
FRETバイオセンサーを利用したリガンドの検出方法
本発明に係わる方法は、リガンド結合蛋白質を利用する多様な種類のFRETバイオセンサーに幅広く適用できる。本発明は、FRET(fluorescence resonance energy transfer、蛍光共鳴エネルギー転移)現象を適用したバイオセンサーを利用したリガンドの検出方法に関し、より具体的にはバイオセンサーを構成するリガンド結合蛋白質(ligand-binding protein)が特定の臨界温度(critical temperature)以上で可逆的なアンフォールディング(reversible unfolding)が現れ、このアンフォールディングのレベルがリガンドの濃度により変わる現象を利用して、前記臨界温度を維持させる条件でバイオセンサーのFRETを測定して、試料中のリガンドを簡単に検出する方法である。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、FRET現象を適用したバイオセンサーを利用したリガンドの検出方法に関し、より具体的にはバイオセンサーを構成するリガンド結合蛋白質が特定の臨界温度(critical temperature)以上で可逆的なアンフォールディング(reversible unfolding)が現れ、このアンフォールディングのレベルがリガンドの濃度により変わる現象を利用して、従来より優秀な効率でリガンド(特に糖)を検出すると共にその濃度を測定する方法に関する。
【背景技術】
【0002】
2002年スタンフォード大学のFrommerは、マルトース測定用FRETバイオセンサーを最初に開発した(Fehr et al., PNAS., 99:9846-9851, 2002)。その後、類似の形態のリボース(Lager et al., FEBS Lett., 553:85, 2003)、グルコース(Fehr et al., J. Biol. Chem., 278:19127-19133, 2003)、スクロース(Ha et al., Appl. Environmicrobiol., 73:7408, 2007)測定用センサー等が持続的に開発されて来た。
【0003】
しかし、初期に開発された前記バイオセンサーは、非常に低いレベルの検出能力を示したため、より正確な測定手段として用いるためには高感度センサーの開発が求められた(Fehr et al. Current Opinion in Plant Biology, 7:345, 2004)。このような努力の一環として、本発明者等は韓国登録特許10−0739529(2007年7月29日)と米国登録特許US7432353(2008年10月7日)では、バイオセンサーを構成している蛋白質ドメインの間のリンカーペプチドを最適化する方法で、マルトース測定用FRETバイオセンサーの検出能力を増加できると提示した。また、理化学研究所のMiyawakiは、蛍光蛋白質を循環置換(circular permutation)させる方法でカルシウム測定用FRETバイオセンサーである「Cameleon」の検出能力を大きく増加させ(Nagai et al., PNAS., 101:10554, 2004)、FrommerはQBP(glutamine-binding protein)に蛍光蛋白質を挿入融合(in-frame fusion)する方法等でFRETバイオセンサーの検出能力を増加させた(Deuschle et al.,Protein Sci., 14:2304, 2006)。しかし、遺伝子操作によるFRETバイオセンサーの改良方法は、多くの試行錯誤と少なくない時間を要し、既に技術的な限界に達しているため、より効率的、かつ簡単な改良方式が求められている。
【0004】
一方、温度変化による蛋白質構造の可逆的変化(reversible change)と安全性(thermal stability)維持を観察する研究は、蛋白質の3次元構造が解明されて以来、蛋白質の折り畳み(folding)過程を予測するための研究と共に、多くの研究者の主な関心事になり、大腸菌由来のPBP(periplasmic-binding protein)は、このような蛋白質構造の熱力学的変化(thermodynamic change)を観察する良いモデルとなって来た。温度によるARBP(arabinose-binding protein)の構造変化をDSC(differential scanning calorimeter)を利用して観察した報告によれば、アラビノースがない条件では、53.5℃で可逆的なアンフォールディングが生じ、1mMのアラビノースが存在する場合には、59℃でアンフォールディングが観察された(Fukuda et al., J. Biol. Chem., 258:13193. 1983)。また、GGBP(glucose/galactose-binding protein)の場合にもグルコースの有無により温度によるアンフォールディング現象が50℃から63℃に増加するという報告があり(Piszczek et al., Biochem. J., 381:97, 2004)、MBPの場合にもマルトースが存在する場合にはpHによりアンフォールディングが発生する温度を8〜15℃まで増加させるという報告があった(Novokhatny et al., Protein Sci., 6:141, 1997)。
【0005】
そこで、本発明者等は、従来のFRETバイオセンサーのリガンド濃度測定能及び検出能を向上させようと鋭意努力した結果、可逆的なアンフォールディング現象が起こる特定臨界温度において融合蛋白質で構成されたバイオセンサーとリガンドを接触させる場合、リガンドを検出して濃度を測定する能力が画期的に改善されることを確認し、本発明を完成することに至った。
【発明の概要】
【0006】
本発明の目的は、従来のFRETバイオセンサーのリガンド検出能及び濃度測定能が改善された新しいリガンドの検出及び濃度測定方法を提供することである。
【0007】
前記目的を達成するために、本発明は、蛍光供与体(fluorescence donor)及び蛍光受容体(fluorescence acceptor)を含む信号発生部(signaling domain)と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部(sensing domain)を含むFRETバイオセンサーを利用したリガンドの検出方法であって、可逆的なアンフォールディング現象が起こり、リガンド結合(リガンドが前記リガンド結合蛋白質に結合すること)によるFRET比(ratio)の変化が最も大きい温度区間である臨界温度でリガンドを含有する試料と接触させることを特徴とするリガンドの検出方法を提供する。
【0008】
さらに本発明は、次の工程を含む、蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンド濃度の測定方法を提供する:
(a)可逆的なアンフォールディングが起こり、リガンドが前記リガンド結合蛋白質に結合することによるFRET比の変化が最も大きい温度区間である臨界温度で、前記FRETバイオセンサーを、リガンドを含有する試料と接触させる工程、及び
(b)前記蛍光供与体と蛍光受容体の発光量比の変化を測定してリガンドの濃度を測定する工程。
本発明の他の特徴及び具現例は、以下の詳細な説明及び添付された特許請求の範囲からより一層明白になる。
【図面の簡単な説明】
【0009】
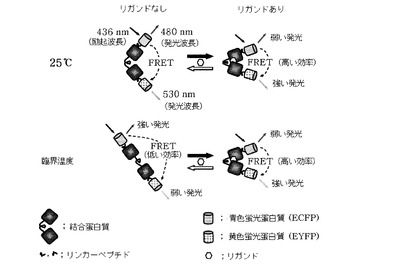
【図1】常温(25℃)と臨界温度でリガンドの有無によるFRETバイオセンサーの構造変化とFRET効率の変化、それによる蛍光蛋白質の発光量差を示した模式図である。
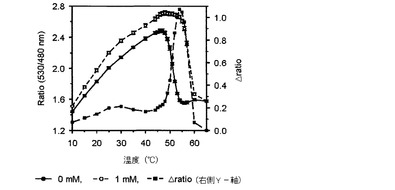
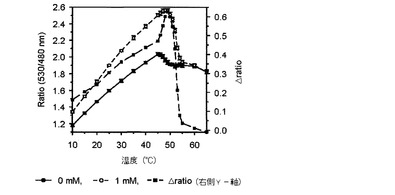
【図2】温度によるマルトースFRETバイオセンサーのratio値とΔratioの変化を示したグラフである。
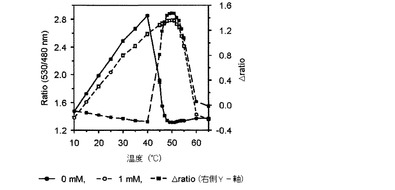
【図3】温度によるグルコースFRETバイオセンサーのratio値とΔratioの変化を示したグラフである。
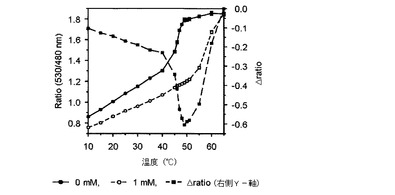
【図4】温度によるアロースFRETバイオセンサーのratio値とΔratioの変化を示したグラフである
【図5】温度によるアラビノースFRETバイオセンサーのratio値とΔratioの変化を示したグラフである。
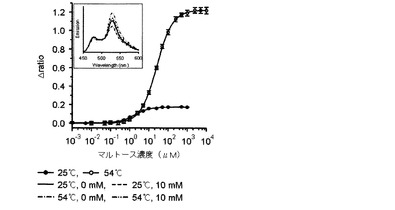
【図6】25℃とΔratio値が最も大きい臨界温度である54℃でリガンドの濃度毎に測定したマルトースFRETバイオセンサーのタイトレーションカーブであり、上段部のスペクトラムは25℃と54℃で各々測定したマルトースFRETバイオセンサーの蛍光スペクトラムである。
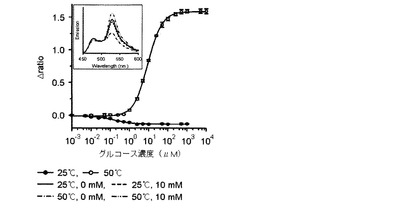
【図7】25℃とΔratio値が最も大きい臨界温度である50℃でリガンドの濃度毎に測定したグルコースFRETバイオセンサーのタイトレーションカーブであり、上段部のスペクトラムは25℃と50℃で各々測定したグルコースFRETバイオセンサーの蛍光スペクトラムである。
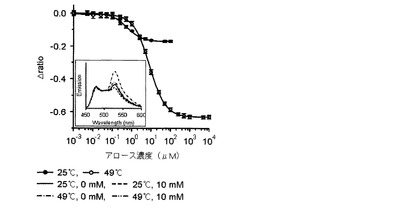
【図8】25℃とΔratio値が最も大きい臨界温度である49℃でリガンドの濃度毎に測定したアロースFRETバイオセンサーのタイトレーションカーブであり、上段部のスペクトラムは25℃と49℃で各々測定したアロースFRETバイオセンサーの蛍光スペクトラムである。
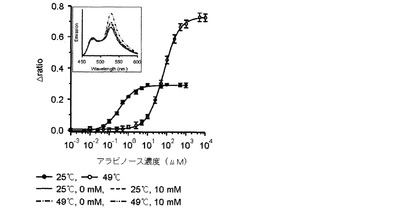
【図9】25℃とΔratio値が最も大きい臨界温度である49℃でリガンドの濃度毎に測定したアラビノースFRETバイオセンサーのタイトレーションカーブである。挿入図は25℃と49℃で各々測定したアラビノースFRETバイオセンサーの蛍光スペクトラムである。
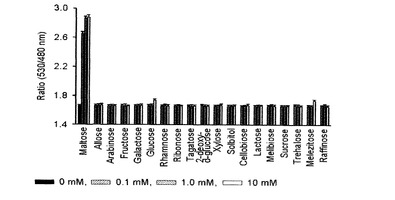
【図10】種々の糖類を対象にマルトースFRETバイオセンサーの特異性を調べたグラフである。
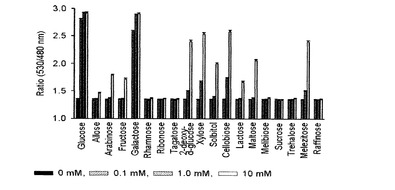
【図11】種々の糖類を対象にグルコースFRETバイオセンサーの特異性を調べたグラフである。
【図12】種々の糖類を対象にアロースFRETバイオセンサーの特異性を調べたグラフである。
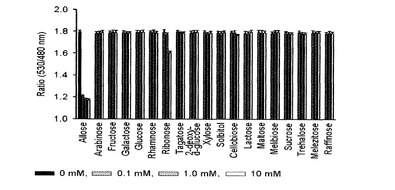
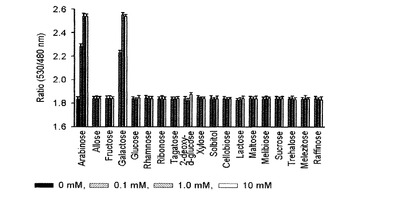
【図13】種々の糖類を対象にアラビノースFRETバイオセンサーの特異性を調べたグラフである。
【発明を実施するための形態】
【0010】
他の方式で定義されない限り、本明細書において使用されたあらゆる技術的・科学的用語は、本発明が属する技術分野に熟練した専門家によって通常理解されるものと同じ意味を有する。通常、本明細書において使用された命名法は、本技術分野において周知であり、しかも汎用されるものである。
本発明の詳細な説明などにおいて使用される主な用語の定義は、下記の通りである。
【0011】
本明細書において「FRET(fluorescence resonance energy transfer)」とは、互いに異なる発光波長帯の二つの蛍光物質の間から発生する非放射性(non-radiative)エネルギー移転現象で、励起(excitation)状態の蛍光供与体(donor)の励起準位エネルギーが蛍光受容体(acceptor)に伝えられて、蛍光受容体から発光(emission)が観察されたり、蛍光供与体の蛍光減少(quenching)が観察される現象である(Lakowicz, J. R. Principles of Fluorescence Spectroscopy, 2nd ed., New York:Plenum Press, 1999)。
【0012】
本明細書において「蛍光供与体」とは、FRET現象で供与体として働く蛍光物質を意味し、「蛍光受容体」とは、FRET現象で受容体として働く蛍光物質を意味する。
【0013】
本明細書において「リガンド結合蛋白質(リガンド−binding protein)」とは、リガンドの結合によって構造的変化(conformational change)を起こす蛋白質の集合体を意味しており、大腸菌由来の細胞膜間結合蛋白質(periplasmic binding protein,PBP)を含む(de Wolf et al., Pharmacol Rev., 52:207, 2000)。
【0014】
本明細書において「リガンド(ligand)」とはリガンド結合蛋白質に結合して構造的な変化を起こす分子で、糖、アミノ酸、蛋白質、脂質、有機酸、金属または金属イオン、酸化物、水酸化物またはそのコンジュゲート(conjugates)、無機イオン、アミンまたはポリアミン及びビタミンのいずれか一つであるが、これに限定されるのではない。
【0015】
本明細書において「試料(sample)」とは、関心があるリガンドを含有したり、含有していると推定されて、分析が行われる組成物を意味し、細胞、水、土壌、空気、食品、廃棄物、動植物器官及び動植物組織のいずれか一つ以上から収集されたものであってもよいが、これに限定されるのではない。この時、前記動植物は人体を含む。
【0016】
本明細書において「臨界温度」とは、FRETバイオセンサーのリガンド結合蛋白質のアンフォールディング(unfolding)が、リガンドの存在の有無によって調節されて、FRETバイオセンサーの検出能及び測定能が向上する温度区間、即ちリガンド結合蛋白質にリガンドの結合の有無によるFRET比の変化が最も大きい温度区間をいう。本発明の実施例3及び4で確認されるようにPBPで構成されたFRETバイオセンサーの場合、49〜54℃の温度区間が検出能及び測定能が向上する「臨界温度」といえる。
【0017】
本発明は一観点において、蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンドの検出方法であって、可逆的なアンフォールディング現象が起こり、リガンド結合(リガンドが前記リガンド結合蛋白質に結合すること)によるFRET比の変化が最も大きい温度区間である臨界温度でリガンドを含有する試料と接触させることを特徴とするリガンドの検出方法に関する。
【0018】
試料中のリガンドの検出は、蛍光供与体及び蛍光受容体の発光量を蛍光分析システム等で測定することによって行われ、蛍光分析システムとしてはフィルター方式及びモノクローム方式の蛍光分光器等を利用することができる。この時、試料中リガンドが存在する場合、前記蛍光供与体と蛍光受容体の発光量変化が感知されるが、これによりリガンドを検出できるようになる。
【0019】
本発明はまた、他の観点において、次の工程を含む、蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンド濃度の測定方法に関する:
(a)可逆的なアンフォールディングが起こり、リガンドが前記リガンド結合蛋白質に結合することによるFRET比の変化が最も大きい温度区間である臨界温度で、前記FRETバイオセンサーをリガンドを含有する試料と接触させる工程、及び
(b)前記蛍光供与体と蛍光受容体の発光量比の変化を測定してリガンドの濃度を測定する工程。
【0020】
前記蛍光供与体及び蛍光受容体の発光量は、蛍光分析システム等で測定され、リガンドの濃度変化が発生する場合、前記二つの蛍光供与体と蛍光受容体の発光量に変化が発生するが、そこで本発明はリガンドの濃度変化測定のために利用される。
【0021】
本発明において、FRETバイオセンサーを構成する融合蛋白質は、信号発生部として蛍光供与体及び蛍光受容体を含み、感知部としてリガンド結合蛋白質を含む形態で、前記蛍光供与体と蛍光受容体は、リガンド結合蛋白質の両末端に結合できる。この時、蛍光供与体または蛍光受容体は、一つ以上のリンカーを利用して前記リガンド結合蛋白質に連結できる。
【0022】
前記リガンド結合蛋白質は、望ましくは本発明の実施例で用いられたMBP(maltose-binding protein)、ALBP(allose-binding protein)、ARBP及びGGBP(galactose/glucose-binding protein)等、大腸菌由来のPBPであることを特徴とするが、リガンドの結合によって構造的変化を起こすリガンド結合蛋白質ならば、これに限定されず本発明による方法及びセンサーで提供できることは明らかである。
【0023】
また、前記バイオセンサーの信号発生部で使われる蛍光供与体及び蛍光受容体の構成は、蛍光供与体の発光スペクトラムと蛍光受容体の吸光スペクトラムが互いに重なってFRETまたは蛍光減少を誘発できるものであれば、いずれのものでもよく、これに前記蛍光供与体として多様な波長の蛍光蛋白質(fluorescent protein)と蛍光顔料(fluorescent dye)、生体発光蛋白質(bioluminescent protein)及び量子ドット(quantum dot)等が利用でき、前記蛍光受容体として前記蛍光供与体と波長が異なる蛍光蛋白質、蛍光顔料及び量子ドット等が用いられる。または、蛍光受容体として前記蛍光供与体の蛍光強度を減少させる消光剤(quencher)と金ナノ粒子(Au-nano particle)等が用いられる。但し、この中でもFRETバイオセンサーを構成する場合、蛍光供与体と蛍光受容体の吸光計数(extinction coefficient)と量子効率(quantum efficiency)、光安全性(photostability)と利用の便宜等を考慮して、蛍光蛋白質であるECFP(enhanced cyan fluorescent protein)とEYFP(enhanced yellow fluorescent protein)を用いることが望ましい。
【0024】
本発明によるリガンドの検出及び濃度測定方法は、蛍光の光学的特性である「FRET」を利用しており、その原理は図1に示されている。FRETは、一般に蛍光供与体から放出される波長が蛍光受容体の吸光スペクトラムと重なり、光子(photon)が出現せず発生するため、共鳴エネルギー転移と呼ばれ、これは蛍光供与体と蛍光受容体との間の長距離双極子相互作用による結果である。FRETのエネルギー転移効率は、蛍光供与体の発光スペクトラムと蛍光受容体の吸光スペクトラムが重なる範囲と蛍光供与体の量子効率、蛍光供与体と蛍光受容体の転移双極子(transition dipoles)の相対方向(relative orientation)、及び蛍光供与体と蛍光受容体との間の距離により変わる。従って、FRETのエネルギー転移効率は、蛍光供与体と蛍光受容体の距離と相対方向により異なるが、Forsterの式によると次のように表現される。
【0025】
[数1]
E=R06/(R6+R06) (1)
前記式(1)において、EはFRET効率を示し、Rは蛍光供与体と蛍光受容体との間の距離であり、蛍光物質により差はあるが通常2〜9nm以内で定義される。また、R0はFRET効率が50%になる蛍光供与体と蛍光受容体との間の距離であり、一般にForster distanceまたはForster radiusと呼ばれる。R0は次の式に表現される。
【0026】
[数2]
R0=0.211[k2n−4QDJ(λ)]1/6 (inÅ) (2)
前記式(2)において、k2は方向係数(orientation factor)であり、通常2/3で計算し、蛍光供与体発光と蛍光受容体吸光の相対方向により0〜4の範囲の値を有する。nは、媒質の屈折率であり、通常25℃の水は〜1.334であり、QDは、蛍光供与体の量子効率である。J(λ)は、蛍光供与体の発光と蛍光受容体の吸光スペクトラム上の重複(overlap)程度であり、M−1cm−1nm4の単位値を有する(Lakowicz, J. R. Principles of Fluorescence Spectroscopy, 2nd ed., New York:Plenum Press, 1999、Patterson et al., Anal. Biochem. 284:438, 2000、 Patterson et al., J. of Cell Sci. 114:837, 2001)。
【0027】
そこで、前記説明したFRETの原理を利用して、本発明者等は韓国登録特許10−0739529号(2007年7月29日)と米国登録特許US7432353号(2008年10月7日)でFRETの蛍光供与体と蛍光受容体として働く蛍光蛋白質であるECFPとEYFPをリガンド結合蛋白質であるPBPの両末端に融合させて、FRETバイオセンサーを構成し、これを利用して各センサーに結合可能なリガンドのアロースとアラビノース、リボース、マルトースを定量的に検出できることを示した。
【0028】
前記FRETバイオセンサーは、ECFP−PBP−EYFPが一つのポリペプチドで構成されて巨大な融合蛋白質として発現され、PBPの概略的な大きさが3×4×6.5nm(Spurlino et al., J. Biol. Chem., 266:5202, 1991)であることを考慮すると、ECFPとEYFPとの間の距離が、概ね5〜6nm程度に位置するようになるため、FRETの発生が可能な距離となる。従って、436nmでECFPを励起させると、ECFPの励起準位エネルギーが、EYFPに伝えられて、ECFPとEYFPの発光を同時に観察することができる(図1参照)。前記FRETバイオセンサーのリガンド結合部位に糖が結合すると、PBPの両末端に融合したECFPとEYFPとの距離と相対方向が変わるようになって、その結果FRET効率の差が生じるため、二つの蛍光蛋白質の発光量比が変わるようになる。従って、二つの蛍光蛋白質の発光量変化を測定することによってリガンドの感知が可能であるが、発光量比の変化は糖濃度に比例するため、定量的な糖濃度の測定が可能になる。
【0029】
それだけではなく、前記式(2)の計算によると、ECFPとEYFPは概ね5nm程度のR0値を有するため(Patterson et al., Anal. Biochem., 284:438, 2000)、ECFPとEYFPとの間の距離が概ね5〜6nm程度と仮定すると、距離または相対的方向の小さい変化が、FRET効率には大きい差を生じさせる。従って、本発明者等はリガンド結合の有無によるFRET効率の差を最大化できると、バイオセンサーの検出能力が大きく向上すると予想した。そこで、本発明では検出能力を最大化するために研究した結果、大腸菌由来のPBPが温度上昇による可逆的なアンフォールディング現象を示し、このような現象はリガンドが存在するとより高い温度でアンフォールディング現象が観察されるとの研究結果に着眼して、従来FRET原理を利用した方法に比べて、リガンド検出能力が増加したリガンドの検出及び濃度測定方法を提供した。
【0030】
即ち、本発明の一実施例では温度変化によるFRETバイオセンサーの蛍光分析を介して、45〜65℃の温度範囲でリガンドの有無によるバイオセンサーの蛍光比が大きく変わることを確認し、より詳しくは49℃〜54℃の温度範囲でセンサーの検出能力である△ratio値が増加することを確認して「臨界温度」区間が存在することを確認した(図2〜図5)。また、本発明が属する技術分野において通常の知識を有する者なら、本発明の臨界温度が結合蛋白質の種類による安全性レベル及び反応液の造成により多様な範囲で変化する可能性があることを理解できる。即ち、低温性または好熱性微生物由来のリガンド結合蛋白質またはこれの基質特異性を改良した結合蛋白質を利用したバイオセンサーは、本発明の臨界温度範囲(49℃〜54℃)より高いか低い温度範囲で、より向上した検出能力を有することもある。特に、蛋白質の構造的堅固性(rigidity)に影響を及ぼす物質、即ち、酸(acid)、塩基(base)、還元剤(reducing agent)、変成剤(denaturant,chaotropic agent)、安定剤(stabilizer)、界面活性剤(surfactant)、乳化剤(emulsifier)、または不活性化剤(detergent)の有無によって臨界温度範囲を調節可能であることは、本発明で期待する通常の知識範囲に属する。
本発明の他の実施形態では、FRETバイオセンサーの検出能力が、25℃で測定した場合より臨界温度において最小2.5倍から最大12倍程度増加することを確認し(図6〜図9)、各センサーの基質に対する特異性も、米国登録特許US7432353号で提示した結果より向上することを確認することができた(図12、図13)。
【実施例】
【0031】
以下、本発明を実施例を挙げて詳述する。これらの実施例は単に本発明をより具体的に説明するためのものであり、本発明の範囲がこれらの実施例に制限されないことは当業界において通常の知識を持った者にとって自明である。
《実施例1:FRETバイオセンサーの製造》
1−1:FRETバイオセンサーのための融合蛋白質製造のための発現ベクターの構築
まず次の式(I)で表される蛋白質を含有するバイオセンサーを提供するために次のように発現ベクターを構築した:
【化1】
ここで、リガンド結合蛋白質であるPBPは、ALBP、ARBP、MBP及びGGBPからなる群から選択され、L1及びL2は各々FP1のC−末端とPBPのN−末端との間、PBPのC−末端とFP2のN−末端との間を連結する2個のアミノ酸で構成されたリンカーペプチドであり、FP1とFP2は、FRETの蛍光供与体と蛍光受容体で、各々ECFPとEYFPで構成される。
【0032】
本発明で用いられたアロースとアラビノース、マルトースを定量的に測定できるFRETバイオセンサーは、本発明の出願人によって先出願された韓国登録特許10−0739529号及び米国登録特許US7432353号で提示されたセンサーと同じであり、詳細には、次の方法によって構築した。
まず、マルトースを定量的に測定するためのマルトースバイオセンサーであるCMY−BIIの発現ベクターは次の方法によって構築した。
【0033】
配列番号1:5’−gatcggatccatggtgagcaagggcgag−3'
配列番号2:5'−gatcaagcttgtacagctcgtccatgc−3'
配列番号3:5'−gatcatatggtgagcaagggcgag−3'
配列番号4:5'−tttaccttcttcgattttcattcgcgacttgtacagctcgtccatgcc−3'
配列番号5:5'−atgaaaatcgaagaaggtaaac−3'
配列番号6:5'−gatcggatcccgagctcgaattagtctg−3'
【0034】
まず、EYFPの遺伝子は、pEYFP−N1ベクター(Clontech,Palo Alto,CA)を鋳型として、各々BamHIとHindIII切断塩基配列が導入された配列番号1と2のプライマーをもちいてPCRを行った。増幅されたEYFP遺伝子は、BamHIとHindIII制限酵素で切断した後、発現ベクターであるpET−21a(Novagen,Madison,WI)の制限酵素認知部位に挿入して、EYFPのC末端に6×His−tagが発現できるベクターであるpEYFP−IIIを構成した。ECFP遺伝子は、pECFPベクター(Clontech,Palo Alto,CA)を鋳型として、NdeIの制限酵素切断塩基配列が導入された配列番号3と、MBPのN末端塩基配列が重なるように作製した配列番号4のプライマーを用いてPCRを行った。同様にMBPの遺伝子は、pMALc2x(NEB,Beverly,MA,USA)を鋳型として、配列番号5と、BamHI制限酵素切断塩基配列が導入された配列番号6のプライマーを用いてPCRを行った。このように増幅された各々のECFPとMBPの遺伝子は互いに重なるように作製されたプライマーによって、重複伸長重合酵素連鎖反応(overlap-extension PCR)が可能であるため、配列番号3と配列番号6のプライマーを利用して、反応液に同量のECFPとMBP遺伝子を添加した後、PCRを行ってECFP−MBP形態の合成遺伝子を得ることができた。前記増幅された合成遺伝子は、NdeIとBamHI制限酵素で切断し、MBP−EYFPの発現ベクターであるpEYFP−IIIの制限酵素認知部位にクローニングする方法で発現ベクターであるpECMY−BIIベクターを構築し、前記方法で構成されたマルトース測定用FRETバイオセンサーをCMY−BIIと命名した。
【0035】
また、アロース測定用バイオセンサーであるCalsBY−QVの発現ベクターを構築するためのECFP遺伝子の増幅は、塩基配列3及び7のプライマーを用いており、ALBP遺伝子は大腸菌MG1655から抽出した染色体遺伝子(chromosomal DNA)を鋳型として、配列番号8と9のプライマーを用いて増幅した。
配列番号7:5'−gacagcatattcggcggccattacttgcttgtacagctcgtccatgc−3'
配列番号8:5'−atggccgccgaatatgctgt−3'
配列番号9:5'−cgcggatcccgattgagtgaccaggatt−3'
【0036】
増幅されたECFP及びALBPの遺伝子は、配列番号3と9のプライマーを利用してECFP−ALBPの合成遺伝子に増幅した。前記ECFP−ALBP遺伝子は、NdeI、BamHI制限酵素認知部位を利用して、pECMY−BII発現ベクターからECFP−MBP遺伝子を取り除いて挿入する方法で、アロース測定用バイオセンサーの発現のためのpECalsBY−QVベクターを構築した。
【0037】
同様に、アラビノース測定用バイオセンサーであるCaraFY−PRの発現ベクターを構築するためのECFP遺伝子の増幅は、塩基配列3と10のプライマーを用いており、ARBPの遺伝子は、大腸菌MG1655から抽出した染色体遺伝子を鋳型として、配列番号11と12のプライマーを用いて増幅した。
配列番号10:5'−ccgagcttcaggttctccatcctaggcttgtacagctcgtccatgc−3'
配列番号11:5'−atggagaacctgaagctcg−3'
配列番号12:5'−cgcggatcccgacttaccgcctaaacctt−3'
【0038】
増幅されたECFPとARBPの遺伝子は、配列番号3と12のプライマーを利用してECFP−ARBPの合成遺伝子で増幅し、pECMY−BII発現ベクターのECFP−MBP遺伝子位置に挿入する方法でpECaraFY−PRを構築した。
また、本発明で用いられたグルコース測定用バイオセンサーであるCmglBY−SSの発現ベクターは下の方法で構築した。
配列番号13:5'−caccaatgcgagtatcagccatcgaagacttgtacagctcgtccatgcc−3'
配列番号14:5'−atggctgatactcgcattggtg−3'
配列番号15:5'−cgcggatcccgatttcttgctgaattcagc−3'
【0039】
まず、ECFP遺伝子はpECFPベクターを鋳型として、配列番号3と、GGBPのN末端塩基配列が重なるように作製した配列番号13の逆方向プライマーを用いてPCRを行った。同様に、GGBPの遺伝子は、大腸菌MG1655から抽出した染色体遺伝子を鋳型として、配列番号14と、BamHI制限酵素切断塩基配列が導入された配列番号15の逆方向プライマーを用いてPCRを行った。前記増幅したECFPとGGBP遺伝子を反応液に共に添加した後、配列番号3と配列番号15のプライマーでPCRを行ってECFP−GGBPの合成遺伝子が得られた。前記増幅されたECFP−GGBP合成遺伝子は、NdeIとBamHI制限酵素で切断し、前記pECMYB−IIのNdeIとBamHI制限酵素認知部位を利用してECFP−MBP遺伝子を取り除いて挿入する方法でpECmglBY−SSベクターを構築し、前記方法で構成されたグルコース測定用FRETバイオセンサーをCmglBY−SSと命名した。
【0040】
1−2:FRETバイオセンサーの製造、純粋分離
実施例1−1で構築したpECalsBY−QV、pECaraFY−PR、pECMY−BII及びpECmglBY−SSが、各々JM109(DE3)に形質転換された大腸菌を50μg/mLのアンピシリンが添加されたLB培地(1%bacto−trypton、0.5%yeast extract、1%NaCl)に接種して、37℃で12時間震盪培養した。
【0041】
前記培養させた大腸菌は、50μg/mLのアンピシリンが添加された1LのLB培地に1%になるよう接種して、37℃で約2時間程度培養させ、O.D.600nmでの吸光度が0.5に到達した時点に、IPTG(isopropyl β-d-thiogalactopyranoside)を0.5mMになるよう添加して、25℃で24時間蛋白質の発現を誘導した。
【0042】
培養を終えた菌株は、6000rpmの速度で遠心分離機(Supra22K,Hanil,Korea)を利用して回収し、20mMリン酸塩緩衝液(pH7.5)に懸濁させ超音波粉砕機で細胞膜を破壊させた。さらに、溶解した菌株は高速遠心分離機で15000rpmで沈殿物を取り除き、上層液だけを0.2μmフィルターでろ過して、以後の精製過程に用いた。
【0043】
蛋白質の精製は、FRETバイオセンサーのC−末端に発現した6×His−tagを利用してFPLC(fast-performance liquid chromatography)に連結した親和性クロマトグラフィーカラムHisTrap(商品名)HP(GE Healthcare,Uppsala,Sweden)を利用して、一次ろ過を行い、陰イオン交換クロマトグラフィーカラムHiTrap(商品名)Q HP(GE Healthcare,Uppsala,Sweden)を利用して、二次精製を行った。精製を終えたFRETセンサーは、20%グリセロールが含まれたPBS緩衝液(pH7.4)に10mg/mLの濃度に濃縮して、−70℃で保管し、下記実施例で用いた。
【0044】
《実施例2:FRETバイオセンサーの蛍光分析方法》
FRETバイオセンサーの蛍光は、各センサー蛋白質を0.5mLのPBS緩衝液(pH7.4)に0.5μMの濃度で同一に調節した条件で、蛍光分析システムのCary Eclipse(Varian Inc.,Mulgrave,Australia)を用いて測定し、436nmで励起させて発生する発光スペクトラムを450nmから600nmまでスキャンして確認した。また、FRET効率を示す指標としては、480nmでのECFP発光とFRETによって発生するEYFPの530nmでの発光量(emission intensity)の比を下記の式(3)に代入した値であるFRET ratioと定義した。
[数3]
ratio=(530nm/480nm) (3)
ratio:EYFPとECFPの発光量比
530nm:FRETによって測定されるEYFPの発光量
480nm:励起光を436nmにした時測定されるECFPの発光量
また、FRETバイオセンサーの検出能力を定義する△ratioは下記の式(4)により決めた。
[数4]
△ratio=ratiomax−ratiomin (4)
△ratio:リガンドの有無によるratioの最大差
ratiomax:リガンドが存在する条件で測定されるratioの比
ratiomin:リガンドがない条件で測定されるratioの比
【0045】
FRETバイオセンサーのリガンドに対するタイトレーションカーブは、リガンドの濃度を1nM〜10mMまで増加させながら測定したratio変化をSigmaplot 10.0(Systat software Inc.,USA)のHill equation、4−parameter方式を適用して、S字形態の曲線(Sigmoidal curve)で表現し、各センサーに対するリガンドの解離常数(dissociation constant)のKdは、S字状の曲線において△ratioが1/2値を示すリガンドの濃度と定めた。また、各センサーを利用して、定量的に測定可能なリガンドの濃度範囲は、△ratio値が10%から90%飽和された範囲以内のリガンド濃度と定義した。
【0046】
《実施例3:FRETバイオセンサーの温度変化による蛍光分析》
温度変化によるFRETバイオセンサーの蛍光分析は、各センサーを0.5mLのPBS緩衝液(pH7.4)に0.5μMの濃度に調節して、リガンドがない条件と飽和濃度の1mMのリガンドが添加された条件でのFRET ratioを測定して、比較分析した。FRETバイオセンサー0.5mLに添加するリガンドの体積は、センサーの過度な希釈を防止するために全体積の1/100に該当する5μLに制限した。温度変化は、10〜65℃区間まで5℃間隔で測定し、ratioの変化が著しい区間である45〜60℃間では1℃間隔で温度を変化させながら測定した。全ての実験群は水分蒸発を防止するために栓が閉じられた蛍光キューベット(cuvette)に入っている状態で実験を行い、キューベットは温度調節装置が連結したペルチエ素子(peltier device)に位置させた状態で目標温度に到達してから3分経過後、蛍光を測定した。
【0047】
その結果、図2乃至図5に示したように、前記実施例1−2で精製したFRETバイオセンサーのリガンドの有無によるFRET ratioの変化が最も大きい臨界温度は、マルトースセンサーは54℃(図2)、グルコースセンサーは50℃(図3)、アロースとアラビノースセンサーは49℃(図4、図5)で若干の差があり、臨界温度±1℃ではFRET ratioの変化が大きくなく、安定した結果を示すことが確認された。
【0048】
また、前記臨界温度でのリガンドに対する各FRETバイオセンサーのタイトレーションカーブを分析した結果、図6乃至9に示したように、マルトースセンサーは1.22(図6)、グルコースセンサーは1.6(図7)、アロースセンサーは−0.62(図8)、アラビノースセンサーは0.72(図9)の△ratio値を示すことが確認された。
【0049】
前記測定された各FRETバイオセンサーの△ratio値を既に25℃で測定した数値と比較すると、マルトースセンサーは7倍(図6)、グルコースセンサーは12倍(図7)、アロースセンサーは3倍(図8)、アラビノースセンサーは2.5倍(図9)以上△ratio値が増加した数値となり、この結果は特定臨界温度で多様な種類のFRETバイオセンサーの検出能力が大きく改善されるという本出願人の主張を直接的に確認する結果である。
前記結果は下記の表1にまとめられた。
【0050】
【表1】
【0051】
一方、各FRETバイオセンサーで測定可能なリガンドの濃度範囲は、生理的(physiological)に有意な濃度範囲と確認され、例えば、血糖濃度を測定するのに重要な血液のグルコースの濃度は70〜200mg/dLであり、これは概ね400〜1100μMに該当する。従って、グルコースセンサーを利用して、血糖濃度を測定する場合には、血液をセンサー体積の1/100となるよう添加するため、4〜11μMの濃度範囲に該当するように、前記濃度範囲はグルコースセンサーのKd値が9.2±0.5μMであるため、最も正確に測定できる濃度区間となる。
【0052】
《実施例4:FRETバイオセンサーのリガンドに対する特異性分析》
FRETバイオセンサーの多様なリガンドに対する特異性分析は、19種の単糖類と多糖類、及び糖アルコール等種々の糖類を対象に測定した。
測定方法は、精製されたFRETバイオセンサーを0.5mLのPBS緩衝液(pH7.4)に0.5μMの濃度に調節して、各リガンドの濃度を100μMと1mM、10mM等になるよう添加して、キューベットを囲むペルチエ素子が前記実施例3で求めた臨界温度に到達した時から3分経過後に蛍光を測定した。
その結果、図10及び12に示したように、マルトースとアロース測定用FRETバイオセンサーは、他の種類の糖とは特異的な結合をしないと確認された。また、25℃で測定した、本発明出願人によって先出願された米国登録特許US7432353号の結果と比較して見ると、アロースFRETバイオセンサーは、高濃度のリボースに対する特異性が減少することが確認できた。
【0053】
一方、アラビノースとグルコースFRETセンサーは、既存の研究結果と同様にガラクトースに非常に親和力が高いことが確認され(Vyas et al., J. Biol. Chem. 266, 5226-5237, 1991、 Fehr et al., J. Biol. Chem. 278, 19127-19133, 2003)、特にグルコースセンサーの場合には、ガラクトース以外にも高濃度で存在する多数の糖類と低いレベルの親和力があると確認された(図11)。しかし、血液に存在する糖類はグルコースを除いて微量に存在するため、グルコースセンサーを血糖測定用に用いても、その他糖類による誤差が大きくないものと判断され、最近の研究によると、ガラクトースに対する親和力が大きく減少したGGBPが報告されたため(Sakaguchi-Mikami et al., Biotechnol. Lett. 30:1453-1460, 2008)、グルコース測定用FRETバイオセンサーの特異性を改善できる余地も十分にある。
【産業上の利用の可能性】
【0054】
以上、説明したように本発明による方法は、バイオセンサーを構成しているリガンド結合蛋白質が、特定の臨界温度以上で可逆的なアンフォールディングが起こり、このアンフォールディングのレベルがリガンド濃度により変わる現象に基づく技術として、前記臨界温度で蛍光を測定する方法でFRETバイオセンサーの検出能力を画期的に増加でき、リガンド結合蛋白質と蛍光蛋白質を利用した全ての種類のFRETバイオセンサーに幅広く適用できるため、活用範囲が広範囲である。
【0055】
以上、本発明の内容の特定の部分を詳述したが、当業界における通常の知識を持った者にとって、このような具体的な記述は単なる好適な実施態様に過ぎず、これにより本発明の範囲が制限されることはないという点は明らかである。よって、本発明の実質的な範囲は特許請求の範囲とこれらの等価物により定義されると言える。
【技術分野】
【0001】
本発明は、FRET現象を適用したバイオセンサーを利用したリガンドの検出方法に関し、より具体的にはバイオセンサーを構成するリガンド結合蛋白質が特定の臨界温度(critical temperature)以上で可逆的なアンフォールディング(reversible unfolding)が現れ、このアンフォールディングのレベルがリガンドの濃度により変わる現象を利用して、従来より優秀な効率でリガンド(特に糖)を検出すると共にその濃度を測定する方法に関する。
【背景技術】
【0002】
2002年スタンフォード大学のFrommerは、マルトース測定用FRETバイオセンサーを最初に開発した(Fehr et al., PNAS., 99:9846-9851, 2002)。その後、類似の形態のリボース(Lager et al., FEBS Lett., 553:85, 2003)、グルコース(Fehr et al., J. Biol. Chem., 278:19127-19133, 2003)、スクロース(Ha et al., Appl. Environmicrobiol., 73:7408, 2007)測定用センサー等が持続的に開発されて来た。
【0003】
しかし、初期に開発された前記バイオセンサーは、非常に低いレベルの検出能力を示したため、より正確な測定手段として用いるためには高感度センサーの開発が求められた(Fehr et al. Current Opinion in Plant Biology, 7:345, 2004)。このような努力の一環として、本発明者等は韓国登録特許10−0739529(2007年7月29日)と米国登録特許US7432353(2008年10月7日)では、バイオセンサーを構成している蛋白質ドメインの間のリンカーペプチドを最適化する方法で、マルトース測定用FRETバイオセンサーの検出能力を増加できると提示した。また、理化学研究所のMiyawakiは、蛍光蛋白質を循環置換(circular permutation)させる方法でカルシウム測定用FRETバイオセンサーである「Cameleon」の検出能力を大きく増加させ(Nagai et al., PNAS., 101:10554, 2004)、FrommerはQBP(glutamine-binding protein)に蛍光蛋白質を挿入融合(in-frame fusion)する方法等でFRETバイオセンサーの検出能力を増加させた(Deuschle et al.,Protein Sci., 14:2304, 2006)。しかし、遺伝子操作によるFRETバイオセンサーの改良方法は、多くの試行錯誤と少なくない時間を要し、既に技術的な限界に達しているため、より効率的、かつ簡単な改良方式が求められている。
【0004】
一方、温度変化による蛋白質構造の可逆的変化(reversible change)と安全性(thermal stability)維持を観察する研究は、蛋白質の3次元構造が解明されて以来、蛋白質の折り畳み(folding)過程を予測するための研究と共に、多くの研究者の主な関心事になり、大腸菌由来のPBP(periplasmic-binding protein)は、このような蛋白質構造の熱力学的変化(thermodynamic change)を観察する良いモデルとなって来た。温度によるARBP(arabinose-binding protein)の構造変化をDSC(differential scanning calorimeter)を利用して観察した報告によれば、アラビノースがない条件では、53.5℃で可逆的なアンフォールディングが生じ、1mMのアラビノースが存在する場合には、59℃でアンフォールディングが観察された(Fukuda et al., J. Biol. Chem., 258:13193. 1983)。また、GGBP(glucose/galactose-binding protein)の場合にもグルコースの有無により温度によるアンフォールディング現象が50℃から63℃に増加するという報告があり(Piszczek et al., Biochem. J., 381:97, 2004)、MBPの場合にもマルトースが存在する場合にはpHによりアンフォールディングが発生する温度を8〜15℃まで増加させるという報告があった(Novokhatny et al., Protein Sci., 6:141, 1997)。
【0005】
そこで、本発明者等は、従来のFRETバイオセンサーのリガンド濃度測定能及び検出能を向上させようと鋭意努力した結果、可逆的なアンフォールディング現象が起こる特定臨界温度において融合蛋白質で構成されたバイオセンサーとリガンドを接触させる場合、リガンドを検出して濃度を測定する能力が画期的に改善されることを確認し、本発明を完成することに至った。
【発明の概要】
【0006】
本発明の目的は、従来のFRETバイオセンサーのリガンド検出能及び濃度測定能が改善された新しいリガンドの検出及び濃度測定方法を提供することである。
【0007】
前記目的を達成するために、本発明は、蛍光供与体(fluorescence donor)及び蛍光受容体(fluorescence acceptor)を含む信号発生部(signaling domain)と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部(sensing domain)を含むFRETバイオセンサーを利用したリガンドの検出方法であって、可逆的なアンフォールディング現象が起こり、リガンド結合(リガンドが前記リガンド結合蛋白質に結合すること)によるFRET比(ratio)の変化が最も大きい温度区間である臨界温度でリガンドを含有する試料と接触させることを特徴とするリガンドの検出方法を提供する。
【0008】
さらに本発明は、次の工程を含む、蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンド濃度の測定方法を提供する:
(a)可逆的なアンフォールディングが起こり、リガンドが前記リガンド結合蛋白質に結合することによるFRET比の変化が最も大きい温度区間である臨界温度で、前記FRETバイオセンサーを、リガンドを含有する試料と接触させる工程、及び
(b)前記蛍光供与体と蛍光受容体の発光量比の変化を測定してリガンドの濃度を測定する工程。
本発明の他の特徴及び具現例は、以下の詳細な説明及び添付された特許請求の範囲からより一層明白になる。
【図面の簡単な説明】
【0009】
【図1】常温(25℃)と臨界温度でリガンドの有無によるFRETバイオセンサーの構造変化とFRET効率の変化、それによる蛍光蛋白質の発光量差を示した模式図である。
【図2】温度によるマルトースFRETバイオセンサーのratio値とΔratioの変化を示したグラフである。
【図3】温度によるグルコースFRETバイオセンサーのratio値とΔratioの変化を示したグラフである。
【図4】温度によるアロースFRETバイオセンサーのratio値とΔratioの変化を示したグラフである
【図5】温度によるアラビノースFRETバイオセンサーのratio値とΔratioの変化を示したグラフである。
【図6】25℃とΔratio値が最も大きい臨界温度である54℃でリガンドの濃度毎に測定したマルトースFRETバイオセンサーのタイトレーションカーブであり、上段部のスペクトラムは25℃と54℃で各々測定したマルトースFRETバイオセンサーの蛍光スペクトラムである。
【図7】25℃とΔratio値が最も大きい臨界温度である50℃でリガンドの濃度毎に測定したグルコースFRETバイオセンサーのタイトレーションカーブであり、上段部のスペクトラムは25℃と50℃で各々測定したグルコースFRETバイオセンサーの蛍光スペクトラムである。
【図8】25℃とΔratio値が最も大きい臨界温度である49℃でリガンドの濃度毎に測定したアロースFRETバイオセンサーのタイトレーションカーブであり、上段部のスペクトラムは25℃と49℃で各々測定したアロースFRETバイオセンサーの蛍光スペクトラムである。
【図9】25℃とΔratio値が最も大きい臨界温度である49℃でリガンドの濃度毎に測定したアラビノースFRETバイオセンサーのタイトレーションカーブである。挿入図は25℃と49℃で各々測定したアラビノースFRETバイオセンサーの蛍光スペクトラムである。
【図10】種々の糖類を対象にマルトースFRETバイオセンサーの特異性を調べたグラフである。
【図11】種々の糖類を対象にグルコースFRETバイオセンサーの特異性を調べたグラフである。
【図12】種々の糖類を対象にアロースFRETバイオセンサーの特異性を調べたグラフである。
【図13】種々の糖類を対象にアラビノースFRETバイオセンサーの特異性を調べたグラフである。
【発明を実施するための形態】
【0010】
他の方式で定義されない限り、本明細書において使用されたあらゆる技術的・科学的用語は、本発明が属する技術分野に熟練した専門家によって通常理解されるものと同じ意味を有する。通常、本明細書において使用された命名法は、本技術分野において周知であり、しかも汎用されるものである。
本発明の詳細な説明などにおいて使用される主な用語の定義は、下記の通りである。
【0011】
本明細書において「FRET(fluorescence resonance energy transfer)」とは、互いに異なる発光波長帯の二つの蛍光物質の間から発生する非放射性(non-radiative)エネルギー移転現象で、励起(excitation)状態の蛍光供与体(donor)の励起準位エネルギーが蛍光受容体(acceptor)に伝えられて、蛍光受容体から発光(emission)が観察されたり、蛍光供与体の蛍光減少(quenching)が観察される現象である(Lakowicz, J. R. Principles of Fluorescence Spectroscopy, 2nd ed., New York:Plenum Press, 1999)。
【0012】
本明細書において「蛍光供与体」とは、FRET現象で供与体として働く蛍光物質を意味し、「蛍光受容体」とは、FRET現象で受容体として働く蛍光物質を意味する。
【0013】
本明細書において「リガンド結合蛋白質(リガンド−binding protein)」とは、リガンドの結合によって構造的変化(conformational change)を起こす蛋白質の集合体を意味しており、大腸菌由来の細胞膜間結合蛋白質(periplasmic binding protein,PBP)を含む(de Wolf et al., Pharmacol Rev., 52:207, 2000)。
【0014】
本明細書において「リガンド(ligand)」とはリガンド結合蛋白質に結合して構造的な変化を起こす分子で、糖、アミノ酸、蛋白質、脂質、有機酸、金属または金属イオン、酸化物、水酸化物またはそのコンジュゲート(conjugates)、無機イオン、アミンまたはポリアミン及びビタミンのいずれか一つであるが、これに限定されるのではない。
【0015】
本明細書において「試料(sample)」とは、関心があるリガンドを含有したり、含有していると推定されて、分析が行われる組成物を意味し、細胞、水、土壌、空気、食品、廃棄物、動植物器官及び動植物組織のいずれか一つ以上から収集されたものであってもよいが、これに限定されるのではない。この時、前記動植物は人体を含む。
【0016】
本明細書において「臨界温度」とは、FRETバイオセンサーのリガンド結合蛋白質のアンフォールディング(unfolding)が、リガンドの存在の有無によって調節されて、FRETバイオセンサーの検出能及び測定能が向上する温度区間、即ちリガンド結合蛋白質にリガンドの結合の有無によるFRET比の変化が最も大きい温度区間をいう。本発明の実施例3及び4で確認されるようにPBPで構成されたFRETバイオセンサーの場合、49〜54℃の温度区間が検出能及び測定能が向上する「臨界温度」といえる。
【0017】
本発明は一観点において、蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンドの検出方法であって、可逆的なアンフォールディング現象が起こり、リガンド結合(リガンドが前記リガンド結合蛋白質に結合すること)によるFRET比の変化が最も大きい温度区間である臨界温度でリガンドを含有する試料と接触させることを特徴とするリガンドの検出方法に関する。
【0018】
試料中のリガンドの検出は、蛍光供与体及び蛍光受容体の発光量を蛍光分析システム等で測定することによって行われ、蛍光分析システムとしてはフィルター方式及びモノクローム方式の蛍光分光器等を利用することができる。この時、試料中リガンドが存在する場合、前記蛍光供与体と蛍光受容体の発光量変化が感知されるが、これによりリガンドを検出できるようになる。
【0019】
本発明はまた、他の観点において、次の工程を含む、蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンド濃度の測定方法に関する:
(a)可逆的なアンフォールディングが起こり、リガンドが前記リガンド結合蛋白質に結合することによるFRET比の変化が最も大きい温度区間である臨界温度で、前記FRETバイオセンサーをリガンドを含有する試料と接触させる工程、及び
(b)前記蛍光供与体と蛍光受容体の発光量比の変化を測定してリガンドの濃度を測定する工程。
【0020】
前記蛍光供与体及び蛍光受容体の発光量は、蛍光分析システム等で測定され、リガンドの濃度変化が発生する場合、前記二つの蛍光供与体と蛍光受容体の発光量に変化が発生するが、そこで本発明はリガンドの濃度変化測定のために利用される。
【0021】
本発明において、FRETバイオセンサーを構成する融合蛋白質は、信号発生部として蛍光供与体及び蛍光受容体を含み、感知部としてリガンド結合蛋白質を含む形態で、前記蛍光供与体と蛍光受容体は、リガンド結合蛋白質の両末端に結合できる。この時、蛍光供与体または蛍光受容体は、一つ以上のリンカーを利用して前記リガンド結合蛋白質に連結できる。
【0022】
前記リガンド結合蛋白質は、望ましくは本発明の実施例で用いられたMBP(maltose-binding protein)、ALBP(allose-binding protein)、ARBP及びGGBP(galactose/glucose-binding protein)等、大腸菌由来のPBPであることを特徴とするが、リガンドの結合によって構造的変化を起こすリガンド結合蛋白質ならば、これに限定されず本発明による方法及びセンサーで提供できることは明らかである。
【0023】
また、前記バイオセンサーの信号発生部で使われる蛍光供与体及び蛍光受容体の構成は、蛍光供与体の発光スペクトラムと蛍光受容体の吸光スペクトラムが互いに重なってFRETまたは蛍光減少を誘発できるものであれば、いずれのものでもよく、これに前記蛍光供与体として多様な波長の蛍光蛋白質(fluorescent protein)と蛍光顔料(fluorescent dye)、生体発光蛋白質(bioluminescent protein)及び量子ドット(quantum dot)等が利用でき、前記蛍光受容体として前記蛍光供与体と波長が異なる蛍光蛋白質、蛍光顔料及び量子ドット等が用いられる。または、蛍光受容体として前記蛍光供与体の蛍光強度を減少させる消光剤(quencher)と金ナノ粒子(Au-nano particle)等が用いられる。但し、この中でもFRETバイオセンサーを構成する場合、蛍光供与体と蛍光受容体の吸光計数(extinction coefficient)と量子効率(quantum efficiency)、光安全性(photostability)と利用の便宜等を考慮して、蛍光蛋白質であるECFP(enhanced cyan fluorescent protein)とEYFP(enhanced yellow fluorescent protein)を用いることが望ましい。
【0024】
本発明によるリガンドの検出及び濃度測定方法は、蛍光の光学的特性である「FRET」を利用しており、その原理は図1に示されている。FRETは、一般に蛍光供与体から放出される波長が蛍光受容体の吸光スペクトラムと重なり、光子(photon)が出現せず発生するため、共鳴エネルギー転移と呼ばれ、これは蛍光供与体と蛍光受容体との間の長距離双極子相互作用による結果である。FRETのエネルギー転移効率は、蛍光供与体の発光スペクトラムと蛍光受容体の吸光スペクトラムが重なる範囲と蛍光供与体の量子効率、蛍光供与体と蛍光受容体の転移双極子(transition dipoles)の相対方向(relative orientation)、及び蛍光供与体と蛍光受容体との間の距離により変わる。従って、FRETのエネルギー転移効率は、蛍光供与体と蛍光受容体の距離と相対方向により異なるが、Forsterの式によると次のように表現される。
【0025】
[数1]
E=R06/(R6+R06) (1)
前記式(1)において、EはFRET効率を示し、Rは蛍光供与体と蛍光受容体との間の距離であり、蛍光物質により差はあるが通常2〜9nm以内で定義される。また、R0はFRET効率が50%になる蛍光供与体と蛍光受容体との間の距離であり、一般にForster distanceまたはForster radiusと呼ばれる。R0は次の式に表現される。
【0026】
[数2]
R0=0.211[k2n−4QDJ(λ)]1/6 (inÅ) (2)
前記式(2)において、k2は方向係数(orientation factor)であり、通常2/3で計算し、蛍光供与体発光と蛍光受容体吸光の相対方向により0〜4の範囲の値を有する。nは、媒質の屈折率であり、通常25℃の水は〜1.334であり、QDは、蛍光供与体の量子効率である。J(λ)は、蛍光供与体の発光と蛍光受容体の吸光スペクトラム上の重複(overlap)程度であり、M−1cm−1nm4の単位値を有する(Lakowicz, J. R. Principles of Fluorescence Spectroscopy, 2nd ed., New York:Plenum Press, 1999、Patterson et al., Anal. Biochem. 284:438, 2000、 Patterson et al., J. of Cell Sci. 114:837, 2001)。
【0027】
そこで、前記説明したFRETの原理を利用して、本発明者等は韓国登録特許10−0739529号(2007年7月29日)と米国登録特許US7432353号(2008年10月7日)でFRETの蛍光供与体と蛍光受容体として働く蛍光蛋白質であるECFPとEYFPをリガンド結合蛋白質であるPBPの両末端に融合させて、FRETバイオセンサーを構成し、これを利用して各センサーに結合可能なリガンドのアロースとアラビノース、リボース、マルトースを定量的に検出できることを示した。
【0028】
前記FRETバイオセンサーは、ECFP−PBP−EYFPが一つのポリペプチドで構成されて巨大な融合蛋白質として発現され、PBPの概略的な大きさが3×4×6.5nm(Spurlino et al., J. Biol. Chem., 266:5202, 1991)であることを考慮すると、ECFPとEYFPとの間の距離が、概ね5〜6nm程度に位置するようになるため、FRETの発生が可能な距離となる。従って、436nmでECFPを励起させると、ECFPの励起準位エネルギーが、EYFPに伝えられて、ECFPとEYFPの発光を同時に観察することができる(図1参照)。前記FRETバイオセンサーのリガンド結合部位に糖が結合すると、PBPの両末端に融合したECFPとEYFPとの距離と相対方向が変わるようになって、その結果FRET効率の差が生じるため、二つの蛍光蛋白質の発光量比が変わるようになる。従って、二つの蛍光蛋白質の発光量変化を測定することによってリガンドの感知が可能であるが、発光量比の変化は糖濃度に比例するため、定量的な糖濃度の測定が可能になる。
【0029】
それだけではなく、前記式(2)の計算によると、ECFPとEYFPは概ね5nm程度のR0値を有するため(Patterson et al., Anal. Biochem., 284:438, 2000)、ECFPとEYFPとの間の距離が概ね5〜6nm程度と仮定すると、距離または相対的方向の小さい変化が、FRET効率には大きい差を生じさせる。従って、本発明者等はリガンド結合の有無によるFRET効率の差を最大化できると、バイオセンサーの検出能力が大きく向上すると予想した。そこで、本発明では検出能力を最大化するために研究した結果、大腸菌由来のPBPが温度上昇による可逆的なアンフォールディング現象を示し、このような現象はリガンドが存在するとより高い温度でアンフォールディング現象が観察されるとの研究結果に着眼して、従来FRET原理を利用した方法に比べて、リガンド検出能力が増加したリガンドの検出及び濃度測定方法を提供した。
【0030】
即ち、本発明の一実施例では温度変化によるFRETバイオセンサーの蛍光分析を介して、45〜65℃の温度範囲でリガンドの有無によるバイオセンサーの蛍光比が大きく変わることを確認し、より詳しくは49℃〜54℃の温度範囲でセンサーの検出能力である△ratio値が増加することを確認して「臨界温度」区間が存在することを確認した(図2〜図5)。また、本発明が属する技術分野において通常の知識を有する者なら、本発明の臨界温度が結合蛋白質の種類による安全性レベル及び反応液の造成により多様な範囲で変化する可能性があることを理解できる。即ち、低温性または好熱性微生物由来のリガンド結合蛋白質またはこれの基質特異性を改良した結合蛋白質を利用したバイオセンサーは、本発明の臨界温度範囲(49℃〜54℃)より高いか低い温度範囲で、より向上した検出能力を有することもある。特に、蛋白質の構造的堅固性(rigidity)に影響を及ぼす物質、即ち、酸(acid)、塩基(base)、還元剤(reducing agent)、変成剤(denaturant,chaotropic agent)、安定剤(stabilizer)、界面活性剤(surfactant)、乳化剤(emulsifier)、または不活性化剤(detergent)の有無によって臨界温度範囲を調節可能であることは、本発明で期待する通常の知識範囲に属する。
本発明の他の実施形態では、FRETバイオセンサーの検出能力が、25℃で測定した場合より臨界温度において最小2.5倍から最大12倍程度増加することを確認し(図6〜図9)、各センサーの基質に対する特異性も、米国登録特許US7432353号で提示した結果より向上することを確認することができた(図12、図13)。
【実施例】
【0031】
以下、本発明を実施例を挙げて詳述する。これらの実施例は単に本発明をより具体的に説明するためのものであり、本発明の範囲がこれらの実施例に制限されないことは当業界において通常の知識を持った者にとって自明である。
《実施例1:FRETバイオセンサーの製造》
1−1:FRETバイオセンサーのための融合蛋白質製造のための発現ベクターの構築
まず次の式(I)で表される蛋白質を含有するバイオセンサーを提供するために次のように発現ベクターを構築した:
【化1】
ここで、リガンド結合蛋白質であるPBPは、ALBP、ARBP、MBP及びGGBPからなる群から選択され、L1及びL2は各々FP1のC−末端とPBPのN−末端との間、PBPのC−末端とFP2のN−末端との間を連結する2個のアミノ酸で構成されたリンカーペプチドであり、FP1とFP2は、FRETの蛍光供与体と蛍光受容体で、各々ECFPとEYFPで構成される。
【0032】
本発明で用いられたアロースとアラビノース、マルトースを定量的に測定できるFRETバイオセンサーは、本発明の出願人によって先出願された韓国登録特許10−0739529号及び米国登録特許US7432353号で提示されたセンサーと同じであり、詳細には、次の方法によって構築した。
まず、マルトースを定量的に測定するためのマルトースバイオセンサーであるCMY−BIIの発現ベクターは次の方法によって構築した。
【0033】
配列番号1:5’−gatcggatccatggtgagcaagggcgag−3'
配列番号2:5'−gatcaagcttgtacagctcgtccatgc−3'
配列番号3:5'−gatcatatggtgagcaagggcgag−3'
配列番号4:5'−tttaccttcttcgattttcattcgcgacttgtacagctcgtccatgcc−3'
配列番号5:5'−atgaaaatcgaagaaggtaaac−3'
配列番号6:5'−gatcggatcccgagctcgaattagtctg−3'
【0034】
まず、EYFPの遺伝子は、pEYFP−N1ベクター(Clontech,Palo Alto,CA)を鋳型として、各々BamHIとHindIII切断塩基配列が導入された配列番号1と2のプライマーをもちいてPCRを行った。増幅されたEYFP遺伝子は、BamHIとHindIII制限酵素で切断した後、発現ベクターであるpET−21a(Novagen,Madison,WI)の制限酵素認知部位に挿入して、EYFPのC末端に6×His−tagが発現できるベクターであるpEYFP−IIIを構成した。ECFP遺伝子は、pECFPベクター(Clontech,Palo Alto,CA)を鋳型として、NdeIの制限酵素切断塩基配列が導入された配列番号3と、MBPのN末端塩基配列が重なるように作製した配列番号4のプライマーを用いてPCRを行った。同様にMBPの遺伝子は、pMALc2x(NEB,Beverly,MA,USA)を鋳型として、配列番号5と、BamHI制限酵素切断塩基配列が導入された配列番号6のプライマーを用いてPCRを行った。このように増幅された各々のECFPとMBPの遺伝子は互いに重なるように作製されたプライマーによって、重複伸長重合酵素連鎖反応(overlap-extension PCR)が可能であるため、配列番号3と配列番号6のプライマーを利用して、反応液に同量のECFPとMBP遺伝子を添加した後、PCRを行ってECFP−MBP形態の合成遺伝子を得ることができた。前記増幅された合成遺伝子は、NdeIとBamHI制限酵素で切断し、MBP−EYFPの発現ベクターであるpEYFP−IIIの制限酵素認知部位にクローニングする方法で発現ベクターであるpECMY−BIIベクターを構築し、前記方法で構成されたマルトース測定用FRETバイオセンサーをCMY−BIIと命名した。
【0035】
また、アロース測定用バイオセンサーであるCalsBY−QVの発現ベクターを構築するためのECFP遺伝子の増幅は、塩基配列3及び7のプライマーを用いており、ALBP遺伝子は大腸菌MG1655から抽出した染色体遺伝子(chromosomal DNA)を鋳型として、配列番号8と9のプライマーを用いて増幅した。
配列番号7:5'−gacagcatattcggcggccattacttgcttgtacagctcgtccatgc−3'
配列番号8:5'−atggccgccgaatatgctgt−3'
配列番号9:5'−cgcggatcccgattgagtgaccaggatt−3'
【0036】
増幅されたECFP及びALBPの遺伝子は、配列番号3と9のプライマーを利用してECFP−ALBPの合成遺伝子に増幅した。前記ECFP−ALBP遺伝子は、NdeI、BamHI制限酵素認知部位を利用して、pECMY−BII発現ベクターからECFP−MBP遺伝子を取り除いて挿入する方法で、アロース測定用バイオセンサーの発現のためのpECalsBY−QVベクターを構築した。
【0037】
同様に、アラビノース測定用バイオセンサーであるCaraFY−PRの発現ベクターを構築するためのECFP遺伝子の増幅は、塩基配列3と10のプライマーを用いており、ARBPの遺伝子は、大腸菌MG1655から抽出した染色体遺伝子を鋳型として、配列番号11と12のプライマーを用いて増幅した。
配列番号10:5'−ccgagcttcaggttctccatcctaggcttgtacagctcgtccatgc−3'
配列番号11:5'−atggagaacctgaagctcg−3'
配列番号12:5'−cgcggatcccgacttaccgcctaaacctt−3'
【0038】
増幅されたECFPとARBPの遺伝子は、配列番号3と12のプライマーを利用してECFP−ARBPの合成遺伝子で増幅し、pECMY−BII発現ベクターのECFP−MBP遺伝子位置に挿入する方法でpECaraFY−PRを構築した。
また、本発明で用いられたグルコース測定用バイオセンサーであるCmglBY−SSの発現ベクターは下の方法で構築した。
配列番号13:5'−caccaatgcgagtatcagccatcgaagacttgtacagctcgtccatgcc−3'
配列番号14:5'−atggctgatactcgcattggtg−3'
配列番号15:5'−cgcggatcccgatttcttgctgaattcagc−3'
【0039】
まず、ECFP遺伝子はpECFPベクターを鋳型として、配列番号3と、GGBPのN末端塩基配列が重なるように作製した配列番号13の逆方向プライマーを用いてPCRを行った。同様に、GGBPの遺伝子は、大腸菌MG1655から抽出した染色体遺伝子を鋳型として、配列番号14と、BamHI制限酵素切断塩基配列が導入された配列番号15の逆方向プライマーを用いてPCRを行った。前記増幅したECFPとGGBP遺伝子を反応液に共に添加した後、配列番号3と配列番号15のプライマーでPCRを行ってECFP−GGBPの合成遺伝子が得られた。前記増幅されたECFP−GGBP合成遺伝子は、NdeIとBamHI制限酵素で切断し、前記pECMYB−IIのNdeIとBamHI制限酵素認知部位を利用してECFP−MBP遺伝子を取り除いて挿入する方法でpECmglBY−SSベクターを構築し、前記方法で構成されたグルコース測定用FRETバイオセンサーをCmglBY−SSと命名した。
【0040】
1−2:FRETバイオセンサーの製造、純粋分離
実施例1−1で構築したpECalsBY−QV、pECaraFY−PR、pECMY−BII及びpECmglBY−SSが、各々JM109(DE3)に形質転換された大腸菌を50μg/mLのアンピシリンが添加されたLB培地(1%bacto−trypton、0.5%yeast extract、1%NaCl)に接種して、37℃で12時間震盪培養した。
【0041】
前記培養させた大腸菌は、50μg/mLのアンピシリンが添加された1LのLB培地に1%になるよう接種して、37℃で約2時間程度培養させ、O.D.600nmでの吸光度が0.5に到達した時点に、IPTG(isopropyl β-d-thiogalactopyranoside)を0.5mMになるよう添加して、25℃で24時間蛋白質の発現を誘導した。
【0042】
培養を終えた菌株は、6000rpmの速度で遠心分離機(Supra22K,Hanil,Korea)を利用して回収し、20mMリン酸塩緩衝液(pH7.5)に懸濁させ超音波粉砕機で細胞膜を破壊させた。さらに、溶解した菌株は高速遠心分離機で15000rpmで沈殿物を取り除き、上層液だけを0.2μmフィルターでろ過して、以後の精製過程に用いた。
【0043】
蛋白質の精製は、FRETバイオセンサーのC−末端に発現した6×His−tagを利用してFPLC(fast-performance liquid chromatography)に連結した親和性クロマトグラフィーカラムHisTrap(商品名)HP(GE Healthcare,Uppsala,Sweden)を利用して、一次ろ過を行い、陰イオン交換クロマトグラフィーカラムHiTrap(商品名)Q HP(GE Healthcare,Uppsala,Sweden)を利用して、二次精製を行った。精製を終えたFRETセンサーは、20%グリセロールが含まれたPBS緩衝液(pH7.4)に10mg/mLの濃度に濃縮して、−70℃で保管し、下記実施例で用いた。
【0044】
《実施例2:FRETバイオセンサーの蛍光分析方法》
FRETバイオセンサーの蛍光は、各センサー蛋白質を0.5mLのPBS緩衝液(pH7.4)に0.5μMの濃度で同一に調節した条件で、蛍光分析システムのCary Eclipse(Varian Inc.,Mulgrave,Australia)を用いて測定し、436nmで励起させて発生する発光スペクトラムを450nmから600nmまでスキャンして確認した。また、FRET効率を示す指標としては、480nmでのECFP発光とFRETによって発生するEYFPの530nmでの発光量(emission intensity)の比を下記の式(3)に代入した値であるFRET ratioと定義した。
[数3]
ratio=(530nm/480nm) (3)
ratio:EYFPとECFPの発光量比
530nm:FRETによって測定されるEYFPの発光量
480nm:励起光を436nmにした時測定されるECFPの発光量
また、FRETバイオセンサーの検出能力を定義する△ratioは下記の式(4)により決めた。
[数4]
△ratio=ratiomax−ratiomin (4)
△ratio:リガンドの有無によるratioの最大差
ratiomax:リガンドが存在する条件で測定されるratioの比
ratiomin:リガンドがない条件で測定されるratioの比
【0045】
FRETバイオセンサーのリガンドに対するタイトレーションカーブは、リガンドの濃度を1nM〜10mMまで増加させながら測定したratio変化をSigmaplot 10.0(Systat software Inc.,USA)のHill equation、4−parameter方式を適用して、S字形態の曲線(Sigmoidal curve)で表現し、各センサーに対するリガンドの解離常数(dissociation constant)のKdは、S字状の曲線において△ratioが1/2値を示すリガンドの濃度と定めた。また、各センサーを利用して、定量的に測定可能なリガンドの濃度範囲は、△ratio値が10%から90%飽和された範囲以内のリガンド濃度と定義した。
【0046】
《実施例3:FRETバイオセンサーの温度変化による蛍光分析》
温度変化によるFRETバイオセンサーの蛍光分析は、各センサーを0.5mLのPBS緩衝液(pH7.4)に0.5μMの濃度に調節して、リガンドがない条件と飽和濃度の1mMのリガンドが添加された条件でのFRET ratioを測定して、比較分析した。FRETバイオセンサー0.5mLに添加するリガンドの体積は、センサーの過度な希釈を防止するために全体積の1/100に該当する5μLに制限した。温度変化は、10〜65℃区間まで5℃間隔で測定し、ratioの変化が著しい区間である45〜60℃間では1℃間隔で温度を変化させながら測定した。全ての実験群は水分蒸発を防止するために栓が閉じられた蛍光キューベット(cuvette)に入っている状態で実験を行い、キューベットは温度調節装置が連結したペルチエ素子(peltier device)に位置させた状態で目標温度に到達してから3分経過後、蛍光を測定した。
【0047】
その結果、図2乃至図5に示したように、前記実施例1−2で精製したFRETバイオセンサーのリガンドの有無によるFRET ratioの変化が最も大きい臨界温度は、マルトースセンサーは54℃(図2)、グルコースセンサーは50℃(図3)、アロースとアラビノースセンサーは49℃(図4、図5)で若干の差があり、臨界温度±1℃ではFRET ratioの変化が大きくなく、安定した結果を示すことが確認された。
【0048】
また、前記臨界温度でのリガンドに対する各FRETバイオセンサーのタイトレーションカーブを分析した結果、図6乃至9に示したように、マルトースセンサーは1.22(図6)、グルコースセンサーは1.6(図7)、アロースセンサーは−0.62(図8)、アラビノースセンサーは0.72(図9)の△ratio値を示すことが確認された。
【0049】
前記測定された各FRETバイオセンサーの△ratio値を既に25℃で測定した数値と比較すると、マルトースセンサーは7倍(図6)、グルコースセンサーは12倍(図7)、アロースセンサーは3倍(図8)、アラビノースセンサーは2.5倍(図9)以上△ratio値が増加した数値となり、この結果は特定臨界温度で多様な種類のFRETバイオセンサーの検出能力が大きく改善されるという本出願人の主張を直接的に確認する結果である。
前記結果は下記の表1にまとめられた。
【0050】
【表1】
【0051】
一方、各FRETバイオセンサーで測定可能なリガンドの濃度範囲は、生理的(physiological)に有意な濃度範囲と確認され、例えば、血糖濃度を測定するのに重要な血液のグルコースの濃度は70〜200mg/dLであり、これは概ね400〜1100μMに該当する。従って、グルコースセンサーを利用して、血糖濃度を測定する場合には、血液をセンサー体積の1/100となるよう添加するため、4〜11μMの濃度範囲に該当するように、前記濃度範囲はグルコースセンサーのKd値が9.2±0.5μMであるため、最も正確に測定できる濃度区間となる。
【0052】
《実施例4:FRETバイオセンサーのリガンドに対する特異性分析》
FRETバイオセンサーの多様なリガンドに対する特異性分析は、19種の単糖類と多糖類、及び糖アルコール等種々の糖類を対象に測定した。
測定方法は、精製されたFRETバイオセンサーを0.5mLのPBS緩衝液(pH7.4)に0.5μMの濃度に調節して、各リガンドの濃度を100μMと1mM、10mM等になるよう添加して、キューベットを囲むペルチエ素子が前記実施例3で求めた臨界温度に到達した時から3分経過後に蛍光を測定した。
その結果、図10及び12に示したように、マルトースとアロース測定用FRETバイオセンサーは、他の種類の糖とは特異的な結合をしないと確認された。また、25℃で測定した、本発明出願人によって先出願された米国登録特許US7432353号の結果と比較して見ると、アロースFRETバイオセンサーは、高濃度のリボースに対する特異性が減少することが確認できた。
【0053】
一方、アラビノースとグルコースFRETセンサーは、既存の研究結果と同様にガラクトースに非常に親和力が高いことが確認され(Vyas et al., J. Biol. Chem. 266, 5226-5237, 1991、 Fehr et al., J. Biol. Chem. 278, 19127-19133, 2003)、特にグルコースセンサーの場合には、ガラクトース以外にも高濃度で存在する多数の糖類と低いレベルの親和力があると確認された(図11)。しかし、血液に存在する糖類はグルコースを除いて微量に存在するため、グルコースセンサーを血糖測定用に用いても、その他糖類による誤差が大きくないものと判断され、最近の研究によると、ガラクトースに対する親和力が大きく減少したGGBPが報告されたため(Sakaguchi-Mikami et al., Biotechnol. Lett. 30:1453-1460, 2008)、グルコース測定用FRETバイオセンサーの特異性を改善できる余地も十分にある。
【産業上の利用の可能性】
【0054】
以上、説明したように本発明による方法は、バイオセンサーを構成しているリガンド結合蛋白質が、特定の臨界温度以上で可逆的なアンフォールディングが起こり、このアンフォールディングのレベルがリガンド濃度により変わる現象に基づく技術として、前記臨界温度で蛍光を測定する方法でFRETバイオセンサーの検出能力を画期的に増加でき、リガンド結合蛋白質と蛍光蛋白質を利用した全ての種類のFRETバイオセンサーに幅広く適用できるため、活用範囲が広範囲である。
【0055】
以上、本発明の内容の特定の部分を詳述したが、当業界における通常の知識を持った者にとって、このような具体的な記述は単なる好適な実施態様に過ぎず、これにより本発明の範囲が制限されることはないという点は明らかである。よって、本発明の実質的な範囲は特許請求の範囲とこれらの等価物により定義されると言える。
【特許請求の範囲】
【請求項1】
蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンドの検出方法であって、可逆的なアンフォールディングが起こり、リガンドの前記リガンド結合蛋白質への結合に伴うFRET比の変化が最も大きい温度区間である臨界温度でリガンドを含有する試料と接触させることを特徴とするリガンドの検出方法。
【請求項2】
次の工程を含む、蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンド濃度の測定方法:
(a)可逆的なアンフォールディングが起こり、リガンドの前記リガンド結合蛋白質への結合に伴うFRET比の変化が最も大きい温度区間である臨界温度で、前記FRETバイオセンサーをリガンドを含有する試料と接触させる工程、及び
(b)前記蛍光供与体と蛍光受容体の発光量比の変化を測定して、リガンドの濃度を測定する工程。
【請求項3】
前記臨界温度に変化を与えるために酸、塩基、還元剤、変成剤(chaotropic agent)、安定剤、界面活性剤、乳化剤及び不活性化剤のいずれか一つ以上を添加した後、変化した臨界温度でリガンドと接触させることを特徴とする請求項1または2に記載の方法。
【請求項4】
前記臨界温度に変化を与えるために前記リガンド結合蛋白質として低温性または好熱性微生物由来のPBPまたはそれの基質特異性を改良したPBPを含むFRETバイオセンサーを利用して変化した臨界温度でリガンドと接触させることを特徴とする請求項1または2に記載の方法。
【請求項5】
前記リガンドは、糖、アミノ酸、蛋白質、脂質、有機酸、金属または金属イオン、酸化物、水酸化物またはそのコンジュゲート、無機イオン、アミンまたはポリアミン及びビタミンからなる群から選択されることを特徴とする請求項1または2に記載の方法。
【請求項6】
前記蛍光供与体は、蛍光蛋白質、蛍光顔料、生体発光蛋白質及び量子ドットからなる群から選択され、前記蛍光受容体は、前記蛍光供与体と波長が異なる蛍光蛋白質、蛍光顔料及び量子ドットからなる群から選択されることを特徴とする請求項1または2に記載の方法。
【請求項7】
前記蛍光供与体は、蛍光蛋白質、蛍光顔料、生体発光蛋白質及び量子ドットからなる群から選択され、前記蛍光受容体は、前記蛍光供与体の蛍光強度を減少させる消光剤または金ナノ粒子であることを特徴とする請求項1または2に記載の方法。
【請求項8】
前記蛍光供与体または蛍光受容体は、一つ以上のリンカーを介して、前記リガンド結合蛋白質に連結することを特徴とする請求項1または2に記載の方法。
【請求項9】
前記リガンド結合蛋白質がPBPである場合、前記臨界温度が49〜54℃であることを特徴とする請求項1または2に記載の方法。
【請求項10】
前記リガンド結合蛋白質がMBPである場合、前記臨界温度が約54℃であることを特徴とする請求項1または2に記載の方法。
【請求項11】
前記リガンド結合蛋白質がGGBPである場合、前記臨界温度が約50℃であることを特徴とする請求項1または2に記載の方法。
【請求項12】
前記リガンド結合蛋白質がALBPまたはARBPである場合、前記臨界温度が約49℃であることを特徴とする請求項1または2に記載の方法。
【請求項1】
蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンドの検出方法であって、可逆的なアンフォールディングが起こり、リガンドの前記リガンド結合蛋白質への結合に伴うFRET比の変化が最も大きい温度区間である臨界温度でリガンドを含有する試料と接触させることを特徴とするリガンドの検出方法。
【請求項2】
次の工程を含む、蛍光供与体及び蛍光受容体を含む信号発生部と前記蛍光供与体及び蛍光受容体を連結するリガンド結合蛋白質を含む感知部を含むFRETバイオセンサーを利用したリガンド濃度の測定方法:
(a)可逆的なアンフォールディングが起こり、リガンドの前記リガンド結合蛋白質への結合に伴うFRET比の変化が最も大きい温度区間である臨界温度で、前記FRETバイオセンサーをリガンドを含有する試料と接触させる工程、及び
(b)前記蛍光供与体と蛍光受容体の発光量比の変化を測定して、リガンドの濃度を測定する工程。
【請求項3】
前記臨界温度に変化を与えるために酸、塩基、還元剤、変成剤(chaotropic agent)、安定剤、界面活性剤、乳化剤及び不活性化剤のいずれか一つ以上を添加した後、変化した臨界温度でリガンドと接触させることを特徴とする請求項1または2に記載の方法。
【請求項4】
前記臨界温度に変化を与えるために前記リガンド結合蛋白質として低温性または好熱性微生物由来のPBPまたはそれの基質特異性を改良したPBPを含むFRETバイオセンサーを利用して変化した臨界温度でリガンドと接触させることを特徴とする請求項1または2に記載の方法。
【請求項5】
前記リガンドは、糖、アミノ酸、蛋白質、脂質、有機酸、金属または金属イオン、酸化物、水酸化物またはそのコンジュゲート、無機イオン、アミンまたはポリアミン及びビタミンからなる群から選択されることを特徴とする請求項1または2に記載の方法。
【請求項6】
前記蛍光供与体は、蛍光蛋白質、蛍光顔料、生体発光蛋白質及び量子ドットからなる群から選択され、前記蛍光受容体は、前記蛍光供与体と波長が異なる蛍光蛋白質、蛍光顔料及び量子ドットからなる群から選択されることを特徴とする請求項1または2に記載の方法。
【請求項7】
前記蛍光供与体は、蛍光蛋白質、蛍光顔料、生体発光蛋白質及び量子ドットからなる群から選択され、前記蛍光受容体は、前記蛍光供与体の蛍光強度を減少させる消光剤または金ナノ粒子であることを特徴とする請求項1または2に記載の方法。
【請求項8】
前記蛍光供与体または蛍光受容体は、一つ以上のリンカーを介して、前記リガンド結合蛋白質に連結することを特徴とする請求項1または2に記載の方法。
【請求項9】
前記リガンド結合蛋白質がPBPである場合、前記臨界温度が49〜54℃であることを特徴とする請求項1または2に記載の方法。
【請求項10】
前記リガンド結合蛋白質がMBPである場合、前記臨界温度が約54℃であることを特徴とする請求項1または2に記載の方法。
【請求項11】
前記リガンド結合蛋白質がGGBPである場合、前記臨界温度が約50℃であることを特徴とする請求項1または2に記載の方法。
【請求項12】
前記リガンド結合蛋白質がALBPまたはARBPである場合、前記臨界温度が約49℃であることを特徴とする請求項1または2に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2012−525582(P2012−525582A)
【公表日】平成24年10月22日(2012.10.22)
【国際特許分類】
【出願番号】特願2012−508395(P2012−508395)
【出願日】平成22年4月27日(2010.4.27)
【国際出願番号】PCT/KR2010/002632
【国際公開番号】WO2010/126272
【国際公開日】平成22年11月4日(2010.11.4)
【出願人】(506272301)コリア リサーチ インスティテュート オブ バイオサイエンス アンド バイオテクノロジー (17)
【Fターム(参考)】
【公表日】平成24年10月22日(2012.10.22)
【国際特許分類】
【出願日】平成22年4月27日(2010.4.27)
【国際出願番号】PCT/KR2010/002632
【国際公開番号】WO2010/126272
【国際公開日】平成22年11月4日(2010.11.4)
【出願人】(506272301)コリア リサーチ インスティテュート オブ バイオサイエンス アンド バイオテクノロジー (17)
【Fターム(参考)】
[ Back to top ]