
Gallibacteriumanatis由来の細胞溶解性RTX毒素
本発明は、動物の健康の分野に関し、特に、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2を含むガリバクテリウム菌種により生じる新たな細菌性家禽疾患の原因因子に関する。本発明は、GtxA(ガリバクテリウム毒素)と名付けた、前記ガリバクテリウム種由来の新規のRTX毒素を提供する。更に、本発明は、GtxAのアミノ酸及びヌクレオチド配列と、不活性化トキソイド又はトキソイドの断片を含むワクチンと、前記疾患を予防するために鳥類を免疫する方法と、鳥類におけるGallibacterium anatis感染を診断する方法とを提供する。
【発明の詳細な説明】
【技術分野】
【0001】
[関連出願の相互参照]
出願、即ち本願において、引用した全ての特許及び非特許文献は、出典を明記することによりその開示内容全体を本願明細書の一部とする。
【0002】
本発明は、動物の健康の分野に関し、特に、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2を含むガリバクテリウム菌種により生じる新たな細菌性家禽疾患の原因因子に関する。本発明は、GtxA(ガリバクテリウム毒素)と名付けた、前記ガリバクテリウム種由来の新規のRTX毒素を提供する。更に、本発明は、GtxAのアミノ酸及びヌクレオチド配列と、不活性化トキソイド又はトキソイドの断片を含むワクチンと、前記疾患を予防するために鳥類を免疫する方法と、鳥類におけるGallibacterium anatis感染を診断する方法とを提供する。
【背景技術】
【0003】
過去10年間、生産性を増加させる集約的養鶏法の結果として、主要な全ての家禽生産国において、疾患の発現が増加してきた。これにより、こうした疾患を抑制する新規の優れたワクチン及び予防接種プログラムの必要性が高まっている。現在、多くの動物は、ウイルス及び細菌を原因とする多数の疾患に対して免疫されている。家禽におけるウイルス性疾患の例には、ニューカッスル病、伝染性気管支炎、トリニューモウイルス、鶏痘、伝染性ファブリキウス嚢病等がある。細菌性疾患の例には、ヘモフィルス・パラガリナルム(上気道)、ボルデテラ・アビウム(上気道)、オルニソバクテリウム・ライノトラキア(下気道)により生じるトリコリーザ、サルモネラ感染症(消化管)、家禽コレラ(敗血症)の原因因子であるパスツレラ・マルトシダ、及び大腸菌感染症がある。
【0004】
産卵家禽の生殖器及び腹膜における炎症は、商用の産卵家禽群において繰り返される問題であり、卵生産量の下落と、死亡率の上昇と、結果的な経済的損失及び動物の生育状態の低下を引き起こす。これらの病変からはトリ病原性大腸菌が単離される場合が多いが、幾つかの研究では、Gallibacterium anatisが単独で、或いは共同病原体として、頻繁に卵巣炎、卵管炎、及び腹膜炎の原因となることが実証されている。更に、トリの敗血症、肝炎、腸炎、及び上気道の病変からG. anatisが単離されている。G. anatisは、産卵雌鶏及び他の鳥類種の上気道及び下部生殖管両方の正常細菌叢に共通する部分であり(Bojesen A.M., Nielsen S.S., Bisgaard M., Prevalence and transmission of haemolytic Gallibacterium species in chicken production systems with different biosecurity levels, Avian Pathol. (2003) 32:503-510)、したがって、日和見病原体と見なすことができる。その病変形成は、特に分子レベルでは、深く研究されておらず、G. anatisが疾患を引き起こす能力の背後にある遺伝子及び機構については殆ど知られていない。G. anatisは、2つの次亜種、β溶血性次亜種ヘモリチカ(haemolytica)と、非溶血性次亜種アナティス(anatis)とに分けられる。赤血球を溶解させる能力は、病原性G. anatis分離株の顕著な表現型である(Christensen H., Bisgaard M., Bojesen A.M., Mutters R., Olsen J.E., Genetic relationships among avian isolates classified as Pasteurella haemolytica, 'Actinobacillus salpingitidis' or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov and description of additional genomospecles within Gallibacterium gen. nov, Int. J. Syst. Evol. Microbiol. (2003) 53:275-287)。ガリバクテリウムは、γプロテオバクテリア科であるパスツレラ科に属するグラム陰性属であり(Christensen et al, 同上)、パスツレラ科の様々な病原性構成員、例えば、ヒトの歯周疾患の原因であるアグリゲイティバクタ・アクチノミセテムコミタンス、ウシ輸送熱の原因物質であるマンヘミア・ヘモリチカ、及びブタ病原体であるアクチノバチルス・プルロニューモニアは、RTX毒素((repeat in toxin)の群に属する溶血素及び白血球毒素を生じる。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Bojesen A.M., Nielsen S.S., Bisgaard M., Prevalence and transmission of haemolytic Gallibacterium species in chicken production systems with different biosecurity levels, Avian Pathol. (2003) 32:503-510)
【非特許文献2】Christensen H., Bisgaard M., Bojesen A.M., Mutters R., Olsen J.E., Genetic relationships among avian isolates classified as Pasteurella haemolytica, 'Actinobacillus salpingitidis' or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov and description of additional genomospecles within Gallibacterium gen. nov, Int. J. Syst. Evol. Microbiol. (2003) 53:275-287)
【発明の概要】
【発明が解決しようとする課題】
【0006】
不活性化又は生存する弱毒化細菌からなるG. anatisワクチンを利用可能である。しかしながら、こうしたワクチンは、この種に由来する分泌溶血性タンパク質に対する保護を与えない。
【0007】
本発明の目的は、G. anatis次亜種ヘモリチカの真核細胞との相互作用を調査すること、及び溶血性表現型の原因となる遺伝子及びタンパク質を特定及び特徴付けすることであった。
【課題を解決するための手段】
【0008】
本発明者は、G. anatisが、病変形成において重要な役割を果たす特質として、トリマクロファージに対する細胞毒性が高いことを見いだした。更に、本発明者は、G. anatis次亜種ヘモリチカにおける白血球毒性及び溶血活性の原因となる新規の種類のRTX毒素の特定及び特徴付けを行った。
【0009】
本発明は、ガリバクテリウム属、最も好ましくは、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される細菌由来のGtxAポリペプチド及びポリヌクレオチドに関する。
【0010】
第1の態様において、本発明は、単離ポリペプチドであって、以下からなる群から選択されるアミノ酸配列を含む、ポリペプチドに関する:
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【0011】
SEQ ID No. 1に記載のアミノ酸配列を有するポリペプチドは、G. anatis由来のRTX毒素である。GtxA(ガリバクテリウム毒素)と名付けたタンパク質は、2026個のアミノ酸(aa)からなる。これは、古典的な膜孔形成RTX毒素のサイズの2倍となる。GtxAのC末端の1000アミノ酸は、パスツレラ科の他の構成員におけるRTX毒素と相同であり、例えば、A. pleuropneumoniae ApxIAとの38%の配列類似性を有する。一方、N末端の略950アミノ酸は、GenBankデータベースにおける有意な一致がなかったが、機能が未知である57アミノ酸の反復を11個含む。
【0012】
GtxA毒素は、トキソイドワクチンとしての用途と、鳥類一般、特に家禽において、G. anatisに対する免疫反応の存在を明らかにする診断用途とを含め、幾つかの有用性を有する。
【0013】
他の態様において、本発明は、単離ポリヌクレオチドであって、以下からなる群から選択される核酸配列を含む、ポリヌクレオチドに関する:
a) SEQ ID No. 4、5、又は6と、
b) SEQ ID No. 4、5、及び6からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも60%である配列変異体と、
c) a)の何れかの少なくとも450個の隣接ヌクレオチドからなる断片であり、選択される配列において指定された任意の核酸が異なる核酸へ変化するのは、このように変化する配列内の核酸が90個以内の場合である断片と、
d) 高ストリンジェンシー下で、SEQ ID No. 4、5、又は6に相補的なポリヌクレオチドとハイブリダイズすることが可能なポリヌクレオチドと、
e) SEQ ID No. 1、2、又は3のポリペプチドをコードするポリヌクレオチドと、
f) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体をコードするポリヌクレオチドと、
g) SEQ ID No. 1、2、又は3の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片をコードするポリヌクレオチド。
【0014】
更に、本発明は、本発明のポリヌクレオチドを含むベクターに関する。
【0015】
他の態様において、本発明は、本発明のポリペプチド、本発明のポリヌクレオチド、及び本発明のベクターの医学的使用に関する。
【0016】
好ましくは、医学的使用は、細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療に関するものである。
【0017】
一態様において、本発明は、細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療用の薬剤を調製するための、本発明のポリペプチド及び/又はポリヌクレオチドの使用に関する。
【0018】
他の態様において、本発明は、本発明のベクターにより形質転換又は形質導入した単離宿主細胞と、本発明の感染性ビリオンを生産可能なパッケージング細胞株とに関する。
【0019】
更に、本発明は、抗体であって、以下からなる群から選択されるアミノ酸配列を有する単離ポリペプチドと特異的に結合可能な抗体に関する。
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【0020】
こうしたGtxAに対する抗体は、診断及び治療の態様において使用することができる。
【0021】
他の態様において、本発明は、本発明の単離ポリペプチドの不活性化の方法に関する。
【0022】
更に、本発明は、本発明の、裸DNA又はベクターとして調製された、単離ポリペプチド又は単離ポリヌクレオチドを、任意に1つ以上の適切なアジュバント、賦形剤、乳化剤、又は担体と共に含むワクチン組成物に関する。
【0023】
別の態様において、本発明は、本明細書に記載したような鳥類種に、本発明のワクチンを投与する方法に関し、前記ワクチンは、筋肉内又は皮下注射、食物又は水などを介した経口投与、エアロゾル、足又は翼の膜部などにおける乱切(らんせつ)、点眼剤、又は卵内投与により投与される。
【0024】
他の態様において、本発明は、診断マーカーとして用いる本発明のポリペプチド又はポリヌクレオチドに関する。
【0025】
本発明は、更に、鳥類種における病原性細菌ガリバクテリウムの感染を診断する方法を提供し、前記方法は、本発明のポリペプチドの検出、前記ポリペプチドに対する抗体の検出、又は本発明のポリヌクレオチドの検出を含む。
【0026】
好ましくは、病原性細菌は、ガリバクテリウム属に由来し、更に好ましくは、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される。
【0027】
更に、本発明のポリペプチドの存在を検出するキットが提供され、前記キットは、前記ポリペプチドと結合可能な少なくとも1つの結合タンパク質を備え、前記結合タンパク質は、固体支持部に結合される。好ましくは、前記結合タンパク質は、抗体である。
【0028】
一態様において、本発明は、本発明のポリペプチドに対する抗体を検出するキットに関し、前記キットは、固体表面に固定された前記ポリペプチドを備える。
【図面の簡単な説明】
【0029】
【図1】G. anatis培養上清の溶血活性及びGtxAの発現。A. G. anatis 12656-12の増殖及び無細胞培養上清の溶血活性。一晩培養した培養物を1:100で希釈し、増殖(600nmでの吸光度により測定した細胞密度)及び無細胞培養上清における溶血活性を記録した。BHIにおいて100倍に希釈した上清の溶血活性を示す。ウェスタンブロッティング(図5B)用の細胞外タンパク質を並行して採取した。実験を3回繰り返し、溶血活性のレベルは変化したが、相対的なパターンは一貫していた。B. ApxI抗血清でのウェスタンブロッティングにより判定したGtxAのレベル。表示した時点で上清を採取し、材料及び方法において説明したように100倍に濃縮し、3〜8%ゲルにおいてSDS-PAGEにより分離した後、ブロッティングを行った。ΔgtxAからの細胞外タンパク質をOD600 = 2で採取した(ΔgtxAと示した列)。WC = 野生型由来の全細胞溶解物: 細胞は、接種5時間後に採取した。サイズマーカーを左側に示す。実験を繰り返し、同じ結果が得られた。
【図2】G. anatis 12656-12野生型(wt)及び同質遺伝子的gtxA突然変異体(ΔgtxA)の溶血活性及び細胞毒性。A. β溶血。5%ウシ血液を含むBHI寒天プレート上に細菌をストリークし、37℃で18時間インキュベートした。B. 生理食塩水(mock)、wt、又はΔgtxAと共に1時間インキュベートした後のHD11細胞の光学顕微鏡検査(倍率100×)。細菌を後期対数期(OD600 = 1)において採取し、感染効率(MOI)を10として添加した。C. LDH活性により定量化した細胞毒性。HD11細胞を、1Bにおいて説明したように細菌と共にインキュベートした。3つの複製ウェルの平均を示しており、棒は、標準誤差(SE)を示す。
【図3】A. G. anatis 12656-12におけるgtxA、gtxC、及びその隣接遺伝子の遺伝子構成。矢印は、オープンリーディングフレームを示す。予測される転写ターミネーターを、gtxCの下流に示す。B. GtxAの構成。Kは、保存されたリジン残基(Lys1484及びLys1607)を示す。グリシン-アスパラギン酸リッチ領域(Gly-Asp-rich region, 位置1640〜1830)に印を付けている。C. GtxAのN末端ドメインにおける15の反復のアライメントであり、アライメントはRaderにより作成した[Heger A., Holm L., Rapid automatic detection and alignment of repeats in protein sequences, Proteins (2000) 41:224-237]。数字は、GtxAにおけるアミノ酸の位置を示す。アミノ酸が反復の50%以上で同一(黒)又は類似(灰色)する位置に印を付けている。
【図4】GtxAを発現する大腸菌の細胞毒性活性。A. 5%ウシ血液及び0.1 mM IPTGを含むLB寒天上で増殖させ、30℃でインキュベートした大腸菌ER2566のβ溶血活性。T1SS: + 又は - は、大腸菌T1SS成分HlyB及びHlyDを発現するプラスミドpLG575の有無を示す。RTX = GtxAのアミノ酸931〜2026であり、N-term = GtxAのアミノ酸1〜949である。B. 様々なバージョンのGtxAを発現する大腸菌ER2566/pLG575による液体溶血アッセイ及びLDH細胞毒性アッセイ。
【図5】大腸菌におけるGtxAの発現。IPTGによる誘導後の大腸菌ER2566に由来する全細胞溶解物(WC)及び細胞外タンパク質(EC)に関するウェスタンブロット。タンパク質は、4〜12%ゲルにおいてSDS-PAGEにより分離し、PVDF膜上にブロットした。ブロットは、ApxI抗血清によりプローブした。B = ブランクであり、サイズマーカーを右側に示している(PageRuler Prestained Protein Ladder Plus(Fermentas))。各列の上方のバンドは、完全長のGtxA(215 kDa)又はRTXドメイン(117 kDa)の予想サイズを有している。サイズの小さなバンドは、分解産物と思われる。
【図6】様々なガリバクテリウム菌株に由来するアミノ酸配列のアライメント。GtxAの位置1133〜1515におけるアミノ酸(SEQ ID No1)を、他のガリバクテリウム菌株に由来する他の毒素のアミノ酸配列に対して整列させている。点は、同一残基を示し、非保存位置を強調して示している。
【図7】様々なG. anatis菌株とG.genomospecies 1 及びG.genomospecies 2 の基準株(それぞれ、CCM5974及びCCM5976)とに由来する培養上清を、OD600 = 0.6において採取し、濾過滅菌した。細胞外タンパク質を方法において説明したように沈殿させ、3〜8% Tris酢酸塩SDSゲル上で分離し、PVDF膜上にブロットし、ApxIA抗血清によりプローブした。GtxA(215kDa)を矢印で示している。大きなバンドは、GtxAに関係しない未確認のタンパク質(図3参照)又はGtxAの修飾バージョンである。M = 分子サイズマーカー(Spectra Multicolor High Range Protein Ladder(Fermentas))であり、サイズを右側に示す(kDa)。
【図8】T1SS突然変異体におけるGtxAのβ溶血活性及び細胞内局在性。A. G. anatis 12656-12及び同質遺伝子的突然変異体のβ溶血。37℃で24時間インキュベート後の12656-12(wt)、gtxA、及びgtxBD突然変異体のストリークを有する血液寒天プレートの写真。ストリークの左部分では細菌を除去し、コロニーの下の溶血を明らかにした。B. GtxAの細胞内局在性。細胞及び上清は、静止期への遷移中に採取した(OD600 = 1.7)。細胞外タンパク質を方法において説明したように沈殿させた。全細胞溶解物(ペレット)及び細胞外タンパク質(上清)を3〜8% Tris酢酸塩SDSゲル上で分離し、PVDF膜上にブロットし、ApxIA抗血清によりプローブした。GtxAを矢印で示している。同一性は、質量分光測定法により検証した。抗血清により、野生型及び突然変異株の両方に存在する略270 kDaの未確認タンパク質が認識された。サイズマーカーを左側に示す(Spectra Multicolor High Range Protein Ladder (Fermentas)。
【図9】G. anatis 12656-12における遺伝子座gtxAC及びgtxEBDの構成。矢印は、オープンリーディングフレーム(ORF)を示す。gtxA及びgtxEBDの検出に使用されるプライマー対の位置は、各ORFの上に小さな矢印により示す。典型的なRTX毒素遺伝子座(Frey & Kuhnert 2002)の構成を比較のために含めている。
【0030】
定義
【0031】
本明細書で使用される「アジュバント」という用語は、投与される免疫原性決定因子/抗原/核酸構築物と混合することにより、前記決定因子に対する免疫反応を高めるか或いは他の形で修飾する物質を示す。
【0032】
本明細書で使用される「対立遺伝子変異体」という用語は、SEQ ID No. 1をコードする遺伝子の代替形態を示す。対立遺伝子は、核酸配列における少なくとも1つの突然変異により生じる場合があり、構造又は機能が変更された或いは変更されない改変mRNA又はポリペプチドを発生させる場合がある。対立遺伝子を発生させる通常の突然変異変化は、一般に、ヌクレオチドの自然欠失、追加、又は置換に起因する。こうした変化のタイプは、それぞれ単独で、或いは他のものと組み合わせて、一定の配列中に1回以上発生する場合がある。
【0033】
本明細書で使用される「抗体」という用語は、免疫グロブリン分子及び免疫グロブリン分子の活性部分を示す。抗体は、例えば、無傷の免疫グロブリン分子又は免疫学的活性を保持する免疫グロブリン分子の断片である。
【0034】
本明細書で使用される「抗原」という用語は、クローン的に分布した免疫受容体(T細胞又はB細胞受容体)に結合可能な物質を示し、通常は、ペプチド、ポリペプチド、又は多量体ポリペプチドを示す。抗原は、好ましくは、免疫反応を引き起こすことが可能である。
【0035】
本明細書で使用される「結合アッセイ」という用語は、任意の2つ以上の分子が互いに共有的又は非共有的に結合して、一方の分子の濃度の測定を可能にする、任意の生物学的又は化学的アッセイを示す。
【0036】
本明細書で使用される「生体試料」という用語は、血清、血漿、全血、唾液、尿、リンパ液、生検、精液、排泄物、涙、汗、乳、脳脊髄液、腹水、滑液から選択される任意の試料を示す。
【0037】
本明細書で使用される「担体」という用語は、抗原が結合して免疫反応の誘導を支援する存在又は化合物を示す。
【0038】
本明細書で定義される「保存的アミノ酸置換」という用語は、1個のアミノ酸が、1つ以上の共通の化学的及び/又は物理的特性を有する別のアミノ酸の代わりとなる置換を示す。アミノ酸は、共有する特性により分類し得る。保存的アミノ酸置換は、アミノ酸の所定の群内にある1個のアミノ酸による、同一群内の別のアミノ酸に対する置換であり、所定の郡内のアミノ酸は、類似又は実質的に類似する特性を示す。
【0039】
本明細書で使用される「検出部分」という用語は、別の分子と結合して検出することが可能な、好ましくはタンパク質であるが限定はされない分子の特定の部分を示す。
【0040】
本明細書で使用される「診断マーカー」という用語は、個体が有する障害を決定するために使用可能なタンパク質等の化合物の特性を示す。
【0041】
本明細書で使用される「障害」という用語は、疾患又は医学的問題を示し、特定の症状及び兆候に関連した、身体機能を害する生物の異常な状態である。侵入微生物等の外的要因により生じる場合があり、或いは内的な機能不全により生じる場合がある。
【0042】
本明細書で使用される「断片」という用語は、核酸又はポリペプチドの非完全長の部分を示す。したがって、断片自体も、それぞれ核酸又はポリペプチドとなる。
【0043】
本明細書で使用される「免疫原性」という用語は、細胞性又は液性となり得る免疫反応を誘発する、抗原又はエピトープ等の特定の物質の能力を示す。
【0044】
本明細書で使用される「薬剤(medicament)」という用語は、予防的、治療的、改善的、又は対症的な使用を対象とした、好ましくはワクチンである任意の化学物質として大まかに定義することが可能な、薬(medicine)又は薬物(medication)とも呼ばれる医薬品を示す。上述したように、本発明の用途には、必ずしも任意の疾患の100%の予防、治療、又は改善だけではなく、部分的な予防、治療、又は改善も含まれることを理解されたい。
【0045】
本明細書で使用される「病原性」という用語は、生物において感染症を発生させる、Gallibacterium anatisといった微生物等の病原体の能力を示す。
【0046】
本明細書で使用される「プラスミド」という用語は、染色体DNAから独立して複製可能な、染色体DNAから分離した一種の染色体外DNA分子を示す。
【0047】
本明細書で使用される「ポリヌクレオチド」という用語は、DNA(デオキシリボ核酸)及びRNA(リボ核酸)等、鎖状に共有結合したヌクレオチドモノマーにより構成された有機高分子を示す。
【0048】
本明細書で使用される「ポリペプチド」という用語は、少なくとも、好ましくは3個以上のアミノ酸を有するペプチドである、タンパク質としても知られる有機化合物を示す。アミノ酸という総称は、天然及び非天然アミノ酸の両方を含み、何れも「D」又は「L」異性体となり得る。
【0049】
本明細書で使用される「予防的治療」という用語は、疾患の治療又は治癒ではなく予防することを目的とした、任意の医療手順を示す。予防するという用語は、絶対的であることを意図しておらず、疾患又は疾患の1つ以上の症状の部分的予防も含む。
【0050】
本明細書で使用される「プロモーター」という用語は、1個以上の近隣の構造遺伝子によりメッセンジャーRNAの転写を開始するためにRNAポリメラーゼが結合するDNA鎖内の結合部位を示す。
【0051】
本明細書で使用される「RTX毒素」(repeats in the structural toxin)という用語は、広範な病原性グラム陰性菌により生成される膜孔形成タンパク質毒素を示す。
【0052】
本明細書で使用される「配列同一性」という用語は、2つの配列間の同一性百分率の決定を示し、数学アルゴリズムを使用して達成可能である。2配列の比較に利用される数学アルゴリズムの好適な非限定的な例は、Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87:2264-2268のアルゴリズムであり、Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877のように修正されたものである。このようなアルゴリズムは、Altschul, et al. (1990) J. Mol. Biol. 215:403-410のBLASTN及びBLASTPプログラムに組み込まれている。
【0053】
同一性を特徴付けるためには、最高次のホモロジー(一致)が得られるように対象配列を整列させる。こうした一般原則に基づき、2つの核酸配列の「同一性百分率」は、米国立バイオテクノロジー情報センター(National Center for Biotechnology Information, NCBI)ウェブサイト(http://www.ncbi.nlm.nih.gov)から入手可能なBLASTNアルゴリズム[Tatiana A. Tatusova, Thomas L. Madden: Blast 2 sequences - a new tool for comparing protein and nucleotide sequences; FEMS Microbiol. Lett. 1999 174 247-250]を使用すると共に、本明細書において提案した初期設定を使用して決定し得る(即ち、Reward for a match = 1; Penalty for a mismatch = -2; Strand option = both strands; Open gap = 5; Extension gap = 2; Penalties gap x_dropoff = 50; Expect = 10; Word size = 11; Filter on)。BLASTNアルゴリズムは、2つの整列させたヌクレオチド配列間の重複範囲における配列同一性%を決定する。Blastは、局所的アライメントであるため、長さの異なる2つの関連配列間において重複範囲内の配列同一性百分率を計算するのに最適である。
【0054】
配列の比較に利用される数学アルゴリズムの他の好適な非限定的な例は、CLUSTAL W(1.7)アライメントアルゴリズムである(Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22:4673-4680.)。CLUSTAL Wは、好ましくはBLOSUM62をスコア行列として使用して、複数の配列アライメントに用いることができる。配列同一性を計算する時、CLUSTAL Wは、参照配列の長さに、アライメントにより形成された任意のギャップを含める。配列同一性は、ギャップを有する整列配列の長さで一致の数を割ることにより計算される。
【0055】
本明細書で使用される「シグナルペプチド」という用語は、局在化ペプチドとも呼ばれる、細胞内のタンパク質の最終的な位置を決定するアミノ酸の短い配列を示す。
【0056】
本明細書で使用される「毒素」という用語は、体組織との接触又はこれによる吸収時に疾患を引き起こすことが可能な、生存細胞又は生物により生産される有毒物質を示す。
【0057】
本明細書で使用される「トキソイド」という用語は、化学処理又は熱処理により毒性は弱化又は抑制されたが、他の特性、例えば免疫原性は維持されている細菌毒素(通常は外毒素)を示す。
【0058】
本明細書で使用される「転写因子」という用語は、特定のDNA配列と結合し、これによりDNAからmRNAへの遺伝情報の転送を制御するタンパク質を示す。
【0059】
本明細書で使用される「ワクチン」という用語は、動物において免疫反応を誘発可能な物質又は組成物を示す。免疫反応は、生物の記憶を誘発し、1次反応ではなく2次反応を受けた感染性因子の宿主生物に対する影響を減じる免疫反応(液性/抗体性及び/又は細胞性)である。
【0060】
本明細書で使用される「ベクター」という用語は、外来遺伝物質を他の細胞へ転送する媒体として使用されるDNA分子を示す。
【発明を実施するための形態】
【0061】
本発明の主目的は、鳥類におけるガリバクテリウム属由来の細菌との細菌感染により生じる任意の疾患の治療及び/又は予防的治療において薬剤として用いる、Gallibacterium anatisに由来する特定のRTX毒素GtxA或いはその免疫原性変異体又は断片を含むワクチン組成物を提供することである。
【0062】
Gallibacterium anatisは、ニワトリ及び他の鳥類種の上気道及び下部生殖管の正常細菌叢の一部である。しかしながら、G. anatisは、病理学的病変からも単離されているため、潜在的病原体であると考えられる。本発明は、非定型の構造と広範な標的細胞を有する新規の病原因子、G. anatis RTX毒素を提供する。
【0063】
G. anatis培養物由来の無細胞濾過滅菌上清は、赤血球及びトリ由来のマクロファージ様細胞(HD11)の両方を溶解させ、1つ以上の外毒素の生成を示唆した。G. anatis 12656-12のゲノム配列において、我々はRTX毒素遺伝子を特定した。コード化タンパク質は、GtxA(Gallibacterium toxin、ガリバクテリウム毒素)と名付けられ、2026個のアミノ酸(aa)により構成されていた。これは、古典的な膜孔形成RTX毒素のサイズの2倍となる。GtxAのC末端の1000アミノ酸は、パスツレラ科の他の構成員におけるRTX毒素と相同であり、例えば、A. pleuropneumoniae ApxIAとの38%の配列類似性を有した。一方、N末端の略950アミノ酸は、GenBankデータベースにおける有意な一致を有していないが、機能が未知である57アミノ酸の反復を11個含む。gtxA及びそのアセチルトランスフェラーゼ活性化因子gtxCを発現する大腸菌は、溶血性及び白血球毒性となった。GtxAの様々な切断バージョンの機能を調査した。C末端RTXドメインは、無傷の毒素よりも低い溶血活性を示し、N末端が、不可欠ではないが、最大の溶血活性に必要であることを示唆した。HD11細胞に対する細胞毒性は、C末端単独では検出されず、新規のN末端反復ドメインが白血球に対する細胞毒性作用にとって不可欠となることを示唆した。
【0064】
G. anatisのgtxAの発現は、ウェスタン及びノーザンブロッティングにより調査した。GtxAは、増殖期に依存する形で細胞外タンパク質画分において検出されたが、細胞随伴タンパク質画分においては検出されず、予測される毒素の分泌と一致した。
【0065】
11の多様な遺伝子型及び表現型のガリバクテリウム菌株を、gtxAの存在及び発現、GtxA分泌レベル、及び培養上清の溶解活性について調査した。gtxAは、広く分布しており、Gallibacterium genomospecies 1及びGallibacterium genomospecies 2(図6、アライメント)を含む、全ての試験株に見られた。発現には菌株間で実質的に幅があり、非病原性非溶血型菌株F149Tは、少量を発現した。上清中のGtxAレベルは、HD11細胞に対する溶血活性及び細胞毒性活性のレベルとある程度の相関を有した。
【0066】
我々は、GtxAがG. anatisの病原性に大きく寄与すると予測する。
【0067】
GtxA
【0068】
これまで説明されていないが、GtxAは、2026個のアミノ酸の大きなポリペプチドであり、215 kDaの分子量を備え、SEQ ID No. 1を有する。GtxAは、C末端及びN末端断片に分割し得る。C末端断片の例は、1,077個のアミノ酸(SEQ ID No. 2)から成り、6つの縦列反復するノナペプチドを特徴とする古典的なRTX毒素に類似する。N末端の例は、949個のアミノ酸(SEQ ID No. 3)から成り、相対的に疎水性であり、他のRTX毒素又はGenBankの他のタンパク質との配列類似性を殆ど共有しない。毒素活性は、GtxAの脂肪酸アシル化を促進する活性化因子GtxCに依存しており、即ち、毒性は、転写後のポリペプチドのアシル化に依存している。非アシル化タンパク質は、毒性を有しない。
【0069】
GtxAは、主に溶血性及び白血球毒性である細胞溶解性の表現型を示す。C末端RTXドメインは、無傷の毒素よりも低い溶血活性を示し、N末端が、不可欠ではないが、最大の溶血活性に必要であることを示す。トリ由来マクロファージ様HD11細胞に対する細胞毒性は、C末端単独では検出されず、新規のN末端反復ドメインが白血球に対する細胞毒性作用にとって不可欠であることを示唆した。
【0070】
異なる地理的領域(デンマーク、チェコ共和国、及びメキシコ)からのガリバクテリウム菌株を、gtxA遺伝子の存在について選別したところ、全ての試験菌株に見られたが、個別の菌株間には発現に実質的な変動が存在した。この変動は、地理的起源には無関係であると思われる。
【0071】
G. anatisは、ニワトリ、産卵雌鶏、及び他の鳥類種の上気道及び下部生殖管の正常細菌叢の一部である。しかしながら、G. anatisは、病理学的病変からも単離されており、G. anatisは、家禽の病変形成において重要な役割を果たすと考えられる。
【0072】
したがって、本発明の目的は、疾患の治療及び/又は予防用の薬剤として用いる、GtxAタンパク質又はその免疫活性ポリペプチド変異体に由来するトキソイドワクチンを提供することである。前記疾患は、温血動物における細菌感染により生じる場合があり、本発明の他の目的は、前記疾患を予防又は治療することである。本発明の他の態様は、疾患の治療及び/又は予防用のGtxAタンパク質をコードするポリヌクレオチド又は免疫活性ポリペプチド変異体をコードするポリヌクレオチドに関する。
【0073】
GtxAポリペプチドは、図6の部分アミノ酸配列のアライメントにより示したように、様々なGallibacterium anatis分離株全体での保存性が高い。そのため、GtxAトキソイドを含むワクチン組成物は、多数の異なるG. anatis分離株に対して、更には、ガリバクテリウム属の関連種に対しても有効となることが予想される。
【0074】
細菌種
【0075】
本発明は、ポリペプチド、ポリヌクレオチド、及びポリヌクレオチド保持発現ベクターに関する。一実施形態において、ポリヌクレオチド及びポリペプチドは、Gallibacterium anatisに由来する。加えて、本発明は、パスツレラ科の細菌、更に好ましくは、ガリバクテリウム属からの細菌、最も好ましくは、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される細菌に由来するGtxAポリペプチド及びポリヌクレオチドを対象とする。
【0076】
GtxAは、他のRTX毒素とは大きく異なることが分かった。本明細書に記載した分子ツールにより、本発明者は、ガリバクテリウム属由来の他のGtxA様毒素のクローニング及びプロービングを可能にした。
【0077】
こうした関連種由来のGtxA様毒素は、SEQ ID NO 1、2、又は3との一定の度合いの配列相同性を示すことが予想され、コード配列は、本発明のポリヌクレオチドに基づくプローブとハイブリダイズすることが予想される。
【0078】
遺伝子組み換え株
【0079】
実施例2において説明されるように、本発明者は、G. anatis gtxA突然変異株を作成した。菌株は、ΔgtxAと命名した。実施例7は、こうしたG. anatis ΔgtxA突然変異体による感染試験を説明しており、野生型微生物に感染させたトリは、通常、野外でのG. anatisへの自然感染により観察される病変に対応する、生殖器系及び腹膜に関与する播種性及び化膿性の炎症を生じた。一方、ΔgtxA突然変異体に感染させたトリは、卵巣に局在する、より軽度の炎症を生じた。したがって、gtxAが、ニワトリにおけるG. anatisの病変形成に実質的に寄与することが実証された。故に、本発明者は、本明細書において上で定義したΔgtxA株を使用して生物への免疫付与が可能であることを実証したと結論付けることができる。このように免疫された鳥類種などの生物は、gtxAを発現する野生型微生物に対する免疫を発生させることができるが、それは、特定の病原性微生物の表面等に存在する非gtxA抗原に対して生成された抗体を介してであり、当該微生物はgtxAの発現及び/又は分泌を無効化した微生物と同じ種である。
【0080】
したがって、一態様において、本発明は、内因性gtxA遺伝子を破壊して、機能性gtxAポリペプチドの発現を無効化したトランスジェニックノックアウト微生物に関し、前記微生物は、非トランスジェニック対照微生物と比較して病原性の減少を示す。
【0081】
一実施形態において、微生物は、Gallibacterium anatisである。
【0082】
他の実施形態において、トランスジェニック微生物は、抗生物質耐性を有していない。
【0083】
GtxAポリペプチド
【0084】
野生型GtxAの1つ、即ち、天然に存在する突然変異のないバージョンのタンパク質は、SEQ ID No. 1において特定される。
【0085】
一実施形態において、本発明は、単離ポリペプチドであって、以下からなる群から選択されるアミノ酸配列を含む、ポリペプチドに関する:
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【0086】
他の野生型GtxAポリペプチドを、他のガリバクテリウム種及びGallibacterium anatisの他の分離株から単離することが可能である。こうしたGtxAは、図6の断片のアライメントに示すように、SEQ ID No. 1、2、及び/又は3との高い度合いの配列同一性を共有することが予想される。
【0087】
好適な実施形態において、本発明は、SEQ ID No. 1と、SEQ ID No. 1に対して少なくとも70%の配列同一性、より好ましくは、GtxA配列との75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性、例えば少なくとも99.5%の配列同一性、例として少なくとも99.9%の配列同一性を備えたGtxAの配列変異体とに関する。
【0088】
他の好適な実施形態において、本発明は、SEQ ID No. 2において定義される、GtxAポリペプチドのC末端ドメインと、SEQ ID No. 2に対して少なくとも70%の配列同一性、より好ましくは、GtxA配列のC末端ドメインとの75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を備えた配列変異体とに関する。
【0089】
他の好適な実施形態において、本発明は、SEQ ID No. 3において定義される、GtxAポリペプチドのN末端ドメインと、SEQ ID No. 3に対して少なくとも70%の配列同一性、より好ましくは、GtxA配列のN末端ドメインとの75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を備えた配列変異体とに関する。
【0090】
完全長のGtxAに加えて、本発明は、GtxAの断片に関する。例えば、C末端GtxAドメイン及びN末端GtxAドメインに関する。加えて、本発明は、こうしたポリペプチドの断片に関する。好適な実施形態において、前記断片は、少なくとも150個の隣接アミノ酸、好ましくは少なくとも200個のアミノ酸、より好ましくは少なくとも300個のアミノ酸、より好ましくは少なくとも500個のアミノ酸、より好ましくは少なくとも750個のアミノ酸、より好ましくは少なくとも1000個のアミノ酸、より好ましくは少なくとも1250個のアミノ酸、より好ましくは少なくとも1500個のアミノ酸、より好ましくは少なくとも1750個のアミノ酸、より好ましくは少なくとも2000個のアミノ酸からなる。
【0091】
GtxA断片は、1つ以上の位置において、野生型GtxA配列と異なっていてよい。好適な実施形態において、前記断片は、30個までのアミノ酸置換、より好ましくは25個までの置換、より好ましくは20個までの置換、より好ましくは15個までの置換、より好ましくは10個までの置換、より好ましくは5個までの置換、例として4個、3個、2個、又は1個の置換を含み得る。
【0092】
本発明が対象とする他の変異体は、保存的アミノ酸置換が生じた、SEQ ID No. 1、2、及び3のポリペプチドの変異体に関する。好適な実施形態において、本発明は、ポリペプチド配列内の任意のアミノ酸が他のアミノ酸に保存的に置換された、SEQ ID No. 1、2、又は3を含む任意のポリペプチドに関する。
【0093】
他の好適な実施形態において、前記配列変異体及び断片は、免疫原性を有する。更に他の好適な実施形態において、前記配列変異体及び断片は、毒性等の生物活性を保持し、前記毒性は、受容者の細胞膜での孔の形成を含み、例えば細胞毒性、例として細胞溶解性細胞毒性、例えば溶血性細胞毒性を含む。
【0094】
本発明は、更に、SEQ ID No. 1、2、及び3のポリペプチドに関し、前記ポリペプチドは、例えば毒性等の生物活性は除去するが、例えば免疫原性等の活性は維持するように特異的に修飾されている。そのため、好適な実施形態において、SEQ ID No. 1、2、及び3のポリペプチドは、好ましくは、熱又は放射線により、更に好ましくは、非アシル化形態で発現させることにより、更に好ましくは、例えばホルムアルデヒド等の化学物質に曝露することにより不活性化される。
【0095】
他の好適な実施形態において、本発明は、シグナルペプチドが異種シグナルペプチドに置き換えられた、SEQ ID No. 1、2、又は3を含む任意のポリペプチドに関する。
【0096】
精製の目的で、本発明には、タグを付けてもよい。好適な実施形態において、SEQ ID No. 1、2、及び3は、親和性タグ、好ましくは、切断可能タグ、例としてポリHisタグ、例えばHAタグ、例としてFLAGタグ、例えばC-mycタグ、例としてHSVタグ、例えばV5タグ、例としてマルトース結合タンパク質タグ、例えばセルロース結合ドメインタグ、例としてBCCPタグ、例えばカルモジュリンタグ、例としてNusタグ、例えばグルタチオン-S-トランスフェラーゼタグ、例として緑色蛍光タンパク質タグ、例えばチオレドキシンタグ、例としてSタグ、例えばStrepタグによりタグ付けし得る。
【0097】
好ましくは、タグは、C末端の一番端など、タンパク質のC末端部分に存在する。より好ましくは、タグは、タグとRTXポリペプチドとの間に挿入されたプロテアーゼ切断部位を有することにより、GtxAポリペプチドから切断可能となる。
【0098】
GtxAポリヌクレオチド
【0099】
本発明により、SEQ ID No. 4、5、及び6において特定のポリヌクレオチド配列が定められる。
【0100】
本発明は、単離ポリヌクレオチドであって、以下からなる群から選択される核酸配列を含む、ポリヌクレオチドに関する:
a) SEQ ID No. 4、5、又は6と、
b) SEQ ID No. 4、5、及び6からなる群から選択されるポリヌクレオチドの配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも60%である配列変異体と、
c) a)の何れかの少なくとも450個の隣接ヌクレオチドからなる断片であり、選択される配列において指定された任意の核酸が異なる核酸へ変化するのは、このように変化する配列内の核酸が90個以内の場合である断片と、
d) 高ストリンジェンシー下で、SEQ ID No. 4、5、又は6に相補的なポリヌクレオチドとハイブリダイズすることが可能なポリヌクレオチドと、
e) SEQ ID No. 1、2、又は3のポリペプチドをコードするポリヌクレオチドと、
f) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体をコードするポリヌクレオチドと、
g) SEQ ID No. 1、2、又は3の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片をコードするポリヌクレオチド。
【0101】
ヌクレオチドプローブと相同DNA又はRNA配列との間のハイブリダイゼーションを決定するための適切な実験条件は、ハイブリダイズするDNA断片又はRNAを含有するフィルタを5×SSC[塩化ナトリウム/クエン酸ナトリウム、Sambrook et al.; Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Lab., Cold Spring Harbor, NY 1989参照]に10分間予め浸漬すること、及び5×SSC、5×デンハルト溶液[Sambrook et al.; 同上参照]、0.5%SDS、及び変性超音波処理サケ精子100 μg/mL [Sambrook et al.; 同上参照]の溶液におけるフィルタのプレハイブリダイゼーションを含み、その後、10 ng/mLの濃度のランダムプライム[Feinberg A P & Vogelstein B; Anal. Biochem. 1983 132 6-13]32P-dCTP標識(比活性度>1×109 cpm/μg)プローブを含有する同じ溶液中で、12時間略45℃でハイブリダイゼーションを行う。その後、フィルタを0.1×SSC、0.5%SDSにおいて、少なくとも60℃(中ストリンジェンシー条件)、好ましくは少なくとも65℃(中/高ストリンジェンシー条件)、より好ましくは少なくとも70℃(高ストリンジェンシー条件)、更により好ましくは少なくとも75℃(超高ストリンジェンシー条件)の温度で、30分間2回洗浄する。こうした条件下でオリゴヌクレオチドプローブがハイブリダイズした分子は、X線フィルムを用いて検出し得る。
【0102】
一実施形態において、核酸分子は、SEQ ID No. 4、5、及び6からなる群から選択される核酸配列と単一のヌクレオチドにおいて異なっている。単一の置換は、サイレント突然変異であってよく、或いは、保存的アミノ酸置換を発生させてもよい。単一の置換又は欠失は、フレームシフト突然変異を発生させてもよい。より好適な実施形態において、本発明は、SEQ ID No. 4のポリヌクレオチドとの配列同一性が少なくとも60%、より好ましくはSEQ ID No. 4との配列同一性が少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%であるポリヌクレオチド配列に関する。
【0103】
他の好適な実施形態において、本発明は、SEQ ID No. 5のポリヌクレオチドとの配列同一性が少なくとも60%、より好ましくはSEQ ID No. 5との配列同一性が少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%であるポリヌクレオチド配列に関する。
【0104】
他の好適な実施形態において、本発明は、SEQ ID No. 6のポリヌクレオチドとの配列同一性が少なくとも60%、より好ましくはSEQ ID No. 6との配列同一性が少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%であるポリヌクレオチド配列に関する。
【0105】
本発明は、更に、SEQ ID No. 1、2、及び3のポリヌクレオチドの断片に関する。好適な実施形態において、前記断片は、少なくとも450個の隣接ヌクレオチド、より好ましくは少なくとも500個の隣接ヌクレオチド、より好ましくは少なくとも600個の隣接ヌクレオチド、より好ましくは少なくとも750個の隣接ヌクレオチド、より好ましくは少なくとも1000個の隣接ヌクレオチド、より好ましくは少なくとも1500個の隣接ヌクレオチド、より好ましくは少なくとも2000個の隣接ヌクレオチド、より好ましくは少なくとも2500個の隣接ヌクレオチド、より好ましくは少なくとも3000個の隣接ヌクレオチド、より好ましくは少なくとも3500個の隣接ヌクレオチド、より好ましくは少なくとも4000個の隣接ヌクレオチド、より好ましくは少なくとも4500個の隣接ヌクレオチド、より好ましくは少なくとも5000個の隣接ヌクレオチド、より好ましくは少なくとも5500個の隣接ヌクレオチド、より好ましくは少なくとも6000個の隣接ヌクレオチドからなる。
【0106】
ポリヌクレオチド断片は、1つ以上の位置において、前記断片が由来する野生型GtxAポリヌクレオチド配列と異なっていてよい。好適な実施形態において、前記断片は、90個までのヌクレオチド置換、より好ましくは80個までの置換、より好ましくは70個までの置換、より好ましくは60個までの置換、より好ましくは50個までの置換、より好ましくは40個までの置換、より好ましくは30個までの置換、より好ましくは20個までの置換、より好ましくは10個までの置換、より好ましくは5個までの置換、例として4個、3個、2個、又は1個の置換を含み得る。
【0107】
本発明は、好ましくは高ストリンジェンシーのハイブリダイズ条件下でSEQ ID No. 4、5、及び6の配列を有するポリヌクレオチドとハイブリダイズすることが可能なポリヌクレオチドに関する。
【0108】
本発明のポリヌクレオチドは、天然に存在する対立遺伝子核酸変異体のヌクレオチド配列を含み得る。
【0109】
本発明のポリヌクレオチドは、更に、SEQ ID No. 4、5、及び6の変異体を含んでよく、前記ポリヌクレオチドは、大腸菌での発現に最適化されている。
【0110】
発現ベクター
【0111】
本発明のポリヌクレオチドは、発現ベクター又はクローニングベクター等、任意の適切なベクター内に含まれてもよい。多数のベクターが利用可能であり、特定の目的に適した任意のベクターを選択し得る。適切な核酸配列を、様々な手順によりベクターに挿入してよく、例えば、DNAは、当技術分野において周知の手法を使用して、適切な制限エンドヌクレアーゼ部位(群)に挿入し得る。本発明に関する核酸配列とは別に、ベクターは、更に、シグナル配列、複製開始点、1つ以上のマーカー遺伝子、エンハンサーエレメント、プロモーター、及び転写終結配列を1つ以上含み得る。ベクターは、更に、エンハンサー、ポリA尾部、リンカー、ポリリンカー、操作リンカー(operative linker)、多重クローニング部位(multiple cloning sites, MCS)、終止コドン、内部リボソーム侵入部位(internal ribosomal entry sites, IRES)、及び組込み用の宿主相同配列といった付加的な配列又は他の定義済みのエレメントを含み得る。ベクターは、好ましくは、発現ベクターであり、適切な細胞においてその発現を指揮する調節核酸配列と適切に操作可能に結合した核酸を含む。
【0112】
好適な実施形態において、本発明のベクターは、真核生物プラスミドベクター等のプラスミドベクターであり、より好ましくは原核生物プラスミドベクターである。
【0113】
別の好適な実施形態において、ベクターは、好ましくはレトロウイルス科に由来するウイルスベクターであり、例としてレンチウイルス、例えばHIV、例としてSIV、例えばEAIV、例としてCIVである。
【0114】
他の好適な実施形態において、ベクターは、限定はされないが、アルファウイルス、アデノウイルス、アデノ随伴ウイルス、バキュロウイルス、HSV、コロナウイルス、ウシパピローマウイルス、Mo-MLVからなる群から選択してよく、好ましくはアデノ随伴ウイルスにしてよい。
【0115】
他の好適な実施形態において、本発明のベクターは、プロモーターを含み、より好ましくは、前記プロモーターは、本発明のポリヌクレオチドと適切に操作可能に結合される。
【0116】
好適な実施形態において、前記プロモーターは、限定はされないが、原核生物プロモーターから選択され、好ましくは、原核生物プロモーターは、他のエレメントを含み、例としてRNAポリメラーゼ結合部位、例えばプリブノーボックス又はその一部、例として-35エレメント又はその一部を含む。本発明の原核生物ベクターは、例えば、大腸菌における、不活性化GtxAの組み換え発現に使用することができる。大腸菌は、GtxC遺伝子を含まないため、大腸菌において発現させたGtxAは、適切にアシル化されず、結果的に無毒性となる。
【0117】
別の好適な実施形態において、前記プロモーターは、限定はされないが、真核生物プロモーターから選択され、好ましくは、真核生物プロモーターは、他のエレメントを含み、例としてRNAポリメラーゼ結合部位、例えばTATAボックス又はその一部、例として任意の真核細胞転写因子のための少なくとも1つの結合部位を含む。
【0118】
本発明のベクターの好適な実施形態は、裸DNAワクチンであり、本発明のポリヌクレオチドと適切に操作可能に結合した真核生物プロモーターを含む。
【0119】
ワクチン
【0120】
一般に、ワクチンは、機能的免疫系を備えた生体検体において免疫反応を誘発可能な物質又は組成物である。組成物は、以下を1つ以上含み得る: 抗原等(例えば、タンパク質、ポリペプチド、ペプチド、核酸等)の「活性成分」、他のエレメントと共に1つ以上の抗原を含む核酸構築物、細胞(例えば、負荷抗原提示細胞(loaded APC)、養子移入用のT細胞)、複合分子(抗体、T細胞受容体(TCRs)、及びMHC複合体等)、担体、アジュバント、及び医薬担体。本発明は、Gallibacterium anatis由来の本発明の単離ポリペプチド、好ましくは前記ポリペプチドの不活性化形態、或いはその免疫原性変異体又は断片を含むワクチン組成物に関する。本明細書で使用される「ワクチン」という用語は、Gallibacterium anatisを起源とする疾患に対する特定の免疫を誘発することを目的とした家畜ワクチンを示す。
【0121】
本発明はワクチン組成物に関し、当該ワクチン組成物は、SEQ ID No. 1、2、又は3のポリペプチド、前記ポリペプチドの不活性化形態、前記ポリペプチドの機能的ホモログ、少なくとも70%の配列同一性を有するポリペプチド、又は前記ポリペプチドの免疫原性活性断片を含む。前記ワクチンは、トキソイドワクチンと呼ばれる。トキソイドワクチンは、毒性を失っているが免疫原性を保持している毒素を使用して標的生物において免疫反応を誘発するワクチンであり、前記標的生物は、同一又は類似の細菌種を起源とする前記毒素又は類似毒素による将来の感染に対して耐性となる。
【0122】
好適な実施形態において、前記ワクチンは、毒性に関して不活性化されているが、免疫原性を維持している不活性化ポリペプチドを含み、好ましくは、熱により、更に好ましくは、ホルムアルデヒド等の化学物質への曝露により、更に好ましくは、非アシル化形態でSEQ ID No. 1のポリペプチドを発現させることにより、不活性化される。
【0123】
他の好適な実施形態において、前記ワクチンは、更に、不活性化されたか又は生存する、弱毒化Gallibacterium anatisを含む。
【0124】
更に他の実施形態において、本発明は、更に、鳥類種に対する病原性を有するウイルス又は微生物由来の少なくとも1つの他の抗原に関し、前記ウイルス又は微生物は、限定はされないが、伝染性気管支炎ウイルス、ニューカッスル病ウイルス、伝染性ファブリキウス嚢病ウイルス、ニワトリ貧血因子、トリレオウイルス、トリニューモウイルス、ニワトリポックスウイルス、トリ脳脊髄炎ウイルス、マイコプラズマ・ガリセプチカム(Mycoplasma gallisepticum)、ヘモフィルス・パラガリナルム(Haemophilus paragallinarum)、パスツレラ・マルトシダ(Pasteurella multocida)、及び大腸菌(Eschericia coli)からなる群から選択される。
【0125】
毒素等の抗原が宿主生物に導入される時、通常、免疫反応を高めるために付加的な成分が使用される。こうした成分は、一般に、アジュバントと呼ばれる。本発明のワクチン組成物は、好ましくは、アジュバント及び/又は担体を含む。アジュバントは、ワクチン組成物との混合がGtxAポリペプチド又はその免疫原性断片に対する免疫反応を高めるか或いは他の形で修飾する任意の物質である。担体は、GtxAポリペプチド又はその免疫原性断片を結び付けることが可能な、抗原の提示を支援する足場構造、例えば、ポリペプチド又は多糖である。
【0126】
したがって、好適な実施形態において、本発明のワクチン組成物は、限定はされないが、フロイント完全及び不完全アジュバント、ビタミンE、非イオン性ブロック重合体、ムラミルジペプチド、Quil A、鉱油及び非鉱物油、野菜油、及びCarbopolの群から選択されるアジュバント及び/又は担体を含む。本発明のワクチンは、Span又はTween等の乳化剤を含み得る。
【0127】
本発明のワクチン組成物は、幾つかの経路、例えば筋肉内又は皮下注射により、例えば食物又は水を介した経口投与により、例としてエアロゾルにより、例えば足又は翼の膜部における乱切により、例として点眼剤により、例えば卵内投与により投与し得る。
【0128】
トリ
【0129】
パスツレラ科の殆どの構成員は、温血動物、好ましくはトリの粘膜において、片利共生生物として生存している。ガリバクテリウム属の構成員は、共生及び病原性菌株を含み、前記病原菌株は、主に、鳥類宿主の気道及び生殖器系において疾患を引き起こす。
【0130】
好適な実施形態において、本発明は、温血動物、より好ましくは、鳥類種、例えば、マガモ属(Anas)、マガン属(Anser)、ハジロ属(Aythya)、ニオイガモ属(Biziura)、コクガン属(Branta)、ハクチョウ属(Cygnus)、アカメカモメ属(Creagrus)、ハシブトアジサシ属(Gelochelidon)、カモメ属(Larus)、ゾウゲカモメ属(Pagophila)、クビワカモメ属(Xemaes)、コウノトリ科(Ciconiidae)、カワラバト属(Columba)、スズメバト属(Columbina)、ミカドバト属(Ducula)、ヒムネバト属(Gallicolumba)、チョウショウバト属(Geopelia)、アメリカウズラバト属(Geotrygon)、カンムリバト属(Goura)、ミヤマバト属(Gymnophaps)、ニュージーランドバト属(Hemiphaga)、アメリカシャコバト属(Leptotila)、ウォンガバト属(Leucosarcia)、オナガバト属(Macropygia)、ハシリバト属(Metriopelia)、レンジャクバト属(Ocyphaps)、シッポウバト属(Oena)、Patagioenas属(Patagioenas)、テリアオバト属(Phapitreron)、ヒメアオバト属(Ptilinopus)、インカバト属(Scardafella)、キジバト属(Streptopelia)、アオバト属(Treron)、アオフバト属(Turtur)、ハジロバト属(Zenaida)、エボシツカツクリ属(Aepypodius)、ヤブツカツクリ属(Alectura)、キジ科(Phasianidae)、ライチョウ亜科(Tetraoninae)、ペリカン科(Pelecanidae)、フラミンゴ科(Phoenicopteridae)、オウム科(Cacatuidae)、ヒインコ科(Loriidae)、インコ科(Psittacidae)、エミュー科(Dromaiidae)、ダーウィンレア属(Pterocnemia)、レア属(Rhea)、ダチョウ科(Struthionidae)、などにおける、細菌感染の治療用としての本発明のポリペプチド、本発明のポリヌクレオチド、及び本発明のベクターに関する。
【0131】
更に好適な実施形態において、本発明のポリペプチド、本発明のポリヌクレオチド、及び本発明のベクターは、カモ、シチメンチョウ、及びニワトリからなる群から選択される鳥類種、より好ましくは産卵雌鶏における、細菌感染の治療において使用される。
【0132】
抗体
【0133】
本発明のポリペプチド又はその免疫原性断片の存在を検出するために、前記ポリペプチド又はその免疫原性断片と特異的に結合することが可能な抗体を生成することが有用である。前記抗体は、前記ポリペプチド上の任意のエピトープと結合し得る。
【0134】
好適な実施形態において、前記抗体は、血清由来のポリクローナル抗体或いはモノクローナル又は組み換え抗体としてよく、前記抗体は、Fv、scFv、Fab、Fab’、又はF(ab)2等の抗体の抗原結合断片、二量体IgA分子又は五価IgM等の多量体形態、アフィボディ(affibodies)、又は二重特異性抗体(diabodies)を含む。
【0135】
好適な実施形態において、本発明は、IgA抗体、最も好ましくは、ニワトリIgA抗体に関する。
【0136】
したがって、好適な実施形態において、本発明は、SEQ ID No. 1、2、又は3のポリペプチドと特異的に結合することが可能な抗体に関する。
【0137】
他の実施形態において、本発明は、SEQ ID No. 1に対して少なくとも70%の配列同一性、より好ましくはSEQ ID No. 1に対して75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を有するポリペプチドと結合することが可能な抗体に関する。
【0138】
更に好適な実施形態において、本発明は、SEQ ID No. 2に対して少なくとも70%の配列同一性、より好ましくはSEQ ID No. 2に対して75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を有するポリペプチドとの結合が可能な抗体に関する。
【0139】
他の好適な実施形態において、本発明は、SEQ ID No. 3に対して少なくとも70%の配列同一性、より好ましくはSEQ ID No. 3に対して75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を有するポリペプチドと結合することが可能な抗体に関する。
【0140】
他の好適な実施形態において、本発明は、SEQ ID No. 1、2、及び3のポリペプチドの何れかの免疫原性断片を含むポリペプチドと結合することが可能な抗体に関し、前記断片は、少なくとも150個の隣接アミノ酸、好ましくは少なくとも200個のアミノ酸、より好ましくは少なくとも300個のアミノ酸、より好ましくは少なくとも500個のアミノ酸、より好ましくは少なくとも750個のアミノ酸、より好ましくは少なくとも1000個のアミノ酸、より好ましくは少なくとも1250個のアミノ酸、より好ましくは少なくとも1500個のアミノ酸、より好ましくは少なくとも1750個のアミノ酸、より好ましくは少なくとも2000個のアミノ酸からなる。
【0141】
更に好適な実施形態において、本発明は、免疫原性断片を含むポリペプチドと結合することが可能な抗体に関し、前記断片は、30個までのアミノ酸置換、より好ましくは25個までの置換、より好ましくは20個までの置換、より好ましくは15個までの置換、より好ましくは10個までの置換、より好ましくは5個までの置換、例として4個、3個、2個、又は1個の置換を含み得る。
【0142】
診断
【0143】
診断試験キットは、本発明に関連する診断の方法を実行するための全ての構成要素を集めたものである。
【0144】
好適な実施形態において、本発明は、ガリバクテリウム属由来の細菌種の存在により生じる細菌感染の兆候を生体試料において検出する試験キットに関する。前記兆候は、SEQ ID No. 1、2、又は3の任意のポリペプチド、又はその機能性変異体の存在、又は前記ポリペプチドの何れかに対する抗体の存在としてよい。
【0145】
好適な実施形態において、前記ポリペプチド又は抗体の存在は、酵素結合免疫吸着測定法(ELISA)により検出し得る。ELISAは、試料中のタンパク質又は他の任意の抗原の存在を検出するために使用される定量的手法である。ELISAでは、未知の量の抗原を表面に付着させ、その後、特異的抗体を表面上に流して、抗原と結合可能にする。この抗体は、酵素と結合しており、最終段階において、当該酵素により何らかの検出可能な信号に転換可能な物質を添加する。
【0146】
幾つかの種類のELISAが存在している:
間接ELISA法
サンドイッチELISA法
競合ELISA法
リバースELISA法
【0147】
更に、化学発光免疫測定法及び解離増強型ランタニド免疫測定法(Dissociation-Enhanced Lanthinide Immunoassays)等、他の免役に基づく測定法を使用して、試料中の前記ポリペプチド又は前記抗体を検出し得る。
【0148】
本発明は、更に、本発明の任意のポリヌクレオチド又はGallibacterium anatis GtxAに特異的な他の特異的DNA又はRNA配列の存在を検出するための診断試験キットに関する。
【0149】
したがって、好適な実施形態において、本発明は、前記ポリヌクレオチドを検出するポリメラーゼ連鎖反応(PCR)又はリアルタイム(RT)-PCR法に関する。PCRは、単一又は数個のDNA片を数桁に渡り増幅し、特定のDNA配列の数千から数百万の複製を生成し、最終的に検出する手法である。当該方法は、DNAの溶解のための反応液の加熱及び冷却並びにDNAの酵素的複製を反復するサイクルからなる温度サイクリングに依存している。標的領域に対して相補的な配列を含むプライマー(短いDNA断片)及びDNAポリメラーゼは、選択的な反復増幅を可能にする主要な成分となる。PCRが進行すると共に、生成されたDNA自体が複製のテンプレートとして使用され、DNAテンプレートを指数関数的に増幅させる連鎖反応が発生する。
【0150】
ポリペプチド、ポリヌクレオチド、及び発現ベクターの医学的使用
【0151】
トキソイドワクチンとしての不活性化毒素は、主に獣医用として、好ましくは家禽群の治療用として、細菌感染の治療及び/又は予防的治療において一般的に使用される。
【0152】
トキソイドワクチンでは、侵入細菌ではなく、侵入細菌により生産される毒素に対抗するように免疫系を条件付けることが目標となる。
【0153】
したがって、本発明のGtxAポリペプチド、GtxAポリペプチドをコードするポリヌクレオチド、又は発現ベクターは、GtxA毒素及び/又は類似する毒素を分泌する細菌により生じた発症状態(pathogenic conditions)の治療及び/又は予防的治療用のトキソイドワクチンを生成するために使用し得る。
【0154】
特定の実施形態において、トリに対して受動免疫をもたらす投与量は、ワクチン1用量当たり略0.25 mLである。他の実施形態において、投与量は、ワクチン1用量当たり約0.4 mLである。更に他の実施形態において、投与量は、ワクチン1用量当たり約0.6 mLである。一実施形態において、投与量は、ワクチン1用量当たり約0.5 mLである。一実施形態において、鳥類は、限定はされないが、カモ、シチメンチョウ、及びニワトリからなる群から選択される。他の好適な実施形態において、鳥類は、産卵雌鶏である。
【0155】
本発明は、更に、他のワクチンを組成物に含めることにより多数の疾患に対して鳥類を保護するために、本発明のワクチンを鳥類に投与する方法を提供する。
【0156】
本発明は、GtxA毒素に対する能動免疫をもたらすための、トリ、好ましくはカモ、シチメンチョウ、及びニワトリ、より好ましくは産卵雌鶏のワクチン接種を含む。ニワトリ及び/又は家禽は、約1日齢又はこれより僅かに高齢となる若年期に(1週齢以内)、1又は2用量のワクチンによりワクチン接種を行う。能動免疫を達成するために適切なワクチン投与量は、約0.05 mL〜約0.5 mLで変化する場合がある。特定の実施形態において、ワクチン投与量は、略0.05 mL〜略0.1 mLである。
【0157】
各トリは、例えば1〜3ヶ月以内に2回等、複数回ワクチン接種を行う必要があると予想される。例えば、産卵雌鶏の生涯に渡る保護には、毎年の再接種を要する場合がある。
【0158】
更に他の実施形態において、本発明の毒素又はその免疫原性断片をトリに投与することが可能である。更に、本発明のワクチンとして使用されるGtxA毒素又はその免疫原性断片の投与量の範囲は、1用量当たり約1μg/kg体重〜約1000μg/kg体重となる場合があり、一例として、1動物に対して1用量当たり約10μg/kg体重〜約100μg/kg体重の範囲となる。
【0159】
【表1】

【0160】
【表2】
【実施例】
【0161】
実施例1: Gallibacterium anatis由来のGtxA、新規のドメイン構成を有する細胞溶解性RTX毒素
【0162】
要約
【0163】
Gallibacterium anatisは、ニワトリ及び他の鳥類種において卵管炎及び腹膜炎の大きな原因となる病原体である。我々は、溶血性のG. anatis次亜種ヘモリチカからの細菌細胞及び無細胞濾過滅菌培養上清がトリ由来マクロファージ様細胞(HD11)に対して高い細胞毒性を有することを見いだした。G. anatis 12656-12のゲノム配列を入手し、合理的手法を用いて2026個のアミノ酸のRTX毒素をコードすると予測される遺伝子を特定し、GtxA(Gallibacterium toxin、ガリバクテリウム毒素)と名付けた。gtxAノックアウト突然変異体の作成により、gtxAがG. anatisの溶血及び白血球毒性活性の原因となることが示された。更に、gtxA及び隣接するアシルトランスフェラーゼgtxCを発現する大腸菌は、細胞溶解性となった。GtxAは、in vitro増殖中に発現され、増殖期に依存する形で細胞外タンパク質画分に局在した。GtxAは、通常とは異なるモジュラー構造を有し、GtxAのC末端の1000個のアミノ酸は、パスツレラ科の他の構成員における古典的な膜孔形成RTX毒素と相同であった。一方、N末端の略950個のアミノ酸は、配列データベースとの有意な一致は殆どなかった。切断GtxAタンパク質の発現により、C末端RTXドメインが完全長の毒素よりも低い溶血活性を有することが実証され、N末端ドメインは最大の溶血活性に必要であることが示唆された。HD11細胞に対する細胞毒性は、C末端単独では検出されず、N末端ドメインが白血球毒性において決定的な役割を果たすことが示唆された。
【0164】
材料及び方法
【0165】
細菌株及び増殖条件
【0166】
Gallibacterium anatis次亜種ヘモリチカ菌株12656-12 Liver (12656-12と呼ぶ)を研究に用いており、この菌株は、敗血症のニワトリの肝臓から最初に単離された[Bojesen A.M., Torpdahl M., Christensen H., Olsen J.E., Bisgaard M., Genetic diversity of Gallibacterium anatis isolates from different chicken flocks, J. Clin. Microbiol. (2003) 41:2737-2740]。G. anatis 12656-12は、閉じたプラスチック袋内で5%クエン酸ウシ血液を補ったブレインハートインフュージョン(BHI)(Oxoid)寒天培地上又はエアレーションしたBHIブロス中において37℃で増殖させた。Anaerogen(Oxoid)を用いて、培養瓶中に嫌気条件を発生させた。大腸菌は、Luria-Bertaniブロス及び寒天培地において増殖させ、培地は、50μg/mLカナマイシン及び20μg/mLクロラムフェニコールにより適宜補った。全ての化学物質は、Sigmaより購入した。
【0167】
バイオインフォマティクス分析
【0168】
G. anatis次亜種ヘモリチカ12656-12 Liverのゲノム配列の草案バージョン(115コンティグ)(A. M. Bojesen、未公開データ)を、454 Life Sciencesからパイロシーケンスに基づく方法を使用して入手した[Margulies M., Egholm M., Altman W.E., Attiya S., Bader J.S., Bemben L.A., et al., Genome sequencing in microfabricated high-density picolitre reactors, Nature (2005) 437:376-380]。遺伝子アノテーションは、オーストラリア、モナシュ大学のビクトリアバイオインフォマティクスコンソーシアムが提供する真核生物用のウェブ上のアノテーションシステム、Wasabiを用いて行った[Bulach D.M., Zuerner R.L., Wilson P., Seemann T., McGrath A., Cullen P.A., Davis J., Johnson M., Kuczek E., Alt D.P., Peterson-Burch B., Coppel R.L., Rood J.I., Davies J.K., Adler B., Genome reduction in Leptospira borgpetersenii reflects limited transmission potential, Proc. Natl. Acad. Sci. USA (2006) 103:14560-14565]。配列類似性の検索は、BLASTP[Altschul S.F., Madden T.L., Schaffer A.A., Zhang J.H., Zhang Z., Miller W., Lipman D.J., Gapped BLAST and PSI-BLAST: a new generation of protein database search programs, Nucleic Acids Res. (1997) 25:3389-3402](データベース:非冗長タンパク質配列(GenBank)及びSwissProt)、FASTA[Pearson W.R., Lipman D.J., Improved tools for biological sequence comparison, Proc. Natl. Acad. Sci. USA (1988) 85:2444-2448]及びSSEARCH(データベース:UniProtKB及びSwissProt)、及びHHpred[Soding J., Biegert A., Lupas A.N., The HHpred interactive server for protein homology detection and structure prediction, Nucleic Acids Res. (2005) 33:W244-W248](データベース:Interpro(2009))を用いて行った。全ての検索は、2009年3月に行った。Transterm[Kingsford C.L., Ayanbule K., Salzberg S.L., Rapid, accurate, computational discovery of Rho-independent transcription terminators illuminates their relationship to DNA uptake, Genome Biol. (2007) 8:R22]を用いて、転写ターミネーターを予測した。
【0169】
溶血アッセイ
【0170】
溶血活性は、以前の記述に従って検査しており[Rowe G.E., Welch R.A., Assays of Hemolytic Toxins, Methods Enzymol. (1994) 235:657-667]、ウシ血液をTN緩衝液(10 mM Tris-HCl, 0.9% NaCl, pH 7.5)中で上部相が無色に見えるまで繰り返し洗浄した。2%(vol/vol)赤血球溶液を、10mM CaCl2で補ったTN緩衝液中で調製した。赤血球は、濾過滅菌細菌培養上清又は細菌と共に、特に記載がない限り、1:1の比において37℃で1時間インキュベートした。非溶解赤血球及び細胞残屑を遠心分離により収集し、放出されたヘモグロビンの量をプレートリーダにおいて540nmで測定した。100%の溶解は、1%triton-X により決定し、バックグラウンドの溶解は、溶血活性の計算前に減算した。熱の影響は、上清を溶血アッセイ前に60℃で30分間インキュベートすることにより調査した。プロテイナーゼKの影響は、上清を溶血アッセイ前に4μg/mLのプロテイナーゼKと共に37℃で30分間インキュベートすることにより調査した。
【0171】
HD11細胞の培養及びLDH細胞毒性アッセイ
【0172】
トリ骨髄細胞のMC29形質転換に由来するマクロファージ様細胞株HD11[Beug H., Vonkirchbach A., Doderlein G., Conscience J.F., Graf T., Chicken Hematopoietic-Cells Transformed by 7 Strains of Defective Avian Leukemia Viruses Display 3 Distinct Phenotypes of Differentiation, Cell (1979) 18:375-390]を、Roswell Park Memorial Institute (RPMI)1640培地+GlutaMAXTM-I + 25 mM HEPES (Gibco)において維持した。培地には、2.5%トリ血清、7.5%ウシ胎仔血清(FBS)、及び25μg/mLゲンタマイシンを補った。細胞は、付着細胞株として37℃で5%CO2雰囲気により培養し、1日又は2日おきに継代培養した。細胞毒性アッセイのために、細胞は、総量100μLの5%FBS添加RPMI中で2×104個ずつ96ウェルプレートに播種した。細胞は、一晩インキュベートし、培地を交換した。生理食塩水(0.9%NaCl)に再懸濁した濾過滅菌培養上清又は細菌を細胞に添加し、1時間インキュベートした。大腸菌は、第2.7節において説明されるように組み換えタンパク質の発現を誘導し、OD600(600nmでの光学密度)を略6×108CFU/mLに相当する1に調節した。G. anatis細胞及び上清は、後期対数期(OD600 = 1)に回収した。G. anatisの懸濁物は、OD600を略4×108CFU/mLに相当する1に調節した。濾過滅菌培養上清を氷上で保存し、採取後30分以内に細胞に添加した。細胞毒性は、LDH細胞毒性アッセイ(Promega)により製造業者の説明に従って決定した。各試料は、3重ウェルにおいて検査し、実験は、少なくとも3回繰り返した。
【0173】
G. anatis gtxA突然変異体の構築
【0174】
gtxAのヌクレオチド140〜1648からなる1508bpの断片を、プライマー4240及び4242によりPCR増幅し、ヌクレオチド3995〜5478からなる1483bpの断片をプライマー4243及び4245により増幅した(プライマー配列は表Iに記載)。2つの断片を制限酵素により消化し、プラスミドpWSK129内の対応制限部位に連結させた[Wang R.F., Kushner S.R., Construction of versatile low-copy-number vectors for cloning, sequencing and gene expression in Escherichia coli, Gene (1991) 100:195-199]。EcoRI消化pUC4-KISSからのゲル精製カナマイシンカセット (Tn903)[Barany F., 2-Codon Insertion Mutagenesis of Plasmid Genes by Using Single-Stranded Hexameric Oligonucleotides, Proc. Natl. Acad. Sci. USA (1985) 82:4202-4206]を2つのPCR断片間のEcoRI部位に連結した。カナマイシン耐性遺伝子をgtxAと同じ転写方向に挿入した。プラスミドDNAは、XhoI及びSalIによる消化により直線化し、カラム精製した。G. anatis 12656-12の自然形質転換受容性(natural competence)を、インフルエンザ菌に関する以前の記述に従ってMIV法により誘発し[Poje G., Redfield R.J., Transformation of Haemophilus influenzae, Methods Mol. Med. (2003) 71:57-70]、G. anatisをBHI中で増殖させてOD600を0.2とし、MIVにおいて1回洗浄し、MIV中で100分間インキュベートした。直鎖DNAを0.5μg DNA/mLの濃度で細胞に添加した。20分後、2体積のBHIを追加し、細菌を1時間インキュベートした後、5μg/mLカナマイシンを含む血液寒天プレート上で形質転換体を選択した。コロニーを再ストリークし、プライマー対39F+kanR、kanF+5334R、及び2871F+3270Rにより欠失を検証した。菌株は、ΔgtxAと命名した。
【0175】
発現プラスミドの構築
【0176】
完全長GtxA、GtxAのN末端ドメイン(アミノ酸1〜949)、及びGtxAのRTXドメイン(アミノ酸931〜2026)を、GtxCと共に又はGtxCなしでコードするプラスミドを、発現ベクターpET28a(Novagen)へのPCR断片のライゲーションにより構築した。PCR断片は、pfx50ポリメラーゼ(Invitrogen)により増幅し、カラム精製し、NcoI及びXhoIにより二重消化し、再度カラム精製するか或いはゲル精製した(断片>6kb)。各構築物に使用したプライマーは、表2に記載している。構築物Nterm+C、即ちgtxCを有するオペロン内のgtxAのヌクレオチド1〜2847は、オーバラップエクステンションによるスプライシング(splicing by overlap extension)を使用して作成し[Horton R.M., Cai Z.L., Ho S.N., Pease L.R., Gene-Splicing by Overlap Extension - Tailor-Made Genes Using the Polymerase Chain-Reaction, Biotechniques (1990) 8:528-535]、プライマーGtxUP-NcoI及びgtxC-r-XhoIをPCRの第二ラウンドにおいて使用した。プラスミドpET28aをNcoI及びXhoIにより二重消化し、アンタークティックホスファターゼ(NEB)により脱リン酸化し、ゲル精製した。ベクター及びPCR断片をモル比1:3で連結し、化学的コンピテント大腸菌Top10F’(Invitrogen)に形質転換し、カナマイシンを含むLB寒天プレート上において選択した。各プラスミドにおけるインサートの配列は、DNAシーケンシング(Macrogen、韓国)により検証した。プラスミドは、大腸菌発現株ER2566(New England Biolabs)に形質転換した。プラスミドpLG575は、HlyAを分泌するT1SSの構成要素である大腸菌hlyB及びhlyDをコードしており[Mackman N., Nicaud J.M., Gray L., Holland I.B., Genetical and functional organisation of the Escherichia coli haemolysin determinant 2001, Mol. Gen. Genet. (1985) 201:282-288]、発現タンパク質の分泌を促進するために導入した。
【0177】
大腸菌における組み換えGtxAタンパク質の発現
【0178】
タンパク質の発現は、30℃でインキュベートした0.1mM IPTGを含む寒天プレート上で誘導した。ブロスでの誘導のために、一晩培養した培養物を1:50で希釈し、培養物のOD600が0.6になるまで37℃で振盪しながらインキュベートした。その後、IPTG(0.2mM)を添加し、誘導を2時間、30℃で維持した。細胞から組み換えタンパク質を放出させるために、細胞を遠心分離によりペレット状にし、元の体積の1/25で0.1M Tris/0.9%NaClに再懸濁させ、45秒間のビーズ破砕(FastPrep)により溶解させ、4℃で遠心沈殿させ、直ちに上清を液体溶血アッセイ及びLDH細胞毒性アッセイに使用した。
【0179】
SDS PAGE及びウェスタンブロット分析
【0180】
培養物500μLを採取し、細胞ペレットを採取時のOD単位当たり10mM Tris 500μLに再懸濁することにより、全細胞タンパク質を取得した。細胞外タンパク質は、濾過滅菌培養上清(低タンパク質結合フィルタ(0.22μm)(Millex(R) GP(Millipore))から調製した。タンパク質は、1体積の氷温96%エタノールの添加により一晩沈殿させ、遠心分離により収集し(13000g、30分間、0℃)、10mM Trisに再懸濁させた(元の体積の1/100)。タンパク質をNuPAGE(R) Novexゲル(Invitrogen)でのSDS-PAGEにより分離させた。ウェスタンブロット分析のために、タンパク質をフッ化ポリビニリデン膜(Invitrogen)にトランスファーした。1次抗体である、A. pleuropneumoniae由来のApxIに対して産出したウサギ抗血清[Schaller A., Kuhn R., Kuhnert P., Nicolet J., Anderson T.J., MacInnes J.I., Seger R.P.A.M., Frey J., Characterization of apxIVA, a new RTX determinant of Actinobacillus pleuropneumoniae, Microbiology (1999) 145:2105-2116]を1:1333の希釈で使用し、Westernbreeze化学発光ウェスタンブロット免疫検出キット(Invitrogen)により製造業者の説明に従って検出した。
【0181】
RNA精製
【0182】
一晩培養した培養物をBHIにおいて1:100で希釈し、37℃でエアレーションと共にインキュベートした。細胞を、OD600が0.17、0.6、2、3の時点で採取し、更に増殖を停止した1時間後、及び24時間のインキュベーション後に採取した。全RNAをRNeasy protect Mini Kit(Qiagen)により単離し、オンカラムDNアーゼ処理を製造業者の説明に従って実行した(Qiagen)。
【0183】
ノーザンブロッティング
【0184】
RNAのブロッティング、プローブ標識([α-32P]-dCTPによる)、及びハイブリダイゼーションを基本的に記述の通りに実行した[Frees D., Chastanet A., Qazi S., Sorensen K., Hill P., Msadek T., Ingmer H., Clp ATPases are required for stress tolerance, intracellular replication and biofilm formation in Staphylococcus aureus, Mol. Microbiol. (2004) 54:1445-1462]。gtxAのRTX側の半分以内にある384bpの断片を、プライマー3487F 5’-GCCTCTACCGCCGTTTCTG-3’及び3874R 5’-GGCTGGCTAATAATTCATCACCTTG-3’によりPCR増幅し、プローブ標識反応のテンプレートとして使用した。
【0185】
結果
【0186】
G. anatisの細胞溶解活性
【0187】
G. anatis次亜種ヘモリチカは、ウシ血液寒天プレート上でβ溶血性を有する[Christensen H., Bisgaard M., Bojesen A.M., Mutters R., Olsen J.E., Genetic relationships among avian isolates classified as Pasteurella haemolytica, 'Actinobacillus salpingitidis' or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov and description of additional genomospecles within Gallibacterium gen. nov, Int. J. Syst. Evol. Microbiol. (2003) 53:275-287]。標的範囲を試験するために、様々な種からの血液により寒天プレート上で菌株12656-12の溶血を調査したところ、細菌は、ウマ、ウシ、ブタ、又はニワトリの血液を含む寒天プレート上で増殖させた時にβ溶血の明瞭なゾーンを発生させ(データを示さず)、Greenham及びHillが報告した広範な標的範囲が確認された[Greenham L.W., Hill T.J., Observations on an Avian Strain of Pasteurella haemolytica, Vet. Rec. (1962) 74:861-863]。細菌は、好気性及び嫌気性の両培養条件下で溶血性を有した。
【0188】
この溶血活性は、分泌された溶血素により生じることが示唆されている[Greenham L.W., Hill T.J., Observations on an Avian Strain of Pasteurella haemolytica, Vet. Rec. (1962) 74:861-863]。これを試験するために、液体溶血アッセイを実施し、様々な増殖段階で採取したG. anatis 12656-12無細胞培養上清を、ウシ赤血球の懸濁液と共にインキュベートし、放出されたヘモグロビンの量を測定した。中期から後期の対数期に由来するG. anatis上清は、赤血球を効率的に溶解させた(図1A)。この活性は、熱(60℃)及びプロテイナーゼKにより不活性化され、カルシウムキレート剤EGTAにより低減されることから(データを示さず)、G. anatisがカルシウム依存性の分泌溶血性タンパク質を生産することが確認された。
【0189】
赤血球の溶解は、宿主の鉄獲得に関与し得るが、しかしながら、他の種類の細胞、例えば、白血球との相互作用は、自然感染中に更に重要な役割を果たし得る。そのため、トリ由来マクロファージ様細胞株HD11を使用して白血球に対するG. anatisの細胞毒性活性を試験した。HD11細胞は、G. anatisへの曝露後、円形化を示し、表面から剥離した(図2B)。細胞毒性は、乳酸脱水素酵素(LDH)細胞毒性アッセイを用いて定量化し、顕著な細胞死が明らかとなった(図2C)。このG. anatisの白血球毒性活性は、この細菌の病変形成において不可欠である可能性が高く、したがって、白血球毒性活性の原因となるタンパク質は、重要な病原因子であると予想される。
【0190】
G. anatisのゲノム配列におけるRTX毒素の特定
【0191】
G. anatisの細胞毒性表現型の原因となる特異タンパク質を特定するために、G. anatis 12656-12のゲノム配列を入手し、潜在的な毒素をコードする配列を検索した。膜孔形成RTX毒素の群に属するタンパク質は、重要な病原因子であり、ガリバクテリウムに関する細菌における溶血性及び白血球毒性活性の原因となることから[Frey J., Kuhnert P., RTX toxins in Pasteurellaceae, Int. J. Med. Microbiol. (2002) 292:149-158]、この種のタンパク質が検索の明確な標的となる。G. anatis 12656-12ゲノム配列に対する様々なRTX毒素(A. pleuropneumoniae由来のApxI及びApxII、A. aggregatibacter由来のLtxA、及び大腸菌由来のHlyAを含む)のアミノ酸配列によるBLAST検索により、2026アミノ酸の推定G. anatis RTX毒素が特定された。このタンパク質をコードする6081ヌクレオチド(nt)のオープンリーディングフレーム(ORF)をgtxAと名付け、gtxはGallibacterium toxin、Aは他のRTXオペロン内での毒素遺伝子の命名に対する類推(analogy)を示す[Frey J., Kuhnert P., RTX toxins in Pasteurellaceae, Int. J. Med. Microbiol. (2002) 292:149-158]。gtxAには、非常に短い5ヌクレオチド(nt)の遺伝子間領域と、163個のアミノ酸の予測タンパク質をコードする492ntのORF(図3A)が続く。このタンパク質は、RTX毒素の活性化に必要なアシルトランスフェラーゼタンパク質との相同性を有し、大腸菌由来のアシルトランスフェラーゼHlyCに対して38%の同一性及び60%の類似性を示した。類推から、遺伝子は、gtxCと名付けた。Rho非依存性転写ターミネーターは、gtxCの下流に発見され、恐らくはgtxA及びgtxCの両方を含む転写単位の終了を示す。gtxA-Cオペロンに隣接する遺伝子は、イノシトール-1-モノホスファターゼ(gtxAの上流のsuhB)及びマンノネートデヒドラターゼ遺伝子(gtxCの下流のuxuA)(図3A)をコードすると予測され、両方とも、GtxACの機能に関与する可能性は低い。
【0192】
興味深いことに、GtxA(2026アミノ酸)は、パスツレラ科の他の構成員及び大腸菌由来のHlyA について記述される「典型的」なRTX毒素(略1000aa[Frey J., Kuhnert P., RTX toxins in Pasteurellaceae, Int. J. Med. Microbiol. (2002) 292:149-158])の2倍の大きさを有する。GtxAのC末端の1000個のアミノ酸は、こうしたRTX毒素と相同であり、この領域は略20%の配列同一性及び35%の配列類似性を有する。このC末端領域は、更に、RTX毒素の幾つかの保存的特徴を含む。大腸菌由来のHlyAは、Lys564及びLys690においてアシル化されており[Stanley P., Packman L.C., Koronakis V., Hughes C., Fatty Acylation of 2 Internal Lysine Residues Required for the Toxic Activity of Escherichia coli Hemolysin, Science (1994) 266:1992-1996]、こうしたリジン及び先行するグリシン残基は、共にGtxAにおいて保存されているため(Lys1484及びLys1607(図3B))、これらはGtxAにおいてGtxCにより仲介されるアシル化部位となる可能性が高い。予測されるアシル化部位(アミノ酸1640〜1830)の下流において、GtxAは、グリシン及びアスパラギン酸塩リッチ領域を有し、これもRTX毒素の保存的特徴である。一方、N末端領域(aa1〜略950)は、利用可能な配列との類似性が限定されており、GenBankデータベースに対するBLASTP検索では、有意なホモログは見つからなかった。しかしながら、aa520〜879の領域は、予測膜タンパク質の未知の機能の保存的ドメイン(COG1511)に対する類似性を有する(E値0.007)。RTXドメインと比較すると、N末端ドメインは、酸性が低く、疎水性アミノ酸、特にセリンを高い割合で含む。2次構造は、主にαヘリックスからなると予測される。
【0193】
N末端ドメインの機能の着想を得るために、配列類似性及び相同性予測用の更に感度の高い検索ツールを用いて、アミノ酸配列のバイオインフォマティクス分析を実行した。FASTA及びSSearchを用いた相同性検索により、アメーバDictyostelium discoide由来の真核生物細胞骨格タンパク質Talin-A及びTalin-B (E値、それぞれ3.4×10-7及び0.0061)及びニワトリ(Gallus gallus) 由来のTalin (E値0.0087)との配列類似性が発見された。更に、相同性検出サーバHHpredでは、Talinに対する相同性が予測された(確率 = 100%)。Talinは、アクチン、ビンキュリン、及びインテグリンの細胞質部分などの、他の一定の範囲のタンパク質と結合する。巨大タンパク質は、複製により生じる反復からなる場合が多く、リピート発見プログラムRadar[16]によるN末端ドメインの調査では、57個のアミノ酸の15の反復が見つかった(図3C)。
【0194】
したがって、GtxAは、2つのドメイン、即ちN末端反復ドメイン及びC末端RTX/細胞溶解素ドメインからなる。
【0195】
GtxAは、GtxCに依存する細胞溶解活性を有する
【0196】
GtxAが細胞溶解性タンパク質であるかを調査するために、gtxAを、予測アシルトランスフェラーゼ遺伝子gtxCと共にクローン化し、非溶血性発現菌株の大腸菌ER2566に導入した。gtxA及びgtxCの発現時、この菌株は、血液寒天プレート上及び液体溶血アッセイにおいて溶血性表現型を呈し(図4)、GtxACが溶血活性を保持することを示した。RTX毒素は、通常、特異的なT1SSにより搬出された細胞外タンパク質である。大腸菌hlyBDによりコードされたT1SSを発現するプラスミド(pLG575)の導入により、溶血ゾーン(図4A)は増加し、イムノブロッティングでは、GtxAの大きな画分が細胞外タンパク質画分に存在することが明らかとなり(図5)、大腸菌分泌系がG. anatis GtxAを分泌可能であることが実証された。HD11細胞に対するGtxAの細胞毒性活性は、LDH放出アッセイにより検査し、gtxACを発現する大腸菌ER2566はHD11細胞に対して毒性を有した。インサートの無いベクターを含む大腸菌(負の対照)は、1時間のインキュベーション後、毒性を示さなかった(図4B)。
【0197】
翻訳後アシル化を必要とすることは、RTX毒素の特徴の1つである。これが非定型のGtxAにも該当するかを評価するために、予測されるアシルトランスフェラーゼGtxC無しで発現させたGtxAの活性を調査した。図4は、GtxAがGtxC不在の状態で発現した場合、赤血球又は白血球に対する細胞溶解活性を有していないことを示している。したがって、非アシル化タンパク質は、不活性であり、翻訳後アシル化は、溶血及び白血球毒性活性の両方に不可欠である。GtxAの分泌は、アシル化の欠如により妨げられず、非アシル化GtxAは、アシル化毒素と同様の量で培養上清内に検出された(図5)。
【0198】
GtxAはG. anatisの細胞毒性活性の原因である
【0199】
G. anatisの溶血及び白血球毒性活性がGtxAを起源とするかを判断するために、gtxAノックアウト突然変異体を構築した。ガリバクテリウムの遺伝子操作用の分子ツールの記述は過去に存在しないが、我々は、G. anatis 12656-12が自然状態でコンピテントであることを発見し、自然形質転換による安定したgtxA突然変異体の構築に、この特質を活用した。結果的に生じた突然変異体では、gtxAの位置1648及び3995間の2347個のヌクレオチドを欠失させ、カナマイシン耐性カセットにより置換させた。
【0200】
野生型とは対照的に、ΔgtxA突然変異体は、血液寒天プレート上(図2A)又は液体溶血アッセイ(データを示さず)において溶血性を有していなかった。更に、ΔgtxAは、HD11細胞に対する細胞毒性を示さなかった(図2B及び2C)。独立して構築した2つのgtxA突然変異体から同一の結果が得られた。したがって、gtxAは、G. anatisの溶血及び白血球毒性活性の原因である。
【0201】
GtxA及びその活性レベルの増殖期依存性
【0202】
G. anatis上清の溶血活性は、増殖期依存性を有し、活性は、後期対数期にピークとなり、静止期への遷移時に降下し、一晩培養した培養物の上清において低くなった(図1A)。このことから、GtxAの発現が同様の増殖期依存性を有するという仮説を立てた。これを調査してGtxAの局在を確証するため、ApxI抗血清によるイムノブロッティングを用いて、培養上清(細胞外タンパク質)及び全細胞溶解物におけるGtxAの量を、増殖中の様々な時点で判定した(図1B)。ApxI抗血清は、完全長GtxA(215kDa)の予測分子量のサイズに対応するバンドを含め、細胞外タンパク質画分内の幾つかのタンパク質を認識した。このバンドは、ΔgtxAには存在せず、バンドがGtxAであることが裏付けられた。溶血活性と同様に、上清中のGtxAの存在及び量は、増殖期依存性を有し(図1B)、GtxAの量は、静止期への遷移時にピークとなり、タンパク質は1日経過した培養物(24時間)では検出されず、A. Pleuropneumoniae ApxI及びApxII [Jarma E., Regassa L.B., Growth phase mediated regulation of the Actinobacillus pleuropneumoniae ApxI and ApxII toxins, Microb. Pathog. (2004) 36:197-203]及びM. haemolytica LktA [Strathdee C.A., Lo R.Y., Regulation of expression of the Pasteurella haemolytica leukotoxin determinant, J. Bacteriol. (1989) 171:5955-5962]について報告されたパターンと同様となった。第二のバンド(>215kDa)は、野生型にも存在するが、ΔgtxAには存在せず、恐らくは翻訳後の修飾により、GtxAが2つの異なる形態で存在し得ることを示唆した。更に2つのバンド(それぞれ65kDa及び>215kDa)は、野生型及び突然変異体の両方において検出されており、GtxAに関連していない可能性が高い。GtxAのサイズのタンパク質は、全細胞溶解物では如何なる時点においても検出されておらず、これは予測される細胞外局在に一致していると共に、GtxAが合成の直後又はそれに関連して分泌されることが裏付けられた。gtxAの転写を調査するために、増殖期全体で採取した細胞からのRNAにより、ノーザンブロッティングを実行した。ブロットにより、gtxAが対数増殖中及び静止期の始めに転写されることが明らかとなったが、静止期に入って2時間の及び一晩培養した培養物では転写物は検出されず、gtxAの転写が静止期中に停止されることが示唆された。結論として、GtxAは、in vitro増殖中に発現され、増殖期依存性の細胞外タンパク質であり、増殖期依存性は、転写調節と、分泌GtxAの蓄積及びその後の分解の間のバランスとに影響される。
【0203】
GtxAのN末端ドメインは完全な細胞溶解活性に必要となる
【0204】
生物情報学的分析では、RTXドメイン及びN末端ドメインの2つの部分からなる他の膜孔形成RTX毒素と比較して、GtxAが非定型の構造を有することが明らかとなった(図3B)。GtxAの細胞溶解活性に対するN末端ドメインの寄与を調査するために、N末端ドメイン(アミノ酸1〜949)及びRTXドメイン(アミノ酸931〜2026)の両方を、大腸菌において別々に発現させ、その溶血及び白血球毒性活性を調査して完全長タンパク質のものと比較した(図4)。RTXドメインをGtxCと共に発現する大腸菌は、血液寒天プレート上及び液体溶血アッセイにおいて、溶血活性を示したことから、RTXドメインは、それ自体が機能性溶血性タンパク質であり、N末端ドメインは、赤血球細胞の溶解に不可欠ではない。しかしながら、RTXドメインは、完全な毒素よりも著しく低い溶血活性を示しており、N末端ドメインが完全な溶血活性に必要であることが示唆された。RTXドメインとHD11細胞との間の相互作用から細胞毒性活性は検出されず、N末端ドメインが白血球毒性に対して不可欠な役割を果たすことが示唆された。イムノブロッティング(図5)は、RTXドメインが発現及び搬出されることを示しており、そのため、活性の違いは、発現レベルの大きな違いによるものではなかった。
【0205】
N末端ドメインのみを発現する大腸菌は、溶血及び細胞毒性活性を有していなかった(図4)。しかしながら、SDS-PAGEでは、この組み換えタンパク質が主に全細胞画分に存在し、細胞外タンパク質画分には少量しか存在しないことが明らかとなった(データを示さず)。そのため、N末端ドメインを発現する細胞の溶解物(FastPrepビーズ破砕又はリゾチーム/超音波処理により生成)の活性を試験したところ、これらは、長期のインキュベーションにもかかわらず、液体溶血アッセイにおいて、或いはトリマクロファージに対して、活性を全く示さなかった。これらの結果は、N末端ドメイン自体が細胞溶解活性を有しないことを示す。
【0206】
実施例2:ガリバクテリウムにおけるgtxAの検出
【0207】
材料及び方法
【0208】
染色体DNAを、DNeasy Kit(Invitrogen)によりガリバクテリウム菌株から精製した。染色体DNAをテンプレート(50ng/50μl反応液)として、標準PCRをプライマーrtxA3368F 5’-CAAACCTAATTCAATCGGATG-3’(SEQ ID NO:24)及びrtxA4625R 5’-TGCTTCAATAATTTTCCATTTTC-3’(SEQ ID NO:25)により行い、gtxAのヌクレオチド3332〜4589を増幅した。PCR条件は、94℃4分間と、94℃30秒、51℃30秒、及び72℃105秒を35サイクルとした。産物は、ゲル電気泳動及びエチジウムブロマイド染色により可視化した。PCR産物は、BigDyeサイクルシーケンシング(Macrogen、韓国)により配列決定した。ヌクレオチド配列は、CLC workbenchにより翻訳及び整列を行った。
【0209】
結果
【0210】
異なるガリバクテリウム菌株からのGtxA毒素のアライメントを図6に提示する。
【0211】
実施例3:配列
【0212】
下記の配列は、本願において言及したアミノ酸及び核酸の配列を示す。
【0213】
【化1】
【化2】
【化3】
【化4−1】
【化4−2】
【化5】
【化6】
【0214】
実施例4:gtxAの配列及び機能は溶血性ガリバクテリウム間で保存されている
【0215】
ガリバクテリウム菌株間でのgtxAヌクレオチド1〜950の配列保存
【0216】
【表3】
【0217】
GtxAは溶血性ガリバクテリウムにより発現及び分泌される
【0218】
材料及び方法
【0219】
タンパク質の単離のために細胞を培養する際には、一晩培養した培養物(16時間)を略1:100に希釈し、OD600を0.03に調整し、細菌を100mLのErlenmeyerフラスコ内の30mL BHIブロスにおいて穏やかなエアレーション(120rpm)下で培養した。細胞及び上清を、後期対数期にOD600 = 0.6(±0.1)で採取した。
【0220】
細胞外タンパク質は、濾過滅菌培養上清(低タンパク質結合フィルタ(0.22μm)(Millex(R) GP(Millipore))から調製した。タンパク質は、1体積の氷温96%エタノールの添加により一晩沈殿させ(-20℃)、遠心分離により収集し(13000g、30分間、0℃)、10mM Trisに再懸濁させた(元の体積の1/100)。タンパク質(13μl)をSDS-PAGEにより、Tris-Acetate SDS Running Buffer (Invitrogen)中のNuPAGE Novex 3-8% Tris-Acetate Mini Gelsにおいて分離させた。ウェスタンブロッティングのために、タンパク質をフッ化ポリビニリデン(PVDF)膜(Invitrogen)にブロットした。ApxI抗血清[24]を1次抗体として使用し、結合したApxI抗体をWesternBreeze Chemiluminescentキット(抗ウサギ)(Invitrogen)により検出した。化学発光信号は、Geliance 600画像化システム(Perkin Elmer Elmer)により捕捉した.
【0221】
GtxAはG. anatisにおける主要な細胞溶解性毒素である
【0222】
8種類の遺伝学的に異なるG. anatis次亜種ヘモリチカ菌株(10672-6、10T4、21K2、24T10、4895、07990、及びAvicor)のgtxAを以前の記述に従って突然変異により不活性化した(Kristensen et al., 2010)。全ての突然変異体が非溶血性であることが判明し(データを示さず)、こうした菌株において、GtxAが溶血活性の原因となることが実証された。この結果は、G. anatisにおける優性な細胞溶解性毒素としてのGtxAの役割を裏付ける。
【0223】
gtxAはG. genomospecies 1及びG. genomospecies 2の溶血活性の原因である
【0224】
G. genomospecies 1及びG. genomospecies 2におけるgtxAの存在から、gtxAがG. genomospecies 1及び2における溶血活性の原因であることが示唆された。G. genomospecies 1及び2型菌株(CCM5974及びCCM5976)においてgtxA突然変異体を構築したところ、これらは非溶血性となり(データを示さず)、これらの種においてもGtxAが溶血活性の原因であることが実証された。
【0225】
実施例5:GtxAの搬出に必要なI型分泌遺伝子座(gtxEBD)の同定及び性質決定
【0226】
GtxAは細胞外タンパク質だが、パスツレラ科の殆どのRTX毒素とは異なり、同族の1型分泌系(type 1 secretion system, T1SS)の成分との共転写は行われない(図9)。T1SSは、3つのタンパク質、即ち、内膜ATPアーゼ、内膜チャネルタンパク質、及び外膜タンパク質の多量体で構成され、大腸菌において、原型のRTX毒素のα溶血素を分泌するT1SSタンパク質は、それぞれHlyB、HlyD、及びTolCと命名されている(Holland et al., Mol Membr Biol 2005, 22: 29-39)。GtxAの搬出を担当するT1SSを特定するために、菌株12656-12のゲノム配列を大腸菌HlyB及びHlyDのアミノ酸配列をクエリとして検索した。この手法により、明白な基質のない予測T1SSオペロンを特定し、GtxA分泌の有望な候補と見なした。このオペロンは、gtxE、gtxB、及びgtxDと名付けた3つの遺伝子を含んでいた(図9)。コード化タンパク質GtxE、GtxB、及びGtxCは、それぞれ、1型分泌系を構成するために必要な3つの構成要素に対応する外膜タンパク質、内膜ATPアーゼ、及び内膜タンパク質になると予測された。
【0227】
オペロンがGtxAの搬出に必要であるかを試験するために、G. anatis 12656-12において、gtxBの大部分(ヌクレオチド位置92から)とgtxDの先頭(最初の123nt)を欠失させることにより突然変異体(ΔgtxBD)を構築した。
【0228】
ΔgtxBDの構築
【0229】
ΔgtxBDの構築のため、2つのPCR断片を生成し、一方(951bp)には、プライマー2870F-XbaI (5’-CTGATCTAGACGCCGTAAATCGCATAATCA-3’) (SEQ ID NO: 26)及び3821R-EcoRI (5’-CGAATTCCCGGCAGAAAAGGTCAACA-3’) (SEQ ID NO: 27)、他方(1233nt)には、6044-SOE (TTTCTGCCGGGAATTCGGCGAATGGTGTGAGAAG-5’) (SEQ ID NO:28)及び7267R-SalI (TCAAGTCGACAAGCCAAAGCCAATACGA-3’) (SEQ ID NO:29)を用いた。プライマー配列の制限部位には下線を付した。2つのPCR断片は、2870f-XbaI及び7267R-SalIによるオーバラップエクステンションPCRによるスプライシングを用いて接続した。結果的に生じた2184ntの断片を、SalI及びXbaIにより消化し、アンタークティックホスファターゼ(Fermentas)によりホスファターゼ処理し、ゲル精製してXbaI/SalI二重消化及びゲル精製したpBluescriptに連結した。EcoRI消化pUC4-KISSからのカナマイシンカセット(Tn903)をゲル精製し、SOEingによる断片内のEcoRI部位に連結した。カナマイシン耐性遺伝子を、gtxBと同じ転写方向に挿入した。プラスミドDNAをSalI及びXbaIによる消化で直線化し、カラム精製した(GFX、Amersham)。G. anatis 12656-12の自然形質転換受容性を、インフルエンザ菌に関する以前の記述に従ってMIV法により誘発した(Poje & Redfield, 2003); 簡潔に言うと、G. anatis 12656-12をBHI中で増殖させてOD600を0.2とし、MIVにおいて1回洗浄し、MIV中で100分間インキュベートした。直鎖DNAを0.5μg DNA/mLの濃度で細胞に添加した。37℃で20分インキュベートした後、2体積のBHIを追加し、細菌を1時間インキュベート(37℃)した後、4μg/mLカナマイシンを含む血液寒天プレート上で形質転換体を選択した。形質転換体を4μg/mLカナマイシンを含むプレート上に再ストリークし、プライマー対TISS2607F (5’-TTTCCTGTAATGCCTGCT-3’) (SEQ ID NO: 30)及びTISS7572R (5’-TTTTGATCGTTCGGGCTT-3’) (SEQ ID NO: 31)によるPCR及びPCR産物の配列決定により欠失を検証した。
【0230】
実施例6: GtxAは全細胞抗原に基づくワクチンの一部となる可能性が低い
【0231】
GtxAは、細胞溶解物中に検出可能なレベルでは存在しておらず、I型分泌gtxEBDが存在しない状態では細胞内に蓄積されない。
【0232】
我々は、GtxAの分泌に必要なI型分泌系遺伝子座(gtxEBD)を特定した。
【0233】
分泌突然変異体ΔgtxBD(実施例5に記載のように構築)において、ΔgtxBDの溶血活性は、野生型(図8A参照)と比較して、大きく減少した。ウェスタンブロッティングでは、gtxBDの欠失により上清中のGtxAの欠如が生じることが明らかとなった(図8B)。こうした観察結果は、gtxEBDがGtxAの搬出を担うことを裏付ける。GtxAは、野生型又はΔgtxBDの全細胞タンパク質内では検出されず、即ち、GtxAは、分泌系が存在しない状態では細胞内部に蓄積されない。同様の観察結果は、A. actinomycetemcomitans (白血球毒素LtxA)及び大腸菌(α溶血素HlyA)におけるRTX毒素分泌不全突然変異体についても得られた。これらの種において、毒素の細胞内蓄積の欠如は、転写の変更によるものではなく、細胞質における毒素の迅速な分解によるものと推測される。
【0234】
実施例7: GtxAを発現できないGallibacterium anatis gtxA突然変異体による感染試験
【0235】
序論
【0236】
細胞溶解性RTX毒素GtxAは、家禽における感染中のG. anatisにとって主要な病原因子とされてきた。これを実証するために、野生型(wt)菌株及び同質遺伝子的ΔgtxA突然変異体による感染中に観察された病変に対するGtxAの寄与を特徴付ける目的で、産卵雌鶏での感染試験を行った
【0237】
材料及び方法
【0238】
合計24羽を研究に含めた。全てのトリを腹腔内経路で感染させ、卵巣の根元に等量のGallibacterium anatisが深く沈着するように試みた。16羽には、G. anatis 12656-12wtを感染させ、一方、16羽には、RTX GtxAを発現できない同質遺伝子的だが非溶血性のΔgtxA突然変異体を感染させた(Kristensen et al., 2010)。
【0239】
結果
【0240】
【表4】
【0241】
wtに感染させたトリは、一般に野外でのG. anatisへの自然感染により観察される病変に対応する、生殖器系及び腹膜に関与する播種性及び化膿性の炎症を生じた。
【0242】
ΔgtxA突然変異体に感染させたトリは、一般に、卵巣に局在する、より軽度の炎症を生じた。幾つかの動物(2/8)では、局在する炎症が強く、感染48時間後に化膿し、一方、8羽のうち2羽は、化膿性卵巣炎及び腹膜炎が感染6日後に観察された。
【0243】
したがって、GtxAがニワトリにおけるG. anatisの病変形成に実質的に寄与すると結論することができる。
【技術分野】
【0001】
[関連出願の相互参照]
出願、即ち本願において、引用した全ての特許及び非特許文献は、出典を明記することによりその開示内容全体を本願明細書の一部とする。
【0002】
本発明は、動物の健康の分野に関し、特に、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2を含むガリバクテリウム菌種により生じる新たな細菌性家禽疾患の原因因子に関する。本発明は、GtxA(ガリバクテリウム毒素)と名付けた、前記ガリバクテリウム種由来の新規のRTX毒素を提供する。更に、本発明は、GtxAのアミノ酸及びヌクレオチド配列と、不活性化トキソイド又はトキソイドの断片を含むワクチンと、前記疾患を予防するために鳥類を免疫する方法と、鳥類におけるGallibacterium anatis感染を診断する方法とを提供する。
【背景技術】
【0003】
過去10年間、生産性を増加させる集約的養鶏法の結果として、主要な全ての家禽生産国において、疾患の発現が増加してきた。これにより、こうした疾患を抑制する新規の優れたワクチン及び予防接種プログラムの必要性が高まっている。現在、多くの動物は、ウイルス及び細菌を原因とする多数の疾患に対して免疫されている。家禽におけるウイルス性疾患の例には、ニューカッスル病、伝染性気管支炎、トリニューモウイルス、鶏痘、伝染性ファブリキウス嚢病等がある。細菌性疾患の例には、ヘモフィルス・パラガリナルム(上気道)、ボルデテラ・アビウム(上気道)、オルニソバクテリウム・ライノトラキア(下気道)により生じるトリコリーザ、サルモネラ感染症(消化管)、家禽コレラ(敗血症)の原因因子であるパスツレラ・マルトシダ、及び大腸菌感染症がある。
【0004】
産卵家禽の生殖器及び腹膜における炎症は、商用の産卵家禽群において繰り返される問題であり、卵生産量の下落と、死亡率の上昇と、結果的な経済的損失及び動物の生育状態の低下を引き起こす。これらの病変からはトリ病原性大腸菌が単離される場合が多いが、幾つかの研究では、Gallibacterium anatisが単独で、或いは共同病原体として、頻繁に卵巣炎、卵管炎、及び腹膜炎の原因となることが実証されている。更に、トリの敗血症、肝炎、腸炎、及び上気道の病変からG. anatisが単離されている。G. anatisは、産卵雌鶏及び他の鳥類種の上気道及び下部生殖管両方の正常細菌叢に共通する部分であり(Bojesen A.M., Nielsen S.S., Bisgaard M., Prevalence and transmission of haemolytic Gallibacterium species in chicken production systems with different biosecurity levels, Avian Pathol. (2003) 32:503-510)、したがって、日和見病原体と見なすことができる。その病変形成は、特に分子レベルでは、深く研究されておらず、G. anatisが疾患を引き起こす能力の背後にある遺伝子及び機構については殆ど知られていない。G. anatisは、2つの次亜種、β溶血性次亜種ヘモリチカ(haemolytica)と、非溶血性次亜種アナティス(anatis)とに分けられる。赤血球を溶解させる能力は、病原性G. anatis分離株の顕著な表現型である(Christensen H., Bisgaard M., Bojesen A.M., Mutters R., Olsen J.E., Genetic relationships among avian isolates classified as Pasteurella haemolytica, 'Actinobacillus salpingitidis' or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov and description of additional genomospecles within Gallibacterium gen. nov, Int. J. Syst. Evol. Microbiol. (2003) 53:275-287)。ガリバクテリウムは、γプロテオバクテリア科であるパスツレラ科に属するグラム陰性属であり(Christensen et al, 同上)、パスツレラ科の様々な病原性構成員、例えば、ヒトの歯周疾患の原因であるアグリゲイティバクタ・アクチノミセテムコミタンス、ウシ輸送熱の原因物質であるマンヘミア・ヘモリチカ、及びブタ病原体であるアクチノバチルス・プルロニューモニアは、RTX毒素((repeat in toxin)の群に属する溶血素及び白血球毒素を生じる。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Bojesen A.M., Nielsen S.S., Bisgaard M., Prevalence and transmission of haemolytic Gallibacterium species in chicken production systems with different biosecurity levels, Avian Pathol. (2003) 32:503-510)
【非特許文献2】Christensen H., Bisgaard M., Bojesen A.M., Mutters R., Olsen J.E., Genetic relationships among avian isolates classified as Pasteurella haemolytica, 'Actinobacillus salpingitidis' or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov and description of additional genomospecles within Gallibacterium gen. nov, Int. J. Syst. Evol. Microbiol. (2003) 53:275-287)
【発明の概要】
【発明が解決しようとする課題】
【0006】
不活性化又は生存する弱毒化細菌からなるG. anatisワクチンを利用可能である。しかしながら、こうしたワクチンは、この種に由来する分泌溶血性タンパク質に対する保護を与えない。
【0007】
本発明の目的は、G. anatis次亜種ヘモリチカの真核細胞との相互作用を調査すること、及び溶血性表現型の原因となる遺伝子及びタンパク質を特定及び特徴付けすることであった。
【課題を解決するための手段】
【0008】
本発明者は、G. anatisが、病変形成において重要な役割を果たす特質として、トリマクロファージに対する細胞毒性が高いことを見いだした。更に、本発明者は、G. anatis次亜種ヘモリチカにおける白血球毒性及び溶血活性の原因となる新規の種類のRTX毒素の特定及び特徴付けを行った。
【0009】
本発明は、ガリバクテリウム属、最も好ましくは、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される細菌由来のGtxAポリペプチド及びポリヌクレオチドに関する。
【0010】
第1の態様において、本発明は、単離ポリペプチドであって、以下からなる群から選択されるアミノ酸配列を含む、ポリペプチドに関する:
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【0011】
SEQ ID No. 1に記載のアミノ酸配列を有するポリペプチドは、G. anatis由来のRTX毒素である。GtxA(ガリバクテリウム毒素)と名付けたタンパク質は、2026個のアミノ酸(aa)からなる。これは、古典的な膜孔形成RTX毒素のサイズの2倍となる。GtxAのC末端の1000アミノ酸は、パスツレラ科の他の構成員におけるRTX毒素と相同であり、例えば、A. pleuropneumoniae ApxIAとの38%の配列類似性を有する。一方、N末端の略950アミノ酸は、GenBankデータベースにおける有意な一致がなかったが、機能が未知である57アミノ酸の反復を11個含む。
【0012】
GtxA毒素は、トキソイドワクチンとしての用途と、鳥類一般、特に家禽において、G. anatisに対する免疫反応の存在を明らかにする診断用途とを含め、幾つかの有用性を有する。
【0013】
他の態様において、本発明は、単離ポリヌクレオチドであって、以下からなる群から選択される核酸配列を含む、ポリヌクレオチドに関する:
a) SEQ ID No. 4、5、又は6と、
b) SEQ ID No. 4、5、及び6からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも60%である配列変異体と、
c) a)の何れかの少なくとも450個の隣接ヌクレオチドからなる断片であり、選択される配列において指定された任意の核酸が異なる核酸へ変化するのは、このように変化する配列内の核酸が90個以内の場合である断片と、
d) 高ストリンジェンシー下で、SEQ ID No. 4、5、又は6に相補的なポリヌクレオチドとハイブリダイズすることが可能なポリヌクレオチドと、
e) SEQ ID No. 1、2、又は3のポリペプチドをコードするポリヌクレオチドと、
f) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体をコードするポリヌクレオチドと、
g) SEQ ID No. 1、2、又は3の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片をコードするポリヌクレオチド。
【0014】
更に、本発明は、本発明のポリヌクレオチドを含むベクターに関する。
【0015】
他の態様において、本発明は、本発明のポリペプチド、本発明のポリヌクレオチド、及び本発明のベクターの医学的使用に関する。
【0016】
好ましくは、医学的使用は、細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療に関するものである。
【0017】
一態様において、本発明は、細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療用の薬剤を調製するための、本発明のポリペプチド及び/又はポリヌクレオチドの使用に関する。
【0018】
他の態様において、本発明は、本発明のベクターにより形質転換又は形質導入した単離宿主細胞と、本発明の感染性ビリオンを生産可能なパッケージング細胞株とに関する。
【0019】
更に、本発明は、抗体であって、以下からなる群から選択されるアミノ酸配列を有する単離ポリペプチドと特異的に結合可能な抗体に関する。
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【0020】
こうしたGtxAに対する抗体は、診断及び治療の態様において使用することができる。
【0021】
他の態様において、本発明は、本発明の単離ポリペプチドの不活性化の方法に関する。
【0022】
更に、本発明は、本発明の、裸DNA又はベクターとして調製された、単離ポリペプチド又は単離ポリヌクレオチドを、任意に1つ以上の適切なアジュバント、賦形剤、乳化剤、又は担体と共に含むワクチン組成物に関する。
【0023】
別の態様において、本発明は、本明細書に記載したような鳥類種に、本発明のワクチンを投与する方法に関し、前記ワクチンは、筋肉内又は皮下注射、食物又は水などを介した経口投与、エアロゾル、足又は翼の膜部などにおける乱切(らんせつ)、点眼剤、又は卵内投与により投与される。
【0024】
他の態様において、本発明は、診断マーカーとして用いる本発明のポリペプチド又はポリヌクレオチドに関する。
【0025】
本発明は、更に、鳥類種における病原性細菌ガリバクテリウムの感染を診断する方法を提供し、前記方法は、本発明のポリペプチドの検出、前記ポリペプチドに対する抗体の検出、又は本発明のポリヌクレオチドの検出を含む。
【0026】
好ましくは、病原性細菌は、ガリバクテリウム属に由来し、更に好ましくは、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される。
【0027】
更に、本発明のポリペプチドの存在を検出するキットが提供され、前記キットは、前記ポリペプチドと結合可能な少なくとも1つの結合タンパク質を備え、前記結合タンパク質は、固体支持部に結合される。好ましくは、前記結合タンパク質は、抗体である。
【0028】
一態様において、本発明は、本発明のポリペプチドに対する抗体を検出するキットに関し、前記キットは、固体表面に固定された前記ポリペプチドを備える。
【図面の簡単な説明】
【0029】
【図1】G. anatis培養上清の溶血活性及びGtxAの発現。A. G. anatis 12656-12の増殖及び無細胞培養上清の溶血活性。一晩培養した培養物を1:100で希釈し、増殖(600nmでの吸光度により測定した細胞密度)及び無細胞培養上清における溶血活性を記録した。BHIにおいて100倍に希釈した上清の溶血活性を示す。ウェスタンブロッティング(図5B)用の細胞外タンパク質を並行して採取した。実験を3回繰り返し、溶血活性のレベルは変化したが、相対的なパターンは一貫していた。B. ApxI抗血清でのウェスタンブロッティングにより判定したGtxAのレベル。表示した時点で上清を採取し、材料及び方法において説明したように100倍に濃縮し、3〜8%ゲルにおいてSDS-PAGEにより分離した後、ブロッティングを行った。ΔgtxAからの細胞外タンパク質をOD600 = 2で採取した(ΔgtxAと示した列)。WC = 野生型由来の全細胞溶解物: 細胞は、接種5時間後に採取した。サイズマーカーを左側に示す。実験を繰り返し、同じ結果が得られた。
【図2】G. anatis 12656-12野生型(wt)及び同質遺伝子的gtxA突然変異体(ΔgtxA)の溶血活性及び細胞毒性。A. β溶血。5%ウシ血液を含むBHI寒天プレート上に細菌をストリークし、37℃で18時間インキュベートした。B. 生理食塩水(mock)、wt、又はΔgtxAと共に1時間インキュベートした後のHD11細胞の光学顕微鏡検査(倍率100×)。細菌を後期対数期(OD600 = 1)において採取し、感染効率(MOI)を10として添加した。C. LDH活性により定量化した細胞毒性。HD11細胞を、1Bにおいて説明したように細菌と共にインキュベートした。3つの複製ウェルの平均を示しており、棒は、標準誤差(SE)を示す。
【図3】A. G. anatis 12656-12におけるgtxA、gtxC、及びその隣接遺伝子の遺伝子構成。矢印は、オープンリーディングフレームを示す。予測される転写ターミネーターを、gtxCの下流に示す。B. GtxAの構成。Kは、保存されたリジン残基(Lys1484及びLys1607)を示す。グリシン-アスパラギン酸リッチ領域(Gly-Asp-rich region, 位置1640〜1830)に印を付けている。C. GtxAのN末端ドメインにおける15の反復のアライメントであり、アライメントはRaderにより作成した[Heger A., Holm L., Rapid automatic detection and alignment of repeats in protein sequences, Proteins (2000) 41:224-237]。数字は、GtxAにおけるアミノ酸の位置を示す。アミノ酸が反復の50%以上で同一(黒)又は類似(灰色)する位置に印を付けている。
【図4】GtxAを発現する大腸菌の細胞毒性活性。A. 5%ウシ血液及び0.1 mM IPTGを含むLB寒天上で増殖させ、30℃でインキュベートした大腸菌ER2566のβ溶血活性。T1SS: + 又は - は、大腸菌T1SS成分HlyB及びHlyDを発現するプラスミドpLG575の有無を示す。RTX = GtxAのアミノ酸931〜2026であり、N-term = GtxAのアミノ酸1〜949である。B. 様々なバージョンのGtxAを発現する大腸菌ER2566/pLG575による液体溶血アッセイ及びLDH細胞毒性アッセイ。
【図5】大腸菌におけるGtxAの発現。IPTGによる誘導後の大腸菌ER2566に由来する全細胞溶解物(WC)及び細胞外タンパク質(EC)に関するウェスタンブロット。タンパク質は、4〜12%ゲルにおいてSDS-PAGEにより分離し、PVDF膜上にブロットした。ブロットは、ApxI抗血清によりプローブした。B = ブランクであり、サイズマーカーを右側に示している(PageRuler Prestained Protein Ladder Plus(Fermentas))。各列の上方のバンドは、完全長のGtxA(215 kDa)又はRTXドメイン(117 kDa)の予想サイズを有している。サイズの小さなバンドは、分解産物と思われる。
【図6】様々なガリバクテリウム菌株に由来するアミノ酸配列のアライメント。GtxAの位置1133〜1515におけるアミノ酸(SEQ ID No1)を、他のガリバクテリウム菌株に由来する他の毒素のアミノ酸配列に対して整列させている。点は、同一残基を示し、非保存位置を強調して示している。
【図7】様々なG. anatis菌株とG.genomospecies 1 及びG.genomospecies 2 の基準株(それぞれ、CCM5974及びCCM5976)とに由来する培養上清を、OD600 = 0.6において採取し、濾過滅菌した。細胞外タンパク質を方法において説明したように沈殿させ、3〜8% Tris酢酸塩SDSゲル上で分離し、PVDF膜上にブロットし、ApxIA抗血清によりプローブした。GtxA(215kDa)を矢印で示している。大きなバンドは、GtxAに関係しない未確認のタンパク質(図3参照)又はGtxAの修飾バージョンである。M = 分子サイズマーカー(Spectra Multicolor High Range Protein Ladder(Fermentas))であり、サイズを右側に示す(kDa)。
【図8】T1SS突然変異体におけるGtxAのβ溶血活性及び細胞内局在性。A. G. anatis 12656-12及び同質遺伝子的突然変異体のβ溶血。37℃で24時間インキュベート後の12656-12(wt)、gtxA、及びgtxBD突然変異体のストリークを有する血液寒天プレートの写真。ストリークの左部分では細菌を除去し、コロニーの下の溶血を明らかにした。B. GtxAの細胞内局在性。細胞及び上清は、静止期への遷移中に採取した(OD600 = 1.7)。細胞外タンパク質を方法において説明したように沈殿させた。全細胞溶解物(ペレット)及び細胞外タンパク質(上清)を3〜8% Tris酢酸塩SDSゲル上で分離し、PVDF膜上にブロットし、ApxIA抗血清によりプローブした。GtxAを矢印で示している。同一性は、質量分光測定法により検証した。抗血清により、野生型及び突然変異株の両方に存在する略270 kDaの未確認タンパク質が認識された。サイズマーカーを左側に示す(Spectra Multicolor High Range Protein Ladder (Fermentas)。
【図9】G. anatis 12656-12における遺伝子座gtxAC及びgtxEBDの構成。矢印は、オープンリーディングフレーム(ORF)を示す。gtxA及びgtxEBDの検出に使用されるプライマー対の位置は、各ORFの上に小さな矢印により示す。典型的なRTX毒素遺伝子座(Frey & Kuhnert 2002)の構成を比較のために含めている。
【0030】
定義
【0031】
本明細書で使用される「アジュバント」という用語は、投与される免疫原性決定因子/抗原/核酸構築物と混合することにより、前記決定因子に対する免疫反応を高めるか或いは他の形で修飾する物質を示す。
【0032】
本明細書で使用される「対立遺伝子変異体」という用語は、SEQ ID No. 1をコードする遺伝子の代替形態を示す。対立遺伝子は、核酸配列における少なくとも1つの突然変異により生じる場合があり、構造又は機能が変更された或いは変更されない改変mRNA又はポリペプチドを発生させる場合がある。対立遺伝子を発生させる通常の突然変異変化は、一般に、ヌクレオチドの自然欠失、追加、又は置換に起因する。こうした変化のタイプは、それぞれ単独で、或いは他のものと組み合わせて、一定の配列中に1回以上発生する場合がある。
【0033】
本明細書で使用される「抗体」という用語は、免疫グロブリン分子及び免疫グロブリン分子の活性部分を示す。抗体は、例えば、無傷の免疫グロブリン分子又は免疫学的活性を保持する免疫グロブリン分子の断片である。
【0034】
本明細書で使用される「抗原」という用語は、クローン的に分布した免疫受容体(T細胞又はB細胞受容体)に結合可能な物質を示し、通常は、ペプチド、ポリペプチド、又は多量体ポリペプチドを示す。抗原は、好ましくは、免疫反応を引き起こすことが可能である。
【0035】
本明細書で使用される「結合アッセイ」という用語は、任意の2つ以上の分子が互いに共有的又は非共有的に結合して、一方の分子の濃度の測定を可能にする、任意の生物学的又は化学的アッセイを示す。
【0036】
本明細書で使用される「生体試料」という用語は、血清、血漿、全血、唾液、尿、リンパ液、生検、精液、排泄物、涙、汗、乳、脳脊髄液、腹水、滑液から選択される任意の試料を示す。
【0037】
本明細書で使用される「担体」という用語は、抗原が結合して免疫反応の誘導を支援する存在又は化合物を示す。
【0038】
本明細書で定義される「保存的アミノ酸置換」という用語は、1個のアミノ酸が、1つ以上の共通の化学的及び/又は物理的特性を有する別のアミノ酸の代わりとなる置換を示す。アミノ酸は、共有する特性により分類し得る。保存的アミノ酸置換は、アミノ酸の所定の群内にある1個のアミノ酸による、同一群内の別のアミノ酸に対する置換であり、所定の郡内のアミノ酸は、類似又は実質的に類似する特性を示す。
【0039】
本明細書で使用される「検出部分」という用語は、別の分子と結合して検出することが可能な、好ましくはタンパク質であるが限定はされない分子の特定の部分を示す。
【0040】
本明細書で使用される「診断マーカー」という用語は、個体が有する障害を決定するために使用可能なタンパク質等の化合物の特性を示す。
【0041】
本明細書で使用される「障害」という用語は、疾患又は医学的問題を示し、特定の症状及び兆候に関連した、身体機能を害する生物の異常な状態である。侵入微生物等の外的要因により生じる場合があり、或いは内的な機能不全により生じる場合がある。
【0042】
本明細書で使用される「断片」という用語は、核酸又はポリペプチドの非完全長の部分を示す。したがって、断片自体も、それぞれ核酸又はポリペプチドとなる。
【0043】
本明細書で使用される「免疫原性」という用語は、細胞性又は液性となり得る免疫反応を誘発する、抗原又はエピトープ等の特定の物質の能力を示す。
【0044】
本明細書で使用される「薬剤(medicament)」という用語は、予防的、治療的、改善的、又は対症的な使用を対象とした、好ましくはワクチンである任意の化学物質として大まかに定義することが可能な、薬(medicine)又は薬物(medication)とも呼ばれる医薬品を示す。上述したように、本発明の用途には、必ずしも任意の疾患の100%の予防、治療、又は改善だけではなく、部分的な予防、治療、又は改善も含まれることを理解されたい。
【0045】
本明細書で使用される「病原性」という用語は、生物において感染症を発生させる、Gallibacterium anatisといった微生物等の病原体の能力を示す。
【0046】
本明細書で使用される「プラスミド」という用語は、染色体DNAから独立して複製可能な、染色体DNAから分離した一種の染色体外DNA分子を示す。
【0047】
本明細書で使用される「ポリヌクレオチド」という用語は、DNA(デオキシリボ核酸)及びRNA(リボ核酸)等、鎖状に共有結合したヌクレオチドモノマーにより構成された有機高分子を示す。
【0048】
本明細書で使用される「ポリペプチド」という用語は、少なくとも、好ましくは3個以上のアミノ酸を有するペプチドである、タンパク質としても知られる有機化合物を示す。アミノ酸という総称は、天然及び非天然アミノ酸の両方を含み、何れも「D」又は「L」異性体となり得る。
【0049】
本明細書で使用される「予防的治療」という用語は、疾患の治療又は治癒ではなく予防することを目的とした、任意の医療手順を示す。予防するという用語は、絶対的であることを意図しておらず、疾患又は疾患の1つ以上の症状の部分的予防も含む。
【0050】
本明細書で使用される「プロモーター」という用語は、1個以上の近隣の構造遺伝子によりメッセンジャーRNAの転写を開始するためにRNAポリメラーゼが結合するDNA鎖内の結合部位を示す。
【0051】
本明細書で使用される「RTX毒素」(repeats in the structural toxin)という用語は、広範な病原性グラム陰性菌により生成される膜孔形成タンパク質毒素を示す。
【0052】
本明細書で使用される「配列同一性」という用語は、2つの配列間の同一性百分率の決定を示し、数学アルゴリズムを使用して達成可能である。2配列の比較に利用される数学アルゴリズムの好適な非限定的な例は、Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87:2264-2268のアルゴリズムであり、Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877のように修正されたものである。このようなアルゴリズムは、Altschul, et al. (1990) J. Mol. Biol. 215:403-410のBLASTN及びBLASTPプログラムに組み込まれている。
【0053】
同一性を特徴付けるためには、最高次のホモロジー(一致)が得られるように対象配列を整列させる。こうした一般原則に基づき、2つの核酸配列の「同一性百分率」は、米国立バイオテクノロジー情報センター(National Center for Biotechnology Information, NCBI)ウェブサイト(http://www.ncbi.nlm.nih.gov)から入手可能なBLASTNアルゴリズム[Tatiana A. Tatusova, Thomas L. Madden: Blast 2 sequences - a new tool for comparing protein and nucleotide sequences; FEMS Microbiol. Lett. 1999 174 247-250]を使用すると共に、本明細書において提案した初期設定を使用して決定し得る(即ち、Reward for a match = 1; Penalty for a mismatch = -2; Strand option = both strands; Open gap = 5; Extension gap = 2; Penalties gap x_dropoff = 50; Expect = 10; Word size = 11; Filter on)。BLASTNアルゴリズムは、2つの整列させたヌクレオチド配列間の重複範囲における配列同一性%を決定する。Blastは、局所的アライメントであるため、長さの異なる2つの関連配列間において重複範囲内の配列同一性百分率を計算するのに最適である。
【0054】
配列の比較に利用される数学アルゴリズムの他の好適な非限定的な例は、CLUSTAL W(1.7)アライメントアルゴリズムである(Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22:4673-4680.)。CLUSTAL Wは、好ましくはBLOSUM62をスコア行列として使用して、複数の配列アライメントに用いることができる。配列同一性を計算する時、CLUSTAL Wは、参照配列の長さに、アライメントにより形成された任意のギャップを含める。配列同一性は、ギャップを有する整列配列の長さで一致の数を割ることにより計算される。
【0055】
本明細書で使用される「シグナルペプチド」という用語は、局在化ペプチドとも呼ばれる、細胞内のタンパク質の最終的な位置を決定するアミノ酸の短い配列を示す。
【0056】
本明細書で使用される「毒素」という用語は、体組織との接触又はこれによる吸収時に疾患を引き起こすことが可能な、生存細胞又は生物により生産される有毒物質を示す。
【0057】
本明細書で使用される「トキソイド」という用語は、化学処理又は熱処理により毒性は弱化又は抑制されたが、他の特性、例えば免疫原性は維持されている細菌毒素(通常は外毒素)を示す。
【0058】
本明細書で使用される「転写因子」という用語は、特定のDNA配列と結合し、これによりDNAからmRNAへの遺伝情報の転送を制御するタンパク質を示す。
【0059】
本明細書で使用される「ワクチン」という用語は、動物において免疫反応を誘発可能な物質又は組成物を示す。免疫反応は、生物の記憶を誘発し、1次反応ではなく2次反応を受けた感染性因子の宿主生物に対する影響を減じる免疫反応(液性/抗体性及び/又は細胞性)である。
【0060】
本明細書で使用される「ベクター」という用語は、外来遺伝物質を他の細胞へ転送する媒体として使用されるDNA分子を示す。
【発明を実施するための形態】
【0061】
本発明の主目的は、鳥類におけるガリバクテリウム属由来の細菌との細菌感染により生じる任意の疾患の治療及び/又は予防的治療において薬剤として用いる、Gallibacterium anatisに由来する特定のRTX毒素GtxA或いはその免疫原性変異体又は断片を含むワクチン組成物を提供することである。
【0062】
Gallibacterium anatisは、ニワトリ及び他の鳥類種の上気道及び下部生殖管の正常細菌叢の一部である。しかしながら、G. anatisは、病理学的病変からも単離されているため、潜在的病原体であると考えられる。本発明は、非定型の構造と広範な標的細胞を有する新規の病原因子、G. anatis RTX毒素を提供する。
【0063】
G. anatis培養物由来の無細胞濾過滅菌上清は、赤血球及びトリ由来のマクロファージ様細胞(HD11)の両方を溶解させ、1つ以上の外毒素の生成を示唆した。G. anatis 12656-12のゲノム配列において、我々はRTX毒素遺伝子を特定した。コード化タンパク質は、GtxA(Gallibacterium toxin、ガリバクテリウム毒素)と名付けられ、2026個のアミノ酸(aa)により構成されていた。これは、古典的な膜孔形成RTX毒素のサイズの2倍となる。GtxAのC末端の1000アミノ酸は、パスツレラ科の他の構成員におけるRTX毒素と相同であり、例えば、A. pleuropneumoniae ApxIAとの38%の配列類似性を有した。一方、N末端の略950アミノ酸は、GenBankデータベースにおける有意な一致を有していないが、機能が未知である57アミノ酸の反復を11個含む。gtxA及びそのアセチルトランスフェラーゼ活性化因子gtxCを発現する大腸菌は、溶血性及び白血球毒性となった。GtxAの様々な切断バージョンの機能を調査した。C末端RTXドメインは、無傷の毒素よりも低い溶血活性を示し、N末端が、不可欠ではないが、最大の溶血活性に必要であることを示唆した。HD11細胞に対する細胞毒性は、C末端単独では検出されず、新規のN末端反復ドメインが白血球に対する細胞毒性作用にとって不可欠となることを示唆した。
【0064】
G. anatisのgtxAの発現は、ウェスタン及びノーザンブロッティングにより調査した。GtxAは、増殖期に依存する形で細胞外タンパク質画分において検出されたが、細胞随伴タンパク質画分においては検出されず、予測される毒素の分泌と一致した。
【0065】
11の多様な遺伝子型及び表現型のガリバクテリウム菌株を、gtxAの存在及び発現、GtxA分泌レベル、及び培養上清の溶解活性について調査した。gtxAは、広く分布しており、Gallibacterium genomospecies 1及びGallibacterium genomospecies 2(図6、アライメント)を含む、全ての試験株に見られた。発現には菌株間で実質的に幅があり、非病原性非溶血型菌株F149Tは、少量を発現した。上清中のGtxAレベルは、HD11細胞に対する溶血活性及び細胞毒性活性のレベルとある程度の相関を有した。
【0066】
我々は、GtxAがG. anatisの病原性に大きく寄与すると予測する。
【0067】
GtxA
【0068】
これまで説明されていないが、GtxAは、2026個のアミノ酸の大きなポリペプチドであり、215 kDaの分子量を備え、SEQ ID No. 1を有する。GtxAは、C末端及びN末端断片に分割し得る。C末端断片の例は、1,077個のアミノ酸(SEQ ID No. 2)から成り、6つの縦列反復するノナペプチドを特徴とする古典的なRTX毒素に類似する。N末端の例は、949個のアミノ酸(SEQ ID No. 3)から成り、相対的に疎水性であり、他のRTX毒素又はGenBankの他のタンパク質との配列類似性を殆ど共有しない。毒素活性は、GtxAの脂肪酸アシル化を促進する活性化因子GtxCに依存しており、即ち、毒性は、転写後のポリペプチドのアシル化に依存している。非アシル化タンパク質は、毒性を有しない。
【0069】
GtxAは、主に溶血性及び白血球毒性である細胞溶解性の表現型を示す。C末端RTXドメインは、無傷の毒素よりも低い溶血活性を示し、N末端が、不可欠ではないが、最大の溶血活性に必要であることを示す。トリ由来マクロファージ様HD11細胞に対する細胞毒性は、C末端単独では検出されず、新規のN末端反復ドメインが白血球に対する細胞毒性作用にとって不可欠であることを示唆した。
【0070】
異なる地理的領域(デンマーク、チェコ共和国、及びメキシコ)からのガリバクテリウム菌株を、gtxA遺伝子の存在について選別したところ、全ての試験菌株に見られたが、個別の菌株間には発現に実質的な変動が存在した。この変動は、地理的起源には無関係であると思われる。
【0071】
G. anatisは、ニワトリ、産卵雌鶏、及び他の鳥類種の上気道及び下部生殖管の正常細菌叢の一部である。しかしながら、G. anatisは、病理学的病変からも単離されており、G. anatisは、家禽の病変形成において重要な役割を果たすと考えられる。
【0072】
したがって、本発明の目的は、疾患の治療及び/又は予防用の薬剤として用いる、GtxAタンパク質又はその免疫活性ポリペプチド変異体に由来するトキソイドワクチンを提供することである。前記疾患は、温血動物における細菌感染により生じる場合があり、本発明の他の目的は、前記疾患を予防又は治療することである。本発明の他の態様は、疾患の治療及び/又は予防用のGtxAタンパク質をコードするポリヌクレオチド又は免疫活性ポリペプチド変異体をコードするポリヌクレオチドに関する。
【0073】
GtxAポリペプチドは、図6の部分アミノ酸配列のアライメントにより示したように、様々なGallibacterium anatis分離株全体での保存性が高い。そのため、GtxAトキソイドを含むワクチン組成物は、多数の異なるG. anatis分離株に対して、更には、ガリバクテリウム属の関連種に対しても有効となることが予想される。
【0074】
細菌種
【0075】
本発明は、ポリペプチド、ポリヌクレオチド、及びポリヌクレオチド保持発現ベクターに関する。一実施形態において、ポリヌクレオチド及びポリペプチドは、Gallibacterium anatisに由来する。加えて、本発明は、パスツレラ科の細菌、更に好ましくは、ガリバクテリウム属からの細菌、最も好ましくは、Gallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される細菌に由来するGtxAポリペプチド及びポリヌクレオチドを対象とする。
【0076】
GtxAは、他のRTX毒素とは大きく異なることが分かった。本明細書に記載した分子ツールにより、本発明者は、ガリバクテリウム属由来の他のGtxA様毒素のクローニング及びプロービングを可能にした。
【0077】
こうした関連種由来のGtxA様毒素は、SEQ ID NO 1、2、又は3との一定の度合いの配列相同性を示すことが予想され、コード配列は、本発明のポリヌクレオチドに基づくプローブとハイブリダイズすることが予想される。
【0078】
遺伝子組み換え株
【0079】
実施例2において説明されるように、本発明者は、G. anatis gtxA突然変異株を作成した。菌株は、ΔgtxAと命名した。実施例7は、こうしたG. anatis ΔgtxA突然変異体による感染試験を説明しており、野生型微生物に感染させたトリは、通常、野外でのG. anatisへの自然感染により観察される病変に対応する、生殖器系及び腹膜に関与する播種性及び化膿性の炎症を生じた。一方、ΔgtxA突然変異体に感染させたトリは、卵巣に局在する、より軽度の炎症を生じた。したがって、gtxAが、ニワトリにおけるG. anatisの病変形成に実質的に寄与することが実証された。故に、本発明者は、本明細書において上で定義したΔgtxA株を使用して生物への免疫付与が可能であることを実証したと結論付けることができる。このように免疫された鳥類種などの生物は、gtxAを発現する野生型微生物に対する免疫を発生させることができるが、それは、特定の病原性微生物の表面等に存在する非gtxA抗原に対して生成された抗体を介してであり、当該微生物はgtxAの発現及び/又は分泌を無効化した微生物と同じ種である。
【0080】
したがって、一態様において、本発明は、内因性gtxA遺伝子を破壊して、機能性gtxAポリペプチドの発現を無効化したトランスジェニックノックアウト微生物に関し、前記微生物は、非トランスジェニック対照微生物と比較して病原性の減少を示す。
【0081】
一実施形態において、微生物は、Gallibacterium anatisである。
【0082】
他の実施形態において、トランスジェニック微生物は、抗生物質耐性を有していない。
【0083】
GtxAポリペプチド
【0084】
野生型GtxAの1つ、即ち、天然に存在する突然変異のないバージョンのタンパク質は、SEQ ID No. 1において特定される。
【0085】
一実施形態において、本発明は、単離ポリペプチドであって、以下からなる群から選択されるアミノ酸配列を含む、ポリペプチドに関する:
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【0086】
他の野生型GtxAポリペプチドを、他のガリバクテリウム種及びGallibacterium anatisの他の分離株から単離することが可能である。こうしたGtxAは、図6の断片のアライメントに示すように、SEQ ID No. 1、2、及び/又は3との高い度合いの配列同一性を共有することが予想される。
【0087】
好適な実施形態において、本発明は、SEQ ID No. 1と、SEQ ID No. 1に対して少なくとも70%の配列同一性、より好ましくは、GtxA配列との75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性、例えば少なくとも99.5%の配列同一性、例として少なくとも99.9%の配列同一性を備えたGtxAの配列変異体とに関する。
【0088】
他の好適な実施形態において、本発明は、SEQ ID No. 2において定義される、GtxAポリペプチドのC末端ドメインと、SEQ ID No. 2に対して少なくとも70%の配列同一性、より好ましくは、GtxA配列のC末端ドメインとの75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を備えた配列変異体とに関する。
【0089】
他の好適な実施形態において、本発明は、SEQ ID No. 3において定義される、GtxAポリペプチドのN末端ドメインと、SEQ ID No. 3に対して少なくとも70%の配列同一性、より好ましくは、GtxA配列のN末端ドメインとの75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を備えた配列変異体とに関する。
【0090】
完全長のGtxAに加えて、本発明は、GtxAの断片に関する。例えば、C末端GtxAドメイン及びN末端GtxAドメインに関する。加えて、本発明は、こうしたポリペプチドの断片に関する。好適な実施形態において、前記断片は、少なくとも150個の隣接アミノ酸、好ましくは少なくとも200個のアミノ酸、より好ましくは少なくとも300個のアミノ酸、より好ましくは少なくとも500個のアミノ酸、より好ましくは少なくとも750個のアミノ酸、より好ましくは少なくとも1000個のアミノ酸、より好ましくは少なくとも1250個のアミノ酸、より好ましくは少なくとも1500個のアミノ酸、より好ましくは少なくとも1750個のアミノ酸、より好ましくは少なくとも2000個のアミノ酸からなる。
【0091】
GtxA断片は、1つ以上の位置において、野生型GtxA配列と異なっていてよい。好適な実施形態において、前記断片は、30個までのアミノ酸置換、より好ましくは25個までの置換、より好ましくは20個までの置換、より好ましくは15個までの置換、より好ましくは10個までの置換、より好ましくは5個までの置換、例として4個、3個、2個、又は1個の置換を含み得る。
【0092】
本発明が対象とする他の変異体は、保存的アミノ酸置換が生じた、SEQ ID No. 1、2、及び3のポリペプチドの変異体に関する。好適な実施形態において、本発明は、ポリペプチド配列内の任意のアミノ酸が他のアミノ酸に保存的に置換された、SEQ ID No. 1、2、又は3を含む任意のポリペプチドに関する。
【0093】
他の好適な実施形態において、前記配列変異体及び断片は、免疫原性を有する。更に他の好適な実施形態において、前記配列変異体及び断片は、毒性等の生物活性を保持し、前記毒性は、受容者の細胞膜での孔の形成を含み、例えば細胞毒性、例として細胞溶解性細胞毒性、例えば溶血性細胞毒性を含む。
【0094】
本発明は、更に、SEQ ID No. 1、2、及び3のポリペプチドに関し、前記ポリペプチドは、例えば毒性等の生物活性は除去するが、例えば免疫原性等の活性は維持するように特異的に修飾されている。そのため、好適な実施形態において、SEQ ID No. 1、2、及び3のポリペプチドは、好ましくは、熱又は放射線により、更に好ましくは、非アシル化形態で発現させることにより、更に好ましくは、例えばホルムアルデヒド等の化学物質に曝露することにより不活性化される。
【0095】
他の好適な実施形態において、本発明は、シグナルペプチドが異種シグナルペプチドに置き換えられた、SEQ ID No. 1、2、又は3を含む任意のポリペプチドに関する。
【0096】
精製の目的で、本発明には、タグを付けてもよい。好適な実施形態において、SEQ ID No. 1、2、及び3は、親和性タグ、好ましくは、切断可能タグ、例としてポリHisタグ、例えばHAタグ、例としてFLAGタグ、例えばC-mycタグ、例としてHSVタグ、例えばV5タグ、例としてマルトース結合タンパク質タグ、例えばセルロース結合ドメインタグ、例としてBCCPタグ、例えばカルモジュリンタグ、例としてNusタグ、例えばグルタチオン-S-トランスフェラーゼタグ、例として緑色蛍光タンパク質タグ、例えばチオレドキシンタグ、例としてSタグ、例えばStrepタグによりタグ付けし得る。
【0097】
好ましくは、タグは、C末端の一番端など、タンパク質のC末端部分に存在する。より好ましくは、タグは、タグとRTXポリペプチドとの間に挿入されたプロテアーゼ切断部位を有することにより、GtxAポリペプチドから切断可能となる。
【0098】
GtxAポリヌクレオチド
【0099】
本発明により、SEQ ID No. 4、5、及び6において特定のポリヌクレオチド配列が定められる。
【0100】
本発明は、単離ポリヌクレオチドであって、以下からなる群から選択される核酸配列を含む、ポリヌクレオチドに関する:
a) SEQ ID No. 4、5、又は6と、
b) SEQ ID No. 4、5、及び6からなる群から選択されるポリヌクレオチドの配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも60%である配列変異体と、
c) a)の何れかの少なくとも450個の隣接ヌクレオチドからなる断片であり、選択される配列において指定された任意の核酸が異なる核酸へ変化するのは、このように変化する配列内の核酸が90個以内の場合である断片と、
d) 高ストリンジェンシー下で、SEQ ID No. 4、5、又は6に相補的なポリヌクレオチドとハイブリダイズすることが可能なポリヌクレオチドと、
e) SEQ ID No. 1、2、又は3のポリペプチドをコードするポリヌクレオチドと、
f) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体をコードするポリヌクレオチドと、
g) SEQ ID No. 1、2、又は3の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片をコードするポリヌクレオチド。
【0101】
ヌクレオチドプローブと相同DNA又はRNA配列との間のハイブリダイゼーションを決定するための適切な実験条件は、ハイブリダイズするDNA断片又はRNAを含有するフィルタを5×SSC[塩化ナトリウム/クエン酸ナトリウム、Sambrook et al.; Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Lab., Cold Spring Harbor, NY 1989参照]に10分間予め浸漬すること、及び5×SSC、5×デンハルト溶液[Sambrook et al.; 同上参照]、0.5%SDS、及び変性超音波処理サケ精子100 μg/mL [Sambrook et al.; 同上参照]の溶液におけるフィルタのプレハイブリダイゼーションを含み、その後、10 ng/mLの濃度のランダムプライム[Feinberg A P & Vogelstein B; Anal. Biochem. 1983 132 6-13]32P-dCTP標識(比活性度>1×109 cpm/μg)プローブを含有する同じ溶液中で、12時間略45℃でハイブリダイゼーションを行う。その後、フィルタを0.1×SSC、0.5%SDSにおいて、少なくとも60℃(中ストリンジェンシー条件)、好ましくは少なくとも65℃(中/高ストリンジェンシー条件)、より好ましくは少なくとも70℃(高ストリンジェンシー条件)、更により好ましくは少なくとも75℃(超高ストリンジェンシー条件)の温度で、30分間2回洗浄する。こうした条件下でオリゴヌクレオチドプローブがハイブリダイズした分子は、X線フィルムを用いて検出し得る。
【0102】
一実施形態において、核酸分子は、SEQ ID No. 4、5、及び6からなる群から選択される核酸配列と単一のヌクレオチドにおいて異なっている。単一の置換は、サイレント突然変異であってよく、或いは、保存的アミノ酸置換を発生させてもよい。単一の置換又は欠失は、フレームシフト突然変異を発生させてもよい。より好適な実施形態において、本発明は、SEQ ID No. 4のポリヌクレオチドとの配列同一性が少なくとも60%、より好ましくはSEQ ID No. 4との配列同一性が少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%であるポリヌクレオチド配列に関する。
【0103】
他の好適な実施形態において、本発明は、SEQ ID No. 5のポリヌクレオチドとの配列同一性が少なくとも60%、より好ましくはSEQ ID No. 5との配列同一性が少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%であるポリヌクレオチド配列に関する。
【0104】
他の好適な実施形態において、本発明は、SEQ ID No. 6のポリヌクレオチドとの配列同一性が少なくとも60%、より好ましくはSEQ ID No. 6との配列同一性が少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%であるポリヌクレオチド配列に関する。
【0105】
本発明は、更に、SEQ ID No. 1、2、及び3のポリヌクレオチドの断片に関する。好適な実施形態において、前記断片は、少なくとも450個の隣接ヌクレオチド、より好ましくは少なくとも500個の隣接ヌクレオチド、より好ましくは少なくとも600個の隣接ヌクレオチド、より好ましくは少なくとも750個の隣接ヌクレオチド、より好ましくは少なくとも1000個の隣接ヌクレオチド、より好ましくは少なくとも1500個の隣接ヌクレオチド、より好ましくは少なくとも2000個の隣接ヌクレオチド、より好ましくは少なくとも2500個の隣接ヌクレオチド、より好ましくは少なくとも3000個の隣接ヌクレオチド、より好ましくは少なくとも3500個の隣接ヌクレオチド、より好ましくは少なくとも4000個の隣接ヌクレオチド、より好ましくは少なくとも4500個の隣接ヌクレオチド、より好ましくは少なくとも5000個の隣接ヌクレオチド、より好ましくは少なくとも5500個の隣接ヌクレオチド、より好ましくは少なくとも6000個の隣接ヌクレオチドからなる。
【0106】
ポリヌクレオチド断片は、1つ以上の位置において、前記断片が由来する野生型GtxAポリヌクレオチド配列と異なっていてよい。好適な実施形態において、前記断片は、90個までのヌクレオチド置換、より好ましくは80個までの置換、より好ましくは70個までの置換、より好ましくは60個までの置換、より好ましくは50個までの置換、より好ましくは40個までの置換、より好ましくは30個までの置換、より好ましくは20個までの置換、より好ましくは10個までの置換、より好ましくは5個までの置換、例として4個、3個、2個、又は1個の置換を含み得る。
【0107】
本発明は、好ましくは高ストリンジェンシーのハイブリダイズ条件下でSEQ ID No. 4、5、及び6の配列を有するポリヌクレオチドとハイブリダイズすることが可能なポリヌクレオチドに関する。
【0108】
本発明のポリヌクレオチドは、天然に存在する対立遺伝子核酸変異体のヌクレオチド配列を含み得る。
【0109】
本発明のポリヌクレオチドは、更に、SEQ ID No. 4、5、及び6の変異体を含んでよく、前記ポリヌクレオチドは、大腸菌での発現に最適化されている。
【0110】
発現ベクター
【0111】
本発明のポリヌクレオチドは、発現ベクター又はクローニングベクター等、任意の適切なベクター内に含まれてもよい。多数のベクターが利用可能であり、特定の目的に適した任意のベクターを選択し得る。適切な核酸配列を、様々な手順によりベクターに挿入してよく、例えば、DNAは、当技術分野において周知の手法を使用して、適切な制限エンドヌクレアーゼ部位(群)に挿入し得る。本発明に関する核酸配列とは別に、ベクターは、更に、シグナル配列、複製開始点、1つ以上のマーカー遺伝子、エンハンサーエレメント、プロモーター、及び転写終結配列を1つ以上含み得る。ベクターは、更に、エンハンサー、ポリA尾部、リンカー、ポリリンカー、操作リンカー(operative linker)、多重クローニング部位(multiple cloning sites, MCS)、終止コドン、内部リボソーム侵入部位(internal ribosomal entry sites, IRES)、及び組込み用の宿主相同配列といった付加的な配列又は他の定義済みのエレメントを含み得る。ベクターは、好ましくは、発現ベクターであり、適切な細胞においてその発現を指揮する調節核酸配列と適切に操作可能に結合した核酸を含む。
【0112】
好適な実施形態において、本発明のベクターは、真核生物プラスミドベクター等のプラスミドベクターであり、より好ましくは原核生物プラスミドベクターである。
【0113】
別の好適な実施形態において、ベクターは、好ましくはレトロウイルス科に由来するウイルスベクターであり、例としてレンチウイルス、例えばHIV、例としてSIV、例えばEAIV、例としてCIVである。
【0114】
他の好適な実施形態において、ベクターは、限定はされないが、アルファウイルス、アデノウイルス、アデノ随伴ウイルス、バキュロウイルス、HSV、コロナウイルス、ウシパピローマウイルス、Mo-MLVからなる群から選択してよく、好ましくはアデノ随伴ウイルスにしてよい。
【0115】
他の好適な実施形態において、本発明のベクターは、プロモーターを含み、より好ましくは、前記プロモーターは、本発明のポリヌクレオチドと適切に操作可能に結合される。
【0116】
好適な実施形態において、前記プロモーターは、限定はされないが、原核生物プロモーターから選択され、好ましくは、原核生物プロモーターは、他のエレメントを含み、例としてRNAポリメラーゼ結合部位、例えばプリブノーボックス又はその一部、例として-35エレメント又はその一部を含む。本発明の原核生物ベクターは、例えば、大腸菌における、不活性化GtxAの組み換え発現に使用することができる。大腸菌は、GtxC遺伝子を含まないため、大腸菌において発現させたGtxAは、適切にアシル化されず、結果的に無毒性となる。
【0117】
別の好適な実施形態において、前記プロモーターは、限定はされないが、真核生物プロモーターから選択され、好ましくは、真核生物プロモーターは、他のエレメントを含み、例としてRNAポリメラーゼ結合部位、例えばTATAボックス又はその一部、例として任意の真核細胞転写因子のための少なくとも1つの結合部位を含む。
【0118】
本発明のベクターの好適な実施形態は、裸DNAワクチンであり、本発明のポリヌクレオチドと適切に操作可能に結合した真核生物プロモーターを含む。
【0119】
ワクチン
【0120】
一般に、ワクチンは、機能的免疫系を備えた生体検体において免疫反応を誘発可能な物質又は組成物である。組成物は、以下を1つ以上含み得る: 抗原等(例えば、タンパク質、ポリペプチド、ペプチド、核酸等)の「活性成分」、他のエレメントと共に1つ以上の抗原を含む核酸構築物、細胞(例えば、負荷抗原提示細胞(loaded APC)、養子移入用のT細胞)、複合分子(抗体、T細胞受容体(TCRs)、及びMHC複合体等)、担体、アジュバント、及び医薬担体。本発明は、Gallibacterium anatis由来の本発明の単離ポリペプチド、好ましくは前記ポリペプチドの不活性化形態、或いはその免疫原性変異体又は断片を含むワクチン組成物に関する。本明細書で使用される「ワクチン」という用語は、Gallibacterium anatisを起源とする疾患に対する特定の免疫を誘発することを目的とした家畜ワクチンを示す。
【0121】
本発明はワクチン組成物に関し、当該ワクチン組成物は、SEQ ID No. 1、2、又は3のポリペプチド、前記ポリペプチドの不活性化形態、前記ポリペプチドの機能的ホモログ、少なくとも70%の配列同一性を有するポリペプチド、又は前記ポリペプチドの免疫原性活性断片を含む。前記ワクチンは、トキソイドワクチンと呼ばれる。トキソイドワクチンは、毒性を失っているが免疫原性を保持している毒素を使用して標的生物において免疫反応を誘発するワクチンであり、前記標的生物は、同一又は類似の細菌種を起源とする前記毒素又は類似毒素による将来の感染に対して耐性となる。
【0122】
好適な実施形態において、前記ワクチンは、毒性に関して不活性化されているが、免疫原性を維持している不活性化ポリペプチドを含み、好ましくは、熱により、更に好ましくは、ホルムアルデヒド等の化学物質への曝露により、更に好ましくは、非アシル化形態でSEQ ID No. 1のポリペプチドを発現させることにより、不活性化される。
【0123】
他の好適な実施形態において、前記ワクチンは、更に、不活性化されたか又は生存する、弱毒化Gallibacterium anatisを含む。
【0124】
更に他の実施形態において、本発明は、更に、鳥類種に対する病原性を有するウイルス又は微生物由来の少なくとも1つの他の抗原に関し、前記ウイルス又は微生物は、限定はされないが、伝染性気管支炎ウイルス、ニューカッスル病ウイルス、伝染性ファブリキウス嚢病ウイルス、ニワトリ貧血因子、トリレオウイルス、トリニューモウイルス、ニワトリポックスウイルス、トリ脳脊髄炎ウイルス、マイコプラズマ・ガリセプチカム(Mycoplasma gallisepticum)、ヘモフィルス・パラガリナルム(Haemophilus paragallinarum)、パスツレラ・マルトシダ(Pasteurella multocida)、及び大腸菌(Eschericia coli)からなる群から選択される。
【0125】
毒素等の抗原が宿主生物に導入される時、通常、免疫反応を高めるために付加的な成分が使用される。こうした成分は、一般に、アジュバントと呼ばれる。本発明のワクチン組成物は、好ましくは、アジュバント及び/又は担体を含む。アジュバントは、ワクチン組成物との混合がGtxAポリペプチド又はその免疫原性断片に対する免疫反応を高めるか或いは他の形で修飾する任意の物質である。担体は、GtxAポリペプチド又はその免疫原性断片を結び付けることが可能な、抗原の提示を支援する足場構造、例えば、ポリペプチド又は多糖である。
【0126】
したがって、好適な実施形態において、本発明のワクチン組成物は、限定はされないが、フロイント完全及び不完全アジュバント、ビタミンE、非イオン性ブロック重合体、ムラミルジペプチド、Quil A、鉱油及び非鉱物油、野菜油、及びCarbopolの群から選択されるアジュバント及び/又は担体を含む。本発明のワクチンは、Span又はTween等の乳化剤を含み得る。
【0127】
本発明のワクチン組成物は、幾つかの経路、例えば筋肉内又は皮下注射により、例えば食物又は水を介した経口投与により、例としてエアロゾルにより、例えば足又は翼の膜部における乱切により、例として点眼剤により、例えば卵内投与により投与し得る。
【0128】
トリ
【0129】
パスツレラ科の殆どの構成員は、温血動物、好ましくはトリの粘膜において、片利共生生物として生存している。ガリバクテリウム属の構成員は、共生及び病原性菌株を含み、前記病原菌株は、主に、鳥類宿主の気道及び生殖器系において疾患を引き起こす。
【0130】
好適な実施形態において、本発明は、温血動物、より好ましくは、鳥類種、例えば、マガモ属(Anas)、マガン属(Anser)、ハジロ属(Aythya)、ニオイガモ属(Biziura)、コクガン属(Branta)、ハクチョウ属(Cygnus)、アカメカモメ属(Creagrus)、ハシブトアジサシ属(Gelochelidon)、カモメ属(Larus)、ゾウゲカモメ属(Pagophila)、クビワカモメ属(Xemaes)、コウノトリ科(Ciconiidae)、カワラバト属(Columba)、スズメバト属(Columbina)、ミカドバト属(Ducula)、ヒムネバト属(Gallicolumba)、チョウショウバト属(Geopelia)、アメリカウズラバト属(Geotrygon)、カンムリバト属(Goura)、ミヤマバト属(Gymnophaps)、ニュージーランドバト属(Hemiphaga)、アメリカシャコバト属(Leptotila)、ウォンガバト属(Leucosarcia)、オナガバト属(Macropygia)、ハシリバト属(Metriopelia)、レンジャクバト属(Ocyphaps)、シッポウバト属(Oena)、Patagioenas属(Patagioenas)、テリアオバト属(Phapitreron)、ヒメアオバト属(Ptilinopus)、インカバト属(Scardafella)、キジバト属(Streptopelia)、アオバト属(Treron)、アオフバト属(Turtur)、ハジロバト属(Zenaida)、エボシツカツクリ属(Aepypodius)、ヤブツカツクリ属(Alectura)、キジ科(Phasianidae)、ライチョウ亜科(Tetraoninae)、ペリカン科(Pelecanidae)、フラミンゴ科(Phoenicopteridae)、オウム科(Cacatuidae)、ヒインコ科(Loriidae)、インコ科(Psittacidae)、エミュー科(Dromaiidae)、ダーウィンレア属(Pterocnemia)、レア属(Rhea)、ダチョウ科(Struthionidae)、などにおける、細菌感染の治療用としての本発明のポリペプチド、本発明のポリヌクレオチド、及び本発明のベクターに関する。
【0131】
更に好適な実施形態において、本発明のポリペプチド、本発明のポリヌクレオチド、及び本発明のベクターは、カモ、シチメンチョウ、及びニワトリからなる群から選択される鳥類種、より好ましくは産卵雌鶏における、細菌感染の治療において使用される。
【0132】
抗体
【0133】
本発明のポリペプチド又はその免疫原性断片の存在を検出するために、前記ポリペプチド又はその免疫原性断片と特異的に結合することが可能な抗体を生成することが有用である。前記抗体は、前記ポリペプチド上の任意のエピトープと結合し得る。
【0134】
好適な実施形態において、前記抗体は、血清由来のポリクローナル抗体或いはモノクローナル又は組み換え抗体としてよく、前記抗体は、Fv、scFv、Fab、Fab’、又はF(ab)2等の抗体の抗原結合断片、二量体IgA分子又は五価IgM等の多量体形態、アフィボディ(affibodies)、又は二重特異性抗体(diabodies)を含む。
【0135】
好適な実施形態において、本発明は、IgA抗体、最も好ましくは、ニワトリIgA抗体に関する。
【0136】
したがって、好適な実施形態において、本発明は、SEQ ID No. 1、2、又は3のポリペプチドと特異的に結合することが可能な抗体に関する。
【0137】
他の実施形態において、本発明は、SEQ ID No. 1に対して少なくとも70%の配列同一性、より好ましくはSEQ ID No. 1に対して75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を有するポリペプチドと結合することが可能な抗体に関する。
【0138】
更に好適な実施形態において、本発明は、SEQ ID No. 2に対して少なくとも70%の配列同一性、より好ましくはSEQ ID No. 2に対して75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を有するポリペプチドとの結合が可能な抗体に関する。
【0139】
他の好適な実施形態において、本発明は、SEQ ID No. 3に対して少なくとも70%の配列同一性、より好ましくはSEQ ID No. 3に対して75%の配列同一性、例えば少なくとも80%の配列同一性、例として少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例として少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例として少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例として少なくとも99%の配列同一性を有するポリペプチドと結合することが可能な抗体に関する。
【0140】
他の好適な実施形態において、本発明は、SEQ ID No. 1、2、及び3のポリペプチドの何れかの免疫原性断片を含むポリペプチドと結合することが可能な抗体に関し、前記断片は、少なくとも150個の隣接アミノ酸、好ましくは少なくとも200個のアミノ酸、より好ましくは少なくとも300個のアミノ酸、より好ましくは少なくとも500個のアミノ酸、より好ましくは少なくとも750個のアミノ酸、より好ましくは少なくとも1000個のアミノ酸、より好ましくは少なくとも1250個のアミノ酸、より好ましくは少なくとも1500個のアミノ酸、より好ましくは少なくとも1750個のアミノ酸、より好ましくは少なくとも2000個のアミノ酸からなる。
【0141】
更に好適な実施形態において、本発明は、免疫原性断片を含むポリペプチドと結合することが可能な抗体に関し、前記断片は、30個までのアミノ酸置換、より好ましくは25個までの置換、より好ましくは20個までの置換、より好ましくは15個までの置換、より好ましくは10個までの置換、より好ましくは5個までの置換、例として4個、3個、2個、又は1個の置換を含み得る。
【0142】
診断
【0143】
診断試験キットは、本発明に関連する診断の方法を実行するための全ての構成要素を集めたものである。
【0144】
好適な実施形態において、本発明は、ガリバクテリウム属由来の細菌種の存在により生じる細菌感染の兆候を生体試料において検出する試験キットに関する。前記兆候は、SEQ ID No. 1、2、又は3の任意のポリペプチド、又はその機能性変異体の存在、又は前記ポリペプチドの何れかに対する抗体の存在としてよい。
【0145】
好適な実施形態において、前記ポリペプチド又は抗体の存在は、酵素結合免疫吸着測定法(ELISA)により検出し得る。ELISAは、試料中のタンパク質又は他の任意の抗原の存在を検出するために使用される定量的手法である。ELISAでは、未知の量の抗原を表面に付着させ、その後、特異的抗体を表面上に流して、抗原と結合可能にする。この抗体は、酵素と結合しており、最終段階において、当該酵素により何らかの検出可能な信号に転換可能な物質を添加する。
【0146】
幾つかの種類のELISAが存在している:
間接ELISA法
サンドイッチELISA法
競合ELISA法
リバースELISA法
【0147】
更に、化学発光免疫測定法及び解離増強型ランタニド免疫測定法(Dissociation-Enhanced Lanthinide Immunoassays)等、他の免役に基づく測定法を使用して、試料中の前記ポリペプチド又は前記抗体を検出し得る。
【0148】
本発明は、更に、本発明の任意のポリヌクレオチド又はGallibacterium anatis GtxAに特異的な他の特異的DNA又はRNA配列の存在を検出するための診断試験キットに関する。
【0149】
したがって、好適な実施形態において、本発明は、前記ポリヌクレオチドを検出するポリメラーゼ連鎖反応(PCR)又はリアルタイム(RT)-PCR法に関する。PCRは、単一又は数個のDNA片を数桁に渡り増幅し、特定のDNA配列の数千から数百万の複製を生成し、最終的に検出する手法である。当該方法は、DNAの溶解のための反応液の加熱及び冷却並びにDNAの酵素的複製を反復するサイクルからなる温度サイクリングに依存している。標的領域に対して相補的な配列を含むプライマー(短いDNA断片)及びDNAポリメラーゼは、選択的な反復増幅を可能にする主要な成分となる。PCRが進行すると共に、生成されたDNA自体が複製のテンプレートとして使用され、DNAテンプレートを指数関数的に増幅させる連鎖反応が発生する。
【0150】
ポリペプチド、ポリヌクレオチド、及び発現ベクターの医学的使用
【0151】
トキソイドワクチンとしての不活性化毒素は、主に獣医用として、好ましくは家禽群の治療用として、細菌感染の治療及び/又は予防的治療において一般的に使用される。
【0152】
トキソイドワクチンでは、侵入細菌ではなく、侵入細菌により生産される毒素に対抗するように免疫系を条件付けることが目標となる。
【0153】
したがって、本発明のGtxAポリペプチド、GtxAポリペプチドをコードするポリヌクレオチド、又は発現ベクターは、GtxA毒素及び/又は類似する毒素を分泌する細菌により生じた発症状態(pathogenic conditions)の治療及び/又は予防的治療用のトキソイドワクチンを生成するために使用し得る。
【0154】
特定の実施形態において、トリに対して受動免疫をもたらす投与量は、ワクチン1用量当たり略0.25 mLである。他の実施形態において、投与量は、ワクチン1用量当たり約0.4 mLである。更に他の実施形態において、投与量は、ワクチン1用量当たり約0.6 mLである。一実施形態において、投与量は、ワクチン1用量当たり約0.5 mLである。一実施形態において、鳥類は、限定はされないが、カモ、シチメンチョウ、及びニワトリからなる群から選択される。他の好適な実施形態において、鳥類は、産卵雌鶏である。
【0155】
本発明は、更に、他のワクチンを組成物に含めることにより多数の疾患に対して鳥類を保護するために、本発明のワクチンを鳥類に投与する方法を提供する。
【0156】
本発明は、GtxA毒素に対する能動免疫をもたらすための、トリ、好ましくはカモ、シチメンチョウ、及びニワトリ、より好ましくは産卵雌鶏のワクチン接種を含む。ニワトリ及び/又は家禽は、約1日齢又はこれより僅かに高齢となる若年期に(1週齢以内)、1又は2用量のワクチンによりワクチン接種を行う。能動免疫を達成するために適切なワクチン投与量は、約0.05 mL〜約0.5 mLで変化する場合がある。特定の実施形態において、ワクチン投与量は、略0.05 mL〜略0.1 mLである。
【0157】
各トリは、例えば1〜3ヶ月以内に2回等、複数回ワクチン接種を行う必要があると予想される。例えば、産卵雌鶏の生涯に渡る保護には、毎年の再接種を要する場合がある。
【0158】
更に他の実施形態において、本発明の毒素又はその免疫原性断片をトリに投与することが可能である。更に、本発明のワクチンとして使用されるGtxA毒素又はその免疫原性断片の投与量の範囲は、1用量当たり約1μg/kg体重〜約1000μg/kg体重となる場合があり、一例として、1動物に対して1用量当たり約10μg/kg体重〜約100μg/kg体重の範囲となる。
【0159】
【表1】
【0160】
【表2】
【実施例】
【0161】
実施例1: Gallibacterium anatis由来のGtxA、新規のドメイン構成を有する細胞溶解性RTX毒素
【0162】
要約
【0163】
Gallibacterium anatisは、ニワトリ及び他の鳥類種において卵管炎及び腹膜炎の大きな原因となる病原体である。我々は、溶血性のG. anatis次亜種ヘモリチカからの細菌細胞及び無細胞濾過滅菌培養上清がトリ由来マクロファージ様細胞(HD11)に対して高い細胞毒性を有することを見いだした。G. anatis 12656-12のゲノム配列を入手し、合理的手法を用いて2026個のアミノ酸のRTX毒素をコードすると予測される遺伝子を特定し、GtxA(Gallibacterium toxin、ガリバクテリウム毒素)と名付けた。gtxAノックアウト突然変異体の作成により、gtxAがG. anatisの溶血及び白血球毒性活性の原因となることが示された。更に、gtxA及び隣接するアシルトランスフェラーゼgtxCを発現する大腸菌は、細胞溶解性となった。GtxAは、in vitro増殖中に発現され、増殖期に依存する形で細胞外タンパク質画分に局在した。GtxAは、通常とは異なるモジュラー構造を有し、GtxAのC末端の1000個のアミノ酸は、パスツレラ科の他の構成員における古典的な膜孔形成RTX毒素と相同であった。一方、N末端の略950個のアミノ酸は、配列データベースとの有意な一致は殆どなかった。切断GtxAタンパク質の発現により、C末端RTXドメインが完全長の毒素よりも低い溶血活性を有することが実証され、N末端ドメインは最大の溶血活性に必要であることが示唆された。HD11細胞に対する細胞毒性は、C末端単独では検出されず、N末端ドメインが白血球毒性において決定的な役割を果たすことが示唆された。
【0164】
材料及び方法
【0165】
細菌株及び増殖条件
【0166】
Gallibacterium anatis次亜種ヘモリチカ菌株12656-12 Liver (12656-12と呼ぶ)を研究に用いており、この菌株は、敗血症のニワトリの肝臓から最初に単離された[Bojesen A.M., Torpdahl M., Christensen H., Olsen J.E., Bisgaard M., Genetic diversity of Gallibacterium anatis isolates from different chicken flocks, J. Clin. Microbiol. (2003) 41:2737-2740]。G. anatis 12656-12は、閉じたプラスチック袋内で5%クエン酸ウシ血液を補ったブレインハートインフュージョン(BHI)(Oxoid)寒天培地上又はエアレーションしたBHIブロス中において37℃で増殖させた。Anaerogen(Oxoid)を用いて、培養瓶中に嫌気条件を発生させた。大腸菌は、Luria-Bertaniブロス及び寒天培地において増殖させ、培地は、50μg/mLカナマイシン及び20μg/mLクロラムフェニコールにより適宜補った。全ての化学物質は、Sigmaより購入した。
【0167】
バイオインフォマティクス分析
【0168】
G. anatis次亜種ヘモリチカ12656-12 Liverのゲノム配列の草案バージョン(115コンティグ)(A. M. Bojesen、未公開データ)を、454 Life Sciencesからパイロシーケンスに基づく方法を使用して入手した[Margulies M., Egholm M., Altman W.E., Attiya S., Bader J.S., Bemben L.A., et al., Genome sequencing in microfabricated high-density picolitre reactors, Nature (2005) 437:376-380]。遺伝子アノテーションは、オーストラリア、モナシュ大学のビクトリアバイオインフォマティクスコンソーシアムが提供する真核生物用のウェブ上のアノテーションシステム、Wasabiを用いて行った[Bulach D.M., Zuerner R.L., Wilson P., Seemann T., McGrath A., Cullen P.A., Davis J., Johnson M., Kuczek E., Alt D.P., Peterson-Burch B., Coppel R.L., Rood J.I., Davies J.K., Adler B., Genome reduction in Leptospira borgpetersenii reflects limited transmission potential, Proc. Natl. Acad. Sci. USA (2006) 103:14560-14565]。配列類似性の検索は、BLASTP[Altschul S.F., Madden T.L., Schaffer A.A., Zhang J.H., Zhang Z., Miller W., Lipman D.J., Gapped BLAST and PSI-BLAST: a new generation of protein database search programs, Nucleic Acids Res. (1997) 25:3389-3402](データベース:非冗長タンパク質配列(GenBank)及びSwissProt)、FASTA[Pearson W.R., Lipman D.J., Improved tools for biological sequence comparison, Proc. Natl. Acad. Sci. USA (1988) 85:2444-2448]及びSSEARCH(データベース:UniProtKB及びSwissProt)、及びHHpred[Soding J., Biegert A., Lupas A.N., The HHpred interactive server for protein homology detection and structure prediction, Nucleic Acids Res. (2005) 33:W244-W248](データベース:Interpro(2009))を用いて行った。全ての検索は、2009年3月に行った。Transterm[Kingsford C.L., Ayanbule K., Salzberg S.L., Rapid, accurate, computational discovery of Rho-independent transcription terminators illuminates their relationship to DNA uptake, Genome Biol. (2007) 8:R22]を用いて、転写ターミネーターを予測した。
【0169】
溶血アッセイ
【0170】
溶血活性は、以前の記述に従って検査しており[Rowe G.E., Welch R.A., Assays of Hemolytic Toxins, Methods Enzymol. (1994) 235:657-667]、ウシ血液をTN緩衝液(10 mM Tris-HCl, 0.9% NaCl, pH 7.5)中で上部相が無色に見えるまで繰り返し洗浄した。2%(vol/vol)赤血球溶液を、10mM CaCl2で補ったTN緩衝液中で調製した。赤血球は、濾過滅菌細菌培養上清又は細菌と共に、特に記載がない限り、1:1の比において37℃で1時間インキュベートした。非溶解赤血球及び細胞残屑を遠心分離により収集し、放出されたヘモグロビンの量をプレートリーダにおいて540nmで測定した。100%の溶解は、1%triton-X により決定し、バックグラウンドの溶解は、溶血活性の計算前に減算した。熱の影響は、上清を溶血アッセイ前に60℃で30分間インキュベートすることにより調査した。プロテイナーゼKの影響は、上清を溶血アッセイ前に4μg/mLのプロテイナーゼKと共に37℃で30分間インキュベートすることにより調査した。
【0171】
HD11細胞の培養及びLDH細胞毒性アッセイ
【0172】
トリ骨髄細胞のMC29形質転換に由来するマクロファージ様細胞株HD11[Beug H., Vonkirchbach A., Doderlein G., Conscience J.F., Graf T., Chicken Hematopoietic-Cells Transformed by 7 Strains of Defective Avian Leukemia Viruses Display 3 Distinct Phenotypes of Differentiation, Cell (1979) 18:375-390]を、Roswell Park Memorial Institute (RPMI)1640培地+GlutaMAXTM-I + 25 mM HEPES (Gibco)において維持した。培地には、2.5%トリ血清、7.5%ウシ胎仔血清(FBS)、及び25μg/mLゲンタマイシンを補った。細胞は、付着細胞株として37℃で5%CO2雰囲気により培養し、1日又は2日おきに継代培養した。細胞毒性アッセイのために、細胞は、総量100μLの5%FBS添加RPMI中で2×104個ずつ96ウェルプレートに播種した。細胞は、一晩インキュベートし、培地を交換した。生理食塩水(0.9%NaCl)に再懸濁した濾過滅菌培養上清又は細菌を細胞に添加し、1時間インキュベートした。大腸菌は、第2.7節において説明されるように組み換えタンパク質の発現を誘導し、OD600(600nmでの光学密度)を略6×108CFU/mLに相当する1に調節した。G. anatis細胞及び上清は、後期対数期(OD600 = 1)に回収した。G. anatisの懸濁物は、OD600を略4×108CFU/mLに相当する1に調節した。濾過滅菌培養上清を氷上で保存し、採取後30分以内に細胞に添加した。細胞毒性は、LDH細胞毒性アッセイ(Promega)により製造業者の説明に従って決定した。各試料は、3重ウェルにおいて検査し、実験は、少なくとも3回繰り返した。
【0173】
G. anatis gtxA突然変異体の構築
【0174】
gtxAのヌクレオチド140〜1648からなる1508bpの断片を、プライマー4240及び4242によりPCR増幅し、ヌクレオチド3995〜5478からなる1483bpの断片をプライマー4243及び4245により増幅した(プライマー配列は表Iに記載)。2つの断片を制限酵素により消化し、プラスミドpWSK129内の対応制限部位に連結させた[Wang R.F., Kushner S.R., Construction of versatile low-copy-number vectors for cloning, sequencing and gene expression in Escherichia coli, Gene (1991) 100:195-199]。EcoRI消化pUC4-KISSからのゲル精製カナマイシンカセット (Tn903)[Barany F., 2-Codon Insertion Mutagenesis of Plasmid Genes by Using Single-Stranded Hexameric Oligonucleotides, Proc. Natl. Acad. Sci. USA (1985) 82:4202-4206]を2つのPCR断片間のEcoRI部位に連結した。カナマイシン耐性遺伝子をgtxAと同じ転写方向に挿入した。プラスミドDNAは、XhoI及びSalIによる消化により直線化し、カラム精製した。G. anatis 12656-12の自然形質転換受容性(natural competence)を、インフルエンザ菌に関する以前の記述に従ってMIV法により誘発し[Poje G., Redfield R.J., Transformation of Haemophilus influenzae, Methods Mol. Med. (2003) 71:57-70]、G. anatisをBHI中で増殖させてOD600を0.2とし、MIVにおいて1回洗浄し、MIV中で100分間インキュベートした。直鎖DNAを0.5μg DNA/mLの濃度で細胞に添加した。20分後、2体積のBHIを追加し、細菌を1時間インキュベートした後、5μg/mLカナマイシンを含む血液寒天プレート上で形質転換体を選択した。コロニーを再ストリークし、プライマー対39F+kanR、kanF+5334R、及び2871F+3270Rにより欠失を検証した。菌株は、ΔgtxAと命名した。
【0175】
発現プラスミドの構築
【0176】
完全長GtxA、GtxAのN末端ドメイン(アミノ酸1〜949)、及びGtxAのRTXドメイン(アミノ酸931〜2026)を、GtxCと共に又はGtxCなしでコードするプラスミドを、発現ベクターpET28a(Novagen)へのPCR断片のライゲーションにより構築した。PCR断片は、pfx50ポリメラーゼ(Invitrogen)により増幅し、カラム精製し、NcoI及びXhoIにより二重消化し、再度カラム精製するか或いはゲル精製した(断片>6kb)。各構築物に使用したプライマーは、表2に記載している。構築物Nterm+C、即ちgtxCを有するオペロン内のgtxAのヌクレオチド1〜2847は、オーバラップエクステンションによるスプライシング(splicing by overlap extension)を使用して作成し[Horton R.M., Cai Z.L., Ho S.N., Pease L.R., Gene-Splicing by Overlap Extension - Tailor-Made Genes Using the Polymerase Chain-Reaction, Biotechniques (1990) 8:528-535]、プライマーGtxUP-NcoI及びgtxC-r-XhoIをPCRの第二ラウンドにおいて使用した。プラスミドpET28aをNcoI及びXhoIにより二重消化し、アンタークティックホスファターゼ(NEB)により脱リン酸化し、ゲル精製した。ベクター及びPCR断片をモル比1:3で連結し、化学的コンピテント大腸菌Top10F’(Invitrogen)に形質転換し、カナマイシンを含むLB寒天プレート上において選択した。各プラスミドにおけるインサートの配列は、DNAシーケンシング(Macrogen、韓国)により検証した。プラスミドは、大腸菌発現株ER2566(New England Biolabs)に形質転換した。プラスミドpLG575は、HlyAを分泌するT1SSの構成要素である大腸菌hlyB及びhlyDをコードしており[Mackman N., Nicaud J.M., Gray L., Holland I.B., Genetical and functional organisation of the Escherichia coli haemolysin determinant 2001, Mol. Gen. Genet. (1985) 201:282-288]、発現タンパク質の分泌を促進するために導入した。
【0177】
大腸菌における組み換えGtxAタンパク質の発現
【0178】
タンパク質の発現は、30℃でインキュベートした0.1mM IPTGを含む寒天プレート上で誘導した。ブロスでの誘導のために、一晩培養した培養物を1:50で希釈し、培養物のOD600が0.6になるまで37℃で振盪しながらインキュベートした。その後、IPTG(0.2mM)を添加し、誘導を2時間、30℃で維持した。細胞から組み換えタンパク質を放出させるために、細胞を遠心分離によりペレット状にし、元の体積の1/25で0.1M Tris/0.9%NaClに再懸濁させ、45秒間のビーズ破砕(FastPrep)により溶解させ、4℃で遠心沈殿させ、直ちに上清を液体溶血アッセイ及びLDH細胞毒性アッセイに使用した。
【0179】
SDS PAGE及びウェスタンブロット分析
【0180】
培養物500μLを採取し、細胞ペレットを採取時のOD単位当たり10mM Tris 500μLに再懸濁することにより、全細胞タンパク質を取得した。細胞外タンパク質は、濾過滅菌培養上清(低タンパク質結合フィルタ(0.22μm)(Millex(R) GP(Millipore))から調製した。タンパク質は、1体積の氷温96%エタノールの添加により一晩沈殿させ、遠心分離により収集し(13000g、30分間、0℃)、10mM Trisに再懸濁させた(元の体積の1/100)。タンパク質をNuPAGE(R) Novexゲル(Invitrogen)でのSDS-PAGEにより分離させた。ウェスタンブロット分析のために、タンパク質をフッ化ポリビニリデン膜(Invitrogen)にトランスファーした。1次抗体である、A. pleuropneumoniae由来のApxIに対して産出したウサギ抗血清[Schaller A., Kuhn R., Kuhnert P., Nicolet J., Anderson T.J., MacInnes J.I., Seger R.P.A.M., Frey J., Characterization of apxIVA, a new RTX determinant of Actinobacillus pleuropneumoniae, Microbiology (1999) 145:2105-2116]を1:1333の希釈で使用し、Westernbreeze化学発光ウェスタンブロット免疫検出キット(Invitrogen)により製造業者の説明に従って検出した。
【0181】
RNA精製
【0182】
一晩培養した培養物をBHIにおいて1:100で希釈し、37℃でエアレーションと共にインキュベートした。細胞を、OD600が0.17、0.6、2、3の時点で採取し、更に増殖を停止した1時間後、及び24時間のインキュベーション後に採取した。全RNAをRNeasy protect Mini Kit(Qiagen)により単離し、オンカラムDNアーゼ処理を製造業者の説明に従って実行した(Qiagen)。
【0183】
ノーザンブロッティング
【0184】
RNAのブロッティング、プローブ標識([α-32P]-dCTPによる)、及びハイブリダイゼーションを基本的に記述の通りに実行した[Frees D., Chastanet A., Qazi S., Sorensen K., Hill P., Msadek T., Ingmer H., Clp ATPases are required for stress tolerance, intracellular replication and biofilm formation in Staphylococcus aureus, Mol. Microbiol. (2004) 54:1445-1462]。gtxAのRTX側の半分以内にある384bpの断片を、プライマー3487F 5’-GCCTCTACCGCCGTTTCTG-3’及び3874R 5’-GGCTGGCTAATAATTCATCACCTTG-3’によりPCR増幅し、プローブ標識反応のテンプレートとして使用した。
【0185】
結果
【0186】
G. anatisの細胞溶解活性
【0187】
G. anatis次亜種ヘモリチカは、ウシ血液寒天プレート上でβ溶血性を有する[Christensen H., Bisgaard M., Bojesen A.M., Mutters R., Olsen J.E., Genetic relationships among avian isolates classified as Pasteurella haemolytica, 'Actinobacillus salpingitidis' or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov and description of additional genomospecles within Gallibacterium gen. nov, Int. J. Syst. Evol. Microbiol. (2003) 53:275-287]。標的範囲を試験するために、様々な種からの血液により寒天プレート上で菌株12656-12の溶血を調査したところ、細菌は、ウマ、ウシ、ブタ、又はニワトリの血液を含む寒天プレート上で増殖させた時にβ溶血の明瞭なゾーンを発生させ(データを示さず)、Greenham及びHillが報告した広範な標的範囲が確認された[Greenham L.W., Hill T.J., Observations on an Avian Strain of Pasteurella haemolytica, Vet. Rec. (1962) 74:861-863]。細菌は、好気性及び嫌気性の両培養条件下で溶血性を有した。
【0188】
この溶血活性は、分泌された溶血素により生じることが示唆されている[Greenham L.W., Hill T.J., Observations on an Avian Strain of Pasteurella haemolytica, Vet. Rec. (1962) 74:861-863]。これを試験するために、液体溶血アッセイを実施し、様々な増殖段階で採取したG. anatis 12656-12無細胞培養上清を、ウシ赤血球の懸濁液と共にインキュベートし、放出されたヘモグロビンの量を測定した。中期から後期の対数期に由来するG. anatis上清は、赤血球を効率的に溶解させた(図1A)。この活性は、熱(60℃)及びプロテイナーゼKにより不活性化され、カルシウムキレート剤EGTAにより低減されることから(データを示さず)、G. anatisがカルシウム依存性の分泌溶血性タンパク質を生産することが確認された。
【0189】
赤血球の溶解は、宿主の鉄獲得に関与し得るが、しかしながら、他の種類の細胞、例えば、白血球との相互作用は、自然感染中に更に重要な役割を果たし得る。そのため、トリ由来マクロファージ様細胞株HD11を使用して白血球に対するG. anatisの細胞毒性活性を試験した。HD11細胞は、G. anatisへの曝露後、円形化を示し、表面から剥離した(図2B)。細胞毒性は、乳酸脱水素酵素(LDH)細胞毒性アッセイを用いて定量化し、顕著な細胞死が明らかとなった(図2C)。このG. anatisの白血球毒性活性は、この細菌の病変形成において不可欠である可能性が高く、したがって、白血球毒性活性の原因となるタンパク質は、重要な病原因子であると予想される。
【0190】
G. anatisのゲノム配列におけるRTX毒素の特定
【0191】
G. anatisの細胞毒性表現型の原因となる特異タンパク質を特定するために、G. anatis 12656-12のゲノム配列を入手し、潜在的な毒素をコードする配列を検索した。膜孔形成RTX毒素の群に属するタンパク質は、重要な病原因子であり、ガリバクテリウムに関する細菌における溶血性及び白血球毒性活性の原因となることから[Frey J., Kuhnert P., RTX toxins in Pasteurellaceae, Int. J. Med. Microbiol. (2002) 292:149-158]、この種のタンパク質が検索の明確な標的となる。G. anatis 12656-12ゲノム配列に対する様々なRTX毒素(A. pleuropneumoniae由来のApxI及びApxII、A. aggregatibacter由来のLtxA、及び大腸菌由来のHlyAを含む)のアミノ酸配列によるBLAST検索により、2026アミノ酸の推定G. anatis RTX毒素が特定された。このタンパク質をコードする6081ヌクレオチド(nt)のオープンリーディングフレーム(ORF)をgtxAと名付け、gtxはGallibacterium toxin、Aは他のRTXオペロン内での毒素遺伝子の命名に対する類推(analogy)を示す[Frey J., Kuhnert P., RTX toxins in Pasteurellaceae, Int. J. Med. Microbiol. (2002) 292:149-158]。gtxAには、非常に短い5ヌクレオチド(nt)の遺伝子間領域と、163個のアミノ酸の予測タンパク質をコードする492ntのORF(図3A)が続く。このタンパク質は、RTX毒素の活性化に必要なアシルトランスフェラーゼタンパク質との相同性を有し、大腸菌由来のアシルトランスフェラーゼHlyCに対して38%の同一性及び60%の類似性を示した。類推から、遺伝子は、gtxCと名付けた。Rho非依存性転写ターミネーターは、gtxCの下流に発見され、恐らくはgtxA及びgtxCの両方を含む転写単位の終了を示す。gtxA-Cオペロンに隣接する遺伝子は、イノシトール-1-モノホスファターゼ(gtxAの上流のsuhB)及びマンノネートデヒドラターゼ遺伝子(gtxCの下流のuxuA)(図3A)をコードすると予測され、両方とも、GtxACの機能に関与する可能性は低い。
【0192】
興味深いことに、GtxA(2026アミノ酸)は、パスツレラ科の他の構成員及び大腸菌由来のHlyA について記述される「典型的」なRTX毒素(略1000aa[Frey J., Kuhnert P., RTX toxins in Pasteurellaceae, Int. J. Med. Microbiol. (2002) 292:149-158])の2倍の大きさを有する。GtxAのC末端の1000個のアミノ酸は、こうしたRTX毒素と相同であり、この領域は略20%の配列同一性及び35%の配列類似性を有する。このC末端領域は、更に、RTX毒素の幾つかの保存的特徴を含む。大腸菌由来のHlyAは、Lys564及びLys690においてアシル化されており[Stanley P., Packman L.C., Koronakis V., Hughes C., Fatty Acylation of 2 Internal Lysine Residues Required for the Toxic Activity of Escherichia coli Hemolysin, Science (1994) 266:1992-1996]、こうしたリジン及び先行するグリシン残基は、共にGtxAにおいて保存されているため(Lys1484及びLys1607(図3B))、これらはGtxAにおいてGtxCにより仲介されるアシル化部位となる可能性が高い。予測されるアシル化部位(アミノ酸1640〜1830)の下流において、GtxAは、グリシン及びアスパラギン酸塩リッチ領域を有し、これもRTX毒素の保存的特徴である。一方、N末端領域(aa1〜略950)は、利用可能な配列との類似性が限定されており、GenBankデータベースに対するBLASTP検索では、有意なホモログは見つからなかった。しかしながら、aa520〜879の領域は、予測膜タンパク質の未知の機能の保存的ドメイン(COG1511)に対する類似性を有する(E値0.007)。RTXドメインと比較すると、N末端ドメインは、酸性が低く、疎水性アミノ酸、特にセリンを高い割合で含む。2次構造は、主にαヘリックスからなると予測される。
【0193】
N末端ドメインの機能の着想を得るために、配列類似性及び相同性予測用の更に感度の高い検索ツールを用いて、アミノ酸配列のバイオインフォマティクス分析を実行した。FASTA及びSSearchを用いた相同性検索により、アメーバDictyostelium discoide由来の真核生物細胞骨格タンパク質Talin-A及びTalin-B (E値、それぞれ3.4×10-7及び0.0061)及びニワトリ(Gallus gallus) 由来のTalin (E値0.0087)との配列類似性が発見された。更に、相同性検出サーバHHpredでは、Talinに対する相同性が予測された(確率 = 100%)。Talinは、アクチン、ビンキュリン、及びインテグリンの細胞質部分などの、他の一定の範囲のタンパク質と結合する。巨大タンパク質は、複製により生じる反復からなる場合が多く、リピート発見プログラムRadar[16]によるN末端ドメインの調査では、57個のアミノ酸の15の反復が見つかった(図3C)。
【0194】
したがって、GtxAは、2つのドメイン、即ちN末端反復ドメイン及びC末端RTX/細胞溶解素ドメインからなる。
【0195】
GtxAは、GtxCに依存する細胞溶解活性を有する
【0196】
GtxAが細胞溶解性タンパク質であるかを調査するために、gtxAを、予測アシルトランスフェラーゼ遺伝子gtxCと共にクローン化し、非溶血性発現菌株の大腸菌ER2566に導入した。gtxA及びgtxCの発現時、この菌株は、血液寒天プレート上及び液体溶血アッセイにおいて溶血性表現型を呈し(図4)、GtxACが溶血活性を保持することを示した。RTX毒素は、通常、特異的なT1SSにより搬出された細胞外タンパク質である。大腸菌hlyBDによりコードされたT1SSを発現するプラスミド(pLG575)の導入により、溶血ゾーン(図4A)は増加し、イムノブロッティングでは、GtxAの大きな画分が細胞外タンパク質画分に存在することが明らかとなり(図5)、大腸菌分泌系がG. anatis GtxAを分泌可能であることが実証された。HD11細胞に対するGtxAの細胞毒性活性は、LDH放出アッセイにより検査し、gtxACを発現する大腸菌ER2566はHD11細胞に対して毒性を有した。インサートの無いベクターを含む大腸菌(負の対照)は、1時間のインキュベーション後、毒性を示さなかった(図4B)。
【0197】
翻訳後アシル化を必要とすることは、RTX毒素の特徴の1つである。これが非定型のGtxAにも該当するかを評価するために、予測されるアシルトランスフェラーゼGtxC無しで発現させたGtxAの活性を調査した。図4は、GtxAがGtxC不在の状態で発現した場合、赤血球又は白血球に対する細胞溶解活性を有していないことを示している。したがって、非アシル化タンパク質は、不活性であり、翻訳後アシル化は、溶血及び白血球毒性活性の両方に不可欠である。GtxAの分泌は、アシル化の欠如により妨げられず、非アシル化GtxAは、アシル化毒素と同様の量で培養上清内に検出された(図5)。
【0198】
GtxAはG. anatisの細胞毒性活性の原因である
【0199】
G. anatisの溶血及び白血球毒性活性がGtxAを起源とするかを判断するために、gtxAノックアウト突然変異体を構築した。ガリバクテリウムの遺伝子操作用の分子ツールの記述は過去に存在しないが、我々は、G. anatis 12656-12が自然状態でコンピテントであることを発見し、自然形質転換による安定したgtxA突然変異体の構築に、この特質を活用した。結果的に生じた突然変異体では、gtxAの位置1648及び3995間の2347個のヌクレオチドを欠失させ、カナマイシン耐性カセットにより置換させた。
【0200】
野生型とは対照的に、ΔgtxA突然変異体は、血液寒天プレート上(図2A)又は液体溶血アッセイ(データを示さず)において溶血性を有していなかった。更に、ΔgtxAは、HD11細胞に対する細胞毒性を示さなかった(図2B及び2C)。独立して構築した2つのgtxA突然変異体から同一の結果が得られた。したがって、gtxAは、G. anatisの溶血及び白血球毒性活性の原因である。
【0201】
GtxA及びその活性レベルの増殖期依存性
【0202】
G. anatis上清の溶血活性は、増殖期依存性を有し、活性は、後期対数期にピークとなり、静止期への遷移時に降下し、一晩培養した培養物の上清において低くなった(図1A)。このことから、GtxAの発現が同様の増殖期依存性を有するという仮説を立てた。これを調査してGtxAの局在を確証するため、ApxI抗血清によるイムノブロッティングを用いて、培養上清(細胞外タンパク質)及び全細胞溶解物におけるGtxAの量を、増殖中の様々な時点で判定した(図1B)。ApxI抗血清は、完全長GtxA(215kDa)の予測分子量のサイズに対応するバンドを含め、細胞外タンパク質画分内の幾つかのタンパク質を認識した。このバンドは、ΔgtxAには存在せず、バンドがGtxAであることが裏付けられた。溶血活性と同様に、上清中のGtxAの存在及び量は、増殖期依存性を有し(図1B)、GtxAの量は、静止期への遷移時にピークとなり、タンパク質は1日経過した培養物(24時間)では検出されず、A. Pleuropneumoniae ApxI及びApxII [Jarma E., Regassa L.B., Growth phase mediated regulation of the Actinobacillus pleuropneumoniae ApxI and ApxII toxins, Microb. Pathog. (2004) 36:197-203]及びM. haemolytica LktA [Strathdee C.A., Lo R.Y., Regulation of expression of the Pasteurella haemolytica leukotoxin determinant, J. Bacteriol. (1989) 171:5955-5962]について報告されたパターンと同様となった。第二のバンド(>215kDa)は、野生型にも存在するが、ΔgtxAには存在せず、恐らくは翻訳後の修飾により、GtxAが2つの異なる形態で存在し得ることを示唆した。更に2つのバンド(それぞれ65kDa及び>215kDa)は、野生型及び突然変異体の両方において検出されており、GtxAに関連していない可能性が高い。GtxAのサイズのタンパク質は、全細胞溶解物では如何なる時点においても検出されておらず、これは予測される細胞外局在に一致していると共に、GtxAが合成の直後又はそれに関連して分泌されることが裏付けられた。gtxAの転写を調査するために、増殖期全体で採取した細胞からのRNAにより、ノーザンブロッティングを実行した。ブロットにより、gtxAが対数増殖中及び静止期の始めに転写されることが明らかとなったが、静止期に入って2時間の及び一晩培養した培養物では転写物は検出されず、gtxAの転写が静止期中に停止されることが示唆された。結論として、GtxAは、in vitro増殖中に発現され、増殖期依存性の細胞外タンパク質であり、増殖期依存性は、転写調節と、分泌GtxAの蓄積及びその後の分解の間のバランスとに影響される。
【0203】
GtxAのN末端ドメインは完全な細胞溶解活性に必要となる
【0204】
生物情報学的分析では、RTXドメイン及びN末端ドメインの2つの部分からなる他の膜孔形成RTX毒素と比較して、GtxAが非定型の構造を有することが明らかとなった(図3B)。GtxAの細胞溶解活性に対するN末端ドメインの寄与を調査するために、N末端ドメイン(アミノ酸1〜949)及びRTXドメイン(アミノ酸931〜2026)の両方を、大腸菌において別々に発現させ、その溶血及び白血球毒性活性を調査して完全長タンパク質のものと比較した(図4)。RTXドメインをGtxCと共に発現する大腸菌は、血液寒天プレート上及び液体溶血アッセイにおいて、溶血活性を示したことから、RTXドメインは、それ自体が機能性溶血性タンパク質であり、N末端ドメインは、赤血球細胞の溶解に不可欠ではない。しかしながら、RTXドメインは、完全な毒素よりも著しく低い溶血活性を示しており、N末端ドメインが完全な溶血活性に必要であることが示唆された。RTXドメインとHD11細胞との間の相互作用から細胞毒性活性は検出されず、N末端ドメインが白血球毒性に対して不可欠な役割を果たすことが示唆された。イムノブロッティング(図5)は、RTXドメインが発現及び搬出されることを示しており、そのため、活性の違いは、発現レベルの大きな違いによるものではなかった。
【0205】
N末端ドメインのみを発現する大腸菌は、溶血及び細胞毒性活性を有していなかった(図4)。しかしながら、SDS-PAGEでは、この組み換えタンパク質が主に全細胞画分に存在し、細胞外タンパク質画分には少量しか存在しないことが明らかとなった(データを示さず)。そのため、N末端ドメインを発現する細胞の溶解物(FastPrepビーズ破砕又はリゾチーム/超音波処理により生成)の活性を試験したところ、これらは、長期のインキュベーションにもかかわらず、液体溶血アッセイにおいて、或いはトリマクロファージに対して、活性を全く示さなかった。これらの結果は、N末端ドメイン自体が細胞溶解活性を有しないことを示す。
【0206】
実施例2:ガリバクテリウムにおけるgtxAの検出
【0207】
材料及び方法
【0208】
染色体DNAを、DNeasy Kit(Invitrogen)によりガリバクテリウム菌株から精製した。染色体DNAをテンプレート(50ng/50μl反応液)として、標準PCRをプライマーrtxA3368F 5’-CAAACCTAATTCAATCGGATG-3’(SEQ ID NO:24)及びrtxA4625R 5’-TGCTTCAATAATTTTCCATTTTC-3’(SEQ ID NO:25)により行い、gtxAのヌクレオチド3332〜4589を増幅した。PCR条件は、94℃4分間と、94℃30秒、51℃30秒、及び72℃105秒を35サイクルとした。産物は、ゲル電気泳動及びエチジウムブロマイド染色により可視化した。PCR産物は、BigDyeサイクルシーケンシング(Macrogen、韓国)により配列決定した。ヌクレオチド配列は、CLC workbenchにより翻訳及び整列を行った。
【0209】
結果
【0210】
異なるガリバクテリウム菌株からのGtxA毒素のアライメントを図6に提示する。
【0211】
実施例3:配列
【0212】
下記の配列は、本願において言及したアミノ酸及び核酸の配列を示す。
【0213】
【化1】
【化2】
【化3】
【化4−1】
【化4−2】
【化5】
【化6】
【0214】
実施例4:gtxAの配列及び機能は溶血性ガリバクテリウム間で保存されている
【0215】
ガリバクテリウム菌株間でのgtxAヌクレオチド1〜950の配列保存
【0216】
【表3】
【0217】
GtxAは溶血性ガリバクテリウムにより発現及び分泌される
【0218】
材料及び方法
【0219】
タンパク質の単離のために細胞を培養する際には、一晩培養した培養物(16時間)を略1:100に希釈し、OD600を0.03に調整し、細菌を100mLのErlenmeyerフラスコ内の30mL BHIブロスにおいて穏やかなエアレーション(120rpm)下で培養した。細胞及び上清を、後期対数期にOD600 = 0.6(±0.1)で採取した。
【0220】
細胞外タンパク質は、濾過滅菌培養上清(低タンパク質結合フィルタ(0.22μm)(Millex(R) GP(Millipore))から調製した。タンパク質は、1体積の氷温96%エタノールの添加により一晩沈殿させ(-20℃)、遠心分離により収集し(13000g、30分間、0℃)、10mM Trisに再懸濁させた(元の体積の1/100)。タンパク質(13μl)をSDS-PAGEにより、Tris-Acetate SDS Running Buffer (Invitrogen)中のNuPAGE Novex 3-8% Tris-Acetate Mini Gelsにおいて分離させた。ウェスタンブロッティングのために、タンパク質をフッ化ポリビニリデン(PVDF)膜(Invitrogen)にブロットした。ApxI抗血清[24]を1次抗体として使用し、結合したApxI抗体をWesternBreeze Chemiluminescentキット(抗ウサギ)(Invitrogen)により検出した。化学発光信号は、Geliance 600画像化システム(Perkin Elmer Elmer)により捕捉した.
【0221】
GtxAはG. anatisにおける主要な細胞溶解性毒素である
【0222】
8種類の遺伝学的に異なるG. anatis次亜種ヘモリチカ菌株(10672-6、10T4、21K2、24T10、4895、07990、及びAvicor)のgtxAを以前の記述に従って突然変異により不活性化した(Kristensen et al., 2010)。全ての突然変異体が非溶血性であることが判明し(データを示さず)、こうした菌株において、GtxAが溶血活性の原因となることが実証された。この結果は、G. anatisにおける優性な細胞溶解性毒素としてのGtxAの役割を裏付ける。
【0223】
gtxAはG. genomospecies 1及びG. genomospecies 2の溶血活性の原因である
【0224】
G. genomospecies 1及びG. genomospecies 2におけるgtxAの存在から、gtxAがG. genomospecies 1及び2における溶血活性の原因であることが示唆された。G. genomospecies 1及び2型菌株(CCM5974及びCCM5976)においてgtxA突然変異体を構築したところ、これらは非溶血性となり(データを示さず)、これらの種においてもGtxAが溶血活性の原因であることが実証された。
【0225】
実施例5:GtxAの搬出に必要なI型分泌遺伝子座(gtxEBD)の同定及び性質決定
【0226】
GtxAは細胞外タンパク質だが、パスツレラ科の殆どのRTX毒素とは異なり、同族の1型分泌系(type 1 secretion system, T1SS)の成分との共転写は行われない(図9)。T1SSは、3つのタンパク質、即ち、内膜ATPアーゼ、内膜チャネルタンパク質、及び外膜タンパク質の多量体で構成され、大腸菌において、原型のRTX毒素のα溶血素を分泌するT1SSタンパク質は、それぞれHlyB、HlyD、及びTolCと命名されている(Holland et al., Mol Membr Biol 2005, 22: 29-39)。GtxAの搬出を担当するT1SSを特定するために、菌株12656-12のゲノム配列を大腸菌HlyB及びHlyDのアミノ酸配列をクエリとして検索した。この手法により、明白な基質のない予測T1SSオペロンを特定し、GtxA分泌の有望な候補と見なした。このオペロンは、gtxE、gtxB、及びgtxDと名付けた3つの遺伝子を含んでいた(図9)。コード化タンパク質GtxE、GtxB、及びGtxCは、それぞれ、1型分泌系を構成するために必要な3つの構成要素に対応する外膜タンパク質、内膜ATPアーゼ、及び内膜タンパク質になると予測された。
【0227】
オペロンがGtxAの搬出に必要であるかを試験するために、G. anatis 12656-12において、gtxBの大部分(ヌクレオチド位置92から)とgtxDの先頭(最初の123nt)を欠失させることにより突然変異体(ΔgtxBD)を構築した。
【0228】
ΔgtxBDの構築
【0229】
ΔgtxBDの構築のため、2つのPCR断片を生成し、一方(951bp)には、プライマー2870F-XbaI (5’-CTGATCTAGACGCCGTAAATCGCATAATCA-3’) (SEQ ID NO: 26)及び3821R-EcoRI (5’-CGAATTCCCGGCAGAAAAGGTCAACA-3’) (SEQ ID NO: 27)、他方(1233nt)には、6044-SOE (TTTCTGCCGGGAATTCGGCGAATGGTGTGAGAAG-5’) (SEQ ID NO:28)及び7267R-SalI (TCAAGTCGACAAGCCAAAGCCAATACGA-3’) (SEQ ID NO:29)を用いた。プライマー配列の制限部位には下線を付した。2つのPCR断片は、2870f-XbaI及び7267R-SalIによるオーバラップエクステンションPCRによるスプライシングを用いて接続した。結果的に生じた2184ntの断片を、SalI及びXbaIにより消化し、アンタークティックホスファターゼ(Fermentas)によりホスファターゼ処理し、ゲル精製してXbaI/SalI二重消化及びゲル精製したpBluescriptに連結した。EcoRI消化pUC4-KISSからのカナマイシンカセット(Tn903)をゲル精製し、SOEingによる断片内のEcoRI部位に連結した。カナマイシン耐性遺伝子を、gtxBと同じ転写方向に挿入した。プラスミドDNAをSalI及びXbaIによる消化で直線化し、カラム精製した(GFX、Amersham)。G. anatis 12656-12の自然形質転換受容性を、インフルエンザ菌に関する以前の記述に従ってMIV法により誘発した(Poje & Redfield, 2003); 簡潔に言うと、G. anatis 12656-12をBHI中で増殖させてOD600を0.2とし、MIVにおいて1回洗浄し、MIV中で100分間インキュベートした。直鎖DNAを0.5μg DNA/mLの濃度で細胞に添加した。37℃で20分インキュベートした後、2体積のBHIを追加し、細菌を1時間インキュベート(37℃)した後、4μg/mLカナマイシンを含む血液寒天プレート上で形質転換体を選択した。形質転換体を4μg/mLカナマイシンを含むプレート上に再ストリークし、プライマー対TISS2607F (5’-TTTCCTGTAATGCCTGCT-3’) (SEQ ID NO: 30)及びTISS7572R (5’-TTTTGATCGTTCGGGCTT-3’) (SEQ ID NO: 31)によるPCR及びPCR産物の配列決定により欠失を検証した。
【0230】
実施例6: GtxAは全細胞抗原に基づくワクチンの一部となる可能性が低い
【0231】
GtxAは、細胞溶解物中に検出可能なレベルでは存在しておらず、I型分泌gtxEBDが存在しない状態では細胞内に蓄積されない。
【0232】
我々は、GtxAの分泌に必要なI型分泌系遺伝子座(gtxEBD)を特定した。
【0233】
分泌突然変異体ΔgtxBD(実施例5に記載のように構築)において、ΔgtxBDの溶血活性は、野生型(図8A参照)と比較して、大きく減少した。ウェスタンブロッティングでは、gtxBDの欠失により上清中のGtxAの欠如が生じることが明らかとなった(図8B)。こうした観察結果は、gtxEBDがGtxAの搬出を担うことを裏付ける。GtxAは、野生型又はΔgtxBDの全細胞タンパク質内では検出されず、即ち、GtxAは、分泌系が存在しない状態では細胞内部に蓄積されない。同様の観察結果は、A. actinomycetemcomitans (白血球毒素LtxA)及び大腸菌(α溶血素HlyA)におけるRTX毒素分泌不全突然変異体についても得られた。これらの種において、毒素の細胞内蓄積の欠如は、転写の変更によるものではなく、細胞質における毒素の迅速な分解によるものと推測される。
【0234】
実施例7: GtxAを発現できないGallibacterium anatis gtxA突然変異体による感染試験
【0235】
序論
【0236】
細胞溶解性RTX毒素GtxAは、家禽における感染中のG. anatisにとって主要な病原因子とされてきた。これを実証するために、野生型(wt)菌株及び同質遺伝子的ΔgtxA突然変異体による感染中に観察された病変に対するGtxAの寄与を特徴付ける目的で、産卵雌鶏での感染試験を行った
【0237】
材料及び方法
【0238】
合計24羽を研究に含めた。全てのトリを腹腔内経路で感染させ、卵巣の根元に等量のGallibacterium anatisが深く沈着するように試みた。16羽には、G. anatis 12656-12wtを感染させ、一方、16羽には、RTX GtxAを発現できない同質遺伝子的だが非溶血性のΔgtxA突然変異体を感染させた(Kristensen et al., 2010)。
【0239】
結果
【0240】
【表4】
【0241】
wtに感染させたトリは、一般に野外でのG. anatisへの自然感染により観察される病変に対応する、生殖器系及び腹膜に関与する播種性及び化膿性の炎症を生じた。
【0242】
ΔgtxA突然変異体に感染させたトリは、一般に、卵巣に局在する、より軽度の炎症を生じた。幾つかの動物(2/8)では、局在する炎症が強く、感染48時間後に化膿し、一方、8羽のうち2羽は、化膿性卵巣炎及び腹膜炎が感染6日後に観察された。
【0243】
したがって、GtxAがニワトリにおけるG. anatisの病変形成に実質的に寄与すると結論することができる。
【特許請求の範囲】
【請求項1】
以下からなる群から選択されるアミノ酸配列を含む、単離ポリペプチド:
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【請求項2】
SEQ ID No. 1、2、及び3からなる群から選択される前記配列の自然発生対立遺伝子変異体である、請求項1記載のポリペプチド。
【請求項3】
前記ポリペプチドは、パスツレラ科由来、より好ましくはガリバクテリウム属由来、より好ましくはGallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される、アミノ酸配列を含む、請求項1記載のポリペプチド。
【請求項4】
前記選択される配列において指定された任意のアミノ酸は、保存的置換をもたらすように変化する、変異ポリペプチドである請求項1記載のポリペプチド。
【請求項5】
シグナルペプチドが、異種シグナルペプチドに置き換えられている、請求項1記載のポリペプチド。
【請求項6】
SEQ ID No. 1との配列同一性が少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 1の配列を有するポリペプチドである、請求項1記載のポリペプチド。
【請求項7】
SEQ ID No. 2との配列同一性が少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 2の配列を有するポリペプチドである、請求項1記載のポリペプチド。
【請求項8】
SEQ ID No. 3との配列同一性が少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 3の配列を有するポリペプチドである、請求項1記載のポリペプチド。
【請求項9】
請求項1記載のポリペプチド、又は少なくとも150個のアミノ酸、好ましくは少なくとも200個のアミノ酸、より好ましくは少なくとも300個のアミノ酸、より好ましくは少なくとも500個のアミノ酸、より好ましくは少なくとも750個のアミノ酸、より好ましくは少なくとも1000個のアミノ酸、より好ましくは少なくとも1250個のアミノ酸、より好ましくは少なくとも1500個のアミノ酸、より好ましくは少なくとも1750個のアミノ酸、最も好ましくは少なくとも2000個のアミノ酸を含む、その断片。
【請求項10】
前記ポリペプチドが、毒性活性を除去するために特異的に修飾されている、請求項1記載のポリペプチド。
【請求項11】
前記変異体が、生物活性を有する、請求項1記載のポリペプチド。
【請求項12】
生物活性が毒性であり、前記毒性が、受容者における孔の形成、より好ましくは細胞毒性、より好ましくは細胞溶解性細胞毒性、更に好ましくは溶血性細胞毒性を含む、請求項11記載のポリペプチド。
【請求項13】
前記変異体が免疫原性を有する、請求項9〜11記載のポリペプチド。
【請求項14】
前記断片が、SEQ ID No. 1と比較して、アミノ酸置換を30個以下、より好ましくは25個以下、より好ましくは20個以下、より好ましくは15個以下、より好ましくは10個以下、より好ましくは5個以下含み、より好ましくはアミノ酸置換を含まない、請求項9記載のポリペプチド。
【請求項15】
更に、ポリHisタグ、GSTタグ、HAタグ、FLAGタグ、C-mycタグ、HSVタグ、V5タグ、マルトース結合タンパク質タグ、セルロース結合ドメインタグ、BCCPタグ、カルモジュリンタグ、Nusタグ、グルタチオン-S-トランスフェラーゼタグ、緑色蛍光タンパク質タグ、チオレドキシンタグ、Sタグ、Strepタグ等の親和性タグを含む、先行請求項の何れかに記載のポリペプチド。
【請求項16】
任意の親和性タグが、切断可能である、請求項15記載のポリペプチド。
【請求項17】
不活性化されている請求項1〜16の何れかに記載のポリペプチド。
【請求項18】
前記ポリペプチドが、熱又は放射線により不活性化されている、請求項17記載のポリペプチド。
【請求項19】
前記ポリペプチドが、化学的に、好ましくはホルムアルデヒドへの曝露により、不活性化される、請求項17記載のポリペプチド。
【請求項20】
前記ポリペプチドが、SEQ ID No. 1の非アシル化形態である、請求項17記載のポリペプチド。
【請求項21】
前記ポリペプチド又はその任意の断片が、免疫原性を有する、請求項17記載のポリペプチド。
【請求項22】
薬剤として使用する先行請求項の何れかに記載のポリペプチド。
【請求項23】
細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療のための請求項22記載の使用。
【請求項24】
前記細菌感染が、温血動物におけるものである、請求項22又は23記載の使用。
【請求項25】
前記温血動物が、鳥類種である、請求項22又は23記載の使用。
【請求項26】
前記鳥類種は、カモ科、より好ましくはマガモ属、マガン属、ハジロ属、ニオイガモ属、コクガン属、又はハクチョウ属、チドリ目、より好ましくはアカメカモメ属、ハシブトアジサシ属、カモメ属、ゾウゲカモメ属、又はクビワカモメ属、コウノトリ目、より好ましくはコウノトリ科、ハト目、より好ましくはカワラバト属、スズメバト属、ミカドバト属、ヒムネバト属、チョウショウバト属、アメリカウズラバト属、カンムリバト属、ミヤマバト属、ニュージーランドバト属、アメリカシャコバト属、ウォンガバト属、オナガバト属、ハシリバト属、レンジャクバト属、シッポウバト属、Patagioenas属、テリアオバト属、ヒメアオバト属、インカバト属、キジバト属、アオバト属、アオフバト属、又はハジロバト属、キジ目、より好ましくはエボシツカツクリ属、ヤブツカツクリ属、キジ科、又はライチョウ亜科、ペリカン目、より好ましくはペリカン科、フラミンゴ目、より好ましくはフラミンゴ科、オウム目、より好ましくはオウム科、ヒインコ科、又はインコ科、ヒクイドリ目、より好ましくはエミュー科、レア目、より好ましくはダーウィンレア属、又はレア属、ダチョウ目、より好まくはダチョウ科の群から選択される、請求項22又は23記載の使用。
【請求項27】
前記鳥類種は、カモ、シチメンチョウ、及びニワトリの群から選択され、好ましくは産卵雌鶏である、請求項22又は23記載の使用。
【請求項28】
以下からなる群から選択される核酸配列を含む、単離ポリヌクレオチド:
a) SEQ ID No. 4、5、又は6と、
b) SEQ ID No. 4、5、及び6からなる群から選択されるヌクレオチド配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも60%である配列変異体と、
c) a)の何れかの少なくとも450個の隣接ヌクレオチドからなる断片であり、選択される配列において指定された任意の核酸が異なる核酸へ変化するのは、このように変化する配列内の核酸が90個以内の場合である断片と、
d) 高ストリンジェンシー下で、SEQ ID No. 4、5、又は6に相補的なポリヌクレオチドとハイブリダイズすることが可能なポリヌクレオチドと、
e) SEQ ID No. 1、2、又は3のポリペプチドをコードするポリヌクレオチドと、
f) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体をコードするポリヌクレオチドと、
g) SEQ ID No. 1、2、又は3の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片をコードするポリヌクレオチド。
【請求項29】
前記ポリヌクレオチドが、自然発生対立遺伝子核酸変異体のヌクレオチド配列を含む、請求項28記載のポリヌクレオチド。
【請求項30】
前記ポリヌクレオチドが、ガリバクテリウム属由来、より好ましくはGallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される、アミノ酸配列をコードする核酸配列を含む、請求項28記載のポリヌクレオチド。
【請求項31】
前記コード化ポリペプチドが、SEQ ID No. 4との配列同一性が少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも85%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 4の配列を有するポリヌクレオチドである、請求項28記載のポリヌクレオチド。
【請求項32】
前記コード化ポリペプチドが、SEQ ID No. 5との配列同一性が少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも85%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 5の配列を有するポリヌクレオチドである、請求項28記載のポリヌクレオチド。
【請求項33】
前記コード化ポリペプチドが、SEQ ID No. 6との配列同一性が少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも85%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 6の配列を有するポリヌクレオチドである、請求項28記載のポリヌクレオチド。
【請求項34】
大腸菌での発現に最適化された、請求項28〜33の何れかに記載のポリヌクレオチド。
【請求項35】
薬剤として使用する、請求項28〜34の何れかに記載のポリヌクレオチド。
【請求項36】
細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療のための請求項35記載の使用。
【請求項37】
前記細菌感染が、温血動物におけるものである、請求項35又は36記載の使用。
【請求項38】
前記温血動物が、鳥類種である、請求項35又は36記載の使用。
【請求項39】
前記鳥類種は、カモ科、より好ましくはマガモ属、マガン属、ハジロ属、ニオイガモ属、コクガン属、又はハクチョウ属、チドリ目、より好ましくはアカメカモメ属、ハシブトアジサシ属、カモメ属、ゾウゲカモメ属、又はクビワカモメ属、コウノトリ目、より好ましくはコウノトリ科、ハト目、より好ましくはカワラバト属、スズメバト属、ミカドバト属、ヒムネバト属、チョウショウバト属、アメリカウズラバト属、カンムリバト属、ミヤマバト属、ニュージーランドバト属、アメリカシャコバト属、ウォンガバト属、オナガバト属、ハシリバト属、レンジャクバト属、シッポウバト属、Patagioenas属、テリアオバト属、ヒメアオバト属、インカバト属、キジバト属、アオバト属、アオフバト属、又はハジロバト属、キジ目、より好ましくはエボシツカツクリ属、ヤブツカツクリ属、キジ科、又はライチョウ亜科、ペリカン目、より好ましくはペリカン科、フラミンゴ目、より好ましくはフラミンゴ科、オウム目、より好ましくはオウム科、ヒインコ科、又はインコ科、ヒクイドリ目、より好ましくはエミュー科、レア目、より好ましくはダーウィンレア属、又はレア属、ダチョウ目、より好まくはダチョウ科の群から選択される、請求項35又は36記載の使用。
【請求項40】
前記鳥類種は、カモ、シチメンチョウ、及びニワトリの群から選択され、好ましくは産卵雌鶏である、請求項35又は36記載の使用。
【請求項41】
請求項28〜34の何れかに記載のポリヌクレオチドを含むベクター。
【請求項42】
更に、前記ポリヌクレオチドと適切に操作可能に連結したプロモーターを含む、請求項41記載のベクター。
【請求項43】
前記プロモーターが、原核生物プロモーターからなる群から選択される、請求項41又は42記載のベクター。
【請求項44】
前記プロモーターが、転写開始点とRNAポリメラーゼの結合部位とを含む、請求項43記載のベクター。
【請求項45】
前記プロモーターが、プリブノーボックス又はその一部を含む、請求項43記載のベクター。
【請求項46】
前記プロモーターが、-35エレメント又はその一部を含む、請求項43記載のベクター。
【請求項47】
前記プロモーターが、真核生物プロモーターからなる群から選択される、請求項41又は42記載のベクター。
【請求項48】
前記プロモーターが、転写開始点とRNAポリメラーゼの結合部位とを含む、請求項47記載のベクター。
【請求項49】
前記プロモーターが、TATAボックスを含む、請求項47記載のベクター。
【請求項50】
前記プロモーターが、任意の真核細胞転写因子の結合部位を少なくとも1つ含む、請求項47記載のベクター。
【請求項51】
前記ベクターが、ウイルスベクター又はプラスミドベクターである、請求項41又は42記載のベクター。
【請求項52】
前記プラスミドベクターが、真核生物プラスミドベクター又は原核生物プラスミドベクターである、請求項51記載のベクター。
【請求項53】
前記ベクターが、レンチウイルス、HIV、SIV、FIV、EAIV、及びCIVを含むレトロウイルス科に由来するベクターからなる群から選択される、請求項41又は42記載のベクター。
【請求項54】
アルファウイルス、アデノウイルス、アデノ随伴ウイルス、バキュロウイルス、HSV、コロナウイルス、ウシパピローマウイルス、Mo-MLVからなる群から選択され、好ましくはアデノ随伴ウイルスである、請求項41又は42記載のベクター。
【請求項55】
薬剤として使用する、請求項41〜54の何れかに記載のベクター。
【請求項56】
細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療のための請求項41〜54の何れかに記載のベクター。
【請求項57】
前記細菌感染が、温血動物におけるものである、請求項55又は56記載の使用。
【請求項58】
前記温血動物が、鳥類種である、請求項55又は56記載の使用。
【請求項59】
前記鳥類種が、カモ科、より好ましくはマガモ属、マガン属、ハジロ属、ニオイガモ属、コクガン属、又はハクチョウ属、チドリ目、より好ましくはアカメカモメ属、ハシブトアジサシ属、カモメ属、ゾウゲカモメ属、又はクビワカモメ属、コウノトリ目、より好ましくはコウノトリ科、ハト目、より好ましくはカワラバト属、スズメバト属、ミカドバト属、ヒムネバト属、チョウショウバト属、アメリカウズラバト属、カンムリバト属、ミヤマバト属、ニュージーランドバト属、アメリカシャコバト属、ウォンガバト属、オナガバト属、ハシリバト属、レンジャクバト属、シッポウバト属、Patagioenas属、テリアオバト属、ヒメアオバト属、インカバト属、キジバト属、アオバト属、アオフバト属、又はハジロバト属、キジ目、より好ましくはエボシツカツクリ属、ヤブツカツクリ属、キジ科、又はライチョウ亜科、ペリカン目、より好ましくはペリカン科、フラミンゴ目、より好ましくはフラミンゴ科、オウム目、より好ましくはオウム科、ヒインコ科、又はインコ科、ヒクイドリ目、より好ましくはエミュー科、レア目、より好ましくはダーウィンレア属、又はレア属、ダチョウ目、より好まくはダチョウ科の群から選択される、請求項55又は56記載の使用。
【請求項60】
前記鳥類種が、カモ、シチメンチョウ、及びニワトリの群から選択され、好ましくは産卵雌鶏である、請求項55又は56記載の使用。
【請求項61】
請求項41〜54の何れかに記載のベクターにより形質転換又は形質導入した単離宿主細胞。
【請求項62】
大腸菌、サッカロミセス菌種、ピキア菌種、アスペルギルス属、Sf9昆虫細胞からなる群から選択され、より好ましくは、CHO、CHO-K1、HEI193T、HEK293、COS、PC12、HiB5、RN33b、又はBHK細胞である、請求項61記載の宿主細胞。
【請求項63】
請求項53又は54の何れかに記載の感染性ビリオンを生産可能なパッケージング細胞株。
【請求項64】
以下からなる群から選択されるアミノ酸配列を有する単離ポリペプチドと特異的に結合可能な抗体:
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【請求項65】
前記抗体が、トリ抗体であり、好ましくはニワトリ抗体である、請求項64記載の抗体。
【請求項66】
前記抗体が、ポリクローナル血清由来抗体又はモノクローナル又は組み換え抗体である、請求項64記載の抗体。
【請求項67】
抗体が、Fv、scFv、Fab、Fab’、又はF(ab)2等の抗体の抗原結合断片、二量体IgA分子又は五価IgM等の多量体形態、アフィボディ、又は二重特異性抗体を含む、請求項64記載の抗体。
【請求項68】
請求項1〜21の何れかに記載の単離ポリペプチドを不活性化する方法であって、熱又は化学物質、好ましくはホルムアルデヒドの使用を含み、或いは非アシル化形態の前記ポリペプチドの発現を含む、方法。
【請求項69】
前記不活性化ポリペプチドが、免疫原性を有する、請求項68記載の方法。
【請求項70】
請求項1〜21の何れかに記載の単離ポリペプチドを含むワクチン組成物。
【請求項71】
前記ポリヌクレオチドが、裸DNA又はベクターとして使用される、請求項28〜34の何れかに記載の単離ポリヌクレオチドを含むワクチン組成物。
【請求項72】
更に、1つ以上の適切なアジュバント、賦形剤、乳化剤、又は担体を含む、請求項70及び71記載のワクチン組成物。
【請求項73】
更に、請求項26の鳥類種に対する病原性を有するウイルス又は微生物由来の他の抗原を少なくとも1つ含む、請求項70及び71記載のワクチン組成物。
【請求項74】
前記ウイルス又は微生物が、伝染性気管支炎ウイルス、ニューカッスル病ウイルス、伝染性ファブリキウス嚢病ウイルス、ニワトリ貧血因子、トリレオウイルス、トリニューモウイルス、ニワトリポックスウイルス、トリ脳脊髄炎ウイルス、マイコプラズマ・ガリセプチカム、ヘモフィルス・パラガリナルム、パスツレラ・マルトシダ、及び大腸菌からなる群から選択される、請求項73記載のワクチン組成物。
【請求項75】
前記ポリペプチドが、SEQ ID No. 1の非アシル化形態と同一である、請求項70記載のワクチン組成物。
【請求項76】
前記ポリペプチド又はその任意の断片が、免疫原性を有する、請求項70記載のワクチン組成物。
【請求項77】
更に、生存する弱毒化Gallibacterium anatisを含む、請求項70及び71記載のワクチン組成物。
【請求項78】
請求項26の鳥類種に対して請求項70〜77の何れかに記載のワクチンを投与する方法であって、前記ワクチンがは、筋肉内又は皮下注射、例えば食物又は水を介した経口投与、エアロゾル、例えば足又は翼の膜部における乱切処理、点眼剤、又は卵内投与により投与される、方法。
【請求項79】
前記鳥類種には、ワクチン接種を少なくとも1回、例として少なくとも2回、例えば少なくとも3回行う、請求項78記載の方法。
【請求項80】
各ワクチン投与量が、1用量当たりのタンパク質を約1μg/kg体重〜約1000μg/kg体重、好ましくは約10μg/kg体重〜約100μg/kg体重含有する、請求項78及び79の何れかに記載の方法。
【請求項81】
ワクチンの各単位投与量が、約0.1mL〜約2mL、例として約0.25mL〜約1mL、例えば略0.5mLである、請求項78及び79の何れかに記載の方法。
【請求項82】
診断マーカーとして使用する、請求項1〜21記載のポリペプチド。
【請求項83】
鳥類種における病原性細菌ガリバクテリウムの感染を診断する方法であって、請求項1〜9の何れかに記載のポリペプチドの検出、前記ポリペプチドに対する抗体の検出、又は請求項28〜33の何れかに記載のポリヌクレオチドの検出を含む方法。
【請求項84】
前記病原性細菌が、ガリバクテリウム属由来であり、より好ましくはGallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される、請求項83記載の方法。
【請求項85】
前記ポリペプチドに対する抗体の検出を含み、好ましくは、前記抗体はIgAである、請求項83記載の方法。
【請求項86】
請求項1〜9記載のポリペプチドを検出するキットであって、前記ポリペプチドと結合可能な少なくとも1つの結合タンパク質を備え、前記結合タンパク質は固体支持部に連結されるキット。
【請求項87】
他の結合タンパク質を備え、1つの結合タンパク質は、検出部分に連結される、請求項86記載のキット。
【請求項88】
前記固体支持部が、微粒子である、請求項86記載のキット。
【請求項89】
前記結合タンパク質が、請求項1〜9の何れかに記載のポリペプチドに対する抗体である、請求項86記載のキット。
【請求項90】
前記ポリペプチドの量を検出可能である、請求項86記載のキット。
【請求項91】
前記1〜9の何れかに記載のポリペプチドに対する抗体を検出するキットであって、固体表面に固定された前記ポリペプチドを備えるキット。
【請求項92】
前記抗体が、IgA型のものである、請求項91記載のキット。
【請求項93】
内因性遺伝子を破壊して、機能性gtxAの発現及び/又は分泌を無効化したトランスジェニックノックアウト微生物であって、同じ種の野生型微生物と比較して病原性の減少を示す微生物。
【請求項94】
前記微生物が、Gallibacterium anatisである、請求項93記載の微生物。
【請求項95】
gtxA遺伝子が、破壊されている、請求項93又は94記載の微生物。
【請求項96】
gtxBD遺伝子が、破壊されている、請求項93又は94記載の微生物。
【請求項1】
以下からなる群から選択されるアミノ酸配列を含む、単離ポリペプチド:
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【請求項2】
SEQ ID No. 1、2、及び3からなる群から選択される前記配列の自然発生対立遺伝子変異体である、請求項1記載のポリペプチド。
【請求項3】
前記ポリペプチドは、パスツレラ科由来、より好ましくはガリバクテリウム属由来、より好ましくはGallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される、アミノ酸配列を含む、請求項1記載のポリペプチド。
【請求項4】
前記選択される配列において指定された任意のアミノ酸は、保存的置換をもたらすように変化する、変異ポリペプチドである請求項1記載のポリペプチド。
【請求項5】
シグナルペプチドが、異種シグナルペプチドに置き換えられている、請求項1記載のポリペプチド。
【請求項6】
SEQ ID No. 1との配列同一性が少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 1の配列を有するポリペプチドである、請求項1記載のポリペプチド。
【請求項7】
SEQ ID No. 2との配列同一性が少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 2の配列を有するポリペプチドである、請求項1記載のポリペプチド。
【請求項8】
SEQ ID No. 3との配列同一性が少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 3の配列を有するポリペプチドである、請求項1記載のポリペプチド。
【請求項9】
請求項1記載のポリペプチド、又は少なくとも150個のアミノ酸、好ましくは少なくとも200個のアミノ酸、より好ましくは少なくとも300個のアミノ酸、より好ましくは少なくとも500個のアミノ酸、より好ましくは少なくとも750個のアミノ酸、より好ましくは少なくとも1000個のアミノ酸、より好ましくは少なくとも1250個のアミノ酸、より好ましくは少なくとも1500個のアミノ酸、より好ましくは少なくとも1750個のアミノ酸、最も好ましくは少なくとも2000個のアミノ酸を含む、その断片。
【請求項10】
前記ポリペプチドが、毒性活性を除去するために特異的に修飾されている、請求項1記載のポリペプチド。
【請求項11】
前記変異体が、生物活性を有する、請求項1記載のポリペプチド。
【請求項12】
生物活性が毒性であり、前記毒性が、受容者における孔の形成、より好ましくは細胞毒性、より好ましくは細胞溶解性細胞毒性、更に好ましくは溶血性細胞毒性を含む、請求項11記載のポリペプチド。
【請求項13】
前記変異体が免疫原性を有する、請求項9〜11記載のポリペプチド。
【請求項14】
前記断片が、SEQ ID No. 1と比較して、アミノ酸置換を30個以下、より好ましくは25個以下、より好ましくは20個以下、より好ましくは15個以下、より好ましくは10個以下、より好ましくは5個以下含み、より好ましくはアミノ酸置換を含まない、請求項9記載のポリペプチド。
【請求項15】
更に、ポリHisタグ、GSTタグ、HAタグ、FLAGタグ、C-mycタグ、HSVタグ、V5タグ、マルトース結合タンパク質タグ、セルロース結合ドメインタグ、BCCPタグ、カルモジュリンタグ、Nusタグ、グルタチオン-S-トランスフェラーゼタグ、緑色蛍光タンパク質タグ、チオレドキシンタグ、Sタグ、Strepタグ等の親和性タグを含む、先行請求項の何れかに記載のポリペプチド。
【請求項16】
任意の親和性タグが、切断可能である、請求項15記載のポリペプチド。
【請求項17】
不活性化されている請求項1〜16の何れかに記載のポリペプチド。
【請求項18】
前記ポリペプチドが、熱又は放射線により不活性化されている、請求項17記載のポリペプチド。
【請求項19】
前記ポリペプチドが、化学的に、好ましくはホルムアルデヒドへの曝露により、不活性化される、請求項17記載のポリペプチド。
【請求項20】
前記ポリペプチドが、SEQ ID No. 1の非アシル化形態である、請求項17記載のポリペプチド。
【請求項21】
前記ポリペプチド又はその任意の断片が、免疫原性を有する、請求項17記載のポリペプチド。
【請求項22】
薬剤として使用する先行請求項の何れかに記載のポリペプチド。
【請求項23】
細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療のための請求項22記載の使用。
【請求項24】
前記細菌感染が、温血動物におけるものである、請求項22又は23記載の使用。
【請求項25】
前記温血動物が、鳥類種である、請求項22又は23記載の使用。
【請求項26】
前記鳥類種は、カモ科、より好ましくはマガモ属、マガン属、ハジロ属、ニオイガモ属、コクガン属、又はハクチョウ属、チドリ目、より好ましくはアカメカモメ属、ハシブトアジサシ属、カモメ属、ゾウゲカモメ属、又はクビワカモメ属、コウノトリ目、より好ましくはコウノトリ科、ハト目、より好ましくはカワラバト属、スズメバト属、ミカドバト属、ヒムネバト属、チョウショウバト属、アメリカウズラバト属、カンムリバト属、ミヤマバト属、ニュージーランドバト属、アメリカシャコバト属、ウォンガバト属、オナガバト属、ハシリバト属、レンジャクバト属、シッポウバト属、Patagioenas属、テリアオバト属、ヒメアオバト属、インカバト属、キジバト属、アオバト属、アオフバト属、又はハジロバト属、キジ目、より好ましくはエボシツカツクリ属、ヤブツカツクリ属、キジ科、又はライチョウ亜科、ペリカン目、より好ましくはペリカン科、フラミンゴ目、より好ましくはフラミンゴ科、オウム目、より好ましくはオウム科、ヒインコ科、又はインコ科、ヒクイドリ目、より好ましくはエミュー科、レア目、より好ましくはダーウィンレア属、又はレア属、ダチョウ目、より好まくはダチョウ科の群から選択される、請求項22又は23記載の使用。
【請求項27】
前記鳥類種は、カモ、シチメンチョウ、及びニワトリの群から選択され、好ましくは産卵雌鶏である、請求項22又は23記載の使用。
【請求項28】
以下からなる群から選択される核酸配列を含む、単離ポリヌクレオチド:
a) SEQ ID No. 4、5、又は6と、
b) SEQ ID No. 4、5、及び6からなる群から選択されるヌクレオチド配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも60%である配列変異体と、
c) a)の何れかの少なくとも450個の隣接ヌクレオチドからなる断片であり、選択される配列において指定された任意の核酸が異なる核酸へ変化するのは、このように変化する配列内の核酸が90個以内の場合である断片と、
d) 高ストリンジェンシー下で、SEQ ID No. 4、5、又は6に相補的なポリヌクレオチドとハイブリダイズすることが可能なポリヌクレオチドと、
e) SEQ ID No. 1、2、又は3のポリペプチドをコードするポリヌクレオチドと、
f) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体をコードするポリヌクレオチドと、
g) SEQ ID No. 1、2、又は3の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片をコードするポリヌクレオチド。
【請求項29】
前記ポリヌクレオチドが、自然発生対立遺伝子核酸変異体のヌクレオチド配列を含む、請求項28記載のポリヌクレオチド。
【請求項30】
前記ポリヌクレオチドが、ガリバクテリウム属由来、より好ましくはGallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される、アミノ酸配列をコードする核酸配列を含む、請求項28記載のポリヌクレオチド。
【請求項31】
前記コード化ポリペプチドが、SEQ ID No. 4との配列同一性が少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも85%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 4の配列を有するポリヌクレオチドである、請求項28記載のポリヌクレオチド。
【請求項32】
前記コード化ポリペプチドが、SEQ ID No. 5との配列同一性が少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも85%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 5の配列を有するポリヌクレオチドである、請求項28記載のポリヌクレオチド。
【請求項33】
前記コード化ポリペプチドが、SEQ ID No. 6との配列同一性が少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも85%、より好ましくは少なくとも98%であり、より好ましくはSEQ ID No. 6の配列を有するポリヌクレオチドである、請求項28記載のポリヌクレオチド。
【請求項34】
大腸菌での発現に最適化された、請求項28〜33の何れかに記載のポリヌクレオチド。
【請求項35】
薬剤として使用する、請求項28〜34の何れかに記載のポリヌクレオチド。
【請求項36】
細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療のための請求項35記載の使用。
【請求項37】
前記細菌感染が、温血動物におけるものである、請求項35又は36記載の使用。
【請求項38】
前記温血動物が、鳥類種である、請求項35又は36記載の使用。
【請求項39】
前記鳥類種は、カモ科、より好ましくはマガモ属、マガン属、ハジロ属、ニオイガモ属、コクガン属、又はハクチョウ属、チドリ目、より好ましくはアカメカモメ属、ハシブトアジサシ属、カモメ属、ゾウゲカモメ属、又はクビワカモメ属、コウノトリ目、より好ましくはコウノトリ科、ハト目、より好ましくはカワラバト属、スズメバト属、ミカドバト属、ヒムネバト属、チョウショウバト属、アメリカウズラバト属、カンムリバト属、ミヤマバト属、ニュージーランドバト属、アメリカシャコバト属、ウォンガバト属、オナガバト属、ハシリバト属、レンジャクバト属、シッポウバト属、Patagioenas属、テリアオバト属、ヒメアオバト属、インカバト属、キジバト属、アオバト属、アオフバト属、又はハジロバト属、キジ目、より好ましくはエボシツカツクリ属、ヤブツカツクリ属、キジ科、又はライチョウ亜科、ペリカン目、より好ましくはペリカン科、フラミンゴ目、より好ましくはフラミンゴ科、オウム目、より好ましくはオウム科、ヒインコ科、又はインコ科、ヒクイドリ目、より好ましくはエミュー科、レア目、より好ましくはダーウィンレア属、又はレア属、ダチョウ目、より好まくはダチョウ科の群から選択される、請求項35又は36記載の使用。
【請求項40】
前記鳥類種は、カモ、シチメンチョウ、及びニワトリの群から選択され、好ましくは産卵雌鶏である、請求項35又は36記載の使用。
【請求項41】
請求項28〜34の何れかに記載のポリヌクレオチドを含むベクター。
【請求項42】
更に、前記ポリヌクレオチドと適切に操作可能に連結したプロモーターを含む、請求項41記載のベクター。
【請求項43】
前記プロモーターが、原核生物プロモーターからなる群から選択される、請求項41又は42記載のベクター。
【請求項44】
前記プロモーターが、転写開始点とRNAポリメラーゼの結合部位とを含む、請求項43記載のベクター。
【請求項45】
前記プロモーターが、プリブノーボックス又はその一部を含む、請求項43記載のベクター。
【請求項46】
前記プロモーターが、-35エレメント又はその一部を含む、請求項43記載のベクター。
【請求項47】
前記プロモーターが、真核生物プロモーターからなる群から選択される、請求項41又は42記載のベクター。
【請求項48】
前記プロモーターが、転写開始点とRNAポリメラーゼの結合部位とを含む、請求項47記載のベクター。
【請求項49】
前記プロモーターが、TATAボックスを含む、請求項47記載のベクター。
【請求項50】
前記プロモーターが、任意の真核細胞転写因子の結合部位を少なくとも1つ含む、請求項47記載のベクター。
【請求項51】
前記ベクターが、ウイルスベクター又はプラスミドベクターである、請求項41又は42記載のベクター。
【請求項52】
前記プラスミドベクターが、真核生物プラスミドベクター又は原核生物プラスミドベクターである、請求項51記載のベクター。
【請求項53】
前記ベクターが、レンチウイルス、HIV、SIV、FIV、EAIV、及びCIVを含むレトロウイルス科に由来するベクターからなる群から選択される、請求項41又は42記載のベクター。
【請求項54】
アルファウイルス、アデノウイルス、アデノ随伴ウイルス、バキュロウイルス、HSV、コロナウイルス、ウシパピローマウイルス、Mo-MLVからなる群から選択され、好ましくはアデノ随伴ウイルスである、請求項41又は42記載のベクター。
【請求項55】
薬剤として使用する、請求項41〜54の何れかに記載のベクター。
【請求項56】
細菌感染により生じた疾患、障害、又は何らかの損傷の治療及び/又は予防的治療のための請求項41〜54の何れかに記載のベクター。
【請求項57】
前記細菌感染が、温血動物におけるものである、請求項55又は56記載の使用。
【請求項58】
前記温血動物が、鳥類種である、請求項55又は56記載の使用。
【請求項59】
前記鳥類種が、カモ科、より好ましくはマガモ属、マガン属、ハジロ属、ニオイガモ属、コクガン属、又はハクチョウ属、チドリ目、より好ましくはアカメカモメ属、ハシブトアジサシ属、カモメ属、ゾウゲカモメ属、又はクビワカモメ属、コウノトリ目、より好ましくはコウノトリ科、ハト目、より好ましくはカワラバト属、スズメバト属、ミカドバト属、ヒムネバト属、チョウショウバト属、アメリカウズラバト属、カンムリバト属、ミヤマバト属、ニュージーランドバト属、アメリカシャコバト属、ウォンガバト属、オナガバト属、ハシリバト属、レンジャクバト属、シッポウバト属、Patagioenas属、テリアオバト属、ヒメアオバト属、インカバト属、キジバト属、アオバト属、アオフバト属、又はハジロバト属、キジ目、より好ましくはエボシツカツクリ属、ヤブツカツクリ属、キジ科、又はライチョウ亜科、ペリカン目、より好ましくはペリカン科、フラミンゴ目、より好ましくはフラミンゴ科、オウム目、より好ましくはオウム科、ヒインコ科、又はインコ科、ヒクイドリ目、より好ましくはエミュー科、レア目、より好ましくはダーウィンレア属、又はレア属、ダチョウ目、より好まくはダチョウ科の群から選択される、請求項55又は56記載の使用。
【請求項60】
前記鳥類種が、カモ、シチメンチョウ、及びニワトリの群から選択され、好ましくは産卵雌鶏である、請求項55又は56記載の使用。
【請求項61】
請求項41〜54の何れかに記載のベクターにより形質転換又は形質導入した単離宿主細胞。
【請求項62】
大腸菌、サッカロミセス菌種、ピキア菌種、アスペルギルス属、Sf9昆虫細胞からなる群から選択され、より好ましくは、CHO、CHO-K1、HEI193T、HEK293、COS、PC12、HiB5、RN33b、又はBHK細胞である、請求項61記載の宿主細胞。
【請求項63】
請求項53又は54の何れかに記載の感染性ビリオンを生産可能なパッケージング細胞株。
【請求項64】
以下からなる群から選択されるアミノ酸配列を有する単離ポリペプチドと特異的に結合可能な抗体:
a) SEQ ID No. 1、2、又は3と、
b) SEQ ID No. 1、2、及び3からなる群から選択されるアミノ酸配列の配列変異体であり、前記SEQ ID No. との配列同一性が少なくとも70%である配列変異体と、
c) a)の何れかの少なくとも150個の隣接アミノ酸からなる断片であり、選択される配列において指定された任意のアミノ酸が異なるアミノ酸へ変化するのは、このように変化する配列内のアミノ酸が30個以内の場合である断片。
【請求項65】
前記抗体が、トリ抗体であり、好ましくはニワトリ抗体である、請求項64記載の抗体。
【請求項66】
前記抗体が、ポリクローナル血清由来抗体又はモノクローナル又は組み換え抗体である、請求項64記載の抗体。
【請求項67】
抗体が、Fv、scFv、Fab、Fab’、又はF(ab)2等の抗体の抗原結合断片、二量体IgA分子又は五価IgM等の多量体形態、アフィボディ、又は二重特異性抗体を含む、請求項64記載の抗体。
【請求項68】
請求項1〜21の何れかに記載の単離ポリペプチドを不活性化する方法であって、熱又は化学物質、好ましくはホルムアルデヒドの使用を含み、或いは非アシル化形態の前記ポリペプチドの発現を含む、方法。
【請求項69】
前記不活性化ポリペプチドが、免疫原性を有する、請求項68記載の方法。
【請求項70】
請求項1〜21の何れかに記載の単離ポリペプチドを含むワクチン組成物。
【請求項71】
前記ポリヌクレオチドが、裸DNA又はベクターとして使用される、請求項28〜34の何れかに記載の単離ポリヌクレオチドを含むワクチン組成物。
【請求項72】
更に、1つ以上の適切なアジュバント、賦形剤、乳化剤、又は担体を含む、請求項70及び71記載のワクチン組成物。
【請求項73】
更に、請求項26の鳥類種に対する病原性を有するウイルス又は微生物由来の他の抗原を少なくとも1つ含む、請求項70及び71記載のワクチン組成物。
【請求項74】
前記ウイルス又は微生物が、伝染性気管支炎ウイルス、ニューカッスル病ウイルス、伝染性ファブリキウス嚢病ウイルス、ニワトリ貧血因子、トリレオウイルス、トリニューモウイルス、ニワトリポックスウイルス、トリ脳脊髄炎ウイルス、マイコプラズマ・ガリセプチカム、ヘモフィルス・パラガリナルム、パスツレラ・マルトシダ、及び大腸菌からなる群から選択される、請求項73記載のワクチン組成物。
【請求項75】
前記ポリペプチドが、SEQ ID No. 1の非アシル化形態と同一である、請求項70記載のワクチン組成物。
【請求項76】
前記ポリペプチド又はその任意の断片が、免疫原性を有する、請求項70記載のワクチン組成物。
【請求項77】
更に、生存する弱毒化Gallibacterium anatisを含む、請求項70及び71記載のワクチン組成物。
【請求項78】
請求項26の鳥類種に対して請求項70〜77の何れかに記載のワクチンを投与する方法であって、前記ワクチンがは、筋肉内又は皮下注射、例えば食物又は水を介した経口投与、エアロゾル、例えば足又は翼の膜部における乱切処理、点眼剤、又は卵内投与により投与される、方法。
【請求項79】
前記鳥類種には、ワクチン接種を少なくとも1回、例として少なくとも2回、例えば少なくとも3回行う、請求項78記載の方法。
【請求項80】
各ワクチン投与量が、1用量当たりのタンパク質を約1μg/kg体重〜約1000μg/kg体重、好ましくは約10μg/kg体重〜約100μg/kg体重含有する、請求項78及び79の何れかに記載の方法。
【請求項81】
ワクチンの各単位投与量が、約0.1mL〜約2mL、例として約0.25mL〜約1mL、例えば略0.5mLである、請求項78及び79の何れかに記載の方法。
【請求項82】
診断マーカーとして使用する、請求項1〜21記載のポリペプチド。
【請求項83】
鳥類種における病原性細菌ガリバクテリウムの感染を診断する方法であって、請求項1〜9の何れかに記載のポリペプチドの検出、前記ポリペプチドに対する抗体の検出、又は請求項28〜33の何れかに記載のポリヌクレオチドの検出を含む方法。
【請求項84】
前記病原性細菌が、ガリバクテリウム属由来であり、より好ましくはGallibacterium anatis、Gallibacterium genomospecies 1、及びGallibacterium genomospecies 2の群から選択される、請求項83記載の方法。
【請求項85】
前記ポリペプチドに対する抗体の検出を含み、好ましくは、前記抗体はIgAである、請求項83記載の方法。
【請求項86】
請求項1〜9記載のポリペプチドを検出するキットであって、前記ポリペプチドと結合可能な少なくとも1つの結合タンパク質を備え、前記結合タンパク質は固体支持部に連結されるキット。
【請求項87】
他の結合タンパク質を備え、1つの結合タンパク質は、検出部分に連結される、請求項86記載のキット。
【請求項88】
前記固体支持部が、微粒子である、請求項86記載のキット。
【請求項89】
前記結合タンパク質が、請求項1〜9の何れかに記載のポリペプチドに対する抗体である、請求項86記載のキット。
【請求項90】
前記ポリペプチドの量を検出可能である、請求項86記載のキット。
【請求項91】
前記1〜9の何れかに記載のポリペプチドに対する抗体を検出するキットであって、固体表面に固定された前記ポリペプチドを備えるキット。
【請求項92】
前記抗体が、IgA型のものである、請求項91記載のキット。
【請求項93】
内因性遺伝子を破壊して、機能性gtxAの発現及び/又は分泌を無効化したトランスジェニックノックアウト微生物であって、同じ種の野生型微生物と比較して病原性の減少を示す微生物。
【請求項94】
前記微生物が、Gallibacterium anatisである、請求項93記載の微生物。
【請求項95】
gtxA遺伝子が、破壊されている、請求項93又は94記載の微生物。
【請求項96】
gtxBD遺伝子が、破壊されている、請求項93又は94記載の微生物。
【図1】

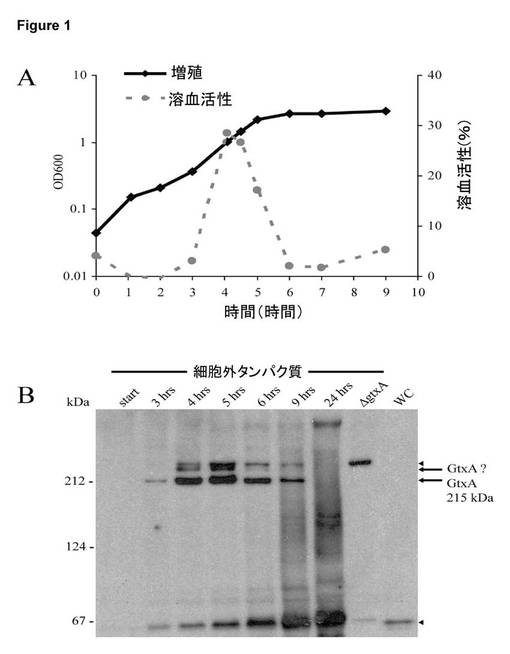
【図2】
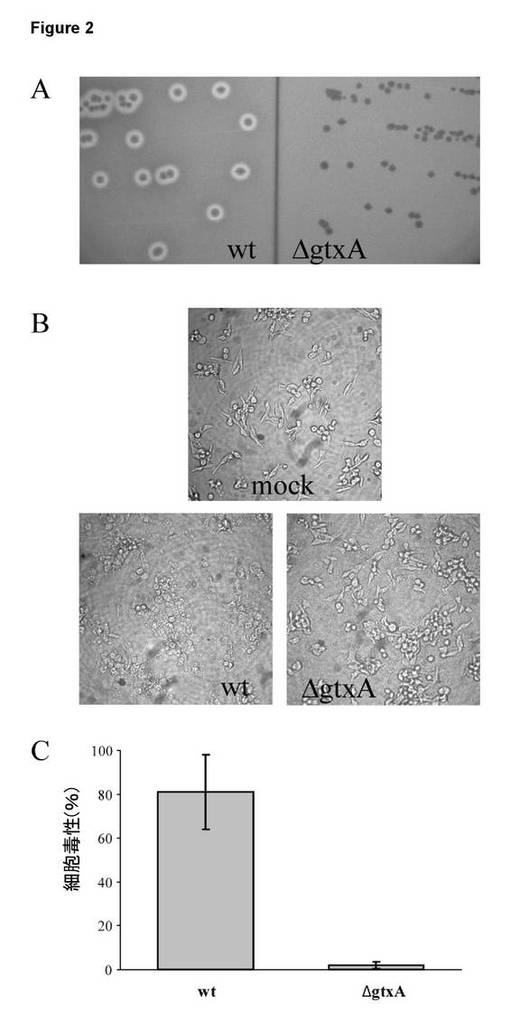
【図3】
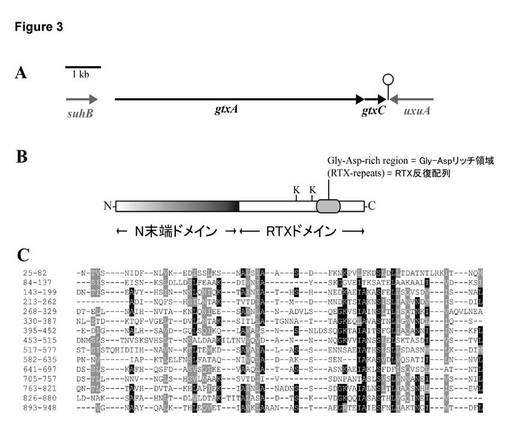
【図4】
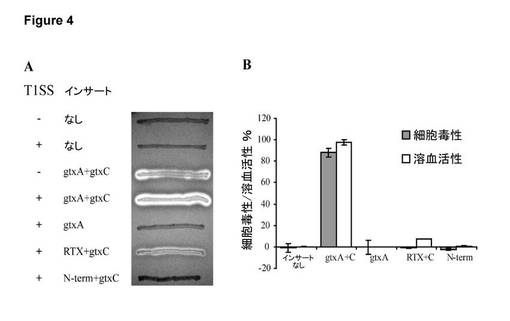
【図5】
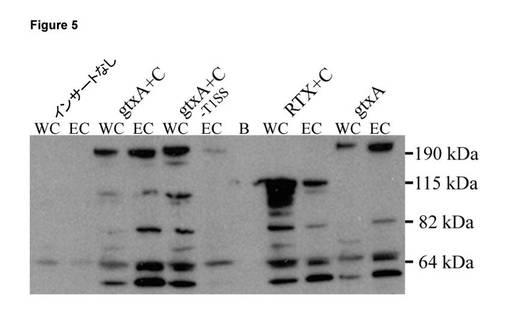
【図6】
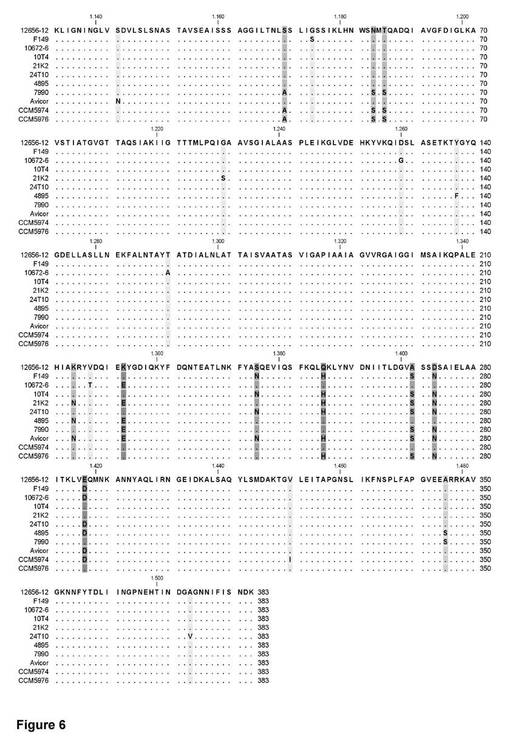
【図7】
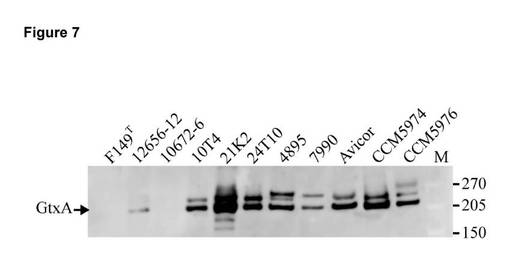
【図8】
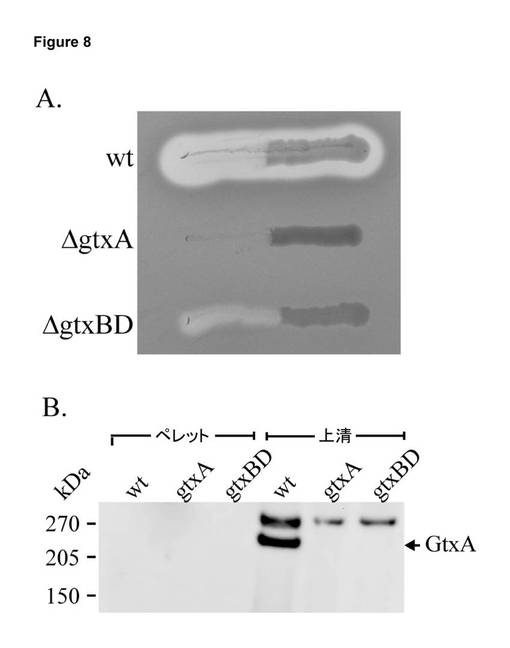
【図9】
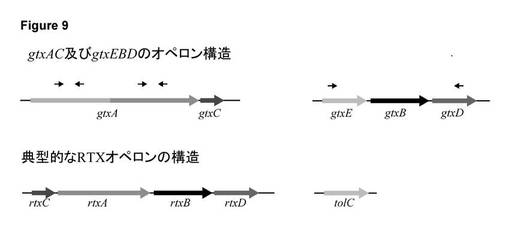
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2013−507103(P2013−507103A)
【公表日】平成25年3月4日(2013.3.4)
【国際特許分類】
【出願番号】特願2012−532457(P2012−532457)
【出願日】平成22年10月7日(2010.10.7)
【国際出願番号】PCT/DK2010/050257
【国際公開番号】WO2011/042026
【国際公開日】平成23年4月14日(2011.4.14)
【出願人】(512092210)ケベンハウンス ウニヴェルジテート(ユニバーシティ オブ コペンハーゲン) (1)
【氏名又は名称原語表記】KOEBENHAVNS UNIVERSITET(UNIVERSITY OF COPENHAGEN)
【Fターム(参考)】
【公表日】平成25年3月4日(2013.3.4)
【国際特許分類】
【出願日】平成22年10月7日(2010.10.7)
【国際出願番号】PCT/DK2010/050257
【国際公開番号】WO2011/042026
【国際公開日】平成23年4月14日(2011.4.14)
【出願人】(512092210)ケベンハウンス ウニヴェルジテート(ユニバーシティ オブ コペンハーゲン) (1)
【氏名又は名称原語表記】KOEBENHAVNS UNIVERSITET(UNIVERSITY OF COPENHAGEN)
【Fターム(参考)】
[ Back to top ]