
GASC1の阻害
本発明により、試験化合物の、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質に結合し、そして任意にその活性を調節する能力を試験する方法が提供される。前記方法には、試験化合物をJumonjiタンパク質のJMJD2サブファミリーのタンパク質、前記タンパク質の補因子、及び任意で脱メチル化の基質と共にインキュベートすることが含まれる。本発明に係る方法を、多数の化合物のスクリーニングに用いて、特定の癌、特に前立腺癌の治療に臨床的に使用する候補化合物となる化合物群を同定することが可能である。スクリーニングアッセイで活性を示さない他の化合物については、候補化合物としての更なる検討から除外できる。従って、本発明の方法は、製薬産業において有用性を有する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、JmjCドメイン含有タンパク質の活性調節剤として作用可能であり、そしてゲノムの不安定性が発症に関与する疾患の予防及び/又は治療に有用な化合物の同定方法に関する。
【背景技術】
【0002】
ヒストンテールのリジン及びアルギニン残基のメチル化は、転写活性が高いクロマチンと低いクロマチンを表す重要なエピジェネティックマーカーである。例えば、ヒストンH3(H3-K9)の第9番リジンのメチル化は、エピジェネティックな要因により不活性なクロマチンと関連している1-3。
【0003】
アセチル化やリン酸化といった他のヒストン修飾とは対照的に、メチル化は、N-CH3結合の高い熱力学的安定性により不可逆的であると考えられてきた。しかしながら、最近において、Shiとその同僚4は、核内のアミンオキシダーゼのホモログであるタンパク質LSD1が、モノメチル化及びジメチル化H3-K4に非常に高い特異性を示すヒストンデメチラーゼであることを同定した。この酵素は、その基質を、アミンオキシダーゼ反応を通じて脱メチル化する。しかしながら、LSD1は、トリメチル化リジンH3-K4を脱メチル化することができず、これは、最も恐らくは、酸化に必要なプロトン化窒素が存在していないことが原因であると考えられる4。本願発明に係る研究を完成する直前に、Tsukada et al.は、Jumonjiタンパク質のFBXL11 (JHDM1a)が、ジメチルH3-K36をFe(II)の存在下でα-ケトグルタレート依存的に特異的に脱メチル化できることを示した5。少なくとも理論上は、FBXL11媒介性の脱メチル化の反応機構は、トリメチルH3-K36を基質として用いることが可能であるが、このような活性は実証できていない5。LSD1及びFBXL11をヒストンデメチラーゼとして同定したことは、メチル基マークの動的な制御を示しており、エピジェネティック研究に重要な軌跡を残すことになるが、彼らは、トリメチル化リジンマークの可逆性の問題は解明していない。
【0004】
SUV39H1ノックアウトマウスの研究で報告されたように、H3-K9のトリメチルバリアントの喪失により、染色体異常が生じ、癌に罹患しやすくなる。故に、このマークを逆行させることが可能な酵素が長年探索されてきたが、その存在が疑問視されていた。後者の見方は、トリメチル化H3-K9が、「非常に安定な遺伝性のクロマチン状態」であるヘテロクロマチンの発生と維持に必要とされるという事実によって、更に強められた。このような酵素とその活性阻害剤の同定により、癌の予防と治療に対する新たなアプローチが提供されるであろう。
【発明の開示】
【0005】
本発明は、試験化合物の、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質に結合し、そして任意にその活性を調節する能力を試験する方法であって、試験化合物をJumonjiタンパク質のJMJD2サブファミリーのタンパク質、前記タンパク質の補因子、及び任意で脱メチル化の基質と共にインキュベートすることを含む方法を提供する。
【0006】
本発明の現時点で好ましい態様においては、前記方法は、以下の工程:
i) 鉄及びα-ケトグルタレートを含む溶液で脱メチル化の基質の懸濁液を調製すること;
ii) 前記試験化合物を、前記懸濁液に添加すること;
iii) ii)で得られた前記懸濁液に、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質を添加すること;
iv) 前記懸濁液をインキュベートすること;並びに
v) 前記基質の脱メチル化の程度を測定すること
を含む。
【0007】
定義
「核酸分子」という用語は、リボ核酸(RNA)若しくはデオキシリボ核酸(DNA)のオリゴマー又はポリマー、又はその模倣体を指す。この用語には、天然の核酸塩基、糖及びヌクレオシド間(バックボーン)の共有結合から構成される分子、並びに同様の機能を有する非天然の核酸塩基、糖及びヌクレオシド間(バックボーン)の共有結合から構成される分子、又はこれらの組合せが含まれる。このような修飾を受けた又は置換された核酸は、例えば、向上した細胞内取り込み、核酸ターゲットへの向上した親和性、並びにヌクレアーぜ及び他の酵素の存在下での増大した安定性等の所望の特性のために天然型よりも好まれる場合があり、本文においては、「核酸類似体」又は「核酸模倣体」の用語で記載される。核酸模倣体の好ましい例には、ペプチド核酸(peptide nucleic acid:PNA)、Locked核酸(Locked Nucleic Acid:LNA)、xylo-LNA、ホスホロチオエート、2'-メトキシ-、2'-メトキシエトキシ-、モルホリノ-及びホスホロアミデートを含有する分子等が挙げられる。
【0008】
「ポリヌクレオチド配列」(例えば、核酸、ポリヌクレオチド、オリゴヌクレオチド等)は、文脈に応じて、ヌクレオチドA、C、T、U、G、若しくは他の天然ヌクレオチド、若しくは人工のヌクレオチド類似体を含むヌクレオチドのポリマーであるか、又は核酸を表す文字列である。いずれかの特定のポリヌクレオチド配列から、所定の核酸又は相補的な核酸のいずれかを決定できる。
【0009】
本発明に関しては、「ポリペプチド」という用語は、完全長タンパク質、オリゴペプチド、短鎖ペプチド及びこれらの断片を含み、アミノ酸残基が共有結合により結合したあらゆる長さのアミノ酸鎖を指す。本明細書及び請求項における全てのポリペプチド配列は、明確な記載がない場合には、慣習的なフォーマットに従い、N末端からC末端の方向へ記載される。
【0010】
任意のアミノ酸ポリマー若しくはヌクレオチドポリマーの番号付けが、選択されたアミノ酸ポリマー若しくは核酸ポリマーの番号に「対応する」又は「関連する」のは、任意のポリマーの成分(例えば、アミノ酸、ヌクレオチド、又は一般的に「残基」と呼ばれるもの)の位置が、その任意のポリマーの成分の実際の番号順の位置ではなく、選択されたアミノ酸ポリマー又はヌクレオチドポリマーの同一又は同等の位置を参照して定められている場合である。従って、例えば、任意のポリペプチド配列の任意のアミノ酸の位置の番号付けは、参照配列として用いられる選択されたポリペプチド配列の同一又は同等のアミノ酸の位置に対応する。
【0011】
「変異体」は、1又は2か所以上のアミノ酸部位において、親ポリペプチドの配列と異なる配列(アミノ酸の欠失、アミノ酸の挿入、及び/又は異なるアミノ酸へのアミノ酸の置換による)を含むポリペプチドである。変異配列は、非天然配列、即ち、自然界に見出されない配列であってもよい。
【0012】
本明細書において、「合成ペプチド」という用語は、in vitroで合成された短鎖ペプチドを含むペプチドを指す。また、この用語には、通常見られないアミノ酸又は非天然アミノ酸を用いた置換により修飾されたペプチド又は短鎖ペプチドも包含される。
【0013】
目的語に付される「天然」とは、その目的語にかかるものが、人間によって人工的に作製されたものとは異なるものとして自然界に見出すことができるという事実を表す。例えば、自然界の供給源から単離可能な生物(ウィルス、細菌、原生動物、昆虫、植物、又は哺乳動物の組織を含む)中に存在し、実験室において人間による意図的な修飾を受けていないポリペプチド又はポリヌクレオチドの配列は、天然のものである。目的語に付される「非天然」とは、その目的語にかかるものが天然のものではなく、即ち、人間により人工的に作製されたものと異なるものとして自然界に見出すことができないことを意味する。
【0014】
「断片」又は「部分配列」とは、所定の配列のいずれかの部分を指す。配列の断片又は部分配列は、その配列自体よりも少なくとも1アミノ酸残基又は1核酸残基は短くなることが理解されるべきである。従って、断片又は部分配列は、それぞれ、より長いアミノ酸配列(例えば、ポリペプチド)又は核酸配列(例えば、ポリヌクレオチド)の一部を含むアミノ酸配列又は核酸配列を指す。
【0015】
「配列同一性」という用語は、同一の長さの2つのアミノ酸配列間又は2つの核酸配列間のホモロジー度の定量的測定値を示す。比較すべき2つの配列が同一の長さにない場合、これらは、可能な限り最適な一致性を示すようにアライメントする必要があり、ギャップが挿入されたり、又はポリペプチド配列若しくはヌクレオチド配列が末端で切断されたりする。配列同一性は、以下の式によって計算可能であり
【数1】
式中、Ndifは、アライメントした際の、2つの配列の同一でない残基の総数であり、Nrefは、一方の配列の残基数である。従って、AGTCAGTCというDNA配列は、AATCAATCというDNA配列と75%の配列同一性を有することになる(Ndif=2及びNref=8)。ギャップについては、具体的な残基の非同一性としてカウントされ、即ち、AGTGTCというDNA配列は、AGTCAGTCというDNA配列と75%の配列同一性を有することになる(Ndif=2及びNref=8)。
【0016】
本発明に係る全てのポリペプチド又はアミノ酸に基づく態様では、1以上の配列間の配列同一性のパーセンテージは、clustalWソフトウェア(http:/www. ebi.ac.uk/clustalW/index. html)により、そのプログラムのデフォルト設定を用いて行われる個々の配列のアライメントに基づく。これらの設定は、以下の通りである:Alignment=3Dfull, Gap Open 10.00, Gap Ext. 0.20, Gap separation Dist. 4, Protein weight matrix: Gonnet。本発明に係るヌクレオチドに基づく態様に関しても、1以上の配列間の配列同一性のパーセンテージは、clustalWソフトウェアをデフォルト設定で用いて行われるアライメントに基づく。ヌクレオチド配列のアライメントにおいては、これらの設定は以下の通りである:Alignment=3Dfull, Gap Open 10.00, Gap Ext. 0.20, Gap separation Dist. 4, DNA weight matrix: identity (IUB)。
【0017】
本文においては、「相補的配列」とは、ストリンジェントな条件下で、本発明の核酸分子にハイブリダイズする核酸配列を指す。「ストリンジェントな条件」は、高いストリンジェンシーを有する一般的な条件を指す。「ストリンジェンシー」という用語は、当該技術分野で良く知られており、核酸のハイブリダイゼーションが行われる条件(温度、イオン強度、及び有機溶媒等の他の化合物の存在)を参照する際に用いられる。「高ストリンジェンシー条件」では、ストリンジェンシーが「弱い」若しくは「低い」条件と比較して、高頻度に相補的な塩基配列を有する核酸フラグメント間でのみ核酸塩基のペア形成が生じる。
【0018】
例えば、高ストリンジェンシーなハイブリダイゼーション条件には、以下が含まれる:(1)洗浄における低イオン強度と高温、例えば、0.1%のドデシル硫酸ナトリウム(SDS)を含む0.015MのNaCl/0.0015Mのクエン酸ナトリウム、pH7.0(0.1xSSC)、50℃;(2)5×Denhardt's溶液(0.1%(重量/体積))高純度ウシ血清アルブミン/0.1%(重量/体積)Ficoll/0.1%(重量/体積)ポリビニルピロリドン)を含む50体積%のホルムアミド、50mMのリン酸ナトリウムバッファー、pH6.5及び5×SSCで42℃でのハイブリダイゼーション;又は(3)50%ホルムアミド、5×SSC、50 mMのリン酸ナトリウム(pH6.8)、0.1%のピロリン酸ナトリウム、5×Denhardt's溶液、超音波処理済みサケ精子DNA(50μg/ml)、0.1%のSDS、及び10%の硫酸デキストラン中で42℃でのハイブリダイゼーション、並びに0.2×SSC及び0.1% SDSで42℃での洗浄。
【0019】
「単離した核酸」という用語は、本発明の核酸が由来する生命体の天然のゲノムでは隣接している(例えば、5'末端で一端が、3'末端で一端が隣接する)両コーディング配列と隣接していない核酸(例えばDNA又はRNA)を指す場合がある。従って、この用語には、例えば、cDNA又はポリメラーゼ連鎖反応(PCR)若しくは制限酵素処理により得られたゲノムDNAの断片が含まれ、このようなcDNA又はゲノムDNA断片が、ベクターに組み込まれているか、これらが元々取り出された生命体と同一若しくは異なる種、例えば、ウィルス等のゲノムに一体化されているか、更なるコーディング配列に連結してキメラポリペプチドをコードするハイブリッド型遺伝子を形成するか、又は他のDNA配列から独立しているかどうかは関係がない。前記DNAは、2本鎖又は1本鎖であってもよく、センス鎖又はアンチセンス鎖であってもよい。
【0020】
「組換えポリヌクレオチド」又は「組換えポリペプチド」とは、それぞれ、複数の供給源である核酸若しくはポリペプチドに由来する核酸配列若しくはアミノ酸配列を含む非天然のポリヌクレオチド又はポリペプチドであり、その供給源である核酸又はポリペプチドは、天然の核酸若しくはポリペプチドであってもよく、又は、それ自体が変異導入若しくは他の種類の修飾を受けたものであってもよい。核酸又はポリペプチドは、人工物であるか若しくは組換え操作を受けている場合、又は人工物であるか若しくは組換え操作を受けたポリペプチド若しくは核酸に由来する場合には、「組換え体」とみなされる場合がある。組換え核酸(例えばDNA又はRNA)は、典型的に一緒に含まれていないか、典型的には互いに関連していないか、又は典型的には互いに分離された状態にある、少なくとも2つの配列セグメントの組合せ(例えば、人工的組合せ)により作製可能である。組換え核酸は、異なる供給源に由来する核酸セグメント及び/又は人工的に合成された核酸セグメントを一緒に連結させるか又は組み合わせることにより形成される核酸分子を含んでいてもよい。「組換えポリペプチド」(又は「組換えタンパク質」)は、しばしばクローニング又は組換え操作された核酸若しくは遺伝子から生じたポリペプチド(又はタンパク質)を指す。異なる核酸配列又はアミノ酸配列が由来する供給源であるポリヌクレオチド又はポリペプチドは、しばしば、相同的であり(即ち、同一又は類似の構造及び/又は機能を有するか、又はこれを有するポリペプチドをコードする)、例えば、しばしば、生命体の異なる単離物、血清型、株、種、又は異なる疾患状態から得られる。
【0021】
「組換え」という用語は、例えば、細胞、ヌクレオチド、ベクター、タンパク質、又はポリペプチドに言及するものとして用いられる場合、典型的には、その細胞、ヌクレオチド又はベクターが、異種の(又は外来性の)核酸の導入若しくは天然の核酸の変更により修飾されていること、又はそのタンパク質若しくはポリペプチドが異種のアミノ酸の導入により修飾されていること、又はその細胞が、その様な修飾を受けた細胞に由来することを示す。組換え細胞は、その細胞の天然(非組換え)型に見出されない核酸配列(例えば、遺伝子)を発現するか、又は、異常に発現しているか、発現が抑制されているか、若しくは全く発現されていない天然の核酸配列(例えば、遺伝子)を発現する。「組換え」という用語は、細胞に言及するものとして用いられる場合、その細胞が異種の核酸を複製するか、又は異種の核酸によりコードされるペプチド若しくはタンパク質を発現することを示す。組換え細胞は、その細胞の天然(非組換え)型に見出されない遺伝子を含んでいてもよい。組換え細胞は、その細胞の天然型に見出される遺伝子を含んでいてもよいが、その場合、その遺伝子は、人工的手段により修飾されてその細胞内に再導入される。前記用語には、その細胞に内在性の核酸であって、その細胞からその核酸を取り出すことなしに修飾された核酸を含む細胞が包含される;このような修飾には、遺伝子置換、部位特異的変異導入及び関連技術により得られるものが含まれる。
【0022】
「組換えにより作製された」という用語は、化学合成手段、核酸セグメントの配列組換えの繰り返し若しくはヌクレオチドの他の種々の作製方法(例えば、シャッフリング)、又は単離された核酸セグメントの、例えば当業者に知られる遺伝子組み換え技術による操作により通常達成される人為的な組換えを指す。「組換え発現された」とは、典型的にはin vitroで組換え核酸を作製し、in vivo、in vitro、又はex vivoで、その組換え核酸を、これを発現又は増幅可能な細胞内へ導入するための技術を指す。
【0023】
ここで用いられる「保存的」とは、(i)その変化が、可能な限り構造的に中性であること、即ち、天然タンパク質と比較して、その変異ペプチド又はポリペプチドの3次構造の変化が最小になるように設計されていることを意味する。構造的に中性であることは、生物活性を保存するために望ましい。当業者に、構造的に中性となる可能性の高い変異を作製するための手引きとなり得る法則が存在する(例えば、(77)及び(78)を参照)。より重要な法則のいくつかには、以下が含まれる:(1)物理化学的に類似する残基、即ち、同義の残基の置換では、置換により挿入されるアミノ酸は、置換されるアミノ酸と同様の構造的役割を果たすことが可能なので、構造変化が起こりにくい;そして(2)進化の過程で保存されている場合、その配列が機能上重要である可能性が示唆されることから、進化の過程で保存されている配列を変化させると、有害な構造上の効果を生じる恐れがある。
【0024】
本明細書の本章又は以下の残りの記載において特に定義されていない限り、ここで使用する全ての科学技術用語は、本発明が属する技術分野の当業者に共通して理解される意味と同様の意味を有する。
【発明を実施するための最良の形態】
【0025】
本発明は、JmjCドメイン含有タンパク質の活性調節剤として作用可能な化合物が、ゲノムの不安定性が発症に関与する疾患の予防及び/又は治療に有用であるという発見に基づく。特に、JmjCドメイン含有タンパク質の調節剤は、癌の予防及び/又は治療に有用である。更に本発明は、Jumonjiタンパク質;ヒストンH3のジメチル化及びトリメチル化リジン9(H3-K9)と相互作用できるタンパク質として、扁平上皮癌1で増幅される遺伝子(GASC1)、の同定に関する。GASC1は、JumonjiファミリーのJMJD2サブファミリーに属する。本発明の発明者らは、JMJD2ファミリーのタンパク質が、トリメチル化及びジメチル化H3-K9を脱メチル化させるヒストンデメチラーゼであることを実証した。更に、本発明者らは、GASC1とJMJD2メンバーの異所発現が、in vivoで、トリメチル化及びジメチル化H3-K9を劇的に減少させ、モノメチル化H3-K9を増加させ、HP1を非局在化させ、ヘテロクロマチンを減少させることも証明した。従って、最初に同定されたヒストントリメチルデメチラーゼであることに加え、JumonjiファミリーのJMJDサブファミリーのタンパク質も、癌の発症に関与している。従って、これらの酵素の触媒活性の阻害剤は、新規な抗ガン治療法の開発のための重要な候補となる。
【0026】
従って、本発明は、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性を調節可能な化合物の同定、スクリーニング、キャラクタリゼーション又は設計の方法に関する。本発明の方法を、多数の化合物のスクリーニングに用いて、特定の癌、特に前立腺癌の治療に臨床的に使用する候補化合物となる化合物群を同定することが可能である。スクリーニングアッセイで活性を示さない他の化合物については、候補化合物としての更なる検討から除外できる。従って、本発明の方法は、製薬産業において有用性を有する。
【0027】
本発明によれば、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質のその基質(ヒストンペプチド、ヒストン又はヌクレオソーム)又は補因子(α-ケトグルタレート又は鉄)への結合を阻害する化合物を、競合結合アッセイにおいて同定できる。また、これとは別に、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質、又はその基質若しくは補因子への試験化合物の結合を直接測定することもできる。後者のタイプのアッセイは、直接結合アッセイと呼ばれる。競合結合アッセイと直接結合アッセイは、両方とも、当該技術分野で一般的に知られるイムノアッセイ及びレセプター結合アッセイで用いられるフォーマットと類似する種々の異なるフォーマットで使用可能である。競合結合アッセイ及び直接結合アッセイを含む結合アッセイの異なるフォーマットの記載については、Basic and Clinical Immunology 7th Edition (D. Stites and A. Terr ed.) 1991; Enzyme Immunoassay, E. T. Maggio, ed., CRC Press, Boca Raton, FIa. (1980);及び"Practice and Theory of Enzyme Immunoassays," P. Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology, Elsevier Science Publishers B. V. Amsterdam (1985) を参照のこと。各参考文献は、本明細書に援用される。
【0028】
例えば、競合結合アッセイでは、試験化合物は、固体表面に結合された結合物質上の特異的な結合部位について、標識された検体と競合する。この種のフォーマットでは、標識された検体は、ヒストンペプチドで標識可能であり、結合物質は、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質であってもよく、これが固相に結合されることになる。これに代わり、標識された検体が、標識されたJMJD2タンパク質であってもよく、結合物質が、固相化されたヒストンペプチドであってもよい。捕捉物質に結合する標識検体の濃度は、結合アッセイにおいて試験化合物が競合する能力と反比例する。試験化合物による標識検体の阻害の量は、その結合アッセイの条件、並びに使用する結合物質、標識検体、及び試験化合物の濃度に依存する。具体的なアッセイ条件のもとでは、その競合結合アッセイにおいて、結合物質への標識検体の結合量が10%以上低下した場合には、試験化合物は、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質への基質(即ち、ヒストンペプチド)又は補因子の結合を阻害できるとみなされる。直接結合アッセイを用いる場合、バックグラウンドのレベルの2倍以上のシグナルが測定される場合には、試験化合物は、JMJD2酵素への基質(即ち、ヒストンペプチド)又は補因子(即ち、α-ケトグルタレート)の結合を阻害しているものとみなされる。
【0029】
競合結合アッセイでは、サンプル化合物は、特異的な結合物質への結合において標識タンパク質と競合する。上述した通り、結合物質を固体表面に結合させることで、結合していない標識タンパク質から結合した標識タンパク質を分離することができる。これに代わり、競合結合アッセイを液相で行い、当該技術分野で知られる種々の技術のいずれかを用いて、結合していない標識タンパク質から結合した標識タンパク質を分離することもできる。分離した後に、結合した標識タンパク質の量を測定する。サンプル中に存在するタンパク質の量は、結合した標識タンパク質の量に反比例する。
【0030】
上記に代わり、相同的結合アッセイ(homogenous binding assay)を行うことも可能であり、このアッセイでは分離工程を要しない。これらの種類の結合アッセイでは、タンパク質上の標識が、タンパク質のその特異的な結合物質への結合により変化する。この標識タンパク質の変化により標識から放出されるシグナルが減少又は増加し、結合測定の終了時に標識を測定することにより、そのタンパク質を検出又は定量することが可能になる。
【0031】
ここに記載される結合アッセイフォーマットは、標識されたアッセイ成分をもちいる。標識は、種々の形態をとることが可能である。標識は、当該技術分野で知られる方法に従って、アッセイの所望の成分に直接的又は間接的に結合できる。多種多様の標識を用いることができる。成分は、種々の方法のいずれによっても標識できる。
【0032】
従来の方法では、3H、125I、35S、14C、又は32Pが組み込まれた放射性標識が用いられる。非放射性標識には、標識した抗体に結合するリガンド、フルオロフォア、化学発光剤、酵素、及び標識されたリガンドの特異的なペア形成剤として機能できる抗体が含まれる。標識の選択は、要求される感度、化合物への結合の容易性、安定性の要求、及び利用可能な機器に依存する。使用可能な種々の標識方法又はシグナル発生システムの総説については、米国特許第4,391,904号(参照により本明細書に組み込まれる)を参照。
【0033】
「ヒストンペプチド」及び「基質」という用語は、JMJD2タンパク質に結合するヒストンの断片を指す。スクリーニングアッセイで用いるJMJD2タンパク質とヒストンタンパク質の製造方法は、当業者に知られている。
【0034】
前記方法は、特に、試験化合物がJumonjiタンパク質のJMJD2サブファミリーのタンパク質に結合し、随意にその活性を調節する調節する能力の試験方法であり、試験化合物を、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質、前記タンパク質の補因子、及び場合により脱メチル化の基質と共にインキュベートすることを含む方法であってもよい。特に、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性を阻害可能な化合物を同定することは、本発明の範囲に含まれる。
【0035】
典型的には、前記方法では、Jumonjiタンパク質のJMJD2サブファミリーのポリペプチド、代表的にはGASC1ポリペプチドを利用する。特に、前記方法では、ヒトGASC1ポリペプチド若しくはこのポリペプチドの断片、又はこれらの変異体を利用する。従って、本発明の重要な態様では、前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質が、以下の:
a)配列番号1に示すアミノ酸配列;
b)配列番号1に示すアミノ酸配列の断片;
c)a)の配列若しくはb)の配列のいずれか、又は両方と少なくとも45%の配列同一性を示すアミノ酸
を含む群から選択される方法が提供される。
【0036】
Jumonjiタンパク質のJMJD2サブファミリーのクローニングされたメンバーには、GASC1(Swiss-Prot accession number Q9H3R0)、並びにそのホモログであるJMJD2a及びJMJD2b(それぞれ、Swiss-Prot accession numbers 075164及び094953)が含まれる。
【0037】
本発明の同程度に重要な態様は、前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質が、以下の:
a)配列番号1(GASC-1(JMJ2Dc))、配列番号2(JMJD2a)、配列番号3(JMJD2b)に示すアミノ酸配列;
b)a)のいずれか1つのアミノ酸配列の断片;
c)a)のいずれか1つの配列及び/又はb)のいずれか1つの配列と少なくとも45%の配列同一性を示すアミノ酸
を含む群から選択される方法に関する。
【0038】
当業者であれば、現時点で知られているJumonjiタンパク質のJMJD2サブファミリーのメンバーは、これらのアミノ酸配列を最適なフィットでアライメントした場合に、約45%の配列同一性を示すことを理解するであろう。従って、本出願では、使用するアミノ酸配列は、Jumonjiタンパク質のJMJD2サブファミリーのいずれのメンバーの配列とも少なくとも45%の同一性を示す。詳細には、前記配列同一性は少なくとも45%、例えば、少なくとも46%、47%、48%、49%、50%、52.5%、55%、57.5%、60%、62.5%、65%、67.5%、70%、72.5%、75%、77.5%、80%、82.5%、85%、87.5%、90%、92.5%、95%、97.5%、98%、98.5%、99%又は99.5%であってもよい。
【0039】
Jumonjiタンパク質のJMJD2サブファミリーの天然ポリペプチドメンバーの機能特性を取り込むためには、前記断片が少なくとも100個のアミノ酸、例えば、少なくとも105、110、115、120、125、130、135、140、145、150、155、160、165、170、175、180、185、190、195、200、210、220、230、240、250、260、270、280、290、300、320、340、360、380、400、420、440、460、480、500、520、540、560、580、600、620、640、660、680、700、750、800、850、900、950、1000個、又は少なくとも1050個のアミノ酸を有することが更に好ましい。
【0040】
好ましくは、前記断片は、少なくとも150個のアミノ酸、例えば、少なくとも151、152、153、154、155、156、157、158、159、160、162、164、166、168、170、172、174、176、178、180、182、184、186、188、190、192、194、196、198又は200個のアミノ酸を有する。
【0041】
本発明は、更に、保存的アミノ酸置換により形成されるアミノ酸配列の類似体も意図している。保存的アミノ酸置換の背後にある原理は、特定のアミノ酸ペアにおいて、一方のアミノ酸を他方で置換した場合にも、そのペプチドの3次構造に最小限の変化しか生じないような相容性の側鎖が存在することである。保存的置換の法則については、以下で説明されている:Bowie et al. Science 247(1990) 1306-1310。本発明の目的の一つは、JMJD2サブファミリータンパク質の基質へ結合する能力を保持するポリペプチド、断片及び変異体を利用することである。いくつかの好ましい態様では、前記ポリペプチド、断片及び変異体は、JMJD2サブファミリータンパク質のα-ケトグルタレート等の補因子へ結合する能力を維持している。
【0042】
必要とされる場合には、前記ポリペプチド、断片及び変異体のそれぞれは、精製された形態又は未精製の形態のいずれかで、例えば、細胞抽出物として、又はこのような抽出物から関連成分を精製することにより提供されてもよい。
【0043】
これに代わり、前記ポリペプチド、断片及び変異体は、組換え発現技術により遺伝子操作で作製し、前記アッセイで使用するために精製することもできる。これに代わり、前記ポリペプチド、断片及び変異体は、細胞ベースのアッセイで用いるために、細胞内で組換え発現することもできる。
【0044】
典型的には、関連成分をコードするポリヌクレオチドは、発現ベクターに組み込まれた状態で提供される。このような発現ベクターは、当該技術分野で通常の技術で組み立てられており、例えば、プラスミドDNA並びに必要とされる可能性のある適切なイニシエーター、プロモーター、エンハンサー及び他の構成成分、例えば、ポリアデニル化シグナルの使用を必要とする場合があり、これらは、完全なタンパク質発現を可能にするため、正しい配向で配置される。適切なベクターは、例えば、本出願の実施例により詳細に記載されるベクターのように、当業者に一瞬で明確に把握されるであろう。プロモーター配列は、選択されるアッセイフォーマットに応じた誘導プロモーター又は構成的プロモーターとなるであろう。
【0045】
Jumonjiタンパク質のJMJD2サブファミリーには、数多くの共通する構造上の特徴が見出される。これらにはJmjN、JmjC、PHD及びTdrの配列が含まれ、これらの1以上は、JMJD2 Jumonjiタンパク質の好ましい特性である。これらの機能ドメインは、そのアミノ酸配列の要となる部位に一定の特定のアミノ酸残基が局在しているという特徴を有する。単に肉眼でタンパク質のアミノ酸を調べただけでは直ちに区別できないかもしれないが、これらのドメインは、SMARTプログラム(http:/smart.embl-heidelberg.de/smart)を用いて決定できる。SMARTプログラムの基礎となるアルゴリズムは、Letunic et al.58とSchultz et al.59に更に詳細に説明されている。
【0046】
従って、本発明の好ましい態様では、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質、並びにポリペプチド、断片及び変異体は、JMJD2aに由来するJmjN、JmjC、PHD及びTUDORのドメインから成る群から選択される1以上のアミノ酸配列を含む:
【化1】
【0047】
本発明の同程度に好ましい態様では、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質並びにポリペプチド、断片及び変異体は、JMJD2bに由来するJmjN、JmjC、PHD及びTUDORのドメインから成る群から選択される1以上のアミノ酸配列を含む:
【化2】
【0048】
現時点で最も好ましい本発明の態様では、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質並びにポリペプチド、断片及び変異体は、JMJD2(GASC1)に由来するJmjN、JmjC、PHD及びTUDORのドメインから成る群から選択される1以上のアミノ酸配列を含む:
【化3】
【0049】
これらのドメインのアミノ酸配列は、その機能を喪失させることなく置換により変更させることが可能なことが理解されるべきである。従って、本発明には、上記の配列のいずれか一つと少なくとも75%の同一性、例えば、上記の配列のいずれかと少なくとも80%、85%、90%、95%、98%、99%の同一性、又は記の配列のいずれかと少なくとも99.5%の同一性を示す1以上のアミノ酸配列を含むJumonjiタンパク質のJMJD2サブファミリーのタンパク質並びにポリペプチド、断片及び変異体が含まれる。
【0050】
Jumonjiタンパク質のJMJD2サブファミリーのタンパク質とその活性の調節剤の候補を、その酵素活性の阻害剤の不存在下でヒストンH3のリジン9を脱メチル化させる条件下で一緒にインキュベートしてもよい。GASC1は、Fe(II)及びα-ケトグルタレートに依存するデメチラーゼであるから、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質は、典型的には、補基質の存在下で基質と接触する。典型的には、α-ケトグルタレート、Fe(II)及びアスコルビン酸が補基質として用いられるが、他の補基質の使用も本発明の範囲に含まれる。
【0051】
当業者はJumonjiタンパク質のJMJD2サブファミリーのタンパク質、並びに上記のポリペプチド、断片及び変異体の活性を測定するにあたって、種々のアプローチを利用可能であり、これらのアプローチには、利用された基質及び/又は補基質の測定並びに形成された生成物及び/又は副生成物の測定が含まれる。従って、本発明の特定の態様では、前記方法は、以下の追加の工程:
a)試験サンプルの以下のいずれかのパラメーターを測定すること:
i)基質のメチル化の状態;
ii)ホルムアルデヒドの放出;
iii)α-ケトグルタレート基質補因子のカルボキシル化の状態;
iv)酸素消費量
b)工程a)で得られた試験サンプルについての数値を、対照サンプルについて得られた数値と比較し、それによって、その試験化合物の、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性を調節する能力を測定すること
を含む。
【0052】
更に、本発明の方法には、上述の要素による下流への効果の測定を組み込むこともできる。特に、GASCの作用は、特定の遺伝子の転写に影響を与える可能性があることから、GASC活性を測定するためにジーンレポーターアッセイを用いることも意図できる。このようなアッセイは、天然状態でJumonjiタンパク質のJMJD2サブファミリーのタンパク質によって抑制されている遺伝子の転写活性の測定に基づくものとなるであろう。更に、メチル化を受けたプロモーターと転写物において結合されるレポーター遺伝子を含む遺伝子操作されたレポーターコンストラクトの使用に基づくin vitroアッセイも意図できる。
【0053】
特定の態様では、これらのレポーターアッセイでは、蛍光レポーターをコードする遺伝子の使用を伴ってもよい;現時点で最も一般的に使用される蛍光レポーターは、緑色蛍光タンパク質(Green Fluorescent Protein:GFP)である。
【0054】
測定は、定量的又は定性的なものでよい。定量的測定及び定性的測定のいずれも、上に示すように、対照の存在下で行うことができる。対照を用いる場合、使用するアッセイ方法にもよるが、以下の工程:
a)Jumonjiタンパク質のJMJD2サブファミリーのタンパク質と脱メチル化の基質を、脱メチル化が可能な条件下でインキュベートすること;
b)試験サンプルの以下のパラメーター:
i)基質のメチル化の状態;
ii)試験サンプル中のホルムアルデヒドの放出;
iii)α-ケトグルタレート基質補因子のカルボキシル化状態
iv)酸素消費量
をモニターすること
により測定を行うことができる。
【0055】
本発明のいくつかの態様では、インキュベート時間と基質のメチル化状態及び/又はホルムアルデヒドの放出を用いて脱メチル化の速度を測定し、これを比較に用いる。
【0056】
GASC1の天然基質がヒストンH3であることから、本発明のいくつかの態様においては、脱メチル化の基質には、ヒストンH3のリジン9に対応するメチル化部位が含まれることが理解されるであろう。本発明の特定の態様では、脱メチル化の基質は、配列番号22に示すアミノ酸配列(TARKSTG)を含むペプチドである。
【0057】
前記アミノ酸配列(配列番号22)のヒストンH3のリジン9に対応するリジン残基はメチル化されている。本発明の大部分の態様において、前記リジン残基は、トリメチル化されていることが好ましいが、前記リジン残基が、ジメチル化又はモノメチル化されていたとしてもアッセイは行うことができる。
【0058】
現時点においては、基質としては、ペプチドを用いることが好ましく、例えば、ヒストンH3のN末端残基6-12、例えば、ヒストンH3の残基5-9、4-10、3-10、2-11、1-12、1-13、1-14、1-15、1-20、1-25、1-30、1-35、1-40、1-45、1-50、1-55、1-60、1-65、1-70、1-75、1-80、1-85、1-90、1-95又は1-100を含む合成ペプチド等を用いることが好ましい。本願においては、ヒストンH3の完全長配列が、配列番号23に示されている。
【0059】
上の脱メチル化基質の説明に従えば、前記基質は、少なくとも6個のアミノ酸残基から成るペプチド、例えば、少なくとも7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、32、34、36、38、40、45、50、60、70、80、90、100、120、140、160、180、200、250、300 350、400、500、600、700、800、900個又は少なくとも1000個のアミノ酸残基から成るペプチドであることが好ましい。80個以下のアミノ酸残基から成るペプチドは、市販の合成サービス又は装置を用いて化学合成により簡便に入手できる。
【0060】
当業者であれば、上記で特定されるようなヒストンH3のN末端残基と少なくとも75%の同一性を示すペプチド、例えば、ヒストンH3のN末端残基と少なくとも77.5%、80%、82.5%、85%、87.5%、90%、92.5%、95%、97.5%、98%、98.5%、99%又は99.5%の同一性を示すペプチドを理解するであろう。前記ペプチドは、上記に詳細に説明されるように、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質に置換が組み込まれたものであってもよい。特に、上記の同一の原理にしたがい、保存的置換が好ましくあろう。
【0061】
本発明の更に好ましい態様では、最小限のものとして、前記基質は、以下から成る群から選択されるアミノ酸配列を含む:
【0062】
現時点で最も好ましい態様では、ヒストンH3のN末端尾部(1-40)を模倣した43個のアミノ酸長の合成ペプチド、ARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPHR(配列番号24)が基質として用いられる。
【0063】
また、バルクヒストン(bulk histone)、合成ペプチド及びヌクレオソームから成る群から選択される基質を用いることが更に好ましい場合もある。
【0064】
上述したように、本発明に係る方法は、基質を用いて行うことができるが、そのメチル化部位は、モノメチル化、ジメチル化、又はトリメチル化されている。
【0065】
上述したように、α-ケトグルタレートは、GASC1の補因子として機能し、従って、本発明に係る方法に必要である。更に、α-ケトグルタレートの類似体を補因子として使用することも本発明の範囲に含まれる。これに加え、アスコルビン酸と、α-ケトグルタレートに加えて、Fe(II)イオンの存在によりもたらされるGASC1触媒活性が、補因子として重要となるであろう。従って、本発明の好ましい態様では、該化合物と、前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質は、Fe(II)イオン及び/又はアスコルビン酸と共にインキュベートされる。アスコルビン酸の存在は不可欠なことではないが、アスコルビン酸により反応系に存在する鉄は適切な形態であるFe(II)に変換されるので、完全な触媒活性を得るためには、アスコルビン酸が必要とされる場合もあることが理解されるべきである。
【0066】
試験化合物がJumonjiタンパク質のJMJD2サブファミリーのタンパク質に直接作用する場合には、無細胞系を用いることもできる。試験系を試験化合物と接触させ、その試験化合物が、前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性を調節する能力を、基質(ヌクレオソーム、ヒストン又はヒストンペプチド)の転換をイムノアッセイを用いて測定することにより測定する。
【0067】
Jumonjiタンパク質のJMJD2サブファミリーのタンパク質のペプチド基質、又はその触媒活性による生成物は、例えば、プレート若しくはビーズ上に固相化し、適切なメチル化部位のメチル化状態又は脱メチル化を、メチル化状態に応じて異なる親和力で前記ペプチドに結合する抗体若しくは他の結合分子を用いて検出できる。従って、トリメチル化、ジメチル化若しくはモノメチル化されたヒストンH3-K9のいずれかを認識する免疫結合パートナーは、試験系の一部を構成し、その活性に対する化合物の作用を迅速に測定することを可能にする。基質特異的抗体及び生成物特異的抗体を含む、このような結合パートナーは市販されている。
【0068】
本発明のアッセイ方法は、in vivoアッセイの形態を取ることも可能である。in vivoでのアッセイは、細胞系、器官系、又は動物全体のアッセイを用いて行うことができる。好ましくは、in vivoアッセイは、細胞系で行われ、例えば、細胞に導入された1又は2種類以上のベクターから関連するポリペプチド若しくはペプチドが発現されるセルラインを用いて行われる。例えば、細胞は、酵母細胞又は哺乳動物由来の細胞であってもよい。
【0069】
更に、本発明との関連で用いられるin vivoアッセイは、単離細胞/初代細胞又は組織切片による場合がある。特定の興味深い態様では、in vivoアッセイは、49歳の日本人女性から放射線照射(腫瘍が隣接構造に浸潤していた)の後に(頚部)食道上部から切除された、あまり分化の進んでいない食道扁平上皮癌から樹立されたヒト扁平上皮癌であるKYSE-150セルラインの使用を要する;増幅されたがん遺伝子c-erb-B(8倍)及びサイクリンD1(4倍)を保持し、ヒトのAST、LDH、MDHのIEFで確認されたようにヌードマウスで腫瘍を発生させると記述される。
【0070】
現時点において好ましくは、メチル化状態の測定は、フルオログラフィーによるホルムアルデヒド-3Hの放出と組み合わせてもよい。ホルムアルデヒド-デヒドロゲナーゼ一体アッセイによるホルムアルデヒド放出を測定するための類似技術も、本発明において意図されている。
【0071】
本発明の好ましい態様では、基質のメチル化状態及び/又はホルムアルデヒドの放出及び/又はα-ケトグルタレート基質補因子のカルボキシル化状態及び/又は酸素消費量は、イムノアッセイ(ELISA、RIA、IRMA、TRIFMA)、免疫沈降、ウェスタンブロッティング、BIAcore、X線結晶解析、溶液NMR、質量分析、分光技術又は蛍光技術により測定される。
【0072】
本発明の現時点で好ましい態様では、基質の利用量及び/又は生成物の形成量の測定は、免疫蛍光分析、好ましくは共焦点免疫染色法により行われる。本発明の現時点で最も好ましい態様では、実施例で説明されるように、ペプチド基質は、ビオチンに結合されている。
【0073】
基質特異的抗体によるアッセイでメチル化状態を測定する場合、適切な対照との比較における免疫結合パートナーの結合レベルの低下は、トリメチル化若しくはジメチル化したH3-K9の低下を意味しており、このことはJumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性の増加に対応している。免疫結合パートナーの結合は、例えば、ウェスタンブロッティング、ELISA、RIA、TRIFMA、免疫沈降又は免疫組織学的手法等、当該技術分野で一般的に知られた方法により評価できる。好ましい技術には、蛍光偏光法、時間分解-蛍光共鳴エネルギー転移法(Time Resolved Fluorescence Resonance Energy Transfer Assay:TR-FRET)、シンチレーション近接アッセイ(Scintillation Proximity Assay)及び「蛍光クエンチング」アッセイ等のハイスループットスクリーニング技術が含まれる。発現物は、容易に検出可能なレポータータンパク質、例えば、以下に限定されるものではないが、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーぜ(CAT)、緑色蛍光タンパク質又はルシフェラーぜ等と連結させることもできる。
【0074】
競合測定では、合成若しくは天然のヒストンペプチドが、標識型又は非標識型で提供される場合がある。抗体を、修飾有り又は無しで用いることもできる。抗体は、その抗体をレポーター分子と、共有結合的又は非共有結合的に連結させて標識できる。検出の容易化に使用可能な適切なレポーター分子又は標識には、放射性ヌクレオチド、酵素、蛍光分子、化学発光剤又は発色剤、並びに、基質、補因子、阻害剤、磁気粒子等が含まれる。キットの抗体又は合成ペプチドは、好ましくは、BSA、チログロブリン、オボアルブミン又はキーホールリンペットヘモシアニン等の適切なタンパク質担持体と結合させることにより、マイクロタイタープレート等の固体表面に固相化させてもよい。
【0075】
本発明の現時点で好ましい態様では、前記方法には、以下の工程:
i) 鉄及びα-ケトグルタレートを含む溶液で請求項8ないし13のいずれか1項に定義される脱メチル化の基質の懸濁液を調製すること;
ii) 前記試験化合物を、前記懸濁液に添加すること;
iii) ii)で得られた前記懸濁液に、請求項2ないし4のいずれか1項に定義されるJumonjiタンパク質のJMJD2サブファミリーのタンパク質を添加すること;
iv) 前記懸濁液を、好ましくは、35〜38℃の温度で10分間以上インキュベートすること;並びに
v) 前記基質の脱メチル化の程度を測定すること
が含まれる。
【0076】
脱メチル化の程度は、リジン9がメチル化されたヒストンH3に特異的な免疫結合パートナーを用いて測定することが更に好ましい。
【0077】
また、前記免疫結合パートナーは、抗トリメチル化ヒストンH3-K9抗体であることが好ましい。また、免疫結合パートナーとして抗ジメチル化ヒストンH3-K9抗体を用いることも、同程度に好ましい。
【0078】
更に好ましい態様では、脱メチル化の程度は、ホルムアルデヒドの放出により評価される。更に、脱メチル化の程度は、酸素消費量を測定することによっても評価できる。
【0079】
これに代わる場合、脱メチル化の程度は、α-ケトグルタレート基質補因子のカルボキシル化状態を測定することにより、評価される。
【0080】
例えば、i)のバッファーは、終濃度で50mMのHepes(pH7.5)、50mMのKCl、4mMのMgCl2、1mMのα-ケトグルタレート、40μMのFeSO4、2mMのアスコルビン酸を含む。実利的な理由により、前記懸濁液は、10-50μlの総量にあることが好ましい。
【0081】
最後に、上記に加えて、反応系のタンパク質成分の分解を最小限に抑えるためには、1又は2種類以上のプロテアーゼ阻害剤を終濃度1-5μg/μlで添加することが有用であろう。例えば、アプロチニンやロイペプチンは、プロテアーゼ阻害剤に含まれるであろう。
【0082】
本発明の種々の局面と、これらの局面の具体的な態様の上記記載に関しては、本発明の一つの局面及び/又は局面の一つの態様と関連して上に記載又は言及されるいずれの特性及び特徴も、記載される本発明の他の全ての局面及び/又は態様に類推して用いるられることが理解されるべきである。
【0083】
本発明の目的又はその特性若しくは特徴の一つについて単数系で言及された場合、その言及は、その目的又はその特性若しくは特徴について複数系で言及していることにもなる。例えば、「1つのペプチド」について言及している場合、1以上のペプチドについて言及していることが理解されるべきである。
【0084】
本明細書を通じて、「含む(comprise)」という単語、及びその変化形である「含む(comprises)」若しくは「含んでいる(comprising)」は、記述する要素、整数若しくは工程、又は要素、整数若しくは工程の群を含むが、いずれかの他の要素、整数若しくは工程、又は要素、整数若しくは工程の群を排除するものではないことを意味することが理解されるべきである。
【実施例】
【0085】
H3K9のトリメチル化バリアントと相互作用するタンパク質の同定を目的とする研究において、ストレプトアビジン-アガロース上に固相化されたビオチン化H3ペプチドを用いてHeLa核タンパク質をアフィニティー精製した。リジン9が非メチル化状態にあるか又はトリメチル化されている、ヒストンH31-40のN末端尾部全体を模倣した合成43-merをin vitro結合実験に用いた。結合したタンパク質をアガロース-マトリックスから溶離し、SDS-PAGEで分離して銀染色し、質量分析により同定した。複数種類のタンパク質では、非メチル化H3K9と比べて、特にトリメチル化H3K9が豊富であった(補足図S1)。
【0086】
これらタンパク質の中から、Jumonjiタンパク質のGASC1を同定した(図1a)。JumonjiドメインのJmjN及び関連するJmjCドメインが存在することから、このタンパク質は、JMJD2c8,9とも呼ばれている(図1a)。Jumonjiタンパク質ファミリーには、α-ケトグルタレート依存的オキシゲナーゼスーパーファミリーに属する多様なタンパク質のグループが含まれる。これらのタンパク質は、細胞周期の進行及び転写調節を含む種々の細胞プロセスを調節している5,10-14。このタンパク質のグループに共通する特徴は、Fe(II)イオンに結合する能力である。更に、これらのタンパク質の多くは、α-ケトグルタレートを補因子としてタンパク質基質をヒドロキシル化する能力を有する15。特に興味深いことに、近年、JmjCドメインを有するタンパク質が、ヒストンデメチラーゼの候補として同定されている16。
【0087】
GASC1は、トリメチル化(me3)H3K9と結合することが分かっているので、GASC1が、このエピジェネティックマークの脱メチル化に関与しているかもしれないという考えは、興味深いものであった。この可能性を試すため、組換え完全長Hisタグ化ヒトGASC1を作製し、バキュロウィルスで感染させた昆虫細胞から精製した(補足図S3)。アフィニティー精製したGASC1を、更にサイズ排除クロマトグラフィーで精製し、溶離した高純度GASC1を、バルクヒストンとインキュベートすることで脱メチル化活性について試験した。図3aに示すように、イムノブロッティング法で評価したところ、GASC1は、非常に効率的にH3K9のトリメチル化を減少させていた。また、GASC1が、関連する生理学的鋳型であるオリゴヌクレオソームを脱メチル化できることも分かった(補足図S4a)。
【0088】
GASC1の脱メチル化活性の特異性を更に分析するため、大量ないし少量のGASC1を示すGASC1のサイズ排除クロマトグラフィーの種々のフラクションをバルクヒストン及びリジンK9若しくはK27がメチル化された合成H3ペプチドとインキュベートし、種々のエピジェネティックマークのメチル化状態を、イムノブロッティング法により評価した(図3b及びS4b)。最も高いGASC1濃度を有するフラクション(F9-F12)では、ジメチル化及びトリメチル化H3K9はヒストンから完全に除去されていた。また、これに付随したモノメチル化(me1)H3K9の増加も認められ、GASC1がトリメチルH3K9をジメチル(me2)型に変換し、更にモノメチルH3K9(me1)に変換することと一致していた。
【0089】
対照的に、ジメチル化H3K4、トリメチル化H3K4、トリメチル化H3K27及びトリメチル化H4K20を含む、試験した他のエピジェネティックマークのレベルは、GASC1処理によって影響を受けていないようであった(図3b及びS4b)。
【0090】
興味深いことに、ジメチル化H3K9は、ヒストンを、少量ないし中程度の量のGASC1(F6、F8、F14及びF16)と反応させた場合に増加したようであった。このことは、ジメチル化H3K9マークの見かけの増加の理由が、脱メチル化反応の初期において、このマークの安定状態のレベルに到達したことによるという可能性を示している。従って、ある程度のジメチルH3K9がGASC1による脱メチル化を通じて失われるが、反応系全体で見れば、少なくとも十分な量のトリメチルH3K9が存在する限りは、GASC1によるトリメチルリジンの脱メチル化により、更なるジメチルH3K9が加わるものと考えられる。
【0091】
図3a及びbの結果と一致して、GASC1の脱メチル化活性は、濃度依存的に変化し、GASC1の熱変性により消失する場合があることから(図S4b、c)、この反応は酵素反応であることが示唆される。更なるin vitro脱メチル化試験を行い、この試験では、純粋な合成トリメチル化H3K9ペプチドをGASC1とインキュベートした。純粋なトリメチル化H3K9とGASC1のインキュベートを30分間行った後では、殆ど全てのトリメチルH3K9が、ジメチル及びモノメチルH3K9に変換されていた(図S4c、d)。
【0092】
GASC1が実際にトリメチル化及びジメチル化H3-K9を脱メチル化することを正確に証明するため、単一又はGASC1の存在下でインキュベートした合成ペプチドの質量分析を行った。この分析により、GASC1とのインキュベート後にH3K9me3ペプチドの大部分(約80%)が、ジメチル化及びモノメチル化H3-K9に変換されたことが示された(図4c-6f)。
【0093】
次に、GASC1に近似するホモログであるJMJD2a及びJMJD2b(図1a、図2b)が、in vivoでH3K9を脱メチル化できるかどうかについて検討した。これを調べるため、完全長のヒトJMJD2a、JMJD2bをクローニングし、バキュロウィルスで感染させた細胞から組換えタンパク質を得た。両ホモログは、バルクヒストンのH3K9me3とH3K9me2を脱メチル化できた(補足図S5)。同様に、GASC1とそのホモログJMJD2Aは、同程度の合成H3K9基質を脱メチル化する能力を有すると考えられた(補足図S4d、e)。これらの結果は、まとめると、トリメチル化及びジメチル化H3K9は両方とも、in vitroでのGASC1とそのホモログの基質であることを強く示唆している。
【0094】
in vivoでの脱メチル化反応の特異性に対する洞察を得るため、ヒトGASC1を、3次元構造が解明された唯一のJumonjiタンパク質である11,15,17-19、FIHの構造上にモデリングした。アライメント(図5及び補足図S2)と一致して、このモデリングでは、ヒスチジンH190、グルタミン酸E192、及びヒスチジンH288の残基が、GASC1の鉄結合用溝の本質的な部分を形成することが示された(図6h、i)。また、in silicoモデルでも、T187とR208の残基が、α-ケトグルタレートの結合に関与しており、GASC1の脱メチル化機能に決定的なものであることが予測された。次にトリメチル化H3ヒストン尾部を、予測されたGASC1構造上にモデリングした。K9がトリメチル化されたH3ヒストン尾部は、GASC1の鉄結合用溝に良好にフィットし、K9メチル基は、メチル化ヒストン尾部とGASC1構造間に立体障害を生じることなく、反応相手の鉄の近傍に位置することができた(図6h)。このin silicoモデルに基づき、H190とE192に変異を導入するだけで、GASC1の鉄結合能が喪失し、従って、その脱メチル化活性が阻害されるであろうことが予測された。
【0095】
上記の仮説を試すため、バルクヒストンを、野生型のGASC1と、ヒスチジンH190 とグルタミン酸E192がそれぞれグリシンとアラニンで置換された変異型GASC1(H190G/E192A)と共にインキュベートすることにより、脱メチル化アッセイを行った。野生型GASC1は、強力な脱メチル化活性を有していたが、変異体GASC1は、検出可能な脱メチル化活性を示さず、予測された鉄結合残基が、GASC1の脱メチル化活性に決定的であることが示された。野生型GASC1と変異型GASC1のサイズ排除クロマトグラフィーのプロファイルは、ほぼ同一であり、前記変異導入が、タンパク質の全体的な構造を失わせなかったことが示された(補足図S6)。
【0096】
次に、H3K9のトリメチルマーク及びジメチルマークのGASC1媒介性脱メチル化の反応機構を検討するため、複数の実験を行った。因子阻害性HIF1(Factor Inhibiting HIF1:FIH)を含むα-ケトグルタレート依存性オキシゲナーゼに関する種々の研究11,17-24では、以下の反応機構が提唱されている:最初に、酵素が、その金属結合モチーフであり、いわゆる表面の3つの分子から成るHXD/EXnH20(GASC1ではHXEXnH)を通じて鉄と結合する。次いで、Fe(II)-酵素の複合体が、補因子であるα-ケトグルタレート(αKG)と結合し、次いで、基質及び酸素と結合する。酸素の結合の後に、αKGの酸化的脱カルボキシル化が起こり、サクシネート、二酸化炭素及びフェリルが生じる。フェリルは、非常に反応性が高い基であり、リジンのメチル基中のC-H結合を酸化して、不安定なカルビノールアミンを形成する場合があり、このカルビノールアミンは、迅速に分解してホルムアルデヒドを生じさせ、そしてリジンはメチル基を奪われることになる(図7a)。
【0097】
cupinスーパーファミリーに属するジオキシゲナーゼは、Fe(II)とα-ケトグルタレートに依存する。更に、FBXL11を含むいくつかのcupinジヒドロゲナーゼは、完全な触媒活性のために更にアスコルビン酸を必要とする5,21,23。アスコルビン酸の作用の様式は、現時点では明確なっていないが、Fe(III)をその活性状態のFe(II)に還元するか、又はフェリル(FeIV=0)中間体が分離して生成された場合に「代理の還元性基質」として機能してジオキシゲナーゼ酵素を「救助する」ことが示唆されている17,22。
【0098】
GASC1媒介性脱メチル化が、上に記載され、図7aに示される反応機構と一致するかどうかを試験するため、脱メチル化反応の推定上の補因子の重要性を検討した。
【0099】
精製したHisタグ化GASC1を、その補因子の存在下又は不存在下で基質としてのバルクヒストンとインキュベートした(図3b)。全ての補因子の存在下ではGASC1はH3K9me3をin vitroで脱メチル化するが、EDTA(鉄をキレート化する)の存在下でのGASC1のインキュベート又は補因子の不存在下でのGASC1のインキュベートでは、脱メチル化は著しく低下していた(図8b)。補因子を添加していないにも関わらず脱メチル化反応を起こさせる能力は、恐らく、組換えGASC1と共に精製された補因子(鉄及びαKG)によるものと考えられる。この理由により、N-オキサリルグリシン、ケルセチン、CoCl2及びNiSO4が、GASC1媒介性のH3K9me3の脱メチル化を阻害する能力を試験することで、上記の補因子の重要性を更に探った。これら全ての化合物が、効果的に脱メチル化反応を阻害できた(図8c及びデータ非開示)。N-オキサリルグリシン(図8d)とケルセチンは、αKG類似体であり、恐らく、GASC1の鉄結合残基からαKGを引き離すことによりGASC1の活性を阻害している。同様に、CoCl2及びNiSO4によるGASC1の阻害には、恐らく、コバルト(II)イオン又はニッケル(II)イオンが競合することにより、GASC1の鉄結合部位からFe(II)が隔離されることが関わっている。脱メチル化反応にアスコルビン酸、αKG及び鉄を必要とすること、並びにニッケル塩及びコバルト塩及びαKG類似体により脱メチル化反応が阻害されることにより、GASC1が、実際にジオキシゲナーゼであることが強く示唆される。
【0100】
更にまた、補因子であるFe(II)及びαKGの存在下では、組換えHisタグ化GASC1は、ホルムアルデヒドを放出し(図8e)、これは、提案される図3aの反応スキームと一致している。これらをまとめると、以上の結果は、GASC1が、鉄とαKGを必要とするヒドロキシル化反応を通じて、in vitroでヒストンH3のトリメチル化及びジメチル化リジンK9を直接脱メチル化し、ホルムアルデヒドとモノメチル化H3K9を生じることを示している。
【0101】
GASC1が、in vitroでジメチル化及びトリメチル化H3K9を脱メチル化できることが立証されたことから、GASC1が、in vivoでヘテロクロマチンの形成/維持を調節できるのかどうかを検討した。この問題に取り組むため、最初に、GASC1の強制発現により、in vivoでH3K9のメチル化を調節できるかどうかを調べた。ヒトの2倍体線維芽細胞を、ヒト野生型GASC1を発現するレトロウィルスで感染させ、H3K9のメチル化状態を、共焦点顕微鏡観察により検証した。GASC1は、核内に局在していたことから、これは、予想上の核内局在シグナル配列が該タンパク質に存在することと一致していた。pBabe−HA−GASC1での感染により、H3K9トリメチルが効果的に減少し、モノメチル化H3K9が増加した(図9a)。対照的に、トリメチル化H3K4とトリメチル化H3K27は、GASC1過剰発現による影響を受けなかった(補足図S7)。
【0102】
次に、ヒト骨肉腫セルラインU2OSを、野生型GASC1を発現するプラスミドと、ヒスチジンH190 とグルタミン酸E192がそれぞれグリシンとアラニンで置換された変異型GASC1(H190G/E192A)を発現するプラスミドでトランスフェクトした。野生型GASC1の異所発現では、in vivoでヒストンH3のK9のトリメチル化とジメチル化から効果的に消失されたが、変異型GASC1では、そうではなかった(図9b)。
【0103】
興味深いことに、GASC1の異所発現では、モノメチル化H3K9が顕著に増加しており、GASC1が、in vivoでトリメチル化H3K9とジメチル化H3K9の両方を脱メチル化することが示唆された。トリメチル化H3K9の減少は、ウェスタンブロッティングによりこのマークの全体的なレベルを評価した場合に、GASC1でトランスフェクトされたU2OS細胞においても明らかであった(図10c)ことから、これらの推定上のFe(II)配位残基の決定的な重要性が確認された。
【0104】
次に、GASC1の近似するホモログであるJMJD2aとJMJD2b(図1a、図2b)も、in vivoでH3K9を脱メチル化できるかどうかを、これらのタンパク質をU2OS細胞内で過剰発現させることにより試験した。GASC1と同様に、JMJD2aとJMJD2bの異所発現により、トリメチル化及びジメチル化H3K9が顕著に減少し、同時にモノメチル化H3K9が増加していた(図10d、e)。これらの結果は、GASC1が、ヒストンH3K9デメチラーゼのファミリーに属していることを示している。
【0105】
ヘテロクロマチンの形成と維持には、トリメチル化及びジメチル化H3K9の存在と並びにHP−1結合が必要とされる25-27。GASC1を異所発現させることにより、H3K9のジメチル化とトリメチル化が全体的に減少したので、HP−1結合と局在性が、GASC1レベルの上昇により影響を受けるかどうかを検討した。NIH 3T3細胞を、HAタグ化GASC1を発現するプラスミドでトランスフェクトし、事前に抽出した細胞内の内在性のHP1βの分布を共焦点顕微鏡観察により確認した。予想した通り、HP1βは、GASC1を過剰発現する細胞内では非局在化しており、H190G/E192A変異体をトランスフェクトした場合には、非局在化は認められなかった(図11f)。同様に、GASC1のホモログであるJMJD2a及びJMJD2bの異所発現では、HP1αが非局在化した(補足図S8)。これらの結果については、GASC1過剰発現細胞内でのクロマチンと結合したHP1α及びHP1γのレベルの低下を示すことにより更に確認した(図11g)。この目的のため、Mycタグ化GASC1をテトラサイクリン制御下で発現させるHEK293細胞を作製した。選択した細胞をテトラサイクリンの存在下又は不存在下で一晩インキュベートしたところ、図11gに示すように、クロマチンと結合したHP1α及びHP1γの量は、GASC1過剰発現細胞内で減少していた。これらの結果をまとめると、GASC1が、ヘテロクロマチンの形成及び維持の調節において、生理学的に重要な役割を果たす可能性が示唆される。
【0106】
近年に至るまで、トリメチル化リジンは、不可逆的な共有結合性のヒストン修飾であると考えられてきており28、ヒストントリメチルデメチラーゼを同定するのは、本報告が最初のものである。我々の結果は、GASC1が、メチル化されたH3K9を特異的に脱メチル化するが、モノメチル化又はジメチル化された基質に特異的なLSD1及びFBXL114,5とは対照的に、GASC1は、in vitro及びin vivoで、H3K9のジメチル種及びトリメチル種の両方を取り除く能力を有することを示している。
【0107】
GASC1は、複数のドメインを含み、恐らく、これらのドメインは、GASC1タンパク質にトリメチル化及びジメチル化H3K9に対する強い特異性を与えるよう作用する(図1a)。JumonjiCドメイン(JmjC)とJumonjiNドメイン(JmjN)に加え、GASC1タンパク質は、2つのplant homeodomain(PHD)並びにtandem Tudor domain(Tdr)を特徴とする。これらのドメインの機能は現時点では明らかとなっていないが、これらはすべて、クロマチンの調節に関与していると考えられている8,10,12,29,30。例えば、53BP1のTudorドメインは、ヒストンH3のリジン79のトリメチル化体に結合することが報告されており31、PHDドメインは、ヌクレオソームへの結合で、Bromoドメインと強調することが実証されている30。どのドメインがGASC1の脱メチル化機能に必要であるのかという洞察を得るためと、どのドメインが基質特異性を決定するのかを解明するために、GASC1タンパク質の変異導入試験を行った。これは、一連のJmJC、JmjN及びPHDドメインに欠失を有するGASC1発現コンストラクトを作成することで行った。これらの結果は、JmjCドメインとJmjNドメインが、GASC1の脱メチル化活性に不可欠であることを示していた(補足図S9)。対照的にPHDドメインの欠失は、GASC1のin vivoでH3K9me3を脱メチル化する能力に影響していなかったと思われた(図S9)。
【0108】
GASC1及びFBXL11の類似する反応機構に照らせば、なぜFBXL11は、その機能を有しないのに、GASC1がトリメチル化ヒストンのリジン基質を脱メチル化する能力を有するのかという疑問が生じるのが妥当である。可能性のある説明としては、鉄/基質結合用溝が、FBXL11ではGASC1よりも小さいことが考えられる。この説明は、実験的な検証を要するが、in silicoモデリングにより支持される。FIHを鋳型としてモデリングした2種類のJumonjiタンパク質のモデルを比較した場合、両者は共に、そのモデルの鉄結合用溝のエッジに位置するループ領域を有するという特徴を有することが判明した。このループ領域は、FBXL11モデルで顕著に長く、これがゆらゆらと動くために、このタンパク質の鉄/基質結合用溝がより小さくなり、結果としてトリメチルリジン基質の収容を阻害しているかもしれないという可能性が考えられる。
【0109】
本報告に記載され、補足図S8bにまとめられる脱メチル化経路は、進化の過程で保存されていると予測される。この関係において、Schizosaccharomyces pombeのJumonjiタンパク質であるEpe1は、ヘテロクロマチン化をアンタゴナイズすることが報告されている10。従って、最近の報告10では、Epe1の強制発現が、ジメチル化H3K9を減少させ、ヘテロクロマチンにアンタゴナイズすると同時にセントロメアと交配型遺伝子座を不安定化させることが証明されている10。このEpe1媒介性表現型は、無傷のJmjCドメインに依存しており、Epe1のヘテロクロマチン化をアンタゴナイズする能力が脱メチル化活性に起因することは証明されていないものの、我々の本データと照らしてこのような役割の仮説を立てることは興味深い。
【0110】
例えば、GASC1等、類似する酵素活性を有する、密接に関連した複数のヒトホモログ(JMJD2a及びJMJD2b、図2b)の存在は、機能の重複性とH3K9トリメチルデメチラーゼ活性の正確な調節の必要性を示唆するものである。ヘテロクロマチンの形成と維持の正確な調節が、適切な生物機能とゲノムの統合性の両方にとって決定的に重要であることから、上記のことは予測されるべきことである。例えば、セントロメアにおけるヘテロクロマチンの形成は、有糸分裂期における染色体の正確な分離に不可欠である32。同様に、マウスのSuv39H1及びSuv39H2(これら遺伝子の産物は、ヘテロクロマチンのH3K9のジメチル化及びトリメチル化に関与する)の欠失は、染色体を不安定化させ、マウスのリンパ腫の誘導において癌遺伝子と共同して機能することが示されている6。従って、GASC1又はそのホモログの異常な活性は、ゲノムを不安定化させ、その結果として癌を生じさせる可能性がある。実際に、GASC1は、元々、食道扁平上皮細胞(oesophageal squamous cell:ESC)の癌で頻繁に増幅される遺伝子として同定されており22、GASC1は、染色体異常を含む種々の種類の癌で過剰発現されている33,34。
【0111】
GASC1の癌への関与に関する更なる支持を得るため、Oncomineデーターベースで、正常組織と腫瘍組織における異なるGASC1発現について探索した。GASC1とそのホモログのJMJD2a及びJMJD2bの発現は、正常組織と比べて前立腺癌で著しく増加していることがわかった(補足図S11)。GASC1は、先に、ESC癌腫由来の複数のセルラインで増幅されていることが示されている33。GASC1遺伝子を増幅させたESCセルライン(KYSE−150)と、より緩やかに発現されているセルライン(KYSE−450)において、H3K9エピジェネティックマークのメチル化状態を試験した。ヒト骨肉腫セルラインU2OSが、対照として含まれた。GASC1増幅の重要性を試験するため、GASC1及び/又はそのホモログに対するsiRNAオリゴでトランスフェクトされた細胞におけるH3K9メチル化状態に対するGASC1ノックダウンによる効果を試験した。細胞をLMP−GASC1でトランスフェクトして、その後、ピューロマイシンを用いて3日間かけてセレクションを行った。GASC1ノックダウンの効果は、RT−QPCRにより評価した(図5a)。ウェスタンブロッティングにより証拠付けられるように、H3K9me3は、KYSE−150細胞内でGASC1ノックダウンに応じて顕著に増大していた(データ非公開)。
【0112】
ジメチル化及びトリメチル化H3K9は、HP1結合に必要であり、ヘテロクロマチン形成に不可欠であり、転写抑制と関連している。更にまた、これらのエピジェネティックマークは、癌から細胞を守る要となる機構である、老化状態にある特定の遺伝子において増加している36-39。従って、GASC1の増幅は、細胞が老化状態になる能力を低下させるか、又はサイレンシング状態にある癌遺伝子の抑制を解除する可能性があることが予測できる(図6d)。従って、GASC1の脱メチル化活性の阻害は、新たな抗腫瘍治療様式を構成する可能性がある。
【0113】
興味深いことに、抗増殖特性及び抗癌特性と関連する植物フラボノイドであるケルセチン(図12b)40-42は、in vitroで効果的にH3K9me3のGASC1媒介性脱メチル化を阻害できた(図12c)。ケルセチンを用いてin vivoでKYSE−150細胞を一晩処理すると、H3K9トリメチル化が驚くほど増大し、このことは、免疫蛍光とウェスタンブロッティングに裏付けられている(図13d及びデータ非開示)。H3K9me3の増大は、増殖速度の顕著な低下、細胞形態の変化、老化関連ヘテロクロマチン病巣(senescence associated heterochromatin foci:SAHF)の出現及びβ−gal染色により特徴付けられる老化状態様の表現型も伴っていた(データ非公開)。興味深いことに、ケルセチン処理は、他のセルライン(低レベル及び中レベルのGASC1を発現する)では、同程度の影響を及ぼさないように思われた。従って、U2OS、WI38及びKYSE−450細胞は、同程度のH3K9me3の増加、又は同様の老化状態様表現型及び増殖阻害を示さなかった。この結果により、JMJD2酵素の発現又は活性の増大とこれに続くH3K9me3/me2の減少により特徴付けられるいくつかの癌が、この化合物による増殖阻害作用に特に高い感受性を示す可能性が示唆される。ケルセチンの生物学的効果は、従来ではトポイソメラーゼIの阻害43、タンパク質と脂質キナーゼシグナル経路の調節44及び活性酸素種の除去45によるものであると考えられてきた。本研究の知見に照らせば、ケルセチンによる抗癌効果は、少なくとも部分的に、そのGASC1及びJMDM2サブファミリーの他のメンバーに対する阻害作用により説明できるという推測が試みられる。
【0114】
以上をまとめると、複数の独立した証拠を用いることで、GASC1が、in vitro及びin vivoの両方で、抑制性のヒストンのジメチル及びトリメチルH3K9マークを直接脱メチル化することが実証された。本知見により、ヒストンのトリメチル化が、可逆性の修飾であることが示された。この知見は、ヒトの疾患、特に癌について、更に深い意味合いを有する可能性がある。
【0115】
材料と方法
脱メチル化アッセイ
バルクヒストン、オリゴヌクレオチド又は合成ヒストンペプチドを、精製したHis-GASC1と、脱メチル化バッファー(50mMのTris(pH8.0)、50mMのKCl、10mMのMgCl2、1mMのα-ケトグルタレート、40μMのFeSO4、2mMのアスコルビン酸、0.01%(w/v)のBSA及び5%(v/v)のグリセロール)中で37℃で反応させた。典型的な反応では、6μgのバルクヒストン又は2μgの修飾ヒストンペプチドを20μgのGASC1と100μlの体積で30分間反応させた。反応混合物を、特異的抗体を用いたウェスタンブロッティングか、又はホルムアルデヒド放出アッセイのいずれかにより分析した。
【0116】
ホルムアルデヒド放出アッセイ
ホルムアルデヒド放出アッセイは、本質的に記載4に沿って行った。全ての反応は、石英製キュベット内で、1反応系当たり200μlの総量で行った。簡潔に言うと、組換えGASC1(典型的には40μg)を150μlの脱メチル化バッファー(上記参照)中で、2mMのNAD+と0.2Uのホルムアルデヒドデヒドロゲナーゼ(FDH)の存在下で、37℃にて5分間インキュベートした。次いで、基質(ヒストンペプチド)を添加して反応を開始させた。340nmの吸光度を、Genesys 10UV Thermospectronic spectrophotometerを用いて0.5mmの間隔を開けて、37℃で測定した(全部で15mm)。
【0117】
質量分析用のH3K9me3ペプチドのGASC1による脱メチル化
6μgの組換えGASC1を、終量90μlのFDHバッファー中で、3μgのH3K9me3ペプチドと37℃で30分間インキュベートした。尿素を終濃度4Mで添加し、混合物を20℃で15分間インキュベートした。同量の1%TFAを添加し、サンプルを、100μlのチップ中にパッキングされた逆相ミニC8カラム(カラム容積20μl)にローディングした。1%TFAで洗浄した後、結合したペプチドを20μl(30%メタノール、25%ギ酸)で溶離させた。溶離した材料の3分の1をSDS-PAGEと、抗H3K9me3及びその後に抗ビオチン抗体を用いたウェスタンブロッティングで分析した(図4c)。残りの材料は、質量分析により分析した(図4d)。
【0118】
質量分析(MS)
溶離物の3分の1を、1%TFA中で、Agilent 1100 Nano HP1C(Palo Alto, CA)を用いて、スプレーエミッター(内径75μm、開口8μm、長さ70 mm;New Objectives、米国)内にパッキングされたC18カラム(Reprosil-Pur C18-AQ 3μm; Dr. Maisch GmbH、アメルブッフ-エントリンゲン、独国)上に注いだ。ペプチドは、10分間でバッファーA(5%アセトニトリル及び0.5%酢酸)からバッファーB(アセトニトリル及び0.5%酢酸)へ0%ないし20%で変化する勾配を用いて、300 nL/分で溶離した。スペクトルをLTQ-FT質量分析器(Thermoelectron、ブレーメン、独国)に記録した。
【0119】
補足的方法
材料
ヒストンH3のN末端尾部(1-40)を模倣する43アミノ酸長の合成ペプチド:
【化4】
このペプチドは、リジンとTtdsスペーサーを介してビオチンに連結された結合用のC末端部のチロシン及びシステインを用いて合成した。この実験で用いたペプチドは、非修飾型であるか、リジン27がトリメチル化されているか、又はリジンK9がモノメチル化、ジメチル化若しくはトリメチル化されており、Jerini, GMBH(独国)から購入した。ホルムアルデヒドデヒドロゲナーゼ(FDH、F1879)及びニコチンアミン及びバルクヒストン(H9250)はSigma社から購入した。本試験で用いた抗体は、以下の通り:抗トリメチル化H3-K9(Upstate 07-523)、抗ジメチル化H3-K9(Upstate 07-212)、抗モノメチル化H3-K9(Abcam Ab9045-50)、抗トリメチルH3-K27(Upstate 07-449)、抗トリメチルH4-K20(Upstate 07-463)、抗トリメチル化H3-K4(Abcam ab8580-50)、抗ジメチル化H3-K4(Upstate 07-030)、抗ヒストンH3(Abcam Ab1791-100)、抗HP1α(Upstate 05-689)、抗HP1γ(upstate 05-690)、抗ビオチン-HRP(Sigma A4541)、抗-His(Upstate 05-531)及び抗HA(CRP Inc. AFC-101P)。
【0120】
セルライン及び組織培養
ヒト食道扁平上皮癌セルラインKYSE-70及び150は、German Collection of Microorganisms and Cell Cultures(ブラオンシュヴァイヒ、独国)から入手した。hTert を発現する2倍体のヒト線維芽細胞(TIG-3)とマウスのエコトロピックレトロウィルスレセプターEcoRを発現するU2OS細胞を実験に用いた。293 TRex-flip-in細胞を基本的に製造元(Invitrogen)による記載に従って用いて、テトラサイクリン誘導性のアミノ末端にmycタグが付いたGASC1を安定に発現するHEK293細胞を作製した。KYSE細胞は、49%のRPMI 1640、2%のウシ胎児血清(FCS)が補充された49%のHam's F12中に5%CO2の下で維持した。他の全ての細胞は、10%のウシ胎児血清(FCS)が補充されたDulbecco's modified Eagle's medium(DMEM)中に5%CO2の下で37℃にて維持した。
【0121】
HeLa核タンパク質のヒストンペプチドプルダウン
HeLa細胞の核抽出物とリジン9がトリメチル化されたヒストンH3ペプチド
【化5】
を用いてプルダウンアッセイを行った。核は、HeLa細胞から調製し、溶解バッファー(50mMのTris(pH7.2)、300mMのNaCl、0.5%のIGEPAL CA-630、1mMのEDTA(pH8.0)、プロテアーゼ阻害剤を含む1mMのPMSF)で溶解させた。溶解物は、遊離ストレブトアビジンビーズを用いて事前除去し(図S1、事前除去1)、次いで、非修飾型H3ヒストンに結合したストレプトアビジンビーズで事前除去した(図S1、事前除去2)。次いで、事前除去した溶解物を、事前にストレプトアビジンビーズと結合させておいた、K9がトリメチル化されたH3ヒストンペプチド(H3K9me3)とインキュベートした。ビーズを十分に洗浄し、続いて、2x LSBバッファー(100mMのTris-HCl(pH6.8)、200mMのDTT、4%のSDS、20%のグリセロール及び0.2%のブロモフェノールブルー)で結合したタンパク質を樹脂から溶離した(図S1、H3K9me3)。タンパク質をSDS-PAGEで分離し、銀染色により可視化した。プルダウンされたH3K9me3を豊富に含むタンパク質バンドを切り出し、ゲルのままトリプシンで処理し、Wilm et al.(Wilmm. (1996) Nature 379, 466-469)の記載に従って、ナノエレクトロスプレータンデム型質量分析器で分析した。
【0122】
免疫蛍光
共焦点免疫染色法を、基本的にSorensen, CS. et al., Nat. Cell. Biol. 7, 195-201 (2005)の記載に従って行った。
【0123】
細胞溶解物の調製
細胞の全溶解物は、同数の細胞を尿素バッファー(1%のSDS、9Mの尿素、25mMのTris-HCl(pH6.8)、1mMのEDTA、0.7mのβ-メルカプトエタノール)に溶解して作製し、5分間沸騰させ、そして超音波処理した。クロマチンの分画化のため、200μlの事前抽出用バッファー(0.5%のIGEPAL-630、5OmMのNaCl、3mMのMgCl2及び300mMのスクロースを含有する2OmMのHepes-KOH(pH7.2))中の同数の細胞を氷上で30分間インキュベートした。100μlを2xLSBバッファー(100mMのTris-HCl(pH6.8)、200mMのDTT、4%のSDS、20%のグリセロール及び0.2%のブロモフェノールブルー)と混合し、沸騰させ、超音波処理して、全溶解物として使用した。残りの100μlは、1300xg、4℃で10分間スピンダウンさせ、1mlの事前抽出用バッファーで洗浄し、100μlの2xLSBバッファーに再懸濁し、沸騰させ、超音波処理し、クロマチン分画として使用した。
【0124】
GASC1及びJMJD2a/bに対するsiRNA及びshRNA
GASC1並びにそのホモログJMJD2a及びJMJD2bに対する低分子干渉RNA(siRNA)オリゴヌクレオチドは、Dharmacon Research, Inc.によって合成された。6ウェルプレート内で、Oligofectamine試薬(Invitrogen)を製造元のプロトコルに従って用いて、細胞(1x105/ウェル)をsiRNAオリゴ(0.3μg/ウェル)でトランスフェクトした。siRNAは、例えば、siRNA retriever;(http://katahdin.cshl.org:9331/homepage/siRNA/RNAi.cgi?type=siRNA)を用いる等、当該技術分野で知られる方法により設計できる。
【0125】
shRNAコンストラクト
短鎖ヘアピンRNAコンストラクトは、Paddison et al.(2005)(Paddison PJ, Clearym, Silva JM, Chang K, Sheth N, Sachidanandam R & Hannon GJ. Cloning of short hairpin RNAs for gene knockdown inmammalian cells Naturemethods pp163-167)の記載に従って、MSCV/LTRmiR30-PIG(LMP)ベクター(Dickens NG et al(2005)"Probing tumor phenotypes using stable and regulated syntheticmicroRNA precursors" Nature Genetics Vol 37, No 11 1289-1295.)内に作製した。
【0126】
「shRNA retriever」(http://katahdin.cshl.org:9331/homepage/siRNA/RNAi.cgi?type=shRNA)を用いて3種類のオリゴヌクレオチドを設計した。以下のオリゴヌクレオチド:
【化6】
を鋳型として用いて150 bpのPCR産物を増幅させ、XhoI及びEcoRIで処理し、そして制限酵素処理したLMPベクター内にライゲーションした。このshRNAコンストラクトは、上記の配列で下線が引かれたGASC1コーディング配列の2117番目の位置の22 bpの配列を標的とする。このshRNAiコンストラクトが、GASC1のmRNAをノックダウンする効率は、リアルタイム定量的RT-PCRで評価したところ、約80%であると評価された。
【0127】
リアルタイム定量的RT-PCR
細胞を、表示するshRNAコンストラクトでトランスフェクトし、簡単なピューロマイシンセレクション(2μg/ml)に付した。Quiagen RNAeasyミニキットを製造元の使用説明書に従って用いて全RNAをトランスフェクトした細胞から作製した。Taqman逆転写キットとポリdTプライマーを製造元の使用説明書に従って用いてcDNAを作製した。このcDNAを鋳型として、GASC1特異的プライマーと共に、Applied biosystems 7700でのリアルタイム定量的PCR反応に用いた。この反応物は、Applied biosystems社製の2xSYBR green反応ミックスを用いて調製した。
【0128】
BrdUの取り込み
siRNAのトランスフェクションから30時間後に、細胞を4ウェルチャンバースライドに分け、BrdUを含む培養培地で15-30分間インキュベートした。
【0129】
ヒトGASC1のcDNAのクローニング
ヒトGASC1及びJMJD2Aの推定上のオープンリーディングフレームをHeLaのcDNAとヒト胎児脳のcDNAライブラリー(Invitrogen、カールスバッド、カリフォルニア州)のcDNAからPCR増幅した。プライマー配列は、請求により入手可能である。PCR産物をゲル精製し、pCR8/GW(Invitrogen)にクローニングし、DNAしーケンシングにより確認した。これらのGateway適合性の導入クローンを用いて、GASC1及びJMJD2AをpCMV-HA、pCMV-myc及びpBabepuroに導入した。標準的な変異導入方法により、GASC1の保存された鉄結合ドメイン(HTE)の、190-192番目のアミノ酸が、それぞれヒスチジン、スレオニン及びグルタミン酸からグリシン、スレオニン及びアラニンに変化した2重変異を作製した。
【0130】
組換えタンパク質
Gatewayを用いてGASC1のcDNA をpCR8/GW からpAcHLT-A(Pharmingen)のGateway修飾型に導入することにより、アミノ末端にヘキサヒスチジンタグが付いた完全長のヒトGASC1バキュロウィルス導入ベクターを作製した。組換えバキュロウィルスは、基本的に、先の文献に記載されたように、GASC1遺伝子を含むバキュロウィルス導入ベクターと、Bsu36IでリニアライズしたBakpak6バキュロウィルスDNAの共トランスフェクションにより作製した。ヒスチジンタグが付いたGASC1とJMJD2Aを発現させ、基本的に先の文献(Christensen et al. Nucleic Acids Research 33, 5458-5470 (2005))に記載されたように、コバルトアフィニティークロマトグラフィーで精製した。溶離した分画を、SDS-PAGEで分析し、選択した分画をサイズ排除クロマトグラフィー(SEC)により更に精製した。簡潔に述べると、SECは、50mM・NaClを含有する25mM・HEPES-KOH(pH7.7)で平衡化させたSuperose 12, 10/300カラム(Pharmacia-Amersham)上で行い、同一のバッファーを用いて、0.3ml/分の流速で溶離した。溶離した物質を、0.5mlの分画で回収し、液体窒素で瞬間冷凍し、-80℃で保存した。全ての工程は、完全にEDTA不含のプロテアーゼ阻害剤(Boehringermannheim、独国)の存在下で、氷上又は4℃で行った。
【0131】
レトロウィルスの形質導入
GASC1とその変異体H190G/E192Aを発現する組換えレトロウィルスを作製するため、GASC1のオープンリーディングフレームを、Gatewayによる組換えによりpBabepuroに導入し、pBabepuro-HA-GASC1とpBabepuro-HA-H190G/E192Aを作製した。Phoenix-Eco 293細胞パッキング用セルラインのトランスフェクションから24-48時間後に、高い力価を有するウィルス粒子が得られた。トランスフェクションは、リン酸カルシウム法により行った。TIG3-hTert-EcoR細胞又はU2OS-EcoR細胞の形質導入は、24時間以内に、ウィルスを含むパッケージングセルラインからの上清を細胞ディッシュに4回添加することにより達成した。形質導入された細胞は、ピューロマイシンの存在下で2-5日間かけて選択した(1μg/μl)。
【0132】
分子モデリング
α-ケトグルタレート、CADペプチド及び鉄と複合体形成したFIHの構造は、1H2Kのアクセス番号でProtein Data Bank(PDB、Brookhaven National Library、アプトン、ニューヨーク州、米国)から入手可能である。ホモロジーモジュールINSIGHT II (2005) (Accelrys、サンディエゴ)を用いて、GASC122-349を鋳型のFIHと共にホモロジーモデリングした。ヒストン尾部(ARTKQTARKSTG)を鋳型のCADペプチドと共にモデリングした。GRID(Version 22、Molecular Discovery Ltd.、オックスフォード、英国)(Goodford, 1985)というプログラムを用いて、GASC1分子の疎水性表面の外形を計算した。続いて、AUTODOCK (Ver. 3.0.5、Scripps Research Institute andmolecular Graphics Laboratory)を用いて、上記のヒストンペプチドを、GASC1の疎水性表面の外形上にドッキングさせた。最後に、前記複合体の構造モデルを、Pymol(http://pymol.sourceforge.net/)で導き出した。
【0133】
補足的方法
材料
ヒストンH3のN末端尾部(1-40)を模倣する43アミノ酸長の合成ペプチド:
【化7】
このペプチドは、リジンとTtdsスペーサーを介してビオチンに連結された結合用のC末端部のチロシン及びシステインを用いて合成した。この実験で用いたペプチドは、非修飾型であるか、リジン27がトリメチル化されているか、又はリジンK9がモノメチル化、ジメチル化若しくはトリメチル化されており、Jerini, GMBH(独国)から購入した。ホルムアルデヒドデヒドロゲナーゼ(FDH、F1879)及びニコチンアミン及びバルクヒストン(H9250)はSigma社から購入した。本試験で用いた抗体は、以下の通り:抗トリメチル化H3-K9(Upstate 07-523)、抗ジメチル化H3-K9(Upstate 07-212)、抗モノメチル化H3-K9(Abcam Ab9045-50)、抗トリメチルH3-K27(Upstate 07-449)、抗トリメチルH4-K20(Upstate 07-463)、抗トリメチル化H3-K4(Abcam ab8580-50)、抗ジメチル化H3-K4(Upstate 07-030)、抗ヒストンH3(Abcam Ab1791-100)、抗HP1α(Upstate 05-689)、抗HP1γ(upstate 05-690)、抗ビオチン-HRP(Sigma A4541)、抗-His(Upstate 05-531)及び抗HA(CRP Inc. AFC-101P)。
【0134】
セルライン及び組織培養
ヒト食道扁平上皮癌セルラインKYSE-70及び150は、German Collection ofmicroorganisms and Cell Cultures(ブラオンシュヴァイヒ、独国)から入手した。hTert を発現する2倍体のヒト線維芽細胞(TIG-3)とマウスのエコトロピックレトロウィルスレセプターEcoRを発現するU2OS細胞を実験に用いた。テトラサイクリン誘導性のアミノ末端にmycタグが付いたGASC1を安定に発現するHEK293細胞を293 TRex-flip-in細胞を基本的に製造元(Invitrogen)による記載に従って用いて作製した。KYSE細胞は、49%のRPMI1640、2%のウシ胎児血清(FCS)が補充された49%のHam's F12中に5%CO2の下で維持した。他の全ての細胞は、10%のウシ胎児血清(FCS)が補充されたDulbecco改変イーグル培地(DMEM)中に5%CO2の下で37℃にて維持した。
【0135】
HeLa核タンパク質のヒストンペプチドプルダウン
HeLa細胞の核抽出物とリジン9がトリメチル化されたヒストンH3ペプチド:
【化8】
を用いてプルダウンアッセイを行った。核は、HeLa細胞から調製し、溶解バッファー(50mMのTris(pH7.2)、300mMのNaCl、0.5%のIGEPAL CA-630、1mMのEDTA(pH8.0)、プロテアーゼ阻害剤を含む1mMのPMSF)で溶解させた。溶解物は、遊離ストレブトアビジンビーズを用いて事前除去し(図S1、事前除去1)、次いで、非修飾型H3ヒストンに結合したストレプトアビジンビーズで事前除去した(図S1、事前除去2)。次いで、事前除去した溶解物を、事前にストレプトアビジンビーズと結合させておいた、K9がトリメチル化されたH3ヒストンペプチド(H3K9me3)とインキュベートした。ビーズを十分に洗浄し、続いて、2x LSBバッファー(100mMのTris-HCl(pH6.8)、200mMのDTT、4%のSDS、20%のグリセロール及び0.2%のブロモフェノールブルー)で結合したタンパク質を樹脂から溶離した(図S1、H3K9me3)。タンパク質をSDS-PAGEで分離し、銀染色により可視化した。プルダウンされたH3K9me3を豊富に含むタンパク質バンドを切り出し、ゲルのままトリプシンで処理し、Wilm et al.(Wilmm. (1996) Nature 379, 466-469)の記載に従って、ナノエレクトロスプレータンデム型質量分析器で分析した。
【0136】
免疫蛍光
共焦点免疫染色法を、基本的にSorensen, CS. et al., Nat. Cell. Biol. 7, 195-201 (2005)の記載に従って行った。
【0137】
細胞溶解物の調製
細胞の全溶解物は、同数の細胞を尿素バッファー(1%のSDS、9Mの尿素、25mMのTris-HCl(pH6.8)、1mMのEDTA、0.7mのβ-メルカプトエタノール)に溶解して作製し、5分間沸騰させ、そして超音波処理した。クロマチンの分画化のため、200μlの事前抽出用バッファー(0.5%のIGEPAL-630、5OmMのNaCl、3mMのMgCl2及び300mMのスクロースを含有する2OmMのHepes-KOH(pH7.2))中の同数の細胞を氷上で30分間インキュベートした。100μlを2xLSBバッファー(100mMのTris-HCl(pH6.8)、200mMのDTT、4%のSDS、20%のグリセロール及び0.2%のブロモフェノールブルー)と混合し、沸騰させ、超音波処理して、全溶解物として使用した。残りの100μlは、1300×g、4℃で10分間スピンダウンさせ、1mlの事前抽出用バッファーで洗浄し、100μlの2xLSBバッファーに再懸濁し、沸騰させ、超音波処理し、クロマチン分画として使用した。
【0138】
GASC1及びJMJD2a/bに対するsiRNA及びshRNA
GASC1並びにそのホモログJMJD2a及びJMJD2bに対する低分子干渉RNA(siRNA)オリゴヌクレオチドは、Dharmacon Research, Inc.によって合成された。6ウェルプレート内で、Oligofectamine試薬(Invitrogen)を製造元のプロトコルに従って用いて、細胞(1x105/ウェル)をsiRNAオリゴ(0.3μg/ウェル)でトランスフェクトした。siRNAは、例えば、siRNA retriever; (http://katahdin.cshl.org:9331/homepage/siRNA/RNAi.cgi?type=siRNA)を用いる等、当該技術分野で知られる方法により設計できる。
【0139】
shRNAコンストラクト
短鎖ヘアピンRNAコンストラクトは、Paddison et al.(2005)(Paddison PJ, Clearym, Silva JM, Chang K, Sheth N, Sachidanandam R & Hannon GJ. Cloning of short hairpin RNAs for gene knockdown inmammalian cells Naturemethods pp163-167)の記載に従って、MSCV/LTRmiR30-PIG (LMP)ベクター(Dickens NG et al(2005)"Probing tumor phenotypes using stable and regulated syntheticmicroRNA precursors" Nature Genetics Vol 37, No 11 1289-1295.)内に作製した。
【0140】
「shRNA retriever」(http://katahdin.cshl.org:9331/homepage/siRNA/RNAi.cgi?type=shRNA)を用いて3種類のオリゴヌクレオチドを設計した。以下のオリゴヌクレオチド:
【化9】
を鋳型として用いて150bpのPCR産物を増幅させ、XhoI及びEcoRIで処理し、そして制限酵素処理したLMPベクター内にライゲーションした。このshRNAコンストラクトは、上記の配列で下線が引かれたGASC1コーディング配列の2117番目の位置の22bpの配列を標的とする。このshRNAiコンストラクトが、GASC1のmRNAをノックダウンする効率は、リアルタイム定量的RT-PCRで評価したところ、約80%であると評価された。
【0141】
リアルタイム定量的RT-PCR
細胞を、表示するshRNAコンストラクトでトランスフェクトし、簡単なピューロマイシンセレクション(2μg/ml)に付した。Quiagen RNAeasyミニキットを製造元の使用説明書に従って用いて全RNAをトランスフェクトした細胞から作製した。Taqman逆転写キットとポリdTプライマーを製造元の使用説明書に従って用いてcDNAを作製した。このcDNAを鋳型として、GASC1特異的プライマーと共に、Applied biosystems 7700でのリアルタイム定量的PCR反応に用いた。この反応物は、Applied biosystems社製の2xSYBR green反応ミックスを用いて調製した。
【0142】
BrdUの取り込み
siRNAのトランスフェクションから30時間後に、細胞を4ウェルチャンバースライドに分け、BrdUを含む培養培地で15-30分間インキュベートした。
【0143】
ヒトGASC1のcDNAのクローニング
ヒトGASC1及びJMJD2Aの推定上のオープンリーディングフレームをHeLaのcDNAとヒト胎児脳のcDNAライブラリー(Invitrogen、カールスバッド、カリフォルニア州)のcDNAからPCR増幅した。プライマー配列は、請求により入手可能である。PCR産物をゲル精製し、pCR8/GW(Invitrogen)にクローニングし、DNAシーケンシングにより確認した。これらのGateway適合性の導入クローンを用いて、GASC1及びJMJD2AをpCMV-HA、pCMV-myc及びpBabepuroに導入した。標準的な変異導入方法により、GASC1の保存された鉄結合ドメイン(HTE)の、190-192番目のアミノ酸が、それぞれヒスチジン、スレオニン及びグルタミン酸からグリシン、スレオニン、アラニンに変化した2重変異を作製した。
【0144】
組換えタンパク質
Gatewayを用いてGASC1のcDNA をpCR8/GW からpAcHLT-A(Pharmingen)のGateway修飾型に導入することにより、アミノ末端にヘキサヒスチジンタグが付いた完全長のヒトGASC1バキュロウィルス導入ベクターを作製した。組換えバキュロウィルスは、基本的に、先の文献に記載されたように、GASC1遺伝子を含むバキュロウィルス導入ベクターと、Bsu36IでリニアライズしたBakpak6バキュロウィルスDNAの共トランスフェクションにより作製した。ヒスチジンタグが付いたGASC1とJMJD2Aを発現させ、基本的に先の文献(Christensen et al. Nucleic Acids Research 33, 5458-5470 (2005))に記載されたように、コバルトアフィニティークロマトグラフィーにより精製した。溶離した分画を、SDS-PAGEで分析し、選択した分画をサイズ排除クロマトグラフィー(SEC)により更に精製した。簡潔に述べると、SECは、50mM・NaClを含有する25mM・HEPES-KOH(pH7.7)で平衡化させたSuperose12、10/300カラム(Pharmacia-Amersham)上で行い、同一のバッファーを用いて、0.3ml/分の流速で溶離した。溶離した物質を、0.5mlの分画で回収し、液体窒素で瞬間冷凍し、-80℃で保存した。全ての工程は、完全にEDTA不含のプロテアーゼ阻害剤(Boehringermannheim、独国)の存在下で、氷上又は4℃で行った。
【0145】
レトロウィルスの形質導入
GASC1とその変異体H190G/E192Aを発現する組換えレトロウィルスを作製するため、GASC1のオープンリーディングフレームを、Gatewayによる組換えによりpBabepuroに導入し、pBabepuro-HA-GASC1とpBabepuro-HA-H190G/E192Aを作製した。Phoenix-Eco 293細胞パッキングセルラインのトランスフェクションから24-48時間後に、高い力価を有するウィルス粒子が得られた。トランスフェクションは、リン酸カルシウム法により行った。TIG3-hTert-EcoR細胞又はU2OS-EcoR細胞の形質導入は、24時間以内に、ウィルスを含むパッケージングセルラインからの上清を細胞ディッシュに4回添加することにより達成した。形質導入された細胞は、ピューロマイシンの存在下で2-5日間かけて選択した(1μg/μl)。
【0146】
分子モデリング
α-ケトグルタレート、CADペプチド及び鉄と複合体形成したFIHの構造は、1H2Kのアクセス番号でProtein Data Bank(PDB、Brookhaven National Library、アプトン、ニューヨーク州、米国)から入手可能である。ホモロジーモジュールINSIGHT II (2005) (Accelrys、サンディエゴ)を用いて、GASC122-349を鋳型のFIHと共にホモロジーモデリングした。ヒストン尾部(ARTKQTARKSTG)を鋳型のCADペプチドと共にモデリングした。GRID(Version 22、Molecular Discovery Ltd.、オックスフォード、英国)(Goodford, 1985)というプログラムを用いて、GASC1分子の疎水性表面の外形を計算した。続いて、AUTODOCK(Ver. 3.0.5、Scripps Research Institute andmolecular Graphics Laboratory)を用いて、上記のヒストンペプチドを、GASC1の疎水性表面の外形上にドッキングさせた。最後に、前記複合体の構造モデルを、Pymol(http://pymol.sourceforge.net/)で導き出した。
【0147】
参考文献
【表1】
【0148】
【表2】
【0149】
【表3】
【0150】
【表4】
【図面の簡単な説明】
【0151】
【図1】JumonjiヒストンデメチラーゼGASC1、FBXL11及びそのヒトホモログ。aは、GASC1、FBXL11、並びにそれらのドメイン:JmjC-Jumonji Cドメイン、JmjN-Jumonji Nドメイン、Tdr-Tudorドメイン、PHD-植物ホメオドメイン、CXXC-CxxCジンクフィンガードメイン、FBX-Fboxドメイン、LR-ロイシンリッチリピートを有するヒトホモログの模式図である。機能ドメインは、SMARTプログラム(http:/smart.embl-heidelberg.de/smart)を用いて決定した。
【図2】bは、ヒトJmjCドメイン含有タンパク質の系統樹である。系統樹は、近接結合(Neighbour Joining:NJ)法によるタンパク質のJmjCドメインに基づいて計算する。この計算に用いた配列を、補足図S2に示す。
【図3】GASC1は、トリメチル化及びジメチル化したヒストンH3K9を脱メチル化する。aは、サイズ排除クロマトグラフィーによる組換えGASC1の精製を表す。0.5mlのフラクションを回収し、10μlの物質をバルクヒストンと37℃で30分間インキュベートした。脱メチル化活性は、トリメチル化H3K9に対するブロッティングにより評価した。bは、GASC1のサイズ排除クロマトグラフィーから選択されたフラクション(F4、F6、F8、F9、FlO、F11、F12、F14、F16)を用いた脱メチル化アッセイを示す。図に示すように、イムノブロッティングによりフラクションの脱メチル化活性を評価した。
【図4】cは、精製したGASC1によるH3K9me3ペプチド(ARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPHRYC-Ttds-K(ビオチン)-NH2)の脱メチル化の質量分析を示す。ペプチドを、GASC1で処理した後(+GASC1)及び未処理のまま(-GASC1)でLTQ-FT上でLC-MSにより分析した。計算値からのモノアイソトピック質量の実測値の偏差値を括弧内に示す。計算上の中性質量は、H3K9me1:5130.885Da、H3K9me2:5144.900Da、H3K9me3:5158.916Daであり、脱メチル化の結果による質量の差異:14.016DaのH3K9(非メチル化)は、観察されなかった。我々のペプチド(分子式C223H390N75O61S2)についてIsoPro 3.0(http://members.aol.com/msmssoft/)により帯電状態+10、解像度50,000(シミュレーションしたスペクトル)でシミュレーションしたところ、個々のピークの多重性は、異なるアイソトープの自然発生により生じている。d、組換えGASC1と、基質として合成トリメチル化H3K9ペプチドを用いた脱メチル化アッセイを示す。ペプチドを、補因子の存在下で6μgのHis-GASC1と共に37℃で30分間インキュベートする。材料の3分の1をウェスタンブロッティングにより解析し、残りを質量分析に用いた(図2e)。
【図5】eは、ヒトJumonjiタンパク質GASC1、JMJD2A、FIH及びFBXL11並びに分裂酵母のJumonjiタンパク質Epe1のJmjCドメインのアライメントを示す。黄色の強調部分は、類似するか又は同一のアミノ酸を表す。黒色及び赤色のアステリスクは、それぞれ鉄の結合とα-ケトグルタレートに寄与すると予測される残基を示す。緑色のアステリスクは、GASC1鉄結合型変異体で変異の生じた残基を示す。矢印は、1h2kのPDB合計入力により定義される、FIHのβ鎖8-15に対応する番号が付された2本鎖β-へリックス鎖を示す。
【図6】fは、トリメチル化H3ヒストン尾部(1-11)(緑色)及びその補因子であるα-ケトグルタレート(赤色)及び鉄(マゼンタ)と複合体形成したGASC1(青色)の予測構造であり、リジンH3K9メチル基は、黄色でマークされている。左側のパネルは、鉄結合残基のHis190、His288、Glu192(白色の棒)の概要であり、右側のパネルは拡大図を示す。gは、バルクヒストンを基質として用い、野生型及び変異型(H190G/E192A)のGASC1の量を変化させながら用いた脱メチル化アッセイを示す。
【図7】GASC1の脱メチル化は、鉄及びα-ケトグルタレートに依存したヒドロキシル化反応を通じて起こる。aは、トリメチル化及びジメチル化H3K9のGASC1媒介性脱メチル化について提唱される反応スキームを示す。1.Fe(II)(マジェンタ)が、表面の3つの金属結合残基His190、Glu192及びHis288(青字)によりGASC1に結合している。2.α-ケトグルタレート(赤色)は、恐らくThr186とLys208の残基を通じて結合している。3.鉄(マジェンタ)は、次いで、分子状態の酸素に結合する。4.次のα-ケトグルタレート(赤色)の酸化的脱カルボキシル化により二酸化炭素、サクシネート(赤色)及びフェリル(ferryl)(FeIV=O、マジェンタ)が生じる。5.フェリル(マジェンタ)は次いで、リジンH3K9のメチル基(緑色)をヒドロキシル化し、ホルムアルデヒドを放出させる。6.最後に、ヒドロキシル化ヒストン尾部(緑色)とサクシネート(赤色)は、GASC1分子を遊離させることが可能であり、新たな脱メチル化サイクルに向けて空いた状態となる。
【図8】bは、合成H3K9me3ペプチドを基質として用いた脱メチル化アッセイを示す。ペプチドを、20μgのHis-GASC1と共に、その補因子であるアスコルビン酸、α-ケトグルタレート及びFe2SO4並びにEDTAの存在下又は不存在下で、37℃にて30分間インキュベートし、ウェスタンブロッティングにより解析した。GASC1の相対脱メチル化活性は、Fujifilm darkbox LAS 3000 readerを用いて定量化し、ローディング対照(α-ビオチン)に対して正規化した。H3K9me3ペプチド(レーン1)を100%に定めた。レーン2-7;EDTA又は提示した補因子の存在下又は不存在下で脱メチル化バッファー中でインキュベートしたH3K9me3ペプチド。cは、組換えGASC1と基質としてバルクヒストンを用いた脱メチル化アッセイを示す。ヒストンを、His-GASC1と共に、補因子(FeSO4及びα-ケトグルタレート)の存在下で、37℃にて30分間、α-ケトグルタレート類似体のN-オキサリルグリシンの存在下でインキュベートした。メチル化状態は、ウェスタンブロッティングにより評価した。dは、Fe(II)活性部位における2-ケトグルタレートとN-オキサリルグリシンの結合様式を示す。eは、GASC1媒介性のH3K9me3脱メチル化によるホルムアルデヒド放出を示す。ホルムアルデヒド放出は、脱メチル化-FDH一体化アッセイ4により測定した。脱メチル化-FDH一体化アッセイは、一定量の酵素GASC1(40μg)を用いたが、基質H3K9me3ペプチドの量を変化させながら行った。ホルムアルデヒド生成は、OD340nmにてNADH精製を測定することでモニターした。基質であるH3K9ペプチド(非メチル化)をネガティブ対照として50μg用いた。
【図9】GASC1の異所発現により、in vivoで、ジメチル化及びトリメチル化H3K9が喪失する。aは、pBabe-GASC1を導入したTig-3細胞の共焦点顕微鏡観察を示す。GASC1を過剰発現するTig-3細胞内でトリメチル化H3K9は喪失する。白色の矢印は、HAタグが付いたGASC1を発現する細胞を示す。bは、免疫蛍光により評価された、GASC1のH190G/E192A変異体を発現するU2OS細胞内でのトリメチル化H3K9を脱メチル化する能力の喪失を示す。白色の矢印は、HAタグ化GASC1を発現するか、又はHAタグ化されたGASC1のH190G/E192A変異体を発現する細胞を示す。
【図10】cは、ウェスタンブロッティングにより評価された、GASC1のH190G/E192A変異体でトランスフェクトされたU2OS細胞内でのトリメチル化H3K9を脱メチル化する能力の喪失を示す。dは、JMJD2aを過剰発現するU2OS細胞内でのトリメチル化及びジメチル化H3K9を脱メチル化する能力の喪失とモノメチル化H3K9の増大を示す。
【図11】fは、HAタグ化GASC1又はHAタグ化されたGASC1のH190G/E192A変異体でトランスフェクトされた3T3細胞の共焦点顕微鏡観察を示す。白色の矢印は、HAタグ化タンパク質を発現する細胞を示す。HP1βは、GASC1を過剰発現する細胞内で非局在化している。gは、テトラサイクリン(Tet)誘導性GASC1発現(Mycタグ化)系を有するHEK293細胞由来の全溶解物及びクロマチン結合フラクションのウェスタンブロッティングを示す。Tetの添加(+)及び省略(-)を、図示する。
【図12】GASC1発現は増殖を促進し、腫瘍発現と正の相関性を有する。aは、GASC1に対するsiRNA又はスクランブルされたsiRNA対照によるKYSE-150及びKYSE-450のトランスフェクションを示す。GASC1のメッセンジャーRNAの相対量は、リアルタイム定量的RT-PCRにより測定した。b、LMP-GASC1又は空のLMPベクターによるKYSE-150のトランスフェクションとその後の3日間のセレクションを示す。GASC1のメッセンジャーRNAの相対量は、リアルタイム定量的RT-PCRにより測定した。cは、GASC1のFe(II)活性部位におけるケルセチンの結合様式を示す。
【図13】dは、組換えGASC1と基質としてバルクヒストンを用いた脱メチル化アッセイを示す。ヒストンを、補因子(FeSO4とα-ケトグルタレート)の存在下でHis-GASC1と共に、可変量のケルセチンの存在化又は不存在下で37℃で30分間インキュベートした。メチル化状態は、ウェスタンブロッティングにより評価した。eは、免疫蛍光での評価による、KYSE-150細胞及びU2OS細胞のH3K9me3メチル化状態に対するケルセチンの効果を示す。
【図14】fは、癌の発症におけるGASC1の関与のモデルを示す。
【図15】(S1)は、HeLa核タンパク質のヒストンペプチドプルダウンを示す。HeLa細胞核抽出物とリジン9がトリメチル化されたヒストンH3ペプチドを用いてプルダウンアッセイを行った。HeLa核抽出物は、遊離ストレブトアビジンビーズを用いて事前除去し(事前除去1)、次いで、非修飾型H3ヒストンに結合したストレプトアビジンビーズで事前除去した(事前除去2)。事前除去した溶解物を、次いで、K9がトリメチル化されたH3ヒストンペプチド(H3K9me3)とインキュベートし、ストレプトアビジンビーズに事前に結合させた。ビーズを洗浄し、次いで結合したタンパク質を溶離させ、SDS-PAGEにより分離して、銀染色により可視化させた。H3K9me3プルダウンで濃縮されたタンパク質のバンドは、次いで、質量分析により同定した。濃縮されたバンドの一つは、Jumonjiタンパク質であるGASC1と同定された(矢印を参照)。やはりプルダウンアッセイで濃縮されたHP1αないしγのタンパク質も同様に示されている。分子量マーカーは左側に示されている。
【図16】(S2)は:ヒトJumonjiタンパク質のJmjCドメインのアライメントを示す。タンパク質は、Clustal Wアルゴリズムを用いたVector NTIによりアライメントさせた。保存されている残基、類似残基及び同一残基のブロックは、グレーで網掛けしてある。括弧内の数字は、そのタンパク質における残基の番号を示す。
【図17】(S2)は:ヒトJumonjiタンパク質のJmjCドメインのアライメントを示す。タンパク質は、Clustal Wアルゴリズムを用いたVector NTIによりアライメントさせた。保存されている残基、類似残基及び同一残基のブロックは、グレーで網掛けしてある。括弧内の数字は、そのタンパク質における残基の番号を示す。
【図18】(S2)は:ヒトJumonjiタンパク質のJmjCドメインのアライメントを示す。タンパク質は、Clustal Wアルゴリズムを用いたVector NTIによりアライメントさせた。保存されている残基、類似残基及び同一残基のブロックは、グレーで網掛けしてある。括弧内の数字は、そのタンパク質における残基の番号を示す。
【図19】(S2)は:ヒトJumonjiタンパク質のJmjCドメインのアライメントを示す。タンパク質は、Clustal Wアルゴリズムを用いたVector NTIによりアライメントさせた。保存されている残基、類似残基及び同一残基のブロックは、グレーで網掛けしてある。括弧内の数字は、そのタンパク質における残基の番号を示す。
【図20】(S3):Hisタグ化組換えGASC1、GASC1(H190G/E192A)変異体並びにJMJD2A及びJMJD2bの精製。コバルト親和性カラムから溶離したフラクション1-4をSDS-PAGEとCoomassie染色に付した。フラクションF2については、サイズ排除クロマトグラフィーにより更に精製して、高純度なタンパク質試料を得た。
【図21】(S4):GASCは、H3K9me3とH3K9me2を脱メチル化する。aは、可変量の組換えGASC1と基質としてオリゴヌクレオソーム又はバルクヒストンを用いた脱メチル化アッセイを示す。bは、組換えGASC1と基質としてバルクヒストンを用いた脱メチル化アッセイを示す。ヒストンを、図示するように可変量のHis-GASC1と共に、その補因子の存在下で、37℃で30分間インキュベートし、ウェスタンブロッティング解析により解析した。60℃で5分間かけて熱失活(失活)させ、GASC1の脱メチル化活性を無力化させた。
【図22】cは、組換えGASC1と基質として合成トリメチル化H3K9ペプチドを用いた脱メチル化アッセイを示す。ペプチドは、10μgのHis-GASC1と共に補因子の存在下で、37℃で30分間インキュベートした。dは、組換えGASC1若しくはJMDM2aと、基質として合成トリメチル化、ジメチル化及びモノメチル化H3-K9ペプチドを用いた脱メチル化アッセイを示す。ペプチドは、8μgのHis-GASC1若しくはHis-JMJD2aと共に補因子の存在下で、37℃で30分間インキュベートし、ウェスタンブロッティング解析により解析した。
【図23】eは、組換えJMDM2aと、基質として合成トリメチル化、ジメチル化及びモノメチル化H3-K9ペプチドを用いた脱メチル化アッセイを示す。ペプチドは、8μgのHis-JMJD2aと共に補因子の存在下で、37℃で30分間インキュベートし、ウェスタンブロッティング解析により解析した。
【図24】(S5):JMJD2A及びJMJD2Bは、H3-K9がトリメチル化及びジメチル化されたヒストンを脱メチル化する。aは、サイズ排除クロマトグラフィーによる組換えJMJD2Aの精製を示す。1mlのフラクションを回収し、10μlの材料をバルクヒストンと共に37℃で30分間インキュベートした。脱メチル化活性は、トリメチル化H3-K9に対するブロッティングにより評価した。b、組換えJMJD2Aと基質としてバルクヒストンを用いた脱メチル化アッセイ。サイズ排除クロマトグラフィーで得られた種々のフラクション(図S5a)をウェスタンブロット解析により解析した。
【図25】cは、サイズ排除クロマトグラフィーによる組換えJMJD2Bの精製。1mlのフラクションを回収し、10μlの材料をバルクヒストンと共に37℃で30分間インキュベートした。脱メチル化活性は、トリメチル化H3-K9に対するブロッティングにより評価した。
【図26】(S6)は、野生型及びH190G/E192A変異型GASC1のサイズ排除クロマトグラフィーを示す。
【図27】(S7)は、α-ケトグルタレート類似体並びにコバルト塩及びニッケル塩は、GASC1の脱メチル化活性を阻害する。組換えGASC1と基質としてバルクヒストンを用いた脱メチル化アッセイ。ヒストンは、His-GASC1と共に、補因子の存在下で、(a)CoCl2若しくは(b)NiSO4のいずれかの存在下又は不存在下で37℃で30分間インキュベートし、ウェスタンブロット解析により解析した。
【図28】(S5):GASC1の異所発現は、H3K4及びH3K27のメチル化に影響しない。aは、免疫蛍光により評価したGASC1を過剰発現するU2OS細胞内のH3K4トリメチル化状態を示す。
【図29】bは、免疫蛍光により評価したGASC1を過剰発現するU2OS及びTig-3細胞内のH3K27トリメチル化状態を示す。
【図30】(S9):ヘテロクロマチンモデリング/可塑性におけるGASC1の推定上の役割。aは、HAタグ化JMJD2a及びJMJD2bでトランスフェクトしたU2OS細胞の共焦点顕微鏡観察を示す。白色の矢印は、HAタグ化GASC1を発現する細胞を示す。JMJD2a及びJMJD2bを過剰発現する細胞では、HP1βが非局在化している。bは、ヘテロクロマチンのモデリング及び可塑性におけるGASC1の役割のモデルを示す。GLP、G9a及びSUV39H1/H2等のH3K9特異的なヒストンメチルトランスフェラーゼ(HMT)のクロマチンに対しての採用により、H3K9マークの進行性のメチル化が生じる。最初に、H3K9は、酵素GLP、G9aによりモノメチル化される可能性がある。SUV39H1/H2等の前進型HMTは、次にジメチル化及びトリメチル化H3K9を生成するように作用し、HP1の結合を可能にする。次に、HP-1結合により、更にHMTが補充され、ヘテロクロマチンが広がることになる。GASC1は、ジメチルマーク及びトリメチルマークを取り除いてHP1結合を妨害することにより、ヘテロクロマチンの拡大と形成に反対する作用を及ぼしている可能性がある。GASC1は、LSD146及びJMJD2a、b等の他のデメチラーゼと協調して、ヘテロクロマチンに反対の作用を及ぼしている可能性がある。
【図31】(S10)は種々のGASC1欠失変異体のH3K9me3脱メチル化活性を示す。aは、本試験で用いた作製されたGASC1欠失変異体を表す図であり、括弧内の数字は、欠失した残基を示す。bは、免疫蛍光で評価した野生型GASC1とGASC欠失変異体の脱メチル化活性を示す。(S11):ヒストンデメチラーゼのJMJD2ファミリーの発現は、前立腺癌腫において上昇している。aは、Lapointe et al (2004) PNAS 101:811-816に記載されるデータを示す。この試験にでは、62種類の初代前立腺腫瘍(PC、61種類の腺癌と1種類の腺様嚢胞癌)、41種類の対応する正常前立腺組織(NP、前立腺の非癌性領域に由来する)、及び9種類の非対応(即ち、異なる被験者の)骨盤リンパ節転移(LNM)が含まれる。26,260種類の異なるヒトの遺伝子を含有するcDNAマイクロアレイ(UniGene clusters)を用いて遺伝子発現プロファイリングを行っている。この試験の病理学的データと臨床データを含む更なる詳細については、Oncomine(www.oncomine.org)又はPNASのウェブサイト(www.PNAS.org)から入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。bは、Yu et al J Clin Oncol (2004) 22:2790-2799に記載されるデータを示す。この試験には、多様な年齢の男性に由来するヒトサンプルが含まれ、前立腺腫瘍(PC、正常な前立腺腺房細胞は全く含まれていない)及び正常な前立腺組織(NP、腫瘍に隣接する部位由来であるが、腫瘍細胞は含まない)が含まれていた。Affymetrix社製(サンタクララ、カリフォルニア州)のU95a、U95b及びU95cのチップセット(37,777種類の遺伝子及び発現配列タグ)を用いて、遺伝子発現プロファイリングを行っている。この試験の更なる詳細は、Oncomineウェブサイト(www.oncomine.org)にて入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。
【図32】(S10)は種々のGASC1欠失変異体のH3K9me3脱メチル化活性を示す。aは、本試験で用いた作製されたGASC1欠失変異体を表す図であり、括弧内の数字は、欠失した残基を示す。bは、免疫蛍光で評価した野生型GASC1とGASC欠失変異体の脱メチル化活性を示す。(S11):ヒストンデメチラーゼのJMJD2ファミリーの発現は、前立腺癌腫において上昇している。aは、Lapointe et al (2004) PNAS 101:811-816に記載されるデータを示す。この試験にでは、62種類の初代前立腺腫瘍(PC、61種類の腺癌と1種類の腺様嚢胞癌)、41種類の対応する正常前立腺組織(NP、前立腺の非癌性領域に由来する)、及び9種類の非対応(即ち、異なる被験者の)骨盤リンパ節転移(LNM)が含まれる。26,260種類の異なるヒトの遺伝子を含有するcDNAマイクロアレイ(UniGene clusters)を用いて遺伝子発現プロファイリングを行っている。この試験の病理学的データと臨床データを含む更なる詳細については、Oncomine(www.oncomine.org)又はPNASのウェブサイト(www.PNAS.org)から入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。bは、Yu et al J Clin Oncol (2004) 22:2790-2799に記載されるデータを示す。この試験には、多様な年齢の男性に由来するヒトサンプルが含まれ、前立腺腫瘍(PC、正常な前立腺腺房細胞は全く含まれていない)及び正常な前立腺組織(NP、腫瘍に隣接する部位由来であるが、腫瘍細胞は含まない)が含まれていた。Affymetrix社製(サンタクララ、カリフォルニア州)のU95a、U95b及びU95cのチップセット(37,777種類の遺伝子及び発現配列タグ)を用いて、遺伝子発現プロファイリングを行っている。この試験の更なる詳細は、Oncomineウェブサイト(www.oncomine.org)にて入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。
【図33】(S10)は種々のGASC1欠失変異体のH3K9me3脱メチル化活性を示す。aは、本試験で用いた作製されたGASC1欠失変異体を表す図であり、括弧内の数字は、欠失した残基を示す。bは、免疫蛍光で評価した野生型GASC1とGASC欠失変異体の脱メチル化活性を示す。(S11):ヒストンデメチラーゼのJMJD2ファミリーの発現は、前立腺癌腫において上昇している。aは、Lapointe et al (2004) PNAS 101:811-816に記載されるデータを示す。この試験にでは、62種類の初代前立腺腫瘍(PC、61種類の腺癌と1種類の腺様嚢胞癌)、41種類の対応する正常前立腺組織(NP、前立腺の非癌性領域に由来する)、及び9種類の非対応(即ち、異なる被験者の)骨盤リンパ節転移(LNM)が含まれる。26,260種類の異なるヒトの遺伝子を含有するcDNAマイクロアレイ(UniGene clusters)を用いて遺伝子発現プロファイリングを行っている。この試験の病理学的データと臨床データを含む更なる詳細については、Oncomine(www.oncomine.org)又はPNASのウェブサイト(www.PNAS.org)から入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。bは、Yu et al J Clin Oncol (2004) 22:2790-2799に記載されるデータを示す。この試験には、多様な年齢の男性に由来するヒトサンプルが含まれ、前立腺腫瘍(PC、正常な前立腺腺房細胞は全く含まれていない)及び正常な前立腺組織(NP、腫瘍に隣接する部位由来であるが、腫瘍細胞は含まない)が含まれていた。Affymetrix社製(サンタクララ、カリフォルニア州)のU95a、U95b及びU95cのチップセット(37,777種類の遺伝子及び発現配列タグ)を用いて、遺伝子発現プロファイリングを行っている。この試験の更なる詳細は、Oncomineウェブサイト(www.oncomine.org)にて入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。
【技術分野】
【0001】
本発明は、JmjCドメイン含有タンパク質の活性調節剤として作用可能であり、そしてゲノムの不安定性が発症に関与する疾患の予防及び/又は治療に有用な化合物の同定方法に関する。
【背景技術】
【0002】
ヒストンテールのリジン及びアルギニン残基のメチル化は、転写活性が高いクロマチンと低いクロマチンを表す重要なエピジェネティックマーカーである。例えば、ヒストンH3(H3-K9)の第9番リジンのメチル化は、エピジェネティックな要因により不活性なクロマチンと関連している1-3。
【0003】
アセチル化やリン酸化といった他のヒストン修飾とは対照的に、メチル化は、N-CH3結合の高い熱力学的安定性により不可逆的であると考えられてきた。しかしながら、最近において、Shiとその同僚4は、核内のアミンオキシダーゼのホモログであるタンパク質LSD1が、モノメチル化及びジメチル化H3-K4に非常に高い特異性を示すヒストンデメチラーゼであることを同定した。この酵素は、その基質を、アミンオキシダーゼ反応を通じて脱メチル化する。しかしながら、LSD1は、トリメチル化リジンH3-K4を脱メチル化することができず、これは、最も恐らくは、酸化に必要なプロトン化窒素が存在していないことが原因であると考えられる4。本願発明に係る研究を完成する直前に、Tsukada et al.は、Jumonjiタンパク質のFBXL11 (JHDM1a)が、ジメチルH3-K36をFe(II)の存在下でα-ケトグルタレート依存的に特異的に脱メチル化できることを示した5。少なくとも理論上は、FBXL11媒介性の脱メチル化の反応機構は、トリメチルH3-K36を基質として用いることが可能であるが、このような活性は実証できていない5。LSD1及びFBXL11をヒストンデメチラーゼとして同定したことは、メチル基マークの動的な制御を示しており、エピジェネティック研究に重要な軌跡を残すことになるが、彼らは、トリメチル化リジンマークの可逆性の問題は解明していない。
【0004】
SUV39H1ノックアウトマウスの研究で報告されたように、H3-K9のトリメチルバリアントの喪失により、染色体異常が生じ、癌に罹患しやすくなる。故に、このマークを逆行させることが可能な酵素が長年探索されてきたが、その存在が疑問視されていた。後者の見方は、トリメチル化H3-K9が、「非常に安定な遺伝性のクロマチン状態」であるヘテロクロマチンの発生と維持に必要とされるという事実によって、更に強められた。このような酵素とその活性阻害剤の同定により、癌の予防と治療に対する新たなアプローチが提供されるであろう。
【発明の開示】
【0005】
本発明は、試験化合物の、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質に結合し、そして任意にその活性を調節する能力を試験する方法であって、試験化合物をJumonjiタンパク質のJMJD2サブファミリーのタンパク質、前記タンパク質の補因子、及び任意で脱メチル化の基質と共にインキュベートすることを含む方法を提供する。
【0006】
本発明の現時点で好ましい態様においては、前記方法は、以下の工程:
i) 鉄及びα-ケトグルタレートを含む溶液で脱メチル化の基質の懸濁液を調製すること;
ii) 前記試験化合物を、前記懸濁液に添加すること;
iii) ii)で得られた前記懸濁液に、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質を添加すること;
iv) 前記懸濁液をインキュベートすること;並びに
v) 前記基質の脱メチル化の程度を測定すること
を含む。
【0007】
定義
「核酸分子」という用語は、リボ核酸(RNA)若しくはデオキシリボ核酸(DNA)のオリゴマー又はポリマー、又はその模倣体を指す。この用語には、天然の核酸塩基、糖及びヌクレオシド間(バックボーン)の共有結合から構成される分子、並びに同様の機能を有する非天然の核酸塩基、糖及びヌクレオシド間(バックボーン)の共有結合から構成される分子、又はこれらの組合せが含まれる。このような修飾を受けた又は置換された核酸は、例えば、向上した細胞内取り込み、核酸ターゲットへの向上した親和性、並びにヌクレアーぜ及び他の酵素の存在下での増大した安定性等の所望の特性のために天然型よりも好まれる場合があり、本文においては、「核酸類似体」又は「核酸模倣体」の用語で記載される。核酸模倣体の好ましい例には、ペプチド核酸(peptide nucleic acid:PNA)、Locked核酸(Locked Nucleic Acid:LNA)、xylo-LNA、ホスホロチオエート、2'-メトキシ-、2'-メトキシエトキシ-、モルホリノ-及びホスホロアミデートを含有する分子等が挙げられる。
【0008】
「ポリヌクレオチド配列」(例えば、核酸、ポリヌクレオチド、オリゴヌクレオチド等)は、文脈に応じて、ヌクレオチドA、C、T、U、G、若しくは他の天然ヌクレオチド、若しくは人工のヌクレオチド類似体を含むヌクレオチドのポリマーであるか、又は核酸を表す文字列である。いずれかの特定のポリヌクレオチド配列から、所定の核酸又は相補的な核酸のいずれかを決定できる。
【0009】
本発明に関しては、「ポリペプチド」という用語は、完全長タンパク質、オリゴペプチド、短鎖ペプチド及びこれらの断片を含み、アミノ酸残基が共有結合により結合したあらゆる長さのアミノ酸鎖を指す。本明細書及び請求項における全てのポリペプチド配列は、明確な記載がない場合には、慣習的なフォーマットに従い、N末端からC末端の方向へ記載される。
【0010】
任意のアミノ酸ポリマー若しくはヌクレオチドポリマーの番号付けが、選択されたアミノ酸ポリマー若しくは核酸ポリマーの番号に「対応する」又は「関連する」のは、任意のポリマーの成分(例えば、アミノ酸、ヌクレオチド、又は一般的に「残基」と呼ばれるもの)の位置が、その任意のポリマーの成分の実際の番号順の位置ではなく、選択されたアミノ酸ポリマー又はヌクレオチドポリマーの同一又は同等の位置を参照して定められている場合である。従って、例えば、任意のポリペプチド配列の任意のアミノ酸の位置の番号付けは、参照配列として用いられる選択されたポリペプチド配列の同一又は同等のアミノ酸の位置に対応する。
【0011】
「変異体」は、1又は2か所以上のアミノ酸部位において、親ポリペプチドの配列と異なる配列(アミノ酸の欠失、アミノ酸の挿入、及び/又は異なるアミノ酸へのアミノ酸の置換による)を含むポリペプチドである。変異配列は、非天然配列、即ち、自然界に見出されない配列であってもよい。
【0012】
本明細書において、「合成ペプチド」という用語は、in vitroで合成された短鎖ペプチドを含むペプチドを指す。また、この用語には、通常見られないアミノ酸又は非天然アミノ酸を用いた置換により修飾されたペプチド又は短鎖ペプチドも包含される。
【0013】
目的語に付される「天然」とは、その目的語にかかるものが、人間によって人工的に作製されたものとは異なるものとして自然界に見出すことができるという事実を表す。例えば、自然界の供給源から単離可能な生物(ウィルス、細菌、原生動物、昆虫、植物、又は哺乳動物の組織を含む)中に存在し、実験室において人間による意図的な修飾を受けていないポリペプチド又はポリヌクレオチドの配列は、天然のものである。目的語に付される「非天然」とは、その目的語にかかるものが天然のものではなく、即ち、人間により人工的に作製されたものと異なるものとして自然界に見出すことができないことを意味する。
【0014】
「断片」又は「部分配列」とは、所定の配列のいずれかの部分を指す。配列の断片又は部分配列は、その配列自体よりも少なくとも1アミノ酸残基又は1核酸残基は短くなることが理解されるべきである。従って、断片又は部分配列は、それぞれ、より長いアミノ酸配列(例えば、ポリペプチド)又は核酸配列(例えば、ポリヌクレオチド)の一部を含むアミノ酸配列又は核酸配列を指す。
【0015】
「配列同一性」という用語は、同一の長さの2つのアミノ酸配列間又は2つの核酸配列間のホモロジー度の定量的測定値を示す。比較すべき2つの配列が同一の長さにない場合、これらは、可能な限り最適な一致性を示すようにアライメントする必要があり、ギャップが挿入されたり、又はポリペプチド配列若しくはヌクレオチド配列が末端で切断されたりする。配列同一性は、以下の式によって計算可能であり
【数1】
式中、Ndifは、アライメントした際の、2つの配列の同一でない残基の総数であり、Nrefは、一方の配列の残基数である。従って、AGTCAGTCというDNA配列は、AATCAATCというDNA配列と75%の配列同一性を有することになる(Ndif=2及びNref=8)。ギャップについては、具体的な残基の非同一性としてカウントされ、即ち、AGTGTCというDNA配列は、AGTCAGTCというDNA配列と75%の配列同一性を有することになる(Ndif=2及びNref=8)。
【0016】
本発明に係る全てのポリペプチド又はアミノ酸に基づく態様では、1以上の配列間の配列同一性のパーセンテージは、clustalWソフトウェア(http:/www. ebi.ac.uk/clustalW/index. html)により、そのプログラムのデフォルト設定を用いて行われる個々の配列のアライメントに基づく。これらの設定は、以下の通りである:Alignment=3Dfull, Gap Open 10.00, Gap Ext. 0.20, Gap separation Dist. 4, Protein weight matrix: Gonnet。本発明に係るヌクレオチドに基づく態様に関しても、1以上の配列間の配列同一性のパーセンテージは、clustalWソフトウェアをデフォルト設定で用いて行われるアライメントに基づく。ヌクレオチド配列のアライメントにおいては、これらの設定は以下の通りである:Alignment=3Dfull, Gap Open 10.00, Gap Ext. 0.20, Gap separation Dist. 4, DNA weight matrix: identity (IUB)。
【0017】
本文においては、「相補的配列」とは、ストリンジェントな条件下で、本発明の核酸分子にハイブリダイズする核酸配列を指す。「ストリンジェントな条件」は、高いストリンジェンシーを有する一般的な条件を指す。「ストリンジェンシー」という用語は、当該技術分野で良く知られており、核酸のハイブリダイゼーションが行われる条件(温度、イオン強度、及び有機溶媒等の他の化合物の存在)を参照する際に用いられる。「高ストリンジェンシー条件」では、ストリンジェンシーが「弱い」若しくは「低い」条件と比較して、高頻度に相補的な塩基配列を有する核酸フラグメント間でのみ核酸塩基のペア形成が生じる。
【0018】
例えば、高ストリンジェンシーなハイブリダイゼーション条件には、以下が含まれる:(1)洗浄における低イオン強度と高温、例えば、0.1%のドデシル硫酸ナトリウム(SDS)を含む0.015MのNaCl/0.0015Mのクエン酸ナトリウム、pH7.0(0.1xSSC)、50℃;(2)5×Denhardt's溶液(0.1%(重量/体積))高純度ウシ血清アルブミン/0.1%(重量/体積)Ficoll/0.1%(重量/体積)ポリビニルピロリドン)を含む50体積%のホルムアミド、50mMのリン酸ナトリウムバッファー、pH6.5及び5×SSCで42℃でのハイブリダイゼーション;又は(3)50%ホルムアミド、5×SSC、50 mMのリン酸ナトリウム(pH6.8)、0.1%のピロリン酸ナトリウム、5×Denhardt's溶液、超音波処理済みサケ精子DNA(50μg/ml)、0.1%のSDS、及び10%の硫酸デキストラン中で42℃でのハイブリダイゼーション、並びに0.2×SSC及び0.1% SDSで42℃での洗浄。
【0019】
「単離した核酸」という用語は、本発明の核酸が由来する生命体の天然のゲノムでは隣接している(例えば、5'末端で一端が、3'末端で一端が隣接する)両コーディング配列と隣接していない核酸(例えばDNA又はRNA)を指す場合がある。従って、この用語には、例えば、cDNA又はポリメラーゼ連鎖反応(PCR)若しくは制限酵素処理により得られたゲノムDNAの断片が含まれ、このようなcDNA又はゲノムDNA断片が、ベクターに組み込まれているか、これらが元々取り出された生命体と同一若しくは異なる種、例えば、ウィルス等のゲノムに一体化されているか、更なるコーディング配列に連結してキメラポリペプチドをコードするハイブリッド型遺伝子を形成するか、又は他のDNA配列から独立しているかどうかは関係がない。前記DNAは、2本鎖又は1本鎖であってもよく、センス鎖又はアンチセンス鎖であってもよい。
【0020】
「組換えポリヌクレオチド」又は「組換えポリペプチド」とは、それぞれ、複数の供給源である核酸若しくはポリペプチドに由来する核酸配列若しくはアミノ酸配列を含む非天然のポリヌクレオチド又はポリペプチドであり、その供給源である核酸又はポリペプチドは、天然の核酸若しくはポリペプチドであってもよく、又は、それ自体が変異導入若しくは他の種類の修飾を受けたものであってもよい。核酸又はポリペプチドは、人工物であるか若しくは組換え操作を受けている場合、又は人工物であるか若しくは組換え操作を受けたポリペプチド若しくは核酸に由来する場合には、「組換え体」とみなされる場合がある。組換え核酸(例えばDNA又はRNA)は、典型的に一緒に含まれていないか、典型的には互いに関連していないか、又は典型的には互いに分離された状態にある、少なくとも2つの配列セグメントの組合せ(例えば、人工的組合せ)により作製可能である。組換え核酸は、異なる供給源に由来する核酸セグメント及び/又は人工的に合成された核酸セグメントを一緒に連結させるか又は組み合わせることにより形成される核酸分子を含んでいてもよい。「組換えポリペプチド」(又は「組換えタンパク質」)は、しばしばクローニング又は組換え操作された核酸若しくは遺伝子から生じたポリペプチド(又はタンパク質)を指す。異なる核酸配列又はアミノ酸配列が由来する供給源であるポリヌクレオチド又はポリペプチドは、しばしば、相同的であり(即ち、同一又は類似の構造及び/又は機能を有するか、又はこれを有するポリペプチドをコードする)、例えば、しばしば、生命体の異なる単離物、血清型、株、種、又は異なる疾患状態から得られる。
【0021】
「組換え」という用語は、例えば、細胞、ヌクレオチド、ベクター、タンパク質、又はポリペプチドに言及するものとして用いられる場合、典型的には、その細胞、ヌクレオチド又はベクターが、異種の(又は外来性の)核酸の導入若しくは天然の核酸の変更により修飾されていること、又はそのタンパク質若しくはポリペプチドが異種のアミノ酸の導入により修飾されていること、又はその細胞が、その様な修飾を受けた細胞に由来することを示す。組換え細胞は、その細胞の天然(非組換え)型に見出されない核酸配列(例えば、遺伝子)を発現するか、又は、異常に発現しているか、発現が抑制されているか、若しくは全く発現されていない天然の核酸配列(例えば、遺伝子)を発現する。「組換え」という用語は、細胞に言及するものとして用いられる場合、その細胞が異種の核酸を複製するか、又は異種の核酸によりコードされるペプチド若しくはタンパク質を発現することを示す。組換え細胞は、その細胞の天然(非組換え)型に見出されない遺伝子を含んでいてもよい。組換え細胞は、その細胞の天然型に見出される遺伝子を含んでいてもよいが、その場合、その遺伝子は、人工的手段により修飾されてその細胞内に再導入される。前記用語には、その細胞に内在性の核酸であって、その細胞からその核酸を取り出すことなしに修飾された核酸を含む細胞が包含される;このような修飾には、遺伝子置換、部位特異的変異導入及び関連技術により得られるものが含まれる。
【0022】
「組換えにより作製された」という用語は、化学合成手段、核酸セグメントの配列組換えの繰り返し若しくはヌクレオチドの他の種々の作製方法(例えば、シャッフリング)、又は単離された核酸セグメントの、例えば当業者に知られる遺伝子組み換え技術による操作により通常達成される人為的な組換えを指す。「組換え発現された」とは、典型的にはin vitroで組換え核酸を作製し、in vivo、in vitro、又はex vivoで、その組換え核酸を、これを発現又は増幅可能な細胞内へ導入するための技術を指す。
【0023】
ここで用いられる「保存的」とは、(i)その変化が、可能な限り構造的に中性であること、即ち、天然タンパク質と比較して、その変異ペプチド又はポリペプチドの3次構造の変化が最小になるように設計されていることを意味する。構造的に中性であることは、生物活性を保存するために望ましい。当業者に、構造的に中性となる可能性の高い変異を作製するための手引きとなり得る法則が存在する(例えば、(77)及び(78)を参照)。より重要な法則のいくつかには、以下が含まれる:(1)物理化学的に類似する残基、即ち、同義の残基の置換では、置換により挿入されるアミノ酸は、置換されるアミノ酸と同様の構造的役割を果たすことが可能なので、構造変化が起こりにくい;そして(2)進化の過程で保存されている場合、その配列が機能上重要である可能性が示唆されることから、進化の過程で保存されている配列を変化させると、有害な構造上の効果を生じる恐れがある。
【0024】
本明細書の本章又は以下の残りの記載において特に定義されていない限り、ここで使用する全ての科学技術用語は、本発明が属する技術分野の当業者に共通して理解される意味と同様の意味を有する。
【発明を実施するための最良の形態】
【0025】
本発明は、JmjCドメイン含有タンパク質の活性調節剤として作用可能な化合物が、ゲノムの不安定性が発症に関与する疾患の予防及び/又は治療に有用であるという発見に基づく。特に、JmjCドメイン含有タンパク質の調節剤は、癌の予防及び/又は治療に有用である。更に本発明は、Jumonjiタンパク質;ヒストンH3のジメチル化及びトリメチル化リジン9(H3-K9)と相互作用できるタンパク質として、扁平上皮癌1で増幅される遺伝子(GASC1)、の同定に関する。GASC1は、JumonjiファミリーのJMJD2サブファミリーに属する。本発明の発明者らは、JMJD2ファミリーのタンパク質が、トリメチル化及びジメチル化H3-K9を脱メチル化させるヒストンデメチラーゼであることを実証した。更に、本発明者らは、GASC1とJMJD2メンバーの異所発現が、in vivoで、トリメチル化及びジメチル化H3-K9を劇的に減少させ、モノメチル化H3-K9を増加させ、HP1を非局在化させ、ヘテロクロマチンを減少させることも証明した。従って、最初に同定されたヒストントリメチルデメチラーゼであることに加え、JumonjiファミリーのJMJDサブファミリーのタンパク質も、癌の発症に関与している。従って、これらの酵素の触媒活性の阻害剤は、新規な抗ガン治療法の開発のための重要な候補となる。
【0026】
従って、本発明は、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性を調節可能な化合物の同定、スクリーニング、キャラクタリゼーション又は設計の方法に関する。本発明の方法を、多数の化合物のスクリーニングに用いて、特定の癌、特に前立腺癌の治療に臨床的に使用する候補化合物となる化合物群を同定することが可能である。スクリーニングアッセイで活性を示さない他の化合物については、候補化合物としての更なる検討から除外できる。従って、本発明の方法は、製薬産業において有用性を有する。
【0027】
本発明によれば、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質のその基質(ヒストンペプチド、ヒストン又はヌクレオソーム)又は補因子(α-ケトグルタレート又は鉄)への結合を阻害する化合物を、競合結合アッセイにおいて同定できる。また、これとは別に、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質、又はその基質若しくは補因子への試験化合物の結合を直接測定することもできる。後者のタイプのアッセイは、直接結合アッセイと呼ばれる。競合結合アッセイと直接結合アッセイは、両方とも、当該技術分野で一般的に知られるイムノアッセイ及びレセプター結合アッセイで用いられるフォーマットと類似する種々の異なるフォーマットで使用可能である。競合結合アッセイ及び直接結合アッセイを含む結合アッセイの異なるフォーマットの記載については、Basic and Clinical Immunology 7th Edition (D. Stites and A. Terr ed.) 1991; Enzyme Immunoassay, E. T. Maggio, ed., CRC Press, Boca Raton, FIa. (1980);及び"Practice and Theory of Enzyme Immunoassays," P. Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology, Elsevier Science Publishers B. V. Amsterdam (1985) を参照のこと。各参考文献は、本明細書に援用される。
【0028】
例えば、競合結合アッセイでは、試験化合物は、固体表面に結合された結合物質上の特異的な結合部位について、標識された検体と競合する。この種のフォーマットでは、標識された検体は、ヒストンペプチドで標識可能であり、結合物質は、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質であってもよく、これが固相に結合されることになる。これに代わり、標識された検体が、標識されたJMJD2タンパク質であってもよく、結合物質が、固相化されたヒストンペプチドであってもよい。捕捉物質に結合する標識検体の濃度は、結合アッセイにおいて試験化合物が競合する能力と反比例する。試験化合物による標識検体の阻害の量は、その結合アッセイの条件、並びに使用する結合物質、標識検体、及び試験化合物の濃度に依存する。具体的なアッセイ条件のもとでは、その競合結合アッセイにおいて、結合物質への標識検体の結合量が10%以上低下した場合には、試験化合物は、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質への基質(即ち、ヒストンペプチド)又は補因子の結合を阻害できるとみなされる。直接結合アッセイを用いる場合、バックグラウンドのレベルの2倍以上のシグナルが測定される場合には、試験化合物は、JMJD2酵素への基質(即ち、ヒストンペプチド)又は補因子(即ち、α-ケトグルタレート)の結合を阻害しているものとみなされる。
【0029】
競合結合アッセイでは、サンプル化合物は、特異的な結合物質への結合において標識タンパク質と競合する。上述した通り、結合物質を固体表面に結合させることで、結合していない標識タンパク質から結合した標識タンパク質を分離することができる。これに代わり、競合結合アッセイを液相で行い、当該技術分野で知られる種々の技術のいずれかを用いて、結合していない標識タンパク質から結合した標識タンパク質を分離することもできる。分離した後に、結合した標識タンパク質の量を測定する。サンプル中に存在するタンパク質の量は、結合した標識タンパク質の量に反比例する。
【0030】
上記に代わり、相同的結合アッセイ(homogenous binding assay)を行うことも可能であり、このアッセイでは分離工程を要しない。これらの種類の結合アッセイでは、タンパク質上の標識が、タンパク質のその特異的な結合物質への結合により変化する。この標識タンパク質の変化により標識から放出されるシグナルが減少又は増加し、結合測定の終了時に標識を測定することにより、そのタンパク質を検出又は定量することが可能になる。
【0031】
ここに記載される結合アッセイフォーマットは、標識されたアッセイ成分をもちいる。標識は、種々の形態をとることが可能である。標識は、当該技術分野で知られる方法に従って、アッセイの所望の成分に直接的又は間接的に結合できる。多種多様の標識を用いることができる。成分は、種々の方法のいずれによっても標識できる。
【0032】
従来の方法では、3H、125I、35S、14C、又は32Pが組み込まれた放射性標識が用いられる。非放射性標識には、標識した抗体に結合するリガンド、フルオロフォア、化学発光剤、酵素、及び標識されたリガンドの特異的なペア形成剤として機能できる抗体が含まれる。標識の選択は、要求される感度、化合物への結合の容易性、安定性の要求、及び利用可能な機器に依存する。使用可能な種々の標識方法又はシグナル発生システムの総説については、米国特許第4,391,904号(参照により本明細書に組み込まれる)を参照。
【0033】
「ヒストンペプチド」及び「基質」という用語は、JMJD2タンパク質に結合するヒストンの断片を指す。スクリーニングアッセイで用いるJMJD2タンパク質とヒストンタンパク質の製造方法は、当業者に知られている。
【0034】
前記方法は、特に、試験化合物がJumonjiタンパク質のJMJD2サブファミリーのタンパク質に結合し、随意にその活性を調節する調節する能力の試験方法であり、試験化合物を、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質、前記タンパク質の補因子、及び場合により脱メチル化の基質と共にインキュベートすることを含む方法であってもよい。特に、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性を阻害可能な化合物を同定することは、本発明の範囲に含まれる。
【0035】
典型的には、前記方法では、Jumonjiタンパク質のJMJD2サブファミリーのポリペプチド、代表的にはGASC1ポリペプチドを利用する。特に、前記方法では、ヒトGASC1ポリペプチド若しくはこのポリペプチドの断片、又はこれらの変異体を利用する。従って、本発明の重要な態様では、前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質が、以下の:
a)配列番号1に示すアミノ酸配列;
b)配列番号1に示すアミノ酸配列の断片;
c)a)の配列若しくはb)の配列のいずれか、又は両方と少なくとも45%の配列同一性を示すアミノ酸
を含む群から選択される方法が提供される。
【0036】
Jumonjiタンパク質のJMJD2サブファミリーのクローニングされたメンバーには、GASC1(Swiss-Prot accession number Q9H3R0)、並びにそのホモログであるJMJD2a及びJMJD2b(それぞれ、Swiss-Prot accession numbers 075164及び094953)が含まれる。
【0037】
本発明の同程度に重要な態様は、前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質が、以下の:
a)配列番号1(GASC-1(JMJ2Dc))、配列番号2(JMJD2a)、配列番号3(JMJD2b)に示すアミノ酸配列;
b)a)のいずれか1つのアミノ酸配列の断片;
c)a)のいずれか1つの配列及び/又はb)のいずれか1つの配列と少なくとも45%の配列同一性を示すアミノ酸
を含む群から選択される方法に関する。
【0038】
当業者であれば、現時点で知られているJumonjiタンパク質のJMJD2サブファミリーのメンバーは、これらのアミノ酸配列を最適なフィットでアライメントした場合に、約45%の配列同一性を示すことを理解するであろう。従って、本出願では、使用するアミノ酸配列は、Jumonjiタンパク質のJMJD2サブファミリーのいずれのメンバーの配列とも少なくとも45%の同一性を示す。詳細には、前記配列同一性は少なくとも45%、例えば、少なくとも46%、47%、48%、49%、50%、52.5%、55%、57.5%、60%、62.5%、65%、67.5%、70%、72.5%、75%、77.5%、80%、82.5%、85%、87.5%、90%、92.5%、95%、97.5%、98%、98.5%、99%又は99.5%であってもよい。
【0039】
Jumonjiタンパク質のJMJD2サブファミリーの天然ポリペプチドメンバーの機能特性を取り込むためには、前記断片が少なくとも100個のアミノ酸、例えば、少なくとも105、110、115、120、125、130、135、140、145、150、155、160、165、170、175、180、185、190、195、200、210、220、230、240、250、260、270、280、290、300、320、340、360、380、400、420、440、460、480、500、520、540、560、580、600、620、640、660、680、700、750、800、850、900、950、1000個、又は少なくとも1050個のアミノ酸を有することが更に好ましい。
【0040】
好ましくは、前記断片は、少なくとも150個のアミノ酸、例えば、少なくとも151、152、153、154、155、156、157、158、159、160、162、164、166、168、170、172、174、176、178、180、182、184、186、188、190、192、194、196、198又は200個のアミノ酸を有する。
【0041】
本発明は、更に、保存的アミノ酸置換により形成されるアミノ酸配列の類似体も意図している。保存的アミノ酸置換の背後にある原理は、特定のアミノ酸ペアにおいて、一方のアミノ酸を他方で置換した場合にも、そのペプチドの3次構造に最小限の変化しか生じないような相容性の側鎖が存在することである。保存的置換の法則については、以下で説明されている:Bowie et al. Science 247(1990) 1306-1310。本発明の目的の一つは、JMJD2サブファミリータンパク質の基質へ結合する能力を保持するポリペプチド、断片及び変異体を利用することである。いくつかの好ましい態様では、前記ポリペプチド、断片及び変異体は、JMJD2サブファミリータンパク質のα-ケトグルタレート等の補因子へ結合する能力を維持している。
【0042】
必要とされる場合には、前記ポリペプチド、断片及び変異体のそれぞれは、精製された形態又は未精製の形態のいずれかで、例えば、細胞抽出物として、又はこのような抽出物から関連成分を精製することにより提供されてもよい。
【0043】
これに代わり、前記ポリペプチド、断片及び変異体は、組換え発現技術により遺伝子操作で作製し、前記アッセイで使用するために精製することもできる。これに代わり、前記ポリペプチド、断片及び変異体は、細胞ベースのアッセイで用いるために、細胞内で組換え発現することもできる。
【0044】
典型的には、関連成分をコードするポリヌクレオチドは、発現ベクターに組み込まれた状態で提供される。このような発現ベクターは、当該技術分野で通常の技術で組み立てられており、例えば、プラスミドDNA並びに必要とされる可能性のある適切なイニシエーター、プロモーター、エンハンサー及び他の構成成分、例えば、ポリアデニル化シグナルの使用を必要とする場合があり、これらは、完全なタンパク質発現を可能にするため、正しい配向で配置される。適切なベクターは、例えば、本出願の実施例により詳細に記載されるベクターのように、当業者に一瞬で明確に把握されるであろう。プロモーター配列は、選択されるアッセイフォーマットに応じた誘導プロモーター又は構成的プロモーターとなるであろう。
【0045】
Jumonjiタンパク質のJMJD2サブファミリーには、数多くの共通する構造上の特徴が見出される。これらにはJmjN、JmjC、PHD及びTdrの配列が含まれ、これらの1以上は、JMJD2 Jumonjiタンパク質の好ましい特性である。これらの機能ドメインは、そのアミノ酸配列の要となる部位に一定の特定のアミノ酸残基が局在しているという特徴を有する。単に肉眼でタンパク質のアミノ酸を調べただけでは直ちに区別できないかもしれないが、これらのドメインは、SMARTプログラム(http:/smart.embl-heidelberg.de/smart)を用いて決定できる。SMARTプログラムの基礎となるアルゴリズムは、Letunic et al.58とSchultz et al.59に更に詳細に説明されている。
【0046】
従って、本発明の好ましい態様では、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質、並びにポリペプチド、断片及び変異体は、JMJD2aに由来するJmjN、JmjC、PHD及びTUDORのドメインから成る群から選択される1以上のアミノ酸配列を含む:
【化1】
【0047】
本発明の同程度に好ましい態様では、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質並びにポリペプチド、断片及び変異体は、JMJD2bに由来するJmjN、JmjC、PHD及びTUDORのドメインから成る群から選択される1以上のアミノ酸配列を含む:
【化2】
【0048】
現時点で最も好ましい本発明の態様では、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質並びにポリペプチド、断片及び変異体は、JMJD2(GASC1)に由来するJmjN、JmjC、PHD及びTUDORのドメインから成る群から選択される1以上のアミノ酸配列を含む:
【化3】
【0049】
これらのドメインのアミノ酸配列は、その機能を喪失させることなく置換により変更させることが可能なことが理解されるべきである。従って、本発明には、上記の配列のいずれか一つと少なくとも75%の同一性、例えば、上記の配列のいずれかと少なくとも80%、85%、90%、95%、98%、99%の同一性、又は記の配列のいずれかと少なくとも99.5%の同一性を示す1以上のアミノ酸配列を含むJumonjiタンパク質のJMJD2サブファミリーのタンパク質並びにポリペプチド、断片及び変異体が含まれる。
【0050】
Jumonjiタンパク質のJMJD2サブファミリーのタンパク質とその活性の調節剤の候補を、その酵素活性の阻害剤の不存在下でヒストンH3のリジン9を脱メチル化させる条件下で一緒にインキュベートしてもよい。GASC1は、Fe(II)及びα-ケトグルタレートに依存するデメチラーゼであるから、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質は、典型的には、補基質の存在下で基質と接触する。典型的には、α-ケトグルタレート、Fe(II)及びアスコルビン酸が補基質として用いられるが、他の補基質の使用も本発明の範囲に含まれる。
【0051】
当業者はJumonjiタンパク質のJMJD2サブファミリーのタンパク質、並びに上記のポリペプチド、断片及び変異体の活性を測定するにあたって、種々のアプローチを利用可能であり、これらのアプローチには、利用された基質及び/又は補基質の測定並びに形成された生成物及び/又は副生成物の測定が含まれる。従って、本発明の特定の態様では、前記方法は、以下の追加の工程:
a)試験サンプルの以下のいずれかのパラメーターを測定すること:
i)基質のメチル化の状態;
ii)ホルムアルデヒドの放出;
iii)α-ケトグルタレート基質補因子のカルボキシル化の状態;
iv)酸素消費量
b)工程a)で得られた試験サンプルについての数値を、対照サンプルについて得られた数値と比較し、それによって、その試験化合物の、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性を調節する能力を測定すること
を含む。
【0052】
更に、本発明の方法には、上述の要素による下流への効果の測定を組み込むこともできる。特に、GASCの作用は、特定の遺伝子の転写に影響を与える可能性があることから、GASC活性を測定するためにジーンレポーターアッセイを用いることも意図できる。このようなアッセイは、天然状態でJumonjiタンパク質のJMJD2サブファミリーのタンパク質によって抑制されている遺伝子の転写活性の測定に基づくものとなるであろう。更に、メチル化を受けたプロモーターと転写物において結合されるレポーター遺伝子を含む遺伝子操作されたレポーターコンストラクトの使用に基づくin vitroアッセイも意図できる。
【0053】
特定の態様では、これらのレポーターアッセイでは、蛍光レポーターをコードする遺伝子の使用を伴ってもよい;現時点で最も一般的に使用される蛍光レポーターは、緑色蛍光タンパク質(Green Fluorescent Protein:GFP)である。
【0054】
測定は、定量的又は定性的なものでよい。定量的測定及び定性的測定のいずれも、上に示すように、対照の存在下で行うことができる。対照を用いる場合、使用するアッセイ方法にもよるが、以下の工程:
a)Jumonjiタンパク質のJMJD2サブファミリーのタンパク質と脱メチル化の基質を、脱メチル化が可能な条件下でインキュベートすること;
b)試験サンプルの以下のパラメーター:
i)基質のメチル化の状態;
ii)試験サンプル中のホルムアルデヒドの放出;
iii)α-ケトグルタレート基質補因子のカルボキシル化状態
iv)酸素消費量
をモニターすること
により測定を行うことができる。
【0055】
本発明のいくつかの態様では、インキュベート時間と基質のメチル化状態及び/又はホルムアルデヒドの放出を用いて脱メチル化の速度を測定し、これを比較に用いる。
【0056】
GASC1の天然基質がヒストンH3であることから、本発明のいくつかの態様においては、脱メチル化の基質には、ヒストンH3のリジン9に対応するメチル化部位が含まれることが理解されるであろう。本発明の特定の態様では、脱メチル化の基質は、配列番号22に示すアミノ酸配列(TARKSTG)を含むペプチドである。
【0057】
前記アミノ酸配列(配列番号22)のヒストンH3のリジン9に対応するリジン残基はメチル化されている。本発明の大部分の態様において、前記リジン残基は、トリメチル化されていることが好ましいが、前記リジン残基が、ジメチル化又はモノメチル化されていたとしてもアッセイは行うことができる。
【0058】
現時点においては、基質としては、ペプチドを用いることが好ましく、例えば、ヒストンH3のN末端残基6-12、例えば、ヒストンH3の残基5-9、4-10、3-10、2-11、1-12、1-13、1-14、1-15、1-20、1-25、1-30、1-35、1-40、1-45、1-50、1-55、1-60、1-65、1-70、1-75、1-80、1-85、1-90、1-95又は1-100を含む合成ペプチド等を用いることが好ましい。本願においては、ヒストンH3の完全長配列が、配列番号23に示されている。
【0059】
上の脱メチル化基質の説明に従えば、前記基質は、少なくとも6個のアミノ酸残基から成るペプチド、例えば、少なくとも7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、32、34、36、38、40、45、50、60、70、80、90、100、120、140、160、180、200、250、300 350、400、500、600、700、800、900個又は少なくとも1000個のアミノ酸残基から成るペプチドであることが好ましい。80個以下のアミノ酸残基から成るペプチドは、市販の合成サービス又は装置を用いて化学合成により簡便に入手できる。
【0060】
当業者であれば、上記で特定されるようなヒストンH3のN末端残基と少なくとも75%の同一性を示すペプチド、例えば、ヒストンH3のN末端残基と少なくとも77.5%、80%、82.5%、85%、87.5%、90%、92.5%、95%、97.5%、98%、98.5%、99%又は99.5%の同一性を示すペプチドを理解するであろう。前記ペプチドは、上記に詳細に説明されるように、Jumonjiタンパク質のJMJD2サブファミリーのタンパク質に置換が組み込まれたものであってもよい。特に、上記の同一の原理にしたがい、保存的置換が好ましくあろう。
【0061】
本発明の更に好ましい態様では、最小限のものとして、前記基質は、以下から成る群から選択されるアミノ酸配列を含む:
【0062】
現時点で最も好ましい態様では、ヒストンH3のN末端尾部(1-40)を模倣した43個のアミノ酸長の合成ペプチド、ARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPHR(配列番号24)が基質として用いられる。
【0063】
また、バルクヒストン(bulk histone)、合成ペプチド及びヌクレオソームから成る群から選択される基質を用いることが更に好ましい場合もある。
【0064】
上述したように、本発明に係る方法は、基質を用いて行うことができるが、そのメチル化部位は、モノメチル化、ジメチル化、又はトリメチル化されている。
【0065】
上述したように、α-ケトグルタレートは、GASC1の補因子として機能し、従って、本発明に係る方法に必要である。更に、α-ケトグルタレートの類似体を補因子として使用することも本発明の範囲に含まれる。これに加え、アスコルビン酸と、α-ケトグルタレートに加えて、Fe(II)イオンの存在によりもたらされるGASC1触媒活性が、補因子として重要となるであろう。従って、本発明の好ましい態様では、該化合物と、前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質は、Fe(II)イオン及び/又はアスコルビン酸と共にインキュベートされる。アスコルビン酸の存在は不可欠なことではないが、アスコルビン酸により反応系に存在する鉄は適切な形態であるFe(II)に変換されるので、完全な触媒活性を得るためには、アスコルビン酸が必要とされる場合もあることが理解されるべきである。
【0066】
試験化合物がJumonjiタンパク質のJMJD2サブファミリーのタンパク質に直接作用する場合には、無細胞系を用いることもできる。試験系を試験化合物と接触させ、その試験化合物が、前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性を調節する能力を、基質(ヌクレオソーム、ヒストン又はヒストンペプチド)の転換をイムノアッセイを用いて測定することにより測定する。
【0067】
Jumonjiタンパク質のJMJD2サブファミリーのタンパク質のペプチド基質、又はその触媒活性による生成物は、例えば、プレート若しくはビーズ上に固相化し、適切なメチル化部位のメチル化状態又は脱メチル化を、メチル化状態に応じて異なる親和力で前記ペプチドに結合する抗体若しくは他の結合分子を用いて検出できる。従って、トリメチル化、ジメチル化若しくはモノメチル化されたヒストンH3-K9のいずれかを認識する免疫結合パートナーは、試験系の一部を構成し、その活性に対する化合物の作用を迅速に測定することを可能にする。基質特異的抗体及び生成物特異的抗体を含む、このような結合パートナーは市販されている。
【0068】
本発明のアッセイ方法は、in vivoアッセイの形態を取ることも可能である。in vivoでのアッセイは、細胞系、器官系、又は動物全体のアッセイを用いて行うことができる。好ましくは、in vivoアッセイは、細胞系で行われ、例えば、細胞に導入された1又は2種類以上のベクターから関連するポリペプチド若しくはペプチドが発現されるセルラインを用いて行われる。例えば、細胞は、酵母細胞又は哺乳動物由来の細胞であってもよい。
【0069】
更に、本発明との関連で用いられるin vivoアッセイは、単離細胞/初代細胞又は組織切片による場合がある。特定の興味深い態様では、in vivoアッセイは、49歳の日本人女性から放射線照射(腫瘍が隣接構造に浸潤していた)の後に(頚部)食道上部から切除された、あまり分化の進んでいない食道扁平上皮癌から樹立されたヒト扁平上皮癌であるKYSE-150セルラインの使用を要する;増幅されたがん遺伝子c-erb-B(8倍)及びサイクリンD1(4倍)を保持し、ヒトのAST、LDH、MDHのIEFで確認されたようにヌードマウスで腫瘍を発生させると記述される。
【0070】
現時点において好ましくは、メチル化状態の測定は、フルオログラフィーによるホルムアルデヒド-3Hの放出と組み合わせてもよい。ホルムアルデヒド-デヒドロゲナーゼ一体アッセイによるホルムアルデヒド放出を測定するための類似技術も、本発明において意図されている。
【0071】
本発明の好ましい態様では、基質のメチル化状態及び/又はホルムアルデヒドの放出及び/又はα-ケトグルタレート基質補因子のカルボキシル化状態及び/又は酸素消費量は、イムノアッセイ(ELISA、RIA、IRMA、TRIFMA)、免疫沈降、ウェスタンブロッティング、BIAcore、X線結晶解析、溶液NMR、質量分析、分光技術又は蛍光技術により測定される。
【0072】
本発明の現時点で好ましい態様では、基質の利用量及び/又は生成物の形成量の測定は、免疫蛍光分析、好ましくは共焦点免疫染色法により行われる。本発明の現時点で最も好ましい態様では、実施例で説明されるように、ペプチド基質は、ビオチンに結合されている。
【0073】
基質特異的抗体によるアッセイでメチル化状態を測定する場合、適切な対照との比較における免疫結合パートナーの結合レベルの低下は、トリメチル化若しくはジメチル化したH3-K9の低下を意味しており、このことはJumonjiタンパク質のJMJD2サブファミリーのタンパク質の活性の増加に対応している。免疫結合パートナーの結合は、例えば、ウェスタンブロッティング、ELISA、RIA、TRIFMA、免疫沈降又は免疫組織学的手法等、当該技術分野で一般的に知られた方法により評価できる。好ましい技術には、蛍光偏光法、時間分解-蛍光共鳴エネルギー転移法(Time Resolved Fluorescence Resonance Energy Transfer Assay:TR-FRET)、シンチレーション近接アッセイ(Scintillation Proximity Assay)及び「蛍光クエンチング」アッセイ等のハイスループットスクリーニング技術が含まれる。発現物は、容易に検出可能なレポータータンパク質、例えば、以下に限定されるものではないが、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーぜ(CAT)、緑色蛍光タンパク質又はルシフェラーぜ等と連結させることもできる。
【0074】
競合測定では、合成若しくは天然のヒストンペプチドが、標識型又は非標識型で提供される場合がある。抗体を、修飾有り又は無しで用いることもできる。抗体は、その抗体をレポーター分子と、共有結合的又は非共有結合的に連結させて標識できる。検出の容易化に使用可能な適切なレポーター分子又は標識には、放射性ヌクレオチド、酵素、蛍光分子、化学発光剤又は発色剤、並びに、基質、補因子、阻害剤、磁気粒子等が含まれる。キットの抗体又は合成ペプチドは、好ましくは、BSA、チログロブリン、オボアルブミン又はキーホールリンペットヘモシアニン等の適切なタンパク質担持体と結合させることにより、マイクロタイタープレート等の固体表面に固相化させてもよい。
【0075】
本発明の現時点で好ましい態様では、前記方法には、以下の工程:
i) 鉄及びα-ケトグルタレートを含む溶液で請求項8ないし13のいずれか1項に定義される脱メチル化の基質の懸濁液を調製すること;
ii) 前記試験化合物を、前記懸濁液に添加すること;
iii) ii)で得られた前記懸濁液に、請求項2ないし4のいずれか1項に定義されるJumonjiタンパク質のJMJD2サブファミリーのタンパク質を添加すること;
iv) 前記懸濁液を、好ましくは、35〜38℃の温度で10分間以上インキュベートすること;並びに
v) 前記基質の脱メチル化の程度を測定すること
が含まれる。
【0076】
脱メチル化の程度は、リジン9がメチル化されたヒストンH3に特異的な免疫結合パートナーを用いて測定することが更に好ましい。
【0077】
また、前記免疫結合パートナーは、抗トリメチル化ヒストンH3-K9抗体であることが好ましい。また、免疫結合パートナーとして抗ジメチル化ヒストンH3-K9抗体を用いることも、同程度に好ましい。
【0078】
更に好ましい態様では、脱メチル化の程度は、ホルムアルデヒドの放出により評価される。更に、脱メチル化の程度は、酸素消費量を測定することによっても評価できる。
【0079】
これに代わる場合、脱メチル化の程度は、α-ケトグルタレート基質補因子のカルボキシル化状態を測定することにより、評価される。
【0080】
例えば、i)のバッファーは、終濃度で50mMのHepes(pH7.5)、50mMのKCl、4mMのMgCl2、1mMのα-ケトグルタレート、40μMのFeSO4、2mMのアスコルビン酸を含む。実利的な理由により、前記懸濁液は、10-50μlの総量にあることが好ましい。
【0081】
最後に、上記に加えて、反応系のタンパク質成分の分解を最小限に抑えるためには、1又は2種類以上のプロテアーゼ阻害剤を終濃度1-5μg/μlで添加することが有用であろう。例えば、アプロチニンやロイペプチンは、プロテアーゼ阻害剤に含まれるであろう。
【0082】
本発明の種々の局面と、これらの局面の具体的な態様の上記記載に関しては、本発明の一つの局面及び/又は局面の一つの態様と関連して上に記載又は言及されるいずれの特性及び特徴も、記載される本発明の他の全ての局面及び/又は態様に類推して用いるられることが理解されるべきである。
【0083】
本発明の目的又はその特性若しくは特徴の一つについて単数系で言及された場合、その言及は、その目的又はその特性若しくは特徴について複数系で言及していることにもなる。例えば、「1つのペプチド」について言及している場合、1以上のペプチドについて言及していることが理解されるべきである。
【0084】
本明細書を通じて、「含む(comprise)」という単語、及びその変化形である「含む(comprises)」若しくは「含んでいる(comprising)」は、記述する要素、整数若しくは工程、又は要素、整数若しくは工程の群を含むが、いずれかの他の要素、整数若しくは工程、又は要素、整数若しくは工程の群を排除するものではないことを意味することが理解されるべきである。
【実施例】
【0085】
H3K9のトリメチル化バリアントと相互作用するタンパク質の同定を目的とする研究において、ストレプトアビジン-アガロース上に固相化されたビオチン化H3ペプチドを用いてHeLa核タンパク質をアフィニティー精製した。リジン9が非メチル化状態にあるか又はトリメチル化されている、ヒストンH31-40のN末端尾部全体を模倣した合成43-merをin vitro結合実験に用いた。結合したタンパク質をアガロース-マトリックスから溶離し、SDS-PAGEで分離して銀染色し、質量分析により同定した。複数種類のタンパク質では、非メチル化H3K9と比べて、特にトリメチル化H3K9が豊富であった(補足図S1)。
【0086】
これらタンパク質の中から、Jumonjiタンパク質のGASC1を同定した(図1a)。JumonjiドメインのJmjN及び関連するJmjCドメインが存在することから、このタンパク質は、JMJD2c8,9とも呼ばれている(図1a)。Jumonjiタンパク質ファミリーには、α-ケトグルタレート依存的オキシゲナーゼスーパーファミリーに属する多様なタンパク質のグループが含まれる。これらのタンパク質は、細胞周期の進行及び転写調節を含む種々の細胞プロセスを調節している5,10-14。このタンパク質のグループに共通する特徴は、Fe(II)イオンに結合する能力である。更に、これらのタンパク質の多くは、α-ケトグルタレートを補因子としてタンパク質基質をヒドロキシル化する能力を有する15。特に興味深いことに、近年、JmjCドメインを有するタンパク質が、ヒストンデメチラーゼの候補として同定されている16。
【0087】
GASC1は、トリメチル化(me3)H3K9と結合することが分かっているので、GASC1が、このエピジェネティックマークの脱メチル化に関与しているかもしれないという考えは、興味深いものであった。この可能性を試すため、組換え完全長Hisタグ化ヒトGASC1を作製し、バキュロウィルスで感染させた昆虫細胞から精製した(補足図S3)。アフィニティー精製したGASC1を、更にサイズ排除クロマトグラフィーで精製し、溶離した高純度GASC1を、バルクヒストンとインキュベートすることで脱メチル化活性について試験した。図3aに示すように、イムノブロッティング法で評価したところ、GASC1は、非常に効率的にH3K9のトリメチル化を減少させていた。また、GASC1が、関連する生理学的鋳型であるオリゴヌクレオソームを脱メチル化できることも分かった(補足図S4a)。
【0088】
GASC1の脱メチル化活性の特異性を更に分析するため、大量ないし少量のGASC1を示すGASC1のサイズ排除クロマトグラフィーの種々のフラクションをバルクヒストン及びリジンK9若しくはK27がメチル化された合成H3ペプチドとインキュベートし、種々のエピジェネティックマークのメチル化状態を、イムノブロッティング法により評価した(図3b及びS4b)。最も高いGASC1濃度を有するフラクション(F9-F12)では、ジメチル化及びトリメチル化H3K9はヒストンから完全に除去されていた。また、これに付随したモノメチル化(me1)H3K9の増加も認められ、GASC1がトリメチルH3K9をジメチル(me2)型に変換し、更にモノメチルH3K9(me1)に変換することと一致していた。
【0089】
対照的に、ジメチル化H3K4、トリメチル化H3K4、トリメチル化H3K27及びトリメチル化H4K20を含む、試験した他のエピジェネティックマークのレベルは、GASC1処理によって影響を受けていないようであった(図3b及びS4b)。
【0090】
興味深いことに、ジメチル化H3K9は、ヒストンを、少量ないし中程度の量のGASC1(F6、F8、F14及びF16)と反応させた場合に増加したようであった。このことは、ジメチル化H3K9マークの見かけの増加の理由が、脱メチル化反応の初期において、このマークの安定状態のレベルに到達したことによるという可能性を示している。従って、ある程度のジメチルH3K9がGASC1による脱メチル化を通じて失われるが、反応系全体で見れば、少なくとも十分な量のトリメチルH3K9が存在する限りは、GASC1によるトリメチルリジンの脱メチル化により、更なるジメチルH3K9が加わるものと考えられる。
【0091】
図3a及びbの結果と一致して、GASC1の脱メチル化活性は、濃度依存的に変化し、GASC1の熱変性により消失する場合があることから(図S4b、c)、この反応は酵素反応であることが示唆される。更なるin vitro脱メチル化試験を行い、この試験では、純粋な合成トリメチル化H3K9ペプチドをGASC1とインキュベートした。純粋なトリメチル化H3K9とGASC1のインキュベートを30分間行った後では、殆ど全てのトリメチルH3K9が、ジメチル及びモノメチルH3K9に変換されていた(図S4c、d)。
【0092】
GASC1が実際にトリメチル化及びジメチル化H3-K9を脱メチル化することを正確に証明するため、単一又はGASC1の存在下でインキュベートした合成ペプチドの質量分析を行った。この分析により、GASC1とのインキュベート後にH3K9me3ペプチドの大部分(約80%)が、ジメチル化及びモノメチル化H3-K9に変換されたことが示された(図4c-6f)。
【0093】
次に、GASC1に近似するホモログであるJMJD2a及びJMJD2b(図1a、図2b)が、in vivoでH3K9を脱メチル化できるかどうかについて検討した。これを調べるため、完全長のヒトJMJD2a、JMJD2bをクローニングし、バキュロウィルスで感染させた細胞から組換えタンパク質を得た。両ホモログは、バルクヒストンのH3K9me3とH3K9me2を脱メチル化できた(補足図S5)。同様に、GASC1とそのホモログJMJD2Aは、同程度の合成H3K9基質を脱メチル化する能力を有すると考えられた(補足図S4d、e)。これらの結果は、まとめると、トリメチル化及びジメチル化H3K9は両方とも、in vitroでのGASC1とそのホモログの基質であることを強く示唆している。
【0094】
in vivoでの脱メチル化反応の特異性に対する洞察を得るため、ヒトGASC1を、3次元構造が解明された唯一のJumonjiタンパク質である11,15,17-19、FIHの構造上にモデリングした。アライメント(図5及び補足図S2)と一致して、このモデリングでは、ヒスチジンH190、グルタミン酸E192、及びヒスチジンH288の残基が、GASC1の鉄結合用溝の本質的な部分を形成することが示された(図6h、i)。また、in silicoモデルでも、T187とR208の残基が、α-ケトグルタレートの結合に関与しており、GASC1の脱メチル化機能に決定的なものであることが予測された。次にトリメチル化H3ヒストン尾部を、予測されたGASC1構造上にモデリングした。K9がトリメチル化されたH3ヒストン尾部は、GASC1の鉄結合用溝に良好にフィットし、K9メチル基は、メチル化ヒストン尾部とGASC1構造間に立体障害を生じることなく、反応相手の鉄の近傍に位置することができた(図6h)。このin silicoモデルに基づき、H190とE192に変異を導入するだけで、GASC1の鉄結合能が喪失し、従って、その脱メチル化活性が阻害されるであろうことが予測された。
【0095】
上記の仮説を試すため、バルクヒストンを、野生型のGASC1と、ヒスチジンH190 とグルタミン酸E192がそれぞれグリシンとアラニンで置換された変異型GASC1(H190G/E192A)と共にインキュベートすることにより、脱メチル化アッセイを行った。野生型GASC1は、強力な脱メチル化活性を有していたが、変異体GASC1は、検出可能な脱メチル化活性を示さず、予測された鉄結合残基が、GASC1の脱メチル化活性に決定的であることが示された。野生型GASC1と変異型GASC1のサイズ排除クロマトグラフィーのプロファイルは、ほぼ同一であり、前記変異導入が、タンパク質の全体的な構造を失わせなかったことが示された(補足図S6)。
【0096】
次に、H3K9のトリメチルマーク及びジメチルマークのGASC1媒介性脱メチル化の反応機構を検討するため、複数の実験を行った。因子阻害性HIF1(Factor Inhibiting HIF1:FIH)を含むα-ケトグルタレート依存性オキシゲナーゼに関する種々の研究11,17-24では、以下の反応機構が提唱されている:最初に、酵素が、その金属結合モチーフであり、いわゆる表面の3つの分子から成るHXD/EXnH20(GASC1ではHXEXnH)を通じて鉄と結合する。次いで、Fe(II)-酵素の複合体が、補因子であるα-ケトグルタレート(αKG)と結合し、次いで、基質及び酸素と結合する。酸素の結合の後に、αKGの酸化的脱カルボキシル化が起こり、サクシネート、二酸化炭素及びフェリルが生じる。フェリルは、非常に反応性が高い基であり、リジンのメチル基中のC-H結合を酸化して、不安定なカルビノールアミンを形成する場合があり、このカルビノールアミンは、迅速に分解してホルムアルデヒドを生じさせ、そしてリジンはメチル基を奪われることになる(図7a)。
【0097】
cupinスーパーファミリーに属するジオキシゲナーゼは、Fe(II)とα-ケトグルタレートに依存する。更に、FBXL11を含むいくつかのcupinジヒドロゲナーゼは、完全な触媒活性のために更にアスコルビン酸を必要とする5,21,23。アスコルビン酸の作用の様式は、現時点では明確なっていないが、Fe(III)をその活性状態のFe(II)に還元するか、又はフェリル(FeIV=0)中間体が分離して生成された場合に「代理の還元性基質」として機能してジオキシゲナーゼ酵素を「救助する」ことが示唆されている17,22。
【0098】
GASC1媒介性脱メチル化が、上に記載され、図7aに示される反応機構と一致するかどうかを試験するため、脱メチル化反応の推定上の補因子の重要性を検討した。
【0099】
精製したHisタグ化GASC1を、その補因子の存在下又は不存在下で基質としてのバルクヒストンとインキュベートした(図3b)。全ての補因子の存在下ではGASC1はH3K9me3をin vitroで脱メチル化するが、EDTA(鉄をキレート化する)の存在下でのGASC1のインキュベート又は補因子の不存在下でのGASC1のインキュベートでは、脱メチル化は著しく低下していた(図8b)。補因子を添加していないにも関わらず脱メチル化反応を起こさせる能力は、恐らく、組換えGASC1と共に精製された補因子(鉄及びαKG)によるものと考えられる。この理由により、N-オキサリルグリシン、ケルセチン、CoCl2及びNiSO4が、GASC1媒介性のH3K9me3の脱メチル化を阻害する能力を試験することで、上記の補因子の重要性を更に探った。これら全ての化合物が、効果的に脱メチル化反応を阻害できた(図8c及びデータ非開示)。N-オキサリルグリシン(図8d)とケルセチンは、αKG類似体であり、恐らく、GASC1の鉄結合残基からαKGを引き離すことによりGASC1の活性を阻害している。同様に、CoCl2及びNiSO4によるGASC1の阻害には、恐らく、コバルト(II)イオン又はニッケル(II)イオンが競合することにより、GASC1の鉄結合部位からFe(II)が隔離されることが関わっている。脱メチル化反応にアスコルビン酸、αKG及び鉄を必要とすること、並びにニッケル塩及びコバルト塩及びαKG類似体により脱メチル化反応が阻害されることにより、GASC1が、実際にジオキシゲナーゼであることが強く示唆される。
【0100】
更にまた、補因子であるFe(II)及びαKGの存在下では、組換えHisタグ化GASC1は、ホルムアルデヒドを放出し(図8e)、これは、提案される図3aの反応スキームと一致している。これらをまとめると、以上の結果は、GASC1が、鉄とαKGを必要とするヒドロキシル化反応を通じて、in vitroでヒストンH3のトリメチル化及びジメチル化リジンK9を直接脱メチル化し、ホルムアルデヒドとモノメチル化H3K9を生じることを示している。
【0101】
GASC1が、in vitroでジメチル化及びトリメチル化H3K9を脱メチル化できることが立証されたことから、GASC1が、in vivoでヘテロクロマチンの形成/維持を調節できるのかどうかを検討した。この問題に取り組むため、最初に、GASC1の強制発現により、in vivoでH3K9のメチル化を調節できるかどうかを調べた。ヒトの2倍体線維芽細胞を、ヒト野生型GASC1を発現するレトロウィルスで感染させ、H3K9のメチル化状態を、共焦点顕微鏡観察により検証した。GASC1は、核内に局在していたことから、これは、予想上の核内局在シグナル配列が該タンパク質に存在することと一致していた。pBabe−HA−GASC1での感染により、H3K9トリメチルが効果的に減少し、モノメチル化H3K9が増加した(図9a)。対照的に、トリメチル化H3K4とトリメチル化H3K27は、GASC1過剰発現による影響を受けなかった(補足図S7)。
【0102】
次に、ヒト骨肉腫セルラインU2OSを、野生型GASC1を発現するプラスミドと、ヒスチジンH190 とグルタミン酸E192がそれぞれグリシンとアラニンで置換された変異型GASC1(H190G/E192A)を発現するプラスミドでトランスフェクトした。野生型GASC1の異所発現では、in vivoでヒストンH3のK9のトリメチル化とジメチル化から効果的に消失されたが、変異型GASC1では、そうではなかった(図9b)。
【0103】
興味深いことに、GASC1の異所発現では、モノメチル化H3K9が顕著に増加しており、GASC1が、in vivoでトリメチル化H3K9とジメチル化H3K9の両方を脱メチル化することが示唆された。トリメチル化H3K9の減少は、ウェスタンブロッティングによりこのマークの全体的なレベルを評価した場合に、GASC1でトランスフェクトされたU2OS細胞においても明らかであった(図10c)ことから、これらの推定上のFe(II)配位残基の決定的な重要性が確認された。
【0104】
次に、GASC1の近似するホモログであるJMJD2aとJMJD2b(図1a、図2b)も、in vivoでH3K9を脱メチル化できるかどうかを、これらのタンパク質をU2OS細胞内で過剰発現させることにより試験した。GASC1と同様に、JMJD2aとJMJD2bの異所発現により、トリメチル化及びジメチル化H3K9が顕著に減少し、同時にモノメチル化H3K9が増加していた(図10d、e)。これらの結果は、GASC1が、ヒストンH3K9デメチラーゼのファミリーに属していることを示している。
【0105】
ヘテロクロマチンの形成と維持には、トリメチル化及びジメチル化H3K9の存在と並びにHP−1結合が必要とされる25-27。GASC1を異所発現させることにより、H3K9のジメチル化とトリメチル化が全体的に減少したので、HP−1結合と局在性が、GASC1レベルの上昇により影響を受けるかどうかを検討した。NIH 3T3細胞を、HAタグ化GASC1を発現するプラスミドでトランスフェクトし、事前に抽出した細胞内の内在性のHP1βの分布を共焦点顕微鏡観察により確認した。予想した通り、HP1βは、GASC1を過剰発現する細胞内では非局在化しており、H190G/E192A変異体をトランスフェクトした場合には、非局在化は認められなかった(図11f)。同様に、GASC1のホモログであるJMJD2a及びJMJD2bの異所発現では、HP1αが非局在化した(補足図S8)。これらの結果については、GASC1過剰発現細胞内でのクロマチンと結合したHP1α及びHP1γのレベルの低下を示すことにより更に確認した(図11g)。この目的のため、Mycタグ化GASC1をテトラサイクリン制御下で発現させるHEK293細胞を作製した。選択した細胞をテトラサイクリンの存在下又は不存在下で一晩インキュベートしたところ、図11gに示すように、クロマチンと結合したHP1α及びHP1γの量は、GASC1過剰発現細胞内で減少していた。これらの結果をまとめると、GASC1が、ヘテロクロマチンの形成及び維持の調節において、生理学的に重要な役割を果たす可能性が示唆される。
【0106】
近年に至るまで、トリメチル化リジンは、不可逆的な共有結合性のヒストン修飾であると考えられてきており28、ヒストントリメチルデメチラーゼを同定するのは、本報告が最初のものである。我々の結果は、GASC1が、メチル化されたH3K9を特異的に脱メチル化するが、モノメチル化又はジメチル化された基質に特異的なLSD1及びFBXL114,5とは対照的に、GASC1は、in vitro及びin vivoで、H3K9のジメチル種及びトリメチル種の両方を取り除く能力を有することを示している。
【0107】
GASC1は、複数のドメインを含み、恐らく、これらのドメインは、GASC1タンパク質にトリメチル化及びジメチル化H3K9に対する強い特異性を与えるよう作用する(図1a)。JumonjiCドメイン(JmjC)とJumonjiNドメイン(JmjN)に加え、GASC1タンパク質は、2つのplant homeodomain(PHD)並びにtandem Tudor domain(Tdr)を特徴とする。これらのドメインの機能は現時点では明らかとなっていないが、これらはすべて、クロマチンの調節に関与していると考えられている8,10,12,29,30。例えば、53BP1のTudorドメインは、ヒストンH3のリジン79のトリメチル化体に結合することが報告されており31、PHDドメインは、ヌクレオソームへの結合で、Bromoドメインと強調することが実証されている30。どのドメインがGASC1の脱メチル化機能に必要であるのかという洞察を得るためと、どのドメインが基質特異性を決定するのかを解明するために、GASC1タンパク質の変異導入試験を行った。これは、一連のJmJC、JmjN及びPHDドメインに欠失を有するGASC1発現コンストラクトを作成することで行った。これらの結果は、JmjCドメインとJmjNドメインが、GASC1の脱メチル化活性に不可欠であることを示していた(補足図S9)。対照的にPHDドメインの欠失は、GASC1のin vivoでH3K9me3を脱メチル化する能力に影響していなかったと思われた(図S9)。
【0108】
GASC1及びFBXL11の類似する反応機構に照らせば、なぜFBXL11は、その機能を有しないのに、GASC1がトリメチル化ヒストンのリジン基質を脱メチル化する能力を有するのかという疑問が生じるのが妥当である。可能性のある説明としては、鉄/基質結合用溝が、FBXL11ではGASC1よりも小さいことが考えられる。この説明は、実験的な検証を要するが、in silicoモデリングにより支持される。FIHを鋳型としてモデリングした2種類のJumonjiタンパク質のモデルを比較した場合、両者は共に、そのモデルの鉄結合用溝のエッジに位置するループ領域を有するという特徴を有することが判明した。このループ領域は、FBXL11モデルで顕著に長く、これがゆらゆらと動くために、このタンパク質の鉄/基質結合用溝がより小さくなり、結果としてトリメチルリジン基質の収容を阻害しているかもしれないという可能性が考えられる。
【0109】
本報告に記載され、補足図S8bにまとめられる脱メチル化経路は、進化の過程で保存されていると予測される。この関係において、Schizosaccharomyces pombeのJumonjiタンパク質であるEpe1は、ヘテロクロマチン化をアンタゴナイズすることが報告されている10。従って、最近の報告10では、Epe1の強制発現が、ジメチル化H3K9を減少させ、ヘテロクロマチンにアンタゴナイズすると同時にセントロメアと交配型遺伝子座を不安定化させることが証明されている10。このEpe1媒介性表現型は、無傷のJmjCドメインに依存しており、Epe1のヘテロクロマチン化をアンタゴナイズする能力が脱メチル化活性に起因することは証明されていないものの、我々の本データと照らしてこのような役割の仮説を立てることは興味深い。
【0110】
例えば、GASC1等、類似する酵素活性を有する、密接に関連した複数のヒトホモログ(JMJD2a及びJMJD2b、図2b)の存在は、機能の重複性とH3K9トリメチルデメチラーゼ活性の正確な調節の必要性を示唆するものである。ヘテロクロマチンの形成と維持の正確な調節が、適切な生物機能とゲノムの統合性の両方にとって決定的に重要であることから、上記のことは予測されるべきことである。例えば、セントロメアにおけるヘテロクロマチンの形成は、有糸分裂期における染色体の正確な分離に不可欠である32。同様に、マウスのSuv39H1及びSuv39H2(これら遺伝子の産物は、ヘテロクロマチンのH3K9のジメチル化及びトリメチル化に関与する)の欠失は、染色体を不安定化させ、マウスのリンパ腫の誘導において癌遺伝子と共同して機能することが示されている6。従って、GASC1又はそのホモログの異常な活性は、ゲノムを不安定化させ、その結果として癌を生じさせる可能性がある。実際に、GASC1は、元々、食道扁平上皮細胞(oesophageal squamous cell:ESC)の癌で頻繁に増幅される遺伝子として同定されており22、GASC1は、染色体異常を含む種々の種類の癌で過剰発現されている33,34。
【0111】
GASC1の癌への関与に関する更なる支持を得るため、Oncomineデーターベースで、正常組織と腫瘍組織における異なるGASC1発現について探索した。GASC1とそのホモログのJMJD2a及びJMJD2bの発現は、正常組織と比べて前立腺癌で著しく増加していることがわかった(補足図S11)。GASC1は、先に、ESC癌腫由来の複数のセルラインで増幅されていることが示されている33。GASC1遺伝子を増幅させたESCセルライン(KYSE−150)と、より緩やかに発現されているセルライン(KYSE−450)において、H3K9エピジェネティックマークのメチル化状態を試験した。ヒト骨肉腫セルラインU2OSが、対照として含まれた。GASC1増幅の重要性を試験するため、GASC1及び/又はそのホモログに対するsiRNAオリゴでトランスフェクトされた細胞におけるH3K9メチル化状態に対するGASC1ノックダウンによる効果を試験した。細胞をLMP−GASC1でトランスフェクトして、その後、ピューロマイシンを用いて3日間かけてセレクションを行った。GASC1ノックダウンの効果は、RT−QPCRにより評価した(図5a)。ウェスタンブロッティングにより証拠付けられるように、H3K9me3は、KYSE−150細胞内でGASC1ノックダウンに応じて顕著に増大していた(データ非公開)。
【0112】
ジメチル化及びトリメチル化H3K9は、HP1結合に必要であり、ヘテロクロマチン形成に不可欠であり、転写抑制と関連している。更にまた、これらのエピジェネティックマークは、癌から細胞を守る要となる機構である、老化状態にある特定の遺伝子において増加している36-39。従って、GASC1の増幅は、細胞が老化状態になる能力を低下させるか、又はサイレンシング状態にある癌遺伝子の抑制を解除する可能性があることが予測できる(図6d)。従って、GASC1の脱メチル化活性の阻害は、新たな抗腫瘍治療様式を構成する可能性がある。
【0113】
興味深いことに、抗増殖特性及び抗癌特性と関連する植物フラボノイドであるケルセチン(図12b)40-42は、in vitroで効果的にH3K9me3のGASC1媒介性脱メチル化を阻害できた(図12c)。ケルセチンを用いてin vivoでKYSE−150細胞を一晩処理すると、H3K9トリメチル化が驚くほど増大し、このことは、免疫蛍光とウェスタンブロッティングに裏付けられている(図13d及びデータ非開示)。H3K9me3の増大は、増殖速度の顕著な低下、細胞形態の変化、老化関連ヘテロクロマチン病巣(senescence associated heterochromatin foci:SAHF)の出現及びβ−gal染色により特徴付けられる老化状態様の表現型も伴っていた(データ非公開)。興味深いことに、ケルセチン処理は、他のセルライン(低レベル及び中レベルのGASC1を発現する)では、同程度の影響を及ぼさないように思われた。従って、U2OS、WI38及びKYSE−450細胞は、同程度のH3K9me3の増加、又は同様の老化状態様表現型及び増殖阻害を示さなかった。この結果により、JMJD2酵素の発現又は活性の増大とこれに続くH3K9me3/me2の減少により特徴付けられるいくつかの癌が、この化合物による増殖阻害作用に特に高い感受性を示す可能性が示唆される。ケルセチンの生物学的効果は、従来ではトポイソメラーゼIの阻害43、タンパク質と脂質キナーゼシグナル経路の調節44及び活性酸素種の除去45によるものであると考えられてきた。本研究の知見に照らせば、ケルセチンによる抗癌効果は、少なくとも部分的に、そのGASC1及びJMDM2サブファミリーの他のメンバーに対する阻害作用により説明できるという推測が試みられる。
【0114】
以上をまとめると、複数の独立した証拠を用いることで、GASC1が、in vitro及びin vivoの両方で、抑制性のヒストンのジメチル及びトリメチルH3K9マークを直接脱メチル化することが実証された。本知見により、ヒストンのトリメチル化が、可逆性の修飾であることが示された。この知見は、ヒトの疾患、特に癌について、更に深い意味合いを有する可能性がある。
【0115】
材料と方法
脱メチル化アッセイ
バルクヒストン、オリゴヌクレオチド又は合成ヒストンペプチドを、精製したHis-GASC1と、脱メチル化バッファー(50mMのTris(pH8.0)、50mMのKCl、10mMのMgCl2、1mMのα-ケトグルタレート、40μMのFeSO4、2mMのアスコルビン酸、0.01%(w/v)のBSA及び5%(v/v)のグリセロール)中で37℃で反応させた。典型的な反応では、6μgのバルクヒストン又は2μgの修飾ヒストンペプチドを20μgのGASC1と100μlの体積で30分間反応させた。反応混合物を、特異的抗体を用いたウェスタンブロッティングか、又はホルムアルデヒド放出アッセイのいずれかにより分析した。
【0116】
ホルムアルデヒド放出アッセイ
ホルムアルデヒド放出アッセイは、本質的に記載4に沿って行った。全ての反応は、石英製キュベット内で、1反応系当たり200μlの総量で行った。簡潔に言うと、組換えGASC1(典型的には40μg)を150μlの脱メチル化バッファー(上記参照)中で、2mMのNAD+と0.2Uのホルムアルデヒドデヒドロゲナーゼ(FDH)の存在下で、37℃にて5分間インキュベートした。次いで、基質(ヒストンペプチド)を添加して反応を開始させた。340nmの吸光度を、Genesys 10UV Thermospectronic spectrophotometerを用いて0.5mmの間隔を開けて、37℃で測定した(全部で15mm)。
【0117】
質量分析用のH3K9me3ペプチドのGASC1による脱メチル化
6μgの組換えGASC1を、終量90μlのFDHバッファー中で、3μgのH3K9me3ペプチドと37℃で30分間インキュベートした。尿素を終濃度4Mで添加し、混合物を20℃で15分間インキュベートした。同量の1%TFAを添加し、サンプルを、100μlのチップ中にパッキングされた逆相ミニC8カラム(カラム容積20μl)にローディングした。1%TFAで洗浄した後、結合したペプチドを20μl(30%メタノール、25%ギ酸)で溶離させた。溶離した材料の3分の1をSDS-PAGEと、抗H3K9me3及びその後に抗ビオチン抗体を用いたウェスタンブロッティングで分析した(図4c)。残りの材料は、質量分析により分析した(図4d)。
【0118】
質量分析(MS)
溶離物の3分の1を、1%TFA中で、Agilent 1100 Nano HP1C(Palo Alto, CA)を用いて、スプレーエミッター(内径75μm、開口8μm、長さ70 mm;New Objectives、米国)内にパッキングされたC18カラム(Reprosil-Pur C18-AQ 3μm; Dr. Maisch GmbH、アメルブッフ-エントリンゲン、独国)上に注いだ。ペプチドは、10分間でバッファーA(5%アセトニトリル及び0.5%酢酸)からバッファーB(アセトニトリル及び0.5%酢酸)へ0%ないし20%で変化する勾配を用いて、300 nL/分で溶離した。スペクトルをLTQ-FT質量分析器(Thermoelectron、ブレーメン、独国)に記録した。
【0119】
補足的方法
材料
ヒストンH3のN末端尾部(1-40)を模倣する43アミノ酸長の合成ペプチド:
【化4】
このペプチドは、リジンとTtdsスペーサーを介してビオチンに連結された結合用のC末端部のチロシン及びシステインを用いて合成した。この実験で用いたペプチドは、非修飾型であるか、リジン27がトリメチル化されているか、又はリジンK9がモノメチル化、ジメチル化若しくはトリメチル化されており、Jerini, GMBH(独国)から購入した。ホルムアルデヒドデヒドロゲナーゼ(FDH、F1879)及びニコチンアミン及びバルクヒストン(H9250)はSigma社から購入した。本試験で用いた抗体は、以下の通り:抗トリメチル化H3-K9(Upstate 07-523)、抗ジメチル化H3-K9(Upstate 07-212)、抗モノメチル化H3-K9(Abcam Ab9045-50)、抗トリメチルH3-K27(Upstate 07-449)、抗トリメチルH4-K20(Upstate 07-463)、抗トリメチル化H3-K4(Abcam ab8580-50)、抗ジメチル化H3-K4(Upstate 07-030)、抗ヒストンH3(Abcam Ab1791-100)、抗HP1α(Upstate 05-689)、抗HP1γ(upstate 05-690)、抗ビオチン-HRP(Sigma A4541)、抗-His(Upstate 05-531)及び抗HA(CRP Inc. AFC-101P)。
【0120】
セルライン及び組織培養
ヒト食道扁平上皮癌セルラインKYSE-70及び150は、German Collection of Microorganisms and Cell Cultures(ブラオンシュヴァイヒ、独国)から入手した。hTert を発現する2倍体のヒト線維芽細胞(TIG-3)とマウスのエコトロピックレトロウィルスレセプターEcoRを発現するU2OS細胞を実験に用いた。293 TRex-flip-in細胞を基本的に製造元(Invitrogen)による記載に従って用いて、テトラサイクリン誘導性のアミノ末端にmycタグが付いたGASC1を安定に発現するHEK293細胞を作製した。KYSE細胞は、49%のRPMI 1640、2%のウシ胎児血清(FCS)が補充された49%のHam's F12中に5%CO2の下で維持した。他の全ての細胞は、10%のウシ胎児血清(FCS)が補充されたDulbecco's modified Eagle's medium(DMEM)中に5%CO2の下で37℃にて維持した。
【0121】
HeLa核タンパク質のヒストンペプチドプルダウン
HeLa細胞の核抽出物とリジン9がトリメチル化されたヒストンH3ペプチド
【化5】
を用いてプルダウンアッセイを行った。核は、HeLa細胞から調製し、溶解バッファー(50mMのTris(pH7.2)、300mMのNaCl、0.5%のIGEPAL CA-630、1mMのEDTA(pH8.0)、プロテアーゼ阻害剤を含む1mMのPMSF)で溶解させた。溶解物は、遊離ストレブトアビジンビーズを用いて事前除去し(図S1、事前除去1)、次いで、非修飾型H3ヒストンに結合したストレプトアビジンビーズで事前除去した(図S1、事前除去2)。次いで、事前除去した溶解物を、事前にストレプトアビジンビーズと結合させておいた、K9がトリメチル化されたH3ヒストンペプチド(H3K9me3)とインキュベートした。ビーズを十分に洗浄し、続いて、2x LSBバッファー(100mMのTris-HCl(pH6.8)、200mMのDTT、4%のSDS、20%のグリセロール及び0.2%のブロモフェノールブルー)で結合したタンパク質を樹脂から溶離した(図S1、H3K9me3)。タンパク質をSDS-PAGEで分離し、銀染色により可視化した。プルダウンされたH3K9me3を豊富に含むタンパク質バンドを切り出し、ゲルのままトリプシンで処理し、Wilm et al.(Wilmm. (1996) Nature 379, 466-469)の記載に従って、ナノエレクトロスプレータンデム型質量分析器で分析した。
【0122】
免疫蛍光
共焦点免疫染色法を、基本的にSorensen, CS. et al., Nat. Cell. Biol. 7, 195-201 (2005)の記載に従って行った。
【0123】
細胞溶解物の調製
細胞の全溶解物は、同数の細胞を尿素バッファー(1%のSDS、9Mの尿素、25mMのTris-HCl(pH6.8)、1mMのEDTA、0.7mのβ-メルカプトエタノール)に溶解して作製し、5分間沸騰させ、そして超音波処理した。クロマチンの分画化のため、200μlの事前抽出用バッファー(0.5%のIGEPAL-630、5OmMのNaCl、3mMのMgCl2及び300mMのスクロースを含有する2OmMのHepes-KOH(pH7.2))中の同数の細胞を氷上で30分間インキュベートした。100μlを2xLSBバッファー(100mMのTris-HCl(pH6.8)、200mMのDTT、4%のSDS、20%のグリセロール及び0.2%のブロモフェノールブルー)と混合し、沸騰させ、超音波処理して、全溶解物として使用した。残りの100μlは、1300xg、4℃で10分間スピンダウンさせ、1mlの事前抽出用バッファーで洗浄し、100μlの2xLSBバッファーに再懸濁し、沸騰させ、超音波処理し、クロマチン分画として使用した。
【0124】
GASC1及びJMJD2a/bに対するsiRNA及びshRNA
GASC1並びにそのホモログJMJD2a及びJMJD2bに対する低分子干渉RNA(siRNA)オリゴヌクレオチドは、Dharmacon Research, Inc.によって合成された。6ウェルプレート内で、Oligofectamine試薬(Invitrogen)を製造元のプロトコルに従って用いて、細胞(1x105/ウェル)をsiRNAオリゴ(0.3μg/ウェル)でトランスフェクトした。siRNAは、例えば、siRNA retriever;(http://katahdin.cshl.org:9331/homepage/siRNA/RNAi.cgi?type=siRNA)を用いる等、当該技術分野で知られる方法により設計できる。
【0125】
shRNAコンストラクト
短鎖ヘアピンRNAコンストラクトは、Paddison et al.(2005)(Paddison PJ, Clearym, Silva JM, Chang K, Sheth N, Sachidanandam R & Hannon GJ. Cloning of short hairpin RNAs for gene knockdown inmammalian cells Naturemethods pp163-167)の記載に従って、MSCV/LTRmiR30-PIG(LMP)ベクター(Dickens NG et al(2005)"Probing tumor phenotypes using stable and regulated syntheticmicroRNA precursors" Nature Genetics Vol 37, No 11 1289-1295.)内に作製した。
【0126】
「shRNA retriever」(http://katahdin.cshl.org:9331/homepage/siRNA/RNAi.cgi?type=shRNA)を用いて3種類のオリゴヌクレオチドを設計した。以下のオリゴヌクレオチド:
【化6】
を鋳型として用いて150 bpのPCR産物を増幅させ、XhoI及びEcoRIで処理し、そして制限酵素処理したLMPベクター内にライゲーションした。このshRNAコンストラクトは、上記の配列で下線が引かれたGASC1コーディング配列の2117番目の位置の22 bpの配列を標的とする。このshRNAiコンストラクトが、GASC1のmRNAをノックダウンする効率は、リアルタイム定量的RT-PCRで評価したところ、約80%であると評価された。
【0127】
リアルタイム定量的RT-PCR
細胞を、表示するshRNAコンストラクトでトランスフェクトし、簡単なピューロマイシンセレクション(2μg/ml)に付した。Quiagen RNAeasyミニキットを製造元の使用説明書に従って用いて全RNAをトランスフェクトした細胞から作製した。Taqman逆転写キットとポリdTプライマーを製造元の使用説明書に従って用いてcDNAを作製した。このcDNAを鋳型として、GASC1特異的プライマーと共に、Applied biosystems 7700でのリアルタイム定量的PCR反応に用いた。この反応物は、Applied biosystems社製の2xSYBR green反応ミックスを用いて調製した。
【0128】
BrdUの取り込み
siRNAのトランスフェクションから30時間後に、細胞を4ウェルチャンバースライドに分け、BrdUを含む培養培地で15-30分間インキュベートした。
【0129】
ヒトGASC1のcDNAのクローニング
ヒトGASC1及びJMJD2Aの推定上のオープンリーディングフレームをHeLaのcDNAとヒト胎児脳のcDNAライブラリー(Invitrogen、カールスバッド、カリフォルニア州)のcDNAからPCR増幅した。プライマー配列は、請求により入手可能である。PCR産物をゲル精製し、pCR8/GW(Invitrogen)にクローニングし、DNAしーケンシングにより確認した。これらのGateway適合性の導入クローンを用いて、GASC1及びJMJD2AをpCMV-HA、pCMV-myc及びpBabepuroに導入した。標準的な変異導入方法により、GASC1の保存された鉄結合ドメイン(HTE)の、190-192番目のアミノ酸が、それぞれヒスチジン、スレオニン及びグルタミン酸からグリシン、スレオニン及びアラニンに変化した2重変異を作製した。
【0130】
組換えタンパク質
Gatewayを用いてGASC1のcDNA をpCR8/GW からpAcHLT-A(Pharmingen)のGateway修飾型に導入することにより、アミノ末端にヘキサヒスチジンタグが付いた完全長のヒトGASC1バキュロウィルス導入ベクターを作製した。組換えバキュロウィルスは、基本的に、先の文献に記載されたように、GASC1遺伝子を含むバキュロウィルス導入ベクターと、Bsu36IでリニアライズしたBakpak6バキュロウィルスDNAの共トランスフェクションにより作製した。ヒスチジンタグが付いたGASC1とJMJD2Aを発現させ、基本的に先の文献(Christensen et al. Nucleic Acids Research 33, 5458-5470 (2005))に記載されたように、コバルトアフィニティークロマトグラフィーで精製した。溶離した分画を、SDS-PAGEで分析し、選択した分画をサイズ排除クロマトグラフィー(SEC)により更に精製した。簡潔に述べると、SECは、50mM・NaClを含有する25mM・HEPES-KOH(pH7.7)で平衡化させたSuperose 12, 10/300カラム(Pharmacia-Amersham)上で行い、同一のバッファーを用いて、0.3ml/分の流速で溶離した。溶離した物質を、0.5mlの分画で回収し、液体窒素で瞬間冷凍し、-80℃で保存した。全ての工程は、完全にEDTA不含のプロテアーゼ阻害剤(Boehringermannheim、独国)の存在下で、氷上又は4℃で行った。
【0131】
レトロウィルスの形質導入
GASC1とその変異体H190G/E192Aを発現する組換えレトロウィルスを作製するため、GASC1のオープンリーディングフレームを、Gatewayによる組換えによりpBabepuroに導入し、pBabepuro-HA-GASC1とpBabepuro-HA-H190G/E192Aを作製した。Phoenix-Eco 293細胞パッキング用セルラインのトランスフェクションから24-48時間後に、高い力価を有するウィルス粒子が得られた。トランスフェクションは、リン酸カルシウム法により行った。TIG3-hTert-EcoR細胞又はU2OS-EcoR細胞の形質導入は、24時間以内に、ウィルスを含むパッケージングセルラインからの上清を細胞ディッシュに4回添加することにより達成した。形質導入された細胞は、ピューロマイシンの存在下で2-5日間かけて選択した(1μg/μl)。
【0132】
分子モデリング
α-ケトグルタレート、CADペプチド及び鉄と複合体形成したFIHの構造は、1H2Kのアクセス番号でProtein Data Bank(PDB、Brookhaven National Library、アプトン、ニューヨーク州、米国)から入手可能である。ホモロジーモジュールINSIGHT II (2005) (Accelrys、サンディエゴ)を用いて、GASC122-349を鋳型のFIHと共にホモロジーモデリングした。ヒストン尾部(ARTKQTARKSTG)を鋳型のCADペプチドと共にモデリングした。GRID(Version 22、Molecular Discovery Ltd.、オックスフォード、英国)(Goodford, 1985)というプログラムを用いて、GASC1分子の疎水性表面の外形を計算した。続いて、AUTODOCK (Ver. 3.0.5、Scripps Research Institute andmolecular Graphics Laboratory)を用いて、上記のヒストンペプチドを、GASC1の疎水性表面の外形上にドッキングさせた。最後に、前記複合体の構造モデルを、Pymol(http://pymol.sourceforge.net/)で導き出した。
【0133】
補足的方法
材料
ヒストンH3のN末端尾部(1-40)を模倣する43アミノ酸長の合成ペプチド:
【化7】
このペプチドは、リジンとTtdsスペーサーを介してビオチンに連結された結合用のC末端部のチロシン及びシステインを用いて合成した。この実験で用いたペプチドは、非修飾型であるか、リジン27がトリメチル化されているか、又はリジンK9がモノメチル化、ジメチル化若しくはトリメチル化されており、Jerini, GMBH(独国)から購入した。ホルムアルデヒドデヒドロゲナーゼ(FDH、F1879)及びニコチンアミン及びバルクヒストン(H9250)はSigma社から購入した。本試験で用いた抗体は、以下の通り:抗トリメチル化H3-K9(Upstate 07-523)、抗ジメチル化H3-K9(Upstate 07-212)、抗モノメチル化H3-K9(Abcam Ab9045-50)、抗トリメチルH3-K27(Upstate 07-449)、抗トリメチルH4-K20(Upstate 07-463)、抗トリメチル化H3-K4(Abcam ab8580-50)、抗ジメチル化H3-K4(Upstate 07-030)、抗ヒストンH3(Abcam Ab1791-100)、抗HP1α(Upstate 05-689)、抗HP1γ(upstate 05-690)、抗ビオチン-HRP(Sigma A4541)、抗-His(Upstate 05-531)及び抗HA(CRP Inc. AFC-101P)。
【0134】
セルライン及び組織培養
ヒト食道扁平上皮癌セルラインKYSE-70及び150は、German Collection ofmicroorganisms and Cell Cultures(ブラオンシュヴァイヒ、独国)から入手した。hTert を発現する2倍体のヒト線維芽細胞(TIG-3)とマウスのエコトロピックレトロウィルスレセプターEcoRを発現するU2OS細胞を実験に用いた。テトラサイクリン誘導性のアミノ末端にmycタグが付いたGASC1を安定に発現するHEK293細胞を293 TRex-flip-in細胞を基本的に製造元(Invitrogen)による記載に従って用いて作製した。KYSE細胞は、49%のRPMI1640、2%のウシ胎児血清(FCS)が補充された49%のHam's F12中に5%CO2の下で維持した。他の全ての細胞は、10%のウシ胎児血清(FCS)が補充されたDulbecco改変イーグル培地(DMEM)中に5%CO2の下で37℃にて維持した。
【0135】
HeLa核タンパク質のヒストンペプチドプルダウン
HeLa細胞の核抽出物とリジン9がトリメチル化されたヒストンH3ペプチド:
【化8】
を用いてプルダウンアッセイを行った。核は、HeLa細胞から調製し、溶解バッファー(50mMのTris(pH7.2)、300mMのNaCl、0.5%のIGEPAL CA-630、1mMのEDTA(pH8.0)、プロテアーゼ阻害剤を含む1mMのPMSF)で溶解させた。溶解物は、遊離ストレブトアビジンビーズを用いて事前除去し(図S1、事前除去1)、次いで、非修飾型H3ヒストンに結合したストレプトアビジンビーズで事前除去した(図S1、事前除去2)。次いで、事前除去した溶解物を、事前にストレプトアビジンビーズと結合させておいた、K9がトリメチル化されたH3ヒストンペプチド(H3K9me3)とインキュベートした。ビーズを十分に洗浄し、続いて、2x LSBバッファー(100mMのTris-HCl(pH6.8)、200mMのDTT、4%のSDS、20%のグリセロール及び0.2%のブロモフェノールブルー)で結合したタンパク質を樹脂から溶離した(図S1、H3K9me3)。タンパク質をSDS-PAGEで分離し、銀染色により可視化した。プルダウンされたH3K9me3を豊富に含むタンパク質バンドを切り出し、ゲルのままトリプシンで処理し、Wilm et al.(Wilmm. (1996) Nature 379, 466-469)の記載に従って、ナノエレクトロスプレータンデム型質量分析器で分析した。
【0136】
免疫蛍光
共焦点免疫染色法を、基本的にSorensen, CS. et al., Nat. Cell. Biol. 7, 195-201 (2005)の記載に従って行った。
【0137】
細胞溶解物の調製
細胞の全溶解物は、同数の細胞を尿素バッファー(1%のSDS、9Mの尿素、25mMのTris-HCl(pH6.8)、1mMのEDTA、0.7mのβ-メルカプトエタノール)に溶解して作製し、5分間沸騰させ、そして超音波処理した。クロマチンの分画化のため、200μlの事前抽出用バッファー(0.5%のIGEPAL-630、5OmMのNaCl、3mMのMgCl2及び300mMのスクロースを含有する2OmMのHepes-KOH(pH7.2))中の同数の細胞を氷上で30分間インキュベートした。100μlを2xLSBバッファー(100mMのTris-HCl(pH6.8)、200mMのDTT、4%のSDS、20%のグリセロール及び0.2%のブロモフェノールブルー)と混合し、沸騰させ、超音波処理して、全溶解物として使用した。残りの100μlは、1300×g、4℃で10分間スピンダウンさせ、1mlの事前抽出用バッファーで洗浄し、100μlの2xLSBバッファーに再懸濁し、沸騰させ、超音波処理し、クロマチン分画として使用した。
【0138】
GASC1及びJMJD2a/bに対するsiRNA及びshRNA
GASC1並びにそのホモログJMJD2a及びJMJD2bに対する低分子干渉RNA(siRNA)オリゴヌクレオチドは、Dharmacon Research, Inc.によって合成された。6ウェルプレート内で、Oligofectamine試薬(Invitrogen)を製造元のプロトコルに従って用いて、細胞(1x105/ウェル)をsiRNAオリゴ(0.3μg/ウェル)でトランスフェクトした。siRNAは、例えば、siRNA retriever; (http://katahdin.cshl.org:9331/homepage/siRNA/RNAi.cgi?type=siRNA)を用いる等、当該技術分野で知られる方法により設計できる。
【0139】
shRNAコンストラクト
短鎖ヘアピンRNAコンストラクトは、Paddison et al.(2005)(Paddison PJ, Clearym, Silva JM, Chang K, Sheth N, Sachidanandam R & Hannon GJ. Cloning of short hairpin RNAs for gene knockdown inmammalian cells Naturemethods pp163-167)の記載に従って、MSCV/LTRmiR30-PIG (LMP)ベクター(Dickens NG et al(2005)"Probing tumor phenotypes using stable and regulated syntheticmicroRNA precursors" Nature Genetics Vol 37, No 11 1289-1295.)内に作製した。
【0140】
「shRNA retriever」(http://katahdin.cshl.org:9331/homepage/siRNA/RNAi.cgi?type=shRNA)を用いて3種類のオリゴヌクレオチドを設計した。以下のオリゴヌクレオチド:
【化9】
を鋳型として用いて150bpのPCR産物を増幅させ、XhoI及びEcoRIで処理し、そして制限酵素処理したLMPベクター内にライゲーションした。このshRNAコンストラクトは、上記の配列で下線が引かれたGASC1コーディング配列の2117番目の位置の22bpの配列を標的とする。このshRNAiコンストラクトが、GASC1のmRNAをノックダウンする効率は、リアルタイム定量的RT-PCRで評価したところ、約80%であると評価された。
【0141】
リアルタイム定量的RT-PCR
細胞を、表示するshRNAコンストラクトでトランスフェクトし、簡単なピューロマイシンセレクション(2μg/ml)に付した。Quiagen RNAeasyミニキットを製造元の使用説明書に従って用いて全RNAをトランスフェクトした細胞から作製した。Taqman逆転写キットとポリdTプライマーを製造元の使用説明書に従って用いてcDNAを作製した。このcDNAを鋳型として、GASC1特異的プライマーと共に、Applied biosystems 7700でのリアルタイム定量的PCR反応に用いた。この反応物は、Applied biosystems社製の2xSYBR green反応ミックスを用いて調製した。
【0142】
BrdUの取り込み
siRNAのトランスフェクションから30時間後に、細胞を4ウェルチャンバースライドに分け、BrdUを含む培養培地で15-30分間インキュベートした。
【0143】
ヒトGASC1のcDNAのクローニング
ヒトGASC1及びJMJD2Aの推定上のオープンリーディングフレームをHeLaのcDNAとヒト胎児脳のcDNAライブラリー(Invitrogen、カールスバッド、カリフォルニア州)のcDNAからPCR増幅した。プライマー配列は、請求により入手可能である。PCR産物をゲル精製し、pCR8/GW(Invitrogen)にクローニングし、DNAシーケンシングにより確認した。これらのGateway適合性の導入クローンを用いて、GASC1及びJMJD2AをpCMV-HA、pCMV-myc及びpBabepuroに導入した。標準的な変異導入方法により、GASC1の保存された鉄結合ドメイン(HTE)の、190-192番目のアミノ酸が、それぞれヒスチジン、スレオニン及びグルタミン酸からグリシン、スレオニン、アラニンに変化した2重変異を作製した。
【0144】
組換えタンパク質
Gatewayを用いてGASC1のcDNA をpCR8/GW からpAcHLT-A(Pharmingen)のGateway修飾型に導入することにより、アミノ末端にヘキサヒスチジンタグが付いた完全長のヒトGASC1バキュロウィルス導入ベクターを作製した。組換えバキュロウィルスは、基本的に、先の文献に記載されたように、GASC1遺伝子を含むバキュロウィルス導入ベクターと、Bsu36IでリニアライズしたBakpak6バキュロウィルスDNAの共トランスフェクションにより作製した。ヒスチジンタグが付いたGASC1とJMJD2Aを発現させ、基本的に先の文献(Christensen et al. Nucleic Acids Research 33, 5458-5470 (2005))に記載されたように、コバルトアフィニティークロマトグラフィーにより精製した。溶離した分画を、SDS-PAGEで分析し、選択した分画をサイズ排除クロマトグラフィー(SEC)により更に精製した。簡潔に述べると、SECは、50mM・NaClを含有する25mM・HEPES-KOH(pH7.7)で平衡化させたSuperose12、10/300カラム(Pharmacia-Amersham)上で行い、同一のバッファーを用いて、0.3ml/分の流速で溶離した。溶離した物質を、0.5mlの分画で回収し、液体窒素で瞬間冷凍し、-80℃で保存した。全ての工程は、完全にEDTA不含のプロテアーゼ阻害剤(Boehringermannheim、独国)の存在下で、氷上又は4℃で行った。
【0145】
レトロウィルスの形質導入
GASC1とその変異体H190G/E192Aを発現する組換えレトロウィルスを作製するため、GASC1のオープンリーディングフレームを、Gatewayによる組換えによりpBabepuroに導入し、pBabepuro-HA-GASC1とpBabepuro-HA-H190G/E192Aを作製した。Phoenix-Eco 293細胞パッキングセルラインのトランスフェクションから24-48時間後に、高い力価を有するウィルス粒子が得られた。トランスフェクションは、リン酸カルシウム法により行った。TIG3-hTert-EcoR細胞又はU2OS-EcoR細胞の形質導入は、24時間以内に、ウィルスを含むパッケージングセルラインからの上清を細胞ディッシュに4回添加することにより達成した。形質導入された細胞は、ピューロマイシンの存在下で2-5日間かけて選択した(1μg/μl)。
【0146】
分子モデリング
α-ケトグルタレート、CADペプチド及び鉄と複合体形成したFIHの構造は、1H2Kのアクセス番号でProtein Data Bank(PDB、Brookhaven National Library、アプトン、ニューヨーク州、米国)から入手可能である。ホモロジーモジュールINSIGHT II (2005) (Accelrys、サンディエゴ)を用いて、GASC122-349を鋳型のFIHと共にホモロジーモデリングした。ヒストン尾部(ARTKQTARKSTG)を鋳型のCADペプチドと共にモデリングした。GRID(Version 22、Molecular Discovery Ltd.、オックスフォード、英国)(Goodford, 1985)というプログラムを用いて、GASC1分子の疎水性表面の外形を計算した。続いて、AUTODOCK(Ver. 3.0.5、Scripps Research Institute andmolecular Graphics Laboratory)を用いて、上記のヒストンペプチドを、GASC1の疎水性表面の外形上にドッキングさせた。最後に、前記複合体の構造モデルを、Pymol(http://pymol.sourceforge.net/)で導き出した。
【0147】
参考文献
【表1】
【0148】
【表2】
【0149】
【表3】
【0150】
【表4】
【図面の簡単な説明】
【0151】
【図1】JumonjiヒストンデメチラーゼGASC1、FBXL11及びそのヒトホモログ。aは、GASC1、FBXL11、並びにそれらのドメイン:JmjC-Jumonji Cドメイン、JmjN-Jumonji Nドメイン、Tdr-Tudorドメイン、PHD-植物ホメオドメイン、CXXC-CxxCジンクフィンガードメイン、FBX-Fboxドメイン、LR-ロイシンリッチリピートを有するヒトホモログの模式図である。機能ドメインは、SMARTプログラム(http:/smart.embl-heidelberg.de/smart)を用いて決定した。
【図2】bは、ヒトJmjCドメイン含有タンパク質の系統樹である。系統樹は、近接結合(Neighbour Joining:NJ)法によるタンパク質のJmjCドメインに基づいて計算する。この計算に用いた配列を、補足図S2に示す。
【図3】GASC1は、トリメチル化及びジメチル化したヒストンH3K9を脱メチル化する。aは、サイズ排除クロマトグラフィーによる組換えGASC1の精製を表す。0.5mlのフラクションを回収し、10μlの物質をバルクヒストンと37℃で30分間インキュベートした。脱メチル化活性は、トリメチル化H3K9に対するブロッティングにより評価した。bは、GASC1のサイズ排除クロマトグラフィーから選択されたフラクション(F4、F6、F8、F9、FlO、F11、F12、F14、F16)を用いた脱メチル化アッセイを示す。図に示すように、イムノブロッティングによりフラクションの脱メチル化活性を評価した。
【図4】cは、精製したGASC1によるH3K9me3ペプチド(ARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPHRYC-Ttds-K(ビオチン)-NH2)の脱メチル化の質量分析を示す。ペプチドを、GASC1で処理した後(+GASC1)及び未処理のまま(-GASC1)でLTQ-FT上でLC-MSにより分析した。計算値からのモノアイソトピック質量の実測値の偏差値を括弧内に示す。計算上の中性質量は、H3K9me1:5130.885Da、H3K9me2:5144.900Da、H3K9me3:5158.916Daであり、脱メチル化の結果による質量の差異:14.016DaのH3K9(非メチル化)は、観察されなかった。我々のペプチド(分子式C223H390N75O61S2)についてIsoPro 3.0(http://members.aol.com/msmssoft/)により帯電状態+10、解像度50,000(シミュレーションしたスペクトル)でシミュレーションしたところ、個々のピークの多重性は、異なるアイソトープの自然発生により生じている。d、組換えGASC1と、基質として合成トリメチル化H3K9ペプチドを用いた脱メチル化アッセイを示す。ペプチドを、補因子の存在下で6μgのHis-GASC1と共に37℃で30分間インキュベートする。材料の3分の1をウェスタンブロッティングにより解析し、残りを質量分析に用いた(図2e)。
【図5】eは、ヒトJumonjiタンパク質GASC1、JMJD2A、FIH及びFBXL11並びに分裂酵母のJumonjiタンパク質Epe1のJmjCドメインのアライメントを示す。黄色の強調部分は、類似するか又は同一のアミノ酸を表す。黒色及び赤色のアステリスクは、それぞれ鉄の結合とα-ケトグルタレートに寄与すると予測される残基を示す。緑色のアステリスクは、GASC1鉄結合型変異体で変異の生じた残基を示す。矢印は、1h2kのPDB合計入力により定義される、FIHのβ鎖8-15に対応する番号が付された2本鎖β-へリックス鎖を示す。
【図6】fは、トリメチル化H3ヒストン尾部(1-11)(緑色)及びその補因子であるα-ケトグルタレート(赤色)及び鉄(マゼンタ)と複合体形成したGASC1(青色)の予測構造であり、リジンH3K9メチル基は、黄色でマークされている。左側のパネルは、鉄結合残基のHis190、His288、Glu192(白色の棒)の概要であり、右側のパネルは拡大図を示す。gは、バルクヒストンを基質として用い、野生型及び変異型(H190G/E192A)のGASC1の量を変化させながら用いた脱メチル化アッセイを示す。
【図7】GASC1の脱メチル化は、鉄及びα-ケトグルタレートに依存したヒドロキシル化反応を通じて起こる。aは、トリメチル化及びジメチル化H3K9のGASC1媒介性脱メチル化について提唱される反応スキームを示す。1.Fe(II)(マジェンタ)が、表面の3つの金属結合残基His190、Glu192及びHis288(青字)によりGASC1に結合している。2.α-ケトグルタレート(赤色)は、恐らくThr186とLys208の残基を通じて結合している。3.鉄(マジェンタ)は、次いで、分子状態の酸素に結合する。4.次のα-ケトグルタレート(赤色)の酸化的脱カルボキシル化により二酸化炭素、サクシネート(赤色)及びフェリル(ferryl)(FeIV=O、マジェンタ)が生じる。5.フェリル(マジェンタ)は次いで、リジンH3K9のメチル基(緑色)をヒドロキシル化し、ホルムアルデヒドを放出させる。6.最後に、ヒドロキシル化ヒストン尾部(緑色)とサクシネート(赤色)は、GASC1分子を遊離させることが可能であり、新たな脱メチル化サイクルに向けて空いた状態となる。
【図8】bは、合成H3K9me3ペプチドを基質として用いた脱メチル化アッセイを示す。ペプチドを、20μgのHis-GASC1と共に、その補因子であるアスコルビン酸、α-ケトグルタレート及びFe2SO4並びにEDTAの存在下又は不存在下で、37℃にて30分間インキュベートし、ウェスタンブロッティングにより解析した。GASC1の相対脱メチル化活性は、Fujifilm darkbox LAS 3000 readerを用いて定量化し、ローディング対照(α-ビオチン)に対して正規化した。H3K9me3ペプチド(レーン1)を100%に定めた。レーン2-7;EDTA又は提示した補因子の存在下又は不存在下で脱メチル化バッファー中でインキュベートしたH3K9me3ペプチド。cは、組換えGASC1と基質としてバルクヒストンを用いた脱メチル化アッセイを示す。ヒストンを、His-GASC1と共に、補因子(FeSO4及びα-ケトグルタレート)の存在下で、37℃にて30分間、α-ケトグルタレート類似体のN-オキサリルグリシンの存在下でインキュベートした。メチル化状態は、ウェスタンブロッティングにより評価した。dは、Fe(II)活性部位における2-ケトグルタレートとN-オキサリルグリシンの結合様式を示す。eは、GASC1媒介性のH3K9me3脱メチル化によるホルムアルデヒド放出を示す。ホルムアルデヒド放出は、脱メチル化-FDH一体化アッセイ4により測定した。脱メチル化-FDH一体化アッセイは、一定量の酵素GASC1(40μg)を用いたが、基質H3K9me3ペプチドの量を変化させながら行った。ホルムアルデヒド生成は、OD340nmにてNADH精製を測定することでモニターした。基質であるH3K9ペプチド(非メチル化)をネガティブ対照として50μg用いた。
【図9】GASC1の異所発現により、in vivoで、ジメチル化及びトリメチル化H3K9が喪失する。aは、pBabe-GASC1を導入したTig-3細胞の共焦点顕微鏡観察を示す。GASC1を過剰発現するTig-3細胞内でトリメチル化H3K9は喪失する。白色の矢印は、HAタグが付いたGASC1を発現する細胞を示す。bは、免疫蛍光により評価された、GASC1のH190G/E192A変異体を発現するU2OS細胞内でのトリメチル化H3K9を脱メチル化する能力の喪失を示す。白色の矢印は、HAタグ化GASC1を発現するか、又はHAタグ化されたGASC1のH190G/E192A変異体を発現する細胞を示す。
【図10】cは、ウェスタンブロッティングにより評価された、GASC1のH190G/E192A変異体でトランスフェクトされたU2OS細胞内でのトリメチル化H3K9を脱メチル化する能力の喪失を示す。dは、JMJD2aを過剰発現するU2OS細胞内でのトリメチル化及びジメチル化H3K9を脱メチル化する能力の喪失とモノメチル化H3K9の増大を示す。
【図11】fは、HAタグ化GASC1又はHAタグ化されたGASC1のH190G/E192A変異体でトランスフェクトされた3T3細胞の共焦点顕微鏡観察を示す。白色の矢印は、HAタグ化タンパク質を発現する細胞を示す。HP1βは、GASC1を過剰発現する細胞内で非局在化している。gは、テトラサイクリン(Tet)誘導性GASC1発現(Mycタグ化)系を有するHEK293細胞由来の全溶解物及びクロマチン結合フラクションのウェスタンブロッティングを示す。Tetの添加(+)及び省略(-)を、図示する。
【図12】GASC1発現は増殖を促進し、腫瘍発現と正の相関性を有する。aは、GASC1に対するsiRNA又はスクランブルされたsiRNA対照によるKYSE-150及びKYSE-450のトランスフェクションを示す。GASC1のメッセンジャーRNAの相対量は、リアルタイム定量的RT-PCRにより測定した。b、LMP-GASC1又は空のLMPベクターによるKYSE-150のトランスフェクションとその後の3日間のセレクションを示す。GASC1のメッセンジャーRNAの相対量は、リアルタイム定量的RT-PCRにより測定した。cは、GASC1のFe(II)活性部位におけるケルセチンの結合様式を示す。
【図13】dは、組換えGASC1と基質としてバルクヒストンを用いた脱メチル化アッセイを示す。ヒストンを、補因子(FeSO4とα-ケトグルタレート)の存在下でHis-GASC1と共に、可変量のケルセチンの存在化又は不存在下で37℃で30分間インキュベートした。メチル化状態は、ウェスタンブロッティングにより評価した。eは、免疫蛍光での評価による、KYSE-150細胞及びU2OS細胞のH3K9me3メチル化状態に対するケルセチンの効果を示す。
【図14】fは、癌の発症におけるGASC1の関与のモデルを示す。
【図15】(S1)は、HeLa核タンパク質のヒストンペプチドプルダウンを示す。HeLa細胞核抽出物とリジン9がトリメチル化されたヒストンH3ペプチドを用いてプルダウンアッセイを行った。HeLa核抽出物は、遊離ストレブトアビジンビーズを用いて事前除去し(事前除去1)、次いで、非修飾型H3ヒストンに結合したストレプトアビジンビーズで事前除去した(事前除去2)。事前除去した溶解物を、次いで、K9がトリメチル化されたH3ヒストンペプチド(H3K9me3)とインキュベートし、ストレプトアビジンビーズに事前に結合させた。ビーズを洗浄し、次いで結合したタンパク質を溶離させ、SDS-PAGEにより分離して、銀染色により可視化させた。H3K9me3プルダウンで濃縮されたタンパク質のバンドは、次いで、質量分析により同定した。濃縮されたバンドの一つは、Jumonjiタンパク質であるGASC1と同定された(矢印を参照)。やはりプルダウンアッセイで濃縮されたHP1αないしγのタンパク質も同様に示されている。分子量マーカーは左側に示されている。
【図16】(S2)は:ヒトJumonjiタンパク質のJmjCドメインのアライメントを示す。タンパク質は、Clustal Wアルゴリズムを用いたVector NTIによりアライメントさせた。保存されている残基、類似残基及び同一残基のブロックは、グレーで網掛けしてある。括弧内の数字は、そのタンパク質における残基の番号を示す。
【図17】(S2)は:ヒトJumonjiタンパク質のJmjCドメインのアライメントを示す。タンパク質は、Clustal Wアルゴリズムを用いたVector NTIによりアライメントさせた。保存されている残基、類似残基及び同一残基のブロックは、グレーで網掛けしてある。括弧内の数字は、そのタンパク質における残基の番号を示す。
【図18】(S2)は:ヒトJumonjiタンパク質のJmjCドメインのアライメントを示す。タンパク質は、Clustal Wアルゴリズムを用いたVector NTIによりアライメントさせた。保存されている残基、類似残基及び同一残基のブロックは、グレーで網掛けしてある。括弧内の数字は、そのタンパク質における残基の番号を示す。
【図19】(S2)は:ヒトJumonjiタンパク質のJmjCドメインのアライメントを示す。タンパク質は、Clustal Wアルゴリズムを用いたVector NTIによりアライメントさせた。保存されている残基、類似残基及び同一残基のブロックは、グレーで網掛けしてある。括弧内の数字は、そのタンパク質における残基の番号を示す。
【図20】(S3):Hisタグ化組換えGASC1、GASC1(H190G/E192A)変異体並びにJMJD2A及びJMJD2bの精製。コバルト親和性カラムから溶離したフラクション1-4をSDS-PAGEとCoomassie染色に付した。フラクションF2については、サイズ排除クロマトグラフィーにより更に精製して、高純度なタンパク質試料を得た。
【図21】(S4):GASCは、H3K9me3とH3K9me2を脱メチル化する。aは、可変量の組換えGASC1と基質としてオリゴヌクレオソーム又はバルクヒストンを用いた脱メチル化アッセイを示す。bは、組換えGASC1と基質としてバルクヒストンを用いた脱メチル化アッセイを示す。ヒストンを、図示するように可変量のHis-GASC1と共に、その補因子の存在下で、37℃で30分間インキュベートし、ウェスタンブロッティング解析により解析した。60℃で5分間かけて熱失活(失活)させ、GASC1の脱メチル化活性を無力化させた。
【図22】cは、組換えGASC1と基質として合成トリメチル化H3K9ペプチドを用いた脱メチル化アッセイを示す。ペプチドは、10μgのHis-GASC1と共に補因子の存在下で、37℃で30分間インキュベートした。dは、組換えGASC1若しくはJMDM2aと、基質として合成トリメチル化、ジメチル化及びモノメチル化H3-K9ペプチドを用いた脱メチル化アッセイを示す。ペプチドは、8μgのHis-GASC1若しくはHis-JMJD2aと共に補因子の存在下で、37℃で30分間インキュベートし、ウェスタンブロッティング解析により解析した。
【図23】eは、組換えJMDM2aと、基質として合成トリメチル化、ジメチル化及びモノメチル化H3-K9ペプチドを用いた脱メチル化アッセイを示す。ペプチドは、8μgのHis-JMJD2aと共に補因子の存在下で、37℃で30分間インキュベートし、ウェスタンブロッティング解析により解析した。
【図24】(S5):JMJD2A及びJMJD2Bは、H3-K9がトリメチル化及びジメチル化されたヒストンを脱メチル化する。aは、サイズ排除クロマトグラフィーによる組換えJMJD2Aの精製を示す。1mlのフラクションを回収し、10μlの材料をバルクヒストンと共に37℃で30分間インキュベートした。脱メチル化活性は、トリメチル化H3-K9に対するブロッティングにより評価した。b、組換えJMJD2Aと基質としてバルクヒストンを用いた脱メチル化アッセイ。サイズ排除クロマトグラフィーで得られた種々のフラクション(図S5a)をウェスタンブロット解析により解析した。
【図25】cは、サイズ排除クロマトグラフィーによる組換えJMJD2Bの精製。1mlのフラクションを回収し、10μlの材料をバルクヒストンと共に37℃で30分間インキュベートした。脱メチル化活性は、トリメチル化H3-K9に対するブロッティングにより評価した。
【図26】(S6)は、野生型及びH190G/E192A変異型GASC1のサイズ排除クロマトグラフィーを示す。
【図27】(S7)は、α-ケトグルタレート類似体並びにコバルト塩及びニッケル塩は、GASC1の脱メチル化活性を阻害する。組換えGASC1と基質としてバルクヒストンを用いた脱メチル化アッセイ。ヒストンは、His-GASC1と共に、補因子の存在下で、(a)CoCl2若しくは(b)NiSO4のいずれかの存在下又は不存在下で37℃で30分間インキュベートし、ウェスタンブロット解析により解析した。
【図28】(S5):GASC1の異所発現は、H3K4及びH3K27のメチル化に影響しない。aは、免疫蛍光により評価したGASC1を過剰発現するU2OS細胞内のH3K4トリメチル化状態を示す。
【図29】bは、免疫蛍光により評価したGASC1を過剰発現するU2OS及びTig-3細胞内のH3K27トリメチル化状態を示す。
【図30】(S9):ヘテロクロマチンモデリング/可塑性におけるGASC1の推定上の役割。aは、HAタグ化JMJD2a及びJMJD2bでトランスフェクトしたU2OS細胞の共焦点顕微鏡観察を示す。白色の矢印は、HAタグ化GASC1を発現する細胞を示す。JMJD2a及びJMJD2bを過剰発現する細胞では、HP1βが非局在化している。bは、ヘテロクロマチンのモデリング及び可塑性におけるGASC1の役割のモデルを示す。GLP、G9a及びSUV39H1/H2等のH3K9特異的なヒストンメチルトランスフェラーゼ(HMT)のクロマチンに対しての採用により、H3K9マークの進行性のメチル化が生じる。最初に、H3K9は、酵素GLP、G9aによりモノメチル化される可能性がある。SUV39H1/H2等の前進型HMTは、次にジメチル化及びトリメチル化H3K9を生成するように作用し、HP1の結合を可能にする。次に、HP-1結合により、更にHMTが補充され、ヘテロクロマチンが広がることになる。GASC1は、ジメチルマーク及びトリメチルマークを取り除いてHP1結合を妨害することにより、ヘテロクロマチンの拡大と形成に反対する作用を及ぼしている可能性がある。GASC1は、LSD146及びJMJD2a、b等の他のデメチラーゼと協調して、ヘテロクロマチンに反対の作用を及ぼしている可能性がある。
【図31】(S10)は種々のGASC1欠失変異体のH3K9me3脱メチル化活性を示す。aは、本試験で用いた作製されたGASC1欠失変異体を表す図であり、括弧内の数字は、欠失した残基を示す。bは、免疫蛍光で評価した野生型GASC1とGASC欠失変異体の脱メチル化活性を示す。(S11):ヒストンデメチラーゼのJMJD2ファミリーの発現は、前立腺癌腫において上昇している。aは、Lapointe et al (2004) PNAS 101:811-816に記載されるデータを示す。この試験にでは、62種類の初代前立腺腫瘍(PC、61種類の腺癌と1種類の腺様嚢胞癌)、41種類の対応する正常前立腺組織(NP、前立腺の非癌性領域に由来する)、及び9種類の非対応(即ち、異なる被験者の)骨盤リンパ節転移(LNM)が含まれる。26,260種類の異なるヒトの遺伝子を含有するcDNAマイクロアレイ(UniGene clusters)を用いて遺伝子発現プロファイリングを行っている。この試験の病理学的データと臨床データを含む更なる詳細については、Oncomine(www.oncomine.org)又はPNASのウェブサイト(www.PNAS.org)から入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。bは、Yu et al J Clin Oncol (2004) 22:2790-2799に記載されるデータを示す。この試験には、多様な年齢の男性に由来するヒトサンプルが含まれ、前立腺腫瘍(PC、正常な前立腺腺房細胞は全く含まれていない)及び正常な前立腺組織(NP、腫瘍に隣接する部位由来であるが、腫瘍細胞は含まない)が含まれていた。Affymetrix社製(サンタクララ、カリフォルニア州)のU95a、U95b及びU95cのチップセット(37,777種類の遺伝子及び発現配列タグ)を用いて、遺伝子発現プロファイリングを行っている。この試験の更なる詳細は、Oncomineウェブサイト(www.oncomine.org)にて入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。
【図32】(S10)は種々のGASC1欠失変異体のH3K9me3脱メチル化活性を示す。aは、本試験で用いた作製されたGASC1欠失変異体を表す図であり、括弧内の数字は、欠失した残基を示す。bは、免疫蛍光で評価した野生型GASC1とGASC欠失変異体の脱メチル化活性を示す。(S11):ヒストンデメチラーゼのJMJD2ファミリーの発現は、前立腺癌腫において上昇している。aは、Lapointe et al (2004) PNAS 101:811-816に記載されるデータを示す。この試験にでは、62種類の初代前立腺腫瘍(PC、61種類の腺癌と1種類の腺様嚢胞癌)、41種類の対応する正常前立腺組織(NP、前立腺の非癌性領域に由来する)、及び9種類の非対応(即ち、異なる被験者の)骨盤リンパ節転移(LNM)が含まれる。26,260種類の異なるヒトの遺伝子を含有するcDNAマイクロアレイ(UniGene clusters)を用いて遺伝子発現プロファイリングを行っている。この試験の病理学的データと臨床データを含む更なる詳細については、Oncomine(www.oncomine.org)又はPNASのウェブサイト(www.PNAS.org)から入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。bは、Yu et al J Clin Oncol (2004) 22:2790-2799に記載されるデータを示す。この試験には、多様な年齢の男性に由来するヒトサンプルが含まれ、前立腺腫瘍(PC、正常な前立腺腺房細胞は全く含まれていない)及び正常な前立腺組織(NP、腫瘍に隣接する部位由来であるが、腫瘍細胞は含まない)が含まれていた。Affymetrix社製(サンタクララ、カリフォルニア州)のU95a、U95b及びU95cのチップセット(37,777種類の遺伝子及び発現配列タグ)を用いて、遺伝子発現プロファイリングを行っている。この試験の更なる詳細は、Oncomineウェブサイト(www.oncomine.org)にて入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。
【図33】(S10)は種々のGASC1欠失変異体のH3K9me3脱メチル化活性を示す。aは、本試験で用いた作製されたGASC1欠失変異体を表す図であり、括弧内の数字は、欠失した残基を示す。bは、免疫蛍光で評価した野生型GASC1とGASC欠失変異体の脱メチル化活性を示す。(S11):ヒストンデメチラーゼのJMJD2ファミリーの発現は、前立腺癌腫において上昇している。aは、Lapointe et al (2004) PNAS 101:811-816に記載されるデータを示す。この試験にでは、62種類の初代前立腺腫瘍(PC、61種類の腺癌と1種類の腺様嚢胞癌)、41種類の対応する正常前立腺組織(NP、前立腺の非癌性領域に由来する)、及び9種類の非対応(即ち、異なる被験者の)骨盤リンパ節転移(LNM)が含まれる。26,260種類の異なるヒトの遺伝子を含有するcDNAマイクロアレイ(UniGene clusters)を用いて遺伝子発現プロファイリングを行っている。この試験の病理学的データと臨床データを含む更なる詳細については、Oncomine(www.oncomine.org)又はPNASのウェブサイト(www.PNAS.org)から入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。bは、Yu et al J Clin Oncol (2004) 22:2790-2799に記載されるデータを示す。この試験には、多様な年齢の男性に由来するヒトサンプルが含まれ、前立腺腫瘍(PC、正常な前立腺腺房細胞は全く含まれていない)及び正常な前立腺組織(NP、腫瘍に隣接する部位由来であるが、腫瘍細胞は含まない)が含まれていた。Affymetrix社製(サンタクララ、カリフォルニア州)のU95a、U95b及びU95cのチップセット(37,777種類の遺伝子及び発現配列タグ)を用いて、遺伝子発現プロファイリングを行っている。この試験の更なる詳細は、Oncomineウェブサイト(www.oncomine.org)にて入手可能である。バーは中央値を示す。Mann-WithneyのU検定(片側検定)のP値を示す。
【特許請求の範囲】
【請求項1】
Jumonjiタンパク質のJMJD2サブファミリーのタンパク質に結合し、そして任意にその活性を調節する試験化合物の能力を試験する方法であって、当該方法が、試験化合物をJumonjiタンパク質のJMJD2サブファミリーのタンパク質、当該タンパク質の補因子、及び任意で脱メチル化の基質とインキュベートすることを含む、前記方法。
【請求項2】
前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質が、以下の:
a)配列番号1に示すアミノ酸配列;
b)配列番号1に示すアミノ酸配列の断片;
c)a)の配列若しくはb)の配列のいずれか、又は両方と少なくとも45%の配列同一性を示すアミノ酸、
を含む群から選択される、請求項1に記載の方法。
【請求項3】
前記Jumonjiタンパク質のJMJD 2サブファミリーのタンパク質が、以下の:
a)配列番号1(GASC-1(JMJD 2c))、配列番号2(JMJD2a)、配列番号3(JMJD2b)に示すアミノ酸配列;
b)a)のいずれか1つのアミノ酸配列の断片;
c)a)のいずれか1つの配列及び/又はb)のいずれか1つの配列と少なくとも75%の配列同一性を示すアミノ酸、
を含む群から選択される、請求項1又は2に記載の方法。
【請求項4】
前記Jumonjiタンパク質のJMJDサブファミリーのタンパク質が、配列番号4、10及び16(JmjN)、配列番号5、11及び17(JmjC)、配列番号6、7、12、13、18及び19(PHD)、並びに配列番号8、9、14、15、20及び21(Tdr/TUDOR)から成る群から選択される、1以上のアミノ酸配列(ドメイン)を含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
前記方法が、以下の追加のステップ:
a)試験サンプルの以下のいずれかのパラメーター:
i)基質のメチル化の状態;
ii)ホルムアルデヒドの放出;
iii)α-ケトグルタレート基質補因子のカルボキシル化の状態;
iv)酸素消費量
をモニターし、
b)ステップa)で得られた試験サンプルについての数値を、対照サンプルについて得られた数値と比較し、それによって、Jumonjiタンパク質のJMJD 2サブファミリーのタンパク質の活性を調節する前記試験化合物の能力を測定すること、
を含む、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
以下のステップ:
a)Jumonjiタンパク質のJMJD2サブファミリーのタンパク質と脱メチル化の基質を、脱メチル化が可能な条件下でインキュベートすること;
b)試験サンプルの以下のいずれかのパラメーター:
i)基質のメチル化の状態;
ii)試験サンプル中のホルムアルデヒドの放出;
iii)α-ケトグルタレート基質補因子のカルボキシル化状態;
iv)酸素消費量
をモニターすること
により、前記対照が提供される、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
前記インキュベートの時間並びに基質のメチル化状態及び/又はホルムアルデヒドの放出を用いて、脱メチル化の速度を測定し、この脱メチル化の速度を比較に用いる、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
前記脱メチル化の基質が、ヒストンH3のリジン9に対応するメチル化された部位を含む、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
前記脱メチル化の基質が、配列番号25に示すアミノ酸配列(TARKSTG)を含むペプチドであり、前記アミノ酸配列中のリジン残基がメチル化されている、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
前記脱メチル化の基質が、少なくとも15個のアミノ酸残基のペプチドである、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記脱メチル化の基質が、少なくとも20個のアミノ酸残基のペプチドである、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記基質が、バルクヒストン、合成ペプチド、及びヌクレオソームから成る群から選択される、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
前記メチル化された部位が、モノメチル化、ジメチル化又はトリメチル化されている、請求項8に記載の方法。
【請求項14】
前記化合物と前記Jumonjiタンパク質のJMJD 2サブファミリーのタンパク質が、Fe(II)イオン、α-ケトグルタレート及び/又はアスコルビン酸とインキュベートされる、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
ステップa)が、イムノブロッティング又は質量分析により行われる、請求項5に記載の方法。
【請求項16】
以下のステップ:
i) 鉄及びα-ケトグルタレートを含む溶液中の請求項8〜13のいずれか1項に記載の脱メチル化の基質の懸濁液を調製し;
ii) 前記試験化合物を、前記懸濁液に添加し;
iii) 上記ii)の懸濁液に、請求項2〜4のいずれか1項に記載のJumonjiタンパク質のJMJD2サブファミリーのタンパク質を添加し;
iv) 前記懸濁液をインキュベートし;そして
v) 前記基質の脱メチル化の程度を測定すること、
を含む、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
前記脱メチル化の程度が、リジン9がメチル化されたヒストンH3に特異的な免疫結合パートナーを用いて評価される、請求項16に記載の方法。
【請求項18】
前記免疫結合パートナーが、抗トリメチル化ヒストンH3-K9抗体である、請求項16又は17に記載の方法。
【請求項19】
前記免疫結合パートナーが、抗ジメチル化ヒストンH3-K9抗体である、請求項16又は17に記載の方法。
【請求項20】
前記脱メチル化の程度が、ホルムアルデヒドの放出を用いて評価される、請求項16に記載の方法。
【請求項21】
前記脱メチル化の程度が、酸素消費量を用いて評価される、請求項16に記載の方法。
【請求項22】
前記脱メチル化の程度が、α-ケトグルタレート基質補因子のカルボキシル化状態を用いて評価される、請求項16に記載の方法。
【請求項1】
Jumonjiタンパク質のJMJD2サブファミリーのタンパク質に結合し、そして任意にその活性を調節する試験化合物の能力を試験する方法であって、当該方法が、試験化合物をJumonjiタンパク質のJMJD2サブファミリーのタンパク質、当該タンパク質の補因子、及び任意で脱メチル化の基質とインキュベートすることを含む、前記方法。
【請求項2】
前記Jumonjiタンパク質のJMJD2サブファミリーのタンパク質が、以下の:
a)配列番号1に示すアミノ酸配列;
b)配列番号1に示すアミノ酸配列の断片;
c)a)の配列若しくはb)の配列のいずれか、又は両方と少なくとも45%の配列同一性を示すアミノ酸、
を含む群から選択される、請求項1に記載の方法。
【請求項3】
前記Jumonjiタンパク質のJMJD 2サブファミリーのタンパク質が、以下の:
a)配列番号1(GASC-1(JMJD 2c))、配列番号2(JMJD2a)、配列番号3(JMJD2b)に示すアミノ酸配列;
b)a)のいずれか1つのアミノ酸配列の断片;
c)a)のいずれか1つの配列及び/又はb)のいずれか1つの配列と少なくとも75%の配列同一性を示すアミノ酸、
を含む群から選択される、請求項1又は2に記載の方法。
【請求項4】
前記Jumonjiタンパク質のJMJDサブファミリーのタンパク質が、配列番号4、10及び16(JmjN)、配列番号5、11及び17(JmjC)、配列番号6、7、12、13、18及び19(PHD)、並びに配列番号8、9、14、15、20及び21(Tdr/TUDOR)から成る群から選択される、1以上のアミノ酸配列(ドメイン)を含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
前記方法が、以下の追加のステップ:
a)試験サンプルの以下のいずれかのパラメーター:
i)基質のメチル化の状態;
ii)ホルムアルデヒドの放出;
iii)α-ケトグルタレート基質補因子のカルボキシル化の状態;
iv)酸素消費量
をモニターし、
b)ステップa)で得られた試験サンプルについての数値を、対照サンプルについて得られた数値と比較し、それによって、Jumonjiタンパク質のJMJD 2サブファミリーのタンパク質の活性を調節する前記試験化合物の能力を測定すること、
を含む、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
以下のステップ:
a)Jumonjiタンパク質のJMJD2サブファミリーのタンパク質と脱メチル化の基質を、脱メチル化が可能な条件下でインキュベートすること;
b)試験サンプルの以下のいずれかのパラメーター:
i)基質のメチル化の状態;
ii)試験サンプル中のホルムアルデヒドの放出;
iii)α-ケトグルタレート基質補因子のカルボキシル化状態;
iv)酸素消費量
をモニターすること
により、前記対照が提供される、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
前記インキュベートの時間並びに基質のメチル化状態及び/又はホルムアルデヒドの放出を用いて、脱メチル化の速度を測定し、この脱メチル化の速度を比較に用いる、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
前記脱メチル化の基質が、ヒストンH3のリジン9に対応するメチル化された部位を含む、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
前記脱メチル化の基質が、配列番号25に示すアミノ酸配列(TARKSTG)を含むペプチドであり、前記アミノ酸配列中のリジン残基がメチル化されている、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
前記脱メチル化の基質が、少なくとも15個のアミノ酸残基のペプチドである、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記脱メチル化の基質が、少なくとも20個のアミノ酸残基のペプチドである、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記基質が、バルクヒストン、合成ペプチド、及びヌクレオソームから成る群から選択される、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
前記メチル化された部位が、モノメチル化、ジメチル化又はトリメチル化されている、請求項8に記載の方法。
【請求項14】
前記化合物と前記Jumonjiタンパク質のJMJD 2サブファミリーのタンパク質が、Fe(II)イオン、α-ケトグルタレート及び/又はアスコルビン酸とインキュベートされる、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
ステップa)が、イムノブロッティング又は質量分析により行われる、請求項5に記載の方法。
【請求項16】
以下のステップ:
i) 鉄及びα-ケトグルタレートを含む溶液中の請求項8〜13のいずれか1項に記載の脱メチル化の基質の懸濁液を調製し;
ii) 前記試験化合物を、前記懸濁液に添加し;
iii) 上記ii)の懸濁液に、請求項2〜4のいずれか1項に記載のJumonjiタンパク質のJMJD2サブファミリーのタンパク質を添加し;
iv) 前記懸濁液をインキュベートし;そして
v) 前記基質の脱メチル化の程度を測定すること、
を含む、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
前記脱メチル化の程度が、リジン9がメチル化されたヒストンH3に特異的な免疫結合パートナーを用いて評価される、請求項16に記載の方法。
【請求項18】
前記免疫結合パートナーが、抗トリメチル化ヒストンH3-K9抗体である、請求項16又は17に記載の方法。
【請求項19】
前記免疫結合パートナーが、抗ジメチル化ヒストンH3-K9抗体である、請求項16又は17に記載の方法。
【請求項20】
前記脱メチル化の程度が、ホルムアルデヒドの放出を用いて評価される、請求項16に記載の方法。
【請求項21】
前記脱メチル化の程度が、酸素消費量を用いて評価される、請求項16に記載の方法。
【請求項22】
前記脱メチル化の程度が、α-ケトグルタレート基質補因子のカルボキシル化状態を用いて評価される、請求項16に記載の方法。
【図1】

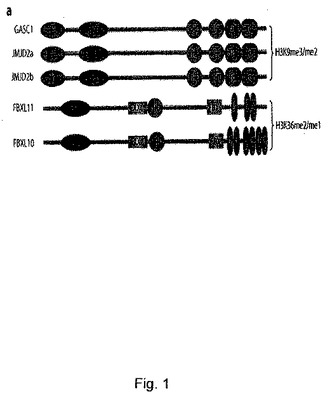
【図2】

【図3】
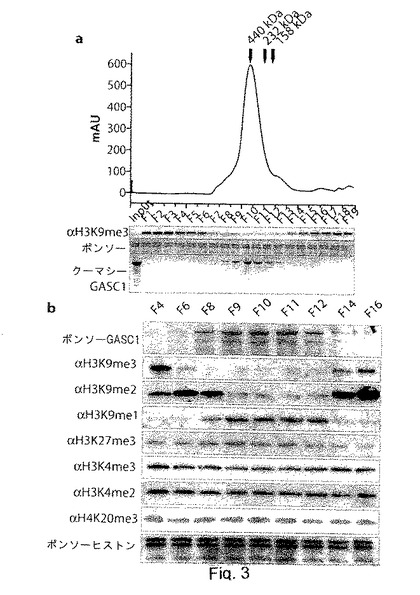
【図4】
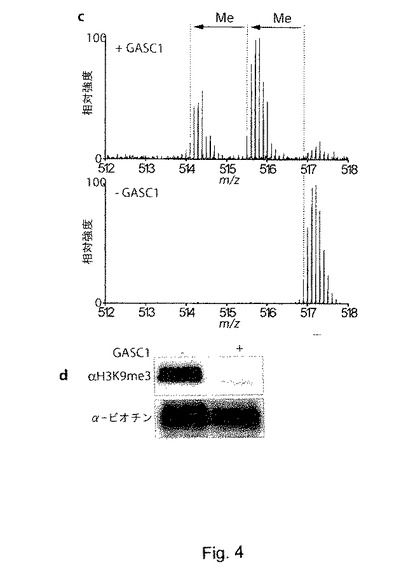
【図5】

【図6】
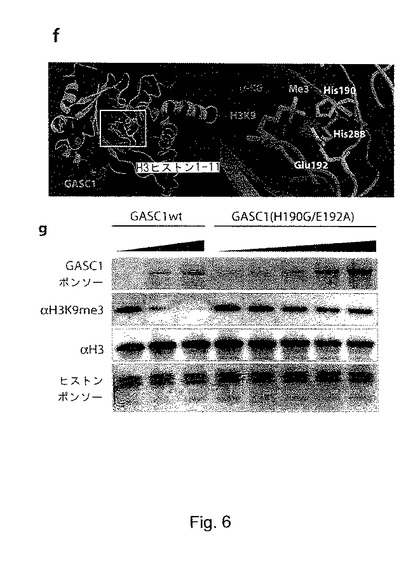
【図7】
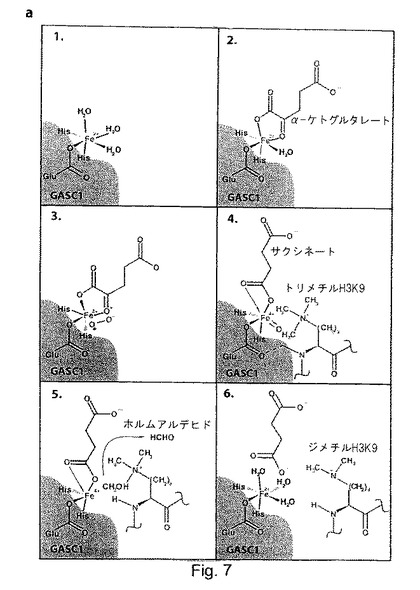
【図8】
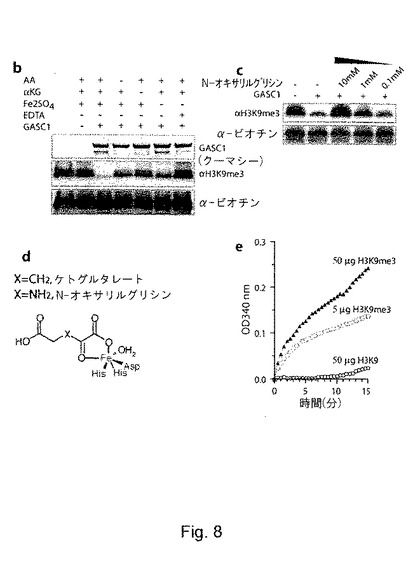
【図9】
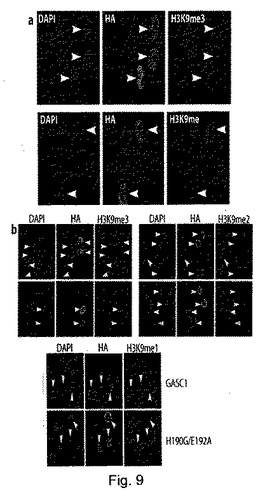
【図10】
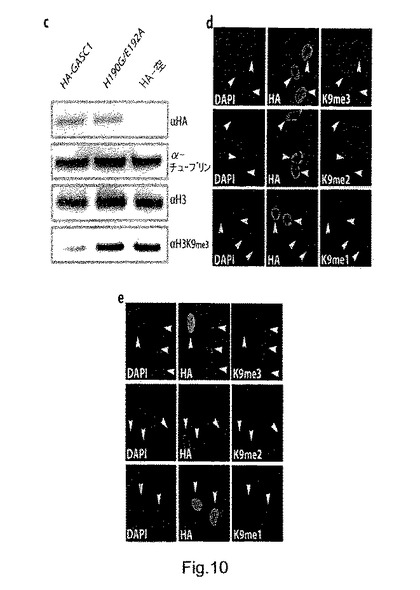
【図11】
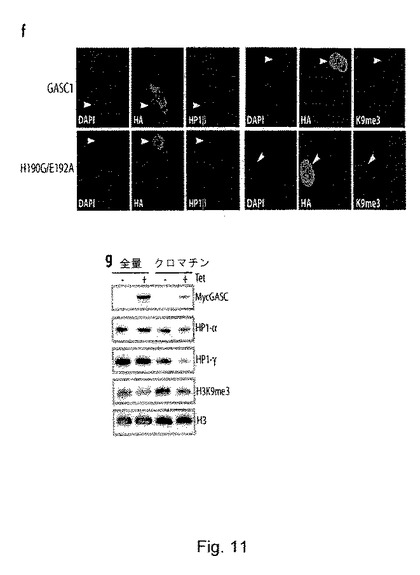
【図12】
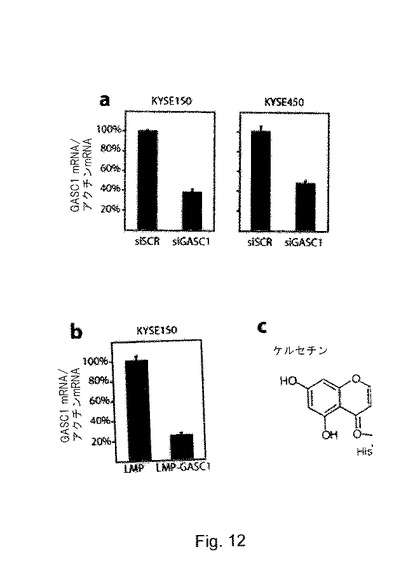
【図13】
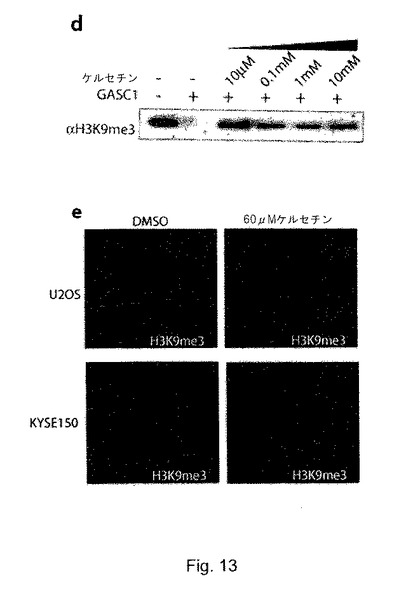
【図14】

【図15】
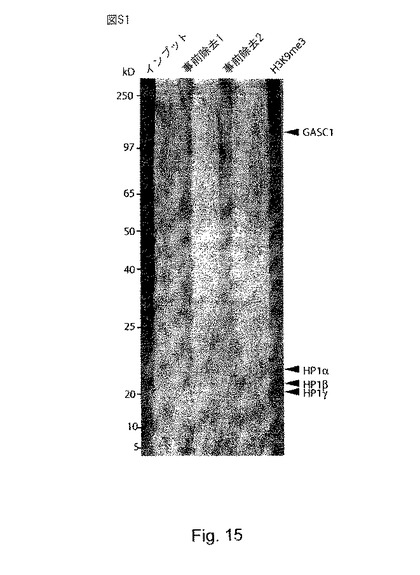
【図16】

【図17】

【図18】

【図19】

【図20】
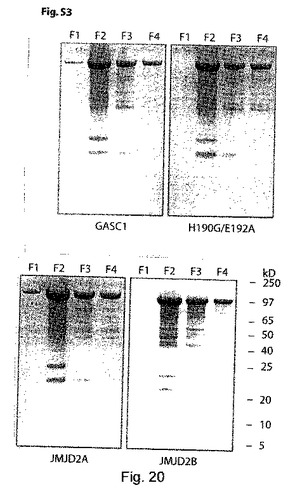
【図21】
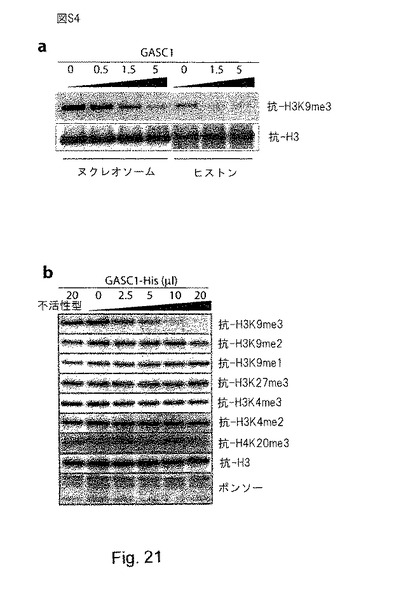
【図22】
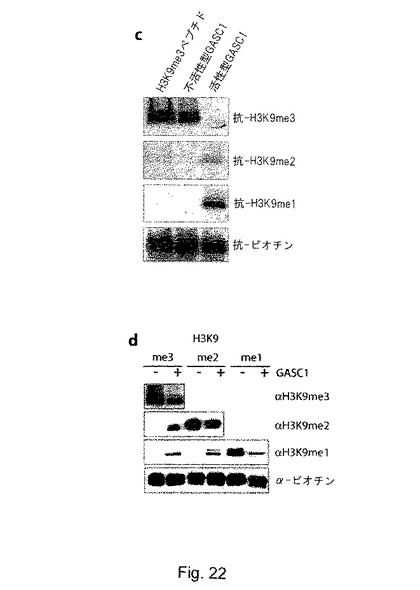
【図23】

【図24】
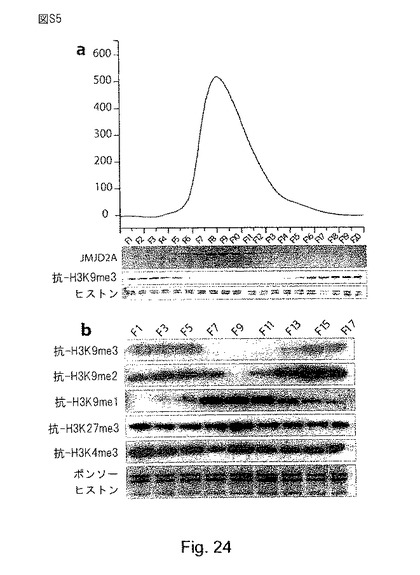
【図25】
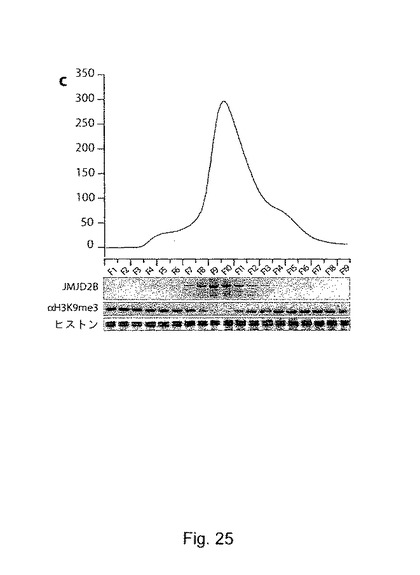
【図26】
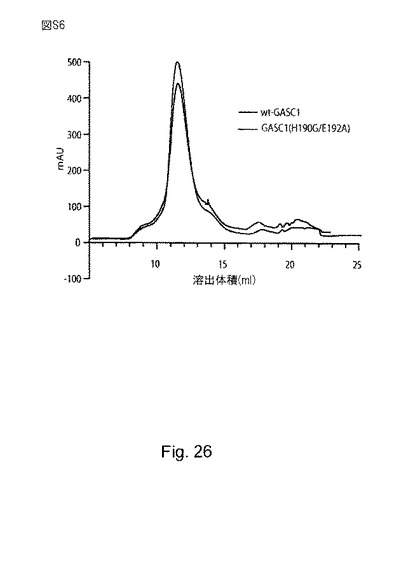
【図27】
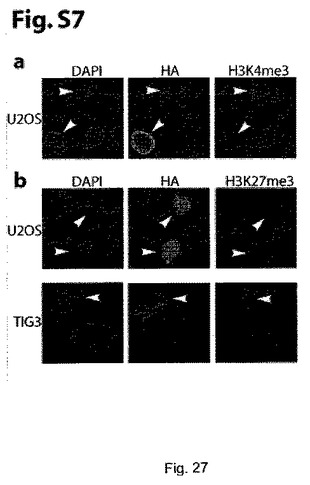
【図28】
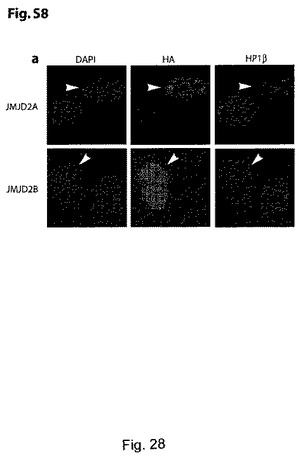
【図29】
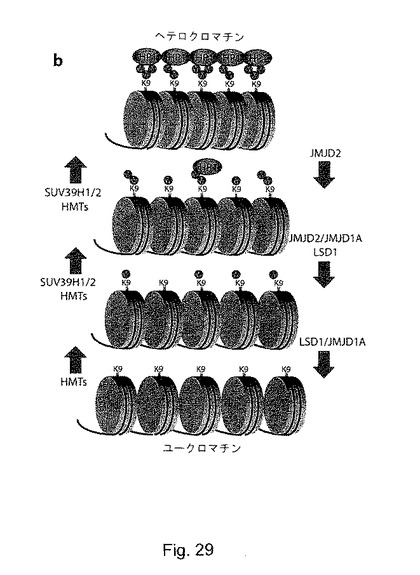
【図30】
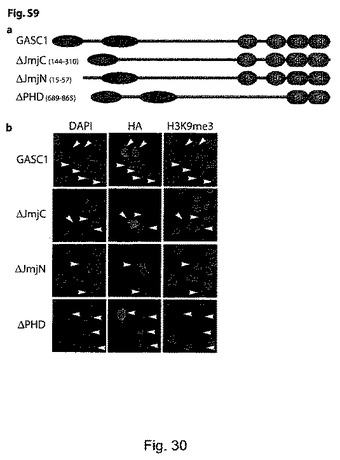
【図31】
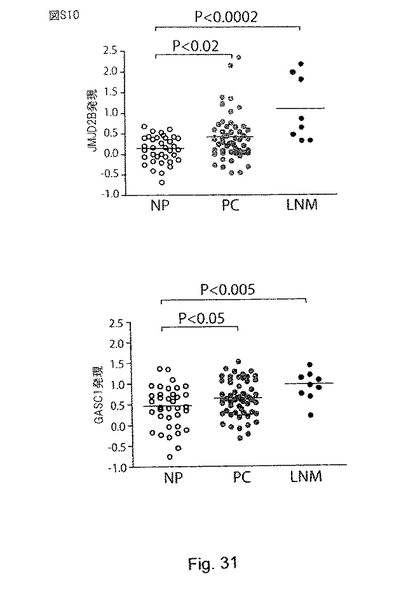
【図32】

【図33】
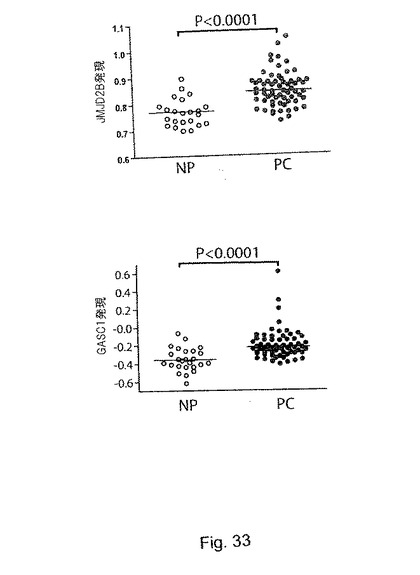
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【公表番号】特表2009−529336(P2009−529336A)
【公表日】平成21年8月20日(2009.8.20)
【国際特許分類】
【出願番号】特願2008−558639(P2008−558639)
【出願日】平成19年3月14日(2007.3.14)
【国際出願番号】PCT/DK2007/000128
【国際公開番号】WO2007/104314
【国際公開日】平成19年9月20日(2007.9.20)
【出願人】(309002994)ケベンハウンス ウニベルシテット (1)
【Fターム(参考)】
【公表日】平成21年8月20日(2009.8.20)
【国際特許分類】
【出願日】平成19年3月14日(2007.3.14)
【国際出願番号】PCT/DK2007/000128
【国際公開番号】WO2007/104314
【国際公開日】平成19年9月20日(2007.9.20)
【出願人】(309002994)ケベンハウンス ウニベルシテット (1)
【Fターム(参考)】
[ Back to top ]