
Gm1遺伝子、G−タンパク質共役型受容体遺伝子及びCRMP2遺伝子が導入されて得られる形質転換体並びにその利用
【課題】CRMP2タンパク質を介したシグナル伝達異常に起因すると思われる疾患、例えば、アルツハイマー病、統合失調症及び軸策損傷等のような疾患症状を改善させるために有用な知識・技術の蓄積・開発が急務であり、これに関連する試験ツール等の開発が期待されている。
【解決手段】Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子が宿主細胞内に導入されて得られることを特徴とする形質転換体等。
【解決手段】Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子が宿主細胞内に導入されて得られることを特徴とする形質転換体等。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Gm1遺伝子、G-タンパク質共役型受容体遺伝子及びCRMP2遺伝子が導入されて得られる形質転換体並びにその利用に関するものである。
【背景技術】
【0002】
Gm1タンパク質は、G−タンパク質αサブユニットの一種であり、G−タンパク質共役受容体(GPCR)刺激による細胞内シグナル伝達に関与していることが知られている。また、Gm1遺伝子は、Golf遺伝子のスプライシングバリアントであり、第1エクソン領域のみ固有の塩基配列を有しており、第2エクソン領域以降はGolf遺伝子と塩基配列を共有している。また、Gm1タンパク質は、NCBIデータベースにおいて「NM_182978」で示されるG−タンパク質αサブユニットの一種であり、guanine nucleotide binding protein(G protein)、alpha activating activity polypeptide、olfactory type(GNAL)、transcript variant 1とも呼ばれることもあり、他のGタンパク質と比較してN末端領域が約80アミノ酸長い特徴的な構造を有している(例えば、特許文献1参照)。
G−タンパク質共役受容体(GPCR)は、7回膜貫通型の受容体であり、G-タンパク質と共役することが知られている(例えば、非特許文献11参照)。
一方、CRMP2タンパク質は、コラプシン応答メディエータタンパク質2(Collapsin Response Mediator Protein 2, CRMP2)は、神経ガイダンス因子であるセマフォリンの情報を媒介する因子として同定されたタンパク質である(例えば、非特許文献1参照)。当該タンパク質は、チューブリン、Numb、Sra−1、TrkB等の分子と直接的又は間接的に結合し、これらの分子を軸策先端へ輸送することにより軸策を伸長させる機能を有することが知られている(非特許文献2、非特許文献3、非特許文献4、非特許文献5)。また、当該タンパク質は、cdk5、GSK3β、Rhoキナーゼ等のキナーゼによりリン酸化されることにより、その活性が調節されること(具体的には、非リン酸化型が活性を有し、キナーゼによりリン酸化されることにより不活化される)が知られている(非特許文献6、非特許文献7)。更にまた、非リン酸化型変異遺伝子を神経細胞に導入すると軸策先端の退縮が損なわれることや、神経細胞でCRMP2タンパク質の生物学的機能を阻害すると軸策の伸長が阻害されること等も報告されている(非特許文献6、非特許文献8)。また、神経損傷後にCRMP2タンパク質を過剰発現させると神経軸索再生が促進されること(例えば、非特許文献9)や、CRMP2タンパク質の変異が統合失調症のリスク要因になっていること(例えば、非特許文献10)等も報告されている。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】特開2004−350672号
【非特許文献】
【0004】
【非特許文献1】Goshimaら, Nature (1995), 376, 509−514
【非特許文献2】Fukata Yら, Nat. Cell Biol., (2002) 4: 583−591
【非特許文献3】Nishimura Tら, Nat. Cell Biol., (2003) 5: 819−826
【非特許文献4】Arimuraら, Nat. Rev. Neurosci., 8: 194−205 (2007)
【非特許文献5】Arimuraら, Developmental Cell, 16, 675−686 (2009)
【非特許文献6】Yoshimura Tら, Cell, 120, 137−149, (2005)
【非特許文献7】Arimura Nら, Mol. Cell. Biol., 25, 9973−9984 (2005)
【非特許文献8】Arimuraら, J.Biol.Chem., 275, 23973−23980 (2000)
【非特許文献9】Suzuki Yら, J. Neurochem., 86, 1042−50, (2003)
【非特許文献10】Nakata Kら, Biol Psychiatry., 53, 571−576, (2003)
【非特許文献11】芳賀、蛋白質核酸酵素 Vol.46, 1764-1771, (2001)
【発明の概要】
【発明が解決しようとする課題】
【0005】
CRMP2タンパク質を介したシグナル伝達異常に起因すると思われる疾患、例えば、アルツハイマー病、統合失調症及び軸策損傷等のような疾患症状を改善させるために有用な知識・技術の蓄積・開発が急務であり、これに関連する試験ツール等の開発が期待されている。
【課題を解決するための手段】
【0006】
本発明者らは、このような状況を鑑み鋭意検討した結果、Gm1タンパク質の生物学的機能の活性化によりCRMP2タンパク質のリン酸化が抑制されること、即ち、CRMP2タンパク質の生物学的機能が活性化されることを新たに見出し、当該知見を利用することにより、本発明に至った。
即ち、本発明は
1.下記(1)、(2)及び(3)の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子が宿主細胞内に導入されて得られることを特徴とする形質転換体(以下、本発明形質転換体と記することもある。);
(1)Gm1タンパク質
(2)G-タンパク質共役型受容体タンパク質
(3)CRMP2タンパク質
2.前記外来遺伝子が、前記の3種類のタンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸であることを特徴とする前項1記載の形質転換体;
3.前項1又は2記載の形質転換体が、前記(1)、(2)及び(3)の3種類のタンパク質をその細胞内で共発現していることを特徴とする形質転換体;
4.(a)被験物質と、前項1、2又は3記載の形質転換体とを接触させる第一工程と、
(b)第一工程後に得られる形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を測定する第二工程と、
(c)第二工程で測定されたリン酸化状態又はそれと相関する指標値を、対照と比較して、前記形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を変動させる被験物質を選択する第三工程と
を含むことを特徴とするGm1タンパク質を介したシグナル伝達調節物質のスクリーニング方法(以下、本発明スクリーニング方法と記すこともある。);
5.第三工程における対照が、被験物質を接触させていない前項1、2又は3記載の形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値であることを特徴とする前項4記載のスクリーニング方法;
6.前項4又は5記載のスクリーニング方法により得られるGm1タンパク質を介したシグナル伝達調節物質;
7.前項6記載のGm1タンパク質を介したシグナル伝達調節物質を有効成分として含有することを特徴とする、CRMP2タンパク質を介したシグナル伝達異常に起因する疾患の治療又は予防剤(以下、本発明薬剤と記すこともある。);
【発明の効果】
【0007】
本発明により、Gm1タンパク質を介したシグナル伝達調節物質のスクリーニング方法及びそれに利用する形質転換体を含む試験ツール等を提供することが可能となる。
【図面の簡単な説明】
【0008】
【図1】図1は、2次元電気泳動サンプルの抗CRMP2抗体でのウエスタンブロットの結果を示す図である。上段図は対照(DMSO)での結果、下段図は被験物質としてRS67333を用いた結果である。アルカリ性側に位置するスポットNo.1は非リン酸化型CRMP2タンパク質のスポットであり、それより酸性側に位置するNo.2−6は、リン酸化型CRMP2タンパク質のスポットである。また、より酸性側に位置するスポットほど、つまりNo.6 ほどリン酸化の程度が高いCRMP2タンパク質のスポットである。
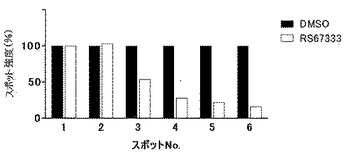
【図2】図2は、図1に示すウエスタンブロットの各スポットのシグナル強度の相対値に基づき、対照(溶媒)と被験物質との間で比較した結果を示す図である。黒棒グラフは対照(DMSO)での結果、白棒グラフは被験物質としてRS67333を用いた結果である。アルカリ性側に位置するスポットNo.1は非リン酸化型CRMP2タンパク質のスポットであり、それより酸性側に位置するNo.2−6は、リン酸化型CRMP2タンパク質のスポットである。また、より酸性側に位置するスポットほど、つまりNo.6 ほどリン酸化の程度が高いCRMP2タンパク質のスポットである。
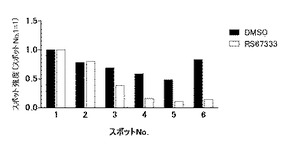
【図3】図3は、図2に示す各スポットのシグナル強度の相対値(パーセンテージ)に基づき、対照(溶媒)と被験物質との間で比較した結果を示す図である。黒棒グラフは対照(DMSO)での結果、白棒グラフは被験物質としてRS67333を用いた結果である。アルカリ性側に位置するスポットNo.1は非リン酸化型CRMP2タンパク質のスポットであり、それより酸性側に位置するNo.2−6は、リン酸化型CRMP2タンパク質のスポットである。また、より酸性側に位置するスポットほど、つまりNo.6 ほどリン酸化の程度が高いCRMP2タンパク質のスポットである。
【発明を実施するための形態】
【0009】
本明細書において、DNAの調製やプラスミドの調製等の遺伝子工学的な手法、タンパク質の抽出やウエスタンブロッティング等のタンパク質工学的な手法は、特に明記しない限り、Molecular Cloning, A Laboratory Manual(T.Maniatis et al., Cold Spring Harbor Laboratory(1982))、新生化学実験講座(日本生化学会編;東京化学同人)等の実験書に記載される方法又はそれに準じた方法により行うことができる。
【0010】
本発明における「Gm1タンパク質」とは、背景技術に記載されるように、G−タンパク質共役受容体(GPCR)刺激による細胞内シグナル伝達に関与していることが知られている、G−タンパク質αサブユニットの一種である。当該タンパク質は、例えば、特開2004−350672号公報等に記載された公知なタンパク質である。
当該タンパク質が有するアミノ酸配列としては、例えば、前記公開公報に記載される配列番号1で示されるアミノ酸配列等を挙げることができる。尚、当該タンパク質は、NCBIデータベースにおいて「NM_182978」で示されるG−タンパク質αサブユニットの一種であり、guanine nucleotide binding protein(G protein)、alpha activating activity polypeptide、olfactory type(GNAL)、transcript variant 1とも呼ばれることもあり、他のGタンパク質と比較してN末端領域が約80アミノ酸長い特徴的な構造を有している。
Gm1遺伝子は、Golf遺伝子のスプライシングバリアントであり、第1エクソン領域のみ固有の塩基配列を有しており、第2エクソン領域以降はGolf遺伝子と塩基配列を共有している。Gm1タンパク質が有するアミノ酸配列をコードする塩基配列としては、例えば、前記公開公報に記載される配列番号2で示される塩基配列等を挙げることができる。
【0011】
本発明における「Gm1タンパク質」は、当該タンパク質の生物学的機能を維持する限り、その一部からなるものを含む。例えば、前記公開公報に記載される配列番号1で示されるアミノ酸配列は、Gタンパク質αサブユニット間で保存されている「GTP結合部位」及び「GTPase活性化部位」のアミノ酸配列と配列同一性が高いアミノ酸配列部分を有する。これらの部分は、前記公開公報に記載される配列番号1で示されるアミノ酸配列におけるアミノ酸番号126〜133、287〜292、353〜359、428〜435の各領域である。これらのアミノ酸領域は、既にGタンパク質αサブユニットとして同定されている「Gs」及び「Golf」における「GTP結合部位」及び「GTPase活性化部位」のアミノ酸配列(NATURE, 117−127 (1991), vol. 349)と一致している。また、配列番号1で示されるアミノ酸配列は、Gタンパク質αサブユニット間で特にGsファミリーに属する「Gs」及び「Golf」で高く保存されている特徴的配列と同じ配列(配列番号1で示されるアミノ酸番号119〜126)を有し、しかもGタンパク質αサブユニット間で保存されているαヘリックス構造をとりうる配列である。
更に、Gm1タンパク質の生物学的機能を喪失すること無く、Gm1タンパク質が有する如何なるアミノ酸残基を何個置換、欠失又は付加できるかの指標は、特開2003−193330に記載された方法等により見出すことができる。前記生物学的機能を喪失しない改変は、例えば、既に同定されている各種のGタンパク質αサブユニットのアミノ酸配列との配列同一性が低い部分について行えばよい。
【0012】
本発明における「CRMP2タンパク質」とは、背景技術に記載されるように、コラプシン応答メディエータタンパク質2(Collapsin Response Mediator Protein 2, CRMP2)は、神経ガイダンス因子であるセマフォリンの情報を媒介する因子として同定されたタンパク質である。当該タンパク質は、例えば、Goshimaら, Nature (1995), 376, 509−514等に記載された公知なタンパク質である。
当該タンパク質が有するアミノ酸配列、及び、当該アミノ酸配列をコードする塩基配列としては、例えば、前記学術文献に記載されるアミノ酸配列、及び、塩基配列等を挙げることができる。尚、当該タンパク質は、NCBIデータベースにおいて「NP_001377」で示されるタンパク質であり、dihydropyrimidinase−like 2、dihydropyrimidinase−like 2、DPYSL2とも呼ばれることもある。
本発明における「G-タンパク質共役型受容体タンパク質」(GPCR)とは、7回膜貫通型の受容体であり、G-タンパク質と共役することが知られている。当該G-タンパク質共役型受容体タンパク質として、例えば、Sevens databaseのホームページ(http://sevens.cbrc.jp/index.php)等に登録されているような受容体を挙げることができ、例えば、ドーパミン受容体、セロトニン受容体及びヒスタミン受容体等を挙げることができ、これらが有するアミノ酸配列、及び、当該アミノ酸配列をコードする塩基配列は、例えば、NCBIのホームページ(http://www.ncbi.nlm.nih.gov/)等より入手することができる。より具体的には、ドーパミンD1受容体の場合には、例えば、NCBIデータベースにおいて「NP_000785.1」で示されるアミノ酸配列、NCBIデータベースにおいて「NM_000794.3」で示される塩基配列等を挙げることができ、またセロトニン4受容体の場合には、例えば、NCBIデータベースにおいて「NP_001035259.1」で示されるアミノ酸配列、NCBIデータベースにおいて「NM_001040169.2」で示される塩基配列等を挙げることができ、またヒスタミンH1受容体の場合には、例えば、NCBIデータベースにおいて「NP_001091683.1」で示されるアミノ酸配列、NCBIデータベースにおいて「NM_001098213.1」で示される塩基配列等を挙げることができる。
【0013】
本発明形質転換体は、下記(1)、(2)及び(3)の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子が宿主細胞内に導入されて得られる。
(1)Gm1タンパク質
(2)G-タンパク質共役型受容体タンパク質
(2)CRMP2タンパク質
【0014】
本発明形質転換体の宿主細胞としては、公知のいかなる宿主細胞でも用いることができ、例えば、大腸菌(例えば、K12等)、バチルス属細菌(例えば、MI114等)等の細菌、酵母(例えば、AH22等)、昆虫細胞(例えば、Sf細胞等)、動物細胞(例えば、COS7細胞等)等を挙げることができる。なかでも、哺乳動物由来の培養細胞(例えば、COS7細胞、CHO細胞、HEK293細胞、Hela細胞、PC12細胞等)を好ましく挙げることができる。
【0015】
本発明形質転換体は、このような宿主細胞に対して、Gm1タンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸、及び、G-タンパク質共役型受容体タンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸、及び、CRMP2タンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸、の全てを外来的に導入するような人為的操作を施すことにより作製することができる。好ましくは、Gm1タンパク質のアミノ酸配列をコードする塩基配列、G-タンパク質共役型受容体タンパク質のアミノ酸配列をコードする塩基配列及びCRMP2タンパク質のアミノ酸配列をコードする塩基配列の全てを含有するプラスミドの形態である核酸を外来的に導入するような人為的操作を施すことにより作製することができる。
前記人為的操作としては、用いられる宿主細胞に応じた通常の遺伝子工学的手法を利用すればよい。具体的には例えば、市販のトランスフェクション試薬を用いて、当該試薬に添付された説明書に記載される方法に従い行う方法を挙げることができる。また、外来遺伝子としてプラスミドの形態である核酸を利用する場合には、リン酸カルシウム法、エレクトロポレーション法、リポフェクション法、DEAEデキストラン法等を挙げることができる。
【0016】
ここで「プラスミド」としては、Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質を宿主細胞内で共発現させることが可能なプラスミドを挙げることができる。このようなプラスミドは、通常の遺伝子工学的手法に従い、Gm1タンパク質のアミノ酸配列をコードする塩基配列を含有する核酸、G-タンパク質共役型受容体タンパク質のアミノ酸配列をコードする塩基配列を含有する核酸、及び、CRMP2タンパク質のアミノ酸配列をコードする塩基配列を含有する核酸、の全てを、公知の発現プラスミドのプロモーター下流の発現可能な位置に連結することにより作製することができる。
ここで「発現プラスミド」としては、例えば、大腸菌を宿主細胞とする場合には、pBR322、pUC12、pUC119、pBluescript等、バチルス属細菌を宿主細胞とする場合には、pUB110、pC194等、酵母を宿主細胞とする場合には、Yip5、Yep24等、昆虫細胞を宿主細胞とする場合には、AcNPV等、動物細胞を宿主細胞とする場合には、pUC18、pcDNA3.1等の公知のプラスミドを挙げることができる。
【0017】
本発明形質転換体における、Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子の宿主細胞内での保持は、一過的に保持されたものであってもよいし、また安定的に保持させたものであってもよい。安定的に保持された形質転換体は、当該外来遺伝子と同時に宿主細胞内に導入された薬剤耐性マーカー等(例えば、発現プラスミドに含有された薬剤耐性マーカー等)を指標にして選択すればよい。
【0018】
本発明形質転換体の細胞内で、Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質を共発現させるには、例えば、本発明形質転換体を培養液の中で培養すればよい。
本発明形質転換体の培養条件は、本発明形質転換体の種類に応じた条件を適宜選択すればよい。
【0019】
例えば、本発明形質転換体が微生物である場合には、微生物の培養に通常使用される液体培地又は平板培地を用いて培養すればよい。培養温度は、微生物の生育可能温度内であればよく、例えば、15〜40℃程度を挙げることができる。培地のpHも、微生物の生育可能範囲であればよく、例えば、pH6〜8程度を挙げることができる。培養時間は、その他の培養条件により異なるが、通常、1〜5日間程度、特に1〜2日間程度にすればよい。温度シフト型やIPTG誘導型等の誘導型の発現プラスミドを用いる場合には、誘導時間は1日間以内、特に数時間程度とすればよい。
【0020】
また形質転換体が昆虫細胞の場合には、昆虫細胞の種類に応じて適した培養条件とすればよい。例えば、FBS及びYeastlateを含むGrace's medium等の昆虫細胞用培地を用いて培養すればよい。培養温度は、25〜35℃程度を挙げることができる。培地のpHも、昆虫細胞の生育可能範囲であればよく、例えば、pH6〜8程度を挙げることができる。培養時間は、その他の培養条件により異なるが、通常、1〜5日間程度、特に2〜3日間程度にすればよい。
発現プラスミドとしてBaculovirus等のウィルスを含む形質転換体の場合は、培養時間は細胞質効果が現れて細胞が死滅する前まで(例えば3〜7日間程度、特に4〜6日間程度)とするのが好ましい。
【0021】
また形質転換体が哺乳動物細胞の場合には、哺乳動物細胞の種類に応じて適した培養条件とすればよい。例えば、FBSを添加したDMEM培地(ニッスイ社製等)を用いて5%CO2存在下、数日毎に新しい培地に交換しながら培養すればよい。哺乳動物細胞がコンフルエントになるまで増殖したら、例えば、トリプシン溶液を加えて個々の細胞に分散させ、得られた細胞の懸濁液を数倍に希釈して新しいシャーレに播種し継代を続ければよい。
培養温度は、36〜38℃程度を挙げることができる。培地のpHも、哺乳動物細胞の生育可能範囲であればよく、例えば、pH6〜8程度を挙げることができる。培養時間は、その他の培養条件により異なるが、通常、2〜5日間程度、特に2〜3日間程度にすればよい。
【0022】
本発明形質転換体の細胞内で、Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質が共発現していることを確認する方法としては、通常のタンパク質工学的方法を利用すればよい。具体的には例えば、本発明形質転換体より調製されたタンパク質抽出液を、抗Gm1タンパク質抗体、抗G-タンパク質共役型受容体タンパク質抗体及び抗CRMP2タンパク質抗体を用いてウエスタンブロットを行う方法等を挙げることができる。
【0023】
本発明スクリーニング方法は、(a)被験物質と、前項1、2又は3記載の形質転換体とを接触させる第一工程と、(b)第一工程後に得られる形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を測定する第二工程と、(c)第二工程で測定されたリン酸化状態又はそれと相関する指標値を、対照と比較して、前記形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を変動させる被験物質を選択する第三工程とを含む。
【0024】
本発明スクリーニング方法において「被験物質」としては、特に限定は無く、例えば、タンパク質(抗体を含む)、ペプチド、非ペプチド性化合物(ヌクレオチド、アミン、糖質、脂質等)、有機低分子化合物、無機低分子化合物、醗酵生産物、細胞抽出液、植物抽出液、動物組織抽出液等を挙げることができる。被験物質の形態としては、特に限定は無く、固体、液体、基剤との混合物、縣濁液又は溶液等を挙げることができる。縣濁液若しくは溶液とする場合、水、pH緩衝液、メチルセルロース溶液、生理食塩水、有機溶媒水溶液(有機溶媒としては通常エタノールやジメチルスルホキシドが用いられる。)等を用いればよい。
被験物質の接触量、接触回数及び接触時間は、例えば、本発明形質転換体に重篤な影響を及ぼさない範囲内とすればよい。
【0025】
本発明スクリーニング方法の第一工程における「接触」は、当業者に汎用されている方法で実施することができる。被験物質を本発明形質転換体に接触する場合、その接触方法には特に限定はなく、例えば、本発明形質転換体を含む培養液・寒天培地等の中に、0.001〜10μM程度の濃度となるように被験物質を添加すればよい。
【0026】
本発明スクリーニング方法の第二工程における「CRMP2タンパク質のリン酸化状態又はそれと相関する指標値を測定する」方法では、タンパク質抽出液の調製、電気泳動法、メンブレンの上への転写、CRMP2タンパク質の検出方法等を用いることができる。これらの方法では、通常の生化学的手法・タンパク質工学的手法等を用いて分析すればよい。例えば、2次元電気泳動法とウエスタンブロット法とを組み合わることにより行えばよい。具体的には例えば、まず被験物質を接触させた本発明形質転換体からタンパク質抽出液を調製する。調製されたタンパク質抽出液を1次元目の電気泳動を行ない、次いで2次元目の電気泳動を行う。分離されたCRMP2タンパク質を適当なメンブレンの上に転写する。次いで、当該メンブレンの上に転写されたCRMP2タンパク質を抗CRMP2タンパク質抗体等で検出する。これら操作の結果、CRMP2タンパク質はリン酸化の程度の違いによって分離される。
非リン酸化型CRMP2は、よりアルカリ性側に検出され、リン酸化の程度が高くなるほど、より酸性側に検出される。尚、は、それ自体公知の通常用いられる方法に従い行うことができる。
また、CRMP2のリン酸化状態を定量する方法としては、公知の定量化法を用いればよいが、例えば、ウエスタンブロットにより検出されたシグナルをイメージアナライザー等にて定量化すればよい。具体的には例えば、本発明形質転換体において2次元電気泳動後のウエスタンブロットにより検出されたCRMP2のスポットの非リン酸化型CRMP2のスポット強度を「1」とし、それぞれのリン酸化型CRMP2のスポット強度を算出する。同様に、対照におけるCRMP2のスポットの強度も、非リン酸化型CRMP2のスポット強度を「1」とし、それぞれのリン酸化型CRMP2のスポット強度を算出する。次いで、対照におけるそれぞれのスポットを100%とし、被検物質を接触させた本発明形質転換体で検出されたCRMP2のそれぞれのスポットのパーセンテージを算出する。被検物質を接触させた本発明形質転換体において検出されたCRMP2のそれぞれのスポットのパーセンテージのいずれかが、例えば70%以下、好ましくは60%以下、更に好ましくは50%以下になる被検物質を、シグナル伝達調節物質の候補物質として選択すればよい。また好ましくは、より高度にリン酸化されたスポットにおいて、スポットのパーセンテージが、例えば70%以下、好ましくは60%以下、更に好ましくは50%以下になる被検物質を、シグナル伝達を活性化する候補物質として選択すればよい。
【0027】
本発明スクリーニング方法の第三工程における対照との比較では、例えば、被験物質を接触させた本発明形質転換体におけるCRMP2タンパク質の第二工程で測定されたリン酸化状態又はそれと相関する指標値を、被験物質を接触させていない本発明形質転換体におけるCRMP2タンパク質の第二工程で測定されたリン酸化状態又はそれと相関する指標値と比較すればよい。ここで「被験物質を接触させていない本発明形質転換体」とは、例えば、溶媒のみを接触させた本発明形質転換体であってもよい。
かかる対照は、対照とするCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を、本発明形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値と併行して測定して求めてもよいし、別途測定して求めてもよい。
例えば、被験物質と接触させた本発明形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値の測定値が、対照とする本発明形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値の測定値よりも低ければ、前記被験物質との接触は、CRMP2タンパク質のリン酸化状態を抑制することによりCRMP2タンパク質を活性化させる効果を有することを意味し、そして当該被験物質はGm1を介したシグナル伝達調節物質として選択することができる。
【0028】
尚、被験物質の代わりに「対照物質」となり得るポジティブコントロール又はネガティブコントロールを用いて本発明スクリーニング方法を実施することにより、場合に応じて上記の対照とすることもできる。
ここで「ポジティブコントロール」とは、CRMP2タンパク質のリン酸化状態を制御する効果を有する任意の物質を表し、また「ネガティブコントロール」としては、被験物質に含まれる溶媒、バックグランドとなる試験系溶液等が挙げられる。
「対照物質」をネガティブコントロールとする場合、被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」が、対照物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」よりも大きければ、当該被験物質は「CRMP2タンパク質のリン酸化状態を抑制する効果」を有すると評価すればよい。一方、被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」が、対照物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」と同程度若しくは小さければ、当該被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」を有さないと評価することができる。
また、対照物質をポジティブコントロールとする場合、被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」と、対照物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」とを比較することにより、被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」を評価すればよい。
このようにして効果を有すると評価された場合には、当該被験物質を、Gm1を介したシグナル伝達調節物質として選択することができる。
【0029】
本発明薬剤は、例えば、上記のようにして選択された「Gm1タンパク質を介したシグナル伝達調節物質」を有効成分として含有する。本発明薬剤は、CRMP2タンパク質を介したシグナル伝達異常に起因する疾患の治療又は予防剤として有用である。
【実施例】
【0030】
以下、本発明の実施例を示してより詳細に説明するが、本発明はこれらの実施例に限定されるものではない。
【0031】
実施例1(本発明スクリーニング方法:Gm1タンパク質を介したシグナル伝達調節物質のスクリーニング)(その1)
(1)Gm1遺伝子の取得
文献(特開2004−350672号)記載の方法に準じた方法を用いて、ヒト脳由来cDNAライブラリー(宝酒造社製)20ngを鋳型に用いて且つ10μMのフォワードプライマーprGm1ATG(5’−ATGGGTCTGTGCTACAGTCTGCGG;配列番号3)、10μMのリバースプライマーprGNAL3’(5’−TCACAAGAGCTCATACTGCTT;配列番号4)及びTAKARA LA Taqポリメラーゼ(TAKARALA Taq with GC Buffer,宝酒造社製)を用いてPCRを行った。PCR条件は、95℃で30秒間、次いで60℃で30秒間、次いで72℃で2分間の保温を1サイクルとしてこれを35サイクル行った。得られたDNA断片をアガロースゲル電気泳動後、当該ゲル上で検出された約1.5kbのバンドを切り出し、pCR2.1−TOPOベクター(インビトロジェン社製)へクローニングしpCR−Gm1を作製した。
(2)pcDNA−Gm1の構築
文献(特開2004−350672号)記載の方法に準じた方法を用いて、実施例1で作製されたプラスミドpCR−Gm1を制限酵素XbaI及びKpnIで二重消化した後、このDNA断片をアガロースゲル電気泳動し、当該ゲル上で検出された約1.5kbのバンドを切り出し、インサートDNAとした。pcDNA3.1(インビトロジェン社製)をXbaI及びKpnIで二重消化したものをベクターとして用い、これにインサートDNAをライゲースを用いて連結することにより、プラスミドpcDNA−Gm1を作製した。
(3)pcDNA−m5HT4の構築
マウス脳より調製したRNA200ngを鋳型に用いて且つ10μMのリバースプライマーm5HT4-3’(5’−CTAAGTATCACTGGGCTGAGCAGCCA;配列番号5)及びSuper Script III逆転写酵素(インビトロジェン社製)を用いて、55℃で50分間、次いで75℃で15分間保温し、逆転写を行った。
得られたcDNA2μlを鋳型に用いて且つ10μMのフォワードプライマーm5HT4-5’(5’−ATGGACAAACTTGATGCTAATGTGAGTT;配列番号6)、10μMのリバースプライマーm5HT4-3’(5’−CTAAGTATCACTGGGCTGAGCAGCCA;配列番号7)及びKOD DNAポリメラーゼ(東洋紡製)を用いてPCRを行った。PCR条件は、95℃で30秒間、次いで60℃で30秒間、次いで68℃で2分間の保温を1サイクルとしてこれを35サイクル行った。得られたDNAをアガロースゲル電気泳動後、当該ゲル上で検出された約1.2kbのバンドを切り出し、pDriveベクター(キアゲン社製)へクローニングしpDr−m5HT4を作製した。次いでプラスミドpDr−m5HT4を制限酵素NheI及びKpnIで二重消化した後、このDNA断片をアガロースゲル電気泳動し、当該ゲル上で検出された約1.2kbのバンドを切り出し、インサートDNAとした。pcDNA3.1(インビトロジェン社製)をNheI及びKpnIで二重消化したものをベクターとして用い、これにインサートDNAをライゲースを用いて連結することにより、プラスミドpcDNA−m5HT4を作製した。
(4)pcDNA−CRMP2の構築
ヒト脳由来cDNAライブラリー(宝酒造社製)20ngを鋳型に用いて且つ10μMのフォワードプライマーhCRMP2(ATG)(5’−CACCATGTCTTATCAGGGGAAGAAAAA;配列番号8)、10μMのリバースプライマーhCRMP2(−STOP)rev(5’−GCCCAGGCTGGTGATGTTGGCACGGCCA;配列番号9)及びKOD DNAポリメラーゼ(東洋紡製)を用いてPCRを行った。PCR条件は、95℃で30秒間、次いで60℃で30秒間、次いで68℃で2分間の保温を1サイクルとしてこれを35サイクル行った。得られたDNAをアガロースゲル電気泳動後、当該ゲル上で検出された約1.7kbのバンドを切り出し、pENTR/D−TOPOベクター(インビトロジェン社製)へクローニングしpENTR−CRMP2を作製した。次いで、4μl pENTR−CRMP2、1μl pcDNA6.2・V5−DEST及び2μl LRクロナーゼII(インビトロジェン社製)を混合し室温で1時間保持することにより、プラスミドpcDNA−CRMP2を作製した。
(5)pcDNA3.1の調製
pcDNA3.1は、インビトロジェン社から販売されている市販品を購入し、これを用いた。
(6)本発明形質転換体の作製およびシグナル伝達調節物質のスクリーニング
COS7細胞(ATCCからの購入品)を10cmシャーレに5×106cells/シャーレで播種し、10%FCSを含むDMEM培地(インビトロジェン社製)中で、37℃、5%CO2条件下にて約24時間培養した。次いで、当該細胞に、上記(2)で構築して得られた「Gm1発現プラスミド(pcDNA−Gm1; 12μg)」、上記(3)で構築して得られた「5HT4発現プラスミド(pcDNA−m5HT4;6μg)」及び、上記(4)で構築して得られた「CRMP2発現プラスミド(pcDNA−CRMP2;6μg)」の全てを、60μlのリポフェクトアミン2000(インビトロジェン社製)と混合し、20分間保持したものをトランスフェクションした。トランスフェクション3時間後に培地を5%透析FCSを含むDMEM培地に全交換し、更に約21時間培養した。
次いで、被験物質RS67333(1-(4-Amino-5-chloro-2-methoxyphenyl)-3-[1-butyl-4-piperidinyl]-1-propanone hydrochloride;C19H29ClN2O2.HCl)を、最終濃度10μMになるように前記で作製された本発明形質転換体である細胞の培養液中に添加した後、得られた培養物を37℃で30分間静置した。
次いで、得られた培養物から、RS67333を含む培地を除去した後、PhosphoProtein Purification kit(キアゲン)を用いて溶解することにより細胞抽出液を得た後、当該細胞抽出液から2−D Clean−Up kit(GEヘルスサイエンス)を用いてタンパク質抽出液(精製タンパク質)を調製した。尚、対照として、RS67333の代わりにDMSO溶媒のみを前記細胞の培養液中に添加した系を設定し、同様にしてタンパク質抽出液(精製タンパク質)を調製した。
得られたタンパク質抽出液(精製タンパク質800μg)を、Immobiline DryStrip(GEヘルスサイエンス)を用いた一次元目の電気泳動に供した。次いで、7.5%SDS−PAGEゲルを用い2次元目の電気泳動を行った。2次元電気泳動後のゲルを、Hybond−P(GEヘルスサイエンス)へ転写した。次いで、当該メンブレンの上に転写されたCRMP2タンパク質を抗CRMP2抗体(C4G)(株式会社免疫生物研究所)を用いてウエスタンブロッティングにより検出した(図1参照)。図1でアルカリ性側に位置するスポットNo.1は、非リン酸化型CRMP2タンパク質のスポットであり、それより酸性側に位置するNo.2−6は、リン酸化型CRMP2タンパク質のスポットである。また、より酸性側に位置するスポットほど、つまりNo.6 ほどリン酸化の程度が高いCRMP2タンパク質のスポットである。
次に、図1に示したウエスタンブロットにより検出されたそれぞれのスポットのシグナル強度を、LAS−4000(富士フィルム)を用い測定した。非リン酸化型CRMP2タンパク質のスポットの強度を「1」とした時のそれぞれリン酸化型CRMP2タンパク質のスポットの強度を算出した(図2参照)。図2に示された結果から明らかな通り、RS67333を接触させた細胞における「より高度にリン酸化されたスポット(スポットNo.3からNo.6)」のシグナル強度が、DMSO溶媒のみを接触させた細胞における「より高度にリン酸化されたスポット(スポットNo.3からNo.6)」のシグナル強度と比較して低下していた。
以上の結果より、RS67333を接触させた細胞において、リン酸化の程度が抑制された結果、より高度にリン酸化された状態のCRMP2タンパク質が減少したことが確認された。
次に、各スポットにおけるシグナルの変化の程度を定量化するために、DMSO溶媒のみを接触させた細胞におけるそれぞれのスポットのシグナル強度の値を100%とした場合の、RS67333を接触させた細胞におけるそれぞれのスポットのシグナル強度の相対値(パーセンテージ)を算出した(図3参照)。その結果、RS67333を接触させた細胞のより高度にリン酸化型されたCRMP2のスポット(スポットNo.3からNo.6)において、スポットのシグナル強度の相対値(パーセンテージ)がそれぞれ、54.1%、27.9%、22.4%及び16.4%であることが確認された。
以上の結果より、RS67333がGm1タンパク質を介するシグナル伝達調節物質であることが判明した。
【0032】
実施例2(本発明スクリーニング方法:Gm1タンパク質を介したシグナル伝達調節物質のスクリーニング)(その2)
(7)pcDNA−GPCRの構築
cDNA2μlを鋳型に用いて且つ10μMのフォワードプライマー、10μMのリバースプライマー及びKOD DNAポリメラーゼ(東洋紡製)を用いてPCRを行う。PCRで得られたDNAをアガロースゲル電気泳動後、当該ゲル上で検出されたバンドを切り出し、pENTR/D−TOPOベクター(インビトロジェン社製)へクローニングしpENTR−GPCRを作製した。次いで、4μl pENTR−GPCR、1μl pcDNA6.2・V5−DEST及び2μl LRクロナーゼII(インビトロジェン社製)を混合し室温で1時間保持することにより、プラスミドpcDNA−GPCRを作製する。
(8)本発明形質転換体の作製およびシグナル伝達調節物質のスクリーニング
COS7細胞(ATCCからの購入品)を6穴プレートに1×106cells/シャーレで播種し、10%FCSを含むDMEM培地(インビトロジェン社製)中で、37℃、5%CO2条件下にて約24時間培養する。次いで、当該細胞に、上記(2)で構築して得られた「Gm1発現プラスミド(pcDNA−Gm1)」、(7)で構築して得られる「GPCR発現プラスミド(pcDNA−GPCR)」及び、上記(4)で構築して得られた「CRMP2発現プラスミド(pcDNA−CRMP2)」の全てを、リポフェクトアミン2000(インビトロジェン社製)と混合し、20分間保持したものをトランスフェクションする。トランスフェクション3時間後に培地を全交換し、更に約21時間培養する。
次いで、各種の被験物質を、最終濃度10μMになるように前記で作製された本発明形質転換体である細胞の培養液中に添加した後、得られた培養物を37℃で30分間静置する。
次いで、得られた培養物から、前記被験物質を含む培地を除去した後、PhosphoProtein Purification kit(キアゲン)を用いて溶解することにより細胞抽出液を得た後、当該細胞抽出液から2−D Clean−Up kit(GEヘルスサイエンス)を用いてタンパク質抽出液(精製タンパク質)を調製する。尚、対照として、前記被験物質の代わりにDMSO溶媒のみを前記細胞の培養液中に添加した系を設定し、同様にしてタンパク質抽出液(精製タンパク質)を調製する。
得られたタンパク質抽出液(精製タンパク質800μg)を、Immobiline DryStrip(GEヘルスサイエンス)を用いた一次元目の電気泳動に供する。次いで、7.5%SDS−PAGEゲルを用い2次元目の電気泳動を行う。2次元電気泳動後のゲルを、Hybond−P(GEヘルスサイエンス)へ転写する。次いで、当該メンブレンの上に転写されたCRMP2タンパク質を抗CRMP2抗体(C4G)(株式会社免疫生物研究所)を用いてウエスタンブロッティングにより検出する。
次に、ウエスタンブロットにより検出されたそれぞれのスポットのシグナル強度を、LAS−4000(富士フィルム)を用い測定する。非リン酸化型CRMP2タンパク質のスポットの強度を「1」とした時のそれぞれリン酸化型CRMP2タンパク質のスポットの強度を算出する。
次に、各スポットにおけるシグナルの変化の程度を定量化するために、DMSO溶媒のみを接触させた細胞におけるそれぞれのスポットのシグナル強度の値を100%とした場合の、前記被験物質を接触させた細胞におけるそれぞれのスポットのシグナル強度の相対値(パーセンテージ)を算出する。その結果、前記被験物質を接触させた細胞のいずれかのリン酸化型CRMP2タンパク質のスポットにおいて、スポットのシグナル強度の相対値(パーセンテージ)が50%以下になる被験物質を、「Gm1タンパク質を介するシグナル伝達調節物質」として選択する。選択された「Gm1タンパク質を介したシグナル伝達調節物質」を有効成分として含有する薬剤(即ち、CRMP2タンパク質を介したシグナル伝達異常に起因する疾患の治療又は予防剤)を調製する。
【産業上の利用可能性】
【0033】
本発明により、Gm1タンパク質を介したシグナル伝達調節物質のスクリーニング方法及びそれに利用する形質転換体を含む試験ツール等を提供することが可能となる。
【配列表フリーテキスト】
【0034】
配列番号3
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号4
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号5
逆転写用PCRのために設計されたオリゴヌクレオチドプライマー
配列番号6
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号7
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号8
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号9
PCRのために設計されたオリゴヌクレオチドプライマー
【技術分野】
【0001】
本発明は、Gm1遺伝子、G-タンパク質共役型受容体遺伝子及びCRMP2遺伝子が導入されて得られる形質転換体並びにその利用に関するものである。
【背景技術】
【0002】
Gm1タンパク質は、G−タンパク質αサブユニットの一種であり、G−タンパク質共役受容体(GPCR)刺激による細胞内シグナル伝達に関与していることが知られている。また、Gm1遺伝子は、Golf遺伝子のスプライシングバリアントであり、第1エクソン領域のみ固有の塩基配列を有しており、第2エクソン領域以降はGolf遺伝子と塩基配列を共有している。また、Gm1タンパク質は、NCBIデータベースにおいて「NM_182978」で示されるG−タンパク質αサブユニットの一種であり、guanine nucleotide binding protein(G protein)、alpha activating activity polypeptide、olfactory type(GNAL)、transcript variant 1とも呼ばれることもあり、他のGタンパク質と比較してN末端領域が約80アミノ酸長い特徴的な構造を有している(例えば、特許文献1参照)。
G−タンパク質共役受容体(GPCR)は、7回膜貫通型の受容体であり、G-タンパク質と共役することが知られている(例えば、非特許文献11参照)。
一方、CRMP2タンパク質は、コラプシン応答メディエータタンパク質2(Collapsin Response Mediator Protein 2, CRMP2)は、神経ガイダンス因子であるセマフォリンの情報を媒介する因子として同定されたタンパク質である(例えば、非特許文献1参照)。当該タンパク質は、チューブリン、Numb、Sra−1、TrkB等の分子と直接的又は間接的に結合し、これらの分子を軸策先端へ輸送することにより軸策を伸長させる機能を有することが知られている(非特許文献2、非特許文献3、非特許文献4、非特許文献5)。また、当該タンパク質は、cdk5、GSK3β、Rhoキナーゼ等のキナーゼによりリン酸化されることにより、その活性が調節されること(具体的には、非リン酸化型が活性を有し、キナーゼによりリン酸化されることにより不活化される)が知られている(非特許文献6、非特許文献7)。更にまた、非リン酸化型変異遺伝子を神経細胞に導入すると軸策先端の退縮が損なわれることや、神経細胞でCRMP2タンパク質の生物学的機能を阻害すると軸策の伸長が阻害されること等も報告されている(非特許文献6、非特許文献8)。また、神経損傷後にCRMP2タンパク質を過剰発現させると神経軸索再生が促進されること(例えば、非特許文献9)や、CRMP2タンパク質の変異が統合失調症のリスク要因になっていること(例えば、非特許文献10)等も報告されている。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】特開2004−350672号
【非特許文献】
【0004】
【非特許文献1】Goshimaら, Nature (1995), 376, 509−514
【非特許文献2】Fukata Yら, Nat. Cell Biol., (2002) 4: 583−591
【非特許文献3】Nishimura Tら, Nat. Cell Biol., (2003) 5: 819−826
【非特許文献4】Arimuraら, Nat. Rev. Neurosci., 8: 194−205 (2007)
【非特許文献5】Arimuraら, Developmental Cell, 16, 675−686 (2009)
【非特許文献6】Yoshimura Tら, Cell, 120, 137−149, (2005)
【非特許文献7】Arimura Nら, Mol. Cell. Biol., 25, 9973−9984 (2005)
【非特許文献8】Arimuraら, J.Biol.Chem., 275, 23973−23980 (2000)
【非特許文献9】Suzuki Yら, J. Neurochem., 86, 1042−50, (2003)
【非特許文献10】Nakata Kら, Biol Psychiatry., 53, 571−576, (2003)
【非特許文献11】芳賀、蛋白質核酸酵素 Vol.46, 1764-1771, (2001)
【発明の概要】
【発明が解決しようとする課題】
【0005】
CRMP2タンパク質を介したシグナル伝達異常に起因すると思われる疾患、例えば、アルツハイマー病、統合失調症及び軸策損傷等のような疾患症状を改善させるために有用な知識・技術の蓄積・開発が急務であり、これに関連する試験ツール等の開発が期待されている。
【課題を解決するための手段】
【0006】
本発明者らは、このような状況を鑑み鋭意検討した結果、Gm1タンパク質の生物学的機能の活性化によりCRMP2タンパク質のリン酸化が抑制されること、即ち、CRMP2タンパク質の生物学的機能が活性化されることを新たに見出し、当該知見を利用することにより、本発明に至った。
即ち、本発明は
1.下記(1)、(2)及び(3)の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子が宿主細胞内に導入されて得られることを特徴とする形質転換体(以下、本発明形質転換体と記することもある。);
(1)Gm1タンパク質
(2)G-タンパク質共役型受容体タンパク質
(3)CRMP2タンパク質
2.前記外来遺伝子が、前記の3種類のタンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸であることを特徴とする前項1記載の形質転換体;
3.前項1又は2記載の形質転換体が、前記(1)、(2)及び(3)の3種類のタンパク質をその細胞内で共発現していることを特徴とする形質転換体;
4.(a)被験物質と、前項1、2又は3記載の形質転換体とを接触させる第一工程と、
(b)第一工程後に得られる形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を測定する第二工程と、
(c)第二工程で測定されたリン酸化状態又はそれと相関する指標値を、対照と比較して、前記形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を変動させる被験物質を選択する第三工程と
を含むことを特徴とするGm1タンパク質を介したシグナル伝達調節物質のスクリーニング方法(以下、本発明スクリーニング方法と記すこともある。);
5.第三工程における対照が、被験物質を接触させていない前項1、2又は3記載の形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値であることを特徴とする前項4記載のスクリーニング方法;
6.前項4又は5記載のスクリーニング方法により得られるGm1タンパク質を介したシグナル伝達調節物質;
7.前項6記載のGm1タンパク質を介したシグナル伝達調節物質を有効成分として含有することを特徴とする、CRMP2タンパク質を介したシグナル伝達異常に起因する疾患の治療又は予防剤(以下、本発明薬剤と記すこともある。);
【発明の効果】
【0007】
本発明により、Gm1タンパク質を介したシグナル伝達調節物質のスクリーニング方法及びそれに利用する形質転換体を含む試験ツール等を提供することが可能となる。
【図面の簡単な説明】
【0008】
【図1】図1は、2次元電気泳動サンプルの抗CRMP2抗体でのウエスタンブロットの結果を示す図である。上段図は対照(DMSO)での結果、下段図は被験物質としてRS67333を用いた結果である。アルカリ性側に位置するスポットNo.1は非リン酸化型CRMP2タンパク質のスポットであり、それより酸性側に位置するNo.2−6は、リン酸化型CRMP2タンパク質のスポットである。また、より酸性側に位置するスポットほど、つまりNo.6 ほどリン酸化の程度が高いCRMP2タンパク質のスポットである。
【図2】図2は、図1に示すウエスタンブロットの各スポットのシグナル強度の相対値に基づき、対照(溶媒)と被験物質との間で比較した結果を示す図である。黒棒グラフは対照(DMSO)での結果、白棒グラフは被験物質としてRS67333を用いた結果である。アルカリ性側に位置するスポットNo.1は非リン酸化型CRMP2タンパク質のスポットであり、それより酸性側に位置するNo.2−6は、リン酸化型CRMP2タンパク質のスポットである。また、より酸性側に位置するスポットほど、つまりNo.6 ほどリン酸化の程度が高いCRMP2タンパク質のスポットである。
【図3】図3は、図2に示す各スポットのシグナル強度の相対値(パーセンテージ)に基づき、対照(溶媒)と被験物質との間で比較した結果を示す図である。黒棒グラフは対照(DMSO)での結果、白棒グラフは被験物質としてRS67333を用いた結果である。アルカリ性側に位置するスポットNo.1は非リン酸化型CRMP2タンパク質のスポットであり、それより酸性側に位置するNo.2−6は、リン酸化型CRMP2タンパク質のスポットである。また、より酸性側に位置するスポットほど、つまりNo.6 ほどリン酸化の程度が高いCRMP2タンパク質のスポットである。
【発明を実施するための形態】
【0009】
本明細書において、DNAの調製やプラスミドの調製等の遺伝子工学的な手法、タンパク質の抽出やウエスタンブロッティング等のタンパク質工学的な手法は、特に明記しない限り、Molecular Cloning, A Laboratory Manual(T.Maniatis et al., Cold Spring Harbor Laboratory(1982))、新生化学実験講座(日本生化学会編;東京化学同人)等の実験書に記載される方法又はそれに準じた方法により行うことができる。
【0010】
本発明における「Gm1タンパク質」とは、背景技術に記載されるように、G−タンパク質共役受容体(GPCR)刺激による細胞内シグナル伝達に関与していることが知られている、G−タンパク質αサブユニットの一種である。当該タンパク質は、例えば、特開2004−350672号公報等に記載された公知なタンパク質である。
当該タンパク質が有するアミノ酸配列としては、例えば、前記公開公報に記載される配列番号1で示されるアミノ酸配列等を挙げることができる。尚、当該タンパク質は、NCBIデータベースにおいて「NM_182978」で示されるG−タンパク質αサブユニットの一種であり、guanine nucleotide binding protein(G protein)、alpha activating activity polypeptide、olfactory type(GNAL)、transcript variant 1とも呼ばれることもあり、他のGタンパク質と比較してN末端領域が約80アミノ酸長い特徴的な構造を有している。
Gm1遺伝子は、Golf遺伝子のスプライシングバリアントであり、第1エクソン領域のみ固有の塩基配列を有しており、第2エクソン領域以降はGolf遺伝子と塩基配列を共有している。Gm1タンパク質が有するアミノ酸配列をコードする塩基配列としては、例えば、前記公開公報に記載される配列番号2で示される塩基配列等を挙げることができる。
【0011】
本発明における「Gm1タンパク質」は、当該タンパク質の生物学的機能を維持する限り、その一部からなるものを含む。例えば、前記公開公報に記載される配列番号1で示されるアミノ酸配列は、Gタンパク質αサブユニット間で保存されている「GTP結合部位」及び「GTPase活性化部位」のアミノ酸配列と配列同一性が高いアミノ酸配列部分を有する。これらの部分は、前記公開公報に記載される配列番号1で示されるアミノ酸配列におけるアミノ酸番号126〜133、287〜292、353〜359、428〜435の各領域である。これらのアミノ酸領域は、既にGタンパク質αサブユニットとして同定されている「Gs」及び「Golf」における「GTP結合部位」及び「GTPase活性化部位」のアミノ酸配列(NATURE, 117−127 (1991), vol. 349)と一致している。また、配列番号1で示されるアミノ酸配列は、Gタンパク質αサブユニット間で特にGsファミリーに属する「Gs」及び「Golf」で高く保存されている特徴的配列と同じ配列(配列番号1で示されるアミノ酸番号119〜126)を有し、しかもGタンパク質αサブユニット間で保存されているαヘリックス構造をとりうる配列である。
更に、Gm1タンパク質の生物学的機能を喪失すること無く、Gm1タンパク質が有する如何なるアミノ酸残基を何個置換、欠失又は付加できるかの指標は、特開2003−193330に記載された方法等により見出すことができる。前記生物学的機能を喪失しない改変は、例えば、既に同定されている各種のGタンパク質αサブユニットのアミノ酸配列との配列同一性が低い部分について行えばよい。
【0012】
本発明における「CRMP2タンパク質」とは、背景技術に記載されるように、コラプシン応答メディエータタンパク質2(Collapsin Response Mediator Protein 2, CRMP2)は、神経ガイダンス因子であるセマフォリンの情報を媒介する因子として同定されたタンパク質である。当該タンパク質は、例えば、Goshimaら, Nature (1995), 376, 509−514等に記載された公知なタンパク質である。
当該タンパク質が有するアミノ酸配列、及び、当該アミノ酸配列をコードする塩基配列としては、例えば、前記学術文献に記載されるアミノ酸配列、及び、塩基配列等を挙げることができる。尚、当該タンパク質は、NCBIデータベースにおいて「NP_001377」で示されるタンパク質であり、dihydropyrimidinase−like 2、dihydropyrimidinase−like 2、DPYSL2とも呼ばれることもある。
本発明における「G-タンパク質共役型受容体タンパク質」(GPCR)とは、7回膜貫通型の受容体であり、G-タンパク質と共役することが知られている。当該G-タンパク質共役型受容体タンパク質として、例えば、Sevens databaseのホームページ(http://sevens.cbrc.jp/index.php)等に登録されているような受容体を挙げることができ、例えば、ドーパミン受容体、セロトニン受容体及びヒスタミン受容体等を挙げることができ、これらが有するアミノ酸配列、及び、当該アミノ酸配列をコードする塩基配列は、例えば、NCBIのホームページ(http://www.ncbi.nlm.nih.gov/)等より入手することができる。より具体的には、ドーパミンD1受容体の場合には、例えば、NCBIデータベースにおいて「NP_000785.1」で示されるアミノ酸配列、NCBIデータベースにおいて「NM_000794.3」で示される塩基配列等を挙げることができ、またセロトニン4受容体の場合には、例えば、NCBIデータベースにおいて「NP_001035259.1」で示されるアミノ酸配列、NCBIデータベースにおいて「NM_001040169.2」で示される塩基配列等を挙げることができ、またヒスタミンH1受容体の場合には、例えば、NCBIデータベースにおいて「NP_001091683.1」で示されるアミノ酸配列、NCBIデータベースにおいて「NM_001098213.1」で示される塩基配列等を挙げることができる。
【0013】
本発明形質転換体は、下記(1)、(2)及び(3)の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子が宿主細胞内に導入されて得られる。
(1)Gm1タンパク質
(2)G-タンパク質共役型受容体タンパク質
(2)CRMP2タンパク質
【0014】
本発明形質転換体の宿主細胞としては、公知のいかなる宿主細胞でも用いることができ、例えば、大腸菌(例えば、K12等)、バチルス属細菌(例えば、MI114等)等の細菌、酵母(例えば、AH22等)、昆虫細胞(例えば、Sf細胞等)、動物細胞(例えば、COS7細胞等)等を挙げることができる。なかでも、哺乳動物由来の培養細胞(例えば、COS7細胞、CHO細胞、HEK293細胞、Hela細胞、PC12細胞等)を好ましく挙げることができる。
【0015】
本発明形質転換体は、このような宿主細胞に対して、Gm1タンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸、及び、G-タンパク質共役型受容体タンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸、及び、CRMP2タンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸、の全てを外来的に導入するような人為的操作を施すことにより作製することができる。好ましくは、Gm1タンパク質のアミノ酸配列をコードする塩基配列、G-タンパク質共役型受容体タンパク質のアミノ酸配列をコードする塩基配列及びCRMP2タンパク質のアミノ酸配列をコードする塩基配列の全てを含有するプラスミドの形態である核酸を外来的に導入するような人為的操作を施すことにより作製することができる。
前記人為的操作としては、用いられる宿主細胞に応じた通常の遺伝子工学的手法を利用すればよい。具体的には例えば、市販のトランスフェクション試薬を用いて、当該試薬に添付された説明書に記載される方法に従い行う方法を挙げることができる。また、外来遺伝子としてプラスミドの形態である核酸を利用する場合には、リン酸カルシウム法、エレクトロポレーション法、リポフェクション法、DEAEデキストラン法等を挙げることができる。
【0016】
ここで「プラスミド」としては、Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質を宿主細胞内で共発現させることが可能なプラスミドを挙げることができる。このようなプラスミドは、通常の遺伝子工学的手法に従い、Gm1タンパク質のアミノ酸配列をコードする塩基配列を含有する核酸、G-タンパク質共役型受容体タンパク質のアミノ酸配列をコードする塩基配列を含有する核酸、及び、CRMP2タンパク質のアミノ酸配列をコードする塩基配列を含有する核酸、の全てを、公知の発現プラスミドのプロモーター下流の発現可能な位置に連結することにより作製することができる。
ここで「発現プラスミド」としては、例えば、大腸菌を宿主細胞とする場合には、pBR322、pUC12、pUC119、pBluescript等、バチルス属細菌を宿主細胞とする場合には、pUB110、pC194等、酵母を宿主細胞とする場合には、Yip5、Yep24等、昆虫細胞を宿主細胞とする場合には、AcNPV等、動物細胞を宿主細胞とする場合には、pUC18、pcDNA3.1等の公知のプラスミドを挙げることができる。
【0017】
本発明形質転換体における、Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子の宿主細胞内での保持は、一過的に保持されたものであってもよいし、また安定的に保持させたものであってもよい。安定的に保持された形質転換体は、当該外来遺伝子と同時に宿主細胞内に導入された薬剤耐性マーカー等(例えば、発現プラスミドに含有された薬剤耐性マーカー等)を指標にして選択すればよい。
【0018】
本発明形質転換体の細胞内で、Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質を共発現させるには、例えば、本発明形質転換体を培養液の中で培養すればよい。
本発明形質転換体の培養条件は、本発明形質転換体の種類に応じた条件を適宜選択すればよい。
【0019】
例えば、本発明形質転換体が微生物である場合には、微生物の培養に通常使用される液体培地又は平板培地を用いて培養すればよい。培養温度は、微生物の生育可能温度内であればよく、例えば、15〜40℃程度を挙げることができる。培地のpHも、微生物の生育可能範囲であればよく、例えば、pH6〜8程度を挙げることができる。培養時間は、その他の培養条件により異なるが、通常、1〜5日間程度、特に1〜2日間程度にすればよい。温度シフト型やIPTG誘導型等の誘導型の発現プラスミドを用いる場合には、誘導時間は1日間以内、特に数時間程度とすればよい。
【0020】
また形質転換体が昆虫細胞の場合には、昆虫細胞の種類に応じて適した培養条件とすればよい。例えば、FBS及びYeastlateを含むGrace's medium等の昆虫細胞用培地を用いて培養すればよい。培養温度は、25〜35℃程度を挙げることができる。培地のpHも、昆虫細胞の生育可能範囲であればよく、例えば、pH6〜8程度を挙げることができる。培養時間は、その他の培養条件により異なるが、通常、1〜5日間程度、特に2〜3日間程度にすればよい。
発現プラスミドとしてBaculovirus等のウィルスを含む形質転換体の場合は、培養時間は細胞質効果が現れて細胞が死滅する前まで(例えば3〜7日間程度、特に4〜6日間程度)とするのが好ましい。
【0021】
また形質転換体が哺乳動物細胞の場合には、哺乳動物細胞の種類に応じて適した培養条件とすればよい。例えば、FBSを添加したDMEM培地(ニッスイ社製等)を用いて5%CO2存在下、数日毎に新しい培地に交換しながら培養すればよい。哺乳動物細胞がコンフルエントになるまで増殖したら、例えば、トリプシン溶液を加えて個々の細胞に分散させ、得られた細胞の懸濁液を数倍に希釈して新しいシャーレに播種し継代を続ければよい。
培養温度は、36〜38℃程度を挙げることができる。培地のpHも、哺乳動物細胞の生育可能範囲であればよく、例えば、pH6〜8程度を挙げることができる。培養時間は、その他の培養条件により異なるが、通常、2〜5日間程度、特に2〜3日間程度にすればよい。
【0022】
本発明形質転換体の細胞内で、Gm1タンパク質、G-タンパク質共役型受容体タンパク質及びCRMP2タンパク質の3種類のタンパク質が共発現していることを確認する方法としては、通常のタンパク質工学的方法を利用すればよい。具体的には例えば、本発明形質転換体より調製されたタンパク質抽出液を、抗Gm1タンパク質抗体、抗G-タンパク質共役型受容体タンパク質抗体及び抗CRMP2タンパク質抗体を用いてウエスタンブロットを行う方法等を挙げることができる。
【0023】
本発明スクリーニング方法は、(a)被験物質と、前項1、2又は3記載の形質転換体とを接触させる第一工程と、(b)第一工程後に得られる形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を測定する第二工程と、(c)第二工程で測定されたリン酸化状態又はそれと相関する指標値を、対照と比較して、前記形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を変動させる被験物質を選択する第三工程とを含む。
【0024】
本発明スクリーニング方法において「被験物質」としては、特に限定は無く、例えば、タンパク質(抗体を含む)、ペプチド、非ペプチド性化合物(ヌクレオチド、アミン、糖質、脂質等)、有機低分子化合物、無機低分子化合物、醗酵生産物、細胞抽出液、植物抽出液、動物組織抽出液等を挙げることができる。被験物質の形態としては、特に限定は無く、固体、液体、基剤との混合物、縣濁液又は溶液等を挙げることができる。縣濁液若しくは溶液とする場合、水、pH緩衝液、メチルセルロース溶液、生理食塩水、有機溶媒水溶液(有機溶媒としては通常エタノールやジメチルスルホキシドが用いられる。)等を用いればよい。
被験物質の接触量、接触回数及び接触時間は、例えば、本発明形質転換体に重篤な影響を及ぼさない範囲内とすればよい。
【0025】
本発明スクリーニング方法の第一工程における「接触」は、当業者に汎用されている方法で実施することができる。被験物質を本発明形質転換体に接触する場合、その接触方法には特に限定はなく、例えば、本発明形質転換体を含む培養液・寒天培地等の中に、0.001〜10μM程度の濃度となるように被験物質を添加すればよい。
【0026】
本発明スクリーニング方法の第二工程における「CRMP2タンパク質のリン酸化状態又はそれと相関する指標値を測定する」方法では、タンパク質抽出液の調製、電気泳動法、メンブレンの上への転写、CRMP2タンパク質の検出方法等を用いることができる。これらの方法では、通常の生化学的手法・タンパク質工学的手法等を用いて分析すればよい。例えば、2次元電気泳動法とウエスタンブロット法とを組み合わることにより行えばよい。具体的には例えば、まず被験物質を接触させた本発明形質転換体からタンパク質抽出液を調製する。調製されたタンパク質抽出液を1次元目の電気泳動を行ない、次いで2次元目の電気泳動を行う。分離されたCRMP2タンパク質を適当なメンブレンの上に転写する。次いで、当該メンブレンの上に転写されたCRMP2タンパク質を抗CRMP2タンパク質抗体等で検出する。これら操作の結果、CRMP2タンパク質はリン酸化の程度の違いによって分離される。
非リン酸化型CRMP2は、よりアルカリ性側に検出され、リン酸化の程度が高くなるほど、より酸性側に検出される。尚、は、それ自体公知の通常用いられる方法に従い行うことができる。
また、CRMP2のリン酸化状態を定量する方法としては、公知の定量化法を用いればよいが、例えば、ウエスタンブロットにより検出されたシグナルをイメージアナライザー等にて定量化すればよい。具体的には例えば、本発明形質転換体において2次元電気泳動後のウエスタンブロットにより検出されたCRMP2のスポットの非リン酸化型CRMP2のスポット強度を「1」とし、それぞれのリン酸化型CRMP2のスポット強度を算出する。同様に、対照におけるCRMP2のスポットの強度も、非リン酸化型CRMP2のスポット強度を「1」とし、それぞれのリン酸化型CRMP2のスポット強度を算出する。次いで、対照におけるそれぞれのスポットを100%とし、被検物質を接触させた本発明形質転換体で検出されたCRMP2のそれぞれのスポットのパーセンテージを算出する。被検物質を接触させた本発明形質転換体において検出されたCRMP2のそれぞれのスポットのパーセンテージのいずれかが、例えば70%以下、好ましくは60%以下、更に好ましくは50%以下になる被検物質を、シグナル伝達調節物質の候補物質として選択すればよい。また好ましくは、より高度にリン酸化されたスポットにおいて、スポットのパーセンテージが、例えば70%以下、好ましくは60%以下、更に好ましくは50%以下になる被検物質を、シグナル伝達を活性化する候補物質として選択すればよい。
【0027】
本発明スクリーニング方法の第三工程における対照との比較では、例えば、被験物質を接触させた本発明形質転換体におけるCRMP2タンパク質の第二工程で測定されたリン酸化状態又はそれと相関する指標値を、被験物質を接触させていない本発明形質転換体におけるCRMP2タンパク質の第二工程で測定されたリン酸化状態又はそれと相関する指標値と比較すればよい。ここで「被験物質を接触させていない本発明形質転換体」とは、例えば、溶媒のみを接触させた本発明形質転換体であってもよい。
かかる対照は、対照とするCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を、本発明形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値と併行して測定して求めてもよいし、別途測定して求めてもよい。
例えば、被験物質と接触させた本発明形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値の測定値が、対照とする本発明形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値の測定値よりも低ければ、前記被験物質との接触は、CRMP2タンパク質のリン酸化状態を抑制することによりCRMP2タンパク質を活性化させる効果を有することを意味し、そして当該被験物質はGm1を介したシグナル伝達調節物質として選択することができる。
【0028】
尚、被験物質の代わりに「対照物質」となり得るポジティブコントロール又はネガティブコントロールを用いて本発明スクリーニング方法を実施することにより、場合に応じて上記の対照とすることもできる。
ここで「ポジティブコントロール」とは、CRMP2タンパク質のリン酸化状態を制御する効果を有する任意の物質を表し、また「ネガティブコントロール」としては、被験物質に含まれる溶媒、バックグランドとなる試験系溶液等が挙げられる。
「対照物質」をネガティブコントロールとする場合、被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」が、対照物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」よりも大きければ、当該被験物質は「CRMP2タンパク質のリン酸化状態を抑制する効果」を有すると評価すればよい。一方、被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」が、対照物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」と同程度若しくは小さければ、当該被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」を有さないと評価することができる。
また、対照物質をポジティブコントロールとする場合、被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」と、対照物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」とを比較することにより、被験物質が有する「CRMP2タンパク質のリン酸化状態を抑制する効果」を評価すればよい。
このようにして効果を有すると評価された場合には、当該被験物質を、Gm1を介したシグナル伝達調節物質として選択することができる。
【0029】
本発明薬剤は、例えば、上記のようにして選択された「Gm1タンパク質を介したシグナル伝達調節物質」を有効成分として含有する。本発明薬剤は、CRMP2タンパク質を介したシグナル伝達異常に起因する疾患の治療又は予防剤として有用である。
【実施例】
【0030】
以下、本発明の実施例を示してより詳細に説明するが、本発明はこれらの実施例に限定されるものではない。
【0031】
実施例1(本発明スクリーニング方法:Gm1タンパク質を介したシグナル伝達調節物質のスクリーニング)(その1)
(1)Gm1遺伝子の取得
文献(特開2004−350672号)記載の方法に準じた方法を用いて、ヒト脳由来cDNAライブラリー(宝酒造社製)20ngを鋳型に用いて且つ10μMのフォワードプライマーprGm1ATG(5’−ATGGGTCTGTGCTACAGTCTGCGG;配列番号3)、10μMのリバースプライマーprGNAL3’(5’−TCACAAGAGCTCATACTGCTT;配列番号4)及びTAKARA LA Taqポリメラーゼ(TAKARALA Taq with GC Buffer,宝酒造社製)を用いてPCRを行った。PCR条件は、95℃で30秒間、次いで60℃で30秒間、次いで72℃で2分間の保温を1サイクルとしてこれを35サイクル行った。得られたDNA断片をアガロースゲル電気泳動後、当該ゲル上で検出された約1.5kbのバンドを切り出し、pCR2.1−TOPOベクター(インビトロジェン社製)へクローニングしpCR−Gm1を作製した。
(2)pcDNA−Gm1の構築
文献(特開2004−350672号)記載の方法に準じた方法を用いて、実施例1で作製されたプラスミドpCR−Gm1を制限酵素XbaI及びKpnIで二重消化した後、このDNA断片をアガロースゲル電気泳動し、当該ゲル上で検出された約1.5kbのバンドを切り出し、インサートDNAとした。pcDNA3.1(インビトロジェン社製)をXbaI及びKpnIで二重消化したものをベクターとして用い、これにインサートDNAをライゲースを用いて連結することにより、プラスミドpcDNA−Gm1を作製した。
(3)pcDNA−m5HT4の構築
マウス脳より調製したRNA200ngを鋳型に用いて且つ10μMのリバースプライマーm5HT4-3’(5’−CTAAGTATCACTGGGCTGAGCAGCCA;配列番号5)及びSuper Script III逆転写酵素(インビトロジェン社製)を用いて、55℃で50分間、次いで75℃で15分間保温し、逆転写を行った。
得られたcDNA2μlを鋳型に用いて且つ10μMのフォワードプライマーm5HT4-5’(5’−ATGGACAAACTTGATGCTAATGTGAGTT;配列番号6)、10μMのリバースプライマーm5HT4-3’(5’−CTAAGTATCACTGGGCTGAGCAGCCA;配列番号7)及びKOD DNAポリメラーゼ(東洋紡製)を用いてPCRを行った。PCR条件は、95℃で30秒間、次いで60℃で30秒間、次いで68℃で2分間の保温を1サイクルとしてこれを35サイクル行った。得られたDNAをアガロースゲル電気泳動後、当該ゲル上で検出された約1.2kbのバンドを切り出し、pDriveベクター(キアゲン社製)へクローニングしpDr−m5HT4を作製した。次いでプラスミドpDr−m5HT4を制限酵素NheI及びKpnIで二重消化した後、このDNA断片をアガロースゲル電気泳動し、当該ゲル上で検出された約1.2kbのバンドを切り出し、インサートDNAとした。pcDNA3.1(インビトロジェン社製)をNheI及びKpnIで二重消化したものをベクターとして用い、これにインサートDNAをライゲースを用いて連結することにより、プラスミドpcDNA−m5HT4を作製した。
(4)pcDNA−CRMP2の構築
ヒト脳由来cDNAライブラリー(宝酒造社製)20ngを鋳型に用いて且つ10μMのフォワードプライマーhCRMP2(ATG)(5’−CACCATGTCTTATCAGGGGAAGAAAAA;配列番号8)、10μMのリバースプライマーhCRMP2(−STOP)rev(5’−GCCCAGGCTGGTGATGTTGGCACGGCCA;配列番号9)及びKOD DNAポリメラーゼ(東洋紡製)を用いてPCRを行った。PCR条件は、95℃で30秒間、次いで60℃で30秒間、次いで68℃で2分間の保温を1サイクルとしてこれを35サイクル行った。得られたDNAをアガロースゲル電気泳動後、当該ゲル上で検出された約1.7kbのバンドを切り出し、pENTR/D−TOPOベクター(インビトロジェン社製)へクローニングしpENTR−CRMP2を作製した。次いで、4μl pENTR−CRMP2、1μl pcDNA6.2・V5−DEST及び2μl LRクロナーゼII(インビトロジェン社製)を混合し室温で1時間保持することにより、プラスミドpcDNA−CRMP2を作製した。
(5)pcDNA3.1の調製
pcDNA3.1は、インビトロジェン社から販売されている市販品を購入し、これを用いた。
(6)本発明形質転換体の作製およびシグナル伝達調節物質のスクリーニング
COS7細胞(ATCCからの購入品)を10cmシャーレに5×106cells/シャーレで播種し、10%FCSを含むDMEM培地(インビトロジェン社製)中で、37℃、5%CO2条件下にて約24時間培養した。次いで、当該細胞に、上記(2)で構築して得られた「Gm1発現プラスミド(pcDNA−Gm1; 12μg)」、上記(3)で構築して得られた「5HT4発現プラスミド(pcDNA−m5HT4;6μg)」及び、上記(4)で構築して得られた「CRMP2発現プラスミド(pcDNA−CRMP2;6μg)」の全てを、60μlのリポフェクトアミン2000(インビトロジェン社製)と混合し、20分間保持したものをトランスフェクションした。トランスフェクション3時間後に培地を5%透析FCSを含むDMEM培地に全交換し、更に約21時間培養した。
次いで、被験物質RS67333(1-(4-Amino-5-chloro-2-methoxyphenyl)-3-[1-butyl-4-piperidinyl]-1-propanone hydrochloride;C19H29ClN2O2.HCl)を、最終濃度10μMになるように前記で作製された本発明形質転換体である細胞の培養液中に添加した後、得られた培養物を37℃で30分間静置した。
次いで、得られた培養物から、RS67333を含む培地を除去した後、PhosphoProtein Purification kit(キアゲン)を用いて溶解することにより細胞抽出液を得た後、当該細胞抽出液から2−D Clean−Up kit(GEヘルスサイエンス)を用いてタンパク質抽出液(精製タンパク質)を調製した。尚、対照として、RS67333の代わりにDMSO溶媒のみを前記細胞の培養液中に添加した系を設定し、同様にしてタンパク質抽出液(精製タンパク質)を調製した。
得られたタンパク質抽出液(精製タンパク質800μg)を、Immobiline DryStrip(GEヘルスサイエンス)を用いた一次元目の電気泳動に供した。次いで、7.5%SDS−PAGEゲルを用い2次元目の電気泳動を行った。2次元電気泳動後のゲルを、Hybond−P(GEヘルスサイエンス)へ転写した。次いで、当該メンブレンの上に転写されたCRMP2タンパク質を抗CRMP2抗体(C4G)(株式会社免疫生物研究所)を用いてウエスタンブロッティングにより検出した(図1参照)。図1でアルカリ性側に位置するスポットNo.1は、非リン酸化型CRMP2タンパク質のスポットであり、それより酸性側に位置するNo.2−6は、リン酸化型CRMP2タンパク質のスポットである。また、より酸性側に位置するスポットほど、つまりNo.6 ほどリン酸化の程度が高いCRMP2タンパク質のスポットである。
次に、図1に示したウエスタンブロットにより検出されたそれぞれのスポットのシグナル強度を、LAS−4000(富士フィルム)を用い測定した。非リン酸化型CRMP2タンパク質のスポットの強度を「1」とした時のそれぞれリン酸化型CRMP2タンパク質のスポットの強度を算出した(図2参照)。図2に示された結果から明らかな通り、RS67333を接触させた細胞における「より高度にリン酸化されたスポット(スポットNo.3からNo.6)」のシグナル強度が、DMSO溶媒のみを接触させた細胞における「より高度にリン酸化されたスポット(スポットNo.3からNo.6)」のシグナル強度と比較して低下していた。
以上の結果より、RS67333を接触させた細胞において、リン酸化の程度が抑制された結果、より高度にリン酸化された状態のCRMP2タンパク質が減少したことが確認された。
次に、各スポットにおけるシグナルの変化の程度を定量化するために、DMSO溶媒のみを接触させた細胞におけるそれぞれのスポットのシグナル強度の値を100%とした場合の、RS67333を接触させた細胞におけるそれぞれのスポットのシグナル強度の相対値(パーセンテージ)を算出した(図3参照)。その結果、RS67333を接触させた細胞のより高度にリン酸化型されたCRMP2のスポット(スポットNo.3からNo.6)において、スポットのシグナル強度の相対値(パーセンテージ)がそれぞれ、54.1%、27.9%、22.4%及び16.4%であることが確認された。
以上の結果より、RS67333がGm1タンパク質を介するシグナル伝達調節物質であることが判明した。
【0032】
実施例2(本発明スクリーニング方法:Gm1タンパク質を介したシグナル伝達調節物質のスクリーニング)(その2)
(7)pcDNA−GPCRの構築
cDNA2μlを鋳型に用いて且つ10μMのフォワードプライマー、10μMのリバースプライマー及びKOD DNAポリメラーゼ(東洋紡製)を用いてPCRを行う。PCRで得られたDNAをアガロースゲル電気泳動後、当該ゲル上で検出されたバンドを切り出し、pENTR/D−TOPOベクター(インビトロジェン社製)へクローニングしpENTR−GPCRを作製した。次いで、4μl pENTR−GPCR、1μl pcDNA6.2・V5−DEST及び2μl LRクロナーゼII(インビトロジェン社製)を混合し室温で1時間保持することにより、プラスミドpcDNA−GPCRを作製する。
(8)本発明形質転換体の作製およびシグナル伝達調節物質のスクリーニング
COS7細胞(ATCCからの購入品)を6穴プレートに1×106cells/シャーレで播種し、10%FCSを含むDMEM培地(インビトロジェン社製)中で、37℃、5%CO2条件下にて約24時間培養する。次いで、当該細胞に、上記(2)で構築して得られた「Gm1発現プラスミド(pcDNA−Gm1)」、(7)で構築して得られる「GPCR発現プラスミド(pcDNA−GPCR)」及び、上記(4)で構築して得られた「CRMP2発現プラスミド(pcDNA−CRMP2)」の全てを、リポフェクトアミン2000(インビトロジェン社製)と混合し、20分間保持したものをトランスフェクションする。トランスフェクション3時間後に培地を全交換し、更に約21時間培養する。
次いで、各種の被験物質を、最終濃度10μMになるように前記で作製された本発明形質転換体である細胞の培養液中に添加した後、得られた培養物を37℃で30分間静置する。
次いで、得られた培養物から、前記被験物質を含む培地を除去した後、PhosphoProtein Purification kit(キアゲン)を用いて溶解することにより細胞抽出液を得た後、当該細胞抽出液から2−D Clean−Up kit(GEヘルスサイエンス)を用いてタンパク質抽出液(精製タンパク質)を調製する。尚、対照として、前記被験物質の代わりにDMSO溶媒のみを前記細胞の培養液中に添加した系を設定し、同様にしてタンパク質抽出液(精製タンパク質)を調製する。
得られたタンパク質抽出液(精製タンパク質800μg)を、Immobiline DryStrip(GEヘルスサイエンス)を用いた一次元目の電気泳動に供する。次いで、7.5%SDS−PAGEゲルを用い2次元目の電気泳動を行う。2次元電気泳動後のゲルを、Hybond−P(GEヘルスサイエンス)へ転写する。次いで、当該メンブレンの上に転写されたCRMP2タンパク質を抗CRMP2抗体(C4G)(株式会社免疫生物研究所)を用いてウエスタンブロッティングにより検出する。
次に、ウエスタンブロットにより検出されたそれぞれのスポットのシグナル強度を、LAS−4000(富士フィルム)を用い測定する。非リン酸化型CRMP2タンパク質のスポットの強度を「1」とした時のそれぞれリン酸化型CRMP2タンパク質のスポットの強度を算出する。
次に、各スポットにおけるシグナルの変化の程度を定量化するために、DMSO溶媒のみを接触させた細胞におけるそれぞれのスポットのシグナル強度の値を100%とした場合の、前記被験物質を接触させた細胞におけるそれぞれのスポットのシグナル強度の相対値(パーセンテージ)を算出する。その結果、前記被験物質を接触させた細胞のいずれかのリン酸化型CRMP2タンパク質のスポットにおいて、スポットのシグナル強度の相対値(パーセンテージ)が50%以下になる被験物質を、「Gm1タンパク質を介するシグナル伝達調節物質」として選択する。選択された「Gm1タンパク質を介したシグナル伝達調節物質」を有効成分として含有する薬剤(即ち、CRMP2タンパク質を介したシグナル伝達異常に起因する疾患の治療又は予防剤)を調製する。
【産業上の利用可能性】
【0033】
本発明により、Gm1タンパク質を介したシグナル伝達調節物質のスクリーニング方法及びそれに利用する形質転換体を含む試験ツール等を提供することが可能となる。
【配列表フリーテキスト】
【0034】
配列番号3
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号4
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号5
逆転写用PCRのために設計されたオリゴヌクレオチドプライマー
配列番号6
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号7
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号8
PCRのために設計されたオリゴヌクレオチドプライマー
配列番号9
PCRのために設計されたオリゴヌクレオチドプライマー
【特許請求の範囲】
【請求項1】
下記(1)、(2)及び(3)の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子が宿主細胞内に導入されて得られることを特徴とする形質転換体。
(1)Gm1タンパク質
(2)G-タンパク質共役型受容体タンパク質
(3)CRMP2タンパク質
【請求項2】
前記外来遺伝子が、前記の3種類のタンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸であることを特徴とする請求項1記載の形質転換体。
【請求項3】
請求項1又は2記載の形質転換体が、前記(1)、(2)及び(3)の3種類のタンパク質をその細胞内で共発現していることを特徴とする形質転換体
【請求項4】
(a)被験物質と、請求項1、2又は3記載の形質転換体とを接触させる第一工程と、
(b)第一工程後に得られる形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を測定する第二工程と、
(c)第二工程で測定されたリン酸化状態又はそれと相関する指標値を、対照と比較して、前記形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を変動させる被験物質を選択する第三工程と
を含むことを特徴とするGm1タンパク質を介したシグナル伝達調節物質のスクリーニング方法。
【請求項5】
第三工程における対照が、被験物質を接触させていない請求項1、2又は3記載の形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値であることを特徴とする請求項4記載のスクリーニング方法。
【請求項6】
請求項4又は5記載のスクリーニング方法により得られるGm1タンパク質を介したシグナル伝達調節物質。
【請求項7】
請求項6記載のGm1タンパク質を介したシグナル伝達調節物質を有効成分として含有することを特徴とする、CRMP2タンパク質を介したシグナル伝達異常に起因する疾患の治療又は予防剤。
【請求項1】
下記(1)、(2)及び(3)の3種類のタンパク質のアミノ酸配列をコードする塩基配列を有する外来遺伝子が宿主細胞内に導入されて得られることを特徴とする形質転換体。
(1)Gm1タンパク質
(2)G-タンパク質共役型受容体タンパク質
(3)CRMP2タンパク質
【請求項2】
前記外来遺伝子が、前記の3種類のタンパク質のアミノ酸配列をコードする塩基配列を含有するプラスミドの形態である核酸であることを特徴とする請求項1記載の形質転換体。
【請求項3】
請求項1又は2記載の形質転換体が、前記(1)、(2)及び(3)の3種類のタンパク質をその細胞内で共発現していることを特徴とする形質転換体
【請求項4】
(a)被験物質と、請求項1、2又は3記載の形質転換体とを接触させる第一工程と、
(b)第一工程後に得られる形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を測定する第二工程と、
(c)第二工程で測定されたリン酸化状態又はそれと相関する指標値を、対照と比較して、前記形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値を変動させる被験物質を選択する第三工程と
を含むことを特徴とするGm1タンパク質を介したシグナル伝達調節物質のスクリーニング方法。
【請求項5】
第三工程における対照が、被験物質を接触させていない請求項1、2又は3記載の形質転換体におけるCRMP2タンパク質のリン酸化状態又はそれと相関する指標値であることを特徴とする請求項4記載のスクリーニング方法。
【請求項6】
請求項4又は5記載のスクリーニング方法により得られるGm1タンパク質を介したシグナル伝達調節物質。
【請求項7】
請求項6記載のGm1タンパク質を介したシグナル伝達調節物質を有効成分として含有することを特徴とする、CRMP2タンパク質を介したシグナル伝達異常に起因する疾患の治療又は予防剤。
【図1】



【図2】
【図3】
【図2】
【図3】
【公開番号】特開2012−165656(P2012−165656A)
【公開日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願番号】特願2011−26902(P2011−26902)
【出願日】平成23年2月10日(2011.2.10)
【出願人】(000002093)住友化学株式会社 (8,981)
【Fターム(参考)】
【公開日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願日】平成23年2月10日(2011.2.10)
【出願人】(000002093)住友化学株式会社 (8,981)
【Fターム(参考)】
[ Back to top ]