
Gタンパク質共役受容体活性化の測定
本発明は、試料中の化合物を検出するための方法およびポリペプチドに関する。詳細には、本発明は、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体を含む無細胞組成物の使用に関する。任意選択で、組成物は、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子も含む。Gタンパク質共役受容体および/または存在する場合アクセサリー分子は、受容体への化合物の結合の検出に使用される生物発光共鳴エネルギー移動(BRET)を可能にする、生物発光タンパク質およびアクセプター分子を一緒に含む。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、試料中の化合物を検出するための方法およびポリペプチドに関する。詳細には、本発明は、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体を含む無細胞組成物の使用に関する。任意選択で、組成物は、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子も含む。Gタンパク質共役受容体および/または存在する場合アクセサリー分子は、生物発光タンパク質およびアクセプター分子を一緒に含み、生物発光タンパク質およびアクセプター分子によって、生物発光共鳴エネルギー移動(BRET)を使用して受容体に結合する化合物を検出することができる。
【背景技術】
【0002】
Gタンパク質共役受容体(GPCR)は、細胞の外側で分子を感知し、内側でシグナル伝達経路を、最終的に細胞応答を活性化する膜貫通受容体のファミリーである。GPCRは、酵母および動物を含む真核生物でしか発見されていない。GPCRと結合しそれを活性化するリガンドには、光感受性化合物、匂い、フェロモン、ホルモン、および神経伝達物質があり、小分子からペプチド、大分子タンパク質までサイズが様々である。Gタンパク質共役受容体は多数の疾患に関与し、治療用薬物の標的でもある。
【0003】
脊椎動物およびC.elegansの嗅覚受容体(OR)は、Gタンパク質共役受容体(GPCR)ファミリーのメンバーである(Buckら、1991;Troemelら、1995)。GPCRは、その7つの膜貫通ドメインを特徴とし、リガンド結合ドメインが膜の細胞外側にあり、Gタンパク質結合ドメインが細胞内側にあると推測される。受容体がリガンドと結合したとき、高次構造変化が受容体中に起こり、ヘテロ三量体Gタンパク質を活性化することを可能にする(Kobilkaら、2007)。次いで、活性化したGタンパク質は、グアニルシクラーゼやホスホリパーゼC経路などのシグナル伝達経路を活性化することができ、シグナルをより高位の処理中枢に伝達する(Gaillardら、2004)。
【0004】
Forster共鳴エネルギー移動、または単に共鳴エネルギー移動(RET)とは、励起状態のドナー分子から基底状態のアクセプター分子へのエネルギーの非放射性移動である(Ghanouniら、2001)。エネルギー移動効率は、ドナーとアクセプターとの距離、スペクトルの重複の程度およびアクセプターとドナーの双極子の相対的配向に依存的である。分子内RETがGPCR活性化のモニタリングに使用されていた以前の場合では、蛍光ドナーおよびアクセプターといった、蛍光共鳴エネルギー移動(FRET)と呼ばれる方法を使用している。ほとんどの場合、蛍光ドナーとアクセプターはどちらも、Aequoria victoria由来の緑色蛍光タンパク質(GFP)の工学的変異体である(Tsien、1998)。最も広く使用されるFRETの対は、ドナーであるシアン蛍光タンパク質(CFP)とアクセプターである黄色蛍光タンパク質(YFP)であり(PistonおよびKremers、2007)、このFRET系は、いくつかのGPCRによる直接的なリガンド結合を定量するのに以前から使用されている(Lohseら、2003および2007;Vilardagaら、2003;Rochaisら、2007;Lisenbeeら、2007)。
【0005】
受容体の活性化をモニタリングする1つの方法では、単一のGPCRを、それぞれ3番目の細胞内ループおよびC末端内の挿入部位において、CFPおよびYFPで二重標識する。436nmの光でCFPを励起すると、480nmでのCFPの放出、および535nmで放出するYFPのFRETを引き起こす。FRETの効率は、ドナーとアクセプターとの距離の6乗によって変化し、このことから、GPCRにおける高次構造変化が極めて感度よく示される。このことは、α2AR、副甲状腺ホルモン受容体(PTHR)、β1−ARおよびセクレチン受容体を用いて、インタクトな細胞において実証された。アゴニストであるノルアドレナリンとα2AR(Lohseら、2003)、副甲状腺ホルモンとPTHR(Vilardagaら、2003)、ノルエピネフリンとβ1−AR(Rochaisら、2007)、およびセクレチンとセクレチン受容体(Lisenbeeら、2007)の相互作用はCFPとYFPとの距離を変化させ、それによってFRETシグナルの変化が生じた。
【0006】
FRET系においてYFPアクセプターをフルオレセインヒ素ヘアピン結合体であるFlAsHと交換すると(Hoffmanら、2005;Nakanishiら、2006)、α2アドレナリン作動性受容体活性化のモニタリングに使用したとき、アゴニスト誘導FRETシグナルがCFP/YFP系と比較して5倍増大した(Nakanishiら、2006)。しかし、FlAsHは、困難な標識化および洗浄手順を伴い、広い調査集団での使用が限定されている。CFP/YFP系は、依然として、分子内の高次構造変化をモニタリングするのに最も頻繁に報告されているFRET系である。FRETに付随する主な不利点は、ドナー蛍光体にエネルギーを与えるのに光源が必要となることである(PistonおよびKremers、2007)。これは、ドナーの励起波長でアクセプターの望ましくない直接励起を引き起こす(「クロストーク」と呼ばれる問題)。
【0007】
生物発光共鳴エネルギー移動(BRET)では、FRETのドナー蛍光体をルシフェラーゼと交換し、アクセプターは任意の適切な蛍光体でよい。基質を添加すると生物発光放出、したがってRETが開始されるので、ルシフェラーゼを使用すると、照明の必要が回避される。BRETの2つの一般的な実行形態は、GFP突然変異体のYFP(BRET1;λem=約530nm)またはGFP2(BRET2;λem=約510nm)のどちらかと結合した、ドナー系であるセレンテラジンh(BRET1;λem=約475nm)またはセレンテラジン400a(Clz400a)基質(BRET2;λem=約395nm)を有するウミシイタケ(Renilla)ルシフェラーゼ(RLuc)を含む。BRET2系は、量子収量を消費して、BRET1系の約55nmと比較して約115nmという、ドナーとアクセプターの放出ピーク間の優れたスペクトル分離をもたらす(PflegerおよびEidneら、2006)。
【0008】
嗅覚受容体でのFRETは、クラスA(a2−アドレナリン作動性および副甲状腺ホルモン、(Vilardagaら、2003))およびクラスB(セクレチン、(Lisenbeeら、2007))GPCRでしか以前には実証されていない。GPCRクラスAに属する哺乳動物ORと異なり、線虫OR(Robertson、1998および2001)は、これらのクラスのどちらにも属さず、進化的および構造的に異なる。さらに、GPCRスーパーファミリーのクラスA内にある、哺乳動物ORを含めた全てのORは、その発現の点では非定型的である。一般に、これらのタンパク質は、化学感覚系列に由来するニューロン以外では機能的に発現できない。哺乳動物および線虫ORの適切な発現にその存在が必要となるいくつかのアクセサリータンパク質が同定されている。
【0009】
したがって、例えば、バイオセンサーで使用するGタンパク質共役受容体と結合する化合物の検出を可能にする適切に感度のよい方法および分子が必要である。
【発明の概要】
【0010】
本発明者らは、驚くべきことに、嗅覚受容体(OR)など、キメラのGタンパク質共役受容体(GPCR)を使用して標的化合物を検出する無細胞の生物発光共鳴エネルギー移動(BRET)が、全細胞を使用したBRETやFRETなど他の検出方法より感度がよいことを見出した。
【0011】
したがって、第1の態様では、本発明は、化合物を検出する方法であって、前記方法が、
i)試料を、
a)脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体と、
b)任意選択で、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子と
を含む無細胞組成物と接触させるステップであって、
Gタンパク質共役受容体が、同じまたは異なる1つまたは複数のサブユニットを含み、Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含むステップと、
ii)ステップi)と同時にまたはそれに連続して、生物発光タンパク質の基質を提供し、生物発光タンパク質が基質を修飾することを可能にするステップと、
iii)ステップii)が生物発光タンパク質とアクセプター分子の間の生物発光共鳴エネルギー移動(BRET)を変動(modulate)させるかどうかを決定するステップと
を含み、化合物がGタンパク質共役受容体と結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化し、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある方法を提供する。
【0012】
一実施形態では、化合物と結合することができるGタンパク質共役受容体のサブユニットは、
i)生物発光タンパク質と、
ii)アクセプター分子と
を含み、化合物がサブユニットと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。例えば、サブユニットは、
i)配列番号13、15、52もしくは54として示されるアミノ酸配列、または
ii)配列番号13、15、52もしくは54のいずれか1つもしくは複数と少なくとも40%同一であるアミノ酸配列
を含んでよい。
【0013】
代替の実施形態では、Gタンパク質共役受容体は、
i)a)生物発光タンパク質と、
b)アクセプター分子と
を含む第1のサブユニットと、
ii)化合物と結合することができる第2のサブユニットと
を含み、化合物が第2のサブユニットと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。
【0014】
さらなる実施形態では、
i)生物発光タンパク質がサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質がサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質がサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子がC末端の一部を形成し、または
vi)アクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質がC末端の一部を形成する。
【0015】
さらに別の代替の実施形態では、Gタンパク質共役受容体は、
i)生物発光タンパク質を含む第1のサブユニットと、
ii)アクセプター分子を含む第2のサブユニットと
を含む。例えば、第1または第2のサブユニットは、
i)配列番号17もしくは19として示されるアミノ酸配列、または
ii)配列番号17および/もしくは19と少なくとも40%同一であるアミノ酸配列
を含んでよい。
【0016】
一実施形態では、
i)生物発光タンパク質が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットの5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットの3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットのC末端の一部を形成し、または
vi)アクセプター分子が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットのC末端の一部を形成する。
【0017】
さらなる代替の実施形態では、
i)Gタンパク質共役受容体が生物発光タンパク質を含むサブユニットを含み、アクセサリー分子がアクセプター分子を含み、または
ii)Gタンパク質共役受容体がアクセプター分子を含むサブユニットを含み、アクセサリー分子が生物発光タンパク質を含む。
【0018】
別の代替の実施形態では、アクセサリー分子は、
i)生物発光タンパク質と、
ii)アクセプター分子と
を含む。
【0019】
さらなる代替の実施形態では、組成物は、少なくとも2つのアクセサリー分子を含み、第1のアクセサリー分子は生物発光タンパク質を含み、第2のアクセサリー分子はアクセプター分子を含む。
【0020】
本発明者らは、FRET−無細胞およびBRET−全細胞検出系と比較したときに本発明がいかに優れているかを発見して特に驚いた。したがって、好ましい実施形態では、本発明の方法は、非生物発光タンパク質をドナー分子として使用し、蛍光共鳴エネルギー移動(FRET)の変動を決定する場合より少なくとも2倍、3倍または4倍感度がよい。さらなる好ましい実施形態では、本発明の方法は、この方法で無細胞組成物中にあるものの代わりにインタクトな細胞中に存在する同じGタンパク質共役受容体を使用する場合より少なくとも10倍、20倍、30倍または40倍強いBRETシグナルをもたらす。
【0021】
クラスAのGPCR、一般にOR、特に線虫ORの非定型的性質を考慮すると、BRETドナーおよびアクセプタータグとともにクラスAのGPCRを含むキメラが、a)構造的完全性を保持し、b)匂い物質による受容体活性化を光学的シグナルに変換することができることは驚くべきことであった。したがって、好ましい実施形態では、Gタンパク質共役受容体はクラスAのGPCRである。さらなる好ましい実施形態では、クラスAのGPCRは嗅覚受容体である。細胞中で発現したときに受容体のN末端が細胞の外側にあり、C末端が細胞の内側にある限り、嗅覚受容体はどんな供給源由来のものでもよい。例として、それだけに限らないが、脊索動物受容体、線虫受容体、またはそのいずれか1つの生物学的に活性な変異体もしくは断片がある。脊索動物受容体の例として、それだけに限らないが、哺乳動物受容体、鳥類受容体および魚類受容体がある。好ましい実施形態では、嗅覚受容体は、線虫受容体、またはその生物学的に活性な変異体もしくは断片である。一実施形態では、線虫受容体は、Caenorhabditis elegans受容体、またはその生物学的に活性な変異体もしくは断片である。
【0022】
一実施形態では、嗅覚受容体は、
i)配列番号1〜6のいずれか1つで示されるアミノ酸配列と、
ii)配列番号1〜6のいずれか1つまたは複数と少なくとも40%同一であるアミノ酸配列と
を含む。
【0023】
別の実施形態では、サブユニットは、2つ以上の異なるGタンパク質共役受容体サブユニットの部分のキメラである。
【0024】
アクセサリー分子は、Gタンパク質共役受容体と直接または間接的に結合する任意の分子でよい。例として、それだけに限らないが、Gタンパク質およびアレスチンがある。
【0025】
BRETの変化の幅に対して化合物の濃度を変化させる効果は、その応答が用量依存的であることを最初に示した。したがって、好ましい実施形態では、BRETのレベルは、試料中の化合物の相対量を示す。
【0026】
生物発光タンパク質の例として、それだけに限らないが、ルシフェラーゼ、β−ガラクトシダーゼ、ラクタマーゼ(lactamase)、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β−グルクロニダーゼまたはβ−グルコシダーゼがある。
【0027】
ルシフェラーゼの例として、それだけに限らないが、ウミシイタケ(Renilla)ルシフェラーゼ、ホタル(Firefly)ルシフェラーゼ、腔腸動物(Coelenterate)ルシフェラーゼ、北アメリカツチボタル(North America glow worm)ルシフェラーゼ、コメツキム(click beetle)シルシフェラーゼ、鉄道虫(railroad worm)ルシフェラーゼ、細菌(bacterial)ルシフェラーゼ、ガウシア(Gaussia)ルシフェラーゼ、エクオリン、ヒカリキノコバエ(Arachnocampa)ルシフェラーゼ、またはそのいずれか1つの生物学的に活性な変異体もしくは断片、またはその2つ以上のキメラがある。
【0028】
基質の例として、それだけに限らないが、甲虫(beetle)ルシフェリン、他のルシフェリン、セレンテラジン、またはセレンテラジンの誘導体がある。また、エクオリンなどの一部の発光タンパク質の場合、基質は、カルシウムイオンなどの補助因子でもよい。
【0029】
一実施形態では、アクセプター分子はタンパク質であり、その例として、それだけに限らないが、緑色蛍光タンパク質(GFP)、GFPの青色蛍光変異体(BFP)、GFPのシアン蛍光変異体(CFP)、GFPの黄色蛍光変異体(YFP)、増強型GFP(EGFP)、増強型CFP(ECFP)、増強型YFP(EYFP)、GFPS65T、エメラルド(Emerald)、トパーズ(Topaz)、GFPuv、不安定化EGFP(dEGFP)、不安定化ECFP(dECFP)、不安定化EYFP(dEYFP)、HcRed、t−HcRed、DsRed、DsRed2、t−ダイマー2、t−ダイマー2(12)、mRFP1、ポシロポリン(pocilloporin)、ウミシイタケ(Renilla)GFP、モンスター(Monster) GFP、paGFP、カエデ(Kaede)タンパク質もしくはフィコビリタンパク質(Phycobiliprotein)、またはそのいずれか1つの生物学的に活性な変異体もしくは断片がある。
【0030】
代替の実施形態では、アクセプター分子は非タンパク質であり、その例として、それだけに限らないが、Alexa Fluor色素、Bodipy色素、Cy色素、フルオレセイン、ダンシル、ウンベリフェロン、蛍光マイクロスフェア、発光マイクロスフェア、蛍光ナノ結晶、マリナブルー、カスケードブルー、カスケードイエロー、パシフィックブルー、オレゴングリーン、テトラメチルローダミン、ローダミン、テキサスレッド、希土類元素キレート、またはその任意の組合せもしくは誘導体がある。
【0031】
別の実施形態では、前記方法は、ステップi)もしくはステップii)と同時にまたはそれに連続して、生物発光タンパク質の補助因子を提供するステップをさらに含む。例として、それだけに限らないが、ATP、マグネシウム、酸素、FMNH2、カルシウム、またはそのいずれか2つ以上の組合せがある。
【0032】
一実施形態では、無細胞組成物は、組換え細胞中でGタンパク質共役受容体を産生し、細胞の膜を破壊することによって得られた。
【0033】
一実施形態では、組換え細胞は、Gタンパク質共役受容体と結合する非内因性タンパク質を産生しない。
【0034】
無細胞組成物を得ることができる細胞は、Gタンパク質共役受容体を発現し、この受容体をその細胞膜に取り込むことができる任意の細胞型である。特に好ましい実施形態では、組換え細胞は酵母細胞である。
【0035】
代替の実施形態では、Gタンパク質共役受容体は、リポソームの脂質二重層中に埋め込まれる。
【0036】
さらなる実施形態では、前記方法は、マイクロ流体工学を使用して行われる。
【0037】
別の態様では、本発明は、化合物を検出するための精製および/または組換えポリペプチドであって、前記ポリペプチドが、
i)Gタンパク質共役受容体のサブユニットと、
ii)生物発光タンパク質および/またはアクセプター分子と
を含み、細胞中で発現したときにサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、ポリペプチドを提供する。
【0038】
一実施形態では、ポリペプチドは、1つまたは複数の異なるGタンパク質共役受容体サブユニット、および任意選択で1つまたは複数の異なるアクセサリー分子を含むGタンパク質共役受容体であるタンパク質複合体の一部を形成する。
【0039】
一実施形態では、
i)生物発光タンパク質がサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質がサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質がサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子がC末端の一部を形成し、または
vi)アクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質がC末端の一部を形成し、または
vii)生物発光タンパク質もしくはアクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、または
viii)生物発光タンパク質もしくはアクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、または
ix)生物発光タンパク質もしくはアクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、または
x)生物発光タンパク質もしくはアクセプター分子がサブユニットのC末端の一部を形成する。
【0040】
さらなる実施形態では、ポリペプチドは、生物発光タンパク質およびアクセプター分子を含み、化合物がポリペプチドと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。
【0041】
代替の実施形態では、ポリペプチドはサブユニットおよび生物発光タンパク質を含み、ポリペプチドはアクセプター分子を含む第2のポリペプチドと直接または間接的に結合し、化合物がポリペプチドおよび/または第2のポリペプチドと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。
【0042】
さらなる代替の実施形態では、ポリペプチドはサブユニットおよびアクセプター分子を含み、ポリペプチドは生物発光タンパク質を含む第2のポリペプチドと直接または間接的に結合し、化合物がポリペプチドおよび/または第2のポリペプチドと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。
【0043】
上記2つの実施形態では、第2のポリペプチドは、(本明細書で定義されるホモもしくはヘテロ二量体または高次の多量体を形成する)Gタンパク質共役受容体サブユニットまたはアクセサリー分子のいずれかを含む。
【0044】
さらなる態様では、本発明は、化合物を検出するための精製および/または組換えポリペプチドであって、前記ポリペプチドが、
i)Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合するアクセサリー分子と、
ii)生物発光タンパク質および/またはアクセプター分子と
を含み、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、ポリペプチドを提供する。
【0045】
熟練した読者なら理解するであろうが、検出の方法に関する上記の実施形態の多くは、本発明のポリペプチドの態様にも当てはまる。
【0046】
本発明のポリペプチドをコードする単離および/または外因性ポリヌクレオチドも提供される。
【0047】
一実施形態では、ポリヌクレオチドは、
i)配列番号7〜12、14、16、18、20、51もしくは53のいずれか1つで示されるヌクレオチド配列、または
ii)配列番号7〜12、14、16、18、20、51もしくは53のいずれか1つまたは複数と少なくとも40%同一であるヌクレオチド配列
を含む。
【0048】
別の態様では、本発明のポリヌクレオチドを含むベクターが提供される。好ましい実施形態では、ポリヌクレオチドは、プロモーターと作動可能に連結する。
【0049】
さらなる態様では、本発明のポリヌクレオチドおよび/または本発明のベクターを含む宿主細胞が提供される。
【0050】
好ましくは、宿主細胞は本発明のポリペプチドを産生し、したがってそれを含む。
【0051】
宿主細胞は任意の細胞型でよい。
【0052】
本発明者らは、酵母の膜における、哺乳動物の嗅覚受容体と進化的に異なる線虫の嗅覚受容体の機能的発現も最初に示す。したがって、好ましい実施形態では、宿主細胞は酵母細胞である。
【0053】
さらなる態様では、本発明は、本発明のポリペプチド、本発明のポリヌクレオチド、本発明のベクター、および/または本発明の宿主細胞を含む組成物を提供する。
【0054】
好ましくは、ポリペプチドは脂質二重層中に埋め込まれている。
【0055】
i)Gタンパク質共役受容体サブユニットを含む本発明のポリペプチドであって、前記ポリペプチドが脂質二重層中に埋め込まれ、ならびに/または
ii)Gタンパク質共役受容体の細胞内ループおよび/もしくはC末端と直接もしくは間接的に結合したアクセサリー分子を含む本発明のポリペプチドであって、Gタンパク質共役受容体が脂質二重層中に埋め込まれ、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、ポリペプチド
を含む無細胞組成物も提供される。
【0056】
好ましくは、脂質二重層は、酵母脂質二重層である。
【0057】
一実施形態では、無細胞組成物は、生物発光タンパク質の基質および/または生物発光タンパク質の補助因子をさらに含む。
【0058】
熟練した読者なら理解するであろうが、検出の方法に関する上記の実施形態の多くは、本発明の無細胞の態様にも当てはまる。
【0059】
さらなる態様では、本発明は、本発明の無細胞組成物を作製する方法であって、本発明の細胞を得るステップと、細胞の膜を破壊するステップとを含む方法を提供する。
【0060】
一実施形態では、細胞を浸透化または溶解する。
【0061】
一実施形態では、細胞をフレンチプレスで溶解する。
【0062】
別の態様では、本発明は、本発明のポリペプチド、本発明の宿主細胞、本発明の組成物および/または本発明の無細胞組成物を含むバイオセンサーを提供する。
【0063】
さらなる態様では、本発明は、Gタンパク質共役受容体と結合する化合物をスクリーニングする方法であって、
i)候補化合物を、
a)脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体と、
b)任意選択で、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子と
を含む無細胞組成物と接触させるステップであって、
Gタンパク質共役受容体が、同じまたは異なる1つまたは複数のサブユニットを含み、Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含むステップと、
ii)ステップi)と同時にまたはそれに連続して、生物発光タンパク質の基質を提供し、生物発光タンパク質が基質を修飾することを可能にするステップと、
iii)ステップii)が生物発光タンパク質とアクセプター分子の間の生物発光共鳴エネルギー移動(BRET)を変動させるかどうかを決定するステップと
を含み、BRETの変動が、化合物がGタンパク質共役受容体と結合することを示し、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記方法を提供する。
【0064】
熟練した読者なら理解するであろうが、検出の方法に関する上記の実施形態の多くは、本発明のスクリーニングの方法の態様にも当てはまる。
【0065】
さらなる態様では、本発明は、本発明のポリペプチド、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明の組成物、本発明の無細胞組成物、および/または本発明のバイオセンサーを含むキットを提供する。
【0066】
明らかとなるように、本発明の1つの態様の好ましい性質および特徴は、本発明の多くの他の態様に適用可能である。
【0067】
本明細書全体にわたって、「含む(comprise)」という語、または「含む(comprises)」や「含んでいる(comprising)」などの変化形は、述べられた要素、整数もしくはステップ、または一群の要素、整数もしくはステップを包含することを意味するが、任意の他の要素、整数もしくはステップ、または一群の要素、整数もしくはステップを除外することを意味しないことが理解されるであろう。
【0068】
以後、下記の非限定的な実施例により、添付図面を参照しながら本発明を説明する。
【図面の簡単な説明】
【0069】
【図1】GFP2およびRLucと融合したODR−10受容体における共鳴エネルギー移動の原理を示す図である。GFP2はGPCRの3番目の細胞内ループ中に、RLucはそのC末端に挿入されている。ジアセチルが結合した結果、高次構造変化が生じ、RLucとGFP2との距離が短く(上部右側)、もしくは長くなり(下部右側)、またはその発色団双極子の相対的配向の変化が生じ得、用量依存的にBRET2シグナルが変化する。
【図2】(A)μM(平均±SEM、n=2)および(B)nM(平均±SEM、n=6)濃度のリガンドまたは水に対するOGORの応答を示す図である(*は、水と比較して有意差がある(P≦0.05)ことを示す)。
【図3】水単独と比較した、(A)μM(平均±SEM、n=2)および(B)nM(平均±SEM、n=6)濃度のリガンドに対するOGORのBRET2比率(%)の変化を示す図である。
【図4】OGORのBRET2応答のジアセチル濃度依存性を示す図である(平均±SEM、n=12)。
【図5】水または1μMのジアセチル水溶液に対するOGOR(平均±SEM、n=3)およびOGOR突然変異体(平均±SEM、n=6)の応答の比較を示す図である。
【図6】水または1μMのジアセチル水溶液に対するOCOY(平均±SEM、n=5)およびOGOR(平均±SEM、n=3)の応答の比較を示す図である。
【図7】様々な濃度のジアセチルを添加した時のOCOYのスペクトルスキャンを示す図である。上部:生データ。下部:標準化した(500nm)データ。
【図8】FRET(平均±SEM、n=5)とBRET2(平均±SEM、n=12)の両方でタグを付けたODR−10の標準化した用量反応曲線を示す図である。
【図9】LB単独と比較した、LB中のOP50細菌上清(LB)またはOP50細菌培養物10μL(平均±SEM、n=3)に対するTGTRの応答を示す図である(*は、LBと比較して有意差がある(P≦0.05)ことを示す)。
【図10】μM(平均±SD、n=3)濃度のリガンドまたは水に対するTGTRの応答を示す図である(*は、水と比較して有意差がある(P≦0.05)ことを示す)。
【図11】ODR−10伝達カセットを示す図である。暗部:ODR−10配列、明部:可変受容体リガンド結合ドメイン。A)カセット中で固定されたN末端、B)N末端可変OR配列。Rルシフェラーゼは410nmで放出するものとして示し、GFP2は515nmで放出するものとして示す。
【図12】共焦点顕微鏡法(Ex488nm、Leica SP2共焦点レーザー走査型顕微鏡)によりodr−10受容体の局在およびタグ付きodr−10の発現レベルを示す、odr−10−Rlucおよびodr−10−GFP2を同時発現した酵母細胞を示す図である。
【図13】生存Invsc1酵母細胞における線虫odr−10の構成的BRET2を示す図である。タグ付きodr−10タンパク質を発現するように、試験した全ての酵母株を15℃で24時間誘導した。10uMのセレンテラジンh(DeepBlueC)を添加し、二重波長マイクロプレート読取り器で、方法において記載するRlucおよびGFP2のフィルター設定を用いて光放出を測定することにより、生存細胞でエネルギー移動測定を行った。データは、2つの独立した実験の平均±StDである。Insvc1/odr−10−RLuc:陰性対照。Invsc1/odr−10−RLuc+Invsc1/odr−10−GFP2:別々の細胞におけるORおよびOG。プレートの読み取りで細胞を一緒に混合した:2つの構築物が別々の細胞で発現したので、BRETは予想されない。Invsc1/odr−10−RLuc;odr−10−GFP2:どちらの構築物も同じ細胞で発現した。BRETが存在し、したがって二量体形成も起こっている。
【図14】免疫沈降によるOdr−10オリゴマーの検出を示す図である。示された受容体を同時発現しているInvsc1酵母細胞の粗製膜を可溶化し、同じ量のルシフェラーゼ活性に調整した。次いで、抗GFP抗体を用いて受容体を免疫沈降し、沈降物におけるルシフェラーゼ活性を決定した。値は、沈降したルシフェラーゼ活性の最大量の%として示す。データは2つの生物学的反復の平均である。
【図15】超遠心分離後の無細胞膜調製物100μLの全細胞アッセイ試料100μLに5μMのセレンテラジン400aを添加した後、BRET2チャネルで測定された光放出量を示す図である。
【図16】全細胞および無細胞アッセイ系を使用してモニタリングした、1μMのジアセチルとのインキュベーション(45分)後のBRET2シグナルを示す図である(平均±S.D.、n=3)。
【発明を実施するための形態】
【0070】
発明の詳細な説明
一般的技術および定義
別段具体的に定義されていない限り、本明細書で使用される全ての科学技術用語は、(例えば、細胞培養、分子遺伝学、バイオセンサー、G共役型タンパク質受容体の生物学、免疫学、免疫組織化学、タンパク質化学、および生化学における)当業者によって一般に理解されているものと同じ意味を有するとみなすものとする。
【0071】
別段示されていない限り、本発明において利用される組換えタンパク質、細胞培養、および免疫学的技術は標準的な手順であり、当業者に周知である。そのような技術は、J.Perbal、A Practical Guide to Molecular Cloning、John Wiley and Sons(1984)、J.Sambrookら、Molecular Cloning:A Laboratory Manual、Cold Spring Harbour Laboratory Press(1989)、T.A.Brown(編)、Essential Molecular Biology:A Practical Approach、第1および2巻、IRL Press(1991)、D.M.GloverおよびB.D.Hames(編)、DNA Cloning:A Practical Approach、第1〜4巻、IRL Press(1995および1996)、ならびにF.M.Ausubelら(編)、Current Protocols in Molecular Biology、Greene Pub.Associates and Wiley−Interscience(1988、現在までの全ての最新版を含む)、Ed HarlowおよびDavid Lane(編)Antibodies:A Laboratory Manual、Cold Spring Harbour Laboratory、(1988)、ならびにJ.E.Coliganら(編)Current Protocols in Immunology、John Wiley & Sons(現在までの全ての最新版を含む)などの出典における文献全体にわたって記載され、説明されている。
【0072】
本明細書において、別段指定されていない限り、「Gタンパク質共役受容体」という用語は、Gタンパク質を介してシグナル伝達する7回膜貫通受容体を指す。この受容体は、単一のサブユニットでもよく、または2つ以上の受容体サブユニットでもよい。2つ以上の受容体サブユニットが存在するとき、それらは同じでもよく、異なってもよく、またはその組合せ(例えば、1つのサブユニットが2つ、別のサブユニットが1つだけ)でもよい。さらに、別段の指定または含意がない限り、「Gタンパク質共役受容体」および「Gタンパク質共役受容体のサブユニット」という用語、またはその変化形は互換的に使用される。
【0073】
本明細書において、「嗅覚受容体」、「嗅受容体」、「OR」という用語またはその変化形は、生物の細胞中に存在するとき、化学感覚的知覚に関与するポリペプチドを指す。一実施形態では、細胞はニューロンである。さらに、「嗅覚受容体」または「嗅受容体」という用語は、匂い物質リガンドと結合し、または匂い物質リガンドと結合するタンパク質複合体の一部を形成し、その結果生理的応答を起こすポリペプチドを指す。
【0074】
本明細書において、「生物発光タンパク質」という用語は、適切な基質に作用し、発光を生じさせることができる任意のタンパク質を指す。
【0075】
本明細書において、「基質」という用語は、生物発光タンパク質とともに、発光を生じさせまたは吸収することができる任意の分子を指す。
【0076】
本明細書において、「生物発光タンパク質が基質を修飾することを可能にする」という語句は、エネルギーを産生する、基質に対する生物発光タンパク質の任意の酵素活性を指す。
【0077】
本明細書において、「アクセプター分子」という用語は、生物発光タンパク質の活性の結果放出されたエネルギーを受容し、光エネルギーとして再び放出することができる任意の化合物を指す。
【0078】
本明細書において、生物発光共鳴エネルギー移動(BRET)とは、生物発光タンパク質ドナーとアクセプター分子の間のエネルギーの非放射性移動に基づく近接アッセイである。
【0079】
本明細書において、「変動させる」もしくは「変動」という用語またはその変化形は、生物発光タンパク質および/またはアクセプター分子の強度および/または放出スペクトルの変化を指す。
【0080】
本明細書において、「空間的位置」という用語は、Gタンパク質共役受容体を含む本明細書で定義されるポリペプチドと化合物が結合した結果変化する、アクセプター分子に対する生物発光タンパク質の三次元での位置付けを指す。
【0081】
本明細書において、「双極子配向」という用語は、三次元空間におけるその配向に対する、生物発光タンパク質および/またはアクセプター分子に関連する双極子モーメントの三次元空間における方向を指す。双極子モーメントは、分子に対して電荷が変化する結果生じるものである。
【0082】
本明細書において、「感度がよい」(more sensitive)という用語は、1つのレポーター系(例えば、BRET)で別のレポーター系(例えば、FRET)に対してリガンドの非結合形態とリガンドの結合形態の間の共鳴エネルギー移動の比率が大きく変化することを指す。Gタンパク質共役受容体サブユニットを含む本発明の単離および/または組換えポリペプチドを含有していること。好ましくは、無細胞組成物は、本発明のポリペプチドを含んでも含まなくても、生細胞を含まない。
【0083】
本明細書において、「接触させる」という用語は、Gタンパク質共役受容体と結合することができるという形で検出される化合物を含む、または含む可能性がある試料を添加することを指す。
【0084】
本明細書において、「脂質二重層」という用語は、内部に共通する疎水性の二重層および2つの親水性の表面を有する、互いに反対に整列した分子の並びの、典型的には両親媒性の2つの層を指す。脂質二重層は、天然に存在するものでもよく、または人工のものでもよい。最も好ましくは、脂質二重層は、Gタンパク質共役受容体を含む本明細書で定義されるポリペプチドが中に挿入される細胞または生体膜、例えば、哺乳動物、昆虫、植物または酵母細胞膜であり、最も好ましくは、酵母細胞膜である。
【0085】
本明細書において、「酵母脂質二重層」という用語は、Gタンパク質共役受容体を含む本明細書で定義されるポリペプチドを発現する酵母細胞に由来する脂質二重層を指す。当業者は、それだけに限らないが、Fus1pタンパク質(TrueheartおよびFink、1989)、Sso1/2p、SncipおよびSec9pタンパク質を含むSNARE複合体(Stropら、2007)、フェロモン受容体Ste2p(Celicら、2003)、および/またはAlr1(Graschopfら、2001)など、膜中に埋め込まれた、酵母中に天然に存在するタンパク質を検出することによって、脂質二重層が酵母に由来するかどうかを容易に決定することができる。
【0086】
本明細書において、「同時にまたは連続して」という用語は、試料を本明細書で定義されるポリペプチドと接触させる前、その間、またはその後に基質を提供できることを意味する。好ましくは、試料と同時またはその後に基質を提供する。
【0087】
「試料」は、検出する化合物を含むことが疑われる任意の物質または組成物でよい。試料の例として、空気、液体、生体材料および土壌がある。試料は、環境もしくは供給源から直接得ることもでき、または本発明の方法を行う前に適切な手順によって少なくとも部分的に精製することもできる。
【0088】
本明細書において、「Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含む」という用語は、生物発光タンパク質が、Gタンパク質共役受容体のサブユニットまたはアクセサリー分子と結合する、好ましくは共有結合する、より好ましくはそれとの融合タンパク質として産生され、アクセプター分子が、Gタンパク質共役受容体のサブユニットまたはアクセサリー分子と結合する、好ましくは共有結合する、より好ましくはそれとの融合タンパク質として産生されることを意味する。生物発光タンパク質およびアクセサリー分子は、同じもしくは異なるGタンパク質共役受容体のサブユニットまたは同じもしくは異なるアクセサリー分子と結合する、好ましくは共有結合する、より好ましくはそれとの融合タンパク質として産生することができる。さらに、生物発光タンパク質およびアクセサリー分子の一方は、Gタンパク質共役受容体のサブユニットと結合する、好ましくは共有結合する、より好ましくはそれとの融合タンパク質として産生することができ、生物発光タンパク質およびアクセサリー分子の他方は、アクセサリー分子と結合することができる。さらに、複数の生物発光タンパク質とアクセサリー分子の対は、多重化して異なるリガンドを検出し、または同じリガンドの検出の感度を増大させる組合せとして存在し得る。
【0089】
本明細書において、「の一部を形成する(forms part of)」という用語は、Gタンパク質共役受容体またはそのサブユニットの特定の領域内に位置する生物発光タンパク質またはアクセプター分子を指す。この用語は、生物発光タンパク質および/またはアクセプター分子が、Gタンパク質共役受容体に付着または結合するが、アミノ酸の連続した鎖を形成しない可能性も含む。一実施形態では、生物発光タンパク質またはアクセプター分子は、Gタンパク質共役受容体の特定の領域と完全に入れ替わる。別の実施形態では、Gタンパク質共役受容体の特定の領域の全部ではないが一部が入れ替わる。さらに別の実施形態では、Gタンパク質共役受容体の特定の領域が入れ替わらない。熟練した受け手なら理解するであろうが、生物発光タンパク質またはアクセプター分子は、本発明のポリペプチドのGタンパク質共役受容体部分が標的化合物と結合してアクセプター分子に対する生物発光タンパク質の位置および/または双極子配向の空間的変化を起こすことができないようには挿入されない。
【0090】
本明細書において、Gタンパク質共役受容体の特定の部分(ドメイン)の「少なくとも大部分」という用語は、特定の領域の少なくとも51%、より好ましくは少なくとも75%、さらにより好ましくは少なくとも90%を指す。
【0091】
「実質的に精製された」または「精製された」とは、その天然の状態で結合する1つまたは複数の脂質、核酸、他のポリペプチド、または他の混入分子から分離しているポリペプチドを意味する。実質的に精製されたポリペプチドは、それと天然に結合する他の構成成分を少なくとも60%、より好ましくは少なくとも75%、より好ましくは少なくとも90%含まないことが好ましい。しかし、現在のところ、本発明のポリペプチドが天然に存在する証拠はない。
【0092】
ポリペプチドとの関連で「組換え」という用語は、その天然の状態と比較して変化した量または変化した速度で、細胞によって、または無細胞発現系で産生されたときのポリペプチドを指す。一実施形態では、細胞は、そのポリペプチドを天然に産生しない細胞である。しかし、細胞は、産生するポリペプチドの量を変化させる、好ましくは増大させる非内因性遺伝子を含む細胞でもよい。本発明の組換えポリペプチドは、それが産生されたトランスジェニック(組換え)細胞、または無細胞発現系の他の構成成分から分離していないポリペプチド、および少なくとも一部の他の構成成分から後で精製して除かれる、そのような細胞または無細胞系で産生されたポリペプチドを含む。
【0093】
「ポリペプチド」および「タンパク質」という用語は一般に互換的に使用され、1本のポリペプチド鎖を指し、非アミノ酸基の付加によって修飾されてもよく、またはされなくてもよい。そのようなポリペプチド鎖は、他のポリペプチドもしくはタンパク質、または補助因子などの他の分子と結合し得ることが理解されるであろう。本明細書において「タンパク質」および「ポリペプチド」という用語は、本明細書に記載のポリペプチドの変異体、突然変異体、生物学的に活性な断片、修飾物、類似体および/または誘導体も含む。
【0094】
ポリペプチドの%同一性は、GAP(NeedlemanおよびWunsch、1970)分析(GCGプログラム)により、ギャップ生成ペナルティー=5、ギャップ伸長ペナルティー=0.3を用いて決定する。質問配列は長さが少なくとも25アミノ酸であり、GAP分析は少なくとも25アミノ酸の領域にわたって2つの配列を整列させる。より好ましくは、質問配列は長さが少なくとも50アミノ酸であり、GAP分析は少なくとも50アミノ酸の領域にわたって2つの配列を整列させる。より好ましくは、質問配列は長さが少なくとも100アミノ酸であり、GAP分析は少なくとも100アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、質問配列は長さが少なくとも250アミノ酸であり、GAP分析は少なくとも250アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、GAP分析はその全長にわたって2つの配列を整列させる。
【0095】
本明細書において、「生物学的に活性な断片」とは、完全長ポリペプチドの定められた活性を維持する、本明細書に記載のポリペプチドの部分である。例えば、Gタンパク質共役受容体の生物学的に活性な断片は、標的化合物と結合して、高次構造変化を起こすことができなければならない。生物学的に活性な断片は、定められた活性を維持する限りどんなサイズでもよい。好ましくは、生物学的に活性な断片は、長さが少なくとも150アミノ酸、より好ましくは少なくとも250アミノ酸である。
【0096】
本明細書において、「生物学的に活性な変異体」とは、1つまたは複数のアミノ酸により天然に存在する分子および/または定められた分子と異なるが、生物学的に活性な断片について上記で定められたような定められた活性を維持する分子である。生物学的に活性な変異体は、天然に存在する分子および/または定められた分子と、典型的には少なくとも50%、より好ましくは少なくとも80%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも97%、さらにより好ましくは少なくとも99%同一である。
【0097】
定められたポリペプチドまたはポリヌクレオチドに関して、上記で示したものより高い%同一性の数字が好ましい実施形態を包含することが理解されるであろう。したがって、適用可能な場合、最小の%同一性の数字に照らして、ポリペプチドまたはポリヌクレオチドが、関連する指定された配列番号と、少なくとも50%、より好ましくは少なくとも60%、より好ましくは少なくとも70%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも91%、より好ましくは少なくとも92%、より好ましくは少なくとも93%、より好ましくは少なくとも94%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%、より好ましくは少なくとも99.1%、より好ましくは少なくとも99.2%、より好ましくは少なくとも99.3%、より好ましくは少なくとも99.4%、より好ましくは少なくとも99.5%、より好ましくは少なくとも99.6%、より好ましくは少なくとも99.7%、より好ましくは少なくとも99.8%、さらにより好ましくは少なくとも99.9%同一であるアミノ酸配列を含むことが好ましい。
【0098】
センスもしくはアンチセンス方向または両方の組合せにおける、一本鎖または二本鎖のDNA、RNA、またはこれらの組合せを含めて、「単離ポリヌクレオチド」とは、その天然の状態で結合または連結するポリヌクレオチド配列から少なくとも部分的に分離しているポリヌクレオチドを意味する。好ましくは、単離ポリヌクレオチドは、それと天然に結合する他の構成成分を少なくとも60%、好ましくは少なくとも75%、最も好ましくは少なくとも90%含まない。
【0099】
ポリヌクレオチドとの関連で、「外因性」という用語は、その天然の状態と比較して変化した量で、細胞または無細胞発現系中に存在するときのポリヌクレオチドを指す。一実施形態では、細胞は、そのポリヌクレオチドを天然に含まない細胞である。しかし、細胞は、コードされたポリペプチドの産生量を変化させる、好ましくは増大させる非内因性ポリヌクレオチドを含む細胞でもよい。本発明の外因性ポリヌクレオチドは、それが存在するトランスジェニック(組換え)細胞、または無細胞発現系の他の構成成分から分離していないポリヌクレオチド、および少なくとも一部の他の構成成分から後で精製して除かれる、そのような細胞または無細胞系で産生されたポリヌクレオチドを含む。
【0100】
さらに、「ポリヌクレオチド」という用語は、本明細書において、「核酸」という用語と互換的に使用される。本発明のポリヌクレオチドは、これとともに提供される分子と比較したときに、ヌクレオチド残基または修飾された残基の欠失、挿入、または置換である1つまたは複数の突然変異を有してよい。突然変異体は、天然に存在するもの(すなわち、天然供給源から単離されたもの)でもよく、または合成のもの(例えば、核酸上で部位特異的突然変異導入を行うことによるもの)でもよい。通常、ポリヌクレオチドの単量体は、ホスホジエステル結合またはその類似物によって連結する。ホスホジエステル結合の類似物には、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホラニリデート(phosphoranilidate)およびホスホラミデートがある。
【0101】
本発明の組成物は、「許容される担体」を含んでよい。そのような許容される担体の例として、水、食塩水、リンガー溶液、デキストロース溶液、ハンクス溶液、および他の生理的に均衡のとれた水性の塩溶液がある。固定油、ゴマ油、オレイン酸エチルやトリグリセリドなどの非水性媒体を使用することもできる。「許容される担体」の正確な性質は、組成物の用途に依存する。本明細書に記載の用途、および組成物における本発明の構成成分の性質を考慮すると、当業者は、特定の用途について適切な「(複数の)許容される担体」を容易に決定することができる。
【0102】
生物発光タンパク質
生物発光タンパク質は、基質を活性化した産物に転換する酵素であり、次いでその産物は緩和するにつれてエネルギーを放出することが当技術分野で理解されている。(基質に対する生物発光タンパク質の活性によって生じた)その活性化した産物は、アクセプター分子に移る、生物発光タンパク質によって生じた発光の供給源である。
【0103】
本発明で使用できるいくつかの異なる生物発光タンパク質がある(例えば、表1を参照)。生物発光タンパク質のサイズを考慮すると、本発明に有用な機能的ポリペプチドを産生できたことは驚きであった。光放出系が知られており、細菌、原生動物、腔腸動物、軟体動物、魚類、ヤスデ類、ハエ、菌類、蠕虫、甲殻類、および甲虫、特にPyrophorus属のコメツキムシ、ならびにPhotinus、Photuris、およびLuciola属のホタルを含めた多数の発光生物から単離されている。生物発光を示すさらなる生物は、WO00/024878、WO99/049019およびViviani(2002)に列挙されている。
【0104】
1つの非常によく知られている例は、特異的な生化学物質であるルシフェリン(天然に存在する蛍光体)がルシフェラーゼ活性を有する酵素によって酸化される、エネルギーを産生する化学反応を触媒するルシフェラーゼとして知られるタンパク質のクラスである(Hastings、1996)。原核生物も真核生物も、細菌、藻類、菌類、昆虫、魚類および他の海洋形態の種を含めて、非常に多様な生物がこのように光エネルギーを放出することができ、それぞれが、他の生物のものと化学的に異なる特異的なルシフェラーゼ活性およびルシフェリンを有する。ルシフェリン/ルシフェラーゼ系は、形態、化学的性質および機能において非常に多様である。したがって、ルシフェラーゼ活性を有する生物発光タンパク質は、様々な供給源からまたは様々な手段によって入手可能である。ルシフェラーゼ活性を有する生物発光タンパク質の例は、米国特許第5,229,285号、第5,219,737号、第5,843,746号、第5,196,524号および第5,670,356号で見つけることができる。最も広く使用されている2つのルシフェラーゼは、(i)ウミシイタケ(Renilla)ルシフェラーゼ(R.reniformis由来)、35kDaのタンパク質、基質としてセレンテラジンを使用し、480nmで光を放出する(Lorenzら、1991);および(ii)ホタルルシフェラーゼ(Photinus pyralis由来)、61kDaのタンパク質、基質としてルシフェリンを使用し、560nmで光を放出する(de Wetら、1987)である。
【0105】
【表1】
【0106】
ガウシア(Gaussia)ルシフェラーゼ(Gaussia princeps由来)は、生化学的アッセイで使用されている(Verhaegenら、2002)。ガウシア(Gaussia)ルシフェラーゼは20kDaのタンパク質であり、セレンテラジンを迅速な反応で酸化し、その結果470nmで明るい光が放出される。
【0107】
本発明に有用なルシフェラーゼは、Anachnocampa種(WO2007/019634)からも特徴付けられている。これらの酵素は、サイズが59kDaであり、放出スペクトルがスペクトルの青色部分に入る発光反応を触媒するATP依存性ルシフェラーゼである。
【0108】
本発明で使用できる代替の非ルシフェラーゼ生物発光タンパク質は、適切な基質に作用して発光シグナルを生じさせる任意の酵素である。そのような酵素の具体例は、β−ガラクトシダーゼ、ラクタマーゼ、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β−グルクロニダーゼおよびβ−グルコシダーゼである。これらの酵素の合成発光基質は当技術分野で周知であり、Tropix Inc.(Bedford、MA、USA)などの企業から市販されている。
【0109】
本発明に有用なペルオキシダーゼの例は、Hushpulianら(2007)によって記載されている。
【0110】
好ましい実施形態では、立体障害による相互作用の阻害を防止するために、低分子量の生物発光タンパク質が使用される。生物発光タンパク質は、好ましくは1本のポリペプチド鎖からなる。また、生物発光タンパク質は、好ましくはオリゴマーまたは凝集物を形成しない。生物発光タンパク質のウミシイタケ(Renilla)ルシフェラーゼ、ガウシア(Gaussia)ルシフェラーゼおよびホタルルシフェラーゼはこれらの基準の全てまたはほとんどを満たす。
【0111】
基質
基質の選択は、生物発光タンパク質によって生じた光の波長および強度に影響を及ぼし得る。
【0112】
広く知られている基質は、刺胞動物、カイアシ類、毛顎動物(chaetgnaths)、有櫛動物、十脚類シュリンプ、アミ類シュリンプ、放散虫類および一部の魚類の分類群に存在するセレンテラジンである(GreerおよびSzalay、2002)。例えばウミシイタケ(Renilla)ルシフェラーゼでは、418〜512nmの間で光を放出するセレンテラジン類似体/誘導体が利用可能である(Inouyeら、1997)。セレンテラジン類似体/誘導体(400A、DeepBlueC)は、ウミシイタケ(Renilla)ルシフェラーゼで光を400nmで放出することが記載されている(WO01/46691)。セレンテラジン類似体/誘導体の他の例は、EnduRenおよびViviRenである。
【0113】
本明細書において、「ルシフェリン」という用語は、生物発光が可能な生物で認められる、光を放出する生物学的色素のクラスを指し、それは酵素ルシフェラーゼの存在下で酸化されて、オキシルシフェリンおよび光の形でのエネルギーを産生する。ルシフェリン、または2−(6−ヒドロキシベンゾチアゾール−2−イル)−2−チアゾリン−4−カルボン酸は、ホタルPhotinus pyralisから初めて単離された。それ以来、様々な異なる生物から、主に海洋、例えば魚類およびイカから様々な形態のルシフェリンが発見され研究されてきたが、多くは陸上生物、例えば、蠕虫、甲虫および様々な他の昆虫において同定されている(Dayら、2004;Viviani、2002)。
【0114】
ルシフェリンには少なくとも5つの一般的な型があり、それぞれが化学的に異なり、広範囲の異なる補助因子を使用する化学的構造的に異なるルシフェラーゼによって触媒される。第1はホタルルシフェリンというホタルルシフェラーゼの基質であり、触媒作用にATPを必要とする(EC1.13.12.7)。第2は細菌ルシフェリンであり、一部のイカおよび魚類でも認められ、長鎖アルデヒドおよび還元型リン酸リボフラビンからなる。細菌ルシフェラーゼはFMNH依存性である。第3は渦鞭毛藻類(dinoflagellate)ルシフェリンであり、夜間の海洋のリン光に関与する生物である渦鞭毛藻類(海洋プランクトン)で認められるテトラピロールクロロフィル(tetrapyrrolic chlorophyll)誘導体である。渦鞭毛藻類ルシフェラーゼは渦鞭毛藻類ルシフェリンの酸化を触媒し、触媒活性のある3つの同一のドメインからなる。第4はイミダゾロピラジンのヴァルグリン(vargulin)であり、特定の貝虫類および深海魚、例えばPorichthysで認められる。最後はセレンテラジン(イミダゾールピラジン)であり、放散虫類、有櫛動物、刺胞動物、イカ、カイアシ類、毛顎動物、魚類およびシュリンプで認められるタンパク質エクオリンの光放出体である。
【0115】
アクセプター分子
本発明で使用できるいくつかの異なるアクセプター分子がある。アクセプター分子は、タンパク質でもよく、または非タンパク質性のものでもよい。タンパク質であるアクセプター分子の例として、それだけに限らないが、緑色蛍光タンパク質(GFP)、GFPの青色蛍光変異体(BFP)、GFPのシアン蛍光変異体(CFP)、GFPの黄色蛍光変異体(YFP)、増強型GFP(EGFP)、増強型CFP(ECFP)、増強型YFP(EYFP)、GFPS65T、エメラルド(Emerald)、トパーズ(Topaz)、GFPuv、不安定化EGFP(dEGFP)、不安定化ECFP(dECFP)、不安定化EYFP(dEYFP)、HcRed、t−HcRed、DsRed、DsRed2、t−ダイマー2、t−ダイマー2(12)、mRFP1、ポシロポリン、ウミシイタケ(Renilla) GFP、モンスター(Monster) GFP、paGFP、カエデ(Kaede)タンパク質もしくはフィコビリタンパク質(Phycobiliprotein)、またはそのいずれか1つの生物学的に活性な変異体もしくは断片がある。タンパク質でないアクセプター分子の例として、それだけに限らないが、Alexa Fluor色素、Bodipy色素、Cy色素、フルオレセイン、ダンシル、ウンベリフェロン、蛍光マイクロスフェア、発光マイクロスフェア、蛍光ナノ結晶、マリナブルー、カスケードブルー、カスケードイエロー、パシフィックブルー、オレゴングリーン、テトラメチルローダミン、ローダミン、テキサスレッド、希土類元素キレート、またはその任意の組合せもしくは誘導体がある。
【0116】
1つの非常によく知られている例は、クラゲAequorea victoria由来の緑色蛍光タンパク質および分子生物学、例えば突然変異導入およびキメラタンパク質技術の適用から生じた多数の他の変異体(GFP)を含む蛍光体の群である(Tsien、1998)。GFPは、その発色団の独特の構成成分に基づいて分類され、各クラスは異なる励起および放出波長を有する:クラス1、中性フェノールとアニオン性フェノレートの野生型混合物:クラス2、フェノレートアニオン:クラス3、中性フェノール:クラス4、積み重なったs電子系を有するフェノレートアニオン:クラス5、インドール:クラス6、イミダゾール:およびクラス7、フェニル。
【0117】
突然変異した、天然に存在するアクセプター分子(変異体)も本発明に有用であり得る。BRETに適する工学的に作製された系の1つの例は、ウミシイタケ(Renilla)ルシフェラーゼとGFPの増強型黄色突然変異体(EYFP)の対であり、1または複数の媒介タンパク質(この場合、Gタンパク質共役受容体)が存在しないと単独では互いにそれほど直接相互作用しない(Xuら、1999)。
【0118】
別の実施形態では、アクセプター分子は蛍光ナノ結晶である。ナノ結晶、または「量子ドット」は、光分解に対する耐性、明るさの向上、非毒性、およびいくつかのプロセスを同時にモニタリングすることを可能にするサイズ依存性の狭い放出スペクトルを含めて、蛍光標識として有機分子を上回るいくつかの利点を有する。さらに、ナノ結晶の吸収スペクトルは最初のピークの上で連続し、全てのサイズ、したがって全ての色を、単一の励起波長を用いて励起することを可能にする。
【0119】
蛍光ナノ結晶は、様々な形でタンパク質に付着させ、または「バイオコンジュゲート」させることができる。例えば、ジヒドロリポ酸(DHLA)または両親媒性ポリマー由来のカルボキシレート基を用いて、「量子ドット」の表面キャップを負に帯電させることができる。直接、または組換えタンパク質と融合した正に帯電したロイシンジッパーペプチドからなる架橋を介して、タンパク質をDHLA−ナノ結晶と静電的にコンジュゲートさせることができる。後者は、意図された標的との特異性によって一次抗体と結合する。あるいは、従来のカルボジイミド化学を用いて、抗体、ストレプトアビジン、または他のタンパク質をナノ結晶のポリアクリレートキャップと共有結合させる。
【0120】
セレン化カドミウム、硫化カドミウム、ヒ化インジウムおよびリン化インジウムを含めたナノ結晶を生成するコロイド法がある。これらの量子ドットは、その量子ドットの体積内にわずか100〜100,000個の原子を含有することができ、直径は原子10〜50個である。一部の量子ドットは、一方の物質が他方の物質に埋まり、大きなバンドギャップを有する小さな領域である。これらはいわゆるコアシェル構造といえ、例えば、コアにCdSe、シェルにZnSを有するもの、またはオルモシル(ormosil)と呼ばれるシリカの特殊な形態に由来するものがある。ドットが大きいと、その蛍光スペクトルは赤く(低エネルギーと)なる。逆に、小さいドットは、青い(高エネルギー)の光を放出する。色合いは、量子ドットのエネルギーレベルに直接関連する。定量的にいうと、蛍光の光のエネルギー(したがって色)を決定するバンドギャップエネルギーは、量子ドットのサイズの2乗に反比例する。大きい量子ドットは、大きいエネルギーレベルを有し、より密集している。このことは、量子ドットが低いエネルギー、すなわちスペクトルの赤い方に近いものを含有する光子を吸収することを可能にしている。
【0121】
代替の実施形態では、アクセプター分子は蛍光マイクロスフェアである。これらは典型的にはポリマーでできており、ポリマーマトリックス中に取り込まれた蛍光分子(例えば、フルオレセインGFPまたはYFP)を含有し、様々な試薬とコンジュゲートさせることができる。蛍光マイクロスフェアは内部標識することもでき、または表面上で標識することもできる。内部標識で、典型的には狭い蛍光放出スペクトルを有する非常に明るく安定した粒子が生成される。内部標識すると、表面の基は、リガンド(例えばタンパク質)とビーズの表面とのコンジュゲートに利用可能なままとなる。内部標識したビーズは、光退色に対して高い耐性を示すので、画像化の用途で広く使用される。
【0122】
カルボキシレート修飾蛍光マイクロスフェアは、1−エチル−3−(3−ジメチルアミノプロピル)カルボジイミド塩酸塩(EDAC)などの水溶性カルボジイミド試薬を使用したタンパク質の共有結合に適している。硫酸蛍光マイクロスフェアは比較的疎水性であり、ほとんどどんなタンパク質も受動的にかつほとんど不可逆的に吸着する。アルデヒド−硫酸蛍光マイクロスフェアは、表面アルデヒド基を付加し、タンパク質と反応するように修飾された硫酸マイクロスフェアである。
【0123】
別の実施形態では、アクセプター分子は発光マイクロスフェアである。これらは典型的にはポリマーでできており、ポリマーマトリックス中に取り込まれた発光分子(例えば、ユーロピウムまたは白金の複合体)を含有し、様々な試薬とコンジュゲートさせることができる。
【0124】
生物発光タンパク質とアクセプター分子の対
BRETに適した対を決定する際に考慮すべき基準は、生物発光タンパク質と比較した、アクセプター分子の相対的な放出/蛍光スペクトルである。生物発光タンパク質の放出スペクトルは、生物発光タンパク質の発光の放出からの光エネルギーが、2つの分子が適切に近接し互いに対して適切な配向にあるときにアクセプター分子を励起することができ、それによってアクセプター分子の蛍光を促進する波長であるように、アクセプター分子の吸光度スペクトルと重複すべきである。例えば、観察可能な放出スペクトルのピークに基づくと、ウミシイタケ(Renilla)ルシフェラーゼ/EGFPの対はウミシイタケ(Renilla)ルシフェラーゼ/EYEFの対ほど良好でないことが実証されている(Xu、1999;Wangら、(1997)、Bioluminescence and Chemiluminescence:Molecular Reporting with Photons、Hastingsら編(Wiley、New York)、pp.419〜422)。潜在的な対を研究するために、選択した生物発光タンパク質およびアクセプター分子を含有するタンパク質融合物を調製し、適当な基質の存在下で試験する。
【0125】
生物発光タンパク質とアクセプター分子が誤って互いに結合しないことも確認すべきである。これは、同じ細胞中で生物発光タンパク質およびアクセプター分子を別々に同時発現させ、次いで発光スペクトルをモニタリングしてBRETが起こるかどうかを決定することにより行うことができる。これは、例えば、Xuら、(1999)の方法を使用して実現することができる。BRETがほとんどまたは全く観察されない場合、選択した生物発光タンパク質とアクセプター分子は適切なBRETの対を形成する。
【0126】
生物発光タンパク質放出は、基質の修飾によって操作することができる。ルシフェラーゼの場合、基質はセレンテラジンである。生物発光タンパク質放出を変化させることの背後にある論理的根拠は、ドナー放出とアクセプター放出の間の分解能を向上させることにある。元のBRET系は、ウミシイタケ(Renilla)ルシフェラーゼをドナーとして、EYFP(またはTopaz)をアクセプターとして、セレンテラジンh誘導体を基質として使用する。BRETアッセイで組み合わせたとき、これらの構成成分は、生物発光タンパク質については475〜480nmで、アクセプター分子については525〜530nmで光を発し、スペクトル分解能は45〜55nmとなる。
【0127】
残念ながら、ウミシイタケ(Renilla)ルシフェラーゼでは実質的にGFP放出と重複する広い放出ピークが生じ、これは系のシグナル対ノイズを低下させる一因となる。ウミシイタケ(Renilla)ルシフェラーゼの基質としてcoel400aを使用する本発明の1つのBRET系は、ドナーとアクセプターの放出波長間の広いスペクトル分解能をもたらす(−105nm)。ウミシイタケ(Renilla)ルシフェラーゼとcoel400aは、390〜400nmで光を発し、この範囲で光を吸収し、505〜508nmで光を再び放出するGFPを調製した。ウミシイタケ(Renilla)ルシフェラーゼとGFPの放出間のスペクトル分解能のこの増大のため、このBRET系は、本発明のポリペプチドの高次構造の小さな変化をモニタリングする優れた生物学的ツールをもたらす。これは、ドナーとアクセプターの波長の差がおよそ51nmである、セレンテラジンh誘導体およびEYFPを使用する以前に記載されている系を上回る著しい向上である。
【0128】
ウミシイタケ(Renilla)ルシフェラーゼ活性の結果(野生型セレンテラジンにより発せられるものとは異なる)様々な波長で光を発するcoel400aを含めて、様々なセレンテラジン誘導体が当技術分野で知られている。当業者は、ドナーの光放出ピークが変化したため、この波長で光を吸収し、それによって効率のよいエネルギー移動を可能にするアクセプター分子を選択する必要があることを理解するであろう。これは、例えばGFPクラス4を、クラス3または1のGFPとなるように変更することによって行うことができる。ドナーの光放出とアクセプターの光吸収ピークとの間のスペクトルの重複は、とりわけ、効率のよいエネルギー移動の1つの条件である。クラス3および1のGFPは、400nmで光を吸収し、505〜511nmで再び放出することが知られている。この結果、およそ111nmのドナーとアクセプターの放出間の波長の差となる。
【0129】
さらなる生物発光タンパク質とアクセプター分子の対の例を表2に示す。
【0130】
【表2】
【0131】
Gタンパク質共役受容体
Gタンパク質共役受容体(GPCR)は、7回膜貫通受容体、7TM受容体、7回らせん(heptahelical)受容体、およびGタンパク質連結受容体(GPLR)としても知られている。GPCRは、細胞の外側で分子を感知し、内側でシグナル伝達経路を、最終的に細胞応答を活性化する膜貫通受容体の大型タンパク質のファミリーである。これらの受容体と結合しそれを活性化するリガンドには、光感受性化合物、匂い、フェロモン、ホルモン、および神経伝達物質があり、小分子からペプチド、大型タンパク質までサイズが様々である。GPCRは多数の疾患に関与するが、現代の全医療用薬物のほぼ半分の標的でもある。GPCRは、配列相同性および機能類似性に基づいて、少なくとも5つのクラスに分類することができる:
クラスAのロドプシン様、
クラスBのセクレチン様、
クラスCの代謝型/フェロモン、
クラスDの菌類フェロモン、および
クラスEのcAMP受容体。
【0132】
クラスAのロドプシン様受容体には、アミン受容体:アセチルコリン、アルファアドレナリン受容体、ベータアドレナリン受容体、ドーパミン、ヒスタミン、セロトニン、オクトパミン、および微量アミン;ペプチド受容体:アンギオテンシン、ボンベシン、ブラジキニン、C5aアナフィラトキシン、Fmet−leu−phe、APJ様、インターロイキン−8、ケモカイン受容体(C−Cケモカイン、C−X−Cケモカイン、BONZO受容体(CXC6R)、C−X3−Cケモカイン、およびXCケモカイン)、CCK受容体、エンドセリン受容体、メラノコルチン受容体、神経ペプチドY受容体、ニューロテンシン受容体、オピオイド受容体、ソマトスタチン受容体、タキキニン受容体(サブスタンスP(NK1)、サブスタンスK(NK2)、ニューロメジンK(NK3)、タキキニン様1、およびタキキニン様2)、バソプレッシン様受容体(バソプレッシン、オキシトシン、およびコノプレッシン)、ガラニン様受容体(ガラニン、アラトスタチン、およびGPCR54)、プロテイナーゼ活性化様受容体(例えば、トロンビン)、オレキシン&神経ペプチドFF、ウロテンシンII受容体、アドレノメデュリン(G10D)受容体、GPR37/エンドセリンB様受容体、ケモカイン受容体様受容体、およびニューロメジンU受容体;ホルモンタンパク質受容体:卵胞刺激ホルモン、ルトロピン−絨毛性ゴナドトロピンホルモン、チロトロピン、およびゴナドトロピン;オプシン(ロドプシン)受容体;嗅受容体;プロスタノイド受容体:プロスタグランジン、プロスタサイクリン、およびトロンボキサン;ヌクレオチド様受容体:アデノシンおよびプリン受容体;カンナビス受容体;血小板活性化因子受容体;ゴナドトロピン放出ホルモン受容体;チロトロピン放出ホルモン&分泌促進物質受容体:チロトロピン放出ホルモン、成長ホルモン分泌促進物質、および成長ホルモン分泌促進物質様;メラトニン受容体;ウイルス受容体;リゾスフィンゴ脂質&LPA(EDG)受容体;ロイコトリエンB4受容体:ロイコトリエンB4受容体BLT1およびロイコトリエンB4受容体BLT2;ならびにクラスAオーファン/他の受容体:血小板ADP&KI01受容体、SREB、Mas原癌遺伝子、RDC1、ORPH、LGR様(ホルモン受容体)、GPR、GPR45様、システイニルロイコトリエン、Mas関連受容体(MRG)、およびGP40様受容体がある。
【0133】
GPCRのクラスB(セクレチン受容体ファミリー)には、ポリペプチドホルモンの受容体(カルシトニン、コルチコトロピン放出因子、胃抑制ペプチド、グルカゴン、グルカゴン様ペプチド−1、−2、成長ホルモン放出ホルモン、副甲状腺ホルモン、PACAP、セクレチン、血管作用性小腸ポリペプチド、利尿ホルモン、EMR1、ラトロフィリン)、形質膜での細胞間相互作用を媒介すると考えられる分子(脳特異的血管新生阻害因子(BAI))ならびにストレス応答および寿命を制御する一群のDrosophilaタンパク質(Methuselah様タンパク質)がある。
【0134】
クラスCの代謝型グルタミン酸/フェロモン受容体には、代謝型グルタミン酸、代謝型グルタミン酸グループI、代謝型グルタミン酸グループII、代謝型グルタミン酸グループIII、他の代謝型グルタミン酸、細胞外カルシウム感知、推定上のフェロモン受容体、GABA−B、GABA−Bサブタイプ1、GABA−Bサブタイプ2、およびオーファンGPRC5受容体がある。
【0135】
GPCRは、視覚、嗅覚、行動および気分の制御、免疫系の活性および炎症の制御、自律神経系の伝達、細胞密度の感知、ならびにその他多くを含めて、多種多様な生理的過程に関与する。不活性なGタンパク質は、その不活性状態で受容体と結合することが知られている。リガンドを認識した後、受容体またはそのサブユニットは高次構造を変化させ、それによってGタンパク質を機械的に活性化し、Gタンパク質は受容体から解離する。受容体はここで別のGタンパク質を活性化することもでき、切り替わって不活性状態に戻ることもできる。受容体分子は、活性と不活性の生物物理的状態の間の高次構造の平衡状態にあると考えられている。リガンドが受容体に結合すると、活性な受容体の状態の方に平衡状態を変化させることができる。本発明のポリペプチドはGタンパク質共役受容体を含み、細胞中で発現したときにその受容体のN末端が細胞の外側にあり、C末端が細胞の内側にある。当業者は、膜貫通タンパク質の配向を検出するのに適した技術を承知している。そのような技術は、それだけに限らないが、結晶解析、NMR研究、モデリング研究ならびに光学または電子顕微鏡標本の界面活性剤浸透化対照と組み合わせた免疫標識のような顕微鏡技術、2つのポリペプチドの断片相補性タギングなどを含む。
【0136】
好ましい実施形態では、Gタンパク質共役受容体はクラスAのGPCRである。さらなる好ましい実施形態では、クラスA(ロドプシン様)GPCRは嗅覚受容体またはアドレナリン作動性受容体であり、より好ましくは嗅覚受容体である。細胞中で発現したときに受容体のN末端が細胞の外側にあり、C末端が細胞の内側にある限り、嗅覚受容体はどんな供給源由来のものでもよい。例として、それだけに限らないが、脊索動物受容体、線虫受容体、またはそのいずれか1つの生物学的に活性な変異体もしくは断片がある。脊索動物受容体の例として、それだけに限らないが、哺乳動物受容体、鳥類受容体および魚類受容体がある。好ましい実施形態では、嗅覚受容体は、線虫受容体、またはその生物学的に活性な変異体もしくは断片である。一実施形態では、線虫受容体は、Caenorhabditis elegans受容体、またはその生物学的に活性な変異体もしくは断片である。本発明のポリペプチドの生成に使用し、かつ/または本発明の方法で使用することができる嗅覚受容体の例は、BuckおよびAxel(1991)、Robertson(1998および2001)、Aloniら(2006)、Feldmesser(2006)、Olenderら(2004aおよびb)、Glusmanら(2000a、2000bおよび2001)、Fuchsら(2001)、PilpelおよびLancet(1999)、Sharonら(1998)、Zozulyaら(2001)、NiimuraおよびNei(2003)、Landerら(2001)、ZhangおよびFirestein(2002)、Youngら(2002)に記載されている。さらに、嗅覚受容体の包括的な一覧がSenseLabのウェブサイトから入手可能である(http://senselab.med.yale.edu)。
【0137】
他の実施形態では、GPCRはクラスBまたはクラスCの受容体であり、これら2つの実施形態のうちクラスCがより好ましい。
【0138】
特に好ましい実施形態では、Gタンパク質共役受容体は7回膜貫通ドメインを含む。
【0139】
生物発光タンパク質は、Gタンパク質共役受容体(または本発明のポリペプチド)の1番目、3番目、5番目の非膜貫通ループ(ドメイン)またはC末端の一部を形成することができる。アクセプター分子も、Gタンパク質共役受容体(または本発明のポリペプチド)の1番目、3番目、5番目の非膜貫通ループ(ドメイン)またはC末端の一部を形成することができる。Gタンパク質共役受容体が発現し、細胞中に存在するときに、これらの領域はそれぞれ細胞内にある。
【0140】
アクセプター分子は、同じ分子の一部(すなわち、同じ1本のポリペプチド鎖)のとき、生物発光タンパク質と同じ領域にある可能性はないが、Gタンパク質共役受容体が二量体または高次の多量体として存在するとき、アクセプター分子は生物発光タンパク質と等しい領域にある可能性がある。例えば、生物発光タンパク質は、受容体の1つのサブユニットのC末端の一部を形成することができ、アクセプター分子は、受容体の別のサブユニットのC末端の一部を形成することができる。この例では、標識が結合したサブユニットは同じでもよく、または異なっていてもよく、例えば2つのサブユニットが同一であり、生物発光タンパク質で標識された一方とアクセプター分子で標識された他方が離れていてもよい。一実施形態では、生物発光タンパク質はGPCRサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子は5番目の非膜貫通ループの一部を形成する。代替の実施形態では、アクセプター分子はGPCRサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質は5番目の非膜貫通ループの一部を形成する。
【0141】
別の実施形態では、生物発光タンパク質はGPCRサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子は3番目の非膜貫通ループの一部を形成する。別の実施形態では、アクセプター分子はGPCRサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質は3番目の非膜貫通ループの一部を形成する。
【0142】
好ましい実施形態では、生物発光タンパク質はGPCRサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子はC末端の一部を形成する。代替の実施形態では、アクセプター分子はGPCRサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質はC末端の一部を形成する。
【0143】
別の実施形態では、Gタンパク質共役受容体は少なくとも2つのサブユニットを含み、生物発光タンパク質が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの5番目の非膜貫通ループの一部を形成する。代替の実施形態では、アクセプター分子は第1のサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質は第2のサブユニットの5番目の非膜貫通ループの一部を形成する。
【0144】
別の実施形態では、Gタンパク質共役受容体は少なくとも2つのサブユニットを含み、生物発光タンパク質が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの3番目の非膜貫通ループの一部を形成する。別の実施形態では、アクセプター分子は第1のサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質は第2のサブユニットの3番目の非膜貫通ループの一部を形成する。
【0145】
別の実施形態では、Gタンパク質共役受容体は少なくとも2つのサブユニットを含み、生物発光タンパク質が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットのC末端の一部を形成する。代替の実施形態では、アクセプター分子は第1のサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質は第2のサブユニットのC末端の一部を形成する。
【0146】
別の実施形態では、Gタンパク質共役受容体は少なくとも2つのサブユニットを含み、ドナーおよびアクセプター分子はそれぞれ第1および第2のサブユニットの同じ部位にある。
【0147】
一実施形態では、生物発光タンパク質またはアクセプター分子は、5番目の膜貫通ドメインの2番目のアミノ酸の後、6番目の膜貫通ドメインの開始の前から2番目のアミノ酸の前に位置する。別の実施形態では、生物発光タンパク質またはアクセプター分子は、5番目の膜貫通ドメインの後のアミノ酸8付近の後、または5番目の膜貫通ドメインの後のアミノ酸22付近の後に位置する。さらなる実施形態では、生物発光タンパク質またはアクセプター分子は、6番目の膜貫通ドメインから約10または12アミノ酸前に挿入される。最も好ましくは、生物発光タンパク質またはアクセプター分子は、3番目の非膜貫通ループ(ドメイン)の中央に位置する。
【0148】
C末端に関しては、天然のC末端の約5〜25アミノ酸が7番目の膜貫通ドメインの末端に残存することが好ましい。好ましくは、生物発光タンパク質またはアクセプター分子は、7番目の膜貫通部の後から約16または20アミノ酸後に挿入される。
【0149】
1番目の非膜貫通ループ(ドメイン)における生物発光タンパク質またはアクセプター分子の位置については、前記標識が1番目の膜貫通ドメインの末端から約2アミノ酸後、および2番目の膜貫通ドメインの開始から約2アミノ酸前に挿入されることが好ましい。最も好ましくは、生物発光タンパク質またはアクセプター分子は、1番目の非膜貫通ループ(ドメイン)の中央に位置する。
【0150】
さらなる実施形態では、生物発光タンパク質は、Gタンパク質共役受容体(または本発明のポリペプチド)のN末端、2番目、4番目または6番目の非膜貫通ループ(ドメイン)の一部を形成することができる。アクセプター分子も、Gタンパク質共役受容体(または本発明のポリペプチド)のN末端、2番目、4番目または6番目の非膜貫通ループ(ドメイン)の一部を形成することができるが、同じ分子の一部のとき、生物発光タンパク質と同じ領域にある可能性はない。Gタンパク質共役受容体が発現し、細胞中に存在するときに、これらの領域はそれぞれ細胞外にある。
【0151】
GPCRは、2つ以上の異なるGPCRの天然に存在しないキメラでもよい。特に、このことは、1つの受容体の部分が、検出する化合物に応じて多種多様なGPCRの他の部分が挿入されたキメラ中に常に存在する伝達カセットを生成することを可能にする。
【0152】
一実施形態では、サブユニットは、第1のGタンパク質共役受容体サブユニットのN末端および1番目の膜貫通ドメインの少なくとも大部分、第2のGタンパク質共役受容体サブユニットの1番目の非膜貫通ループの少なくとも大部分から5番目の膜貫通ドメインの少なくとも大部分まで、ならびに第1のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む。
【0153】
別の実施形態では、サブユニットは、第1のGタンパク質共役受容体サブユニットのN末端から5番目の膜貫通ドメインの少なくとも大部分まで、および第2のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む。
【0154】
当業者は、Gタンパク質共役物のN末端、膜貫通ドメイン、非膜貫通ループ(ドメイン)およびC末端を容易に決定することができる。例えば、様々なバイオインフォマティクスの手法を使用して、そのアミノ酸配列およびGタンパク質共役受容体の既知の膜貫通ドメインとの類似性に基づいて、タンパク質における膜貫通ドメインの位置およびトポロジーを決定することができる。アラインメントおよびアミノ酸配列比較は、例えば、BLASTプログラムまたはCLUSTAL Wプログラムを使用することにより、当技術分野で日常的に行われる。既知の膜貫通ドメイン含有タンパク質とのアラインメントに基づいて、当業者は膜貫通ドメインの位置を予測することができる。さらに、一部の膜貫通タンパク質の三次元構造が知られており、例えば、7回膜貫通Gタンパク質共役ロドプシン光受容体構造がX線結晶解析によって解析されている。そのような3D構造の分析およびそれとの比較に基づいて、他の膜タンパク質における膜貫通ドメインの位置およびトポロジーを予測することができる。タンパク質における膜貫通ドメインの位置およびトポロジーの予測に利用可能な多数のプログラムも存在する。例えば、膜貫通タンパク質のセグメントを予測するTMpred(HofmannおよびStoffel、1993);膜タンパク質のトポロジーを予測するTopPred(von Heijneら、1992);単一および複数の配列から二次構造を予測するPREDATOR(FrishmanおよびArgos、1997);複数整列させた配列からタンパク質の膜貫通領域を予測するTMAP(PerssonおよびArgos、1994);ならびに単一の配列から膜貫通領域を予測するALOM2(Klienら、1984)の1つまたは組合せを使用することができる。
【0155】
標準的な命名法に従って、膜貫通ドメインおよび非膜貫通ループ(ドメイン)の番号付けは、ポリペプチドのN末端に応じて行う。
【0156】
天然に存在するGタンパク質共役受容体のアミノ酸配列突然変異体/変異体は、コードするポリヌクレオチド中に適当なヌクレオチド変化を導入することにより、または所望のポリペプチドのin vitro合成により調製することができる。そのような突然変異体には、例えば、アミノ酸配列内の残基の欠失、挿入または置換物がある。最終的なポリペプチド産物が所望の特徴を有することを条件として、欠失、挿入および置換の組合せを行って最終的な構築物に到達することができる。
【0157】
突然変異(変異)ポリペプチドは、当技術分野で知られている任意の技術を使用して調製することができる。例えば、本明細書に記載のポリヌクレオチドをin vitro突然変異導入にかけることができる。そのようなin vitro突然変異導入技術には、適切なベクターへのポリヌクレオチドのサブクローニング、E.coli XL−1red(Stratagene)などの「突然変異誘発」株へのベクターの形質転換、および適切な数の世代にわたる形質転換細菌の増殖が含まれ得る。別の例では、Harayama(1998)によって大枠が記載されている通りに、Gタンパク質共役受容体をコードするポリヌクレオチドをDNAシャッフリング技術にかける。本明細書に記載の技術を使用して突然変異/変異DNAに由来する産物を容易にスクリーニングして、それが本発明の方法に有用であるかを決定することができる。
【0158】
アミノ酸配列突然変異体の設計において、突然変異部位の位置および突然変異の性質は、改変する(複数の)特徴に依存する。突然変異の部位は、例えば、(1)最初に保存的アミノ酸を選択して置換し、次いで得られた結果に応じて選択を急進的にして置換すること、(2)標的残基を欠失させること、または(3)位置する部位の隣に他の残基を挿入することによって、個々にまたは連続して改変することができる。
【0159】
アミノ酸配列の欠失は、一般に約1〜15個の残基であり、より好ましくは約1〜10個の残基であり、典型的には約1〜5個の連続した残基に及ぶ。
【0160】
置換突然変異体は、Gタンパク質共役受容体における少なくとも1つのアミノ酸残基が除去され、その場所に異なる残基が挿入される。置換突然変異導入で最も興味深い部位は、機能にとって重要なものとして同定された部位を含む。他の興味深い部位は、様々な株または種から得られた特定の残基が同一であるものである。これらの位置は、生物活性にとって重要である可能性がある。これらの部位、特に少なくとも3つの他の同一に保存されている部位の配列に入るものは、比較的保存的な形で置換することが好ましい。そのような保存的置換を表3に示す。
【0161】
さらに、所望される場合、置換物または付加物として非天然アミノ酸または化学的アミノ酸類似体を本明細書に記載のポリペプチドに導入することができる。そのようなアミノ酸には、それだけに限らないが、一般的なアミノ酸のD−異性体、2,4−ジアミノ酪酸、α−アミノイソ酪酸、4−アミノ酪酸、2−アミノ酪酸、6−アミノヘキサン酸、2−アミノイソ酪酸、3−アミノプロピオン酸、オルニチン、ノルロイシン、ノルバリン、ヒドロキシプロリン、サルコシン、シトルリン、ホモシトルリン、システイン酸、t−ブチルグリシン、t−ブチルアラニン、フェニルグリシン、シクロヘキシルアラニン、β−アラニン、フルオロアミノ酸、β−メチルアミノ酸、Cα−メチルアミノ酸、Nα−メチルアミノ酸などのデザイナーアミノ酸、および一般のアミノ酸類似体がある。
【0162】
例えば、ビオチン化、ベンジル化、グリコシル化、アセチル化、リン酸化、アミド化、既知の保護/遮断基による誘導体化、タンパク質分解性切断、抗体分子または他の細胞性リガンドとの連結などにより、合成の間またはその後に異なった形で修飾されたポリペプチドも、本発明の範囲に含まれる。これらの修飾物は、ポリペプチドの安定性および/または生物活性を増大させる働きをし得る。
【0163】
【表3】
【0164】
本明細書に記載のポリペプチドは、組換えポリペプチドの産生および回収、ならびにポリペプチドの化学合成を含めた様々な形で生成することができる。一実施形態では、ポリペプチドを発現することができる細胞をポリペプチドの産生に有効な条件下で培養し、そのポリペプチドを回収することにより、本発明の単離ポリペプチドを生成する。培養するのに好ましい細胞は、本発明の組換え細胞である。有効な培養条件には、それだけに限らないが、有効な培地、バイオリアクター、ポリペプチド産生を可能にする温度、pHおよび酸素条件がある。有効な培地とは、細胞を培養して本発明のポリペプチドを産生する任意の培地を指す。典型的には、そのような培地は、同化できる炭素、窒素およびリン酸源、ならびに適当な塩、ミネラル、金属およびビタミンなどの他の栄養素を有する水性培地を含む。従来の発酵バイオリアクター、振盪フラスコ、試験管、マイクロタイター皿、およびペトリ皿の中で本発明の細胞を培養することができる。組換え細胞に適した温度、pHおよび酸素含量で培養を実施することができる。そのような培養条件は、当業者の専門知識の範囲内にある。
【0165】
アクセサリー分子
アクセサリー分子は、G共役型タンパク質受容体のシグナル伝達に関与するタンパク質である。例として、Gタンパク質およびアレスチンがある。任意の標識アクセサリー分子は、Gタンパク質共役受容体(および/または本明細書で定義されるポリペプチド)と直接または間接的に結合し、化合物とGタンパク質共役受容体が結合した後、2つの標識の相対的位置が空間的に変化して、BRETの検出を可能にする限り、本発明の方法で使用することができる。
【0166】
当業者は、当技術分野で標準的な技術を使用して、特定のGタンパク質共役受容体と結合するアクセサリー分子を容易に同定することができる。
【0167】
Gタンパク質共役受容体について上記で記載した突然変異体(変異体)の詳細は、アクセサリー分子にも当てはまる。
【0168】
生物発光タンパク質またはアクセプター分子は、アクセサリー分子が依然として受容体と直接または間接的に結合することができ、対象とする化合物と結合した後、BRETの変化を検出することができる限り、アクセサリー分子中のどこにでも位置することができる。一実施形態では、生物発光タンパク質またはアクセプター分子は、アクセサリー分子のC末端またはN末端に位置する。GPCRとの複合体の形成に少なくとも1つの他のアクセサリー分子を必要とする場合、アクセサリー分子は受容体と間接的に結合し、例えば、第1のアクセサリー分子は受容体と結合し、第2のアクセサリー分子は、第1のアクセサリー分子と直接結合することによって受容体と間接的に結合する。
【0169】
Gタンパク質
Gタンパク質は、細胞内シグナル伝達のセカンドメッセンジャーカスケードに関与するタンパク質のファミリーである。Gタンパク質は、不活性なグアノシン二リン酸(GDP)結合状態と活性なグアノシン三リン酸(GTP)結合状態とを入れ替える「分子スイッチ」として機能する。最終的に、Gタンパク質はシグナル伝達ネットワークのカスケードを開始することにより下流の細胞過程を制御するHofmannら、2009;OldhamおよびHamm、2008)。
【0170】
2つの異なるGタンパク質のファミリー:Gタンパク質共役受容体によって活性化され、アルファ(α)、ベータ(β)およびガンマ(γ)サブユニットで構成される、時折「大型」Gタンパク質と称されるヘテロ三量体Gタンパク質;ならびに低分子量GTPアーゼのRasスーパーファミリーに属する「小型」Gタンパク質(20〜25kDa)がある。これらのタンパク質は、ヘテロ三量体Gタンパク質で認められるアルファ(α)サブユニットと相同であり、やはりGTPおよびGDPと結合し、シグナル伝達に関与する。形質膜と結合するために、多数のGタンパク質が共有結合により脂質で修飾され、例えば、ヘテロ三量体Gタンパク質サブユニットはミリストイル化、パルミトイル化、またはプレニル化されていることがあり、一方で小型Gタンパク質はプレニル化されていることがある。
【0171】
熟練した受け手なら気づくであろうが、既知のGタンパク質が多数ある。線虫においていくつかのヘテロ三量体Gタンパク質が同定されており、例えばC.elegansは21個のGα、2個のGβおよび2個のGγ遺伝子を有する(Jansenら、1999;Cuppenら、2003)。配列類似性に基づいて、哺乳動物Gαサブユニットは、4つのファミリー:Gs、Gi/o、GqおよびG12に分けられている(Nevesら、2002)。C.elegansは各哺乳動物ファミリーの1つのオルソログを発現する:GSA−1(Gs)、GOA−1(Gi/o)、EGL−30(Gq)およびGPA−12(G12)。残りのC.elegansαサブユニット(GPA−1〜11、GPA−13〜17およびODR−3)は、分類を可能にするほど十分な相同性を有さない。保存されるGαサブユニットは、GPA−12を除いて幅広く発現されるが、新たなGα遺伝子のうち14個が化学感覚ニューロンのサブセット中で発現される。
【0172】
GβサブユニットGPB−1ならびにGγサブユニットGPC−2は、古典的なGタンパク質ヘテロ三量体中でαサブユニットと一緒に機能すると思われる。残りのGβサブユニットGPB−2は特定のRGSタンパク質の機能を制御すると考えられるが、残りのGγサブユニットGPC−1は、化学感覚において制限された役割を有する。したがって、C.elegansにおけるほとんどのGタンパク質経路の機能的差異はαサブユニットにある。
【0173】
アレスチン
アレスチンは、Gタンパク質シグナル伝達の終結に重要なタンパク質の1つである。活性化したGPCRはGタンパク質共役受容体キナーゼ(GRK)によって特異的にリン酸化され、次いでアレスチンと結合して受容体/Gタンパク質相互作用が妨げられ、その結果後のシグナル伝達が抑えられる。アレスチンは、アゴニストで活性化されているものを含むGPCRと結合し、GRKによってリン酸化されていないものよりされているものと堅く結合する。
【0174】
GPCRの分類をもたらす、細胞内のアレスチン輸送の2つの異なるパターンが以下のように描写されている:アレスチンが細胞表面で受容体と相互作用するが、エンドサイトーシスで小胞中に取り込まれず、したがって受容体との一過性の相互作用を示すクラスA、ならびにR−アレスチンおよび受容体が細胞膜からエンドサイトーシス小胞へと一緒に輸送されるクラスB。これら2つのクラスの受容体は、異なるアレスチンアイソフォームに対する親和性に関しても異なる。さらに、クラスA受容体は、R−アレスチン2と優先的に結合するが、クラスB受容体は、等しい親和性でβ−アレスチン1およびH−アレスチン2と結合する。
【0175】
R−アレスチンの結合により受容体がその同族Gタンパク質から脱共役し、下流のセカンドメッセンジャー分子を介したGPCRシグナル伝達の抑制または脱感作を引き起こす。
【0176】
Gタンパク質シグナルを終結する一方で、アレスチンの結合は、GPCRからの新たなシグナル伝達を開始することができる。例えば、R−アレスチンは、Srcなどの非受容体チロシンキナーゼに、内在化している受容体とのシグナル伝達複合体を形成させるアダプターとして働く。H−アレスチンは、ASK−MKK4−JNK3やRAF−MEK−ERK1/2などのMAPKモジュールの、GPCRによって制御された足場として機能する。さらに、アレスチンはクラスリン、AP2複合体のβ−アダプチンサブユニット2やArf−6などのエンドサイトーシス機構のタンパク質と相互作用し、したがってクラスリン被覆小胞を介して受容体の内在化を促進する。
【0177】
線虫アレスチンの例は、C.elegans由来のARR−1であり、これはHSNニューロンおよび様々な化学感覚ニューロンを含めた神経系で主として発現し、可溶性および揮発性匂い物質の検出に関与する。20を超える推定上のアレスチンが線虫C.elegansにおいて現在同定されている。そのようなアレスチンは神経系全体を通じて発現し、脊椎動物非視覚性アレスチンと同様に受容体内在化を支援し、視覚/感覚性サブタイプと同様に嗅覚および視覚にも関与する。
【0178】
組換えベクター
本発明の一実施形態は、宿主細胞中にポリヌクレオチド分子を送達することができる任意のベクター中に挿入された、本明細書に記載のポリペプチドをコードする少なくとも1つの単離/外因性ポリヌクレオチドを含む組換えベクターを含む。そのようなベクターは、異種のポリヌクレオチド配列を含有し、この配列は本発明のポリヌクレオチド分子の隣に天然には認められないポリヌクレオチド配列であり、好ましくは、その(複数の)ポリヌクレオチド分子が由来する種以外の種に由来する。ベクターはRNAでもまたはDNAでもよく、原核生物性でもまたは真核生物性でもよく、典型的には(米国特許第5,792,294号に記載のような)トランスポゾン、ウイルスまたはプラスミドである。
【0179】
組換えベクターの1つの型は、発現ベクターと作動可能に連結する(複数の)ポリヌクレオチドを含む。作動可能に連結するという語句は、分子が宿主細胞に形質転換されたときに発現することができるような形で発現ベクター中にポリヌクレオチド分子を挿入することを指す。本明細書において、発現ベクターは、宿主細胞を形質転換し、特定のポリヌクレオチド分子の発現を行うことができるDNAまたはRNAベクターである。好ましくは、発現ベクターは、宿主細胞内で複製することもできる。発現ベクターは原核生物性でもまたは真核生物性でもよく、典型的にはウイルスまたはプラスミドである。発現ベクターは、細菌、菌類、内部寄生生物、節足動物、動物、および植物細胞を含めた組換え細胞中で機能する(すなわち、遺伝子発現を誘導する)任意のベクターを含む。本発明のベクターを使用して無細胞発現系でポリペプチドを産生することもでき、そのような系は当技術分野で周知である。
【0180】
本明細書において「作動可能に連結する」とは、2つ以上の核酸(例えばDNA)セグメント間の機能的関係を指す。典型的には、それは、転写される配列に対する転写制御エレメントの機能的関係を指す。例えば、プロモーターは、適当な宿主細胞および/または無細胞発現系中でコード配列の転写を刺激または調節する場合、本明細書で定義されるポリヌクレオチドなどのコード配列と作動可能に連結する。一般に、転写される配列と作動可能に連結しているプロモーター転写制御エレメントは、転写される配列と物理的に連続し、すなわちそれはシス作用性である。しかし、エンハンサーなど一部の転写制御エレメントは、転写を亢進するコード配列と物理的に連続し、または近接して位置する必要はない。
【0181】
具体的には、本発明の発現ベクターは、転写調節配列、翻訳調節配列、複製起点や、組換え細胞に適合し本発明のポリヌクレオチド分子の発現を調節する他の制御配列などの制御配列を含有する。具体的には、本発明の組換え分子は、転写調節配列を含む。転写調節配列は、転写の開始、延長、および終結を調節する配列である。特に重要な転写調節配列は、プロモーター、エンハンサー、オペレーターやレプレッサー配列など、転写開始を調節するものである。適切な転写調節配列は、本発明の組換え細胞の少なくとも1つの中で機能することができる任意の転写調節配列を含む。様々なそのような転写調節配列が当業者に知られている。好ましい転写調節配列には、それだけに限らないが、tac、lac、trp、trc、oxy−pro、omp/lpp、rrnB、バクテリオファージラムダ、バクテリオファージT7、T7lac、バクテリオファージT3、バクテリオファージSP6、バクテリオファージSP01、メタロチオネイン、α接合因子、Pichiaアルコールオキシダーゼ、アルファウイルスサブゲノムプロモーター(シンドビズウイルスサブゲノムプロモーターなど)、抗生物質耐性遺伝子、バキュロウイルス、Heliothis zea昆虫ウイルス、ワクシニアウイルス、ヘルペスウイルス、アライグマポックスウイルス、他のポックスウイルス、アデノウイルス、サイトメガロウイルス(中間初期プロモーターなど)、シミアンウイルス40、レトロウイルス、アクチン、レトロウイルス長末端反復配列、ラウス肉腫ウイルス、熱ショック、リン酸および硝酸転写調節配列、ならびに原核または真核細胞中で遺伝子発現を調節することができる他の配列など、細菌、酵母、節足動物、線虫、植物または動物細胞中で機能するものがある。
【0182】
宿主細胞
本発明の別の実施形態は、本明細書に記載の1つもしくは複数の組換え分子で形質転換された宿主細胞またはその子孫細胞を含む。細胞へのポリヌクレオチド分子の形質転換は、ポリヌクレオチド分子を細胞中に挿入できる任意の方法で行うことができる。形質転換の技術には、それだけに限らないが、トランスフェクション、エレクトロポレーション、マイクロインジェクション、リポフェクション、吸着、およびプロトプラスト融合がある。組換え細胞は、単細胞のままでもよく、または成長して組織、臓器もしくは多細胞生物になってもよい。本発明の形質転換されたポリヌクレオチド分子は、染色体外にあるままでもよく、または発現する能力が保持される形で、形質転換される(すなわち組換え)細胞の染色体内の1つまたは複数の部位に組み込むこともできる。
【0183】
形質転換するのに適した宿主細胞は、本発明のポリヌクレオチドで形質転換することができる任意の細胞を含む。本発明の宿主細胞は、本明細書に記載のポリペプチドを内因的に(すなわち天然に)産生することもでき、または本明細書に記載の少なくとも1つのポリヌクレオチド分子で形質転換した後にそのようなポリペプチドを産生することもできる。本発明の宿主細胞は、本明細書で定義される少なくとも1つのタンパク質を産生することができる任意の細胞でよく、細菌、菌類(酵母を含む)、寄生生物、線虫、節足動物、動物および植物細胞を含む。宿主細胞の例として、Salmonella、Escherichia、Bacillus、Listeria、Saccharomyces、Spodoptera、Mycobacteria、Trichoplusia、BHK(仔ハムスター腎)細胞、MDCK細胞、CRFK細胞、CV−1細胞、COS(例えばCOS−7)細胞、およびVero細胞がある。宿主細胞のさらなる例は、E.coli K−12誘導体を含むE.coli;Salmonella typhi;弱毒化株を含むSalmonella typhimurium;Spodoptera frugiperda;Trichoplusia ni;および非腫瘍原性マウス筋原細胞のG8細胞(例えば、ATCC CRL 1246)である。特に好ましい宿主細胞は、それだけに限らないが、Saccharomyces種、特にSaccharomyces cerevisiae、Candida種、Hansenula種やPichia種などの酵母細胞である。
【0184】
組換えDNA技術を使用して、例えば、宿主細胞内のポリヌクレオチド分子のコピー数、そのポリヌクレオチド分子が転写される効率、得られた転写物が翻訳される効率、および翻訳後修飾の効率を操作することにより、形質転換されたポリヌクレオチド分子の発現を向上させることができる。本発明のポリヌクレオチド分子の発現の増大に有用な組換え技術には、それだけに限らないが、高コピー数のプラスミドへのポリヌクレオチド分子の作動的連結、1つまたは複数の宿主細胞染色体へのポリヌクレオチド分子の組み込み、プラスミドへのベクター安定性配列の付加、転写調節シグナル(例えば、プロモーター、オペレーター、エンハンサー)の置換または改変、翻訳調節シグナル(例えば、リボソーム結合部位、シャイン−ダルガルノ配列)の置換または改変、宿主細胞のコドン使用に対応するような本発明のポリヌクレオチド分子の改変、および転写物を不安定にする配列の欠失がある。
【0185】
無細胞組成物
細胞から無細胞組成物を調製する方法は当技術分野で周知であり、それには、凍結融解サイクルの反復、粉砕、超音波処理デバイスにおける超音波での細胞の処理、ホモジナイズ処理、フレンチプレスの使用、界面活性剤および/または酵素の添加、ガラスビーズ溶解、分画遠心、様々な勾配媒体を使用するいくつかの密度勾配の手順がある。これらの技術は当業者によく知られており、例えば「Current Protocols in Protein Science」;John E.Caligan;Ben M.Dunn;Hidde L.Ploegh;David W.Speicher;Paul T.Wingfield;Wiley and Sons)で詳細に論じられている。
【0186】
細胞膜抽出物の単離または調製では、これらの方法の組合せが通常使用される(RiederおよびEmr、2001)。一般に、機械的手段により、または界面活性剤を使用して細胞を溶解し、例えば、米国特許第7,393,934号に概略が述べられているように、分画遠心により膜分画を単離する。Moore(1994)に概略が述べられている方法が一般的に使用される。そのような方法の例は、米国特許第7,320,875号で見つけることができる。他の方法、例えば水性二相アフィニティー分配も膜分画の調製に利用可能である(PerssonおよびJergil、1994)。
【0187】
細胞膜分画の精製に利用可能な多数の市販のキット、例えば、QiagenのQproteome形質膜タンパク質キット(Qproteome Plasma Membrane Protein Kit)もある。細胞を低張性緩衝液中でインキュベートし、それを膨張させる。弱い界面活性剤を添加し、得られた細胞懸濁液を機械的破壊によってホモジナイズする。インタクトな細胞、細胞片、核および主要な細胞内小器官を遠心分離によって除去する。得られた上清は、細胞質タンパク質およびミクロソーム、ゴルジ小胞、ならびに形質膜を含有する。細胞膜上の分子に特異的なリガンドを上清に添加する。リガンドは細胞膜小胞と結合し、リガンドと結合する磁性ビーズを使用してリガンド−小胞複合体を沈降させる。洗浄後、天然の条件下で形質膜小胞を溶出させ、リガンドはビーズと結合したままである。
【0188】
酵母細胞では、細胞膜の調製に一般的に使用される方法は、細胞をスフェロプラストにすることである。これは高収量をもたらし、形質膜の大規模単離に申し分なく適している。一般に、そのような方法では、スフェロプラストの負に帯電した表面を高密度のカチオン性シリカビーズで被覆し、その膜を細胞の他のどんな膜の細胞内小器官よりも高密度にする必要がある。洗浄手順で、過剰なカチオン性ビーズを除去し、その後ポリアクリル酸を添加して、ビーズ上の遊離カチオン性基を遮断する。その後、EGTA含有溶解緩衝液中で、手作業でホモジナイズすることにより、被覆したスフェロプラストを溶解して膜構成成分の凝集を防止する。スフェロプラスト溶解物の遠心分離により、重い形質膜−マイクロビーズ集合物をペレットにし、細胞内にある膜の細胞内小器官を上清中に残す。
【0189】
本発明に使用することができるGタンパク質共役受容体関連ポリペプチドを含む無細胞組成物の生成のさらなる例は、Kaiserら(2008)に記載されている。
【0190】
特定の実施形態では、GPCRを含む本発明のポリペプチドは、リポソーム調製物の脂質二重層中に埋め込まれる。本明細書において、「リポソーム」という用語は、両親媒性のリン脂質、例えばホスファチジルエタノールアミンおよびコレステロールの二重層を含む閉じた小胞を指す。リポソームは、(ホスファチジルエタノールアミンのような)脂質鎖が混合した天然由来のリン脂質、またはDOPE(ジオレオイルホスファチジルエタノールアミン)のような純粋な表面活性物質成分から構成され得る。
【0191】
本明細書に記載のポリペプチドを含有するリポソームは、水中で、例えば超音波処理により、タンパク質を発現する細胞のリン脂質膜を破壊することによって作製することができる。リン脂質は再び集合して、水溶液の中心部を含有するリポソームの球体になる。剪断速度が低いと多数の層を有する多重膜リポソームが生じる。高速剪断超音波処理を続けると、より小さい単層のリポソームが形成され、これは本発明の適用により適している。配向および到達性が最適である最適なリポソームサイズにする実験は、当業者の日常的な能力の範囲内である。
【0192】
本明細書に記載のポリペプチドを生成し、界面活性剤中で解離させ、その後脂質の存在下で膜を再構成し、界面活性剤を除去する膜再構成技術によってそのポリペプチドをリポソーム中に取り込むこともできる。結合相手(化合物)との、リポソームに入っているポリペプチドの配向および到達性が確実に最適になるように、リポソーム組成物を調整する。最適なリポソーム組成物にするそのような調整は、当技術分野で日常的である。
【0193】
BRETのアッセイ
好ましい実施形態では、生物発光タンパク質とアクセプター分子の間で起こるエネルギー移動は、特定の波長を選択する光学フィルターを使用して測定された放出(1つはアクセプター分子の放出、もう1つは生物発光タンパク質の放出)から計算された比率として示される(方程式1を参照)。
Ea/Ed=BRET比率 (1)
上式で、Eaはアクセプター分子の放出強度として定義され(放出光はアクセプターの放出に適合した特異的なフィルターを使用して選択される)、Edは生物発光タンパク質の放出強度として定義される(放出光は生物発光タンパク質の放出に適合した特異的なフィルターを使用して選択される)。
【0194】
光学フィルターは、BRETに適した波長識別を可能にする任意の型のフィルターでよいことが、当業者には容易に理解されるはずである。例えば、本発明に従って使用される光学フィルターは、干渉フィルター、ロングパスフィルター、ショートパスフィルターなどであり得る。フィルターを通過する波長の強度(通常はカウント毎秒(CPS)または相対発光単位(RLU))は、光電子増倍管(PMT)またはCCDカメラを使用して定量することができる。その後、定量したシグナルを使用してBRET比率を計算し、エネルギー移動効率を表す。アクセプター放出の強度が増大するにつれて、BRET比率は増大する。
【0195】
一般に、ドナー放出強度に対するアクセプター放出強度の比率が決定され(方程式1を参照)、これは、エネルギー移動効率を反映する任意の単位で表された数である。この比率は、エネルギー移動効率の増大につれて増大する(Xuら、1999を参照)。
【0196】
エネルギー移動効率は、アクセプター放出強度に対するドナー放出強度の逆比率を使用して表すこともできる(方程式2を参照)。この場合、エネルギー移動効率が増大するにつれて比率は低下する。この計算を行う前に、バックグラウンドの光および基質の自己発光の存在について放出強度を補正する。この補正は、基質を含有するが、生物発光タンパク質、アクセプター分子または本発明のポリペプチドを含有しない対照試料から適当な波長で測定された放出強度を減算することにより一般に行われる。
Ed/Ea=BRET比率 (2)
上式で、EaおよびEdは上記で定義した通りである。
【0197】
生物発光タンパク質およびアクセプター分子の放出の光強度は、分光蛍光光度計など単色光分光器ベースの機器、電荷結合素子(CCD)カメラまたはダイオードアレイ検出器を使用して定量することもできる。分光蛍光光度計を使用して、基質の添加後に生物発光タンパク質とアクセプター分子の放出ピークがどちらも検出されるように放出スキャンを行う。ピーク下の面積は相対的な光強度を表し、上記で概略を述べた通り、これを比率の計算に使用する。同じ試料から生物発光タンパク質およびアクセプター分子の光を測定することができる任意の機器を使用して、本発明のBRET系をモニタリングすることができる。
【0198】
代替的な実施形態では、単独のアクセプター分子の放出がBRETの有効な検出および/または定量に適している。この場合、エネルギー移動効率は、アクセプター放出強度のみを使用して表される。エネルギー移動を測定するために、比率の計算を行わずにアクセプター放出強度を使用できることが、当業者には容易に明らかとなるであろう。これは、理想的には、アクセプター分子が、生物発光タンパク質から移動した光を吸収する場合にのみ光を放出することに起因する。この場合、光フィルターが1つだけ必要である。
【0199】
関連する実施形態では、単独の生物発光タンパク質の放出がBRETの有効な検出および/または定量に適している。この場合、エネルギー移動効率は、生物発光タンパク質放出強度のみを使用して計算される。エネルギー移動を測定するために、比率の計算を行わずにドナー放出強度を使用できることが、当業者には容易に明らかとなるであろう。これは、アクセプター分子が、生物発光タンパク質から移動した光を吸収するので、生物発光タンパク質から検出可能な放出が対応して低下することに起因する。この場合、光フィルターが1つだけ必要である。
【0200】
代替的な実施形態では、エネルギー移動効率は、測定に1つの光学フィルターしか必要としない比率的測定を使用して表される。この場合、適当な光学フィルターを使用してドナーまたはアクセプターの光強度を決定し、フィルターを使用せずに試料の別の測定を行う(オープンスペクトルの強度)。この後者の測定では、(全ての波長について)光の総放出量を定量する。次いで、方程式3または4を使用して比率の計算を行う。方程式3では、アクセプターの光学フィルターのみを必要とする。方程式4では、ドナーの光学フィルターのみを必要とする。
Ea/Eo−Ea=BRET比率または=Eo−Ea/Ea (3)
Eo−Ed/Ed=BRET比率または=Ed/Eo−Ed (4)
上式で、EaおよびEdは上記で定義した通りであり、Eoは、全ての波長を組み合わせた(オープンスペクトル)放出強度として定義される。
【0201】
方程式1〜4からさらなる方程式を導き出せることが、当業者には容易に明らかであるはずである。例えば、そのように導き出された式の1つでは、生物発光タンパク質および/またはアクセプター分子の放出波長で存在するバックグラウンドの光について補正する。
【0202】
BRETアッセイを行う際に、BRETCountを使用して、各ウェルから光放出を決定することができる。BRETCount機器は改変型のTopCountであり、TopCountは、Packard Instrument(Meriden、CT)から販売されているマイクロタイタープレートシンチレーションおよび発光計数器である。バックグラウンドノイズを除去するために同時に2つの光電子増倍管(PMT)を利用する古典的な計数器と異なり、TopCountは、ノイズ低減に単一のPMT技術および時間分解パルス計数を使用して、標準の不透明なマイクロタイタープレートでの計数を可能にする。不透明なマイクロタイタープレートの使用は、光学的なクロストークをごくわずかなレベルまで低減することができる。TopCountには、1、2、6または12個の試料の同時読み取りを可能にするそれぞれ1、2、6および12個の検出器(PMT)を含めて、様々なフォーマットがある。BRETCountの他に、他の市販の機器:Victor2(Wallac、Finland(Perkin Elmer Life Sciences))およびFusion(Packard Instrument、Meriden)でBRETを行うことが可能である。少なくともアクセプター分子の放出、および好ましくは2つの波長(アクセプター分子および生物発光タンパク質)またはそれ以上を検出できる読取り器を使用して、BRETを行うことができる。
【0203】
本発明の一実施形態では、マイクロ流体工学デバイスを使用してBRETを検出する。好都合なことに、マイクロ流体工学デバイスは、マイクロ流体工学デバイスの試料貯蔵所に移すのに、一般に約50μL以下の試料のアリコートしか必要としない。これは、手作業でまたはシリンジ、キャピラリーなどを介した圧気注入によって行われる。
【0204】
マイクロ流体工学を使用した自動化発光バイオチップデバイスを使用して、全ての必要なBRET反応ステップを行うことができる。複数の手作業での操作ステップを回避し、実験を行う際の人間の時間および労力を低減するため、マイクロ流体バイオチッププラットホームでBRET反応を自動化することは望ましい。マイクロ流体工学デバイスは、パターン化されたマイクロチャネルを有する内蔵型の使い捨てバイオチップ、ならびに複数の試料、試薬、および基質を貯蔵するための貯蔵手段を有する区画を含有し得る。区画からマイクロチャネルを介して反応部位へと試料、または試薬の少なくとも1つ、次いで発光基質を連続して移すステップを自動化することができる。次いで発光基質がドナー分子と反応して発光し、それが光学検出器によって検出される。発光を検出するマイクロ流体工学デバイスの例が米国特許第6,949,377号に記載されている。
【0205】
使用
本発明を使用して、多種多様な化合物、特に匂い物質を検出することができる。典型的には、匂い物質は、少なくとも1つの生物の化学感覚的嗅覚受容体によって検出することができる揮発性の有機もしくは無機化合物または無機ガスとなる。これらは、アミンおよび/またはスルフヒドリル含有化合物、カルボン酸、アルコール、アルデヒド、アルカン、アルケン、芳香族化合物、エステル、テルペンまたはテルペン誘導体、エーテル、CO2など、ならびにこれらの性質の組合せを有する化合物を含み得る。
【0206】
匂い物質は、ヒトにとって価値のあるまたは興味深いいくつかの生物学的または化学的状態を示し得る。そのように示される状態には、以下が含まれ得る:
・食料および飲料、医薬品または関連材料の感覚的訴え、品質または安全性。
・ヒトまたは動物の健康、栄養または運動状態。
・病原体を含めた危険な物質の存在の有無。
・産業プロセスの進行または状態。
・環境汚染または状態。
・香料、芳香剤または他の化粧品の感覚的訴え、品質または安全性。
【0207】
特に好ましい実施形態では、化合物は、本明細書に記載のポリペプチドの受容体部分とのみ結合する。
【0208】
特に好ましい実施形態では、嗅覚受容体を含む本明細書で定義されるポリペプチドをバイオセンサーで使用して、対象とする(複数の)化合物を検出する。バイオセンサーとは、生物学的な構成成分を物理化学的な検出器構成成分と組み合わせた、分析対象を検出するためのデバイスである。それは、典型的には3つの部分からなり、第1はGタンパク質共役受容体を含む本明細書で定義される少なくとも1つのポリペプチドである。第2に、変換器または検出器エレメントを必要とし、これは物理化学的に(例えば、光学的、電気化学的に)働き、化合物とポリペプチドの相互作用から生じるシグナルを、容易に測定および定量できる別のシグナルに変換する(すなわち、変換器)。第3に、関連の電子またはシグナル処理装置を必要とし、ユーザーフレンドリーに相互作用の結果を提示する。BRETを含むバイオセンサーの例は、Charestら(2005)に記載されている。
【0209】
別の実施形態では、スクリーニング方法を薬物の発見および/または開発に使用する。より具体的には、受容体は、潜在的な治療薬の標的である。したがって、この実施形態では、受容体が、それだけに限らないが、アドレナリン作動性受容体、セロトニン受容体、ドーパミン受容体、代謝型/グルタミン酸受容体、GABA受容体、鋤鼻受容体(vomeronasal receptor)、味覚受容体やセクレチン様受容体などの臨床的に重要な分子であることが好ましい。
【0210】
Gタンパク質共役受容体と相互作用することが可能な分子を同定し、特徴付け、スクリーニングする方法も本発明の範囲内にあり、96ウェル、384ウェル、1536ウェル(および他)の市販のプレートを使用して実施される、当技術分野で知られているいわゆるハイスループットスクリーニング方法および類似した手法(Spencer、1998;Oldenburg、1998;Milligan、1999)を含む。本発明に従って使用されるさらなる方法は、それだけに限らないが、(とりわけPope、1999に記載の)ハイスループットスクリーニングにおける均等蛍光読取り(homogenous fluorescence readout)を含む。
【0211】
当業者なら気づくであろうが、本発明を多重化することもできる。この系では、異なる化合物と結合する、異なるGタンパク質共役受容体を含む2つ以上のポリペプチドを提供する。異なる波長で放出して異なる標的化合物の検出および定量を可能にするように、異なる各Gタンパク質共役受容体を異なる生物発光タンパク質および/またはアクセプター分子で標識する。
【実施例】
【0212】
[実施例1]
二重標識BRET ODR−10(OGPR)構築物
材料および方法
二重標識BRET OR構築物の設計および構築
3番目の細胞内ループ(IC3)およびC末端に挿入されたBRET2構成成分を用いてBRET2受容体構築物を作製した。(1)IC3の中央にあるGFP2、およびC末端にあるRLuc(h);(2)IC3の中央にあるRLuc(h)およびC末端にあるGFP2の付いたORを用いて2つのBRET構築物を作製した。The Biology WorkbenchにあるTMAPという、膜貫通セグメントを予測するウェブベースのツール(http://seqtool.sdsc.edu)を使用して、3番目の細胞内ループ(IC3)の位置を決定した。ODR−10では、IC3の中央はアミノ酸240〜241である。OGORと称されるこの構築物のアミノ酸配列は、配列番号15として示される(コードするヌクレオチド配列は配列番号16として示される)。
【0213】
ODR−10は、superscriptII(Invitrogen)を使用して、標準的な技術を用いて調製したC.elegans cDNAから増幅した。PCR条件は、94℃で2分を行い、94℃で15秒、59℃で30秒、68℃で70秒を30サイクル、最後の伸長ステップが68℃で5分であった。標準のPfx50 PCR成分を、プライマーODR−10Xba1F 5’−AGTCTAGAATGTCGGGAGAATTGTGGATTA−3’(配列番号21)およびODR−10−attb1−R 5’−GGGGACAAGTTTGTACAAAAAAGCAGGCTTCATCACGTCGGAACTTGAG−3’(配列番号22)とともに使用した。増幅したcDNAをpGEM−T(Promega)にクローニングし、配列を決定した。
【0214】
BRET構成成分のGFP2およびウミシイタケ(Renilla)ルシフェラーゼRLuc(h)は、市販のプラスミドpGFP2−MCS−Rluc(h)(PerkinElmer;Lot#6310051−6D1)から入手した。
【0215】
構築物の個々の断片をPCR増幅することによってBRET構築物を構築した(プライマーおよびアニーリング温度については表4を参照)。Pfx50(商標)標準成分(Invitrogen)を用いてPCRを行い、条件は、94℃で2分を行い、94℃で15秒、59℃で30秒、68℃で70秒を30サイクル、最後の伸長ステップが68℃で5分であった。構築物の別々の構成成分を、重複するプライマーを使用して増幅した。
【0216】
【表4】
【0217】
次いで、構成成分を精製し、対の反応に入れて94℃で変性し、52℃で(重複するプライマー部位で)アニールし、68℃で5分伸長して必要な対にした。次いでこれを、その対のほとんど5’および3’のプライマーを使用するPCRの鋳型として使用した。完全長の構築物が作製されるまでこれを反復した。次いでこれらをpGEM−Tにクローニングし、配列を決定した。ApaLIを使用してエラーのないクローンをpDONR201(Invitrogen)にサブクローニングした。次いでGateway(登録商標)技術を使用してこれらをpYES−DEST52(Invitrogen)にサブクローニングして(attB部位は元の5’および3’プライマーに含まれていた)、その後S.cerevisiaeで発現させた。
【0218】
OGOR突然変異体(H110Y)の構築
二本鎖鋳型としてpDONR201ベクター中のOGOR(上記参照)を用いて部位特異的突然変異導入を実施した。標準的なプロトコールに従ってStratagene Quickchange部位特異的突然変異導入キットを使用して、以下のプライマー:5’−CACCAGTTTTGTTGTCTCAGGAGTTTATTTTGTTTATCGATATTTTGCAACTT−3’(WT)(配列番号43)および5’−AAGTTGCAAAATATCGATAAACAAAATAAACTCCTGAGACAACAAAACTGGTG−3’(H110Yアンチセンス)(配列番号44)を使用してヒスチジン110からチロシンへの(H110Y)突然変異を行った。
【0219】
Pfx50(商標)標準成分(Invitrogen)を用いてPCRを行い、条件は、94℃で2分を行い、94℃で30秒、55℃で1分、68℃で6.8分を16サイクルであった。次いで、配列決定の後、Gateway(登録商標)技術を使用してOGOR突然変異体をpYES−DEST52(Invitrogen)にサブクローニングして(5’および3’のattB部位は元のOGOR配列に含まれていた)、その後S.cerevisiaeで発現させた。
【0220】
溶液調製
20%(w/v)グルコース − グルコース20gを脱イオン水100mLに添加し、溶解するまでホットプレート(50℃)上で加熱し、溶液をフィルター精製(0.2μmフィルター)する。
【0221】
20%(w/v)ラフィノース − グルコース20gを脱イオン水100mLに添加し、溶解するまでホットプレート(50℃)上で加熱し、溶液をフィルター精製(0.2μmフィルター)する。
【0222】
20%(w/v)ガラクトース − ガラクトース20gを脱イオン水100mLに添加し、溶解するまでホットプレート(50℃)上で加熱し、溶液をフィルター精製(0.2μmフィルター)する。
【0223】
YPD培地 − 細菌用ペプトン20g、酵母エキス10gを脱イオン水1Lに添加する。15分間オートクレーブをかける。
【0224】
YPDプレート − 細菌用ペプトン20g、酵母エキス10gおよび寒天20gを脱イオン水1Lに添加する。15分間オートクレーブをかける。オートクレーブした後、20%グルコースを20mL添加し、寒天プレートに注ぐ。
【0225】
Yeast Synthetic Drop−Out Media without Uracil(SCMM−U)培地− アミノ酸不含酵母窒素塩基(yeast nitrogen base without amino acids)(製品番号Y 0626、Sigma)6.7gおよびウラシル不含酵母合成離脱培地補充物(yeast synthetic drop-out media supplement without uracil)(製品番号Y 1501)1.92gを脱イオン水1Lに添加する。15分間オートクレーブをかける。
【0226】
SCMM−Uプレート − アミノ酸不含酵母窒素塩基(製品番号Y 0626、Sigma)1.34g、ウラシル不含酵母合成離脱培地補充物(製品番号Y 1501、Sigma)0.384gおよび寒天4gを脱イオン水180mLに添加する。15分間オートクレーブをかける。オートクレーブした後、20%グルコースを20mL添加し、寒天プレートに注ぐ。
【0227】
1×リン酸ナトリウム緩衝液(1×PBS、pH7.4) − リン酸水素二ナトリウム82.33g(0.58M)、リン酸ナトリウム一塩基酸23.45g(0.17M)、および塩化ナトリウム(NaCl)40gを脱イオン水中で溶解することにより10×ストック溶液を調製した。ストック溶液を脱イオン水で1×PBSに希釈した。
【0228】
セレンテラジン400a − 純エタノール10.20mL中でセレンテラジン400a(Clz400a、Biosynth)1mgを溶解して250μMストック溶液を作製する。40または400μLのセレンテラジン400aのアリコートをピペットで微量遠心管に入れ、次いでSpeedVac(登録商標)Plus SC110A(Savant)を用いてドライダウンする。−80℃でアリコートを貯蔵する。試料を100または1000μLの無水エタノール(それぞれ40または400μLのアリコート)で再構成して、50μMの濃度を得る。
【0229】
匂い物質調製物 − 匂い物質のジアセチル、ベンズアルデヒドおよびオクタナールの1%ストック溶液をそれぞれの溶媒である水、エタノールおよびDMSOで作製した。匂い物質を水で段階希釈して、所望の終濃度を得た。匂い物質を添加していない溶媒の同じ希釈物も調製した。
【0230】
プラスミド形質転換プロトコール
酵母形質転換キット(YEAST−1、Sigma)を使用して、酵母形質転換を実施した。YPDプレートをINVSC1(Invitrogen)S.cervesiae株で画線し、28℃で2〜3日間インキュベートした。YPDプレートからInVSC1細胞1ループ分を掻き取って微量遠心管に入れる。形質転換緩衝液(製品コードT 0809)0.5mL中で細胞を再懸濁し、5秒間遠心する。上清を除去し、緩衝液50〜100μLを管に残す。10mg/mLサケ精巣DNA(製品コードD 9156)10μLを管に添加する。研究するpYES−DEST52−ORプラスミドDNA1μgを添加し、10秒間ボルテックスをかける。PLATE緩衝液(製品コード8966)600μLを添加し、ボルテックスをかける。室温で4時間インキュベートする。42℃熱ブロックで試料に15分間熱ショックをかける。微量遠心機で3秒間遠心し、上清を除去する。滅菌水500μL中で細胞を再懸濁する。SCMM−Uプレートに100μLを播く。コロニーが現れるまで28℃で2〜3日間インキュベートする。
【0231】
発現プロトコール
pYES−DEST52−OR構築物を含有する単一コロニーを、2%グルコースを含有するSCMM−U培地15mLに接種する。振盪(200rpm)しながら28℃で一晩増殖させる。
【0232】
一晩培養物のOD600を決定した。誘導培地(SCMM−U、2%ガラクトース、2%ラフィノース)30mL中で0.4のOD600を得るのに必要な一晩培養物の量を取り出し、1500×g、5分間で細胞をペレットにした。誘導培地1mL中で細胞を再懸濁し、それを誘導培地29mLに接種した。振盪(200rpm)しながら15℃で72時間増殖させる。フラスコから培養物を取り出し、1500×gで5分間、4℃で細胞を遠心分離する。上清をデカントし、滅菌水1mL中で細胞を再懸濁する。滅菌した微量遠心管に細胞を移し、試料を10,000×gで1分間遠心分離する。上清を除去し、膜単離を実施する。
【0233】
膜調製
1500×g(4℃)で5分間の遠心分離によって細胞を回収し、水で洗浄し、1000mg/Lグルコース、36mg/Lピルビン酸ナトリウム、カルシウムおよびマグネシウム(Invitrogen)を含有するダルベッコリン酸緩衝食塩水(D−PBS)または1×PBS4mL中で再懸濁した。フレンチプレス(約18000psi)によって細胞を溶解し、9300×g(4℃)で15分間の遠心分離によって可溶性タンパク質分画を単離した。この後、13.15mLポリカーボネート厚肉遠心管に可溶性タンパク質分画を移し、4℃で1時間、40,000rpm(Beckman Coulter L−80超遠心分離機)で超遠心分離をかけた。上清をデカントし、D−PBDまたは1×PBS1mL中で膜ペレットを再懸濁し、可溶化するまで4℃で一晩放置した。
【0234】
スペクトル測定
プレートを読み取るSpectraMax M2分光蛍光光度計(Molecular Devices)を用いて、全てのスペクトルスキャンを記録した。96ウェルプレート(Perkin−Elmer)を使用して、100μLの膜調製物試料のアリコートをスキャンした。蛍光スペクトルスキャンは、励起波長420nmで、455nm放出カットオフフィルターを使用して、450から600nmまで記録した。
【0235】
同時二重放出検出
POLARstar OPTIMAマイクロプレート読取り器(BMG LabTech)を用いて、同時二重放出RET測定を実施した。同時放出測定ではBRET2フィルターセット(410〜80/515〜30)を使用し、各チャネルについてセットのそれぞれのゲインを用いた(3300/4095)。
【0236】
RET分析プロトコール
試料濃度の標準化
BRET2タグ付き受容体試料の相対的濃度を評価するために、スペクトルスキャンから試料の蛍光強度を決定し、OGORの蛍光強度によって試料の量を標準化した。GFP2の強度を濃度に転換するために、精製したGFP2タンパク質を様々な濃度でアッセイして、蛍光強度に対する濃度に関して較正グラフを作成した。コバルトアフィニティークロマトグラフィーを使用して、供給された使用説明書に従って(BD Talon(BD Biosciences、Clontech))GFP2タンパク質を精製した。最良適合の方程式の線を再編成すると(y=271x+233.31)、蛍光強度が濃度に転換される。最終体積100μLで5または10nMの概算濃度をアッセイに使用した。
【0237】
選択性
96ウェルプレート(Perkin−Elmer)でRET分析を実施し、特定の二重タグ付き受容体タンパク質(OGOR)を1μMの特定の各匂い物質とともに45分間インキュベートした。最終体積は100μLであり、10μLの匂い物質、受容体タンパク質試料(「試料濃度の標準化」の節を参照)、D−PBS(または1×PBS)および5μLのClz400a(5μM)を含んでいた。D−PBS(または1×PBS)の試料をアッセイすることによって、バックグラウンドシグナルを記録した。インキュベーション時間の間、96ウェルプレートウェルをTopseal(商標)−A(Packard)で密封し、各試料の領域を個々に密封した。非匂い物質試料(水、ピルビン酸、クエン酸、乳酸)を調製し、最初にアッセイした。この後、ドラフト中で匂い物質溶液を調製し、以下の順序でアッセイした:2−ブタノン、2,3−ペンタンジオン、2,3−ブタンジオールおよびジアセチル。
【0238】
BRET2測定では、45分のインキュベーション時間後にClz400a基質(5μM)を添加し、5秒間に対して0.50sの積分時間(integration time)を使用してシグナルを記録した。Clz400aを添加した後、シグナルを記録する間ウェルを再度密封して、試料の交差汚染を低減した。試料をウェルに添加した順にアッセイを連続して実施した。
【0239】
BRET2比率は、((受容体試料の515nmでの放出)−(D−PBD(または1×PBS)の515nmでの放出)/(受容体試料の410nmでの放出)−(D−PBDの410nmでの放出)(または1×PBS)のように測定された放出比率として計算する。
【0240】
用量反応曲線
GFP2の強度によって、OGORの濃度を10nMに標準化した。本文中に示したように、濃度範囲にわたって匂い物質の濃度を変化させることにより、用量反応曲線を構築した。常に最低の濃度を最初に、最高の濃度を最後にアッセイした。
【0241】
Windows(登録商標) XP用GraphPad Prismを使用した非線形回帰分析により、S字状の用量反応曲線を適合させ、EC50を計算した。
【0242】
結果
リガンド応答および選択性
生成した構築物は、嗅覚受容体の3番目の細胞内ループ内およびC末端に挿入されたBRET2ドナーおよびアクセプターを有する(図1)。ODR−10によってジアセチルが結合すると、ドナーおよびアクセプター構成成分が離れた場合にBRET2比率が増大し、またはこれらの構成成分が互いに近づいた場合にシグナルが増大する。
【0243】
OGORを10nMまたは1μMの以下のリガンド:水、ピルビン酸、クエン酸、乳酸、2−ブタノン、2,3−ペンタンジオン、2,3−ブタンジオールおよびジアセチルとともにインキュベートした後、BRET2シグナルを記録した(図2aおよびb)。BRET2シグナルの最大の変化が、OGORをμM濃度のジアセチルとともにインキュベートした時に起こり、溶媒(水)の応答と比較してシグナルの37.5%の差が生じた(図3a)。これは、無細胞アッセイのフォーマットで二重標識BRET2タグ付き受容体を使用してリガンド結合をモニタリングした最初の例である。
【0244】
nMとμMどちらの濃度のジアセチルをOGORに添加した時でも、BRET2比率の変化は、対照応答(水)に対して有意差があった(P>0.05)(図3aおよびb)。ジアセチル刺激に対するOGORの応答は、nMとμM両方の濃度で試験した他の全てのリガンドと比較して、対照応答に対して有意差があった唯一の応答であった。これらの結果から、ODR−10がジアセチルと選択的に結合することが確認される。ジアセチルは、ODR−10を発現するHEK293細胞においてCa2+上昇応答を生じさせる唯一の揮発性化合物であることが以前に確認されていた(Zhangら、1997)。
【0245】
37.5%という、リガンド結合に対するBRET2応答の最大の変化は、α2A受容体による10μMのノルエピネフリンの結合時(約5%)(Lohseら、2003)およびPTHRによる1μMのPTHの結合時(約20%)(Vilardagaら、2003)にFRETシグナルで観察された変化より実質的に大きい。ジアセチルの添加時のBRET2シグナルの低下から、BRET2構成成分が匂い物質の結合時に離れることが示唆される。これは、分子内変化のモニタリングについて、このBRET2伝達系が、FRET伝達系と比較して優れた感度を示す最初の実証である。
【0246】
用量応答
OGORのBRET2シグナルの変化の幅に対する匂い物質の濃度変化の効果(図4)は、用量依存的応答の最初の実証をもたらす。EC50値は3.55fMジアセチルと計算され、これは0.31千兆分率(ppq)に等しい。ヒト細胞で発現され、カルシウム画像化技術によってモニタリングされたとき、ジアセチルに対するODR−10の見かけの親和性は、2.3μM(約0.2百万分率(ppm))と計算され、EC50値は走化性の結果と一致する(Zhangら、1997)。
【0247】
ここで示した無細胞アッセイは、全細胞アッセイ、および蛍光検出(Liら、2009)や炎イオン化検出を伴うガスクロマトグラフィー技術(GC−FID)(Macciolaら、2008)など既存の化学的検出系と比較して、ジアセチル定量について9桁を超えて感度が高い。
【0248】
陰性対照−OGOR突然変異(H110Y)
ジアセチルに対するOGOR応答の理想的な陰性対照は、OGOR受容体の場合と同じ距離離れてBRET2構成成分を置くが、その受容体はジアセチル自体に応答しない。ODR−10の3番目の膜貫通残基にある110位でヒスチジンをチロシンに交換すると、ジアセチルに対する走化性応答を強く欠損する(Troemelら、1995;Senguptaら、1996)。
【0249】
OGOR構築物にヒスチジン110からチロシンへの(H110Y)突然変異を導入すると、μM濃度のジアセチルに対する応答が、OGORについて32.4%から4.1%に低減した(図5)。ジアセチルに対する応答のこの低下は、溶媒(水)の応答とは有意差がなく(P<0.05)、この突然変異から機能喪失が生じたことが示唆された。このことから、ジアセチルに対するOGORの応答が、ODR−10受容体とジアセチルの相互作用に起因し、BRET2構成成分自体との非特異的な相互作用に起因しないことが推測される。
【0250】
[実施例2]
二重標識FRET ODR−10(OCOY)と二重標識BRET2 ODR−10(OGOR)の比較
GPCR活性化のモニタリングについてBRET2系とFRET系の感度を比較するため、ODR−10受容体にタグを付けるのに使用したBRET2構成成分をFRET構成成分と交換し、その結果3番目の細胞内ループにCFPが、C末端にYFPが挿入された(OCOY)。CFP(mCFP)(配列番号45)およびYFP(mシトリン)(配列番号46)誘導体はどちらも単量体であり、酵母発現についてコドンを最適化した(EUROSCARF)(それぞれ配列番号47および48)。
【0251】
材料および方法
Genscriptにより、KpnIおよびXhoI制限部位に挟まれたOCOY配列を合成した。そのOCOY配列を、pENTR11ベクターのKpnIおよびXhoI部位に挿入した。次いで、Gateway(登録商標)技術を使用してこれらをpYES−DEST52(Invitrogen)に組み換えて、その後S.cerevisiaeで発現させた。OCOYコード配列は配列番号49として示されるが、そのアミノ酸配列は配列番号50として示される。
【0252】
OCOY発現
OGOR発現と同じ発現プロトコールを、InVSC1におけるOCOYの発現に使用した。
【0253】
濃度の標準化
二重タグ付き受容体試料の相対的濃度を評価するために、スペクトルスキャンから試料の蛍光強度を決定し、OCOYの蛍光強度によって試料の量を標準化した。YFPの強度(530nm)は、OCOYについて721.32a.u.に標準化された。50μLのOCOYをピペットで適当なウェルに入れ、1×PBSで体積を100μLにした。
【0254】
結果
リガンド応答
μM濃度のジアセチルに応答して、OCOYのFRET比率(520nmでの強度/480nmでの強度)は、OGORでの32.4%の低減と比較して、7.6%低減した(図6)。OCOYの応答は、溶媒(水)の応答に対して有意差がある(P<0.05)ことが示された。OGORは、ジアセチル結合のモニタリングについて、OCOYより4倍を超えて感度が高いことが示された。
【0255】
BRET2系の、最大のRET効率の50%に対応するRETプローブ分離であるForster距離(R0)が7.5nmであると最近決定され、遺伝的にコードされた任意のRET対について最大のR0値が決定された(Dacresら、2009)。FRET系のR0値は4.8nmであると以前に決定され(Eversら、2006)、BRET2系が、FRET系(2.4〜7.2nm)より大きな距離範囲(3.8〜11.3nm)でプローブを置くことができることが示唆される。OGORで測定された移動効率は、ジアセチル(1μM)の添加時64.3%から47%に変化し、6.8nmから7.6nmへの距離の変化が示唆される。FRET系など、Forster距離が4.8nmの系は、ジアセチルの添加時にこの距離の変化で11.3%から6.0%への移動効率の変化しか示さない。このことから、BRET2系が、検出感度の点から、この距離範囲におけるこの距離の変化の測定により適していることが確認される。
【0256】
OCOY用量反応曲線
ジアセチルに対するOCOYの応答が溶媒(水)の応答に対して有意差があったことを確認した後、用量反応をプロットした。このデータを得るために、様々な濃度のジアセチルを添加してスペクトルスキャンを記録した(図7)。ジアセチルの濃度が1×10−21Mから1×10−10Mに増大すると、YFPの放出強度は低下し、CFPの放出強度は付随して増大した(図7)。OCOYの応答を、標準化されたOGORの反応曲線に対してスケーリングすると(図8)、BRET2検出が、ODR−10によるジアセチル結合のモニタリングについてFRET検出より感度が高いことが確認された。FRET検出系で計算されたEC50対数値は−16.43±0.9741M(平均±SEM、n=5)であったが、これは、BRET2系で計算されたEC50対数値−14.43±0.5468(平均±SEM、n=12)に対して有意差がない(P<0.05)。このことから、ジアセチルに対するODR−10の親和性がどちらの検出系でも統計的に類似していることが実証され、ジアセチルがODR−10受容体と特異的に相互作用し、RET構成成分自体とは相互作用しないことが推測される。
【0257】
[実施例3]
二重タグ付きStr112(TGTR)
材料および方法
Genscriptにより、NcoIおよびNotI制限部位に挟まれたTGTR配列を合成した。そのTGTR配列を、pENTR11ベクターのNcoIおよびNotI部位に挿入した。次いで、Gateway(登録商標)技術を使用してこれらをpYES−DEST52(Invitrogen)に組み換えて、その後S.cerevisiaeで発現させた。
【0258】
TGTR発現
OGOR発現と同じ発現プロトコールを、InVSC1におけるTGTRの発現に使用した。TGTRコード配列は配列番号51として示されるが、そのアミノ酸配列は配列番号52として示される。
【0259】
RET分析プロトコール
OGOR選択性研究で記載したのと同じRET分析プロトコール。10nMのTGTRで、1μMのリガンドのピルビン酸、クエン酸、酢酸エチル、アセトインおよびジアセチルをアッセイした。
【0260】
細菌抽出物アッセイの調製
LB中で一晩、37℃で振盪(200rpm)しながら細菌株OP50を増殖させた。E.coli株OP50は、実験室において線虫成長用食物源として通常使用される(Brenner、1974)。OP50細菌培養物を使用して2つのアッセイを行った。第1のアッセイでは、10μLの細菌培養物または10μLのLBを10nMのTGTRに添加した。第2のアッセイは、1mLの細菌培養物を微量遠心管に入れ、10000×gで1分間遠心分離することを必要とした。新鮮な微量遠心管にデカントで入れた後の10μLの上清、または10μLのLBを10nMのTGTRに添加した。細菌試料を添加した後、OGORのRET分析と同じプロトコールに従った。
【0261】
結果
OP50細菌上清またはLB中の細菌培養物10μLの添加に対するTGTRの応答は、LB単独に対して有意差があった(P≦0.05)(図9)。増殖の間にOP50から放出された複雑な匂い物質混合物に対してStr112受容体を刺激すると、BRET2比率が増大し、このことから、BRET2構成成分が匂い物質の結合時に互いに近づくことが示唆される。
【0262】
いくつかのリガンドを個々に試験し(図10)、ピルビン酸とアセトインでどちらも水に対するTGTRの応答と比較して有意に高いBRET2シグナルが生じた(P≦0.05)。このことから、リガンド結合時にBRET2構成成分が互いに近づくことが確認される。アセトインに応答したBRET2シグナルの20.8%の増大は、ノルエピネフリンに対するFRETタグ付きα2Aアドレナリン作動性受容体の応答(Lohseら、2003)より4倍大きい。これは、Str112線虫受容体による匂い物質結合の最初の実証であり、BRET2タグ付きStr112による匂い物質結合をモニタリングした最初の例でもある。
【0263】
[実施例4]
受容体キメラ
BRETタグの付いたODR−10のN末端(aa1〜32)およびIC3からC末端(aa225〜aa339)がマルチクローニング部位を挟むように、ODR−10伝達カセットを工学的に作製する(図11)。キメラ受容体を酵母で発現させる。ORのリガンド結合領域をシャッフルして「インフレーム」でこのカセットに入れ、これらのキメラ受容体を使用して、BRETによるリガンド結合を評価することができる。
【0264】
[実施例5]
嗅受容体のオリゴマー形成
C末端においてRlucで標識されたOdr10(配列番号17)およびC末端においてGFP2で標識されたOdr10(配列番号19)を酵母で同時発現させて、受容体のオリゴマー形成およびBRETの変動を示した。
【0265】
材料および方法
形質転換体の培養およびタグ付き遺伝子の誘導条件
形質転換された酵母細胞のコロニーを、適当な栄養素(ウラシル(U)、トリプトファン(T)など)を欠く酵母合成離脱培地(SCMM、Sigma)中で増殖させて、2%グルコースを用いて一晩、28℃、190rpmで振盪しながらプラスミド含有細胞について選択した;SCMM−U−Tは、Invsc1に対してpYesDest52−Odr10−RlucおよびpDestpESC−Odr10−GFP2を同時発現させ(Invsc1/OR/OG)、SCMM−Uは、Invsc1に対してそれぞれ単独のpVV214−odr10−Rluc(Invsc1/OR)および単独のpYesDest52 odr10−GFP2(Invsc1/OG)を発現させる。
【0266】
一晩培養後、OD600を決定し、細胞を誘導培地(培養培地のようなグルコースの代わりに2%ガラクトースおよび1%ラフィノースを有する)中で懸濁して0.4の最終的なOD600を得た。この誘導培地を15℃で振盪しながら24時間インキュベートして融合受容体発現を誘導した。次いで培養物をペレットにし、冷やしたリン酸緩衝食塩水(PBS)でそれを2回洗浄した。細胞ペレットは−80℃で貯蔵することができ、BRET2アッセイの直前にPBS緩衝液中で懸濁した。
【0267】
BRET2アッセイ用酵母細胞の定量
最初に、SpectraMax M2分光蛍光光度計(Molecular Devices)を使用して、白色96ウェルマイクロプレート(Perkin Elmer)中で直接Ex420nmでGFP2を励起することによるGFP強度(1ウェル当たり100μl中約3000RLUまたはc.p.s)によってInvsc1/OR/OGを定量した。ルシフェリン天然セレンテラジン(終濃度5μM)を96ウェル白色マイクロプレート中の総体積100μlの細胞に添加し、直ちに発光シグナルを測定する(Polarstar Optima、BMGLABTECH)ことによって、定量したInvsc1/OR/OG細胞のウミシイタケ(Renilla)ルシフェラーゼ活性を決定した。
【0268】
Invsc1/OR/OGと類似したGFP強度を有するように、Invsc1/OGを標準化した。Invsc1/OR/OGと類似したRluc活性を有するように、Invsc1/ORを標準化した。Invsc1/OR/OGと同じ細胞密度を有するように、陰性対照(宿主株)Invsc1を定量した。
【0269】
マイクロプレートBRET2細胞ベースアッセイ
IssadおよびJockers(2006)からアッセイ手順を改変し、96ウェル白色マイクロプレート中で、総体積100μlで、各試料について2回反復してそれを行った。3つの生物学的に独立したアッセイを行い、データをプールする。終濃度10μMでセレンテラジンh DeepBlueC基質(Biosynth AG)を添加し、二重波長発光/蛍光測定マイクロプレート読取り器(BMGLabtech)を用いて読み取りを行った。以下の最適化されたフィルター設定を使用して、Rlucの光放出(410±80nm)およびGFP2の光放出(515±30nm)を測定した。同時発現したRlucおよびGFP2融合タンパク質の515nm/410nmでの放出、ならびにRluc融合タンパク質単独の515nm/410nmでの放出の差として、BRET比率を定義した。結果をミリBRET単位で表し、1ミリBRETは1000を乗じたBRET比率の値に対応する(Ayoub、2002)。同じ設定での、RlucまたはGFP2融合物を発現しない対照細胞(宿主株Invsc1)の照明の結果、細胞の自己蛍光が検出される。BRET比率の値を計算する前に、試験した全ての試料から自己蛍光を減算した。
【0270】
粗製膜調製、可溶化、および免疫沈降
適切に誘導された(GFP2レベルが宿主株Invsc1細胞のバックグラウンドに対して2倍以下であった)酵母細胞を、氷冷した緩衝液A(75mM トリス−HCl、pH7.4、12mM MgCl2および2mM EDTA)で3回洗浄し、緩衝液B(緩衝液A+プロテアーゼ阻害剤カクテル、コンプリートMini EDTA−フリー(Roche Applied Science))中で最終的なペレットを懸濁した。次いでフレンチプレスを使用して酵母細胞を破壊し、それを10,000×g、4℃で15分間遠心分離する。130000×gで上清を1時間超遠心分離し、冷やした緩衝液B中でペレットを懸濁した。
【0271】
Ayoubら(2002)に記載の方法に従って、膜タンパク質の可溶化および免疫沈降の手順を改変した。粗製膜を0.5%ジギトニン(Sigma)で可溶化し、4℃でこの混合物を3時間撹拌した。18,000×g、4℃で30分間遠心分離することにより、可溶化されていない膜タンパク質を除去した。冷やした緩衝液Bを使用してジギトニン濃度を0.2%に調整し、ポリクローナル抗GFP抗体(Sapphire Bioscience、カタログ番号120−29000)を添加して最終希釈率を1:1000にした。穏やかにかき混ぜながら、免疫複合体形成を4℃で12時間進行させ、プロテインA−アガロース懸濁物(Sigma)を添加して最終体積を1:10(アガロース懸濁物:試料)にし、穏やかにかき混ぜながら4℃でさらに6時間インキュベートした。5000×gで1分間遠心分離した後、上清をデカントし、冷やした緩衝液C(緩衝液B+0.2%ジギトニン)0.5mlでアガロースビーズを5回洗浄した。基質としてセレンテラジンを使用して(Biosynth AG、Switzerland)、発光測定器(BMGLabtech)でルシフェラーゼ活性を評価することにより、沈降物を分析した。
【0272】
結果
酵母細胞における線虫odr−10の視覚化 − 共焦点顕微鏡法
24時間の誘導後にodr−10−Rlucおよびodr−10−GFP2を同時発現した酵母株Invsc1の共焦点画像から、GFP2融合タンパク質がほとんど細胞の膜上でのみ発現したことが明らかとなり、タグ付きodr−10の過剰発現は観察されなかった。細胞の約32%がタグ付きodr−10タンパク質を発現した。誘導時間を48時間まで延長すると、共焦点画像は、細胞中でのGFP2の過剰発現を示し、凝集したGFP2粒子が細胞の細胞質ゾル中で観察された(図12)。
【0273】
BRET2による生存酵母細胞における構成的odr−10オリゴマーの検出
BRET2実験の結果から、odr−10−Rlucおよびodr−10−GFP2を同時発現する酵母細胞で著しいエネルギー移動が観察されたことが示され(図13)、odr−10がホモオリゴマーを形成することが示唆される。半分の酵母細胞がodr−10−Rlucを発現した試料およびあと半分の酵母細胞がodr−10−GFP2を発現した試料では、著しいエネルギー移動は得られなかった。
【0274】
免疫沈降による生存酵母細胞における構成的Odr−10オリゴマーの検出
受容体−Rlucおよび−GFP2融合物の対の発現、その後の抗緑色蛍光タンパク質抗体を使用した沈降、および免疫共沈降したルシフェラーゼ活性の測定によって、生存酵母細胞におけるOdr−10のオリゴマー形成が確認された。図14で示された結果から、Odr−10が細胞中でオリゴマーを形成することが示唆される。免疫共沈降が受容体に非依存的な、GFP2とRlucとの相互作用から生じ得る可能性を除外するために、(1)odr−10−Rluc/GFP2、(2)受容体−GFP2または受容体−Rlucのどちらかを発現した可溶化膜の混合物を使用した免疫共沈降も実施した。同じ細胞中でのodr−10−GFP2およびodr−10−Rlucの同時発現と比較してルシフェラーゼ活性の20%未満が回復したこれらの試料の結果から、同時発現するodr−10−GFP2およびodr−10−Rlucの融合タンパク質間の相互作用が特異的であったことが示唆される。
【0275】
[実施例6]
BRET2タグ付きマウスα2Aアドレナリン作動性受容体
そのN末端がAla250の後にあり、そのC末端がArg372の隣にある3番目の細胞内ループ中にGFP2が挿入されるように、マウスα2Aアドレナリン作動性受容体(配列番号56)を工学的に作製した。受容体のC末端中のVal461にRlucを融合させた(アミノ酸配列は配列番号54として、コードするオープンリーディングフレームは配列番号53として示される)。
【0276】
二重タグ付きアドレナリン作動性受容体を酵母で発現させる。これらの受容体を使用して、BRET2比率の変化を介してマウスα2Aアドレナリン作動性受容体によるリガンド結合をモニタリングすることができる。
【0277】
[実施例7]
全細胞対無細胞BRETアッセイ
本発明の無細胞アッセイ系を全細胞アッセイ系と比較すると、RLucチャネルで測定したときの光放出量の点から、全細胞アッセイと比較して無細胞アッセイで、BRET2シグナルの強度が40倍を超えて大きかったことが実証される(図15)。1μMのジアセチルのアッセイの結果、全細胞アッセイを使用するとBRET2シグナルの有意な変化はなかったが(P≦0.05)、無細胞アッセイ系を使用すると52%の有意な低下(P≦0.05)が観察された(図16)。
【0278】
広く記載されている本発明の趣旨または範囲を逸脱せずに、具体的な実施形態に示されている本発明に多数の変更および/または改変をなし得ることが当業者に理解されるであろう。したがって、本実施形態は、全ての点において例示的であり制限的ではないとみなされるべきである。
【0279】
本出願は、その内容全体が参照により本明細書に組み込まれる、2009年1月29日に出願されたUS61/148,271の優先権を主張するものである。
【0280】
本明細書で論じ、かつ/または言及した全ての刊行物は、その全体が本明細書に組み込まれる。
【0281】
本明細書に含まれている、文書、行為、材料、デバイス、物品などのどんな論述も、本発明の場面を提供する目的のものに過ぎない。それは、これらの事項のいずれかまたは全てが従来技術の基礎の一部を形成し、または本出願の各請求項の優先日前から存在する、本発明に関連する分野における共通の一般知識であったことを承認するものとみなすべきでない。
【0282】
(参考文献)
【配列表フリーテキスト】
【0283】
配列番号1−C.elegans Odr10受容体。
配列番号2−C.elegans Str112受容体。
配列番号3−C.elegans Str113受容体。
配列番号4−C.elegans Str114受容体。
配列番号5−C.elegans Str115受容体。
配列番号6−C.elegans Str116受容体。
配列番号7−C.elegans Odr10受容体をコードするORF。
配列番号8−C.elegans Str112受容体をコードするORF。
配列番号9−C.elegans Str113受容体をコードするORF。
配列番号10−C.elegans Str114受容体をコードするORF。
配列番号11−C.elegans Str115受容体をコードするORF。
配列番号12−C.elegans Str116受容体をコードするORF。
配列番号13−C.elegans Odr10受容体、3番目の細胞内ループ(5番目の非膜貫通ループ)中に挿入されたRLucおよびC末端に挿入されたGFPを含む本発明のポリペプチド。
配列番号14−C.elegans Odr10受容体、3番目の細胞内ループ(5番目の非膜貫通ループ)中に挿入されたRLucおよびC末端に挿入されたGFPを含む本発明のポリペプチドをコードするオープンリーディングフレーム。
配列番号15−C.elegans Odr10受容体、3番目の細胞内ループ(5番目の非膜貫通ループ)中に挿入されたGFPおよびC末端に挿入されたRLucを含む本発明のポリペプチド。
配列番号16−C.elegans Odr10受容体、3番目の細胞内ループ(5番目の非膜貫通ループ)中に挿入されたGFPおよびC末端に挿入されたRLucを含む本発明のポリペプチドをコードするオープンリーディングフレーム。
配列番号17−C.elegans Odr10受容体およびC末端に挿入されたRLucを含む本発明のポリペプチド。
配列番号18−C.elegans Odr10受容体、C末端に挿入されたRLucを含む本発明のポリペプチドをコードするオープンリーディングフレーム。
配列番号19−C.elegans Odr10受容体およびC末端に挿入されたGFP2を含む本発明のポリペプチド。
配列番号20−C.elegans Odr10受容体、C末端に挿入されたGFP2を含む本発明のポリペプチドをコードするオープンリーディングフレーム。
配列番号21〜44−オリゴヌクレオチドプライマー。
配列番号45−mシトリン誘導体。
配列番号46−mCFP誘導体。
配列番号47−mシトリン誘導体をコードするオープンリーディングフレーム。
配列番号48−mCFP誘導体をコードするオープンリーディングフレーム。
配列番号49−Odr10FRET二重標識融合タンパク質をコードするオープンリーディングフレーム。
配列番号50−Odr10FRET二重標識融合タンパク質。
配列番号51−Str112BRET二重標識融合タンパク質をコードするオープンリーディングフレーム。
配列番号52−Str112BRET二重標識融合タンパク質。
配列番号53−マウスα2Aアドレナリン作動性受容体BRET二重標識融合タンパク質をコードするオープンリーディングフレーム。
配列番号54−マウスα2Aアドレナリン作動性受容体BRET二重標識融合タンパク質。
配列番号55−マウスα2Aアドレナリン作動性受容体をコードするオープンリーディングフレーム。
配列番号56−マウスα2Aアドレナリン作動性受容体。
【技術分野】
【0001】
本発明は、試料中の化合物を検出するための方法およびポリペプチドに関する。詳細には、本発明は、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体を含む無細胞組成物の使用に関する。任意選択で、組成物は、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子も含む。Gタンパク質共役受容体および/または存在する場合アクセサリー分子は、生物発光タンパク質およびアクセプター分子を一緒に含み、生物発光タンパク質およびアクセプター分子によって、生物発光共鳴エネルギー移動(BRET)を使用して受容体に結合する化合物を検出することができる。
【背景技術】
【0002】
Gタンパク質共役受容体(GPCR)は、細胞の外側で分子を感知し、内側でシグナル伝達経路を、最終的に細胞応答を活性化する膜貫通受容体のファミリーである。GPCRは、酵母および動物を含む真核生物でしか発見されていない。GPCRと結合しそれを活性化するリガンドには、光感受性化合物、匂い、フェロモン、ホルモン、および神経伝達物質があり、小分子からペプチド、大分子タンパク質までサイズが様々である。Gタンパク質共役受容体は多数の疾患に関与し、治療用薬物の標的でもある。
【0003】
脊椎動物およびC.elegansの嗅覚受容体(OR)は、Gタンパク質共役受容体(GPCR)ファミリーのメンバーである(Buckら、1991;Troemelら、1995)。GPCRは、その7つの膜貫通ドメインを特徴とし、リガンド結合ドメインが膜の細胞外側にあり、Gタンパク質結合ドメインが細胞内側にあると推測される。受容体がリガンドと結合したとき、高次構造変化が受容体中に起こり、ヘテロ三量体Gタンパク質を活性化することを可能にする(Kobilkaら、2007)。次いで、活性化したGタンパク質は、グアニルシクラーゼやホスホリパーゼC経路などのシグナル伝達経路を活性化することができ、シグナルをより高位の処理中枢に伝達する(Gaillardら、2004)。
【0004】
Forster共鳴エネルギー移動、または単に共鳴エネルギー移動(RET)とは、励起状態のドナー分子から基底状態のアクセプター分子へのエネルギーの非放射性移動である(Ghanouniら、2001)。エネルギー移動効率は、ドナーとアクセプターとの距離、スペクトルの重複の程度およびアクセプターとドナーの双極子の相対的配向に依存的である。分子内RETがGPCR活性化のモニタリングに使用されていた以前の場合では、蛍光ドナーおよびアクセプターといった、蛍光共鳴エネルギー移動(FRET)と呼ばれる方法を使用している。ほとんどの場合、蛍光ドナーとアクセプターはどちらも、Aequoria victoria由来の緑色蛍光タンパク質(GFP)の工学的変異体である(Tsien、1998)。最も広く使用されるFRETの対は、ドナーであるシアン蛍光タンパク質(CFP)とアクセプターである黄色蛍光タンパク質(YFP)であり(PistonおよびKremers、2007)、このFRET系は、いくつかのGPCRによる直接的なリガンド結合を定量するのに以前から使用されている(Lohseら、2003および2007;Vilardagaら、2003;Rochaisら、2007;Lisenbeeら、2007)。
【0005】
受容体の活性化をモニタリングする1つの方法では、単一のGPCRを、それぞれ3番目の細胞内ループおよびC末端内の挿入部位において、CFPおよびYFPで二重標識する。436nmの光でCFPを励起すると、480nmでのCFPの放出、および535nmで放出するYFPのFRETを引き起こす。FRETの効率は、ドナーとアクセプターとの距離の6乗によって変化し、このことから、GPCRにおける高次構造変化が極めて感度よく示される。このことは、α2AR、副甲状腺ホルモン受容体(PTHR)、β1−ARおよびセクレチン受容体を用いて、インタクトな細胞において実証された。アゴニストであるノルアドレナリンとα2AR(Lohseら、2003)、副甲状腺ホルモンとPTHR(Vilardagaら、2003)、ノルエピネフリンとβ1−AR(Rochaisら、2007)、およびセクレチンとセクレチン受容体(Lisenbeeら、2007)の相互作用はCFPとYFPとの距離を変化させ、それによってFRETシグナルの変化が生じた。
【0006】
FRET系においてYFPアクセプターをフルオレセインヒ素ヘアピン結合体であるFlAsHと交換すると(Hoffmanら、2005;Nakanishiら、2006)、α2アドレナリン作動性受容体活性化のモニタリングに使用したとき、アゴニスト誘導FRETシグナルがCFP/YFP系と比較して5倍増大した(Nakanishiら、2006)。しかし、FlAsHは、困難な標識化および洗浄手順を伴い、広い調査集団での使用が限定されている。CFP/YFP系は、依然として、分子内の高次構造変化をモニタリングするのに最も頻繁に報告されているFRET系である。FRETに付随する主な不利点は、ドナー蛍光体にエネルギーを与えるのに光源が必要となることである(PistonおよびKremers、2007)。これは、ドナーの励起波長でアクセプターの望ましくない直接励起を引き起こす(「クロストーク」と呼ばれる問題)。
【0007】
生物発光共鳴エネルギー移動(BRET)では、FRETのドナー蛍光体をルシフェラーゼと交換し、アクセプターは任意の適切な蛍光体でよい。基質を添加すると生物発光放出、したがってRETが開始されるので、ルシフェラーゼを使用すると、照明の必要が回避される。BRETの2つの一般的な実行形態は、GFP突然変異体のYFP(BRET1;λem=約530nm)またはGFP2(BRET2;λem=約510nm)のどちらかと結合した、ドナー系であるセレンテラジンh(BRET1;λem=約475nm)またはセレンテラジン400a(Clz400a)基質(BRET2;λem=約395nm)を有するウミシイタケ(Renilla)ルシフェラーゼ(RLuc)を含む。BRET2系は、量子収量を消費して、BRET1系の約55nmと比較して約115nmという、ドナーとアクセプターの放出ピーク間の優れたスペクトル分離をもたらす(PflegerおよびEidneら、2006)。
【0008】
嗅覚受容体でのFRETは、クラスA(a2−アドレナリン作動性および副甲状腺ホルモン、(Vilardagaら、2003))およびクラスB(セクレチン、(Lisenbeeら、2007))GPCRでしか以前には実証されていない。GPCRクラスAに属する哺乳動物ORと異なり、線虫OR(Robertson、1998および2001)は、これらのクラスのどちらにも属さず、進化的および構造的に異なる。さらに、GPCRスーパーファミリーのクラスA内にある、哺乳動物ORを含めた全てのORは、その発現の点では非定型的である。一般に、これらのタンパク質は、化学感覚系列に由来するニューロン以外では機能的に発現できない。哺乳動物および線虫ORの適切な発現にその存在が必要となるいくつかのアクセサリータンパク質が同定されている。
【0009】
したがって、例えば、バイオセンサーで使用するGタンパク質共役受容体と結合する化合物の検出を可能にする適切に感度のよい方法および分子が必要である。
【発明の概要】
【0010】
本発明者らは、驚くべきことに、嗅覚受容体(OR)など、キメラのGタンパク質共役受容体(GPCR)を使用して標的化合物を検出する無細胞の生物発光共鳴エネルギー移動(BRET)が、全細胞を使用したBRETやFRETなど他の検出方法より感度がよいことを見出した。
【0011】
したがって、第1の態様では、本発明は、化合物を検出する方法であって、前記方法が、
i)試料を、
a)脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体と、
b)任意選択で、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子と
を含む無細胞組成物と接触させるステップであって、
Gタンパク質共役受容体が、同じまたは異なる1つまたは複数のサブユニットを含み、Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含むステップと、
ii)ステップi)と同時にまたはそれに連続して、生物発光タンパク質の基質を提供し、生物発光タンパク質が基質を修飾することを可能にするステップと、
iii)ステップii)が生物発光タンパク質とアクセプター分子の間の生物発光共鳴エネルギー移動(BRET)を変動(modulate)させるかどうかを決定するステップと
を含み、化合物がGタンパク質共役受容体と結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化し、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある方法を提供する。
【0012】
一実施形態では、化合物と結合することができるGタンパク質共役受容体のサブユニットは、
i)生物発光タンパク質と、
ii)アクセプター分子と
を含み、化合物がサブユニットと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。例えば、サブユニットは、
i)配列番号13、15、52もしくは54として示されるアミノ酸配列、または
ii)配列番号13、15、52もしくは54のいずれか1つもしくは複数と少なくとも40%同一であるアミノ酸配列
を含んでよい。
【0013】
代替の実施形態では、Gタンパク質共役受容体は、
i)a)生物発光タンパク質と、
b)アクセプター分子と
を含む第1のサブユニットと、
ii)化合物と結合することができる第2のサブユニットと
を含み、化合物が第2のサブユニットと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。
【0014】
さらなる実施形態では、
i)生物発光タンパク質がサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質がサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質がサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子がC末端の一部を形成し、または
vi)アクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質がC末端の一部を形成する。
【0015】
さらに別の代替の実施形態では、Gタンパク質共役受容体は、
i)生物発光タンパク質を含む第1のサブユニットと、
ii)アクセプター分子を含む第2のサブユニットと
を含む。例えば、第1または第2のサブユニットは、
i)配列番号17もしくは19として示されるアミノ酸配列、または
ii)配列番号17および/もしくは19と少なくとも40%同一であるアミノ酸配列
を含んでよい。
【0016】
一実施形態では、
i)生物発光タンパク質が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットの5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットの3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットのC末端の一部を形成し、または
vi)アクセプター分子が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットのC末端の一部を形成する。
【0017】
さらなる代替の実施形態では、
i)Gタンパク質共役受容体が生物発光タンパク質を含むサブユニットを含み、アクセサリー分子がアクセプター分子を含み、または
ii)Gタンパク質共役受容体がアクセプター分子を含むサブユニットを含み、アクセサリー分子が生物発光タンパク質を含む。
【0018】
別の代替の実施形態では、アクセサリー分子は、
i)生物発光タンパク質と、
ii)アクセプター分子と
を含む。
【0019】
さらなる代替の実施形態では、組成物は、少なくとも2つのアクセサリー分子を含み、第1のアクセサリー分子は生物発光タンパク質を含み、第2のアクセサリー分子はアクセプター分子を含む。
【0020】
本発明者らは、FRET−無細胞およびBRET−全細胞検出系と比較したときに本発明がいかに優れているかを発見して特に驚いた。したがって、好ましい実施形態では、本発明の方法は、非生物発光タンパク質をドナー分子として使用し、蛍光共鳴エネルギー移動(FRET)の変動を決定する場合より少なくとも2倍、3倍または4倍感度がよい。さらなる好ましい実施形態では、本発明の方法は、この方法で無細胞組成物中にあるものの代わりにインタクトな細胞中に存在する同じGタンパク質共役受容体を使用する場合より少なくとも10倍、20倍、30倍または40倍強いBRETシグナルをもたらす。
【0021】
クラスAのGPCR、一般にOR、特に線虫ORの非定型的性質を考慮すると、BRETドナーおよびアクセプタータグとともにクラスAのGPCRを含むキメラが、a)構造的完全性を保持し、b)匂い物質による受容体活性化を光学的シグナルに変換することができることは驚くべきことであった。したがって、好ましい実施形態では、Gタンパク質共役受容体はクラスAのGPCRである。さらなる好ましい実施形態では、クラスAのGPCRは嗅覚受容体である。細胞中で発現したときに受容体のN末端が細胞の外側にあり、C末端が細胞の内側にある限り、嗅覚受容体はどんな供給源由来のものでもよい。例として、それだけに限らないが、脊索動物受容体、線虫受容体、またはそのいずれか1つの生物学的に活性な変異体もしくは断片がある。脊索動物受容体の例として、それだけに限らないが、哺乳動物受容体、鳥類受容体および魚類受容体がある。好ましい実施形態では、嗅覚受容体は、線虫受容体、またはその生物学的に活性な変異体もしくは断片である。一実施形態では、線虫受容体は、Caenorhabditis elegans受容体、またはその生物学的に活性な変異体もしくは断片である。
【0022】
一実施形態では、嗅覚受容体は、
i)配列番号1〜6のいずれか1つで示されるアミノ酸配列と、
ii)配列番号1〜6のいずれか1つまたは複数と少なくとも40%同一であるアミノ酸配列と
を含む。
【0023】
別の実施形態では、サブユニットは、2つ以上の異なるGタンパク質共役受容体サブユニットの部分のキメラである。
【0024】
アクセサリー分子は、Gタンパク質共役受容体と直接または間接的に結合する任意の分子でよい。例として、それだけに限らないが、Gタンパク質およびアレスチンがある。
【0025】
BRETの変化の幅に対して化合物の濃度を変化させる効果は、その応答が用量依存的であることを最初に示した。したがって、好ましい実施形態では、BRETのレベルは、試料中の化合物の相対量を示す。
【0026】
生物発光タンパク質の例として、それだけに限らないが、ルシフェラーゼ、β−ガラクトシダーゼ、ラクタマーゼ(lactamase)、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β−グルクロニダーゼまたはβ−グルコシダーゼがある。
【0027】
ルシフェラーゼの例として、それだけに限らないが、ウミシイタケ(Renilla)ルシフェラーゼ、ホタル(Firefly)ルシフェラーゼ、腔腸動物(Coelenterate)ルシフェラーゼ、北アメリカツチボタル(North America glow worm)ルシフェラーゼ、コメツキム(click beetle)シルシフェラーゼ、鉄道虫(railroad worm)ルシフェラーゼ、細菌(bacterial)ルシフェラーゼ、ガウシア(Gaussia)ルシフェラーゼ、エクオリン、ヒカリキノコバエ(Arachnocampa)ルシフェラーゼ、またはそのいずれか1つの生物学的に活性な変異体もしくは断片、またはその2つ以上のキメラがある。
【0028】
基質の例として、それだけに限らないが、甲虫(beetle)ルシフェリン、他のルシフェリン、セレンテラジン、またはセレンテラジンの誘導体がある。また、エクオリンなどの一部の発光タンパク質の場合、基質は、カルシウムイオンなどの補助因子でもよい。
【0029】
一実施形態では、アクセプター分子はタンパク質であり、その例として、それだけに限らないが、緑色蛍光タンパク質(GFP)、GFPの青色蛍光変異体(BFP)、GFPのシアン蛍光変異体(CFP)、GFPの黄色蛍光変異体(YFP)、増強型GFP(EGFP)、増強型CFP(ECFP)、増強型YFP(EYFP)、GFPS65T、エメラルド(Emerald)、トパーズ(Topaz)、GFPuv、不安定化EGFP(dEGFP)、不安定化ECFP(dECFP)、不安定化EYFP(dEYFP)、HcRed、t−HcRed、DsRed、DsRed2、t−ダイマー2、t−ダイマー2(12)、mRFP1、ポシロポリン(pocilloporin)、ウミシイタケ(Renilla)GFP、モンスター(Monster) GFP、paGFP、カエデ(Kaede)タンパク質もしくはフィコビリタンパク質(Phycobiliprotein)、またはそのいずれか1つの生物学的に活性な変異体もしくは断片がある。
【0030】
代替の実施形態では、アクセプター分子は非タンパク質であり、その例として、それだけに限らないが、Alexa Fluor色素、Bodipy色素、Cy色素、フルオレセイン、ダンシル、ウンベリフェロン、蛍光マイクロスフェア、発光マイクロスフェア、蛍光ナノ結晶、マリナブルー、カスケードブルー、カスケードイエロー、パシフィックブルー、オレゴングリーン、テトラメチルローダミン、ローダミン、テキサスレッド、希土類元素キレート、またはその任意の組合せもしくは誘導体がある。
【0031】
別の実施形態では、前記方法は、ステップi)もしくはステップii)と同時にまたはそれに連続して、生物発光タンパク質の補助因子を提供するステップをさらに含む。例として、それだけに限らないが、ATP、マグネシウム、酸素、FMNH2、カルシウム、またはそのいずれか2つ以上の組合せがある。
【0032】
一実施形態では、無細胞組成物は、組換え細胞中でGタンパク質共役受容体を産生し、細胞の膜を破壊することによって得られた。
【0033】
一実施形態では、組換え細胞は、Gタンパク質共役受容体と結合する非内因性タンパク質を産生しない。
【0034】
無細胞組成物を得ることができる細胞は、Gタンパク質共役受容体を発現し、この受容体をその細胞膜に取り込むことができる任意の細胞型である。特に好ましい実施形態では、組換え細胞は酵母細胞である。
【0035】
代替の実施形態では、Gタンパク質共役受容体は、リポソームの脂質二重層中に埋め込まれる。
【0036】
さらなる実施形態では、前記方法は、マイクロ流体工学を使用して行われる。
【0037】
別の態様では、本発明は、化合物を検出するための精製および/または組換えポリペプチドであって、前記ポリペプチドが、
i)Gタンパク質共役受容体のサブユニットと、
ii)生物発光タンパク質および/またはアクセプター分子と
を含み、細胞中で発現したときにサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、ポリペプチドを提供する。
【0038】
一実施形態では、ポリペプチドは、1つまたは複数の異なるGタンパク質共役受容体サブユニット、および任意選択で1つまたは複数の異なるアクセサリー分子を含むGタンパク質共役受容体であるタンパク質複合体の一部を形成する。
【0039】
一実施形態では、
i)生物発光タンパク質がサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質がサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質がサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子がC末端の一部を形成し、または
vi)アクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質がC末端の一部を形成し、または
vii)生物発光タンパク質もしくはアクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、または
viii)生物発光タンパク質もしくはアクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、または
ix)生物発光タンパク質もしくはアクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、または
x)生物発光タンパク質もしくはアクセプター分子がサブユニットのC末端の一部を形成する。
【0040】
さらなる実施形態では、ポリペプチドは、生物発光タンパク質およびアクセプター分子を含み、化合物がポリペプチドと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。
【0041】
代替の実施形態では、ポリペプチドはサブユニットおよび生物発光タンパク質を含み、ポリペプチドはアクセプター分子を含む第2のポリペプチドと直接または間接的に結合し、化合物がポリペプチドおよび/または第2のポリペプチドと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。
【0042】
さらなる代替の実施形態では、ポリペプチドはサブユニットおよびアクセプター分子を含み、ポリペプチドは生物発光タンパク質を含む第2のポリペプチドと直接または間接的に結合し、化合物がポリペプチドおよび/または第2のポリペプチドと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する。
【0043】
上記2つの実施形態では、第2のポリペプチドは、(本明細書で定義されるホモもしくはヘテロ二量体または高次の多量体を形成する)Gタンパク質共役受容体サブユニットまたはアクセサリー分子のいずれかを含む。
【0044】
さらなる態様では、本発明は、化合物を検出するための精製および/または組換えポリペプチドであって、前記ポリペプチドが、
i)Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合するアクセサリー分子と、
ii)生物発光タンパク質および/またはアクセプター分子と
を含み、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、ポリペプチドを提供する。
【0045】
熟練した読者なら理解するであろうが、検出の方法に関する上記の実施形態の多くは、本発明のポリペプチドの態様にも当てはまる。
【0046】
本発明のポリペプチドをコードする単離および/または外因性ポリヌクレオチドも提供される。
【0047】
一実施形態では、ポリヌクレオチドは、
i)配列番号7〜12、14、16、18、20、51もしくは53のいずれか1つで示されるヌクレオチド配列、または
ii)配列番号7〜12、14、16、18、20、51もしくは53のいずれか1つまたは複数と少なくとも40%同一であるヌクレオチド配列
を含む。
【0048】
別の態様では、本発明のポリヌクレオチドを含むベクターが提供される。好ましい実施形態では、ポリヌクレオチドは、プロモーターと作動可能に連結する。
【0049】
さらなる態様では、本発明のポリヌクレオチドおよび/または本発明のベクターを含む宿主細胞が提供される。
【0050】
好ましくは、宿主細胞は本発明のポリペプチドを産生し、したがってそれを含む。
【0051】
宿主細胞は任意の細胞型でよい。
【0052】
本発明者らは、酵母の膜における、哺乳動物の嗅覚受容体と進化的に異なる線虫の嗅覚受容体の機能的発現も最初に示す。したがって、好ましい実施形態では、宿主細胞は酵母細胞である。
【0053】
さらなる態様では、本発明は、本発明のポリペプチド、本発明のポリヌクレオチド、本発明のベクター、および/または本発明の宿主細胞を含む組成物を提供する。
【0054】
好ましくは、ポリペプチドは脂質二重層中に埋め込まれている。
【0055】
i)Gタンパク質共役受容体サブユニットを含む本発明のポリペプチドであって、前記ポリペプチドが脂質二重層中に埋め込まれ、ならびに/または
ii)Gタンパク質共役受容体の細胞内ループおよび/もしくはC末端と直接もしくは間接的に結合したアクセサリー分子を含む本発明のポリペプチドであって、Gタンパク質共役受容体が脂質二重層中に埋め込まれ、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、ポリペプチド
を含む無細胞組成物も提供される。
【0056】
好ましくは、脂質二重層は、酵母脂質二重層である。
【0057】
一実施形態では、無細胞組成物は、生物発光タンパク質の基質および/または生物発光タンパク質の補助因子をさらに含む。
【0058】
熟練した読者なら理解するであろうが、検出の方法に関する上記の実施形態の多くは、本発明の無細胞の態様にも当てはまる。
【0059】
さらなる態様では、本発明は、本発明の無細胞組成物を作製する方法であって、本発明の細胞を得るステップと、細胞の膜を破壊するステップとを含む方法を提供する。
【0060】
一実施形態では、細胞を浸透化または溶解する。
【0061】
一実施形態では、細胞をフレンチプレスで溶解する。
【0062】
別の態様では、本発明は、本発明のポリペプチド、本発明の宿主細胞、本発明の組成物および/または本発明の無細胞組成物を含むバイオセンサーを提供する。
【0063】
さらなる態様では、本発明は、Gタンパク質共役受容体と結合する化合物をスクリーニングする方法であって、
i)候補化合物を、
a)脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体と、
b)任意選択で、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子と
を含む無細胞組成物と接触させるステップであって、
Gタンパク質共役受容体が、同じまたは異なる1つまたは複数のサブユニットを含み、Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含むステップと、
ii)ステップi)と同時にまたはそれに連続して、生物発光タンパク質の基質を提供し、生物発光タンパク質が基質を修飾することを可能にするステップと、
iii)ステップii)が生物発光タンパク質とアクセプター分子の間の生物発光共鳴エネルギー移動(BRET)を変動させるかどうかを決定するステップと
を含み、BRETの変動が、化合物がGタンパク質共役受容体と結合することを示し、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記方法を提供する。
【0064】
熟練した読者なら理解するであろうが、検出の方法に関する上記の実施形態の多くは、本発明のスクリーニングの方法の態様にも当てはまる。
【0065】
さらなる態様では、本発明は、本発明のポリペプチド、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明の組成物、本発明の無細胞組成物、および/または本発明のバイオセンサーを含むキットを提供する。
【0066】
明らかとなるように、本発明の1つの態様の好ましい性質および特徴は、本発明の多くの他の態様に適用可能である。
【0067】
本明細書全体にわたって、「含む(comprise)」という語、または「含む(comprises)」や「含んでいる(comprising)」などの変化形は、述べられた要素、整数もしくはステップ、または一群の要素、整数もしくはステップを包含することを意味するが、任意の他の要素、整数もしくはステップ、または一群の要素、整数もしくはステップを除外することを意味しないことが理解されるであろう。
【0068】
以後、下記の非限定的な実施例により、添付図面を参照しながら本発明を説明する。
【図面の簡単な説明】
【0069】
【図1】GFP2およびRLucと融合したODR−10受容体における共鳴エネルギー移動の原理を示す図である。GFP2はGPCRの3番目の細胞内ループ中に、RLucはそのC末端に挿入されている。ジアセチルが結合した結果、高次構造変化が生じ、RLucとGFP2との距離が短く(上部右側)、もしくは長くなり(下部右側)、またはその発色団双極子の相対的配向の変化が生じ得、用量依存的にBRET2シグナルが変化する。
【図2】(A)μM(平均±SEM、n=2)および(B)nM(平均±SEM、n=6)濃度のリガンドまたは水に対するOGORの応答を示す図である(*は、水と比較して有意差がある(P≦0.05)ことを示す)。
【図3】水単独と比較した、(A)μM(平均±SEM、n=2)および(B)nM(平均±SEM、n=6)濃度のリガンドに対するOGORのBRET2比率(%)の変化を示す図である。
【図4】OGORのBRET2応答のジアセチル濃度依存性を示す図である(平均±SEM、n=12)。
【図5】水または1μMのジアセチル水溶液に対するOGOR(平均±SEM、n=3)およびOGOR突然変異体(平均±SEM、n=6)の応答の比較を示す図である。
【図6】水または1μMのジアセチル水溶液に対するOCOY(平均±SEM、n=5)およびOGOR(平均±SEM、n=3)の応答の比較を示す図である。
【図7】様々な濃度のジアセチルを添加した時のOCOYのスペクトルスキャンを示す図である。上部:生データ。下部:標準化した(500nm)データ。
【図8】FRET(平均±SEM、n=5)とBRET2(平均±SEM、n=12)の両方でタグを付けたODR−10の標準化した用量反応曲線を示す図である。
【図9】LB単独と比較した、LB中のOP50細菌上清(LB)またはOP50細菌培養物10μL(平均±SEM、n=3)に対するTGTRの応答を示す図である(*は、LBと比較して有意差がある(P≦0.05)ことを示す)。
【図10】μM(平均±SD、n=3)濃度のリガンドまたは水に対するTGTRの応答を示す図である(*は、水と比較して有意差がある(P≦0.05)ことを示す)。
【図11】ODR−10伝達カセットを示す図である。暗部:ODR−10配列、明部:可変受容体リガンド結合ドメイン。A)カセット中で固定されたN末端、B)N末端可変OR配列。Rルシフェラーゼは410nmで放出するものとして示し、GFP2は515nmで放出するものとして示す。
【図12】共焦点顕微鏡法(Ex488nm、Leica SP2共焦点レーザー走査型顕微鏡)によりodr−10受容体の局在およびタグ付きodr−10の発現レベルを示す、odr−10−Rlucおよびodr−10−GFP2を同時発現した酵母細胞を示す図である。
【図13】生存Invsc1酵母細胞における線虫odr−10の構成的BRET2を示す図である。タグ付きodr−10タンパク質を発現するように、試験した全ての酵母株を15℃で24時間誘導した。10uMのセレンテラジンh(DeepBlueC)を添加し、二重波長マイクロプレート読取り器で、方法において記載するRlucおよびGFP2のフィルター設定を用いて光放出を測定することにより、生存細胞でエネルギー移動測定を行った。データは、2つの独立した実験の平均±StDである。Insvc1/odr−10−RLuc:陰性対照。Invsc1/odr−10−RLuc+Invsc1/odr−10−GFP2:別々の細胞におけるORおよびOG。プレートの読み取りで細胞を一緒に混合した:2つの構築物が別々の細胞で発現したので、BRETは予想されない。Invsc1/odr−10−RLuc;odr−10−GFP2:どちらの構築物も同じ細胞で発現した。BRETが存在し、したがって二量体形成も起こっている。
【図14】免疫沈降によるOdr−10オリゴマーの検出を示す図である。示された受容体を同時発現しているInvsc1酵母細胞の粗製膜を可溶化し、同じ量のルシフェラーゼ活性に調整した。次いで、抗GFP抗体を用いて受容体を免疫沈降し、沈降物におけるルシフェラーゼ活性を決定した。値は、沈降したルシフェラーゼ活性の最大量の%として示す。データは2つの生物学的反復の平均である。
【図15】超遠心分離後の無細胞膜調製物100μLの全細胞アッセイ試料100μLに5μMのセレンテラジン400aを添加した後、BRET2チャネルで測定された光放出量を示す図である。
【図16】全細胞および無細胞アッセイ系を使用してモニタリングした、1μMのジアセチルとのインキュベーション(45分)後のBRET2シグナルを示す図である(平均±S.D.、n=3)。
【発明を実施するための形態】
【0070】
発明の詳細な説明
一般的技術および定義
別段具体的に定義されていない限り、本明細書で使用される全ての科学技術用語は、(例えば、細胞培養、分子遺伝学、バイオセンサー、G共役型タンパク質受容体の生物学、免疫学、免疫組織化学、タンパク質化学、および生化学における)当業者によって一般に理解されているものと同じ意味を有するとみなすものとする。
【0071】
別段示されていない限り、本発明において利用される組換えタンパク質、細胞培養、および免疫学的技術は標準的な手順であり、当業者に周知である。そのような技術は、J.Perbal、A Practical Guide to Molecular Cloning、John Wiley and Sons(1984)、J.Sambrookら、Molecular Cloning:A Laboratory Manual、Cold Spring Harbour Laboratory Press(1989)、T.A.Brown(編)、Essential Molecular Biology:A Practical Approach、第1および2巻、IRL Press(1991)、D.M.GloverおよびB.D.Hames(編)、DNA Cloning:A Practical Approach、第1〜4巻、IRL Press(1995および1996)、ならびにF.M.Ausubelら(編)、Current Protocols in Molecular Biology、Greene Pub.Associates and Wiley−Interscience(1988、現在までの全ての最新版を含む)、Ed HarlowおよびDavid Lane(編)Antibodies:A Laboratory Manual、Cold Spring Harbour Laboratory、(1988)、ならびにJ.E.Coliganら(編)Current Protocols in Immunology、John Wiley & Sons(現在までの全ての最新版を含む)などの出典における文献全体にわたって記載され、説明されている。
【0072】
本明細書において、別段指定されていない限り、「Gタンパク質共役受容体」という用語は、Gタンパク質を介してシグナル伝達する7回膜貫通受容体を指す。この受容体は、単一のサブユニットでもよく、または2つ以上の受容体サブユニットでもよい。2つ以上の受容体サブユニットが存在するとき、それらは同じでもよく、異なってもよく、またはその組合せ(例えば、1つのサブユニットが2つ、別のサブユニットが1つだけ)でもよい。さらに、別段の指定または含意がない限り、「Gタンパク質共役受容体」および「Gタンパク質共役受容体のサブユニット」という用語、またはその変化形は互換的に使用される。
【0073】
本明細書において、「嗅覚受容体」、「嗅受容体」、「OR」という用語またはその変化形は、生物の細胞中に存在するとき、化学感覚的知覚に関与するポリペプチドを指す。一実施形態では、細胞はニューロンである。さらに、「嗅覚受容体」または「嗅受容体」という用語は、匂い物質リガンドと結合し、または匂い物質リガンドと結合するタンパク質複合体の一部を形成し、その結果生理的応答を起こすポリペプチドを指す。
【0074】
本明細書において、「生物発光タンパク質」という用語は、適切な基質に作用し、発光を生じさせることができる任意のタンパク質を指す。
【0075】
本明細書において、「基質」という用語は、生物発光タンパク質とともに、発光を生じさせまたは吸収することができる任意の分子を指す。
【0076】
本明細書において、「生物発光タンパク質が基質を修飾することを可能にする」という語句は、エネルギーを産生する、基質に対する生物発光タンパク質の任意の酵素活性を指す。
【0077】
本明細書において、「アクセプター分子」という用語は、生物発光タンパク質の活性の結果放出されたエネルギーを受容し、光エネルギーとして再び放出することができる任意の化合物を指す。
【0078】
本明細書において、生物発光共鳴エネルギー移動(BRET)とは、生物発光タンパク質ドナーとアクセプター分子の間のエネルギーの非放射性移動に基づく近接アッセイである。
【0079】
本明細書において、「変動させる」もしくは「変動」という用語またはその変化形は、生物発光タンパク質および/またはアクセプター分子の強度および/または放出スペクトルの変化を指す。
【0080】
本明細書において、「空間的位置」という用語は、Gタンパク質共役受容体を含む本明細書で定義されるポリペプチドと化合物が結合した結果変化する、アクセプター分子に対する生物発光タンパク質の三次元での位置付けを指す。
【0081】
本明細書において、「双極子配向」という用語は、三次元空間におけるその配向に対する、生物発光タンパク質および/またはアクセプター分子に関連する双極子モーメントの三次元空間における方向を指す。双極子モーメントは、分子に対して電荷が変化する結果生じるものである。
【0082】
本明細書において、「感度がよい」(more sensitive)という用語は、1つのレポーター系(例えば、BRET)で別のレポーター系(例えば、FRET)に対してリガンドの非結合形態とリガンドの結合形態の間の共鳴エネルギー移動の比率が大きく変化することを指す。Gタンパク質共役受容体サブユニットを含む本発明の単離および/または組換えポリペプチドを含有していること。好ましくは、無細胞組成物は、本発明のポリペプチドを含んでも含まなくても、生細胞を含まない。
【0083】
本明細書において、「接触させる」という用語は、Gタンパク質共役受容体と結合することができるという形で検出される化合物を含む、または含む可能性がある試料を添加することを指す。
【0084】
本明細書において、「脂質二重層」という用語は、内部に共通する疎水性の二重層および2つの親水性の表面を有する、互いに反対に整列した分子の並びの、典型的には両親媒性の2つの層を指す。脂質二重層は、天然に存在するものでもよく、または人工のものでもよい。最も好ましくは、脂質二重層は、Gタンパク質共役受容体を含む本明細書で定義されるポリペプチドが中に挿入される細胞または生体膜、例えば、哺乳動物、昆虫、植物または酵母細胞膜であり、最も好ましくは、酵母細胞膜である。
【0085】
本明細書において、「酵母脂質二重層」という用語は、Gタンパク質共役受容体を含む本明細書で定義されるポリペプチドを発現する酵母細胞に由来する脂質二重層を指す。当業者は、それだけに限らないが、Fus1pタンパク質(TrueheartおよびFink、1989)、Sso1/2p、SncipおよびSec9pタンパク質を含むSNARE複合体(Stropら、2007)、フェロモン受容体Ste2p(Celicら、2003)、および/またはAlr1(Graschopfら、2001)など、膜中に埋め込まれた、酵母中に天然に存在するタンパク質を検出することによって、脂質二重層が酵母に由来するかどうかを容易に決定することができる。
【0086】
本明細書において、「同時にまたは連続して」という用語は、試料を本明細書で定義されるポリペプチドと接触させる前、その間、またはその後に基質を提供できることを意味する。好ましくは、試料と同時またはその後に基質を提供する。
【0087】
「試料」は、検出する化合物を含むことが疑われる任意の物質または組成物でよい。試料の例として、空気、液体、生体材料および土壌がある。試料は、環境もしくは供給源から直接得ることもでき、または本発明の方法を行う前に適切な手順によって少なくとも部分的に精製することもできる。
【0088】
本明細書において、「Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含む」という用語は、生物発光タンパク質が、Gタンパク質共役受容体のサブユニットまたはアクセサリー分子と結合する、好ましくは共有結合する、より好ましくはそれとの融合タンパク質として産生され、アクセプター分子が、Gタンパク質共役受容体のサブユニットまたはアクセサリー分子と結合する、好ましくは共有結合する、より好ましくはそれとの融合タンパク質として産生されることを意味する。生物発光タンパク質およびアクセサリー分子は、同じもしくは異なるGタンパク質共役受容体のサブユニットまたは同じもしくは異なるアクセサリー分子と結合する、好ましくは共有結合する、より好ましくはそれとの融合タンパク質として産生することができる。さらに、生物発光タンパク質およびアクセサリー分子の一方は、Gタンパク質共役受容体のサブユニットと結合する、好ましくは共有結合する、より好ましくはそれとの融合タンパク質として産生することができ、生物発光タンパク質およびアクセサリー分子の他方は、アクセサリー分子と結合することができる。さらに、複数の生物発光タンパク質とアクセサリー分子の対は、多重化して異なるリガンドを検出し、または同じリガンドの検出の感度を増大させる組合せとして存在し得る。
【0089】
本明細書において、「の一部を形成する(forms part of)」という用語は、Gタンパク質共役受容体またはそのサブユニットの特定の領域内に位置する生物発光タンパク質またはアクセプター分子を指す。この用語は、生物発光タンパク質および/またはアクセプター分子が、Gタンパク質共役受容体に付着または結合するが、アミノ酸の連続した鎖を形成しない可能性も含む。一実施形態では、生物発光タンパク質またはアクセプター分子は、Gタンパク質共役受容体の特定の領域と完全に入れ替わる。別の実施形態では、Gタンパク質共役受容体の特定の領域の全部ではないが一部が入れ替わる。さらに別の実施形態では、Gタンパク質共役受容体の特定の領域が入れ替わらない。熟練した受け手なら理解するであろうが、生物発光タンパク質またはアクセプター分子は、本発明のポリペプチドのGタンパク質共役受容体部分が標的化合物と結合してアクセプター分子に対する生物発光タンパク質の位置および/または双極子配向の空間的変化を起こすことができないようには挿入されない。
【0090】
本明細書において、Gタンパク質共役受容体の特定の部分(ドメイン)の「少なくとも大部分」という用語は、特定の領域の少なくとも51%、より好ましくは少なくとも75%、さらにより好ましくは少なくとも90%を指す。
【0091】
「実質的に精製された」または「精製された」とは、その天然の状態で結合する1つまたは複数の脂質、核酸、他のポリペプチド、または他の混入分子から分離しているポリペプチドを意味する。実質的に精製されたポリペプチドは、それと天然に結合する他の構成成分を少なくとも60%、より好ましくは少なくとも75%、より好ましくは少なくとも90%含まないことが好ましい。しかし、現在のところ、本発明のポリペプチドが天然に存在する証拠はない。
【0092】
ポリペプチドとの関連で「組換え」という用語は、その天然の状態と比較して変化した量または変化した速度で、細胞によって、または無細胞発現系で産生されたときのポリペプチドを指す。一実施形態では、細胞は、そのポリペプチドを天然に産生しない細胞である。しかし、細胞は、産生するポリペプチドの量を変化させる、好ましくは増大させる非内因性遺伝子を含む細胞でもよい。本発明の組換えポリペプチドは、それが産生されたトランスジェニック(組換え)細胞、または無細胞発現系の他の構成成分から分離していないポリペプチド、および少なくとも一部の他の構成成分から後で精製して除かれる、そのような細胞または無細胞系で産生されたポリペプチドを含む。
【0093】
「ポリペプチド」および「タンパク質」という用語は一般に互換的に使用され、1本のポリペプチド鎖を指し、非アミノ酸基の付加によって修飾されてもよく、またはされなくてもよい。そのようなポリペプチド鎖は、他のポリペプチドもしくはタンパク質、または補助因子などの他の分子と結合し得ることが理解されるであろう。本明細書において「タンパク質」および「ポリペプチド」という用語は、本明細書に記載のポリペプチドの変異体、突然変異体、生物学的に活性な断片、修飾物、類似体および/または誘導体も含む。
【0094】
ポリペプチドの%同一性は、GAP(NeedlemanおよびWunsch、1970)分析(GCGプログラム)により、ギャップ生成ペナルティー=5、ギャップ伸長ペナルティー=0.3を用いて決定する。質問配列は長さが少なくとも25アミノ酸であり、GAP分析は少なくとも25アミノ酸の領域にわたって2つの配列を整列させる。より好ましくは、質問配列は長さが少なくとも50アミノ酸であり、GAP分析は少なくとも50アミノ酸の領域にわたって2つの配列を整列させる。より好ましくは、質問配列は長さが少なくとも100アミノ酸であり、GAP分析は少なくとも100アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、質問配列は長さが少なくとも250アミノ酸であり、GAP分析は少なくとも250アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、GAP分析はその全長にわたって2つの配列を整列させる。
【0095】
本明細書において、「生物学的に活性な断片」とは、完全長ポリペプチドの定められた活性を維持する、本明細書に記載のポリペプチドの部分である。例えば、Gタンパク質共役受容体の生物学的に活性な断片は、標的化合物と結合して、高次構造変化を起こすことができなければならない。生物学的に活性な断片は、定められた活性を維持する限りどんなサイズでもよい。好ましくは、生物学的に活性な断片は、長さが少なくとも150アミノ酸、より好ましくは少なくとも250アミノ酸である。
【0096】
本明細書において、「生物学的に活性な変異体」とは、1つまたは複数のアミノ酸により天然に存在する分子および/または定められた分子と異なるが、生物学的に活性な断片について上記で定められたような定められた活性を維持する分子である。生物学的に活性な変異体は、天然に存在する分子および/または定められた分子と、典型的には少なくとも50%、より好ましくは少なくとも80%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも97%、さらにより好ましくは少なくとも99%同一である。
【0097】
定められたポリペプチドまたはポリヌクレオチドに関して、上記で示したものより高い%同一性の数字が好ましい実施形態を包含することが理解されるであろう。したがって、適用可能な場合、最小の%同一性の数字に照らして、ポリペプチドまたはポリヌクレオチドが、関連する指定された配列番号と、少なくとも50%、より好ましくは少なくとも60%、より好ましくは少なくとも70%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも91%、より好ましくは少なくとも92%、より好ましくは少なくとも93%、より好ましくは少なくとも94%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%、より好ましくは少なくとも99.1%、より好ましくは少なくとも99.2%、より好ましくは少なくとも99.3%、より好ましくは少なくとも99.4%、より好ましくは少なくとも99.5%、より好ましくは少なくとも99.6%、より好ましくは少なくとも99.7%、より好ましくは少なくとも99.8%、さらにより好ましくは少なくとも99.9%同一であるアミノ酸配列を含むことが好ましい。
【0098】
センスもしくはアンチセンス方向または両方の組合せにおける、一本鎖または二本鎖のDNA、RNA、またはこれらの組合せを含めて、「単離ポリヌクレオチド」とは、その天然の状態で結合または連結するポリヌクレオチド配列から少なくとも部分的に分離しているポリヌクレオチドを意味する。好ましくは、単離ポリヌクレオチドは、それと天然に結合する他の構成成分を少なくとも60%、好ましくは少なくとも75%、最も好ましくは少なくとも90%含まない。
【0099】
ポリヌクレオチドとの関連で、「外因性」という用語は、その天然の状態と比較して変化した量で、細胞または無細胞発現系中に存在するときのポリヌクレオチドを指す。一実施形態では、細胞は、そのポリヌクレオチドを天然に含まない細胞である。しかし、細胞は、コードされたポリペプチドの産生量を変化させる、好ましくは増大させる非内因性ポリヌクレオチドを含む細胞でもよい。本発明の外因性ポリヌクレオチドは、それが存在するトランスジェニック(組換え)細胞、または無細胞発現系の他の構成成分から分離していないポリヌクレオチド、および少なくとも一部の他の構成成分から後で精製して除かれる、そのような細胞または無細胞系で産生されたポリヌクレオチドを含む。
【0100】
さらに、「ポリヌクレオチド」という用語は、本明細書において、「核酸」という用語と互換的に使用される。本発明のポリヌクレオチドは、これとともに提供される分子と比較したときに、ヌクレオチド残基または修飾された残基の欠失、挿入、または置換である1つまたは複数の突然変異を有してよい。突然変異体は、天然に存在するもの(すなわち、天然供給源から単離されたもの)でもよく、または合成のもの(例えば、核酸上で部位特異的突然変異導入を行うことによるもの)でもよい。通常、ポリヌクレオチドの単量体は、ホスホジエステル結合またはその類似物によって連結する。ホスホジエステル結合の類似物には、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホラニリデート(phosphoranilidate)およびホスホラミデートがある。
【0101】
本発明の組成物は、「許容される担体」を含んでよい。そのような許容される担体の例として、水、食塩水、リンガー溶液、デキストロース溶液、ハンクス溶液、および他の生理的に均衡のとれた水性の塩溶液がある。固定油、ゴマ油、オレイン酸エチルやトリグリセリドなどの非水性媒体を使用することもできる。「許容される担体」の正確な性質は、組成物の用途に依存する。本明細書に記載の用途、および組成物における本発明の構成成分の性質を考慮すると、当業者は、特定の用途について適切な「(複数の)許容される担体」を容易に決定することができる。
【0102】
生物発光タンパク質
生物発光タンパク質は、基質を活性化した産物に転換する酵素であり、次いでその産物は緩和するにつれてエネルギーを放出することが当技術分野で理解されている。(基質に対する生物発光タンパク質の活性によって生じた)その活性化した産物は、アクセプター分子に移る、生物発光タンパク質によって生じた発光の供給源である。
【0103】
本発明で使用できるいくつかの異なる生物発光タンパク質がある(例えば、表1を参照)。生物発光タンパク質のサイズを考慮すると、本発明に有用な機能的ポリペプチドを産生できたことは驚きであった。光放出系が知られており、細菌、原生動物、腔腸動物、軟体動物、魚類、ヤスデ類、ハエ、菌類、蠕虫、甲殻類、および甲虫、特にPyrophorus属のコメツキムシ、ならびにPhotinus、Photuris、およびLuciola属のホタルを含めた多数の発光生物から単離されている。生物発光を示すさらなる生物は、WO00/024878、WO99/049019およびViviani(2002)に列挙されている。
【0104】
1つの非常によく知られている例は、特異的な生化学物質であるルシフェリン(天然に存在する蛍光体)がルシフェラーゼ活性を有する酵素によって酸化される、エネルギーを産生する化学反応を触媒するルシフェラーゼとして知られるタンパク質のクラスである(Hastings、1996)。原核生物も真核生物も、細菌、藻類、菌類、昆虫、魚類および他の海洋形態の種を含めて、非常に多様な生物がこのように光エネルギーを放出することができ、それぞれが、他の生物のものと化学的に異なる特異的なルシフェラーゼ活性およびルシフェリンを有する。ルシフェリン/ルシフェラーゼ系は、形態、化学的性質および機能において非常に多様である。したがって、ルシフェラーゼ活性を有する生物発光タンパク質は、様々な供給源からまたは様々な手段によって入手可能である。ルシフェラーゼ活性を有する生物発光タンパク質の例は、米国特許第5,229,285号、第5,219,737号、第5,843,746号、第5,196,524号および第5,670,356号で見つけることができる。最も広く使用されている2つのルシフェラーゼは、(i)ウミシイタケ(Renilla)ルシフェラーゼ(R.reniformis由来)、35kDaのタンパク質、基質としてセレンテラジンを使用し、480nmで光を放出する(Lorenzら、1991);および(ii)ホタルルシフェラーゼ(Photinus pyralis由来)、61kDaのタンパク質、基質としてルシフェリンを使用し、560nmで光を放出する(de Wetら、1987)である。
【0105】
【表1】
【0106】
ガウシア(Gaussia)ルシフェラーゼ(Gaussia princeps由来)は、生化学的アッセイで使用されている(Verhaegenら、2002)。ガウシア(Gaussia)ルシフェラーゼは20kDaのタンパク質であり、セレンテラジンを迅速な反応で酸化し、その結果470nmで明るい光が放出される。
【0107】
本発明に有用なルシフェラーゼは、Anachnocampa種(WO2007/019634)からも特徴付けられている。これらの酵素は、サイズが59kDaであり、放出スペクトルがスペクトルの青色部分に入る発光反応を触媒するATP依存性ルシフェラーゼである。
【0108】
本発明で使用できる代替の非ルシフェラーゼ生物発光タンパク質は、適切な基質に作用して発光シグナルを生じさせる任意の酵素である。そのような酵素の具体例は、β−ガラクトシダーゼ、ラクタマーゼ、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β−グルクロニダーゼおよびβ−グルコシダーゼである。これらの酵素の合成発光基質は当技術分野で周知であり、Tropix Inc.(Bedford、MA、USA)などの企業から市販されている。
【0109】
本発明に有用なペルオキシダーゼの例は、Hushpulianら(2007)によって記載されている。
【0110】
好ましい実施形態では、立体障害による相互作用の阻害を防止するために、低分子量の生物発光タンパク質が使用される。生物発光タンパク質は、好ましくは1本のポリペプチド鎖からなる。また、生物発光タンパク質は、好ましくはオリゴマーまたは凝集物を形成しない。生物発光タンパク質のウミシイタケ(Renilla)ルシフェラーゼ、ガウシア(Gaussia)ルシフェラーゼおよびホタルルシフェラーゼはこれらの基準の全てまたはほとんどを満たす。
【0111】
基質
基質の選択は、生物発光タンパク質によって生じた光の波長および強度に影響を及ぼし得る。
【0112】
広く知られている基質は、刺胞動物、カイアシ類、毛顎動物(chaetgnaths)、有櫛動物、十脚類シュリンプ、アミ類シュリンプ、放散虫類および一部の魚類の分類群に存在するセレンテラジンである(GreerおよびSzalay、2002)。例えばウミシイタケ(Renilla)ルシフェラーゼでは、418〜512nmの間で光を放出するセレンテラジン類似体/誘導体が利用可能である(Inouyeら、1997)。セレンテラジン類似体/誘導体(400A、DeepBlueC)は、ウミシイタケ(Renilla)ルシフェラーゼで光を400nmで放出することが記載されている(WO01/46691)。セレンテラジン類似体/誘導体の他の例は、EnduRenおよびViviRenである。
【0113】
本明細書において、「ルシフェリン」という用語は、生物発光が可能な生物で認められる、光を放出する生物学的色素のクラスを指し、それは酵素ルシフェラーゼの存在下で酸化されて、オキシルシフェリンおよび光の形でのエネルギーを産生する。ルシフェリン、または2−(6−ヒドロキシベンゾチアゾール−2−イル)−2−チアゾリン−4−カルボン酸は、ホタルPhotinus pyralisから初めて単離された。それ以来、様々な異なる生物から、主に海洋、例えば魚類およびイカから様々な形態のルシフェリンが発見され研究されてきたが、多くは陸上生物、例えば、蠕虫、甲虫および様々な他の昆虫において同定されている(Dayら、2004;Viviani、2002)。
【0114】
ルシフェリンには少なくとも5つの一般的な型があり、それぞれが化学的に異なり、広範囲の異なる補助因子を使用する化学的構造的に異なるルシフェラーゼによって触媒される。第1はホタルルシフェリンというホタルルシフェラーゼの基質であり、触媒作用にATPを必要とする(EC1.13.12.7)。第2は細菌ルシフェリンであり、一部のイカおよび魚類でも認められ、長鎖アルデヒドおよび還元型リン酸リボフラビンからなる。細菌ルシフェラーゼはFMNH依存性である。第3は渦鞭毛藻類(dinoflagellate)ルシフェリンであり、夜間の海洋のリン光に関与する生物である渦鞭毛藻類(海洋プランクトン)で認められるテトラピロールクロロフィル(tetrapyrrolic chlorophyll)誘導体である。渦鞭毛藻類ルシフェラーゼは渦鞭毛藻類ルシフェリンの酸化を触媒し、触媒活性のある3つの同一のドメインからなる。第4はイミダゾロピラジンのヴァルグリン(vargulin)であり、特定の貝虫類および深海魚、例えばPorichthysで認められる。最後はセレンテラジン(イミダゾールピラジン)であり、放散虫類、有櫛動物、刺胞動物、イカ、カイアシ類、毛顎動物、魚類およびシュリンプで認められるタンパク質エクオリンの光放出体である。
【0115】
アクセプター分子
本発明で使用できるいくつかの異なるアクセプター分子がある。アクセプター分子は、タンパク質でもよく、または非タンパク質性のものでもよい。タンパク質であるアクセプター分子の例として、それだけに限らないが、緑色蛍光タンパク質(GFP)、GFPの青色蛍光変異体(BFP)、GFPのシアン蛍光変異体(CFP)、GFPの黄色蛍光変異体(YFP)、増強型GFP(EGFP)、増強型CFP(ECFP)、増強型YFP(EYFP)、GFPS65T、エメラルド(Emerald)、トパーズ(Topaz)、GFPuv、不安定化EGFP(dEGFP)、不安定化ECFP(dECFP)、不安定化EYFP(dEYFP)、HcRed、t−HcRed、DsRed、DsRed2、t−ダイマー2、t−ダイマー2(12)、mRFP1、ポシロポリン、ウミシイタケ(Renilla) GFP、モンスター(Monster) GFP、paGFP、カエデ(Kaede)タンパク質もしくはフィコビリタンパク質(Phycobiliprotein)、またはそのいずれか1つの生物学的に活性な変異体もしくは断片がある。タンパク質でないアクセプター分子の例として、それだけに限らないが、Alexa Fluor色素、Bodipy色素、Cy色素、フルオレセイン、ダンシル、ウンベリフェロン、蛍光マイクロスフェア、発光マイクロスフェア、蛍光ナノ結晶、マリナブルー、カスケードブルー、カスケードイエロー、パシフィックブルー、オレゴングリーン、テトラメチルローダミン、ローダミン、テキサスレッド、希土類元素キレート、またはその任意の組合せもしくは誘導体がある。
【0116】
1つの非常によく知られている例は、クラゲAequorea victoria由来の緑色蛍光タンパク質および分子生物学、例えば突然変異導入およびキメラタンパク質技術の適用から生じた多数の他の変異体(GFP)を含む蛍光体の群である(Tsien、1998)。GFPは、その発色団の独特の構成成分に基づいて分類され、各クラスは異なる励起および放出波長を有する:クラス1、中性フェノールとアニオン性フェノレートの野生型混合物:クラス2、フェノレートアニオン:クラス3、中性フェノール:クラス4、積み重なったs電子系を有するフェノレートアニオン:クラス5、インドール:クラス6、イミダゾール:およびクラス7、フェニル。
【0117】
突然変異した、天然に存在するアクセプター分子(変異体)も本発明に有用であり得る。BRETに適する工学的に作製された系の1つの例は、ウミシイタケ(Renilla)ルシフェラーゼとGFPの増強型黄色突然変異体(EYFP)の対であり、1または複数の媒介タンパク質(この場合、Gタンパク質共役受容体)が存在しないと単独では互いにそれほど直接相互作用しない(Xuら、1999)。
【0118】
別の実施形態では、アクセプター分子は蛍光ナノ結晶である。ナノ結晶、または「量子ドット」は、光分解に対する耐性、明るさの向上、非毒性、およびいくつかのプロセスを同時にモニタリングすることを可能にするサイズ依存性の狭い放出スペクトルを含めて、蛍光標識として有機分子を上回るいくつかの利点を有する。さらに、ナノ結晶の吸収スペクトルは最初のピークの上で連続し、全てのサイズ、したがって全ての色を、単一の励起波長を用いて励起することを可能にする。
【0119】
蛍光ナノ結晶は、様々な形でタンパク質に付着させ、または「バイオコンジュゲート」させることができる。例えば、ジヒドロリポ酸(DHLA)または両親媒性ポリマー由来のカルボキシレート基を用いて、「量子ドット」の表面キャップを負に帯電させることができる。直接、または組換えタンパク質と融合した正に帯電したロイシンジッパーペプチドからなる架橋を介して、タンパク質をDHLA−ナノ結晶と静電的にコンジュゲートさせることができる。後者は、意図された標的との特異性によって一次抗体と結合する。あるいは、従来のカルボジイミド化学を用いて、抗体、ストレプトアビジン、または他のタンパク質をナノ結晶のポリアクリレートキャップと共有結合させる。
【0120】
セレン化カドミウム、硫化カドミウム、ヒ化インジウムおよびリン化インジウムを含めたナノ結晶を生成するコロイド法がある。これらの量子ドットは、その量子ドットの体積内にわずか100〜100,000個の原子を含有することができ、直径は原子10〜50個である。一部の量子ドットは、一方の物質が他方の物質に埋まり、大きなバンドギャップを有する小さな領域である。これらはいわゆるコアシェル構造といえ、例えば、コアにCdSe、シェルにZnSを有するもの、またはオルモシル(ormosil)と呼ばれるシリカの特殊な形態に由来するものがある。ドットが大きいと、その蛍光スペクトルは赤く(低エネルギーと)なる。逆に、小さいドットは、青い(高エネルギー)の光を放出する。色合いは、量子ドットのエネルギーレベルに直接関連する。定量的にいうと、蛍光の光のエネルギー(したがって色)を決定するバンドギャップエネルギーは、量子ドットのサイズの2乗に反比例する。大きい量子ドットは、大きいエネルギーレベルを有し、より密集している。このことは、量子ドットが低いエネルギー、すなわちスペクトルの赤い方に近いものを含有する光子を吸収することを可能にしている。
【0121】
代替の実施形態では、アクセプター分子は蛍光マイクロスフェアである。これらは典型的にはポリマーでできており、ポリマーマトリックス中に取り込まれた蛍光分子(例えば、フルオレセインGFPまたはYFP)を含有し、様々な試薬とコンジュゲートさせることができる。蛍光マイクロスフェアは内部標識することもでき、または表面上で標識することもできる。内部標識で、典型的には狭い蛍光放出スペクトルを有する非常に明るく安定した粒子が生成される。内部標識すると、表面の基は、リガンド(例えばタンパク質)とビーズの表面とのコンジュゲートに利用可能なままとなる。内部標識したビーズは、光退色に対して高い耐性を示すので、画像化の用途で広く使用される。
【0122】
カルボキシレート修飾蛍光マイクロスフェアは、1−エチル−3−(3−ジメチルアミノプロピル)カルボジイミド塩酸塩(EDAC)などの水溶性カルボジイミド試薬を使用したタンパク質の共有結合に適している。硫酸蛍光マイクロスフェアは比較的疎水性であり、ほとんどどんなタンパク質も受動的にかつほとんど不可逆的に吸着する。アルデヒド−硫酸蛍光マイクロスフェアは、表面アルデヒド基を付加し、タンパク質と反応するように修飾された硫酸マイクロスフェアである。
【0123】
別の実施形態では、アクセプター分子は発光マイクロスフェアである。これらは典型的にはポリマーでできており、ポリマーマトリックス中に取り込まれた発光分子(例えば、ユーロピウムまたは白金の複合体)を含有し、様々な試薬とコンジュゲートさせることができる。
【0124】
生物発光タンパク質とアクセプター分子の対
BRETに適した対を決定する際に考慮すべき基準は、生物発光タンパク質と比較した、アクセプター分子の相対的な放出/蛍光スペクトルである。生物発光タンパク質の放出スペクトルは、生物発光タンパク質の発光の放出からの光エネルギーが、2つの分子が適切に近接し互いに対して適切な配向にあるときにアクセプター分子を励起することができ、それによってアクセプター分子の蛍光を促進する波長であるように、アクセプター分子の吸光度スペクトルと重複すべきである。例えば、観察可能な放出スペクトルのピークに基づくと、ウミシイタケ(Renilla)ルシフェラーゼ/EGFPの対はウミシイタケ(Renilla)ルシフェラーゼ/EYEFの対ほど良好でないことが実証されている(Xu、1999;Wangら、(1997)、Bioluminescence and Chemiluminescence:Molecular Reporting with Photons、Hastingsら編(Wiley、New York)、pp.419〜422)。潜在的な対を研究するために、選択した生物発光タンパク質およびアクセプター分子を含有するタンパク質融合物を調製し、適当な基質の存在下で試験する。
【0125】
生物発光タンパク質とアクセプター分子が誤って互いに結合しないことも確認すべきである。これは、同じ細胞中で生物発光タンパク質およびアクセプター分子を別々に同時発現させ、次いで発光スペクトルをモニタリングしてBRETが起こるかどうかを決定することにより行うことができる。これは、例えば、Xuら、(1999)の方法を使用して実現することができる。BRETがほとんどまたは全く観察されない場合、選択した生物発光タンパク質とアクセプター分子は適切なBRETの対を形成する。
【0126】
生物発光タンパク質放出は、基質の修飾によって操作することができる。ルシフェラーゼの場合、基質はセレンテラジンである。生物発光タンパク質放出を変化させることの背後にある論理的根拠は、ドナー放出とアクセプター放出の間の分解能を向上させることにある。元のBRET系は、ウミシイタケ(Renilla)ルシフェラーゼをドナーとして、EYFP(またはTopaz)をアクセプターとして、セレンテラジンh誘導体を基質として使用する。BRETアッセイで組み合わせたとき、これらの構成成分は、生物発光タンパク質については475〜480nmで、アクセプター分子については525〜530nmで光を発し、スペクトル分解能は45〜55nmとなる。
【0127】
残念ながら、ウミシイタケ(Renilla)ルシフェラーゼでは実質的にGFP放出と重複する広い放出ピークが生じ、これは系のシグナル対ノイズを低下させる一因となる。ウミシイタケ(Renilla)ルシフェラーゼの基質としてcoel400aを使用する本発明の1つのBRET系は、ドナーとアクセプターの放出波長間の広いスペクトル分解能をもたらす(−105nm)。ウミシイタケ(Renilla)ルシフェラーゼとcoel400aは、390〜400nmで光を発し、この範囲で光を吸収し、505〜508nmで光を再び放出するGFPを調製した。ウミシイタケ(Renilla)ルシフェラーゼとGFPの放出間のスペクトル分解能のこの増大のため、このBRET系は、本発明のポリペプチドの高次構造の小さな変化をモニタリングする優れた生物学的ツールをもたらす。これは、ドナーとアクセプターの波長の差がおよそ51nmである、セレンテラジンh誘導体およびEYFPを使用する以前に記載されている系を上回る著しい向上である。
【0128】
ウミシイタケ(Renilla)ルシフェラーゼ活性の結果(野生型セレンテラジンにより発せられるものとは異なる)様々な波長で光を発するcoel400aを含めて、様々なセレンテラジン誘導体が当技術分野で知られている。当業者は、ドナーの光放出ピークが変化したため、この波長で光を吸収し、それによって効率のよいエネルギー移動を可能にするアクセプター分子を選択する必要があることを理解するであろう。これは、例えばGFPクラス4を、クラス3または1のGFPとなるように変更することによって行うことができる。ドナーの光放出とアクセプターの光吸収ピークとの間のスペクトルの重複は、とりわけ、効率のよいエネルギー移動の1つの条件である。クラス3および1のGFPは、400nmで光を吸収し、505〜511nmで再び放出することが知られている。この結果、およそ111nmのドナーとアクセプターの放出間の波長の差となる。
【0129】
さらなる生物発光タンパク質とアクセプター分子の対の例を表2に示す。
【0130】
【表2】
【0131】
Gタンパク質共役受容体
Gタンパク質共役受容体(GPCR)は、7回膜貫通受容体、7TM受容体、7回らせん(heptahelical)受容体、およびGタンパク質連結受容体(GPLR)としても知られている。GPCRは、細胞の外側で分子を感知し、内側でシグナル伝達経路を、最終的に細胞応答を活性化する膜貫通受容体の大型タンパク質のファミリーである。これらの受容体と結合しそれを活性化するリガンドには、光感受性化合物、匂い、フェロモン、ホルモン、および神経伝達物質があり、小分子からペプチド、大型タンパク質までサイズが様々である。GPCRは多数の疾患に関与するが、現代の全医療用薬物のほぼ半分の標的でもある。GPCRは、配列相同性および機能類似性に基づいて、少なくとも5つのクラスに分類することができる:
クラスAのロドプシン様、
クラスBのセクレチン様、
クラスCの代謝型/フェロモン、
クラスDの菌類フェロモン、および
クラスEのcAMP受容体。
【0132】
クラスAのロドプシン様受容体には、アミン受容体:アセチルコリン、アルファアドレナリン受容体、ベータアドレナリン受容体、ドーパミン、ヒスタミン、セロトニン、オクトパミン、および微量アミン;ペプチド受容体:アンギオテンシン、ボンベシン、ブラジキニン、C5aアナフィラトキシン、Fmet−leu−phe、APJ様、インターロイキン−8、ケモカイン受容体(C−Cケモカイン、C−X−Cケモカイン、BONZO受容体(CXC6R)、C−X3−Cケモカイン、およびXCケモカイン)、CCK受容体、エンドセリン受容体、メラノコルチン受容体、神経ペプチドY受容体、ニューロテンシン受容体、オピオイド受容体、ソマトスタチン受容体、タキキニン受容体(サブスタンスP(NK1)、サブスタンスK(NK2)、ニューロメジンK(NK3)、タキキニン様1、およびタキキニン様2)、バソプレッシン様受容体(バソプレッシン、オキシトシン、およびコノプレッシン)、ガラニン様受容体(ガラニン、アラトスタチン、およびGPCR54)、プロテイナーゼ活性化様受容体(例えば、トロンビン)、オレキシン&神経ペプチドFF、ウロテンシンII受容体、アドレノメデュリン(G10D)受容体、GPR37/エンドセリンB様受容体、ケモカイン受容体様受容体、およびニューロメジンU受容体;ホルモンタンパク質受容体:卵胞刺激ホルモン、ルトロピン−絨毛性ゴナドトロピンホルモン、チロトロピン、およびゴナドトロピン;オプシン(ロドプシン)受容体;嗅受容体;プロスタノイド受容体:プロスタグランジン、プロスタサイクリン、およびトロンボキサン;ヌクレオチド様受容体:アデノシンおよびプリン受容体;カンナビス受容体;血小板活性化因子受容体;ゴナドトロピン放出ホルモン受容体;チロトロピン放出ホルモン&分泌促進物質受容体:チロトロピン放出ホルモン、成長ホルモン分泌促進物質、および成長ホルモン分泌促進物質様;メラトニン受容体;ウイルス受容体;リゾスフィンゴ脂質&LPA(EDG)受容体;ロイコトリエンB4受容体:ロイコトリエンB4受容体BLT1およびロイコトリエンB4受容体BLT2;ならびにクラスAオーファン/他の受容体:血小板ADP&KI01受容体、SREB、Mas原癌遺伝子、RDC1、ORPH、LGR様(ホルモン受容体)、GPR、GPR45様、システイニルロイコトリエン、Mas関連受容体(MRG)、およびGP40様受容体がある。
【0133】
GPCRのクラスB(セクレチン受容体ファミリー)には、ポリペプチドホルモンの受容体(カルシトニン、コルチコトロピン放出因子、胃抑制ペプチド、グルカゴン、グルカゴン様ペプチド−1、−2、成長ホルモン放出ホルモン、副甲状腺ホルモン、PACAP、セクレチン、血管作用性小腸ポリペプチド、利尿ホルモン、EMR1、ラトロフィリン)、形質膜での細胞間相互作用を媒介すると考えられる分子(脳特異的血管新生阻害因子(BAI))ならびにストレス応答および寿命を制御する一群のDrosophilaタンパク質(Methuselah様タンパク質)がある。
【0134】
クラスCの代謝型グルタミン酸/フェロモン受容体には、代謝型グルタミン酸、代謝型グルタミン酸グループI、代謝型グルタミン酸グループII、代謝型グルタミン酸グループIII、他の代謝型グルタミン酸、細胞外カルシウム感知、推定上のフェロモン受容体、GABA−B、GABA−Bサブタイプ1、GABA−Bサブタイプ2、およびオーファンGPRC5受容体がある。
【0135】
GPCRは、視覚、嗅覚、行動および気分の制御、免疫系の活性および炎症の制御、自律神経系の伝達、細胞密度の感知、ならびにその他多くを含めて、多種多様な生理的過程に関与する。不活性なGタンパク質は、その不活性状態で受容体と結合することが知られている。リガンドを認識した後、受容体またはそのサブユニットは高次構造を変化させ、それによってGタンパク質を機械的に活性化し、Gタンパク質は受容体から解離する。受容体はここで別のGタンパク質を活性化することもでき、切り替わって不活性状態に戻ることもできる。受容体分子は、活性と不活性の生物物理的状態の間の高次構造の平衡状態にあると考えられている。リガンドが受容体に結合すると、活性な受容体の状態の方に平衡状態を変化させることができる。本発明のポリペプチドはGタンパク質共役受容体を含み、細胞中で発現したときにその受容体のN末端が細胞の外側にあり、C末端が細胞の内側にある。当業者は、膜貫通タンパク質の配向を検出するのに適した技術を承知している。そのような技術は、それだけに限らないが、結晶解析、NMR研究、モデリング研究ならびに光学または電子顕微鏡標本の界面活性剤浸透化対照と組み合わせた免疫標識のような顕微鏡技術、2つのポリペプチドの断片相補性タギングなどを含む。
【0136】
好ましい実施形態では、Gタンパク質共役受容体はクラスAのGPCRである。さらなる好ましい実施形態では、クラスA(ロドプシン様)GPCRは嗅覚受容体またはアドレナリン作動性受容体であり、より好ましくは嗅覚受容体である。細胞中で発現したときに受容体のN末端が細胞の外側にあり、C末端が細胞の内側にある限り、嗅覚受容体はどんな供給源由来のものでもよい。例として、それだけに限らないが、脊索動物受容体、線虫受容体、またはそのいずれか1つの生物学的に活性な変異体もしくは断片がある。脊索動物受容体の例として、それだけに限らないが、哺乳動物受容体、鳥類受容体および魚類受容体がある。好ましい実施形態では、嗅覚受容体は、線虫受容体、またはその生物学的に活性な変異体もしくは断片である。一実施形態では、線虫受容体は、Caenorhabditis elegans受容体、またはその生物学的に活性な変異体もしくは断片である。本発明のポリペプチドの生成に使用し、かつ/または本発明の方法で使用することができる嗅覚受容体の例は、BuckおよびAxel(1991)、Robertson(1998および2001)、Aloniら(2006)、Feldmesser(2006)、Olenderら(2004aおよびb)、Glusmanら(2000a、2000bおよび2001)、Fuchsら(2001)、PilpelおよびLancet(1999)、Sharonら(1998)、Zozulyaら(2001)、NiimuraおよびNei(2003)、Landerら(2001)、ZhangおよびFirestein(2002)、Youngら(2002)に記載されている。さらに、嗅覚受容体の包括的な一覧がSenseLabのウェブサイトから入手可能である(http://senselab.med.yale.edu)。
【0137】
他の実施形態では、GPCRはクラスBまたはクラスCの受容体であり、これら2つの実施形態のうちクラスCがより好ましい。
【0138】
特に好ましい実施形態では、Gタンパク質共役受容体は7回膜貫通ドメインを含む。
【0139】
生物発光タンパク質は、Gタンパク質共役受容体(または本発明のポリペプチド)の1番目、3番目、5番目の非膜貫通ループ(ドメイン)またはC末端の一部を形成することができる。アクセプター分子も、Gタンパク質共役受容体(または本発明のポリペプチド)の1番目、3番目、5番目の非膜貫通ループ(ドメイン)またはC末端の一部を形成することができる。Gタンパク質共役受容体が発現し、細胞中に存在するときに、これらの領域はそれぞれ細胞内にある。
【0140】
アクセプター分子は、同じ分子の一部(すなわち、同じ1本のポリペプチド鎖)のとき、生物発光タンパク質と同じ領域にある可能性はないが、Gタンパク質共役受容体が二量体または高次の多量体として存在するとき、アクセプター分子は生物発光タンパク質と等しい領域にある可能性がある。例えば、生物発光タンパク質は、受容体の1つのサブユニットのC末端の一部を形成することができ、アクセプター分子は、受容体の別のサブユニットのC末端の一部を形成することができる。この例では、標識が結合したサブユニットは同じでもよく、または異なっていてもよく、例えば2つのサブユニットが同一であり、生物発光タンパク質で標識された一方とアクセプター分子で標識された他方が離れていてもよい。一実施形態では、生物発光タンパク質はGPCRサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子は5番目の非膜貫通ループの一部を形成する。代替の実施形態では、アクセプター分子はGPCRサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質は5番目の非膜貫通ループの一部を形成する。
【0141】
別の実施形態では、生物発光タンパク質はGPCRサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子は3番目の非膜貫通ループの一部を形成する。別の実施形態では、アクセプター分子はGPCRサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質は3番目の非膜貫通ループの一部を形成する。
【0142】
好ましい実施形態では、生物発光タンパク質はGPCRサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子はC末端の一部を形成する。代替の実施形態では、アクセプター分子はGPCRサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質はC末端の一部を形成する。
【0143】
別の実施形態では、Gタンパク質共役受容体は少なくとも2つのサブユニットを含み、生物発光タンパク質が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの5番目の非膜貫通ループの一部を形成する。代替の実施形態では、アクセプター分子は第1のサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質は第2のサブユニットの5番目の非膜貫通ループの一部を形成する。
【0144】
別の実施形態では、Gタンパク質共役受容体は少なくとも2つのサブユニットを含み、生物発光タンパク質が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの3番目の非膜貫通ループの一部を形成する。別の実施形態では、アクセプター分子は第1のサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質は第2のサブユニットの3番目の非膜貫通ループの一部を形成する。
【0145】
別の実施形態では、Gタンパク質共役受容体は少なくとも2つのサブユニットを含み、生物発光タンパク質が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットのC末端の一部を形成する。代替の実施形態では、アクセプター分子は第1のサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質は第2のサブユニットのC末端の一部を形成する。
【0146】
別の実施形態では、Gタンパク質共役受容体は少なくとも2つのサブユニットを含み、ドナーおよびアクセプター分子はそれぞれ第1および第2のサブユニットの同じ部位にある。
【0147】
一実施形態では、生物発光タンパク質またはアクセプター分子は、5番目の膜貫通ドメインの2番目のアミノ酸の後、6番目の膜貫通ドメインの開始の前から2番目のアミノ酸の前に位置する。別の実施形態では、生物発光タンパク質またはアクセプター分子は、5番目の膜貫通ドメインの後のアミノ酸8付近の後、または5番目の膜貫通ドメインの後のアミノ酸22付近の後に位置する。さらなる実施形態では、生物発光タンパク質またはアクセプター分子は、6番目の膜貫通ドメインから約10または12アミノ酸前に挿入される。最も好ましくは、生物発光タンパク質またはアクセプター分子は、3番目の非膜貫通ループ(ドメイン)の中央に位置する。
【0148】
C末端に関しては、天然のC末端の約5〜25アミノ酸が7番目の膜貫通ドメインの末端に残存することが好ましい。好ましくは、生物発光タンパク質またはアクセプター分子は、7番目の膜貫通部の後から約16または20アミノ酸後に挿入される。
【0149】
1番目の非膜貫通ループ(ドメイン)における生物発光タンパク質またはアクセプター分子の位置については、前記標識が1番目の膜貫通ドメインの末端から約2アミノ酸後、および2番目の膜貫通ドメインの開始から約2アミノ酸前に挿入されることが好ましい。最も好ましくは、生物発光タンパク質またはアクセプター分子は、1番目の非膜貫通ループ(ドメイン)の中央に位置する。
【0150】
さらなる実施形態では、生物発光タンパク質は、Gタンパク質共役受容体(または本発明のポリペプチド)のN末端、2番目、4番目または6番目の非膜貫通ループ(ドメイン)の一部を形成することができる。アクセプター分子も、Gタンパク質共役受容体(または本発明のポリペプチド)のN末端、2番目、4番目または6番目の非膜貫通ループ(ドメイン)の一部を形成することができるが、同じ分子の一部のとき、生物発光タンパク質と同じ領域にある可能性はない。Gタンパク質共役受容体が発現し、細胞中に存在するときに、これらの領域はそれぞれ細胞外にある。
【0151】
GPCRは、2つ以上の異なるGPCRの天然に存在しないキメラでもよい。特に、このことは、1つの受容体の部分が、検出する化合物に応じて多種多様なGPCRの他の部分が挿入されたキメラ中に常に存在する伝達カセットを生成することを可能にする。
【0152】
一実施形態では、サブユニットは、第1のGタンパク質共役受容体サブユニットのN末端および1番目の膜貫通ドメインの少なくとも大部分、第2のGタンパク質共役受容体サブユニットの1番目の非膜貫通ループの少なくとも大部分から5番目の膜貫通ドメインの少なくとも大部分まで、ならびに第1のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む。
【0153】
別の実施形態では、サブユニットは、第1のGタンパク質共役受容体サブユニットのN末端から5番目の膜貫通ドメインの少なくとも大部分まで、および第2のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む。
【0154】
当業者は、Gタンパク質共役物のN末端、膜貫通ドメイン、非膜貫通ループ(ドメイン)およびC末端を容易に決定することができる。例えば、様々なバイオインフォマティクスの手法を使用して、そのアミノ酸配列およびGタンパク質共役受容体の既知の膜貫通ドメインとの類似性に基づいて、タンパク質における膜貫通ドメインの位置およびトポロジーを決定することができる。アラインメントおよびアミノ酸配列比較は、例えば、BLASTプログラムまたはCLUSTAL Wプログラムを使用することにより、当技術分野で日常的に行われる。既知の膜貫通ドメイン含有タンパク質とのアラインメントに基づいて、当業者は膜貫通ドメインの位置を予測することができる。さらに、一部の膜貫通タンパク質の三次元構造が知られており、例えば、7回膜貫通Gタンパク質共役ロドプシン光受容体構造がX線結晶解析によって解析されている。そのような3D構造の分析およびそれとの比較に基づいて、他の膜タンパク質における膜貫通ドメインの位置およびトポロジーを予測することができる。タンパク質における膜貫通ドメインの位置およびトポロジーの予測に利用可能な多数のプログラムも存在する。例えば、膜貫通タンパク質のセグメントを予測するTMpred(HofmannおよびStoffel、1993);膜タンパク質のトポロジーを予測するTopPred(von Heijneら、1992);単一および複数の配列から二次構造を予測するPREDATOR(FrishmanおよびArgos、1997);複数整列させた配列からタンパク質の膜貫通領域を予測するTMAP(PerssonおよびArgos、1994);ならびに単一の配列から膜貫通領域を予測するALOM2(Klienら、1984)の1つまたは組合せを使用することができる。
【0155】
標準的な命名法に従って、膜貫通ドメインおよび非膜貫通ループ(ドメイン)の番号付けは、ポリペプチドのN末端に応じて行う。
【0156】
天然に存在するGタンパク質共役受容体のアミノ酸配列突然変異体/変異体は、コードするポリヌクレオチド中に適当なヌクレオチド変化を導入することにより、または所望のポリペプチドのin vitro合成により調製することができる。そのような突然変異体には、例えば、アミノ酸配列内の残基の欠失、挿入または置換物がある。最終的なポリペプチド産物が所望の特徴を有することを条件として、欠失、挿入および置換の組合せを行って最終的な構築物に到達することができる。
【0157】
突然変異(変異)ポリペプチドは、当技術分野で知られている任意の技術を使用して調製することができる。例えば、本明細書に記載のポリヌクレオチドをin vitro突然変異導入にかけることができる。そのようなin vitro突然変異導入技術には、適切なベクターへのポリヌクレオチドのサブクローニング、E.coli XL−1red(Stratagene)などの「突然変異誘発」株へのベクターの形質転換、および適切な数の世代にわたる形質転換細菌の増殖が含まれ得る。別の例では、Harayama(1998)によって大枠が記載されている通りに、Gタンパク質共役受容体をコードするポリヌクレオチドをDNAシャッフリング技術にかける。本明細書に記載の技術を使用して突然変異/変異DNAに由来する産物を容易にスクリーニングして、それが本発明の方法に有用であるかを決定することができる。
【0158】
アミノ酸配列突然変異体の設計において、突然変異部位の位置および突然変異の性質は、改変する(複数の)特徴に依存する。突然変異の部位は、例えば、(1)最初に保存的アミノ酸を選択して置換し、次いで得られた結果に応じて選択を急進的にして置換すること、(2)標的残基を欠失させること、または(3)位置する部位の隣に他の残基を挿入することによって、個々にまたは連続して改変することができる。
【0159】
アミノ酸配列の欠失は、一般に約1〜15個の残基であり、より好ましくは約1〜10個の残基であり、典型的には約1〜5個の連続した残基に及ぶ。
【0160】
置換突然変異体は、Gタンパク質共役受容体における少なくとも1つのアミノ酸残基が除去され、その場所に異なる残基が挿入される。置換突然変異導入で最も興味深い部位は、機能にとって重要なものとして同定された部位を含む。他の興味深い部位は、様々な株または種から得られた特定の残基が同一であるものである。これらの位置は、生物活性にとって重要である可能性がある。これらの部位、特に少なくとも3つの他の同一に保存されている部位の配列に入るものは、比較的保存的な形で置換することが好ましい。そのような保存的置換を表3に示す。
【0161】
さらに、所望される場合、置換物または付加物として非天然アミノ酸または化学的アミノ酸類似体を本明細書に記載のポリペプチドに導入することができる。そのようなアミノ酸には、それだけに限らないが、一般的なアミノ酸のD−異性体、2,4−ジアミノ酪酸、α−アミノイソ酪酸、4−アミノ酪酸、2−アミノ酪酸、6−アミノヘキサン酸、2−アミノイソ酪酸、3−アミノプロピオン酸、オルニチン、ノルロイシン、ノルバリン、ヒドロキシプロリン、サルコシン、シトルリン、ホモシトルリン、システイン酸、t−ブチルグリシン、t−ブチルアラニン、フェニルグリシン、シクロヘキシルアラニン、β−アラニン、フルオロアミノ酸、β−メチルアミノ酸、Cα−メチルアミノ酸、Nα−メチルアミノ酸などのデザイナーアミノ酸、および一般のアミノ酸類似体がある。
【0162】
例えば、ビオチン化、ベンジル化、グリコシル化、アセチル化、リン酸化、アミド化、既知の保護/遮断基による誘導体化、タンパク質分解性切断、抗体分子または他の細胞性リガンドとの連結などにより、合成の間またはその後に異なった形で修飾されたポリペプチドも、本発明の範囲に含まれる。これらの修飾物は、ポリペプチドの安定性および/または生物活性を増大させる働きをし得る。
【0163】
【表3】
【0164】
本明細書に記載のポリペプチドは、組換えポリペプチドの産生および回収、ならびにポリペプチドの化学合成を含めた様々な形で生成することができる。一実施形態では、ポリペプチドを発現することができる細胞をポリペプチドの産生に有効な条件下で培養し、そのポリペプチドを回収することにより、本発明の単離ポリペプチドを生成する。培養するのに好ましい細胞は、本発明の組換え細胞である。有効な培養条件には、それだけに限らないが、有効な培地、バイオリアクター、ポリペプチド産生を可能にする温度、pHおよび酸素条件がある。有効な培地とは、細胞を培養して本発明のポリペプチドを産生する任意の培地を指す。典型的には、そのような培地は、同化できる炭素、窒素およびリン酸源、ならびに適当な塩、ミネラル、金属およびビタミンなどの他の栄養素を有する水性培地を含む。従来の発酵バイオリアクター、振盪フラスコ、試験管、マイクロタイター皿、およびペトリ皿の中で本発明の細胞を培養することができる。組換え細胞に適した温度、pHおよび酸素含量で培養を実施することができる。そのような培養条件は、当業者の専門知識の範囲内にある。
【0165】
アクセサリー分子
アクセサリー分子は、G共役型タンパク質受容体のシグナル伝達に関与するタンパク質である。例として、Gタンパク質およびアレスチンがある。任意の標識アクセサリー分子は、Gタンパク質共役受容体(および/または本明細書で定義されるポリペプチド)と直接または間接的に結合し、化合物とGタンパク質共役受容体が結合した後、2つの標識の相対的位置が空間的に変化して、BRETの検出を可能にする限り、本発明の方法で使用することができる。
【0166】
当業者は、当技術分野で標準的な技術を使用して、特定のGタンパク質共役受容体と結合するアクセサリー分子を容易に同定することができる。
【0167】
Gタンパク質共役受容体について上記で記載した突然変異体(変異体)の詳細は、アクセサリー分子にも当てはまる。
【0168】
生物発光タンパク質またはアクセプター分子は、アクセサリー分子が依然として受容体と直接または間接的に結合することができ、対象とする化合物と結合した後、BRETの変化を検出することができる限り、アクセサリー分子中のどこにでも位置することができる。一実施形態では、生物発光タンパク質またはアクセプター分子は、アクセサリー分子のC末端またはN末端に位置する。GPCRとの複合体の形成に少なくとも1つの他のアクセサリー分子を必要とする場合、アクセサリー分子は受容体と間接的に結合し、例えば、第1のアクセサリー分子は受容体と結合し、第2のアクセサリー分子は、第1のアクセサリー分子と直接結合することによって受容体と間接的に結合する。
【0169】
Gタンパク質
Gタンパク質は、細胞内シグナル伝達のセカンドメッセンジャーカスケードに関与するタンパク質のファミリーである。Gタンパク質は、不活性なグアノシン二リン酸(GDP)結合状態と活性なグアノシン三リン酸(GTP)結合状態とを入れ替える「分子スイッチ」として機能する。最終的に、Gタンパク質はシグナル伝達ネットワークのカスケードを開始することにより下流の細胞過程を制御するHofmannら、2009;OldhamおよびHamm、2008)。
【0170】
2つの異なるGタンパク質のファミリー:Gタンパク質共役受容体によって活性化され、アルファ(α)、ベータ(β)およびガンマ(γ)サブユニットで構成される、時折「大型」Gタンパク質と称されるヘテロ三量体Gタンパク質;ならびに低分子量GTPアーゼのRasスーパーファミリーに属する「小型」Gタンパク質(20〜25kDa)がある。これらのタンパク質は、ヘテロ三量体Gタンパク質で認められるアルファ(α)サブユニットと相同であり、やはりGTPおよびGDPと結合し、シグナル伝達に関与する。形質膜と結合するために、多数のGタンパク質が共有結合により脂質で修飾され、例えば、ヘテロ三量体Gタンパク質サブユニットはミリストイル化、パルミトイル化、またはプレニル化されていることがあり、一方で小型Gタンパク質はプレニル化されていることがある。
【0171】
熟練した受け手なら気づくであろうが、既知のGタンパク質が多数ある。線虫においていくつかのヘテロ三量体Gタンパク質が同定されており、例えばC.elegansは21個のGα、2個のGβおよび2個のGγ遺伝子を有する(Jansenら、1999;Cuppenら、2003)。配列類似性に基づいて、哺乳動物Gαサブユニットは、4つのファミリー:Gs、Gi/o、GqおよびG12に分けられている(Nevesら、2002)。C.elegansは各哺乳動物ファミリーの1つのオルソログを発現する:GSA−1(Gs)、GOA−1(Gi/o)、EGL−30(Gq)およびGPA−12(G12)。残りのC.elegansαサブユニット(GPA−1〜11、GPA−13〜17およびODR−3)は、分類を可能にするほど十分な相同性を有さない。保存されるGαサブユニットは、GPA−12を除いて幅広く発現されるが、新たなGα遺伝子のうち14個が化学感覚ニューロンのサブセット中で発現される。
【0172】
GβサブユニットGPB−1ならびにGγサブユニットGPC−2は、古典的なGタンパク質ヘテロ三量体中でαサブユニットと一緒に機能すると思われる。残りのGβサブユニットGPB−2は特定のRGSタンパク質の機能を制御すると考えられるが、残りのGγサブユニットGPC−1は、化学感覚において制限された役割を有する。したがって、C.elegansにおけるほとんどのGタンパク質経路の機能的差異はαサブユニットにある。
【0173】
アレスチン
アレスチンは、Gタンパク質シグナル伝達の終結に重要なタンパク質の1つである。活性化したGPCRはGタンパク質共役受容体キナーゼ(GRK)によって特異的にリン酸化され、次いでアレスチンと結合して受容体/Gタンパク質相互作用が妨げられ、その結果後のシグナル伝達が抑えられる。アレスチンは、アゴニストで活性化されているものを含むGPCRと結合し、GRKによってリン酸化されていないものよりされているものと堅く結合する。
【0174】
GPCRの分類をもたらす、細胞内のアレスチン輸送の2つの異なるパターンが以下のように描写されている:アレスチンが細胞表面で受容体と相互作用するが、エンドサイトーシスで小胞中に取り込まれず、したがって受容体との一過性の相互作用を示すクラスA、ならびにR−アレスチンおよび受容体が細胞膜からエンドサイトーシス小胞へと一緒に輸送されるクラスB。これら2つのクラスの受容体は、異なるアレスチンアイソフォームに対する親和性に関しても異なる。さらに、クラスA受容体は、R−アレスチン2と優先的に結合するが、クラスB受容体は、等しい親和性でβ−アレスチン1およびH−アレスチン2と結合する。
【0175】
R−アレスチンの結合により受容体がその同族Gタンパク質から脱共役し、下流のセカンドメッセンジャー分子を介したGPCRシグナル伝達の抑制または脱感作を引き起こす。
【0176】
Gタンパク質シグナルを終結する一方で、アレスチンの結合は、GPCRからの新たなシグナル伝達を開始することができる。例えば、R−アレスチンは、Srcなどの非受容体チロシンキナーゼに、内在化している受容体とのシグナル伝達複合体を形成させるアダプターとして働く。H−アレスチンは、ASK−MKK4−JNK3やRAF−MEK−ERK1/2などのMAPKモジュールの、GPCRによって制御された足場として機能する。さらに、アレスチンはクラスリン、AP2複合体のβ−アダプチンサブユニット2やArf−6などのエンドサイトーシス機構のタンパク質と相互作用し、したがってクラスリン被覆小胞を介して受容体の内在化を促進する。
【0177】
線虫アレスチンの例は、C.elegans由来のARR−1であり、これはHSNニューロンおよび様々な化学感覚ニューロンを含めた神経系で主として発現し、可溶性および揮発性匂い物質の検出に関与する。20を超える推定上のアレスチンが線虫C.elegansにおいて現在同定されている。そのようなアレスチンは神経系全体を通じて発現し、脊椎動物非視覚性アレスチンと同様に受容体内在化を支援し、視覚/感覚性サブタイプと同様に嗅覚および視覚にも関与する。
【0178】
組換えベクター
本発明の一実施形態は、宿主細胞中にポリヌクレオチド分子を送達することができる任意のベクター中に挿入された、本明細書に記載のポリペプチドをコードする少なくとも1つの単離/外因性ポリヌクレオチドを含む組換えベクターを含む。そのようなベクターは、異種のポリヌクレオチド配列を含有し、この配列は本発明のポリヌクレオチド分子の隣に天然には認められないポリヌクレオチド配列であり、好ましくは、その(複数の)ポリヌクレオチド分子が由来する種以外の種に由来する。ベクターはRNAでもまたはDNAでもよく、原核生物性でもまたは真核生物性でもよく、典型的には(米国特許第5,792,294号に記載のような)トランスポゾン、ウイルスまたはプラスミドである。
【0179】
組換えベクターの1つの型は、発現ベクターと作動可能に連結する(複数の)ポリヌクレオチドを含む。作動可能に連結するという語句は、分子が宿主細胞に形質転換されたときに発現することができるような形で発現ベクター中にポリヌクレオチド分子を挿入することを指す。本明細書において、発現ベクターは、宿主細胞を形質転換し、特定のポリヌクレオチド分子の発現を行うことができるDNAまたはRNAベクターである。好ましくは、発現ベクターは、宿主細胞内で複製することもできる。発現ベクターは原核生物性でもまたは真核生物性でもよく、典型的にはウイルスまたはプラスミドである。発現ベクターは、細菌、菌類、内部寄生生物、節足動物、動物、および植物細胞を含めた組換え細胞中で機能する(すなわち、遺伝子発現を誘導する)任意のベクターを含む。本発明のベクターを使用して無細胞発現系でポリペプチドを産生することもでき、そのような系は当技術分野で周知である。
【0180】
本明細書において「作動可能に連結する」とは、2つ以上の核酸(例えばDNA)セグメント間の機能的関係を指す。典型的には、それは、転写される配列に対する転写制御エレメントの機能的関係を指す。例えば、プロモーターは、適当な宿主細胞および/または無細胞発現系中でコード配列の転写を刺激または調節する場合、本明細書で定義されるポリヌクレオチドなどのコード配列と作動可能に連結する。一般に、転写される配列と作動可能に連結しているプロモーター転写制御エレメントは、転写される配列と物理的に連続し、すなわちそれはシス作用性である。しかし、エンハンサーなど一部の転写制御エレメントは、転写を亢進するコード配列と物理的に連続し、または近接して位置する必要はない。
【0181】
具体的には、本発明の発現ベクターは、転写調節配列、翻訳調節配列、複製起点や、組換え細胞に適合し本発明のポリヌクレオチド分子の発現を調節する他の制御配列などの制御配列を含有する。具体的には、本発明の組換え分子は、転写調節配列を含む。転写調節配列は、転写の開始、延長、および終結を調節する配列である。特に重要な転写調節配列は、プロモーター、エンハンサー、オペレーターやレプレッサー配列など、転写開始を調節するものである。適切な転写調節配列は、本発明の組換え細胞の少なくとも1つの中で機能することができる任意の転写調節配列を含む。様々なそのような転写調節配列が当業者に知られている。好ましい転写調節配列には、それだけに限らないが、tac、lac、trp、trc、oxy−pro、omp/lpp、rrnB、バクテリオファージラムダ、バクテリオファージT7、T7lac、バクテリオファージT3、バクテリオファージSP6、バクテリオファージSP01、メタロチオネイン、α接合因子、Pichiaアルコールオキシダーゼ、アルファウイルスサブゲノムプロモーター(シンドビズウイルスサブゲノムプロモーターなど)、抗生物質耐性遺伝子、バキュロウイルス、Heliothis zea昆虫ウイルス、ワクシニアウイルス、ヘルペスウイルス、アライグマポックスウイルス、他のポックスウイルス、アデノウイルス、サイトメガロウイルス(中間初期プロモーターなど)、シミアンウイルス40、レトロウイルス、アクチン、レトロウイルス長末端反復配列、ラウス肉腫ウイルス、熱ショック、リン酸および硝酸転写調節配列、ならびに原核または真核細胞中で遺伝子発現を調節することができる他の配列など、細菌、酵母、節足動物、線虫、植物または動物細胞中で機能するものがある。
【0182】
宿主細胞
本発明の別の実施形態は、本明細書に記載の1つもしくは複数の組換え分子で形質転換された宿主細胞またはその子孫細胞を含む。細胞へのポリヌクレオチド分子の形質転換は、ポリヌクレオチド分子を細胞中に挿入できる任意の方法で行うことができる。形質転換の技術には、それだけに限らないが、トランスフェクション、エレクトロポレーション、マイクロインジェクション、リポフェクション、吸着、およびプロトプラスト融合がある。組換え細胞は、単細胞のままでもよく、または成長して組織、臓器もしくは多細胞生物になってもよい。本発明の形質転換されたポリヌクレオチド分子は、染色体外にあるままでもよく、または発現する能力が保持される形で、形質転換される(すなわち組換え)細胞の染色体内の1つまたは複数の部位に組み込むこともできる。
【0183】
形質転換するのに適した宿主細胞は、本発明のポリヌクレオチドで形質転換することができる任意の細胞を含む。本発明の宿主細胞は、本明細書に記載のポリペプチドを内因的に(すなわち天然に)産生することもでき、または本明細書に記載の少なくとも1つのポリヌクレオチド分子で形質転換した後にそのようなポリペプチドを産生することもできる。本発明の宿主細胞は、本明細書で定義される少なくとも1つのタンパク質を産生することができる任意の細胞でよく、細菌、菌類(酵母を含む)、寄生生物、線虫、節足動物、動物および植物細胞を含む。宿主細胞の例として、Salmonella、Escherichia、Bacillus、Listeria、Saccharomyces、Spodoptera、Mycobacteria、Trichoplusia、BHK(仔ハムスター腎)細胞、MDCK細胞、CRFK細胞、CV−1細胞、COS(例えばCOS−7)細胞、およびVero細胞がある。宿主細胞のさらなる例は、E.coli K−12誘導体を含むE.coli;Salmonella typhi;弱毒化株を含むSalmonella typhimurium;Spodoptera frugiperda;Trichoplusia ni;および非腫瘍原性マウス筋原細胞のG8細胞(例えば、ATCC CRL 1246)である。特に好ましい宿主細胞は、それだけに限らないが、Saccharomyces種、特にSaccharomyces cerevisiae、Candida種、Hansenula種やPichia種などの酵母細胞である。
【0184】
組換えDNA技術を使用して、例えば、宿主細胞内のポリヌクレオチド分子のコピー数、そのポリヌクレオチド分子が転写される効率、得られた転写物が翻訳される効率、および翻訳後修飾の効率を操作することにより、形質転換されたポリヌクレオチド分子の発現を向上させることができる。本発明のポリヌクレオチド分子の発現の増大に有用な組換え技術には、それだけに限らないが、高コピー数のプラスミドへのポリヌクレオチド分子の作動的連結、1つまたは複数の宿主細胞染色体へのポリヌクレオチド分子の組み込み、プラスミドへのベクター安定性配列の付加、転写調節シグナル(例えば、プロモーター、オペレーター、エンハンサー)の置換または改変、翻訳調節シグナル(例えば、リボソーム結合部位、シャイン−ダルガルノ配列)の置換または改変、宿主細胞のコドン使用に対応するような本発明のポリヌクレオチド分子の改変、および転写物を不安定にする配列の欠失がある。
【0185】
無細胞組成物
細胞から無細胞組成物を調製する方法は当技術分野で周知であり、それには、凍結融解サイクルの反復、粉砕、超音波処理デバイスにおける超音波での細胞の処理、ホモジナイズ処理、フレンチプレスの使用、界面活性剤および/または酵素の添加、ガラスビーズ溶解、分画遠心、様々な勾配媒体を使用するいくつかの密度勾配の手順がある。これらの技術は当業者によく知られており、例えば「Current Protocols in Protein Science」;John E.Caligan;Ben M.Dunn;Hidde L.Ploegh;David W.Speicher;Paul T.Wingfield;Wiley and Sons)で詳細に論じられている。
【0186】
細胞膜抽出物の単離または調製では、これらの方法の組合せが通常使用される(RiederおよびEmr、2001)。一般に、機械的手段により、または界面活性剤を使用して細胞を溶解し、例えば、米国特許第7,393,934号に概略が述べられているように、分画遠心により膜分画を単離する。Moore(1994)に概略が述べられている方法が一般的に使用される。そのような方法の例は、米国特許第7,320,875号で見つけることができる。他の方法、例えば水性二相アフィニティー分配も膜分画の調製に利用可能である(PerssonおよびJergil、1994)。
【0187】
細胞膜分画の精製に利用可能な多数の市販のキット、例えば、QiagenのQproteome形質膜タンパク質キット(Qproteome Plasma Membrane Protein Kit)もある。細胞を低張性緩衝液中でインキュベートし、それを膨張させる。弱い界面活性剤を添加し、得られた細胞懸濁液を機械的破壊によってホモジナイズする。インタクトな細胞、細胞片、核および主要な細胞内小器官を遠心分離によって除去する。得られた上清は、細胞質タンパク質およびミクロソーム、ゴルジ小胞、ならびに形質膜を含有する。細胞膜上の分子に特異的なリガンドを上清に添加する。リガンドは細胞膜小胞と結合し、リガンドと結合する磁性ビーズを使用してリガンド−小胞複合体を沈降させる。洗浄後、天然の条件下で形質膜小胞を溶出させ、リガンドはビーズと結合したままである。
【0188】
酵母細胞では、細胞膜の調製に一般的に使用される方法は、細胞をスフェロプラストにすることである。これは高収量をもたらし、形質膜の大規模単離に申し分なく適している。一般に、そのような方法では、スフェロプラストの負に帯電した表面を高密度のカチオン性シリカビーズで被覆し、その膜を細胞の他のどんな膜の細胞内小器官よりも高密度にする必要がある。洗浄手順で、過剰なカチオン性ビーズを除去し、その後ポリアクリル酸を添加して、ビーズ上の遊離カチオン性基を遮断する。その後、EGTA含有溶解緩衝液中で、手作業でホモジナイズすることにより、被覆したスフェロプラストを溶解して膜構成成分の凝集を防止する。スフェロプラスト溶解物の遠心分離により、重い形質膜−マイクロビーズ集合物をペレットにし、細胞内にある膜の細胞内小器官を上清中に残す。
【0189】
本発明に使用することができるGタンパク質共役受容体関連ポリペプチドを含む無細胞組成物の生成のさらなる例は、Kaiserら(2008)に記載されている。
【0190】
特定の実施形態では、GPCRを含む本発明のポリペプチドは、リポソーム調製物の脂質二重層中に埋め込まれる。本明細書において、「リポソーム」という用語は、両親媒性のリン脂質、例えばホスファチジルエタノールアミンおよびコレステロールの二重層を含む閉じた小胞を指す。リポソームは、(ホスファチジルエタノールアミンのような)脂質鎖が混合した天然由来のリン脂質、またはDOPE(ジオレオイルホスファチジルエタノールアミン)のような純粋な表面活性物質成分から構成され得る。
【0191】
本明細書に記載のポリペプチドを含有するリポソームは、水中で、例えば超音波処理により、タンパク質を発現する細胞のリン脂質膜を破壊することによって作製することができる。リン脂質は再び集合して、水溶液の中心部を含有するリポソームの球体になる。剪断速度が低いと多数の層を有する多重膜リポソームが生じる。高速剪断超音波処理を続けると、より小さい単層のリポソームが形成され、これは本発明の適用により適している。配向および到達性が最適である最適なリポソームサイズにする実験は、当業者の日常的な能力の範囲内である。
【0192】
本明細書に記載のポリペプチドを生成し、界面活性剤中で解離させ、その後脂質の存在下で膜を再構成し、界面活性剤を除去する膜再構成技術によってそのポリペプチドをリポソーム中に取り込むこともできる。結合相手(化合物)との、リポソームに入っているポリペプチドの配向および到達性が確実に最適になるように、リポソーム組成物を調整する。最適なリポソーム組成物にするそのような調整は、当技術分野で日常的である。
【0193】
BRETのアッセイ
好ましい実施形態では、生物発光タンパク質とアクセプター分子の間で起こるエネルギー移動は、特定の波長を選択する光学フィルターを使用して測定された放出(1つはアクセプター分子の放出、もう1つは生物発光タンパク質の放出)から計算された比率として示される(方程式1を参照)。
Ea/Ed=BRET比率 (1)
上式で、Eaはアクセプター分子の放出強度として定義され(放出光はアクセプターの放出に適合した特異的なフィルターを使用して選択される)、Edは生物発光タンパク質の放出強度として定義される(放出光は生物発光タンパク質の放出に適合した特異的なフィルターを使用して選択される)。
【0194】
光学フィルターは、BRETに適した波長識別を可能にする任意の型のフィルターでよいことが、当業者には容易に理解されるはずである。例えば、本発明に従って使用される光学フィルターは、干渉フィルター、ロングパスフィルター、ショートパスフィルターなどであり得る。フィルターを通過する波長の強度(通常はカウント毎秒(CPS)または相対発光単位(RLU))は、光電子増倍管(PMT)またはCCDカメラを使用して定量することができる。その後、定量したシグナルを使用してBRET比率を計算し、エネルギー移動効率を表す。アクセプター放出の強度が増大するにつれて、BRET比率は増大する。
【0195】
一般に、ドナー放出強度に対するアクセプター放出強度の比率が決定され(方程式1を参照)、これは、エネルギー移動効率を反映する任意の単位で表された数である。この比率は、エネルギー移動効率の増大につれて増大する(Xuら、1999を参照)。
【0196】
エネルギー移動効率は、アクセプター放出強度に対するドナー放出強度の逆比率を使用して表すこともできる(方程式2を参照)。この場合、エネルギー移動効率が増大するにつれて比率は低下する。この計算を行う前に、バックグラウンドの光および基質の自己発光の存在について放出強度を補正する。この補正は、基質を含有するが、生物発光タンパク質、アクセプター分子または本発明のポリペプチドを含有しない対照試料から適当な波長で測定された放出強度を減算することにより一般に行われる。
Ed/Ea=BRET比率 (2)
上式で、EaおよびEdは上記で定義した通りである。
【0197】
生物発光タンパク質およびアクセプター分子の放出の光強度は、分光蛍光光度計など単色光分光器ベースの機器、電荷結合素子(CCD)カメラまたはダイオードアレイ検出器を使用して定量することもできる。分光蛍光光度計を使用して、基質の添加後に生物発光タンパク質とアクセプター分子の放出ピークがどちらも検出されるように放出スキャンを行う。ピーク下の面積は相対的な光強度を表し、上記で概略を述べた通り、これを比率の計算に使用する。同じ試料から生物発光タンパク質およびアクセプター分子の光を測定することができる任意の機器を使用して、本発明のBRET系をモニタリングすることができる。
【0198】
代替的な実施形態では、単独のアクセプター分子の放出がBRETの有効な検出および/または定量に適している。この場合、エネルギー移動効率は、アクセプター放出強度のみを使用して表される。エネルギー移動を測定するために、比率の計算を行わずにアクセプター放出強度を使用できることが、当業者には容易に明らかとなるであろう。これは、理想的には、アクセプター分子が、生物発光タンパク質から移動した光を吸収する場合にのみ光を放出することに起因する。この場合、光フィルターが1つだけ必要である。
【0199】
関連する実施形態では、単独の生物発光タンパク質の放出がBRETの有効な検出および/または定量に適している。この場合、エネルギー移動効率は、生物発光タンパク質放出強度のみを使用して計算される。エネルギー移動を測定するために、比率の計算を行わずにドナー放出強度を使用できることが、当業者には容易に明らかとなるであろう。これは、アクセプター分子が、生物発光タンパク質から移動した光を吸収するので、生物発光タンパク質から検出可能な放出が対応して低下することに起因する。この場合、光フィルターが1つだけ必要である。
【0200】
代替的な実施形態では、エネルギー移動効率は、測定に1つの光学フィルターしか必要としない比率的測定を使用して表される。この場合、適当な光学フィルターを使用してドナーまたはアクセプターの光強度を決定し、フィルターを使用せずに試料の別の測定を行う(オープンスペクトルの強度)。この後者の測定では、(全ての波長について)光の総放出量を定量する。次いで、方程式3または4を使用して比率の計算を行う。方程式3では、アクセプターの光学フィルターのみを必要とする。方程式4では、ドナーの光学フィルターのみを必要とする。
Ea/Eo−Ea=BRET比率または=Eo−Ea/Ea (3)
Eo−Ed/Ed=BRET比率または=Ed/Eo−Ed (4)
上式で、EaおよびEdは上記で定義した通りであり、Eoは、全ての波長を組み合わせた(オープンスペクトル)放出強度として定義される。
【0201】
方程式1〜4からさらなる方程式を導き出せることが、当業者には容易に明らかであるはずである。例えば、そのように導き出された式の1つでは、生物発光タンパク質および/またはアクセプター分子の放出波長で存在するバックグラウンドの光について補正する。
【0202】
BRETアッセイを行う際に、BRETCountを使用して、各ウェルから光放出を決定することができる。BRETCount機器は改変型のTopCountであり、TopCountは、Packard Instrument(Meriden、CT)から販売されているマイクロタイタープレートシンチレーションおよび発光計数器である。バックグラウンドノイズを除去するために同時に2つの光電子増倍管(PMT)を利用する古典的な計数器と異なり、TopCountは、ノイズ低減に単一のPMT技術および時間分解パルス計数を使用して、標準の不透明なマイクロタイタープレートでの計数を可能にする。不透明なマイクロタイタープレートの使用は、光学的なクロストークをごくわずかなレベルまで低減することができる。TopCountには、1、2、6または12個の試料の同時読み取りを可能にするそれぞれ1、2、6および12個の検出器(PMT)を含めて、様々なフォーマットがある。BRETCountの他に、他の市販の機器:Victor2(Wallac、Finland(Perkin Elmer Life Sciences))およびFusion(Packard Instrument、Meriden)でBRETを行うことが可能である。少なくともアクセプター分子の放出、および好ましくは2つの波長(アクセプター分子および生物発光タンパク質)またはそれ以上を検出できる読取り器を使用して、BRETを行うことができる。
【0203】
本発明の一実施形態では、マイクロ流体工学デバイスを使用してBRETを検出する。好都合なことに、マイクロ流体工学デバイスは、マイクロ流体工学デバイスの試料貯蔵所に移すのに、一般に約50μL以下の試料のアリコートしか必要としない。これは、手作業でまたはシリンジ、キャピラリーなどを介した圧気注入によって行われる。
【0204】
マイクロ流体工学を使用した自動化発光バイオチップデバイスを使用して、全ての必要なBRET反応ステップを行うことができる。複数の手作業での操作ステップを回避し、実験を行う際の人間の時間および労力を低減するため、マイクロ流体バイオチッププラットホームでBRET反応を自動化することは望ましい。マイクロ流体工学デバイスは、パターン化されたマイクロチャネルを有する内蔵型の使い捨てバイオチップ、ならびに複数の試料、試薬、および基質を貯蔵するための貯蔵手段を有する区画を含有し得る。区画からマイクロチャネルを介して反応部位へと試料、または試薬の少なくとも1つ、次いで発光基質を連続して移すステップを自動化することができる。次いで発光基質がドナー分子と反応して発光し、それが光学検出器によって検出される。発光を検出するマイクロ流体工学デバイスの例が米国特許第6,949,377号に記載されている。
【0205】
使用
本発明を使用して、多種多様な化合物、特に匂い物質を検出することができる。典型的には、匂い物質は、少なくとも1つの生物の化学感覚的嗅覚受容体によって検出することができる揮発性の有機もしくは無機化合物または無機ガスとなる。これらは、アミンおよび/またはスルフヒドリル含有化合物、カルボン酸、アルコール、アルデヒド、アルカン、アルケン、芳香族化合物、エステル、テルペンまたはテルペン誘導体、エーテル、CO2など、ならびにこれらの性質の組合せを有する化合物を含み得る。
【0206】
匂い物質は、ヒトにとって価値のあるまたは興味深いいくつかの生物学的または化学的状態を示し得る。そのように示される状態には、以下が含まれ得る:
・食料および飲料、医薬品または関連材料の感覚的訴え、品質または安全性。
・ヒトまたは動物の健康、栄養または運動状態。
・病原体を含めた危険な物質の存在の有無。
・産業プロセスの進行または状態。
・環境汚染または状態。
・香料、芳香剤または他の化粧品の感覚的訴え、品質または安全性。
【0207】
特に好ましい実施形態では、化合物は、本明細書に記載のポリペプチドの受容体部分とのみ結合する。
【0208】
特に好ましい実施形態では、嗅覚受容体を含む本明細書で定義されるポリペプチドをバイオセンサーで使用して、対象とする(複数の)化合物を検出する。バイオセンサーとは、生物学的な構成成分を物理化学的な検出器構成成分と組み合わせた、分析対象を検出するためのデバイスである。それは、典型的には3つの部分からなり、第1はGタンパク質共役受容体を含む本明細書で定義される少なくとも1つのポリペプチドである。第2に、変換器または検出器エレメントを必要とし、これは物理化学的に(例えば、光学的、電気化学的に)働き、化合物とポリペプチドの相互作用から生じるシグナルを、容易に測定および定量できる別のシグナルに変換する(すなわち、変換器)。第3に、関連の電子またはシグナル処理装置を必要とし、ユーザーフレンドリーに相互作用の結果を提示する。BRETを含むバイオセンサーの例は、Charestら(2005)に記載されている。
【0209】
別の実施形態では、スクリーニング方法を薬物の発見および/または開発に使用する。より具体的には、受容体は、潜在的な治療薬の標的である。したがって、この実施形態では、受容体が、それだけに限らないが、アドレナリン作動性受容体、セロトニン受容体、ドーパミン受容体、代謝型/グルタミン酸受容体、GABA受容体、鋤鼻受容体(vomeronasal receptor)、味覚受容体やセクレチン様受容体などの臨床的に重要な分子であることが好ましい。
【0210】
Gタンパク質共役受容体と相互作用することが可能な分子を同定し、特徴付け、スクリーニングする方法も本発明の範囲内にあり、96ウェル、384ウェル、1536ウェル(および他)の市販のプレートを使用して実施される、当技術分野で知られているいわゆるハイスループットスクリーニング方法および類似した手法(Spencer、1998;Oldenburg、1998;Milligan、1999)を含む。本発明に従って使用されるさらなる方法は、それだけに限らないが、(とりわけPope、1999に記載の)ハイスループットスクリーニングにおける均等蛍光読取り(homogenous fluorescence readout)を含む。
【0211】
当業者なら気づくであろうが、本発明を多重化することもできる。この系では、異なる化合物と結合する、異なるGタンパク質共役受容体を含む2つ以上のポリペプチドを提供する。異なる波長で放出して異なる標的化合物の検出および定量を可能にするように、異なる各Gタンパク質共役受容体を異なる生物発光タンパク質および/またはアクセプター分子で標識する。
【実施例】
【0212】
[実施例1]
二重標識BRET ODR−10(OGPR)構築物
材料および方法
二重標識BRET OR構築物の設計および構築
3番目の細胞内ループ(IC3)およびC末端に挿入されたBRET2構成成分を用いてBRET2受容体構築物を作製した。(1)IC3の中央にあるGFP2、およびC末端にあるRLuc(h);(2)IC3の中央にあるRLuc(h)およびC末端にあるGFP2の付いたORを用いて2つのBRET構築物を作製した。The Biology WorkbenchにあるTMAPという、膜貫通セグメントを予測するウェブベースのツール(http://seqtool.sdsc.edu)を使用して、3番目の細胞内ループ(IC3)の位置を決定した。ODR−10では、IC3の中央はアミノ酸240〜241である。OGORと称されるこの構築物のアミノ酸配列は、配列番号15として示される(コードするヌクレオチド配列は配列番号16として示される)。
【0213】
ODR−10は、superscriptII(Invitrogen)を使用して、標準的な技術を用いて調製したC.elegans cDNAから増幅した。PCR条件は、94℃で2分を行い、94℃で15秒、59℃で30秒、68℃で70秒を30サイクル、最後の伸長ステップが68℃で5分であった。標準のPfx50 PCR成分を、プライマーODR−10Xba1F 5’−AGTCTAGAATGTCGGGAGAATTGTGGATTA−3’(配列番号21)およびODR−10−attb1−R 5’−GGGGACAAGTTTGTACAAAAAAGCAGGCTTCATCACGTCGGAACTTGAG−3’(配列番号22)とともに使用した。増幅したcDNAをpGEM−T(Promega)にクローニングし、配列を決定した。
【0214】
BRET構成成分のGFP2およびウミシイタケ(Renilla)ルシフェラーゼRLuc(h)は、市販のプラスミドpGFP2−MCS−Rluc(h)(PerkinElmer;Lot#6310051−6D1)から入手した。
【0215】
構築物の個々の断片をPCR増幅することによってBRET構築物を構築した(プライマーおよびアニーリング温度については表4を参照)。Pfx50(商標)標準成分(Invitrogen)を用いてPCRを行い、条件は、94℃で2分を行い、94℃で15秒、59℃で30秒、68℃で70秒を30サイクル、最後の伸長ステップが68℃で5分であった。構築物の別々の構成成分を、重複するプライマーを使用して増幅した。
【0216】
【表4】
【0217】
次いで、構成成分を精製し、対の反応に入れて94℃で変性し、52℃で(重複するプライマー部位で)アニールし、68℃で5分伸長して必要な対にした。次いでこれを、その対のほとんど5’および3’のプライマーを使用するPCRの鋳型として使用した。完全長の構築物が作製されるまでこれを反復した。次いでこれらをpGEM−Tにクローニングし、配列を決定した。ApaLIを使用してエラーのないクローンをpDONR201(Invitrogen)にサブクローニングした。次いでGateway(登録商標)技術を使用してこれらをpYES−DEST52(Invitrogen)にサブクローニングして(attB部位は元の5’および3’プライマーに含まれていた)、その後S.cerevisiaeで発現させた。
【0218】
OGOR突然変異体(H110Y)の構築
二本鎖鋳型としてpDONR201ベクター中のOGOR(上記参照)を用いて部位特異的突然変異導入を実施した。標準的なプロトコールに従ってStratagene Quickchange部位特異的突然変異導入キットを使用して、以下のプライマー:5’−CACCAGTTTTGTTGTCTCAGGAGTTTATTTTGTTTATCGATATTTTGCAACTT−3’(WT)(配列番号43)および5’−AAGTTGCAAAATATCGATAAACAAAATAAACTCCTGAGACAACAAAACTGGTG−3’(H110Yアンチセンス)(配列番号44)を使用してヒスチジン110からチロシンへの(H110Y)突然変異を行った。
【0219】
Pfx50(商標)標準成分(Invitrogen)を用いてPCRを行い、条件は、94℃で2分を行い、94℃で30秒、55℃で1分、68℃で6.8分を16サイクルであった。次いで、配列決定の後、Gateway(登録商標)技術を使用してOGOR突然変異体をpYES−DEST52(Invitrogen)にサブクローニングして(5’および3’のattB部位は元のOGOR配列に含まれていた)、その後S.cerevisiaeで発現させた。
【0220】
溶液調製
20%(w/v)グルコース − グルコース20gを脱イオン水100mLに添加し、溶解するまでホットプレート(50℃)上で加熱し、溶液をフィルター精製(0.2μmフィルター)する。
【0221】
20%(w/v)ラフィノース − グルコース20gを脱イオン水100mLに添加し、溶解するまでホットプレート(50℃)上で加熱し、溶液をフィルター精製(0.2μmフィルター)する。
【0222】
20%(w/v)ガラクトース − ガラクトース20gを脱イオン水100mLに添加し、溶解するまでホットプレート(50℃)上で加熱し、溶液をフィルター精製(0.2μmフィルター)する。
【0223】
YPD培地 − 細菌用ペプトン20g、酵母エキス10gを脱イオン水1Lに添加する。15分間オートクレーブをかける。
【0224】
YPDプレート − 細菌用ペプトン20g、酵母エキス10gおよび寒天20gを脱イオン水1Lに添加する。15分間オートクレーブをかける。オートクレーブした後、20%グルコースを20mL添加し、寒天プレートに注ぐ。
【0225】
Yeast Synthetic Drop−Out Media without Uracil(SCMM−U)培地− アミノ酸不含酵母窒素塩基(yeast nitrogen base without amino acids)(製品番号Y 0626、Sigma)6.7gおよびウラシル不含酵母合成離脱培地補充物(yeast synthetic drop-out media supplement without uracil)(製品番号Y 1501)1.92gを脱イオン水1Lに添加する。15分間オートクレーブをかける。
【0226】
SCMM−Uプレート − アミノ酸不含酵母窒素塩基(製品番号Y 0626、Sigma)1.34g、ウラシル不含酵母合成離脱培地補充物(製品番号Y 1501、Sigma)0.384gおよび寒天4gを脱イオン水180mLに添加する。15分間オートクレーブをかける。オートクレーブした後、20%グルコースを20mL添加し、寒天プレートに注ぐ。
【0227】
1×リン酸ナトリウム緩衝液(1×PBS、pH7.4) − リン酸水素二ナトリウム82.33g(0.58M)、リン酸ナトリウム一塩基酸23.45g(0.17M)、および塩化ナトリウム(NaCl)40gを脱イオン水中で溶解することにより10×ストック溶液を調製した。ストック溶液を脱イオン水で1×PBSに希釈した。
【0228】
セレンテラジン400a − 純エタノール10.20mL中でセレンテラジン400a(Clz400a、Biosynth)1mgを溶解して250μMストック溶液を作製する。40または400μLのセレンテラジン400aのアリコートをピペットで微量遠心管に入れ、次いでSpeedVac(登録商標)Plus SC110A(Savant)を用いてドライダウンする。−80℃でアリコートを貯蔵する。試料を100または1000μLの無水エタノール(それぞれ40または400μLのアリコート)で再構成して、50μMの濃度を得る。
【0229】
匂い物質調製物 − 匂い物質のジアセチル、ベンズアルデヒドおよびオクタナールの1%ストック溶液をそれぞれの溶媒である水、エタノールおよびDMSOで作製した。匂い物質を水で段階希釈して、所望の終濃度を得た。匂い物質を添加していない溶媒の同じ希釈物も調製した。
【0230】
プラスミド形質転換プロトコール
酵母形質転換キット(YEAST−1、Sigma)を使用して、酵母形質転換を実施した。YPDプレートをINVSC1(Invitrogen)S.cervesiae株で画線し、28℃で2〜3日間インキュベートした。YPDプレートからInVSC1細胞1ループ分を掻き取って微量遠心管に入れる。形質転換緩衝液(製品コードT 0809)0.5mL中で細胞を再懸濁し、5秒間遠心する。上清を除去し、緩衝液50〜100μLを管に残す。10mg/mLサケ精巣DNA(製品コードD 9156)10μLを管に添加する。研究するpYES−DEST52−ORプラスミドDNA1μgを添加し、10秒間ボルテックスをかける。PLATE緩衝液(製品コード8966)600μLを添加し、ボルテックスをかける。室温で4時間インキュベートする。42℃熱ブロックで試料に15分間熱ショックをかける。微量遠心機で3秒間遠心し、上清を除去する。滅菌水500μL中で細胞を再懸濁する。SCMM−Uプレートに100μLを播く。コロニーが現れるまで28℃で2〜3日間インキュベートする。
【0231】
発現プロトコール
pYES−DEST52−OR構築物を含有する単一コロニーを、2%グルコースを含有するSCMM−U培地15mLに接種する。振盪(200rpm)しながら28℃で一晩増殖させる。
【0232】
一晩培養物のOD600を決定した。誘導培地(SCMM−U、2%ガラクトース、2%ラフィノース)30mL中で0.4のOD600を得るのに必要な一晩培養物の量を取り出し、1500×g、5分間で細胞をペレットにした。誘導培地1mL中で細胞を再懸濁し、それを誘導培地29mLに接種した。振盪(200rpm)しながら15℃で72時間増殖させる。フラスコから培養物を取り出し、1500×gで5分間、4℃で細胞を遠心分離する。上清をデカントし、滅菌水1mL中で細胞を再懸濁する。滅菌した微量遠心管に細胞を移し、試料を10,000×gで1分間遠心分離する。上清を除去し、膜単離を実施する。
【0233】
膜調製
1500×g(4℃)で5分間の遠心分離によって細胞を回収し、水で洗浄し、1000mg/Lグルコース、36mg/Lピルビン酸ナトリウム、カルシウムおよびマグネシウム(Invitrogen)を含有するダルベッコリン酸緩衝食塩水(D−PBS)または1×PBS4mL中で再懸濁した。フレンチプレス(約18000psi)によって細胞を溶解し、9300×g(4℃)で15分間の遠心分離によって可溶性タンパク質分画を単離した。この後、13.15mLポリカーボネート厚肉遠心管に可溶性タンパク質分画を移し、4℃で1時間、40,000rpm(Beckman Coulter L−80超遠心分離機)で超遠心分離をかけた。上清をデカントし、D−PBDまたは1×PBS1mL中で膜ペレットを再懸濁し、可溶化するまで4℃で一晩放置した。
【0234】
スペクトル測定
プレートを読み取るSpectraMax M2分光蛍光光度計(Molecular Devices)を用いて、全てのスペクトルスキャンを記録した。96ウェルプレート(Perkin−Elmer)を使用して、100μLの膜調製物試料のアリコートをスキャンした。蛍光スペクトルスキャンは、励起波長420nmで、455nm放出カットオフフィルターを使用して、450から600nmまで記録した。
【0235】
同時二重放出検出
POLARstar OPTIMAマイクロプレート読取り器(BMG LabTech)を用いて、同時二重放出RET測定を実施した。同時放出測定ではBRET2フィルターセット(410〜80/515〜30)を使用し、各チャネルについてセットのそれぞれのゲインを用いた(3300/4095)。
【0236】
RET分析プロトコール
試料濃度の標準化
BRET2タグ付き受容体試料の相対的濃度を評価するために、スペクトルスキャンから試料の蛍光強度を決定し、OGORの蛍光強度によって試料の量を標準化した。GFP2の強度を濃度に転換するために、精製したGFP2タンパク質を様々な濃度でアッセイして、蛍光強度に対する濃度に関して較正グラフを作成した。コバルトアフィニティークロマトグラフィーを使用して、供給された使用説明書に従って(BD Talon(BD Biosciences、Clontech))GFP2タンパク質を精製した。最良適合の方程式の線を再編成すると(y=271x+233.31)、蛍光強度が濃度に転換される。最終体積100μLで5または10nMの概算濃度をアッセイに使用した。
【0237】
選択性
96ウェルプレート(Perkin−Elmer)でRET分析を実施し、特定の二重タグ付き受容体タンパク質(OGOR)を1μMの特定の各匂い物質とともに45分間インキュベートした。最終体積は100μLであり、10μLの匂い物質、受容体タンパク質試料(「試料濃度の標準化」の節を参照)、D−PBS(または1×PBS)および5μLのClz400a(5μM)を含んでいた。D−PBS(または1×PBS)の試料をアッセイすることによって、バックグラウンドシグナルを記録した。インキュベーション時間の間、96ウェルプレートウェルをTopseal(商標)−A(Packard)で密封し、各試料の領域を個々に密封した。非匂い物質試料(水、ピルビン酸、クエン酸、乳酸)を調製し、最初にアッセイした。この後、ドラフト中で匂い物質溶液を調製し、以下の順序でアッセイした:2−ブタノン、2,3−ペンタンジオン、2,3−ブタンジオールおよびジアセチル。
【0238】
BRET2測定では、45分のインキュベーション時間後にClz400a基質(5μM)を添加し、5秒間に対して0.50sの積分時間(integration time)を使用してシグナルを記録した。Clz400aを添加した後、シグナルを記録する間ウェルを再度密封して、試料の交差汚染を低減した。試料をウェルに添加した順にアッセイを連続して実施した。
【0239】
BRET2比率は、((受容体試料の515nmでの放出)−(D−PBD(または1×PBS)の515nmでの放出)/(受容体試料の410nmでの放出)−(D−PBDの410nmでの放出)(または1×PBS)のように測定された放出比率として計算する。
【0240】
用量反応曲線
GFP2の強度によって、OGORの濃度を10nMに標準化した。本文中に示したように、濃度範囲にわたって匂い物質の濃度を変化させることにより、用量反応曲線を構築した。常に最低の濃度を最初に、最高の濃度を最後にアッセイした。
【0241】
Windows(登録商標) XP用GraphPad Prismを使用した非線形回帰分析により、S字状の用量反応曲線を適合させ、EC50を計算した。
【0242】
結果
リガンド応答および選択性
生成した構築物は、嗅覚受容体の3番目の細胞内ループ内およびC末端に挿入されたBRET2ドナーおよびアクセプターを有する(図1)。ODR−10によってジアセチルが結合すると、ドナーおよびアクセプター構成成分が離れた場合にBRET2比率が増大し、またはこれらの構成成分が互いに近づいた場合にシグナルが増大する。
【0243】
OGORを10nMまたは1μMの以下のリガンド:水、ピルビン酸、クエン酸、乳酸、2−ブタノン、2,3−ペンタンジオン、2,3−ブタンジオールおよびジアセチルとともにインキュベートした後、BRET2シグナルを記録した(図2aおよびb)。BRET2シグナルの最大の変化が、OGORをμM濃度のジアセチルとともにインキュベートした時に起こり、溶媒(水)の応答と比較してシグナルの37.5%の差が生じた(図3a)。これは、無細胞アッセイのフォーマットで二重標識BRET2タグ付き受容体を使用してリガンド結合をモニタリングした最初の例である。
【0244】
nMとμMどちらの濃度のジアセチルをOGORに添加した時でも、BRET2比率の変化は、対照応答(水)に対して有意差があった(P>0.05)(図3aおよびb)。ジアセチル刺激に対するOGORの応答は、nMとμM両方の濃度で試験した他の全てのリガンドと比較して、対照応答に対して有意差があった唯一の応答であった。これらの結果から、ODR−10がジアセチルと選択的に結合することが確認される。ジアセチルは、ODR−10を発現するHEK293細胞においてCa2+上昇応答を生じさせる唯一の揮発性化合物であることが以前に確認されていた(Zhangら、1997)。
【0245】
37.5%という、リガンド結合に対するBRET2応答の最大の変化は、α2A受容体による10μMのノルエピネフリンの結合時(約5%)(Lohseら、2003)およびPTHRによる1μMのPTHの結合時(約20%)(Vilardagaら、2003)にFRETシグナルで観察された変化より実質的に大きい。ジアセチルの添加時のBRET2シグナルの低下から、BRET2構成成分が匂い物質の結合時に離れることが示唆される。これは、分子内変化のモニタリングについて、このBRET2伝達系が、FRET伝達系と比較して優れた感度を示す最初の実証である。
【0246】
用量応答
OGORのBRET2シグナルの変化の幅に対する匂い物質の濃度変化の効果(図4)は、用量依存的応答の最初の実証をもたらす。EC50値は3.55fMジアセチルと計算され、これは0.31千兆分率(ppq)に等しい。ヒト細胞で発現され、カルシウム画像化技術によってモニタリングされたとき、ジアセチルに対するODR−10の見かけの親和性は、2.3μM(約0.2百万分率(ppm))と計算され、EC50値は走化性の結果と一致する(Zhangら、1997)。
【0247】
ここで示した無細胞アッセイは、全細胞アッセイ、および蛍光検出(Liら、2009)や炎イオン化検出を伴うガスクロマトグラフィー技術(GC−FID)(Macciolaら、2008)など既存の化学的検出系と比較して、ジアセチル定量について9桁を超えて感度が高い。
【0248】
陰性対照−OGOR突然変異(H110Y)
ジアセチルに対するOGOR応答の理想的な陰性対照は、OGOR受容体の場合と同じ距離離れてBRET2構成成分を置くが、その受容体はジアセチル自体に応答しない。ODR−10の3番目の膜貫通残基にある110位でヒスチジンをチロシンに交換すると、ジアセチルに対する走化性応答を強く欠損する(Troemelら、1995;Senguptaら、1996)。
【0249】
OGOR構築物にヒスチジン110からチロシンへの(H110Y)突然変異を導入すると、μM濃度のジアセチルに対する応答が、OGORについて32.4%から4.1%に低減した(図5)。ジアセチルに対する応答のこの低下は、溶媒(水)の応答とは有意差がなく(P<0.05)、この突然変異から機能喪失が生じたことが示唆された。このことから、ジアセチルに対するOGORの応答が、ODR−10受容体とジアセチルの相互作用に起因し、BRET2構成成分自体との非特異的な相互作用に起因しないことが推測される。
【0250】
[実施例2]
二重標識FRET ODR−10(OCOY)と二重標識BRET2 ODR−10(OGOR)の比較
GPCR活性化のモニタリングについてBRET2系とFRET系の感度を比較するため、ODR−10受容体にタグを付けるのに使用したBRET2構成成分をFRET構成成分と交換し、その結果3番目の細胞内ループにCFPが、C末端にYFPが挿入された(OCOY)。CFP(mCFP)(配列番号45)およびYFP(mシトリン)(配列番号46)誘導体はどちらも単量体であり、酵母発現についてコドンを最適化した(EUROSCARF)(それぞれ配列番号47および48)。
【0251】
材料および方法
Genscriptにより、KpnIおよびXhoI制限部位に挟まれたOCOY配列を合成した。そのOCOY配列を、pENTR11ベクターのKpnIおよびXhoI部位に挿入した。次いで、Gateway(登録商標)技術を使用してこれらをpYES−DEST52(Invitrogen)に組み換えて、その後S.cerevisiaeで発現させた。OCOYコード配列は配列番号49として示されるが、そのアミノ酸配列は配列番号50として示される。
【0252】
OCOY発現
OGOR発現と同じ発現プロトコールを、InVSC1におけるOCOYの発現に使用した。
【0253】
濃度の標準化
二重タグ付き受容体試料の相対的濃度を評価するために、スペクトルスキャンから試料の蛍光強度を決定し、OCOYの蛍光強度によって試料の量を標準化した。YFPの強度(530nm)は、OCOYについて721.32a.u.に標準化された。50μLのOCOYをピペットで適当なウェルに入れ、1×PBSで体積を100μLにした。
【0254】
結果
リガンド応答
μM濃度のジアセチルに応答して、OCOYのFRET比率(520nmでの強度/480nmでの強度)は、OGORでの32.4%の低減と比較して、7.6%低減した(図6)。OCOYの応答は、溶媒(水)の応答に対して有意差がある(P<0.05)ことが示された。OGORは、ジアセチル結合のモニタリングについて、OCOYより4倍を超えて感度が高いことが示された。
【0255】
BRET2系の、最大のRET効率の50%に対応するRETプローブ分離であるForster距離(R0)が7.5nmであると最近決定され、遺伝的にコードされた任意のRET対について最大のR0値が決定された(Dacresら、2009)。FRET系のR0値は4.8nmであると以前に決定され(Eversら、2006)、BRET2系が、FRET系(2.4〜7.2nm)より大きな距離範囲(3.8〜11.3nm)でプローブを置くことができることが示唆される。OGORで測定された移動効率は、ジアセチル(1μM)の添加時64.3%から47%に変化し、6.8nmから7.6nmへの距離の変化が示唆される。FRET系など、Forster距離が4.8nmの系は、ジアセチルの添加時にこの距離の変化で11.3%から6.0%への移動効率の変化しか示さない。このことから、BRET2系が、検出感度の点から、この距離範囲におけるこの距離の変化の測定により適していることが確認される。
【0256】
OCOY用量反応曲線
ジアセチルに対するOCOYの応答が溶媒(水)の応答に対して有意差があったことを確認した後、用量反応をプロットした。このデータを得るために、様々な濃度のジアセチルを添加してスペクトルスキャンを記録した(図7)。ジアセチルの濃度が1×10−21Mから1×10−10Mに増大すると、YFPの放出強度は低下し、CFPの放出強度は付随して増大した(図7)。OCOYの応答を、標準化されたOGORの反応曲線に対してスケーリングすると(図8)、BRET2検出が、ODR−10によるジアセチル結合のモニタリングについてFRET検出より感度が高いことが確認された。FRET検出系で計算されたEC50対数値は−16.43±0.9741M(平均±SEM、n=5)であったが、これは、BRET2系で計算されたEC50対数値−14.43±0.5468(平均±SEM、n=12)に対して有意差がない(P<0.05)。このことから、ジアセチルに対するODR−10の親和性がどちらの検出系でも統計的に類似していることが実証され、ジアセチルがODR−10受容体と特異的に相互作用し、RET構成成分自体とは相互作用しないことが推測される。
【0257】
[実施例3]
二重タグ付きStr112(TGTR)
材料および方法
Genscriptにより、NcoIおよびNotI制限部位に挟まれたTGTR配列を合成した。そのTGTR配列を、pENTR11ベクターのNcoIおよびNotI部位に挿入した。次いで、Gateway(登録商標)技術を使用してこれらをpYES−DEST52(Invitrogen)に組み換えて、その後S.cerevisiaeで発現させた。
【0258】
TGTR発現
OGOR発現と同じ発現プロトコールを、InVSC1におけるTGTRの発現に使用した。TGTRコード配列は配列番号51として示されるが、そのアミノ酸配列は配列番号52として示される。
【0259】
RET分析プロトコール
OGOR選択性研究で記載したのと同じRET分析プロトコール。10nMのTGTRで、1μMのリガンドのピルビン酸、クエン酸、酢酸エチル、アセトインおよびジアセチルをアッセイした。
【0260】
細菌抽出物アッセイの調製
LB中で一晩、37℃で振盪(200rpm)しながら細菌株OP50を増殖させた。E.coli株OP50は、実験室において線虫成長用食物源として通常使用される(Brenner、1974)。OP50細菌培養物を使用して2つのアッセイを行った。第1のアッセイでは、10μLの細菌培養物または10μLのLBを10nMのTGTRに添加した。第2のアッセイは、1mLの細菌培養物を微量遠心管に入れ、10000×gで1分間遠心分離することを必要とした。新鮮な微量遠心管にデカントで入れた後の10μLの上清、または10μLのLBを10nMのTGTRに添加した。細菌試料を添加した後、OGORのRET分析と同じプロトコールに従った。
【0261】
結果
OP50細菌上清またはLB中の細菌培養物10μLの添加に対するTGTRの応答は、LB単独に対して有意差があった(P≦0.05)(図9)。増殖の間にOP50から放出された複雑な匂い物質混合物に対してStr112受容体を刺激すると、BRET2比率が増大し、このことから、BRET2構成成分が匂い物質の結合時に互いに近づくことが示唆される。
【0262】
いくつかのリガンドを個々に試験し(図10)、ピルビン酸とアセトインでどちらも水に対するTGTRの応答と比較して有意に高いBRET2シグナルが生じた(P≦0.05)。このことから、リガンド結合時にBRET2構成成分が互いに近づくことが確認される。アセトインに応答したBRET2シグナルの20.8%の増大は、ノルエピネフリンに対するFRETタグ付きα2Aアドレナリン作動性受容体の応答(Lohseら、2003)より4倍大きい。これは、Str112線虫受容体による匂い物質結合の最初の実証であり、BRET2タグ付きStr112による匂い物質結合をモニタリングした最初の例でもある。
【0263】
[実施例4]
受容体キメラ
BRETタグの付いたODR−10のN末端(aa1〜32)およびIC3からC末端(aa225〜aa339)がマルチクローニング部位を挟むように、ODR−10伝達カセットを工学的に作製する(図11)。キメラ受容体を酵母で発現させる。ORのリガンド結合領域をシャッフルして「インフレーム」でこのカセットに入れ、これらのキメラ受容体を使用して、BRETによるリガンド結合を評価することができる。
【0264】
[実施例5]
嗅受容体のオリゴマー形成
C末端においてRlucで標識されたOdr10(配列番号17)およびC末端においてGFP2で標識されたOdr10(配列番号19)を酵母で同時発現させて、受容体のオリゴマー形成およびBRETの変動を示した。
【0265】
材料および方法
形質転換体の培養およびタグ付き遺伝子の誘導条件
形質転換された酵母細胞のコロニーを、適当な栄養素(ウラシル(U)、トリプトファン(T)など)を欠く酵母合成離脱培地(SCMM、Sigma)中で増殖させて、2%グルコースを用いて一晩、28℃、190rpmで振盪しながらプラスミド含有細胞について選択した;SCMM−U−Tは、Invsc1に対してpYesDest52−Odr10−RlucおよびpDestpESC−Odr10−GFP2を同時発現させ(Invsc1/OR/OG)、SCMM−Uは、Invsc1に対してそれぞれ単独のpVV214−odr10−Rluc(Invsc1/OR)および単独のpYesDest52 odr10−GFP2(Invsc1/OG)を発現させる。
【0266】
一晩培養後、OD600を決定し、細胞を誘導培地(培養培地のようなグルコースの代わりに2%ガラクトースおよび1%ラフィノースを有する)中で懸濁して0.4の最終的なOD600を得た。この誘導培地を15℃で振盪しながら24時間インキュベートして融合受容体発現を誘導した。次いで培養物をペレットにし、冷やしたリン酸緩衝食塩水(PBS)でそれを2回洗浄した。細胞ペレットは−80℃で貯蔵することができ、BRET2アッセイの直前にPBS緩衝液中で懸濁した。
【0267】
BRET2アッセイ用酵母細胞の定量
最初に、SpectraMax M2分光蛍光光度計(Molecular Devices)を使用して、白色96ウェルマイクロプレート(Perkin Elmer)中で直接Ex420nmでGFP2を励起することによるGFP強度(1ウェル当たり100μl中約3000RLUまたはc.p.s)によってInvsc1/OR/OGを定量した。ルシフェリン天然セレンテラジン(終濃度5μM)を96ウェル白色マイクロプレート中の総体積100μlの細胞に添加し、直ちに発光シグナルを測定する(Polarstar Optima、BMGLABTECH)ことによって、定量したInvsc1/OR/OG細胞のウミシイタケ(Renilla)ルシフェラーゼ活性を決定した。
【0268】
Invsc1/OR/OGと類似したGFP強度を有するように、Invsc1/OGを標準化した。Invsc1/OR/OGと類似したRluc活性を有するように、Invsc1/ORを標準化した。Invsc1/OR/OGと同じ細胞密度を有するように、陰性対照(宿主株)Invsc1を定量した。
【0269】
マイクロプレートBRET2細胞ベースアッセイ
IssadおよびJockers(2006)からアッセイ手順を改変し、96ウェル白色マイクロプレート中で、総体積100μlで、各試料について2回反復してそれを行った。3つの生物学的に独立したアッセイを行い、データをプールする。終濃度10μMでセレンテラジンh DeepBlueC基質(Biosynth AG)を添加し、二重波長発光/蛍光測定マイクロプレート読取り器(BMGLabtech)を用いて読み取りを行った。以下の最適化されたフィルター設定を使用して、Rlucの光放出(410±80nm)およびGFP2の光放出(515±30nm)を測定した。同時発現したRlucおよびGFP2融合タンパク質の515nm/410nmでの放出、ならびにRluc融合タンパク質単独の515nm/410nmでの放出の差として、BRET比率を定義した。結果をミリBRET単位で表し、1ミリBRETは1000を乗じたBRET比率の値に対応する(Ayoub、2002)。同じ設定での、RlucまたはGFP2融合物を発現しない対照細胞(宿主株Invsc1)の照明の結果、細胞の自己蛍光が検出される。BRET比率の値を計算する前に、試験した全ての試料から自己蛍光を減算した。
【0270】
粗製膜調製、可溶化、および免疫沈降
適切に誘導された(GFP2レベルが宿主株Invsc1細胞のバックグラウンドに対して2倍以下であった)酵母細胞を、氷冷した緩衝液A(75mM トリス−HCl、pH7.4、12mM MgCl2および2mM EDTA)で3回洗浄し、緩衝液B(緩衝液A+プロテアーゼ阻害剤カクテル、コンプリートMini EDTA−フリー(Roche Applied Science))中で最終的なペレットを懸濁した。次いでフレンチプレスを使用して酵母細胞を破壊し、それを10,000×g、4℃で15分間遠心分離する。130000×gで上清を1時間超遠心分離し、冷やした緩衝液B中でペレットを懸濁した。
【0271】
Ayoubら(2002)に記載の方法に従って、膜タンパク質の可溶化および免疫沈降の手順を改変した。粗製膜を0.5%ジギトニン(Sigma)で可溶化し、4℃でこの混合物を3時間撹拌した。18,000×g、4℃で30分間遠心分離することにより、可溶化されていない膜タンパク質を除去した。冷やした緩衝液Bを使用してジギトニン濃度を0.2%に調整し、ポリクローナル抗GFP抗体(Sapphire Bioscience、カタログ番号120−29000)を添加して最終希釈率を1:1000にした。穏やかにかき混ぜながら、免疫複合体形成を4℃で12時間進行させ、プロテインA−アガロース懸濁物(Sigma)を添加して最終体積を1:10(アガロース懸濁物:試料)にし、穏やかにかき混ぜながら4℃でさらに6時間インキュベートした。5000×gで1分間遠心分離した後、上清をデカントし、冷やした緩衝液C(緩衝液B+0.2%ジギトニン)0.5mlでアガロースビーズを5回洗浄した。基質としてセレンテラジンを使用して(Biosynth AG、Switzerland)、発光測定器(BMGLabtech)でルシフェラーゼ活性を評価することにより、沈降物を分析した。
【0272】
結果
酵母細胞における線虫odr−10の視覚化 − 共焦点顕微鏡法
24時間の誘導後にodr−10−Rlucおよびodr−10−GFP2を同時発現した酵母株Invsc1の共焦点画像から、GFP2融合タンパク質がほとんど細胞の膜上でのみ発現したことが明らかとなり、タグ付きodr−10の過剰発現は観察されなかった。細胞の約32%がタグ付きodr−10タンパク質を発現した。誘導時間を48時間まで延長すると、共焦点画像は、細胞中でのGFP2の過剰発現を示し、凝集したGFP2粒子が細胞の細胞質ゾル中で観察された(図12)。
【0273】
BRET2による生存酵母細胞における構成的odr−10オリゴマーの検出
BRET2実験の結果から、odr−10−Rlucおよびodr−10−GFP2を同時発現する酵母細胞で著しいエネルギー移動が観察されたことが示され(図13)、odr−10がホモオリゴマーを形成することが示唆される。半分の酵母細胞がodr−10−Rlucを発現した試料およびあと半分の酵母細胞がodr−10−GFP2を発現した試料では、著しいエネルギー移動は得られなかった。
【0274】
免疫沈降による生存酵母細胞における構成的Odr−10オリゴマーの検出
受容体−Rlucおよび−GFP2融合物の対の発現、その後の抗緑色蛍光タンパク質抗体を使用した沈降、および免疫共沈降したルシフェラーゼ活性の測定によって、生存酵母細胞におけるOdr−10のオリゴマー形成が確認された。図14で示された結果から、Odr−10が細胞中でオリゴマーを形成することが示唆される。免疫共沈降が受容体に非依存的な、GFP2とRlucとの相互作用から生じ得る可能性を除外するために、(1)odr−10−Rluc/GFP2、(2)受容体−GFP2または受容体−Rlucのどちらかを発現した可溶化膜の混合物を使用した免疫共沈降も実施した。同じ細胞中でのodr−10−GFP2およびodr−10−Rlucの同時発現と比較してルシフェラーゼ活性の20%未満が回復したこれらの試料の結果から、同時発現するodr−10−GFP2およびodr−10−Rlucの融合タンパク質間の相互作用が特異的であったことが示唆される。
【0275】
[実施例6]
BRET2タグ付きマウスα2Aアドレナリン作動性受容体
そのN末端がAla250の後にあり、そのC末端がArg372の隣にある3番目の細胞内ループ中にGFP2が挿入されるように、マウスα2Aアドレナリン作動性受容体(配列番号56)を工学的に作製した。受容体のC末端中のVal461にRlucを融合させた(アミノ酸配列は配列番号54として、コードするオープンリーディングフレームは配列番号53として示される)。
【0276】
二重タグ付きアドレナリン作動性受容体を酵母で発現させる。これらの受容体を使用して、BRET2比率の変化を介してマウスα2Aアドレナリン作動性受容体によるリガンド結合をモニタリングすることができる。
【0277】
[実施例7]
全細胞対無細胞BRETアッセイ
本発明の無細胞アッセイ系を全細胞アッセイ系と比較すると、RLucチャネルで測定したときの光放出量の点から、全細胞アッセイと比較して無細胞アッセイで、BRET2シグナルの強度が40倍を超えて大きかったことが実証される(図15)。1μMのジアセチルのアッセイの結果、全細胞アッセイを使用するとBRET2シグナルの有意な変化はなかったが(P≦0.05)、無細胞アッセイ系を使用すると52%の有意な低下(P≦0.05)が観察された(図16)。
【0278】
広く記載されている本発明の趣旨または範囲を逸脱せずに、具体的な実施形態に示されている本発明に多数の変更および/または改変をなし得ることが当業者に理解されるであろう。したがって、本実施形態は、全ての点において例示的であり制限的ではないとみなされるべきである。
【0279】
本出願は、その内容全体が参照により本明細書に組み込まれる、2009年1月29日に出願されたUS61/148,271の優先権を主張するものである。
【0280】
本明細書で論じ、かつ/または言及した全ての刊行物は、その全体が本明細書に組み込まれる。
【0281】
本明細書に含まれている、文書、行為、材料、デバイス、物品などのどんな論述も、本発明の場面を提供する目的のものに過ぎない。それは、これらの事項のいずれかまたは全てが従来技術の基礎の一部を形成し、または本出願の各請求項の優先日前から存在する、本発明に関連する分野における共通の一般知識であったことを承認するものとみなすべきでない。
【0282】
(参考文献)
【配列表フリーテキスト】
【0283】
配列番号1−C.elegans Odr10受容体。
配列番号2−C.elegans Str112受容体。
配列番号3−C.elegans Str113受容体。
配列番号4−C.elegans Str114受容体。
配列番号5−C.elegans Str115受容体。
配列番号6−C.elegans Str116受容体。
配列番号7−C.elegans Odr10受容体をコードするORF。
配列番号8−C.elegans Str112受容体をコードするORF。
配列番号9−C.elegans Str113受容体をコードするORF。
配列番号10−C.elegans Str114受容体をコードするORF。
配列番号11−C.elegans Str115受容体をコードするORF。
配列番号12−C.elegans Str116受容体をコードするORF。
配列番号13−C.elegans Odr10受容体、3番目の細胞内ループ(5番目の非膜貫通ループ)中に挿入されたRLucおよびC末端に挿入されたGFPを含む本発明のポリペプチド。
配列番号14−C.elegans Odr10受容体、3番目の細胞内ループ(5番目の非膜貫通ループ)中に挿入されたRLucおよびC末端に挿入されたGFPを含む本発明のポリペプチドをコードするオープンリーディングフレーム。
配列番号15−C.elegans Odr10受容体、3番目の細胞内ループ(5番目の非膜貫通ループ)中に挿入されたGFPおよびC末端に挿入されたRLucを含む本発明のポリペプチド。
配列番号16−C.elegans Odr10受容体、3番目の細胞内ループ(5番目の非膜貫通ループ)中に挿入されたGFPおよびC末端に挿入されたRLucを含む本発明のポリペプチドをコードするオープンリーディングフレーム。
配列番号17−C.elegans Odr10受容体およびC末端に挿入されたRLucを含む本発明のポリペプチド。
配列番号18−C.elegans Odr10受容体、C末端に挿入されたRLucを含む本発明のポリペプチドをコードするオープンリーディングフレーム。
配列番号19−C.elegans Odr10受容体およびC末端に挿入されたGFP2を含む本発明のポリペプチド。
配列番号20−C.elegans Odr10受容体、C末端に挿入されたGFP2を含む本発明のポリペプチドをコードするオープンリーディングフレーム。
配列番号21〜44−オリゴヌクレオチドプライマー。
配列番号45−mシトリン誘導体。
配列番号46−mCFP誘導体。
配列番号47−mシトリン誘導体をコードするオープンリーディングフレーム。
配列番号48−mCFP誘導体をコードするオープンリーディングフレーム。
配列番号49−Odr10FRET二重標識融合タンパク質をコードするオープンリーディングフレーム。
配列番号50−Odr10FRET二重標識融合タンパク質。
配列番号51−Str112BRET二重標識融合タンパク質をコードするオープンリーディングフレーム。
配列番号52−Str112BRET二重標識融合タンパク質。
配列番号53−マウスα2Aアドレナリン作動性受容体BRET二重標識融合タンパク質をコードするオープンリーディングフレーム。
配列番号54−マウスα2Aアドレナリン作動性受容体BRET二重標識融合タンパク質。
配列番号55−マウスα2Aアドレナリン作動性受容体をコードするオープンリーディングフレーム。
配列番号56−マウスα2Aアドレナリン作動性受容体。
【特許請求の範囲】
【請求項1】
化合物を検出する方法であって、前記方法が、
i)試料を、
a)脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体と、
b)任意選択で、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子と
を含む無細胞組成物と接触させるステップであって、
Gタンパク質共役受容体が、同じまたは異なる1つまたは複数のサブユニットを含み、Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含むステップと、
ii)ステップi)と同時にまたはそれに連続して、生物発光タンパク質の基質を提供し、生物発光タンパク質が基質を修飾することを可能にするステップと、
iii)ステップii)が生物発光タンパク質とアクセプター分子の間の生物発光共鳴エネルギー移動(BRET)を変動させるかどうかを決定するステップと
を含み、化合物がGタンパク質共役受容体と結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化し、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記方法。
【請求項2】
化合物と結合することができるGタンパク質共役受容体のサブユニットが、
i)生物発光タンパク質と、
ii)アクセプター分子と
を含み、化合物がサブユニットと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する、請求項1に記載の方法。
【請求項3】
Gタンパク質共役受容体が、
i)a)生物発光タンパク質と、
b)アクセプター分子と
を含む第1のサブユニットと、
ii)化合物と結合することができる第2のサブユニットと
を含み、化合物が第2のサブユニットと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する、請求項1に記載の方法。
【請求項4】
i)生物発光タンパク質がサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質がサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質がサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子がC末端の一部を形成し、または
vi)アクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質がC末端の一部を形成する、
請求項2または請求項3に記載の方法。
【請求項5】
Gタンパク質共役受容体が、
i)生物発光タンパク質を含む第1のサブユニットと、
ii)アクセプター分子を含む第2のサブユニットと
を含む、請求項1に記載の方法。
【請求項6】
i)生物発光タンパク質が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットの5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットの3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットのC末端の一部を形成し、または
vi)アクセプター分子が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットのC末端の一部を形成する、
請求項5に記載の方法。
【請求項7】
i)Gタンパク質共役受容体が生物発光タンパク質を含むサブユニットを含み、アクセサリー分子がアクセプター分子を含み、または
ii)Gタンパク質共役受容体がアクセプター分子を含むサブユニットを含み、アクセサリー分子が生物発光タンパク質を含む、
請求項1に記載の方法。
【請求項8】
アクセサリー分子が、
i)生物発光タンパク質と、
ii)アクセプター分子と
を含む、請求項1に記載の方法。
【請求項9】
組成物が少なくとも2つのアクセサリー分子を含み、第1のアクセサリー分子が生物発光タンパク質を含み、第2のアクセサリー分子がアクセプター分子を含む、請求項1に記載の方法。
【請求項10】
Gタンパク質共役受容体が嗅覚受容体である、請求項1から9のいずれか一項に記載の方法。
【請求項11】
嗅覚受容体が、脊索動物受容体、線虫受容体、またはそのいずれか1つの生物学的に活性な変異体もしくは断片である、請求項10に記載の方法。
【請求項12】
嗅覚受容体が、
i)配列番号1〜6のいずれか1つで示されるアミノ酸配列と、
ii)配列番号1〜6のいずれか1つまたは複数と少なくとも40%同一であるアミノ酸配列と
を含む、請求項11に記載の方法。
【請求項13】
サブユニットが、2つ以上の異なるGタンパク質共役受容体サブユニットの部分のキメラである、請求項1から12のいずれか一項に記載の方法。
【請求項14】
サブユニットが、第1のGタンパク質共役受容体サブユニットのN末端および1番目の膜貫通ドメインの少なくとも大部分、第2のGタンパク質共役受容体サブユニットの1番目の非膜貫通ループの少なくとも大部分から5番目の膜貫通ドメインの少なくとも大部分まで、ならびに第1のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む、請求項13に記載の方法。
【請求項15】
サブユニットが、第1のGタンパク質共役受容体サブユニットのN末端から5番目の膜貫通ドメインの少なくとも大部分まで、および第2のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む、請求項13に記載の方法。
【請求項16】
アクセサリー分子がGタンパク質またはアレスチンである、請求項1から15のいずれか一項に記載の方法。
【請求項17】
BRETのレベルが、試料中の化合物の相対量を示す、請求項1から16のいずれか一項に記載の方法。
【請求項18】
生物発光タンパク質が、ルシフェラーゼ、β−ガラクトシダーゼ、ラクタマーゼ、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β−グルクロニダーゼまたはβ−グルコシダーゼである、請求項1から17のいずれか一項に記載の方法。
【請求項19】
ルシフェラーゼが、ウミシイタケ(Renilla)ルシフェラーゼ、ホタルルシフェラーゼ、腔腸動物ルシフェラーゼ、北アメリカツチボタルルシフェラーゼ、コメツキムシルシフェラーゼ、鉄道虫ルシフェラーゼ、細菌ルシフェラーゼ、ガウシア(Gaussia)ルシフェラーゼ、エクオリン、ヒカリキノコバエ(Arachnocampa)ルシフェラーゼ、またはそのいずれか1つの生物学的に活性な変異体もしくは断片、またはその2つ以上のキメラである、請求項18に記載の方法。
【請求項20】
基質が、ルシフェリン、カルシウム、セレンテラジン、またはセレンテラジンの誘導体である、請求項1から19のいずれか一項に記載の方法。
【請求項21】
アクセプター分子がタンパク質である、請求項1から20のいずれか一項に記載の方法。
【請求項22】
アクセプター分子が、緑色蛍光タンパク質(GFP)、GFPの青色蛍光変異体(BFP)、GFPのシアン蛍光変異体(CFP)、GFPの黄色蛍光変異体(YFP)、増強型GFP(EGFP)、増強型CFP(ECFP)、増強型YFP(EYFP)、GFPS65T、エメラルド(Emerald)、トパーズ(Topaz)、GFPuv、不安定化EGFP(dEGFP)、不安定化ECFP(dECFP)、不安定化EYFP(dEYFP)、HcRed、t−HcRed、DsRed、DsRed2、t−ダイマー2、t−ダイマー2(12)、mRFP1、ポシロポリン、ウミシイタケ(Renilla) GFP、モンスター(Monster) GFP、paGFP、カエデ(Kaede)タンパク質もしくはフィコビリタンパク質、またはそのいずれか1つの生物学的に活性な変異体もしくは断片である、請求項21に記載の方法。
【請求項23】
アクセプター分子が非タンパク質である、請求項1から20のいずれか一項に記載の方法。
【請求項24】
アクセプター分子が、Alexa Fluor色素、Bodipy色素、Cy色素、フルオレセイン、ダンシル、ウンベリフェロン、蛍光マイクロスフェア、発光マイクロスフェア、蛍光ナノ結晶、マリナブルー、カスケードブルー、カスケードイエロー、パシフィックブルー、オレゴングリーン、テトラメチルローダミン、ローダミン、テキサスレッド、希土類元素キレート、またはその任意の組合せもしくは誘導体である、請求項23に記載の方法。
【請求項25】
ステップi)もしくはステップii)と同時にまたはそれに連続して、生物発光タンパク質の補助因子を提供するステップをさらに含む、請求項1から24のいずれか一項に記載の方法。
【請求項26】
補助因子が、ATP、マグネシウム、酸素、FMNH2、カルシウム、またはそのいずれか2つ以上の組合せである、請求項25に記載の方法。
【請求項27】
無細胞組成物が、組換え細胞中でGタンパク質共役受容体を産生し、細胞の膜を破壊することによって得られた、請求項1から26のいずれか一項に記載の方法。
【請求項28】
組換え細胞が、Gタンパク質共役受容体と結合する非内因性タンパク質を産生しない、請求項27に記載の方法。
【請求項29】
組換え細胞が酵母細胞である、請求項27または請求項28に記載の方法。
【請求項30】
Gタンパク質共役受容体が、リポソームの脂質二重層中に埋め込まれる、請求項1から29のいずれか一項に記載の方法。
【請求項31】
マイクロ流体工学を使用して行われる、請求項1から30のいずれか一項に記載の方法。
【請求項32】
化合物を検出するための精製および/または組換えポリペプチドであって、前記ポリペプチドが、
i)Gタンパク質共役受容体のサブユニットと、
ii)生物発光タンパク質および/またはアクセプター分子と
を含み、細胞中で発現したときにサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記ポリペプチド。
【請求項33】
i)生物発光タンパク質がサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質がサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質がサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子がC末端の一部を形成し、または
vi)アクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質がC末端の一部を形成し、または
vii)生物発光タンパク質もしくはアクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、または
viii)生物発光タンパク質もしくはアクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、または
ix)生物発光タンパク質もしくはアクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、または
x)生物発光タンパク質もしくはアクセプター分子がサブユニットのC末端の一部を形成する、
請求項32に記載のポリペプチド。
【請求項34】
生物発光タンパク質およびアクセプター分子を含み、化合物がポリペプチドと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する、請求項32に記載のポリペプチド。
【請求項35】
化合物を検出するための精製および/または組換えポリペプチドであって、前記ポリペプチドが、
i)Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合するアクセサリー分子と、
ii)生物発光タンパク質および/またはアクセプター分子と
を含み、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記ポリペプチド。
【請求項36】
Gタンパク質共役受容体が嗅覚受容体である、請求項32から35のいずれか一項に記載のポリペプチド。
【請求項37】
嗅覚受容体が、脊索動物受容体、線虫受容体、またはそのいずれか1つの生物学的に活性な変異体もしくは断片である、請求項36に記載のポリペプチド。
【請求項38】
嗅覚受容体が、
i)配列番号1〜6のいずれか1つで示されるアミノ酸配列と、
ii)配列番号1〜6のいずれか1つまたは複数と少なくとも40%同一であるアミノ酸配列と
を含む、請求項37に記載のポリペプチド。
【請求項39】
サブユニットが、2つ以上の異なるGタンパク質共役受容体サブユニットの部分のキメラである、請求項32から38のいずれか一項に記載のポリペプチド。
【請求項40】
サブユニットが、第1のGタンパク質共役受容体サブユニットのN末端および1番目の膜貫通ドメインの少なくとも大部分、第2のGタンパク質共役受容体サブユニットの1番目の非膜貫通ループの少なくとも大部分から5番目の膜貫通ドメインの少なくとも大部分まで、ならびに第1のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む、請求項39に記載のポリペプチド。
【請求項41】
サブユニットが、第1のGタンパク質共役受容体サブユニットのN末端から5番目の膜貫通ドメインの少なくとも大部分まで、および第2のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む、請求項39に記載のポリペプチド。
【請求項42】
請求項32から41のいずれか一項に記載のポリペプチドをコードする単離および/または外因性ポリヌクレオチド。
【請求項43】
請求項42に記載のポリヌクレオチドを含むベクター。
【請求項44】
ポリヌクレオチドが、プロモーターと作動可能に連結する、請求項43に記載のベクター。
【請求項45】
請求項42に記載のポリヌクレオチドおよび/または請求項43もしくは請求項44に記載のベクターを含む宿主細胞。
【請求項46】
請求項32から41のいずれか一項に記載のポリペプチドをさらに含む、請求項45に記載の宿主細胞。
【請求項47】
酵母細胞である、請求項45または請求項46に記載の宿主細胞。
【請求項48】
請求項32から41のいずれか一項に記載のポリペプチド、請求項42に記載のポリヌクレオチド、請求項43もしくは請求項44に記載のベクター、および/または請求項45から47のいずれか一項に記載の宿主細胞を含む組成物。
【請求項49】
ポリペプチドが脂質二重層中に埋め込まれている、請求項48に記載の組成物。
【請求項50】
i)脂質二重層中に埋め込まれている、請求項32から34もしくは36から41のいずれか一項に記載のポリペプチド、ならびに/または
ii)Gタンパク質共役受容体が脂質二重層中に埋め込まれ、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、Gタンパク質共役受容体の細胞内ループおよび/もしくはC末端と直接もしくは間接的に結合した請求項35に記載のポリペプチド
を含む無細胞組成物。
【請求項51】
脂質二重層が酵母脂質二重層である、請求項50に記載の無細胞組成物。
【請求項52】
生物発光タンパク質の基質および/または生物発光タンパク質の補助因子をさらに含む、請求項50または請求項51に記載の無細胞組成物。
【請求項53】
請求項50から52のいずれか一項に記載の無細胞組成物を作製する方法であって、前記方法が、請求項45から47のいずれか一項に記載の細胞を得るステップと、細胞の膜を破壊するステップとを含む、前記方法。
【請求項54】
細胞を浸透化または溶解する、請求項53に記載の方法。
【請求項55】
細胞をフレンチプレスで溶解する、請求項54に記載の方法。
【請求項56】
請求項32から41のいずれか一項に記載のポリペプチド、請求項45から47のいずれか一項に記載の宿主細胞、請求項48もしくは請求項49に記載の組成物、および/または請求項50から52のいずれか一項に記載の無細胞組成物を含むバイオセンサー。
【請求項57】
Gタンパク質共役受容体と結合する化合物をスクリーニングする方法であって、前記方法が、
i)候補化合物を、
a)脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体と、
b)任意選択で、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子と
を含む無細胞組成物と接触させるステップであって、
Gタンパク質共役受容体が、同じまたは異なる1つまたは複数のサブユニットを含み、Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含むステップと、
ii)ステップi)と同時にまたはそれに連続して、生物発光タンパク質の基質を提供し、生物発光タンパク質が基質を修飾することを可能にするステップと、
iii)ステップii)が生物発光タンパク質とアクセプター分子の間の生物発光共鳴エネルギー移動(BRET)を変動させるかどうかを決定するステップと
を含み、BRETの変動が、化合物がGタンパク質共役受容体と結合することを示し、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記方法。
【請求項58】
請求項32から41のいずれか一項に記載のポリペプチド、請求項42に記載のポリヌクレオチド、請求項43もしくは請求項44に記載のベクター、請求項45から47のいずれか一項に記載の宿主細胞、請求項48もしくは請求項49に記載の組成物、請求項50から52のいずれか一項に記載の無細胞組成物、および/または請求項56に記載のバイオセンサーを含むキット。
【請求項1】
化合物を検出する方法であって、前記方法が、
i)試料を、
a)脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体と、
b)任意選択で、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子と
を含む無細胞組成物と接触させるステップであって、
Gタンパク質共役受容体が、同じまたは異なる1つまたは複数のサブユニットを含み、Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含むステップと、
ii)ステップi)と同時にまたはそれに連続して、生物発光タンパク質の基質を提供し、生物発光タンパク質が基質を修飾することを可能にするステップと、
iii)ステップii)が生物発光タンパク質とアクセプター分子の間の生物発光共鳴エネルギー移動(BRET)を変動させるかどうかを決定するステップと
を含み、化合物がGタンパク質共役受容体と結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化し、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記方法。
【請求項2】
化合物と結合することができるGタンパク質共役受容体のサブユニットが、
i)生物発光タンパク質と、
ii)アクセプター分子と
を含み、化合物がサブユニットと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する、請求項1に記載の方法。
【請求項3】
Gタンパク質共役受容体が、
i)a)生物発光タンパク質と、
b)アクセプター分子と
を含む第1のサブユニットと、
ii)化合物と結合することができる第2のサブユニットと
を含み、化合物が第2のサブユニットと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する、請求項1に記載の方法。
【請求項4】
i)生物発光タンパク質がサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質がサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質がサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子がC末端の一部を形成し、または
vi)アクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質がC末端の一部を形成する、
請求項2または請求項3に記載の方法。
【請求項5】
Gタンパク質共役受容体が、
i)生物発光タンパク質を含む第1のサブユニットと、
ii)アクセプター分子を含む第2のサブユニットと
を含む、請求項1に記載の方法。
【請求項6】
i)生物発光タンパク質が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子が第1のサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットの5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットの3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子が第1のサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットの3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子が第2のサブユニットのC末端の一部を形成し、または
vi)アクセプター分子が第1のサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質が第2のサブユニットのC末端の一部を形成する、
請求項5に記載の方法。
【請求項7】
i)Gタンパク質共役受容体が生物発光タンパク質を含むサブユニットを含み、アクセサリー分子がアクセプター分子を含み、または
ii)Gタンパク質共役受容体がアクセプター分子を含むサブユニットを含み、アクセサリー分子が生物発光タンパク質を含む、
請求項1に記載の方法。
【請求項8】
アクセサリー分子が、
i)生物発光タンパク質と、
ii)アクセプター分子と
を含む、請求項1に記載の方法。
【請求項9】
組成物が少なくとも2つのアクセサリー分子を含み、第1のアクセサリー分子が生物発光タンパク質を含み、第2のアクセサリー分子がアクセプター分子を含む、請求項1に記載の方法。
【請求項10】
Gタンパク質共役受容体が嗅覚受容体である、請求項1から9のいずれか一項に記載の方法。
【請求項11】
嗅覚受容体が、脊索動物受容体、線虫受容体、またはそのいずれか1つの生物学的に活性な変異体もしくは断片である、請求項10に記載の方法。
【請求項12】
嗅覚受容体が、
i)配列番号1〜6のいずれか1つで示されるアミノ酸配列と、
ii)配列番号1〜6のいずれか1つまたは複数と少なくとも40%同一であるアミノ酸配列と
を含む、請求項11に記載の方法。
【請求項13】
サブユニットが、2つ以上の異なるGタンパク質共役受容体サブユニットの部分のキメラである、請求項1から12のいずれか一項に記載の方法。
【請求項14】
サブユニットが、第1のGタンパク質共役受容体サブユニットのN末端および1番目の膜貫通ドメインの少なくとも大部分、第2のGタンパク質共役受容体サブユニットの1番目の非膜貫通ループの少なくとも大部分から5番目の膜貫通ドメインの少なくとも大部分まで、ならびに第1のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む、請求項13に記載の方法。
【請求項15】
サブユニットが、第1のGタンパク質共役受容体サブユニットのN末端から5番目の膜貫通ドメインの少なくとも大部分まで、および第2のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む、請求項13に記載の方法。
【請求項16】
アクセサリー分子がGタンパク質またはアレスチンである、請求項1から15のいずれか一項に記載の方法。
【請求項17】
BRETのレベルが、試料中の化合物の相対量を示す、請求項1から16のいずれか一項に記載の方法。
【請求項18】
生物発光タンパク質が、ルシフェラーゼ、β−ガラクトシダーゼ、ラクタマーゼ、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β−グルクロニダーゼまたはβ−グルコシダーゼである、請求項1から17のいずれか一項に記載の方法。
【請求項19】
ルシフェラーゼが、ウミシイタケ(Renilla)ルシフェラーゼ、ホタルルシフェラーゼ、腔腸動物ルシフェラーゼ、北アメリカツチボタルルシフェラーゼ、コメツキムシルシフェラーゼ、鉄道虫ルシフェラーゼ、細菌ルシフェラーゼ、ガウシア(Gaussia)ルシフェラーゼ、エクオリン、ヒカリキノコバエ(Arachnocampa)ルシフェラーゼ、またはそのいずれか1つの生物学的に活性な変異体もしくは断片、またはその2つ以上のキメラである、請求項18に記載の方法。
【請求項20】
基質が、ルシフェリン、カルシウム、セレンテラジン、またはセレンテラジンの誘導体である、請求項1から19のいずれか一項に記載の方法。
【請求項21】
アクセプター分子がタンパク質である、請求項1から20のいずれか一項に記載の方法。
【請求項22】
アクセプター分子が、緑色蛍光タンパク質(GFP)、GFPの青色蛍光変異体(BFP)、GFPのシアン蛍光変異体(CFP)、GFPの黄色蛍光変異体(YFP)、増強型GFP(EGFP)、増強型CFP(ECFP)、増強型YFP(EYFP)、GFPS65T、エメラルド(Emerald)、トパーズ(Topaz)、GFPuv、不安定化EGFP(dEGFP)、不安定化ECFP(dECFP)、不安定化EYFP(dEYFP)、HcRed、t−HcRed、DsRed、DsRed2、t−ダイマー2、t−ダイマー2(12)、mRFP1、ポシロポリン、ウミシイタケ(Renilla) GFP、モンスター(Monster) GFP、paGFP、カエデ(Kaede)タンパク質もしくはフィコビリタンパク質、またはそのいずれか1つの生物学的に活性な変異体もしくは断片である、請求項21に記載の方法。
【請求項23】
アクセプター分子が非タンパク質である、請求項1から20のいずれか一項に記載の方法。
【請求項24】
アクセプター分子が、Alexa Fluor色素、Bodipy色素、Cy色素、フルオレセイン、ダンシル、ウンベリフェロン、蛍光マイクロスフェア、発光マイクロスフェア、蛍光ナノ結晶、マリナブルー、カスケードブルー、カスケードイエロー、パシフィックブルー、オレゴングリーン、テトラメチルローダミン、ローダミン、テキサスレッド、希土類元素キレート、またはその任意の組合せもしくは誘導体である、請求項23に記載の方法。
【請求項25】
ステップi)もしくはステップii)と同時にまたはそれに連続して、生物発光タンパク質の補助因子を提供するステップをさらに含む、請求項1から24のいずれか一項に記載の方法。
【請求項26】
補助因子が、ATP、マグネシウム、酸素、FMNH2、カルシウム、またはそのいずれか2つ以上の組合せである、請求項25に記載の方法。
【請求項27】
無細胞組成物が、組換え細胞中でGタンパク質共役受容体を産生し、細胞の膜を破壊することによって得られた、請求項1から26のいずれか一項に記載の方法。
【請求項28】
組換え細胞が、Gタンパク質共役受容体と結合する非内因性タンパク質を産生しない、請求項27に記載の方法。
【請求項29】
組換え細胞が酵母細胞である、請求項27または請求項28に記載の方法。
【請求項30】
Gタンパク質共役受容体が、リポソームの脂質二重層中に埋め込まれる、請求項1から29のいずれか一項に記載の方法。
【請求項31】
マイクロ流体工学を使用して行われる、請求項1から30のいずれか一項に記載の方法。
【請求項32】
化合物を検出するための精製および/または組換えポリペプチドであって、前記ポリペプチドが、
i)Gタンパク質共役受容体のサブユニットと、
ii)生物発光タンパク質および/またはアクセプター分子と
を含み、細胞中で発現したときにサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記ポリペプチド。
【請求項33】
i)生物発光タンパク質がサブユニットの3番目の非膜貫通ループの一部を形成し、アクセプター分子が5番目の非膜貫通ループの一部を形成し、または
ii)アクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、生物発光タンパク質が5番目の非膜貫通ループの一部を形成し、または
iii)生物発光タンパク質がサブユニットの1番目の非膜貫通ループの一部を形成し、アクセプター分子が3番目の非膜貫通ループの一部を形成し、または
iv)アクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、生物発光タンパク質が3番目の非膜貫通ループの一部を形成し、または
v)生物発光タンパク質がサブユニットの5番目の非膜貫通ループの一部を形成し、アクセプター分子がC末端の一部を形成し、または
vi)アクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、生物発光タンパク質がC末端の一部を形成し、または
vii)生物発光タンパク質もしくはアクセプター分子がサブユニットの1番目の非膜貫通ループの一部を形成し、または
viii)生物発光タンパク質もしくはアクセプター分子がサブユニットの3番目の非膜貫通ループの一部を形成し、または
ix)生物発光タンパク質もしくはアクセプター分子がサブユニットの5番目の非膜貫通ループの一部を形成し、または
x)生物発光タンパク質もしくはアクセプター分子がサブユニットのC末端の一部を形成する、
請求項32に記載のポリペプチド。
【請求項34】
生物発光タンパク質およびアクセプター分子を含み、化合物がポリペプチドと結合したときにアクセプター分子に対する生物発光タンパク質の空間的位置および/または双極子配向が変化する、請求項32に記載のポリペプチド。
【請求項35】
化合物を検出するための精製および/または組換えポリペプチドであって、前記ポリペプチドが、
i)Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合するアクセサリー分子と、
ii)生物発光タンパク質および/またはアクセプター分子と
を含み、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記ポリペプチド。
【請求項36】
Gタンパク質共役受容体が嗅覚受容体である、請求項32から35のいずれか一項に記載のポリペプチド。
【請求項37】
嗅覚受容体が、脊索動物受容体、線虫受容体、またはそのいずれか1つの生物学的に活性な変異体もしくは断片である、請求項36に記載のポリペプチド。
【請求項38】
嗅覚受容体が、
i)配列番号1〜6のいずれか1つで示されるアミノ酸配列と、
ii)配列番号1〜6のいずれか1つまたは複数と少なくとも40%同一であるアミノ酸配列と
を含む、請求項37に記載のポリペプチド。
【請求項39】
サブユニットが、2つ以上の異なるGタンパク質共役受容体サブユニットの部分のキメラである、請求項32から38のいずれか一項に記載のポリペプチド。
【請求項40】
サブユニットが、第1のGタンパク質共役受容体サブユニットのN末端および1番目の膜貫通ドメインの少なくとも大部分、第2のGタンパク質共役受容体サブユニットの1番目の非膜貫通ループの少なくとも大部分から5番目の膜貫通ドメインの少なくとも大部分まで、ならびに第1のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む、請求項39に記載のポリペプチド。
【請求項41】
サブユニットが、第1のGタンパク質共役受容体サブユニットのN末端から5番目の膜貫通ドメインの少なくとも大部分まで、および第2のGタンパク質共役受容体サブユニットの5番目の非膜貫通ループの少なくとも大部分からC末端までを含む、請求項39に記載のポリペプチド。
【請求項42】
請求項32から41のいずれか一項に記載のポリペプチドをコードする単離および/または外因性ポリヌクレオチド。
【請求項43】
請求項42に記載のポリヌクレオチドを含むベクター。
【請求項44】
ポリヌクレオチドが、プロモーターと作動可能に連結する、請求項43に記載のベクター。
【請求項45】
請求項42に記載のポリヌクレオチドおよび/または請求項43もしくは請求項44に記載のベクターを含む宿主細胞。
【請求項46】
請求項32から41のいずれか一項に記載のポリペプチドをさらに含む、請求項45に記載の宿主細胞。
【請求項47】
酵母細胞である、請求項45または請求項46に記載の宿主細胞。
【請求項48】
請求項32から41のいずれか一項に記載のポリペプチド、請求項42に記載のポリヌクレオチド、請求項43もしくは請求項44に記載のベクター、および/または請求項45から47のいずれか一項に記載の宿主細胞を含む組成物。
【請求項49】
ポリペプチドが脂質二重層中に埋め込まれている、請求項48に記載の組成物。
【請求項50】
i)脂質二重層中に埋め込まれている、請求項32から34もしくは36から41のいずれか一項に記載のポリペプチド、ならびに/または
ii)Gタンパク質共役受容体が脂質二重層中に埋め込まれ、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、Gタンパク質共役受容体の細胞内ループおよび/もしくはC末端と直接もしくは間接的に結合した請求項35に記載のポリペプチド
を含む無細胞組成物。
【請求項51】
脂質二重層が酵母脂質二重層である、請求項50に記載の無細胞組成物。
【請求項52】
生物発光タンパク質の基質および/または生物発光タンパク質の補助因子をさらに含む、請求項50または請求項51に記載の無細胞組成物。
【請求項53】
請求項50から52のいずれか一項に記載の無細胞組成物を作製する方法であって、前記方法が、請求項45から47のいずれか一項に記載の細胞を得るステップと、細胞の膜を破壊するステップとを含む、前記方法。
【請求項54】
細胞を浸透化または溶解する、請求項53に記載の方法。
【請求項55】
細胞をフレンチプレスで溶解する、請求項54に記載の方法。
【請求項56】
請求項32から41のいずれか一項に記載のポリペプチド、請求項45から47のいずれか一項に記載の宿主細胞、請求項48もしくは請求項49に記載の組成物、および/または請求項50から52のいずれか一項に記載の無細胞組成物を含むバイオセンサー。
【請求項57】
Gタンパク質共役受容体と結合する化合物をスクリーニングする方法であって、前記方法が、
i)候補化合物を、
a)脂質二重層中に埋め込まれ、化合物と結合することができる少なくとも1つのGタンパク質共役受容体と、
b)任意選択で、Gタンパク質共役受容体の細胞内ループおよび/またはC末端と直接または間接的に結合する少なくとも1つのアクセサリー分子と
を含む無細胞組成物と接触させるステップであって、
Gタンパク質共役受容体が、同じまたは異なる1つまたは複数のサブユニットを含み、Gタンパク質共役受容体および/または存在する場合アクセサリー分子が、生物発光タンパク質およびアクセプター分子を一緒に含むステップと、
ii)ステップi)と同時にまたはそれに連続して、生物発光タンパク質の基質を提供し、生物発光タンパク質が基質を修飾することを可能にするステップと、
iii)ステップii)が生物発光タンパク質とアクセプター分子の間の生物発光共鳴エネルギー移動(BRET)を変動させるかどうかを決定するステップと
を含み、BRETの変動が、化合物がGタンパク質共役受容体と結合することを示し、細胞中で発現したときにGタンパク質共役受容体またはそのサブユニットのN末端が細胞の外側にあり、C末端が細胞の内側にある、前記方法。
【請求項58】
請求項32から41のいずれか一項に記載のポリペプチド、請求項42に記載のポリヌクレオチド、請求項43もしくは請求項44に記載のベクター、請求項45から47のいずれか一項に記載の宿主細胞、請求項48もしくは請求項49に記載の組成物、請求項50から52のいずれか一項に記載の無細胞組成物、および/または請求項56に記載のバイオセンサーを含むキット。
【図1】

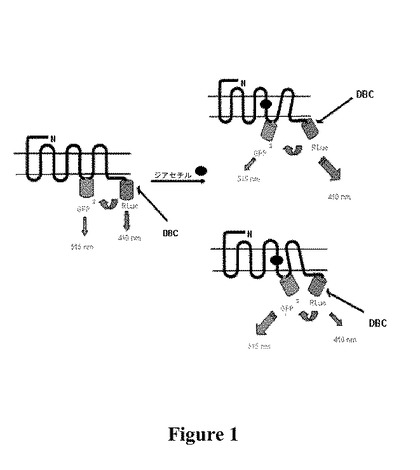
【図2】
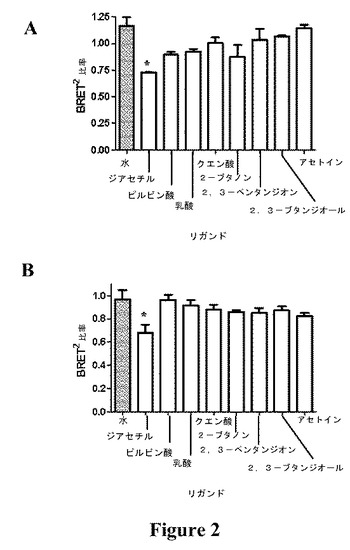
【図3】
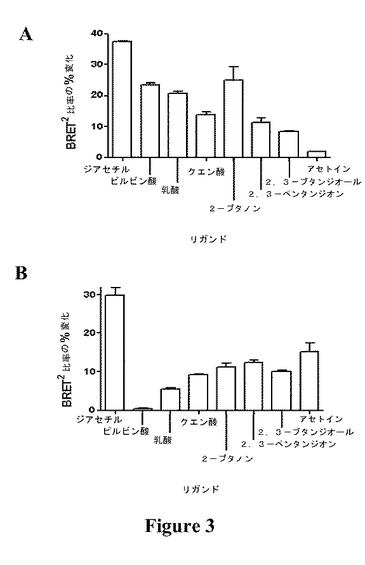
【図4】
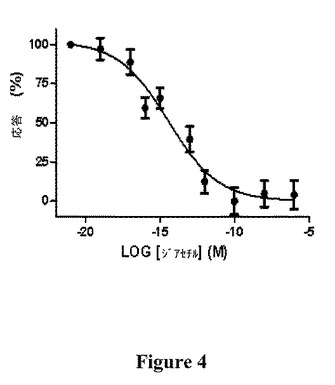
【図5】
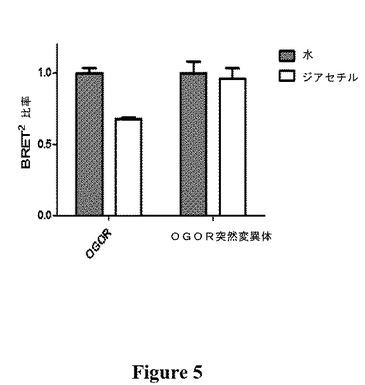
【図6】
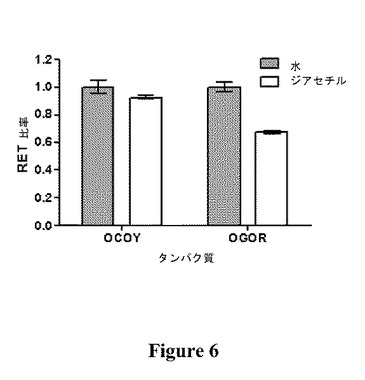
【図7】
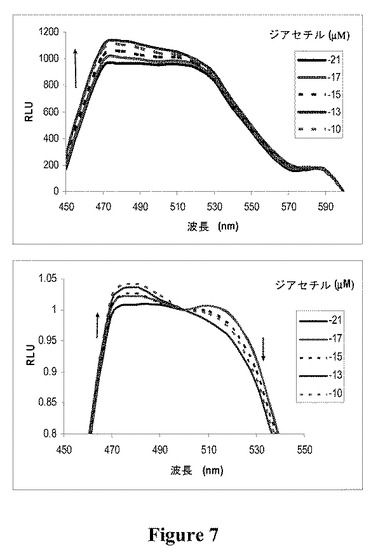
【図8】
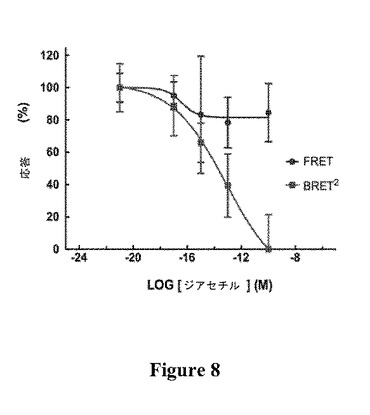
【図9】
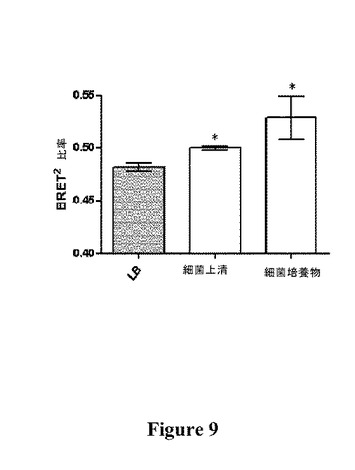
【図10】
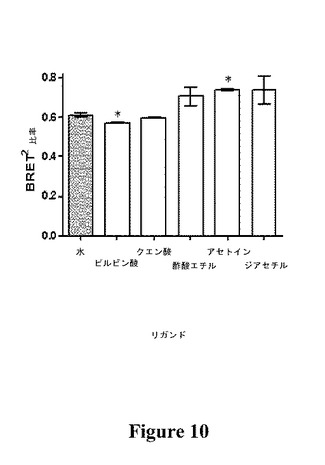
【図11】
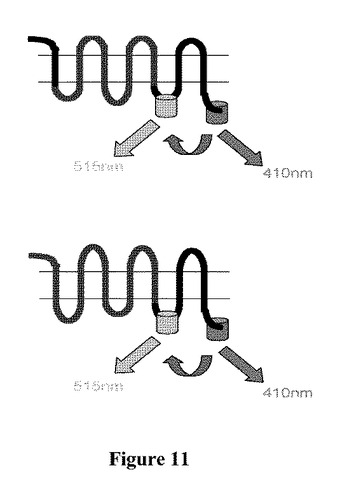
【図12】

【図13】
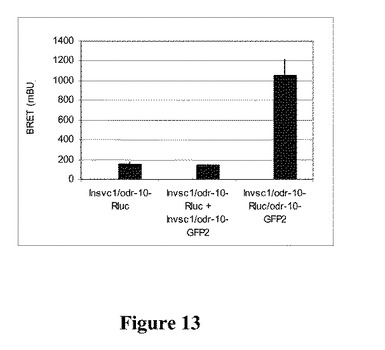
【図14】
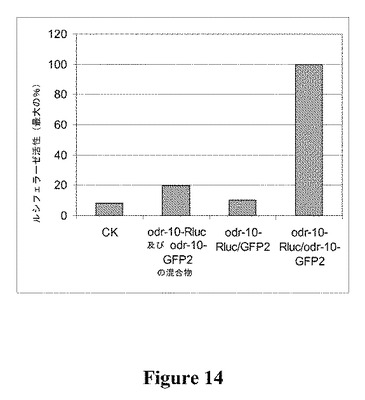
【図15】
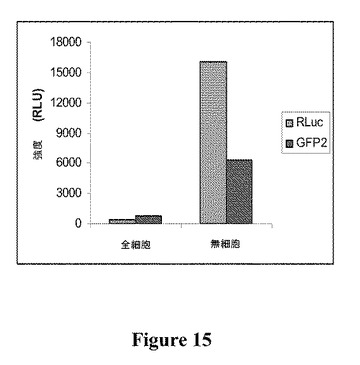
【図16】
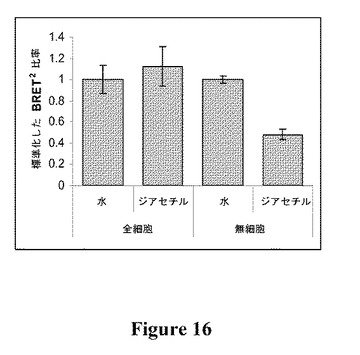
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公表番号】特表2012−516429(P2012−516429A)
【公表日】平成24年7月19日(2012.7.19)
【国際特許分類】
【出願番号】特願2011−546532(P2011−546532)
【出願日】平成22年1月28日(2010.1.28)
【国際出願番号】PCT/AU2010/000080
【国際公開番号】WO2010/085844
【国際公開日】平成22年8月5日(2010.8.5)
【出願人】(305039998)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガニゼイション (92)
【Fターム(参考)】
【公表日】平成24年7月19日(2012.7.19)
【国際特許分類】
【出願日】平成22年1月28日(2010.1.28)
【国際出願番号】PCT/AU2010/000080
【国際公開番号】WO2010/085844
【国際公開日】平成22年8月5日(2010.8.5)
【出願人】(305039998)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガニゼイション (92)
【Fターム(参考)】
[ Back to top ]