
GSK3β阻害効果に基づく抗がん剤の評価方法
【課題】グリコーゲン合成酵素キナーゼ3β(GSK3β)のがん細胞の生存・増殖への関与を明らかにし、がんの治療・診断のための新たな手段を提供する。
【解決手段】グリコーゲン合成酵素キナーゼ3β(GSK3β)の第216番目チロシン残基のリン酸化により活性化される前記酵素の活性に対する阻害効果を指標として、被験物質の抗がん剤としての効果を評価する方法。
【解決手段】グリコーゲン合成酵素キナーゼ3β(GSK3β)の第216番目チロシン残基のリン酸化により活性化される前記酵素の活性に対する阻害効果を指標として、被験物質の抗がん剤としての効果を評価する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、第216番目チロシン残基がリン酸化されたグリコーゲン合成酵素キナーゼ3β(GSK3β)の活性を阻害すること、またはGSK3βの第216番目チロシン残基のリン酸化を阻害することによりがん細胞の生存や増殖を抑制する方法、ならびに前記阻害効果に基づく新規抗がん剤の開発および抗がん剤の評価方法に関する。
【背景技術】
【0002】
近年、大腸がんの発生・進展の分子機構においてβ-カテニンの安定化による活性化が、rasがん遺伝子産物のがん化作用とともに有力な新規がん化シグナルとして注目されている(非特許文献1および2参照)。β-カテニンは細胞増殖にかかわるWntシグナル伝達系の中心的な実行因子であり、そのがん化作用はこれまでin vitroの実験系やトランスジェニック動物モデルを用いた多数の研究により支持されている(非特許文献3参照)。
【0003】
発明者らは、最近、腫瘍細胞核内におけるβ-カテニンの集積様式の違いから、ヒト大腸癌には2種類のβ-カテニン活性化パターンがあることを見出した(非特許文献4参照)。つまり、40-50%の大腸がん症例ではβ-カテニンの核内集積(NA)、すなわち活性化が腫瘍全体に認められる(NAd)のに対し、10-15%の症例ではその集積(活性化)が腫瘍浸潤先進部に限局して検出される(NAinv)。そして、このβ-カテニンのNAinv型活性化が大腸癌の病期の進行と術後の再発に有意に相関する。この結果は、大腸癌におけるβ-カテニンのがん化シグナル活性化パターンが、腫瘍の転移・再発や患者の術後生存率の信頼性の高い予測因子になり得ることを示唆する(非特許文献4および5参照)。
【0004】
グリコーゲン合成酵素キナーゼ3β(以下、「GSK3β」と記載する)は、このWnt/β-カテニンシグナル系の制御因子群(非特許文献2および3参照)の1つであり、生理的条件下の細胞においてβ-カテニンをリン酸化して、APCがん抑制蛋白質やアキシン(いずれもGSK3βの基質)などから構成されるユビキチン分解系に誘導する。このことは、GSK3β活性と大腸癌の仮説的接点として、GSK3βの第9セリン残基リン酸化に伴う不活性化が標的基質であるβ-カテニンをユビキチン分解系から回避させ、がん化シグナルの活性化に導くことを示唆する。
【0005】
GSK3βは様々な細胞内シグナル伝達を媒介する多機能セリン・スレオニンキナーゼ(リン酸化酵素)であり、エネルギー代謝、転写調節、細胞増殖・生存等を含む広範な細胞調節過程を制御する(非特許文献6−9参照)。つまり、Wnt/β-カテニンシグナルの制御は本酵素の多彩な機能の1つにすぎない。GSK3βは細胞内シグナル伝達を媒介する多くの蛋白質リン酸化酵素とは異なり、正常細胞において活性化型として存在し、種々の刺激によってその活性が抑制的に制御されている。GSK3βの基質には、c-Jun、c-Myc等の多様ながん化関連転写因子(非特許文献10および11参照)や、β-カテニン、Gli蛋白質等の原がん遺伝子産物 (非特許文献12および13参照)が含まれ、これら基質の不活性化を介して細胞の腫瘍性形質転換や腫瘍発生を妨げる。また、GSK3βはNF-κB細胞不死化シグナルを介して細胞の生存に本質的な役割も果たしている(非特許文献15および16参照)。こうしたGSK3βの機能から、GSK3β遺伝子を細胞の異常増殖マーカーとして利用することを報告した特許出願もある(特許文献1参照)。
【0006】
最近、GSK3βがインスリン非依存性糖尿病(non-insulin-dependent diabetes mellitus [NIDDM]:2型糖尿病)やアルツハイマー病をはじめとする神経変性疾患の発症にも関与していることがわかってきた(非特許文献14参照)。そのため、GSK3β阻害剤を神経変性疾患や2型糖尿病やがん等の治療に応用する特許出願もなされているが(特許文献2および3参照)、いずれも特定化合物のGSK3β阻害効果と既知の知見から当該化合物のこれら疾患への適用を予測するものにすぎず、とくにがんについては実証されていない。
【0007】
上記のとおり、GSK3βはWnt/β-カテニンシグナル伝達の負の制御因子であることから、GSK3βを介したシグナル伝達制御機能の異常ががんにおけるβ-カテニンシグナル活性化パターンに関連しているかどうかを明らかにすることは重要な課題である。一方、本酵素遺伝子のノックアウト動物の解析により、GSK3βはNF-κB経路を介した細胞の生存と増殖を維持する作用が報告されている(非特許文献12および13参照)。また、前立腺癌由来細胞株において、GSK3β阻害がTRAIL(tumor necrosis factor-related apoptosis-inducing ligand)による薬剤感受性を亢進させたり、細胞増殖を抑制する報告もある(非特許文献17、18および19参照)。このように、細胞生存・増殖やがん化にかかわる基幹的な細胞内シグナルに対して相反する作用が提唱されていることから、GSK3βがWntシグナルとは異なる独自の分子メカニズムを介してがん細胞に影響を及ぼしている可能性があるのではないかと想定される。しかしながら、これまでGSK3βは細胞の腫瘍性形質転換や腫瘍発生に対し抑制的に作用するという仮説から、GSK3β阻害剤には潜在的発がん性が予測されている。それゆえ、GSK3βとがんの発生・進展との関与については報告が少なく、詳細な検討がなされてはいなかった。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特表2004−528837号公報
【特許文献2】特表2004−504838号公報
【特許文献3】特表2004−507545号公報
【非特許文献】
【0009】
【非特許文献1】Jass JR, Whitehall VLJ, Young J, Leggett BA. Emerging concepts in colorectal neoplasia. Gastroenterology 2002; 123(3): 862-76.
【非特許文献2】Wong NA, Pignatelli M. β-catenin- a linchpin in colorectal carcinogenesis. Am J Pathol 2002; 160(2): 389-401.
【非特許文献3】Polakis P. The oncogenic activation of beta-catenin. Curr Opin Genet Dev 1999; 9(1): 15-21.
【非特許文献4】Ougolkov AV, Yamashita K, Mai M, Minamoto T. Oncogenic β-catenin and MMP-7 (matrilysin) cosegregate in late-stage clinical colon cancer. Gastroenterology 2002; 122(1): 60-71.
【非特許文献5】Zhang B, Ougolkov A, Yamashita K, Takahashi Y, Mai M, Minamoto T. β-catenin and ras oncogenes detect most human colorectal cancers. Clin Cancer Res 2003; 9(8): 3073-9.
【非特許文献6】Manoukian AS, Woodgett JR. Role of glycogen synthase kinase-3 in cancer: regulation by Wnts and other signaling pathways. Adv Cancer Res 2002; 84: 203-29.
【非特許文献7】Doble BW, Woodgett JR. GSK-3: tricks of the trade for a multi-tasking kinase. J Cell Sci 2003; 116(Pt 7): 1175-86.
【非特許文献8】Harwood AJ. Regulation of GSK-3; a cellular multiprocessor. Cell 2001; 105(7): 821-4.
【非特許文献9】Kim L, Harwood A, Kimmel AR. Receptor-dependent and tyrosine phosphatase-mediated inhibition of GSK3 regulates cell fate choice. Dev Cell 2002; 3(4): 523-32.
【非特許文献10】Morton S, Davis RJ, McLaren A, Cohen P. A reinvestigation of the multiple phosphorylation of the transcription factor c-Jun. EMBO J 2003; 22(15): 3876-86.
【非特許文献11】Gregory MA, Qi Y, Hann SR. Phosphorylation by glycogen kinase-3 controls c-myc proteolysis and subnuclear localisation. J Biol Chem 2003; 278(51): 51606-12.
【非特許文献12】Price MA, Kalderon D. Proteolysis of the Hedgehog signaling effector Cubitus interruptus requires phosphorylation by glycogen synthase kinase 3 and casein kinase 1. Cell 2002; 108(6): 823-35.
【非特許文献13】Jia J, Amanai K, Wang G, Tang J, Wang B, Jiang J. Shaggy/GSK3 antagonizes Hedgehog signalling by regulating cubitus interruptus. Nature 2002; 416(6880): 548-52.
【非特許文献14】Cohen P, Goedert M. GSK3 inhibitors: development and therapeutic potential. Nat Rev Drug Discov 2004; 3(6): 479-87.
【非特許文献15】Hoeflich KP, Luo J, Rubie EA, Tsao MS, Jin O, Woodgett JR. Requirement for glycogen synthase kinase-3β in cell survival and NF-κB activation. Nature 2000; 406(6791): 86-90.
【非特許文献16】Schwabe RF, Brenner DA. Role of glycogen synthase kinase-3 in TNF-α-induced NF-κB activation and apoptosis in hepatocytes. Am J Physiol Gastrointest Liver Physiol 2002; 283(1): G204-11.
【非特許文献17】Liao X, Zhang L, Thrasher JB, Du J, Li B. Glycogen synthase kinase-3β suppression eliminates tumor necrosis factor-related apoptosis-inducing ligand resistance in prostate cancer. Mol Cancer Ther 2003; 2(11): 1215-1222.
【非特許文献18】Wang L, Lin H-K, Hu Y-C, Xie S, Yang L, Chang C. Suppression of androgen receptor-mediated transactivation and cell growth by the Glycogen synthase kinase 3β in prostate cells. J Biol Chem 2004; 279(31): 32444-32452.
【非特許文献19】Mazor M, Kawano Y, Zhu H, Waxman J, Kypta RM. Inhibition of glycogen synthase kinase-3 represses androgen receptor activity and prostate cancer cell. Oncogene 2004; 23(47): 7882-7892
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明は、グリコーゲン合成酵素キナーゼ3β(GSK3β)とがんの発生・進展との関与を明らかにし、がんの治療・診断のための新たな手段を提供することにある。
【0011】
発明者らは、上記課題を検討するために、大腸癌細胞株および同一大腸がん患者の非がん組織と癌組織におけるGSK3βの発現と第9セリン残基リン酸化解析を通じた酵素活性化の程度を比較解析した。さらに、GSK3βの発現と活性をそれぞれ転写調節的ならびに薬理学的に修飾することにより大腸癌細胞の増殖に及ぼす本酵素の影響を研究した。
【0012】
その結果、大部分の大腸癌組織や大腸癌細胞株においてそれぞれの非がん粘膜や細胞(HEK293)に比べてGSK3βとその第216チロシン残基がリン酸化された活性化型phospho-GSK3βTyr216分画の発現が亢進し、一方、第9セリン残基がリン酸化された不活性化型phospho-GSK3βSer9分画の発現はほとんど認められなかった。また、in vitroにおいて大腸癌細胞株や神経膠芽腫細胞株等のがん細胞の生存と増殖は低分子量タイプのGSK3β阻害剤の投与により用量(濃度)依存的に抑制され、同様にGSK3βに対するRNA干渉(RNAi)によっても抑制された。さらに、大腸がんモデル動物においてもGSK3β阻害剤投与による腫瘍組織の用量依存的縮小が観察された。このことは、従来の知見とは異なり、GSK3βの発現や活性ががん(とくに大腸がん)の治療や予防の新たな標的となりうることを提示している。
【課題を解決するための手段】
【0013】
すなわち本発明は、GSK3βの発現や第216番目チロシン残基リン酸化による活性化阻害を通じて本酵素活性を抑制することにより、がん細胞の生存や増殖を抑制する方法に関する。
【0014】
前記方法は、式(I)または(II)の構造を有する化合物もしくはその薬理学的に許容される塩をがん細胞に作用させることによって実施できる。
【0015】
【化1】

【0016】
(ここで、Q1は-(CH2)n-Ar-O-(nは1〜5の整数、Arは置換または非置換アリール)であり、Q2はハロゲン、アルキル、ヒドロキシ、アルコキシ、アリールオキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、モノアルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルまたはアルキルスルホニルから選ばれる基である)
【0017】
【化2】
【0018】
(ここで、R1は水素、アルキル、アラルキル、アルコキシアルキル、ヒドロキシアルキル、ハロアルキル、アミノアルキル、モノアルキルアミノアルキル、ジアルキルアミノアルキル、アジドアルキル、アシルアミノアルキル、アルキルスルホニルアミノアルキル、アリールスルホニルアミノアルキル、メルカプトアルキル、アルキルチオアルキル、グルコピラノシル、カルボキシアルキル、アルコキシカルボニルアルキル、アミノカルボニルアルキル、ヒドロキシアルキルチオアルキル、メルカプトアルキルチオアルキル、アリールチオアルキルまたはカルボキシアルキルチオアルキルであり、
R2は水素、アルキル、アラルキル、アルコキシアルキル、ヒドロキシアルキル、ハロアルキル、アミノアルキル、モノアルキルアミノアルキル、ジアルキルアミノアルキル、アシルアミノアルキル、アルキルスルホニルアミノアルキル、アリールスルホニルアミノアルキル、メルカプトアルキル、アルキルチオアルキル、カルボキシアルキル、アルコキシカルボニルアルキル、アミノカルボニルアルキル、アルキルチオまたはアルキルスルフィニルであり、
R3はハロゲン、アルキル、ヒドロキシ、アルコキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、アルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルおよびアルキルスルホニルからなる群より選ばれる1または2以上の置換基で置換されたフェニル基であり、
R4、R5、R6およびR7は、互いに独立して、水素、ハロゲン、アルキル、ヒドロキシ、アルコキシ、アリールオキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、モノアルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルまたはアルキルスルホニルである)
【0019】
前記化合物の好ましい一例としては、例えば、下式に示されるN-(4-メトキシベンジル)-N'-(5-ニトロ-1,3-チアゾール-2-イル)ウレアまたは3-(2,4-ジクロロフェニル)-4-(1-メチル-1H-インドール-3-イル)-1H-ピロール-2,5-ジオンを挙げることができる。
【0020】
【化3】
【0021】
本発明はまた、上記した化合物もしくはその薬理学的に許容される塩を有効成分として含む、がんの治療用医薬組成物も提供する。本明細書では、前記医薬組成物の大腸がん、胃がん、膵がん、肝細胞がん、および神経膠芽腫細胞に対する効果や、ヌードマウス移植大腸癌に対する薬理効果を示したが、本発明の医薬組成物の効果はこれに限られるものでなく、あらゆるがんに対してその効果が期待される。
【0022】
本発明はまた、GSK3βの第216番目チロシン残基のリン酸化による本酵素の活性化に対する阻害効果を指標として、被験物質の抗がん剤としての効果を評価する方法を提供する。
【0023】
前記方法は、たとえば、以下の工程により実施できる:
1)細胞を被験物質の添加または非添加条件下で培養する;
2)上記細胞内における第216チロシン残基リン酸化GSK3β蛋白質分画の発現量を検出するか、あるいはGSK3β酵素活性を検出・測定する
3)被験物質の添加または非添加条件下における、上記の酵素発現量あるいは活性の相違に基づいて、該被験物質の抗がん剤としての効果を評価する。
【0024】
前記方法において、細胞はがん患者から単離された細胞または癌細胞株、例えば、SW480, SW620, HT29, LoVo, HCT116, SW48およびRKO等の大腸癌細胞株やA172, T98G, U251, U87等の神経膠芽腫細胞株等を用いることができる。
【0025】
本発明の方法は、また、以下の工程によっても実施できる:
1)がんモデル動物を被験物質の投与または非投与の条件下で飼育する。
2)上記動物の血液または細胞中における第216チロシン残基リン酸化GSK3β蛋白質分画の発現量を検出する。
3)被験物質の投与または非投与条件下における、上記発現量の相違と移植腫瘍の増殖の変化に基づいて、該被験物質の抗がん剤としての効果を評価する。
【0026】
前記方法において、動物は齧歯類、特にマウスが好ましい。
また、本発明の方法において、蛋白質の発現量は、ウエスタンブロット法、ドットブロット法、スロットブロット法、ELISA法、およびRIA法から選ばれるいずれか一つの方法によって検出することができる。
【発明の効果】
【0027】
本発明によれば、GSK3β抑制効果に基づく新規の抗がん剤の提供とそのスクリーニングが可能になる。また、GSK3βレベルを指標とするがんの早期診断が可能になる。
【図面の簡単な説明】
【0028】
【図1】図1は、大腸癌細胞株とヒト胎児腎細胞 HEK293における、(A)ウエスタンブロット法による総GSK3βとphospho-GSK3βSer9(不活性化型)およびphospho-GSK3βTyr216(活性化型)の発現をそれぞれ特異的抗体(BiosciencesおよびCell Signaling Technology)を用いて解析した結果、(B) GSK3β活性を合成ヒトβ-カテニン蛋白を基質にして、in vitro kinase assayにより検出した (Adler V, et al. Proc Natl Acad Sci USA 1997;94:1686-91に記載されている検出方法を用いた) 結果、(C) GSK3β(シグナル)の上流で活性化されるプロテインキナーゼB(PKB/Akt)の活性化を抗phospho-AktThr308抗体や抗phospho-AktSer473抗体(Cell Signaling Technology)を用いて検出した結果。
【図2】図2は、HEK293細胞(上4枚の図)とSW480大腸癌細胞株(下4枚の図)における総GSK3β(赤色蛍光)とphospho-GSK3βSer9(緑色蛍光)の二重免疫蛍光細胞化学染色による発現と局在の観察結果を示す。図の尺度線は100μmに相当し、画像はすべて同一倍率で撮影された。
【図3】図3は、大腸がん症例の正常粘膜および腫瘍組織における、総GSK3β、phospho-GSK3βSer9(不活性化型)およびphospho-GSK3βTyr216(活性化型)分画の発現レベルを検出した結果を示す。各パネル上段の数字は患者ID番号(表2と対応)。N:非がん(正常)粘膜組織;T:癌組織。
【図4】図4は、AR-A014418とSB-216763のHEK293細胞および各種大腸癌細胞株のin vitroにおける生存、増殖とアポトーシスにおよぼす影響(効果)を解析した結果を示す。A, B, E: 0日目から特定時間(24, 48, 72および96時間)経過した時点でMTSアッセイにより計測した各ウェルの生細胞数。C, D:抗PCNA(proliferating cell nuclear antigen)抗体による免疫蛍光法とHoechstにより2重染色し、蛍光顕微鏡で観察したときの、全観察細胞に占める増殖細胞核(PCNA陽性)(D)あるいは、アポトーシスを生じている核(C)の比率。
【図5】図5は、GSK3βを標的とするsiRNAのHEK293細胞(上)とSW480大腸癌細胞(下)のin vitroでの細胞生存率におよぼす効果を示す。
【図6】図6は、大腸癌細胞移植動物モデルに対するGSK3β阻害剤投与による治療実験プロトコールの概略を示している。
【図7】図7は、異なる用量のGSK3β阻害剤(AR-A014418あるいはSB-216763)、あるいはDMSO投与開始して5週間後における大腸癌細胞移植マウスの写真である。
【図8】図8は、大腸癌細胞移植マウスにおいて、異なる用量のGSK3β阻害剤(AR-A014418あるいはSB-216763)、あるいはDMSOを投与開始後の移植腫瘍体積の経時的変化を示すグラフである。
【図9】図9は、各種の神経膠芽腫細胞株(T98G、A172、U251、U87)の細胞生存におよぼす、異なる濃度のGSK3β阻害剤(AR-A014418あるいはSB-216763)、あるいはDMSOの作用を経時的に示すグラフである。
【図10】図10は、各種の神経膠芽腫細胞株(A172、T98G、U251、U87)および非がん細胞株(HEK293)に対するGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響を示す。(A)は、AR-A014418処理後96時間の生存腫瘍細胞数を示すグラフである。(B)は、AR-A014418あるいはSB-216763処理後の腫瘍細胞におけるアポトーシス誘導を示すグラフである。(C)は、AR-A014418あるいはSB-216763処理後の増殖期細胞分画(bromodeoxy-uridine [BrdU]取り込み量)を示すグラフである。
【図11】図11は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)の細胞老化に対する、それぞれ25μMのGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響をDMSOの場合と比較して、β-galactosidase活性の指標であるx-gal染色により調べた結果を示す。
【図12】図12は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)をそれぞれ25μMのGSK3β阻害剤(AR-A014418あるいはSB-216763)あるいはDMSOにより処理して72時間後の老化細胞比率の変化を、x-gal染色により調べた結果を示す。
【図13】図13は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)における、human telomerase reverse transcriptase (hTERT)の発現に対するGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響を調べた結果である。
【図14】図14は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)における、telomerase活性に対するGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響を調べた結果である。
【図15】図15は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)における、(A)telomerase活性や(B)hTERT mRNAの発現に対するGSK3β発現阻害の影響を、siRNAトランスフェクション(小分子干渉RNA導入)により調べた結果を示す。
【図16】図16は、本発明に係る非放射性試験管内GSK3β酵素活性検出法(Non-RI in vitro kinase assay)の原理と概要を示す。
【図17】図17は、非放射性試験管内GSK3β酵素活性検出法により各種細胞株におけるGSK3β酵素活性を検出・比較した結果を示す。
【図18】図18は、HEK293細胞と各種の培養癌細胞株:SW480, HCT116, HT29(大腸癌);MKN-28, NKPS, TMK-1, NUGC-4(胃癌);MIA-PaCa-2, Capan-1, BxPC-3(膵臓癌);HEP-G2(肝細胞癌)におけるGSK3β酵素活性を非放射性試験管内GSK3β酵素活性検出法により検出した結果を示している。P,C:同一細胞からの抽出蛋白質に対して抗GSK3β抗体により免疫沈降した検体(P)と、同抗体の代わりに非特異的マウスIgGを添加して同様に解析した検体(陰性対照)(C)。
【図19】図19は、大腸癌細胞株(SW480とHCT116)から免疫沈降したGSK3β酵素活性に対する、それぞれ25μMのGSK3β阻害剤(AR-A014418あるいはSB-216763)と陰性対照であるDMSOの阻害効果を、非放射性試験管内GSK3β酵素活性検出法を応用して検証した結果を示している。
【0029】
本明細書は、本願の優先権の基礎である特願2005−133号の明細書に記載された内容を包含する。
【発明を実施するための形態】
【0030】
以下、本発明について詳細に説明する。
1.グリコーゲン合成酵素キナーゼ3β阻害剤によるがんの治療
本発明は、グリコーゲン合成酵素キナーゼ3β(以下、「GSK3β」と記載することもある)阻害剤の大腸がん治療用医薬組成物としての利用を提供する。
1.1 GSK3β
GSK3βはβ-カテニン原がん遺伝子産物を含む一連の細胞調節蛋白質をリン酸化し不活性化する抑制因子として作用する多機能蛋白質リン酸化酵素である。発明者らは、(1)大腸がん患者の癌組織や大腸癌細胞株で特異的なGSK3βの発現亢進が見られること、(2)GSK3β阻害剤の投与により用量依存的に大腸癌細胞株の生存と増殖が抑制され、アポトーシスが誘導されるること、(3)大腸癌細胞株の増殖がGSK3βに対するsiRNAによって抑制されることを初めて見出し、GSK3β阻害剤が抗がん剤として有用であること、また、GSK3β抑制効果が抗がん剤評価の指標として有用であることを実証した。
【0031】
本発明に係るGSK3βにはヒトGSK3βのほか、マウスGSK3βなど、そのオーソログも含まれるが、本発明の目的においてはヒトGSK3βが最も好ましい。GSK3β遺伝子やGSK3β蛋白質の配列情報は公知であり、公共データベースであるGenBankより容易に入手することができる。例えば、マウスGSK3β遺伝子と対応するGSK3β蛋白質のアミノ酸配列は、Accession No. BC060743およびAAH60743.1、あるいはBC006936およびAAH06936.1として開示されている。また、ヒトGSK3β遺伝子と対応するGSK3β蛋白質のアミノ酸配列は、Accession No.NM_002093およびNP_002084として開示されている。もちろん、これらの配列は一例であって、本発明のGSK3β遺伝子やGSK3β蛋白質はこれら配列に限定されない。
【0032】
1.2 GSK3β阻害剤
本明細書において、GSK3β阻害剤とは、GSK3βの機能を物理的あるいは化学的に阻害する物質を意味する。こうしたGSK3β阻害剤については、既に多くの化合物が同定されているが、その構造と作用メカニズムは多岐にわたる(Meijer L, Flajolet M, Greengard P. Trends in Pharmacol Sci 2004; 25 (9): 471-480 参照)。
【0033】
こうしたGSK3β阻害剤の構造と製造方法は、例えば、以下の特許出願あるいは登録特許に詳細に記載されている:特表2005-536517記載のピラゾロピリミジン、特表2005-535593記載のチアゾリジンジチオン誘導体、特表2005-534713記載のジヒドロピラゾロピリジン化合物、特表2005-533066記載のアゾールメチリデンシアニド誘導体、特表2005-531609記載のシスインドリル−マレイミド誘導体、特表2005-526835記載の2−置換−1,3−チアゾール化合物、特表2005-526814記載のモルホリン誘導体、特表2005-519087記載の1−[アルキル]、1−[(ヘテロアリール)アルキル]および1−[(アリール)アルキル]−7−(ピリミジン−4−イル)−イミダゾ[1,2−a]ピリミジン−5(1H)−オン誘導体、特表2005-519086記載のヘテロアリール置換2−ピリジニルおよび2−ピリミジニル−6,7,8,9−テトラヒドロピリミド[1,2−a]ピリミジン−4−オン誘導体、特表2005-516961記載のオキシインドール誘導体、特表2005-516960記載のオキシインドール誘導体、特表2005-516948記載のキナゾリン類、特表2005-513082記載のイソキノリン類、特表2005-509641記載のヒドロキシアルキルインドロカルバゾール化合物、特表2005-505515記載のアリールアミン、特表2005-504810記載および特表2005-504809記載の置換2−ピリジニル−6,7,8,9−テトラヒドロピリミド[1,2−a]ピリミジン−4−オンおよび7−ピリジニル−2,3−ジヒドロイミダゾ[1,2−a]ピリミジン−5(1H)オン誘導体、特表2005-501800記載のジヒドロピラゾロピリジン化合物、特表2004-53831記載の2,4−ジアミノチアゾール誘導体、特表2004-536111記載の4−(4−メトキシベンジル)−N′−(5−ニトロ−1,3−チアゾール−2−イル)尿素、特表2004-536110記載の複素環式アミン、特表2004-527486記載のピリミジン化合物、特表2004-522777記載の2−アリルアミノ−ピリミジン、特表2004-516297記載のトリアジン類、特表2004-515499記載のインダゾリル置換ピロリン化合物、特表2004-505078記載の3−インドリル−4−フェニル−1H−ピロール−2,5−ジオン誘導体、特表2003-528095記載の2−アミノ−3−(アルキル)−ピリミドン誘導体、特表2003-527303記載のピリミジンまたはピリジン類、特表2003-516974記載のピラジン誘導体、特表2002-527419記載のピロール−2,5−ジオン類、再表01/081345記載の芳香族アミド化合物、および特表平01-233281記載のピロール類等。当業者であれば、これらの記載に基づいて、所望のGSK3β阻害剤を製造し、使用することができるであろう。
【0034】
GSK3βの活性は種々の段階:(a) 転写後修飾(転写後の第9セリンと第216チロシン残基のリン酸化による制御)、(b) タンパク複合体との相互作用、(c) 基質プライミング、(d) 細胞内分布、で制御されており、上記したGSK3β阻害剤はそのいずれかの段階でGSK3βに作用するものと思われる(Meijer L, Flajolet M, Greengard P. Trends Pharmacol Sci 2004;25 (9), 471-480 参照)。
【0035】
発明者らは、2つの代表的GSK3β阻害剤(AR-A014418およびSB-216763)について、これらががんに対して特異的な抑制作用を有することを、in vivoおよびin vitroの両面で実証した。また、これらのGSK3β阻害剤が、がん細胞において転写後修飾(リン酸化)を受けたGSK3βの活性を阻害しうることを確認した。
【0036】
(1) AR-A014418
AR-A014418は下記の式で示されるチアゾール誘導体である(CAS NO: 487021-52-3、WO2003004478、特表2004-536111号参照)。
【0037】
【化4】
【0038】
AR-A014418は、試薬としてCalbiochem等により販売されており、一般に入手可能である。
【0039】
上記化合物に限定されず、下記の一般式(I)で示されるチアゾール誘導体もまた、同様のGSK3β阻害作用を有する限り、AR-A014418と同様、本発明に係るがん細胞の増殖抑制やがんの治療用医薬組成物として利用可能である。
【0040】
【化5】
【0041】
ここで、Q1は-(CH2)n-Ar-O-(nは1〜5の整数、Arは置換または非置換アリール)であり、Q2はハロゲン、アルキル、ヒドロキシ、アルコキシ、アリールオキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、モノアルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルまたはアルキルスルホニルから選ばれる基である。
【0042】
(2) SB-216763
SB-216763は、化学名を3-(2,4-Dichlorophenyl)-4-(1-methyl-1H-indol-3-yl)-1H-pyrrole-2,5-dioneと称し、下記の式で示されるピロール誘導体である(EP328026、特表01-233281号 実施例45参照)。
【0043】
【化6】
【0044】
SB-216763もまた、試薬としてSigma-Aldrich等により販売されており、一般に入手可能である。
【0045】
なお上記化合物に限定されず、下記の一般式(II)で示されるピロール類もまた、同様のGSK3β阻害作用を有する限り、SB-216763と同様、本発明に係るがん細胞の増殖抑制やがんの治療用医薬組成物として利用可能である。
【0046】
【化7】
【0047】
(ここで、R1は水素、アルキル、アラルキル、アルコキシアルキル、ヒドロキシアルキル、ハロアルキル、アミノアルキル、モノアルキルアミノアルキル、ジアルキルアミノアルキル、アジドアルキル、アシルアミノアルキル、アルキルスルホニルアミノアルキル、アリールスルホニルアミノアルキル、メルカプトアルキル、アルキルチオアルキル、グルコピラノシル、カルボキシアルキル、アルコキシカルボニルアルキル、アミノカルボニルアルキル、ヒドロキシアルキルチオアルキル、メルカプトアルキルチオアルキル、アリールチオアルキルまたはカルボキシアルキルチオアルキルであり、
R2は水素、アルキル、アラルキル、アルコキシアルキル、ヒドロキシアルキル、ハロアルキル、アミノアルキル、モノアルキルアミノアルキル、ジアルキルアミノアルキル、アシルアミノアルキル、アルキルスルホニルアミノアルキル、アリールスルホニルアミノアルキル、メルカプトアルキル、アルキルチオアルキル、カルボキシアルキル、アルコキシカルボニルアルキル、アミノカルボニルアルキル、アルキルチオまたはアルキルスルフィニルであり、
R3はハロゲン、アルキル、ヒドロキシ、アルコキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、アルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルおよびアルキルスルホニルからなる群より選ばれる1または2以上の置換基で置換されたフェニル基であり、
R4、R5、R6およびR7は、互いに独立して、水素、ハロゲン、アルキル、ヒドロキシ、アルコキシ、アリールオキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、モノアルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルまたはアルキルスルホニルである)
【0048】
さらに上記した化合物の薬学的に許容しうる塩、例えば塩基性の酸付加塩、無機もしくは有機の酸またはアルカリ金属塩、アルカリ土類金属塩等もまた、本発明に係るがん細胞の増殖抑制や医薬組成物に利用可能である。
【0049】
1.3 GSK3βのリン酸化とがんの治療
GSK3βの生理的活性はリン酸化(第9セリン残基、不活性化型;第216チロシン残基、活性化型)により調節されている。大腸癌細胞や組織においてGSK3βの発現と活性はβ-カテニン活性化(核内集積)とは関係なく亢進し、非がん細胞や組織で検証された第9セリン残基のリン酸化(不活性化型)による酵素活性制御は認められなかった。一方で、癌細胞や癌組織特異的に第216チロシン残基がリン酸化されたGSK3β(活性化型)分画が高頻度に発現していた。
【0050】
培養細胞のGSK3β活性を薬理学的阻害剤(AR-A014418、 SB-216763)を用いて抑制すると、阻害剤の用量依存的に癌細胞のアポトーシスが誘導され、増殖は阻止されるが、非がん細胞株(HEK293)ではこうした効果は認められなかった。RNA干渉法でGSK3β発現をノックダウンしても、薬理的阻害剤の効果と同様に癌細胞の生存と増殖が特異的に抑制された。以上の結果より、GSK3β活性の増強、特に第216チロシン残基リン酸化GSK3β(活性化型)分画の高発現が、癌細胞の生存・増殖を維持・促進するというがん化のあらたなメカニズムが明らかとなった。
すなわち、第216チロシン残基のリン酸化により活性化されているGSK3β活性を薬理学的ならびに転写調節的技法を駆使して修飾することにより、がんの治療や予防が可能になることがはじめて示された。
【0051】
2.被験物質の抗がん剤としての効果を評価する方法
本発明は、GSK3β遺伝子、またはGSK3β蛋白質に対する阻害効果、とくに第216番目チロシン残基のリン酸化により活性化されている本酵素活性に対する阻害効果を指標として、被験物質の抗がん剤としての効果を評価する方法を提供する。
【0052】
前記方法は、1つの被験物質について、その投与(添加)および非投与条件下における、阻害効果を比較評価するものであってもよいし、2つ以上の被験物質についての同様な効果の比較評価であってもよい。あるいは、前記遺伝子や蛋白質の発現量や活性と抗腫瘍効果との相関関係が経験的に確立されれば、その関係に基づいて、比較対照なしに絶対評価するものであってもよい。
【0053】
本発明の評価方法において、GSK3β遺伝子やGSK3β蛋白質に対する阻害効果は、GSK3β遺伝子やGSK3β蛋白質の発現量を指標として評価してもよいし、GSK3β遺伝子やGSK3β蛋白質の活性(安定性含む)を指標として評価しても良い。また、評価系はin vivo系であってもよいし、in vitro系であってもよい。以下、本発明の評価方法の具体的な実施方法について説明する。
【0054】
2.1 培養細胞
本発明の評価方法で用いられる細胞は、GSK3β遺伝子を高発現する哺乳動物細胞であればとくに限定されない。好ましくは哺乳動物由来の癌細胞(好ましくは、初代培養細胞)や樹立された癌細胞株が好ましい。前記哺乳動物としては、ヒト、マウス、ラットまたはハムスター等が好ましく、ヒトまたはマウスがより好ましい。また、GSK3β遺伝子をそのプロモーター領域とともに導入し、該遺伝子を高発現するように組換えた細胞など、人為的に形質転換された細胞を使用してもよい。
【0055】
細胞は、被検物質の添加または非添加条件下で培養する。培養方法は特に限定されず、当該細胞に適した培養方法を適宜選択すればよい。培養細胞への被検物質の添加方法や添加量も特に限定されず、上記細胞の培養前あるいは培養期間中の適当な時点で、被検物質を培養培地に添加し一定期間培養する。被験物質添加後の培養期間もとくに限定されないが、好ましくは30分〜24時間である。
【0056】
2.2 がんモデル動物
本発明の方法で用いられるがんモデル動物は、非ヒト哺乳動物であれば特に限定されないが、齧歯類が好ましく、マウスが特に好ましい。そのようながんモデル動物は市販のものであっても、公知の方法にしたがって作製されたものでもよい。例えば、大腸がんモデルマウスは、既報(Gupta RA., et al. Activation of nuclear hormone receptor peroxisome proliferator-activated receptor-δ accelerates intestinal adenoma growth. Nat Med 2004; 10(3): 245-247)に従って作製することができる。
【0057】
前記動物は、被験物質の投与または非投与条件下で適当な期間飼育を行う。動物への被験物質の投与量は特に限定されず、被験物質の性状や動物の体重に合わせて、適宜用量を設定すればよい。また、動物への被験物質の投与方法および投与期間も特に限定されず、被験物質の性状に合わせて、適宜その投与経路と投与期間を設定すればよい。
【0058】
被験物質の投与または非投与条件下で飼育された前記動物の血液または細胞を単離し、該血液または細胞中のGSK3β遺伝子や蛋白質の発現量を検出する。検出対象とする細胞としては、患部、すなわち癌組織細胞が好ましい。
【0059】
2.3 GSK3β遺伝子の検出
GSK3β遺伝子の検出方法としては、例えば、単離された血液または細胞からまず全RNAを抽出し、該全RNA中におけるmRNAの発現量を検出する方法を挙げることができる。
【0060】
全RNA中のGSK3β遺伝子の発現量は、得られた全RNAよりcRNAまたはcDNAを調製し、これを適当な標識化合物でラベルすることにより、そのシグナル強度として検出することができる。遺伝子の発現量の検出方法は特に限定されず、以下に説明する公知の方法から適宜選択して用いればよい。
【0061】
遺伝子チップ、アレイ、メンブレンフィルター等の固相化試料を用いた解析では、検出対象である遺伝子が固相化されている、市販のGeneChip(Affimetrix)、インテリジーン(宝酒造)、フィルター製マイクロアレイ アトラスシステム(クローンテック)を用いて簡便に実施できる。あるいは、公知の方法(Lipshutz RJ, et al. Nat Genet 1999; 21 (suppl) 20-24)に基づいて作製されたGSK3β固相化試料を用いて解析してもよい。
【0062】
RT-PCR法やその1つであるリアルタイムPCR(TaqMan PCR)法は微量なRNAを高感度かつ定量的に検出できるという点で本発明の評価方法に好適である。リアルタイムPCR(TaqMan PCR)法では、5’端を蛍光色素(レポーター)で、3’端を蛍光色素(クエンチャー)で標識した、目的遺伝子の特定領域にハイブリダイズするオリゴヌクレオチドプローブが使用される。該プローブは、通常の状態ではクエンチャーによってレポーターの蛍光が抑制されている。この蛍光プローブを目的遺伝子に完全にハイブリダイズさせた状態で、その外側からTaq DNAポリメラーゼを用いてPCRを行う。Taq DNAポリメラーゼによる伸長反応が進むと、そのエキソヌクレアーゼ活性により蛍光プローブが5’端から加水分解され、レポーター色素が遊離し、蛍光を発する。リアルタイムPCR法は、この蛍光強度をリアルタイムでモニタリングすることにより、鋳型DNAの初期量を正確に定量することができる。
【0063】
上記以外に、遺伝子発現量を解析する方法としては、例えば、サブトラクション法(Sive HL, John T. St. Nucleic Acids Res 1988; 16: 10937、Wang Z, Brown DD. Proc Natl Acad Sci U. S. A. 1991; 88: 11505-11509)、ディファレンシャル・ディスプレイ法(Liang P, Pardee AB. Science 1992; 257, 967-971、Liang P, Averboukh L, Keyomarsi K, Sager R, Pardee AB. Cancer Res 1992; 52: 6966-6968)、ディファレンシャル・ハイブリダイゼーション法(John T. St, Davis RW. Cell 1979; 16: 443-452)、また、適当なプローブを用いたクロスハイブリダイゼーション法(”Molecular Cloning, A Laboratory Manual”Maniatis, T., Fritsch, E.F., Sambrook, J. (1982) Cold Spring Harbor Laboratory Press)等を挙げることができる。
【0064】
上記方法のほか、GSK3β遺伝子のプロモーター支配下に、該プロモーター活性の検出を可能にする遺伝子(以下「レポーター遺伝子」と記載する。)を利用して、間接的にGSK3β遺伝子やGSK3β蛋白質の発現を検出することもできる。レポーター遺伝子としては、例えば、クロラムフェニコールアセチルトランスフェラーゼ、ホタルルシフェラーゼ、β−ガラクトシダーゼ、分泌型アルカリホスファターゼ、緑色蛍光蛋白質(green-fluorescent protein)等を利用することができる。
【0065】
以上のような、一過的な遺伝子導入法を利用した試験方法とは別に、レポーター遺伝子、およびGSK3β遺伝子の発現ベクターで、宿主細胞を二重に形質転換した細胞を利用した試験方法も採択可能である。この場合には、pIND(インビトロジェン社製)やpTet-On(クロンテック社製)等の発現ベクターを利用して、GSK3β遺伝子の発現を誘導する条件下で該レポーター遺伝子の発現が促進されるような細胞株を樹立することが必要となる。得られた細胞株を、GSK3β遺伝子の発現が誘導される条件下におくと、該遺伝子の発現依存的にレポーター遺伝子の転写が促進される。したがって、レポーター遺伝子の発現が可能な条件下において、培地中に任意の被検物質を添加した場合と添加しない場合でのレポーター遺伝子の発現量変化をみれば、GSK3βの発現量が評価できる。
【0066】
2.4 GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の検出
GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の発現量は、該蛋白質に特異的に結合する抗体を用いて検出することができる。抗体を利用した蛋白質の検出方法は特に限定されないが、ウエスタンブロット法、ドットブロット法、スロットブロット法、ELISA法、およびRIA法から選ばれるいずれか一つの方法であることが好ましい。
【0067】
検体である血液または細胞(細胞抽出液として使用する)は、必要に応じて高速遠心を行うことにより不溶性の物質を除去した後、以下のようにELISA/RIA用試料やウエスタンブロット用試料として調製する。
【0068】
ELISA/RIA用試料は、例えば、回収した血清をそのまま使用するか、緩衝液で適宜希釈したものを用いる。ウエスタンブロット用(電気泳動用)試料は、例えば、細胞抽出液をそのまま使用するか、緩衝液で適宜希釈して、SDS−ポリアクリルアミドゲル電気泳動用の2−メルカプトエタノールを含むサンプル緩衝液(シグマ−アルドリッチ社製等)と混合したものを用いる。ドット/スロットブロット用試料は、例えば、回収した細胞抽出液そのもの、または緩衝液で適宜希釈したものを、ブロッティング装置を使用するなどして、直接メンブレンへ吸着させたものを用いる。
【0069】
本工程で用いられる「GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)に特異的に結合する抗体(以下、「抗GSK3β抗体」と記載する。)」は、公知の方法にしたがって調製してもよいし、市販のものを用いてもよい。抗体は、常法により、抗原となるGSK3β蛋白質、あるいはそのアミノ酸配列から選択される任意のポリペプチドを用いて動物を免疫し、該動物生体内に産生される抗体を採取、精製することによって得ることができる。また、公知の方法(例えば、Kohler and Milstein, Nature 256, 495-497, 1975、Kennet, R. ed., Monoclonal Antibody p.365-367, 1980, Prenum Press, N.Y.)にしたがって、抗GSK3β抗体を産生する抗体産生細胞とミエローマ細胞とを融合させることによりハイブリドーマを樹立し、これよりモノクローナル抗体を得ることもできる。
【0070】
抗体作製用の抗原としては、GSK3β蛋白質またはその少なくとも6個の連続した部分アミノ酸配列からなるポリペプチド、あるいはこれらに任意のアミノ酸配列や担体(例えば、N末端付加するキーホールリンペットヘモシアニン)が付加された誘導体を挙げることができる。
【0071】
前記抗原ポリペプチドは、GSK3β蛋白質を遺伝子操作により宿主細胞に産生させることによって得ることができる。具体的には、GSK3β遺伝子を発現可能なベクターを作製し、これを宿主細胞に導入して該遺伝子を発現させればよい。
【0072】
抗GSK3β抗体は、それを直接標識するか、または該抗体を一次抗体とし、該一次抗体を特異的に認識する(抗体を作製した動物由来の抗体を認識する)標識二次抗体と協同で検出に用いられる。
【0073】
前記標識の種類として好ましいものは、酵素(アルカリホスファターゼまたは西洋ワサビペルオキシダーゼ)またはビオチン(ただし二次抗体のビオチンにさらに酵素標識ストレプトアビジンを結合させる操作が加わる)であるが、これらに限定されない。標識二次抗体(または標識ストレプトアビジン)としては、予め標識された抗体(またはストレプトアビジン)が、各種市販されている。なお、RIAの場合は125I等の放射性同位元素で標識された抗体を用い、測定は液体シンチレーションカウンター等を用いて行う。
【0074】
これら標識された酵素の活性を検出することにより、抗原の発現量が測定される。アルカリホスファターゼまたは西洋ワサビペルオキシダーゼで標識する場合、これら酵素の触媒により発色する基質や発光する基質が市販されている。
【0075】
発色する基質を用いた場合、ウエスタンブロット法やドット/スロットブロット法を利用すれば、目視で検出できる。ELISA法では、市販のマイクロプレートリーダーを用いて各ウェルの吸光度(測定波長は基質により異なる)を測定し、定量することが好ましい。また上述の抗体作製に使用した抗原の希釈系列を調製し、これを標準抗原試料として他の試料と同時に検出操作を行い、標準抗原濃度と測定値をプロットした標準曲線を作成することにより、他の試料中の抗原濃度を定量することも可能である。
【0076】
一方、発光する基質を使用した場合は、ウエスタンブロット法やドット/スロットブロット法においては、X線フィルムまたはイメージングプレートを用いたオートラジオグラフィーや、インスタントカメラを用いた写真撮影により検出することができる。また、デンシトメトリーやモレキュラー・イメージャーFxシステム(バイオラッド社製)等を利用した定量も可能である。さらに、ELISA法で発光基質を用いる場合は、発光マイクロプレートリーダー(例えば、バイオラッド社製等)を用いて酵素活性を測定する。
【0077】
2.5 被験物質の評価
被験物質の投与(添加)または非投与(非添加)における、GSK3β遺伝子またはGSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の発現量の相違に基づいて、該被験物質の抗がん剤としての効果を評価する。
【0078】
すなわち、被験物質の投与条件下で非投与条件下よりもGSK3β遺伝子またはGSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の発現量が有意に減少している場合、該被験物質は抗がん剤として有用であると評価できる。ここで、「有意に減少している」とは、例えば、被験物質の投与および非投与条件下でのGSK3β遺伝子またはGSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の発現量に統計的有意差(p<0.05)があることを意味する。
【0079】
2.6 非放射性試験管内GSK3β酵素活性検出法(Non-RI in vitro kinase assay)
GSK3βの生理的活性はリン酸化(第9セリン残基、不活性化型;第216チロシン残基、活性化型)により調節されているので、各細胞におけるGSK3β活性はin vitro kinase assayあるいは、第216チロシン残基リン酸化ペプチド特異抗体による免疫ブロッティングのいずれかの手法により検出あるいは推定することができる。また第216チロシン残基リン酸化GSK3β蛋白質分画の発現量は、GSK3βの酵素活性を相対的(あるいは、間接的)に反映しているに過ぎないため、GSK3β活性を直接、検出・測定する手法は、本発明におけるGSK3β活性阻害効果の評価方法において重要といえる。発明者らは、このGSK3βの活性測定の新たな手段として、非放射性試験管内GSK3β酵素活性検出法を開発した。
【0080】
本検出法の概要は、図16に示すとおりである。まず、一定量の細胞抽出液に抗GSK3β特異的抗体を加え、GSK3β蛋白質を免疫沈降させる(図16のS)。コントロールとして、上記細胞抽出液に同濃度の非特異的IgG(抗GSK3β抗体を作成した動物種で、たとえばマウスIgG)を加える(図16のC)。遠心後、上清の細胞抽出液を除去し、免疫沈降物をリン酸緩衝液とリン酸化反応用緩衝液で十分に洗浄する。ついで、リン酸化反応用緩衝液の存在下で、組換えヒトβカテニン蛋白質(基質)と(非放射性)ATPを加え、リン酸化反応を行う。その後、この反応液にSDS電気泳動用バッファー加えて加熱・変性したものをウエスタンブロッティングにより解析し、GSK3β認識配列に特異的な抗リン酸化βカテニンペプチド抗体によりリン酸化組み換えヒトβカテニン蛋白質を検出する。この手法により検出されるβカテニン蛋白質のリン酸化の程度(ウエスタンブロッティングのシグナル強度)から、当該細胞(抽出液中)のGSK3β活性を計測することができる。組換えヒトβカテニン蛋白質をそのヒスチジン基を介してビーズ(resins)等に固定化すれば、ウエスタンブロッティングを用いることなく、on-resins colorimetric assayも可能となる。
【0081】
3.抗がん剤評価用キット
本発明の方法を実施するためのキットとして、例えば、以下のa)〜f)の少なくとも一つ以上を含むキットを挙げることができる。
a)GSK3β遺伝子またはGSK3β遺伝子を特異的に増幅するための、15〜30塩基長の連続したオリゴヌクレオチドプライマー
b)GSK3β遺伝子またはGSK3β遺伝子に特異的に結合し、該遺伝子を検出するための20〜1500塩基長の連続したポリヌクレオチドプローブ
c)上記b)記載のポリヌクレオチドプローブが固定された固相化試料
d)GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)またはそのGSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)に特異的に結合し、該蛋白質を検出するための抗体
e)上記d)記載の抗体に特異的に結合しうる二次抗体
f)GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)
【0082】
前記a)記載のプライマーは、上記遺伝子の塩基配列に基づき、市販のプライマー設計ソフトを用いる等、常法にしたがい容易に設計し、増幅することができる。
【0083】
前記b)記載のプローブは、ノーザンハイブリダイゼーション法であれば、20塩基長程度の1本鎖オリゴヌクレオチドか2本鎖DNAが好適に用いられる。マイクロアレイであれば、100〜1500塩基長程度の2本鎖DNA、または20〜100塩基長程度の1本鎖オリゴヌクレオチドが好適に用いられる。Affimetrix社のGene Chipシステムであれば25塩基長程度の1本鎖オリゴがよい。これらは、特にGSK3β遺伝子の配列特異性が高い部分に特異的にハイブリダイズするプローブとして設計することが好ましい。これらのプローブや前述のプライマーは、適当な標識によりラベル(例えば、酵素標識、放射性標識、蛍光標識等)されていてもよく、またビオチン、リン酸、アミン等により修飾されていてもよい。
【0084】
前記c)記載の固相化試料は、前記b)記載のプローブをガラス板、ナイロンメンブレン、マイクロビーズ、シリコンチップ等の固相に固定することにより作製される。
【0085】
前記d)およびe)記載の抗体やf)の蛋白質は、2.4に記載した方法により作製することができる。抗体は、適当な標識によりラベル(例えば、酵素標識、放射性標識、蛍光標識等)されていてもよいし、ビオチン等により適当に修飾されていてもよい。
【0086】
本発明のキットは上記した構成要素のほか、必要に応じて、ハイブリダイゼーション、プローブの標識、あるいはラベル体の検出のための試薬、反応用緩衝液、酵素基質等、本発明の評価方法に実施に必要な他の要素を含んでいてもよい。
【0087】
4.がんの診断方法
本発明者らは、ヒトGSK3β遺伝子やGSK3β蛋白質は、がん(特に大腸がん)患者の病変組織で特異的に活性化されていることを確認した。したがって、被験者から単離された検体中の、これら遺伝子や蛋白質の発現量を測定すれば、該被験者のがんの発症や病態を予測することができる。遺伝子や蛋白質の発現量の測定は、基本的に2で記載した方法にしたがって実施することができる。
【実施例】
【0088】
<実施例1> 各種細胞株および大腸がん患者の腫瘍におけるGSK3β活性とGSK3β阻害剤の影響
1.対象と方法
細胞株:
7種類の大腸癌細胞株(表1:SW480, SW620, HT29, LoVo, HCT116, SW48およびRKO)およびヒト胎児腎細胞株(HEK293)はAmerican Type Culture Collection (ATCC,Manassas,VA)から購入し、推奨プロトコールに従って培養・維持した。各細胞株はコンフルエントになる前の指数関数的増殖期に採取し、リン酸緩衝液(PBS)で洗浄・遠心して−80℃で保管した。GSK3βとphospho-GSK3βSer9の免疫細胞学的染色を行うために、カバーグラス上で培養した各細胞は4%パラフォルムアルデヒドで固定した。
【0089】
大腸がん症例と組織検体:
対象患者は、2002年および2003年に金沢大学がん研究所病院で大腸がんが切除された20例の患者(男性13名、女性7名:年齢は44-93歳(平均73歳))であり、本研究に登録されることを同意し、インフォームドコンセントを提出した者である。各患者の新鮮手術標本から採取した1組の正常粘膜と腫瘍組織検体は、液体窒素を用いてすみやかに凍結し、-80℃で保管した。手術標本は10%中性緩衝ホルマリンで固定し、組織病理学的検索に供して腫瘍の性状やTNM分類に準拠した病期を決定した。各腫瘍の組織学的特性を代表するパラフィン切片は免疫組織化学的に解析し、β-カテニンの活性化を判定した。
【0090】
ウエスタンブロッティング解析:
凍結保存してあった手術標本検体や培養細胞集塊(ペレット)から細胞蛋白質を、蛋白質分解酵素阻害剤と脱リン酸酵素阻害剤(いずれもSigma-Aldrich)を混入した溶解液(CelLyticTM-MT,Sigma-Aldrich)により抽出した。各検体の蛋白質濃度はCoomasie Protein Assay Reagent (Pierce)を用いてBradford法により測定した。
【0091】
抽出された蛋白質100 μgをSDS-ポリアクリルアミド(10%)ゲル電気泳動(SDS-PAGE)で分離し、ニトロセルロース膜(Amersham)に転写後、それぞれ1000倍に希釈したウサギポリクローナル抗体(Cell Signaling TechnologyおよびBD Biosciences)を反応させ、化学発光検出試薬(ECL(登録商標),Amersham)で可視化することにより、第9セリン残基がリン酸化されたGSK3β分画(phospho-GSK3βSer9)および第216チロシン残基がリン酸化された分画(phospho-GSK3βTyr216)をそれぞれ検出した。ついで、ラットGSK3βのN-末端側第1-160アミノ酸残基に対して作成されたマウスモノクローナル抗体(2500倍希釈,BD Transduction Laboratories)を反応させ、同様にして(ヒト)総GSK3βを検出した。
【0092】
GSK3β(シグナル)の上流で活性化される代表的キナーゼとして、プロテインキナーゼB(PKB、あるいはAkt)の活性化をphospho-AktThr308やphospho-AktSer473に対する抗体(それぞれ1000倍希釈、Cell Signaling Technology)を用いて、同様に検出した。
【0093】
GSK3βのin vitro kinase assay:
それぞれの細胞株から抽出した1mgの蛋白質から、免疫沈降法によりGSK3βを調整し、その酵素活性を組換えヒトβ-カテニン蛋白質を基質にして、32P標識ATPを用い、既報(Adler V, et al. Proc Natl Acad Sci USA 1997;94:1686-91)にしたがってin vitro kinase assayにより検出した。β-カテニン蛋白質の放射性リン酸化シグナルはオートラジオグラフィーにより検出した。
【0094】
免疫蛍光細胞化学的染色:
HEK293細胞と各大腸癌細胞株をカバーグラス上で培養し、免疫蛍光染色により総GSK3βとphospho-GSK3βSer9を検出した。一次抗体はウエスタンブロット法で用いた抗体(100倍希釈)を使用し、二次抗体としてそれぞれ200倍希釈したCy3標識抗マウスIgG (H+L)抗体およびFITC標識抗ウサギIgG (H+L)抗体(いずれもJackson ImmunoResearch)を使用した。また、核染色にはHoechst 33342(Hoechst)を使用した。免疫蛍光染色のすべての手法は既報(Shakoori A, et al. Biochem Biophys Res Commun 2003; 312(3): 850-7)に準じて行った。染色された細胞はCCDデジタルカメラ(オリンパスDP70, Olympus)に接続した蛍光顕微鏡(オリンパスAX80, Olympus)により観察・記録した。
【0095】
GSK3βSer9とK-ras遺伝子における突然変異の検出:
大腸癌培養細胞株と手術標本から採取した正常粘膜および腫瘍組織からゲノムDNAをプロティナーゼK消化とフェノール・クロロフォルム処理により抽出・精製した。GSK3βの第9セリン残基をコードするコドン9を含むエクソン1を上流プライマー(5’-ATTCGCGAAGAGAGTGATCAT-3’:配列番号1)と下流プライマー(5’-CACTGCTAACTTTCATGCTGC-3’:配列番号2)を用いて94℃ 1分、55℃ 1分、72℃ 1分の35サイクルプログラムとそれに続く72℃ 10分間伸長反応によりPCR増幅した。
【0096】
GSK3βコドン9(TCC)の突然変異はPCR産物の制限酵素Mnl I(認識配列:CCTC[N]7;New England Biolabs)消化による制限酵素断片長多型(PCR-RFLP)により解析した。この制限酵素は110塩基対の野生型コドン9のPCR産物をそれぞれ52と58塩基対の断片に切断し、一方、コドン9の第1あるいは第2塩基に変異を有するPCR産物は切断できない。K-ras遺伝子コドン12と13の変異は既報(Zhang B, et al. Clin Cancer Res 2003; 9(8): 3073-9.)にしたがい、それぞれ制限酵素Mva I(Takara)とBgl I(TOYOBO)を用いて、ミスマッチプライマーによるPCR産物のPCR-RFLP解析により検出した。
【0097】
β-カテニン活性化の免疫組織化学的解析:
各症例において、大腸原発腫瘍の主要な組織学的特徴を代表する組織切片をアビジン-ビオチン複合体法を用いて免疫組織化学的に検索し、既報(Ougolkov AV, et al. Gastroenterology 2002; 122(1): 60-71)にしたがい、β-カテニンがん化シグナル活性化を判定した。抗β-カテニン抗体(BD Transduction Laboratories)は100倍希釈して使用した。原発腫瘍におけるβ-カテニンの発現は発明者らが最近、定義した3種類の特徴的パターン:細胞膜発現型(M)、正常の腺窩と同じ発現を示すもの;び慢性核内集積型(NAd)、β-カテニンの核内集積を示す癌細胞が腫瘍全体に認められるもの;腫瘍浸潤先進部核内集積型(NAinv)、β-カテニンの核内集積を示す癌細胞が腫瘍浸潤先進部(腫瘍の浸潤最深部で間質と接する部分)に限局して認められ、それ以外の腫瘍部分における癌細胞はM型のβ-カテニン発現を示すもの、にしたがって分類した。
【0098】
β-カテニンは核内に移行すると例外を除いて造腫瘍活性型であるという根拠(Wong NA, et al. Am J Pathol 2002; 160(2): 389-401., Polakis P. Curr Opin Genet Dev 1999; 9(1): 15-21.)に基づいて、本研究ではβ-カテニンの核内集積はその造腫瘍活性を示すものと見做した。経験豊富な2名の研究者(AV,BZ)が対象腫瘍におけるGSK3βの活性に関する情報を知らされない状態で、それぞれ個別に腫瘍におけるβ-カテニン活性化パターンを判定した。そして、両者の判定はすべての腫瘍について一致した。
【0099】
細胞増殖解析:
各大腸癌細胞株とHEK293細胞を96穴培養プレートの各ウェルに3 x 103個播種し、対応する培地中で一晩、培養した(この状態が0日)。既知のGSK3β阻害剤であるAR-A014418(N-(4-methoxybenzyl)-N'-(5-nitro-1,3-thiazol-2-yl)urea, Calbiochem)とSB-216763(3-(2,4-dichlorophenyl)-4-(1-methyl-1H-indol-3-yl)-1H-pyrrole-2,5-dione, Sigma-Aldrich)はジメチルスルフォキシド(DMSO)に溶解した。第0日に各細胞株の培地を交換し、種々の濃度(5, 10, 25, 50μM)になるようにAR-A014418あるいはSB-216763を添加した。対照として、細胞を含まないウェルに同濃度のAR-A014418あるいはSB-216763を添加したものと、細胞を含むウェルにDMSOのみを添加したものを同時に解析した。
【0100】
0日目から特定時間(24, 48, 72および96時間)経過した時点で各ウェルの生細胞数をMTS*生存細胞測定キット(CellTiter 96TM Aqueous, Promega Corp)を用いて、同社のプロトコールにしたがって計測した。すなわち、各ウェルに20μLの直前に調製したMTS混合液を添加し、5% CO2の湿潤条件下において37℃で3時間反応させ、490nm波長の吸光度をTHERMOmaxマイクロプレートリーダー(Molecular Devices)を用いて計測した。各細胞は、それぞれの濃度について6ウェルずつ測定を行い、その平均吸光度を生細胞数として算出した。
*3-[4,5-dimethylthiazol-2-yl]-5-[3-carboxy-methoxyphenyl]-2H-tetrazolium inner salt
【0101】
siRNAトランスフェクション(小分子干渉RNA導入)実験:
ヒトGSK3βを標的とする4種類の2本鎖siRNAを混合したもの(siRNA SMARTpoolTM GSK-3β)と陰性対照のsiRNA(siGLOTM RISK-Free siRNA)はDharmacon RNA Tchnologies社(Lafayette, CO)により合成・供給された。それぞれ1 x 105個のHEK293細胞と各大腸癌細胞株を6穴細胞培養プレートに播き、1晩培養した。siRNA導入効果が最適となるような条件を検定するために、指定されたプロトコールに準拠して、約50%コンフルエントの細胞にTransIT(登録商標)-TKOトランスフェクション試薬(Mirus Corporation)を用いて異なる濃度のGSK3β特異的siRNAあるいは同濃度の陰性対照siRNAを導入した。siRNA導入のGSK3β発現に対する効果を調べるために、異なる濃度のそれぞれのsiRNAを導入して4日後に収集した細胞から蛋白質を抽出し、GSK3αとGSK3βをともに認識する抗ヒトGSK3抗体(1,000倍希釈;Upstate Biotechnology)を用いてウエスタンブロット法により解析した。
【0102】
この予備実験により決定されたsiRNAの最適条件において、GSK3β標的siRNA導入の細胞増殖に及ぼす効果を前項と同様にして細胞増殖解析法により解析した。すなわち、3 x 103個のHEK293細胞と各大腸癌細胞株を96穴細胞培養プレートの各ウェルに播き、対応する培地中で一晩、培養した。その後、最適濃度のGSK3β siRNAあるいは陰性対照siRNAを各細胞に導入した。当日から起算して指定時間(24, 48, 72および96時間)後に各ウェル中の生細胞数をMTS細胞増殖解析キット(Promega)を用いて計測した。
【0103】
2.結果
2.1 各種細胞株におけるGSK3β活性
本研究で用いた大腸癌細胞株はいずれもHEK293細胞に比べて高いGSK3βの発現レベルを示した。全ての大腸癌細胞株において、GSK3β特異的抗体によるウエスタンブロット法により検出可能な不活性化型phosph-GSK3βSer9分画の発現は認められなかったが、逆に活性化型のphospho-GSK3βTyr216分画の発現はより高かった(図1A)。また、in vitro kinase assayにより、これらの大腸癌細胞株におけるGSK3βの酵素活性が確認された(図1B)。これらの結果から、全ての大腸癌細胞株においてGSK3βは恒常的に活性化状態にあることが示された。
【0104】
大腸癌細胞株で認められた結果とは対照的に、HEK293細胞ではphospho-GSK3βSer9分画の発現が検出された。なお、この細胞ではβ-カテニンが安定した低発現を示すよう生理的に制御されていることを発明者らはこれまでに証明している(Spiegelman VS, et al., Mol Cell 2000; 5(5): 877-82, Ougolkov A, et al., J Natl Cancer Inst 2004; 96(15): 1161-70.)。
【0105】
免疫蛍光細胞化学的染色により、GSK3βはHEK293細胞とすべての大腸癌細胞の細胞質に局在し、一方、phospho-GSK3βSer9はHEK293細胞にのみ検出された(図2)。これらの細胞株における総GSK3βとphospho-GSK3βSer9の発現レベルはウエスタンブロット法の結果(図1A)と一致していた。以上の結果より、β-カテニンがAPC不活性化(SW480,SW620, LoVo)あるいはβ-カテニン遺伝子(CTNNB1)変異(HCT116、SW48)のいずれかによってGSK3βによるリン酸化から回避されて活性化されている大腸癌細胞株(表1)において、(総)GSK3βの過剰発現とその不活性化型であるphosph-GSK3βSer9分画の低発現、および活性化型であるphospho-GSK3βTyr216分画のが高発現していることが示された。
【0106】
【表1】
【0107】
2.2 大腸がん患者の腫瘍組織におけるGSK3β活性
樹立された培養細胞株は長期間の細胞発育が可能になるよう適応し、そして実際の生体における同種の腫瘍の特性を必ずしも反映しなくなっていることがあるので、20例の大腸がん患者から採取した正常大腸粘膜と腫瘍組織におけるGSK3βの発現と第9セリン残基および第216チロシン残基のリン酸化を解析した。この解析で得られた結果をそれぞれの腫瘍におけるβ-カテニン活性化の有無やそのパターンと、臨床的ならびに病理組織学的因子と比較検討した。その結果、図3に示すように、ほとんどの症例において腫瘍におけるGSK3βの発現とphospho-GSK3βTyr216分画は同一症例の正常粘膜におけるものに比べて高いレベルであった。phospho-GSK3βSer9分画は非腫瘍性粘膜において高頻度に高いレベルで検出されたが、症例番号74と79を除いてほとんどの症例の腫瘍においてこの不活性化型酵素分画はわずかに検出されるのみかあるいはまったく検出されなかった(表2)。
【0108】
大腸癌におけるβ-カテニンの過剰発現と核内集積の相関を示してきた発明者らのこれまでの報告(前掲)に基づいて、前記した方法で、β-カテニンの活性化はその腫瘍核内集積に基づいて判定した。その結果、β-カテニン活性化は13例(65%)の腫瘍に検出され、そのうち7例はNAd型、6例はNAinv型であった。大腸癌細胞株における観察所見と同様に、これらの結果はGSK3βの抑制的制御を欠落した過剰発現は大腸癌における普遍的な特性であり、β-カテニン活性化の有無あるいは活性化パターン(NAd,NAinv)とは無関係であることを示している(表2)。腫瘍におけるGSK3βの発現や活性と大腸がん症例の腫瘍病期や転移なども含めた臨床的特徴との間には特定の相関は認められなかった(表2)。
【0109】
【表2】
【0110】
2.3 GSK3β活性を制御する因子
大腸癌におけるGSK3βの発現やその第9セリン残基や第216チロシン残基のリン酸化を介した不活性化を制御する可能性のある上流因子(シグナル)について検討した。GSK3βを不活性化する上流のキナーゼ(リン酸化酵素)のうち最もよく知られているものはPKB/Aktであり、この酵素はリン酸イノシチド3-キナーゼ(PI3K)依存的リン酸化により活性化される。PI3K活性化はインスリンおよびrasシグナルの下流に生じるもので、とくに後者のシグナルは大腸癌においてK-ras遺伝子の活性化突然変異によって高頻度に活性化されている(Adjei AA. J Natl Cancer Inst 2001; 93(14): 1062-74, Minamoto T, et al. Expert Rev Mol Diag 2002; 2(6): 565-75.)。これらのがん化シグナルネットワークを考慮して、大腸癌における活性化型PKB/Akt(phospho-AktThr308,phospho-AktSer473)とK-ras原がん遺伝子の既知のホットスポット変異(コドン12,13)を解析し、これらの因子のいずれかがGSK3βの発現やその第9セリン残基や第216チロシン残基のリン酸化に影響しているかどうかを検討した。
【0111】
いずれかの活性化型PKB/Akt分画は7種類の大腸癌細胞株のうち6種類に、また、20例の大腸がん症例のうち13例の腫瘍において検出された(図1C,表1および2)。K-ras遺伝子の突然変異による活性化は6種類の大腸癌細胞株のうち4種類に検出され(表1)、そして20例の大腸がん症例ではそのうち5例の腫瘍において検出された(表2)。対象とした大腸癌細胞株と20例の大腸がん症例の腫瘍において、GSK3βの活性とPKB/Aktのリン酸化による活性化あるいはK-rasの突然変異による活性化との間には明らかな相関は認められなかった。これは大腸癌細胞株や大腸がん症例の腫瘍においてphospho-GSK3βSer9が高頻度に欠落していることに起因する(表1,2)。この結果は、RasあるいはPKB/Akt以外の未知のシグナルを介する上流因子あるいは分子機構が大腸癌におけるGSK3βの発現亢進あるいはその不活性化を抑制していることを示している。
【0112】
最近、GSK3βの第9セリン残基を別のアミノ酸に置換するような突然変異を同遺伝子のコドン9に人為的に導入することによりGSK3βは第9セリン残基リン酸化から回避され、恒常的に活性化されることが報告されている(Wang L, et al., J Biol Chem 2004; 279(31): 32444-52.)。今回、大腸癌細胞株あるいは大腸がん症例の腫瘍組織検体のDNAから増幅したPCR産物をPCR-RFLP法や塩基配列決定法により調べたが、GSK3βコドン9に変異は検出されなかった(データ提示なし)。このことは、GSK3βの第9セリン残基リン酸化を阻害するような遺伝子変異が大腸癌におけるGSK3β活性化の推測的(あるいは候補)機構ではないことを示す。
【0113】
2.4 各種細胞株に対するGSK3β阻害剤の影響
実験的にGSK3βの活性や発現を抑制することが大腸癌細胞株のin vitroにおける生存や増殖に影響を及ぼすかどうかを検討した。大腸癌細胞株のSW480とHCT116を種々の濃度のAR-A014418あるいはSB-216763により処理したところ、これらの細胞の生存(率)は各阻害剤の用量(濃度)依存的に減少した(図4A)。GSK3β阻害剤の同様の効果は他の大腸癌細胞株についても観察された(図4B,E)。しかし、これらの阻害剤の細胞生存抑制効果はHEK293細胞では観察されなかった(図4A)。GSK3β阻害剤を作用させた大腸癌細胞ではアポトーシス(Hoechst染色で観察・計測)の頻度は増加し、細胞増殖率(増殖細胞核抗原:PCNA標識率により計測)は低下したが、HEK293細胞ではこれらの変化は軽微であった(図4C, D)。
【0114】
さらに、大腸癌におけるGSK3βの発現増強の役割を解明するために、個々の大腸癌細胞の生存に及ぼすGSK3βを標的としたRNA干渉(RNAi)の効果を観察した。GSK3βに特異的な4種類のsiRNA(最終濃度はそれぞれ25nM)を混合して細胞に導入したところ、HEK293細胞と大腸癌細胞SW480におけるGSK3β蛋白質の発現は特異的に減少した(図5)。この選択的なGSK3βの発現抑制は、GSK3αやアクチンの発現がGSK3β特異的siRNAあるいは陰性対照siRNAのいずれの導入によっても影響されないことでも裏付けられた(図5)。注目すべきことに、GSK3βに対するRNAiは大腸癌細胞SW480やHEK293細胞におけるGSK3βの発現を同程度に抑制したのに対し、大腸癌細胞SW480の生存は抑制し、HEK293細胞の増殖生存には影響しなかった(図5)。この結果は大腸癌細胞において過剰発現し活性化されているGSK3βの特異的がん化作用をさらに支持するものであった。
【0115】
3.まとめ
種々の大腸癌細胞株や臨床的大腸癌におけるGSK3βの発現亢進と活性化、およびGSK3β阻害剤の大腸癌細胞増殖に対する顕著な抑制効果を示した今回の結果は、GSK3βが既知のがん化シグナルに対して抑制的に作用し、それゆえGSK3β阻害剤には潜在的発がん性が予測されるという従来の知見を覆すものであった。すなわち、本研究により、癌細胞において第9セリン残基のリン酸化の欠如と第216チロシン残基のリン酸化を介して恒常的に活性化されているGSK3βの活性や発現を薬理学的ならびに転写調節的技法を駆使して修飾することにより、がんの新たな治療や予防が可能になることが示された。
【0116】
<実施例2> 大腸癌細胞移植動物モデルに対するGSK3β阻害剤投与の効果
GSK3β阻害(剤)のin vitroにおける制がん効果を明らかにする目的で、以下の動物実験を実施した。
1.対象と方法
6週齢のヌードマウス(BALB/cA Jcl, nu/nu,日本クレア)の左腋窩の皮下に107個の大腸がん細胞SW480を23G(ゲージ)の注射針を用いて移植した。癌細胞を移植して2週間後に腫瘍生着が確認されたマウスを、AR-A014418投与群(N=12)、SB-216763投与群(N=12)、DMSO投与(コントロール)群、(N=5)の3群に分けた。さらに、AR-A014418投与群とSB-216763投与群をそれぞれ1mg/kg(培養細胞実験の5μMに相当)投与群(N=4)、2mg/kg(培養細胞実験の10μMに相当)投与群および5mg/kg(培養細胞実験の25μMに相当)投与群の3群、合計6群に分けて、治療実験を行った(表3,図6)。DMSOに溶解した3種類の濃度のAR-A014418(283μg/mL=0.35mM; 567μg/mL=0.7mM; 1419μg/mL=1.75mM)あるいはSB-216763(235μg/mL=0.35mM; 471μg/mL=0.7mM; 1177μg/mL=1.75mM)をそれぞれ0.2mlずつ、DMSO投与群には同量のDMSOを週3回(隔日)、腹腔内に注入した。
【0117】
【表3】
【0118】
これらのGSK3β阻害剤投与量は、マウスの体液量を体重(20g程度)の70%と想定して、培養細胞に使用した培養液中の濃度と同レベルになるように算出した。すなわち、1mg/kg、2mg/kgおよび5mg/kgの阻害剤投与量はマウス体液量に換算して、それぞれ、細胞培養液中の阻害剤濃度の5μM、10μMおよび25μMに相当する。
【0119】
各群のマウスにDMSOあるいはそれぞれの用量の阻害剤の投与期間に毎週、体重、食餌摂取量と移植腫瘍の大きさ(長径と短径)を計測した。DMSOあるいは各阻害剤を投与して5週間後に、エーテル深麻酔によりマウスを安楽死させ、移植腫瘍と主要臓器(肺、肝、膵、腎、脾)を摘出した(図6)。腫瘍の体積は、以下の計算式により算出した。
腫瘍体積(cm3)=0.5 x 長径(cm) x [短径(cm)]2
【0120】
2.結果
図7は、治療実験開始5週間後、各群(4匹ずつ)の代表的な固体を示したものである。対照(DMSO投与)群の移植腫瘍に比べて、GSK3β阻害剤を投与されたいずれの群でも左腋窩の皮下移植腫瘍の増殖が抑制されていることが確認された。図8は、異なる用量の各阻害剤投与後の腫瘍体積の変化を経時的に示したものである。このグラフから、移植腫瘍の増殖は、対照群の腫瘍に比べてGSK3β阻害剤の投与用量に依存して有意に抑制されていることが確認された。なお、対照群を含めていずれの群でも、食餌摂取量や体重変化などの観点から、明らかな有害事象は認められなかった。
【0121】
この実施例により、in vivoにおけるGSK3β阻害(剤)の抗腫瘍効果が初めて検証された。また、短期間の実験ではあるが、GSK3β阻害剤の毒性あるいは有害事象はないものと考えられる。
【0122】
<実施例3> 神経膠芽腫細胞に対するGSK3β阻害剤の作用
GSK3β阻害(剤)の制がん効果が大腸癌に限定されず、それ以外のがん種、とくに難治性がんに対しても有効かどうかを明らかにすることは、本酵素の阻害に基づくがん治療法の開発には重要な課題である。そこで、大腸癌以外の難治性がん(胃癌、膵癌、肝細胞癌、脳腫瘍など)由来の細胞について実施している実験のうち、難治性がんの一例として神経膠芽腫を対象にした成果を記載する。
【0123】
1.対象と方法
神経膠芽腫(glioblastoma)は、極めて分化度が低く悪性度が最も高い神経膠腫で、原発性脳腫瘍の約9%を占める。実際の腫瘍は手術で完全摘出できないことが多く、既存の抗がん剤治療や放射線照射治療に対しても耐性を示し、極めて難治性である。このように、本腫瘍に対する有効な治療法がないことから、その生命予後は最近の20−30年間、改善されていない実情であり、新しい治療法の開発が切望されている。
【0124】
本実施例では、4種類の神経膠芽腫細胞株A172、T98G、U251およびU87(American Type Culture Collection [ATCC,Manassas,VA]から購入した)と、ヒト胎児腎細胞 HEK293を対象にした。これらの細胞株におけるGSK3βとそのリン酸化分画の発現を<実施例1>に準じて解析した。また、既知のGSK3β阻害剤(AR-A014418あるいはSB-216763)の、腫瘍細胞の生存、増殖やアポトーシスなどへの作用を、大腸がん細胞の場合と同様に解析した。
【0125】
各細胞は一晩培養した後、それぞれDMSOに溶解したAR-A014418あるいはSB-216763を所定の濃度(5μM, 10μM, 25μMまたは40μM, 50μM)となるように添加した培地で培養し、生存細胞数とアポトーシスの頻度を経時的に測定した。コントロールとして、同量のDMSOを添加して培養した細胞についても、その生存細胞数とアポトーシスの頻度を測定した。なお、<実施例1>とは異なり、生存細胞数の計測にはWST-8**アッセイキット(Cell Counting Kit-8,和光,大阪)とBio-Rad社(Hercules, CA)製のマイクロプレートリーダーを使用した。また、増殖期細胞分画はbromodeoxy uridine(BrdU)を取り込む細胞数をCell Proliferation ELISA BrdU Kit(Roche, Panzberg, Germany)を用いて測定し、腫瘍細胞の増殖に及ぼすGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響を解析した。
**WST-8: 4-[3-(4-iodophenyl)-2-(4-nitrophenyl)-2H5-tetrazolio]-1,3-benzene disulfonate
【0126】
2.結果
<実施例1>と同様にウエスタンブロット法により解析した結果、4種類の神経膠芽腫細胞株 A172、T98G、U251およびU87のいずれにも、GSK3βとその第216チロシン残基がリン酸化された活性化型分画が検出された(データの提示なし)。現在、これらの神経膠芽腫細胞におけるGSK3βの酵素活性を非放射性試験管内酵素活性検出法(後述の実施例5を参照)により検証している。
【0127】
大腸癌細胞の場合と同様に、AR-A014418およびSB-216763で処置した神経膠芽腫細胞はいずれも、DMSOで処理した場合(コントロール)に比較して、阻害剤の濃度依存的に生存細胞数が減少し、アポトーシスが顕著に誘導された(図9,10A,10B)。一方、HEK293細胞では、GSK3β阻害剤を培地に加えることによりアポトーシスが軽度に誘導されるものの、DMSOで処理した場合(コントロール)との有意差は認められなかった(図10B)。また、これらの神経膠芽腫細胞では、GSK3β阻害剤処理によって、その増殖が抑制された(図10C)。
【0128】
以上の結果より、大腸がんのほかに神経膠芽腫においても(活性化型)GSK3βが腫瘍細胞の生存、増殖を支持し、アポトーシスを阻止していることが明らかにされた。そして、(活性化型)GSK3βは大腸癌以外のがん種に対しても治療標的であることが示唆された。
【0129】
<実施例4> がん細胞の不死化に対するGSK3β阻害の影響
GSK3β阻害(剤)の制がん効果の分子および細胞レベルのメカニズムを明らかにすることは重要である。そこで、がんに共通する重要な生物学的特性の1つである細胞の不死化(immortalization)に対するGSK3β阻害(剤)の作用を、細胞の老化とテロメア(telomere)維持機構に着目して解析した。
【0130】
1.対象と方法
大腸癌細胞および非がん細胞をそれぞれGSK3β阻害剤あるいは特異的RNA干渉により処理し、細胞老化や細胞不死化にかかわる細胞レベル、分子レベルでの変化を解析した。
【0131】
<実施例1>にしたがって、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(ヒト胎児腎細胞 HEK293)を25μMのGSK3β阻害剤(AR-A014418あるいはSB-216763)、あるいはGSK3β特異的RNA干渉で処理した。対照として、同じ細胞に対してDMSO投与あるいは非特異的RNA干渉を行った。そして、GSK3βを阻害した細胞と対応する対照細胞における老化細胞の頻度を比較・解析した。また、これらの細胞に対して、細胞不死化の分子指標であるhuman telomerase reverse transcriptase(hTERT)mRNAの発現およびtelomerase活性の変化を経時的に比較解析した。
【0132】
老化細胞は、β-galactosidase活性の指標であるx-gal染色陽性の細胞として判定した。具体的には、本染色のためカバーグラス上で培養した細胞をリン酸緩衝液(PBS)で洗浄し、クエン酸・リン酸固定液(0.1Mクエン酸,0.2M Na2HPO4, pH 6.0)により室温で5分間固定した。ついで、細胞をPBSで2回洗浄し、x-gal染色液(1mg/mL x-gal [Sigma-Aldrich], 5mM K3Fe[CN]6, 2mM MgCl2, 150mM NaCl, 30mM クエン酸・リン酸緩衝液,5mM K4Fe[CN]6)を37℃で20時間反応させた。染色した細胞をPBSで洗浄後、4%パラフォルムアルデヒドで再固定し、光学顕微鏡観察によりx-gal染色陽性細胞を同定した。老化細胞の頻度は強拡大で5視野観察し、各視野における全細胞数(約100個の細胞)に占めるx-gal染色陽性細胞の頻度の平均値として計測・算出した。
【0133】
hTERT mRNAの発現は、対象細胞からtotal RNAを抽出し、RT-PCR法により解析した。RT-PCRには上流primer(5’-CGGAAGAGTGTCTGGAGCAA-3’: 配列番号3)と下流primer(5’-GGATGAAGCGGAGTCTGGA-3’: 配列番号4)を用い、(15秒@94℃,10秒@55℃,9秒@72℃) x 36ないし38サイクルのRT-PCRプログラムによりhTERT遺伝子を増幅した。telomeraseの活性は市販のキット(TRAPEZE(R) Telomerase Detection Kit, Chemicon International)を用いて、推奨プロトコールに準拠して検出した。GSK3β阻害細胞と対照細胞におけるhTERTの発現レベルとtelomerase活性は、電気泳動上のシグナルをdensitometer (NIH Image Software)により計測・定量し、比較解析した。
【0134】
2.結果
25μMのGSK3β阻害剤で72時間処理することにより、大腸癌細胞株ではいずれも、AR-A014418、SB-216763、DMSO処理した対照細胞に比べてx-gal染色陽性細胞として同定される老化細胞の頻度が有意に増加した。一方、HEK293細胞では、いずれかのGSK3β阻害剤処理した細胞とDMSO処理細胞の老化細胞の頻度に有意差はみとめられなかった(図11,12)。
【0135】
つぎに、hTERTの発現とtelomerase活性に対するGSK3β阻害の効果を検討した。その結果、大腸癌細胞株では、AR-A014418、あるいはSB-216763処理の72ないし96時間後にhTERT発現およびtelomerase活性が抑制された。一方、HEK293細胞では、GSK3β阻害剤によるhTERTの発現変化あるいはtelomerase活性の抑制は認められなかった(図13,14)。また、これらの細胞において特異的RNA干渉によりGSK3βの発現を減弱させることにより、GSK3β阻害剤の場合と同様に大腸癌細胞ではhTERT発現およびtelomerase活性が抑制されたが、HEK293細胞では変化が認められなかった(図15)。
【0136】
以上の結果より、癌細胞の不死化の阻止と老化の促進が、GSK3β阻害による制がん効果の分子・細胞レベルのメカニズムの1つである可能性が示唆された。
【0137】
<実施例5> 非放射性試験管内GSK3β酵素活性検出法(Non-RI in vitro kinase assay)の開発
第216チロシン残基リン酸化GSK3β蛋白質分画の発現量は、GSK3βの酵素活性を相対的(あるいは、間接的)に反映しているに過ぎないため、本酵素活性を直接、検出・測定する手法は、既存の物質のGSK3β活性阻害効果の評価のために重要である。また、本酵素(活性)が発病あるいは発症にかかわる疾患(2型糖尿病、アルツハイマー病、がんなど)の診断法の開発とともに、これらの疾患に対する薬剤開発の観点から、本酵素活性の阻害効果を有する可能性のある物質のスクリーニングや同定にも極めて有用である。GSK3βに限らず、蛋白質リン酸化酵素の活性測定の従来法では放射性同位元素(32Pなど)を標識したATPが必要とされることが、その実施上の制約となっている。そこで発明者らは、放射性同位元素を必要としない簡便なGSK3βの活性測定の新しい手法として、非放射性試験管内GSK3β酵素活性検出法を開発した。
【0138】
1.原理
本検出法の原理について、図16を参照して概説する。まず、細胞抽出液に抗GSK3β特異的抗体とビーズ(resins)を加え、GSK3β蛋白質を免疫沈降させる(図中S)。陰性対照(コントロール)として、同じ細胞の抽出液に同濃度の非特異的マウスIgGを加える(図中C)。ついで、kinase buffer存在下で、組換えヒトβ-catenin蛋白質(基質)と非放射性ATP(図中P)を加え、リン酸化反応を行う。その後、この反応液をウエスタンブロット法により解析し、抗リン酸化β-cateninペプチド抗体によりリン酸化ヒトβ-catenin蛋白質を検出する。組換えヒトβ−catenin蛋白質をそのヒスチジン基を介してビーズ(resins)等に固定化すれば、ウエスタンブロット法を用いることなく、on-resins colorimetric assayも可能である。
【0139】
2.対象と方法
本法の開発にはHEK293細胞と各種の培養癌細胞株:SW480, HCT116, HT29(大腸癌);MKN-28, NKPS, TMK-1, NUGC-4(胃癌);MIA-PaCa-2, Capan-1, BxPC-3(膵臓癌);HEP-G2(肝細胞癌)を使用した。これらの細胞はいずれもAmerican Type Culture Collection(ATCC,Manassas,VA)から購入し、それぞれ推奨の培地と培養条件により培養した。
【0140】
<実施例1>の方法にしたがって、細胞溶解用緩衝液(0.2M Tris-HCl pH 7.5, 1.5M NaCl, 10mM EDTA, 10mM EGTA, 10% Triton X-100, 25mM sodium pyrophosphate, 10mM β-glyceophosphate, 10mM Na3VO4, Protease inhibitors cocktail [Sigma-Aldrich])を用いて、それぞれの細胞から蛋白質を抽出した。細胞由来の蛋白質1mg(1000μg)から、定法にしたがってマウスモノクローナル抗GSK3β抗体(BD Transduction Laboratories)と20μLのProtein A/G Agarose resins(Calbiochem)によりGSK3βを免疫沈降して、resin上に精製した(以後、GSK3βと記載する)。免疫沈降の陰性対照には、抗GSK3β抗体の代わりに非特異的マウスIgGを使用した。細胞由来のGSK3βをリン酸緩衝液(PBS)で2回、ついでリン酸化反応用緩衝液(kinase buffer)(0.25M Tris-HCl pH 7.5, 10mM β-glyceophosphate, 20mM dithiothreitol [DTT], 1mM Na3VO4, 0.1M MgCl2)を用いて2回洗浄した。この細胞由来のGSK3βをkinase bufferに懸濁し、10mM ATP(非放射性)2μLと4μgの組換えヒトβ-catenin蛋白質(基質)を加えて計50μLとした。このkinase反応溶液を温水槽中(30℃)で30分間反応させた後、等量の2xSDS電気泳動用サンプルバッファーを添加して95℃で5分間過熱してリン酸化反応を停止した。
【0141】
この反応液上清をSDSポリアクリルアミド電気泳動で分離し、<実施例1>と同様に、ウエスタンブロット法により目的とする以下の蛋白質を解析した。GSK3βによるリン酸化の基質として用いた組換えヒトβ-catenin蛋白質のリン酸化は、リン酸化β-cateninペプチド(p-β-cateninSer33/37Thr41)特異抗体(Cell Signaling)により検出した。ついで、反応液中のGSK3βと組換えヒトβ-catenin蛋白質を同様にウエスタンブロット法により検出した。これにより、それぞれの細胞におけるGSK3βの発現と相対的な活性の比較も可能になる。
【0142】
この手法をさらに、GSK3β阻害剤の酵素活性阻害効果の判定に応用した。具体的には、kinase反応液中に最終濃度が25μMになるようにSB-216763あるいはAR-A014418を添加し、これらの反応液を同様にウエスタンブロット法により解析し、阻害剤の代わりにDMSOだけを添加した場合のβ-cateninリン酸化と比較した。
【0143】
2.結果
本検出法を用いて、まず、各種大腸癌細胞株(SW480、HT29、HCT116)および非がん細胞株(HEK293)を解析したところ、いずれの細胞においてもGSK3βの酵素活性が検出された(図17)。リン酸化β-catenin のシグナル強度から、HEK293細胞に比べて大腸癌細胞のGSK3β活性は高く、この結果はウエスタンブロット法によるphospho-GSK3βTyr216分画の検出結果(図1A)と対応していると判断された。
【0144】
つぎに、大腸癌を含めた種々の消化器癌細胞を対象にして、同様に解析した。その結果、大腸癌以外に、胃癌、膵癌および肝細胞癌の細胞株にもGSK3βの酵素活性が特異的に検出された。とくに、膵癌細胞(BxPC-3)と肝細胞癌(HEP-G2)に強いGSK3β活性が検出された(図18)。この解析により、それぞれの細胞種におけるGSK3β活性とともに、その総発現量を定量的に比較することが可能であり、同一細胞においては本酵素の発現量と活性に相関がみられた(図18)。
【0145】
さらに、本検出法を用いて、既知のGSK3β阻害剤の効果検証を試みた。その結果、2種類の大腸癌細胞株(SW480, HCT116)における GSK3βの活性は25μMのSB-216763あるいは同濃度のAR-A-014418により完全に阻害されることが確認された(図19)。
【0146】
一連の結果より、この非放射性試験管内GSK3β酵素活性検出法の有用性が検証された。また、この検出法をon resins colorimetric assay法に深化させることにより、本酵素を指標にする疾患診断法の開発や、新しいGSK3β活性阻害物質のスクリーニング、ならびに本酵素阻害手法の開発に応用可能であることが示唆された。
【0147】
<実施例6>胃癌、膵癌および肝細胞癌の細胞株におけるGSK3β阻害の作用
<実施例3>と同様の目的で、大腸癌以外の消化器系臓器の癌腫、とくに難治性の消化器癌から樹立された培養細胞株を対象にして、GSK3βの発現と活性を解析した。<実施例5>と図18AおよびBに示したように、解析対象にした胃癌細胞株 MKN-28、NKPS、TMK-1、NUGC-4、膵癌細胞株MIAPaCa-2、Capan1、BxPC-3、および肝細胞癌HEP-G2のうち、MKN-28とMIAPaCa-2以外のすべての細胞株に明瞭なGSK3βの発現と活性が認められた。
【0148】
本解析の結果に基づいて、これらの癌細胞の生存、増殖およびアポトーシスに対するGSK3β阻害(小分子阻害剤、およびRNA干渉法)の作用を<実施例1>および<実施例3>の方法により検討する。前記解析結果にしたがい、それぞれの臓器由来でGSK3βの高い活性を示す癌細胞株(たとえば、NUGC-4、BxPC-3、HEP-G2)についてがん移植動物モデルを作成し、<実施例2>にしたがい、in vivoにおけるこれらの難治性消化器癌細胞に対するGSK3β阻害剤の(腫瘍増殖抑制)効果を検討する。
【0149】
本明細書中で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書中にとり入れるものとする。
【産業上の利用可能性】
【0150】
本発明は、がん治療や予防のための新規戦略の開発とがんの分子疫学的研究の展開に貢献するものと期待される。
【配列表フリーテキスト】
【0151】
配列番号1−人工配列の説明:プライマー
配列番号2−人工配列の説明:プライマー
配列番号3−人工配列の説明:プライマー
配列番号4−人工配列の説明:プライマー
【技術分野】
【0001】
本発明は、第216番目チロシン残基がリン酸化されたグリコーゲン合成酵素キナーゼ3β(GSK3β)の活性を阻害すること、またはGSK3βの第216番目チロシン残基のリン酸化を阻害することによりがん細胞の生存や増殖を抑制する方法、ならびに前記阻害効果に基づく新規抗がん剤の開発および抗がん剤の評価方法に関する。
【背景技術】
【0002】
近年、大腸がんの発生・進展の分子機構においてβ-カテニンの安定化による活性化が、rasがん遺伝子産物のがん化作用とともに有力な新規がん化シグナルとして注目されている(非特許文献1および2参照)。β-カテニンは細胞増殖にかかわるWntシグナル伝達系の中心的な実行因子であり、そのがん化作用はこれまでin vitroの実験系やトランスジェニック動物モデルを用いた多数の研究により支持されている(非特許文献3参照)。
【0003】
発明者らは、最近、腫瘍細胞核内におけるβ-カテニンの集積様式の違いから、ヒト大腸癌には2種類のβ-カテニン活性化パターンがあることを見出した(非特許文献4参照)。つまり、40-50%の大腸がん症例ではβ-カテニンの核内集積(NA)、すなわち活性化が腫瘍全体に認められる(NAd)のに対し、10-15%の症例ではその集積(活性化)が腫瘍浸潤先進部に限局して検出される(NAinv)。そして、このβ-カテニンのNAinv型活性化が大腸癌の病期の進行と術後の再発に有意に相関する。この結果は、大腸癌におけるβ-カテニンのがん化シグナル活性化パターンが、腫瘍の転移・再発や患者の術後生存率の信頼性の高い予測因子になり得ることを示唆する(非特許文献4および5参照)。
【0004】
グリコーゲン合成酵素キナーゼ3β(以下、「GSK3β」と記載する)は、このWnt/β-カテニンシグナル系の制御因子群(非特許文献2および3参照)の1つであり、生理的条件下の細胞においてβ-カテニンをリン酸化して、APCがん抑制蛋白質やアキシン(いずれもGSK3βの基質)などから構成されるユビキチン分解系に誘導する。このことは、GSK3β活性と大腸癌の仮説的接点として、GSK3βの第9セリン残基リン酸化に伴う不活性化が標的基質であるβ-カテニンをユビキチン分解系から回避させ、がん化シグナルの活性化に導くことを示唆する。
【0005】
GSK3βは様々な細胞内シグナル伝達を媒介する多機能セリン・スレオニンキナーゼ(リン酸化酵素)であり、エネルギー代謝、転写調節、細胞増殖・生存等を含む広範な細胞調節過程を制御する(非特許文献6−9参照)。つまり、Wnt/β-カテニンシグナルの制御は本酵素の多彩な機能の1つにすぎない。GSK3βは細胞内シグナル伝達を媒介する多くの蛋白質リン酸化酵素とは異なり、正常細胞において活性化型として存在し、種々の刺激によってその活性が抑制的に制御されている。GSK3βの基質には、c-Jun、c-Myc等の多様ながん化関連転写因子(非特許文献10および11参照)や、β-カテニン、Gli蛋白質等の原がん遺伝子産物 (非特許文献12および13参照)が含まれ、これら基質の不活性化を介して細胞の腫瘍性形質転換や腫瘍発生を妨げる。また、GSK3βはNF-κB細胞不死化シグナルを介して細胞の生存に本質的な役割も果たしている(非特許文献15および16参照)。こうしたGSK3βの機能から、GSK3β遺伝子を細胞の異常増殖マーカーとして利用することを報告した特許出願もある(特許文献1参照)。
【0006】
最近、GSK3βがインスリン非依存性糖尿病(non-insulin-dependent diabetes mellitus [NIDDM]:2型糖尿病)やアルツハイマー病をはじめとする神経変性疾患の発症にも関与していることがわかってきた(非特許文献14参照)。そのため、GSK3β阻害剤を神経変性疾患や2型糖尿病やがん等の治療に応用する特許出願もなされているが(特許文献2および3参照)、いずれも特定化合物のGSK3β阻害効果と既知の知見から当該化合物のこれら疾患への適用を予測するものにすぎず、とくにがんについては実証されていない。
【0007】
上記のとおり、GSK3βはWnt/β-カテニンシグナル伝達の負の制御因子であることから、GSK3βを介したシグナル伝達制御機能の異常ががんにおけるβ-カテニンシグナル活性化パターンに関連しているかどうかを明らかにすることは重要な課題である。一方、本酵素遺伝子のノックアウト動物の解析により、GSK3βはNF-κB経路を介した細胞の生存と増殖を維持する作用が報告されている(非特許文献12および13参照)。また、前立腺癌由来細胞株において、GSK3β阻害がTRAIL(tumor necrosis factor-related apoptosis-inducing ligand)による薬剤感受性を亢進させたり、細胞増殖を抑制する報告もある(非特許文献17、18および19参照)。このように、細胞生存・増殖やがん化にかかわる基幹的な細胞内シグナルに対して相反する作用が提唱されていることから、GSK3βがWntシグナルとは異なる独自の分子メカニズムを介してがん細胞に影響を及ぼしている可能性があるのではないかと想定される。しかしながら、これまでGSK3βは細胞の腫瘍性形質転換や腫瘍発生に対し抑制的に作用するという仮説から、GSK3β阻害剤には潜在的発がん性が予測されている。それゆえ、GSK3βとがんの発生・進展との関与については報告が少なく、詳細な検討がなされてはいなかった。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特表2004−528837号公報
【特許文献2】特表2004−504838号公報
【特許文献3】特表2004−507545号公報
【非特許文献】
【0009】
【非特許文献1】Jass JR, Whitehall VLJ, Young J, Leggett BA. Emerging concepts in colorectal neoplasia. Gastroenterology 2002; 123(3): 862-76.
【非特許文献2】Wong NA, Pignatelli M. β-catenin- a linchpin in colorectal carcinogenesis. Am J Pathol 2002; 160(2): 389-401.
【非特許文献3】Polakis P. The oncogenic activation of beta-catenin. Curr Opin Genet Dev 1999; 9(1): 15-21.
【非特許文献4】Ougolkov AV, Yamashita K, Mai M, Minamoto T. Oncogenic β-catenin and MMP-7 (matrilysin) cosegregate in late-stage clinical colon cancer. Gastroenterology 2002; 122(1): 60-71.
【非特許文献5】Zhang B, Ougolkov A, Yamashita K, Takahashi Y, Mai M, Minamoto T. β-catenin and ras oncogenes detect most human colorectal cancers. Clin Cancer Res 2003; 9(8): 3073-9.
【非特許文献6】Manoukian AS, Woodgett JR. Role of glycogen synthase kinase-3 in cancer: regulation by Wnts and other signaling pathways. Adv Cancer Res 2002; 84: 203-29.
【非特許文献7】Doble BW, Woodgett JR. GSK-3: tricks of the trade for a multi-tasking kinase. J Cell Sci 2003; 116(Pt 7): 1175-86.
【非特許文献8】Harwood AJ. Regulation of GSK-3; a cellular multiprocessor. Cell 2001; 105(7): 821-4.
【非特許文献9】Kim L, Harwood A, Kimmel AR. Receptor-dependent and tyrosine phosphatase-mediated inhibition of GSK3 regulates cell fate choice. Dev Cell 2002; 3(4): 523-32.
【非特許文献10】Morton S, Davis RJ, McLaren A, Cohen P. A reinvestigation of the multiple phosphorylation of the transcription factor c-Jun. EMBO J 2003; 22(15): 3876-86.
【非特許文献11】Gregory MA, Qi Y, Hann SR. Phosphorylation by glycogen kinase-3 controls c-myc proteolysis and subnuclear localisation. J Biol Chem 2003; 278(51): 51606-12.
【非特許文献12】Price MA, Kalderon D. Proteolysis of the Hedgehog signaling effector Cubitus interruptus requires phosphorylation by glycogen synthase kinase 3 and casein kinase 1. Cell 2002; 108(6): 823-35.
【非特許文献13】Jia J, Amanai K, Wang G, Tang J, Wang B, Jiang J. Shaggy/GSK3 antagonizes Hedgehog signalling by regulating cubitus interruptus. Nature 2002; 416(6880): 548-52.
【非特許文献14】Cohen P, Goedert M. GSK3 inhibitors: development and therapeutic potential. Nat Rev Drug Discov 2004; 3(6): 479-87.
【非特許文献15】Hoeflich KP, Luo J, Rubie EA, Tsao MS, Jin O, Woodgett JR. Requirement for glycogen synthase kinase-3β in cell survival and NF-κB activation. Nature 2000; 406(6791): 86-90.
【非特許文献16】Schwabe RF, Brenner DA. Role of glycogen synthase kinase-3 in TNF-α-induced NF-κB activation and apoptosis in hepatocytes. Am J Physiol Gastrointest Liver Physiol 2002; 283(1): G204-11.
【非特許文献17】Liao X, Zhang L, Thrasher JB, Du J, Li B. Glycogen synthase kinase-3β suppression eliminates tumor necrosis factor-related apoptosis-inducing ligand resistance in prostate cancer. Mol Cancer Ther 2003; 2(11): 1215-1222.
【非特許文献18】Wang L, Lin H-K, Hu Y-C, Xie S, Yang L, Chang C. Suppression of androgen receptor-mediated transactivation and cell growth by the Glycogen synthase kinase 3β in prostate cells. J Biol Chem 2004; 279(31): 32444-32452.
【非特許文献19】Mazor M, Kawano Y, Zhu H, Waxman J, Kypta RM. Inhibition of glycogen synthase kinase-3 represses androgen receptor activity and prostate cancer cell. Oncogene 2004; 23(47): 7882-7892
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明は、グリコーゲン合成酵素キナーゼ3β(GSK3β)とがんの発生・進展との関与を明らかにし、がんの治療・診断のための新たな手段を提供することにある。
【0011】
発明者らは、上記課題を検討するために、大腸癌細胞株および同一大腸がん患者の非がん組織と癌組織におけるGSK3βの発現と第9セリン残基リン酸化解析を通じた酵素活性化の程度を比較解析した。さらに、GSK3βの発現と活性をそれぞれ転写調節的ならびに薬理学的に修飾することにより大腸癌細胞の増殖に及ぼす本酵素の影響を研究した。
【0012】
その結果、大部分の大腸癌組織や大腸癌細胞株においてそれぞれの非がん粘膜や細胞(HEK293)に比べてGSK3βとその第216チロシン残基がリン酸化された活性化型phospho-GSK3βTyr216分画の発現が亢進し、一方、第9セリン残基がリン酸化された不活性化型phospho-GSK3βSer9分画の発現はほとんど認められなかった。また、in vitroにおいて大腸癌細胞株や神経膠芽腫細胞株等のがん細胞の生存と増殖は低分子量タイプのGSK3β阻害剤の投与により用量(濃度)依存的に抑制され、同様にGSK3βに対するRNA干渉(RNAi)によっても抑制された。さらに、大腸がんモデル動物においてもGSK3β阻害剤投与による腫瘍組織の用量依存的縮小が観察された。このことは、従来の知見とは異なり、GSK3βの発現や活性ががん(とくに大腸がん)の治療や予防の新たな標的となりうることを提示している。
【課題を解決するための手段】
【0013】
すなわち本発明は、GSK3βの発現や第216番目チロシン残基リン酸化による活性化阻害を通じて本酵素活性を抑制することにより、がん細胞の生存や増殖を抑制する方法に関する。
【0014】
前記方法は、式(I)または(II)の構造を有する化合物もしくはその薬理学的に許容される塩をがん細胞に作用させることによって実施できる。
【0015】
【化1】
【0016】
(ここで、Q1は-(CH2)n-Ar-O-(nは1〜5の整数、Arは置換または非置換アリール)であり、Q2はハロゲン、アルキル、ヒドロキシ、アルコキシ、アリールオキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、モノアルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルまたはアルキルスルホニルから選ばれる基である)
【0017】
【化2】
【0018】
(ここで、R1は水素、アルキル、アラルキル、アルコキシアルキル、ヒドロキシアルキル、ハロアルキル、アミノアルキル、モノアルキルアミノアルキル、ジアルキルアミノアルキル、アジドアルキル、アシルアミノアルキル、アルキルスルホニルアミノアルキル、アリールスルホニルアミノアルキル、メルカプトアルキル、アルキルチオアルキル、グルコピラノシル、カルボキシアルキル、アルコキシカルボニルアルキル、アミノカルボニルアルキル、ヒドロキシアルキルチオアルキル、メルカプトアルキルチオアルキル、アリールチオアルキルまたはカルボキシアルキルチオアルキルであり、
R2は水素、アルキル、アラルキル、アルコキシアルキル、ヒドロキシアルキル、ハロアルキル、アミノアルキル、モノアルキルアミノアルキル、ジアルキルアミノアルキル、アシルアミノアルキル、アルキルスルホニルアミノアルキル、アリールスルホニルアミノアルキル、メルカプトアルキル、アルキルチオアルキル、カルボキシアルキル、アルコキシカルボニルアルキル、アミノカルボニルアルキル、アルキルチオまたはアルキルスルフィニルであり、
R3はハロゲン、アルキル、ヒドロキシ、アルコキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、アルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルおよびアルキルスルホニルからなる群より選ばれる1または2以上の置換基で置換されたフェニル基であり、
R4、R5、R6およびR7は、互いに独立して、水素、ハロゲン、アルキル、ヒドロキシ、アルコキシ、アリールオキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、モノアルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルまたはアルキルスルホニルである)
【0019】
前記化合物の好ましい一例としては、例えば、下式に示されるN-(4-メトキシベンジル)-N'-(5-ニトロ-1,3-チアゾール-2-イル)ウレアまたは3-(2,4-ジクロロフェニル)-4-(1-メチル-1H-インドール-3-イル)-1H-ピロール-2,5-ジオンを挙げることができる。
【0020】
【化3】
【0021】
本発明はまた、上記した化合物もしくはその薬理学的に許容される塩を有効成分として含む、がんの治療用医薬組成物も提供する。本明細書では、前記医薬組成物の大腸がん、胃がん、膵がん、肝細胞がん、および神経膠芽腫細胞に対する効果や、ヌードマウス移植大腸癌に対する薬理効果を示したが、本発明の医薬組成物の効果はこれに限られるものでなく、あらゆるがんに対してその効果が期待される。
【0022】
本発明はまた、GSK3βの第216番目チロシン残基のリン酸化による本酵素の活性化に対する阻害効果を指標として、被験物質の抗がん剤としての効果を評価する方法を提供する。
【0023】
前記方法は、たとえば、以下の工程により実施できる:
1)細胞を被験物質の添加または非添加条件下で培養する;
2)上記細胞内における第216チロシン残基リン酸化GSK3β蛋白質分画の発現量を検出するか、あるいはGSK3β酵素活性を検出・測定する
3)被験物質の添加または非添加条件下における、上記の酵素発現量あるいは活性の相違に基づいて、該被験物質の抗がん剤としての効果を評価する。
【0024】
前記方法において、細胞はがん患者から単離された細胞または癌細胞株、例えば、SW480, SW620, HT29, LoVo, HCT116, SW48およびRKO等の大腸癌細胞株やA172, T98G, U251, U87等の神経膠芽腫細胞株等を用いることができる。
【0025】
本発明の方法は、また、以下の工程によっても実施できる:
1)がんモデル動物を被験物質の投与または非投与の条件下で飼育する。
2)上記動物の血液または細胞中における第216チロシン残基リン酸化GSK3β蛋白質分画の発現量を検出する。
3)被験物質の投与または非投与条件下における、上記発現量の相違と移植腫瘍の増殖の変化に基づいて、該被験物質の抗がん剤としての効果を評価する。
【0026】
前記方法において、動物は齧歯類、特にマウスが好ましい。
また、本発明の方法において、蛋白質の発現量は、ウエスタンブロット法、ドットブロット法、スロットブロット法、ELISA法、およびRIA法から選ばれるいずれか一つの方法によって検出することができる。
【発明の効果】
【0027】
本発明によれば、GSK3β抑制効果に基づく新規の抗がん剤の提供とそのスクリーニングが可能になる。また、GSK3βレベルを指標とするがんの早期診断が可能になる。
【図面の簡単な説明】
【0028】
【図1】図1は、大腸癌細胞株とヒト胎児腎細胞 HEK293における、(A)ウエスタンブロット法による総GSK3βとphospho-GSK3βSer9(不活性化型)およびphospho-GSK3βTyr216(活性化型)の発現をそれぞれ特異的抗体(BiosciencesおよびCell Signaling Technology)を用いて解析した結果、(B) GSK3β活性を合成ヒトβ-カテニン蛋白を基質にして、in vitro kinase assayにより検出した (Adler V, et al. Proc Natl Acad Sci USA 1997;94:1686-91に記載されている検出方法を用いた) 結果、(C) GSK3β(シグナル)の上流で活性化されるプロテインキナーゼB(PKB/Akt)の活性化を抗phospho-AktThr308抗体や抗phospho-AktSer473抗体(Cell Signaling Technology)を用いて検出した結果。
【図2】図2は、HEK293細胞(上4枚の図)とSW480大腸癌細胞株(下4枚の図)における総GSK3β(赤色蛍光)とphospho-GSK3βSer9(緑色蛍光)の二重免疫蛍光細胞化学染色による発現と局在の観察結果を示す。図の尺度線は100μmに相当し、画像はすべて同一倍率で撮影された。
【図3】図3は、大腸がん症例の正常粘膜および腫瘍組織における、総GSK3β、phospho-GSK3βSer9(不活性化型)およびphospho-GSK3βTyr216(活性化型)分画の発現レベルを検出した結果を示す。各パネル上段の数字は患者ID番号(表2と対応)。N:非がん(正常)粘膜組織;T:癌組織。
【図4】図4は、AR-A014418とSB-216763のHEK293細胞および各種大腸癌細胞株のin vitroにおける生存、増殖とアポトーシスにおよぼす影響(効果)を解析した結果を示す。A, B, E: 0日目から特定時間(24, 48, 72および96時間)経過した時点でMTSアッセイにより計測した各ウェルの生細胞数。C, D:抗PCNA(proliferating cell nuclear antigen)抗体による免疫蛍光法とHoechstにより2重染色し、蛍光顕微鏡で観察したときの、全観察細胞に占める増殖細胞核(PCNA陽性)(D)あるいは、アポトーシスを生じている核(C)の比率。
【図5】図5は、GSK3βを標的とするsiRNAのHEK293細胞(上)とSW480大腸癌細胞(下)のin vitroでの細胞生存率におよぼす効果を示す。
【図6】図6は、大腸癌細胞移植動物モデルに対するGSK3β阻害剤投与による治療実験プロトコールの概略を示している。
【図7】図7は、異なる用量のGSK3β阻害剤(AR-A014418あるいはSB-216763)、あるいはDMSO投与開始して5週間後における大腸癌細胞移植マウスの写真である。
【図8】図8は、大腸癌細胞移植マウスにおいて、異なる用量のGSK3β阻害剤(AR-A014418あるいはSB-216763)、あるいはDMSOを投与開始後の移植腫瘍体積の経時的変化を示すグラフである。
【図9】図9は、各種の神経膠芽腫細胞株(T98G、A172、U251、U87)の細胞生存におよぼす、異なる濃度のGSK3β阻害剤(AR-A014418あるいはSB-216763)、あるいはDMSOの作用を経時的に示すグラフである。
【図10】図10は、各種の神経膠芽腫細胞株(A172、T98G、U251、U87)および非がん細胞株(HEK293)に対するGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響を示す。(A)は、AR-A014418処理後96時間の生存腫瘍細胞数を示すグラフである。(B)は、AR-A014418あるいはSB-216763処理後の腫瘍細胞におけるアポトーシス誘導を示すグラフである。(C)は、AR-A014418あるいはSB-216763処理後の増殖期細胞分画(bromodeoxy-uridine [BrdU]取り込み量)を示すグラフである。
【図11】図11は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)の細胞老化に対する、それぞれ25μMのGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響をDMSOの場合と比較して、β-galactosidase活性の指標であるx-gal染色により調べた結果を示す。
【図12】図12は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)をそれぞれ25μMのGSK3β阻害剤(AR-A014418あるいはSB-216763)あるいはDMSOにより処理して72時間後の老化細胞比率の変化を、x-gal染色により調べた結果を示す。
【図13】図13は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)における、human telomerase reverse transcriptase (hTERT)の発現に対するGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響を調べた結果である。
【図14】図14は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)における、telomerase活性に対するGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響を調べた結果である。
【図15】図15は、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(HEK293)における、(A)telomerase活性や(B)hTERT mRNAの発現に対するGSK3β発現阻害の影響を、siRNAトランスフェクション(小分子干渉RNA導入)により調べた結果を示す。
【図16】図16は、本発明に係る非放射性試験管内GSK3β酵素活性検出法(Non-RI in vitro kinase assay)の原理と概要を示す。
【図17】図17は、非放射性試験管内GSK3β酵素活性検出法により各種細胞株におけるGSK3β酵素活性を検出・比較した結果を示す。
【図18】図18は、HEK293細胞と各種の培養癌細胞株:SW480, HCT116, HT29(大腸癌);MKN-28, NKPS, TMK-1, NUGC-4(胃癌);MIA-PaCa-2, Capan-1, BxPC-3(膵臓癌);HEP-G2(肝細胞癌)におけるGSK3β酵素活性を非放射性試験管内GSK3β酵素活性検出法により検出した結果を示している。P,C:同一細胞からの抽出蛋白質に対して抗GSK3β抗体により免疫沈降した検体(P)と、同抗体の代わりに非特異的マウスIgGを添加して同様に解析した検体(陰性対照)(C)。
【図19】図19は、大腸癌細胞株(SW480とHCT116)から免疫沈降したGSK3β酵素活性に対する、それぞれ25μMのGSK3β阻害剤(AR-A014418あるいはSB-216763)と陰性対照であるDMSOの阻害効果を、非放射性試験管内GSK3β酵素活性検出法を応用して検証した結果を示している。
【0029】
本明細書は、本願の優先権の基礎である特願2005−133号の明細書に記載された内容を包含する。
【発明を実施するための形態】
【0030】
以下、本発明について詳細に説明する。
1.グリコーゲン合成酵素キナーゼ3β阻害剤によるがんの治療
本発明は、グリコーゲン合成酵素キナーゼ3β(以下、「GSK3β」と記載することもある)阻害剤の大腸がん治療用医薬組成物としての利用を提供する。
1.1 GSK3β
GSK3βはβ-カテニン原がん遺伝子産物を含む一連の細胞調節蛋白質をリン酸化し不活性化する抑制因子として作用する多機能蛋白質リン酸化酵素である。発明者らは、(1)大腸がん患者の癌組織や大腸癌細胞株で特異的なGSK3βの発現亢進が見られること、(2)GSK3β阻害剤の投与により用量依存的に大腸癌細胞株の生存と増殖が抑制され、アポトーシスが誘導されるること、(3)大腸癌細胞株の増殖がGSK3βに対するsiRNAによって抑制されることを初めて見出し、GSK3β阻害剤が抗がん剤として有用であること、また、GSK3β抑制効果が抗がん剤評価の指標として有用であることを実証した。
【0031】
本発明に係るGSK3βにはヒトGSK3βのほか、マウスGSK3βなど、そのオーソログも含まれるが、本発明の目的においてはヒトGSK3βが最も好ましい。GSK3β遺伝子やGSK3β蛋白質の配列情報は公知であり、公共データベースであるGenBankより容易に入手することができる。例えば、マウスGSK3β遺伝子と対応するGSK3β蛋白質のアミノ酸配列は、Accession No. BC060743およびAAH60743.1、あるいはBC006936およびAAH06936.1として開示されている。また、ヒトGSK3β遺伝子と対応するGSK3β蛋白質のアミノ酸配列は、Accession No.NM_002093およびNP_002084として開示されている。もちろん、これらの配列は一例であって、本発明のGSK3β遺伝子やGSK3β蛋白質はこれら配列に限定されない。
【0032】
1.2 GSK3β阻害剤
本明細書において、GSK3β阻害剤とは、GSK3βの機能を物理的あるいは化学的に阻害する物質を意味する。こうしたGSK3β阻害剤については、既に多くの化合物が同定されているが、その構造と作用メカニズムは多岐にわたる(Meijer L, Flajolet M, Greengard P. Trends in Pharmacol Sci 2004; 25 (9): 471-480 参照)。
【0033】
こうしたGSK3β阻害剤の構造と製造方法は、例えば、以下の特許出願あるいは登録特許に詳細に記載されている:特表2005-536517記載のピラゾロピリミジン、特表2005-535593記載のチアゾリジンジチオン誘導体、特表2005-534713記載のジヒドロピラゾロピリジン化合物、特表2005-533066記載のアゾールメチリデンシアニド誘導体、特表2005-531609記載のシスインドリル−マレイミド誘導体、特表2005-526835記載の2−置換−1,3−チアゾール化合物、特表2005-526814記載のモルホリン誘導体、特表2005-519087記載の1−[アルキル]、1−[(ヘテロアリール)アルキル]および1−[(アリール)アルキル]−7−(ピリミジン−4−イル)−イミダゾ[1,2−a]ピリミジン−5(1H)−オン誘導体、特表2005-519086記載のヘテロアリール置換2−ピリジニルおよび2−ピリミジニル−6,7,8,9−テトラヒドロピリミド[1,2−a]ピリミジン−4−オン誘導体、特表2005-516961記載のオキシインドール誘導体、特表2005-516960記載のオキシインドール誘導体、特表2005-516948記載のキナゾリン類、特表2005-513082記載のイソキノリン類、特表2005-509641記載のヒドロキシアルキルインドロカルバゾール化合物、特表2005-505515記載のアリールアミン、特表2005-504810記載および特表2005-504809記載の置換2−ピリジニル−6,7,8,9−テトラヒドロピリミド[1,2−a]ピリミジン−4−オンおよび7−ピリジニル−2,3−ジヒドロイミダゾ[1,2−a]ピリミジン−5(1H)オン誘導体、特表2005-501800記載のジヒドロピラゾロピリジン化合物、特表2004-53831記載の2,4−ジアミノチアゾール誘導体、特表2004-536111記載の4−(4−メトキシベンジル)−N′−(5−ニトロ−1,3−チアゾール−2−イル)尿素、特表2004-536110記載の複素環式アミン、特表2004-527486記載のピリミジン化合物、特表2004-522777記載の2−アリルアミノ−ピリミジン、特表2004-516297記載のトリアジン類、特表2004-515499記載のインダゾリル置換ピロリン化合物、特表2004-505078記載の3−インドリル−4−フェニル−1H−ピロール−2,5−ジオン誘導体、特表2003-528095記載の2−アミノ−3−(アルキル)−ピリミドン誘導体、特表2003-527303記載のピリミジンまたはピリジン類、特表2003-516974記載のピラジン誘導体、特表2002-527419記載のピロール−2,5−ジオン類、再表01/081345記載の芳香族アミド化合物、および特表平01-233281記載のピロール類等。当業者であれば、これらの記載に基づいて、所望のGSK3β阻害剤を製造し、使用することができるであろう。
【0034】
GSK3βの活性は種々の段階:(a) 転写後修飾(転写後の第9セリンと第216チロシン残基のリン酸化による制御)、(b) タンパク複合体との相互作用、(c) 基質プライミング、(d) 細胞内分布、で制御されており、上記したGSK3β阻害剤はそのいずれかの段階でGSK3βに作用するものと思われる(Meijer L, Flajolet M, Greengard P. Trends Pharmacol Sci 2004;25 (9), 471-480 参照)。
【0035】
発明者らは、2つの代表的GSK3β阻害剤(AR-A014418およびSB-216763)について、これらががんに対して特異的な抑制作用を有することを、in vivoおよびin vitroの両面で実証した。また、これらのGSK3β阻害剤が、がん細胞において転写後修飾(リン酸化)を受けたGSK3βの活性を阻害しうることを確認した。
【0036】
(1) AR-A014418
AR-A014418は下記の式で示されるチアゾール誘導体である(CAS NO: 487021-52-3、WO2003004478、特表2004-536111号参照)。
【0037】
【化4】
【0038】
AR-A014418は、試薬としてCalbiochem等により販売されており、一般に入手可能である。
【0039】
上記化合物に限定されず、下記の一般式(I)で示されるチアゾール誘導体もまた、同様のGSK3β阻害作用を有する限り、AR-A014418と同様、本発明に係るがん細胞の増殖抑制やがんの治療用医薬組成物として利用可能である。
【0040】
【化5】
【0041】
ここで、Q1は-(CH2)n-Ar-O-(nは1〜5の整数、Arは置換または非置換アリール)であり、Q2はハロゲン、アルキル、ヒドロキシ、アルコキシ、アリールオキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、モノアルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルまたはアルキルスルホニルから選ばれる基である。
【0042】
(2) SB-216763
SB-216763は、化学名を3-(2,4-Dichlorophenyl)-4-(1-methyl-1H-indol-3-yl)-1H-pyrrole-2,5-dioneと称し、下記の式で示されるピロール誘導体である(EP328026、特表01-233281号 実施例45参照)。
【0043】
【化6】
【0044】
SB-216763もまた、試薬としてSigma-Aldrich等により販売されており、一般に入手可能である。
【0045】
なお上記化合物に限定されず、下記の一般式(II)で示されるピロール類もまた、同様のGSK3β阻害作用を有する限り、SB-216763と同様、本発明に係るがん細胞の増殖抑制やがんの治療用医薬組成物として利用可能である。
【0046】
【化7】
【0047】
(ここで、R1は水素、アルキル、アラルキル、アルコキシアルキル、ヒドロキシアルキル、ハロアルキル、アミノアルキル、モノアルキルアミノアルキル、ジアルキルアミノアルキル、アジドアルキル、アシルアミノアルキル、アルキルスルホニルアミノアルキル、アリールスルホニルアミノアルキル、メルカプトアルキル、アルキルチオアルキル、グルコピラノシル、カルボキシアルキル、アルコキシカルボニルアルキル、アミノカルボニルアルキル、ヒドロキシアルキルチオアルキル、メルカプトアルキルチオアルキル、アリールチオアルキルまたはカルボキシアルキルチオアルキルであり、
R2は水素、アルキル、アラルキル、アルコキシアルキル、ヒドロキシアルキル、ハロアルキル、アミノアルキル、モノアルキルアミノアルキル、ジアルキルアミノアルキル、アシルアミノアルキル、アルキルスルホニルアミノアルキル、アリールスルホニルアミノアルキル、メルカプトアルキル、アルキルチオアルキル、カルボキシアルキル、アルコキシカルボニルアルキル、アミノカルボニルアルキル、アルキルチオまたはアルキルスルフィニルであり、
R3はハロゲン、アルキル、ヒドロキシ、アルコキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、アルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルおよびアルキルスルホニルからなる群より選ばれる1または2以上の置換基で置換されたフェニル基であり、
R4、R5、R6およびR7は、互いに独立して、水素、ハロゲン、アルキル、ヒドロキシ、アルコキシ、アリールオキシ、ハロアルキル、ニトロ、アミノ、アシルアミノ、モノアルキルアミノ、ジアルキルアミノ、アルキルチオ、アルキルスルフィニルまたはアルキルスルホニルである)
【0048】
さらに上記した化合物の薬学的に許容しうる塩、例えば塩基性の酸付加塩、無機もしくは有機の酸またはアルカリ金属塩、アルカリ土類金属塩等もまた、本発明に係るがん細胞の増殖抑制や医薬組成物に利用可能である。
【0049】
1.3 GSK3βのリン酸化とがんの治療
GSK3βの生理的活性はリン酸化(第9セリン残基、不活性化型;第216チロシン残基、活性化型)により調節されている。大腸癌細胞や組織においてGSK3βの発現と活性はβ-カテニン活性化(核内集積)とは関係なく亢進し、非がん細胞や組織で検証された第9セリン残基のリン酸化(不活性化型)による酵素活性制御は認められなかった。一方で、癌細胞や癌組織特異的に第216チロシン残基がリン酸化されたGSK3β(活性化型)分画が高頻度に発現していた。
【0050】
培養細胞のGSK3β活性を薬理学的阻害剤(AR-A014418、 SB-216763)を用いて抑制すると、阻害剤の用量依存的に癌細胞のアポトーシスが誘導され、増殖は阻止されるが、非がん細胞株(HEK293)ではこうした効果は認められなかった。RNA干渉法でGSK3β発現をノックダウンしても、薬理的阻害剤の効果と同様に癌細胞の生存と増殖が特異的に抑制された。以上の結果より、GSK3β活性の増強、特に第216チロシン残基リン酸化GSK3β(活性化型)分画の高発現が、癌細胞の生存・増殖を維持・促進するというがん化のあらたなメカニズムが明らかとなった。
すなわち、第216チロシン残基のリン酸化により活性化されているGSK3β活性を薬理学的ならびに転写調節的技法を駆使して修飾することにより、がんの治療や予防が可能になることがはじめて示された。
【0051】
2.被験物質の抗がん剤としての効果を評価する方法
本発明は、GSK3β遺伝子、またはGSK3β蛋白質に対する阻害効果、とくに第216番目チロシン残基のリン酸化により活性化されている本酵素活性に対する阻害効果を指標として、被験物質の抗がん剤としての効果を評価する方法を提供する。
【0052】
前記方法は、1つの被験物質について、その投与(添加)および非投与条件下における、阻害効果を比較評価するものであってもよいし、2つ以上の被験物質についての同様な効果の比較評価であってもよい。あるいは、前記遺伝子や蛋白質の発現量や活性と抗腫瘍効果との相関関係が経験的に確立されれば、その関係に基づいて、比較対照なしに絶対評価するものであってもよい。
【0053】
本発明の評価方法において、GSK3β遺伝子やGSK3β蛋白質に対する阻害効果は、GSK3β遺伝子やGSK3β蛋白質の発現量を指標として評価してもよいし、GSK3β遺伝子やGSK3β蛋白質の活性(安定性含む)を指標として評価しても良い。また、評価系はin vivo系であってもよいし、in vitro系であってもよい。以下、本発明の評価方法の具体的な実施方法について説明する。
【0054】
2.1 培養細胞
本発明の評価方法で用いられる細胞は、GSK3β遺伝子を高発現する哺乳動物細胞であればとくに限定されない。好ましくは哺乳動物由来の癌細胞(好ましくは、初代培養細胞)や樹立された癌細胞株が好ましい。前記哺乳動物としては、ヒト、マウス、ラットまたはハムスター等が好ましく、ヒトまたはマウスがより好ましい。また、GSK3β遺伝子をそのプロモーター領域とともに導入し、該遺伝子を高発現するように組換えた細胞など、人為的に形質転換された細胞を使用してもよい。
【0055】
細胞は、被検物質の添加または非添加条件下で培養する。培養方法は特に限定されず、当該細胞に適した培養方法を適宜選択すればよい。培養細胞への被検物質の添加方法や添加量も特に限定されず、上記細胞の培養前あるいは培養期間中の適当な時点で、被検物質を培養培地に添加し一定期間培養する。被験物質添加後の培養期間もとくに限定されないが、好ましくは30分〜24時間である。
【0056】
2.2 がんモデル動物
本発明の方法で用いられるがんモデル動物は、非ヒト哺乳動物であれば特に限定されないが、齧歯類が好ましく、マウスが特に好ましい。そのようながんモデル動物は市販のものであっても、公知の方法にしたがって作製されたものでもよい。例えば、大腸がんモデルマウスは、既報(Gupta RA., et al. Activation of nuclear hormone receptor peroxisome proliferator-activated receptor-δ accelerates intestinal adenoma growth. Nat Med 2004; 10(3): 245-247)に従って作製することができる。
【0057】
前記動物は、被験物質の投与または非投与条件下で適当な期間飼育を行う。動物への被験物質の投与量は特に限定されず、被験物質の性状や動物の体重に合わせて、適宜用量を設定すればよい。また、動物への被験物質の投与方法および投与期間も特に限定されず、被験物質の性状に合わせて、適宜その投与経路と投与期間を設定すればよい。
【0058】
被験物質の投与または非投与条件下で飼育された前記動物の血液または細胞を単離し、該血液または細胞中のGSK3β遺伝子や蛋白質の発現量を検出する。検出対象とする細胞としては、患部、すなわち癌組織細胞が好ましい。
【0059】
2.3 GSK3β遺伝子の検出
GSK3β遺伝子の検出方法としては、例えば、単離された血液または細胞からまず全RNAを抽出し、該全RNA中におけるmRNAの発現量を検出する方法を挙げることができる。
【0060】
全RNA中のGSK3β遺伝子の発現量は、得られた全RNAよりcRNAまたはcDNAを調製し、これを適当な標識化合物でラベルすることにより、そのシグナル強度として検出することができる。遺伝子の発現量の検出方法は特に限定されず、以下に説明する公知の方法から適宜選択して用いればよい。
【0061】
遺伝子チップ、アレイ、メンブレンフィルター等の固相化試料を用いた解析では、検出対象である遺伝子が固相化されている、市販のGeneChip(Affimetrix)、インテリジーン(宝酒造)、フィルター製マイクロアレイ アトラスシステム(クローンテック)を用いて簡便に実施できる。あるいは、公知の方法(Lipshutz RJ, et al. Nat Genet 1999; 21 (suppl) 20-24)に基づいて作製されたGSK3β固相化試料を用いて解析してもよい。
【0062】
RT-PCR法やその1つであるリアルタイムPCR(TaqMan PCR)法は微量なRNAを高感度かつ定量的に検出できるという点で本発明の評価方法に好適である。リアルタイムPCR(TaqMan PCR)法では、5’端を蛍光色素(レポーター)で、3’端を蛍光色素(クエンチャー)で標識した、目的遺伝子の特定領域にハイブリダイズするオリゴヌクレオチドプローブが使用される。該プローブは、通常の状態ではクエンチャーによってレポーターの蛍光が抑制されている。この蛍光プローブを目的遺伝子に完全にハイブリダイズさせた状態で、その外側からTaq DNAポリメラーゼを用いてPCRを行う。Taq DNAポリメラーゼによる伸長反応が進むと、そのエキソヌクレアーゼ活性により蛍光プローブが5’端から加水分解され、レポーター色素が遊離し、蛍光を発する。リアルタイムPCR法は、この蛍光強度をリアルタイムでモニタリングすることにより、鋳型DNAの初期量を正確に定量することができる。
【0063】
上記以外に、遺伝子発現量を解析する方法としては、例えば、サブトラクション法(Sive HL, John T. St. Nucleic Acids Res 1988; 16: 10937、Wang Z, Brown DD. Proc Natl Acad Sci U. S. A. 1991; 88: 11505-11509)、ディファレンシャル・ディスプレイ法(Liang P, Pardee AB. Science 1992; 257, 967-971、Liang P, Averboukh L, Keyomarsi K, Sager R, Pardee AB. Cancer Res 1992; 52: 6966-6968)、ディファレンシャル・ハイブリダイゼーション法(John T. St, Davis RW. Cell 1979; 16: 443-452)、また、適当なプローブを用いたクロスハイブリダイゼーション法(”Molecular Cloning, A Laboratory Manual”Maniatis, T., Fritsch, E.F., Sambrook, J. (1982) Cold Spring Harbor Laboratory Press)等を挙げることができる。
【0064】
上記方法のほか、GSK3β遺伝子のプロモーター支配下に、該プロモーター活性の検出を可能にする遺伝子(以下「レポーター遺伝子」と記載する。)を利用して、間接的にGSK3β遺伝子やGSK3β蛋白質の発現を検出することもできる。レポーター遺伝子としては、例えば、クロラムフェニコールアセチルトランスフェラーゼ、ホタルルシフェラーゼ、β−ガラクトシダーゼ、分泌型アルカリホスファターゼ、緑色蛍光蛋白質(green-fluorescent protein)等を利用することができる。
【0065】
以上のような、一過的な遺伝子導入法を利用した試験方法とは別に、レポーター遺伝子、およびGSK3β遺伝子の発現ベクターで、宿主細胞を二重に形質転換した細胞を利用した試験方法も採択可能である。この場合には、pIND(インビトロジェン社製)やpTet-On(クロンテック社製)等の発現ベクターを利用して、GSK3β遺伝子の発現を誘導する条件下で該レポーター遺伝子の発現が促進されるような細胞株を樹立することが必要となる。得られた細胞株を、GSK3β遺伝子の発現が誘導される条件下におくと、該遺伝子の発現依存的にレポーター遺伝子の転写が促進される。したがって、レポーター遺伝子の発現が可能な条件下において、培地中に任意の被検物質を添加した場合と添加しない場合でのレポーター遺伝子の発現量変化をみれば、GSK3βの発現量が評価できる。
【0066】
2.4 GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の検出
GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の発現量は、該蛋白質に特異的に結合する抗体を用いて検出することができる。抗体を利用した蛋白質の検出方法は特に限定されないが、ウエスタンブロット法、ドットブロット法、スロットブロット法、ELISA法、およびRIA法から選ばれるいずれか一つの方法であることが好ましい。
【0067】
検体である血液または細胞(細胞抽出液として使用する)は、必要に応じて高速遠心を行うことにより不溶性の物質を除去した後、以下のようにELISA/RIA用試料やウエスタンブロット用試料として調製する。
【0068】
ELISA/RIA用試料は、例えば、回収した血清をそのまま使用するか、緩衝液で適宜希釈したものを用いる。ウエスタンブロット用(電気泳動用)試料は、例えば、細胞抽出液をそのまま使用するか、緩衝液で適宜希釈して、SDS−ポリアクリルアミドゲル電気泳動用の2−メルカプトエタノールを含むサンプル緩衝液(シグマ−アルドリッチ社製等)と混合したものを用いる。ドット/スロットブロット用試料は、例えば、回収した細胞抽出液そのもの、または緩衝液で適宜希釈したものを、ブロッティング装置を使用するなどして、直接メンブレンへ吸着させたものを用いる。
【0069】
本工程で用いられる「GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)に特異的に結合する抗体(以下、「抗GSK3β抗体」と記載する。)」は、公知の方法にしたがって調製してもよいし、市販のものを用いてもよい。抗体は、常法により、抗原となるGSK3β蛋白質、あるいはそのアミノ酸配列から選択される任意のポリペプチドを用いて動物を免疫し、該動物生体内に産生される抗体を採取、精製することによって得ることができる。また、公知の方法(例えば、Kohler and Milstein, Nature 256, 495-497, 1975、Kennet, R. ed., Monoclonal Antibody p.365-367, 1980, Prenum Press, N.Y.)にしたがって、抗GSK3β抗体を産生する抗体産生細胞とミエローマ細胞とを融合させることによりハイブリドーマを樹立し、これよりモノクローナル抗体を得ることもできる。
【0070】
抗体作製用の抗原としては、GSK3β蛋白質またはその少なくとも6個の連続した部分アミノ酸配列からなるポリペプチド、あるいはこれらに任意のアミノ酸配列や担体(例えば、N末端付加するキーホールリンペットヘモシアニン)が付加された誘導体を挙げることができる。
【0071】
前記抗原ポリペプチドは、GSK3β蛋白質を遺伝子操作により宿主細胞に産生させることによって得ることができる。具体的には、GSK3β遺伝子を発現可能なベクターを作製し、これを宿主細胞に導入して該遺伝子を発現させればよい。
【0072】
抗GSK3β抗体は、それを直接標識するか、または該抗体を一次抗体とし、該一次抗体を特異的に認識する(抗体を作製した動物由来の抗体を認識する)標識二次抗体と協同で検出に用いられる。
【0073】
前記標識の種類として好ましいものは、酵素(アルカリホスファターゼまたは西洋ワサビペルオキシダーゼ)またはビオチン(ただし二次抗体のビオチンにさらに酵素標識ストレプトアビジンを結合させる操作が加わる)であるが、これらに限定されない。標識二次抗体(または標識ストレプトアビジン)としては、予め標識された抗体(またはストレプトアビジン)が、各種市販されている。なお、RIAの場合は125I等の放射性同位元素で標識された抗体を用い、測定は液体シンチレーションカウンター等を用いて行う。
【0074】
これら標識された酵素の活性を検出することにより、抗原の発現量が測定される。アルカリホスファターゼまたは西洋ワサビペルオキシダーゼで標識する場合、これら酵素の触媒により発色する基質や発光する基質が市販されている。
【0075】
発色する基質を用いた場合、ウエスタンブロット法やドット/スロットブロット法を利用すれば、目視で検出できる。ELISA法では、市販のマイクロプレートリーダーを用いて各ウェルの吸光度(測定波長は基質により異なる)を測定し、定量することが好ましい。また上述の抗体作製に使用した抗原の希釈系列を調製し、これを標準抗原試料として他の試料と同時に検出操作を行い、標準抗原濃度と測定値をプロットした標準曲線を作成することにより、他の試料中の抗原濃度を定量することも可能である。
【0076】
一方、発光する基質を使用した場合は、ウエスタンブロット法やドット/スロットブロット法においては、X線フィルムまたはイメージングプレートを用いたオートラジオグラフィーや、インスタントカメラを用いた写真撮影により検出することができる。また、デンシトメトリーやモレキュラー・イメージャーFxシステム(バイオラッド社製)等を利用した定量も可能である。さらに、ELISA法で発光基質を用いる場合は、発光マイクロプレートリーダー(例えば、バイオラッド社製等)を用いて酵素活性を測定する。
【0077】
2.5 被験物質の評価
被験物質の投与(添加)または非投与(非添加)における、GSK3β遺伝子またはGSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の発現量の相違に基づいて、該被験物質の抗がん剤としての効果を評価する。
【0078】
すなわち、被験物質の投与条件下で非投与条件下よりもGSK3β遺伝子またはGSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の発現量が有意に減少している場合、該被験物質は抗がん剤として有用であると評価できる。ここで、「有意に減少している」とは、例えば、被験物質の投与および非投与条件下でのGSK3β遺伝子またはGSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)の発現量に統計的有意差(p<0.05)があることを意味する。
【0079】
2.6 非放射性試験管内GSK3β酵素活性検出法(Non-RI in vitro kinase assay)
GSK3βの生理的活性はリン酸化(第9セリン残基、不活性化型;第216チロシン残基、活性化型)により調節されているので、各細胞におけるGSK3β活性はin vitro kinase assayあるいは、第216チロシン残基リン酸化ペプチド特異抗体による免疫ブロッティングのいずれかの手法により検出あるいは推定することができる。また第216チロシン残基リン酸化GSK3β蛋白質分画の発現量は、GSK3βの酵素活性を相対的(あるいは、間接的)に反映しているに過ぎないため、GSK3β活性を直接、検出・測定する手法は、本発明におけるGSK3β活性阻害効果の評価方法において重要といえる。発明者らは、このGSK3βの活性測定の新たな手段として、非放射性試験管内GSK3β酵素活性検出法を開発した。
【0080】
本検出法の概要は、図16に示すとおりである。まず、一定量の細胞抽出液に抗GSK3β特異的抗体を加え、GSK3β蛋白質を免疫沈降させる(図16のS)。コントロールとして、上記細胞抽出液に同濃度の非特異的IgG(抗GSK3β抗体を作成した動物種で、たとえばマウスIgG)を加える(図16のC)。遠心後、上清の細胞抽出液を除去し、免疫沈降物をリン酸緩衝液とリン酸化反応用緩衝液で十分に洗浄する。ついで、リン酸化反応用緩衝液の存在下で、組換えヒトβカテニン蛋白質(基質)と(非放射性)ATPを加え、リン酸化反応を行う。その後、この反応液にSDS電気泳動用バッファー加えて加熱・変性したものをウエスタンブロッティングにより解析し、GSK3β認識配列に特異的な抗リン酸化βカテニンペプチド抗体によりリン酸化組み換えヒトβカテニン蛋白質を検出する。この手法により検出されるβカテニン蛋白質のリン酸化の程度(ウエスタンブロッティングのシグナル強度)から、当該細胞(抽出液中)のGSK3β活性を計測することができる。組換えヒトβカテニン蛋白質をそのヒスチジン基を介してビーズ(resins)等に固定化すれば、ウエスタンブロッティングを用いることなく、on-resins colorimetric assayも可能となる。
【0081】
3.抗がん剤評価用キット
本発明の方法を実施するためのキットとして、例えば、以下のa)〜f)の少なくとも一つ以上を含むキットを挙げることができる。
a)GSK3β遺伝子またはGSK3β遺伝子を特異的に増幅するための、15〜30塩基長の連続したオリゴヌクレオチドプライマー
b)GSK3β遺伝子またはGSK3β遺伝子に特異的に結合し、該遺伝子を検出するための20〜1500塩基長の連続したポリヌクレオチドプローブ
c)上記b)記載のポリヌクレオチドプローブが固定された固相化試料
d)GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)またはそのGSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)に特異的に結合し、該蛋白質を検出するための抗体
e)上記d)記載の抗体に特異的に結合しうる二次抗体
f)GSK3β蛋白質(第216チロシン残基リン酸化GSK3β蛋白質)
【0082】
前記a)記載のプライマーは、上記遺伝子の塩基配列に基づき、市販のプライマー設計ソフトを用いる等、常法にしたがい容易に設計し、増幅することができる。
【0083】
前記b)記載のプローブは、ノーザンハイブリダイゼーション法であれば、20塩基長程度の1本鎖オリゴヌクレオチドか2本鎖DNAが好適に用いられる。マイクロアレイであれば、100〜1500塩基長程度の2本鎖DNA、または20〜100塩基長程度の1本鎖オリゴヌクレオチドが好適に用いられる。Affimetrix社のGene Chipシステムであれば25塩基長程度の1本鎖オリゴがよい。これらは、特にGSK3β遺伝子の配列特異性が高い部分に特異的にハイブリダイズするプローブとして設計することが好ましい。これらのプローブや前述のプライマーは、適当な標識によりラベル(例えば、酵素標識、放射性標識、蛍光標識等)されていてもよく、またビオチン、リン酸、アミン等により修飾されていてもよい。
【0084】
前記c)記載の固相化試料は、前記b)記載のプローブをガラス板、ナイロンメンブレン、マイクロビーズ、シリコンチップ等の固相に固定することにより作製される。
【0085】
前記d)およびe)記載の抗体やf)の蛋白質は、2.4に記載した方法により作製することができる。抗体は、適当な標識によりラベル(例えば、酵素標識、放射性標識、蛍光標識等)されていてもよいし、ビオチン等により適当に修飾されていてもよい。
【0086】
本発明のキットは上記した構成要素のほか、必要に応じて、ハイブリダイゼーション、プローブの標識、あるいはラベル体の検出のための試薬、反応用緩衝液、酵素基質等、本発明の評価方法に実施に必要な他の要素を含んでいてもよい。
【0087】
4.がんの診断方法
本発明者らは、ヒトGSK3β遺伝子やGSK3β蛋白質は、がん(特に大腸がん)患者の病変組織で特異的に活性化されていることを確認した。したがって、被験者から単離された検体中の、これら遺伝子や蛋白質の発現量を測定すれば、該被験者のがんの発症や病態を予測することができる。遺伝子や蛋白質の発現量の測定は、基本的に2で記載した方法にしたがって実施することができる。
【実施例】
【0088】
<実施例1> 各種細胞株および大腸がん患者の腫瘍におけるGSK3β活性とGSK3β阻害剤の影響
1.対象と方法
細胞株:
7種類の大腸癌細胞株(表1:SW480, SW620, HT29, LoVo, HCT116, SW48およびRKO)およびヒト胎児腎細胞株(HEK293)はAmerican Type Culture Collection (ATCC,Manassas,VA)から購入し、推奨プロトコールに従って培養・維持した。各細胞株はコンフルエントになる前の指数関数的増殖期に採取し、リン酸緩衝液(PBS)で洗浄・遠心して−80℃で保管した。GSK3βとphospho-GSK3βSer9の免疫細胞学的染色を行うために、カバーグラス上で培養した各細胞は4%パラフォルムアルデヒドで固定した。
【0089】
大腸がん症例と組織検体:
対象患者は、2002年および2003年に金沢大学がん研究所病院で大腸がんが切除された20例の患者(男性13名、女性7名:年齢は44-93歳(平均73歳))であり、本研究に登録されることを同意し、インフォームドコンセントを提出した者である。各患者の新鮮手術標本から採取した1組の正常粘膜と腫瘍組織検体は、液体窒素を用いてすみやかに凍結し、-80℃で保管した。手術標本は10%中性緩衝ホルマリンで固定し、組織病理学的検索に供して腫瘍の性状やTNM分類に準拠した病期を決定した。各腫瘍の組織学的特性を代表するパラフィン切片は免疫組織化学的に解析し、β-カテニンの活性化を判定した。
【0090】
ウエスタンブロッティング解析:
凍結保存してあった手術標本検体や培養細胞集塊(ペレット)から細胞蛋白質を、蛋白質分解酵素阻害剤と脱リン酸酵素阻害剤(いずれもSigma-Aldrich)を混入した溶解液(CelLyticTM-MT,Sigma-Aldrich)により抽出した。各検体の蛋白質濃度はCoomasie Protein Assay Reagent (Pierce)を用いてBradford法により測定した。
【0091】
抽出された蛋白質100 μgをSDS-ポリアクリルアミド(10%)ゲル電気泳動(SDS-PAGE)で分離し、ニトロセルロース膜(Amersham)に転写後、それぞれ1000倍に希釈したウサギポリクローナル抗体(Cell Signaling TechnologyおよびBD Biosciences)を反応させ、化学発光検出試薬(ECL(登録商標),Amersham)で可視化することにより、第9セリン残基がリン酸化されたGSK3β分画(phospho-GSK3βSer9)および第216チロシン残基がリン酸化された分画(phospho-GSK3βTyr216)をそれぞれ検出した。ついで、ラットGSK3βのN-末端側第1-160アミノ酸残基に対して作成されたマウスモノクローナル抗体(2500倍希釈,BD Transduction Laboratories)を反応させ、同様にして(ヒト)総GSK3βを検出した。
【0092】
GSK3β(シグナル)の上流で活性化される代表的キナーゼとして、プロテインキナーゼB(PKB、あるいはAkt)の活性化をphospho-AktThr308やphospho-AktSer473に対する抗体(それぞれ1000倍希釈、Cell Signaling Technology)を用いて、同様に検出した。
【0093】
GSK3βのin vitro kinase assay:
それぞれの細胞株から抽出した1mgの蛋白質から、免疫沈降法によりGSK3βを調整し、その酵素活性を組換えヒトβ-カテニン蛋白質を基質にして、32P標識ATPを用い、既報(Adler V, et al. Proc Natl Acad Sci USA 1997;94:1686-91)にしたがってin vitro kinase assayにより検出した。β-カテニン蛋白質の放射性リン酸化シグナルはオートラジオグラフィーにより検出した。
【0094】
免疫蛍光細胞化学的染色:
HEK293細胞と各大腸癌細胞株をカバーグラス上で培養し、免疫蛍光染色により総GSK3βとphospho-GSK3βSer9を検出した。一次抗体はウエスタンブロット法で用いた抗体(100倍希釈)を使用し、二次抗体としてそれぞれ200倍希釈したCy3標識抗マウスIgG (H+L)抗体およびFITC標識抗ウサギIgG (H+L)抗体(いずれもJackson ImmunoResearch)を使用した。また、核染色にはHoechst 33342(Hoechst)を使用した。免疫蛍光染色のすべての手法は既報(Shakoori A, et al. Biochem Biophys Res Commun 2003; 312(3): 850-7)に準じて行った。染色された細胞はCCDデジタルカメラ(オリンパスDP70, Olympus)に接続した蛍光顕微鏡(オリンパスAX80, Olympus)により観察・記録した。
【0095】
GSK3βSer9とK-ras遺伝子における突然変異の検出:
大腸癌培養細胞株と手術標本から採取した正常粘膜および腫瘍組織からゲノムDNAをプロティナーゼK消化とフェノール・クロロフォルム処理により抽出・精製した。GSK3βの第9セリン残基をコードするコドン9を含むエクソン1を上流プライマー(5’-ATTCGCGAAGAGAGTGATCAT-3’:配列番号1)と下流プライマー(5’-CACTGCTAACTTTCATGCTGC-3’:配列番号2)を用いて94℃ 1分、55℃ 1分、72℃ 1分の35サイクルプログラムとそれに続く72℃ 10分間伸長反応によりPCR増幅した。
【0096】
GSK3βコドン9(TCC)の突然変異はPCR産物の制限酵素Mnl I(認識配列:CCTC[N]7;New England Biolabs)消化による制限酵素断片長多型(PCR-RFLP)により解析した。この制限酵素は110塩基対の野生型コドン9のPCR産物をそれぞれ52と58塩基対の断片に切断し、一方、コドン9の第1あるいは第2塩基に変異を有するPCR産物は切断できない。K-ras遺伝子コドン12と13の変異は既報(Zhang B, et al. Clin Cancer Res 2003; 9(8): 3073-9.)にしたがい、それぞれ制限酵素Mva I(Takara)とBgl I(TOYOBO)を用いて、ミスマッチプライマーによるPCR産物のPCR-RFLP解析により検出した。
【0097】
β-カテニン活性化の免疫組織化学的解析:
各症例において、大腸原発腫瘍の主要な組織学的特徴を代表する組織切片をアビジン-ビオチン複合体法を用いて免疫組織化学的に検索し、既報(Ougolkov AV, et al. Gastroenterology 2002; 122(1): 60-71)にしたがい、β-カテニンがん化シグナル活性化を判定した。抗β-カテニン抗体(BD Transduction Laboratories)は100倍希釈して使用した。原発腫瘍におけるβ-カテニンの発現は発明者らが最近、定義した3種類の特徴的パターン:細胞膜発現型(M)、正常の腺窩と同じ発現を示すもの;び慢性核内集積型(NAd)、β-カテニンの核内集積を示す癌細胞が腫瘍全体に認められるもの;腫瘍浸潤先進部核内集積型(NAinv)、β-カテニンの核内集積を示す癌細胞が腫瘍浸潤先進部(腫瘍の浸潤最深部で間質と接する部分)に限局して認められ、それ以外の腫瘍部分における癌細胞はM型のβ-カテニン発現を示すもの、にしたがって分類した。
【0098】
β-カテニンは核内に移行すると例外を除いて造腫瘍活性型であるという根拠(Wong NA, et al. Am J Pathol 2002; 160(2): 389-401., Polakis P. Curr Opin Genet Dev 1999; 9(1): 15-21.)に基づいて、本研究ではβ-カテニンの核内集積はその造腫瘍活性を示すものと見做した。経験豊富な2名の研究者(AV,BZ)が対象腫瘍におけるGSK3βの活性に関する情報を知らされない状態で、それぞれ個別に腫瘍におけるβ-カテニン活性化パターンを判定した。そして、両者の判定はすべての腫瘍について一致した。
【0099】
細胞増殖解析:
各大腸癌細胞株とHEK293細胞を96穴培養プレートの各ウェルに3 x 103個播種し、対応する培地中で一晩、培養した(この状態が0日)。既知のGSK3β阻害剤であるAR-A014418(N-(4-methoxybenzyl)-N'-(5-nitro-1,3-thiazol-2-yl)urea, Calbiochem)とSB-216763(3-(2,4-dichlorophenyl)-4-(1-methyl-1H-indol-3-yl)-1H-pyrrole-2,5-dione, Sigma-Aldrich)はジメチルスルフォキシド(DMSO)に溶解した。第0日に各細胞株の培地を交換し、種々の濃度(5, 10, 25, 50μM)になるようにAR-A014418あるいはSB-216763を添加した。対照として、細胞を含まないウェルに同濃度のAR-A014418あるいはSB-216763を添加したものと、細胞を含むウェルにDMSOのみを添加したものを同時に解析した。
【0100】
0日目から特定時間(24, 48, 72および96時間)経過した時点で各ウェルの生細胞数をMTS*生存細胞測定キット(CellTiter 96TM Aqueous, Promega Corp)を用いて、同社のプロトコールにしたがって計測した。すなわち、各ウェルに20μLの直前に調製したMTS混合液を添加し、5% CO2の湿潤条件下において37℃で3時間反応させ、490nm波長の吸光度をTHERMOmaxマイクロプレートリーダー(Molecular Devices)を用いて計測した。各細胞は、それぞれの濃度について6ウェルずつ測定を行い、その平均吸光度を生細胞数として算出した。
*3-[4,5-dimethylthiazol-2-yl]-5-[3-carboxy-methoxyphenyl]-2H-tetrazolium inner salt
【0101】
siRNAトランスフェクション(小分子干渉RNA導入)実験:
ヒトGSK3βを標的とする4種類の2本鎖siRNAを混合したもの(siRNA SMARTpoolTM GSK-3β)と陰性対照のsiRNA(siGLOTM RISK-Free siRNA)はDharmacon RNA Tchnologies社(Lafayette, CO)により合成・供給された。それぞれ1 x 105個のHEK293細胞と各大腸癌細胞株を6穴細胞培養プレートに播き、1晩培養した。siRNA導入効果が最適となるような条件を検定するために、指定されたプロトコールに準拠して、約50%コンフルエントの細胞にTransIT(登録商標)-TKOトランスフェクション試薬(Mirus Corporation)を用いて異なる濃度のGSK3β特異的siRNAあるいは同濃度の陰性対照siRNAを導入した。siRNA導入のGSK3β発現に対する効果を調べるために、異なる濃度のそれぞれのsiRNAを導入して4日後に収集した細胞から蛋白質を抽出し、GSK3αとGSK3βをともに認識する抗ヒトGSK3抗体(1,000倍希釈;Upstate Biotechnology)を用いてウエスタンブロット法により解析した。
【0102】
この予備実験により決定されたsiRNAの最適条件において、GSK3β標的siRNA導入の細胞増殖に及ぼす効果を前項と同様にして細胞増殖解析法により解析した。すなわち、3 x 103個のHEK293細胞と各大腸癌細胞株を96穴細胞培養プレートの各ウェルに播き、対応する培地中で一晩、培養した。その後、最適濃度のGSK3β siRNAあるいは陰性対照siRNAを各細胞に導入した。当日から起算して指定時間(24, 48, 72および96時間)後に各ウェル中の生細胞数をMTS細胞増殖解析キット(Promega)を用いて計測した。
【0103】
2.結果
2.1 各種細胞株におけるGSK3β活性
本研究で用いた大腸癌細胞株はいずれもHEK293細胞に比べて高いGSK3βの発現レベルを示した。全ての大腸癌細胞株において、GSK3β特異的抗体によるウエスタンブロット法により検出可能な不活性化型phosph-GSK3βSer9分画の発現は認められなかったが、逆に活性化型のphospho-GSK3βTyr216分画の発現はより高かった(図1A)。また、in vitro kinase assayにより、これらの大腸癌細胞株におけるGSK3βの酵素活性が確認された(図1B)。これらの結果から、全ての大腸癌細胞株においてGSK3βは恒常的に活性化状態にあることが示された。
【0104】
大腸癌細胞株で認められた結果とは対照的に、HEK293細胞ではphospho-GSK3βSer9分画の発現が検出された。なお、この細胞ではβ-カテニンが安定した低発現を示すよう生理的に制御されていることを発明者らはこれまでに証明している(Spiegelman VS, et al., Mol Cell 2000; 5(5): 877-82, Ougolkov A, et al., J Natl Cancer Inst 2004; 96(15): 1161-70.)。
【0105】
免疫蛍光細胞化学的染色により、GSK3βはHEK293細胞とすべての大腸癌細胞の細胞質に局在し、一方、phospho-GSK3βSer9はHEK293細胞にのみ検出された(図2)。これらの細胞株における総GSK3βとphospho-GSK3βSer9の発現レベルはウエスタンブロット法の結果(図1A)と一致していた。以上の結果より、β-カテニンがAPC不活性化(SW480,SW620, LoVo)あるいはβ-カテニン遺伝子(CTNNB1)変異(HCT116、SW48)のいずれかによってGSK3βによるリン酸化から回避されて活性化されている大腸癌細胞株(表1)において、(総)GSK3βの過剰発現とその不活性化型であるphosph-GSK3βSer9分画の低発現、および活性化型であるphospho-GSK3βTyr216分画のが高発現していることが示された。
【0106】
【表1】
【0107】
2.2 大腸がん患者の腫瘍組織におけるGSK3β活性
樹立された培養細胞株は長期間の細胞発育が可能になるよう適応し、そして実際の生体における同種の腫瘍の特性を必ずしも反映しなくなっていることがあるので、20例の大腸がん患者から採取した正常大腸粘膜と腫瘍組織におけるGSK3βの発現と第9セリン残基および第216チロシン残基のリン酸化を解析した。この解析で得られた結果をそれぞれの腫瘍におけるβ-カテニン活性化の有無やそのパターンと、臨床的ならびに病理組織学的因子と比較検討した。その結果、図3に示すように、ほとんどの症例において腫瘍におけるGSK3βの発現とphospho-GSK3βTyr216分画は同一症例の正常粘膜におけるものに比べて高いレベルであった。phospho-GSK3βSer9分画は非腫瘍性粘膜において高頻度に高いレベルで検出されたが、症例番号74と79を除いてほとんどの症例の腫瘍においてこの不活性化型酵素分画はわずかに検出されるのみかあるいはまったく検出されなかった(表2)。
【0108】
大腸癌におけるβ-カテニンの過剰発現と核内集積の相関を示してきた発明者らのこれまでの報告(前掲)に基づいて、前記した方法で、β-カテニンの活性化はその腫瘍核内集積に基づいて判定した。その結果、β-カテニン活性化は13例(65%)の腫瘍に検出され、そのうち7例はNAd型、6例はNAinv型であった。大腸癌細胞株における観察所見と同様に、これらの結果はGSK3βの抑制的制御を欠落した過剰発現は大腸癌における普遍的な特性であり、β-カテニン活性化の有無あるいは活性化パターン(NAd,NAinv)とは無関係であることを示している(表2)。腫瘍におけるGSK3βの発現や活性と大腸がん症例の腫瘍病期や転移なども含めた臨床的特徴との間には特定の相関は認められなかった(表2)。
【0109】
【表2】
【0110】
2.3 GSK3β活性を制御する因子
大腸癌におけるGSK3βの発現やその第9セリン残基や第216チロシン残基のリン酸化を介した不活性化を制御する可能性のある上流因子(シグナル)について検討した。GSK3βを不活性化する上流のキナーゼ(リン酸化酵素)のうち最もよく知られているものはPKB/Aktであり、この酵素はリン酸イノシチド3-キナーゼ(PI3K)依存的リン酸化により活性化される。PI3K活性化はインスリンおよびrasシグナルの下流に生じるもので、とくに後者のシグナルは大腸癌においてK-ras遺伝子の活性化突然変異によって高頻度に活性化されている(Adjei AA. J Natl Cancer Inst 2001; 93(14): 1062-74, Minamoto T, et al. Expert Rev Mol Diag 2002; 2(6): 565-75.)。これらのがん化シグナルネットワークを考慮して、大腸癌における活性化型PKB/Akt(phospho-AktThr308,phospho-AktSer473)とK-ras原がん遺伝子の既知のホットスポット変異(コドン12,13)を解析し、これらの因子のいずれかがGSK3βの発現やその第9セリン残基や第216チロシン残基のリン酸化に影響しているかどうかを検討した。
【0111】
いずれかの活性化型PKB/Akt分画は7種類の大腸癌細胞株のうち6種類に、また、20例の大腸がん症例のうち13例の腫瘍において検出された(図1C,表1および2)。K-ras遺伝子の突然変異による活性化は6種類の大腸癌細胞株のうち4種類に検出され(表1)、そして20例の大腸がん症例ではそのうち5例の腫瘍において検出された(表2)。対象とした大腸癌細胞株と20例の大腸がん症例の腫瘍において、GSK3βの活性とPKB/Aktのリン酸化による活性化あるいはK-rasの突然変異による活性化との間には明らかな相関は認められなかった。これは大腸癌細胞株や大腸がん症例の腫瘍においてphospho-GSK3βSer9が高頻度に欠落していることに起因する(表1,2)。この結果は、RasあるいはPKB/Akt以外の未知のシグナルを介する上流因子あるいは分子機構が大腸癌におけるGSK3βの発現亢進あるいはその不活性化を抑制していることを示している。
【0112】
最近、GSK3βの第9セリン残基を別のアミノ酸に置換するような突然変異を同遺伝子のコドン9に人為的に導入することによりGSK3βは第9セリン残基リン酸化から回避され、恒常的に活性化されることが報告されている(Wang L, et al., J Biol Chem 2004; 279(31): 32444-52.)。今回、大腸癌細胞株あるいは大腸がん症例の腫瘍組織検体のDNAから増幅したPCR産物をPCR-RFLP法や塩基配列決定法により調べたが、GSK3βコドン9に変異は検出されなかった(データ提示なし)。このことは、GSK3βの第9セリン残基リン酸化を阻害するような遺伝子変異が大腸癌におけるGSK3β活性化の推測的(あるいは候補)機構ではないことを示す。
【0113】
2.4 各種細胞株に対するGSK3β阻害剤の影響
実験的にGSK3βの活性や発現を抑制することが大腸癌細胞株のin vitroにおける生存や増殖に影響を及ぼすかどうかを検討した。大腸癌細胞株のSW480とHCT116を種々の濃度のAR-A014418あるいはSB-216763により処理したところ、これらの細胞の生存(率)は各阻害剤の用量(濃度)依存的に減少した(図4A)。GSK3β阻害剤の同様の効果は他の大腸癌細胞株についても観察された(図4B,E)。しかし、これらの阻害剤の細胞生存抑制効果はHEK293細胞では観察されなかった(図4A)。GSK3β阻害剤を作用させた大腸癌細胞ではアポトーシス(Hoechst染色で観察・計測)の頻度は増加し、細胞増殖率(増殖細胞核抗原:PCNA標識率により計測)は低下したが、HEK293細胞ではこれらの変化は軽微であった(図4C, D)。
【0114】
さらに、大腸癌におけるGSK3βの発現増強の役割を解明するために、個々の大腸癌細胞の生存に及ぼすGSK3βを標的としたRNA干渉(RNAi)の効果を観察した。GSK3βに特異的な4種類のsiRNA(最終濃度はそれぞれ25nM)を混合して細胞に導入したところ、HEK293細胞と大腸癌細胞SW480におけるGSK3β蛋白質の発現は特異的に減少した(図5)。この選択的なGSK3βの発現抑制は、GSK3αやアクチンの発現がGSK3β特異的siRNAあるいは陰性対照siRNAのいずれの導入によっても影響されないことでも裏付けられた(図5)。注目すべきことに、GSK3βに対するRNAiは大腸癌細胞SW480やHEK293細胞におけるGSK3βの発現を同程度に抑制したのに対し、大腸癌細胞SW480の生存は抑制し、HEK293細胞の増殖生存には影響しなかった(図5)。この結果は大腸癌細胞において過剰発現し活性化されているGSK3βの特異的がん化作用をさらに支持するものであった。
【0115】
3.まとめ
種々の大腸癌細胞株や臨床的大腸癌におけるGSK3βの発現亢進と活性化、およびGSK3β阻害剤の大腸癌細胞増殖に対する顕著な抑制効果を示した今回の結果は、GSK3βが既知のがん化シグナルに対して抑制的に作用し、それゆえGSK3β阻害剤には潜在的発がん性が予測されるという従来の知見を覆すものであった。すなわち、本研究により、癌細胞において第9セリン残基のリン酸化の欠如と第216チロシン残基のリン酸化を介して恒常的に活性化されているGSK3βの活性や発現を薬理学的ならびに転写調節的技法を駆使して修飾することにより、がんの新たな治療や予防が可能になることが示された。
【0116】
<実施例2> 大腸癌細胞移植動物モデルに対するGSK3β阻害剤投与の効果
GSK3β阻害(剤)のin vitroにおける制がん効果を明らかにする目的で、以下の動物実験を実施した。
1.対象と方法
6週齢のヌードマウス(BALB/cA Jcl, nu/nu,日本クレア)の左腋窩の皮下に107個の大腸がん細胞SW480を23G(ゲージ)の注射針を用いて移植した。癌細胞を移植して2週間後に腫瘍生着が確認されたマウスを、AR-A014418投与群(N=12)、SB-216763投与群(N=12)、DMSO投与(コントロール)群、(N=5)の3群に分けた。さらに、AR-A014418投与群とSB-216763投与群をそれぞれ1mg/kg(培養細胞実験の5μMに相当)投与群(N=4)、2mg/kg(培養細胞実験の10μMに相当)投与群および5mg/kg(培養細胞実験の25μMに相当)投与群の3群、合計6群に分けて、治療実験を行った(表3,図6)。DMSOに溶解した3種類の濃度のAR-A014418(283μg/mL=0.35mM; 567μg/mL=0.7mM; 1419μg/mL=1.75mM)あるいはSB-216763(235μg/mL=0.35mM; 471μg/mL=0.7mM; 1177μg/mL=1.75mM)をそれぞれ0.2mlずつ、DMSO投与群には同量のDMSOを週3回(隔日)、腹腔内に注入した。
【0117】
【表3】
【0118】
これらのGSK3β阻害剤投与量は、マウスの体液量を体重(20g程度)の70%と想定して、培養細胞に使用した培養液中の濃度と同レベルになるように算出した。すなわち、1mg/kg、2mg/kgおよび5mg/kgの阻害剤投与量はマウス体液量に換算して、それぞれ、細胞培養液中の阻害剤濃度の5μM、10μMおよび25μMに相当する。
【0119】
各群のマウスにDMSOあるいはそれぞれの用量の阻害剤の投与期間に毎週、体重、食餌摂取量と移植腫瘍の大きさ(長径と短径)を計測した。DMSOあるいは各阻害剤を投与して5週間後に、エーテル深麻酔によりマウスを安楽死させ、移植腫瘍と主要臓器(肺、肝、膵、腎、脾)を摘出した(図6)。腫瘍の体積は、以下の計算式により算出した。
腫瘍体積(cm3)=0.5 x 長径(cm) x [短径(cm)]2
【0120】
2.結果
図7は、治療実験開始5週間後、各群(4匹ずつ)の代表的な固体を示したものである。対照(DMSO投与)群の移植腫瘍に比べて、GSK3β阻害剤を投与されたいずれの群でも左腋窩の皮下移植腫瘍の増殖が抑制されていることが確認された。図8は、異なる用量の各阻害剤投与後の腫瘍体積の変化を経時的に示したものである。このグラフから、移植腫瘍の増殖は、対照群の腫瘍に比べてGSK3β阻害剤の投与用量に依存して有意に抑制されていることが確認された。なお、対照群を含めていずれの群でも、食餌摂取量や体重変化などの観点から、明らかな有害事象は認められなかった。
【0121】
この実施例により、in vivoにおけるGSK3β阻害(剤)の抗腫瘍効果が初めて検証された。また、短期間の実験ではあるが、GSK3β阻害剤の毒性あるいは有害事象はないものと考えられる。
【0122】
<実施例3> 神経膠芽腫細胞に対するGSK3β阻害剤の作用
GSK3β阻害(剤)の制がん効果が大腸癌に限定されず、それ以外のがん種、とくに難治性がんに対しても有効かどうかを明らかにすることは、本酵素の阻害に基づくがん治療法の開発には重要な課題である。そこで、大腸癌以外の難治性がん(胃癌、膵癌、肝細胞癌、脳腫瘍など)由来の細胞について実施している実験のうち、難治性がんの一例として神経膠芽腫を対象にした成果を記載する。
【0123】
1.対象と方法
神経膠芽腫(glioblastoma)は、極めて分化度が低く悪性度が最も高い神経膠腫で、原発性脳腫瘍の約9%を占める。実際の腫瘍は手術で完全摘出できないことが多く、既存の抗がん剤治療や放射線照射治療に対しても耐性を示し、極めて難治性である。このように、本腫瘍に対する有効な治療法がないことから、その生命予後は最近の20−30年間、改善されていない実情であり、新しい治療法の開発が切望されている。
【0124】
本実施例では、4種類の神経膠芽腫細胞株A172、T98G、U251およびU87(American Type Culture Collection [ATCC,Manassas,VA]から購入した)と、ヒト胎児腎細胞 HEK293を対象にした。これらの細胞株におけるGSK3βとそのリン酸化分画の発現を<実施例1>に準じて解析した。また、既知のGSK3β阻害剤(AR-A014418あるいはSB-216763)の、腫瘍細胞の生存、増殖やアポトーシスなどへの作用を、大腸がん細胞の場合と同様に解析した。
【0125】
各細胞は一晩培養した後、それぞれDMSOに溶解したAR-A014418あるいはSB-216763を所定の濃度(5μM, 10μM, 25μMまたは40μM, 50μM)となるように添加した培地で培養し、生存細胞数とアポトーシスの頻度を経時的に測定した。コントロールとして、同量のDMSOを添加して培養した細胞についても、その生存細胞数とアポトーシスの頻度を測定した。なお、<実施例1>とは異なり、生存細胞数の計測にはWST-8**アッセイキット(Cell Counting Kit-8,和光,大阪)とBio-Rad社(Hercules, CA)製のマイクロプレートリーダーを使用した。また、増殖期細胞分画はbromodeoxy uridine(BrdU)を取り込む細胞数をCell Proliferation ELISA BrdU Kit(Roche, Panzberg, Germany)を用いて測定し、腫瘍細胞の増殖に及ぼすGSK3β阻害剤(AR-A014418あるいはSB-216763)の影響を解析した。
**WST-8: 4-[3-(4-iodophenyl)-2-(4-nitrophenyl)-2H5-tetrazolio]-1,3-benzene disulfonate
【0126】
2.結果
<実施例1>と同様にウエスタンブロット法により解析した結果、4種類の神経膠芽腫細胞株 A172、T98G、U251およびU87のいずれにも、GSK3βとその第216チロシン残基がリン酸化された活性化型分画が検出された(データの提示なし)。現在、これらの神経膠芽腫細胞におけるGSK3βの酵素活性を非放射性試験管内酵素活性検出法(後述の実施例5を参照)により検証している。
【0127】
大腸癌細胞の場合と同様に、AR-A014418およびSB-216763で処置した神経膠芽腫細胞はいずれも、DMSOで処理した場合(コントロール)に比較して、阻害剤の濃度依存的に生存細胞数が減少し、アポトーシスが顕著に誘導された(図9,10A,10B)。一方、HEK293細胞では、GSK3β阻害剤を培地に加えることによりアポトーシスが軽度に誘導されるものの、DMSOで処理した場合(コントロール)との有意差は認められなかった(図10B)。また、これらの神経膠芽腫細胞では、GSK3β阻害剤処理によって、その増殖が抑制された(図10C)。
【0128】
以上の結果より、大腸がんのほかに神経膠芽腫においても(活性化型)GSK3βが腫瘍細胞の生存、増殖を支持し、アポトーシスを阻止していることが明らかにされた。そして、(活性化型)GSK3βは大腸癌以外のがん種に対しても治療標的であることが示唆された。
【0129】
<実施例4> がん細胞の不死化に対するGSK3β阻害の影響
GSK3β阻害(剤)の制がん効果の分子および細胞レベルのメカニズムを明らかにすることは重要である。そこで、がんに共通する重要な生物学的特性の1つである細胞の不死化(immortalization)に対するGSK3β阻害(剤)の作用を、細胞の老化とテロメア(telomere)維持機構に着目して解析した。
【0130】
1.対象と方法
大腸癌細胞および非がん細胞をそれぞれGSK3β阻害剤あるいは特異的RNA干渉により処理し、細胞老化や細胞不死化にかかわる細胞レベル、分子レベルでの変化を解析した。
【0131】
<実施例1>にしたがって、各大腸癌細胞株(SW480、HCT116)および非がん細胞株(ヒト胎児腎細胞 HEK293)を25μMのGSK3β阻害剤(AR-A014418あるいはSB-216763)、あるいはGSK3β特異的RNA干渉で処理した。対照として、同じ細胞に対してDMSO投与あるいは非特異的RNA干渉を行った。そして、GSK3βを阻害した細胞と対応する対照細胞における老化細胞の頻度を比較・解析した。また、これらの細胞に対して、細胞不死化の分子指標であるhuman telomerase reverse transcriptase(hTERT)mRNAの発現およびtelomerase活性の変化を経時的に比較解析した。
【0132】
老化細胞は、β-galactosidase活性の指標であるx-gal染色陽性の細胞として判定した。具体的には、本染色のためカバーグラス上で培養した細胞をリン酸緩衝液(PBS)で洗浄し、クエン酸・リン酸固定液(0.1Mクエン酸,0.2M Na2HPO4, pH 6.0)により室温で5分間固定した。ついで、細胞をPBSで2回洗浄し、x-gal染色液(1mg/mL x-gal [Sigma-Aldrich], 5mM K3Fe[CN]6, 2mM MgCl2, 150mM NaCl, 30mM クエン酸・リン酸緩衝液,5mM K4Fe[CN]6)を37℃で20時間反応させた。染色した細胞をPBSで洗浄後、4%パラフォルムアルデヒドで再固定し、光学顕微鏡観察によりx-gal染色陽性細胞を同定した。老化細胞の頻度は強拡大で5視野観察し、各視野における全細胞数(約100個の細胞)に占めるx-gal染色陽性細胞の頻度の平均値として計測・算出した。
【0133】
hTERT mRNAの発現は、対象細胞からtotal RNAを抽出し、RT-PCR法により解析した。RT-PCRには上流primer(5’-CGGAAGAGTGTCTGGAGCAA-3’: 配列番号3)と下流primer(5’-GGATGAAGCGGAGTCTGGA-3’: 配列番号4)を用い、(15秒@94℃,10秒@55℃,9秒@72℃) x 36ないし38サイクルのRT-PCRプログラムによりhTERT遺伝子を増幅した。telomeraseの活性は市販のキット(TRAPEZE(R) Telomerase Detection Kit, Chemicon International)を用いて、推奨プロトコールに準拠して検出した。GSK3β阻害細胞と対照細胞におけるhTERTの発現レベルとtelomerase活性は、電気泳動上のシグナルをdensitometer (NIH Image Software)により計測・定量し、比較解析した。
【0134】
2.結果
25μMのGSK3β阻害剤で72時間処理することにより、大腸癌細胞株ではいずれも、AR-A014418、SB-216763、DMSO処理した対照細胞に比べてx-gal染色陽性細胞として同定される老化細胞の頻度が有意に増加した。一方、HEK293細胞では、いずれかのGSK3β阻害剤処理した細胞とDMSO処理細胞の老化細胞の頻度に有意差はみとめられなかった(図11,12)。
【0135】
つぎに、hTERTの発現とtelomerase活性に対するGSK3β阻害の効果を検討した。その結果、大腸癌細胞株では、AR-A014418、あるいはSB-216763処理の72ないし96時間後にhTERT発現およびtelomerase活性が抑制された。一方、HEK293細胞では、GSK3β阻害剤によるhTERTの発現変化あるいはtelomerase活性の抑制は認められなかった(図13,14)。また、これらの細胞において特異的RNA干渉によりGSK3βの発現を減弱させることにより、GSK3β阻害剤の場合と同様に大腸癌細胞ではhTERT発現およびtelomerase活性が抑制されたが、HEK293細胞では変化が認められなかった(図15)。
【0136】
以上の結果より、癌細胞の不死化の阻止と老化の促進が、GSK3β阻害による制がん効果の分子・細胞レベルのメカニズムの1つである可能性が示唆された。
【0137】
<実施例5> 非放射性試験管内GSK3β酵素活性検出法(Non-RI in vitro kinase assay)の開発
第216チロシン残基リン酸化GSK3β蛋白質分画の発現量は、GSK3βの酵素活性を相対的(あるいは、間接的)に反映しているに過ぎないため、本酵素活性を直接、検出・測定する手法は、既存の物質のGSK3β活性阻害効果の評価のために重要である。また、本酵素(活性)が発病あるいは発症にかかわる疾患(2型糖尿病、アルツハイマー病、がんなど)の診断法の開発とともに、これらの疾患に対する薬剤開発の観点から、本酵素活性の阻害効果を有する可能性のある物質のスクリーニングや同定にも極めて有用である。GSK3βに限らず、蛋白質リン酸化酵素の活性測定の従来法では放射性同位元素(32Pなど)を標識したATPが必要とされることが、その実施上の制約となっている。そこで発明者らは、放射性同位元素を必要としない簡便なGSK3βの活性測定の新しい手法として、非放射性試験管内GSK3β酵素活性検出法を開発した。
【0138】
1.原理
本検出法の原理について、図16を参照して概説する。まず、細胞抽出液に抗GSK3β特異的抗体とビーズ(resins)を加え、GSK3β蛋白質を免疫沈降させる(図中S)。陰性対照(コントロール)として、同じ細胞の抽出液に同濃度の非特異的マウスIgGを加える(図中C)。ついで、kinase buffer存在下で、組換えヒトβ-catenin蛋白質(基質)と非放射性ATP(図中P)を加え、リン酸化反応を行う。その後、この反応液をウエスタンブロット法により解析し、抗リン酸化β-cateninペプチド抗体によりリン酸化ヒトβ-catenin蛋白質を検出する。組換えヒトβ−catenin蛋白質をそのヒスチジン基を介してビーズ(resins)等に固定化すれば、ウエスタンブロット法を用いることなく、on-resins colorimetric assayも可能である。
【0139】
2.対象と方法
本法の開発にはHEK293細胞と各種の培養癌細胞株:SW480, HCT116, HT29(大腸癌);MKN-28, NKPS, TMK-1, NUGC-4(胃癌);MIA-PaCa-2, Capan-1, BxPC-3(膵臓癌);HEP-G2(肝細胞癌)を使用した。これらの細胞はいずれもAmerican Type Culture Collection(ATCC,Manassas,VA)から購入し、それぞれ推奨の培地と培養条件により培養した。
【0140】
<実施例1>の方法にしたがって、細胞溶解用緩衝液(0.2M Tris-HCl pH 7.5, 1.5M NaCl, 10mM EDTA, 10mM EGTA, 10% Triton X-100, 25mM sodium pyrophosphate, 10mM β-glyceophosphate, 10mM Na3VO4, Protease inhibitors cocktail [Sigma-Aldrich])を用いて、それぞれの細胞から蛋白質を抽出した。細胞由来の蛋白質1mg(1000μg)から、定法にしたがってマウスモノクローナル抗GSK3β抗体(BD Transduction Laboratories)と20μLのProtein A/G Agarose resins(Calbiochem)によりGSK3βを免疫沈降して、resin上に精製した(以後、GSK3βと記載する)。免疫沈降の陰性対照には、抗GSK3β抗体の代わりに非特異的マウスIgGを使用した。細胞由来のGSK3βをリン酸緩衝液(PBS)で2回、ついでリン酸化反応用緩衝液(kinase buffer)(0.25M Tris-HCl pH 7.5, 10mM β-glyceophosphate, 20mM dithiothreitol [DTT], 1mM Na3VO4, 0.1M MgCl2)を用いて2回洗浄した。この細胞由来のGSK3βをkinase bufferに懸濁し、10mM ATP(非放射性)2μLと4μgの組換えヒトβ-catenin蛋白質(基質)を加えて計50μLとした。このkinase反応溶液を温水槽中(30℃)で30分間反応させた後、等量の2xSDS電気泳動用サンプルバッファーを添加して95℃で5分間過熱してリン酸化反応を停止した。
【0141】
この反応液上清をSDSポリアクリルアミド電気泳動で分離し、<実施例1>と同様に、ウエスタンブロット法により目的とする以下の蛋白質を解析した。GSK3βによるリン酸化の基質として用いた組換えヒトβ-catenin蛋白質のリン酸化は、リン酸化β-cateninペプチド(p-β-cateninSer33/37Thr41)特異抗体(Cell Signaling)により検出した。ついで、反応液中のGSK3βと組換えヒトβ-catenin蛋白質を同様にウエスタンブロット法により検出した。これにより、それぞれの細胞におけるGSK3βの発現と相対的な活性の比較も可能になる。
【0142】
この手法をさらに、GSK3β阻害剤の酵素活性阻害効果の判定に応用した。具体的には、kinase反応液中に最終濃度が25μMになるようにSB-216763あるいはAR-A014418を添加し、これらの反応液を同様にウエスタンブロット法により解析し、阻害剤の代わりにDMSOだけを添加した場合のβ-cateninリン酸化と比較した。
【0143】
2.結果
本検出法を用いて、まず、各種大腸癌細胞株(SW480、HT29、HCT116)および非がん細胞株(HEK293)を解析したところ、いずれの細胞においてもGSK3βの酵素活性が検出された(図17)。リン酸化β-catenin のシグナル強度から、HEK293細胞に比べて大腸癌細胞のGSK3β活性は高く、この結果はウエスタンブロット法によるphospho-GSK3βTyr216分画の検出結果(図1A)と対応していると判断された。
【0144】
つぎに、大腸癌を含めた種々の消化器癌細胞を対象にして、同様に解析した。その結果、大腸癌以外に、胃癌、膵癌および肝細胞癌の細胞株にもGSK3βの酵素活性が特異的に検出された。とくに、膵癌細胞(BxPC-3)と肝細胞癌(HEP-G2)に強いGSK3β活性が検出された(図18)。この解析により、それぞれの細胞種におけるGSK3β活性とともに、その総発現量を定量的に比較することが可能であり、同一細胞においては本酵素の発現量と活性に相関がみられた(図18)。
【0145】
さらに、本検出法を用いて、既知のGSK3β阻害剤の効果検証を試みた。その結果、2種類の大腸癌細胞株(SW480, HCT116)における GSK3βの活性は25μMのSB-216763あるいは同濃度のAR-A-014418により完全に阻害されることが確認された(図19)。
【0146】
一連の結果より、この非放射性試験管内GSK3β酵素活性検出法の有用性が検証された。また、この検出法をon resins colorimetric assay法に深化させることにより、本酵素を指標にする疾患診断法の開発や、新しいGSK3β活性阻害物質のスクリーニング、ならびに本酵素阻害手法の開発に応用可能であることが示唆された。
【0147】
<実施例6>胃癌、膵癌および肝細胞癌の細胞株におけるGSK3β阻害の作用
<実施例3>と同様の目的で、大腸癌以外の消化器系臓器の癌腫、とくに難治性の消化器癌から樹立された培養細胞株を対象にして、GSK3βの発現と活性を解析した。<実施例5>と図18AおよびBに示したように、解析対象にした胃癌細胞株 MKN-28、NKPS、TMK-1、NUGC-4、膵癌細胞株MIAPaCa-2、Capan1、BxPC-3、および肝細胞癌HEP-G2のうち、MKN-28とMIAPaCa-2以外のすべての細胞株に明瞭なGSK3βの発現と活性が認められた。
【0148】
本解析の結果に基づいて、これらの癌細胞の生存、増殖およびアポトーシスに対するGSK3β阻害(小分子阻害剤、およびRNA干渉法)の作用を<実施例1>および<実施例3>の方法により検討する。前記解析結果にしたがい、それぞれの臓器由来でGSK3βの高い活性を示す癌細胞株(たとえば、NUGC-4、BxPC-3、HEP-G2)についてがん移植動物モデルを作成し、<実施例2>にしたがい、in vivoにおけるこれらの難治性消化器癌細胞に対するGSK3β阻害剤の(腫瘍増殖抑制)効果を検討する。
【0149】
本明細書中で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書中にとり入れるものとする。
【産業上の利用可能性】
【0150】
本発明は、がん治療や予防のための新規戦略の開発とがんの分子疫学的研究の展開に貢献するものと期待される。
【配列表フリーテキスト】
【0151】
配列番号1−人工配列の説明:プライマー
配列番号2−人工配列の説明:プライマー
配列番号3−人工配列の説明:プライマー
配列番号4−人工配列の説明:プライマー
【特許請求の範囲】
【請求項1】
グリコーゲン合成酵素キナーゼ3β(GSK3β)の第216番目チロシン残基のリン酸化により活性化される前記酵素の活性に対する阻害効果を指標として、被験物質の抗がん剤としての効果を評価する方法。
【請求項2】
下記の工程を含む、請求項1に記載の方法:
1)細胞を被験物質の添加または非添加条件下で培養する。
2)上記細胞内におけるGSK3β蛋白質の第216チロシン残基リン酸化の程度を検出するか、あるいはGSK3βの酵素活性を検出・測定する。
3)被験物質の添加または非添加条件下における、上記のリン酸化の程度、あるいはGSK3βの酵素活性の相違に基づいて、該被験物質の抗がん剤としての効果を評価する。
【請求項3】
細胞ががん患者から単離された細胞または癌細胞株である、請求項2に記載の方法。
【請求項4】
下記の工程を含む、請求項1に記載の方法:
1)がんモデル動物を被験物質の投与または非投与の条件下で飼育する。
2)上記動物の血液または細胞中におけるGSK3β蛋白質の第216チロシン残基リン酸化の程度を検出するか、あるいはGSK3βの酵素活性を検出する。
3)被験物質の投与または非投与条件下における、上記のリン酸化の程度、あるいはGSK3βの酵素活性の相違と移植腫瘍の増殖の変化に基づいて、該被験物質の抗がん剤としての効果を評価する。
【請求項5】
動物がマウスである、請求項4に記載の方法。
【請求項6】
GSK3β蛋白質の酵素活性が非放射性試験管内酵素活性検出法により検出されること、あるいは、前記の第216チロシン残基リン酸化の程度がウエスタンブロット法、ドットブロット法、スロットブロット法、ELISA法、およびRIA法から選ばれるいずれか一つの方法によって検出されることを特徴とする、請求項2〜5のいずれか1項に記載の方法。
【請求項1】
グリコーゲン合成酵素キナーゼ3β(GSK3β)の第216番目チロシン残基のリン酸化により活性化される前記酵素の活性に対する阻害効果を指標として、被験物質の抗がん剤としての効果を評価する方法。
【請求項2】
下記の工程を含む、請求項1に記載の方法:
1)細胞を被験物質の添加または非添加条件下で培養する。
2)上記細胞内におけるGSK3β蛋白質の第216チロシン残基リン酸化の程度を検出するか、あるいはGSK3βの酵素活性を検出・測定する。
3)被験物質の添加または非添加条件下における、上記のリン酸化の程度、あるいはGSK3βの酵素活性の相違に基づいて、該被験物質の抗がん剤としての効果を評価する。
【請求項3】
細胞ががん患者から単離された細胞または癌細胞株である、請求項2に記載の方法。
【請求項4】
下記の工程を含む、請求項1に記載の方法:
1)がんモデル動物を被験物質の投与または非投与の条件下で飼育する。
2)上記動物の血液または細胞中におけるGSK3β蛋白質の第216チロシン残基リン酸化の程度を検出するか、あるいはGSK3βの酵素活性を検出する。
3)被験物質の投与または非投与条件下における、上記のリン酸化の程度、あるいはGSK3βの酵素活性の相違と移植腫瘍の増殖の変化に基づいて、該被験物質の抗がん剤としての効果を評価する。
【請求項5】
動物がマウスである、請求項4に記載の方法。
【請求項6】
GSK3β蛋白質の酵素活性が非放射性試験管内酵素活性検出法により検出されること、あるいは、前記の第216チロシン残基リン酸化の程度がウエスタンブロット法、ドットブロット法、スロットブロット法、ELISA法、およびRIA法から選ばれるいずれか一つの方法によって検出されることを特徴とする、請求項2〜5のいずれか1項に記載の方法。
【図1】

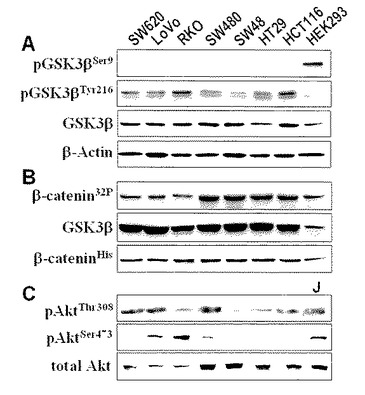
【図2】
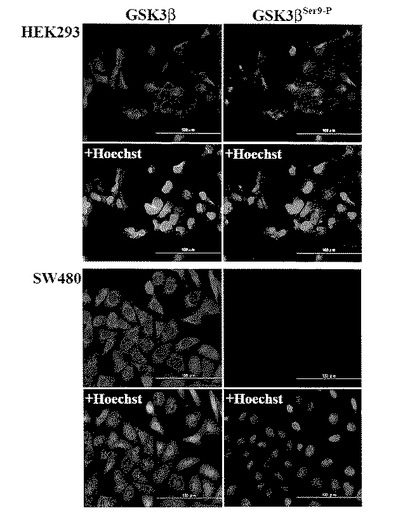
【図3】
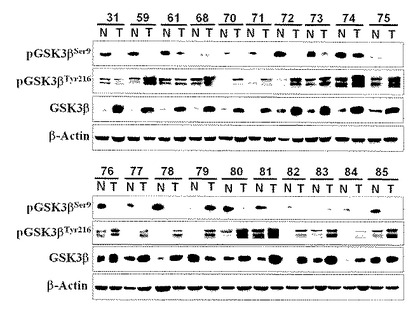
【図4】
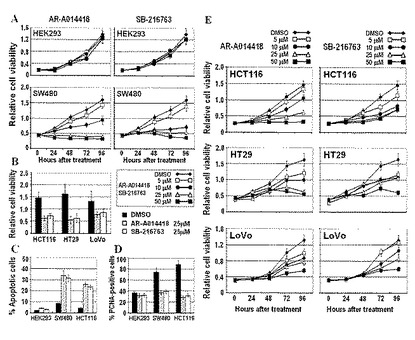
【図5】
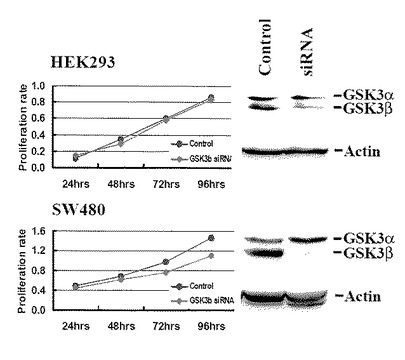
【図6】
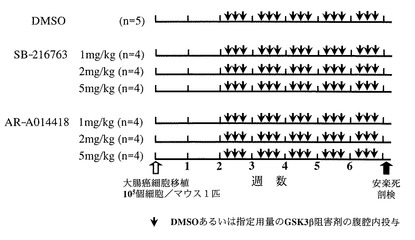
【図7】
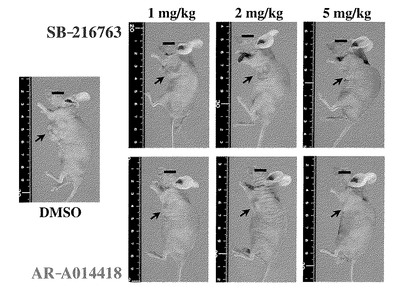
【図8】
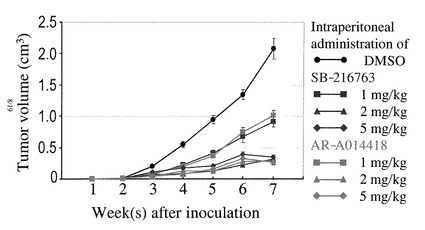
【図9】
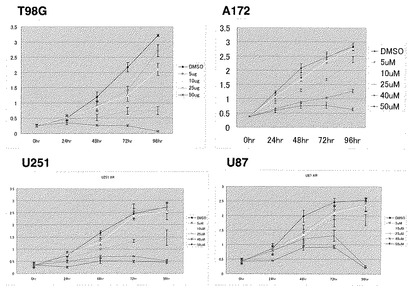
【図10】
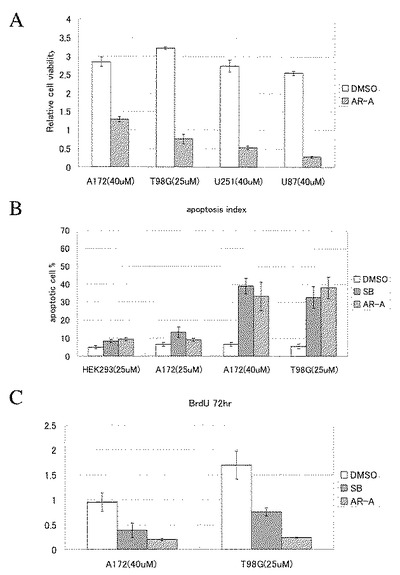
【図11】
【図12】
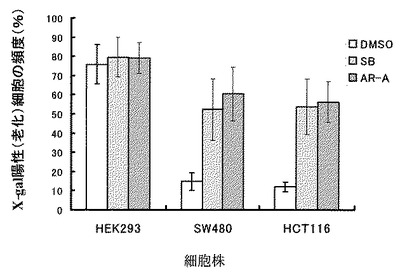
【図13】
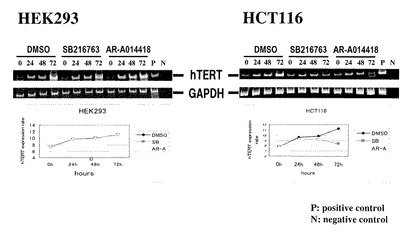
【図14】
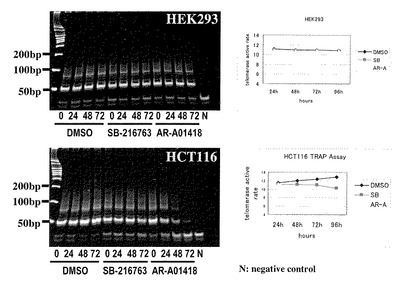
【図15】
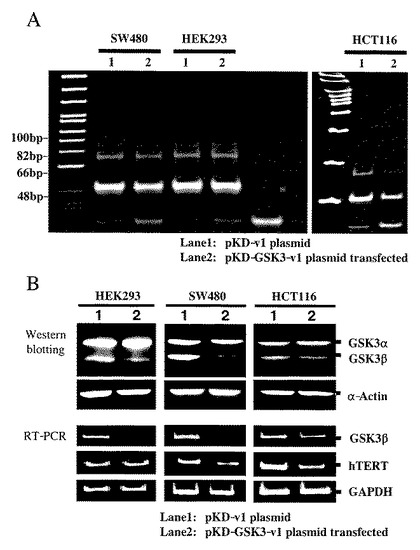
【図16】
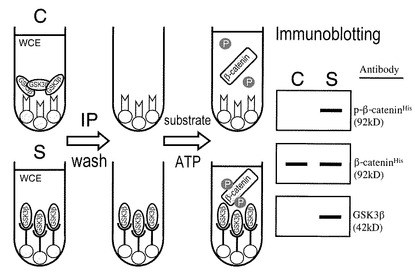
【図17】
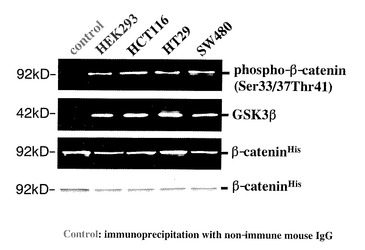
【図18】
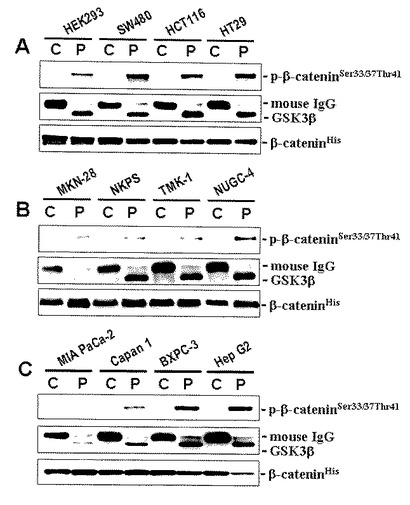
【図19】
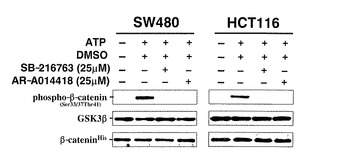
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【公開番号】特開2012−187105(P2012−187105A)
【公開日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願番号】特願2012−91933(P2012−91933)
【出願日】平成24年4月13日(2012.4.13)
【分割の表示】特願2006−550915(P2006−550915)の分割
【原出願日】平成18年1月4日(2006.1.4)
【出願人】(504160781)国立大学法人金沢大学 (282)
【Fターム(参考)】
【公開日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願日】平成24年4月13日(2012.4.13)
【分割の表示】特願2006−550915(P2006−550915)の分割
【原出願日】平成18年1月4日(2006.1.4)
【出願人】(504160781)国立大学法人金沢大学 (282)
【Fターム(参考)】
[ Back to top ]