
Fcレセプター細胞外領域を含むポリペプチド及びその製造方法
【課題】工業生産規模において、Fcレセプター細胞外領域を含むポリペプチド、特に、FcγRI細胞外領域を含むポリペプチドを生産する微生物の提供。
【解決手段】FcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、特定の塩基配列を含むポリヌクレオチド、又は特定の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。該ポリヌクレオチドが挿入された組換えベクター。該ベクターにより形質転換された形質転換体。
【解決手段】FcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、特定の塩基配列を含むポリヌクレオチド、又は特定の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。該ポリヌクレオチドが挿入された組換えベクター。該ベクターにより形質転換された形質転換体。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Fcレセプター細胞外領域を含むポリペプチド及びその製造方法、並びに該ポリペプチドの利用方法に関する。
【背景技術】
【0002】
Fcレセプターは、免疫グロブリン分子のFc領域に結合する一群の分子である。個々の分子は、免疫グロブリンスーパーファミリーに属する認識ドメインによって、単一の、あるいは同じグループの免疫グロブリンイソタイプをFcレセプター上の認識ドメインによって認識している。これによって、一定の免疫応答においてどのアクセサリー細胞が動因されるかが決まってくる(非特許文献1)。Fcレセプターは、さらにいくつかのサブタイプに分類することができ、IgG(免疫グロブリンG)に対するレセプターとしては、FcγRI、FcγRIIa、FcγRIIb及びFcγR3の存在が報告されている(非特許文献1)。上記レセプターの中でも、FcγRIとIgGとの親和性は高く、その平衡解離定数(Kd)は10−8 M以下であることが報告されている(非特許文献2)。
【0003】
FcγRIはIgG(免疫グロブリンG)に対するレセプターであり、単球及びマクロファージ上に構成的に発現され、好中球および好酸球上においては誘導的に発現される。FcγRIは、細胞外領域、膜貫通領域、細胞質内領域に区分され、IgGとの結合は、IgGのFc領域とFcγRIの細胞外領域で起こり、その後細胞質へとシグナルが伝達される。FcγRIはIgGとの結合に直接関わる分子量約42,000のα鎖と、γ鎖の2種類のサブユニットによって構成されており、γ鎖は細胞膜と細胞外領域の境界にあるシステインを介した共有結合によりホモダイマーを形成している(非特許文献1)。
【0004】
近年になり、Fcレセプターの予想外の免疫抑制的な生物学的特性は、特に、自己免疫疾患または自己免疫症候群、移植物の拒絶および悪性リンパ増殖の領域において医薬として注目を浴びつつある。また、FcγR1の機能である抗体の吸着能は各種抗体精製用クロマトグラフィーゲルの捕捉機能タンパク質としても利用することができる。
FcγRIα鎖のアミノ酸配列および遺伝子配列は、Janet等により明らかにされ(非特許文献2)、その後、遺伝子組換え技術により、大腸菌(非特許文献、特許文献1)あるいは動物細胞を利用することによる、FcγRIの発現が報告された。しかしながら、大腸菌を利用した発現系においてはFcγRIの細胞外領域タンパク質の発現量が極めて低く、工業生産規模において生産することは困難である。また、動物細胞を用いた系では、大腸菌を用いた系よりも高い発現量を実現できることが報告(非特許文献3)されているが、培養に時間がかかるため、生産性は必ずしも高くない。
【0005】
【特許文献1】特表2004−530419号公報
【非特許文献1】J.V.Ravetch等, Annu.Rev.Immunol., 9, 457頁, 1991年
【非特許文献2】J.M.Allen等, Science, 243, 378頁, 1989年
【非特許文献3】A.Paetz等, Biochem.Biophys.Res.Commun., 338, 1811頁, 2005年
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は工業生産規模において、Fcレセプター細胞外領域を含むポリペプチド、特に、FcγRI細胞外領域を含むポリペプチドを生産する微生物を作製することである。さらには、新規FcγRI細胞外領域を含むポリペプチドの生産系を利用してFcγRI細胞外領域を含むポリペプチドの生産を行い、該FcγRI細胞外領域を含むポリペプチドを提供することにある。
【課題を解決するための手段】
【0007】
本願発明者らは、上記課題を解決すべく鋭意検討した結果、ヒトFcγRI遺伝子から転写されるmRNAのコドンを、大腸菌宿主において利用頻度が高いコドンにするべく該遺伝子の配列を変換し、変換したレセプター遺伝子が挿入された組換えベクターに導入することにより、大腸菌においてFcγRIの発現量が向上することを見出した。
すなわち、本願発明者らは、ヒトFcレセプターの一種類であるFcγRIの遺伝子配列を見直し、大腸菌で該レセプターを発現させるべく大腸菌が容易に翻訳するコドンに対応するようにDNA配列を変換し、さらに、対応する変換コドンからなるオリゴヌクレオチドを合成後、PCR法により全長FcγRI遺伝子を作製し、該遺伝子が挿入された発現プラスミドベクターを大腸菌に導入した結果、大腸菌宿主においてFcγRI細胞外領域タンパク質の発現量が飛躍的に向上することを見出し、本発明を完成するに至った。
【0008】
すなわち、本発明は、FcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、配列番号:2に記載の塩基配列を含むポリヌクレオチド、又は配列番号:2に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチドを提供する。
また、本発明は、FcγRI細胞外領域を含むポリペプチドからシグナル配列を除去した領域をコードするポリヌクレオチドであって、配列番号:3に記載の塩基配列を含むポリヌクレオチド、又は配列番号:3に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチドを提供する。
また、本発明は、シグナル配列を除去した、FcγRI細胞外領域を含むポリペプチドのN−末端側にメチオニンが付加されたポリペプチドをコードするポリヌクレオチドであって、配列番号:4に記載の塩基配列を含むポリヌクレオチド、又は配列番号:4に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチドを提供する。
【0009】
また、本発明は、配列番号2〜4のいずれか1に記載の塩基配列を含むポリヌクレオチドによりコードされるポリペプチドを提供する。
また、本発明は、配列番号2〜4のいずれか1に記載の塩基配列を含むポリヌクレオチドに、発現制御配列が機能的に付加されたポリヌクレオチドを提供する。
また、本発明は、上記のいずれかのポリヌクレオチドが挿入された組換えベクターを提供する。
また、本発明は、上記のいずれかのポリヌクレオチド、又は上記組換えベクターにより形質転換された形質転換体を提供する。
また、本発明は、上記形質転換体を培養する工程を含む、FcγRI細胞外領域を含むポリペプチドの製造方法を提供する。
また、本発明は、上記製造方法によって得られたポリペプチドを、担体上に固定してなる、抗体精製用吸着剤を提供する。
また、本発明は、上記抗体精製用吸着剤に抗体を吸着させる方法を提供する。
【発明の効果】
【0010】
ヒトFcγRI細胞外領域を含むポリペプチドをコードする遺伝子コドンを、大腸菌型にすることによって、FcγRI細胞外領域を含むポリペプチドを、大腸菌に大量に発現させることが可能となる。この発現系は、工業的規模におけるFcγRI細胞外領域を含むポリペプチドの生産に有用なものである。また、このようにして得られた、FcγRI細胞外領域を含むポリペプチドは、抗体アフィニティークロマトゲルの捕捉基として利用でき、すなわち、抗体精製用吸着剤として利用することができる。
【発明を実施するための最良の形態】
【0011】
以下、本発明について詳細に説明する。
まず、FcγRIタンパク質をコードする遺伝子について説明すると、FcγRI遺伝子は配列番号:1に記載の約2.2Kベースの塩基配列を含むDNAである。このDNAがコードするタンパク質の機能構造は、は公的データベース(European Bioinformatics Institute、 URL:http://www.ebi.ac.uk/、 アクセス日:2006年3月2日)に公表され公知であり、構造上のドメイン、あるいはシグナル配列、細胞膜貫通領域等のアミノ酸配列の決定は容易である。
【0012】
次に、本発明のポリヌクレオチドについて説明する。
本発明のポリヌクレオチドは、ヒトFcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、その遺伝子情報のコドンを、適当な宿主細胞のコドン使用頻度に最適化したポリヌクレオチドである。すなわち、ヒトFcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドが挿入された組換えベクターを宿主細胞に形質転換して得られた形質転換体を培養してポリペプチドを生産せる場合に、その宿主細胞が容易に翻訳することができるコドンに対応することができるように、ポリヌクレオトドのコドンを変換することによって、効率よく目的のポリペプチドの生産をすることができるようにしたものである。本明細書において、本発明を説明する場合、大腸菌を宿主細胞として用いた場合について説明するが、本発明は、宿主細胞として大腸菌を用いる場合に限定されず、他の宿主細胞の場合についても適用される。すなわち、他の宿主細胞を用いる場合は、その宿主細胞の最適なコドンについてゲノム解析することによって決定し、他の宿主細胞においても適用することが可能である。
【0013】
本発明のポリヌクレオチドは、以下の(a)、(b)又は(c)で表わされるポリヌクレオチドである。
(a)FcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、配列番号:2に記載の塩基配列を含むポリヌクレオチド、又は配列番号:2に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
(b)FcγRI細胞外領域を含むポリペプチドからシグナル配列を除去した領域をコードするポリヌクレオチドであって、配列番号:3に記載の塩基配列を含むポリヌクレオチド、又は配列番号:3に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
(c)シグナル配列を除去した、FcγRI細胞外領域を含むポリペプチドのN−末端側にメチオニンが付加されたポリペプチドをコードするポリヌクレオチドであって、配列番号:4に記載の塩基配列を含むポリヌクレオチド、又は配列番号:4に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
【0014】
(a)のポリヌクレオチドについて説明する。(a)のポリヌクレオチドは、ヒトFcγRI細胞該領域を含むポリペプチドをコードする遺伝子であり、その遺伝子情報のコドンを、ヒト型から大腸菌型のコドンとなるように変換した、すなわち大腸菌のコドン使用頻度に最適化したポリヌクレオチドである。
言い換えると、(a)のポリヌクレオチドは、ヒトFcγRI細胞外領域をコードする遺伝子DNAから転写されるmRNA中において存在する大腸菌レアコドン(rare codon)を、コードするアミノ酸を同一のまま、大腸菌コドンで利用頻度が高いコドンに変換したポリヌクレオチド配列である。なお、レアコドンとは、その宿主におけるコドンの使用頻度が少ないものと定義することができる。宿主における使用頻度は、ゲノム遺伝子の解析結果を参考にすることで推測することが可能であり、大腸菌においては、アミノ酸であるArgコドンのAGA、AGG、CGG、CGA、IleコドンのAUA、LeuコドンのCUA、GlyコドンのGGA、ProコドンのCCCを例示することができる。コドンの使用頻度の解析は公的データベース(URL:http://www.kazusa.or.jp/codon/、アクセス日:2006年3月2日)を利用することによっても可能である。
【0015】
コドンの変換は、対応するDNA配列の塩基配列を変えることにより可能である。DNA配列の塩基置換を行なうには、たとえば、Site−directed mutagenesis法など公知の変異導入法を利用することができる。好ましくは、合成オリゴヌクレオチドとPCRを組合わせたDNAWorks法を例示することができる(Nucleic Acid Res., 30, e43頁, 2002年)。この方法によれば、アミノ酸配列から数十塩基からなるオリゴヌクレオチド群を合成し、PCRにより合成オリゴヌクレオチドをアッセンブリーさせることによって完全長の遺伝子を合成することができる。
遺伝子すべてのレアコドンを変換することも可能であり、部分的に変換することも可能である。mRNAの発現量を定量することによって適宜変換割合を変えることができる。
【0016】
(b)のポリヌクレオチドは、(a)のポリヌクレオチドから、シグナル配列をコードする部分を除去したものであり、すなわち、FcγRI細胞外領域を含むポリペプチドからシグナル配列を除去した領域をコードするポリヌクレオチドである。
なお、シグナル配列とは、細胞内で発現したタンパク質が細胞膜を通過し、細胞外において分泌するための領域であり、通常、該タンパク質のN−末端側に存在しており、細胞膜通過後、特定のプロテアーゼ酵素によって切断される。シグナル配列としては、配列番号1のアミノ酸番号1から15、あるいは1から20のペプチド配列を例示することができる。FcγRI細胞外領域は、さらに、シグナル配列を除去し、IgGと相互作用する部位のみでも機能的には十分である。
【0017】
(c)のポリペプチドは、シグナル配列を除去した、FcγRI細胞外領域を含むポリペプチドのN−末端側にメチオニンが付加されたポリペプチドをコードするポリヌクレオチドである。
【0018】
本発明において、ポリペプチドとは、天然に存在するポリヌクレオチドであってもよいし、また、人工的に合成されたヌクレオチドであってもよい。
【0019】
(a)、(b)及び(c)のポリヌクレオチドは、それぞれ、配列番号:2、配列番号:3及び配列番号:4に記載の塩基配列を含むポリヌクレオチドに加え、例えば、配列番号:2、配列番号:3及び配列番号:4に記載の塩基配列を含むポリヌクレオチドがコードするポリペプチドにおいて、生物学的活性の同一性が損なわれない範囲内において、そのポリペプチド鎖に対する、アミノ酸配列中の一部(好ましくは1〜20個、更に好ましくは1〜10個程度、最も好ましくは1〜3個)のアミノ酸が欠失、置換又は付加したアミノ酸配列からなるポリペプチドをコードするようなポリヌクレオチドをも含む。この場合も、生物学的活性の同一性とは、FcγRIのイムノグロブリンに対する結合性を意味する。ここで、イムノグロブリンとは、好ましくはIgGであり、更に好ましくはIgG1である。
当業者であれば、(a)、(b)及び(c)のポリペプチドに、部位特異的突然変異法などを用いて、適宜、置換、欠失、及び/又は付加変異を導入することができる。
【0020】
上記(a)、(b)及び(c)のポリヌクレオチドは、上記(a)、(b)、(c)の配列に対して実質的に相同なポリヌクレオチド配列をも含む。実質的に相同であるとは、二つの核酸に関して、下記の配列比較アルゴリズムの一つを用いて、または、手動整列および目視試験により測定し、最大対応で整列化させた場合に、75%を超え、好ましくは85%を超え、より好ましくは95%を超えるDNA同一性を有する配列または部分配列を意味する。なお、この定義は、その相補体が試験配列にハイブリダイズする配列も意味することは言うまでもない。配列比較のために、典型的には一つの配列が基準配列として働き、それに対して試験配列を比較する。配列比較アルゴリズムを用いる場合は、試験配列および基準配列をコンピュータに入力し、必要があれば部分座標を指定し、配列アルゴリズムプログラムのパラメータを指定する。パラメータはデフォルト値を用いてもよく、または、代わりのパラメータを指定することもできる。次いで、配列比較アルゴリズムが、プログラムのパラメータに基づき、基準配列に対する試験配列の配列同一性の割合を計算する。比較のための配列整列方法は当該技術分野において公知である。比較のための、最適配列は、例えば、局所相同性アルゴリズム(Smith等, Adv.Appl.Math., 2, 482頁, 1981年)、相同性整列化アルゴリズム(Needlman等, J.Mol.Biol., 48, 443頁, 1970年)、類似性検索法(Person等, Proc.Natl.Acd.Sci.USA, 85, 2444頁, 1988年)を例示することができる。
【0021】
有用なアルゴリズムの一例はPileup法であり、連続性のペアワイズアライメントを用いて関連配列群から多重配列を作製し、関係と配列同一性の割合を示すことができる。配列同一性の割合および配列類似性を求めるのに適したアルゴリズムのもう一例はBLASTアルゴリズムである(Altschul等, J.Mol. iol., 215, 403頁, 1990年)。
【0022】
また、本発明のポリヌクレオチドには、上記(a)、(b)、(c)のポリヌクレオチドと、ストリンジェントな条件下でハイブリダイズするDNA配列が含まれるものである。ストリンジェントな条件下でハイブリダイズするとは、プローブが核酸混合物中で、その標的配列にはハイブリダイズするが、他の核酸にはハイブリダイズしない条件を意味する。ストリンジェントな条件は配列依存性があり、異なる環境下では異なることになる。より長い配列では、特異的に、より高温でハイブリダイズする。一般に、高度にストリンジェントな条件は、既定のイオン強度およびpHで特異的配列の熱融解温度より約5〜10℃低くなるように選択される。低ストリンジェントな条件は、一般的には融解温度より約15〜30℃低くなるように選択される。融解温度は、既定のイオン強度、pH、核酸配列で標的核酸に対して相補的なプローブにおいて50%が平衡状態で占める温度である。
【0023】
具体的には、ストリンジェントな条件下でハイブリダイズできるポリヌクレオチドとは、配列番号:1に記載中の任意の少なくとも20個、好ましくは少なくとも30個、例えば40、60または100個の連続した配列を有するポリヌクレオチドをプローブとし、例えばECL direct nucleic acid labeling and detection system(Amersham Pharmacia Biotech社製)を用いて、マニュアルに記載の条件(例えば、wash:42℃、0.5×SSCを含むprimary wash buffer)においてハイブリダイズするポリヌクレオチドを指す。より具体的な「ストリンジェントな条件」とは、例えば、通常、42℃、2×SSC、0.1%SDSの条件であり、好ましくは50℃、2×SSC、0.1%SDSの条件であり、さらに好ましくは、65℃、0.1×SSCおよび0.1%SDSの条件であるが、これらの条件に特に制限されない。ハイブリダイゼーションのストリンジェンシーに影響する要素としてはこれら温度、塩濃度に加えプローブ濃度、プローブの長さ、反応時間を含む複数の要素が考えられ、当業者であればこれら要素を適宜選択することで最適なストリンジェンシーを実現することが可能である。ハイブリダイゼーション法の詳細な手順については、「Molecular Cloning, A Laboratory Manual 2nd ed.」(Cold Spring Harbor Press(1989)、特にSection9.47-9.58)、「Current Protocols in Molecular Biology」(John Wiley&Sons(1987-1997)、特にSection6.3-6.4)、「DNA Cloning 1: Core Techniques, A Practical Approach 2nd ed.」(Oxford University(1995)、特にSection2.10)等を参照することができる。
【0024】
本発明は、上述した、(a)、(b)及び(c)のポリヌクレオチドによりコードされるポリペプチドを含む。このようなポリペプチドとしては、例えば、配列番号:1に記載のアミノ酸配列からなるポリペプチド、及びがいアミノ酸配列中の一部(好ましくは1〜20個、更に好ましくは1〜10個程度、最も好ましくは1〜3個)のアミノ酸が欠失、置換又は付加したアミノ酸配列からなるポリペプチドが挙げられる。
【0025】
本発明は、本発明のポリヌクレオチドが挿入された組換えベクターを提供する。上記ポリヌクレオチドを公知の発現ベクターに挿入することにより、FcγRI細胞外領域を含むポリペプチドを発現するベクター(組換えベクター)が提供される。また、この組換えベクターで形質転換した形質転換体を培養することにより、FcγRI細胞外領域を含むポリペプチドを形質転換体から得ることができる。すなわち、本発明は、本発明のポリヌクレオチドが挿入された組換えベクターにより形質転換された形質転換体を培養し、その培養物から、FcγRI細胞外領域を含むポリペプチドを回収することを含む、FcγRI細胞外領域を含むポリペプチドの製造方法が提供される。
【0026】
本発明のポリヌクレオチドは、そのまま用いて適当な細胞に形質転換してもよいが、通常は、適当な公知の発現ベクターに挿入される。発現ベクターへの挿入は、その発現ベクターの適当な位置に遺伝子工学的に挿入すればよい。適当な位置とは、発現ベクターの複製機能、所望の抗生物質マーカー、又は伝達性に関する領域を破壊しないような位置を意味する。本発明のポリヌクレオチドは、発現制御配列が機能的に付加されたものであることが好ましい。発現制御配列が機能的に付加されるとは、例えば、プロモーター、エンハンサー、オペレーター、インデューサー等が付加されることであり、このような配列が付加されることによって、ポリペプチドの宿主細胞内での発現が可能となる。
【0027】
本発明において、組換えベクターを形質転換する宿主細胞としては、微生物が好ましく、原核細胞が更に好ましく用いられる。原核細胞の中でも、大腸菌が好ましいが、本発明のおいては宿主細胞を限定するものでなく、遺伝子工学において用いられる宿主細胞については全て用いられるものとする。利用可能な微生物としては、例えば以下のような微生物が挙げられる。
【0028】
大腸菌(エシェリヒア(Escherichia)属)
バチルス(Bacillus)属
シュードモナス(Pseudomonas)属
セラチア(Serratia)属
ブレビバクテリウム(Brevibacterium)属
コリネバクテリイウム(Corynebacterium)属
ストレプトコッカス(Streptococcus)属
ラクトバチルス(Lactobacillus)属など宿主ベクター系の開発されている細菌
ロドコッカス(Rhodococcus)属
ストレプトマイセス(Streptomyces)属など宿主ベクター系の開発されている放線菌
サッカロマイセス(Saccharomyces)属
クライベロマイセス(Kluyveromyces)属
シゾサッカロマイセス(Schizosaccharomyces)属
チゴサッカロマイセス(Zygosaccharomyces)属
ヤロウイア(Yarrowia)属
トリコスポロン(Trichosporon)属
ロドスポリジウム(Rhodosporidium)属
ピキア(Pichia)属
キャンディダ(Candida)属などの宿主ベクター系の開発されている酵母
ノイロスポラ(Neurospora)属
アスペルギルス(Aspergillus)属
セファロスポリウム(Cephalosporium)属
トリコデルマ(Trichoderma)属などの宿主ベクター系の開発されているカビ
【0029】
以下、宿主として大腸菌を用いる場合について説明する。発現ベクターにポリヌクレオチドを挿入する場合、上記ポリヌクレオチドをプロモータ機能を有するDNA配列を付加させることが好ましい。プロモータとは、タンパク質のコード領域又は機能的RNAの発現を制御することが可能なDNA配列を意味し、大腸菌の場合、lacプロモータ、trcプロモータ、T7プロモータ等を例示することができる。その中でも、好ましくは転写活性が強いT7プロモータである。
【0030】
プラスミドベクター(発現ベクター)としては、形質転換する細胞内で安定に存在し、複製することができる特性を有するものであれば特に制限なく用いることができる。このようなプラスミドベクターとしては、大腸菌の形質転換に利用されるpUC系、pBR系、pET系、広宿主域(Broad−Host−Range)プラスミドベクター等、あるいは対象細胞内で複製可能なプラスミドベクターとのシャトルベクターを挙げることができる。例えば、成書(Barbara E. Funnell, PLASMID BIOLOGY,545〜566頁, ASM press)に記載されているものが挙げられる。さらに、形質転換対象となる細胞が保有するプラスミドを利用しても良い。また、後述するようなベクターを用いることもできる。
【0031】
宿主細胞としては、大腸菌を用いる場合、例えば、K12系のJM109株を用いることが好ましい。さらに好ましくは、T7プロモータ配列が利用できるBL21(DE3)株などを例示することができる。
また、本発明の宿主細胞としては上記大腸菌等を変異処理することにより誘導される大腸菌変異株を利用することもできる。変異処理はニトロソグアニジン、メタンスルホン酸エチル、紫外線、放射線等の当業者では周知の変異処理剤を利用して行えばよい。
【0032】
大腸菌等の微生物への外来遺伝子の導入および発現のための手順乃至方法は、本発明において下記したところ以外のものにおいても、遺伝子工学の分野により慣用されているものを含み、その手法乃至方法(たとえば、“Plasmid transformation of Escherichia coli and other bacteria”, Method in Enzymology, 204, 63〜113頁, 1991年, Academic Press)に準じて実施すればよい。具体的にはヒートショック法、エレクトロポレーション法等を挙げることができる。
【0033】
本発明のFcγRI細胞外領域を含むポリペプチドの製造方法は、本発明のポリペプチドが挿入された組換えベクターにより形質転換された形質転換体を培養する工程を含む。上記方法は、具体的には、上記形質転換体を培養し、その培養物から、FcγRI細胞外領域を含むポリペプチドを回収することを含む。
【0034】
本発明のFcγRIを生産するために用いられる形質転換体(大腸菌)は、当該技術分野で周知であり、かつ、選択した宿主細胞の培養に好適な培地で増殖させる。好適な培地の例としては必要な栄養源を補ったLB(ルリアブロス)培地を挙げることができる。好ましい実施態様においては、組換えベクターで形質転換体の増殖を選択的に可能にするために、培地に、組換えベクターの構成を基にした選抜剤を含有させることが好ましい。例えば、カナマイシン耐性遺伝子を発現する細胞の増殖のためにカナマイシンが培地に添加される。培地には、炭素、窒素および無機塩供給源の他に、適当な栄養源を加えてもよい。所望により、グルタチオン、システイン、シスタミン、チオグリコレート、ジチオスレイトールからなる群から選択される一種類以上の還元剤を含んでも良い。大腸菌増殖における培養温度は、20℃〜40℃、より好ましくは25℃〜35℃であり、より好ましくは約30℃である。培地のpHは、大腸菌培養においては約6.8〜7.4であり、より好ましくは7.0である。
【0035】
本発明の組換えベクターに誘導性のプロモータを用いる場合には、良好なタンパク質発現が起こるような条件下で誘導する。誘導剤としてはIPTGを例示することができ、培養液の濁度(660 nmにおける吸光度)を測定し、約0.5〜1.0の増殖期間に適当量のIPTGを添加し、引き続き培養することによりタンパク質の発現を誘導することができる。IPTGの添加濃度は0.1〜1.0mMであり、好ましくは0.5mM程度である。IPTG誘導に関する種々の条件は当該技術分野においては周知である。
【0036】
培養液から、本発明のFcγRI細胞外領域を含むポリペプチドを回収するには、発現の形態によって適宜抽出方法を選択すればよい。培養上清に発現する場合は菌体を遠心分離操作によって分離し、得られる培養上清からFcγRI細胞外領域を含むポリペプチドを抽出すればよい。一方、細胞内にFcγRI細胞外領域を含むポリペプチドが発現する場合には、遠心分離操作により菌体を集め、酵素処理剤や界面活性剤等を添加することにより菌体を破砕し、FcγRI細胞外領域を含むポリペプチドを抽出することができる。抽出タンパク質の中からFcγRI細胞外領域を含むポリペプチドを分離・精製するには、当該技術分野において公知の方法を用いることができる。例えば、液体クロマトグラフィーを利用することができる。液体クロマトグラフィーとしては、イオン交換クロマト、疎水性相互作用クロマト、ゲルろ過、アフィニティークロマトなどを挙げることができる。これらのクロマトを組み合わせて精製操作を行うことによって、高純度なFcγRI細胞外領域を含むポリペプチドを調製することができる。
【0037】
次に、本発明の抗体精製用吸着剤について説明する。
本発明の抗体精製用吸着剤は、上述したポリペプチドの製造方法によって得られたポリペプチドを、担体上に固定してなる。
用いられる担体としては、ポリペプチドを化学的又は物理的に固定化することのできるものであれば、特に制限なく用いることができ、クロマトグラフィー用担体を用いることができる。このような担体としては、例えば、セルロース、アガロース等の多糖類、ガラス、セラミックス、又はポリプロピレン、塩化ビニル、ポリスチレン等のプラスチック素材等が挙げられる。このような担体は市販されているものを用いることができ、例えば、Novagen社製、His・Bind purificationキット(登録商標)中のHis・Bind Resinゲル、商品名「HiTrap NHS−Activated HP」(Amersham Biosciences社製)、商品名「トシル基活性化ダイナビーズM−280」(Dynal社製)等を用いることができる。
【0038】
なお、ポリペプチドを担体に固定化する際に、ペプチドの末端に担体との競合に利用できる適当なスペーサーを挿入したポリペプチドを用いることが好ましい。スペーサーとしては、例えば、アミノ酸、ペプチド、炭素数2〜8個程度の主鎖を有糸、好ましくは水酸基等の親水性の官能基を有するもの(例えば、両末端に活性基を有するポリビニルアルコールなど)、糖鎖等が挙げられる。上記アミノ酸としては、例えば、ヒスチジン、アルギニン、リシン、アスパラギン、グルタミン、システイン、アスパラギン酸、グルタミン酸等が例示でき、ヒスチジンが複数個(通常、2〜10個)連続して結合した、ポリヒスチジンが好ましく用いられる。このようなポリヒスチジンをスペーサーとして用いる場合、担体とポリペプチドとの結合はNiキレートとの親和性により結合する。アミノ酸がスペーサーである場合は、スペーサーを挿入したペプチドを連続した1個のポリペプチドとして合成することができる。遺伝子組み換え法によりポリペプチドを産生させる場合、ベクターとして、組換タンパク質の末端にヒスチジンタグを付加させるようなベクターに、ポリヌクレオチドを挿入し、それを適当な宿主に形質転換させ、ポリペプチドを産生させることができる。このようなベクターとしては、市販されているものを用いることができ、例えば、ノバジェン社製のpET28b+等が挙げられる。
【0039】
また、本発明は、上記抗体精製用吸着剤に抗体を吸着させる方法を提供する。また、上記抗体精製用吸着剤は、例えば、抗体の精製分離に用いることができる。すなわち、本発明は、上記抗体精製用吸着剤を用いた、抗体の精製方法を提供する。
抗体精製用吸着剤に抗体を吸着させるには、例えば、上記抗体精製用吸着剤を、例えばカラム等につめ、ここに、抗体を含む試料を中性条件下に導入する。これにより、試料中の抗体が、抗体精製用吸着剤に固定化されているポリペプチドと反応して吸着される。
また、抗体精製用吸着剤を、抗体の精製分離に用いる場合は以下の通りに実施する。抗体が吸着した抗体精製用吸着剤から抗体を精製分離するには、カラム中に、塩濃度の高い、例えばNaCl、KCl、Na2SO4を含む緩衝液を流すことにより、又はカラム中のpHを変化させることにより実施する。
【実施例】
【0040】
以下、本発明の実施の形態について詳細に説明する。なお、本発明は、これらの実施例のみに限定されるものではなく、発明の要旨を逸脱しない範囲で、任意に変更が可能であることは言うまでもない。
(実施例1)FcレセプターをコードするDNA配列の設計
配列番号:1のFcγRIの細胞外領域のアミノ酸配列を基にDNAworks法(Nucleic Acid Res., 30, e43頁, 2002年)を用いて、コドンを大腸菌型に変換した。該方法によりコドンを変換したDNA配列とヒト型コドンをアライメントした結果を図1に示す。図1に示す通り、アミノ酸配列は変わらずに、DNAの配列変換が行われ、そのDNA配列間の類似性は75%であった。
【0041】
(実施例2)FcレセプターをコードするDNA配列の作製
ヒト型コドンから大腸菌型コドンに変換したDNA配列を作製するために52種類のオリゴヌクレオチドを合成した。合成したオリゴヌクレオチドを配列番号5から56に示す。次いで、二段階のPCRを行い、オリゴヌクレオチドから完全長のFcγRIをコードするDNAを作製した。一段階目のPCRの反応液は表1に示す通りであり、反応条件は94℃、5分の熱処理後、94℃、30秒間の第一ステップ、62℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、次いで、72℃、7分の第四ステップである。表1中のDNAミックスとは、52種類の合成した50 pmol/μlのオリゴヌクレオチドをそれぞれ一定量サンプリングし混合した溶液を意味する。
【0042】
第二段階目のPCRは一段階目のPCRの反応液を用いて、表2の反応液組成で行った。PCRプライマーの配列は、配列番号5(5’−ATGTGGTTTCTGACCACGCTGTTGCTGTGGGTGCCGGT−3’)と配列番号56(5’−GGTCGCGCCCTGCGGCTCCTTACGATGCAC−3’)のオリゴヌクレオチドを用いた。反応条件は94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップである。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。 次いで、目的バンドを抽出(QIAquick Gel extraction kit:キアゲン社)後、抽出DNAの5’末端をリン酸化(TaKaRa BKL Kit:タカラバイオ社)し、制限酵素Sma Iで消化したpUC19プラスミドベクターに挿入し、大腸菌JM109株(タカラバイオ社)を形質転換し調製した。これをpECFcRとした。図2に構造を示す。
【0043】
【表1】

【0044】
【表2】
【0045】
(実施例3)Fcレセプター発現プラスミドベクターの作製
FcγRIの細胞外領域を発現させるために、pECFcRをテンプレートにし配列番号57(5’−CCCATGGCTGTGATTACGCTGCAACCACCG−3’:アンダーライン部分(CCATGG)は制限酵素NcoIサイト)と配列番号 58(5’−CCAACCTTGACCGGGGTCGGCAGTTGAAGACC−3’ :アンダーライン部分(AACCTT)は制限酵素HindIIIサイト) のオリゴヌクレオチドをプライマーにしてPCRを行い、細胞外領域をコードするDNAを増幅した。反応条件は94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップである。反応液組成を表3に示す。
【0046】
【表3】
【0047】
反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。次いで、QIAquick Gel extraction kit(キアゲン社)を用いて、目的バンドを抽出した後、増幅DNAを制限酵素NcoIとXhoIにより消化し、これらの制限酵素を用いて事前に消化したpET28b+(ノバジェン社)と、Ligation Kit Ver2(タカラバイオ社)を用いてライゲーションした。次いで、50μg/mlの抗生物質カナマイシンを添加したLB寒天培地により大腸菌JM109を形質転換し調製した。これをpETECFcRとした。図3に構造を示す。得られたpETECFcRは、ヒト型コドンを用いているものである。
【0048】
上記と並行して、FcγRIの細胞外領域をコードするDNAを用いた発現ベクターも作製した。挿入断片の作製はPCRで行い、Human cDNAcloneTC119841プラスミドベクター(Origene社)をテンプレートとし、配列番号59(5’−CCCATGGCAGTGATCACTTTGCAGCCTCC−3’ :アンダーライン部分(CCATGG)は制限酵素NcoIサイト)と配列番号60(5’−GCTCGAGGACAGGAGTTGGTAACTGGAGGCC−3’:アンダーライン部分(CTCGAG)は制限酵素XhoIサイト)のオリゴヌクレオチドをPCRプライマーとした。PCRは94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップである。反応液組成を表4に示す。
【0049】
【表4】
【0050】
反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。次いで、QIAquick Gel extraction kit(キアゲン社)を用いて、目的バンドを抽出した後、増幅DNAを制限酵素NcoIとXhoIにより消化し、これらの制限酵素を用いて事前に消化したpET28a+(ノバジェン社)と、Ligation Kit Ver2(タカラバイオ社)を用いてライゲーションした。次いで、50μg/mlの抗生物質カナマイシンを添加したLB寒天培地により大腸菌JM109を形質転換し調製した。これをpETHUFcRとした。図4に構造を示す。
【0051】
(実施例4)配列の確認
実施例3において作製したpETECFcR およびpETHUFcRプラスミドベクターに挿入したDNAの配列をチェーンターミネータ法に基づくBig Dye Terminator Cycle Sequencing FS read Reaction kit(登録商標)(PEアプライドバイオシステム社)を用いてサイクルシークエンス反応に供し、全自動DNAシークエンサーABI Prism 3700 DNA analyzer(登録商標)(PEアプライドバイオシステム社)にて解析した。なお、配列番号61(5’−TAATACGACTCACTATAGGG−3’)に示すオリゴヌクレオチドをシークエンス用プライマーとして使用した。解析の結果、pETECFcR およびpETHUFcRに挿入したDNAの塩基配列は設計通りであることが確認された。pETECFcRに挿入したDNAによってコードされるFcγR1タンパク質のアミノ酸配列を配列番号62に、pETHUFcRに挿入したDNAによってコードされるアミノ酸配列を配列番号63に示す。
【0052】
(実施例5)Fcレセプターの発現誘導
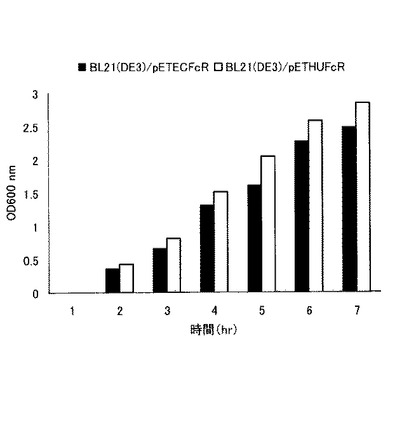
pETECFcRプラスミドベクターを50μg/mlの抗生物質カナマイシンを添加したLB寒天培地により大腸菌BL21(DE3)(タカラバイオ社)に形質転換した。得られた形質転換体を、BL21(DE3)/pETECFcRと命名した。pETHUFcRプラスミドベクターについても同様に操作を行い、形質転換体を得、BL21(DE3)/pETHUFcRと命名した。
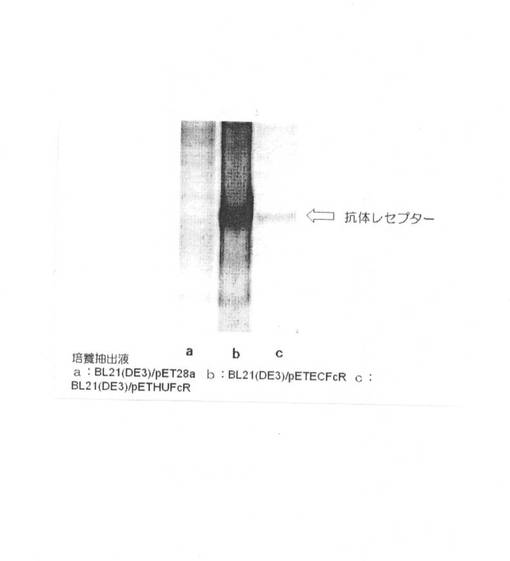
得られた形質転換体について、37℃で18時間培養した後、出現した任意のコロニーをピックアップし50μg/mlの抗生物質カナマイシンを添加したLB液体培地に接種した。37℃で培養し、OD600 nmの値が約0.5になったところで0.5mMのIPTG(isopropyl−β−D−thiogalactopyranoside、タカラバイオ社製)を添加した。IPTGを添加した後も培養を続け、一時間おきに培地をサンプリングし、それぞれの600nmにおける吸光度を測定した。結果を図5に示す。図5において、■はBL21(DE3)/pETECFcRについての結果を示し、□はBL21(DE3)/pETHUFcRについての結果を示す。または、X軸(横軸)は時間(単位はhr)を示し、Y軸(縦軸)はOD600nmの吸光度(単位は任意)を示す。図5に示すように、両者において増殖には特に差異は認められなかった。
【0053】
次いで、吸光度を測定した培地について、それぞれの形質転換体からタンパク質溶液を調製した。調製はBugBuster protein extraction Kit(登録商標)(ノバジェン社)を用いて行い、調製したタンパク質溶液をSDS−PAGEに供した。電気泳動像の結果を図6に示す。結果は、IPTG添加4時間後の結果を示す。図6において、aのレーンは、BL21(DE3)/pET28a(ベクターのみ)を形質転換したもの、bのレーンは、BL21(DE3)/pETECFcRについての結果を示し、cのレーンは、BL21(DE3)/pETHUFcRについての結果を示す。矢印の位置は、抗体レセプターの泳動位置を示す。図6に示す通り、発現するタンパク質のアミノ酸配列は同一であるものの、両者で発現量が異なることがわかった。すなわち、BL21(DE3)/pETECFcRの方が、BL21(DE3)/pETHUFcRに比べ、増殖は同程度であるにもかかわらず、発現量が多く、この結果は、宿主である大腸菌に合わせたコドンではヒト型コドンに比べて有意に発現量を増加させることに成功したことを示す。
【0054】
(実施例6)Fcレセプターのリフォールディングおよび抗体結合活性評価
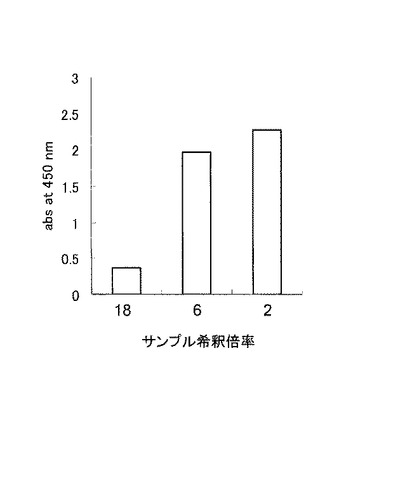
実施例5において、BL21(DE3)/pETECFcRからは、FcγRIタンパク質は大腸菌不溶性画分としてのみ得られたため、リフォールディング操作により再活性化を行った。リフォールディングは、Refolding CAキット(タカラバイオ社)を用いて行った。
抗体結合活性はELISA法により行った。先ず、抗体である10 μg/mlのガンマーグロブリン(化学及血清療法研究所製)を、マイクロプレートに固定し(4℃、18時間)、StartingBlock Blocking Buffers(登録商標)(PIERCE社)によりブロッキングした。次いで、実施例5で調製したタンパク質溶液を段階希釈し、ELISA反応に供した。反応は30℃、2時間で行った。次いで、0.2%(w/v)Tween20、150 mM NaClを含むTris−HCl緩衝液(pH8.0)で洗浄し、His−probe(H−15)HRP抗体(Santa Cruz Biotechnology社)を添加した。30℃で2時間反応させた後、先の緩衝液で洗浄し、TMB Peroxidase Substrate(KPL社)を添加し450 nmの吸光度を測定した。結果を図7に示す。図中、X軸(横軸)はサンプルの希釈倍率を示し、Y軸(縦軸)は、OD450nmの吸光度(単位は任意)を示す。図7に示す通り、サンプル6倍希釈した場合は、吸光度の減少はあまり観察されなかったが、18倍に希釈した場合、吸光度はかなり減少した。
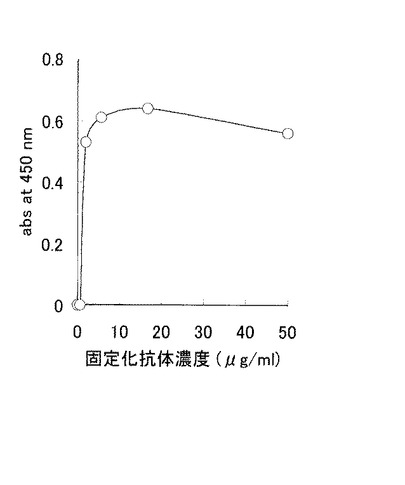
【0055】
また、固定化時の抗体濃度を50 μg/mlから段階的に希釈したELISA反応も行った。ここでは、添加サンプルであるタンパク質調製溶液を一定量添加し反応させた。結果を図8に示す。図8に示す通り、固定化抗体濃度を上げることにより吸光度は上昇した。これらの結果より、大腸菌不溶性画分に発現させたFcγRI細胞外領域を含むポリペプチドをリフォールディングさせることにより特異的な抗体結合活性が有意に発現することが確認された。
【0056】
(実施例7)Fcレセプターの抗体吸着能評価
実施例6で得られた、FcγR1細胞外領域を含むポリペプチドは、配列番号:62及び配列番号:63から明らかなように、C末端にヒスチジンが6個連続した配列を有しているので、Niキレートの親和性により、担体と結合させることができる。実施例6で得られたポリペプチド100μgを、Novagen社製のHis・Bind Purificationキット(登録商標)中のHis・Bind Resinゲル500μLに固定化した。具体的には、実施例で得られたポリペプチド100μg、及び上記ゲル500μLをカラムに充填し、純水で洗浄した後、キット中のNiチャージ緩衝液(50mM NiSO4)を用いてNiをゲルに担持させた。
【0057】
次いで、抗体のゲルへの非特異吸着を防ぐために20mMのイミダゾール、500mMのNaCl、10%グリセロール、0.001%の界面活性剤(SB3−14)を含むTris−HCl緩衝液(pH8.0)により十分平衡化した。平衡化終了後のゲルに、平衡化緩衝液で希釈した100μg/mlのガンマグロブリンを2.0ml添加した。平衡化緩衝液で洗浄後、500mMのイミダゾール、500mMのNaCl、10%グリセロール、0.001%界面活性剤(SB3−14)を含むTris−HCl緩衝液(pH8.0)で溶出させた。
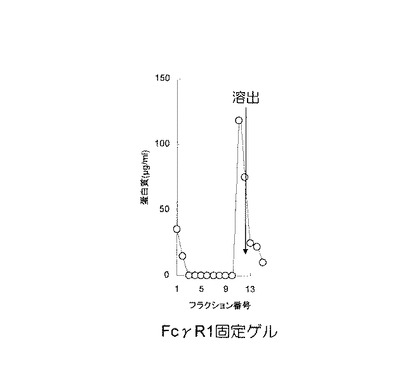
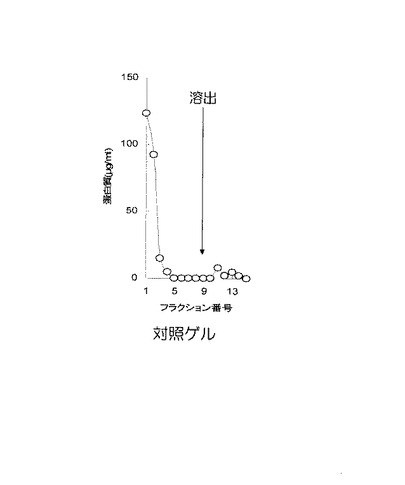
【0058】
なお、対照実験として、ゲルにFcγRI細胞外領域を含むポリペプチドを吸着しない区、すなわち、担体のみをカラムに充填した実験を行った。カラムから溶出した溶出液を集め、溶出液中のガンマグロブリン量を、バイオ・ラド社のプロテインアッセイキットを用い、イムノグロブリンを標準タンパク質として定量した。FcγRI細胞外領域を含むポリペプチドを固定化したゲルを用いた結果を図9に、対照実験の結果を図10に示す。
【0059】
図9及び図10において、X軸(横軸)はフラクション番号を示し、Y軸(縦軸)は溶出されたタンパク質の濃度(単位はμg/ml)を示し、矢印の位置は、溶出液を添加した位置を示す。図9に示す通り、FcγRI細胞外領域を含むポリペプチドを固定化したゲルにおいては、平衡化緩衝液で洗浄中にガンマグロブリンの溶出は認められず、溶出画分(図9におけるフラクション番号10〜15)に、ガンマグロブリンが検出された。図10に示すように、対照実験においては、ガンマグロブリンはゲルに吸着されることなく、フロースルー画分(図10におけるフラクション番号1〜3)に溶出された。
上記結果より、本発明の方法により得られた、FcガンマRI細胞外領域を含むポリペプチドを利用し、抗体を精製するための抗体精製用吸着剤を得ることができ、また、その抗体精製用吸着剤に抗体を吸着させ、その後精製する方法が提供された。
【図面の簡単な説明】
【0060】
【図1−1】抗体レセプターFcγR1のヒトおよび大腸菌間の遺伝子配列を比較した図である。図中、EcoliFcgR1が大腸菌型コドンに変換したDNA配列の配列、HumanFcGR1がヒトのFcγR1の配列である。
【図1−2】抗体レセプターFcγR1のヒトおよび大腸菌間の遺伝子配列を比較した図である。図中、EcoliFcgR1が大腸菌型コドンに変換したDNA配列の配列、HumanFcGR1がヒトのFcγR1の配列である。
【図2】プラスミドベクターpECFcRの構造を示す図である。
【図3】プラスミドベクターpETECFcRの構造を示す図である。
【図4】プラスミドベクターpETHUFcRの構造を示す図である。
【図5】実施例5におけるFcレセプターの発現誘導を示す図である。
【図6】抗体レセプターFcγR1細胞外領域を含むポリペプチドの発現量を比較した電気泳動像である。
【図7】実施例6におけるFcレセプターの抗体結合活性を示す図である。
【図8】実施例6におけるFcレセプターの抗体結合活性を示す図である。
【図9】実施例7におけるFcレセプターの抗体吸着能を示す図である。
【図10】実施例7におけるFcレセプターの抗体吸着能の対照実験を示す図である。
【技術分野】
【0001】
本発明は、Fcレセプター細胞外領域を含むポリペプチド及びその製造方法、並びに該ポリペプチドの利用方法に関する。
【背景技術】
【0002】
Fcレセプターは、免疫グロブリン分子のFc領域に結合する一群の分子である。個々の分子は、免疫グロブリンスーパーファミリーに属する認識ドメインによって、単一の、あるいは同じグループの免疫グロブリンイソタイプをFcレセプター上の認識ドメインによって認識している。これによって、一定の免疫応答においてどのアクセサリー細胞が動因されるかが決まってくる(非特許文献1)。Fcレセプターは、さらにいくつかのサブタイプに分類することができ、IgG(免疫グロブリンG)に対するレセプターとしては、FcγRI、FcγRIIa、FcγRIIb及びFcγR3の存在が報告されている(非特許文献1)。上記レセプターの中でも、FcγRIとIgGとの親和性は高く、その平衡解離定数(Kd)は10−8 M以下であることが報告されている(非特許文献2)。
【0003】
FcγRIはIgG(免疫グロブリンG)に対するレセプターであり、単球及びマクロファージ上に構成的に発現され、好中球および好酸球上においては誘導的に発現される。FcγRIは、細胞外領域、膜貫通領域、細胞質内領域に区分され、IgGとの結合は、IgGのFc領域とFcγRIの細胞外領域で起こり、その後細胞質へとシグナルが伝達される。FcγRIはIgGとの結合に直接関わる分子量約42,000のα鎖と、γ鎖の2種類のサブユニットによって構成されており、γ鎖は細胞膜と細胞外領域の境界にあるシステインを介した共有結合によりホモダイマーを形成している(非特許文献1)。
【0004】
近年になり、Fcレセプターの予想外の免疫抑制的な生物学的特性は、特に、自己免疫疾患または自己免疫症候群、移植物の拒絶および悪性リンパ増殖の領域において医薬として注目を浴びつつある。また、FcγR1の機能である抗体の吸着能は各種抗体精製用クロマトグラフィーゲルの捕捉機能タンパク質としても利用することができる。
FcγRIα鎖のアミノ酸配列および遺伝子配列は、Janet等により明らかにされ(非特許文献2)、その後、遺伝子組換え技術により、大腸菌(非特許文献、特許文献1)あるいは動物細胞を利用することによる、FcγRIの発現が報告された。しかしながら、大腸菌を利用した発現系においてはFcγRIの細胞外領域タンパク質の発現量が極めて低く、工業生産規模において生産することは困難である。また、動物細胞を用いた系では、大腸菌を用いた系よりも高い発現量を実現できることが報告(非特許文献3)されているが、培養に時間がかかるため、生産性は必ずしも高くない。
【0005】
【特許文献1】特表2004−530419号公報
【非特許文献1】J.V.Ravetch等, Annu.Rev.Immunol., 9, 457頁, 1991年
【非特許文献2】J.M.Allen等, Science, 243, 378頁, 1989年
【非特許文献3】A.Paetz等, Biochem.Biophys.Res.Commun., 338, 1811頁, 2005年
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は工業生産規模において、Fcレセプター細胞外領域を含むポリペプチド、特に、FcγRI細胞外領域を含むポリペプチドを生産する微生物を作製することである。さらには、新規FcγRI細胞外領域を含むポリペプチドの生産系を利用してFcγRI細胞外領域を含むポリペプチドの生産を行い、該FcγRI細胞外領域を含むポリペプチドを提供することにある。
【課題を解決するための手段】
【0007】
本願発明者らは、上記課題を解決すべく鋭意検討した結果、ヒトFcγRI遺伝子から転写されるmRNAのコドンを、大腸菌宿主において利用頻度が高いコドンにするべく該遺伝子の配列を変換し、変換したレセプター遺伝子が挿入された組換えベクターに導入することにより、大腸菌においてFcγRIの発現量が向上することを見出した。
すなわち、本願発明者らは、ヒトFcレセプターの一種類であるFcγRIの遺伝子配列を見直し、大腸菌で該レセプターを発現させるべく大腸菌が容易に翻訳するコドンに対応するようにDNA配列を変換し、さらに、対応する変換コドンからなるオリゴヌクレオチドを合成後、PCR法により全長FcγRI遺伝子を作製し、該遺伝子が挿入された発現プラスミドベクターを大腸菌に導入した結果、大腸菌宿主においてFcγRI細胞外領域タンパク質の発現量が飛躍的に向上することを見出し、本発明を完成するに至った。
【0008】
すなわち、本発明は、FcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、配列番号:2に記載の塩基配列を含むポリヌクレオチド、又は配列番号:2に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチドを提供する。
また、本発明は、FcγRI細胞外領域を含むポリペプチドからシグナル配列を除去した領域をコードするポリヌクレオチドであって、配列番号:3に記載の塩基配列を含むポリヌクレオチド、又は配列番号:3に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチドを提供する。
また、本発明は、シグナル配列を除去した、FcγRI細胞外領域を含むポリペプチドのN−末端側にメチオニンが付加されたポリペプチドをコードするポリヌクレオチドであって、配列番号:4に記載の塩基配列を含むポリヌクレオチド、又は配列番号:4に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチドを提供する。
【0009】
また、本発明は、配列番号2〜4のいずれか1に記載の塩基配列を含むポリヌクレオチドによりコードされるポリペプチドを提供する。
また、本発明は、配列番号2〜4のいずれか1に記載の塩基配列を含むポリヌクレオチドに、発現制御配列が機能的に付加されたポリヌクレオチドを提供する。
また、本発明は、上記のいずれかのポリヌクレオチドが挿入された組換えベクターを提供する。
また、本発明は、上記のいずれかのポリヌクレオチド、又は上記組換えベクターにより形質転換された形質転換体を提供する。
また、本発明は、上記形質転換体を培養する工程を含む、FcγRI細胞外領域を含むポリペプチドの製造方法を提供する。
また、本発明は、上記製造方法によって得られたポリペプチドを、担体上に固定してなる、抗体精製用吸着剤を提供する。
また、本発明は、上記抗体精製用吸着剤に抗体を吸着させる方法を提供する。
【発明の効果】
【0010】
ヒトFcγRI細胞外領域を含むポリペプチドをコードする遺伝子コドンを、大腸菌型にすることによって、FcγRI細胞外領域を含むポリペプチドを、大腸菌に大量に発現させることが可能となる。この発現系は、工業的規模におけるFcγRI細胞外領域を含むポリペプチドの生産に有用なものである。また、このようにして得られた、FcγRI細胞外領域を含むポリペプチドは、抗体アフィニティークロマトゲルの捕捉基として利用でき、すなわち、抗体精製用吸着剤として利用することができる。
【発明を実施するための最良の形態】
【0011】
以下、本発明について詳細に説明する。
まず、FcγRIタンパク質をコードする遺伝子について説明すると、FcγRI遺伝子は配列番号:1に記載の約2.2Kベースの塩基配列を含むDNAである。このDNAがコードするタンパク質の機能構造は、は公的データベース(European Bioinformatics Institute、 URL:http://www.ebi.ac.uk/、 アクセス日:2006年3月2日)に公表され公知であり、構造上のドメイン、あるいはシグナル配列、細胞膜貫通領域等のアミノ酸配列の決定は容易である。
【0012】
次に、本発明のポリヌクレオチドについて説明する。
本発明のポリヌクレオチドは、ヒトFcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、その遺伝子情報のコドンを、適当な宿主細胞のコドン使用頻度に最適化したポリヌクレオチドである。すなわち、ヒトFcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドが挿入された組換えベクターを宿主細胞に形質転換して得られた形質転換体を培養してポリペプチドを生産せる場合に、その宿主細胞が容易に翻訳することができるコドンに対応することができるように、ポリヌクレオトドのコドンを変換することによって、効率よく目的のポリペプチドの生産をすることができるようにしたものである。本明細書において、本発明を説明する場合、大腸菌を宿主細胞として用いた場合について説明するが、本発明は、宿主細胞として大腸菌を用いる場合に限定されず、他の宿主細胞の場合についても適用される。すなわち、他の宿主細胞を用いる場合は、その宿主細胞の最適なコドンについてゲノム解析することによって決定し、他の宿主細胞においても適用することが可能である。
【0013】
本発明のポリヌクレオチドは、以下の(a)、(b)又は(c)で表わされるポリヌクレオチドである。
(a)FcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、配列番号:2に記載の塩基配列を含むポリヌクレオチド、又は配列番号:2に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
(b)FcγRI細胞外領域を含むポリペプチドからシグナル配列を除去した領域をコードするポリヌクレオチドであって、配列番号:3に記載の塩基配列を含むポリヌクレオチド、又は配列番号:3に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
(c)シグナル配列を除去した、FcγRI細胞外領域を含むポリペプチドのN−末端側にメチオニンが付加されたポリペプチドをコードするポリヌクレオチドであって、配列番号:4に記載の塩基配列を含むポリヌクレオチド、又は配列番号:4に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
【0014】
(a)のポリヌクレオチドについて説明する。(a)のポリヌクレオチドは、ヒトFcγRI細胞該領域を含むポリペプチドをコードする遺伝子であり、その遺伝子情報のコドンを、ヒト型から大腸菌型のコドンとなるように変換した、すなわち大腸菌のコドン使用頻度に最適化したポリヌクレオチドである。
言い換えると、(a)のポリヌクレオチドは、ヒトFcγRI細胞外領域をコードする遺伝子DNAから転写されるmRNA中において存在する大腸菌レアコドン(rare codon)を、コードするアミノ酸を同一のまま、大腸菌コドンで利用頻度が高いコドンに変換したポリヌクレオチド配列である。なお、レアコドンとは、その宿主におけるコドンの使用頻度が少ないものと定義することができる。宿主における使用頻度は、ゲノム遺伝子の解析結果を参考にすることで推測することが可能であり、大腸菌においては、アミノ酸であるArgコドンのAGA、AGG、CGG、CGA、IleコドンのAUA、LeuコドンのCUA、GlyコドンのGGA、ProコドンのCCCを例示することができる。コドンの使用頻度の解析は公的データベース(URL:http://www.kazusa.or.jp/codon/、アクセス日:2006年3月2日)を利用することによっても可能である。
【0015】
コドンの変換は、対応するDNA配列の塩基配列を変えることにより可能である。DNA配列の塩基置換を行なうには、たとえば、Site−directed mutagenesis法など公知の変異導入法を利用することができる。好ましくは、合成オリゴヌクレオチドとPCRを組合わせたDNAWorks法を例示することができる(Nucleic Acid Res., 30, e43頁, 2002年)。この方法によれば、アミノ酸配列から数十塩基からなるオリゴヌクレオチド群を合成し、PCRにより合成オリゴヌクレオチドをアッセンブリーさせることによって完全長の遺伝子を合成することができる。
遺伝子すべてのレアコドンを変換することも可能であり、部分的に変換することも可能である。mRNAの発現量を定量することによって適宜変換割合を変えることができる。
【0016】
(b)のポリヌクレオチドは、(a)のポリヌクレオチドから、シグナル配列をコードする部分を除去したものであり、すなわち、FcγRI細胞外領域を含むポリペプチドからシグナル配列を除去した領域をコードするポリヌクレオチドである。
なお、シグナル配列とは、細胞内で発現したタンパク質が細胞膜を通過し、細胞外において分泌するための領域であり、通常、該タンパク質のN−末端側に存在しており、細胞膜通過後、特定のプロテアーゼ酵素によって切断される。シグナル配列としては、配列番号1のアミノ酸番号1から15、あるいは1から20のペプチド配列を例示することができる。FcγRI細胞外領域は、さらに、シグナル配列を除去し、IgGと相互作用する部位のみでも機能的には十分である。
【0017】
(c)のポリペプチドは、シグナル配列を除去した、FcγRI細胞外領域を含むポリペプチドのN−末端側にメチオニンが付加されたポリペプチドをコードするポリヌクレオチドである。
【0018】
本発明において、ポリペプチドとは、天然に存在するポリヌクレオチドであってもよいし、また、人工的に合成されたヌクレオチドであってもよい。
【0019】
(a)、(b)及び(c)のポリヌクレオチドは、それぞれ、配列番号:2、配列番号:3及び配列番号:4に記載の塩基配列を含むポリヌクレオチドに加え、例えば、配列番号:2、配列番号:3及び配列番号:4に記載の塩基配列を含むポリヌクレオチドがコードするポリペプチドにおいて、生物学的活性の同一性が損なわれない範囲内において、そのポリペプチド鎖に対する、アミノ酸配列中の一部(好ましくは1〜20個、更に好ましくは1〜10個程度、最も好ましくは1〜3個)のアミノ酸が欠失、置換又は付加したアミノ酸配列からなるポリペプチドをコードするようなポリヌクレオチドをも含む。この場合も、生物学的活性の同一性とは、FcγRIのイムノグロブリンに対する結合性を意味する。ここで、イムノグロブリンとは、好ましくはIgGであり、更に好ましくはIgG1である。
当業者であれば、(a)、(b)及び(c)のポリペプチドに、部位特異的突然変異法などを用いて、適宜、置換、欠失、及び/又は付加変異を導入することができる。
【0020】
上記(a)、(b)及び(c)のポリヌクレオチドは、上記(a)、(b)、(c)の配列に対して実質的に相同なポリヌクレオチド配列をも含む。実質的に相同であるとは、二つの核酸に関して、下記の配列比較アルゴリズムの一つを用いて、または、手動整列および目視試験により測定し、最大対応で整列化させた場合に、75%を超え、好ましくは85%を超え、より好ましくは95%を超えるDNA同一性を有する配列または部分配列を意味する。なお、この定義は、その相補体が試験配列にハイブリダイズする配列も意味することは言うまでもない。配列比較のために、典型的には一つの配列が基準配列として働き、それに対して試験配列を比較する。配列比較アルゴリズムを用いる場合は、試験配列および基準配列をコンピュータに入力し、必要があれば部分座標を指定し、配列アルゴリズムプログラムのパラメータを指定する。パラメータはデフォルト値を用いてもよく、または、代わりのパラメータを指定することもできる。次いで、配列比較アルゴリズムが、プログラムのパラメータに基づき、基準配列に対する試験配列の配列同一性の割合を計算する。比較のための配列整列方法は当該技術分野において公知である。比較のための、最適配列は、例えば、局所相同性アルゴリズム(Smith等, Adv.Appl.Math., 2, 482頁, 1981年)、相同性整列化アルゴリズム(Needlman等, J.Mol.Biol., 48, 443頁, 1970年)、類似性検索法(Person等, Proc.Natl.Acd.Sci.USA, 85, 2444頁, 1988年)を例示することができる。
【0021】
有用なアルゴリズムの一例はPileup法であり、連続性のペアワイズアライメントを用いて関連配列群から多重配列を作製し、関係と配列同一性の割合を示すことができる。配列同一性の割合および配列類似性を求めるのに適したアルゴリズムのもう一例はBLASTアルゴリズムである(Altschul等, J.Mol. iol., 215, 403頁, 1990年)。
【0022】
また、本発明のポリヌクレオチドには、上記(a)、(b)、(c)のポリヌクレオチドと、ストリンジェントな条件下でハイブリダイズするDNA配列が含まれるものである。ストリンジェントな条件下でハイブリダイズするとは、プローブが核酸混合物中で、その標的配列にはハイブリダイズするが、他の核酸にはハイブリダイズしない条件を意味する。ストリンジェントな条件は配列依存性があり、異なる環境下では異なることになる。より長い配列では、特異的に、より高温でハイブリダイズする。一般に、高度にストリンジェントな条件は、既定のイオン強度およびpHで特異的配列の熱融解温度より約5〜10℃低くなるように選択される。低ストリンジェントな条件は、一般的には融解温度より約15〜30℃低くなるように選択される。融解温度は、既定のイオン強度、pH、核酸配列で標的核酸に対して相補的なプローブにおいて50%が平衡状態で占める温度である。
【0023】
具体的には、ストリンジェントな条件下でハイブリダイズできるポリヌクレオチドとは、配列番号:1に記載中の任意の少なくとも20個、好ましくは少なくとも30個、例えば40、60または100個の連続した配列を有するポリヌクレオチドをプローブとし、例えばECL direct nucleic acid labeling and detection system(Amersham Pharmacia Biotech社製)を用いて、マニュアルに記載の条件(例えば、wash:42℃、0.5×SSCを含むprimary wash buffer)においてハイブリダイズするポリヌクレオチドを指す。より具体的な「ストリンジェントな条件」とは、例えば、通常、42℃、2×SSC、0.1%SDSの条件であり、好ましくは50℃、2×SSC、0.1%SDSの条件であり、さらに好ましくは、65℃、0.1×SSCおよび0.1%SDSの条件であるが、これらの条件に特に制限されない。ハイブリダイゼーションのストリンジェンシーに影響する要素としてはこれら温度、塩濃度に加えプローブ濃度、プローブの長さ、反応時間を含む複数の要素が考えられ、当業者であればこれら要素を適宜選択することで最適なストリンジェンシーを実現することが可能である。ハイブリダイゼーション法の詳細な手順については、「Molecular Cloning, A Laboratory Manual 2nd ed.」(Cold Spring Harbor Press(1989)、特にSection9.47-9.58)、「Current Protocols in Molecular Biology」(John Wiley&Sons(1987-1997)、特にSection6.3-6.4)、「DNA Cloning 1: Core Techniques, A Practical Approach 2nd ed.」(Oxford University(1995)、特にSection2.10)等を参照することができる。
【0024】
本発明は、上述した、(a)、(b)及び(c)のポリヌクレオチドによりコードされるポリペプチドを含む。このようなポリペプチドとしては、例えば、配列番号:1に記載のアミノ酸配列からなるポリペプチド、及びがいアミノ酸配列中の一部(好ましくは1〜20個、更に好ましくは1〜10個程度、最も好ましくは1〜3個)のアミノ酸が欠失、置換又は付加したアミノ酸配列からなるポリペプチドが挙げられる。
【0025】
本発明は、本発明のポリヌクレオチドが挿入された組換えベクターを提供する。上記ポリヌクレオチドを公知の発現ベクターに挿入することにより、FcγRI細胞外領域を含むポリペプチドを発現するベクター(組換えベクター)が提供される。また、この組換えベクターで形質転換した形質転換体を培養することにより、FcγRI細胞外領域を含むポリペプチドを形質転換体から得ることができる。すなわち、本発明は、本発明のポリヌクレオチドが挿入された組換えベクターにより形質転換された形質転換体を培養し、その培養物から、FcγRI細胞外領域を含むポリペプチドを回収することを含む、FcγRI細胞外領域を含むポリペプチドの製造方法が提供される。
【0026】
本発明のポリヌクレオチドは、そのまま用いて適当な細胞に形質転換してもよいが、通常は、適当な公知の発現ベクターに挿入される。発現ベクターへの挿入は、その発現ベクターの適当な位置に遺伝子工学的に挿入すればよい。適当な位置とは、発現ベクターの複製機能、所望の抗生物質マーカー、又は伝達性に関する領域を破壊しないような位置を意味する。本発明のポリヌクレオチドは、発現制御配列が機能的に付加されたものであることが好ましい。発現制御配列が機能的に付加されるとは、例えば、プロモーター、エンハンサー、オペレーター、インデューサー等が付加されることであり、このような配列が付加されることによって、ポリペプチドの宿主細胞内での発現が可能となる。
【0027】
本発明において、組換えベクターを形質転換する宿主細胞としては、微生物が好ましく、原核細胞が更に好ましく用いられる。原核細胞の中でも、大腸菌が好ましいが、本発明のおいては宿主細胞を限定するものでなく、遺伝子工学において用いられる宿主細胞については全て用いられるものとする。利用可能な微生物としては、例えば以下のような微生物が挙げられる。
【0028】
大腸菌(エシェリヒア(Escherichia)属)
バチルス(Bacillus)属
シュードモナス(Pseudomonas)属
セラチア(Serratia)属
ブレビバクテリウム(Brevibacterium)属
コリネバクテリイウム(Corynebacterium)属
ストレプトコッカス(Streptococcus)属
ラクトバチルス(Lactobacillus)属など宿主ベクター系の開発されている細菌
ロドコッカス(Rhodococcus)属
ストレプトマイセス(Streptomyces)属など宿主ベクター系の開発されている放線菌
サッカロマイセス(Saccharomyces)属
クライベロマイセス(Kluyveromyces)属
シゾサッカロマイセス(Schizosaccharomyces)属
チゴサッカロマイセス(Zygosaccharomyces)属
ヤロウイア(Yarrowia)属
トリコスポロン(Trichosporon)属
ロドスポリジウム(Rhodosporidium)属
ピキア(Pichia)属
キャンディダ(Candida)属などの宿主ベクター系の開発されている酵母
ノイロスポラ(Neurospora)属
アスペルギルス(Aspergillus)属
セファロスポリウム(Cephalosporium)属
トリコデルマ(Trichoderma)属などの宿主ベクター系の開発されているカビ
【0029】
以下、宿主として大腸菌を用いる場合について説明する。発現ベクターにポリヌクレオチドを挿入する場合、上記ポリヌクレオチドをプロモータ機能を有するDNA配列を付加させることが好ましい。プロモータとは、タンパク質のコード領域又は機能的RNAの発現を制御することが可能なDNA配列を意味し、大腸菌の場合、lacプロモータ、trcプロモータ、T7プロモータ等を例示することができる。その中でも、好ましくは転写活性が強いT7プロモータである。
【0030】
プラスミドベクター(発現ベクター)としては、形質転換する細胞内で安定に存在し、複製することができる特性を有するものであれば特に制限なく用いることができる。このようなプラスミドベクターとしては、大腸菌の形質転換に利用されるpUC系、pBR系、pET系、広宿主域(Broad−Host−Range)プラスミドベクター等、あるいは対象細胞内で複製可能なプラスミドベクターとのシャトルベクターを挙げることができる。例えば、成書(Barbara E. Funnell, PLASMID BIOLOGY,545〜566頁, ASM press)に記載されているものが挙げられる。さらに、形質転換対象となる細胞が保有するプラスミドを利用しても良い。また、後述するようなベクターを用いることもできる。
【0031】
宿主細胞としては、大腸菌を用いる場合、例えば、K12系のJM109株を用いることが好ましい。さらに好ましくは、T7プロモータ配列が利用できるBL21(DE3)株などを例示することができる。
また、本発明の宿主細胞としては上記大腸菌等を変異処理することにより誘導される大腸菌変異株を利用することもできる。変異処理はニトロソグアニジン、メタンスルホン酸エチル、紫外線、放射線等の当業者では周知の変異処理剤を利用して行えばよい。
【0032】
大腸菌等の微生物への外来遺伝子の導入および発現のための手順乃至方法は、本発明において下記したところ以外のものにおいても、遺伝子工学の分野により慣用されているものを含み、その手法乃至方法(たとえば、“Plasmid transformation of Escherichia coli and other bacteria”, Method in Enzymology, 204, 63〜113頁, 1991年, Academic Press)に準じて実施すればよい。具体的にはヒートショック法、エレクトロポレーション法等を挙げることができる。
【0033】
本発明のFcγRI細胞外領域を含むポリペプチドの製造方法は、本発明のポリペプチドが挿入された組換えベクターにより形質転換された形質転換体を培養する工程を含む。上記方法は、具体的には、上記形質転換体を培養し、その培養物から、FcγRI細胞外領域を含むポリペプチドを回収することを含む。
【0034】
本発明のFcγRIを生産するために用いられる形質転換体(大腸菌)は、当該技術分野で周知であり、かつ、選択した宿主細胞の培養に好適な培地で増殖させる。好適な培地の例としては必要な栄養源を補ったLB(ルリアブロス)培地を挙げることができる。好ましい実施態様においては、組換えベクターで形質転換体の増殖を選択的に可能にするために、培地に、組換えベクターの構成を基にした選抜剤を含有させることが好ましい。例えば、カナマイシン耐性遺伝子を発現する細胞の増殖のためにカナマイシンが培地に添加される。培地には、炭素、窒素および無機塩供給源の他に、適当な栄養源を加えてもよい。所望により、グルタチオン、システイン、シスタミン、チオグリコレート、ジチオスレイトールからなる群から選択される一種類以上の還元剤を含んでも良い。大腸菌増殖における培養温度は、20℃〜40℃、より好ましくは25℃〜35℃であり、より好ましくは約30℃である。培地のpHは、大腸菌培養においては約6.8〜7.4であり、より好ましくは7.0である。
【0035】
本発明の組換えベクターに誘導性のプロモータを用いる場合には、良好なタンパク質発現が起こるような条件下で誘導する。誘導剤としてはIPTGを例示することができ、培養液の濁度(660 nmにおける吸光度)を測定し、約0.5〜1.0の増殖期間に適当量のIPTGを添加し、引き続き培養することによりタンパク質の発現を誘導することができる。IPTGの添加濃度は0.1〜1.0mMであり、好ましくは0.5mM程度である。IPTG誘導に関する種々の条件は当該技術分野においては周知である。
【0036】
培養液から、本発明のFcγRI細胞外領域を含むポリペプチドを回収するには、発現の形態によって適宜抽出方法を選択すればよい。培養上清に発現する場合は菌体を遠心分離操作によって分離し、得られる培養上清からFcγRI細胞外領域を含むポリペプチドを抽出すればよい。一方、細胞内にFcγRI細胞外領域を含むポリペプチドが発現する場合には、遠心分離操作により菌体を集め、酵素処理剤や界面活性剤等を添加することにより菌体を破砕し、FcγRI細胞外領域を含むポリペプチドを抽出することができる。抽出タンパク質の中からFcγRI細胞外領域を含むポリペプチドを分離・精製するには、当該技術分野において公知の方法を用いることができる。例えば、液体クロマトグラフィーを利用することができる。液体クロマトグラフィーとしては、イオン交換クロマト、疎水性相互作用クロマト、ゲルろ過、アフィニティークロマトなどを挙げることができる。これらのクロマトを組み合わせて精製操作を行うことによって、高純度なFcγRI細胞外領域を含むポリペプチドを調製することができる。
【0037】
次に、本発明の抗体精製用吸着剤について説明する。
本発明の抗体精製用吸着剤は、上述したポリペプチドの製造方法によって得られたポリペプチドを、担体上に固定してなる。
用いられる担体としては、ポリペプチドを化学的又は物理的に固定化することのできるものであれば、特に制限なく用いることができ、クロマトグラフィー用担体を用いることができる。このような担体としては、例えば、セルロース、アガロース等の多糖類、ガラス、セラミックス、又はポリプロピレン、塩化ビニル、ポリスチレン等のプラスチック素材等が挙げられる。このような担体は市販されているものを用いることができ、例えば、Novagen社製、His・Bind purificationキット(登録商標)中のHis・Bind Resinゲル、商品名「HiTrap NHS−Activated HP」(Amersham Biosciences社製)、商品名「トシル基活性化ダイナビーズM−280」(Dynal社製)等を用いることができる。
【0038】
なお、ポリペプチドを担体に固定化する際に、ペプチドの末端に担体との競合に利用できる適当なスペーサーを挿入したポリペプチドを用いることが好ましい。スペーサーとしては、例えば、アミノ酸、ペプチド、炭素数2〜8個程度の主鎖を有糸、好ましくは水酸基等の親水性の官能基を有するもの(例えば、両末端に活性基を有するポリビニルアルコールなど)、糖鎖等が挙げられる。上記アミノ酸としては、例えば、ヒスチジン、アルギニン、リシン、アスパラギン、グルタミン、システイン、アスパラギン酸、グルタミン酸等が例示でき、ヒスチジンが複数個(通常、2〜10個)連続して結合した、ポリヒスチジンが好ましく用いられる。このようなポリヒスチジンをスペーサーとして用いる場合、担体とポリペプチドとの結合はNiキレートとの親和性により結合する。アミノ酸がスペーサーである場合は、スペーサーを挿入したペプチドを連続した1個のポリペプチドとして合成することができる。遺伝子組み換え法によりポリペプチドを産生させる場合、ベクターとして、組換タンパク質の末端にヒスチジンタグを付加させるようなベクターに、ポリヌクレオチドを挿入し、それを適当な宿主に形質転換させ、ポリペプチドを産生させることができる。このようなベクターとしては、市販されているものを用いることができ、例えば、ノバジェン社製のpET28b+等が挙げられる。
【0039】
また、本発明は、上記抗体精製用吸着剤に抗体を吸着させる方法を提供する。また、上記抗体精製用吸着剤は、例えば、抗体の精製分離に用いることができる。すなわち、本発明は、上記抗体精製用吸着剤を用いた、抗体の精製方法を提供する。
抗体精製用吸着剤に抗体を吸着させるには、例えば、上記抗体精製用吸着剤を、例えばカラム等につめ、ここに、抗体を含む試料を中性条件下に導入する。これにより、試料中の抗体が、抗体精製用吸着剤に固定化されているポリペプチドと反応して吸着される。
また、抗体精製用吸着剤を、抗体の精製分離に用いる場合は以下の通りに実施する。抗体が吸着した抗体精製用吸着剤から抗体を精製分離するには、カラム中に、塩濃度の高い、例えばNaCl、KCl、Na2SO4を含む緩衝液を流すことにより、又はカラム中のpHを変化させることにより実施する。
【実施例】
【0040】
以下、本発明の実施の形態について詳細に説明する。なお、本発明は、これらの実施例のみに限定されるものではなく、発明の要旨を逸脱しない範囲で、任意に変更が可能であることは言うまでもない。
(実施例1)FcレセプターをコードするDNA配列の設計
配列番号:1のFcγRIの細胞外領域のアミノ酸配列を基にDNAworks法(Nucleic Acid Res., 30, e43頁, 2002年)を用いて、コドンを大腸菌型に変換した。該方法によりコドンを変換したDNA配列とヒト型コドンをアライメントした結果を図1に示す。図1に示す通り、アミノ酸配列は変わらずに、DNAの配列変換が行われ、そのDNA配列間の類似性は75%であった。
【0041】
(実施例2)FcレセプターをコードするDNA配列の作製
ヒト型コドンから大腸菌型コドンに変換したDNA配列を作製するために52種類のオリゴヌクレオチドを合成した。合成したオリゴヌクレオチドを配列番号5から56に示す。次いで、二段階のPCRを行い、オリゴヌクレオチドから完全長のFcγRIをコードするDNAを作製した。一段階目のPCRの反応液は表1に示す通りであり、反応条件は94℃、5分の熱処理後、94℃、30秒間の第一ステップ、62℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、次いで、72℃、7分の第四ステップである。表1中のDNAミックスとは、52種類の合成した50 pmol/μlのオリゴヌクレオチドをそれぞれ一定量サンプリングし混合した溶液を意味する。
【0042】
第二段階目のPCRは一段階目のPCRの反応液を用いて、表2の反応液組成で行った。PCRプライマーの配列は、配列番号5(5’−ATGTGGTTTCTGACCACGCTGTTGCTGTGGGTGCCGGT−3’)と配列番号56(5’−GGTCGCGCCCTGCGGCTCCTTACGATGCAC−3’)のオリゴヌクレオチドを用いた。反応条件は94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップである。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。 次いで、目的バンドを抽出(QIAquick Gel extraction kit:キアゲン社)後、抽出DNAの5’末端をリン酸化(TaKaRa BKL Kit:タカラバイオ社)し、制限酵素Sma Iで消化したpUC19プラスミドベクターに挿入し、大腸菌JM109株(タカラバイオ社)を形質転換し調製した。これをpECFcRとした。図2に構造を示す。
【0043】
【表1】
【0044】
【表2】
【0045】
(実施例3)Fcレセプター発現プラスミドベクターの作製
FcγRIの細胞外領域を発現させるために、pECFcRをテンプレートにし配列番号57(5’−CCCATGGCTGTGATTACGCTGCAACCACCG−3’:アンダーライン部分(CCATGG)は制限酵素NcoIサイト)と配列番号 58(5’−CCAACCTTGACCGGGGTCGGCAGTTGAAGACC−3’ :アンダーライン部分(AACCTT)は制限酵素HindIIIサイト) のオリゴヌクレオチドをプライマーにしてPCRを行い、細胞外領域をコードするDNAを増幅した。反応条件は94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップである。反応液組成を表3に示す。
【0046】
【表3】
【0047】
反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。次いで、QIAquick Gel extraction kit(キアゲン社)を用いて、目的バンドを抽出した後、増幅DNAを制限酵素NcoIとXhoIにより消化し、これらの制限酵素を用いて事前に消化したpET28b+(ノバジェン社)と、Ligation Kit Ver2(タカラバイオ社)を用いてライゲーションした。次いで、50μg/mlの抗生物質カナマイシンを添加したLB寒天培地により大腸菌JM109を形質転換し調製した。これをpETECFcRとした。図3に構造を示す。得られたpETECFcRは、ヒト型コドンを用いているものである。
【0048】
上記と並行して、FcγRIの細胞外領域をコードするDNAを用いた発現ベクターも作製した。挿入断片の作製はPCRで行い、Human cDNAcloneTC119841プラスミドベクター(Origene社)をテンプレートとし、配列番号59(5’−CCCATGGCAGTGATCACTTTGCAGCCTCC−3’ :アンダーライン部分(CCATGG)は制限酵素NcoIサイト)と配列番号60(5’−GCTCGAGGACAGGAGTTGGTAACTGGAGGCC−3’:アンダーライン部分(CTCGAG)は制限酵素XhoIサイト)のオリゴヌクレオチドをPCRプライマーとした。PCRは94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップである。反応液組成を表4に示す。
【0049】
【表4】
【0050】
反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。次いで、QIAquick Gel extraction kit(キアゲン社)を用いて、目的バンドを抽出した後、増幅DNAを制限酵素NcoIとXhoIにより消化し、これらの制限酵素を用いて事前に消化したpET28a+(ノバジェン社)と、Ligation Kit Ver2(タカラバイオ社)を用いてライゲーションした。次いで、50μg/mlの抗生物質カナマイシンを添加したLB寒天培地により大腸菌JM109を形質転換し調製した。これをpETHUFcRとした。図4に構造を示す。
【0051】
(実施例4)配列の確認
実施例3において作製したpETECFcR およびpETHUFcRプラスミドベクターに挿入したDNAの配列をチェーンターミネータ法に基づくBig Dye Terminator Cycle Sequencing FS read Reaction kit(登録商標)(PEアプライドバイオシステム社)を用いてサイクルシークエンス反応に供し、全自動DNAシークエンサーABI Prism 3700 DNA analyzer(登録商標)(PEアプライドバイオシステム社)にて解析した。なお、配列番号61(5’−TAATACGACTCACTATAGGG−3’)に示すオリゴヌクレオチドをシークエンス用プライマーとして使用した。解析の結果、pETECFcR およびpETHUFcRに挿入したDNAの塩基配列は設計通りであることが確認された。pETECFcRに挿入したDNAによってコードされるFcγR1タンパク質のアミノ酸配列を配列番号62に、pETHUFcRに挿入したDNAによってコードされるアミノ酸配列を配列番号63に示す。
【0052】
(実施例5)Fcレセプターの発現誘導
pETECFcRプラスミドベクターを50μg/mlの抗生物質カナマイシンを添加したLB寒天培地により大腸菌BL21(DE3)(タカラバイオ社)に形質転換した。得られた形質転換体を、BL21(DE3)/pETECFcRと命名した。pETHUFcRプラスミドベクターについても同様に操作を行い、形質転換体を得、BL21(DE3)/pETHUFcRと命名した。
得られた形質転換体について、37℃で18時間培養した後、出現した任意のコロニーをピックアップし50μg/mlの抗生物質カナマイシンを添加したLB液体培地に接種した。37℃で培養し、OD600 nmの値が約0.5になったところで0.5mMのIPTG(isopropyl−β−D−thiogalactopyranoside、タカラバイオ社製)を添加した。IPTGを添加した後も培養を続け、一時間おきに培地をサンプリングし、それぞれの600nmにおける吸光度を測定した。結果を図5に示す。図5において、■はBL21(DE3)/pETECFcRについての結果を示し、□はBL21(DE3)/pETHUFcRについての結果を示す。または、X軸(横軸)は時間(単位はhr)を示し、Y軸(縦軸)はOD600nmの吸光度(単位は任意)を示す。図5に示すように、両者において増殖には特に差異は認められなかった。
【0053】
次いで、吸光度を測定した培地について、それぞれの形質転換体からタンパク質溶液を調製した。調製はBugBuster protein extraction Kit(登録商標)(ノバジェン社)を用いて行い、調製したタンパク質溶液をSDS−PAGEに供した。電気泳動像の結果を図6に示す。結果は、IPTG添加4時間後の結果を示す。図6において、aのレーンは、BL21(DE3)/pET28a(ベクターのみ)を形質転換したもの、bのレーンは、BL21(DE3)/pETECFcRについての結果を示し、cのレーンは、BL21(DE3)/pETHUFcRについての結果を示す。矢印の位置は、抗体レセプターの泳動位置を示す。図6に示す通り、発現するタンパク質のアミノ酸配列は同一であるものの、両者で発現量が異なることがわかった。すなわち、BL21(DE3)/pETECFcRの方が、BL21(DE3)/pETHUFcRに比べ、増殖は同程度であるにもかかわらず、発現量が多く、この結果は、宿主である大腸菌に合わせたコドンではヒト型コドンに比べて有意に発現量を増加させることに成功したことを示す。
【0054】
(実施例6)Fcレセプターのリフォールディングおよび抗体結合活性評価
実施例5において、BL21(DE3)/pETECFcRからは、FcγRIタンパク質は大腸菌不溶性画分としてのみ得られたため、リフォールディング操作により再活性化を行った。リフォールディングは、Refolding CAキット(タカラバイオ社)を用いて行った。
抗体結合活性はELISA法により行った。先ず、抗体である10 μg/mlのガンマーグロブリン(化学及血清療法研究所製)を、マイクロプレートに固定し(4℃、18時間)、StartingBlock Blocking Buffers(登録商標)(PIERCE社)によりブロッキングした。次いで、実施例5で調製したタンパク質溶液を段階希釈し、ELISA反応に供した。反応は30℃、2時間で行った。次いで、0.2%(w/v)Tween20、150 mM NaClを含むTris−HCl緩衝液(pH8.0)で洗浄し、His−probe(H−15)HRP抗体(Santa Cruz Biotechnology社)を添加した。30℃で2時間反応させた後、先の緩衝液で洗浄し、TMB Peroxidase Substrate(KPL社)を添加し450 nmの吸光度を測定した。結果を図7に示す。図中、X軸(横軸)はサンプルの希釈倍率を示し、Y軸(縦軸)は、OD450nmの吸光度(単位は任意)を示す。図7に示す通り、サンプル6倍希釈した場合は、吸光度の減少はあまり観察されなかったが、18倍に希釈した場合、吸光度はかなり減少した。
【0055】
また、固定化時の抗体濃度を50 μg/mlから段階的に希釈したELISA反応も行った。ここでは、添加サンプルであるタンパク質調製溶液を一定量添加し反応させた。結果を図8に示す。図8に示す通り、固定化抗体濃度を上げることにより吸光度は上昇した。これらの結果より、大腸菌不溶性画分に発現させたFcγRI細胞外領域を含むポリペプチドをリフォールディングさせることにより特異的な抗体結合活性が有意に発現することが確認された。
【0056】
(実施例7)Fcレセプターの抗体吸着能評価
実施例6で得られた、FcγR1細胞外領域を含むポリペプチドは、配列番号:62及び配列番号:63から明らかなように、C末端にヒスチジンが6個連続した配列を有しているので、Niキレートの親和性により、担体と結合させることができる。実施例6で得られたポリペプチド100μgを、Novagen社製のHis・Bind Purificationキット(登録商標)中のHis・Bind Resinゲル500μLに固定化した。具体的には、実施例で得られたポリペプチド100μg、及び上記ゲル500μLをカラムに充填し、純水で洗浄した後、キット中のNiチャージ緩衝液(50mM NiSO4)を用いてNiをゲルに担持させた。
【0057】
次いで、抗体のゲルへの非特異吸着を防ぐために20mMのイミダゾール、500mMのNaCl、10%グリセロール、0.001%の界面活性剤(SB3−14)を含むTris−HCl緩衝液(pH8.0)により十分平衡化した。平衡化終了後のゲルに、平衡化緩衝液で希釈した100μg/mlのガンマグロブリンを2.0ml添加した。平衡化緩衝液で洗浄後、500mMのイミダゾール、500mMのNaCl、10%グリセロール、0.001%界面活性剤(SB3−14)を含むTris−HCl緩衝液(pH8.0)で溶出させた。
【0058】
なお、対照実験として、ゲルにFcγRI細胞外領域を含むポリペプチドを吸着しない区、すなわち、担体のみをカラムに充填した実験を行った。カラムから溶出した溶出液を集め、溶出液中のガンマグロブリン量を、バイオ・ラド社のプロテインアッセイキットを用い、イムノグロブリンを標準タンパク質として定量した。FcγRI細胞外領域を含むポリペプチドを固定化したゲルを用いた結果を図9に、対照実験の結果を図10に示す。
【0059】
図9及び図10において、X軸(横軸)はフラクション番号を示し、Y軸(縦軸)は溶出されたタンパク質の濃度(単位はμg/ml)を示し、矢印の位置は、溶出液を添加した位置を示す。図9に示す通り、FcγRI細胞外領域を含むポリペプチドを固定化したゲルにおいては、平衡化緩衝液で洗浄中にガンマグロブリンの溶出は認められず、溶出画分(図9におけるフラクション番号10〜15)に、ガンマグロブリンが検出された。図10に示すように、対照実験においては、ガンマグロブリンはゲルに吸着されることなく、フロースルー画分(図10におけるフラクション番号1〜3)に溶出された。
上記結果より、本発明の方法により得られた、FcガンマRI細胞外領域を含むポリペプチドを利用し、抗体を精製するための抗体精製用吸着剤を得ることができ、また、その抗体精製用吸着剤に抗体を吸着させ、その後精製する方法が提供された。
【図面の簡単な説明】
【0060】
【図1−1】抗体レセプターFcγR1のヒトおよび大腸菌間の遺伝子配列を比較した図である。図中、EcoliFcgR1が大腸菌型コドンに変換したDNA配列の配列、HumanFcGR1がヒトのFcγR1の配列である。
【図1−2】抗体レセプターFcγR1のヒトおよび大腸菌間の遺伝子配列を比較した図である。図中、EcoliFcgR1が大腸菌型コドンに変換したDNA配列の配列、HumanFcGR1がヒトのFcγR1の配列である。
【図2】プラスミドベクターpECFcRの構造を示す図である。
【図3】プラスミドベクターpETECFcRの構造を示す図である。
【図4】プラスミドベクターpETHUFcRの構造を示す図である。
【図5】実施例5におけるFcレセプターの発現誘導を示す図である。
【図6】抗体レセプターFcγR1細胞外領域を含むポリペプチドの発現量を比較した電気泳動像である。
【図7】実施例6におけるFcレセプターの抗体結合活性を示す図である。
【図8】実施例6におけるFcレセプターの抗体結合活性を示す図である。
【図9】実施例7におけるFcレセプターの抗体吸着能を示す図である。
【図10】実施例7におけるFcレセプターの抗体吸着能の対照実験を示す図である。
【特許請求の範囲】
【請求項1】
FcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、配列番号:2に記載の塩基配列を含むポリヌクレオチド、又は配列番号:2に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
【請求項2】
FcγRI細胞外領域を含むポリペプチドからシグナル配列を除去した領域をコードするポリヌクレオチドであって、配列番号:3に記載の塩基配列を含むポリヌクレオチド、又は配列番号:3に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
【請求項3】
シグナル配列を除去した、FcγRI細胞外領域を含むポリペプチドのN−末端側にメチオニンが付加されたポリペプチドをコードするポリヌクレオチドであって、配列番号:4に記載の塩基配列を含むポリヌクレオチド、又は配列番号:4に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
【請求項4】
配列番号2〜4のいずれか1に記載の塩基配列を含むポリヌクレオチドに、発現制御配列が機能的に付加されたポリヌクレオチド。
【請求項5】
配列番号2〜4のいずれか1に記載の塩基配列を含むポリヌクレオチドによりコードされるポリペプチド。
【請求項6】
請求項1〜4のいずれか1項に記載のポリヌクレオチドが挿入された組換えベクター。
【請求項7】
請求項1〜4のいずれか1項に記載のポリヌクレオチド、又は請求項6に記載の組換えベクターにより形質転換された形質転換体。
【請求項8】
形質転換体が原核細胞である、請求項7に記載の形質転換体。
【請求項9】
請求項7又は8に記載の形質転換体を培養する工程を含む、FcγRI細胞外領域を含むポリペプチドの製造方法。
【請求項10】
請求項9に記載の製造方法によって得られたポリペプチドを、担体上に固定してなる、抗体精製用吸着剤。
【請求項11】
担体がクロマトグラフィー用担体である、請求項10に記載の抗体精製用吸着剤。
【請求項12】
請求項10又は11に記載の抗体精製用吸着剤に抗体を吸着させる方法。
【請求項1】
FcγRI細胞外領域を含むポリペプチドをコードするポリヌクレオチドであって、配列番号:2に記載の塩基配列を含むポリヌクレオチド、又は配列番号:2に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
【請求項2】
FcγRI細胞外領域を含むポリペプチドからシグナル配列を除去した領域をコードするポリヌクレオチドであって、配列番号:3に記載の塩基配列を含むポリヌクレオチド、又は配列番号:3に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
【請求項3】
シグナル配列を除去した、FcγRI細胞外領域を含むポリペプチドのN−末端側にメチオニンが付加されたポリペプチドをコードするポリヌクレオチドであって、配列番号:4に記載の塩基配列を含むポリヌクレオチド、又は配列番号:4に記載の塩基配列を含むポリヌクレオチドと実質的に相同的なポリヌクレオチド。
【請求項4】
配列番号2〜4のいずれか1に記載の塩基配列を含むポリヌクレオチドに、発現制御配列が機能的に付加されたポリヌクレオチド。
【請求項5】
配列番号2〜4のいずれか1に記載の塩基配列を含むポリヌクレオチドによりコードされるポリペプチド。
【請求項6】
請求項1〜4のいずれか1項に記載のポリヌクレオチドが挿入された組換えベクター。
【請求項7】
請求項1〜4のいずれか1項に記載のポリヌクレオチド、又は請求項6に記載の組換えベクターにより形質転換された形質転換体。
【請求項8】
形質転換体が原核細胞である、請求項7に記載の形質転換体。
【請求項9】
請求項7又は8に記載の形質転換体を培養する工程を含む、FcγRI細胞外領域を含むポリペプチドの製造方法。
【請求項10】
請求項9に記載の製造方法によって得られたポリペプチドを、担体上に固定してなる、抗体精製用吸着剤。
【請求項11】
担体がクロマトグラフィー用担体である、請求項10に記載の抗体精製用吸着剤。
【請求項12】
請求項10又は11に記載の抗体精製用吸着剤に抗体を吸着させる方法。
【図2】


【図3】
【図4】
【図5】
【図7】
【図8】
【図9】
【図10】
【図1−1】

【図1−2】

【図6】
【図3】
【図4】
【図5】
【図7】
【図8】
【図9】
【図10】
【図1−1】
【図1−2】
【図6】
【公開番号】特開2008−245580(P2008−245580A)
【公開日】平成20年10月16日(2008.10.16)
【国際特許分類】
【出願番号】特願2007−91281(P2007−91281)
【出願日】平成19年3月30日(2007.3.30)
【出願人】(000003300)東ソー株式会社 (1,901)
【出願人】(000173762)財団法人相模中央化学研究所 (151)
【Fターム(参考)】
【公開日】平成20年10月16日(2008.10.16)
【国際特許分類】
【出願日】平成19年3月30日(2007.3.30)
【出願人】(000003300)東ソー株式会社 (1,901)
【出願人】(000173762)財団法人相模中央化学研究所 (151)
【Fターム(参考)】
[ Back to top ]