
GLP−1融合タンパク質
【課題】本発明は、血漿半減期が短い化合物を送達することに付随する課題を克服するものである。
【解決手段】本発明は、グルカゴン様ペプチド1の生体内半減期を延ばす効果を持つタンパク質に融合されたグルカゴン様ペプチド1化合物に関する。これらの融合タンパク質は、インスリン非依存性糖尿病および他の様々な状態の処置に使用することができる。
【解決手段】本発明は、グルカゴン様ペプチド1の生体内半減期を延ばす効果を持つタンパク質に融合されたグルカゴン様ペプチド1化合物に関する。これらの融合タンパク質は、インスリン非依存性糖尿病および他の様々な状態の処置に使用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、グルカゴン様ペプチドの生体内半減期を延ばす効果を持つタンパク質に融合されたグルカゴン様ペプチド(その類似体および誘導体を含む)に関する。これらの融合タンパク質は、インスリン非依存性糖尿病および他の様々な状態の処置に使用することができる。
【背景技術】
【0002】
グルカゴン様ペプチド1(GLP-1)は食物摂取に反応して腸のL細胞が分泌する37アミノ酸ペプチドである。これは、インスリン分泌を刺激すること(インスリン分泌促進作用)により、細胞によるグルコースの取込みおよび血清グルコースレベルの低下を引き起こすことがわかっている[例えば非特許文献1を参照されたい]。しかしGLP-1は活性に乏しい。次に、第6位と第7位の間で内因的に切断されることにより、生物学的に活性でより強力なGLP-1(7-37)OHペプチドが生成する。数多くのGLP-1類似体および誘導体が知られているが、本明細書ではそれらを「GLP-1化合物」と呼ぶ。これらのGLP-1類似体には、アメリカドクトカゲの毒液中に見いだされるペプチドであるエクセンディン(Exendin)類が含まれる。エクセンディン類は、天然GLP-1に対して配列相同性を持ち、GLP-1受容体を結合して、GLP-1(7-37)OHに起因するとされている数多くの活性を担うシグナル伝達カスケードを開始させることができる。
【0003】
GLP-1化合物は生理学的に重要な種々の活性を持っている。例えば、GLP-1はインスリン放出を刺激し、グルカゴン分泌を低下させ、胃内容排出を阻害し、グルコース利用を増進することが明らかにされている[非特許文献2、非特許文献3、非特許文献4]。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Mojsov, S., (1992) Int. J. Peptide Protein Research, 40:333-343
【非特許文献2】Nauck, M.A.ら (1993) Diabetologia 36:741-744
【非特許文献3】Gutniak, M.ら (1992) New England J. of Med. 326:1316-1322
【非特許文献4】Nauck, M.A.ら (1993) J. Clin. Invest. 91:301-307
【発明の概要】
【発明が解決しようとする課題】
【0005】
GLP-1はインスリン非依存性糖尿病(NIDDM)の処置として最も有望視されている。NIDDMに関係するインスリン抵抗性を処置するために、数多くの経口薬が市場に出ている。しかし疾患の進行に伴って、患者は、インスリンの放出を刺激する処置に移行しなければならず、そしてついにはインスリンの注射を含む処置に移行しなければならなくなる。しかし、現在用いられているインスリン放出刺激薬は、実際にインスリンを投与した場合と同様に、低血糖を引き起こしうる。しかしGLP-1活性は血中グルコースレベルによって制御される。レベルが一定の閾レベルまで低下すると、GLP-1は活性でなくなる。したがって、GLP-1を使用する処置は、低血糖の危険を伴わない。
【0006】
しかし、GLP-1ペプチドはクリアランスが速く半減期が短いので、GLP-1ペプチドを使用する治療法の有用性には限界があった。例えばGLP-1(7-37)の血清半減期は3〜5分しかない。GLP-1(7-36)アミドは、皮下投与した場合、約50分の作用時間を持つ。内在性プロテアーゼ切断に対して耐性な類似体および誘導体でも、反復投与を24時間以上避けることができるほど長い半減期は持たない。治療薬の速いクリアランスは、反復投与が必要になるので、薬剤の血中レベルを長時間にわたって高く維持することが望まれる場合には不都合である。また、過去の処置方法で経口薬の服用しか経験していない糖尿病患者にとって、持効性化合物は特に重要である。これらの患者は、薬剤を複数回注射する必要がある方法に移行するのに、しばしば極めて苦しい思いをする。
【課題を解決するための手段】
【0007】
本発明は、血漿半減期が短い化合物を送達することに付随する課題を克服するものである。本発明の化合物は、循環半減期が長い別のタンパク質(免疫グロブリンのFc部分またはアルブミンなど)に融合されたGLP-1化合物を包含する。
【0008】
小さい治療ペプチドは、わずかな構造変化でも安定性および/または生物学的活性に影響が及びうるので、一般に、操作することが難しい。これは、現在開発中のGLP-1化合物には、特に当てはまることであった。例えばGLP-1(7-37)OHには、αヘリックスを主とする構造からβシートを主とする構造へのコンフォメーション変化を起こす傾向がある。このβシート型は凝集物をもたらし、その凝集物は不活性であると考えられている。したがって、増加した半減期を持つ、生物学的に活性なGLP-1融合タンパク質を開発できることは、驚くべきことだった。これは、GLP-1(7-37)OHの単独での取り扱いが困難であることと、結合している小さなGLP-1ペプチドと比較して融合パートナーのサイズが大きいことを考えると、とりわけ予想外なことだった。
【0009】
本発明の化合物には、N末端およびC末端を持つ第1ポリペプチドがN末端およびC末端を持つ第2のポリペプチドに融合してなる異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)ヒトアルブミン、
b)ヒトアルブミン類似体、および
c)ヒトアルブミンの断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端に融合している異種融合タンパク質が包含される。
【0010】
本発明の化合物には、N末端およびC末端を持つ第1ポリペプチドがN末端およびC末端を持つ第2ポリペプチドに融合してなる異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)ヒトアルブミン、
b)ヒトアルブミン類似体、および
c)ヒトアルブミンの断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端にペプチドリンカーを介して融合している異種融合タンパク質も包含される。ペプチドリンカーは、
a)グリシンリッチペプチド、
b)[Gly-Gly-Gly-Gly-Ser]nという配列(式中、nは1、2、3、4、5または6である)を持つペプチド、および
c)[Gly-Gly-Gly-Gly-Ser]3という配列を持つペプチド、
からなる群より選択することが好ましい。
【0011】
他にも、本発明の化合物には、N末端およびC末端を持つ第1ポリペプチドがN末端およびC末端を持つ第2ポリペプチドに融合してなる異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)免疫グロブリンのFc部分、
b)免疫グロブリンのFc部分の類似体、および
c)免疫グロブリンのFc部分の断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端に融合している異種融合タンパク質も含まれる。GLP-1化合物はペプチドリンカーを介して第2ポリペプチドに融合していてもよい。ペプチドリンカーは、
a)グリシンリッチペプチド、
b)[Gly-Gly-Gly-Gly-Ser]nという配列(式中、nは1、2、3、4、5または6である)を持つペプチド、および
c)[Gly-Gly-Gly-Gly-Ser]3という配列を持つペプチド、
からなる群より選択することが好ましい。
【0012】
異種融合タンパク質の一部であるGLP-1化合物は、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン(Exendin)4中の対応するアミノ酸と相違するアミノ酸を、6個を超えて含まないことが、一般に好ましい。GLP-1化合物は、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、5個を超えて含まないことが、さらに好ましい。GLP-1化合物は、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、4個、3個、または2個を超えて含まないことが、最も好ましい。異種融合タンパク質の一部であるGLP-1化合物は、好ましくは、8位にグリシンまたはバリンを持つ。
【0013】
本発明は、本明細書に記載の異種融合タンパク質をコードするポリヌクレオチド、これらのポリヌクレオチドを含むベクター、および本明細書に記載のベクターでトランスフェクトまたは形質転換された宿主細胞も包含する。また、異種融合タンパク質を製造する方法であって、検出可能な量の異種融合タンパク質が発現される条件で、本明細書に記載のポリヌクレオチドを転写し翻訳する工程を含む方法も包含される。
【0014】
本発明は、血中グルコースレベルの正常化を必要とする哺乳動物の血中グルコースレベルを正常化する方法であって、治療有効量の本明細書に記載の異種融合タンパク質を投与することを含む方法も包含する。
【0015】
添付の図面を参照して本発明をさらに詳しく説明する。
【図面の簡単な説明】
【0016】
【図1】ヒンジ領域、CH2ドメインおよびCH3ドメインを含むIgG1 Fcアミノ酸配列示す図である。
【図2−1】ヒト血清アルブミンアミノ酸配列を示す図である。
【図2−2】ヒト血清アルブミンアミノ酸配列を示す図である。
【図3】(A)IgG1-FcおよびGLP-1-Fc融合タンパク質の分子量を示すSDS-PAGEゲルおよび同じゲルの免疫ブロットを示す図である(レーン1、MW標品;レーン2、精製Fc;レーン3、疑似トランスフェクトした培地;レーン4、 Val8-GLP-1-Fc;レーン5、エクセンディン4-Fc)。(B)ヒトHSAおよびGLP-1-HSA融合タンパク質の分子量を示すSDS-PAGEゲルおよび同じゲルの免疫ブロットを示す図である(レーン1、MW標品;レーン2、精製HSA;レーン3、疑似トランスフェクトした培地;レーン4、Val8-GLP-1-HSA;レーン5、Val8-GLP-1-[Gly-Gly-Gly-Gly-Ser]3-HSA;レーン6、エクセンディン4-HSA;レーン7、エクセンディン4-[Gly-Gly-Gly-Gly-Ser]3-HSA)。
【図4】精製Fc、アルブミンおよびGLP-1融合タンパク質のSDS-PAGEゲルを示す図である(レーン1、MW標品;レーン2、精製Fc;レーン3、Val8-GLP-1-Fc;レーン4、エクセンディン4-Fc、レーン5、MW標品;レーン6、Val8-GLP-1-HSA;レーン7、エクセンディン4-HSA;レーン8、エクセンディン4-[Gly-Gly-Gly-Gly-Ser]3-HSA)。
【図5】図1に示すFc領域を含む発現クローニングベクターを示す図である。
【図6】図2に示すアルブミン配列を含む発現クローニングベクターを示す図である。
【図7】図2に示すアルブミン配列の5'にインフレームで融合した15アミノ酸リンカーをコードするDNAを含む発現クローニングベクターを示す図である。
【図8】GLP-1融合タンパク質の試験管内用量反応活性を示す図である。
【図9】GLP-1 FcおよびHSA融合タンパク質の薬物動態を示す図である。
【図10−1】2頭の正常絶食イヌにおけるエクセンディン-Fcに対するグルコース動態反応を示す図である。
【図10−2】2頭の正常絶食イヌにおけるエクセンディン-Fcに対するグルコース動態反応を示す図である。
【図11−1】2頭の正常絶食イヌにおけるエクセンディン-Fcに対するインスリン分泌促進反応を示す図である。
【図11−2】2頭の正常絶食イヌにおけるエクセンディン-Fcに対するインスリン分泌促進反応を示す図である。
【図12】ヒトIgG1 Fc領域をコードするDNA配列を示す図である。
【図13−1】ヒトアルブミンタンパク質をコードするDNA配列を示す図である。
【図13−2】ヒトアルブミンタンパク質をコードするDNA配列を示す図である。
【発明を実施するための形態】
【0017】
本発明の異種融合タンパク質は、ヒトアルブミン、ヒトアルブミン類似体、ヒトアルブミン断片、免疫グロブリンのFc部分、免疫グロブリンのFc部分の類似体、または免疫グロブリンのFc部分の断片に融合されたGLP-1化合物を含む。GLP-1化合物のC末端は、アルブミンまたはFcタンパク質のN末端に直接融合するか、またはペプチドリンカーを介して融合することができる。これらの異種融合タンパク質は生物学的に活性であり、天然GLP-1と比較して増加した半減期を持つ。
【0018】
異種融合タンパク質の一部を構成するGLP-1化合物は、膵臓のβ細胞上のGLP-1受容体に結合することによってインスリン分泌促進活性を示すことができるように天然のGLP-1(7-37)OHに対して十分な相同性を持つ、約25〜約39個の天然または非天然アミノ酸を有するポリペプチドを含むことが好ましい。GLP-1化合物は、典型的には、GLP-1(7-37)OH、GLP-1(7-37)OHの類似体、GLP-1(7-37)OHの断片、またはGLP-1(7-37)OH類似体の断片のアミノ酸配列を持つポリペプチドを含む。GLP-1(7-37)OHは配列番号1のアミノ酸配列:
【化1】

(配列番号1)
を持つ。
【0019】
当技術分野の慣例により、GLP-1(7-37)OHのアミノ末端には残基番号7が割り当てられ、カルボキシ末端には番号37が割り当てられている。配列番号1に示すように、ポリペプチド中の他のアミノ酸には連続した番号が与えられる。例えば12位はフェニルアラニンであり、22位はグリシンである。
【0020】
GLP-1化合物には「GLP-1断片」も包含される。GLP-1断片は、GLP-1(7-37)OHまたはその類似体もしくは誘導体のN末端および/またはC末端から1つまたは複数のアミノ酸を切り落とした後に得られるポリペプチドである。GLP-1(7-37)OHの記述に使用される命名法はGLP-1断片にも適用することができる。例えば、GLP-1(9-36)OHは、N末端から2つのアミノ酸を切り落とし、C末端から1つのアミノ酸を切り落とすことによって得られるGLP-1断片を表す。断片中のアミノ酸は、GLP-1(7-37)OH中の対応するアミノ酸と同じ番号で表示される。例えば、GLP-1(7-37)OHの場合と同様に、GLP-1(9-36)OH中のN末端グルタミン酸は9位にあり、12位はフェニルアラニンで占められ、22位はグリシンで占められている。GLP-1(7-36)OHの場合は、GLP-1(7-37)OHの37位のグリシンが欠失している。
【0021】
GLP-1化合物には、GLP-1(7-37)OHまたはその断片もしくは類似体のN末端および/またはC末端に1つまたは複数のアミノ酸が付加されているポリペプチドも包含される。このタイプのGLP-1化合物は約39個までのアミノ酸を持つことが好ましい。「伸長型」GLP-1化合物中のアミノ酸は、GLP-1(7-37)OH中の対応するアミノ酸と同じ番号で表示される。例えば、GLP-1(7-37)OHのN末端に2つのアミノ酸を付加することによって得られるGLP-1化合物のN末端アミノ酸は5位にあり、GLP-1(7-37)OHのC末端に1つのアミノ酸を付加することによって得られるGLP-1化合物のC末端アミノ酸は38位にある。したがって、これら「伸長型」GLP-1化合物のどちらにおいても、GLP-1(7-37)OHの場合と同様に、12位はフェニルアラニンで占められ、22位はグリシンで占められる。伸長型GLP-1化合物のアミノ酸1〜6は、好ましくは、GLP-1(1-37)OHの対応する位置にあるアミノ酸と同じであるか、またはその保存的置換である。伸長型GLP-1化合物のアミノ酸38〜45は、好ましくは、グルカゴンまたはエクセンディン4の対応する位置にあるアミノ酸と同じであるか、またはその保存的置換である。
【0022】
本発明のGLP-1化合物は「GLP-1類似体」を包含する。GLP-1類似体は、その類似体がインスリン分泌促進活性を持つように、GLP-1(7-37)OHまたはGLP-1(7-37)OHの断片に対して十分な相同性を持つ。好ましくは、GLP-1類似体は、1、2、3、4または5個のアミノ酸がGLP-1(7-37)OHまたはGLP1(7-37)OHの断片の対応する位置にあるアミノ酸とは異なるように改変されている、GLP-1(7-37)OHまたはその断片のアミノ酸配列を持つ。GLP-1化合物を指定するために本明細書で使用する命名法では、置換アミノ酸およびその位置を親構造の前に示す。例えばGlu22-GLP-1(7-37)OHは、GLP-1(7-37)OHの22位に通常見いだされるグリシンがグルタミン酸で置き換えられているGLP-1化合物を表し、Val8-Glu22-GLP-1(7-37)OHは、GLP-1(7-37)OHの8位に通常見いだされるアラニンおよび22位に通常見いだされるグリシンがそれぞれバリンおよびグルタミン酸で置き換えられているGLP-1化合物を表す。
【0023】
本発明のGLP-1化合物には「GLP-1誘導体」も包含される。GLP-1誘導体は、GLP-1またはGLP-1類似体のアミノ酸配列を持つが、さらにそのアミノ酸側鎖基、α炭素原子、末端アミノ基、または末端カルボン酸基の1つまたは複数が化学修飾されている分子であると定義される。化学修飾には、例えば化学部分の付加、新しい結合の生成、および化学部分の除去などが含まれるが、これらに限るわけではない。アミノ酸側鎖基での修飾には、例えばリジンε-アミノ基のアシル化、アルギニン、ヒスチジンまたはリジンのN-アルキル化、グルタミン酸またはアスパラギン酸カルボン酸基のアルキル化、およびグルタミンまたはアスパラギンの脱アミド化などがあるが、これらに限るわけではない。末端アミノ基の修飾には、例えばデスアミノ、N-低級アルキル、N-ジ-低級アルキル、およびN-アシル修飾などがあるが、これらに限るわけではない。末端カルボキシ基の修飾には、例えばアミド、低級アルキルアミド、ジアルキルアミド、および低級アルキルエステル修飾などがあるが、これらに限るわけではない。低級アルキルはC1〜C4アルキルである。さらに、1つまたは複数の側鎖基または末端基を、通常の技術を有するタンパク質化学者に知られている保護基で保護することもできる。アミノ酸のα炭素はモノメチル化またはジメチル化することができる。
【0024】
GLP-1化合物は、そのGLP-1化合物自体がGLP-1受容体を結合し、当該受容体を介したシグナリングを誘発することができる限り、どれでも本発明の異種融合タンパク質の一部とすることができる。GLP-1受容体結合およびシグナル伝達は、EP619,322および米国特許第5,120,712号にそれぞれ記載されているような試験管内アッセイを使って評価することができる。
【0025】
当技術分野では数多くの活性なGLP-1断片、類似体および誘導体が知られており、これらの類似体および誘導体はいずれも、本発明の異種融合タンパク質の一部とすることができる。新規GLP-1類似体ならびに当技術分野で知られているGLP-1類似体および誘導体の例をいくつか挙げる。
【0026】
当技術分野で知られているGLP-1類似体およびGLP-1断片には、例えばGLP-1(7-34)およびGLP-1(7-35)、GLP-1(7-36)、Gln9-GLP-1(7-37)、D-Gln9-GLP-1(7-37)、Thr16-Lys18-GLP-1(7-37)、およびLys18-GLP-1(7-37)などがある。GLP-1(7-34)およびGLP-1(7-35)などのGLP-1類似体は米国特許第5,118,666号に開示されている。インスリン分泌促進性を持つ生物学的プロセシングを受けた形態のGLP-1、例えばGLP-1(7-36)なども知られている。生物学的に活性な他の既知のGLP-1化合物は、Hoffmannらの米国特許第5,977,071号、Buckleyらの米国特許第5,545,618号、およびAdelhorstら, J. Biol. Chem. 269:6275 (1994)に開示されている。
【0027】
好ましいGLP-1類似体群は式I(配列番号2)のGLP-1類似体から構成される:
【化2】
式I(配列番号2)
[式中、
8位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
9位のXaaは、Glu、Asp、またはLysであり、
11位のXaaは、Thr、Ala、Gly、Ser、Leu、Ile、Val、Glu、Asp、またはLysであり、
14位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
16位のXaaは、Val、Ala、Gly、Ser、Thr、Leu、Ile、Tyr、Glu、Asp、Trp、またはLysであり、
17位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
18位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、Trp、Tyr、またはLysであり、
19位のXaaは、Tyr、Phe、Trp、Glu、Asp、Gln、またはLysであり、
20位のXaaは、Leu、Ala、Gly、Ser、Thr、Ile、Val、Glu、Asp、Met、Trp、Tyr、またはLysであり、
21位のXaaは、Glu、Asp、またはLysであり、
22位のXaaは、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
23位のXaaは、Gln、Asn、Arg、Glu、Asp、またはLysであり、
24位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Arg、Glu、Asp、またはLysであり、
25位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
26位のXaaは、Lys、Arg、Gln、Glu、Asp、またはHisであり、
27位のXaaは、Leu、Glu、Asp、またはLysであり、
30位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
31位のXaaは、Trp、Phe、Tyr、Glu、Asp、またはLysであり、
32位のXaaは、Leu、Gly、Ala、Ser、Thr、Ile、Val、Glu、Asp、またはLysであり、
33位のXaaは、Val、Gly、Ala、Ser、Thr、Leu、Ile、Glu、Asp、またはLysであり、
34位のXaaは、Asn、Lys、Arg、Glu、Asp、またはHisであり、
35位のXaaは、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
36位のXaaは、Gly、Arg、Lys、Glu、Asp、またはHisであり、
37位のXaaは、Pro、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、もしくはLysであるか、または欠失しており、
38位のXaaは、Ser、Arg、Lys、Glu、Asp、もしくはHisであるか、または欠失しており、
39位のXaaは、Ser、Arg、Lys、Glu、Asp、もしくはHisであるか、または欠失しており、
40位のXaaは、Gly、Asp、Glu、もしくはLysであるか、または欠失しており、
41位のXaaは、Ala、Phe、Trp、Tyr、Glu、Asp、もしくはLysであるか、または欠失しており、
42位のXaaは、Ser、Pro、Lys、Glu、もしくはAspであるか、または欠失しており、
43位のXaaは、Ser、Pro、Glu、Asp、もしくはLysであるか、または欠失しており、
44位のXaaは、Gly、Pro、Glu、Asp、もしくはLysであるか、または欠失しており、
45位のXaaは、Ala、Ser、Val、Glu、Asp、もしくはLysであるか、または欠失している。
ただし、37、38、39、40、41、42、43、または44位のアミノ酸が欠失している場合は、そのアミノ酸の下流にある各アミノ酸も欠失しているものとする]。
【0028】
式IのGLP-1化合物は、GLP-1(7-37)OHまたはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、6個を超えて含まないことが好ましい。GLP-1(7-37)OHまたはエクセンディン4中の対応するアミノ酸と相違するアミノ酸は5個を超えないことがより好ましい。GLP-1(7-37)OHまたはエクセンディン4中の対応するアミノ酸と相違するアミノ酸は4個を超えないことがさらに好ましい。
【0029】
本発明のGLP-1化合物は、式Iの誘導体、例えばそのC-1-6-エステル、またはアミド、またはC-1-6-アルキルアミド、またはC-1-6-ジアルキルアミドなどを包含する。WO99/43706には式IのGLP-1化合物の誘導体が記載されており、この公開明細書は参考文献としてそのまま本明細書の一部を構成する。本発明は、WO99/43706に記載されているように誘導体化された式Iの化合物、および誘導体化されていない式Iの化合物を包含する。
【0030】
もう一つの好ましいGLP-1化合物群は、式II(配列番号3)のGLP-1類似体から構成される:
【化3】
式II(配列番号3)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、Val、Leu、Ile、Ser、またはThrであり、
9位のXaaは、Thr、Ser、Arg、Lys、Trp、Phe、Tyr、Glu、またはHisであり、
11位のXaaは、Asp、Glu、Arg、Thr、Ala、Lys、またはHisであり、
12位のXaaは、His、Trp、Phe、またはTyrであり、
16位のXaaは、Leu、Ser、Thr、Trp、His、Phe、Asp、Val、Tyr、Glu、またはAlaであり、
18位のXaaは、His、Pro、Asp、Glu、Arg、Ser、Ala、またはLysであり、
22位のXaaは、Gly、Asp、Glu、Gln、Asn、Lys、Arg、またはCysであり、
23位のXaaは、His、Asp、Lys、Glu、Gln、またはArgであり、
24位のXaaは、Glu、Arg、Ala、またはLysであり、
26位のXaaは、Trp、Tyr、Phe、Asp、Lys、Glu、またはHisであり、
27位のXaaは、Ala、Glu、His、Phe、Tyr、Trp、Arg、またはLysであり、
30位のXaaは、Ala、Glu、Asp、Ser、またはHisであり、
31位のXaaは、Asp、Glu、Ser、Thr、Arg、Trp、またはLysであり、
33位のXaaは、Asp、Arg、Val、Lys、Ala、Gly、またはGluであり、
34位のXaaは、Glu、Lys、またはAspであり、
35位のXaaは、Thr、Ser、Lys、Arg、Trp、Tyr、Phe、Asp、Gly、Pro、His、またはGluであり、
36位のXaaは、Thr、Ser、Asp、Trp、Tyr、Phe、Arg、Glu、またはHisであり、
37位のRは、Lys、Arg、Thr、Ser、Glu、Asp、Trp、Tyr、Phe、His、Gly、もしくはGly-Proであるか、または欠失している]。
【0031】
もう一つの好ましいGLP-1化合物群は、式III(配列番号4)のGLP-1類似体から構成される:
【化4】
式III(配列番号4)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、Val、Leu、Ile、Ser、またはThrであり、
11位のXaaは、Asp、Glu、Arg、Thr、Ala、Lys、またはHisであり、
12位のXaaは、His、Trp、Phe、またはTyrであり、
16位のXaaは、Leu、Ser、Thr、Trp、His、Phe、Asp、Val、Glu、またはAlaであり、
22位のXaaは、Gly、Asp、Glu、Gln、Asn、Lys、Arg、またはCysであり、
23位のXaaは、His、Asp、Lys、Glu、またはGlnであり、
24位のXaaは、Glu、His、Ala、またはLysであり、
25位のXaaは、Asp、Lys、Glu、またはHisであり、
27位のXaaは、Ala、Glu、His、Phe、Tyr、Trp、Arg、またはLysであり、
30位のXaaは、Ala、Glu、Asp、Ser、またはHisであり、
33位のXaaは、Asp、Arg、Val、Lys、Ala、Gly、またはGluであり、
34位のXaaは、Glu、Lys、またはAspであり、
35位のXaaは、Thr、Ser、Lys、Arg、Trp、Tyr、Phe、Asp、Gly、Pro、His、またはGluであり、
36位のXaaは、Arg、Glu、またはHisであり、
37位のRは、Lys、Arg、Thr、Ser、Glu、Asp、Trp、Tyr、Phe、His、Gly、もしくはGly-Proであるか、または欠失している]。
【0032】
もう一つの好ましいGLP-1化合物群は、式IV(配列番号5)のGLP-1類似体から構成される:
【化5】
式IV(配列番号5)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、Val、Leu、Ile、Ser、Met、またはThrであり、
12位のXaaは、His、Trp、Phe、またはTyrであり、
16位のXaaは、Leu、Ser、Thr、Trp、His、Phe、Asp、Val、Glu、またはAlaであり、
22位のXaaは、Gly、Asp、Glu、Gln、Asn、Lys、Arg、またはCysであり、
23位のXaaは、His、Asp、Lys、Glu、またはGlnであり、
26位のXaaは、Asp、Lys、Glu、またはHisであり、
30位のXaaは、Ala、Glu、Asp、Ser、またはHisであり、
35位のXaaは、Thr、Ser、Lys、Arg、Trp、Tyr、Phe、Asp、Gly、Pro、His、またはGluであり、
37位のRは、Lys、Arg、Thr、Ser、Glu、Asp、Trp、Tyr、Phe、His、Gly、もしくはGly-Proであるか、または欠失している]。
【0033】
もう一つの好ましいGLP-1群は、式V(配列番号6)のGLP-1類似体から構成される:
【化6】
式V(配列番号6)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、Val、Leu、Ile、Ser、またはThrであり、
22位のXaaは、Gly、Asp、Glu、Gln、Asn、Lys、Arg、またはCysであり、
23位のXaaは、His、Asp、Lys、Glu、またはGlnであり、
24位のXaaは、Ala、Glu、His、Phe、Tyr、Trp、Arg、またはLysであり、
30位のXaaは、Ala、Glu、Asp、Ser、またはHisであり、
37位のRは、Lys、Arg、Thr、Ser、Glu、Asp、Trp、Tyr、Phe、His、Gly、もしくはGly-Proであるか、または欠失している]。
【0034】
式I、II、III、IVおよびVの好ましいGLP-1化合物は、8位にアラニン以外のアミノ酸を持つGLP-1類似体またはGLP-1類似体の断片を含む(8位類似体)。これらの8位類似体は、さらに9、11、12、16、18、22、23、24、26、27、30、31、33、34、35、36、および37位にも、天然GLP-1(7-37)OHの対応するアミノ酸と比較して、1つまたは複数の変異を持つことが好ましい。また、これらの類似体は、天然のGLP-1(7-37)OHまたはGLP-1(7-36)OH中の対応するアミノ酸と比較して、6個以下の変異を持つことが好ましい。より好ましい類似体は、天然のGLP-1(7-37)OHもしくはGLP-1(7-36)OH中の対応するアミノ酸と比較して5個以下の変異を持つか、または天然のGLP-1(7-37)OHもしくはGLP-1(7-36)OH中の対応するアミノ酸と比較して4個以下の変異を持つ。これらの類似体は、天然のGLP-1(7-37)OHまたはGLP-1(7-36)OH中の対応するアミノ酸と比較して3個以下の変異を持つことが、さらに好ましい。これらの類似体は、天然のGLP-1(7-37)OH中の対応するアミノ酸と比較して2個以下の変異を持つことが、最も好ましい。
【0035】
式II、III、IVおよびVの化合物は、凝集して不溶型を生成する傾向が低下していることがわかった。比較的小さいGLP-1ペプチドが、はるかに大きいタンパク質に融合されているにもかかわらず、活性なコンフォメーションを維持しなければならない融合タンパク質にあっては、これも重要なことである。本発明の融合タンパク質に包含される式II、III、IVおよびVの好ましいGLP-1化合物は、22位のグリシンと、好ましくは8位のアラニンが、別のアミノ酸で置き換えられているGLP-1類似体またはGLP-1類似体の断片を含む。
【0036】
22位がアスパラギン酸、グルタミン酸、アルギニンまたはリジンである場合、8位は好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニンまたはメチオニンであり、より好ましくはバリンまたはグリシンである。22位がシステイン酸などのスルホン酸である場合は、8位は好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニンまたはメチオニンであり、より好ましくはバリンまたはグリシンである。
【0037】
他の好ましいGLP-1化合物として、8位のアミノ酸が好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンであり、30位がグルタミン酸、アスパラギン酸、セリン、またはヒスチジン、より好ましくはグルタミン酸である点以外は、GLP-1(7-37)OHの配列を持つ、式IV(配列番号5)のGLP-1類似体が挙げられる。
【0038】
他の好ましいGLP-1化合物として、8位のアミノ酸が好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンであり、37位がヒスチジン、リジン、アルギニン、スレオニン、セリン、グルタミン酸、アスパラギン酸、トリプトファン、チロシン、フェニルアラニン、より好ましくはヒスチジンである点以外は、GLP-1(7-37)OHの配列を持つ、式IV(配列番号5)のGLP-1類似体が挙げられる。
【0039】
他の好ましいGLP-1化合物として、8位のアミノ酸が好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンであり、22位がグルタミン酸、リジン、アスパラギン酸、またはアルギニン、より好ましくはグルタミン酸またはリジンであり、23位がリジン、アルギニン、グルタミン酸、アスパラギン酸およびヒスチジン、より好ましくはリジンまたはグルタミン酸である点以外は、GLP-1(7-37)OHの配列を持つ、式IV(配列番号5)のGLP-1類似体が挙げられる。
【0040】
他の好ましいGLP-1化合物として、8位のアミノ酸が好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンであり、22位がグルタミン酸、リジン、アスパラギン酸、またはアルギニン、より好ましくはグルタミン酸またはリジンであり、27位がアラニン、リジン、アルギニン、トリプトファン、チロシン、フェニルアラニン、またはヒスチジン、より好ましくはアラニンである点以外は、GLP-1(7-37)OHの配列を持つ、式V(配列番号6)のGLP-1類似体が挙げられる。
【0041】
他の好ましいGLP-1化合物として、8位のアミノ酸ならびに9位、11位、12位、16位、18位、22位、23位、24位、26位、27位、30位、31位、33位、34位、35位、36位、および37位からなる群より選択される1、2または3個のアミノ酸が、天然GLP-1(7-37)OHの対応する位置にあるアミノ酸とは相違する点以外は、GLP-1(7-37)OHの配列を持つ、式IIのGLP-1類似体が挙げられる。
【0042】
他の好ましい式IIのGLP-1化合物には、以下に挙げるものがある:Val8-GLP-1(7-37)OH、Gly8-GLP-1(7-37)OH、Glu22-GLP-1(7-37)OH、Asp22-GLP-1(7-37)OH、Arg22-GLP-1(7-37)OH、Lys22-GLP-1(7-37)OH、Cys22-GLP-1(7-37)OH、Val8-Glu22-GLP-1(7-37)OH、Val8-Asp22-GLP-1(7-37)OH、Val8-Arg22-GLP-1(7-37)OH、Val8-Lys22-GLP-1(7-37)OH、Val8-Cys22-GLP-1(7-37)OH、Gly8-Glu22-GLP-1(7-37)OH、Gly8-Asp22-GLP-1(7-37)OH、Gly8-Arg22-GLP-1(7-37)OH、Gly8-Lys22-GLP-1(7-37)OH、Gly8-Cys22-GLP-1(7-37)OH、Glu22-GLP-1(7-36)OH、Asp22-GLP-1(7-36)OH、Arg22-GLP-1(7-36)OH、Lys22-GLP-1(7-36)OH、Cys22-GLP-1(7-36)OH、Val8-Glu22-GLP-1(7-36)OH、Val8-Asp22-GLP-1(7-36)OH、Val8-Arg22-GLP-1(7-36)OH、Val8-Lys22-GLP-1(7-36)OH、Val8-Cys22-GLP-1(7-36)OH、Gly8-Glu22-GLP-1(7-36)OH、Gly8-Asp22-GLP-1(7-36)OH、Gly8-Arg22-GLP-1(7-36)OH、Gly8-Lys22-GLP-1(7-36)OH、Gly8-Cys22-GLP-1(7-36)OH、Lys23-GLP-1(7-37)OH、Val8-Lys23-GLP-1(7-37)OH、Gly8-Lys23-GLP-1(7-37)OH、His24-GLP-1(7-37)OH、Val8-His24-GLP-1(7-37)OH、Gly8-His24-GLP-1(7-37)OH、Lys24-GLP-1(7-37)OH、Val8-Lys24-GLP-1(7-37)OH、Gly8-Lys23-GLP-1(7-37)OH、Glu30-GLP-1(7-37)OH、Val8-Glu30-GLP-1(7-37)OH、Gly8-Glu30-GLP-1(7-37)OH、Asp30-GLP-1(7-37)OH、Val8-Asp30-GLP-1(7-37)OH、Gly8-Asp30-GLP-1(7-37)OH、Gln30-GLP-1(7-37)OH、Val8-Gln30-GLP-1(7-37)OH、Gly8-Gln30-GLP-1(7-37)OH、Tyr30-GLP-1(7-37)OH、Val8-Tyr30-GLP-1(7-37)OH、Gly8-Tyr30-GLP-1(7-37)OH、Ser30-GLP-1(7-37)OH、Val8-Ser30-GLP-1(7-37)OH、Gly8-Ser30-GLP-1(7-37)OH、His30-GLP-1(7-37)OH、Val8-His30-GLP-1(7-37)OH、Gly8-His30-GLP-1(7-37)OH、Glu34-GLP-1(7-37)OH、Val8-Glu34-GLP-1(7-37)OH、Gly8-Glu34-GLP-1(7-37)OH、Ala34-GLP-1(7-37)OH、Val8-Ala34-GLP-1(7-37)OH、Gly8-Ala34-GLP-1(7-37)OH、Gly34-GLP-1(7-37)OH、Val8-Gly34-GLP-1(7-37)OH、Gly8-Gly34-GLP-1(7-37)OH、Ala35-GLP-1(7-37)OH、Val8-Ala35-GLP-1(7-37)OH、Gly8-Ala35-GLP-1(7-37)OH、Lys35-GLP-1(7-37)OH、Val8-Lys35-GLP-1(7-37)OH、Gly8-Lys35-GLP-1(7-37)OH、His35-GLP-1(7-37)OH Val8-His35-GLP-1(7-37)OH、Gly8-His35-GLP-1(7-37)OH、Pro35-GLP-1(7-37)OH、Val8-Pro35-GLP-1(7-37)OH、Gly8-Pro35-GLP-1(7-37)OH、Glu35-GLP-1(7-37)OH、Val8-Glu35-GLP-1(7-37)OH、Gly8-Glu35-GLP-1(7-37)OH、Val8-Ala27-GLP-1(7-37)OH、Val8-His37-GLP-1(7-37)OH、Val8-Glu22-Lys23-GLP-1(7-37)OH、Val8-Glu22-Glu23-GLP-1(7-37)OH、Val8-Glu22-Ala27-GLP-1(7-37)OH、Val8-Gly34-Lys35-GLP-1(7-37)OH、Val8-His37-GLP-1(7-37)OH、Gly8-His37-GLP-1(7-37)OH、Val8-Glu22-Ala27-GLP-1(7-37)OH、Gly8-Glu22-Ala27-GLP-1(7-37)OH、Val8-Lys22-Glu23-GLP-1(7-37)OH、およびGly8-Lys22-Glu23-GLP-1(7-37)OH。
【0043】
本発明での使用に好ましいもう一つのGLP-1類似体および誘導体群は、式VI(配列番号7)の分子から構成される:
R1-X-Glu-Gly10-Thr-Phe-Thr-Ser-Asp15-Val-Ser-Ser-Tyr-Leu20-Y -Gly-Gln-Ala-Ala25-Lys-Z-Phe-Ile-Ala30-Trp-Leu-Val-Lys-Gly35-Arg-R2
式VI(配列番号7)
[式中、R1は、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジン、およびα-メチルヒスチジンであり、XはAla、Gly、Val、Thr、Ile、およびα-メチル-Alaからなる群より選択され、YはGlu、Gln、Ala、Thr、Ser、およびGlyからなる群より選択され、ZはGlu、Gln、Ala、Thr、Ser、およびGlyからなる群より選択され、R2はGly-OHである]。
【0044】
本発明での使用に好ましいもう一つのGLP-1化合物群はWO91/11457に開示されており、本質的に、以下の(a)〜(e)からなる群より選択される少なくとも1つの改変を持つ、GLP-1(7-34)、GLP-1(7-35)、GLP-1(7-36)、もしくはGLP-1(7-37)、またはそのアミド型、およびその医薬的に許容できる塩からなる:
(a)グリシン、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、アルギニン、またはD-リジンによる26位および/または34位のリジンの置換、またはグリシン、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、リジン、またはD-アルギニンによる36位のアルギニンの置換、
(b)耐酸化性アミノ酸による31位のトリプトファンの置換、
(c)次に挙げる置換の少なくとも1つ:チロシンによる16位のバリンの置換、リジンによる18位のセリンの置換、アスパラギン酸による21位のグルタミン酸の置換、セリンによる22位のグリシンの置換、アルギニンによる23位のグルタミンの置換、アルギニンによる24位のアラニンの置換、およびグルタミンによる26位のリジンの置換、
(d)次に挙げる置換の少なくとも1つ:グリシン、セリン、またはシステインによる8位のアラニンの置換、アスパラギン酸、グリシン、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、またはフェニルアラニンによる9位のグルタミン酸の置換、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、またはフェニルアラニンによる10位のグリシンの置換、およびグルタミン酸による15位のアスパラギン酸の置換、
(e)グリシン、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、またはフェニルアラニン、またはヒスチジンのD-もしくはN-アシル化もしくはアルキル化型による7位のヒスチジンの置換、
ただし、(a)、(b)、(d)、および(e)の置換において、置換アミノ酸は所望によりD型であってもよく、7位の置換アミノ酸は所望によりN-アシル化型またはN-アルキル化型であってもよい。
【0045】
酵素ジペプチジル-ペプチダーゼIV(DPP IV)は、投与されたGLP-1に観察される迅速な生体内失活の原因であると考え得るので[例えばMentlein, R.ら, Eur. J. Biochem., 214:829-835 (1993)参照]、融合タンパク質の形でDPP IVの活性から保護されるGLP-1類似体および誘導体は好ましく、GLP-1化合物がGly8-GLP-1(7-37)OH、Val8-GLP-1(7-37)OH、α-メチル-Ala8-GLP-1(7-37)OH、またはGly8-Gln21-GLP-1(7-37)OHである融合タンパク質は、さらに好ましい。
【0046】
本発明での使用に好ましいもう一つのGLP-1化合物群は、参考文献として特に本明細書の一部を構成する米国特許第5,512,549号にクレームされている式VII(配列番号8)の化合物からなる。
【化7】
式VII(配列番号8)
[式中、R1は、4-イミダゾプロピオニル、4-イミダゾアセチル、または4-イミダゾ-α,α-ジメチルアセチルからなる群より選択され、R2は、C6-C10非分岐アシルからなる群から選択されるか、または存在せず、R3はGly-OHまたはNH2からなる群より選択され、XaaはLysまたはArgである]。
【0047】
本発明での使用に、より好ましい式IVの化合物は、XaaがArgであり、R2がC6-C10非分岐アシルである式IVの化合物である。本発明での使用に、さらに好ましい式IVの化合物は、XaaがArgであり、R2がC6-C10非分岐アシルであり、R3がGly-OHである化合物である。本発明での使用に著しく好ましい他の式IVの化合物は、XaaがArgであり、R2がC6-C10非分岐アシルであり、R3がGly-OHであり、R1が4-イミダゾプロピオニルである式VIの化合物である。本発明での使用に特に好ましい式IVの化合物は、XaaがArgであり、R2がC8非分岐アシルであり、R3がGly-OHであり、R1が4-イミダゾプロピオニルである式IVの化合物である。
【0048】
GLP-1化合物は、好ましくは、8位がアラニン以外のアミノ酸である主鎖を持つGLP-1類似体(8位類似体)を含む。また主鎖は、7位にL-ヒスチジン、D-ヒスチジン、またはデスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジン、もしくはα-メチルヒスチジンなどの修飾型ヒスチジンを含んでもよい。これらの8位類似体は、さらに12、16、18、19、20、22、25、27、30、33、および37位にも、天然GLP-1(7-37)OHの対応するアミノ酸と比較して、1つまたは複数の変異を持つことが好ましい。これらの8位類似体は、さらに16、18、22、25および33位にも、天然GLP-1(7-37)OHの対応するアミノ酸と比較して、1つまたは複数の変異を持つことが、より好ましい。
【0049】
好ましい一態様として、GLP-1類似体は、12位のアミノ酸がトリプトファンまたはチロシンからなる群より選択される、GLP-1(7-37)OHである。12位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。12位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。
【0050】
もう一つの好ましい態様として、GLP-1類似体は、16位のアミノ酸がトリプトファン、イソロイシン、ロイシン、フェニルアラニン、またはチロシンからなる群より選択される、GLP-1(7-37)である。16位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。16位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。16位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。16位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0051】
もう一つの好ましい態様として、GLP-1類似体は、18位のアミノ酸がトリプトファン、チロシン、フェニルアラニン、リジン、ロイシン、またはイソロイシン、好ましくはトリプトファン、チロシン、およびイソロイシンからなる群より選択される、GLP-1(7-37)OHである。18位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。18位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。18位および8位での置換に加えて、30位のアミノ酸が、グルタミン酸で置換されていることも、好ましい。18位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0052】
もう一つの好ましい態様として、GLP-1類似体は、19位のアミノ酸がトリプトファンまたはフェニルアラニンからなる群より選択される(好ましくはトリプトファンである)、GLP-1(7-37)OHである。19位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。19位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。19位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。19位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0053】
もう一つの好ましい態様として、GLP-1類似体は、20位のアミノ酸がフェニルアラニン、チロシン、またはトリプトファンである、GLP-1(7-37)OHである。20位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。20位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。20位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。20位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0054】
もう一つの好ましい態様として、GLP-1類似体は、25位のアミノ酸がバリン、イソロイシン、およびロイシンからなる群より選択される(好ましくはバリンである)、GLP-1(7-37)OHである。25位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。25位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。25位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。25位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0055】
もう一つの好ましい態様として、GLP-1類似体は、27位のアミノ酸がイソロイシンまたはアラニンからなる群より選択される、GLP-1(7-37)OHである。27位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。27位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。27位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。27位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0056】
もう一つの好ましい態様として、GLP-1類似体は、33位のアミノ酸がイソロイシンである、GLP-1(7-37)OHである。33位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。33位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。33位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。33位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0057】
本GLP-1化合物は以下の位置の1つまたは複数が改変されている:8、12、16、18、19、20、22、25、27、30、33、および37。これらのGLP-1化合物は、GLP-1(7-37)OHと比較して増加した効力を示し、式IX(配列番号12)のアミノ酸配列を含む:
Xaa7-Xaa8-Glu-Gly-Thr-Xaa12-Thr-Ser-Asp-Xaa16-Ser-Xaa18-Xaa19-Xaa20-Glu-Xaa22-Gln-Ala-Xaa25-Lys-Xaa27-Phe-Ile-Xaa30-Trp-Leu-Xaa33-Lys-Gly-Arg-Xaa37
式IX(配列番号12)
[式中、
Xaa7は、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジン、またはα-メチルヒスチジンであり、
Xaa8は、Ala、Gly、Val、Leu、Ile、Ser、またはThrであり、
Xaa12は、Phe、Trp、またはTyrであり、
Xaa16は、Val、Trp、Ile、Leu、Phe、またはTyrであり、
Xaa18は、Ser、Trp、Tyr、Phe、Lys、Ile、Leu、Valであり、
Xaa19は、Tyr、Trp、またはPheであり、
Xaa20は、Leu、Phe、Tyr、またはTrpであり、
Xaa22は、Gly、Glu、Asp、またはLysであり、
Xaa25は、Ala、Val、Ile、またはLeuであり、
Xaa27は、Glu、Ile、またはAlaであり、
Xaa30は、AlaまたはGluであり、
Xaa33は、Val、またはIleであり、
Xaa37は、Gly、His、もしくはNH2であるか、または存在しない]。
【0058】
式IXの好ましいGLP-1化合物としては、例えば、GLP-1(7-37)OH、GLP-1(7-36)-NH2、Gly8-GLP-1(7-37)OH、Gly8-GLP-1(7-36)NH2、Val8-GLP-1(7-37)OH、Val8-GLP-1(7-36)NH2、Leu8-GLP-1(7-37)OH、Leu8-GLP-1(7-36)NH2、Ile8-GLP-1(7-37)OH、Ile8-GLP-1(7-36)NH2、Ser8-GLP-1(7-37)OH、Ser8-GLP-1(7-36)NH2、Thr8-GLP-1(7-37)OH、Thr8-GLP-1(7-36)NH2、Val8-Tyr12-GLP-1(7-37)OH、Val8-Tyr12-GLP-1(7-36)NH2、Val8-Tyr16-GLP-1(7-37)OH、Val8-Tyr16-GLP-1(7-36)NH2、Val8-Glu22-GLP-1(7-37)OH、Val8-Glu22-GLP-1(7-36)NH2、Gly8-Glu22-GLP-1(7-37)OH、Gly8-Glu22-GLP-1(7-36)NH2、Val8-Asp22-GLP-1(7-37)OH、Val8-Asp22-GLP-1(7-36)NH2、Gly8-Asp22-GLP-1(7-37)OH、Gly8-Asp22-GLP-1(7-36)NH2、Val8-Lys22-GLP-1(7-37)OH、Val8-Lys22-GLP-1(7-36)NH2、Gly8-Lys22-GLP-1(7-37)OH、Gly8-Lys22-GLP-1(7-36)NH2、Leu8-Glu22-GLP-1(7-37)OH、Leu8-Glu22-GLP-1(7-36)NH2、Ile8-Glu22-GLP-1(7-37)OH、Ile8-Glu22-GLP-1(7-36)NH2、Leu8-Asp22-GLP-1(7-37)OH、Leu8-Asp22-GLP-1(7-36)NH2、Ile8-Asp22-GLP-1(7-37)OH、Ile8-Asp22-GLP-1(7-36)NH2、Leu8-Lys22-GLP-1(7-37)OH、Leu8-Lys22-GLP-1(7-36)NH2、Ile8-Lys22-GLP-1(7-37)OH、Ile8-Lys22-GLP-1(7-36)NH2、Ser8-Glu22-GLP-1(7-37)OH、Ser8-Glu22-GLP-1(7-36)NH2、Thr8-Glu22-GLP-1(7-37)OH、Thr8-Glu22-GLP-1(7-36)NH2、Ser8-Asp22-GLP-1(7-37)OH、Ser8-Asp22-GLP-1(7-36)NH2、Thr8-Asp22-GLP-1(7-37)OH、Thr8-Asp22-GLP-1(7-36)NH2、Ser8-Lys22-GLP-1(7-37)OH、Ser8-Lys22-GLP-1(7-36)NH2、Thr8-Lys22-GLP-1(7-37)OH、Thr8-Lys22-GLP-1(7-36)NH2、Glu22-GLP-1(7-37)OH、Glu22-GLP-1(7-36)NH2、Asp22-GLP-1(7-37)OH、Asp22-GLP-1(7-36)NH2、Lys22-GLP-1(7-37)OH、Lys22-GLP-1(7-36)NH2、Val8-Ala27-GLP-1(7-37)OH、Val8-Glu22-Ala27-GLP-1(7-37)OH、Val8-Glu30-GLP-1(7-37)OH、Val8-Glu30-GLP-1(7-36)NH2、Gly8-Glu30-GLP-1(7-37)OH、Gly8-Glu30-GLP-1(7-36)NH2、Leu8-Glu30-GLP-1(7-37)OH、Leu8-Glu30-GLP-1(7-36)NH2、Ile8-Glu30-GLP-1(7-37)OH、Ile8-Glu30-GLP-1(7-36)NH2、Ser8-Glu30-GLP-1(7-37)OH、Ser8-Glu30-GLP-1(7-36)NH2、Thr8-Glu30-GLP-1(7-37)OH、Thr8-Glu30-GLP-1(7-36)NH2、Val8-His37-GLP-1(7-37)OH、Val8-His37-GLP-1(7-36)NH2、Gly8-His37-GLP-1(7-37)OH、Gly8-His37-GLP-1(7-36)NH2、Leu8-His37-GLP-1(7-37)OH、Leu8-His37-GLP-1(7-36)NH2、Ile8-His37-GLP-1(7-37)OH、Ile8-His37-GLP-1(7-36)NH2、Ser8-His37-GLP-1(7-37)OH、Ser8-His37-GLP-1(7-36)NH2、Thr8-His37-GLP-1(7-37)OH、Thr8-His37-GLP-1(7-36)NH2などが挙げられる。
【0059】
複数の置換を持つ式IXの好ましいGLP-1化合物としては、例えば、8位がバリンまたはグリシンであり、22位がグルタミン酸であり、16位がチロシン、ロイシンまたはトリプトファンであり、18位がチロシン、トリプトファン、またはイソロイシンであり、25位がバリンであり、33位がイソロイシンであるGLP-1(7-37)OHが挙げられる。他の好ましいGLP-1化合物には、以下の化合物が含まれる:Val8-Tyr16-GLP-1(7-37)OH、Val8-Tyr12-Glu22-GLP-1(7-37)OH、Val8-Tyr16-Phe19-GLP-1(7-37)OH、Val8-Tyr16-Glu22-GLP-1(7-37)OH、Val8-Trp16-Glu22-GLP-1(7-37)OH、Val8-Leu16-Glu22-GLP-1(7-37)OH、Val8-Ile16-Glu22-GLP-1(7-37)OH、Val8-Phe16-Glu22-GLP-1(7-37)OH、Val8-Trp18-Glu22-GLP-1(7-37)OH、Val8-Tyr18-Glu22-GLP-1(7-37)OH、Val8-Phe18-Glu22-GLP-1(7-37)OH、およびVal8-Ile18-Glu22-GLP-1(7-37)OH。
【0060】
本発明のGLP-1化合物には、エクセンディン化合物も包含される。エクセンディン3およびエクセンディン4は、ドクトカゲ科トカゲ毒から最初に単離された生物学的に活性なペプチドであり、GLP-1受容体に結合して、哺乳類旁細胞におけるcAMP依存的H+生成を刺激することが明らかにされている。エクセンディン3およびエクセンディン4はどちらも、GLP-1に対して約53%相同な39アミノ酸ペプチドである。これらはGLP-1活性の強力なアゴニストとして作用する。注目すべきことに、エクセンディン(9-39アミノ酸)として知られるエクセンディンのN末端切断型誘導体は、エクセンディン3、エクセンディン4およびGLP-1の阻害剤である。
【0061】
エクセンディン化合物は、典型的には、エクセンディン3、エクセンディン4、またはその類似体もしくは断片のアミノ酸配列を持つポリペプチドを含む。エクセンディン3およびエクセンディン4は米国特許第5,424,286号に開示されている。
【0062】
エクセンディン3は配列番号9のアミノ酸配列:
【化8】
(配列番号9)
を持つ。
【0063】
エクセンディン4は配列番号10のアミノ酸配列:
【化9】
(配列番号10)
を持つ。
【0064】
GLP-1化合物には、エクセンディンまたはエクセンディン類似体のN末端および/またはC末端から1つまたは複数のアミノ酸を切り落とした後に得られるポリペプチドであるエクセンディン断片も含まれる。さらに、GLP-1化合物には、エクセンディンまたはその断片のN末端および/またはC末端に1つまたは複数のアミノ酸が付加されているエクセンディンポリペプチドも含まれる。このタイプのエクセンディン化合物は約45個までのアミノ酸を持つ。
【0065】
GLP-1化合物には「エクセンディン類似体」も包含される。エクセンディン類似体は、その類似体がインスリン分泌促進活性を持つように、エクセンディン4、エクセンディン3、またはその断片に対して十分な相同性を持つ。エクセンディン断片および/または類似体の活性は、EP619,322および米国特許第5,120,712号に記載されているような試験管内アッセイを使って評価することができる。
【0066】
好ましくは、エクセンディン類似体は、1、2、3、4または5個のアミノ酸がエクセンディン4またはエクセンディン4の断片の対応する位置にあるアミノ酸とは異なるように改変されている、エクセンディン4またはその断片のアミノ酸配列を持つ。エクセンディン化合物を指定するために本明細書で使用する命名法では、置換アミノ酸およびその位置を親構造の前に示す。Val8-エクセンディン4は、エクセンディン4の8位に通常見いだされるグリシンがバリンで置き換えられているエクセンディン化合物を表す。
【0067】
もう一つの好ましいGLP-1化合物群は、式VIII(配列番号11)のGLP-1/エクセンディン4類似体から構成される。
【化10】
式VIII(配列番号11)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、またはValであり、
16位のXaaは、LeuまたはValであり、
18位のXaaは、LysまたはSerであり、
19位のXaaは、GlnまたはTyrであり、
20位のXaaは、MetまたはLeuであり、
22位のXaaは、GluまたはGlnであり、
23位のXaaは、Glu、またはGlnであり、
25位のXaaは、ValまたはAlaであり、
26位のXaaは、ArgまたはLysであり、
27位のXaaは、LeuまたはGluであり、
30位のXaaは、GluまたはAlaであり、
33位のXaaは、ValまたはLysであり、
34位のXaaは、AsnまたはLysであり、
36位のXaaは、GlyまたはArgであり、
37位のRは、Gly、Pro、もしくはPro-Ser-Ser-Gly-Ala-Pro-Pro-Pro-Serであるか、または存在しない]。
この群に含まれる18種類の分子種の活性を表6に記載する。
【0068】
本発明にとって有用な他のエクセンディン類似体は、PCT特許公開WO99/25728(Beeleyら)、WO99/25727(Beeleyら)、WO98/05351(Youngら)、WO99/40788(Youngら)、WO99/07404(Beeleyら)、およびWO99/43708(Knudsenら)に記載されている。
【0069】
本発明のGLP-1融合タンパク質はグリコシル化部位を含みうる。グリコシル化は、タンパク質の特定部位に糖部分が付加される化学修飾である。タンパク質のグリコシル化は、成熟タンパク質の正しい電荷、コンフォメーション、および安定性の確保に役割を果たし、そのタンパク質を細胞表面に導いて、最終的にはそのタンパク質を分泌させることができる。最も重要なことに、グリコシル化は、多くのタンパク質で、その生体内クリアランス速度を左右する。糖鎖はO結合型でもN結合型でもよい。一般に、O結合型糖鎖はセリンおよびスレオニンのヒドロキシル基酸素に付加され、N結合型糖鎖はアスパラギンのアミド窒素に付加される。N-グリコシル化のコンセンサス配列はAsn X1 X2[X1はProを除く任意のアミノ酸であり、X2はSerまたはThrである]である。
【0070】
GLP-1化合物は生体内では一般にグリコシル化されない。しかし、興味深いことに、Fc配列に融合されたC末端伸長部を持つGLP-1化合物を含む本発明のGLP-1融合タンパク質は、C末端伸長部の最後のセリン(SSGAPPPS*)と、FcのN末端領域の11位にあるスレオニン(AEPKSCDKTHT*CPPC...)でグリコシル化される。
【0071】
異種Fc融合タンパク質:
上述のGLP-1化合物は免疫グロブリンのFc部分に直接融合するか、またはペプチドリンカーを介して融合することができる。
【0072】
免疫グロブリンは、ジスルフィド結合でつながったポリペプチド鎖を含み、典型的には2本の軽鎖と2本の重鎖を持つ分子である。各鎖では、1つのドメイン(V)が、その分子の抗体特異性に依存して可変的なアミノ酸配列を持つ。他のドメイン(C)は同じクラスの分子に共通するかなり一定した配列を持つ。
【0073】
本明細書で使用する「免疫グロブリンのFc部分」という用語は、免疫学の分野でこの用語に通常与えられている意味を持つ。具体的に述べると、この用語は、抗体から2つの抗原結合領域(Fab断片)を除去することによって得られる抗体断片を表す。Fab断片を除去する一方法は、免疫グロブリンをパパインプロテアーゼで消化することである。したがってFc部分は、非共有結合相互作用およびジスルフィド結合によって会合した両重鎖由来のほぼ同サイズの定常領域断片から形成される。Fc部分はヒンジ領域を含み、CH2およびCH3ドメインから抗体のC末端までにわたることができる。ヒトおよびマウス免疫グロブリンの代表的なヒンジ領域は、Borrebaeck, C.A.K.編「Antibody Engineering, A Practical Guide」(W.H. Freeman and Co., 1992)に記載されており、その内容は参考文献として本明細書の一部を構成する。Fc部分はさらに1つまたは複数のグリコシル化部位を含む場合もある。ヒンジ領域、CH2およびCH3ドメインを含み、82位に1つのN-グリコシル化部位を持つ代表的Fc部分のアミノ酸配列を図1に示す。
【0074】
ヒト免疫グロブリンFc領域には、異なるエフェクターおよび薬物動態特性を持つIgG、IgA、IgM、IgD、およびIgEの5種類がある。IgGは血清中で最も豊富な免疫グロブリンである。また、IgGは血清中で、免疫グロブリンのなかで最も長い半減期(23日)も持っている。IgGは他の免疫グロブリンとは異なり、Fc受容体への結合後に、効率よく再循環する。IgGには4つのサブクラスG1、G2、G3、およびG4があり、それぞれに異なるエフェクター機能を持っている。G1、G2、およびG3は、C1qを結合し補体を結合することができるが、G4はできない。G3はG1より効率よくC1qを結合することができるが、G1の方が補体による細胞溶解をより効果的に媒介する。G2の補体結合効率は非常に低い。IgG中のC1q結合部位は、CH2ドメインのカルボキシ末端領域にある。
【0075】
どのIgGサブクラスもFc受容体(CD16、CD32、CD64)に結合する能力を持つが、G1およびG3はG2およびG4よりも効果的である。IgGのFc受容体結合領域は、ヒンジ領域と、CH2ドメインのカルボキシ末端領域との両方に位置する残基によって形成される。
【0076】
IgAは、単量体型と、J鎖でつながれた二量体型の、両方で存在することができる。IgAは血清中に2番目に豊富なIgであるが、半減期は6日しかない。IgAは3つのエフェクター機能を持っている。これは、マクロファージおよび好酸球上のIgA特異的受容体に結合して、それぞれ食作用および脱顆粒を引き起こす。またこれは、未知の代替経路によって補体を結合することもできる。
【0077】
IgMは、J鎖でつながれた五量体またはJ鎖でつながれた六量体として発現する。IgMは5日の血清半減期を持つ。これは、そのCH3ドメイン中にある結合部位を介してC1qに弱く結合する。IgDは血清中で3日の半減期を持つ。このIgに起因すると考えられるエフェクター機能ははっきりしていない。IgEは単量体型Igであり、2.5日の血清半減期を持つ。IgEは2つのFc受容体に結合して、脱顆粒を引き起こし、炎症誘発因子の放出をもたらす。
【0078】
所望する生体内効果に応じて、本発明の異種融合タンパク質は、上述したイソタイプをどれでも含むことができ、補体および/またはFc受容体結合機能が改変されている突然変異型Fc領域を含んでもよい。したがって本発明の異種融合タンパク質は、GLP-1化合物に融合された免疫グロブリンのFc部分全体、免疫グロブリンのFc部分の断片、またはその類似体を含むことができる。
【0079】
本発明の融合タンパク質は、一本鎖タンパク質からなるか、または多鎖ポリペプチドとして存在することができる。2以上のFc融合タンパク質を、それらがFc領域間に自然に形成されるジスルフィド結合を介して相互作用するような形で、製造することができる。これらの多量体はGLP-1化合物に関して均一であってもよいし、融合タンパク質のFc部分のN末端に異なるGLP-1化合物が融合されていてもよい。
【0080】
融合タンパク質の最終構造がどうであれ、FcまたはFc様領域は、そのN末端に融合されたGLP-1化合物の生体内血漿半減期を延長する役割を果たさなければならない。また、融合されたGLP-1化合物は、なんらかの生物学的活性を保っていなければならない。半減期の増加は、融合タンパク質の半減期をGLP-1化合物単独の半減期と比較する実施例7に記載の方法を使って証明することができる。生物学的活性は当技術分野で知られている試験管内法および生体内法によって決定することができる。代表的な生物学的アッセイを実施例6、8、および9に記載する。
【0081】
タンパク質分解によって製造されるIgGのFc領域は、無傷のIgG分子と同じ生体内半減期を持ち、Fab断片は迅速に分解されるので、半減期の延長に関係する配列はCH2および/またはCH3ドメイン中に存在すると考えられる。また、高親和性Fc受容体もC1qも結合しないIgG変異体の異化速度は、親野生型抗体のクリアランス速度と識別できないことが、文献で明らかにされているが、これは、異化部位がFc受容体結合またはC1q結合に関与する部位とは異なることを示している[Wawrzynczakら, (1992) Molecular Immunology 29:221]。ネズミIgG1 Fc領域を使った部位特異的突然変異誘発研究により、IgG1 Fc領域のうち異化速度を制御する部位は、CH2-CH3ドメイン境界に位置することが示唆された。
【0082】
これらの研究に基づいて、融合タンパク質の半減期が最適化されるように、Fc領域の異化部位を改変することができる。本発明の異種融合タンパク質に使用するFc領域は、IgG1またはIgG4 Fc領域に由来することが好ましい。Fc領域はIgG4であるかまたはIgG4に由来することが、さらに好ましい。IgG Fc領域は、好ましくは、ヒンジ領域を含めてCH2領域とCH3領域の両方を含む。
【0083】
異種アルブミン融合タンパク質:
上述のGLP-1化合物はアルブミンまたはその類似体、断片、もしくは誘導体に直接融合するか、またはペプチドリンカーを介して融合することができる。
【0084】
本発明の融合タンパク質の一部を構成するアルブミンタンパク質は、一般的には、どの種からクローン化されたアルブミンに由来してもよい。しかし、融合タンパク質がヒトで免疫原性を示す危険を低下させるには、ヒトアルブミンならびにその断片および類似体が好ましい。ヒト血清アルブミン(HSA)は、66,500の分子式量を持つ585アミノ酸の単一非グリコシル化ポリペプチド鎖からなる。ヒトHSAのアミノ酸配列を図2に示す[Melounら (1975) FEBS Letters 58:136;Behrensら (1975) Fed. Proc. 34:591;Lawnら (1981) Nucleic Acids Research 9:6102-6114;Minghettiら (1986) J. Biol. Chem. 261:6747参照]。アルブミンには、様々な多型変異体ならびに類似体および断片が記載されている[Weitkampら, (1973) Ann. Hum. Genet. 37:219参照]。例えばEP322,094には、種々の短縮型HSAが開示されている。これらの断片には、例えばHSA(1-373)、HSA(1-388)、HSA(1-389)、HSA(1-369)、およびHSA(1-419)、ならびに1-369と1-419の間の断片が含まれる。EP399,666には、例えばHSA(1-177)およびHSA(1-200)ならびにHSA(1-177)とHSA(1-200)の間の断片を含むアルブミン断片が開示されている。
【0085】
本発明の異種融合タンパク質は、当該融合タンパク質が生物学的に活性であり、かつGLP-1化合物単独よりも長い血漿半減期を持つような、任意のアルブミンタンパク質(断片、類似体、および誘導体を含む)にカップリングされたGLP-1化合物を包含すると理解される。したがって、融合タンパク質のアルブミン部分が天然ヒトアルブミンに匹敵する血漿半減期を持つ必要は、必ずしもない。長い半減期を持つ断片、類似体、および誘導体、または天然ヒトアルブミンと対象GLP-1化合物との中間の半減期を持つ断片、類似体、および誘導体は、知られているか、または作製することができる。
【0086】
本発明の異種融合タンパク質には、融合タンパク質のGLP-1化合物部分および/またはFcもしくはアルブミン部分に保存的アミノ酸置換を持つタンパク質が包含される。「保存的置換」は、同じ正味の電荷を持ち、ほぼ同じサイズおよび形状を持つ別のアミノ酸による、あるアミノ酸の置換である。脂肪族アミノ酸側鎖または置換脂肪族アミノ酸側鎖を持つアミノ酸は、その側鎖中の炭素およびヘテロ原子の総数の差がだいたい4以下であれば、ほぼ同じサイズを持つ。またこれらは、その側鎖中の分枝数の差が1以下であれば、ほぼ同じ形状を持つ。側鎖中にフェニル基または置換フェニル基を持つアミノ酸はほぼ同じサイズおよび形状を持つとみなされる。特に別段の条件を記す場合を除いて、保存的置換は、好ましくは、天然アミノ酸を使ってなされる。
【0087】
しかし「アミノ酸」という用語は、ここでは広い意味で使用され、天然のアミノ酸だけでなく、アミノ酸類似体および誘導体を含む非天然アミノ酸を包含する。後者にはアミノ酸部分を含む分子が包含される。この広い定義からみて、本明細書にいうアミノ酸に、例えば天然の蛋白原性L-アミノ酸;D-アミノ酸;アミノ酸類似体および誘導体などの化学修飾アミノ酸;ノルロイシン、β-アラニン、オルニチン、GABAなどの天然の非蛋白原性アミノ酸;およびアミノ酸に特有であることが当技術分野で知られている性質を持つ化学合成化合物などが含まれることは、当業者には理解されるだろう。本明細書で使用する「蛋白原性」という用語は、そのアミノ酸が細胞内で代謝経路によりペプチド、ポリペプチド、またはタンパク質に組み込まれうることを示す。
【0088】
合成非天然アミノ酸、置換アミノ酸、または1もしくは複数のD-アミノ酸を含む非天然アミノ酸を、本発明の異種融合タンパク質に組み込むと、いくつかの点で有利になる場合がある。D-アミノ酸含有ペプチド等は、対応するL-アミノ酸含有ペプチドと比較して、試験管内または生体内で、増加した安定性を示す。したがって、D-アミノ酸を組み込んだペプチド等の構築は、細胞内安定性の向上が望ましい時または必要な時には、特に有用な場合がある。より具体的に述べると、D-ペプチド等は、必要に応じて、内因性ペプチダーゼおよびプロテアーゼに対して耐性を示すことより、その分子の生物学的利用能を向上させ、生体内での寿命を引き延ばす。また、D-ペプチド等は、Tヘルパー細胞に対して主要組織適合複合体クラスII拘束性の提示を行なうためのプロセシングを効率よく受けることができず、したがって丸ごとの生物における体液性免疫反応を誘導する可能性が少ない。
【0089】
置換用のアミノ酸を選択する際には、本発明に包含される様々なポリペプチドの構造/機能解析の他にも、考慮することができる因子が数多くある。そのような改変を行なう際に考慮することができる一つの因子は、アミノ酸のハイドロパシー指標である。タンパク質に相互作用的な生物学的機能を付与する際のハイドロパシーアミノ酸指標の重要性は、KyteおよびDoolittle(1982, J. Mol. Biol., 157:105-132)によって論じられている。アミノ酸の相対的ハイドロパシー特性が、結果として得られるタンパク質の二次構造に寄与することは、一般に認められている。これは、結果として、タンパク質と、例えば酵素、基質、受容体、リガンド、DNA、抗体、抗原などの分子との相互作用に影響する。各アミノ酸には、その疎水性および電荷特徴に基づいて、ハイドロパシー指標が以下のように割り当てられている:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(-0.4);スレオニン(-0.7);セリン(-0.8);トリプトファン(-0.9);チロシン(-1.3);プロリン(-1.6);ヒスチジン(-3.2);グルタミン酸/グルタミン/アスパラギン酸/アスパラギン(-3.5);リジン(-3.9);およびアルギニン(-4.5)。
【0090】
当技術分野では知られているように、特定のアミノ酸を、類似するハイドロパシー指標またはハイドロパシースコアを持つ他のアミノ酸の代わりに、ペプチド、ポリペプチド、またはタンパク質中に使用して、結果として、類似する生物学的活性を持つか、または改善された生物学的活性を持つペプチド等を、製造することができる。そのような改変を行なう際には、±2以内のハイドロパシー指標を持つアミノ酸同士を置換することが好ましい。より好ましい置換は、アミノ酸が±1以内のハイドロパシー指標を持つような置換である。最も好ましい置換は、アミノ酸が±0.5以内のハイドロパシー指標を持つような置換である。
【0091】
類似するアミノ酸は親水性に基づいて置換することもできる。米国特許第4,554,101号には、隣接アミノ酸の親水性によって支配される、タンパク質の最大局所平均親水性は、そのタンパク質の生物学的性質と相関関係にあることが開示されている。アミノ酸には以下の親水性値が割り当てられている:アルギニン/リジン(+3.0);アスパラギン酸/グルタミン酸(+3.0±1);セリン(+0.3);アスパラギン/グルタミン(+0.2);グリシン(0);スレオニン(-0.4);プロリン(-0.5±1);アラニン/ヒスチジン(-0.5);システイン(-1.0);メチオニン(-1.3);バリン(-1.5);ロイシン/イソロイシン(-1.8);チロシン(-2.3);フェニルアラニン(-2.5);およびトリプトファン(-3.4)。したがって、ペプチド、ポリペプチド、またはタンパク質中のあるアミノ酸を、類似する親水性スコアを持つ別のアミノ酸で置換して、結果として、類似する生物学的活性を持っている、すなわち正しい生物学的機能をまだ保っているペプチド等を、製造することができる。そのような改変を行なう際には、好ましくは、±2以内のハイドロパシー指標を持つアミノ酸同士を置換し、±1以内のアミノ酸同士はより好ましく、±0.5以内のアミノ酸同士は最も好ましい。
【0092】
上に略述したように、本発明の融合タンパク質中のアミノ酸置換は、アミノ酸側鎖置換基の相対的類似性、例えばそれらの疎水性、親水性、電荷、サイズなどに基づいて行なうことができる。また、二次構造傾向に基づいて置換を行なうこともできる。例えば、らせんアミノ酸は、そのらせん構造を保つであろうアミノ酸で置換することができる。本ペプチド等にサイレントな変化をもたらす保存的アミノ酸変異になるように上述の様々な特徴を考慮した代表的置換は、当該天然アミノ酸が属するクラスの他の構成要素から選択することができる。アミノ酸は以下の4群に分類することができる:(1)酸性アミノ酸、(2)塩基性アミノ酸、(3)中性極性アミノ酸、および(4)中性無極性アミノ酸。
【0093】
本発明の異種融合タンパク質を製造するための一般的方法
本発明の異種融合タンパク質は様々な方法で製造することができるが、なかでも組換え法は好ましい。ここに開示し特許請求する本発明のために、下記の一般的な分子生物学用語および略号を、以下に定義する。本願で使用する用語および略号は、別段の指定がない限り、それぞれ通常の意味を持つ。例えば「℃」は摂氏度を表し、「mmol」はミリモルを表す。「mg」はミリグラムを表し、「μg」はマイクログラムを表す。「mlまたはmL」はミリリットルを表し、「μlまたはμL」はマイクロリットルを表す。アミノ酸略号は37 C.F.R.§1.822(b)(2)(1994)に記載されているとおりである。
【0094】
本明細書で使用する「塩基対」または「bp」はDNAまたはRNAを表す。略号A、C、G、およびTは、DNA分子においては、それぞれデオキシリボヌクレオシドである(デオキシ)アデノシン、(デオキシ)シチジン、(デオキシ)グアノシン、およびチミジンの5'-一リン酸型に対応する。略号U、C、G、およびAは、RNA分子においては、リボヌクレオシドであるウリジン、シチジン、グアノシン、およびアデノシンの5'-一リン酸型に対応する。二本鎖DNAの場合、塩基対はAとTまたはCとGの組合せを表しうる。DNA/RNAの場合、ヘテロ二重鎖塩基対はAとUまたはCとGの組合せを表しうる(下記「相補性」の定義を参照されたい)。
【0095】
DNAの「消化」または「制限(消化)」は、DNA中の特定配列だけに作用する制限酵素(「配列特異的エンドヌクレアーゼ」)によるDNAの触媒的切断を表す。本明細書で使用した種々の制限酵素は市販されており、その反応条件、補因子、および他の必要条件は、当業者に知られているであろうとおりに使用した。特定の制限酵素に適した緩衝液および基質量は、製造者によって指定されるか、または文献中に容易に見いだすことができる。
【0096】
「ライゲーション(連結)」は、2つの二本鎖核酸断片間にリン酸ジエステル結合を形成させる過程を表す。別段の定めがない限り、ライゲーションは、既知の緩衝液および条件により、T4 DNAリガーゼなどのDNAリガーゼを使って達成することができる。
【0097】
「プラスミド」は、染色体外にある(通常は)自己複製性の遺伝要素を表す。プラスミドは一般に小文字「p」と、それに続く文字および/または数字によって指定される。本明細書での出発プラスミドは市販されているか、公に無制限に入手することができるか、または公表された手順に従って、入手可能なプラスミドから構築することができる。また、本明細書に記載のプラスミドと等価なプラスミドが当技術分野では知られており、それらは当業者には明らかだろう。
【0098】
本明細書にいう「組換えDNAクローニングベクター」は、1つもしくは複数の追加DNAセグメントを付加することができる、またはそのようなDNAセグメントが既に付加されている、自律的に複製する任意の物質(例えばプラスミドおよびファージ)を表す。
【0099】
本明細書にいう「組換えDNA発現ベクター」は、挿入されたDNAの転写を制御するプロモーターが組み込まれている任意の組換えDNAクローニングベクターを表す。
【0100】
「転写」は、DNAのヌクレオチド配列に含まれる情報が相補的RNA配列に移される過程を表す。
【0101】
「トランスフェクション」は、宿主細胞による発現ベクターの取り込みを表し、何らかのコード配列が実際に発現されるかどうかを問わない。当業者には、例えばリン酸カルシウム共沈法、リポソームトランスフェクション、およびエレクトロポレーションなど、数多くのトランスフェクション法が知られている。一般にトランスフェクションの成功は、そのベクターの作動を示す何らかのしるしがその宿主細胞内で起こる場合に認識される。
【0102】
「形質転換」は、当該DNAが染色体外要素として、または染色体組込みによって、複製することができるように、DNAをある生物に導入することをいう。細菌および真核細胞宿主を形質転換する方法は当技術分野ではよく知られており、例えば核注入法、プロトプラスト融合、または塩化カルシウムを使ったカルシウム処理による方法など、それらの方法の多くは、J. Sambrookら「Molecular Cloning: A Laboratory Manual」(1989)に要約されている。一般に、DNAを酵母に導入する場合には、形質転換という用語は、トランスフェクションという用語と相対する用語として使用される。
【0103】
本明細書にいう「翻訳」は、メッセンジャーRNA(mRNA)の遺伝情報を使ってポリペプチド鎖を指定し、その合成を指図する過程を表す。
【0104】
「ベクター」は、遺伝子操作において細胞のトランスフェクションおよび/または形質転換に使用される核酸化合物であって、適当な制御配列と組み合わせた場合にトランスフェクションおよび/または形質転換される宿主細胞に特定の性質を付与する適当なタンパク質分子に対応するポリヌクレオチド配列を持つ核酸化合物を表す。プラスミド、ウイルス、およびバクテリオファージは適切なベクターである。人工ベクターは、異なる供給源に由来するDNA分子を制限酵素およびリガーゼで切断し、つなぎ合わせることによって構築される。本明細書で使用する「ベクター」という用語は、組換えDNAクローニングベクターおよび組換えDNA発現ベクターを包含する。
【0105】
本明細書にいう「相補的」または「相補性」は、二本鎖核酸中で水素結合によって会合する塩基(プリンおよびピリミジン)の対を表す。以下の塩基対は相補的である:グアニンとシトシン;アデニンとチミン;およびアデニンとウラシル。
【0106】
本明細書にいう「ハイブリダイゼーション」は、核酸の鎖が相補的な鎖と塩基対形成によって結合する過程を表す。同一ではないが極めて類似している2本の相補的核酸のハイブリダイゼーションに使用される条件は、それら2本の鎖の相補性の程度および鎖の長さによって相違する。そのような技術および条件は当業者には周知である。
【0107】
「単離されたアミノ酸配列」は、どんな方法で構築または合成されるにせよ、天然の配列とは異なる位置にある任意のアミノ酸を表す。
【0108】
「単離されたDNA化合物」は、どんな方法で構築または合成されるにせよ、ゲノムDNA中でのそれ本来の位置とは異なる位置にある任意のDNA配列を表す。
【0109】
「単離された核酸化合物」は、どんな方法で構築または合成されるにせよ、それ本来の位置とは異なる位置にある任意のRNAまたはDNA配列を表す。
【0110】
「プライマー」は、酵素的または合成的伸長の開始基質として機能する核酸断片を表す。
【0111】
「プロモーター」は、DNAまたはRNAの転写を指図するDNA配列を表す。
【0112】
「プローブ」は、別の核酸化合物とハイブリダイズする核酸化合物またはその断片を表す。
【0113】
当業者は、ハイブリダイゼーション反応の「ストリンジェンシー」を容易に決定することができ、ハイブリダイゼーション反応の「ストリンジェンシー」は一般的には、プローブ長、洗浄温度、および塩濃度に依存する実験的計算の結果である。一般に、長いプローブほど適正なアニーリングに高い温度を必要とし、短いプローブは必要とする温度が低い。ハイブリダイゼーションは、一般に、相補的な鎖がそれらの融解温度未満の環境に存在する時の、変性したDNAの再アニール能力に依存する。プローブとハイブリダイズ可能な配列との間に所望する相同性の程度が高いほど、使用することができる相対温度は高くなる。そのため、高い相対温度ほど、反応を、よりストリンジェントにする傾向を持ち、低い温度ほどその傾向が少ないということになる。ハイブリダイゼーション反応のストリンジェンシーに関するさらなる詳細と説明については、Ausubelら「Current Protocols in Molecular Biology」(Wiley Interscience Publishers, 1995)を参照されたい。
【0114】
本明細書で定義する「ストリンジェントな条件」または「高ストリンジェンシー条件」は、(1)洗浄に低イオン強度および高温を使用する条件、例えば15mM塩化ナトリウム/1.5mMクエン酸ナトリウム/0.1%ドデシル硫酸ナトリウムを50℃で使用する条件、(2)ハイブリダイゼーション時にホルムアミドなどの変性剤を使用する条件、例えば50%(v/v)ホルムアミドおよび0.1%ウシ血清アルブミン/0.1%フィコール/0.1%ポリビニルピロリドン/50mMリン酸ナトリウム緩衝液(pH6.5)および750mM塩化ナトリウム/75mMクエン酸ナトリウムを42℃で使用する条件、または(3)50%ホルムアミド、5×SSC(750mM塩化ナトリウム、75mMクエン酸ナトリウム)、50mMリン酸ナトリウム(pH6.8)、0.1%ピロリン酸ナトリウム、5×デンハート液、超音波処理サケ精子DNA(50μg/ml)、0.1%SDS、および10%硫酸デキストランを42℃で使用し、0.2×SSC(30mM塩化ナトリウム/3mMクエン酸ナトリウム)および50%ホルムアミド中、42℃および55℃で洗浄した後、EDTAを含む0.1×SSCからなる高ストリンジェンシー洗浄を55℃で行なう条件で識別しうる。
【0115】
「中等度にストリンジェントな条件」は、Sambrookら「Molecular Cloning: A Laboratory Manual」(ニューヨーク, Cold Spring Harbor Press, (1989))に記載されているように識別することができ、上述した条件よりストリンジェンシーの低い洗浄溶液およびハイブリダイゼーション条件(例えば温度、イオン強度、およびSDSのパーセント)の使用が含まれる。中等度にストリンジェントな条件の一例は、20%ホルムアミド、5×SSC(750mM塩化ナトリウム、75mMクエン酸ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20mg/mL変性剪断サケ精子DNAを含む溶液中、37℃で終夜インキュベートした後、フィルターを1×SSC中、約37〜50℃で洗浄することである。プローブ長などの因子を考慮して必要に応じて温度、イオン強度などを調節する方法は、当業者にはわかるだろう。
【0116】
「PCR」は、熱安定性DNAポリメラーゼを利用する周知のポリメラーゼ連鎖反応を表す。
【0117】
「リーダー配列」は、目的とする所望のポリペプチドを生成させるために、酵素的または化学的に除去することができるアミノ酸の配列を表す。
【0118】
「分泌シグナル配列」は、より大きいポリペプチドのN末端領域に一般に存在し、そのポリペプチドと小胞体などの細胞膜コンパートメントションとの会合および形質膜を通したそのポリペプチドの分泌を開始させる機能を果たす、アミノ酸の配列を表す。
【0119】
本発明の異種融合タンパク質をコードするDNAの構築
野生型アルブミンおよび免疫グロブリンタンパク質は様々な供給源から入手することができる。例えばこれらのタンパク質は、目的とするmRNAを検出可能なレベルに発現させる組織または細胞から作製されたcDNAライブラリーから得ることができる。目的とするタンパク質の公表されたDNA配列またはタンパク質配列を使って設計されたプローブで、上記ライブラリーをスクリーニングすることができる。例えば免疫グロブリン軽鎖または重鎖定常領域は、Adamsら (1980) Biochemistry 19:2711-2719;Goughetら (1980) Biochemistry 19:2702-2710;Dolbyら (1980) Proc. Natl. Acad. Sci. USA 77:6027-6031;Riceら (1982) Proc. Natl. Acad. Sci. USA 79:7862-7862;Falknerら (1982) Nature 298:286-288;およびMorrisonら (1984) Ann. Rev. Immunol. 2:239-256に記載されている。アルブミンのタンパク質配列およびDNA配列を開示している文献には、例えばMelounら (1975) FEBS Letters 58:136;Behrensら (1975) Fed. Proc. 34:591;Lawnら (1981) Nucleic Acids Research 9:6102-6114;およびMinghettiら (1986) J. Biol. Chem. 261:6747などがある。
【0120】
選択したプローブによるcDNAライブラリーまたはゲノムライブラリーのスクリーニングは、Sambrookら「Molecular Cloning: A Laboratory Manual」(Cold Spring Harbor Laboratory Press, ニューヨーク (1989))に記載されているような標準的方法を使って行なうことができる。アルブミンまたは免疫グロブリンタンパク質をコードする遺伝子を単離するもう一つの手段は、PCR法を使用することである[Sambrookら, 前掲;Dieffenbachら「PCR Primer: A Laboratory Manual」Cold Spring Harbor Laboratory Press, ニューヨーク (1995)]。PCRプライマーは公表された配列に基づいて設計することができる。
【0121】
一般に、特定の種からクローン化された完全長野生型配列は、融合タンパク質の一部であるGLP-1化合物の血漿半減期を延ばす能力を保っている類似体、断片、および誘導体を作製するためのテンプレートとして役立ちうる。融合タンパク質がヒトで潜在的免疫原性を示す危険を低下させるために、本発明の異種融合タンパク質のFc部分およびアルブミン部分は、天然ヒト配列に由来することが好ましい。
【0122】
特に、本発明に包含される融合タンパク質の免疫グロブリン部分は、免疫グロブリンのFc断片だけを含むことが好ましい。特定のエフェクター機能を所望するかどうかと、融合タンパク質の構造的特徴とに応じて、Fc部分は、CH2およびCH3ドメインに加えてヒンジ領域を含むことができ、またはそれらの他の何らかの組合せを含むことができる。これらのFc断片は、その断片の所望する末端に対応する配列にハイブリダイズするように設計されたプライマーにより、PCR技術を使って作製することができる。同様に、アルブミンの断片が望ましい場合は、アルブミン内部配列に相補的なPCRプライマーを設計することができる。発現ベクターへのクローニングを容易にするための制限酵素部位が生成するようにPCRプライマーを設計することもできる。
【0123】
本発明のGLP-1化合物をコードするDNAは、上述した方法のようなクローニング法および化学合成DNAを含む多種多様な方法で作製することができる。コードされるペプチドの長さが短いことを考えると、化学合成は魅力的な方法であるだろう。GLP-1のアミノ酸配列は、プレプログルカゴン遺伝子の配列と共に、公表されている[Lopezら (1983) Proc. Natl. Acad. Sci., USA 80:5485-5489;Bellら (1983) Nature, 302:716-718;Heinrich, G.ら (1984) Endocrinol, 115:2176-2181;Ghiglione, M.ら (1984) Diabetologia 27:599-600]。したがって、天然GLP-1化合物およびその断片をPCRするためのプライマーを設計することができる。
【0124】
次に、GLP-1化合物をコードするDNAを、アルブミンまたはFcタンパク質をコードするDNAにインフレームで連結することにより、融合タンパク質をコードする遺伝子を構築することができる。GLP-1化合物をコードする遺伝子と、アルブミンまたはFcタンパク質をコードする遺伝子は、リンカーペプチドをコードするDNAを介してインフレームでつなぎ合わせることもできる。
【0125】
本発明の異種融合タンパク質の生体内での機能および安定性は、小さいペプチドリンカーを加えて、潜在的に望ましくないドメイン相互作用を防ぐことにより、最適化することができる。これらのリンカーは潜在的には任意の長さを持ち、アミノ酸の任意の組合せからなりうるが、その長さは、望ましくないドメイン相互作用を防ぎそして/または生物学的活性および/または安定性を最適化するのに必要な長さを超えないことが好ましい。一般に、リンカーは、極端にかさ高い側鎖を持つアミノ酸や、有意な二次構造を導入しそうなアミノ酸を含まないべきである。リンカーは、セリン-グリシンリッチであると共に、長さは30アミノ酸未満であることが好ましい。リンカーの長さは20アミノ酸を超えないことがより好ましい。リンカーの長さは15アミノ酸を超えないことがさらに好ましい。好ましいリンカーは、Gly-Gly-Gly-Gly-Serという配列の反復を含む。この配列は2〜6回反復することが好ましい。この配列は3〜4回反復することがさらに好ましい。
【0126】
野生型GLP-1、アルブミン、およびFcポリペプチドをコードするDNAならびにその断片は、ライゲーションに先立って、または融合タンパク質全体をコードするcDNAの形で、突然変異させることができる。当技術分野では様々な突然変異導入技術がよく知られている。例えば突然変異導入PCR法では、対応するタンパク質中の特定のアミノ酸配列を変化させる目的で、鎖オーバーラップ伸長を利用して、特定の塩基突然変異を生成する。このPCR突然変異導入法では4つのプライマー、すなわちフォワード方向の2つ(プライマーAおよびC)とリバース方向の2つ(プライマーBおよびD)を使用する必要がある。突然変異した遺伝子は、2つの異なる段階で野生型テンプレートから増幅される。第1反応では、AからBへの反応とCからDへの反応とを個別に行なって、遺伝子を半分ずつ増幅する。ここに、BおよびCプライマーは、突然変異させようとする遺伝子の領域を標的としている。これらのプライマーを標的領域と整列させた場合、それらは、変化させようとしている塩基に関してミスマッチを持つ。AからBまでおよびCからDまでの反応が完了したら、反応生成物を単離し、それらを混合して、AからDまでの反応のテンプレートとして使用する。そうすると、この反応により、完全な突然変異型産物が得られる。
【0127】
融合タンパク質全体をコードする遺伝子を作製したら、それを適当な発現ベクターにクローニングすることができる。本発明のGLP-1融合タンパク質を製造するために使用することができる具体的戦略を実施例1に説明する。
【0128】
本発明の異種融合タンパク質を組換え発現させるための一般的方法
本明細書に記載の異種融合タンパク質産生用発現ベクターまたはクローニングベクターで宿主細胞をトランスフェクトまたは形質転換し、プロモーターを誘導するため、形質転換体を選択するため、または所望の配列をコードする遺伝子を増幅するために適宜改良された通常の栄養培地で培養する。培地、温度、pHなどの培養条件は、当業者であれば、はなはだしい実験を行なわなくても選択することができる。細胞培養の生産性を最大にするための原理、手順、および実用的技術は、一般に、M. Butler編「Mammalian Cell Biotechnology: A Practical Approach」(IRL Press, 1991)およびSambrookら(前掲)に記載されている。トランスフェクションの方法は、例えばCaPO4およびエレクトロポレーションなど、当業者に知られている。哺乳類細胞宿主系形質転換の一般的側面は、米国特許第4,399,216号に記載されている。酵母への形質転換は通例、van Solingenら, J Bact. 130(2): 946-7(1977)およびHsiaoら, Proc. Natl. Acad. Sci. USA 76(8):3829-33(1979)の方法に従って行なわれる。しかし、細胞にDNAを導入する他の方法、例えば核マイクロインジェクション、エレクトロポレーション、無傷細胞との細菌プロトプラスト融合、またはポリブレンもしくはポリオミチン(polyomithine)などのポリカチオンも、使用することができる。哺乳類細胞を形質転換するための様々な技術については、Keownら, Methods in Enzymology 185: 527-37(1990)およびMansourら, Nature 336(6197): 348-52(1988)を参照されたい。
【0129】
本明細書に記載するベクター中の核酸(例えばDNA)をクローニングまたは発現させるのに適した宿主細胞には、原核生物、酵母、または高等真核細胞が含まれる。適切な原核生物には、グラム陰性生物またはグラム陽性生物などの真正細菌、例えば大腸菌などの腸内細菌が含まれるが、これらに限るわけではない。例えば大腸菌K12株MM294(ATCC 3 1.446)、大腸菌X1 776(ATCC 3 1.537)、大腸菌W3 110株(ATCC 27.325)およびK5 772株(ATCC 53.635)など、様々な大腸菌株を公に入手することができる。他の適切な原核宿主細胞には、エシェリキア(例えば大腸菌)、エンテロバクター、エルウィニア、クレブシエラ、プロテウス、サルモネラ(例えばネズミチフス菌)、セラチア(例えば霊菌)、および赤痢菌などの腸内細菌、ならびに枯草菌やバチルス・リケニフォルミス(例えば1989年4月12日に公開されたDD266,710に開示されているバチルス・リケニフォルミス 41P)、緑膿菌などのシュードモナス、およびストレプトミセスなどが含まれる。これらの例は限定ではなく説明のための実例である。W3110株は、組換えDNA産物醗酵用の一般的な宿主株であるので、特に好ましい宿主または親宿主の一つである。宿主細胞が分泌するタンパク質加水分解酵素は極微量であることが好ましい。例えばW3110株を改変して、その宿主に内在するタンパク質をコードする遺伝子に、遺伝的突然変異をもたらしてもよい。そのような宿主の例には、完全遺伝子型ronAを持つ大腸菌W3110 1A2株、完全遺伝子型tonA ptr3を持つ大腸菌W3110 9E4株、完全遺伝子型tonA, ptr3 phoA E15 (argF-lac) I69 degP ompT /can'を持つ大腸菌W3110 27C7株(ATCC 55,244)、非カナマイシン耐性degP欠失突然変異を持つ37D6株である大腸菌W3110 40B4株、および1990年8月7日発行の米国特許第4,946,783号に開示されている突然変異型ぺリプラスムプロテアーゼを持つ大腸菌株がある。あるいは、生体内クローニング法、例えばPCRまたは他の核酸ポリメラーゼ反応なども適切である。
【0130】
原核生物に加えて、糸状菌または酵母などの真核微生物も、融合タンパク質ベクターにとって適切なクローニング宿主または発現宿主である。サッカロミセス・セレビシェは広く使用されている下等真核宿主微生物である。他にも、シゾサッカロミセス・ポンベ[BeachおよびNurse, Nature 290:140-3 (1981)、1995年5月2日公開のEP 139,383)、クルイベロミセス宿主[米国特許第4,943,529号、Fleerら,Bio/Technology 9(10):968-75 (1991)]、例えばクルイベロミセス・ラクチス(MW98-8C、CBS683、CBS4574)[de Louvencourtら, J. Bacteriol., 154(2):737-42 (1983)]、クルイベロミセス・フラギリス(ATCC 12,424)、クルイベロミセス・ブルガリカス(ATCC 16,045)、クルイベロミセス・ウィッカーラミイ(ATCC 24,178)、クルイベロミセス・ワルティイ(ATCC 56,500)、クルイベロミセス・ドロソフィラルム(ATCC 36,906)[Van den Bergら, Bio/Technology 8(2):135-9 (1990)]、クルイベロミセス・サーモトレランス、およびクルイベロミセス・マルキシアヌス、ヤロウィア(EP 402,226)、ピキア・パストリス(EP 183,070)[Sreekrishnaら, J. Basic Microbiol. 28(4):265-78 (1988)]、カンジダ、トリコデルマ・リーシア(EP 244,234)、ニューロスポラ・クラッサ[Caseら, Proc. Natl. Acad. Sci. USA 76(10):5259-5263 (1979)]、シュワンニオミセス、例えばシュワンニオミセス・オシデンツリス(1990年10月31日公開のEP 394,538)、例えばニューロスポラ、ペニシリウム、トリポクラジウム(1991年1月10日公開のWO 91/00357)、およびアスペルギルス宿主(例えばアスペルギルス・ニデュランス[Ballanceら, Biochem. Biophys. Res. Comm. 112(1):284-9 (1983)、Tilburnら, Gene 26(2-3):205-21 (1983)、Yeltonら, Proc. Natl. Acad. Sci. USA 81(5):1470-4 (1984)]およびアスペルギルス・ニガー[KellyおよびHynes, EMBO J. 4(2):475-9 (1985)])などの糸状菌などがある。メチロトローフ酵母は、ハンゼヌラ、カンジダ、クロエケラ、ピキア、サッカロミセス、トルロプシス、およびロドトルイアからなる属群から選択される。この種類の酵母の具体的な種の一覧は、C. Antony「The Biochemistry of Methylotrophs」269(1982)に記載されている。
【0131】
本発明の融合タンパク質の発現に適した宿主細胞は多細胞生物から得られる。無脊椎動物細胞の例には、ドロソフィラS2およびスポドプテラSp、スポドプテラhigh5などの昆虫細胞ならびに植物細胞が含まれる。有用な哺乳類宿主細胞株の例には、チャイニーズハムスター卵巣(CHO)細胞およびCOS細胞が含まれる。より具体的な例としては、SV40で形質転換されたサル腎臓CV1株(COS-7、ATCC CRL 1651)、ヒト胎児腎臓株[293細胞または懸濁培養で成長するようにサブクローニングされた293細胞;Grahamら, J. Gen. Virol. 36(1):59 (1977)]、チャイニーズハムスター卵巣細胞/-DHFR[CHO、UrlaubおよびChasin, Proc. Natl. Acad. Sci. USA 77(7):4216-20 (1980)]、マウスセルトリ細胞[TM4, Mather, Biol. Reprod. 23(1):243-52 (1980)]、ヒト肺細胞(W138, ATCC CCL 75)、ヒト肝臓細胞(Hep G2, HB 8065)、およびマウス乳癌細胞(MMT 060562, ATCC CCL51)などが挙げられる。適切な宿主細胞の選択は、技術上可能であると考えられる。
【0132】
本発明の融合タンパク質は直接的に組換え生産するか、または成熟融合タンパク質のN末端に特異的切断部位をもたらすシグナル配列または他の追加配列を持つタンパク質として組換え生産することができる。一般に、シグナル配列はベクターの一成分であってもよいし、ベクターに挿入される融合タンパク質コードDNAの一部であってもよい。シグナル配列は、例えばアルカリホスファターゼ、ペニシリナーゼ、lpp、または耐熱性エンテロトキシンIIリーダーの群から選択される原核シグナル配列とすることができる。酵母分泌の場合は、シグナル配列を、例えば酵母インベルターゼリーダー、アルファ因子リーダー(例えばサッカロミセスおよびクルイベロミセスのa因子リーダー;後者は米国特許第5,010,182号に記述されている)、または酸性ホスファターゼリーダー、カンジダ・アルビカンス・グルコアミラーゼリーダー(EP 362,179)またはWO 90/13646に記載のシグナルなどにすることができる。哺乳類細胞発現の場合は、同じ種または近縁種の分泌ポリペプチドに由来する哺乳類シグナル配列ならびにウイルス分泌リーダーを使って、タンパク質の分泌を指示することができる。
【0133】
発現ベクターおよびクローニングベクターはどちらも、1つまたは複数の選択した宿主細胞内でそのベクターの複製を可能にする核酸配列を含有する。そのような配列は、種々の細菌、酵母およびウイルスについてよく知られている。プラスミドpBR322由来の複製起点はほとんどのグラム陰性菌に適し、2μプラスミド起点は酵母に適し、種々のウイルス起点(SV40、ポリオーマ、アデノウイルス、VSVまたはBPV)は哺乳類細胞におけるベクターのクローニングに役立つ。
【0134】
発現ベクターおよびクローニングベクターは典型的には選択遺伝子(選択可能マーカーともいう)を含有する。典型的な選択遺伝子は、(a)抗生物質または他の毒素、例えばアンピシリン、ネオマイシン、メトトレキセート、またはテトラサイクリンなどに対する耐性を付与するタンパク質をコードするか、(b)独立栄養不全を補うタンパク質をコードするか、または(c)天然培地からは利用できない必須栄養素を供給するタンパク質をコードする(例えばバチルスの場合、D-アラニンラセマーゼをコードする遺伝子など)。
【0135】
適切な哺乳類細胞用選択マーカーの一例は、融合タンパク質コード核酸を取り込む能力を持つ細胞の同定を可能にするもの、例えばDHFRまたはチミジンキナーゼなどである。野生型DHFRを使用する場合に適切な宿主細胞は、UrlaubおよびChasin, Proc. Natl. Acad. Sci. USA, 77(7):4216-20(1980)に記述されているように作出し増殖されるDHFR活性欠損CHO細胞株である。酵母での使用に適した選択遺伝子は、酵母プラスミドYRp7中に存在するtrp1遺伝子である[Stinchcombら, Nature 282(5734):39-43 (1979)、Kingsmanら, Gene 7(2):141-52 (1979)、またはTschemperら, Gene 10(2):157-66 (1980)]。trp1遺伝子は、トリプトファン中で増殖する能力を欠く酵母の突然変異株(例えばATCC番号44076またはPEPC1[Jones, Genetics 85:23-33 (1977)]用の選択マーカーになる。
【0136】
発現ベクターおよびクローニングベクターは、通常、mRNA合成を指示するために融合タンパク質コード核酸配列に作動可能に連結されたプロモーターを含有する。種々の潜在的宿主細胞によって認識されるプロモーターはよく知られている。原核宿主での使用に適したプロモーターには、β-ラクタマーゼおよびラクトースプロモーター系[Changら, Nature 275(5681):617-24 (1978)、Goeddelら, Nature 281(5732):544-8 (1979)]、アルカリホスファターゼ、トリプトファン(up)プロモーター系[Goeddel, Nucleic Acids Res. 8(18):4057-74 (1980)、1981年9月30日公開のEP 36,776)、およびtatプロモーターなどのハイブリッドプロモーター[deBoerら, Proc. Natl. Acad. Sci. USA 80(1):21-25 (1983)]などがある。細菌系用のプロモーターは、融合タンパク質をコードするDNAに作動可能に連結されたシャイン・ダルガノ(S.D.)配列も含有するだろう。
【0137】
酵母宿主での使用に適した促進配列の例としては、3-ホスホグリセリン酸キナーゼ[Hitzemanら, J. Biol. Chem. 255(24):12073 (1980)]または他の解糖系酵素[Hessら, J. Adv. Enzyme Reg. 7:149 (1968)、およびHolland, Biochemistry 17(23):4900-7 (1978)]、例えばエノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース-6-リン酸イソメラーゼ、3-ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼ、およびグルコキナーゼなどのプロモーターが挙げられる。
【0138】
生育条件によって転写が制御されるという追加の利点を持つ誘導性プロモーターである他の酵母プロモーターは、アルコールデヒドロゲナーゼ2、イソチトクロームC、酸性ホスファターゼ、窒素代謝に関係する分解酵素、メタロチオネイン、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、ならびにマルトースおよびガラクトース資化を担う酵素のプロモーター領域である。酵母発現用の適切なベクターおよびプロモーターはEP 73,657にさらに詳しく記載されている。哺乳類宿主細胞におけるベクターからの融合タンパク質コードmRNAの転写は、例えば、ポリオーマウイルス、鶏痘ウイルス、アデノウイルス(アデノウイルス2など)、ウシ乳頭腫ウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよびシミアンウイルス40(SV40)などのウイルスのゲノムから得られるプロモーター、アクチンプロモーターまたは免疫グロブリンプロモーターなどの異種哺乳類プロモーター、および熱ショックプロモーターから得られるプロモーターによって制御しうる(ただし、それらのプロモーターは当該宿主細胞系に適合するものとする)。
【0139】
融合タンパク質をコードするポリヌクレオチドの高等真核生物による転写は、ベクターにエンハンサー配列を挿入することによって増進させうる。エンハンサーは通常約10〜300bpのDNAのシス作用要素であって、プロモーターに作用してその転写を増進する。現在では、哺乳類遺伝子(グロビン、エステラーゼ、アルブミン、α-フェトプロテインおよびインスリン)に由来する多くのエンハンサー配列が知られている。しかし、典型的には、真核細胞ウイルス由来のエンハンサーが使用されるだろう。複製起点の後期側(bp100-270)にあるSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後期側にあるポリオーマエンハンサー、およびアデノウイルスエンハンサーなどが、その例である。エンハンサーをベクターに接合する際は、融合タンパク質コード配列に対して5'側に接合してもよいし、3'側に接合してもよいが、好ましくはエンハンサーをプロモーターの5'側に置く。
【0140】
真核宿主細胞(酵母、カビ、昆虫、植物、動物、ヒト、または他の多細胞生物に由来する有核細胞)で使用される発現ベクターは、転写の終結およびmRNAの安定化に必要な配列も含有するだろう。そのような配列は、真核細胞またはウイルスのDNAまたはcDNAの5'非翻訳領域および時には3'非翻訳領域から、一般に入手することができる。これらの領域は、融合タンパク質をコードするmRNAの非翻訳部分にポリアデニル化された断片として転写されるヌクレオチドセグメントを含有する。
【0141】
様々な形態の融合タンパク質を培養培地または宿主細胞溶解物から回収することができる。膜結合型の場合は、適切な界面活性剤溶液(例えばトリトンX100)を使用するか、酵素的切断によって、膜から遊離させることができる。融合タンパク質の発現に使用した細胞は、例えば凍結融解サイクリング、超音波処理、機械的破壊、または細胞溶解剤など、様々な物理的または化学的手段によって破壊することができる。
【0142】
本発明の異種融合タンパク質の精製:
本発明の異種融合タンパク質を適当な宿主細胞中で発現させたら、その類似体を単離し、精製することができる。以下の方法は、適切な精製法の代表例である:カルボキシメチルセルロースでの分画、セファデックスG-75などのゲル濾過、DEAEまたはMono-Qなどの陰イオン交換樹脂、CMまたはMono-Sなどの陽イオン交換体、IgGなどの夾雑物を除去するためのプロテインAセファロース、エピトープタグ付のポリペプチドを結合するための金属キレートカラム、逆相HPLC、クロマトフォーカシング、シリカゲル、エタノール沈殿、および硫酸アンモニウム沈殿。。
【0143】
様々なタンパク質精製方法を使用することができ、そのような方法は当技術分野で知られていると共に、例えばDeutscher, Methods in Enzymology 182:83-9(1990)およびScopes「Protein Purification: Principles and Practice」Springer-Verlag, ニューヨーク(1982)などに記載されている。選択される精製工程は、使用する製造方法および製造される融合タンパク質の性質に依存するだろう。例えばFc断片を含む融合タンパク質は、プロテインAまたはプロテインGアフィニティーマトリックスを使って効果的に精製することができる。低pHまたは高pH緩衝液を使って、アフィニティーマトリックスから融合タンパク質を溶出させることができる。穏やかな溶出条件は、融合タンパク質の不可逆的変性を防止するのに役立つだろう。イミダゾール含有緩衝液を使用することもできる。実施例3に本発明融合タンパク質の精製手順の成功例をいくつか説明する。
【0144】
本発明の異種融合タンパク質の特徴づけ:
本発明の融合タンパク質を特徴づける方法は数多く存在する。これらの方法には、一例として、SDS-PAGEとタンパク質染色法または抗IgGまたは抗HSA抗体を用いた免疫ブロット法との併用が含まれる。他の方法には、例えばマトリックス支援レーザー脱離/イオン化質量分析法(MALDI-MS)、液体クロマトグラフィー/質量分析法、等電点フォーカシング、分析用イオン交換、クロマトフォーカシング、および円二色性などがある。SDS-PAGEと免疫ブロット法との併用ならびに質量分析法を用いて、いくつかの代表的な異種融合タンパク質を特徴づけた(実施例4および5ならびに図3および4参照)。
【0145】
例えば表3(実施例5参照)に、いくつかの代表的な融合タンパク質の計算分子量と、質量分析法によって決定された質量とを例示する。また、図3および図4に、SDS PAGEで決定したいくつかの代表的な融合タンパク質の分子量を例示する。試験した異種融合タンパク質は全て、一過性に発現、分泌させた。また、Igκシグナル配列を切断して、正しいN末端を持つタンパク質を得た。
【0146】
また表3は、質量分析法によって決定した分子量が予想より大きい場合があることも示している。これはFc部分のグリコシル化およびC末端伸長の結果である。融合タンパク質の酵素消化に続いて、逆相HPLCおよび質量分析を行なう事により、糖鎖部分を含むペプチド画分を同定することができる。次に、これらの画分をN末端アミノ酸配列決定にかけて、潜在的グリコシル化部位を同定することができる。例えばエクセンディン4-Fc(配列番号29)の特徴づけにより、39位のセリンおよび50位のスレオニンがO結合型グリコシル化を、また122位のアスパラギンがN結合型グリコシル化を受けることがわかる。
【0147】
いくつかの代表的なGLP-1融合タンパク質を活性についても調べた。試験管内および生体内でGLP-1活性を検出する方法は数多く存在する(実施例6、7、8および9参照)。表4(実施例6)にいくつかのGLP-1融合物に付随するGLP-1受容体活性を例示する。数字は、Val8-GLP-1(7-37)OHに付随する活性に対する相対値である。試験した融合タンパク質は全てGLP-1受容体活性を持っていた。低い試験管内活性は、必ずしも、生体内での効果が弱いことを示さない。これらの融合タンパク質の半減期はかなり増加しているので、弱い試験管内活性は一般に弱い生体内活性を予言するものではない。図7および実施例7は、本発明の融合タンパク質に付随する延長された半減期を例示している。例えば、Val8-GLP-1-Fcはサルで約45時間の半減期を持ち、Val8-GLP-1-HSAはサルで約87時間の半減期を持ち、Gly8-Glu22-GLP-1-CEx-リンカー-IgG1はイヌでIV投与後に約55時間の半減期を持ち、Gly8-Glu22-GLP-1-CEx-リンカー-IgG1はイヌでSC投与後に約38時間の半減期を持った。
【0148】
本発明の組成物:
治療タンパク質組成物には物理的安定性も不可欠な特徴である。GLP-1化合物は加工中に構造変化が起こるために製造および製剤がとりわけ困難だった。例えば一部のGLP-1化合物には凝集しようとする一般的傾向がある。また、一部のGLP-1化合物は可溶性かつ活性なαヘリックス型から不溶性で潜在的に不活性なβシート型に変換することが明らかにされている。IgGのFc領域またはアルブミンなどの大きいタンパク質へのGLP-1化合物の融合は、GLP-1化合物の半減期を増大させるだけでなく、GLP-1化合物の物理的安定性およびコンフォメーションの安定性にも寄与する。例えば、PBS中のVal8-GLP-1-リンカー-HSAは、37℃で約30日まで安定である。
【0149】
本発明の異種融合タンパク質は、1つまたは複数の賦形剤と共に製剤化することができる。本発明の活性な融合タンパク質は、医薬的に許容できる緩衝液と混合し、許容できる安定性および腸管外投与などの投与が可能なpHが得られるように、pHを調節することができる。
【0150】
所望により、1つまたは複数の医薬的に許容できる抗微生物剤を加えてもよい。メタクレゾールおよびフェノールは、好ましい医薬的に許容できる微生物剤である。イオン強度または張性を調節するために、1つまたは複数の医薬的に許容できる塩を加えてもよい。製剤の等張性をさらに調節するために、1つまたは複数の賦形剤を加えてもよい。グリセリンは等張性調節賦形剤の一例である。医薬的に許容できるとは、ヒトまたは他の動物への投与に適切しており、有毒な要素または望ましくない夾雑物を含まず、製剤中の活性化合物の活性を妨害しないことを意味する。
【0151】
本発明では、本発明の異種融合タンパク質の医薬的に許容できる塩型を使用することができる。酸付加塩を形成させるためによく使用される酸は、塩酸、臭化水素酸、ヨウ化水素酸、硫酸、リン酸などの無機酸、およびp-トルエンスルホン酸、メタンスルホン酸、シュウ酸、p-ブロモフェニルスルホン酸、炭酸、コハク酸、クエン酸、安息香酸、酢酸などの有機酸である。好ましい酸付加塩は、塩酸および臭化水素酸などの鉱酸によって形成されるものである。
【0152】
塩基付加塩には、無機塩基、例えばアンモニウムまたはアルカリもしくはアルカリ土類金属の水酸化物、炭酸塩、炭酸水素塩などから誘導されるものが含まれる。したがって、本発明の塩の製造に役立つそのような塩には、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、炭酸カリウムなどが含まれる。
【0153】
組成物の投与:
投与は、通常の技術を持つ医師に有効であることが知られているどの経路で行なってもよい。末梢、腸管外投与はそのような方法の一つである。腸管外投与は医学的文献では滅菌注射器または他の何らかの機械的装置、例えば注入ポンプなどによる、身体への剤形の注射であると一般に理解されている。末梢腸管外経路には静脈内、筋肉内、皮下、および腹腔内投与経路が含まれる。
【0154】
本発明の異種融合タンパク質は、非腸管外経路である経口、直腸、鼻腔、または下気道経路での投与にも馴染みうる。これらの非腸管外経路のうち、下気道経路および経口経路は好ましい。
【0155】
本発明の融合タンパク質は、広範囲にわたる多様な疾患および状態の処置に使用することができる。本発明の融合タンパク質は主に、「GLP-1受容体」と呼ばれる受容体に作用することによって、その生物学的効果を発揮する。したがって、GLP-1受容体刺激またはGLP-1化合物の投与に有利な反応を示す疾患および/または状態を持つ対象は、本発明のGLP-1融合タンパク質で処置することができる。これらの対象を、「GLP-1化合物による処置を必要としている」または「GLP-1受容体刺激を必要としている」という。これには、インスリン非依存性糖尿病、インスリン依存性糖尿病、脳卒中(WO00/16797参照)、心筋梗塞(WO98/08531参照)、肥満(WO98/19698参照)、手術後の異化変化(米国特許第6,006,753号)、機能性消化不良および過敏性腸症候群(WO99/64060参照)を持つ対象が含まれる。また、GLP-1化合物による予防的処置を必要とする対象、例えばインスリン非依存性糖尿病を発症する危険がある対象(WO00/07617参照)も含まれる。耐糖能障害または空腹時グルコース異常を持つ対象、当該対象の身長および体格での正常体重より約25%高い体重を持つ対象、部分膵切除を受けた対象、片親または両親がインスリン非依存性糖尿病である対象、妊娠糖尿病の病歴がある対象、および急性または慢性膵炎の病歴がある対象は、インスリン非依存性糖尿病を発症する危険がある。
【0156】
GLP-1化合物の「有効量」は、GLP-1受容体刺激を必要としている対象に投与したときに、許容できない副作用を引き起こすことなく、望ましい治療効果および/または予防効果をもたらす量である。「望ましい治療効果」には以下に挙げる効果の1つまたは複数が含まれる:1)疾患または状態に関係する症状の改善、2)疾患または状態に関係する症状の発症の遅延、3)処置しない場合と比較した生存期間の延長、および4)処置しない場合と比較した生活の質の向上。例えば糖尿病の処置に関してGLP-1化合物の「有効量」とは、処置しない場合よりも血中グルコース濃度がよりよく管理され、よって網膜症、ニューロパシーまたは腎疾患などの糖尿病合併症の発症が遅延するような量である。糖尿病の予防に関してGLP-1化合物の「有効量」とは、スルホニル尿素、チアゾリジンジオン、インスリンおよび/またはビスグアニジンなどの抗低血糖薬による処置を必要とする高血中グルコースレベルの発生を、処置しない場合と比較して遅延させるような量である。
【0157】
患者の血中グルコースを正常化するのに有効な融合タンパク質の用量は、例えば当該対象の性別、体重および年齢、血中グルコース調節不能の重症度、投与経路および生物学的利用能、融合タンパク質の薬物動態プロフィール、効力、および製剤などを含む数多くの因子に依存するだろう。
【0158】
本発明は、アルブミンタンパク質、アルブミン断片、アルブミン類似体、Fcタンパク質、Fc断片、またはFc類似体に融合されているために改善された生化学的特性および生物物理学的特性を持つGLP-1化合物を包含する。これらの異種タンパク質は、宿主細胞中でうまく発現させ、GLP-1受容体の活性化に関係するシグナリング活性を保ち、長くなった半減期を持つことができる。
【実施例】
【0159】
本発明をさらに詳しく説明するために、以下に実施例を記載する。本発明の範囲は単に以下の実施例からなるとみなしてはならない。ここに記載する特定の試薬、装置、および方法が単なる例示であって、決して本発明の限定を意図するものでないことは、当業者には理解されるだろう。
【0160】
実施例1:異種融合タンパク質をコードするDNAの構築
実施例1a
Val8-GLP-1(7-37)-FcをコードするDNAの構築:
ヒトIgG1のFc部分をcDNAライブラリーから単離した。これは全ヒンジ領域とCH2およびCH3ドメインとを含む。このヒトIgG1のFc部分の696塩基対を含む断片を、哺乳類発現ベクターpJB02のNheI部位およびEco47III部位にサブクローニングして、pJB02/Fcを作製した(図5参照)。Val8-GLP-1(7-37)に融合されたIgκ分泌シグナル配列をコードするDNAを、4本のオーバーラップした相補的オリゴヌクレオチド、すなわち
5'-CTAGCCACCATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACCAGTG-3'[配列番号12]
5'-GAGGGCACCTTCACCTCCGACGTGTCCTCCTATCTGGAGGGCCAGGCCGCCAAGGAGTTCATCGCCTGGCTGGTGAAGGGAAGAGGC-3'[配列番号13]
5'-TGAAGGTGCCCTCCACGTGGTCACCAGTGGAACCTGGAACCCAGAGCAGCAGTACCCATAGCAGGAGTGTGTCTGTCTCCATGGTGG-3'[配列番号14]
5'-GCCTCTTCCCTTCACCAGCCAGGCGATGAACTCCTTGGCGGCCTGGCCCTCCAGATAGGAGGACACGTCGGAGG-3'[配列番号15]
の試験管内ハイブリダイゼーションによって作製した。
【0161】
ハイブリダイゼーション反応は、等量の各オリゴヌクレオチド(各オリゴの最終濃度は1pm/μl)を使って行なった。オリゴヌクレオチドの混合物をライゲーション緩衝液(50mMトリス-HCl、pH7.5、10mM MgCl2、10mM DTT、1mM ATP、25μg/mlウシ血清アルブミン)中、100℃で5分間加熱した後、少なくとも2時間かけて30℃まで冷却した。
【0162】
得られたハイブリダイゼーション産物を、NheIおよびEco47IIIで消化しておいたpJB02/Fcベクター骨格に、室温で2時間、または16℃で一晩かけて連結した。そのライゲーション産物を使ってコンピテントXL-1Blue細胞(Stratagene)を形質転換した。クローンをNcoIで消化することにより、組換えプラスミドを、ペプチドコード配列(コザック配列とシグナルペプチドの最初のMetをコードするもの)の存在についてスクリーニングし、配列決定した。得られた発現プラスミドはトランスフェクションアッセイに使用したが、このプラスミドをpJB02-V8-GLP-1-Fcと名付けた(図5)。
【0163】
実施例1b
Val8-GLP-1(7-37)-HSAをコードするDNAの構築:
プラスミドHSA/pcDNA3.1GSをInvitrogenから購入し(カタログ番号H-M12523M-pcDNA3.1/GS)、ヒト血清アルブミン(HSA)をコードするcDNAを単離するためのテンプレートとして使用した。HSA cDNAはPCRを使って調製した。この際、リーダー配列および6アミノ酸プロペプチドをコードするDNAを5'末端から除去した。さらに停止コドンをHSAコード配列の3'末端に直接付加した。最後に、クローニングが容易になるように、5'末端および3'末端に制限酵素部位を作った。Invitorogenから購入した元のベクター中に存在するHSA DNA配列は、天然ヒト配列と比較して、遺伝子の3'領域(667位)に一塩基変異を含んでいた。この変異により、AspではなくAsnのコドンになる。そこで、上述した鎖オーバーラップPCR突然変異導入法を使って、この位置がAspをコードするように、コドンを改変した。得られたHSAコードDNAをpJB02のNheI部位およびHindIII部位にクローニングしてpJB02-HSA(図6)を作製した。
【0164】
Val8-GLP-1(7-37)配列に融合されたIgκリーダー配列を実施例1aで述べたように作製した。このDNAをpJB02-HSAのNheI部位およびFspI部位に連結してpJB02-Val8-GLP-1-HSAを作製した。
【0165】
実施例1c
Val8-GLP-1(7-37)-リンカー-HSAをコードするDNAの構築:
ベクターpJB02-HSAを実施例1bで述べたように作製した。リンカー配列[GGGGS]3をコードするDNAを、HSAコードDNAの5'末端にインフレームで連結してpJB02-リンカー-HSA(図7)を作製した。Igκリーダー配列をコードし、Val8-GLP-1(7-37)配列およびリンカー配列の5'部分に融合されたDNAを、実施例1aで述べたように作製した。このDNAをpJB02のNheIおよびBspEIに連結してpJB02- Val8-GLP-1-リンカー-HSAを作製した。
【0166】
実施例1d
エクセンディン4-FcをコードするDNAの構築:
プラスミドpJB02/Fcを実施例1aで述べたように作製した。エクセンディン4に融合されたIgκシグナル配列をコードするDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CTAGCCACCATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACCGGTCAC-3'[配列番号16]
5'-GGAGAGGGAACCTTCACCAGCGACCTGAGCAAGCAGATGGAGGAGGAGGCCGTGAGACTG-3'[配列番号17]
5'-TTCATCGAGTGGCTGAAGAACGGAGGACCAAGCAGCGGAGCCCCTCCTCCTAGC-3'[配列番号18]
5'-GAACCTGGAACCCAGAGCAGCAGTACCCATAGCAGGAGTGTGTCTGTCTCCATGGTGG-3'[配列番号19]
5'-CTCCTCCTCCATCTGCTTGCTCAGGTCGCTGGTGAAGGTTCCCTCTCCGTGACCGGTG-3'[配列番号20]
5'-GCTAGGAGGAGGGGCTCCGCTGCTTGGTCCTCCGTTCTTCAGCCACTCGATGAACAGTCTCACGGC-3'[配列番号21]。
【0167】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、実施例1aで述べたように、NheIおよびEco47IIIで消化しておいたpJB02ベクターに連結して、pJB02-エクセンディン4-Fcを作製した。
【0168】
実施例1e
エクセンディン4-HSAをコードするDNAの構築:
プラスミドpJB02-HSAを実施例1bで述べたように作製した。エクセンディン4に融合されたIgκシグナル配列をコードするDNAを、オーバーラップした相補的オリゴヌクレオチド(実施例1dに記載したものと同じもの)の試験管内ハイブリダイゼーションによって作製した。ハイブリダイゼーション反応も上述したとおりに行なった。DNAをpJB02-HSA中のユニークなNheI部位およびFspI部位にクローニングしてpJB02-エクセンディン4-HSAを作製した。
【0169】
実施例1f
エクセンディン4-リンカー-HSAをコードするDNAの構築:
プラスミドpJB02-リンカー-HSAを実施例1cで述べたように構築した。エクセンディン4およびリンカー配列の5'部分に融合されたIgκシグナルをコードするDNAを実施例1dで述べたように作製した。このDNAをpJB02-リンカー-HSAのユニークなNheI部位およびBspEI部位にクローニングしてpJB02-エクセンディン4-リンカー-HSAを作製した。
【0170】
実施例1g
Val8-GLP-1/C-Ex-FcをコードするDNAの構築:
プラスミドpJB02-エクセンディン4-Fcを実施例1dで述べたように作製した。エクセンディン4コードDNAを、AgeIおよびEco47IIIで、ベクターから切り出した。Val8-GLP-1/C-ExコードDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CCGGTCACGTGGAGGGCACCTTCACCTCCGACGTGTCCTCCTATCTGGAGGGCCAGGCCGCCA-3'[配列番号22]
5'-AGGAATTCATCGCCTGGCTGGTGAAGGGCCGGGGCAGCAGCGGAGCCCCTCCTCCTAGC-3'[配列番号23]
5'-CTCCAGATAGGAGGACACGTCGGAGGTGAAGGTGCCCTCCACGTGA-3'[配列番号24]
5'-GCTAGGAGGAGGGGCTCCGCTGCTGCCCCGGCCCTTCACCAGCCAGGCGATGAATTCCTTGGCGGCCTGGCC-3'[配列番号25]。
【0171】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、エクセンディン4の代わりに、pJB02-エクセンディン4-Fc発現ベクターに連結して、pJB02-Val8-GLP-1/C-Ex-Fcを作製した。
【0172】
実施例1h
Val8-Glu22-GLP-1-FcをコードするDNAの構築:
プラスミドpJB02-エクセンディン4-Fcを実施例1dで述べたように作製した。エクセンディン4コードDNAを、AgeIおよびEco47IIIで、ベクターから切り出した。Val8-Glu22-GLP-1コードDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CCGGTCACGTGGAGGGCACCTTCACCTCCGACGTGTCCTCCTATCTCGAGGAGCAGGCCGCCA-3'[配列番号26]
5'-AGGAGTTCATCGCCTGGCTGGTGAAGGGCCGGGGC-3'[配列番号27]
5'-GCCCCGGCCCTTCACCAGCCAGGCGATGAACTCCTTGGCGGCCTGCTC-3'[配列番号28]
5'-CTCGAGATAGGAGGACACGTCGGAGGTGAAGGTGCCCTCCACGTGA-3'[配列番号29]。
【0173】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、エクセンディン4の代わりに、pJB02-エクセンディン4-Fc発現ベクターに連結して、pJB02-Val8-Glu22-GLP-1-Fcを作製した。
【0174】
実施例1i
Val8-Glu22GLP-1/C-Ex-FcをコードするDNAの構築:
プラスミドpJB02-エクセンディン4-Fcを実施例1dで述べたように製造した。エクセンディン4コードDNAを、AgeIおよびEco47IIIで、ベクターから切り出した。Val8-Glu22GLP-1/C-ExコードDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CCGGTCACGTGGAGGGCACCTTCACCTCCGACGTGTCCTCCTATCTCGAGGAGCAGGCCGCCA-3'[配列番号30]
5'-AGGAATTCATCGCCTGGCTGGTGAAGGGCCGGGGCAGCAGCGGAGCCCCTCCTCCTAGC-3'[配列番号31]
5'-CTCGAGATAGGAGGACACGTCGGAGGTGAAGGTGCCCTCCACGTGA-3'[配列番号32]
5'-GCTAGGAGGAGGGGCTCCGCTGCTGCCCCGGCCCTTCACCAGCCAGGCGATGAATTCCTTGGCGGCCTGCTC-3'[配列番号33]。
【0175】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、エクセンディン4の代わりに、pJB02-エクセンディン4-Fc発現ベクターに連結して、pJB02-Val8-Glu22-GLP-1/C-Ex-Fcを作製した。
【0176】
実施例1j
Gly8-GLP-1-FcをコードするDNAの構築:
プラスミドpJB02-エクセンディン4-Fcを実施例1dで述べたように製造した。エクセンディン4コードDNAを、AgeIおよびEco47IIIで、ベクターから切り出した。Gly8-GLP-1コードDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CCGGTCACGGCGAGGGCACCTTCACTAGTGACGTGTCCTCCTATCTGGAGGGCCAGGCCGCCA-3'[配列番号34]
5'-AGGAGTTCATCGCCTGGCTGGTGAAGGGCCGGGGC-3'[配列番号35]
5'-CTCCAGATAGGAGGACACGTCACTAGTGAAGGTGCCCTCGCCGTGA-3'[配列番号36]
5'-GCCCCGGCCCTTCACCAGCCAGGCGATGAACTCCTTGGCGGCCTGGCC-3'[配列番号37]。
【0177】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、エクセンディン4の代わりに、pJB02-エクセンディン4-Fc発現ベクターに連結して、pJB02-Gly8-GLP-1-Fcを作製した。
【0178】
実施例2:異種融合タンパク質の発現
HEK293EBNA細胞(付着細胞および浮遊細胞の両方)を一過性にトランスフェクトすることにより、実施例1のDNAコンストラクトがコードする融合タンパク質の発現を行なった。細胞を計数し、トランスフェクションの24時間前に播種した。FuGene(商標)6トランスフェクション試薬(Roche Molecular Biochemicals、カタログ番号1814443)とOptiMEM(Gibco/BRL)とを混合し、室温で5分間インキュベートした時点でDNAを加えることによってトランスフェクションカクテルを調製し、そのカクテルをさらに15分間インキュベートした。トランスフェクション直前に、新しい成長培地をプレートに加えた。トランスフェクションのさらなる詳細を表1および表2に示す。
【0179】
293EBNA細胞の一過性トランスフェクションに使用した試薬
【表1】
【0180】
培地組成
【表2】
【0181】
小規模トランスフェクション(35〜10mm容器)の場合は、トランスフェクションの24時間後に細胞をPBSで洗浄して収穫培地に切り替え、数日間にわたって24時間毎に培地を収集し、置換した。大規模トランスフェクション(700cm2ローラーボトル)の場合は、トランスフェクションの48時間後にローラーボトルをPBSで洗浄し、収穫培地に換えた。少なくとも連続する10日間にわたって24時間毎に培地を収集し、交換した。以降のタンパク質生成には、常に、10回分の収穫物だけを使用した。
【0182】
実施例3:異種融合タンパク質の精製
実施例3a
Val8-GLP-1-Fcの精製:
大規模トランスフェクションによって得た約4.5リットルの調整培地(融合タンパク質発現レベル約20μg/ml)を、CUNOフィルターシステムを使ってろ過し、10Kフィルターメンブレンを装着したProFluxタンジェンシャルフロー濾過システムを使って250mlに濃縮した。Val8-GLP-1-Fcを、1×PBS(pH7.4)中の5ml HiTrapプロテインAカラムに、2ml/分の流速で捕捉し、50mMクエン酸(pH3.3)で溶出させた。4mlの1×PBSと100μlの1Mトリス(pH8)が入っているチューブに、各画分(1ml)を収集した。
【0183】
SDS-PAGEおよびZorbax C8での逆相HPLCによって融合タンパク質を含むことが確認された画分をプールして、1×PBS(pH7.4)中のSuperdex 75 60/60カラムに10ml/分の流速で適用した。陽性画分(20ml/チューブ)を集めて、プールした。次に、プールした画分を3ml/分の流速で0.1%TFA水溶液でのC4逆相クロマトグラフィーにかけた。70分で5%B(0.1%TFA/アセトニトリル)から100%Bに至る勾配を使ってVal8-GLP-1-Fcを溶出させた。溶出画分(3ml/チューブ)を集めた。減圧乾燥によってアセトニトリルを除去し、1mlのH2Oを加えた。精製試料(約32ml)を4リットルの1×PBS(pH7.4)に対して2回透析した。
【0184】
次に、透析した試料を、MILLEX-GV 0.22μmフィルターユニットを使って濾過し、280nmでの吸収を使って濃度を決定した。
【0185】
実施例3b
Val8-GLP-1-HSAまたはVal8-GLP-1-リンカー-HSAの精製:
約6.5リットルの調整培地(融合タンパク質発現レベル約10μg/ml)を、CUNOフィルターシステムを使ってろ過し、10Kフィルターメンブレンを装着したProFluxタンジェンシャルフロー濾過システムを使って380mlに濃縮した。
【0186】
融合タンパク質を、20mMトリス(ph7.4)中の50ml Fast Flow Qカラム(Pharmacia)に、5ml/分の流速で捕捉した。10CVで0%→50% 20mMトリス(pH7.4)、1M NaClの後、2CVで100%Bへの勾配を使って、タンパク質を溶出させた。
【0187】
融合タンパク質を含む画分をプールし、5ml/分の流速で、0.1%TFA水溶液でのC4逆相クロマトグラフィーにかけた。120分で20%B(0.1%TFA/アセトニトリル)から90%Bへの勾配を使って、融合タンパク質を溶出させた。各画分(3.5ml/チューブ)を集めた。減圧乾燥によってアセトニトリルを除去した。
【0188】
プールした試料約9mlを1×PBS(pH7.4)で40mlに希釈し、4リットルの1×PBS(pH7.4)に対して一晩透析した。試料を濾過し、280nmでの吸収によって濃度を決定した。
【0189】
実施例3c
エクセンディン4-Fcの精製:
約4リットルの調整培地(融合タンパク質発現レベル約8μg/ml)を、CUNOフィルターシステムを使ってろ過し、30Kフィルターメンブレンを装着したProFluxタンジェンシャルフロー濾過システムを使って250mlに濃縮した。
【0190】
エクセンディン4-Fcを、1×PBS(pH7.4)中の5ml HiTrapプロテインAカラムに、2ml/分の流速で捕捉し、50mMクエン酸(pH3.3)で溶出させた。融合タンパク質を含む画分をプールし、濾過し、4リットルの1×PBSに対して一晩透析した。次に、透析した試料を、1×PBS(pH7.4)、0.5M NaCl中のSuperdex 75 60/60カラムに、10ml/分の流速で適用した。融合タンパク質を含む画分(20ml/チューブ)を集め、プールし、約1mg/mlに濃縮した。次に、濃縮した試料をMILLEX-GV 0.22μmフィルターユニットを使って濾過した。
【0191】
実施例3d
エクセンディン4-HSAおよびエクセンディン4-リンカー-HSAの精製
約1.1リットルの調整培地(融合タンパク質発現レベル約6μg/ml)を、CUNOフィルターシステムを使って濾過し、30Kフィルターメンブレンを装着したProFluxタンジェンシャルフロー濾過システムを使って175mlに濃縮した。
【0192】
20mMトリス(pH7.4)中の5ml HiTrap Q-セファロースカラムを使って、2ml/分の流速で融合タンパク質を捕捉した。12CVで0%→50% 20mMトリス(pH7.4)、1M NaClの後、4CVで100%Bへの勾配を使って、タンパク質を溶出させた。
【0193】
融合タンパク質を含む画分をプールし、5ml/分の流速で、0.1%TFA水溶液でのC4逆相クロマトグラフィーにかけた。70分で10%B(0.1%TFA/アセトニトリル)から100%Bへの勾配を使って、融合タンパク質を溶出させた。融合タンパク質を含む画分(10ml/チューブ)を集めた。減圧乾燥によってアセトニトリルを除去した。
【0194】
プールした試料約8mlを4リットルの1×PBS(pH7.4)に対して一晩透析した。試料を濾過し、280nmでの吸収によって濃度を決定した。次に、透析した試料を、1×PBS(pH7.4)、0.5M NaCl中のSuperdex 200 26/60カラムに、2ml/分の流速で適用した。融合タンパク質を含む画分(3ml/チューブ)を集め、プールし、濃縮し、濾過した。
【0195】
実施例4:SDS PAGEによる融合タンパク質の特徴づけ
SDS-PAGEを行った後、免疫ブロット法を実施することにより、様々な融合タンパク質発現ベクターでトランスフェクトした細胞から得られる精製融合タンパク質と調整培地の両方を分析した。SDS-PAGEは、Novex 16%トリス-グリシン成形済みゲル(EC6498)、泳動緩衝液(10×、LC2675)、および試料緩衝液(L2676)を使って、Novex Powerease 500システムで行なった。充填に先だって、試料を50mM DTTで還元し、95℃で3〜5分間加熱した。
【0196】
SDS-PAGEゲルでの泳動を行なった後、水および転写緩衝液(20%エタノールを含む1×トリス-グリシンSeprabuff(Owl Scientific、カタログ番号ER26-S))を使って、ゲルからSDSを洗い流した。Novex転写装置を、PVDFメンブレン(BioRad、カタログ番号162-0174)およびニトロセルロースメンブレン(BioRad、カタログ番号1703965または1703932)と共に使用した。転写は室温にて30〜35Vで90分間行なった。0.1%Tween-20(Sigma、カタログ番号P-7949)および5%ミルク(BioRad、カタログ番号170-6404)を含む1×PBS中、4℃で1〜12時間、メンブレンをブロッキングした。抗体を1×PBS+5%ミルクに希釈し、ブロットをこれらの溶液中、4℃で1〜2時間インキュベートする。インキュベーションの合間に、ブロットを、室温の1×PBSおよび0.2%Tween-20で、5分間ずつ4回洗浄する。PBSは、GIBCO 10×PBS(カタログ番号70011)から、最終組成が1mM第一リン酸カリウム、3mM第二リン酸ナトリウム、153mM塩化ナトリウム、pH7.4になるように調製するか、またはSigmaのPBSポーチ(カタログ番号1000-3)から、120mM NaCl、2.7mM KClおよび10mMリン酸塩、25℃でpH7.4になるように調製した。
【0197】
一次抗体は、ポリクローナルヤギ抗IgG1またはウサギ抗HSAとした。二次抗体は抗ヤギIgG HRPまたは抗ウサギIgG HRPとした。二次抗体は1:5000に希釈した。ブロットの発色にはECLシステム(Amersham Pharmacia Biotech、カタログ番号RN2108およびカタログ番号RPN1674H)を使用した。
【0198】
図3Aは、精製Fcタンパク質を、pJB02-Val8-GLP-1-Fcトランスフェクト細胞およびpJB02-エクセンディン4-Fcトランスフェクト細胞から得られる調整培地と比較した図である。移動度の減少は、融合タンパク質のGLP-1部分によるサイズの増加と合致している。図3Bも同様に、精製HSAを、pJB02-Val8-GLP-1-HSA、pJB02-Val8-GLP-1-リンカー-HSA、pJB02-エクセンディン4-HSA、またはpJB02-エクセンディン4-リンカー-HSAでトランスフェクトした細胞から得られる調整培地と比較した図である。図4に精製融合タンパク質調製物を示す。
【0199】
実施例5:質量分析法を使った融合タンパク質の特徴づけ
実験は全て、タイムラグフォーカシング電子機器(Time Lag Focuing electronics)、リフレクトロン(Reflectron)(0〜8000Daペプチド範囲の分析に使用)、リニア検出器(Linear detector)(高分子量/高シグナル分析時に使用)、および後段加速検出器(Post Acceleratio Detector)(すなわちP.A.D、高分子量/極低シグナル分析に使用)を装着したMicromass TofSpec-2E質量分析計で行なった。装置の実効飛行行路長はリニアモードで1.2メートル、リフレクトロンモードで2.3メートルである。リニアおよびリフレクトロンモード検出用に2つのデュアルマイクロチャンネルプレート検出器を備える。使用するレーザーは、毎秒5回のレーザー照射で337nmで作動するLaser Science Inc. VSL-337i窒素レーザーである。データは全て、2GHz8ビット内蔵ディジタイザを使って取得し、1スペクトルにつき50回までのレーザー照射を平均した。
【0200】
問題にしているGLP-1融合タンパク質を分析する場合は、装置をリニアモードで操作した。リニア検出器は、MALDI-ToF-MS装置の飛行管を縦断するイオンを検出する装置である。この検出器は、イオン存在度を経時的に測定し、変換用ディジタイザにシグナルを送信する。ディジタイザは、質量分析器からのシグナルをコンピュータに転送できるようにするアナログ-ディジタル変換器であり、コンピュータに転送されたシグナルは、そこで便利なm/zスペクトルに再構築される。
【0201】
再結晶飽和シナピン酸溶液(50/50 Acn/H2Oおよび0.1%TFAに希釈)をイオン化マトリックスとして利用した。シナピン酸は、10kDaを超えるタンパク質に適したマトリックスである。分析した試料の質量が正確に決定されるように、内部および外部較正ファイル用に、適当な質量基準タンパク質を使用した。試料は全て1:2の試料:マトリックス希釈液を使って分析した。装置は以下のリニア検出器条件で初期設定した:
ソース電圧(Source Voltage):20.0keV
抽出電圧(Extraction Voltage):20.0keV
収束電圧(Focus Voltage):16.0keV
リニア検出器:3.7keV
P.A.D.:(オフライン)
パルス電圧(Pulse Voltage):3.0keV
レーザーコース(Laser Coarse):50
レーザーファイン(Laser Fine):50。
【0202】
最適な信号/雑音比と最も高い解像度が得られるように、これらの設定を(必要に応じて)変更した。表3に様々なGLP-1融合タンパク質の特徴を示す。
【0203】
【表3】
CExはC末端伸長を表し、Ser-Ser-Gly-Ala-Pro-Pro-Pro-Serという配列を含む。
リンカーはGly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Serである。
【0204】
実施例6:異種融合タンパク質の活性:
本発明融合タンパク質のGLP-1受容体活性化能力を、GelfandらのEP619,322および米国特許第5,120,712にそれぞれ記載されているような、試験管内アッセイを使って評価した。Val8-GLP-1(7-37)OHの活性と比較した、これらの化合物の活性を、表4に報告する。図8に、Val8-GLP-1およびエクセンディン4融合タンパク質に関する試験管内用量反応曲線を図示する。さらに、Fcまたはアルブミンタンパク質と融合させて生物学的に活性な融合タンパク質を製造することができる多くのGLP-1類似体の試験管内活性を、表5aおよび表5bに示す。これらの活性はGLP-1(7-37)OHと比較されている。
【0205】
GLP-1融合タンパク質の試験管内活性
【表4】
CExはC末端伸長を表し、Ser-Ser-Gly-Ala-Pro-Pro-Pro-Serという配列を含む。
リンカーはGly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Serである。
C2はSer-Ser-Gly-Ala-Ser-Ser-Gly-Alaである。
【0206】
表3および表4に記載した融合タンパク質のアミノ酸配列を配列番号13〜31に示す。
【0207】
Val8-GLP-1-ヒト血清アルブミンアミノ酸配列は配列番号13によって表される。
1 HVEGTFTSDV SSYLEGQAAK EFIAWLVKGR GDAHKSEVAH RFKDLGEENF KALVLIAFAQ
61 YLQQCPFEDH VKLVNEVTEF AKTCVADESA ENCDKSLHTL FGDKLCTVAT LRETYGEMAD
121 CCAKQEPERN ECFLQHKDDN PNLPRLVRPE VDVMCTAFHD NEETFLKKYL YEIARRHPYF
181 YAPELLFFAK RYKAAFTECC QAADKAACLL PKLDELRDEG KASSAKQRLK CASLQKFGER
241 AFKAWAVARL SQRFPKAEFA EVSKLVTDLT KVHTECCHGD LLECADDRAD LAKYICENQD
301 SISSKLKECC EKPLLEKSHC IAEVENDEMP ADLPSLAADF VESKDVCKNY AEAKDVFLGM
361 FLYEYARRHP DYSVVLLLRL AKTYETTLEK CCAAADPHEC YAKVFDEFKP LVEEPQNLIK
421 QNCELFEQLG EYKFQNALLV RYTKKVPQVS TPTLVEVSRN LGKVGSKCCK HPEAKRMPCA
481 EDYLSVVLNQ LCVLHEKTPV SDRVTKCCTE SLVNRRPCFS ALEVDETYVP KEFNAETFTF
541 HADICTLSEK ERQIKKQTAL VELVKHKPKA TKEQLKAVMD DFAAFVEKCC KADDKETCFA
601 EEGKKLVAAS QAALGL[配列番号13]
【0208】
Val8-GLP-1-リンカー-ヒト血清アルブミンアミノ酸配列は配列番号14によって表される。
1 HVEGTFTSDV SSYLEGQAAK EFIAWLVKGR GGGGGSGGGG SGGGGSDAHK SEVAHRFKDL
61 GEENFKALVL IAFAQYLQQC PFEDHVKLVN EVTEFAKTCV ADESAENCDK SLHTLFGDKL
121 CTVATLRETY GEMADCCAKQ EPERNECFLQ HKDDNPNLPR LVRPEVDVMC TAFHDNEETF
181 LKKYLYEIAR RHPYFYAPEL LFFAKRYKAA FTECCQAADK AACLLPKLDE LRDEGKASSA
241 KQRLKCASLQ KFGERAFKAW AVARLSQRFP KAEFAEVSKL VTDLTKVHTE CCHGDLLECA
301 DDRADLAKYI CENQDSISSK LKECCEKPLL EKSHCIAEVE NDEMPADLPS LAADFVESKD
361 VCKNYAEAKD VFLGMFLYEY ARRHPDYSVV LLLRLAKTYE TTLEKCCAAA DPHECYAKVF
421 DEFKPLVEEP QNLIKQNCEL FEQLGEYKFQ NALLVRYTKK VPQVSTPTLV EVSRNLGKVG
481 SKCCKHPEAK RMPCAEDYLS VVLNQLCVLH EKTPVSDRVT KCCTESLVNR RPCFSALEVD
541 ETYVPKEFNA ETFTFHADIC TLSEKERQIK KQTALVELVK HKPKATKEQL KAVMDDFAAF
601 VEKCCKADDK ETCFAEEGKK LVAASQAALG L[配列番号14]
【0209】
Gly8-Glu22-GLP-1-CEx-リンカー-ヒト血清アルブミンアミノ酸配列は配列番号15によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSG GGGGSGGGGS GGGGSDAHKS
61 EVAHRFKDLG EENFKALVLI AFAQYLQQCP FEDHVKLVNE VTEFAKTCVA DESAENCDKS
121 LHTLFGDKLC TVATLRETYG EMADCCAKQE PERNECFLQH KDDNPNLPRL VRPEVDVMCT
181 AFHDNEETFL KKYLYEIARR HPYFYAPELL FFAKRYKAAF TECCQAADKA ACLLPKLDEL
241 RDEGKASSAK QRLKCASLQK FGERAFKAWA VARLSQRFPK AEFAEVSKLV TDLTKVHTEC
301 CHGDLLECAD DRADLAKYIC ENQDSISSKL KECCEKPLLE KSHCIAEVEN DEMPADLPSL
361 AADFVESKDV CKNYAEAKDV FLGMFLYEYA RRHPDYSVVL LLRLAKTYET TLEKCCAAAD
421 PHECYAKVFD EFKPLVEEPQ NLIKQNCELF EQLGEYKFQN ALLVRYTKKV PQVSTPTLVE
481 VSRNLGKVGS KCCKHPEAKR MPCAEDYLSV VLNQLCVLHE KTPVSDRVTK CCTESLVNRR
541 PCFSALEVDE TYVPKEFNAE TFTFHADICT LSEKERQIKK QTALVELVKH KPKATKEQLK
601 AVMDDFAAFV EKCCKADDKE TCFAEEGKKL VAASQAALGL[配列番号15]
【0210】
エクセンディン4-ヒト血清アルブミンアミノ酸配列は配列番号16によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGAPPPSD AHKSEVAHRF KDLGEENFKA
61 LVLIAFAQYL QQCPFEDHVK LVNEVTEFAK TCVADESAEN CDKSLHTLFG DKLCTVATLR
121 ETYGEMADCC AKQEPERNEC FLQHKDDNPN LPRLVRPEVD VMCTAFHDNE ETFLKKYLYE
181 IARRHPYFYA PELLFFAKRY KAAFTECCQA ADKAACLLPK LDELRDEGKA SSAKQRLKCA
241 SLQKFGERAF KAWAVARLSQ RFPKAEFAEV SKLVTDLTKV HTECCHGDLL ECADDRADLA
301 KYICENQDSI SSKLKECCEK PLLEKSHCIA EVENDEMPAD LPSLAADFVE SKDVCKNYAE
361 AKDVFLGMFL YEYARRHPDY SVVLLLRLAK TYETTLEKCC AAADPHECYA KVFDEFKPLV
421 EEPQNLIKQN CELFEQLGEY KFQNALLVRY TKKVPQVSTP TLVEVSRNLG KVGSKCCKHP
481 EAKRMPCAED YLSVVLNQLC VLHEKTPVSD RVTKCCTESL VNRRPCFSAL EVDETYVPKE
541 FNAETFTFHA DICTLSEKER QIKKQTALVE LVKHKPKATK EQLKAVMDDF AAFVEKCCKA
601 DDKETCFAEE GKKLVAASQA ALGL[配列番号16]
【0211】
エクセンディン4-リンカー-ヒト血清アルブミンアミノ酸配列は配列番号17によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGAPPPSG GGGGSGGGGS GGGGSDAHKS
61 EVAHRFKDLG EENFKALVLI AFAQYLQQCP FEDHVKLVNE VTEFAKTCVA DESAENCDKS
121 LHTLFGDKLC TVATLRETYG EMADCCAKQE PERNECFLQH KDDNPNLPRL VRPEVDVMCT
181 AFHDNEETFL KKYLYEIARR HPYFYAPELL FFAKRYKAAF TECCQAADKA ACLLPKLDEL
241 RDEGKASSAK QRLKCASLQK FGERAFKAWA VARLSQRFPK AEFAEVSKLV TDLTKVHTEC
301 CHGDLLECAD DRADLAKYIC ENQDSISSKL KECCEKPLLE KSHCIAEVEN DEMPADLPSL
361 AADFVESKDV CKNYAEAKDV FLGMFLYEYA RRHPDYSVVL LLRLAKTYET TLEKCCAAAD
421 PHECYAKVFD EFKPLVEEPQ NLIKQNCELF EQLGEYKFQN ALLVRYTKKV PQVSTPTLVE
481 VSRNLGKVGS KCCKHPEAKR MPCAEDYLSV VLNQLCVLHE KTPVSDRVTK CCTESLVNRR
541 PCFSALEVDE TYVPKEFNAE TFTFHADICT LSEKERQIKK QTALVELVKH KPKATKEQLK
601 AVMDDFAAFV EKCCKADDKE TCFAEEGKKL VAASQAALGL[配列番号17]
【0212】
Val8-GLP-1-IgG1アミノ酸配列は配列番号18によって表される。
1 HVEGTFTSDV SSYLEGQAAK EFIAWLVKGR GAEPKSCDKT HTCPPCPAPE LLGGPSVFLF
61 PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV
121 SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SREEMTKNQV
181 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF
241 SCSVMHEALH NHYTQKSLSL SPGK[配列番号18]
【0213】
Val8-GLP-1-Cex-IgG1アミノ酸配列は配列番号19によって表される。
1 HVEGTFTSDV SSYLEGQAAK EFIAWLVKGR GSSGAPPPSA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号19]
【0214】
Val8-Glu22GLP-1-IgG1アミノ酸配列は配列番号20によって表される。
1 HVEGTFTSDV SSYLEEQAAK EFIAWLVKGR GAEPKSCDKT HTCPPCPAPE LLGGPSVFLF
61 PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV
121 SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SREEMTKNQV
181 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF
241 SCSVMHEALH NHYTQKSLSL SPGK[配列番号20]
【0215】
Val8-Glu22GLP-1-CEx-IgG1アミノ酸配列は配列番号21によって表される。
1 HVEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号21]
【0216】
Gly8-Glu22GLP-1-C2-IgG1アミノ酸配列は配列番号22によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGASSGAA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号22]
【0217】
Gly8-Glu22GLP-1-CEx-リンカー-IgG1アミノ酸配列は配列番号23によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSG GGGSGGGGSG GGGSAEPKSC
61 DKTHTCPPCP APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
121 GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK
181 GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS
241 DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK[配列番号23]
【0218】
Gly8-Glu22GLP-1-CEx-リンカー-IgG4アミノ酸配列は配列番号24によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSG GGGSGGGGSG GGGSAESKYG
61 PPCPSCPAPE FLGGPSVFLF PPKPKDTLMI SRTPEVTCVV VDVSQEDPEV QFNWYVDGVE
121 VHNAKTKPRE EQFNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKGLPSSIE KTISKAKGQP
181 REPQVYTLPP SQEEMTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS
241 FFLYSRLTVD KSRWQEGNVF SCSVMHEALH NHYTQKSLSL SLGK[配列番号24]
【0219】
Gly8-Glu22GLP-1-CEx-2リンカー-IgG1アミノ酸配列は配列番号25によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSG GGGSGGGGSG GGGSGGGGSG
61 GGGSGGGGSA EPKSCDKTHT CPPCPAPELL GGPSVFLFPP KPKDTLMISR TPEVTCVVVD
121 VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN
181 KALPAPIEKT ISKAKGQPRE PQVYTLPPSR EEMTKNQVSL TCLVKGFYPS DIAVEWESNG
241 QPENNYKTTP PVLDSDGSFF LYSKLTVDKS RWQQGNVFSC SVMHEALHNH YTQKSLSLSP
301 GK[配列番号25]
【0220】
Gly8-Glu22GLP-1-2リンカー-IgG1アミノ酸配列は配列番号26によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GGGGGSGGGG SGGGGSGGGG SGGGGSGGGG
61 SAEPKSCDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV
121 KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE
181 KTISKAKGQP REPQVYTLPP SREEMTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT
241 TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK
[配列番号26]
【0221】
Gly8-Glu22GLP-1-2CEx-IgG1アミノ酸配列は配列番号27によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSS SGAPPPSAEP KSCDKTHTCP
61 PCPAPELLGG PSVFLFPPKP KDTLMISRTP EVTCVVVDVS HEDPEVKFNW YVDGVEVHNA
121 KTKPREEQYN STYRVVSVLT VLHQDWLNGK EYKCKVSNKA LPAPIEKTIS KAKGQPREPQ
181 VYTLPPSREE MTKNQVSLTC LVKGFYPSDI AVEWESNGQP ENNYKTTPPV LDSDGSFFLY
241 SKLTVDKSRW QQGNVFSCSV MHEALHNHYT QKSLSLSPGK[配列番号27]
【0222】
Gly8-Glu22-Val25-Ile33GLP-1-CEx-リンカー-IgG1アミノ酸配列は配列番号28によって表される。
1 HGEGTFTSDV SSYLEEQAVK EFIAWLIKGR GSSGAPPPSG GGGSGGGGSG GGGSAEPKSC
61 DKTHTCPPCP APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
121 GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK
181 GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS
241 DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK[配列番号28]
【0223】
エクセンディン4-IgG1アミノ酸配列は配列番号29によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGAPPPSA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号29]
【0224】
エクセンディン4-C2-IgG1アミノ酸配列は配列番号30によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGASSGAA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号30]
【0225】
エクセンディン4-リンカー-IgG1アミノ酸配列は配列番号31によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGAPPPSG GGGSGGGGSG GGGSAEPKSC
61 DKTHTCPPCP APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
121 GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK
181 GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS
241 DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK[配列番号31]
【0226】
試験管内GLP-1類似体活性
【表5a−1】
【表5a−2】
【0227】
試験管内GLP-1類似体活性
【表5b−1】
【表5b−2】
【0228】
GLP/エクセンディン類似体の試験管内活性
【表6】
【0229】
実施例7:Val8-GLP-1-IgG1およびVal8-GLP-1-HSAの生体内薬物動態:
Val8-GLP-1-IgG1およびVal8-GLP-1-HSAの薬物動態研究をカニクイザルで行なった。サルに5.6ナノモル/kgの精製Val8-GLP-1-IgG1またはVal8-GLP-1-HSAを投与した。化合物は静脈内ボーラス投与として投与した。投与前および投与の0.083、0.25、0.5、1、4、8、12、24、48、72、96、120、144、168、および216時間後に、EDTAを含むチューブに血液を採取した。免疫反応性Val8-GLP-1の血漿濃度を、Val8-GLP-1(7-37)のN末端(7-16)領域に対して主たる特異性を持つポリクローナル抗血清を利用するラジオイムノアッセイを使って決定した。図9は、2頭のカニクイザルに単回静脈内投与した後のVal8-GLP-1-FcおよびVal8-GLP-1-リンカー-HSAの血漿濃度を表す。Fc融合タンパク質は約45時間の半減期を持ち、アルブミン融合タンパク質は約87時間の半減期を持っていた。
【0230】
実施例8:エクセンディン4-IgG1の生体内薬物動態:
カニューレを留置した正常雄ビーグル犬2頭を一晩絶食後に調べた。動脈および静脈血管アクセス口にアクセスして、橈側皮静脈にカテーテルを経皮的に挿入し、確保した。動物をケージに入れ、カテーテルを回り継手/テザー系にとりつけた。融合タンパク質エクセンディン4-IgG1(11.8μM)を含む溶液を、橈側皮静脈カテーテルを通して静脈内注射した(1.0nmol/kg)。次に、10mlの食塩水でカテーテルを洗浄した。2時間後に高血糖(150mg/dl)クランプを開始し、これを3時間続けた。この5時間の全体にわたって動脈血試料を採取し、融合タンパク質、グルコース、およびインスリンの血漿濃度の決定に使用した。
【0231】
この研究の結果を、両動物に食塩水を皮下ボーラス投与し、3時間後に3時間高血糖(150mg/dl)クランプ法を使って調べた過去の同様の研究で得られた結果と比較した。
【0232】
どちらの研究でも、血漿グルコース濃度はBeckmanグルコース分析機を使って決定した。血漿インスリン濃度は、Linco Research, Inc.の従業員がその研究室で開発されたRIAキットを使って決定した。データを図10および図11に示す。
【0233】
実施例9:Gly8-Glu22-GLP-1-CEx-リンカー-IgG1の生体内薬物動態:
3頭の正常雄ビーグル犬からなる2群に、0.1mg/kgのGly8-Glu22-GLP-1-CEx-リンカー-IgG1を皮下(SC)または静脈内(IV)投与した。IV群とSC群の両方について、投与の30分前から投与の216時間後まで集めた試料で、Gly8-Glu22-GLP-1-CEx-リンカー-IgG1免疫反応性の血漿濃度を、ラジオイムノアッセイによって決定した。次に、これらの濃度を使って、本明細書に記載の薬物動態パラメータを決定した。IV投与されたGly8-Glu22-GLP-1-CEx-リンカー-IgG1の平均消失半減期は約55時間であり、全身クリアランスは1.5mL/時/kgだった。SC投与されたGly8-Glu22-GLP-1-CEx-リンカー-IgG1の平均消失半減期は約38時間だった。
【技術分野】
【0001】
本発明は、グルカゴン様ペプチドの生体内半減期を延ばす効果を持つタンパク質に融合されたグルカゴン様ペプチド(その類似体および誘導体を含む)に関する。これらの融合タンパク質は、インスリン非依存性糖尿病および他の様々な状態の処置に使用することができる。
【背景技術】
【0002】
グルカゴン様ペプチド1(GLP-1)は食物摂取に反応して腸のL細胞が分泌する37アミノ酸ペプチドである。これは、インスリン分泌を刺激すること(インスリン分泌促進作用)により、細胞によるグルコースの取込みおよび血清グルコースレベルの低下を引き起こすことがわかっている[例えば非特許文献1を参照されたい]。しかしGLP-1は活性に乏しい。次に、第6位と第7位の間で内因的に切断されることにより、生物学的に活性でより強力なGLP-1(7-37)OHペプチドが生成する。数多くのGLP-1類似体および誘導体が知られているが、本明細書ではそれらを「GLP-1化合物」と呼ぶ。これらのGLP-1類似体には、アメリカドクトカゲの毒液中に見いだされるペプチドであるエクセンディン(Exendin)類が含まれる。エクセンディン類は、天然GLP-1に対して配列相同性を持ち、GLP-1受容体を結合して、GLP-1(7-37)OHに起因するとされている数多くの活性を担うシグナル伝達カスケードを開始させることができる。
【0003】
GLP-1化合物は生理学的に重要な種々の活性を持っている。例えば、GLP-1はインスリン放出を刺激し、グルカゴン分泌を低下させ、胃内容排出を阻害し、グルコース利用を増進することが明らかにされている[非特許文献2、非特許文献3、非特許文献4]。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Mojsov, S., (1992) Int. J. Peptide Protein Research, 40:333-343
【非特許文献2】Nauck, M.A.ら (1993) Diabetologia 36:741-744
【非特許文献3】Gutniak, M.ら (1992) New England J. of Med. 326:1316-1322
【非特許文献4】Nauck, M.A.ら (1993) J. Clin. Invest. 91:301-307
【発明の概要】
【発明が解決しようとする課題】
【0005】
GLP-1はインスリン非依存性糖尿病(NIDDM)の処置として最も有望視されている。NIDDMに関係するインスリン抵抗性を処置するために、数多くの経口薬が市場に出ている。しかし疾患の進行に伴って、患者は、インスリンの放出を刺激する処置に移行しなければならず、そしてついにはインスリンの注射を含む処置に移行しなければならなくなる。しかし、現在用いられているインスリン放出刺激薬は、実際にインスリンを投与した場合と同様に、低血糖を引き起こしうる。しかしGLP-1活性は血中グルコースレベルによって制御される。レベルが一定の閾レベルまで低下すると、GLP-1は活性でなくなる。したがって、GLP-1を使用する処置は、低血糖の危険を伴わない。
【0006】
しかし、GLP-1ペプチドはクリアランスが速く半減期が短いので、GLP-1ペプチドを使用する治療法の有用性には限界があった。例えばGLP-1(7-37)の血清半減期は3〜5分しかない。GLP-1(7-36)アミドは、皮下投与した場合、約50分の作用時間を持つ。内在性プロテアーゼ切断に対して耐性な類似体および誘導体でも、反復投与を24時間以上避けることができるほど長い半減期は持たない。治療薬の速いクリアランスは、反復投与が必要になるので、薬剤の血中レベルを長時間にわたって高く維持することが望まれる場合には不都合である。また、過去の処置方法で経口薬の服用しか経験していない糖尿病患者にとって、持効性化合物は特に重要である。これらの患者は、薬剤を複数回注射する必要がある方法に移行するのに、しばしば極めて苦しい思いをする。
【課題を解決するための手段】
【0007】
本発明は、血漿半減期が短い化合物を送達することに付随する課題を克服するものである。本発明の化合物は、循環半減期が長い別のタンパク質(免疫グロブリンのFc部分またはアルブミンなど)に融合されたGLP-1化合物を包含する。
【0008】
小さい治療ペプチドは、わずかな構造変化でも安定性および/または生物学的活性に影響が及びうるので、一般に、操作することが難しい。これは、現在開発中のGLP-1化合物には、特に当てはまることであった。例えばGLP-1(7-37)OHには、αヘリックスを主とする構造からβシートを主とする構造へのコンフォメーション変化を起こす傾向がある。このβシート型は凝集物をもたらし、その凝集物は不活性であると考えられている。したがって、増加した半減期を持つ、生物学的に活性なGLP-1融合タンパク質を開発できることは、驚くべきことだった。これは、GLP-1(7-37)OHの単独での取り扱いが困難であることと、結合している小さなGLP-1ペプチドと比較して融合パートナーのサイズが大きいことを考えると、とりわけ予想外なことだった。
【0009】
本発明の化合物には、N末端およびC末端を持つ第1ポリペプチドがN末端およびC末端を持つ第2のポリペプチドに融合してなる異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)ヒトアルブミン、
b)ヒトアルブミン類似体、および
c)ヒトアルブミンの断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端に融合している異種融合タンパク質が包含される。
【0010】
本発明の化合物には、N末端およびC末端を持つ第1ポリペプチドがN末端およびC末端を持つ第2ポリペプチドに融合してなる異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)ヒトアルブミン、
b)ヒトアルブミン類似体、および
c)ヒトアルブミンの断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端にペプチドリンカーを介して融合している異種融合タンパク質も包含される。ペプチドリンカーは、
a)グリシンリッチペプチド、
b)[Gly-Gly-Gly-Gly-Ser]nという配列(式中、nは1、2、3、4、5または6である)を持つペプチド、および
c)[Gly-Gly-Gly-Gly-Ser]3という配列を持つペプチド、
からなる群より選択することが好ましい。
【0011】
他にも、本発明の化合物には、N末端およびC末端を持つ第1ポリペプチドがN末端およびC末端を持つ第2ポリペプチドに融合してなる異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)免疫グロブリンのFc部分、
b)免疫グロブリンのFc部分の類似体、および
c)免疫グロブリンのFc部分の断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端に融合している異種融合タンパク質も含まれる。GLP-1化合物はペプチドリンカーを介して第2ポリペプチドに融合していてもよい。ペプチドリンカーは、
a)グリシンリッチペプチド、
b)[Gly-Gly-Gly-Gly-Ser]nという配列(式中、nは1、2、3、4、5または6である)を持つペプチド、および
c)[Gly-Gly-Gly-Gly-Ser]3という配列を持つペプチド、
からなる群より選択することが好ましい。
【0012】
異種融合タンパク質の一部であるGLP-1化合物は、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン(Exendin)4中の対応するアミノ酸と相違するアミノ酸を、6個を超えて含まないことが、一般に好ましい。GLP-1化合物は、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、5個を超えて含まないことが、さらに好ましい。GLP-1化合物は、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、4個、3個、または2個を超えて含まないことが、最も好ましい。異種融合タンパク質の一部であるGLP-1化合物は、好ましくは、8位にグリシンまたはバリンを持つ。
【0013】
本発明は、本明細書に記載の異種融合タンパク質をコードするポリヌクレオチド、これらのポリヌクレオチドを含むベクター、および本明細書に記載のベクターでトランスフェクトまたは形質転換された宿主細胞も包含する。また、異種融合タンパク質を製造する方法であって、検出可能な量の異種融合タンパク質が発現される条件で、本明細書に記載のポリヌクレオチドを転写し翻訳する工程を含む方法も包含される。
【0014】
本発明は、血中グルコースレベルの正常化を必要とする哺乳動物の血中グルコースレベルを正常化する方法であって、治療有効量の本明細書に記載の異種融合タンパク質を投与することを含む方法も包含する。
【0015】
添付の図面を参照して本発明をさらに詳しく説明する。
【図面の簡単な説明】
【0016】
【図1】ヒンジ領域、CH2ドメインおよびCH3ドメインを含むIgG1 Fcアミノ酸配列示す図である。
【図2−1】ヒト血清アルブミンアミノ酸配列を示す図である。
【図2−2】ヒト血清アルブミンアミノ酸配列を示す図である。
【図3】(A)IgG1-FcおよびGLP-1-Fc融合タンパク質の分子量を示すSDS-PAGEゲルおよび同じゲルの免疫ブロットを示す図である(レーン1、MW標品;レーン2、精製Fc;レーン3、疑似トランスフェクトした培地;レーン4、 Val8-GLP-1-Fc;レーン5、エクセンディン4-Fc)。(B)ヒトHSAおよびGLP-1-HSA融合タンパク質の分子量を示すSDS-PAGEゲルおよび同じゲルの免疫ブロットを示す図である(レーン1、MW標品;レーン2、精製HSA;レーン3、疑似トランスフェクトした培地;レーン4、Val8-GLP-1-HSA;レーン5、Val8-GLP-1-[Gly-Gly-Gly-Gly-Ser]3-HSA;レーン6、エクセンディン4-HSA;レーン7、エクセンディン4-[Gly-Gly-Gly-Gly-Ser]3-HSA)。
【図4】精製Fc、アルブミンおよびGLP-1融合タンパク質のSDS-PAGEゲルを示す図である(レーン1、MW標品;レーン2、精製Fc;レーン3、Val8-GLP-1-Fc;レーン4、エクセンディン4-Fc、レーン5、MW標品;レーン6、Val8-GLP-1-HSA;レーン7、エクセンディン4-HSA;レーン8、エクセンディン4-[Gly-Gly-Gly-Gly-Ser]3-HSA)。
【図5】図1に示すFc領域を含む発現クローニングベクターを示す図である。
【図6】図2に示すアルブミン配列を含む発現クローニングベクターを示す図である。
【図7】図2に示すアルブミン配列の5'にインフレームで融合した15アミノ酸リンカーをコードするDNAを含む発現クローニングベクターを示す図である。
【図8】GLP-1融合タンパク質の試験管内用量反応活性を示す図である。
【図9】GLP-1 FcおよびHSA融合タンパク質の薬物動態を示す図である。
【図10−1】2頭の正常絶食イヌにおけるエクセンディン-Fcに対するグルコース動態反応を示す図である。
【図10−2】2頭の正常絶食イヌにおけるエクセンディン-Fcに対するグルコース動態反応を示す図である。
【図11−1】2頭の正常絶食イヌにおけるエクセンディン-Fcに対するインスリン分泌促進反応を示す図である。
【図11−2】2頭の正常絶食イヌにおけるエクセンディン-Fcに対するインスリン分泌促進反応を示す図である。
【図12】ヒトIgG1 Fc領域をコードするDNA配列を示す図である。
【図13−1】ヒトアルブミンタンパク質をコードするDNA配列を示す図である。
【図13−2】ヒトアルブミンタンパク質をコードするDNA配列を示す図である。
【発明を実施するための形態】
【0017】
本発明の異種融合タンパク質は、ヒトアルブミン、ヒトアルブミン類似体、ヒトアルブミン断片、免疫グロブリンのFc部分、免疫グロブリンのFc部分の類似体、または免疫グロブリンのFc部分の断片に融合されたGLP-1化合物を含む。GLP-1化合物のC末端は、アルブミンまたはFcタンパク質のN末端に直接融合するか、またはペプチドリンカーを介して融合することができる。これらの異種融合タンパク質は生物学的に活性であり、天然GLP-1と比較して増加した半減期を持つ。
【0018】
異種融合タンパク質の一部を構成するGLP-1化合物は、膵臓のβ細胞上のGLP-1受容体に結合することによってインスリン分泌促進活性を示すことができるように天然のGLP-1(7-37)OHに対して十分な相同性を持つ、約25〜約39個の天然または非天然アミノ酸を有するポリペプチドを含むことが好ましい。GLP-1化合物は、典型的には、GLP-1(7-37)OH、GLP-1(7-37)OHの類似体、GLP-1(7-37)OHの断片、またはGLP-1(7-37)OH類似体の断片のアミノ酸配列を持つポリペプチドを含む。GLP-1(7-37)OHは配列番号1のアミノ酸配列:
【化1】
(配列番号1)
を持つ。
【0019】
当技術分野の慣例により、GLP-1(7-37)OHのアミノ末端には残基番号7が割り当てられ、カルボキシ末端には番号37が割り当てられている。配列番号1に示すように、ポリペプチド中の他のアミノ酸には連続した番号が与えられる。例えば12位はフェニルアラニンであり、22位はグリシンである。
【0020】
GLP-1化合物には「GLP-1断片」も包含される。GLP-1断片は、GLP-1(7-37)OHまたはその類似体もしくは誘導体のN末端および/またはC末端から1つまたは複数のアミノ酸を切り落とした後に得られるポリペプチドである。GLP-1(7-37)OHの記述に使用される命名法はGLP-1断片にも適用することができる。例えば、GLP-1(9-36)OHは、N末端から2つのアミノ酸を切り落とし、C末端から1つのアミノ酸を切り落とすことによって得られるGLP-1断片を表す。断片中のアミノ酸は、GLP-1(7-37)OH中の対応するアミノ酸と同じ番号で表示される。例えば、GLP-1(7-37)OHの場合と同様に、GLP-1(9-36)OH中のN末端グルタミン酸は9位にあり、12位はフェニルアラニンで占められ、22位はグリシンで占められている。GLP-1(7-36)OHの場合は、GLP-1(7-37)OHの37位のグリシンが欠失している。
【0021】
GLP-1化合物には、GLP-1(7-37)OHまたはその断片もしくは類似体のN末端および/またはC末端に1つまたは複数のアミノ酸が付加されているポリペプチドも包含される。このタイプのGLP-1化合物は約39個までのアミノ酸を持つことが好ましい。「伸長型」GLP-1化合物中のアミノ酸は、GLP-1(7-37)OH中の対応するアミノ酸と同じ番号で表示される。例えば、GLP-1(7-37)OHのN末端に2つのアミノ酸を付加することによって得られるGLP-1化合物のN末端アミノ酸は5位にあり、GLP-1(7-37)OHのC末端に1つのアミノ酸を付加することによって得られるGLP-1化合物のC末端アミノ酸は38位にある。したがって、これら「伸長型」GLP-1化合物のどちらにおいても、GLP-1(7-37)OHの場合と同様に、12位はフェニルアラニンで占められ、22位はグリシンで占められる。伸長型GLP-1化合物のアミノ酸1〜6は、好ましくは、GLP-1(1-37)OHの対応する位置にあるアミノ酸と同じであるか、またはその保存的置換である。伸長型GLP-1化合物のアミノ酸38〜45は、好ましくは、グルカゴンまたはエクセンディン4の対応する位置にあるアミノ酸と同じであるか、またはその保存的置換である。
【0022】
本発明のGLP-1化合物は「GLP-1類似体」を包含する。GLP-1類似体は、その類似体がインスリン分泌促進活性を持つように、GLP-1(7-37)OHまたはGLP-1(7-37)OHの断片に対して十分な相同性を持つ。好ましくは、GLP-1類似体は、1、2、3、4または5個のアミノ酸がGLP-1(7-37)OHまたはGLP1(7-37)OHの断片の対応する位置にあるアミノ酸とは異なるように改変されている、GLP-1(7-37)OHまたはその断片のアミノ酸配列を持つ。GLP-1化合物を指定するために本明細書で使用する命名法では、置換アミノ酸およびその位置を親構造の前に示す。例えばGlu22-GLP-1(7-37)OHは、GLP-1(7-37)OHの22位に通常見いだされるグリシンがグルタミン酸で置き換えられているGLP-1化合物を表し、Val8-Glu22-GLP-1(7-37)OHは、GLP-1(7-37)OHの8位に通常見いだされるアラニンおよび22位に通常見いだされるグリシンがそれぞれバリンおよびグルタミン酸で置き換えられているGLP-1化合物を表す。
【0023】
本発明のGLP-1化合物には「GLP-1誘導体」も包含される。GLP-1誘導体は、GLP-1またはGLP-1類似体のアミノ酸配列を持つが、さらにそのアミノ酸側鎖基、α炭素原子、末端アミノ基、または末端カルボン酸基の1つまたは複数が化学修飾されている分子であると定義される。化学修飾には、例えば化学部分の付加、新しい結合の生成、および化学部分の除去などが含まれるが、これらに限るわけではない。アミノ酸側鎖基での修飾には、例えばリジンε-アミノ基のアシル化、アルギニン、ヒスチジンまたはリジンのN-アルキル化、グルタミン酸またはアスパラギン酸カルボン酸基のアルキル化、およびグルタミンまたはアスパラギンの脱アミド化などがあるが、これらに限るわけではない。末端アミノ基の修飾には、例えばデスアミノ、N-低級アルキル、N-ジ-低級アルキル、およびN-アシル修飾などがあるが、これらに限るわけではない。末端カルボキシ基の修飾には、例えばアミド、低級アルキルアミド、ジアルキルアミド、および低級アルキルエステル修飾などがあるが、これらに限るわけではない。低級アルキルはC1〜C4アルキルである。さらに、1つまたは複数の側鎖基または末端基を、通常の技術を有するタンパク質化学者に知られている保護基で保護することもできる。アミノ酸のα炭素はモノメチル化またはジメチル化することができる。
【0024】
GLP-1化合物は、そのGLP-1化合物自体がGLP-1受容体を結合し、当該受容体を介したシグナリングを誘発することができる限り、どれでも本発明の異種融合タンパク質の一部とすることができる。GLP-1受容体結合およびシグナル伝達は、EP619,322および米国特許第5,120,712号にそれぞれ記載されているような試験管内アッセイを使って評価することができる。
【0025】
当技術分野では数多くの活性なGLP-1断片、類似体および誘導体が知られており、これらの類似体および誘導体はいずれも、本発明の異種融合タンパク質の一部とすることができる。新規GLP-1類似体ならびに当技術分野で知られているGLP-1類似体および誘導体の例をいくつか挙げる。
【0026】
当技術分野で知られているGLP-1類似体およびGLP-1断片には、例えばGLP-1(7-34)およびGLP-1(7-35)、GLP-1(7-36)、Gln9-GLP-1(7-37)、D-Gln9-GLP-1(7-37)、Thr16-Lys18-GLP-1(7-37)、およびLys18-GLP-1(7-37)などがある。GLP-1(7-34)およびGLP-1(7-35)などのGLP-1類似体は米国特許第5,118,666号に開示されている。インスリン分泌促進性を持つ生物学的プロセシングを受けた形態のGLP-1、例えばGLP-1(7-36)なども知られている。生物学的に活性な他の既知のGLP-1化合物は、Hoffmannらの米国特許第5,977,071号、Buckleyらの米国特許第5,545,618号、およびAdelhorstら, J. Biol. Chem. 269:6275 (1994)に開示されている。
【0027】
好ましいGLP-1類似体群は式I(配列番号2)のGLP-1類似体から構成される:
【化2】
式I(配列番号2)
[式中、
8位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
9位のXaaは、Glu、Asp、またはLysであり、
11位のXaaは、Thr、Ala、Gly、Ser、Leu、Ile、Val、Glu、Asp、またはLysであり、
14位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
16位のXaaは、Val、Ala、Gly、Ser、Thr、Leu、Ile、Tyr、Glu、Asp、Trp、またはLysであり、
17位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
18位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、Trp、Tyr、またはLysであり、
19位のXaaは、Tyr、Phe、Trp、Glu、Asp、Gln、またはLysであり、
20位のXaaは、Leu、Ala、Gly、Ser、Thr、Ile、Val、Glu、Asp、Met、Trp、Tyr、またはLysであり、
21位のXaaは、Glu、Asp、またはLysであり、
22位のXaaは、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
23位のXaaは、Gln、Asn、Arg、Glu、Asp、またはLysであり、
24位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Arg、Glu、Asp、またはLysであり、
25位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
26位のXaaは、Lys、Arg、Gln、Glu、Asp、またはHisであり、
27位のXaaは、Leu、Glu、Asp、またはLysであり、
30位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
31位のXaaは、Trp、Phe、Tyr、Glu、Asp、またはLysであり、
32位のXaaは、Leu、Gly、Ala、Ser、Thr、Ile、Val、Glu、Asp、またはLysであり、
33位のXaaは、Val、Gly、Ala、Ser、Thr、Leu、Ile、Glu、Asp、またはLysであり、
34位のXaaは、Asn、Lys、Arg、Glu、Asp、またはHisであり、
35位のXaaは、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
36位のXaaは、Gly、Arg、Lys、Glu、Asp、またはHisであり、
37位のXaaは、Pro、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、もしくはLysであるか、または欠失しており、
38位のXaaは、Ser、Arg、Lys、Glu、Asp、もしくはHisであるか、または欠失しており、
39位のXaaは、Ser、Arg、Lys、Glu、Asp、もしくはHisであるか、または欠失しており、
40位のXaaは、Gly、Asp、Glu、もしくはLysであるか、または欠失しており、
41位のXaaは、Ala、Phe、Trp、Tyr、Glu、Asp、もしくはLysであるか、または欠失しており、
42位のXaaは、Ser、Pro、Lys、Glu、もしくはAspであるか、または欠失しており、
43位のXaaは、Ser、Pro、Glu、Asp、もしくはLysであるか、または欠失しており、
44位のXaaは、Gly、Pro、Glu、Asp、もしくはLysであるか、または欠失しており、
45位のXaaは、Ala、Ser、Val、Glu、Asp、もしくはLysであるか、または欠失している。
ただし、37、38、39、40、41、42、43、または44位のアミノ酸が欠失している場合は、そのアミノ酸の下流にある各アミノ酸も欠失しているものとする]。
【0028】
式IのGLP-1化合物は、GLP-1(7-37)OHまたはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、6個を超えて含まないことが好ましい。GLP-1(7-37)OHまたはエクセンディン4中の対応するアミノ酸と相違するアミノ酸は5個を超えないことがより好ましい。GLP-1(7-37)OHまたはエクセンディン4中の対応するアミノ酸と相違するアミノ酸は4個を超えないことがさらに好ましい。
【0029】
本発明のGLP-1化合物は、式Iの誘導体、例えばそのC-1-6-エステル、またはアミド、またはC-1-6-アルキルアミド、またはC-1-6-ジアルキルアミドなどを包含する。WO99/43706には式IのGLP-1化合物の誘導体が記載されており、この公開明細書は参考文献としてそのまま本明細書の一部を構成する。本発明は、WO99/43706に記載されているように誘導体化された式Iの化合物、および誘導体化されていない式Iの化合物を包含する。
【0030】
もう一つの好ましいGLP-1化合物群は、式II(配列番号3)のGLP-1類似体から構成される:
【化3】
式II(配列番号3)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、Val、Leu、Ile、Ser、またはThrであり、
9位のXaaは、Thr、Ser、Arg、Lys、Trp、Phe、Tyr、Glu、またはHisであり、
11位のXaaは、Asp、Glu、Arg、Thr、Ala、Lys、またはHisであり、
12位のXaaは、His、Trp、Phe、またはTyrであり、
16位のXaaは、Leu、Ser、Thr、Trp、His、Phe、Asp、Val、Tyr、Glu、またはAlaであり、
18位のXaaは、His、Pro、Asp、Glu、Arg、Ser、Ala、またはLysであり、
22位のXaaは、Gly、Asp、Glu、Gln、Asn、Lys、Arg、またはCysであり、
23位のXaaは、His、Asp、Lys、Glu、Gln、またはArgであり、
24位のXaaは、Glu、Arg、Ala、またはLysであり、
26位のXaaは、Trp、Tyr、Phe、Asp、Lys、Glu、またはHisであり、
27位のXaaは、Ala、Glu、His、Phe、Tyr、Trp、Arg、またはLysであり、
30位のXaaは、Ala、Glu、Asp、Ser、またはHisであり、
31位のXaaは、Asp、Glu、Ser、Thr、Arg、Trp、またはLysであり、
33位のXaaは、Asp、Arg、Val、Lys、Ala、Gly、またはGluであり、
34位のXaaは、Glu、Lys、またはAspであり、
35位のXaaは、Thr、Ser、Lys、Arg、Trp、Tyr、Phe、Asp、Gly、Pro、His、またはGluであり、
36位のXaaは、Thr、Ser、Asp、Trp、Tyr、Phe、Arg、Glu、またはHisであり、
37位のRは、Lys、Arg、Thr、Ser、Glu、Asp、Trp、Tyr、Phe、His、Gly、もしくはGly-Proであるか、または欠失している]。
【0031】
もう一つの好ましいGLP-1化合物群は、式III(配列番号4)のGLP-1類似体から構成される:
【化4】
式III(配列番号4)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、Val、Leu、Ile、Ser、またはThrであり、
11位のXaaは、Asp、Glu、Arg、Thr、Ala、Lys、またはHisであり、
12位のXaaは、His、Trp、Phe、またはTyrであり、
16位のXaaは、Leu、Ser、Thr、Trp、His、Phe、Asp、Val、Glu、またはAlaであり、
22位のXaaは、Gly、Asp、Glu、Gln、Asn、Lys、Arg、またはCysであり、
23位のXaaは、His、Asp、Lys、Glu、またはGlnであり、
24位のXaaは、Glu、His、Ala、またはLysであり、
25位のXaaは、Asp、Lys、Glu、またはHisであり、
27位のXaaは、Ala、Glu、His、Phe、Tyr、Trp、Arg、またはLysであり、
30位のXaaは、Ala、Glu、Asp、Ser、またはHisであり、
33位のXaaは、Asp、Arg、Val、Lys、Ala、Gly、またはGluであり、
34位のXaaは、Glu、Lys、またはAspであり、
35位のXaaは、Thr、Ser、Lys、Arg、Trp、Tyr、Phe、Asp、Gly、Pro、His、またはGluであり、
36位のXaaは、Arg、Glu、またはHisであり、
37位のRは、Lys、Arg、Thr、Ser、Glu、Asp、Trp、Tyr、Phe、His、Gly、もしくはGly-Proであるか、または欠失している]。
【0032】
もう一つの好ましいGLP-1化合物群は、式IV(配列番号5)のGLP-1類似体から構成される:
【化5】
式IV(配列番号5)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、Val、Leu、Ile、Ser、Met、またはThrであり、
12位のXaaは、His、Trp、Phe、またはTyrであり、
16位のXaaは、Leu、Ser、Thr、Trp、His、Phe、Asp、Val、Glu、またはAlaであり、
22位のXaaは、Gly、Asp、Glu、Gln、Asn、Lys、Arg、またはCysであり、
23位のXaaは、His、Asp、Lys、Glu、またはGlnであり、
26位のXaaは、Asp、Lys、Glu、またはHisであり、
30位のXaaは、Ala、Glu、Asp、Ser、またはHisであり、
35位のXaaは、Thr、Ser、Lys、Arg、Trp、Tyr、Phe、Asp、Gly、Pro、His、またはGluであり、
37位のRは、Lys、Arg、Thr、Ser、Glu、Asp、Trp、Tyr、Phe、His、Gly、もしくはGly-Proであるか、または欠失している]。
【0033】
もう一つの好ましいGLP-1群は、式V(配列番号6)のGLP-1類似体から構成される:
【化6】
式V(配列番号6)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、Val、Leu、Ile、Ser、またはThrであり、
22位のXaaは、Gly、Asp、Glu、Gln、Asn、Lys、Arg、またはCysであり、
23位のXaaは、His、Asp、Lys、Glu、またはGlnであり、
24位のXaaは、Ala、Glu、His、Phe、Tyr、Trp、Arg、またはLysであり、
30位のXaaは、Ala、Glu、Asp、Ser、またはHisであり、
37位のRは、Lys、Arg、Thr、Ser、Glu、Asp、Trp、Tyr、Phe、His、Gly、もしくはGly-Proであるか、または欠失している]。
【0034】
式I、II、III、IVおよびVの好ましいGLP-1化合物は、8位にアラニン以外のアミノ酸を持つGLP-1類似体またはGLP-1類似体の断片を含む(8位類似体)。これらの8位類似体は、さらに9、11、12、16、18、22、23、24、26、27、30、31、33、34、35、36、および37位にも、天然GLP-1(7-37)OHの対応するアミノ酸と比較して、1つまたは複数の変異を持つことが好ましい。また、これらの類似体は、天然のGLP-1(7-37)OHまたはGLP-1(7-36)OH中の対応するアミノ酸と比較して、6個以下の変異を持つことが好ましい。より好ましい類似体は、天然のGLP-1(7-37)OHもしくはGLP-1(7-36)OH中の対応するアミノ酸と比較して5個以下の変異を持つか、または天然のGLP-1(7-37)OHもしくはGLP-1(7-36)OH中の対応するアミノ酸と比較して4個以下の変異を持つ。これらの類似体は、天然のGLP-1(7-37)OHまたはGLP-1(7-36)OH中の対応するアミノ酸と比較して3個以下の変異を持つことが、さらに好ましい。これらの類似体は、天然のGLP-1(7-37)OH中の対応するアミノ酸と比較して2個以下の変異を持つことが、最も好ましい。
【0035】
式II、III、IVおよびVの化合物は、凝集して不溶型を生成する傾向が低下していることがわかった。比較的小さいGLP-1ペプチドが、はるかに大きいタンパク質に融合されているにもかかわらず、活性なコンフォメーションを維持しなければならない融合タンパク質にあっては、これも重要なことである。本発明の融合タンパク質に包含される式II、III、IVおよびVの好ましいGLP-1化合物は、22位のグリシンと、好ましくは8位のアラニンが、別のアミノ酸で置き換えられているGLP-1類似体またはGLP-1類似体の断片を含む。
【0036】
22位がアスパラギン酸、グルタミン酸、アルギニンまたはリジンである場合、8位は好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニンまたはメチオニンであり、より好ましくはバリンまたはグリシンである。22位がシステイン酸などのスルホン酸である場合は、8位は好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニンまたはメチオニンであり、より好ましくはバリンまたはグリシンである。
【0037】
他の好ましいGLP-1化合物として、8位のアミノ酸が好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンであり、30位がグルタミン酸、アスパラギン酸、セリン、またはヒスチジン、より好ましくはグルタミン酸である点以外は、GLP-1(7-37)OHの配列を持つ、式IV(配列番号5)のGLP-1類似体が挙げられる。
【0038】
他の好ましいGLP-1化合物として、8位のアミノ酸が好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンであり、37位がヒスチジン、リジン、アルギニン、スレオニン、セリン、グルタミン酸、アスパラギン酸、トリプトファン、チロシン、フェニルアラニン、より好ましくはヒスチジンである点以外は、GLP-1(7-37)OHの配列を持つ、式IV(配列番号5)のGLP-1類似体が挙げられる。
【0039】
他の好ましいGLP-1化合物として、8位のアミノ酸が好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンであり、22位がグルタミン酸、リジン、アスパラギン酸、またはアルギニン、より好ましくはグルタミン酸またはリジンであり、23位がリジン、アルギニン、グルタミン酸、アスパラギン酸およびヒスチジン、より好ましくはリジンまたはグルタミン酸である点以外は、GLP-1(7-37)OHの配列を持つ、式IV(配列番号5)のGLP-1類似体が挙げられる。
【0040】
他の好ましいGLP-1化合物として、8位のアミノ酸が好ましくはグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンであり、22位がグルタミン酸、リジン、アスパラギン酸、またはアルギニン、より好ましくはグルタミン酸またはリジンであり、27位がアラニン、リジン、アルギニン、トリプトファン、チロシン、フェニルアラニン、またはヒスチジン、より好ましくはアラニンである点以外は、GLP-1(7-37)OHの配列を持つ、式V(配列番号6)のGLP-1類似体が挙げられる。
【0041】
他の好ましいGLP-1化合物として、8位のアミノ酸ならびに9位、11位、12位、16位、18位、22位、23位、24位、26位、27位、30位、31位、33位、34位、35位、36位、および37位からなる群より選択される1、2または3個のアミノ酸が、天然GLP-1(7-37)OHの対応する位置にあるアミノ酸とは相違する点以外は、GLP-1(7-37)OHの配列を持つ、式IIのGLP-1類似体が挙げられる。
【0042】
他の好ましい式IIのGLP-1化合物には、以下に挙げるものがある:Val8-GLP-1(7-37)OH、Gly8-GLP-1(7-37)OH、Glu22-GLP-1(7-37)OH、Asp22-GLP-1(7-37)OH、Arg22-GLP-1(7-37)OH、Lys22-GLP-1(7-37)OH、Cys22-GLP-1(7-37)OH、Val8-Glu22-GLP-1(7-37)OH、Val8-Asp22-GLP-1(7-37)OH、Val8-Arg22-GLP-1(7-37)OH、Val8-Lys22-GLP-1(7-37)OH、Val8-Cys22-GLP-1(7-37)OH、Gly8-Glu22-GLP-1(7-37)OH、Gly8-Asp22-GLP-1(7-37)OH、Gly8-Arg22-GLP-1(7-37)OH、Gly8-Lys22-GLP-1(7-37)OH、Gly8-Cys22-GLP-1(7-37)OH、Glu22-GLP-1(7-36)OH、Asp22-GLP-1(7-36)OH、Arg22-GLP-1(7-36)OH、Lys22-GLP-1(7-36)OH、Cys22-GLP-1(7-36)OH、Val8-Glu22-GLP-1(7-36)OH、Val8-Asp22-GLP-1(7-36)OH、Val8-Arg22-GLP-1(7-36)OH、Val8-Lys22-GLP-1(7-36)OH、Val8-Cys22-GLP-1(7-36)OH、Gly8-Glu22-GLP-1(7-36)OH、Gly8-Asp22-GLP-1(7-36)OH、Gly8-Arg22-GLP-1(7-36)OH、Gly8-Lys22-GLP-1(7-36)OH、Gly8-Cys22-GLP-1(7-36)OH、Lys23-GLP-1(7-37)OH、Val8-Lys23-GLP-1(7-37)OH、Gly8-Lys23-GLP-1(7-37)OH、His24-GLP-1(7-37)OH、Val8-His24-GLP-1(7-37)OH、Gly8-His24-GLP-1(7-37)OH、Lys24-GLP-1(7-37)OH、Val8-Lys24-GLP-1(7-37)OH、Gly8-Lys23-GLP-1(7-37)OH、Glu30-GLP-1(7-37)OH、Val8-Glu30-GLP-1(7-37)OH、Gly8-Glu30-GLP-1(7-37)OH、Asp30-GLP-1(7-37)OH、Val8-Asp30-GLP-1(7-37)OH、Gly8-Asp30-GLP-1(7-37)OH、Gln30-GLP-1(7-37)OH、Val8-Gln30-GLP-1(7-37)OH、Gly8-Gln30-GLP-1(7-37)OH、Tyr30-GLP-1(7-37)OH、Val8-Tyr30-GLP-1(7-37)OH、Gly8-Tyr30-GLP-1(7-37)OH、Ser30-GLP-1(7-37)OH、Val8-Ser30-GLP-1(7-37)OH、Gly8-Ser30-GLP-1(7-37)OH、His30-GLP-1(7-37)OH、Val8-His30-GLP-1(7-37)OH、Gly8-His30-GLP-1(7-37)OH、Glu34-GLP-1(7-37)OH、Val8-Glu34-GLP-1(7-37)OH、Gly8-Glu34-GLP-1(7-37)OH、Ala34-GLP-1(7-37)OH、Val8-Ala34-GLP-1(7-37)OH、Gly8-Ala34-GLP-1(7-37)OH、Gly34-GLP-1(7-37)OH、Val8-Gly34-GLP-1(7-37)OH、Gly8-Gly34-GLP-1(7-37)OH、Ala35-GLP-1(7-37)OH、Val8-Ala35-GLP-1(7-37)OH、Gly8-Ala35-GLP-1(7-37)OH、Lys35-GLP-1(7-37)OH、Val8-Lys35-GLP-1(7-37)OH、Gly8-Lys35-GLP-1(7-37)OH、His35-GLP-1(7-37)OH Val8-His35-GLP-1(7-37)OH、Gly8-His35-GLP-1(7-37)OH、Pro35-GLP-1(7-37)OH、Val8-Pro35-GLP-1(7-37)OH、Gly8-Pro35-GLP-1(7-37)OH、Glu35-GLP-1(7-37)OH、Val8-Glu35-GLP-1(7-37)OH、Gly8-Glu35-GLP-1(7-37)OH、Val8-Ala27-GLP-1(7-37)OH、Val8-His37-GLP-1(7-37)OH、Val8-Glu22-Lys23-GLP-1(7-37)OH、Val8-Glu22-Glu23-GLP-1(7-37)OH、Val8-Glu22-Ala27-GLP-1(7-37)OH、Val8-Gly34-Lys35-GLP-1(7-37)OH、Val8-His37-GLP-1(7-37)OH、Gly8-His37-GLP-1(7-37)OH、Val8-Glu22-Ala27-GLP-1(7-37)OH、Gly8-Glu22-Ala27-GLP-1(7-37)OH、Val8-Lys22-Glu23-GLP-1(7-37)OH、およびGly8-Lys22-Glu23-GLP-1(7-37)OH。
【0043】
本発明での使用に好ましいもう一つのGLP-1類似体および誘導体群は、式VI(配列番号7)の分子から構成される:
R1-X-Glu-Gly10-Thr-Phe-Thr-Ser-Asp15-Val-Ser-Ser-Tyr-Leu20-Y -Gly-Gln-Ala-Ala25-Lys-Z-Phe-Ile-Ala30-Trp-Leu-Val-Lys-Gly35-Arg-R2
式VI(配列番号7)
[式中、R1は、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジン、およびα-メチルヒスチジンであり、XはAla、Gly、Val、Thr、Ile、およびα-メチル-Alaからなる群より選択され、YはGlu、Gln、Ala、Thr、Ser、およびGlyからなる群より選択され、ZはGlu、Gln、Ala、Thr、Ser、およびGlyからなる群より選択され、R2はGly-OHである]。
【0044】
本発明での使用に好ましいもう一つのGLP-1化合物群はWO91/11457に開示されており、本質的に、以下の(a)〜(e)からなる群より選択される少なくとも1つの改変を持つ、GLP-1(7-34)、GLP-1(7-35)、GLP-1(7-36)、もしくはGLP-1(7-37)、またはそのアミド型、およびその医薬的に許容できる塩からなる:
(a)グリシン、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、アルギニン、またはD-リジンによる26位および/または34位のリジンの置換、またはグリシン、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、リジン、またはD-アルギニンによる36位のアルギニンの置換、
(b)耐酸化性アミノ酸による31位のトリプトファンの置換、
(c)次に挙げる置換の少なくとも1つ:チロシンによる16位のバリンの置換、リジンによる18位のセリンの置換、アスパラギン酸による21位のグルタミン酸の置換、セリンによる22位のグリシンの置換、アルギニンによる23位のグルタミンの置換、アルギニンによる24位のアラニンの置換、およびグルタミンによる26位のリジンの置換、
(d)次に挙げる置換の少なくとも1つ:グリシン、セリン、またはシステインによる8位のアラニンの置換、アスパラギン酸、グリシン、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、またはフェニルアラニンによる9位のグルタミン酸の置換、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、またはフェニルアラニンによる10位のグリシンの置換、およびグルタミン酸による15位のアスパラギン酸の置換、
(e)グリシン、セリン、システイン、スレオニン、アスパラギン、グルタミン、チロシン、アラニン、バリン、イソロイシン、ロイシン、メチオニン、またはフェニルアラニン、またはヒスチジンのD-もしくはN-アシル化もしくはアルキル化型による7位のヒスチジンの置換、
ただし、(a)、(b)、(d)、および(e)の置換において、置換アミノ酸は所望によりD型であってもよく、7位の置換アミノ酸は所望によりN-アシル化型またはN-アルキル化型であってもよい。
【0045】
酵素ジペプチジル-ペプチダーゼIV(DPP IV)は、投与されたGLP-1に観察される迅速な生体内失活の原因であると考え得るので[例えばMentlein, R.ら, Eur. J. Biochem., 214:829-835 (1993)参照]、融合タンパク質の形でDPP IVの活性から保護されるGLP-1類似体および誘導体は好ましく、GLP-1化合物がGly8-GLP-1(7-37)OH、Val8-GLP-1(7-37)OH、α-メチル-Ala8-GLP-1(7-37)OH、またはGly8-Gln21-GLP-1(7-37)OHである融合タンパク質は、さらに好ましい。
【0046】
本発明での使用に好ましいもう一つのGLP-1化合物群は、参考文献として特に本明細書の一部を構成する米国特許第5,512,549号にクレームされている式VII(配列番号8)の化合物からなる。
【化7】
式VII(配列番号8)
[式中、R1は、4-イミダゾプロピオニル、4-イミダゾアセチル、または4-イミダゾ-α,α-ジメチルアセチルからなる群より選択され、R2は、C6-C10非分岐アシルからなる群から選択されるか、または存在せず、R3はGly-OHまたはNH2からなる群より選択され、XaaはLysまたはArgである]。
【0047】
本発明での使用に、より好ましい式IVの化合物は、XaaがArgであり、R2がC6-C10非分岐アシルである式IVの化合物である。本発明での使用に、さらに好ましい式IVの化合物は、XaaがArgであり、R2がC6-C10非分岐アシルであり、R3がGly-OHである化合物である。本発明での使用に著しく好ましい他の式IVの化合物は、XaaがArgであり、R2がC6-C10非分岐アシルであり、R3がGly-OHであり、R1が4-イミダゾプロピオニルである式VIの化合物である。本発明での使用に特に好ましい式IVの化合物は、XaaがArgであり、R2がC8非分岐アシルであり、R3がGly-OHであり、R1が4-イミダゾプロピオニルである式IVの化合物である。
【0048】
GLP-1化合物は、好ましくは、8位がアラニン以外のアミノ酸である主鎖を持つGLP-1類似体(8位類似体)を含む。また主鎖は、7位にL-ヒスチジン、D-ヒスチジン、またはデスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジン、もしくはα-メチルヒスチジンなどの修飾型ヒスチジンを含んでもよい。これらの8位類似体は、さらに12、16、18、19、20、22、25、27、30、33、および37位にも、天然GLP-1(7-37)OHの対応するアミノ酸と比較して、1つまたは複数の変異を持つことが好ましい。これらの8位類似体は、さらに16、18、22、25および33位にも、天然GLP-1(7-37)OHの対応するアミノ酸と比較して、1つまたは複数の変異を持つことが、より好ましい。
【0049】
好ましい一態様として、GLP-1類似体は、12位のアミノ酸がトリプトファンまたはチロシンからなる群より選択される、GLP-1(7-37)OHである。12位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。12位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。
【0050】
もう一つの好ましい態様として、GLP-1類似体は、16位のアミノ酸がトリプトファン、イソロイシン、ロイシン、フェニルアラニン、またはチロシンからなる群より選択される、GLP-1(7-37)である。16位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。16位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。16位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。16位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0051】
もう一つの好ましい態様として、GLP-1類似体は、18位のアミノ酸がトリプトファン、チロシン、フェニルアラニン、リジン、ロイシン、またはイソロイシン、好ましくはトリプトファン、チロシン、およびイソロイシンからなる群より選択される、GLP-1(7-37)OHである。18位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。18位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。18位および8位での置換に加えて、30位のアミノ酸が、グルタミン酸で置換されていることも、好ましい。18位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0052】
もう一つの好ましい態様として、GLP-1類似体は、19位のアミノ酸がトリプトファンまたはフェニルアラニンからなる群より選択される(好ましくはトリプトファンである)、GLP-1(7-37)OHである。19位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。19位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。19位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。19位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0053】
もう一つの好ましい態様として、GLP-1類似体は、20位のアミノ酸がフェニルアラニン、チロシン、またはトリプトファンである、GLP-1(7-37)OHである。20位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。20位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。20位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。20位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0054】
もう一つの好ましい態様として、GLP-1類似体は、25位のアミノ酸がバリン、イソロイシン、およびロイシンからなる群より選択される(好ましくはバリンである)、GLP-1(7-37)OHである。25位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。25位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。25位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。25位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0055】
もう一つの好ましい態様として、GLP-1類似体は、27位のアミノ酸がイソロイシンまたはアラニンからなる群より選択される、GLP-1(7-37)OHである。27位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。27位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。27位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。27位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0056】
もう一つの好ましい態様として、GLP-1類似体は、33位のアミノ酸がイソロイシンである、GLP-1(7-37)OHである。33位での置換に加えて、8位のアミノ酸がグリシン、バリン、ロイシン、イソロイシン、セリン、スレオニン、またはメチオニン、より好ましくはバリンまたはグリシンで置換されていると、より好ましい。33位および8位での置換に加えて、22位のアミノ酸がグルタミン酸で置換されていると、さらに好ましい。33位および8位での置換に加えて、30位のアミノ酸がグルタミン酸で置換されていることも、好ましい。33位および8位での置換に加えて、37位のアミノ酸がヒスチジンで置換されていることも、好ましい。
【0057】
本GLP-1化合物は以下の位置の1つまたは複数が改変されている:8、12、16、18、19、20、22、25、27、30、33、および37。これらのGLP-1化合物は、GLP-1(7-37)OHと比較して増加した効力を示し、式IX(配列番号12)のアミノ酸配列を含む:
Xaa7-Xaa8-Glu-Gly-Thr-Xaa12-Thr-Ser-Asp-Xaa16-Ser-Xaa18-Xaa19-Xaa20-Glu-Xaa22-Gln-Ala-Xaa25-Lys-Xaa27-Phe-Ile-Xaa30-Trp-Leu-Xaa33-Lys-Gly-Arg-Xaa37
式IX(配列番号12)
[式中、
Xaa7は、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジン、またはα-メチルヒスチジンであり、
Xaa8は、Ala、Gly、Val、Leu、Ile、Ser、またはThrであり、
Xaa12は、Phe、Trp、またはTyrであり、
Xaa16は、Val、Trp、Ile、Leu、Phe、またはTyrであり、
Xaa18は、Ser、Trp、Tyr、Phe、Lys、Ile、Leu、Valであり、
Xaa19は、Tyr、Trp、またはPheであり、
Xaa20は、Leu、Phe、Tyr、またはTrpであり、
Xaa22は、Gly、Glu、Asp、またはLysであり、
Xaa25は、Ala、Val、Ile、またはLeuであり、
Xaa27は、Glu、Ile、またはAlaであり、
Xaa30は、AlaまたはGluであり、
Xaa33は、Val、またはIleであり、
Xaa37は、Gly、His、もしくはNH2であるか、または存在しない]。
【0058】
式IXの好ましいGLP-1化合物としては、例えば、GLP-1(7-37)OH、GLP-1(7-36)-NH2、Gly8-GLP-1(7-37)OH、Gly8-GLP-1(7-36)NH2、Val8-GLP-1(7-37)OH、Val8-GLP-1(7-36)NH2、Leu8-GLP-1(7-37)OH、Leu8-GLP-1(7-36)NH2、Ile8-GLP-1(7-37)OH、Ile8-GLP-1(7-36)NH2、Ser8-GLP-1(7-37)OH、Ser8-GLP-1(7-36)NH2、Thr8-GLP-1(7-37)OH、Thr8-GLP-1(7-36)NH2、Val8-Tyr12-GLP-1(7-37)OH、Val8-Tyr12-GLP-1(7-36)NH2、Val8-Tyr16-GLP-1(7-37)OH、Val8-Tyr16-GLP-1(7-36)NH2、Val8-Glu22-GLP-1(7-37)OH、Val8-Glu22-GLP-1(7-36)NH2、Gly8-Glu22-GLP-1(7-37)OH、Gly8-Glu22-GLP-1(7-36)NH2、Val8-Asp22-GLP-1(7-37)OH、Val8-Asp22-GLP-1(7-36)NH2、Gly8-Asp22-GLP-1(7-37)OH、Gly8-Asp22-GLP-1(7-36)NH2、Val8-Lys22-GLP-1(7-37)OH、Val8-Lys22-GLP-1(7-36)NH2、Gly8-Lys22-GLP-1(7-37)OH、Gly8-Lys22-GLP-1(7-36)NH2、Leu8-Glu22-GLP-1(7-37)OH、Leu8-Glu22-GLP-1(7-36)NH2、Ile8-Glu22-GLP-1(7-37)OH、Ile8-Glu22-GLP-1(7-36)NH2、Leu8-Asp22-GLP-1(7-37)OH、Leu8-Asp22-GLP-1(7-36)NH2、Ile8-Asp22-GLP-1(7-37)OH、Ile8-Asp22-GLP-1(7-36)NH2、Leu8-Lys22-GLP-1(7-37)OH、Leu8-Lys22-GLP-1(7-36)NH2、Ile8-Lys22-GLP-1(7-37)OH、Ile8-Lys22-GLP-1(7-36)NH2、Ser8-Glu22-GLP-1(7-37)OH、Ser8-Glu22-GLP-1(7-36)NH2、Thr8-Glu22-GLP-1(7-37)OH、Thr8-Glu22-GLP-1(7-36)NH2、Ser8-Asp22-GLP-1(7-37)OH、Ser8-Asp22-GLP-1(7-36)NH2、Thr8-Asp22-GLP-1(7-37)OH、Thr8-Asp22-GLP-1(7-36)NH2、Ser8-Lys22-GLP-1(7-37)OH、Ser8-Lys22-GLP-1(7-36)NH2、Thr8-Lys22-GLP-1(7-37)OH、Thr8-Lys22-GLP-1(7-36)NH2、Glu22-GLP-1(7-37)OH、Glu22-GLP-1(7-36)NH2、Asp22-GLP-1(7-37)OH、Asp22-GLP-1(7-36)NH2、Lys22-GLP-1(7-37)OH、Lys22-GLP-1(7-36)NH2、Val8-Ala27-GLP-1(7-37)OH、Val8-Glu22-Ala27-GLP-1(7-37)OH、Val8-Glu30-GLP-1(7-37)OH、Val8-Glu30-GLP-1(7-36)NH2、Gly8-Glu30-GLP-1(7-37)OH、Gly8-Glu30-GLP-1(7-36)NH2、Leu8-Glu30-GLP-1(7-37)OH、Leu8-Glu30-GLP-1(7-36)NH2、Ile8-Glu30-GLP-1(7-37)OH、Ile8-Glu30-GLP-1(7-36)NH2、Ser8-Glu30-GLP-1(7-37)OH、Ser8-Glu30-GLP-1(7-36)NH2、Thr8-Glu30-GLP-1(7-37)OH、Thr8-Glu30-GLP-1(7-36)NH2、Val8-His37-GLP-1(7-37)OH、Val8-His37-GLP-1(7-36)NH2、Gly8-His37-GLP-1(7-37)OH、Gly8-His37-GLP-1(7-36)NH2、Leu8-His37-GLP-1(7-37)OH、Leu8-His37-GLP-1(7-36)NH2、Ile8-His37-GLP-1(7-37)OH、Ile8-His37-GLP-1(7-36)NH2、Ser8-His37-GLP-1(7-37)OH、Ser8-His37-GLP-1(7-36)NH2、Thr8-His37-GLP-1(7-37)OH、Thr8-His37-GLP-1(7-36)NH2などが挙げられる。
【0059】
複数の置換を持つ式IXの好ましいGLP-1化合物としては、例えば、8位がバリンまたはグリシンであり、22位がグルタミン酸であり、16位がチロシン、ロイシンまたはトリプトファンであり、18位がチロシン、トリプトファン、またはイソロイシンであり、25位がバリンであり、33位がイソロイシンであるGLP-1(7-37)OHが挙げられる。他の好ましいGLP-1化合物には、以下の化合物が含まれる:Val8-Tyr16-GLP-1(7-37)OH、Val8-Tyr12-Glu22-GLP-1(7-37)OH、Val8-Tyr16-Phe19-GLP-1(7-37)OH、Val8-Tyr16-Glu22-GLP-1(7-37)OH、Val8-Trp16-Glu22-GLP-1(7-37)OH、Val8-Leu16-Glu22-GLP-1(7-37)OH、Val8-Ile16-Glu22-GLP-1(7-37)OH、Val8-Phe16-Glu22-GLP-1(7-37)OH、Val8-Trp18-Glu22-GLP-1(7-37)OH、Val8-Tyr18-Glu22-GLP-1(7-37)OH、Val8-Phe18-Glu22-GLP-1(7-37)OH、およびVal8-Ile18-Glu22-GLP-1(7-37)OH。
【0060】
本発明のGLP-1化合物には、エクセンディン化合物も包含される。エクセンディン3およびエクセンディン4は、ドクトカゲ科トカゲ毒から最初に単離された生物学的に活性なペプチドであり、GLP-1受容体に結合して、哺乳類旁細胞におけるcAMP依存的H+生成を刺激することが明らかにされている。エクセンディン3およびエクセンディン4はどちらも、GLP-1に対して約53%相同な39アミノ酸ペプチドである。これらはGLP-1活性の強力なアゴニストとして作用する。注目すべきことに、エクセンディン(9-39アミノ酸)として知られるエクセンディンのN末端切断型誘導体は、エクセンディン3、エクセンディン4およびGLP-1の阻害剤である。
【0061】
エクセンディン化合物は、典型的には、エクセンディン3、エクセンディン4、またはその類似体もしくは断片のアミノ酸配列を持つポリペプチドを含む。エクセンディン3およびエクセンディン4は米国特許第5,424,286号に開示されている。
【0062】
エクセンディン3は配列番号9のアミノ酸配列:
【化8】
(配列番号9)
を持つ。
【0063】
エクセンディン4は配列番号10のアミノ酸配列:
【化9】
(配列番号10)
を持つ。
【0064】
GLP-1化合物には、エクセンディンまたはエクセンディン類似体のN末端および/またはC末端から1つまたは複数のアミノ酸を切り落とした後に得られるポリペプチドであるエクセンディン断片も含まれる。さらに、GLP-1化合物には、エクセンディンまたはその断片のN末端および/またはC末端に1つまたは複数のアミノ酸が付加されているエクセンディンポリペプチドも含まれる。このタイプのエクセンディン化合物は約45個までのアミノ酸を持つ。
【0065】
GLP-1化合物には「エクセンディン類似体」も包含される。エクセンディン類似体は、その類似体がインスリン分泌促進活性を持つように、エクセンディン4、エクセンディン3、またはその断片に対して十分な相同性を持つ。エクセンディン断片および/または類似体の活性は、EP619,322および米国特許第5,120,712号に記載されているような試験管内アッセイを使って評価することができる。
【0066】
好ましくは、エクセンディン類似体は、1、2、3、4または5個のアミノ酸がエクセンディン4またはエクセンディン4の断片の対応する位置にあるアミノ酸とは異なるように改変されている、エクセンディン4またはその断片のアミノ酸配列を持つ。エクセンディン化合物を指定するために本明細書で使用する命名法では、置換アミノ酸およびその位置を親構造の前に示す。Val8-エクセンディン4は、エクセンディン4の8位に通常見いだされるグリシンがバリンで置き換えられているエクセンディン化合物を表す。
【0067】
もう一つの好ましいGLP-1化合物群は、式VIII(配列番号11)のGLP-1/エクセンディン4類似体から構成される。
【化10】
式VIII(配列番号11)
[式中、
7位のXaaは、L-ヒスチジン、D-ヒスチジン、デスアミノヒスチジン、2-アミノヒスチジン、β-ヒドロキシヒスチジン、ホモヒスチジン、α-フルオロメチルヒスチジンまたはα-メチルヒスチジンであり、
8位のXaaは、Gly、Ala、またはValであり、
16位のXaaは、LeuまたはValであり、
18位のXaaは、LysまたはSerであり、
19位のXaaは、GlnまたはTyrであり、
20位のXaaは、MetまたはLeuであり、
22位のXaaは、GluまたはGlnであり、
23位のXaaは、Glu、またはGlnであり、
25位のXaaは、ValまたはAlaであり、
26位のXaaは、ArgまたはLysであり、
27位のXaaは、LeuまたはGluであり、
30位のXaaは、GluまたはAlaであり、
33位のXaaは、ValまたはLysであり、
34位のXaaは、AsnまたはLysであり、
36位のXaaは、GlyまたはArgであり、
37位のRは、Gly、Pro、もしくはPro-Ser-Ser-Gly-Ala-Pro-Pro-Pro-Serであるか、または存在しない]。
この群に含まれる18種類の分子種の活性を表6に記載する。
【0068】
本発明にとって有用な他のエクセンディン類似体は、PCT特許公開WO99/25728(Beeleyら)、WO99/25727(Beeleyら)、WO98/05351(Youngら)、WO99/40788(Youngら)、WO99/07404(Beeleyら)、およびWO99/43708(Knudsenら)に記載されている。
【0069】
本発明のGLP-1融合タンパク質はグリコシル化部位を含みうる。グリコシル化は、タンパク質の特定部位に糖部分が付加される化学修飾である。タンパク質のグリコシル化は、成熟タンパク質の正しい電荷、コンフォメーション、および安定性の確保に役割を果たし、そのタンパク質を細胞表面に導いて、最終的にはそのタンパク質を分泌させることができる。最も重要なことに、グリコシル化は、多くのタンパク質で、その生体内クリアランス速度を左右する。糖鎖はO結合型でもN結合型でもよい。一般に、O結合型糖鎖はセリンおよびスレオニンのヒドロキシル基酸素に付加され、N結合型糖鎖はアスパラギンのアミド窒素に付加される。N-グリコシル化のコンセンサス配列はAsn X1 X2[X1はProを除く任意のアミノ酸であり、X2はSerまたはThrである]である。
【0070】
GLP-1化合物は生体内では一般にグリコシル化されない。しかし、興味深いことに、Fc配列に融合されたC末端伸長部を持つGLP-1化合物を含む本発明のGLP-1融合タンパク質は、C末端伸長部の最後のセリン(SSGAPPPS*)と、FcのN末端領域の11位にあるスレオニン(AEPKSCDKTHT*CPPC...)でグリコシル化される。
【0071】
異種Fc融合タンパク質:
上述のGLP-1化合物は免疫グロブリンのFc部分に直接融合するか、またはペプチドリンカーを介して融合することができる。
【0072】
免疫グロブリンは、ジスルフィド結合でつながったポリペプチド鎖を含み、典型的には2本の軽鎖と2本の重鎖を持つ分子である。各鎖では、1つのドメイン(V)が、その分子の抗体特異性に依存して可変的なアミノ酸配列を持つ。他のドメイン(C)は同じクラスの分子に共通するかなり一定した配列を持つ。
【0073】
本明細書で使用する「免疫グロブリンのFc部分」という用語は、免疫学の分野でこの用語に通常与えられている意味を持つ。具体的に述べると、この用語は、抗体から2つの抗原結合領域(Fab断片)を除去することによって得られる抗体断片を表す。Fab断片を除去する一方法は、免疫グロブリンをパパインプロテアーゼで消化することである。したがってFc部分は、非共有結合相互作用およびジスルフィド結合によって会合した両重鎖由来のほぼ同サイズの定常領域断片から形成される。Fc部分はヒンジ領域を含み、CH2およびCH3ドメインから抗体のC末端までにわたることができる。ヒトおよびマウス免疫グロブリンの代表的なヒンジ領域は、Borrebaeck, C.A.K.編「Antibody Engineering, A Practical Guide」(W.H. Freeman and Co., 1992)に記載されており、その内容は参考文献として本明細書の一部を構成する。Fc部分はさらに1つまたは複数のグリコシル化部位を含む場合もある。ヒンジ領域、CH2およびCH3ドメインを含み、82位に1つのN-グリコシル化部位を持つ代表的Fc部分のアミノ酸配列を図1に示す。
【0074】
ヒト免疫グロブリンFc領域には、異なるエフェクターおよび薬物動態特性を持つIgG、IgA、IgM、IgD、およびIgEの5種類がある。IgGは血清中で最も豊富な免疫グロブリンである。また、IgGは血清中で、免疫グロブリンのなかで最も長い半減期(23日)も持っている。IgGは他の免疫グロブリンとは異なり、Fc受容体への結合後に、効率よく再循環する。IgGには4つのサブクラスG1、G2、G3、およびG4があり、それぞれに異なるエフェクター機能を持っている。G1、G2、およびG3は、C1qを結合し補体を結合することができるが、G4はできない。G3はG1より効率よくC1qを結合することができるが、G1の方が補体による細胞溶解をより効果的に媒介する。G2の補体結合効率は非常に低い。IgG中のC1q結合部位は、CH2ドメインのカルボキシ末端領域にある。
【0075】
どのIgGサブクラスもFc受容体(CD16、CD32、CD64)に結合する能力を持つが、G1およびG3はG2およびG4よりも効果的である。IgGのFc受容体結合領域は、ヒンジ領域と、CH2ドメインのカルボキシ末端領域との両方に位置する残基によって形成される。
【0076】
IgAは、単量体型と、J鎖でつながれた二量体型の、両方で存在することができる。IgAは血清中に2番目に豊富なIgであるが、半減期は6日しかない。IgAは3つのエフェクター機能を持っている。これは、マクロファージおよび好酸球上のIgA特異的受容体に結合して、それぞれ食作用および脱顆粒を引き起こす。またこれは、未知の代替経路によって補体を結合することもできる。
【0077】
IgMは、J鎖でつながれた五量体またはJ鎖でつながれた六量体として発現する。IgMは5日の血清半減期を持つ。これは、そのCH3ドメイン中にある結合部位を介してC1qに弱く結合する。IgDは血清中で3日の半減期を持つ。このIgに起因すると考えられるエフェクター機能ははっきりしていない。IgEは単量体型Igであり、2.5日の血清半減期を持つ。IgEは2つのFc受容体に結合して、脱顆粒を引き起こし、炎症誘発因子の放出をもたらす。
【0078】
所望する生体内効果に応じて、本発明の異種融合タンパク質は、上述したイソタイプをどれでも含むことができ、補体および/またはFc受容体結合機能が改変されている突然変異型Fc領域を含んでもよい。したがって本発明の異種融合タンパク質は、GLP-1化合物に融合された免疫グロブリンのFc部分全体、免疫グロブリンのFc部分の断片、またはその類似体を含むことができる。
【0079】
本発明の融合タンパク質は、一本鎖タンパク質からなるか、または多鎖ポリペプチドとして存在することができる。2以上のFc融合タンパク質を、それらがFc領域間に自然に形成されるジスルフィド結合を介して相互作用するような形で、製造することができる。これらの多量体はGLP-1化合物に関して均一であってもよいし、融合タンパク質のFc部分のN末端に異なるGLP-1化合物が融合されていてもよい。
【0080】
融合タンパク質の最終構造がどうであれ、FcまたはFc様領域は、そのN末端に融合されたGLP-1化合物の生体内血漿半減期を延長する役割を果たさなければならない。また、融合されたGLP-1化合物は、なんらかの生物学的活性を保っていなければならない。半減期の増加は、融合タンパク質の半減期をGLP-1化合物単独の半減期と比較する実施例7に記載の方法を使って証明することができる。生物学的活性は当技術分野で知られている試験管内法および生体内法によって決定することができる。代表的な生物学的アッセイを実施例6、8、および9に記載する。
【0081】
タンパク質分解によって製造されるIgGのFc領域は、無傷のIgG分子と同じ生体内半減期を持ち、Fab断片は迅速に分解されるので、半減期の延長に関係する配列はCH2および/またはCH3ドメイン中に存在すると考えられる。また、高親和性Fc受容体もC1qも結合しないIgG変異体の異化速度は、親野生型抗体のクリアランス速度と識別できないことが、文献で明らかにされているが、これは、異化部位がFc受容体結合またはC1q結合に関与する部位とは異なることを示している[Wawrzynczakら, (1992) Molecular Immunology 29:221]。ネズミIgG1 Fc領域を使った部位特異的突然変異誘発研究により、IgG1 Fc領域のうち異化速度を制御する部位は、CH2-CH3ドメイン境界に位置することが示唆された。
【0082】
これらの研究に基づいて、融合タンパク質の半減期が最適化されるように、Fc領域の異化部位を改変することができる。本発明の異種融合タンパク質に使用するFc領域は、IgG1またはIgG4 Fc領域に由来することが好ましい。Fc領域はIgG4であるかまたはIgG4に由来することが、さらに好ましい。IgG Fc領域は、好ましくは、ヒンジ領域を含めてCH2領域とCH3領域の両方を含む。
【0083】
異種アルブミン融合タンパク質:
上述のGLP-1化合物はアルブミンまたはその類似体、断片、もしくは誘導体に直接融合するか、またはペプチドリンカーを介して融合することができる。
【0084】
本発明の融合タンパク質の一部を構成するアルブミンタンパク質は、一般的には、どの種からクローン化されたアルブミンに由来してもよい。しかし、融合タンパク質がヒトで免疫原性を示す危険を低下させるには、ヒトアルブミンならびにその断片および類似体が好ましい。ヒト血清アルブミン(HSA)は、66,500の分子式量を持つ585アミノ酸の単一非グリコシル化ポリペプチド鎖からなる。ヒトHSAのアミノ酸配列を図2に示す[Melounら (1975) FEBS Letters 58:136;Behrensら (1975) Fed. Proc. 34:591;Lawnら (1981) Nucleic Acids Research 9:6102-6114;Minghettiら (1986) J. Biol. Chem. 261:6747参照]。アルブミンには、様々な多型変異体ならびに類似体および断片が記載されている[Weitkampら, (1973) Ann. Hum. Genet. 37:219参照]。例えばEP322,094には、種々の短縮型HSAが開示されている。これらの断片には、例えばHSA(1-373)、HSA(1-388)、HSA(1-389)、HSA(1-369)、およびHSA(1-419)、ならびに1-369と1-419の間の断片が含まれる。EP399,666には、例えばHSA(1-177)およびHSA(1-200)ならびにHSA(1-177)とHSA(1-200)の間の断片を含むアルブミン断片が開示されている。
【0085】
本発明の異種融合タンパク質は、当該融合タンパク質が生物学的に活性であり、かつGLP-1化合物単独よりも長い血漿半減期を持つような、任意のアルブミンタンパク質(断片、類似体、および誘導体を含む)にカップリングされたGLP-1化合物を包含すると理解される。したがって、融合タンパク質のアルブミン部分が天然ヒトアルブミンに匹敵する血漿半減期を持つ必要は、必ずしもない。長い半減期を持つ断片、類似体、および誘導体、または天然ヒトアルブミンと対象GLP-1化合物との中間の半減期を持つ断片、類似体、および誘導体は、知られているか、または作製することができる。
【0086】
本発明の異種融合タンパク質には、融合タンパク質のGLP-1化合物部分および/またはFcもしくはアルブミン部分に保存的アミノ酸置換を持つタンパク質が包含される。「保存的置換」は、同じ正味の電荷を持ち、ほぼ同じサイズおよび形状を持つ別のアミノ酸による、あるアミノ酸の置換である。脂肪族アミノ酸側鎖または置換脂肪族アミノ酸側鎖を持つアミノ酸は、その側鎖中の炭素およびヘテロ原子の総数の差がだいたい4以下であれば、ほぼ同じサイズを持つ。またこれらは、その側鎖中の分枝数の差が1以下であれば、ほぼ同じ形状を持つ。側鎖中にフェニル基または置換フェニル基を持つアミノ酸はほぼ同じサイズおよび形状を持つとみなされる。特に別段の条件を記す場合を除いて、保存的置換は、好ましくは、天然アミノ酸を使ってなされる。
【0087】
しかし「アミノ酸」という用語は、ここでは広い意味で使用され、天然のアミノ酸だけでなく、アミノ酸類似体および誘導体を含む非天然アミノ酸を包含する。後者にはアミノ酸部分を含む分子が包含される。この広い定義からみて、本明細書にいうアミノ酸に、例えば天然の蛋白原性L-アミノ酸;D-アミノ酸;アミノ酸類似体および誘導体などの化学修飾アミノ酸;ノルロイシン、β-アラニン、オルニチン、GABAなどの天然の非蛋白原性アミノ酸;およびアミノ酸に特有であることが当技術分野で知られている性質を持つ化学合成化合物などが含まれることは、当業者には理解されるだろう。本明細書で使用する「蛋白原性」という用語は、そのアミノ酸が細胞内で代謝経路によりペプチド、ポリペプチド、またはタンパク質に組み込まれうることを示す。
【0088】
合成非天然アミノ酸、置換アミノ酸、または1もしくは複数のD-アミノ酸を含む非天然アミノ酸を、本発明の異種融合タンパク質に組み込むと、いくつかの点で有利になる場合がある。D-アミノ酸含有ペプチド等は、対応するL-アミノ酸含有ペプチドと比較して、試験管内または生体内で、増加した安定性を示す。したがって、D-アミノ酸を組み込んだペプチド等の構築は、細胞内安定性の向上が望ましい時または必要な時には、特に有用な場合がある。より具体的に述べると、D-ペプチド等は、必要に応じて、内因性ペプチダーゼおよびプロテアーゼに対して耐性を示すことより、その分子の生物学的利用能を向上させ、生体内での寿命を引き延ばす。また、D-ペプチド等は、Tヘルパー細胞に対して主要組織適合複合体クラスII拘束性の提示を行なうためのプロセシングを効率よく受けることができず、したがって丸ごとの生物における体液性免疫反応を誘導する可能性が少ない。
【0089】
置換用のアミノ酸を選択する際には、本発明に包含される様々なポリペプチドの構造/機能解析の他にも、考慮することができる因子が数多くある。そのような改変を行なう際に考慮することができる一つの因子は、アミノ酸のハイドロパシー指標である。タンパク質に相互作用的な生物学的機能を付与する際のハイドロパシーアミノ酸指標の重要性は、KyteおよびDoolittle(1982, J. Mol. Biol., 157:105-132)によって論じられている。アミノ酸の相対的ハイドロパシー特性が、結果として得られるタンパク質の二次構造に寄与することは、一般に認められている。これは、結果として、タンパク質と、例えば酵素、基質、受容体、リガンド、DNA、抗体、抗原などの分子との相互作用に影響する。各アミノ酸には、その疎水性および電荷特徴に基づいて、ハイドロパシー指標が以下のように割り当てられている:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(-0.4);スレオニン(-0.7);セリン(-0.8);トリプトファン(-0.9);チロシン(-1.3);プロリン(-1.6);ヒスチジン(-3.2);グルタミン酸/グルタミン/アスパラギン酸/アスパラギン(-3.5);リジン(-3.9);およびアルギニン(-4.5)。
【0090】
当技術分野では知られているように、特定のアミノ酸を、類似するハイドロパシー指標またはハイドロパシースコアを持つ他のアミノ酸の代わりに、ペプチド、ポリペプチド、またはタンパク質中に使用して、結果として、類似する生物学的活性を持つか、または改善された生物学的活性を持つペプチド等を、製造することができる。そのような改変を行なう際には、±2以内のハイドロパシー指標を持つアミノ酸同士を置換することが好ましい。より好ましい置換は、アミノ酸が±1以内のハイドロパシー指標を持つような置換である。最も好ましい置換は、アミノ酸が±0.5以内のハイドロパシー指標を持つような置換である。
【0091】
類似するアミノ酸は親水性に基づいて置換することもできる。米国特許第4,554,101号には、隣接アミノ酸の親水性によって支配される、タンパク質の最大局所平均親水性は、そのタンパク質の生物学的性質と相関関係にあることが開示されている。アミノ酸には以下の親水性値が割り当てられている:アルギニン/リジン(+3.0);アスパラギン酸/グルタミン酸(+3.0±1);セリン(+0.3);アスパラギン/グルタミン(+0.2);グリシン(0);スレオニン(-0.4);プロリン(-0.5±1);アラニン/ヒスチジン(-0.5);システイン(-1.0);メチオニン(-1.3);バリン(-1.5);ロイシン/イソロイシン(-1.8);チロシン(-2.3);フェニルアラニン(-2.5);およびトリプトファン(-3.4)。したがって、ペプチド、ポリペプチド、またはタンパク質中のあるアミノ酸を、類似する親水性スコアを持つ別のアミノ酸で置換して、結果として、類似する生物学的活性を持っている、すなわち正しい生物学的機能をまだ保っているペプチド等を、製造することができる。そのような改変を行なう際には、好ましくは、±2以内のハイドロパシー指標を持つアミノ酸同士を置換し、±1以内のアミノ酸同士はより好ましく、±0.5以内のアミノ酸同士は最も好ましい。
【0092】
上に略述したように、本発明の融合タンパク質中のアミノ酸置換は、アミノ酸側鎖置換基の相対的類似性、例えばそれらの疎水性、親水性、電荷、サイズなどに基づいて行なうことができる。また、二次構造傾向に基づいて置換を行なうこともできる。例えば、らせんアミノ酸は、そのらせん構造を保つであろうアミノ酸で置換することができる。本ペプチド等にサイレントな変化をもたらす保存的アミノ酸変異になるように上述の様々な特徴を考慮した代表的置換は、当該天然アミノ酸が属するクラスの他の構成要素から選択することができる。アミノ酸は以下の4群に分類することができる:(1)酸性アミノ酸、(2)塩基性アミノ酸、(3)中性極性アミノ酸、および(4)中性無極性アミノ酸。
【0093】
本発明の異種融合タンパク質を製造するための一般的方法
本発明の異種融合タンパク質は様々な方法で製造することができるが、なかでも組換え法は好ましい。ここに開示し特許請求する本発明のために、下記の一般的な分子生物学用語および略号を、以下に定義する。本願で使用する用語および略号は、別段の指定がない限り、それぞれ通常の意味を持つ。例えば「℃」は摂氏度を表し、「mmol」はミリモルを表す。「mg」はミリグラムを表し、「μg」はマイクログラムを表す。「mlまたはmL」はミリリットルを表し、「μlまたはμL」はマイクロリットルを表す。アミノ酸略号は37 C.F.R.§1.822(b)(2)(1994)に記載されているとおりである。
【0094】
本明細書で使用する「塩基対」または「bp」はDNAまたはRNAを表す。略号A、C、G、およびTは、DNA分子においては、それぞれデオキシリボヌクレオシドである(デオキシ)アデノシン、(デオキシ)シチジン、(デオキシ)グアノシン、およびチミジンの5'-一リン酸型に対応する。略号U、C、G、およびAは、RNA分子においては、リボヌクレオシドであるウリジン、シチジン、グアノシン、およびアデノシンの5'-一リン酸型に対応する。二本鎖DNAの場合、塩基対はAとTまたはCとGの組合せを表しうる。DNA/RNAの場合、ヘテロ二重鎖塩基対はAとUまたはCとGの組合せを表しうる(下記「相補性」の定義を参照されたい)。
【0095】
DNAの「消化」または「制限(消化)」は、DNA中の特定配列だけに作用する制限酵素(「配列特異的エンドヌクレアーゼ」)によるDNAの触媒的切断を表す。本明細書で使用した種々の制限酵素は市販されており、その反応条件、補因子、および他の必要条件は、当業者に知られているであろうとおりに使用した。特定の制限酵素に適した緩衝液および基質量は、製造者によって指定されるか、または文献中に容易に見いだすことができる。
【0096】
「ライゲーション(連結)」は、2つの二本鎖核酸断片間にリン酸ジエステル結合を形成させる過程を表す。別段の定めがない限り、ライゲーションは、既知の緩衝液および条件により、T4 DNAリガーゼなどのDNAリガーゼを使って達成することができる。
【0097】
「プラスミド」は、染色体外にある(通常は)自己複製性の遺伝要素を表す。プラスミドは一般に小文字「p」と、それに続く文字および/または数字によって指定される。本明細書での出発プラスミドは市販されているか、公に無制限に入手することができるか、または公表された手順に従って、入手可能なプラスミドから構築することができる。また、本明細書に記載のプラスミドと等価なプラスミドが当技術分野では知られており、それらは当業者には明らかだろう。
【0098】
本明細書にいう「組換えDNAクローニングベクター」は、1つもしくは複数の追加DNAセグメントを付加することができる、またはそのようなDNAセグメントが既に付加されている、自律的に複製する任意の物質(例えばプラスミドおよびファージ)を表す。
【0099】
本明細書にいう「組換えDNA発現ベクター」は、挿入されたDNAの転写を制御するプロモーターが組み込まれている任意の組換えDNAクローニングベクターを表す。
【0100】
「転写」は、DNAのヌクレオチド配列に含まれる情報が相補的RNA配列に移される過程を表す。
【0101】
「トランスフェクション」は、宿主細胞による発現ベクターの取り込みを表し、何らかのコード配列が実際に発現されるかどうかを問わない。当業者には、例えばリン酸カルシウム共沈法、リポソームトランスフェクション、およびエレクトロポレーションなど、数多くのトランスフェクション法が知られている。一般にトランスフェクションの成功は、そのベクターの作動を示す何らかのしるしがその宿主細胞内で起こる場合に認識される。
【0102】
「形質転換」は、当該DNAが染色体外要素として、または染色体組込みによって、複製することができるように、DNAをある生物に導入することをいう。細菌および真核細胞宿主を形質転換する方法は当技術分野ではよく知られており、例えば核注入法、プロトプラスト融合、または塩化カルシウムを使ったカルシウム処理による方法など、それらの方法の多くは、J. Sambrookら「Molecular Cloning: A Laboratory Manual」(1989)に要約されている。一般に、DNAを酵母に導入する場合には、形質転換という用語は、トランスフェクションという用語と相対する用語として使用される。
【0103】
本明細書にいう「翻訳」は、メッセンジャーRNA(mRNA)の遺伝情報を使ってポリペプチド鎖を指定し、その合成を指図する過程を表す。
【0104】
「ベクター」は、遺伝子操作において細胞のトランスフェクションおよび/または形質転換に使用される核酸化合物であって、適当な制御配列と組み合わせた場合にトランスフェクションおよび/または形質転換される宿主細胞に特定の性質を付与する適当なタンパク質分子に対応するポリヌクレオチド配列を持つ核酸化合物を表す。プラスミド、ウイルス、およびバクテリオファージは適切なベクターである。人工ベクターは、異なる供給源に由来するDNA分子を制限酵素およびリガーゼで切断し、つなぎ合わせることによって構築される。本明細書で使用する「ベクター」という用語は、組換えDNAクローニングベクターおよび組換えDNA発現ベクターを包含する。
【0105】
本明細書にいう「相補的」または「相補性」は、二本鎖核酸中で水素結合によって会合する塩基(プリンおよびピリミジン)の対を表す。以下の塩基対は相補的である:グアニンとシトシン;アデニンとチミン;およびアデニンとウラシル。
【0106】
本明細書にいう「ハイブリダイゼーション」は、核酸の鎖が相補的な鎖と塩基対形成によって結合する過程を表す。同一ではないが極めて類似している2本の相補的核酸のハイブリダイゼーションに使用される条件は、それら2本の鎖の相補性の程度および鎖の長さによって相違する。そのような技術および条件は当業者には周知である。
【0107】
「単離されたアミノ酸配列」は、どんな方法で構築または合成されるにせよ、天然の配列とは異なる位置にある任意のアミノ酸を表す。
【0108】
「単離されたDNA化合物」は、どんな方法で構築または合成されるにせよ、ゲノムDNA中でのそれ本来の位置とは異なる位置にある任意のDNA配列を表す。
【0109】
「単離された核酸化合物」は、どんな方法で構築または合成されるにせよ、それ本来の位置とは異なる位置にある任意のRNAまたはDNA配列を表す。
【0110】
「プライマー」は、酵素的または合成的伸長の開始基質として機能する核酸断片を表す。
【0111】
「プロモーター」は、DNAまたはRNAの転写を指図するDNA配列を表す。
【0112】
「プローブ」は、別の核酸化合物とハイブリダイズする核酸化合物またはその断片を表す。
【0113】
当業者は、ハイブリダイゼーション反応の「ストリンジェンシー」を容易に決定することができ、ハイブリダイゼーション反応の「ストリンジェンシー」は一般的には、プローブ長、洗浄温度、および塩濃度に依存する実験的計算の結果である。一般に、長いプローブほど適正なアニーリングに高い温度を必要とし、短いプローブは必要とする温度が低い。ハイブリダイゼーションは、一般に、相補的な鎖がそれらの融解温度未満の環境に存在する時の、変性したDNAの再アニール能力に依存する。プローブとハイブリダイズ可能な配列との間に所望する相同性の程度が高いほど、使用することができる相対温度は高くなる。そのため、高い相対温度ほど、反応を、よりストリンジェントにする傾向を持ち、低い温度ほどその傾向が少ないということになる。ハイブリダイゼーション反応のストリンジェンシーに関するさらなる詳細と説明については、Ausubelら「Current Protocols in Molecular Biology」(Wiley Interscience Publishers, 1995)を参照されたい。
【0114】
本明細書で定義する「ストリンジェントな条件」または「高ストリンジェンシー条件」は、(1)洗浄に低イオン強度および高温を使用する条件、例えば15mM塩化ナトリウム/1.5mMクエン酸ナトリウム/0.1%ドデシル硫酸ナトリウムを50℃で使用する条件、(2)ハイブリダイゼーション時にホルムアミドなどの変性剤を使用する条件、例えば50%(v/v)ホルムアミドおよび0.1%ウシ血清アルブミン/0.1%フィコール/0.1%ポリビニルピロリドン/50mMリン酸ナトリウム緩衝液(pH6.5)および750mM塩化ナトリウム/75mMクエン酸ナトリウムを42℃で使用する条件、または(3)50%ホルムアミド、5×SSC(750mM塩化ナトリウム、75mMクエン酸ナトリウム)、50mMリン酸ナトリウム(pH6.8)、0.1%ピロリン酸ナトリウム、5×デンハート液、超音波処理サケ精子DNA(50μg/ml)、0.1%SDS、および10%硫酸デキストランを42℃で使用し、0.2×SSC(30mM塩化ナトリウム/3mMクエン酸ナトリウム)および50%ホルムアミド中、42℃および55℃で洗浄した後、EDTAを含む0.1×SSCからなる高ストリンジェンシー洗浄を55℃で行なう条件で識別しうる。
【0115】
「中等度にストリンジェントな条件」は、Sambrookら「Molecular Cloning: A Laboratory Manual」(ニューヨーク, Cold Spring Harbor Press, (1989))に記載されているように識別することができ、上述した条件よりストリンジェンシーの低い洗浄溶液およびハイブリダイゼーション条件(例えば温度、イオン強度、およびSDSのパーセント)の使用が含まれる。中等度にストリンジェントな条件の一例は、20%ホルムアミド、5×SSC(750mM塩化ナトリウム、75mMクエン酸ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20mg/mL変性剪断サケ精子DNAを含む溶液中、37℃で終夜インキュベートした後、フィルターを1×SSC中、約37〜50℃で洗浄することである。プローブ長などの因子を考慮して必要に応じて温度、イオン強度などを調節する方法は、当業者にはわかるだろう。
【0116】
「PCR」は、熱安定性DNAポリメラーゼを利用する周知のポリメラーゼ連鎖反応を表す。
【0117】
「リーダー配列」は、目的とする所望のポリペプチドを生成させるために、酵素的または化学的に除去することができるアミノ酸の配列を表す。
【0118】
「分泌シグナル配列」は、より大きいポリペプチドのN末端領域に一般に存在し、そのポリペプチドと小胞体などの細胞膜コンパートメントションとの会合および形質膜を通したそのポリペプチドの分泌を開始させる機能を果たす、アミノ酸の配列を表す。
【0119】
本発明の異種融合タンパク質をコードするDNAの構築
野生型アルブミンおよび免疫グロブリンタンパク質は様々な供給源から入手することができる。例えばこれらのタンパク質は、目的とするmRNAを検出可能なレベルに発現させる組織または細胞から作製されたcDNAライブラリーから得ることができる。目的とするタンパク質の公表されたDNA配列またはタンパク質配列を使って設計されたプローブで、上記ライブラリーをスクリーニングすることができる。例えば免疫グロブリン軽鎖または重鎖定常領域は、Adamsら (1980) Biochemistry 19:2711-2719;Goughetら (1980) Biochemistry 19:2702-2710;Dolbyら (1980) Proc. Natl. Acad. Sci. USA 77:6027-6031;Riceら (1982) Proc. Natl. Acad. Sci. USA 79:7862-7862;Falknerら (1982) Nature 298:286-288;およびMorrisonら (1984) Ann. Rev. Immunol. 2:239-256に記載されている。アルブミンのタンパク質配列およびDNA配列を開示している文献には、例えばMelounら (1975) FEBS Letters 58:136;Behrensら (1975) Fed. Proc. 34:591;Lawnら (1981) Nucleic Acids Research 9:6102-6114;およびMinghettiら (1986) J. Biol. Chem. 261:6747などがある。
【0120】
選択したプローブによるcDNAライブラリーまたはゲノムライブラリーのスクリーニングは、Sambrookら「Molecular Cloning: A Laboratory Manual」(Cold Spring Harbor Laboratory Press, ニューヨーク (1989))に記載されているような標準的方法を使って行なうことができる。アルブミンまたは免疫グロブリンタンパク質をコードする遺伝子を単離するもう一つの手段は、PCR法を使用することである[Sambrookら, 前掲;Dieffenbachら「PCR Primer: A Laboratory Manual」Cold Spring Harbor Laboratory Press, ニューヨーク (1995)]。PCRプライマーは公表された配列に基づいて設計することができる。
【0121】
一般に、特定の種からクローン化された完全長野生型配列は、融合タンパク質の一部であるGLP-1化合物の血漿半減期を延ばす能力を保っている類似体、断片、および誘導体を作製するためのテンプレートとして役立ちうる。融合タンパク質がヒトで潜在的免疫原性を示す危険を低下させるために、本発明の異種融合タンパク質のFc部分およびアルブミン部分は、天然ヒト配列に由来することが好ましい。
【0122】
特に、本発明に包含される融合タンパク質の免疫グロブリン部分は、免疫グロブリンのFc断片だけを含むことが好ましい。特定のエフェクター機能を所望するかどうかと、融合タンパク質の構造的特徴とに応じて、Fc部分は、CH2およびCH3ドメインに加えてヒンジ領域を含むことができ、またはそれらの他の何らかの組合せを含むことができる。これらのFc断片は、その断片の所望する末端に対応する配列にハイブリダイズするように設計されたプライマーにより、PCR技術を使って作製することができる。同様に、アルブミンの断片が望ましい場合は、アルブミン内部配列に相補的なPCRプライマーを設計することができる。発現ベクターへのクローニングを容易にするための制限酵素部位が生成するようにPCRプライマーを設計することもできる。
【0123】
本発明のGLP-1化合物をコードするDNAは、上述した方法のようなクローニング法および化学合成DNAを含む多種多様な方法で作製することができる。コードされるペプチドの長さが短いことを考えると、化学合成は魅力的な方法であるだろう。GLP-1のアミノ酸配列は、プレプログルカゴン遺伝子の配列と共に、公表されている[Lopezら (1983) Proc. Natl. Acad. Sci., USA 80:5485-5489;Bellら (1983) Nature, 302:716-718;Heinrich, G.ら (1984) Endocrinol, 115:2176-2181;Ghiglione, M.ら (1984) Diabetologia 27:599-600]。したがって、天然GLP-1化合物およびその断片をPCRするためのプライマーを設計することができる。
【0124】
次に、GLP-1化合物をコードするDNAを、アルブミンまたはFcタンパク質をコードするDNAにインフレームで連結することにより、融合タンパク質をコードする遺伝子を構築することができる。GLP-1化合物をコードする遺伝子と、アルブミンまたはFcタンパク質をコードする遺伝子は、リンカーペプチドをコードするDNAを介してインフレームでつなぎ合わせることもできる。
【0125】
本発明の異種融合タンパク質の生体内での機能および安定性は、小さいペプチドリンカーを加えて、潜在的に望ましくないドメイン相互作用を防ぐことにより、最適化することができる。これらのリンカーは潜在的には任意の長さを持ち、アミノ酸の任意の組合せからなりうるが、その長さは、望ましくないドメイン相互作用を防ぎそして/または生物学的活性および/または安定性を最適化するのに必要な長さを超えないことが好ましい。一般に、リンカーは、極端にかさ高い側鎖を持つアミノ酸や、有意な二次構造を導入しそうなアミノ酸を含まないべきである。リンカーは、セリン-グリシンリッチであると共に、長さは30アミノ酸未満であることが好ましい。リンカーの長さは20アミノ酸を超えないことがより好ましい。リンカーの長さは15アミノ酸を超えないことがさらに好ましい。好ましいリンカーは、Gly-Gly-Gly-Gly-Serという配列の反復を含む。この配列は2〜6回反復することが好ましい。この配列は3〜4回反復することがさらに好ましい。
【0126】
野生型GLP-1、アルブミン、およびFcポリペプチドをコードするDNAならびにその断片は、ライゲーションに先立って、または融合タンパク質全体をコードするcDNAの形で、突然変異させることができる。当技術分野では様々な突然変異導入技術がよく知られている。例えば突然変異導入PCR法では、対応するタンパク質中の特定のアミノ酸配列を変化させる目的で、鎖オーバーラップ伸長を利用して、特定の塩基突然変異を生成する。このPCR突然変異導入法では4つのプライマー、すなわちフォワード方向の2つ(プライマーAおよびC)とリバース方向の2つ(プライマーBおよびD)を使用する必要がある。突然変異した遺伝子は、2つの異なる段階で野生型テンプレートから増幅される。第1反応では、AからBへの反応とCからDへの反応とを個別に行なって、遺伝子を半分ずつ増幅する。ここに、BおよびCプライマーは、突然変異させようとする遺伝子の領域を標的としている。これらのプライマーを標的領域と整列させた場合、それらは、変化させようとしている塩基に関してミスマッチを持つ。AからBまでおよびCからDまでの反応が完了したら、反応生成物を単離し、それらを混合して、AからDまでの反応のテンプレートとして使用する。そうすると、この反応により、完全な突然変異型産物が得られる。
【0127】
融合タンパク質全体をコードする遺伝子を作製したら、それを適当な発現ベクターにクローニングすることができる。本発明のGLP-1融合タンパク質を製造するために使用することができる具体的戦略を実施例1に説明する。
【0128】
本発明の異種融合タンパク質を組換え発現させるための一般的方法
本明細書に記載の異種融合タンパク質産生用発現ベクターまたはクローニングベクターで宿主細胞をトランスフェクトまたは形質転換し、プロモーターを誘導するため、形質転換体を選択するため、または所望の配列をコードする遺伝子を増幅するために適宜改良された通常の栄養培地で培養する。培地、温度、pHなどの培養条件は、当業者であれば、はなはだしい実験を行なわなくても選択することができる。細胞培養の生産性を最大にするための原理、手順、および実用的技術は、一般に、M. Butler編「Mammalian Cell Biotechnology: A Practical Approach」(IRL Press, 1991)およびSambrookら(前掲)に記載されている。トランスフェクションの方法は、例えばCaPO4およびエレクトロポレーションなど、当業者に知られている。哺乳類細胞宿主系形質転換の一般的側面は、米国特許第4,399,216号に記載されている。酵母への形質転換は通例、van Solingenら, J Bact. 130(2): 946-7(1977)およびHsiaoら, Proc. Natl. Acad. Sci. USA 76(8):3829-33(1979)の方法に従って行なわれる。しかし、細胞にDNAを導入する他の方法、例えば核マイクロインジェクション、エレクトロポレーション、無傷細胞との細菌プロトプラスト融合、またはポリブレンもしくはポリオミチン(polyomithine)などのポリカチオンも、使用することができる。哺乳類細胞を形質転換するための様々な技術については、Keownら, Methods in Enzymology 185: 527-37(1990)およびMansourら, Nature 336(6197): 348-52(1988)を参照されたい。
【0129】
本明細書に記載するベクター中の核酸(例えばDNA)をクローニングまたは発現させるのに適した宿主細胞には、原核生物、酵母、または高等真核細胞が含まれる。適切な原核生物には、グラム陰性生物またはグラム陽性生物などの真正細菌、例えば大腸菌などの腸内細菌が含まれるが、これらに限るわけではない。例えば大腸菌K12株MM294(ATCC 3 1.446)、大腸菌X1 776(ATCC 3 1.537)、大腸菌W3 110株(ATCC 27.325)およびK5 772株(ATCC 53.635)など、様々な大腸菌株を公に入手することができる。他の適切な原核宿主細胞には、エシェリキア(例えば大腸菌)、エンテロバクター、エルウィニア、クレブシエラ、プロテウス、サルモネラ(例えばネズミチフス菌)、セラチア(例えば霊菌)、および赤痢菌などの腸内細菌、ならびに枯草菌やバチルス・リケニフォルミス(例えば1989年4月12日に公開されたDD266,710に開示されているバチルス・リケニフォルミス 41P)、緑膿菌などのシュードモナス、およびストレプトミセスなどが含まれる。これらの例は限定ではなく説明のための実例である。W3110株は、組換えDNA産物醗酵用の一般的な宿主株であるので、特に好ましい宿主または親宿主の一つである。宿主細胞が分泌するタンパク質加水分解酵素は極微量であることが好ましい。例えばW3110株を改変して、その宿主に内在するタンパク質をコードする遺伝子に、遺伝的突然変異をもたらしてもよい。そのような宿主の例には、完全遺伝子型ronAを持つ大腸菌W3110 1A2株、完全遺伝子型tonA ptr3を持つ大腸菌W3110 9E4株、完全遺伝子型tonA, ptr3 phoA E15 (argF-lac) I69 degP ompT /can'を持つ大腸菌W3110 27C7株(ATCC 55,244)、非カナマイシン耐性degP欠失突然変異を持つ37D6株である大腸菌W3110 40B4株、および1990年8月7日発行の米国特許第4,946,783号に開示されている突然変異型ぺリプラスムプロテアーゼを持つ大腸菌株がある。あるいは、生体内クローニング法、例えばPCRまたは他の核酸ポリメラーゼ反応なども適切である。
【0130】
原核生物に加えて、糸状菌または酵母などの真核微生物も、融合タンパク質ベクターにとって適切なクローニング宿主または発現宿主である。サッカロミセス・セレビシェは広く使用されている下等真核宿主微生物である。他にも、シゾサッカロミセス・ポンベ[BeachおよびNurse, Nature 290:140-3 (1981)、1995年5月2日公開のEP 139,383)、クルイベロミセス宿主[米国特許第4,943,529号、Fleerら,Bio/Technology 9(10):968-75 (1991)]、例えばクルイベロミセス・ラクチス(MW98-8C、CBS683、CBS4574)[de Louvencourtら, J. Bacteriol., 154(2):737-42 (1983)]、クルイベロミセス・フラギリス(ATCC 12,424)、クルイベロミセス・ブルガリカス(ATCC 16,045)、クルイベロミセス・ウィッカーラミイ(ATCC 24,178)、クルイベロミセス・ワルティイ(ATCC 56,500)、クルイベロミセス・ドロソフィラルム(ATCC 36,906)[Van den Bergら, Bio/Technology 8(2):135-9 (1990)]、クルイベロミセス・サーモトレランス、およびクルイベロミセス・マルキシアヌス、ヤロウィア(EP 402,226)、ピキア・パストリス(EP 183,070)[Sreekrishnaら, J. Basic Microbiol. 28(4):265-78 (1988)]、カンジダ、トリコデルマ・リーシア(EP 244,234)、ニューロスポラ・クラッサ[Caseら, Proc. Natl. Acad. Sci. USA 76(10):5259-5263 (1979)]、シュワンニオミセス、例えばシュワンニオミセス・オシデンツリス(1990年10月31日公開のEP 394,538)、例えばニューロスポラ、ペニシリウム、トリポクラジウム(1991年1月10日公開のWO 91/00357)、およびアスペルギルス宿主(例えばアスペルギルス・ニデュランス[Ballanceら, Biochem. Biophys. Res. Comm. 112(1):284-9 (1983)、Tilburnら, Gene 26(2-3):205-21 (1983)、Yeltonら, Proc. Natl. Acad. Sci. USA 81(5):1470-4 (1984)]およびアスペルギルス・ニガー[KellyおよびHynes, EMBO J. 4(2):475-9 (1985)])などの糸状菌などがある。メチロトローフ酵母は、ハンゼヌラ、カンジダ、クロエケラ、ピキア、サッカロミセス、トルロプシス、およびロドトルイアからなる属群から選択される。この種類の酵母の具体的な種の一覧は、C. Antony「The Biochemistry of Methylotrophs」269(1982)に記載されている。
【0131】
本発明の融合タンパク質の発現に適した宿主細胞は多細胞生物から得られる。無脊椎動物細胞の例には、ドロソフィラS2およびスポドプテラSp、スポドプテラhigh5などの昆虫細胞ならびに植物細胞が含まれる。有用な哺乳類宿主細胞株の例には、チャイニーズハムスター卵巣(CHO)細胞およびCOS細胞が含まれる。より具体的な例としては、SV40で形質転換されたサル腎臓CV1株(COS-7、ATCC CRL 1651)、ヒト胎児腎臓株[293細胞または懸濁培養で成長するようにサブクローニングされた293細胞;Grahamら, J. Gen. Virol. 36(1):59 (1977)]、チャイニーズハムスター卵巣細胞/-DHFR[CHO、UrlaubおよびChasin, Proc. Natl. Acad. Sci. USA 77(7):4216-20 (1980)]、マウスセルトリ細胞[TM4, Mather, Biol. Reprod. 23(1):243-52 (1980)]、ヒト肺細胞(W138, ATCC CCL 75)、ヒト肝臓細胞(Hep G2, HB 8065)、およびマウス乳癌細胞(MMT 060562, ATCC CCL51)などが挙げられる。適切な宿主細胞の選択は、技術上可能であると考えられる。
【0132】
本発明の融合タンパク質は直接的に組換え生産するか、または成熟融合タンパク質のN末端に特異的切断部位をもたらすシグナル配列または他の追加配列を持つタンパク質として組換え生産することができる。一般に、シグナル配列はベクターの一成分であってもよいし、ベクターに挿入される融合タンパク質コードDNAの一部であってもよい。シグナル配列は、例えばアルカリホスファターゼ、ペニシリナーゼ、lpp、または耐熱性エンテロトキシンIIリーダーの群から選択される原核シグナル配列とすることができる。酵母分泌の場合は、シグナル配列を、例えば酵母インベルターゼリーダー、アルファ因子リーダー(例えばサッカロミセスおよびクルイベロミセスのa因子リーダー;後者は米国特許第5,010,182号に記述されている)、または酸性ホスファターゼリーダー、カンジダ・アルビカンス・グルコアミラーゼリーダー(EP 362,179)またはWO 90/13646に記載のシグナルなどにすることができる。哺乳類細胞発現の場合は、同じ種または近縁種の分泌ポリペプチドに由来する哺乳類シグナル配列ならびにウイルス分泌リーダーを使って、タンパク質の分泌を指示することができる。
【0133】
発現ベクターおよびクローニングベクターはどちらも、1つまたは複数の選択した宿主細胞内でそのベクターの複製を可能にする核酸配列を含有する。そのような配列は、種々の細菌、酵母およびウイルスについてよく知られている。プラスミドpBR322由来の複製起点はほとんどのグラム陰性菌に適し、2μプラスミド起点は酵母に適し、種々のウイルス起点(SV40、ポリオーマ、アデノウイルス、VSVまたはBPV)は哺乳類細胞におけるベクターのクローニングに役立つ。
【0134】
発現ベクターおよびクローニングベクターは典型的には選択遺伝子(選択可能マーカーともいう)を含有する。典型的な選択遺伝子は、(a)抗生物質または他の毒素、例えばアンピシリン、ネオマイシン、メトトレキセート、またはテトラサイクリンなどに対する耐性を付与するタンパク質をコードするか、(b)独立栄養不全を補うタンパク質をコードするか、または(c)天然培地からは利用できない必須栄養素を供給するタンパク質をコードする(例えばバチルスの場合、D-アラニンラセマーゼをコードする遺伝子など)。
【0135】
適切な哺乳類細胞用選択マーカーの一例は、融合タンパク質コード核酸を取り込む能力を持つ細胞の同定を可能にするもの、例えばDHFRまたはチミジンキナーゼなどである。野生型DHFRを使用する場合に適切な宿主細胞は、UrlaubおよびChasin, Proc. Natl. Acad. Sci. USA, 77(7):4216-20(1980)に記述されているように作出し増殖されるDHFR活性欠損CHO細胞株である。酵母での使用に適した選択遺伝子は、酵母プラスミドYRp7中に存在するtrp1遺伝子である[Stinchcombら, Nature 282(5734):39-43 (1979)、Kingsmanら, Gene 7(2):141-52 (1979)、またはTschemperら, Gene 10(2):157-66 (1980)]。trp1遺伝子は、トリプトファン中で増殖する能力を欠く酵母の突然変異株(例えばATCC番号44076またはPEPC1[Jones, Genetics 85:23-33 (1977)]用の選択マーカーになる。
【0136】
発現ベクターおよびクローニングベクターは、通常、mRNA合成を指示するために融合タンパク質コード核酸配列に作動可能に連結されたプロモーターを含有する。種々の潜在的宿主細胞によって認識されるプロモーターはよく知られている。原核宿主での使用に適したプロモーターには、β-ラクタマーゼおよびラクトースプロモーター系[Changら, Nature 275(5681):617-24 (1978)、Goeddelら, Nature 281(5732):544-8 (1979)]、アルカリホスファターゼ、トリプトファン(up)プロモーター系[Goeddel, Nucleic Acids Res. 8(18):4057-74 (1980)、1981年9月30日公開のEP 36,776)、およびtatプロモーターなどのハイブリッドプロモーター[deBoerら, Proc. Natl. Acad. Sci. USA 80(1):21-25 (1983)]などがある。細菌系用のプロモーターは、融合タンパク質をコードするDNAに作動可能に連結されたシャイン・ダルガノ(S.D.)配列も含有するだろう。
【0137】
酵母宿主での使用に適した促進配列の例としては、3-ホスホグリセリン酸キナーゼ[Hitzemanら, J. Biol. Chem. 255(24):12073 (1980)]または他の解糖系酵素[Hessら, J. Adv. Enzyme Reg. 7:149 (1968)、およびHolland, Biochemistry 17(23):4900-7 (1978)]、例えばエノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース-6-リン酸イソメラーゼ、3-ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼ、およびグルコキナーゼなどのプロモーターが挙げられる。
【0138】
生育条件によって転写が制御されるという追加の利点を持つ誘導性プロモーターである他の酵母プロモーターは、アルコールデヒドロゲナーゼ2、イソチトクロームC、酸性ホスファターゼ、窒素代謝に関係する分解酵素、メタロチオネイン、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、ならびにマルトースおよびガラクトース資化を担う酵素のプロモーター領域である。酵母発現用の適切なベクターおよびプロモーターはEP 73,657にさらに詳しく記載されている。哺乳類宿主細胞におけるベクターからの融合タンパク質コードmRNAの転写は、例えば、ポリオーマウイルス、鶏痘ウイルス、アデノウイルス(アデノウイルス2など)、ウシ乳頭腫ウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよびシミアンウイルス40(SV40)などのウイルスのゲノムから得られるプロモーター、アクチンプロモーターまたは免疫グロブリンプロモーターなどの異種哺乳類プロモーター、および熱ショックプロモーターから得られるプロモーターによって制御しうる(ただし、それらのプロモーターは当該宿主細胞系に適合するものとする)。
【0139】
融合タンパク質をコードするポリヌクレオチドの高等真核生物による転写は、ベクターにエンハンサー配列を挿入することによって増進させうる。エンハンサーは通常約10〜300bpのDNAのシス作用要素であって、プロモーターに作用してその転写を増進する。現在では、哺乳類遺伝子(グロビン、エステラーゼ、アルブミン、α-フェトプロテインおよびインスリン)に由来する多くのエンハンサー配列が知られている。しかし、典型的には、真核細胞ウイルス由来のエンハンサーが使用されるだろう。複製起点の後期側(bp100-270)にあるSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後期側にあるポリオーマエンハンサー、およびアデノウイルスエンハンサーなどが、その例である。エンハンサーをベクターに接合する際は、融合タンパク質コード配列に対して5'側に接合してもよいし、3'側に接合してもよいが、好ましくはエンハンサーをプロモーターの5'側に置く。
【0140】
真核宿主細胞(酵母、カビ、昆虫、植物、動物、ヒト、または他の多細胞生物に由来する有核細胞)で使用される発現ベクターは、転写の終結およびmRNAの安定化に必要な配列も含有するだろう。そのような配列は、真核細胞またはウイルスのDNAまたはcDNAの5'非翻訳領域および時には3'非翻訳領域から、一般に入手することができる。これらの領域は、融合タンパク質をコードするmRNAの非翻訳部分にポリアデニル化された断片として転写されるヌクレオチドセグメントを含有する。
【0141】
様々な形態の融合タンパク質を培養培地または宿主細胞溶解物から回収することができる。膜結合型の場合は、適切な界面活性剤溶液(例えばトリトンX100)を使用するか、酵素的切断によって、膜から遊離させることができる。融合タンパク質の発現に使用した細胞は、例えば凍結融解サイクリング、超音波処理、機械的破壊、または細胞溶解剤など、様々な物理的または化学的手段によって破壊することができる。
【0142】
本発明の異種融合タンパク質の精製:
本発明の異種融合タンパク質を適当な宿主細胞中で発現させたら、その類似体を単離し、精製することができる。以下の方法は、適切な精製法の代表例である:カルボキシメチルセルロースでの分画、セファデックスG-75などのゲル濾過、DEAEまたはMono-Qなどの陰イオン交換樹脂、CMまたはMono-Sなどの陽イオン交換体、IgGなどの夾雑物を除去するためのプロテインAセファロース、エピトープタグ付のポリペプチドを結合するための金属キレートカラム、逆相HPLC、クロマトフォーカシング、シリカゲル、エタノール沈殿、および硫酸アンモニウム沈殿。。
【0143】
様々なタンパク質精製方法を使用することができ、そのような方法は当技術分野で知られていると共に、例えばDeutscher, Methods in Enzymology 182:83-9(1990)およびScopes「Protein Purification: Principles and Practice」Springer-Verlag, ニューヨーク(1982)などに記載されている。選択される精製工程は、使用する製造方法および製造される融合タンパク質の性質に依存するだろう。例えばFc断片を含む融合タンパク質は、プロテインAまたはプロテインGアフィニティーマトリックスを使って効果的に精製することができる。低pHまたは高pH緩衝液を使って、アフィニティーマトリックスから融合タンパク質を溶出させることができる。穏やかな溶出条件は、融合タンパク質の不可逆的変性を防止するのに役立つだろう。イミダゾール含有緩衝液を使用することもできる。実施例3に本発明融合タンパク質の精製手順の成功例をいくつか説明する。
【0144】
本発明の異種融合タンパク質の特徴づけ:
本発明の融合タンパク質を特徴づける方法は数多く存在する。これらの方法には、一例として、SDS-PAGEとタンパク質染色法または抗IgGまたは抗HSA抗体を用いた免疫ブロット法との併用が含まれる。他の方法には、例えばマトリックス支援レーザー脱離/イオン化質量分析法(MALDI-MS)、液体クロマトグラフィー/質量分析法、等電点フォーカシング、分析用イオン交換、クロマトフォーカシング、および円二色性などがある。SDS-PAGEと免疫ブロット法との併用ならびに質量分析法を用いて、いくつかの代表的な異種融合タンパク質を特徴づけた(実施例4および5ならびに図3および4参照)。
【0145】
例えば表3(実施例5参照)に、いくつかの代表的な融合タンパク質の計算分子量と、質量分析法によって決定された質量とを例示する。また、図3および図4に、SDS PAGEで決定したいくつかの代表的な融合タンパク質の分子量を例示する。試験した異種融合タンパク質は全て、一過性に発現、分泌させた。また、Igκシグナル配列を切断して、正しいN末端を持つタンパク質を得た。
【0146】
また表3は、質量分析法によって決定した分子量が予想より大きい場合があることも示している。これはFc部分のグリコシル化およびC末端伸長の結果である。融合タンパク質の酵素消化に続いて、逆相HPLCおよび質量分析を行なう事により、糖鎖部分を含むペプチド画分を同定することができる。次に、これらの画分をN末端アミノ酸配列決定にかけて、潜在的グリコシル化部位を同定することができる。例えばエクセンディン4-Fc(配列番号29)の特徴づけにより、39位のセリンおよび50位のスレオニンがO結合型グリコシル化を、また122位のアスパラギンがN結合型グリコシル化を受けることがわかる。
【0147】
いくつかの代表的なGLP-1融合タンパク質を活性についても調べた。試験管内および生体内でGLP-1活性を検出する方法は数多く存在する(実施例6、7、8および9参照)。表4(実施例6)にいくつかのGLP-1融合物に付随するGLP-1受容体活性を例示する。数字は、Val8-GLP-1(7-37)OHに付随する活性に対する相対値である。試験した融合タンパク質は全てGLP-1受容体活性を持っていた。低い試験管内活性は、必ずしも、生体内での効果が弱いことを示さない。これらの融合タンパク質の半減期はかなり増加しているので、弱い試験管内活性は一般に弱い生体内活性を予言するものではない。図7および実施例7は、本発明の融合タンパク質に付随する延長された半減期を例示している。例えば、Val8-GLP-1-Fcはサルで約45時間の半減期を持ち、Val8-GLP-1-HSAはサルで約87時間の半減期を持ち、Gly8-Glu22-GLP-1-CEx-リンカー-IgG1はイヌでIV投与後に約55時間の半減期を持ち、Gly8-Glu22-GLP-1-CEx-リンカー-IgG1はイヌでSC投与後に約38時間の半減期を持った。
【0148】
本発明の組成物:
治療タンパク質組成物には物理的安定性も不可欠な特徴である。GLP-1化合物は加工中に構造変化が起こるために製造および製剤がとりわけ困難だった。例えば一部のGLP-1化合物には凝集しようとする一般的傾向がある。また、一部のGLP-1化合物は可溶性かつ活性なαヘリックス型から不溶性で潜在的に不活性なβシート型に変換することが明らかにされている。IgGのFc領域またはアルブミンなどの大きいタンパク質へのGLP-1化合物の融合は、GLP-1化合物の半減期を増大させるだけでなく、GLP-1化合物の物理的安定性およびコンフォメーションの安定性にも寄与する。例えば、PBS中のVal8-GLP-1-リンカー-HSAは、37℃で約30日まで安定である。
【0149】
本発明の異種融合タンパク質は、1つまたは複数の賦形剤と共に製剤化することができる。本発明の活性な融合タンパク質は、医薬的に許容できる緩衝液と混合し、許容できる安定性および腸管外投与などの投与が可能なpHが得られるように、pHを調節することができる。
【0150】
所望により、1つまたは複数の医薬的に許容できる抗微生物剤を加えてもよい。メタクレゾールおよびフェノールは、好ましい医薬的に許容できる微生物剤である。イオン強度または張性を調節するために、1つまたは複数の医薬的に許容できる塩を加えてもよい。製剤の等張性をさらに調節するために、1つまたは複数の賦形剤を加えてもよい。グリセリンは等張性調節賦形剤の一例である。医薬的に許容できるとは、ヒトまたは他の動物への投与に適切しており、有毒な要素または望ましくない夾雑物を含まず、製剤中の活性化合物の活性を妨害しないことを意味する。
【0151】
本発明では、本発明の異種融合タンパク質の医薬的に許容できる塩型を使用することができる。酸付加塩を形成させるためによく使用される酸は、塩酸、臭化水素酸、ヨウ化水素酸、硫酸、リン酸などの無機酸、およびp-トルエンスルホン酸、メタンスルホン酸、シュウ酸、p-ブロモフェニルスルホン酸、炭酸、コハク酸、クエン酸、安息香酸、酢酸などの有機酸である。好ましい酸付加塩は、塩酸および臭化水素酸などの鉱酸によって形成されるものである。
【0152】
塩基付加塩には、無機塩基、例えばアンモニウムまたはアルカリもしくはアルカリ土類金属の水酸化物、炭酸塩、炭酸水素塩などから誘導されるものが含まれる。したがって、本発明の塩の製造に役立つそのような塩には、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、炭酸カリウムなどが含まれる。
【0153】
組成物の投与:
投与は、通常の技術を持つ医師に有効であることが知られているどの経路で行なってもよい。末梢、腸管外投与はそのような方法の一つである。腸管外投与は医学的文献では滅菌注射器または他の何らかの機械的装置、例えば注入ポンプなどによる、身体への剤形の注射であると一般に理解されている。末梢腸管外経路には静脈内、筋肉内、皮下、および腹腔内投与経路が含まれる。
【0154】
本発明の異種融合タンパク質は、非腸管外経路である経口、直腸、鼻腔、または下気道経路での投与にも馴染みうる。これらの非腸管外経路のうち、下気道経路および経口経路は好ましい。
【0155】
本発明の融合タンパク質は、広範囲にわたる多様な疾患および状態の処置に使用することができる。本発明の融合タンパク質は主に、「GLP-1受容体」と呼ばれる受容体に作用することによって、その生物学的効果を発揮する。したがって、GLP-1受容体刺激またはGLP-1化合物の投与に有利な反応を示す疾患および/または状態を持つ対象は、本発明のGLP-1融合タンパク質で処置することができる。これらの対象を、「GLP-1化合物による処置を必要としている」または「GLP-1受容体刺激を必要としている」という。これには、インスリン非依存性糖尿病、インスリン依存性糖尿病、脳卒中(WO00/16797参照)、心筋梗塞(WO98/08531参照)、肥満(WO98/19698参照)、手術後の異化変化(米国特許第6,006,753号)、機能性消化不良および過敏性腸症候群(WO99/64060参照)を持つ対象が含まれる。また、GLP-1化合物による予防的処置を必要とする対象、例えばインスリン非依存性糖尿病を発症する危険がある対象(WO00/07617参照)も含まれる。耐糖能障害または空腹時グルコース異常を持つ対象、当該対象の身長および体格での正常体重より約25%高い体重を持つ対象、部分膵切除を受けた対象、片親または両親がインスリン非依存性糖尿病である対象、妊娠糖尿病の病歴がある対象、および急性または慢性膵炎の病歴がある対象は、インスリン非依存性糖尿病を発症する危険がある。
【0156】
GLP-1化合物の「有効量」は、GLP-1受容体刺激を必要としている対象に投与したときに、許容できない副作用を引き起こすことなく、望ましい治療効果および/または予防効果をもたらす量である。「望ましい治療効果」には以下に挙げる効果の1つまたは複数が含まれる:1)疾患または状態に関係する症状の改善、2)疾患または状態に関係する症状の発症の遅延、3)処置しない場合と比較した生存期間の延長、および4)処置しない場合と比較した生活の質の向上。例えば糖尿病の処置に関してGLP-1化合物の「有効量」とは、処置しない場合よりも血中グルコース濃度がよりよく管理され、よって網膜症、ニューロパシーまたは腎疾患などの糖尿病合併症の発症が遅延するような量である。糖尿病の予防に関してGLP-1化合物の「有効量」とは、スルホニル尿素、チアゾリジンジオン、インスリンおよび/またはビスグアニジンなどの抗低血糖薬による処置を必要とする高血中グルコースレベルの発生を、処置しない場合と比較して遅延させるような量である。
【0157】
患者の血中グルコースを正常化するのに有効な融合タンパク質の用量は、例えば当該対象の性別、体重および年齢、血中グルコース調節不能の重症度、投与経路および生物学的利用能、融合タンパク質の薬物動態プロフィール、効力、および製剤などを含む数多くの因子に依存するだろう。
【0158】
本発明は、アルブミンタンパク質、アルブミン断片、アルブミン類似体、Fcタンパク質、Fc断片、またはFc類似体に融合されているために改善された生化学的特性および生物物理学的特性を持つGLP-1化合物を包含する。これらの異種タンパク質は、宿主細胞中でうまく発現させ、GLP-1受容体の活性化に関係するシグナリング活性を保ち、長くなった半減期を持つことができる。
【実施例】
【0159】
本発明をさらに詳しく説明するために、以下に実施例を記載する。本発明の範囲は単に以下の実施例からなるとみなしてはならない。ここに記載する特定の試薬、装置、および方法が単なる例示であって、決して本発明の限定を意図するものでないことは、当業者には理解されるだろう。
【0160】
実施例1:異種融合タンパク質をコードするDNAの構築
実施例1a
Val8-GLP-1(7-37)-FcをコードするDNAの構築:
ヒトIgG1のFc部分をcDNAライブラリーから単離した。これは全ヒンジ領域とCH2およびCH3ドメインとを含む。このヒトIgG1のFc部分の696塩基対を含む断片を、哺乳類発現ベクターpJB02のNheI部位およびEco47III部位にサブクローニングして、pJB02/Fcを作製した(図5参照)。Val8-GLP-1(7-37)に融合されたIgκ分泌シグナル配列をコードするDNAを、4本のオーバーラップした相補的オリゴヌクレオチド、すなわち
5'-CTAGCCACCATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACCAGTG-3'[配列番号12]
5'-GAGGGCACCTTCACCTCCGACGTGTCCTCCTATCTGGAGGGCCAGGCCGCCAAGGAGTTCATCGCCTGGCTGGTGAAGGGAAGAGGC-3'[配列番号13]
5'-TGAAGGTGCCCTCCACGTGGTCACCAGTGGAACCTGGAACCCAGAGCAGCAGTACCCATAGCAGGAGTGTGTCTGTCTCCATGGTGG-3'[配列番号14]
5'-GCCTCTTCCCTTCACCAGCCAGGCGATGAACTCCTTGGCGGCCTGGCCCTCCAGATAGGAGGACACGTCGGAGG-3'[配列番号15]
の試験管内ハイブリダイゼーションによって作製した。
【0161】
ハイブリダイゼーション反応は、等量の各オリゴヌクレオチド(各オリゴの最終濃度は1pm/μl)を使って行なった。オリゴヌクレオチドの混合物をライゲーション緩衝液(50mMトリス-HCl、pH7.5、10mM MgCl2、10mM DTT、1mM ATP、25μg/mlウシ血清アルブミン)中、100℃で5分間加熱した後、少なくとも2時間かけて30℃まで冷却した。
【0162】
得られたハイブリダイゼーション産物を、NheIおよびEco47IIIで消化しておいたpJB02/Fcベクター骨格に、室温で2時間、または16℃で一晩かけて連結した。そのライゲーション産物を使ってコンピテントXL-1Blue細胞(Stratagene)を形質転換した。クローンをNcoIで消化することにより、組換えプラスミドを、ペプチドコード配列(コザック配列とシグナルペプチドの最初のMetをコードするもの)の存在についてスクリーニングし、配列決定した。得られた発現プラスミドはトランスフェクションアッセイに使用したが、このプラスミドをpJB02-V8-GLP-1-Fcと名付けた(図5)。
【0163】
実施例1b
Val8-GLP-1(7-37)-HSAをコードするDNAの構築:
プラスミドHSA/pcDNA3.1GSをInvitrogenから購入し(カタログ番号H-M12523M-pcDNA3.1/GS)、ヒト血清アルブミン(HSA)をコードするcDNAを単離するためのテンプレートとして使用した。HSA cDNAはPCRを使って調製した。この際、リーダー配列および6アミノ酸プロペプチドをコードするDNAを5'末端から除去した。さらに停止コドンをHSAコード配列の3'末端に直接付加した。最後に、クローニングが容易になるように、5'末端および3'末端に制限酵素部位を作った。Invitorogenから購入した元のベクター中に存在するHSA DNA配列は、天然ヒト配列と比較して、遺伝子の3'領域(667位)に一塩基変異を含んでいた。この変異により、AspではなくAsnのコドンになる。そこで、上述した鎖オーバーラップPCR突然変異導入法を使って、この位置がAspをコードするように、コドンを改変した。得られたHSAコードDNAをpJB02のNheI部位およびHindIII部位にクローニングしてpJB02-HSA(図6)を作製した。
【0164】
Val8-GLP-1(7-37)配列に融合されたIgκリーダー配列を実施例1aで述べたように作製した。このDNAをpJB02-HSAのNheI部位およびFspI部位に連結してpJB02-Val8-GLP-1-HSAを作製した。
【0165】
実施例1c
Val8-GLP-1(7-37)-リンカー-HSAをコードするDNAの構築:
ベクターpJB02-HSAを実施例1bで述べたように作製した。リンカー配列[GGGGS]3をコードするDNAを、HSAコードDNAの5'末端にインフレームで連結してpJB02-リンカー-HSA(図7)を作製した。Igκリーダー配列をコードし、Val8-GLP-1(7-37)配列およびリンカー配列の5'部分に融合されたDNAを、実施例1aで述べたように作製した。このDNAをpJB02のNheIおよびBspEIに連結してpJB02- Val8-GLP-1-リンカー-HSAを作製した。
【0166】
実施例1d
エクセンディン4-FcをコードするDNAの構築:
プラスミドpJB02/Fcを実施例1aで述べたように作製した。エクセンディン4に融合されたIgκシグナル配列をコードするDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CTAGCCACCATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACCGGTCAC-3'[配列番号16]
5'-GGAGAGGGAACCTTCACCAGCGACCTGAGCAAGCAGATGGAGGAGGAGGCCGTGAGACTG-3'[配列番号17]
5'-TTCATCGAGTGGCTGAAGAACGGAGGACCAAGCAGCGGAGCCCCTCCTCCTAGC-3'[配列番号18]
5'-GAACCTGGAACCCAGAGCAGCAGTACCCATAGCAGGAGTGTGTCTGTCTCCATGGTGG-3'[配列番号19]
5'-CTCCTCCTCCATCTGCTTGCTCAGGTCGCTGGTGAAGGTTCCCTCTCCGTGACCGGTG-3'[配列番号20]
5'-GCTAGGAGGAGGGGCTCCGCTGCTTGGTCCTCCGTTCTTCAGCCACTCGATGAACAGTCTCACGGC-3'[配列番号21]。
【0167】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、実施例1aで述べたように、NheIおよびEco47IIIで消化しておいたpJB02ベクターに連結して、pJB02-エクセンディン4-Fcを作製した。
【0168】
実施例1e
エクセンディン4-HSAをコードするDNAの構築:
プラスミドpJB02-HSAを実施例1bで述べたように作製した。エクセンディン4に融合されたIgκシグナル配列をコードするDNAを、オーバーラップした相補的オリゴヌクレオチド(実施例1dに記載したものと同じもの)の試験管内ハイブリダイゼーションによって作製した。ハイブリダイゼーション反応も上述したとおりに行なった。DNAをpJB02-HSA中のユニークなNheI部位およびFspI部位にクローニングしてpJB02-エクセンディン4-HSAを作製した。
【0169】
実施例1f
エクセンディン4-リンカー-HSAをコードするDNAの構築:
プラスミドpJB02-リンカー-HSAを実施例1cで述べたように構築した。エクセンディン4およびリンカー配列の5'部分に融合されたIgκシグナルをコードするDNAを実施例1dで述べたように作製した。このDNAをpJB02-リンカー-HSAのユニークなNheI部位およびBspEI部位にクローニングしてpJB02-エクセンディン4-リンカー-HSAを作製した。
【0170】
実施例1g
Val8-GLP-1/C-Ex-FcをコードするDNAの構築:
プラスミドpJB02-エクセンディン4-Fcを実施例1dで述べたように作製した。エクセンディン4コードDNAを、AgeIおよびEco47IIIで、ベクターから切り出した。Val8-GLP-1/C-ExコードDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CCGGTCACGTGGAGGGCACCTTCACCTCCGACGTGTCCTCCTATCTGGAGGGCCAGGCCGCCA-3'[配列番号22]
5'-AGGAATTCATCGCCTGGCTGGTGAAGGGCCGGGGCAGCAGCGGAGCCCCTCCTCCTAGC-3'[配列番号23]
5'-CTCCAGATAGGAGGACACGTCGGAGGTGAAGGTGCCCTCCACGTGA-3'[配列番号24]
5'-GCTAGGAGGAGGGGCTCCGCTGCTGCCCCGGCCCTTCACCAGCCAGGCGATGAATTCCTTGGCGGCCTGGCC-3'[配列番号25]。
【0171】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、エクセンディン4の代わりに、pJB02-エクセンディン4-Fc発現ベクターに連結して、pJB02-Val8-GLP-1/C-Ex-Fcを作製した。
【0172】
実施例1h
Val8-Glu22-GLP-1-FcをコードするDNAの構築:
プラスミドpJB02-エクセンディン4-Fcを実施例1dで述べたように作製した。エクセンディン4コードDNAを、AgeIおよびEco47IIIで、ベクターから切り出した。Val8-Glu22-GLP-1コードDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CCGGTCACGTGGAGGGCACCTTCACCTCCGACGTGTCCTCCTATCTCGAGGAGCAGGCCGCCA-3'[配列番号26]
5'-AGGAGTTCATCGCCTGGCTGGTGAAGGGCCGGGGC-3'[配列番号27]
5'-GCCCCGGCCCTTCACCAGCCAGGCGATGAACTCCTTGGCGGCCTGCTC-3'[配列番号28]
5'-CTCGAGATAGGAGGACACGTCGGAGGTGAAGGTGCCCTCCACGTGA-3'[配列番号29]。
【0173】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、エクセンディン4の代わりに、pJB02-エクセンディン4-Fc発現ベクターに連結して、pJB02-Val8-Glu22-GLP-1-Fcを作製した。
【0174】
実施例1i
Val8-Glu22GLP-1/C-Ex-FcをコードするDNAの構築:
プラスミドpJB02-エクセンディン4-Fcを実施例1dで述べたように製造した。エクセンディン4コードDNAを、AgeIおよびEco47IIIで、ベクターから切り出した。Val8-Glu22GLP-1/C-ExコードDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CCGGTCACGTGGAGGGCACCTTCACCTCCGACGTGTCCTCCTATCTCGAGGAGCAGGCCGCCA-3'[配列番号30]
5'-AGGAATTCATCGCCTGGCTGGTGAAGGGCCGGGGCAGCAGCGGAGCCCCTCCTCCTAGC-3'[配列番号31]
5'-CTCGAGATAGGAGGACACGTCGGAGGTGAAGGTGCCCTCCACGTGA-3'[配列番号32]
5'-GCTAGGAGGAGGGGCTCCGCTGCTGCCCCGGCCCTTCACCAGCCAGGCGATGAATTCCTTGGCGGCCTGCTC-3'[配列番号33]。
【0175】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、エクセンディン4の代わりに、pJB02-エクセンディン4-Fc発現ベクターに連結して、pJB02-Val8-Glu22-GLP-1/C-Ex-Fcを作製した。
【0176】
実施例1j
Gly8-GLP-1-FcをコードするDNAの構築:
プラスミドpJB02-エクセンディン4-Fcを実施例1dで述べたように製造した。エクセンディン4コードDNAを、AgeIおよびEco47IIIで、ベクターから切り出した。Gly8-GLP-1コードDNAを、以下のオーバーラップした相補的オリゴヌクレオチドの試験管内ハイブリダイゼーションによって作製した:
5'-CCGGTCACGGCGAGGGCACCTTCACTAGTGACGTGTCCTCCTATCTGGAGGGCCAGGCCGCCA-3'[配列番号34]
5'-AGGAGTTCATCGCCTGGCTGGTGAAGGGCCGGGGC-3'[配列番号35]
5'-CTCCAGATAGGAGGACACGTCACTAGTGAAGGTGCCCTCGCCGTGA-3'[配列番号36]
5'-GCCCCGGCCCTTCACCAGCCAGGCGATGAACTCCTTGGCGGCCTGGCC-3'[配列番号37]。
【0177】
ハイブリダイゼーション反応は実施例1aで述べたように行なった。ハイブリダイズした産物を、エクセンディン4の代わりに、pJB02-エクセンディン4-Fc発現ベクターに連結して、pJB02-Gly8-GLP-1-Fcを作製した。
【0178】
実施例2:異種融合タンパク質の発現
HEK293EBNA細胞(付着細胞および浮遊細胞の両方)を一過性にトランスフェクトすることにより、実施例1のDNAコンストラクトがコードする融合タンパク質の発現を行なった。細胞を計数し、トランスフェクションの24時間前に播種した。FuGene(商標)6トランスフェクション試薬(Roche Molecular Biochemicals、カタログ番号1814443)とOptiMEM(Gibco/BRL)とを混合し、室温で5分間インキュベートした時点でDNAを加えることによってトランスフェクションカクテルを調製し、そのカクテルをさらに15分間インキュベートした。トランスフェクション直前に、新しい成長培地をプレートに加えた。トランスフェクションのさらなる詳細を表1および表2に示す。
【0179】
293EBNA細胞の一過性トランスフェクションに使用した試薬
【表1】
【0180】
培地組成
【表2】
【0181】
小規模トランスフェクション(35〜10mm容器)の場合は、トランスフェクションの24時間後に細胞をPBSで洗浄して収穫培地に切り替え、数日間にわたって24時間毎に培地を収集し、置換した。大規模トランスフェクション(700cm2ローラーボトル)の場合は、トランスフェクションの48時間後にローラーボトルをPBSで洗浄し、収穫培地に換えた。少なくとも連続する10日間にわたって24時間毎に培地を収集し、交換した。以降のタンパク質生成には、常に、10回分の収穫物だけを使用した。
【0182】
実施例3:異種融合タンパク質の精製
実施例3a
Val8-GLP-1-Fcの精製:
大規模トランスフェクションによって得た約4.5リットルの調整培地(融合タンパク質発現レベル約20μg/ml)を、CUNOフィルターシステムを使ってろ過し、10Kフィルターメンブレンを装着したProFluxタンジェンシャルフロー濾過システムを使って250mlに濃縮した。Val8-GLP-1-Fcを、1×PBS(pH7.4)中の5ml HiTrapプロテインAカラムに、2ml/分の流速で捕捉し、50mMクエン酸(pH3.3)で溶出させた。4mlの1×PBSと100μlの1Mトリス(pH8)が入っているチューブに、各画分(1ml)を収集した。
【0183】
SDS-PAGEおよびZorbax C8での逆相HPLCによって融合タンパク質を含むことが確認された画分をプールして、1×PBS(pH7.4)中のSuperdex 75 60/60カラムに10ml/分の流速で適用した。陽性画分(20ml/チューブ)を集めて、プールした。次に、プールした画分を3ml/分の流速で0.1%TFA水溶液でのC4逆相クロマトグラフィーにかけた。70分で5%B(0.1%TFA/アセトニトリル)から100%Bに至る勾配を使ってVal8-GLP-1-Fcを溶出させた。溶出画分(3ml/チューブ)を集めた。減圧乾燥によってアセトニトリルを除去し、1mlのH2Oを加えた。精製試料(約32ml)を4リットルの1×PBS(pH7.4)に対して2回透析した。
【0184】
次に、透析した試料を、MILLEX-GV 0.22μmフィルターユニットを使って濾過し、280nmでの吸収を使って濃度を決定した。
【0185】
実施例3b
Val8-GLP-1-HSAまたはVal8-GLP-1-リンカー-HSAの精製:
約6.5リットルの調整培地(融合タンパク質発現レベル約10μg/ml)を、CUNOフィルターシステムを使ってろ過し、10Kフィルターメンブレンを装着したProFluxタンジェンシャルフロー濾過システムを使って380mlに濃縮した。
【0186】
融合タンパク質を、20mMトリス(ph7.4)中の50ml Fast Flow Qカラム(Pharmacia)に、5ml/分の流速で捕捉した。10CVで0%→50% 20mMトリス(pH7.4)、1M NaClの後、2CVで100%Bへの勾配を使って、タンパク質を溶出させた。
【0187】
融合タンパク質を含む画分をプールし、5ml/分の流速で、0.1%TFA水溶液でのC4逆相クロマトグラフィーにかけた。120分で20%B(0.1%TFA/アセトニトリル)から90%Bへの勾配を使って、融合タンパク質を溶出させた。各画分(3.5ml/チューブ)を集めた。減圧乾燥によってアセトニトリルを除去した。
【0188】
プールした試料約9mlを1×PBS(pH7.4)で40mlに希釈し、4リットルの1×PBS(pH7.4)に対して一晩透析した。試料を濾過し、280nmでの吸収によって濃度を決定した。
【0189】
実施例3c
エクセンディン4-Fcの精製:
約4リットルの調整培地(融合タンパク質発現レベル約8μg/ml)を、CUNOフィルターシステムを使ってろ過し、30Kフィルターメンブレンを装着したProFluxタンジェンシャルフロー濾過システムを使って250mlに濃縮した。
【0190】
エクセンディン4-Fcを、1×PBS(pH7.4)中の5ml HiTrapプロテインAカラムに、2ml/分の流速で捕捉し、50mMクエン酸(pH3.3)で溶出させた。融合タンパク質を含む画分をプールし、濾過し、4リットルの1×PBSに対して一晩透析した。次に、透析した試料を、1×PBS(pH7.4)、0.5M NaCl中のSuperdex 75 60/60カラムに、10ml/分の流速で適用した。融合タンパク質を含む画分(20ml/チューブ)を集め、プールし、約1mg/mlに濃縮した。次に、濃縮した試料をMILLEX-GV 0.22μmフィルターユニットを使って濾過した。
【0191】
実施例3d
エクセンディン4-HSAおよびエクセンディン4-リンカー-HSAの精製
約1.1リットルの調整培地(融合タンパク質発現レベル約6μg/ml)を、CUNOフィルターシステムを使って濾過し、30Kフィルターメンブレンを装着したProFluxタンジェンシャルフロー濾過システムを使って175mlに濃縮した。
【0192】
20mMトリス(pH7.4)中の5ml HiTrap Q-セファロースカラムを使って、2ml/分の流速で融合タンパク質を捕捉した。12CVで0%→50% 20mMトリス(pH7.4)、1M NaClの後、4CVで100%Bへの勾配を使って、タンパク質を溶出させた。
【0193】
融合タンパク質を含む画分をプールし、5ml/分の流速で、0.1%TFA水溶液でのC4逆相クロマトグラフィーにかけた。70分で10%B(0.1%TFA/アセトニトリル)から100%Bへの勾配を使って、融合タンパク質を溶出させた。融合タンパク質を含む画分(10ml/チューブ)を集めた。減圧乾燥によってアセトニトリルを除去した。
【0194】
プールした試料約8mlを4リットルの1×PBS(pH7.4)に対して一晩透析した。試料を濾過し、280nmでの吸収によって濃度を決定した。次に、透析した試料を、1×PBS(pH7.4)、0.5M NaCl中のSuperdex 200 26/60カラムに、2ml/分の流速で適用した。融合タンパク質を含む画分(3ml/チューブ)を集め、プールし、濃縮し、濾過した。
【0195】
実施例4:SDS PAGEによる融合タンパク質の特徴づけ
SDS-PAGEを行った後、免疫ブロット法を実施することにより、様々な融合タンパク質発現ベクターでトランスフェクトした細胞から得られる精製融合タンパク質と調整培地の両方を分析した。SDS-PAGEは、Novex 16%トリス-グリシン成形済みゲル(EC6498)、泳動緩衝液(10×、LC2675)、および試料緩衝液(L2676)を使って、Novex Powerease 500システムで行なった。充填に先だって、試料を50mM DTTで還元し、95℃で3〜5分間加熱した。
【0196】
SDS-PAGEゲルでの泳動を行なった後、水および転写緩衝液(20%エタノールを含む1×トリス-グリシンSeprabuff(Owl Scientific、カタログ番号ER26-S))を使って、ゲルからSDSを洗い流した。Novex転写装置を、PVDFメンブレン(BioRad、カタログ番号162-0174)およびニトロセルロースメンブレン(BioRad、カタログ番号1703965または1703932)と共に使用した。転写は室温にて30〜35Vで90分間行なった。0.1%Tween-20(Sigma、カタログ番号P-7949)および5%ミルク(BioRad、カタログ番号170-6404)を含む1×PBS中、4℃で1〜12時間、メンブレンをブロッキングした。抗体を1×PBS+5%ミルクに希釈し、ブロットをこれらの溶液中、4℃で1〜2時間インキュベートする。インキュベーションの合間に、ブロットを、室温の1×PBSおよび0.2%Tween-20で、5分間ずつ4回洗浄する。PBSは、GIBCO 10×PBS(カタログ番号70011)から、最終組成が1mM第一リン酸カリウム、3mM第二リン酸ナトリウム、153mM塩化ナトリウム、pH7.4になるように調製するか、またはSigmaのPBSポーチ(カタログ番号1000-3)から、120mM NaCl、2.7mM KClおよび10mMリン酸塩、25℃でpH7.4になるように調製した。
【0197】
一次抗体は、ポリクローナルヤギ抗IgG1またはウサギ抗HSAとした。二次抗体は抗ヤギIgG HRPまたは抗ウサギIgG HRPとした。二次抗体は1:5000に希釈した。ブロットの発色にはECLシステム(Amersham Pharmacia Biotech、カタログ番号RN2108およびカタログ番号RPN1674H)を使用した。
【0198】
図3Aは、精製Fcタンパク質を、pJB02-Val8-GLP-1-Fcトランスフェクト細胞およびpJB02-エクセンディン4-Fcトランスフェクト細胞から得られる調整培地と比較した図である。移動度の減少は、融合タンパク質のGLP-1部分によるサイズの増加と合致している。図3Bも同様に、精製HSAを、pJB02-Val8-GLP-1-HSA、pJB02-Val8-GLP-1-リンカー-HSA、pJB02-エクセンディン4-HSA、またはpJB02-エクセンディン4-リンカー-HSAでトランスフェクトした細胞から得られる調整培地と比較した図である。図4に精製融合タンパク質調製物を示す。
【0199】
実施例5:質量分析法を使った融合タンパク質の特徴づけ
実験は全て、タイムラグフォーカシング電子機器(Time Lag Focuing electronics)、リフレクトロン(Reflectron)(0〜8000Daペプチド範囲の分析に使用)、リニア検出器(Linear detector)(高分子量/高シグナル分析時に使用)、および後段加速検出器(Post Acceleratio Detector)(すなわちP.A.D、高分子量/極低シグナル分析に使用)を装着したMicromass TofSpec-2E質量分析計で行なった。装置の実効飛行行路長はリニアモードで1.2メートル、リフレクトロンモードで2.3メートルである。リニアおよびリフレクトロンモード検出用に2つのデュアルマイクロチャンネルプレート検出器を備える。使用するレーザーは、毎秒5回のレーザー照射で337nmで作動するLaser Science Inc. VSL-337i窒素レーザーである。データは全て、2GHz8ビット内蔵ディジタイザを使って取得し、1スペクトルにつき50回までのレーザー照射を平均した。
【0200】
問題にしているGLP-1融合タンパク質を分析する場合は、装置をリニアモードで操作した。リニア検出器は、MALDI-ToF-MS装置の飛行管を縦断するイオンを検出する装置である。この検出器は、イオン存在度を経時的に測定し、変換用ディジタイザにシグナルを送信する。ディジタイザは、質量分析器からのシグナルをコンピュータに転送できるようにするアナログ-ディジタル変換器であり、コンピュータに転送されたシグナルは、そこで便利なm/zスペクトルに再構築される。
【0201】
再結晶飽和シナピン酸溶液(50/50 Acn/H2Oおよび0.1%TFAに希釈)をイオン化マトリックスとして利用した。シナピン酸は、10kDaを超えるタンパク質に適したマトリックスである。分析した試料の質量が正確に決定されるように、内部および外部較正ファイル用に、適当な質量基準タンパク質を使用した。試料は全て1:2の試料:マトリックス希釈液を使って分析した。装置は以下のリニア検出器条件で初期設定した:
ソース電圧(Source Voltage):20.0keV
抽出電圧(Extraction Voltage):20.0keV
収束電圧(Focus Voltage):16.0keV
リニア検出器:3.7keV
P.A.D.:(オフライン)
パルス電圧(Pulse Voltage):3.0keV
レーザーコース(Laser Coarse):50
レーザーファイン(Laser Fine):50。
【0202】
最適な信号/雑音比と最も高い解像度が得られるように、これらの設定を(必要に応じて)変更した。表3に様々なGLP-1融合タンパク質の特徴を示す。
【0203】
【表3】
CExはC末端伸長を表し、Ser-Ser-Gly-Ala-Pro-Pro-Pro-Serという配列を含む。
リンカーはGly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Serである。
【0204】
実施例6:異種融合タンパク質の活性:
本発明融合タンパク質のGLP-1受容体活性化能力を、GelfandらのEP619,322および米国特許第5,120,712にそれぞれ記載されているような、試験管内アッセイを使って評価した。Val8-GLP-1(7-37)OHの活性と比較した、これらの化合物の活性を、表4に報告する。図8に、Val8-GLP-1およびエクセンディン4融合タンパク質に関する試験管内用量反応曲線を図示する。さらに、Fcまたはアルブミンタンパク質と融合させて生物学的に活性な融合タンパク質を製造することができる多くのGLP-1類似体の試験管内活性を、表5aおよび表5bに示す。これらの活性はGLP-1(7-37)OHと比較されている。
【0205】
GLP-1融合タンパク質の試験管内活性
【表4】
CExはC末端伸長を表し、Ser-Ser-Gly-Ala-Pro-Pro-Pro-Serという配列を含む。
リンカーはGly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Ser-Gly-Gly-Gly-Gly-Serである。
C2はSer-Ser-Gly-Ala-Ser-Ser-Gly-Alaである。
【0206】
表3および表4に記載した融合タンパク質のアミノ酸配列を配列番号13〜31に示す。
【0207】
Val8-GLP-1-ヒト血清アルブミンアミノ酸配列は配列番号13によって表される。
1 HVEGTFTSDV SSYLEGQAAK EFIAWLVKGR GDAHKSEVAH RFKDLGEENF KALVLIAFAQ
61 YLQQCPFEDH VKLVNEVTEF AKTCVADESA ENCDKSLHTL FGDKLCTVAT LRETYGEMAD
121 CCAKQEPERN ECFLQHKDDN PNLPRLVRPE VDVMCTAFHD NEETFLKKYL YEIARRHPYF
181 YAPELLFFAK RYKAAFTECC QAADKAACLL PKLDELRDEG KASSAKQRLK CASLQKFGER
241 AFKAWAVARL SQRFPKAEFA EVSKLVTDLT KVHTECCHGD LLECADDRAD LAKYICENQD
301 SISSKLKECC EKPLLEKSHC IAEVENDEMP ADLPSLAADF VESKDVCKNY AEAKDVFLGM
361 FLYEYARRHP DYSVVLLLRL AKTYETTLEK CCAAADPHEC YAKVFDEFKP LVEEPQNLIK
421 QNCELFEQLG EYKFQNALLV RYTKKVPQVS TPTLVEVSRN LGKVGSKCCK HPEAKRMPCA
481 EDYLSVVLNQ LCVLHEKTPV SDRVTKCCTE SLVNRRPCFS ALEVDETYVP KEFNAETFTF
541 HADICTLSEK ERQIKKQTAL VELVKHKPKA TKEQLKAVMD DFAAFVEKCC KADDKETCFA
601 EEGKKLVAAS QAALGL[配列番号13]
【0208】
Val8-GLP-1-リンカー-ヒト血清アルブミンアミノ酸配列は配列番号14によって表される。
1 HVEGTFTSDV SSYLEGQAAK EFIAWLVKGR GGGGGSGGGG SGGGGSDAHK SEVAHRFKDL
61 GEENFKALVL IAFAQYLQQC PFEDHVKLVN EVTEFAKTCV ADESAENCDK SLHTLFGDKL
121 CTVATLRETY GEMADCCAKQ EPERNECFLQ HKDDNPNLPR LVRPEVDVMC TAFHDNEETF
181 LKKYLYEIAR RHPYFYAPEL LFFAKRYKAA FTECCQAADK AACLLPKLDE LRDEGKASSA
241 KQRLKCASLQ KFGERAFKAW AVARLSQRFP KAEFAEVSKL VTDLTKVHTE CCHGDLLECA
301 DDRADLAKYI CENQDSISSK LKECCEKPLL EKSHCIAEVE NDEMPADLPS LAADFVESKD
361 VCKNYAEAKD VFLGMFLYEY ARRHPDYSVV LLLRLAKTYE TTLEKCCAAA DPHECYAKVF
421 DEFKPLVEEP QNLIKQNCEL FEQLGEYKFQ NALLVRYTKK VPQVSTPTLV EVSRNLGKVG
481 SKCCKHPEAK RMPCAEDYLS VVLNQLCVLH EKTPVSDRVT KCCTESLVNR RPCFSALEVD
541 ETYVPKEFNA ETFTFHADIC TLSEKERQIK KQTALVELVK HKPKATKEQL KAVMDDFAAF
601 VEKCCKADDK ETCFAEEGKK LVAASQAALG L[配列番号14]
【0209】
Gly8-Glu22-GLP-1-CEx-リンカー-ヒト血清アルブミンアミノ酸配列は配列番号15によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSG GGGGSGGGGS GGGGSDAHKS
61 EVAHRFKDLG EENFKALVLI AFAQYLQQCP FEDHVKLVNE VTEFAKTCVA DESAENCDKS
121 LHTLFGDKLC TVATLRETYG EMADCCAKQE PERNECFLQH KDDNPNLPRL VRPEVDVMCT
181 AFHDNEETFL KKYLYEIARR HPYFYAPELL FFAKRYKAAF TECCQAADKA ACLLPKLDEL
241 RDEGKASSAK QRLKCASLQK FGERAFKAWA VARLSQRFPK AEFAEVSKLV TDLTKVHTEC
301 CHGDLLECAD DRADLAKYIC ENQDSISSKL KECCEKPLLE KSHCIAEVEN DEMPADLPSL
361 AADFVESKDV CKNYAEAKDV FLGMFLYEYA RRHPDYSVVL LLRLAKTYET TLEKCCAAAD
421 PHECYAKVFD EFKPLVEEPQ NLIKQNCELF EQLGEYKFQN ALLVRYTKKV PQVSTPTLVE
481 VSRNLGKVGS KCCKHPEAKR MPCAEDYLSV VLNQLCVLHE KTPVSDRVTK CCTESLVNRR
541 PCFSALEVDE TYVPKEFNAE TFTFHADICT LSEKERQIKK QTALVELVKH KPKATKEQLK
601 AVMDDFAAFV EKCCKADDKE TCFAEEGKKL VAASQAALGL[配列番号15]
【0210】
エクセンディン4-ヒト血清アルブミンアミノ酸配列は配列番号16によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGAPPPSD AHKSEVAHRF KDLGEENFKA
61 LVLIAFAQYL QQCPFEDHVK LVNEVTEFAK TCVADESAEN CDKSLHTLFG DKLCTVATLR
121 ETYGEMADCC AKQEPERNEC FLQHKDDNPN LPRLVRPEVD VMCTAFHDNE ETFLKKYLYE
181 IARRHPYFYA PELLFFAKRY KAAFTECCQA ADKAACLLPK LDELRDEGKA SSAKQRLKCA
241 SLQKFGERAF KAWAVARLSQ RFPKAEFAEV SKLVTDLTKV HTECCHGDLL ECADDRADLA
301 KYICENQDSI SSKLKECCEK PLLEKSHCIA EVENDEMPAD LPSLAADFVE SKDVCKNYAE
361 AKDVFLGMFL YEYARRHPDY SVVLLLRLAK TYETTLEKCC AAADPHECYA KVFDEFKPLV
421 EEPQNLIKQN CELFEQLGEY KFQNALLVRY TKKVPQVSTP TLVEVSRNLG KVGSKCCKHP
481 EAKRMPCAED YLSVVLNQLC VLHEKTPVSD RVTKCCTESL VNRRPCFSAL EVDETYVPKE
541 FNAETFTFHA DICTLSEKER QIKKQTALVE LVKHKPKATK EQLKAVMDDF AAFVEKCCKA
601 DDKETCFAEE GKKLVAASQA ALGL[配列番号16]
【0211】
エクセンディン4-リンカー-ヒト血清アルブミンアミノ酸配列は配列番号17によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGAPPPSG GGGGSGGGGS GGGGSDAHKS
61 EVAHRFKDLG EENFKALVLI AFAQYLQQCP FEDHVKLVNE VTEFAKTCVA DESAENCDKS
121 LHTLFGDKLC TVATLRETYG EMADCCAKQE PERNECFLQH KDDNPNLPRL VRPEVDVMCT
181 AFHDNEETFL KKYLYEIARR HPYFYAPELL FFAKRYKAAF TECCQAADKA ACLLPKLDEL
241 RDEGKASSAK QRLKCASLQK FGERAFKAWA VARLSQRFPK AEFAEVSKLV TDLTKVHTEC
301 CHGDLLECAD DRADLAKYIC ENQDSISSKL KECCEKPLLE KSHCIAEVEN DEMPADLPSL
361 AADFVESKDV CKNYAEAKDV FLGMFLYEYA RRHPDYSVVL LLRLAKTYET TLEKCCAAAD
421 PHECYAKVFD EFKPLVEEPQ NLIKQNCELF EQLGEYKFQN ALLVRYTKKV PQVSTPTLVE
481 VSRNLGKVGS KCCKHPEAKR MPCAEDYLSV VLNQLCVLHE KTPVSDRVTK CCTESLVNRR
541 PCFSALEVDE TYVPKEFNAE TFTFHADICT LSEKERQIKK QTALVELVKH KPKATKEQLK
601 AVMDDFAAFV EKCCKADDKE TCFAEEGKKL VAASQAALGL[配列番号17]
【0212】
Val8-GLP-1-IgG1アミノ酸配列は配列番号18によって表される。
1 HVEGTFTSDV SSYLEGQAAK EFIAWLVKGR GAEPKSCDKT HTCPPCPAPE LLGGPSVFLF
61 PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV
121 SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SREEMTKNQV
181 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF
241 SCSVMHEALH NHYTQKSLSL SPGK[配列番号18]
【0213】
Val8-GLP-1-Cex-IgG1アミノ酸配列は配列番号19によって表される。
1 HVEGTFTSDV SSYLEGQAAK EFIAWLVKGR GSSGAPPPSA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号19]
【0214】
Val8-Glu22GLP-1-IgG1アミノ酸配列は配列番号20によって表される。
1 HVEGTFTSDV SSYLEEQAAK EFIAWLVKGR GAEPKSCDKT HTCPPCPAPE LLGGPSVFLF
61 PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV
121 SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SREEMTKNQV
181 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF
241 SCSVMHEALH NHYTQKSLSL SPGK[配列番号20]
【0215】
Val8-Glu22GLP-1-CEx-IgG1アミノ酸配列は配列番号21によって表される。
1 HVEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号21]
【0216】
Gly8-Glu22GLP-1-C2-IgG1アミノ酸配列は配列番号22によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGASSGAA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号22]
【0217】
Gly8-Glu22GLP-1-CEx-リンカー-IgG1アミノ酸配列は配列番号23によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSG GGGSGGGGSG GGGSAEPKSC
61 DKTHTCPPCP APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
121 GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK
181 GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS
241 DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK[配列番号23]
【0218】
Gly8-Glu22GLP-1-CEx-リンカー-IgG4アミノ酸配列は配列番号24によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSG GGGSGGGGSG GGGSAESKYG
61 PPCPSCPAPE FLGGPSVFLF PPKPKDTLMI SRTPEVTCVV VDVSQEDPEV QFNWYVDGVE
121 VHNAKTKPRE EQFNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKGLPSSIE KTISKAKGQP
181 REPQVYTLPP SQEEMTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS
241 FFLYSRLTVD KSRWQEGNVF SCSVMHEALH NHYTQKSLSL SLGK[配列番号24]
【0219】
Gly8-Glu22GLP-1-CEx-2リンカー-IgG1アミノ酸配列は配列番号25によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSG GGGSGGGGSG GGGSGGGGSG
61 GGGSGGGGSA EPKSCDKTHT CPPCPAPELL GGPSVFLFPP KPKDTLMISR TPEVTCVVVD
121 VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN
181 KALPAPIEKT ISKAKGQPRE PQVYTLPPSR EEMTKNQVSL TCLVKGFYPS DIAVEWESNG
241 QPENNYKTTP PVLDSDGSFF LYSKLTVDKS RWQQGNVFSC SVMHEALHNH YTQKSLSLSP
301 GK[配列番号25]
【0220】
Gly8-Glu22GLP-1-2リンカー-IgG1アミノ酸配列は配列番号26によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GGGGGSGGGG SGGGGSGGGG SGGGGSGGGG
61 SAEPKSCDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV
121 KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE
181 KTISKAKGQP REPQVYTLPP SREEMTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT
241 TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK
[配列番号26]
【0221】
Gly8-Glu22GLP-1-2CEx-IgG1アミノ酸配列は配列番号27によって表される。
1 HGEGTFTSDV SSYLEEQAAK EFIAWLVKGR GSSGAPPPSS SGAPPPSAEP KSCDKTHTCP
61 PCPAPELLGG PSVFLFPPKP KDTLMISRTP EVTCVVVDVS HEDPEVKFNW YVDGVEVHNA
121 KTKPREEQYN STYRVVSVLT VLHQDWLNGK EYKCKVSNKA LPAPIEKTIS KAKGQPREPQ
181 VYTLPPSREE MTKNQVSLTC LVKGFYPSDI AVEWESNGQP ENNYKTTPPV LDSDGSFFLY
241 SKLTVDKSRW QQGNVFSCSV MHEALHNHYT QKSLSLSPGK[配列番号27]
【0222】
Gly8-Glu22-Val25-Ile33GLP-1-CEx-リンカー-IgG1アミノ酸配列は配列番号28によって表される。
1 HGEGTFTSDV SSYLEEQAVK EFIAWLIKGR GSSGAPPPSG GGGSGGGGSG GGGSAEPKSC
61 DKTHTCPPCP APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
121 GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK
181 GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS
241 DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK[配列番号28]
【0223】
エクセンディン4-IgG1アミノ酸配列は配列番号29によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGAPPPSA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号29]
【0224】
エクセンディン4-C2-IgG1アミノ酸配列は配列番号30によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGASSGAA EPKSCDKTHT CPPCPAPELL
61 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
121 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
181 EEMTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF LYSKLTVDKS
241 RWQQGNVFSC SVMHEALHNH YTQKSLSLSP GK[配列番号30]
【0225】
エクセンディン4-リンカー-IgG1アミノ酸配列は配列番号31によって表される。
1 HGEGTFTSDL SKQMEEEAVR LFIEWLKNGG PSSGAPPPSG GGGSGGGGSG GGGSAEPKSC
61 DKTHTCPPCP APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
121 GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK
181 GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS
241 DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK[配列番号31]
【0226】
試験管内GLP-1類似体活性
【表5a−1】
【表5a−2】
【0227】
試験管内GLP-1類似体活性
【表5b−1】
【表5b−2】
【0228】
GLP/エクセンディン類似体の試験管内活性
【表6】
【0229】
実施例7:Val8-GLP-1-IgG1およびVal8-GLP-1-HSAの生体内薬物動態:
Val8-GLP-1-IgG1およびVal8-GLP-1-HSAの薬物動態研究をカニクイザルで行なった。サルに5.6ナノモル/kgの精製Val8-GLP-1-IgG1またはVal8-GLP-1-HSAを投与した。化合物は静脈内ボーラス投与として投与した。投与前および投与の0.083、0.25、0.5、1、4、8、12、24、48、72、96、120、144、168、および216時間後に、EDTAを含むチューブに血液を採取した。免疫反応性Val8-GLP-1の血漿濃度を、Val8-GLP-1(7-37)のN末端(7-16)領域に対して主たる特異性を持つポリクローナル抗血清を利用するラジオイムノアッセイを使って決定した。図9は、2頭のカニクイザルに単回静脈内投与した後のVal8-GLP-1-FcおよびVal8-GLP-1-リンカー-HSAの血漿濃度を表す。Fc融合タンパク質は約45時間の半減期を持ち、アルブミン融合タンパク質は約87時間の半減期を持っていた。
【0230】
実施例8:エクセンディン4-IgG1の生体内薬物動態:
カニューレを留置した正常雄ビーグル犬2頭を一晩絶食後に調べた。動脈および静脈血管アクセス口にアクセスして、橈側皮静脈にカテーテルを経皮的に挿入し、確保した。動物をケージに入れ、カテーテルを回り継手/テザー系にとりつけた。融合タンパク質エクセンディン4-IgG1(11.8μM)を含む溶液を、橈側皮静脈カテーテルを通して静脈内注射した(1.0nmol/kg)。次に、10mlの食塩水でカテーテルを洗浄した。2時間後に高血糖(150mg/dl)クランプを開始し、これを3時間続けた。この5時間の全体にわたって動脈血試料を採取し、融合タンパク質、グルコース、およびインスリンの血漿濃度の決定に使用した。
【0231】
この研究の結果を、両動物に食塩水を皮下ボーラス投与し、3時間後に3時間高血糖(150mg/dl)クランプ法を使って調べた過去の同様の研究で得られた結果と比較した。
【0232】
どちらの研究でも、血漿グルコース濃度はBeckmanグルコース分析機を使って決定した。血漿インスリン濃度は、Linco Research, Inc.の従業員がその研究室で開発されたRIAキットを使って決定した。データを図10および図11に示す。
【0233】
実施例9:Gly8-Glu22-GLP-1-CEx-リンカー-IgG1の生体内薬物動態:
3頭の正常雄ビーグル犬からなる2群に、0.1mg/kgのGly8-Glu22-GLP-1-CEx-リンカー-IgG1を皮下(SC)または静脈内(IV)投与した。IV群とSC群の両方について、投与の30分前から投与の216時間後まで集めた試料で、Gly8-Glu22-GLP-1-CEx-リンカー-IgG1免疫反応性の血漿濃度を、ラジオイムノアッセイによって決定した。次に、これらの濃度を使って、本明細書に記載の薬物動態パラメータを決定した。IV投与されたGly8-Glu22-GLP-1-CEx-リンカー-IgG1の平均消失半減期は約55時間であり、全身クリアランスは1.5mL/時/kgだった。SC投与されたGly8-Glu22-GLP-1-CEx-リンカー-IgG1の平均消失半減期は約38時間だった。
【特許請求の範囲】
【請求項1】
N末端およびC末端を持つ第1ポリペプチドと、第1ポリペプチドに融合しているN末端およびC末端を持つ第2ポリペプチドを有する異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)ヒトアルブミン、
b)ヒトアルブミン類似体、および
c)ヒトアルブミンの断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端に融合している異種融合タンパク質。
【請求項2】
N末端およびC末端を持つ第1ポリペプチドと、第1ポリペプチドに融合しているN末端およびC末端を持つ第2ポリペプチドを有する異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)ヒトアルブミン、
b)ヒトアルブミン類似体、および
c)ヒトアルブミンの断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端にペプチドリンカーを介して融合している異種融合タンパク質。
【請求項3】
ペプチドリンカーが、
a)グリシンリッチペプチド、
b)配列:[Gly-Gly-Gly-Gly-Ser]n(式中、nは1、2、3、4、5または6である)を持つペプチド、および
c)配列:[Gly-Gly-Gly-Gly-Ser]3を持つペプチド、
からなる群より選択される、請求項2記載の異種融合タンパク質。
【請求項4】
GLP-1化合物が、以下の式1[配列番号2]の配列を含む、請求項1、2、または3記載の異種融合タンパク質:
【化1】
式I(配列番号2)
[式中、
8位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
9位のXaaは、Glu、Asp、またはLysであり、
11位のXaaは、Thr、Ala、Gly、Ser、Leu、Ile、Val、Glu、Asp、またはLysであり、
14位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
16位のXaaは、Val、Ala、Gly、Ser、Thr、Leu、Ile、Tyr、Glu、Asp、Trp、またはLysであり、
17位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
18位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、Trp、Tyr、またはLysであり、
19位のXaaは、Tyr、Phe、Trp、Glu、Asp、Gln、またはLysであり、
20位のXaaは、Leu、Ala、Gly、Ser、Thr、Ile、Val、Glu、Asp、Met、Trp、Tyr、またはLysであり、
21位のXaaは、Glu、Asp、またはLysであり、
22位のXaaは、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
23位のXaaは、Gln、Asn、Arg、Glu、Asp、またはLysであり、
24位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Arg、Glu、Asp、またはLysであり、
25位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
26位のXaaは、Lys、Arg、Gln、Glu、Asp、またはHisであり、
27位のXaaは、Leu、Glu、Asp、またはLysであり、
30位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
31位のXaaは、Trp、Phe、Tyr、Glu、Asp、またはLysであり、
32位のXaaは、Leu、Gly、Ala、Ser、Thr、Ile、Val、Glu、Asp、またはLysであり、
33位のXaaは、Val、Gly、Ala、Ser、Thr、Leu、Ile、Glu、Asp、またはLysであり、
34位のXaaは、Asn、Lys、Arg、Glu、Asp、またはHisであり、
35位のXaaは、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
36位のXaaは、Gly、Arg、Lys、Glu、Asp、またはHisであり、
37位のXaaは、Pro、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、もしくはLysであるか、または欠失しており、
38位のXaaは、Ser、Arg、Lys、Glu、Asp、もしくはHisであるか、または欠失しており、
39位のXaaは、Ser、Arg、Lys、Glu、Asp、もしくはHisであるか、または欠失しており、
40位のXaaは、Gly、Asp、Glu、もしくはLysであるか、または欠失しており、
41位のXaaは、Ala、Phe、Trp、Tyr、Glu、Asp、もしくはLysであるか、または欠失しており、
42位のXaaは、Ser、Pro、Lys、Glu、もしくはAspであるか、または欠失しており、
43位のXaaは、Ser、Pro、Glu、Asp、もしくはLysであるか、または欠失しており、
44位のXaaは、Gly、Pro、Glu、Asp、もしくはLysであるか、または欠失しており、
45位のXaaは、Ala、Ser、Val、Glu、Asp、もしくはLysであるか、または欠失しており、
ただし、37、38、39、40、41、42、43、または44位のアミノ酸が欠失している場合は、そのアミノ酸の下流にある各アミノ酸も欠失している]。
【請求項5】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、6個を超えて含まない、請求項4記載の異種融合タンパク質。
【請求項6】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、5個を超えて含まない、請求項5記載の異種融合タンパク質。
【請求項7】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、4個を超えて含まない、請求項6記載の異種融合タンパク質。
【請求項8】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、3個を超えて含まない、請求項7記載の異種融合タンパク質。
【請求項9】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、2個を超えて含まない、請求項8記載の異種融合タンパク質。
【請求項10】
インスリン非依存性糖尿病患者処置用医薬を製造するための、請求項1〜9のいずれか一項に記載の異種融合タンパク質の使用。
【請求項11】
肥満患者処置用医薬を製造するための、請求項1〜9のいずれか一項に記載の異種融合タンパク質の使用。
【請求項1】
N末端およびC末端を持つ第1ポリペプチドと、第1ポリペプチドに融合しているN末端およびC末端を持つ第2ポリペプチドを有する異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)ヒトアルブミン、
b)ヒトアルブミン類似体、および
c)ヒトアルブミンの断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端に融合している異種融合タンパク質。
【請求項2】
N末端およびC末端を持つ第1ポリペプチドと、第1ポリペプチドに融合しているN末端およびC末端を持つ第2ポリペプチドを有する異種融合タンパク質であって、第1ポリペプチドがGLP-1化合物であり、第2ポリペプチドが、
a)ヒトアルブミン、
b)ヒトアルブミン類似体、および
c)ヒトアルブミンの断片
からなる群より選択され、かつ第1ポリペプチドのC末端が第2ポリペプチドのN末端にペプチドリンカーを介して融合している異種融合タンパク質。
【請求項3】
ペプチドリンカーが、
a)グリシンリッチペプチド、
b)配列:[Gly-Gly-Gly-Gly-Ser]n(式中、nは1、2、3、4、5または6である)を持つペプチド、および
c)配列:[Gly-Gly-Gly-Gly-Ser]3を持つペプチド、
からなる群より選択される、請求項2記載の異種融合タンパク質。
【請求項4】
GLP-1化合物が、以下の式1[配列番号2]の配列を含む、請求項1、2、または3記載の異種融合タンパク質:
【化1】
式I(配列番号2)
[式中、
8位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
9位のXaaは、Glu、Asp、またはLysであり、
11位のXaaは、Thr、Ala、Gly、Ser、Leu、Ile、Val、Glu、Asp、またはLysであり、
14位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
16位のXaaは、Val、Ala、Gly、Ser、Thr、Leu、Ile、Tyr、Glu、Asp、Trp、またはLysであり、
17位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
18位のXaaは、Ser、Ala、Gly、Thr、Leu、Ile、Val、Glu、Asp、Trp、Tyr、またはLysであり、
19位のXaaは、Tyr、Phe、Trp、Glu、Asp、Gln、またはLysであり、
20位のXaaは、Leu、Ala、Gly、Ser、Thr、Ile、Val、Glu、Asp、Met、Trp、Tyr、またはLysであり、
21位のXaaは、Glu、Asp、またはLysであり、
22位のXaaは、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
23位のXaaは、Gln、Asn、Arg、Glu、Asp、またはLysであり、
24位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Arg、Glu、Asp、またはLysであり、
25位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
26位のXaaは、Lys、Arg、Gln、Glu、Asp、またはHisであり、
27位のXaaは、Leu、Glu、Asp、またはLysであり、
30位のXaaは、Ala、Gly、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
31位のXaaは、Trp、Phe、Tyr、Glu、Asp、またはLysであり、
32位のXaaは、Leu、Gly、Ala、Ser、Thr、Ile、Val、Glu、Asp、またはLysであり、
33位のXaaは、Val、Gly、Ala、Ser、Thr、Leu、Ile、Glu、Asp、またはLysであり、
34位のXaaは、Asn、Lys、Arg、Glu、Asp、またはHisであり、
35位のXaaは、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、またはLysであり、
36位のXaaは、Gly、Arg、Lys、Glu、Asp、またはHisであり、
37位のXaaは、Pro、Gly、Ala、Ser、Thr、Leu、Ile、Val、Glu、Asp、もしくはLysであるか、または欠失しており、
38位のXaaは、Ser、Arg、Lys、Glu、Asp、もしくはHisであるか、または欠失しており、
39位のXaaは、Ser、Arg、Lys、Glu、Asp、もしくはHisであるか、または欠失しており、
40位のXaaは、Gly、Asp、Glu、もしくはLysであるか、または欠失しており、
41位のXaaは、Ala、Phe、Trp、Tyr、Glu、Asp、もしくはLysであるか、または欠失しており、
42位のXaaは、Ser、Pro、Lys、Glu、もしくはAspであるか、または欠失しており、
43位のXaaは、Ser、Pro、Glu、Asp、もしくはLysであるか、または欠失しており、
44位のXaaは、Gly、Pro、Glu、Asp、もしくはLysであるか、または欠失しており、
45位のXaaは、Ala、Ser、Val、Glu、Asp、もしくはLysであるか、または欠失しており、
ただし、37、38、39、40、41、42、43、または44位のアミノ酸が欠失している場合は、そのアミノ酸の下流にある各アミノ酸も欠失している]。
【請求項5】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、6個を超えて含まない、請求項4記載の異種融合タンパク質。
【請求項6】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、5個を超えて含まない、請求項5記載の異種融合タンパク質。
【請求項7】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、4個を超えて含まない、請求項6記載の異種融合タンパク質。
【請求項8】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、3個を超えて含まない、請求項7記載の異種融合タンパク質。
【請求項9】
GLP-1化合物が、GLP-1(7-37)OH、GLP-1(7-36)OH、またはエクセンディン4中の対応するアミノ酸と相違するアミノ酸を、2個を超えて含まない、請求項8記載の異種融合タンパク質。
【請求項10】
インスリン非依存性糖尿病患者処置用医薬を製造するための、請求項1〜9のいずれか一項に記載の異種融合タンパク質の使用。
【請求項11】
肥満患者処置用医薬を製造するための、請求項1〜9のいずれか一項に記載の異種融合タンパク質の使用。
【図1】


【図2−2】

【図5】
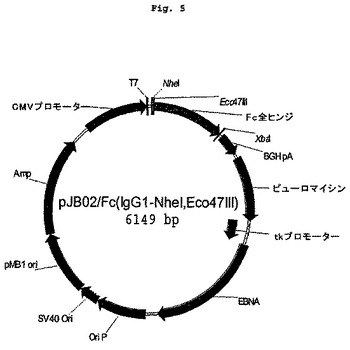
【図6】
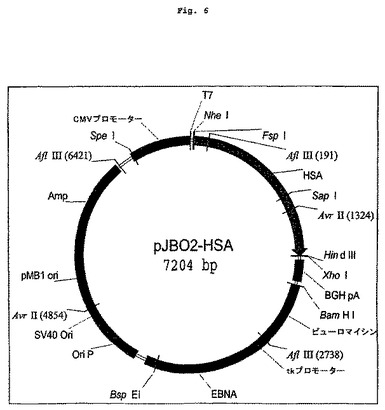
【図7】
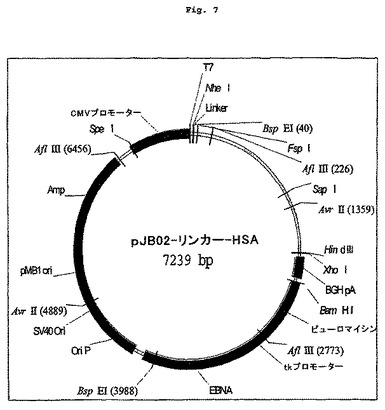
【図8】
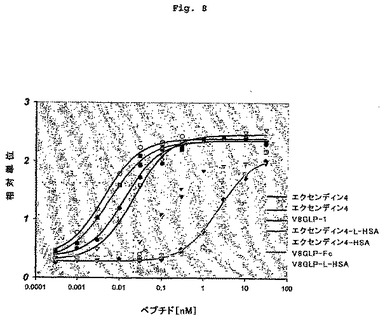
【図9】
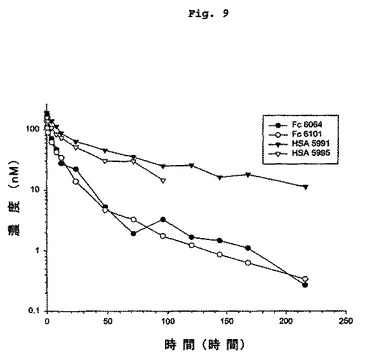
【図10−1】
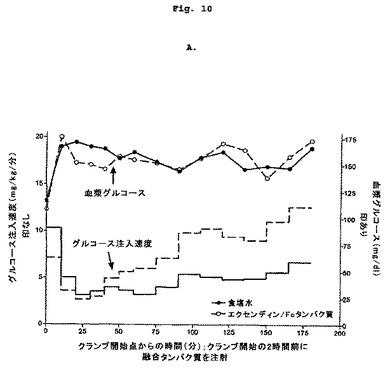
【図10−2】
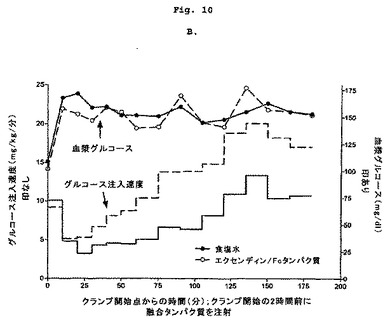
【図11−1】
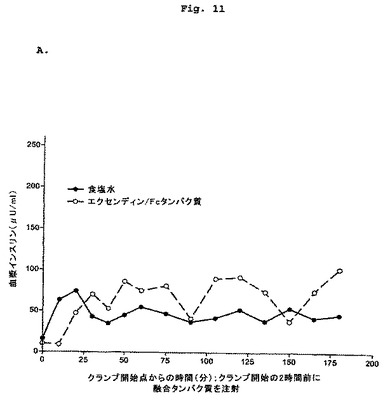
【図11−2】
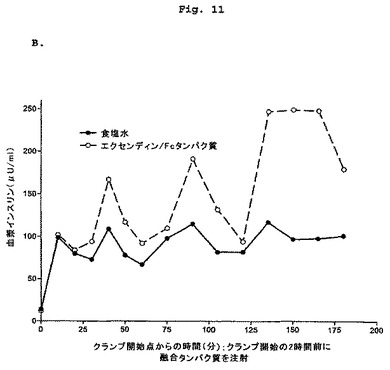
【図12】

【図13−2】

【図2−1】

【図3】
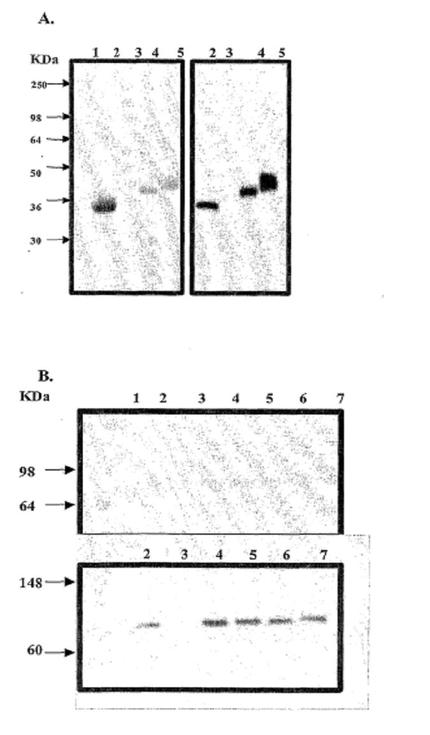
【図4】
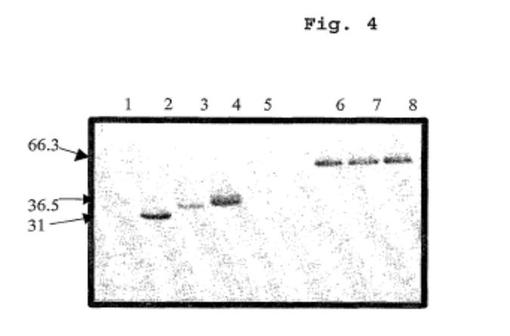
【図13−1】

【図2−2】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10−1】
【図10−2】
【図11−1】
【図11−2】
【図12】
【図13−2】
【図2−1】
【図3】
【図4】
【図13−1】
【公開番号】特開2011−6447(P2011−6447A)
【公開日】平成23年1月13日(2011.1.13)
【国際特許分類】
【出願番号】特願2010−185227(P2010−185227)
【出願日】平成22年8月20日(2010.8.20)
【分割の表示】特願2002−547963(P2002−547963)の分割
【原出願日】平成13年11月29日(2001.11.29)
【出願人】(594197872)イーライ リリー アンド カンパニー (301)
【Fターム(参考)】
【公開日】平成23年1月13日(2011.1.13)
【国際特許分類】
【出願日】平成22年8月20日(2010.8.20)
【分割の表示】特願2002−547963(P2002−547963)の分割
【原出願日】平成13年11月29日(2001.11.29)
【出願人】(594197872)イーライ リリー アンド カンパニー (301)
【Fターム(参考)】
[ Back to top ]