
GAG結合蛋白質
【課題】ケモカイン、及び白血球上のそれらの高親和性受容体の相互作用の阻害のための方法とツール及び炎症性疾患の治療処置の提供。
【解決手段】GAG結合部位を、特性決定された蛋白質に導入する方法であって、工程:−構造保持に必須でない蛋白質中の領域を同定すること;−塩基性アミノ酸を上記部位中に導入するか、及び/又は、少なくとも一つの巨大な及び/又は酸性のアミノ酸を上記部位において欠失させることを含み、GAG結合部位はKd≦10μM、好ましくは≦1μM、さらに好ましくは≦0.1μMのGAG結合親和性を有する方法。
【解決手段】GAG結合部位を、特性決定された蛋白質に導入する方法であって、工程:−構造保持に必須でない蛋白質中の領域を同定すること;−塩基性アミノ酸を上記部位中に導入するか、及び/又は、少なくとも一つの巨大な及び/又は酸性のアミノ酸を上記部位において欠失させることを含み、GAG結合部位はKd≦10μM、好ましくは≦1μM、さらに好ましくは≦0.1μMのGAG結合親和性を有する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ケモカインと白血球上のそれらの高親和性受容体の相互作用の阻害のための方法とツール及び炎症性疾患の治療処置のための方法に関する。
【背景技術】
【0002】
ケモカインは、化学誘引物質サイトカインを起源とし、実際は50より多いメンバーを含み、そして小さくて誘導可能な、そして分泌される、低分子量の蛋白質(それらのモノマー形態において6−12kDa)のファミリーを表し、免疫監視機構及び炎症のプロセスの間に決定的な役割を担う。免疫及び炎症におけるそれらの機能に依存して、それらは、2つのクラスに区別することができる。多くの異なる組織細胞により、並びに細菌の毒素及び炎症性サイトカイン、例えばIL−1,TNF及びインターフェロンに応答して白血球を移入する(immigrating)ことにより、炎症性ケモカインは生産される。それらの
主な機能は、宿主の防御のため及び炎症のプロセスにおいて白血球を補充することである。ホーミングケモカインは、他方、リンパ組織の規定されたエリアにおいて構成的に発現される。それらは、免疫系において白血球及び星状細胞の往来及びホーミングを指示する。これらのケモカインは、BCA−1,SDF−1又はSLCにより例示されたとおり、成熟、分化、活性化のコンテクストにおいて白血球の再配置及び再循環を制御し、そして二次リンパ器官内でそれらの正確なホーミングを確保する(ensure)。
【0003】
多数の見本にも拘わらず、ケモカインは顕著に類似の構造的折り畳み(folds)を示す
が、配列相同性は20から70パーセントの間で変化する。ケモカインは、大まかに、70−130アミノ酸からなり、4つの保存されたシステイン残基を伴う。システインは2つのジスルフィド結合を形成し(Cys 1→Cys 3,Cys 2→Cys 4)、それらの特徴的な三次元構造に必須である。走化性ケモカインは、短いアミノ末端ドメイン(3−10アミノ酸)、第1システイン残基、β鎖並びに第2及び第4システイン残基の間に見いだされる接続ループ、並びに20−60アミノ酸のカルボキシル末端αヘリックスからなる。蛋白質のコアは、うまく整列された構造を有するが、N−及びC−末端部分は乱れている。分泌蛋白質のように、それらは、20−25アミノ酸のリーダー配列を伴って合成されて、放出の前に分割されて、はずれる。
【0004】
ケモカインは成熟蛋白質内のそれらのシステイン残基の相対的な位置に基づいて4つのファミリーに副次的に分けられる。αケモカインサブファミリーにおいては、4つのシステインのうちの最初の2つが単一アミノ酸(CXC)として分離され、一方βケモカインにおいては対応するシステイン残基が互いに隣接し合う(CC)。αケモカインは、さらに、N末端にELR配列を含むことにより好中球に走化性であるもの(IL−8、例えば)、及びELRモチーフを欠きリンパ球に作用するもの(I−TAC、例えば)に分類することができる。構造上、βケモカインは2つのファミリーに副次的に分類することができ、単球−化学誘引性蛋白質エオタキシンファミリーであって5つの単球化学誘引性蛋白質(MCP)と互いに約65パーセント同一のエオタキシンを含むものと、それ以外のβケモカインである。CXC−ファミリーの場合のように、CC−残基の上流のN−末端アミノ酸が、ケモカインの生物活性及び白血球選択性に必須の成分である。βケモカインは、一般に、好中球には作用せず、しかし単球、好酸球、好塩基球及びリンパ球を可変性選択性をもって誘引する。
【0005】
ほんのわずかなケモカインがCC−又はCXC−ファミリーに適合しない。リンフォタクチンは、これまで、その初期構造において4つの特徴的なシステインの代わりにたった2つしか示さない唯一のケモカインであり、即ち、γ−又はC−ケモカインとして分類さ
れる。他方、この分類を締めくくることにより、フラクタルカイン(fractalkine)は最
初の2つのシステインを隔てる3つのアミノ酸を有するδ−又はCXXXC−サブファミリーの唯一の代表である。リンフォタキシン及びフラクタルカインはどちらもT−細胞及びナチュラルキラー細胞の走化性を誘導する。
【0006】
ケモカインは、特定の細胞表面、標的細胞上の7つの膜貫通スパニング(7TM)G−プロテイン共役受容体に結合することにより、細胞の移動(migration)及び活性化を誘
導する。18のケモカイン受容体がこれまでにクローン化されており、6つのCXC、10のCC、1つのCX3C及び1つのXC受容体を含む。ケモカイン受容体は特定の細胞に限定され(例えば、CXCR1は好中球に限定される)るが、その他は、より広く発現される(例えば、CCR2は単球、T細胞、ナチュラルキラー細胞及び好塩基球上で発現される)。ケモカインに類似して、上記受容体は特定の細胞上で構成的に発現されるが、いくつかは誘導可能である。それらのいくつかはダウン制御さえもされ得て、細胞を特定のケモカインに対して非応答性にするが、その他に対しての応答性は残す。ほとんどの受容体が1つより多いケモカインを認識し、逆もまた同じで、しかしながら認識は対応するサブファミリーのケモカインに限定される(表1を参照)。
【0007】
【表1】

【0008】
ケモカインはそれらの受容体への相互作用のための主要な2つの部位を有し、一つはアミノ末端ドメイン中にあり、そして他方は第2と第3システイン残基の間に伸びるバックボーンの露出されたループ内にある。両部位はジスルフィド結合により近い位置を持つ。受容体は最初にドッキングドメインとして機能するらしいループ領域内の結合部位を認識
する。この相互作用はケモカインの移動性(mobility)を制限し、即ち、アミノ末端ドメインの正確な位置付けを容易にする。それらの受容体に対して有効に今まで通り結合するがシグナルを生じない変異体ケモカインを用いて、研究が実施された。これらの変異体は例えば、IL−8,RANTES及びMCP−1のN末端内のアミノ酸欠失又は修飾により得られた。
【0009】
複数の細胞内シグナリング経路が、受容体活性化後に、ケモカイン結合の結果として生じる。ケモカインは、2種類の非シグナリング分子とも相互作用する。一つはDARC受容体であって、赤血球上及び内皮細胞上で発現されてCC−ケモカイン並びにCXC−ケモカインに結合することによりそれらが循環されるのを阻止する。第2の種類は、硫酸ヘパラングリコサミノグリカン(GAGs)であり、プロテオグリカンの一部であって、ケモカインの共同受容体として機能する。それらは、ホーミング組織(例えば、内皮細胞)の表面上でケモカインを捕捉して提示することにより、局所的濃度勾配を確立する。炎症性応答、例えば慢性関節リウマチにおいては、セレクチン媒介性プロセスにおいて内皮上の白血球ローリングが、細胞表面のプロテオグリカンにより提示されるケモカインと接触する。それにより、白血球インテグリンが活性化されるようになり、安定した接着及び溢出を導く。補充された白血球は、局所炎症性ケモカインにより活性化されて、ケモカインの局所性高濃度のためにさらなるケモカインシグナリングに対して鈍感になるかもしれない。組織血流ケモカイン勾配を保持するために、DARC受容体は過剰のケモカインのための下水だめ(sink)として機能する。
【0010】
硫酸ヘパラン(HS)グリコサミノグリカンは、グリコサミノグリカンの側鎖に共有結合したコア蛋白質からなり、ほとんどの哺乳類細胞及び組織に見いだされる。蛋白質部分は細胞膜中又は細胞外マトリックス内のプロテオグリカンの位置決めを決定するが、グリコサミノグリカン成分は様々な細胞外リガンド、例えば成長因子、ケモカイン及び接着分子との相互作用を媒介する。プロテオグリカンの生合成は以前に広く総説された。細胞表面プロテオグリカンの主要なグループは、膜貫通蛋白質のシンデカンファミリー(哺乳類においては4つのメンバー)及びグリコシルホスファチジルイノシトール(GPI)テイルにより細胞膜に結合される蛋白質のグリピカン(哺乳類においては6つのメンバー)である。グリピカンは、神経系において、腎臓において、広範囲に、そして少ない程度では骨格筋及び平滑筋において発現されるが、シンデカン−1は上皮細胞において主要なHSPGであり、シンデカン−2は繊維芽細胞及び内皮細胞において支配的であり、そしてシンデカン−3は神経細胞において富裕であり、そしてシンデカン−4は広範囲に発現される。特定のセリン上のテトラサッカライド結合領域を通してシンデカンコア蛋白質に添加されるGAG鎖の大多数は、HS鎖である。膜貫通及びサイトプラズムドメインとは対照的に、特定のシンデカンタイプの細胞外ドメインのアミノ酸配列は異なる種の間では保存されていないが、GAG鎖の数及び位置は高度に保存されている。GAGの構造は、しかしながら、種特異的であり、さらに、HSPG発現組織の性質に依存する。
【0011】
硫酸ヘパラン(HS)は直鎖状ポリサッカライドのグリコサミノグリカン(GAG)ファミリーのもっとも豊富な蛋白質であり、ヘパリン、硫酸コンドロイチン、硫酸デルマタン及び硫酸ケラタンも含む。天然のHSはD−グルコサミン(GlcNAc)及びD−グルクロン酸(GlcA)から構成される20−100のジサッカライドユニットの直鎖により特徴付けされ、N−及びO−硫酸化(グルコサミンの6−O及び3−O硫酸化及びウロン酸の2−O硫酸化)並びにβ−D−グルクロン酸のα−L−イズロン酸(IdoA)へのエピマー化(epimerisation)を含むように修飾され得る。
【0012】
低硫酸化の領域により分離されるN−及びO−硫酸化糖残基のクラスターは、多数の蛋白質結合及びHSの制御特性に主に必須であると推測される。HSの硫酸基の塩基性アミノ酸への静電相互作用に加えて、ファンデルワース相互作用及び疎水性相互作用も蛋白質
結合に関与すると考えられる。さらに、コンフォメーション上柔軟なイズロン酸残基の存在は、蛋白質へのGAG結合に好都合のようである。他の因子、例えば蛋白質結合部位の間の間隔をあけることも、蛋白質−GAG結合相互作用において決定的な役割を担う:例えば、HSにより誘導される−インターフェロンの二量体化は、低硫酸化の7kDa領域により分離される2つの蛋白質結合配列を伴うGAG鎖を必要とする。いくつかのリガンドの完全な生物活性には付加的な配列がときどき必要とされ:FGF−2シグナルトランスダクションをサポートするためには、HSがFGF受容体と相互作用すると想像される最少結合配列並びに追加の残基の両方を有さねばならない。
【0013】
ヘパリン結合蛋白質は、しばしば、塩基性アミノ酸残基のクラスターからなる。リジン、アルギニン、アスパラギン、ヒスチジン及びグルタミンは、しばしば、GAG上の硫酸基及びカルボキシル基との静電接触に関与する。塩基性アミノ酸の間隔(spacing)は、
ときどき蛋白質3−D構造により決定されるが、GAG結合特異性及び親和性を制御すると想定される。当該リガンドの生物活性はHS−蛋白質相互作用のキネティクスによっても影響され得る。成長因子拡散の面積を減少させることは、GAG鎖の長い繰り返しの特性並びにそれらの相対的に早い蛋白質結合のオン及びオフの速度が理想的に適合される、示唆されたHSPG機能の一つである。いくつかの場合、熱力学よりもキネティクスが、HS−蛋白質結合の生理機能を推進する。ほとんどのHSリガンド機能は、明確な長さ及び構造のGAG配列を必要とする。マスト細胞により生産されるヘアピンは、構造上、硫酸ヘパランに類似しているが、相対的に小さな程度の構造多様性を伴って均一に高度な硫酸化をもたらす高いレベルの重合後修飾により特徴付けされる。即ち、硫酸ヘパラン中の高度に修飾されたブロックは、ときどき、「ヘアピン様」と呼ばれる。このため、ヘアピンはそのまま蛋白質の生物物理研究のための完璧なHS類似体として使用可能であり、さらには、大量に利用できるので、HSよりも経費がかからない。異なる細胞種が異なるグリコサミノグリカン構造を有するプロテオグリカンを合成することが示されており、その構造は病理の間、発生の間、又は細胞外シグナル、例えば成長因子に応答して変化する。HSPGsのこの構造多様性は、高い結合融通性を導き、プロテオグリカンの多大な重要性を強調する。
【0014】
硫酸ヘパランプロテオグリカンはFGFシグナリングに必須であるとの証明以来、幾人かの研究者がケモカイン活性を促進させるためのケモカイン−GAG結合の重要性を示そうと実行した。第1に、これまでに研究されたほとんど全てのケモカインはインビトロにおいてHSを結合するらしく、これはこれらの蛋白質の根本的な特性を表すことを示唆する。第2に、インビボにおいてTリンパ球がCC−ケモカインをグリコサミノグリカンとの複合体として分泌するとの発見は、この形態の相互作用が生理学上関連していることを示す。さらに、ケモカインとHSの対合(association)は内皮表面を横切る濃度勾配を
安定化させるのを助け、それにより白血球を移動させるための方向性の情報を提供することが知られている。HSは蛋白質分解からケモカインを保護すること及びそれらのオリゴマー化を誘導すること、即ち、G−共役シグナリング受容体の付近で局所的高濃度を促進するとも考えられる。オリゴマー化の機能的な関連性は、しかしながら、議論が残されているが、全てのケモカインはマルチマー化のための明確な構造上の基礎を有する。β−シートの対合を通したダイマー化はCXC−ファミリー(例えば、IL−8)の全てのケモカインにおいて観察されており、それらのN−末端鎖を通してダイマー化するCC−ケモカインファミリー(例えば、RANTES)のほとんどのメンバーとは対照的である。
【0015】
豊富なデータがケモカインの相互作用の阻害及び低分子量化合物による白血球上のそれらの親和性受容体に関して蓄積した。しかしながら、このアプローチによる炎症性疾患の治療処置におけるブレイクスルーはない。
【0016】
インターロイキン−8(IL−8)は慢性及び急性の炎症の間、好中球誘引(attracti
on)に関与する重要分子である。これまでにIL−8の作用をブロックするためにいくつかのアプローチが企てられたが、例えば、グルココルチコイド、ビタミンD3,シクロスポリンA,形質転換成長因子β、インターフェロン等によるIL−8生産の阻害に始まるが、それら全てはIL−8のmRNAの生産レベルにおいてIl−8活性を阻害する。以前使用されたさらなるアプローチは、特異的アンタゴニストとして作用するために白血球上の受容体に対するか又はIL−8自身に対する特異的抗体を用いることによりIL−8のその受容体への結合を阻害すること、即ちIL−8活性を阻害することである。
【発明の概要】
【0017】
本発明の目的は、よって、白血球上のケモカイン/受容体の相互作用の阻害又は妨害のための別の戦略を提供することである。特に、そのような戦略により、IL−9,RANTES又はMCP−1の作用を標的にすべきである。
【図面の簡単な説明】
【0018】
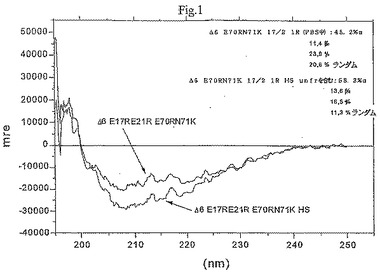
【図1】図1はCDスペクトルである。
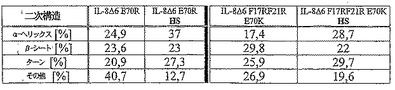
【図2】図2は様々な変異体の二次構造コンテンツを示す。
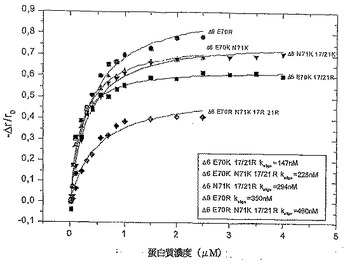
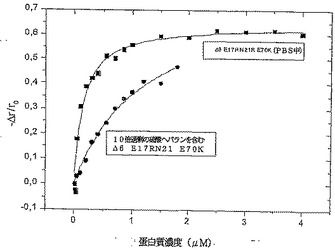
【図3】図3は様々な変異体の蛍光異方性試験の結果のグラフである。
【図4】図4は様々な変異体の蛍光異方性試験の結果のグラフである。
【図5】図5は恒温蛍光滴定の結果のグラフである。
【図6】図6は様々な変異体のアンフォールド実験の結果のグラフである。
【図7】図7はIL−8変異体の走化性指数を示す。
【図8】図8はRANTES走化性アッセイの結果を示す。
【発明を実施するための形態】
【0019】
本発明の主題は、よって、新規なGAG結合蛋白質並びにGAG−共同受容体に対して(野生型よりも)高い親和性を示す別のGAG結合蛋白質を生産する方法を提供することである。そのような修飾されたGAG結合蛋白質は野生型のGAG結合蛋白質との競合因子として作用することができ、そして野生型GAG結合蛋白質の活性を阻害する点ではダウン制御することができるが、しかしながら当業界において使用される公知の組換え蛋白質を用いたときに生じる副作用がない。本発明による分子は上記の不都合をもたない。本修飾GAG結合蛋白質は様々な治療用用途のためのドラッグにおいて使用することができ、特に、ケモカインの場合、当業界において使用される公知の組換え蛋白質を用いたときに生じる公知の不都合なしに炎症性疾患を治療するためのドラッグにおいて使用することができる。本発明によるGAG結合部位の修飾はこの部位に対する結合事象上にその活性が基づく全ての蛋白質、特にGAG部位によるケモカイン、のための広く適用可能な戦略であることが判明した。高いGAG結合親和性を有する本発明による好ましい分子は、それらの生物学上の効果、特にGAG部位に関する野生型分子とのそれらの競合によりそれらの抗炎症活性に関して特に有利であることが証明された。
【0020】
よって、本発明は、GAG結合部位を、特性決定された蛋白質に導入する方法を提供するが、工程:
―構造保持に必須でない蛋白質中の領域を同定すること
―塩基性アミノ酸を上記部位中に導入するか、及び/又は、少なくとも一つの巨大な及び/又は酸性のアミノ酸を上記部位において欠失させること
を含み、GAG結合部位はKd≦10μM、好ましくは≦1μM、さらに好ましくは≦0.1μMのGAG結合親和性を有する。塩基性アミノ酸を上記部位中に導入するか、及び/又は、少なくとも一つの巨大な及び/又は酸性のアミノ酸を上記部位において欠失させることにより、新規で改良された「人工の」GAG結合部位が上記蛋白質に導入される。これは、上記修飾の前にはGAG結合活性を示さなかった蛋白質へのGAG結合部位の新規で完璧な導入を含む。これは、既にGAG結合活性を示す蛋白質へのGAG結合部位の
導入も含む。新規なGAG結合部位は、GAG結合親和性を示さない蛋白質の領域並びにGAG結合親和性を示す蛋白質の領域へ導入することができる。しかしながら、本発明のもっとも好ましい態様によれば、与えられたGAG結合蛋白質のGAG結合親和性の修飾が提供され、上記修飾された蛋白質のGAG結合能力が野生型蛋白質に比べて増加する。本発明は、GAG結合部位を蛋白質に導入する方法、修飾されたGAG結合蛋白質、並びに単離されたDNA分子、ベクター、組換え細胞、薬学組成物、及び上記修飾された蛋白質の使用法に関する。
【0021】
用語「少なくとも一つの塩基性アミノ酸を導入する」は、追加のアミノ酸の導入並びにアミノ酸の置換に関する。主要な目的は、塩基性アミノ酸、好ましくはArg,Lys,His,Asn及び/又はGlnの上記部位における全アミノ酸に対する相対量を増加させることであり、それにより、結果のGAG結合部位が好ましくは少なくとも3つの塩基性アミノ酸、さらに好ましくは4、もっとも好ましくは5つのアミノ酸を含むべきである。
【0022】
GAG結合部位は、好ましくは、溶媒露出位置、即ち、ループにある。これは、有効な修飾を保証する。
蛋白質の領域が構造の保持に必須であるか否かは、例えば、当業者には知られている特定のプログラムとともにコンピュータの方法により試験することができる。蛋白質の修飾の後に、コンフォメーションの安定性をインシリコにて試験することが好ましい。
【0023】
用語「巨大な」アミノ酸は、長いか又は立体構造干渉性の側鎖を有するアミノ酸を意味する;これらは特にTrp,Ile,Leu,Phe,Tyrである。酸性アミノ酸は、特に、Glu及びAspである。好ましくは、結果のGAG結合部位が巨大かつ酸性のアミノ酸を含まないことであり、すべての巨大なかつ酸性のアミノ酸が除去されていることを意味する。
【0024】
GAG結合親和性を測定する−本出願の保護の範囲のため−解離定数Kdを通して。一つの可能性は、リガンド結合における構造変化によるあらゆる与えられた蛋白質の解離定数(Kd)を測定することである。さまざまな技術が当業者には知られており、例えば、恒温蛍光滴定、恒温滴定熱量測定、表面プラズモン共鳴、ゲル移動度アッセイ、放射性標識されたGAGリガンドを用いた競合実験により間接的に行う。さらなる可能性は、これも当業者には知られているコンピュータによる方法によりによる計算により結合領域を予測することであり、いくつかのプログラムを使用してよい。
【0025】
蛋白質中にGAG結合部位を導入するためのプロトコルは、例えば以下のとおりである:
・全体の構造保持に必須ではなくかつGAG結合に適しているかもしれない蛋白質の領域を同定する
・何れかの位置において塩基性Arg,Lys,His,Asn及び/又はGln残基を導入すること(置換又は挿入)により新規のGAG結合部位をデザインする
・インシリコにおいて結果の変異蛋白質のコンフォメーションの安定性をチェックする
・野生型蛋白質のcDNAをクローン化する(あるいは:cDNAを購入する)
・これをPCR−補助変異導入法のための鋳型として使用することによりアミノ酸配列に上記の変化を導入する
・変異体遺伝子を適当な発現系にサブクローン化する(生物学上必要とされる翻訳後修飾に依存して原核生物又は真核生物)
・インビトロにおいて変異体蛋白質の発現、精製及び特性決定
・導入されたGAG結合親和性KdGAG(変異体)<10μMの基準。
【0026】
新規のGAG結合部位を有する上記の操作された蛋白質の例は、例えば、IgGのFc並びに以下のとおりに修飾された補体因子C3及びC4である。
【0027】
【化1】
【0028】
本発明のさらなる側面は、上記の発明の方法により得ることが可能な蛋白質である。発明の蛋白質は、よって、−野生型の蛋白質に比較すると−上記のとおり新規なGAG結合部位を含み、よって、天然GAG結合蛋白質との競合因子として作用することになるが、特に発明の蛋白質のGAG結合親和性が極めて高い、例えばKd≦10μMだからからである。
【0029】
本発明のさらなる側面は修飾されたGAG結合蛋白質であり、上記の蛋白質の中のGAG結合領域が少なくとも一つのアミノ酸の置換、挿入、及び/又は欠失により修飾されることにより、上記GAG結合領域の塩基性アミノ酸の相対量を増加させるか、及び/又はGAG結合領域の巨大及び/又は酸性のアミノ酸の量を増加させるか減少させるが、好ましくは溶剤露出位置においてであり、そして上記蛋白質のGAG結合親和性が各々の野生型蛋白質のGAG結合親和性に比較して増加する。
【0030】
驚きをもって示されたことは、GAG結合領域中の塩基性アミノ酸、特にArg,Lys,His,Asn及び/又はGlnの相対領域を増加させることにより、修飾されたGAG結合蛋白質が野生型蛋白質に比較して増加したGAG結合親和性を示すことであり、特に塩基性アミノ酸の相対量を溶剤露出位置において増加させた場合であるが、蛋白質表面上の陽性荷電したエリアが結合親和性を増強することが示されたからである。好ましくは、3つの塩基性アミノ酸、さらに好ましくは4、もっとも好ましくは5つのアミノ酸がGAG結合領域中に存在する。
【0031】
用語「GAG結合蛋白質」は、GAG共同受容体に結合するあらゆる蛋白質を意味する。蛋白質がGAG共同受容体に結合するか否かは、上記のとおりに公知のプロトコルの助けを借りて試験することができる。Hilemanら(BioEssays 20(1998),156−167)は、グリコサミノグリカン結合する蛋白質内のコンセンサス部位を開示する。この文献内に開示された情報は、本発明のための開始情報としても有用である。用語「蛋白質」は、本発明により提供される分子が少なくとも80アミノ酸の長さであることを明らかにする。これは、それらを、本抗炎症戦略のために適した候補にするために要求される。GAG結合部位と相互作用し、かつその相互作用のための生理学上又は病理学上関連するより小さな分子は知られていないし、本発明に関係しない。好ましくは、本発明による分子は、少なくとも90、少なくとも100、少なくとも120、少なくとも150、少なくとも200、少なくとも300、少なくとも400又は少なくとも500のアミノ酸残基から構成される。
【0032】
本発明の範囲において、用語「GAG結合領域」は、恒温蛍光滴定により測定されたところによれば、100μM以下、好ましくは50μM以下、さらに好ましくは20μM以下の解離定数(Kd−値)にてGAGに結合する領域として定義される(以下の実施例を参照)。
【0033】
本出願において言及される如何なる修飾も、公知の生化学法、例えば部位特異的変異導入法を用いて実施することができる。GAG結合部位の分子はクローン化はもちろん従来
技術であり(例えば、WO96/34965 A,WO02/07935 A,Jaya
raman et al.(FEBS Letters 482(2000),154−158),WO02/20715 A,Yang et al.(J.Cell.Biochem.56(1994),455−468)、GAG領域の分子シャッフリング又は
デノボ合成が記載されていることも注目すべきである;Butcher et al.,(FEBS Letters 4009(1997),183−187)(人工ペプチドに関し、蛋白質ではない);Junno−Oue et al,(J.Virol.75(2001),12439−12445)デノボ合成)を参照)。
【0034】
GAG結合領域は、置換、挿入及び/又は欠失により修飾することができる。これは、非塩基性アミノ酸が塩基性アミノ酸により置換されてよいこと、塩基性アミノ酸がGAG結合領域に挿入されてよいこと又は非塩基性アミノ酸が欠失してよいことを意味する。さらに、GAG結合を干渉するアミノ酸、好ましくはすべての干渉するアミノ酸結合が欠失する。そのようなアミノ酸は特に上記のとおり巨大なアミノ酸又は酸性アミノ酸であり、例えばGlu及びAspである。アミノ酸がGAG結合を干渉するか否かは、例えば数学の方法又はコンピュータの方法により試験してよい。これらの修飾の如何なる結果も、上記GAG結合領域中の塩基性アミノ酸の相対量が増加することであり、「相対」はGAG結合領域中のすべてのアミノ酸の数に比較したGAG結合領域中の塩基性アミノ酸の量を意味する。さらに、立体構造上又は静電上GAG結合を干渉するアミノ酸を欠失させる。
【0035】
アミノ酸が溶剤露出位置中に存在するか否かは、例えば、上記のとおり蛋白質の公知の三次元構造の助けを借りてか又はコンピュータ法の助けを借りて決定することができる。
上記の修飾の蛋白質のGAG結合親和性が各野生型蛋白質のGAG結合親和性に比較して増加しているか否かは、上記のとおり、例えば、解離定数を測定する蛍光滴定実験の助けを借りて測定することができる。改良されたGAG結合親和性に関する基準は、Kd(変異体)<Kd(野生型)であり、好ましくは少なくとも100%である。特別に改良されて修飾のされた蛋白質は、野生型のKdに比較して、少なくとも5倍、好ましくは少なくとも10倍、さらに好ましくは少なくとも100倍高いGAG結合親和性を有する。増加したGAG結合親和性は、よって、好ましくは10μM以下、好ましくは10μM以下、さらに好ましくは0.1μM以下のKdを示す。
【0036】
GAG結合親和性を増加させることにより、修飾された蛋白質は、特異的アンタゴニストになり、そしてGAG結合に関して野生型のGAG結合蛋白質と競合することになる。
好ましくは、Arg,Lys,及びHisからなる群から選択される少なくとも一つのアミノ酸が上記GAG結合領域に挿入される。これらのアミノ酸は、GAG結合領域に容易に挿入されるが、用語「挿入される」は、挿入それ自体並びにあらゆる非塩基性アミノ酸をアルギニン、リジン又はヒスチジンで置換することに関する。もちろん、一つより覆い塩基性アミノ酸を挿入することが可能であり、それにより同じ塩基性アミノ酸が挿入されるか又は上記アミノ酸2つ又は3つの組み合わせでもよい。
【0037】
さらに好ましくは、上記蛋白質がケモカイン、好ましくはIL−8,RANTES又はMCP−1である。ケモカインは共同受容体との相互作用の部位を有することが知られており、このケモカイン結合はしばしば上記のとおりさらなる受容体活性化の条件である。ケモカインはしばしば炎症性疾患において見いだされるから、ケモカイン受容体活性化をブロックすることに主な興味がある。そのようなケモカインは、好ましくは、IL−8,RANTES又はMCP−1であり、よく特性決定された分子であって、GAG結合領域がよく知られている分子である(例えば、Lortat−Jacob et al.,PNAS 99(3)(2002),1229−1234を参照)。これらのケモカイン中のGAG結合領域の中の塩基性アミノ酸の量を増加させることにより、それらの結合親和性が増加し、よって野生型蛋白質の濃度に関しての修飾された蛋白質の濃度に依存して、
野生型ケモカインはほとんど又は全く結合しなくなる。
【0038】
有利な側面によれば、上記GAG結合領域はC末端のαヘリックスである。典型的な化学的モノマーがC末端のαヘリックスによりオーバーレイされた三重鎖逆平行βシートの周囲に組織化される。ケモカイン中のこのC末端αヘリックスがGAG結合に関与する主要な部分であり、そのため、塩基性アミノ酸の量を増加させるためにこのC末端αヘリックス中の修飾が増加したGAG結合親和性を有する修飾ケモカインをもたらすことが示された。
【0039】
有利なことには、IL−8中の17、21、70、及び/又は71位が、Arg,Lys,His,Asn及び/又はGlnにより置換されている。ここでは、これらの前記の部位のたった一つが修飾されることがあり得る。しかしながら、これらの部位の一つより多く並びに全ても修飾されてよく、全ての修飾がArg又はLys又はHis又はAsn又はGln又はそれらの混合の何れかであってよい。IL−8においては、これらの位置がIL−8のGAG結合親和性を大いに増加させることが示されていることから、これらの位置は修飾に特に適している。
【0040】
好ましくは、増加した結合親和性は、硫酸ヘパラン及び/又はヘパリンに対しての増加した結合親和性である。硫酸ヘパランは直鎖状のポリサッカライドのGAGファミリーのもっとも豊富なメンバーであり、ヘパリンも含む。ヘパリンは硫酸ヘパランに構造上極めて類似しており、相対的に小さい程度の構造多様性の、均一に高度な硫酸化をもたらす高いレベルの重合後修飾により特徴付けられる。よって、硫酸ヘパラン中の高度に修飾されたブロックは、ときに、ヘパリン様と呼ばれ、そしてヘパリンは蛋白質の生物物理研究のための硫酸ヘパラン類似体として使用することができる。何れにせよ、硫酸ヘパランとヘパリンは共に特に適している。
【0041】
さらに好ましくは、さらに生物学上活性な領域が修飾されることにより、上記蛋白質のさらなる生物学上の活性を阻害するか又はダウン制御する。このさらなる生物学上の活性は、ほとんどのGAG結合蛋白質、例えばケモカインに関して知られている。これは、受容体、例えば7TM受容体に対する結合領域になる。用語「さらなる(further)」は、
GAG結合領域ではないが、しかし他の分子、細胞又は受容体に結合するか及び/又はそれらを活性化する、生物学上活性な領域を規定する。このさらなる生物学上活性な領域を修飾することにより、この蛋白質のさらなる生物学上の活性が阻害されるか又はダウン制御され、そしてそれにより野生型蛋白質よりも強いアンタゴニストである修飾された蛋白質が提供される。これは、他方で、GAG結合親和性が野生型GAG結合蛋白質よりも高いことを意味し、修飾された蛋白質は野生型蛋白質に代わりGAGに対して大部分結合する。一方、蛋白質がGAGに結合したときに主に起こる野生型蛋白質のさらなる活性は阻害されるか又はダウン制御されるが、修飾された蛋白質がこの特異的な活性を実行しないか又は低い程度でこの活性を実行するからである。この修飾された蛋白質を用いて、当業界の状況において記載されるとおり、他の組換え蛋白質から知られる副作用を示さない、野生型GAG結合蛋白質に関して有効なアンタゴニストが提供される。このさらなる生物学上の活性領域は、例えば、受容体競合アッセイによりインビトロにおいて測定することができる(蛍光標識されたwtケモカインを用いる、カルシウム流入、及び細胞移動(天然白血球又は7TM安定にトランスフェクトされた細胞系)を用いて)。そのようなさらなる生物学上活性な領域の例は、さらなる受容体結合部位に加えて(成長因子ファミリーにおけるように)、酵素部位(ヒドロラーゼ、リアーゼ、スルフォトランスフェラーゼ、N−デアセチラーゼ、及びコポリメラーゼにおけるように)、蛋白質相互作用部位(アンチトロンビンIIIにおけるように)、及び膜結合ドメイン(ヘルペスシンプレックスウイルスgD蛋白質におけるように)である。二重に修飾された蛋白質のこの好ましい態様により、よって、優性(GAG結合に関して)陰性(受容体に関して)変異体が提供され
、本発明の対象物のセットに関して特に有利である。
【0042】
さらに好ましくは、上記のさらなる生物学上の活性領域が、欠失、挿入、及び/又は置換により、好ましくはアラニン、立体構造上及び/又は静電上類似の残基により修飾される。もちろん、上記のさらなる生物学上活性な領域中で少なくとも一つのアミノ酸を欠失させるか又は挿入するか又は置換するかの何れかが可能である。しかしながら、これらの修飾の少なくとも2つ又はそれら全部3つの組み合わせを提供することも可能である。与えられたアミノ酸をアラニン又は立体構造上/静電上類似の残基で修飾することにより−「類似」は置換されるアミノ酸に類似なことを意味する−修飾された蛋白質は、全くか又はほとんど立体構造上/静電上修飾されない。これは特に有利であるが、何故ならば、修飾された蛋白質の他の活性、特にGAG結合領域に対する親和性が変化しないからである。
【0043】
有利には、上記蛋白質がケモカインであり、そして上記のさらなる生物学上の活性が白血球活性化である。上記のとおり、ケモカインは慢性及び急性炎症の間、白血球誘引に関与する。よって、白血球活性化を阻害するか又はダウン制御することにより、炎症が減少するか又は阻害され、この特定の修飾された蛋白質を、炎症性疾患を研究し、診断し、そして治療するための重要なツールにする。
【0044】
有利な側面によれば、上記蛋白質がIL−8であり、そしてさらなる生物学上活性な領域が最初の10個のN末端アミノ酸内に位置する。最初のN末端のアミノ酸は白血球活性化に関与し、特に、Glu−4,Leu−5及びArg−6は受容体結合及び活性化に必須であることが分かっている。よって、これら3つ又は最初の10個のN末端アミノ酸全てさえ置換するか又は欠失させることにより、受容体結合及び活性化を阻害するか又はダウン制御することができる。
【0045】
さらなる有利な蛋白質は、最初の6個のN末端アミノ酸が欠失したIL−8変異体である。上記のとおり、この変異体は全くか又はほとんど白血球に結合して白血球を活性化せず、その結果、炎症性疾患を研究し、診断し、そして治療するのに適している。
【0046】
好ましくは、上記蛋白質がdel6F17RE70KN71R,del6F17RE70RN71K及びdel6E70KN71Kからなる群から選択されるIL−8変異体である。これらの変異体が特に有利であることが示されたのは、最初の6個のN末端アミノ酸の欠失が受容体結合及び活性化を阻害するか又はダウン制御するからである。さらに、17及び21位の2つのフェニルアラニンが、受容体ののちの活性化を促進するためにそのN末端細胞外ドメイン上で受容体と最初に接触することがわかった。あらゆる好中球接触を阻止するために、これらの2つのアミノ酸17及び21を交換するが、それらは塩基性アミノ酸に交換され、何故ならば、それらが蛋白質の三次元モデル上で観察され得るように、C末端のαヘリックスのGAG結合モチーフに極めて近接しているからである。17及び/又は21位をアルギニン又はリジンに交換することにより、GAG結合親和性は、よって、増加する。
【0047】
本発明のさらなる側面は、上記の発明の蛋白質をコードする単離されたポリ核酸分子である。当該ポリ核酸はDNA又はRNAであってよい。それにより、発明の修飾された蛋白質を導く修飾がDNA又はRNAレベルで実施される。この発明の単離されたポリ核酸分子は、診断方法並びに遺伝子治療及び発明の修飾された蛋白質の大規模な生産に適している。
【0048】
さらに好ましくは、単離されたポリ核酸分子は、ストリンジェントな条件下で上において規定された発明の単離されたポリ核酸分子にハイブリダイズする。ハイブリダイゼーシ
ョン条件に依存して、相補二重鎖が2つのDNA又はRNA分子間に形成され、完全にマッチするか又はミスマッチ塩基を含むかの何れかによる(Sambrook et al.,Molecular Cloning:A laboratory manual,第2版、Cold Spring Harbor,N.Y.1989)。約50ヌクレオチドを超える長さのプローブは、25から30%間でのミスマッチ塩基をもってよい。小さなプローブは少数のミスマッチしかもたない。ミスマッチ塩基対を含む二重鎖を形成する標的及びプローブの傾向は、ハイブリダイゼーション条件のストリンジェンシーにより制御され、それ自身は因子、例えばハイブリダイゼーションバッファー中の塩濃度又はホルムアルデヒド濃度、ハイブリダイゼーションの温度及びハイブリダイゼーション後の洗浄条件の函数である。ハイブリッド二重鎖条件の形成において起こるよく知られた原理を適用することにより、所望のストリンジェンシーを有する条件が、当業者により、ハイブリダイゼーションバッファー、温度及び洗浄条件の様々から選択することにより達成され得る。即ち、完全にマッチしたか又は部分的にミスマッチの二重鎖の何れかの検出を許容する条件が選択可能である。二重鎖の融解温度(Tm)は適切なハイブリダイゼーション条件を選択するために有用である。200ヌクレオチドの長さを超えるポリヌクレオチド分子のためのストリンジェントなハイブリダイゼーション条件は、典型的には、予測される二重鎖の融解温度の15−25℃低い温度においてハイブリダイズさせることを含む。長いプローブよりも安定性の劣る二重鎖を形成させる30ヌクレオチドを超えるオリゴヌクレオチドに関しては、ストリンジェントハイブリダイゼーションは通常Tmより5から10℃低い温度にてハイブリダイズさせることにより達成される。核酸二重鎖のTmは、核酸中に含まれるG+Cパーセントに基づいた式を用いて計算することができ、鎖の長さを参酌し、例えば、式Tm=81.5−16.6(log[Na+]))+0.41(%G+C)−(600/N)であり、式中、Nは鎖の長さである。
【0049】
本発明のさらなる側面は、上で規定された本発明の単離されたDNA分子を含むベクターに関する。当該ベクターは、有効なトランスフェクション並びに蛋白質の有効な発現に必要な全ての制御要素を含む。そのようなベクターは、当業界でよく知られており、この目的のために如何なる適切なベクターも選択することができる。
【0050】
本発明のさらなる側面は、上で規定された本発明のベクターにより安定にトランスフェクトされた組換え細胞に関する。そのような組換え細胞並びにそれからの子孫細胞は上記ベクターを含む。それにより、構成的又は上記ベクターの活性化に際して、上記修飾蛋白質を発現する細胞系が提供される。
【0051】
本発明のさらなる側面は、上で規定された本発明による蛋白質、ポリ核酸又はベクター及び薬学上受容可能な担体を含むことを特徴とする薬学組成物に関する。もちろん、当該薬学組成物は、薬学組成物中に通常存在する追加の物質、例えば、塩、バッファー、乳化剤、発色剤、等をさらに含んでよい。
【0052】
本発明のさらなる側面は、上で規定された本発明による修飾蛋白質、ポリ核酸又はベクターの、各野生型蛋白質の生物活性を阻害するか又は抑圧するための方法における使用に関する。上で言及されたとおり、修飾蛋白質は、公知の組換え蛋白質を用いて起こる副作用が発明の修飾蛋白質を用いて起こらないようなアンタゴニストとして作用する。ケモカインの場合、これは特に炎症性反応に関与する生物活性である。
【0053】
よって、本発明による修飾蛋白質、ポリ核酸又はベクターのさらなる使用は、炎症性の症状の治療のための医薬を製造するための方法における。特に、修飾蛋白質がケモカインの場合、それは副作用の内アンタゴニストとして作用して、炎症性症状の治療に特に適する。よって、本出願のさらなる側面は、本発明による単離された修飾蛋白質、本発明による単離されたポリ核酸分子又はベクター又は本発明による薬学組成物を患者に投与するた
めの方法でもある。
【0054】
好ましくは、炎症性の症状は、慢性関節リウマチ、乾癬、骨関節症、喘息、アルツハイマー疾患、及び多発性硬化症からなる群から選択される。ケモカインを通した活性化は本発明による修飾蛋白質により阻害され得るから、上で言及された炎症性の症状が阻止されるか又は治療されるようにして、炎症性反応が阻害されるか又はダウン制御され得る。
【0055】
本発明は、発明を限定するためのものではない以下の実施例及び図面の助けを借りてさらに詳細に説明されるが、図1はCDスペクトルであり;図2は様々な変異体の二次構造コンテンツを示し;図3及び4は様々な変異体の蛍光異方性試験の結果のグラフであり;図5は恒温蛍光滴定の結果のグラフであり;図6は様々な変異体のアンフォールド実験の結果のグラフであり;図7はIL−8変異体の走化性指数を示し;そして図8はRANTES走化性アッセイの結果を示す。
【実施例】
【0056】
実施例1:組換えIL−8遺伝子の生成及び変異体のクローン化
ポリメラーゼ鎖反応(PCR)技術を用いることにより、センス及びアンチセンス変異導入プライマーを用いて変異を導入して変異体に関しての所望のcDNAsを生成した。wtIL−8に関するcDNAを含む合成プラスミドを鋳型として用い、Clontech Advantage(登録商標)2ポリメラーゼミックスをDNAポリメラーゼとして適用し、そしてPCR反応をエッペンドルフマスターグラディエントサイクラーを用いて実施した。用いられた変異導入プライマーは以下の表に要約され、フォワードプライマーから始める(5’から3’):
・CACC ATG TGT CAG TGT ATA AAG ACA TAC TCC(変異Δ6のためのプライマー)
・CACC ATG TGT CAG TGT ATA AAG ACA TAC TCC AAA CCT AGG CAC CCC AAA AGG ATA(変異Δ6 F17R F21Rのためのプライマー)
リバースプライマーは(5’から3’):
・TTA TGA ATT CCT AGC CCT CTT(変異E70Rのためのプライマー)
・TTA TGA ATT CTT AGC CCT CTT(変異E70Kのためのプライマー)
・TTA TGA CTT CTC AGC CCT CTT(変異N71Kのためのプライマー)
・TTA TGA CTT CTT AGC CCT CTT(変異E70K N71Kのためのプライマー)
・TTA TGA CTT CCT AGC CCT CTT(変異E70R N71Kのためのプライマー)
・TTA TGA CCT CTT AGC CCT CTT(変異E70K N71Rのためのプライマー)
・TTA TGA CCT CCT AGC CCT CTT(変異E70R N71Rのためのプライマー)。
【0057】
PCR産物は精製し、pCR(登録商標)T7/NT−TOPO(登録商標)TA(インビトロジェン)にクローン化して、そしてTOP10Fコンピテントエシェリヒアコリ(インビトロジェン)を形質転換した。次の工程として、配列の確認を二重鎖DNA配列決定によりABI PRISM CE1シークエンサーにより実施した。
実施例2:組換え蛋白質の発現及び精製
配列が確認されたら、発現のために、構築物によりカルシウム−コンピテントBL21
(DE3)エシェリヒアコリを形質転換した。細胞を撹拌しながら、100μg/mlアンピシリンを含む1lのLennox Broth(シグマ)中で37℃においてOD600が0.8に到達するまで生育させた。蛋白質発現の誘導は、イソプロピル−β−D−チオガラクトピラノシド(IPTG)を最終濃度1mMまで加えることにより達成した。4時間後に、細胞を6000gにおいて20分間遠心分離することにより回収した。細胞沈殿物を次に、20mM TRIS/HCl,50mM NaCl,pH8を含むバッファーに懸濁し、100ワットにて5x20秒音波処理し、そして最後に再び10,000gにて20分間遠心分離した。組換えIL−8蛋白質の主要フラクションが封入体(inclusion body)中に見いだされたので、さらなる精製のために変性条件を選択した。そうして、細胞沈殿物を6M Gua/HcL及び50mM MES,pH6.5を含むバッファーに懸濁した。懸濁液を次に4℃において4時間撹拌して、次に、50mM MES,pH6.5に対する透析工程に続いた。結果の懸濁液を次に遠心分離し、そして濾過して強カチオン交換カラム(ファルマシアバイオテックのSP Sepharose(登録商標))に負荷した。50mM MESバッファー,pH6.5中の0M−1M NaClからの直線勾配により、60分かけて溶出を達成した。所望の蛋白質を含むフラクションの凍結乾燥後に、第2の精製工程をC18カラムを用いた逆相HPLCにより実施した。この場合、10%−90%アセトニトリルの非直線勾配を選択することにより、所望の蛋白質を溶出した。上記と同じ条件下で同じカチオン交換カラムにより、変性蛋白質のリフォールディングを最後に達成した。
【0058】
蛋白質を、次に、精製度及び同一性に関して、最初の場合は銀染色、そして第2においてはウエスタンブロット分析により、IL−8に対する特異的モノクローナル抗体を用いてチェックした。円偏光二色性(CD)測定により、蛋白質のリフォールディングも確認した。
実施例3:変異体の生物物理特性決定
3.1円偏光二色性測定及び分析
硫酸ヘパラン(HS)の存在及び不在下で上記変異体蛋白質の二次構造変化を調査するために、CD分光法を実施した。測定値は、Jasco J−710旋光分散計(spectropolarimeter)上で195−250nmの範囲にわたり記録し、そして0.1cmの経路(path)の長さのセルを用いた。5μMの濃度による蛋白質溶液のスペクトルを1秒のレスポンス時間、0.2nmの工程解析、50nm/分の速度、1nmのバンド幅及び20mdegの感度にて記録した。3つのスキャンを平均化してスムーズなスペクトルを生じさせた。蛋白質のスペクトルを、次に、バッファー自体又はバッファー/BSの何れかのCD−シグナルに関してバックグラウンド補正した。プログラムSELCONを用いて、HS存在及び不在下での蛋白質の二次構造分析を最後に実施した。
【0059】
非常に多くのアミノ酸が新規な組み合わせにおいて交換されたため、円偏光二色性により結果の二次構造変化の寸法を見いだそうと試みた。
導入された変異に依存して異なる構造が得られた。何の構造も示さなかった一つの発現変異体(変異Δ6 F17R F21R E70K N71R)を除いて、全ての変異体が測定可能なα−ヘリックス、β−シート及びループを呈した。IL−8wtに比較して、一つの変異体のみ(変異Δ6 E70R)がほぼ類似の構造を示したが、その他は主にそれらのα−ヘリックスに差異があり、全体の構造の17.2%から45.2%の範囲であった。にも拘らず、この事実は、IL−8wtの全体の構造が蛋白質配列内での多くの変化にも拘わらず保持されたことを示唆する。これは、以前には予測され得なかった。硫酸ヘパランオリゴサッカライドのみがIL−8wt二次構造に影響し得てヘパリンは不可能であったという既に見いだされた事実から、未分画の硫酸ヘパランにより誘導された効果に注意が引き付けられた。試験された変異体全てがHS結合に際して構造変化を示し、複合体形成の証拠として観察され得る。
【0060】
導入された変異及び硫酸ヘパラン添加に際しての構造変化を証明するために、得られたいくつかのデータを上及び下のグラフにおいて要約する。
3.2蛍光測定
濃度及びリガンド依存性の第4の構造変化を研究するため、蛍光分光法を実施した。その高い感度のために、ナノグラム量の蛋白質しか必要とせず、蛍光技術は所望の調査を実施するための選択の方法であった。測定は、Perkin−Elmer(ビーカンズフィールド、英国)LS50B分光計を用いて企てられた。
3.3蛍光異方性(Anisotropy)
外来発色団bisANSによりもたらされるケモカインの濃度依存性の蛍光異方性を記録することにより、上記変異体のダイマー化定数を発見することを目指した。測定はPBS中で実施し、高濃度で開始し(4μMまでの蛋白質)、徐々に希釈した。各データ点に関して、507nmにおいて記録された異方性シグナル(r)を60秒間かけて平均化した。
【0061】
IL−8のオリゴマー化は、蛋白質GAG結合特性に対して関連性をもって影響することが報告されていた。モノマー濃度における設定により、IL−8結合サイズはダイマー濃度よりも1000倍強固にオリゴサッカライドを限定した(defined)。よって、蛍光
異方性により、IL−8変異体のオリゴマー化特性を調査した。IL−8固有のフルオロフォア(Trp57)は全ての変異体に関して十分な感度がないため、外来のフルオロフォアbis−ANSをこれらの測定に用いた。再び、二次構造に関しては既に注目されたとおり、変異体Δ6 E70Rは、オリゴマー化定数においてもIL−8wt(koligo=379nM)に比較して高い類似度を示した(koligo=350nM)。高いkoligo(koligo=460nM)の変異体は、よって、ダイマー化が不完全であり、Δ6 F17RF21RE70RN21Kであった。以前の研究は、β−シートが主にダイマー化プロセスに関与していることを明らかにしたが、事実、この変異体の二次構造に関する結果と相関しており、たった11.4%のβ−シートの極めて低い割り当て(share)を示した。もっとも低いkoligo(koligo=147nM)の変異体
はΔ6 F17RF21R E70Kであることがわかったが、調査された全変異体のβ−シート構造のもっとも高い割り当て(29.8%)を再び示した。硫酸ヘパラン付加の衝撃も観察された。IL−8wtに関しては、硫酸ヘパランがオリゴマー化定数のはるかに高いレベルへのシフト(koligo=1.075nM)を誘導したが、これは調査されたIL−8変異体に関しても観察された。Δ6 F17RF21R E70Kは0.147μMから1.162μMにシフトし、そして変異体Δ6 E70Rは硫酸ヘパラン存在下において0.35μMから1.505μMにシフトした。得られた結果のいくつかを図3及び4に示すが、図3はケモカイン濃度におけるPBS中のIL−8変異体の蛍光異方性の依存度を示し、そして図4はHSの存在(10倍過剰)及び不在((pc=10xy過剰)蛋白質濃度)下でのケモカイン濃度におけるPBS中のΔ6 F17RF21R
E70Kの蛍光異方性の依存度を示す。
3.4恒温蛍光滴定(IFT)実験
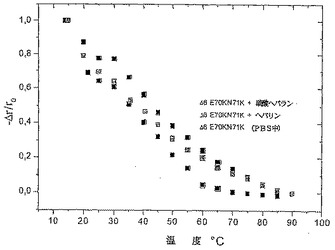
解離定数(Kd値)はリガンドの蛋白質に対する結合親和性に関する測定値であり、よって、リガンド結合に際して蛋白質の蛍光放射特性の濃度依存性変化(蛍光クエンチング)がKdの測定に用いられた。これらの変異体は、提案された(proposed)GAG結合部位においてか又はその近くに位置する固有のトリプトファン発色団を含むためにリガンド結合に際して構造的な変化に感受性であるべきだから、IFT実験はこの種の調査に適しているようであった。蛍光強度滴定をPBS中で700nMの蛋白質濃度を用いて実施した。各GAGリガンドのアリコートの付加及び60秒の平衡期間の後に、282nMにおける励起に際しての蛋白質溶液の放射を300−400nmの範囲にわたり記録した。
【0062】
未分画のヘパリン及び硫酸ヘパランの結合を調査した。十分な感度を保証するために、変異体をダイマー濃度に設定した。GAG結合に際したTrp57蛍光強度のクエンチン
グが25−35%の範囲内で記録された。リガンド結合の顕著な改善が、特にヘパリン結合に関して観察された。IL−8wt(Kd=37μM)に比較して、Δ6 F17RN71R E70K(Kd=14nM)及びΔ6 F17RF21R N71K(Kd=14.6nM)は、2600倍良好な結合を示し、そしてΔ6 E70K N71K(Kd=74nM)は1760倍良好な結合を示した。良好な結果は硫酸ヘパランに関しても得られた。Δ6 F17RN71R E70Kに関しては、107nMのKdが観察され、Δ6 F17RF21R N71Kに関してはKdが95nMであり、そして変異体Δ6
E70K N71Kは34nMのKdを示した。IL−8wtが4.2μMのKdにて結合するように、変異体に関して見いだされたKdsは結合に関しての異例の改善を表したので、図5を参照されたい。
3.5アンフォールド実験
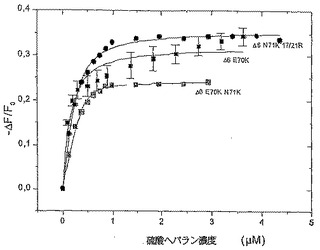
蛋白質の安定性及びこの安定性がGAGリガンド結合に際して変化するのか否かについての情報を得るため、アンフォールド実験を企画した。上記のとおり、蛍光技術は第4の構造変化を観察するのに極めて感度がよく、よって蛋白質の熱構造変化を調査するためのえり抜きの方法である。測定はIFTに関して記載されたとおりに企画し、リガンド濃度は変化させずに温度を変化させた。蛋白質の構造は0.7μMの濃度において15−85℃の温度において10倍過剰の硫酸ヘパラン及びヘパリンの不在下及び存在下において観察された。
【0063】
蛋白質の放射の最大は340nmから357nmの範囲であり、値は溶剤露出トリプトファン残基に関して典型的である。アンフォールド実験を15℃において開始し、変異体の放射の最大は340nm−351nmの間で変化した。その放射最大が340nmにおいて観察されたIL−8wtに比較して、これはわずかに高い値を意味する。温度の増加と共に、放射最大の強度が低下し、最大値の高いか又は低い波長へのシフトの何れかを伴った。Δ6 E70R及びΔ6 E70K N71Kの放射最大は、352.5nm−357nm及び343nm−345nmにシフトし、Trp57残基の溶剤への温度増加を伴うさらなる暴露に関して典型的であるが、おもしろいことにΔ6 F17RN71R E70K及びΔ6 F17RF21R E70Rは青いシフトを示し、350nm−343nm及び著しくないが350nm−348nmの範囲であった(図6を参照)。温度をゆっくりと低下させることにより、アンフォールドのプロセスが波長シフト及び強度の変化に関しては部分的に可逆性であった。5倍過剰の硫酸ヘパランの付加は蛋白質の安定性の増加を導いたが、おそらくは複合体形成のためである。これは、一方では融解温度の高い温度へのシフトにより観察できたし、他方では温度増加に際しての放射最大の著しくは顕著でないシフトにより観察できた。
実施例4:優性−陰性IL−8変異体の受容体『陰性』機能の細胞に基づくアッセイ
好中球誘引に関してのIL−8変異体の損なわれた受容体機能を特性決定するために、IL−8変異体に応答した好中球のトランスフィルターに基づく走化性を、5μmのPVP−無しのポリカーボネート膜を備えたマイクロ走化性チェンバー内でアッセイした。
細胞調製:
簡単に言えば、好中球フラクションを新たに回収されたヒト血液から調製した。これは、6%デキストラン溶液のヘパリン処理された血液への添加(2:1)により行い、次に、45分間沈降のために放置した。上清の透明な溶液を回収し、そしてHBSS w/o
Ca及びMgにより2回洗浄した。細胞を数え、そして最後にHBSSにより2Mio/ml細胞懸濁液にて希釈して、数えられた細胞の60%のみが好中球であることを考慮した。
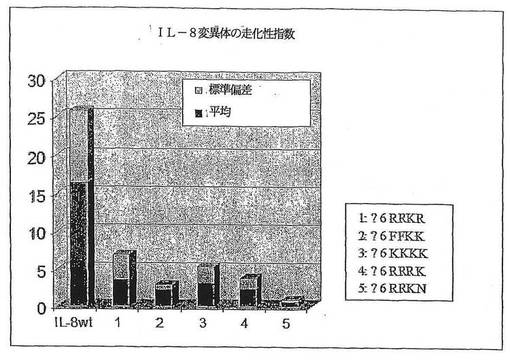
走化性アッセイ:
IL−8変異体を10μg/ml、1μg/ml及び0.1μg/mlの濃度に希釈し、そしてチェンバーの底部区画へ同じものを3通りにして置いた(ウエルあたり26μl)。新たに調製された好中球を上部チェンバー内に接種し(ウエルあたり50μl)、そして30分間、37℃において5%のCO2加湿インキュベーター中でインキュベートし
た。インキュベーション後に、チェンバーを解体し、フィルターの上部側を洗浄し、そしてふき取り(wiped off)、そして底部側に付着した細胞をメタノールで固定してHem
acolor溶液(メルク)により染色した。細胞を、次に、400xの倍率にて、ウエルあたり4つのランダムに選択された顕微鏡の視野の中で数えた。最後に、3つの個別の実験の平均をケモカイン濃度に対してプロットした。図7において、様々なIL−8変異体の走化性指数を示す。予測されたとおり、全ての変異体が顕著に低下した受容体結合活性を示した。
実施例5:組換えRANTES遺伝子の生成、発現、変異体の生物物理特性決定及び活性の特性決定
優性な陰性「GAG−マスキング」ケモカイン変異体のコンセプトを、移植拒絶のようなタイプIVの高感受性反応、アトピー性皮膚炎並びに関節炎、進行性糸球体腎炎及び炎症性肺疾患のような他の炎症性傷害に関与するケモカインであるRANTESにも用いた。
【0064】
wt蛋白質を開始メチオニン残基に導入することにより、受容体結合活性が損なわれた。エシェリヒアコリ内でのwt RANTESの発現は、このメチオニン残基の保有を導き、wt RANTESを単球移動の有力な阻害剤にし、いわゆるMet−RANTESである。GAG結合親和性を増強する別の変異をPCRに基づく部位特異的変異導入法により導入した。
【0065】
これらの9つのRANTES変異体がこれまでにクローン化され、発現されて、精製された、Met−RANTES A22K,Met−RANTES H23K,Met−RANTES T43K,Met−RANTES N46R,Met−RANTES N46K,Met−RANTES Q48K,Met−RANTES A22K/N46R,Met−RANTES V49R/E66S及びMet−RANTES 15LSLA18 V49R/E66S。
【0066】
恒温蛍光滴定実験を実施することにより、サイズで規定されたヘパリンに関するRANTESの相対的親和性定数(Kd値)を測定した。表に見られるとおり、全てのRANTES変異蛋白質はこのヘパリンに関して高い親和性を示し、Met−RANTES A22K,Met−RANTES H23K,Met−RANTES T43K及びMet−RANTES A22K/N46Rがもっとも有望な結果を示した。
【0067】
Kd(nM)
野生型Rantes 456.2± 8.5
Met-Rantes V49R/E66S 345.5±21.7
Rantes 15LSLA18 V49R/E66S 297.3±14.1
Rantes N46R 367.7±11.7
Rantes N46K 257.4±10.2
Rantes H23K 202.5±12.8
Rantes Q48K 383.4±39.6
Rantes T43K 139.2±30.1
Rantes A22K 202.1± 9.8
Rantes A22K/N46R 164.0±16.6
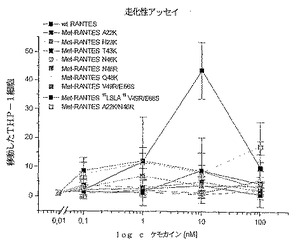
RANTES走化性アッセイ
RANTES変異体により指示された細胞移動を、5μmのPVP−コートされたポリカーボネート膜を備えた48ウエルBoydenチェンバーシステムを用いて調査した。RANTES及びRANTES変異体の20mM HEPES pH7.3及び1mg/mlのBSAを含むRPMI 1640の希釈液を3通りにしてチェンバーの底部ウエル内に置いた。同じ培地中の2x106/ml細胞の50μlのTHP−1細胞懸濁液(セ
ルカルチャーのヨーロッパコレクションからの前単球細胞系)を上部ウエル中に置いた。37℃における5%CO2加湿の2時間のインキュベーションの後に、フィルターの上部表面をHBSS溶液で洗浄した。移動した細胞をメタノールにより固定して、Hemacolor溶液(メルク)により染色した。400xの倍率にて、ウエルあたり5つを数え、3つの個別の実験の平均を図8においてケモカイン濃度に対してプロットした。エラーバーは3つの実験の平均の標準誤差を表す。再び、IL−8変異体の場合のように、全てのRANTES変異体が顕著に低下した受容体結合活性を示した。
実施例6:GAG結合領域を有する蛋白質
バイオインフォマティックス及びプロテオームにより、GAG結合蛋白質をそれらのGAG結合領域と共に特性決定した。以下の表2及び3において、ケモカインとそれらのGAG結合領域(表2)を示し、そして他の蛋白質の例もそれらのGAG結合領域と共に提供する(表3)。
【0068】
【表2】
【0069】
【表3】
【0070】
【表4】
【0071】
【表5】
【0072】
【表6】
【0073】
【表7】
【0074】
【表8】
【0075】
【表9】
【0076】
【表10】
【0077】
【表11】
【0078】
【表12】
【0079】
【表13】
【0080】
【表14】
【0081】
【表15】
【0082】
【表16】
【0083】
【表17】
【0084】
【表18】
【0085】
【表19】
【0086】
【表20】
【0087】
【表21】
【0088】
【表22】
【0089】
【表23】
【0090】
【表24】
【0091】
【表25】
【0092】
【表26】
【0093】
【表27】
【0094】
【表28】
【0095】
【表29】
【0096】
【表30】
【技術分野】
【0001】
本発明は、ケモカインと白血球上のそれらの高親和性受容体の相互作用の阻害のための方法とツール及び炎症性疾患の治療処置のための方法に関する。
【背景技術】
【0002】
ケモカインは、化学誘引物質サイトカインを起源とし、実際は50より多いメンバーを含み、そして小さくて誘導可能な、そして分泌される、低分子量の蛋白質(それらのモノマー形態において6−12kDa)のファミリーを表し、免疫監視機構及び炎症のプロセスの間に決定的な役割を担う。免疫及び炎症におけるそれらの機能に依存して、それらは、2つのクラスに区別することができる。多くの異なる組織細胞により、並びに細菌の毒素及び炎症性サイトカイン、例えばIL−1,TNF及びインターフェロンに応答して白血球を移入する(immigrating)ことにより、炎症性ケモカインは生産される。それらの
主な機能は、宿主の防御のため及び炎症のプロセスにおいて白血球を補充することである。ホーミングケモカインは、他方、リンパ組織の規定されたエリアにおいて構成的に発現される。それらは、免疫系において白血球及び星状細胞の往来及びホーミングを指示する。これらのケモカインは、BCA−1,SDF−1又はSLCにより例示されたとおり、成熟、分化、活性化のコンテクストにおいて白血球の再配置及び再循環を制御し、そして二次リンパ器官内でそれらの正確なホーミングを確保する(ensure)。
【0003】
多数の見本にも拘わらず、ケモカインは顕著に類似の構造的折り畳み(folds)を示す
が、配列相同性は20から70パーセントの間で変化する。ケモカインは、大まかに、70−130アミノ酸からなり、4つの保存されたシステイン残基を伴う。システインは2つのジスルフィド結合を形成し(Cys 1→Cys 3,Cys 2→Cys 4)、それらの特徴的な三次元構造に必須である。走化性ケモカインは、短いアミノ末端ドメイン(3−10アミノ酸)、第1システイン残基、β鎖並びに第2及び第4システイン残基の間に見いだされる接続ループ、並びに20−60アミノ酸のカルボキシル末端αヘリックスからなる。蛋白質のコアは、うまく整列された構造を有するが、N−及びC−末端部分は乱れている。分泌蛋白質のように、それらは、20−25アミノ酸のリーダー配列を伴って合成されて、放出の前に分割されて、はずれる。
【0004】
ケモカインは成熟蛋白質内のそれらのシステイン残基の相対的な位置に基づいて4つのファミリーに副次的に分けられる。αケモカインサブファミリーにおいては、4つのシステインのうちの最初の2つが単一アミノ酸(CXC)として分離され、一方βケモカインにおいては対応するシステイン残基が互いに隣接し合う(CC)。αケモカインは、さらに、N末端にELR配列を含むことにより好中球に走化性であるもの(IL−8、例えば)、及びELRモチーフを欠きリンパ球に作用するもの(I−TAC、例えば)に分類することができる。構造上、βケモカインは2つのファミリーに副次的に分類することができ、単球−化学誘引性蛋白質エオタキシンファミリーであって5つの単球化学誘引性蛋白質(MCP)と互いに約65パーセント同一のエオタキシンを含むものと、それ以外のβケモカインである。CXC−ファミリーの場合のように、CC−残基の上流のN−末端アミノ酸が、ケモカインの生物活性及び白血球選択性に必須の成分である。βケモカインは、一般に、好中球には作用せず、しかし単球、好酸球、好塩基球及びリンパ球を可変性選択性をもって誘引する。
【0005】
ほんのわずかなケモカインがCC−又はCXC−ファミリーに適合しない。リンフォタクチンは、これまで、その初期構造において4つの特徴的なシステインの代わりにたった2つしか示さない唯一のケモカインであり、即ち、γ−又はC−ケモカインとして分類さ
れる。他方、この分類を締めくくることにより、フラクタルカイン(fractalkine)は最
初の2つのシステインを隔てる3つのアミノ酸を有するδ−又はCXXXC−サブファミリーの唯一の代表である。リンフォタキシン及びフラクタルカインはどちらもT−細胞及びナチュラルキラー細胞の走化性を誘導する。
【0006】
ケモカインは、特定の細胞表面、標的細胞上の7つの膜貫通スパニング(7TM)G−プロテイン共役受容体に結合することにより、細胞の移動(migration)及び活性化を誘
導する。18のケモカイン受容体がこれまでにクローン化されており、6つのCXC、10のCC、1つのCX3C及び1つのXC受容体を含む。ケモカイン受容体は特定の細胞に限定され(例えば、CXCR1は好中球に限定される)るが、その他は、より広く発現される(例えば、CCR2は単球、T細胞、ナチュラルキラー細胞及び好塩基球上で発現される)。ケモカインに類似して、上記受容体は特定の細胞上で構成的に発現されるが、いくつかは誘導可能である。それらのいくつかはダウン制御さえもされ得て、細胞を特定のケモカインに対して非応答性にするが、その他に対しての応答性は残す。ほとんどの受容体が1つより多いケモカインを認識し、逆もまた同じで、しかしながら認識は対応するサブファミリーのケモカインに限定される(表1を参照)。
【0007】
【表1】
【0008】
ケモカインはそれらの受容体への相互作用のための主要な2つの部位を有し、一つはアミノ末端ドメイン中にあり、そして他方は第2と第3システイン残基の間に伸びるバックボーンの露出されたループ内にある。両部位はジスルフィド結合により近い位置を持つ。受容体は最初にドッキングドメインとして機能するらしいループ領域内の結合部位を認識
する。この相互作用はケモカインの移動性(mobility)を制限し、即ち、アミノ末端ドメインの正確な位置付けを容易にする。それらの受容体に対して有効に今まで通り結合するがシグナルを生じない変異体ケモカインを用いて、研究が実施された。これらの変異体は例えば、IL−8,RANTES及びMCP−1のN末端内のアミノ酸欠失又は修飾により得られた。
【0009】
複数の細胞内シグナリング経路が、受容体活性化後に、ケモカイン結合の結果として生じる。ケモカインは、2種類の非シグナリング分子とも相互作用する。一つはDARC受容体であって、赤血球上及び内皮細胞上で発現されてCC−ケモカイン並びにCXC−ケモカインに結合することによりそれらが循環されるのを阻止する。第2の種類は、硫酸ヘパラングリコサミノグリカン(GAGs)であり、プロテオグリカンの一部であって、ケモカインの共同受容体として機能する。それらは、ホーミング組織(例えば、内皮細胞)の表面上でケモカインを捕捉して提示することにより、局所的濃度勾配を確立する。炎症性応答、例えば慢性関節リウマチにおいては、セレクチン媒介性プロセスにおいて内皮上の白血球ローリングが、細胞表面のプロテオグリカンにより提示されるケモカインと接触する。それにより、白血球インテグリンが活性化されるようになり、安定した接着及び溢出を導く。補充された白血球は、局所炎症性ケモカインにより活性化されて、ケモカインの局所性高濃度のためにさらなるケモカインシグナリングに対して鈍感になるかもしれない。組織血流ケモカイン勾配を保持するために、DARC受容体は過剰のケモカインのための下水だめ(sink)として機能する。
【0010】
硫酸ヘパラン(HS)グリコサミノグリカンは、グリコサミノグリカンの側鎖に共有結合したコア蛋白質からなり、ほとんどの哺乳類細胞及び組織に見いだされる。蛋白質部分は細胞膜中又は細胞外マトリックス内のプロテオグリカンの位置決めを決定するが、グリコサミノグリカン成分は様々な細胞外リガンド、例えば成長因子、ケモカイン及び接着分子との相互作用を媒介する。プロテオグリカンの生合成は以前に広く総説された。細胞表面プロテオグリカンの主要なグループは、膜貫通蛋白質のシンデカンファミリー(哺乳類においては4つのメンバー)及びグリコシルホスファチジルイノシトール(GPI)テイルにより細胞膜に結合される蛋白質のグリピカン(哺乳類においては6つのメンバー)である。グリピカンは、神経系において、腎臓において、広範囲に、そして少ない程度では骨格筋及び平滑筋において発現されるが、シンデカン−1は上皮細胞において主要なHSPGであり、シンデカン−2は繊維芽細胞及び内皮細胞において支配的であり、そしてシンデカン−3は神経細胞において富裕であり、そしてシンデカン−4は広範囲に発現される。特定のセリン上のテトラサッカライド結合領域を通してシンデカンコア蛋白質に添加されるGAG鎖の大多数は、HS鎖である。膜貫通及びサイトプラズムドメインとは対照的に、特定のシンデカンタイプの細胞外ドメインのアミノ酸配列は異なる種の間では保存されていないが、GAG鎖の数及び位置は高度に保存されている。GAGの構造は、しかしながら、種特異的であり、さらに、HSPG発現組織の性質に依存する。
【0011】
硫酸ヘパラン(HS)は直鎖状ポリサッカライドのグリコサミノグリカン(GAG)ファミリーのもっとも豊富な蛋白質であり、ヘパリン、硫酸コンドロイチン、硫酸デルマタン及び硫酸ケラタンも含む。天然のHSはD−グルコサミン(GlcNAc)及びD−グルクロン酸(GlcA)から構成される20−100のジサッカライドユニットの直鎖により特徴付けされ、N−及びO−硫酸化(グルコサミンの6−O及び3−O硫酸化及びウロン酸の2−O硫酸化)並びにβ−D−グルクロン酸のα−L−イズロン酸(IdoA)へのエピマー化(epimerisation)を含むように修飾され得る。
【0012】
低硫酸化の領域により分離されるN−及びO−硫酸化糖残基のクラスターは、多数の蛋白質結合及びHSの制御特性に主に必須であると推測される。HSの硫酸基の塩基性アミノ酸への静電相互作用に加えて、ファンデルワース相互作用及び疎水性相互作用も蛋白質
結合に関与すると考えられる。さらに、コンフォメーション上柔軟なイズロン酸残基の存在は、蛋白質へのGAG結合に好都合のようである。他の因子、例えば蛋白質結合部位の間の間隔をあけることも、蛋白質−GAG結合相互作用において決定的な役割を担う:例えば、HSにより誘導される−インターフェロンの二量体化は、低硫酸化の7kDa領域により分離される2つの蛋白質結合配列を伴うGAG鎖を必要とする。いくつかのリガンドの完全な生物活性には付加的な配列がときどき必要とされ:FGF−2シグナルトランスダクションをサポートするためには、HSがFGF受容体と相互作用すると想像される最少結合配列並びに追加の残基の両方を有さねばならない。
【0013】
ヘパリン結合蛋白質は、しばしば、塩基性アミノ酸残基のクラスターからなる。リジン、アルギニン、アスパラギン、ヒスチジン及びグルタミンは、しばしば、GAG上の硫酸基及びカルボキシル基との静電接触に関与する。塩基性アミノ酸の間隔(spacing)は、
ときどき蛋白質3−D構造により決定されるが、GAG結合特異性及び親和性を制御すると想定される。当該リガンドの生物活性はHS−蛋白質相互作用のキネティクスによっても影響され得る。成長因子拡散の面積を減少させることは、GAG鎖の長い繰り返しの特性並びにそれらの相対的に早い蛋白質結合のオン及びオフの速度が理想的に適合される、示唆されたHSPG機能の一つである。いくつかの場合、熱力学よりもキネティクスが、HS−蛋白質結合の生理機能を推進する。ほとんどのHSリガンド機能は、明確な長さ及び構造のGAG配列を必要とする。マスト細胞により生産されるヘアピンは、構造上、硫酸ヘパランに類似しているが、相対的に小さな程度の構造多様性を伴って均一に高度な硫酸化をもたらす高いレベルの重合後修飾により特徴付けされる。即ち、硫酸ヘパラン中の高度に修飾されたブロックは、ときどき、「ヘアピン様」と呼ばれる。このため、ヘアピンはそのまま蛋白質の生物物理研究のための完璧なHS類似体として使用可能であり、さらには、大量に利用できるので、HSよりも経費がかからない。異なる細胞種が異なるグリコサミノグリカン構造を有するプロテオグリカンを合成することが示されており、その構造は病理の間、発生の間、又は細胞外シグナル、例えば成長因子に応答して変化する。HSPGsのこの構造多様性は、高い結合融通性を導き、プロテオグリカンの多大な重要性を強調する。
【0014】
硫酸ヘパランプロテオグリカンはFGFシグナリングに必須であるとの証明以来、幾人かの研究者がケモカイン活性を促進させるためのケモカイン−GAG結合の重要性を示そうと実行した。第1に、これまでに研究されたほとんど全てのケモカインはインビトロにおいてHSを結合するらしく、これはこれらの蛋白質の根本的な特性を表すことを示唆する。第2に、インビボにおいてTリンパ球がCC−ケモカインをグリコサミノグリカンとの複合体として分泌するとの発見は、この形態の相互作用が生理学上関連していることを示す。さらに、ケモカインとHSの対合(association)は内皮表面を横切る濃度勾配を
安定化させるのを助け、それにより白血球を移動させるための方向性の情報を提供することが知られている。HSは蛋白質分解からケモカインを保護すること及びそれらのオリゴマー化を誘導すること、即ち、G−共役シグナリング受容体の付近で局所的高濃度を促進するとも考えられる。オリゴマー化の機能的な関連性は、しかしながら、議論が残されているが、全てのケモカインはマルチマー化のための明確な構造上の基礎を有する。β−シートの対合を通したダイマー化はCXC−ファミリー(例えば、IL−8)の全てのケモカインにおいて観察されており、それらのN−末端鎖を通してダイマー化するCC−ケモカインファミリー(例えば、RANTES)のほとんどのメンバーとは対照的である。
【0015】
豊富なデータがケモカインの相互作用の阻害及び低分子量化合物による白血球上のそれらの親和性受容体に関して蓄積した。しかしながら、このアプローチによる炎症性疾患の治療処置におけるブレイクスルーはない。
【0016】
インターロイキン−8(IL−8)は慢性及び急性の炎症の間、好中球誘引(attracti
on)に関与する重要分子である。これまでにIL−8の作用をブロックするためにいくつかのアプローチが企てられたが、例えば、グルココルチコイド、ビタミンD3,シクロスポリンA,形質転換成長因子β、インターフェロン等によるIL−8生産の阻害に始まるが、それら全てはIL−8のmRNAの生産レベルにおいてIl−8活性を阻害する。以前使用されたさらなるアプローチは、特異的アンタゴニストとして作用するために白血球上の受容体に対するか又はIL−8自身に対する特異的抗体を用いることによりIL−8のその受容体への結合を阻害すること、即ちIL−8活性を阻害することである。
【発明の概要】
【0017】
本発明の目的は、よって、白血球上のケモカイン/受容体の相互作用の阻害又は妨害のための別の戦略を提供することである。特に、そのような戦略により、IL−9,RANTES又はMCP−1の作用を標的にすべきである。
【図面の簡単な説明】
【0018】
【図1】図1はCDスペクトルである。
【図2】図2は様々な変異体の二次構造コンテンツを示す。
【図3】図3は様々な変異体の蛍光異方性試験の結果のグラフである。
【図4】図4は様々な変異体の蛍光異方性試験の結果のグラフである。
【図5】図5は恒温蛍光滴定の結果のグラフである。
【図6】図6は様々な変異体のアンフォールド実験の結果のグラフである。
【図7】図7はIL−8変異体の走化性指数を示す。
【図8】図8はRANTES走化性アッセイの結果を示す。
【発明を実施するための形態】
【0019】
本発明の主題は、よって、新規なGAG結合蛋白質並びにGAG−共同受容体に対して(野生型よりも)高い親和性を示す別のGAG結合蛋白質を生産する方法を提供することである。そのような修飾されたGAG結合蛋白質は野生型のGAG結合蛋白質との競合因子として作用することができ、そして野生型GAG結合蛋白質の活性を阻害する点ではダウン制御することができるが、しかしながら当業界において使用される公知の組換え蛋白質を用いたときに生じる副作用がない。本発明による分子は上記の不都合をもたない。本修飾GAG結合蛋白質は様々な治療用用途のためのドラッグにおいて使用することができ、特に、ケモカインの場合、当業界において使用される公知の組換え蛋白質を用いたときに生じる公知の不都合なしに炎症性疾患を治療するためのドラッグにおいて使用することができる。本発明によるGAG結合部位の修飾はこの部位に対する結合事象上にその活性が基づく全ての蛋白質、特にGAG部位によるケモカイン、のための広く適用可能な戦略であることが判明した。高いGAG結合親和性を有する本発明による好ましい分子は、それらの生物学上の効果、特にGAG部位に関する野生型分子とのそれらの競合によりそれらの抗炎症活性に関して特に有利であることが証明された。
【0020】
よって、本発明は、GAG結合部位を、特性決定された蛋白質に導入する方法を提供するが、工程:
―構造保持に必須でない蛋白質中の領域を同定すること
―塩基性アミノ酸を上記部位中に導入するか、及び/又は、少なくとも一つの巨大な及び/又は酸性のアミノ酸を上記部位において欠失させること
を含み、GAG結合部位はKd≦10μM、好ましくは≦1μM、さらに好ましくは≦0.1μMのGAG結合親和性を有する。塩基性アミノ酸を上記部位中に導入するか、及び/又は、少なくとも一つの巨大な及び/又は酸性のアミノ酸を上記部位において欠失させることにより、新規で改良された「人工の」GAG結合部位が上記蛋白質に導入される。これは、上記修飾の前にはGAG結合活性を示さなかった蛋白質へのGAG結合部位の新規で完璧な導入を含む。これは、既にGAG結合活性を示す蛋白質へのGAG結合部位の
導入も含む。新規なGAG結合部位は、GAG結合親和性を示さない蛋白質の領域並びにGAG結合親和性を示す蛋白質の領域へ導入することができる。しかしながら、本発明のもっとも好ましい態様によれば、与えられたGAG結合蛋白質のGAG結合親和性の修飾が提供され、上記修飾された蛋白質のGAG結合能力が野生型蛋白質に比べて増加する。本発明は、GAG結合部位を蛋白質に導入する方法、修飾されたGAG結合蛋白質、並びに単離されたDNA分子、ベクター、組換え細胞、薬学組成物、及び上記修飾された蛋白質の使用法に関する。
【0021】
用語「少なくとも一つの塩基性アミノ酸を導入する」は、追加のアミノ酸の導入並びにアミノ酸の置換に関する。主要な目的は、塩基性アミノ酸、好ましくはArg,Lys,His,Asn及び/又はGlnの上記部位における全アミノ酸に対する相対量を増加させることであり、それにより、結果のGAG結合部位が好ましくは少なくとも3つの塩基性アミノ酸、さらに好ましくは4、もっとも好ましくは5つのアミノ酸を含むべきである。
【0022】
GAG結合部位は、好ましくは、溶媒露出位置、即ち、ループにある。これは、有効な修飾を保証する。
蛋白質の領域が構造の保持に必須であるか否かは、例えば、当業者には知られている特定のプログラムとともにコンピュータの方法により試験することができる。蛋白質の修飾の後に、コンフォメーションの安定性をインシリコにて試験することが好ましい。
【0023】
用語「巨大な」アミノ酸は、長いか又は立体構造干渉性の側鎖を有するアミノ酸を意味する;これらは特にTrp,Ile,Leu,Phe,Tyrである。酸性アミノ酸は、特に、Glu及びAspである。好ましくは、結果のGAG結合部位が巨大かつ酸性のアミノ酸を含まないことであり、すべての巨大なかつ酸性のアミノ酸が除去されていることを意味する。
【0024】
GAG結合親和性を測定する−本出願の保護の範囲のため−解離定数Kdを通して。一つの可能性は、リガンド結合における構造変化によるあらゆる与えられた蛋白質の解離定数(Kd)を測定することである。さまざまな技術が当業者には知られており、例えば、恒温蛍光滴定、恒温滴定熱量測定、表面プラズモン共鳴、ゲル移動度アッセイ、放射性標識されたGAGリガンドを用いた競合実験により間接的に行う。さらなる可能性は、これも当業者には知られているコンピュータによる方法によりによる計算により結合領域を予測することであり、いくつかのプログラムを使用してよい。
【0025】
蛋白質中にGAG結合部位を導入するためのプロトコルは、例えば以下のとおりである:
・全体の構造保持に必須ではなくかつGAG結合に適しているかもしれない蛋白質の領域を同定する
・何れかの位置において塩基性Arg,Lys,His,Asn及び/又はGln残基を導入すること(置換又は挿入)により新規のGAG結合部位をデザインする
・インシリコにおいて結果の変異蛋白質のコンフォメーションの安定性をチェックする
・野生型蛋白質のcDNAをクローン化する(あるいは:cDNAを購入する)
・これをPCR−補助変異導入法のための鋳型として使用することによりアミノ酸配列に上記の変化を導入する
・変異体遺伝子を適当な発現系にサブクローン化する(生物学上必要とされる翻訳後修飾に依存して原核生物又は真核生物)
・インビトロにおいて変異体蛋白質の発現、精製及び特性決定
・導入されたGAG結合親和性KdGAG(変異体)<10μMの基準。
【0026】
新規のGAG結合部位を有する上記の操作された蛋白質の例は、例えば、IgGのFc並びに以下のとおりに修飾された補体因子C3及びC4である。
【0027】
【化1】
【0028】
本発明のさらなる側面は、上記の発明の方法により得ることが可能な蛋白質である。発明の蛋白質は、よって、−野生型の蛋白質に比較すると−上記のとおり新規なGAG結合部位を含み、よって、天然GAG結合蛋白質との競合因子として作用することになるが、特に発明の蛋白質のGAG結合親和性が極めて高い、例えばKd≦10μMだからからである。
【0029】
本発明のさらなる側面は修飾されたGAG結合蛋白質であり、上記の蛋白質の中のGAG結合領域が少なくとも一つのアミノ酸の置換、挿入、及び/又は欠失により修飾されることにより、上記GAG結合領域の塩基性アミノ酸の相対量を増加させるか、及び/又はGAG結合領域の巨大及び/又は酸性のアミノ酸の量を増加させるか減少させるが、好ましくは溶剤露出位置においてであり、そして上記蛋白質のGAG結合親和性が各々の野生型蛋白質のGAG結合親和性に比較して増加する。
【0030】
驚きをもって示されたことは、GAG結合領域中の塩基性アミノ酸、特にArg,Lys,His,Asn及び/又はGlnの相対領域を増加させることにより、修飾されたGAG結合蛋白質が野生型蛋白質に比較して増加したGAG結合親和性を示すことであり、特に塩基性アミノ酸の相対量を溶剤露出位置において増加させた場合であるが、蛋白質表面上の陽性荷電したエリアが結合親和性を増強することが示されたからである。好ましくは、3つの塩基性アミノ酸、さらに好ましくは4、もっとも好ましくは5つのアミノ酸がGAG結合領域中に存在する。
【0031】
用語「GAG結合蛋白質」は、GAG共同受容体に結合するあらゆる蛋白質を意味する。蛋白質がGAG共同受容体に結合するか否かは、上記のとおりに公知のプロトコルの助けを借りて試験することができる。Hilemanら(BioEssays 20(1998),156−167)は、グリコサミノグリカン結合する蛋白質内のコンセンサス部位を開示する。この文献内に開示された情報は、本発明のための開始情報としても有用である。用語「蛋白質」は、本発明により提供される分子が少なくとも80アミノ酸の長さであることを明らかにする。これは、それらを、本抗炎症戦略のために適した候補にするために要求される。GAG結合部位と相互作用し、かつその相互作用のための生理学上又は病理学上関連するより小さな分子は知られていないし、本発明に関係しない。好ましくは、本発明による分子は、少なくとも90、少なくとも100、少なくとも120、少なくとも150、少なくとも200、少なくとも300、少なくとも400又は少なくとも500のアミノ酸残基から構成される。
【0032】
本発明の範囲において、用語「GAG結合領域」は、恒温蛍光滴定により測定されたところによれば、100μM以下、好ましくは50μM以下、さらに好ましくは20μM以下の解離定数(Kd−値)にてGAGに結合する領域として定義される(以下の実施例を参照)。
【0033】
本出願において言及される如何なる修飾も、公知の生化学法、例えば部位特異的変異導入法を用いて実施することができる。GAG結合部位の分子はクローン化はもちろん従来
技術であり(例えば、WO96/34965 A,WO02/07935 A,Jaya
raman et al.(FEBS Letters 482(2000),154−158),WO02/20715 A,Yang et al.(J.Cell.Biochem.56(1994),455−468)、GAG領域の分子シャッフリング又は
デノボ合成が記載されていることも注目すべきである;Butcher et al.,(FEBS Letters 4009(1997),183−187)(人工ペプチドに関し、蛋白質ではない);Junno−Oue et al,(J.Virol.75(2001),12439−12445)デノボ合成)を参照)。
【0034】
GAG結合領域は、置換、挿入及び/又は欠失により修飾することができる。これは、非塩基性アミノ酸が塩基性アミノ酸により置換されてよいこと、塩基性アミノ酸がGAG結合領域に挿入されてよいこと又は非塩基性アミノ酸が欠失してよいことを意味する。さらに、GAG結合を干渉するアミノ酸、好ましくはすべての干渉するアミノ酸結合が欠失する。そのようなアミノ酸は特に上記のとおり巨大なアミノ酸又は酸性アミノ酸であり、例えばGlu及びAspである。アミノ酸がGAG結合を干渉するか否かは、例えば数学の方法又はコンピュータの方法により試験してよい。これらの修飾の如何なる結果も、上記GAG結合領域中の塩基性アミノ酸の相対量が増加することであり、「相対」はGAG結合領域中のすべてのアミノ酸の数に比較したGAG結合領域中の塩基性アミノ酸の量を意味する。さらに、立体構造上又は静電上GAG結合を干渉するアミノ酸を欠失させる。
【0035】
アミノ酸が溶剤露出位置中に存在するか否かは、例えば、上記のとおり蛋白質の公知の三次元構造の助けを借りてか又はコンピュータ法の助けを借りて決定することができる。
上記の修飾の蛋白質のGAG結合親和性が各野生型蛋白質のGAG結合親和性に比較して増加しているか否かは、上記のとおり、例えば、解離定数を測定する蛍光滴定実験の助けを借りて測定することができる。改良されたGAG結合親和性に関する基準は、Kd(変異体)<Kd(野生型)であり、好ましくは少なくとも100%である。特別に改良されて修飾のされた蛋白質は、野生型のKdに比較して、少なくとも5倍、好ましくは少なくとも10倍、さらに好ましくは少なくとも100倍高いGAG結合親和性を有する。増加したGAG結合親和性は、よって、好ましくは10μM以下、好ましくは10μM以下、さらに好ましくは0.1μM以下のKdを示す。
【0036】
GAG結合親和性を増加させることにより、修飾された蛋白質は、特異的アンタゴニストになり、そしてGAG結合に関して野生型のGAG結合蛋白質と競合することになる。
好ましくは、Arg,Lys,及びHisからなる群から選択される少なくとも一つのアミノ酸が上記GAG結合領域に挿入される。これらのアミノ酸は、GAG結合領域に容易に挿入されるが、用語「挿入される」は、挿入それ自体並びにあらゆる非塩基性アミノ酸をアルギニン、リジン又はヒスチジンで置換することに関する。もちろん、一つより覆い塩基性アミノ酸を挿入することが可能であり、それにより同じ塩基性アミノ酸が挿入されるか又は上記アミノ酸2つ又は3つの組み合わせでもよい。
【0037】
さらに好ましくは、上記蛋白質がケモカイン、好ましくはIL−8,RANTES又はMCP−1である。ケモカインは共同受容体との相互作用の部位を有することが知られており、このケモカイン結合はしばしば上記のとおりさらなる受容体活性化の条件である。ケモカインはしばしば炎症性疾患において見いだされるから、ケモカイン受容体活性化をブロックすることに主な興味がある。そのようなケモカインは、好ましくは、IL−8,RANTES又はMCP−1であり、よく特性決定された分子であって、GAG結合領域がよく知られている分子である(例えば、Lortat−Jacob et al.,PNAS 99(3)(2002),1229−1234を参照)。これらのケモカイン中のGAG結合領域の中の塩基性アミノ酸の量を増加させることにより、それらの結合親和性が増加し、よって野生型蛋白質の濃度に関しての修飾された蛋白質の濃度に依存して、
野生型ケモカインはほとんど又は全く結合しなくなる。
【0038】
有利な側面によれば、上記GAG結合領域はC末端のαヘリックスである。典型的な化学的モノマーがC末端のαヘリックスによりオーバーレイされた三重鎖逆平行βシートの周囲に組織化される。ケモカイン中のこのC末端αヘリックスがGAG結合に関与する主要な部分であり、そのため、塩基性アミノ酸の量を増加させるためにこのC末端αヘリックス中の修飾が増加したGAG結合親和性を有する修飾ケモカインをもたらすことが示された。
【0039】
有利なことには、IL−8中の17、21、70、及び/又は71位が、Arg,Lys,His,Asn及び/又はGlnにより置換されている。ここでは、これらの前記の部位のたった一つが修飾されることがあり得る。しかしながら、これらの部位の一つより多く並びに全ても修飾されてよく、全ての修飾がArg又はLys又はHis又はAsn又はGln又はそれらの混合の何れかであってよい。IL−8においては、これらの位置がIL−8のGAG結合親和性を大いに増加させることが示されていることから、これらの位置は修飾に特に適している。
【0040】
好ましくは、増加した結合親和性は、硫酸ヘパラン及び/又はヘパリンに対しての増加した結合親和性である。硫酸ヘパランは直鎖状のポリサッカライドのGAGファミリーのもっとも豊富なメンバーであり、ヘパリンも含む。ヘパリンは硫酸ヘパランに構造上極めて類似しており、相対的に小さい程度の構造多様性の、均一に高度な硫酸化をもたらす高いレベルの重合後修飾により特徴付けられる。よって、硫酸ヘパラン中の高度に修飾されたブロックは、ときに、ヘパリン様と呼ばれ、そしてヘパリンは蛋白質の生物物理研究のための硫酸ヘパラン類似体として使用することができる。何れにせよ、硫酸ヘパランとヘパリンは共に特に適している。
【0041】
さらに好ましくは、さらに生物学上活性な領域が修飾されることにより、上記蛋白質のさらなる生物学上の活性を阻害するか又はダウン制御する。このさらなる生物学上の活性は、ほとんどのGAG結合蛋白質、例えばケモカインに関して知られている。これは、受容体、例えば7TM受容体に対する結合領域になる。用語「さらなる(further)」は、
GAG結合領域ではないが、しかし他の分子、細胞又は受容体に結合するか及び/又はそれらを活性化する、生物学上活性な領域を規定する。このさらなる生物学上活性な領域を修飾することにより、この蛋白質のさらなる生物学上の活性が阻害されるか又はダウン制御され、そしてそれにより野生型蛋白質よりも強いアンタゴニストである修飾された蛋白質が提供される。これは、他方で、GAG結合親和性が野生型GAG結合蛋白質よりも高いことを意味し、修飾された蛋白質は野生型蛋白質に代わりGAGに対して大部分結合する。一方、蛋白質がGAGに結合したときに主に起こる野生型蛋白質のさらなる活性は阻害されるか又はダウン制御されるが、修飾された蛋白質がこの特異的な活性を実行しないか又は低い程度でこの活性を実行するからである。この修飾された蛋白質を用いて、当業界の状況において記載されるとおり、他の組換え蛋白質から知られる副作用を示さない、野生型GAG結合蛋白質に関して有効なアンタゴニストが提供される。このさらなる生物学上の活性領域は、例えば、受容体競合アッセイによりインビトロにおいて測定することができる(蛍光標識されたwtケモカインを用いる、カルシウム流入、及び細胞移動(天然白血球又は7TM安定にトランスフェクトされた細胞系)を用いて)。そのようなさらなる生物学上活性な領域の例は、さらなる受容体結合部位に加えて(成長因子ファミリーにおけるように)、酵素部位(ヒドロラーゼ、リアーゼ、スルフォトランスフェラーゼ、N−デアセチラーゼ、及びコポリメラーゼにおけるように)、蛋白質相互作用部位(アンチトロンビンIIIにおけるように)、及び膜結合ドメイン(ヘルペスシンプレックスウイルスgD蛋白質におけるように)である。二重に修飾された蛋白質のこの好ましい態様により、よって、優性(GAG結合に関して)陰性(受容体に関して)変異体が提供され
、本発明の対象物のセットに関して特に有利である。
【0042】
さらに好ましくは、上記のさらなる生物学上の活性領域が、欠失、挿入、及び/又は置換により、好ましくはアラニン、立体構造上及び/又は静電上類似の残基により修飾される。もちろん、上記のさらなる生物学上活性な領域中で少なくとも一つのアミノ酸を欠失させるか又は挿入するか又は置換するかの何れかが可能である。しかしながら、これらの修飾の少なくとも2つ又はそれら全部3つの組み合わせを提供することも可能である。与えられたアミノ酸をアラニン又は立体構造上/静電上類似の残基で修飾することにより−「類似」は置換されるアミノ酸に類似なことを意味する−修飾された蛋白質は、全くか又はほとんど立体構造上/静電上修飾されない。これは特に有利であるが、何故ならば、修飾された蛋白質の他の活性、特にGAG結合領域に対する親和性が変化しないからである。
【0043】
有利には、上記蛋白質がケモカインであり、そして上記のさらなる生物学上の活性が白血球活性化である。上記のとおり、ケモカインは慢性及び急性炎症の間、白血球誘引に関与する。よって、白血球活性化を阻害するか又はダウン制御することにより、炎症が減少するか又は阻害され、この特定の修飾された蛋白質を、炎症性疾患を研究し、診断し、そして治療するための重要なツールにする。
【0044】
有利な側面によれば、上記蛋白質がIL−8であり、そしてさらなる生物学上活性な領域が最初の10個のN末端アミノ酸内に位置する。最初のN末端のアミノ酸は白血球活性化に関与し、特に、Glu−4,Leu−5及びArg−6は受容体結合及び活性化に必須であることが分かっている。よって、これら3つ又は最初の10個のN末端アミノ酸全てさえ置換するか又は欠失させることにより、受容体結合及び活性化を阻害するか又はダウン制御することができる。
【0045】
さらなる有利な蛋白質は、最初の6個のN末端アミノ酸が欠失したIL−8変異体である。上記のとおり、この変異体は全くか又はほとんど白血球に結合して白血球を活性化せず、その結果、炎症性疾患を研究し、診断し、そして治療するのに適している。
【0046】
好ましくは、上記蛋白質がdel6F17RE70KN71R,del6F17RE70RN71K及びdel6E70KN71Kからなる群から選択されるIL−8変異体である。これらの変異体が特に有利であることが示されたのは、最初の6個のN末端アミノ酸の欠失が受容体結合及び活性化を阻害するか又はダウン制御するからである。さらに、17及び21位の2つのフェニルアラニンが、受容体ののちの活性化を促進するためにそのN末端細胞外ドメイン上で受容体と最初に接触することがわかった。あらゆる好中球接触を阻止するために、これらの2つのアミノ酸17及び21を交換するが、それらは塩基性アミノ酸に交換され、何故ならば、それらが蛋白質の三次元モデル上で観察され得るように、C末端のαヘリックスのGAG結合モチーフに極めて近接しているからである。17及び/又は21位をアルギニン又はリジンに交換することにより、GAG結合親和性は、よって、増加する。
【0047】
本発明のさらなる側面は、上記の発明の蛋白質をコードする単離されたポリ核酸分子である。当該ポリ核酸はDNA又はRNAであってよい。それにより、発明の修飾された蛋白質を導く修飾がDNA又はRNAレベルで実施される。この発明の単離されたポリ核酸分子は、診断方法並びに遺伝子治療及び発明の修飾された蛋白質の大規模な生産に適している。
【0048】
さらに好ましくは、単離されたポリ核酸分子は、ストリンジェントな条件下で上において規定された発明の単離されたポリ核酸分子にハイブリダイズする。ハイブリダイゼーシ
ョン条件に依存して、相補二重鎖が2つのDNA又はRNA分子間に形成され、完全にマッチするか又はミスマッチ塩基を含むかの何れかによる(Sambrook et al.,Molecular Cloning:A laboratory manual,第2版、Cold Spring Harbor,N.Y.1989)。約50ヌクレオチドを超える長さのプローブは、25から30%間でのミスマッチ塩基をもってよい。小さなプローブは少数のミスマッチしかもたない。ミスマッチ塩基対を含む二重鎖を形成する標的及びプローブの傾向は、ハイブリダイゼーション条件のストリンジェンシーにより制御され、それ自身は因子、例えばハイブリダイゼーションバッファー中の塩濃度又はホルムアルデヒド濃度、ハイブリダイゼーションの温度及びハイブリダイゼーション後の洗浄条件の函数である。ハイブリッド二重鎖条件の形成において起こるよく知られた原理を適用することにより、所望のストリンジェンシーを有する条件が、当業者により、ハイブリダイゼーションバッファー、温度及び洗浄条件の様々から選択することにより達成され得る。即ち、完全にマッチしたか又は部分的にミスマッチの二重鎖の何れかの検出を許容する条件が選択可能である。二重鎖の融解温度(Tm)は適切なハイブリダイゼーション条件を選択するために有用である。200ヌクレオチドの長さを超えるポリヌクレオチド分子のためのストリンジェントなハイブリダイゼーション条件は、典型的には、予測される二重鎖の融解温度の15−25℃低い温度においてハイブリダイズさせることを含む。長いプローブよりも安定性の劣る二重鎖を形成させる30ヌクレオチドを超えるオリゴヌクレオチドに関しては、ストリンジェントハイブリダイゼーションは通常Tmより5から10℃低い温度にてハイブリダイズさせることにより達成される。核酸二重鎖のTmは、核酸中に含まれるG+Cパーセントに基づいた式を用いて計算することができ、鎖の長さを参酌し、例えば、式Tm=81.5−16.6(log[Na+]))+0.41(%G+C)−(600/N)であり、式中、Nは鎖の長さである。
【0049】
本発明のさらなる側面は、上で規定された本発明の単離されたDNA分子を含むベクターに関する。当該ベクターは、有効なトランスフェクション並びに蛋白質の有効な発現に必要な全ての制御要素を含む。そのようなベクターは、当業界でよく知られており、この目的のために如何なる適切なベクターも選択することができる。
【0050】
本発明のさらなる側面は、上で規定された本発明のベクターにより安定にトランスフェクトされた組換え細胞に関する。そのような組換え細胞並びにそれからの子孫細胞は上記ベクターを含む。それにより、構成的又は上記ベクターの活性化に際して、上記修飾蛋白質を発現する細胞系が提供される。
【0051】
本発明のさらなる側面は、上で規定された本発明による蛋白質、ポリ核酸又はベクター及び薬学上受容可能な担体を含むことを特徴とする薬学組成物に関する。もちろん、当該薬学組成物は、薬学組成物中に通常存在する追加の物質、例えば、塩、バッファー、乳化剤、発色剤、等をさらに含んでよい。
【0052】
本発明のさらなる側面は、上で規定された本発明による修飾蛋白質、ポリ核酸又はベクターの、各野生型蛋白質の生物活性を阻害するか又は抑圧するための方法における使用に関する。上で言及されたとおり、修飾蛋白質は、公知の組換え蛋白質を用いて起こる副作用が発明の修飾蛋白質を用いて起こらないようなアンタゴニストとして作用する。ケモカインの場合、これは特に炎症性反応に関与する生物活性である。
【0053】
よって、本発明による修飾蛋白質、ポリ核酸又はベクターのさらなる使用は、炎症性の症状の治療のための医薬を製造するための方法における。特に、修飾蛋白質がケモカインの場合、それは副作用の内アンタゴニストとして作用して、炎症性症状の治療に特に適する。よって、本出願のさらなる側面は、本発明による単離された修飾蛋白質、本発明による単離されたポリ核酸分子又はベクター又は本発明による薬学組成物を患者に投与するた
めの方法でもある。
【0054】
好ましくは、炎症性の症状は、慢性関節リウマチ、乾癬、骨関節症、喘息、アルツハイマー疾患、及び多発性硬化症からなる群から選択される。ケモカインを通した活性化は本発明による修飾蛋白質により阻害され得るから、上で言及された炎症性の症状が阻止されるか又は治療されるようにして、炎症性反応が阻害されるか又はダウン制御され得る。
【0055】
本発明は、発明を限定するためのものではない以下の実施例及び図面の助けを借りてさらに詳細に説明されるが、図1はCDスペクトルであり;図2は様々な変異体の二次構造コンテンツを示し;図3及び4は様々な変異体の蛍光異方性試験の結果のグラフであり;図5は恒温蛍光滴定の結果のグラフであり;図6は様々な変異体のアンフォールド実験の結果のグラフであり;図7はIL−8変異体の走化性指数を示し;そして図8はRANTES走化性アッセイの結果を示す。
【実施例】
【0056】
実施例1:組換えIL−8遺伝子の生成及び変異体のクローン化
ポリメラーゼ鎖反応(PCR)技術を用いることにより、センス及びアンチセンス変異導入プライマーを用いて変異を導入して変異体に関しての所望のcDNAsを生成した。wtIL−8に関するcDNAを含む合成プラスミドを鋳型として用い、Clontech Advantage(登録商標)2ポリメラーゼミックスをDNAポリメラーゼとして適用し、そしてPCR反応をエッペンドルフマスターグラディエントサイクラーを用いて実施した。用いられた変異導入プライマーは以下の表に要約され、フォワードプライマーから始める(5’から3’):
・CACC ATG TGT CAG TGT ATA AAG ACA TAC TCC(変異Δ6のためのプライマー)
・CACC ATG TGT CAG TGT ATA AAG ACA TAC TCC AAA CCT AGG CAC CCC AAA AGG ATA(変異Δ6 F17R F21Rのためのプライマー)
リバースプライマーは(5’から3’):
・TTA TGA ATT CCT AGC CCT CTT(変異E70Rのためのプライマー)
・TTA TGA ATT CTT AGC CCT CTT(変異E70Kのためのプライマー)
・TTA TGA CTT CTC AGC CCT CTT(変異N71Kのためのプライマー)
・TTA TGA CTT CTT AGC CCT CTT(変異E70K N71Kのためのプライマー)
・TTA TGA CTT CCT AGC CCT CTT(変異E70R N71Kのためのプライマー)
・TTA TGA CCT CTT AGC CCT CTT(変異E70K N71Rのためのプライマー)
・TTA TGA CCT CCT AGC CCT CTT(変異E70R N71Rのためのプライマー)。
【0057】
PCR産物は精製し、pCR(登録商標)T7/NT−TOPO(登録商標)TA(インビトロジェン)にクローン化して、そしてTOP10Fコンピテントエシェリヒアコリ(インビトロジェン)を形質転換した。次の工程として、配列の確認を二重鎖DNA配列決定によりABI PRISM CE1シークエンサーにより実施した。
実施例2:組換え蛋白質の発現及び精製
配列が確認されたら、発現のために、構築物によりカルシウム−コンピテントBL21
(DE3)エシェリヒアコリを形質転換した。細胞を撹拌しながら、100μg/mlアンピシリンを含む1lのLennox Broth(シグマ)中で37℃においてOD600が0.8に到達するまで生育させた。蛋白質発現の誘導は、イソプロピル−β−D−チオガラクトピラノシド(IPTG)を最終濃度1mMまで加えることにより達成した。4時間後に、細胞を6000gにおいて20分間遠心分離することにより回収した。細胞沈殿物を次に、20mM TRIS/HCl,50mM NaCl,pH8を含むバッファーに懸濁し、100ワットにて5x20秒音波処理し、そして最後に再び10,000gにて20分間遠心分離した。組換えIL−8蛋白質の主要フラクションが封入体(inclusion body)中に見いだされたので、さらなる精製のために変性条件を選択した。そうして、細胞沈殿物を6M Gua/HcL及び50mM MES,pH6.5を含むバッファーに懸濁した。懸濁液を次に4℃において4時間撹拌して、次に、50mM MES,pH6.5に対する透析工程に続いた。結果の懸濁液を次に遠心分離し、そして濾過して強カチオン交換カラム(ファルマシアバイオテックのSP Sepharose(登録商標))に負荷した。50mM MESバッファー,pH6.5中の0M−1M NaClからの直線勾配により、60分かけて溶出を達成した。所望の蛋白質を含むフラクションの凍結乾燥後に、第2の精製工程をC18カラムを用いた逆相HPLCにより実施した。この場合、10%−90%アセトニトリルの非直線勾配を選択することにより、所望の蛋白質を溶出した。上記と同じ条件下で同じカチオン交換カラムにより、変性蛋白質のリフォールディングを最後に達成した。
【0058】
蛋白質を、次に、精製度及び同一性に関して、最初の場合は銀染色、そして第2においてはウエスタンブロット分析により、IL−8に対する特異的モノクローナル抗体を用いてチェックした。円偏光二色性(CD)測定により、蛋白質のリフォールディングも確認した。
実施例3:変異体の生物物理特性決定
3.1円偏光二色性測定及び分析
硫酸ヘパラン(HS)の存在及び不在下で上記変異体蛋白質の二次構造変化を調査するために、CD分光法を実施した。測定値は、Jasco J−710旋光分散計(spectropolarimeter)上で195−250nmの範囲にわたり記録し、そして0.1cmの経路(path)の長さのセルを用いた。5μMの濃度による蛋白質溶液のスペクトルを1秒のレスポンス時間、0.2nmの工程解析、50nm/分の速度、1nmのバンド幅及び20mdegの感度にて記録した。3つのスキャンを平均化してスムーズなスペクトルを生じさせた。蛋白質のスペクトルを、次に、バッファー自体又はバッファー/BSの何れかのCD−シグナルに関してバックグラウンド補正した。プログラムSELCONを用いて、HS存在及び不在下での蛋白質の二次構造分析を最後に実施した。
【0059】
非常に多くのアミノ酸が新規な組み合わせにおいて交換されたため、円偏光二色性により結果の二次構造変化の寸法を見いだそうと試みた。
導入された変異に依存して異なる構造が得られた。何の構造も示さなかった一つの発現変異体(変異Δ6 F17R F21R E70K N71R)を除いて、全ての変異体が測定可能なα−ヘリックス、β−シート及びループを呈した。IL−8wtに比較して、一つの変異体のみ(変異Δ6 E70R)がほぼ類似の構造を示したが、その他は主にそれらのα−ヘリックスに差異があり、全体の構造の17.2%から45.2%の範囲であった。にも拘らず、この事実は、IL−8wtの全体の構造が蛋白質配列内での多くの変化にも拘わらず保持されたことを示唆する。これは、以前には予測され得なかった。硫酸ヘパランオリゴサッカライドのみがIL−8wt二次構造に影響し得てヘパリンは不可能であったという既に見いだされた事実から、未分画の硫酸ヘパランにより誘導された効果に注意が引き付けられた。試験された変異体全てがHS結合に際して構造変化を示し、複合体形成の証拠として観察され得る。
【0060】
導入された変異及び硫酸ヘパラン添加に際しての構造変化を証明するために、得られたいくつかのデータを上及び下のグラフにおいて要約する。
3.2蛍光測定
濃度及びリガンド依存性の第4の構造変化を研究するため、蛍光分光法を実施した。その高い感度のために、ナノグラム量の蛋白質しか必要とせず、蛍光技術は所望の調査を実施するための選択の方法であった。測定は、Perkin−Elmer(ビーカンズフィールド、英国)LS50B分光計を用いて企てられた。
3.3蛍光異方性(Anisotropy)
外来発色団bisANSによりもたらされるケモカインの濃度依存性の蛍光異方性を記録することにより、上記変異体のダイマー化定数を発見することを目指した。測定はPBS中で実施し、高濃度で開始し(4μMまでの蛋白質)、徐々に希釈した。各データ点に関して、507nmにおいて記録された異方性シグナル(r)を60秒間かけて平均化した。
【0061】
IL−8のオリゴマー化は、蛋白質GAG結合特性に対して関連性をもって影響することが報告されていた。モノマー濃度における設定により、IL−8結合サイズはダイマー濃度よりも1000倍強固にオリゴサッカライドを限定した(defined)。よって、蛍光
異方性により、IL−8変異体のオリゴマー化特性を調査した。IL−8固有のフルオロフォア(Trp57)は全ての変異体に関して十分な感度がないため、外来のフルオロフォアbis−ANSをこれらの測定に用いた。再び、二次構造に関しては既に注目されたとおり、変異体Δ6 E70Rは、オリゴマー化定数においてもIL−8wt(koligo=379nM)に比較して高い類似度を示した(koligo=350nM)。高いkoligo(koligo=460nM)の変異体は、よって、ダイマー化が不完全であり、Δ6 F17RF21RE70RN21Kであった。以前の研究は、β−シートが主にダイマー化プロセスに関与していることを明らかにしたが、事実、この変異体の二次構造に関する結果と相関しており、たった11.4%のβ−シートの極めて低い割り当て(share)を示した。もっとも低いkoligo(koligo=147nM)の変異体
はΔ6 F17RF21R E70Kであることがわかったが、調査された全変異体のβ−シート構造のもっとも高い割り当て(29.8%)を再び示した。硫酸ヘパラン付加の衝撃も観察された。IL−8wtに関しては、硫酸ヘパランがオリゴマー化定数のはるかに高いレベルへのシフト(koligo=1.075nM)を誘導したが、これは調査されたIL−8変異体に関しても観察された。Δ6 F17RF21R E70Kは0.147μMから1.162μMにシフトし、そして変異体Δ6 E70Rは硫酸ヘパラン存在下において0.35μMから1.505μMにシフトした。得られた結果のいくつかを図3及び4に示すが、図3はケモカイン濃度におけるPBS中のIL−8変異体の蛍光異方性の依存度を示し、そして図4はHSの存在(10倍過剰)及び不在((pc=10xy過剰)蛋白質濃度)下でのケモカイン濃度におけるPBS中のΔ6 F17RF21R
E70Kの蛍光異方性の依存度を示す。
3.4恒温蛍光滴定(IFT)実験
解離定数(Kd値)はリガンドの蛋白質に対する結合親和性に関する測定値であり、よって、リガンド結合に際して蛋白質の蛍光放射特性の濃度依存性変化(蛍光クエンチング)がKdの測定に用いられた。これらの変異体は、提案された(proposed)GAG結合部位においてか又はその近くに位置する固有のトリプトファン発色団を含むためにリガンド結合に際して構造的な変化に感受性であるべきだから、IFT実験はこの種の調査に適しているようであった。蛍光強度滴定をPBS中で700nMの蛋白質濃度を用いて実施した。各GAGリガンドのアリコートの付加及び60秒の平衡期間の後に、282nMにおける励起に際しての蛋白質溶液の放射を300−400nmの範囲にわたり記録した。
【0062】
未分画のヘパリン及び硫酸ヘパランの結合を調査した。十分な感度を保証するために、変異体をダイマー濃度に設定した。GAG結合に際したTrp57蛍光強度のクエンチン
グが25−35%の範囲内で記録された。リガンド結合の顕著な改善が、特にヘパリン結合に関して観察された。IL−8wt(Kd=37μM)に比較して、Δ6 F17RN71R E70K(Kd=14nM)及びΔ6 F17RF21R N71K(Kd=14.6nM)は、2600倍良好な結合を示し、そしてΔ6 E70K N71K(Kd=74nM)は1760倍良好な結合を示した。良好な結果は硫酸ヘパランに関しても得られた。Δ6 F17RN71R E70Kに関しては、107nMのKdが観察され、Δ6 F17RF21R N71Kに関してはKdが95nMであり、そして変異体Δ6
E70K N71Kは34nMのKdを示した。IL−8wtが4.2μMのKdにて結合するように、変異体に関して見いだされたKdsは結合に関しての異例の改善を表したので、図5を参照されたい。
3.5アンフォールド実験
蛋白質の安定性及びこの安定性がGAGリガンド結合に際して変化するのか否かについての情報を得るため、アンフォールド実験を企画した。上記のとおり、蛍光技術は第4の構造変化を観察するのに極めて感度がよく、よって蛋白質の熱構造変化を調査するためのえり抜きの方法である。測定はIFTに関して記載されたとおりに企画し、リガンド濃度は変化させずに温度を変化させた。蛋白質の構造は0.7μMの濃度において15−85℃の温度において10倍過剰の硫酸ヘパラン及びヘパリンの不在下及び存在下において観察された。
【0063】
蛋白質の放射の最大は340nmから357nmの範囲であり、値は溶剤露出トリプトファン残基に関して典型的である。アンフォールド実験を15℃において開始し、変異体の放射の最大は340nm−351nmの間で変化した。その放射最大が340nmにおいて観察されたIL−8wtに比較して、これはわずかに高い値を意味する。温度の増加と共に、放射最大の強度が低下し、最大値の高いか又は低い波長へのシフトの何れかを伴った。Δ6 E70R及びΔ6 E70K N71Kの放射最大は、352.5nm−357nm及び343nm−345nmにシフトし、Trp57残基の溶剤への温度増加を伴うさらなる暴露に関して典型的であるが、おもしろいことにΔ6 F17RN71R E70K及びΔ6 F17RF21R E70Rは青いシフトを示し、350nm−343nm及び著しくないが350nm−348nmの範囲であった(図6を参照)。温度をゆっくりと低下させることにより、アンフォールドのプロセスが波長シフト及び強度の変化に関しては部分的に可逆性であった。5倍過剰の硫酸ヘパランの付加は蛋白質の安定性の増加を導いたが、おそらくは複合体形成のためである。これは、一方では融解温度の高い温度へのシフトにより観察できたし、他方では温度増加に際しての放射最大の著しくは顕著でないシフトにより観察できた。
実施例4:優性−陰性IL−8変異体の受容体『陰性』機能の細胞に基づくアッセイ
好中球誘引に関してのIL−8変異体の損なわれた受容体機能を特性決定するために、IL−8変異体に応答した好中球のトランスフィルターに基づく走化性を、5μmのPVP−無しのポリカーボネート膜を備えたマイクロ走化性チェンバー内でアッセイした。
細胞調製:
簡単に言えば、好中球フラクションを新たに回収されたヒト血液から調製した。これは、6%デキストラン溶液のヘパリン処理された血液への添加(2:1)により行い、次に、45分間沈降のために放置した。上清の透明な溶液を回収し、そしてHBSS w/o
Ca及びMgにより2回洗浄した。細胞を数え、そして最後にHBSSにより2Mio/ml細胞懸濁液にて希釈して、数えられた細胞の60%のみが好中球であることを考慮した。
走化性アッセイ:
IL−8変異体を10μg/ml、1μg/ml及び0.1μg/mlの濃度に希釈し、そしてチェンバーの底部区画へ同じものを3通りにして置いた(ウエルあたり26μl)。新たに調製された好中球を上部チェンバー内に接種し(ウエルあたり50μl)、そして30分間、37℃において5%のCO2加湿インキュベーター中でインキュベートし
た。インキュベーション後に、チェンバーを解体し、フィルターの上部側を洗浄し、そしてふき取り(wiped off)、そして底部側に付着した細胞をメタノールで固定してHem
acolor溶液(メルク)により染色した。細胞を、次に、400xの倍率にて、ウエルあたり4つのランダムに選択された顕微鏡の視野の中で数えた。最後に、3つの個別の実験の平均をケモカイン濃度に対してプロットした。図7において、様々なIL−8変異体の走化性指数を示す。予測されたとおり、全ての変異体が顕著に低下した受容体結合活性を示した。
実施例5:組換えRANTES遺伝子の生成、発現、変異体の生物物理特性決定及び活性の特性決定
優性な陰性「GAG−マスキング」ケモカイン変異体のコンセプトを、移植拒絶のようなタイプIVの高感受性反応、アトピー性皮膚炎並びに関節炎、進行性糸球体腎炎及び炎症性肺疾患のような他の炎症性傷害に関与するケモカインであるRANTESにも用いた。
【0064】
wt蛋白質を開始メチオニン残基に導入することにより、受容体結合活性が損なわれた。エシェリヒアコリ内でのwt RANTESの発現は、このメチオニン残基の保有を導き、wt RANTESを単球移動の有力な阻害剤にし、いわゆるMet−RANTESである。GAG結合親和性を増強する別の変異をPCRに基づく部位特異的変異導入法により導入した。
【0065】
これらの9つのRANTES変異体がこれまでにクローン化され、発現されて、精製された、Met−RANTES A22K,Met−RANTES H23K,Met−RANTES T43K,Met−RANTES N46R,Met−RANTES N46K,Met−RANTES Q48K,Met−RANTES A22K/N46R,Met−RANTES V49R/E66S及びMet−RANTES 15LSLA18 V49R/E66S。
【0066】
恒温蛍光滴定実験を実施することにより、サイズで規定されたヘパリンに関するRANTESの相対的親和性定数(Kd値)を測定した。表に見られるとおり、全てのRANTES変異蛋白質はこのヘパリンに関して高い親和性を示し、Met−RANTES A22K,Met−RANTES H23K,Met−RANTES T43K及びMet−RANTES A22K/N46Rがもっとも有望な結果を示した。
【0067】
Kd(nM)
野生型Rantes 456.2± 8.5
Met-Rantes V49R/E66S 345.5±21.7
Rantes 15LSLA18 V49R/E66S 297.3±14.1
Rantes N46R 367.7±11.7
Rantes N46K 257.4±10.2
Rantes H23K 202.5±12.8
Rantes Q48K 383.4±39.6
Rantes T43K 139.2±30.1
Rantes A22K 202.1± 9.8
Rantes A22K/N46R 164.0±16.6
RANTES走化性アッセイ
RANTES変異体により指示された細胞移動を、5μmのPVP−コートされたポリカーボネート膜を備えた48ウエルBoydenチェンバーシステムを用いて調査した。RANTES及びRANTES変異体の20mM HEPES pH7.3及び1mg/mlのBSAを含むRPMI 1640の希釈液を3通りにしてチェンバーの底部ウエル内に置いた。同じ培地中の2x106/ml細胞の50μlのTHP−1細胞懸濁液(セ
ルカルチャーのヨーロッパコレクションからの前単球細胞系)を上部ウエル中に置いた。37℃における5%CO2加湿の2時間のインキュベーションの後に、フィルターの上部表面をHBSS溶液で洗浄した。移動した細胞をメタノールにより固定して、Hemacolor溶液(メルク)により染色した。400xの倍率にて、ウエルあたり5つを数え、3つの個別の実験の平均を図8においてケモカイン濃度に対してプロットした。エラーバーは3つの実験の平均の標準誤差を表す。再び、IL−8変異体の場合のように、全てのRANTES変異体が顕著に低下した受容体結合活性を示した。
実施例6:GAG結合領域を有する蛋白質
バイオインフォマティックス及びプロテオームにより、GAG結合蛋白質をそれらのGAG結合領域と共に特性決定した。以下の表2及び3において、ケモカインとそれらのGAG結合領域(表2)を示し、そして他の蛋白質の例もそれらのGAG結合領域と共に提供する(表3)。
【0068】
【表2】
【0069】
【表3】
【0070】
【表4】
【0071】
【表5】
【0072】
【表6】
【0073】
【表7】
【0074】
【表8】
【0075】
【表9】
【0076】
【表10】
【0077】
【表11】
【0078】
【表12】
【0079】
【表13】
【0080】
【表14】
【0081】
【表15】
【0082】
【表16】
【0083】
【表17】
【0084】
【表18】
【0085】
【表19】
【0086】
【表20】
【0087】
【表21】
【0088】
【表22】
【0089】
【表23】
【0090】
【表24】
【0091】
【表25】
【0092】
【表26】
【0093】
【表27】
【0094】
【表28】
【0095】
【表29】
【0096】
【表30】
【特許請求の範囲】
【請求項1】
GAG結合部位を、特性決定された蛋白質に導入する方法であって、工程:
・構造保持に必須でない蛋白質中の領域を同定すること
・塩基性アミノ酸を上記部位中に導入するか、及び/又は、少なくとも一つの巨大な及び/又は酸性のアミノ酸を上記部位において欠失させること
を含み、
GAG結合部位はKd≦10μM、好ましくは≦1μM、さらに好ましくは≦0.1μMのGAG結合親和性を有する、
方法。
【請求項2】
請求項1記載の方法により得ることができる蛋白質。
【請求項3】
修飾されたGAG結合蛋白質であって、当該蛋白質内のGAG結合領域が少なくとも一つのアミノ酸の置換、挿入、及び/又は欠失により修飾されることにより、上記GAG結合領域中の塩基性アミノ酸の相対量を増加させるか、及び/又は上記GAG結合量中、好ましくは溶剤露出位置において巨大か及び/又は酸性のアミノ酸を減少させることを特徴とし、そして当該蛋白質のGAG結合親和性が各野生型蛋白質のGAG結合親和性に比較して増加していることを特徴とする、修飾されたGAG結合蛋白質。
【請求項4】
Arg,Lys,及びHisからなる群から選択される少なくとも一つの塩基性アミノ酸がGAG結合領域に挿入されていることを特徴とする、請求項2又は3記載の蛋白質。
【請求項5】
蛋白質がケモカイン、好ましくはIL−8,RANTES又はMCP−1であることを特徴とする、請求項2乃至4の何れか1項記載の蛋白質。
【請求項6】
GAG結合領域がC末端αヘリックスであることを特徴とする、請求項5記載の蛋白質。
【請求項7】
IL−8の17、21、70、及び/又は71位がArg,Lys,His,Asn及び/又はGlnにより置換されていることを特徴とする、請求項6記載の蛋白質。
【請求項8】
増加したGAG結合親和性が硫酸ヘパラン及び/又はヘパリンに対する増加した結合親和性であることを特徴とする、請求項2乃至7の何れか1項記載の蛋白質。
【請求項9】
さらなる生物学上活性な領域が修飾されることにより上記蛋白質のさらなる生物学上の活性を阻害するか及び/又はダウン制御することを特徴とする、請求項2乃至8の何れか1項記載の蛋白質。
【請求項10】
さらなる生物学上活性な領域が、欠失、挿入、及び/又は置換により、好ましくはアラニン、立体構造上及び/又は静電上類似の残基により修飾されていることを特徴とする、請求項9記載の蛋白質。
【請求項11】
蛋白質がケモカインであり、そしてさらなる生物学上の活性が白血球活性化であることを特徴とする、請求項9又は10記載の蛋白質。
【請求項12】
蛋白質がIL−8であり、そしてさらなる生物学上活性な領域が最初の10個のN末端アミノ酸の中に位置することを特徴とする、請求項11記載の蛋白質。
【請求項13】
蛋白質がIL−8であり、そして最初の6個のN末端アミノ酸が欠失していることを特
徴とする、請求項12記載の蛋白質。
【請求項14】
蛋白質が、del6F17RE70KN71R,del6F17RE70RN71K及びdel6E70KN71Kからなる群から選択されるIL−8であることを特徴とする、請求項12記載の蛋白質。
【請求項15】
請求項2乃至14の何れか1項に記載された蛋白質をコードすることを特徴とする、単離されたポリ核酸分子。
【請求項16】
ストリンジェントな条件下で請求項15記載のDNA分子にハイブリダイズすることを特徴とする、単離されたポリ核酸分子。
【請求項17】
請求項15又は16記載の単離されたDNA分子を含むことを特徴とする、ベクター。
【請求項18】
請求項17記載のベクターにより安定にトランスフェクトされていることを特徴とする、組換え細胞。
【請求項19】
請求項2乃至14の何れか1項記載の蛋白質、請求項15又は16記載のポリ核酸又は請求項17記載のベクター及び薬学上受容可能な担体を含むことを特徴とする、薬学組成物。
【請求項20】
請求項2乃至14の何れか1項記載の蛋白質、請求項15又は16記載のポリ核酸又は請求項17記載のベクターの、各野生型蛋白質の生物活性を阻害するか又は抑圧するための方法における、使用。
【請求項21】
請求項2乃至14の何れか1項記載の蛋白質、請求項15又は16記載のポリ核酸又は請求項17記載のベクターの、炎症性の症状の治療のための医薬を製造するための方法における、使用。
【請求項22】
炎症性症状が、慢性関節リウマチ、乾癬、骨関節症、喘息、アルツハイマー疾患、及び多発性硬化症からなる群から選択されることを特徴とする、請求項21記載の使用。
【請求項1】
GAG結合部位を、特性決定された蛋白質に導入する方法であって、工程:
・構造保持に必須でない蛋白質中の領域を同定すること
・塩基性アミノ酸を上記部位中に導入するか、及び/又は、少なくとも一つの巨大な及び/又は酸性のアミノ酸を上記部位において欠失させること
を含み、
GAG結合部位はKd≦10μM、好ましくは≦1μM、さらに好ましくは≦0.1μMのGAG結合親和性を有する、
方法。
【請求項2】
請求項1記載の方法により得ることができる蛋白質。
【請求項3】
修飾されたGAG結合蛋白質であって、当該蛋白質内のGAG結合領域が少なくとも一つのアミノ酸の置換、挿入、及び/又は欠失により修飾されることにより、上記GAG結合領域中の塩基性アミノ酸の相対量を増加させるか、及び/又は上記GAG結合量中、好ましくは溶剤露出位置において巨大か及び/又は酸性のアミノ酸を減少させることを特徴とし、そして当該蛋白質のGAG結合親和性が各野生型蛋白質のGAG結合親和性に比較して増加していることを特徴とする、修飾されたGAG結合蛋白質。
【請求項4】
Arg,Lys,及びHisからなる群から選択される少なくとも一つの塩基性アミノ酸がGAG結合領域に挿入されていることを特徴とする、請求項2又は3記載の蛋白質。
【請求項5】
蛋白質がケモカイン、好ましくはIL−8,RANTES又はMCP−1であることを特徴とする、請求項2乃至4の何れか1項記載の蛋白質。
【請求項6】
GAG結合領域がC末端αヘリックスであることを特徴とする、請求項5記載の蛋白質。
【請求項7】
IL−8の17、21、70、及び/又は71位がArg,Lys,His,Asn及び/又はGlnにより置換されていることを特徴とする、請求項6記載の蛋白質。
【請求項8】
増加したGAG結合親和性が硫酸ヘパラン及び/又はヘパリンに対する増加した結合親和性であることを特徴とする、請求項2乃至7の何れか1項記載の蛋白質。
【請求項9】
さらなる生物学上活性な領域が修飾されることにより上記蛋白質のさらなる生物学上の活性を阻害するか及び/又はダウン制御することを特徴とする、請求項2乃至8の何れか1項記載の蛋白質。
【請求項10】
さらなる生物学上活性な領域が、欠失、挿入、及び/又は置換により、好ましくはアラニン、立体構造上及び/又は静電上類似の残基により修飾されていることを特徴とする、請求項9記載の蛋白質。
【請求項11】
蛋白質がケモカインであり、そしてさらなる生物学上の活性が白血球活性化であることを特徴とする、請求項9又は10記載の蛋白質。
【請求項12】
蛋白質がIL−8であり、そしてさらなる生物学上活性な領域が最初の10個のN末端アミノ酸の中に位置することを特徴とする、請求項11記載の蛋白質。
【請求項13】
蛋白質がIL−8であり、そして最初の6個のN末端アミノ酸が欠失していることを特
徴とする、請求項12記載の蛋白質。
【請求項14】
蛋白質が、del6F17RE70KN71R,del6F17RE70RN71K及びdel6E70KN71Kからなる群から選択されるIL−8であることを特徴とする、請求項12記載の蛋白質。
【請求項15】
請求項2乃至14の何れか1項に記載された蛋白質をコードすることを特徴とする、単離されたポリ核酸分子。
【請求項16】
ストリンジェントな条件下で請求項15記載のDNA分子にハイブリダイズすることを特徴とする、単離されたポリ核酸分子。
【請求項17】
請求項15又は16記載の単離されたDNA分子を含むことを特徴とする、ベクター。
【請求項18】
請求項17記載のベクターにより安定にトランスフェクトされていることを特徴とする、組換え細胞。
【請求項19】
請求項2乃至14の何れか1項記載の蛋白質、請求項15又は16記載のポリ核酸又は請求項17記載のベクター及び薬学上受容可能な担体を含むことを特徴とする、薬学組成物。
【請求項20】
請求項2乃至14の何れか1項記載の蛋白質、請求項15又は16記載のポリ核酸又は請求項17記載のベクターの、各野生型蛋白質の生物活性を阻害するか又は抑圧するための方法における、使用。
【請求項21】
請求項2乃至14の何れか1項記載の蛋白質、請求項15又は16記載のポリ核酸又は請求項17記載のベクターの、炎症性の症状の治療のための医薬を製造するための方法における、使用。
【請求項22】
炎症性症状が、慢性関節リウマチ、乾癬、骨関節症、喘息、アルツハイマー疾患、及び多発性硬化症からなる群から選択されることを特徴とする、請求項21記載の使用。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図8】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図8】
【図7】
【公開番号】特開2012−165747(P2012−165747A)
【公開日】平成24年9月6日(2012.9.6)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−86483(P2012−86483)
【出願日】平成24年4月5日(2012.4.5)
【分割の表示】特願2012−40123(P2012−40123)の分割
【原出願日】平成16年12月2日(2004.12.2)
【出願人】(506191046)プロタフィン・ビオテヒノロギー・アクチェンゲゼルシャフト (3)
【Fターム(参考)】
【公開日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願番号】特願2012−86483(P2012−86483)
【出願日】平成24年4月5日(2012.4.5)
【分割の表示】特願2012−40123(P2012−40123)の分割
【原出願日】平成16年12月2日(2004.12.2)
【出願人】(506191046)プロタフィン・ビオテヒノロギー・アクチェンゲゼルシャフト (3)
【Fターム(参考)】
[ Back to top ]