
HCVを遺伝子型分類するためのヘテロ二本鎖トラッキングアッセイ(HTA)
【課題】配列決定を必要とすることなくサブタイプ内遺伝的分岐をまた与え得る、HCV遺伝子型分類およびサブタイプ分類のための正確な信頼できるアッセイを開発すること。
【解決手段】HCVを遺伝子型分類するための、ハイブリダイゼーションに基づいて2つ以上のウイルスゲノム間の遺伝的関係を決定する方法である、ヘテロ二本鎖トラッキングアッセイ(HTA)が開示される。HCVを遺伝子型分類するためのHTAは、HCVサブタイプ(1a、1b、2a、2b、および3a)に対するE1のコアのカルボキシ末端および一部に由来する、一本鎖プローブを用いて開発された。HTAは、HCVのサブタイプ分類についてRFLPよりも正確であり、そして新規の変異体を同定する能力を有し、そして疫学研究に有用である。
【解決手段】HCVを遺伝子型分類するための、ハイブリダイゼーションに基づいて2つ以上のウイルスゲノム間の遺伝的関係を決定する方法である、ヘテロ二本鎖トラッキングアッセイ(HTA)が開示される。HCVを遺伝子型分類するためのHTAは、HCVサブタイプ(1a、1b、2a、2b、および3a)に対するE1のコアのカルボキシ末端および一部に由来する、一本鎖プローブを用いて開発された。HTAは、HCVのサブタイプ分類についてRFLPよりも正確であり、そして新規の変異体を同定する能力を有し、そして疫学研究に有用である。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、C型肝炎ウイルス(HCV)の遺伝子型分類に関する。特に、本発明は、好ま
しくはHCVのコアおよびエンベロープ領域由来の特異的プライマー、ならびにヘテロ二本
鎖移動度またはトラッキングアッセイ(次にこれは特異的プライマーを利用する)を用いたHCVの遺伝子型を決定するための方法に関する。
【背景技術】
【0002】
(発明の背景)
ウイルス性肝炎は、A、B、C、D、およびE型肝炎として知られる5つの異なるウイルスにより引き起こされることが知られている。HAVはRNAウイルスであり、そして長期の臨床的徴候を導かない。HBVは、DNAウイルスである。HDVは、HBVの非存在下で細胞を感染させ得ない依存性ウイルスである。HEVは、飲料水媒介性ウイルスである。HCVは最初に同定され、そして非A非B肝炎NANBHの原因として特徴付けられた(Houghtonら、特許文献
1および特許文献2)。これは、HCVの同定における免疫学的試薬として有用な多くの一
般的および特異的なポリペプチドの開示を導いた。例えば、非特許文献1;非特許文献2、および非特許文献3を参照のこと。
【0003】
HCVは一本鎖RNAウイルスであり、ペスティウイルスおよびフラビウイルスの遠縁にあたり、そして輸血関連肝炎の非常に大多数の、および世界中の市中(community acquired
)非A非B肝炎のほとんどの場合の原因因子である。HCVゲノムは、単一の長いオープン
リーディングフレーム(ORF)に隣接する5'および3'非コード(NC)領域からなる。このORFは、アミノ末端に3つの構造タンパク質およびカルボキシ末端に6つの非構造(NS)タンパク質をコードする。構造タンパク質は、ヌクレオキャプシド(コア;C)タンパク質ならびに2つの糖蛋白質であるエンベロープ1(E1)およびエンベロープ2(E2)から表される。非構造タンパク質は、NS2、NS3、NS4a、NS4b、NS5a、NS5bと呼ばれる。5'NCRは
、HCVゲノムの最も高度に保存された部分であるが、2つのエンベロープタンパク質(E1
およびE2)の配列は、異なるHCV単離物間で高度に可変的である。最も高い程度の変異はE2内の領域で観察されており、現在一般に超可変領域1(HVR1)またはH2HVと呼ばれてい
る。HVR2と呼ばれる第2の可変領域もまた、単離物のサブセットに存在する。代表的に、HCVの遺伝的不均一性は、2つの上位準種(quasispecies)および遺伝子型で分類されて
いる。本明細書中で使用する用語「準種」は、感染個体内のHCV集団の遺伝的不均一性を
いう。本明細書中で使用する用語「遺伝子型」および「サブタイプ(subtype)」は、異
なるHCV単離物間で観察された遺伝的不均一性をいう。約9.4kb RNA分子のポジティブ鎖である、HCVゲノムの核酸配列変異の分析は、遺伝的可変性が重要なウイルス学的および
臨床学的意味合いに関連することを示唆する。
【0004】
HCVの原型単離物は、特許文献1および特許文献2において特徴づけられた。本明細書
中で使用する用語「HCV」は、新たに単離されたNANBHウイルス種を含む。用語「HCV-1」
は、上記の公報に記載されたウイルスをいう。
【0005】
HCVの最初の同定から、少なくとも6つの異なる主要なウイルス型が同定されており(
全長ゲノムが報告されており)、そして1、2、3、4、5、および6型と呼ばれている。これらの型には、多くのサブタイプが含まれる。患者が感染するウイルス型は、臨床的予後に影響し得、そしてまた種々の処置に応答する。非特許文献4を参照のこと。HCV感
染のほとんどの重篤な臨床結果が肝細胞ガン腫であることを考慮すると、患者がどのHCV
型(単数または複数)に感染するかを決定し得ることは有用である。従って、配列決定を必要とすることなくサブタイプ内(intra-subtype)遺伝的分岐をまた与え得る、HCV遺伝子型分類およびサブタイプ分類のための正確な信頼できるアッセイを開発することは特に重要である。完全なウイルスゲノムを用いた理想的なヌクレオチド配列に基づく系は実用的でないので、異なる領域の分析に基づいたHCV遺伝子型分類のためのいくつかの分類が
提案されている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】欧州特許第388,232号明細書
【特許文献2】欧州特許第318,216号明細書
【非特許文献】
【0007】
【非特許文献1】Chooら、Science,1989年,第244巻,pp.359-262
【非特許文献2】Kuoら、Science,1989年,第244巻,pp.362-364
【非特許文献3】Houghtonら、Hepatology,1991年,第14巻,pp.381-388
【非特許文献4】Yoshiokeら、Hepatology,1992年,第16巻,pp.293-299
【発明の概要】
【発明が解決しようとする課題】
【0008】
配列決定を必要とすることなくサブタイプ内遺伝的分岐をまた与え得る、HCV遺伝子型
分類およびサブタイプ分類のための正確な信頼できるアッセイを開発すること。
【課題を解決するための手段】
【0009】
本発明は、例えば、上記課題を解決するために、以下の手段を提供する。
(項目1)配列番号1に記載の配列からなるオリゴヌクレオチド。
(項目2)配列番号2に記載の配列からなるオリゴヌクレオチド。
(項目3)配列番号3に記載の配列からなるオリゴヌクレオチド。
(項目4)配列番号4に記載の配列からなるオリゴヌクレオチド。
(項目5)配列番号5に記載の配列からなるオリゴヌクレオチド。
(項目6)配列番号6に記載の配列からなるオリゴヌクレオチド。
(項目7)配列番号7に記載の配列からなるオリゴヌクレオチド。
(項目8)配列番号8に記載の配列からなるオリゴヌクレオチド。
(項目9)配列番号9に記載の配列からなるオリゴヌクレオチド。
(項目10)センスプライマーが配列番号1からなり、そしてアンチセンスプライマーが、配列番号2、配列番号3、配列番号4、および配列番号5からなる群より選択される、PCRプライマー対。
(項目11)アンチセンスプライマーが配列番号6からなり、そしてセンスプライマーが、配列番号7、配列番号8、および配列番号9からなる群より選択される、PCRプライマ
ー対。
(項目12)HCV株のHCV遺伝子型を決定する方法であって、以下の工程:
(a)該HCV株を、1段階以上のPCRに供する工程であって、ここで該1段階以上のPCRが
、HCVゲノムのコアまたはE1領域由来のセンスプローブおよびHCVゲノムのコアまたはE1領域由来のアンチセンスプローブを利用する、工程;
(b)工程(a)で得られた増幅産物の混合物と、既知のHCV遺伝子型のDNAまたはRNAフ
ラグメントとを変性および再アニールすることにより、ヘテロ二本鎖を形成する工程;
(c)サイズにより分離する系において該ヘテロ二本鎖の移動度と、既知遺伝子型のDNA
またはRNAフラグメントのホモ二本鎖の移動度とを比較して、該HCV株の遺伝子型を決定する工程
を包含する、方法。
(項目13)項目12に記載の方法であって、ここで前記HCV株が、2段階のPCRに供され、第1のセットのプライマーが、前記HCVゲノムのコアまたはE1領域由来のユニバーサル
センスプローブおよび前記HCVゲノムのコアまたはE1領域由来の型特異的アンチセンスプ
ローブを含み、そして第2のセットのPCRプライマーが、前記HCVゲノムのコアまたはE1領域由来のユニバーサルアンチセンスプローブおよび前記HCVゲノムのコアまたはE1領域由
来の型特異的センスプローブを含む、方法。
(項目14)項目12に記載の方法であって、ここで前記第1のセットのPCRプライマー
が項目10に記載のプライマーであり、そして前記第2のセットのPCRプライマーが項目
11に記載のプライマーである、方法。
(項目15)前記既知の遺伝子型のDNAまたはRNAフラグメントがDNAプローブを含む、項
目12に記載の方法。
(項目16)前記プローブが一本鎖である、項目15に記載の方法。
(項目17)前記DNAプローブが放射標識される、項目16に記載の方法。
(項目18)前記一本鎖DNAプローブがPCR増幅により得られる、項目16に記載の方法。(項目19)項目18に記載の方法であって、ここで前記DNAプローブが、前記第1の段
階に項目10に記載のプライマーおよび前記第2の段階に項目11に記載のプライマーを利用する2段階PCR増幅により得られる、方法。
(項目20)前記HCV株が前記ヘテロ二本鎖を形成する混合物中に過剰に存在する、項目
12に記載の方法。
(項目21)HCV株の薬物治療に対する応答を、該HCV株に感染した患者から予測する方法であって、既知のHCV遺伝子型の該薬物治療に対する感受性を決定する工程、項目12に
記載の方法により該HCV株の該HCV遺伝子型を決定する工程、および該薬物治療前の該株の該HCV遺伝子型と、該薬物治療に対する既知のHCV遺伝子型の該感受性とを比較する工程を包含する、方法。
(項目22)HCV株の治療ワクチンに対する応答を、該HCV株に感染した患者から予測する方法であって、既知のHCV遺伝子型の該治療ワクチンに対する感受性を決定する工程、項
目12に記載の方法により該HCV株の該HCV遺伝子型を決定する工程、および該治療ワクチンの投与前の該株の該HCV遺伝子型と、該治療ワクチンに対する既知のHCV遺伝子型の該感受性とを比較する工程を包含する、方法。
(項目23)所定のサンプル集団に対する予防ワクチン組成物の適性を予測する方法であって、該予防ワクチンの遺伝子型を決定する工程、項目12に記載の方法により該サンプル集団における既知のHCV遺伝子型の優勢を決定する工程、および該予防ワクチンの該集
団サンプルへの投与前に、該予防ワクチン株の該HCV遺伝子型と決定された優勢遺伝子型
とを比較する工程を包含する、方法。
【0010】
(発明の要旨)
本発明は、ヘテロ二本鎖トラッキングアッセイ(HTA)に基づいた、HCV遺伝子型分類およびサブタイプ内変異の特徴付けのためのプライマーおよび方法を含む。好ましいプローブ/プライマーは、HCVのコアのカルボキシ末端およびE1領域の一部由来の一本鎖である。
【0011】
HTAは、2つ以上のウイルスゲノム間の遺伝的関係を決定する方法に基づいたハイブリ
ダイゼーションである。この方法の基礎は、分岐テンプレートから同時増幅された関連DNA産物が、ランダムに再アニールして、中性ポリアクリルアミドゲルのようなサイズに基
づいて分子を分離するように設計された系において、減少した移動度で移動するヘテロ二本鎖を形成することであり、HTAは、HIV-1を遺伝子型分類するために、そして患者および集団におけるHIV-1のインビボ進化を追随するために最初に使用された。例えば、Delwartら(1993)Science 262:1757-1261およびDelwartら(1994)J.Virol.68:6722-6883
を参照のこと。
【0012】
本発明の1つの局面は、HCVを遺伝子型分類するための方法であり、この方法は、部分
的に相補的なDNA鎖またはRNA鎖を変性および再アニールする工程、ならびにサイズに基づいて分子を分離するように設計された系において(例えば、ポリアクリルアミドまたはMDEゲルを介する電気泳動に従うことにより)DNAヘテロ二本鎖の電気泳動移動度を書き留めることにより配列変異を検出する工程を含む。
【0013】
本発明の別の局面は、HCVゲノムのコアおよびE1領域から選択された、遺伝子型分類において使用されるプローブに関する。
【0014】
本発明の別の局面は、異なる既知の遺伝子型の薬物治療に対する感受性を決定すること、患者を感染させるHCV株の遺伝子型を決定すること、およびその遺伝子型とその薬物治
療感受性とを比較して、患者の薬物治療に対する応答を予測することにより、HCV株に感
染した患者の薬物治療に対する応答を予測する方法に関する。
【0015】
本発明の別の局面は、治療ワクチンに関し、そしてHCV株に感染した患者の遺伝子型を
決定すること、および同じ遺伝子型の治療ワクチンを投与することにより、どの治療ワクチンが利用されるべきかを予測することに関する。
【0016】
本発明の別の局面は、予防ワクチンに関し、そして同様のサンプルにおいて優勢な遺伝子型を決定すること、および集団サンプルに対して優勢な遺伝子型であると思われる遺伝子型の予防ワクチンを投与することにより、どのワクチンが特定の集団サンプルに投与されるべきか予測することに関する。
【0017】
本発明の別の局面は、本発明の方法を用いて新規のHCV遺伝子型を発見する能力に関す
る。
【図面の簡単な説明】
【0018】
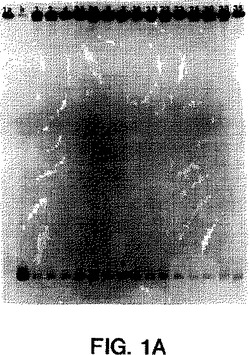
【図1A】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
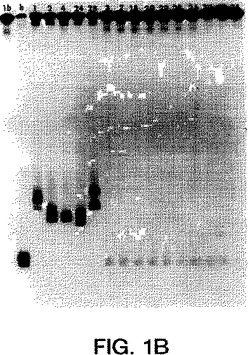
【図1B】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
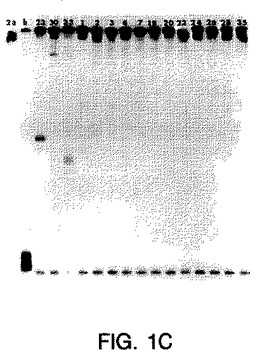
【図1C】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
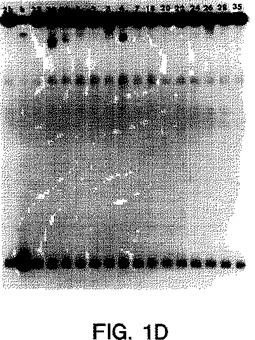
【図1D】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
【図1E】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
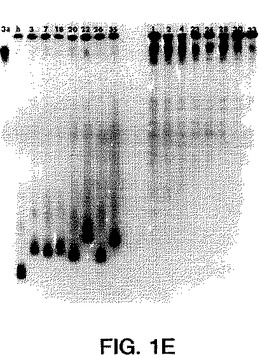
【図2A】図2A〜2Cは、本明細書中に記載の透析患者由来の推定1型(nt625〜930)、2型(nt583〜915)、または3型(nt558〜834)単離物の配列決定により得られた部分的E1配列と、ゲノムの同じ領域上での1a型(HCV-1)(Chooら、PNAS(1991)88:2451-2455、全てのヌクレオチド「nt」はこの論文に従った名称である)、1b型(HCV-J)(Katoら、PNAS(1990)87:9524-2528)、2a型(HC-J6)(Okamotoら、Virol.(1992)188:331-341)、2b型(HC-J8)(Okamotoら、Virol.(1992)188:331-341)、2c型(Bukhら、PNAS(1993)90:8234-8239)、および3a型(NZL-1)(Sakamotoら)J.Gen.Virol.(1994)75:1761-1768について公開された遺伝子型配列とを比較することにより形成された系統樹(すなわち各部分的E1ヌクレオチド配列の関連性を示す系統発生樹)である。
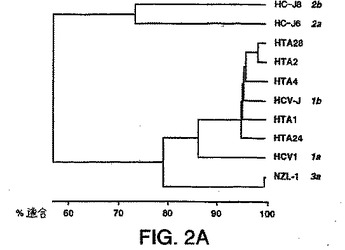
【図2B】図2A〜2Cは、本明細書中に記載の透析患者由来の推定1型(nt625〜930)、2型(nt583〜915)、または3型(nt558〜834)単離物の配列決定により得られた部分的E1配列と、ゲノムの同じ領域上での1a型(HCV-1)(Chooら、PNAS(1991)88:2451-2455、全てのヌクレオチド「nt」はこの論文に従った名称である)、1b型(HCV-J)(Katoら、PNAS(1990)87:9524-2528)、2a型(HC-J6)(Okamotoら、Virol.(1992)188:331-341)、2b型(HC-J8)(Okamotoら、Virol.(1992)188:331-341)、2c型(Bukhら、PNAS(1993)90:8234-8239)、および3a型(NZL-1)(Sakamotoら)J.Gen.Virol.(1994)75:1761-1768について公開された遺伝子型配列とを比較することにより形成された系統樹(すなわち各部分的E1ヌクレオチド配列の関連性を示す系統発生樹)である。
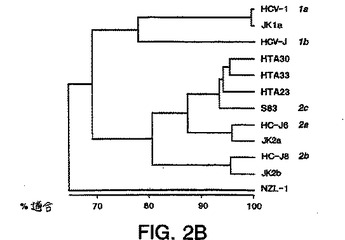
【図2C】図2A〜2Cは、本明細書中に記載の透析患者由来の推定1型(nt625〜930)、2型(nt583〜915)、または3型(nt558〜834)単離物の配列決定により得られた部分的E1配列と、ゲノムの同じ領域上での1a型(HCV-1)(Chooら、PNAS(1991)88:2451-2455、全てのヌクレオチド「nt」はこの論文に従った名称である)、1b型(HCV-J)(Katoら、PNAS(1990)87:9524-2528)、2a型(HC-J6)(Okamotoら、Virol.(1992)188:331-341)、2b型(HC-J8)(Okamotoら、Virol.(1992)188:331-341)、2c型(Bukhら、PNAS(1993)90:8234-8239)、および3a型(NZL-1)(Sakamotoら)J.Gen.Virol.(1994)75:1761-1768について公開された遺伝子型配列とを比較することにより形成された系統樹(すなわち各部分的E1ヌクレオチド配列の関連性を示す系統発生樹)である。
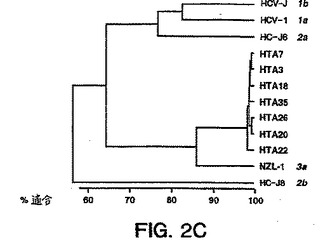
【図2D】図2Dは、直接配列決定により得られた単離物23、30、および33の部分的5'UTR配列のいずれかと、ゲノムの同じ領域についての公開された1、2、および3型(nt-274〜nt-81)との比較により形成された系統樹(系統発生樹)である。
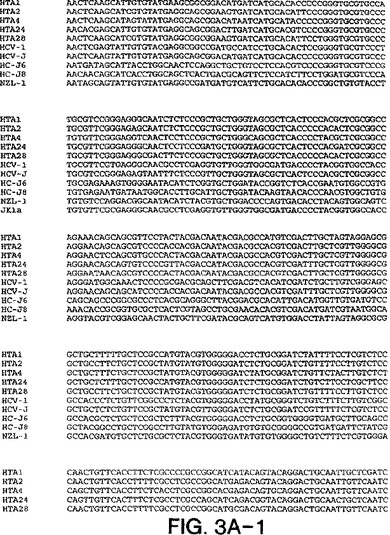
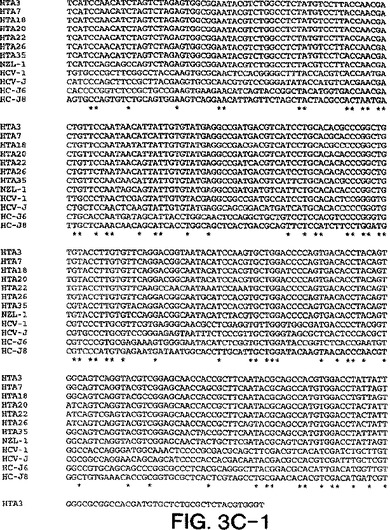
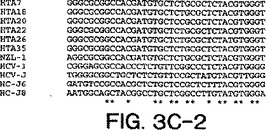
【図3A−1】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
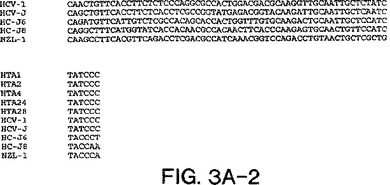
【図3A−2】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
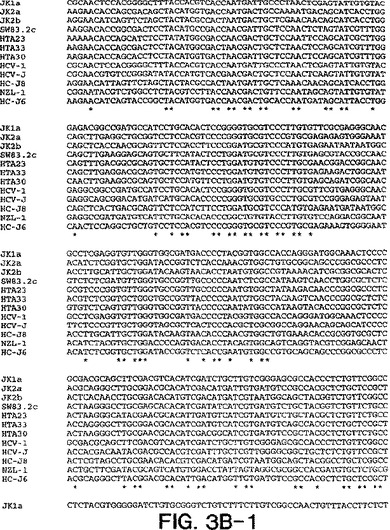
【図3B−1】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
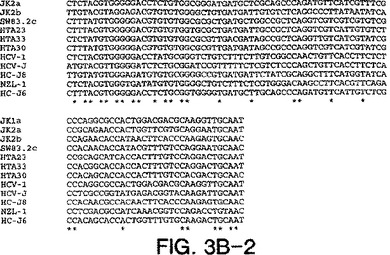
【図3B−2】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3C−1】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3C−2】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3D】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【発明を実施するための形態】
【0019】
(発明の説明)
本発明の実施は、他に示されない限り、当該技術の範囲内の分子生物学、微生物学、組換えDNA、ポリペプチド合成および核酸合成、ならびに免疫学の従来技術を使用する。こ
のような技術は、文献中で十分に説明される。例えば、Sambrookら,MOLECULAR CLONING;A LABORATORY MANUAL,第2版(1989);DNA CLONING,第IおよびII巻(D.N Glover編、1985);OLIGONUCLEOTIDE SYNTHESIS(M.J.Gait編,1984);NUCLEIC ACID HYBRIDIZATION(B.D.HamesおよびS.J.Higgins編,1984);TRANSCRIPTION AND TRANSLATION(B.D.HamesおよびS.J.Higgins編,1984);ANIMAL CELL CULTURE(R.I.Freshney編,1986);IMMOBILIZED CELLS AND ENZYMES(IRL Press,1986);B.Perbal、A PRACTICAL GUIDE TO MOLECULAR CLONING(1984);シリーズMETHODS IN ENZYMOLOGY(Academic Press,Inc.);GENE TRANSFER VECTORS FOR MAMMALIAN CELLS(J.H.MillerおよびM.P.Calos編,1987,Cold Spring Harbor Laboratory)、Methods in Enzymology第154巻および第155巻(それぞれ、WuおよびGrossman、ならび
にWu編)、MayerおよびWalker編(1987),IMMUNOCHEMICAL METHODS IN CELL AND MOLECULAR BIOLOGY(Academic Press,London)、Scopes,(1987),PROTEIN PURIFICATION:PRINCIPLES AND PRACTICE,第2版(Springer-Verlag,N.Y.)、ならびにHANDBOOK OF EXPERIMENTAL IMMUNOLOGY,第I-IV巻(D.M.WeirおよびC.C.Blackwell編
1986)を参照のこと。
【0020】
ヌクレオチドおよびアミノ酸の標準的な略語が本明細書中で使用される。本明細書中に引用された全ての刊行物、特許、および特許出願が参考として援用される。
【0021】
本明細書中で使用する用語「組換えポリヌクレオチド」は、その起源または操作により、(1)天然に関連するポリヌクレオチドの全てもしくは一部に関連しないか、(2)天然に連結するポリヌクレオチド以外のポリヌクレオチドに連結するか、または(3)天然に生じない、ゲノム、cDNA、半合成、もしくは合成起源のポリヌクレオチドを意図する。
【0022】
本明細書中に使用する用語「ポリヌクレオチド」は、リボヌクレオチドまたはデオキシリボヌクレオチドのいずれかの、任意の長さのヌクレオチドのポリマー形態をいう。この用語は、分子の一次構造のみをいう。従って、この用語は、二本鎖および一本鎖DNAおよ
びRNAを含む。これは、既知の型の修飾(例えば、当該分野で公知の標識、メチル化、「キャップ」、1つ以上の天然に生じるヌクレオチドのアナログによる置換)、ヌクレオチド内修飾(例えば、非荷電結合(例えば、メチルホスホネート、ホスホトリエステル、ホスホアミデート、カルバメート等)による修飾および荷電結合(例えば、ホスホロチオエート、ホスホロジチオエート等)による修飾、ペンダント部分(例えば、タンパク質(例えば、ヌクレアーゼ、毒素、抗体、シグナルペプチド、ポリ-L-リジン等を含む))を含
む修飾、インターカレーター(例えば、アクリジン、ソラレン等)による修飾、キレーター(例えば、金属、放射性金属、ホウ素、酸化金属等)を含む修飾、アルキル化剤を含む修飾、修飾結合(例えば、αアノマー核酸等)による修飾)、ならびにポリヌクレオチドの非修飾形態をも含む。
【0023】
「PCR」は、本明細書中でポリメラーゼ連鎖反応(PCR)技術を意味し、これはMullis(米国特許第4,683,195号(Mullisら)および同第4,683,202号、本明細書中に参考として援用される)により開示される。PCR技術において、所望の配列の反対の末端に適合す
る短いオリゴヌクレオチドプライマーが調製される。プライマー間の配列は既知である必要はない。DNA(またはRNA)サンプルは抽出され、そして変性される(好ましくは、熱により)。次いで、オリゴヌクレオチドプライマーを、dNTPおよびポリメラーゼ(好ましくは、熱安定性のTaqポリメラーゼ)と共にモル過剰で添加する。DNAは複製され、次いで再度変性される。これは、それぞれのプライマーで始まる2つの「長い産物」および2つの最初の鎖(二本鎖DNA分子あたり)を生じる。次いで、反応混合物は、重合条件に戻され
(例えば、温度を低下させるか、変性剤を不活化するか、またはより多くのポリメラーゼを添加することにより)、そして第2のサイクルが開始される。第2のサイクルは、2つの最初の鎖、サイクル1由来の2つの長い産物、2つの新たな長い産物(最初の鎖から複製された)、および長い産物から複製された2つの「短い産物」を提供する。短い産物は、各末端でプライマーを有する標的配列(センスまたはアンチセンス)の配列を有する。さらなる各サイクルで、さらなる2つの長い産物が生成され、そして短い産物の数は、前のサイクルの終わりに残った長い産物および短い産物の数に等しい。従って、短い産物の数は、各サイクルで指数的に増殖する。特定の分析配列のこの増殖は、極端に少ない量のDNA検出を可能にする。
【0024】
本明細書中で使用する用語「3SR」は、欧州特許公開第373,960号(1990年6月20日に
公開)に記載されたような「自立(self-sustained)配列複製」としても知られる標的核酸増幅法をいう。
【0025】
本明細書中で使用する用語「LCR」は、Barany,Proc.Natl.Acad.Sci.(USA)(1991)88:189-193により記載されるような「リガーゼ連鎖反応」としても知られる標的核酸増幅法をいう。
【0026】
「オープンリーディングフレーム」(ORF)は、ポリペプチドをコードするポリヌクレ
オチド配列の領域であり;この領域は、コード配列の一部または総コード配列を示し得る。
【0027】
「コード配列」は、適切な調節配列の制御下に置かれた場合、通常mRNAを介してポリペプチドに翻訳されるポリヌクレオチド配列である。コード配列の境界は、5'末端での翻訳開始コドンおよび3'末端での翻訳停止コドンにより決定される。コード配列は、cDNAおよび組換えポリヌクレオチド配列を含み得るが、これらに限定されない。
【0028】
本明細書中で使用する用語「ポリペプチド」は、アミノ酸のポリマーをいい、そして特定の長さの産物をいわず;従って、ペプチド、オリゴペプチド、およびタンパク質がポリペプチドの定義内に含まれる。この用語はまた、ポリペプチドの発現後修飾(例えば、グリコシル化、アセチル化、リン酸化など)をいわないか、または除外する。例えば、1つ以上のアミノ酸アナログ(例えば、非天然アミノ酸等を含む)を含むポリペプチド、置換された結合を有するポリペプチド、ならびに天然および非天然の両方の当該分野で公知の他の修飾が、定義内に含まれる。
【0029】
示された核酸配列「に由来する」ポリペプチドまたはアミノ酸配列は、その配列にコードされるポリペプチドまたはその一部(ここで、その一部は、少なくとも3〜5アミノ酸、そしてより好ましくは少なくとも8〜10アミノ酸、そしてさらにより好ましくは少なくとも11〜15アミノ酸からなるか、またはその配列にコードされるポリペプチドと免疫学的に同定可能である)のアミノ酸配列と同一の、アミノ酸配列を有するポリペプチドをいう。この用語はまた、示された核酸配列から発現されるポリペプチドを含む。
【0030】
タンパク質は、そのタンパク質に特異的な抗体(モノクローナルまたはポリクローナルのいずれか)を生成するために使用され得る。これらの抗体を生成するための方法は、当該分野で公知である。
【0031】
「組換え宿主細胞」、「宿主細胞」、「細胞」、「細胞培養物」、および他のこのような用語は、組換えベクターまたは他の転移DNAのレシピエントとして使用され得るかもし
くは使用されてきた、そして形質転換された最初の細胞の子孫を含む、例えば、微生物、昆虫細胞、および哺乳動物細胞を意味する。単一の親細胞の子孫は、自然、偶発的、または計画的変異により、形態またはゲノムもしくは総DNA相補物において最初の親細胞と必
ずしも完全に同一ではあり得ないことが理解される。哺乳動物宿主細胞の例として、チャイニーズハムスター卵巣(CHO)およびサル腎臓(COS)細胞が挙げられる。
【0032】
「cDNA」は、mRNAの相補鎖にハイブリダイズする相補的mRNA配列を意味する。
【0033】
「精製」および「単離」は、ポリペプチドまたはヌクレオチド配列をいう場合に、示された分子が、同じ型の他の生物学的巨大分子の実質的な非存在下で存在することを意味する。本明細書中で使用する用語「精製」は、存在する同じ型の生物学的巨大分子の、好ましくは少なくとも75重量%、より好ましくは少なくとも85重量%、なおより好ましくは少なくとも95重量%、そして最も好ましくは少なくとも98重量%を意味する(しかし、水、緩衝液、および他の低分子(特に、1000未満の分子量を有する分子)は存在し得る)。
【0034】
「薬学的に受容可能なキャリア」は、それ自体が組成物を受ける個体に有害な抗体の産生を誘導しない、任意の薬学的キャリアを意味する。適切なキャリアは、代表的には、大きな緩徐に代謝される巨大分子(例えば、タンパク質、多糖、ポリ乳酸、ポリグリコール酸、ポリマーアミノ酸、アミノ酸コポリマー;および不活性ウイルス粒子)である。このようなキャリアは、当業者に周知である。
【0035】
治療組成物は、代表的には、薬学的に受容可能なビヒクル(例えば、水、生理食塩水、グリセロール、エタノール等)を含む。さらに、補助物質(例えば、湿潤剤または乳化剤、pH緩衝化物質等)がこのようなビヒクルに存在し得る。
【0036】
代表的には、治療組成物は、液体の溶液または懸濁液のいずれかのような注射可能物質として調製される;注射前の液体ビヒクル中の溶液または懸濁液に適した固体形態もまた調製され得る。調製物はまた、増強したアジュバント効果のために乳化され得るか、またはリポソーム内にカプセル化され得る。
【0037】
異なるHCV遺伝子型が、異なる病原性ならびに別々の地理的分布を有し得、そして感染
した患者において異なる血清学的プロフィールを部分的に誘発し得ることが、証拠によって示される。Cammarotaら、J.Clin.Microb.(1995)33:2781-2784を参照のこと。本発明は、HCVを検出し、そして異なる型のHCVによる感染を同定する方法を含む。本発明は、HCVを遺伝子型分類すること、HCVの新規の遺伝子型を発見する能力、および薬物治療に対する応答を予測する能力についてウイルス集団を評価することを含む。本発明はまた、HCVの遺伝子型分類における使用のためのプローブを含む。
【0038】
HCVを遺伝子型分類する方法として、HCVゲノムのコア/E1領域由来のプローブ/プライマーを利用する、ヘテロ二本鎖トラッキングまたは移動度アッセイが挙げられるが、これらに限定されない。HCV遺伝子型間の証明された抗原的差異は、血液ドナースクリーニング
およびIFN処置に対する応答の予測においてのみならず、異なる国におけるHCVに対する候補ワクチンの示された組成物、治療ワクチンの選択、ならびに新規の遺伝子型の同定においてもまた有用性を有する。ゲノムの異なる領域の分析(例えば、RFLP)に基づいた、集団を感染させる主要な遺伝子型を同定するための他の方法が提案されている。5'非コード領域(NCR)から同時増幅された配列のRFCPを用いてHCVを遺伝子型分類する議論については、Davidsonら、J.Gen Virol.(1995)76:1197-1204を参照のこと。
【0039】
既知の核酸に基づく遺伝子型分類法は、サブタイプ特異的RT-PCR(逆転写酵素-PCR)プライマー(Okamoto(1992)J.Gen Virol 73:673-679を参照のこと)米国特許第5,427,909号;(2)特異的プローブ(G.Marteenら、Lineプローブアッセイ);(3)制限部位多型性(ヌクレオチド配列(nt)の機能)、または(4)遺伝子型を決定するための直接的配列を必要とする。5'NC配列のRFLPによる分析は、実施しやすいが、全てのHCV遺
伝子型を正確に予測せず、そしていくつかのサブタイプは誤分類され得る。例えば、制限酵素により認識される1aと1bとの間の配列における変化は絶対的でなく、そして1aおよび1b、ならびに2aおよび2b以外の配列は誤分類される。例えば、1c型は1a型と思われ、2c型は2a型または2b型のいずれかであると思われる。Cammarotaら、J.Clin.Microb.,(1995)33:2781-2784を参照のこと。この理由のため、RFLPは、「エスケープ」種、新規の
分岐種、または疫学的傾向を検出し得ない。RFLPのような分類方法は、HCV配列不均一性
に関して回収される急速に増大する情報に適応するために、恐らく連続的に改変されなければならない。
【0040】
上記のように、核酸に基づく遺伝子型分類法を使用する場合、型もしくはサブタイプのいずれかの結果、または「分類不可能な」結果である陰性が得られる。例えば、遺伝子型特異的PCRにより分類されないままであった単離物が、コアおよびNS5遺伝子から得られたPCR増幅物の配列分析に基づいてサブタイプ2cに分類された、Cammarotaら、J.Clin.Microb.(1995)33:2781-2784を参照のこと。この問題は、コアのC末端/E1の中央2/3におけるRT-PCRプライマーを選択することによりHCV遺伝子型を決定する、目下請求される本
発明を用いることによって避けられる。さらに、単離物のサブタイプは、本発明のHCV遺
伝子型分類を用いて正確に決定され得、そして単離物は、約30%分岐未満のものでさえも検出され得、これは配列決定なしで新規のサブタイプの特徴付けを可能にする。
【0041】
(ヘテロ二本鎖トラッキングまたは移動度アッセイ)
本発明におけるHCV遺伝子型を決定する方法は、複雑な準種における少数の変異体を利
用する。1つのこのような技術は、二本鎖トラッキングアッセイ(HTA)である。HIVを用いた使用について当該分野で周知のHTA(例えば、Delwartら、J.Virol.(1994)68:6672-6683;Delwartら、Science(1993)262:1257-1261;Delwartら、PCR Methods and
Applications 4:S202-S216(1995)Cold Springs Harbor);およびDelwartら、Heteroduprex Mobility Analysis HIV-1 env Subtyping Kit Protocol Version 3(これらの各々は、その全体が参考として本明細書中に援用される)を参照のこと)は、配列が感染個体の末梢血単核細胞からネスティッドPCRにより増幅された場合、分岐テン
プレートから同時増幅された関連DNA産物が、ランダムに再アニールして、中性ポリアク
リルアミドゲルにおいて減少した移動度で移動するヘテロ二本鎖を形成し得たという観察に起因する。これらの技術を用いて、複数のウイルスDNAテンプレート分子間の遺伝的関
係が確立され得る。
【0042】
HTAは、特に、標識プローブとして第1のPCR産物を利用する。これは放射性であり得、異なる供給源(すなわち、分類が所望される供給源)由来の過剰量(ドライバー(driver))の非標識PCR産物と混合される。次いで、プローブ配列は、ドライバーと完全にヘテ
ロ二本鎖にさせられ、そしてサイズに基づいて分離される。得られたポリアクリルアミドゲルの例のためのオートラジオグラムは、これらのヘテロ二本鎖のみを明らかにし、そして研究下での2つのウイルス集団間の関係の視覚的な表示を提供する。ヘテロ二本鎖が異なる移動度で移動するという事実は、非適合(mismatched)および非対合(unpaired)ヌクレオチドの鎖特異的組成物がそれらの移動度に影響することを示す。
【0043】
次いで、Delwartらに記載される指数方程式を使用して、既知の配列の遺伝子の対様(pairwise)分析からの実験データに適合する曲線を記載する。本発明において、この方程
式を使用して、プローブの既知の遺伝子型と患者サンプルの未知の遺伝子型との間の遺伝的距離を評価する。
【0044】
(HTAにおける使用のためのプライマー)
E1またはコア領域がHCV不均一性を研究するための最良の領域であり得ることが決定さ
れ、従って、E1領域は、本発明におけるプライマーのための選択となった。本発明のゲノムの最も不均一な領域である部分的E1配列、ならびにより長いフラグメント(すなわち、400nt)(1000ntほどの長さであり得るが)の使用は、サブタイプ/型間で交差ハイブリダイズしないプローブの設計を可能にし、従って正確な遺伝子型分類を可能にする。不均一な領域を隣接させることにより、センスおよびアンチセンスプライマーの保存nt配列が同定された。好ましくは、第1回のPCRのためのユニバーサルセンスプライマーと型特異的
アンチセンスプライマーとの組合せ、ならびに第2回のためのユニバーサルアンチセンスプライマーと型特異的センスプライマーとの組合せが利用された。PCRは2回である必要
はなく、そしてプライマーは、上記の組合せに限定されない。しかし、好ましい組合せは、一本鎖プローブの調製を可能にし、そしてPCRプライマー組合せの数を最小にする。
【0045】
好ましいプローブは、コアおよびE1領域における配列である。この広範な遺伝子型のついての配列は公開され、そして少なくとも12個の異なる遺伝子型およびサブタイプ:I/1a、II/1b、III/2a、IV/2b、2c、3a、4a、4b、4c、4d、5a、6aに分類されている。同じ遺伝子型のHCV単離物間のE1遺伝子のヌクレオチド配列同一性は、88.0%〜99.1%の範囲で
あるが、異なる遺伝子型のHCV単離物のものは、53.5%〜78.6%の範囲である。中性ポ
リアクリルアミドゲルにおけるヘテロ二本鎖の良好な区別のための変異の程度は、快適には、分岐テンプレートが恐らく再アニールして、それらが同じサブタイプである場合にヘテロ二本鎖を形成するような、3〜20%の範囲内である。この理由のために、一本鎖32p
標識DNAプローブが、ヘテロ二本鎖の形成が不可能な場合、ss-DNAプローブが再アニール
し、そしてホモ二本鎖バンドを恐らく形成し得ないように使用された。直接的配列決定をすることなく、本発明は、サブタイプの確実な同定だけでなく、同じサブタイプ内の遺伝的関係もまた迅速に与え得る。例えば、分析された遺伝子型(すなわち、1a、1b、2a、2b、3a)は、異なるサブタイプ間で重複を示さなかった。
【0046】
さらに、単離物の約30%分岐がゲル上で視覚化され得るので、新規のサブタイプは視覚化され得、そして集団中の単離物の分布は特徴付けられ得、そして集団または個々の単離物の後に、疫学的研究における集団または個体が続き得る。
【0047】
(HCV遺伝子型分類キット)
HCVの遺伝子型を決定するためのキットは、本発明の範囲内にある。Delwartら、Heteroduprex Mobility Analysis HIV-1 env Subtyping Kit Protocol Version 3においてHIVについて記載されるように、このようなキットは、特異的なプライマーを含む。
好ましいプライマーは、HCVゲノムのコアおよびE1領域由来である。2段階のPCRが所望される場合、第1回のプライマーは、例えば、ユニバーサルセンスプローブ(好ましくは、HCVゲノムのコア/E1領域に位置する)を含み得る。1つのこのようなユニバーサルプライマーは、HCV-1のヌクレオチド508〜529に位置し、そして表1に示される。ユニバーサル
プライマーと対となるのは、好ましくはまたHCVゲノムのコア/E1領域に位置し得る、型特異的アンチセンスプライマーであり得る。これらのプライマーの例は、HCVゲノムの1型
、2a型、2b型、および3a型についてはヌクレオチド1032〜1012由来であり、そしてまた表1に示される。
【0048】
第2回のPCRが所望される場合、第2回のプライマーは、同様に、HCVゲノムのコア/E1
領域由来である。好ましい第2回のプライマーは、HCV-1ゲノムのヌクレオチド978〜958
に由来するユニバーサルアンチセンスプライマーを含み得、このプライマーは表1に示される。さらに、第2回のプライマーは、コア/E1領域由来の型特異的センスプライマーを
含み得る。好ましい第2回の型特異的センスプライマーは、HCVゲノム1型、2型、また
は3型のヌクレオチド536〜557由来であり、表1に示される。
【0049】
第1回または第2回のプライマーは、ウイルス濃度が十分に高い場合、第2回のPCRを
使用することなくウイルスRNAを増幅するのに十分であり得る。すなわちネスティッドPCRは必ずしも必要とされず、必要とされるものは100倍過剰量のプローブ中のPCR産物である。
【0050】
本発明のHCV遺伝子型分類キットはまた、新規のサブタイプがこのキットにおける使用
のために発見および評価されるにつれて変化し得る、サブタイプ参照を含む。単一の参照に対する比較が明確な結果をいつも提供するとは限らないので、所定のサブタイプ由来の1つより多い参照の使用は推奨される。
【0051】
前述の議論および以下の実施例は、本発明を例示するのみであり、当業者は、本発明が他の方法で実行され得、そして本発明が請求の範囲の参照によってのみ定義されることを理解する。
【実施例】
【0052】
(実施例1)
(患者サンプル)
規則的な血液透析を受けている35人の血液透析患者を研究した:平均年齢64.8±13歳
の20人男性(57%)および15人女性(43%)。血清サンプルを、1995年8月に回収し、アリコートに分け、そして-80℃で保存した。26人の患者が抗HCV ELISA陽性であり、そし
て9人が抗HCV ELISA陰性であった。26人のELISA陽性のうち25人はまたRIBA III陽性であったが、1人は不確定であった。9人のELISA陰性は、全てRIBA III陰性であった。15人の患者がHCV-RNA 5'UTRおよびE1 PCR陽性であった。15個の5'NCR産物の直接的配列決定により、5人の患者が1型;3人が2型;そして7人の患者が3型に帰された。
【0053】
(実施例2)
(cDNAおよびPCR)
HCV-RNAを、Stratagene RNA Isolation KitからのStratagene試薬を用いて、少なくとも2つの異なる回数で抽出した(ChomezynskyおよびSacchi法)。
【0054】
RNAを20μlの血漿から抽出し、これを100pmolのPCRプライマーを用いて25μlのcDNA混
合物(BRL cDNA合成キット、8085SB)中で逆転写させた。cDNA混合物を5分間煮沸し、氷上ですばやく冷却し、そしてPerkin PCRキット(N801-0055)説明書に従って最終濃度でPCR cDNA試薬に添加した。40回のPCRサイクル(94℃10秒、55℃30秒、および72℃30秒)を行った。10μlの第1のPCR反応混合物を、ネスティッドPCRプライマーを含む第2のPCR反応混合物に添加し、そして上記のように40サイクル増幅した。
【0055】
最初の抽出物を、Shimizuら、PNAS(1992)5477-5481に以前に記載された5'NCRに特異
的なプライマーを用いたネスティッドPCR反応に使用し、そしてこの産物を直接配列決定
し、そしてRFLPに使用した。同じ抽出物由来のRNAを、コア/E1プライマーを用いたHTAに
使用した。第2のRNA抽出を、RFLPおよび/またはHTAのために行って、結果を確認した。HTAに使用したプライマーを表1に示す。これらのE1産物を得るために使用したPCRプライ
マーのネスティッド対は、1、2a、2b、および3a型で異なった。第1回の増幅のためのユニバーサルセンスプローブは、Chooら、PNAS、1991の5'〜3'nt508〜529、アミノ酸170〜176に対応するが、第2回の増幅のためのユニバーサルアンチセンスプライマーは、Chooら、PNAS、1991のnt978〜958、アミノ酸320〜326に対応する。
【0056】
ssDNA DNAプローブをHTAにおける使用のために調製した場合、ネスティッドPCRのためのプライマーの1つをビオチン化した。例えば、表1における配列番号6を参照のこと。
【0057】
【表1】

(実施例3)
(HTA)
一本鎖プローブを、プライマー320Aの1つをビオチン化した以外は、上記のような同じPCRプライマーを用いた既知の遺伝子型のHCV ELISAおよびRIBA陽性血清のRT-PCRにより
調製した。ssDNAプローブを、Heng PanおよびEric Delwartのプロトコルに従って、Dynabeads M-280 Streptavidinを用いて作製した。非ビオチン化一本鎖を、磁気ビーズ/ストレプトアビジンカラムから溶出した。プローブを、異なる遺伝子型の20ngのssDNAから
作製し、そしてT4ポリヌクレオチドキナーゼ(Gibco BRL)および100μCiの32P ATPを
用いて末端標識し、次いでカラム精製した。キナーゼプローブを、Pharmacia Bio Sepharoseカラムを用いて32P ATPから分離した。32P標識一本鎖プローブを、100倍過剰量の
ドライバーと混合し、そしてPCR産物を、患者サンプルまたはコントロール血清/血漿から生成した。ハイブリダイゼーションは2×SSCであった。混合物を、94℃のヒートブロッ
クに3分間置いた。次いで、それらを55℃のヒートブロックに少なくとも2時間移した。全反応容量を、1mm厚の6%ポリアクリルアミドMDEゲル(Baker)にロードし、そして500Vで16時間電気泳動した。ゲルを80℃で濾紙上で真空乾燥し、そしてX線フィルムに暴露した。各サンプルの遺伝子型を、Delwart法に基づいて決定した。表2は、HTAを用いることにより決定した遺伝子型結果を示す。
【0058】
図1A〜1Eは、表1における一本鎖プローブの各々を示すオートラジオグラムであり、すなわち遺伝子型1a、1b、2a、2b、3aについて既知の特異的なプローブは、それぞれ図1A〜E中である(MDEゲルの左端のレーンを参照のこと)。ホモ二本鎖(h)(由来した二本鎖RT-PCR産物形態に対するssプローブ)を、プローブに隣接して示す。プローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物もまた、各図の適切なレーンとして示される。
【0059】
図1A〜1Eにおいて示され得るように、1型ssサブタイププローブは各1型サブタイプに特異的であり、そして他のサブタイプ1b、2a、2b、3aに交差ハイブリダイズしなかった(2a、2bは示さず)。3a型ssサブタイプ特異的プローブもまたサブタイプ3aに特異的であり、そして1a、2c、または2a、2s単離物に交差ハイブリダイズしなかった(データは示さず)。ssサブタイプ2プローブは、互いに交差ハイブリダイズしないが(データは示さず)、サブタイプ2c単離物に交差ハイブリダイズした;しかし、ホモ二本鎖と2c単離物との間の距離は高度の分岐を示し、これは患者23、30、および33が異なるサブタイプを有することを示唆した。血清23、30、および33中のウイルスは、部分的E1の配列決定によりサブタイプ2cに最も密接に関連することが確認されたが(図2bを参照のこと)、51UTR配列決定
によっては不明瞭であった。(図2Dを参照のこと)。
【0060】
単離物23、30、および33は、2aプローブとハイブリダイズするが、30および33のみが2bプローブにハイブリダイズする。ゲルはまた、単離物30が2bより2aに密接に関連することを示す。従って、3つの全ての血清は明らかに2型非a非bサブタイプであるが、それらは、全てが等しく2aおよび2b型と分岐しているとは限らない。図1Bおよび1Dにおいて示されるように、患者4は、1b型および非a非b型サブタイプに同時感染しているらしい。
【0061】
1bプローブは患者(JK16)に由来し、そしてホモ二本鎖レーン(h)に反映される2つのウイルスゲノムを有するらしく、従って各1b患者は2つのバンドを有する。
【0062】
ssプローブ3aは、3a型個体(JK3a)からの1つのRT-PCR産物のプラスミドクローンに由来した(図1E、レーンhを参照のこと)。従って、レーン22における複数のバンドは、最も恐らくは、この患者における2つの密接に関連したウイルスを反映する。
【0063】
非常にしばしば、患者が独特のウイルス単離物を有すると思われた。患者3および18が、同一のまたは高度に関連したウイルス単離物を有することは可能である。同様に、MDE
ゲルでのバンドの同時移動に基づいて、患者20および26は、同じ3a型ウイルス単離物を有し、そして患者2および4は、同じ1b型単離物を有した。
【0064】
図2a〜2cは、部分的E1ヌクレオチド配列の各々の遺伝的関係を示す系統発生樹(系統樹)を示す。これらの系統樹を、Weinerら、J.Virol.67:4365-4368頁(1993)により記
載されたように、計算手段を有するコンピューターソフトウェアプログラムGene Works
Unweighted Pair Group Methodsを使用することによる、互いに対するヌクレオチド配列の対様連続的アラインメントにより構築した。図2a〜2cにおける系統樹を、ゲノムの同じ領域上での1a型(HCV-1)(Chooら、PNAS 1991);1b(HCV-J)(Katoら);2a(HC-J6)(Okamotoら(1992));2b(HC-J8)(Okamotoら、1992);2c(Bukhら、PNAS、1993)、および3a(NZL-1)(Sakamotoら、1994)について公開された遺伝子型配列に対す
る配列分析により決定されたように、透析患者由来の推定1型(nt625〜93)、2型(nt583〜915)、または3型(nt558〜834)単離物の部分的E1配列を比較することにより形成
した。
【0065】
図2Dは、ゲノムの同じ領域について、単離物23、30、および33の部分的5'UTR配列のい
ずれかと、公開された1、2、および3型(nt-274〜-81)遺伝子型配列とを比較するこ
とにより、上記のように形成した系統樹である。
【0066】
RFLPおよびHTAの結果を比較し、そして表2に示す。
【0067】
【表2】
図3a〜3dにおける部分的E1配列は、表2において与えられたHTAサブタイプ指定を確証
し、そして患者23、30、および33が2cと最も密接に関連し、33が2cと最も離れて(18.6
%分岐)関連することを決定的に示す。
【0068】
ScrFI(Davidsonら、J.Gen.Virol.(1995)76:1197-1204を参照のこと)を用いたRFLP結果は、23、30、および33を2a型と誤って指定した。この誤った指定は図2Dにおいて
反映され、これはコンピューターが、5'UTR nt配列に基づいても、ゲノムのこの領域に
おける不十分なnt分岐に起因して正確にHCV 2cをサブタイプ分類しなかったことを示す
。
【0069】
コアおよびエンベロープ領域に対するプライマーを利用する本発明のHTAは、HCVゲノムの3レベルの特徴付けを可能にした。第1は、RT-PCRプライマーの選択における型特異性であった。第2は、コア/E1領域における、400ntを超える領域由来のプライマーの選択に基づくサブタイプ特異性であり、これは、サブタイププローブ(例えば、1および3、2a、2b)間の交差ハイブリダイゼーションの欠如、ならびに遺伝子型間の差異を最大にする高度の不均一性(交差ハイブリダイゼーションの欠如)をもたらした。最後に、単離物特異性を、図1.E〜1.Eにおいて例示されるようなホモ二本鎖からの距離により決定した。他の遺伝子型分類方法は、単離物差異を分析する能力を有さない。
【0070】
【表3−1】
【0071】
【表3−2】
【0072】
【表3−3】
【0073】
【表3−4】
【0074】
【表3−5】
【0075】
【表3−6】
【0076】
【表3−7】
【0077】
【表3−8】
【0078】
【表3−9】
【0079】
【表3−10】
【0080】
【表3−11】
【0081】
【表3−12】
【0082】
【表3−13】
【0083】
【表3−14】
【0084】
【表3−15】
【0085】
【表3−16】
【0086】
【表3−17】
【0087】
【表3−18】
【0088】
【表3−19】
【0089】
【表3−20】
【0090】
【表3−21】
【0091】
【表3−22】
【0092】
【表3−23】
【0093】
【表3−24】
【0094】
【表3−25】
【技術分野】
【0001】
(発明の分野)
本発明は、C型肝炎ウイルス(HCV)の遺伝子型分類に関する。特に、本発明は、好ま
しくはHCVのコアおよびエンベロープ領域由来の特異的プライマー、ならびにヘテロ二本
鎖移動度またはトラッキングアッセイ(次にこれは特異的プライマーを利用する)を用いたHCVの遺伝子型を決定するための方法に関する。
【背景技術】
【0002】
(発明の背景)
ウイルス性肝炎は、A、B、C、D、およびE型肝炎として知られる5つの異なるウイルスにより引き起こされることが知られている。HAVはRNAウイルスであり、そして長期の臨床的徴候を導かない。HBVは、DNAウイルスである。HDVは、HBVの非存在下で細胞を感染させ得ない依存性ウイルスである。HEVは、飲料水媒介性ウイルスである。HCVは最初に同定され、そして非A非B肝炎NANBHの原因として特徴付けられた(Houghtonら、特許文献
1および特許文献2)。これは、HCVの同定における免疫学的試薬として有用な多くの一
般的および特異的なポリペプチドの開示を導いた。例えば、非特許文献1;非特許文献2、および非特許文献3を参照のこと。
【0003】
HCVは一本鎖RNAウイルスであり、ペスティウイルスおよびフラビウイルスの遠縁にあたり、そして輸血関連肝炎の非常に大多数の、および世界中の市中(community acquired
)非A非B肝炎のほとんどの場合の原因因子である。HCVゲノムは、単一の長いオープン
リーディングフレーム(ORF)に隣接する5'および3'非コード(NC)領域からなる。このORFは、アミノ末端に3つの構造タンパク質およびカルボキシ末端に6つの非構造(NS)タンパク質をコードする。構造タンパク質は、ヌクレオキャプシド(コア;C)タンパク質ならびに2つの糖蛋白質であるエンベロープ1(E1)およびエンベロープ2(E2)から表される。非構造タンパク質は、NS2、NS3、NS4a、NS4b、NS5a、NS5bと呼ばれる。5'NCRは
、HCVゲノムの最も高度に保存された部分であるが、2つのエンベロープタンパク質(E1
およびE2)の配列は、異なるHCV単離物間で高度に可変的である。最も高い程度の変異はE2内の領域で観察されており、現在一般に超可変領域1(HVR1)またはH2HVと呼ばれてい
る。HVR2と呼ばれる第2の可変領域もまた、単離物のサブセットに存在する。代表的に、HCVの遺伝的不均一性は、2つの上位準種(quasispecies)および遺伝子型で分類されて
いる。本明細書中で使用する用語「準種」は、感染個体内のHCV集団の遺伝的不均一性を
いう。本明細書中で使用する用語「遺伝子型」および「サブタイプ(subtype)」は、異
なるHCV単離物間で観察された遺伝的不均一性をいう。約9.4kb RNA分子のポジティブ鎖である、HCVゲノムの核酸配列変異の分析は、遺伝的可変性が重要なウイルス学的および
臨床学的意味合いに関連することを示唆する。
【0004】
HCVの原型単離物は、特許文献1および特許文献2において特徴づけられた。本明細書
中で使用する用語「HCV」は、新たに単離されたNANBHウイルス種を含む。用語「HCV-1」
は、上記の公報に記載されたウイルスをいう。
【0005】
HCVの最初の同定から、少なくとも6つの異なる主要なウイルス型が同定されており(
全長ゲノムが報告されており)、そして1、2、3、4、5、および6型と呼ばれている。これらの型には、多くのサブタイプが含まれる。患者が感染するウイルス型は、臨床的予後に影響し得、そしてまた種々の処置に応答する。非特許文献4を参照のこと。HCV感
染のほとんどの重篤な臨床結果が肝細胞ガン腫であることを考慮すると、患者がどのHCV
型(単数または複数)に感染するかを決定し得ることは有用である。従って、配列決定を必要とすることなくサブタイプ内(intra-subtype)遺伝的分岐をまた与え得る、HCV遺伝子型分類およびサブタイプ分類のための正確な信頼できるアッセイを開発することは特に重要である。完全なウイルスゲノムを用いた理想的なヌクレオチド配列に基づく系は実用的でないので、異なる領域の分析に基づいたHCV遺伝子型分類のためのいくつかの分類が
提案されている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】欧州特許第388,232号明細書
【特許文献2】欧州特許第318,216号明細書
【非特許文献】
【0007】
【非特許文献1】Chooら、Science,1989年,第244巻,pp.359-262
【非特許文献2】Kuoら、Science,1989年,第244巻,pp.362-364
【非特許文献3】Houghtonら、Hepatology,1991年,第14巻,pp.381-388
【非特許文献4】Yoshiokeら、Hepatology,1992年,第16巻,pp.293-299
【発明の概要】
【発明が解決しようとする課題】
【0008】
配列決定を必要とすることなくサブタイプ内遺伝的分岐をまた与え得る、HCV遺伝子型
分類およびサブタイプ分類のための正確な信頼できるアッセイを開発すること。
【課題を解決するための手段】
【0009】
本発明は、例えば、上記課題を解決するために、以下の手段を提供する。
(項目1)配列番号1に記載の配列からなるオリゴヌクレオチド。
(項目2)配列番号2に記載の配列からなるオリゴヌクレオチド。
(項目3)配列番号3に記載の配列からなるオリゴヌクレオチド。
(項目4)配列番号4に記載の配列からなるオリゴヌクレオチド。
(項目5)配列番号5に記載の配列からなるオリゴヌクレオチド。
(項目6)配列番号6に記載の配列からなるオリゴヌクレオチド。
(項目7)配列番号7に記載の配列からなるオリゴヌクレオチド。
(項目8)配列番号8に記載の配列からなるオリゴヌクレオチド。
(項目9)配列番号9に記載の配列からなるオリゴヌクレオチド。
(項目10)センスプライマーが配列番号1からなり、そしてアンチセンスプライマーが、配列番号2、配列番号3、配列番号4、および配列番号5からなる群より選択される、PCRプライマー対。
(項目11)アンチセンスプライマーが配列番号6からなり、そしてセンスプライマーが、配列番号7、配列番号8、および配列番号9からなる群より選択される、PCRプライマ
ー対。
(項目12)HCV株のHCV遺伝子型を決定する方法であって、以下の工程:
(a)該HCV株を、1段階以上のPCRに供する工程であって、ここで該1段階以上のPCRが
、HCVゲノムのコアまたはE1領域由来のセンスプローブおよびHCVゲノムのコアまたはE1領域由来のアンチセンスプローブを利用する、工程;
(b)工程(a)で得られた増幅産物の混合物と、既知のHCV遺伝子型のDNAまたはRNAフ
ラグメントとを変性および再アニールすることにより、ヘテロ二本鎖を形成する工程;
(c)サイズにより分離する系において該ヘテロ二本鎖の移動度と、既知遺伝子型のDNA
またはRNAフラグメントのホモ二本鎖の移動度とを比較して、該HCV株の遺伝子型を決定する工程
を包含する、方法。
(項目13)項目12に記載の方法であって、ここで前記HCV株が、2段階のPCRに供され、第1のセットのプライマーが、前記HCVゲノムのコアまたはE1領域由来のユニバーサル
センスプローブおよび前記HCVゲノムのコアまたはE1領域由来の型特異的アンチセンスプ
ローブを含み、そして第2のセットのPCRプライマーが、前記HCVゲノムのコアまたはE1領域由来のユニバーサルアンチセンスプローブおよび前記HCVゲノムのコアまたはE1領域由
来の型特異的センスプローブを含む、方法。
(項目14)項目12に記載の方法であって、ここで前記第1のセットのPCRプライマー
が項目10に記載のプライマーであり、そして前記第2のセットのPCRプライマーが項目
11に記載のプライマーである、方法。
(項目15)前記既知の遺伝子型のDNAまたはRNAフラグメントがDNAプローブを含む、項
目12に記載の方法。
(項目16)前記プローブが一本鎖である、項目15に記載の方法。
(項目17)前記DNAプローブが放射標識される、項目16に記載の方法。
(項目18)前記一本鎖DNAプローブがPCR増幅により得られる、項目16に記載の方法。(項目19)項目18に記載の方法であって、ここで前記DNAプローブが、前記第1の段
階に項目10に記載のプライマーおよび前記第2の段階に項目11に記載のプライマーを利用する2段階PCR増幅により得られる、方法。
(項目20)前記HCV株が前記ヘテロ二本鎖を形成する混合物中に過剰に存在する、項目
12に記載の方法。
(項目21)HCV株の薬物治療に対する応答を、該HCV株に感染した患者から予測する方法であって、既知のHCV遺伝子型の該薬物治療に対する感受性を決定する工程、項目12に
記載の方法により該HCV株の該HCV遺伝子型を決定する工程、および該薬物治療前の該株の該HCV遺伝子型と、該薬物治療に対する既知のHCV遺伝子型の該感受性とを比較する工程を包含する、方法。
(項目22)HCV株の治療ワクチンに対する応答を、該HCV株に感染した患者から予測する方法であって、既知のHCV遺伝子型の該治療ワクチンに対する感受性を決定する工程、項
目12に記載の方法により該HCV株の該HCV遺伝子型を決定する工程、および該治療ワクチンの投与前の該株の該HCV遺伝子型と、該治療ワクチンに対する既知のHCV遺伝子型の該感受性とを比較する工程を包含する、方法。
(項目23)所定のサンプル集団に対する予防ワクチン組成物の適性を予測する方法であって、該予防ワクチンの遺伝子型を決定する工程、項目12に記載の方法により該サンプル集団における既知のHCV遺伝子型の優勢を決定する工程、および該予防ワクチンの該集
団サンプルへの投与前に、該予防ワクチン株の該HCV遺伝子型と決定された優勢遺伝子型
とを比較する工程を包含する、方法。
【0010】
(発明の要旨)
本発明は、ヘテロ二本鎖トラッキングアッセイ(HTA)に基づいた、HCV遺伝子型分類およびサブタイプ内変異の特徴付けのためのプライマーおよび方法を含む。好ましいプローブ/プライマーは、HCVのコアのカルボキシ末端およびE1領域の一部由来の一本鎖である。
【0011】
HTAは、2つ以上のウイルスゲノム間の遺伝的関係を決定する方法に基づいたハイブリ
ダイゼーションである。この方法の基礎は、分岐テンプレートから同時増幅された関連DNA産物が、ランダムに再アニールして、中性ポリアクリルアミドゲルのようなサイズに基
づいて分子を分離するように設計された系において、減少した移動度で移動するヘテロ二本鎖を形成することであり、HTAは、HIV-1を遺伝子型分類するために、そして患者および集団におけるHIV-1のインビボ進化を追随するために最初に使用された。例えば、Delwartら(1993)Science 262:1757-1261およびDelwartら(1994)J.Virol.68:6722-6883
を参照のこと。
【0012】
本発明の1つの局面は、HCVを遺伝子型分類するための方法であり、この方法は、部分
的に相補的なDNA鎖またはRNA鎖を変性および再アニールする工程、ならびにサイズに基づいて分子を分離するように設計された系において(例えば、ポリアクリルアミドまたはMDEゲルを介する電気泳動に従うことにより)DNAヘテロ二本鎖の電気泳動移動度を書き留めることにより配列変異を検出する工程を含む。
【0013】
本発明の別の局面は、HCVゲノムのコアおよびE1領域から選択された、遺伝子型分類において使用されるプローブに関する。
【0014】
本発明の別の局面は、異なる既知の遺伝子型の薬物治療に対する感受性を決定すること、患者を感染させるHCV株の遺伝子型を決定すること、およびその遺伝子型とその薬物治
療感受性とを比較して、患者の薬物治療に対する応答を予測することにより、HCV株に感
染した患者の薬物治療に対する応答を予測する方法に関する。
【0015】
本発明の別の局面は、治療ワクチンに関し、そしてHCV株に感染した患者の遺伝子型を
決定すること、および同じ遺伝子型の治療ワクチンを投与することにより、どの治療ワクチンが利用されるべきかを予測することに関する。
【0016】
本発明の別の局面は、予防ワクチンに関し、そして同様のサンプルにおいて優勢な遺伝子型を決定すること、および集団サンプルに対して優勢な遺伝子型であると思われる遺伝子型の予防ワクチンを投与することにより、どのワクチンが特定の集団サンプルに投与されるべきか予測することに関する。
【0017】
本発明の別の局面は、本発明の方法を用いて新規のHCV遺伝子型を発見する能力に関す
る。
【図面の簡単な説明】
【0018】
【図1A】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
【図1B】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
【図1C】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
【図1D】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
【図1E】図1A〜1Eは、サンプルのホモ二本鎖およびヘテロ二本鎖が既知の遺伝子型のプローブで分類されたことを示すオートラジオグラムである(ssプローブは、図1A〜1EにおいてそれぞれMDEゲルの左端のレーンにある、遺伝子型1a、1b、2a、2b、3aのプローブである)。ホモ二本鎖(h)(由来する既知の遺伝子型の二本鎖RT-PCR産物に対するssプローブ)は、プローブに隣接して示される。ssプローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物のヘテロ二本鎖は、各図の適切なレーンにおいて上記のように示される。
【図2A】図2A〜2Cは、本明細書中に記載の透析患者由来の推定1型(nt625〜930)、2型(nt583〜915)、または3型(nt558〜834)単離物の配列決定により得られた部分的E1配列と、ゲノムの同じ領域上での1a型(HCV-1)(Chooら、PNAS(1991)88:2451-2455、全てのヌクレオチド「nt」はこの論文に従った名称である)、1b型(HCV-J)(Katoら、PNAS(1990)87:9524-2528)、2a型(HC-J6)(Okamotoら、Virol.(1992)188:331-341)、2b型(HC-J8)(Okamotoら、Virol.(1992)188:331-341)、2c型(Bukhら、PNAS(1993)90:8234-8239)、および3a型(NZL-1)(Sakamotoら)J.Gen.Virol.(1994)75:1761-1768について公開された遺伝子型配列とを比較することにより形成された系統樹(すなわち各部分的E1ヌクレオチド配列の関連性を示す系統発生樹)である。
【図2B】図2A〜2Cは、本明細書中に記載の透析患者由来の推定1型(nt625〜930)、2型(nt583〜915)、または3型(nt558〜834)単離物の配列決定により得られた部分的E1配列と、ゲノムの同じ領域上での1a型(HCV-1)(Chooら、PNAS(1991)88:2451-2455、全てのヌクレオチド「nt」はこの論文に従った名称である)、1b型(HCV-J)(Katoら、PNAS(1990)87:9524-2528)、2a型(HC-J6)(Okamotoら、Virol.(1992)188:331-341)、2b型(HC-J8)(Okamotoら、Virol.(1992)188:331-341)、2c型(Bukhら、PNAS(1993)90:8234-8239)、および3a型(NZL-1)(Sakamotoら)J.Gen.Virol.(1994)75:1761-1768について公開された遺伝子型配列とを比較することにより形成された系統樹(すなわち各部分的E1ヌクレオチド配列の関連性を示す系統発生樹)である。
【図2C】図2A〜2Cは、本明細書中に記載の透析患者由来の推定1型(nt625〜930)、2型(nt583〜915)、または3型(nt558〜834)単離物の配列決定により得られた部分的E1配列と、ゲノムの同じ領域上での1a型(HCV-1)(Chooら、PNAS(1991)88:2451-2455、全てのヌクレオチド「nt」はこの論文に従った名称である)、1b型(HCV-J)(Katoら、PNAS(1990)87:9524-2528)、2a型(HC-J6)(Okamotoら、Virol.(1992)188:331-341)、2b型(HC-J8)(Okamotoら、Virol.(1992)188:331-341)、2c型(Bukhら、PNAS(1993)90:8234-8239)、および3a型(NZL-1)(Sakamotoら)J.Gen.Virol.(1994)75:1761-1768について公開された遺伝子型配列とを比較することにより形成された系統樹(すなわち各部分的E1ヌクレオチド配列の関連性を示す系統発生樹)である。
【図2D】図2Dは、直接配列決定により得られた単離物23、30、および33の部分的5'UTR配列のいずれかと、ゲノムの同じ領域についての公開された1、2、および3型(nt-274〜nt-81)との比較により形成された系統樹(系統発生樹)である。
【図3A−1】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3A−2】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3B−1】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3B−2】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3C−1】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3C−2】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【図3D】図3A〜3Dは、図2A〜2Dにおいて示された系統樹のヌクレオチド配列を示す。
【発明を実施するための形態】
【0019】
(発明の説明)
本発明の実施は、他に示されない限り、当該技術の範囲内の分子生物学、微生物学、組換えDNA、ポリペプチド合成および核酸合成、ならびに免疫学の従来技術を使用する。こ
のような技術は、文献中で十分に説明される。例えば、Sambrookら,MOLECULAR CLONING;A LABORATORY MANUAL,第2版(1989);DNA CLONING,第IおよびII巻(D.N Glover編、1985);OLIGONUCLEOTIDE SYNTHESIS(M.J.Gait編,1984);NUCLEIC ACID HYBRIDIZATION(B.D.HamesおよびS.J.Higgins編,1984);TRANSCRIPTION AND TRANSLATION(B.D.HamesおよびS.J.Higgins編,1984);ANIMAL CELL CULTURE(R.I.Freshney編,1986);IMMOBILIZED CELLS AND ENZYMES(IRL Press,1986);B.Perbal、A PRACTICAL GUIDE TO MOLECULAR CLONING(1984);シリーズMETHODS IN ENZYMOLOGY(Academic Press,Inc.);GENE TRANSFER VECTORS FOR MAMMALIAN CELLS(J.H.MillerおよびM.P.Calos編,1987,Cold Spring Harbor Laboratory)、Methods in Enzymology第154巻および第155巻(それぞれ、WuおよびGrossman、ならび
にWu編)、MayerおよびWalker編(1987),IMMUNOCHEMICAL METHODS IN CELL AND MOLECULAR BIOLOGY(Academic Press,London)、Scopes,(1987),PROTEIN PURIFICATION:PRINCIPLES AND PRACTICE,第2版(Springer-Verlag,N.Y.)、ならびにHANDBOOK OF EXPERIMENTAL IMMUNOLOGY,第I-IV巻(D.M.WeirおよびC.C.Blackwell編
1986)を参照のこと。
【0020】
ヌクレオチドおよびアミノ酸の標準的な略語が本明細書中で使用される。本明細書中に引用された全ての刊行物、特許、および特許出願が参考として援用される。
【0021】
本明細書中で使用する用語「組換えポリヌクレオチド」は、その起源または操作により、(1)天然に関連するポリヌクレオチドの全てもしくは一部に関連しないか、(2)天然に連結するポリヌクレオチド以外のポリヌクレオチドに連結するか、または(3)天然に生じない、ゲノム、cDNA、半合成、もしくは合成起源のポリヌクレオチドを意図する。
【0022】
本明細書中に使用する用語「ポリヌクレオチド」は、リボヌクレオチドまたはデオキシリボヌクレオチドのいずれかの、任意の長さのヌクレオチドのポリマー形態をいう。この用語は、分子の一次構造のみをいう。従って、この用語は、二本鎖および一本鎖DNAおよ
びRNAを含む。これは、既知の型の修飾(例えば、当該分野で公知の標識、メチル化、「キャップ」、1つ以上の天然に生じるヌクレオチドのアナログによる置換)、ヌクレオチド内修飾(例えば、非荷電結合(例えば、メチルホスホネート、ホスホトリエステル、ホスホアミデート、カルバメート等)による修飾および荷電結合(例えば、ホスホロチオエート、ホスホロジチオエート等)による修飾、ペンダント部分(例えば、タンパク質(例えば、ヌクレアーゼ、毒素、抗体、シグナルペプチド、ポリ-L-リジン等を含む))を含
む修飾、インターカレーター(例えば、アクリジン、ソラレン等)による修飾、キレーター(例えば、金属、放射性金属、ホウ素、酸化金属等)を含む修飾、アルキル化剤を含む修飾、修飾結合(例えば、αアノマー核酸等)による修飾)、ならびにポリヌクレオチドの非修飾形態をも含む。
【0023】
「PCR」は、本明細書中でポリメラーゼ連鎖反応(PCR)技術を意味し、これはMullis(米国特許第4,683,195号(Mullisら)および同第4,683,202号、本明細書中に参考として援用される)により開示される。PCR技術において、所望の配列の反対の末端に適合す
る短いオリゴヌクレオチドプライマーが調製される。プライマー間の配列は既知である必要はない。DNA(またはRNA)サンプルは抽出され、そして変性される(好ましくは、熱により)。次いで、オリゴヌクレオチドプライマーを、dNTPおよびポリメラーゼ(好ましくは、熱安定性のTaqポリメラーゼ)と共にモル過剰で添加する。DNAは複製され、次いで再度変性される。これは、それぞれのプライマーで始まる2つの「長い産物」および2つの最初の鎖(二本鎖DNA分子あたり)を生じる。次いで、反応混合物は、重合条件に戻され
(例えば、温度を低下させるか、変性剤を不活化するか、またはより多くのポリメラーゼを添加することにより)、そして第2のサイクルが開始される。第2のサイクルは、2つの最初の鎖、サイクル1由来の2つの長い産物、2つの新たな長い産物(最初の鎖から複製された)、および長い産物から複製された2つの「短い産物」を提供する。短い産物は、各末端でプライマーを有する標的配列(センスまたはアンチセンス)の配列を有する。さらなる各サイクルで、さらなる2つの長い産物が生成され、そして短い産物の数は、前のサイクルの終わりに残った長い産物および短い産物の数に等しい。従って、短い産物の数は、各サイクルで指数的に増殖する。特定の分析配列のこの増殖は、極端に少ない量のDNA検出を可能にする。
【0024】
本明細書中で使用する用語「3SR」は、欧州特許公開第373,960号(1990年6月20日に
公開)に記載されたような「自立(self-sustained)配列複製」としても知られる標的核酸増幅法をいう。
【0025】
本明細書中で使用する用語「LCR」は、Barany,Proc.Natl.Acad.Sci.(USA)(1991)88:189-193により記載されるような「リガーゼ連鎖反応」としても知られる標的核酸増幅法をいう。
【0026】
「オープンリーディングフレーム」(ORF)は、ポリペプチドをコードするポリヌクレ
オチド配列の領域であり;この領域は、コード配列の一部または総コード配列を示し得る。
【0027】
「コード配列」は、適切な調節配列の制御下に置かれた場合、通常mRNAを介してポリペプチドに翻訳されるポリヌクレオチド配列である。コード配列の境界は、5'末端での翻訳開始コドンおよび3'末端での翻訳停止コドンにより決定される。コード配列は、cDNAおよび組換えポリヌクレオチド配列を含み得るが、これらに限定されない。
【0028】
本明細書中で使用する用語「ポリペプチド」は、アミノ酸のポリマーをいい、そして特定の長さの産物をいわず;従って、ペプチド、オリゴペプチド、およびタンパク質がポリペプチドの定義内に含まれる。この用語はまた、ポリペプチドの発現後修飾(例えば、グリコシル化、アセチル化、リン酸化など)をいわないか、または除外する。例えば、1つ以上のアミノ酸アナログ(例えば、非天然アミノ酸等を含む)を含むポリペプチド、置換された結合を有するポリペプチド、ならびに天然および非天然の両方の当該分野で公知の他の修飾が、定義内に含まれる。
【0029】
示された核酸配列「に由来する」ポリペプチドまたはアミノ酸配列は、その配列にコードされるポリペプチドまたはその一部(ここで、その一部は、少なくとも3〜5アミノ酸、そしてより好ましくは少なくとも8〜10アミノ酸、そしてさらにより好ましくは少なくとも11〜15アミノ酸からなるか、またはその配列にコードされるポリペプチドと免疫学的に同定可能である)のアミノ酸配列と同一の、アミノ酸配列を有するポリペプチドをいう。この用語はまた、示された核酸配列から発現されるポリペプチドを含む。
【0030】
タンパク質は、そのタンパク質に特異的な抗体(モノクローナルまたはポリクローナルのいずれか)を生成するために使用され得る。これらの抗体を生成するための方法は、当該分野で公知である。
【0031】
「組換え宿主細胞」、「宿主細胞」、「細胞」、「細胞培養物」、および他のこのような用語は、組換えベクターまたは他の転移DNAのレシピエントとして使用され得るかもし
くは使用されてきた、そして形質転換された最初の細胞の子孫を含む、例えば、微生物、昆虫細胞、および哺乳動物細胞を意味する。単一の親細胞の子孫は、自然、偶発的、または計画的変異により、形態またはゲノムもしくは総DNA相補物において最初の親細胞と必
ずしも完全に同一ではあり得ないことが理解される。哺乳動物宿主細胞の例として、チャイニーズハムスター卵巣(CHO)およびサル腎臓(COS)細胞が挙げられる。
【0032】
「cDNA」は、mRNAの相補鎖にハイブリダイズする相補的mRNA配列を意味する。
【0033】
「精製」および「単離」は、ポリペプチドまたはヌクレオチド配列をいう場合に、示された分子が、同じ型の他の生物学的巨大分子の実質的な非存在下で存在することを意味する。本明細書中で使用する用語「精製」は、存在する同じ型の生物学的巨大分子の、好ましくは少なくとも75重量%、より好ましくは少なくとも85重量%、なおより好ましくは少なくとも95重量%、そして最も好ましくは少なくとも98重量%を意味する(しかし、水、緩衝液、および他の低分子(特に、1000未満の分子量を有する分子)は存在し得る)。
【0034】
「薬学的に受容可能なキャリア」は、それ自体が組成物を受ける個体に有害な抗体の産生を誘導しない、任意の薬学的キャリアを意味する。適切なキャリアは、代表的には、大きな緩徐に代謝される巨大分子(例えば、タンパク質、多糖、ポリ乳酸、ポリグリコール酸、ポリマーアミノ酸、アミノ酸コポリマー;および不活性ウイルス粒子)である。このようなキャリアは、当業者に周知である。
【0035】
治療組成物は、代表的には、薬学的に受容可能なビヒクル(例えば、水、生理食塩水、グリセロール、エタノール等)を含む。さらに、補助物質(例えば、湿潤剤または乳化剤、pH緩衝化物質等)がこのようなビヒクルに存在し得る。
【0036】
代表的には、治療組成物は、液体の溶液または懸濁液のいずれかのような注射可能物質として調製される;注射前の液体ビヒクル中の溶液または懸濁液に適した固体形態もまた調製され得る。調製物はまた、増強したアジュバント効果のために乳化され得るか、またはリポソーム内にカプセル化され得る。
【0037】
異なるHCV遺伝子型が、異なる病原性ならびに別々の地理的分布を有し得、そして感染
した患者において異なる血清学的プロフィールを部分的に誘発し得ることが、証拠によって示される。Cammarotaら、J.Clin.Microb.(1995)33:2781-2784を参照のこと。本発明は、HCVを検出し、そして異なる型のHCVによる感染を同定する方法を含む。本発明は、HCVを遺伝子型分類すること、HCVの新規の遺伝子型を発見する能力、および薬物治療に対する応答を予測する能力についてウイルス集団を評価することを含む。本発明はまた、HCVの遺伝子型分類における使用のためのプローブを含む。
【0038】
HCVを遺伝子型分類する方法として、HCVゲノムのコア/E1領域由来のプローブ/プライマーを利用する、ヘテロ二本鎖トラッキングまたは移動度アッセイが挙げられるが、これらに限定されない。HCV遺伝子型間の証明された抗原的差異は、血液ドナースクリーニング
およびIFN処置に対する応答の予測においてのみならず、異なる国におけるHCVに対する候補ワクチンの示された組成物、治療ワクチンの選択、ならびに新規の遺伝子型の同定においてもまた有用性を有する。ゲノムの異なる領域の分析(例えば、RFLP)に基づいた、集団を感染させる主要な遺伝子型を同定するための他の方法が提案されている。5'非コード領域(NCR)から同時増幅された配列のRFCPを用いてHCVを遺伝子型分類する議論については、Davidsonら、J.Gen Virol.(1995)76:1197-1204を参照のこと。
【0039】
既知の核酸に基づく遺伝子型分類法は、サブタイプ特異的RT-PCR(逆転写酵素-PCR)プライマー(Okamoto(1992)J.Gen Virol 73:673-679を参照のこと)米国特許第5,427,909号;(2)特異的プローブ(G.Marteenら、Lineプローブアッセイ);(3)制限部位多型性(ヌクレオチド配列(nt)の機能)、または(4)遺伝子型を決定するための直接的配列を必要とする。5'NC配列のRFLPによる分析は、実施しやすいが、全てのHCV遺
伝子型を正確に予測せず、そしていくつかのサブタイプは誤分類され得る。例えば、制限酵素により認識される1aと1bとの間の配列における変化は絶対的でなく、そして1aおよび1b、ならびに2aおよび2b以外の配列は誤分類される。例えば、1c型は1a型と思われ、2c型は2a型または2b型のいずれかであると思われる。Cammarotaら、J.Clin.Microb.,(1995)33:2781-2784を参照のこと。この理由のため、RFLPは、「エスケープ」種、新規の
分岐種、または疫学的傾向を検出し得ない。RFLPのような分類方法は、HCV配列不均一性
に関して回収される急速に増大する情報に適応するために、恐らく連続的に改変されなければならない。
【0040】
上記のように、核酸に基づく遺伝子型分類法を使用する場合、型もしくはサブタイプのいずれかの結果、または「分類不可能な」結果である陰性が得られる。例えば、遺伝子型特異的PCRにより分類されないままであった単離物が、コアおよびNS5遺伝子から得られたPCR増幅物の配列分析に基づいてサブタイプ2cに分類された、Cammarotaら、J.Clin.Microb.(1995)33:2781-2784を参照のこと。この問題は、コアのC末端/E1の中央2/3におけるRT-PCRプライマーを選択することによりHCV遺伝子型を決定する、目下請求される本
発明を用いることによって避けられる。さらに、単離物のサブタイプは、本発明のHCV遺
伝子型分類を用いて正確に決定され得、そして単離物は、約30%分岐未満のものでさえも検出され得、これは配列決定なしで新規のサブタイプの特徴付けを可能にする。
【0041】
(ヘテロ二本鎖トラッキングまたは移動度アッセイ)
本発明におけるHCV遺伝子型を決定する方法は、複雑な準種における少数の変異体を利
用する。1つのこのような技術は、二本鎖トラッキングアッセイ(HTA)である。HIVを用いた使用について当該分野で周知のHTA(例えば、Delwartら、J.Virol.(1994)68:6672-6683;Delwartら、Science(1993)262:1257-1261;Delwartら、PCR Methods and
Applications 4:S202-S216(1995)Cold Springs Harbor);およびDelwartら、Heteroduprex Mobility Analysis HIV-1 env Subtyping Kit Protocol Version 3(これらの各々は、その全体が参考として本明細書中に援用される)を参照のこと)は、配列が感染個体の末梢血単核細胞からネスティッドPCRにより増幅された場合、分岐テン
プレートから同時増幅された関連DNA産物が、ランダムに再アニールして、中性ポリアク
リルアミドゲルにおいて減少した移動度で移動するヘテロ二本鎖を形成し得たという観察に起因する。これらの技術を用いて、複数のウイルスDNAテンプレート分子間の遺伝的関
係が確立され得る。
【0042】
HTAは、特に、標識プローブとして第1のPCR産物を利用する。これは放射性であり得、異なる供給源(すなわち、分類が所望される供給源)由来の過剰量(ドライバー(driver))の非標識PCR産物と混合される。次いで、プローブ配列は、ドライバーと完全にヘテ
ロ二本鎖にさせられ、そしてサイズに基づいて分離される。得られたポリアクリルアミドゲルの例のためのオートラジオグラムは、これらのヘテロ二本鎖のみを明らかにし、そして研究下での2つのウイルス集団間の関係の視覚的な表示を提供する。ヘテロ二本鎖が異なる移動度で移動するという事実は、非適合(mismatched)および非対合(unpaired)ヌクレオチドの鎖特異的組成物がそれらの移動度に影響することを示す。
【0043】
次いで、Delwartらに記載される指数方程式を使用して、既知の配列の遺伝子の対様(pairwise)分析からの実験データに適合する曲線を記載する。本発明において、この方程
式を使用して、プローブの既知の遺伝子型と患者サンプルの未知の遺伝子型との間の遺伝的距離を評価する。
【0044】
(HTAにおける使用のためのプライマー)
E1またはコア領域がHCV不均一性を研究するための最良の領域であり得ることが決定さ
れ、従って、E1領域は、本発明におけるプライマーのための選択となった。本発明のゲノムの最も不均一な領域である部分的E1配列、ならびにより長いフラグメント(すなわち、400nt)(1000ntほどの長さであり得るが)の使用は、サブタイプ/型間で交差ハイブリダイズしないプローブの設計を可能にし、従って正確な遺伝子型分類を可能にする。不均一な領域を隣接させることにより、センスおよびアンチセンスプライマーの保存nt配列が同定された。好ましくは、第1回のPCRのためのユニバーサルセンスプライマーと型特異的
アンチセンスプライマーとの組合せ、ならびに第2回のためのユニバーサルアンチセンスプライマーと型特異的センスプライマーとの組合せが利用された。PCRは2回である必要
はなく、そしてプライマーは、上記の組合せに限定されない。しかし、好ましい組合せは、一本鎖プローブの調製を可能にし、そしてPCRプライマー組合せの数を最小にする。
【0045】
好ましいプローブは、コアおよびE1領域における配列である。この広範な遺伝子型のついての配列は公開され、そして少なくとも12個の異なる遺伝子型およびサブタイプ:I/1a、II/1b、III/2a、IV/2b、2c、3a、4a、4b、4c、4d、5a、6aに分類されている。同じ遺伝子型のHCV単離物間のE1遺伝子のヌクレオチド配列同一性は、88.0%〜99.1%の範囲で
あるが、異なる遺伝子型のHCV単離物のものは、53.5%〜78.6%の範囲である。中性ポ
リアクリルアミドゲルにおけるヘテロ二本鎖の良好な区別のための変異の程度は、快適には、分岐テンプレートが恐らく再アニールして、それらが同じサブタイプである場合にヘテロ二本鎖を形成するような、3〜20%の範囲内である。この理由のために、一本鎖32p
標識DNAプローブが、ヘテロ二本鎖の形成が不可能な場合、ss-DNAプローブが再アニール
し、そしてホモ二本鎖バンドを恐らく形成し得ないように使用された。直接的配列決定をすることなく、本発明は、サブタイプの確実な同定だけでなく、同じサブタイプ内の遺伝的関係もまた迅速に与え得る。例えば、分析された遺伝子型(すなわち、1a、1b、2a、2b、3a)は、異なるサブタイプ間で重複を示さなかった。
【0046】
さらに、単離物の約30%分岐がゲル上で視覚化され得るので、新規のサブタイプは視覚化され得、そして集団中の単離物の分布は特徴付けられ得、そして集団または個々の単離物の後に、疫学的研究における集団または個体が続き得る。
【0047】
(HCV遺伝子型分類キット)
HCVの遺伝子型を決定するためのキットは、本発明の範囲内にある。Delwartら、Heteroduprex Mobility Analysis HIV-1 env Subtyping Kit Protocol Version 3においてHIVについて記載されるように、このようなキットは、特異的なプライマーを含む。
好ましいプライマーは、HCVゲノムのコアおよびE1領域由来である。2段階のPCRが所望される場合、第1回のプライマーは、例えば、ユニバーサルセンスプローブ(好ましくは、HCVゲノムのコア/E1領域に位置する)を含み得る。1つのこのようなユニバーサルプライマーは、HCV-1のヌクレオチド508〜529に位置し、そして表1に示される。ユニバーサル
プライマーと対となるのは、好ましくはまたHCVゲノムのコア/E1領域に位置し得る、型特異的アンチセンスプライマーであり得る。これらのプライマーの例は、HCVゲノムの1型
、2a型、2b型、および3a型についてはヌクレオチド1032〜1012由来であり、そしてまた表1に示される。
【0048】
第2回のPCRが所望される場合、第2回のプライマーは、同様に、HCVゲノムのコア/E1
領域由来である。好ましい第2回のプライマーは、HCV-1ゲノムのヌクレオチド978〜958
に由来するユニバーサルアンチセンスプライマーを含み得、このプライマーは表1に示される。さらに、第2回のプライマーは、コア/E1領域由来の型特異的センスプライマーを
含み得る。好ましい第2回の型特異的センスプライマーは、HCVゲノム1型、2型、また
は3型のヌクレオチド536〜557由来であり、表1に示される。
【0049】
第1回または第2回のプライマーは、ウイルス濃度が十分に高い場合、第2回のPCRを
使用することなくウイルスRNAを増幅するのに十分であり得る。すなわちネスティッドPCRは必ずしも必要とされず、必要とされるものは100倍過剰量のプローブ中のPCR産物である。
【0050】
本発明のHCV遺伝子型分類キットはまた、新規のサブタイプがこのキットにおける使用
のために発見および評価されるにつれて変化し得る、サブタイプ参照を含む。単一の参照に対する比較が明確な結果をいつも提供するとは限らないので、所定のサブタイプ由来の1つより多い参照の使用は推奨される。
【0051】
前述の議論および以下の実施例は、本発明を例示するのみであり、当業者は、本発明が他の方法で実行され得、そして本発明が請求の範囲の参照によってのみ定義されることを理解する。
【実施例】
【0052】
(実施例1)
(患者サンプル)
規則的な血液透析を受けている35人の血液透析患者を研究した:平均年齢64.8±13歳
の20人男性(57%)および15人女性(43%)。血清サンプルを、1995年8月に回収し、アリコートに分け、そして-80℃で保存した。26人の患者が抗HCV ELISA陽性であり、そし
て9人が抗HCV ELISA陰性であった。26人のELISA陽性のうち25人はまたRIBA III陽性であったが、1人は不確定であった。9人のELISA陰性は、全てRIBA III陰性であった。15人の患者がHCV-RNA 5'UTRおよびE1 PCR陽性であった。15個の5'NCR産物の直接的配列決定により、5人の患者が1型;3人が2型;そして7人の患者が3型に帰された。
【0053】
(実施例2)
(cDNAおよびPCR)
HCV-RNAを、Stratagene RNA Isolation KitからのStratagene試薬を用いて、少なくとも2つの異なる回数で抽出した(ChomezynskyおよびSacchi法)。
【0054】
RNAを20μlの血漿から抽出し、これを100pmolのPCRプライマーを用いて25μlのcDNA混
合物(BRL cDNA合成キット、8085SB)中で逆転写させた。cDNA混合物を5分間煮沸し、氷上ですばやく冷却し、そしてPerkin PCRキット(N801-0055)説明書に従って最終濃度でPCR cDNA試薬に添加した。40回のPCRサイクル(94℃10秒、55℃30秒、および72℃30秒)を行った。10μlの第1のPCR反応混合物を、ネスティッドPCRプライマーを含む第2のPCR反応混合物に添加し、そして上記のように40サイクル増幅した。
【0055】
最初の抽出物を、Shimizuら、PNAS(1992)5477-5481に以前に記載された5'NCRに特異
的なプライマーを用いたネスティッドPCR反応に使用し、そしてこの産物を直接配列決定
し、そしてRFLPに使用した。同じ抽出物由来のRNAを、コア/E1プライマーを用いたHTAに
使用した。第2のRNA抽出を、RFLPおよび/またはHTAのために行って、結果を確認した。HTAに使用したプライマーを表1に示す。これらのE1産物を得るために使用したPCRプライ
マーのネスティッド対は、1、2a、2b、および3a型で異なった。第1回の増幅のためのユニバーサルセンスプローブは、Chooら、PNAS、1991の5'〜3'nt508〜529、アミノ酸170〜176に対応するが、第2回の増幅のためのユニバーサルアンチセンスプライマーは、Chooら、PNAS、1991のnt978〜958、アミノ酸320〜326に対応する。
【0056】
ssDNA DNAプローブをHTAにおける使用のために調製した場合、ネスティッドPCRのためのプライマーの1つをビオチン化した。例えば、表1における配列番号6を参照のこと。
【0057】
【表1】
(実施例3)
(HTA)
一本鎖プローブを、プライマー320Aの1つをビオチン化した以外は、上記のような同じPCRプライマーを用いた既知の遺伝子型のHCV ELISAおよびRIBA陽性血清のRT-PCRにより
調製した。ssDNAプローブを、Heng PanおよびEric Delwartのプロトコルに従って、Dynabeads M-280 Streptavidinを用いて作製した。非ビオチン化一本鎖を、磁気ビーズ/ストレプトアビジンカラムから溶出した。プローブを、異なる遺伝子型の20ngのssDNAから
作製し、そしてT4ポリヌクレオチドキナーゼ(Gibco BRL)および100μCiの32P ATPを
用いて末端標識し、次いでカラム精製した。キナーゼプローブを、Pharmacia Bio Sepharoseカラムを用いて32P ATPから分離した。32P標識一本鎖プローブを、100倍過剰量の
ドライバーと混合し、そしてPCR産物を、患者サンプルまたはコントロール血清/血漿から生成した。ハイブリダイゼーションは2×SSCであった。混合物を、94℃のヒートブロッ
クに3分間置いた。次いで、それらを55℃のヒートブロックに少なくとも2時間移した。全反応容量を、1mm厚の6%ポリアクリルアミドMDEゲル(Baker)にロードし、そして500Vで16時間電気泳動した。ゲルを80℃で濾紙上で真空乾燥し、そしてX線フィルムに暴露した。各サンプルの遺伝子型を、Delwart法に基づいて決定した。表2は、HTAを用いることにより決定した遺伝子型結果を示す。
【0058】
図1A〜1Eは、表1における一本鎖プローブの各々を示すオートラジオグラムであり、すなわち遺伝子型1a、1b、2a、2b、3aについて既知の特異的なプローブは、それぞれ図1A〜E中である(MDEゲルの左端のレーンを参照のこと)。ホモ二本鎖(h)(由来した二本鎖RT-PCR産物形態に対するssプローブ)を、プローブに隣接して示す。プローブにハイブリダイズした15人の透析患者(番号1、2、3、4、7、18、20、22、23、24、26、28、30、33、35)由来のRT-PCR産物もまた、各図の適切なレーンとして示される。
【0059】
図1A〜1Eにおいて示され得るように、1型ssサブタイププローブは各1型サブタイプに特異的であり、そして他のサブタイプ1b、2a、2b、3aに交差ハイブリダイズしなかった(2a、2bは示さず)。3a型ssサブタイプ特異的プローブもまたサブタイプ3aに特異的であり、そして1a、2c、または2a、2s単離物に交差ハイブリダイズしなかった(データは示さず)。ssサブタイプ2プローブは、互いに交差ハイブリダイズしないが(データは示さず)、サブタイプ2c単離物に交差ハイブリダイズした;しかし、ホモ二本鎖と2c単離物との間の距離は高度の分岐を示し、これは患者23、30、および33が異なるサブタイプを有することを示唆した。血清23、30、および33中のウイルスは、部分的E1の配列決定によりサブタイプ2cに最も密接に関連することが確認されたが(図2bを参照のこと)、51UTR配列決定
によっては不明瞭であった。(図2Dを参照のこと)。
【0060】
単離物23、30、および33は、2aプローブとハイブリダイズするが、30および33のみが2bプローブにハイブリダイズする。ゲルはまた、単離物30が2bより2aに密接に関連することを示す。従って、3つの全ての血清は明らかに2型非a非bサブタイプであるが、それらは、全てが等しく2aおよび2b型と分岐しているとは限らない。図1Bおよび1Dにおいて示されるように、患者4は、1b型および非a非b型サブタイプに同時感染しているらしい。
【0061】
1bプローブは患者(JK16)に由来し、そしてホモ二本鎖レーン(h)に反映される2つのウイルスゲノムを有するらしく、従って各1b患者は2つのバンドを有する。
【0062】
ssプローブ3aは、3a型個体(JK3a)からの1つのRT-PCR産物のプラスミドクローンに由来した(図1E、レーンhを参照のこと)。従って、レーン22における複数のバンドは、最も恐らくは、この患者における2つの密接に関連したウイルスを反映する。
【0063】
非常にしばしば、患者が独特のウイルス単離物を有すると思われた。患者3および18が、同一のまたは高度に関連したウイルス単離物を有することは可能である。同様に、MDE
ゲルでのバンドの同時移動に基づいて、患者20および26は、同じ3a型ウイルス単離物を有し、そして患者2および4は、同じ1b型単離物を有した。
【0064】
図2a〜2cは、部分的E1ヌクレオチド配列の各々の遺伝的関係を示す系統発生樹(系統樹)を示す。これらの系統樹を、Weinerら、J.Virol.67:4365-4368頁(1993)により記
載されたように、計算手段を有するコンピューターソフトウェアプログラムGene Works
Unweighted Pair Group Methodsを使用することによる、互いに対するヌクレオチド配列の対様連続的アラインメントにより構築した。図2a〜2cにおける系統樹を、ゲノムの同じ領域上での1a型(HCV-1)(Chooら、PNAS 1991);1b(HCV-J)(Katoら);2a(HC-J6)(Okamotoら(1992));2b(HC-J8)(Okamotoら、1992);2c(Bukhら、PNAS、1993)、および3a(NZL-1)(Sakamotoら、1994)について公開された遺伝子型配列に対す
る配列分析により決定されたように、透析患者由来の推定1型(nt625〜93)、2型(nt583〜915)、または3型(nt558〜834)単離物の部分的E1配列を比較することにより形成
した。
【0065】
図2Dは、ゲノムの同じ領域について、単離物23、30、および33の部分的5'UTR配列のい
ずれかと、公開された1、2、および3型(nt-274〜-81)遺伝子型配列とを比較するこ
とにより、上記のように形成した系統樹である。
【0066】
RFLPおよびHTAの結果を比較し、そして表2に示す。
【0067】
【表2】
図3a〜3dにおける部分的E1配列は、表2において与えられたHTAサブタイプ指定を確証
し、そして患者23、30、および33が2cと最も密接に関連し、33が2cと最も離れて(18.6
%分岐)関連することを決定的に示す。
【0068】
ScrFI(Davidsonら、J.Gen.Virol.(1995)76:1197-1204を参照のこと)を用いたRFLP結果は、23、30、および33を2a型と誤って指定した。この誤った指定は図2Dにおいて
反映され、これはコンピューターが、5'UTR nt配列に基づいても、ゲノムのこの領域に
おける不十分なnt分岐に起因して正確にHCV 2cをサブタイプ分類しなかったことを示す
。
【0069】
コアおよびエンベロープ領域に対するプライマーを利用する本発明のHTAは、HCVゲノムの3レベルの特徴付けを可能にした。第1は、RT-PCRプライマーの選択における型特異性であった。第2は、コア/E1領域における、400ntを超える領域由来のプライマーの選択に基づくサブタイプ特異性であり、これは、サブタイププローブ(例えば、1および3、2a、2b)間の交差ハイブリダイゼーションの欠如、ならびに遺伝子型間の差異を最大にする高度の不均一性(交差ハイブリダイゼーションの欠如)をもたらした。最後に、単離物特異性を、図1.E〜1.Eにおいて例示されるようなホモ二本鎖からの距離により決定した。他の遺伝子型分類方法は、単離物差異を分析する能力を有さない。
【0070】
【表3−1】
【0071】
【表3−2】
【0072】
【表3−3】
【0073】
【表3−4】
【0074】
【表3−5】
【0075】
【表3−6】
【0076】
【表3−7】
【0077】
【表3−8】
【0078】
【表3−9】
【0079】
【表3−10】
【0080】
【表3−11】
【0081】
【表3−12】
【0082】
【表3−13】
【0083】
【表3−14】
【0084】
【表3−15】
【0085】
【表3−16】
【0086】
【表3−17】
【0087】
【表3−18】
【0088】
【表3−19】
【0089】
【表3−20】
【0090】
【表3−21】
【0091】
【表3−22】
【0092】
【表3−23】
【0093】
【表3−24】
【0094】
【表3−25】
【特許請求の範囲】
【請求項1】
本願明細書に記載された発明。
【請求項1】
本願明細書に記載された発明。
【図1A】

【図1B】
【図1C】
【図1D】
【図1E】
【図2A】
【図2B】
【図2C】
【図2D】

【図3A−1】
【図3A−2】
【図3B−1】
【図3B−2】
【図3C−1】
【図3C−2】
【図3D】

【図1B】
【図1C】
【図1D】
【図1E】
【図2A】
【図2B】
【図2C】
【図2D】
【図3A−1】
【図3A−2】
【図3B−1】
【図3B−2】
【図3C−1】
【図3C−2】
【図3D】
【公開番号】特開2012−80890(P2012−80890A)
【公開日】平成24年4月26日(2012.4.26)
【国際特許分類】
【出願番号】特願2011−256583(P2011−256583)
【出願日】平成23年11月24日(2011.11.24)
【分割の表示】特願2008−105151(P2008−105151)の分割
【原出願日】平成9年4月11日(1997.4.11)
【出願人】(591076811)ノバルティス バクシンズ アンド ダイアグノスティックス,インコーポレーテッド (265)
【Fターム(参考)】
【公開日】平成24年4月26日(2012.4.26)
【国際特許分類】
【出願日】平成23年11月24日(2011.11.24)
【分割の表示】特願2008−105151(P2008−105151)の分割
【原出願日】平成9年4月11日(1997.4.11)
【出願人】(591076811)ノバルティス バクシンズ アンド ダイアグノスティックス,インコーポレーテッド (265)
【Fターム(参考)】
[ Back to top ]