
Gタンパク質共役受容体シグナルによるダイオキシン作用の軽減
【課題】生体内に既存している情報伝達システムを利用するダイオキシン作用の阻害、軽減を可能にする手段を提供する。
【解決手段】Gタンパク質とダイオキシン作用に関係するAIPが相互作用しており、AIPと相互作用するGα13の過剰発現、Gα13を活性化するリゾホスファチジン酸刺激によりAhレセプターを介する遺伝子発現が抑制されること見出したことに基づいて、Gタンパク質とAIPの相互作用を促進する手段を含む医薬又は食品、ダイオキシン作用の阻害、軽減に有効な候補物質の選択方法、Gタンパク質共役型受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物の同定方法、ダイオキシン作用の阻害剤、ダイオキシン作用に起因する疾患の予防治療剤又は予防治療方法、Gタンパク質共役型受容体、ダイオキシン作用への制御試験系からなる。
【解決手段】Gタンパク質とダイオキシン作用に関係するAIPが相互作用しており、AIPと相互作用するGα13の過剰発現、Gα13を活性化するリゾホスファチジン酸刺激によりAhレセプターを介する遺伝子発現が抑制されること見出したことに基づいて、Gタンパク質とAIPの相互作用を促進する手段を含む医薬又は食品、ダイオキシン作用の阻害、軽減に有効な候補物質の選択方法、Gタンパク質共役型受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物の同定方法、ダイオキシン作用の阻害剤、ダイオキシン作用に起因する疾患の予防治療剤又は予防治療方法、Gタンパク質共役型受容体、ダイオキシン作用への制御試験系からなる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アリル炭化水素受容体結合タンパク質(arylhydrocarbon receptor interacting protein以下AIPと略称する)とアリル炭化水素受容体(arylhydrocarbon receptor、以下Ahレセプターと略称する)の結合を抑制することにより、ダイオキシン類が結合するAhレセプターを介する作用を抑制する医薬、食品及び方法に関する。より詳しくは、GTP結合タンパク質(Gタンパク質)等のAIPと相互作用する物質によりAhレセプターの安定性や細胞内局在に影響を与えて、ダイオキシン類の作用を軽減する技術に関する。
【背景技術】
【0002】
ダイオキシンは有機塩素化合物を焼却炉で燃焼する際などに発生する。ダイオキシンの産生を根絶することは根本的な廃棄物の減少が図られない限り困難である。中国、および開発途上国などにおける世界的な規模でのダイオキシンの環境汚染が懸念されており、ダイオキシンの毒性の軽減を目的とする新しい取り組みが将来の課題として残されている。
【0003】
ダイオキシンは、急性毒性、発がん性、催奇形性などの毒性を有するとともに、極微量でホルモンや成長因子などの働きを乱す環境ホルモンとして作用する。ダイオキシンの有機塩素化合物は、化学的に極めて安定であり、大気中、土壌、海水など自然界に一度拡散すると分解されることなく蓄積される。また、ダイオキシンは生体内に取り込まれると肝臓、脂肪組織などに蓄積され、体外へ排出されにくい性質を持っている。従来よりダイオキシンの産生および除去に関する技術の開発は盛んに行われてきたが、ダイオキシンの作用発現機構に他の細胞情報伝達系を用いて影響を与えて、ダイオキシンの作用を軽減する技術は報告されていなかった。
【0004】
以下に本明細書において引用した文献を列記する。
【非特許文献1】Gohla A., Harhammer R., Schultz G. (1998) J Biol Chem. 273, 4653-4659
【非特許文献2】Yamaguchi Y., Katoh H., Yasui H., Aoki J., Nakamura K., Negishi M.(2000) J Neurochem. 2000 75, 708-717
【非特許文献3】Katoh H., Aoki J., Yamaguchi Y., Kitano Y.,Ichikawa A., Negishi M.(1998) J Biol Chem. 273, 28700-28707
【非特許文献4】Voyno-Yasenetskaya T., Conklin B.R., Gilbert R.L., Hooley R., Bourne H.R., Barber D.L.(1994) J Biol Chem. 269, 4721-4724
【非特許文献5】Berestetskaya Y.V., Faure M.P., Ichijo H., Voyno-YasenetskayaT.A.(1998) J Biol Chem. 273, 27816-27823.
【非特許文献6】Yuan J., Slice L.W., Rozengurt E. (2001) J Biol Chem. 276, 38619-38627
【非特許文献7】Voyno-Yasenetskaya T.A., Faure M.P., Ahn N.G., Bourne H.R.(1996) JBiol Chem. 271, 21081-21087
【非特許文献8】Kozasa T., Jiang X., Hart M.J., Sternweis P.M., Singer W.D.,Gilman A.G., Bollag G., Sternweis P.C.(1998) Science 280, 2109-2111
【非特許文献9】Vaiskunaite R., Adarichev V., Furthmayr H., Kozasa T., GudkovA., Voyno-Yasenetskaya T.A. (2000) J Biol Chem. 275, 26206-26212
【非特許文献10】Meigs T.E., Fedor-Chaiken M., Kaplan D.D., Brackenbury R.,Casey P.J.(2001) Proc. Natl. Acad. Sci. U S A. 98, 519-524
【非特許文献11】Kozak, M. (1999) Gene, 234, 187-208
【非特許文献12】Prickett K.S., Amberg D.C., Hopp T.P.(1989) Bio Techniques, 7, 580-589
【非特許文献13】Vertes A.A. et al.(1994) Molecular Microbiol., 11, 739-46
【非特許文献14】Ulmer (1983) Science, 219, 666-671
【非特許文献15】Tuschl T.(2002) Nature Biotechnology, 20, 446-448
【非特許文献16】Brummelkamp T.R., Bernards R., Agami R.(2002) Science, 296, 550-553
【非特許文献17】Andersson S., Davis D.L., Dahlback H., Jornvall H., RussellD.W.(1989) J Biol. Chem., 264, 8222-8229
【非特許文献18】Kobayashi A., Numayama-Tsuruta K., Sogawa K., Fujii-KuriyamaY.(1997) J Biochem. 122, 703-710
【非特許文献19】Mimura J., Ema M., Sogawa K., Fujii-Kuriyama Y. (1999) Gene Dev., 13, 20-25
【発明の開示】
【発明が解決しようとする課題】
【0005】
従来、ダイオキシンが体内に一度摂取されてしまった後に、その作用を抑制するための技術的な解決法は考案されていなかった。したがって、本発明は生体内に既存している情報伝達システムを利用するダイオキシン作用の阻害、軽減を可能にする手段を提供することを課題とする。
【課題を解決するための手段】
【0006】
上記課題を解決すべく本発明者らは鋭意努力し、哺乳動物のGタンパク質遺伝子を細胞に導入して組換え体Gタンパク質を産生する系を用いて、それと相互作用する細胞内タンパク質を網羅的に探索した。その結果、Gタンパク質と相互作用する分子として、新たにダイオキシン作用に関係するAIPを見出した。次に、Ahレセプターを介するダイオキシン作用に対するGタンパク質シグナルの影響を検討した。その結果、AIPと相互作用するGα13の過剰発現、Gα13を活性化するリゾホスファチジン酸(lysophosphatidic acid、以下LPAと略称することがある)刺激によりAhレセプターを介する遺伝子発現が抑制されることが判明した。さらに、Gタンパク質シグナルの活性化によりAhレセプターの細胞内局在が変化することを見出し、本発明を完成した。
【0007】
すなわち本発明は、
1.GTP結合タンパク質(以下、Gタンパク質)とアリル炭化水素受容体結合タンパク質(以下、AIP)の相互作用を促進する手段を含む医薬又は食品、
2.用途が、ダイオキシン作用の阻害、軽減である前項1の医薬又は食品、
3.手段が、Gタンパク質の増加による前項1又は2の医薬又は食品、
4.Gタンパク質の増加が、Gタンパク質遺伝子を担持するベクターの投与、又は活性型Gタンパク質自体の投与若しくは摂取である前項3の医薬又は食品、
5.手段が、Gタンパク質の活性化による前項1又は2の医薬又は食品、
6.Gタンパク質を活性化する手段が、Gタンパク質共役受容体シグナル伝達系への作用である前項5の医薬又は食品、
7.作用が、Gタンパク質共役受容体シグナル伝達系に対する活性化剤の投与又は摂取による前項6の医薬又は食品、
8.活性化剤が、リゾホスファチジン酸である前項7の医薬又は食品、
9.ダイオキシン作用に影響するメカニズムが、アリル炭化水素受容体(以下、Ahレセプター)とダイオキシンの結合の阻害である前項2〜8の何れか1項に記載の医薬又は食品、
10.Gタンパク質が、Gタンパク質αサブユニット13(Gα13)である前項1〜9の何れか1項に記載の医薬又は食品、
11.以下の少なくとも1の方法を選択するダイオキシン作用の阻害、軽減に有効な候補物質の選択方法;
1)Gタンパク質とAIPの相互作用の促進の可否
2)Gタンパク質の発現の促進の可否
3)Gタンパク質遺伝子の転写促進の可否
4)Gタンパク質の活性化の可否
5)Gタンパク質共役受容体への作用の可否
12.Gタンパク質共役受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物の同定方法であって、Gタンパク質とAIPの相互作用を可能にする系で、候補化合物をこの系に接触させ、次いでGタンパク質とAIPの相互作用により生じるシグナルおよび/またはマーカーを使用する系を用いて、該シグナルおよび/またはマーカーの存在若しくは不存在および/または変化を検出することにより、該化合物がGタンパク質とAIPとの結合を阻害するか否かを決定することを特徴とする同定方法、
13.Gタンパク質がGα13である前項11又は12に記載の方法、
14.前項13に記載の同定方法により同定された化合物を含有するダイオキシン作用の阻害剤、
15.前項1〜10の何れか1項に記載の医薬若しくは食品からなる、又は前項14に記載の阻害剤を含む、ダイオキシン作用に起因する疾患の予防治療剤又は予防治療方法、
16.ダイオキシン作用に基づく疾患が、肝臓毒性、胸腺の萎縮、皮膚がん、又は子宮内膜症である前項15に記載の予防治療剤又は予防治療方法、
17.Gタンパク質共役受容体、Gタンパク質の優性活性型変異体および/または優性抑制型変異体を用いることを特徴とするダイオキシン作用への制御試験系、
18.Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する方法、
19.Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する手段を含む医薬又は食品、
に関する。
【発明の効果】
【0008】
本発明により、Gタンパク質共役受容体シグナルに働きかける薬剤を利用した、ダイオキシン作用の阻害および軽減法の開発が可能となった。Gタンパク質共役受容体はヒトゲノム全3万種類の遺伝子の中で1000種類にもおよぶ大きなファミリーを形成している。また、組織、臓器特異的な発現を示すGタンパク質共役受容体が数多く存在しており、既に臨床上使用されている薬剤の40%近くがGタンパク質共役受容体を標的としている。したがって、本発明により生体内でダイオキシンの毒性作用が見られる、肝臓、胸腺、皮膚、子宮内膜に特異的に発現しているGタンパク質共役受容体を標的とする薬剤を検索し、その薬剤の投与によりダイオキシンの効果だけを軽減する理想的な薬剤が見出される可能性がある。
【発明を実施するための最良の形態】
【0009】
以下、本発明について発明の実施の態様をさらに詳しく説明する。以下の詳細な説明は例示であり、説明のためのものに過ぎず、本発明を何ら限定するものではない。
本明細書においては単離された若しくは合成の完全長蛋白質;単離された若しくは合成の完全長ポリペプチド;または単離された若しくは合成の完全長オリゴペプチドを意味する総称的用語として「ポリペプチド」という用語を使用することがある。ここで蛋白質、ポリペプチド若しくはオリゴペプチドはペプチド結合または修飾されたペプチド結合により互いに結合している2個以上のアミノ酸を含むものである。
【0010】
Gタンパク質は、細胞膜に存在するGタンパク質共役受容体が受け取ったホルモンや神経伝達物質などの刺激を細胞内に伝えるトランスデューサーであり、α、β、γサブユニットからなるヘテロ三量体を形成する。Gタンパク質は不活性化状態では、三量体を形成し、αサブユニット(Gα)はGDPと結合している。受容体にリガンドが結合すると、Gαに結合していたGDPが解離し、細胞内に豊富に存在するGTPが結合してGタンパク質が活性化される。Gタンパク質が活性化されるとGαとβγサブユニット(Gβγ)が解離し、それぞれが下流のエフェクタータンパク質にシグナルを伝える。Gαに結合したGTPがGDPに加水分解されることによって、Gタンパク質は再び不活性化状態にもどり、Gαβγの三量体を形成する。
【0011】
三量体Gタンパク質はGαのアミノ酸配列によって4つのファミリー(Gαs、Gαi、Gαq、Gα12)に分類される。このなかでGα12ファミリーにはGα12とGα13が存在し、どちらも広範な組織に発現している。Gα12やGα13の活性化によってアクチンストレスファイバーの形成(非特許文献1)や、神経伝達物質の放出に関わるCa2+依存性エクソサイトーシスの抑制(非特許文献2)、神経突起の退縮(非特許文献3)、Na+/H+交換因子の活性化(非特許文献4)、アポトーシスの誘導(非特許文献5)、PKDの活性化(非特許文献6)、JNKの活性化(非特許文献7)が起こる。このような細胞応答に至るシグナル伝達経路を明らかにするために、直接相互作用する分子の同定が行われ、Gα13のGTP加水分解反応を促進するRGSドメインとRhoのGTP/GDP交換反応を促進するDHドメインをもったp115RhoGEFが同定された(非特許文献8)。他にもGα13と相互作用する分子として、radixin(非特許文献9)やcadherin(非特許文献10)などが同定されている。
【0012】
ダイオキシンは、一般に、75種類の異性体を持つポリ塩化ジベンゾパラジオキシン(PCDD)および135種類の異性体を持つポリ塩化ジベンゾフラン(PCDF)の総称をいう。ダイオキシンによる肝臓毒性、胸腺の萎縮、皮膚がん、子宮内膜症の発生には、Ahレセプターが関与している。Ahレセプターはベンツピレン、3−メチルコラントレンのような多環性芳香族炭化水素化合物とダイオキシンのようなハロゲン化芳香族炭化水素化合物をリガンドとする受容体型転写因子である。Ahレセプターはリガンドがない状態では熱ショックタンパク質90(heat shock protein 90、Hsp90)、AIP(またはXAP2、ARA9とも呼ばれる)と複合体を形成し、細胞質に存在する。リガンドがAhレセプターに結合するとAhレセプターは複合体から離れ、核内へ移行する。Ahレセプターは核内でアリル炭化水素受容体核輸送体(arylhydrocarbon receptor nuclear translocator、以下Arntと略称する)と2量体を形成する。その2量体はシトクロームP450遺伝子(CYP1A1)のプロモーター上に存在する異物応答配列(xenobiotic responsive element、以下XRE略称する)に結合する。
【0013】
本発明においては、Gタンパク質と相互作用する細胞内タンパク質の探索を行い、ダイオキシン作用にかかわるAIP がGタンパク質と相互作用することを初めて見出した。したがって、本発明の一態様はGタンパク質とAIPの相互作用を促進する手段を含む医薬又は食品である。また、本発明においてはGタンパク質シグナルがAhレセプターよる遺伝子発現を抑制することを見出した。このことから、Gタンパク質はAIPのAhレセプターとの結合を阻害することで、Ahレセプターに結合するリガンドの作用を抑える可能性が示唆された。Ahレセプターに結合するリガンドとしてダイオキシンが好適に挙げられる。したがって、本発明の医薬又は食品の用途の一つはダイオキシン作用の阻害、軽減である。
【0014】
本明細書において、「Gタンパク質とAIPの相互作用」とは、Gタンパク質とAIPがある様式により互いに作用を及ぼすことをいう。「ある様式」とは、結合、物理学的接触、近接等を含むものであり、結果として互いに作用を及ぼし得る様式であればいずれのものであってもよい。
【0015】
Gタンパク質とAIPの相互作用を促進する手段は、Gタンパク質の増加により達成可能である。Gタンパク質の増加は、具体的には、遺伝子導入手段によって、機能するように連結されたプロモーターを有するGタンパク質遺伝子を担持するベクターを標的組織へ投与することにより達成することができる。自己増殖ベクターに遺伝子を担持する手段によっても本発明の目的は達成可能である。
【0016】
遺伝子導入手段は、望ましくはインテグレーションさせる方法、例えばリポソーム法、リン酸カルシウム法、エレクトロポレーション法、レトロウイルス法等によるのが好ましい。しかし一過性に発現させるリポソーム法、リン酸カルシウム法、エレクトロポレーション法、ウイルスベクター法、アテロコラーゲン法等の方法であってもよい。用いるベクターは特に限定されず、公知のプラスミド、ファージ、コスミド、BAC、YAC、組換えウイルス、トランスポゾン等、通常の組み換え実験によって挿入DNA断片を導入することが可能な全ての組換えベクターに適用することができる。ベクターは、当然に自体公知の組合せが好適であるプロモーター、エンハンサーと共に構築することができる。例えば、通常宿主に適したプロモーターが挿入されている市販のタンパク質発現ベクターを用いることができる。具体的には、ZAPExpress(Stratagene)、pSVK3(Amersham Bioscience)、pEGFP-C1(Clontech)、アテロコラーゲン等が挙げられる。
【0017】
ベクターへのタンパク質発現遺伝子の挿入は、該遺伝子又はこれを含むDNA断片をベクター中のプロモーターの下流にプロモーターの制御下に配置されるように連結して行う。また、プロモーターとタンパク質発現遺伝子との間にコザック配列(非特許文献11)を挿入したり、タンパク質発現遺伝子の下流にタグとなるポリペプチドをコードするDNAを挿入した構造を有するベクターも好ましく用いられる。タグとなるポリペプチドとしては特に制限はないが、例えば、FLAGタグ(非特許文献12)等が挙げられる。プロモーターを連結したGタンパク質発現遺伝子を標的細胞の染色体中に直接挿入する相同組換え技術、あるいはトランスポゾンや挿入配列(非特許文献13)等を用いて発現させることができる。
【0018】
Gタンパク質の増加は上記のように得られたGタンパク質を投与又は摂取することにより達成可能である。Gタンパク質は、天然に存在するものであってよく、Gタンパク質に変異を導入したものであってもよい。具体的には、活性型Gタンパク質であることが望ましい。また、Gタンパク質とAIPの相互作用部位を含むGタンパク質の部分ポリペプチドであってもよい。Gタンパク質とAIPの相互作用部位のアミノ酸配列からなるポリペプチドはかかるポリペプチドとして好適である。
【0019】
ポリペプチドに欠失、置換、付加または挿入等の変異を導入する手段は自体公知であり、例えばウルマー(Ulmer)の技術(非特許文献14)を利用できる。このような変異の導入において、当該ポリペプチドの基本的な性質(物性、機能または免疫学的活性等)を変化させないという観点から、例えば、同族アミノ酸(極性アミノ酸、非極性アミノ酸、疎水性アミノ酸、親水性アミノ酸、陽性荷電アミノ酸、陰性荷電アミノ酸および芳香族アミノ酸等)の間での相互置換は容易に想定される。さらに、これら利用できるポリペプチドは、その構成アミノ基またはカルボキシル基等を、例えばアミド化修飾する等、機能の著しい変更を伴わない程度に改変が可能である。
【0020】
ポリペプチドは、常法に従って製造できる。具体的には、通常の液相法および固相法によるポリペプチド合成法を挙げることができるが、これらに限らず公知の方法が広く利用可能である。または市販のポリペプチド合成装置を用いて製造可能である。あるいは遺伝子工学的手法により取得することもできる。例えば目的とするポリペプチドをコードする遺伝子を宿主細胞中で発現できる組換えDNA(発現ベクター)を作成し、これを適当な宿主細胞、例えば大腸菌にトランスフェクションして形質転換した後に該形質転換体を培養し、次いで得られる培養物から目的とするポリペプチドを回収することにより製造可能である。
【0021】
Gタンパク質とAIPの相互作用を促進する手段は、Gタンパク質の活性化により達成可能である。前記Gタンパク質の活性化はGタンパク質共役受容体シグナル伝達系に作用することにより達成可能であり、具体的には、Gタンパク質共役受容体シグナル伝達系に対する活性化剤の投与又は摂取によることができる。Gタンパク質共役受容体シグナル伝達系に対する活性化剤の好適な例として、リゾホスファチジン酸が挙げられる。
【0022】
Ahレセプターはリガンドが無い状態ではAIPと複合体を形成し細胞質に存在するが、Gタンパク質がAIPに結合するとAhレセプターは複合体から離れ、核内へ移行する。本発明においては、Gタンパク質シグナルがダイオキシン受容体であるAhレセプターの細胞内局在に影響を与えることを見出した。このことから、Gタンパク質がAIPと結合することによりAhレセプターの細胞内局在が変化することにより、ダイオキシンとAhレセプターの結合が阻害される可能性が考えられる。したがって、本発明の一態様はダイオキシン作用に影響するメカニズムがAhレセプターとダイオキシンの結合の阻害である医薬又は食品である。 本発明に用いるGタンパク質は、好ましくはGαi以外のGαであり、より好ましくはGα13である。
【0023】
ダイオキシン作用の阻害、軽減に有効な候補物質の選択方法は以下の方法から選択することができる;1)Gタンパク質とAIPの相互作用の促進の可否、2)Gタンパク質の発現の促進の可否、3)Gタンパク質遺伝子の転写促進の可否、4)Gタンパク質の活性化の可否、5)Gタンパク質共役受容体への作用の可否、である。1)〜5)の方法においては、1)Gタンパク質とAIPの相互作用、2)Gタンパク質の発現、3)Gタンパク質遺伝子の転写、4)Gタンパク質の活性化、5)Gタンパク質共役受容体への作用を可能にする条件を選択し、Gタンパク質とAIPを前記候補物質と接触させ、1)Gタンパク質とAIPの相互作用、2)Gタンパク質の発現、3)Gタンパク質遺伝子の転写、4)Gタンパク質の活性化、5)Gタンパク質共役受容体への作用を検出することができるシグナルおよび/またはマーカーを使用する系を用いて、このシグナルおよび/またはマーカーの存在若しくは不存在または変化を検出することにより、ダイオキシン作用の阻害、軽減に有効な候補物質を選択することができる。1)Gタンパク質とAIPの相互作用、2)Gタンパク質の発現、3)Gタンパク質遺伝子の転写、4)Gタンパク質の活性化、または5)Gタンパク質共役受容体への作用を可能にする条件はインビトロのものであってよく、インビボのものであってもよい。例えば、Gタンパク質とAIPを共発現させた細胞を用いることもできる。
【0024】
Gタンパク質共役受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物の同定方法は、自体公知のスクリーニングシステムを利用して構築可能である。例えば、Gタンパク質とAIPの相互作用を可能にする系で、調べようとする化合物(候補化合物)をこの系に接触させ、次いでGタンパク質とAIPの相互作用により生じるシグナルおよび/またはマーカーを使用する系を用いて、このシグナルおよび/またはマーカーの存在若しくは不存在または変化を検出することにより、Gタンパク質共役受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物を同定することができる。Gタンパク質とAIPの相互作用を可能にする系は、インビトロのものであってよく、インビボのものであってもよい。例えば、Gタンパク質とAIPを共発現させた細胞を用いることができる。
【0025】
前記候補物質又は候補化合物としては、例えば化学ライブラリーや天然物由来の化合物、またはGタンパク質、AIPの一次構造や立体構造に基づいてドラッグデザインして得られた化合物等が挙げられる。あるいは、AIPとGタンパク質との相互作用部位のアミノ酸配列からなるポリペプチドの構造に基づいてドラッグデザインして得られた化合物等も候補物質又は候補化合物として好適である。
【0026】
上記同定方法で得られた化合物は、ダイオキシン作用の阻害剤に利用可能である。本発明の医薬若しくは食品、または上記同定方法で得られた化合物を含む阻害剤は、ダイオキシン作用に基づく疾患の予防治療剤又は予防治療方法に利用可能である。かかる疾患としては、肝臓毒性、胸腺の萎縮、皮膚がん、又は子宮内膜症等を挙げることができる。
【0027】
本発明においては、Gタンパク質共役受容体シグナルによりダイオキシンによる遺伝子発現が抑制されることが見出された。このことから、Gタンパク質共役受容体、Gタンパク質の優性活性型変異体および/または優性抑制型変異体を用いてダイオキシン作用への制御試験系を構築することができる。前記制御試験系においては、ダイオキシン作用を制御する可能性のある候補物質をGタンパク質共役受容体、Gタンパク質の優性活性型変異体および/または優性抑制型変異体と接触させ、ダイオキシンによる遺伝子発現により生じるシグナルおよび/またはマーカーを使用する系を用いて、該シグナルおよび/またはマーカーの存在若しくは不存在または変化を検出することにより、前記候補物質がダイオキシン作用を制御するか否かを判定する。前記制御試験系は、インビトロのものであってよく、インビボのものであってもよい。例えば、Gタンパク質共役受容体、Gタンパク質の優性活性型受容体および/または優性抑制型変異体を共発現させた細胞を用いることができる。
【0028】
本発明においては、AIPとGタンパク質の相互作用によりAhレセプターの転写活性化能が抑制された。このことから、AIPにGタンパク質が相互作用したことにより、Ahレセプターと結合できるAIP分子の数が減少し、Ahレセプターの転写活性化能が抑制された可能性が示唆された。したがって、Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する方法が構築可能である。また、本発明の一態様は、Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する手段を含む医薬又は食品である。具体的には、AIPに相互作用する物質をAIPと相互作用させ、Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターの転写活性化能の促進又は抑制を達成することができる。好ましいAIPに相互作用する物質はGタンパク質であり、より好ましくはGαi以外のGαであり、さらに好ましくはGα13である。
【0029】
Ahレセプターと結合できるAIP分子の数を減少させることにより、Ahレセプターによる転写活性化能を抑制する手法の一つとして、RNAi法が挙げられる。詳しくは、RNAi法によりAIPの生理活性を制御することにより、AHレセプターと結合できるAIP分子の数を減少させる。RNAi法は機能を阻害しようとする遺伝子の特定領域と相同なセンスRNAとアンチセンスRNAからなる二本鎖RNAが標的遺伝子のmRNAの相同部分を干渉・破壊する方法である。二本鎖RNAはRNAiの目的に適した配列を有する領域から選択する。具体例として、AIPのAhレセプターとの結合部位を含む領域から当該配列を選択することができる。一般的には、二本鎖RNAの配列として開始コドンの50から100ヌクレオチド下流の領域を選択する。通常、RNAi法には、塩基数が21〜23塩基であるセンスRNAおよびアンチセンスRNAの一対の組み合わせからなる二本鎖RNAが必要である。
【0030】
前記二本鎖RNAを構成するオリゴヌクレオチドは、公知の方法によって作製することができる。例えば化学的に合成することができるし、あるいは天然の核酸を制限酵素などによって切断し、上記のような塩基配列で構成されるように改変し、あるいは連結することも可能である。具体的には、オリゴヌクレオチド合成装置等を用いて合成することができる。また、siRNA合成キットが既に市販されており(ニッポンジーンテク社等)、これらを利用して二本鎖RNAを合成することもできる。前記二本鎖RNAを細胞に導入するには、公知の方法を用いることができる。例えば、リン酸カルシウム法、リポフェクチン法などが挙げられる。前記二本鎖RNAは、適当なベクターを用いて細胞内で恒常的に発現することもできる。例えば、非特許文献15および16に開示されたsiRNA発現専用ベクターを利用すればよい。さらに、当該ベクターに二本鎖RNAを組み込む方法は、自体公知の方法を適用することができる。例えば二本鎖RNAに適当なリンカーをライゲーションし、これを目的に適したベクターのマルチクローニングサイトへ挿入することによって、所望のベクターを得ることもできる。
【0031】
本明細書においてシグナルとは、そのもの自体がその物理的または化学的性質により直接検出され得るものを指し、マーカーとは、そのものの物理的または生物学的性質を指標として間接的に検出され得るものを指す。シグナルおよび/またはマーカーとしては、ルフェラーゼ、蛍光蛋白質(GFP、YFP、BFP等)、および放射性同位体等、レポーター遺伝子、例えばクロラムフェニコールアセチルトランスフェラーゼ遺伝子等、または検出用のエピトープタグ、例えば6×Hisタグ等、公知のものが利用できる。これらシグナルまたはマーカーの検出方法は当業者には周知のものである。
【0032】
本発明において使用するGタンパク質、Gタンパク質の優性活性型変異体および/または優性抑制型変異体、Gタンパク質共役受容体、AIPは、これらを遺伝子工学的手法で発現させた細胞、無細胞系合成産物、化学合成産物、または該細胞や生体試料から調製したものであってよく、これらからさらに精製されたものであってもよい。また、Gタンパク質とAIPの相互作用、およびこれら蛋白質の機能に影響がなければ、N末端側やC末端側に別の蛋白質やポリペプチド、例えばβ−ガラクトシダーゼ、IgG等の免疫グロブリンFc断片、Hisタグ、Mycタグ、HAタグ、FLAGタグ、またはXpressタグ等のタグペプチド類を、直接的にまたはリンカーペプチド等を介して間接的に、遺伝子工学的手法等を用いて付加したものであってもよい。
【0033】
本発明の医薬は、予防又は治療の目的に応じて、散剤、顆粒剤、錠剤、カプセル剤、腸溶剤、液剤、注射剤(液剤、懸濁剤)などの各種の形態に、常法にしたがって調製することができる。本発明の医薬は、通常は1種または複数の医薬用担体を用いて医薬組成物として製造することが好ましい。本発明の医薬中に含まれる有効成分の量は、広範囲から適宜選択されるが、通常約0.00001〜70重量%、好ましくは0.0001〜5重量%程度の範囲とするのが適当である。
【0034】
本発明の医薬を投与するときには、該医薬を単独で使用してもよく、あるいは目的の疾患の防止、治療または改善に必要な他の化合物または医薬と共に使用してもよい。投与経路は、全身投与または局所投与のいずれも選択することができる。この場合、疾患、症状等に応じた適当な投与経路を選択する。例えば、非経口経路として、通常の静脈内投与、動脈内投与のほか、皮下、皮内、筋肉内等への投与を挙げることができる。あるいは経口による投与も可能である。さらに、経粘膜投与または経皮投与も可能である。
【0035】
本発明の医薬の投与形態としては、各種の形態が目的に応じて選択でき、その代表的なものとしては、錠剤、丸剤、散剤、粉末剤、細粒剤、顆粒剤、カプセル剤等の固体投与形態や、水溶液製剤、エタノール溶液製剤、懸濁剤、脂肪乳剤、リポソーム製剤、シクロデキストリン等の包接体、シロップ、エリキシル等の液剤投与形態が含まれる。これらは更に投与経路に応じて経口剤、非経口剤(点滴剤、注射剤)、経鼻剤、吸入剤、経膣剤、坐剤、舌下剤、点眼剤、点耳剤、軟膏剤、クリーム剤、経皮吸収剤、経粘膜吸収剤等に分類され、それぞれ通常の方法に従い、調合、成形、調製することができる。
【0036】
本発明の食品は、例えば、塊状、液状(液体、粘稠流体、ペースト等)、ゼリー状、粉末状、顆粒状などの各種形態に、常法に従って調製することができる。かかる形態の具体的な例としては、飲料(ドリンク剤、清涼飲料水、ジュース、茶類など)、粉末飲料(粉末ジュース、粉末スープなど)、菓子類(クッキー、ビスケット、シリアル、チュアブル錠剤、チューイングガム、キャンディー、グミ、ウェハース、せんべいなど)、調味料(ドレッシング、ソース、粉末調味料など)、主食製品(パン、麺類、餅など)などが挙げられる。また、本発明の食品は、一般食品以外にも、病者用食品、高齢者用食品、特定保健用食品などの特別用途食品や、医薬部外品、機能性食品、健康食品などの目的に幅広く使用することができる。
【0037】
本発明の医薬又は食品には、本発明の効果を損なわない範囲であれば、必要に応じて、通常医薬や食品に配合される成分、例えば、栄養素(他の蛋白質、脂質、炭水化物、ミネラル、ビタミンなど)、動植物成分(粉砕物、脱脂物、抽出エキス、抽出末など)、賦形剤(二糖類、糖アルコール、多糖類、水溶性高分子など)、増量剤、甘味料、香味剤、着色剤、防腐剤、乳化剤、界面活性剤(可溶化剤、泡安定化剤、消泡剤など)、ハイドロトロープ剤(アルコール類、多価アルコール類など)などを配合することができる。本発明の医薬又は食品におけるこれら成分の配合量は特に限定されず、剤型、形態、用途等に応じて適宜設定することができ、例えば医薬又は食品の全重量に基づいて、0.1〜99.9重量%程度とすることができる。
【0038】
本発明の医薬又は食品の投与量又は摂取量については、本発明の効果が得られるものであれば特に限定されるものではなく、含有される成分の有効性、投与形態、投与経路、疾患の種類、対象の性質(体重、年齢、病状および他の医薬の使用の有無等)、および担当医師の判断等に応じて適宜選択される。本発明の医薬又は食品は、1日1〜数回に分けて投与又は摂取することができ、数日または数週間に1回の割合で間欠的に投与又は摂取してもよい。
【0039】
以下、本発明を実施例に基づき具体的に説明するが、本発明は下記の実施例に限定されない。
【実施例】
【0040】
実施例1
(酵母ツーハイブリッド法を用いた新規Gα13結合タンパク質の探索)
Gα13を介したシグナル伝達経路を解明するために、酵母ツーハイブリッド法を用いて、新規Gα13結合タンパク質の探索を行った。
【0041】
<方法>
1.ベイト側プラスミド(pGBKT7-Gα13QL)の構築
酵母ツーハイブリッド法によるスクリーニングはMATCHMAKER Two-hybrid System 3(Clontech)を用いて行った。ヒトGα13(GenbankAF493902、5729847)のcDNAはヒト293T細胞のRNAを鋳型とした逆転写酵素反応により作成し、翻訳領域の両端にEcoRIサイトを持つプライマー(配列番号1および2)で増幅した。そのDNA断片を哺乳動物細胞発現用プラスミドpCMV5(非特許文献17)のEcoRIサイトに挿入した。226番目のグルタミンをロイシンへの置換はQuickChangeキット(Stratagene)を用いて行った。活性型変異体Gα13QLをコードするcDNAをpCMV5-Gα13QLよりEcoRIで切り出し、GAL4結合領域(GAL4BD)を含むベイト側プラスミドpGBKT7のEcoRIサイトに挿入し、pGBKT7-Gα13QLを作成した。
【0042】
Forward primer:5'-CCGGAATTCATGGCGGACTTCCTGCCG-3'(配列番号1)
Reverse primer:5'-CCGGAATTCTCACTGTAGCATAAGCTGCTTGAGG-3'(配列番号2)
【0043】
2.酵母コンピテントセルの作成と形質転換
酵母の形質転換はYeast Transformation System(Clontech)を用い、酢酸リチウム法によって行った。
酵母(AH109)をYPDA寒天培地で30℃、2〜3日間培養した。培地上に生じた2mmのコロニーを6〜7個掻きとり、YPDA液体培地 50mlに移し、30℃、250rpmにて18時間振盪培養した。定常期(OD600>1.5)まで培養した細胞をYPDA液体培地 300mlに移し、さらに30℃、250rpmにて3時間振盪培養した。細胞を室温、1000×gにて5分間遠心し、細胞を回収した。滅菌水 40mlで細胞を懸濁した後、再び室温、1000×gにて5分間遠心し、細胞を回収した。1×TE/LiAc(10mM Tris-HCl(pH7.5)、1mM EDTA、0.1mM 酢酸リチウム) 1.5mlで細胞を懸濁したものをコンピテントセルとし、トランスフォーメーションに用いた。
【0044】
次に、酵母の発現プラスミド 200ng、ニシン精巣キャリアーDNA(94℃で20分加熱処理済)100ngとコンピテントセル 100μlを混ぜ合わせた。その後、PEG/LiAc(10mM Tris-HCl(pH 7.5)、1mM EDTA、1M 酢酸リチウム、40% PEG)600μlを添加し、ボルテックスで攪拌した。30℃で30分間インキュベートした後、DMSO 70μlを加えて穏やかに混ぜた。42℃にて15分間インキュベートした後、氷上で1〜2分間冷却した。室温、14000×gにて5秒間遠心し、ペレットを回収した。TE 500μlで2回洗浄した後、TE 500μlに懸濁し、培地上に細胞をプレーティングした。
【0045】
3.ライブラリーのスクリーニング
ライブラリーのスクリーニングにはPretransformed MATCHMAKER mouse brain cDNA library(Clontech)を使用した。これはGAL4活性化ドメイン(GAL4AD)をコードする酵母発現ベクター(pACT2)にマウス脳cDNAが挿入されたプラスミドが既に酵母(Y187)に導入されたものである。この酵母とベイト側プラスミドを導入した酵母(AH109)を30℃、40rpmにて25時間インキュベートして接合させた酵母をSD(-Leu/-Try/-His/Ade)選択培地で培養し、生育した陽性クローンについてさらにα−ガラクトシダーゼアッセイを行った。
【0046】
<結果>
Gα13の活性型変異体をベイトとし、マウス脳cDNAライブラリーのスクリーニングを行った結果、8.8×106個のクローンから一次選択により、20クローンを単離した。さらにα−ガラクトシダーゼアッセイによる二次選択により、11クローンを単離した。これらのクローンについて、シーケンスの解析を行った。このうち、Ahレセプターと結合し、そのシグナルに影響を及ぼす分子として報告されているAIPの遺伝子を含むプラスミドを、再度Gα13発現プラスミドとともに酵母(AH109)に導入し、選択培地における細胞の増殖の確認を行った。
【0047】
実施例2
(293細胞内におけるGα13とAIPの相互作用の確認)
哺乳動物細胞におけるGα13とAIPの相互作用を確認するために、ヒト胎児腎由来HEK293細胞にGα13とAIPを強制発現させ、免疫沈降法とウエスタンブロッティングを行い、相互作用の検討を行った。
AIP全長、AIPのN末端側、AIPのC末端側をコードするcDNAを発現用プラスミドに挿入し、これらのプラスミドをHEK293細胞に導入した発現を確認するとともに、Gα13と一緒に沈降するかどうか検討した。
【0048】
<方法>
1.哺乳動物細胞発現プラスミド(pCMV5-2HA-AIP)の構築
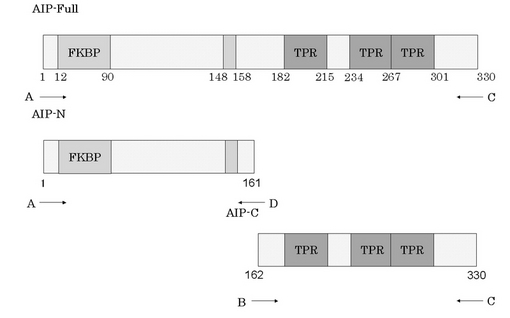
酵母ツーハイブリッド法によって、Gα13をベイトとして単離したクローンはN末端側13アミノ酸欠いたAIP(AIPΔ13)のcDNAを含むものであった。このクローンを鋳型とし、PCR法によりAIP全長(1-993:AIP-Full)、N末側断片(1-483:AIP-N)、C末側断片(484-993:AIP-C)を作製した(図1)。
【0049】
使用したプライマーの配列を以下に示す。
5'AIP-Full/5'-AIP-N:5'-GAAGATCTATGGCGGATCTCATCGCAAGACTCCGAGAGGACGGGATCCAAAAGCGTGTGA-3'(配列番号3)
5'AIP-C:5'-GAAGATCTACCTATCAACAGGACCC-3'(配列番号4)
3'AIP-Full/3'AIP-C:5'-GAAGATCTTCAGTGGGAAAAGATGCC-3'(配列番号5)
3'AIP-N:5'-GAAGATCTTCAGCCAGGACTCTCTAC-3'(配列番号6)
【0050】
293細胞内におけるGα13とAIPの相互作用を確認するために、HAタグのついた哺乳動物細胞発現ベクターpCMV5-2HAに上記のAIP遺伝子を挿入したプラスミドを構築した。AIP-Full、AIP-N、AIP-CはそれぞれのPCR産物の両端をBglIIで切断し、ベクターのBglIIサイトに挿入し、pCMV5-2HA-AIP-Full、pCMV5-2HA-AIP-N、pCMV5-2HA-AIP-Cを作製した。ライゲーションは、LigationHigh(TOYOBO)15μl、DNA溶液 15μl(インサートDNA 100ng、ベクタープラスミドDNA 50ng を含む)を16℃にて30分間インキュベートして行った。
【0051】
2.細胞培養とトランスフェクション
ヒト胎児腎由来HEK293細胞は10% ウシ胎児血清(FBS)と100μg/ml カナマイシンを含むダルベッコ変法イーグル培地(DMEM)を使用し、5% CO2存在下、37℃で培養した。トランスフェクションはリン酸カルシウム法によって行った。細胞は6cmディッシュで培養し、サブコンフルエントに達したものを用いた。トランスフェクションを行う前に、10% FBSを含むDMEM 4mlに交換した。HAタグのついたAIP(pCMV5-2HA-AIP-N 3μg、pCMV5-2HA-AIP-C 1μg、pCMV5-2HA-AIP-Full 1μg)、FLAGタグのついたGα13QL(pCMV5-FLAG-Gα13QL 2μg)にpCMV5を加え合計で5μgになるようにDNA溶液を調製した。さらに2.5M CaCl2・2H2O 50μlと水を合計で500μlになるように添加し、混合した。このDNA溶液を2×HeBS(50mM HEPES-NaOH(pH 7.0)、280mM NaCl、1.5mM Na2PO4) 500μlに加えて、よく攪拌した。室温で20分放置した後、HEK293細胞培養液に滴下した。細胞を5% CO2 存在下、37℃にて18時間培養後、培養液を取り除き3mlの培地(100μg/ml カナマイシンを含むDMEM)に交換し、さらに24時間培養した。
【0052】
3.細胞抽出液(ライセート)の調製
HEK293細胞に遺伝子導入後、5% CO2 存在下、37℃にて36時間培養した細胞からライセートを調製した。細胞の培養液を取り除いた後、ディッシュを氷上で冷やしながらLysisBuffer(20mM HEPES-NaOH(pH 7.5)、3mM MgCl2・6H2O、100mM NaCl、1mM NaVO4、10mM NaF、20mM β-Glycerophosphate、1mM PMSF、1mM DTT、1μM leupeptin、1mM EGTA、0.5% LB-PX)を600μM加えた。セルスクレーパーで細胞を掻き取り、1.5mlチューブに回収した。チューブを氷上で10分間放置し、細胞を融解させた。4℃、14000×rpmにて10分間遠心分離を行い、上清を回収した。
【0053】
4.免疫沈降
細胞抽出液 300μlに30% Protein G Sepharose(Amasham Bioscience) 30μl、抗FLAG抗体(シグマ)1μgを加え、ローテーターで4℃にて8時間旋回させた。4℃、2000×rpmにて3分間遠心分離を行い、上清を取り除いた。Lysis Buffer 300μl で3回洗浄した後、30μlの2×Sample Buffer(0.1M Tris-HCl(pH6.8)、4% SDS、20% グリセロール、0.06M DTT)を加え、95℃にて3分間煮沸した。室温、14000×rpmにて10秒間遠心分離し、上清をSDS-PAGEによって分離した。
【0054】
5.ウエスタンブロッティング
SDS-PAGEによりサンプルを分離した後、PVDF膜に100Vで1時間転写した。5% スキムミルクにPVDF膜を30分浸しブロッキングした。PBST(138mM NaCl、2.68mM KCl、8.10mM Na2HPO4・12H2O、1.47mM KH2PO4、0.1% Tween20)で500倍に希釈した一次抗体と室温で1時間反応させた後、PBSTで3回洗浄した。二次抗体はペルオキシダーゼ標識した抗マウスIgG抗体を5% スキムミルクで4000倍に希釈したものを用い、室温にて30分間反応させた。PBSTによって3回洗浄した後、ECL Western Blotting Detection System(Amersham Bioscience)により検出した。
【0055】
<結果>
図2に示すようにAIP全長、AIPのC末端側はGα13と共沈するが、AIPのN末端側はGα13と共沈しないことがわかった。このことから、AIPはそのC末端側を介してGα13と相互作用することが示された。
【0056】
実施例3
(レポーターアッセイ)
実施例2の結果から哺乳動物細胞内でGα13とAIPが相互作用することが確認できたので、Gα13がAIPの機能に影響を及ぼす可能性が示唆された。図3に示すようにAIPは細胞質に存在するAhレセプターと結合し、レセプターを安定化し、リガンドとの結合を促進することで、Ahレセプターを介した転写活性を上昇すると考えられている。このことからGα13とAIPの相互作用がAhレセプターの機能に影響を及ぼす可能性が考えられた。Gα13によるAhレセプターを介した転写活性の変化について検討するためにレポーターアッセイを行った。内在性のAhレセプターが存在しないことが報告されているCOS-7細胞内に、AhレセプターとArnt、AhレセプターのDNA認識配列であるXREをホタルルシフェラーゼ遺伝子の上流に組み込んだレポータープラスミドと、内部コントロールとしてウミシイタケルシフェラーゼ遺伝子の上流にヒトポリペプチド鎖延長因子(elongation factor)のプロモーターを組み込んだコントロールプラスミドを導入しレポーターアッセイを行った。
【0057】
<方法>
1.プラスミド
活性型Gα13QLプラスミドの他に、Ahレセプター発現プラスミド(pBOS-HA-AhR)と、Arnt発現プラスミド(pBOS-GST-Arnt)(非特許文献18)、ルシフェラーゼ遺伝子の上流にXRE配列を組み込んだレポータープラスミド(pX4Tk-Luc)(非特許文献19)を使用した。pBOS-HA-AhRはpBSK-mAhR(非特許文献19)からHindIII/XbaIで切り出したマウスAhレセプターcDNA断片を平滑末端にした後、pBOST7HAのSmaIサイトに挿入した。
【0058】
2.細胞培養とトランスフェクション
実験には、アフリカミドリザル腎臓由来COS-7細胞を用い、実施例2におけるHEK293細胞と同様の条件(10% FBS、100μg/ml カナマイシンを含むDMEMにおいて、5% CO2存在下、37℃)で培養し、リン酸カルシウム法でトランスフェクションを行った。24時間培養後、培養液を取り除き、1μM 3-Methylcholanthrene(3-MC)や10μM リゾホスファチジン酸(LPA)を含む無血清培地 3mlに交換し、さらに24時間培養した。
【0059】
3.細胞ライセートの調製
遺伝子導入後、48時間培養した細胞を回収し、細胞ライセートを調製した。細胞ライセートの調製にはDual-Luciferase Reporter Assay(Promega)に含まれるPassive Lysis Bufferを用いた。まず、細胞の培養液を取り除き、PBS(138mM NaCl、2.68mM KCl、8.10mM Na2HPO4・12H2O、1.47mM KH2PO4)を添加し、洗浄した。洗浄後、PBSを完全に取り除き、1×Passive Lysis Buffer存在下で細胞を回収し、1.5mlチューブに移した。回収した細胞ライセートを液体窒素で1〜2回凍結融解させ、細胞を完全に融解させた。
【0060】
4.レポーターアッセイおよび化学発光測定
レポーターアッセイはDual-Luciferase Reporter Assay(Promega)を用いて行った。また、化学発光測定はプレートリーダーMithras LB940(BELTHOLD)を用いて行った。96穴プレートに細胞ライセート 10μlにLuciferase Assay Reagent II(ホタルルシフェラーゼの基質含む)を50μl加えた。2秒後から、10秒間、ホタルルシフェラーゼ活性を測定した。測定後、Stop&GloReagent(コントロールのウミシイタケルシフェラーゼの基質を含む)を50μl加えた。再び2秒待った後、10秒間ウミイシイタケルシフェラーゼ活性を測定した。2種類の異なる酵素により触媒されて生じる発光の強度を上記のように測定し、ウミシイタケルシフェラーゼによる発光強度を用いてサンプル間の遺伝子導入効率の違いを補正してホタルルシフェラーゼ活性を評価した。
【0061】
<結果>
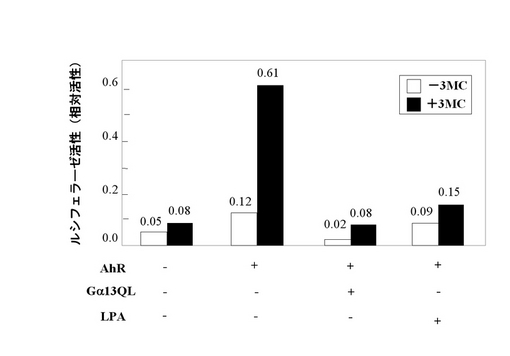
図4に示すように、Ahレセプターの発現プラスミドとArntの発現プラスミドをトランスフェクションした細胞において、Ahレセプターのリガンドである3-Methylcholantherene(3-MC)依存的にホタルルシフェラーゼ活性の上昇が見られた。AIPとの相互作用が確認されたGα13の活性型変異体(Gα13QL)を共発現させるとAhレセプターを介したルシフェラーゼ活性が低下した。また、リゾホスファチジン酸(LPA)によっても、Ahレセプターを介したルシフェラーゼ活性が抑制された。この結果から、Gα13がAhレセプターシグナルを負に制御することが示された。
【0062】
実施例4
(Ahレセプターの細胞内局在の検討)
これまでの実験から、Gα13はAIPと相互作用し、Ahレセプターシグナルを負に制御する可能性が示唆された。また、AIPはAhレセプターの細胞内局在に関わることから、Gα13とAIPの相互作用がAhレセプターの細胞内局在に影響を及ぼす可能性が考えられた。そこで、Gα13の活性化がAhレセプターの細胞内局在に及ぼす影響について、免疫染色法を用いて検討を行った。COS-7細胞内にHAタグのついたAhレセプターとArnt、Gα13QLを過剰発現させ、3-MC依存的なAhレセプターの細胞内局在の変化について調べた。
【0063】
<方法>
1.細胞培養とトランスフェクション
細胞はレポーターアッセイに用いたCOS-7細胞をカバーガラス上において実施例2におけるHEK293細胞と同様の条件(10% FBS、100μg/ml カナマイシンを含むDMEMにおいて、5% CO2存在下、37℃)で培養を行った。細胞にpBOS-HA-AhR 4μg、pBOS-GST-Arnt 1μg、pCMV5-Gα13QL 1μgをリン酸カルシウム法によってトランスフェクションを行った。24時間培養後、培地を3-MCやLPAを含む無血清培地に交換し、さらに24時間培養した。
【0064】
2.免疫染色法
トランスフェクションした細胞が付着したカバーガラスを4% パラホルムアルデヒドに20分間浸し、細胞を固定した。PBSで細胞を洗浄した後、10% FBSに浸してブロッキングを行った。PBSで200倍に希釈した抗HA抗体(Roche Diagnostics、12CA5)と室温で1時間反応させた。PBSで3回洗浄した後、ブロッキングバッファー(10% FBS)で1000倍に希釈したAlexa488標識二次抗体と室温で1時間反応させた。Hoechstで核を染色した後、100% グリセロールと共にマウントした。顕微鏡はAxioplan2imaging(ZEISS)を用いて観察を行った。
【0065】
<結果>
図5に示すように、Gα13QLを共発現させていない細胞では、Ahレセプターが3-MC依存的に細胞質から核へ移行することが確認できた。しかしGα13QLを共発現させた細胞では3-MC刺激をしていない細胞においても、Ahレセプターが核および核周辺に存在し、3-MC刺激後もAhレセプターの局在は変化しなかった。
また、3-MCと同時にLPAも培養液に加え、内在性Gα13活性化時におけるAhレセプターの細胞内局在の変化について調べた結果、Gα13QLを共発現させた時と同様に、3-MC依存的なAhレセプターの細胞内局在の変化は見られなかった。
【産業上の利用可能性】
【0066】
Ahレセプターを介するシグナルは、薬物代謝酵素の一つであるシトクロームP450の発現を誘導して、生体内に取り込まれた異物を細胞内で代謝して体外に排出するための機構としても働いている。また、Gタンパク質シグナルは常に生体内で作動して恒常性を維持しているシステムである。そこで、Gタンパク質受容体のアンタゴニストの投与で、Ahレセプターシグナルが逆に亢進して、薬物代謝活性が上昇することも期待される。Ahレセプターシグナルを評価系としてGタンパク質共役受容体のアゴニスト、アンタゴニストを検討することにより、新たな薬剤が見出される可能性がある。
【図面の簡単な説明】
【0067】
【図1】実験に用いたプラスミドのコンストラクトを示す。AIP全長(AIP-Full:1−330)、AIP N末端側(AIP-N:1−161)、AIP C末端側(AIP-C:162−330)をそれぞれAIPΔ13を鋳型としPCRによって作製した。(実施例2)
【図2】293細胞内においてAIPはC末端側を介してGα13と相互作用することを示す。FLAGタグのついたGα13QLとHAタグのついたAIP全長(Full)、AIPのN末端側(N)、AIPのC末端側(C)を293細胞内に共発現させ、細胞抽出液に抗FLAG抗体を加え免疫沈降を行い、得られたサンプルを抗FLAG抗体と抗HA抗体によりウエスタンブロッティングを行った。(実施例2)
【図3】ダイオキシンによるAhレセプターを介したシトクロームP450の遺伝子発現機構を示す図である。Ahレセプターはリガンドがない状態ではHsp90、AIP(またはXAP2、ARA9とも呼ばれる)と複合体を形成し、細胞質に存在する。リガンドがAhレセプターに結合すると複合体から離れ、核内へ移行する。Ahレセプターは核内でArntと2量体を形成する。その2量体はシトクロームP450遺伝子(CYP1A1)のプロモーター上に存在するXREに結合する。
【図4】Gα13の活性化がAhレセプターの転写活性に及ぼす影響を示す。COS-7細胞にコントロールベクター(pEF-RL)とレポータープラスミド(pXRE-Luc)、Ahレセプター(AhR)、Arnt、Gα13QLの遺伝子を導入後、24時間培養した。その後、1μM 3-MC、10μM LPAを加え、さらに24時間培養を行い、レポーターアッセイを行った。(実施例3)
【図5】Gα13の活性化がAhレセプターの細胞内局在に及ぼす影響を示す。COS-7細胞にHAタグのついたAhレセプター、Arnt、Gα13QLの遺伝子を導入した24時間後に1μM 3-MC、10μM LPAを加え、さらに24時間培養した。遺伝子導入後48時間で細胞をパラホルムアルデヒドで固定し、抗HA抗体で免疫染色を行った。(実施例4)
【技術分野】
【0001】
本発明は、アリル炭化水素受容体結合タンパク質(arylhydrocarbon receptor interacting protein以下AIPと略称する)とアリル炭化水素受容体(arylhydrocarbon receptor、以下Ahレセプターと略称する)の結合を抑制することにより、ダイオキシン類が結合するAhレセプターを介する作用を抑制する医薬、食品及び方法に関する。より詳しくは、GTP結合タンパク質(Gタンパク質)等のAIPと相互作用する物質によりAhレセプターの安定性や細胞内局在に影響を与えて、ダイオキシン類の作用を軽減する技術に関する。
【背景技術】
【0002】
ダイオキシンは有機塩素化合物を焼却炉で燃焼する際などに発生する。ダイオキシンの産生を根絶することは根本的な廃棄物の減少が図られない限り困難である。中国、および開発途上国などにおける世界的な規模でのダイオキシンの環境汚染が懸念されており、ダイオキシンの毒性の軽減を目的とする新しい取り組みが将来の課題として残されている。
【0003】
ダイオキシンは、急性毒性、発がん性、催奇形性などの毒性を有するとともに、極微量でホルモンや成長因子などの働きを乱す環境ホルモンとして作用する。ダイオキシンの有機塩素化合物は、化学的に極めて安定であり、大気中、土壌、海水など自然界に一度拡散すると分解されることなく蓄積される。また、ダイオキシンは生体内に取り込まれると肝臓、脂肪組織などに蓄積され、体外へ排出されにくい性質を持っている。従来よりダイオキシンの産生および除去に関する技術の開発は盛んに行われてきたが、ダイオキシンの作用発現機構に他の細胞情報伝達系を用いて影響を与えて、ダイオキシンの作用を軽減する技術は報告されていなかった。
【0004】
以下に本明細書において引用した文献を列記する。
【非特許文献1】Gohla A., Harhammer R., Schultz G. (1998) J Biol Chem. 273, 4653-4659
【非特許文献2】Yamaguchi Y., Katoh H., Yasui H., Aoki J., Nakamura K., Negishi M.(2000) J Neurochem. 2000 75, 708-717
【非特許文献3】Katoh H., Aoki J., Yamaguchi Y., Kitano Y.,Ichikawa A., Negishi M.(1998) J Biol Chem. 273, 28700-28707
【非特許文献4】Voyno-Yasenetskaya T., Conklin B.R., Gilbert R.L., Hooley R., Bourne H.R., Barber D.L.(1994) J Biol Chem. 269, 4721-4724
【非特許文献5】Berestetskaya Y.V., Faure M.P., Ichijo H., Voyno-YasenetskayaT.A.(1998) J Biol Chem. 273, 27816-27823.
【非特許文献6】Yuan J., Slice L.W., Rozengurt E. (2001) J Biol Chem. 276, 38619-38627
【非特許文献7】Voyno-Yasenetskaya T.A., Faure M.P., Ahn N.G., Bourne H.R.(1996) JBiol Chem. 271, 21081-21087
【非特許文献8】Kozasa T., Jiang X., Hart M.J., Sternweis P.M., Singer W.D.,Gilman A.G., Bollag G., Sternweis P.C.(1998) Science 280, 2109-2111
【非特許文献9】Vaiskunaite R., Adarichev V., Furthmayr H., Kozasa T., GudkovA., Voyno-Yasenetskaya T.A. (2000) J Biol Chem. 275, 26206-26212
【非特許文献10】Meigs T.E., Fedor-Chaiken M., Kaplan D.D., Brackenbury R.,Casey P.J.(2001) Proc. Natl. Acad. Sci. U S A. 98, 519-524
【非特許文献11】Kozak, M. (1999) Gene, 234, 187-208
【非特許文献12】Prickett K.S., Amberg D.C., Hopp T.P.(1989) Bio Techniques, 7, 580-589
【非特許文献13】Vertes A.A. et al.(1994) Molecular Microbiol., 11, 739-46
【非特許文献14】Ulmer (1983) Science, 219, 666-671
【非特許文献15】Tuschl T.(2002) Nature Biotechnology, 20, 446-448
【非特許文献16】Brummelkamp T.R., Bernards R., Agami R.(2002) Science, 296, 550-553
【非特許文献17】Andersson S., Davis D.L., Dahlback H., Jornvall H., RussellD.W.(1989) J Biol. Chem., 264, 8222-8229
【非特許文献18】Kobayashi A., Numayama-Tsuruta K., Sogawa K., Fujii-KuriyamaY.(1997) J Biochem. 122, 703-710
【非特許文献19】Mimura J., Ema M., Sogawa K., Fujii-Kuriyama Y. (1999) Gene Dev., 13, 20-25
【発明の開示】
【発明が解決しようとする課題】
【0005】
従来、ダイオキシンが体内に一度摂取されてしまった後に、その作用を抑制するための技術的な解決法は考案されていなかった。したがって、本発明は生体内に既存している情報伝達システムを利用するダイオキシン作用の阻害、軽減を可能にする手段を提供することを課題とする。
【課題を解決するための手段】
【0006】
上記課題を解決すべく本発明者らは鋭意努力し、哺乳動物のGタンパク質遺伝子を細胞に導入して組換え体Gタンパク質を産生する系を用いて、それと相互作用する細胞内タンパク質を網羅的に探索した。その結果、Gタンパク質と相互作用する分子として、新たにダイオキシン作用に関係するAIPを見出した。次に、Ahレセプターを介するダイオキシン作用に対するGタンパク質シグナルの影響を検討した。その結果、AIPと相互作用するGα13の過剰発現、Gα13を活性化するリゾホスファチジン酸(lysophosphatidic acid、以下LPAと略称することがある)刺激によりAhレセプターを介する遺伝子発現が抑制されることが判明した。さらに、Gタンパク質シグナルの活性化によりAhレセプターの細胞内局在が変化することを見出し、本発明を完成した。
【0007】
すなわち本発明は、
1.GTP結合タンパク質(以下、Gタンパク質)とアリル炭化水素受容体結合タンパク質(以下、AIP)の相互作用を促進する手段を含む医薬又は食品、
2.用途が、ダイオキシン作用の阻害、軽減である前項1の医薬又は食品、
3.手段が、Gタンパク質の増加による前項1又は2の医薬又は食品、
4.Gタンパク質の増加が、Gタンパク質遺伝子を担持するベクターの投与、又は活性型Gタンパク質自体の投与若しくは摂取である前項3の医薬又は食品、
5.手段が、Gタンパク質の活性化による前項1又は2の医薬又は食品、
6.Gタンパク質を活性化する手段が、Gタンパク質共役受容体シグナル伝達系への作用である前項5の医薬又は食品、
7.作用が、Gタンパク質共役受容体シグナル伝達系に対する活性化剤の投与又は摂取による前項6の医薬又は食品、
8.活性化剤が、リゾホスファチジン酸である前項7の医薬又は食品、
9.ダイオキシン作用に影響するメカニズムが、アリル炭化水素受容体(以下、Ahレセプター)とダイオキシンの結合の阻害である前項2〜8の何れか1項に記載の医薬又は食品、
10.Gタンパク質が、Gタンパク質αサブユニット13(Gα13)である前項1〜9の何れか1項に記載の医薬又は食品、
11.以下の少なくとも1の方法を選択するダイオキシン作用の阻害、軽減に有効な候補物質の選択方法;
1)Gタンパク質とAIPの相互作用の促進の可否
2)Gタンパク質の発現の促進の可否
3)Gタンパク質遺伝子の転写促進の可否
4)Gタンパク質の活性化の可否
5)Gタンパク質共役受容体への作用の可否
12.Gタンパク質共役受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物の同定方法であって、Gタンパク質とAIPの相互作用を可能にする系で、候補化合物をこの系に接触させ、次いでGタンパク質とAIPの相互作用により生じるシグナルおよび/またはマーカーを使用する系を用いて、該シグナルおよび/またはマーカーの存在若しくは不存在および/または変化を検出することにより、該化合物がGタンパク質とAIPとの結合を阻害するか否かを決定することを特徴とする同定方法、
13.Gタンパク質がGα13である前項11又は12に記載の方法、
14.前項13に記載の同定方法により同定された化合物を含有するダイオキシン作用の阻害剤、
15.前項1〜10の何れか1項に記載の医薬若しくは食品からなる、又は前項14に記載の阻害剤を含む、ダイオキシン作用に起因する疾患の予防治療剤又は予防治療方法、
16.ダイオキシン作用に基づく疾患が、肝臓毒性、胸腺の萎縮、皮膚がん、又は子宮内膜症である前項15に記載の予防治療剤又は予防治療方法、
17.Gタンパク質共役受容体、Gタンパク質の優性活性型変異体および/または優性抑制型変異体を用いることを特徴とするダイオキシン作用への制御試験系、
18.Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する方法、
19.Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する手段を含む医薬又は食品、
に関する。
【発明の効果】
【0008】
本発明により、Gタンパク質共役受容体シグナルに働きかける薬剤を利用した、ダイオキシン作用の阻害および軽減法の開発が可能となった。Gタンパク質共役受容体はヒトゲノム全3万種類の遺伝子の中で1000種類にもおよぶ大きなファミリーを形成している。また、組織、臓器特異的な発現を示すGタンパク質共役受容体が数多く存在しており、既に臨床上使用されている薬剤の40%近くがGタンパク質共役受容体を標的としている。したがって、本発明により生体内でダイオキシンの毒性作用が見られる、肝臓、胸腺、皮膚、子宮内膜に特異的に発現しているGタンパク質共役受容体を標的とする薬剤を検索し、その薬剤の投与によりダイオキシンの効果だけを軽減する理想的な薬剤が見出される可能性がある。
【発明を実施するための最良の形態】
【0009】
以下、本発明について発明の実施の態様をさらに詳しく説明する。以下の詳細な説明は例示であり、説明のためのものに過ぎず、本発明を何ら限定するものではない。
本明細書においては単離された若しくは合成の完全長蛋白質;単離された若しくは合成の完全長ポリペプチド;または単離された若しくは合成の完全長オリゴペプチドを意味する総称的用語として「ポリペプチド」という用語を使用することがある。ここで蛋白質、ポリペプチド若しくはオリゴペプチドはペプチド結合または修飾されたペプチド結合により互いに結合している2個以上のアミノ酸を含むものである。
【0010】
Gタンパク質は、細胞膜に存在するGタンパク質共役受容体が受け取ったホルモンや神経伝達物質などの刺激を細胞内に伝えるトランスデューサーであり、α、β、γサブユニットからなるヘテロ三量体を形成する。Gタンパク質は不活性化状態では、三量体を形成し、αサブユニット(Gα)はGDPと結合している。受容体にリガンドが結合すると、Gαに結合していたGDPが解離し、細胞内に豊富に存在するGTPが結合してGタンパク質が活性化される。Gタンパク質が活性化されるとGαとβγサブユニット(Gβγ)が解離し、それぞれが下流のエフェクタータンパク質にシグナルを伝える。Gαに結合したGTPがGDPに加水分解されることによって、Gタンパク質は再び不活性化状態にもどり、Gαβγの三量体を形成する。
【0011】
三量体Gタンパク質はGαのアミノ酸配列によって4つのファミリー(Gαs、Gαi、Gαq、Gα12)に分類される。このなかでGα12ファミリーにはGα12とGα13が存在し、どちらも広範な組織に発現している。Gα12やGα13の活性化によってアクチンストレスファイバーの形成(非特許文献1)や、神経伝達物質の放出に関わるCa2+依存性エクソサイトーシスの抑制(非特許文献2)、神経突起の退縮(非特許文献3)、Na+/H+交換因子の活性化(非特許文献4)、アポトーシスの誘導(非特許文献5)、PKDの活性化(非特許文献6)、JNKの活性化(非特許文献7)が起こる。このような細胞応答に至るシグナル伝達経路を明らかにするために、直接相互作用する分子の同定が行われ、Gα13のGTP加水分解反応を促進するRGSドメインとRhoのGTP/GDP交換反応を促進するDHドメインをもったp115RhoGEFが同定された(非特許文献8)。他にもGα13と相互作用する分子として、radixin(非特許文献9)やcadherin(非特許文献10)などが同定されている。
【0012】
ダイオキシンは、一般に、75種類の異性体を持つポリ塩化ジベンゾパラジオキシン(PCDD)および135種類の異性体を持つポリ塩化ジベンゾフラン(PCDF)の総称をいう。ダイオキシンによる肝臓毒性、胸腺の萎縮、皮膚がん、子宮内膜症の発生には、Ahレセプターが関与している。Ahレセプターはベンツピレン、3−メチルコラントレンのような多環性芳香族炭化水素化合物とダイオキシンのようなハロゲン化芳香族炭化水素化合物をリガンドとする受容体型転写因子である。Ahレセプターはリガンドがない状態では熱ショックタンパク質90(heat shock protein 90、Hsp90)、AIP(またはXAP2、ARA9とも呼ばれる)と複合体を形成し、細胞質に存在する。リガンドがAhレセプターに結合するとAhレセプターは複合体から離れ、核内へ移行する。Ahレセプターは核内でアリル炭化水素受容体核輸送体(arylhydrocarbon receptor nuclear translocator、以下Arntと略称する)と2量体を形成する。その2量体はシトクロームP450遺伝子(CYP1A1)のプロモーター上に存在する異物応答配列(xenobiotic responsive element、以下XRE略称する)に結合する。
【0013】
本発明においては、Gタンパク質と相互作用する細胞内タンパク質の探索を行い、ダイオキシン作用にかかわるAIP がGタンパク質と相互作用することを初めて見出した。したがって、本発明の一態様はGタンパク質とAIPの相互作用を促進する手段を含む医薬又は食品である。また、本発明においてはGタンパク質シグナルがAhレセプターよる遺伝子発現を抑制することを見出した。このことから、Gタンパク質はAIPのAhレセプターとの結合を阻害することで、Ahレセプターに結合するリガンドの作用を抑える可能性が示唆された。Ahレセプターに結合するリガンドとしてダイオキシンが好適に挙げられる。したがって、本発明の医薬又は食品の用途の一つはダイオキシン作用の阻害、軽減である。
【0014】
本明細書において、「Gタンパク質とAIPの相互作用」とは、Gタンパク質とAIPがある様式により互いに作用を及ぼすことをいう。「ある様式」とは、結合、物理学的接触、近接等を含むものであり、結果として互いに作用を及ぼし得る様式であればいずれのものであってもよい。
【0015】
Gタンパク質とAIPの相互作用を促進する手段は、Gタンパク質の増加により達成可能である。Gタンパク質の増加は、具体的には、遺伝子導入手段によって、機能するように連結されたプロモーターを有するGタンパク質遺伝子を担持するベクターを標的組織へ投与することにより達成することができる。自己増殖ベクターに遺伝子を担持する手段によっても本発明の目的は達成可能である。
【0016】
遺伝子導入手段は、望ましくはインテグレーションさせる方法、例えばリポソーム法、リン酸カルシウム法、エレクトロポレーション法、レトロウイルス法等によるのが好ましい。しかし一過性に発現させるリポソーム法、リン酸カルシウム法、エレクトロポレーション法、ウイルスベクター法、アテロコラーゲン法等の方法であってもよい。用いるベクターは特に限定されず、公知のプラスミド、ファージ、コスミド、BAC、YAC、組換えウイルス、トランスポゾン等、通常の組み換え実験によって挿入DNA断片を導入することが可能な全ての組換えベクターに適用することができる。ベクターは、当然に自体公知の組合せが好適であるプロモーター、エンハンサーと共に構築することができる。例えば、通常宿主に適したプロモーターが挿入されている市販のタンパク質発現ベクターを用いることができる。具体的には、ZAPExpress(Stratagene)、pSVK3(Amersham Bioscience)、pEGFP-C1(Clontech)、アテロコラーゲン等が挙げられる。
【0017】
ベクターへのタンパク質発現遺伝子の挿入は、該遺伝子又はこれを含むDNA断片をベクター中のプロモーターの下流にプロモーターの制御下に配置されるように連結して行う。また、プロモーターとタンパク質発現遺伝子との間にコザック配列(非特許文献11)を挿入したり、タンパク質発現遺伝子の下流にタグとなるポリペプチドをコードするDNAを挿入した構造を有するベクターも好ましく用いられる。タグとなるポリペプチドとしては特に制限はないが、例えば、FLAGタグ(非特許文献12)等が挙げられる。プロモーターを連結したGタンパク質発現遺伝子を標的細胞の染色体中に直接挿入する相同組換え技術、あるいはトランスポゾンや挿入配列(非特許文献13)等を用いて発現させることができる。
【0018】
Gタンパク質の増加は上記のように得られたGタンパク質を投与又は摂取することにより達成可能である。Gタンパク質は、天然に存在するものであってよく、Gタンパク質に変異を導入したものであってもよい。具体的には、活性型Gタンパク質であることが望ましい。また、Gタンパク質とAIPの相互作用部位を含むGタンパク質の部分ポリペプチドであってもよい。Gタンパク質とAIPの相互作用部位のアミノ酸配列からなるポリペプチドはかかるポリペプチドとして好適である。
【0019】
ポリペプチドに欠失、置換、付加または挿入等の変異を導入する手段は自体公知であり、例えばウルマー(Ulmer)の技術(非特許文献14)を利用できる。このような変異の導入において、当該ポリペプチドの基本的な性質(物性、機能または免疫学的活性等)を変化させないという観点から、例えば、同族アミノ酸(極性アミノ酸、非極性アミノ酸、疎水性アミノ酸、親水性アミノ酸、陽性荷電アミノ酸、陰性荷電アミノ酸および芳香族アミノ酸等)の間での相互置換は容易に想定される。さらに、これら利用できるポリペプチドは、その構成アミノ基またはカルボキシル基等を、例えばアミド化修飾する等、機能の著しい変更を伴わない程度に改変が可能である。
【0020】
ポリペプチドは、常法に従って製造できる。具体的には、通常の液相法および固相法によるポリペプチド合成法を挙げることができるが、これらに限らず公知の方法が広く利用可能である。または市販のポリペプチド合成装置を用いて製造可能である。あるいは遺伝子工学的手法により取得することもできる。例えば目的とするポリペプチドをコードする遺伝子を宿主細胞中で発現できる組換えDNA(発現ベクター)を作成し、これを適当な宿主細胞、例えば大腸菌にトランスフェクションして形質転換した後に該形質転換体を培養し、次いで得られる培養物から目的とするポリペプチドを回収することにより製造可能である。
【0021】
Gタンパク質とAIPの相互作用を促進する手段は、Gタンパク質の活性化により達成可能である。前記Gタンパク質の活性化はGタンパク質共役受容体シグナル伝達系に作用することにより達成可能であり、具体的には、Gタンパク質共役受容体シグナル伝達系に対する活性化剤の投与又は摂取によることができる。Gタンパク質共役受容体シグナル伝達系に対する活性化剤の好適な例として、リゾホスファチジン酸が挙げられる。
【0022】
Ahレセプターはリガンドが無い状態ではAIPと複合体を形成し細胞質に存在するが、Gタンパク質がAIPに結合するとAhレセプターは複合体から離れ、核内へ移行する。本発明においては、Gタンパク質シグナルがダイオキシン受容体であるAhレセプターの細胞内局在に影響を与えることを見出した。このことから、Gタンパク質がAIPと結合することによりAhレセプターの細胞内局在が変化することにより、ダイオキシンとAhレセプターの結合が阻害される可能性が考えられる。したがって、本発明の一態様はダイオキシン作用に影響するメカニズムがAhレセプターとダイオキシンの結合の阻害である医薬又は食品である。 本発明に用いるGタンパク質は、好ましくはGαi以外のGαであり、より好ましくはGα13である。
【0023】
ダイオキシン作用の阻害、軽減に有効な候補物質の選択方法は以下の方法から選択することができる;1)Gタンパク質とAIPの相互作用の促進の可否、2)Gタンパク質の発現の促進の可否、3)Gタンパク質遺伝子の転写促進の可否、4)Gタンパク質の活性化の可否、5)Gタンパク質共役受容体への作用の可否、である。1)〜5)の方法においては、1)Gタンパク質とAIPの相互作用、2)Gタンパク質の発現、3)Gタンパク質遺伝子の転写、4)Gタンパク質の活性化、5)Gタンパク質共役受容体への作用を可能にする条件を選択し、Gタンパク質とAIPを前記候補物質と接触させ、1)Gタンパク質とAIPの相互作用、2)Gタンパク質の発現、3)Gタンパク質遺伝子の転写、4)Gタンパク質の活性化、5)Gタンパク質共役受容体への作用を検出することができるシグナルおよび/またはマーカーを使用する系を用いて、このシグナルおよび/またはマーカーの存在若しくは不存在または変化を検出することにより、ダイオキシン作用の阻害、軽減に有効な候補物質を選択することができる。1)Gタンパク質とAIPの相互作用、2)Gタンパク質の発現、3)Gタンパク質遺伝子の転写、4)Gタンパク質の活性化、または5)Gタンパク質共役受容体への作用を可能にする条件はインビトロのものであってよく、インビボのものであってもよい。例えば、Gタンパク質とAIPを共発現させた細胞を用いることもできる。
【0024】
Gタンパク質共役受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物の同定方法は、自体公知のスクリーニングシステムを利用して構築可能である。例えば、Gタンパク質とAIPの相互作用を可能にする系で、調べようとする化合物(候補化合物)をこの系に接触させ、次いでGタンパク質とAIPの相互作用により生じるシグナルおよび/またはマーカーを使用する系を用いて、このシグナルおよび/またはマーカーの存在若しくは不存在または変化を検出することにより、Gタンパク質共役受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物を同定することができる。Gタンパク質とAIPの相互作用を可能にする系は、インビトロのものであってよく、インビボのものであってもよい。例えば、Gタンパク質とAIPを共発現させた細胞を用いることができる。
【0025】
前記候補物質又は候補化合物としては、例えば化学ライブラリーや天然物由来の化合物、またはGタンパク質、AIPの一次構造や立体構造に基づいてドラッグデザインして得られた化合物等が挙げられる。あるいは、AIPとGタンパク質との相互作用部位のアミノ酸配列からなるポリペプチドの構造に基づいてドラッグデザインして得られた化合物等も候補物質又は候補化合物として好適である。
【0026】
上記同定方法で得られた化合物は、ダイオキシン作用の阻害剤に利用可能である。本発明の医薬若しくは食品、または上記同定方法で得られた化合物を含む阻害剤は、ダイオキシン作用に基づく疾患の予防治療剤又は予防治療方法に利用可能である。かかる疾患としては、肝臓毒性、胸腺の萎縮、皮膚がん、又は子宮内膜症等を挙げることができる。
【0027】
本発明においては、Gタンパク質共役受容体シグナルによりダイオキシンによる遺伝子発現が抑制されることが見出された。このことから、Gタンパク質共役受容体、Gタンパク質の優性活性型変異体および/または優性抑制型変異体を用いてダイオキシン作用への制御試験系を構築することができる。前記制御試験系においては、ダイオキシン作用を制御する可能性のある候補物質をGタンパク質共役受容体、Gタンパク質の優性活性型変異体および/または優性抑制型変異体と接触させ、ダイオキシンによる遺伝子発現により生じるシグナルおよび/またはマーカーを使用する系を用いて、該シグナルおよび/またはマーカーの存在若しくは不存在または変化を検出することにより、前記候補物質がダイオキシン作用を制御するか否かを判定する。前記制御試験系は、インビトロのものであってよく、インビボのものであってもよい。例えば、Gタンパク質共役受容体、Gタンパク質の優性活性型受容体および/または優性抑制型変異体を共発現させた細胞を用いることができる。
【0028】
本発明においては、AIPとGタンパク質の相互作用によりAhレセプターの転写活性化能が抑制された。このことから、AIPにGタンパク質が相互作用したことにより、Ahレセプターと結合できるAIP分子の数が減少し、Ahレセプターの転写活性化能が抑制された可能性が示唆された。したがって、Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する方法が構築可能である。また、本発明の一態様は、Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する手段を含む医薬又は食品である。具体的には、AIPに相互作用する物質をAIPと相互作用させ、Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターの転写活性化能の促進又は抑制を達成することができる。好ましいAIPに相互作用する物質はGタンパク質であり、より好ましくはGαi以外のGαであり、さらに好ましくはGα13である。
【0029】
Ahレセプターと結合できるAIP分子の数を減少させることにより、Ahレセプターによる転写活性化能を抑制する手法の一つとして、RNAi法が挙げられる。詳しくは、RNAi法によりAIPの生理活性を制御することにより、AHレセプターと結合できるAIP分子の数を減少させる。RNAi法は機能を阻害しようとする遺伝子の特定領域と相同なセンスRNAとアンチセンスRNAからなる二本鎖RNAが標的遺伝子のmRNAの相同部分を干渉・破壊する方法である。二本鎖RNAはRNAiの目的に適した配列を有する領域から選択する。具体例として、AIPのAhレセプターとの結合部位を含む領域から当該配列を選択することができる。一般的には、二本鎖RNAの配列として開始コドンの50から100ヌクレオチド下流の領域を選択する。通常、RNAi法には、塩基数が21〜23塩基であるセンスRNAおよびアンチセンスRNAの一対の組み合わせからなる二本鎖RNAが必要である。
【0030】
前記二本鎖RNAを構成するオリゴヌクレオチドは、公知の方法によって作製することができる。例えば化学的に合成することができるし、あるいは天然の核酸を制限酵素などによって切断し、上記のような塩基配列で構成されるように改変し、あるいは連結することも可能である。具体的には、オリゴヌクレオチド合成装置等を用いて合成することができる。また、siRNA合成キットが既に市販されており(ニッポンジーンテク社等)、これらを利用して二本鎖RNAを合成することもできる。前記二本鎖RNAを細胞に導入するには、公知の方法を用いることができる。例えば、リン酸カルシウム法、リポフェクチン法などが挙げられる。前記二本鎖RNAは、適当なベクターを用いて細胞内で恒常的に発現することもできる。例えば、非特許文献15および16に開示されたsiRNA発現専用ベクターを利用すればよい。さらに、当該ベクターに二本鎖RNAを組み込む方法は、自体公知の方法を適用することができる。例えば二本鎖RNAに適当なリンカーをライゲーションし、これを目的に適したベクターのマルチクローニングサイトへ挿入することによって、所望のベクターを得ることもできる。
【0031】
本明細書においてシグナルとは、そのもの自体がその物理的または化学的性質により直接検出され得るものを指し、マーカーとは、そのものの物理的または生物学的性質を指標として間接的に検出され得るものを指す。シグナルおよび/またはマーカーとしては、ルフェラーゼ、蛍光蛋白質(GFP、YFP、BFP等)、および放射性同位体等、レポーター遺伝子、例えばクロラムフェニコールアセチルトランスフェラーゼ遺伝子等、または検出用のエピトープタグ、例えば6×Hisタグ等、公知のものが利用できる。これらシグナルまたはマーカーの検出方法は当業者には周知のものである。
【0032】
本発明において使用するGタンパク質、Gタンパク質の優性活性型変異体および/または優性抑制型変異体、Gタンパク質共役受容体、AIPは、これらを遺伝子工学的手法で発現させた細胞、無細胞系合成産物、化学合成産物、または該細胞や生体試料から調製したものであってよく、これらからさらに精製されたものであってもよい。また、Gタンパク質とAIPの相互作用、およびこれら蛋白質の機能に影響がなければ、N末端側やC末端側に別の蛋白質やポリペプチド、例えばβ−ガラクトシダーゼ、IgG等の免疫グロブリンFc断片、Hisタグ、Mycタグ、HAタグ、FLAGタグ、またはXpressタグ等のタグペプチド類を、直接的にまたはリンカーペプチド等を介して間接的に、遺伝子工学的手法等を用いて付加したものであってもよい。
【0033】
本発明の医薬は、予防又は治療の目的に応じて、散剤、顆粒剤、錠剤、カプセル剤、腸溶剤、液剤、注射剤(液剤、懸濁剤)などの各種の形態に、常法にしたがって調製することができる。本発明の医薬は、通常は1種または複数の医薬用担体を用いて医薬組成物として製造することが好ましい。本発明の医薬中に含まれる有効成分の量は、広範囲から適宜選択されるが、通常約0.00001〜70重量%、好ましくは0.0001〜5重量%程度の範囲とするのが適当である。
【0034】
本発明の医薬を投与するときには、該医薬を単独で使用してもよく、あるいは目的の疾患の防止、治療または改善に必要な他の化合物または医薬と共に使用してもよい。投与経路は、全身投与または局所投与のいずれも選択することができる。この場合、疾患、症状等に応じた適当な投与経路を選択する。例えば、非経口経路として、通常の静脈内投与、動脈内投与のほか、皮下、皮内、筋肉内等への投与を挙げることができる。あるいは経口による投与も可能である。さらに、経粘膜投与または経皮投与も可能である。
【0035】
本発明の医薬の投与形態としては、各種の形態が目的に応じて選択でき、その代表的なものとしては、錠剤、丸剤、散剤、粉末剤、細粒剤、顆粒剤、カプセル剤等の固体投与形態や、水溶液製剤、エタノール溶液製剤、懸濁剤、脂肪乳剤、リポソーム製剤、シクロデキストリン等の包接体、シロップ、エリキシル等の液剤投与形態が含まれる。これらは更に投与経路に応じて経口剤、非経口剤(点滴剤、注射剤)、経鼻剤、吸入剤、経膣剤、坐剤、舌下剤、点眼剤、点耳剤、軟膏剤、クリーム剤、経皮吸収剤、経粘膜吸収剤等に分類され、それぞれ通常の方法に従い、調合、成形、調製することができる。
【0036】
本発明の食品は、例えば、塊状、液状(液体、粘稠流体、ペースト等)、ゼリー状、粉末状、顆粒状などの各種形態に、常法に従って調製することができる。かかる形態の具体的な例としては、飲料(ドリンク剤、清涼飲料水、ジュース、茶類など)、粉末飲料(粉末ジュース、粉末スープなど)、菓子類(クッキー、ビスケット、シリアル、チュアブル錠剤、チューイングガム、キャンディー、グミ、ウェハース、せんべいなど)、調味料(ドレッシング、ソース、粉末調味料など)、主食製品(パン、麺類、餅など)などが挙げられる。また、本発明の食品は、一般食品以外にも、病者用食品、高齢者用食品、特定保健用食品などの特別用途食品や、医薬部外品、機能性食品、健康食品などの目的に幅広く使用することができる。
【0037】
本発明の医薬又は食品には、本発明の効果を損なわない範囲であれば、必要に応じて、通常医薬や食品に配合される成分、例えば、栄養素(他の蛋白質、脂質、炭水化物、ミネラル、ビタミンなど)、動植物成分(粉砕物、脱脂物、抽出エキス、抽出末など)、賦形剤(二糖類、糖アルコール、多糖類、水溶性高分子など)、増量剤、甘味料、香味剤、着色剤、防腐剤、乳化剤、界面活性剤(可溶化剤、泡安定化剤、消泡剤など)、ハイドロトロープ剤(アルコール類、多価アルコール類など)などを配合することができる。本発明の医薬又は食品におけるこれら成分の配合量は特に限定されず、剤型、形態、用途等に応じて適宜設定することができ、例えば医薬又は食品の全重量に基づいて、0.1〜99.9重量%程度とすることができる。
【0038】
本発明の医薬又は食品の投与量又は摂取量については、本発明の効果が得られるものであれば特に限定されるものではなく、含有される成分の有効性、投与形態、投与経路、疾患の種類、対象の性質(体重、年齢、病状および他の医薬の使用の有無等)、および担当医師の判断等に応じて適宜選択される。本発明の医薬又は食品は、1日1〜数回に分けて投与又は摂取することができ、数日または数週間に1回の割合で間欠的に投与又は摂取してもよい。
【0039】
以下、本発明を実施例に基づき具体的に説明するが、本発明は下記の実施例に限定されない。
【実施例】
【0040】
実施例1
(酵母ツーハイブリッド法を用いた新規Gα13結合タンパク質の探索)
Gα13を介したシグナル伝達経路を解明するために、酵母ツーハイブリッド法を用いて、新規Gα13結合タンパク質の探索を行った。
【0041】
<方法>
1.ベイト側プラスミド(pGBKT7-Gα13QL)の構築
酵母ツーハイブリッド法によるスクリーニングはMATCHMAKER Two-hybrid System 3(Clontech)を用いて行った。ヒトGα13(GenbankAF493902、5729847)のcDNAはヒト293T細胞のRNAを鋳型とした逆転写酵素反応により作成し、翻訳領域の両端にEcoRIサイトを持つプライマー(配列番号1および2)で増幅した。そのDNA断片を哺乳動物細胞発現用プラスミドpCMV5(非特許文献17)のEcoRIサイトに挿入した。226番目のグルタミンをロイシンへの置換はQuickChangeキット(Stratagene)を用いて行った。活性型変異体Gα13QLをコードするcDNAをpCMV5-Gα13QLよりEcoRIで切り出し、GAL4結合領域(GAL4BD)を含むベイト側プラスミドpGBKT7のEcoRIサイトに挿入し、pGBKT7-Gα13QLを作成した。
【0042】
Forward primer:5'-CCGGAATTCATGGCGGACTTCCTGCCG-3'(配列番号1)
Reverse primer:5'-CCGGAATTCTCACTGTAGCATAAGCTGCTTGAGG-3'(配列番号2)
【0043】
2.酵母コンピテントセルの作成と形質転換
酵母の形質転換はYeast Transformation System(Clontech)を用い、酢酸リチウム法によって行った。
酵母(AH109)をYPDA寒天培地で30℃、2〜3日間培養した。培地上に生じた2mmのコロニーを6〜7個掻きとり、YPDA液体培地 50mlに移し、30℃、250rpmにて18時間振盪培養した。定常期(OD600>1.5)まで培養した細胞をYPDA液体培地 300mlに移し、さらに30℃、250rpmにて3時間振盪培養した。細胞を室温、1000×gにて5分間遠心し、細胞を回収した。滅菌水 40mlで細胞を懸濁した後、再び室温、1000×gにて5分間遠心し、細胞を回収した。1×TE/LiAc(10mM Tris-HCl(pH7.5)、1mM EDTA、0.1mM 酢酸リチウム) 1.5mlで細胞を懸濁したものをコンピテントセルとし、トランスフォーメーションに用いた。
【0044】
次に、酵母の発現プラスミド 200ng、ニシン精巣キャリアーDNA(94℃で20分加熱処理済)100ngとコンピテントセル 100μlを混ぜ合わせた。その後、PEG/LiAc(10mM Tris-HCl(pH 7.5)、1mM EDTA、1M 酢酸リチウム、40% PEG)600μlを添加し、ボルテックスで攪拌した。30℃で30分間インキュベートした後、DMSO 70μlを加えて穏やかに混ぜた。42℃にて15分間インキュベートした後、氷上で1〜2分間冷却した。室温、14000×gにて5秒間遠心し、ペレットを回収した。TE 500μlで2回洗浄した後、TE 500μlに懸濁し、培地上に細胞をプレーティングした。
【0045】
3.ライブラリーのスクリーニング
ライブラリーのスクリーニングにはPretransformed MATCHMAKER mouse brain cDNA library(Clontech)を使用した。これはGAL4活性化ドメイン(GAL4AD)をコードする酵母発現ベクター(pACT2)にマウス脳cDNAが挿入されたプラスミドが既に酵母(Y187)に導入されたものである。この酵母とベイト側プラスミドを導入した酵母(AH109)を30℃、40rpmにて25時間インキュベートして接合させた酵母をSD(-Leu/-Try/-His/Ade)選択培地で培養し、生育した陽性クローンについてさらにα−ガラクトシダーゼアッセイを行った。
【0046】
<結果>
Gα13の活性型変異体をベイトとし、マウス脳cDNAライブラリーのスクリーニングを行った結果、8.8×106個のクローンから一次選択により、20クローンを単離した。さらにα−ガラクトシダーゼアッセイによる二次選択により、11クローンを単離した。これらのクローンについて、シーケンスの解析を行った。このうち、Ahレセプターと結合し、そのシグナルに影響を及ぼす分子として報告されているAIPの遺伝子を含むプラスミドを、再度Gα13発現プラスミドとともに酵母(AH109)に導入し、選択培地における細胞の増殖の確認を行った。
【0047】
実施例2
(293細胞内におけるGα13とAIPの相互作用の確認)
哺乳動物細胞におけるGα13とAIPの相互作用を確認するために、ヒト胎児腎由来HEK293細胞にGα13とAIPを強制発現させ、免疫沈降法とウエスタンブロッティングを行い、相互作用の検討を行った。
AIP全長、AIPのN末端側、AIPのC末端側をコードするcDNAを発現用プラスミドに挿入し、これらのプラスミドをHEK293細胞に導入した発現を確認するとともに、Gα13と一緒に沈降するかどうか検討した。
【0048】
<方法>
1.哺乳動物細胞発現プラスミド(pCMV5-2HA-AIP)の構築
酵母ツーハイブリッド法によって、Gα13をベイトとして単離したクローンはN末端側13アミノ酸欠いたAIP(AIPΔ13)のcDNAを含むものであった。このクローンを鋳型とし、PCR法によりAIP全長(1-993:AIP-Full)、N末側断片(1-483:AIP-N)、C末側断片(484-993:AIP-C)を作製した(図1)。
【0049】
使用したプライマーの配列を以下に示す。
5'AIP-Full/5'-AIP-N:5'-GAAGATCTATGGCGGATCTCATCGCAAGACTCCGAGAGGACGGGATCCAAAAGCGTGTGA-3'(配列番号3)
5'AIP-C:5'-GAAGATCTACCTATCAACAGGACCC-3'(配列番号4)
3'AIP-Full/3'AIP-C:5'-GAAGATCTTCAGTGGGAAAAGATGCC-3'(配列番号5)
3'AIP-N:5'-GAAGATCTTCAGCCAGGACTCTCTAC-3'(配列番号6)
【0050】
293細胞内におけるGα13とAIPの相互作用を確認するために、HAタグのついた哺乳動物細胞発現ベクターpCMV5-2HAに上記のAIP遺伝子を挿入したプラスミドを構築した。AIP-Full、AIP-N、AIP-CはそれぞれのPCR産物の両端をBglIIで切断し、ベクターのBglIIサイトに挿入し、pCMV5-2HA-AIP-Full、pCMV5-2HA-AIP-N、pCMV5-2HA-AIP-Cを作製した。ライゲーションは、LigationHigh(TOYOBO)15μl、DNA溶液 15μl(インサートDNA 100ng、ベクタープラスミドDNA 50ng を含む)を16℃にて30分間インキュベートして行った。
【0051】
2.細胞培養とトランスフェクション
ヒト胎児腎由来HEK293細胞は10% ウシ胎児血清(FBS)と100μg/ml カナマイシンを含むダルベッコ変法イーグル培地(DMEM)を使用し、5% CO2存在下、37℃で培養した。トランスフェクションはリン酸カルシウム法によって行った。細胞は6cmディッシュで培養し、サブコンフルエントに達したものを用いた。トランスフェクションを行う前に、10% FBSを含むDMEM 4mlに交換した。HAタグのついたAIP(pCMV5-2HA-AIP-N 3μg、pCMV5-2HA-AIP-C 1μg、pCMV5-2HA-AIP-Full 1μg)、FLAGタグのついたGα13QL(pCMV5-FLAG-Gα13QL 2μg)にpCMV5を加え合計で5μgになるようにDNA溶液を調製した。さらに2.5M CaCl2・2H2O 50μlと水を合計で500μlになるように添加し、混合した。このDNA溶液を2×HeBS(50mM HEPES-NaOH(pH 7.0)、280mM NaCl、1.5mM Na2PO4) 500μlに加えて、よく攪拌した。室温で20分放置した後、HEK293細胞培養液に滴下した。細胞を5% CO2 存在下、37℃にて18時間培養後、培養液を取り除き3mlの培地(100μg/ml カナマイシンを含むDMEM)に交換し、さらに24時間培養した。
【0052】
3.細胞抽出液(ライセート)の調製
HEK293細胞に遺伝子導入後、5% CO2 存在下、37℃にて36時間培養した細胞からライセートを調製した。細胞の培養液を取り除いた後、ディッシュを氷上で冷やしながらLysisBuffer(20mM HEPES-NaOH(pH 7.5)、3mM MgCl2・6H2O、100mM NaCl、1mM NaVO4、10mM NaF、20mM β-Glycerophosphate、1mM PMSF、1mM DTT、1μM leupeptin、1mM EGTA、0.5% LB-PX)を600μM加えた。セルスクレーパーで細胞を掻き取り、1.5mlチューブに回収した。チューブを氷上で10分間放置し、細胞を融解させた。4℃、14000×rpmにて10分間遠心分離を行い、上清を回収した。
【0053】
4.免疫沈降
細胞抽出液 300μlに30% Protein G Sepharose(Amasham Bioscience) 30μl、抗FLAG抗体(シグマ)1μgを加え、ローテーターで4℃にて8時間旋回させた。4℃、2000×rpmにて3分間遠心分離を行い、上清を取り除いた。Lysis Buffer 300μl で3回洗浄した後、30μlの2×Sample Buffer(0.1M Tris-HCl(pH6.8)、4% SDS、20% グリセロール、0.06M DTT)を加え、95℃にて3分間煮沸した。室温、14000×rpmにて10秒間遠心分離し、上清をSDS-PAGEによって分離した。
【0054】
5.ウエスタンブロッティング
SDS-PAGEによりサンプルを分離した後、PVDF膜に100Vで1時間転写した。5% スキムミルクにPVDF膜を30分浸しブロッキングした。PBST(138mM NaCl、2.68mM KCl、8.10mM Na2HPO4・12H2O、1.47mM KH2PO4、0.1% Tween20)で500倍に希釈した一次抗体と室温で1時間反応させた後、PBSTで3回洗浄した。二次抗体はペルオキシダーゼ標識した抗マウスIgG抗体を5% スキムミルクで4000倍に希釈したものを用い、室温にて30分間反応させた。PBSTによって3回洗浄した後、ECL Western Blotting Detection System(Amersham Bioscience)により検出した。
【0055】
<結果>
図2に示すようにAIP全長、AIPのC末端側はGα13と共沈するが、AIPのN末端側はGα13と共沈しないことがわかった。このことから、AIPはそのC末端側を介してGα13と相互作用することが示された。
【0056】
実施例3
(レポーターアッセイ)
実施例2の結果から哺乳動物細胞内でGα13とAIPが相互作用することが確認できたので、Gα13がAIPの機能に影響を及ぼす可能性が示唆された。図3に示すようにAIPは細胞質に存在するAhレセプターと結合し、レセプターを安定化し、リガンドとの結合を促進することで、Ahレセプターを介した転写活性を上昇すると考えられている。このことからGα13とAIPの相互作用がAhレセプターの機能に影響を及ぼす可能性が考えられた。Gα13によるAhレセプターを介した転写活性の変化について検討するためにレポーターアッセイを行った。内在性のAhレセプターが存在しないことが報告されているCOS-7細胞内に、AhレセプターとArnt、AhレセプターのDNA認識配列であるXREをホタルルシフェラーゼ遺伝子の上流に組み込んだレポータープラスミドと、内部コントロールとしてウミシイタケルシフェラーゼ遺伝子の上流にヒトポリペプチド鎖延長因子(elongation factor)のプロモーターを組み込んだコントロールプラスミドを導入しレポーターアッセイを行った。
【0057】
<方法>
1.プラスミド
活性型Gα13QLプラスミドの他に、Ahレセプター発現プラスミド(pBOS-HA-AhR)と、Arnt発現プラスミド(pBOS-GST-Arnt)(非特許文献18)、ルシフェラーゼ遺伝子の上流にXRE配列を組み込んだレポータープラスミド(pX4Tk-Luc)(非特許文献19)を使用した。pBOS-HA-AhRはpBSK-mAhR(非特許文献19)からHindIII/XbaIで切り出したマウスAhレセプターcDNA断片を平滑末端にした後、pBOST7HAのSmaIサイトに挿入した。
【0058】
2.細胞培養とトランスフェクション
実験には、アフリカミドリザル腎臓由来COS-7細胞を用い、実施例2におけるHEK293細胞と同様の条件(10% FBS、100μg/ml カナマイシンを含むDMEMにおいて、5% CO2存在下、37℃)で培養し、リン酸カルシウム法でトランスフェクションを行った。24時間培養後、培養液を取り除き、1μM 3-Methylcholanthrene(3-MC)や10μM リゾホスファチジン酸(LPA)を含む無血清培地 3mlに交換し、さらに24時間培養した。
【0059】
3.細胞ライセートの調製
遺伝子導入後、48時間培養した細胞を回収し、細胞ライセートを調製した。細胞ライセートの調製にはDual-Luciferase Reporter Assay(Promega)に含まれるPassive Lysis Bufferを用いた。まず、細胞の培養液を取り除き、PBS(138mM NaCl、2.68mM KCl、8.10mM Na2HPO4・12H2O、1.47mM KH2PO4)を添加し、洗浄した。洗浄後、PBSを完全に取り除き、1×Passive Lysis Buffer存在下で細胞を回収し、1.5mlチューブに移した。回収した細胞ライセートを液体窒素で1〜2回凍結融解させ、細胞を完全に融解させた。
【0060】
4.レポーターアッセイおよび化学発光測定
レポーターアッセイはDual-Luciferase Reporter Assay(Promega)を用いて行った。また、化学発光測定はプレートリーダーMithras LB940(BELTHOLD)を用いて行った。96穴プレートに細胞ライセート 10μlにLuciferase Assay Reagent II(ホタルルシフェラーゼの基質含む)を50μl加えた。2秒後から、10秒間、ホタルルシフェラーゼ活性を測定した。測定後、Stop&GloReagent(コントロールのウミシイタケルシフェラーゼの基質を含む)を50μl加えた。再び2秒待った後、10秒間ウミイシイタケルシフェラーゼ活性を測定した。2種類の異なる酵素により触媒されて生じる発光の強度を上記のように測定し、ウミシイタケルシフェラーゼによる発光強度を用いてサンプル間の遺伝子導入効率の違いを補正してホタルルシフェラーゼ活性を評価した。
【0061】
<結果>
図4に示すように、Ahレセプターの発現プラスミドとArntの発現プラスミドをトランスフェクションした細胞において、Ahレセプターのリガンドである3-Methylcholantherene(3-MC)依存的にホタルルシフェラーゼ活性の上昇が見られた。AIPとの相互作用が確認されたGα13の活性型変異体(Gα13QL)を共発現させるとAhレセプターを介したルシフェラーゼ活性が低下した。また、リゾホスファチジン酸(LPA)によっても、Ahレセプターを介したルシフェラーゼ活性が抑制された。この結果から、Gα13がAhレセプターシグナルを負に制御することが示された。
【0062】
実施例4
(Ahレセプターの細胞内局在の検討)
これまでの実験から、Gα13はAIPと相互作用し、Ahレセプターシグナルを負に制御する可能性が示唆された。また、AIPはAhレセプターの細胞内局在に関わることから、Gα13とAIPの相互作用がAhレセプターの細胞内局在に影響を及ぼす可能性が考えられた。そこで、Gα13の活性化がAhレセプターの細胞内局在に及ぼす影響について、免疫染色法を用いて検討を行った。COS-7細胞内にHAタグのついたAhレセプターとArnt、Gα13QLを過剰発現させ、3-MC依存的なAhレセプターの細胞内局在の変化について調べた。
【0063】
<方法>
1.細胞培養とトランスフェクション
細胞はレポーターアッセイに用いたCOS-7細胞をカバーガラス上において実施例2におけるHEK293細胞と同様の条件(10% FBS、100μg/ml カナマイシンを含むDMEMにおいて、5% CO2存在下、37℃)で培養を行った。細胞にpBOS-HA-AhR 4μg、pBOS-GST-Arnt 1μg、pCMV5-Gα13QL 1μgをリン酸カルシウム法によってトランスフェクションを行った。24時間培養後、培地を3-MCやLPAを含む無血清培地に交換し、さらに24時間培養した。
【0064】
2.免疫染色法
トランスフェクションした細胞が付着したカバーガラスを4% パラホルムアルデヒドに20分間浸し、細胞を固定した。PBSで細胞を洗浄した後、10% FBSに浸してブロッキングを行った。PBSで200倍に希釈した抗HA抗体(Roche Diagnostics、12CA5)と室温で1時間反応させた。PBSで3回洗浄した後、ブロッキングバッファー(10% FBS)で1000倍に希釈したAlexa488標識二次抗体と室温で1時間反応させた。Hoechstで核を染色した後、100% グリセロールと共にマウントした。顕微鏡はAxioplan2imaging(ZEISS)を用いて観察を行った。
【0065】
<結果>
図5に示すように、Gα13QLを共発現させていない細胞では、Ahレセプターが3-MC依存的に細胞質から核へ移行することが確認できた。しかしGα13QLを共発現させた細胞では3-MC刺激をしていない細胞においても、Ahレセプターが核および核周辺に存在し、3-MC刺激後もAhレセプターの局在は変化しなかった。
また、3-MCと同時にLPAも培養液に加え、内在性Gα13活性化時におけるAhレセプターの細胞内局在の変化について調べた結果、Gα13QLを共発現させた時と同様に、3-MC依存的なAhレセプターの細胞内局在の変化は見られなかった。
【産業上の利用可能性】
【0066】
Ahレセプターを介するシグナルは、薬物代謝酵素の一つであるシトクロームP450の発現を誘導して、生体内に取り込まれた異物を細胞内で代謝して体外に排出するための機構としても働いている。また、Gタンパク質シグナルは常に生体内で作動して恒常性を維持しているシステムである。そこで、Gタンパク質受容体のアンタゴニストの投与で、Ahレセプターシグナルが逆に亢進して、薬物代謝活性が上昇することも期待される。Ahレセプターシグナルを評価系としてGタンパク質共役受容体のアゴニスト、アンタゴニストを検討することにより、新たな薬剤が見出される可能性がある。
【図面の簡単な説明】
【0067】
【図1】実験に用いたプラスミドのコンストラクトを示す。AIP全長(AIP-Full:1−330)、AIP N末端側(AIP-N:1−161)、AIP C末端側(AIP-C:162−330)をそれぞれAIPΔ13を鋳型としPCRによって作製した。(実施例2)
【図2】293細胞内においてAIPはC末端側を介してGα13と相互作用することを示す。FLAGタグのついたGα13QLとHAタグのついたAIP全長(Full)、AIPのN末端側(N)、AIPのC末端側(C)を293細胞内に共発現させ、細胞抽出液に抗FLAG抗体を加え免疫沈降を行い、得られたサンプルを抗FLAG抗体と抗HA抗体によりウエスタンブロッティングを行った。(実施例2)
【図3】ダイオキシンによるAhレセプターを介したシトクロームP450の遺伝子発現機構を示す図である。Ahレセプターはリガンドがない状態ではHsp90、AIP(またはXAP2、ARA9とも呼ばれる)と複合体を形成し、細胞質に存在する。リガンドがAhレセプターに結合すると複合体から離れ、核内へ移行する。Ahレセプターは核内でArntと2量体を形成する。その2量体はシトクロームP450遺伝子(CYP1A1)のプロモーター上に存在するXREに結合する。
【図4】Gα13の活性化がAhレセプターの転写活性に及ぼす影響を示す。COS-7細胞にコントロールベクター(pEF-RL)とレポータープラスミド(pXRE-Luc)、Ahレセプター(AhR)、Arnt、Gα13QLの遺伝子を導入後、24時間培養した。その後、1μM 3-MC、10μM LPAを加え、さらに24時間培養を行い、レポーターアッセイを行った。(実施例3)
【図5】Gα13の活性化がAhレセプターの細胞内局在に及ぼす影響を示す。COS-7細胞にHAタグのついたAhレセプター、Arnt、Gα13QLの遺伝子を導入した24時間後に1μM 3-MC、10μM LPAを加え、さらに24時間培養した。遺伝子導入後48時間で細胞をパラホルムアルデヒドで固定し、抗HA抗体で免疫染色を行った。(実施例4)
【特許請求の範囲】
【請求項1】
GTP結合タンパク質(以下、Gタンパク質)とアリル炭化水素受容体結合タンパク質(以下、AIP)の相互作用を促進する手段を含む医薬又は食品。
【請求項2】
用途が、ダイオキシン作用の阻害、軽減である請求項1の医薬又は食品。
【請求項3】
手段が、Gタンパク質の増加による請求項1又は2の医薬又は食品。
【請求項4】
Gタンパク質の増加が、Gタンパク質遺伝子を担持するベクターの投与、又は活性型Gタンパク質自体の投与若しくは摂取である請求項3の医薬又は食品。
【請求項5】
手段が、Gタンパク質の活性化による請求項1又は2の医薬又は食品。
【請求項6】
Gタンパク質を活性化する手段が、Gタンパク質共役受容体シグナル伝達系への作用である請求項5の医薬又は食品。
【請求項7】
作用が、Gタンパク質共役受容体シグナル伝達系に対する活性化剤の投与又は摂取による請求項6の医薬又は食品。
【請求項8】
活性化剤が、リゾホスファチジン酸である請求項7の医薬又は食品。
【請求項9】
ダイオキシン作用に影響するメカニズムが、アリル炭化水素受容体(以下、Ahレセプター)とダイオキシンの結合の阻害である請求項2〜8の何れか1項に記載の医薬又は食品。
【請求項10】
Gタンパク質が、Gタンパク質αサブユニット13(Gα13)である請求項1〜9の何れか1項に記載の医薬又は食品。
【請求項11】
以下の少なくとも1の方法を選択するダイオキシン作用の阻害、軽減に有効な候補物質の選択方法;
1)Gタンパク質とAIPの相互作用の促進の可否
2)Gタンパク質の発現の促進の可否
3)Gタンパク質遺伝子の転写促進の可否
4)Gタンパク質の活性化の可否
5)Gタンパク質共役受容体への作用の可否
【請求項12】
Gタンパク質共役受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物の同定方法であって、Gタンパク質とAIPの相互作用を可能にする系で、候補化合物をこの系に接触させ、次いでGタンパク質とAIPの相互作用により生じるシグナルおよび/またはマーカーを使用する系を用いて、該シグナルおよび/またはマーカーの存在若しくは不存在および/または変化を検出することにより、該化合物がGタンパク質とAIPとの結合を阻害するか否かを決定することを特徴とする同定方法。
【請求項13】
Gタンパク質がGα13である請求項11又は12に記載の方法。
【請求項14】
請求項13に記載の同定方法により同定された化合物を含有するダイオキシン作用の阻害剤。
【請求項15】
請求項1〜10の何れか1項に記載の医薬若しくは食品からなる、又は請求項14に記載の阻害剤を含む、ダイオキシン作用に起因する疾患の予防治療剤又は予防治療方法。
【請求項16】
ダイオキシン作用に基づく疾患が、肝臓毒性、胸腺の萎縮、皮膚がん、又は子宮内膜症である請求項15に記載の予防治療剤又は予防治療方法。
【請求項17】
Gタンパク質共役受容体、Gタンパク質の優性活性型変異体および/または優性抑制型変異体を用いることを特徴とするダイオキシン作用への制御試験系。
【請求項18】
Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する方法。
【請求項19】
Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する手段を含む医薬又は食品。
【請求項1】
GTP結合タンパク質(以下、Gタンパク質)とアリル炭化水素受容体結合タンパク質(以下、AIP)の相互作用を促進する手段を含む医薬又は食品。
【請求項2】
用途が、ダイオキシン作用の阻害、軽減である請求項1の医薬又は食品。
【請求項3】
手段が、Gタンパク質の増加による請求項1又は2の医薬又は食品。
【請求項4】
Gタンパク質の増加が、Gタンパク質遺伝子を担持するベクターの投与、又は活性型Gタンパク質自体の投与若しくは摂取である請求項3の医薬又は食品。
【請求項5】
手段が、Gタンパク質の活性化による請求項1又は2の医薬又は食品。
【請求項6】
Gタンパク質を活性化する手段が、Gタンパク質共役受容体シグナル伝達系への作用である請求項5の医薬又は食品。
【請求項7】
作用が、Gタンパク質共役受容体シグナル伝達系に対する活性化剤の投与又は摂取による請求項6の医薬又は食品。
【請求項8】
活性化剤が、リゾホスファチジン酸である請求項7の医薬又は食品。
【請求項9】
ダイオキシン作用に影響するメカニズムが、アリル炭化水素受容体(以下、Ahレセプター)とダイオキシンの結合の阻害である請求項2〜8の何れか1項に記載の医薬又は食品。
【請求項10】
Gタンパク質が、Gタンパク質αサブユニット13(Gα13)である請求項1〜9の何れか1項に記載の医薬又は食品。
【請求項11】
以下の少なくとも1の方法を選択するダイオキシン作用の阻害、軽減に有効な候補物質の選択方法;
1)Gタンパク質とAIPの相互作用の促進の可否
2)Gタンパク質の発現の促進の可否
3)Gタンパク質遺伝子の転写促進の可否
4)Gタンパク質の活性化の可否
5)Gタンパク質共役受容体への作用の可否
【請求項12】
Gタンパク質共役受容体シグナルの活性化によるダイオキシン作用の阻害を制御する化合物の同定方法であって、Gタンパク質とAIPの相互作用を可能にする系で、候補化合物をこの系に接触させ、次いでGタンパク質とAIPの相互作用により生じるシグナルおよび/またはマーカーを使用する系を用いて、該シグナルおよび/またはマーカーの存在若しくは不存在および/または変化を検出することにより、該化合物がGタンパク質とAIPとの結合を阻害するか否かを決定することを特徴とする同定方法。
【請求項13】
Gタンパク質がGα13である請求項11又は12に記載の方法。
【請求項14】
請求項13に記載の同定方法により同定された化合物を含有するダイオキシン作用の阻害剤。
【請求項15】
請求項1〜10の何れか1項に記載の医薬若しくは食品からなる、又は請求項14に記載の阻害剤を含む、ダイオキシン作用に起因する疾患の予防治療剤又は予防治療方法。
【請求項16】
ダイオキシン作用に基づく疾患が、肝臓毒性、胸腺の萎縮、皮膚がん、又は子宮内膜症である請求項15に記載の予防治療剤又は予防治療方法。
【請求項17】
Gタンパク質共役受容体、Gタンパク質の優性活性型変異体および/または優性抑制型変異体を用いることを特徴とするダイオキシン作用への制御試験系。
【請求項18】
Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する方法。
【請求項19】
Ahレセプターと結合できるAIP分子の数を増加又は減少させることにより、Ahレセプターによる転写活性化能を促進又は抑制する手段を含む医薬又は食品。
【図1】


【図2】

【図3】

【図4】
【図5】

【図2】
【図3】
【図4】
【図5】
【公開番号】特開2006−143690(P2006−143690A)
【公開日】平成18年6月8日(2006.6.8)
【国際特許分類】
【出願番号】特願2004−339219(P2004−339219)
【出願日】平成16年11月24日(2004.11.24)
【出願人】(504143441)国立大学法人 奈良先端科学技術大学院大学 (226)
【Fターム(参考)】
【公開日】平成18年6月8日(2006.6.8)
【国際特許分類】
【出願日】平成16年11月24日(2004.11.24)
【出願人】(504143441)国立大学法人 奈良先端科学技術大学院大学 (226)
【Fターム(参考)】
[ Back to top ]