
G−CSF類似体組成物及び方法
【課題】顆粒球コロニー刺激因子(G−CSF)類似体、該類似体を含む組成物及び関連組成物の提供。
【解決手段】第1の化学的部分を介して第2の化学的部分が間接的に結合した、特定な配列からなるアミノ酸配列を含む改変されたG−CSFポリペプチドを製造する方法であって、(a)in vitroで第1の化学的部分を外部ループに結合させる工程および、(b)第1の化学的部分を介して第2の化学的部分を間接的に外部ループに結合させる工程を含み、当該第1の化学的部分が多糖であり、第2の化学的部分がポリエチレングリコールであり、特定な配列からなるアミノ酸配列のN末端メチオニンが存在し、または存在しておらず、当該改変G−CSFがG−CSF受容体に結合する生物活性を維持している前記方法。
【解決手段】第1の化学的部分を介して第2の化学的部分が間接的に結合した、特定な配列からなるアミノ酸配列を含む改変されたG−CSFポリペプチドを製造する方法であって、(a)in vitroで第1の化学的部分を外部ループに結合させる工程および、(b)第1の化学的部分を介して第2の化学的部分を間接的に外部ループに結合させる工程を含み、当該第1の化学的部分が多糖であり、第2の化学的部分がポリエチレングリコールであり、特定な配列からなるアミノ酸配列のN末端メチオニンが存在し、または存在しておらず、当該改変G−CSFがG−CSF受容体に結合する生物活性を維持している前記方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は顆粒球コロニー刺激因子(“G−CSF”)類似体、前記類似体を含む組成物、及び関連組成物に関する。別の態様によると、本発明は本発明の類似体をコードする核酸又は関連核酸、関連宿主細胞及びベクターに関する。別の態様によると、本発明はG−CSF及びその類似体の三次元構造を表現するためのコンピュータープログラム及び装置に関する。別の態様によると、本発明はG−CSF類似体及び関連組成物を合理的に設計するための方法に関する。更に別の態様によると、本発明は本発明のG−CSF類似体を使用する治療方法に関する。
【背景技術】
【0002】
造血は、骨髄微小環境内の細胞と成長因子との2つの系により制御される。成長因子はコロニー刺激因子とも呼称され、前駆細胞を刺激して増殖させ、分化性血球のコロニーを形成する。本明細書中でG−CSFと呼称する顆粒球コロニー刺激因子はこれらの因子の1種であり、好中球の成長及び発生を優先的に刺激するので、好中球減少症に使用できると予想される。Welteら,PNAS−USA 82: 1526−1530(1985); Souzaら,Science 232: 61−65(1986)及びGabrilove,J.Seminars in Hematology 26: (2)1−14(1989)。
【0003】
ヒトでは血漿中に内因性G−CSFが検出可能である。Jonesら,Bailliere’s Clinical Hematology 2(1): 83−111(1989)。G−CSFは繊維芽細胞、マクロファージ、T細胞トロホブラスト、内皮細胞及び上皮細胞により産生され、17番染色体に位置する4個のエキソンと5個のイントロンから構成される単一コピー遺伝子の発現産物である。この遺伝子座の転写により生成されるmRNA種は示差的プロセッシングを受け、177アミノ酸のタンパク質をコードする形態と174アミノ酸のタンパク質をコードする形態との2形態のG−CSF mRNAをもたらし(Nagataら,EMBO J 5:575−581(1986))、174アミノ酸からなる形態は最大のin vivo生物比活性を有することが判明した。G−CSFは種交差反応性であるため、ヒトG−CSFを例えばマウス、イヌ又はサル等の別の哺乳動物に投与すると、持続的な好中球白血球増加症が誘発される。Mooreら,PNAS−USA 84: 7134−7138(1987)。
【0004】
ヒトG−CSFは多数の起源から取得及び精製できる。天然ヒトG−CSF(nhG−CSF)は培養ヒト腫瘍細胞系の上清から単離することができる。組換えDNA技術の発達(例えば参考資料として本明細書の一部とする米国特許第4,810,643号(Souza)参照)により、真核宿主細胞発現産物としてグリコシル化形態のG−CSF及び原核宿主細胞発現産物として非グリコシル化形態のG−CSFを商業規模の量で生産することが可能になった。
【0005】
G−CSFは、好中球の増加が有益であるような症状の治療に有用であることが判明した。例えば癌患者にとってG−CSFは、化学療法又は放射線療法に起因する造血欠損を補償するために好中球産生を選択的に刺激する手段として有益である。この他に、細菌代謝物により主に誘発される種々の感染症及び関連症状(例えば敗血症)の治療にも使用できる。更に例えば骨髄移植の目的で培養細胞を成長又は増殖させるために、G−CSFを単独使用又は他の化合物(例えば他のサイトカイン)と併用しても有用である。
【0006】
G−CSFの細胞代謝過程であるシグナル変換は現在、十分に解明されていない。個々の前駆細胞内に変化を誘発すると思われる細胞表面レセプターにG−CSFが結合し、細胞分化に至ると考えられる。
【0007】
種々の改変G−CSFが報告されている。一般に、薬剤を設計するためには、ある種の構造効果を及ぼすように特定の改変を加えることが知られている。例えば、1個のシステインを欠失させると、未改変状態では通常はジスルフィド結合を介して折り畳まれている分子が解放される。この他に、タンパク質の機能を変えるために、アミノ酸を付加、欠失又は置換させる方法も知られている。
【0008】
組換えヒトG−CSF突然変異体は製造されているが、その製造方法は全体構造/機能関係情報を考慮していない。例えば、Cys18の突然変異及び生化学的修飾が報告されている。Kugaら,Biochem.Biophy.Res.Comm 159: 103−111(1989); Luら,Arch.Biochem.Biophys.268: 81−92(1989)。
【0009】
(上記)米国特許第4,810,643号(発明の名称「多能性顆粒球コロニー刺激因子の製造」)には、G−CSFのポリペプチド類似体及びペプチドフラグメントが広く開示されている。特定のG−CSF類似体としては、(174アミノ酸種又はこれにN末端メチオニンを付加した175アミノ酸種の)17、36、42、64及び74位のシステインを別のアミノ酸(例えばセリン)で置換した類似体や、最初の(N末端)位置にアラニンを有するG−CSFが開示されている。
【0010】
ヨーロッパ特許第0335423号(発明の名称「改変ヒトG−CSF」)は、hG−CSF活性を有するポリペプチドにおける少なくとも1個のアミノ酸の修飾を開示しているらしい。
【0011】
ヨーロッパ特許第0272703号(発明の名称「新規ポリペプチド」)は、N末端又はその「近傍」のアミノ酸を置換又は欠失させたG−CSF誘導体を開示しているらしい。
【0012】
ヨーロッパ特許第0459630号(発明の名称「ポリペプチド」)は、天然に存在するG−CSFの生物学的特性の少なくとも1種と5mg/mlで少なくとも35%の溶液安定性とを有する天然に存在するG−CSFの誘導体を開示している模様であり、該誘導体は、少なくとも天然配列のCys17がSer17残基で置換され、天然配列のAsp27がSer27残基で置換されている。
【0013】
ヨーロッパ特許第0256843号(発明の名称「G−CSFとそのムテインの発現及びその使用」)は、タンパク質のアミノ酸配列を変えずに組換え宿主におけるタンパク質の発現を強化するようにN末端を修飾した、G−CSFをコードする改変DNA配列を開示しているらしい。
【0014】
ヨーロッパ特許第0243153号(発明の名称「ヒトG−CSFタンパク質発現」)は、少なくとも1個の酵母KEX2プロテアーゼプロセッシング部位を不活性化することによりG−CSFを修飾すると、酵母を使用した組換え生産収率を増加できると開示しているらしい。
【0015】
Shawの米国特許第4,904,584号(発明の名称「ポリペプチドの部位特異的均質改変」)はリジン改変タンパク質を開示しているらしい。
【0016】
WO/9012874号は、タンパク質のシステイン改変変異体を開示しているらしい。
【0017】
オーストラリア特許出願第AU−A−10948/92号(発明の名称「組換えタンパク質の活性化の改善」)は、原核発現後の分子の折り畳みの助長を目的としてG−CSF分子の各末端にアミノ酸を付加することを開示しているらしい。
【0018】
オーストラリア特許出願第AU−A−76380/91号(発明の名称「顆粒球コロニー刺激因子(G−CSF)のムテイン」)は、174アミノ酸を有するG−CSFの50〜56位及び177アミノ酸を有するG−CSFの53〜59位の配列Leu−Gly−His−Ser−Leu−Gly−Ile、及び/又は174アミノ酸を有する成熟G−CSFの43、79、156及び170位もしくは177アミノ酸を有する成熟G−CSFの46、82、159及び173位の4個のヒスチジン残基の少なくとも1個における顆粒球刺激因子G−CSFのムテインを開示しているらしい。
【0019】
英国特許第2213821号(発明の名称「合成ヒト顆粒球コロニー刺激因子遺伝子」)は、選択された領域のカセット突然変異誘発を助長するための制限部位と、所望の発現系への遺伝子の組み込みを助長するためのフランキング制限部位とを含む合成G−CSFコーディング核酸配列を開示しているらしい。
【0020】
G−CSFは、ある程度まで結晶化されたと報告されており(例えばヨーロッパ特許第344796号)、G−CSFの全体構造は推定されているが、粗レベルに止まっている。Bazan,Immunology Today 11: 350−354 (1990); Parryら,J.Molecular Recognition 8: 107−110(1988)。G−CSFの全体構造は未だ報告されておらず、また、G−CSF類似体の体系的設計には分子の全体構造と機能の関係の体系的研究が必須であるが、このような研究も報告されていない。従って、G−CSF類似体のこの体系的設計方法及び該方法により得られる組成物が必要とされている。
【先行技術文献】
【特許文献】
【0021】
【特許文献1】米国特許第4,810,643号明細書
【特許文献2】欧州特許第0335423号明細書
【特許文献3】欧州特許第0272703号明細書
【特許文献4】欧州特許第0459630号明細書
【特許文献5】欧州特許第0256843号明細書
【特許文献6】欧州特許第0243153号明細書
【特許文献7】米国特許第4,904,584号明細書
【特許文献8】国際公開第90/12874号パンフレット
【特許文献9】豪国特許出願第10948/92号明細書
【特許文献10】豪国特許出願第76380/91号明細書
【特許文献11】英国特許第2213821号明細書
【特許文献12】欧州特許第344796号明細書
【非特許文献】
【0022】
【非特許文献1】Welteら,PNAS−USA 82: 1526−1530(1985)
【非特許文献2】Souzaら,Science 232: 61−65(1986)
【非特許文献3】Gabrilove,J.Seminars in Hematology 26: (2)1−14(1989)
【非特許文献4】Jonesら,Bailliere’s Clinical Hematology 2(1): 83−111(1989)
【非特許文献5】Nagataら,EMBO J 5:575−581(1986)
【非特許文献6】Mooreら,PNAS−USA 84: 7134−7138(1987)
【非特許文献7】Kugaら,Biochem.Biophy.Res.Comm 159: 103−111(1989)
【非特許文献8】Luら,Arch.Biochem.Biophys.268: 81−92(1989)
【非特許文献9】Bazan,Immunology Today 11: 350−354 (1990)
【非特許文献10】Parryら,J.Molecular Recognition 8: 107−110(1988)
【発明の概要】
【0023】
(発明の要約)
本発明により、G−CSFの三次元構造は原子レベルまで決定されるに至った。この三次元構造から、G−CSF分子の組成がどのように変化すると構造変化を生じるかをかなり確実に予想することができる。これらの構造特性を生物活性と相関させると、G−CSF類似体を設計及び製造することができる。
【0024】
G−CSFの三次元構造については他にも考察されている(Bazan,Immunology Today 11: 350−354(1990); Parryら,J.Molecular Recognition 8: 107−110(1988))が、推定構造が不正確(Parryら,前出)及び/又は推定構造から構成部分と構造の相関の詳細が得られないという理由により、これらの考察はG−CSF類似体を製造しようと意図する人々には何ら役立たなかった。三次元構造を原子レベルまで決定するという本発明の構造分析は従来の最も完全な分析よりも遥かにすぐれており、G−CSF類似体を設計及び製造しようとする人々に重要な情報を提供する。例えば、本発明の三次元構造分析から、厳密な疎水性及び親水性領域が決定された。
【0025】
相対疎水性は分子の安定性に直接関係するので重要である。一般に、水性環境中に存在する生物分子は外部が親水性であり、内部が疎水性であり、熱力学の第2法則によると、これは最低エネルギー状態であり、安定性を提供する。G−CSFの内部コアは疎水性であり且つ外部領域は親水性であると予想することができるが、特定の疎水性又は親水性領域を知る術はなかった。本発明により提供された疎水性/親水性領域の知識に基づき、G−CSF分子のどのような変化が分子の全体構造に影響するかをかなり確実に予想することができる。
【0026】
一般に、G−CSF全体構造を変化させずに生物活性を変化させるような類似体を設計するために、疎水性及び親水性領域の位置の知識を使用することができる(本明細書中では「生物活性」なる用語は機能を表すために最も広義に使用する)。生物活性を構造に相関させることができる。構造が変化せず且つ突然変異が生物活性に影響しないならば、突然変異は生物学的機能をもたない。他方、構造が変化せず且つ突然変異が生物活性に影響するならば、残基(又は原子)は少なくとも1種の生物学的機能に不可欠である。本発明の実施例のいくつかは全体構造を変えずに生物学的機能を変化させるように意図した。
【0027】
構造と生物活性との相関に基づき、本発明の1態様はG−CSF類似体に関する。これらの類似体は、非常に少数のアミノ酸残基をG−CSFアミノ酸配列と相異させるか又は改変した分子である。改変は1個以上のアミノ酸残基の付加、置換又は欠失であり得る。改変は、アミノ酸自体の類似体(例えばペプチド類似体)又は改変部分(例えば改変側基)を有するアミノ酸の付加又は置換を含み得る。比較の基準として使用されるG−CSFはヒト、動物又は組換え核酸技術起源であり得る(本明細書中の実施例は付加的N末端メチオニル残基を有するヒトG−CSFの174アミノ酸種の組換え製造に基づく)。類似体は天然ヒトG−CSF分子と異なる機能を有してもよいし、同一機能を示してもよいし、同一機能を別程度で示してもよい。例えば、類似体は改変前よりも生物活性が高くても低くてもよいし、貯蔵寿命を延ばしてもよいし、安定性を低下させてもよいし、調合し易くしてもよいし、他の成分と結合しにくくしてもよい。類似体は造血活性をもたなくてもよく、従って、(例えばG−CSFの過産生の場合等のように)G−CSF効果に対するアンタゴニストとして有用であり得る。本明細書中では本発明の類似体を便宜上タンパク質又はペプチドと呼称する場合もあるが、本発明はペプチド類似体又は化学的に修飾されたペプチド等の他の型の分子も包含する。
【0028】
別の態様によると、本発明はG−CSF類似体を活性成分として含有する関連組成物に関する。本明細書中で使用する「関連組成物」なる用語は、G−CSF類似体の存在を確認後に得られる組成物(例えば検出可能なラベルで標識したG−CSF類似体、関連レセプター又は医薬組成物)を意味する。少なくとも1個のポリエチレングリコール分子を結合した類似体等の化学的に修飾したG−CSF類似体も関連組成物とみなされる。
【0029】
例えば、検出可能なラベル(例えば蛍光、化学発光又は放射性分子)を結合したG−CSF類似体を製造することができる。
【0030】
別の例は公知材料を使用して公知技術により調剤することが可能な医薬組成物である。例えば、参考資料として本明細書の一部とするRemington’s Pharmaceutical Science, 第18版(1990,Mack Publishing Co., Easton, Pennsylvania 18042)pp.1435−1712参照。一般に、組成は投与法、安定性、製法等の種々の因子及び他の因子に依存する。G−CSF類似体は注射又は吸入による肺投与により投与することができる。本発明のG−CSF類似体組成物には腸溶性剤形も使用可能であり、従って経口投与が有効であり得る。G−CSF類似体をリポソーム又は他の送達用マイクロキャリヤーに組み込んでもよいし、ジェル又は他の徐放用組成物に配合してもよい。好適組成物は組成物の用途により異なるが、一般に天然G−CSFの生物活性の少なくとも1種を有するG−CSF類似体の好適医薬組成物は皮下注射又は吸入による肺投与用に製造した組成物であり、各投与型毎の特定組成は類似体の特性に依存する。
【0031】
関連組成物の別の例は本発明の類似体のレセプターである。本明細書中で使用する「レセプター」なる用語は、本発明の類似体分子と選択的に結合する部分を意味する。例えば、抗体又はそのフラグメント又は「組換え抗体」(Huseら,Science 246:1275(1989))をレセプターとして使用することができる。(本発明は結合特異的レセプターを包含するが)選択的結合は特異的結合のみを意味するものではないものの、どちらかといえば結合はランダムではない。レセプターは細胞表面上又は細胞内もしくは細胞外に位置し、本発明の類似体の生物活性を提供、抑制又は局在化するように機能し得る。レセプター結合は類似体自体と間接的に関係する活性カスケードの始動メカニズムでもあり得る。本発明は更に前記レセプターをコードする核酸、該核酸を含むベクター及び該核酸を含む宿主細胞も包含する。
【0032】
関連組成物の別の例は、化学的部分を結合させたG−CSF類似体である。一般に、化学修飾はタンパク質の生物活性又は抗原性を変えるか、又は他の特性を変えることができ、これらの因子は当業者に考慮されよう。上述のように、このような化学的部分の1例はポリエチレングリコールである。修飾は親水性もしくは疎水性ポリマー分子、脂肪酸分子、又は多糖分子の1種以上の付加を含み得る。化学修飾剤の例としては、ポリエチレングリコール、アルキルポリエチレングリコール、DI−ポリ(アミノ酸)、ポリビニルピロリドン、ポリビニルアルコール、ピランコポリマー、酢酸/アシル化、プロピオン酸、パルミチン酸、ステアリン酸、デキストラン、カルボキシメチルセルロース、プルラン又はアガロースが挙げられる。Francis,Focus on Growth Factors 3: 4−10(May 1992)(Mediscript,Mountview Court, Friern Barnet Lane, London N20 OLD,英国刊)参照。更に、化学修飾は付加的タンパク質もしくはその部分、細胞毒性物質又は抗体の使用を含み得る。化学修飾はレシチンも使用し得る。
【0033】
別の態様によると、本発明は前記類似体をコードする核酸に関する。核酸はDNA又はRNA又はその誘導体であり、典型的には適当な調節配列を含むファージ又はプラスミド等のベクターにクローニングされ、このベクターで発現される。核酸は、例えば診断又は予後の目的で(例えば放射性、化学発光又は蛍光ラベルを使用して)標識してもよい。核酸配列は、例えば細菌発現に好適なコドンを含むなどして発現に最適化することができる。核酸とその相補鎖及び、所望の類似体をコードするのを妨げないようなその改変体も本発明に含まれる。
【0034】
別の態様によると、本発明は本発明の類似体をコードする上記核酸を含む宿主細胞に関する。宿主細胞は真核細胞でも原核細胞でもよく、発現系は糖基の結合(グリコシル化)(又は阻止)、分子の適正な折り畳み、組換え発現に付随するリーダー配列もしくは他の因子の付加又は欠失に関する追加の段階を含み得る。
【0035】
別の態様によると、本発明はこのような核酸配列の発現の型又は量を抑制又は変更するように機能するアンチセンス核酸に関する。これらの核酸は公知方法により製造することができる。
【0036】
本発明の別の態様によると、本発明の類似体をコードする核酸は、例えば類似体をコードする配列を含むベクターを受容体に導入し、類似体組成物を必要とする受容体の体内で核酸自体を発現させることにより、遺伝子療法の目的で使用することができる。まず最初にベクターを細胞等のキャリヤーに導入後、キャリヤーを受容体に導入する。このような発現は局在化してもよいし、全身でもよい。他のキャリヤーとしては、受容体への遺伝子移入を媒介するように機能し得る非天然に存在するキャリヤー(例えばリポソーム又は他のマイクロキャリヤーもしくは粒子)が挙げられる。
【0037】
本発明は更に、G−CSF又は類似体の三次元構造を表現(例えば視覚表示)するためのコンピュータープログラム及び、G−CSF分子の各成分の存在と全体構造内の原子レベルまでの該成分の厳密な位置を表現するコンピュータープログラムを提供する。このようなプログラムの1例を以下に説明する。分子の三次元構造を表現するためのコンピュータープログラムは多数のものが現在市販されている。一般に、これらのプログラムは分子の三次元構造の座標(即ちx、y及びz軸に沿うG−CSF分子の各原子の数値帰属)の入力手段、前記座標の表現手段(例えば視覚表示)、前記座標の修正手段、及びこのような修正座標を有する分子の画像の表現手段を提供する。結晶情報即ち、G−CSF分子の結晶分析から得られた三次元空間におけるG−CSF分子の原子の位置の座標をプログラムすると、G−CSF三次元構造の表現(例えば視覚表示)のためのコンピュータープログラムを作成することができる。従って、G−CSF類似体三次元構造の表現用コンピュータープログラムも提供される。San Diego,Californiaに所在のBiosymから市販されているコンピュータープログラムInsight II,version 4に図5に示すような座標を入力すると好適である。好適表現手段としては、Silicon Graphics 320 VGXコンピューターでG−CSF分子又はその類似体を立体的に観察することが可能なCrystal Eyes眼鏡(Silicon Graphicsの市販品)を着用する。また、本発明のG−CSF結晶座標及び回折データは、米国、Upton,New York 119723に所在のProtein Data Bank,Chemistry Department,Brookhaven National Laboratoryに寄託されている。これらのデータを使用してG−CSF分子又はその類似体の三次元構造の表現用の別のコンピュータープログラムを作成することもできる。従って、本発明の別の態様は、G−CSF分子の三次元構造の表現用コンピュータープログラムである。G−CSF分子の三次元構造の視覚表示用コンピュータープログラムも提供され、更に、該プログラムはこのような視覚表示を変更するための手段を有する。このようなコンピュータープログラムの表現に有用な装置、特にG−CSF分子又はその類似体の前記三次元構造のコンピューター画像の視覚表示に有用な装置と、前記コンピュータープログラム及び装置を製造するための手段も提供される。
【0038】
G−CSF分子上の被改変特定部位を選択することができ、この改変がG−CSF分子の全体構造に及ぼす影響を容易に確認することができるので、コンピュータープログラムはG−CSF類似体を製造するために有用である。前記被改変部位の選択は、G−CSF類似体の所望の生物特性に依存する。前記G−CSF分子(r−met−hu−G−CSF)をランダムに改変したならば、17520通りもの置換が考えられ、複数の置換、付加又は欠失を有する類似体の数は更に多い。分子の組成と相関させながら三次元構造を観察することにより、改変部位をランダムに選択する必要がなくなり、改変部位を合理的に決定することができる。
【0039】
上述のように、各成分の原子レベルまでの位置決定を含めてG−CSFの三次元構造が確認されたので、どの部分がG−CSF分子の全体構造を維持するために必要であるかに関する情報を得られるようになった。従って、本発明のG−CSF類似体の製造時にG−CSF分子の全体構造を維持すべきか、又は本発明のG−CSF類似体の製造時にG−CSF分子の全体構造を変更すべきか(及びどのように変更すべきか)を選択することができる。場合により、このような類似体を製造後、このような類似体が所望の特性を有するか否かを試験することができる。
【0040】
例えば、非改変天然又は組換えG−CSF分子の全体構造を維持しようとすることができる。全体構造を図2、3及び4に示し、以下に詳細に説明する。全体構造の維持により、天然G−CSFの造血能を有する類似体に必要な特性であるレセプター結合を確保することができる(レセプター結合が存在しないならば、類似体の存在からシグナル変換は行われない)。あるクラスのG−CSF類似体は天然又は組換え(非改変)G−CSF分子の三次元コア構造を有しながら、好中球を選択的に刺激する能力の増加等の別の特性も有すると予想される。別のクラスのG−CSF類似体は別の全体構造を有する類似体であり、G−CSF類似体分子とG−CSFレセプターの結合能が低く、非改変天然又は組換えG−CSFに比較して好中球を選択的に刺激する能力が低い。
【0041】
例えば、こうしてG−CSF分子の内部領域内のどの部分が疎水性であるかが分かり、これに対応してG−CSF分子の外部のどの分子が親水性であるかが分かる。好ましくは本発明のように原子レベルまで全体三次元構造を認識できなければ、この疎水性内部領域内のどの改変が分子の全体構造コンフォーメーションの変化をもたらすかを予測することはできない。全体構造が変化すると、例えばレセプター結合の欠損等の機能的変化が生じ、従って、非改変G−CSFに認められるような生物活性の低下が生じ得る。従って、別のクラスのG−CSF類似体は、(非改変)天然又は組換えG−CSFと同一の疎水性を有するG−CSF類似体である。より特定的には、別のクラスのG−CSF類似体は、(非改変)天然又は組換えG−CSFの疎水性部分と同一の疎水性部分をその内部コアの4螺旋束の内側に有しており、且つ前記非改変天然又は組換えG−CSFとは異なる組成を有する。
【0042】
別の例は、G−CSF分子の内部コア(螺旋)と結合する構造である外部ループに関する。アミノ酸残基の空間位置に関する情報を含む三次元構造から、特定のループにおける特定の変化は全体的コンフォーメーション変化をもたらさないと予想することができる。従って、本発明の別のクラスのG−CSF類似体は、改変外部ループを有しており且つ(非改変)天然又は組換えG−CSFと同一の全体構造を有する。より特定的には、本発明の別のクラスのG−CSF類似体は改変外部ループを有しており、該ループは本明細書中のループ及び螺旋の表記に従い、螺旋A及びBの間、螺旋B及びCの間、螺旋C及びDの間、螺旋D及びAの間に存在するループから選択される。より特定的には、前記ループ、好ましくはABループ及び/又はCDループは該ループを安定化させることにより分子の半減期を増加させるように改変されている。このような安定化は、前記ループの全部又は一部をG−CSF(又は類似体)分子のコアに存在するα螺旋束の一部と結合することにより得られる。このような結合は、βシート、塩架橋、ジスルフィド結合、疎水性相互作用又は当業者に使用可能な他の結合手段を介して実施され、このような結合手段は前記外部ループを安定化するように機能する。例えば、ABループを分子の内部領域内の螺旋の1つと結合することによりAB又はCDループを安定化させることができる。
【0043】
N末端は内部螺旋の構造安定性に影響しないので、N末端を改変してもG−CSF分子の全体構造は変化せず、外部ループは改変には好適であるが、同一の一般論がN末端にも当てはまる。
【0044】
また、(非改変)天然又は組換えG−CSFではこのような外部ループは比較的フレキシブルであり、レセプター結合を妨げない傾向があるので、このような外部ループは化学的修飾部位となり得る。従って、更に化学的部分を直接結合(又は化学的結合手段として機能する別の化学的部分を介して間接結合)する余地がある。化学的部分は、G−CSF分子の1種以上の機能の変更に使用可能な種々の部分から選択することができる。例えば、外部ループはポリエチレングリコール分子等のような血清半減期を増加させるように機能する1種以上のポリマーを添加するための部位を提供し得る。このようなポリエチレングリコール分子を加え、ポリエチレングリコール部分と結合可能な反応性側基を有する付加的リジンを含むように前記ループを改変させることができる。他のクラスの化学的部分も1個以上のループに結合することができ、このような化学的部分の非限定的な例としては、他の生物活性分子(例えばレセプター)、他の治療用タンパク質(例えばハイブリッド分子をもたらす他の造血因子)、又は細胞毒性物質(例えばジフテリア毒素)が挙げられる。当然これらの例に留まることなく、所望の化学的部分を有する当業者は、該所望の部分を所望の外部ループに結合することができる。従って、別のクラスの本発明のG−CSF類似体は、少なくとも1個のポリエチレングリコール分子等の化学的部分を付加する少なくとも1つの改変を外部ループに有する類似体を含む。
【0045】
欠失(例えば分子の分解のためにタンパク質により認識される部位の欠失)も外部ループに有効であり得る。こうして、一方ではG−CSFレセプター結合及びシグナル変換能(即ち好中球の突然変異を選択的に刺激する能力)を有する分子の半減期を増加することができる。従って、別のクラスの本発明のG−CSF類似体は、プロテアーゼによる前記類似体の代謝回転を低下させる少なくとも1つの改変を外部ループに有する類似体を含む。この改変に好適なループはABループ及びCDループである。(以下に詳細に説明する)外部ループに存在するアミノ酸残基の一部を欠失させることにより短縮G−CSF分子を製造することができ、該短縮G−CSF分子は製造又は生物学的機能に付加的利点を有する。
【0046】
別の例は、相互に近接するアミノ酸残基間の相対電荷に関する。上述のように、G−CSF分子は比較的密に充填された4螺旋束を含む。螺旋上の面のいくつかは他の螺旋に向き合う。螺旋が別の螺旋に向き合う点(例えば残基)において、相互に向き合う2つのアミノ酸分子は同一電荷を有することができ、従って、相互に反発しあい、分子全体を不安定にする。これは、反発が生じないようにアミノ酸部分の一方又は両方の電荷を(逆電荷又は中性電荷に)変えることによりこれを解決することができる。従って、別のクラスのG−CSF類似体は、電荷位置等の表面相互作用による不安定を修正するように改変されたG−CSF類似体を含む。
【0047】
別の態様によると、本発明はG−CSF類似体及び関連組成物の設計方法並びに該方法の産物に関する。前記方法の終産物は上記G−CSF類似体又は関連組成物であり得る。例えば、本明細書中に開示する実施例は、(a)G−CSF分子の成分(即ち化学的部分)の改変がG−CSF構造に及ぼす作用及び(b)構造変化が生物学的機能に及ぼす作用を立証する。従って、本質的に本発明の別の態様は、
(a)G−CSF分子の各アミノ酸残基又は各アミノ酸残基の各原子等の化学的部分を相関させたG−CSF分子の三次元構造を含む情報を観察する段階と、
(b)前記情報からG−CSF分子上の被改変部位を選択する段階と、
(c)このような改変を有するG−CSF類似体分子を製造する段階と、
(d)任意に、このようなG−CSF類似体分子が所望の特性を有するか否かを試験する段階と
を含むG−CSF類似体の製造方法である。
【0048】
本発明のコンピュータープログラムは、コンピューターに基づくG−CSF類似体の製造方法に使用することができる。従って、本発明の別の態様は、
(a)G−CSF分子の各アミノ酸残基又は各アミノ酸残基の各原子等の化学的部分と相関させたG−CSF分子の三次元構造のコンピューター表現を提供する段階と、
(b)前記情報からG−CSF分子上の被改変部位を選択する段階と、
(c)このような改変を有するG−CSF分子を製造する段階と、
(d)任意に、このようなG−CSF分子が所望の特性を有するか否かを試験する段階と
を含む、コンピューターに基づくG−CSF類似体の製造方法である。
【0049】
より特定的には本発明は、
(a)(i)G−CSF分子の座標を三次元空間に表示し且つ(ii)前記G−CSF表現の変更及びその観察のための情報を入力できるようにプログラムされたコンピューターを介してG−CSF分子の三次元構造を観察する段階と、
(b)前記G−CSF分子の前記視覚画像上の被改変部位を選択する段階と、
(c)前記コンピューターに前記改変のための情報を入力する段階と、
(d)前記コンピューターを介して前記改変G−CSF分子の三次元構造を観察する段階と、
(e)任意に段階(a)〜(e)を繰り返す段階と、
(f)前記改変を有するG−CSF類似体を製造する段階と、
(g)任意に前記G−CSF類似体が所望の特性を有するか否かを試験する段階と
を含む、G−CSF類似体の製造方法を提供する。
【0050】
別の態様によると、本発明は本発明のG−CSF類似体及び関連組成物の使用方法、並びに造血障害治療において単独又は他の造血因子もしくは薬剤と併用して哺乳動物を治療又は予防する方法に関する。G−CSF類似体を設計する1態様は、非改変G−CSFが有するとして知られている特性を増加又は変更することを目的とすると意図される。
【0051】
例えば、本発明の類似体は強化又は変更された活性を有するので、G−CSFが(例えば)好中球減少症の治療に有用である場合には本発明の組成物及び方法は同様にこのような治療に使用することができる。
【0052】
別の例は、特に造血障害の治療で他の因子と併用した場合により有効に相互作用することを目的としたG−CSFの改変である。このような併用の1例は、初期作用性造血因子(即ち造血カスケードの初期に比較的未分化の細胞に対して作用する因子)の使用及び後期作用造血因子、例えばG−CSF又はその類似体(G−CSFは好中球の選択刺激においてCFU−GM系に作用する)の同時又は順次使用である。本発明の方法及び組成物は、造血因子のこのような組み合わせ又は「カクテル」を使用する療法で有用である。
【0053】
本発明の組成物及び方法は、白血球減少症、骨髄性白血病、重度慢性好中球減少症、再生不能性貧血、糖原病、粘膜炎及び他の骨髄不全症状の治療にも有用である。本発明の組成物及び方法は、化学療法又は放射線療法に起因する造血欠損の治療にも有用であり得る。例えば骨髄移植の成功又は移植における末梢前駆血球の使用は、本発明の組成物(遺伝子療法の場合にはタンパク質又は核酸)及び方法を適用することにより増進することができる。本発明の組成物及び方法は、創傷治癒、火傷治療、菌血症、敗血症、真菌感染、心内膜炎、骨腎盂炎、腹部外傷に関連する感染、抗生物質に反応しない感染、肺炎等の感染症の治療にも有用であり、本発明の組成物及び方法を細菌炎症の治療に利用することもできる。更に、本発明の組成物及び方法は白血病細胞分化能の報告(Welteら,PNAS−USA 82: 1526−1530(1985))に基づき白血病の治療にも有用であり得る。この他に、腫瘍細胞に結合するレセプター(例えば抗体)の任意存在下で本発明の組成物及び方法を使用して腫瘍個体を治療することもできる。治療用途に関する研究論文については、いずれも参考資料として本明細書の一部とするLieshhkeとBurgess,N.Engl.J.Med.327: 28−34及び99−106(1992)を参照されたい。
【0054】
本発明の組成物及び方法は、他の部分の製造の中間体として使用するのにも有用であり、例えば、G−CSFは他の造血因子の生成に影響を及ぼすことが報告されており、(確認されたならば)この機能は本発明の組成物及び/又は方法により増進又は変更することができる。
【0055】
本発明のG−CSF類似体に関連する組成物(例えばレセプター)は、G−CSF又は類似体の活性を阻止するアンタゴニストとして作用するのにも有用である。非改変G−CSF又はG−CSF類似体の活性の一部又は全部を有する組成物を得ることができ、1種以上の化学的部分を付加してこのようなG−CSF又は類似体の1種以上の特性を改変することができる。三次元コンフォーメーションの知識に基づき、所望の効果に達するためにこのような化学修飾の最良の幾何学的位置を予測することができる。
【0056】
化学修飾の一般目的としては、半減期の改善(例えば腎性クリアランス、免疫クリアランス又は細胞クリアランスの減少)、生物活性の変更(例えば酵素特性の変更、有機溶媒中の生物活性又は活性の分離)、毒性の低下(例えば毒性エピトープの隠蔽、区画化及び選択的生物分布)、免疫反応性の改変(免疫原性の低下、抗原性又はアジュバント作用の低下)、又は物性の低下(例えば溶解度の増加、熱安定性の改善、機械的安定性の改善又はコンフォーメーション安定化)が挙げられる。Francis,Focus on Growth Factors 3: 4−10(May 1992)(Mediscript,Mountview Court,Friern Barnet Lane,London N20 OLD,英国刊)参照。
【0057】
以下、実施例により本発明を非限定的に説明する。当業者は種々の変形及び変更が可能であり、請求の範囲は本発明の範囲に該当するこのような全等価態様を包含する。
【0058】
(発明の詳細な説明)
本発明はG−CSFの三次元構造の知見に基づく。この三次元構造は立体的に観察するためのコンピュータープログラムにより表現された。これを立体的に観察することにより、構造−機能関係を認識し、G−CSF類似体を設計及び製造した。
G−CSFの三次元全体構造
構造を確認するために使用したG−CSFは、細菌発現に付随する付加的N末端メチオニン残基を有する非グリコシル化174アミノ酸種であった。このG−CSFのDNA及びアミノ酸配列を図1に示す。
【0059】
全体として、G−CSFの三次元構造は主に螺旋状であり、175個の残基のうちの103個は4α螺旋束を形成する。他の二次構造は最初の2つの長い螺旋の間のループに存在し、4残基310螺旋の直後には6残基α螺旋が存在する。図2に示すように、全体構造を他のタンパク質、即ち成長ホルモン(Abdel−Meguidら,PNAS−USA 84: 6434(1987)及びVosら,Science 255: 305−312(1992))、顆粒球マクロファージコロニー刺激因子(Diederichsら,Science 254: 1779−1782(1991))、インターフェロン−β(Sendaら,EMBO J.11: (3193−3201))、インターロイキン−2(McKay Science 257: 1673−1677(1992))、及びインターロイキン−4(Powersら,Science 256: 1673−1677(1992)、及びSmithら,J.Mol.Biol.224: 899−904(1992))に報告されている構造と比較した。アミノ酸配列は類似していないにも拘わらず、これらの成長因子間には構造類似性が存在する。
【0060】
構造情報はG−CSF生化学の相関であり、これは以下のように要約することができる(配列位置1はN末端とする)。
【0061】
【表1】

【0062】
抗体結合試験から収集したこの生化学情報(Laytonら,Biochemistry 266: 23815−23823(1991)参照)を三次元構造に重ね合わせてG−CSF類似体を設計した。これらのG−CSF類似体の設計、製造及び試験を下記実施例1に説明する。
【図面の簡単な説明】
【0063】
【図1】付加的N末端メチオニンを有するG−CSFの174アミノ酸種のアミノ酸配列図である(配列番号1)(配列番号2)。
【図2A】G−CSFの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2B】hGHの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2C】pGHの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2D】GM−CSFの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2E】INF−Bの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。螺旋の元の分類を括弧内に示す。
【図2F】IL−2の結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2G】IL−4の結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図3】G−CSFの三次元構造の「リボン図」である。螺旋Aはアミノ酸残基11〜39(上記図1による番号付け)、螺旋Bはアミノ酸残基72〜91、螺旋Cはアミノ酸残基100〜123、螺旋Dはアミノ酸残基143〜173である。比較的短い310螺旋はアミノ酸残基45〜48に位置し、α螺旋はアミノ酸残基48〜53に位置する。残基93〜95は左回り螺旋のほぼ1回転を形成する。
【図4】G−CSFの三次元構造の「バレル図」である。G−CSFの三次元構造の全体円筒体とその向きを種々の濃淡で示す。数字は図1によるアミノ酸残基位置を示す。
【図5A】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5B】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5C】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5D】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5E】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5F】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5G】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5H】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5I】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5J】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5K】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5L】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5M】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5N】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5O】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5P】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5Q】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5R】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5S】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5T】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5U】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5V】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5W】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5X】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5Y】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5Z】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5AA】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5BB】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5CC】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5DD】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5EE】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5FF】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5GG】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5HH】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5II】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5JJ】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5KK】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5LL】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5MM】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5NN】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5OO】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5PP】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5QQ】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5RR】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5SS】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5TT】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5UU】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5VV】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5WW】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5XX】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5YY】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5ZZ】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5AAA】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5BBB】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5CCC】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5DDD】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5EEE】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5FFF】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図6】結晶化に関与するパラメーターに関して結晶化マトリックスの最適化に使用されるストラテジーを示す。結晶化マトリックスは24穴組織培養プレートのウェル中の結晶化溶液の成分(塩、緩衝液及び沈殿剤)の最終濃度に対応する。これらの濃度は、適量のストック溶液をマイクロタイタープレートのウェルにピペット分注することにより生成される。マトリックスを設計するために、結晶学者は成分の高低濃度を決定する。これらの高低濃度を行(例えばA1〜A6、B1〜B6、C1〜C6又はD1〜D6)又は全トレー(A1〜D6)に沿ってピペット分注することができる。前者方法は制限された数のウェルに沿って単一成分の結晶成長の再現性を調べるために有用であり、後者方法は初期スクリーニングで有用である。結晶化マトリックスの数段の最適化段階の結果を3つのプレートの図により示す。ウェルの色が濃いほど正の結晶化結果を表し、最終段階ではX線品質結晶であり、初期段階では約0.05mm未満の寸法の油滴、顆粒状沈殿又は小結晶であり得る。Aは最初のウェル(A1)と最後のウェル(D6)の間の濃度範囲が広く且つウェル間の濃度増加が((濃度A1)−(濃度D6))/23)として計算される1パラメーターの初期スクリーンを示す。Bは、濃度の結晶化マトリックス最適化の後期段階でA1とD6の間の差が縮まり、プレート当たりに形成される結晶が増加していることを示す。Cはプレートの大部分のウェルに高品質結晶が存在するマトリックス最適化の最終段階を示す。
【実施例1】
【0064】
本実施例は結晶質G−CSFの製造、コンピューター生成画像による組換えヒトG−CSFの三次元構造の可視化、特定部位突然変異誘発又は核酸増幅法を使用した類似体の製造、G−CSF類似体を分析するために使用した生物アッセイ及びHPLC分析、並びに全体構造/機能関係の決定について説明する。全引用文献は参考資料として本明細書の一部とする。
【0065】
A.自動結晶化の使用
組換えヒト顆粒球コロニー刺激因子(r−hu−G−CSF)の三次元構造が必要であることと、大量の精製タンパク質が入手可能であることから、不完全階乗サンプリング及び接種による結晶成長法を実施した。JancarikとKim,J.Appl.Crystallogr.24: 409(1991)に記載されている不完全階乗結晶化の実施から出発して、油滴及び複屈折凝集体を生成する溶液条件を確認した。また、自動ピペッティングシステムのソフトウェア及びハードウェアを改造して毎日約400の異なる結晶化条件を設定した。Weber,J.Appl.Crystallogr.20: 366−373(1987)。この手順により、r−hu−G−CSF結晶を生成する結晶化溶液を調製した。
【0066】
結晶の寸法、再現性及び品質は、「核生成開始単位」の数を接種溶液の連続希釈により推定する接種法により改善された。これらの方法により、2.0mmのr−hu−G−CSF結晶を再現可能に成長させることができた。これらの結晶の空間群はP212121であり、セル寸法はa=90Å、b=110Å及びc=49Åであり、2.0Åの分解能まで回折する。
【0067】
1.全体的手法
新規タンパク質の結晶化条件を探索するために、CarterとCarter,J.Biol.Chem.254: 122219−12223(1979)は不完全階乗法を提案した。同氏らによると、一般に有望であるとみなされるランダムに選択された多数の結晶化条件のサンプリングにより、タンパク質結晶化をもたらす好適な試薬組み合わせが得られる。このアイデアは、JancarikとKim,J.Appl.Crystallogr.24: 409(1991)により実現され、同氏らはpH範囲、塩及び沈殿剤を考慮した初期結晶化試験のための32種の溶液を記載した。本発明では同氏らの作業を拡張し、70種までの溶液を記載する。溶液調製の人的労力及び誤差を最小限にするために、自動ピペッティング機にあわせて手法をプログラムした。
【0068】
Weberの連続自動格子探索法(SAGS),J.Cryst,Growth 90: 318−324(1988)に従い、ロボットシステムを使用して温度、pH、塩及び沈殿剤の結晶化条件を連続的に最適化する一連の溶液を調製した。結晶を再現可能に成長させることが可能な溶液が決定されると、結晶の品質を著しく改善する接種技術が開発された。これらの方法を組み合わせると、数百種の回折品質結晶(少なくとも約2.5Åに回折し、好ましくは少なくとも部分的に2Å未満、より好ましくは約1Åに回折する部分を有する結晶)が数日で生成された。
【0069】
一般に、結晶化を所望する任意のタンパク質と共に使用され得る結晶化方法は、
(a)(i)異なる塩濃度を各々有する塩溶液のアリコート又は(ii)異なる沈殿剤濃度を各々有する沈殿剤溶液のアリコートと所望のタンパク質の水性アリコートとを一定範囲のpHの任意存在下で結合する段階と、
(b)こうして結合したアリコートの予備結晶形成を観察し、予備結晶形態の生成に有効な前記塩もしくは沈殿剤組み合わせ及び前記pHを選択するか、又は予備結晶形態が生成されない場合には、タンパク質の前記水性アリコートのタンパク質出発濃度を増加させる段階と、
(c)前記塩又は前記沈殿剤濃度を選択後、前記選択された濃度の存在下で前に選択しなかった前記溶液を用いて段階(a)を繰り返す段階と、
(d)所望の品質の結晶が得られるまで段階(b)及び段階(a)を繰り返す段階と
を含む。
【0070】
上記方法は任意に自動化することができ、時間と労力を著しく節約することができる。好適タンパク質出発濃度は10mg/ml〜20mg/mlであるが、この出発濃度はタンパク質により異なる(下記G−CSFは33mg/mlを使用して分析した)。分析を開始するために好適な塩溶液範囲は0〜2.5M(NaCl)である。好適沈殿剤はポリエチレングリコール8000であるが、他の沈殿剤としては、有機溶媒(例えばエタノール)、分子量500〜20,000のポリエチレングリコール分子、及び当業者に公知の他の沈殿剤を挙げることができる。好適pH範囲はpH4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5及び9.0である。予備結晶化形態は、油状物、複屈折沈殿、小結晶(<約0.05mm)、中結晶(約0.5〜.5mm)及び大結晶(>約0.5mm)を含む。結晶構造を検出するまでの好適待機時間は48時間であるが、週に一度の観察でも好適であり、一般には約1カ月後に別のタンパク質濃度を使用する(一般にタンパク質濃度を増加させる)。自動化はAccuflexシステムを改造すると好適である。好適自動化パラメーターを以下に説明する。
【0071】
一般に、濃度10mg/ml〜20mg/mlのタンパク質を0〜2.5MのNaCl溶液と組み合わせ、上記濃度範囲の存在下で(別々に)このような各組み合わせを実施した。予備結晶化構造が観察されたら、所望の結晶品質に達するまで前記塩濃度及びpH範囲を別実験で最適化させる。次いで、pHレベルを変えながら沈殿剤濃度も最適化させる。両者が最適化されたら、最適条件を同時に実施し、所望の結果を得る(これを図6に示す)。
【0072】
a.自動ピペッティングシステムの実施
標準シリアルインターフェースを介してASCIIコードを送信するパーソナルコンピューターにより制御されるAccuflexピペッティングシステム(ICN Pharmaceuticals,Costa Mesa,CA)により、液滴及び液溜溶液を調製した。ピペッターは回転バルブにより6種の異なる溶液をサンプリングし、xy座標系内の並進を制御可能なプレートにこれらの溶液をピペット分注する。座標系の鉛直成分は液体を分配及び回収することが可能な注射器を操作する。
【0073】
Accuflexのソフトウェアは、CoxとWeber,J.Appl.Crystallogr.20: 366−373(1987)により提案されているようなSAGS法を利用した。この方法は、他の2つのパラメーターを変化させながら、pH及び沈殿剤濃度という2つの主要な結晶化パラメーターを体系的に変化させるものである。これらのコンセプトに基づきながら、本発明で使用するソフトウェアは自動格子探索ストラテジーで使用される結晶化溶液の設計及び実現により高いフレキシビリティーを提供する。このフレキシビリティーの結果として、本発明のソフトウェアはより多数の異なる溶液を生成することができた。これは、下記不完全階乗法の実施に不可欠である。
【0074】
自動格子探索ストラテジーの速度及び設計を改善するために、Accuflexピペッティングシステムはソフトウェア及びハードウェアの改造が必要であった。ハードウェアの改造により、懸垂液滴(hanging drop)実験用トレーと、放置液滴(sitting drop)実験用トレーとの2種の異なるマイクロタイタートレーと、24種の付加的緩衝液、塩及び沈殿剤溶液を保持するPlexglasトレーを使用できるようになった。これらの付加溶液により、調査可能な結晶化条件の格子を拡大した。
【0075】
ハードウェア改造を利用するために、ピペッティングソフトウェアを2つのサブルーチンに書き込んだ。結晶学者は一方のサブルーチンにより成分の濃度に基づいて結晶化溶液のマトリックスを設計することができ、第2のサブルーチンにより適正容量の溶液を結晶化トレーにピペット分注するコンピューターコードにこれらの濃度を翻訳することができる。濃度マトリックスは2種のプログラムのどちらかを使用して作成することができる。第1のプログラム(Thousand Oaks,CAに所在のAmgen Inc.の市販品MRF)は結晶学者により供給されるストック溶液濃度のリストに関するものであり、指定濃度に達するために必要なピペット分注容量を計算する。より好適な第2の方法は、沈殿剤の勾配又はpHを改善するために使用可能なスプレッドシートプログラム(Lotus)を組み込む。これらのプログラムにより生成される濃度マトリックスは制御プログラム(Accuflexピペッターに元々内蔵されているプログラムの改造であり、Thousand Oaks,CAに所在のAmgen,Inc.の市販品SUX)により翻訳され、ウェルはこれに従って充填される。
【0076】
b.不完全階乗法の実施
改造ピペッティングシステムは種々の溶液を調製するのに便利であるため、拡張不完全階乗法の実施を改善することができた。3つの変数から1つの因子の96のランダムな組み合わせを含むリストを作成するプログラムINFAC(Carterら,J.Cryst.Growth 90: 60−73(1988))を使用して「ランダム」な成分を有する結晶化溶液の新しい組合わせを展開した。即座に沈殿したカルシウムとリン酸の組み合わせを除くと、沈殿剤、塩及び緩衝液の70種の異なる組み合わせが残った。自動ピペッターを使用してこれらの組み合わせを調製し、1週間インキュベートした。混合物を検査し、沈殿を形成した溶液をより低い成分濃度で再び調製した。全ウェルから沈殿が除去されるまでこれを繰り返した。
【0077】
c.r−hu−G−CSFの結晶化
X線品質結晶を生成する溶液を見いだすために数種の異なる結晶化ストラテジーを使用した。これらのストラテジーは、不完全階乗法の使用、連続自動格子探索(SAGS)を使用した結晶化条件の最適化、接種技術の実施、及び数百の品質結晶を一晩で生成する結晶生成手順の開発を含むものであった。特に指定しない限り、スクリーニング及びr−hu−G−CSF結晶の生成は懸垂液滴蒸気拡散法を使用した。Afinsenら,Physical principles of protein crystallization.:Eisenberg(編),Advances in Protein Chemistry 41: 1−33(1991)。
【0078】
r−hu−G−CSFの結晶化条件の初期スクリーニングはJancarikとKim,J.Appl.Crystallogr.24: 409(1991)の不完全階乗法を使用し、「予備結晶化」結果をもたらす数種の溶液を調製した。これらの結果は複屈折沈殿、油状物及び非常に小さい結晶(<0.05mm)を含むものであった。これらの予備結晶化溶液をその後、体系的スクリーニングの出発点として使用した。
【0079】
スクリーニングプロセスには結晶化マトリックスの開発が必要であった。これらのマトリックスは結晶化溶液中の成分の濃度に対応し、IBM−PCに基づくスプレッドシートLotus(商標)を使用して生成し、改造Accuflexピペッティングシステムを用いて実施した。マトリックスの設計におけるストラテジーは、他の条件(例えばpH及び沈殿剤濃度)を一定に維持しながら1つの結晶化条件(例えば塩濃度)を変化させることであった。スクリーニングの開始時に変更条件の濃度範囲は広かったが、マイクロタイタートレー中の全ウェルが同一結晶化結果を生じるまで濃度を連続的に最適化した。これらの結果を結晶、複屈折沈殿、顆粒状沈殿、油滴及び非晶質塊の分類により評点した。ある結晶化パラメーターの濃度が少なくとも沈殿を生成しなかった場合には、沈殿が形成されるまでこのパラメーターの濃度を増加させた。各トレーを調製後、少なくとも2日間撹乱せずに静置した後、結晶成長を検査した。この初期スクリーニング後、トレーを週に一度の割合で検査した。
【0080】
このスクリーニングプロセスから、pH及び沈殿剤が同一でありながら塩(MgCl,LiSO4)が異なり、小(0.1×0.05×0.05mm)結晶を生成した2種の別々の溶液を確認した。これらの結果に基づき、他の結晶化パラメーターを一定に維持しながらLiSO4に対してMgClの濃度を変化させた濃度マトリックスの新しい組合わせを作成した。この実験系の結果、約3週間で回折品質結晶(>約0.5mm)を生成する溶液が確認された。この結晶化成長溶液(100mM Mes pH5.8,380mM MgCl2,220mM LiSO4及び8%PEG8k)を見いだすために、約300mgのタンパク質を消費した約8,000の条件をスクリーニングした。
【0081】
結晶の寸法は液滴当たりの結晶形成数に依存した。典型的には平均寸法(1.0×0.7×0.7mm)を有する3〜5個の結晶が形成された。接種(下記参照)を実施したか否かに依存して、等しい空間群(P212121)及び単位セル寸法a=90.2,b=110.2,c=49.5を有する2種の形態が得られた。接種を実施しない場合には、r−hu−G−CSF結晶は1つの長い平坦な表面と丸みを帯びた縁部を有していた。
【0082】
接種を使用した場合には、4〜6時間以内に鋭角面を有する結晶(0.05×0.05×0.05mm)が液滴内に観察された。24時間以内に結晶は(0.7×0.7×0.7mmまで)成長し、液滴中の結晶形成数に依存して2mmを越えるまで成長し続けた。
【0083】
d.接種及び核生成開始部位の決定
本発明の結晶接種方法は、(ここでは最適結晶成長条件を決定後に)使用した各個ウェル中の核生成開始単位数を確定する。この方法は、「種晶」の数が結晶の品質に影響し、分解能に影響するという点で有利である。本発明の接種は更に、接種を実施しない場合には成長に約3週間を要するが、接種を実施すると、G−CSF結晶は約3日で成長するという利点がある。
【0084】
ある生成成長系列(手法の項参照のこと)では、確定された小結晶(<0.01×0.01×0.01mm)のシャワーが一晩で生成された。前に使用したピペット先端を再使用した以外は上記と同様の結晶化条件に従った。恐らく、使用済み先端に形成されており、結晶の核生成部位を提供する小核生成単位により結晶シャワー効果が生じたと予想される。標準生成成長条件下で結晶シャワーを含む少量(0.5μl)の液滴を新しい液滴に加えると、一晩で結晶のシャワーが生成された。この方法を使用して結晶シャワーを含む液滴のトレー数個を作成し、これを「種晶ストック」と名付けた。
【0085】
「種晶ストック」液滴内に含まれる核生成開始単位(NIU)の数を推定し、r−hu−G−CSF結晶の再現性及び品質を改善するように試みた。「種晶ストック」中のNIUの数を決定するために、96穴マイクロタイタープレートに沿って液滴のアリコートを連続希釈した。マイクロタイタープレートは、等容量のr−hu−G−CSF(33mg/ml)と(上記)結晶成長溶液を含有する溶液50μlを各ウェルに加えることにより調製した。「種晶ストック」のアリコート(3μl)をマイクロタイタープレートの第1のウェルに移した。ウェル内の溶液を混合した後、3μlをマイクロタイタープレートの行に沿って次のウェルに移した。マイクロタイタープレートの各行を同様に調製し、トレーをプラスチックテープで密閉した。一晩でマイクロタイタープレートのウェルの底に小結晶が形成されたので、ウェル中の結晶の数を元の「種晶ストック」の希釈液に相関させた。大きい単結晶を生成するために、「種晶ストック」液滴を新鮮なCGS中に適宜希釈した後、NIUを含有するこの溶液のアリコートを液滴に移した。
【0086】
結晶化条件が一旦最適化されたら、CGS及びr−hu−G−CSF(33mg/ml)各3mlを混合して5個のトレー(各24穴)を作成する生成法で結晶を成長させた。この方法は、リットル量の最適化結晶化溶液を調製し、この溶液をタンパク質と混合し、懸垂液滴又は放置液滴トレーにタンパク質/結晶化溶液を加えるものであった。この方法では典型的には約5日間で100〜300の品質結晶(>0.5mm)が得られた。
【0087】
e.実験法
材料
図1に示すアミノ酸配列と比活性1.0±0.6×108U/mg(pH4.0の10mM酢酸緩衝液中で細胞マイトジェン生成アッセイにより測定)を有する濃度約3mg/mlのr−hu−G−CSFから出発して結晶学的情報を得た。YM10フィルターを使用して75psiでAmicon濃縮器を使用して溶液を濃縮した。溶液を典型的には4℃で10倍に濃縮し、数カ月貯蔵した。
【0088】
初期スクリーニング
懸垂液滴を使用して蒸気拡散平衡によりX線分析に適した結晶を得た。予備スクリーニングのために、(上記のように調製した)33mg/mlタンパク質溶液7μlを等容量のウェル溶液と混合し、シリコンガラスプレートに載せ、Linbro組織培養プレート(McLean,Vaに所在のFlow Laboratories)を使用してウェル溶液上に懸垂させた。全ピペッティングはAccuflexピペッターを使用して実施したが、ウェル溶液の生成後はトレーを自動ピペッターから取り外し、卓上振盪機で少なくとも10分間十分に混合した。次にLinbroトレーを再びピペッターに取り付け、ウェル溶液及びタンパク質溶液をシリコンカバースリップに加えた。次にカバースリップを裏返し、ウェル溶液1mlをシリコン油脂で覆い、密封した。
【0089】
自動結晶化システムの成分は以下の通りである。PC−DOSコンピューターシステムを使用して、その成分の濃度に基づいて結晶化溶液のマトリックスを設計した。これらのマトリックスは(上記)MRF又はLotusスプレッドシートのいずれかを用いて作成した。これらのプログラムの終産物はデータファイルである。このファイルは、マトリックスに示された濃度を得るために適した容量のストック溶液をピペットするためにSUXプログラムにより必要とされる情報を含む。SUXプログラム情報をシリアルI/Oポートに送り、ストック溶液に対するバルブの位置、回収した後にマイクロタイタープレートのウェルにピペット分注すべき溶液の量及び各ウェルのXY位置(各ウェルの行列)をAccuflexピペッティングシステムに指令するためにこの情報を使用した。充填中の注射器のZ位置(高さ)と、異なる溶液の充填間に注射器を洗浄するためにシステムを停止させるドレインの位置とを含む付加情報をピペッターに送信した。24穴マイクロタイタープレート(Linbro又はCryschem)及びカバースリップホルダーをプレートに載せ、プレートをXY面内で移動させた。プレートの移動により、ウェルにピペット分注するようにピペッターに注射器を位置決めさせた。プレートの移動により、カバースリップ及びバイアルも位置決めし、これらから溶液を抽出した。ピペット分注前に、Linbroマイクロタイタープレートはウェルの縁部の周囲に油脂薄膜を塗布しておいた。結晶化溶液をウェル内で調製してから、カバースリップに移す前に、マイクロタイタープレートをピペッティングシステムから取り外し、溶液を10分間卓上振盪機で混合した。混合後、ウェル溶液をカバースリップ(懸垂液滴プロトコルの場合)又はウェルの中央ポスト(放置液滴プロトコルの場合)に移した。タンパク質をバイアルから抽出し、ウェル溶液を含有するカバースリップ液滴(又はポスト)に加えた。プラスチックテープをCryschemプレートの頂部に張り付けてウェルを密封した。
【0090】
生成成長
結晶化条件が最適化されたら、「生成」法を使用して結晶成長を実施した。100mM Mes(pH5.8)、380mM MgCl2、220mM LiSO4及び8%PEG 8Kを含有する結晶化溶液1リットルを調製した。エッペンドルフ注射器ピペッターを使用してこの溶液の1mlアリコートをLinbroプレートのウェルの各々にピペット分注した。この溶液50%及びG−CSF(33mg/ml)50%を含有する混合溶液を調製し、シリコンカバースリップにピペット分注した。これらの液滴の典型量は50〜100μlであり、これらの液滴の寸法が大きいため、カバースリップのめくれ上がり及び液滴のウェル懸垂に細心の注意を払った。
【0091】
データ収集
R−AXIS(Houston,TXに所在のMolecular Structure,Corp.)イメージングプレートデテクターで収集した2.2Åデータに対してX−PLOR(Bruniger,X−PLOR version 3.0,結晶分析システム及びNMR,New Have CTに所在のイエール大学)を用いて構造を最適化させた。
【0092】
f.考察
有効な組換えヒト治療剤として、r−hu−G−CSFは大量生産され、構造分析のためにグラムレベルが入手可能になっている。他の該当タンパク質が入手可能になるにつれ、本発明の結晶化方法は他の用途にも利用可能である。この方法は、大量のタンパク質(>約200mg)を有する任意の結晶学的プロジェクトに適用することができる。当業者に認識される通り、本発明の材料及び手法は変形可能であり、等価材料及び手法が他のタンパク質の結晶化に使用可能である。
【0093】
B.G−CSFの三次元構造を可視化するためのコンピュータープログラム
添付図に示すような図はG−CSFの三次元構造を可視化するために有用であるが、分子を立体的観察を可能にするコンピュータープログラムが好適であると予想される。この立体観察は当業者には「バーチャルリアリティー」とも呼称され、高分子構造から原子レベルまで広範囲の分解能で全角度から構造を三次元形態で可視化することができる。本発明のコンピュータープログラムにより、例えば分子を回転させることにより、分子の観察角度の透視図を変更することもできる。本発明のプログラムは更に変更に応答するので、例えば完全アミノ酸残基を含む1個以上の原子画像を削除、追加もしくは置換えることができ、又は存在する基もしくは置換された基に化学的部分を付加することができ、構造変化を可視化することができる。
【0094】
他のコンピューターに基づくシステムを使用することもでき、その要素は、(a)例えばG−CSFの三次元構造の直交座標又は他の数値帰属座標等の情報を入力するための手段と、(b)前記座標を表現するための手段、例えば三次元構造を観察してこのような三次元構造をアミノ酸組成等のG−CSF分子の組成と相関できるようにする視覚手段と、(c)任意要素として、前記三次元構造の画像が改変組成を表示するように、表現されたG−CSF分子の組成を改変する情報を入力するための手段である。
【0095】
使用される好適コンピュータープログラムの座標を図5に示す。好適コンピュータープログラムはSan Diego,CAに所在のBiosymの市販品Insight II,version 4である。粗結晶学的構造については、回折データの観察強度(F−obs)及び直交座標も米国,Upton,New York 119723に所在のProtein Data Bank,Chemistry Department,Brookhaven National Laboratoryに寄託されており、参考資料として本明細書の一部とする。
【0096】
座標をInsight IIプログラムに入力すると、コンピュータースクリーン上に三次元G−CSF分子像を容易に表示することができる。表示に好適なコンピューターシステムはSilicon Graphics 320 VGX(San Diego,CA)である。立体観察の際には、G−CSF分子を三次元で立体的に観察することが可能な眼鏡(Crystal Eyes,Silicon Graphics)を装着し、分子を回転させて分子設計を想像することができる。
【0097】
従って、本発明はコンピューターを用いてG−CSF類似体を設計又は製造する方法を提供し、該方法は、
(a)G−CSF分子の部分の組成の表示、好ましくは各アミノ酸の三次元配置の表示、より好ましくはG−CSF分子の各原子の三次元位置の表示を含めてG−CSF分子の三次元構造を表示するための手段を前記コンピューターに備える段階と、
(b)前記表示を観察する段階と、
(c)前記分子の組成又は部分の位置の改変のための部位を前記表示上で選択する段階と、
(d)前記改変を有するG−CSF類似体を製造する段階とを含む。
【0098】
改変は、終産物G−CSF類似体の所望の構造特性に基づいて選択することができ、このような設計には以下に詳述する要素を考慮する。このような要素とは、特にG−CSF分子の螺旋構造の内側の残基であって、このような残基を改変すると、分子の内部コアの全体構造が変化し、レセプター結合を妨げ得る疎水性アミノ酸残基の位置及び組成と、改変しても、G−CSF分子の全体構造には影響しない外部ループ構造の位置及び組成を含む。
【0099】
図2A〜4は、種々の方法で全体的三次元コンフォーメーションを示す。トポロジー図、リボン図、及びバレル図はいずれもG−CSFのコンフォーメーションの特徴を示す。
【0100】
図2A〜Gは、G−CSFと他の分子との比較を示す。アーキテクチャーは類似しているが、これらの成長因子はそのループの局所コンフォーメーション及び螺旋束の幾何学的配置が異なる。他方、アミノ酸配列の相違にも拘わらず、これらの分子の全6種間には2つの長いクロスオーバー結合を含むアップ−アップ−ダウン−ダウントポロジーが保存されている。
【0101】
図3は組換えヒトG−CSFの二次構造を詳細に示す。このリボン図は螺旋の向きとその相対位置を示す。
【0102】
図4は組換えヒトG−CSFのコンフォーメーションを別の方法で示す。この「バレル」図は組換えヒトG−CSFの全体的なアーキテクチャーを示す。
【0103】
C.M13突然変異誘発を用いた類似体の調製
本実施例は、PCT出願公開第WO85/00817号(Souzaら,1985年2月28日公開、参考資料として本明細書の一部とする)に記載の方法に従って、1本鎖バクテリオファージM13を用いる部位特異的突然変異誘発技術を使用したG−CSF類似体の調製に関する。この方法は主に非突然変異誘発配列の1本鎖核酸鋳型を使用し、配列中に所望の改変を含むより小さいオリゴヌクレオチドを前記鋳型に結合することからなる。非同一配列をハイブリダイズさせるようなハイブリダイゼーション条件を使用し、残りの配列は最初の鋳型と同一になるように充填する。その結果、2本の鎖の一方が所望の改変を含むような2本鎖分子が生成される。この突然変異誘発した1本鎖を分離し、その相補鎖の鋳型として使用する。こうして所望の改変を含む2本鎖分子を生成する。
【0104】
使用した改変前のG−CSF核酸配列を図1に示し、突然変異誘発した核酸を含むオリゴヌクレオチドを表2に示す。本明細書中でアミノ酸残基及びヌクレオチドに使用した略記は慣用通りであり、Stryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y., N.Y.1988,裏内表紙を参照されたい。
【0105】
改変前のG−CSF核酸配列をまず最初にベクターM13mp21に挿入した。次に改変前のG−CSF配列を含む1本鎖ファージM13mp21からのDNAを単離し、水に再懸濁した。各反応につきこのDNA200ngをリン酸化オリゴヌクレオチド1.5pmolと混合し(表2)、0.1M Tris,0.01M MgCl2,0.005M DTT,0.1mM ATP(pH8.0)に懸濁した。65℃まで加熱し、室温までゆっくりと冷却することによりDNAをアニールした。
【0106】
冷却後、ATP、dATP、dCTP、dGTP、TTP各0.5mM、T4 DNAリガーゼ1単位及び大腸菌ポリメラーゼ1のKlenowフラグメント1単位を、0.1M Tris,0.025M NaCl,0.01M MgCl2,0.01M DTT(pH7.5)中のアニールしたDNA1単位に加えた。
【0107】
こうして2本鎖になった閉環状DNAをそれ以上精製せずに使用して大腸菌にトランスフェクトした。プラークをニトロセルロースフィルターで釣り上げた後、P32で末端標識した1本鎖DNAとフィルターを1時間55〜60℃でハイブリダイズさせることによりプラークをスクリーニングした。ハイブリダイゼーション後、オリゴの融点(A−Tでは2℃、G−Cでは4℃)よりも0〜3℃低い温度でフィルターを洗浄し、突然変異配列を含むファージを有するプラークに対応するオートラジオグラフィーシグナルを選択的に発生させた。陽性クローンを配列決定により確認した。
【0108】
M13突然変異誘発法により製造した各G−CSF類似体毎に使用したオリゴヌクレオチドを下表に示す。命名法は、改変前のアミノ酸の残基及び位置(例えば17位リジン)と、置換したアミノ酸の残基及び位置(例えばアルギニン17)を示す。2個以上の残基が置換されている場合には、位置を指定する上付き数字をカンマで区切るか、又はセミコロンで残基を区切る。置換でなく欠失の場合にはその旨明記した。M13に基づく突然変異誘発に使用したオリゴヌクレオチド配列を次の欄に示す。これらのオリゴヌクレオチドは合成により製造したが、その製法は限定的ではなく、任意の核酸合成法及び/又は装置を使用することができる。オリゴ長も示す。上述のように、これらのオリゴを1本鎖ファージベクターと接触させた後、単一ヌクレオチドを加え、G−CSF類似体核酸配列を完成させた。
【0109】
【表2】
【0110】
D.DNA増幅を使用したG−CSF類似体の調製
本実施例は、DNA増幅技術を用いたG−CSF類似体の製造方法に関する。主に、各類似体をコードするDNAを別個の2部分として増幅して結合した後、完全配列を増幅した。改変前のG−CSF DNAのどの場所に所望の改変を加えるべきかに依存して、内部プライマーを使用して改変を組み込み、2つの別個の増幅部分を生成した。例えば、所望の類似体DNAの5’末端を増幅するためには、増幅すべき領域の一端には(改変前のG−CSF DNAの上流のプラスミドの配列に相補的な)5’フランキングプライマーを使用し、他方の末端をプライムするためには、改変前のDNAとハイブリダイズすることが可能でありながら所望の改変を含み得る内部プライマーを使用した。得られた増幅領域は5’フランキングプライマーから内部プライマーに及ぶ領域であっだ。(改変前のG−CSF DNAの下流のプラスミドの配列に相補的な)3’フランキングプライマーと所期突然変異の領域に相補的な内部プライマーを使用して3’末端も同様に処理した。(内部プライマーの位置に依存して同一寸法でもよいし、同一寸法でなくてもよい)2つの「半部分」を増幅後、2つの「半部分」を結合した。結合後、5’フランキングプライマーと3’フランキングプライマーを使用して所望の改変を含む完全配列を増幅した。
【0111】
2つ以上の改変が所望される場合には、内部プライマーに改変を組み込むように上記プロセスを変更するか、又は別の内部プライマーを使用してプロセスを繰り返す。あるいは、核酸配列に改変をもたらすための他の方法、例えば上記のようなファージに基づく突然変異誘発技術と遺伝子増幅法を併用してもよい。2つ以上の改変を有する類似体を製造する方法の例を以下に記載する。
【0112】
下記G−CSF類似体を生成するために使用した鋳型DNAは、図1の配列に(G−CSFコーディング領域を含むプラスミドからの)所定のフランキング領域を加えたものであった。これらのフランキング領域を以下に説明するように5’及び3’フランキングプライマーとして使用した。増幅反応は、10mM Tris−HCl、1.5mM MgCl2、50mM KCl、0.1mg/mlゼラチン(pH8.3)を含有する40μl容量中で20℃で実施した。40μl反応溶液は更に各dNTP 0.1mM、各プライマー10pmol及び鋳型DNA1ngを含有するものとした。各増幅は15サイクル繰り返した。各サイクルは94℃で0.5分間、50℃で0.5分間、72℃で0.75分間から構成した。フランキングプライマーは20ヌクレオチド長、内部プライマーは20〜25ヌクレオチド長であった。こうして、所望のG−CSF類似体の前部又は後部をコードする2本鎖DNAの多重コピーを生成した。
【0113】
2つの「半部分」を結合するために、2つの反応物の各々の40分の1を第3のDNA増幅反応で結合した。突然変異を有するこれらの2部分の末端は相補的であるので、2部分を内部プライマー位置でアニールさせ、重合サイクル後、完全長DNA配列を得た。こうしてアニール後、5’及び3’フランキングプライマーを使用して完全類似体を増幅した。この増幅プロセスは上述のように15サイクル繰り返した。
【0114】
完全増幅類似体DNA配列をXbaI及びXhoI制限エンドヌクレアーゼで切断し、ベクターに挿入するための付着末端を生成した。切断したDNAをプラスミドベクターに挿入し、このベクターを使用して大腸菌を形質転換させた。形質転換細胞を50μg/mlのカナマイシンで攻撃し、30℃でインキュベートした。G−CSF類似体タンパク質の生成を全細胞溶解物のポリアクリルアミドゲル電気泳動により確認した。生成単離物から精製したプラスミドのDNA配列分析により所望の突然変異の存在を確認した。次に培養物を増殖させ、細胞を回収し、G−CSF類似体を以下のように精製した。
【0115】
下表3は遺伝子増幅を使用して製造した各類似体に使用した特定プライマーを示す。
【0116】
【表3】
【0117】
1.二重突然変異の構築
G−CSF類似体Gln12.21→Glu12.21を製造するために、DNA増幅を2回別々に実施し、2つのDNA突然変異体を製造した。使用した鋳型DNAは図1の配列に(G−CSFコーディング領域を含むプラスミドからの)所定のフランキング領域を加えたものである。厳密な配列は下記の通りである。Perkin Elmer/Cetus DNA熱サイクラーを使用して2つのDNA増幅反応の各々を実施した。40μl反応混合物は、1×PCR緩衝液(Cetus)、4dXTP(Cetus)各0.2mM、各プライマーオリゴヌクレオチド50pmol、(プラスミドベクター上の)G−CSF鋳型DNA2ng及びTaqポリメラーゼ(Cetus)1単位から構成した。増幅プロセスは30サイクル実施した。各サイクルは94℃で1分間、50℃で2分間、72℃で3分間から構成した。
【0118】
DNA増幅“A”は下記オリゴヌクレオチド:
【0119】
【化1】
を使用した。
【0120】
DNA増幅“B”は下記オリゴヌクレオチド:
【0121】
【化2】
を使用した。
【0122】
DNA増幅“A”後に得られた109塩基対2本鎖DNA産物からDNA突然変異Gln12→Glu12を含む64塩基対XbaI−HindIII DNAフラグメントを切り出し、単離した。DNA増幅“B”後に得られた509塩基対2本鎖DNA産物からDNA突然変異Gln21→Glu21を含む197塩基対HindIII−BsmI DNAフラグメントを切り出し、単離した。
【0123】
“A”及び“B”フラグメントを4.8kbp XbaI−BsmI DNAプラスミドベクターフラグメントと連結した。連結混合物は等モルDNA制限フラグメント、連結緩衝液(25mM Tris−HCl(pH7.8),10mM MgCl2,2mM DTT,0.5mM rATP及び100μg/ml BSA)及びT4 DNAリガーゼから構成し、14℃で一晩インキュベートした。その後、Bio Rad Gene Pulsar装置(BioRad,Richmond,CA)を使用するエレクトロポレーションにより連結DNAを大腸菌FM5に形質転換した。クローンを単離し、DNA配列決定によりプラスミド構築物が2つの突然変異を含むことを確認した。この「中間」ベクターは更に193塩基対BsmI−BsmI DNAフラグメントを欠失していた。最終プラスミドベクターは中間ベクターから2kbp SstI−BamHI DNAフラグメント、プラスミドベクターから2.8kbp SstI−EcoRI DNAフラグメント、及びプラスミドベクターから360bp BamHI−EcoRI DNAフラグメントを切り出し、単離することにより得たDNAフラグメントの(上記のような)連結及び形質転換により構築した。DNA配列決定により最終構築物がG−CSF遺伝子を含むことを確認した。下記のように培養物を増殖し、細胞を回収し、G−CSF類似体を精製した。
【0124】
上述のように、突然変異誘発法の任意の組み合わせを使用して、1つ以上の改変を有するG−CSF類似体核酸(及び発現産物)を製造することができる。M13に基づく突然変異誘発及び遺伝子増幅に基づく突然変異誘発を使用した上記2例はその実例である。
【0125】
E.G−CSF類似体DNAの発現
次にG−CSF類似体DNAをプラスミドベクターに挿入し、大腸菌株FM5(ATCC#53911)を形質転換させるために使用した。プラスミド上及び細菌宿主細胞中に含まれる本発明のG−CSF類似体DNAは、Rockville,MDに所在のAmerican Type Culture Collectionから入手可能であり、その受託番号は以下に示す。
【0126】
トリプトン10g、酵母エキス5g及びNaCl 5gを含有するブロスで密度A600=0.5に達するまで1リットル培養物を30℃で増殖させ、前記密度に達したら42℃まで迅速に加熱した。フラスコを3時間振盪し続けた。
【0127】
他の原核又は真核宿主細胞も使用可能であり、例えば他の細菌細胞、株もしくは種、哺乳動物培養細胞(COS、CHO又は他の型)、昆虫細胞もしくは多細胞器官もしくは生物、又は植物細胞もしくは多細胞器官もしくは生物を使用することができ、当業者は適切な宿主に想到しよう。本発明のG−CSF類似体及び関連組成物は、例えば固相ペプチド合成法又は他の化学的製法により合成することもできる。他のクローニング及び発現系は当業者に自明である。
【0128】
F.G−CSF類似体タンパク質の精製
遠心分離(10,000×G,20分間,4℃)により細胞を回収した。ペレット(通常は5g)を1mM DTT30mlに再懸濁し、10,000psiのフレンチプレスセルに3回通した。破壊した細胞懸濁液を10,000gで30分間遠心分離し、上清を除去し、ペレットを水30〜40mlに再懸濁した。これを10,000×Gで30分間再遠心分離し、このペレットを2% Sarkosyl及び50mM Tris(pH8)25mlに溶解させた。硫酸銅を濃度40μMまで加え、混合物を少なくとも15時間15〜25℃で撹拌した。次いで混合物を20,000×Gで30分間遠心分離した。得られた可溶化タンパク質混合物を13.3mM Tris(pH7.7)で4倍に希釈し、Sarkosylを除去した後、20mM Tris(pH7.7)で平衡化したDEAE−セルロース(Whatman DE−52)カラムに上清を充填した。カラム充填及び同一緩衝液でカラムを洗浄後、類似体を20mM Tris/NaCl(下記のように類似体に依存して35mM〜100mM)(pH7.7)で溶離した。類似体のほとんどはDEAEカラムからの溶出液を50%酢酸でpH5.4に調整し、必要に応じて(適正な導電率を得るために)5mM酢酸ナトリウム(pH5.4)で希釈した。次に20mM酢酸ナトリウム(pH5.4)で平衡化したCM−セファロースカラムに溶液を充填した。次いで280nmの吸光度がほぼ0になるまでカラムを20mM NaAc(pH5.4)で洗浄した。次いでG−CSF類似体を下表4に記載する濃度の酢酸ナトリウム/NaClで溶離した。CM−セファロースカラムに充填しなかった類似体のDEAEカラム溶出液を10mM NaAc(pH4.0)緩衝液で直接透析した。次いで精製G−CSF類似体をin vitro分析のために適宜単離した。類似体の溶離に使用した塩濃度は上述のように変化させた。DEAEセルロースカラム及びCM−セファロースカラムの濃度を下表に示す
【0129】
【表4】
*167欠失のデータは得られなかった。
**これらの類似体ではDEAEセルロースカラムを単独で精製に使用した。
【0130】
上記精製法は例示であり、当業者には自明の通り、発明のG−CSF類似体を得るために他の手段も使用可能である。
【0131】
G.生物アッセイ
本発明のG−CSF類似体をどの方法で製造したかに関係なく、類似体の生物活性をアッセイした。3H−チミジンアッセイを実施して細胞分裂度を確認した。他の生物アッセイを使用して所望の活性を確認することもできる。マウスWEHI−3B(D+)白血病細胞系に末端分化を誘導する能力をアッセイする等の生物アッセイによりG−CSF活性を得ることもできる。Nicolaら,Blood 54: 614−27(1979)参照。生物活性を確認するために他のin vitroアッセイも使用できる。Nicola,Annu.Rev.Biochem.58: 45−77(1989)参照。一般に、生物活性試験では(非改変G−CSFと比較した)生物活性の増加又は低下、(非改変G−CSFと比較した)生物活性の相違、レセプターアフィニティー分析又は血清半減期分析等の所望の結果を分析すべきである。これらの例に止まることなく、当業者は所望の最終結果の試験に有用な他のアッセイにも想到しよう。
【0132】
常法を使用して3H−チミジンアッセイを実施した。雌Balb Cマウスを殺して骨髄を取り出した。骨髄細胞を成長培地に短時間懸濁し、遠心分離し、再懸濁した。約10,000個の細胞を含む160μlアリコートを96穴マイクロタイタープレートの各ウェルに加えた。(上記のように調製した)精製G−CSF類似体のサンプルを各ウェルに加え、68時間インキュベートした。3H−チミジンをウェルに加え、更に5時間インキュベートした。5時間のインキュベーション時間後、細胞を回収し、濾過し、十分に濯いだ。シンチレーション液を収容するバイアルにフィルターを加えた。β線を計数した(LKBβプレートシンチレーションカウンター)。標準及び類似体を3回ずつ分析し、標準曲線よりも実質的に上又は下に位置するサンプルを適正な希釈度で再アッセイした。ここに報告する結果は非改変組換えヒトG−CSF標準結果に対する3回の類似体データの平均である。
【0133】
H.HPLC分析
類似体の精製サンプルで高圧液体クロマトグラフィーを実施した。逆相HPLCカラム上のピークの位置は2種のタンパク質間の構造類似を決定的に示すものではないが、類似の保持時間を有する類似体はHPLCカラムに対して非改変分子と同一型の疎水性相互作用を有し得る。これは全体構造の類似を示す1面である。
【0134】
類似体及び非改変組換えヒトG−CSFのサンプルを逆相(0.46×25cm)Vydac 214TP54カラム(Separations Group,Inc.Hesperia,CA)で分析した。類似体がカラム中でどのように挙動したかに依存して0.1mg/ml〜5mg/mlの最終濃度まで20mM酢酸及び40mM NaCl緩衝溶液(pH5.2)中で精製類似体G−CSFサンプルを調製した。1%イソプロパノール、52.8%アセトニトリル及び0.38%トリフルオロ酢酸(TFA)を含有する水溶液で予め平衡化しておいたHPLCカラムに(濃度に依存して)種々の量を充填した。サンプルを0.86%/分のアセトニトリル及び0.002%TFAの勾配にかけた。
【0135】
I.結果
上記生物アッセイ及びHPLC分析の結果を以下に示す。生物活性は3回のデータの平均であり、対照標準(非改変G−CSF)の百分率として示す。相対HPLCピーク位置は対照標準(非改変G−CSF)ピークに対する類似体G−CSFの位置である。+又は−の記号は、類似体HPLCピークが対照標準ピークよりも(何分)先又は後に現れるかを示す。変異体の全部について相対HPLCピークを分析した訳ではなく、分析したもののみを下表に示す。上記のように調製した本発明の類似体をコードする核酸を含む大腸菌宿主細胞のAmerican Type Collection番号も示す。
【0136】
【表5】
【0137】
1.構造−機能関係の確認
本発明の類似体を設計するために使用した第1段階は、G−CSF分子の構造完全性にどの部分が必要であるかを決定することであった。これはアミノ酸残基レベルで実施したが、原子レベルも分析に利用できる。構造完全性に必要な残基を修飾すると、G−CSF分子の全体構造が変化する。これは製造しようとする類似体に依存して望ましい場合と望ましくない場合がある。本実施例は、G−CSFレセプター(本項で以下に記載する「G−CSFレセプター」は造血細胞に存在する天然G−CSFレセプターを意味する)への類似体のG−CSFレセプター結合を維持する目的で、G−CSF分子の全体構造完全性を維持するように設計した。G−CSFレセプター結合は上記生物アッセイにより決定されるような少なくとも1種の生物活性に必要な段階であると仮定され、本試験により確認された。
【0138】
図面に示すように、G−CSF(ここでは組換えヒトmet−G−CSF)は45Å×30Å×24Åの全体寸法を有する左巻き螺旋を有する逆平行4α螺旋束である。螺旋束内の4つの螺旋は螺旋A、B、C及びDと呼称され、その結合ループはAB、BC及びCDループとして知られる。螺旋交差角度は−167.5°〜−159.4°である。螺旋A、B及びCは直鎖であり、螺旋Dは(組換えヒトmet−G−CSFの)Gly150及びSer160に2種の構造特徴を含む。全体としてG−CSF分子は外部ループにより直列に結合された4つの螺旋からなる束である。この構造情報を公知機能情報と相関させた。(1位メチオニンを含む)残基47、23、24、20、21、44、53、113、110、28及び114を修飾することができ、生物活性に及ぼす作用は相当のものであることが判明した。
【0139】
生物活性を低下させた単一突然変異の多くは30Åの間隔で4螺旋束の異なった面に位置するG−CSFの2つの領域の周囲に集中していた。一方の領域はA螺旋とD螺旋の相互作用をもたらす。これは更に下記非改変分子における塩橋の存在により確認される。
【0140】
【表6】
【0141】
ここに報告する距離は図5A〜FFFに示すような分子Aの距離である(3つのG−CSF分子は共に結晶化し、A、B及びCと命名した)。螺旋A及びDの間には塩橋ウェブが存在し、螺旋A構造を安定化させるように機能するので、G−CSF分子の全体構造に影響する。
【0142】
残基Glu20、Arg23及びLys24の周囲の領域はA螺旋の親水性面(残基20〜37)に存在する。20及び23位の非帯電アラニン残基で残基を置換すると、類似のHPLC保持時間が得られ、構造の類似性が予想される。(本アッセイにより示されるように)これらの部位を改変すると、生物活性が変化した。Lys24を置換した場合には生物活性は変化したが、他の2種の改変のように類似のHPLC保持時間は得られなかった。
【0143】
改変により生物活性が低下した第2の部位はAB螺旋である。47位のグルタミンをアラニンで置換する(上記類似体19)と、(チミジン取り込みアッセイにおいて)生物活性がゼロに低下した。AB螺旋はアミノ及びカルボキシ末端を除いて主に疎水性であり、310螺旋1回転を含む。各末端には2個のヒスチジン(His44及びHis56)と、His44との塩架橋を形成することが可能な残基46の付加的グルタミンとが存在する。類似体のフーリエ変換赤外スペクトログラフィー分析(FTIR)によると、この類似体は非改変組換えG−CSF分子に構造的に類似する。更に試験した結果、この類似体は非改変組換え分子と同一条件下では結晶化しないことが判明した。
【0144】
カルボキシ末端の改変(Gln174、Arg167及びArg170)は生物活性にほとんど影響しなかった。他方、最後の8個の残基(167〜175)が欠失すると、生物活性は低下した。これらの結果から、欠失は全体構造を不安定にし、突然変異体がG−CSFレセプターと適正に結合するのを妨げる(その結果、シグナル変換の開始を妨げる)と予想される。
【0145】
一般に、G−CSF内部コア(外部ループを欠く内部4螺旋束)では疎水性内部残基は構造完全性に必須である。例えば螺旋Aにおいて内部疎水性残基は(メチオニンを1位として)Phe14、Cys18、Val22、Ile25、Ile32及びLeu36である。他の疎水性残基(同様にmetを1位とする)は螺旋BのAla72、Leu76、Leu79、Leu83、Tyr86、Leu90、Leu93であり、螺旋CのLeu104、Leu107、Val111、Ala114、Ile118、Met122であり、螺旋DのVal154、Val158、Phe161、Val164、Val168、Leu172である。
【0146】
本発明により製造したG−CSF類似体からの上記生物活性データによると、外部ループの改変はG−CSF全体構造にほとんど影響しない。類似体製造のための好適ループはABループ及びCDループである。ループは螺旋に比較して比較的にフレキシブルな構造である。ループは該分子のタンパク分解により分解され得る。該分子を使用する目的は生物攻撃に対する応答を発生すること、即ち好中球を選択的に刺激することであるので、G−CSFはin vivoで比較的迅速に作用する。G−CSF代謝回転も比較的迅速である。ループのフレキシビリティーはプロテアーゼが該分子と結合する「手」を提供し、該分子を不活性化する。プロテアーゼ分解を阻止しながら(非改変G−CSFの全体構造の保持を介して)生物活性を低下させないようにループを改変することができる。
【0147】
この現象は恐らくG−CSF分子に限定されず、図2に示すような類似の公知全体構造を有する他の分子にも共通すると思われる。例えばhGH、インターフェロンB、IL−2、GM−CSF及びIL−4の外部ループを改変しても全体構造の変化は最小に止まる。GM−CSF分子上の外部ループはG−CSF分子上に存在する外部ループほどフレキシブルではないので、GM−CSFはその広い生物活性に一致して血清寿命が長いと予想される。従って、β−シート構造から外部ループを解放することにより、(G−CSFと同様に)ループをよりフレキシブルにし、分子をプロテアーゼ分解に対してより感受性にする(その結果、回転率を増加する)ようにGM−CSFの外部ループを改変することができる。
【0148】
これらの外部ループの改変は、内部螺旋の1つ以上と結合することによりループを安定化することにより実施し得る。βシート、塩橋、ジスルフィド結合又は疎水性相互作用の形成等の結合手段が当業者に公知であり、他の手段も使用可能である。更に、1個以上のアミノ酸残基又はその部分等の1個以上の部分を欠失させ、短縮分子を製造し、こうして外部ループの所定の部分を除去することもできる。
【0149】
従って、外部ループ、好ましくはABループ(r−hu−met G−CSFのアミノ酸58〜72)又はCDループ(r−hu−met−G−CSFのアミノ酸119〜145)、これらのループより好ましくはないがアミノ末端(アミノ酸1〜10)の改変により、G−CSF G−CSFレセプター結合を除去せずに生物機能を変えることができる。例えば、(1)例えばプロテアーゼがG−CSF分子に作用する能力を低下させることにより又は、上記のようにG−CSF分子の所定の特徴を変化させる(例えば血清もしくは他の半減期を増加させるか又は抗原性を低下させる)ように機能する1個以上のポリエチレングリコール分子もしくは経口製剤用腸溶性コーティング等の化学修飾をG−CSF分子に加えることにより、G−CSF分子の半減期を増加させる(又は例えば経口剤形を製造する)こともできるし、(2)G−CSF G−CSFレセプター輸送メカニズムを介する細胞侵入によりシグナル変換を行う別のサイトカイン又は別のタンパク質等の別のタンパク質の一部又は全部とG−CSFとを結合するなどしてハイブリッド分子を作成することもできるし、(3)例えば(非改変G−CSF分子に比較して)好中球を選択的に刺激する能力等の生物活性を増加させることができる。このような改変は上記例に限定されない。
【0150】
上記データから観察される別の特徴は、安定化表面相互作用が生物活性に影響し得ることである。これは類似体23と40を比較すると明白である。類似体23は28位の中性帯電アラニン残基を同位の帯電アスパラギン残基で置換したものであり、この置換の結果、(本明細書に開示のチミジン取り込みアッセイにより測定されるように)生物活性は50%増加した。28位のアスパラギン残基は113位のアスパラギン残基との表面相互作用を有しており、両者の残基は負に帯電しているので、(同様の帯電部分の排斥により)所定量の不安定が存在する。他方、113位のアスパラギンを中性帯電アラニンで置換すると、(本アッセイシステムでは)生物活性はゼロまで低下する。このことから明らかなように、113位のアスパラギンは生物活性に重要であり、28位のアスパラギンを除去すると、113位のアスパラギンが有する作用が増加する。
【0151】
上記のように製造した類似体及びG−CSF構造に基づいてG−CSFレセプター結合に必要なドメインも決定した。G−CSFレセプター結合ドメインは(メチオニンを1位として)残基11〜57(A及びAB螺旋間)及び100〜118(B及びC螺旋間)に位置する。外部ループ構造を改変し、レセプター結合領域を無傷のままにすることにより、G−CSFレセプターと結合することが可能な短縮分子を製造し、好中球を選択的に刺激するためのシグナル変換を開始することもできる。
【0152】
生物活性に必須であり、恐らくG−CSFレセプター結合又はシグナル変換にも必須であると予想される残基を同定した。二次構造の2つの異なる領域には2つの別個の部位が位置する。ここで「部位A」なる部位は、螺旋束の他の2つの構成員間の塩橋接触により限定される螺旋上に位置する。第2部位「部位B」は比較的フレキシブルな螺旋AB上に位置する。AB螺旋はカルボキシ及びアミノ末端における残基の種類及び位置の理由から、局所pH変化の影響を受け易い。このフレキシブル螺旋の機能的重要性は、G−CSFレセプターとの結合時にコンフォーメンションを誘導適合できるという点にある。更に、直接突然変異及び間接相対タンパク質構造分析により確認されるように、D螺旋の延長部分もG−CSFレセプター結合ドメインである。r−hu−met−G−CSFのカルボキシ末端を欠失させると、hGHの場合と同様に活性が低下する(CunninghamとWells,Science 244: 1081−1084(1989)参照)。類似のトポロジーを有すると予想されるIL−6及びGM−CSF等の類似構造を有するサイトカインもD螺旋のカルボキシ末端に沿ってその生物活性の中心を有する(Bazan,Immunology Today 11: 350−354(1990)参照)。
【0153】
G−CSFとhGHとのG−CSFレセプター結合決定基の構造及び位置を比較すると、両者分子は類似のシグナル変換手段を有すると予想される。hGHでは2つの別個のG−CSFレセプター結合部位が確認されている(De Vosら,Science 255: 306−32(1991))。これらの結合部位の一方(部位Iと呼称する)は螺旋1及び2と螺旋4の間の結合領域であるhGHの螺旋1の露出面上の残基により形成される。第2の結合部位(部位IIと呼称する)は螺旋1と螺旋3の表面残基により形成される。
【0154】
G−CSFに確認されたG−CSFレセプター結合決定基はhGHに確認されると同一の相対位置に位置する。AB螺旋上の螺旋A及びB間の結合領域に位置するG−CSFレセプター結合部位(部位A)は、hGHの螺旋の小部分(残基38〜47)に報告されている位置と類似する。G−CSFのAB螺旋における単一点突然変異は、(本アッセイで確認されるように)生物活性を著しく低下させるので、G−CSFレセプター−リガンド界面に作用するとも思われる。G−CSFレセプターの結合は、この領域の310螺旋を不安定にし、リガンド/G−CSFレセプター複合体の結合エネルギーを改善するコンフォーメーション変化をもたらす。
【0155】
hGHレセプター複合体において、螺旋束の第1の螺旋はhGHレセプターを二量体化するために必要な残基を結合部位の両方に与える。G−CSFの対応する螺旋(螺旋A)の突然変異分析の結果、生物活性に必要な3個の残基が確認された。これらの3個の残基のうちでGlu20及びArg24は螺旋Cに向かう螺旋束の1つの表面に位置し、(非対称単位における3分子のうちの2分子に存在する)Arg23の側鎖は螺旋Dに向かう束の表面に向いている。これらの生物学的に重要な残基の側鎖の位置は、hGHの場合と同様にG−CSFが螺旋A及び螺旋C間の界面に沿って第2のG−CSFレセプター結合部位を有することを示す。hGH分子とは対照的に、最初の11個の残基の欠失は生物活性にほとんど影響しないので、G−CSFのアミノ末端は制限された生物機能を有する。
【0156】
上述のように(例えば図2参照)、G−CSFは他のサイトカインとトポロジー類似性を有する。構造と上記生物化学試験との相関、突然変異分析及びhGHレセプター複合体の特定残基の直接比較によると、G−CSFは2つのレセプター結合部位を有する。部位Aは螺旋A及びDの界面に沿って存在し、小さいAB螺旋中に残基を含む。部位BもA螺旋中に残基を含むが、螺旋A及びCの間の界面に沿って位置する。G−CSFとhGHとの間の生物学的に重要な残基の構造と相対位置の保存は、共通のシグナル変換方法でレセプターが2地点で結合されることを示す1つの指標である。従って、G−CSFレセプター結合部位(残基20〜57及び145〜175)のいずれか一方の改変により、改変G−CSFレセプター結合ドメインを有するG−CSF類似体を製造できるとみなされる。
【0157】
G−CSFタンパク質の三次元構造の知見と組成の相関により、G−CSF類似体の体系的合理的製造方法が可能になる。上記実施例から明らかなように、構造のコア内の側鎖の寸法及び極性の制限は、全体構造が変化する前に分子がどの程度の変化に耐え得るかを決定する。
【技術分野】
【0001】
本発明は顆粒球コロニー刺激因子(“G−CSF”)類似体、前記類似体を含む組成物、及び関連組成物に関する。別の態様によると、本発明は本発明の類似体をコードする核酸又は関連核酸、関連宿主細胞及びベクターに関する。別の態様によると、本発明はG−CSF及びその類似体の三次元構造を表現するためのコンピュータープログラム及び装置に関する。別の態様によると、本発明はG−CSF類似体及び関連組成物を合理的に設計するための方法に関する。更に別の態様によると、本発明は本発明のG−CSF類似体を使用する治療方法に関する。
【背景技術】
【0002】
造血は、骨髄微小環境内の細胞と成長因子との2つの系により制御される。成長因子はコロニー刺激因子とも呼称され、前駆細胞を刺激して増殖させ、分化性血球のコロニーを形成する。本明細書中でG−CSFと呼称する顆粒球コロニー刺激因子はこれらの因子の1種であり、好中球の成長及び発生を優先的に刺激するので、好中球減少症に使用できると予想される。Welteら,PNAS−USA 82: 1526−1530(1985); Souzaら,Science 232: 61−65(1986)及びGabrilove,J.Seminars in Hematology 26: (2)1−14(1989)。
【0003】
ヒトでは血漿中に内因性G−CSFが検出可能である。Jonesら,Bailliere’s Clinical Hematology 2(1): 83−111(1989)。G−CSFは繊維芽細胞、マクロファージ、T細胞トロホブラスト、内皮細胞及び上皮細胞により産生され、17番染色体に位置する4個のエキソンと5個のイントロンから構成される単一コピー遺伝子の発現産物である。この遺伝子座の転写により生成されるmRNA種は示差的プロセッシングを受け、177アミノ酸のタンパク質をコードする形態と174アミノ酸のタンパク質をコードする形態との2形態のG−CSF mRNAをもたらし(Nagataら,EMBO J 5:575−581(1986))、174アミノ酸からなる形態は最大のin vivo生物比活性を有することが判明した。G−CSFは種交差反応性であるため、ヒトG−CSFを例えばマウス、イヌ又はサル等の別の哺乳動物に投与すると、持続的な好中球白血球増加症が誘発される。Mooreら,PNAS−USA 84: 7134−7138(1987)。
【0004】
ヒトG−CSFは多数の起源から取得及び精製できる。天然ヒトG−CSF(nhG−CSF)は培養ヒト腫瘍細胞系の上清から単離することができる。組換えDNA技術の発達(例えば参考資料として本明細書の一部とする米国特許第4,810,643号(Souza)参照)により、真核宿主細胞発現産物としてグリコシル化形態のG−CSF及び原核宿主細胞発現産物として非グリコシル化形態のG−CSFを商業規模の量で生産することが可能になった。
【0005】
G−CSFは、好中球の増加が有益であるような症状の治療に有用であることが判明した。例えば癌患者にとってG−CSFは、化学療法又は放射線療法に起因する造血欠損を補償するために好中球産生を選択的に刺激する手段として有益である。この他に、細菌代謝物により主に誘発される種々の感染症及び関連症状(例えば敗血症)の治療にも使用できる。更に例えば骨髄移植の目的で培養細胞を成長又は増殖させるために、G−CSFを単独使用又は他の化合物(例えば他のサイトカイン)と併用しても有用である。
【0006】
G−CSFの細胞代謝過程であるシグナル変換は現在、十分に解明されていない。個々の前駆細胞内に変化を誘発すると思われる細胞表面レセプターにG−CSFが結合し、細胞分化に至ると考えられる。
【0007】
種々の改変G−CSFが報告されている。一般に、薬剤を設計するためには、ある種の構造効果を及ぼすように特定の改変を加えることが知られている。例えば、1個のシステインを欠失させると、未改変状態では通常はジスルフィド結合を介して折り畳まれている分子が解放される。この他に、タンパク質の機能を変えるために、アミノ酸を付加、欠失又は置換させる方法も知られている。
【0008】
組換えヒトG−CSF突然変異体は製造されているが、その製造方法は全体構造/機能関係情報を考慮していない。例えば、Cys18の突然変異及び生化学的修飾が報告されている。Kugaら,Biochem.Biophy.Res.Comm 159: 103−111(1989); Luら,Arch.Biochem.Biophys.268: 81−92(1989)。
【0009】
(上記)米国特許第4,810,643号(発明の名称「多能性顆粒球コロニー刺激因子の製造」)には、G−CSFのポリペプチド類似体及びペプチドフラグメントが広く開示されている。特定のG−CSF類似体としては、(174アミノ酸種又はこれにN末端メチオニンを付加した175アミノ酸種の)17、36、42、64及び74位のシステインを別のアミノ酸(例えばセリン)で置換した類似体や、最初の(N末端)位置にアラニンを有するG−CSFが開示されている。
【0010】
ヨーロッパ特許第0335423号(発明の名称「改変ヒトG−CSF」)は、hG−CSF活性を有するポリペプチドにおける少なくとも1個のアミノ酸の修飾を開示しているらしい。
【0011】
ヨーロッパ特許第0272703号(発明の名称「新規ポリペプチド」)は、N末端又はその「近傍」のアミノ酸を置換又は欠失させたG−CSF誘導体を開示しているらしい。
【0012】
ヨーロッパ特許第0459630号(発明の名称「ポリペプチド」)は、天然に存在するG−CSFの生物学的特性の少なくとも1種と5mg/mlで少なくとも35%の溶液安定性とを有する天然に存在するG−CSFの誘導体を開示している模様であり、該誘導体は、少なくとも天然配列のCys17がSer17残基で置換され、天然配列のAsp27がSer27残基で置換されている。
【0013】
ヨーロッパ特許第0256843号(発明の名称「G−CSFとそのムテインの発現及びその使用」)は、タンパク質のアミノ酸配列を変えずに組換え宿主におけるタンパク質の発現を強化するようにN末端を修飾した、G−CSFをコードする改変DNA配列を開示しているらしい。
【0014】
ヨーロッパ特許第0243153号(発明の名称「ヒトG−CSFタンパク質発現」)は、少なくとも1個の酵母KEX2プロテアーゼプロセッシング部位を不活性化することによりG−CSFを修飾すると、酵母を使用した組換え生産収率を増加できると開示しているらしい。
【0015】
Shawの米国特許第4,904,584号(発明の名称「ポリペプチドの部位特異的均質改変」)はリジン改変タンパク質を開示しているらしい。
【0016】
WO/9012874号は、タンパク質のシステイン改変変異体を開示しているらしい。
【0017】
オーストラリア特許出願第AU−A−10948/92号(発明の名称「組換えタンパク質の活性化の改善」)は、原核発現後の分子の折り畳みの助長を目的としてG−CSF分子の各末端にアミノ酸を付加することを開示しているらしい。
【0018】
オーストラリア特許出願第AU−A−76380/91号(発明の名称「顆粒球コロニー刺激因子(G−CSF)のムテイン」)は、174アミノ酸を有するG−CSFの50〜56位及び177アミノ酸を有するG−CSFの53〜59位の配列Leu−Gly−His−Ser−Leu−Gly−Ile、及び/又は174アミノ酸を有する成熟G−CSFの43、79、156及び170位もしくは177アミノ酸を有する成熟G−CSFの46、82、159及び173位の4個のヒスチジン残基の少なくとも1個における顆粒球刺激因子G−CSFのムテインを開示しているらしい。
【0019】
英国特許第2213821号(発明の名称「合成ヒト顆粒球コロニー刺激因子遺伝子」)は、選択された領域のカセット突然変異誘発を助長するための制限部位と、所望の発現系への遺伝子の組み込みを助長するためのフランキング制限部位とを含む合成G−CSFコーディング核酸配列を開示しているらしい。
【0020】
G−CSFは、ある程度まで結晶化されたと報告されており(例えばヨーロッパ特許第344796号)、G−CSFの全体構造は推定されているが、粗レベルに止まっている。Bazan,Immunology Today 11: 350−354 (1990); Parryら,J.Molecular Recognition 8: 107−110(1988)。G−CSFの全体構造は未だ報告されておらず、また、G−CSF類似体の体系的設計には分子の全体構造と機能の関係の体系的研究が必須であるが、このような研究も報告されていない。従って、G−CSF類似体のこの体系的設計方法及び該方法により得られる組成物が必要とされている。
【先行技術文献】
【特許文献】
【0021】
【特許文献1】米国特許第4,810,643号明細書
【特許文献2】欧州特許第0335423号明細書
【特許文献3】欧州特許第0272703号明細書
【特許文献4】欧州特許第0459630号明細書
【特許文献5】欧州特許第0256843号明細書
【特許文献6】欧州特許第0243153号明細書
【特許文献7】米国特許第4,904,584号明細書
【特許文献8】国際公開第90/12874号パンフレット
【特許文献9】豪国特許出願第10948/92号明細書
【特許文献10】豪国特許出願第76380/91号明細書
【特許文献11】英国特許第2213821号明細書
【特許文献12】欧州特許第344796号明細書
【非特許文献】
【0022】
【非特許文献1】Welteら,PNAS−USA 82: 1526−1530(1985)
【非特許文献2】Souzaら,Science 232: 61−65(1986)
【非特許文献3】Gabrilove,J.Seminars in Hematology 26: (2)1−14(1989)
【非特許文献4】Jonesら,Bailliere’s Clinical Hematology 2(1): 83−111(1989)
【非特許文献5】Nagataら,EMBO J 5:575−581(1986)
【非特許文献6】Mooreら,PNAS−USA 84: 7134−7138(1987)
【非特許文献7】Kugaら,Biochem.Biophy.Res.Comm 159: 103−111(1989)
【非特許文献8】Luら,Arch.Biochem.Biophys.268: 81−92(1989)
【非特許文献9】Bazan,Immunology Today 11: 350−354 (1990)
【非特許文献10】Parryら,J.Molecular Recognition 8: 107−110(1988)
【発明の概要】
【0023】
(発明の要約)
本発明により、G−CSFの三次元構造は原子レベルまで決定されるに至った。この三次元構造から、G−CSF分子の組成がどのように変化すると構造変化を生じるかをかなり確実に予想することができる。これらの構造特性を生物活性と相関させると、G−CSF類似体を設計及び製造することができる。
【0024】
G−CSFの三次元構造については他にも考察されている(Bazan,Immunology Today 11: 350−354(1990); Parryら,J.Molecular Recognition 8: 107−110(1988))が、推定構造が不正確(Parryら,前出)及び/又は推定構造から構成部分と構造の相関の詳細が得られないという理由により、これらの考察はG−CSF類似体を製造しようと意図する人々には何ら役立たなかった。三次元構造を原子レベルまで決定するという本発明の構造分析は従来の最も完全な分析よりも遥かにすぐれており、G−CSF類似体を設計及び製造しようとする人々に重要な情報を提供する。例えば、本発明の三次元構造分析から、厳密な疎水性及び親水性領域が決定された。
【0025】
相対疎水性は分子の安定性に直接関係するので重要である。一般に、水性環境中に存在する生物分子は外部が親水性であり、内部が疎水性であり、熱力学の第2法則によると、これは最低エネルギー状態であり、安定性を提供する。G−CSFの内部コアは疎水性であり且つ外部領域は親水性であると予想することができるが、特定の疎水性又は親水性領域を知る術はなかった。本発明により提供された疎水性/親水性領域の知識に基づき、G−CSF分子のどのような変化が分子の全体構造に影響するかをかなり確実に予想することができる。
【0026】
一般に、G−CSF全体構造を変化させずに生物活性を変化させるような類似体を設計するために、疎水性及び親水性領域の位置の知識を使用することができる(本明細書中では「生物活性」なる用語は機能を表すために最も広義に使用する)。生物活性を構造に相関させることができる。構造が変化せず且つ突然変異が生物活性に影響しないならば、突然変異は生物学的機能をもたない。他方、構造が変化せず且つ突然変異が生物活性に影響するならば、残基(又は原子)は少なくとも1種の生物学的機能に不可欠である。本発明の実施例のいくつかは全体構造を変えずに生物学的機能を変化させるように意図した。
【0027】
構造と生物活性との相関に基づき、本発明の1態様はG−CSF類似体に関する。これらの類似体は、非常に少数のアミノ酸残基をG−CSFアミノ酸配列と相異させるか又は改変した分子である。改変は1個以上のアミノ酸残基の付加、置換又は欠失であり得る。改変は、アミノ酸自体の類似体(例えばペプチド類似体)又は改変部分(例えば改変側基)を有するアミノ酸の付加又は置換を含み得る。比較の基準として使用されるG−CSFはヒト、動物又は組換え核酸技術起源であり得る(本明細書中の実施例は付加的N末端メチオニル残基を有するヒトG−CSFの174アミノ酸種の組換え製造に基づく)。類似体は天然ヒトG−CSF分子と異なる機能を有してもよいし、同一機能を示してもよいし、同一機能を別程度で示してもよい。例えば、類似体は改変前よりも生物活性が高くても低くてもよいし、貯蔵寿命を延ばしてもよいし、安定性を低下させてもよいし、調合し易くしてもよいし、他の成分と結合しにくくしてもよい。類似体は造血活性をもたなくてもよく、従って、(例えばG−CSFの過産生の場合等のように)G−CSF効果に対するアンタゴニストとして有用であり得る。本明細書中では本発明の類似体を便宜上タンパク質又はペプチドと呼称する場合もあるが、本発明はペプチド類似体又は化学的に修飾されたペプチド等の他の型の分子も包含する。
【0028】
別の態様によると、本発明はG−CSF類似体を活性成分として含有する関連組成物に関する。本明細書中で使用する「関連組成物」なる用語は、G−CSF類似体の存在を確認後に得られる組成物(例えば検出可能なラベルで標識したG−CSF類似体、関連レセプター又は医薬組成物)を意味する。少なくとも1個のポリエチレングリコール分子を結合した類似体等の化学的に修飾したG−CSF類似体も関連組成物とみなされる。
【0029】
例えば、検出可能なラベル(例えば蛍光、化学発光又は放射性分子)を結合したG−CSF類似体を製造することができる。
【0030】
別の例は公知材料を使用して公知技術により調剤することが可能な医薬組成物である。例えば、参考資料として本明細書の一部とするRemington’s Pharmaceutical Science, 第18版(1990,Mack Publishing Co., Easton, Pennsylvania 18042)pp.1435−1712参照。一般に、組成は投与法、安定性、製法等の種々の因子及び他の因子に依存する。G−CSF類似体は注射又は吸入による肺投与により投与することができる。本発明のG−CSF類似体組成物には腸溶性剤形も使用可能であり、従って経口投与が有効であり得る。G−CSF類似体をリポソーム又は他の送達用マイクロキャリヤーに組み込んでもよいし、ジェル又は他の徐放用組成物に配合してもよい。好適組成物は組成物の用途により異なるが、一般に天然G−CSFの生物活性の少なくとも1種を有するG−CSF類似体の好適医薬組成物は皮下注射又は吸入による肺投与用に製造した組成物であり、各投与型毎の特定組成は類似体の特性に依存する。
【0031】
関連組成物の別の例は本発明の類似体のレセプターである。本明細書中で使用する「レセプター」なる用語は、本発明の類似体分子と選択的に結合する部分を意味する。例えば、抗体又はそのフラグメント又は「組換え抗体」(Huseら,Science 246:1275(1989))をレセプターとして使用することができる。(本発明は結合特異的レセプターを包含するが)選択的結合は特異的結合のみを意味するものではないものの、どちらかといえば結合はランダムではない。レセプターは細胞表面上又は細胞内もしくは細胞外に位置し、本発明の類似体の生物活性を提供、抑制又は局在化するように機能し得る。レセプター結合は類似体自体と間接的に関係する活性カスケードの始動メカニズムでもあり得る。本発明は更に前記レセプターをコードする核酸、該核酸を含むベクター及び該核酸を含む宿主細胞も包含する。
【0032】
関連組成物の別の例は、化学的部分を結合させたG−CSF類似体である。一般に、化学修飾はタンパク質の生物活性又は抗原性を変えるか、又は他の特性を変えることができ、これらの因子は当業者に考慮されよう。上述のように、このような化学的部分の1例はポリエチレングリコールである。修飾は親水性もしくは疎水性ポリマー分子、脂肪酸分子、又は多糖分子の1種以上の付加を含み得る。化学修飾剤の例としては、ポリエチレングリコール、アルキルポリエチレングリコール、DI−ポリ(アミノ酸)、ポリビニルピロリドン、ポリビニルアルコール、ピランコポリマー、酢酸/アシル化、プロピオン酸、パルミチン酸、ステアリン酸、デキストラン、カルボキシメチルセルロース、プルラン又はアガロースが挙げられる。Francis,Focus on Growth Factors 3: 4−10(May 1992)(Mediscript,Mountview Court, Friern Barnet Lane, London N20 OLD,英国刊)参照。更に、化学修飾は付加的タンパク質もしくはその部分、細胞毒性物質又は抗体の使用を含み得る。化学修飾はレシチンも使用し得る。
【0033】
別の態様によると、本発明は前記類似体をコードする核酸に関する。核酸はDNA又はRNA又はその誘導体であり、典型的には適当な調節配列を含むファージ又はプラスミド等のベクターにクローニングされ、このベクターで発現される。核酸は、例えば診断又は予後の目的で(例えば放射性、化学発光又は蛍光ラベルを使用して)標識してもよい。核酸配列は、例えば細菌発現に好適なコドンを含むなどして発現に最適化することができる。核酸とその相補鎖及び、所望の類似体をコードするのを妨げないようなその改変体も本発明に含まれる。
【0034】
別の態様によると、本発明は本発明の類似体をコードする上記核酸を含む宿主細胞に関する。宿主細胞は真核細胞でも原核細胞でもよく、発現系は糖基の結合(グリコシル化)(又は阻止)、分子の適正な折り畳み、組換え発現に付随するリーダー配列もしくは他の因子の付加又は欠失に関する追加の段階を含み得る。
【0035】
別の態様によると、本発明はこのような核酸配列の発現の型又は量を抑制又は変更するように機能するアンチセンス核酸に関する。これらの核酸は公知方法により製造することができる。
【0036】
本発明の別の態様によると、本発明の類似体をコードする核酸は、例えば類似体をコードする配列を含むベクターを受容体に導入し、類似体組成物を必要とする受容体の体内で核酸自体を発現させることにより、遺伝子療法の目的で使用することができる。まず最初にベクターを細胞等のキャリヤーに導入後、キャリヤーを受容体に導入する。このような発現は局在化してもよいし、全身でもよい。他のキャリヤーとしては、受容体への遺伝子移入を媒介するように機能し得る非天然に存在するキャリヤー(例えばリポソーム又は他のマイクロキャリヤーもしくは粒子)が挙げられる。
【0037】
本発明は更に、G−CSF又は類似体の三次元構造を表現(例えば視覚表示)するためのコンピュータープログラム及び、G−CSF分子の各成分の存在と全体構造内の原子レベルまでの該成分の厳密な位置を表現するコンピュータープログラムを提供する。このようなプログラムの1例を以下に説明する。分子の三次元構造を表現するためのコンピュータープログラムは多数のものが現在市販されている。一般に、これらのプログラムは分子の三次元構造の座標(即ちx、y及びz軸に沿うG−CSF分子の各原子の数値帰属)の入力手段、前記座標の表現手段(例えば視覚表示)、前記座標の修正手段、及びこのような修正座標を有する分子の画像の表現手段を提供する。結晶情報即ち、G−CSF分子の結晶分析から得られた三次元空間におけるG−CSF分子の原子の位置の座標をプログラムすると、G−CSF三次元構造の表現(例えば視覚表示)のためのコンピュータープログラムを作成することができる。従って、G−CSF類似体三次元構造の表現用コンピュータープログラムも提供される。San Diego,Californiaに所在のBiosymから市販されているコンピュータープログラムInsight II,version 4に図5に示すような座標を入力すると好適である。好適表現手段としては、Silicon Graphics 320 VGXコンピューターでG−CSF分子又はその類似体を立体的に観察することが可能なCrystal Eyes眼鏡(Silicon Graphicsの市販品)を着用する。また、本発明のG−CSF結晶座標及び回折データは、米国、Upton,New York 119723に所在のProtein Data Bank,Chemistry Department,Brookhaven National Laboratoryに寄託されている。これらのデータを使用してG−CSF分子又はその類似体の三次元構造の表現用の別のコンピュータープログラムを作成することもできる。従って、本発明の別の態様は、G−CSF分子の三次元構造の表現用コンピュータープログラムである。G−CSF分子の三次元構造の視覚表示用コンピュータープログラムも提供され、更に、該プログラムはこのような視覚表示を変更するための手段を有する。このようなコンピュータープログラムの表現に有用な装置、特にG−CSF分子又はその類似体の前記三次元構造のコンピューター画像の視覚表示に有用な装置と、前記コンピュータープログラム及び装置を製造するための手段も提供される。
【0038】
G−CSF分子上の被改変特定部位を選択することができ、この改変がG−CSF分子の全体構造に及ぼす影響を容易に確認することができるので、コンピュータープログラムはG−CSF類似体を製造するために有用である。前記被改変部位の選択は、G−CSF類似体の所望の生物特性に依存する。前記G−CSF分子(r−met−hu−G−CSF)をランダムに改変したならば、17520通りもの置換が考えられ、複数の置換、付加又は欠失を有する類似体の数は更に多い。分子の組成と相関させながら三次元構造を観察することにより、改変部位をランダムに選択する必要がなくなり、改変部位を合理的に決定することができる。
【0039】
上述のように、各成分の原子レベルまでの位置決定を含めてG−CSFの三次元構造が確認されたので、どの部分がG−CSF分子の全体構造を維持するために必要であるかに関する情報を得られるようになった。従って、本発明のG−CSF類似体の製造時にG−CSF分子の全体構造を維持すべきか、又は本発明のG−CSF類似体の製造時にG−CSF分子の全体構造を変更すべきか(及びどのように変更すべきか)を選択することができる。場合により、このような類似体を製造後、このような類似体が所望の特性を有するか否かを試験することができる。
【0040】
例えば、非改変天然又は組換えG−CSF分子の全体構造を維持しようとすることができる。全体構造を図2、3及び4に示し、以下に詳細に説明する。全体構造の維持により、天然G−CSFの造血能を有する類似体に必要な特性であるレセプター結合を確保することができる(レセプター結合が存在しないならば、類似体の存在からシグナル変換は行われない)。あるクラスのG−CSF類似体は天然又は組換え(非改変)G−CSF分子の三次元コア構造を有しながら、好中球を選択的に刺激する能力の増加等の別の特性も有すると予想される。別のクラスのG−CSF類似体は別の全体構造を有する類似体であり、G−CSF類似体分子とG−CSFレセプターの結合能が低く、非改変天然又は組換えG−CSFに比較して好中球を選択的に刺激する能力が低い。
【0041】
例えば、こうしてG−CSF分子の内部領域内のどの部分が疎水性であるかが分かり、これに対応してG−CSF分子の外部のどの分子が親水性であるかが分かる。好ましくは本発明のように原子レベルまで全体三次元構造を認識できなければ、この疎水性内部領域内のどの改変が分子の全体構造コンフォーメーションの変化をもたらすかを予測することはできない。全体構造が変化すると、例えばレセプター結合の欠損等の機能的変化が生じ、従って、非改変G−CSFに認められるような生物活性の低下が生じ得る。従って、別のクラスのG−CSF類似体は、(非改変)天然又は組換えG−CSFと同一の疎水性を有するG−CSF類似体である。より特定的には、別のクラスのG−CSF類似体は、(非改変)天然又は組換えG−CSFの疎水性部分と同一の疎水性部分をその内部コアの4螺旋束の内側に有しており、且つ前記非改変天然又は組換えG−CSFとは異なる組成を有する。
【0042】
別の例は、G−CSF分子の内部コア(螺旋)と結合する構造である外部ループに関する。アミノ酸残基の空間位置に関する情報を含む三次元構造から、特定のループにおける特定の変化は全体的コンフォーメーション変化をもたらさないと予想することができる。従って、本発明の別のクラスのG−CSF類似体は、改変外部ループを有しており且つ(非改変)天然又は組換えG−CSFと同一の全体構造を有する。より特定的には、本発明の別のクラスのG−CSF類似体は改変外部ループを有しており、該ループは本明細書中のループ及び螺旋の表記に従い、螺旋A及びBの間、螺旋B及びCの間、螺旋C及びDの間、螺旋D及びAの間に存在するループから選択される。より特定的には、前記ループ、好ましくはABループ及び/又はCDループは該ループを安定化させることにより分子の半減期を増加させるように改変されている。このような安定化は、前記ループの全部又は一部をG−CSF(又は類似体)分子のコアに存在するα螺旋束の一部と結合することにより得られる。このような結合は、βシート、塩架橋、ジスルフィド結合、疎水性相互作用又は当業者に使用可能な他の結合手段を介して実施され、このような結合手段は前記外部ループを安定化するように機能する。例えば、ABループを分子の内部領域内の螺旋の1つと結合することによりAB又はCDループを安定化させることができる。
【0043】
N末端は内部螺旋の構造安定性に影響しないので、N末端を改変してもG−CSF分子の全体構造は変化せず、外部ループは改変には好適であるが、同一の一般論がN末端にも当てはまる。
【0044】
また、(非改変)天然又は組換えG−CSFではこのような外部ループは比較的フレキシブルであり、レセプター結合を妨げない傾向があるので、このような外部ループは化学的修飾部位となり得る。従って、更に化学的部分を直接結合(又は化学的結合手段として機能する別の化学的部分を介して間接結合)する余地がある。化学的部分は、G−CSF分子の1種以上の機能の変更に使用可能な種々の部分から選択することができる。例えば、外部ループはポリエチレングリコール分子等のような血清半減期を増加させるように機能する1種以上のポリマーを添加するための部位を提供し得る。このようなポリエチレングリコール分子を加え、ポリエチレングリコール部分と結合可能な反応性側基を有する付加的リジンを含むように前記ループを改変させることができる。他のクラスの化学的部分も1個以上のループに結合することができ、このような化学的部分の非限定的な例としては、他の生物活性分子(例えばレセプター)、他の治療用タンパク質(例えばハイブリッド分子をもたらす他の造血因子)、又は細胞毒性物質(例えばジフテリア毒素)が挙げられる。当然これらの例に留まることなく、所望の化学的部分を有する当業者は、該所望の部分を所望の外部ループに結合することができる。従って、別のクラスの本発明のG−CSF類似体は、少なくとも1個のポリエチレングリコール分子等の化学的部分を付加する少なくとも1つの改変を外部ループに有する類似体を含む。
【0045】
欠失(例えば分子の分解のためにタンパク質により認識される部位の欠失)も外部ループに有効であり得る。こうして、一方ではG−CSFレセプター結合及びシグナル変換能(即ち好中球の突然変異を選択的に刺激する能力)を有する分子の半減期を増加することができる。従って、別のクラスの本発明のG−CSF類似体は、プロテアーゼによる前記類似体の代謝回転を低下させる少なくとも1つの改変を外部ループに有する類似体を含む。この改変に好適なループはABループ及びCDループである。(以下に詳細に説明する)外部ループに存在するアミノ酸残基の一部を欠失させることにより短縮G−CSF分子を製造することができ、該短縮G−CSF分子は製造又は生物学的機能に付加的利点を有する。
【0046】
別の例は、相互に近接するアミノ酸残基間の相対電荷に関する。上述のように、G−CSF分子は比較的密に充填された4螺旋束を含む。螺旋上の面のいくつかは他の螺旋に向き合う。螺旋が別の螺旋に向き合う点(例えば残基)において、相互に向き合う2つのアミノ酸分子は同一電荷を有することができ、従って、相互に反発しあい、分子全体を不安定にする。これは、反発が生じないようにアミノ酸部分の一方又は両方の電荷を(逆電荷又は中性電荷に)変えることによりこれを解決することができる。従って、別のクラスのG−CSF類似体は、電荷位置等の表面相互作用による不安定を修正するように改変されたG−CSF類似体を含む。
【0047】
別の態様によると、本発明はG−CSF類似体及び関連組成物の設計方法並びに該方法の産物に関する。前記方法の終産物は上記G−CSF類似体又は関連組成物であり得る。例えば、本明細書中に開示する実施例は、(a)G−CSF分子の成分(即ち化学的部分)の改変がG−CSF構造に及ぼす作用及び(b)構造変化が生物学的機能に及ぼす作用を立証する。従って、本質的に本発明の別の態様は、
(a)G−CSF分子の各アミノ酸残基又は各アミノ酸残基の各原子等の化学的部分を相関させたG−CSF分子の三次元構造を含む情報を観察する段階と、
(b)前記情報からG−CSF分子上の被改変部位を選択する段階と、
(c)このような改変を有するG−CSF類似体分子を製造する段階と、
(d)任意に、このようなG−CSF類似体分子が所望の特性を有するか否かを試験する段階と
を含むG−CSF類似体の製造方法である。
【0048】
本発明のコンピュータープログラムは、コンピューターに基づくG−CSF類似体の製造方法に使用することができる。従って、本発明の別の態様は、
(a)G−CSF分子の各アミノ酸残基又は各アミノ酸残基の各原子等の化学的部分と相関させたG−CSF分子の三次元構造のコンピューター表現を提供する段階と、
(b)前記情報からG−CSF分子上の被改変部位を選択する段階と、
(c)このような改変を有するG−CSF分子を製造する段階と、
(d)任意に、このようなG−CSF分子が所望の特性を有するか否かを試験する段階と
を含む、コンピューターに基づくG−CSF類似体の製造方法である。
【0049】
より特定的には本発明は、
(a)(i)G−CSF分子の座標を三次元空間に表示し且つ(ii)前記G−CSF表現の変更及びその観察のための情報を入力できるようにプログラムされたコンピューターを介してG−CSF分子の三次元構造を観察する段階と、
(b)前記G−CSF分子の前記視覚画像上の被改変部位を選択する段階と、
(c)前記コンピューターに前記改変のための情報を入力する段階と、
(d)前記コンピューターを介して前記改変G−CSF分子の三次元構造を観察する段階と、
(e)任意に段階(a)〜(e)を繰り返す段階と、
(f)前記改変を有するG−CSF類似体を製造する段階と、
(g)任意に前記G−CSF類似体が所望の特性を有するか否かを試験する段階と
を含む、G−CSF類似体の製造方法を提供する。
【0050】
別の態様によると、本発明は本発明のG−CSF類似体及び関連組成物の使用方法、並びに造血障害治療において単独又は他の造血因子もしくは薬剤と併用して哺乳動物を治療又は予防する方法に関する。G−CSF類似体を設計する1態様は、非改変G−CSFが有するとして知られている特性を増加又は変更することを目的とすると意図される。
【0051】
例えば、本発明の類似体は強化又は変更された活性を有するので、G−CSFが(例えば)好中球減少症の治療に有用である場合には本発明の組成物及び方法は同様にこのような治療に使用することができる。
【0052】
別の例は、特に造血障害の治療で他の因子と併用した場合により有効に相互作用することを目的としたG−CSFの改変である。このような併用の1例は、初期作用性造血因子(即ち造血カスケードの初期に比較的未分化の細胞に対して作用する因子)の使用及び後期作用造血因子、例えばG−CSF又はその類似体(G−CSFは好中球の選択刺激においてCFU−GM系に作用する)の同時又は順次使用である。本発明の方法及び組成物は、造血因子のこのような組み合わせ又は「カクテル」を使用する療法で有用である。
【0053】
本発明の組成物及び方法は、白血球減少症、骨髄性白血病、重度慢性好中球減少症、再生不能性貧血、糖原病、粘膜炎及び他の骨髄不全症状の治療にも有用である。本発明の組成物及び方法は、化学療法又は放射線療法に起因する造血欠損の治療にも有用であり得る。例えば骨髄移植の成功又は移植における末梢前駆血球の使用は、本発明の組成物(遺伝子療法の場合にはタンパク質又は核酸)及び方法を適用することにより増進することができる。本発明の組成物及び方法は、創傷治癒、火傷治療、菌血症、敗血症、真菌感染、心内膜炎、骨腎盂炎、腹部外傷に関連する感染、抗生物質に反応しない感染、肺炎等の感染症の治療にも有用であり、本発明の組成物及び方法を細菌炎症の治療に利用することもできる。更に、本発明の組成物及び方法は白血病細胞分化能の報告(Welteら,PNAS−USA 82: 1526−1530(1985))に基づき白血病の治療にも有用であり得る。この他に、腫瘍細胞に結合するレセプター(例えば抗体)の任意存在下で本発明の組成物及び方法を使用して腫瘍個体を治療することもできる。治療用途に関する研究論文については、いずれも参考資料として本明細書の一部とするLieshhkeとBurgess,N.Engl.J.Med.327: 28−34及び99−106(1992)を参照されたい。
【0054】
本発明の組成物及び方法は、他の部分の製造の中間体として使用するのにも有用であり、例えば、G−CSFは他の造血因子の生成に影響を及ぼすことが報告されており、(確認されたならば)この機能は本発明の組成物及び/又は方法により増進又は変更することができる。
【0055】
本発明のG−CSF類似体に関連する組成物(例えばレセプター)は、G−CSF又は類似体の活性を阻止するアンタゴニストとして作用するのにも有用である。非改変G−CSF又はG−CSF類似体の活性の一部又は全部を有する組成物を得ることができ、1種以上の化学的部分を付加してこのようなG−CSF又は類似体の1種以上の特性を改変することができる。三次元コンフォーメーションの知識に基づき、所望の効果に達するためにこのような化学修飾の最良の幾何学的位置を予測することができる。
【0056】
化学修飾の一般目的としては、半減期の改善(例えば腎性クリアランス、免疫クリアランス又は細胞クリアランスの減少)、生物活性の変更(例えば酵素特性の変更、有機溶媒中の生物活性又は活性の分離)、毒性の低下(例えば毒性エピトープの隠蔽、区画化及び選択的生物分布)、免疫反応性の改変(免疫原性の低下、抗原性又はアジュバント作用の低下)、又は物性の低下(例えば溶解度の増加、熱安定性の改善、機械的安定性の改善又はコンフォーメーション安定化)が挙げられる。Francis,Focus on Growth Factors 3: 4−10(May 1992)(Mediscript,Mountview Court,Friern Barnet Lane,London N20 OLD,英国刊)参照。
【0057】
以下、実施例により本発明を非限定的に説明する。当業者は種々の変形及び変更が可能であり、請求の範囲は本発明の範囲に該当するこのような全等価態様を包含する。
【0058】
(発明の詳細な説明)
本発明はG−CSFの三次元構造の知見に基づく。この三次元構造は立体的に観察するためのコンピュータープログラムにより表現された。これを立体的に観察することにより、構造−機能関係を認識し、G−CSF類似体を設計及び製造した。
G−CSFの三次元全体構造
構造を確認するために使用したG−CSFは、細菌発現に付随する付加的N末端メチオニン残基を有する非グリコシル化174アミノ酸種であった。このG−CSFのDNA及びアミノ酸配列を図1に示す。
【0059】
全体として、G−CSFの三次元構造は主に螺旋状であり、175個の残基のうちの103個は4α螺旋束を形成する。他の二次構造は最初の2つの長い螺旋の間のループに存在し、4残基310螺旋の直後には6残基α螺旋が存在する。図2に示すように、全体構造を他のタンパク質、即ち成長ホルモン(Abdel−Meguidら,PNAS−USA 84: 6434(1987)及びVosら,Science 255: 305−312(1992))、顆粒球マクロファージコロニー刺激因子(Diederichsら,Science 254: 1779−1782(1991))、インターフェロン−β(Sendaら,EMBO J.11: (3193−3201))、インターロイキン−2(McKay Science 257: 1673−1677(1992))、及びインターロイキン−4(Powersら,Science 256: 1673−1677(1992)、及びSmithら,J.Mol.Biol.224: 899−904(1992))に報告されている構造と比較した。アミノ酸配列は類似していないにも拘わらず、これらの成長因子間には構造類似性が存在する。
【0060】
構造情報はG−CSF生化学の相関であり、これは以下のように要約することができる(配列位置1はN末端とする)。
【0061】
【表1】
【0062】
抗体結合試験から収集したこの生化学情報(Laytonら,Biochemistry 266: 23815−23823(1991)参照)を三次元構造に重ね合わせてG−CSF類似体を設計した。これらのG−CSF類似体の設計、製造及び試験を下記実施例1に説明する。
【図面の簡単な説明】
【0063】
【図1】付加的N末端メチオニンを有するG−CSFの174アミノ酸種のアミノ酸配列図である(配列番号1)(配列番号2)。
【図2A】G−CSFの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2B】hGHの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2C】pGHの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2D】GM−CSFの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2E】INF−Bの結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。螺旋の元の分類を括弧内に示す。
【図2F】IL−2の結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図2G】IL−4の結晶構造のトポロジー図である。これらの図は引用文献を参考にした。二次構造元素の長さは残基の数に比例して表した。A、B、C及びD螺旋はG−CSFについて本発明で使用する方式に従って分類した。
【図3】G−CSFの三次元構造の「リボン図」である。螺旋Aはアミノ酸残基11〜39(上記図1による番号付け)、螺旋Bはアミノ酸残基72〜91、螺旋Cはアミノ酸残基100〜123、螺旋Dはアミノ酸残基143〜173である。比較的短い310螺旋はアミノ酸残基45〜48に位置し、α螺旋はアミノ酸残基48〜53に位置する。残基93〜95は左回り螺旋のほぼ1回転を形成する。
【図4】G−CSFの三次元構造の「バレル図」である。G−CSFの三次元構造の全体円筒体とその向きを種々の濃淡で示す。数字は図1によるアミノ酸残基位置を示す。
【図5A】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5B】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5C】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5D】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5E】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5F】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5G】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5H】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5I】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5J】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5K】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5L】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5M】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5N】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5O】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5P】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5Q】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5R】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5S】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5T】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5U】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5V】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5W】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5X】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5Y】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5Z】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5AA】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5BB】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5CC】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5DD】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5EE】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5FF】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5GG】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5HH】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5II】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5JJ】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5KK】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5LL】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5MM】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5NN】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5OO】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5PP】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5QQ】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5RR】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5SS】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5TT】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5UU】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5VV】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5WW】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5XX】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5YY】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5ZZ】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5AAA】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5BBB】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5CCC】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5DDD】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5EEE】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図5FFF】G−CSFの三次元構造のコンピューターによる視覚画像を生成するために使用される座標のリストである。座標は以下の通りである。各列は別個フィールドに対応する。(i)(左側から)フィールド1は原子、(ii)フィールド2は帰属原子の番号、(iii)フィールド3は原子名(周期表標準命名法により、CBは炭素原子β、CGは炭素原子γ等である)、(iv)フィールド4は残基型(例えばStryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y.1988、裏内表紙に示されるようなアミノ酸の三文字命名法による)、(v)フィールド5〜7は原子のx軸、y軸及びz軸位置、(vi)フィールド8(多くの場合には“1.00”)は前記位置における占有度、(vii)フィールド9はB因子、(viii)フィールド10は分子表示を示す。G−CSFの3分子(a、b及びc)は1単位として相互に結晶した。a、b又はcの表示はどの座標がどの分子に由来するかを示す。文字の後に記載した数字(1、2又は3)は、帰属アミノ酸残基位置を示し、分子Aは10〜175位を割り当てられ、分子Bは210〜375位を割り当てられ、分子Cは410〜575位を割り当てられている。これらの位置は、相互に結晶した3分子がオーバーラップしないように指定した。(W表示は水を表す)。
【図6】結晶化に関与するパラメーターに関して結晶化マトリックスの最適化に使用されるストラテジーを示す。結晶化マトリックスは24穴組織培養プレートのウェル中の結晶化溶液の成分(塩、緩衝液及び沈殿剤)の最終濃度に対応する。これらの濃度は、適量のストック溶液をマイクロタイタープレートのウェルにピペット分注することにより生成される。マトリックスを設計するために、結晶学者は成分の高低濃度を決定する。これらの高低濃度を行(例えばA1〜A6、B1〜B6、C1〜C6又はD1〜D6)又は全トレー(A1〜D6)に沿ってピペット分注することができる。前者方法は制限された数のウェルに沿って単一成分の結晶成長の再現性を調べるために有用であり、後者方法は初期スクリーニングで有用である。結晶化マトリックスの数段の最適化段階の結果を3つのプレートの図により示す。ウェルの色が濃いほど正の結晶化結果を表し、最終段階ではX線品質結晶であり、初期段階では約0.05mm未満の寸法の油滴、顆粒状沈殿又は小結晶であり得る。Aは最初のウェル(A1)と最後のウェル(D6)の間の濃度範囲が広く且つウェル間の濃度増加が((濃度A1)−(濃度D6))/23)として計算される1パラメーターの初期スクリーンを示す。Bは、濃度の結晶化マトリックス最適化の後期段階でA1とD6の間の差が縮まり、プレート当たりに形成される結晶が増加していることを示す。Cはプレートの大部分のウェルに高品質結晶が存在するマトリックス最適化の最終段階を示す。
【実施例1】
【0064】
本実施例は結晶質G−CSFの製造、コンピューター生成画像による組換えヒトG−CSFの三次元構造の可視化、特定部位突然変異誘発又は核酸増幅法を使用した類似体の製造、G−CSF類似体を分析するために使用した生物アッセイ及びHPLC分析、並びに全体構造/機能関係の決定について説明する。全引用文献は参考資料として本明細書の一部とする。
【0065】
A.自動結晶化の使用
組換えヒト顆粒球コロニー刺激因子(r−hu−G−CSF)の三次元構造が必要であることと、大量の精製タンパク質が入手可能であることから、不完全階乗サンプリング及び接種による結晶成長法を実施した。JancarikとKim,J.Appl.Crystallogr.24: 409(1991)に記載されている不完全階乗結晶化の実施から出発して、油滴及び複屈折凝集体を生成する溶液条件を確認した。また、自動ピペッティングシステムのソフトウェア及びハードウェアを改造して毎日約400の異なる結晶化条件を設定した。Weber,J.Appl.Crystallogr.20: 366−373(1987)。この手順により、r−hu−G−CSF結晶を生成する結晶化溶液を調製した。
【0066】
結晶の寸法、再現性及び品質は、「核生成開始単位」の数を接種溶液の連続希釈により推定する接種法により改善された。これらの方法により、2.0mmのr−hu−G−CSF結晶を再現可能に成長させることができた。これらの結晶の空間群はP212121であり、セル寸法はa=90Å、b=110Å及びc=49Åであり、2.0Åの分解能まで回折する。
【0067】
1.全体的手法
新規タンパク質の結晶化条件を探索するために、CarterとCarter,J.Biol.Chem.254: 122219−12223(1979)は不完全階乗法を提案した。同氏らによると、一般に有望であるとみなされるランダムに選択された多数の結晶化条件のサンプリングにより、タンパク質結晶化をもたらす好適な試薬組み合わせが得られる。このアイデアは、JancarikとKim,J.Appl.Crystallogr.24: 409(1991)により実現され、同氏らはpH範囲、塩及び沈殿剤を考慮した初期結晶化試験のための32種の溶液を記載した。本発明では同氏らの作業を拡張し、70種までの溶液を記載する。溶液調製の人的労力及び誤差を最小限にするために、自動ピペッティング機にあわせて手法をプログラムした。
【0068】
Weberの連続自動格子探索法(SAGS),J.Cryst,Growth 90: 318−324(1988)に従い、ロボットシステムを使用して温度、pH、塩及び沈殿剤の結晶化条件を連続的に最適化する一連の溶液を調製した。結晶を再現可能に成長させることが可能な溶液が決定されると、結晶の品質を著しく改善する接種技術が開発された。これらの方法を組み合わせると、数百種の回折品質結晶(少なくとも約2.5Åに回折し、好ましくは少なくとも部分的に2Å未満、より好ましくは約1Åに回折する部分を有する結晶)が数日で生成された。
【0069】
一般に、結晶化を所望する任意のタンパク質と共に使用され得る結晶化方法は、
(a)(i)異なる塩濃度を各々有する塩溶液のアリコート又は(ii)異なる沈殿剤濃度を各々有する沈殿剤溶液のアリコートと所望のタンパク質の水性アリコートとを一定範囲のpHの任意存在下で結合する段階と、
(b)こうして結合したアリコートの予備結晶形成を観察し、予備結晶形態の生成に有効な前記塩もしくは沈殿剤組み合わせ及び前記pHを選択するか、又は予備結晶形態が生成されない場合には、タンパク質の前記水性アリコートのタンパク質出発濃度を増加させる段階と、
(c)前記塩又は前記沈殿剤濃度を選択後、前記選択された濃度の存在下で前に選択しなかった前記溶液を用いて段階(a)を繰り返す段階と、
(d)所望の品質の結晶が得られるまで段階(b)及び段階(a)を繰り返す段階と
を含む。
【0070】
上記方法は任意に自動化することができ、時間と労力を著しく節約することができる。好適タンパク質出発濃度は10mg/ml〜20mg/mlであるが、この出発濃度はタンパク質により異なる(下記G−CSFは33mg/mlを使用して分析した)。分析を開始するために好適な塩溶液範囲は0〜2.5M(NaCl)である。好適沈殿剤はポリエチレングリコール8000であるが、他の沈殿剤としては、有機溶媒(例えばエタノール)、分子量500〜20,000のポリエチレングリコール分子、及び当業者に公知の他の沈殿剤を挙げることができる。好適pH範囲はpH4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5及び9.0である。予備結晶化形態は、油状物、複屈折沈殿、小結晶(<約0.05mm)、中結晶(約0.5〜.5mm)及び大結晶(>約0.5mm)を含む。結晶構造を検出するまでの好適待機時間は48時間であるが、週に一度の観察でも好適であり、一般には約1カ月後に別のタンパク質濃度を使用する(一般にタンパク質濃度を増加させる)。自動化はAccuflexシステムを改造すると好適である。好適自動化パラメーターを以下に説明する。
【0071】
一般に、濃度10mg/ml〜20mg/mlのタンパク質を0〜2.5MのNaCl溶液と組み合わせ、上記濃度範囲の存在下で(別々に)このような各組み合わせを実施した。予備結晶化構造が観察されたら、所望の結晶品質に達するまで前記塩濃度及びpH範囲を別実験で最適化させる。次いで、pHレベルを変えながら沈殿剤濃度も最適化させる。両者が最適化されたら、最適条件を同時に実施し、所望の結果を得る(これを図6に示す)。
【0072】
a.自動ピペッティングシステムの実施
標準シリアルインターフェースを介してASCIIコードを送信するパーソナルコンピューターにより制御されるAccuflexピペッティングシステム(ICN Pharmaceuticals,Costa Mesa,CA)により、液滴及び液溜溶液を調製した。ピペッターは回転バルブにより6種の異なる溶液をサンプリングし、xy座標系内の並進を制御可能なプレートにこれらの溶液をピペット分注する。座標系の鉛直成分は液体を分配及び回収することが可能な注射器を操作する。
【0073】
Accuflexのソフトウェアは、CoxとWeber,J.Appl.Crystallogr.20: 366−373(1987)により提案されているようなSAGS法を利用した。この方法は、他の2つのパラメーターを変化させながら、pH及び沈殿剤濃度という2つの主要な結晶化パラメーターを体系的に変化させるものである。これらのコンセプトに基づきながら、本発明で使用するソフトウェアは自動格子探索ストラテジーで使用される結晶化溶液の設計及び実現により高いフレキシビリティーを提供する。このフレキシビリティーの結果として、本発明のソフトウェアはより多数の異なる溶液を生成することができた。これは、下記不完全階乗法の実施に不可欠である。
【0074】
自動格子探索ストラテジーの速度及び設計を改善するために、Accuflexピペッティングシステムはソフトウェア及びハードウェアの改造が必要であった。ハードウェアの改造により、懸垂液滴(hanging drop)実験用トレーと、放置液滴(sitting drop)実験用トレーとの2種の異なるマイクロタイタートレーと、24種の付加的緩衝液、塩及び沈殿剤溶液を保持するPlexglasトレーを使用できるようになった。これらの付加溶液により、調査可能な結晶化条件の格子を拡大した。
【0075】
ハードウェア改造を利用するために、ピペッティングソフトウェアを2つのサブルーチンに書き込んだ。結晶学者は一方のサブルーチンにより成分の濃度に基づいて結晶化溶液のマトリックスを設計することができ、第2のサブルーチンにより適正容量の溶液を結晶化トレーにピペット分注するコンピューターコードにこれらの濃度を翻訳することができる。濃度マトリックスは2種のプログラムのどちらかを使用して作成することができる。第1のプログラム(Thousand Oaks,CAに所在のAmgen Inc.の市販品MRF)は結晶学者により供給されるストック溶液濃度のリストに関するものであり、指定濃度に達するために必要なピペット分注容量を計算する。より好適な第2の方法は、沈殿剤の勾配又はpHを改善するために使用可能なスプレッドシートプログラム(Lotus)を組み込む。これらのプログラムにより生成される濃度マトリックスは制御プログラム(Accuflexピペッターに元々内蔵されているプログラムの改造であり、Thousand Oaks,CAに所在のAmgen,Inc.の市販品SUX)により翻訳され、ウェルはこれに従って充填される。
【0076】
b.不完全階乗法の実施
改造ピペッティングシステムは種々の溶液を調製するのに便利であるため、拡張不完全階乗法の実施を改善することができた。3つの変数から1つの因子の96のランダムな組み合わせを含むリストを作成するプログラムINFAC(Carterら,J.Cryst.Growth 90: 60−73(1988))を使用して「ランダム」な成分を有する結晶化溶液の新しい組合わせを展開した。即座に沈殿したカルシウムとリン酸の組み合わせを除くと、沈殿剤、塩及び緩衝液の70種の異なる組み合わせが残った。自動ピペッターを使用してこれらの組み合わせを調製し、1週間インキュベートした。混合物を検査し、沈殿を形成した溶液をより低い成分濃度で再び調製した。全ウェルから沈殿が除去されるまでこれを繰り返した。
【0077】
c.r−hu−G−CSFの結晶化
X線品質結晶を生成する溶液を見いだすために数種の異なる結晶化ストラテジーを使用した。これらのストラテジーは、不完全階乗法の使用、連続自動格子探索(SAGS)を使用した結晶化条件の最適化、接種技術の実施、及び数百の品質結晶を一晩で生成する結晶生成手順の開発を含むものであった。特に指定しない限り、スクリーニング及びr−hu−G−CSF結晶の生成は懸垂液滴蒸気拡散法を使用した。Afinsenら,Physical principles of protein crystallization.:Eisenberg(編),Advances in Protein Chemistry 41: 1−33(1991)。
【0078】
r−hu−G−CSFの結晶化条件の初期スクリーニングはJancarikとKim,J.Appl.Crystallogr.24: 409(1991)の不完全階乗法を使用し、「予備結晶化」結果をもたらす数種の溶液を調製した。これらの結果は複屈折沈殿、油状物及び非常に小さい結晶(<0.05mm)を含むものであった。これらの予備結晶化溶液をその後、体系的スクリーニングの出発点として使用した。
【0079】
スクリーニングプロセスには結晶化マトリックスの開発が必要であった。これらのマトリックスは結晶化溶液中の成分の濃度に対応し、IBM−PCに基づくスプレッドシートLotus(商標)を使用して生成し、改造Accuflexピペッティングシステムを用いて実施した。マトリックスの設計におけるストラテジーは、他の条件(例えばpH及び沈殿剤濃度)を一定に維持しながら1つの結晶化条件(例えば塩濃度)を変化させることであった。スクリーニングの開始時に変更条件の濃度範囲は広かったが、マイクロタイタートレー中の全ウェルが同一結晶化結果を生じるまで濃度を連続的に最適化した。これらの結果を結晶、複屈折沈殿、顆粒状沈殿、油滴及び非晶質塊の分類により評点した。ある結晶化パラメーターの濃度が少なくとも沈殿を生成しなかった場合には、沈殿が形成されるまでこのパラメーターの濃度を増加させた。各トレーを調製後、少なくとも2日間撹乱せずに静置した後、結晶成長を検査した。この初期スクリーニング後、トレーを週に一度の割合で検査した。
【0080】
このスクリーニングプロセスから、pH及び沈殿剤が同一でありながら塩(MgCl,LiSO4)が異なり、小(0.1×0.05×0.05mm)結晶を生成した2種の別々の溶液を確認した。これらの結果に基づき、他の結晶化パラメーターを一定に維持しながらLiSO4に対してMgClの濃度を変化させた濃度マトリックスの新しい組合わせを作成した。この実験系の結果、約3週間で回折品質結晶(>約0.5mm)を生成する溶液が確認された。この結晶化成長溶液(100mM Mes pH5.8,380mM MgCl2,220mM LiSO4及び8%PEG8k)を見いだすために、約300mgのタンパク質を消費した約8,000の条件をスクリーニングした。
【0081】
結晶の寸法は液滴当たりの結晶形成数に依存した。典型的には平均寸法(1.0×0.7×0.7mm)を有する3〜5個の結晶が形成された。接種(下記参照)を実施したか否かに依存して、等しい空間群(P212121)及び単位セル寸法a=90.2,b=110.2,c=49.5を有する2種の形態が得られた。接種を実施しない場合には、r−hu−G−CSF結晶は1つの長い平坦な表面と丸みを帯びた縁部を有していた。
【0082】
接種を使用した場合には、4〜6時間以内に鋭角面を有する結晶(0.05×0.05×0.05mm)が液滴内に観察された。24時間以内に結晶は(0.7×0.7×0.7mmまで)成長し、液滴中の結晶形成数に依存して2mmを越えるまで成長し続けた。
【0083】
d.接種及び核生成開始部位の決定
本発明の結晶接種方法は、(ここでは最適結晶成長条件を決定後に)使用した各個ウェル中の核生成開始単位数を確定する。この方法は、「種晶」の数が結晶の品質に影響し、分解能に影響するという点で有利である。本発明の接種は更に、接種を実施しない場合には成長に約3週間を要するが、接種を実施すると、G−CSF結晶は約3日で成長するという利点がある。
【0084】
ある生成成長系列(手法の項参照のこと)では、確定された小結晶(<0.01×0.01×0.01mm)のシャワーが一晩で生成された。前に使用したピペット先端を再使用した以外は上記と同様の結晶化条件に従った。恐らく、使用済み先端に形成されており、結晶の核生成部位を提供する小核生成単位により結晶シャワー効果が生じたと予想される。標準生成成長条件下で結晶シャワーを含む少量(0.5μl)の液滴を新しい液滴に加えると、一晩で結晶のシャワーが生成された。この方法を使用して結晶シャワーを含む液滴のトレー数個を作成し、これを「種晶ストック」と名付けた。
【0085】
「種晶ストック」液滴内に含まれる核生成開始単位(NIU)の数を推定し、r−hu−G−CSF結晶の再現性及び品質を改善するように試みた。「種晶ストック」中のNIUの数を決定するために、96穴マイクロタイタープレートに沿って液滴のアリコートを連続希釈した。マイクロタイタープレートは、等容量のr−hu−G−CSF(33mg/ml)と(上記)結晶成長溶液を含有する溶液50μlを各ウェルに加えることにより調製した。「種晶ストック」のアリコート(3μl)をマイクロタイタープレートの第1のウェルに移した。ウェル内の溶液を混合した後、3μlをマイクロタイタープレートの行に沿って次のウェルに移した。マイクロタイタープレートの各行を同様に調製し、トレーをプラスチックテープで密閉した。一晩でマイクロタイタープレートのウェルの底に小結晶が形成されたので、ウェル中の結晶の数を元の「種晶ストック」の希釈液に相関させた。大きい単結晶を生成するために、「種晶ストック」液滴を新鮮なCGS中に適宜希釈した後、NIUを含有するこの溶液のアリコートを液滴に移した。
【0086】
結晶化条件が一旦最適化されたら、CGS及びr−hu−G−CSF(33mg/ml)各3mlを混合して5個のトレー(各24穴)を作成する生成法で結晶を成長させた。この方法は、リットル量の最適化結晶化溶液を調製し、この溶液をタンパク質と混合し、懸垂液滴又は放置液滴トレーにタンパク質/結晶化溶液を加えるものであった。この方法では典型的には約5日間で100〜300の品質結晶(>0.5mm)が得られた。
【0087】
e.実験法
材料
図1に示すアミノ酸配列と比活性1.0±0.6×108U/mg(pH4.0の10mM酢酸緩衝液中で細胞マイトジェン生成アッセイにより測定)を有する濃度約3mg/mlのr−hu−G−CSFから出発して結晶学的情報を得た。YM10フィルターを使用して75psiでAmicon濃縮器を使用して溶液を濃縮した。溶液を典型的には4℃で10倍に濃縮し、数カ月貯蔵した。
【0088】
初期スクリーニング
懸垂液滴を使用して蒸気拡散平衡によりX線分析に適した結晶を得た。予備スクリーニングのために、(上記のように調製した)33mg/mlタンパク質溶液7μlを等容量のウェル溶液と混合し、シリコンガラスプレートに載せ、Linbro組織培養プレート(McLean,Vaに所在のFlow Laboratories)を使用してウェル溶液上に懸垂させた。全ピペッティングはAccuflexピペッターを使用して実施したが、ウェル溶液の生成後はトレーを自動ピペッターから取り外し、卓上振盪機で少なくとも10分間十分に混合した。次にLinbroトレーを再びピペッターに取り付け、ウェル溶液及びタンパク質溶液をシリコンカバースリップに加えた。次にカバースリップを裏返し、ウェル溶液1mlをシリコン油脂で覆い、密封した。
【0089】
自動結晶化システムの成分は以下の通りである。PC−DOSコンピューターシステムを使用して、その成分の濃度に基づいて結晶化溶液のマトリックスを設計した。これらのマトリックスは(上記)MRF又はLotusスプレッドシートのいずれかを用いて作成した。これらのプログラムの終産物はデータファイルである。このファイルは、マトリックスに示された濃度を得るために適した容量のストック溶液をピペットするためにSUXプログラムにより必要とされる情報を含む。SUXプログラム情報をシリアルI/Oポートに送り、ストック溶液に対するバルブの位置、回収した後にマイクロタイタープレートのウェルにピペット分注すべき溶液の量及び各ウェルのXY位置(各ウェルの行列)をAccuflexピペッティングシステムに指令するためにこの情報を使用した。充填中の注射器のZ位置(高さ)と、異なる溶液の充填間に注射器を洗浄するためにシステムを停止させるドレインの位置とを含む付加情報をピペッターに送信した。24穴マイクロタイタープレート(Linbro又はCryschem)及びカバースリップホルダーをプレートに載せ、プレートをXY面内で移動させた。プレートの移動により、ウェルにピペット分注するようにピペッターに注射器を位置決めさせた。プレートの移動により、カバースリップ及びバイアルも位置決めし、これらから溶液を抽出した。ピペット分注前に、Linbroマイクロタイタープレートはウェルの縁部の周囲に油脂薄膜を塗布しておいた。結晶化溶液をウェル内で調製してから、カバースリップに移す前に、マイクロタイタープレートをピペッティングシステムから取り外し、溶液を10分間卓上振盪機で混合した。混合後、ウェル溶液をカバースリップ(懸垂液滴プロトコルの場合)又はウェルの中央ポスト(放置液滴プロトコルの場合)に移した。タンパク質をバイアルから抽出し、ウェル溶液を含有するカバースリップ液滴(又はポスト)に加えた。プラスチックテープをCryschemプレートの頂部に張り付けてウェルを密封した。
【0090】
生成成長
結晶化条件が最適化されたら、「生成」法を使用して結晶成長を実施した。100mM Mes(pH5.8)、380mM MgCl2、220mM LiSO4及び8%PEG 8Kを含有する結晶化溶液1リットルを調製した。エッペンドルフ注射器ピペッターを使用してこの溶液の1mlアリコートをLinbroプレートのウェルの各々にピペット分注した。この溶液50%及びG−CSF(33mg/ml)50%を含有する混合溶液を調製し、シリコンカバースリップにピペット分注した。これらの液滴の典型量は50〜100μlであり、これらの液滴の寸法が大きいため、カバースリップのめくれ上がり及び液滴のウェル懸垂に細心の注意を払った。
【0091】
データ収集
R−AXIS(Houston,TXに所在のMolecular Structure,Corp.)イメージングプレートデテクターで収集した2.2Åデータに対してX−PLOR(Bruniger,X−PLOR version 3.0,結晶分析システム及びNMR,New Have CTに所在のイエール大学)を用いて構造を最適化させた。
【0092】
f.考察
有効な組換えヒト治療剤として、r−hu−G−CSFは大量生産され、構造分析のためにグラムレベルが入手可能になっている。他の該当タンパク質が入手可能になるにつれ、本発明の結晶化方法は他の用途にも利用可能である。この方法は、大量のタンパク質(>約200mg)を有する任意の結晶学的プロジェクトに適用することができる。当業者に認識される通り、本発明の材料及び手法は変形可能であり、等価材料及び手法が他のタンパク質の結晶化に使用可能である。
【0093】
B.G−CSFの三次元構造を可視化するためのコンピュータープログラム
添付図に示すような図はG−CSFの三次元構造を可視化するために有用であるが、分子を立体的観察を可能にするコンピュータープログラムが好適であると予想される。この立体観察は当業者には「バーチャルリアリティー」とも呼称され、高分子構造から原子レベルまで広範囲の分解能で全角度から構造を三次元形態で可視化することができる。本発明のコンピュータープログラムにより、例えば分子を回転させることにより、分子の観察角度の透視図を変更することもできる。本発明のプログラムは更に変更に応答するので、例えば完全アミノ酸残基を含む1個以上の原子画像を削除、追加もしくは置換えることができ、又は存在する基もしくは置換された基に化学的部分を付加することができ、構造変化を可視化することができる。
【0094】
他のコンピューターに基づくシステムを使用することもでき、その要素は、(a)例えばG−CSFの三次元構造の直交座標又は他の数値帰属座標等の情報を入力するための手段と、(b)前記座標を表現するための手段、例えば三次元構造を観察してこのような三次元構造をアミノ酸組成等のG−CSF分子の組成と相関できるようにする視覚手段と、(c)任意要素として、前記三次元構造の画像が改変組成を表示するように、表現されたG−CSF分子の組成を改変する情報を入力するための手段である。
【0095】
使用される好適コンピュータープログラムの座標を図5に示す。好適コンピュータープログラムはSan Diego,CAに所在のBiosymの市販品Insight II,version 4である。粗結晶学的構造については、回折データの観察強度(F−obs)及び直交座標も米国,Upton,New York 119723に所在のProtein Data Bank,Chemistry Department,Brookhaven National Laboratoryに寄託されており、参考資料として本明細書の一部とする。
【0096】
座標をInsight IIプログラムに入力すると、コンピュータースクリーン上に三次元G−CSF分子像を容易に表示することができる。表示に好適なコンピューターシステムはSilicon Graphics 320 VGX(San Diego,CA)である。立体観察の際には、G−CSF分子を三次元で立体的に観察することが可能な眼鏡(Crystal Eyes,Silicon Graphics)を装着し、分子を回転させて分子設計を想像することができる。
【0097】
従って、本発明はコンピューターを用いてG−CSF類似体を設計又は製造する方法を提供し、該方法は、
(a)G−CSF分子の部分の組成の表示、好ましくは各アミノ酸の三次元配置の表示、より好ましくはG−CSF分子の各原子の三次元位置の表示を含めてG−CSF分子の三次元構造を表示するための手段を前記コンピューターに備える段階と、
(b)前記表示を観察する段階と、
(c)前記分子の組成又は部分の位置の改変のための部位を前記表示上で選択する段階と、
(d)前記改変を有するG−CSF類似体を製造する段階とを含む。
【0098】
改変は、終産物G−CSF類似体の所望の構造特性に基づいて選択することができ、このような設計には以下に詳述する要素を考慮する。このような要素とは、特にG−CSF分子の螺旋構造の内側の残基であって、このような残基を改変すると、分子の内部コアの全体構造が変化し、レセプター結合を妨げ得る疎水性アミノ酸残基の位置及び組成と、改変しても、G−CSF分子の全体構造には影響しない外部ループ構造の位置及び組成を含む。
【0099】
図2A〜4は、種々の方法で全体的三次元コンフォーメーションを示す。トポロジー図、リボン図、及びバレル図はいずれもG−CSFのコンフォーメーションの特徴を示す。
【0100】
図2A〜Gは、G−CSFと他の分子との比較を示す。アーキテクチャーは類似しているが、これらの成長因子はそのループの局所コンフォーメーション及び螺旋束の幾何学的配置が異なる。他方、アミノ酸配列の相違にも拘わらず、これらの分子の全6種間には2つの長いクロスオーバー結合を含むアップ−アップ−ダウン−ダウントポロジーが保存されている。
【0101】
図3は組換えヒトG−CSFの二次構造を詳細に示す。このリボン図は螺旋の向きとその相対位置を示す。
【0102】
図4は組換えヒトG−CSFのコンフォーメーションを別の方法で示す。この「バレル」図は組換えヒトG−CSFの全体的なアーキテクチャーを示す。
【0103】
C.M13突然変異誘発を用いた類似体の調製
本実施例は、PCT出願公開第WO85/00817号(Souzaら,1985年2月28日公開、参考資料として本明細書の一部とする)に記載の方法に従って、1本鎖バクテリオファージM13を用いる部位特異的突然変異誘発技術を使用したG−CSF類似体の調製に関する。この方法は主に非突然変異誘発配列の1本鎖核酸鋳型を使用し、配列中に所望の改変を含むより小さいオリゴヌクレオチドを前記鋳型に結合することからなる。非同一配列をハイブリダイズさせるようなハイブリダイゼーション条件を使用し、残りの配列は最初の鋳型と同一になるように充填する。その結果、2本の鎖の一方が所望の改変を含むような2本鎖分子が生成される。この突然変異誘発した1本鎖を分離し、その相補鎖の鋳型として使用する。こうして所望の改変を含む2本鎖分子を生成する。
【0104】
使用した改変前のG−CSF核酸配列を図1に示し、突然変異誘発した核酸を含むオリゴヌクレオチドを表2に示す。本明細書中でアミノ酸残基及びヌクレオチドに使用した略記は慣用通りであり、Stryer,Biochemistry,第3版,W.H.Freeman and Company,N.Y., N.Y.1988,裏内表紙を参照されたい。
【0105】
改変前のG−CSF核酸配列をまず最初にベクターM13mp21に挿入した。次に改変前のG−CSF配列を含む1本鎖ファージM13mp21からのDNAを単離し、水に再懸濁した。各反応につきこのDNA200ngをリン酸化オリゴヌクレオチド1.5pmolと混合し(表2)、0.1M Tris,0.01M MgCl2,0.005M DTT,0.1mM ATP(pH8.0)に懸濁した。65℃まで加熱し、室温までゆっくりと冷却することによりDNAをアニールした。
【0106】
冷却後、ATP、dATP、dCTP、dGTP、TTP各0.5mM、T4 DNAリガーゼ1単位及び大腸菌ポリメラーゼ1のKlenowフラグメント1単位を、0.1M Tris,0.025M NaCl,0.01M MgCl2,0.01M DTT(pH7.5)中のアニールしたDNA1単位に加えた。
【0107】
こうして2本鎖になった閉環状DNAをそれ以上精製せずに使用して大腸菌にトランスフェクトした。プラークをニトロセルロースフィルターで釣り上げた後、P32で末端標識した1本鎖DNAとフィルターを1時間55〜60℃でハイブリダイズさせることによりプラークをスクリーニングした。ハイブリダイゼーション後、オリゴの融点(A−Tでは2℃、G−Cでは4℃)よりも0〜3℃低い温度でフィルターを洗浄し、突然変異配列を含むファージを有するプラークに対応するオートラジオグラフィーシグナルを選択的に発生させた。陽性クローンを配列決定により確認した。
【0108】
M13突然変異誘発法により製造した各G−CSF類似体毎に使用したオリゴヌクレオチドを下表に示す。命名法は、改変前のアミノ酸の残基及び位置(例えば17位リジン)と、置換したアミノ酸の残基及び位置(例えばアルギニン17)を示す。2個以上の残基が置換されている場合には、位置を指定する上付き数字をカンマで区切るか、又はセミコロンで残基を区切る。置換でなく欠失の場合にはその旨明記した。M13に基づく突然変異誘発に使用したオリゴヌクレオチド配列を次の欄に示す。これらのオリゴヌクレオチドは合成により製造したが、その製法は限定的ではなく、任意の核酸合成法及び/又は装置を使用することができる。オリゴ長も示す。上述のように、これらのオリゴを1本鎖ファージベクターと接触させた後、単一ヌクレオチドを加え、G−CSF類似体核酸配列を完成させた。
【0109】
【表2】
【0110】
D.DNA増幅を使用したG−CSF類似体の調製
本実施例は、DNA増幅技術を用いたG−CSF類似体の製造方法に関する。主に、各類似体をコードするDNAを別個の2部分として増幅して結合した後、完全配列を増幅した。改変前のG−CSF DNAのどの場所に所望の改変を加えるべきかに依存して、内部プライマーを使用して改変を組み込み、2つの別個の増幅部分を生成した。例えば、所望の類似体DNAの5’末端を増幅するためには、増幅すべき領域の一端には(改変前のG−CSF DNAの上流のプラスミドの配列に相補的な)5’フランキングプライマーを使用し、他方の末端をプライムするためには、改変前のDNAとハイブリダイズすることが可能でありながら所望の改変を含み得る内部プライマーを使用した。得られた増幅領域は5’フランキングプライマーから内部プライマーに及ぶ領域であっだ。(改変前のG−CSF DNAの下流のプラスミドの配列に相補的な)3’フランキングプライマーと所期突然変異の領域に相補的な内部プライマーを使用して3’末端も同様に処理した。(内部プライマーの位置に依存して同一寸法でもよいし、同一寸法でなくてもよい)2つの「半部分」を増幅後、2つの「半部分」を結合した。結合後、5’フランキングプライマーと3’フランキングプライマーを使用して所望の改変を含む完全配列を増幅した。
【0111】
2つ以上の改変が所望される場合には、内部プライマーに改変を組み込むように上記プロセスを変更するか、又は別の内部プライマーを使用してプロセスを繰り返す。あるいは、核酸配列に改変をもたらすための他の方法、例えば上記のようなファージに基づく突然変異誘発技術と遺伝子増幅法を併用してもよい。2つ以上の改変を有する類似体を製造する方法の例を以下に記載する。
【0112】
下記G−CSF類似体を生成するために使用した鋳型DNAは、図1の配列に(G−CSFコーディング領域を含むプラスミドからの)所定のフランキング領域を加えたものであった。これらのフランキング領域を以下に説明するように5’及び3’フランキングプライマーとして使用した。増幅反応は、10mM Tris−HCl、1.5mM MgCl2、50mM KCl、0.1mg/mlゼラチン(pH8.3)を含有する40μl容量中で20℃で実施した。40μl反応溶液は更に各dNTP 0.1mM、各プライマー10pmol及び鋳型DNA1ngを含有するものとした。各増幅は15サイクル繰り返した。各サイクルは94℃で0.5分間、50℃で0.5分間、72℃で0.75分間から構成した。フランキングプライマーは20ヌクレオチド長、内部プライマーは20〜25ヌクレオチド長であった。こうして、所望のG−CSF類似体の前部又は後部をコードする2本鎖DNAの多重コピーを生成した。
【0113】
2つの「半部分」を結合するために、2つの反応物の各々の40分の1を第3のDNA増幅反応で結合した。突然変異を有するこれらの2部分の末端は相補的であるので、2部分を内部プライマー位置でアニールさせ、重合サイクル後、完全長DNA配列を得た。こうしてアニール後、5’及び3’フランキングプライマーを使用して完全類似体を増幅した。この増幅プロセスは上述のように15サイクル繰り返した。
【0114】
完全増幅類似体DNA配列をXbaI及びXhoI制限エンドヌクレアーゼで切断し、ベクターに挿入するための付着末端を生成した。切断したDNAをプラスミドベクターに挿入し、このベクターを使用して大腸菌を形質転換させた。形質転換細胞を50μg/mlのカナマイシンで攻撃し、30℃でインキュベートした。G−CSF類似体タンパク質の生成を全細胞溶解物のポリアクリルアミドゲル電気泳動により確認した。生成単離物から精製したプラスミドのDNA配列分析により所望の突然変異の存在を確認した。次に培養物を増殖させ、細胞を回収し、G−CSF類似体を以下のように精製した。
【0115】
下表3は遺伝子増幅を使用して製造した各類似体に使用した特定プライマーを示す。
【0116】
【表3】
【0117】
1.二重突然変異の構築
G−CSF類似体Gln12.21→Glu12.21を製造するために、DNA増幅を2回別々に実施し、2つのDNA突然変異体を製造した。使用した鋳型DNAは図1の配列に(G−CSFコーディング領域を含むプラスミドからの)所定のフランキング領域を加えたものである。厳密な配列は下記の通りである。Perkin Elmer/Cetus DNA熱サイクラーを使用して2つのDNA増幅反応の各々を実施した。40μl反応混合物は、1×PCR緩衝液(Cetus)、4dXTP(Cetus)各0.2mM、各プライマーオリゴヌクレオチド50pmol、(プラスミドベクター上の)G−CSF鋳型DNA2ng及びTaqポリメラーゼ(Cetus)1単位から構成した。増幅プロセスは30サイクル実施した。各サイクルは94℃で1分間、50℃で2分間、72℃で3分間から構成した。
【0118】
DNA増幅“A”は下記オリゴヌクレオチド:
【0119】
【化1】
を使用した。
【0120】
DNA増幅“B”は下記オリゴヌクレオチド:
【0121】
【化2】
を使用した。
【0122】
DNA増幅“A”後に得られた109塩基対2本鎖DNA産物からDNA突然変異Gln12→Glu12を含む64塩基対XbaI−HindIII DNAフラグメントを切り出し、単離した。DNA増幅“B”後に得られた509塩基対2本鎖DNA産物からDNA突然変異Gln21→Glu21を含む197塩基対HindIII−BsmI DNAフラグメントを切り出し、単離した。
【0123】
“A”及び“B”フラグメントを4.8kbp XbaI−BsmI DNAプラスミドベクターフラグメントと連結した。連結混合物は等モルDNA制限フラグメント、連結緩衝液(25mM Tris−HCl(pH7.8),10mM MgCl2,2mM DTT,0.5mM rATP及び100μg/ml BSA)及びT4 DNAリガーゼから構成し、14℃で一晩インキュベートした。その後、Bio Rad Gene Pulsar装置(BioRad,Richmond,CA)を使用するエレクトロポレーションにより連結DNAを大腸菌FM5に形質転換した。クローンを単離し、DNA配列決定によりプラスミド構築物が2つの突然変異を含むことを確認した。この「中間」ベクターは更に193塩基対BsmI−BsmI DNAフラグメントを欠失していた。最終プラスミドベクターは中間ベクターから2kbp SstI−BamHI DNAフラグメント、プラスミドベクターから2.8kbp SstI−EcoRI DNAフラグメント、及びプラスミドベクターから360bp BamHI−EcoRI DNAフラグメントを切り出し、単離することにより得たDNAフラグメントの(上記のような)連結及び形質転換により構築した。DNA配列決定により最終構築物がG−CSF遺伝子を含むことを確認した。下記のように培養物を増殖し、細胞を回収し、G−CSF類似体を精製した。
【0124】
上述のように、突然変異誘発法の任意の組み合わせを使用して、1つ以上の改変を有するG−CSF類似体核酸(及び発現産物)を製造することができる。M13に基づく突然変異誘発及び遺伝子増幅に基づく突然変異誘発を使用した上記2例はその実例である。
【0125】
E.G−CSF類似体DNAの発現
次にG−CSF類似体DNAをプラスミドベクターに挿入し、大腸菌株FM5(ATCC#53911)を形質転換させるために使用した。プラスミド上及び細菌宿主細胞中に含まれる本発明のG−CSF類似体DNAは、Rockville,MDに所在のAmerican Type Culture Collectionから入手可能であり、その受託番号は以下に示す。
【0126】
トリプトン10g、酵母エキス5g及びNaCl 5gを含有するブロスで密度A600=0.5に達するまで1リットル培養物を30℃で増殖させ、前記密度に達したら42℃まで迅速に加熱した。フラスコを3時間振盪し続けた。
【0127】
他の原核又は真核宿主細胞も使用可能であり、例えば他の細菌細胞、株もしくは種、哺乳動物培養細胞(COS、CHO又は他の型)、昆虫細胞もしくは多細胞器官もしくは生物、又は植物細胞もしくは多細胞器官もしくは生物を使用することができ、当業者は適切な宿主に想到しよう。本発明のG−CSF類似体及び関連組成物は、例えば固相ペプチド合成法又は他の化学的製法により合成することもできる。他のクローニング及び発現系は当業者に自明である。
【0128】
F.G−CSF類似体タンパク質の精製
遠心分離(10,000×G,20分間,4℃)により細胞を回収した。ペレット(通常は5g)を1mM DTT30mlに再懸濁し、10,000psiのフレンチプレスセルに3回通した。破壊した細胞懸濁液を10,000gで30分間遠心分離し、上清を除去し、ペレットを水30〜40mlに再懸濁した。これを10,000×Gで30分間再遠心分離し、このペレットを2% Sarkosyl及び50mM Tris(pH8)25mlに溶解させた。硫酸銅を濃度40μMまで加え、混合物を少なくとも15時間15〜25℃で撹拌した。次いで混合物を20,000×Gで30分間遠心分離した。得られた可溶化タンパク質混合物を13.3mM Tris(pH7.7)で4倍に希釈し、Sarkosylを除去した後、20mM Tris(pH7.7)で平衡化したDEAE−セルロース(Whatman DE−52)カラムに上清を充填した。カラム充填及び同一緩衝液でカラムを洗浄後、類似体を20mM Tris/NaCl(下記のように類似体に依存して35mM〜100mM)(pH7.7)で溶離した。類似体のほとんどはDEAEカラムからの溶出液を50%酢酸でpH5.4に調整し、必要に応じて(適正な導電率を得るために)5mM酢酸ナトリウム(pH5.4)で希釈した。次に20mM酢酸ナトリウム(pH5.4)で平衡化したCM−セファロースカラムに溶液を充填した。次いで280nmの吸光度がほぼ0になるまでカラムを20mM NaAc(pH5.4)で洗浄した。次いでG−CSF類似体を下表4に記載する濃度の酢酸ナトリウム/NaClで溶離した。CM−セファロースカラムに充填しなかった類似体のDEAEカラム溶出液を10mM NaAc(pH4.0)緩衝液で直接透析した。次いで精製G−CSF類似体をin vitro分析のために適宜単離した。類似体の溶離に使用した塩濃度は上述のように変化させた。DEAEセルロースカラム及びCM−セファロースカラムの濃度を下表に示す
【0129】
【表4】
*167欠失のデータは得られなかった。
**これらの類似体ではDEAEセルロースカラムを単独で精製に使用した。
【0130】
上記精製法は例示であり、当業者には自明の通り、発明のG−CSF類似体を得るために他の手段も使用可能である。
【0131】
G.生物アッセイ
本発明のG−CSF類似体をどの方法で製造したかに関係なく、類似体の生物活性をアッセイした。3H−チミジンアッセイを実施して細胞分裂度を確認した。他の生物アッセイを使用して所望の活性を確認することもできる。マウスWEHI−3B(D+)白血病細胞系に末端分化を誘導する能力をアッセイする等の生物アッセイによりG−CSF活性を得ることもできる。Nicolaら,Blood 54: 614−27(1979)参照。生物活性を確認するために他のin vitroアッセイも使用できる。Nicola,Annu.Rev.Biochem.58: 45−77(1989)参照。一般に、生物活性試験では(非改変G−CSFと比較した)生物活性の増加又は低下、(非改変G−CSFと比較した)生物活性の相違、レセプターアフィニティー分析又は血清半減期分析等の所望の結果を分析すべきである。これらの例に止まることなく、当業者は所望の最終結果の試験に有用な他のアッセイにも想到しよう。
【0132】
常法を使用して3H−チミジンアッセイを実施した。雌Balb Cマウスを殺して骨髄を取り出した。骨髄細胞を成長培地に短時間懸濁し、遠心分離し、再懸濁した。約10,000個の細胞を含む160μlアリコートを96穴マイクロタイタープレートの各ウェルに加えた。(上記のように調製した)精製G−CSF類似体のサンプルを各ウェルに加え、68時間インキュベートした。3H−チミジンをウェルに加え、更に5時間インキュベートした。5時間のインキュベーション時間後、細胞を回収し、濾過し、十分に濯いだ。シンチレーション液を収容するバイアルにフィルターを加えた。β線を計数した(LKBβプレートシンチレーションカウンター)。標準及び類似体を3回ずつ分析し、標準曲線よりも実質的に上又は下に位置するサンプルを適正な希釈度で再アッセイした。ここに報告する結果は非改変組換えヒトG−CSF標準結果に対する3回の類似体データの平均である。
【0133】
H.HPLC分析
類似体の精製サンプルで高圧液体クロマトグラフィーを実施した。逆相HPLCカラム上のピークの位置は2種のタンパク質間の構造類似を決定的に示すものではないが、類似の保持時間を有する類似体はHPLCカラムに対して非改変分子と同一型の疎水性相互作用を有し得る。これは全体構造の類似を示す1面である。
【0134】
類似体及び非改変組換えヒトG−CSFのサンプルを逆相(0.46×25cm)Vydac 214TP54カラム(Separations Group,Inc.Hesperia,CA)で分析した。類似体がカラム中でどのように挙動したかに依存して0.1mg/ml〜5mg/mlの最終濃度まで20mM酢酸及び40mM NaCl緩衝溶液(pH5.2)中で精製類似体G−CSFサンプルを調製した。1%イソプロパノール、52.8%アセトニトリル及び0.38%トリフルオロ酢酸(TFA)を含有する水溶液で予め平衡化しておいたHPLCカラムに(濃度に依存して)種々の量を充填した。サンプルを0.86%/分のアセトニトリル及び0.002%TFAの勾配にかけた。
【0135】
I.結果
上記生物アッセイ及びHPLC分析の結果を以下に示す。生物活性は3回のデータの平均であり、対照標準(非改変G−CSF)の百分率として示す。相対HPLCピーク位置は対照標準(非改変G−CSF)ピークに対する類似体G−CSFの位置である。+又は−の記号は、類似体HPLCピークが対照標準ピークよりも(何分)先又は後に現れるかを示す。変異体の全部について相対HPLCピークを分析した訳ではなく、分析したもののみを下表に示す。上記のように調製した本発明の類似体をコードする核酸を含む大腸菌宿主細胞のAmerican Type Collection番号も示す。
【0136】
【表5】
【0137】
1.構造−機能関係の確認
本発明の類似体を設計するために使用した第1段階は、G−CSF分子の構造完全性にどの部分が必要であるかを決定することであった。これはアミノ酸残基レベルで実施したが、原子レベルも分析に利用できる。構造完全性に必要な残基を修飾すると、G−CSF分子の全体構造が変化する。これは製造しようとする類似体に依存して望ましい場合と望ましくない場合がある。本実施例は、G−CSFレセプター(本項で以下に記載する「G−CSFレセプター」は造血細胞に存在する天然G−CSFレセプターを意味する)への類似体のG−CSFレセプター結合を維持する目的で、G−CSF分子の全体構造完全性を維持するように設計した。G−CSFレセプター結合は上記生物アッセイにより決定されるような少なくとも1種の生物活性に必要な段階であると仮定され、本試験により確認された。
【0138】
図面に示すように、G−CSF(ここでは組換えヒトmet−G−CSF)は45Å×30Å×24Åの全体寸法を有する左巻き螺旋を有する逆平行4α螺旋束である。螺旋束内の4つの螺旋は螺旋A、B、C及びDと呼称され、その結合ループはAB、BC及びCDループとして知られる。螺旋交差角度は−167.5°〜−159.4°である。螺旋A、B及びCは直鎖であり、螺旋Dは(組換えヒトmet−G−CSFの)Gly150及びSer160に2種の構造特徴を含む。全体としてG−CSF分子は外部ループにより直列に結合された4つの螺旋からなる束である。この構造情報を公知機能情報と相関させた。(1位メチオニンを含む)残基47、23、24、20、21、44、53、113、110、28及び114を修飾することができ、生物活性に及ぼす作用は相当のものであることが判明した。
【0139】
生物活性を低下させた単一突然変異の多くは30Åの間隔で4螺旋束の異なった面に位置するG−CSFの2つの領域の周囲に集中していた。一方の領域はA螺旋とD螺旋の相互作用をもたらす。これは更に下記非改変分子における塩橋の存在により確認される。
【0140】
【表6】
【0141】
ここに報告する距離は図5A〜FFFに示すような分子Aの距離である(3つのG−CSF分子は共に結晶化し、A、B及びCと命名した)。螺旋A及びDの間には塩橋ウェブが存在し、螺旋A構造を安定化させるように機能するので、G−CSF分子の全体構造に影響する。
【0142】
残基Glu20、Arg23及びLys24の周囲の領域はA螺旋の親水性面(残基20〜37)に存在する。20及び23位の非帯電アラニン残基で残基を置換すると、類似のHPLC保持時間が得られ、構造の類似性が予想される。(本アッセイにより示されるように)これらの部位を改変すると、生物活性が変化した。Lys24を置換した場合には生物活性は変化したが、他の2種の改変のように類似のHPLC保持時間は得られなかった。
【0143】
改変により生物活性が低下した第2の部位はAB螺旋である。47位のグルタミンをアラニンで置換する(上記類似体19)と、(チミジン取り込みアッセイにおいて)生物活性がゼロに低下した。AB螺旋はアミノ及びカルボキシ末端を除いて主に疎水性であり、310螺旋1回転を含む。各末端には2個のヒスチジン(His44及びHis56)と、His44との塩架橋を形成することが可能な残基46の付加的グルタミンとが存在する。類似体のフーリエ変換赤外スペクトログラフィー分析(FTIR)によると、この類似体は非改変組換えG−CSF分子に構造的に類似する。更に試験した結果、この類似体は非改変組換え分子と同一条件下では結晶化しないことが判明した。
【0144】
カルボキシ末端の改変(Gln174、Arg167及びArg170)は生物活性にほとんど影響しなかった。他方、最後の8個の残基(167〜175)が欠失すると、生物活性は低下した。これらの結果から、欠失は全体構造を不安定にし、突然変異体がG−CSFレセプターと適正に結合するのを妨げる(その結果、シグナル変換の開始を妨げる)と予想される。
【0145】
一般に、G−CSF内部コア(外部ループを欠く内部4螺旋束)では疎水性内部残基は構造完全性に必須である。例えば螺旋Aにおいて内部疎水性残基は(メチオニンを1位として)Phe14、Cys18、Val22、Ile25、Ile32及びLeu36である。他の疎水性残基(同様にmetを1位とする)は螺旋BのAla72、Leu76、Leu79、Leu83、Tyr86、Leu90、Leu93であり、螺旋CのLeu104、Leu107、Val111、Ala114、Ile118、Met122であり、螺旋DのVal154、Val158、Phe161、Val164、Val168、Leu172である。
【0146】
本発明により製造したG−CSF類似体からの上記生物活性データによると、外部ループの改変はG−CSF全体構造にほとんど影響しない。類似体製造のための好適ループはABループ及びCDループである。ループは螺旋に比較して比較的にフレキシブルな構造である。ループは該分子のタンパク分解により分解され得る。該分子を使用する目的は生物攻撃に対する応答を発生すること、即ち好中球を選択的に刺激することであるので、G−CSFはin vivoで比較的迅速に作用する。G−CSF代謝回転も比較的迅速である。ループのフレキシビリティーはプロテアーゼが該分子と結合する「手」を提供し、該分子を不活性化する。プロテアーゼ分解を阻止しながら(非改変G−CSFの全体構造の保持を介して)生物活性を低下させないようにループを改変することができる。
【0147】
この現象は恐らくG−CSF分子に限定されず、図2に示すような類似の公知全体構造を有する他の分子にも共通すると思われる。例えばhGH、インターフェロンB、IL−2、GM−CSF及びIL−4の外部ループを改変しても全体構造の変化は最小に止まる。GM−CSF分子上の外部ループはG−CSF分子上に存在する外部ループほどフレキシブルではないので、GM−CSFはその広い生物活性に一致して血清寿命が長いと予想される。従って、β−シート構造から外部ループを解放することにより、(G−CSFと同様に)ループをよりフレキシブルにし、分子をプロテアーゼ分解に対してより感受性にする(その結果、回転率を増加する)ようにGM−CSFの外部ループを改変することができる。
【0148】
これらの外部ループの改変は、内部螺旋の1つ以上と結合することによりループを安定化することにより実施し得る。βシート、塩橋、ジスルフィド結合又は疎水性相互作用の形成等の結合手段が当業者に公知であり、他の手段も使用可能である。更に、1個以上のアミノ酸残基又はその部分等の1個以上の部分を欠失させ、短縮分子を製造し、こうして外部ループの所定の部分を除去することもできる。
【0149】
従って、外部ループ、好ましくはABループ(r−hu−met G−CSFのアミノ酸58〜72)又はCDループ(r−hu−met−G−CSFのアミノ酸119〜145)、これらのループより好ましくはないがアミノ末端(アミノ酸1〜10)の改変により、G−CSF G−CSFレセプター結合を除去せずに生物機能を変えることができる。例えば、(1)例えばプロテアーゼがG−CSF分子に作用する能力を低下させることにより又は、上記のようにG−CSF分子の所定の特徴を変化させる(例えば血清もしくは他の半減期を増加させるか又は抗原性を低下させる)ように機能する1個以上のポリエチレングリコール分子もしくは経口製剤用腸溶性コーティング等の化学修飾をG−CSF分子に加えることにより、G−CSF分子の半減期を増加させる(又は例えば経口剤形を製造する)こともできるし、(2)G−CSF G−CSFレセプター輸送メカニズムを介する細胞侵入によりシグナル変換を行う別のサイトカイン又は別のタンパク質等の別のタンパク質の一部又は全部とG−CSFとを結合するなどしてハイブリッド分子を作成することもできるし、(3)例えば(非改変G−CSF分子に比較して)好中球を選択的に刺激する能力等の生物活性を増加させることができる。このような改変は上記例に限定されない。
【0150】
上記データから観察される別の特徴は、安定化表面相互作用が生物活性に影響し得ることである。これは類似体23と40を比較すると明白である。類似体23は28位の中性帯電アラニン残基を同位の帯電アスパラギン残基で置換したものであり、この置換の結果、(本明細書に開示のチミジン取り込みアッセイにより測定されるように)生物活性は50%増加した。28位のアスパラギン残基は113位のアスパラギン残基との表面相互作用を有しており、両者の残基は負に帯電しているので、(同様の帯電部分の排斥により)所定量の不安定が存在する。他方、113位のアスパラギンを中性帯電アラニンで置換すると、(本アッセイシステムでは)生物活性はゼロまで低下する。このことから明らかなように、113位のアスパラギンは生物活性に重要であり、28位のアスパラギンを除去すると、113位のアスパラギンが有する作用が増加する。
【0151】
上記のように製造した類似体及びG−CSF構造に基づいてG−CSFレセプター結合に必要なドメインも決定した。G−CSFレセプター結合ドメインは(メチオニンを1位として)残基11〜57(A及びAB螺旋間)及び100〜118(B及びC螺旋間)に位置する。外部ループ構造を改変し、レセプター結合領域を無傷のままにすることにより、G−CSFレセプターと結合することが可能な短縮分子を製造し、好中球を選択的に刺激するためのシグナル変換を開始することもできる。
【0152】
生物活性に必須であり、恐らくG−CSFレセプター結合又はシグナル変換にも必須であると予想される残基を同定した。二次構造の2つの異なる領域には2つの別個の部位が位置する。ここで「部位A」なる部位は、螺旋束の他の2つの構成員間の塩橋接触により限定される螺旋上に位置する。第2部位「部位B」は比較的フレキシブルな螺旋AB上に位置する。AB螺旋はカルボキシ及びアミノ末端における残基の種類及び位置の理由から、局所pH変化の影響を受け易い。このフレキシブル螺旋の機能的重要性は、G−CSFレセプターとの結合時にコンフォーメンションを誘導適合できるという点にある。更に、直接突然変異及び間接相対タンパク質構造分析により確認されるように、D螺旋の延長部分もG−CSFレセプター結合ドメインである。r−hu−met−G−CSFのカルボキシ末端を欠失させると、hGHの場合と同様に活性が低下する(CunninghamとWells,Science 244: 1081−1084(1989)参照)。類似のトポロジーを有すると予想されるIL−6及びGM−CSF等の類似構造を有するサイトカインもD螺旋のカルボキシ末端に沿ってその生物活性の中心を有する(Bazan,Immunology Today 11: 350−354(1990)参照)。
【0153】
G−CSFとhGHとのG−CSFレセプター結合決定基の構造及び位置を比較すると、両者分子は類似のシグナル変換手段を有すると予想される。hGHでは2つの別個のG−CSFレセプター結合部位が確認されている(De Vosら,Science 255: 306−32(1991))。これらの結合部位の一方(部位Iと呼称する)は螺旋1及び2と螺旋4の間の結合領域であるhGHの螺旋1の露出面上の残基により形成される。第2の結合部位(部位IIと呼称する)は螺旋1と螺旋3の表面残基により形成される。
【0154】
G−CSFに確認されたG−CSFレセプター結合決定基はhGHに確認されると同一の相対位置に位置する。AB螺旋上の螺旋A及びB間の結合領域に位置するG−CSFレセプター結合部位(部位A)は、hGHの螺旋の小部分(残基38〜47)に報告されている位置と類似する。G−CSFのAB螺旋における単一点突然変異は、(本アッセイで確認されるように)生物活性を著しく低下させるので、G−CSFレセプター−リガンド界面に作用するとも思われる。G−CSFレセプターの結合は、この領域の310螺旋を不安定にし、リガンド/G−CSFレセプター複合体の結合エネルギーを改善するコンフォーメーション変化をもたらす。
【0155】
hGHレセプター複合体において、螺旋束の第1の螺旋はhGHレセプターを二量体化するために必要な残基を結合部位の両方に与える。G−CSFの対応する螺旋(螺旋A)の突然変異分析の結果、生物活性に必要な3個の残基が確認された。これらの3個の残基のうちでGlu20及びArg24は螺旋Cに向かう螺旋束の1つの表面に位置し、(非対称単位における3分子のうちの2分子に存在する)Arg23の側鎖は螺旋Dに向かう束の表面に向いている。これらの生物学的に重要な残基の側鎖の位置は、hGHの場合と同様にG−CSFが螺旋A及び螺旋C間の界面に沿って第2のG−CSFレセプター結合部位を有することを示す。hGH分子とは対照的に、最初の11個の残基の欠失は生物活性にほとんど影響しないので、G−CSFのアミノ末端は制限された生物機能を有する。
【0156】
上述のように(例えば図2参照)、G−CSFは他のサイトカインとトポロジー類似性を有する。構造と上記生物化学試験との相関、突然変異分析及びhGHレセプター複合体の特定残基の直接比較によると、G−CSFは2つのレセプター結合部位を有する。部位Aは螺旋A及びDの界面に沿って存在し、小さいAB螺旋中に残基を含む。部位BもA螺旋中に残基を含むが、螺旋A及びCの間の界面に沿って位置する。G−CSFとhGHとの間の生物学的に重要な残基の構造と相対位置の保存は、共通のシグナル変換方法でレセプターが2地点で結合されることを示す1つの指標である。従って、G−CSFレセプター結合部位(残基20〜57及び145〜175)のいずれか一方の改変により、改変G−CSFレセプター結合ドメインを有するG−CSF類似体を製造できるとみなされる。
【0157】
G−CSFタンパク質の三次元構造の知見と組成の相関により、G−CSF類似体の体系的合理的製造方法が可能になる。上記実施例から明らかなように、構造のコア内の側鎖の寸法及び極性の制限は、全体構造が変化する前に分子がどの程度の変化に耐え得るかを決定する。
【特許請求の範囲】
【請求項1】
第1の化学的部分を介して第2の化学的部分が間接的に結合した、配列番号2に示すアミノ酸配列を含む改変されたG−CSFポリペプチドを製造する方法であって、
(a)in vitroで第1の化学的部分を外部ループに結合させる工程および
(b)第1の化学的部分を介して第2の化学的部分を間接的に外部ループに結合させる工程を含み、当該第1の化学的部分が多糖であり、第2の化学的部分がポリエチレングリコールであり、配列番号2のN末端メチオニンが存在し、または存在しておらず、当該改変G−CSFがG−CSF受容体に結合する生物活性を維持していることを特徴とする前記方法。
【請求項2】
外部ループがCDループである請求項1に記載の方法。
【請求項3】
外部ループがABループである請求項1に記載の方法。
【請求項4】
外部ループがBCループである請求項1に記載の方法。
【請求項1】
第1の化学的部分を介して第2の化学的部分が間接的に結合した、配列番号2に示すアミノ酸配列を含む改変されたG−CSFポリペプチドを製造する方法であって、
(a)in vitroで第1の化学的部分を外部ループに結合させる工程および
(b)第1の化学的部分を介して第2の化学的部分を間接的に外部ループに結合させる工程を含み、当該第1の化学的部分が多糖であり、第2の化学的部分がポリエチレングリコールであり、配列番号2のN末端メチオニンが存在し、または存在しておらず、当該改変G−CSFがG−CSF受容体に結合する生物活性を維持していることを特徴とする前記方法。
【請求項2】
外部ループがCDループである請求項1に記載の方法。
【請求項3】
外部ループがABループである請求項1に記載の方法。
【請求項4】
外部ループがBCループである請求項1に記載の方法。
【図1】


【図2A】
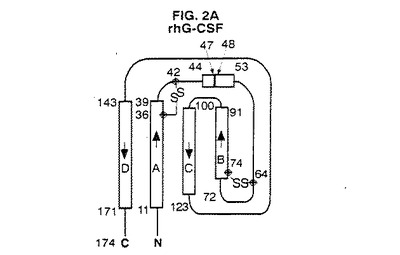
【図2B】
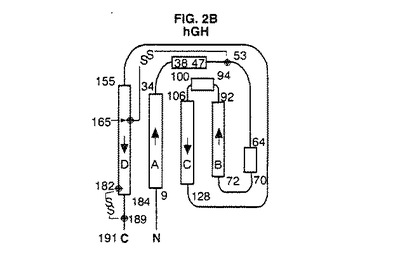
【図2C】
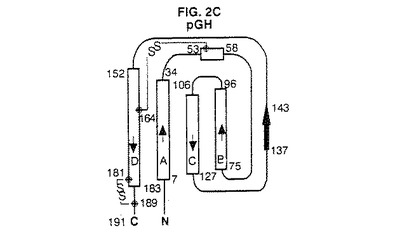
【図2D】
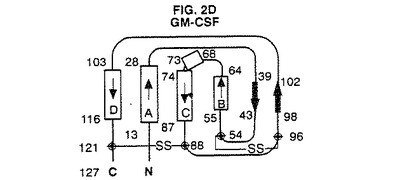
【図2E】
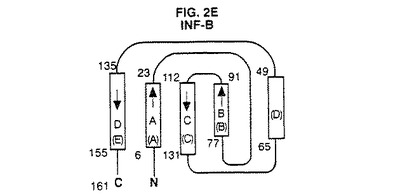
【図2F】
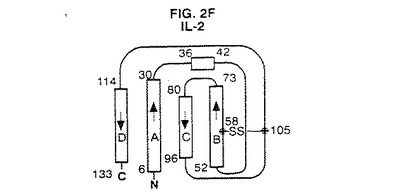
【図2G】
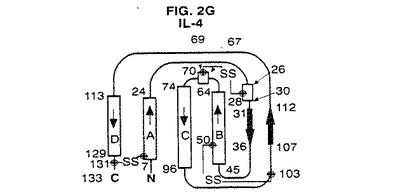
【図3】
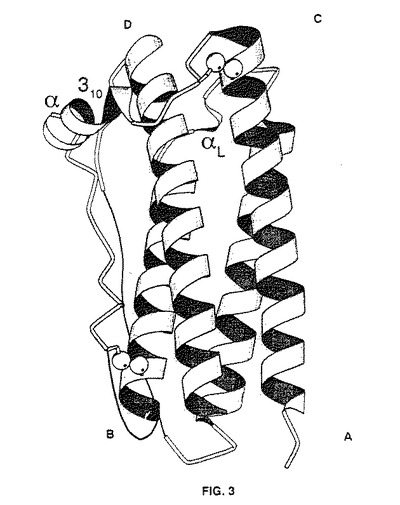
【図4】
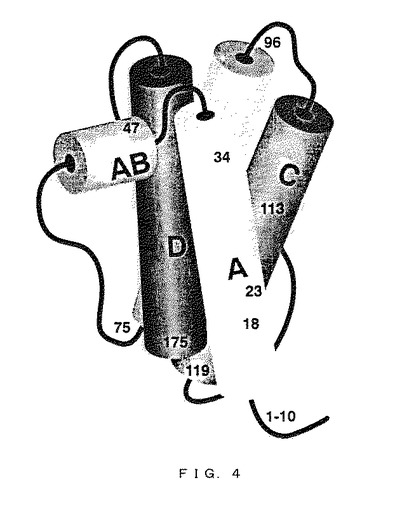
【図5A】

【図5B】

【図5C】

【図5D】

【図5E】

【図5F】

【図5G】

【図5H】

【図5I】

【図5J】

【図5K】

【図5L】

【図5M】

【図5N】

【図5O】

【図5P】

【図5Q】

【図5R】

【図5S】

【図5T】

【図5U】

【図5V】

【図5W】

【図5X】

【図5Y】

【図5Z】

【図5AA】

【図5BB】

【図5CC】

【図5DD】

【図5EE】

【図5FF】

【図5GG】

【図5HH】

【図5II】

【図5JJ】

【図5KK】

【図5LL】

【図5MM】

【図5NN】

【図5OO】

【図5PP】

【図5QQ】
【図5RR】

【図5SS】

【図5TT】

【図5UU】

【図5VV】

【図5WW】

【図5XX】

【図5YY】

【図5ZZ】

【図5AAA】

【図5BBB】

【図5CCC】

【図5DDD】

【図5EEE】

【図5FFF】

【図6】
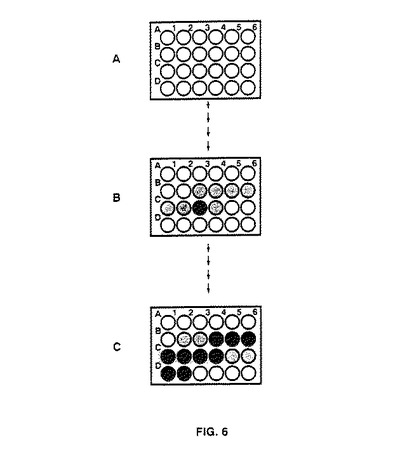
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図5I】
【図5J】
【図5K】
【図5L】
【図5M】
【図5N】
【図5O】
【図5P】
【図5Q】
【図5R】
【図5S】
【図5T】
【図5U】
【図5V】
【図5W】
【図5X】
【図5Y】
【図5Z】
【図5AA】
【図5BB】
【図5CC】
【図5DD】
【図5EE】
【図5FF】
【図5GG】
【図5HH】
【図5II】
【図5JJ】
【図5KK】
【図5LL】
【図5MM】
【図5NN】
【図5OO】
【図5PP】
【図5QQ】
【図5RR】
【図5SS】
【図5TT】
【図5UU】
【図5VV】
【図5WW】
【図5XX】
【図5YY】
【図5ZZ】
【図5AAA】
【図5BBB】
【図5CCC】
【図5DDD】
【図5EEE】
【図5FFF】
【図6】
【公開番号】特開2011−207885(P2011−207885A)
【公開日】平成23年10月20日(2011.10.20)
【国際特許分類】
【出願番号】特願2011−87129(P2011−87129)
【出願日】平成23年4月11日(2011.4.11)
【分割の表示】特願2005−333254(P2005−333254)の分割
【原出願日】平成6年1月25日(1994.1.25)
【出願人】(500049716)アムジエン・インコーポレーテツド (242)
【Fターム(参考)】
【公開日】平成23年10月20日(2011.10.20)
【国際特許分類】
【出願日】平成23年4月11日(2011.4.11)
【分割の表示】特願2005−333254(P2005−333254)の分割
【原出願日】平成6年1月25日(1994.1.25)
【出願人】(500049716)アムジエン・インコーポレーテツド (242)
【Fターム(参考)】
[ Back to top ]