
H.ピロリのflbA遺伝子のクローニング及び特徴づけ、並びに非鞭毛(Aflagellate)株の産生
【課題】本発明は、ヘリコバクター ピロリの鞭毛蛋白質の生合成制御に関わるヌクレオチド配列、該配列がコードする蛋白質、非鞭毛細菌株、及びH.ピロリによる感染を検出するための方法、またはそのような感染を予防するための方法を提供すること。
【解決手段】感染患者の血清による認識には影響を与えないが、特にカンピロバクター(Campylobacter )ファミリの細菌(例としてはCampylobacter jejuni)に関しての「偽陽性」型の反応を避けることが可能な、H. ピロリ株の修飾を行う。得られた修飾済み細菌を、免疫組成物または組成物を構築するにあたってワクチン接種に用いる。
【解決手段】感染患者の血清による認識には影響を与えないが、特にカンピロバクター(Campylobacter )ファミリの細菌(例としてはCampylobacter jejuni)に関しての「偽陽性」型の反応を避けることが可能な、H. ピロリ株の修飾を行う。得られた修飾済み細菌を、免疫組成物または組成物を構築するにあたってワクチン接種に用いる。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[発明の属する技術分野]
本発明は、現在までのところヒトの胃粘膜表面上に特異的に見られることがわかっている、グラム陰性細菌であるヘリコバクター・ピロリ(H. ピロリとも命名されている)に関する。
【0002】
[従来の技術]
多くの細菌と共通して、H. ピロリもまた、酸性p Hの培養液に対しては感受性であるが、生理学的濃度の尿素存在下においては、酸性度を許容することが可能である(Marshallら、1990、Gastroenterol. 99: 697- 702)。尿素を加水分解して二酸化炭素とアンモニアにし、細菌の微小環境下に放出することで、H. ピロリのウレアーゼは、胃の酸性環境下においても該細菌を生存させることを可能にする。動物モデルを使った最近の研究により、ウレアーゼは消化器官の粘膜でのコロニー形成における重要な因子であることが示唆された(Eaton ら、1991、Infect. Immun. 59: 2470- 2475)。該ウレアーゼはまた、直接的もしくは間接的に消化器粘膜に損傷を引き起こすことが疑われている。
【0003】
現在は、ヘリコバクター・ピロリ(H. ピロリ)はアントラル・ガストリテス(antral gastrites)の病因であると認識されていて、潰瘍の発達に必要な共因子の一つであることがわかっている。更に、胃癌の発達に、H. ピロリの存在が関連している可能性があることがわかっている。
【0004】
[発明が解決しようとする課題]
ヘリコバクター・ピロリ種の細菌による試験管内感染を検出する感受性、特異的な新規の方法を開発するために、発明者らは当該細菌の移動性を制御するシステムに興味を有している。
【0005】
[課題を解決するための手段]
この目的を見据え、発明者らは、感染患者の血清による認識には影響を与えないが、特にカンピロバクター(Campylobacter )ファミリの細菌(例としてはCampylobacter jejuni)に関しての「偽陽性」型の反応を避けることが可能な、H. ピロリ株の異なる修飾に興味を持っている。
【0006】
更に発明者らは、必要ならば、免疫組成物または組成物を構築するにあたって、得られた修飾済み細菌をワクチン接種に用いることが可能であると述べている。
【0007】
[発明の実施の形態]
第一段階において発明者らは、H. ピロリの鞭毛の生合成の制御、及びその結果として当該細菌の移動性の制御に関わるflbA遺伝子を同定し、単離した。鞭毛の生合成は、フラジェリンA及びBの合成と鞘(sheath)の生合成とを具備している。flbA遺伝子はフラジェリンA及びBの生合成、並びにフラジェリンを含む鞘の生合成の両方を制御する。発明者らは、flbA遺伝子が該細菌の固着蛋白質(「フック」とも呼ばれる)の生合成を制御する点においても重要であるということを確立している。
【0008】
故に本発明は、ヘリコバクター・ピロリ(Helicobacter pylori) の鞭毛タンパク質の生合成を調節すると共に、高厳密度の条件下において、下記の配列を有する二つのオリゴヌクレオチドを用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブとハイブリダイズすることができるか、
OLFlbA−1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2: GAAATCTTCATACTGGCAGCTCCAGTC
或いは、高厳密度の条件下において、これらのオリゴヌクレオチドとハイブリダイズすることができるヌクレオチド配列に関する。
【0009】
上記の配列は、ヘリコバクター・ピロリ(Helicobacter pylori) の鞭毛タンパク質の生合成を調節するflbA遺伝子のヌクレオチド配列であって、このヌクレオチド配列は、H. ピロリ株の染色体DNAを含むゲノムライブラリーを、下記の配列を有する二つのオリゴヌクレオチドを用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブか、
OLFlbA−1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2: GAAATCTTCATACTGGCAGCTCCAGTC
或いは、高厳密度の条件下において、これらのオリゴヌクレオチドとハイブリダイズすることができるプローブを用いてスクリーニングする工程と、前記プローブとハイブリダイズするDNA配列を回収する工程と、得られたDNA配列をプラスミドタイプの適切なベクター中にサブクローニングし、次いで、高厳密度の条件下において、オリゴヌクレオチドOLFlbA−1およびOLFlbA−2を用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブとハイブリダイズする修飾ベクターを選別する工程と、上記プローブとハイブリダイズするプラスミドベクター中に含まれているDNAフラグメントの配列決定を行い、これらフラグメントに含まれるオープンリーディングフレームを決定する工程とによって得られる。
【0010】
有益には、これらのDNA断片を使って、約2169ヌクレオチドからなるオープンリーディングフレームに対応した、flbA遺伝子のコード配列が再構築される。
【0011】
H. ピロリの染色体DNAを含むゲノムライブラリは、どんなH. ピロリの株からも得ることができる。コスミドライブラリもまた、H. ピロリの染色体DNAから調製することが可能である。
【0012】
このライブラリを構築するのに使う株の例は、N6株であり、これは1992年6月26日にNCIMB40512としてNCIMBに寄託された。
【0013】
H. ピロリDNAライブラリ中に存在する検索すべきのDNAに対してハイブリダイズさせる為の、プローブの調製に使った二つのオリゴヌクレオチドプライマは、Lcr D/ FlbFファミリの種々の蛋白質の保存領域から選択した。
【0014】
二つのオリゴヌクレオチドプライマOLFb A- 1及びOLFb A- 2は、プローブとして利用でき、以下の配列
ATG CCA GGA AAG CAA ATG GCG ATT GAT
GCG GAT TTA AAT TCA GGA CTT ATT GAT
GAT AAG GAA GCT AAA AAA CGG CGC GCC
GCT CTA AGC CAA GAA GCG GAT TTT TAT
GGT GCG ATG GAT GGC GCG TCT AAA TTT
を有する130塩基対の断片を増幅させることが可能である。
【0015】
上記した高厳密度の条件は以下のとおりである:ハイブリダイゼーションは42℃で、0. 1%SDSを含んだ2 X SSC緩衝液中の50%のホルムアミド(1 X SSCは、0. 15M Na Cl に15m Mのクエン酸ナトリウムを加えたものでp Hは7. 0)の存在下で行った。洗浄は68℃で行い、例としては2 X SSCプラス0. 1%SDSを使って1時間の間に2回行った。
【0016】
本発明に関しての特に興味深いヌクレオチド配列は、図3、4に記載のヌクレオチド配列、または高厳密度において上記した配列とハイブリダイズするヌクレオチド配列に対応したflbA遺伝子の配列である。
【0017】
本発明の別の態様によると、本出願の主題である該ヌクレオチド配列は、図3、4に記載のヌクレオチド配列であって、図3、4に示したアミノ酸配列を有するタンパク質、またはH. ピロリの鞭毛タンパク質の生合成に関して上記配列と同じ調節特性をもったアミノ酸配列を有するタンパク質をコードすることで特徴付けられる。
【0018】
本発明はまた、ヌクレオチド配列であって、該配列は前述の定義に対応し、更に、flbA遺伝子がもはやホスト細胞内で発現せず、またはホスト細胞内でのflbA遺伝子の発現がH. ピロリの鞭毛の生合成を可能としないように、また適切な場合には、H. ピロリのフックタンパク質の合成ができないように、欠失、置換または挿入によって修飾されているヌクレオチド配列に関する。
【0019】
本発明のヌクレオチド配列に対する修飾は、不可逆であって、特に、この配列がこのようにして修飾されたヌクレオチド配列で形質転換した細菌内に存在する、flbA遺伝子と組み換えを起こした時に、不可逆であるものでなくてはならない。この組み換えは例として、二重交差型である。好ましくはヌクレオチド配列の該修飾は、(この修飾配列によって、与えられたH. ピロリ株内のそれに対応した正常なflbA遺伝子の断片を置換した後に)隣り合う遺伝子群の機能に関して、実質的な修飾を起こさせるべきではない。
【0020】
本発明の範囲に更に含まれるものとしては、上記した基準に合致したflbA遺伝子のフラグメントを構成するヌクレオチド配列がある。例としては、本発明の主題である断片は少なくとも6ヌクレオチド、好ましくは、100ヌクレオチドではないにしろ、少なくとも50ヌクレオチドのフラグメントからなる。
【0021】
このような断片が選択される理由は、例えばそのflbA遺伝子の特異的な特徴のため、もしくはLcr D/ FlbFファミリの蛋白質をコードしたいくつかの遺伝子の保存領域に属するためである。
【0022】
別の態様において本発明はまた、遺伝子中に存在する制限部位を境界として持つflbA遺伝子断片に向けられている。これらの部位のうちいくつかを例として、図2で定義する。
【0023】
本発明に記載の別の断片は、flbA遺伝子のどこかの領域に由来し、好ましくは制限部位を含むか、または制限部位を含むことができる、少なくとも1000塩基対の断片である。
【0024】
本発明における他のヌクレオチド配列は例として、上記した様なヌクレオチド配列を含み、該配列はマーカー、例えば抗生物質に対する耐性遺伝子、または1994年5月20日に出願のFR9406202に記載された様な重金属に対する耐性遺伝子を含むカセットの挿入によって修飾されていることを特徴とする組み換え核酸がある。
【0025】
それゆえに、カナマイシンに対する抵抗性用のカセットを挿入することが可能である。この状況においては、種々の技術を利用することが可能であり、参考文献として特に、Labigne A. ら(J.of Bacteriology, 170, 1988, 1704- 1708)と、Labigne A.らの論文(Res.Microbiol. 1992, 143, 15- 26)を参照されたい。
【0026】
本発明はまた、前述で定義したヌクレオチド配列由来の特異的なオリゴヌクレオチドであって、以下の配列
OLFlb- 1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlb- 2: GAAATCTTCATACTGGCAGCTCCAGTC
OLFlb- 7: CGGGATCCGTGGTTACTAATGGTTCTAC
OLFlb- 8: CGGGATCCTCATGGCCTCTTCAGAGACC
のうちどれか一つを有することで特徴付けられる特異的なヌクレオチド配列に関するものである。
【0027】
別の態様において本発明は、前述のように定義された様なヌクレオチド配列によってコードされることを特徴とする、H. ピロリのFlbA蛋白質のアミノ酸配列に関するものである。
【0028】
H. ピロリのFlbA蛋白質の特異的なアミノ酸配列は図3、4に記載されている。
【0029】
それゆえに本発明の範囲内において、flbA遺伝子及びこの遺伝子により発現される蛋白質は、特にワクチン接種において利用する免疫学的組成物もしくは組成物にとって興味深い。
【0030】
本発明はまた、ヘリコバクター・ピロリ菌株であって、塩基またはヌクレオチド断片の置換、付加および/または欠失によって、H. ピロリの鞭毛タンパク質の生合成の調節に関与する、上記のflbA遺伝子のヌクレオチド配列に生じた突然変異によってもたらされる、非鞭毛フェノタイプを有することを特徴とする菌株に向けられたものである。
【0031】
flbA遺伝子の関するこの修飾は、非鞭毛フェノタイプの株を得ることを可能とし、すなわち該株はFlaA蛋白質もFlaB蛋白質も発現しなくなり、更に好ましくは鞘蛋白質も発現しなくなる。
【0032】
この菌株に関する一つの態様において、得られた該株は更にH. ピロリのフック蛋白質を欠損している。
【0033】
好ましくは、上記した基準に見合う菌株であって、N6株から得られたものであり、1992年6月26日に、NCIMB受付番号第40512号でNCIMBに寄託されたものであることを特徴とする。
【0034】
例として本発明は、H. ピロリの組み換え非鞭毛株であって、N6flbA-と命名され、1995年6月30日に、NCIMB受付番号第40747号でNCIMBに寄託されたH. ピロリの非鞭毛株に関する。
【0035】
このようなH. ピロリの非鞭毛株は、血清学における利用と、その結果として試験管内でのH. ピロリの感染の検出にとって特に興味深い。有益には、これらの株は組み換え型である。
【0036】
特にこれらの株は、H. ピロリによる感染を特異的且つ感度の良い方法で検出するのに有利である。言い換えれば、本発明は有益なことに、例えばサルモネラ(Salmonerlla) またはカンピロバクタ(Campylobacter) 等の菌株による「偽陽性」の結果を避けながら、H. ピロリによる感染を特異的に検出せしめる。
【0037】
このように定義された、非鞭毛型のH. ピロリ株が仮に、たとえばワクチン接種組成物の調製に使われる用に、他の適用が考えられるならば、組み換え体の非鞭毛菌株であって、例えば弱められたウレアーゼを産生するか、或いはもはやウレアーゼを産生しないように更に突然変異されており、この突然変異は、例えばureA遺伝子、ureB遺伝子、ureC遺伝子、ureD遺伝子、ureE遺伝子、ureF遺伝子、ureG遺伝子、ureH遺伝子、およびureI遺伝子の中から選択される一以上のヌクレオチドの突然変異からなることを特徴とする、第二の修飾もしくは変異を有する菌株を調製することは興味深い。ウレアーゼの構造遺伝子群である、ureA、ureB、ureC、ureDは、論文に発表されている(Labigne ら、1991、J.of Bacteriol. 173: 1920- 1931). その他の遺伝子群は特許出願EP 0610322に記載されている。
【0038】
本発明の菌株はそのままで、もしくは抽出液として利用でき、更に本発明は特に、以前記載された株から得た細菌の全抽出物に関するものである。
【0039】
そのようなバクテリア抽出物は、n-オクチルグルコシドで抽出することができる。この場合に使う調製技術は、Lelwala-Guruge J. (Scan. J. Infectt.Dis.1992、24:457- 465)によって記述されたものである。
【0040】
別のバクテリア抽出物も記述された技術により、PBS(Bazaillou M.ら、Clin.Diagn. Lab. Immuno., 1994, 1: 310- 317による方法)もしくはグリシン(Aguirre P.M., Eur.J.Clin.Microbiol.Infect.Dis., 1992, 11: 634- 639による方法) を使った抽出により得ることができる。
【0041】
本願の範囲において本発明は、患者由来の生物学的液体のサンプル( 特に血清サンプル)におけるH. ピロリによる感染をインビトロで検出するための組成物であって、活性成分として本発明の菌株または、上記のバクテリア抽出物を含有したものに関する。
【0042】
用いる生物学的サンプルはどのようなタイプのものであってもよく、また特に、どのようなタイプの生物学的液体サンプルでもよく、例としては血清、唾液、尿があげられる。
【0043】
同様にして検出に用いる方法は、免疫学的な反応を含む、どのようなタイプのものでもよく特に抗原/抗体タイプのものがよい。例としてはウェスタンブロットやELISAなどの技術を使った使用があげられる。
本発明はまた、患者由来の生物学的液体のサンプル(特に血清サンプル)におけるH. ピロリによる感染をインビトロで検出するための方法であって、前記被検サンプルを、本発明の菌株、または上記のバクテリア抽出物と接触させる工程と、前記菌株と、前記被検サンプル中に存在し且つH. ピロリに向けられる抗体との間の免疫反応を検出する工程とを具備する方法に関する。
【0044】
例としては、生物学的サンプルにおけるH. ピロリによる感染のインビトロでの検出は、
天然の、もしくは組み換え体の蛋白質、非鞭毛株、またはバクテリア抽出物、特にNOG(n-オクチルグルコシド)を使ったN6flbA株の抽出物(例としては、抽出物の量は3μg/ml、抗原の量は2μg/ml)などの検出に使う抗原でプレートを覆う工程と、ネガティブ及びポジティブのコントロールの範囲で使い(ポジティブコントロールは異なる希釈度で用いられる)、100分の1に希釈された患者の血清を平行して試験する(供せられる容量は、100μl )工程と、インキュベーションの工程が例として、37℃、1時間で行われ、その後の洗浄を連続的に幾度か行い、次にインキュベーション(例としては37℃で1時間)をモノクローナル接合体(conjugate) (ペルオキシダーゼでラベルしたヒトIgGタイプ)と行い、該接合体が異なる希釈度で用いられ(例としては抗原の場合は1/ 32, 000で、菌抽出物の場合は1/ 64, 000)、供せられる容量が100μl である工程と、モノクローナル接合体とのインキュベーション後にいくつかの異なる洗浄を行い(例えば例4)、酵素反応を「OPDと基質」を使い、30分間遮光して行わせることを満たすことで実行できる。次に酵素反応は、硫酸を加えて終了させて、光学濃度であるO Dを492nm/ 620nmで読む。
【0045】
本発明は更に、H. ピロリに対する抗体を得るための免疫原組成物であって、活性成分として、本発明の菌株、または上記のバクテリア抽出物を含有することを特徴とする免疫原組成物に向けられている。
【0046】
本発明の一つの特異的な態様において、H. ピロリに対する抗体を得るための免疫原組成物は、FlbA蛋白質由来のアミノ酸配列を含んでいることを特徴とする。
【0047】
本発明の範囲に更に含まれるものとしては、H. ピロリによる感染に対して防御する抗体を得るためのワクチン接種組成物であって、活性成分として本発明の菌株または、上記のバクテリア抽出物を含有することを特徴とする組成物がある。
【0048】
H. ピロリによる感染に対して防御する抗体を得るための別のワクチン接種組成物は、活性成分としてウレアーゼタイプの抗原、特にureA、ureB、ureC、またはureD遺伝子によってコードされ、上記に定義したアミノ酸を有する蛋白質を有することを特徴とする。
【0049】
本発明は上記したアミノ酸配列に向けられるモノクローナル抗体またはポリクローナル血清に関する。この抗体は、それ自体は知られた方法によって得られ、特に、動物を選択した抗原で免役し、次に抗体を合成し、合成された抗体を回収し、その中からH. ピロリを特異的に認識するものを選択するか、または前もって免役しておいた動物の脾臓細胞をミエローマ細胞と融合させてハイブリドーマを調製し、次にこのハイブリドーマを培養して、選択したH. ピロリ抗原に対しての特異的認識をもとに選択したモノクローナル抗体を得ることにより得られる。
【0050】
本発明の他のモノクローナル抗体、またはポリクローナル血清は、上記したように非鞭毛性H. ピロリに向けられている。
【0051】
本発明は更に、生物学的サンプルにおけるH. ピロリによる感染をインビトロで検出するための組成物であって、活性成分として、本発明の非鞭毛性のフェノタイプのH. ピロリに対して得られるモノクローナル抗体、またはポリクロナール血清を含有する組成物に関する。
【0052】
本発明はまた、薬物の活性成分として、本発明によるアミノ酸配列をコードしたヌクレオチド配列であって、動物またはヒトで免役学的反応を誘導することができるヌクレオチド配列に関する。薬物としてヌクレオチド配列を使用する技術は、Donneyら(1995; Nature Medic. 1( 6), pp 583- 587)に記載されている。
【0053】
例I
flbA遺伝子の同定及び非鞭毛株の調製
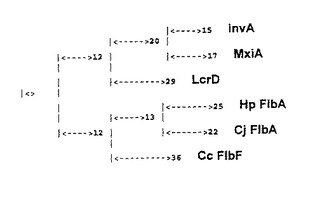
細菌の移動度の発現を制御する機能を担うことが知られている蛋白質の中で興味のあるものは、最近同定されたLcrD/FlbFファミリに属する蛋白質であり、その中にはイェルシニア属の細菌のLcrD蛋白質(6)、サルモネラのInvA蛋白質(2)、シゲラのMxiA(1)、カウロバクター クレセンタスのFlbF(7)、及びカンピロバクター ジェジュニのLfbA(4)がある。LcrD、InvA、及びMxia蛋白質は、該蛋白質を発現する細菌の病原性に関わる蛋白質の制御及び/または分泌に関わり、一方カウロバクター クレセンタスのFlbF蛋白質、及びカンピロバクター ジェジュニのFlbAは鞭毛の生合成の制御に関わり、それゆえに移動度に関わる。現在までに知られたLcr D/ FlbAファミリの相同体は、特にこの蛋白質のN末端部分が非常によく保存された領域を有しており、それゆえにこの2つの保存領域(イェルシニアのLcr D蛋白質の151から156番目のアミノ酸であるMPGKQM、及びLcr Dの189から195番目のアミノ酸であるMDGAMKF)を使って2つの縮退したオリゴヌクレオチド(L OLFlbA- 1及びOLFlbA- 2、表1)を限定することが可能であったが、該ヌクレオチドは、ヘリコバクター・ピロリの染色体DNA上で行う遺伝子増幅におけるヌクレオチドプライマとして合成されて使用される。このようにして、130塩基対(bp)の断片を増幅することが可能であり、そのヌクレオチド配列の決定により、該断片がLcrD/ FlbFファミリの蛋白質に非常に相同である蛋白質の一部をコードしていることが証明された。この増大された断片は次に、放射性のラベルがなされH. ピロリのコスミドベクタのスクリーニングに使われた。
【0054】
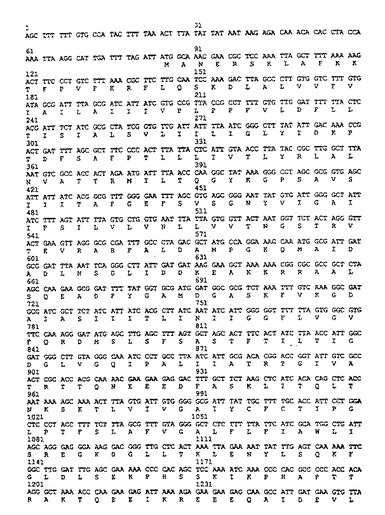
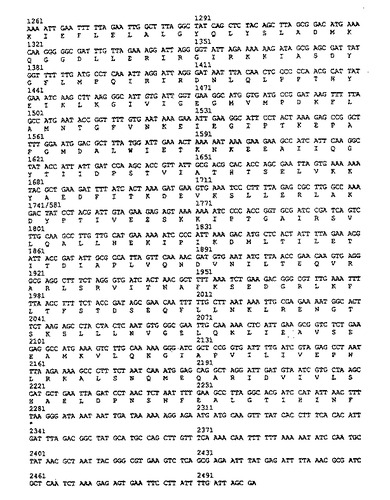
この断片は図3、4に示した配列の575から707の間のヌクレオチドに対応した配列である。
【0055】
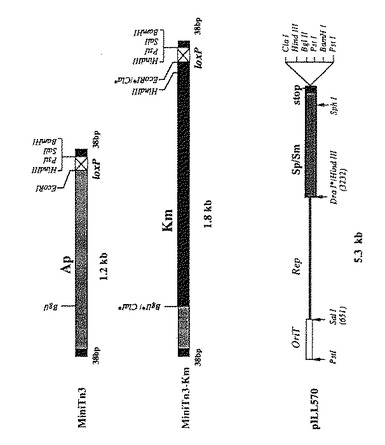
ゲノムライブラリのコスミドのうちの一つは、H. ピロリのLcrD/ FlbF相同体をコードしているものとして同定され、Sau3Aによる部分消化を行って、2から5(キロ塩基対)の大きさをもつ挿入断片を含む、ベクタpILL570中のコスミドのミニライブラリ(200サブクローン)を構築した。ベクタpILL570はLabinge A. らの論文(パスツール研究所/Elsevier Paris1992.Res. Microbiol, 1992, 143, 15- 26)に記載されている。このベクタの制限地図は、図1に示してある。上記の200クローンを130塩基対のプローブにハイブリダイズすると、プラスミドpSUS39およびpSUS207をもつクローンが陽性を示した。これら二つの組み換えプラスミドの線形制限地図は、図2に示してあり、これらのクローンの挿入部がそれぞれ重なり合った配列をゆすることを示した。これらの二つの挿入部のヌクレオチド配列を決定して、これら導入部にはflbA遺伝子がそのままは含まれないことを明かにした。カンピロバクタ ジェジュニのFlbA遺伝子に対する相同体であるために、H. ピロリのflbAと命名される当該遺伝子は、2196塩基対のオープンリーディングフレームに対応し、80. 1キロダルトンの計算上の分子量をもつ蛋白質をコードしている。flbAのヌクレオチド配列及びFlbAのアミノ酸配列を図3、4に示してある。プロモータ及びターミネータに特徴のコンセンサス配列は、該オープンリーディングフレームの上流及び下流には検出さていない。
【0056】
FlbA蛋白質は、共に移動度に関わるカンピロバクタ ジェジュニのFlbA蛋白質及びカウロバクタ クレセンタスのFlbF蛋白質と類似点を呈しているが(それぞれ51. 7%、40. 4%)、この類似の割合は、移動度に関わらないLcr D/ FlbF蛋白質ファミリとの比較ではより低く、イェルシニアのLcrDとは32. 8%の相同性、シゲラのMxiAとは30. 5%の相同性、そしてサルモネラのInvAとは29. 3%の相同性である。これらの蛋白質の、H. ピロリのFlbAに対するマルチプル整列(multiple alighnment )を図5、6に示してある。LcrD/ FlbFファミリの相同体で最も保存された領域は蛋白質のN末端部分に位置している。
【0057】
移動度に関わる蛋白質(FlbA及びFlbF)の系統学的進化、及び病原性に関わる蛋白質の発現及び/または分泌の制御に関わる蛋白質の系統学的進化は、系統樹を使って示してある(図7)。二つの別個の分岐が見られる:H. ピロリのFlbAが、鞭毛の生合成に関わる制御蛋白質に対応する分岐に属することは、はっきりとしている。
【0058】
FlbA蛋白質の合成を欠損している、H. ピロリのアイソジェニック変異体の構築と特徴づけ
プラスミドpSUS39の1600塩基対の断片を、それぞれの5’末端にBamHI制限部を有するオリゴヌクレオチドOLFlbA- 1及びOLFlbA-7を使って増幅した(表 )。この断片の中央部には、ユニークなHind III制限部位があり、この断片を、pUC19の誘導体であってそのマルチプルクローニング部位のHind III部位を取り除いた、pSUS33へクローニングした。pSUS33を得るために、プラスミドpUC19をHindIIIで消化し、その結果得られる粘着末端をクレノー酵素及びT4DNAポリメラーゼで処理して平滑末端にし、その結果の断片をT4DNAポリメラーゼで再連結して、pSUS33を得るために大腸菌DH5αへ挿入した。上記の1600塩基対の断片をpSUS33へ挿入して得た組み換えプラスミドをpSUS40と命名し、HindIIIで線状化し、その末端を平滑にして、プラスミドpILL600(LabingeA.ら1988、J.Bact. 170, 1704- 1708)由来のSmaIカナマイシンカセットをこのユニークな部位へクローニングして、プラスミドpSUS42を得た。プラスミドpSUS42を次に、電気穿孔法によりH. ピロリのN6株へ導入した。電気穿孔法は、Ferrero R.L.ら(Journal of Bacteriology, July,1992, pp4212- 4217,Vol174, No 13)が記載の方法に添って行った。次にカナマイシン(25μg/ml)を含んだ選択培地で選択した形質転換体に関して、遺伝型及び表現型の特徴づけを行った。図8は変異体作成にあたって従った手順を示している。遺伝子増幅及びサザンハイブリダイゼーションによるこれらの変異体の遺伝型分析により、カナマイシン抵抗性形質転換体のゲノムがflbA遺伝子の真ん中に、該抵抗性遺伝子を含んでいることと、それゆえに二重交差による、野生型のflbA遺伝子の不活型flbA- Kmへの対立遺伝子置換が、pSUS33ベクタのヌクレオチド配列の喪失を伴って起きたこととが示された。H. ピロリのflbA- 変異体の表現型分析により、該変異体は移動度を有しないこと、更に電子顕微鏡による分析により、鞭毛要素の完全な欠失及び鞭毛の鞘の欠失があることを示した。H. ピロリの鞭毛全体の蛋白質に向けられた抗体を使ったイミュノブロット実験(ウェスタンブロット)(図9)は、鞭毛のサブユニットであるFlaA及びFlaBに対応したペプチドのバンドが欠失していることと、また同様に最近O'Tooleと共同研究者(5)によってフック蛋白質(もしくは固定蛋白質)または鞭毛と同定された蛋白質(5)で、見かけ分子量が90キロダルトンのポリペプチドに対応したバンドも欠失していることを示した。
【0059】
全体としてみると、これらの結果はH. ピロリのFlbA蛋白質は鞭毛の構造全体の合成に必須であり、この蛋白質をコードした遺伝子の不活化は、該構造の構成要素の輸送の阻害ではなくて、鞭毛形成にくわわるいかなる構造の合成をも完全に停止する結果となることを示唆している。
【0060】
【表1】

例II
調べたH. ピロリの血清学モデル
1)HspAmalE: 47. 5k Da (HspA=13kDa)の組み換え蛋白質
保存されていた血清の、母集団Iと呼ばれるものにおいて41%の感受性、96%の特異性が得られた。
【0061】
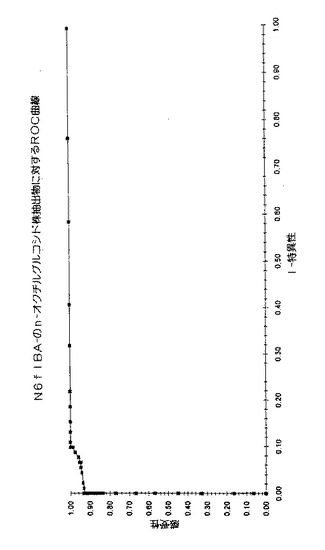
2)N6flbA−:ヘリコバクター・ピロリの非鞭毛株3種類の抽出法を行った(n-オクチルグルコシド、PBS、グリシン)。さしあたり、n-オクチルグルコシドによる抽出が最もよいことがわかった。
【0062】
3)−N6: 野生型株に対応している。抽出はn-オクチルグルコシドで行った。
【0063】
血清の第二の母集団も使った(母集団II)。この母集団は、臨床的、内視鏡的、組織学的、細菌学的、及び解剖病理学的(anatomopathological)見地からよく記録された数百の血清からなっている。本調査において異なるモデル系の出来栄えを調べるために使ったのが、この母集団IIであった。5つの異なる母集団を試験した。
【0064】
試験した5つの血清は、
300の通常の血清(FNTS)
WHITTAKER血清学で陽性であった18の血清(CBMS)
母集団II血清と名付けられた92の、よく記録された血清
細菌学的、解剖病理学的(anatomopathological )見地からよく記録され、また母集団Iと名付けられた、87の血清
以下の交差反応を示す23の血清
抗レジオネラ(Legionella)陽性の10の血清
抗クラミジア(Chlamydia)陽性の10の血清
抗カンピロバクタ(Campylobacter)陽性の3つの血清
文献で効果が認められている2つの競合するキットをパラレルに使って試験した。
【0065】
2つの市販のキット
Cobas Core(ROCHE)
Pylori Stat(WHITTAKER)
結果
通常の血清(FNTS)( 図10から14、表2、3)
以下のモデルを通して得た300の血清
Hsp A malE
N6 flBA
N6
疫学的な調査によると、フランスでは20から25%の血清陽性率(sero-prevalences)となっている。300人の血液提供者の血清の分布を調べ、CBMSライブラリを使って以前に定義された域値(WHITTAKER血清学)を確証する為に異なる域値に対する陽性の支配度を計算した。
【0066】
この研究により、同じ血清陽性率を使って異なる試験を比較することができる。
【0067】
最初の43の血清は、以下のモデルを通して得た
Cobas Core(Roche)
Pylori Stat(WHITTAKER)
JLF血清学として知られる血清学(数種類の水溶性バクテリア抽出物に基づくELISA試験)
結果は任意の単位に基づいて、異なる域値に対して表してある;陽性の結果は1、陰性の結果は0と表す。
【0068】
異なる試験によって、これら43の血清を比較して、以下のことが判明する:非鞭毛株であるN6flbA- 及びCobas Core試験( Roche)では、20%のオーダーに匹敵する血清陽性率となる。
【0069】
HspAでは、非常に低い血清陽性率(7%)となり、これは引き続く結果の観点からは、感受性の欠失を示唆している。
【0070】
引き続く結果を考慮すると、JLF血清学は、血清陽性率がわずかに14%であるので非常に感度がよいことがわかる。
【0071】
Pylori Stat(Whittaker) 試験では、高い血清陽性率(29%)となるが、これは特異性の欠如、または域値が低すぎることを示している。
【0072】
【表2】
【表3】
WHITTAKER血清学で陽性の血清(CBMS)(表4)
Pylori Stat試験(Whittaker)で陽性であった血清はわずか3つであった。この3つの血清は、他の試験では確認できなかった。
【0073】
この結果は特異性の欠如によるものであると思われる。もし、現在の市場で最も良いCobas Core試験(Roche)を参照するならば、我々の異なるモデルをCobas Coreに対して比較することが可能である。
【0074】
非鞭毛N6flbA- 株はCobas Coreと完全に一致する。
【0075】
Cobas Coreで陰性であった上記の3つの血清は、N6flbA- でも陰性であった。
【0076】
Cobas Coreで陽性であった15の血清は、N6flbA- でも陽性であった。
【0077】
野生型N6株は非鞭毛株と同じ結果となった。
【0078】
9つのCobas Core血清がHspAで陰性であるので、HspAもまた感受性がない。
【0079】
Cobas Coreで陰性である上記の3つの血清は、HspAでも陰性である。
【0080】
【表4】
母集団IIの血清
92の血清を選択し、3つのグループへ分類した:
34:消化不良患者、内視鏡または組織学での潰瘍との診断(十二指腸、胃)、培養及び/または解剖病理学によりヘリコバクター・ピロリが存在すること、迅速な尿素試験も実行、このグループはHp+/U+と命名。
【0081】
27:消化不良患者、内視鏡または組織学での潰瘍との差動的診断(十二指腸、胃)、培養及び/または解剖病理学によりヘリコバクター・ピロリが存在すること、迅速な尿素試験も実行、このグループはHp+/U−と命名。
【0082】
31:消化不良のまたは消化不良でない患者、内視鏡及び組織学では正常な胃十二指腸、培養及び/または解剖病理学によりヘリコバクター・ピロリが存在しないこと、迅速な尿素試験も実行、このグループはHp−と命名.
臨床、内視鏡、組織学、細菌学、及び解剖病理学な知見を、それぞれの患者に対して示した。
【0083】
この良く記録された集団により、感受性及び特異性の基準の定義がなされる。
【0084】
Hp A: 母集団Iに見られるように、実質的な感受性の欠如がいぜんとして見られる。
【0085】
感受性は59%で特異性は100%である。
【0086】
N6flbA:n- オクチルグルコシド抽出物に対して100%の感受性が確認でき、特異性は90%である。
【0087】
この結果は、RocheのCobas Core試験の結果で得られたもの(感受性が98%で特異性が94%)と匹敵する。
【0088】
N6: 母集団IIにおいて、野生型株は非鞭毛株に完全に匹敵する。
【0089】
陰性の31の血清は野生型に対しては陽性である。鞭毛による交差反応は、母集団IIに関しては全く見られなかった。
【0090】
【表5】
【表6】
【表7】
【表8】
【表9】
【表10】
【表11】
【表12】
【表13】
【表14】
【表15】
血清学の位置づけ
血清学を2つのレベルで位置づけた。
【0091】
高感度血清学:心窩部痛をうったえる若年患者に存在する細菌を検出するためのもの。
【0092】
この血清学で陰性となった場合は、患者は内視鏡の検査または生検を受けることはなく、他の痛みの原因となるものを調べる。
【0093】
危険特異的血清学:これはヘリコバクター・ピロリによる感染での危険性、すなわち潰瘍、癌、または胃リンパ腫(MAL Tリンパ腫)を示すこと含む。
【0094】
疑いのある危険性に特異的な分子を使うか
または危険特異的な域値を使う(域値は危険のある患者では、危険のない患者よりも高い)。
【0095】
この特異的な血清学を使って、一般の母集団をスクリーニングにかけ、それによってヘリコバクター・ピロリに関わり、また症状がないために検出できない様な癌及びリンパ腫を検出する(痛みを訴えている患者のみが胃腸科にかかる。)
感受性の問題に対する応答は良い。
【0096】
【表16】
【表17】
【表18】
胃炎の強度と抗体レベルとの間の相関関係
胃炎は3つの変数で定義される:
萎縮(Gの後の最初の数字で表現);強度は1から4で表す。
【0097】
中性好性多核細胞及び単球の侵入に対応する全体の炎症(Gの後の二番目の数字で表現);強度は1から3で表す。
【0098】
中性好性多核細胞の数に対応した活性(Gの後の三番目の数字で表現);強度は0から3までで表す。ろ胞状のものはFと表現する。
【0099】
通常は、以下の相関関係が見られる:
活性はヘリコバクター・ピロリと非常に良く相関する。
【0100】
炎症はヘリコバクター・ピロリと相関する。
【0101】
それぞれのグループで見られる力価の平均は、故に上記の3つの変数とその強度とを使って計算される。
【0102】
結果の評価:
t 試験を使って2つのデータの平均が顕著に異なるか、または5%の危険度でそうでないかを証明することが可能となる。
【0103】
t 試験が基づく仮定が、変数の等価性であり、F試験(Fisher試験)で示されている。
【0104】
変数の中には等しくないものがあるので、よってそれぞれを全ての平均値と比較することはできない。
【0105】
可能であるならば平均を比較することで、異なるグループの間の差が顕著であるかそうでないかを示すことができる。
【0106】
顕著な差:HspAに対する「2」及び「3」、と「炎症」グループのNOGとの間の平均。
【0107】
顕著でない差:活性に関しては、異なる強度レベル間での顕著な差は見られなかった:
HspA: 0と2、0と3、1と2、1と3、及び2と3との間には顕著な差はない
NPG: 0と1、0と2、1と2、1と3、及び2と3との間には顕著な差はない.
しかしながら、力価が胃炎の強度に依存して増大する傾向は見られる:
萎縮に関して:HspA及び非鞭毛株のNOG抽出のは平均値は、レベルが1から2、及び2から3へ移ると、二倍になる。
【0108】
炎症に関して:レベルが2から3になると、平均が二倍になる。
【0109】
それぞれのグループにおける数は、統計的に顕著な差であると結論するには、相対的に少ない(それぞれ30より少ない)。
【0110】
【表19】
【表20】
交差反応性を呈する血清
2つのタイプの血清を用いた。
20の血清(10の抗- レジオネラ+ 、10の抗- クラミジア+ ):これら3つの細菌はそれぞれが非常に似た熱ショック蛋白質を有しているため、HspAに対して交差反応することができる
3つの抗- クラミジア陽性抗体:非鞭毛株であるN6flbAでは消出する、鞭毛株N6との交差反応性を示すため。抗−カンピロバクタ陽性血清を得ることは非常に難しい;これが、わずかに3つの血清しか得られない理由である。
【0111】
HspAは、10の抗- レジオネラ陽性抗体とも、10の抗- クラミジア陽性抗体とも、交差反応性を全く示さない。
【0112】
これらの血清のうちいくつかは、鞭毛株及び非鞭毛株の両方について、抗- ヘリコバクター・ピロリ抗体に関する陽性力価を有しているが、これらの血清の臨床的意義はわかっていない。
【0113】
【表21】
結論
HspA malE
感受性が欠損しているために、この分子自体を利用することはできないが、他の分子と結合しているならば興味深いであろう。
【0114】
しかしながら、異なる細菌間で実質的に保存された熱ショック蛋白質による交差反応の危険性をはらんでいる。
【0115】
N6flbA-
この非鞭毛変異体は非常に興味深いことがわかってきている;母集団IIで判明した感受性及び特異性は、非常に好ましい効果を示している。
【0116】
N6
さしあたっては、鞭毛株が興味深い。しかしながら鞭毛に関連した交差反応性に関してはわずかしか調べられていないが、これはカンピロバクタ血清学に関して良く考証されている血清を得ることが難しいためである。
【0117】
JLF試験
ヘリコバクター・ピロリの幾つかの株についての水溶性(PBS)抽出に基づいた血清学的試験を開発した。
【0118】
非鞭毛株変異体のNOG抽出を使って母集団Iの血清を試験した。
【0119】
細菌学的及び解剖病理学的見地からのみ考証された87の血清を、非鞭毛細菌抽出を使って試験した。
【0120】
培養が陽性であるか、または解剖病理学的試験及び迅速な尿素試験で陽性であれば、血清は陽性である。
【0121】
3つの試験(培養、解剖病理学、及び迅速な尿素試験)で陰性であれば血清は陰性である。
【0122】
感受性は90. 3%(28/ 31)で、特異性は71. 4%(40/ 56)であった。
【0123】
はじめの試験であやまって陽性とされた16の血清のうち、9つはJLF血清学またはJLFウェスタンブロット、もしくはその両方により陽性であった。
【0124】
はじめの試験であやまって陰性とされた3つの血清全ては、JLF血清学またはJLFウェスタンブロットで陰性であり、一つは両方で陰性であった。
【0125】
【表22】
【表23】
技法
プレートは以下のものでコートした:
2μg/mlのHspA抗原
3μg/mlの、NflbA及びN6のNO G抽出
範囲
5つの範囲でのネガティブコントロール
ポジティブコントロールは4つの希釈度で使用
患者血清
1/ 100希釈
容量:100μl
インキュベーション 37℃1時間
3回洗浄
モノクローナル接合体(IgG toxo)
使用濃度
HspAに対しては1/ 32000
N6flbA- に対しては1/ 64000
N6に対しては1/ 56000
容量 100μl
接合体のインキュベーション:37℃、1時間
4回の洗浄
OPD+ 基質による酵素反応の進行
遮光で30分間
硫酸による酵素反応終結
492nm/ 620nmでのOD測定
ODの、任意の単位(AU)への変換
【表24】
【表25】
【表26】
【表27】
【表28】
【表29】
【表30】
参考文献
1. Andrews,G.P., Maurelli,A.T.: mxiAof Shigella flexneri 2a, which facilitates export of invasion plasmid antigens encodes a homolog of the low−calcium−response protein LcrD of Yersinia pestis. Infect. Immun. 60: 3287- 3295( 1992).
2. Galan,J.E., Ginocchio,C., Costeas, P.:Molecular and functional characterization of the Salmonella invasion gene invA:homology of InvA to menbers of a new protein family. J.Bacteriol. 174: 4338- 4349( 1992).
3. Leying,H., Suebaum, S., Geis, G.,Haas,R.:Cloning and genetic characterization of a Helicobacter pylori flagellin gene. Mol.Microbiol.6:2563-2874( 1993).
4. O’Toole,P.W., Kostrzynska, M., Trust,T.J.:Non−mobile mutants of Helicobacter pilori and Helicobacter mustelae defective in flagellar hook production. Mol. Microbiol. 14: 691- 703(1994).
5.Plano,G.V., Barve,S.S., Straley,S.C.: LcrD, a menbrane −bound regulator of the Yersinia pestis low−calcium response. J.Bacteriol. 173: 7293- 7303( 1991).
6. Ramakrishnaan, G., Zhao,J−L., Newton,A.:The cell cycle−regulated geneflbF of Caulobacter crescentus is homologous to a virulence locus of Yersinia pestis. J.Bacteriol. 173: 7283- 7292( 1991).
7.Suerbaum, S., Josenhans,C., Labigne,A.:Cloning and genetic characterization of the Helicobacter pylori and Helicobacter mustelae flb flagellin genes and constructionof H.pylorifla− And flB− negative mutantsby electroporation−mediated allelic exchange. J.Bacteriol., 175: 3278- 3288( 1993).
【図面の簡単な説明】
【0126】
【図1】プラスミドpILL570、およびカナマイシン抵抗性遺伝子カセットを含んだ小型トランスポゾンTn 3の制限地図。
【図2】組み換えプラスミドpSUS39及びpSUS207の線形制限地図。表示の番号は制限断片の長さに対応していて、塩基対で表示されている。H:HindIII、Bg:BglII。星印は、クローニング中に制限部位が修飾されて、対応していた制限酵素で認識されなくなったことを示す。
【図3】H. ピロリのflbAのヌクレオチド配列及び、一文字表示による推定のアミノ酸配列。
【図4】H. ピロリのflbAのヌクレオチド配列及び、一文字表示による推定のアミノ酸配列。
【図5】H. ピロリのFlbA蛋白質と、他の5つのLcr D/ FlbFファミリとのマルチプル整列(multiple alignment)。Cj FlbA:カンピロバクタ ジェジュニ( Campylobacter jejuni) FlbA; Cc FlbF:カウロバクタ クレセントゥス( Caulobacter creacentus) FlbF; YpLcsD: イェルシニア ペスティス( Yersiniapestis)LcrD; StlnvA: サルモネラ ティフィムリウム( Salmonera typhimurium) InvA; SfMxiA: シゲラフレクスネリ( Shigella flexneri)MxiA。星印はLcrD/ FlbFファミリの相同体全てにおいて保存されているアミノ酸の位置を示し、点は相同体6つのうち少なくとも5つにおいて保存されているアミノ酸の位置を示す;縮退したオリゴヌクレオチドの合成に用いられた、保存アミノ酸配列には下線が引いてある。これらの相同体のN末端領域においての保存の度合いにたいして注意するべきであり、これはC末端の疎水性領域の多様性の度合いと対照的である。
【図6】H. ピロリのFlbA蛋白質と、他の5つのLcrD/ FlbFファミリとのマルチプル整列(multiple alignment)。Cj FlbA:カンピロバクタ ジェジュニ( Campylobacter jejuni) FlbA; Cc FlbF: カウロバクタ クレセントゥス( Caulobactercreacentus) FlbF; YpLcs D: イェルシニア ペスティス( Yersinia pestis) LcrD; StlnvA: サルモネラ ティフィムリウム( Salmonera typhimurium) InvA; SfMxiA: シゲラ フレクスネリ( Shigella flexneri) MxiA。星印はLcrD/ FlbFファミリの相同体全てにおいて保存されているアミノ酸の位置を示し、点は相同体6つのうち少なくとも5つにおいて保存されているアミノ酸の位置を示す;縮退したオリゴヌクレオチドの合成に用いられた、保存アミノ酸配列には下線が引いてある。これらの相同体のN末端領域においての保存の度合いにたいして注意するべきであり、これはC末端の疎水性領域の多様性の度合いと対照的である。
【図7】Lcr D/ FlbFファミリに属する6つの蛋白質の系統樹を描いた図。移動度の発現制御に関わる蛋白質、すなわちH. ピロリのFlbA( Hp FlbA) 及びカンピロバクタ ジェジュニのFlbA( Cj FlbA) 、並びにカウロバクタ クレセントゥス( Caulobacter creacentus) のFlbF( Cc FlbF) は、病原性蛋白質( サルモネラのInvA、シゲラのMxiA、及びイェルシニアのLcrD) の分泌に関わる蛋白質とはかけ離れたブランチを形成する。表示の番号は、相対的な進化の距離を表す。
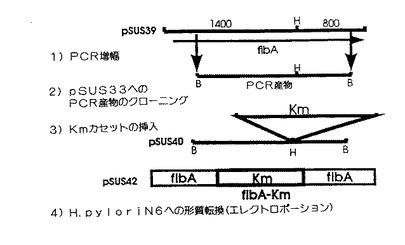
【図8】H. ピロリの同質遺伝子的変異体株N6、すなわちFlbA蛋白質をコードした遺伝子が、カナマイシン抵抗性をコードした遺伝子の挿入によって不活化された変異体を構築する際に従った方法を描いた図。
【図9】H. ピロリから精製した鞭毛に対して特異的なAK 179抗血清( 3) を使って行った、N6flbA- 変異体由来の蛋白質の免疫ブロット(ウェスタンブロット);1:N6- flbA変異体、2:flaA/ flaB二重変異体、3: flaB( 8) 変異体、4:fla A( 8) 変異体、5:野生型N6株。
【図10】H. ピロリに対して行った血清学の結果比較。
【図11】H. ピロリに対して行った血清学の結果比較。
【図12】H. ピロリに対して行った血清学の結果比較。
【図13】H. ピロリに対して行った血清学の結果比較。
【図14】H. ピロリに対して行った血清学の結果比較。
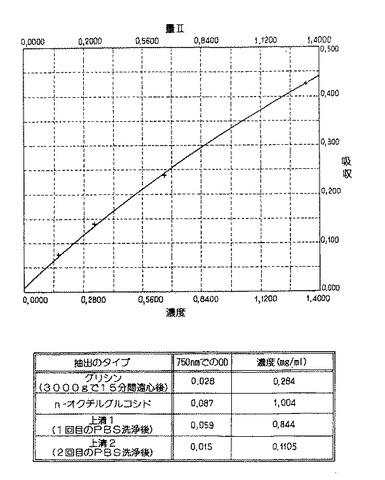
【図15】非鞭毛株N6flbA- を使った抽出:グリシン、PBS、またはNOGを使って行った抽出。いかのデータに基づいて曲線を描いた。 Net Abs 計算したSTD# 濃度 750. 0 濃度 差 1 0. 0000 0. 0020 - 0. 008 0. 0080 2 0. 1660 0. 0760 0. 1721 -0. 006 3 0. 3300 0. 1400 0. 3459 -0. 016 4 0. 6650 0. 2390 0. 6474 0. 0176 5 1. 3300 0. 4280 1. 3336 -0. 004係数:P 2= 2. 0324, P 1= 2. 2753, P 0= 0平均:- 1. 0356E- 07, 標準偏差:0. 0130
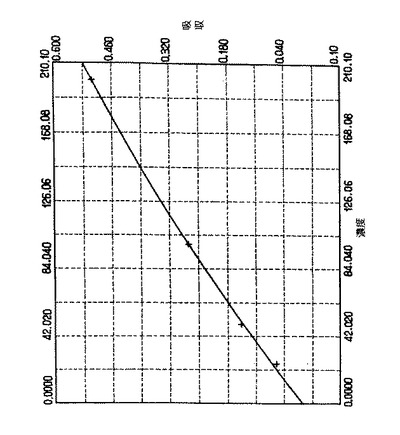
【図16】非鞭毛株N6flbA- を使った抽出:グリシン、PBS、またはNOGを使って行った抽出。ミニメソッド( BioRad) の蛋白質アッセイ。グリシン:1/ 2希釈、グルコシド:1/ 10希釈、上清1:1/ 4希釈、上清2:希釈せず。以下のデータに基づいて曲線を描いた。 Net Abs 計算したSTD# 濃度 750. 0 濃度 差 1 0. 0000 -0. 003 1. 5398 -1. 540 2 25. 000 0. 0600 21. 861 3. 1392 3 50. 000 0. 1470 51. 810 -1. 810 4 100. 00 0. 2750 99. 855 0. 1454 5 200. 00 0. 5090 199. 94 0. 0636係数:P 2= 144. 63, P 1= 314. 31,P0= 2. 4815
【図17】非鞭毛株N6flbA- を使った抽出:グリシン、PBS、またはNOGを使って行った抽出。
【発明の詳細な説明】
【0001】
[発明の属する技術分野]
本発明は、現在までのところヒトの胃粘膜表面上に特異的に見られることがわかっている、グラム陰性細菌であるヘリコバクター・ピロリ(H. ピロリとも命名されている)に関する。
【0002】
[従来の技術]
多くの細菌と共通して、H. ピロリもまた、酸性p Hの培養液に対しては感受性であるが、生理学的濃度の尿素存在下においては、酸性度を許容することが可能である(Marshallら、1990、Gastroenterol. 99: 697- 702)。尿素を加水分解して二酸化炭素とアンモニアにし、細菌の微小環境下に放出することで、H. ピロリのウレアーゼは、胃の酸性環境下においても該細菌を生存させることを可能にする。動物モデルを使った最近の研究により、ウレアーゼは消化器官の粘膜でのコロニー形成における重要な因子であることが示唆された(Eaton ら、1991、Infect. Immun. 59: 2470- 2475)。該ウレアーゼはまた、直接的もしくは間接的に消化器粘膜に損傷を引き起こすことが疑われている。
【0003】
現在は、ヘリコバクター・ピロリ(H. ピロリ)はアントラル・ガストリテス(antral gastrites)の病因であると認識されていて、潰瘍の発達に必要な共因子の一つであることがわかっている。更に、胃癌の発達に、H. ピロリの存在が関連している可能性があることがわかっている。
【0004】
[発明が解決しようとする課題]
ヘリコバクター・ピロリ種の細菌による試験管内感染を検出する感受性、特異的な新規の方法を開発するために、発明者らは当該細菌の移動性を制御するシステムに興味を有している。
【0005】
[課題を解決するための手段]
この目的を見据え、発明者らは、感染患者の血清による認識には影響を与えないが、特にカンピロバクター(Campylobacter )ファミリの細菌(例としてはCampylobacter jejuni)に関しての「偽陽性」型の反応を避けることが可能な、H. ピロリ株の異なる修飾に興味を持っている。
【0006】
更に発明者らは、必要ならば、免疫組成物または組成物を構築するにあたって、得られた修飾済み細菌をワクチン接種に用いることが可能であると述べている。
【0007】
[発明の実施の形態]
第一段階において発明者らは、H. ピロリの鞭毛の生合成の制御、及びその結果として当該細菌の移動性の制御に関わるflbA遺伝子を同定し、単離した。鞭毛の生合成は、フラジェリンA及びBの合成と鞘(sheath)の生合成とを具備している。flbA遺伝子はフラジェリンA及びBの生合成、並びにフラジェリンを含む鞘の生合成の両方を制御する。発明者らは、flbA遺伝子が該細菌の固着蛋白質(「フック」とも呼ばれる)の生合成を制御する点においても重要であるということを確立している。
【0008】
故に本発明は、ヘリコバクター・ピロリ(Helicobacter pylori) の鞭毛タンパク質の生合成を調節すると共に、高厳密度の条件下において、下記の配列を有する二つのオリゴヌクレオチドを用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブとハイブリダイズすることができるか、
OLFlbA−1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2: GAAATCTTCATACTGGCAGCTCCAGTC
或いは、高厳密度の条件下において、これらのオリゴヌクレオチドとハイブリダイズすることができるヌクレオチド配列に関する。
【0009】
上記の配列は、ヘリコバクター・ピロリ(Helicobacter pylori) の鞭毛タンパク質の生合成を調節するflbA遺伝子のヌクレオチド配列であって、このヌクレオチド配列は、H. ピロリ株の染色体DNAを含むゲノムライブラリーを、下記の配列を有する二つのオリゴヌクレオチドを用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブか、
OLFlbA−1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2: GAAATCTTCATACTGGCAGCTCCAGTC
或いは、高厳密度の条件下において、これらのオリゴヌクレオチドとハイブリダイズすることができるプローブを用いてスクリーニングする工程と、前記プローブとハイブリダイズするDNA配列を回収する工程と、得られたDNA配列をプラスミドタイプの適切なベクター中にサブクローニングし、次いで、高厳密度の条件下において、オリゴヌクレオチドOLFlbA−1およびOLFlbA−2を用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブとハイブリダイズする修飾ベクターを選別する工程と、上記プローブとハイブリダイズするプラスミドベクター中に含まれているDNAフラグメントの配列決定を行い、これらフラグメントに含まれるオープンリーディングフレームを決定する工程とによって得られる。
【0010】
有益には、これらのDNA断片を使って、約2169ヌクレオチドからなるオープンリーディングフレームに対応した、flbA遺伝子のコード配列が再構築される。
【0011】
H. ピロリの染色体DNAを含むゲノムライブラリは、どんなH. ピロリの株からも得ることができる。コスミドライブラリもまた、H. ピロリの染色体DNAから調製することが可能である。
【0012】
このライブラリを構築するのに使う株の例は、N6株であり、これは1992年6月26日にNCIMB40512としてNCIMBに寄託された。
【0013】
H. ピロリDNAライブラリ中に存在する検索すべきのDNAに対してハイブリダイズさせる為の、プローブの調製に使った二つのオリゴヌクレオチドプライマは、Lcr D/ FlbFファミリの種々の蛋白質の保存領域から選択した。
【0014】
二つのオリゴヌクレオチドプライマOLFb A- 1及びOLFb A- 2は、プローブとして利用でき、以下の配列
ATG CCA GGA AAG CAA ATG GCG ATT GAT
GCG GAT TTA AAT TCA GGA CTT ATT GAT
GAT AAG GAA GCT AAA AAA CGG CGC GCC
GCT CTA AGC CAA GAA GCG GAT TTT TAT
GGT GCG ATG GAT GGC GCG TCT AAA TTT
を有する130塩基対の断片を増幅させることが可能である。
【0015】
上記した高厳密度の条件は以下のとおりである:ハイブリダイゼーションは42℃で、0. 1%SDSを含んだ2 X SSC緩衝液中の50%のホルムアミド(1 X SSCは、0. 15M Na Cl に15m Mのクエン酸ナトリウムを加えたものでp Hは7. 0)の存在下で行った。洗浄は68℃で行い、例としては2 X SSCプラス0. 1%SDSを使って1時間の間に2回行った。
【0016】
本発明に関しての特に興味深いヌクレオチド配列は、図3、4に記載のヌクレオチド配列、または高厳密度において上記した配列とハイブリダイズするヌクレオチド配列に対応したflbA遺伝子の配列である。
【0017】
本発明の別の態様によると、本出願の主題である該ヌクレオチド配列は、図3、4に記載のヌクレオチド配列であって、図3、4に示したアミノ酸配列を有するタンパク質、またはH. ピロリの鞭毛タンパク質の生合成に関して上記配列と同じ調節特性をもったアミノ酸配列を有するタンパク質をコードすることで特徴付けられる。
【0018】
本発明はまた、ヌクレオチド配列であって、該配列は前述の定義に対応し、更に、flbA遺伝子がもはやホスト細胞内で発現せず、またはホスト細胞内でのflbA遺伝子の発現がH. ピロリの鞭毛の生合成を可能としないように、また適切な場合には、H. ピロリのフックタンパク質の合成ができないように、欠失、置換または挿入によって修飾されているヌクレオチド配列に関する。
【0019】
本発明のヌクレオチド配列に対する修飾は、不可逆であって、特に、この配列がこのようにして修飾されたヌクレオチド配列で形質転換した細菌内に存在する、flbA遺伝子と組み換えを起こした時に、不可逆であるものでなくてはならない。この組み換えは例として、二重交差型である。好ましくはヌクレオチド配列の該修飾は、(この修飾配列によって、与えられたH. ピロリ株内のそれに対応した正常なflbA遺伝子の断片を置換した後に)隣り合う遺伝子群の機能に関して、実質的な修飾を起こさせるべきではない。
【0020】
本発明の範囲に更に含まれるものとしては、上記した基準に合致したflbA遺伝子のフラグメントを構成するヌクレオチド配列がある。例としては、本発明の主題である断片は少なくとも6ヌクレオチド、好ましくは、100ヌクレオチドではないにしろ、少なくとも50ヌクレオチドのフラグメントからなる。
【0021】
このような断片が選択される理由は、例えばそのflbA遺伝子の特異的な特徴のため、もしくはLcr D/ FlbFファミリの蛋白質をコードしたいくつかの遺伝子の保存領域に属するためである。
【0022】
別の態様において本発明はまた、遺伝子中に存在する制限部位を境界として持つflbA遺伝子断片に向けられている。これらの部位のうちいくつかを例として、図2で定義する。
【0023】
本発明に記載の別の断片は、flbA遺伝子のどこかの領域に由来し、好ましくは制限部位を含むか、または制限部位を含むことができる、少なくとも1000塩基対の断片である。
【0024】
本発明における他のヌクレオチド配列は例として、上記した様なヌクレオチド配列を含み、該配列はマーカー、例えば抗生物質に対する耐性遺伝子、または1994年5月20日に出願のFR9406202に記載された様な重金属に対する耐性遺伝子を含むカセットの挿入によって修飾されていることを特徴とする組み換え核酸がある。
【0025】
それゆえに、カナマイシンに対する抵抗性用のカセットを挿入することが可能である。この状況においては、種々の技術を利用することが可能であり、参考文献として特に、Labigne A. ら(J.of Bacteriology, 170, 1988, 1704- 1708)と、Labigne A.らの論文(Res.Microbiol. 1992, 143, 15- 26)を参照されたい。
【0026】
本発明はまた、前述で定義したヌクレオチド配列由来の特異的なオリゴヌクレオチドであって、以下の配列
OLFlb- 1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlb- 2: GAAATCTTCATACTGGCAGCTCCAGTC
OLFlb- 7: CGGGATCCGTGGTTACTAATGGTTCTAC
OLFlb- 8: CGGGATCCTCATGGCCTCTTCAGAGACC
のうちどれか一つを有することで特徴付けられる特異的なヌクレオチド配列に関するものである。
【0027】
別の態様において本発明は、前述のように定義された様なヌクレオチド配列によってコードされることを特徴とする、H. ピロリのFlbA蛋白質のアミノ酸配列に関するものである。
【0028】
H. ピロリのFlbA蛋白質の特異的なアミノ酸配列は図3、4に記載されている。
【0029】
それゆえに本発明の範囲内において、flbA遺伝子及びこの遺伝子により発現される蛋白質は、特にワクチン接種において利用する免疫学的組成物もしくは組成物にとって興味深い。
【0030】
本発明はまた、ヘリコバクター・ピロリ菌株であって、塩基またはヌクレオチド断片の置換、付加および/または欠失によって、H. ピロリの鞭毛タンパク質の生合成の調節に関与する、上記のflbA遺伝子のヌクレオチド配列に生じた突然変異によってもたらされる、非鞭毛フェノタイプを有することを特徴とする菌株に向けられたものである。
【0031】
flbA遺伝子の関するこの修飾は、非鞭毛フェノタイプの株を得ることを可能とし、すなわち該株はFlaA蛋白質もFlaB蛋白質も発現しなくなり、更に好ましくは鞘蛋白質も発現しなくなる。
【0032】
この菌株に関する一つの態様において、得られた該株は更にH. ピロリのフック蛋白質を欠損している。
【0033】
好ましくは、上記した基準に見合う菌株であって、N6株から得られたものであり、1992年6月26日に、NCIMB受付番号第40512号でNCIMBに寄託されたものであることを特徴とする。
【0034】
例として本発明は、H. ピロリの組み換え非鞭毛株であって、N6flbA-と命名され、1995年6月30日に、NCIMB受付番号第40747号でNCIMBに寄託されたH. ピロリの非鞭毛株に関する。
【0035】
このようなH. ピロリの非鞭毛株は、血清学における利用と、その結果として試験管内でのH. ピロリの感染の検出にとって特に興味深い。有益には、これらの株は組み換え型である。
【0036】
特にこれらの株は、H. ピロリによる感染を特異的且つ感度の良い方法で検出するのに有利である。言い換えれば、本発明は有益なことに、例えばサルモネラ(Salmonerlla) またはカンピロバクタ(Campylobacter) 等の菌株による「偽陽性」の結果を避けながら、H. ピロリによる感染を特異的に検出せしめる。
【0037】
このように定義された、非鞭毛型のH. ピロリ株が仮に、たとえばワクチン接種組成物の調製に使われる用に、他の適用が考えられるならば、組み換え体の非鞭毛菌株であって、例えば弱められたウレアーゼを産生するか、或いはもはやウレアーゼを産生しないように更に突然変異されており、この突然変異は、例えばureA遺伝子、ureB遺伝子、ureC遺伝子、ureD遺伝子、ureE遺伝子、ureF遺伝子、ureG遺伝子、ureH遺伝子、およびureI遺伝子の中から選択される一以上のヌクレオチドの突然変異からなることを特徴とする、第二の修飾もしくは変異を有する菌株を調製することは興味深い。ウレアーゼの構造遺伝子群である、ureA、ureB、ureC、ureDは、論文に発表されている(Labigne ら、1991、J.of Bacteriol. 173: 1920- 1931). その他の遺伝子群は特許出願EP 0610322に記載されている。
【0038】
本発明の菌株はそのままで、もしくは抽出液として利用でき、更に本発明は特に、以前記載された株から得た細菌の全抽出物に関するものである。
【0039】
そのようなバクテリア抽出物は、n-オクチルグルコシドで抽出することができる。この場合に使う調製技術は、Lelwala-Guruge J. (Scan. J. Infectt.Dis.1992、24:457- 465)によって記述されたものである。
【0040】
別のバクテリア抽出物も記述された技術により、PBS(Bazaillou M.ら、Clin.Diagn. Lab. Immuno., 1994, 1: 310- 317による方法)もしくはグリシン(Aguirre P.M., Eur.J.Clin.Microbiol.Infect.Dis., 1992, 11: 634- 639による方法) を使った抽出により得ることができる。
【0041】
本願の範囲において本発明は、患者由来の生物学的液体のサンプル( 特に血清サンプル)におけるH. ピロリによる感染をインビトロで検出するための組成物であって、活性成分として本発明の菌株または、上記のバクテリア抽出物を含有したものに関する。
【0042】
用いる生物学的サンプルはどのようなタイプのものであってもよく、また特に、どのようなタイプの生物学的液体サンプルでもよく、例としては血清、唾液、尿があげられる。
【0043】
同様にして検出に用いる方法は、免疫学的な反応を含む、どのようなタイプのものでもよく特に抗原/抗体タイプのものがよい。例としてはウェスタンブロットやELISAなどの技術を使った使用があげられる。
本発明はまた、患者由来の生物学的液体のサンプル(特に血清サンプル)におけるH. ピロリによる感染をインビトロで検出するための方法であって、前記被検サンプルを、本発明の菌株、または上記のバクテリア抽出物と接触させる工程と、前記菌株と、前記被検サンプル中に存在し且つH. ピロリに向けられる抗体との間の免疫反応を検出する工程とを具備する方法に関する。
【0044】
例としては、生物学的サンプルにおけるH. ピロリによる感染のインビトロでの検出は、
天然の、もしくは組み換え体の蛋白質、非鞭毛株、またはバクテリア抽出物、特にNOG(n-オクチルグルコシド)を使ったN6flbA株の抽出物(例としては、抽出物の量は3μg/ml、抗原の量は2μg/ml)などの検出に使う抗原でプレートを覆う工程と、ネガティブ及びポジティブのコントロールの範囲で使い(ポジティブコントロールは異なる希釈度で用いられる)、100分の1に希釈された患者の血清を平行して試験する(供せられる容量は、100μl )工程と、インキュベーションの工程が例として、37℃、1時間で行われ、その後の洗浄を連続的に幾度か行い、次にインキュベーション(例としては37℃で1時間)をモノクローナル接合体(conjugate) (ペルオキシダーゼでラベルしたヒトIgGタイプ)と行い、該接合体が異なる希釈度で用いられ(例としては抗原の場合は1/ 32, 000で、菌抽出物の場合は1/ 64, 000)、供せられる容量が100μl である工程と、モノクローナル接合体とのインキュベーション後にいくつかの異なる洗浄を行い(例えば例4)、酵素反応を「OPDと基質」を使い、30分間遮光して行わせることを満たすことで実行できる。次に酵素反応は、硫酸を加えて終了させて、光学濃度であるO Dを492nm/ 620nmで読む。
【0045】
本発明は更に、H. ピロリに対する抗体を得るための免疫原組成物であって、活性成分として、本発明の菌株、または上記のバクテリア抽出物を含有することを特徴とする免疫原組成物に向けられている。
【0046】
本発明の一つの特異的な態様において、H. ピロリに対する抗体を得るための免疫原組成物は、FlbA蛋白質由来のアミノ酸配列を含んでいることを特徴とする。
【0047】
本発明の範囲に更に含まれるものとしては、H. ピロリによる感染に対して防御する抗体を得るためのワクチン接種組成物であって、活性成分として本発明の菌株または、上記のバクテリア抽出物を含有することを特徴とする組成物がある。
【0048】
H. ピロリによる感染に対して防御する抗体を得るための別のワクチン接種組成物は、活性成分としてウレアーゼタイプの抗原、特にureA、ureB、ureC、またはureD遺伝子によってコードされ、上記に定義したアミノ酸を有する蛋白質を有することを特徴とする。
【0049】
本発明は上記したアミノ酸配列に向けられるモノクローナル抗体またはポリクローナル血清に関する。この抗体は、それ自体は知られた方法によって得られ、特に、動物を選択した抗原で免役し、次に抗体を合成し、合成された抗体を回収し、その中からH. ピロリを特異的に認識するものを選択するか、または前もって免役しておいた動物の脾臓細胞をミエローマ細胞と融合させてハイブリドーマを調製し、次にこのハイブリドーマを培養して、選択したH. ピロリ抗原に対しての特異的認識をもとに選択したモノクローナル抗体を得ることにより得られる。
【0050】
本発明の他のモノクローナル抗体、またはポリクローナル血清は、上記したように非鞭毛性H. ピロリに向けられている。
【0051】
本発明は更に、生物学的サンプルにおけるH. ピロリによる感染をインビトロで検出するための組成物であって、活性成分として、本発明の非鞭毛性のフェノタイプのH. ピロリに対して得られるモノクローナル抗体、またはポリクロナール血清を含有する組成物に関する。
【0052】
本発明はまた、薬物の活性成分として、本発明によるアミノ酸配列をコードしたヌクレオチド配列であって、動物またはヒトで免役学的反応を誘導することができるヌクレオチド配列に関する。薬物としてヌクレオチド配列を使用する技術は、Donneyら(1995; Nature Medic. 1( 6), pp 583- 587)に記載されている。
【0053】
例I
flbA遺伝子の同定及び非鞭毛株の調製
細菌の移動度の発現を制御する機能を担うことが知られている蛋白質の中で興味のあるものは、最近同定されたLcrD/FlbFファミリに属する蛋白質であり、その中にはイェルシニア属の細菌のLcrD蛋白質(6)、サルモネラのInvA蛋白質(2)、シゲラのMxiA(1)、カウロバクター クレセンタスのFlbF(7)、及びカンピロバクター ジェジュニのLfbA(4)がある。LcrD、InvA、及びMxia蛋白質は、該蛋白質を発現する細菌の病原性に関わる蛋白質の制御及び/または分泌に関わり、一方カウロバクター クレセンタスのFlbF蛋白質、及びカンピロバクター ジェジュニのFlbAは鞭毛の生合成の制御に関わり、それゆえに移動度に関わる。現在までに知られたLcr D/ FlbAファミリの相同体は、特にこの蛋白質のN末端部分が非常によく保存された領域を有しており、それゆえにこの2つの保存領域(イェルシニアのLcr D蛋白質の151から156番目のアミノ酸であるMPGKQM、及びLcr Dの189から195番目のアミノ酸であるMDGAMKF)を使って2つの縮退したオリゴヌクレオチド(L OLFlbA- 1及びOLFlbA- 2、表1)を限定することが可能であったが、該ヌクレオチドは、ヘリコバクター・ピロリの染色体DNA上で行う遺伝子増幅におけるヌクレオチドプライマとして合成されて使用される。このようにして、130塩基対(bp)の断片を増幅することが可能であり、そのヌクレオチド配列の決定により、該断片がLcrD/ FlbFファミリの蛋白質に非常に相同である蛋白質の一部をコードしていることが証明された。この増大された断片は次に、放射性のラベルがなされH. ピロリのコスミドベクタのスクリーニングに使われた。
【0054】
この断片は図3、4に示した配列の575から707の間のヌクレオチドに対応した配列である。
【0055】
ゲノムライブラリのコスミドのうちの一つは、H. ピロリのLcrD/ FlbF相同体をコードしているものとして同定され、Sau3Aによる部分消化を行って、2から5(キロ塩基対)の大きさをもつ挿入断片を含む、ベクタpILL570中のコスミドのミニライブラリ(200サブクローン)を構築した。ベクタpILL570はLabinge A. らの論文(パスツール研究所/Elsevier Paris1992.Res. Microbiol, 1992, 143, 15- 26)に記載されている。このベクタの制限地図は、図1に示してある。上記の200クローンを130塩基対のプローブにハイブリダイズすると、プラスミドpSUS39およびpSUS207をもつクローンが陽性を示した。これら二つの組み換えプラスミドの線形制限地図は、図2に示してあり、これらのクローンの挿入部がそれぞれ重なり合った配列をゆすることを示した。これらの二つの挿入部のヌクレオチド配列を決定して、これら導入部にはflbA遺伝子がそのままは含まれないことを明かにした。カンピロバクタ ジェジュニのFlbA遺伝子に対する相同体であるために、H. ピロリのflbAと命名される当該遺伝子は、2196塩基対のオープンリーディングフレームに対応し、80. 1キロダルトンの計算上の分子量をもつ蛋白質をコードしている。flbAのヌクレオチド配列及びFlbAのアミノ酸配列を図3、4に示してある。プロモータ及びターミネータに特徴のコンセンサス配列は、該オープンリーディングフレームの上流及び下流には検出さていない。
【0056】
FlbA蛋白質は、共に移動度に関わるカンピロバクタ ジェジュニのFlbA蛋白質及びカウロバクタ クレセンタスのFlbF蛋白質と類似点を呈しているが(それぞれ51. 7%、40. 4%)、この類似の割合は、移動度に関わらないLcr D/ FlbF蛋白質ファミリとの比較ではより低く、イェルシニアのLcrDとは32. 8%の相同性、シゲラのMxiAとは30. 5%の相同性、そしてサルモネラのInvAとは29. 3%の相同性である。これらの蛋白質の、H. ピロリのFlbAに対するマルチプル整列(multiple alighnment )を図5、6に示してある。LcrD/ FlbFファミリの相同体で最も保存された領域は蛋白質のN末端部分に位置している。
【0057】
移動度に関わる蛋白質(FlbA及びFlbF)の系統学的進化、及び病原性に関わる蛋白質の発現及び/または分泌の制御に関わる蛋白質の系統学的進化は、系統樹を使って示してある(図7)。二つの別個の分岐が見られる:H. ピロリのFlbAが、鞭毛の生合成に関わる制御蛋白質に対応する分岐に属することは、はっきりとしている。
【0058】
FlbA蛋白質の合成を欠損している、H. ピロリのアイソジェニック変異体の構築と特徴づけ
プラスミドpSUS39の1600塩基対の断片を、それぞれの5’末端にBamHI制限部を有するオリゴヌクレオチドOLFlbA- 1及びOLFlbA-7を使って増幅した(表 )。この断片の中央部には、ユニークなHind III制限部位があり、この断片を、pUC19の誘導体であってそのマルチプルクローニング部位のHind III部位を取り除いた、pSUS33へクローニングした。pSUS33を得るために、プラスミドpUC19をHindIIIで消化し、その結果得られる粘着末端をクレノー酵素及びT4DNAポリメラーゼで処理して平滑末端にし、その結果の断片をT4DNAポリメラーゼで再連結して、pSUS33を得るために大腸菌DH5αへ挿入した。上記の1600塩基対の断片をpSUS33へ挿入して得た組み換えプラスミドをpSUS40と命名し、HindIIIで線状化し、その末端を平滑にして、プラスミドpILL600(LabingeA.ら1988、J.Bact. 170, 1704- 1708)由来のSmaIカナマイシンカセットをこのユニークな部位へクローニングして、プラスミドpSUS42を得た。プラスミドpSUS42を次に、電気穿孔法によりH. ピロリのN6株へ導入した。電気穿孔法は、Ferrero R.L.ら(Journal of Bacteriology, July,1992, pp4212- 4217,Vol174, No 13)が記載の方法に添って行った。次にカナマイシン(25μg/ml)を含んだ選択培地で選択した形質転換体に関して、遺伝型及び表現型の特徴づけを行った。図8は変異体作成にあたって従った手順を示している。遺伝子増幅及びサザンハイブリダイゼーションによるこれらの変異体の遺伝型分析により、カナマイシン抵抗性形質転換体のゲノムがflbA遺伝子の真ん中に、該抵抗性遺伝子を含んでいることと、それゆえに二重交差による、野生型のflbA遺伝子の不活型flbA- Kmへの対立遺伝子置換が、pSUS33ベクタのヌクレオチド配列の喪失を伴って起きたこととが示された。H. ピロリのflbA- 変異体の表現型分析により、該変異体は移動度を有しないこと、更に電子顕微鏡による分析により、鞭毛要素の完全な欠失及び鞭毛の鞘の欠失があることを示した。H. ピロリの鞭毛全体の蛋白質に向けられた抗体を使ったイミュノブロット実験(ウェスタンブロット)(図9)は、鞭毛のサブユニットであるFlaA及びFlaBに対応したペプチドのバンドが欠失していることと、また同様に最近O'Tooleと共同研究者(5)によってフック蛋白質(もしくは固定蛋白質)または鞭毛と同定された蛋白質(5)で、見かけ分子量が90キロダルトンのポリペプチドに対応したバンドも欠失していることを示した。
【0059】
全体としてみると、これらの結果はH. ピロリのFlbA蛋白質は鞭毛の構造全体の合成に必須であり、この蛋白質をコードした遺伝子の不活化は、該構造の構成要素の輸送の阻害ではなくて、鞭毛形成にくわわるいかなる構造の合成をも完全に停止する結果となることを示唆している。
【0060】
【表1】
例II
調べたH. ピロリの血清学モデル
1)HspAmalE: 47. 5k Da (HspA=13kDa)の組み換え蛋白質
保存されていた血清の、母集団Iと呼ばれるものにおいて41%の感受性、96%の特異性が得られた。
【0061】
2)N6flbA−:ヘリコバクター・ピロリの非鞭毛株3種類の抽出法を行った(n-オクチルグルコシド、PBS、グリシン)。さしあたり、n-オクチルグルコシドによる抽出が最もよいことがわかった。
【0062】
3)−N6: 野生型株に対応している。抽出はn-オクチルグルコシドで行った。
【0063】
血清の第二の母集団も使った(母集団II)。この母集団は、臨床的、内視鏡的、組織学的、細菌学的、及び解剖病理学的(anatomopathological)見地からよく記録された数百の血清からなっている。本調査において異なるモデル系の出来栄えを調べるために使ったのが、この母集団IIであった。5つの異なる母集団を試験した。
【0064】
試験した5つの血清は、
300の通常の血清(FNTS)
WHITTAKER血清学で陽性であった18の血清(CBMS)
母集団II血清と名付けられた92の、よく記録された血清
細菌学的、解剖病理学的(anatomopathological )見地からよく記録され、また母集団Iと名付けられた、87の血清
以下の交差反応を示す23の血清
抗レジオネラ(Legionella)陽性の10の血清
抗クラミジア(Chlamydia)陽性の10の血清
抗カンピロバクタ(Campylobacter)陽性の3つの血清
文献で効果が認められている2つの競合するキットをパラレルに使って試験した。
【0065】
2つの市販のキット
Cobas Core(ROCHE)
Pylori Stat(WHITTAKER)
結果
通常の血清(FNTS)( 図10から14、表2、3)
以下のモデルを通して得た300の血清
Hsp A malE
N6 flBA
N6
疫学的な調査によると、フランスでは20から25%の血清陽性率(sero-prevalences)となっている。300人の血液提供者の血清の分布を調べ、CBMSライブラリを使って以前に定義された域値(WHITTAKER血清学)を確証する為に異なる域値に対する陽性の支配度を計算した。
【0066】
この研究により、同じ血清陽性率を使って異なる試験を比較することができる。
【0067】
最初の43の血清は、以下のモデルを通して得た
Cobas Core(Roche)
Pylori Stat(WHITTAKER)
JLF血清学として知られる血清学(数種類の水溶性バクテリア抽出物に基づくELISA試験)
結果は任意の単位に基づいて、異なる域値に対して表してある;陽性の結果は1、陰性の結果は0と表す。
【0068】
異なる試験によって、これら43の血清を比較して、以下のことが判明する:非鞭毛株であるN6flbA- 及びCobas Core試験( Roche)では、20%のオーダーに匹敵する血清陽性率となる。
【0069】
HspAでは、非常に低い血清陽性率(7%)となり、これは引き続く結果の観点からは、感受性の欠失を示唆している。
【0070】
引き続く結果を考慮すると、JLF血清学は、血清陽性率がわずかに14%であるので非常に感度がよいことがわかる。
【0071】
Pylori Stat(Whittaker) 試験では、高い血清陽性率(29%)となるが、これは特異性の欠如、または域値が低すぎることを示している。
【0072】
【表2】
【表3】
WHITTAKER血清学で陽性の血清(CBMS)(表4)
Pylori Stat試験(Whittaker)で陽性であった血清はわずか3つであった。この3つの血清は、他の試験では確認できなかった。
【0073】
この結果は特異性の欠如によるものであると思われる。もし、現在の市場で最も良いCobas Core試験(Roche)を参照するならば、我々の異なるモデルをCobas Coreに対して比較することが可能である。
【0074】
非鞭毛N6flbA- 株はCobas Coreと完全に一致する。
【0075】
Cobas Coreで陰性であった上記の3つの血清は、N6flbA- でも陰性であった。
【0076】
Cobas Coreで陽性であった15の血清は、N6flbA- でも陽性であった。
【0077】
野生型N6株は非鞭毛株と同じ結果となった。
【0078】
9つのCobas Core血清がHspAで陰性であるので、HspAもまた感受性がない。
【0079】
Cobas Coreで陰性である上記の3つの血清は、HspAでも陰性である。
【0080】
【表4】
母集団IIの血清
92の血清を選択し、3つのグループへ分類した:
34:消化不良患者、内視鏡または組織学での潰瘍との診断(十二指腸、胃)、培養及び/または解剖病理学によりヘリコバクター・ピロリが存在すること、迅速な尿素試験も実行、このグループはHp+/U+と命名。
【0081】
27:消化不良患者、内視鏡または組織学での潰瘍との差動的診断(十二指腸、胃)、培養及び/または解剖病理学によりヘリコバクター・ピロリが存在すること、迅速な尿素試験も実行、このグループはHp+/U−と命名。
【0082】
31:消化不良のまたは消化不良でない患者、内視鏡及び組織学では正常な胃十二指腸、培養及び/または解剖病理学によりヘリコバクター・ピロリが存在しないこと、迅速な尿素試験も実行、このグループはHp−と命名.
臨床、内視鏡、組織学、細菌学、及び解剖病理学な知見を、それぞれの患者に対して示した。
【0083】
この良く記録された集団により、感受性及び特異性の基準の定義がなされる。
【0084】
Hp A: 母集団Iに見られるように、実質的な感受性の欠如がいぜんとして見られる。
【0085】
感受性は59%で特異性は100%である。
【0086】
N6flbA:n- オクチルグルコシド抽出物に対して100%の感受性が確認でき、特異性は90%である。
【0087】
この結果は、RocheのCobas Core試験の結果で得られたもの(感受性が98%で特異性が94%)と匹敵する。
【0088】
N6: 母集団IIにおいて、野生型株は非鞭毛株に完全に匹敵する。
【0089】
陰性の31の血清は野生型に対しては陽性である。鞭毛による交差反応は、母集団IIに関しては全く見られなかった。
【0090】
【表5】
【表6】
【表7】
【表8】
【表9】
【表10】
【表11】
【表12】
【表13】
【表14】
【表15】
血清学の位置づけ
血清学を2つのレベルで位置づけた。
【0091】
高感度血清学:心窩部痛をうったえる若年患者に存在する細菌を検出するためのもの。
【0092】
この血清学で陰性となった場合は、患者は内視鏡の検査または生検を受けることはなく、他の痛みの原因となるものを調べる。
【0093】
危険特異的血清学:これはヘリコバクター・ピロリによる感染での危険性、すなわち潰瘍、癌、または胃リンパ腫(MAL Tリンパ腫)を示すこと含む。
【0094】
疑いのある危険性に特異的な分子を使うか
または危険特異的な域値を使う(域値は危険のある患者では、危険のない患者よりも高い)。
【0095】
この特異的な血清学を使って、一般の母集団をスクリーニングにかけ、それによってヘリコバクター・ピロリに関わり、また症状がないために検出できない様な癌及びリンパ腫を検出する(痛みを訴えている患者のみが胃腸科にかかる。)
感受性の問題に対する応答は良い。
【0096】
【表16】
【表17】
【表18】
胃炎の強度と抗体レベルとの間の相関関係
胃炎は3つの変数で定義される:
萎縮(Gの後の最初の数字で表現);強度は1から4で表す。
【0097】
中性好性多核細胞及び単球の侵入に対応する全体の炎症(Gの後の二番目の数字で表現);強度は1から3で表す。
【0098】
中性好性多核細胞の数に対応した活性(Gの後の三番目の数字で表現);強度は0から3までで表す。ろ胞状のものはFと表現する。
【0099】
通常は、以下の相関関係が見られる:
活性はヘリコバクター・ピロリと非常に良く相関する。
【0100】
炎症はヘリコバクター・ピロリと相関する。
【0101】
それぞれのグループで見られる力価の平均は、故に上記の3つの変数とその強度とを使って計算される。
【0102】
結果の評価:
t 試験を使って2つのデータの平均が顕著に異なるか、または5%の危険度でそうでないかを証明することが可能となる。
【0103】
t 試験が基づく仮定が、変数の等価性であり、F試験(Fisher試験)で示されている。
【0104】
変数の中には等しくないものがあるので、よってそれぞれを全ての平均値と比較することはできない。
【0105】
可能であるならば平均を比較することで、異なるグループの間の差が顕著であるかそうでないかを示すことができる。
【0106】
顕著な差:HspAに対する「2」及び「3」、と「炎症」グループのNOGとの間の平均。
【0107】
顕著でない差:活性に関しては、異なる強度レベル間での顕著な差は見られなかった:
HspA: 0と2、0と3、1と2、1と3、及び2と3との間には顕著な差はない
NPG: 0と1、0と2、1と2、1と3、及び2と3との間には顕著な差はない.
しかしながら、力価が胃炎の強度に依存して増大する傾向は見られる:
萎縮に関して:HspA及び非鞭毛株のNOG抽出のは平均値は、レベルが1から2、及び2から3へ移ると、二倍になる。
【0108】
炎症に関して:レベルが2から3になると、平均が二倍になる。
【0109】
それぞれのグループにおける数は、統計的に顕著な差であると結論するには、相対的に少ない(それぞれ30より少ない)。
【0110】
【表19】
【表20】
交差反応性を呈する血清
2つのタイプの血清を用いた。
20の血清(10の抗- レジオネラ+ 、10の抗- クラミジア+ ):これら3つの細菌はそれぞれが非常に似た熱ショック蛋白質を有しているため、HspAに対して交差反応することができる
3つの抗- クラミジア陽性抗体:非鞭毛株であるN6flbAでは消出する、鞭毛株N6との交差反応性を示すため。抗−カンピロバクタ陽性血清を得ることは非常に難しい;これが、わずかに3つの血清しか得られない理由である。
【0111】
HspAは、10の抗- レジオネラ陽性抗体とも、10の抗- クラミジア陽性抗体とも、交差反応性を全く示さない。
【0112】
これらの血清のうちいくつかは、鞭毛株及び非鞭毛株の両方について、抗- ヘリコバクター・ピロリ抗体に関する陽性力価を有しているが、これらの血清の臨床的意義はわかっていない。
【0113】
【表21】
結論
HspA malE
感受性が欠損しているために、この分子自体を利用することはできないが、他の分子と結合しているならば興味深いであろう。
【0114】
しかしながら、異なる細菌間で実質的に保存された熱ショック蛋白質による交差反応の危険性をはらんでいる。
【0115】
N6flbA-
この非鞭毛変異体は非常に興味深いことがわかってきている;母集団IIで判明した感受性及び特異性は、非常に好ましい効果を示している。
【0116】
N6
さしあたっては、鞭毛株が興味深い。しかしながら鞭毛に関連した交差反応性に関してはわずかしか調べられていないが、これはカンピロバクタ血清学に関して良く考証されている血清を得ることが難しいためである。
【0117】
JLF試験
ヘリコバクター・ピロリの幾つかの株についての水溶性(PBS)抽出に基づいた血清学的試験を開発した。
【0118】
非鞭毛株変異体のNOG抽出を使って母集団Iの血清を試験した。
【0119】
細菌学的及び解剖病理学的見地からのみ考証された87の血清を、非鞭毛細菌抽出を使って試験した。
【0120】
培養が陽性であるか、または解剖病理学的試験及び迅速な尿素試験で陽性であれば、血清は陽性である。
【0121】
3つの試験(培養、解剖病理学、及び迅速な尿素試験)で陰性であれば血清は陰性である。
【0122】
感受性は90. 3%(28/ 31)で、特異性は71. 4%(40/ 56)であった。
【0123】
はじめの試験であやまって陽性とされた16の血清のうち、9つはJLF血清学またはJLFウェスタンブロット、もしくはその両方により陽性であった。
【0124】
はじめの試験であやまって陰性とされた3つの血清全ては、JLF血清学またはJLFウェスタンブロットで陰性であり、一つは両方で陰性であった。
【0125】
【表22】
【表23】
技法
プレートは以下のものでコートした:
2μg/mlのHspA抗原
3μg/mlの、NflbA及びN6のNO G抽出
範囲
5つの範囲でのネガティブコントロール
ポジティブコントロールは4つの希釈度で使用
患者血清
1/ 100希釈
容量:100μl
インキュベーション 37℃1時間
3回洗浄
モノクローナル接合体(IgG toxo)
使用濃度
HspAに対しては1/ 32000
N6flbA- に対しては1/ 64000
N6に対しては1/ 56000
容量 100μl
接合体のインキュベーション:37℃、1時間
4回の洗浄
OPD+ 基質による酵素反応の進行
遮光で30分間
硫酸による酵素反応終結
492nm/ 620nmでのOD測定
ODの、任意の単位(AU)への変換
【表24】
【表25】
【表26】
【表27】
【表28】
【表29】
【表30】
参考文献
1. Andrews,G.P., Maurelli,A.T.: mxiAof Shigella flexneri 2a, which facilitates export of invasion plasmid antigens encodes a homolog of the low−calcium−response protein LcrD of Yersinia pestis. Infect. Immun. 60: 3287- 3295( 1992).
2. Galan,J.E., Ginocchio,C., Costeas, P.:Molecular and functional characterization of the Salmonella invasion gene invA:homology of InvA to menbers of a new protein family. J.Bacteriol. 174: 4338- 4349( 1992).
3. Leying,H., Suebaum, S., Geis, G.,Haas,R.:Cloning and genetic characterization of a Helicobacter pylori flagellin gene. Mol.Microbiol.6:2563-2874( 1993).
4. O’Toole,P.W., Kostrzynska, M., Trust,T.J.:Non−mobile mutants of Helicobacter pilori and Helicobacter mustelae defective in flagellar hook production. Mol. Microbiol. 14: 691- 703(1994).
5.Plano,G.V., Barve,S.S., Straley,S.C.: LcrD, a menbrane −bound regulator of the Yersinia pestis low−calcium response. J.Bacteriol. 173: 7293- 7303( 1991).
6. Ramakrishnaan, G., Zhao,J−L., Newton,A.:The cell cycle−regulated geneflbF of Caulobacter crescentus is homologous to a virulence locus of Yersinia pestis. J.Bacteriol. 173: 7283- 7292( 1991).
7.Suerbaum, S., Josenhans,C., Labigne,A.:Cloning and genetic characterization of the Helicobacter pylori and Helicobacter mustelae flb flagellin genes and constructionof H.pylorifla− And flB− negative mutantsby electroporation−mediated allelic exchange. J.Bacteriol., 175: 3278- 3288( 1993).
【図面の簡単な説明】
【0126】
【図1】プラスミドpILL570、およびカナマイシン抵抗性遺伝子カセットを含んだ小型トランスポゾンTn 3の制限地図。
【図2】組み換えプラスミドpSUS39及びpSUS207の線形制限地図。表示の番号は制限断片の長さに対応していて、塩基対で表示されている。H:HindIII、Bg:BglII。星印は、クローニング中に制限部位が修飾されて、対応していた制限酵素で認識されなくなったことを示す。
【図3】H. ピロリのflbAのヌクレオチド配列及び、一文字表示による推定のアミノ酸配列。
【図4】H. ピロリのflbAのヌクレオチド配列及び、一文字表示による推定のアミノ酸配列。
【図5】H. ピロリのFlbA蛋白質と、他の5つのLcr D/ FlbFファミリとのマルチプル整列(multiple alignment)。Cj FlbA:カンピロバクタ ジェジュニ( Campylobacter jejuni) FlbA; Cc FlbF:カウロバクタ クレセントゥス( Caulobacter creacentus) FlbF; YpLcsD: イェルシニア ペスティス( Yersiniapestis)LcrD; StlnvA: サルモネラ ティフィムリウム( Salmonera typhimurium) InvA; SfMxiA: シゲラフレクスネリ( Shigella flexneri)MxiA。星印はLcrD/ FlbFファミリの相同体全てにおいて保存されているアミノ酸の位置を示し、点は相同体6つのうち少なくとも5つにおいて保存されているアミノ酸の位置を示す;縮退したオリゴヌクレオチドの合成に用いられた、保存アミノ酸配列には下線が引いてある。これらの相同体のN末端領域においての保存の度合いにたいして注意するべきであり、これはC末端の疎水性領域の多様性の度合いと対照的である。
【図6】H. ピロリのFlbA蛋白質と、他の5つのLcrD/ FlbFファミリとのマルチプル整列(multiple alignment)。Cj FlbA:カンピロバクタ ジェジュニ( Campylobacter jejuni) FlbA; Cc FlbF: カウロバクタ クレセントゥス( Caulobactercreacentus) FlbF; YpLcs D: イェルシニア ペスティス( Yersinia pestis) LcrD; StlnvA: サルモネラ ティフィムリウム( Salmonera typhimurium) InvA; SfMxiA: シゲラ フレクスネリ( Shigella flexneri) MxiA。星印はLcrD/ FlbFファミリの相同体全てにおいて保存されているアミノ酸の位置を示し、点は相同体6つのうち少なくとも5つにおいて保存されているアミノ酸の位置を示す;縮退したオリゴヌクレオチドの合成に用いられた、保存アミノ酸配列には下線が引いてある。これらの相同体のN末端領域においての保存の度合いにたいして注意するべきであり、これはC末端の疎水性領域の多様性の度合いと対照的である。
【図7】Lcr D/ FlbFファミリに属する6つの蛋白質の系統樹を描いた図。移動度の発現制御に関わる蛋白質、すなわちH. ピロリのFlbA( Hp FlbA) 及びカンピロバクタ ジェジュニのFlbA( Cj FlbA) 、並びにカウロバクタ クレセントゥス( Caulobacter creacentus) のFlbF( Cc FlbF) は、病原性蛋白質( サルモネラのInvA、シゲラのMxiA、及びイェルシニアのLcrD) の分泌に関わる蛋白質とはかけ離れたブランチを形成する。表示の番号は、相対的な進化の距離を表す。
【図8】H. ピロリの同質遺伝子的変異体株N6、すなわちFlbA蛋白質をコードした遺伝子が、カナマイシン抵抗性をコードした遺伝子の挿入によって不活化された変異体を構築する際に従った方法を描いた図。
【図9】H. ピロリから精製した鞭毛に対して特異的なAK 179抗血清( 3) を使って行った、N6flbA- 変異体由来の蛋白質の免疫ブロット(ウェスタンブロット);1:N6- flbA変異体、2:flaA/ flaB二重変異体、3: flaB( 8) 変異体、4:fla A( 8) 変異体、5:野生型N6株。
【図10】H. ピロリに対して行った血清学の結果比較。
【図11】H. ピロリに対して行った血清学の結果比較。
【図12】H. ピロリに対して行った血清学の結果比較。
【図13】H. ピロリに対して行った血清学の結果比較。
【図14】H. ピロリに対して行った血清学の結果比較。
【図15】非鞭毛株N6flbA- を使った抽出:グリシン、PBS、またはNOGを使って行った抽出。いかのデータに基づいて曲線を描いた。 Net Abs 計算したSTD# 濃度 750. 0 濃度 差 1 0. 0000 0. 0020 - 0. 008 0. 0080 2 0. 1660 0. 0760 0. 1721 -0. 006 3 0. 3300 0. 1400 0. 3459 -0. 016 4 0. 6650 0. 2390 0. 6474 0. 0176 5 1. 3300 0. 4280 1. 3336 -0. 004係数:P 2= 2. 0324, P 1= 2. 2753, P 0= 0平均:- 1. 0356E- 07, 標準偏差:0. 0130
【図16】非鞭毛株N6flbA- を使った抽出:グリシン、PBS、またはNOGを使って行った抽出。ミニメソッド( BioRad) の蛋白質アッセイ。グリシン:1/ 2希釈、グルコシド:1/ 10希釈、上清1:1/ 4希釈、上清2:希釈せず。以下のデータに基づいて曲線を描いた。 Net Abs 計算したSTD# 濃度 750. 0 濃度 差 1 0. 0000 -0. 003 1. 5398 -1. 540 2 25. 000 0. 0600 21. 861 3. 1392 3 50. 000 0. 1470 51. 810 -1. 810 4 100. 00 0. 2750 99. 855 0. 1454 5 200. 00 0. 5090 199. 94 0. 0636係数:P 2= 144. 63, P 1= 314. 31,P0= 2. 4815
【図17】非鞭毛株N6flbA- を使った抽出:グリシン、PBS、またはNOGを使って行った抽出。
【特許請求の範囲】
【請求項1】
ヘリコバクター・ピロリ(Helicobacter pylori)の鞭毛タンパク質の生合成を調節すると共に、高厳密度の条件下において、下記の配列を有する二つのオリゴヌクレオチドを用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブとハイブリダイズすることができるか、
OLFlbA−1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2: GAAATCTTCATACTGGCAGCTCCAGTC
或いは、高厳密度の条件下において、これらのオリゴヌクレオチドとハイブリダイズすることができるヌクレオチド配列。
【請求項2】
ヘリコバクター・ピロリ(Helicobacter pylori)の鞭毛タンパク質の生合成を調節するflbA遺伝子のヌクレオチド配列であって、このヌクレオチド配列は、H. ピロリ株の染色体DNAを含むゲノムライブラリーを、下記の配列を有する二つのオリゴヌクレオチドを用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブか、
OLFlbA−1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2: GAAATCTTCATACTGGCAGCTCCAGTC
或いは、高厳密度の条件下において、これらのオリゴヌクレオチドとハイブリダイズすることができるプローブを用いてスリーニングする工程と、前記プローブとハイブリダイズするDNA配列を回収する工程と、得られたDNA配列をプラスミドタイプの適切なベクター中にサブクローニングし、次いで、高厳密度の条件下において、オリゴヌクレオチドOLFlbA−1およびOLFlbA−2を用いて増幅されたH.ピロリ由来のヌクレオチドフラグメントに対応するプローブとハイブリダイズする修飾ベクターを選別する工程と、上記プローブとハイブリダイズするプラスミドベクター中に含まれているDNAフラグメントの配列決定を行い、これらフラグメントに含まれるオープンリーディングフレームを決定する工程とによって得られるヌクレオチド配列。
【請求項3】
請求項1または2に記載のヌクレオチド配列であって、図3、4に示したヌクレオチド配列を有することを特徴とするヌクレオチド配列。
【請求項4】
請求項1〜3の何れか1項に記載のヌクレオチド配列であって、図3、4に示したアミノ酸配列を有するタンパク質、またはH.ピロリの鞭毛タンパク質の生合成に関して上記配列と同じ調節特性をもったアミノ酸配列を有するタンパク質をコードすることを特徴とするヌクレオチド配列。
【請求項5】
請求項1〜4の何れか1項に記載のヌクレオチド配列であって、該配列は、flbA遺伝子がもはやホスト細胞内で発現せず、またはホスト細胞内でのflbA遺伝子の発現がH. ピロリの鞭毛の生合成を可能としないように、また適切な場合には、H. ピロリのフックタンパク質の合成ができないように、欠失、置換または挿入によって修飾されているヌクレオチド配列。
【請求項6】
flbA遺伝子のフラグメントに対応する請求項1〜4の何れか1項に記載のヌクレオチド配列であって、flbA遺伝子に由来し、好ましくはflbA遺伝子の配列に存在する制限酵素部位で切断された、少なくとも6ヌクレオチド、好ましくは少なくとも100ヌクレオチドのフラグメントであることを特徴とするヌクレオチド配列。
【請求項7】
請求項1〜6の何れか1項のヌクレオチド配列を含み、該配列はマーカー、例えば抗生物質に対する耐性遺伝子または重金属に対する耐性遺伝子を含むカセットの挿入によって修飾されていることを特徴とする組み換え核酸。
【請求項8】
請求項7に記載の組み換え核酸であって、請求項1〜5の何れか1項に記載の前記ヌクレオチド配列が、カナマイシンに対する耐性カセットの挿入によって修飾されていることを特徴とする組み換え核酸。
【請求項9】
請求項1〜3の何れか1項に記載の配列に対して特異的であることと、下記の配列の一つを有することとを特徴とするオリゴヌクレオチド。
OLFlbA−1:ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2:GAAATCTTCATACTGGCAGCTCCAGTC
OLFlbA−7:CGGGATCCGGGTTACTAATGGTTCTAC
OLFlbA−8:CGGGATCCTCATGGCCTCTTCAGAGACC
【請求項10】
請求項1および2の何れか1項に記載のヌクレオチド配列によってコードされることを特徴とする、H. ピロリのFlbAタンパク質のアミノ酸配列。
【請求項11】
H. ピロリのFlbAタンパク質であり、図3、4に示した配列を有すること、またはFlbAタンパク質に対する抗体によって認識される該蛋白質のフラグメントであることを特徴とするアミノ酸配列。
【請求項12】
ヘリコバクター・ピロリ菌株であって、塩基またはヌクレオチドフラグメントの置換、付加および/または欠失によって、H. ピロリの鞭毛タンパク質の生合成の調節に関与するflbA遺伝子の請求項1〜3の何れか1項に記載のヌクレオチド配列に生じた突然変異によってもたらされる、非鞭毛フェノタイプを有することを特徴とする菌株。
【請求項13】
請求項12に記載の菌株であって、更に、H. ピロリのフックタンパク質が欠失していることを特徴とする菌株。
【請求項14】
請求項12または13に記載の組み換え菌株であって、N6株から得られたものであり、1992年6月26日に、NCIMB受付番号第40512号でNCIMBに寄託されたものであることを特徴とする菌株。
【請求項15】
請求項12または14に記載の組み換え菌株であって、N6flbA−株であり、1995年6月30日に、NCIMB受付番号第40747号でNCIMBに寄託されたものであることを特徴とする菌株。
【請求項16】
請求項12〜15の何れか1項に記載の菌株であって、弱められたウレアーゼを産生するか、或いはもはやウレアーゼを産生しないように更に突然変異されており、この突然変異は、例えばureA遺伝子ureB遺伝子、ureC遺伝子、ureD遺伝子、ureE遺伝子、ureF遺伝子、ureG遺伝子、ureH遺伝子、およびureI遺伝子の中から選択される一以上のヌクレオチドの突然変異からなることを特徴とする菌株。
【請求項17】
請求項12〜15の何れか1項に記載の菌株の抽出物であることを特徴とするバクテリア抽出物。
【請求項18】
請求項17に記載のバクテリア抽出物であって、n−オクチルグルコシドで抽出した後に得られることを特徴とするバクテリア抽出物。
【請求項19】
請求項17に記載のバクテリア抽出物であって、PBSまたはグリシンで抽出した後に得られるバクテリア抽出物。
【請求項20】
患者由来の生物学的液体のサンプル(特に血清サンプル)におけるH. ピロリによる感染をインビトロで検出するための組成物であって、活性成分として、請求項12〜15の何れか1項の菌株、または請求項17〜19の何れか1項のバクテリア抽出物を含有することを特徴とする組成物。
【請求項21】
患者由来の生物学的液体のサンプル(特に血清サンプル)におけるH. ピロリによる感染をインビトロで検出するための方法であって、前記被検サンプルを、請求項12〜15の何れか1項の菌株、または請求項17〜19の何れか1項のバクテリア抽出物と接触させる工程と、前記菌株と、前記被検サンプル中に存在し且つH. ピロリに向けられる抗体との間の免疫反応を検出する工程とを具備する方法。
【請求項22】
H. ピロリに対する抗体を得るための免疫原組成物であって、活性成分として、請求項12〜16の何れか1項の菌株、または請求項17〜19の何れか1項のバクテリア抽出物を含有することを特徴とする組成物。
【請求項23】
H. ピロリに対する抗体を得るための免疫原組成物であって、請求項10または11の何れか1項に記載のアミノ酸配列を含有する組成物。
【請求項24】
H. ピロリによる感染に対する防護的抗体を得るためのワクチン接種組成物であって、活性成分として、請求項12〜16の何れか1項の菌株、または請求項17〜19の何れか1項のバクテリア抽出物を含有することを特徴とする組成物。
【請求項25】
H. ピロリによる感染に対する抗体を得るためのワクチン接種組成物であって、活性成分として、ウレアーゼタイプである抗原またはH.ピロリのウレアーゼ活性に関与する抗原、特にureA遺伝子、ureB遺伝子、ureC遺伝子、またはureD遺伝子によってコードされる抗原と、請求項10または11の何れかのアミノ酸配列を有するタンパクとを含有することを特徴とする組成物。
【請求項26】
請求項10または11の何れかに記載のアミノ酸配列に向けられる、モノクローナル抗体またはポリクローナル血清。
【請求項27】
請求項12〜15の何れか1項に記載のH. ピロリ株に向けられる、モノクローナル抗体またはポリクローナル血清。
【請求項28】
生物学的サンプルにおけるH. ピロリによる感染をインビトロで検出するための組成物であって、活性成分として、請求項12〜15の何れか1項の非鞭毛フェノタイプを有するH. ピロリに対して得られるモノクローナル抗体またはポリクローナル血清を含有する組成物。
【請求項29】
H. ピロリに対する抗体取得に用いる免疫原組成物を調製するための、請求項1〜9の何れか1項に記載のヌクレオチド配列の使用。
【請求項30】
H. ピロリに感染した患者の抗体を診断するためのキットであって、請求項15〜19の何れか1項に記載のバクテリア抽出物と、抗原/抗体タイプの反応を明示するために必要とされる試薬とを含むキット。
【請求項1】
ヘリコバクター・ピロリ(Helicobacter pylori)の鞭毛タンパク質の生合成を調節すると共に、高厳密度の条件下において、下記の配列を有する二つのオリゴヌクレオチドを用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブとハイブリダイズすることができるか、
OLFlbA−1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2: GAAATCTTCATACTGGCAGCTCCAGTC
或いは、高厳密度の条件下において、これらのオリゴヌクレオチドとハイブリダイズすることができるヌクレオチド配列。
【請求項2】
ヘリコバクター・ピロリ(Helicobacter pylori)の鞭毛タンパク質の生合成を調節するflbA遺伝子のヌクレオチド配列であって、このヌクレオチド配列は、H. ピロリ株の染色体DNAを含むゲノムライブラリーを、下記の配列を有する二つのオリゴヌクレオチドを用いて増幅されたH. ピロリ由来のヌクレオチドフラグメントに対応するプローブか、
OLFlbA−1: ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2: GAAATCTTCATACTGGCAGCTCCAGTC
或いは、高厳密度の条件下において、これらのオリゴヌクレオチドとハイブリダイズすることができるプローブを用いてスリーニングする工程と、前記プローブとハイブリダイズするDNA配列を回収する工程と、得られたDNA配列をプラスミドタイプの適切なベクター中にサブクローニングし、次いで、高厳密度の条件下において、オリゴヌクレオチドOLFlbA−1およびOLFlbA−2を用いて増幅されたH.ピロリ由来のヌクレオチドフラグメントに対応するプローブとハイブリダイズする修飾ベクターを選別する工程と、上記プローブとハイブリダイズするプラスミドベクター中に含まれているDNAフラグメントの配列決定を行い、これらフラグメントに含まれるオープンリーディングフレームを決定する工程とによって得られるヌクレオチド配列。
【請求項3】
請求項1または2に記載のヌクレオチド配列であって、図3、4に示したヌクレオチド配列を有することを特徴とするヌクレオチド配列。
【請求項4】
請求項1〜3の何れか1項に記載のヌクレオチド配列であって、図3、4に示したアミノ酸配列を有するタンパク質、またはH.ピロリの鞭毛タンパク質の生合成に関して上記配列と同じ調節特性をもったアミノ酸配列を有するタンパク質をコードすることを特徴とするヌクレオチド配列。
【請求項5】
請求項1〜4の何れか1項に記載のヌクレオチド配列であって、該配列は、flbA遺伝子がもはやホスト細胞内で発現せず、またはホスト細胞内でのflbA遺伝子の発現がH. ピロリの鞭毛の生合成を可能としないように、また適切な場合には、H. ピロリのフックタンパク質の合成ができないように、欠失、置換または挿入によって修飾されているヌクレオチド配列。
【請求項6】
flbA遺伝子のフラグメントに対応する請求項1〜4の何れか1項に記載のヌクレオチド配列であって、flbA遺伝子に由来し、好ましくはflbA遺伝子の配列に存在する制限酵素部位で切断された、少なくとも6ヌクレオチド、好ましくは少なくとも100ヌクレオチドのフラグメントであることを特徴とするヌクレオチド配列。
【請求項7】
請求項1〜6の何れか1項のヌクレオチド配列を含み、該配列はマーカー、例えば抗生物質に対する耐性遺伝子または重金属に対する耐性遺伝子を含むカセットの挿入によって修飾されていることを特徴とする組み換え核酸。
【請求項8】
請求項7に記載の組み換え核酸であって、請求項1〜5の何れか1項に記載の前記ヌクレオチド配列が、カナマイシンに対する耐性カセットの挿入によって修飾されていることを特徴とする組み換え核酸。
【請求項9】
請求項1〜3の何れか1項に記載の配列に対して特異的であることと、下記の配列の一つを有することとを特徴とするオリゴヌクレオチド。
OLFlbA−1:ATGCCTCGAGGTCGAAAAGCAAGATG
OLFlbA−2:GAAATCTTCATACTGGCAGCTCCAGTC
OLFlbA−7:CGGGATCCGGGTTACTAATGGTTCTAC
OLFlbA−8:CGGGATCCTCATGGCCTCTTCAGAGACC
【請求項10】
請求項1および2の何れか1項に記載のヌクレオチド配列によってコードされることを特徴とする、H. ピロリのFlbAタンパク質のアミノ酸配列。
【請求項11】
H. ピロリのFlbAタンパク質であり、図3、4に示した配列を有すること、またはFlbAタンパク質に対する抗体によって認識される該蛋白質のフラグメントであることを特徴とするアミノ酸配列。
【請求項12】
ヘリコバクター・ピロリ菌株であって、塩基またはヌクレオチドフラグメントの置換、付加および/または欠失によって、H. ピロリの鞭毛タンパク質の生合成の調節に関与するflbA遺伝子の請求項1〜3の何れか1項に記載のヌクレオチド配列に生じた突然変異によってもたらされる、非鞭毛フェノタイプを有することを特徴とする菌株。
【請求項13】
請求項12に記載の菌株であって、更に、H. ピロリのフックタンパク質が欠失していることを特徴とする菌株。
【請求項14】
請求項12または13に記載の組み換え菌株であって、N6株から得られたものであり、1992年6月26日に、NCIMB受付番号第40512号でNCIMBに寄託されたものであることを特徴とする菌株。
【請求項15】
請求項12または14に記載の組み換え菌株であって、N6flbA−株であり、1995年6月30日に、NCIMB受付番号第40747号でNCIMBに寄託されたものであることを特徴とする菌株。
【請求項16】
請求項12〜15の何れか1項に記載の菌株であって、弱められたウレアーゼを産生するか、或いはもはやウレアーゼを産生しないように更に突然変異されており、この突然変異は、例えばureA遺伝子ureB遺伝子、ureC遺伝子、ureD遺伝子、ureE遺伝子、ureF遺伝子、ureG遺伝子、ureH遺伝子、およびureI遺伝子の中から選択される一以上のヌクレオチドの突然変異からなることを特徴とする菌株。
【請求項17】
請求項12〜15の何れか1項に記載の菌株の抽出物であることを特徴とするバクテリア抽出物。
【請求項18】
請求項17に記載のバクテリア抽出物であって、n−オクチルグルコシドで抽出した後に得られることを特徴とするバクテリア抽出物。
【請求項19】
請求項17に記載のバクテリア抽出物であって、PBSまたはグリシンで抽出した後に得られるバクテリア抽出物。
【請求項20】
患者由来の生物学的液体のサンプル(特に血清サンプル)におけるH. ピロリによる感染をインビトロで検出するための組成物であって、活性成分として、請求項12〜15の何れか1項の菌株、または請求項17〜19の何れか1項のバクテリア抽出物を含有することを特徴とする組成物。
【請求項21】
患者由来の生物学的液体のサンプル(特に血清サンプル)におけるH. ピロリによる感染をインビトロで検出するための方法であって、前記被検サンプルを、請求項12〜15の何れか1項の菌株、または請求項17〜19の何れか1項のバクテリア抽出物と接触させる工程と、前記菌株と、前記被検サンプル中に存在し且つH. ピロリに向けられる抗体との間の免疫反応を検出する工程とを具備する方法。
【請求項22】
H. ピロリに対する抗体を得るための免疫原組成物であって、活性成分として、請求項12〜16の何れか1項の菌株、または請求項17〜19の何れか1項のバクテリア抽出物を含有することを特徴とする組成物。
【請求項23】
H. ピロリに対する抗体を得るための免疫原組成物であって、請求項10または11の何れか1項に記載のアミノ酸配列を含有する組成物。
【請求項24】
H. ピロリによる感染に対する防護的抗体を得るためのワクチン接種組成物であって、活性成分として、請求項12〜16の何れか1項の菌株、または請求項17〜19の何れか1項のバクテリア抽出物を含有することを特徴とする組成物。
【請求項25】
H. ピロリによる感染に対する抗体を得るためのワクチン接種組成物であって、活性成分として、ウレアーゼタイプである抗原またはH.ピロリのウレアーゼ活性に関与する抗原、特にureA遺伝子、ureB遺伝子、ureC遺伝子、またはureD遺伝子によってコードされる抗原と、請求項10または11の何れかのアミノ酸配列を有するタンパクとを含有することを特徴とする組成物。
【請求項26】
請求項10または11の何れかに記載のアミノ酸配列に向けられる、モノクローナル抗体またはポリクローナル血清。
【請求項27】
請求項12〜15の何れか1項に記載のH. ピロリ株に向けられる、モノクローナル抗体またはポリクローナル血清。
【請求項28】
生物学的サンプルにおけるH. ピロリによる感染をインビトロで検出するための組成物であって、活性成分として、請求項12〜15の何れか1項の非鞭毛フェノタイプを有するH. ピロリに対して得られるモノクローナル抗体またはポリクローナル血清を含有する組成物。
【請求項29】
H. ピロリに対する抗体取得に用いる免疫原組成物を調製するための、請求項1〜9の何れか1項に記載のヌクレオチド配列の使用。
【請求項30】
H. ピロリに感染した患者の抗体を診断するためのキットであって、請求項15〜19の何れか1項に記載のバクテリア抽出物と、抗原/抗体タイプの反応を明示するために必要とされる試薬とを含むキット。
【図1】

【図2】

【図3】
【図4】
【図5】

【図6】

【図7】
【図8】
【図9】

【図10】

【図11】

【図12】

【図13】

【図14】
【図15】
【図16】
【図17】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公開番号】特開2009−131288(P2009−131288A)
【公開日】平成21年6月18日(2009.6.18)
【国際特許分類】
【出願番号】特願2009−66575(P2009−66575)
【出願日】平成21年3月18日(2009.3.18)
【分割の表示】特願平8−175168の分割
【原出願日】平成8年7月4日(1996.7.4)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(596005872)アンスティテュ・ナシオナル・デゥ・ラ・サンテ・エ・デゥ・ラ・ルシェルシュ・メディカル (8)
【Fターム(参考)】
【公開日】平成21年6月18日(2009.6.18)
【国際特許分類】
【出願日】平成21年3月18日(2009.3.18)
【分割の表示】特願平8−175168の分割
【原出願日】平成8年7月4日(1996.7.4)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(596005872)アンスティテュ・ナシオナル・デゥ・ラ・サンテ・エ・デゥ・ラ・ルシェルシュ・メディカル (8)
【Fターム(参考)】
[ Back to top ]