
Fasシグナルを介した幹細胞の分化制御方法
【課題】幹細胞の分化状態を制御する方法を提供する。
【解決手段】幹細胞のFasシグナルの活性を制御することを含む、幹細胞の分化制御方法。
【解決手段】幹細胞のFasシグナルの活性を制御することを含む、幹細胞の分化制御方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Fasシグナルを介して幹細胞の分化を制御する方法に関する。
【背景技術】
【0002】
胚性幹細胞(ES細胞)や誘導多能性幹細胞(iPS細胞)をはじめとする幹細胞は、生体に存在する様々な細胞に分化する能力を有し、細胞移植が有効な治療方法として期待される疾病、例えば、白血病、パーキンソン病等の治療に用いることが期待されている。
幹細胞を適切に維持・管理するためには、幹細胞の未分化性を維持した状態を保ちながら継代培養することが必要になる。したがって、幹細胞の未分化性を、確実にしかも簡便に維持するための手法の開発が望まれている。
一方、幹細胞を分化させる際には、より迅速に目的の分化状態へと方向付けすることが要求される。したがって、幹細胞の未分化性をより迅速に解除するための手法の開発も望まれている。
【0003】
糖鎖は、細胞の生存に必須であるばかりでなく、がん、免疫、発生・分化や感染症などにおいて特に重要な役割を果たしていることが報告されている。発生分化の過程においては、その分化程度や状態に応じて糖鎖を合成する糖転移酵素遺伝子の発現パターンが変化し、合成される糖鎖構造も変化することが知られている。しかし、糖鎖に硫酸基を付加する硫酸基転移酵素の活性が幹細胞の分化に与える影響についてはこれまで知られていない。
【0004】
Fasシグナルはアポトーシスを引き起こすシグナル経路として知られており、アポトーシスは細胞の分化に関与することが知られている。
しかし、幹細胞におけるFasシグナルと分化状態との関係についてこれまで報告はなく、また、幹細胞におけるFasシグナルと硫酸基転移酵素との関係についても知られていない。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、幹細胞の分化状態を制御する方法の提供を課題とする。
【課題を解決するための手段】
【0006】
本発明者らは、上記課題の解決のために鋭意検討を行った。その結果、Fasシグナルの活性を制御することにより、幹細胞の分化状態を制御できることを見い出した。また、本発明者らは、硫酸基転移酵素の活性を制御することでFasシグナルの活性を制御することができ、これにより幹細胞の分化状態をも制御できることを見い出した。本発明はこれらの知見に基づき完成するに至ったものである。
【0007】
本発明の課題は下記の手段により達成された。
[1]幹細胞のFasシグナルの活性を制御することを含む、幹細胞の分化制御方法。
[2]幹細胞中で硫酸基転移酵素の活性を制御し、これにより該幹細胞のFasシグナルの活性を制御する、[1]に記載の方法。
[3]硫酸基転移酵素がヘパラン硫酸硫酸基転移酵素である、[2]に記載の方法。
[4]ヘパラン硫酸硫酸基転移酵素がヘパラン硫酸3位硫酸基転移酵素である、[3]に記載の方法。
[5]ヘパラン硫酸3位硫酸基転移酵素が下記(A)〜(C)のいずれかのアミノ酸配列を有する、[4]に記載の方法:
(A)配列番号2のアミノ酸配列、
(B)配列番号2のアミノ酸配列において1もしくは数個のアミノ酸が置換、欠失、挿入又は付加されたアミノ酸配列、
(C)配列番号2のアミノ酸配列と80%以上の同一性を有するアミノ酸配列。
[6]幹細胞のFasシグナルの活性を抑制し、これにより幹細胞の未分化性を維持する、[1]〜[5]のいずれかに記載の方法。
[7]幹細胞のFasシグナルを活性化し、これにより幹細胞の未分化性を解除する、[1]〜[5]のいずれかに記載の方法。
[8]幹細胞中で硫酸基転移酵素の活性を抑制して該幹細胞のFasシグナルを抑制し、これにより幹細胞の未分化性を維持する、[6]に記載の方法。
[9]硫酸基転移酵素をコードする遺伝子の発現を抑制する核酸を幹細胞に導入し、該幹細胞中で該遺伝子の発現を抑制することで硫酸基転移酵素の活性を抑制する、[8]に記載の方法。
[10]幹細胞中で硫酸基転移酵素の活性を高めて該幹細胞のFasシグナルを活性化し、これにより幹細胞の未分化性を解除する、[7]に記載の方法。
[11]硫酸基転移酵素をコードする遺伝子を幹細胞に導入し、該幹細胞中で該遺伝子を過剰発現させることで硫酸基転移酵素の活性を高める、[10]に記載の方法。
[12]幹細胞にFasシグナル阻害剤を投与して該幹細胞のFasシグナルを抑制し、これにより幹細胞の未分化性を維持する、[6]に記載の方法。
[13]Fasシグナル阻害剤が、配列番号39のアミノ酸配列を有するペプチドを含む、[12]に記載の方法。
[14]幹細胞が哺乳類に由来する、[1]〜[13]のいずれかに記載の方法。
[15]幹細胞がES細胞である、[14]に記載の方法。
[16][1]〜[15]のいずれかに記載の方法で分化制御された細胞。
【発明の効果】
【0008】
本発明によれば、幹細胞を所望の分化状態に制御することができる。
【図面の簡単な説明】
【0009】
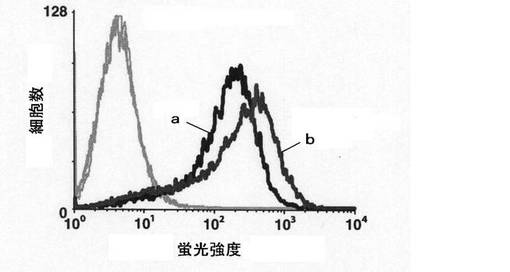
【図1】mHS3OST−5過剰発現細胞において、グルコサミン残基の3位に硫酸基が転移したHSの細胞表面における存在量をFACS分析により測定した結果を示す図である。図1中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
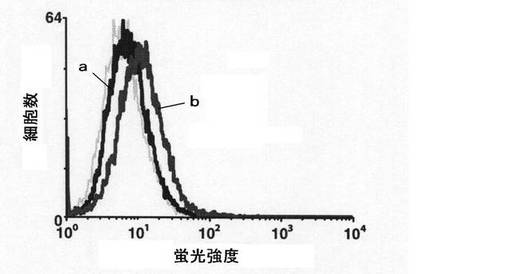
【図2】mHS3OST−5過剰発現細胞において、細胞表面に存在するFasの量をFACS分析により測定した結果を示す図である。図2中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図3】mHS3OST−5過剰発現細胞における脂質ラフト中のFasの存在量をイムノブロットにより調べた結果を示す写真である。図3中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。各画分の番号は実施例で使用した遠心管の最上層から数えた層の順番を示す。
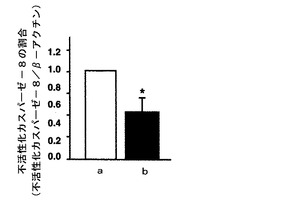
【図4】mHS3OST−5過剰発現細胞における不活性型カスパーゼ−8の量をイムノブロットにより調べた結果を示す図である。図4中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
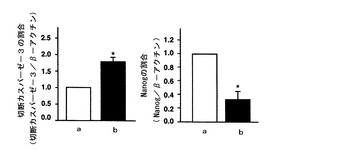
【図5】mHS3OST−5過剰発現細胞における切断カスパーゼ−3及びNanogの量をイムノブロットにより調べた結果を示す図である。図5中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
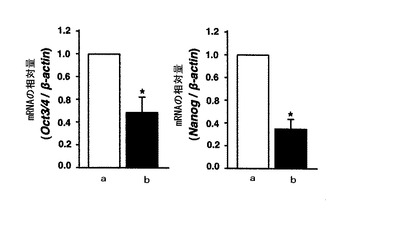
【図6】mHS3OST−5過剰発現細胞における未分化マーカー(Oct3/4及びNanog)のmRNA量をリアルタイムPCRにより調べた結果を示す図である。図6中、aはコントロール細胞の結果を示し、bはHS3OST−5過剰発現細胞の結果を示す。
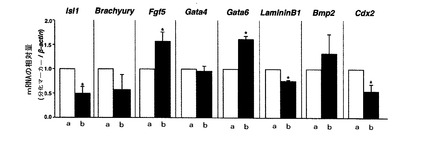
【図7】mHS3OST−5過剰発現細胞における分化マーカー(Isl1、Brachyury、Fgf5、Gata4、Gata6、LamininB1、Bmp2及びCdx2)のmRNA量をリアルタイムPCRにより調べた結果を示す図である。図7中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図8】mHS3OST−5過剰発現細胞におけるHSとFasとの結合機序をオーバーレイアッセイにより調べた結果を示す写真である。図7中、AはHS4C3(抗へパラン硫酸抗体:グルコサミン残基の3位が硫酸化された硫酸化構造を特異的に認識する抗体)を反応させた後にHRP標識抗マウスIgG抗体を用いて発色させた結果を示し、BはGST融合Fasを反応させた後にHRP標識抗GST抗体を用いて発色させた結果を示し、CはGST融合FasとHS4C3との混合物を反応させた後にHRP標識抗GST抗体を用いて発色させた結果を示し、DはGST融合変異導入Fasを反応させた後にHRP標識抗GST抗体を用いて発色させた結果を示す。Eはβ−アクチンの量をイムノブロットにより調べた結果を示す。また、図7中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
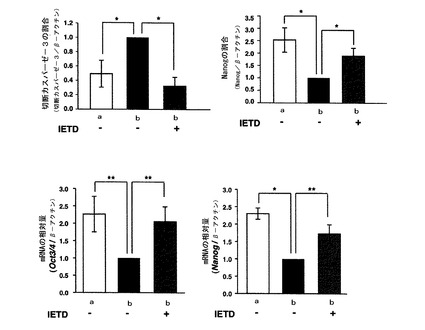
【図9】mHS3OST−5過剰発現細胞において、IETDの添加によりFasシグナルを阻害したときの分化状態の変化をマーカー発現量を指標に調べた結果を示す図である。図8中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図10】R1細胞の分化に対するFasシグナル阻害剤(IETD)の影響を調べた結果を示す図である。
【発明を実施するための形態】
【0010】
以下、本発明について、その好ましい実施態様に基づき詳細に説明する。
【0011】
本発明は、幹細胞中においてFasシグナルの活性を制御することにより該幹細胞の分化状態を制御する方法である。上記Fasシグナルの活性制御手段に特に制限はないが、幹細胞中の硫酸基転移酵素活性の制御や、幹細胞へのFasシグナル阻害剤の投与により行うことが好ましい。本発明において「分化状態を制御する」とは、未分化性を維持することを含む概念である。また、本発明において「Fas」とはFas受容体を意味する。
【0012】
本発明に用いる幹細胞に特に制限はなく、分化万能性、分化多能性、分化単一性のいずれの分化能を有する幹細胞であってもよいが、分化万能性を有する幹細胞であることが好ましい。本発明に用いる幹細胞の具体例として、例えば、胚性幹細胞(ES細胞)、誘導多能性幹細胞(iPS細胞)、胚性生殖幹細胞、核移植ES細胞、神経幹細胞、造血幹細胞、間葉系幹細胞、肝幹細胞、皮膚幹細胞、筋幹細胞等が挙げられるが、ES細胞又はiPS細胞であることが好ましく、ES細胞であることがより好ましい。上記幹細胞の由来に特に制限はなく、哺乳類、鳥類等に由来するものを用いることができるが、哺乳類に由来する幹細胞を好適に用いることができる。前記哺乳類としては、例えば、ヒト、サル、ウシ、ヤギ、ヒツジ、ブタ、ウマ、ウサギ、ラット、マウス、モルモット等が挙げられる。
上記幹細胞は通常の方法で調製することができる。また、寄託機関等に寄託されているセルライン等の分譲により得ることもできる。
【0013】
続いて、幹細胞のFasシグナルの活性を制御する手段について説明する。
【0014】
本発明において、幹細胞のFasシグナルは、幹細胞中で硫酸基転移酵素活性の制御することで行うことができる。
本発明における硫酸基転移酵素活性の制御とは、硫酸基転移酵素の活性を高めること及び抑制することの両概念を含む。
本発明において活性が制御されうる硫酸基転移酵素は、硫酸基供与体から糖鎖に硫酸基を転移する酵素である。上記硫酸基転移酵素は、その活性を高めることでFasシグナルを所望のレベルまで活性化させることができ、また、その活性を低下させるとFasシグナルを所望のレベルまで不活性化できる酵素である。上記硫酸基転移酵素は硫酸基供与体からヘパラン硫酸(HS)に硫酸基を転移する硫酸基転移酵素であることが好ましく、硫酸基供与体からヘパラン硫酸のグルコサミン残基の3位のヒドロキシル基に硫酸基を転移する活性を有する硫酸基転移酵素(ヘパラン硫酸3位硫酸基転移酵素(HS 3−O−sulfotransferase)、以下、HS3OSTと呼ぶことがある。)であることがより好ましい。
【0015】
上記HS3OSTは、配列番号2のアミノ酸配列又は配列番号2のアミノ酸配列と70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%以上の同一性(identity)を有することが好ましい。上記「アミノ酸配列の同一性(%)」とは、比較する2つのアミノ酸配列を必要に応じて間隙を導入して整列(アラインメント)させ、得られる最大のアミノ酸配列の同一性(%)をいう。アミノ酸配列の同一性を決定する目的のためのアラインメントは、通常の方法を用いて行うことができ、例えばBLASTのような公に入手可能なコンピュータソフトウエアや、DNASIS Pro(日立ソフトウェアエンジニアリング社製)並びにGENETYX(ゼネティックス社製)等の市販のソフトウェアを使用することもできる。
【0016】
また、上記HS3OSTは、配列番号2のアミノ酸配列において1もしくは数個のアミノ酸が置換、欠失、挿入又は付加されたアミノ酸配列を有するものであってもよい。上記「1もしくは数個」とは、1〜50個の整数であることが好ましく、1〜30個の整数であることが好ましく、1〜20個の整数であることがより好ましく、1〜15個の整数であることがさらに好ましく、1〜10個の整数であることがさらに好ましい。
【0017】
上記HS3OSTは、配列番号1の塩基配列又は配列番号1の塩基配列と80%以上、好ましくは90%以上、より好ましくは95%以上、さらに好ましくは97%以上の同一性を有するDNAにコードされるものであることが好ましい。上記「塩基配列の同一性(%)」とは、比較する2つの塩基配列を必要に応じて間隙を導入して整列(アラインメント)させ、得られる最大の塩基配列の同一性(%)をいう。塩基配列の同一性を決定する目的のためのアラインメントは、通常の方法を用いて行うことができ、例えばBLASTのような公に入手可能なコンピュータソフトウエアや、DNASIS Pro(日立ソフトウェアエンジニアリング社製)並びにGENETYX(ゼネティックス社製)等の市販のソフトウェアを使用することもできる。
【0018】
また、上記HS3OSTは、配列番号1の塩基配列と相補的な塩基配列からなるDNAにストリンジェントな条件下でハイブリダイズするDNAにコードされるものであってもよい。本発明において「ストリンジェントな条件」とは、塩基配列の同一性が高いDNA同士、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%以上の同一性を有するDNA同士がハイブリダイズし、それより同一性が低いDNA同士がハイブリダイズしない条件、あるいは通常のサザンハイブリダイゼーションの洗浄条件である60℃、1×SSC、0.1%SDS、好ましくは、60℃、0.1×SSC、0.1%SDS、より好ましくは、65℃、0.1×SSC、0.1%SDS、さらに好ましくは68℃、0.1×SSC、0.1%SDSに相当する温度及び塩濃度で、1回、より好ましくは2〜3回洗浄する条件が挙げられる。上記ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、公知の方法で行うことができる。
【0019】
本発明において活性が制御される硫酸基転移酵素は1種類であっても2種以上であってもよい。
本発明において活性が制御される硫酸基転移酵素は、使用する幹細胞の由来と同一の由来であることが好ましいが、異なっていてもいてよい。例えば、使用する幹細胞がヒト由来のES細胞を用いる場合には、該幹細胞中でヒト由来の硫酸基転移酵素の活性を制御することが好ましいが、該幹細胞中で硫酸基転移酵素の活性を高める際には、ヒト以外に由来する硫酸基転移酵素を導入することで、該幹細胞とは由来の異なる硫酸基転移酵素の活性を高めてもよい。
硫酸基転移反応における上記硫酸基供与体は、通常、3’ホスホアデノシン5’ホスホ硫酸(PAPS)が利用される。
【0020】
幹細胞中における硫酸基転移酵素活性の制御は通常の方法により行うことができる。硫酸基転移酵素活性を抑制する場合には、RNA干渉(RNA interference、以下RNAiと記す)を用いる方法、アンチセンス核酸を用いる方法、所望の硫酸基転移酵素遺伝子の転写産物を特異的に開裂するリボザイム活性を有する核酸を用いる方法、又は幹細胞のゲノムDNAに存在する硫酸基転移酵素遺伝子を変性・破壊する方法のような通常の遺伝子発現抑制手法を用いることが好ましい。より確実に、しかも簡便に硫酸基転移酵素活性を抑制する観点から、RNAiを用いる方法で制御することがより好ましい。
【0021】
RNAiに用いる核酸は、硫酸基転移酵素をコードする遺伝子の少なくとも一部と完全な相同性を有することが好ましいが、必ずしも完全同一である必要はない。
【0022】
上記RNAiに用いる核酸としてsiRNA(短鎖二本鎖RNA)を好適に用いることができる。siRNAの設計は、制御対象である硫酸基転移酵素の任意の塩基配列領域を標的として通常の方法で行うことができる。すなわち、所望の硫酸基転移酵素の一部の領域を標的とし、該領域と同一の塩基配列を有するsiRNAを通常の方法により作製することができる。
【0023】
また、siRNAは、標的配列に対して1又は数個の塩基の置換、欠失、挿入又は付加を有するものであっても、上述の糖鎖関連遺伝子の発現を抑制する機能を有する限り使用することができる。上記「1又は数個」とは、1〜10個の整数、好ましくは1〜5個の整数を表わす。
【0024】
さらに、これらのsiRNAの相補鎖とストリンジェントな条件下においてハイブリダイズしうる核酸も、本発明の核酸に包含される。この場合の「ストリンジェントな条件」として好ましくは60℃、1×SSC、0.1%SDS、より好ましくは60℃、0.1×SSC、0.1%SDSに相当する温度及び塩濃度で、1回、より好ましくは2〜3回洗浄する条件が挙げられる。
【0025】
ここで、siRNAには、ショートヘアピンRNA(shRNA:short hairpin RNA)のような、分子内において二本鎖RNA構造を形成し得る分子も含む。
【0026】
また、siRNAは、必ずしも標的配列に対する一組の二本鎖RNAである必要はなく、標的配列を含んだ領域に対する複数組の二本鎖RNAの混合物であってもよい。例えば、標的となる糖鎖関連遺伝子のmRNA全長又は全長に近い領域に対応する長鎖二本鎖RNAを予めDICER酵素で分解し、その分解産物を本発明の核酸として用いることができる。
【0027】
上述したRNAを発現し得る発現ベクターを幹細胞に導入することで、所望のRNAiの効果を得ることができる。かかる発現ベクターは、当業者であれば、一般的な遺伝子工学技術により適宜作製することができる。発現ベクターとして、pBAsiやpiGENEといったNakedプラスミドベクター、アデノウイルスベクター、レトロウイルスベクター等を好適に用いることができる。
【0028】
一方、幹細胞中で硫酸基転移酵素活性を高める場合には、硫酸基転移酵素の発現を促進させる機能を有する核酸や、所望の硫酸基転移酵素をコードする遺伝子を所望のプロモーターの下流に連結した発現ベクターを幹細胞に導入することにより該幹細胞中で硫酸基転移酵素を発現させることができ、これにより該幹細胞中で硫酸基転移酵素の活性を高めることができる。
また、硫酸基転移酵素を含有する培地中で幹細胞を培養することで、該幹細胞中で硫酸基転移酵素の活性を高めることもできる。
【0029】
上述した硫酸基転移酵素活性の制御は、幹細胞におけるFasシグナルの活性を制御するための手段である。Fasシグナルは、アポトーシスを誘起するシグナル経路として知られている。リボソームで翻訳されたFasはゴルジ体に移行し、ゴルジ体に移行したFasはゴルジ体において所定の糖鎖と結合する。適切に糖鎖が結合したFasは脂質ラフト(膜マイクロドメインの一種であり、スフィンゴ脂質とコレステロールとに富む細胞膜領域を意味する。)にリクルートされ、これが引き金となってFasシグナルが活性化されると考えられている。
ES細胞等の幹細胞では、通常、Fasシグナルは活性化されていない。このような幹細胞において、硫酸基転移酵素の活性を高めると、Fasシグナルが活性化し、未分化性の解除が引き起こされる。そのメカニズムとして以下のことが考えられる。
【0030】
特定の硫酸基転移酵素の活性が高まると、該硫酸基転移酵素が硫酸基供与体から硫酸基を特定の糖鎖に転移する速度が上昇し、該硫酸基転移酵素により硫酸基が転移された糖鎖の数が増大する。この硫酸基が転移された糖鎖はFasとの結合能が増大し、Fasに結合しやすくなる。これによりゴルジ体で糖鎖がFasに結合するようになると、糖鎖が結合したFasがゴルジ体から細胞膜の脂質ラフトに移行してFasシグナルカスケードが活性化される。
Fasシグナルが活性化すると、その下流に位置するカスパーゼ−8やカスパーゼ−3が活性化され、これにより幹細胞中でNanog等の未分化性維持因子が分解され、幹細胞の未分化性を解除する。また、同時に、Fasシグナルが活性化すると、未分化性維持因子のmRNAレベルが低下すると同時に特定の分化因子のmRNAレベルが上昇し、幹細胞の未分化性の解除を促進する。
逆に、幹細胞中において特定の硫酸基転移酵素の活性を抑制しておけばFasの脂質ラフトへの移行が抑制される。その結果、Fasシグナルの活性化を防ぐことができるため、幹細胞の未分化性を維持することが可能になる。
【0031】
また、本発明において、Fasシグナルの活性は、Fasシグナル阻害剤によっても制御することができる。Fasシグナル阻害剤によるFasシグナルの活性制御は、Fasシグナルの活性抑制である。
Fasシグナル阻害剤は、幹細胞中でFasシグナルを阻害しうるものであれば特に制限はないが、配列番号39のアミノ酸配列(IETD)を有するペプチド又は該ペプチドの誘導体を含む分子を好適に用いることができる。配列番号39のアミノ酸配列は、カスパーゼ−8に認識される配列であるため、上記ペプチド又はその誘導体が存在すると、カスパーゼ−8が本来の働きができなくなる。カスパーゼ−8はFasシグナルの下流で働く酵素であるため、カスパーゼ−8の働きが抑制されると、結果としてFasシグナルの活性も抑制される。
【0032】
Fasシグナル阻害剤の幹細胞への投与は、例えば、Fasシグナル阻害剤を幹細胞の培養液中に含有させておくことで行うことができる。
【0033】
本発明の方法は、幹細胞の分化を制御するために用いることができる。本発明の方法によれば、簡便に幹細胞の分化を制御・管理することができ、再生医療の実現に貢献することができる。
【0034】
本発明の細胞は、上記本発明の分化制御方法により分化が制御された細胞である。ここで、「分化が制御された」とは、未分化性が維持された状態を含む概念である。該細胞には、未分化性が維持されている細胞、分化開始前の細胞、分化過程の細胞、分化した細胞のいずれもが含まれる。本発明の細胞には、本発明の分化制御方法によって幹細胞から分化した組織や器官に含まれる細胞をも含む。
【0035】
以下、本発明を実施例に基づきさらに詳細に説明するが、本発明はこれに限定されるものではない。
【実施例】
【0036】
調製例1 硫酸基転移酵素の活性が高められた幹細胞の調製
マウス由来ヘパラン硫酸3位硫酸基転移酵素−5(mHS3OST−5)の発現ベクターは、pCAGIPuroベクター(東京大学大学院理学系研究科、程久美子准教授より供与)を用いて、J.Biol.Chem 2006;281:10945〜10953に記載の方法に基づき調製した。調製した発現ベクターは、mHS3OST−5の全長をコードする配列番号1の塩基配列を含み、配列番号2のアミノ酸配列からなる蛋白質を発現するように設計された。
【0037】
1000U/mLのleukemia inhibitory factor(LIF)を含むESC培地(15質量%FBS(Hyclone社製)、1%ペニシリン/ストレプトマイシン(Gibco社製)、0.1mM 2−メルカプトエタノール(Gibco社製)、0.1mM非必須アミノ酸(Gibco社製)を含有するDMEM培地)を用いて、10μg/mLのマイトマイシンCで不活化したマウス胎児繊維芽細胞(MEFs)上で、マウスES細胞であるR1セルライン(Proc.Natl.Acad.Sci.USA 90(1993)8424−8428に記載、自然科学研究機構 生理学研究所 分子神経生理研究部門 より供与)を維持した。
上記R1細胞を回収し、1×106個の細胞をゼラチンコートしたフィーダーフリーの60mm培養皿(Iwaki社製)に移し、LIFを含むESC培地で16時間培養した。培養後の細胞に、Lipofectamine 2000(Invitrogen社製)を用いて、上記で調製したHS3OST−5を含むCAGIPuroベクター2μgを導入した。その後1日間培養し、2μg/mLのピューロマイシン(シグマ社製)の存在下で24時間セレクションを行った。セレクションされた細胞を、mHS3OST−5の活性が高められたマウスES細胞(以下、mHS3OST−5過剰発現細胞ということがある。)として後述する実験に用いた。
なお、以下で使用されるコントロール細胞は、HS3OST−5を含まない空のpCAGIPuroベクターを上記と同様の方法によりR1細胞に導入したものである。
【0038】
試験例1 FACS分析
mHS3OST−5は、ヘパラン硫酸のグルコサミン残基の3位のヒドロキシル基に硫酸基を転移する。したがって、ヘパラン硫酸のグルコサミン残基の3位の硫酸化を含む硫酸化構造に特異的に結合する抗体(HS4C3)が認識するエピトープを増加させると考えられる。そこで、mHS3OST−5過剰発現細胞について、細胞表面に存在するHS4C3認識エピトープの量をFACS分析により調べた。
具体的には、遺伝子導入後2日目のHS3OST−5過剰発現細胞をFACS緩衝液(0.5質量%BSA、0.1質量%アジ化ナトリウムを含むPBS)中でVesicular Stomatitis Virus(VSV)標識したHS4C3(Department of Biochemistry, Nijmegen Center for Molecular Life Sciences, Radboud University Nijmegen Medical Centre, Nijmegen, The Netherlands Toin H. Van Kuppevelt教授より供与 (J Biol Chem
2006; 281:4654-4662))と共にインキュベートし、洗浄後、細胞懸濁液をマウス由来抗VSVグリコプロテイン抗体(IgG、シグマ社製)と共にFACS緩衝液中でインキュベートした。続いてFITC標識抗マウスIgG抗体(シグマ社製)を用いて標識した。その後、細胞をFACSria Cell Sorter(ベクトンディッキンソン社製)に通してFACS分析を行った。
結果を図1に示す。
【0039】
図1の結果から、mHS3OST−5過剰発現細胞において細胞の蛍光強度が高く、細胞表面のHS4C3認識エピトープが増加していることがわかる。
【0040】
続いて、mHS3OST−5過剰発現細胞において細胞表面へとリクルートされたFasの量をFACS分析により調べた。使用する抗体をFITC標識抗Fas抗体(商品名:Jo2、ベクトンディッキンソン社製)としたこと以外は上記と同様にFACS分析を行った。
結果を図2に示す。
【0041】
図2に示すように、mHS3OST−5過剰発現細胞において蛍光強度が高く、細胞表面に存在するFasの量が増加していることがわかった。
【0042】
上述した図1及び図2の結果は、幹細胞においてHS3OSTの活性を高めることで、ヘパラン硫酸のグルコサミン酸基の3位の硫酸化が促進され、これによりFasにヘパラン硫酸が結合しやすくなり、その結果Fasが細胞表面へと移動したことを示す。
【0043】
試験例2 脂質ラフト中のFasの存在量
細胞表面の存在するFasは、脂質ラフトに組み込まれることで、Fasシグナルを活性化する。そこで、mHS3OST−5過剰発現細胞において細胞表面に存在するFasが脂質ラフトに存在しているか否かをイムノブロッットにより調べた。
【0044】
脂質ラフトは、J.Immunol 1999;162:7217−7223及びCell Death Differ 2002;9:329−339の記載に基づき調製した。
具体的には、遺伝子導入後2日目の細胞を0.6mLの細胞溶解液(50mM Tris−HCl pH7.4、150mM NaCl、1%TritonX−100、プロテアーゼインヒビター)を用いて氷上でホモジナイズし(tight Dounce homogenizerを用いて50回)、抽出物をタンパク質1mg当たり60質量%Optiprep(シグマ社製)1.4mLと混合して42.5質量%のOptiprep溶液を調製した。これを遠心管(ベックマンコールター社製)に移して、5mLの30質量%Optiprep溶液と、50mMのTris−HCl pH7.4及び150mMのNaClを含有する3mLの5%Optiprepを積層した。これをSW41Tiローター(ベックマンコールター社製)を用いて4℃で16時間、30,000rpmで遠心することで溶液を9層に分断した。遠心管の上から3〜5層目が脂質ラフトを含みうる画分であり、上から7〜9層目は脂質ラフトを含まない画分である。
【0045】
上記各画分を10%SDS−PAGEにかけた後、PVDFメンブレン(ミリポア社製)に転写した。この転写後のメンブレンをブロッキングした後、マウス由来抗Fas抗体(IgG、BD biosciences 社製)を反応させた。洗浄後、西洋ワサビペルオキシダーゼ(HRP)標識抗マウスIgG抗体(Cell Signaling Technology社製)を2次抗体として反応させ、さらに洗浄後、ECL Plus試薬(GEヘルスケア社製)によりHRPを発色させた。
結果を図3に示す。
【0046】
図3の結果から、mHS3OST−5過剰発現細胞において脂質ラフトに存在するFasの量が増加していることがわかる。この結果は、mHS3OST−5過剰発現細胞においてFasシグナルが活性化していることを強く示唆するものである。
【0047】
試験例3 不活性化カスパーゼ−8の存在量
カスパーゼ−8はFasシグナルの下流で働く。そこで、mHS3OST−5過剰発現細胞におけるカスパーゼ−8の状態を調べるために、不活性化カスパーゼ−8の量をイムノブロットにより調べた。
具体的には、遺伝子導入後2日目のmHS3OST−5過剰発現細胞を細胞溶解液(50mM Tris−HCl pH7.4、150mM NaCl、1質量%TritonX−100、1mM Na3VO4、10mM NaF、プロテアーゼインヒビター)で溶解し、10%SDS−PAGEにかけた後、上記と同様にPVDFメンブレンに転写し、プライマリー抗体としてラビット由来抗不活性化カスパーゼ−8抗体(IgG、Cell Signaling Technology社製)を、2次抗体としてHRP標識抗ラビットIgG抗体を用いて不活性化カスパーゼ8の量を調べた。
得られたバンド強度をNIHimageで測定した結果を図4に示す。なお、図4の値はβ−アクチンのバンド強度により補正した値である。
【0048】
図4に示すように、mHS3OST−5過剰発現細胞では不活性化カスパーゼ−8の量が減少していることから、活性化カスパーゼ−8が増加していることを示す。すなわち、mHS3OST−5過剰発現細胞では、Fasシグナルが活性化されていることが示された。
【0049】
試験例4 切断カスパーゼ−3及びNanogの存在量
カスパーゼ−3はFasシグナルの下流で機能するタンパク質であり、カスパーゼ−8の下流で働くことが知られている。また、カスパーゼ−3は切断されることで活性化されることが知られている。そこで、切断カスパーゼ−3及びさらにその下流で機能する未分化性維持因子であるNanogについて、遺伝子導入後2日目のmHS3OST−5過剰発現細胞における存在量をイムノブロットにより調べた。
プライマリー抗体として上記のラビット由来抗不活性化カスパーゼ−8抗体に代えてラビット由来抗切断カスパーゼ−3抗体(Cell Signaling Technology社製)又はラビット由来抗Nanog抗体(ReproCELL社製)を用いた以外は上記と同様の方法により、mHS3OST−5過剰発現細胞における切断カスパーゼ−3及びNanogの存在量を調べた。
結果を図5に示す。
【0050】
図5に示すように、HS3OST−5過剰発現細胞において切断カスパーゼ−3の量が増加しており、カスパーゼ−3の活性が増加していることがわかる。また、これに伴いNanogの量が減少しており、切断カスパーゼ−3によりNanogが分解されたことが示唆された。Nanogは幹細胞の未分化性維持因子である。したがって、この結果は、mHS3OST−5過剰発現細胞において未分化性が解除されたことを示す。
【0051】
試験例5 Oct3/4とNanogのmRNAレベル
遺伝子導入後4日目のmHS3OST−5過剰発現細胞において、未分化性マーカーとして知られるOct3/4及びNanogのmRNAの発現量をリアルタイムPCRにより調べた。各mRNAの発現量は、下記表1に示す配列番号の塩基配列からなるプライマーセット及びプローブを用い、ABI PRISM 7700 Sequence Detection System(アプライドバイオシステムズ社製)を用いて調べた。
【0052】
【表1】

【0053】
結果を図6に示す。
図6の結果から、mHS3OST−5過剰発現細胞においていずれの未分化性マーカーのmRNAレベルも顕著に低下していることがわかる。これにより、mHS3OST−5過剰発現細胞において、タンパク質レベルでの分解のみならず、転写レベルにおいても未分化性維持に必要な因子の発現が抑制されていることが示された。
【0054】
試験例6 分化マーカーのmRNAレベル
遺伝子導入後4日目のmHS3OST−5過剰発現細胞において、各種分化マーカーについて、mRNAの発現量を調べた。各mRNAの発現量は、下記表2に示す配列番号の塩基配列からなるプライマー及びプローブを用い、ABI PRISM 7700 Sequence Detection System(アプライドバイオシステムズ社製)を用いて行った。
【0055】
【表2】
【0056】
結果を図7に示す。
図7に示すように、各種分化マーカーにおいてもその発現量の変化が認められ、特に、神経外胚葉マーカーであるIsl1、遠位内胚葉マーカーであるLamininB1、及び栄養膜マーカーであるCdx2におけるmRNAレベルの低下、並びに原始外胚葉マーカーであるFgf5及び原始内胚葉マーカーであるGata6のmRNAレベルの上昇がコントロール細胞に比べて有意であった。
これらの結果もまた、mHS3OST−5過剰発現細胞において未分化性が解除されていること示す。
【0057】
試験例7 HS中のmHS4C3認識エピトープの増加とFasとの結合能
mHS3OST−5過剰発現細胞においてHSとFasとの結合能が高まっていることをバインディングアッセイにより調べた。
具体的には、遺伝子導入後2日目のmHS3OST−5過剰発現細胞の細胞溶解液を8%SDS−PAGEにかけた後、PVDFメンブレンに転写し、これをブロッキング液(5%non−fatドライミルクを含むPBS pH6.5)でブロッキングし、さらにGST(グルタチオンセファローストランスフェラーゼ)融合Fas、GST融合変異導入Fas、又はGST融合FasとHS4C3との混合物を10nMの濃度で反応させた。その後、メンブレンを洗浄液(0.1%Tween20を含むPBS pH6.5)で洗浄し、HRP標識抗GST抗体(Santa Cruz社製)を室温で1時間反応させ後、HRPをECL Plus試薬を用いて発色させた。
GST融合Fas、GST融合変異導入Fasは、Fas又は変異導入FasをGSTとの融合タンパク質として大腸菌BL21株で常法により発現させ、これらをグルタチオンセファロース4B樹脂(GEヘルスケア社製)を用いて精製することで得た。
上記変異導入Fasは、HSとの結合に寄与することが予測された正電荷アミノ酸領域(配列番号36;KLRRRVH)を配列番号37(ALAAAVA)のアミノ酸配列に置き換えた構造をもつ。
結果を図8に示す。
【0058】
図8Aは、転写後のPVDFにHS4C3を反応させ、さらにHRP標識2次抗体を用いて発色させた結果を示す。mHS3OST−5過剰発現細胞において強いバンド強度が認められることから、mHS3OST−5過剰発現細胞においてグルコサミン残基の3位が硫酸化されたHSが増加していることがわかる。
図8Bは、転写後のPVDFにGST標識Fasを反応させ、HRP標識抗GST抗体で検出たものである。mHS3OST−5過剰発現細胞においてFasの結合量が高いことから、mHS3OST−5過剰発現細胞においてFas結合物質が増加していることがわかる。
図8Cは、転写後のPVDFにGST標識FasとHS4C3の混合物を反応させ、HRP標識抗GST抗体で検出したものである。この場合にはバンドが確認できないことから、FasのHSとの結合部位がHS4C3と競合していることがわかる。すなわち、FasがHSのグルコサミン残基の3位が硫酸化された部分を特異的に認識して結合していることがわかる。
図8Dは、転写後のPVDFにGST標識変異導入Fasを反応させたものである。変異導入Fasでは未変異Fasに比べてバンド強度が弱いことから、Fasの配列番号36のアミノ酸配列領域がHSとの結合に主要な役割を果たしていることがわかる。
なお、図8Eは、コントロールとしてβアクチンの量を調べた結果である。
【0059】
試験例8 Fasシグナル阻害による未分化性の回復
IETD(Ac-IETD-CHO)はカスパーゼ−8の働きを抑制する薬剤であり、Fasシグナルカスケードを遮断する薬剤である。そこで、IETDによりmHS3OST−5過剰発現細胞において引き起こされている未分化性の解除を脱することができるか、すなわち、再び未分化状態に戻りうるか調べた。
具体的には、HS3OST−5過剰発現細胞を10μM濃度のIETD(PEPTIDE INSTITUTE, INC社製)の存在下で24時間培養し、細胞を回収して溶解し、イムノブロットにより切断カスパーゼ−3とNanogのタンパク質量を、リアルタイムPCRによりOct3/とNanogのmRNA量をそれぞれ上述した方法により調べた。
結果を図9に示す。
【0060】
図9の結果から、IETD存在下で培養したHS3OST−5過剰発現細胞において、切断カスパーゼ−3のタンパク質量が低下し、これに伴いNanogのタンパク質量が増加していることがわかった。これらはいずれもコントロール細胞と同等のレベルまで回復していることもわかる。
また、IETDの存在下で培養したHS3OST−5過剰発現細胞では、Oct3/4及びNanogのmRNAの発現量もコントロール細胞と同レベルにまで回復していることがわかる。
これらの結果は、一度未分化性が解除された細胞においてさえ、Fasシグナルを阻害すれば、再び未分化性を回復しうることを示す。また、このことから、幹細胞においてFasシグナルを活性化しうる硫酸基転移酵素の活性を抑制しておけば、Fasシグナルの抑制状態が維持され、その結果幹細胞の未分化性を維持できることもわかる。
【0061】
試験例9 Fasシグナル阻害による未分化性の維持
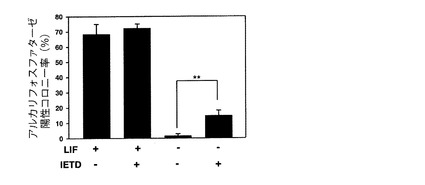
ES細胞ではアルカリフォスファターゼ(ALP)が活性化している。したがって、ES細胞をALP染色し、陽性コロニー数を評価することにより、未分化性を維持したES細胞の評価を行うことができる。本試験では、ES細胞をFasシグナル阻害ペプチドである上記IETDで処理した場合のES細胞の分化状態をALP活性を指標として評価した。
【0062】
具体的には、6cmディッシュに1×104のES細胞(R1細胞)をまき、LIF及び10μM Ac-IETD-CHOの存在下又は非存在下で、5日間培養した。培養後、ホルマリン/メタノールで細胞を固定し、洗浄後、5-bromo-4-chloro-3-indolyl phosphate-nitroblue tetrazolium (Nacalai Tesque)で染色を行った。染色された(陽性)コロニーは顕微鏡下でカウントした。結果を図10に示す。
上記ES細胞の未分化性の維持に必要なLIFを除いて分化させた細胞では、ALP陽性コロニー率が低下していた。しかし、Ac-IETD-CHOを加え、Fasシグナルをブロックした細胞では、ALP陽性コロニー率が顕著に増加した。このことから、Fasシグナル阻害剤には、ES細胞の未分化性を維持する作用があることがわかった。
【0063】
調製例2 硫酸基転移酵素の活性が抑制された幹細胞の調製
mHS3OST−5を標的としたショートヘアピンRNAを発現するための発現プラスミドをpSUPER.retro.puro(OligoEngine社製)を用いて構築した。具体的には、配列番号38の塩基配列を有するDNAをpSUPER.retro.puroのBamHIとHindIIIサイトの間に挿入した。レトロウィルスを産生するために、上記のコンストラクトをウィルスパッケージング細胞(PLAT−E)に感染させ、このウィルスを含むPLAT−E培養液を8μg/mLPolybrene(シグマ社製)と混合した後、R1細胞と共に24時間インキュベートした。こうしてR1細胞にウィルスを感染させた後、R1細胞をLIFを含むESC培地に移し、2μg/mLのピューロマイシンの存在下で5〜7日間培養することで、mHS3OST−5の活性が抑制されたマウスES細胞(以下、mHS3OST−5抑制細胞という。)を得た。
【0064】
mHS3OST−5抑制細胞はヘパラン硫酸のグルコサミン残基の3位のOH基に対する硫酸基の転移反応が抑制されているため、ゴルジ体においてFasが糖鎖と適切に結合することができない。したがって、Fasが脂質ラフトに移動することができず、Fasシグナルの活性が抑制されている。
【0065】
mHS3OST−5以外の硫酸基転移酵素であっても、糖鎖のFasに対する親和性を制御することができるものであれば、幹細胞中で該硫酸基転移酵素の活性を制御することでFasシグナルの活性を制御することができ、mHS3OST−5と同様のメカニズムにより幹細胞の分化を制御することができる。
【0066】
以上の結果、幹細胞中でFasシグナルの活性を制御することにより、該幹細胞の分化を制御できることが示された。より具体的には、Fasシグナルの活性を抑制することにより、幹細胞の未分化性を維持することができ、逆にFasシグナルを活性化すれば、幹細胞の未分化性が解除できることが示された。
【技術分野】
【0001】
本発明は、Fasシグナルを介して幹細胞の分化を制御する方法に関する。
【背景技術】
【0002】
胚性幹細胞(ES細胞)や誘導多能性幹細胞(iPS細胞)をはじめとする幹細胞は、生体に存在する様々な細胞に分化する能力を有し、細胞移植が有効な治療方法として期待される疾病、例えば、白血病、パーキンソン病等の治療に用いることが期待されている。
幹細胞を適切に維持・管理するためには、幹細胞の未分化性を維持した状態を保ちながら継代培養することが必要になる。したがって、幹細胞の未分化性を、確実にしかも簡便に維持するための手法の開発が望まれている。
一方、幹細胞を分化させる際には、より迅速に目的の分化状態へと方向付けすることが要求される。したがって、幹細胞の未分化性をより迅速に解除するための手法の開発も望まれている。
【0003】
糖鎖は、細胞の生存に必須であるばかりでなく、がん、免疫、発生・分化や感染症などにおいて特に重要な役割を果たしていることが報告されている。発生分化の過程においては、その分化程度や状態に応じて糖鎖を合成する糖転移酵素遺伝子の発現パターンが変化し、合成される糖鎖構造も変化することが知られている。しかし、糖鎖に硫酸基を付加する硫酸基転移酵素の活性が幹細胞の分化に与える影響についてはこれまで知られていない。
【0004】
Fasシグナルはアポトーシスを引き起こすシグナル経路として知られており、アポトーシスは細胞の分化に関与することが知られている。
しかし、幹細胞におけるFasシグナルと分化状態との関係についてこれまで報告はなく、また、幹細胞におけるFasシグナルと硫酸基転移酵素との関係についても知られていない。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、幹細胞の分化状態を制御する方法の提供を課題とする。
【課題を解決するための手段】
【0006】
本発明者らは、上記課題の解決のために鋭意検討を行った。その結果、Fasシグナルの活性を制御することにより、幹細胞の分化状態を制御できることを見い出した。また、本発明者らは、硫酸基転移酵素の活性を制御することでFasシグナルの活性を制御することができ、これにより幹細胞の分化状態をも制御できることを見い出した。本発明はこれらの知見に基づき完成するに至ったものである。
【0007】
本発明の課題は下記の手段により達成された。
[1]幹細胞のFasシグナルの活性を制御することを含む、幹細胞の分化制御方法。
[2]幹細胞中で硫酸基転移酵素の活性を制御し、これにより該幹細胞のFasシグナルの活性を制御する、[1]に記載の方法。
[3]硫酸基転移酵素がヘパラン硫酸硫酸基転移酵素である、[2]に記載の方法。
[4]ヘパラン硫酸硫酸基転移酵素がヘパラン硫酸3位硫酸基転移酵素である、[3]に記載の方法。
[5]ヘパラン硫酸3位硫酸基転移酵素が下記(A)〜(C)のいずれかのアミノ酸配列を有する、[4]に記載の方法:
(A)配列番号2のアミノ酸配列、
(B)配列番号2のアミノ酸配列において1もしくは数個のアミノ酸が置換、欠失、挿入又は付加されたアミノ酸配列、
(C)配列番号2のアミノ酸配列と80%以上の同一性を有するアミノ酸配列。
[6]幹細胞のFasシグナルの活性を抑制し、これにより幹細胞の未分化性を維持する、[1]〜[5]のいずれかに記載の方法。
[7]幹細胞のFasシグナルを活性化し、これにより幹細胞の未分化性を解除する、[1]〜[5]のいずれかに記載の方法。
[8]幹細胞中で硫酸基転移酵素の活性を抑制して該幹細胞のFasシグナルを抑制し、これにより幹細胞の未分化性を維持する、[6]に記載の方法。
[9]硫酸基転移酵素をコードする遺伝子の発現を抑制する核酸を幹細胞に導入し、該幹細胞中で該遺伝子の発現を抑制することで硫酸基転移酵素の活性を抑制する、[8]に記載の方法。
[10]幹細胞中で硫酸基転移酵素の活性を高めて該幹細胞のFasシグナルを活性化し、これにより幹細胞の未分化性を解除する、[7]に記載の方法。
[11]硫酸基転移酵素をコードする遺伝子を幹細胞に導入し、該幹細胞中で該遺伝子を過剰発現させることで硫酸基転移酵素の活性を高める、[10]に記載の方法。
[12]幹細胞にFasシグナル阻害剤を投与して該幹細胞のFasシグナルを抑制し、これにより幹細胞の未分化性を維持する、[6]に記載の方法。
[13]Fasシグナル阻害剤が、配列番号39のアミノ酸配列を有するペプチドを含む、[12]に記載の方法。
[14]幹細胞が哺乳類に由来する、[1]〜[13]のいずれかに記載の方法。
[15]幹細胞がES細胞である、[14]に記載の方法。
[16][1]〜[15]のいずれかに記載の方法で分化制御された細胞。
【発明の効果】
【0008】
本発明によれば、幹細胞を所望の分化状態に制御することができる。
【図面の簡単な説明】
【0009】
【図1】mHS3OST−5過剰発現細胞において、グルコサミン残基の3位に硫酸基が転移したHSの細胞表面における存在量をFACS分析により測定した結果を示す図である。図1中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図2】mHS3OST−5過剰発現細胞において、細胞表面に存在するFasの量をFACS分析により測定した結果を示す図である。図2中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図3】mHS3OST−5過剰発現細胞における脂質ラフト中のFasの存在量をイムノブロットにより調べた結果を示す写真である。図3中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。各画分の番号は実施例で使用した遠心管の最上層から数えた層の順番を示す。
【図4】mHS3OST−5過剰発現細胞における不活性型カスパーゼ−8の量をイムノブロットにより調べた結果を示す図である。図4中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図5】mHS3OST−5過剰発現細胞における切断カスパーゼ−3及びNanogの量をイムノブロットにより調べた結果を示す図である。図5中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図6】mHS3OST−5過剰発現細胞における未分化マーカー(Oct3/4及びNanog)のmRNA量をリアルタイムPCRにより調べた結果を示す図である。図6中、aはコントロール細胞の結果を示し、bはHS3OST−5過剰発現細胞の結果を示す。
【図7】mHS3OST−5過剰発現細胞における分化マーカー(Isl1、Brachyury、Fgf5、Gata4、Gata6、LamininB1、Bmp2及びCdx2)のmRNA量をリアルタイムPCRにより調べた結果を示す図である。図7中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
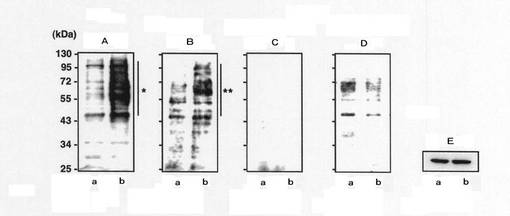
【図8】mHS3OST−5過剰発現細胞におけるHSとFasとの結合機序をオーバーレイアッセイにより調べた結果を示す写真である。図7中、AはHS4C3(抗へパラン硫酸抗体:グルコサミン残基の3位が硫酸化された硫酸化構造を特異的に認識する抗体)を反応させた後にHRP標識抗マウスIgG抗体を用いて発色させた結果を示し、BはGST融合Fasを反応させた後にHRP標識抗GST抗体を用いて発色させた結果を示し、CはGST融合FasとHS4C3との混合物を反応させた後にHRP標識抗GST抗体を用いて発色させた結果を示し、DはGST融合変異導入Fasを反応させた後にHRP標識抗GST抗体を用いて発色させた結果を示す。Eはβ−アクチンの量をイムノブロットにより調べた結果を示す。また、図7中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図9】mHS3OST−5過剰発現細胞において、IETDの添加によりFasシグナルを阻害したときの分化状態の変化をマーカー発現量を指標に調べた結果を示す図である。図8中、aはコントロール細胞の結果を示し、bはmHS3OST−5過剰発現細胞の結果を示す。
【図10】R1細胞の分化に対するFasシグナル阻害剤(IETD)の影響を調べた結果を示す図である。
【発明を実施するための形態】
【0010】
以下、本発明について、その好ましい実施態様に基づき詳細に説明する。
【0011】
本発明は、幹細胞中においてFasシグナルの活性を制御することにより該幹細胞の分化状態を制御する方法である。上記Fasシグナルの活性制御手段に特に制限はないが、幹細胞中の硫酸基転移酵素活性の制御や、幹細胞へのFasシグナル阻害剤の投与により行うことが好ましい。本発明において「分化状態を制御する」とは、未分化性を維持することを含む概念である。また、本発明において「Fas」とはFas受容体を意味する。
【0012】
本発明に用いる幹細胞に特に制限はなく、分化万能性、分化多能性、分化単一性のいずれの分化能を有する幹細胞であってもよいが、分化万能性を有する幹細胞であることが好ましい。本発明に用いる幹細胞の具体例として、例えば、胚性幹細胞(ES細胞)、誘導多能性幹細胞(iPS細胞)、胚性生殖幹細胞、核移植ES細胞、神経幹細胞、造血幹細胞、間葉系幹細胞、肝幹細胞、皮膚幹細胞、筋幹細胞等が挙げられるが、ES細胞又はiPS細胞であることが好ましく、ES細胞であることがより好ましい。上記幹細胞の由来に特に制限はなく、哺乳類、鳥類等に由来するものを用いることができるが、哺乳類に由来する幹細胞を好適に用いることができる。前記哺乳類としては、例えば、ヒト、サル、ウシ、ヤギ、ヒツジ、ブタ、ウマ、ウサギ、ラット、マウス、モルモット等が挙げられる。
上記幹細胞は通常の方法で調製することができる。また、寄託機関等に寄託されているセルライン等の分譲により得ることもできる。
【0013】
続いて、幹細胞のFasシグナルの活性を制御する手段について説明する。
【0014】
本発明において、幹細胞のFasシグナルは、幹細胞中で硫酸基転移酵素活性の制御することで行うことができる。
本発明における硫酸基転移酵素活性の制御とは、硫酸基転移酵素の活性を高めること及び抑制することの両概念を含む。
本発明において活性が制御されうる硫酸基転移酵素は、硫酸基供与体から糖鎖に硫酸基を転移する酵素である。上記硫酸基転移酵素は、その活性を高めることでFasシグナルを所望のレベルまで活性化させることができ、また、その活性を低下させるとFasシグナルを所望のレベルまで不活性化できる酵素である。上記硫酸基転移酵素は硫酸基供与体からヘパラン硫酸(HS)に硫酸基を転移する硫酸基転移酵素であることが好ましく、硫酸基供与体からヘパラン硫酸のグルコサミン残基の3位のヒドロキシル基に硫酸基を転移する活性を有する硫酸基転移酵素(ヘパラン硫酸3位硫酸基転移酵素(HS 3−O−sulfotransferase)、以下、HS3OSTと呼ぶことがある。)であることがより好ましい。
【0015】
上記HS3OSTは、配列番号2のアミノ酸配列又は配列番号2のアミノ酸配列と70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%以上の同一性(identity)を有することが好ましい。上記「アミノ酸配列の同一性(%)」とは、比較する2つのアミノ酸配列を必要に応じて間隙を導入して整列(アラインメント)させ、得られる最大のアミノ酸配列の同一性(%)をいう。アミノ酸配列の同一性を決定する目的のためのアラインメントは、通常の方法を用いて行うことができ、例えばBLASTのような公に入手可能なコンピュータソフトウエアや、DNASIS Pro(日立ソフトウェアエンジニアリング社製)並びにGENETYX(ゼネティックス社製)等の市販のソフトウェアを使用することもできる。
【0016】
また、上記HS3OSTは、配列番号2のアミノ酸配列において1もしくは数個のアミノ酸が置換、欠失、挿入又は付加されたアミノ酸配列を有するものであってもよい。上記「1もしくは数個」とは、1〜50個の整数であることが好ましく、1〜30個の整数であることが好ましく、1〜20個の整数であることがより好ましく、1〜15個の整数であることがさらに好ましく、1〜10個の整数であることがさらに好ましい。
【0017】
上記HS3OSTは、配列番号1の塩基配列又は配列番号1の塩基配列と80%以上、好ましくは90%以上、より好ましくは95%以上、さらに好ましくは97%以上の同一性を有するDNAにコードされるものであることが好ましい。上記「塩基配列の同一性(%)」とは、比較する2つの塩基配列を必要に応じて間隙を導入して整列(アラインメント)させ、得られる最大の塩基配列の同一性(%)をいう。塩基配列の同一性を決定する目的のためのアラインメントは、通常の方法を用いて行うことができ、例えばBLASTのような公に入手可能なコンピュータソフトウエアや、DNASIS Pro(日立ソフトウェアエンジニアリング社製)並びにGENETYX(ゼネティックス社製)等の市販のソフトウェアを使用することもできる。
【0018】
また、上記HS3OSTは、配列番号1の塩基配列と相補的な塩基配列からなるDNAにストリンジェントな条件下でハイブリダイズするDNAにコードされるものであってもよい。本発明において「ストリンジェントな条件」とは、塩基配列の同一性が高いDNA同士、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%以上の同一性を有するDNA同士がハイブリダイズし、それより同一性が低いDNA同士がハイブリダイズしない条件、あるいは通常のサザンハイブリダイゼーションの洗浄条件である60℃、1×SSC、0.1%SDS、好ましくは、60℃、0.1×SSC、0.1%SDS、より好ましくは、65℃、0.1×SSC、0.1%SDS、さらに好ましくは68℃、0.1×SSC、0.1%SDSに相当する温度及び塩濃度で、1回、より好ましくは2〜3回洗浄する条件が挙げられる。上記ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、公知の方法で行うことができる。
【0019】
本発明において活性が制御される硫酸基転移酵素は1種類であっても2種以上であってもよい。
本発明において活性が制御される硫酸基転移酵素は、使用する幹細胞の由来と同一の由来であることが好ましいが、異なっていてもいてよい。例えば、使用する幹細胞がヒト由来のES細胞を用いる場合には、該幹細胞中でヒト由来の硫酸基転移酵素の活性を制御することが好ましいが、該幹細胞中で硫酸基転移酵素の活性を高める際には、ヒト以外に由来する硫酸基転移酵素を導入することで、該幹細胞とは由来の異なる硫酸基転移酵素の活性を高めてもよい。
硫酸基転移反応における上記硫酸基供与体は、通常、3’ホスホアデノシン5’ホスホ硫酸(PAPS)が利用される。
【0020】
幹細胞中における硫酸基転移酵素活性の制御は通常の方法により行うことができる。硫酸基転移酵素活性を抑制する場合には、RNA干渉(RNA interference、以下RNAiと記す)を用いる方法、アンチセンス核酸を用いる方法、所望の硫酸基転移酵素遺伝子の転写産物を特異的に開裂するリボザイム活性を有する核酸を用いる方法、又は幹細胞のゲノムDNAに存在する硫酸基転移酵素遺伝子を変性・破壊する方法のような通常の遺伝子発現抑制手法を用いることが好ましい。より確実に、しかも簡便に硫酸基転移酵素活性を抑制する観点から、RNAiを用いる方法で制御することがより好ましい。
【0021】
RNAiに用いる核酸は、硫酸基転移酵素をコードする遺伝子の少なくとも一部と完全な相同性を有することが好ましいが、必ずしも完全同一である必要はない。
【0022】
上記RNAiに用いる核酸としてsiRNA(短鎖二本鎖RNA)を好適に用いることができる。siRNAの設計は、制御対象である硫酸基転移酵素の任意の塩基配列領域を標的として通常の方法で行うことができる。すなわち、所望の硫酸基転移酵素の一部の領域を標的とし、該領域と同一の塩基配列を有するsiRNAを通常の方法により作製することができる。
【0023】
また、siRNAは、標的配列に対して1又は数個の塩基の置換、欠失、挿入又は付加を有するものであっても、上述の糖鎖関連遺伝子の発現を抑制する機能を有する限り使用することができる。上記「1又は数個」とは、1〜10個の整数、好ましくは1〜5個の整数を表わす。
【0024】
さらに、これらのsiRNAの相補鎖とストリンジェントな条件下においてハイブリダイズしうる核酸も、本発明の核酸に包含される。この場合の「ストリンジェントな条件」として好ましくは60℃、1×SSC、0.1%SDS、より好ましくは60℃、0.1×SSC、0.1%SDSに相当する温度及び塩濃度で、1回、より好ましくは2〜3回洗浄する条件が挙げられる。
【0025】
ここで、siRNAには、ショートヘアピンRNA(shRNA:short hairpin RNA)のような、分子内において二本鎖RNA構造を形成し得る分子も含む。
【0026】
また、siRNAは、必ずしも標的配列に対する一組の二本鎖RNAである必要はなく、標的配列を含んだ領域に対する複数組の二本鎖RNAの混合物であってもよい。例えば、標的となる糖鎖関連遺伝子のmRNA全長又は全長に近い領域に対応する長鎖二本鎖RNAを予めDICER酵素で分解し、その分解産物を本発明の核酸として用いることができる。
【0027】
上述したRNAを発現し得る発現ベクターを幹細胞に導入することで、所望のRNAiの効果を得ることができる。かかる発現ベクターは、当業者であれば、一般的な遺伝子工学技術により適宜作製することができる。発現ベクターとして、pBAsiやpiGENEといったNakedプラスミドベクター、アデノウイルスベクター、レトロウイルスベクター等を好適に用いることができる。
【0028】
一方、幹細胞中で硫酸基転移酵素活性を高める場合には、硫酸基転移酵素の発現を促進させる機能を有する核酸や、所望の硫酸基転移酵素をコードする遺伝子を所望のプロモーターの下流に連結した発現ベクターを幹細胞に導入することにより該幹細胞中で硫酸基転移酵素を発現させることができ、これにより該幹細胞中で硫酸基転移酵素の活性を高めることができる。
また、硫酸基転移酵素を含有する培地中で幹細胞を培養することで、該幹細胞中で硫酸基転移酵素の活性を高めることもできる。
【0029】
上述した硫酸基転移酵素活性の制御は、幹細胞におけるFasシグナルの活性を制御するための手段である。Fasシグナルは、アポトーシスを誘起するシグナル経路として知られている。リボソームで翻訳されたFasはゴルジ体に移行し、ゴルジ体に移行したFasはゴルジ体において所定の糖鎖と結合する。適切に糖鎖が結合したFasは脂質ラフト(膜マイクロドメインの一種であり、スフィンゴ脂質とコレステロールとに富む細胞膜領域を意味する。)にリクルートされ、これが引き金となってFasシグナルが活性化されると考えられている。
ES細胞等の幹細胞では、通常、Fasシグナルは活性化されていない。このような幹細胞において、硫酸基転移酵素の活性を高めると、Fasシグナルが活性化し、未分化性の解除が引き起こされる。そのメカニズムとして以下のことが考えられる。
【0030】
特定の硫酸基転移酵素の活性が高まると、該硫酸基転移酵素が硫酸基供与体から硫酸基を特定の糖鎖に転移する速度が上昇し、該硫酸基転移酵素により硫酸基が転移された糖鎖の数が増大する。この硫酸基が転移された糖鎖はFasとの結合能が増大し、Fasに結合しやすくなる。これによりゴルジ体で糖鎖がFasに結合するようになると、糖鎖が結合したFasがゴルジ体から細胞膜の脂質ラフトに移行してFasシグナルカスケードが活性化される。
Fasシグナルが活性化すると、その下流に位置するカスパーゼ−8やカスパーゼ−3が活性化され、これにより幹細胞中でNanog等の未分化性維持因子が分解され、幹細胞の未分化性を解除する。また、同時に、Fasシグナルが活性化すると、未分化性維持因子のmRNAレベルが低下すると同時に特定の分化因子のmRNAレベルが上昇し、幹細胞の未分化性の解除を促進する。
逆に、幹細胞中において特定の硫酸基転移酵素の活性を抑制しておけばFasの脂質ラフトへの移行が抑制される。その結果、Fasシグナルの活性化を防ぐことができるため、幹細胞の未分化性を維持することが可能になる。
【0031】
また、本発明において、Fasシグナルの活性は、Fasシグナル阻害剤によっても制御することができる。Fasシグナル阻害剤によるFasシグナルの活性制御は、Fasシグナルの活性抑制である。
Fasシグナル阻害剤は、幹細胞中でFasシグナルを阻害しうるものであれば特に制限はないが、配列番号39のアミノ酸配列(IETD)を有するペプチド又は該ペプチドの誘導体を含む分子を好適に用いることができる。配列番号39のアミノ酸配列は、カスパーゼ−8に認識される配列であるため、上記ペプチド又はその誘導体が存在すると、カスパーゼ−8が本来の働きができなくなる。カスパーゼ−8はFasシグナルの下流で働く酵素であるため、カスパーゼ−8の働きが抑制されると、結果としてFasシグナルの活性も抑制される。
【0032】
Fasシグナル阻害剤の幹細胞への投与は、例えば、Fasシグナル阻害剤を幹細胞の培養液中に含有させておくことで行うことができる。
【0033】
本発明の方法は、幹細胞の分化を制御するために用いることができる。本発明の方法によれば、簡便に幹細胞の分化を制御・管理することができ、再生医療の実現に貢献することができる。
【0034】
本発明の細胞は、上記本発明の分化制御方法により分化が制御された細胞である。ここで、「分化が制御された」とは、未分化性が維持された状態を含む概念である。該細胞には、未分化性が維持されている細胞、分化開始前の細胞、分化過程の細胞、分化した細胞のいずれもが含まれる。本発明の細胞には、本発明の分化制御方法によって幹細胞から分化した組織や器官に含まれる細胞をも含む。
【0035】
以下、本発明を実施例に基づきさらに詳細に説明するが、本発明はこれに限定されるものではない。
【実施例】
【0036】
調製例1 硫酸基転移酵素の活性が高められた幹細胞の調製
マウス由来ヘパラン硫酸3位硫酸基転移酵素−5(mHS3OST−5)の発現ベクターは、pCAGIPuroベクター(東京大学大学院理学系研究科、程久美子准教授より供与)を用いて、J.Biol.Chem 2006;281:10945〜10953に記載の方法に基づき調製した。調製した発現ベクターは、mHS3OST−5の全長をコードする配列番号1の塩基配列を含み、配列番号2のアミノ酸配列からなる蛋白質を発現するように設計された。
【0037】
1000U/mLのleukemia inhibitory factor(LIF)を含むESC培地(15質量%FBS(Hyclone社製)、1%ペニシリン/ストレプトマイシン(Gibco社製)、0.1mM 2−メルカプトエタノール(Gibco社製)、0.1mM非必須アミノ酸(Gibco社製)を含有するDMEM培地)を用いて、10μg/mLのマイトマイシンCで不活化したマウス胎児繊維芽細胞(MEFs)上で、マウスES細胞であるR1セルライン(Proc.Natl.Acad.Sci.USA 90(1993)8424−8428に記載、自然科学研究機構 生理学研究所 分子神経生理研究部門 より供与)を維持した。
上記R1細胞を回収し、1×106個の細胞をゼラチンコートしたフィーダーフリーの60mm培養皿(Iwaki社製)に移し、LIFを含むESC培地で16時間培養した。培養後の細胞に、Lipofectamine 2000(Invitrogen社製)を用いて、上記で調製したHS3OST−5を含むCAGIPuroベクター2μgを導入した。その後1日間培養し、2μg/mLのピューロマイシン(シグマ社製)の存在下で24時間セレクションを行った。セレクションされた細胞を、mHS3OST−5の活性が高められたマウスES細胞(以下、mHS3OST−5過剰発現細胞ということがある。)として後述する実験に用いた。
なお、以下で使用されるコントロール細胞は、HS3OST−5を含まない空のpCAGIPuroベクターを上記と同様の方法によりR1細胞に導入したものである。
【0038】
試験例1 FACS分析
mHS3OST−5は、ヘパラン硫酸のグルコサミン残基の3位のヒドロキシル基に硫酸基を転移する。したがって、ヘパラン硫酸のグルコサミン残基の3位の硫酸化を含む硫酸化構造に特異的に結合する抗体(HS4C3)が認識するエピトープを増加させると考えられる。そこで、mHS3OST−5過剰発現細胞について、細胞表面に存在するHS4C3認識エピトープの量をFACS分析により調べた。
具体的には、遺伝子導入後2日目のHS3OST−5過剰発現細胞をFACS緩衝液(0.5質量%BSA、0.1質量%アジ化ナトリウムを含むPBS)中でVesicular Stomatitis Virus(VSV)標識したHS4C3(Department of Biochemistry, Nijmegen Center for Molecular Life Sciences, Radboud University Nijmegen Medical Centre, Nijmegen, The Netherlands Toin H. Van Kuppevelt教授より供与 (J Biol Chem
2006; 281:4654-4662))と共にインキュベートし、洗浄後、細胞懸濁液をマウス由来抗VSVグリコプロテイン抗体(IgG、シグマ社製)と共にFACS緩衝液中でインキュベートした。続いてFITC標識抗マウスIgG抗体(シグマ社製)を用いて標識した。その後、細胞をFACSria Cell Sorter(ベクトンディッキンソン社製)に通してFACS分析を行った。
結果を図1に示す。
【0039】
図1の結果から、mHS3OST−5過剰発現細胞において細胞の蛍光強度が高く、細胞表面のHS4C3認識エピトープが増加していることがわかる。
【0040】
続いて、mHS3OST−5過剰発現細胞において細胞表面へとリクルートされたFasの量をFACS分析により調べた。使用する抗体をFITC標識抗Fas抗体(商品名:Jo2、ベクトンディッキンソン社製)としたこと以外は上記と同様にFACS分析を行った。
結果を図2に示す。
【0041】
図2に示すように、mHS3OST−5過剰発現細胞において蛍光強度が高く、細胞表面に存在するFasの量が増加していることがわかった。
【0042】
上述した図1及び図2の結果は、幹細胞においてHS3OSTの活性を高めることで、ヘパラン硫酸のグルコサミン酸基の3位の硫酸化が促進され、これによりFasにヘパラン硫酸が結合しやすくなり、その結果Fasが細胞表面へと移動したことを示す。
【0043】
試験例2 脂質ラフト中のFasの存在量
細胞表面の存在するFasは、脂質ラフトに組み込まれることで、Fasシグナルを活性化する。そこで、mHS3OST−5過剰発現細胞において細胞表面に存在するFasが脂質ラフトに存在しているか否かをイムノブロッットにより調べた。
【0044】
脂質ラフトは、J.Immunol 1999;162:7217−7223及びCell Death Differ 2002;9:329−339の記載に基づき調製した。
具体的には、遺伝子導入後2日目の細胞を0.6mLの細胞溶解液(50mM Tris−HCl pH7.4、150mM NaCl、1%TritonX−100、プロテアーゼインヒビター)を用いて氷上でホモジナイズし(tight Dounce homogenizerを用いて50回)、抽出物をタンパク質1mg当たり60質量%Optiprep(シグマ社製)1.4mLと混合して42.5質量%のOptiprep溶液を調製した。これを遠心管(ベックマンコールター社製)に移して、5mLの30質量%Optiprep溶液と、50mMのTris−HCl pH7.4及び150mMのNaClを含有する3mLの5%Optiprepを積層した。これをSW41Tiローター(ベックマンコールター社製)を用いて4℃で16時間、30,000rpmで遠心することで溶液を9層に分断した。遠心管の上から3〜5層目が脂質ラフトを含みうる画分であり、上から7〜9層目は脂質ラフトを含まない画分である。
【0045】
上記各画分を10%SDS−PAGEにかけた後、PVDFメンブレン(ミリポア社製)に転写した。この転写後のメンブレンをブロッキングした後、マウス由来抗Fas抗体(IgG、BD biosciences 社製)を反応させた。洗浄後、西洋ワサビペルオキシダーゼ(HRP)標識抗マウスIgG抗体(Cell Signaling Technology社製)を2次抗体として反応させ、さらに洗浄後、ECL Plus試薬(GEヘルスケア社製)によりHRPを発色させた。
結果を図3に示す。
【0046】
図3の結果から、mHS3OST−5過剰発現細胞において脂質ラフトに存在するFasの量が増加していることがわかる。この結果は、mHS3OST−5過剰発現細胞においてFasシグナルが活性化していることを強く示唆するものである。
【0047】
試験例3 不活性化カスパーゼ−8の存在量
カスパーゼ−8はFasシグナルの下流で働く。そこで、mHS3OST−5過剰発現細胞におけるカスパーゼ−8の状態を調べるために、不活性化カスパーゼ−8の量をイムノブロットにより調べた。
具体的には、遺伝子導入後2日目のmHS3OST−5過剰発現細胞を細胞溶解液(50mM Tris−HCl pH7.4、150mM NaCl、1質量%TritonX−100、1mM Na3VO4、10mM NaF、プロテアーゼインヒビター)で溶解し、10%SDS−PAGEにかけた後、上記と同様にPVDFメンブレンに転写し、プライマリー抗体としてラビット由来抗不活性化カスパーゼ−8抗体(IgG、Cell Signaling Technology社製)を、2次抗体としてHRP標識抗ラビットIgG抗体を用いて不活性化カスパーゼ8の量を調べた。
得られたバンド強度をNIHimageで測定した結果を図4に示す。なお、図4の値はβ−アクチンのバンド強度により補正した値である。
【0048】
図4に示すように、mHS3OST−5過剰発現細胞では不活性化カスパーゼ−8の量が減少していることから、活性化カスパーゼ−8が増加していることを示す。すなわち、mHS3OST−5過剰発現細胞では、Fasシグナルが活性化されていることが示された。
【0049】
試験例4 切断カスパーゼ−3及びNanogの存在量
カスパーゼ−3はFasシグナルの下流で機能するタンパク質であり、カスパーゼ−8の下流で働くことが知られている。また、カスパーゼ−3は切断されることで活性化されることが知られている。そこで、切断カスパーゼ−3及びさらにその下流で機能する未分化性維持因子であるNanogについて、遺伝子導入後2日目のmHS3OST−5過剰発現細胞における存在量をイムノブロットにより調べた。
プライマリー抗体として上記のラビット由来抗不活性化カスパーゼ−8抗体に代えてラビット由来抗切断カスパーゼ−3抗体(Cell Signaling Technology社製)又はラビット由来抗Nanog抗体(ReproCELL社製)を用いた以外は上記と同様の方法により、mHS3OST−5過剰発現細胞における切断カスパーゼ−3及びNanogの存在量を調べた。
結果を図5に示す。
【0050】
図5に示すように、HS3OST−5過剰発現細胞において切断カスパーゼ−3の量が増加しており、カスパーゼ−3の活性が増加していることがわかる。また、これに伴いNanogの量が減少しており、切断カスパーゼ−3によりNanogが分解されたことが示唆された。Nanogは幹細胞の未分化性維持因子である。したがって、この結果は、mHS3OST−5過剰発現細胞において未分化性が解除されたことを示す。
【0051】
試験例5 Oct3/4とNanogのmRNAレベル
遺伝子導入後4日目のmHS3OST−5過剰発現細胞において、未分化性マーカーとして知られるOct3/4及びNanogのmRNAの発現量をリアルタイムPCRにより調べた。各mRNAの発現量は、下記表1に示す配列番号の塩基配列からなるプライマーセット及びプローブを用い、ABI PRISM 7700 Sequence Detection System(アプライドバイオシステムズ社製)を用いて調べた。
【0052】
【表1】
【0053】
結果を図6に示す。
図6の結果から、mHS3OST−5過剰発現細胞においていずれの未分化性マーカーのmRNAレベルも顕著に低下していることがわかる。これにより、mHS3OST−5過剰発現細胞において、タンパク質レベルでの分解のみならず、転写レベルにおいても未分化性維持に必要な因子の発現が抑制されていることが示された。
【0054】
試験例6 分化マーカーのmRNAレベル
遺伝子導入後4日目のmHS3OST−5過剰発現細胞において、各種分化マーカーについて、mRNAの発現量を調べた。各mRNAの発現量は、下記表2に示す配列番号の塩基配列からなるプライマー及びプローブを用い、ABI PRISM 7700 Sequence Detection System(アプライドバイオシステムズ社製)を用いて行った。
【0055】
【表2】
【0056】
結果を図7に示す。
図7に示すように、各種分化マーカーにおいてもその発現量の変化が認められ、特に、神経外胚葉マーカーであるIsl1、遠位内胚葉マーカーであるLamininB1、及び栄養膜マーカーであるCdx2におけるmRNAレベルの低下、並びに原始外胚葉マーカーであるFgf5及び原始内胚葉マーカーであるGata6のmRNAレベルの上昇がコントロール細胞に比べて有意であった。
これらの結果もまた、mHS3OST−5過剰発現細胞において未分化性が解除されていること示す。
【0057】
試験例7 HS中のmHS4C3認識エピトープの増加とFasとの結合能
mHS3OST−5過剰発現細胞においてHSとFasとの結合能が高まっていることをバインディングアッセイにより調べた。
具体的には、遺伝子導入後2日目のmHS3OST−5過剰発現細胞の細胞溶解液を8%SDS−PAGEにかけた後、PVDFメンブレンに転写し、これをブロッキング液(5%non−fatドライミルクを含むPBS pH6.5)でブロッキングし、さらにGST(グルタチオンセファローストランスフェラーゼ)融合Fas、GST融合変異導入Fas、又はGST融合FasとHS4C3との混合物を10nMの濃度で反応させた。その後、メンブレンを洗浄液(0.1%Tween20を含むPBS pH6.5)で洗浄し、HRP標識抗GST抗体(Santa Cruz社製)を室温で1時間反応させ後、HRPをECL Plus試薬を用いて発色させた。
GST融合Fas、GST融合変異導入Fasは、Fas又は変異導入FasをGSTとの融合タンパク質として大腸菌BL21株で常法により発現させ、これらをグルタチオンセファロース4B樹脂(GEヘルスケア社製)を用いて精製することで得た。
上記変異導入Fasは、HSとの結合に寄与することが予測された正電荷アミノ酸領域(配列番号36;KLRRRVH)を配列番号37(ALAAAVA)のアミノ酸配列に置き換えた構造をもつ。
結果を図8に示す。
【0058】
図8Aは、転写後のPVDFにHS4C3を反応させ、さらにHRP標識2次抗体を用いて発色させた結果を示す。mHS3OST−5過剰発現細胞において強いバンド強度が認められることから、mHS3OST−5過剰発現細胞においてグルコサミン残基の3位が硫酸化されたHSが増加していることがわかる。
図8Bは、転写後のPVDFにGST標識Fasを反応させ、HRP標識抗GST抗体で検出たものである。mHS3OST−5過剰発現細胞においてFasの結合量が高いことから、mHS3OST−5過剰発現細胞においてFas結合物質が増加していることがわかる。
図8Cは、転写後のPVDFにGST標識FasとHS4C3の混合物を反応させ、HRP標識抗GST抗体で検出したものである。この場合にはバンドが確認できないことから、FasのHSとの結合部位がHS4C3と競合していることがわかる。すなわち、FasがHSのグルコサミン残基の3位が硫酸化された部分を特異的に認識して結合していることがわかる。
図8Dは、転写後のPVDFにGST標識変異導入Fasを反応させたものである。変異導入Fasでは未変異Fasに比べてバンド強度が弱いことから、Fasの配列番号36のアミノ酸配列領域がHSとの結合に主要な役割を果たしていることがわかる。
なお、図8Eは、コントロールとしてβアクチンの量を調べた結果である。
【0059】
試験例8 Fasシグナル阻害による未分化性の回復
IETD(Ac-IETD-CHO)はカスパーゼ−8の働きを抑制する薬剤であり、Fasシグナルカスケードを遮断する薬剤である。そこで、IETDによりmHS3OST−5過剰発現細胞において引き起こされている未分化性の解除を脱することができるか、すなわち、再び未分化状態に戻りうるか調べた。
具体的には、HS3OST−5過剰発現細胞を10μM濃度のIETD(PEPTIDE INSTITUTE, INC社製)の存在下で24時間培養し、細胞を回収して溶解し、イムノブロットにより切断カスパーゼ−3とNanogのタンパク質量を、リアルタイムPCRによりOct3/とNanogのmRNA量をそれぞれ上述した方法により調べた。
結果を図9に示す。
【0060】
図9の結果から、IETD存在下で培養したHS3OST−5過剰発現細胞において、切断カスパーゼ−3のタンパク質量が低下し、これに伴いNanogのタンパク質量が増加していることがわかった。これらはいずれもコントロール細胞と同等のレベルまで回復していることもわかる。
また、IETDの存在下で培養したHS3OST−5過剰発現細胞では、Oct3/4及びNanogのmRNAの発現量もコントロール細胞と同レベルにまで回復していることがわかる。
これらの結果は、一度未分化性が解除された細胞においてさえ、Fasシグナルを阻害すれば、再び未分化性を回復しうることを示す。また、このことから、幹細胞においてFasシグナルを活性化しうる硫酸基転移酵素の活性を抑制しておけば、Fasシグナルの抑制状態が維持され、その結果幹細胞の未分化性を維持できることもわかる。
【0061】
試験例9 Fasシグナル阻害による未分化性の維持
ES細胞ではアルカリフォスファターゼ(ALP)が活性化している。したがって、ES細胞をALP染色し、陽性コロニー数を評価することにより、未分化性を維持したES細胞の評価を行うことができる。本試験では、ES細胞をFasシグナル阻害ペプチドである上記IETDで処理した場合のES細胞の分化状態をALP活性を指標として評価した。
【0062】
具体的には、6cmディッシュに1×104のES細胞(R1細胞)をまき、LIF及び10μM Ac-IETD-CHOの存在下又は非存在下で、5日間培養した。培養後、ホルマリン/メタノールで細胞を固定し、洗浄後、5-bromo-4-chloro-3-indolyl phosphate-nitroblue tetrazolium (Nacalai Tesque)で染色を行った。染色された(陽性)コロニーは顕微鏡下でカウントした。結果を図10に示す。
上記ES細胞の未分化性の維持に必要なLIFを除いて分化させた細胞では、ALP陽性コロニー率が低下していた。しかし、Ac-IETD-CHOを加え、Fasシグナルをブロックした細胞では、ALP陽性コロニー率が顕著に増加した。このことから、Fasシグナル阻害剤には、ES細胞の未分化性を維持する作用があることがわかった。
【0063】
調製例2 硫酸基転移酵素の活性が抑制された幹細胞の調製
mHS3OST−5を標的としたショートヘアピンRNAを発現するための発現プラスミドをpSUPER.retro.puro(OligoEngine社製)を用いて構築した。具体的には、配列番号38の塩基配列を有するDNAをpSUPER.retro.puroのBamHIとHindIIIサイトの間に挿入した。レトロウィルスを産生するために、上記のコンストラクトをウィルスパッケージング細胞(PLAT−E)に感染させ、このウィルスを含むPLAT−E培養液を8μg/mLPolybrene(シグマ社製)と混合した後、R1細胞と共に24時間インキュベートした。こうしてR1細胞にウィルスを感染させた後、R1細胞をLIFを含むESC培地に移し、2μg/mLのピューロマイシンの存在下で5〜7日間培養することで、mHS3OST−5の活性が抑制されたマウスES細胞(以下、mHS3OST−5抑制細胞という。)を得た。
【0064】
mHS3OST−5抑制細胞はヘパラン硫酸のグルコサミン残基の3位のOH基に対する硫酸基の転移反応が抑制されているため、ゴルジ体においてFasが糖鎖と適切に結合することができない。したがって、Fasが脂質ラフトに移動することができず、Fasシグナルの活性が抑制されている。
【0065】
mHS3OST−5以外の硫酸基転移酵素であっても、糖鎖のFasに対する親和性を制御することができるものであれば、幹細胞中で該硫酸基転移酵素の活性を制御することでFasシグナルの活性を制御することができ、mHS3OST−5と同様のメカニズムにより幹細胞の分化を制御することができる。
【0066】
以上の結果、幹細胞中でFasシグナルの活性を制御することにより、該幹細胞の分化を制御できることが示された。より具体的には、Fasシグナルの活性を抑制することにより、幹細胞の未分化性を維持することができ、逆にFasシグナルを活性化すれば、幹細胞の未分化性が解除できることが示された。
【特許請求の範囲】
【請求項1】
幹細胞のFasシグナルの活性を制御することを含む、幹細胞の分化制御方法。
【請求項2】
幹細胞中で硫酸基転移酵素の活性を制御し、これにより該幹細胞のFasシグナルの活性を制御する、請求項1に記載の方法。
【請求項3】
硫酸基転移酵素がヘパラン硫酸硫酸基転移酵素である、請求項2に記載の方法。
【請求項4】
ヘパラン硫酸硫酸基転移酵素がヘパラン硫酸3位硫酸基転移酵素である、請求項3に記載の方法。
【請求項5】
ヘパラン硫酸3位硫酸基転移酵素が下記(A)〜(C)のいずれかのアミノ酸配列を有する、請求項4に記載の方法:
(A)配列番号2のアミノ酸配列、
(B)配列番号2のアミノ酸配列において1もしくは数個のアミノ酸が置換、欠失、挿入又は付加されたアミノ酸配列、
(C)配列番号2のアミノ酸配列と80%以上の同一性を有するアミノ酸配列。
【請求項6】
幹細胞のFasシグナルの活性を抑制し、これにより幹細胞の未分化性を維持する、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
幹細胞のFasシグナルを活性化し、これにより幹細胞の未分化性を解除する、請求項1〜5のいずれか1項に記載の方法。
【請求項8】
幹細胞中で硫酸基転移酵素の活性を抑制して該幹細胞のFasシグナルを抑制し、これにより幹細胞の未分化性を維持する、請求項6に記載の方法。
【請求項9】
硫酸基転移酵素をコードする遺伝子の発現を抑制する核酸を幹細胞に導入し、該幹細胞中で該遺伝子の発現を抑制することで硫酸基転移酵素の活性を抑制する、請求項8に記載の方法。
【請求項10】
幹細胞中で硫酸基転移酵素の活性を高めて該幹細胞のFasシグナルを活性化し、これにより幹細胞の未分化性を解除する、請求項7に記載の方法。
【請求項11】
硫酸基転移酵素をコードする遺伝子を幹細胞に導入し、該幹細胞中で該遺伝子を過剰発現させることで硫酸基転移酵素の活性を高める、請求項10に記載の方法。
【請求項12】
幹細胞にFasシグナル阻害剤を投与して該幹細胞のFasシグナルを抑制し、これにより幹細胞の未分化性を維持する、請求項6に記載の方法。
【請求項13】
Fasシグナル阻害剤が、配列番号39のアミノ酸配列を有するペプチドを含む、請求項12に記載の方法。
【請求項14】
幹細胞が哺乳類に由来する、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
幹細胞がES細胞である、請求項14に記載の方法。
【請求項16】
請求項1〜15のいずれか1項に記載の方法で分化制御された細胞。
【請求項1】
幹細胞のFasシグナルの活性を制御することを含む、幹細胞の分化制御方法。
【請求項2】
幹細胞中で硫酸基転移酵素の活性を制御し、これにより該幹細胞のFasシグナルの活性を制御する、請求項1に記載の方法。
【請求項3】
硫酸基転移酵素がヘパラン硫酸硫酸基転移酵素である、請求項2に記載の方法。
【請求項4】
ヘパラン硫酸硫酸基転移酵素がヘパラン硫酸3位硫酸基転移酵素である、請求項3に記載の方法。
【請求項5】
ヘパラン硫酸3位硫酸基転移酵素が下記(A)〜(C)のいずれかのアミノ酸配列を有する、請求項4に記載の方法:
(A)配列番号2のアミノ酸配列、
(B)配列番号2のアミノ酸配列において1もしくは数個のアミノ酸が置換、欠失、挿入又は付加されたアミノ酸配列、
(C)配列番号2のアミノ酸配列と80%以上の同一性を有するアミノ酸配列。
【請求項6】
幹細胞のFasシグナルの活性を抑制し、これにより幹細胞の未分化性を維持する、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
幹細胞のFasシグナルを活性化し、これにより幹細胞の未分化性を解除する、請求項1〜5のいずれか1項に記載の方法。
【請求項8】
幹細胞中で硫酸基転移酵素の活性を抑制して該幹細胞のFasシグナルを抑制し、これにより幹細胞の未分化性を維持する、請求項6に記載の方法。
【請求項9】
硫酸基転移酵素をコードする遺伝子の発現を抑制する核酸を幹細胞に導入し、該幹細胞中で該遺伝子の発現を抑制することで硫酸基転移酵素の活性を抑制する、請求項8に記載の方法。
【請求項10】
幹細胞中で硫酸基転移酵素の活性を高めて該幹細胞のFasシグナルを活性化し、これにより幹細胞の未分化性を解除する、請求項7に記載の方法。
【請求項11】
硫酸基転移酵素をコードする遺伝子を幹細胞に導入し、該幹細胞中で該遺伝子を過剰発現させることで硫酸基転移酵素の活性を高める、請求項10に記載の方法。
【請求項12】
幹細胞にFasシグナル阻害剤を投与して該幹細胞のFasシグナルを抑制し、これにより幹細胞の未分化性を維持する、請求項6に記載の方法。
【請求項13】
Fasシグナル阻害剤が、配列番号39のアミノ酸配列を有するペプチドを含む、請求項12に記載の方法。
【請求項14】
幹細胞が哺乳類に由来する、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
幹細胞がES細胞である、請求項14に記載の方法。
【請求項16】
請求項1〜15のいずれか1項に記載の方法で分化制御された細胞。
【図4】

【図5】
【図6】
【図7】
【図9】
【図10】
【図1】
【図2】
【図3】

【図8】
【図5】
【図6】
【図7】
【図9】
【図10】
【図1】
【図2】
【図3】
【図8】
【公開番号】特開2012−161247(P2012−161247A)
【公開日】平成24年8月30日(2012.8.30)
【国際特許分類】
【出願番号】特願2011−21846(P2011−21846)
【出願日】平成23年2月3日(2011.2.3)
【出願人】(598123138)学校法人 創価大学 (49)
【Fターム(参考)】
【公開日】平成24年8月30日(2012.8.30)
【国際特許分類】
【出願日】平成23年2月3日(2011.2.3)
【出願人】(598123138)学校法人 創価大学 (49)
【Fターム(参考)】
[ Back to top ]