
HER3結合ポリペプチド
本発明は、HER3結合モチーフ(BM)を含むHER3結合ポリペプチドを提供し、該モチーフは、
i) EX2X3X4A X6X7EIW X11LPNL X16X17X18QX20 X21AFIX25 X26LX28D、及び
ii) i)において定義される配列に対して少なくとも90%の同一性を有するアミノ酸配列
から選択されるアミノ酸配列からなり、ここで該ポリペプチドは、HER3の細胞外ドメインに結合する。HER3及びHER2に対して又はHER3及びEGFRに対して結合親和性を有し、かつ本明細書で定義されるHER3結合ポリペプチドとHER2結合ポリペプチド又はEGFR結合ポリペプチドとを含む、二重特異性リガンドも提供される。

i) EX2X3X4A X6X7EIW X11LPNL X16X17X18QX20 X21AFIX25 X26LX28D、及び
ii) i)において定義される配列に対して少なくとも90%の同一性を有するアミノ酸配列
から選択されるアミノ酸配列からなり、ここで該ポリペプチドは、HER3の細胞外ドメインに結合する。HER3及びHER2に対して又はHER3及びEGFRに対して結合親和性を有し、かつ本明細書で定義されるHER3結合ポリペプチドとHER2結合ポリペプチド又はEGFR結合ポリペプチドとを含む、二重特異性リガンドも提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒト上皮成長因子受容体3(本明細書ではHER3と呼ぶ)に結合するポリペプチド並びに画像化及び治療における上記ポリペプチドの使用に関する。本発明はまた、HER3及びHER2、又はHER3及びEGFRの両方に対する結合親和性を有する二重特異性リガンドに関する。
【背景技術】
【0002】
EGFR(ErbB1又はHER1)、ErbB2(HER2)、ErbB3(ERBB3又はHER3)及びErbB4(HER4)を含む膜貫通チロシンキナーゼ受容体の上皮増殖因子ファミリーは、細胞内シグナル伝達経路の複雑なネットワークを通した重要な細胞機能(例えば細胞増殖、生存、分化及び遊走)の調節に関与する。今日では、これらの受容体の異常な発現及びシグナル伝達がいくつかの型のがんの発生及び進行に関連するということが十分に確立されており、それによりこれらの受容体は新規ながん治療の開発のための重要な標的となっている。これまでに、EGFR及びHER2受容体が最も広く研究されており、いくつかの標的の薬剤はヒトがんの診断又は処置について行政当局により認可されている。HER3はチロシンキナーゼ活性を欠いており、従ってそのシグナル伝達機能を発揮するために他のErbB受容体と(例えばHER2と)ヘテロ二量体を形成しなければならない(非特許文献1)。HER2−HER3ヘテロ二量体は潜在的に発がん性の単位であると考えられており(非特許文献2)、そしてHER3はHER2の好ましいヘテロ二量化パートナーとして近年注目が高まっている。HER2との二量化はPI3K/Akt経路の活性化並びに腫瘍細胞生存及び増殖の促進をもたらす(非特許文献3)。HER3は、乳がん(非特許文献4)、卵巣がん及び膀胱がん(非特許文献5)を含む多数のヒトのがんにおいて発現され、そしていくつかの肺がん(非特許文献6)及び前立腺がん(非特許文献7)を含む他のがん(非特許文献8)におけるシグナル伝達において重要な役割を果たす。さらに、高レベルの受容体発現は、HER2を過剰発現する患者と比較して有意に短い生存期間と関連するので、HER3発現は予後的な価値を有する(非特許文献9、非特許文献10)。実際HER3は、ErbB受容体−PI3Kシグナル伝達軸のリガンド誘導活性化における主要なノードであると提案されてきた(非特許文献11)。
【0003】
より最近に認可された治療の比較的大部分はEGFRに向けられたものであり、そしてHER2受容体はモノクローナル抗体に基づく。この新しいクラスの生物学的薬剤のがん治療における成功の背後にある理由の1つは、低分子化学薬を使用して以前には達成できなかった新しく独特な作用機構を抗体が提供するということである。より伝統的なアゴニスト(例えば細胞表面受容体の刺激)及びアンタゴニスト(例えば天然のタンパク質−タンパク質相互作用のブロック)の治療効果に加えて、抗体はまた、様々な範囲の免疫学的防御機構を特異的にがん性細胞に向けるために、さらには種々の画像化又は治療用抱合体(例えば化学療法薬、放射性核種及び毒素)の特異的送達を達成するための標的化剤として使用され得る。ErbBファミリーの十分に研究されたEGFR及びHER2受容体メンバーと対照的に、抗HER3抗体の使用に関しては比較的少ない報告しかない。
【0004】
しかしUllrich及び共同研究者らは、抗HER3モノクローナル抗体が乳がんの細胞モデルにおいてHER3仲介シグナル伝達を阻害するということを報告しており(非特許文献12)、そして現在では2つのモノクローナル抗HER3抗体が第1相臨床試験にある(AMG 888、非特許文献13、及びMM−121、Schoeberlら、上記)。
【0005】
しかし、全長モノクローナル抗体を使用するいくつかの成功したがん治療研究が報告されているが、このクラスの薬剤は固形腫瘍を標的とするために(診断的なペイロードの目的でも治療的なペイロードの目的でもなく)常に最適というわけではない。治療効果は、腫瘍全体の薬物の有効な分布に依存し、そして分子画像化は、腫瘍取り込みと周囲の正常組織との間の高い比率に依存する。腫瘍浸透率(血管外漏出を含む)は分子の大きさと逆の関係があるので、比較的大きなIgG分子は本質的に乏しい組織分布及び浸透能を有する。さらに、分子画像化については、抗体の非常に長いインビボでの半減期が比較的高い血液シグナルを生じ、それにより比較的少ない腫瘍対血液コントラストを生じる。
【0006】
HER3はEGFファミリーの他のメンバーと同じ腫瘍細胞で発現され得るので、HER3及びEGFファミリーの別のメンバーを標的にする二重特異性分子の製造は、近年いくらかの興味を集めている。このような二重特異性分子は、例えば分子画像化適用における標的化及びHER3と腫瘍上に発現される別の抗原との同時標的化のための標的化ビヒクルとして利用され得る。
【0007】
しかし、2つの別々の標的に結合する二重特異性モノクローナル抗体を製造することは複雑である。4つの必要なポリペプチド鎖をコードする遺伝子が細胞において産生される場合、10もの異なる組み合わせが可能であり、そして1つの組み合わせのみが望まれる二重特異性抗体を示す。従って、二重特異的結合という構想は、抗体を使用して十分に探査されていない。代わりに、抗体フラグメント及び他の結合分子が二重特異性分子(二重特異性Z変異体を含む)を作製するためにより広く利用されてきた(非特許文献14)。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Citri et al,Exp Cell Res 284(1):54−65(2003)
【非特許文献2】Holbro et al,Proc Natl Acad Sci 100(15):8933−8(2003)
【非特許文献3】Hynes et al,Nat Rev Cancer 5(5):341−54(2005)
【非特許文献4】Bobrow et al,Eur J Cancer 33(11):1846−50(1997)
【非特許文献5】Rajkumar et al,Clin Mol Pathol 49(4)(1996)
【非特許文献6】Engelman et al,Proc Natl Acad Sci 102(10):3788−93(2005)
【非特許文献7】Gregory et al,Clin Cancer Res 11(5):1704−12(2005)
【非特許文献8】Stove et al,Clin Exp Metastasis 21(8):665−84(2004)
【非特許文献9】Tanner et al,J Clin Oncol 24(26):4317−23(2006)
【非特許文献10】Reschke et al,Clin Cancer Res 14(16):5188−97(2008)
【非特許文献11】Schoeberl et al,Sci Signal 2(77):ra 31(2009)
【非特許文献12】van der Horst et al,Int J Cancer 115(4):519−27(2005)
【非特許文献13】Baselga et al,Nat Rev Cancer 9(7):463−75(2009)
【非特許文献14】Friedman et al,Biotechnol Appl Biochem 54:121−31(2009)
【発明の概要】
【発明が解決しようとする課題】
【0009】
発明の説明
例えばHER3を発現する細胞の標的化、このようなHER3を発現する細胞の分子画像化及びHER3関連状態の処置において使用され得る新規なHER3結合剤を提供することが本発明の目的である。
【0010】
本発明のさらなる目的は、HER2及びHER3の両方を発現する病変に対する高い特異性を有する標的化剤を提供することである。
【課題を解決するための手段】
【0011】
一局面によれば、HER3結合モチーフ(BM)を含むHER3結合ポリペプチドが提供され、該モチーフは、
i) EX2X3X4A X6X7EIW X11LPNL X16X17X18QX20 X21AFIX25 X26LX28D、
[ここで、互いに独立して、
X2はR、K、L、W及びVから選択され;
X3はR、H、K、M、S、W、Y及びVから選択され;
X4はA、R、N、D、Q、E、G、H、I、L、K、M、S、T、W及びVから選択され;
X6はA、R、Q、M、S、T及びYから選択され;
X7はA、E、G、H、K、F、S、T、W、Y及びVから選択され;
X11はA、N、D、Q、E、I、L及びTから選択され;
X16はN及びTから選択され;
X17はA、R、N、Q、K、P、S、T及びVから選択され;
X18はA、R、N、E、F、D、Q、G、H、I、L、K、S、T、W、Y及びVから選択され;
X20はA、R及びKから選択され;
X21はA、R、N、G、H、S及びVから選択され;
X25はA、R、E、G、I、K、S、T及びVから選択され;
X26はS及びKから選択され;
X28はA、D、Q、E、F、P、S、T、W及びYから選択される];
及び
ii) i)において定義された配列に対して少なくとも90%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列からなり、
ここでポリペプチドはHER3の細胞外ドメイン(HER3−ECD)に結合する。
【0012】
本明細書において記載されるポリペプチドは、HER3によく結合するという点でHER3に対する良好な結合親和性を示す。配列に関連したHER3結合ポリペプチドのクラスの上記の定義は、選択実験においてHER3とのそれらの相互作用について選択された親骨格の多数の無作為なポリペプチド変異体の分析に基づく。同定されたHER3結合モチーフ又は「BM」は、親骨格の結合領域に相当し、この領域は3ヘリックスバンドルタンパク質ドメイン(three−helical bundle protein domain)内の2つのアルファヘリックスを構成する。親骨格において、2つのBMヘリックスの変更されたアミノ酸残基は、抗体の定常Fc部分との相互作用のための結合表面を構成する。結合表面の無作為な変更及びその後の変異体の選択により、結合表面のFc相互作用能がHER3との相互作用の能力で置き換えられた。
【0013】
当業者に理解されるように、本明細書に定義されるポリペプチドのHER3結合能のように、ポリペプチドの機能はポリペプチドの三次構造に依存する。従って、その三次構造及び機能に大きな影響を与えずにポリペプチドのアミノ酸配列に小さな変化を生成することが可能である。従って一実施態様において、本ポリペプチドは、結果として生じた配列がi)により定義されるクラスに属する配列に対して少なくとも90%同一である、例えばi)により定義されるクラスに属する配列に対して少なくとも93%同一、例えば少なくとも97%同一であるように改変された、i)BMの変異体を含む。いくつかの実施態様において、小さな変化は本明細書において開示されるHER3結合ポリペプチドの配列の全ての位置において作製され得る。他の実施態様において、小さな変化は、骨格アミノ酸残基とも示される不変位置においてのみ作製され得る。このような場合において、変化は可変位置、すなわち「X」で示される位置(例えば上記の定義されたBMのX2、X3、X4、X6、X7、X11、X16、X17、X18、X20、X21、X25、X26及びX28)において許容されていない。例えば、アミノ酸残基の特定の機能による分類(例えば疎水性、親水性、極性など)に属するアミノ酸残基は、同じ機能グループからの別のアミノ酸残基と交換され得るということが可能である。
【0014】
本明細書及び添付の特許請求の範囲を通じて使用される用語「%の同一性(同一性%)」は以下のようにして算出される。CLUSTAL Wアルゴリズム(Thompson、J.D.、Higgins、D.G.and Gibson、T.J.、Nucleic Acids Research、22: 4673−4680(1994))を使用して問い合わせ配列を対象配列と整列させる。整列された配列の最短のものに対応する範囲にわたって比較を行う。整列された配列の最短のものは、いくつかの場合には標的配列であるかもしれない(例えば29アミノ酸残基のHER3結合モチーフ)。他の場合、問い合わせ配列が整列された配列の最短のものを構成するかもしれない。問い合わせ配列は、例えば少なくとも10アミノ酸残基、例えば少なくとも20アミノ酸残基からなるものであり得る。各位置におけるアミノ酸残基を比較し、そして標的配列において同一の対応を有する問い合わせ配列中の位置のパーセントを同一性%として報告する。
【0015】
本明細書及び添付の特許請求の範囲を通じて使用される用語「%の類似性(類似性%)」は以下のやり方で計算される。配列アラインメント及び比較を基本的に同一性%の計算に関して記載したように行う。しかし「類似性」は以下のように解釈されるべきである。2つのアミノ酸残基は、それらがアミノ酸残基の同じグループに属する場合に類似と見なされる。アミノ酸残基のグループの非限定的な例は、アミノ酸残基Ala、Val、Phe、Pro、Leu、Ile、Trp、Met及びCysを含む疎水性グループ;アミノ酸残基Lys、Arg及びHisを含む塩基性グループ;アミノ酸残基Glu及びAspを含む酸性グループ;非荷電アミノ酸残基Gln、Asn、Ser、Thr及びTyrを含む親水性グループ;並びにアミノ酸残基Glyを含む天然グループである。従って、各位置におけるアミノ酸残基を比較し、そして標的配列中に類似した対応を有する問い合わせ配列中の位置のパーセントを類似性%として報告する。
【0016】
この説明全体を通して、本発明に従う代替の実施態様は、特定の同一性パーセントの代わりに、対応する類似性パーセントを満たす。他の代替の実施態様は、特定の同一性パーセントに加えて、各配列についての好ましい同一性パーセントの群から選択される別のより高い類似性パーセント満たす。例えば、配列は別の配列に対して70%の類似性であってもよく;又はそれは別の配列に対して70%の同一性であってもよく;又は別の配列に対して70%の同一性及び90%の類似性であってもよい。
【0017】
上に開示されるポリペプチドの一実施態様において、X2はR、K及びWから選択される。
【0018】
上に開示されるポリペプチドのさらなる実施態様において、X3はR、K、S及びYから選択される。
【0019】
上に開示されるポリペプチドのさらなる実施態様において、X4はA、R、N、Q、H、I、L、K、S、T及びVから選択される。
【0020】
上に開示されるポリペプチドのさらなる実施態様において、X6はA、S、T及びYから選択される。
【0021】
上に開示されるポリペプチドのさらなる実施態様において、X7はA、G、F、W、Y及びVから選択される。
【0022】
上に開示されるポリペプチドのさらなる実施態様において、X11はQ及びEから選択される。
【0023】
上に開示されるポリペプチドのさらなる実施態様において、X17はR、Q、T及びVから選択される。
【0024】
上に開示されるポリペプチドのさらなる実施態様において、X18はR、Q、G、H、I、L、S、T、W、Y及びVから選択される。
【0025】
上に開示されるポリペプチドのさらなる実施態様において、X20はR及びKから選択される。
【0026】
上に開示されるポリペプチドのさらなる実施態様において、X21はA及びGから選択される。
【0027】
上に開示されるポリペプチドのさらなる実施態様において、X25はA、R、G、K及びSから選択される。
【0028】
上に開示されるポリペプチドのさらなる実施態様において、X28はA、Q、E、F、S、W及びYから選択される。
【0029】
上に開示されるポリペプチドのさらなる実施態様において、X2はR及びKから選択される。
【0030】
上に開示されるポリペプチドのさらなる実施態様において、X3はK及びYから選択される。
【0031】
上に開示されるポリペプチドのさらなる実施態様において、X4はA、R、N、Q、H、L、K、S、T及びVから選択される。
【0032】
上に開示されるポリペプチドのさらなる実施態様において、X6はT及びYから選択される。
【0033】
上に開示されるポリペプチドのさらなる実施態様において、X7はA、G、F、Y及びVから選択される。
【0034】
上に開示されるポリペプチドのさらなる実施態様において、X17はR、Q及びVから選択される。
【0035】
上に開示されるポリペプチドのさらなる実施態様において、X18はR、Q、I、L、T、Y及びVから選択される。
【0036】
上に開示されるポリペプチドのさらなる実施態様において、X25はA、G、K及びSから選択される。
【0037】
上に開示されるポリペプチドのさらなる実施態様において、X28はA、Q、E、F、S及びWから選択される。
【0038】
上で開示されるHER3結合ポリペプチドの一実施態様において、該ポリペプチドはHER3結合モチーフ(BM)を含み、このモチーフは、
i) EX2X3X4A YX7EIW X11LPNL X16X17X18QX20 X21AFIX25 X26LX28D、
[ここで、互いに独立して、
X2はK、R及びWから選択され;
X3はK、R、及びYから選択され;
X4はA、D、E、G、H、I、K、L、M、N、Q、R、S、T、V及びWから選択され;
X7はA、E、F、G、H、K、S、T、V、W及びYから選択され;
X11はE及びQから選択され;
X16はN及びTから選択され;
X17はR、T及びVから選択され;
X18はR、T及びVから選択され;
X20はK及びRから選択され;
X21はA及びGから選択され;
X25はA、G、K、R及びSから選択され;
X26はS及びKから選択され;
X28はA、D、E、F、P、Q、S、T、W及びYから選択される];
及び
ii) i)において定義される配列に対して少なくとも93%の同一性を有するアミノ酸配列、から選択されるアミノ酸配列からなり、ここで該ポリペプチドは、HER3の細胞外ドメインに結合する。
【0039】
上に開示されるポリペプチドのさらなる実施態様において、X2X3はKY、RY、WK及びWRから選択される。
【0040】
上に開示されるポリペプチドのさらなる実施態様において、X11はQである。
【0041】
上に開示されるポリペプチドのさらなる実施態様において、X17X18はRV、RT、VR及びRRから選択される。
【0042】
上に開示されるポリペプチドのさらなる実施態様において、X20はKである。
【0043】
上に開示されるポリペプチドのさらなる実施態様において、X21はAである。
【0044】
上に開示されるポリペプチドのさらなる実施態様において、X2X3はKY及びRYから選択される。
【0045】
上に開示されるポリペプチドのさらなる実施態様において、X4はK、L、S及びTから選択される。
【0046】
上に開示されるポリペプチドのさらなる実施態様において、X7はF、G及びYから選択される。
【0047】
上に開示されるポリペプチドのさらなる実施態様において、X17X18はRV、RT及びVRから選択される。
【0048】
上に開示されるポリペプチドのさらなる実施態様において、X25はA、G及びSから選択される。
【0049】
上で開示されるポリペプチドのなおさらなる実施態様において、アミノ酸配列i)は、以下の条件I、II、III及びIV:
I) X2X3はKY及びRYから選択される;
II) X11はQである;
III) X20はKである;
IV) X21はAである、
の少なくとも1つを満たす。
【0050】
アミノ酸配列i)は、例えば4つの条件I、II、III及びIVのうち少なくとも2つ、例えば少なくとも3つ、又は全てを満たし得る。
【0051】
以下の実験項に詳細に記載されるように、HER3結合変異体の選択は、個々のHER3結合モチーフ(BM)配列の同定をもたらした。これらの配列は、この局面に従うHER3結合ポリペプチドの個々の実施態様を構成する。個々のHER3結合モチーフの配列を図1に、そして配列番号1〜334として示す。この局面のいくつかの実施態様において、BM配列i)は、配列番号1〜配列番号66のいずれか1つから選択され、例えば配列番号22〜66のいずれか1つから、例えば配列番号23〜25、配列番号27〜28、配列番号32、配列番号35〜36、配列番号40、配列番号42、配列番号44〜45、配列番号53〜54及び配列番号56から選択される。
【0052】
特定の実施態様において、HER3結合モチーフ(BM)はこのようにして3ヘリックスバンドルタンパク質ドメインの一部を形成する。例えば、BMは該3ヘリックスバンドルタンパク質ドメイン内で相互接続ループを有する2つのアルファヘリックスを本質的に構成し得る。これらの実施態様において、本発明のHER3結合ポリペプチドは:
i) K−[BM]−DPSQS XaXbLLXc EAKKL NDXdQ;
[ここで
[BM]は上で定義されたHER3結合モチーフであり;
XaはA及びSから選択され;
XbはN及びEから選択され;
XcはA、S及びCから選択され;
XdはA及びSから選択される];
及び
ii) 上で定義される配列のいずれか1つに対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含み得る。上記アミノ酸配列は、上で定義された配列のいずれか1つに対して少なくとも82%、例えば少なくとも84%、例えば少なくとも86%、例えば少なくとも88%、例えば少なくとも90%、例えば少なくとも92%、例えば少なくとも94%、例えば少なくとも96%、例えば少なくとも98%の同一性を有し得る。
【0053】
上で定義されたHER3結合ポリペプチドの一実施態様において、XaはAであり;XbはNであり;XcはAであり、そしてXdはAである。
【0054】
上で定義されたHER3結合ポリペプチドのさらなる実施態様において、XaはAであり;XbはNであり;XcはCであり、そしてXdはAである。
【0055】
上で定義されたHER3結合ポリペプチドのさらなる実施態様において、XaはSであり;XbはEであり;XcはSであり、そしてXdはSである。
【0056】
上で定義されたHER3結合ポリペプチドのさらなる実施態様において、XaはSであり;XbはEであり;XcはCであり、そしてXdはSである。
【0057】
なおさらなる実施態様において、上で定義されるHER3結合ポリペプチドのアミノ酸配列は、配列番号335〜668から選択され、特に配列番号335〜400から、例えば配列番号356〜400から、例えば配列番号357〜359、配列番号361〜362、配列番号366、配列番号369〜370、配列番号374、配列番号376、配列番号378〜379、配列番号387〜388及び配列番号390から選択される。
【0058】
特定の実施態様において、上記3ヘリックスバンドルタンパク質ドメインは細菌受容体タンパク質のドメインから選択される。このようなドメインの非限定的な例は、黄色ブドウ球菌(Staphylococcus aureus)由来のプロテインAの5つの異なる3ヘリックスドメイン、例えばドメインB、及びその誘導体である。いくつかの実施態様において、3ヘリックスバンドルタンパク質ドメインはプロテインZの変異体であり、これはブドウ球菌プロテインAの上記ドメインBから誘導される。
【0059】
従って、さらなる実施態様において:
i) YAK−[BM]−DPSQS SELLXc EAKKL NDSQA P;
[ここで[BM]は上で定義されたHER3結合モチーフであり、そして
XcはS及びCから選択される];及び
ii) i)において定義される配列のいずれか1つに対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含むHER3結合ポリペプチドが提供される。
【0060】
あるいは:
i) FNK−[BM]−DPSQS ANLLXc EAKKL NDAQA P;
[ここで[BM]は上で定義されたHER3結合モチーフであり、そして
XcはA及びCから選択される];及び
ii) 上記i)において定義された配列のいずれか1つに対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含むHER3結合ポリペプチドが提供される。
【0061】
上で考察したように、上記のアミノ酸配列と比較して小さな変化を含むポリペプチドもまた本出願の範囲内である。従って、いくつかの実施態様において、上で定義されたHER3結合ポリペプチドは、例えば、本明細書中に記載される配列に対して、少なくとも82%、少なくとも83%、少なくとも85%、少なくとも87%、少なくとも89%、少なくとも91%、少なくとも92%、少なくとも94%、少なくとも96%又は少なくとも98%同一である配列を有し得る。
【0062】
いくつかの実施態様において及び以下の実験項において開示されるように、HER3結合モチーフは58アミノ酸のポリペプチドの一部を形成し得る。このようなポリペプチドは、例えば配列番号669〜1002のいずれか1つから選択される配列、特に配列番号669〜734から選択される配列、例えば配列番号690〜734から、例えば配列番号691〜693、配列番号695〜696、配列番号700、配列番号703〜704、配列番号708、配列番号710、配列番号712〜713、配列番号721、配列番号722及び配列番号724から選択される配列を含み得る。
【0063】
本明細書に記載されるいずれかの局面に従うHER3結合ポリペプチドは、相互作用のKD値が多くとも1×10-6M、又は例えば多くとも1×10-7M、例えば多くとも1×10-8MであるようにHER3に結合し得る。本明細書において使用される用語「HER3結合」及び「HER3に対する結合親和性」は、例えばBiacore機器(GE Healthcare)におけるような表面プラズモン共鳴技術の使用により試験され得るポリペプチドの特性を指す。HER3結合親和性は、例えばHER3をBiacore機器のセンサーチップに固定し、そして試験しようとするポリペプチドを含有するサンプルをチップ上に通す実験において試験され得る。あるいは以下の実施例に記載されるように、HER3結合親和性は細胞上(on−cell)実験で試験され得る。細胞上実験は、例えば様々な濃度の標識されたHER3を試験しようとするポリペプチドを提示している細胞とともにインキュベートすることにより行われ得る。その後、細胞集団を蛍光細胞分析分離システム(FACS)におけるようなフローサイトメーターで分析し得、そして平均蛍光強度データを決定し得る。続いて平均蛍光をHER3濃度に対してプロットし、そして見かけの平衡解離定数KDを決定するために1部位結合モデルにフィッティングし得る。KD測定において使用されるHER3又はそのフラグメントは、例えば配列番号1007で表されるアミノ酸配列(HER3−ECD)を含み得る。
【0064】
以下の実施例において説明されるように、HER3結合変異体の選択は、例えばタンパク質骨格の未処理の変異体の選択のためのファージディスプレイ、続いて場合により親和性成熟HER3結合変異体の選択のための親和性成熟及び細胞ディスプレイにより達成され得る。しかし、ファージベースでも、細胞ベースでもその他でも、いずれの選択システムもHER3結合ポリペプチドの選択に使用され得ることが理解される。以下の実施例において、グラム陽性細菌スタフィロコッカス・カルノーサス(Staphylococcus carnosus)での組み換えタンパク質の細胞表面ディスプレイのための発現系を使用した(Jonsson et al、Biotechnol Applied Biochem 54:93−103(2009))。しかし、ファージ粒子の大きさを比較して、より大きなサイズの細胞は一般的にハイスループットスクリーニング及び細胞提示タンパク質ライブラリーからの選択のための蛍光細胞分析分離(FACS)の使用を可能にする。
【0065】
IgG分子と比較して、本発明のHER3結合ポリペプチドは比較的小さく、これにより、HER3結合ポリペプチドを広範囲の適用において有用にするだけでなく、固相ペプチド合成を使用した製造が可能になり得る。固相合成経路は、例えば分子画像化における放射標識に使用されるキレート剤のような非生物学的基の部位特異的結合を容易にする。
【0066】
従って、当業者には当然のことながら、様々な修飾及び/又は付加が、本発明の範囲から逸脱することなくポリペプチドを特定の適用に合わせるために、本明細書に開示されるいずれかの局面にしたがってなされ得る。例えば、いずれかの局面に従うHER3結合ポリペプチドは、さらなるC末端アミノ酸及び/又はN末端アミノ酸を含み得る。このようなポリペプチドは、ポリペプチド鎖におけるかなり最初の方及び/又は最後の方の位置に、すなわちN末端及び/又はC末端にさらなるアミノ酸残基を有するポリペプチドとして理解されるべきである。従って、HER3結合ポリペプチドは、いずれかの適切な数のさらなるアミノ酸残基、例えば少なくとも1つの付加的なアミノ酸残基を含み得る。付加的アミノ酸残基はそれぞれ、例えばポリペプチドの製造、精製、インビボ若しくはインビトロでの安定化、カップリング、又は検出を改善するために、独立して又は集合的に付加され得る。このような付加的アミノ酸残基は、化学的カップリングの目的のために付加された1つ又はそれ以上のアミノ酸残基を含み得る。この一例はシステイン残基の付加である。このような付加的アミノ酸残基はまた、ポリペプチドの精製又は検出のための「タグ」、例えばタグに対して特異的な抗体との相互作用のため、又はHis6タグの場合には固定化金属アフィニティークロマトグラフィー(IMAC)のための、His6タグ又は「myc」(c−myc)タグ又は「FLAG」タグを生じ得る。
【0067】
上で考察されたさらなるアミノ酸を、化学的連結を用いて(公知の有機化学方法を使用して)、又は融合タンパク質としてのHER3結合ポリペプチドの発現のような他の手段を用いてHER3結合ポリペプチドにカップリングさせ得る。
【0068】
上で考察されたさらなるアミノ酸は、例えば1つ又はそれ以上のポリペプチドドメインを含み得る。さらなるポリペプチドドメインは、HER3結合ポリペプチドに別の機能、例えば別の結合機能、又は酵素的機能、又は毒素機能(例えばイムノトキシン)、又は蛍光シグナル機能、又はその組み合わせを与え得る。
【0069】
いくつかの実施態様において、さらなるポリペプチドドメインは、インビボでHER3結合ポリペプチドの半減期を増加させる半減期延長部分を含み得る。当業者に理解されるように、増加した、又は延長された半減期は、特定の分子の血液からの遅延されたクリアランスを意味する。半減期延長部分は、例えば血清アルブミンへのインビボでの結合を可能にするペプチド又はタンパク質を含み得る。特に、半減期延長部分はアルブミン結合部分であり得る。アルブミン結合部分は、例えば天然に存在するポリペプチド、又はそのアルブミン結合フラグメント、又は改変ポリペプチドからなるものであってもよい。改変ポリペプチドは、新しいか又は増強された特性、例えばアルブミンのような分子に対する結合親和性を付与する目的で、それをタンパク質工学技術、例えば部位特異的又は無作為化されたアプローチでの変異及び変更を受けさせることにより、天然に存在する出発ポリペプチドから誘導され得る。
【0070】
このような改変アルブミン結合ポリペプチドは、例えばアルブミンに対するその特異的結合親和性について選択された、タンパク質骨格の変異体であってもよい。特定の実施態様において、タンパク質骨格は、連鎖球菌プロテインGのドメイン又はその誘導体、例えば連鎖球菌株G148由来のプロテインGのドメインGA1、ドメインGA2及びドメインGA3、特にドメインGA3から選択され得る。従って、HER3結合ポリペプチドの一実施態様において、さらなるアミノ酸は、連鎖球菌プロテインGのアルブミン結合ドメイン(ABD)、又はその誘導体を含む。本発明のHER3結合ポリペプチドにおけるさらなるポリペプチドドメインとして含まれ得るアルブミン結合ドメインの一例は、配列番号1005で示される。適切なアルブミン結合ドメインの他の例はWO2009/016043に開示される。このようなABD延長ポリペプチドはインビボで血清アルブミンに結合し、そしてそのより長い半減期から利益を得、これによりポリペプチド自体の正味の半減期が増加する(例えばWO91/01743を参照のこと)。
【0071】
本発明に従うHER3結合ポリペプチドが上で定義されるアルブミン結合部分を含む場合、HER3結合(bindnig)ポリペプチドの全体の大きさは比較的小さい。例えばヒト対象のような哺乳動物対象に投与された場合、HER3結合ポリペプチドのアルブミン結合部分は、血清アルブミンと非共有結合で結合し、そしてそれによりポリペプチドは減少した腎クリアランス及び上皮細胞での増加した再循環から利益が得られ得る。しかし組織浸透は、血清アルブミンの血管外漏出特性に起因して速いままである。さらに、半減期延長部分を含むHER3結合ポリペプチドは、対応する半減期延長部分の無いポリペプチドと比較して、インビボで延長された半減期を示すだけでなく、インビボで減少した免疫応答を示す(例えばWO2005/097202を参照のこと)。
【0072】
さらなるポリペプチドドメインは、同じ結合機能を有するHER3結合ポリペプチドを生じ得る。従って、さらなる実施態様において、そのアミノ酸配列は同じであっても異なっていてもよい少なくとも2つのHER3結合ポリペプチド単量体単位を含むHER3結合ポリペプチドが提供される。ポリペプチドの多量体形態は、それぞれがHER3結合モチーフを有し、かつそれぞれが多量体内の「単量体」を形成する、適切な数のドメインを含み得る。これらのドメインは全て同じアミノ酸配列を有していてもよいが、あるいはそれらが異なるアミノ酸配列を有していてもよい。特に、本発明のHER3結合ポリペプチドは、ホモ二量体又はヘテロ二量体を形成し得る。
【0073】
上記のさらなるポリペプチドドメイン(単数又は複数)は、公知の有機化学方法を使用して共有結合カップリングによりHER3結合ポリペプチドに結合され得る。あるいは、さらなるポリペプチドドメイン(単数又は複数)を含むHER3結合ポリペプチドを1つ又はそれ以上の融合ポリペプチドとして、例えばポリペプチドのの組み換え発現のための系において発現させてもよいし、直接的又はリンカー(例えばアミノ酸リンカー)を介して、いずれかの他のやり方で結合させてもよい。
【0074】
HER3結合ポリペプチドの一実施態様において、本ポリペプチドは天然に存在するHER3リガンドと同じエピトープに結合するか、又はそれに十分に接近して天然に存在するHER3リガンド(例えばニューレグリン−1及び2)のHER3への結合をブロックする。従ってHER3結合ポリペプチドは、HER3遮断薬として、かつ/又はインビボでのHER3媒介細胞シグナル伝達を阻害するために使用され得る。
【0075】
さらなる局面において、上記のHER3結合ポリペプチドをコードするポリヌクレオチドが提供される。このようなポリヌクレオチドを含む発現ベクターは、例えば宿主細胞における発現によりHER3結合ポリペプチドの生産を可能にし得る。
【0076】
HER3結合ポリペプチドは、様々な医療、獣医学及び診断適用における従来の抗体又は低分子量物質に対する代替として使用され得る。当業者は、本発明のポリペプチドが、試薬のHER3に対する親和性を必要とするいずれかの方法において有用であり得るということを理解するだろう。従って、HER3結合ポリペプチドは、このような方法における検出試薬、捕捉試薬又は分離試薬として使用され得る。さらに、HER3結合ポリペプチドは、例えば哺乳動物対象における細胞表面でのHER3発現を特徴とする疾患の存在を明らかにし、診断し、そして試験するために診断法において使用され得る。従って、HER3発現は、診断上、本発明のポリペプチドの使用によりインビボでモニタリングされ得る。診断モニタリングは、例えば分子画像化によるHER3発現の判断のための診断用核種の送達により達成され得る。本発明に従うHER3結合ポリペプチドはまた、それ自身だけで治療剤若しくは診断剤として、又はHER3タンパク質に対する直接的(例えば、イットリウム−90、ルテチウム−177及びトリウム−227のような放射性核種、マイタンシノイドのような毒素を含む毒性分子)若しくは間接的(例えばがんワクチン、免疫賦活性分子)な効果を有する他の治療剤若しくは診断剤を標的とするための手段として有用であり得る。治療効果は、受容体相互作用をブロックすることにより、あるいは腫瘍細胞へ有効なペイロード(payload)を送達するための標的としてHER3結合ポリペプチドを使用することにより、潜在的に媒介され得る。
【0077】
さらなる局面において、上で定義されたHER3結合ポリペプチド;HER2結合ポリペプチド;及びHER3結合ポリペプチドとHER2結合ポリペプチドとを連結するための連結部分を含む、HER3及びHER2に対する結合親和性を有するリガンドが提供される。このようなリガンドは、2つの異なる結合機能を含む二重特異性ヘテロ二量体の例として解釈されるべきであり:1つは上で開示されるポリペプチドによりもたらされるHER3に対する結合機能であり、そして1つはHER2結合ポリペプチドによりもたらされるHER2に対する結合機能である。当然のことながら、本発明に従うHER3結合ポリペプチドは、リガンド中に含まれ得る。2つの結合機能は、2つの別々の単量体単位としてリガンドに含まれる。
【0078】
このような本発明に従う二重特異性リガンドは、HER3及びHER2の両方が関与するがんの検出、特徴付け及び/又は診断において有用であり得る。検出、特徴付け及び/又は診断は、例えばガンマ線若しくは陽電子放出核種を使用する分子画像により、又は近赤外蛍光プローブを使用することにより達成され得る。上記のように、HER3はHER2についての好ましいヘテロ二量化パートナーであり、そしてHER2の過剰発現を特徴とする腫瘍の増殖を促進するために重要である。HER2は公知のリガンドがなく、リガンド誘導シグナル伝達に関してHERファミリーの他の受容体に依存すると現在では考えられている。しかし本発明に従う二重特異性リガンドは、診断のためだけでなく、標的化治療ペイロード適用にも使用され得る。ペイロード適用において使用される化合物としては、とりわけ、放射性核種、プロドラッグ酵素、サイトカイン、化学毒性分子、毒素及び光増感剤が挙げられる。標的化ペイロード適用については、正常組織と疾患組織との間の高度な識別が非常に重要である。
【0079】
2つの異なる受容体に結合する二重特異性リガンドに含まれる両方の結合ポリペプチドの親和性を微調整することにより、リガンドは両方の受容体を同時に発現している病変にのみ優先的に位置し、そして蓄積し得る。従って、HER2及びHER3結合機能は、それらの個々の標的に対して2つの同等に強力な結合機能を有するリガンドをもたらし得る。従って、HER3結合ポリペプチドのHER3に対する結合親和性は、HER2結合ポリペプチドのHER2に対する結合親和性とほぼ同位であり得る。一実施態様において、HER2結合ポリペプチドは、相互作用のKD値が多くとも1×10-6M、例えば多くとも1×10-7M、例えば多くとも1×10-8MであるようにHER2に結合し得る。別の実施態様において、HER2結合ポリペプチドのHER2に対する結合は、対応するHER3結合ポリペプチドのHER3に対する結合と同程度に強力であるかより強力であり得る。従って、HER2結合ポリペプチドとHER2との間の相互作用のKD値は、多くてHER3結合ポリペプチドとHER3との間の相互作用のKD値である。HER3結合ポリペプチドとHER3との間の相互作用のKD値は、HER2結合ポリペプチドとHER2との間の相互作用のKD値の例えば5倍、10倍又は100倍であり得る。
【0080】
一実施態様において、二重特異性リガンドのHER2結合ポリペプチドは改変ポリペプチドである。HER2結合ポリペプチドは、HER2に対するその特異的結合親和性について選択された、タンパク質骨格の変異体である改変ポリペプチドであり得る。タンパク質骨格の非限定的な例は、ブドウ球菌プロテインA、並びにそのドメイン及び誘導体からなる群より選択される。いくつかの実施態様において、HER2結合ポリペプチドは、黄色ブドウ球菌(Staphylococcus aureus)由来のプロテインAのドメインBから誘導された改変タンパク質である。この改変タンパク質は、特にプロテインZ誘導体を含んでいてもよく、これは上記プロテインAのドメインBから誘導されたプロテインZの変異体である。
【0081】
リガンドのいくつかの実施態様において、改変HER2結合タンパク質は、
i) 配列番号1003に対応する、
YAKEM RNAYW EIALL PNLTN QQKRA FIRKL YDDPS QSSEL LSEAK KLNDS Q;及び
ii) i)において定義される配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む。
【0082】
例えば1から45個までのアミノ酸を含むペプチドリンカーによりHER3結合ポリペプチド及びHER2結合ポリペプチドが連結され得る。連結部分は特にHER3結合ポリペプチドとHER2結合ポリペプチドのN末端及び/又はC末端のカップリングのために配置され得る。このようにして生じる連結は、C末端とC末端との連結、C末端とN末端との連結又はN末端とC末端の連結から選択され得る。連結部分のHER3結合ポリペプチド及びHER2結合ポリペプチドへのカップリングは、例えば化学的結合(公知の有機化学方法を使用する)を用いて、又は融合タンパク質としてのリガンドの発現のようないずれかの他の手段により達成され得る。
【0083】
リガンドは、上記のように、インビボでのリガンドの半減期を延長するための半減期延長部分をさらに含み得る。半減期の延長は、多数の種々の半減期延長部分により達成され得、そしてこのような部分の非限定的な例は、アルブミン、HER3結合ポリペプチドに関連して上で定義されたようなアルブミン結合部分、IgGからのFc部分、及びポリエチレングリコール(PEG)のようなポリマー化合物から選択される。哺乳動物血清において最も豊富なタンパク質である血清アルブミンへのリガンドのカップリングは、インビボでのリガンドの半減期の延長をもたらし得る。
【0084】
特に、アルブミン結合部分は、
i) 配列番号1005で示される、
LAEAK VLANR ELDKY GVSDF YKRLI NKAKT VEGVE ALKLH ILAAL P;及び
ii) i)において定義された配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含むアルブミン結合部分のような、連鎖球菌プロテインGのドメインGA3から誘導された改変タンパク質であり得る。
【0085】
半減期延長部分は、例えば公知の有機化学方法を使用して共有結合カップリングによりリガンドの残部に接続されても、他のやり方で直接、又は多数のアミノ酸を含むリンカーを介して結合されてもよい。半減期延長部分がアルブミン又はアルブミン結合部分により表される場合、リガンドはそれ自体1つ又はそれ以上の融合ポリペプチドとしてポリペプチドの組み換え発現のための系において発現されてもよい。一実施態様において、半減期延長部分は、リガンドに存在するシステイン残基を介してリガンドの残部に結合する。特に、半減期延長部分は、結合モチーフから離れた部位に導入された、HER3又はHER2結合ポリペプチドのシステイン残基に結合され得る。別の実施態様において、半減期延長部分は連結部分を介してリガンドの残部に結合する。
【0086】
特定の実施態様において、連結部分は上で定義された半減期延長部分を含む。従って、アルブミン、アルブミン結合部分又はPEGは例えばHER3結合ポリペプチドをHER2結合ポリペプチドに接続するために使用され得る。
【0087】
いくつかの実施態様において、本発明に従うリガンドは、さらなるC末端又はN末端アミノ酸を含み得る。このようなさらなるアミノ酸は、HER3結合ポリペプチドに関して上で記載したように、例えばポリペプチドの生産、精製、インビボ若しくはインビトロでの安定化、カップリング又は検出を改善するために、個別に、又は集合的に付加され得る。特定の実施態様において、さらなるアミノ酸は1つ又はそれ以上のポリペプチドドメインを構成し得る。このようなさらなるポリペプチドドメインの機能は、既にリガンドに含まれている機能と同じであっても異なっていてもよい。さらなるポリペプチドドメインは、例えば別のHER3結合機能及び/又は別のHER2結合機能を有するリガンドを生じ得る。従って、本発明に従う二重特異性リガンドは、HER3結合機能の多量体及び/又はHER2結合機能の多量体を含み得る。多量体結合機能を含む二重特異性リガンドの一例は、2つのHER3結合ポリペプチド及び2つのHER2結合ポリペプチドを含むリガンドである。
【0088】
上で定義されたリガンドは、一実施態様において、インビボでのHER2及びHER3の二量化を防止し得る。より詳細には、リガンドは、例えば哺乳動物対象の細胞表面で発現されたHER2及び/又はHER3に結合することにより、対象におけるHER3仲介シグナル伝達を阻害し得る。リガンドの二重特異性に起因して、これは、それらがもはやシグナル伝達を行うことができないような位置において2つの受容体に結合してブロックし得る。当業者には当然のことながら、従って本発明のリガンドは、細胞表面上のHER2及び/又はHER3の発現、特に過剰発現に関連する状態の処置のために利用され得る。
【0089】
しかしHER3はEGFRともヘテロ二量体を形成し得、それ故PI3K/Akt経路を活性化する活性シグナル伝達単位を形成する(Engelman et al、Proc Natl Acad Sci 102(10):3788−93(2005))。実際、HER3は、明らかにErbBリガンドの上方調節のために非小細胞肺がん(NSCLC)において過剰発現されるが(Fujimoto et al、Cancer Res 65(24):11478−85(2005))、一方でHER3は正常肺上皮で発現されない。従って、関連する局面において、上で定義されたHER3結合ポリペプチド;EGFR結合ポリペプチド;及びHER3結合ポリペプチドとEGFR結合ポリペプチドとを連結するための連結部分を含む、HER3及びEGFRに対する結合親和性を有するリガンドが提供される。当然のことながら、本発明の上記の局面において記載される特徴は、本発明のこの局面にも適用可能である。特に、このような二重特異性リガンドは、HER3及びEGFRを標的にし得、そしてHER3及びHER2に対する二重特異性リガンド結合に関して上で記載した作用様式と類似しているか又は同じ作用様式で作用する。HER3及びEGFRに対する二重特異性リガンド結合の例は、
i) 配列番号1004に相当する、
YAKEM WIAWE EIRNL PNLNG WQMTA FIAKL LDDPS QSSEL LSEAK KLNDS Q;及び
ii) i)において定義される配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるEGFR結合ポリペプチドを含み得る。
【0090】
HER3及びEGFR結合リガンドにおいて使用され得るEGFR結合ポリペプチドの他の例はWO2007/065635に開示される。
【0091】
他の実施態様において、本発明に従うHER3結合ポリペプチド又は二重特異性リガンドは、治療剤又は診断剤に、インビボ及びインビトロの両方で、HER3を発現する細胞、特にHER3を過剰発現する細胞を標的にさせる際に使用される。上で考察したように、本発明のリガンドは、HER3及び/若しくはHER2、又はHER3及び/若しくはEGFRを発現するか又は過剰発現する細胞を標的にする際に利用され得る。従って関連する局面において、本明細書において定義されるHER3結合ポリペプチド、又は上で定義されるリガンド、及び治療剤の組み合わせが提供される。治療剤の非限定的な例は、細胞傷害性ペイロード、例えば放射性核種、低分子量薬物又は毒素から選択される。このような治療剤は、化学的に結合されるか、そうでなければHER3結合ポリペプチド又はリガンドに結合される。
【0092】
本発明に従う組み合わせ、HER3結合ポリペプチド又はリガンドはさらに、治療における、例えばHER3関連状態、例えばがん疾患、例えば結腸がん、子宮内膜がん、胃がん、神経膠腫、乳がん、膵臓がん、頭頸部扁平上皮がん、肺がん、黒色種、髄芽腫、神経上皮腫、卵巣がん、パジェット病、乳頭様甲状腺がん、前立腺がん、皮膚扁平上皮がん、移行上皮がん及び前庭神経鞘腫から選択される、哺乳動物(例えばヒト対象)におけるHER3関連状態の処置のための、医薬として使用され得る。従って、HER3関連状態、例えば上述のHER3関連状態の処置のための医薬の製造のための本発明のHER3結合ポリペプチド又はリガンドの使用もまた、本発明の範囲内である。
【0093】
関連する局面において、HER3結合ポリペプチド、又は上記のリガンド若しくは組み合わせを、それを必要とする哺乳動物に投与することを含む、HER3関連状態の処置方法が提供される。結果として、本処置方法において、対象は、本発明に従うHER3結合ポリペプチド、リガンド又は組み合わせで処置される。この方法のより特定の実施態様において、HER3結合ポリペプチド、リガンド又は組み合わせの、対象の細胞表面で発現されたHER3に対する結合は、受容体活性化を阻害する。この結合は、例えばHER3仲介シグナル伝達を阻害し得る。本処置方法の一実施態様において、上記HER3関連状態は、がん疾患、例えば結腸がん、子宮内膜がん、胃がん、神経膠腫、乳がん、膵臓がん、頭頸部扁平上皮がん、肺がん、黒色種、髄芽腫、神経上皮腫、卵巣がん、パジェット病、乳頭様甲状腺がん、前立腺がん、皮膚扁平上皮がん、移行上皮がん及び前庭神経鞘腫から選択される。
【0094】
別の実施態様において、本発明のリガンドを、それを必要とする哺乳動物対象に投与することを含む、HER3及び/又はHER2関連状態の処置方法が提供される。
【0095】
当業者に理解されるように、そして上で考察したように、本発明に従うHER3結合ポリペプチド又はリガンドは、HER3又はHER3及び/若しくはHER2関連状態の診断のような診断において使用され得る。本発明に従うHER3結合ポリペプチドは、診断剤と別々に使用されても組み合わせて使用されてもよい。このような組み合わせは、HER3関連状態の診断において、例えばHER3を過剰発現する細胞の分子画像化において使用され得る。
【0096】
本発明はここで以下の非限定的な実施例によりさらに説明される。
【図面の簡単な説明】
【0097】
【図1−1】本発明のHER3結合ポリペプチドに含まれるHER3結合モチーフの例(配列番号1〜334)、本発明に従う49量体HER3結合ポリペプチドの例(配列番号335〜668)及び本発明に従う58量体HER3結合ポリペプチドの例(配列番号669〜1002)、本発明に従うリガンドに含まれるHER2結合ポリペプチドの例(配列番号1003)、本発明のリガンドの一実施態様において含まれ得るようなアルブミン結合部分の例(配列番号1005)、本発明に従うリガンドの一実施態様において含まれ得るEGFR結合ポリペプチドの例(配列番号1004)のアミノ酸配列表、さらにはプロテインZの配列(配列番号1006)、並びにHER3の細胞外ドメイン(配列番号1007)、HER2の細胞外ドメイン(配列番号1008)及びHER4の細胞外ドメイン(配列番号1009)の配列の表である。
【図1−2】図1−1の続きである。
【図1−3】図1−2の続きである。
【図1−4】図1−3の続きである。
【図1−5】図1−4の続きである。
【図1−6】図1−5の続きである。
【図1−7】図1−6の続きである。
【図1−8】図1−7の続きである。
【図1−9】図1−8の続きである。
【図1−10】図1−9の続きである。
【図1−11】図1−10の続きである。
【図1−12】図1−11の続きである。
【図1−13】図1−12の続きである。
【図1−14】図1−13の続きである。
【図1−15】図1−14の続きである。
【図1−16】図1−15の続きである。
【図1−17】図1−16の続きである。
【図1−18】図1−17の続きである。
【図1−19】図1−18の続きである。
【図1−20】図1−19の続きである。
【図1−21】図1−20の続きである。
【図1−22】図1−21の続きである。
【図1−23】図1−22の続きである。
【図1−24】図1−23の続きである。
【図1−25】図1−24の続きである。
【図1−26】図1−25の続きである。
【図1−27】図1−26の続きである。
【図1−28】図1−27の続きである。
【図1−29】図1−28の続きである。
【図1−30】図1−29の続きである。
【図1−31】図1−30の続きである。
【図1−32】図1−31の続きである。
【図1−33】図1−32の続きである。
【図1−34】図1−33の続きである。
【図1−35】図1−35の続きである。
【図1−36】図1−35の続きである。
【図1−37】図1−36の続きである。
【図1−38】図1−37の続きである。
【図1−39】図1−38の続きである。
【図1−40】図1−39の続きである。
【図1−41】図1−40の続きである。
【図1−42】図1−41の続きである。
【図2】HSA結合ドメインABDへの融合として発現された個々のHER3結合Z変異体からの周辺質フラクションを使用して行われたドットブロットアッセイの結果を示す。21のコントロールタンパク質に対する潜在的バックグラウンド結合、さらにはHER3−ECD及びHER3−Fcへの結合を分析した。タンパク質をメンブレン上に3列で以下の順序にてドットを打った; 列1: HSA、IgG、IgM、IgA、アルファ−2マクログロブリン、フィブリノゲン、ホロ−トランスフェリン、アルファ−1−抗トリプシン、 列2:C3、ハプトグロブリン(haptoglobulin)、アルファ−1酸性糖タンパク質、アルファ−1抗キモトリプシン、C4、マウスIgEカッパ、ヘモペキシン、トランスサイレチン、 列3:ストレプトアビジン、ニュートラアビジン、HER2−ECD、HER3−Fc、HER4−Fc、HER3−ECD及びHER4−ECD。 画像は、A) Z01748、B) Z01813、C) Z01814、D) Z01815、E) Z02009、F) Z01821、G) Z02010、H) Z01824、I) Z01825、J) Z01817、K) Z01818、L) Z01820、M) Z01826、N) Z01828、O) Z02011、P) Z01830、及びQ)ネガティブコントロールとしての、ABDのみを発現するpAY01247からの生成物を示す。
【図3】実施例2に記載される免疫蛍光実験からの画像を示す。HER3ポジティブAU566細胞(A〜D)並びにHER2ポジティブ及びHER3ネガティブのSKOV−3細胞(E〜F)をHER3特異的Z変異体又は以下に従うコントロールで染色した:A) His6−(Z01814)2−cys、B) His6−(Z01820)2−cys、C) 抗HER3抗体(ポジティブコントロール)、D)(Z01154)2(ネガティブコントロールZ変異体)、E) His6−(Z01814)2−cys、F) His6−(Z01820)2−cys。
【図4】S.カルノーサス(S.carnosus)(B)の表面での本発明のHER3結合ポリペプチドの細胞ディスプレイのために使用したブドウ球菌ディスプレイベクターの略図である。ベクターpSCZ1(A)は:i)リパーゼ遺伝子(関連するスタフィロコッカス・ヒイカス(Staphylococcus hyicus)の表面で発現される)からのプロモーター及び分泌シグナル(S)、ii) 同じリパーゼ遺伝子からのプロペプチド(PP)をコードする遺伝子フラグメント、(組み換えタンパク質の効率的な転座に重要であることが実証されている)、iii)細胞壁固定領域(XM)、(ブドウ球菌プロテインA由来)、iv)標的結合シグナルの表面発現レベル正規化をもたらすために導入されるブドウ球菌プロテインG由来のアルブミン結合タンパク質(ABP、Samuelson et al、J Bacteriology 177(6):1470−1476、(1995))、v) ブドウ球菌における安定した複製及び発現のためのブドウ球菌複製起点及びクロラムフェニコールアセチルトランスフェラーゼ遺伝子(Cmlr)、並びにvi)E.coliでのサブクローニング作業を容易にする、E.coliのための複製起点(OriE)さらにはβ−ラクタマーゼ遺伝子(Bla)を含む。
【図5】Sc:ZHER3LIBのフローサイトメトリー分離からの結果として密度プロットを示す。これらのプロットは、x軸に表面発現レベルに対応するFL−4チャネル蛍光強度(HSA結合によりモニタリング)、及びy軸にHER3結合に対応するFL−1チャネル蛍光を示す。密度プロットは、各プロットにおいて四角で輪郭を描いたゲーティングにおいて使用された領域と共に、それぞれ1、2、3及び4回目のフローサイトメトリー分離前のブドウ球菌ライブラリーを示す。
【図6−1】細胞ベースのフローサイトメトリーアッセイのヒストグラム図を示し、ここではアルブミン結合タンパク質(ABP)への融合でZ変異体を提示するFACSで単離した細胞を、蛍光標識したHER3−Fc及び蛍光標識したHSAと共にインキュベートした。HER3結合シグナル(MFI、FL−1)とHSA結合シグナル(MFI、FL−4)との比率がy軸にある。全てのFL−1/FL−4比は;A) Z05405のFL−1/FL−4比、及びB〜D) 第一世代バインダー(binder)(Z01753)のFL−1/FL−4比を用いて正規化される。45の個々のクローンを4つの別々のアッセイ(A〜D)において分析し、ここでZ05405及びZ05409を比較のために全てのアッセイに含めた。A)はZ05403〜Z05415の細胞ベースのフローサイトメトリーアッセイ結果を示し、B)はZ05416〜Z05426細胞ベースのフローサイトメトリーアッセイ結果を示し、C)はZ05427〜Z05434の細胞ベースのフローサイトメトリーアッセイ結果を示し、そしてD)はZ05435〜Z05447の細胞ベースのフローサイトメトリーアッセイ結果を示す。
【図6−2】細胞ベースのフローサイトメトリーアッセイのヒストグラム図を示し、ここではアルブミン結合タンパク質(ABP)への融合でZ変異体を提示するFACSで単離した細胞を、蛍光標識したHER3−Fc及び蛍光標識したHSAと共にインキュベートした。HER3結合シグナル(MFI、FL−1)とHSA結合シグナル(MFI、FL−4)との比率がy軸にある。全てのFL−1/FL−4比は;A) Z05405のFL−1/FL−4比、及びB〜D) 第一世代バインダー(binder)(Z01753)のFL−1/FL−4比を用いて正規化される。45の個々のクローンを4つの別々のアッセイ(A〜D)において分析し、ここでZ05405及びZ05409を比較のために全てのアッセイに含めた。A)はZ05403〜Z05415の細胞ベースのフローサイトメトリーアッセイ結果を示し、B)はZ05416〜Z05426細胞ベースのフローサイトメトリーアッセイ結果を示し、C)はZ05427〜Z05434の細胞ベースのフローサイトメトリーアッセイ結果を示し、そしてD)はZ05435〜Z05447の細胞ベースのフローサイトメトリーアッセイ結果を示す。
【図7A】4つの異なる濃度での4つの異なるZ変異体の表面プラズモン共鳴(SPR)動力学的分析からのセンサーグラムを示す。各センサーグラムにおける一番上の曲線は最も高い濃度の注入されたZ変異体に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は3番目に高い濃度に対応するなどである。46、15、1.5及び1.5nMの濃度のZ変異体Z05405(A)、57、19、6.3及び1.9nMの濃度のZ05413(B)、67、22、7.5及び2.2nMの濃度のZ05416(C)、並びに59、20、6.5及び2nMの濃度のZ05417(D)を、ヒトHER3−Fcを固定した表面上に注入し、そしてkon及びkoffに基づいて解離定数を決定した。
【図7B】4つの異なる濃度での4つの異なるZ変異体の表面プラズモン共鳴(SPR)動力学的分析からのセンサーグラムを示す。各センサーグラムにおける一番上の曲線は最も高い濃度の注入されたZ変異体に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は3番目に高い濃度に対応するなどである。46、15、1.5及び1.5nMの濃度のZ変異体Z05405(A)、57、19、6.3及び1.9nMの濃度のZ05413(B)、67、22、7.5及び2.2nMの濃度のZ05416(C)、並びに59、20、6.5及び2nMの濃度のZ05417(D)を、ヒトHER3−Fcを固定した表面上に注入し、そしてkon及びkoffに基づいて解離定数を決定した。
【図7C】4つの異なる濃度での4つの異なるZ変異体の表面プラズモン共鳴(SPR)動力学的分析からのセンサーグラムを示す。各センサーグラムにおける一番上の曲線は最も高い濃度の注入されたZ変異体に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は3番目に高い濃度に対応するなどである。46、15、1.5及び1.5nMの濃度のZ変異体Z05405(A)、57、19、6.3及び1.9nMの濃度のZ05413(B)、67、22、7.5及び2.2nMの濃度のZ05416(C)、並びに59、20、6.5及び2nMの濃度のZ05417(D)を、ヒトHER3−Fcを固定した表面上に注入し、そしてkon及びkoffに基づいて解離定数を決定した。
【図7D】4つの異なる濃度での4つの異なるZ変異体の表面プラズモン共鳴(SPR)動力学的分析からのセンサーグラムを示す。各センサーグラムにおける一番上の曲線は最も高い濃度の注入されたZ変異体に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は3番目に高い濃度に対応するなどである。46、15、1.5及び1.5nMの濃度のZ変異体Z05405(A)、57、19、6.3及び1.9nMの濃度のZ05413(B)、67、22、7.5及び2.2nMの濃度のZ05416(C)、並びに59、20、6.5及び2nMの濃度のZ05417(D)を、ヒトHER3−Fcを固定した表面上に注入し、そしてkon及びkoffに基づいて解離定数を決定した。
【図8A】Z05417のヘレグリンとのSPR競合分析からのセンサーグラムを示す。A)はヒトヘレグリンを固定した表面上での、単独及び40倍モル濃度過剰のZ05417と共にプレインキュベートした、HER3−Fcの注入から得られたセンサーグラムを示す。B)は、ヒトHER3−Fcを固定化した表面上でのヘレグリンとのZ05417の同時注入から得られたセンサーグラムを示す。Z05417を、5つの異なる濃度、10nM、5nM、1nM、0.5nM及び0nMで二回注入した。一番上の曲線は最も高い濃度の注入されたZ05417に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は三番目に高い濃度に対応するなどである。
【図8B】Z05417のヘレグリンとのSPR競合分析からのセンサーグラムを示す。A)はヒトヘレグリンを固定した表面上での、単独及び40倍モル濃度過剰のZ05417と共にプレインキュベートした、HER3−Fcの注入から得られたセンサーグラムを示す。B)は、ヒトHER3−Fcを固定化した表面上でのヘレグリンとのZ05417の同時注入から得られたセンサーグラムを示す。Z05417を、5つの異なる濃度、10nM、5nM、1nM、0.5nM及び0nMで二回注入した。一番上の曲線は最も高い濃度の注入されたZ05417に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は三番目に高い濃度に対応するなどである。
【図9】HER3結合Z変異体の存在しない場合及び存在下でのHER3リン酸化のパーセントに関する図表を示す。MCF−7細胞を、Z変異体の存在しない場合(100%リン酸化、塗りつぶしたバー)、並びにHER3結合Z変異体Z05416及びZ05417の存在下で、並びにネガティブコントロールZ01155の存在下で(taqポリメラーゼ結合Z変異体、縞模様のバー)又は培地のみを用いて(白いバー)、5nMヘレグリンで刺激した。この図表は、ヘレグリン誘導HER3リン酸化(100%)と比較した、Z変異体又は培地の存在下でのHER3リン酸化のパーセントを示す。
【実施例】
【0098】
実施例1:HER3結合ポリペプチドの選択及びスクリーニング
材料及び方法
HER3及びHSAの標識: 組み換えヒトHER3/Fcキメラ(R&D Systems、#348−RB−050)(本明細書ではHER3−Fcと示される)のビオチン化を、Biotin−XXマイクロスケールタンパク質標識キット(Invitrogen、#B30010)を使用して供給業者の推奨にしたがって行った。HER3の細胞外ドメイン(配列番号1007)(本明細書ではHER3−ECDと示される)を、EZ−LinkTM−Sulfo−NHS−LC−LC−Biotin(Pierce、#21338)を使用して供給業者の推奨にしたがってビオチン化した。ヒト血清アルブミン(HSA; Sigma、#A−3782)を、Alexa Fluor(R) 647スクシンイミジルエステル(Invitrogen、#A20006)を使用して供給業者の推奨にしたがって蛍光標識した。タンパク質緩衝液をPBS(10mMホスフェート、137mM NaCl、2.68mM KCl、pH7.4)又は過剰のビオチン若しくはフルオロフォアを除去するために0.1%Pluronic(R)F108 NF界面活性剤(PBSP; BASF Corporation、#30085231)を補充したPBSに変更した。
HER3結合ポリペプチドのファージディスプレイ選択: Groenwall et al、J Biotechnol 128:162−183(2007)に記載されるように、ファージミドpAffi1/pAY00065において構築されたバクテリオファージで提示されるZ分子のランダム変異体のライブラリー(Zlib2002と示される)を、HER3−ECD−結合ポリペプチドの選択のために使用した。ファージミドライブラリーから、さらには選択ラウンドの間に、ファージストックの製造を以前に記載された手順(Nord et al、Nat Biotech 15:772−777(1997); Hansson et al、Immunotechnology 4:237−252(1999))にしたがってヘルパーファージM13K07(New England Biolabs、#NO315S)を使用して行った。大腸菌(Escherichia coli)アンバー抑圧遺伝子系統RR1ΔM15(Ruether、Nucleic Acids Res 10:5765−5772、1982)を全てのクローニング手順のために、そしてファージ産生のための宿主として使用した。
【0099】
ファージ選択を、溶液でビオチン化HER3タンパク質及びストレプトアビジン被覆常磁性ビーズ(Dynabeads(R) M−280ストレプトアビジン、Dynal #112.06)を、結合したファージの捕捉のために使用して行った。さらに、ファージ選択を、固相においてMaxiSorp immunotubes(Nunc)に固定化されたHER3−ECDを使用して行った。ファージ粒子の非特異的結合を、全てのチューブ及びビーズをPBST 0.1−ゼラチン(0.1%Tween−20及び0.1%ゼラチンを補充したPBS)で予備処理することにより最少にした。選択を4サイクルで行った。全ての選択をPBST 0.1−ゼラチン中でかつ体積1mlで行った。
【0100】
溶液で行われた選択については、4サイクルの選択を室温(RT)にて異なる濃度のビオチン化組み換えHER3−ECDを標的タンパク質として使用して行った。サイクル1及び2で使用したファージストックを、ストレプトアビジン結合ファージを除去するために0.1mgストレプトアビジン被覆ビーズとともに1時間プレインキュベートした。サイクル1において、Zlib2002ライブラリーからのファージ粒子を、100nMのビオチン化HER3−ECDと共にPBST 0.1−ゼラチン中で2時間インキュベートした。それに続く選択サイクルを2つの選択トラックに分割し、そのために標的濃度をサイクル2について50(トラック1)又は20nM(トラック2)まで、サイクル3について20(トラック1)又は10nM(トラック2)まで、そしてサイクル4について20(トラック1)又は5nM(トラック2)まで下げた。結合したファージをストレプトアビジン被覆M−280 Dynabeads(R)を用いて捕捉し、ビーズ1ミリグラムあたりHER3−ECD4μgを固定化させた。
【0101】
固相での選択については、immunotubeに1ml HER3−ECD(50mM炭酸ナトリウム(sodium carbonat)緩衝液中10μg/ml、pH 9.6)を1.5時間室温にて固定化させ、その後3ml PBST 0.1−ゼラチンを用いて1時間室温にてブロックした。ファージストックをこのチューブに加え、そして2時間インキュベートした。
【0102】
両方の選択ストラテジーのために、PBST 0.1−ゼラチンでの洗浄回数を、選択ストリンジェンシーを高めるためにラウンド間で増やした。従って、洗浄をサイクル1において2回、サイクル2において3回、サイクル3において6回、そしてサイクル4において12回行った。結合したファージ粒子を0.1Mグリシン−HCl(pH2.2)で溶出し、続いてすぐに1M Tris−HCl(pH8)で中和した。ファージ含有溶出液を対数期RR1ΔM15細胞に感染させるために使用し、そしてファージミド粒子を、ヘルパーファージM13K07を使用して感染した細胞からレスキューした。選択過程を、各選択ラウンドの前及び標的タンパク質からの溶出後にファージストックの滴定によりモニタリングした。ファージ溶液の段階希釈を、対数期RR1ΔM15細胞の感染のために使用した。
【0103】
Z変異体のELISAスクリーニング: Z変異体分子が実際にHER3と相互作用するかどうかを試験するために、2つの異なるELISAを行い、一方は標的タンパク質としてビオチン化HER3−ECDを使用し、そして2つ目はHER3−Fcを使用した。上で記載したように製造された単一コロニーを、100μg/mlアンピシリン及び0.1mMイソプロピル−β−D−1−チオガラクトピラノシド(IPTG)を補充した1ml TSB−YE培地(30g/l TSB、5g/l酵母抽出物)中でディープウェルプレート(Nunc #278752)において播種することによりZ変異体を生成した。ネガティブコントロールとして、インスリン結合Z変異体(Z00801)を播種し、そして各プレートにおいて増殖させた。これらのプレートを18〜24時間37℃でインキュベートした。インキュベーション後に、−20℃での貯蔵のために15%グリセロールを含む96ウェルプレートに各培養物を少量移すことにより複製のプレートを作製した。残りの細胞を遠心分離によりペレット化し、300μl PBST 0.05(0.05% Tween 20を補充したPBS)中に再懸濁し、そして細胞の周辺質部分を放出させるために−80℃で凍結した。凍結したサンプルを水浴で解凍し、そして細胞を遠心分離によりペレット化した。周辺質上清は、連鎖球菌株G148由来のプロテインGのGA3からのアルブミン結合ドメインへの融合物としてのZ変異体(AQHDEALE−[Z#####]−VDYV−[ABD]−YVPGとして表される)を含有していた(Groenwall et al、前出)。Z#####は個々のZ変異体を指す。
【0104】
ハーフエリア96ウェルELISAプレート(Costar #3690)を、6μg/mlヒト血清アルブミン(HSA、Sigma #A3782)を含有する50μl/ウェルのコーティング緩衝液(50mM炭酸ナトリウム、pH9.6)で被覆し、そして終夜(ON)インキュベートした。HSA溶液を捨てて、2%脱脂粉乳溶液(Semper AB)を補充したPBST 0.1 100μlを用いてウェルを1時間室温にてブロックした。ブロッキング溶液を廃棄し、そして周辺質溶液50μlを各ウェルに加え、そして1.5時間室温でゆっくりと震盪しながらインキュベートした。上清を捨てて、ウェルをPBST 0.05で4回洗浄した。PBST 0.05中0.5μg/mlの濃度のHER3−Fc 50μl又はPBS中1μg/mlの濃度のビオチン化HER3−ECDを各ウェルに加えた。プレートを1.5時間室温でインキュベートし、続いてPBST 0.05で4回洗浄した。HER3−Fcプレートにおいて、西洋ワサビペルオキシダーゼ(HRP)で標識されPBST0.05で1:4000に希釈されたヒトFcに対する抗体(DAKO Cytomation、#P0214)をウェルに加え、そして1時間室温でインキュベートした。ビオチン化HER3−ECDを含むプレートにおいて、HRPと結合したストレプトアビジン(DAKO、#P0397)を各ウェルに加えてPBST 0.05で1:5000に希釈した。上で記載したように洗浄した後、50μl ImmunoPure TMB基質(Pierce #34021)をウェルに加え、そしてプレートを製造者の推奨に従って処理した。ブロッキングから読み取りまでの全ての工程を、Tecan Genesis Freedom 200ロボット(Tecan Group LTD)で行った。ウェルの吸光度を450nmでELISAリーダーTecan Ultra 384(Tecan)で読み取り、そしてMagellan v.5.0ソフトウェア(Tecan)を用いて評価した。
【0105】
配列決定: ELISAスクリーニングに基づいて、陽性と考えられる全てのクローンの一部を配列決定のために選別した。PCRフラグメントを、標準的なPCRプログラム並びにプライマーAFFI−21(5’−tgcttccggctcgtatgttgtgtg)及びAFFI−22(5'−cggaaccagagccaccaccgg)を使用して単一コロニーから増幅した。増幅されたフラグメントの配列決定を、製造者のプロトコルに従って使用されるビオチン化オリゴヌクレオチドAFFI−72(5'−ビオチン−cggaaccagagccaccaccgg)及びBigDye(R)ターミネーターv3.1サイクルシークエンスキット(Applied Biosystems)を使用して行った。配列決定反応混合物を、磁気ストレプトアビジン被覆ビーズへの結合により、Magnatrix 8000(Magnetic Biosolution)を使用して精製し、そしてABI PRISM(R)3100 Genetic Analyzer(PE Applied Biosystems)で分析した。配列決定結果をインポートし、そしてALD LIMS NautilusTM 2003 R2 B3ソフトウェア(Thermo Electronics Corp.)を用いて分析した。
【0106】
ドットブロット分析: 生成されたHER3結合Z変異体を、特異性についてドットブロット分析によりアルファ−2マクログロブリン(MP biomedicals/Cappel、#55833)、アルファ−1酸性糖タンパク質(RDI、#RDI−SCP−153−1)、アルファ−1抗キモトリプシン(RDI、#RDI−SCP−159−0)、アルファ−1−抗トリプシン(RDI、#RDI−SCP−165−5)、補体C3(RDI、#RDI−SCP−150−0)、補体C4(RDI、#RDI−SCP−151−0)、フィブリノゲン(Enzyme research、#031015E)、ハプトグロブリン(RDI、#RDI−SCP−119−1)、ヘモペキシン(Agilent)、ホロ−トランスフェリン(Sigma、#T0665)、ヒトIgA(Bethyl、#P80−102)、マウスIgEカッパ(BD、#55079)、ヒトIgG(Sigma、#G4386)、ヒトIgM(Sigma、#I8260)、ヒト血清アルブミン(HSA、Sigma、#A3782)、ニュートラアビジン(Pierce、#31000)、ストレプトアビジン(Pierce、#21122)、トランスサイレチン(Sigma、#P1742)、HER2−ECD(配列番号1008)、HER3−ECD(配列番号1007)、HER3−Fc(R&D Systems)、HER4−ECD(配列番号1009)、HER4−Fc(R&D Systems)に対して試験した。
【0107】
可溶性タンパク質を含有する周辺質上清を得るために、Z変異体をELISAスクリーニングで記載された方法と本質的に同じ方法で製造した。上清を0.45μmメンブレンを使用してろ過した。
【0108】
ニトロセルロースメンブレン(Invitrogen)に各タンパク質1μlを、それぞれ0.29mg/ml及び0.14mg/mlの濃度を有していたHER3−ECD及びHER4−ECDを除いて0.25mg/mlの濃度で用いてドットを打った。これらのメンブレンを0.5%カゼイン(ブロッキング液)を補充したPBST0.1中で終夜4℃でブロックした。溶液を除去した後、これらのメンブレンを1時間、0.5%カゼインを補充した周辺質上清と共にインキュベートした。これらのメンブレンをPBST 0.1中で非常に短く3回、そして4×5分洗浄した。Z変異体を、PBSTブロッキング液中で1:5000に希釈された全てのZ変異体(社内で製造)に共通のエピトープに対するABD融合超免疫ポリクローナルウサギIgを用いて1時間室温でインキュベートすることにより検出した。上記のように洗浄した後、ブロッキング溶液で1:5000希釈された、HRPと複合体化されたウサギIgGに対する抗体(DAKO Cytomation、#P0448)をメンブレンに加え、続いてこれらのメンブレンを1時間室温でインキュベートした。メンブレンを洗浄し、PBSですすぎ、そしてSupersignal(Pierce #34075)と共に浸漬した。発光をChemiImager 5500(Alpha Innotech Corp.)を用いて撮影した。
【0109】
Z変異体のサブクローニング: 二量体Z変異体をpAffi1/pAY00065ベクターから増幅した。異なるプライマー対を使用してPCRを行い、そして得られた遺伝子フラグメントを精製し、そしてリガーゼ緩衝液中でハイブリダイズさせた。ハイブリダイズした遺伝子フラグメントをpAY00430ベクターでサブクローニングし、N末端His6タグ及びC末端システインを生じた(His6−(Z#####)2−Cys)。HER3結合Z変異体をダイマーとしてサブクローニングし、そして発現ベクターによりコードされる構築物は、MGSSHHHHHHLQ−[Z#####][Z#####]−VDCであった。3つの部分のライゲーションを、両方の挿入フラグメントをベクター中に同じ工程で挿入するために使用した。ハイブリダイズした遺伝子フラグメント及びAccI−消化して脱リン酸化した発現ベクターをリガーゼ緩衝液中で連結し、エレクトロコンピテント(electrocompetent)E.coli TOP10細胞中にエレクトロポレーション処理した。形質転換細胞を、50μg/mlのカナマイシンを補充したTBABプレート(30g/lトリプトース(tryptose)血液寒天基礎培地)上に広げ、続いて37℃で終夜インキュベートした。コロニーをPCRを使用してスクリーニングし、そしてPCRフラグメントの長さをアガロースゲルで確認した。配列を確認するために、上記のように配列決定を行った。プラスミドDNAストックを配列決定したクローンから調製し、そして−80℃に置いた。さらに、E.coli BL21(DE3)細胞をエレクトロポレーションによりプラスミドを用いて形質転換した。
【0110】
結果
HER3結合ポリペプチドのファージディスプレイ選択: 4ラウンドのファージディスプレイ選択を、ビオチン化又は非ビオチン化ヒトHER3−ECDに対して行った。4回の選択サイクルを、2つの異なる標的濃度でビオチン化標的を含む溶液で、又は非ビオチン化標的を含む固相で行った。溶液では、ファージ粒子標的複合体がストレプトアビジン被覆ビーズに捕捉された。各選択サイクルについて、洗浄回数を増やした。ファージ粒子力価及び収率を、各選択サイクル後に計算した。ファージ粒子収率(選択された(out)ファージ粒子/導入した(in)ファージ粒子)は、3回目又は最後のサイクルで増加し、標的結合クローンの濃縮を示していた。
【0111】
Z変異体のELISAスクリーニング: 4サイクルの選択後に得られたクローンを96ウェルプレートで作製し、そしてHER3−ECD及びHER3−Fc−結合活性についてELISAでスクリーニングした。各選択トラックから合計で93のクローンをスクリーニングした。吸光度測定は、選択方法と無関係に、約90%のクローンがHER3に対して陽性(ネガティブコントロールのシグナルの2倍の応答として定義される)であることを示した。インスリンに結合するZ変異体分子を、ネガティブ及びポジティブコントロール実験において使用した。ポジティブコントロールをビオチン化インスリンを用いて検出し、そしてネガティブコントロールについては標的タンパク質を除いた。
【0112】
配列決定: ELISAスクリーニングにおいてHER3−Fcに対して様々な(variying)シグナル吸光度値を有することに基づいて選択されたクローンについて配列決定を行い、従って配列決定のための多くの異なる代表的なバインダーを得た。合計で135のHER3ポジティブクローンを配列決定した。クローンの半分は合計23の新しいクローンを生じるいくつかのコピーにおいて見出され、それらのうち2つはバックグラウンドバインダーと同一であった。各変異体に固有の識別番号#####を付与し、そして個々の変異体をZ#####と呼ぶ。58量体Z変異体のアミノ酸配列を図1及び配列表に配列番号669〜689として載せる。これらのZ変異体の推定HER3結合モチーフを図1及び配列表に配列番号1〜21として載せる。これらの各Z変異体内で完全3ヘリックスバンドルを構成すると予測される49量体ポリペプチドのアミノ酸配列を図1及び配列表に配列番号335〜355として載せる。
【0113】
ドットブロット分析: 配列決定された独特なHER3結合ポリペプチド(AQHDEALE−[Z#####]−VDYV−[ABD]−YVPGの形態)のうち16の特異性をドットブロット分析を使用して試験した。23の異なるタンパク質をニトロセルロースメンブレン上にブロットした。これらの23のタンパク質には、16の高度に豊富なヒト血清タンパク質、HER2−ECD、HER3−ECD及びHER3−Fc並びにHER4−ECD及びHER4−Fcが含まれていた。Z01824以外の全てのZ変異体融合タンパク質はHSAと同様にHER3−ECD及びHER3−Fcに特異的に結合した。従って、試験したZ変異体の特異性は十分なものであった(図2)。
【0114】
Z変異体のサブクローニング: 16の独特なクローンを二量体として発現ベクターpAY004308でサブクローニングするために選択した。クローニングは、15の二量体のHER3結合Z変異体を生じた(Z01748、Z01749、Z01751、Z01753、Z01814、Z01815、Z01817、Z01820、Z01821、Z01826、Z01828、Z01830、Z02009、Z02010及びZ02011の二量体)。
【0115】
実施例2: Z変異体の産生及び特徴付け
材料及び方法
タンパク質発現及び精製:実施例1においてサブクローニングした形質転換したE.coli BL21(DE3)培養物を、TSB−YEで約1の光学密度まで増殖させた。次いで1M IPTGを最終濃度0.5mMまで加えることによりタンパク質発現を誘導した。培養物を誘導の5時間後に遠心分離により採取した。上清を廃棄し、そして細胞ペレットを集めて−20℃で貯蔵した。
【0116】
HER3結合Z変異体を、1.5ml Ni−NTA Superflow Columns(Qiagen)で変性条件下で細胞ペレットから精製し、そしてバッファーをPBSに交換してPD−10カラム(GE Healthcare)を使用した。精製したZ変異体をアリコートに分けて−80℃で貯蔵した。
【0117】
タンパク質特徴付け: 精製されたZ変異体(His6−(Z#####)2−Cys形態)の濃度を280nmにおける吸光度測定により決定した。純度を10ウェル4〜12% NuPAGETMゲル(Invitrogen)でのSDS−PAGE分析でクマシーブルー染色を使用して見積もった。精製したZ変異体のアイデンティティを確認し、分子量を決定するために、LC/MS分析をAgilent1100LC/MSDシステム(Agilent Technologies)で行った。
【0118】
CD分析: 精製したZ変異体を解凍し、そしてPBS中0.5mg/mlに希釈した。それぞれの希釈したZ変異体について、250〜195nmでのCDスペクトルを20℃で得た。さらに、温度可変測定(VTM)を、融点(Tm)を決定するために行った。VTMにおいて、5℃/分の温度勾配で温度を20から90℃に上げながら吸光度を220nmで測定した。CD測定を、光路長1mmを有するセルを使用してJasco J−810分光偏光計(Jasco Scandinavia AB)で行った。
【0119】
蛍光免疫染色: HER3ポジティブヒト乳腺細胞株AU565(ATCC、#CRL−2351)及びHER3ネガティブヒト卵巣がん細胞株SKOV−3(Ecacc、#91091004)を供給業者に推奨されるように培養した。アッセイ前日に、25000個の細胞をマルチウェルスライド(スライドあたり8ウェル、Histolab #ER−201−1B)の各ウェルに加え、そして終夜CO2インキュベーターで板状形態を呈するように増殖させた。パスツールピペットを用いてスライド上に液滴を加えることによりこれらのスライドをPBSで穏やかに洗浄した。
【0120】
2つの実験を行った。1つ目の実験では、AU565細胞を11の異なるHER3特異的Z変異体で染色した;His6−(Z01748)2−Cys、His6−(Z01749)2−Cys、His6−(Z01751)2−Cys、His6−(Z01753)2−Cys、His6−(Z01814)2−Cys、His6−(Z01815)2−Cys、His6−(Z01817)2−Cys、His6−(Z01820)2−Cys、His6−(Z01828)2−Cys、His6−(Z01830)2−Cys及びHis6−(Z02011)2−Cys。2つ目の実験は、His6−(Z01814)2−Cys及びHis6−(Z01820)2−Cysを用いて、そして(Z01154)2をネガティブコントロールZ変異体として用いて行った。染色を、HER3ポジティブAU565及びHER3ネガティブSKOV−3細胞を用いてスライド上で行った。R&D Systemsからの抗HER3抗体(#AF234)をポジティブコントロールとして両方の実験に
含めた。
【0121】
両方の実験において、Z変異体をPBS中15μg/mlの濃度でウェルに加え、そして試薬の全ての希釈はPBS中で行い、全ての洗浄も同様にした。1時間室温でのインキュベーション後、マルチウェルスライドを上記のように穏やかに洗浄し、そしてヤギ抗Affibody分子Ig(Affibody AB、#20.1000.01.0005)を各ウェルに5μg/mlの濃度で加えた。45分室温でのインキュベーション後、スライドを上記のように穏やかに洗浄し、そして5μg/mlのニワトリ抗ヤギAlexa Fluor488(Invitrogen、#A21467)を各ウェルに加えた。さらに45分のインキュベーション後、スライドを穏やかに洗浄した。ポジティブコントロール、ヤギ抗HER3抗体を15μg/mlの濃度で、続いて5μg/mlのニワトリ抗ヤギAlexa Fluor488を使用した。染色が完了した後、PBS中3%ホルムアルデヒド(Sigma、#1365)中で10分間室温で固定した。次いでスライドを2回すすぎ、乾燥し、そしてDAPI(4',6−ジアミジン−2−フェニルインドール、Vector laboratories、#H1200)を含有する退色防止液を用いて封入した。生体画像化用のビデオカメラを備えたLEICA DMLA顕微鏡を使用して染色を記録した。
【0122】
フローサイトメトリーを使用した細胞上親和性順位付け: Z変異体Z01751、Z01753、Z01814及びZ01820をコードする全長遺伝子配列を、それらの対応するpAY00430ベクター構築物からPCR増幅し、そして制限酵素XhoI及びSalI(New England Biolabs)で予め消化したブドウ球菌ディスプレイベクターpSCZ1(Kronqvist et al、Protein Eng Des Sel 21:247−255(2008))に結合させた。E.coli株RR1ΔM15をプラスミド構築及び製造のための宿主として使用し、そして以前に記載されたプロトコル(Loefblom et al、J Appl Microbiol 102:736−747(2007))に従ってこれらの構築物をエレクトロコンピテントなスタフィロコッカス・カルノーサス(Staphylococcus carnosus)TM300(Goetz、Soc Appl Bacteriol Symp Ser.19:49−53(1990))に形質転換した。独立して4つの異なるZ変異体を提示しているブドウ球菌細胞を、20μg/mlクロラムフェニコールを補充したTSB−YE 10mlに播種し、そして終夜37℃及び150rpmで増殖させた。培養物から、106個の細胞を1ml PBSPで洗浄した。これらの細胞を遠心分離(3500xg、4℃、6分)によりペレット化し、そして異なる濃度のビオチン化HER3−Fc(5、20、50及び100nM;ビオチン化については実施例1を参照のこと)を含有するPBSP100μl中に再懸濁した。穏やかに混合しながら室温で1時間インキュベーションすることにより平衡結合に達した。細胞を氷冷PBSP 1mlで1回洗浄し、続いて氷上で1.25μg/ml Alexa Fluor(R)488複合体化ストレパビジン(strepavidin)(Invitrogen、#S32354)及び225nM Alexa Fluor(R)647複合体化HSA(実施例1を参照のこと)を含有する氷冷PBSP 100μl中で40分間インキュベートした。氷冷PBSP1mlで1回洗浄した後、細胞をフローサイトメトリー分析の前に氷冷PBSP300μlに再懸濁した。平均蛍光強度(MFI)を、FACS Vantage SE(BD Biosciences)フローサイトメーターを使用して測定した。
【0123】
結果
タンパク質発現及び精製: 15の二量体Z変異体分子(His6−(Z#####)2−Cysの形態)は許容しうる産生レベルの可溶性タンパク質を生じ、そして産生されたバッチの純度は、基準を満たさなかった2つのZ変異体(His6−(Z01826)2−Cys及びHis6−(Z02009)2−Cys)を除いて、SDS−PAGE分析により90%を超えると見積もられた。LC/MS分析により、全ての純粋なZ変異体分子について正確な分子量が確認された。
CD分析: CDスペクトルは、Z変異体分子がα−ヘリックス構造を20℃で有していることを示した。この結果は融点(Tm)を決定した温度可変測定(表1)でも確認された。
【0124】
【表1】
【0125】
蛍光免疫染色: 蛍光免疫顕微鏡検査法を、ヒトがん細胞において発現される天然HER3に対する特異性及び結合活性の特徴付けのために使用した。実施例1のELISAにおいてポジティブシグナルを示す11のクローンを、N末端His6−タグ及びC末端システインに融合された二量体として製造した。各Z変異体をHER3ポジティブAU565細胞(ヒト乳房乳腺細胞株)でインキュベートした。抗HER3抗体をポジティブコントロールとして使用した。染色手順の後に細胞を固定した。11のバインダーのうち、6つは膜結合染色パターンを生じた。バックグラウンドはいくつかのZ変異体について高く、そしていくつかの場合にはそれらが特異的であるか否かを判別することが困難であった。2つのHER3特異的分子、His6−(Z01814)2−Cys及びHis6−(Z01820)2−Cysを、HER3ポジティブAU565細胞並びにHER2ポジティブ及びHER3ネガティブSKOV−3細胞(ヒト卵巣がん細胞株)の両方に対するさらなる分析のために選択した。明確な細胞膜染色がAU565で得られたがSKOV−3細胞では得られず、HER2よりもHER3に対するZ変異体の選択性を示した(図3)。
【0126】
フローサイトメトリーを使用した細胞上親和性順位付け: HER3結合分子をさらに特徴付けし、そして細菌細胞での機能発現を確認するために、蛍光免疫染色においてHER3に対する特異性を示す4つのHER3結合分子を、ブドウ球菌細胞ディスプレイ及びフローサイトメトリーを使用する細胞上親和性順位付け実験のために使用した。4つの変異体の単量体構築物を、その後のブドウ球菌宿主への形質転換のためにブドウ球菌ディスプレイベクターにサブクローニングした。4つのZ変異体を提示しているブドウ球菌細胞をそれぞれ、5〜100nMの範囲に及ぶ4つの濃度のビオチン化HER3−Fcとともにインキュベートした。フローサイトメトリーを使用して細胞を分析し、4つ全ての変異体についてクローン間での相対的親和性において有意な差異はなく、細胞表面上での効果的な発現及びHER3への特異的結合が明らかとなった(データは示されていない)。ビオチン化HER2を実験においてネガティブコントロールとして使用して交差特異性は観察されず、実施例1における周辺質上清を用いて行われたドットブロット及びこの実施例における蛍光免疫染色の結果を裏付けた。
【0127】
実施例3: HER3結合Z変異体の成熟ライブラリーの設計及び構築
この実施例において、成熟ライブラリーを構築した。このライブラリーをHER3結合ポリペプチドの選択のために使用した。成熟ライブラリーからの選択は通常、増加した親和性を有するバインダーを生じると期待される(Orlova et al、Cancer Res 66(8):4339−48(2006)。しかしモノヌクレオチド及び縮重コドンを使用する伝統的なオリゴヌクレオチド合成技術は、ブドウ球菌系統がアンバー停止コドンを抑制することができないために設計を制限し得る。この研究において、無作為化二本鎖リンカーをSlonomics(R)技術により代わりに生成し、これはその後に構成される二本鎖DNAのライゲーション及び制限を使用したトリヌクレオチドビルディングブロックの無作為化セットの組み込みを可能にする。
【0128】
材料及び方法
ライブラリー設計: ライブラリーは、実施例1及び2に記載されるHER3結合Z変異体の配列に基づくものであった。新しいライブラリーにおいて、Z分子骨格における13の可変位置は、配列番号1〜21において定義されるZ変異体配列の結合モチーフに基づいたストラテジーにしたがって、特定のアミノ酸残基に偏っていた。制限部位XhoI及びNheIが隣接した、HER3結合ポリペプチドのヘリックス1及び2において部分的に無作為化された位置をコードする131bpの二本鎖リンカー、例えば5’−GAA NNN NNN NNN GCG NNN NNN GAG ATC TGG NNN TTA CCT AAC TTA AAC NNN NNN CAA NNN NNN GCC TTC ATC NNN AGT TTA NNN GAT GAC CCA AGC CAA AGC GCT AAC TT−3’(無作為化コドンはNNNと示される)の混合物を含有するSlonoMax(R)ライブラリーをSloning BioTechnology GmbHに発注した。13の可変Z位置についての新しいライブラリーにおけるアミノ酸残基の理論的分布を表3に示す:
【0129】
【表2】
【0130】
ライブラリー構築及びクローニング: Phusion DNAポリメラーゼ(Finnzymes、#F530L)を使用して8サイクルのPCRの間にライブラリーを増幅し、そしてプールした生成物をQIAquick PCR精製キット(QIAGEN)を用いて供給業者の推奨にしたがって精製した。無作為化ライブラリーフラグメントの精製したプールを制限酵素XhoI及びNheI(New England Biolabs)で消化し、そして2%アガロースゲルでの分取ゲル電気泳動を使用して精製した。E.coli系統RR1ΔM15を、Jetstar Maxi Kit(Genomed、#220020)を用いたブドウ球菌ディスプレイベクターpSCZ1(図4A)のプラスミド産生のための宿主系統として使用した。
【0131】
このベクターを同じ酵素XhoI及びNheIで消化し、そして1%アガロースゲルでの分取ゲル電気泳動を使用して精製した。pSCZ1と無作為化ライブラリーフラグメントとのライゲーションを、1:4のベクター対フラグメントのモル比のT4 DNAリガーゼ(New England Biolabs、#M0202T)を使用して行った。ライゲーション混合物を、QIAquick PCR精製キットを使用して供給業者の推奨に従って、エレクトロコンピテントなE.coli DH5α細胞(Invitrogen、#18263−012)への形質転換前に精製した。形質転換後、さらには終夜の増殖後に直接プレーティングした個々のクローンを、配列確認のためにBigDyeサーモサイクルシークエンシング(Thermo Cycle Sequencing)反応及びABI Prism 3700機器(PE Applied Biosystems)を使用してPCR増幅した。プラスミドをJetstar Maxi Kitを使用したE.coliの終夜培養から製造し、そしてエレクトロコンピテントなS.カルノーサス(S.carnosus)に以前に記載されたように(Loefblom et al、前出)形質転換した。ブドウ球菌提示ライブラリーを本明細書以下ではSc:ZHER3LIBと示す。
【0132】
ライブラリー品質分析: Sc:ZHER3LIBのアリコート(少なくともライブラリーサイズの10倍、すなわち1.3×108より多い)を、20μg/mlクロラムフェニコールを含む100ml TSB−YEに播種し、そして終夜37℃及び150rpmで増殖させた。16時間後、107個の細胞を1ml PBSPで1回洗浄した。細胞を遠心分離(3500xg、4℃、6分)でペレット化し、そして225nM Alexa Fluor(R)647結合体化HSAを含有するPBSPに再懸濁し、そして1時間室温にて暗所でインキュベートした。1ml氷冷PBSPで1回洗浄した後、フローサイトメトリー分析の前に細胞を300μl氷冷PBSPに再懸濁した。平均蛍光強度(MFI)を、FACS Vantage SE(BD Biosciences)フローサイトメーターを使用して測定した。
【0133】
結果
ライブラリー構築及びクローニング: 新しいライブラリーを、確認された結合特性(実施例1及び2)を有する一組のHER3結合Z変異体(配列番号669〜689)に基づいて設計した。設計されたライブラリーの理論的なサイズは7.4×108のZ変異体であった。DNAフラグメントのライブラリーをブドウ球菌発現ベクター中にクローニングし、そしてS.カルノーサス(S.carnosus)に形質転換して約1.3×107の個々のクローンを含有する細胞提示ライブラリーを生成した。個々のライブラリーメンバーの配列分析により、理論的設計に従ったコドンの分布、並びに予期しないコドン、多重挿入及びフレームシフトの低い比率が示された。
ライブラリー品質分析: Z変異体成熟ライブラリーが細菌表面に機能的に提示されたことを確認するために、ライブラリーからのブドウ球菌細胞を蛍光標識したHSAと共にインキュベートし、そしてフローサイトメトリーを使用して分析した。結果は、ライブラリーの約72%が細胞表面で機能的ABP融合を有する全長タンパク質を発現したことを示した(データは示されていない)。従ってHER3結合ポリペプチドの成熟ライブラリーは首尾よく構築された。
【0134】
実施例4: ブドウ球菌表面ディスプレイライブラリーからのZ変異体の選択、スクリーニング及び特徴付け
材料及び方法
細胞標識及びFACSを使用したブドウ球菌細胞選別: Sc:ZHER3LIBのアリコート(ライブラリーサイズの少なくとも10倍、すなわち1.3×108より多い)を、20μg/mlクロラムフェニコールを含むTSB−YE 100mlに播種し、そして終夜37℃及び150rpmで増殖させた。16時間後、細胞(その後のサンプリング数の少なくとも4倍)を1ml PBSPで洗浄した。細胞を遠心分離(3500×g、4℃、6分)でペレット化し、そしてビオチン化HER3−Fcを含有するPBSP中に再懸濁し、そして室温で穏やかに混合しながら2時間インキュベートして平衡結合に到達させた。その後細胞を氷冷PBSPで洗浄し、続いて1.25μg/ml Alexa Fluor(R)488結合体化ストレプトアビジン(Invitrogen)及び225nM Alexa Fluor(R)647結合体化HSAを含有する1ml PBSP中で1時間氷上で暗所にてインキュベートした。氷冷PBSP1mlでの最終洗浄工程の後、細胞を選別前に氷冷PBSPに再懸濁した。細胞をFACSVantage SE(BD Biosciences)フローサイトメーターを使用して選別した。ソートゲートは 最も高いAlexa Fluor(R)488対Alexa Fluor(R)647蛍光強度比を示すZ変異体提示細胞の上部(top)フラクション(典型的に0.1%)を選別するように設定した。細胞を直接0.5ml TSB−YE培地中に選別し、そしてその後10μg/mlクロラムフェニコールを含有するTSBに播種し、そして37℃で16時間、標識及びFACSの次のラウンドのため、増殖により単離された細胞を増幅するためにインキュベートした。この手順を4回繰り返した。
【0135】
配列決定: 個々のブドウ球菌クローンの配列決定を、実施例3のライブラリー構築において記載した細胞選別サイクル3及び4の後に行った。
【0136】
細胞上親和性順位付け及びKD決定: HER3−Fcをビオチン化し、そして実施例1において記載されるようにHSAをAlexa Fluor(R)647と結合体化した。異なるZ変異体を提示しているブドウ球菌細胞をそれぞれ10ml TSB−YE及び20μg/mlクロラムフェニコールに播種し、そして終夜37℃及び150rpmで増殖させた。培養物から、106個の細胞を1ml PBSPで洗浄した。細胞を遠心分離(3500×g、4℃、6分)でペレット化し、そして推定KDに及ぶ異なる濃度のビオチン化HER3−Fc(Z05405、Z05413、Z05416及びZ05417について135、90、27、9、2.7、0.9、0.45、0.22、0.11及び0.054nM;Z01820について670、337、110、45、18、6.7、2.2、1.1、0.37及び0.27nM)を含有するPBSPに再懸濁した。穏やかに混合しながら室温で1時間インキュベーションすることにより平衡結合に達した。細胞を氷冷PBSP1mlで洗浄し、続いて1.25μg/mlAlexa Fluor(R)488結合体化ストレプトアビジン(Invitrogen)、そして順位付け実験では225nM Alexa Fluor(R)647結合体化HSAを含有する氷冷PBSP100μl中で氷上で40分間インキュベートした。氷冷PBSP1mlで洗浄した後、フローサイトメトリー分析前に細胞を氷冷PBSP300μlに再懸濁した。平均蛍光強度(MFI)をFACS Vantage SE(BD Biosciences)フローサイトメーターを使用して測定した。
【0137】
結果
改善されたZ変異体の単離のためのフローサイトメトリー選別: 成熟HER3結合Z変異体の単離のために、ブドウ球菌ライブラリーを、細胞増殖による増幅の交互のラウンドを含むラウンドの蛍光細胞分析分離にかけた。
【0138】
手短には、細胞をビオチン化HER3−Fcと共に実施例1及び2からのZ変異体の推定KDと比較して約10倍低い濃度でインキュベートした。その後細胞を洗浄し、そしてその後の細胞に結合したHER3の蛍光媒介検出のための蛍光標識したストレプトアビジン、さらには表面発現レベルのモニタリングのための蛍光標識したHSAと共にインキュベートした。結合したHER3の解離速度を小さくするために、二次的試薬及びHSAのインキュベーションを氷上で行った。さらに洗浄した後、標識された細胞ライブラリーをフローサイトメーターでスクリーニング及び選別した。標的濃度、選別パラメーター及びソートゲートに関する選択ストリンジェンシーは、各選別ラウンドとともに増加し、そして典型的には、最も高い標的結合対表面発現比を示すライブラリーの上位0.1%を、増幅及びその後の選別ラウンドのためにゲーティングして単離した。フローサイトメーターでのライブラリーの標的結合特性の可視化により、各選別ラウンドでのHER3ポジティブクローンの濃縮そして最終ラウンドでは本質的にHER3ポジティブクローンのみであることが明らかとなった(図5)。FACSの4ラウンドまでの後、単離された細胞を個々の候補の配列決定及び特徴付けのために半固形培地に広げた。
配列決定: 576の個々のクローンを配列決定し、結果として443の配列が可読であり、そのうち45のクローンは同じ選別ラウンドにおいて1回以上出現した(配列番号690〜734)。各変異体に固有の識別番号#####を付与し、そして個々の変異体をZ#####と呼ぶ。58量体Z変異体のアミノ酸配列を図1及び配列表に配列番号690〜1002として載せる。これらのZ変異体の推定HER3結合モチーフを図1及び配列表に配列番号22〜334として載せる。これらの各Z変異体内で完全3ヘリックスバンドルを構成すると予測される49量体ポリペプチドのアミノ酸配列を図1及び配列表に配列番号356〜668として載せる。
【0139】
細胞上親和性順位付け: 配列決定結果に基づいて1回より多く出現した45のクローンを、FER3に対する親和性に関して変異体を順位付けするために全細胞ELISAアッセイにかけた。ブドウ球菌細胞集団をフローサイトメトリーを使用して分析し、そしてブドウ球菌細胞において表面提示された実施例1及び2からの1つのZ変異体(Z01753)を比較のために分析に含めた。全ての45の候補がアッセイにおいてHER3結合に関してポジティブであり、さらに、43は実施例1及び2からのZ変異体と比較してより高いシグナルを示した(図6A〜D)。アッセイにおいて最も高いシグナルは配列決定後に最も頻繁に出現した4つのクローンで見られ、フローサイトメトリー選別におけるZ変異体の定量的な単離が示された。
【0140】
KD決定: 見掛けの平衡解離定数(KD)を、4つのZ変異(図4B)並びに実施例1及び2からの1つのHER3結合Z変異体(Z01820)について細胞で決定した。4つの変異体及びZ01820を提示しているブドウ球菌細胞を、推定KDに及ぶ様々な濃度の標識HER3−Fcでインキュベートした。細胞集団をフローサイトメーターで分析し、続いて平均蛍光強度データをHER3−Fc濃度に対してプロットし、そしてKDを決定するために1部位結合モデルにフィッティングした。全ての4つの成熟Z変異体は低いナノモルの見掛けのKDを示した。最も強いバインダーは、実施例1及び2からのZ変異体と比較してHER3−Fcに対する親和性において15倍の改善を示した(表3)。
【0141】
【表3】
【0142】
実施例5:Z変異体の表面プラズモン共鳴KD決定
材料及び方法
表面プラズモン共鳴分析: 表面プラズモン共鳴(SPR)分析をHER3結合Z変異体Z05405、Z05413、Z05416及びZ05417についてBIAcoreTM 2000機器(GE Healthcare)で行った。ヒトHER3−Fc(R&D Systems)及びマウスHER3−Fc(R&D systems)をCM−5センサーチップ(GE Healthcare)上でNHS/ECDアミンカップリング化学により固定化した。固定化は10mM NaAc(pH4.5)中で30μl/分の流量で、そして10μg/mlの濃度で行い、4000RUの固定化レベルを目標とした。ランニングバッファーとしてPBSPを20μl/分(別の記載がなければ)の流量で、そして再生のために5mM NaOHを使用した。
【0143】
動力学の決定のために、4つの異なる濃度(1.5〜67nM、PBSP中で希釈)の4つのHER3結合Z変異体250μlを、CM−5チップ表面上に固定化したHER3−Fc上に注入した。5mM NaOH 15μlの4回の注入及びランニングバッファを用いた徹底的な洗浄により表面を再生した。各サンプルを二つ組で測定し、そしてブランク表面からの応答を各濃度での各Z変異体の応答から差し引いた。平均会合速度(kon)及び解離速度(koff)に基づいてKD値を決定するために、得られたセンサーグラム(sensorgrams)を1部位結合モデルにフィッティングした。
【0144】
対照実験: HER3−FcのFc部分に対する親和性について選択されたのではないことを確認するために、各Z変異体(50nM)を、各サンプル100μl(上記の濃度で)をバイオセンサーチップ表面に固定化されたHER3−Fc上に注入する前に、10倍モル濃度過剰のヒトポリクローナルIgG(500nM)と共に1時間室温でインキュベートした。さらなる対照実験において、センサーチップ表面に固定化されたHER3−ECD上にZ変異体を注入した。
【0145】
結果
表面プラズモン共鳴分析: 4つのZ変異体Z05405、Z05413、Z05416及びZ05417の解離平衡定数(KD)をSPR技術により決定した。1部位結合モデルに対する非線形回帰を使用してZ変異体の会合速度及び解離速度から親和性を決定し、Z05405について1.61nM、Z05413について0.78nM、Z05416について0.78nM、そしてZ05417について0.69nMの解離定数を得た(図7、表4)。表4において、全ての値は同日に行われた各濃度の二つ組の平均である。
【0146】
【表4】
【0147】
対照実験: HER3結合Z変異体の分析はヒトIgG Fc領域に融合されたHER3を用いて行ったので、Z変異体がHER3−FcのFc部分に対する親和性について選択されたのではないことを確認するために2つの対照実験を行った。IgGと共にプレインキュベートしたZ変異体とZ変異体のみを含有するサンプルとの間に応答の差異は見られなかった(データは示していない)。さらに、HER3−ECDを有する表面上に注入されたZ変異体は、全ての4つの変異体に対して特異的結合を示した(データは示していない)。総合すると、これら2つの実験は、FcがHER3−Fc上のZ変異体の結合部位の一部ではないということを明確に示す。
【0148】
実施例6: ヘレグリン及びHER3結合Z変異体についての競合アッセイ
この実施例において、HER3結合Z変異体がリガンドヘレグリンのHER3に対する結合をブロックする能力を競合アッセイにおいてSPRにより調べた。
【0149】
材料及び方法
天然リガンドヘレグリン(HRG1−β1、EGF様ドメイン、R&D Systems)と4つのHER3特異的Z変異体Z05405、Z05413、Z05416及びZ05417との間の競合を、SPRを使用して2つの異なるアッセイで分析した。
【0150】
第一のアッセイにおいて、2.5nM HER3−Fcを、100nMの4つのHER3特異的Z変異体の各々(40倍モル濃度過剰)とともに1時間室温でプレインキュベートした。その後、HER3−Fc及びZ変異体の各混合物75μlを、ヘレグリンが固定化されたセンサーチップ表面上に注入した。さらに、Z変異体とプレインキュベートしていないHER3−Fcを、ヘレグリンが固定化されたチップ表面上に注入した。
【0151】
第二のアッセイにおいて、5つの異なる濃度(0〜10nM)の各Z変異体100μlを、HER3が固定されたセンサーチップ表面上に注入し、続いてすぐに250nMヘレグリン100μlを注入した(BIAcore COINJECTコマンドを使用)。全てのサンプルを二つ組で注入し、そしてHER2特異的Z変異体を、同じ設定を使用してネガティブコントロールとして注入した。5mM NaOHの2回の注入及びランニングバッファでの徹底的な洗浄で表面を再生させた。
【0152】
結果
HER3特異的Z変異体Z05405、Z05413、Z05416及びZ05417の天然HER3リガンドヘレグリンとのHER3に対する競合をSPR技術により調べた。
【0153】
チップ表面上に固定化されたヘレグリンを使用した第一のアッセイの結果は、Z変異体とプレインキュベートしていないHER3−Fcを注入した場合と比較してZ変異体とともにプレインキュベートしたHER3−Fcを注入した場合に、応答レベルのほとんど完全な減少を示した(図8A)。
【0154】
チップ表面上に固定化されたHER3−Fcを使用した第二のアッセイの結果は、HER3受容体とその天然リガンドとの間の相互作用と濃度依存的に競合することを示した(図8B)。同じパターンがマウスHER3−Fcについて観察された(データは示していない)。ネガティブコントロールとヘレグリンとの間の競合はヒト又はマウスHER3のいずれについても観察することができなかった(データは示していない)。
【0155】
これらの結果は、HER3特異的Z変異体が天然リガンドヘレグリンと同じHER3上の結合部位と相互作用し、それ故リガンドと受容体との間の結合とインビトロで競合し得るということを示した。この効果は、今後の治療的なインビボ適用において利用されるかもしれない。
【0156】
実施例7: インビトロでのヘレグリン誘導HER3リン酸化の阻害
この実施例では、HER3結合Z変異体のヘレグリン誘導HER3リン酸化を妨害する能力を細胞アッセイで調べた。
【0157】
材料及び方法
刺激: MCF−7(ACC115、DSMZ)乳がん細胞を、L−グルタミン(BE17−605E、Cambrex)、非必須アミノ酸(BE13−114E、Cambrex)、ピルビン酸ナトリウム(BE13−115E、Cambrex)及び10%ウシ胎児血清(FBS、10108−165、Gibco)を補充したRPMI培地(BE12−167F、Lonza)で培養した。刺激の前日に、1×106の細胞を6mmペトリ皿(430168、BD Biosciences)に上記の培地5ml中で播種した。
【0158】
刺激の1時間前に、培地をRPMI+L−グルタミン+2%透析FBS(26400−036、Gibco)からなる培地に変更した。HER3のリン酸化を5nMヘレグリン(HRG1−β、396−HB、R&D systems)を用いて誘導した。それぞれN末端hisタグ及びC末端システインを含むHER3結合Z変異体Z05416及びZ05417(His6−Z05416−cys及びHis6−Z05417−cys)を、10又は100倍モル濃度過剰のヘレグリンと同時に加えた。taqポリメラーゼ特異的Z変異体(His6−Z01155−cys)をネガティブコントロールとして使用して、そして上記と同じ手順に従って加えた。
【0159】
10分間37℃でインキュベーションした後、ペトリ皿を氷上に置き、続いて氷冷PBSで洗浄することにより細胞プロセスを停止させた。洗浄後、1mM活性化オルトバナデート(450243、Sigma)を含有するPBS 2mlを各皿に加え、そして細胞を細胞スクレーパーを使用して剥離した。細胞溶液を10mlのチューブに移し、そして細胞を1000rpmでの3分間の予冷遠心分離で回転させることによりペレット化した。乾燥ペレットを氷冷溶解緩衝液100μl(1% NP−40(Sigma I3021)、20mM Tris(pH8.0)、137mM NaCl、10%グリセロール、2mM EDTA、1mM活性化ナトリウムオルトバナデート)に溶解して細胞を溶解させた。溶解物を回転させて4℃にて30分間インキュベートし、そして13000rpmで15分間遠心分離した。上清を集めてさらなる分析まで−80℃で保存した。
【0160】
pHER3−ELISA: リン酸化HER3タンパク質(pHER3)の細胞溶解物中の存在を、Duo−Set IC phospo ErbB3サンドイッチELISA(DYC−1769、R&D Systems)を使用して分析した。
【0161】
96ウェルハーフエリアプレート(Costar 3690)を終夜50μl/ウェルのPBS中に希釈された2μg/mlの捕捉抗体で被覆した。次の日、プレートを自動化ELISA洗浄機でPBS+0.05%Tween(PBST)で4回洗浄した。1% BSAを含有するPBS(100μl/ウェル)で2時間室温にてウェルをブロックし、そして以前に記載したように洗浄した。
【0162】
IC Diluent #12(1% NP−40、20mM Tris(pH8.0)、137mM NaCl、10%グリセロール、2mM EDTA、及び1mM活性化ナトリウムオルトバナデート)で2回又は4回希釈した50μl/ウェルの細胞溶解物を加えた。製造者により提供されたポジティブコントロールもウェルに加えた。続いてプレートを2時間インキュベートした。
【0163】
洗浄後、製造者のプロトコルにしたがってIC diluent #14(20mM Tris、137mM NaCl、0.05% Tween 20、0.1%BSA、pH7.2−7.4)で希釈した50μl/ウェルの検出抗体を加え、そしてプレートをさらに2時間インキュベートした。インキュベーション後に、プレートを洗浄し、そして50μl/ウェルのTMB(Immunopure、Termo Fisher Scientific)を加えた。20分後に50μl/ウェルの2M H2SO4を加えることにより反応を停止させ、そして吸光度を450nmでVictor3 ELISAプレートリーダーを使用して測定した。
【0164】
結果
HER3特異的Z変異体Z05416及びZ05417を、ネガティブコントロール(taqポリメラーゼ特異的Z01155)と一緒に、HER3のヘレグリン誘導リン酸化をブロックするそれらの能力について評価した。pHER3の相対含有量(%)を、ヘレグリン刺激培養物(ヘレグリン刺激培養物から得られたOD値)に対するブロックされた培養物(Z変異体を含有する培養物から得られたOD値)の関係として決定した。図9は、HER3結合変異体Z05416及びZ05417が用量依存的様式でヘレグリン誘導リン酸化を阻害し、一方でtaqポリメラーゼ結合変異体Z01155はいずれの効果も有していなかったことを示す。
【0165】
実施例8: HER3及びHER2に結合する二重特異性Z変異体
背景の項に記載したように、HER3はHER2にとって好ましいヘテロ二量体化パートナーであり、そしてHER2はリガンド誘導シグナル伝達に関してHERファミリーの他の受容体に依存している。二重特異性HER3及びHER2標的化リガンド(例えばHER3−HER2結合Z変異体)は、両方の受容体を発現している腫瘍において受容体シグナル促進腫瘍成長をブロックするかもしれない。
【0166】
場合により様々なリンカー長を含む二重特異性分子のための種々の分子構築物を考慮した。構築物の例は、HER2結合ポリペプチドに対してHER3結合ポリペプチドをN末端又はC末端に位置づけることである。リンカーが2つの結合ポリペプチドの間に導入される場合、それは1〜60アミノ酸残基、例えば1〜45アミノ酸を含み得る。さらに、二重特異性構築物は、半減期延長部分、例えばアルブミン融合、アルブミン結合部分又はPEGのような分子へのカップリングを分子のN末端若しくはC末端部分又はリンカー領域内に備えていてもよい。あるいは、半減期延長部分はそれ自体で結合ポリペプチド間のリンカー、又はスペーサーとして利用され得る。
【0167】
材料及び方法
二量体二重特異性Z変異体分子を、Friedman et al、Biotechnol Appl Biochem、54(2):121−31(2009)に以前に報告されたように操作した。FriedmanらはHERファミリーの2つのメンバーに対して特異的な四量体Z変異体の製造について記載しているが、HER3及びHER2二重特異性Z変異体の構築は、原理上は同じプロトコルに従う。当業者は、Friedmanらに基づく手順を適用する方法を用意に理解するだろう。HER3及びHER2に結合するヘテロ二量体Z変異体の構築については、Jonssonらにより記載される手順(前出)に従うことにより、2つの結合ポリペプチドの間に40アミノ酸のペプチドリンカーを使用してHER3結合変異体をコードする遺伝子フラグメントを、HER2結合Z変異体をコードする遺伝子フラグメントとインフレームで遺伝子融合した。
【0168】
二重特異性タンパク質をコードする遺伝子を、HER3−HER2結合Z変異体をコードするベクターのクローニングにより構築した。HER3結合Z変異体は、配列番号669〜1002から選択され、特に配列番号669〜734から選択される配列、例えば配列番号690〜734から、例えば配列番号691〜693、配列番号695〜696、配列番号700、配列番号703〜704、配列番号708、配列番号710、配列番号712〜713、配列番号721、配列番号722及び配列番号724から選択され、そしてHER2結合Z変異体は配列番号1003から選択される。HER2及びHER3結合Z変異体は、異なる受容体に対して異なる親和性を有し得る。構築物は同じ受容体に対する様々な親和性を有するZ変異体をコードするベクターから作製されてもよい。二重特異性HER3−HER2結合Z変異体をコードするベクターを、適切なエンドヌクレアーゼ(例えばSfiI、AccI、PstI、BamHI、New England Biolabs)を使用して切断し、そしてHER3−HER2特異的Z変異体をコードする遺伝子フラグメントを回収し、そしてライゲーションして二重特異性分子をコードする遺伝子を形成した。HER3及びHER2結合ポリペプチドは遺伝子フラグメント内で個々に単量体単位を形成しても多量体単位、例えば二量体、三量体又は四量体を形成してもよい。
【0169】
ペプチドリンカーの導入のために、リンカー領域をコードする遺伝子フラグメント(例えばGGGS又はGGGGSGGGGLVGLGSGGGGS)の倍数を異なるリンカー長を有する構築物を作製するために導入する。リンカー領域はアルブミン遺伝子又はアルブミン結合ドメイン(ABD)のための遺伝子も含有し得る。ここでは、ABD遺伝子をPCRを使用して増幅し、そしてリンカーコード単位を導入する。この後、BamHI制限消化し、そしてHER結合ポリペプチド間で二量体HER3−HER2二重特異性遺伝子中にライゲーションする。これは本質的に、Jonssonら(前出)により三量体TNF構築物の作製について記載されたように行われるが、Jonssonにおいて記載される中心TNF分子をABDに置き換える。あるいは、HER3−HER2結合Z変異体をコードする遺伝子フラグメントを、種々の形式で二重特異性分子をクローニングするるための所望のリンカー領域及び制限部位も導入する適切なPCRプライマーを使用して増幅する。二重特異性分子の種々の形式は、例えばN末端若しくはC末端の位置づけ、結合Z変異体間の様々な長さのリンカー及び/又は中間のABD遺伝子である。所望の二重特異性分子をコードする遺伝子の構築についての別の可能性は、単に完全合成遺伝子をそのようなサービスを提供している企業(例えばGeneart AG)から購入することである。
【0170】
結果として得られた、HER3結合Z変異体、HER2結合Z変異体、ペプチドリンカー及び場合によりABDをコードする遺伝子フラグメントを、発現ベクターにライゲーションし、発現細胞(例えばE.coli)に形質転換する。タンパク質を例えば実施例2に記載されるように発現させる。さらなるhis−tag及び/又は標識目的のための独特のシステインをコードする発現ベクターを使用し、そして発現されたタンパク質を例えば実施例2に記載されるように精製する。タンパク質発現及び精製に使用される方法は、当業者に公知の標準的な方法である。
【0171】
次いで得られた二重特異性タンパク質を、Friedmanら(前出)に記載されるように、バイオセンサー分析(Biacore、GE Healthcare)を使用することによりインビトロでの同時結合について試験する。ヒトHER3−Fc(hHER3−Fc)及びヒトHER2−Fc(hHER2−Fc、両方ともR&D Systemsから)を、アミンカップリングによりCM5センサーチップ(GE Healthcare)のカルボキシル化デキストラン層上に製造者の指示に従って固定化した。別のフローセル表面を活性化し、そして参照表面として不活性化した。二重特異性HER3−HER2分子をランニングバッファ、HBS(10mM Hepes、150mM NaCl、3.4mM EDTA及び0.005%界面活性剤P20、pH7.4)中で希釈し、その後結合分析を25℃で行った。第一の実験において、二重特異性HER3−HER2分子を125nMから2μMの範囲に及ぶ濃度で全ての表面に30μl/分の流量で注入した。HER3結合Z変異体及びHER2結合Z変異体もまたコントロールとして注入した。各注入後に、10mM HCl 10mlを注入することによりフローセルを再生した。
【0172】
第二の実験において、5μM HER3−HER2分子を、hHER3−Fc又はhHER2−Fc(R&D Systems)がアミンカップリングによりCM5センサーチップのカルボキシル化デキストラン層上に固定化されたフローセル表面上に注入した。1分の短い解離時間の後、115nM hHER3−Fc及びhHER2−Fc(ランニングバッファで希釈)を注入し、そして二重特異性分子のそれらの標的両方に同時に結合する能力をモニタリングした。
【0173】
蛍光標識した二重特異性HER3−HER2分子を、HER3を発現しHER2を発現していない細胞、又はHER2を発現しHER3を発現していない細胞のいずれかとインキュベートすることにより、HER3−HER2分子の生存細胞での結合能力を、蛍光顕微鏡及びフローサイトメトリー分析を使用してFriedmanら(前出)に記載されるようにモニタリングした。それぞれの受容体に対する異なる親和性を有する異なる構築物が、細胞での異なる受容体発現レベルを区別するかどうかを調べるために特異性の差異を同時に調べた。
【0174】
細胞に対する結合二重特異性Z変異体の生物学的効果を、細胞培養物を0.05nMから5000nMまでの範囲にわたる様々な量の二重特異性Z変異体に曝露し、そして細胞増殖速度及び生存を分析し、そして重要な細胞シグナル伝達経路のリン酸化パターンをウェスタンブロット分析を使用して、HER2特異的Z変異体についてEkerljungらにより(Tumour Biol 27(4):201−10(2006)及びBiochem Biophys Res Commun 377(2):489−94(2008)において)記載されるように分析することにより試験した。試験することができる細胞株としては、MDA−MB−361、NCI−N87、CALU−3及びSKOV−3が挙げられる。またHER3形質導入BT−474及びZR75細胞を使用することもできる。
【0175】
インビボでのHER3−HER2 Z変異体の効果を試験するために、二重特異性構築物を腫瘍標的化及び腫瘍における生物学的効果について評価した。二重特異性HER3−HER2結合Z変異体を以前に記載されたように(Ahlgren et al、Bioconjug Chem 19(1):235−43(2008))放射標識し、そしてインビトロでの細胞取り込み、保持及び内部移行について、記載された方法(Steffen et al、Cancer Biother Radiopharm 20(3):239−48(2005))を使用して特徴付けした。HER3及びHER2受容体を発現している異種移植された腫瘍(例えばMDA−MB−361、NCI−N87、CALU−3及びSKOV−3)を有する動物を、体内分布研究において放射標識したHER3−HER2結合Z変異体を注射し、そして腫瘍取り込み及び他の器官との対比についてモニタリングした。半減期延長部分を有していない構築物については、異種移植されたマウスにおける放射能の分布を、典型的には放射標識二重特異性HER3−HER2結合Z変異体の注射後1、2、4、8、24、48及び72時間の時点で評価した。半減期延長部分を有する構築物については、放射能の分布を典型的には放射標識された二重特異性HER3−HER2結合Z変異体の注射後1、4、12、24、48、72、168及び332時間の時点で、本質的にTolmachevらにより記載されるように(Cancer Res 67(6):2773−82(2007))評価した。放射標識二重特異性Z変異体の取り込みを、HER2及び/又はHER3結合Z変異体単独の取り込みと比較した。二重特異性Z変異体が、単一特異性の相当するものと比較した場合にインビボで腫瘍において優れた取り込み及び保持及び/又は特異性を有する場合、それらの二重特異性変異体は標的化ペイロード治療に適切であり得る。このような治療は、二重特異性分子に融合又は結合体化された潜在的なエフェクター機能、例えば放射性核種、毒性化学分子、毒素、サイトカイン又は光増感剤を利用するだろう。
【0176】
二重特異性Z変異体を、インビボでの内因性の効果についてさらに特徴付けした。マウスに、両方の受容体を発現している腫瘍細胞、又はコントロールとして、受容体の一方のみを発現しているか受容体をどちらも発現していない腫瘍細胞を移植し、そして二重特異性Z変異体の単回又は複数回の注射を行う。腫瘍成長を経時的に、そして注射緩衝液のみで活性化合物(すなわち二重特異性HER3−HER2結合分子)を注射されていないビヒクル群を含むコントロール群に対してモニタリングした、
【0177】
実施例9:HER3及びEGFRに対する二重特異性を有するZ変異体
導入部に記載したように、HER3はHER2にについての好ましいヘテロ二量化パートナーである。しかし、特定の条件では、EGF受容体(EGFR又はHER1)に対する重要な二量化パートナーでもある。二重特異性HER3及びEGFR標的化リガンドを使用することにより、両方の受容体を発現している腫瘍が標的にされ、そして腫瘍促進受容体シグナル伝達がブロックされ得る。
【0178】
二重特異性HER3及びEGFR結合Z変異体は、本質的に上記実施例8に記載されるように操作される。得られた構築物を、HER3及びEGFR受容体を発現する細胞株(例えばA431細胞)において、実施例8で二重特異性HER3及びHER2結合Z変異体について記載される方法と類似の方法で試験することができる。
【技術分野】
【0001】
本発明は、ヒト上皮成長因子受容体3(本明細書ではHER3と呼ぶ)に結合するポリペプチド並びに画像化及び治療における上記ポリペプチドの使用に関する。本発明はまた、HER3及びHER2、又はHER3及びEGFRの両方に対する結合親和性を有する二重特異性リガンドに関する。
【背景技術】
【0002】
EGFR(ErbB1又はHER1)、ErbB2(HER2)、ErbB3(ERBB3又はHER3)及びErbB4(HER4)を含む膜貫通チロシンキナーゼ受容体の上皮増殖因子ファミリーは、細胞内シグナル伝達経路の複雑なネットワークを通した重要な細胞機能(例えば細胞増殖、生存、分化及び遊走)の調節に関与する。今日では、これらの受容体の異常な発現及びシグナル伝達がいくつかの型のがんの発生及び進行に関連するということが十分に確立されており、それによりこれらの受容体は新規ながん治療の開発のための重要な標的となっている。これまでに、EGFR及びHER2受容体が最も広く研究されており、いくつかの標的の薬剤はヒトがんの診断又は処置について行政当局により認可されている。HER3はチロシンキナーゼ活性を欠いており、従ってそのシグナル伝達機能を発揮するために他のErbB受容体と(例えばHER2と)ヘテロ二量体を形成しなければならない(非特許文献1)。HER2−HER3ヘテロ二量体は潜在的に発がん性の単位であると考えられており(非特許文献2)、そしてHER3はHER2の好ましいヘテロ二量化パートナーとして近年注目が高まっている。HER2との二量化はPI3K/Akt経路の活性化並びに腫瘍細胞生存及び増殖の促進をもたらす(非特許文献3)。HER3は、乳がん(非特許文献4)、卵巣がん及び膀胱がん(非特許文献5)を含む多数のヒトのがんにおいて発現され、そしていくつかの肺がん(非特許文献6)及び前立腺がん(非特許文献7)を含む他のがん(非特許文献8)におけるシグナル伝達において重要な役割を果たす。さらに、高レベルの受容体発現は、HER2を過剰発現する患者と比較して有意に短い生存期間と関連するので、HER3発現は予後的な価値を有する(非特許文献9、非特許文献10)。実際HER3は、ErbB受容体−PI3Kシグナル伝達軸のリガンド誘導活性化における主要なノードであると提案されてきた(非特許文献11)。
【0003】
より最近に認可された治療の比較的大部分はEGFRに向けられたものであり、そしてHER2受容体はモノクローナル抗体に基づく。この新しいクラスの生物学的薬剤のがん治療における成功の背後にある理由の1つは、低分子化学薬を使用して以前には達成できなかった新しく独特な作用機構を抗体が提供するということである。より伝統的なアゴニスト(例えば細胞表面受容体の刺激)及びアンタゴニスト(例えば天然のタンパク質−タンパク質相互作用のブロック)の治療効果に加えて、抗体はまた、様々な範囲の免疫学的防御機構を特異的にがん性細胞に向けるために、さらには種々の画像化又は治療用抱合体(例えば化学療法薬、放射性核種及び毒素)の特異的送達を達成するための標的化剤として使用され得る。ErbBファミリーの十分に研究されたEGFR及びHER2受容体メンバーと対照的に、抗HER3抗体の使用に関しては比較的少ない報告しかない。
【0004】
しかしUllrich及び共同研究者らは、抗HER3モノクローナル抗体が乳がんの細胞モデルにおいてHER3仲介シグナル伝達を阻害するということを報告しており(非特許文献12)、そして現在では2つのモノクローナル抗HER3抗体が第1相臨床試験にある(AMG 888、非特許文献13、及びMM−121、Schoeberlら、上記)。
【0005】
しかし、全長モノクローナル抗体を使用するいくつかの成功したがん治療研究が報告されているが、このクラスの薬剤は固形腫瘍を標的とするために(診断的なペイロードの目的でも治療的なペイロードの目的でもなく)常に最適というわけではない。治療効果は、腫瘍全体の薬物の有効な分布に依存し、そして分子画像化は、腫瘍取り込みと周囲の正常組織との間の高い比率に依存する。腫瘍浸透率(血管外漏出を含む)は分子の大きさと逆の関係があるので、比較的大きなIgG分子は本質的に乏しい組織分布及び浸透能を有する。さらに、分子画像化については、抗体の非常に長いインビボでの半減期が比較的高い血液シグナルを生じ、それにより比較的少ない腫瘍対血液コントラストを生じる。
【0006】
HER3はEGFファミリーの他のメンバーと同じ腫瘍細胞で発現され得るので、HER3及びEGFファミリーの別のメンバーを標的にする二重特異性分子の製造は、近年いくらかの興味を集めている。このような二重特異性分子は、例えば分子画像化適用における標的化及びHER3と腫瘍上に発現される別の抗原との同時標的化のための標的化ビヒクルとして利用され得る。
【0007】
しかし、2つの別々の標的に結合する二重特異性モノクローナル抗体を製造することは複雑である。4つの必要なポリペプチド鎖をコードする遺伝子が細胞において産生される場合、10もの異なる組み合わせが可能であり、そして1つの組み合わせのみが望まれる二重特異性抗体を示す。従って、二重特異的結合という構想は、抗体を使用して十分に探査されていない。代わりに、抗体フラグメント及び他の結合分子が二重特異性分子(二重特異性Z変異体を含む)を作製するためにより広く利用されてきた(非特許文献14)。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Citri et al,Exp Cell Res 284(1):54−65(2003)
【非特許文献2】Holbro et al,Proc Natl Acad Sci 100(15):8933−8(2003)
【非特許文献3】Hynes et al,Nat Rev Cancer 5(5):341−54(2005)
【非特許文献4】Bobrow et al,Eur J Cancer 33(11):1846−50(1997)
【非特許文献5】Rajkumar et al,Clin Mol Pathol 49(4)(1996)
【非特許文献6】Engelman et al,Proc Natl Acad Sci 102(10):3788−93(2005)
【非特許文献7】Gregory et al,Clin Cancer Res 11(5):1704−12(2005)
【非特許文献8】Stove et al,Clin Exp Metastasis 21(8):665−84(2004)
【非特許文献9】Tanner et al,J Clin Oncol 24(26):4317−23(2006)
【非特許文献10】Reschke et al,Clin Cancer Res 14(16):5188−97(2008)
【非特許文献11】Schoeberl et al,Sci Signal 2(77):ra 31(2009)
【非特許文献12】van der Horst et al,Int J Cancer 115(4):519−27(2005)
【非特許文献13】Baselga et al,Nat Rev Cancer 9(7):463−75(2009)
【非特許文献14】Friedman et al,Biotechnol Appl Biochem 54:121−31(2009)
【発明の概要】
【発明が解決しようとする課題】
【0009】
発明の説明
例えばHER3を発現する細胞の標的化、このようなHER3を発現する細胞の分子画像化及びHER3関連状態の処置において使用され得る新規なHER3結合剤を提供することが本発明の目的である。
【0010】
本発明のさらなる目的は、HER2及びHER3の両方を発現する病変に対する高い特異性を有する標的化剤を提供することである。
【課題を解決するための手段】
【0011】
一局面によれば、HER3結合モチーフ(BM)を含むHER3結合ポリペプチドが提供され、該モチーフは、
i) EX2X3X4A X6X7EIW X11LPNL X16X17X18QX20 X21AFIX25 X26LX28D、
[ここで、互いに独立して、
X2はR、K、L、W及びVから選択され;
X3はR、H、K、M、S、W、Y及びVから選択され;
X4はA、R、N、D、Q、E、G、H、I、L、K、M、S、T、W及びVから選択され;
X6はA、R、Q、M、S、T及びYから選択され;
X7はA、E、G、H、K、F、S、T、W、Y及びVから選択され;
X11はA、N、D、Q、E、I、L及びTから選択され;
X16はN及びTから選択され;
X17はA、R、N、Q、K、P、S、T及びVから選択され;
X18はA、R、N、E、F、D、Q、G、H、I、L、K、S、T、W、Y及びVから選択され;
X20はA、R及びKから選択され;
X21はA、R、N、G、H、S及びVから選択され;
X25はA、R、E、G、I、K、S、T及びVから選択され;
X26はS及びKから選択され;
X28はA、D、Q、E、F、P、S、T、W及びYから選択される];
及び
ii) i)において定義された配列に対して少なくとも90%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列からなり、
ここでポリペプチドはHER3の細胞外ドメイン(HER3−ECD)に結合する。
【0012】
本明細書において記載されるポリペプチドは、HER3によく結合するという点でHER3に対する良好な結合親和性を示す。配列に関連したHER3結合ポリペプチドのクラスの上記の定義は、選択実験においてHER3とのそれらの相互作用について選択された親骨格の多数の無作為なポリペプチド変異体の分析に基づく。同定されたHER3結合モチーフ又は「BM」は、親骨格の結合領域に相当し、この領域は3ヘリックスバンドルタンパク質ドメイン(three−helical bundle protein domain)内の2つのアルファヘリックスを構成する。親骨格において、2つのBMヘリックスの変更されたアミノ酸残基は、抗体の定常Fc部分との相互作用のための結合表面を構成する。結合表面の無作為な変更及びその後の変異体の選択により、結合表面のFc相互作用能がHER3との相互作用の能力で置き換えられた。
【0013】
当業者に理解されるように、本明細書に定義されるポリペプチドのHER3結合能のように、ポリペプチドの機能はポリペプチドの三次構造に依存する。従って、その三次構造及び機能に大きな影響を与えずにポリペプチドのアミノ酸配列に小さな変化を生成することが可能である。従って一実施態様において、本ポリペプチドは、結果として生じた配列がi)により定義されるクラスに属する配列に対して少なくとも90%同一である、例えばi)により定義されるクラスに属する配列に対して少なくとも93%同一、例えば少なくとも97%同一であるように改変された、i)BMの変異体を含む。いくつかの実施態様において、小さな変化は本明細書において開示されるHER3結合ポリペプチドの配列の全ての位置において作製され得る。他の実施態様において、小さな変化は、骨格アミノ酸残基とも示される不変位置においてのみ作製され得る。このような場合において、変化は可変位置、すなわち「X」で示される位置(例えば上記の定義されたBMのX2、X3、X4、X6、X7、X11、X16、X17、X18、X20、X21、X25、X26及びX28)において許容されていない。例えば、アミノ酸残基の特定の機能による分類(例えば疎水性、親水性、極性など)に属するアミノ酸残基は、同じ機能グループからの別のアミノ酸残基と交換され得るということが可能である。
【0014】
本明細書及び添付の特許請求の範囲を通じて使用される用語「%の同一性(同一性%)」は以下のようにして算出される。CLUSTAL Wアルゴリズム(Thompson、J.D.、Higgins、D.G.and Gibson、T.J.、Nucleic Acids Research、22: 4673−4680(1994))を使用して問い合わせ配列を対象配列と整列させる。整列された配列の最短のものに対応する範囲にわたって比較を行う。整列された配列の最短のものは、いくつかの場合には標的配列であるかもしれない(例えば29アミノ酸残基のHER3結合モチーフ)。他の場合、問い合わせ配列が整列された配列の最短のものを構成するかもしれない。問い合わせ配列は、例えば少なくとも10アミノ酸残基、例えば少なくとも20アミノ酸残基からなるものであり得る。各位置におけるアミノ酸残基を比較し、そして標的配列において同一の対応を有する問い合わせ配列中の位置のパーセントを同一性%として報告する。
【0015】
本明細書及び添付の特許請求の範囲を通じて使用される用語「%の類似性(類似性%)」は以下のやり方で計算される。配列アラインメント及び比較を基本的に同一性%の計算に関して記載したように行う。しかし「類似性」は以下のように解釈されるべきである。2つのアミノ酸残基は、それらがアミノ酸残基の同じグループに属する場合に類似と見なされる。アミノ酸残基のグループの非限定的な例は、アミノ酸残基Ala、Val、Phe、Pro、Leu、Ile、Trp、Met及びCysを含む疎水性グループ;アミノ酸残基Lys、Arg及びHisを含む塩基性グループ;アミノ酸残基Glu及びAspを含む酸性グループ;非荷電アミノ酸残基Gln、Asn、Ser、Thr及びTyrを含む親水性グループ;並びにアミノ酸残基Glyを含む天然グループである。従って、各位置におけるアミノ酸残基を比較し、そして標的配列中に類似した対応を有する問い合わせ配列中の位置のパーセントを類似性%として報告する。
【0016】
この説明全体を通して、本発明に従う代替の実施態様は、特定の同一性パーセントの代わりに、対応する類似性パーセントを満たす。他の代替の実施態様は、特定の同一性パーセントに加えて、各配列についての好ましい同一性パーセントの群から選択される別のより高い類似性パーセント満たす。例えば、配列は別の配列に対して70%の類似性であってもよく;又はそれは別の配列に対して70%の同一性であってもよく;又は別の配列に対して70%の同一性及び90%の類似性であってもよい。
【0017】
上に開示されるポリペプチドの一実施態様において、X2はR、K及びWから選択される。
【0018】
上に開示されるポリペプチドのさらなる実施態様において、X3はR、K、S及びYから選択される。
【0019】
上に開示されるポリペプチドのさらなる実施態様において、X4はA、R、N、Q、H、I、L、K、S、T及びVから選択される。
【0020】
上に開示されるポリペプチドのさらなる実施態様において、X6はA、S、T及びYから選択される。
【0021】
上に開示されるポリペプチドのさらなる実施態様において、X7はA、G、F、W、Y及びVから選択される。
【0022】
上に開示されるポリペプチドのさらなる実施態様において、X11はQ及びEから選択される。
【0023】
上に開示されるポリペプチドのさらなる実施態様において、X17はR、Q、T及びVから選択される。
【0024】
上に開示されるポリペプチドのさらなる実施態様において、X18はR、Q、G、H、I、L、S、T、W、Y及びVから選択される。
【0025】
上に開示されるポリペプチドのさらなる実施態様において、X20はR及びKから選択される。
【0026】
上に開示されるポリペプチドのさらなる実施態様において、X21はA及びGから選択される。
【0027】
上に開示されるポリペプチドのさらなる実施態様において、X25はA、R、G、K及びSから選択される。
【0028】
上に開示されるポリペプチドのさらなる実施態様において、X28はA、Q、E、F、S、W及びYから選択される。
【0029】
上に開示されるポリペプチドのさらなる実施態様において、X2はR及びKから選択される。
【0030】
上に開示されるポリペプチドのさらなる実施態様において、X3はK及びYから選択される。
【0031】
上に開示されるポリペプチドのさらなる実施態様において、X4はA、R、N、Q、H、L、K、S、T及びVから選択される。
【0032】
上に開示されるポリペプチドのさらなる実施態様において、X6はT及びYから選択される。
【0033】
上に開示されるポリペプチドのさらなる実施態様において、X7はA、G、F、Y及びVから選択される。
【0034】
上に開示されるポリペプチドのさらなる実施態様において、X17はR、Q及びVから選択される。
【0035】
上に開示されるポリペプチドのさらなる実施態様において、X18はR、Q、I、L、T、Y及びVから選択される。
【0036】
上に開示されるポリペプチドのさらなる実施態様において、X25はA、G、K及びSから選択される。
【0037】
上に開示されるポリペプチドのさらなる実施態様において、X28はA、Q、E、F、S及びWから選択される。
【0038】
上で開示されるHER3結合ポリペプチドの一実施態様において、該ポリペプチドはHER3結合モチーフ(BM)を含み、このモチーフは、
i) EX2X3X4A YX7EIW X11LPNL X16X17X18QX20 X21AFIX25 X26LX28D、
[ここで、互いに独立して、
X2はK、R及びWから選択され;
X3はK、R、及びYから選択され;
X4はA、D、E、G、H、I、K、L、M、N、Q、R、S、T、V及びWから選択され;
X7はA、E、F、G、H、K、S、T、V、W及びYから選択され;
X11はE及びQから選択され;
X16はN及びTから選択され;
X17はR、T及びVから選択され;
X18はR、T及びVから選択され;
X20はK及びRから選択され;
X21はA及びGから選択され;
X25はA、G、K、R及びSから選択され;
X26はS及びKから選択され;
X28はA、D、E、F、P、Q、S、T、W及びYから選択される];
及び
ii) i)において定義される配列に対して少なくとも93%の同一性を有するアミノ酸配列、から選択されるアミノ酸配列からなり、ここで該ポリペプチドは、HER3の細胞外ドメインに結合する。
【0039】
上に開示されるポリペプチドのさらなる実施態様において、X2X3はKY、RY、WK及びWRから選択される。
【0040】
上に開示されるポリペプチドのさらなる実施態様において、X11はQである。
【0041】
上に開示されるポリペプチドのさらなる実施態様において、X17X18はRV、RT、VR及びRRから選択される。
【0042】
上に開示されるポリペプチドのさらなる実施態様において、X20はKである。
【0043】
上に開示されるポリペプチドのさらなる実施態様において、X21はAである。
【0044】
上に開示されるポリペプチドのさらなる実施態様において、X2X3はKY及びRYから選択される。
【0045】
上に開示されるポリペプチドのさらなる実施態様において、X4はK、L、S及びTから選択される。
【0046】
上に開示されるポリペプチドのさらなる実施態様において、X7はF、G及びYから選択される。
【0047】
上に開示されるポリペプチドのさらなる実施態様において、X17X18はRV、RT及びVRから選択される。
【0048】
上に開示されるポリペプチドのさらなる実施態様において、X25はA、G及びSから選択される。
【0049】
上で開示されるポリペプチドのなおさらなる実施態様において、アミノ酸配列i)は、以下の条件I、II、III及びIV:
I) X2X3はKY及びRYから選択される;
II) X11はQである;
III) X20はKである;
IV) X21はAである、
の少なくとも1つを満たす。
【0050】
アミノ酸配列i)は、例えば4つの条件I、II、III及びIVのうち少なくとも2つ、例えば少なくとも3つ、又は全てを満たし得る。
【0051】
以下の実験項に詳細に記載されるように、HER3結合変異体の選択は、個々のHER3結合モチーフ(BM)配列の同定をもたらした。これらの配列は、この局面に従うHER3結合ポリペプチドの個々の実施態様を構成する。個々のHER3結合モチーフの配列を図1に、そして配列番号1〜334として示す。この局面のいくつかの実施態様において、BM配列i)は、配列番号1〜配列番号66のいずれか1つから選択され、例えば配列番号22〜66のいずれか1つから、例えば配列番号23〜25、配列番号27〜28、配列番号32、配列番号35〜36、配列番号40、配列番号42、配列番号44〜45、配列番号53〜54及び配列番号56から選択される。
【0052】
特定の実施態様において、HER3結合モチーフ(BM)はこのようにして3ヘリックスバンドルタンパク質ドメインの一部を形成する。例えば、BMは該3ヘリックスバンドルタンパク質ドメイン内で相互接続ループを有する2つのアルファヘリックスを本質的に構成し得る。これらの実施態様において、本発明のHER3結合ポリペプチドは:
i) K−[BM]−DPSQS XaXbLLXc EAKKL NDXdQ;
[ここで
[BM]は上で定義されたHER3結合モチーフであり;
XaはA及びSから選択され;
XbはN及びEから選択され;
XcはA、S及びCから選択され;
XdはA及びSから選択される];
及び
ii) 上で定義される配列のいずれか1つに対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含み得る。上記アミノ酸配列は、上で定義された配列のいずれか1つに対して少なくとも82%、例えば少なくとも84%、例えば少なくとも86%、例えば少なくとも88%、例えば少なくとも90%、例えば少なくとも92%、例えば少なくとも94%、例えば少なくとも96%、例えば少なくとも98%の同一性を有し得る。
【0053】
上で定義されたHER3結合ポリペプチドの一実施態様において、XaはAであり;XbはNであり;XcはAであり、そしてXdはAである。
【0054】
上で定義されたHER3結合ポリペプチドのさらなる実施態様において、XaはAであり;XbはNであり;XcはCであり、そしてXdはAである。
【0055】
上で定義されたHER3結合ポリペプチドのさらなる実施態様において、XaはSであり;XbはEであり;XcはSであり、そしてXdはSである。
【0056】
上で定義されたHER3結合ポリペプチドのさらなる実施態様において、XaはSであり;XbはEであり;XcはCであり、そしてXdはSである。
【0057】
なおさらなる実施態様において、上で定義されるHER3結合ポリペプチドのアミノ酸配列は、配列番号335〜668から選択され、特に配列番号335〜400から、例えば配列番号356〜400から、例えば配列番号357〜359、配列番号361〜362、配列番号366、配列番号369〜370、配列番号374、配列番号376、配列番号378〜379、配列番号387〜388及び配列番号390から選択される。
【0058】
特定の実施態様において、上記3ヘリックスバンドルタンパク質ドメインは細菌受容体タンパク質のドメインから選択される。このようなドメインの非限定的な例は、黄色ブドウ球菌(Staphylococcus aureus)由来のプロテインAの5つの異なる3ヘリックスドメイン、例えばドメインB、及びその誘導体である。いくつかの実施態様において、3ヘリックスバンドルタンパク質ドメインはプロテインZの変異体であり、これはブドウ球菌プロテインAの上記ドメインBから誘導される。
【0059】
従って、さらなる実施態様において:
i) YAK−[BM]−DPSQS SELLXc EAKKL NDSQA P;
[ここで[BM]は上で定義されたHER3結合モチーフであり、そして
XcはS及びCから選択される];及び
ii) i)において定義される配列のいずれか1つに対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含むHER3結合ポリペプチドが提供される。
【0060】
あるいは:
i) FNK−[BM]−DPSQS ANLLXc EAKKL NDAQA P;
[ここで[BM]は上で定義されたHER3結合モチーフであり、そして
XcはA及びCから選択される];及び
ii) 上記i)において定義された配列のいずれか1つに対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含むHER3結合ポリペプチドが提供される。
【0061】
上で考察したように、上記のアミノ酸配列と比較して小さな変化を含むポリペプチドもまた本出願の範囲内である。従って、いくつかの実施態様において、上で定義されたHER3結合ポリペプチドは、例えば、本明細書中に記載される配列に対して、少なくとも82%、少なくとも83%、少なくとも85%、少なくとも87%、少なくとも89%、少なくとも91%、少なくとも92%、少なくとも94%、少なくとも96%又は少なくとも98%同一である配列を有し得る。
【0062】
いくつかの実施態様において及び以下の実験項において開示されるように、HER3結合モチーフは58アミノ酸のポリペプチドの一部を形成し得る。このようなポリペプチドは、例えば配列番号669〜1002のいずれか1つから選択される配列、特に配列番号669〜734から選択される配列、例えば配列番号690〜734から、例えば配列番号691〜693、配列番号695〜696、配列番号700、配列番号703〜704、配列番号708、配列番号710、配列番号712〜713、配列番号721、配列番号722及び配列番号724から選択される配列を含み得る。
【0063】
本明細書に記載されるいずれかの局面に従うHER3結合ポリペプチドは、相互作用のKD値が多くとも1×10-6M、又は例えば多くとも1×10-7M、例えば多くとも1×10-8MであるようにHER3に結合し得る。本明細書において使用される用語「HER3結合」及び「HER3に対する結合親和性」は、例えばBiacore機器(GE Healthcare)におけるような表面プラズモン共鳴技術の使用により試験され得るポリペプチドの特性を指す。HER3結合親和性は、例えばHER3をBiacore機器のセンサーチップに固定し、そして試験しようとするポリペプチドを含有するサンプルをチップ上に通す実験において試験され得る。あるいは以下の実施例に記載されるように、HER3結合親和性は細胞上(on−cell)実験で試験され得る。細胞上実験は、例えば様々な濃度の標識されたHER3を試験しようとするポリペプチドを提示している細胞とともにインキュベートすることにより行われ得る。その後、細胞集団を蛍光細胞分析分離システム(FACS)におけるようなフローサイトメーターで分析し得、そして平均蛍光強度データを決定し得る。続いて平均蛍光をHER3濃度に対してプロットし、そして見かけの平衡解離定数KDを決定するために1部位結合モデルにフィッティングし得る。KD測定において使用されるHER3又はそのフラグメントは、例えば配列番号1007で表されるアミノ酸配列(HER3−ECD)を含み得る。
【0064】
以下の実施例において説明されるように、HER3結合変異体の選択は、例えばタンパク質骨格の未処理の変異体の選択のためのファージディスプレイ、続いて場合により親和性成熟HER3結合変異体の選択のための親和性成熟及び細胞ディスプレイにより達成され得る。しかし、ファージベースでも、細胞ベースでもその他でも、いずれの選択システムもHER3結合ポリペプチドの選択に使用され得ることが理解される。以下の実施例において、グラム陽性細菌スタフィロコッカス・カルノーサス(Staphylococcus carnosus)での組み換えタンパク質の細胞表面ディスプレイのための発現系を使用した(Jonsson et al、Biotechnol Applied Biochem 54:93−103(2009))。しかし、ファージ粒子の大きさを比較して、より大きなサイズの細胞は一般的にハイスループットスクリーニング及び細胞提示タンパク質ライブラリーからの選択のための蛍光細胞分析分離(FACS)の使用を可能にする。
【0065】
IgG分子と比較して、本発明のHER3結合ポリペプチドは比較的小さく、これにより、HER3結合ポリペプチドを広範囲の適用において有用にするだけでなく、固相ペプチド合成を使用した製造が可能になり得る。固相合成経路は、例えば分子画像化における放射標識に使用されるキレート剤のような非生物学的基の部位特異的結合を容易にする。
【0066】
従って、当業者には当然のことながら、様々な修飾及び/又は付加が、本発明の範囲から逸脱することなくポリペプチドを特定の適用に合わせるために、本明細書に開示されるいずれかの局面にしたがってなされ得る。例えば、いずれかの局面に従うHER3結合ポリペプチドは、さらなるC末端アミノ酸及び/又はN末端アミノ酸を含み得る。このようなポリペプチドは、ポリペプチド鎖におけるかなり最初の方及び/又は最後の方の位置に、すなわちN末端及び/又はC末端にさらなるアミノ酸残基を有するポリペプチドとして理解されるべきである。従って、HER3結合ポリペプチドは、いずれかの適切な数のさらなるアミノ酸残基、例えば少なくとも1つの付加的なアミノ酸残基を含み得る。付加的アミノ酸残基はそれぞれ、例えばポリペプチドの製造、精製、インビボ若しくはインビトロでの安定化、カップリング、又は検出を改善するために、独立して又は集合的に付加され得る。このような付加的アミノ酸残基は、化学的カップリングの目的のために付加された1つ又はそれ以上のアミノ酸残基を含み得る。この一例はシステイン残基の付加である。このような付加的アミノ酸残基はまた、ポリペプチドの精製又は検出のための「タグ」、例えばタグに対して特異的な抗体との相互作用のため、又はHis6タグの場合には固定化金属アフィニティークロマトグラフィー(IMAC)のための、His6タグ又は「myc」(c−myc)タグ又は「FLAG」タグを生じ得る。
【0067】
上で考察されたさらなるアミノ酸を、化学的連結を用いて(公知の有機化学方法を使用して)、又は融合タンパク質としてのHER3結合ポリペプチドの発現のような他の手段を用いてHER3結合ポリペプチドにカップリングさせ得る。
【0068】
上で考察されたさらなるアミノ酸は、例えば1つ又はそれ以上のポリペプチドドメインを含み得る。さらなるポリペプチドドメインは、HER3結合ポリペプチドに別の機能、例えば別の結合機能、又は酵素的機能、又は毒素機能(例えばイムノトキシン)、又は蛍光シグナル機能、又はその組み合わせを与え得る。
【0069】
いくつかの実施態様において、さらなるポリペプチドドメインは、インビボでHER3結合ポリペプチドの半減期を増加させる半減期延長部分を含み得る。当業者に理解されるように、増加した、又は延長された半減期は、特定の分子の血液からの遅延されたクリアランスを意味する。半減期延長部分は、例えば血清アルブミンへのインビボでの結合を可能にするペプチド又はタンパク質を含み得る。特に、半減期延長部分はアルブミン結合部分であり得る。アルブミン結合部分は、例えば天然に存在するポリペプチド、又はそのアルブミン結合フラグメント、又は改変ポリペプチドからなるものであってもよい。改変ポリペプチドは、新しいか又は増強された特性、例えばアルブミンのような分子に対する結合親和性を付与する目的で、それをタンパク質工学技術、例えば部位特異的又は無作為化されたアプローチでの変異及び変更を受けさせることにより、天然に存在する出発ポリペプチドから誘導され得る。
【0070】
このような改変アルブミン結合ポリペプチドは、例えばアルブミンに対するその特異的結合親和性について選択された、タンパク質骨格の変異体であってもよい。特定の実施態様において、タンパク質骨格は、連鎖球菌プロテインGのドメイン又はその誘導体、例えば連鎖球菌株G148由来のプロテインGのドメインGA1、ドメインGA2及びドメインGA3、特にドメインGA3から選択され得る。従って、HER3結合ポリペプチドの一実施態様において、さらなるアミノ酸は、連鎖球菌プロテインGのアルブミン結合ドメイン(ABD)、又はその誘導体を含む。本発明のHER3結合ポリペプチドにおけるさらなるポリペプチドドメインとして含まれ得るアルブミン結合ドメインの一例は、配列番号1005で示される。適切なアルブミン結合ドメインの他の例はWO2009/016043に開示される。このようなABD延長ポリペプチドはインビボで血清アルブミンに結合し、そしてそのより長い半減期から利益を得、これによりポリペプチド自体の正味の半減期が増加する(例えばWO91/01743を参照のこと)。
【0071】
本発明に従うHER3結合ポリペプチドが上で定義されるアルブミン結合部分を含む場合、HER3結合(bindnig)ポリペプチドの全体の大きさは比較的小さい。例えばヒト対象のような哺乳動物対象に投与された場合、HER3結合ポリペプチドのアルブミン結合部分は、血清アルブミンと非共有結合で結合し、そしてそれによりポリペプチドは減少した腎クリアランス及び上皮細胞での増加した再循環から利益が得られ得る。しかし組織浸透は、血清アルブミンの血管外漏出特性に起因して速いままである。さらに、半減期延長部分を含むHER3結合ポリペプチドは、対応する半減期延長部分の無いポリペプチドと比較して、インビボで延長された半減期を示すだけでなく、インビボで減少した免疫応答を示す(例えばWO2005/097202を参照のこと)。
【0072】
さらなるポリペプチドドメインは、同じ結合機能を有するHER3結合ポリペプチドを生じ得る。従って、さらなる実施態様において、そのアミノ酸配列は同じであっても異なっていてもよい少なくとも2つのHER3結合ポリペプチド単量体単位を含むHER3結合ポリペプチドが提供される。ポリペプチドの多量体形態は、それぞれがHER3結合モチーフを有し、かつそれぞれが多量体内の「単量体」を形成する、適切な数のドメインを含み得る。これらのドメインは全て同じアミノ酸配列を有していてもよいが、あるいはそれらが異なるアミノ酸配列を有していてもよい。特に、本発明のHER3結合ポリペプチドは、ホモ二量体又はヘテロ二量体を形成し得る。
【0073】
上記のさらなるポリペプチドドメイン(単数又は複数)は、公知の有機化学方法を使用して共有結合カップリングによりHER3結合ポリペプチドに結合され得る。あるいは、さらなるポリペプチドドメイン(単数又は複数)を含むHER3結合ポリペプチドを1つ又はそれ以上の融合ポリペプチドとして、例えばポリペプチドのの組み換え発現のための系において発現させてもよいし、直接的又はリンカー(例えばアミノ酸リンカー)を介して、いずれかの他のやり方で結合させてもよい。
【0074】
HER3結合ポリペプチドの一実施態様において、本ポリペプチドは天然に存在するHER3リガンドと同じエピトープに結合するか、又はそれに十分に接近して天然に存在するHER3リガンド(例えばニューレグリン−1及び2)のHER3への結合をブロックする。従ってHER3結合ポリペプチドは、HER3遮断薬として、かつ/又はインビボでのHER3媒介細胞シグナル伝達を阻害するために使用され得る。
【0075】
さらなる局面において、上記のHER3結合ポリペプチドをコードするポリヌクレオチドが提供される。このようなポリヌクレオチドを含む発現ベクターは、例えば宿主細胞における発現によりHER3結合ポリペプチドの生産を可能にし得る。
【0076】
HER3結合ポリペプチドは、様々な医療、獣医学及び診断適用における従来の抗体又は低分子量物質に対する代替として使用され得る。当業者は、本発明のポリペプチドが、試薬のHER3に対する親和性を必要とするいずれかの方法において有用であり得るということを理解するだろう。従って、HER3結合ポリペプチドは、このような方法における検出試薬、捕捉試薬又は分離試薬として使用され得る。さらに、HER3結合ポリペプチドは、例えば哺乳動物対象における細胞表面でのHER3発現を特徴とする疾患の存在を明らかにし、診断し、そして試験するために診断法において使用され得る。従って、HER3発現は、診断上、本発明のポリペプチドの使用によりインビボでモニタリングされ得る。診断モニタリングは、例えば分子画像化によるHER3発現の判断のための診断用核種の送達により達成され得る。本発明に従うHER3結合ポリペプチドはまた、それ自身だけで治療剤若しくは診断剤として、又はHER3タンパク質に対する直接的(例えば、イットリウム−90、ルテチウム−177及びトリウム−227のような放射性核種、マイタンシノイドのような毒素を含む毒性分子)若しくは間接的(例えばがんワクチン、免疫賦活性分子)な効果を有する他の治療剤若しくは診断剤を標的とするための手段として有用であり得る。治療効果は、受容体相互作用をブロックすることにより、あるいは腫瘍細胞へ有効なペイロード(payload)を送達するための標的としてHER3結合ポリペプチドを使用することにより、潜在的に媒介され得る。
【0077】
さらなる局面において、上で定義されたHER3結合ポリペプチド;HER2結合ポリペプチド;及びHER3結合ポリペプチドとHER2結合ポリペプチドとを連結するための連結部分を含む、HER3及びHER2に対する結合親和性を有するリガンドが提供される。このようなリガンドは、2つの異なる結合機能を含む二重特異性ヘテロ二量体の例として解釈されるべきであり:1つは上で開示されるポリペプチドによりもたらされるHER3に対する結合機能であり、そして1つはHER2結合ポリペプチドによりもたらされるHER2に対する結合機能である。当然のことながら、本発明に従うHER3結合ポリペプチドは、リガンド中に含まれ得る。2つの結合機能は、2つの別々の単量体単位としてリガンドに含まれる。
【0078】
このような本発明に従う二重特異性リガンドは、HER3及びHER2の両方が関与するがんの検出、特徴付け及び/又は診断において有用であり得る。検出、特徴付け及び/又は診断は、例えばガンマ線若しくは陽電子放出核種を使用する分子画像により、又は近赤外蛍光プローブを使用することにより達成され得る。上記のように、HER3はHER2についての好ましいヘテロ二量化パートナーであり、そしてHER2の過剰発現を特徴とする腫瘍の増殖を促進するために重要である。HER2は公知のリガンドがなく、リガンド誘導シグナル伝達に関してHERファミリーの他の受容体に依存すると現在では考えられている。しかし本発明に従う二重特異性リガンドは、診断のためだけでなく、標的化治療ペイロード適用にも使用され得る。ペイロード適用において使用される化合物としては、とりわけ、放射性核種、プロドラッグ酵素、サイトカイン、化学毒性分子、毒素及び光増感剤が挙げられる。標的化ペイロード適用については、正常組織と疾患組織との間の高度な識別が非常に重要である。
【0079】
2つの異なる受容体に結合する二重特異性リガンドに含まれる両方の結合ポリペプチドの親和性を微調整することにより、リガンドは両方の受容体を同時に発現している病変にのみ優先的に位置し、そして蓄積し得る。従って、HER2及びHER3結合機能は、それらの個々の標的に対して2つの同等に強力な結合機能を有するリガンドをもたらし得る。従って、HER3結合ポリペプチドのHER3に対する結合親和性は、HER2結合ポリペプチドのHER2に対する結合親和性とほぼ同位であり得る。一実施態様において、HER2結合ポリペプチドは、相互作用のKD値が多くとも1×10-6M、例えば多くとも1×10-7M、例えば多くとも1×10-8MであるようにHER2に結合し得る。別の実施態様において、HER2結合ポリペプチドのHER2に対する結合は、対応するHER3結合ポリペプチドのHER3に対する結合と同程度に強力であるかより強力であり得る。従って、HER2結合ポリペプチドとHER2との間の相互作用のKD値は、多くてHER3結合ポリペプチドとHER3との間の相互作用のKD値である。HER3結合ポリペプチドとHER3との間の相互作用のKD値は、HER2結合ポリペプチドとHER2との間の相互作用のKD値の例えば5倍、10倍又は100倍であり得る。
【0080】
一実施態様において、二重特異性リガンドのHER2結合ポリペプチドは改変ポリペプチドである。HER2結合ポリペプチドは、HER2に対するその特異的結合親和性について選択された、タンパク質骨格の変異体である改変ポリペプチドであり得る。タンパク質骨格の非限定的な例は、ブドウ球菌プロテインA、並びにそのドメイン及び誘導体からなる群より選択される。いくつかの実施態様において、HER2結合ポリペプチドは、黄色ブドウ球菌(Staphylococcus aureus)由来のプロテインAのドメインBから誘導された改変タンパク質である。この改変タンパク質は、特にプロテインZ誘導体を含んでいてもよく、これは上記プロテインAのドメインBから誘導されたプロテインZの変異体である。
【0081】
リガンドのいくつかの実施態様において、改変HER2結合タンパク質は、
i) 配列番号1003に対応する、
YAKEM RNAYW EIALL PNLTN QQKRA FIRKL YDDPS QSSEL LSEAK KLNDS Q;及び
ii) i)において定義される配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む。
【0082】
例えば1から45個までのアミノ酸を含むペプチドリンカーによりHER3結合ポリペプチド及びHER2結合ポリペプチドが連結され得る。連結部分は特にHER3結合ポリペプチドとHER2結合ポリペプチドのN末端及び/又はC末端のカップリングのために配置され得る。このようにして生じる連結は、C末端とC末端との連結、C末端とN末端との連結又はN末端とC末端の連結から選択され得る。連結部分のHER3結合ポリペプチド及びHER2結合ポリペプチドへのカップリングは、例えば化学的結合(公知の有機化学方法を使用する)を用いて、又は融合タンパク質としてのリガンドの発現のようないずれかの他の手段により達成され得る。
【0083】
リガンドは、上記のように、インビボでのリガンドの半減期を延長するための半減期延長部分をさらに含み得る。半減期の延長は、多数の種々の半減期延長部分により達成され得、そしてこのような部分の非限定的な例は、アルブミン、HER3結合ポリペプチドに関連して上で定義されたようなアルブミン結合部分、IgGからのFc部分、及びポリエチレングリコール(PEG)のようなポリマー化合物から選択される。哺乳動物血清において最も豊富なタンパク質である血清アルブミンへのリガンドのカップリングは、インビボでのリガンドの半減期の延長をもたらし得る。
【0084】
特に、アルブミン結合部分は、
i) 配列番号1005で示される、
LAEAK VLANR ELDKY GVSDF YKRLI NKAKT VEGVE ALKLH ILAAL P;及び
ii) i)において定義された配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含むアルブミン結合部分のような、連鎖球菌プロテインGのドメインGA3から誘導された改変タンパク質であり得る。
【0085】
半減期延長部分は、例えば公知の有機化学方法を使用して共有結合カップリングによりリガンドの残部に接続されても、他のやり方で直接、又は多数のアミノ酸を含むリンカーを介して結合されてもよい。半減期延長部分がアルブミン又はアルブミン結合部分により表される場合、リガンドはそれ自体1つ又はそれ以上の融合ポリペプチドとしてポリペプチドの組み換え発現のための系において発現されてもよい。一実施態様において、半減期延長部分は、リガンドに存在するシステイン残基を介してリガンドの残部に結合する。特に、半減期延長部分は、結合モチーフから離れた部位に導入された、HER3又はHER2結合ポリペプチドのシステイン残基に結合され得る。別の実施態様において、半減期延長部分は連結部分を介してリガンドの残部に結合する。
【0086】
特定の実施態様において、連結部分は上で定義された半減期延長部分を含む。従って、アルブミン、アルブミン結合部分又はPEGは例えばHER3結合ポリペプチドをHER2結合ポリペプチドに接続するために使用され得る。
【0087】
いくつかの実施態様において、本発明に従うリガンドは、さらなるC末端又はN末端アミノ酸を含み得る。このようなさらなるアミノ酸は、HER3結合ポリペプチドに関して上で記載したように、例えばポリペプチドの生産、精製、インビボ若しくはインビトロでの安定化、カップリング又は検出を改善するために、個別に、又は集合的に付加され得る。特定の実施態様において、さらなるアミノ酸は1つ又はそれ以上のポリペプチドドメインを構成し得る。このようなさらなるポリペプチドドメインの機能は、既にリガンドに含まれている機能と同じであっても異なっていてもよい。さらなるポリペプチドドメインは、例えば別のHER3結合機能及び/又は別のHER2結合機能を有するリガンドを生じ得る。従って、本発明に従う二重特異性リガンドは、HER3結合機能の多量体及び/又はHER2結合機能の多量体を含み得る。多量体結合機能を含む二重特異性リガンドの一例は、2つのHER3結合ポリペプチド及び2つのHER2結合ポリペプチドを含むリガンドである。
【0088】
上で定義されたリガンドは、一実施態様において、インビボでのHER2及びHER3の二量化を防止し得る。より詳細には、リガンドは、例えば哺乳動物対象の細胞表面で発現されたHER2及び/又はHER3に結合することにより、対象におけるHER3仲介シグナル伝達を阻害し得る。リガンドの二重特異性に起因して、これは、それらがもはやシグナル伝達を行うことができないような位置において2つの受容体に結合してブロックし得る。当業者には当然のことながら、従って本発明のリガンドは、細胞表面上のHER2及び/又はHER3の発現、特に過剰発現に関連する状態の処置のために利用され得る。
【0089】
しかしHER3はEGFRともヘテロ二量体を形成し得、それ故PI3K/Akt経路を活性化する活性シグナル伝達単位を形成する(Engelman et al、Proc Natl Acad Sci 102(10):3788−93(2005))。実際、HER3は、明らかにErbBリガンドの上方調節のために非小細胞肺がん(NSCLC)において過剰発現されるが(Fujimoto et al、Cancer Res 65(24):11478−85(2005))、一方でHER3は正常肺上皮で発現されない。従って、関連する局面において、上で定義されたHER3結合ポリペプチド;EGFR結合ポリペプチド;及びHER3結合ポリペプチドとEGFR結合ポリペプチドとを連結するための連結部分を含む、HER3及びEGFRに対する結合親和性を有するリガンドが提供される。当然のことながら、本発明の上記の局面において記載される特徴は、本発明のこの局面にも適用可能である。特に、このような二重特異性リガンドは、HER3及びEGFRを標的にし得、そしてHER3及びHER2に対する二重特異性リガンド結合に関して上で記載した作用様式と類似しているか又は同じ作用様式で作用する。HER3及びEGFRに対する二重特異性リガンド結合の例は、
i) 配列番号1004に相当する、
YAKEM WIAWE EIRNL PNLNG WQMTA FIAKL LDDPS QSSEL LSEAK KLNDS Q;及び
ii) i)において定義される配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるEGFR結合ポリペプチドを含み得る。
【0090】
HER3及びEGFR結合リガンドにおいて使用され得るEGFR結合ポリペプチドの他の例はWO2007/065635に開示される。
【0091】
他の実施態様において、本発明に従うHER3結合ポリペプチド又は二重特異性リガンドは、治療剤又は診断剤に、インビボ及びインビトロの両方で、HER3を発現する細胞、特にHER3を過剰発現する細胞を標的にさせる際に使用される。上で考察したように、本発明のリガンドは、HER3及び/若しくはHER2、又はHER3及び/若しくはEGFRを発現するか又は過剰発現する細胞を標的にする際に利用され得る。従って関連する局面において、本明細書において定義されるHER3結合ポリペプチド、又は上で定義されるリガンド、及び治療剤の組み合わせが提供される。治療剤の非限定的な例は、細胞傷害性ペイロード、例えば放射性核種、低分子量薬物又は毒素から選択される。このような治療剤は、化学的に結合されるか、そうでなければHER3結合ポリペプチド又はリガンドに結合される。
【0092】
本発明に従う組み合わせ、HER3結合ポリペプチド又はリガンドはさらに、治療における、例えばHER3関連状態、例えばがん疾患、例えば結腸がん、子宮内膜がん、胃がん、神経膠腫、乳がん、膵臓がん、頭頸部扁平上皮がん、肺がん、黒色種、髄芽腫、神経上皮腫、卵巣がん、パジェット病、乳頭様甲状腺がん、前立腺がん、皮膚扁平上皮がん、移行上皮がん及び前庭神経鞘腫から選択される、哺乳動物(例えばヒト対象)におけるHER3関連状態の処置のための、医薬として使用され得る。従って、HER3関連状態、例えば上述のHER3関連状態の処置のための医薬の製造のための本発明のHER3結合ポリペプチド又はリガンドの使用もまた、本発明の範囲内である。
【0093】
関連する局面において、HER3結合ポリペプチド、又は上記のリガンド若しくは組み合わせを、それを必要とする哺乳動物に投与することを含む、HER3関連状態の処置方法が提供される。結果として、本処置方法において、対象は、本発明に従うHER3結合ポリペプチド、リガンド又は組み合わせで処置される。この方法のより特定の実施態様において、HER3結合ポリペプチド、リガンド又は組み合わせの、対象の細胞表面で発現されたHER3に対する結合は、受容体活性化を阻害する。この結合は、例えばHER3仲介シグナル伝達を阻害し得る。本処置方法の一実施態様において、上記HER3関連状態は、がん疾患、例えば結腸がん、子宮内膜がん、胃がん、神経膠腫、乳がん、膵臓がん、頭頸部扁平上皮がん、肺がん、黒色種、髄芽腫、神経上皮腫、卵巣がん、パジェット病、乳頭様甲状腺がん、前立腺がん、皮膚扁平上皮がん、移行上皮がん及び前庭神経鞘腫から選択される。
【0094】
別の実施態様において、本発明のリガンドを、それを必要とする哺乳動物対象に投与することを含む、HER3及び/又はHER2関連状態の処置方法が提供される。
【0095】
当業者に理解されるように、そして上で考察したように、本発明に従うHER3結合ポリペプチド又はリガンドは、HER3又はHER3及び/若しくはHER2関連状態の診断のような診断において使用され得る。本発明に従うHER3結合ポリペプチドは、診断剤と別々に使用されても組み合わせて使用されてもよい。このような組み合わせは、HER3関連状態の診断において、例えばHER3を過剰発現する細胞の分子画像化において使用され得る。
【0096】
本発明はここで以下の非限定的な実施例によりさらに説明される。
【図面の簡単な説明】
【0097】
【図1−1】本発明のHER3結合ポリペプチドに含まれるHER3結合モチーフの例(配列番号1〜334)、本発明に従う49量体HER3結合ポリペプチドの例(配列番号335〜668)及び本発明に従う58量体HER3結合ポリペプチドの例(配列番号669〜1002)、本発明に従うリガンドに含まれるHER2結合ポリペプチドの例(配列番号1003)、本発明のリガンドの一実施態様において含まれ得るようなアルブミン結合部分の例(配列番号1005)、本発明に従うリガンドの一実施態様において含まれ得るEGFR結合ポリペプチドの例(配列番号1004)のアミノ酸配列表、さらにはプロテインZの配列(配列番号1006)、並びにHER3の細胞外ドメイン(配列番号1007)、HER2の細胞外ドメイン(配列番号1008)及びHER4の細胞外ドメイン(配列番号1009)の配列の表である。
【図1−2】図1−1の続きである。
【図1−3】図1−2の続きである。
【図1−4】図1−3の続きである。
【図1−5】図1−4の続きである。
【図1−6】図1−5の続きである。
【図1−7】図1−6の続きである。
【図1−8】図1−7の続きである。
【図1−9】図1−8の続きである。
【図1−10】図1−9の続きである。
【図1−11】図1−10の続きである。
【図1−12】図1−11の続きである。
【図1−13】図1−12の続きである。
【図1−14】図1−13の続きである。
【図1−15】図1−14の続きである。
【図1−16】図1−15の続きである。
【図1−17】図1−16の続きである。
【図1−18】図1−17の続きである。
【図1−19】図1−18の続きである。
【図1−20】図1−19の続きである。
【図1−21】図1−20の続きである。
【図1−22】図1−21の続きである。
【図1−23】図1−22の続きである。
【図1−24】図1−23の続きである。
【図1−25】図1−24の続きである。
【図1−26】図1−25の続きである。
【図1−27】図1−26の続きである。
【図1−28】図1−27の続きである。
【図1−29】図1−28の続きである。
【図1−30】図1−29の続きである。
【図1−31】図1−30の続きである。
【図1−32】図1−31の続きである。
【図1−33】図1−32の続きである。
【図1−34】図1−33の続きである。
【図1−35】図1−35の続きである。
【図1−36】図1−35の続きである。
【図1−37】図1−36の続きである。
【図1−38】図1−37の続きである。
【図1−39】図1−38の続きである。
【図1−40】図1−39の続きである。
【図1−41】図1−40の続きである。
【図1−42】図1−41の続きである。
【図2】HSA結合ドメインABDへの融合として発現された個々のHER3結合Z変異体からの周辺質フラクションを使用して行われたドットブロットアッセイの結果を示す。21のコントロールタンパク質に対する潜在的バックグラウンド結合、さらにはHER3−ECD及びHER3−Fcへの結合を分析した。タンパク質をメンブレン上に3列で以下の順序にてドットを打った; 列1: HSA、IgG、IgM、IgA、アルファ−2マクログロブリン、フィブリノゲン、ホロ−トランスフェリン、アルファ−1−抗トリプシン、 列2:C3、ハプトグロブリン(haptoglobulin)、アルファ−1酸性糖タンパク質、アルファ−1抗キモトリプシン、C4、マウスIgEカッパ、ヘモペキシン、トランスサイレチン、 列3:ストレプトアビジン、ニュートラアビジン、HER2−ECD、HER3−Fc、HER4−Fc、HER3−ECD及びHER4−ECD。 画像は、A) Z01748、B) Z01813、C) Z01814、D) Z01815、E) Z02009、F) Z01821、G) Z02010、H) Z01824、I) Z01825、J) Z01817、K) Z01818、L) Z01820、M) Z01826、N) Z01828、O) Z02011、P) Z01830、及びQ)ネガティブコントロールとしての、ABDのみを発現するpAY01247からの生成物を示す。
【図3】実施例2に記載される免疫蛍光実験からの画像を示す。HER3ポジティブAU566細胞(A〜D)並びにHER2ポジティブ及びHER3ネガティブのSKOV−3細胞(E〜F)をHER3特異的Z変異体又は以下に従うコントロールで染色した:A) His6−(Z01814)2−cys、B) His6−(Z01820)2−cys、C) 抗HER3抗体(ポジティブコントロール)、D)(Z01154)2(ネガティブコントロールZ変異体)、E) His6−(Z01814)2−cys、F) His6−(Z01820)2−cys。
【図4】S.カルノーサス(S.carnosus)(B)の表面での本発明のHER3結合ポリペプチドの細胞ディスプレイのために使用したブドウ球菌ディスプレイベクターの略図である。ベクターpSCZ1(A)は:i)リパーゼ遺伝子(関連するスタフィロコッカス・ヒイカス(Staphylococcus hyicus)の表面で発現される)からのプロモーター及び分泌シグナル(S)、ii) 同じリパーゼ遺伝子からのプロペプチド(PP)をコードする遺伝子フラグメント、(組み換えタンパク質の効率的な転座に重要であることが実証されている)、iii)細胞壁固定領域(XM)、(ブドウ球菌プロテインA由来)、iv)標的結合シグナルの表面発現レベル正規化をもたらすために導入されるブドウ球菌プロテインG由来のアルブミン結合タンパク質(ABP、Samuelson et al、J Bacteriology 177(6):1470−1476、(1995))、v) ブドウ球菌における安定した複製及び発現のためのブドウ球菌複製起点及びクロラムフェニコールアセチルトランスフェラーゼ遺伝子(Cmlr)、並びにvi)E.coliでのサブクローニング作業を容易にする、E.coliのための複製起点(OriE)さらにはβ−ラクタマーゼ遺伝子(Bla)を含む。
【図5】Sc:ZHER3LIBのフローサイトメトリー分離からの結果として密度プロットを示す。これらのプロットは、x軸に表面発現レベルに対応するFL−4チャネル蛍光強度(HSA結合によりモニタリング)、及びy軸にHER3結合に対応するFL−1チャネル蛍光を示す。密度プロットは、各プロットにおいて四角で輪郭を描いたゲーティングにおいて使用された領域と共に、それぞれ1、2、3及び4回目のフローサイトメトリー分離前のブドウ球菌ライブラリーを示す。
【図6−1】細胞ベースのフローサイトメトリーアッセイのヒストグラム図を示し、ここではアルブミン結合タンパク質(ABP)への融合でZ変異体を提示するFACSで単離した細胞を、蛍光標識したHER3−Fc及び蛍光標識したHSAと共にインキュベートした。HER3結合シグナル(MFI、FL−1)とHSA結合シグナル(MFI、FL−4)との比率がy軸にある。全てのFL−1/FL−4比は;A) Z05405のFL−1/FL−4比、及びB〜D) 第一世代バインダー(binder)(Z01753)のFL−1/FL−4比を用いて正規化される。45の個々のクローンを4つの別々のアッセイ(A〜D)において分析し、ここでZ05405及びZ05409を比較のために全てのアッセイに含めた。A)はZ05403〜Z05415の細胞ベースのフローサイトメトリーアッセイ結果を示し、B)はZ05416〜Z05426細胞ベースのフローサイトメトリーアッセイ結果を示し、C)はZ05427〜Z05434の細胞ベースのフローサイトメトリーアッセイ結果を示し、そしてD)はZ05435〜Z05447の細胞ベースのフローサイトメトリーアッセイ結果を示す。
【図6−2】細胞ベースのフローサイトメトリーアッセイのヒストグラム図を示し、ここではアルブミン結合タンパク質(ABP)への融合でZ変異体を提示するFACSで単離した細胞を、蛍光標識したHER3−Fc及び蛍光標識したHSAと共にインキュベートした。HER3結合シグナル(MFI、FL−1)とHSA結合シグナル(MFI、FL−4)との比率がy軸にある。全てのFL−1/FL−4比は;A) Z05405のFL−1/FL−4比、及びB〜D) 第一世代バインダー(binder)(Z01753)のFL−1/FL−4比を用いて正規化される。45の個々のクローンを4つの別々のアッセイ(A〜D)において分析し、ここでZ05405及びZ05409を比較のために全てのアッセイに含めた。A)はZ05403〜Z05415の細胞ベースのフローサイトメトリーアッセイ結果を示し、B)はZ05416〜Z05426細胞ベースのフローサイトメトリーアッセイ結果を示し、C)はZ05427〜Z05434の細胞ベースのフローサイトメトリーアッセイ結果を示し、そしてD)はZ05435〜Z05447の細胞ベースのフローサイトメトリーアッセイ結果を示す。
【図7A】4つの異なる濃度での4つの異なるZ変異体の表面プラズモン共鳴(SPR)動力学的分析からのセンサーグラムを示す。各センサーグラムにおける一番上の曲線は最も高い濃度の注入されたZ変異体に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は3番目に高い濃度に対応するなどである。46、15、1.5及び1.5nMの濃度のZ変異体Z05405(A)、57、19、6.3及び1.9nMの濃度のZ05413(B)、67、22、7.5及び2.2nMの濃度のZ05416(C)、並びに59、20、6.5及び2nMの濃度のZ05417(D)を、ヒトHER3−Fcを固定した表面上に注入し、そしてkon及びkoffに基づいて解離定数を決定した。
【図7B】4つの異なる濃度での4つの異なるZ変異体の表面プラズモン共鳴(SPR)動力学的分析からのセンサーグラムを示す。各センサーグラムにおける一番上の曲線は最も高い濃度の注入されたZ変異体に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は3番目に高い濃度に対応するなどである。46、15、1.5及び1.5nMの濃度のZ変異体Z05405(A)、57、19、6.3及び1.9nMの濃度のZ05413(B)、67、22、7.5及び2.2nMの濃度のZ05416(C)、並びに59、20、6.5及び2nMの濃度のZ05417(D)を、ヒトHER3−Fcを固定した表面上に注入し、そしてkon及びkoffに基づいて解離定数を決定した。
【図7C】4つの異なる濃度での4つの異なるZ変異体の表面プラズモン共鳴(SPR)動力学的分析からのセンサーグラムを示す。各センサーグラムにおける一番上の曲線は最も高い濃度の注入されたZ変異体に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は3番目に高い濃度に対応するなどである。46、15、1.5及び1.5nMの濃度のZ変異体Z05405(A)、57、19、6.3及び1.9nMの濃度のZ05413(B)、67、22、7.5及び2.2nMの濃度のZ05416(C)、並びに59、20、6.5及び2nMの濃度のZ05417(D)を、ヒトHER3−Fcを固定した表面上に注入し、そしてkon及びkoffに基づいて解離定数を決定した。
【図7D】4つの異なる濃度での4つの異なるZ変異体の表面プラズモン共鳴(SPR)動力学的分析からのセンサーグラムを示す。各センサーグラムにおける一番上の曲線は最も高い濃度の注入されたZ変異体に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は3番目に高い濃度に対応するなどである。46、15、1.5及び1.5nMの濃度のZ変異体Z05405(A)、57、19、6.3及び1.9nMの濃度のZ05413(B)、67、22、7.5及び2.2nMの濃度のZ05416(C)、並びに59、20、6.5及び2nMの濃度のZ05417(D)を、ヒトHER3−Fcを固定した表面上に注入し、そしてkon及びkoffに基づいて解離定数を決定した。
【図8A】Z05417のヘレグリンとのSPR競合分析からのセンサーグラムを示す。A)はヒトヘレグリンを固定した表面上での、単独及び40倍モル濃度過剰のZ05417と共にプレインキュベートした、HER3−Fcの注入から得られたセンサーグラムを示す。B)は、ヒトHER3−Fcを固定化した表面上でのヘレグリンとのZ05417の同時注入から得られたセンサーグラムを示す。Z05417を、5つの異なる濃度、10nM、5nM、1nM、0.5nM及び0nMで二回注入した。一番上の曲線は最も高い濃度の注入されたZ05417に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は三番目に高い濃度に対応するなどである。
【図8B】Z05417のヘレグリンとのSPR競合分析からのセンサーグラムを示す。A)はヒトヘレグリンを固定した表面上での、単独及び40倍モル濃度過剰のZ05417と共にプレインキュベートした、HER3−Fcの注入から得られたセンサーグラムを示す。B)は、ヒトHER3−Fcを固定化した表面上でのヘレグリンとのZ05417の同時注入から得られたセンサーグラムを示す。Z05417を、5つの異なる濃度、10nM、5nM、1nM、0.5nM及び0nMで二回注入した。一番上の曲線は最も高い濃度の注入されたZ05417に対応し、上から二番目の曲線は二番目に高い濃度に対応し、上から三番目の曲線は三番目に高い濃度に対応するなどである。
【図9】HER3結合Z変異体の存在しない場合及び存在下でのHER3リン酸化のパーセントに関する図表を示す。MCF−7細胞を、Z変異体の存在しない場合(100%リン酸化、塗りつぶしたバー)、並びにHER3結合Z変異体Z05416及びZ05417の存在下で、並びにネガティブコントロールZ01155の存在下で(taqポリメラーゼ結合Z変異体、縞模様のバー)又は培地のみを用いて(白いバー)、5nMヘレグリンで刺激した。この図表は、ヘレグリン誘導HER3リン酸化(100%)と比較した、Z変異体又は培地の存在下でのHER3リン酸化のパーセントを示す。
【実施例】
【0098】
実施例1:HER3結合ポリペプチドの選択及びスクリーニング
材料及び方法
HER3及びHSAの標識: 組み換えヒトHER3/Fcキメラ(R&D Systems、#348−RB−050)(本明細書ではHER3−Fcと示される)のビオチン化を、Biotin−XXマイクロスケールタンパク質標識キット(Invitrogen、#B30010)を使用して供給業者の推奨にしたがって行った。HER3の細胞外ドメイン(配列番号1007)(本明細書ではHER3−ECDと示される)を、EZ−LinkTM−Sulfo−NHS−LC−LC−Biotin(Pierce、#21338)を使用して供給業者の推奨にしたがってビオチン化した。ヒト血清アルブミン(HSA; Sigma、#A−3782)を、Alexa Fluor(R) 647スクシンイミジルエステル(Invitrogen、#A20006)を使用して供給業者の推奨にしたがって蛍光標識した。タンパク質緩衝液をPBS(10mMホスフェート、137mM NaCl、2.68mM KCl、pH7.4)又は過剰のビオチン若しくはフルオロフォアを除去するために0.1%Pluronic(R)F108 NF界面活性剤(PBSP; BASF Corporation、#30085231)を補充したPBSに変更した。
HER3結合ポリペプチドのファージディスプレイ選択: Groenwall et al、J Biotechnol 128:162−183(2007)に記載されるように、ファージミドpAffi1/pAY00065において構築されたバクテリオファージで提示されるZ分子のランダム変異体のライブラリー(Zlib2002と示される)を、HER3−ECD−結合ポリペプチドの選択のために使用した。ファージミドライブラリーから、さらには選択ラウンドの間に、ファージストックの製造を以前に記載された手順(Nord et al、Nat Biotech 15:772−777(1997); Hansson et al、Immunotechnology 4:237−252(1999))にしたがってヘルパーファージM13K07(New England Biolabs、#NO315S)を使用して行った。大腸菌(Escherichia coli)アンバー抑圧遺伝子系統RR1ΔM15(Ruether、Nucleic Acids Res 10:5765−5772、1982)を全てのクローニング手順のために、そしてファージ産生のための宿主として使用した。
【0099】
ファージ選択を、溶液でビオチン化HER3タンパク質及びストレプトアビジン被覆常磁性ビーズ(Dynabeads(R) M−280ストレプトアビジン、Dynal #112.06)を、結合したファージの捕捉のために使用して行った。さらに、ファージ選択を、固相においてMaxiSorp immunotubes(Nunc)に固定化されたHER3−ECDを使用して行った。ファージ粒子の非特異的結合を、全てのチューブ及びビーズをPBST 0.1−ゼラチン(0.1%Tween−20及び0.1%ゼラチンを補充したPBS)で予備処理することにより最少にした。選択を4サイクルで行った。全ての選択をPBST 0.1−ゼラチン中でかつ体積1mlで行った。
【0100】
溶液で行われた選択については、4サイクルの選択を室温(RT)にて異なる濃度のビオチン化組み換えHER3−ECDを標的タンパク質として使用して行った。サイクル1及び2で使用したファージストックを、ストレプトアビジン結合ファージを除去するために0.1mgストレプトアビジン被覆ビーズとともに1時間プレインキュベートした。サイクル1において、Zlib2002ライブラリーからのファージ粒子を、100nMのビオチン化HER3−ECDと共にPBST 0.1−ゼラチン中で2時間インキュベートした。それに続く選択サイクルを2つの選択トラックに分割し、そのために標的濃度をサイクル2について50(トラック1)又は20nM(トラック2)まで、サイクル3について20(トラック1)又は10nM(トラック2)まで、そしてサイクル4について20(トラック1)又は5nM(トラック2)まで下げた。結合したファージをストレプトアビジン被覆M−280 Dynabeads(R)を用いて捕捉し、ビーズ1ミリグラムあたりHER3−ECD4μgを固定化させた。
【0101】
固相での選択については、immunotubeに1ml HER3−ECD(50mM炭酸ナトリウム(sodium carbonat)緩衝液中10μg/ml、pH 9.6)を1.5時間室温にて固定化させ、その後3ml PBST 0.1−ゼラチンを用いて1時間室温にてブロックした。ファージストックをこのチューブに加え、そして2時間インキュベートした。
【0102】
両方の選択ストラテジーのために、PBST 0.1−ゼラチンでの洗浄回数を、選択ストリンジェンシーを高めるためにラウンド間で増やした。従って、洗浄をサイクル1において2回、サイクル2において3回、サイクル3において6回、そしてサイクル4において12回行った。結合したファージ粒子を0.1Mグリシン−HCl(pH2.2)で溶出し、続いてすぐに1M Tris−HCl(pH8)で中和した。ファージ含有溶出液を対数期RR1ΔM15細胞に感染させるために使用し、そしてファージミド粒子を、ヘルパーファージM13K07を使用して感染した細胞からレスキューした。選択過程を、各選択ラウンドの前及び標的タンパク質からの溶出後にファージストックの滴定によりモニタリングした。ファージ溶液の段階希釈を、対数期RR1ΔM15細胞の感染のために使用した。
【0103】
Z変異体のELISAスクリーニング: Z変異体分子が実際にHER3と相互作用するかどうかを試験するために、2つの異なるELISAを行い、一方は標的タンパク質としてビオチン化HER3−ECDを使用し、そして2つ目はHER3−Fcを使用した。上で記載したように製造された単一コロニーを、100μg/mlアンピシリン及び0.1mMイソプロピル−β−D−1−チオガラクトピラノシド(IPTG)を補充した1ml TSB−YE培地(30g/l TSB、5g/l酵母抽出物)中でディープウェルプレート(Nunc #278752)において播種することによりZ変異体を生成した。ネガティブコントロールとして、インスリン結合Z変異体(Z00801)を播種し、そして各プレートにおいて増殖させた。これらのプレートを18〜24時間37℃でインキュベートした。インキュベーション後に、−20℃での貯蔵のために15%グリセロールを含む96ウェルプレートに各培養物を少量移すことにより複製のプレートを作製した。残りの細胞を遠心分離によりペレット化し、300μl PBST 0.05(0.05% Tween 20を補充したPBS)中に再懸濁し、そして細胞の周辺質部分を放出させるために−80℃で凍結した。凍結したサンプルを水浴で解凍し、そして細胞を遠心分離によりペレット化した。周辺質上清は、連鎖球菌株G148由来のプロテインGのGA3からのアルブミン結合ドメインへの融合物としてのZ変異体(AQHDEALE−[Z#####]−VDYV−[ABD]−YVPGとして表される)を含有していた(Groenwall et al、前出)。Z#####は個々のZ変異体を指す。
【0104】
ハーフエリア96ウェルELISAプレート(Costar #3690)を、6μg/mlヒト血清アルブミン(HSA、Sigma #A3782)を含有する50μl/ウェルのコーティング緩衝液(50mM炭酸ナトリウム、pH9.6)で被覆し、そして終夜(ON)インキュベートした。HSA溶液を捨てて、2%脱脂粉乳溶液(Semper AB)を補充したPBST 0.1 100μlを用いてウェルを1時間室温にてブロックした。ブロッキング溶液を廃棄し、そして周辺質溶液50μlを各ウェルに加え、そして1.5時間室温でゆっくりと震盪しながらインキュベートした。上清を捨てて、ウェルをPBST 0.05で4回洗浄した。PBST 0.05中0.5μg/mlの濃度のHER3−Fc 50μl又はPBS中1μg/mlの濃度のビオチン化HER3−ECDを各ウェルに加えた。プレートを1.5時間室温でインキュベートし、続いてPBST 0.05で4回洗浄した。HER3−Fcプレートにおいて、西洋ワサビペルオキシダーゼ(HRP)で標識されPBST0.05で1:4000に希釈されたヒトFcに対する抗体(DAKO Cytomation、#P0214)をウェルに加え、そして1時間室温でインキュベートした。ビオチン化HER3−ECDを含むプレートにおいて、HRPと結合したストレプトアビジン(DAKO、#P0397)を各ウェルに加えてPBST 0.05で1:5000に希釈した。上で記載したように洗浄した後、50μl ImmunoPure TMB基質(Pierce #34021)をウェルに加え、そしてプレートを製造者の推奨に従って処理した。ブロッキングから読み取りまでの全ての工程を、Tecan Genesis Freedom 200ロボット(Tecan Group LTD)で行った。ウェルの吸光度を450nmでELISAリーダーTecan Ultra 384(Tecan)で読み取り、そしてMagellan v.5.0ソフトウェア(Tecan)を用いて評価した。
【0105】
配列決定: ELISAスクリーニングに基づいて、陽性と考えられる全てのクローンの一部を配列決定のために選別した。PCRフラグメントを、標準的なPCRプログラム並びにプライマーAFFI−21(5’−tgcttccggctcgtatgttgtgtg)及びAFFI−22(5'−cggaaccagagccaccaccgg)を使用して単一コロニーから増幅した。増幅されたフラグメントの配列決定を、製造者のプロトコルに従って使用されるビオチン化オリゴヌクレオチドAFFI−72(5'−ビオチン−cggaaccagagccaccaccgg)及びBigDye(R)ターミネーターv3.1サイクルシークエンスキット(Applied Biosystems)を使用して行った。配列決定反応混合物を、磁気ストレプトアビジン被覆ビーズへの結合により、Magnatrix 8000(Magnetic Biosolution)を使用して精製し、そしてABI PRISM(R)3100 Genetic Analyzer(PE Applied Biosystems)で分析した。配列決定結果をインポートし、そしてALD LIMS NautilusTM 2003 R2 B3ソフトウェア(Thermo Electronics Corp.)を用いて分析した。
【0106】
ドットブロット分析: 生成されたHER3結合Z変異体を、特異性についてドットブロット分析によりアルファ−2マクログロブリン(MP biomedicals/Cappel、#55833)、アルファ−1酸性糖タンパク質(RDI、#RDI−SCP−153−1)、アルファ−1抗キモトリプシン(RDI、#RDI−SCP−159−0)、アルファ−1−抗トリプシン(RDI、#RDI−SCP−165−5)、補体C3(RDI、#RDI−SCP−150−0)、補体C4(RDI、#RDI−SCP−151−0)、フィブリノゲン(Enzyme research、#031015E)、ハプトグロブリン(RDI、#RDI−SCP−119−1)、ヘモペキシン(Agilent)、ホロ−トランスフェリン(Sigma、#T0665)、ヒトIgA(Bethyl、#P80−102)、マウスIgEカッパ(BD、#55079)、ヒトIgG(Sigma、#G4386)、ヒトIgM(Sigma、#I8260)、ヒト血清アルブミン(HSA、Sigma、#A3782)、ニュートラアビジン(Pierce、#31000)、ストレプトアビジン(Pierce、#21122)、トランスサイレチン(Sigma、#P1742)、HER2−ECD(配列番号1008)、HER3−ECD(配列番号1007)、HER3−Fc(R&D Systems)、HER4−ECD(配列番号1009)、HER4−Fc(R&D Systems)に対して試験した。
【0107】
可溶性タンパク質を含有する周辺質上清を得るために、Z変異体をELISAスクリーニングで記載された方法と本質的に同じ方法で製造した。上清を0.45μmメンブレンを使用してろ過した。
【0108】
ニトロセルロースメンブレン(Invitrogen)に各タンパク質1μlを、それぞれ0.29mg/ml及び0.14mg/mlの濃度を有していたHER3−ECD及びHER4−ECDを除いて0.25mg/mlの濃度で用いてドットを打った。これらのメンブレンを0.5%カゼイン(ブロッキング液)を補充したPBST0.1中で終夜4℃でブロックした。溶液を除去した後、これらのメンブレンを1時間、0.5%カゼインを補充した周辺質上清と共にインキュベートした。これらのメンブレンをPBST 0.1中で非常に短く3回、そして4×5分洗浄した。Z変異体を、PBSTブロッキング液中で1:5000に希釈された全てのZ変異体(社内で製造)に共通のエピトープに対するABD融合超免疫ポリクローナルウサギIgを用いて1時間室温でインキュベートすることにより検出した。上記のように洗浄した後、ブロッキング溶液で1:5000希釈された、HRPと複合体化されたウサギIgGに対する抗体(DAKO Cytomation、#P0448)をメンブレンに加え、続いてこれらのメンブレンを1時間室温でインキュベートした。メンブレンを洗浄し、PBSですすぎ、そしてSupersignal(Pierce #34075)と共に浸漬した。発光をChemiImager 5500(Alpha Innotech Corp.)を用いて撮影した。
【0109】
Z変異体のサブクローニング: 二量体Z変異体をpAffi1/pAY00065ベクターから増幅した。異なるプライマー対を使用してPCRを行い、そして得られた遺伝子フラグメントを精製し、そしてリガーゼ緩衝液中でハイブリダイズさせた。ハイブリダイズした遺伝子フラグメントをpAY00430ベクターでサブクローニングし、N末端His6タグ及びC末端システインを生じた(His6−(Z#####)2−Cys)。HER3結合Z変異体をダイマーとしてサブクローニングし、そして発現ベクターによりコードされる構築物は、MGSSHHHHHHLQ−[Z#####][Z#####]−VDCであった。3つの部分のライゲーションを、両方の挿入フラグメントをベクター中に同じ工程で挿入するために使用した。ハイブリダイズした遺伝子フラグメント及びAccI−消化して脱リン酸化した発現ベクターをリガーゼ緩衝液中で連結し、エレクトロコンピテント(electrocompetent)E.coli TOP10細胞中にエレクトロポレーション処理した。形質転換細胞を、50μg/mlのカナマイシンを補充したTBABプレート(30g/lトリプトース(tryptose)血液寒天基礎培地)上に広げ、続いて37℃で終夜インキュベートした。コロニーをPCRを使用してスクリーニングし、そしてPCRフラグメントの長さをアガロースゲルで確認した。配列を確認するために、上記のように配列決定を行った。プラスミドDNAストックを配列決定したクローンから調製し、そして−80℃に置いた。さらに、E.coli BL21(DE3)細胞をエレクトロポレーションによりプラスミドを用いて形質転換した。
【0110】
結果
HER3結合ポリペプチドのファージディスプレイ選択: 4ラウンドのファージディスプレイ選択を、ビオチン化又は非ビオチン化ヒトHER3−ECDに対して行った。4回の選択サイクルを、2つの異なる標的濃度でビオチン化標的を含む溶液で、又は非ビオチン化標的を含む固相で行った。溶液では、ファージ粒子標的複合体がストレプトアビジン被覆ビーズに捕捉された。各選択サイクルについて、洗浄回数を増やした。ファージ粒子力価及び収率を、各選択サイクル後に計算した。ファージ粒子収率(選択された(out)ファージ粒子/導入した(in)ファージ粒子)は、3回目又は最後のサイクルで増加し、標的結合クローンの濃縮を示していた。
【0111】
Z変異体のELISAスクリーニング: 4サイクルの選択後に得られたクローンを96ウェルプレートで作製し、そしてHER3−ECD及びHER3−Fc−結合活性についてELISAでスクリーニングした。各選択トラックから合計で93のクローンをスクリーニングした。吸光度測定は、選択方法と無関係に、約90%のクローンがHER3に対して陽性(ネガティブコントロールのシグナルの2倍の応答として定義される)であることを示した。インスリンに結合するZ変異体分子を、ネガティブ及びポジティブコントロール実験において使用した。ポジティブコントロールをビオチン化インスリンを用いて検出し、そしてネガティブコントロールについては標的タンパク質を除いた。
【0112】
配列決定: ELISAスクリーニングにおいてHER3−Fcに対して様々な(variying)シグナル吸光度値を有することに基づいて選択されたクローンについて配列決定を行い、従って配列決定のための多くの異なる代表的なバインダーを得た。合計で135のHER3ポジティブクローンを配列決定した。クローンの半分は合計23の新しいクローンを生じるいくつかのコピーにおいて見出され、それらのうち2つはバックグラウンドバインダーと同一であった。各変異体に固有の識別番号#####を付与し、そして個々の変異体をZ#####と呼ぶ。58量体Z変異体のアミノ酸配列を図1及び配列表に配列番号669〜689として載せる。これらのZ変異体の推定HER3結合モチーフを図1及び配列表に配列番号1〜21として載せる。これらの各Z変異体内で完全3ヘリックスバンドルを構成すると予測される49量体ポリペプチドのアミノ酸配列を図1及び配列表に配列番号335〜355として載せる。
【0113】
ドットブロット分析: 配列決定された独特なHER3結合ポリペプチド(AQHDEALE−[Z#####]−VDYV−[ABD]−YVPGの形態)のうち16の特異性をドットブロット分析を使用して試験した。23の異なるタンパク質をニトロセルロースメンブレン上にブロットした。これらの23のタンパク質には、16の高度に豊富なヒト血清タンパク質、HER2−ECD、HER3−ECD及びHER3−Fc並びにHER4−ECD及びHER4−Fcが含まれていた。Z01824以外の全てのZ変異体融合タンパク質はHSAと同様にHER3−ECD及びHER3−Fcに特異的に結合した。従って、試験したZ変異体の特異性は十分なものであった(図2)。
【0114】
Z変異体のサブクローニング: 16の独特なクローンを二量体として発現ベクターpAY004308でサブクローニングするために選択した。クローニングは、15の二量体のHER3結合Z変異体を生じた(Z01748、Z01749、Z01751、Z01753、Z01814、Z01815、Z01817、Z01820、Z01821、Z01826、Z01828、Z01830、Z02009、Z02010及びZ02011の二量体)。
【0115】
実施例2: Z変異体の産生及び特徴付け
材料及び方法
タンパク質発現及び精製:実施例1においてサブクローニングした形質転換したE.coli BL21(DE3)培養物を、TSB−YEで約1の光学密度まで増殖させた。次いで1M IPTGを最終濃度0.5mMまで加えることによりタンパク質発現を誘導した。培養物を誘導の5時間後に遠心分離により採取した。上清を廃棄し、そして細胞ペレットを集めて−20℃で貯蔵した。
【0116】
HER3結合Z変異体を、1.5ml Ni−NTA Superflow Columns(Qiagen)で変性条件下で細胞ペレットから精製し、そしてバッファーをPBSに交換してPD−10カラム(GE Healthcare)を使用した。精製したZ変異体をアリコートに分けて−80℃で貯蔵した。
【0117】
タンパク質特徴付け: 精製されたZ変異体(His6−(Z#####)2−Cys形態)の濃度を280nmにおける吸光度測定により決定した。純度を10ウェル4〜12% NuPAGETMゲル(Invitrogen)でのSDS−PAGE分析でクマシーブルー染色を使用して見積もった。精製したZ変異体のアイデンティティを確認し、分子量を決定するために、LC/MS分析をAgilent1100LC/MSDシステム(Agilent Technologies)で行った。
【0118】
CD分析: 精製したZ変異体を解凍し、そしてPBS中0.5mg/mlに希釈した。それぞれの希釈したZ変異体について、250〜195nmでのCDスペクトルを20℃で得た。さらに、温度可変測定(VTM)を、融点(Tm)を決定するために行った。VTMにおいて、5℃/分の温度勾配で温度を20から90℃に上げながら吸光度を220nmで測定した。CD測定を、光路長1mmを有するセルを使用してJasco J−810分光偏光計(Jasco Scandinavia AB)で行った。
【0119】
蛍光免疫染色: HER3ポジティブヒト乳腺細胞株AU565(ATCC、#CRL−2351)及びHER3ネガティブヒト卵巣がん細胞株SKOV−3(Ecacc、#91091004)を供給業者に推奨されるように培養した。アッセイ前日に、25000個の細胞をマルチウェルスライド(スライドあたり8ウェル、Histolab #ER−201−1B)の各ウェルに加え、そして終夜CO2インキュベーターで板状形態を呈するように増殖させた。パスツールピペットを用いてスライド上に液滴を加えることによりこれらのスライドをPBSで穏やかに洗浄した。
【0120】
2つの実験を行った。1つ目の実験では、AU565細胞を11の異なるHER3特異的Z変異体で染色した;His6−(Z01748)2−Cys、His6−(Z01749)2−Cys、His6−(Z01751)2−Cys、His6−(Z01753)2−Cys、His6−(Z01814)2−Cys、His6−(Z01815)2−Cys、His6−(Z01817)2−Cys、His6−(Z01820)2−Cys、His6−(Z01828)2−Cys、His6−(Z01830)2−Cys及びHis6−(Z02011)2−Cys。2つ目の実験は、His6−(Z01814)2−Cys及びHis6−(Z01820)2−Cysを用いて、そして(Z01154)2をネガティブコントロールZ変異体として用いて行った。染色を、HER3ポジティブAU565及びHER3ネガティブSKOV−3細胞を用いてスライド上で行った。R&D Systemsからの抗HER3抗体(#AF234)をポジティブコントロールとして両方の実験に
含めた。
【0121】
両方の実験において、Z変異体をPBS中15μg/mlの濃度でウェルに加え、そして試薬の全ての希釈はPBS中で行い、全ての洗浄も同様にした。1時間室温でのインキュベーション後、マルチウェルスライドを上記のように穏やかに洗浄し、そしてヤギ抗Affibody分子Ig(Affibody AB、#20.1000.01.0005)を各ウェルに5μg/mlの濃度で加えた。45分室温でのインキュベーション後、スライドを上記のように穏やかに洗浄し、そして5μg/mlのニワトリ抗ヤギAlexa Fluor488(Invitrogen、#A21467)を各ウェルに加えた。さらに45分のインキュベーション後、スライドを穏やかに洗浄した。ポジティブコントロール、ヤギ抗HER3抗体を15μg/mlの濃度で、続いて5μg/mlのニワトリ抗ヤギAlexa Fluor488を使用した。染色が完了した後、PBS中3%ホルムアルデヒド(Sigma、#1365)中で10分間室温で固定した。次いでスライドを2回すすぎ、乾燥し、そしてDAPI(4',6−ジアミジン−2−フェニルインドール、Vector laboratories、#H1200)を含有する退色防止液を用いて封入した。生体画像化用のビデオカメラを備えたLEICA DMLA顕微鏡を使用して染色を記録した。
【0122】
フローサイトメトリーを使用した細胞上親和性順位付け: Z変異体Z01751、Z01753、Z01814及びZ01820をコードする全長遺伝子配列を、それらの対応するpAY00430ベクター構築物からPCR増幅し、そして制限酵素XhoI及びSalI(New England Biolabs)で予め消化したブドウ球菌ディスプレイベクターpSCZ1(Kronqvist et al、Protein Eng Des Sel 21:247−255(2008))に結合させた。E.coli株RR1ΔM15をプラスミド構築及び製造のための宿主として使用し、そして以前に記載されたプロトコル(Loefblom et al、J Appl Microbiol 102:736−747(2007))に従ってこれらの構築物をエレクトロコンピテントなスタフィロコッカス・カルノーサス(Staphylococcus carnosus)TM300(Goetz、Soc Appl Bacteriol Symp Ser.19:49−53(1990))に形質転換した。独立して4つの異なるZ変異体を提示しているブドウ球菌細胞を、20μg/mlクロラムフェニコールを補充したTSB−YE 10mlに播種し、そして終夜37℃及び150rpmで増殖させた。培養物から、106個の細胞を1ml PBSPで洗浄した。これらの細胞を遠心分離(3500xg、4℃、6分)によりペレット化し、そして異なる濃度のビオチン化HER3−Fc(5、20、50及び100nM;ビオチン化については実施例1を参照のこと)を含有するPBSP100μl中に再懸濁した。穏やかに混合しながら室温で1時間インキュベーションすることにより平衡結合に達した。細胞を氷冷PBSP 1mlで1回洗浄し、続いて氷上で1.25μg/ml Alexa Fluor(R)488複合体化ストレパビジン(strepavidin)(Invitrogen、#S32354)及び225nM Alexa Fluor(R)647複合体化HSA(実施例1を参照のこと)を含有する氷冷PBSP 100μl中で40分間インキュベートした。氷冷PBSP1mlで1回洗浄した後、細胞をフローサイトメトリー分析の前に氷冷PBSP300μlに再懸濁した。平均蛍光強度(MFI)を、FACS Vantage SE(BD Biosciences)フローサイトメーターを使用して測定した。
【0123】
結果
タンパク質発現及び精製: 15の二量体Z変異体分子(His6−(Z#####)2−Cysの形態)は許容しうる産生レベルの可溶性タンパク質を生じ、そして産生されたバッチの純度は、基準を満たさなかった2つのZ変異体(His6−(Z01826)2−Cys及びHis6−(Z02009)2−Cys)を除いて、SDS−PAGE分析により90%を超えると見積もられた。LC/MS分析により、全ての純粋なZ変異体分子について正確な分子量が確認された。
CD分析: CDスペクトルは、Z変異体分子がα−ヘリックス構造を20℃で有していることを示した。この結果は融点(Tm)を決定した温度可変測定(表1)でも確認された。
【0124】
【表1】
【0125】
蛍光免疫染色: 蛍光免疫顕微鏡検査法を、ヒトがん細胞において発現される天然HER3に対する特異性及び結合活性の特徴付けのために使用した。実施例1のELISAにおいてポジティブシグナルを示す11のクローンを、N末端His6−タグ及びC末端システインに融合された二量体として製造した。各Z変異体をHER3ポジティブAU565細胞(ヒト乳房乳腺細胞株)でインキュベートした。抗HER3抗体をポジティブコントロールとして使用した。染色手順の後に細胞を固定した。11のバインダーのうち、6つは膜結合染色パターンを生じた。バックグラウンドはいくつかのZ変異体について高く、そしていくつかの場合にはそれらが特異的であるか否かを判別することが困難であった。2つのHER3特異的分子、His6−(Z01814)2−Cys及びHis6−(Z01820)2−Cysを、HER3ポジティブAU565細胞並びにHER2ポジティブ及びHER3ネガティブSKOV−3細胞(ヒト卵巣がん細胞株)の両方に対するさらなる分析のために選択した。明確な細胞膜染色がAU565で得られたがSKOV−3細胞では得られず、HER2よりもHER3に対するZ変異体の選択性を示した(図3)。
【0126】
フローサイトメトリーを使用した細胞上親和性順位付け: HER3結合分子をさらに特徴付けし、そして細菌細胞での機能発現を確認するために、蛍光免疫染色においてHER3に対する特異性を示す4つのHER3結合分子を、ブドウ球菌細胞ディスプレイ及びフローサイトメトリーを使用する細胞上親和性順位付け実験のために使用した。4つの変異体の単量体構築物を、その後のブドウ球菌宿主への形質転換のためにブドウ球菌ディスプレイベクターにサブクローニングした。4つのZ変異体を提示しているブドウ球菌細胞をそれぞれ、5〜100nMの範囲に及ぶ4つの濃度のビオチン化HER3−Fcとともにインキュベートした。フローサイトメトリーを使用して細胞を分析し、4つ全ての変異体についてクローン間での相対的親和性において有意な差異はなく、細胞表面上での効果的な発現及びHER3への特異的結合が明らかとなった(データは示されていない)。ビオチン化HER2を実験においてネガティブコントロールとして使用して交差特異性は観察されず、実施例1における周辺質上清を用いて行われたドットブロット及びこの実施例における蛍光免疫染色の結果を裏付けた。
【0127】
実施例3: HER3結合Z変異体の成熟ライブラリーの設計及び構築
この実施例において、成熟ライブラリーを構築した。このライブラリーをHER3結合ポリペプチドの選択のために使用した。成熟ライブラリーからの選択は通常、増加した親和性を有するバインダーを生じると期待される(Orlova et al、Cancer Res 66(8):4339−48(2006)。しかしモノヌクレオチド及び縮重コドンを使用する伝統的なオリゴヌクレオチド合成技術は、ブドウ球菌系統がアンバー停止コドンを抑制することができないために設計を制限し得る。この研究において、無作為化二本鎖リンカーをSlonomics(R)技術により代わりに生成し、これはその後に構成される二本鎖DNAのライゲーション及び制限を使用したトリヌクレオチドビルディングブロックの無作為化セットの組み込みを可能にする。
【0128】
材料及び方法
ライブラリー設計: ライブラリーは、実施例1及び2に記載されるHER3結合Z変異体の配列に基づくものであった。新しいライブラリーにおいて、Z分子骨格における13の可変位置は、配列番号1〜21において定義されるZ変異体配列の結合モチーフに基づいたストラテジーにしたがって、特定のアミノ酸残基に偏っていた。制限部位XhoI及びNheIが隣接した、HER3結合ポリペプチドのヘリックス1及び2において部分的に無作為化された位置をコードする131bpの二本鎖リンカー、例えば5’−GAA NNN NNN NNN GCG NNN NNN GAG ATC TGG NNN TTA CCT AAC TTA AAC NNN NNN CAA NNN NNN GCC TTC ATC NNN AGT TTA NNN GAT GAC CCA AGC CAA AGC GCT AAC TT−3’(無作為化コドンはNNNと示される)の混合物を含有するSlonoMax(R)ライブラリーをSloning BioTechnology GmbHに発注した。13の可変Z位置についての新しいライブラリーにおけるアミノ酸残基の理論的分布を表3に示す:
【0129】
【表2】
【0130】
ライブラリー構築及びクローニング: Phusion DNAポリメラーゼ(Finnzymes、#F530L)を使用して8サイクルのPCRの間にライブラリーを増幅し、そしてプールした生成物をQIAquick PCR精製キット(QIAGEN)を用いて供給業者の推奨にしたがって精製した。無作為化ライブラリーフラグメントの精製したプールを制限酵素XhoI及びNheI(New England Biolabs)で消化し、そして2%アガロースゲルでの分取ゲル電気泳動を使用して精製した。E.coli系統RR1ΔM15を、Jetstar Maxi Kit(Genomed、#220020)を用いたブドウ球菌ディスプレイベクターpSCZ1(図4A)のプラスミド産生のための宿主系統として使用した。
【0131】
このベクターを同じ酵素XhoI及びNheIで消化し、そして1%アガロースゲルでの分取ゲル電気泳動を使用して精製した。pSCZ1と無作為化ライブラリーフラグメントとのライゲーションを、1:4のベクター対フラグメントのモル比のT4 DNAリガーゼ(New England Biolabs、#M0202T)を使用して行った。ライゲーション混合物を、QIAquick PCR精製キットを使用して供給業者の推奨に従って、エレクトロコンピテントなE.coli DH5α細胞(Invitrogen、#18263−012)への形質転換前に精製した。形質転換後、さらには終夜の増殖後に直接プレーティングした個々のクローンを、配列確認のためにBigDyeサーモサイクルシークエンシング(Thermo Cycle Sequencing)反応及びABI Prism 3700機器(PE Applied Biosystems)を使用してPCR増幅した。プラスミドをJetstar Maxi Kitを使用したE.coliの終夜培養から製造し、そしてエレクトロコンピテントなS.カルノーサス(S.carnosus)に以前に記載されたように(Loefblom et al、前出)形質転換した。ブドウ球菌提示ライブラリーを本明細書以下ではSc:ZHER3LIBと示す。
【0132】
ライブラリー品質分析: Sc:ZHER3LIBのアリコート(少なくともライブラリーサイズの10倍、すなわち1.3×108より多い)を、20μg/mlクロラムフェニコールを含む100ml TSB−YEに播種し、そして終夜37℃及び150rpmで増殖させた。16時間後、107個の細胞を1ml PBSPで1回洗浄した。細胞を遠心分離(3500xg、4℃、6分)でペレット化し、そして225nM Alexa Fluor(R)647結合体化HSAを含有するPBSPに再懸濁し、そして1時間室温にて暗所でインキュベートした。1ml氷冷PBSPで1回洗浄した後、フローサイトメトリー分析の前に細胞を300μl氷冷PBSPに再懸濁した。平均蛍光強度(MFI)を、FACS Vantage SE(BD Biosciences)フローサイトメーターを使用して測定した。
【0133】
結果
ライブラリー構築及びクローニング: 新しいライブラリーを、確認された結合特性(実施例1及び2)を有する一組のHER3結合Z変異体(配列番号669〜689)に基づいて設計した。設計されたライブラリーの理論的なサイズは7.4×108のZ変異体であった。DNAフラグメントのライブラリーをブドウ球菌発現ベクター中にクローニングし、そしてS.カルノーサス(S.carnosus)に形質転換して約1.3×107の個々のクローンを含有する細胞提示ライブラリーを生成した。個々のライブラリーメンバーの配列分析により、理論的設計に従ったコドンの分布、並びに予期しないコドン、多重挿入及びフレームシフトの低い比率が示された。
ライブラリー品質分析: Z変異体成熟ライブラリーが細菌表面に機能的に提示されたことを確認するために、ライブラリーからのブドウ球菌細胞を蛍光標識したHSAと共にインキュベートし、そしてフローサイトメトリーを使用して分析した。結果は、ライブラリーの約72%が細胞表面で機能的ABP融合を有する全長タンパク質を発現したことを示した(データは示されていない)。従ってHER3結合ポリペプチドの成熟ライブラリーは首尾よく構築された。
【0134】
実施例4: ブドウ球菌表面ディスプレイライブラリーからのZ変異体の選択、スクリーニング及び特徴付け
材料及び方法
細胞標識及びFACSを使用したブドウ球菌細胞選別: Sc:ZHER3LIBのアリコート(ライブラリーサイズの少なくとも10倍、すなわち1.3×108より多い)を、20μg/mlクロラムフェニコールを含むTSB−YE 100mlに播種し、そして終夜37℃及び150rpmで増殖させた。16時間後、細胞(その後のサンプリング数の少なくとも4倍)を1ml PBSPで洗浄した。細胞を遠心分離(3500×g、4℃、6分)でペレット化し、そしてビオチン化HER3−Fcを含有するPBSP中に再懸濁し、そして室温で穏やかに混合しながら2時間インキュベートして平衡結合に到達させた。その後細胞を氷冷PBSPで洗浄し、続いて1.25μg/ml Alexa Fluor(R)488結合体化ストレプトアビジン(Invitrogen)及び225nM Alexa Fluor(R)647結合体化HSAを含有する1ml PBSP中で1時間氷上で暗所にてインキュベートした。氷冷PBSP1mlでの最終洗浄工程の後、細胞を選別前に氷冷PBSPに再懸濁した。細胞をFACSVantage SE(BD Biosciences)フローサイトメーターを使用して選別した。ソートゲートは 最も高いAlexa Fluor(R)488対Alexa Fluor(R)647蛍光強度比を示すZ変異体提示細胞の上部(top)フラクション(典型的に0.1%)を選別するように設定した。細胞を直接0.5ml TSB−YE培地中に選別し、そしてその後10μg/mlクロラムフェニコールを含有するTSBに播種し、そして37℃で16時間、標識及びFACSの次のラウンドのため、増殖により単離された細胞を増幅するためにインキュベートした。この手順を4回繰り返した。
【0135】
配列決定: 個々のブドウ球菌クローンの配列決定を、実施例3のライブラリー構築において記載した細胞選別サイクル3及び4の後に行った。
【0136】
細胞上親和性順位付け及びKD決定: HER3−Fcをビオチン化し、そして実施例1において記載されるようにHSAをAlexa Fluor(R)647と結合体化した。異なるZ変異体を提示しているブドウ球菌細胞をそれぞれ10ml TSB−YE及び20μg/mlクロラムフェニコールに播種し、そして終夜37℃及び150rpmで増殖させた。培養物から、106個の細胞を1ml PBSPで洗浄した。細胞を遠心分離(3500×g、4℃、6分)でペレット化し、そして推定KDに及ぶ異なる濃度のビオチン化HER3−Fc(Z05405、Z05413、Z05416及びZ05417について135、90、27、9、2.7、0.9、0.45、0.22、0.11及び0.054nM;Z01820について670、337、110、45、18、6.7、2.2、1.1、0.37及び0.27nM)を含有するPBSPに再懸濁した。穏やかに混合しながら室温で1時間インキュベーションすることにより平衡結合に達した。細胞を氷冷PBSP1mlで洗浄し、続いて1.25μg/mlAlexa Fluor(R)488結合体化ストレプトアビジン(Invitrogen)、そして順位付け実験では225nM Alexa Fluor(R)647結合体化HSAを含有する氷冷PBSP100μl中で氷上で40分間インキュベートした。氷冷PBSP1mlで洗浄した後、フローサイトメトリー分析前に細胞を氷冷PBSP300μlに再懸濁した。平均蛍光強度(MFI)をFACS Vantage SE(BD Biosciences)フローサイトメーターを使用して測定した。
【0137】
結果
改善されたZ変異体の単離のためのフローサイトメトリー選別: 成熟HER3結合Z変異体の単離のために、ブドウ球菌ライブラリーを、細胞増殖による増幅の交互のラウンドを含むラウンドの蛍光細胞分析分離にかけた。
【0138】
手短には、細胞をビオチン化HER3−Fcと共に実施例1及び2からのZ変異体の推定KDと比較して約10倍低い濃度でインキュベートした。その後細胞を洗浄し、そしてその後の細胞に結合したHER3の蛍光媒介検出のための蛍光標識したストレプトアビジン、さらには表面発現レベルのモニタリングのための蛍光標識したHSAと共にインキュベートした。結合したHER3の解離速度を小さくするために、二次的試薬及びHSAのインキュベーションを氷上で行った。さらに洗浄した後、標識された細胞ライブラリーをフローサイトメーターでスクリーニング及び選別した。標的濃度、選別パラメーター及びソートゲートに関する選択ストリンジェンシーは、各選別ラウンドとともに増加し、そして典型的には、最も高い標的結合対表面発現比を示すライブラリーの上位0.1%を、増幅及びその後の選別ラウンドのためにゲーティングして単離した。フローサイトメーターでのライブラリーの標的結合特性の可視化により、各選別ラウンドでのHER3ポジティブクローンの濃縮そして最終ラウンドでは本質的にHER3ポジティブクローンのみであることが明らかとなった(図5)。FACSの4ラウンドまでの後、単離された細胞を個々の候補の配列決定及び特徴付けのために半固形培地に広げた。
配列決定: 576の個々のクローンを配列決定し、結果として443の配列が可読であり、そのうち45のクローンは同じ選別ラウンドにおいて1回以上出現した(配列番号690〜734)。各変異体に固有の識別番号#####を付与し、そして個々の変異体をZ#####と呼ぶ。58量体Z変異体のアミノ酸配列を図1及び配列表に配列番号690〜1002として載せる。これらのZ変異体の推定HER3結合モチーフを図1及び配列表に配列番号22〜334として載せる。これらの各Z変異体内で完全3ヘリックスバンドルを構成すると予測される49量体ポリペプチドのアミノ酸配列を図1及び配列表に配列番号356〜668として載せる。
【0139】
細胞上親和性順位付け: 配列決定結果に基づいて1回より多く出現した45のクローンを、FER3に対する親和性に関して変異体を順位付けするために全細胞ELISAアッセイにかけた。ブドウ球菌細胞集団をフローサイトメトリーを使用して分析し、そしてブドウ球菌細胞において表面提示された実施例1及び2からの1つのZ変異体(Z01753)を比較のために分析に含めた。全ての45の候補がアッセイにおいてHER3結合に関してポジティブであり、さらに、43は実施例1及び2からのZ変異体と比較してより高いシグナルを示した(図6A〜D)。アッセイにおいて最も高いシグナルは配列決定後に最も頻繁に出現した4つのクローンで見られ、フローサイトメトリー選別におけるZ変異体の定量的な単離が示された。
【0140】
KD決定: 見掛けの平衡解離定数(KD)を、4つのZ変異(図4B)並びに実施例1及び2からの1つのHER3結合Z変異体(Z01820)について細胞で決定した。4つの変異体及びZ01820を提示しているブドウ球菌細胞を、推定KDに及ぶ様々な濃度の標識HER3−Fcでインキュベートした。細胞集団をフローサイトメーターで分析し、続いて平均蛍光強度データをHER3−Fc濃度に対してプロットし、そしてKDを決定するために1部位結合モデルにフィッティングした。全ての4つの成熟Z変異体は低いナノモルの見掛けのKDを示した。最も強いバインダーは、実施例1及び2からのZ変異体と比較してHER3−Fcに対する親和性において15倍の改善を示した(表3)。
【0141】
【表3】
【0142】
実施例5:Z変異体の表面プラズモン共鳴KD決定
材料及び方法
表面プラズモン共鳴分析: 表面プラズモン共鳴(SPR)分析をHER3結合Z変異体Z05405、Z05413、Z05416及びZ05417についてBIAcoreTM 2000機器(GE Healthcare)で行った。ヒトHER3−Fc(R&D Systems)及びマウスHER3−Fc(R&D systems)をCM−5センサーチップ(GE Healthcare)上でNHS/ECDアミンカップリング化学により固定化した。固定化は10mM NaAc(pH4.5)中で30μl/分の流量で、そして10μg/mlの濃度で行い、4000RUの固定化レベルを目標とした。ランニングバッファーとしてPBSPを20μl/分(別の記載がなければ)の流量で、そして再生のために5mM NaOHを使用した。
【0143】
動力学の決定のために、4つの異なる濃度(1.5〜67nM、PBSP中で希釈)の4つのHER3結合Z変異体250μlを、CM−5チップ表面上に固定化したHER3−Fc上に注入した。5mM NaOH 15μlの4回の注入及びランニングバッファを用いた徹底的な洗浄により表面を再生した。各サンプルを二つ組で測定し、そしてブランク表面からの応答を各濃度での各Z変異体の応答から差し引いた。平均会合速度(kon)及び解離速度(koff)に基づいてKD値を決定するために、得られたセンサーグラム(sensorgrams)を1部位結合モデルにフィッティングした。
【0144】
対照実験: HER3−FcのFc部分に対する親和性について選択されたのではないことを確認するために、各Z変異体(50nM)を、各サンプル100μl(上記の濃度で)をバイオセンサーチップ表面に固定化されたHER3−Fc上に注入する前に、10倍モル濃度過剰のヒトポリクローナルIgG(500nM)と共に1時間室温でインキュベートした。さらなる対照実験において、センサーチップ表面に固定化されたHER3−ECD上にZ変異体を注入した。
【0145】
結果
表面プラズモン共鳴分析: 4つのZ変異体Z05405、Z05413、Z05416及びZ05417の解離平衡定数(KD)をSPR技術により決定した。1部位結合モデルに対する非線形回帰を使用してZ変異体の会合速度及び解離速度から親和性を決定し、Z05405について1.61nM、Z05413について0.78nM、Z05416について0.78nM、そしてZ05417について0.69nMの解離定数を得た(図7、表4)。表4において、全ての値は同日に行われた各濃度の二つ組の平均である。
【0146】
【表4】
【0147】
対照実験: HER3結合Z変異体の分析はヒトIgG Fc領域に融合されたHER3を用いて行ったので、Z変異体がHER3−FcのFc部分に対する親和性について選択されたのではないことを確認するために2つの対照実験を行った。IgGと共にプレインキュベートしたZ変異体とZ変異体のみを含有するサンプルとの間に応答の差異は見られなかった(データは示していない)。さらに、HER3−ECDを有する表面上に注入されたZ変異体は、全ての4つの変異体に対して特異的結合を示した(データは示していない)。総合すると、これら2つの実験は、FcがHER3−Fc上のZ変異体の結合部位の一部ではないということを明確に示す。
【0148】
実施例6: ヘレグリン及びHER3結合Z変異体についての競合アッセイ
この実施例において、HER3結合Z変異体がリガンドヘレグリンのHER3に対する結合をブロックする能力を競合アッセイにおいてSPRにより調べた。
【0149】
材料及び方法
天然リガンドヘレグリン(HRG1−β1、EGF様ドメイン、R&D Systems)と4つのHER3特異的Z変異体Z05405、Z05413、Z05416及びZ05417との間の競合を、SPRを使用して2つの異なるアッセイで分析した。
【0150】
第一のアッセイにおいて、2.5nM HER3−Fcを、100nMの4つのHER3特異的Z変異体の各々(40倍モル濃度過剰)とともに1時間室温でプレインキュベートした。その後、HER3−Fc及びZ変異体の各混合物75μlを、ヘレグリンが固定化されたセンサーチップ表面上に注入した。さらに、Z変異体とプレインキュベートしていないHER3−Fcを、ヘレグリンが固定化されたチップ表面上に注入した。
【0151】
第二のアッセイにおいて、5つの異なる濃度(0〜10nM)の各Z変異体100μlを、HER3が固定されたセンサーチップ表面上に注入し、続いてすぐに250nMヘレグリン100μlを注入した(BIAcore COINJECTコマンドを使用)。全てのサンプルを二つ組で注入し、そしてHER2特異的Z変異体を、同じ設定を使用してネガティブコントロールとして注入した。5mM NaOHの2回の注入及びランニングバッファでの徹底的な洗浄で表面を再生させた。
【0152】
結果
HER3特異的Z変異体Z05405、Z05413、Z05416及びZ05417の天然HER3リガンドヘレグリンとのHER3に対する競合をSPR技術により調べた。
【0153】
チップ表面上に固定化されたヘレグリンを使用した第一のアッセイの結果は、Z変異体とプレインキュベートしていないHER3−Fcを注入した場合と比較してZ変異体とともにプレインキュベートしたHER3−Fcを注入した場合に、応答レベルのほとんど完全な減少を示した(図8A)。
【0154】
チップ表面上に固定化されたHER3−Fcを使用した第二のアッセイの結果は、HER3受容体とその天然リガンドとの間の相互作用と濃度依存的に競合することを示した(図8B)。同じパターンがマウスHER3−Fcについて観察された(データは示していない)。ネガティブコントロールとヘレグリンとの間の競合はヒト又はマウスHER3のいずれについても観察することができなかった(データは示していない)。
【0155】
これらの結果は、HER3特異的Z変異体が天然リガンドヘレグリンと同じHER3上の結合部位と相互作用し、それ故リガンドと受容体との間の結合とインビトロで競合し得るということを示した。この効果は、今後の治療的なインビボ適用において利用されるかもしれない。
【0156】
実施例7: インビトロでのヘレグリン誘導HER3リン酸化の阻害
この実施例では、HER3結合Z変異体のヘレグリン誘導HER3リン酸化を妨害する能力を細胞アッセイで調べた。
【0157】
材料及び方法
刺激: MCF−7(ACC115、DSMZ)乳がん細胞を、L−グルタミン(BE17−605E、Cambrex)、非必須アミノ酸(BE13−114E、Cambrex)、ピルビン酸ナトリウム(BE13−115E、Cambrex)及び10%ウシ胎児血清(FBS、10108−165、Gibco)を補充したRPMI培地(BE12−167F、Lonza)で培養した。刺激の前日に、1×106の細胞を6mmペトリ皿(430168、BD Biosciences)に上記の培地5ml中で播種した。
【0158】
刺激の1時間前に、培地をRPMI+L−グルタミン+2%透析FBS(26400−036、Gibco)からなる培地に変更した。HER3のリン酸化を5nMヘレグリン(HRG1−β、396−HB、R&D systems)を用いて誘導した。それぞれN末端hisタグ及びC末端システインを含むHER3結合Z変異体Z05416及びZ05417(His6−Z05416−cys及びHis6−Z05417−cys)を、10又は100倍モル濃度過剰のヘレグリンと同時に加えた。taqポリメラーゼ特異的Z変異体(His6−Z01155−cys)をネガティブコントロールとして使用して、そして上記と同じ手順に従って加えた。
【0159】
10分間37℃でインキュベーションした後、ペトリ皿を氷上に置き、続いて氷冷PBSで洗浄することにより細胞プロセスを停止させた。洗浄後、1mM活性化オルトバナデート(450243、Sigma)を含有するPBS 2mlを各皿に加え、そして細胞を細胞スクレーパーを使用して剥離した。細胞溶液を10mlのチューブに移し、そして細胞を1000rpmでの3分間の予冷遠心分離で回転させることによりペレット化した。乾燥ペレットを氷冷溶解緩衝液100μl(1% NP−40(Sigma I3021)、20mM Tris(pH8.0)、137mM NaCl、10%グリセロール、2mM EDTA、1mM活性化ナトリウムオルトバナデート)に溶解して細胞を溶解させた。溶解物を回転させて4℃にて30分間インキュベートし、そして13000rpmで15分間遠心分離した。上清を集めてさらなる分析まで−80℃で保存した。
【0160】
pHER3−ELISA: リン酸化HER3タンパク質(pHER3)の細胞溶解物中の存在を、Duo−Set IC phospo ErbB3サンドイッチELISA(DYC−1769、R&D Systems)を使用して分析した。
【0161】
96ウェルハーフエリアプレート(Costar 3690)を終夜50μl/ウェルのPBS中に希釈された2μg/mlの捕捉抗体で被覆した。次の日、プレートを自動化ELISA洗浄機でPBS+0.05%Tween(PBST)で4回洗浄した。1% BSAを含有するPBS(100μl/ウェル)で2時間室温にてウェルをブロックし、そして以前に記載したように洗浄した。
【0162】
IC Diluent #12(1% NP−40、20mM Tris(pH8.0)、137mM NaCl、10%グリセロール、2mM EDTA、及び1mM活性化ナトリウムオルトバナデート)で2回又は4回希釈した50μl/ウェルの細胞溶解物を加えた。製造者により提供されたポジティブコントロールもウェルに加えた。続いてプレートを2時間インキュベートした。
【0163】
洗浄後、製造者のプロトコルにしたがってIC diluent #14(20mM Tris、137mM NaCl、0.05% Tween 20、0.1%BSA、pH7.2−7.4)で希釈した50μl/ウェルの検出抗体を加え、そしてプレートをさらに2時間インキュベートした。インキュベーション後に、プレートを洗浄し、そして50μl/ウェルのTMB(Immunopure、Termo Fisher Scientific)を加えた。20分後に50μl/ウェルの2M H2SO4を加えることにより反応を停止させ、そして吸光度を450nmでVictor3 ELISAプレートリーダーを使用して測定した。
【0164】
結果
HER3特異的Z変異体Z05416及びZ05417を、ネガティブコントロール(taqポリメラーゼ特異的Z01155)と一緒に、HER3のヘレグリン誘導リン酸化をブロックするそれらの能力について評価した。pHER3の相対含有量(%)を、ヘレグリン刺激培養物(ヘレグリン刺激培養物から得られたOD値)に対するブロックされた培養物(Z変異体を含有する培養物から得られたOD値)の関係として決定した。図9は、HER3結合変異体Z05416及びZ05417が用量依存的様式でヘレグリン誘導リン酸化を阻害し、一方でtaqポリメラーゼ結合変異体Z01155はいずれの効果も有していなかったことを示す。
【0165】
実施例8: HER3及びHER2に結合する二重特異性Z変異体
背景の項に記載したように、HER3はHER2にとって好ましいヘテロ二量体化パートナーであり、そしてHER2はリガンド誘導シグナル伝達に関してHERファミリーの他の受容体に依存している。二重特異性HER3及びHER2標的化リガンド(例えばHER3−HER2結合Z変異体)は、両方の受容体を発現している腫瘍において受容体シグナル促進腫瘍成長をブロックするかもしれない。
【0166】
場合により様々なリンカー長を含む二重特異性分子のための種々の分子構築物を考慮した。構築物の例は、HER2結合ポリペプチドに対してHER3結合ポリペプチドをN末端又はC末端に位置づけることである。リンカーが2つの結合ポリペプチドの間に導入される場合、それは1〜60アミノ酸残基、例えば1〜45アミノ酸を含み得る。さらに、二重特異性構築物は、半減期延長部分、例えばアルブミン融合、アルブミン結合部分又はPEGのような分子へのカップリングを分子のN末端若しくはC末端部分又はリンカー領域内に備えていてもよい。あるいは、半減期延長部分はそれ自体で結合ポリペプチド間のリンカー、又はスペーサーとして利用され得る。
【0167】
材料及び方法
二量体二重特異性Z変異体分子を、Friedman et al、Biotechnol Appl Biochem、54(2):121−31(2009)に以前に報告されたように操作した。FriedmanらはHERファミリーの2つのメンバーに対して特異的な四量体Z変異体の製造について記載しているが、HER3及びHER2二重特異性Z変異体の構築は、原理上は同じプロトコルに従う。当業者は、Friedmanらに基づく手順を適用する方法を用意に理解するだろう。HER3及びHER2に結合するヘテロ二量体Z変異体の構築については、Jonssonらにより記載される手順(前出)に従うことにより、2つの結合ポリペプチドの間に40アミノ酸のペプチドリンカーを使用してHER3結合変異体をコードする遺伝子フラグメントを、HER2結合Z変異体をコードする遺伝子フラグメントとインフレームで遺伝子融合した。
【0168】
二重特異性タンパク質をコードする遺伝子を、HER3−HER2結合Z変異体をコードするベクターのクローニングにより構築した。HER3結合Z変異体は、配列番号669〜1002から選択され、特に配列番号669〜734から選択される配列、例えば配列番号690〜734から、例えば配列番号691〜693、配列番号695〜696、配列番号700、配列番号703〜704、配列番号708、配列番号710、配列番号712〜713、配列番号721、配列番号722及び配列番号724から選択され、そしてHER2結合Z変異体は配列番号1003から選択される。HER2及びHER3結合Z変異体は、異なる受容体に対して異なる親和性を有し得る。構築物は同じ受容体に対する様々な親和性を有するZ変異体をコードするベクターから作製されてもよい。二重特異性HER3−HER2結合Z変異体をコードするベクターを、適切なエンドヌクレアーゼ(例えばSfiI、AccI、PstI、BamHI、New England Biolabs)を使用して切断し、そしてHER3−HER2特異的Z変異体をコードする遺伝子フラグメントを回収し、そしてライゲーションして二重特異性分子をコードする遺伝子を形成した。HER3及びHER2結合ポリペプチドは遺伝子フラグメント内で個々に単量体単位を形成しても多量体単位、例えば二量体、三量体又は四量体を形成してもよい。
【0169】
ペプチドリンカーの導入のために、リンカー領域をコードする遺伝子フラグメント(例えばGGGS又はGGGGSGGGGLVGLGSGGGGS)の倍数を異なるリンカー長を有する構築物を作製するために導入する。リンカー領域はアルブミン遺伝子又はアルブミン結合ドメイン(ABD)のための遺伝子も含有し得る。ここでは、ABD遺伝子をPCRを使用して増幅し、そしてリンカーコード単位を導入する。この後、BamHI制限消化し、そしてHER結合ポリペプチド間で二量体HER3−HER2二重特異性遺伝子中にライゲーションする。これは本質的に、Jonssonら(前出)により三量体TNF構築物の作製について記載されたように行われるが、Jonssonにおいて記載される中心TNF分子をABDに置き換える。あるいは、HER3−HER2結合Z変異体をコードする遺伝子フラグメントを、種々の形式で二重特異性分子をクローニングするるための所望のリンカー領域及び制限部位も導入する適切なPCRプライマーを使用して増幅する。二重特異性分子の種々の形式は、例えばN末端若しくはC末端の位置づけ、結合Z変異体間の様々な長さのリンカー及び/又は中間のABD遺伝子である。所望の二重特異性分子をコードする遺伝子の構築についての別の可能性は、単に完全合成遺伝子をそのようなサービスを提供している企業(例えばGeneart AG)から購入することである。
【0170】
結果として得られた、HER3結合Z変異体、HER2結合Z変異体、ペプチドリンカー及び場合によりABDをコードする遺伝子フラグメントを、発現ベクターにライゲーションし、発現細胞(例えばE.coli)に形質転換する。タンパク質を例えば実施例2に記載されるように発現させる。さらなるhis−tag及び/又は標識目的のための独特のシステインをコードする発現ベクターを使用し、そして発現されたタンパク質を例えば実施例2に記載されるように精製する。タンパク質発現及び精製に使用される方法は、当業者に公知の標準的な方法である。
【0171】
次いで得られた二重特異性タンパク質を、Friedmanら(前出)に記載されるように、バイオセンサー分析(Biacore、GE Healthcare)を使用することによりインビトロでの同時結合について試験する。ヒトHER3−Fc(hHER3−Fc)及びヒトHER2−Fc(hHER2−Fc、両方ともR&D Systemsから)を、アミンカップリングによりCM5センサーチップ(GE Healthcare)のカルボキシル化デキストラン層上に製造者の指示に従って固定化した。別のフローセル表面を活性化し、そして参照表面として不活性化した。二重特異性HER3−HER2分子をランニングバッファ、HBS(10mM Hepes、150mM NaCl、3.4mM EDTA及び0.005%界面活性剤P20、pH7.4)中で希釈し、その後結合分析を25℃で行った。第一の実験において、二重特異性HER3−HER2分子を125nMから2μMの範囲に及ぶ濃度で全ての表面に30μl/分の流量で注入した。HER3結合Z変異体及びHER2結合Z変異体もまたコントロールとして注入した。各注入後に、10mM HCl 10mlを注入することによりフローセルを再生した。
【0172】
第二の実験において、5μM HER3−HER2分子を、hHER3−Fc又はhHER2−Fc(R&D Systems)がアミンカップリングによりCM5センサーチップのカルボキシル化デキストラン層上に固定化されたフローセル表面上に注入した。1分の短い解離時間の後、115nM hHER3−Fc及びhHER2−Fc(ランニングバッファで希釈)を注入し、そして二重特異性分子のそれらの標的両方に同時に結合する能力をモニタリングした。
【0173】
蛍光標識した二重特異性HER3−HER2分子を、HER3を発現しHER2を発現していない細胞、又はHER2を発現しHER3を発現していない細胞のいずれかとインキュベートすることにより、HER3−HER2分子の生存細胞での結合能力を、蛍光顕微鏡及びフローサイトメトリー分析を使用してFriedmanら(前出)に記載されるようにモニタリングした。それぞれの受容体に対する異なる親和性を有する異なる構築物が、細胞での異なる受容体発現レベルを区別するかどうかを調べるために特異性の差異を同時に調べた。
【0174】
細胞に対する結合二重特異性Z変異体の生物学的効果を、細胞培養物を0.05nMから5000nMまでの範囲にわたる様々な量の二重特異性Z変異体に曝露し、そして細胞増殖速度及び生存を分析し、そして重要な細胞シグナル伝達経路のリン酸化パターンをウェスタンブロット分析を使用して、HER2特異的Z変異体についてEkerljungらにより(Tumour Biol 27(4):201−10(2006)及びBiochem Biophys Res Commun 377(2):489−94(2008)において)記載されるように分析することにより試験した。試験することができる細胞株としては、MDA−MB−361、NCI−N87、CALU−3及びSKOV−3が挙げられる。またHER3形質導入BT−474及びZR75細胞を使用することもできる。
【0175】
インビボでのHER3−HER2 Z変異体の効果を試験するために、二重特異性構築物を腫瘍標的化及び腫瘍における生物学的効果について評価した。二重特異性HER3−HER2結合Z変異体を以前に記載されたように(Ahlgren et al、Bioconjug Chem 19(1):235−43(2008))放射標識し、そしてインビトロでの細胞取り込み、保持及び内部移行について、記載された方法(Steffen et al、Cancer Biother Radiopharm 20(3):239−48(2005))を使用して特徴付けした。HER3及びHER2受容体を発現している異種移植された腫瘍(例えばMDA−MB−361、NCI−N87、CALU−3及びSKOV−3)を有する動物を、体内分布研究において放射標識したHER3−HER2結合Z変異体を注射し、そして腫瘍取り込み及び他の器官との対比についてモニタリングした。半減期延長部分を有していない構築物については、異種移植されたマウスにおける放射能の分布を、典型的には放射標識二重特異性HER3−HER2結合Z変異体の注射後1、2、4、8、24、48及び72時間の時点で評価した。半減期延長部分を有する構築物については、放射能の分布を典型的には放射標識された二重特異性HER3−HER2結合Z変異体の注射後1、4、12、24、48、72、168及び332時間の時点で、本質的にTolmachevらにより記載されるように(Cancer Res 67(6):2773−82(2007))評価した。放射標識二重特異性Z変異体の取り込みを、HER2及び/又はHER3結合Z変異体単独の取り込みと比較した。二重特異性Z変異体が、単一特異性の相当するものと比較した場合にインビボで腫瘍において優れた取り込み及び保持及び/又は特異性を有する場合、それらの二重特異性変異体は標的化ペイロード治療に適切であり得る。このような治療は、二重特異性分子に融合又は結合体化された潜在的なエフェクター機能、例えば放射性核種、毒性化学分子、毒素、サイトカイン又は光増感剤を利用するだろう。
【0176】
二重特異性Z変異体を、インビボでの内因性の効果についてさらに特徴付けした。マウスに、両方の受容体を発現している腫瘍細胞、又はコントロールとして、受容体の一方のみを発現しているか受容体をどちらも発現していない腫瘍細胞を移植し、そして二重特異性Z変異体の単回又は複数回の注射を行う。腫瘍成長を経時的に、そして注射緩衝液のみで活性化合物(すなわち二重特異性HER3−HER2結合分子)を注射されていないビヒクル群を含むコントロール群に対してモニタリングした、
【0177】
実施例9:HER3及びEGFRに対する二重特異性を有するZ変異体
導入部に記載したように、HER3はHER2にについての好ましいヘテロ二量化パートナーである。しかし、特定の条件では、EGF受容体(EGFR又はHER1)に対する重要な二量化パートナーでもある。二重特異性HER3及びEGFR標的化リガンドを使用することにより、両方の受容体を発現している腫瘍が標的にされ、そして腫瘍促進受容体シグナル伝達がブロックされ得る。
【0178】
二重特異性HER3及びEGFR結合Z変異体は、本質的に上記実施例8に記載されるように操作される。得られた構築物を、HER3及びEGFR受容体を発現する細胞株(例えばA431細胞)において、実施例8で二重特異性HER3及びHER2結合Z変異体について記載される方法と類似の方法で試験することができる。
【特許請求の範囲】
【請求項1】
HER3結合モチーフ(BM)を含むHER3結合ポリペプチドであって、該モチーフは、
i) EX2X3X4A YX7EIW X11LPNL X16X17X18QX20 X21AFIX25 X26LX28D、
[ここで、互いに独立して、
X2はK、R及びWから選択され;
X3はK、R、及びYから選択され;
X4は、A、D、E、G、H、I、K、L、M、N、Q、R、S、T、V及びWから選択され;
X7はA、E、F、G、H、K、S、T、V、W及びYから選択され;
X11はE及びQから選択され;
X16はN及びTから選択され;
X17はR、T及びVから選択され;
X18はR、T及びVから選択され;
X20はK及びRから選択され;
X21はA及びGから選択され;
X25はA、G、K、R及びSから選択され;
X26はS及びKから選択され;
X28はA、D、E、F、P、Q、S、T、W及びYから選択される];
及び
ii) i)において定義された配列に対して少なくとも93%の同一性を有するアミノ酸配列[ここで該ポリペプチドは、HER3の細胞外ドメインに結合する]、
から選択されるアミノ酸配列からなる、上記HER3結合ポリペプチド。
【請求項2】
X2X3はKY、RY、WK及びWRから選択される、請求項1に記載のHER3結合ポリペプチド。
【請求項3】
X4はA、H、I、K、L、N、Q、R、S、T及びVから選択される、請求項1又は2に記載のHER3結合ポリペプチド。
【請求項4】
X7はA、F、G、V、W及びYから選択される、請求項1〜3のいずれか1項に記載のHER3結合ポリペプチド。
【請求項5】
X11はQである、請求項1〜4のいずれか1項に記載のHER3結合ポリペプチド。
【請求項6】
X17X18はRV、RT、VR及びRRから選択される、請求項1〜5のいずれか1項に記載のHER3結合ポリペプチド。
【請求項7】
X20はKである、請求項1〜6のいずれか1項に記載のHER3結合ポリペプチド。
【請求項8】
X21はAである、請求項1〜7のいずれか1項に記載のHER3結合ポリペプチド。
【請求項9】
X25はA、G、K及びSから選択される、請求項1〜8のいずれか1項に記載のHER3結合ポリペプチド。
【請求項10】
X28はA、E、F、Q、S、W及びYから選択される、請求項1〜9のいずれか1項に記載のHER3結合ポリペプチド。
【請求項11】
X2X3はKY及びRYから選択される、請求項2に記載のHER3結合ポリペプチド。
【請求項12】
X4はK、L、S及びTから選択される、請求項3に記載のHER3結合ポリペプチド。
【請求項13】
X7はF、G及びYから選択される、請求項4に記載のHER3結合ポリペプチド。
【請求項14】
X17X18はRV、RT及びVRから選択される、請求項6に記載のHER3結合ポリペプチド。
【請求項15】
X25はA、G及びSから選択される、請求項9に記載のHER3結合ポリペプチド。
【請求項16】
X28はA、E、F、Q、S及びWから選択される、請求項10に記載のHER3結合ポリペプチド。
【請求項17】
アミノ酸配列i)が、以下の4つの条件I、II、III及びIV:
I) X2X3はKY及びRYから選択される;
II) X11はQである;
III) X20はKである;
IV) X21はAである、
のうち少なくとも1つを満たす、請求項1〜16のいずれか1項に記載のHER3結合ポリペプチド。
【請求項18】
4つの条件I、II、III及びIVのうち少なくとも2つを満たす、請求項17に記載のHER3結合ポリペプチド。
【請求項19】
4つの条件I、II、III及びIVのうち少なくとも3つを満たす、請求項18に記載のHER3結合ポリペプチド。
【請求項20】
4つの条件I、II、III及びIVの全てを満たす、請求項19に記載のHER3結合ポリペプチド。
【請求項21】
アミノ酸配列が配列番号1〜334のいずれか1つから選択される、請求項1〜20のいずれか1項に記載のHER3結合ポリペプチド。
【請求項22】
アミノ酸配列が配列番号1〜66のいずれか1つから選択される、請求項21に記載のHER3結合ポリペプチド
【請求項23】
アミノ酸配列が、配列番号23〜25、配列番号27〜28、配列番号32、配列番号35〜36、配列番号40、配列番号42、配列番号44〜45、配列番号53〜54及び配列番号56のいずれか1つから選択される、請求項22に記載のHER3結合ポリペプチド。
【請求項24】
前記HER3結合モチーフが、3ヘリックスバンドルタンパク質ドメインの一部を形成する、請求項1〜23のいずれか1項に記載のHER3結合ポリペプチド。
【請求項25】
前記HER3結合モチーフが、前記3ヘリックスバンドルタンパク質ドメイン内で、相互接続ループを有する2つのアルファヘリックスの一部を本質的に形成する、請求項24に記載のHER3結合ポリペプチド。
【請求項26】
前記3ヘリックスバンドルタンパク質ドメインが、黄色ブドウ球菌由来のプロテインAのドメイン及びその誘導体から選択される、請求項24又は25に記載のHER3結合ポリペプチド。
【請求項27】
i) K−[BM]−DPSQS XaXbLLXc EAKKL NDXdQ;
[ここで、
[BM]は、請求項1〜23のいずれか1項において定義されるHER3結合モチーフであり;
XaはA及びSから選択され;
XbはN及びEから選択され;
XcはA、S及びCから選択され;
XdはA及びSから選択される]
及び
ii) 上で定義された配列のいずれか1つに対して少なくとも94%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項1〜26のいずれか1項に記載のHER3結合ポリペプチド。
【請求項28】
XaはAであり;XbはNであり;XcはAであり、そしてXdはAである、請求項27に記載のHER3結合ポリペプチド。
【請求項29】
XaはAであり;XbはNであり;XcはCであり、そしてXdはAである、請求項27に記載のHER3結合ポリペプチド。
【請求項30】
XaはSであり;XbはEであり;XcはSであり、そしてXdはSである、請求項27に記載のHER3結合ポリペプチド。
【請求項31】
XaはSであり;XbはEであり;XcはCであり、そしてXdはSである、請求項27に記載のHER3結合ポリペプチド。
【請求項32】
アミノ酸配列が配列番号335〜668のいずれか1つから選択される、請求項27〜31のいずれか1項に記載のHER3結合ポリペプチド。
【請求項33】
アミノ酸配列が、配列番号335〜400のいずれか1つから選択される、請求項32に記載のHER3結合ポリペプチド。
【請求項34】
アミノ酸配列が、配列番号357〜359、配列番号361〜362、配列番号366、配列番号369〜370、配列番号374、配列番号376、配列番号378〜379、配列番号387〜388及び配列番号390のいずか1つから選択される、請求項33に記載のHER3結合ポリペプチド。
【請求項35】
i) YAK−[BM]−DPSQS SELLXc EAKKL NDSQA P;
[ここで[BM]は請求項1〜23のいずれか1項において定義されたHER3結合モチーフであり、そしてXcはS及びCから選択される];及び
ii) 上で定義される配列のいずれか1つに対して少なくとも94%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項1〜26のいずれか1項に記載のHER3結合ポリペプチド。
【請求項36】
i) FNK−[BM]−DPSQS ANLLXc EAKKL NDAQA P;
[ここで[BM]は請求項1〜23のいずれか1項において定義されたHER3結合モチーフであり、そしてXcはA及びCから選択される];及び
ii) 上で定義された配列のいずれか1つに対して少なくとも94%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項1〜26のいずれか1項に記載のHER3結合ポリペプチド。
【請求項37】
アミノ酸配列が配列番号669〜1002から選択される、請求項1〜26のいずれか1項に記載のHER3結合ポリペプチド。
【請求項38】
アミノ酸配列が配列番号669〜734から選択される、請求項37に記載のHER3結合ポリペプチド。
【請求項39】
アミノ酸配列が、配列番号691〜693、配列番号695〜696、配列番号700、配列番号703〜704、配列番号708、配列番号710、配列番号712〜713、配列番号721、配列番号722及び配列番号724から選択される、請求項38に記載のHER3結合ポリペプチド。
【請求項40】
相互作用のKD値が多くとも1×10-6M、例えば多くとも1×10-7M、例えば多くとも1×10-8MであるようにHER3結合ポリペプチドがHER3に結合する、請求項1〜39のいずれか1項に記載のHER3結合ポリペプチド。
【請求項41】
さらなるC末端アミノ酸及び/又はN末端アミノ酸を含む、請求項1〜40のいずれか1項に記載のHER3結合ポリペプチド。
【請求項42】
さらなるC末端アミノ酸及び/又はN末端アミノ酸の各々が、ポリペプチドの生産、精製、インビボ若しくはインビトロでの安定化、カップリング又は検出を改善する、請求項41に記載のHER3結合ポリペプチド。
【請求項43】
アミノ酸配列が同じであっても異なっていてもよい、少なくとも2つのHER3結合ポリペプチド単量体単位を含む、多量体形態である、請求項1〜42のいずれか1項に記載のHER3結合ポリペプチド。
【請求項44】
二量体形態である、請求項43に記載のHER3結合ポリペプチド。
【請求項45】
天然に存在するHER3リガンドのHER3への結合をブロックする、請求項1〜44のいずれか1項に記載のHER3結合ポリペプチド。
【請求項46】
インビボでHER3仲介シグナル伝達を阻害する、請求項45に記載のHER3結合ポリペプチド。
【請求項47】
請求項1〜46のいずれか1項に記載のポリペプチドをコードするポリヌクレオチド。
【請求項48】
請求項47に記載のポリヌクレオチドを含む発現ベクター。
【請求項49】
請求項48に記載の発現ベクターを含む宿主細胞。
【請求項50】
請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド;HER2結合ポリペプチド;及びHER3結合ポリペプチドとHER2結合ポリペプチドとを連結するための連結部分を含む、HER3及びHER2に対する結合親和性を有するリガンド。
【請求項51】
HER2結合ポリペプチドが、相互作用のKD値が多くとも1×10-6M、例えば多くとも1×10-7M、例えば多くとも1×10-8MであるようにHER2に結合する、請求項50に記載のリガンド。
【請求項52】
HER2結合ポリペプチドが、黄色ブドウ球菌由来のプロテインAのドメインBから誘導された改変タンパク質である、請求項50又は51に記載のリガンド。
【請求項53】
改変タンパク質がプロテインZ誘導体を含む、請求項52に記載のリガンド。
【請求項54】
改変タンパク質が、
i) YAKEM RNAYW EIALL PNLTN QQKRA FIRKL YDDPS QSSEL LSEAK KLNDS Q;及び
ii) i)において定義される配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項53に記載のリガンド。
【請求項55】
連結部分が1から最大で45個までのアミノ酸を含むペプチドである、請求項50〜54のいずれか1項に記載のリガンド。
【請求項56】
インビボでリガンド半減期を延長させるための半減期延長部分をさらに含む、請求項50〜55のいずれか1項に記載のリガンド。
【請求項57】
半減期延長部分がアルブミンである、請求項56に記載のリガンド。
【請求項58】
半減期延長部分がアルブミン結合部分である、請求項56に記載のリガンド。
【請求項59】
アルブミン結合部分が、連鎖球菌プロテインGのドメインGA3から誘導された改変タンパク質である、請求項58に記載のリガンド。
【請求項60】
アルブミン結合部分が、
i) LAEAK VLANR ELDKY GVSDF YKRLI NKAKT VEGVE ALKLH ILAAL P;及び
ii) i)において定義された配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項59に記載のリガンド。
【請求項61】
半減期延長部分がポリエチレングリコールである、請求項56に記載のリガンド。
【請求項62】
半減期延長部分が、リガンド中に存在する少なくとも1つのシステイン残基を介してリガンドの残部に結合する、請求項56〜61のいずれか1項に記載のリガンド。
【請求項63】
半減期延長部分が連結部分を介してリガンドの残部に結合する、請求項56〜62のいずれか1項に記載のリガンド。
【請求項64】
連結部分が、請求項56〜60のいずれか1項において定義される半減期延長部分を含む、請求項50〜62のいずれか1項に記載のリガンド。
【請求項65】
融合ポリペプチドである、請求項50〜60又は64のいずれか1項に記載のリガンド。
【請求項66】
インビボでHER2及びHER3の二量化を防止する、請求項50〜65のいずれか1項に記載のリガンド。
【請求項67】
インビボでHER3仲介シグナル伝達を阻害する、請求項66に記載のリガンド。
【請求項68】
請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド;EGFR結合ポリペプチド;及びHER3結合ポリペプチドをEGFR結合ポリペプチドと連結するための連結部分を含む、HER3及びEGFRに対する結合親和性を有するリガンド。
【請求項69】
請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド又は請求項50〜68のいずれか1項に記載のリガンドと治療剤との組み合わせ。
【請求項70】
治療における使用のための、請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド、請求項50〜68のいずれか1項に記載のリガンド又は請求項69に記載の組み合わせ。
【請求項71】
HER3関連状態の処置のための、請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド、請求項50〜68のいずれか1項に記載のリガンド又は請求項69に記載の組み合わせ。
【請求項72】
がん疾患、例えば結腸がん、子宮内膜がん、胃がん、神経膠腫、乳がん、膵臓がん、頭頸部扁平上皮がん、肺がん、黒色種、髄芽腫、神経上皮腫、卵巣がん、パジェット病、乳頭様甲状腺がん、前立腺がん、皮膚扁平上皮がん、移行上皮がん及び前庭神経鞘腫から選択されるHER3関連状態の処置のための、請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド、請求項50〜68のいずれか1項に記載のリガンド又は請求項69に記載の組み合わせ。
【請求項73】
請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド、請求項50〜68のいずれか1項に記載のリガンド又は請求項69に記載の組み合わせを、それを必要とする哺乳動物対象に投与する工程を含む、HER3関連状態の処置方法。
【請求項74】
HER3結合ポリペプチド、リガンド又は組み合わせの、対象の細胞表面で発現されたHER3への結合が、HER3仲介シグナル伝達を阻害する、請求項73に記載の処置方法。
【請求項75】
HER3関連状態が、がん疾患、例えば結腸がん、子宮内膜がん、胃がん、神経膠腫、乳がん、膵臓がん、頭頸部扁平上皮がん、肺がん、黒色種、髄芽腫、神経上皮腫、卵巣がん、パジェット病、乳頭様甲状腺がん、前立腺がん、皮膚扁平上皮がん、移行上皮がん及び前庭神経鞘腫から選択される、請求項73又は74に記載の方法。
【請求項1】
HER3結合モチーフ(BM)を含むHER3結合ポリペプチドであって、該モチーフは、
i) EX2X3X4A YX7EIW X11LPNL X16X17X18QX20 X21AFIX25 X26LX28D、
[ここで、互いに独立して、
X2はK、R及びWから選択され;
X3はK、R、及びYから選択され;
X4は、A、D、E、G、H、I、K、L、M、N、Q、R、S、T、V及びWから選択され;
X7はA、E、F、G、H、K、S、T、V、W及びYから選択され;
X11はE及びQから選択され;
X16はN及びTから選択され;
X17はR、T及びVから選択され;
X18はR、T及びVから選択され;
X20はK及びRから選択され;
X21はA及びGから選択され;
X25はA、G、K、R及びSから選択され;
X26はS及びKから選択され;
X28はA、D、E、F、P、Q、S、T、W及びYから選択される];
及び
ii) i)において定義された配列に対して少なくとも93%の同一性を有するアミノ酸配列[ここで該ポリペプチドは、HER3の細胞外ドメインに結合する]、
から選択されるアミノ酸配列からなる、上記HER3結合ポリペプチド。
【請求項2】
X2X3はKY、RY、WK及びWRから選択される、請求項1に記載のHER3結合ポリペプチド。
【請求項3】
X4はA、H、I、K、L、N、Q、R、S、T及びVから選択される、請求項1又は2に記載のHER3結合ポリペプチド。
【請求項4】
X7はA、F、G、V、W及びYから選択される、請求項1〜3のいずれか1項に記載のHER3結合ポリペプチド。
【請求項5】
X11はQである、請求項1〜4のいずれか1項に記載のHER3結合ポリペプチド。
【請求項6】
X17X18はRV、RT、VR及びRRから選択される、請求項1〜5のいずれか1項に記載のHER3結合ポリペプチド。
【請求項7】
X20はKである、請求項1〜6のいずれか1項に記載のHER3結合ポリペプチド。
【請求項8】
X21はAである、請求項1〜7のいずれか1項に記載のHER3結合ポリペプチド。
【請求項9】
X25はA、G、K及びSから選択される、請求項1〜8のいずれか1項に記載のHER3結合ポリペプチド。
【請求項10】
X28はA、E、F、Q、S、W及びYから選択される、請求項1〜9のいずれか1項に記載のHER3結合ポリペプチド。
【請求項11】
X2X3はKY及びRYから選択される、請求項2に記載のHER3結合ポリペプチド。
【請求項12】
X4はK、L、S及びTから選択される、請求項3に記載のHER3結合ポリペプチド。
【請求項13】
X7はF、G及びYから選択される、請求項4に記載のHER3結合ポリペプチド。
【請求項14】
X17X18はRV、RT及びVRから選択される、請求項6に記載のHER3結合ポリペプチド。
【請求項15】
X25はA、G及びSから選択される、請求項9に記載のHER3結合ポリペプチド。
【請求項16】
X28はA、E、F、Q、S及びWから選択される、請求項10に記載のHER3結合ポリペプチド。
【請求項17】
アミノ酸配列i)が、以下の4つの条件I、II、III及びIV:
I) X2X3はKY及びRYから選択される;
II) X11はQである;
III) X20はKである;
IV) X21はAである、
のうち少なくとも1つを満たす、請求項1〜16のいずれか1項に記載のHER3結合ポリペプチド。
【請求項18】
4つの条件I、II、III及びIVのうち少なくとも2つを満たす、請求項17に記載のHER3結合ポリペプチド。
【請求項19】
4つの条件I、II、III及びIVのうち少なくとも3つを満たす、請求項18に記載のHER3結合ポリペプチド。
【請求項20】
4つの条件I、II、III及びIVの全てを満たす、請求項19に記載のHER3結合ポリペプチド。
【請求項21】
アミノ酸配列が配列番号1〜334のいずれか1つから選択される、請求項1〜20のいずれか1項に記載のHER3結合ポリペプチド。
【請求項22】
アミノ酸配列が配列番号1〜66のいずれか1つから選択される、請求項21に記載のHER3結合ポリペプチド
【請求項23】
アミノ酸配列が、配列番号23〜25、配列番号27〜28、配列番号32、配列番号35〜36、配列番号40、配列番号42、配列番号44〜45、配列番号53〜54及び配列番号56のいずれか1つから選択される、請求項22に記載のHER3結合ポリペプチド。
【請求項24】
前記HER3結合モチーフが、3ヘリックスバンドルタンパク質ドメインの一部を形成する、請求項1〜23のいずれか1項に記載のHER3結合ポリペプチド。
【請求項25】
前記HER3結合モチーフが、前記3ヘリックスバンドルタンパク質ドメイン内で、相互接続ループを有する2つのアルファヘリックスの一部を本質的に形成する、請求項24に記載のHER3結合ポリペプチド。
【請求項26】
前記3ヘリックスバンドルタンパク質ドメインが、黄色ブドウ球菌由来のプロテインAのドメイン及びその誘導体から選択される、請求項24又は25に記載のHER3結合ポリペプチド。
【請求項27】
i) K−[BM]−DPSQS XaXbLLXc EAKKL NDXdQ;
[ここで、
[BM]は、請求項1〜23のいずれか1項において定義されるHER3結合モチーフであり;
XaはA及びSから選択され;
XbはN及びEから選択され;
XcはA、S及びCから選択され;
XdはA及びSから選択される]
及び
ii) 上で定義された配列のいずれか1つに対して少なくとも94%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項1〜26のいずれか1項に記載のHER3結合ポリペプチド。
【請求項28】
XaはAであり;XbはNであり;XcはAであり、そしてXdはAである、請求項27に記載のHER3結合ポリペプチド。
【請求項29】
XaはAであり;XbはNであり;XcはCであり、そしてXdはAである、請求項27に記載のHER3結合ポリペプチド。
【請求項30】
XaはSであり;XbはEであり;XcはSであり、そしてXdはSである、請求項27に記載のHER3結合ポリペプチド。
【請求項31】
XaはSであり;XbはEであり;XcはCであり、そしてXdはSである、請求項27に記載のHER3結合ポリペプチド。
【請求項32】
アミノ酸配列が配列番号335〜668のいずれか1つから選択される、請求項27〜31のいずれか1項に記載のHER3結合ポリペプチド。
【請求項33】
アミノ酸配列が、配列番号335〜400のいずれか1つから選択される、請求項32に記載のHER3結合ポリペプチド。
【請求項34】
アミノ酸配列が、配列番号357〜359、配列番号361〜362、配列番号366、配列番号369〜370、配列番号374、配列番号376、配列番号378〜379、配列番号387〜388及び配列番号390のいずか1つから選択される、請求項33に記載のHER3結合ポリペプチド。
【請求項35】
i) YAK−[BM]−DPSQS SELLXc EAKKL NDSQA P;
[ここで[BM]は請求項1〜23のいずれか1項において定義されたHER3結合モチーフであり、そしてXcはS及びCから選択される];及び
ii) 上で定義される配列のいずれか1つに対して少なくとも94%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項1〜26のいずれか1項に記載のHER3結合ポリペプチド。
【請求項36】
i) FNK−[BM]−DPSQS ANLLXc EAKKL NDAQA P;
[ここで[BM]は請求項1〜23のいずれか1項において定義されたHER3結合モチーフであり、そしてXcはA及びCから選択される];及び
ii) 上で定義された配列のいずれか1つに対して少なくとも94%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項1〜26のいずれか1項に記載のHER3結合ポリペプチド。
【請求項37】
アミノ酸配列が配列番号669〜1002から選択される、請求項1〜26のいずれか1項に記載のHER3結合ポリペプチド。
【請求項38】
アミノ酸配列が配列番号669〜734から選択される、請求項37に記載のHER3結合ポリペプチド。
【請求項39】
アミノ酸配列が、配列番号691〜693、配列番号695〜696、配列番号700、配列番号703〜704、配列番号708、配列番号710、配列番号712〜713、配列番号721、配列番号722及び配列番号724から選択される、請求項38に記載のHER3結合ポリペプチド。
【請求項40】
相互作用のKD値が多くとも1×10-6M、例えば多くとも1×10-7M、例えば多くとも1×10-8MであるようにHER3結合ポリペプチドがHER3に結合する、請求項1〜39のいずれか1項に記載のHER3結合ポリペプチド。
【請求項41】
さらなるC末端アミノ酸及び/又はN末端アミノ酸を含む、請求項1〜40のいずれか1項に記載のHER3結合ポリペプチド。
【請求項42】
さらなるC末端アミノ酸及び/又はN末端アミノ酸の各々が、ポリペプチドの生産、精製、インビボ若しくはインビトロでの安定化、カップリング又は検出を改善する、請求項41に記載のHER3結合ポリペプチド。
【請求項43】
アミノ酸配列が同じであっても異なっていてもよい、少なくとも2つのHER3結合ポリペプチド単量体単位を含む、多量体形態である、請求項1〜42のいずれか1項に記載のHER3結合ポリペプチド。
【請求項44】
二量体形態である、請求項43に記載のHER3結合ポリペプチド。
【請求項45】
天然に存在するHER3リガンドのHER3への結合をブロックする、請求項1〜44のいずれか1項に記載のHER3結合ポリペプチド。
【請求項46】
インビボでHER3仲介シグナル伝達を阻害する、請求項45に記載のHER3結合ポリペプチド。
【請求項47】
請求項1〜46のいずれか1項に記載のポリペプチドをコードするポリヌクレオチド。
【請求項48】
請求項47に記載のポリヌクレオチドを含む発現ベクター。
【請求項49】
請求項48に記載の発現ベクターを含む宿主細胞。
【請求項50】
請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド;HER2結合ポリペプチド;及びHER3結合ポリペプチドとHER2結合ポリペプチドとを連結するための連結部分を含む、HER3及びHER2に対する結合親和性を有するリガンド。
【請求項51】
HER2結合ポリペプチドが、相互作用のKD値が多くとも1×10-6M、例えば多くとも1×10-7M、例えば多くとも1×10-8MであるようにHER2に結合する、請求項50に記載のリガンド。
【請求項52】
HER2結合ポリペプチドが、黄色ブドウ球菌由来のプロテインAのドメインBから誘導された改変タンパク質である、請求項50又は51に記載のリガンド。
【請求項53】
改変タンパク質がプロテインZ誘導体を含む、請求項52に記載のリガンド。
【請求項54】
改変タンパク質が、
i) YAKEM RNAYW EIALL PNLTN QQKRA FIRKL YDDPS QSSEL LSEAK KLNDS Q;及び
ii) i)において定義される配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項53に記載のリガンド。
【請求項55】
連結部分が1から最大で45個までのアミノ酸を含むペプチドである、請求項50〜54のいずれか1項に記載のリガンド。
【請求項56】
インビボでリガンド半減期を延長させるための半減期延長部分をさらに含む、請求項50〜55のいずれか1項に記載のリガンド。
【請求項57】
半減期延長部分がアルブミンである、請求項56に記載のリガンド。
【請求項58】
半減期延長部分がアルブミン結合部分である、請求項56に記載のリガンド。
【請求項59】
アルブミン結合部分が、連鎖球菌プロテインGのドメインGA3から誘導された改変タンパク質である、請求項58に記載のリガンド。
【請求項60】
アルブミン結合部分が、
i) LAEAK VLANR ELDKY GVSDF YKRLI NKAKT VEGVE ALKLH ILAAL P;及び
ii) i)において定義された配列に対して少なくとも80%の同一性を有するアミノ酸配列、
から選択されるアミノ酸配列を含む、請求項59に記載のリガンド。
【請求項61】
半減期延長部分がポリエチレングリコールである、請求項56に記載のリガンド。
【請求項62】
半減期延長部分が、リガンド中に存在する少なくとも1つのシステイン残基を介してリガンドの残部に結合する、請求項56〜61のいずれか1項に記載のリガンド。
【請求項63】
半減期延長部分が連結部分を介してリガンドの残部に結合する、請求項56〜62のいずれか1項に記載のリガンド。
【請求項64】
連結部分が、請求項56〜60のいずれか1項において定義される半減期延長部分を含む、請求項50〜62のいずれか1項に記載のリガンド。
【請求項65】
融合ポリペプチドである、請求項50〜60又は64のいずれか1項に記載のリガンド。
【請求項66】
インビボでHER2及びHER3の二量化を防止する、請求項50〜65のいずれか1項に記載のリガンド。
【請求項67】
インビボでHER3仲介シグナル伝達を阻害する、請求項66に記載のリガンド。
【請求項68】
請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド;EGFR結合ポリペプチド;及びHER3結合ポリペプチドをEGFR結合ポリペプチドと連結するための連結部分を含む、HER3及びEGFRに対する結合親和性を有するリガンド。
【請求項69】
請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド又は請求項50〜68のいずれか1項に記載のリガンドと治療剤との組み合わせ。
【請求項70】
治療における使用のための、請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド、請求項50〜68のいずれか1項に記載のリガンド又は請求項69に記載の組み合わせ。
【請求項71】
HER3関連状態の処置のための、請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド、請求項50〜68のいずれか1項に記載のリガンド又は請求項69に記載の組み合わせ。
【請求項72】
がん疾患、例えば結腸がん、子宮内膜がん、胃がん、神経膠腫、乳がん、膵臓がん、頭頸部扁平上皮がん、肺がん、黒色種、髄芽腫、神経上皮腫、卵巣がん、パジェット病、乳頭様甲状腺がん、前立腺がん、皮膚扁平上皮がん、移行上皮がん及び前庭神経鞘腫から選択されるHER3関連状態の処置のための、請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド、請求項50〜68のいずれか1項に記載のリガンド又は請求項69に記載の組み合わせ。
【請求項73】
請求項1〜46のいずれか1項に記載のHER3結合ポリペプチド、請求項50〜68のいずれか1項に記載のリガンド又は請求項69に記載の組み合わせを、それを必要とする哺乳動物対象に投与する工程を含む、HER3関連状態の処置方法。
【請求項74】
HER3結合ポリペプチド、リガンド又は組み合わせの、対象の細胞表面で発現されたHER3への結合が、HER3仲介シグナル伝達を阻害する、請求項73に記載の処置方法。
【請求項75】
HER3関連状態が、がん疾患、例えば結腸がん、子宮内膜がん、胃がん、神経膠腫、乳がん、膵臓がん、頭頸部扁平上皮がん、肺がん、黒色種、髄芽腫、神経上皮腫、卵巣がん、パジェット病、乳頭様甲状腺がん、前立腺がん、皮膚扁平上皮がん、移行上皮がん及び前庭神経鞘腫から選択される、請求項73又は74に記載の方法。
【図1−1】

【図1−2】

【図1−3】

【図1−4】

【図1−5】

【図1−6】

【図1−7】

【図1−8】

【図1−9】

【図1−10】

【図1−11】

【図1−12】

【図1−13】

【図1−14】

【図1−15】

【図1−16】

【図1−17】

【図1−18】

【図1−19】

【図1−20】

【図1−21】

【図1−22】

【図1−23】

【図1−24】

【図1−25】

【図1−26】

【図1−27】

【図1−28】

【図1−29】

【図1−30】

【図1−31】

【図1−32】

【図1−33】

【図1−34】

【図1−35】

【図1−36】

【図1−37】

【図1−38】

【図1−39】

【図1−40】

【図1−41】

【図1−42】

【図2】
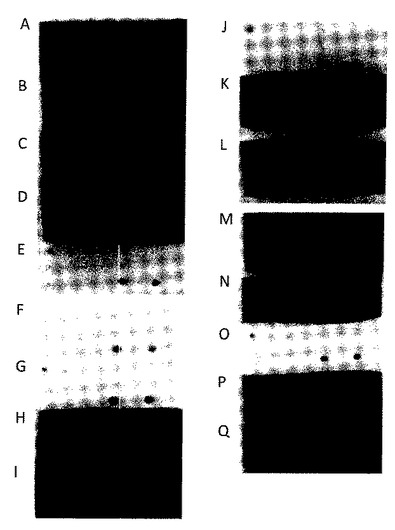
【図3】

【図4】
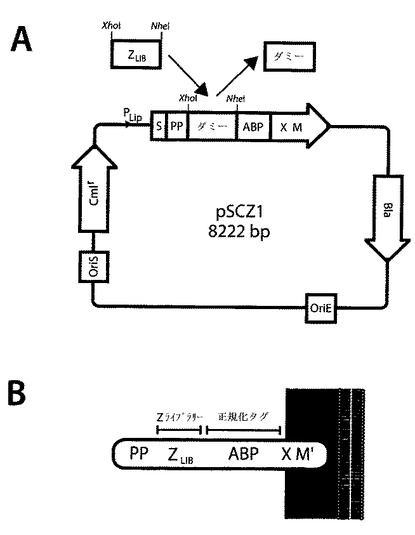
【図5】
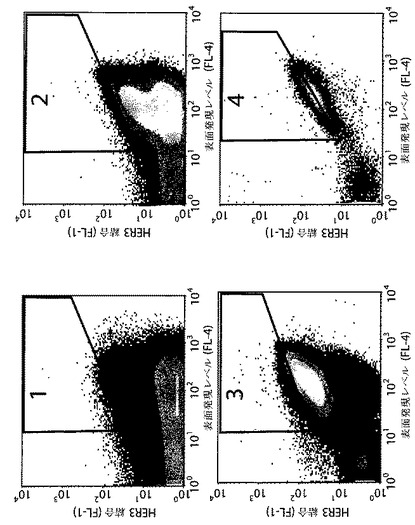
【図6−1】
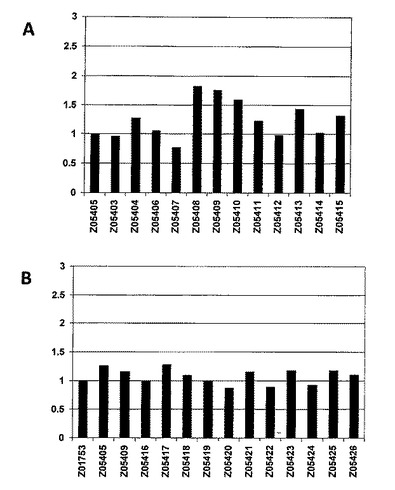
【図6−2】
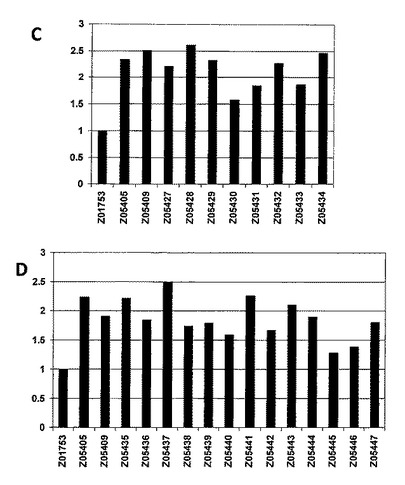
【図7A】
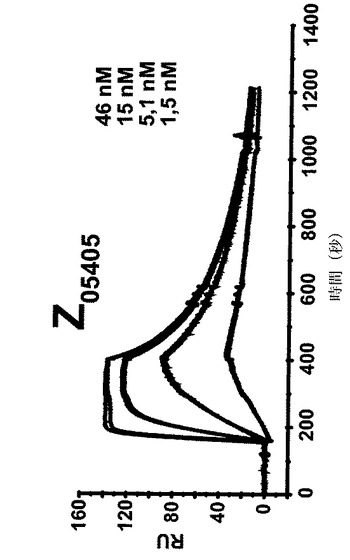
【図7B】
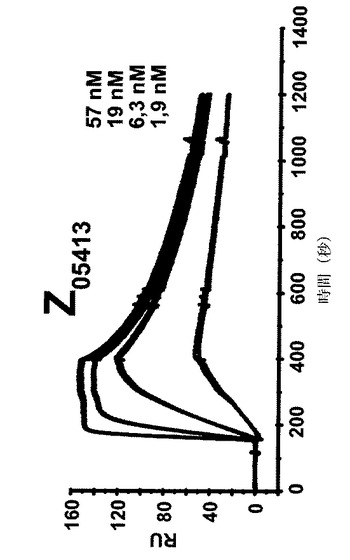
【図7C】
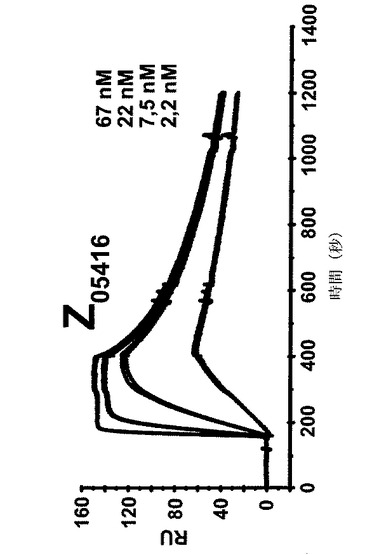
【図7D】
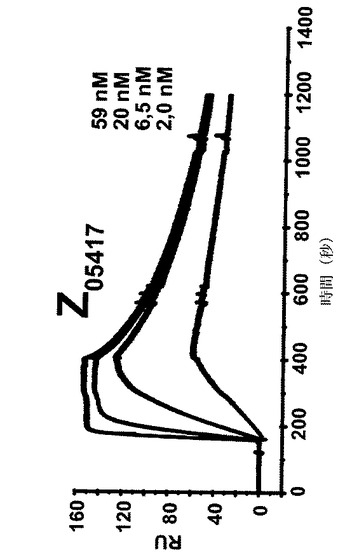
【図8A】

【図8B】
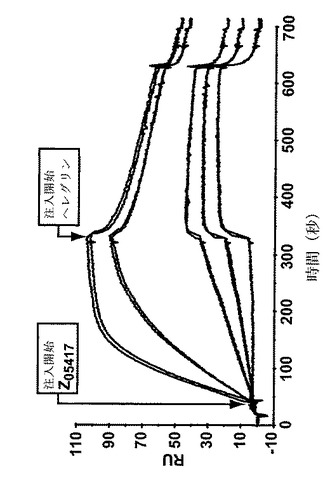
【図9】
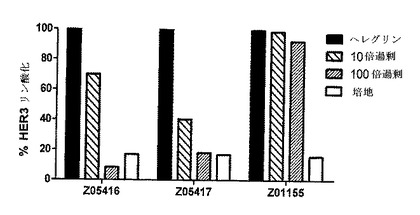
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図1−6】
【図1−7】
【図1−8】
【図1−9】
【図1−10】
【図1−11】
【図1−12】
【図1−13】
【図1−14】
【図1−15】
【図1−16】
【図1−17】
【図1−18】
【図1−19】
【図1−20】
【図1−21】
【図1−22】
【図1−23】
【図1−24】
【図1−25】
【図1−26】
【図1−27】
【図1−28】
【図1−29】
【図1−30】
【図1−31】
【図1−32】
【図1−33】
【図1−34】
【図1−35】
【図1−36】
【図1−37】
【図1−38】
【図1−39】
【図1−40】
【図1−41】
【図1−42】
【図2】
【図3】
【図4】
【図5】
【図6−1】
【図6−2】
【図7A】
【図7B】
【図7C】
【図7D】
【図8A】
【図8B】
【図9】
【公表番号】特表2013−509861(P2013−509861A)
【公表日】平成25年3月21日(2013.3.21)
【国際特許分類】
【出願番号】特願2012−536757(P2012−536757)
【出願日】平成22年10月27日(2010.10.27)
【国際出願番号】PCT/SE2010/051164
【国際公開番号】WO2011/056124
【国際公開日】平成23年5月12日(2011.5.12)
【出願人】(307015286)アフィボディ・アーベー (9)
【Fターム(参考)】
【公表日】平成25年3月21日(2013.3.21)
【国際特許分類】
【出願日】平成22年10月27日(2010.10.27)
【国際出願番号】PCT/SE2010/051164
【国際公開番号】WO2011/056124
【国際公開日】平成23年5月12日(2011.5.12)
【出願人】(307015286)アフィボディ・アーベー (9)
【Fターム(参考)】
[ Back to top ]