも用いられうる。
【0038】
あるいはプロテアーゼ活性の可視的検出は、システイン誘導体による金コロイドの色の変化に基づく新規アッセイを通じて行われうる(Guarise, C.他、Proc. Natl. Acad. Sci. 103:3978〜82頁、2006)。このアッセイにおいて、末端がアセチル化されたシステインを2つ持つペプチドがナノメーターサイズの金コロイドの凝集を妨げることで可視的な色のシフトを誘導する能力と、かかるアセチル化システイン末端を1つだけ有するペプチドの色の変化を誘導する能力の欠如との対比が用いられる。次いで、かかるアッセイのための本発明の基質は以下のように表される。
AcNH-Cys(S-Ac)-X1-X2-Cys(S-Ac)-OHまたは
AcNH-Cys(S-Ac)-X1-X2-X3-Cys(S-Ac)-OH
ここでX1、X2およびX3は上で定義されたものである。
【0039】
このアッセイにおいて、基質が試料に添加され、次いで試料および基質の全量が金ナノ粒子の懸濁液に添加される。金ナノ粒子の色が変わる場合、インタクトな基質はなお利用可能であり、これは基質が切断されていないことを意味し、B.アンスラシスが試料中に存在しないことを示す。色が変化しないことは切断された基質および炭疽菌の存在を示す。
【0040】
本発明の検出方法はB.アンスラシスの検出に適しているが、実施例中で数種の試料において示されるように、他の微生物も検出されうる。最も好ましくは、炭疽の疑いがある患者由来の試料が用いられる。かかる試料は血液、血清もしくは血漿試料であることができ、または例えば尿、唾液、リンパ液などの他の体液由来であってもよい。
【0041】
あるいは試料は土壌、水または空気などの環境資源から採取されうる。後者の場合、好ましくは空気を通して微生物を収集するフィルターが用いられる。言うまでもなく、あらゆる疑いのある粉末(郵送された封筒中の粉末など)、炭疽菌で汚染されうる食品または微生物が通常増殖される培地から試料を採取することも可能である。試料は採取後、本発明の基質と直接的に反応させることができ、蛍光または他の可視的なシグナルがB.アンスラシスが存在する試料において検出されうる。このようにして炭疽菌の存在の迅速診断がなされうる。
【0042】
さらにジペプチドの切断に関与するB.アンスラシス由来の酵素活性はPXO1プラスミド上に属すると思われる。このプラスミドは致死因子タンパク質などの炭疽菌の有毒遺伝子も抱えることから、本発明の検出方法はそれらの有害なB.アンスラシス菌株に対しても特異的である。それゆえに本発明の検出方法はB.アンスラシスPXO1+株を同定することもできる。
【0043】
B.アンスラシスの検出および診断のためのFRET基質を開発する中で、別の基質であるBikKam21がP.エルギノーサの検出において特異的であると思われた(図3J)。この基質の作用メカニズムをより理解するため、BikKam20からBikKam24(Table 1(表4))で表示されるBikKam21アナログが設計された。グリシン1つの追加または除去(BikKam20およびBikKam22)は、P.エルギノーサによる基質切断の増強をもたらした(図6A〜C)。一方、基質配列からもっとグリシンを除去した場合(BikKam23)、有意な活性の低下が観察された(図6D)。グリシンを1つだけ有するBikKam21アナログ(BikKam24)はP.エルギノーサ活性を呈さなかった(図6E)。
【0044】
P.エルギノーサ基質の切断はおそらくLasAによるものだが、これはP.エルギノーサの毒性において重要な酵素である。LasA以外に、BikKam20およびBikKam21のペプチド配列は細菌酵素であるリゾスタフィンによっても認識される。したがってBikKam21基質およびそのアナログの特異性がリゾスタフィン(Sigma)およびストレプトコッカス・エクイ・ズーエピデミカス(Streptococcus equi zooepidemicus)のようなリゾスタフィン産生細菌の培養上清を用いて調べられた。BikKam20およびBikKam21はリゾスタフィンにより切断された。基質を3つのグリシンに短くするとP.エルギノーサ活性の上昇およびリゾスタフィン活性の低下がもたらされた(図6C)。BikKam23はP.エルギノーサに対する活性がより低かったが、リゾスタフィンによって切断されなかった(図6D)。
【0045】
記述された基質のいくつかが精製されたリゾスタフィンによって切断された場合を除いて、リゾスタフィン産生細菌はいずれもBikKam基質を切断できなかった(図6)。結論として、FRET基質であるBikKam21からBikKam23はP.エルギノーサの検出および診断において用いられうるが、BikKam23はリゾスタフィンと相互作用しないことから最も好ましい。
【0046】
(実施例)
(実施例1)
材料と方法
細菌
Brain Heart Infusion(BHI)培地または70%ヒト血清(HuS)中、適切な増殖温度で細菌を一晩増殖させた。翌日、細菌を10分間10000rpmでの遠心分離によりペレット化した。上清を0.22uMフィルター(MilliPore)での濾過により滅菌した。質量分析の解析ならびにインビトロおよびエクスビボの結果の確認のため、B.アンスラシス感染を受けたマウスの血清をWaters Sep-Pak classic cartridge WAT051910 C18カラムで濾過した。
【0047】
試薬
FRETペプチドはPepScan(オランダ)により合成した。同一性および純度を質量分析および逆相HPLCにより確認した。炭疽菌致死因子(LF)はList Laboratoriesから購入した。サーモリシン、クロストリジウム由来コラゲナーゼおよびリゾスタフィンはSigmaから購入した。ビブリオ由来コラゲナーゼ/ディスパーゼはRocheから購入した。LF阻害剤12155はPyxis(オランダ)により合成した。9つの試験ペプチドを合成し、FITCおよびDABCYLと反応させて、以下の試薬を形成した。
【0048】
【表4】
【0049】
FRETアッセイ
アッセイ反応は、黒色ウェルで底部が透明の96ウェルプレート(Corning)中で行った。酵素アッセイのため、上述の試薬各々10uMを37℃でサーモリシン(1ug)、クロストリジウム由来コラゲナーゼ(10ug)、ビブリオ由来コラゲナーゼ/ディスパーゼ(5ug)またはLF(1ug)の存在下でインキュベートした。細菌上清中の酵素活性は、16uMのFRETペプチドを100uLの濾過された上清とともに37℃でインキュベートすることにより決定した。濾過されたBHI培地または70% HuSを対照として用いた。プレートは10分間隔でCytofluor 4000(Applied Biosystems)上で、485nmフィルターを用いた励起および530nmフィルターを用いた発光で読み取った。結果を図1、2および3に示す。
【0050】
質量分析
BikKam1のSepPak画分(Waters Sep-pak classic cartridge C18)を1)BHI中でB.アンスラシスを添加しておよび添加しないで、2)感染および対照マウスの血清とともにインキュベートしたものについて、[FITC]-Ahx-Leu-D-Leuおよび[dabcyl]-K(NH2)の存在を分析した。
【0051】
これらの試料はES-LC-MS/MSで以下の条件下で分析した。
【0052】
【表5】
【0053】
0.6ml/分の流速はLC-packingsスプリッターにより、およそ40μl/分に減速した。
カラム:PepMap C18;3μm;15cmx1mm サイズ:10 en 50μl
【0054】
[dabcyl]-K(NH2)の二価分子イオン(MH22+199.1)からコーン電圧20V、衝突エネルギー10〜11eVで生成したイオンのスペクトル(MS/MS)を記録した。[FITC]-Ahx-Leu-D-Leuの二価分子イオン(MH22+374.2)からのコーン電圧20〜25V、衝突エネルギー13eVでのスペクトルを記録した。
【0055】
アルゴンガス圧力は10-4mBarであった。
【0056】
図4の結果は非感染マウス(図4A)と比べてB.アンスラシス感染マウスにおいて追加ピークが観察されることを示しており、本実施例においては[FITC]-Ahx-Leu-D-Leuを表している。
【0057】
(実施例2)
材料と方法
B.セレウスおよびB.グロビギ細胞を5mlのBHI培地中で37℃で一晩増殖させた。翌日、細菌をBHI中で1:10に希釈して0.01MのBikKam1(PepScan、オランダ)存在下で増殖させた。増殖中、細胞試料を1時間間隔で採取した。細胞試料をPBSで2回洗浄し、20uLのPBSで再構成してガラススライド上にスポットした。陰性対照として、BikKam1を添加せずに16時間増殖させた細菌を用いた。蛍光顕微鏡検査はLeitz Orthoplan顕微鏡で1000倍の倍率を用いて行った。
【0058】
結果
2時間のインキュベーション後、蛍光のBikKam1断片をB.セレウスの細胞質中で検出できた(図5B)。4時間時では、蛍光のBikKam1断片は細胞質中に加えて、新たに分裂した側の細胞壁(側壁)中にも存在していた(図5C)。最終的に16時間のインキュベーション後、全てのBikKam1断片がB.セレウスの細胞質から側壁に移動した(図5D)。細菌B.グロビギはBikKam1基質を切断できないことから、陰性対照として用いた。予想された通り、BikKam1存在下で培養したB.グロビギは蛍光細菌にならなかった。また0時間時およびBikKam1を添加せずに増殖させた試料において、蛍光細菌を検出できなかった(図5A)。これらの結果は前述のFRETアッセイ、細菌酵素の検出に加えて、BikKam1基質を用いると細菌の可視化も可能であることを示す。
【特許請求の範囲】
【請求項1】
微生物、より具体的にはB.アンスラシスを検出するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、アミノ酸X1およびX2、またはX1、X2およびX3からなるジまたはトリペプチドに連結した一組の分子マーカーを含み、前記アミノ酸の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい、基質。
【請求項2】
D-アミノ酸が、中性アミノ酸、好ましくはDLeu、DVal、DIle、DGlyまたはDAla、より好ましくはDLeu、DValまたはDIleである、請求項1に記載の基質。
【請求項3】
Lアミノ酸1つまたはLアミノ酸2つのうち1つがLeuである、請求項1または2に記載の基質。
【請求項4】
前記一組の分子マーカーが、蛍光標識、好ましくはFITC(フルオレセイン-5-イソチオシアネート)および前記蛍光標識の消光物質、好ましくはDABCYL(4-((-4-(ジメチルアミノ)-フェニル)-アゾ)-安息香酸)を含む、請求項1から3のいずれか一項に記載のバチルス・アンスラシスを蛍光検出するための基質。
【請求項5】
リンカーがアミノヘキソン酸(FITC用)およびリジン(DABCYL用)である、請求項4に記載の基質。
【請求項6】
FITC-Ahx-X1-X2-X3-Lys-DABCYL(ここでX1、X2およびX3は請求項1に定義されており、X3は存在しなくてもよく、Lysはリジンを表し、Ahxはアミノヘキソン酸を表す)である、請求項5に記載の基質。
【請求項7】
次の表1からなる群から選択される、請求項1から6のいずれか一項に記載の基質。
【表1】
【請求項8】
前記一組の分子マーカーが末端アセチル化システインを2つ含み、リンカーが存在しない、請求項1から3のいずれか一項に記載のバチルス・アンスラシスを検出するための基質。
【請求項9】
AcNH-Cys(S-Ac)-X1-X2-X3-Cys(S-Ac)-OH(ここでX1、X2およびX3は請求項3に定義されており、X3は存在しなくてもよく、Cys(S-Ac)はチオールアセチル化システインを表し、AcNHはアセチル化アミノ末端を表す)である、請求項8に記載の基質。
【請求項10】
次の表2からなる群から選択される、請求項8または9に記載の基質。
【表2】
【請求項11】
試料中の微生物、特にバチルス・アンスラシスを検出するための方法であって、請求項1から10のいずれか一項に記載の基質を試料に添加する段階と、蛍光または色のシフトを検出する段階とを含む方法。
【請求項12】
蛍光シグナルが微生物細胞において細胞内に位置する、請求項11に記載の方法。
【請求項13】
試料が、体液、粉末、水、食品、培地または任意の他の生物学的マトリックスから選択される、請求項11または12に記載の方法。
【請求項14】
試料中のバチルス・アンスラシスPXO1+を同定するための方法であって、請求項1から10のいずれか一項に記載の基質を試料に添加する段階と、蛍光、色のシフトもしくは特異的な質量スペクトルを検出するまたはキャピラリー電気泳動により検出を達成する段階とを含む方法。
【請求項15】
バチルス・アンスラシスを検出および診断するための、請求項1から10のいずれか一項に記載の基質の使用。
【請求項16】
微生物、より具体的にはP.エルギノーサを検出するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、グリシンアミノ酸からなるトリ、テトラまたはペンタペプチドに連結した一組の分子マーカーを含む、基質。
【請求項17】
前記一組の分子マーカーが、蛍光標識、好ましくはFITC(フルオレセイン-5-イソチオシアネート)および前記蛍光標識の消光物質、好ましくはDABCYL(4-((-4-(ジメチルアミノ)-フェニル)-アゾ)-安息香酸)を含む、請求項16に記載の基質。
【請求項18】
リンカーがアミノヘキソン酸(FITC用)およびリジン(DABCYL用)である、請求項17に記載の基質。
【請求項19】
次の表3からなる群から選択される、請求項16に記載の基質。
【表3】
【請求項20】
試料中の微生物、特にシュードモナス・エルギノーサを検出するための方法であって、請求項16から19のいずれか一項に記載の基質を試料に添加する段階と、蛍光または色のシフトを検出する段階とを含む方法。
【請求項21】
蛍光シグナルが微生物細胞において細胞内に位置する、請求項20に記載の方法。
【請求項22】
試料が、体液、粉末、水、食品、培地または任意の他の生物学的マトリックスから選択される、請求項20または21に記載の方法。
【請求項23】
シュードモナス・エルギノーサを検出および診断するための、請求項16から19のいずれか一項に記載の基質の使用。
【請求項1】
微生物、より具体的にはB.アンスラシスを検出するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、アミノ酸X1およびX2、またはX1、X2およびX3からなるジまたはトリペプチドに連結した一組の分子マーカーを含み、前記アミノ酸の1つ、例えばX1がD-アミノ酸であり、残りのアミノ酸、例えばX2およびX3が任意のD-またはL-アミノ酸であってもよい、基質。
【請求項2】
D-アミノ酸が、中性アミノ酸、好ましくはDLeu、DVal、DIle、DGlyまたはDAla、より好ましくはDLeu、DValまたはDIleである、請求項1に記載の基質。
【請求項3】
Lアミノ酸1つまたはLアミノ酸2つのうち1つがLeuである、請求項1または2に記載の基質。
【請求項4】
前記一組の分子マーカーが、蛍光標識、好ましくはFITC(フルオレセイン-5-イソチオシアネート)および前記蛍光標識の消光物質、好ましくはDABCYL(4-((-4-(ジメチルアミノ)-フェニル)-アゾ)-安息香酸)を含む、請求項1から3のいずれか一項に記載のバチルス・アンスラシスを蛍光検出するための基質。
【請求項5】
リンカーがアミノヘキソン酸(FITC用)およびリジン(DABCYL用)である、請求項4に記載の基質。
【請求項6】
FITC-Ahx-X1-X2-X3-Lys-DABCYL(ここでX1、X2およびX3は請求項1に定義されており、X3は存在しなくてもよく、Lysはリジンを表し、Ahxはアミノヘキソン酸を表す)である、請求項5に記載の基質。
【請求項7】
次の表1からなる群から選択される、請求項1から6のいずれか一項に記載の基質。
【表1】
【請求項8】
前記一組の分子マーカーが末端アセチル化システインを2つ含み、リンカーが存在しない、請求項1から3のいずれか一項に記載のバチルス・アンスラシスを検出するための基質。
【請求項9】
AcNH-Cys(S-Ac)-X1-X2-X3-Cys(S-Ac)-OH(ここでX1、X2およびX3は請求項3に定義されており、X3は存在しなくてもよく、Cys(S-Ac)はチオールアセチル化システインを表し、AcNHはアセチル化アミノ末端を表す)である、請求項8に記載の基質。
【請求項10】
次の表2からなる群から選択される、請求項8または9に記載の基質。
【表2】
【請求項11】
試料中の微生物、特にバチルス・アンスラシスを検出するための方法であって、請求項1から10のいずれか一項に記載の基質を試料に添加する段階と、蛍光または色のシフトを検出する段階とを含む方法。
【請求項12】
蛍光シグナルが微生物細胞において細胞内に位置する、請求項11に記載の方法。
【請求項13】
試料が、体液、粉末、水、食品、培地または任意の他の生物学的マトリックスから選択される、請求項11または12に記載の方法。
【請求項14】
試料中のバチルス・アンスラシスPXO1+を同定するための方法であって、請求項1から10のいずれか一項に記載の基質を試料に添加する段階と、蛍光、色のシフトもしくは特異的な質量スペクトルを検出するまたはキャピラリー電気泳動により検出を達成する段階とを含む方法。
【請求項15】
バチルス・アンスラシスを検出および診断するための、請求項1から10のいずれか一項に記載の基質の使用。
【請求項16】
微生物、より具体的にはP.エルギノーサを検出するための基質であって、任意選択でリンカー分子またはリンカー部分を用いて、グリシンアミノ酸からなるトリ、テトラまたはペンタペプチドに連結した一組の分子マーカーを含む、基質。
【請求項17】
前記一組の分子マーカーが、蛍光標識、好ましくはFITC(フルオレセイン-5-イソチオシアネート)および前記蛍光標識の消光物質、好ましくはDABCYL(4-((-4-(ジメチルアミノ)-フェニル)-アゾ)-安息香酸)を含む、請求項16に記載の基質。
【請求項18】
リンカーがアミノヘキソン酸(FITC用)およびリジン(DABCYL用)である、請求項17に記載の基質。
【請求項19】
次の表3からなる群から選択される、請求項16に記載の基質。
【表3】
【請求項20】
試料中の微生物、特にシュードモナス・エルギノーサを検出するための方法であって、請求項16から19のいずれか一項に記載の基質を試料に添加する段階と、蛍光または色のシフトを検出する段階とを含む方法。
【請求項21】
蛍光シグナルが微生物細胞において細胞内に位置する、請求項20に記載の方法。
【請求項22】
試料が、体液、粉末、水、食品、培地または任意の他の生物学的マトリックスから選択される、請求項20または21に記載の方法。
【請求項23】
シュードモナス・エルギノーサを検出および診断するための、請求項16から19のいずれか一項に記載の基質の使用。
【図1A】

【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図1I】
【図1J】
【図2】
【図3AB】
【図3CD】
【図3EF】
【図3GH】
【図3IJ】
【図4A】
【図4B】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図5】

【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図1I】
【図1J】
【図2】
【図3AB】
【図3CD】
【図3EF】
【図3GH】
【図3IJ】
【図4A】
【図4B】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図5】
【公表番号】特表2012−509079(P2012−509079A)
【公表日】平成24年4月19日(2012.4.19)
【国際特許分類】
【出願番号】特願2011−537382(P2011−537382)
【出願日】平成21年11月19日(2009.11.19)
【国際出願番号】PCT/NL2009/050702
【国際公開番号】WO2010/059051
【国際公開日】平成22年5月27日(2010.5.27)
【出願人】(511095850)ネーデルランドセ・オルガニサティ・フォール・トゥーヘパスト−ナトゥールウェテンスハッペライク・オンデルズーク・テーエヌオー (16)
【Fターム(参考)】
【公表日】平成24年4月19日(2012.4.19)
【国際特許分類】
【出願日】平成21年11月19日(2009.11.19)
【国際出願番号】PCT/NL2009/050702
【国際公開番号】WO2010/059051
【国際公開日】平成22年5月27日(2010.5.27)
【出願人】(511095850)ネーデルランドセ・オルガニサティ・フォール・トゥーヘパスト−ナトゥールウェテンスハッペライク・オンデルズーク・テーエヌオー (16)
【Fターム(参考)】
[ Back to top ]