
Fc融合タンパク質の血清半減期を増加させるための組成物および方法
増加した血清半減期を有する糖改変体Fc融合タンパク質が本発明において提供される。1つ以上の非内因性グリコシル化部位を導入することによってFc融合タンパク質の血清半減期を増加させるための方法もまた提供される。本発明は、免疫グロブリンFcドメインおよび少なくとも1つの異種ポリペプチドドメインを含む融合タンパク質の血清半減期を延長するための方法を提供する。ある特定の実施形態において、それらの方法は、最初のFc融合タンパク質の異種部分をコードする核酸を、1つ以上のさらなるN結合型グリコシル化部位をコードするように改変することによって、最初のFc融合タンパク質と比べて延長された血清半減期を有する改変されたFc融合タンパク質をコードする改変された核酸を調製する工程を包含する。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願は、2009年12月2日に出願された米国仮出願番号第61/266,095号、および2010年4月23日に出願された同第61/327,582号の利益を主張する。上記米国仮出願の全教示は、参考として本明細書に援用される。
【背景技術】
【0002】
背景
天然状態または組換え的に生成されたときの治療用のタンパク質またはペプチドは、多くの場合、短い安定期間または短い血清半減期を示す不安定な分子である。さらに、これらの分子は、処方されるとき、特に、診断および治療の目的のために水溶液として処方されるとき、極度に不安定であることが多い。タンパク質性の治療用分子のインビボまたはインビトロにおける安定性を拡大するかまたは促進する実用的な溶液はほとんど無い。多くの治療薬、特に、ペプチド薬は、インビボでの不十分な血清半減期に悩まされている。これにより、治療効果に必要な血清レベルを維持するために、高頻度および/もしくはより高用量でのそのような治療薬の投与、または徐放製剤の使用を余儀なくされる。薬物の高頻度の全身投与は、かなりの負の副作用を伴う。例えば、高頻度の、例えば、毎日の、全身注射は、被験体にとってかなりの苦痛の種であり、投与関連感染症の高リスクの原因となり、特に、その治療薬が静脈内投与されるべきであるとき、入院または高頻度の来院を必要とし得る。そのうえ、長期間の処置における毎日の静脈内注射は、血管穿刺が繰り返されることによって引き起こされるかなりの組織瘢痕および血管病理ももたらし得る。同様の問題が、治療薬の高頻度の全身投与のすべてについて、例えば、糖尿病患者に対するインスリンの投与、または多発性硬化症に罹患している患者におけるインターフェロン薬物の投与について、知られている。これらの因子のすべてが、患者の服薬率を低下させ、医療制度に対するコストを増加させる。
【0003】
医薬品の血清半減期を改変するために可能性のある1つの解決方法は、半減期を増加させ得る薬剤分子に共有結合的に付着することである。以前に、ポリペプチドに対するポリエチレングリコールまたは「PEG」などのポリマーの付着によって、それらの血清半減期が増加し得ることが示された。しかしながら、ポリマーの付着によって、薬物活性が低下し得る。不完全または不均一な付着によって、異なる特性を有する化合物の混合集団がもたらされる。さらに、そのような改変に起因する半減期の変化は、予測不可能である。例えば、種々のポリエチレングリコールと、IL−8、G−CSFおよびIL−1raとの結合体化によって、様々な活性および半減期を有する分子が生成された(非特許文献1)。IL−8とPEG20との結合体化は、その半減期を変化させなかったが、IL−1raとPEG20との結合体化は、半減期を7倍近く増加させた。さらに、IL−8/PEG20結合体は、天然タンパク質よりも10〜20倍有効性が低い。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】GaertnerおよびOfford,Bioconjugate Chem.(1996),7:38−44
【発明の概要】
【発明が解決しようとする課題】
【0005】
したがって、生物学的に活性な分子の生物学的機能を著しく減少させずにその分子の血清半減期を増加することができる方法が、大いに望ましいだろう。
【課題を解決するための手段】
【0006】
要旨
一つには、本開示は、免疫グロブリンFcドメインおよび少なくとも1つの異種ポリペプチドドメインを含む融合タンパク質の血清半減期を延長するための方法を提供する。ある特定の実施形態において、それらの方法は、最初のFc融合タンパク質の異種部分をコードする核酸を、1つ以上のさらなるN結合型グリコシル化部位をコードするように改変することによって、最初のFc融合タンパク質と比べて延長された血清半減期を有する改変されたFc融合タンパク質をコードする改変された核酸を調製する工程を包含する。いくつかの実施形態において、本発明の改変された核酸は、適当な細胞培養物において発現されるとき、最初のFc融合タンパク質の血清半減期よりも少なくとも10%長い血清半減期を有する改変されたFc融合タンパク質をコードし得る。いくつかの実施形態において、本発明の改変されたFc融合タンパク質は、改変されていないFc融合タンパク質と比べて、実質的に同じまたは高いインビボ生物学的活性を有する。ある特定の実施形態において、さらなる(すなわち、導入される)グリコシル化部位の1つ以上におけるグリコシル化は、そのさらなるグリコシル化部位を欠く融合ポリペプチドの血清半減期と比べて、改変されたFc融合タンパク質の半減期(例えば、インビトロ、インビボにおける血清半減期)を少なくとも10%、20%、30%、40%、50%、75%、100%、125%、150%、175%、200%、225%もしくは250%またはそれ以上増加させる。ある特定の実施形態において、改変されたFc融合タンパク質、およびさらなるグリコシル化部位を欠く融合ポリペプチドの血清半減期は、比較するために、同じ動物モデルまたは動物種において測定される。例示的な実施形態において、血清半減期は、本明細書中に記載されるような薬物動態学的ラットアッセイまたは薬物動態学的サルアッセイにおいて測定される。
【0007】
いくつかの実施形態において、本開示は、a)少なくとも1つのさらなるN結合型グリコシル化部位を導入するように改変された核酸を、哺乳動物または哺乳動物様のグリコシル化を提供する細胞培養物において発現させること、およびb)その細胞培養物から改変されたFc融合タンパク質を収集することによって、最初のFc融合タンパク質と比べて延長された半減期を有する改変されたFc融合タンパク質を調製するための方法を提供する。Fc融合タンパク質は、細胞培養物からタンパク質を得るのに適した任意の手法を用いて、粗画分、部分的に精製された画分または高度に精製された画分として収集され得る。ある特定の態様において、改変された核酸は、シアル酸を含むN結合型糖成分を産生する細胞株において発現される。ある特定の態様において、改変された核酸は、哺乳動物細胞株によって発現され、その細胞株としては、CHO細胞株、NSO細胞株、COS細胞株またはHEK236細胞株が挙げられるがこれらに限定されない。他の態様において、改変された核酸は、哺乳動物または哺乳動物様のグリコシル化を提供するように操作された非哺乳動物細胞によって発現され、その細胞としては、遺伝的に操作された真菌細胞、昆虫細胞または植物細胞が挙げられるがこれらに限定されない。ある特定の態様において、精製は、改変されたFc融合タンパク質をプロテインAに曝露し、そのプロテインAに結合した改変されたFc融合タンパク質を収集する工程を含み得る。好ましい実施形態において、本開示の方法によって生成されるFc融合タンパク質は、患者に投与するために処方される。
【0008】
いくつかの実施形態において、本開示は、本明細書中に記載される方法のいずれかに従って調製される改変された核酸を含む細胞株を提供する。本発明の細胞株は、哺乳動物細胞株(例えば、CHO細胞株、NSO細胞株、COS細胞株またはHEK236細胞株)、または哺乳動物もしくは哺乳動物様のグリコシル化を提供するように遺伝的に改変された非哺乳動物細胞株(例えば、真菌細胞、昆虫細胞または植物細胞)を含み得る。
【0009】
ある特定の実施形態において、本開示は、増加された安定性および/または血清半減期を特徴とする糖改変体Fc融合タンパク質、ならびに増加された半減期を有する融合タンパク質を生成するための方法を提供する。本発明のFc融合タンパク質としては、免疫グロブリンFcドメインおよび少なくとも1つの異種ポリペプチドドメインを含むポリペプチドが挙げられるがこれに限定されない。そのFc融合タンパク質は、少なくとも1つの非内因性N結合型グリコシル化部位を導入するように免疫グロブリンFcドメインの外側で改変され、それにより、導入されるグリコシル化部位を欠く融合タンパク質の半減期(例えば、インビトロ、インビボまたは血清半減期)と比べて、改変された融合タンパク質の血清半減期が増加する。ある特定の実施形態において、本発明の融合タンパク質は、その融合物の血清半減期を、導入されるグリコシル化部位を欠く融合タンパク質の血清半減期(serum half−live)と比べて少なくとも10%、20%、30%、40%、50%、75%、100%、125%、150%、175%、200%、225%もしくは250%またはそれ以上増加させる、少なくとも1つの非内因性の、すなわち導入される、N結合型グリコシル化部位を含む。ある特定の実施形態において、改変されたFc融合タンパク質、およびさらなるグリコシル化部位を欠く融合ポリペプチドの血清半減期は、比較のために同じ動物モデルまたは動物種において測定される。例示的な実施形態において、血清半減期は、本明細書中に記載されるような薬物動態学的ラットアッセイまたは薬物動態学的サルアッセイにおいて測定される。
【0010】
本発明のFc融合タンパク質は、1つのN結合型グリコシル化部位を導入するために、1つ以上のアミノ酸残基の付加、欠失または置換によって改変され得る。ある特定の実施形態において、最初のFc融合タンパク質の異種部分は、各50、55、60、65、70、75、80、85、90、95 100、105 110、115、120、130、140または150アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む。ある特定の実施形態において、最初のFc融合タンパク質の異種部分は、各60、70、80、90、100、110、120または125アミノ酸あたり1個より少ないN結合型グリコシル化部位を含む。ある特定の実施形態において、N結合型グリコシル化に付着された、改変されたFc融合タンパク質の異種部分の各アミノ酸は、N結合型グリコシル化によって改変された他の任意のアミノ酸と少なくとも15、20、30、40、50、60、70、80、90または100アミノ酸離れている。ある特定の実施形態において、さらなるN結合型グリコシル化部位は、改変されたFc融合タンパク質のN末端、C末端、またはN末端とC末端の両方の10、20、30、40または50アミノ酸以内に存在しない。
【0011】
好ましい実施形態において、本発明のFc融合タンパク質は、表面に露出したアミノ酸配列(すなわち、可溶性ループドメイン)によって接続されており、かつαヘリックスまたはβシートに組み込まれない、少なくとも2つの構造的に異なるドメインを含む。好ましい実施形態において、さらなるN結合型グリコシル化部位は、表面に露出しているアミノ酸配列内に位置する。好ましい実施形態において、さらなるN結合型グリコシル化部位は、二次構造エレメント、例えば、αヘリックスまたはβシートを有するタンパク質の領域に組み込まれない。本発明において有用な異種部分には、タンパク質の機能的ドメイン(例えば、酵素ドメインもしくは触媒ドメインまたはリガンド結合ドメイン)が含まれ得る。いくつかの実施形態において、本開示のFc融合タンパク質は、リンカードメインをさらに含む。いくつかの実施形態において、導入されるグリコシル化部位のうちの少なくとも1つは、リンカードメイン内に配置される。本発明の融合ポリペプチドの異種ドメインは、好ましくは、高分子量であり、少なくとも25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、105または110kDaである。
【0012】
特定の実施形態において、異種部分は、細胞レセプター(例えば、膜貫通レセプター)の細胞外(すなわち、可溶性)ドメイン、ならびにその任意の改変体(変異体、フラグメントおよびペプチド模倣型を含む)を含む。好ましい実施形態において、異種部分(heterologous potion)には、膜貫通レセプターのリガンド結合ドメインが含まれる。本開示の方法は、付着される任意のさらなる糖成分が、リガンド結合ドメインを実質的に干渉しないような(例えば、導入されるグリコシル化部位を欠くFc融合タンパク質のリガンド結合性と比べて2、3、5、10または15倍未満の結合活性の減少)その異種の位置において1つ以上のN結合型グリコシル化部位を導入するために使用され得る。ゆえに、いくつかの態様において、導入されるN結合型グリコシル化部位の1つ以上は、リガンド結合ドメインのアミノ酸配列が改変されないような位置、および/またはリガンド境界面を実質的に干渉しないと予測される位置に導入される。ある特定の実施形態において、Fc融合タンパク質は、最初のFc融合タンパク質のIC50(すなわち、最大半量阻害濃度)よりも2、3、5または10倍以下だけ低いIC50を有する。
【0013】
ある特定の実施形態において、Fc融合タンパク質は、神経成長因子/腫瘍壊死因子レセプターファミリーのメンバーから選択される異種部分を含む。特定の実施形態において、細胞外レセプタードメインは、腫瘍壊死因子2型レセプター(すなわち、TNFR2)の一部分を含む。好ましい実施形態において、本開示は、1マイクロモル濃度未満または100、10もしくは1ナノモル濃度未満のKDでTNF−αに結合するTNFR2融合タンパク質を提供する。いくつかの実施形態において、本発明のTNFR2融合タンパク質は、ヒトTNFR2前駆体タンパク質(すなわち、配列番号1)のD47、Q48、A50、E155またはG253位に配置される1つ以上の改変されたアミノ酸を有するTNFR2細胞外ドメインを含む。これらの残基は、TNFR2(すなわち、配列番号2)のプロセシングされた細胞外のドメインのそれぞれD25、Q26、A28、E133およびG231位に対応する。好ましい改変には、TNFR2ポリペプチドに1つ以上のN結合型グリコシル化部位を導入する、アミノ酸の付加、置換および/または欠失が含まれ、例えば、配列番号2のアミノ酸配列に対するアミノ酸置換Q26N、A28S、D25N、E133N、D25N、G231Nが挙げられる。いくつかの実施形態において、1つ以上のN結合型グリコシル化部位を導入するために、1つより多い改変(例えば、付加、欠失および/または置換)がTNFR2融合タンパク質に対して行われ、その改変としては、例えば、配列番号2のアミノ酸配列に対する、Q26N/A28S;D25N/E133N;D25N/G231N;Q26N/A28S/E133N;Q26N/A28S/G231N;またはE133N/G231N置換を含むTNFR2改変体が挙げられる。本発明のTNFR2−Fc融合タンパク質は、本明細書中に開示されるもののいずれかであり得る(例えば、配列番号1、2、5、6、7もしくは8から選択されるアミノ酸配列を有するポリペプチドを含むか、または配列番号1、2、5、6、7もしくは8から選択されるアミノ酸配列と少なくとも80%、85%、90%、95%、97%もしくは99%同一であるアミノ酸配列を含む、融合タンパク質)。TNFR2−Fc融合タンパク質は、TNFR2−Fc融合タンパク質および薬学的に許容され得るキャリアを含む薬学的調製物として処方され得、ここで、その薬学的調製物は、哺乳動物への投与に適するように、発熱性材料を実質的に含まない。本組成物は、サイズ排除クロマトグラフィーによって評価されるとき、他のポリペプチド構成要素に対して少なくとも95%純粋であり得、必要に応じて本組成物は、少なくとも98%純粋である。
【図面の簡単な説明】
【0014】
本特許または本出願ファイルは、カラーで仕上げられた少なくとも1つの図面を含む。カラーの図面を含む本特許または本特許出願公報の写しは、請求され、必要な手数料の支払いがなされたときに、当局によって提供され得る。
【図1】図1は、TNFR2−h(1)Fcポリペプチドのアミノ酸配列(配列番号5)を示している。リンカーなしで免疫グロブリンFcドメインに融合されたヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質(h(1)Fc改変体と命名される)が示されている。免疫グロブリンFc領域に一重下線が引かれており、天然に存在するN結合型グリコシル化部位に二重下線が引かれている。
【図2】図2は、Q48N/A50S改変体TNFR2−h(1)Fcポリペプチドのアミノ酸配列(配列番号6)を示している。リンカーなしでh(1)Fc免疫グロブリンFcドメインに融合されたヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質が示されている。天然に存在するTNFR2−Fc細胞外ドメインが、N結合型グリコシル化部位を導入するために48および50位で改変された。免疫グロブリンFc領域に一重下線が引かれており、置換されたアミノ酸に二重下線が引かれている。
【図3】図3は、改変体(D47N/E155N)TNFR2−h(1)Fcポリペプチドのアミノ酸配列(配列番号7)を示している。リンカーなしでh(1)Fc免疫グロブリンFcドメインに融合されたヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質が示されている。天然に存在するTNFR2−Fc細胞外ドメインが、N結合型グリコシル化部位を導入するために47および155位で改変された。免疫グロブリンFc領域に一重下線が引かれており、置換されたアミノ酸に二重下線が引かれている。
【図4】図4は、改変体(D47N/G253N)TNFR2−h(1)Fcポリペプチドのアミノ酸配列(配列番号8)を示している。リンカーなしでh(1)Fc免疫グロブリンFcドメインに融合されたヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質が示されている。天然に存在するTNFR2−Fc細胞外ドメインが、N結合型グリコシル化部位を導入するために47および253位で改変された。免疫グロブリンFc領域に一重下線が引かれており、置換されたアミノ酸に二重下線が引かれている。
【図5】図5は、天然のリーダー配列を有する、TNFR2−h(1)Fcポリペプチド(すなわち、配列番号5)をコードする核酸配列(配列番号9)を示している。
【図6】図6は、TNFとTNFR2レセプターの細胞外ドメインとの結合境界面を示しているリボン図である。TNFは、左側の緑色のリボン構造であり、TNFR2レセプターは、右側の青色のリボン構造である。N結合型グリコシル化部位を導入するために操作された部位は、円で示されている。単一変異は、青色の円によって表示されており、二重変異は、緑色の円によって表示されている。リガンド結合活性を有意に減少させた変異は、TNF結合部位の近位に配置されており、赤色の円として示されている。
【図7】図7は、強調されている非内因性グリコシル化部位を導入するための、例示的なアミノ酸改変を有するTNFR2の細胞外ドメインの配列を示している。強調された各領域について、非内因性グリコシル化部位を導入し得る改変の特定の例が示されている。シグナルペプチドが、一重下線で示されており、内因性グリコシル化部位が、二重下線で示されている。
【図8】図8は、TNFR2構造内のベータ鎖の予測される位置(矢印)を示している。ナンバリングは、天然のTNFR2前駆体配列(配列番号1)に基づく。TNFR1(PDB ID:1EXT)、サイトカイン応答変更因子E(CrmE;PDB ID:2UWI)およびCD134(OX40;PDB ID:2HEY)との配列比較および構造相同性によって、二次構造を推測した。
【図9】図9は、天然のリーダー配列を有する改変体TNFR2−h(2)Fcポリペプチドのアミノ酸配列(配列番号16)を示している。4アミノ酸リンカーを介して例示的なヒトFcドメイン(配列番号14)に融合された天然のヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質が示されている。Fcドメインが、一重下線で示されており、リーダー配列およびリンカー配列に二重下線が引かれている。
【図10】図10は、TNFR2−h(2)Fcポリペプチド(すなわち、配列番号16)をコードするヌクレオチド配列(配列番号17)を示している。Fcドメインをコードする配列が、一重下線で示されており、リーダーおよびリンカーをコードする配列に二重下線が引かれている。
【図11】図11は、ラットコラーゲン誘発関節炎モデルにおける炎症のマーカーとしての、足の体積に対する糖改変体Q48N/A50S TNFR2−h(1)Fcの作用を示している。0および7日目にコラーゲンを投与し、被験物質での処置を6日目に開始した。示されているデータは、平均値±SEMである。TNFR2−h(1)Fcと同様に、Q48N/A50S TNFR2−h(1)Fcでの処置は、この研究の後半のビヒクル処置ラットにおいて観察された足の腫脹を防止した。
【図12】図12は、ラットコラーゲン誘発関節炎モデルにおける骨質に対する糖改変体Q48N/A50S TNFR2−h(1)Fcの作用を示している。21日目の研究完了後のマイクロコンピュータ断層撮影(マイクロCT)によって、エキソビボにおいて足根骨の像を得た。TNFR2−h(1)Fcと同様に、Q48N/A50S TNFR2−h(1)Fcでの処置は、ビヒクル処置ラットにおいて観察された骨侵食を防止した。
【発明を実施するための形態】
【0015】
詳細な説明
1.概要
ある特定の態様において、本開示は、適切な標的タンパク質の適切な位置に少なくとも1つの非内因性N結合型グリコシル化部位を導入することによって、Fc融合タンパク質の血清半減期が延長され得るという驚くべき発見に関する。したがって、本開示は、そのポリペプチド上のN結合型グリコシル化部位の数を増加させることによって、特定の治療用ポリペプチドであるFc融合タンパク質の血清半減期を増加させるための方法を提供する。本明細書中で証明されるように、本開示の方法は、リガンド結合ドメインを含むレセプターの細胞外ドメインの一部分を含むFc融合タンパク質の血清半減期を増加させるために使用された。特定の例において、本開示は、それぞれの融合タンパク質の改変されていない形態と比べて増加された半減期を特徴とする改変されたTNFR2−Fc融合タンパク質を提供する。本開示を任意の特定の作用機序に限定しないが、Fc融合タンパク質の非Fc部分(「異種部分」)は、その患者の体内滞留時間中に様々な細胞内および細胞外の環境に曝露されると提唱される。これらの種々の条件によって、その異種部分の一部分が、プロテアーゼに対する基質、またはタンパク質の改変もしくは分解のプロセスを開始する他の酵素もしくは分子に対する基質として脆弱になる可能性がある。したがって、本開示は、Fc融合タンパク質の血清半減期が、異種部分を分解するように作用する物質に有意に影響され、その異種部分をそのような物質から保護する傾向がある改変が、より長い血清半減期をもたらし得るという新規の提案を提供する。本明細書中に記載されるように、1つ以上のN結合型グリコシル化部位の付加によって、異種ドメインの脆弱な部分を望まれない分解または変更から保護するため、および場合によっては、そのような分子の望ましい構造上のコンフォメーションを安定化するための、単一工程の生体適合性の系を提供する。
【0016】
最初のFc融合タンパク質の異種ドメイン内のどの位置がさらなるN結合型グリコシル化部位に最も適用できるかを演繹的に予測することは困難であり得、何の制約または指針もないときは、任意の1つの異種ドメインが、理論上は、1つ以上のさらなるN結合型部位のほぼ無限の組み合わせで改変され得る。種々の最初のFc融合タンパク質を用いた研究を通して、本出願人は、改変された分子の生物学的活性(特に、インビボ生物学的活性)を保ちつつ、最初のFc融合タンパク質と比べて延長された血清半減期を示す改変されたFc融合タンパク質に到達するために、妥当な部位数での最初のFc融合タンパク質の改変を可能にする様々な指針を見出した。本開示全体の範囲を限定することを望むものではないが、本出願人は、以下の原理の1つ以上を用いることにより、半減期が延長された活性なFc融合タンパク質を生成する実行可能なアプローチが提供されることを見出した:(1)タンパク質構造解析によって決定されるか経験的方法によって決定されるかに関係なく、N結合型グリコシル化部位は、Fc融合タンパク質の異種部分の表面に露出した位置、特に、規定のタンパク質構造エレメント(例えば、アルファヘリックスまたはベータシート)内に含まれない位置に配置されることが有利であり得る;(2)N結合型グリコシル化部位は、改変または分解に供された場合にタンパク質の構造または機能の実質的な混乱を引き起こし得るFc融合タンパク質の異種部分内の位置に配置されることが有利であり得る(例えば、多くのタンパク質は、切断された場合にタンパク質全体およびその活性の混乱をほとんどまたはまったく引き起こさない非構造領域をN末端に有し、したがって、そのような位置に配置されるN結合型グリコシル化部位は、血清半減期に対して比較的小さい作用しか有さない可能性があるが、対照的に、構造エレメント間に配置される表面露出領域は、高露出と、望ましくない分解に供された場合のそのタンパク質に対する重大な結果との組み合わせを示すので、構造エレメント間の表面露出アミノ酸は、N結合型グリコシル化部位の導入にとって望ましい位置に相当する);(3)N結合型グリコシル化部位は、最初のFc融合タンパク質の重要な機能的部位(例えば、リガンド結合表面または触媒部位)を干渉しない位置に配置され得、これは、その機能的部位自体の外部の位置を選択すること、および/または任意の新しいN結合型糖成分がその機能的部位に突き出ないであろう位置を選択することによって達成され得る;(4)N結合型グリコシル化部位の付加によって達成される作用の程度は、すでに最初のFc融合タンパク質に存在するN結合型糖の密度と反比例し得るので、そのアプローチは、比較的低レベルのグリコシル化(例えば、60、70、80、90、100、110、125アミノ酸またはそれ以上の異種部分あたり1個未満のN結合型部位)を有する異種部分を有する最初のFc融合タンパク質を用いるとき、最も有効であり得る;(5)重度にグリコシル化された異種部分と軽度にグリコシル化された異種部分の両方において、達成される作用の程度は、N結合型グリコシル化部位間の間隔に比例し得る(これは、1つ以上のさらなるN結合型グリコシル化部位が、比較的規則正しい間隔、かつ既存のN結合型グリコシル化部位または他のさらなるN結合型グリコシル化部位から離れた(例えば、N結合型グリコシル化部位のN残基間が10、15、20、25、30、35または40アミノ酸を超える)間隔で配置されることが有利であり得ることを意味し、重度にグリコシル化されたタンパク質でさえ、糖が密集する場合、十分に間隔が空けられたN結合型部位の付加によって、血清半減期の有意な増加が促進され得る)。上記原理の1つ以上を用いることにより、最初のFc融合タンパク質と比べて延長された血清半減期を示す単一の改変されたFc融合タンパク質を設計することが可能であり得るが、単一の最初のFc融合タンパク質に基づいて複数の改変されたFc融合タンパク質を生成すること、これらの変更された形態を試験すること、次いで、増加された半減期と保持された生物学的活性との最善の組み合わせを示す改変された形態の組み合わせを試験することが有益であることが多い。
【0017】
ある特定の態様において、本開示は、そのポリペプチド上のN結合型グリコシル化部位の数を増加させることによって、特定の治療用ポリペプチドであるFc融合タンパク質の血清半減期を増加させるための方法に関する。本明細書中で証明されるように、本開示の方法は、リガンド結合ドメインを含むレセプターの細胞外ドメインの一部分を含むFc融合タンパク質の血清半減期を増加させるために使用された。特定の例において、本開示は、それぞれの融合タンパク質の改変されていない形態と比べて増加された半減期を特徴とする改変されたTNFR2−Fc融合タンパク質を提供する。その機序に関係なく、さらなるグリコシル化部位の導入が、高分子量の生物製剤、特に、Fc融合タンパク質の半減期を延ばす有効な方法であることは、本明細書中に提示されるデータから明らかである。
【0018】
特定の実施形態において、本開示は、増加された血清半減期を特徴とする改変されたTNFR2−Fc糖改変体融合タンパク質を提供する。腫瘍壊死因子(TNF)は、正常な炎症応答および免疫応答に関わる、天然に存在するサイトカインである。腫瘍壊死因子−α(TNFα)および腫瘍壊死因子−β(TNFβ)は、相同的な多機能性サイトカインである。これらのポリペプチドの構造的および機能的な特徴が非常に類似していることにより、それらは、腫瘍壊死因子または「TNF」と集合的に記述される。一般にTNFに帰されている活性としては:他のサイトカイン(IL−1、IL−6、GM−CSFおよびIL−10を含む)の放出、ケモカインの誘導、接着分子の増加、血管の成長、組織破壊酵素の放出、およびT細胞の活性化が挙げられる。例えば、Feldmannら、1997,Adv.Immunol.,64:283−350,Nawrothら、1986,J.Exp.Med.,163:1363−1375;Moserら、1989,J.Clin.Invest,83:444−455;Shinguら、1993,Clin.Exp.Immunol.94:145−149;MacNaulら、1992,Matrix Suppl.,1:198−199;およびAhmadzadehら、1990,Clin.Exp.Rheumatol.8:387−391を参照のこと。これらの活性のすべてが、炎症応答を増強するように働き得る。
【0019】
TNFは、組織の損傷をもたらす炎症促進性作用(例えば、血管内皮細胞に対して凝血促進活性を誘導すること(Poberら、J.Immunol.136:1680(1986))、好中球およびリンパ球の接着を増加させること(Poberら、J.Immunol.138:3319(1987))ならびにマクロファージ、好中球および血管内皮細胞からの血小板活性化因子の放出を刺激すること(Camussiら、J.Exp.Med.166:1390(1987)))を引き起こす。TNFはまた、感染症(Ceramiら、Immunol.Today 9:28(1988))、免疫障害、腫瘍性の病態(Oliffら、Cell 50:555(1987))、自己免疫性の病態および移植片対宿主病態(Piguetら、J.Exp.Med.166:1280(1987))にも関連する。うっ血性心不全、炎症性腸疾患(クローン病を含む)、関節炎および喘息が、そのようなTNF関連障害の中に入る。
【0020】
特に、TNFは、発熱、倦怠感、食欲不振および悪液質を含む、グラム陰性敗血症およびエンドトキシンショックにおいて中心的役割を果たす(Michieら、Br.J.Surg.76:670−671(1989);Debetsら、Second Vienna Shock Forum,p.463−466(1989);Simpsonら、Crit.Care Clin.5:27−47(1989);Waageら、Lancet 1:355−357(1987);Hammerleら、Second Vienna Shock Forum p.715−718(1989);Debetsら、Crit.Care Med.17:489−497(1989);Calandraら、J.Infect.Dis.161:982−987(1990);Revhaugら、Arch.Surg.123:162−170(1988))。
【0021】
TNFは、TNF応答性細胞上の特異的な細胞表面レセプターとの相互作用を介して、その生物学的作用を開始する。細胞表面腫瘍壊死因子レセプター(TNFR)には、p75(または2型)およびp55(または1型)と命名された異なる2つの形態がある(Smithら、1990,Science 248:1019−1023;Loetscherら、1990,Cell 61:351−359)。TNFR 1型およびTNFR 2型の各々は、TNFαとTNFβの両方に結合する。TNFの生物学的活性は、いずれかの細胞表面TNFRへの結合に依存する。1型レセプター(TNF−R55、TNF−RIまたはTNFR−βとも呼ばれる)は、TNF−αの細胞傷害性、抗ウイルス性および増殖性の活性をもたらすシグナルを伝達すると示されている55kdの糖タンパク質である。p75レセプター(TNF−R75、TNFR2またはTNFR−αとも呼ばれる)は、細胞傷害性および増殖性のシグナル、ならびにGM−CSFの分泌をもたらすシグナルを伝達すると示されている75kDaの糖タンパク質である。
【0022】
TNFアンタゴニスト(例えば、可溶性TNFRおよび抗TNF抗体)は、TNF活性を阻止して、TNFに応答したIL−1、GM−CSF、IL−6、IL−8、接着分子および組織破壊の減少を引き起こすと証明されている(Feldmannら、1997)。ハムスター抗マウスTNF抗体を利用したTNF拮抗作用の影響が、DBA/1マウスにおけるII型コラーゲン関節炎のモデルにおいて試験された(Williamsら、1992,Proc.Natl.Acad.Sci.USA,89:9784−9788)。疾患の発症後に開始された処置によって、足蹠の腫脹、臨床スコア、および関節破壊の組織病理学が改善された。他の研究も、抗体(Thorbeckeら、1992,Proc.Natl.Acad.Sci.USA,89:7375−7379)またはTNFR構築物(Husbyら、1988,J.Autoimmun.1:363−71;Tettaら、1990,Ann.Rheum.Dis.49:665−667;Wooleyら、1993,J.Immunol.151:6602−6607;Piguetら、1992,Immunology 77:510−514)のいずれかを用いて同様の結果を得ている。
【0023】
3つの特異的なTNFアンタゴニストが、現在、FDAによって承認されている:エタネルセプト(Enbrel(登録商標))、インフリキシマブ(Remicade(登録商標))およびアダリムマブ(Humira(登録商標))。これらの薬物の1つ以上が、関節リウマチ、若年性関節リウマチ、乾癬、乾癬性関節炎、強直性脊椎炎および炎症性腸疾患(クローン病または潰瘍性大腸炎)の処置について承認されている。
【0024】
ヒトIgG1のFc部分に連結された可溶性ヒトTNFR(p75)の組換えバージョン(sTNFR(p75):Fc,Enbrel(登録商標),Immunex)の臨床試験から、その投与によって、RA疾患の活動性の有意かつ迅速な減少がもたらされたことが示された(Morelandら、1997,N.Eng.J.Med.,337:141−147)。さらに、Enbrel(登録商標)に対する小児科の臨床試験からの安全性データは、この薬物が、一般に、若年性関節リウマチ(JRA)を有する患者によって十分許容されることを示唆している(Garrisonら、1998,Am.College of Rheumatology meeting,Nov.9,1998,abstract 584)。
【0025】
上で述べたように、Enbrel(登録商標)は、ヒトIgG1のFc部分に連結されたヒト75キロダルトン(p75)TNFRの細胞外リガンド結合部分からなる二量体の融合タンパク質である。Enbrel(登録商標)のFc構成要素は、CH2ドメイン、CH3ドメインおよびヒンジ領域を含むが、IgG1のCH1ドメインを含まない。Enbrel(登録商標)は、チャイニーズハムスター卵巣(CHO)哺乳動物細胞発現系において産生される。それは、934アミノ酸からなり、およそ150キロダルトンの見かけの分子量を有する(Smithら、1990,Science 248:1019−1023;Mohlerら、1993,J.Immunol.151:1548−1561;米国特許第5,395,760号(Immunex Corporation,Seattle,Wash.);米国特許第5,605,690号(Immunex Corporation,Seattle,Wash.)。
【0026】
Enbrel(登録商標)は、現在、1つ以上の疾患修飾性抗リウマチ薬(DMARD)に対して不十分な反応を有する患者における中等度から重度の活動性関節リウマチの徴候および症状の減少について示唆されている。Enbrel(登録商標)は、メトトレキサート単独に十分に反応しない患者において、メトトレキサートと併用して使用され得る。Enbrel(登録商標)は、1つ以上のDMARDに対して不十分な反応を有する患者における中等度から重度の活動性多関節型若年性関節リウマチの徴候および症状の減少についても示唆されている(1999年5月28日)。Enbrel(登録商標)は、皮下注射として、1週間に2回、25mgでRA患者に投与される。
【0027】
現在、ENBREL調製物を用いた処置は、1週間に2回皮下に投与されるが、これは、高価であり、患者にとって不快かつ不便である。したがって、本開示は、導入されるグリコシル化部位を欠く可溶性TNFレセプターの血清半減期と比べて改変されたポリペプチドの血清半減期を増加させる少なくとも1つの非内因性N結合型グリコシル化部位を導入するように改変されたTNF結合レセプター(例えば、Enbrel(登録商標))の可溶性(例えば、細胞外)部分を含むTNFアンタゴニストを提供する。
【0028】
本明細書において使用される用語は、概して、当該分野における、本発明の文脈内における、および各用語が使用される特定の文脈における、それらの通常の意味を有する。ある特定の用語が、本発明の組成物および方法、ならびにそれらをどのように作製し使用するかを説明する際に、当業者にさらなる指針を提供するために下記または本明細書の他の箇所で論じられる。用語の任意の使用の範囲または意味は、その用語が使用される特定の文脈から明らかになるだろう。
【0029】
「約」および「およそ」は、測定の性質または精度を考慮に入れると、通常、測定された量について許容され得る誤差の程度を意味するものとする。代表的には、例示的な誤差の程度は、所与の値または所与の値の範囲の20パーセント(%)以内、好ましくは、10%以内、より好ましくは、5%以内である。
【0030】
あるいは、および生体系において特に、用語「約」および「およそ」は、1桁以内の値、好ましくは、所与の値の5倍以内の値、より好ましくは、2倍以内の値を意味し得る。
【0031】
本発明の方法は、配列を互いに比較する工程(野生型配列と1つ以上の変異体(配列改変体)との比較を含む)を包含し得る。そのような比較は、代表的には、例えば、当該分野で周知の配列アラインメントプログラムおよび/またはアルゴリズム(いくつかの例を挙げれば、例えば、BLAST、FASTAおよびMEGALIGN)を用いた、ポリマー配列のアラインメントを含む。当業者は、そのようなアラインメントでは、変異が残基の挿入または欠失を含む場合、配列アラインメントは、挿入されたまたは欠失された残基を含まないポリマー配列内に「ギャップ」(代表的には、ダッシュまたは「A」によって表される)を導入し得ることを容易に認識し得る。
【0032】
そのすべての文法的な形および綴りバリエーションにおける「相同」とは、「共通の進化的起源」を有する2つのタンパク質(同じ生物種内の上科由来のタンパク質ならびに異なる生物種由来の相同タンパク質を含む)間の関係性のことを指す。そのようなタンパク質(およびそれらをコードする核酸)は、同一性パーセントに関するものであるか、または特定の残基もしくはモチーフおよび保存された位置の存在によるものであるかに関係なく、それらの配列類似性によって反映される配列相同性を有する。
【0033】
そのすべての文法上の形態における用語「配列類似性」とは、共通の進化的起源を共有するかもしれないし共有しないかもしれない、核酸配列間またはアミノ酸配列間の同一性または対応の程度のことを指す。
【0034】
しかしながら、通常の使用法および本願において、用語「相同」は、「高度に」などの副詞で修飾されるとき、配列類似性のことを指し得、共通の進化的起源に関係することもあるし、関係しないこともある。
【0035】
用語「薬物動態学的特性」とは、生理活性物質(例えば、小分子、ポリペプチド薬など)の吸収、分布、代謝および排出のことを指す。
【0036】
2.糖改変体Fc融合タンパク質
異種ポリペプチドに連結された免疫グロブリン分子由来のFcドメインを有する糖改変体融合タンパク質が、本明細書中に提供される。その糖改変体融合タンパク質は、融合タンパク質のFcドメインの外側に少なくとも1つの非内因性N結合型グリコシル化部位を含む。そのFcドメインは、直接、またはポリペプチドリンカーを介して間接的に、異種ポリペプチドに連結され得る。非内因性グリコシル化部位は、異種ポリペプチド、リンカーまたはその両方に導入され得る。
【0037】
「非内因性」、「導入される」または「新規の」グリコシル化部位とは、そのポリペプチドの改変されていないバージョンに存在しないグリコシル化部位のことを指す。したがって、本明細書中に記載される糖改変体融合タンパク質は、融合タンパク質の改変されていないバージョンに存在するグリコシル化部位の数と比べて、少なくとも1つのさらなるN結合型グリコシル化部位を有する。例えば、融合タンパク質の改変されていないバージョンが、Fcドメインの外側に2つのグリコシル化部位を有し、1つのN結合型グリコシル化部位が、異種ポリペプチドに導入される場合、その糖改変体融合タンパク質は、2つの天然のグリコシル化部位および1つの導入されたグリコシル化部位を含む3つのN結合型グリコシル化部位を有することになる。天然に存在する改変体または人工の改変体もまた、グリコシル化部位の数を増加させる本発明の方法に従って改変され得るので、グリコシル化部位の数を増加させるための融合タンパク質の改変は、特定の「野生型」アミノ酸配列に限定されないことが理解されるべきである。
【0038】
いくつかのタンパク質が多数の糖側鎖を担持し得ることは、周知である。O結合型グリコシル化部位間の距離は、わずか1つおきのアミノ酸であり得る(例えば、Kolset & Tveit(2008)Cell.Mol.Life Sci 65:1073−1085およびKianiら(2002)Cell Research 12(1):19−32を参照のこと)。N結合型グリコシル化部位については、部位間の距離は、わずか3、4、5または6アミノ酸であり得る(例えば、Lundinら(2007)FEBS Letters 581:5601−5604(2007);Apweilerら(1991)Biochimica et Biophysica Acta 1473:4−8(これらの各々の全内容が本明細書中で参考として援用される)を参照のこと)。したがって、ある特定の実施形態において、少なくとも約1、2、3、4、5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95もしくは100またはそれ以上のN結合型グリコシル化部位が、本明細書中に記載される融合タンパク質に付加され得る。ある特定の実施形態において、本開示の糖改変体融合タンパク質は、各10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、125、150または200アミノ酸あたり少なくとも1つのグリコシル化されたアミノ酸を含む。ある特定の実施形態において、N結合型グリコシル化された各アミノ酸は、他の任意のN結合型、O結合型、またはN結合型およびO結合型のグリコシル化されたアミノ酸と少なくとも10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95もしくは100またはそれ以上のアミノ酸だけ離れている。
【0039】
本明細書中で使用されるとき、「グリコシル化部位」は、糖付着コンセンサス配列(すなわち、糖、(単糖、オリゴ糖または多糖のいずれか)をアミノ酸配列に付着するためのコンセンサス配列として作用する、ある範囲のアミノ酸)のことを意味し得るか、または糖成分が共有結合的に連結される実際のアミノ酸残基のことを意味し得る。その糖成分は、単糖(単糖分子)、オリゴ糖または多糖であり得る。
【0040】
N結合型グリコシル化部位は、糖成分を付加するように、タンパク質のアミノ酸配列を改変することによって、または融合タンパク質におけるアミノ酸残基を化学的に改変することによって、融合タンパク質に導入され得る。好ましい実施形態において、本明細書中に記載される融合タンパク質は、コンセンサス配列NXT/S(アスパラギン−X−セリン/トレオニン)(ここで、Xは、プロリン以外の任意のアミノ酸である)を有する少なくとも1つのN結合型グリコシル化部位を導入するように(例えば、指定のアミノ酸の挿入、欠失または置換によって)改変される。タンパク質上の炭水化物部分の数を増加させる別の手段は、ポリペプチドへのグリコシドの化学的または酵素的な結合によるものである。例えば、用いられる結合の様式に依存して、糖成分は、(a)アルギニンおよびヒスチジン;(b)遊離カルボキシル基;(c)遊離スルフヒドリル基(例えば、システインのもの);(d)遊離ヒドロキシル基(例えば、セリン、トレオニンまたはヒドロキシプロリンのもの);(e)芳香族残基(例えば、フェニルアラニン、チロシンまたはトリプトファンのもの);または(f)グルタミンのアミド基に付着され得る。例えば、WO87/05330およびAplin and Wriston(1981)CRC Crit.Rev.Biochem.,pp.259−306(本明細書中で参考として援用される)を参照のこと。例示的な実施形態において、N結合型グリカンを生成するために、糖成分は、アルギニン残基に結合される。
【0041】
例示的な実施形態において、糖成分は、宿主細胞において融合タンパク質を発現させることによって細胞機構を用いて、導入されたグリコシル化部位に付加される。一般に、融合タンパク質は、適切なグリコシル化を提供する哺乳動物細胞株(例えば、HEK293またはCHO細胞株)または他の哺乳動物発現細胞株において発現され得る。
【0042】
ある特定の実施形態において、糖成分は、哺乳動物様のグリコシル化をもたらすように操作された非哺乳動物宿主細胞(例えば、酵母、細菌または昆虫細胞)を用いて、導入されたグリコシル化部位に付加され得る。ヒトにおける糖タンパク質のプロセシングを模倣する一連の酵素反応を行う遺伝的に改変されたグリコシル化経路を有する細胞株が開発されている。これらの操作された細胞において発現される組換えタンパク質は、実質的に同一でないにせよ、ヒトの対応物に類似の糖タンパク質(例えば、哺乳動物様のグリコシル化)をもたらす。
【0043】
発現されるペプチドのグリコシル化プロファイルを変更するように宿主細胞を遺伝的に改変するための手法は、周知である。例えば、Altmannら(1999,Glycoconjugate J.16:109−123)、Ailorら(2000,Glycobiology 10(8):837−847)、Jarvisら(In vitrogen Conference,March,1999,abstract)、Hollister and Jarvis(2001,Glycobiology 11(1):1−9)およびPalacpacら(1999,PNAS USA 96:4697)、Jarvisら(1998.Curr.Opin.Biotechnol.9:528−533)、Gemgross(米国特許公開番号20020137134)(これらのすべてが、昆虫細胞または植物細胞をグリコシルトランスフェラーゼ遺伝子でトランスフェクトすることによって、昆虫細胞発現系または植物細胞発現系を「ヒト化する(hummanize)」手法を開示している)を参照のこと。
【0044】
原核生物系において発現されるペプチドのグリコシル化プロファイルを遺伝的に変更する手法も存在する。インビボにおいてオリゴ糖を産生する、細菌Neisseria meningitidisおよびAzorhizobium由来の様々なグリコシルトランスフェラーゼで大腸菌が操作されている(Bettlerら、1999,Glycoconj.J.16:205−212)。Neisseria meningitidisのβ1,3Nアセチルグルコサミニルトランスフェラーゼ(acetyl glucoaminyltransferase)lgta遺伝子を過剰発現するように遺伝的に操作された大腸菌は、外来性ラクトースを効率的にグリコシル化する(Priemら、2002,Glycobiology 12:235−240)。
【0045】
真菌細胞は、外来性グリコシルトランスフェラーゼを産生するようにも遺伝的に改変されている(Yoshidaら、1999,Glycobiology,9(1):53−58;Kalsnerら、1995,Glycoconj.J.12:360−370;Schwientek and Ernst,1994,Gene 145(2):299−303;Chibaら、1995,Biochem J.308:405−409)。
【0046】
ある特定の実施形態において、本開示のFc融合タンパク質を発現する宿主細胞は、1つ以上の外来性グリコシルトランスフェラーゼ酵素および/または1つ以上の外来性グリコシダーゼ酵素を発現する真核細胞または原核細胞であり得、ここで、その宿主細胞内での組換え糖ペプチドの発現によって、「ヒト」グリカン構造を有する組換え糖ペプチドが産生される。
【0047】
いくつかの実施形態において、上記細胞において有用な異種グリコシルトランスフェラーゼ酵素は、例えば、Taniguchiら(2002,Handbook of Glycosyltransferases and Related Genes,Springer,N.Y.)において入手可能なグリコシルトランスフェラーゼファミリーのリストに含められた任意の公知のグリコシルトランスフェラーゼ酵素からなる群より選択され得る。
【0048】
いくつかの実施形態において、宿主細胞は、宿主細胞内での組換え糖ペプチドの発現によって、「ヒト」グリカン構造を有する組換え糖ペプチドが産生されるように、1つ以上の内因性グリコシルトランスフェラーゼ酵素および/または1つ以上の内因性グリコシダーゼ酵素が不活性化された、真核細胞または原核細胞であり得る。
【0049】
いくつかの実施形態において、宿主細胞は、異種のグリコシルトランスフェラーゼ酵素および/またはグリコシダーゼ酵素を発現し得、同時に、1つ以上の内因性のグリコシルトランスフェラーゼ酵素および/またはグリコシダーゼ酵素が不活性化されている。内因性のグリコシルトランスフェラーゼ酵素および/またはグリコシダーゼ酵素は、当業者に公知の任意の手法を用いて不活性化され得、その手法としては、アンチセンス法、および宿主細胞のゲノムに核酸を挿入することを含む手法が挙げられるがこれらに限定されない。
【0050】
例示的な実施形態において、本明細書中に記載される糖改変体融合タンパク質は、その融合タンパク質の改変されていない形態と比べて高い安定性および/または増加された血清半減期を有する。例示的な実施形態において、糖改変体融合タンパク質の血清半減期は、その融合タンパク質の改変されていない形態と比べて、少なくとも10%、15%、20%、25%、30%、40%、50%、55%、60%、70%、75%、80%、85%、90%、95%、100%、125%、150%、200%、250%もしくは300%またはそれ以上増加される。ある特定の実施形態において、改変されたFc融合タンパク質およびさらなるグリコシル化部位を欠く融合ポリペプチドの血清半減期は、比較のために、同じ動物モデルまたは動物種において測定される。例示的な実施形態において、血清半減期は、本明細書中に記載されるような薬物動態学的ラットアッセイまたは薬物動態学的サルアッセイにおいて測定される(例えば、実施例4および5を参照のこと)。
【0051】
例示的な実施形態において、1つ以上の非内因性グリコシル化部位の導入は、融合タンパク質の異種ポリペプチド部分の1つ以上の生物学的活性に有意に影響しない。例えば、融合タンパク質の異種ポリペプチド部分の1つ以上の生物学的活性は、10倍、5倍、3倍、2.5倍、2倍、1.5倍、1倍、0.5倍未満またはそれより少なく、影響され得る。異種ポリペプチド部分の生物学的活性の例は、本明細書中にさらに記載されるが、例えば、タンパク質間相互作用、リガンド結合、生理学的活性などが挙げられる。
【0052】
例示的な実施形態において、1つ以上の非内因性グリコシル化部位が、Fc融合タンパク質の異種部分に導入される。Fc融合タンパク質の異種部分は、触媒ドメインまたはリガンド結合ドメインを含み得る。本明細書中で使用されるとき、「リガンド結合ドメイン」とは、別の分子、例えば、リガンドと相互作用するタンパク質上の領域のことを指す。例えば、リガンド結合ドメインとは、抗体によって結合されるタンパク質上の領域、抗原に結合する抗体上の領域、リガンドに結合するレセプター上の領域、レセプターに結合するリガンド上の領域、第2のポリペプチドに結合する第1のポリペプチド上の領域、小分子に結合するポリペプチド上の領域などのことを指し得る。例示的な実施形態において、リガンド結合ドメインは、リガンドに結合する膜貫通レセプタータンパク質の細胞外ドメイン内の領域である。本明細書中で使用されるとき、「触媒ドメイン」とは、機能活性を有する領域のことを指す。触媒ドメインの例としては、例えば、キナーゼドメイン、ホスファターゼドメイン、プロテアーゼドメインが挙げられる。
【0053】
例示的な実施形態において、1つ以上の非内因性グリコシル化部位は、可撓性または非構造である、表面に露出した領域(例えば、αヘリックスおよび/またはβシートを接続するループ領域)に導入される。通常、タンパク質は、二次構造および三次構造を有するポリペプチド鎖である。ポリペプチドの二次構造は、αヘリックスおよびβシートと呼ばれる2つの一般的な構造ドメインへのポリペプチド鎖のフォールディングを含む。タンパク質の三次構造は、様々な長さおよび不規則な形状のループ領域によって接続される、αヘリックスおよびβシートである二次構造の組み合わせからなる。一般に、二次構造エレメントの組み合わせによって、安定な疎水性コアが形成され、ループ領域は、タンパク質の表面に提示される。ループ領域は、溶媒に曝露されるので、それらは、一般に、内部の構造的に安定な二次構造ドメイン(例えば、αヘリックスおよびβシートドメイン)よりも、様々な反応性分子およびタンパク質酵素(例えば、プロテアーゼ)による求核攻撃/ペプチド結合切断を受けやすい。
【0054】
ある特定の実施形態において、融合タンパク質の異種ポリペプチド部分における1つ以上の天然に存在するグリコシル化部位(例えば、改変されていない形態のタンパク質におけるグリコシル化部位)を除去することが望ましい場合がある。例えば、N結合型グリコシル化部位の間隔を変更するために、天然に存在するグリコシル化部位を除去し、次いで、所望の位置に2つの非内因性グリコシル化部位を導入することが望ましい場合がある。さらに、融合タンパク質に存在する天然に存在するO結合型グリコシル化部位の1つ以上を除去することが望ましい場合がある。
【0055】
N結合型およびO結合型のグリコシル化部位は、コンセンサスアミノ酸配列を削除することによって、またはアミノ酸残基から糖成分を化学的もしくは酵素的に切断することによって、除去され得る。例えば、N結合型グリコシル化部位は、N結合型グリコシル化認識部位(すなわち、NXT/S)の1番目もしくは3番目のアミノ酸の位置の一方もしくは両方におけるアミノ酸の置換もしくは欠失、および/またはそのトリペプチド配列の2番目の位置のアミノ酸の欠失によって除去され得る。O結合型グリコシル化部位は、1つ以上のセリンもしくはトレオニン残基の付加、欠失もしくは置換、および/または下記に示されるようなO結合型コンセンサス配列の破壊によって除去され得る。あるいは、ポリペプチド上に存在する1つ以上の天然に存在する炭水化物部分の除去が、化学的および/または酵素的に達成され得る。化学的な脱グリコシル化は、例えば、グリコシル化されたポリペプチドをトリフルオロメタンスルホン酸化合物または等価な化合物に曝露することを含み得る。この処理は、アミノ酸配列がインタクトなままで、連結糖(N−アセチルグルコサミンまたはN−アセチルガラクトサミン)を除くほとんどまたはすべての糖の切断をもたらす。化学的な脱グリコシル化は、Hakimuddinら(1987)Arch.Biochem.Biophys.259:52およびEdgeら(1981)Anal.Biochem.118:131によってさらに説明されている。グリコシル化されたポリペプチド上の炭水化物部分の酵素的切断は、Thotakuraら(1987)Meth.Enzymol.138:350によって説明されているような様々なエンドグリコシダーゼおよびエキソグリコシダーゼの使用によって達成され得る。
【0056】
ある特定の実施形態において、1つ以上の非内因性N結合型グリコシル化部位に加えて1つ以上のO結合型グリコシル化部位を導入することが望ましい場合がある。O結合型グリコシル化部位は、O結合型グリコシル化コンセンサス配列をポリペプチドに導入することによって、タンパク質に付加され得る。例示的なO結合型コンセンサス配列としては、例えば、CXXGGT/S−C、NSTE/DA、NITQS、QSlQS、0/E−FI1RZK−V、C−E/D−SNおよびGGSC−K/Rが挙げられる。あるいは、O結合型糖成分は、ポリペプチド内のアミノ酸を化学的に改変することによって導入され得る。例えば、WO87/05330およびAplin and Wriston(1981)CRC Crit.Rev.Biochem.,pp.259−306(本明細書中で参考として援用される)を参照のこと。
【0057】
ある特定の実施形態において、本明細書中に記載される糖改変体ポリペプチドは、さらなるドメイン(例えば、リーダー配列、リンカーおよび/または精製/同定タグ)を含み得る。
【0058】
いくつかの実施形態において、融合タンパク質は、シグナル配列、例えば、ミツバチメリチンリーダー(HBML):MKFLVNVALVFMVVYISYIYA(配列番号10);組織プラスミノゲンアクチベーター(TPA)リーダー:MDAMKRGLCCVLLLCGAVFVSP(配列番号11);または(iii)天然のリーダー配列を含み得る。
【0059】
本開示の融合タンパク質は、必要に応じてリンカードメインを含み得る。ある特定の実施形態において、融合タンパク質は、Fcドメインと異種ポリペプチドドメインとの間に位置する比較的非構造のリンカーを含む。この非構造リンカーは、1、2、3、4もしくは5アミノ酸の人工配列、または二次構造を比較的含まない5〜10、15、20、30、50もしくはそれ以上の長さのアミノ酸であり得る。リンカーは、グリシン残基およびプロリン残基に富んでいることがあり、例えば、トレオニン/セリンおよびグリシンの単一配列またはトレオニン/セリンおよびグリシンの反復配列(例えば、TG3(配列番号12)またはSG3(配列番号13)のシングレットまたは反復)を含み得る。ある特定の実施形態において、導入されるグリコシル化部位の1つ以上が、リンカードメイン内に配置され得る。
【0060】
本開示の融合タンパク質は、その融合タンパク質の単離および/または検出を容易にするドメインを含み得る。そのようなドメインの周知の例としては、ポリヒスチジン、Glu−Glu、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、マルトース結合タンパク質(MBP)またはヒト血清アルブミンが挙げられるがこれらに限定されない。融合ドメインは、所望の特性を付与するように選択され得る。例えば、いくつかの融合ドメインが、アフィニティークロマトグラフィーによる融合タンパク質の単離にとって特に有用である。親和性精製のために、アフィニティークロマトグラフィーにとって適切なマトリックス(例えば、グルタチオン、アミラーゼおよびニッケルまたはコバルトに結合体化された樹脂)が使用される。そのようなマトリックスの多くは、「キット」の形態で入手可能である(例えば、Pharmacia GST精製システム、および(HIS6)融合パートナーを用いたとき有用であるQIAexpress(商標)システム(Qiagen))。別の例として、融合ドメインは、ポリペプチドの検出を容易にするように選択され得る。そのような検出ドメインの例としては、様々な蛍光タンパク質(例えば、GFP)、ならびに特異的な抗体が入手可能な通常短いペプチド配列である「エピトープタグ」が挙げられる。特異的なモノクローナル抗体が容易に入手可能な周知のエピトープタグとしては、FLAG、インフルエンザウイルス赤血球凝集素(HA)およびc−mycタグが挙げられる。場合によっては、融合ドメインは、適切なプロテアーゼが融合タンパク質を部分的に消化することによってそこから組換えタンパク質が遊離されるのを可能にするプロテアーゼ切断部位(例えば、Xa因子またはトロンビンに対するもの)を有する。次いで、遊離されたタンパク質は、その後のクロマトグラフィー分離によって、融合ドメインから単離され得る。
【0061】
融合タンパク質の種々のエレメントが、所望の機能性と一致する任意の様式で配列され得ることが理解される。例えば、Fcドメインが、異種ポリペプチドドメインに対してC末端に配置され得るか、あるいは、異種ポリペプチドドメインが、Fcドメインに対してC末端に配置され得る。そのFcドメインおよび異種ドメインは、融合タンパク質において隣接している必要はなく、さらなるドメインまたはアミノ酸配列が、いずれかのドメインに対してC末端もしくはN末端に、またはそれらのドメイン間に含められ得る。
【0062】
ある特定の実施形態において、糖改変体融合タンパク質は:PEG化されたアミノ酸、ファルネシル化されたアミノ酸、アセチル化されたアミノ酸、ビオチン化されたアミノ酸、脂質部分に結合体化されたアミノ酸、および有機誘導体化剤に結合体化されたアミノ酸から選択される1つ以上のさらなる改変されたアミノ酸残基を含み得る。そのような翻訳後修飾の結果として、本明細書中に記載される融合タンパク質は、非アミノ酸エレメント(例えば、ポリエチレングリコール、脂質およびホスフェート)を含み得る。種々の細胞(例えば、CHO、HeLa、MDCK、293、W138、NIH−3T3またはHEK293)は、そのような翻訳後の活性に対する特定の細胞機構および特徴的な機構を有し、本発明のポリペプチドの正確な改変およびプロセシングを確実にするように選択され得る。
【0063】
本開示の融合タンパク質は、通常、少なくとも30、40、50、60、70、80、90、100もしくは110kDaまたはそれ以上のサイズである高分子量である。
【0064】
例示的な実施形態において、本開示は、Fcドメインの外側に少なくとも1つの非内因性N結合型グリコシル化部位を有するTNFR2−Fc糖改変体融合タンパク質を提供する。ある特定の実施形態において、本発明の糖改変体融合タンパク質は、配列番号18、21、22、23または24から選択されるアミノ酸配列と少なくとも75%同一であるアミノ酸配列を含み得る。ある特定の場合において、糖改変体融合タンパク質は、配列番号18、21、22、23または24から選択されるアミノ酸配列と少なくとも80%、85%、90%、95%、97%、98%、99%または100%同一であるアミノ酸配列を有する。
【0065】
ある特定の実施形態において、本明細書中に記載される糖改変体Fc融合タンパク質は、ActRIIbレセプターの細胞外ドメインおよび/または抗体の可変領域を含まない。
【0066】
Fcドメイン
本明細書中に記載される糖改変体融合タンパク質は、異種ポリペプチドに融合された免疫グロブリン重鎖Fcドメインを含む。免疫グロブリンのFc部分との融合は、広範囲のタンパク質に対して望ましい薬物動態学的特性を付与すると知られている。本明細書中で使用される用語「Fc領域」または「Fcドメイン」とは、天然配列Fc領域および改変体Fc領域を含む、免疫グロブリン重鎖のC末端領域のことを指す。免疫グロブリン重鎖のFc領域の境界は、様々であり得るが、ヒトIgG重鎖Fc領域は、通常、Cys226位のアミノ酸残基またはPro230から免疫グロブリンのカルボキシル末端に及ぶと規定される。Fc領域のC末端のリジン(EUナンバリングシステムによれば残基447)は、例えば、抗体の重鎖をコードする核酸を組換え的に操作するとき、除去され得る。別段示されない限り、本明細書において免疫グロブリン重鎖内の残基のナンバリングは、Kabatら、Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,MD(1991)(本明細書中で参考として明確に援用される)におけるようなEUインデックスのナンバリングである。「KabatにおけるようなEUインデックス」とは、ヒトIgG1 EU抗体の残基のナンバリングのことを指す。
【0067】
「天然配列Fc領域」は、天然に見られるFc領域のアミノ酸配列と同一のアミノ酸配列を含む。天然配列ヒトFc領域には、天然配列ヒトIgG1 Fc領域(非AおよびAアロタイプ);天然配列ヒトIgG2 Fc領域;天然配列ヒトIgG3 Fc領域;および天然配列ヒトIgG4 Fc領域、ならびにそれらの天然に存在する改変体が含まれる。
【0068】
「改変体Fc領域」は、少なくとも1つのアミノ酸の改変、好ましくは、1つ以上のアミノ酸置換のせいで、天然配列Fc領域のアミノ酸配列と異なるアミノ酸配列を含む。好ましくは、改変体Fc領域は、天然配列Fc領域または親ポリペプチドのFc領域と比較して、少なくとも1つのアミノ酸置換(例えば、天然配列Fc領域または親ポリペプチドのFc領域における、約1〜約10個のアミノ酸の置換、好ましくは、約1〜約5個のアミノ酸の置換)を有する。本明細書中の改変体Fc領域は、好ましくは、天然配列Fc領域および/または親ポリペプチドのFc領域と少なくとも約80%の相同性、最も好ましくは、それらと少なくとも約90%の相同性、より好ましくは、それらと少なくとも約95%の相同性を有し得る。
【0069】
好ましい実施形態において、本明細書中に記載されるFc融合タンパク質は、免疫グロブリン(immunoglobuline)FcドメインのC末端部分のN末端に融合された異種部分を有する。好ましくは、パパイン切断部位のすぐ上流のヒンジ領域から開始する配列(例えば、重鎖定常領域の1番目の残基を114または他の免疫グロブリンの類似部位と考えて)が融合に使用される。1つの実施形態において、異種ドメインは、IgG1、IgG2またはIgG3重鎖の、ヒンジ領域およびCH2およびCH3、またはCH1、ヒンジ、CH2およびCH3ドメインに融合される。いくつかの実施形態において、融合が行われる正確な部位は重大ではなく、最適な部位が日常的な実験によって決定され得る。
【0070】
ヒトFcドメインについては、ヒトIgG1およびIgG3免疫グロブリン配列を使用することが好ましい。IgG1を使用する主な利点は、IgG1融合タンパク質が、固定化されたプロテインA上で効率的に精製され得る点である。対照的に、IgG3融合タンパク質の精製には、それほど著しく多用途でない基材であるプロテインGが必要である。しかしながら、免疫グロブリンの他の構造的および機能的な特性が、特定の構築に対するIg融合パートナーを選択する際に考慮されるべきである。例えば、IgG3ヒンジは、より長く、かつより可撓性であるので、IgG1に融合されたときに正しく折り畳まれないかまたは機能しない可能性があるより大きな異種部分を受け入れることができる。別の考慮すべき点は、結合価であり得る;IgGイムノアドヘシンは、二価のホモ二量体であるのに対し、IgAおよびIgMのようなIgサブタイプはそれぞれ、基本的なIgホモ二量体単位の二量体または五量体の構造を生じ得る。
【0071】
インビボで適用するために設計されたFc融合タンパク質については、薬物動態学的特性およびFc領域によって特定されるエフェクター機能が、同様に重要である。IgG1、IgG2およびIgG4のすべてが、21日というインビボ半減期を有するが、補体系を活性化する際のそれらの相対的な力価は異なる。IgG4は、補体を活性化せず、IgG2は、補体活性化がIgG1よりも有意に弱い。さらに、IgG1とは異なり、IgG2は、単核細胞上または好中球上のFcレセプターに結合しない。IgG3は、補体活性化にとって最適であるが、そのインビボ半減期は、他のIgGアイソタイプのおよそ3分の1である。
【0072】
ヒト治療薬として使用するために設計されるFc融合タンパク質に対する別の重要な考慮すべき点は、特定のアイソタイプのアロタイプ改変体の数である。一般に、血清学的に定義されるアロタイプがより少ないIgGアイソタイプが好ましい。例えば、IgG1は、4つの血清学的に定義されるアロタイプ部位しか有さず、そのうちの2つ(GImおよび2)は、Fc領域内に配置されており;これらの部位のうちの1つであるGlmlは、免疫原性でない。対照的に、IgG3には、血清学的に定義されるアロタイプが12個存在し、そのすべてが、Fc領域に存在し;これらの部位のうちの3つ(G3m5、11および21)だけが、免疫原性でない1つのアロタイプを有する。したがって、IgG3融合タンパク質の潜在的な免疫原性は、IgG1融合タンパク質のそれよりも高い。
【0073】
ある特定の実施形態において、Fc融合タンパク質において使用されるFcドメインは、野生型Fcドメインと比較して1つ以上の変更を含み得る。それにもかかわらず、これらのFcドメインは、それらの野生型対応物と比較して、治療的有用性に必要な実質的に同じ特徴を保持し得る。例えばWO99/51642に記載されているような、例えば、変更された(すなわち、改善されたまたは減少された)CIq結合および/または補体依存性細胞傷害(CDC)をもたらし得るある特定の変更がFc領域内において行われ得ると考えられる。Fc領域改変体の他の例に関係する、Duncan & Winter Nature 322:738−40(1988);米国特許第5,648,260号;米国特許第5,624,821号;およびWO94/29351も参照のこと。WO00/42072(Presta)およびWO2004/056312(Lowman)には、FcRに対する結合性が改善されたまたは減少された抗体改変体が記載されている。これらの特許公報の内容は、本明細書中で明確に参考として援用される。Shieldsら、J.Biol.Chem.9(2):6591−6604(2001)もまた参照のこと。増加された半減期、および母体のIgGを胎児に移行させるのに関与する新生児Fcレセプター(FcRn)(Guyerら、J.Immunol.117:587(1976)およびKimら、J.Immunol.24:249(1994))に対する改善された結合性を有する抗体は、US2005/0014934A1(Hintonら)に記載されている。これらの抗体は、FcRnへのFc領域の結合を改善する1つ以上の置換をその抗体中に有するFc領域(reg on)を含む。変更されたFc領域アミノ酸配列および増加されたまたは減少されたCIq結合能を有するポリペプチド改変体は、米国特許第6,194,551号および国際公開WO99/51642に記載されている。Idusogieら、J.Immunol.164:4178−4184(2000)もまた参照のこと。これらの内容は、本明細書中で明確に参考として援用される。
【0074】
例示的なFcドメインを下に示す。
【0075】
【化1】
都合のよいリンカー(TG3)とFcドメインとが組み合わされた例を下に示す。
【0076】
【化2】
必要に応じて、Fcドメインは、残基(例えば、Asp−265、リジン322およびAsn−434)に1つ以上の変異を有する。ある特定の場合において、これらの変異の1つ以上(例えば、Asp−265変異)を有する変異体Fcドメインは、野生型Fcドメインと比べて、Fcγレセプターに結合する能力が低下している。他の場合では、これらの変異の1つ以上(例えば、Asn−434変異)を有する変異体Fcドメインは、野生型Fcドメインと比べて、MHCクラスI関連Fcレセプター(FcRN)に結合する能力が増加している。
【0077】
異種ポリペプチド
本明細書中に記載される糖改変体融合タンパク質は、Fcドメインに直接または間接的に融合された異種ポリペプチド部分も含む。その異種ポリペプチド部分は、任意のポリペプチドであり得る。例示的な実施形態において、異種ポリペプチド部分は、少なくとも10、15、20、25、30、35、40、50、60、70、80、90もしくは100kDaまたはそれ以上の分子量を有する。
【0078】
異種ポリペプチド部分は、治療用タンパク質またはそのフラグメント(例えば、成長因子、酵素、血清酵素、内分泌因子(例えば、GLP1)、骨形成タンパク質および可溶性レセプターフラグメント)であり得る。例示的な異種ポリペプチドとしては、成長因子(例えば、肝細胞成長因子(HGF)、神経成長因子(NGF)、上皮成長因子(EGF)、線維芽細胞成長因子(FGF)、トランスフォーミング成長因子(例えば、TGF−アルファ、TGF−ベータ、TGF−ベータ2、TGF−ベータ3)、血管内皮成長因子(VEGF;例えば、VEGF−2))、インターフェロン(例えば、INF−アルファ、INF−ベータ)およびインスリンが挙げられる。他の例示的な異種ポリペプチドとしては、アルファ−ガラクトシダーゼ(例えば、Fabrazyme(商標))などの酵素が挙げられる。他の例示的な異種ポリペプチドとしては、骨形成タンパク質(BMP)、エリトロポイエチン(EPO)、ミオスタチンおよび腫瘍壊死因子(例えば、TNF−α)が挙げられる。他の例示的な異種ポリペプチドとしては、膜貫通レセプターの細胞外ドメイン(細胞レセプターの任意の天然に存在する細胞外ドメイン、ならびにその任意の改変体(変異体、フラグメントおよびペプチド模倣型を含む)を含む)が挙げられる。
【0079】
例示的な実施形態において、異種ポリペプチド部分は、TNF/NGFファミリーのレセプター由来のレセプターの細胞外ドメインである。可溶性レセプターポリペプチドの例としては、例えば、本開示の配列番号2、5、6、7または8が挙げられる。好ましい実施形態において、少なくとも1つの非内因性N結合型グリコシル化部位を有する糖改変体融合タンパク質に含められる細胞外レセプタードメインは、天然に存在するレセプターのリガンドに結合する能力を有する。ある特定の実施形態において、非内因性N結合型グリコシル化部位が、細胞外レセプタードメインのリガンド結合ポケットの外側に導入される。好ましくは、導入されるグリコシル化部位のグリコシル化は、レセプタータンパク質と可溶性リガンドとの結合に有意に影響しない。例示的な実施形態において、導入されるグリコシル化部位(glycosylation sited)のグリコシル化は、細胞外レセプタードメインとそのリガンドとの結合を、10倍、5倍、3倍、2.5倍、2倍、1.5倍、1倍、0.5倍またはそれ未満より大きく減少させない。好ましくは、少なくとも1つの非内因性N結合型グリコシル化部位を含む可溶性レセプタードメインは、1μM未満または100、10もしくは1nM未満の解離定数で特定のリガンドに結合し得る。
【0080】
1つの実施形態において、本明細書中に記載される糖改変体融合タンパク質は、神経成長因子(NGF)/腫瘍壊死因子(TNF)レセプターファミリー由来のレセプターの細胞外ドメインを含む異種ポリペプチド部分を含む。例えば、M.Lotzら、J.Leukocyte Biology,60:1−7(1990)を参照のこと。そのNGF/TNFレセプターファミリーには、例えば、神経成長因子レセプター(NGFR)、TNFR1(またはTNFR55)、TNFR2(またはTNFR75)、TNFレセプター関連タンパク質(TNFRrp)、CD40、Hodgkin抗原CD30、T細胞抗原CD27、Fas/APO−1、OX−40および4−1BB/ILAが含まれる。例示的な実施形態において、本明細書中に記載される融合タンパク質は、少なくとも1つの非内因性N結合型グリコシル化部位を有するTNFR2の細胞外ドメインを含む。
【0081】
本明細書中で使用されるとき、用語「TNFR2」とは、任意の種由来の腫瘍壊死因子レセプター2型(TNFR2)タンパク質のファミリー、および突然変異誘発または他の改変によるそのようなTNFR2タンパク質に由来する改変体のことを指す。TNFR2ファミリーのメンバーは、一般に、システインリッチ領域を含むリガンド結合細胞外ドメイン、膜貫通ドメイン、および細胞内シグナル伝達活性を有すると予測される細胞質ドメインから構成される膜貫通タンパク質である。
【0082】
用語「TNFR2ポリペプチド」には、TNFR2ファミリーメンバーの任意の天然に存在するポリペプチド、ならびに有用な活性を有するその任意の改変体(変異体、フラグメント、融合物およびペプチド模倣型を含む)を含むポリペプチドが含まれる。例えば、TNFR2ポリペプチドには、TNFR2ポリペプチドの配列と少なくとも約80%同一、好ましくは、少なくとも85%、90%、95%、97%、99%またはそれ以上同一の配列を有する任意の公知のTNFR2の配列に由来するポリペプチドが含まれる。例えば、本発明のTNFR2ポリペプチドは、TNFαまたはTNFβに結合し得、TNFR2タンパク質および/またはTNFαもしくはTNFβの機能を阻害し得る。TNFR2ポリペプチドの例としては、ヒトTNFR2前駆体ポリペプチド(配列番号1)および可溶性ヒトTNFR2ポリペプチド(例えば、配列番号2、5、6、7または8)が挙げられる。
【0083】
ヒトTNFR2前駆体タンパク質配列は、以下のとおりである:
【0084】
【化3】
シグナルペプチドに一重下線が引かれている;細胞外ドメインは太字であり、潜在的なN結合型グリコシル化部位には二重下線が引かれている。
【0085】
プロセシングされたヒトTNFR2可溶性(細胞外)ポリペプチド配列は、以下のとおりである:
【0086】
【化4】
天然に存在するN結合型グリコシル化部位に下線が引かれている。本明細書中で報告されるように、アミノ末端のロイシンは、削除され得、アミノ末端にはプロリン残基が残っている。
【0087】
ヒトTRFR2前駆体タンパク質をコードする核酸配列は、以下のとおりである(GenbankエントリーNM_001066.2):
【0088】
【化5】
【0089】
【化6】
ヒトTNFR2可溶性(細胞外)ポリペプチドをコードする核酸配列は、以下のとおりである:
【0090】
【化7】
本開示は、1つ以上のN結合型グリコシル化部位(NXS/T)の付加によって、改変されていないTNFR2−Fcタンパク質と比べてTNFR2−Fc融合タンパク質の血清半減期が増加されることを証明する。N結合型グリコシル化部位は、既存のSもしくはTに対して、正しい位置にNを導入することによって、または既存のNに対して、正しい位置にSもしくはTを導入することによって、最小労力で導入され得る。TNFR2における非内因性NXS/T配列の導入に特に適した部位としては、例えば、配列番号2について、アミノ酸25〜27(DQT)、26〜28(QIA)、133〜135(ETS)および231〜233(GST)が挙げられる。例えば、N結合型グリコシル化部位を導入し得る望ましい変更は:D25N、Q26N、A28S、E133NおよびG231Nである。場合によっては、いくつかのアミノ酸が、非内因性のNXS/T配列を導入するように改変され得る。例えば、Q26N/A28Sの二重置換は、配列番号2の残基26〜28において、新規のN結合型グリコシル化部位を導入するために必要なN残基とS残基の両方を提供する。さらに、TNFR2−Fcポリペプチド上の1つより多い領域が、複数のN結合型グリコシル化部位を導入するように改変され得る。例えば、D25N/E133Nの二重置換は、配列番号2の残基25〜27および133〜135において2つのN結合型グリコシル化部位を導入し、D25N/G231Nの二重置換は、配列番号2の残基25〜27および231〜233において2つのN結合型グリコシル化部位を導入する。他の例示的なTNFR2改変体としては、例えば、E133N/G231N、Q26N/A28S/E133NおよびQ26N/A28S/G231Nが挙げられる。グリコシル化されると予測される任意のSが、グリコシル化によってもたらされる保護に起因して、免疫原性部位を作り出すことなくTに変更され得る。同様に、グリコシル化されると予測される任意のTが、Sに変更され得る。
【0091】
3.糖改変体融合タンパク質をコードする核酸
ある特定の態様において、本発明は、本発明の糖改変体融合タンパク質(本明細書中に開示されるフラグメントおよび機能的改変体を含む)のいずれかをコードする単離された核酸および/または組換え核酸を提供する。その主題の核酸は、一本鎖または二本鎖であり得る。そのような核酸は、DNA分子またはRNA分子であり得る。これらの核酸は、例えば、本発明の糖改変体融合タンパク質を作製するための方法において、または直接的な治療薬として(例えば、遺伝子治療アプローチにおいて)、使用され得る。
【0092】
例示的な実施形態において、本発明の核酸は、TNFR2−Fc糖改変体融合タンパク質をコードする。例えば、配列番号3は、天然に存在するヒトTNFR2前駆体ポリペプチドをコードし、配列番号4は、TNFR2のプロセシングされた細胞外ドメインをコードする。
【0093】
ある特定の実施形態において、本発明の糖改変体融合タンパク質をコードする主題の核酸は、配列番号3、4、9または17の改変体である核酸を含むとさらに理解される。改変体ヌクレオチド配列は、1つ以上のヌクレオチドの置換、付加または欠失によって異なる配列(例えば、対立遺伝子改変体)を含む。例示的な実施形態において、その改変体核酸は、コードされたタンパク質内に1つ以上の非内因性N結合型グリコシル化部位を導入する核酸配列改変を含む。
【0094】
ある特定の実施形態において、本発明は、配列番号3、4、9または17と少なくとも80%、85%、90%、95%、97%、98%、99%または100%同一であり、かつコードされたタンパク質内に非内因性N結合型グリコシル化部位を導入する1つ以上の配列改変を含む、単離された核酸配列または組換え核酸配列を提供する。当業者は、コードされたタンパク質内に1つ以上の非内因性N結合型グリコシル化部位を導入する、配列番号3、4、9または17に相補的な核酸配列および配列番号3、4、9または17の改変体もまた本発明の範囲内であることを認識するだろう。さらなる実施形態において、本発明の核酸配列は、単離され得、組換えであり得、および/もしくは異種ヌクレオチド配列と融合され得るか、またはDNAライブラリー中のものであり得る。
【0095】
他の実施形態において、本発明の核酸はまた、配列番号3、4、9もしくは17において指定されるヌクレオチド配列、配列番号3、4、9もしくは17の相補配列、またはそれらのフラグメントに高度にストリンジェントな条件下でハイブリダイズするヌクレオチド配列も含む。そのようなヌクレオチド配列は、1つ以上の非内因性N結合型グリコシル化部位を有する糖改変体タンパク質融合物をコードする。当業者は、DNAハイブリダイゼーションを促進する適切なストリンジェンシー条件が変動し得ることを容易に理解するだろう。当業者は、DNAハイブリダイゼーションを促進する適切なストリンジェンシー条件が変動し得ることを容易に理解するだろう。例えば、約45℃の6.0×塩化ナトリウム/クエン酸ナトリウム(SSC)におけるハイブリダイゼーションに続く、50℃の2.0×SSCでの洗浄が行われ得る。例えば、洗浄工程における塩濃度は、50℃の約2.0×SSCの低ストリンジェンシーから50℃の約0.2×SSCの高ストリンジェンシーより選択され得る。さらに、洗浄工程における温度は、室温である約22℃での低ストリンジェンシー条件から約65℃の高ストリンジェンシー条件に上昇され得る。温度と塩の両方が、変動し得るか、または温度もしくは塩濃度が一定に保持され得る一方で、他方の変数が変更される。1つの実施形態において、本発明は、室温の6×SSCに続く室温の2×SSCでの洗浄の低ストリンジェンシー条件下でハイブリダイズする核酸を提供する。
【0096】
遺伝暗号の縮重に起因して配列番号3、4または9に示されるような核酸と異なり、かつコードされるタンパク質に1つ以上の非内因性N結合型グリコシル化部位を導入する、単離された核酸改変体もまた、本発明の範囲内である。例えば、いくつかのアミノ酸が、1つより多いトリプレットによって指定される。同じアミノ酸を特定するコドン、すなわちシノニム(例えば、CAUおよびCACは、ヒスチジンに対するシノニムである)は、そのタンパク質のアミノ酸配列に影響しない「サイレント」変異をもたらし得る。しかしながら、その主題のタンパク質のアミノ酸配列を変更させるDNA配列多型が哺乳動物細胞間に存在し得ると予想される。当業者は、特定のタンパク質をコードする核酸の1つ以上のヌクレオチド(ヌクレオチドの約3〜5%まで)におけるこれらのバリエーションが、天然の対立遺伝子バリエーションに起因して所与の種の個体間に存在し得ることを認識するだろう。任意のおよびすべてのそのようなヌクレオチドバリエーションおよびその結果得られるアミノ酸多型が、本発明の範囲内である。
【0097】
ある特定の実施形態において、本発明の組換え核酸は、発現構築物内の1つ以上の制御ヌクレオチド配列に作動可能に連結され得る。制御ヌクレオチド配列は、一般に、発現のために使用される宿主細胞にとって適切なものであり得る。数多くのタイプの適切な発現ベクターおよび適当な制御配列が、様々な宿主細胞について当該分野で公知である。代表的には、前記1つ以上の制御ヌクレオチド配列としては、プロモーター配列、リーダーまたはシグナル配列、リボソーム結合部位、転写開始配列および転写終結配列、翻訳開始配列および翻訳終結配列、ならびにエンハンサーまたはアクチベーター配列が挙げられ得るがこれらに限定されない。当該分野で公知であるような構成的プロモーターまたは誘導性プロモーターが、本発明によって企図される。それらのプロモーターは、天然に存在するプロモーター、または2つ以上のプロモーターのエレメントを組み合わせたハイブリッドプロモーターであり得る。発現構築物は、エピソーム(例えば、プラスミド)上において細胞内に存在し得るか、または発現構築物は、染色体に挿入され得る。好ましい実施形態において、発現ベクターは、形質転換された宿主細胞の選択を可能にする選択可能なマーカー遺伝子を含む。選択可能なマーカー遺伝子は、当該分野で周知であり、使用される宿主細胞によって異なる。
【0098】
ある特定の実施形態において、主題の核酸は、本発明の糖改変体融合タンパク質をコードし、かつ少なくとも1つの制御配列に作動可能に連結された、ヌクレオチド配列を含む発現ベクターとして提供される。制御配列は、当該分野において認められたものであり、本発明のポリペプチドの発現を指示するように選択される。したがって、制御配列という用語には、プロモーター、エンハンサーおよび他の発現調節エレメントが含まれる。例示的な制御配列は、Goeddel;Gene Expression Technology:Methods in Enzymology,Academic Press,San Diego,Calif.(1990)に記載されている。例えば、作動可能に連結されたときDNA配列の発現を調節する多種多様な発現調節配列のいずれかが、糖改変体融合タンパク質をコードするDNA配列を発現させるためにこれらのベクターにおいて使用され得る。そのような有用な発現調節配列としては、例えば、SV40の初期および後期プロモーター、tetプロモーター、アデノウイルスまたはサイトメガロウイルスの最初期プロモーター、RSVプロモーター、lac系、trp系、TACまたはTRC系、発現がT7 RNAポリメラーゼによって指示されるT7プロモーター、ファージラムダの主要なオペレーターおよびプロモーター領域、fdコートタンパク質に対する調節領域、3−ホスホグリセリン酸キナーゼまたは他の糖分解酵素に対するプロモーター、酸ホスファターゼ(例えば、Pho5)のプロモーター、酵母アルファ接合因子のプロモーター、バキュロウイルス系の多角体プロモーター、および原核細胞もしくは真核細胞またはそれらのウイルスの遺伝子の発現を調節すると知られている他の配列、ならびにそれらの様々な組み合わせが挙げられる。発現ベクターのデザインは、形質転換される宿主細胞の選択および/または発現が望まれるタンパク質のタイプのような因子に依存し得ることが理解されるべきである。さらに、ベクターのコピー数、そのコピー数およびそのベクターによってコードされる他の任意のタンパク質(例えば、抗生物質マーカー)の発現を調節する能力もまた考慮されるべきである。
【0099】
本発明の組換え核酸は、クローニングされた遺伝子またはその一部分を、原核細胞、真核細胞(酵母、鳥類、昆虫または哺乳動物)またはその両方における発現に適したベクターにライゲートすることによって作製され得る。本発明の組換え糖改変体融合タンパク質を生成するための発現ビヒクルには、プラスミドおよび他のベクターが含まれる。例えば、適当なベクターには、以下のタイプのプラスミドが含まれる:大腸菌などの原核細胞における発現用の、pBR322由来プラスミド、pEMBL由来プラスミド、pEX由来プラスミド、pBTac由来プラスミドおよびpUC由来プラスミド。
【0100】
いくつかの哺乳動物発現ベクターは、細菌内でのベクターの増殖を促す原核生物の配列と、真核細胞内で発現される1つ以上の真核生物の転写単位の両方を含む。pcDNAI/amp、pcDNAI/neo、pRc/CMV、pSV2gpt、pSV2neo、pSV2−dhfr、pTk2、pRSVneo、pMSG、pSVT7、pko−neoおよびpHyg由来ベクターが、真核細胞のトランスフェクションに適した哺乳動物発現ベクターの例である。これらのベクターのうちのいくつかは、原核細胞と真核細胞の両方における複製および薬物耐性選択を容易にするために、pBR322などの細菌のプラスミド由来の配列で改変される。あるいは、ウイルスの誘導体(例えば、ウシパピローマウイルス(BPV−1)またはエプスタイン・バーウイルス(pHEBo、pREP由来およびp205))が、真核細胞内でのタンパク質の一過性の発現のために使用され得る。他のウイルス(レトロウイルスを含む)の発現系の例は、下記の遺伝子治療送達系の説明において見出され得る。プラスミドの調製および宿主生物の形質転換において使用される様々な方法が、当該分野で周知である。原核細胞と真核細胞の両方に適した他の適した発現系、ならびに一般的な組換え手順については、Molecular Cloning A Laboratory Manual,3rd Ed.,Sambrook,FritschおよびManiatis編(Cold Spring Harbor Laboratory Press,2001)を参照のこと。場合によっては、バキュロウイルス発現系を使用することによって組換え糖改変体融合タンパク質を発現することが望ましい場合がある。そのようなバキュロウイルス発現系の例としては、pVL由来ベクター(例えば、pVL1392、pVL1393およびpVL941)、pAcUW由来ベクター(例えば、pAcUW1)およびpBlueBac由来ベクター(例えば、β−gal含有pBlueBacIII)が挙げられる。
【0101】
好ましい実施形態において、Pcmv−Scriptベクター(Stratagene,La Jolla,Calif.)、pcDNA4ベクター(Invitrogen,Carlsbad,Calif.)およびpCI−neoベクター(Promega,Madison,Wis.)などのベクターが、CHO細胞において本発明の主題の糖改変体融合タンパク質を産生するために設計され得る。明らかであるように、その主題の遺伝子構築物は、精製にむけて、培養において増殖する細胞内でその主題の糖改変体融合タンパク質の発現を引き起こすために、例えば、タンパク質(融合タンパク質または改変体タンパク質を含む)を産生するために、使用され得る。
【0102】
本開示はまた、本発明の主題の糖改変体タンパク質融合物の1つ以上に対するコード配列を含む組換え遺伝子でトランスフェクトされた宿主細胞に関係する。その宿主細胞は、任意の原核細胞または真核細胞であり得る。例えば、本発明の糖改変体融合タンパク質は、大腸菌などの細菌細胞、昆虫細胞(例えば、バキュロウイルス発現系を使用する)、酵母または哺乳動物細胞において発現され得る。他の適当な宿主細胞は、当業者に公知である。好ましくは、その宿主細胞は、発現されるタンパク質に対して哺乳動物のグリコシル化パターンをもたらし得る哺乳動物宿主細胞(例えば、CHOまたはBHK細胞株)であり得る。
【0103】
したがって、本発明はさらに、本発明の主題の糖改変体融合タンパク質を生成する方法に関係する。例えば、Fc融合タンパク質をコードする発現ベクターでトランスフェクトされた宿主細胞が、Fc融合タンパク質の発現が生じるのを可能にする適切な条件下で培養され得る。そのFc融合タンパク質は、分泌され得、そして細胞とFc融合タンパク質を含む培地との混合物から単離され得る。あるいは、そのFc融合タンパク質は、細胞質に保持され得るか、または膜画分中、および収集されて溶解されてタンパク質が単離される細胞中に保持され得る。細胞培養物は、宿主細胞、培地および他の副産物を含む。細胞培養に適した培地は、当該分野で周知である。主題のFc融合タンパク質は、タンパク質を精製するための当該分野で公知の手法(イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、限外濾過、電気泳動、Fc融合タンパク質の特定のエピトープに特異的な抗体を用いる免疫親和性精製、およびFc融合タンパク質に融合されたドメインに結合する物質を用いる親和性精製(例えば、プロテインAカラムが、Fc融合タンパク質を精製するために使用され得る)を含む)を用いて、細胞培養液、宿主細胞またはその両方から単離され得る。好ましい実施形態において、Fc融合タンパク質は、その精製を容易にするさらなるドメインを含む。好ましい実施形態において、精製は、例えば、以下のうちの3つ以上を任意の順序で含む一連のカラムクロマトグラフィー工程によって達成され得る:プロテインAクロマトグラフィー、Qセファロースクロマトグラフィー、フェニルセファロースクロマトグラフィー、サイズ排除クロマトグラフィーおよび陽イオン交換クロマトグラフィー。その精製は、ウイルス濾過および緩衝液交換によって完了され得る。本発明の糖改変体融合タンパク質は、サイズ排除クロマトグラフィーによって測定されるとき>98%の純度に、およびSDS PAGEによって測定されるとき>95%に精製され得る。
【0104】
別の実施形態において、本発明の組換え糖改変体Fc融合タンパク質の所望の部分のN末端におけるポリ−(His)/エンテロキナーゼ切断部位配列などの精製リーダー配列をコードする融合遺伝子のおかげで、Ni2+金属樹脂を用いるアフィニティークロマトグラフィーによる、発現された融合タンパク質の精製が可能になり得る。次いで、その精製リーダー配列が、その後、エンテロキナーゼでの処理によって除去されることにより、本発明の精製された糖改変体融合タンパク質が提供され得る(例えば、Hochuliら(1987)J.Chromatography 411:177;およびJanknechtら、PNAS USA 88:8972を参照のこと)。
【0105】
融合遺伝子を作製するための手法は、周知である。本質的には、種々のポリペプチド配列をコードする様々なDNAフラグメントの連結が、従来の手法(平滑末端化されたまたは粘着末端化された末端をライゲーションに使用すること、適切な末端をもたらす制限酵素消化、必要に応じて付着末端を埋めること、望ましくない連結を回避するためのアルカリホスファターゼ処理、および酵素的ライゲーション)に従って行われる。別の実施形態において、融合遺伝子は、自動化DNA合成装置を含む従来の手法によって合成され得る。あるいは、後にアニールしてキメラ遺伝子配列を生成し得る2つの連続した遺伝子フラグメント間に相補的なオーバーハングを生じるアンカープライマーを用いて、遺伝子フラグメントのPCR増幅が行われ得る(例えば、Current Protocols in Molecular Biology,eds.Ausubelら、John Wiley & Sons:1992を参照のこと)。
【0106】
4.血清半減期を増加させる方法
特定の実施形態において、本発明は、Fcドメインの外側に1つ以上の非内因性N結合型グリコシル化部位を導入するように融合タンパク質を改変することによって、Fc融合タンパク質の安定性および/または半減期(例えば、インビトロ、インビボまたは血清半減期)を増加させるための方法に関する。例えば、そのような方法は、1つ以上の非内因性N結合型グリコシル化部位を導入するようにFc融合タンパク質の配列を改変する工程、および糖改変体Fc融合タンパク質を産生する適当な細胞(例えば、チャイニーズハムスター卵巣(CHO)細胞またはヒト細胞)において前記改変されたポリペプチドをコードする核酸を発現させる工程を包含し得る。そのような方法は:a)改変されたポリペプチドの発現に適した条件下で細胞を培養する工程(ここで、前記細胞は、改変されたポリペプチド発現構築物で形質転換されている);およびb)そのように発現された改変されたタンパク質を収集する工程を包含し得る。精製は、例えば、以下のうちの1、2もしくは3つまたはそれ以上を任意の順序で含む一連の精製工程によって達成され得る:プロテインAクロマトグラフィー(chromatograph)、陰イオン交換クロマトグラフィー(chromotography)(例えば、Qセファロース)、疎水性相互作用クロマトグラフィー(例えば、フェニルセファロース)、サイズ排除クロマトグラフィーおよび陽イオン交換クロマトグラフィー。本開示のそのようなポリペプチドは、さらに液体または固体(例えば、凍結乾燥された形態)として処方され得る。本明細書中に記載される任意の糖改変体融合タンパク質が、前記方法を用いて生成され得る。
【0107】
好ましい実施形態において、導入されたグリコシル化部位の1つ以上におけるグリコシル化によって、改変された融合タンパク質の半減期(例えば、インビトロ、インビボ、血清半減期)が、導入されるグリコシル化部位を欠く融合タンパク質の血清半減期と比べて少なくとも10%、25%、50%、75%、100%、125%、150%、175%、200%、225%もしくは250%またはそれ以上増加する。ある特定の実施形態において、本開示の方法は、本開示の融合タンパク質の各10、20、30、40、50、60、70、80、90、100、110、120、130、140、150、160、170、180、190または200アミノ酸あたり少なくとも1つのグリコシル化されたアミノ酸を導入するために使用され得る。ある特定の実施形態において、本開示の方法は、N結合型グリコシル化、O結合型グリコシル化またはその両方によって改変された各アミノ酸間の間隙が少なくとも10、20、30、40、50、60、70、80、90もしくは100アミノ酸またはそれ以上を維持するようにグリコシル化部位を導入するために使用され得る。
【0108】
ある特定の実施形態において、本開示は、少なくとも1つの異種ポリペプチドドメインおよび免疫グロブリンFcドメインを含む融合タンパク質の半減期を延長するための方法を提供する。例えば、そのような方法は、a)1つ以上の非内因性N結合型グリコシル化部位を導入するように免疫グロブリンFcドメインの外側でFc融合タンパク質を改変する工程、およびb)導入されたグリコシル化部位のうちの1つがグリコシル化されるようにFc融合タンパク質を発現させる工程を包含し得る。本明細書中に開示される方法は、例えば、リガンド結合ドメインを含むレセプターの細胞外ドメインの一部分(例えば、可溶性部分)を含むFc融合タンパク質の半減期を延長するために使用され得る。ある特定の実施形態において、その方法は、Fc融合タンパク質のリガンド結合ドメインの外側にグリコシル化部位を導入するために使用される。好ましい実施形態において、グリコシル化部位を導入するための方法は、可溶性リガンドに対する融合タンパク質のレセプター部分の結合に有意に影響せず、例えば、リガンドの結合は、導入されるグリコシル化部位を欠くFc融合タンパク質に対するリガンドの結合と比べて2、3、5、10または15倍未満だけ影響される。いくつかの実施形態において、本開示の方法は、リンカードメインを含む融合タンパク質に1つ以上のグリコシル化部位を導入するために使用される。ある特定の実施形態において、本開示の方法は、リンカードメイン内に少なくとも1つのグリコシル化部位を導入するために使用され得る。一般に、本明細書中に開示される方法は、高分子量、例えば、少なくとも25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、105または110kDaである高分子のタンパク質である、異種ドメインを有する融合タンパク質の半減期を増加させるために使用される。
【0109】
5.スクリーニングアッセイ
ある特定の態様において、本発明は、少なくとも1つの生物学的活性を維持し、かつ高い安定性および/または増加された血清半減期を有する、糖改変体Fc融合タンパク質を同定するためのスクリーニングアッセイを提供する。糖改変体融合タンパク質は、生物学的活性を調節する能力を査定するため、それらの安定性を査定するため、および/またはインビボもしくはインビトロにおいてそれらの血清半減期を査定するために試験され得る。糖改変体融合タンパク質は、例えば、動物モデルにおいて試験され得る。
【0110】
本発明の糖改変体融合タンパク質の生物学的活性についてスクリーニングするアプローチは数多く存在し、例えば、糖改変体レセプター−Fc融合タンパク質をアッセイするときは、改変されたレセプターがリガンドに結合する能力が測定され得るか、またはその糖改変体融合タンパク質がリガンド−レセプターシグナル伝達を乱す能力もまたアッセイされ得る。ある特定の実施形態において、改変されていないタンパク質の少なくとも1つの生物学的活性を保持する糖改変体(例えば、選択された細胞株に対する、リガンドまたはレセプターが媒介する作用を混乱させるレセプター改変体)を同定する糖改変体融合タンパク質のハイスループットスクリーニングが行われ得る。
【0111】
様々なアッセイ形式が十分であるが、それにもかかわらず、本開示に照らして、本明細書中にはっきりと記載されないものも、当業者によって理解されるだろう。本明細書中に記載されるように、本発明の糖改変体融合タンパク質は、前記融合タンパク質をコードする核酸の対応するフラグメントから組換え的に生成されたポリペプチドをスクリーニングすることによって得ることができる。さらに、フラグメントは、従来のMerrifield固相f−Mocまたはt−Boc化学などの当該分野で公知の手法を用いて化学的に合成され得る。
【0112】
本発明の機能的に活性な糖改変体融合タンパク質は、前記融合タンパク質をコードする、対応する変異誘発された核酸から組換え的に生成された改変された融合タンパク質のライブラリーをスクリーニングすることによって得ることができる。それらの糖改変体は、改変されていない融合タンパク質の生物学的活性、例えば、糖改変体レセプター融合物が様々な細胞レセプタータンパク質(例えば、TNFR2レセプタータンパク質)のアンタゴニスト(インヒビター)として作用する能力、および/またはリガンド−レセプター結合によって媒介される細胞内のシグナル伝達を少なくとも保持するものを同定するために、生成され、試験され得る。
【0113】
機能的な糖改変体は、安定性および/または血清半減期を増加させるように本発明の融合タンパク質のアミノ酸配列を改変することによって作製され得る。そのような改変された糖改変体タンパク質は、生物学的活性(例えば、リガンド結合性)を保持するように選択されるとき、改変されていない融合タンパク質の機能的等価物と考えられる。改変された糖改変体融合タンパク質はまた、例えば、1つ以上のN結合型グリコシル化部位を導入するアミノ酸の置換、欠失もしくは付加、および/または他の配列改変(例えば、保存的置換)によって作製され得る。例えば、イソロイシンまたはバリンによるロイシン、グルタミン酸によるアスパラギン酸、セリンによるトレオニンの個別の置換、または構造的に関係があるアミノ酸によるアミノ酸の類似の置換(例えば、保存的変異)が、その結果生じる分子の生物学的活性に対して大きな作用を有さないであろうと予想されることは妥当である。保存的置換は、側鎖に関するアミノ酸のファミリー内で行われる置換である。本発明の融合タンパク質のアミノ酸配列の変化が機能的ホモログをもたらすか否かは、改変体融合タンパク質が、改変されていない融合タンパク質と類似の様式で生物学的活性を示す能力を査定することによって容易に判定され得る。例えば、改変体レセプターFc融合タンパク質(例えば、TNFR2−Fc融合タンパク質)は、特異的なリガンド(例えば、TNFαまたはTNFβ)に結合する能力、レセプターポリペプチドへのリガンドの結合を妨害する能力、またはレセプターへのリガンドの結合によって引き起こされるシグナル伝達を干渉する能力についてスクリーニングされ得る。
【0114】
糖改変体融合タンパク質のコンビナトリアルライブラリーは、各々が潜在的なポリペプチド配列の少なくとも一部分を含む融合タンパク質のライブラリーをコードする遺伝子の縮重ライブラリーを経由して作製され得る。例えば、合成オリゴヌクレオチドの混合物が、潜在的なポリペプチドヌクレオチド配列の縮重セットが個別のポリペプチドとして、あるいは、より大きな融合タンパク質のセットとして(例えば、ファージディスプレイの場合)発現可能であるように、遺伝子配列に酵素的にライゲートされ得る。
【0115】
潜在的な糖改変体のライブラリーを縮重オリゴヌクレオチド配列から生成し得る方法が多く存在する。縮重遺伝子配列の化学的合成が、自動DNA合成装置において行われ得、次いで、その合成遺伝子が、発現用の適切なベクターにライゲートされ得る。縮重オリゴヌクレオチドの合成法は、当該分野で周知である(例えば、Narang,SA(1983)Tetrahedron 39:3;Itakuraら(1981)Recombinant DNA,Proc.3rd Cleveland Sympos.Macromolecules,ed.AG Walton,Amsterdam:Elsevier pp 273−289;Itakuraら(1984)Annu.Rev.Biochem.53:323;Itakuraら(1984)Science 198:1056;Ikeら(1983)Nucleic Acid Res.11:477を参照のこと)。そのような手法は、他のタンパク質の指向性進化において使用されている(例えば、Scottら(1990)Science 249:386−390;Robertsら(1992)PNAS USA 89:2429−2433;Devlinら(1990)Science 249:404−406;Cwirlaら(1990)PNAS USA 87:6378−6382;ならびに米国特許第5,223,409号、同第5,198,346号および同第5,096,815号を参照のこと)。
【0116】
点変異および切断によって作製されるコンビナトリアルライブラリーの遺伝子産物をスクリーニングするため、および、もっと詳しく言うなら、ある特定の特性を有する遺伝子産物についてcDNAライブラリーをスクリーニングするための、広範囲の手法が当該分野で公知である。そのような手法は、一般に、本発明の糖改変体融合タンパク質のコンビナトリアル突然変異誘発によって作製された遺伝子ライブラリーの迅速なスクリーニングに適用可能であり得る。大きな遺伝子ライブラリーをスクリーニングするために最も広く使用されている手法は、代表的には、複製可能な発現ベクターに遺伝子ライブラリーをクローニングすること、得られたベクターのライブラリーで適切な細胞を形質転換すること、および所望の活性の検出が遺伝子(その産物が検出される)をコードするベクターの比較的簡便な単離を容易にする条件下でそのコンビナトリアル遺伝子を発現させることを含む。
【0117】
好ましいアッセイには、レセプター−リガンド結合アッセイおよびリガンド媒介性細胞シグナル伝達アッセイが含まれる。本発明の糖改変体レセプター−Fc融合タンパク質とリガンドとの複合体の形成は、様々な手法によって検出され得る。例えば、複合体の形成の調節は、イムノアッセイまたはクロマトグラフィー検出によって、例えば、検出可能に標識されたタンパク質(例えば、放射性標識された(例えば、32P、35S、14Cまたは3H)、蛍光標識された(例えば、FITC)、または酵素的に標識された、レセプターポリペプチドまたはリガンド)を用いて定量され得る。
【0118】
ある特定の実施形態において、本発明は、糖改変体レセプター−Fc融合タンパク質とその結合リガンドとの相互作用の程度を直接または間接的に測定する際の、蛍光偏光アッセイおよび蛍光共鳴エネルギー転移(FRET)アッセイの使用を企図する。さらに、他の検出様式(例えば、光導波路(PCT公開WO96/26432および米国特許第5,677,196号)、表面プラズモン共鳴(SPR)、表面電荷センサーおよび表面力センサーに基づく検出様式)が、本発明の多くの実施形態と適合可能である。
【0119】
さらに、本発明は、糖改変体Fc融合タンパク質とその結合パートナーとの相互作用を乱すかまたは増強する物質を同定するための「2ハイブリッドアッセイ」としても知られる相互作用捕捉アッセイの使用を企図する。例えば、米国特許第5,283,317号;Zervosら(1993)Cell 72:223−232;Maduraら(1993)J Biol Chem 268:12046−12054;Bartelら(1993)Biotechniques 14:920−924;およびIwabuchiら(1993)Oncogene 8:1693−1696)を参照のこと。あるいは、そのようなタンパク質間相互作用は、光架橋、放射性標識されたリガンドの結合、およびアフィニティークロマトグラフィー(Jakoby WBら、1974,Methods in Enzymology 46:1)を含むインビトロでの生化学的方法を用いてタンパク質レベルで同定され得る。ある特定の場合において、糖改変体融合タンパク質は、機序に基づいたアッセイ(例えば、リガンドまたはレセプターポリペプチドに結合する糖改変体融合タンパク質を検出するアッセイ)においてスクリーニングされ得る。これは、固相または流体相の結合事象を含み得る。あるいは、リガンドまたはレセプターポリペプチドをコードする遺伝子が、レポーター系(例えば、β−ガラクトシダーゼ、ルシフェラーゼまたは緑色蛍光タンパク質)とともに細胞にトランスフェクトされ得、必要に応じてハイスループットスクリーニングによって糖改変体ライブラリーに対してスクリーニングされ得るか、または糖改変体ライブラリーの個別のメンバーを用いてスクリーニングされ得る。機序に基づいた他の結合アッセイ、例えば、自由エネルギーの変化を検出する結合アッセイが使用され得る。結合アッセイは、ウェル、ビーズもしくはチップに固定された標的、または固定化された抗体によって捕捉される標的、またはキャピラリー電気泳動によって分離される標的を用いて、行われ得る。その結合したタンパク質は、通常、比色定量または蛍光または表面プラズモン共鳴を用いて、検出され得る。
【0120】
6.例示的な治療上の使用
様々な実施形態において、本発明の糖改変体融合タンパク質で処置される患者または本発明の糖改変体融合タンパク質での処置の候補患者は、哺乳動物(例えば、げっ歯類および霊長類)、特に、ヒト患者であり得る。ある特定の実施形態において、本開示は、増加された半減期を特徴とする改変された可溶性レセプター−Fc融合物を提供する。レセプター−Fc融合物は、本明細書中で要約される様々な障害および状態を処置する際の使用について先に記載された。
【0121】
本明細書中で使用されるとき、障害または状態を「防止する」治療薬とは、統計的サンプルにおいて、処置されるサンプルにおけるその障害または状態の発生を未処置のコントロールサンプルと比べて減少させるか、あるいは未処置のコントロールサンプルと比べてその障害もしくは状態の発生を遅延させるかまたはその1つ以上の症状の重症度を低下させる、治療薬のことを指す。本明細書中で使用される用語「処置する」は、指名された状態の予防、またはその状態がいったん確立されたらその回復もしくは排除を含む。どちらにしても、防止または処置は、医師または他の医療提供者によって提供される診断、およびその治療薬の投与の意図される結果において、識別され得る。
【0122】
いくつかの実施形態において、本開示は、増加された血清半減期を有するTNFR2レセプターの細胞外ドメインを含む糖改変体融合タンパク質を提供する。そのようなTNFR2−Fc糖改変体融合物は、TNFαが介在する様々な障害または状態(例えば、急性および慢性の免疫性および自己免疫性の病態(例えば、全身性エリテマトーデス(SLE)、関節リウマチ、若年性慢性関節炎、甲状腺炎(thyroidosis)、移植片対宿主病、強皮症、真性糖尿病、グレーヴズ病)、脊椎関節症、感染症に関連する障害(例えば、敗血症症候群、悪液質、急性または慢性の細菌感染に起因する循環虚脱およびショック、急性および慢性の寄生虫病ならびに/または細菌、ウイルスもしくは真菌の感染症)、慢性の炎症性の病態(例えば、強皮症、サルコイドーシス、慢性炎症性腸疾患、潰瘍性大腸炎およびクローン病の病態)、血管の炎症性の病態(例えば、播種性血管内凝固、アテローム性動脈硬化症および川崎病の病態)および神経変性疾患(例えば、多発性硬化症、急性横断性脊髄炎、錐体外路系疾患、ハンチントン舞踏病および老人性舞踏病;パーキンソン病、進行性核上性麻痺(Progressive supranucleo palsy)、脊髄性運動失調症、フリードライヒ運動失調症、メンセル(Mencel)病、デジェリーヌ・トーマス病、シャイ・ドレーガー(Shi−Drager)病、マシャド・ジョセフ病、レフサム病、無βリポタンパク質血症(abetalipoprotemia)、毛細血管拡張、ミトコンドリア多系統障害、筋萎縮性側索硬化症、乳児性脊髄性筋萎縮症、若年性脊髄性筋萎縮症、アルツハイマー病、ダウン症候群、びまん性レヴィー小体病、ウェルニッケ・コルサコフ症候群、クロイツフェルト・ヤコブ病、亜急性硬化性全脳炎、ハラーフォルデン・シュパッツ(Hallerrorden−Spatz)病、ならびに拳闘家痴呆)が挙げられる)を防止するためまたは処置するために使用され得る。
【0123】
7.薬学的組成物
ある特定の実施形態において、本明細書中に記載される糖改変体融合タンパク質は、薬学的に許容され得るキャリアとともに処方される。例えば、本開示の融合タンパク質は、単独で、または薬学的製剤(治療用組成物)の構成要素として、投与され得る。主題の化合物は、ヒトまたは獣医学において使用するために好都合な任意の方法で投与用に処方され得る。適切な投薬レジメンが、主治医によって決定され得、その治療用タンパク質の改変されていないバージョンに対する標準的な投薬レジメンによって形成され得る。
【0124】
ある特定の実施形態において、本発明の治療方法は、本組成物を全身的に、または埋没物もしくはデバイスとして局所的に、投与する工程を包含する。投与されるとき、本発明において使用するための治療用組成物は、当然のことながら、発熱物質を含まない生理的に許容され得る形態である。上に記載されたような組成物に必要に応じて含められ得る、本明細書中に記載される融合タンパク質以外の治療上有用な薬剤は、本発明の方法において、その主題の融合タンパク質と同時にまたは連続的に投与され得る。
【0125】
代表的には、本明細書中に記載される融合タンパク質は、非経口的に(parentally)投与され得る。非経口投与に適した薬学的組成物は、1つ以上の薬学的に許容され得る滅菌された等張性の水性もしくは非水性の溶液、分散液、懸濁液もしくはエマルジョン、または滅菌された注射可能な溶液もしくは分散液に使用直前に再構成され得る滅菌された散剤とともに、1つ以上の本開示の融合タンパク質を含み得、その薬学的組成物は、酸化防止剤、緩衝剤、静菌剤、意図されるレシピエントの血液とその製剤を等張性にする溶質、または懸濁剤もしくは増粘剤を含み得る。本発明の薬学的組成物において使用され得る適当な水性および非水性のキャリアの例としては、水、エタノール、ポリオール類(例えば、グリセロール、プロピレングリコール、ポリエチレングリコールなど)およびそれらの適当な混合物、オリーブ油などの植物油、ならびにオレイン酸エチルなどの注射可能な有機エステルが挙げられる。例えば、レシチンなどのコーティング材料を使用することによって、分散液の場合は、必要な粒径を維持することによって、および界面活性物質を使用することによって、適切な流動性が維持され得る。
【0126】
さらに、本組成物は、標的組織部位(例えば、骨、筋肉、循環器系など)への送達用の形態として、被包され得るか、または注射され得る。ある特定の実施形態において、本発明の組成物は、1つ以上の治療用化合物(例えば、糖改変体融合タンパク質)を標的組織部位(例えば、骨、筋肉、循環器系)に送達することができる、発生中の組織に構造を提供することができる、および体内に最適に再吸収されることができる、マトリックスを含み得る。例えば、そのマトリックスは、本開示の融合タンパク質の緩効性を提供し得る。そのようなマトリックスは、他の埋め込まれる医学的適用のために現在使用されている材料から形成され得る。
【0127】
マトリックス材の選択は、生体適合性、生分解性、力学的特性、美容上の外観および境界面特性に基づく。主題の組成物の特定の適用法が、適切な製剤を規定し得る。本組成物に対する潜在的なマトリックスは、生分解性かつ化学的に定義された硫酸カルシウム、リン酸三カルシウム、ヒドロキシアパタイト、ポリ乳酸およびポリ無水物であり得る。他の潜在的な材料は、生分解性かつ生物学的に十分定義されているもの(例えば、骨または皮膚コラーゲン)である。さらなるマトリックスは、純粋なタンパク質または細胞外マトリックス構成要素を含む。他の潜在的なマトリックスは、生分解性でない化学的に定義されたもの(例えば、焼結ヒドロキシアパタイト、バイオガラス、アルミネートまたは他のセラミックス)である。マトリックスは、上で述べられたタイプの材料(例えば、ポリ乳酸およびヒドロキシアパタイトまたはコラーゲンおよびリン酸三カルシウム)のいずれかの組み合わせを含み得る。バイオセラミックスは、組成(例えば、カルシウム−アルミネート−ホスフェート)、ならびにポアサイズ、粒径、粒子の形状および生分解性を変化させる処理において変化され得る。
【0128】
ある特定の実施形態において、融合タンパク質は、経口投与のために、例えば、カプセル、カシェ剤、丸剤、錠剤、ロゼンジ(風味がつけられた基剤、通常、スクロースおよびアカシアまたはトラガントを用いる)、散剤、顆粒剤の形態で、または水性もしくは非水性の液体中の溶液もしくは懸濁液として、または水中油型もしくは油中水型の液体エマルジョンとして、またはエリキシル剤もしくはシロップ剤として、またはトローチ剤(不活性な基剤(例えば、ゼラチンおよびグリセリン、またはスクロースおよびアカシア)を用いる)として、および/または口腔洗浄薬などとして、処方され得、それらの各々は、活性成分として所定量の薬剤を含む。ある薬剤は、ボーラス、舐剤またはペーストとしても投与され得る。
【0129】
経口投与用の固体剤形(例えば、カプセル、錠剤、丸剤、糖衣錠および散剤顆粒剤)において、本発明の1つ以上の融合タンパク質は、1つ以上の薬学的に許容され得るキャリア(例えば、クエン酸ナトリウムまたはリン酸二カルシウム)、および/または以下のうちのいずれかと混合され得る:(1)充填剤または増量剤(例えば、デンプン、ラクトース、スクロース、グルコース、マンニトールおよび/またはケイ酸);(2)結合剤(例えば、カルボキシメチルセルロース、アルギネート、ゼラチン、ポリビニルピロリドン、スクロースおよび/またはアカシア);(3)吸湿剤(例えば、グリセロール);(4)崩壊剤(例えば、寒天、炭酸カルシウム、ジャガイモまたはタピオカのデンプン、アルギン酸、ある特定のシリケート、および炭酸ナトリウム);(5)溶解遅延剤(例えば、パラフィン);(6)吸収促進物質(例えば、四級アンモニウム化合物);(7)湿潤剤(例えば、セチルアルコールおよびモノステアリン酸グリセロール);(8)吸収剤(例えば、カオリンおよびベントナイト粘土)(9)潤滑剤(例えば、タルク、ステアリン酸カルシウム、ステアリン酸マグネシウム、固体ポリエチレングリコール、ラウリル硫酸ナトリウムおよびそれらの混合物);および(10)着色剤。カプセル、錠剤および丸剤の場合、薬学的組成物は、緩衝剤も含み得る。類似のタイプの固体組成物は、ラクトースまたは乳糖のような賦形剤、ならびに高分子量ポリエチレングリコールを用いて、軟および硬充填ゼラチンカプセルにおいて充填剤としても使用され得る。
【0130】
経口投与用の液体剤形には、薬学的に許容され得るエマルジョン、マイクロエマルジョン、溶液、懸濁液、シロップ剤およびエリキシル剤が含まれる。その液体剤形は、活性成分に加えて、当該分野において通常使用される不活性な希釈剤(例えば、水または他の溶媒)、可溶化剤および乳化剤(例えば、エチルアルコール、イソプロピルアルコール、炭酸エチル、酢酸エチル、ベンジルアルコール、安息香酸ベンジル、プロピレングリコール、1,3−ブチレングリコール、油(特に、綿実油、ラッカセイ油、トウモロコシ油、胚芽油、オリーブ油、ヒマシ油およびゴマ油)、グリセロール、テトラヒドロフリルアルコール、ポリエチレングリコールおよびソルビタンの脂肪酸エステル)、ならびにそれらの混合物を含み得る。経口用組成物は、不活性な希釈剤のほかにも、佐剤(例えば、湿潤剤、乳化剤および懸濁剤、甘味剤、香味剤、着色剤、芳香剤および保存剤)も含み得る。
【0131】
懸濁液は、融合タンパク質に加えて、懸濁剤(例えば、エトキシ化イソステアリルアルコール、ポリオキシエチレンソルビトールおよびソルビタンエステル、微結晶性セルロース、メタ水酸化アルミニウム、ベントナイト、寒天およびトラガント、ならびにそれらの混合物)を含み得る。
【0132】
本発明の組成物は、佐剤(例えば、保存剤、湿潤剤、乳化剤および分散剤)も含み得る。様々な抗菌剤および抗真菌剤、例えば、パラベン、クロロブタノール、フェノールソルビン酸などを含めることによって、微生物の作用の防止が確実にされ得る。等張剤(例えば、糖、塩化ナトリウムなど)を組成物に含めることも望ましい場合がある。さらに、注射可能な薬学的形態の長時間の吸収は、モノステアリン酸アルミニウムおよびゼラチンなどの吸収を遅延させる物質を含めることによってもたらされ得る。
【実施例】
【0133】
例示
本発明は、本明細書中で広く説明されてきたが、以下の実施例を参照することによってより容易に理解され得、その実施例は、ある特定の実施形態および本発明の実施形態を説明する目的のためだけに含められるものであって、本発明を限定することを目的としていない。
【0134】
実施例1:糖改変体TNFR2−Fc融合タンパク質の発現
本出願人は、様々なTNFR2−Fc融合タンパク質(各々がさらなるN結合型グリコシル化部位を含む)を発現するためのベクターを構築した。コントロールTNFR2−Fc融合タンパク質(すなわち、基礎的な、すなわち「最初の」改変されていない形態のTNFR2−Fc)は、介在リンカーなしでヒトIgG1 Fcドメインに融合されたヒトTNFR2の細胞外ドメインを有する。その最初のTNFR2−Fc融合タンパク質の配列を図1に示し(配列番号5)、コード核酸(リーダー配列を含む)を図5に示す(配列番号8)。図1に示される融合タンパク質は、TNFR2−h(1)Fcとさらに完全に命名される。TNFR2−h(2)Fcと命名された改変体は、図9(配列番号16)に示されるように代替リンカー/境界配列を有し、図10に示されるヌクレオチド配列(配列番号17)によってコードされる。タンパク質構築物を、哺乳動物細胞において発現させるためにpAID4ベクター(Pearsallら、PNAS 105(2008):7082−7087)内にクローニングした。
【0135】
最初の実験にむけて、TNFR2−Fcの糖改変体をHEK293細胞において一過性に発現させた。簡潔には、500mlスピナーにおいて、HEK293T細胞を、250mlの体積のFreestyle(Invitrogen)培地中で6×105細胞/mlに調整し、一晩生育した。翌日、これらの細胞を、0.5μg/mlの最終DNA濃度のDNA:PEI(1:1)複合体で処理した。4時間後、250mlの培地を加え、細胞を7日間生育した。その細胞を遠沈することによって培養上清を収集し、濃縮した。
【0136】
改変体を、様々な手法(例えば、プロテインAカラム(Mab Select(商標),GE Healthcare LifeSciences,USA)が挙げられる)を用いて精製し、低pH(3.0)グリシン緩衝液で溶出した後、サイズ排除クロマトグラフィーに供した。リン酸緩衝食塩水またはTris緩衝食塩水に対してタンパク質を透析した。
【0137】
図5に提供されるタンパク質配列は、「LPA・・・」のN末端の配列を示しているが、各タンパク質のかなりの部分が、そのN末端のロイシンが除去されることにより、「PAQ・・・」というN末端の配列がもたらされることを示した。したがって、本明細書中に開示されるいずれのTNFR2−Fc分子も、1つまたは2つのアミノ酸がN末端から除去されていることがある。
【0138】
使用するにあたり、3つの異なるリーダー配列が考えられた:
【0139】
【化8】
さらなる精製は、例えば、以下のうちの3つ以上を任意の順序で含む一連のカラムクロマトグラフィー工程によって達成され得る:Qセファロースクロマトグラフィー、フェニルセファロースクロマトグラフィー、ヒドロキシアパタイトクロマトグラフィーおよび陽イオン交換クロマトグラフィー。精製は、ウイルス濾過および緩衝液交換によって完了され得る。
【0140】
実施例2:糖改変体TNFR2−Fc融合タンパク質の設計
さらなるN結合型グリコシル化部位を導入するための位置を、TNFR2の細胞外ドメインとそのリガンドであるTNF(結晶座標は公的に入手可能)との共結晶構造の検討、および本明細書中に記載される原理の適用によって、選択した。半減期を延長しつつ、そのタンパク質のTNFアンタゴニスト活性を保存すると予測される位置におけるような変更を選択し、その変更を図7に示す。アミノ酸のナンバリングが、図7に示されるような天然のプロセシングされていないTNFR2アミノ酸配列に基づくことに注意されたい。以下の位置のうちの1つにさらなるN結合型糖成分を有するように、変更されたTNFR2−Fc融合タンパク質を設計した:47、48、62、114、155、202、203、222および253。天然のTNFR2−Fcは、171位および193位にN結合型糖成分を有すること(これは、最初のTNFR2−Fc融合タンパク質の異種(heterolgous)ドメインが、1:117.5というN結合型糖成分とアミノ酸との比を有することを意味する)に注意されたい。さらなる糖成分の予測される位置を、TNFR2−TNF共結晶構造上にマッピングし、これらを図6に示す。図8は、関連するタンパク質との配列比較および構造上の相同性によって推測される、TNFR2細胞外ドメインにおけるベータ鎖二次構造の予測される位置を示している。新しいN結合型部位の各々は、構造エレメントに対して内側に位置された。例えば、47位および48位に導入される部位は、第1のベータシート構造エレメントに対して内側である。単一のさらなるN結合型グリコシル化部位を有するタンパク質を作製し、試験した後、組み合わせ変異体を作製し、試験した。
【0141】
実施例3:TNFR2−Fc糖改変体タンパク質の試験
改変された分子の機能性、およびその分子の薬物動態学的特性に対する改変の影響を査定する一連のアッセイにおいて、TNFR2−h(1)Fcの糖改変体を試験した。
【0142】
無細胞生化学的アッセイにおいてリガンド結合性を査定するために、Biacore(登録商標)3000バイオセンサー装置を使用した。簡潔には、TNFR2−h(1)Fc改変体をフローセルに充填し、次いで、TNFに曝露した。動態パラメータ(kaおよびkd)を評価することによって、解離定数(KD)を算出した。
【0143】
リガンド阻害を査定するために、本質的にはKhabarら、Immunol.Lett.46(1995):107−110に記載されているようなTNFシグナル伝達についての細胞ベースアッセイを使用した。簡潔には、WEHI細胞(ATCC)を、TNF−アルファ(R&D Systems,Minneapolis,Minn.)およびアクチノマイシンDの存在下において培養した。TNF−アルファは、これらの細胞においてアポトーシスを引き起こし、溶解速度をA490nmにおいて検出した。TNF−アルファアンタゴニストが存在することにより、IC50の検出を可能にするシグナルが減少する。
【0144】
上記の分子の薬物動態学的特性を査定するために、標準的な手法を用いてSprague−Dawleyラットにおいて研究を行った。簡潔には、順化期間後に、TNFR2−h(1)Fc分子を動物に投与し、最初の投与の6、10、24、32、48、72、96、144、216および312時間後に尾に切り目を入れることによって出血させた。TNFR2−Fc分子の存在を、ヒトFc部分に対するELISAによって検出した。
【0145】
そのデータを、天然のTNFR2−h(1)Fc分子から得られたデータに対して正規化することにより、実験ごとのばらつきを最小にした。正規された値を下に示す。望ましい質を示すタンパク質を灰色かつ下線で示している。
【0146】
【表1】
*D47Nタンパク質は、インビボでの試験を可能にするのに十分な量で発現されなかった。
**N結合型糖を62位または114位に配置する変異によって、リガンド結合の完全な破壊が引き起こされたことから、他の特性を評価しなかった。
【0147】
上に提示されたデータは、野生型分子のリガンド結合および拮抗作用と類似のリガンド結合および拮抗作用を保持しつつ、単一のさらなるN結合型部位を含む改変体のほぼ半数(9つ中4つ)が、実質的に改善された血清半減期を示したことを示唆している。血清半減期に対する異種間物差し法の影響のため、ラットにおける20〜30%の血清半減期の変化は、ヒト患者に対して投薬レジメンの意味ある改善を提供するはずである。興味深いことに、組み合わせ変異は、一般に、血清半減期またはタンパク質活性を減少させるとみられたことから、この分子では、グリコシル化密度のさらなる増加が、もはや有益でなかったことが示唆される。
【0148】
これらのデータは、本明細書中に提供される教示を用いることにより、延長された血清半減期を有するFc融合分子を効率的に作製することができることを証明している。
【0149】
実施例4:薬物動態学的ラットアッセイ
糖改変体分子の薬物動態学的特性は、標準的な手法を用いてSprague−Dawleyラットにおいて測定され得る。簡潔には、順化期間後、糖改変体分子を8匹の動物の皮下に(SC)投与し、最初の投与の6、10、24、32、48、72、96、144、216および312時間後に尾に切り目を入れることによって出血させる。糖改変体分子の存在は、標的タンパク質に適した任意の方法を用いて検出され得る。例えば、糖改変体Fc融合タンパク質は、ヒトFc部分に対するELISAを用いて検出され得る。各個別の動物について、血清排出半減期(T1/2)を算出する。平均T1/2および標準偏差を糖改変体分子について算出する。同じ用量で投与される適切なコントロール分子(例えば、導入されるN結合型グリコシル化部位を含まないFc融合タンパク質)に対する平均T1/2および標準偏差がまだ決定されていない場合、それらを決定するために同様の研究が行われ得る。スチューデントのt検定などの適切な統計的検定を用いて、平均T1/2を解析する。糖改変体分子の平均T1/2は、適切なコントロール分子の平均T1/2と比較され得る。
【0150】
前述の方法に従って、ラットにおいてTNFR2−h(1)Fc、Q48N/A50S TNFR2−h(1)FcおよびQ48N/A50S TNFR2−h(2)Fcの薬物動態学的特性を比較する研究を行った。この研究では、5mg/kgという単回用量の皮下投与の28日後までタンパク質濃度が測定された(構築物1つあたりn=3匹のラット)。
【0151】
【表2】
TNFR2−h(1)Fcと比較したところ、糖改変体Q48N/A50S TNFR2−h(1)FcおよびQ48N/A50S TNFR2−h(2)Fcの血清半減期は、それぞれ50%および80%長いことが見出された。著しいことに、Q48N/A50S TNFR2−h(2)Fcに対する最高血清濃度(Cmax)は、Q48N/A50S TNFR2−h(1)Fcよりもほぼ5倍高かった(20.8μg/ml対4.4μg/ml,p<0.05)。これらの結果は、TNFR2−Fc糖改変体のリンカー/境界領域の改変(配列番号16と配列番号5とを比較のこと)によって、改善された薬物動態学的特性を有する改変体(この実施例におけるQ48N/A50S TNFR2−h(2)Fc)がもたらされ得ることを証明している。したがって、配列番号18として提供されるリンカー−hFc配列は、特に、本明細書中に開示される任意の糖改変体との組み合わせにおいて有用であり得る。
【0152】
実施例5:薬物動態学的サルアッセイ
糖改変体分子の薬物動態学的特性は、標準的な手法を用いてカニクイザルモデルにおいて行われ得る。簡潔には、4匹の雄および4匹の雌のカニクイザルを選択する。それらの動物は、この手順の当初は実験的にナイーブであり、およそ2〜6歳であり、少なくとも2.5kgの体重である。標準的な食餌を動物に与え、処置全体にわたってそれらの動物を標準的な手順に従って収容する。皮下(SC)投与または静脈内(IV)投与によって糖改変体分子を動物に投薬する。22日間にわたって、以下の投与後の時点において大腿静脈穿刺によって血液サンプルを収集する:5分後、15分後、30分後、1時間後、2時間後、4時間後、8時間後、12時間後、24時間後、34時間後、48時間後、58時間後、72時間後、120時間後、168時間後、240時間後、336時間後、408時間後および504時間後。投薬前の血液サンプルも収集する。血液を収集した後、そのサンプルを氷上に保管し、遠心分離によって血清を単離する。血清サンプル中の糖改変体分子の存在は、標的タンパク質に適した任意の方法を用いて検出され得る。例えば、糖改変体Fc融合タンパク質は、ヒトFc部分に対するELISAを用いて検出され得る。各個別の動物について、血清排出半減期(T1/2)を算出する。平均T1/2および標準偏差を糖改変体分子について算出する。同じ用量で投与される適切なコントロール分子(例えば、導入されるN結合型グリコシル化部位を含まないFc融合タンパク質)に対する平均T1/2および標準偏差がまだ決定されていない場合、それらを決定するために同様の研究が行われ得る。スチューデントのt検定などの適切な統計的検定を用いて、平均T1/2を解析する。糖改変体分子の平均T1/2は、適切なコントロール分子の平均T1/2と比較され得る。
【0153】
実施例6:ラットコラーゲン誘発関節炎モデルにおけるQ48N/A50S TNFR2−h(1)Fcの作用
糖改変体Q48N/A50S TNFR2−h(1)Fcを、ラットにおけるコラーゲン誘発関節炎に対する有効性について評価した。ウシコラーゲンII型(Elastin Products,カタログ番号CN276)を0.01M酢酸およびフロイント不完全アジュバント(Difco,カタログ番号263910)に1mg/mlの濃度に溶解し、研究の0および7日目に、雌Lewisラット(150〜200g)の尾の底部に皮内注射(2mg/kg)によって投与した。6日目から開始して、3mg/kgのQ48N/A50S TNFR2−h(1)Fc、3mg/kgのTNFR2−h(1)Fc、またはビヒクルでラットを1週間に3回皮下処置した。足の体積は、ベースラインおよび研究の全体にわたる複数の時点においてプレチスモメトリーによって測定したのに対し、骨質は、研究の終わりに(エキソビボで)マイクロコンピュータ断層撮影(マイクロ−CT)によって査定した。
【0154】
足の腫脹をコラーゲン誘発炎症に対するマーカーとして使用した。予想通り、ビヒクルで処置されたラットにおける足の体積は、研究の途中で増加し、終わりまで上昇したままだった。この足の腫脹は、ポジティブコントロールであるTNFR2−h(1)Fcで処置されたラットと同程度に、糖改変体Q48N/A50S TNFR2−h(1)Fcで処置されたラットにおいて効果的に阻害された(図11)。さらに、マイクロ−CTによって査定された骨質は、ビヒクルで処置されたコントロールと比べて、これらの両方の処置群において同様に改善されたことから(図12)、抗炎症性の有効性のさらなる証拠がもたらされた。これらのデータは、糖改変体Q48N/A50S TNFR2−h(1)Fcが、その置換されていない対応物であるTNFR2−h(1)Fcの抗炎症性の有効性と等価な抗炎症性の有効性をインビボにおいて有することを証明している。
【0155】
まとめると、前述の結果は、TNFR2−h(1)Fcのある特定の糖改変体、例えば、Q48N/A50S TNFR2−h(1)Fcが、TNFR2−h(1)Fc自体と比べて、増加された血清半減期および低下しない抗炎症性の有効性を有することを示唆している。
【0156】
参考としての援用
本明細書中で言及されるすべての刊行物および特許は、各個別の刊行物または特許が参考として援用されると明確かつ個別に示されたかのように、それらの全体が参考として援用される。
【0157】
主題の特定の実施形態を論じてきたが、上記の明細書は、例示的なものであって、限定的なものではない。この明細書および下記の請求項を検討すると、多くのバリエーションが当業者に明らかになるだろう。本発明の完全な範囲は、等価物の完全な範囲に加えて請求項、およびそのようなバリエーションに加えて本明細書を参照することにより決定されるべきである。
【技術分野】
【0001】
関連出願
本出願は、2009年12月2日に出願された米国仮出願番号第61/266,095号、および2010年4月23日に出願された同第61/327,582号の利益を主張する。上記米国仮出願の全教示は、参考として本明細書に援用される。
【背景技術】
【0002】
背景
天然状態または組換え的に生成されたときの治療用のタンパク質またはペプチドは、多くの場合、短い安定期間または短い血清半減期を示す不安定な分子である。さらに、これらの分子は、処方されるとき、特に、診断および治療の目的のために水溶液として処方されるとき、極度に不安定であることが多い。タンパク質性の治療用分子のインビボまたはインビトロにおける安定性を拡大するかまたは促進する実用的な溶液はほとんど無い。多くの治療薬、特に、ペプチド薬は、インビボでの不十分な血清半減期に悩まされている。これにより、治療効果に必要な血清レベルを維持するために、高頻度および/もしくはより高用量でのそのような治療薬の投与、または徐放製剤の使用を余儀なくされる。薬物の高頻度の全身投与は、かなりの負の副作用を伴う。例えば、高頻度の、例えば、毎日の、全身注射は、被験体にとってかなりの苦痛の種であり、投与関連感染症の高リスクの原因となり、特に、その治療薬が静脈内投与されるべきであるとき、入院または高頻度の来院を必要とし得る。そのうえ、長期間の処置における毎日の静脈内注射は、血管穿刺が繰り返されることによって引き起こされるかなりの組織瘢痕および血管病理ももたらし得る。同様の問題が、治療薬の高頻度の全身投与のすべてについて、例えば、糖尿病患者に対するインスリンの投与、または多発性硬化症に罹患している患者におけるインターフェロン薬物の投与について、知られている。これらの因子のすべてが、患者の服薬率を低下させ、医療制度に対するコストを増加させる。
【0003】
医薬品の血清半減期を改変するために可能性のある1つの解決方法は、半減期を増加させ得る薬剤分子に共有結合的に付着することである。以前に、ポリペプチドに対するポリエチレングリコールまたは「PEG」などのポリマーの付着によって、それらの血清半減期が増加し得ることが示された。しかしながら、ポリマーの付着によって、薬物活性が低下し得る。不完全または不均一な付着によって、異なる特性を有する化合物の混合集団がもたらされる。さらに、そのような改変に起因する半減期の変化は、予測不可能である。例えば、種々のポリエチレングリコールと、IL−8、G−CSFおよびIL−1raとの結合体化によって、様々な活性および半減期を有する分子が生成された(非特許文献1)。IL−8とPEG20との結合体化は、その半減期を変化させなかったが、IL−1raとPEG20との結合体化は、半減期を7倍近く増加させた。さらに、IL−8/PEG20結合体は、天然タンパク質よりも10〜20倍有効性が低い。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】GaertnerおよびOfford,Bioconjugate Chem.(1996),7:38−44
【発明の概要】
【発明が解決しようとする課題】
【0005】
したがって、生物学的に活性な分子の生物学的機能を著しく減少させずにその分子の血清半減期を増加することができる方法が、大いに望ましいだろう。
【課題を解決するための手段】
【0006】
要旨
一つには、本開示は、免疫グロブリンFcドメインおよび少なくとも1つの異種ポリペプチドドメインを含む融合タンパク質の血清半減期を延長するための方法を提供する。ある特定の実施形態において、それらの方法は、最初のFc融合タンパク質の異種部分をコードする核酸を、1つ以上のさらなるN結合型グリコシル化部位をコードするように改変することによって、最初のFc融合タンパク質と比べて延長された血清半減期を有する改変されたFc融合タンパク質をコードする改変された核酸を調製する工程を包含する。いくつかの実施形態において、本発明の改変された核酸は、適当な細胞培養物において発現されるとき、最初のFc融合タンパク質の血清半減期よりも少なくとも10%長い血清半減期を有する改変されたFc融合タンパク質をコードし得る。いくつかの実施形態において、本発明の改変されたFc融合タンパク質は、改変されていないFc融合タンパク質と比べて、実質的に同じまたは高いインビボ生物学的活性を有する。ある特定の実施形態において、さらなる(すなわち、導入される)グリコシル化部位の1つ以上におけるグリコシル化は、そのさらなるグリコシル化部位を欠く融合ポリペプチドの血清半減期と比べて、改変されたFc融合タンパク質の半減期(例えば、インビトロ、インビボにおける血清半減期)を少なくとも10%、20%、30%、40%、50%、75%、100%、125%、150%、175%、200%、225%もしくは250%またはそれ以上増加させる。ある特定の実施形態において、改変されたFc融合タンパク質、およびさらなるグリコシル化部位を欠く融合ポリペプチドの血清半減期は、比較するために、同じ動物モデルまたは動物種において測定される。例示的な実施形態において、血清半減期は、本明細書中に記載されるような薬物動態学的ラットアッセイまたは薬物動態学的サルアッセイにおいて測定される。
【0007】
いくつかの実施形態において、本開示は、a)少なくとも1つのさらなるN結合型グリコシル化部位を導入するように改変された核酸を、哺乳動物または哺乳動物様のグリコシル化を提供する細胞培養物において発現させること、およびb)その細胞培養物から改変されたFc融合タンパク質を収集することによって、最初のFc融合タンパク質と比べて延長された半減期を有する改変されたFc融合タンパク質を調製するための方法を提供する。Fc融合タンパク質は、細胞培養物からタンパク質を得るのに適した任意の手法を用いて、粗画分、部分的に精製された画分または高度に精製された画分として収集され得る。ある特定の態様において、改変された核酸は、シアル酸を含むN結合型糖成分を産生する細胞株において発現される。ある特定の態様において、改変された核酸は、哺乳動物細胞株によって発現され、その細胞株としては、CHO細胞株、NSO細胞株、COS細胞株またはHEK236細胞株が挙げられるがこれらに限定されない。他の態様において、改変された核酸は、哺乳動物または哺乳動物様のグリコシル化を提供するように操作された非哺乳動物細胞によって発現され、その細胞としては、遺伝的に操作された真菌細胞、昆虫細胞または植物細胞が挙げられるがこれらに限定されない。ある特定の態様において、精製は、改変されたFc融合タンパク質をプロテインAに曝露し、そのプロテインAに結合した改変されたFc融合タンパク質を収集する工程を含み得る。好ましい実施形態において、本開示の方法によって生成されるFc融合タンパク質は、患者に投与するために処方される。
【0008】
いくつかの実施形態において、本開示は、本明細書中に記載される方法のいずれかに従って調製される改変された核酸を含む細胞株を提供する。本発明の細胞株は、哺乳動物細胞株(例えば、CHO細胞株、NSO細胞株、COS細胞株またはHEK236細胞株)、または哺乳動物もしくは哺乳動物様のグリコシル化を提供するように遺伝的に改変された非哺乳動物細胞株(例えば、真菌細胞、昆虫細胞または植物細胞)を含み得る。
【0009】
ある特定の実施形態において、本開示は、増加された安定性および/または血清半減期を特徴とする糖改変体Fc融合タンパク質、ならびに増加された半減期を有する融合タンパク質を生成するための方法を提供する。本発明のFc融合タンパク質としては、免疫グロブリンFcドメインおよび少なくとも1つの異種ポリペプチドドメインを含むポリペプチドが挙げられるがこれに限定されない。そのFc融合タンパク質は、少なくとも1つの非内因性N結合型グリコシル化部位を導入するように免疫グロブリンFcドメインの外側で改変され、それにより、導入されるグリコシル化部位を欠く融合タンパク質の半減期(例えば、インビトロ、インビボまたは血清半減期)と比べて、改変された融合タンパク質の血清半減期が増加する。ある特定の実施形態において、本発明の融合タンパク質は、その融合物の血清半減期を、導入されるグリコシル化部位を欠く融合タンパク質の血清半減期(serum half−live)と比べて少なくとも10%、20%、30%、40%、50%、75%、100%、125%、150%、175%、200%、225%もしくは250%またはそれ以上増加させる、少なくとも1つの非内因性の、すなわち導入される、N結合型グリコシル化部位を含む。ある特定の実施形態において、改変されたFc融合タンパク質、およびさらなるグリコシル化部位を欠く融合ポリペプチドの血清半減期は、比較のために同じ動物モデルまたは動物種において測定される。例示的な実施形態において、血清半減期は、本明細書中に記載されるような薬物動態学的ラットアッセイまたは薬物動態学的サルアッセイにおいて測定される。
【0010】
本発明のFc融合タンパク質は、1つのN結合型グリコシル化部位を導入するために、1つ以上のアミノ酸残基の付加、欠失または置換によって改変され得る。ある特定の実施形態において、最初のFc融合タンパク質の異種部分は、各50、55、60、65、70、75、80、85、90、95 100、105 110、115、120、130、140または150アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む。ある特定の実施形態において、最初のFc融合タンパク質の異種部分は、各60、70、80、90、100、110、120または125アミノ酸あたり1個より少ないN結合型グリコシル化部位を含む。ある特定の実施形態において、N結合型グリコシル化に付着された、改変されたFc融合タンパク質の異種部分の各アミノ酸は、N結合型グリコシル化によって改変された他の任意のアミノ酸と少なくとも15、20、30、40、50、60、70、80、90または100アミノ酸離れている。ある特定の実施形態において、さらなるN結合型グリコシル化部位は、改変されたFc融合タンパク質のN末端、C末端、またはN末端とC末端の両方の10、20、30、40または50アミノ酸以内に存在しない。
【0011】
好ましい実施形態において、本発明のFc融合タンパク質は、表面に露出したアミノ酸配列(すなわち、可溶性ループドメイン)によって接続されており、かつαヘリックスまたはβシートに組み込まれない、少なくとも2つの構造的に異なるドメインを含む。好ましい実施形態において、さらなるN結合型グリコシル化部位は、表面に露出しているアミノ酸配列内に位置する。好ましい実施形態において、さらなるN結合型グリコシル化部位は、二次構造エレメント、例えば、αヘリックスまたはβシートを有するタンパク質の領域に組み込まれない。本発明において有用な異種部分には、タンパク質の機能的ドメイン(例えば、酵素ドメインもしくは触媒ドメインまたはリガンド結合ドメイン)が含まれ得る。いくつかの実施形態において、本開示のFc融合タンパク質は、リンカードメインをさらに含む。いくつかの実施形態において、導入されるグリコシル化部位のうちの少なくとも1つは、リンカードメイン内に配置される。本発明の融合ポリペプチドの異種ドメインは、好ましくは、高分子量であり、少なくとも25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、105または110kDaである。
【0012】
特定の実施形態において、異種部分は、細胞レセプター(例えば、膜貫通レセプター)の細胞外(すなわち、可溶性)ドメイン、ならびにその任意の改変体(変異体、フラグメントおよびペプチド模倣型を含む)を含む。好ましい実施形態において、異種部分(heterologous potion)には、膜貫通レセプターのリガンド結合ドメインが含まれる。本開示の方法は、付着される任意のさらなる糖成分が、リガンド結合ドメインを実質的に干渉しないような(例えば、導入されるグリコシル化部位を欠くFc融合タンパク質のリガンド結合性と比べて2、3、5、10または15倍未満の結合活性の減少)その異種の位置において1つ以上のN結合型グリコシル化部位を導入するために使用され得る。ゆえに、いくつかの態様において、導入されるN結合型グリコシル化部位の1つ以上は、リガンド結合ドメインのアミノ酸配列が改変されないような位置、および/またはリガンド境界面を実質的に干渉しないと予測される位置に導入される。ある特定の実施形態において、Fc融合タンパク質は、最初のFc融合タンパク質のIC50(すなわち、最大半量阻害濃度)よりも2、3、5または10倍以下だけ低いIC50を有する。
【0013】
ある特定の実施形態において、Fc融合タンパク質は、神経成長因子/腫瘍壊死因子レセプターファミリーのメンバーから選択される異種部分を含む。特定の実施形態において、細胞外レセプタードメインは、腫瘍壊死因子2型レセプター(すなわち、TNFR2)の一部分を含む。好ましい実施形態において、本開示は、1マイクロモル濃度未満または100、10もしくは1ナノモル濃度未満のKDでTNF−αに結合するTNFR2融合タンパク質を提供する。いくつかの実施形態において、本発明のTNFR2融合タンパク質は、ヒトTNFR2前駆体タンパク質(すなわち、配列番号1)のD47、Q48、A50、E155またはG253位に配置される1つ以上の改変されたアミノ酸を有するTNFR2細胞外ドメインを含む。これらの残基は、TNFR2(すなわち、配列番号2)のプロセシングされた細胞外のドメインのそれぞれD25、Q26、A28、E133およびG231位に対応する。好ましい改変には、TNFR2ポリペプチドに1つ以上のN結合型グリコシル化部位を導入する、アミノ酸の付加、置換および/または欠失が含まれ、例えば、配列番号2のアミノ酸配列に対するアミノ酸置換Q26N、A28S、D25N、E133N、D25N、G231Nが挙げられる。いくつかの実施形態において、1つ以上のN結合型グリコシル化部位を導入するために、1つより多い改変(例えば、付加、欠失および/または置換)がTNFR2融合タンパク質に対して行われ、その改変としては、例えば、配列番号2のアミノ酸配列に対する、Q26N/A28S;D25N/E133N;D25N/G231N;Q26N/A28S/E133N;Q26N/A28S/G231N;またはE133N/G231N置換を含むTNFR2改変体が挙げられる。本発明のTNFR2−Fc融合タンパク質は、本明細書中に開示されるもののいずれかであり得る(例えば、配列番号1、2、5、6、7もしくは8から選択されるアミノ酸配列を有するポリペプチドを含むか、または配列番号1、2、5、6、7もしくは8から選択されるアミノ酸配列と少なくとも80%、85%、90%、95%、97%もしくは99%同一であるアミノ酸配列を含む、融合タンパク質)。TNFR2−Fc融合タンパク質は、TNFR2−Fc融合タンパク質および薬学的に許容され得るキャリアを含む薬学的調製物として処方され得、ここで、その薬学的調製物は、哺乳動物への投与に適するように、発熱性材料を実質的に含まない。本組成物は、サイズ排除クロマトグラフィーによって評価されるとき、他のポリペプチド構成要素に対して少なくとも95%純粋であり得、必要に応じて本組成物は、少なくとも98%純粋である。
【図面の簡単な説明】
【0014】
本特許または本出願ファイルは、カラーで仕上げられた少なくとも1つの図面を含む。カラーの図面を含む本特許または本特許出願公報の写しは、請求され、必要な手数料の支払いがなされたときに、当局によって提供され得る。
【図1】図1は、TNFR2−h(1)Fcポリペプチドのアミノ酸配列(配列番号5)を示している。リンカーなしで免疫グロブリンFcドメインに融合されたヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質(h(1)Fc改変体と命名される)が示されている。免疫グロブリンFc領域に一重下線が引かれており、天然に存在するN結合型グリコシル化部位に二重下線が引かれている。
【図2】図2は、Q48N/A50S改変体TNFR2−h(1)Fcポリペプチドのアミノ酸配列(配列番号6)を示している。リンカーなしでh(1)Fc免疫グロブリンFcドメインに融合されたヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質が示されている。天然に存在するTNFR2−Fc細胞外ドメインが、N結合型グリコシル化部位を導入するために48および50位で改変された。免疫グロブリンFc領域に一重下線が引かれており、置換されたアミノ酸に二重下線が引かれている。
【図3】図3は、改変体(D47N/E155N)TNFR2−h(1)Fcポリペプチドのアミノ酸配列(配列番号7)を示している。リンカーなしでh(1)Fc免疫グロブリンFcドメインに融合されたヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質が示されている。天然に存在するTNFR2−Fc細胞外ドメインが、N結合型グリコシル化部位を導入するために47および155位で改変された。免疫グロブリンFc領域に一重下線が引かれており、置換されたアミノ酸に二重下線が引かれている。
【図4】図4は、改変体(D47N/G253N)TNFR2−h(1)Fcポリペプチドのアミノ酸配列(配列番号8)を示している。リンカーなしでh(1)Fc免疫グロブリンFcドメインに融合されたヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質が示されている。天然に存在するTNFR2−Fc細胞外ドメインが、N結合型グリコシル化部位を導入するために47および253位で改変された。免疫グロブリンFc領域に一重下線が引かれており、置換されたアミノ酸に二重下線が引かれている。
【図5】図5は、天然のリーダー配列を有する、TNFR2−h(1)Fcポリペプチド(すなわち、配列番号5)をコードする核酸配列(配列番号9)を示している。
【図6】図6は、TNFとTNFR2レセプターの細胞外ドメインとの結合境界面を示しているリボン図である。TNFは、左側の緑色のリボン構造であり、TNFR2レセプターは、右側の青色のリボン構造である。N結合型グリコシル化部位を導入するために操作された部位は、円で示されている。単一変異は、青色の円によって表示されており、二重変異は、緑色の円によって表示されている。リガンド結合活性を有意に減少させた変異は、TNF結合部位の近位に配置されており、赤色の円として示されている。
【図7】図7は、強調されている非内因性グリコシル化部位を導入するための、例示的なアミノ酸改変を有するTNFR2の細胞外ドメインの配列を示している。強調された各領域について、非内因性グリコシル化部位を導入し得る改変の特定の例が示されている。シグナルペプチドが、一重下線で示されており、内因性グリコシル化部位が、二重下線で示されている。
【図8】図8は、TNFR2構造内のベータ鎖の予測される位置(矢印)を示している。ナンバリングは、天然のTNFR2前駆体配列(配列番号1)に基づく。TNFR1(PDB ID:1EXT)、サイトカイン応答変更因子E(CrmE;PDB ID:2UWI)およびCD134(OX40;PDB ID:2HEY)との配列比較および構造相同性によって、二次構造を推測した。
【図9】図9は、天然のリーダー配列を有する改変体TNFR2−h(2)Fcポリペプチドのアミノ酸配列(配列番号16)を示している。4アミノ酸リンカーを介して例示的なヒトFcドメイン(配列番号14)に融合された天然のヒトTNFR2の細胞外ドメインを有する可溶性TNFR2融合タンパク質が示されている。Fcドメインが、一重下線で示されており、リーダー配列およびリンカー配列に二重下線が引かれている。
【図10】図10は、TNFR2−h(2)Fcポリペプチド(すなわち、配列番号16)をコードするヌクレオチド配列(配列番号17)を示している。Fcドメインをコードする配列が、一重下線で示されており、リーダーおよびリンカーをコードする配列に二重下線が引かれている。
【図11】図11は、ラットコラーゲン誘発関節炎モデルにおける炎症のマーカーとしての、足の体積に対する糖改変体Q48N/A50S TNFR2−h(1)Fcの作用を示している。0および7日目にコラーゲンを投与し、被験物質での処置を6日目に開始した。示されているデータは、平均値±SEMである。TNFR2−h(1)Fcと同様に、Q48N/A50S TNFR2−h(1)Fcでの処置は、この研究の後半のビヒクル処置ラットにおいて観察された足の腫脹を防止した。
【図12】図12は、ラットコラーゲン誘発関節炎モデルにおける骨質に対する糖改変体Q48N/A50S TNFR2−h(1)Fcの作用を示している。21日目の研究完了後のマイクロコンピュータ断層撮影(マイクロCT)によって、エキソビボにおいて足根骨の像を得た。TNFR2−h(1)Fcと同様に、Q48N/A50S TNFR2−h(1)Fcでの処置は、ビヒクル処置ラットにおいて観察された骨侵食を防止した。
【発明を実施するための形態】
【0015】
詳細な説明
1.概要
ある特定の態様において、本開示は、適切な標的タンパク質の適切な位置に少なくとも1つの非内因性N結合型グリコシル化部位を導入することによって、Fc融合タンパク質の血清半減期が延長され得るという驚くべき発見に関する。したがって、本開示は、そのポリペプチド上のN結合型グリコシル化部位の数を増加させることによって、特定の治療用ポリペプチドであるFc融合タンパク質の血清半減期を増加させるための方法を提供する。本明細書中で証明されるように、本開示の方法は、リガンド結合ドメインを含むレセプターの細胞外ドメインの一部分を含むFc融合タンパク質の血清半減期を増加させるために使用された。特定の例において、本開示は、それぞれの融合タンパク質の改変されていない形態と比べて増加された半減期を特徴とする改変されたTNFR2−Fc融合タンパク質を提供する。本開示を任意の特定の作用機序に限定しないが、Fc融合タンパク質の非Fc部分(「異種部分」)は、その患者の体内滞留時間中に様々な細胞内および細胞外の環境に曝露されると提唱される。これらの種々の条件によって、その異種部分の一部分が、プロテアーゼに対する基質、またはタンパク質の改変もしくは分解のプロセスを開始する他の酵素もしくは分子に対する基質として脆弱になる可能性がある。したがって、本開示は、Fc融合タンパク質の血清半減期が、異種部分を分解するように作用する物質に有意に影響され、その異種部分をそのような物質から保護する傾向がある改変が、より長い血清半減期をもたらし得るという新規の提案を提供する。本明細書中に記載されるように、1つ以上のN結合型グリコシル化部位の付加によって、異種ドメインの脆弱な部分を望まれない分解または変更から保護するため、および場合によっては、そのような分子の望ましい構造上のコンフォメーションを安定化するための、単一工程の生体適合性の系を提供する。
【0016】
最初のFc融合タンパク質の異種ドメイン内のどの位置がさらなるN結合型グリコシル化部位に最も適用できるかを演繹的に予測することは困難であり得、何の制約または指針もないときは、任意の1つの異種ドメインが、理論上は、1つ以上のさらなるN結合型部位のほぼ無限の組み合わせで改変され得る。種々の最初のFc融合タンパク質を用いた研究を通して、本出願人は、改変された分子の生物学的活性(特に、インビボ生物学的活性)を保ちつつ、最初のFc融合タンパク質と比べて延長された血清半減期を示す改変されたFc融合タンパク質に到達するために、妥当な部位数での最初のFc融合タンパク質の改変を可能にする様々な指針を見出した。本開示全体の範囲を限定することを望むものではないが、本出願人は、以下の原理の1つ以上を用いることにより、半減期が延長された活性なFc融合タンパク質を生成する実行可能なアプローチが提供されることを見出した:(1)タンパク質構造解析によって決定されるか経験的方法によって決定されるかに関係なく、N結合型グリコシル化部位は、Fc融合タンパク質の異種部分の表面に露出した位置、特に、規定のタンパク質構造エレメント(例えば、アルファヘリックスまたはベータシート)内に含まれない位置に配置されることが有利であり得る;(2)N結合型グリコシル化部位は、改変または分解に供された場合にタンパク質の構造または機能の実質的な混乱を引き起こし得るFc融合タンパク質の異種部分内の位置に配置されることが有利であり得る(例えば、多くのタンパク質は、切断された場合にタンパク質全体およびその活性の混乱をほとんどまたはまったく引き起こさない非構造領域をN末端に有し、したがって、そのような位置に配置されるN結合型グリコシル化部位は、血清半減期に対して比較的小さい作用しか有さない可能性があるが、対照的に、構造エレメント間に配置される表面露出領域は、高露出と、望ましくない分解に供された場合のそのタンパク質に対する重大な結果との組み合わせを示すので、構造エレメント間の表面露出アミノ酸は、N結合型グリコシル化部位の導入にとって望ましい位置に相当する);(3)N結合型グリコシル化部位は、最初のFc融合タンパク質の重要な機能的部位(例えば、リガンド結合表面または触媒部位)を干渉しない位置に配置され得、これは、その機能的部位自体の外部の位置を選択すること、および/または任意の新しいN結合型糖成分がその機能的部位に突き出ないであろう位置を選択することによって達成され得る;(4)N結合型グリコシル化部位の付加によって達成される作用の程度は、すでに最初のFc融合タンパク質に存在するN結合型糖の密度と反比例し得るので、そのアプローチは、比較的低レベルのグリコシル化(例えば、60、70、80、90、100、110、125アミノ酸またはそれ以上の異種部分あたり1個未満のN結合型部位)を有する異種部分を有する最初のFc融合タンパク質を用いるとき、最も有効であり得る;(5)重度にグリコシル化された異種部分と軽度にグリコシル化された異種部分の両方において、達成される作用の程度は、N結合型グリコシル化部位間の間隔に比例し得る(これは、1つ以上のさらなるN結合型グリコシル化部位が、比較的規則正しい間隔、かつ既存のN結合型グリコシル化部位または他のさらなるN結合型グリコシル化部位から離れた(例えば、N結合型グリコシル化部位のN残基間が10、15、20、25、30、35または40アミノ酸を超える)間隔で配置されることが有利であり得ることを意味し、重度にグリコシル化されたタンパク質でさえ、糖が密集する場合、十分に間隔が空けられたN結合型部位の付加によって、血清半減期の有意な増加が促進され得る)。上記原理の1つ以上を用いることにより、最初のFc融合タンパク質と比べて延長された血清半減期を示す単一の改変されたFc融合タンパク質を設計することが可能であり得るが、単一の最初のFc融合タンパク質に基づいて複数の改変されたFc融合タンパク質を生成すること、これらの変更された形態を試験すること、次いで、増加された半減期と保持された生物学的活性との最善の組み合わせを示す改変された形態の組み合わせを試験することが有益であることが多い。
【0017】
ある特定の態様において、本開示は、そのポリペプチド上のN結合型グリコシル化部位の数を増加させることによって、特定の治療用ポリペプチドであるFc融合タンパク質の血清半減期を増加させるための方法に関する。本明細書中で証明されるように、本開示の方法は、リガンド結合ドメインを含むレセプターの細胞外ドメインの一部分を含むFc融合タンパク質の血清半減期を増加させるために使用された。特定の例において、本開示は、それぞれの融合タンパク質の改変されていない形態と比べて増加された半減期を特徴とする改変されたTNFR2−Fc融合タンパク質を提供する。その機序に関係なく、さらなるグリコシル化部位の導入が、高分子量の生物製剤、特に、Fc融合タンパク質の半減期を延ばす有効な方法であることは、本明細書中に提示されるデータから明らかである。
【0018】
特定の実施形態において、本開示は、増加された血清半減期を特徴とする改変されたTNFR2−Fc糖改変体融合タンパク質を提供する。腫瘍壊死因子(TNF)は、正常な炎症応答および免疫応答に関わる、天然に存在するサイトカインである。腫瘍壊死因子−α(TNFα)および腫瘍壊死因子−β(TNFβ)は、相同的な多機能性サイトカインである。これらのポリペプチドの構造的および機能的な特徴が非常に類似していることにより、それらは、腫瘍壊死因子または「TNF」と集合的に記述される。一般にTNFに帰されている活性としては:他のサイトカイン(IL−1、IL−6、GM−CSFおよびIL−10を含む)の放出、ケモカインの誘導、接着分子の増加、血管の成長、組織破壊酵素の放出、およびT細胞の活性化が挙げられる。例えば、Feldmannら、1997,Adv.Immunol.,64:283−350,Nawrothら、1986,J.Exp.Med.,163:1363−1375;Moserら、1989,J.Clin.Invest,83:444−455;Shinguら、1993,Clin.Exp.Immunol.94:145−149;MacNaulら、1992,Matrix Suppl.,1:198−199;およびAhmadzadehら、1990,Clin.Exp.Rheumatol.8:387−391を参照のこと。これらの活性のすべてが、炎症応答を増強するように働き得る。
【0019】
TNFは、組織の損傷をもたらす炎症促進性作用(例えば、血管内皮細胞に対して凝血促進活性を誘導すること(Poberら、J.Immunol.136:1680(1986))、好中球およびリンパ球の接着を増加させること(Poberら、J.Immunol.138:3319(1987))ならびにマクロファージ、好中球および血管内皮細胞からの血小板活性化因子の放出を刺激すること(Camussiら、J.Exp.Med.166:1390(1987)))を引き起こす。TNFはまた、感染症(Ceramiら、Immunol.Today 9:28(1988))、免疫障害、腫瘍性の病態(Oliffら、Cell 50:555(1987))、自己免疫性の病態および移植片対宿主病態(Piguetら、J.Exp.Med.166:1280(1987))にも関連する。うっ血性心不全、炎症性腸疾患(クローン病を含む)、関節炎および喘息が、そのようなTNF関連障害の中に入る。
【0020】
特に、TNFは、発熱、倦怠感、食欲不振および悪液質を含む、グラム陰性敗血症およびエンドトキシンショックにおいて中心的役割を果たす(Michieら、Br.J.Surg.76:670−671(1989);Debetsら、Second Vienna Shock Forum,p.463−466(1989);Simpsonら、Crit.Care Clin.5:27−47(1989);Waageら、Lancet 1:355−357(1987);Hammerleら、Second Vienna Shock Forum p.715−718(1989);Debetsら、Crit.Care Med.17:489−497(1989);Calandraら、J.Infect.Dis.161:982−987(1990);Revhaugら、Arch.Surg.123:162−170(1988))。
【0021】
TNFは、TNF応答性細胞上の特異的な細胞表面レセプターとの相互作用を介して、その生物学的作用を開始する。細胞表面腫瘍壊死因子レセプター(TNFR)には、p75(または2型)およびp55(または1型)と命名された異なる2つの形態がある(Smithら、1990,Science 248:1019−1023;Loetscherら、1990,Cell 61:351−359)。TNFR 1型およびTNFR 2型の各々は、TNFαとTNFβの両方に結合する。TNFの生物学的活性は、いずれかの細胞表面TNFRへの結合に依存する。1型レセプター(TNF−R55、TNF−RIまたはTNFR−βとも呼ばれる)は、TNF−αの細胞傷害性、抗ウイルス性および増殖性の活性をもたらすシグナルを伝達すると示されている55kdの糖タンパク質である。p75レセプター(TNF−R75、TNFR2またはTNFR−αとも呼ばれる)は、細胞傷害性および増殖性のシグナル、ならびにGM−CSFの分泌をもたらすシグナルを伝達すると示されている75kDaの糖タンパク質である。
【0022】
TNFアンタゴニスト(例えば、可溶性TNFRおよび抗TNF抗体)は、TNF活性を阻止して、TNFに応答したIL−1、GM−CSF、IL−6、IL−8、接着分子および組織破壊の減少を引き起こすと証明されている(Feldmannら、1997)。ハムスター抗マウスTNF抗体を利用したTNF拮抗作用の影響が、DBA/1マウスにおけるII型コラーゲン関節炎のモデルにおいて試験された(Williamsら、1992,Proc.Natl.Acad.Sci.USA,89:9784−9788)。疾患の発症後に開始された処置によって、足蹠の腫脹、臨床スコア、および関節破壊の組織病理学が改善された。他の研究も、抗体(Thorbeckeら、1992,Proc.Natl.Acad.Sci.USA,89:7375−7379)またはTNFR構築物(Husbyら、1988,J.Autoimmun.1:363−71;Tettaら、1990,Ann.Rheum.Dis.49:665−667;Wooleyら、1993,J.Immunol.151:6602−6607;Piguetら、1992,Immunology 77:510−514)のいずれかを用いて同様の結果を得ている。
【0023】
3つの特異的なTNFアンタゴニストが、現在、FDAによって承認されている:エタネルセプト(Enbrel(登録商標))、インフリキシマブ(Remicade(登録商標))およびアダリムマブ(Humira(登録商標))。これらの薬物の1つ以上が、関節リウマチ、若年性関節リウマチ、乾癬、乾癬性関節炎、強直性脊椎炎および炎症性腸疾患(クローン病または潰瘍性大腸炎)の処置について承認されている。
【0024】
ヒトIgG1のFc部分に連結された可溶性ヒトTNFR(p75)の組換えバージョン(sTNFR(p75):Fc,Enbrel(登録商標),Immunex)の臨床試験から、その投与によって、RA疾患の活動性の有意かつ迅速な減少がもたらされたことが示された(Morelandら、1997,N.Eng.J.Med.,337:141−147)。さらに、Enbrel(登録商標)に対する小児科の臨床試験からの安全性データは、この薬物が、一般に、若年性関節リウマチ(JRA)を有する患者によって十分許容されることを示唆している(Garrisonら、1998,Am.College of Rheumatology meeting,Nov.9,1998,abstract 584)。
【0025】
上で述べたように、Enbrel(登録商標)は、ヒトIgG1のFc部分に連結されたヒト75キロダルトン(p75)TNFRの細胞外リガンド結合部分からなる二量体の融合タンパク質である。Enbrel(登録商標)のFc構成要素は、CH2ドメイン、CH3ドメインおよびヒンジ領域を含むが、IgG1のCH1ドメインを含まない。Enbrel(登録商標)は、チャイニーズハムスター卵巣(CHO)哺乳動物細胞発現系において産生される。それは、934アミノ酸からなり、およそ150キロダルトンの見かけの分子量を有する(Smithら、1990,Science 248:1019−1023;Mohlerら、1993,J.Immunol.151:1548−1561;米国特許第5,395,760号(Immunex Corporation,Seattle,Wash.);米国特許第5,605,690号(Immunex Corporation,Seattle,Wash.)。
【0026】
Enbrel(登録商標)は、現在、1つ以上の疾患修飾性抗リウマチ薬(DMARD)に対して不十分な反応を有する患者における中等度から重度の活動性関節リウマチの徴候および症状の減少について示唆されている。Enbrel(登録商標)は、メトトレキサート単独に十分に反応しない患者において、メトトレキサートと併用して使用され得る。Enbrel(登録商標)は、1つ以上のDMARDに対して不十分な反応を有する患者における中等度から重度の活動性多関節型若年性関節リウマチの徴候および症状の減少についても示唆されている(1999年5月28日)。Enbrel(登録商標)は、皮下注射として、1週間に2回、25mgでRA患者に投与される。
【0027】
現在、ENBREL調製物を用いた処置は、1週間に2回皮下に投与されるが、これは、高価であり、患者にとって不快かつ不便である。したがって、本開示は、導入されるグリコシル化部位を欠く可溶性TNFレセプターの血清半減期と比べて改変されたポリペプチドの血清半減期を増加させる少なくとも1つの非内因性N結合型グリコシル化部位を導入するように改変されたTNF結合レセプター(例えば、Enbrel(登録商標))の可溶性(例えば、細胞外)部分を含むTNFアンタゴニストを提供する。
【0028】
本明細書において使用される用語は、概して、当該分野における、本発明の文脈内における、および各用語が使用される特定の文脈における、それらの通常の意味を有する。ある特定の用語が、本発明の組成物および方法、ならびにそれらをどのように作製し使用するかを説明する際に、当業者にさらなる指針を提供するために下記または本明細書の他の箇所で論じられる。用語の任意の使用の範囲または意味は、その用語が使用される特定の文脈から明らかになるだろう。
【0029】
「約」および「およそ」は、測定の性質または精度を考慮に入れると、通常、測定された量について許容され得る誤差の程度を意味するものとする。代表的には、例示的な誤差の程度は、所与の値または所与の値の範囲の20パーセント(%)以内、好ましくは、10%以内、より好ましくは、5%以内である。
【0030】
あるいは、および生体系において特に、用語「約」および「およそ」は、1桁以内の値、好ましくは、所与の値の5倍以内の値、より好ましくは、2倍以内の値を意味し得る。
【0031】
本発明の方法は、配列を互いに比較する工程(野生型配列と1つ以上の変異体(配列改変体)との比較を含む)を包含し得る。そのような比較は、代表的には、例えば、当該分野で周知の配列アラインメントプログラムおよび/またはアルゴリズム(いくつかの例を挙げれば、例えば、BLAST、FASTAおよびMEGALIGN)を用いた、ポリマー配列のアラインメントを含む。当業者は、そのようなアラインメントでは、変異が残基の挿入または欠失を含む場合、配列アラインメントは、挿入されたまたは欠失された残基を含まないポリマー配列内に「ギャップ」(代表的には、ダッシュまたは「A」によって表される)を導入し得ることを容易に認識し得る。
【0032】
そのすべての文法的な形および綴りバリエーションにおける「相同」とは、「共通の進化的起源」を有する2つのタンパク質(同じ生物種内の上科由来のタンパク質ならびに異なる生物種由来の相同タンパク質を含む)間の関係性のことを指す。そのようなタンパク質(およびそれらをコードする核酸)は、同一性パーセントに関するものであるか、または特定の残基もしくはモチーフおよび保存された位置の存在によるものであるかに関係なく、それらの配列類似性によって反映される配列相同性を有する。
【0033】
そのすべての文法上の形態における用語「配列類似性」とは、共通の進化的起源を共有するかもしれないし共有しないかもしれない、核酸配列間またはアミノ酸配列間の同一性または対応の程度のことを指す。
【0034】
しかしながら、通常の使用法および本願において、用語「相同」は、「高度に」などの副詞で修飾されるとき、配列類似性のことを指し得、共通の進化的起源に関係することもあるし、関係しないこともある。
【0035】
用語「薬物動態学的特性」とは、生理活性物質(例えば、小分子、ポリペプチド薬など)の吸収、分布、代謝および排出のことを指す。
【0036】
2.糖改変体Fc融合タンパク質
異種ポリペプチドに連結された免疫グロブリン分子由来のFcドメインを有する糖改変体融合タンパク質が、本明細書中に提供される。その糖改変体融合タンパク質は、融合タンパク質のFcドメインの外側に少なくとも1つの非内因性N結合型グリコシル化部位を含む。そのFcドメインは、直接、またはポリペプチドリンカーを介して間接的に、異種ポリペプチドに連結され得る。非内因性グリコシル化部位は、異種ポリペプチド、リンカーまたはその両方に導入され得る。
【0037】
「非内因性」、「導入される」または「新規の」グリコシル化部位とは、そのポリペプチドの改変されていないバージョンに存在しないグリコシル化部位のことを指す。したがって、本明細書中に記載される糖改変体融合タンパク質は、融合タンパク質の改変されていないバージョンに存在するグリコシル化部位の数と比べて、少なくとも1つのさらなるN結合型グリコシル化部位を有する。例えば、融合タンパク質の改変されていないバージョンが、Fcドメインの外側に2つのグリコシル化部位を有し、1つのN結合型グリコシル化部位が、異種ポリペプチドに導入される場合、その糖改変体融合タンパク質は、2つの天然のグリコシル化部位および1つの導入されたグリコシル化部位を含む3つのN結合型グリコシル化部位を有することになる。天然に存在する改変体または人工の改変体もまた、グリコシル化部位の数を増加させる本発明の方法に従って改変され得るので、グリコシル化部位の数を増加させるための融合タンパク質の改変は、特定の「野生型」アミノ酸配列に限定されないことが理解されるべきである。
【0038】
いくつかのタンパク質が多数の糖側鎖を担持し得ることは、周知である。O結合型グリコシル化部位間の距離は、わずか1つおきのアミノ酸であり得る(例えば、Kolset & Tveit(2008)Cell.Mol.Life Sci 65:1073−1085およびKianiら(2002)Cell Research 12(1):19−32を参照のこと)。N結合型グリコシル化部位については、部位間の距離は、わずか3、4、5または6アミノ酸であり得る(例えば、Lundinら(2007)FEBS Letters 581:5601−5604(2007);Apweilerら(1991)Biochimica et Biophysica Acta 1473:4−8(これらの各々の全内容が本明細書中で参考として援用される)を参照のこと)。したがって、ある特定の実施形態において、少なくとも約1、2、3、4、5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95もしくは100またはそれ以上のN結合型グリコシル化部位が、本明細書中に記載される融合タンパク質に付加され得る。ある特定の実施形態において、本開示の糖改変体融合タンパク質は、各10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、125、150または200アミノ酸あたり少なくとも1つのグリコシル化されたアミノ酸を含む。ある特定の実施形態において、N結合型グリコシル化された各アミノ酸は、他の任意のN結合型、O結合型、またはN結合型およびO結合型のグリコシル化されたアミノ酸と少なくとも10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95もしくは100またはそれ以上のアミノ酸だけ離れている。
【0039】
本明細書中で使用されるとき、「グリコシル化部位」は、糖付着コンセンサス配列(すなわち、糖、(単糖、オリゴ糖または多糖のいずれか)をアミノ酸配列に付着するためのコンセンサス配列として作用する、ある範囲のアミノ酸)のことを意味し得るか、または糖成分が共有結合的に連結される実際のアミノ酸残基のことを意味し得る。その糖成分は、単糖(単糖分子)、オリゴ糖または多糖であり得る。
【0040】
N結合型グリコシル化部位は、糖成分を付加するように、タンパク質のアミノ酸配列を改変することによって、または融合タンパク質におけるアミノ酸残基を化学的に改変することによって、融合タンパク質に導入され得る。好ましい実施形態において、本明細書中に記載される融合タンパク質は、コンセンサス配列NXT/S(アスパラギン−X−セリン/トレオニン)(ここで、Xは、プロリン以外の任意のアミノ酸である)を有する少なくとも1つのN結合型グリコシル化部位を導入するように(例えば、指定のアミノ酸の挿入、欠失または置換によって)改変される。タンパク質上の炭水化物部分の数を増加させる別の手段は、ポリペプチドへのグリコシドの化学的または酵素的な結合によるものである。例えば、用いられる結合の様式に依存して、糖成分は、(a)アルギニンおよびヒスチジン;(b)遊離カルボキシル基;(c)遊離スルフヒドリル基(例えば、システインのもの);(d)遊離ヒドロキシル基(例えば、セリン、トレオニンまたはヒドロキシプロリンのもの);(e)芳香族残基(例えば、フェニルアラニン、チロシンまたはトリプトファンのもの);または(f)グルタミンのアミド基に付着され得る。例えば、WO87/05330およびAplin and Wriston(1981)CRC Crit.Rev.Biochem.,pp.259−306(本明細書中で参考として援用される)を参照のこと。例示的な実施形態において、N結合型グリカンを生成するために、糖成分は、アルギニン残基に結合される。
【0041】
例示的な実施形態において、糖成分は、宿主細胞において融合タンパク質を発現させることによって細胞機構を用いて、導入されたグリコシル化部位に付加される。一般に、融合タンパク質は、適切なグリコシル化を提供する哺乳動物細胞株(例えば、HEK293またはCHO細胞株)または他の哺乳動物発現細胞株において発現され得る。
【0042】
ある特定の実施形態において、糖成分は、哺乳動物様のグリコシル化をもたらすように操作された非哺乳動物宿主細胞(例えば、酵母、細菌または昆虫細胞)を用いて、導入されたグリコシル化部位に付加され得る。ヒトにおける糖タンパク質のプロセシングを模倣する一連の酵素反応を行う遺伝的に改変されたグリコシル化経路を有する細胞株が開発されている。これらの操作された細胞において発現される組換えタンパク質は、実質的に同一でないにせよ、ヒトの対応物に類似の糖タンパク質(例えば、哺乳動物様のグリコシル化)をもたらす。
【0043】
発現されるペプチドのグリコシル化プロファイルを変更するように宿主細胞を遺伝的に改変するための手法は、周知である。例えば、Altmannら(1999,Glycoconjugate J.16:109−123)、Ailorら(2000,Glycobiology 10(8):837−847)、Jarvisら(In vitrogen Conference,March,1999,abstract)、Hollister and Jarvis(2001,Glycobiology 11(1):1−9)およびPalacpacら(1999,PNAS USA 96:4697)、Jarvisら(1998.Curr.Opin.Biotechnol.9:528−533)、Gemgross(米国特許公開番号20020137134)(これらのすべてが、昆虫細胞または植物細胞をグリコシルトランスフェラーゼ遺伝子でトランスフェクトすることによって、昆虫細胞発現系または植物細胞発現系を「ヒト化する(hummanize)」手法を開示している)を参照のこと。
【0044】
原核生物系において発現されるペプチドのグリコシル化プロファイルを遺伝的に変更する手法も存在する。インビボにおいてオリゴ糖を産生する、細菌Neisseria meningitidisおよびAzorhizobium由来の様々なグリコシルトランスフェラーゼで大腸菌が操作されている(Bettlerら、1999,Glycoconj.J.16:205−212)。Neisseria meningitidisのβ1,3Nアセチルグルコサミニルトランスフェラーゼ(acetyl glucoaminyltransferase)lgta遺伝子を過剰発現するように遺伝的に操作された大腸菌は、外来性ラクトースを効率的にグリコシル化する(Priemら、2002,Glycobiology 12:235−240)。
【0045】
真菌細胞は、外来性グリコシルトランスフェラーゼを産生するようにも遺伝的に改変されている(Yoshidaら、1999,Glycobiology,9(1):53−58;Kalsnerら、1995,Glycoconj.J.12:360−370;Schwientek and Ernst,1994,Gene 145(2):299−303;Chibaら、1995,Biochem J.308:405−409)。
【0046】
ある特定の実施形態において、本開示のFc融合タンパク質を発現する宿主細胞は、1つ以上の外来性グリコシルトランスフェラーゼ酵素および/または1つ以上の外来性グリコシダーゼ酵素を発現する真核細胞または原核細胞であり得、ここで、その宿主細胞内での組換え糖ペプチドの発現によって、「ヒト」グリカン構造を有する組換え糖ペプチドが産生される。
【0047】
いくつかの実施形態において、上記細胞において有用な異種グリコシルトランスフェラーゼ酵素は、例えば、Taniguchiら(2002,Handbook of Glycosyltransferases and Related Genes,Springer,N.Y.)において入手可能なグリコシルトランスフェラーゼファミリーのリストに含められた任意の公知のグリコシルトランスフェラーゼ酵素からなる群より選択され得る。
【0048】
いくつかの実施形態において、宿主細胞は、宿主細胞内での組換え糖ペプチドの発現によって、「ヒト」グリカン構造を有する組換え糖ペプチドが産生されるように、1つ以上の内因性グリコシルトランスフェラーゼ酵素および/または1つ以上の内因性グリコシダーゼ酵素が不活性化された、真核細胞または原核細胞であり得る。
【0049】
いくつかの実施形態において、宿主細胞は、異種のグリコシルトランスフェラーゼ酵素および/またはグリコシダーゼ酵素を発現し得、同時に、1つ以上の内因性のグリコシルトランスフェラーゼ酵素および/またはグリコシダーゼ酵素が不活性化されている。内因性のグリコシルトランスフェラーゼ酵素および/またはグリコシダーゼ酵素は、当業者に公知の任意の手法を用いて不活性化され得、その手法としては、アンチセンス法、および宿主細胞のゲノムに核酸を挿入することを含む手法が挙げられるがこれらに限定されない。
【0050】
例示的な実施形態において、本明細書中に記載される糖改変体融合タンパク質は、その融合タンパク質の改変されていない形態と比べて高い安定性および/または増加された血清半減期を有する。例示的な実施形態において、糖改変体融合タンパク質の血清半減期は、その融合タンパク質の改変されていない形態と比べて、少なくとも10%、15%、20%、25%、30%、40%、50%、55%、60%、70%、75%、80%、85%、90%、95%、100%、125%、150%、200%、250%もしくは300%またはそれ以上増加される。ある特定の実施形態において、改変されたFc融合タンパク質およびさらなるグリコシル化部位を欠く融合ポリペプチドの血清半減期は、比較のために、同じ動物モデルまたは動物種において測定される。例示的な実施形態において、血清半減期は、本明細書中に記載されるような薬物動態学的ラットアッセイまたは薬物動態学的サルアッセイにおいて測定される(例えば、実施例4および5を参照のこと)。
【0051】
例示的な実施形態において、1つ以上の非内因性グリコシル化部位の導入は、融合タンパク質の異種ポリペプチド部分の1つ以上の生物学的活性に有意に影響しない。例えば、融合タンパク質の異種ポリペプチド部分の1つ以上の生物学的活性は、10倍、5倍、3倍、2.5倍、2倍、1.5倍、1倍、0.5倍未満またはそれより少なく、影響され得る。異種ポリペプチド部分の生物学的活性の例は、本明細書中にさらに記載されるが、例えば、タンパク質間相互作用、リガンド結合、生理学的活性などが挙げられる。
【0052】
例示的な実施形態において、1つ以上の非内因性グリコシル化部位が、Fc融合タンパク質の異種部分に導入される。Fc融合タンパク質の異種部分は、触媒ドメインまたはリガンド結合ドメインを含み得る。本明細書中で使用されるとき、「リガンド結合ドメイン」とは、別の分子、例えば、リガンドと相互作用するタンパク質上の領域のことを指す。例えば、リガンド結合ドメインとは、抗体によって結合されるタンパク質上の領域、抗原に結合する抗体上の領域、リガンドに結合するレセプター上の領域、レセプターに結合するリガンド上の領域、第2のポリペプチドに結合する第1のポリペプチド上の領域、小分子に結合するポリペプチド上の領域などのことを指し得る。例示的な実施形態において、リガンド結合ドメインは、リガンドに結合する膜貫通レセプタータンパク質の細胞外ドメイン内の領域である。本明細書中で使用されるとき、「触媒ドメイン」とは、機能活性を有する領域のことを指す。触媒ドメインの例としては、例えば、キナーゼドメイン、ホスファターゼドメイン、プロテアーゼドメインが挙げられる。
【0053】
例示的な実施形態において、1つ以上の非内因性グリコシル化部位は、可撓性または非構造である、表面に露出した領域(例えば、αヘリックスおよび/またはβシートを接続するループ領域)に導入される。通常、タンパク質は、二次構造および三次構造を有するポリペプチド鎖である。ポリペプチドの二次構造は、αヘリックスおよびβシートと呼ばれる2つの一般的な構造ドメインへのポリペプチド鎖のフォールディングを含む。タンパク質の三次構造は、様々な長さおよび不規則な形状のループ領域によって接続される、αヘリックスおよびβシートである二次構造の組み合わせからなる。一般に、二次構造エレメントの組み合わせによって、安定な疎水性コアが形成され、ループ領域は、タンパク質の表面に提示される。ループ領域は、溶媒に曝露されるので、それらは、一般に、内部の構造的に安定な二次構造ドメイン(例えば、αヘリックスおよびβシートドメイン)よりも、様々な反応性分子およびタンパク質酵素(例えば、プロテアーゼ)による求核攻撃/ペプチド結合切断を受けやすい。
【0054】
ある特定の実施形態において、融合タンパク質の異種ポリペプチド部分における1つ以上の天然に存在するグリコシル化部位(例えば、改変されていない形態のタンパク質におけるグリコシル化部位)を除去することが望ましい場合がある。例えば、N結合型グリコシル化部位の間隔を変更するために、天然に存在するグリコシル化部位を除去し、次いで、所望の位置に2つの非内因性グリコシル化部位を導入することが望ましい場合がある。さらに、融合タンパク質に存在する天然に存在するO結合型グリコシル化部位の1つ以上を除去することが望ましい場合がある。
【0055】
N結合型およびO結合型のグリコシル化部位は、コンセンサスアミノ酸配列を削除することによって、またはアミノ酸残基から糖成分を化学的もしくは酵素的に切断することによって、除去され得る。例えば、N結合型グリコシル化部位は、N結合型グリコシル化認識部位(すなわち、NXT/S)の1番目もしくは3番目のアミノ酸の位置の一方もしくは両方におけるアミノ酸の置換もしくは欠失、および/またはそのトリペプチド配列の2番目の位置のアミノ酸の欠失によって除去され得る。O結合型グリコシル化部位は、1つ以上のセリンもしくはトレオニン残基の付加、欠失もしくは置換、および/または下記に示されるようなO結合型コンセンサス配列の破壊によって除去され得る。あるいは、ポリペプチド上に存在する1つ以上の天然に存在する炭水化物部分の除去が、化学的および/または酵素的に達成され得る。化学的な脱グリコシル化は、例えば、グリコシル化されたポリペプチドをトリフルオロメタンスルホン酸化合物または等価な化合物に曝露することを含み得る。この処理は、アミノ酸配列がインタクトなままで、連結糖(N−アセチルグルコサミンまたはN−アセチルガラクトサミン)を除くほとんどまたはすべての糖の切断をもたらす。化学的な脱グリコシル化は、Hakimuddinら(1987)Arch.Biochem.Biophys.259:52およびEdgeら(1981)Anal.Biochem.118:131によってさらに説明されている。グリコシル化されたポリペプチド上の炭水化物部分の酵素的切断は、Thotakuraら(1987)Meth.Enzymol.138:350によって説明されているような様々なエンドグリコシダーゼおよびエキソグリコシダーゼの使用によって達成され得る。
【0056】
ある特定の実施形態において、1つ以上の非内因性N結合型グリコシル化部位に加えて1つ以上のO結合型グリコシル化部位を導入することが望ましい場合がある。O結合型グリコシル化部位は、O結合型グリコシル化コンセンサス配列をポリペプチドに導入することによって、タンパク質に付加され得る。例示的なO結合型コンセンサス配列としては、例えば、CXXGGT/S−C、NSTE/DA、NITQS、QSlQS、0/E−FI1RZK−V、C−E/D−SNおよびGGSC−K/Rが挙げられる。あるいは、O結合型糖成分は、ポリペプチド内のアミノ酸を化学的に改変することによって導入され得る。例えば、WO87/05330およびAplin and Wriston(1981)CRC Crit.Rev.Biochem.,pp.259−306(本明細書中で参考として援用される)を参照のこと。
【0057】
ある特定の実施形態において、本明細書中に記載される糖改変体ポリペプチドは、さらなるドメイン(例えば、リーダー配列、リンカーおよび/または精製/同定タグ)を含み得る。
【0058】
いくつかの実施形態において、融合タンパク質は、シグナル配列、例えば、ミツバチメリチンリーダー(HBML):MKFLVNVALVFMVVYISYIYA(配列番号10);組織プラスミノゲンアクチベーター(TPA)リーダー:MDAMKRGLCCVLLLCGAVFVSP(配列番号11);または(iii)天然のリーダー配列を含み得る。
【0059】
本開示の融合タンパク質は、必要に応じてリンカードメインを含み得る。ある特定の実施形態において、融合タンパク質は、Fcドメインと異種ポリペプチドドメインとの間に位置する比較的非構造のリンカーを含む。この非構造リンカーは、1、2、3、4もしくは5アミノ酸の人工配列、または二次構造を比較的含まない5〜10、15、20、30、50もしくはそれ以上の長さのアミノ酸であり得る。リンカーは、グリシン残基およびプロリン残基に富んでいることがあり、例えば、トレオニン/セリンおよびグリシンの単一配列またはトレオニン/セリンおよびグリシンの反復配列(例えば、TG3(配列番号12)またはSG3(配列番号13)のシングレットまたは反復)を含み得る。ある特定の実施形態において、導入されるグリコシル化部位の1つ以上が、リンカードメイン内に配置され得る。
【0060】
本開示の融合タンパク質は、その融合タンパク質の単離および/または検出を容易にするドメインを含み得る。そのようなドメインの周知の例としては、ポリヒスチジン、Glu−Glu、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、マルトース結合タンパク質(MBP)またはヒト血清アルブミンが挙げられるがこれらに限定されない。融合ドメインは、所望の特性を付与するように選択され得る。例えば、いくつかの融合ドメインが、アフィニティークロマトグラフィーによる融合タンパク質の単離にとって特に有用である。親和性精製のために、アフィニティークロマトグラフィーにとって適切なマトリックス(例えば、グルタチオン、アミラーゼおよびニッケルまたはコバルトに結合体化された樹脂)が使用される。そのようなマトリックスの多くは、「キット」の形態で入手可能である(例えば、Pharmacia GST精製システム、および(HIS6)融合パートナーを用いたとき有用であるQIAexpress(商標)システム(Qiagen))。別の例として、融合ドメインは、ポリペプチドの検出を容易にするように選択され得る。そのような検出ドメインの例としては、様々な蛍光タンパク質(例えば、GFP)、ならびに特異的な抗体が入手可能な通常短いペプチド配列である「エピトープタグ」が挙げられる。特異的なモノクローナル抗体が容易に入手可能な周知のエピトープタグとしては、FLAG、インフルエンザウイルス赤血球凝集素(HA)およびc−mycタグが挙げられる。場合によっては、融合ドメインは、適切なプロテアーゼが融合タンパク質を部分的に消化することによってそこから組換えタンパク質が遊離されるのを可能にするプロテアーゼ切断部位(例えば、Xa因子またはトロンビンに対するもの)を有する。次いで、遊離されたタンパク質は、その後のクロマトグラフィー分離によって、融合ドメインから単離され得る。
【0061】
融合タンパク質の種々のエレメントが、所望の機能性と一致する任意の様式で配列され得ることが理解される。例えば、Fcドメインが、異種ポリペプチドドメインに対してC末端に配置され得るか、あるいは、異種ポリペプチドドメインが、Fcドメインに対してC末端に配置され得る。そのFcドメインおよび異種ドメインは、融合タンパク質において隣接している必要はなく、さらなるドメインまたはアミノ酸配列が、いずれかのドメインに対してC末端もしくはN末端に、またはそれらのドメイン間に含められ得る。
【0062】
ある特定の実施形態において、糖改変体融合タンパク質は:PEG化されたアミノ酸、ファルネシル化されたアミノ酸、アセチル化されたアミノ酸、ビオチン化されたアミノ酸、脂質部分に結合体化されたアミノ酸、および有機誘導体化剤に結合体化されたアミノ酸から選択される1つ以上のさらなる改変されたアミノ酸残基を含み得る。そのような翻訳後修飾の結果として、本明細書中に記載される融合タンパク質は、非アミノ酸エレメント(例えば、ポリエチレングリコール、脂質およびホスフェート)を含み得る。種々の細胞(例えば、CHO、HeLa、MDCK、293、W138、NIH−3T3またはHEK293)は、そのような翻訳後の活性に対する特定の細胞機構および特徴的な機構を有し、本発明のポリペプチドの正確な改変およびプロセシングを確実にするように選択され得る。
【0063】
本開示の融合タンパク質は、通常、少なくとも30、40、50、60、70、80、90、100もしくは110kDaまたはそれ以上のサイズである高分子量である。
【0064】
例示的な実施形態において、本開示は、Fcドメインの外側に少なくとも1つの非内因性N結合型グリコシル化部位を有するTNFR2−Fc糖改変体融合タンパク質を提供する。ある特定の実施形態において、本発明の糖改変体融合タンパク質は、配列番号18、21、22、23または24から選択されるアミノ酸配列と少なくとも75%同一であるアミノ酸配列を含み得る。ある特定の場合において、糖改変体融合タンパク質は、配列番号18、21、22、23または24から選択されるアミノ酸配列と少なくとも80%、85%、90%、95%、97%、98%、99%または100%同一であるアミノ酸配列を有する。
【0065】
ある特定の実施形態において、本明細書中に記載される糖改変体Fc融合タンパク質は、ActRIIbレセプターの細胞外ドメインおよび/または抗体の可変領域を含まない。
【0066】
Fcドメイン
本明細書中に記載される糖改変体融合タンパク質は、異種ポリペプチドに融合された免疫グロブリン重鎖Fcドメインを含む。免疫グロブリンのFc部分との融合は、広範囲のタンパク質に対して望ましい薬物動態学的特性を付与すると知られている。本明細書中で使用される用語「Fc領域」または「Fcドメイン」とは、天然配列Fc領域および改変体Fc領域を含む、免疫グロブリン重鎖のC末端領域のことを指す。免疫グロブリン重鎖のFc領域の境界は、様々であり得るが、ヒトIgG重鎖Fc領域は、通常、Cys226位のアミノ酸残基またはPro230から免疫グロブリンのカルボキシル末端に及ぶと規定される。Fc領域のC末端のリジン(EUナンバリングシステムによれば残基447)は、例えば、抗体の重鎖をコードする核酸を組換え的に操作するとき、除去され得る。別段示されない限り、本明細書において免疫グロブリン重鎖内の残基のナンバリングは、Kabatら、Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,MD(1991)(本明細書中で参考として明確に援用される)におけるようなEUインデックスのナンバリングである。「KabatにおけるようなEUインデックス」とは、ヒトIgG1 EU抗体の残基のナンバリングのことを指す。
【0067】
「天然配列Fc領域」は、天然に見られるFc領域のアミノ酸配列と同一のアミノ酸配列を含む。天然配列ヒトFc領域には、天然配列ヒトIgG1 Fc領域(非AおよびAアロタイプ);天然配列ヒトIgG2 Fc領域;天然配列ヒトIgG3 Fc領域;および天然配列ヒトIgG4 Fc領域、ならびにそれらの天然に存在する改変体が含まれる。
【0068】
「改変体Fc領域」は、少なくとも1つのアミノ酸の改変、好ましくは、1つ以上のアミノ酸置換のせいで、天然配列Fc領域のアミノ酸配列と異なるアミノ酸配列を含む。好ましくは、改変体Fc領域は、天然配列Fc領域または親ポリペプチドのFc領域と比較して、少なくとも1つのアミノ酸置換(例えば、天然配列Fc領域または親ポリペプチドのFc領域における、約1〜約10個のアミノ酸の置換、好ましくは、約1〜約5個のアミノ酸の置換)を有する。本明細書中の改変体Fc領域は、好ましくは、天然配列Fc領域および/または親ポリペプチドのFc領域と少なくとも約80%の相同性、最も好ましくは、それらと少なくとも約90%の相同性、より好ましくは、それらと少なくとも約95%の相同性を有し得る。
【0069】
好ましい実施形態において、本明細書中に記載されるFc融合タンパク質は、免疫グロブリン(immunoglobuline)FcドメインのC末端部分のN末端に融合された異種部分を有する。好ましくは、パパイン切断部位のすぐ上流のヒンジ領域から開始する配列(例えば、重鎖定常領域の1番目の残基を114または他の免疫グロブリンの類似部位と考えて)が融合に使用される。1つの実施形態において、異種ドメインは、IgG1、IgG2またはIgG3重鎖の、ヒンジ領域およびCH2およびCH3、またはCH1、ヒンジ、CH2およびCH3ドメインに融合される。いくつかの実施形態において、融合が行われる正確な部位は重大ではなく、最適な部位が日常的な実験によって決定され得る。
【0070】
ヒトFcドメインについては、ヒトIgG1およびIgG3免疫グロブリン配列を使用することが好ましい。IgG1を使用する主な利点は、IgG1融合タンパク質が、固定化されたプロテインA上で効率的に精製され得る点である。対照的に、IgG3融合タンパク質の精製には、それほど著しく多用途でない基材であるプロテインGが必要である。しかしながら、免疫グロブリンの他の構造的および機能的な特性が、特定の構築に対するIg融合パートナーを選択する際に考慮されるべきである。例えば、IgG3ヒンジは、より長く、かつより可撓性であるので、IgG1に融合されたときに正しく折り畳まれないかまたは機能しない可能性があるより大きな異種部分を受け入れることができる。別の考慮すべき点は、結合価であり得る;IgGイムノアドヘシンは、二価のホモ二量体であるのに対し、IgAおよびIgMのようなIgサブタイプはそれぞれ、基本的なIgホモ二量体単位の二量体または五量体の構造を生じ得る。
【0071】
インビボで適用するために設計されたFc融合タンパク質については、薬物動態学的特性およびFc領域によって特定されるエフェクター機能が、同様に重要である。IgG1、IgG2およびIgG4のすべてが、21日というインビボ半減期を有するが、補体系を活性化する際のそれらの相対的な力価は異なる。IgG4は、補体を活性化せず、IgG2は、補体活性化がIgG1よりも有意に弱い。さらに、IgG1とは異なり、IgG2は、単核細胞上または好中球上のFcレセプターに結合しない。IgG3は、補体活性化にとって最適であるが、そのインビボ半減期は、他のIgGアイソタイプのおよそ3分の1である。
【0072】
ヒト治療薬として使用するために設計されるFc融合タンパク質に対する別の重要な考慮すべき点は、特定のアイソタイプのアロタイプ改変体の数である。一般に、血清学的に定義されるアロタイプがより少ないIgGアイソタイプが好ましい。例えば、IgG1は、4つの血清学的に定義されるアロタイプ部位しか有さず、そのうちの2つ(GImおよび2)は、Fc領域内に配置されており;これらの部位のうちの1つであるGlmlは、免疫原性でない。対照的に、IgG3には、血清学的に定義されるアロタイプが12個存在し、そのすべてが、Fc領域に存在し;これらの部位のうちの3つ(G3m5、11および21)だけが、免疫原性でない1つのアロタイプを有する。したがって、IgG3融合タンパク質の潜在的な免疫原性は、IgG1融合タンパク質のそれよりも高い。
【0073】
ある特定の実施形態において、Fc融合タンパク質において使用されるFcドメインは、野生型Fcドメインと比較して1つ以上の変更を含み得る。それにもかかわらず、これらのFcドメインは、それらの野生型対応物と比較して、治療的有用性に必要な実質的に同じ特徴を保持し得る。例えばWO99/51642に記載されているような、例えば、変更された(すなわち、改善されたまたは減少された)CIq結合および/または補体依存性細胞傷害(CDC)をもたらし得るある特定の変更がFc領域内において行われ得ると考えられる。Fc領域改変体の他の例に関係する、Duncan & Winter Nature 322:738−40(1988);米国特許第5,648,260号;米国特許第5,624,821号;およびWO94/29351も参照のこと。WO00/42072(Presta)およびWO2004/056312(Lowman)には、FcRに対する結合性が改善されたまたは減少された抗体改変体が記載されている。これらの特許公報の内容は、本明細書中で明確に参考として援用される。Shieldsら、J.Biol.Chem.9(2):6591−6604(2001)もまた参照のこと。増加された半減期、および母体のIgGを胎児に移行させるのに関与する新生児Fcレセプター(FcRn)(Guyerら、J.Immunol.117:587(1976)およびKimら、J.Immunol.24:249(1994))に対する改善された結合性を有する抗体は、US2005/0014934A1(Hintonら)に記載されている。これらの抗体は、FcRnへのFc領域の結合を改善する1つ以上の置換をその抗体中に有するFc領域(reg on)を含む。変更されたFc領域アミノ酸配列および増加されたまたは減少されたCIq結合能を有するポリペプチド改変体は、米国特許第6,194,551号および国際公開WO99/51642に記載されている。Idusogieら、J.Immunol.164:4178−4184(2000)もまた参照のこと。これらの内容は、本明細書中で明確に参考として援用される。
【0074】
例示的なFcドメインを下に示す。
【0075】
【化1】
都合のよいリンカー(TG3)とFcドメインとが組み合わされた例を下に示す。
【0076】
【化2】
必要に応じて、Fcドメインは、残基(例えば、Asp−265、リジン322およびAsn−434)に1つ以上の変異を有する。ある特定の場合において、これらの変異の1つ以上(例えば、Asp−265変異)を有する変異体Fcドメインは、野生型Fcドメインと比べて、Fcγレセプターに結合する能力が低下している。他の場合では、これらの変異の1つ以上(例えば、Asn−434変異)を有する変異体Fcドメインは、野生型Fcドメインと比べて、MHCクラスI関連Fcレセプター(FcRN)に結合する能力が増加している。
【0077】
異種ポリペプチド
本明細書中に記載される糖改変体融合タンパク質は、Fcドメインに直接または間接的に融合された異種ポリペプチド部分も含む。その異種ポリペプチド部分は、任意のポリペプチドであり得る。例示的な実施形態において、異種ポリペプチド部分は、少なくとも10、15、20、25、30、35、40、50、60、70、80、90もしくは100kDaまたはそれ以上の分子量を有する。
【0078】
異種ポリペプチド部分は、治療用タンパク質またはそのフラグメント(例えば、成長因子、酵素、血清酵素、内分泌因子(例えば、GLP1)、骨形成タンパク質および可溶性レセプターフラグメント)であり得る。例示的な異種ポリペプチドとしては、成長因子(例えば、肝細胞成長因子(HGF)、神経成長因子(NGF)、上皮成長因子(EGF)、線維芽細胞成長因子(FGF)、トランスフォーミング成長因子(例えば、TGF−アルファ、TGF−ベータ、TGF−ベータ2、TGF−ベータ3)、血管内皮成長因子(VEGF;例えば、VEGF−2))、インターフェロン(例えば、INF−アルファ、INF−ベータ)およびインスリンが挙げられる。他の例示的な異種ポリペプチドとしては、アルファ−ガラクトシダーゼ(例えば、Fabrazyme(商標))などの酵素が挙げられる。他の例示的な異種ポリペプチドとしては、骨形成タンパク質(BMP)、エリトロポイエチン(EPO)、ミオスタチンおよび腫瘍壊死因子(例えば、TNF−α)が挙げられる。他の例示的な異種ポリペプチドとしては、膜貫通レセプターの細胞外ドメイン(細胞レセプターの任意の天然に存在する細胞外ドメイン、ならびにその任意の改変体(変異体、フラグメントおよびペプチド模倣型を含む)を含む)が挙げられる。
【0079】
例示的な実施形態において、異種ポリペプチド部分は、TNF/NGFファミリーのレセプター由来のレセプターの細胞外ドメインである。可溶性レセプターポリペプチドの例としては、例えば、本開示の配列番号2、5、6、7または8が挙げられる。好ましい実施形態において、少なくとも1つの非内因性N結合型グリコシル化部位を有する糖改変体融合タンパク質に含められる細胞外レセプタードメインは、天然に存在するレセプターのリガンドに結合する能力を有する。ある特定の実施形態において、非内因性N結合型グリコシル化部位が、細胞外レセプタードメインのリガンド結合ポケットの外側に導入される。好ましくは、導入されるグリコシル化部位のグリコシル化は、レセプタータンパク質と可溶性リガンドとの結合に有意に影響しない。例示的な実施形態において、導入されるグリコシル化部位(glycosylation sited)のグリコシル化は、細胞外レセプタードメインとそのリガンドとの結合を、10倍、5倍、3倍、2.5倍、2倍、1.5倍、1倍、0.5倍またはそれ未満より大きく減少させない。好ましくは、少なくとも1つの非内因性N結合型グリコシル化部位を含む可溶性レセプタードメインは、1μM未満または100、10もしくは1nM未満の解離定数で特定のリガンドに結合し得る。
【0080】
1つの実施形態において、本明細書中に記載される糖改変体融合タンパク質は、神経成長因子(NGF)/腫瘍壊死因子(TNF)レセプターファミリー由来のレセプターの細胞外ドメインを含む異種ポリペプチド部分を含む。例えば、M.Lotzら、J.Leukocyte Biology,60:1−7(1990)を参照のこと。そのNGF/TNFレセプターファミリーには、例えば、神経成長因子レセプター(NGFR)、TNFR1(またはTNFR55)、TNFR2(またはTNFR75)、TNFレセプター関連タンパク質(TNFRrp)、CD40、Hodgkin抗原CD30、T細胞抗原CD27、Fas/APO−1、OX−40および4−1BB/ILAが含まれる。例示的な実施形態において、本明細書中に記載される融合タンパク質は、少なくとも1つの非内因性N結合型グリコシル化部位を有するTNFR2の細胞外ドメインを含む。
【0081】
本明細書中で使用されるとき、用語「TNFR2」とは、任意の種由来の腫瘍壊死因子レセプター2型(TNFR2)タンパク質のファミリー、および突然変異誘発または他の改変によるそのようなTNFR2タンパク質に由来する改変体のことを指す。TNFR2ファミリーのメンバーは、一般に、システインリッチ領域を含むリガンド結合細胞外ドメイン、膜貫通ドメイン、および細胞内シグナル伝達活性を有すると予測される細胞質ドメインから構成される膜貫通タンパク質である。
【0082】
用語「TNFR2ポリペプチド」には、TNFR2ファミリーメンバーの任意の天然に存在するポリペプチド、ならびに有用な活性を有するその任意の改変体(変異体、フラグメント、融合物およびペプチド模倣型を含む)を含むポリペプチドが含まれる。例えば、TNFR2ポリペプチドには、TNFR2ポリペプチドの配列と少なくとも約80%同一、好ましくは、少なくとも85%、90%、95%、97%、99%またはそれ以上同一の配列を有する任意の公知のTNFR2の配列に由来するポリペプチドが含まれる。例えば、本発明のTNFR2ポリペプチドは、TNFαまたはTNFβに結合し得、TNFR2タンパク質および/またはTNFαもしくはTNFβの機能を阻害し得る。TNFR2ポリペプチドの例としては、ヒトTNFR2前駆体ポリペプチド(配列番号1)および可溶性ヒトTNFR2ポリペプチド(例えば、配列番号2、5、6、7または8)が挙げられる。
【0083】
ヒトTNFR2前駆体タンパク質配列は、以下のとおりである:
【0084】
【化3】
シグナルペプチドに一重下線が引かれている;細胞外ドメインは太字であり、潜在的なN結合型グリコシル化部位には二重下線が引かれている。
【0085】
プロセシングされたヒトTNFR2可溶性(細胞外)ポリペプチド配列は、以下のとおりである:
【0086】
【化4】
天然に存在するN結合型グリコシル化部位に下線が引かれている。本明細書中で報告されるように、アミノ末端のロイシンは、削除され得、アミノ末端にはプロリン残基が残っている。
【0087】
ヒトTRFR2前駆体タンパク質をコードする核酸配列は、以下のとおりである(GenbankエントリーNM_001066.2):
【0088】
【化5】
【0089】
【化6】
ヒトTNFR2可溶性(細胞外)ポリペプチドをコードする核酸配列は、以下のとおりである:
【0090】
【化7】
本開示は、1つ以上のN結合型グリコシル化部位(NXS/T)の付加によって、改変されていないTNFR2−Fcタンパク質と比べてTNFR2−Fc融合タンパク質の血清半減期が増加されることを証明する。N結合型グリコシル化部位は、既存のSもしくはTに対して、正しい位置にNを導入することによって、または既存のNに対して、正しい位置にSもしくはTを導入することによって、最小労力で導入され得る。TNFR2における非内因性NXS/T配列の導入に特に適した部位としては、例えば、配列番号2について、アミノ酸25〜27(DQT)、26〜28(QIA)、133〜135(ETS)および231〜233(GST)が挙げられる。例えば、N結合型グリコシル化部位を導入し得る望ましい変更は:D25N、Q26N、A28S、E133NおよびG231Nである。場合によっては、いくつかのアミノ酸が、非内因性のNXS/T配列を導入するように改変され得る。例えば、Q26N/A28Sの二重置換は、配列番号2の残基26〜28において、新規のN結合型グリコシル化部位を導入するために必要なN残基とS残基の両方を提供する。さらに、TNFR2−Fcポリペプチド上の1つより多い領域が、複数のN結合型グリコシル化部位を導入するように改変され得る。例えば、D25N/E133Nの二重置換は、配列番号2の残基25〜27および133〜135において2つのN結合型グリコシル化部位を導入し、D25N/G231Nの二重置換は、配列番号2の残基25〜27および231〜233において2つのN結合型グリコシル化部位を導入する。他の例示的なTNFR2改変体としては、例えば、E133N/G231N、Q26N/A28S/E133NおよびQ26N/A28S/G231Nが挙げられる。グリコシル化されると予測される任意のSが、グリコシル化によってもたらされる保護に起因して、免疫原性部位を作り出すことなくTに変更され得る。同様に、グリコシル化されると予測される任意のTが、Sに変更され得る。
【0091】
3.糖改変体融合タンパク質をコードする核酸
ある特定の態様において、本発明は、本発明の糖改変体融合タンパク質(本明細書中に開示されるフラグメントおよび機能的改変体を含む)のいずれかをコードする単離された核酸および/または組換え核酸を提供する。その主題の核酸は、一本鎖または二本鎖であり得る。そのような核酸は、DNA分子またはRNA分子であり得る。これらの核酸は、例えば、本発明の糖改変体融合タンパク質を作製するための方法において、または直接的な治療薬として(例えば、遺伝子治療アプローチにおいて)、使用され得る。
【0092】
例示的な実施形態において、本発明の核酸は、TNFR2−Fc糖改変体融合タンパク質をコードする。例えば、配列番号3は、天然に存在するヒトTNFR2前駆体ポリペプチドをコードし、配列番号4は、TNFR2のプロセシングされた細胞外ドメインをコードする。
【0093】
ある特定の実施形態において、本発明の糖改変体融合タンパク質をコードする主題の核酸は、配列番号3、4、9または17の改変体である核酸を含むとさらに理解される。改変体ヌクレオチド配列は、1つ以上のヌクレオチドの置換、付加または欠失によって異なる配列(例えば、対立遺伝子改変体)を含む。例示的な実施形態において、その改変体核酸は、コードされたタンパク質内に1つ以上の非内因性N結合型グリコシル化部位を導入する核酸配列改変を含む。
【0094】
ある特定の実施形態において、本発明は、配列番号3、4、9または17と少なくとも80%、85%、90%、95%、97%、98%、99%または100%同一であり、かつコードされたタンパク質内に非内因性N結合型グリコシル化部位を導入する1つ以上の配列改変を含む、単離された核酸配列または組換え核酸配列を提供する。当業者は、コードされたタンパク質内に1つ以上の非内因性N結合型グリコシル化部位を導入する、配列番号3、4、9または17に相補的な核酸配列および配列番号3、4、9または17の改変体もまた本発明の範囲内であることを認識するだろう。さらなる実施形態において、本発明の核酸配列は、単離され得、組換えであり得、および/もしくは異種ヌクレオチド配列と融合され得るか、またはDNAライブラリー中のものであり得る。
【0095】
他の実施形態において、本発明の核酸はまた、配列番号3、4、9もしくは17において指定されるヌクレオチド配列、配列番号3、4、9もしくは17の相補配列、またはそれらのフラグメントに高度にストリンジェントな条件下でハイブリダイズするヌクレオチド配列も含む。そのようなヌクレオチド配列は、1つ以上の非内因性N結合型グリコシル化部位を有する糖改変体タンパク質融合物をコードする。当業者は、DNAハイブリダイゼーションを促進する適切なストリンジェンシー条件が変動し得ることを容易に理解するだろう。当業者は、DNAハイブリダイゼーションを促進する適切なストリンジェンシー条件が変動し得ることを容易に理解するだろう。例えば、約45℃の6.0×塩化ナトリウム/クエン酸ナトリウム(SSC)におけるハイブリダイゼーションに続く、50℃の2.0×SSCでの洗浄が行われ得る。例えば、洗浄工程における塩濃度は、50℃の約2.0×SSCの低ストリンジェンシーから50℃の約0.2×SSCの高ストリンジェンシーより選択され得る。さらに、洗浄工程における温度は、室温である約22℃での低ストリンジェンシー条件から約65℃の高ストリンジェンシー条件に上昇され得る。温度と塩の両方が、変動し得るか、または温度もしくは塩濃度が一定に保持され得る一方で、他方の変数が変更される。1つの実施形態において、本発明は、室温の6×SSCに続く室温の2×SSCでの洗浄の低ストリンジェンシー条件下でハイブリダイズする核酸を提供する。
【0096】
遺伝暗号の縮重に起因して配列番号3、4または9に示されるような核酸と異なり、かつコードされるタンパク質に1つ以上の非内因性N結合型グリコシル化部位を導入する、単離された核酸改変体もまた、本発明の範囲内である。例えば、いくつかのアミノ酸が、1つより多いトリプレットによって指定される。同じアミノ酸を特定するコドン、すなわちシノニム(例えば、CAUおよびCACは、ヒスチジンに対するシノニムである)は、そのタンパク質のアミノ酸配列に影響しない「サイレント」変異をもたらし得る。しかしながら、その主題のタンパク質のアミノ酸配列を変更させるDNA配列多型が哺乳動物細胞間に存在し得ると予想される。当業者は、特定のタンパク質をコードする核酸の1つ以上のヌクレオチド(ヌクレオチドの約3〜5%まで)におけるこれらのバリエーションが、天然の対立遺伝子バリエーションに起因して所与の種の個体間に存在し得ることを認識するだろう。任意のおよびすべてのそのようなヌクレオチドバリエーションおよびその結果得られるアミノ酸多型が、本発明の範囲内である。
【0097】
ある特定の実施形態において、本発明の組換え核酸は、発現構築物内の1つ以上の制御ヌクレオチド配列に作動可能に連結され得る。制御ヌクレオチド配列は、一般に、発現のために使用される宿主細胞にとって適切なものであり得る。数多くのタイプの適切な発現ベクターおよび適当な制御配列が、様々な宿主細胞について当該分野で公知である。代表的には、前記1つ以上の制御ヌクレオチド配列としては、プロモーター配列、リーダーまたはシグナル配列、リボソーム結合部位、転写開始配列および転写終結配列、翻訳開始配列および翻訳終結配列、ならびにエンハンサーまたはアクチベーター配列が挙げられ得るがこれらに限定されない。当該分野で公知であるような構成的プロモーターまたは誘導性プロモーターが、本発明によって企図される。それらのプロモーターは、天然に存在するプロモーター、または2つ以上のプロモーターのエレメントを組み合わせたハイブリッドプロモーターであり得る。発現構築物は、エピソーム(例えば、プラスミド)上において細胞内に存在し得るか、または発現構築物は、染色体に挿入され得る。好ましい実施形態において、発現ベクターは、形質転換された宿主細胞の選択を可能にする選択可能なマーカー遺伝子を含む。選択可能なマーカー遺伝子は、当該分野で周知であり、使用される宿主細胞によって異なる。
【0098】
ある特定の実施形態において、主題の核酸は、本発明の糖改変体融合タンパク質をコードし、かつ少なくとも1つの制御配列に作動可能に連結された、ヌクレオチド配列を含む発現ベクターとして提供される。制御配列は、当該分野において認められたものであり、本発明のポリペプチドの発現を指示するように選択される。したがって、制御配列という用語には、プロモーター、エンハンサーおよび他の発現調節エレメントが含まれる。例示的な制御配列は、Goeddel;Gene Expression Technology:Methods in Enzymology,Academic Press,San Diego,Calif.(1990)に記載されている。例えば、作動可能に連結されたときDNA配列の発現を調節する多種多様な発現調節配列のいずれかが、糖改変体融合タンパク質をコードするDNA配列を発現させるためにこれらのベクターにおいて使用され得る。そのような有用な発現調節配列としては、例えば、SV40の初期および後期プロモーター、tetプロモーター、アデノウイルスまたはサイトメガロウイルスの最初期プロモーター、RSVプロモーター、lac系、trp系、TACまたはTRC系、発現がT7 RNAポリメラーゼによって指示されるT7プロモーター、ファージラムダの主要なオペレーターおよびプロモーター領域、fdコートタンパク質に対する調節領域、3−ホスホグリセリン酸キナーゼまたは他の糖分解酵素に対するプロモーター、酸ホスファターゼ(例えば、Pho5)のプロモーター、酵母アルファ接合因子のプロモーター、バキュロウイルス系の多角体プロモーター、および原核細胞もしくは真核細胞またはそれらのウイルスの遺伝子の発現を調節すると知られている他の配列、ならびにそれらの様々な組み合わせが挙げられる。発現ベクターのデザインは、形質転換される宿主細胞の選択および/または発現が望まれるタンパク質のタイプのような因子に依存し得ることが理解されるべきである。さらに、ベクターのコピー数、そのコピー数およびそのベクターによってコードされる他の任意のタンパク質(例えば、抗生物質マーカー)の発現を調節する能力もまた考慮されるべきである。
【0099】
本発明の組換え核酸は、クローニングされた遺伝子またはその一部分を、原核細胞、真核細胞(酵母、鳥類、昆虫または哺乳動物)またはその両方における発現に適したベクターにライゲートすることによって作製され得る。本発明の組換え糖改変体融合タンパク質を生成するための発現ビヒクルには、プラスミドおよび他のベクターが含まれる。例えば、適当なベクターには、以下のタイプのプラスミドが含まれる:大腸菌などの原核細胞における発現用の、pBR322由来プラスミド、pEMBL由来プラスミド、pEX由来プラスミド、pBTac由来プラスミドおよびpUC由来プラスミド。
【0100】
いくつかの哺乳動物発現ベクターは、細菌内でのベクターの増殖を促す原核生物の配列と、真核細胞内で発現される1つ以上の真核生物の転写単位の両方を含む。pcDNAI/amp、pcDNAI/neo、pRc/CMV、pSV2gpt、pSV2neo、pSV2−dhfr、pTk2、pRSVneo、pMSG、pSVT7、pko−neoおよびpHyg由来ベクターが、真核細胞のトランスフェクションに適した哺乳動物発現ベクターの例である。これらのベクターのうちのいくつかは、原核細胞と真核細胞の両方における複製および薬物耐性選択を容易にするために、pBR322などの細菌のプラスミド由来の配列で改変される。あるいは、ウイルスの誘導体(例えば、ウシパピローマウイルス(BPV−1)またはエプスタイン・バーウイルス(pHEBo、pREP由来およびp205))が、真核細胞内でのタンパク質の一過性の発現のために使用され得る。他のウイルス(レトロウイルスを含む)の発現系の例は、下記の遺伝子治療送達系の説明において見出され得る。プラスミドの調製および宿主生物の形質転換において使用される様々な方法が、当該分野で周知である。原核細胞と真核細胞の両方に適した他の適した発現系、ならびに一般的な組換え手順については、Molecular Cloning A Laboratory Manual,3rd Ed.,Sambrook,FritschおよびManiatis編(Cold Spring Harbor Laboratory Press,2001)を参照のこと。場合によっては、バキュロウイルス発現系を使用することによって組換え糖改変体融合タンパク質を発現することが望ましい場合がある。そのようなバキュロウイルス発現系の例としては、pVL由来ベクター(例えば、pVL1392、pVL1393およびpVL941)、pAcUW由来ベクター(例えば、pAcUW1)およびpBlueBac由来ベクター(例えば、β−gal含有pBlueBacIII)が挙げられる。
【0101】
好ましい実施形態において、Pcmv−Scriptベクター(Stratagene,La Jolla,Calif.)、pcDNA4ベクター(Invitrogen,Carlsbad,Calif.)およびpCI−neoベクター(Promega,Madison,Wis.)などのベクターが、CHO細胞において本発明の主題の糖改変体融合タンパク質を産生するために設計され得る。明らかであるように、その主題の遺伝子構築物は、精製にむけて、培養において増殖する細胞内でその主題の糖改変体融合タンパク質の発現を引き起こすために、例えば、タンパク質(融合タンパク質または改変体タンパク質を含む)を産生するために、使用され得る。
【0102】
本開示はまた、本発明の主題の糖改変体タンパク質融合物の1つ以上に対するコード配列を含む組換え遺伝子でトランスフェクトされた宿主細胞に関係する。その宿主細胞は、任意の原核細胞または真核細胞であり得る。例えば、本発明の糖改変体融合タンパク質は、大腸菌などの細菌細胞、昆虫細胞(例えば、バキュロウイルス発現系を使用する)、酵母または哺乳動物細胞において発現され得る。他の適当な宿主細胞は、当業者に公知である。好ましくは、その宿主細胞は、発現されるタンパク質に対して哺乳動物のグリコシル化パターンをもたらし得る哺乳動物宿主細胞(例えば、CHOまたはBHK細胞株)であり得る。
【0103】
したがって、本発明はさらに、本発明の主題の糖改変体融合タンパク質を生成する方法に関係する。例えば、Fc融合タンパク質をコードする発現ベクターでトランスフェクトされた宿主細胞が、Fc融合タンパク質の発現が生じるのを可能にする適切な条件下で培養され得る。そのFc融合タンパク質は、分泌され得、そして細胞とFc融合タンパク質を含む培地との混合物から単離され得る。あるいは、そのFc融合タンパク質は、細胞質に保持され得るか、または膜画分中、および収集されて溶解されてタンパク質が単離される細胞中に保持され得る。細胞培養物は、宿主細胞、培地および他の副産物を含む。細胞培養に適した培地は、当該分野で周知である。主題のFc融合タンパク質は、タンパク質を精製するための当該分野で公知の手法(イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、限外濾過、電気泳動、Fc融合タンパク質の特定のエピトープに特異的な抗体を用いる免疫親和性精製、およびFc融合タンパク質に融合されたドメインに結合する物質を用いる親和性精製(例えば、プロテインAカラムが、Fc融合タンパク質を精製するために使用され得る)を含む)を用いて、細胞培養液、宿主細胞またはその両方から単離され得る。好ましい実施形態において、Fc融合タンパク質は、その精製を容易にするさらなるドメインを含む。好ましい実施形態において、精製は、例えば、以下のうちの3つ以上を任意の順序で含む一連のカラムクロマトグラフィー工程によって達成され得る:プロテインAクロマトグラフィー、Qセファロースクロマトグラフィー、フェニルセファロースクロマトグラフィー、サイズ排除クロマトグラフィーおよび陽イオン交換クロマトグラフィー。その精製は、ウイルス濾過および緩衝液交換によって完了され得る。本発明の糖改変体融合タンパク質は、サイズ排除クロマトグラフィーによって測定されるとき>98%の純度に、およびSDS PAGEによって測定されるとき>95%に精製され得る。
【0104】
別の実施形態において、本発明の組換え糖改変体Fc融合タンパク質の所望の部分のN末端におけるポリ−(His)/エンテロキナーゼ切断部位配列などの精製リーダー配列をコードする融合遺伝子のおかげで、Ni2+金属樹脂を用いるアフィニティークロマトグラフィーによる、発現された融合タンパク質の精製が可能になり得る。次いで、その精製リーダー配列が、その後、エンテロキナーゼでの処理によって除去されることにより、本発明の精製された糖改変体融合タンパク質が提供され得る(例えば、Hochuliら(1987)J.Chromatography 411:177;およびJanknechtら、PNAS USA 88:8972を参照のこと)。
【0105】
融合遺伝子を作製するための手法は、周知である。本質的には、種々のポリペプチド配列をコードする様々なDNAフラグメントの連結が、従来の手法(平滑末端化されたまたは粘着末端化された末端をライゲーションに使用すること、適切な末端をもたらす制限酵素消化、必要に応じて付着末端を埋めること、望ましくない連結を回避するためのアルカリホスファターゼ処理、および酵素的ライゲーション)に従って行われる。別の実施形態において、融合遺伝子は、自動化DNA合成装置を含む従来の手法によって合成され得る。あるいは、後にアニールしてキメラ遺伝子配列を生成し得る2つの連続した遺伝子フラグメント間に相補的なオーバーハングを生じるアンカープライマーを用いて、遺伝子フラグメントのPCR増幅が行われ得る(例えば、Current Protocols in Molecular Biology,eds.Ausubelら、John Wiley & Sons:1992を参照のこと)。
【0106】
4.血清半減期を増加させる方法
特定の実施形態において、本発明は、Fcドメインの外側に1つ以上の非内因性N結合型グリコシル化部位を導入するように融合タンパク質を改変することによって、Fc融合タンパク質の安定性および/または半減期(例えば、インビトロ、インビボまたは血清半減期)を増加させるための方法に関する。例えば、そのような方法は、1つ以上の非内因性N結合型グリコシル化部位を導入するようにFc融合タンパク質の配列を改変する工程、および糖改変体Fc融合タンパク質を産生する適当な細胞(例えば、チャイニーズハムスター卵巣(CHO)細胞またはヒト細胞)において前記改変されたポリペプチドをコードする核酸を発現させる工程を包含し得る。そのような方法は:a)改変されたポリペプチドの発現に適した条件下で細胞を培養する工程(ここで、前記細胞は、改変されたポリペプチド発現構築物で形質転換されている);およびb)そのように発現された改変されたタンパク質を収集する工程を包含し得る。精製は、例えば、以下のうちの1、2もしくは3つまたはそれ以上を任意の順序で含む一連の精製工程によって達成され得る:プロテインAクロマトグラフィー(chromatograph)、陰イオン交換クロマトグラフィー(chromotography)(例えば、Qセファロース)、疎水性相互作用クロマトグラフィー(例えば、フェニルセファロース)、サイズ排除クロマトグラフィーおよび陽イオン交換クロマトグラフィー。本開示のそのようなポリペプチドは、さらに液体または固体(例えば、凍結乾燥された形態)として処方され得る。本明細書中に記載される任意の糖改変体融合タンパク質が、前記方法を用いて生成され得る。
【0107】
好ましい実施形態において、導入されたグリコシル化部位の1つ以上におけるグリコシル化によって、改変された融合タンパク質の半減期(例えば、インビトロ、インビボ、血清半減期)が、導入されるグリコシル化部位を欠く融合タンパク質の血清半減期と比べて少なくとも10%、25%、50%、75%、100%、125%、150%、175%、200%、225%もしくは250%またはそれ以上増加する。ある特定の実施形態において、本開示の方法は、本開示の融合タンパク質の各10、20、30、40、50、60、70、80、90、100、110、120、130、140、150、160、170、180、190または200アミノ酸あたり少なくとも1つのグリコシル化されたアミノ酸を導入するために使用され得る。ある特定の実施形態において、本開示の方法は、N結合型グリコシル化、O結合型グリコシル化またはその両方によって改変された各アミノ酸間の間隙が少なくとも10、20、30、40、50、60、70、80、90もしくは100アミノ酸またはそれ以上を維持するようにグリコシル化部位を導入するために使用され得る。
【0108】
ある特定の実施形態において、本開示は、少なくとも1つの異種ポリペプチドドメインおよび免疫グロブリンFcドメインを含む融合タンパク質の半減期を延長するための方法を提供する。例えば、そのような方法は、a)1つ以上の非内因性N結合型グリコシル化部位を導入するように免疫グロブリンFcドメインの外側でFc融合タンパク質を改変する工程、およびb)導入されたグリコシル化部位のうちの1つがグリコシル化されるようにFc融合タンパク質を発現させる工程を包含し得る。本明細書中に開示される方法は、例えば、リガンド結合ドメインを含むレセプターの細胞外ドメインの一部分(例えば、可溶性部分)を含むFc融合タンパク質の半減期を延長するために使用され得る。ある特定の実施形態において、その方法は、Fc融合タンパク質のリガンド結合ドメインの外側にグリコシル化部位を導入するために使用される。好ましい実施形態において、グリコシル化部位を導入するための方法は、可溶性リガンドに対する融合タンパク質のレセプター部分の結合に有意に影響せず、例えば、リガンドの結合は、導入されるグリコシル化部位を欠くFc融合タンパク質に対するリガンドの結合と比べて2、3、5、10または15倍未満だけ影響される。いくつかの実施形態において、本開示の方法は、リンカードメインを含む融合タンパク質に1つ以上のグリコシル化部位を導入するために使用される。ある特定の実施形態において、本開示の方法は、リンカードメイン内に少なくとも1つのグリコシル化部位を導入するために使用され得る。一般に、本明細書中に開示される方法は、高分子量、例えば、少なくとも25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100、105または110kDaである高分子のタンパク質である、異種ドメインを有する融合タンパク質の半減期を増加させるために使用される。
【0109】
5.スクリーニングアッセイ
ある特定の態様において、本発明は、少なくとも1つの生物学的活性を維持し、かつ高い安定性および/または増加された血清半減期を有する、糖改変体Fc融合タンパク質を同定するためのスクリーニングアッセイを提供する。糖改変体融合タンパク質は、生物学的活性を調節する能力を査定するため、それらの安定性を査定するため、および/またはインビボもしくはインビトロにおいてそれらの血清半減期を査定するために試験され得る。糖改変体融合タンパク質は、例えば、動物モデルにおいて試験され得る。
【0110】
本発明の糖改変体融合タンパク質の生物学的活性についてスクリーニングするアプローチは数多く存在し、例えば、糖改変体レセプター−Fc融合タンパク質をアッセイするときは、改変されたレセプターがリガンドに結合する能力が測定され得るか、またはその糖改変体融合タンパク質がリガンド−レセプターシグナル伝達を乱す能力もまたアッセイされ得る。ある特定の実施形態において、改変されていないタンパク質の少なくとも1つの生物学的活性を保持する糖改変体(例えば、選択された細胞株に対する、リガンドまたはレセプターが媒介する作用を混乱させるレセプター改変体)を同定する糖改変体融合タンパク質のハイスループットスクリーニングが行われ得る。
【0111】
様々なアッセイ形式が十分であるが、それにもかかわらず、本開示に照らして、本明細書中にはっきりと記載されないものも、当業者によって理解されるだろう。本明細書中に記載されるように、本発明の糖改変体融合タンパク質は、前記融合タンパク質をコードする核酸の対応するフラグメントから組換え的に生成されたポリペプチドをスクリーニングすることによって得ることができる。さらに、フラグメントは、従来のMerrifield固相f−Mocまたはt−Boc化学などの当該分野で公知の手法を用いて化学的に合成され得る。
【0112】
本発明の機能的に活性な糖改変体融合タンパク質は、前記融合タンパク質をコードする、対応する変異誘発された核酸から組換え的に生成された改変された融合タンパク質のライブラリーをスクリーニングすることによって得ることができる。それらの糖改変体は、改変されていない融合タンパク質の生物学的活性、例えば、糖改変体レセプター融合物が様々な細胞レセプタータンパク質(例えば、TNFR2レセプタータンパク質)のアンタゴニスト(インヒビター)として作用する能力、および/またはリガンド−レセプター結合によって媒介される細胞内のシグナル伝達を少なくとも保持するものを同定するために、生成され、試験され得る。
【0113】
機能的な糖改変体は、安定性および/または血清半減期を増加させるように本発明の融合タンパク質のアミノ酸配列を改変することによって作製され得る。そのような改変された糖改変体タンパク質は、生物学的活性(例えば、リガンド結合性)を保持するように選択されるとき、改変されていない融合タンパク質の機能的等価物と考えられる。改変された糖改変体融合タンパク質はまた、例えば、1つ以上のN結合型グリコシル化部位を導入するアミノ酸の置換、欠失もしくは付加、および/または他の配列改変(例えば、保存的置換)によって作製され得る。例えば、イソロイシンまたはバリンによるロイシン、グルタミン酸によるアスパラギン酸、セリンによるトレオニンの個別の置換、または構造的に関係があるアミノ酸によるアミノ酸の類似の置換(例えば、保存的変異)が、その結果生じる分子の生物学的活性に対して大きな作用を有さないであろうと予想されることは妥当である。保存的置換は、側鎖に関するアミノ酸のファミリー内で行われる置換である。本発明の融合タンパク質のアミノ酸配列の変化が機能的ホモログをもたらすか否かは、改変体融合タンパク質が、改変されていない融合タンパク質と類似の様式で生物学的活性を示す能力を査定することによって容易に判定され得る。例えば、改変体レセプターFc融合タンパク質(例えば、TNFR2−Fc融合タンパク質)は、特異的なリガンド(例えば、TNFαまたはTNFβ)に結合する能力、レセプターポリペプチドへのリガンドの結合を妨害する能力、またはレセプターへのリガンドの結合によって引き起こされるシグナル伝達を干渉する能力についてスクリーニングされ得る。
【0114】
糖改変体融合タンパク質のコンビナトリアルライブラリーは、各々が潜在的なポリペプチド配列の少なくとも一部分を含む融合タンパク質のライブラリーをコードする遺伝子の縮重ライブラリーを経由して作製され得る。例えば、合成オリゴヌクレオチドの混合物が、潜在的なポリペプチドヌクレオチド配列の縮重セットが個別のポリペプチドとして、あるいは、より大きな融合タンパク質のセットとして(例えば、ファージディスプレイの場合)発現可能であるように、遺伝子配列に酵素的にライゲートされ得る。
【0115】
潜在的な糖改変体のライブラリーを縮重オリゴヌクレオチド配列から生成し得る方法が多く存在する。縮重遺伝子配列の化学的合成が、自動DNA合成装置において行われ得、次いで、その合成遺伝子が、発現用の適切なベクターにライゲートされ得る。縮重オリゴヌクレオチドの合成法は、当該分野で周知である(例えば、Narang,SA(1983)Tetrahedron 39:3;Itakuraら(1981)Recombinant DNA,Proc.3rd Cleveland Sympos.Macromolecules,ed.AG Walton,Amsterdam:Elsevier pp 273−289;Itakuraら(1984)Annu.Rev.Biochem.53:323;Itakuraら(1984)Science 198:1056;Ikeら(1983)Nucleic Acid Res.11:477を参照のこと)。そのような手法は、他のタンパク質の指向性進化において使用されている(例えば、Scottら(1990)Science 249:386−390;Robertsら(1992)PNAS USA 89:2429−2433;Devlinら(1990)Science 249:404−406;Cwirlaら(1990)PNAS USA 87:6378−6382;ならびに米国特許第5,223,409号、同第5,198,346号および同第5,096,815号を参照のこと)。
【0116】
点変異および切断によって作製されるコンビナトリアルライブラリーの遺伝子産物をスクリーニングするため、および、もっと詳しく言うなら、ある特定の特性を有する遺伝子産物についてcDNAライブラリーをスクリーニングするための、広範囲の手法が当該分野で公知である。そのような手法は、一般に、本発明の糖改変体融合タンパク質のコンビナトリアル突然変異誘発によって作製された遺伝子ライブラリーの迅速なスクリーニングに適用可能であり得る。大きな遺伝子ライブラリーをスクリーニングするために最も広く使用されている手法は、代表的には、複製可能な発現ベクターに遺伝子ライブラリーをクローニングすること、得られたベクターのライブラリーで適切な細胞を形質転換すること、および所望の活性の検出が遺伝子(その産物が検出される)をコードするベクターの比較的簡便な単離を容易にする条件下でそのコンビナトリアル遺伝子を発現させることを含む。
【0117】
好ましいアッセイには、レセプター−リガンド結合アッセイおよびリガンド媒介性細胞シグナル伝達アッセイが含まれる。本発明の糖改変体レセプター−Fc融合タンパク質とリガンドとの複合体の形成は、様々な手法によって検出され得る。例えば、複合体の形成の調節は、イムノアッセイまたはクロマトグラフィー検出によって、例えば、検出可能に標識されたタンパク質(例えば、放射性標識された(例えば、32P、35S、14Cまたは3H)、蛍光標識された(例えば、FITC)、または酵素的に標識された、レセプターポリペプチドまたはリガンド)を用いて定量され得る。
【0118】
ある特定の実施形態において、本発明は、糖改変体レセプター−Fc融合タンパク質とその結合リガンドとの相互作用の程度を直接または間接的に測定する際の、蛍光偏光アッセイおよび蛍光共鳴エネルギー転移(FRET)アッセイの使用を企図する。さらに、他の検出様式(例えば、光導波路(PCT公開WO96/26432および米国特許第5,677,196号)、表面プラズモン共鳴(SPR)、表面電荷センサーおよび表面力センサーに基づく検出様式)が、本発明の多くの実施形態と適合可能である。
【0119】
さらに、本発明は、糖改変体Fc融合タンパク質とその結合パートナーとの相互作用を乱すかまたは増強する物質を同定するための「2ハイブリッドアッセイ」としても知られる相互作用捕捉アッセイの使用を企図する。例えば、米国特許第5,283,317号;Zervosら(1993)Cell 72:223−232;Maduraら(1993)J Biol Chem 268:12046−12054;Bartelら(1993)Biotechniques 14:920−924;およびIwabuchiら(1993)Oncogene 8:1693−1696)を参照のこと。あるいは、そのようなタンパク質間相互作用は、光架橋、放射性標識されたリガンドの結合、およびアフィニティークロマトグラフィー(Jakoby WBら、1974,Methods in Enzymology 46:1)を含むインビトロでの生化学的方法を用いてタンパク質レベルで同定され得る。ある特定の場合において、糖改変体融合タンパク質は、機序に基づいたアッセイ(例えば、リガンドまたはレセプターポリペプチドに結合する糖改変体融合タンパク質を検出するアッセイ)においてスクリーニングされ得る。これは、固相または流体相の結合事象を含み得る。あるいは、リガンドまたはレセプターポリペプチドをコードする遺伝子が、レポーター系(例えば、β−ガラクトシダーゼ、ルシフェラーゼまたは緑色蛍光タンパク質)とともに細胞にトランスフェクトされ得、必要に応じてハイスループットスクリーニングによって糖改変体ライブラリーに対してスクリーニングされ得るか、または糖改変体ライブラリーの個別のメンバーを用いてスクリーニングされ得る。機序に基づいた他の結合アッセイ、例えば、自由エネルギーの変化を検出する結合アッセイが使用され得る。結合アッセイは、ウェル、ビーズもしくはチップに固定された標的、または固定化された抗体によって捕捉される標的、またはキャピラリー電気泳動によって分離される標的を用いて、行われ得る。その結合したタンパク質は、通常、比色定量または蛍光または表面プラズモン共鳴を用いて、検出され得る。
【0120】
6.例示的な治療上の使用
様々な実施形態において、本発明の糖改変体融合タンパク質で処置される患者または本発明の糖改変体融合タンパク質での処置の候補患者は、哺乳動物(例えば、げっ歯類および霊長類)、特に、ヒト患者であり得る。ある特定の実施形態において、本開示は、増加された半減期を特徴とする改変された可溶性レセプター−Fc融合物を提供する。レセプター−Fc融合物は、本明細書中で要約される様々な障害および状態を処置する際の使用について先に記載された。
【0121】
本明細書中で使用されるとき、障害または状態を「防止する」治療薬とは、統計的サンプルにおいて、処置されるサンプルにおけるその障害または状態の発生を未処置のコントロールサンプルと比べて減少させるか、あるいは未処置のコントロールサンプルと比べてその障害もしくは状態の発生を遅延させるかまたはその1つ以上の症状の重症度を低下させる、治療薬のことを指す。本明細書中で使用される用語「処置する」は、指名された状態の予防、またはその状態がいったん確立されたらその回復もしくは排除を含む。どちらにしても、防止または処置は、医師または他の医療提供者によって提供される診断、およびその治療薬の投与の意図される結果において、識別され得る。
【0122】
いくつかの実施形態において、本開示は、増加された血清半減期を有するTNFR2レセプターの細胞外ドメインを含む糖改変体融合タンパク質を提供する。そのようなTNFR2−Fc糖改変体融合物は、TNFαが介在する様々な障害または状態(例えば、急性および慢性の免疫性および自己免疫性の病態(例えば、全身性エリテマトーデス(SLE)、関節リウマチ、若年性慢性関節炎、甲状腺炎(thyroidosis)、移植片対宿主病、強皮症、真性糖尿病、グレーヴズ病)、脊椎関節症、感染症に関連する障害(例えば、敗血症症候群、悪液質、急性または慢性の細菌感染に起因する循環虚脱およびショック、急性および慢性の寄生虫病ならびに/または細菌、ウイルスもしくは真菌の感染症)、慢性の炎症性の病態(例えば、強皮症、サルコイドーシス、慢性炎症性腸疾患、潰瘍性大腸炎およびクローン病の病態)、血管の炎症性の病態(例えば、播種性血管内凝固、アテローム性動脈硬化症および川崎病の病態)および神経変性疾患(例えば、多発性硬化症、急性横断性脊髄炎、錐体外路系疾患、ハンチントン舞踏病および老人性舞踏病;パーキンソン病、進行性核上性麻痺(Progressive supranucleo palsy)、脊髄性運動失調症、フリードライヒ運動失調症、メンセル(Mencel)病、デジェリーヌ・トーマス病、シャイ・ドレーガー(Shi−Drager)病、マシャド・ジョセフ病、レフサム病、無βリポタンパク質血症(abetalipoprotemia)、毛細血管拡張、ミトコンドリア多系統障害、筋萎縮性側索硬化症、乳児性脊髄性筋萎縮症、若年性脊髄性筋萎縮症、アルツハイマー病、ダウン症候群、びまん性レヴィー小体病、ウェルニッケ・コルサコフ症候群、クロイツフェルト・ヤコブ病、亜急性硬化性全脳炎、ハラーフォルデン・シュパッツ(Hallerrorden−Spatz)病、ならびに拳闘家痴呆)が挙げられる)を防止するためまたは処置するために使用され得る。
【0123】
7.薬学的組成物
ある特定の実施形態において、本明細書中に記載される糖改変体融合タンパク質は、薬学的に許容され得るキャリアとともに処方される。例えば、本開示の融合タンパク質は、単独で、または薬学的製剤(治療用組成物)の構成要素として、投与され得る。主題の化合物は、ヒトまたは獣医学において使用するために好都合な任意の方法で投与用に処方され得る。適切な投薬レジメンが、主治医によって決定され得、その治療用タンパク質の改変されていないバージョンに対する標準的な投薬レジメンによって形成され得る。
【0124】
ある特定の実施形態において、本発明の治療方法は、本組成物を全身的に、または埋没物もしくはデバイスとして局所的に、投与する工程を包含する。投与されるとき、本発明において使用するための治療用組成物は、当然のことながら、発熱物質を含まない生理的に許容され得る形態である。上に記載されたような組成物に必要に応じて含められ得る、本明細書中に記載される融合タンパク質以外の治療上有用な薬剤は、本発明の方法において、その主題の融合タンパク質と同時にまたは連続的に投与され得る。
【0125】
代表的には、本明細書中に記載される融合タンパク質は、非経口的に(parentally)投与され得る。非経口投与に適した薬学的組成物は、1つ以上の薬学的に許容され得る滅菌された等張性の水性もしくは非水性の溶液、分散液、懸濁液もしくはエマルジョン、または滅菌された注射可能な溶液もしくは分散液に使用直前に再構成され得る滅菌された散剤とともに、1つ以上の本開示の融合タンパク質を含み得、その薬学的組成物は、酸化防止剤、緩衝剤、静菌剤、意図されるレシピエントの血液とその製剤を等張性にする溶質、または懸濁剤もしくは増粘剤を含み得る。本発明の薬学的組成物において使用され得る適当な水性および非水性のキャリアの例としては、水、エタノール、ポリオール類(例えば、グリセロール、プロピレングリコール、ポリエチレングリコールなど)およびそれらの適当な混合物、オリーブ油などの植物油、ならびにオレイン酸エチルなどの注射可能な有機エステルが挙げられる。例えば、レシチンなどのコーティング材料を使用することによって、分散液の場合は、必要な粒径を維持することによって、および界面活性物質を使用することによって、適切な流動性が維持され得る。
【0126】
さらに、本組成物は、標的組織部位(例えば、骨、筋肉、循環器系など)への送達用の形態として、被包され得るか、または注射され得る。ある特定の実施形態において、本発明の組成物は、1つ以上の治療用化合物(例えば、糖改変体融合タンパク質)を標的組織部位(例えば、骨、筋肉、循環器系)に送達することができる、発生中の組織に構造を提供することができる、および体内に最適に再吸収されることができる、マトリックスを含み得る。例えば、そのマトリックスは、本開示の融合タンパク質の緩効性を提供し得る。そのようなマトリックスは、他の埋め込まれる医学的適用のために現在使用されている材料から形成され得る。
【0127】
マトリックス材の選択は、生体適合性、生分解性、力学的特性、美容上の外観および境界面特性に基づく。主題の組成物の特定の適用法が、適切な製剤を規定し得る。本組成物に対する潜在的なマトリックスは、生分解性かつ化学的に定義された硫酸カルシウム、リン酸三カルシウム、ヒドロキシアパタイト、ポリ乳酸およびポリ無水物であり得る。他の潜在的な材料は、生分解性かつ生物学的に十分定義されているもの(例えば、骨または皮膚コラーゲン)である。さらなるマトリックスは、純粋なタンパク質または細胞外マトリックス構成要素を含む。他の潜在的なマトリックスは、生分解性でない化学的に定義されたもの(例えば、焼結ヒドロキシアパタイト、バイオガラス、アルミネートまたは他のセラミックス)である。マトリックスは、上で述べられたタイプの材料(例えば、ポリ乳酸およびヒドロキシアパタイトまたはコラーゲンおよびリン酸三カルシウム)のいずれかの組み合わせを含み得る。バイオセラミックスは、組成(例えば、カルシウム−アルミネート−ホスフェート)、ならびにポアサイズ、粒径、粒子の形状および生分解性を変化させる処理において変化され得る。
【0128】
ある特定の実施形態において、融合タンパク質は、経口投与のために、例えば、カプセル、カシェ剤、丸剤、錠剤、ロゼンジ(風味がつけられた基剤、通常、スクロースおよびアカシアまたはトラガントを用いる)、散剤、顆粒剤の形態で、または水性もしくは非水性の液体中の溶液もしくは懸濁液として、または水中油型もしくは油中水型の液体エマルジョンとして、またはエリキシル剤もしくはシロップ剤として、またはトローチ剤(不活性な基剤(例えば、ゼラチンおよびグリセリン、またはスクロースおよびアカシア)を用いる)として、および/または口腔洗浄薬などとして、処方され得、それらの各々は、活性成分として所定量の薬剤を含む。ある薬剤は、ボーラス、舐剤またはペーストとしても投与され得る。
【0129】
経口投与用の固体剤形(例えば、カプセル、錠剤、丸剤、糖衣錠および散剤顆粒剤)において、本発明の1つ以上の融合タンパク質は、1つ以上の薬学的に許容され得るキャリア(例えば、クエン酸ナトリウムまたはリン酸二カルシウム)、および/または以下のうちのいずれかと混合され得る:(1)充填剤または増量剤(例えば、デンプン、ラクトース、スクロース、グルコース、マンニトールおよび/またはケイ酸);(2)結合剤(例えば、カルボキシメチルセルロース、アルギネート、ゼラチン、ポリビニルピロリドン、スクロースおよび/またはアカシア);(3)吸湿剤(例えば、グリセロール);(4)崩壊剤(例えば、寒天、炭酸カルシウム、ジャガイモまたはタピオカのデンプン、アルギン酸、ある特定のシリケート、および炭酸ナトリウム);(5)溶解遅延剤(例えば、パラフィン);(6)吸収促進物質(例えば、四級アンモニウム化合物);(7)湿潤剤(例えば、セチルアルコールおよびモノステアリン酸グリセロール);(8)吸収剤(例えば、カオリンおよびベントナイト粘土)(9)潤滑剤(例えば、タルク、ステアリン酸カルシウム、ステアリン酸マグネシウム、固体ポリエチレングリコール、ラウリル硫酸ナトリウムおよびそれらの混合物);および(10)着色剤。カプセル、錠剤および丸剤の場合、薬学的組成物は、緩衝剤も含み得る。類似のタイプの固体組成物は、ラクトースまたは乳糖のような賦形剤、ならびに高分子量ポリエチレングリコールを用いて、軟および硬充填ゼラチンカプセルにおいて充填剤としても使用され得る。
【0130】
経口投与用の液体剤形には、薬学的に許容され得るエマルジョン、マイクロエマルジョン、溶液、懸濁液、シロップ剤およびエリキシル剤が含まれる。その液体剤形は、活性成分に加えて、当該分野において通常使用される不活性な希釈剤(例えば、水または他の溶媒)、可溶化剤および乳化剤(例えば、エチルアルコール、イソプロピルアルコール、炭酸エチル、酢酸エチル、ベンジルアルコール、安息香酸ベンジル、プロピレングリコール、1,3−ブチレングリコール、油(特に、綿実油、ラッカセイ油、トウモロコシ油、胚芽油、オリーブ油、ヒマシ油およびゴマ油)、グリセロール、テトラヒドロフリルアルコール、ポリエチレングリコールおよびソルビタンの脂肪酸エステル)、ならびにそれらの混合物を含み得る。経口用組成物は、不活性な希釈剤のほかにも、佐剤(例えば、湿潤剤、乳化剤および懸濁剤、甘味剤、香味剤、着色剤、芳香剤および保存剤)も含み得る。
【0131】
懸濁液は、融合タンパク質に加えて、懸濁剤(例えば、エトキシ化イソステアリルアルコール、ポリオキシエチレンソルビトールおよびソルビタンエステル、微結晶性セルロース、メタ水酸化アルミニウム、ベントナイト、寒天およびトラガント、ならびにそれらの混合物)を含み得る。
【0132】
本発明の組成物は、佐剤(例えば、保存剤、湿潤剤、乳化剤および分散剤)も含み得る。様々な抗菌剤および抗真菌剤、例えば、パラベン、クロロブタノール、フェノールソルビン酸などを含めることによって、微生物の作用の防止が確実にされ得る。等張剤(例えば、糖、塩化ナトリウムなど)を組成物に含めることも望ましい場合がある。さらに、注射可能な薬学的形態の長時間の吸収は、モノステアリン酸アルミニウムおよびゼラチンなどの吸収を遅延させる物質を含めることによってもたらされ得る。
【実施例】
【0133】
例示
本発明は、本明細書中で広く説明されてきたが、以下の実施例を参照することによってより容易に理解され得、その実施例は、ある特定の実施形態および本発明の実施形態を説明する目的のためだけに含められるものであって、本発明を限定することを目的としていない。
【0134】
実施例1:糖改変体TNFR2−Fc融合タンパク質の発現
本出願人は、様々なTNFR2−Fc融合タンパク質(各々がさらなるN結合型グリコシル化部位を含む)を発現するためのベクターを構築した。コントロールTNFR2−Fc融合タンパク質(すなわち、基礎的な、すなわち「最初の」改変されていない形態のTNFR2−Fc)は、介在リンカーなしでヒトIgG1 Fcドメインに融合されたヒトTNFR2の細胞外ドメインを有する。その最初のTNFR2−Fc融合タンパク質の配列を図1に示し(配列番号5)、コード核酸(リーダー配列を含む)を図5に示す(配列番号8)。図1に示される融合タンパク質は、TNFR2−h(1)Fcとさらに完全に命名される。TNFR2−h(2)Fcと命名された改変体は、図9(配列番号16)に示されるように代替リンカー/境界配列を有し、図10に示されるヌクレオチド配列(配列番号17)によってコードされる。タンパク質構築物を、哺乳動物細胞において発現させるためにpAID4ベクター(Pearsallら、PNAS 105(2008):7082−7087)内にクローニングした。
【0135】
最初の実験にむけて、TNFR2−Fcの糖改変体をHEK293細胞において一過性に発現させた。簡潔には、500mlスピナーにおいて、HEK293T細胞を、250mlの体積のFreestyle(Invitrogen)培地中で6×105細胞/mlに調整し、一晩生育した。翌日、これらの細胞を、0.5μg/mlの最終DNA濃度のDNA:PEI(1:1)複合体で処理した。4時間後、250mlの培地を加え、細胞を7日間生育した。その細胞を遠沈することによって培養上清を収集し、濃縮した。
【0136】
改変体を、様々な手法(例えば、プロテインAカラム(Mab Select(商標),GE Healthcare LifeSciences,USA)が挙げられる)を用いて精製し、低pH(3.0)グリシン緩衝液で溶出した後、サイズ排除クロマトグラフィーに供した。リン酸緩衝食塩水またはTris緩衝食塩水に対してタンパク質を透析した。
【0137】
図5に提供されるタンパク質配列は、「LPA・・・」のN末端の配列を示しているが、各タンパク質のかなりの部分が、そのN末端のロイシンが除去されることにより、「PAQ・・・」というN末端の配列がもたらされることを示した。したがって、本明細書中に開示されるいずれのTNFR2−Fc分子も、1つまたは2つのアミノ酸がN末端から除去されていることがある。
【0138】
使用するにあたり、3つの異なるリーダー配列が考えられた:
【0139】
【化8】
さらなる精製は、例えば、以下のうちの3つ以上を任意の順序で含む一連のカラムクロマトグラフィー工程によって達成され得る:Qセファロースクロマトグラフィー、フェニルセファロースクロマトグラフィー、ヒドロキシアパタイトクロマトグラフィーおよび陽イオン交換クロマトグラフィー。精製は、ウイルス濾過および緩衝液交換によって完了され得る。
【0140】
実施例2:糖改変体TNFR2−Fc融合タンパク質の設計
さらなるN結合型グリコシル化部位を導入するための位置を、TNFR2の細胞外ドメインとそのリガンドであるTNF(結晶座標は公的に入手可能)との共結晶構造の検討、および本明細書中に記載される原理の適用によって、選択した。半減期を延長しつつ、そのタンパク質のTNFアンタゴニスト活性を保存すると予測される位置におけるような変更を選択し、その変更を図7に示す。アミノ酸のナンバリングが、図7に示されるような天然のプロセシングされていないTNFR2アミノ酸配列に基づくことに注意されたい。以下の位置のうちの1つにさらなるN結合型糖成分を有するように、変更されたTNFR2−Fc融合タンパク質を設計した:47、48、62、114、155、202、203、222および253。天然のTNFR2−Fcは、171位および193位にN結合型糖成分を有すること(これは、最初のTNFR2−Fc融合タンパク質の異種(heterolgous)ドメインが、1:117.5というN結合型糖成分とアミノ酸との比を有することを意味する)に注意されたい。さらなる糖成分の予測される位置を、TNFR2−TNF共結晶構造上にマッピングし、これらを図6に示す。図8は、関連するタンパク質との配列比較および構造上の相同性によって推測される、TNFR2細胞外ドメインにおけるベータ鎖二次構造の予測される位置を示している。新しいN結合型部位の各々は、構造エレメントに対して内側に位置された。例えば、47位および48位に導入される部位は、第1のベータシート構造エレメントに対して内側である。単一のさらなるN結合型グリコシル化部位を有するタンパク質を作製し、試験した後、組み合わせ変異体を作製し、試験した。
【0141】
実施例3:TNFR2−Fc糖改変体タンパク質の試験
改変された分子の機能性、およびその分子の薬物動態学的特性に対する改変の影響を査定する一連のアッセイにおいて、TNFR2−h(1)Fcの糖改変体を試験した。
【0142】
無細胞生化学的アッセイにおいてリガンド結合性を査定するために、Biacore(登録商標)3000バイオセンサー装置を使用した。簡潔には、TNFR2−h(1)Fc改変体をフローセルに充填し、次いで、TNFに曝露した。動態パラメータ(kaおよびkd)を評価することによって、解離定数(KD)を算出した。
【0143】
リガンド阻害を査定するために、本質的にはKhabarら、Immunol.Lett.46(1995):107−110に記載されているようなTNFシグナル伝達についての細胞ベースアッセイを使用した。簡潔には、WEHI細胞(ATCC)を、TNF−アルファ(R&D Systems,Minneapolis,Minn.)およびアクチノマイシンDの存在下において培養した。TNF−アルファは、これらの細胞においてアポトーシスを引き起こし、溶解速度をA490nmにおいて検出した。TNF−アルファアンタゴニストが存在することにより、IC50の検出を可能にするシグナルが減少する。
【0144】
上記の分子の薬物動態学的特性を査定するために、標準的な手法を用いてSprague−Dawleyラットにおいて研究を行った。簡潔には、順化期間後に、TNFR2−h(1)Fc分子を動物に投与し、最初の投与の6、10、24、32、48、72、96、144、216および312時間後に尾に切り目を入れることによって出血させた。TNFR2−Fc分子の存在を、ヒトFc部分に対するELISAによって検出した。
【0145】
そのデータを、天然のTNFR2−h(1)Fc分子から得られたデータに対して正規化することにより、実験ごとのばらつきを最小にした。正規された値を下に示す。望ましい質を示すタンパク質を灰色かつ下線で示している。
【0146】
【表1】
*D47Nタンパク質は、インビボでの試験を可能にするのに十分な量で発現されなかった。
**N結合型糖を62位または114位に配置する変異によって、リガンド結合の完全な破壊が引き起こされたことから、他の特性を評価しなかった。
【0147】
上に提示されたデータは、野生型分子のリガンド結合および拮抗作用と類似のリガンド結合および拮抗作用を保持しつつ、単一のさらなるN結合型部位を含む改変体のほぼ半数(9つ中4つ)が、実質的に改善された血清半減期を示したことを示唆している。血清半減期に対する異種間物差し法の影響のため、ラットにおける20〜30%の血清半減期の変化は、ヒト患者に対して投薬レジメンの意味ある改善を提供するはずである。興味深いことに、組み合わせ変異は、一般に、血清半減期またはタンパク質活性を減少させるとみられたことから、この分子では、グリコシル化密度のさらなる増加が、もはや有益でなかったことが示唆される。
【0148】
これらのデータは、本明細書中に提供される教示を用いることにより、延長された血清半減期を有するFc融合分子を効率的に作製することができることを証明している。
【0149】
実施例4:薬物動態学的ラットアッセイ
糖改変体分子の薬物動態学的特性は、標準的な手法を用いてSprague−Dawleyラットにおいて測定され得る。簡潔には、順化期間後、糖改変体分子を8匹の動物の皮下に(SC)投与し、最初の投与の6、10、24、32、48、72、96、144、216および312時間後に尾に切り目を入れることによって出血させる。糖改変体分子の存在は、標的タンパク質に適した任意の方法を用いて検出され得る。例えば、糖改変体Fc融合タンパク質は、ヒトFc部分に対するELISAを用いて検出され得る。各個別の動物について、血清排出半減期(T1/2)を算出する。平均T1/2および標準偏差を糖改変体分子について算出する。同じ用量で投与される適切なコントロール分子(例えば、導入されるN結合型グリコシル化部位を含まないFc融合タンパク質)に対する平均T1/2および標準偏差がまだ決定されていない場合、それらを決定するために同様の研究が行われ得る。スチューデントのt検定などの適切な統計的検定を用いて、平均T1/2を解析する。糖改変体分子の平均T1/2は、適切なコントロール分子の平均T1/2と比較され得る。
【0150】
前述の方法に従って、ラットにおいてTNFR2−h(1)Fc、Q48N/A50S TNFR2−h(1)FcおよびQ48N/A50S TNFR2−h(2)Fcの薬物動態学的特性を比較する研究を行った。この研究では、5mg/kgという単回用量の皮下投与の28日後までタンパク質濃度が測定された(構築物1つあたりn=3匹のラット)。
【0151】
【表2】
TNFR2−h(1)Fcと比較したところ、糖改変体Q48N/A50S TNFR2−h(1)FcおよびQ48N/A50S TNFR2−h(2)Fcの血清半減期は、それぞれ50%および80%長いことが見出された。著しいことに、Q48N/A50S TNFR2−h(2)Fcに対する最高血清濃度(Cmax)は、Q48N/A50S TNFR2−h(1)Fcよりもほぼ5倍高かった(20.8μg/ml対4.4μg/ml,p<0.05)。これらの結果は、TNFR2−Fc糖改変体のリンカー/境界領域の改変(配列番号16と配列番号5とを比較のこと)によって、改善された薬物動態学的特性を有する改変体(この実施例におけるQ48N/A50S TNFR2−h(2)Fc)がもたらされ得ることを証明している。したがって、配列番号18として提供されるリンカー−hFc配列は、特に、本明細書中に開示される任意の糖改変体との組み合わせにおいて有用であり得る。
【0152】
実施例5:薬物動態学的サルアッセイ
糖改変体分子の薬物動態学的特性は、標準的な手法を用いてカニクイザルモデルにおいて行われ得る。簡潔には、4匹の雄および4匹の雌のカニクイザルを選択する。それらの動物は、この手順の当初は実験的にナイーブであり、およそ2〜6歳であり、少なくとも2.5kgの体重である。標準的な食餌を動物に与え、処置全体にわたってそれらの動物を標準的な手順に従って収容する。皮下(SC)投与または静脈内(IV)投与によって糖改変体分子を動物に投薬する。22日間にわたって、以下の投与後の時点において大腿静脈穿刺によって血液サンプルを収集する:5分後、15分後、30分後、1時間後、2時間後、4時間後、8時間後、12時間後、24時間後、34時間後、48時間後、58時間後、72時間後、120時間後、168時間後、240時間後、336時間後、408時間後および504時間後。投薬前の血液サンプルも収集する。血液を収集した後、そのサンプルを氷上に保管し、遠心分離によって血清を単離する。血清サンプル中の糖改変体分子の存在は、標的タンパク質に適した任意の方法を用いて検出され得る。例えば、糖改変体Fc融合タンパク質は、ヒトFc部分に対するELISAを用いて検出され得る。各個別の動物について、血清排出半減期(T1/2)を算出する。平均T1/2および標準偏差を糖改変体分子について算出する。同じ用量で投与される適切なコントロール分子(例えば、導入されるN結合型グリコシル化部位を含まないFc融合タンパク質)に対する平均T1/2および標準偏差がまだ決定されていない場合、それらを決定するために同様の研究が行われ得る。スチューデントのt検定などの適切な統計的検定を用いて、平均T1/2を解析する。糖改変体分子の平均T1/2は、適切なコントロール分子の平均T1/2と比較され得る。
【0153】
実施例6:ラットコラーゲン誘発関節炎モデルにおけるQ48N/A50S TNFR2−h(1)Fcの作用
糖改変体Q48N/A50S TNFR2−h(1)Fcを、ラットにおけるコラーゲン誘発関節炎に対する有効性について評価した。ウシコラーゲンII型(Elastin Products,カタログ番号CN276)を0.01M酢酸およびフロイント不完全アジュバント(Difco,カタログ番号263910)に1mg/mlの濃度に溶解し、研究の0および7日目に、雌Lewisラット(150〜200g)の尾の底部に皮内注射(2mg/kg)によって投与した。6日目から開始して、3mg/kgのQ48N/A50S TNFR2−h(1)Fc、3mg/kgのTNFR2−h(1)Fc、またはビヒクルでラットを1週間に3回皮下処置した。足の体積は、ベースラインおよび研究の全体にわたる複数の時点においてプレチスモメトリーによって測定したのに対し、骨質は、研究の終わりに(エキソビボで)マイクロコンピュータ断層撮影(マイクロ−CT)によって査定した。
【0154】
足の腫脹をコラーゲン誘発炎症に対するマーカーとして使用した。予想通り、ビヒクルで処置されたラットにおける足の体積は、研究の途中で増加し、終わりまで上昇したままだった。この足の腫脹は、ポジティブコントロールであるTNFR2−h(1)Fcで処置されたラットと同程度に、糖改変体Q48N/A50S TNFR2−h(1)Fcで処置されたラットにおいて効果的に阻害された(図11)。さらに、マイクロ−CTによって査定された骨質は、ビヒクルで処置されたコントロールと比べて、これらの両方の処置群において同様に改善されたことから(図12)、抗炎症性の有効性のさらなる証拠がもたらされた。これらのデータは、糖改変体Q48N/A50S TNFR2−h(1)Fcが、その置換されていない対応物であるTNFR2−h(1)Fcの抗炎症性の有効性と等価な抗炎症性の有効性をインビボにおいて有することを証明している。
【0155】
まとめると、前述の結果は、TNFR2−h(1)Fcのある特定の糖改変体、例えば、Q48N/A50S TNFR2−h(1)Fcが、TNFR2−h(1)Fc自体と比べて、増加された血清半減期および低下しない抗炎症性の有効性を有することを示唆している。
【0156】
参考としての援用
本明細書中で言及されるすべての刊行物および特許は、各個別の刊行物または特許が参考として援用されると明確かつ個別に示されたかのように、それらの全体が参考として援用される。
【0157】
主題の特定の実施形態を論じてきたが、上記の明細書は、例示的なものであって、限定的なものではない。この明細書および下記の請求項を検討すると、多くのバリエーションが当業者に明らかになるだろう。本発明の完全な範囲は、等価物の完全な範囲に加えて請求項、およびそのようなバリエーションに加えて本明細書を参照することにより決定されるべきである。
【特許請求の範囲】
【請求項1】
改変されたFc融合タンパク質をコードする改変された核酸を調製するための方法であって、該改変されたFc融合タンパク質は、最初のFc融合タンパク質と比べて延長された血清半減期を有し、該改変されたFc融合タンパク質および該最初のFc融合タンパク質の各々は、Fc部分および異種部分を含み、該方法は、該最初のFc融合タンパク質の該異種部分をコードする核酸を、1つ以上のさらなるN結合型グリコシル化部位をコードするように改変する工程を包含し、ここで、(a)該改変された核酸は、適当な細胞培養物において発現されるとき、薬物動態学的サルアッセイにおいて測定される該最初のFc融合タンパク質の血清半減期よりも少なくとも10%長い血清半減期を有する改変されたFc融合タンパク質をコードし、かつ(b)該改変されたFc融合タンパク質は、改変されていないFc融合タンパク質と比べて、実質的に同じかまたは高いインビボ生物学的活性を有する、方法。
【請求項2】
前記1つ以上のさらなるN結合型グリコシル化部位が、前記異種部分の1つ以上のアミノ酸配列に導入され、該アミノ酸配列が、表面に露出している、請求項1に記載の方法。
【請求項3】
前記1つ以上のさらなるN結合型グリコシル化部位が、前記異種部分の1つ以上のアミノ酸配列に導入され、該アミノ酸配列が、表面に露出しており、かつβシートまたはαヘリックスに組み込まれていない、請求項2に記載の方法。
【請求項4】
前記異種部分が、リガンド結合ドメインを含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
前記リガンド結合ドメインが、膜貫通レセプターの細胞外ドメインに由来する、請求項4に記載の方法。
【請求項6】
前記1つ以上のさらなるN結合型グリコシル化部位が、前記異種部分内の位置に導入され、ここで、任意のさらなる糖成分が前記リガンド結合ドメインを実質的に干渉しない、請求項4または5に記載の方法。
【請求項7】
前記1つ以上のさらなるN結合型グリコシル化部位が、前記リガンド結合ドメインのアミノ酸配列が改変されない位置に導入される、請求項4〜6のいずれか1項に記載の方法。
【請求項8】
前記1つ以上のさらなるN結合型グリコシル化部位は、該さらなるN結合型グリコシル化部位に付着される任意の糖成分がリガンド結合境界面を実質的に干渉しないと予測される位置に導入される、請求項4〜7のいずれか1項に記載の方法。
【請求項9】
前記改変されたFc融合タンパク質が、前記最初のFc融合タンパク質の最大半量阻害濃度(IC50)の1/2以下のIC50でリガンドに結合する、請求項4〜8のいずれか1項に記載の方法。
【請求項10】
前記最初のFc融合タンパク質が、少なくとも1つのグリコシル化部位が導入されるように、少なくとも1つのアミノ酸残基の付加または欠失によって改変される、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記最初のFc融合タンパク質が、少なくとも1つのグリコシル化部位が導入されるように、少なくとも1つのアミノ酸残基の置換によって改変される、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記最初のFc融合タンパク質の前記異種部分が、少なくとも2つの構造的に異なるαヘリックスドメインおよび/またはβシートドメインを含み、該ドメインは、表面に露出している非構造ポリペプチド領域によって接続されている、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
さらなるN結合型グリコシル化部位が、前記非構造ペプチド領域内に位置する、請求項12に記載の方法。
【請求項14】
前記最初のFc融合タンパク質の前記異種部分が、各90アミノ酸あたり1つより少ないN結合型グリコシル化部位を含む、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
前記最初のFc融合タンパク質の前記異種部分が、各125アミノ酸あたり1つより少ないN結合型グリコシル化部位を含む、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
前記改変されたFc融合タンパク質の前記異種部分が、各90アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
前記改変されたFc融合タンパク質の前記異種部分が、各65アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む、請求項1〜16のいずれか1項に記載の方法。
【請求項18】
前記改変されたFc融合タンパク質の前記異種部分の各アミノ酸であって、N結合型グリコシル化が付着している、各アミノ酸は、N結合型グリコシル化によって改変された他の任意のアミノ酸と少なくとも20アミノ酸離れている、請求項1〜17のいずれか1項に記載の方法。
【請求項19】
前記最初のFc融合タンパク質が、前記Fc部分と前記異種部分との間にポリペプチドリンカーをさらに含み、該ポリペプチドリンカーがN結合型グリコシル化部位を含まない、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
前記改変されたFc融合タンパク質が、前記リンカー内に位置するN結合型グリコシル化部位を含む、請求項1〜18のいずれか1項に記載の方法。
【請求項21】
前記異種部分がTNFR2レセプターの細胞外ドメインを含み、該細胞外ドメインがリガンド結合ドメインを含む、請求項1〜20のいずれか1項に記載の方法。
【請求項22】
前記TNFR2レセプターの前記細胞外ドメインが、配列番号2と少なくとも90%同一であるアミノ酸配列を含む、請求項21に記載の方法。
【請求項23】
改変されたFc融合タンパク質を調製するための方法であって、該改変されたFc融合タンパク質は、最初のFc融合タンパク質と比べて延長された血清半減期を有し、該方法は:
(a)請求項1から22のいずれかに記載の方法に従って調製される改変された核酸を、哺乳動物または哺乳動物様のグリコシル化を提供する細胞培養物において発現させる工程;および
(b)該細胞培養物から該改変されたFc融合タンパク質を収集する工程
を包含する、方法。
【請求項24】
前記改変されたFc融合タンパク質を精製する工程をさらに包含する、請求項23に記載の方法。
【請求項25】
前記改変されたFc融合タンパク質を精製する工程が、該改変されたFc融合タンパク質をプロテインAに曝露する工程、および該プロテインAに結合した該改変されたFc融合タンパク質を収集する工程を包含する、請求項24に記載の方法。
【請求項26】
患者に投与するために、前記改変されたFc融合タンパク質を処方する工程をさらに包含する、請求項23〜25のいずれか1項に記載の方法。
【請求項27】
前記改変された核酸が、哺乳動物細胞株によって発現される、請求項23に記載の方法。
【請求項28】
前記哺乳動物細胞株が、CHO細胞株、NSO細胞株、COS細胞株およびHEK236細胞株からなる群より選択される、請求項27に記載の方法。
【請求項29】
前記改変された核酸が、シアル酸を含むN結合型糖成分を産生する細胞株において発現される、請求項23、27または28のいずれか1項に記載の方法。
【請求項30】
前記改変された核酸が、哺乳動物または哺乳動物様のグリコシル化を提供するように操作された非哺乳動物細胞によって発現される、請求項23または29に記載の方法。
【請求項31】
前記細胞株が、真菌細胞、昆虫細胞または植物細胞に由来する、請求項30に記載の方法。
【請求項32】
請求項1〜22のいずれかに記載の方法に従って調製される改変された核酸を含む、細胞株。
【請求項33】
請求項23〜32のいずれか1項に記載の方法に従って調製される改変されたFc融合タンパク質。
【請求項34】
免疫グロブリンFcドメインおよび少なくとも1つの異種ポリペプチドドメインを含む融合タンパク質であって、ここで、該融合タンパク質は、少なくとも1つの非内因性N結合型グリコシル化部位が導入されるように、該免疫グロブリンFcドメインの外側で改変されており、ここで、該導入された1つ以上のグリコシル化部位におけるグリコシル化によって、薬物動態学的サルアッセイにおいて測定される該改変された融合タンパク質の血清半減期が、導入されたグリコシル化部位を欠く融合タンパク質の血清半減期と比べて少なくとも10%増加する、融合タンパク質。
【請求項35】
前記異種ポリペプチドドメインが、レセプターの細胞外ドメインの少なくとも一部分を含み、該一部分が、リガンド結合ドメインを含む、請求項34に記載の融合タンパク質。
【請求項36】
前記グリコシル化部位が、リガンド結合ポケットの外側の前記細胞外ドメインに導入される、請求項35に記載の融合タンパク質。
【請求項37】
前記導入された1つ以上のグリコシル化部位におけるグリコシル化が、前記レセプターのリガンド結合活性に3倍を超えて影響しない、請求項35または36に記載の融合タンパク質。
【請求項38】
前記融合タンパク質が、少なくとも1つのグリコシル化部位が導入されるように、少なくとも1つのアミノ酸残基の付加または欠失によって改変されている、請求項34〜37のいずれか1項に記載の融合タンパク質。
【請求項39】
前記融合タンパク質が、少なくとも1つのグリコシル化部位が導入されるように、少なくとも1つのアミノ酸残基の置換によって改変されている、請求項34〜38のいずれか1項に記載の融合タンパク質。
【請求項40】
前記異種ポリペプチドドメインの分子量が、少なくとも25kDaである、請求項34〜39のいずれか1項に記載の融合タンパク質。
【請求項41】
前記導入された1つ以上のグリコシル化部位におけるグリコシル化によって、前記融合タンパク質の血清半減期が、導入されたグリコシル化部位を欠く融合タンパク質の血清半減期と比べて少なくとも20%増加する、請求項34〜40のいずれか1項に記載の融合タンパク質。
【請求項42】
改変されていない異種ポリペプチドドメインが、各90アミノ酸あたり1つより少ないN結合型グリコシル化部位を含む、請求項34〜41のいずれか1項に記載の融合タンパク質。
【請求項43】
改変されていない異種ポリペプチドドメインが、各125アミノ酸あたり1つより少ないN結合型グリコシル化部位を含む、請求項34〜41のいずれか1項に記載の融合タンパク質。
【請求項44】
前記改変された異種ポリペプチドが、各90アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む、請求項34〜41のいずれか1項に記載の融合タンパク質。
【請求項45】
前記改変された異種ポリペプチドが、各65アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む、請求項34〜41のいずれか1項に記載の融合タンパク質。
【請求項46】
N結合型グリコシル化によって改変された各アミノ酸が、N結合型グリコシル化によって改変された他の任意のアミノ酸と少なくとも20アミノ酸離れている、請求項34〜45のいずれか1項に記載の融合タンパク質。
【請求項47】
前記融合タンパク質が、前記Fcドメインと前記異種ポリペプチドドメインとの間にポリペプチドリンカーをさらに含む、請求項34〜46のいずれか1項に記載の融合タンパク質。
【請求項48】
前記導入されたグリコシル化部位の少なくとも1つが、前記リンカー内に配置されている、請求項47に記載の融合タンパク質。
【請求項49】
前記Fc融合タンパク質の前記異種部分が、少なくとも2つの構造的に異なるαヘリックスドメインおよび/またはβシートドメインを含み、該ドメインは、表面に露出した非構造ポリペプチド領域によって接続されている、請求項34〜48のいずれか1項に記載の融合タンパク質。
【請求項50】
導入されるグリコシル化部位が、前記非構造ペプチド領域内に位置する、請求項49に記載の融合タンパク質。
【請求項51】
前記レセプターが、神経成長因子/腫瘍壊死因子レセプターファミリーのメンバーである、請求項35〜50に記載の融合タンパク質。
【請求項52】
前記レセプターが、TNFR2レセプターである、請求項51に記載の融合タンパク質。
【請求項53】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のQ26、D25、A28、E133またはG231から選択される1つ以上のアミノ酸残基においてアミノ酸置換を含む、請求項52に記載の融合タンパク質。
【請求項54】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸26におけるQからNへの置換、および配列番号2のアミノ酸28におけるAからSへの置換を含む、請求項53に記載の融合タンパク質。
【請求項55】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸25におけるDからNへの置換、および配列番号2のアミノ酸133におけるEからNへの置換を含む、請求項53に記載の融合タンパク質。
【請求項56】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸25におけるDからNへの置換、および配列番号2のアミノ酸231におけるGからNへの置換を含む、請求項53に記載の融合タンパク質。
【請求項57】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸配列と少なくとも90%同一である、請求項53〜56のいずれか1項に記載の融合タンパク質。
【請求項58】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸配列と少なくとも95%同一である、請求項57に記載の融合タンパク質。
【請求項59】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸配列と少なくとも99%同一である、請求項58に記載の融合タンパク質。
【請求項60】
前記融合タンパク質が:
a)配列番号5のアミノ酸13〜257を含むポリペプチド;
b)配列番号6のアミノ酸13〜257を含むポリペプチド;
c)配列番号7のアミノ酸13〜257を含むポリペプチド;および
d)配列番号8のアミノ酸13〜257を含むポリペプチド
から選択される異種ドメインを含む、請求項53に記載の融合タンパク質。
【請求項61】
前記融合タンパク質が、リンカーおよびFc部分を含み、該リンカーおよびFc部分が、配列番号18のアミノ酸配列を有する、請求項60に記載の融合タンパク質。
【請求項62】
請求項34〜61のいずれか1項に記載の融合タンパク質および薬学的に許容され得るキャリアを含む薬学的調製物であって、該調製物は、哺乳動物への投与に適するように、発熱性材料を実質的に含まない、薬学的調製物。
【請求項1】
改変されたFc融合タンパク質をコードする改変された核酸を調製するための方法であって、該改変されたFc融合タンパク質は、最初のFc融合タンパク質と比べて延長された血清半減期を有し、該改変されたFc融合タンパク質および該最初のFc融合タンパク質の各々は、Fc部分および異種部分を含み、該方法は、該最初のFc融合タンパク質の該異種部分をコードする核酸を、1つ以上のさらなるN結合型グリコシル化部位をコードするように改変する工程を包含し、ここで、(a)該改変された核酸は、適当な細胞培養物において発現されるとき、薬物動態学的サルアッセイにおいて測定される該最初のFc融合タンパク質の血清半減期よりも少なくとも10%長い血清半減期を有する改変されたFc融合タンパク質をコードし、かつ(b)該改変されたFc融合タンパク質は、改変されていないFc融合タンパク質と比べて、実質的に同じかまたは高いインビボ生物学的活性を有する、方法。
【請求項2】
前記1つ以上のさらなるN結合型グリコシル化部位が、前記異種部分の1つ以上のアミノ酸配列に導入され、該アミノ酸配列が、表面に露出している、請求項1に記載の方法。
【請求項3】
前記1つ以上のさらなるN結合型グリコシル化部位が、前記異種部分の1つ以上のアミノ酸配列に導入され、該アミノ酸配列が、表面に露出しており、かつβシートまたはαヘリックスに組み込まれていない、請求項2に記載の方法。
【請求項4】
前記異種部分が、リガンド結合ドメインを含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
前記リガンド結合ドメインが、膜貫通レセプターの細胞外ドメインに由来する、請求項4に記載の方法。
【請求項6】
前記1つ以上のさらなるN結合型グリコシル化部位が、前記異種部分内の位置に導入され、ここで、任意のさらなる糖成分が前記リガンド結合ドメインを実質的に干渉しない、請求項4または5に記載の方法。
【請求項7】
前記1つ以上のさらなるN結合型グリコシル化部位が、前記リガンド結合ドメインのアミノ酸配列が改変されない位置に導入される、請求項4〜6のいずれか1項に記載の方法。
【請求項8】
前記1つ以上のさらなるN結合型グリコシル化部位は、該さらなるN結合型グリコシル化部位に付着される任意の糖成分がリガンド結合境界面を実質的に干渉しないと予測される位置に導入される、請求項4〜7のいずれか1項に記載の方法。
【請求項9】
前記改変されたFc融合タンパク質が、前記最初のFc融合タンパク質の最大半量阻害濃度(IC50)の1/2以下のIC50でリガンドに結合する、請求項4〜8のいずれか1項に記載の方法。
【請求項10】
前記最初のFc融合タンパク質が、少なくとも1つのグリコシル化部位が導入されるように、少なくとも1つのアミノ酸残基の付加または欠失によって改変される、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記最初のFc融合タンパク質が、少なくとも1つのグリコシル化部位が導入されるように、少なくとも1つのアミノ酸残基の置換によって改変される、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記最初のFc融合タンパク質の前記異種部分が、少なくとも2つの構造的に異なるαヘリックスドメインおよび/またはβシートドメインを含み、該ドメインは、表面に露出している非構造ポリペプチド領域によって接続されている、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
さらなるN結合型グリコシル化部位が、前記非構造ペプチド領域内に位置する、請求項12に記載の方法。
【請求項14】
前記最初のFc融合タンパク質の前記異種部分が、各90アミノ酸あたり1つより少ないN結合型グリコシル化部位を含む、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
前記最初のFc融合タンパク質の前記異種部分が、各125アミノ酸あたり1つより少ないN結合型グリコシル化部位を含む、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
前記改変されたFc融合タンパク質の前記異種部分が、各90アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
前記改変されたFc融合タンパク質の前記異種部分が、各65アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む、請求項1〜16のいずれか1項に記載の方法。
【請求項18】
前記改変されたFc融合タンパク質の前記異種部分の各アミノ酸であって、N結合型グリコシル化が付着している、各アミノ酸は、N結合型グリコシル化によって改変された他の任意のアミノ酸と少なくとも20アミノ酸離れている、請求項1〜17のいずれか1項に記載の方法。
【請求項19】
前記最初のFc融合タンパク質が、前記Fc部分と前記異種部分との間にポリペプチドリンカーをさらに含み、該ポリペプチドリンカーがN結合型グリコシル化部位を含まない、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
前記改変されたFc融合タンパク質が、前記リンカー内に位置するN結合型グリコシル化部位を含む、請求項1〜18のいずれか1項に記載の方法。
【請求項21】
前記異種部分がTNFR2レセプターの細胞外ドメインを含み、該細胞外ドメインがリガンド結合ドメインを含む、請求項1〜20のいずれか1項に記載の方法。
【請求項22】
前記TNFR2レセプターの前記細胞外ドメインが、配列番号2と少なくとも90%同一であるアミノ酸配列を含む、請求項21に記載の方法。
【請求項23】
改変されたFc融合タンパク質を調製するための方法であって、該改変されたFc融合タンパク質は、最初のFc融合タンパク質と比べて延長された血清半減期を有し、該方法は:
(a)請求項1から22のいずれかに記載の方法に従って調製される改変された核酸を、哺乳動物または哺乳動物様のグリコシル化を提供する細胞培養物において発現させる工程;および
(b)該細胞培養物から該改変されたFc融合タンパク質を収集する工程
を包含する、方法。
【請求項24】
前記改変されたFc融合タンパク質を精製する工程をさらに包含する、請求項23に記載の方法。
【請求項25】
前記改変されたFc融合タンパク質を精製する工程が、該改変されたFc融合タンパク質をプロテインAに曝露する工程、および該プロテインAに結合した該改変されたFc融合タンパク質を収集する工程を包含する、請求項24に記載の方法。
【請求項26】
患者に投与するために、前記改変されたFc融合タンパク質を処方する工程をさらに包含する、請求項23〜25のいずれか1項に記載の方法。
【請求項27】
前記改変された核酸が、哺乳動物細胞株によって発現される、請求項23に記載の方法。
【請求項28】
前記哺乳動物細胞株が、CHO細胞株、NSO細胞株、COS細胞株およびHEK236細胞株からなる群より選択される、請求項27に記載の方法。
【請求項29】
前記改変された核酸が、シアル酸を含むN結合型糖成分を産生する細胞株において発現される、請求項23、27または28のいずれか1項に記載の方法。
【請求項30】
前記改変された核酸が、哺乳動物または哺乳動物様のグリコシル化を提供するように操作された非哺乳動物細胞によって発現される、請求項23または29に記載の方法。
【請求項31】
前記細胞株が、真菌細胞、昆虫細胞または植物細胞に由来する、請求項30に記載の方法。
【請求項32】
請求項1〜22のいずれかに記載の方法に従って調製される改変された核酸を含む、細胞株。
【請求項33】
請求項23〜32のいずれか1項に記載の方法に従って調製される改変されたFc融合タンパク質。
【請求項34】
免疫グロブリンFcドメインおよび少なくとも1つの異種ポリペプチドドメインを含む融合タンパク質であって、ここで、該融合タンパク質は、少なくとも1つの非内因性N結合型グリコシル化部位が導入されるように、該免疫グロブリンFcドメインの外側で改変されており、ここで、該導入された1つ以上のグリコシル化部位におけるグリコシル化によって、薬物動態学的サルアッセイにおいて測定される該改変された融合タンパク質の血清半減期が、導入されたグリコシル化部位を欠く融合タンパク質の血清半減期と比べて少なくとも10%増加する、融合タンパク質。
【請求項35】
前記異種ポリペプチドドメインが、レセプターの細胞外ドメインの少なくとも一部分を含み、該一部分が、リガンド結合ドメインを含む、請求項34に記載の融合タンパク質。
【請求項36】
前記グリコシル化部位が、リガンド結合ポケットの外側の前記細胞外ドメインに導入される、請求項35に記載の融合タンパク質。
【請求項37】
前記導入された1つ以上のグリコシル化部位におけるグリコシル化が、前記レセプターのリガンド結合活性に3倍を超えて影響しない、請求項35または36に記載の融合タンパク質。
【請求項38】
前記融合タンパク質が、少なくとも1つのグリコシル化部位が導入されるように、少なくとも1つのアミノ酸残基の付加または欠失によって改変されている、請求項34〜37のいずれか1項に記載の融合タンパク質。
【請求項39】
前記融合タンパク質が、少なくとも1つのグリコシル化部位が導入されるように、少なくとも1つのアミノ酸残基の置換によって改変されている、請求項34〜38のいずれか1項に記載の融合タンパク質。
【請求項40】
前記異種ポリペプチドドメインの分子量が、少なくとも25kDaである、請求項34〜39のいずれか1項に記載の融合タンパク質。
【請求項41】
前記導入された1つ以上のグリコシル化部位におけるグリコシル化によって、前記融合タンパク質の血清半減期が、導入されたグリコシル化部位を欠く融合タンパク質の血清半減期と比べて少なくとも20%増加する、請求項34〜40のいずれか1項に記載の融合タンパク質。
【請求項42】
改変されていない異種ポリペプチドドメインが、各90アミノ酸あたり1つより少ないN結合型グリコシル化部位を含む、請求項34〜41のいずれか1項に記載の融合タンパク質。
【請求項43】
改変されていない異種ポリペプチドドメインが、各125アミノ酸あたり1つより少ないN結合型グリコシル化部位を含む、請求項34〜41のいずれか1項に記載の融合タンパク質。
【請求項44】
前記改変された異種ポリペプチドが、各90アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む、請求項34〜41のいずれか1項に記載の融合タンパク質。
【請求項45】
前記改変された異種ポリペプチドが、各65アミノ酸あたり少なくとも1つのN結合型グリコシル化部位を含む、請求項34〜41のいずれか1項に記載の融合タンパク質。
【請求項46】
N結合型グリコシル化によって改変された各アミノ酸が、N結合型グリコシル化によって改変された他の任意のアミノ酸と少なくとも20アミノ酸離れている、請求項34〜45のいずれか1項に記載の融合タンパク質。
【請求項47】
前記融合タンパク質が、前記Fcドメインと前記異種ポリペプチドドメインとの間にポリペプチドリンカーをさらに含む、請求項34〜46のいずれか1項に記載の融合タンパク質。
【請求項48】
前記導入されたグリコシル化部位の少なくとも1つが、前記リンカー内に配置されている、請求項47に記載の融合タンパク質。
【請求項49】
前記Fc融合タンパク質の前記異種部分が、少なくとも2つの構造的に異なるαヘリックスドメインおよび/またはβシートドメインを含み、該ドメインは、表面に露出した非構造ポリペプチド領域によって接続されている、請求項34〜48のいずれか1項に記載の融合タンパク質。
【請求項50】
導入されるグリコシル化部位が、前記非構造ペプチド領域内に位置する、請求項49に記載の融合タンパク質。
【請求項51】
前記レセプターが、神経成長因子/腫瘍壊死因子レセプターファミリーのメンバーである、請求項35〜50に記載の融合タンパク質。
【請求項52】
前記レセプターが、TNFR2レセプターである、請求項51に記載の融合タンパク質。
【請求項53】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のQ26、D25、A28、E133またはG231から選択される1つ以上のアミノ酸残基においてアミノ酸置換を含む、請求項52に記載の融合タンパク質。
【請求項54】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸26におけるQからNへの置換、および配列番号2のアミノ酸28におけるAからSへの置換を含む、請求項53に記載の融合タンパク質。
【請求項55】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸25におけるDからNへの置換、および配列番号2のアミノ酸133におけるEからNへの置換を含む、請求項53に記載の融合タンパク質。
【請求項56】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸25におけるDからNへの置換、および配列番号2のアミノ酸231におけるGからNへの置換を含む、請求項53に記載の融合タンパク質。
【請求項57】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸配列と少なくとも90%同一である、請求項53〜56のいずれか1項に記載の融合タンパク質。
【請求項58】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸配列と少なくとも95%同一である、請求項57に記載の融合タンパク質。
【請求項59】
前記TNFR2レセプターの細胞外ドメインが、配列番号2のアミノ酸配列と少なくとも99%同一である、請求項58に記載の融合タンパク質。
【請求項60】
前記融合タンパク質が:
a)配列番号5のアミノ酸13〜257を含むポリペプチド;
b)配列番号6のアミノ酸13〜257を含むポリペプチド;
c)配列番号7のアミノ酸13〜257を含むポリペプチド;および
d)配列番号8のアミノ酸13〜257を含むポリペプチド
から選択される異種ドメインを含む、請求項53に記載の融合タンパク質。
【請求項61】
前記融合タンパク質が、リンカーおよびFc部分を含み、該リンカーおよびFc部分が、配列番号18のアミノ酸配列を有する、請求項60に記載の融合タンパク質。
【請求項62】
請求項34〜61のいずれか1項に記載の融合タンパク質および薬学的に許容され得るキャリアを含む薬学的調製物であって、該調製物は、哺乳動物への投与に適するように、発熱性材料を実質的に含まない、薬学的調製物。
【図1】


【図2】

【図3】

【図4】

【図5】

【図6】
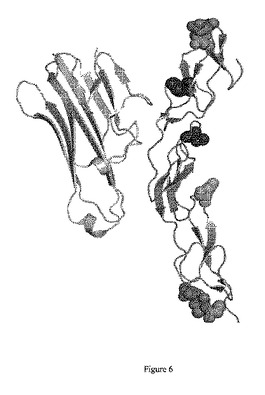
【図7】

【図8】

【図9】

【図10】
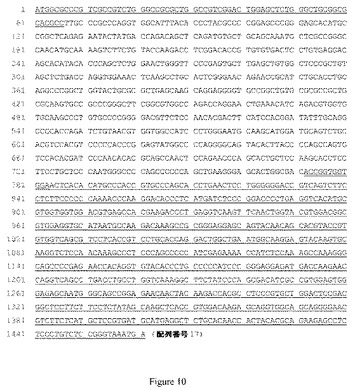
【図11】
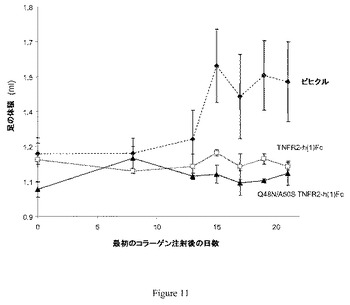
【図12】
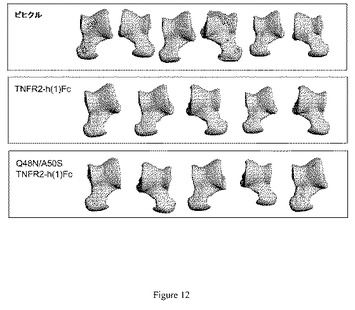
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2013−512674(P2013−512674A)
【公表日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2012−542194(P2012−542194)
【出願日】平成22年12月2日(2010.12.2)
【国際出願番号】PCT/US2010/058765
【国際公開番号】WO2011/068993
【国際公開日】平成23年6月9日(2011.6.9)
【出願人】(509125475)アクセルロン ファーマ, インコーポレイテッド (21)
【Fターム(参考)】
【公表日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成22年12月2日(2010.12.2)
【国際出願番号】PCT/US2010/058765
【国際公開番号】WO2011/068993
【国際公開日】平成23年6月9日(2011.6.9)
【出願人】(509125475)アクセルロン ファーマ, インコーポレイテッド (21)
【Fターム(参考)】
[ Back to top ]