
HIV感染を阻害する方法および組成物
本発明はHIV感染を阻害する薬剤を同定するための新規方法を提供する。抗HIV薬を、イソペプチダーゼT(IsoT)の生物学的活性、例えばそのイソペプチダーゼ活性またはウイルスタンパク質R(Vpr)のような他の分子へのその結合を調節する能力について、試験化合物をスクリーニングすることによって同定する。かかるIsoTモジュレーターを、さらにHIV感染またはHIV複製の指標である活性を阻害する活性について試験することができる。これらの新規な抗HIV薬はHIV感染およびHIV感染に関するもしくはHIV感染によってもたらされる状態の、予防または処置に有用である。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本願は米国仮出願第60/673,623(出願日:2005年4月21日)に基づき合衆国法典第35巻、119条(e)の優先権を主張する。先の出願の開示はその全体をあらゆる目的のために引用し、本明細書の一部とする。
【0002】
発明分野
本発明は、一般的にHIV感染を阻害する化合物を同定するための方法と、当該化合物の治療的に関する。より具体的には、本発明はイソペプチダーゼT(IsoT)の阻害剤をスクリーニングすること、そしてかかる阻害剤をHIVの処置または予防のために使用することに関する。
【背景技術】
【0003】
発明の背景
ヒト免疫不全ウイルス(HIV)はレトロウイルス科に属するレンチウイルスである。性的接触および妊娠期間中のHIVの感染は世界中のAIDS症例のほぼ90%を占めると概算された。この感染は、感染個体の体液、例えば精液、膣分泌物または血液に曝露されたとき、性器の粘膜または胎盤を通じてHIVが通過して引き起こされる。のこりのAIDS症例はHIVに汚染された血液の輸血、静脈内薬剤使用者間の針の共有、侵襲処置におけるHIV汚染体液への事故的曝露、そして感染ウイルスが受容ヒト組織へ直接接触することができる他の状況のためである。
【発明の開示】
【発明が解決しようとする課題】
【0004】
現在利用可能なHIV感染およびAIDSの処置用薬剤は満足のいくものではない。HIVを処置するための一般薬剤、例えばAZTまたはHIVプロテアーゼ阻害剤の毒性または望ましくない副作用は、有効医薬濃度で使用したときそれらの抗ウイルス活性と相容れない。したがって、AIDSおよびHIV感染を予防および処置するためのよりよい他の化合物に対するこの分野における要求は未だに存在する。本発明は、このおよび他の要求に対処する。
【課題を解決するための手段】
【0005】
発明の要約
ある態様において、本発明はHIV感染を阻害する薬剤を同定するための方法を提供する。当該方法は、イソペプチダーゼT(IsoT)分子の生物学的活性を試験化合物の存在下でアッセイして、IsoT分子の生物学的活性を阻害する化合物を同定することを必要とする。これらの方法は、同定した化合物のHIV感染阻害能をさらに試験することを含むことができる。ある方法では、アッセイする生物学的活性はIsoT分子のイソペプチダーゼ活性または他の分子(例えばユビキタンまたはVpr)とのその結合である。また、IsoT分子をコード化する遺伝子の発現であってもよい。典型的には、スクリーニングに使用するIsoT分子は哺乳類細胞に由来する。好ましい態様において、ヒトIsoT分子を使用する。
【0006】
これらのスクリーニング法のいくつかにおいて、同定した化合物によるHIV−1感染の阻害能を、当該化合物で処理した人工HIV許容細胞におけるHIV複製と、当該化合物で処理しなかった対照細胞におけるHIV複製を比較することによって試験する。これらの方法のいくつかにおいて、使用するHIV許容細胞はHeLa−T4−βGal HIV細胞である。ある方法において、HIV複製をp24抗原ELISAアッセイまたは逆転写酵素活性アッセイによってモニターする。
【0007】
本発明の他の方法において、同定されたIsoT調節化合物によるHIV感染の阻害能を、当該化合物で処理した宿主細胞における偽ウイルス生産と、当該化合物で処理しなかった対照宿主細胞における偽ウイルス生産を比較することによって試験する。使用される宿主細胞は293T HEK細胞である。いくつかの方法では、宿主細胞を、細胞においてHIV偽ウイルスを生産する偽ウイルスプラスミドでトランスフェクトする。
【0008】
関連する局面において、本発明はHIV感染を阻害する薬剤を同定する方法を提供する。これらの方法には、第1に試験化合物をスクリーニングしてIsoTイソペプチダーゼ活性またはVprとのその結合を下方制御するIsoT調節化合物を同定すること、次に同定したIsoT調節化合物のHIV感染阻害能について試験することを含む。典型的には、スクリーニングに使用するIsoT分子は哺乳類細胞に由来する。好ましい態様において、スクリーニングに使用するIsoT分子はヒトIsoTである。
【0009】
これらの方法のいくつかにおいて、IsoT調節化合物によるHIV−1感染の阻害能を、当該化合物で処理したHIV許容細胞におけるHIV複製と、当該化合物で処理しなかった対照細胞におけるHIV複製を比較することによって試験する。いくつかの方法において、使用されるHIV許容細胞はHeLa−T4−βGal HIV細胞である。いくつかのスクリーニング法において、HIV複製をp24抗原ELISAアッセイまたは逆転写酵素活性アッセイによってモニターする。
【0010】
他の方法において、IsoT調節化合物によるHIV感染の阻害能を、当該化合物で処理した宿主細胞における偽ウイルス生産と、当該化合物で処理しなかった対照宿主細胞における偽ウイルス生産を比較することによって試験する。使用される宿主細胞は293T HEK細胞である。いくつかの方法では、宿主細胞を、当該細胞においてHIV偽ウイルスを生産する偽ウイルスプラスミドでトランスフェクトする。
【0011】
他の態様において、本発明は対象のHIV感染を処置する方法を提供する。かかる方法は、HIV感染を有する対象に、有効量のIsoT調節化合物を含む医薬組成物を投与することを必要とする。IsoT調節化合物はIsoTイソペプチダーゼ活性、VprとのIsoT結合またはIsoTをコード化する遺伝子の発現を下方制御することができる。いくつかの方法において、使用するIsoT調節化合物はヒトIsoTを下方制御することができる。いくつかの方法において、使用するIsoT調節化合物はインビトロでIsoT発現細胞におけるHIV複製を阻害することができる。これらの治療法において、IsoT調節化合物を既知の抗HIV薬剤と共に投与することがある。いくつかの方法において、IsoT阻害薬剤はIsoT活性を特異的に阻害する本発明によって同定された化合物である。他の方法において、IsoT阻害薬剤はIsoTの発現または細胞レベルを特異的に下方制御する核酸分子、例えばsiRNAまたはアンチセンスDNAである。
【0012】
本発明の性質および利点の更なる理解を、明細書および特許請求の範囲の残りの部分を参照することによって理解することができる。
【0013】
図面の簡単な説明
図1A−1BはVprのIsoTへの結合を示す:(A)概略図およびIsoTプレイクローンの位置を酵母−2−ハイブリッドを全長IsoT cDNAと比較して同定した。プレイクローンは2個のUBAドメインを有するC末端227アミノ酸を含む。本明細書で使用する全長IsoT cDNAクローンを膵臓由来のcDNAライブラリから増幅した。C335の相対位置(活性部位システイン)を示す。(B)VprとIsoTの特異的相互作用を示すために、β−ガラクトシダーゼフィルターリフトアッセイを使用して酵母−2−ハイブリッドをアッセイする。VP16は空ベクターである。
【0014】
図2A−2Cは野生型IsoT過剰発現がHIV−1ウイルスの力価を増加させるが、活性部位変異はそうではないことを示す:(A)全長(FL)野生型IsoTの過剰発現がトランスフェクション後24および48時間で偽ウイルス過剰発現を増加させる。293T細胞をIsoTをコード化する第4cDNAの存在下(+)または非存在下(−)において3個の偽ウイルスプラスミドとコトランスフェクトした。レポーターウイルス上清をトランスフェクション後24または48時間で回収し、293T標的細胞におけるレポーター活性についてアッセイした。細胞溶解物を平行してウェスタン分析で、IsoT特異的抗体を使用して内因性IsoT発現ならびにIsoT過剰発現について分析した。1列目はプラスミドでトランスフェクトしていない、トランスフェクション試薬のみのものである。(B)293T細胞を(A)のとおりに、第4プラスミドとしてVIP3(プラスミド骨格)、FL−野生型、PL−C末端227、またはFL−C335Aでコトランスフェクトした。トランスフェクション後30時間に回収したウイルス上清由来のルシフェラーゼレポーター活性を293T標的細胞においてアッセイした。(C)HIV力価に対する用量依存関数効果。FL−WTおよびFL−C335Aを力価判定し、平行して第4プラスミドとして偽プラスミドとトランスフェクトした。
【0015】
図3A−3BはVpr(A30L)ウイルスの減少した出芽が感染性ではなく減少した粒子生産と関連することを示す。293T細胞を偽ウイルスDNA発現WTおよびVprの変異型とトランスフェクトした。(A)WTまたは核局在化変異体W54Rではなく、A30L Vpr取り込み変異体は、293T標的細胞における感染性によって測定するとウイルス力価を減少した。(B)p24レベルを標的293T細胞を感染するために使用した偽ウイルスインプットについて、(A)から標準化(9.38ng)した。
【0016】
図4A−4BはRNA干渉によるIsoTまたはVprの欠乏が293T細胞における偽ウイルス生産を阻害することを示す。(A)293T細胞をHIV Tat、IsoT、VprまたはルシフェラーゼGL3(陰性対照)を標的とする示したsiRNAでトランスフェクトした。細胞を一晩培養し、ウェルあたり2:0.05:0.5または100ng:2.5ng:25ngのベクターRNA:デルタプサイ:envの比で、3プラスミド偽ウイルス系のcDNAトランスフェクションに供した。3つの複写ウェルを条件毎に行った。ウイルス上清を48時間後に回収し、標的293T細胞に感染させるために使用した。これらの形質導入細胞におけるルシフェラーゼ活性を48時間後に測定し、Aに示す。(B)293T細胞をHIV Tat、IsoT、VprまたはルシフェラーゼGL3に対するsiRNAとトランスフェクトした。72時間後細胞生存率をCellTiterGloを使用し、蛍光を測定して決定した。
【0017】
図5はIsoTまたはVprの欠乏がHeLa−CD4−β−gal細胞におけるHIV−1 IIIb複製を阻害することを示す。細胞をTat、IsoT、Vpr、ルシフェラーゼGL2(陰性対照)、またはPolR2A(細胞毒性についての陽性対照)を標的とする示したsiRNAとトランスフェクトした。24時間培養後、細胞をHIV−1 IIIbに感染させる。β−ガラクトシダーゼ活性を、感染の測定として感染後72時間で測定した。細胞毒性をsiRNAトランスフェクト非感染細胞について平行して測定した。データを陰性対照GL2 siRNAトランスフェクト細胞によって発生したシグナルの割合として表現する。
【発明を実施するための最良の形態】
【0018】
詳細な説明
I.概説
本発明は一部において、IsoTがHIV感染に関与するという本発明者らの発見に基づく。ヒトIsoTは、遊離K48結合ポリユビキチン鎖のモノマーへの分解に関与する93kDaの亜鉛結合脱ユビキチン化酵素である。本発明者らはIsoTがHIV−1ウイルスのウイルス性タンパク質R(Vpr)の結合パートナーであることを見出した。Vprは、単球およびマクロファージにおけるHIVウイルス複製に必須であり、T細胞およびT細胞株におけるウイルス複製を増加させる96アミノ酸、14kDa付属タンパク質である(例えば、Cohen et al., J Virol 64:30973099, 1990;Paxton et al., J Virol 67:72297237, 1993;およびAndersen et al., Curr HIV Res. 3:43-51, 2005参照)。本発明者らはまた、活性部位変異体ではなく野生型IsoTの過剰発現が生産細胞由来のウイルス力価を上昇させることを見出した。さらに、低分子干渉RNAを用いたIsoTレベルの欠乏がHEK293T生産細胞におけるウイルス力価を減少させ、そして実験室株HIV−1(IIIb)およびHeLa−T4−βGalにおけるHIV−1 BaL、ならびにU373 HIV−1インジケーター細胞株それぞれの感染性を減少させることを発見した。これらのデータはIsoTが宿主細胞からのHIV−1放出の効果を上昇させるのに重要な役割を果たしていることを示す。
【0019】
これらの発見により、本発明はHIV感染を阻害する新規薬剤をスクリーニングする方法を提供する。試験化合物を、第1にIsoT分子の生物学的活性、例えばその発現、そのVprとの相互作用、またはそのイソペプチダーゼ活性を調節する能力についてスクリーニングする。したがって同定された薬剤を、典型的にはさらにHIV感染またはHIV感染を示す活性を調節する能力について試験する。様々なIsoT分子をスクリーニングアッセイにおいて使用することができる。例えば、ヒト、ウサギ、ラットまたはマウス由来のIsoTを調節剤のスクリーニングのために使用することができる。いくつかの好ましい態様において、ヒトIsoT分子を使用する。本発明に開示したスクリーニング法の有用性を例示するために、本発明者らはヒトIsoTの阻害剤のために低分子化合物のハイスループットスクリーニングを行った。下記実施例に詳述するとおり、IsoTを阻害することによる抗ウイルス活性を有する化合物がスクリーニングによって同定された。
【0020】
本発明はまた、治療使用を含む。IsoT活性の薬理学的阻害はHIV感染に関連する状態の処置または予防の新規アプローチを提供する。典型的には、アプローチには本発明によって同定することができるIsoT阻害剤(例えばsiRNAまたは低分子有機化合物)を対象に投与することを含む。
【0021】
以下の項は本発明の組成物を製造し、使用するための、そして本発明の方法を実施するための指標を提供する。
【0022】
II.定義
他に定義がない限り、本明細書で使用されている全ての技術および科学用語は本発明が属する分野における当業者が通常理解するとおりの意味を有する。下記文献は当業者に本発明において使用される多くの用語の一般的な定義を提供する: Oxford Dictionary of Biochemistry and Molecular Biology, Smith et al. (eds.), Oxford University Press (revised ed., 2000); Dictionary of Microbiology and Molecular Biology, Singleton et al. (Eds.), John Wiley & Sons (3rd ed., 2002);および A Dictionary of Biology (Oxford Paperback Reference), Martin and Hine (Eds.), Oxford University Press (4th ed., 2000)。さらに、下記定義を本発明の実施において読者を補助するために提供する。
【0023】
「試験薬剤」または「試験化合物」なる用語には、あらゆる物質、分子、要素、化合物、実体、またはそれらの組合せを含む。限定されないが、例えばタンパク質、ポリペプチド、小有機分子、ポリサッカライド、ポリヌクレオチド等が含まれる。天然物、合成化合物または化学化合物または2種以上の物質の組合せであり得る。特に記載がない限り、「薬剤」、「物質」および「化合物」なる用語は相互に交換可能に使用することができる。
【0024】
本明細書において「アナログ」なる用語は、参照分子と構造的に類似しているが参照分子の特定の置換基を別の置換基で置換することによって標的化および制御方法が修正されている分子を意味するものとして使用する。参照分子と比較して、アナログは同一の、類似のまたは改善された有用性を示すものと、当業者には予期される。アナログの合成およびスクリーニングは改善された特徴(例えば標的分子に対する高い結合親和性)を有する既知の化合物の変異体を同定するための薬学化学において周知の方法である。
【0025】
HIVはレトロウイルスのファミリーであるヒト免疫不全ウイルス(HIV)を意味する。限定されないが、これらのウイルスにはHIV−I、HIV−II、HIV−III(HTLV−II、LAV−1、LAV−2としても知られる)等が含まれる。本明細書において使用するとき、HIVはHIVファミリーにおけるあらゆる株、形態、サブタイプおよび変異体であり得る。
【0026】
2個の核酸配列またはアミノ酸配列の文脈中での「同一」または「配列同一」なる用語は、特定の比較窓にわたって最大対応を並べたとき、2個の配列における残基が同じであることを意味する。比較のための配列のアラインメント法は、当業者に既知である。比較のための配列の最適なアラインメントを、Smith and Waterman (1981) Adv. Appl. Math. 2:482の局所ホモロジーアルゴリズムによって; Needleman and Wunsch (1970) J. Mol. Biol. 48:443のアラインメントアルゴリズムによって; Pearson and Lipman (1988) Proc. Nat. Acad. Sci U.S.A. 85:2444の同一性の調査法によって;by computerized implementations of these algorithms これらのアルゴリズムのコンピュータによる実行(限定されないが、Intelligentics, Mountain View, CAによるPC/GeneプログラムのCLUSTAL;およびWisconsin Genetics Software Package, Genetics Computer Group (GCG), 575 Science Dr., Madison, Wis., U.S.A.におけるGAP、BESTFIT、BLAST、FASTA、またはTFASTAを含む)によって、行うことができる。アラインメントは、検査およびマニュアルアラインメントによってしばしば行うこともできる。
【0027】
「実質的に同一の」核酸またはアミノ酸配列なる用語は、上記プログラム(例えばBLAST)を標準的なパラメータを用いて使用して標準配列と比較したとき、少なくとも90%、好ましくは少なくとも95%、より好ましくは少なくとも98%、そしてもっとも好ましくは少なくとも99%以上の配列同一性を有する配列を含む核酸またはアミノ酸を意味する。好ましくは、実質的同一性は少なくとも約50残基長である配列領域にわたって、より好ましくは少なくとも約100残基の領域にわたって存在し、そしてもっとも好ましくは配列は少なくとも約150残基にわたって実質的に同一である。もっとも好ましい態様において、配列はコーディング領域の全長にわたって実質的に同一である。
【0028】
IsoT分子の生物学的活性に関する「調節」なる用語は、IsoTの細胞レベル、細胞内局在性または他の生物学的活性(例えばそのイソペプチダーゼ活性)における変化を意味する。IsoT活性の調節は上方制御(すなわち、活性化または刺激)あるいは下方制御(すなわち阻害または抑制)であってよい。例えば、調節は細胞レベル、あるいはIsoTの酵素的修飾(例えばリン酸化)、結合特性(例えばVprまたは基質への結合)、またはIsoTタンパク質の他の生物学的、機能的または免疫学的特徴における変化を引き起こし得る。活性の変化は例えば、IsoTコード化遺伝子の発現、IsoTタンパク質をコード化するmRNAの安定性、翻訳効率の増加もしくは減少、またはIsoT酵素の他の生物学的活性(例えばそのイソペプチダーゼ活性)の変化によって引き起こされ得る。IsoTモジュレーターの作用の形態は、直接、例えばIsoTタンパク質またはIsoTタンパク質をコード化する遺伝子に結合することによっていても良い。変化はまた、関節、例えばIsoTを調節する他の分子と結合および/または修飾(例えば酵素的に)することによっていても良い。
【0029】
「対象」なる用語は、哺乳類、とりわけヒトを意味する。これは他の非ヒト動物、例えばウシ、ウマ、ヒツジ、ブタ、ネコ、イヌ、マウス、ラット、ウサギ、モルモット、サルを含む。
【0030】
IsoTのような分子の「変異体」は、分子全体またはそのフラグメントと構造および生物学的活性が実質的に同一の分子を意味する。したがって、2個の分子が類似の活性を有するならば、一方の分子の構成または2次、3次もしくは4次構造が他方のものと同一でなくとも、あるいはアミノ酸残基の配列が同一でなくとも、それらは本明細書で使用する用語としての変異体であると考える。
【0031】
III.スクリーニングスキーム
本発明によると、HIV感染の新規な阻害剤を、第1に試験化合物をIsoTの生物学的活性を調節(例えば阻害)する能力についてスクリーニングすることによって同定する。スクリーニングアッセイにおいてモニターするIsoTの生物学的活性は、そのイソペプチダーゼ活性であってもよい。それは他の分子、例えばVprまたはユビキチンへのその結合であってもよい。生物学的活性はまた、IsoT発現またはその細胞レベルであり得る。IsoTの生物学的活性を調節する試験化合物を同定した後、それらを典型的にはさらに、HIV感染を調節する、あるいはHIV感染もしくはHIV複製の指標である活性を調節する能力について試験する。この工程は、IsoTの生物学的活性を調節することによって第1の工程で同定された化合物が実際にHIV感染を制御(例えば阻害)することができることを確認するために行われる。
【0032】
当業者に周知の様々な生化学および分子生物学の技術またはアッセイをIsoTモジュレーターをスクリーニングするために使用することができる。かかる技術は、例えばHandbook of Drug Screening, Seethala et al. (eds.), Marcel Dekker (1st ed., 2001);High Throughput Screening: Methods and Protocols (Methods in Molecular Biology, 190), Janzen (ed.), Humana Press (1st ed., 2002); Current Protocols in Immunology, Coligan et al. (Ed.), John Wiley & Sons Inc (2002); Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press (3rd ed., 2001); および Brent et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (ringbou ed., 2003)に記載されている。
【0033】
様々な種由来のIsoTを、本発明のIsoTモジュレーターのスクリーニングにおいて使用することができる。好ましくは、哺乳類細胞由来のIsoT分子を使用する。例えば、ヒトIsoT(受託番号 NM_003481 および NP_003472)がクローン化され、文献、例えば Falquet et al., FEBS Lett. 376: 233-237; 1995; Wilkinson et al., Biochemistry 34:14535-14546, 1995; and Lacombe et al., FEBS Lett. 531:469-474, 2002に記載された。他の例には、チンパンジーIsoT(受託番号 XM_526393 および XP_526393)およびマウスIsoT(受託番号 BC066993 および AAH66993)が含まれる。あらゆるこれらのIsoT配列または実質的にその配列と同一の配列を、本発明のIsoTモジュレーターを同定するためのスクリーニングアッセイに使用することができる。下記実施例に例示したとおり、IsoTポリヌクレオチドのクローニングおよび発現、ならびにIsoTポリペプチドの精製を、通常行われている方法および技術で行うことができる。かかる方法および技術は例えば、Sambrook et al., supra;および Brent et al., supraに記載されている。IsoTを得る具体的な方法は、文献、例えばGabriel et al., Biochemistry 41:13755 -13766, 2002にも提供されている。
【0034】
無傷のIsoT分子または無傷のIsoT分子をコード化する核酸に加えて、IsoTフラグメント(例えば触媒ドメインまたはVpr結合ドメイン)、アナログまたは機能的誘導体も使用することができる。これらのアッセイにおいて使用することができるIsoTフラグメントは通常、IsoT分子の生物学的活性(例えばそのイソペプチダーゼ活性またはVprとの結合)の1種以上を保持している。上記のとおり、異なる種由来のIsoTは既に配列決定され、十分に特徴付けられている。したがって、それらのフラグメント、アナログ、誘導体または融合タンパク質を当業者に周知の方法を使用して容易に得ることができる。例えば、IsoTの機能的誘導体をタンパク質分解的切断によって天然に発生した、あるいは組み換えによって発現したタンパク質から、当業者に既知の常套の精製方法によって製造することができる。あるいは、機能的誘導体をそのイソペプチダーゼ活性を保持しているIsoTのフラグメントのみを発現することによって、組み換えDNA技術により製造することができる。
【0035】
IV.試験化合物
本発明の方法でスクリーニングすることができる試験化合物には、ポリペプチド、βターン模倣薬、ポリサッカライド、リン脂質、ホルモン、プロスタグランジン、ステロイド、芳香族化合物、ヘテロ環式化合物、ベンゾジアゼピン、オリゴマーN−置換グリシン、オリゴカルバメート、ポリペプチド、サッカライド、脂肪酸、ステロイド、プリン類、ピリミジン類、誘導体、構造アナログまたはそれらの組合せが含まれる。試験化合物は合成分子であることもあるし、天然分子であることもある。
【0036】
試験化合物を、合成または天然化合物のライブラリーを含む広い種類のソースから得る。コンビナトリアルライブラリーを段階的方法で合成することができる多くのタイプの化合物のために製造することができる。化合物の大きなコンビナトリアルライブラリーをWO 95/12608、WO 93/06121、WO 94/08051、WO 95/35503およびWO 95/30642に記載のコード化合成ライブラリー(ESL)法によって構成することができる。ペプチドライブラリーをファージディスプレイ法(例えばDevlin, WO 91/18980参照)によって製造することもできる。バクテリア、菌、植物および動物抽出物の形態の天然化合物のライブラリーを商業的ソースから得るか、あるいは実際に収集することができる。既知の薬理学的薬剤は、構造アナログを製造するためのアシル化、アルキル化、エステル化、アミド化のような指向性またはランダム化学修飾に供することができる。
【0037】
他の化合物またはペプチドのコンビナトリアルライブラリーを任意の位置で配列優先または固定なしで十分にランダム化することができる。あるいは、ライブラリーを偏向させることができる、すなわち配列中のいくつかの位置が一定であるか、あるいは限定された数の可能性から選択される。例えば、いくつかの場合において、ヌクレオチドまたはアミノ酸残基を、例えば疎水性アミノ酸、親水性残基、立体的に偏向した(小または大)残基、システインの製造の方向、架橋、SH−3ドメインに関するプロリン、セリン、スレオニン、チロシンまたはリン酸化部位に関するヒスチジン、またはプリンのような明らかなクラス内でランダム化する。
【0038】
試験化合物は、天然に発生するタンパク質またはそれらのフラグメントであり得る。かかる試験化合物は天然ソース、例えば細胞または組織溶解物から得ることができる。ポリペプチド薬剤のライブラリーを、例えば商業的に入手可能なcDNAライブラリーから製造するか、あるいは常套の方法で製造することができる。試験化合物はまた、約5〜約30個、好ましくは約5〜約20個、とりわけ好ましくは約7〜約15個のアミノ酸のペプチドのようなペプチドであり得る。ペプチドは天然に発生したタンパク質の消化物、ランダムペプチドまたは「偏向した」ランダムペプチドであってもよい。いくつかの方法において、試験化合物はポリペプチドまたはタンパク質である。
【0039】
試験化合物は核酸でもあり得る。核酸試験化合物は天然に発生した核酸、ランダム核酸または「偏向した」ランダム核酸であってもよい。例えば、原核または真核ゲノムの消化物、を上記タンパク質と同様に使用することができる。
【0040】
好ましい態様において、試験化合物は低分子、例えば約500または1000未満の分子量の分子である。好ましくはハイスループットアッセイを適用し、かかる低分子をスクリーニングするために使用することができる。いくつかの方法において、上記低分子試験化合物のコンビナトリアルライブラリーを容易に使用してIsoTの低分子モジュレーターをスクリーニングすることができる。かかるスクリーニングには多くのアッセイ、例えばSchultz et al., Bioorg Med Chem Lett 8:2409-2414, 1998; Weller et al., Mol Divers. 3:61-70, 1997; Fernandes et al., Curr Opin Chem Biol 2:597-603, 1998;およびSittampalam et al., Curr Opin Chem Biol 1:384-91, 1997に記載のものを使用することができる。
【0041】
本発明の方法でスクリーニングする試験化合物のライブラリーはまた、IsoTポリペプチド、それらのフラグメントまたはアナログの構造的研究に基づき製造することができる。かかる構造的研究はIsoポリペプチドとより結合しやすい試験化合物を同定することができる。IsoTポリペプチドの3次元構造を多くの方法、例えば結晶構造および分子モデリングで研究することができる。X線結晶解析を使用したタンパク質の構造研究法は文献に周知である。Physical Bio-chemistry, Van Holde, K. E. (Prentice-Hall, New Jersey 1971), pp. 221-239, および Physical Chemistry with Applications to the Life Sciences, D. Eisenberg & D. C. Crothers (Benjamin Cummings, Menlo Park 1979)参照。IsoTポリペプチド構造のコンピューターモデリングは、IsoTモジュレーターをスクリーニングするための試験化合物を設計する他の方法を提供する。分子モデリング法は文献、例えばU.S. Patent No. 5,612,894、表題“System and method for molecular modeling utilizing a sensitivity factor”およびU.S. Patent No. 5,583,973、表題“Molecular modeling method and system”に記載されている。さらに、タンパク質構造も中性子回折および核磁気共鳴(NMR)によって決定することができる。例えばPhysical Chemistry, 4th Ed. Moore, W. J. (Prentice-Hall, New Jersey 1972), およびNMR of Proteins and Nucleic Acids, K. Wuthrich (Wiley-Interscience, New York 1986)参照。
【0042】
本発明のモジュレーターにはIsoTポリペプチドに特異的に結合する抗体も含むことができる。かかる抗体はモノクローナルまたはポリクローナルであり得る。かかる抗体は当業者に周知の方法を使用して製造することができる。例えば、非ヒト、例えばマウスもしくはラットモノクローナル抗体の製造を、例えばIsoT ポリペプチドまたはそのフラグメントで動物を免疫することによって行うことができる(Harlow & Lane, Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York, 1988参照)。かかる免疫源は天然ソースから、ペプチド合成または組換発現によって得ることができる。
【0043】
マウス抗体のヒト化形態を、非ヒト抗体のCDR領域をヒト定常領域と組み換えDNA技術によって結合することによって製造することができる。Queen et al., Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989) および WO 90/07861参照。ヒト抗体をファージディスプレー法を使用して得ることができる。例えばDower et al., WO 91/17271; McCafferty et al., WO 92/01047参照。これらの方法において、ファージのライブラリーが形成され、そこではメンバーは外部表面に異なる抗体を提示する。抗体はFvまたはFabフラグメントとして通常提示される。所望の特異性を有するファージ提示抗体を本発明のIsoTポリペプチドに対する親和性濃縮によって選択する。
【0044】
IsoTポリペプチドに対するヒト抗体を、少なくともヒト免疫グロブリン遺伝子座および不活性内因性免疫グロブリン遺伝子座のセグメントをコード化する導入遺伝子を有する非ヒトトランスジェニック哺乳類から製造することもできる。例えばLonberg et al., WO93/12227 (1993); Kucherlapati, WO 91/10741 (1991)参照。ヒト抗体を競合的結合実験またはその他によって選択して、特定のマウス抗体と同じエピトープ特異性を有することができる。かかる抗体はとりわけ、マウス抗体の有用な機能的特徴を共有する可能性がある。ヒトポリクローナル抗体はまた、免疫源で免疫したヒト由来の血清の形態で提供され得る。所望により、かかるポリクローナル抗体はIsoTポリペプチドまたはそのフラグメントを使用した親和性精製によって濃縮することができる。
【0045】
V.IsoTモジュレーターのための試験化合物スクリーニング
HIV感染を阻害する新規化合物を同定するために、本明細書に記載のとおり、試験化合物を第1にIsoTの生物学的活性を調節する能力についてスクリーニングする。好ましい態様において、試験化合物はIsoTのイソペプチダーゼ活性を調節(例えば阻害)する能力について試験する。IsoTの酵素活性を調節(例えば阻害)する化合物を多くのアッセイ形式で同定することができる。例えば、下記実施例に示すとおり、スクリーニングは精製IsoT酵素およびユビキチン−7−アミド−4−メチルクマリン(methlcoumarin)(Ub−AMC)を基質として使用して生化学アッセイを行うことができる。試験化合物のIsoTのイソペプチダーゼ活性に対する効果を当業者に既知の、例えばLacombe et al., FEBS Lett. 20:531:469-74, 2002;および Gabriel et al., Biochemistry. 19:41:13755-66, 2002のIsoT活性アッセイを使用してモニターすることができる。例えば、IsoTの酵素活性をユビキチンダイマー基質でアッセイすることができる。IsoTによる基質の切断は、Lacombe et al., FEBS Lett. 20:531:469-74, 2002に記載のとおり、電気泳動およびクーマシー染色によって分析することができる。分枝および直線ユビキチンダイマー基質を文献、例えばGabriel et al., Biochemistry 41:13755 -13766, 2002に記載のとおり製造することができる。ユビキチンダイマーの他に、他の基質もIsoT酵素活性のモジュレーターをスクリーニングするために使用することができる。例えば多くのアミノメチルクマリン(AMC)ベースのペプチド基質を、IsoT活性をモニターするために使用することができる(Stein et al., Biochemistry 34:12616-12623, 1995)。これらの基質(例えばZ-Leu-Arg-Gly-Gly-AMC または ユビキチン−アルデヒド)は、IsoTイソペプチダーゼの反応速度論およびIsoT活性のユビキチンアルデヒドによる阻害を研究するために文献において使用されている(Stein et al., Biochemistry 34:12616-12623, 1995;および Melandri et al., Biochemistry 35: 12893 -12900, 1996)。
【0046】
他の態様において、試験化合物を第1にIsoTのVprまたはユビキチンのような他の分子との結合を調節する活性についてスクリーニングすることができる。Vprはクローン化され、下記実施例および文献、例えばAmini et al., J Biol Chem. 279:46046-56, 2004; Iijima et al., Virol. 327:249-61, 2004; Yao et al., Retrovirology. 1:21-31, 2004; および Muthumani et al., Int Immunol. 17:103-16, 2005に既知の方法を使用して発現される。多くのアッセイおよび方法をIsoTポリペプチドのVprまたはユビキチンとの結合に対する試験化合物の効果を試験するために使用することができる。これらにはタンパク質と他の化合物(他のタンパク質を含む)の相互作用を研究するために当業者に一般的に知られている方法、例えば標識インビトロタンパク質−タンパク質結合アッセイ、免疫沈降法、GSTプルダウンアッセイ、酵母または哺乳類2種ハイブリッドスクリーニングおよび蛍光消光または蛍光偏光が含まれる。さらに、IsoTの他の分子(例えばユビキチン)との結合を試験するための具体的な方法は、下記実施例および文献(Amini et al., J. Biol. Chem., 279:46046-46056, 2004;および Yao et al., Retrovirology. 1:21-31, 2004参照)に提供される。
【0047】
ある態様において、IsoTとVpr結合の物理的相互作用を下記実施例に例示のような共免疫沈降法によって分析することができる。標識または分子タグIsoT(例えばFLAG−IsoT)およびVpr(例えばMyc−Vpr)を宿主細胞、例えば293T細胞において共発現させることができる。IsoTのVprとの結合を細胞溶解物を抗FLAG抗体で免疫沈降し、その後ウェスタンブロット分析することによって試験することができる。同様に、IsoTのユビキチンとの結合も当業者に既知の方法で分析することができる。例えば、IsoT−ユビキチン結合をタンデムKClおよびウレア直線グラジエントを用いるユビキチン−セファロースカラムを使用して試験することができる。この方法はLacombe et al. (FEBS Lett. 20:531:469-74, 2002) およびGabriel et al. (Biochemistry 41:13755 -13766, 2002)に記載されている。
【0048】
他の方法において、試験化合物をIsoTの発現または細胞レベル(転写、翻訳または翻訳後修飾)を調節する活性についてアッセイする。周知の様々な生化学および分子生物学技術をIsoT遺伝子の発現またはIsoTポリペプチドの細胞レベルを試験するために使用することができる。かかる技術は例えば、Sambrook et al., supra;および Brent et al., supraに記載されている。ある態様において、IsoT分子の内因性レベルを細胞が通常発現しているIsoTにおいて直接モニターすることができる。ある態様において、IsoT分子の発言または細胞レベルをクローンcDNAまたはIsoTをコード化するゲノム配列を使用して発現系において試験することができる。
【0049】
あるいは、IsoT遺伝子の発現の調節を細胞ベースの系において、発現ベクターの培養細胞株への短期または安定トランスフェクションによって試験することができる。レポーター遺伝子と作動可能なように結合しているIsoT遺伝子の転写制御配列(例えばプロモーター)を有するアッセイベクターを、プロモーター活性のアッセイのためにあらゆる哺乳類宿主細胞株にトランスフェクトすることができる。レポーター遺伝子と作動可能なように結合しているIsoT遺伝子(またはIsoT遺伝子の転写制御因子)を含む構成物を、分子生物学において通常実施されている技術および方法のみを使用して製造することができる(例えばSambrook et al. および Brent et al., supra参照)。細胞培養、トランスフェクションおよびレポーター遺伝子アッセイの一般的な方法は、文献、例えばBrent et al., supra; および Transfection Guide, Promega Corporation, Madison, WI (1998)に記載されている。あらゆる容易にトランスフェクト可能な哺乳類細胞株をIsoTプロモーター機能をアッセイするために、またはIsoTを発現するために使用することができる。例えばCHO、COS、HCT116、HEK293、MCF−7、およびHepG2は全て好適な細胞株である。
【0050】
適当な宿主細胞に挿入したとき、発現ベクターにおける転写制御因子は宿主RNAポリメラーゼによるレポーター遺伝子の転写を誘導する。レポーター遺伝子は典型的には、宿主細胞には本来存在しない容易にアッセイされる酵素活性を有するポリペプチドをコード化する。真核細胞プロモーターの典型的なレポーターポリペプチドには例えば、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、ホタルもしくはRenillaルシフェラーゼ、β−ガラクトシダーゼ、β−グルクロニダーゼ、アルカリホスファターゼおよび緑蛍光タンパク質(GFP)が含まれる。
【0051】
一般的に、スクリーニングで同定されたIsoT−調節化合物はIsoTの生物学的活性(例えばそのイソペプチダーゼ活性またはVprとの結合)を顕著に改変することができる。調節の程度を同じ化合物の対照タンパク質(例えばルシフェラーゼまたはタンパク質キナーゼ)に対する活性と、あるいは対照化合物の同じIsoT分子に対する活性と比較して測定する。例えば、IsoT発現またはIsoTイソペプチダーゼ活性を阻害するIsoT調節化合物は典型的には、対照酵素についてのIC50よりも少なくとも2、5、10、25、50、100、500または1000倍低いIsoT分子(例えばヒトIsoT)についてのIC50(最大阻害の50%を引き起こす有効濃度)を有する。IsoT分子の同じ活性を阻害する陰性対照化合物のIC50よりもIsoTのIC50は少なくとも5、10、25、50、100、500または1000倍低いと定義することもできる。同様に、調節をIsoTのVprとの結合で測定するならば、同じ程度の阻害を達成するための対照化合物の濃度よりも5、10、25、50または100倍低い濃度でのIsoT調節化合物が存在することで、通常IsoT−Vpr結合活性を予防または阻害することができる。本明細書において使用するとき、IsoTの他の分子への結合の阻害(例えばVprまたはユビキチン多量体)はIsoTのその分子への結合親和性を少なくとも2倍、好ましくは少なくとも5倍、より好ましくは少なくとも10、20、50または100倍減少させることを意味する。
【0052】
VI.HIV阻害化合物のスクリーニング
HIVの新規阻害剤を同定するために、上記IsoTモジュレーターを典型的にはさらに、それらのHIV感染に対する阻害作用を確認するために試験する。典型的には、化合物をHIV感染またはHIV複製の指標である活性を調節する能力についてスクリーニングする。多くのアッセイおよび方法をIsoT調節化合物のHIV阻害活性を試験するために使用することができる。ある方法では、IsoT調節化合物のHIV感染に対する潜在的な阻害活性を、当業者が通常実施している方法を使用して、インビトロでの培養細胞のHIV感染に対するそれらの効果を試験することによって試験することができる。例えば下記実施例に記載のように、化合物の抗ウイルス活性をHIVウイルスに感染する前に化合物と接触させたHeLaCD4βgal細胞においてウイルス複製をモニターすることによって評価することができる。化合物を、Seddiki et al., AIDS Res Hum Retroviruses. 15:381-90, 1999に記載のように第1マクロファージ培養物のHIV感染について試験することもできる。それらを、Fujii et al., J Vet Med Sci. 66:115-21, 2004に記載のように他のT細胞および単球細胞株のHIV感染について試験することもできる。HIV感染をモニターするためのさらなるインビトロ系は文献に記載されている。例えばLi et al., Pediatr Res. 54:282-8, 2003; Steinberg et al., Virol. 193:524-7, 1993; Hansen et al., Antiviral Res. 16:233-42, 1991;および Piedimonte et al., AIDS Res Hum Retroviruses. 6:251-60, 1990参照。
【0053】
これらのアッセイにおいて、細胞のHIV感染を形態学的に、例えば顕微鏡細胞変異効果アッセイ(例えばFujii et al., J Vet Med Sci. 66:115-21, 2004参照)によってモニターすることができる。酵素的に、例えば細胞培養物の上清におけるHIV逆転写(RT)活性をアッセイすることによって評価することもできる。かかるアッセイは文献、例えばReynolds et al., Proc Natl Acad Sci U S A. 100:1615-20, 2003;および Li et al., Pediatr Res. 54:282-8, 2003に記載されている。他のアッセイではHIV感染をウイルス核酸またはウイルス抗原の定量的集積によってモニターする。例えば、Winters et al. (PCR Methods Appl. 1:257-62, 1992)はHIV感染細胞培養物からHIV gag RNAおよびDNAをアッセイする方法を記載した。Vanitharani et al.はウイルスp24抗原の生産を測定するHIV感染アッセイを記載した(Virology 289:334-42, 2001)。ウイルス複製はまた、例えばChargelegue et al., J Virol Methods. 38(3):323-32, 1992;およびKlein et al., J Virol Methods. 107(2):169-75, 2003に記載のとおり、p24抗原ELISAアッセイによってインビトロでモニターすることができる。全てのこれらのアッセイを、本発明のIsoT調節化合物の抗HIV活性を評価するために使用し、そして改変することができる。
【0054】
ある方法において、IsoT調節化合物のHIV感染に対する潜在的な阻害効果をHIV複製が許容状態である人工レポーター細胞において試験することができる。これらの細胞において、HIV転写制御因子、例えばHIV−LTRの制御下でのレポーター遺伝子の発現を試験することによって、HIV感染および複製をモニターする。かかる細胞の一つの例がHeLa−T4−βGal HIVレポーター細胞である。下記実施例に説明のとおり、IsoT調節化合物で処理した後HeLa−T4−βGalレポーター細胞をHIV−IIIbで感染させるることができる。β−ガラクトシダーゼ活性を測定することによってモニターすると、化合物処理細胞のウイルス感染を化合物で処理しなかった対照細胞のものと比較することができる。IsoT調節化合物で処理した細胞のウイルス力価が減少しており、あるいは感染が減少していることで、当該化合物が実際にHIV感染またはウイルス複製を阻害することができると確認する。
【0055】
本明細書で試験したHela−T4−βGal細胞に加えて、多くの類似のレポーターアッセイも文献に記載されている。例えばGervaix et al. (Proc Natl Acad Sci USA. 94:4653-8, 1997)はヒト化緑蛍光タンパク質(GFP)をコード化するプラスミドをHIV−I LTRプロモーターの制御下で発現する安定なT細胞株を開発した。HIV−Iの感染によって、非感染細胞と比較して100倍〜1000倍の感染細胞の蛍光の増加が観察される。これらのアッセイ系のいずれかを本発明においてIsoT調節化合物のHIV感染に対する効果をリアルタイムで観察するために使用することができる。これらのインビトロ系は感染細胞の時間にわたる定量と、化合物の感受性の測定を行うことができる。
【0056】
他の方法において、IsoT調節化合物のHIV複製に対する効果を当該化合物で処理した細胞におけるHIV−1偽ウイルスの生産を試験することによって試験することができる。細胞はIsoTを内因的にまたは外因的に発現し得る。IsoT−コード化構造を宿主細胞に、とりわけIsoTを内因的に発現していない宿主細胞にトランスフェクトすることができる。下記実施例に記載のとおり、HIV−1偽ウイルスの生産を、レポーター遺伝子(例えばルシフェラーゼ遺伝子)をコード化するプサイ−陽性RNA、全構造タンパク質をコード化するデルタプサイパッケージング構造、ならびに制御または修飾タンパク質、例えばTat、Rev、VprおよびVif、ならびにVSV−gエンベロープ発現プラスミドで生産細胞(例えば293T HEK細胞)をトランスフェクトすることによって得ることができる。生産細胞で生産される偽ウイルスはレポーター遺伝子のみをコード化する。標的細胞を生産細胞由来の上清中の偽ウイルスでトランスフェクトした後、レポーター遺伝子を下記逆転写および標的細胞ゲノムへの集積によって発現する。
【0057】
HIV複製の阻害剤をスクリーニングするために、生産宿主細胞を偽ウイルスプラスミドのトランスフェクションの前、同時、または後にIsoT調節化合物で処理することができる。好ましくは、化合物を偽ウイルスプラスミドのトランスフェクション前に宿主細胞に投与し、アッセイプロセス全体で存在させる。生産された偽ウイルスの力価を生産細胞由来の上清における偽ウイルスで標的細胞を感染させること、そして標的細胞におけるレポーターの活性(例えばルシフェラーゼ活性)を評価することによってモニターすることができる。対照として化合物で処理しなかった生産細胞由来の上清で感染させた標的細胞におけるレポーター活性も測定する。IsoT調節化合物がウイルス出芽に対する阻害効果を有するならば、化合物で処理した生産細胞由来の上清と接触させた標的細胞は対照細胞と比較して減少したレポーター活性を有する。
【0058】
VII.治療適用
(i)IsoTイソペプチダーゼ活性;(ii)IsoTのVprとの結合、または(iii)IsoTコード化遺伝子の発現を下方制御することによって、上記IsoT調節化合物は本発明の有用な治療適用を提供する。それらはHIVに感染した細胞におけるHIV複製を阻害するために容易に使用することができる。それらはまた、HIV感染ならびにHIV感染に関連する疾患または状態(例えばAIDS)を有する対象を予防的または治療的に処置するために有用である。本発明のIsoT調節化合物での処置に適しているHIV感染には、あらゆるレトロウイルスのHIVファミリー(例えばHIV−1またはHIV−2)による対象、とりわけヒト対象の感染を含む。IsoT調節化合物は、レトロウイルスのHIVファミリーのあらゆるメンバーのキャリアーである対象の処置に有用である。それらは活性AIDSで診断される対象の処置に有用であり得る。化合物はまた、かかる対象におけるAIDS関連状態の処置または予防に有用である。HIV感染を有するとは診断されていないがHIVによる感染の危険があると考えられる対象も、本発明のIsoT調節化合物での処置に適している。
【0059】
本発明の治療適用に使用することができるIsoT阻害化合物には、下記実施例に記載のような本発明の方法によって同定され得る低分子有機化合物が含まれる。一度同定されると、かかる化合物を有機化学の通常使用される方法を用いて容易に合成することができ、そして/または商業的供給者から容易に入手することができる。本発明の治療法を実施するために好適なIsoTモジュレーターには、IsoT細胞レベルまたはその生物学的活性(例えばその酵素活性)を特異的に調節することができる他の化合物も含む。かかる化合物には、IsoTポリペプチドを特異的に認識するアンタゴニストまたはアゴニスト抗体が含まれる。それらにはまた、低分子干渉RNA(siRNA)、マイクロRNA(miRNA)および合成ヘアピンRNA(shRNA)、アンチセンス核酸、ならびに相補DNA(cDNA)が含まれる。例えば、下記実施例に記載のIsoT特異的またはVpr特異的siRNAを使用することができる。さらに、治療的適用は、上記スクリーニング方法によって同定することができる他のIsoT阻害剤を使用することができる。
【0060】
いずれかのAIDS関連状態を有する対象は本発明のIsoT調節化合物で処置するのに適している。かかる状態にはAIDS関連複合(ARC)、進行性全身性リンパ節腫(PGL)、抗HIV抗体陽性状態およびHIV陽性状態、AIDS関連神経学的状態(例えば認知症または熱帯性麻痺)、カポジ肉腫、血小板減少症紫斑病および関連する日和見感染、例えばニューモシスティス・カリニ肺炎、マイコバクテリア結核、食道カンジダ症、脳トキソプラズマ症、CMV網膜炎、HIV関連脳障害、HIV関連消耗症候群等が含まれる。
【0061】
インビボでHIV感染およびAIDSの進行を測定するための標準的な方法を、対象が本発明のIsoT調節化合物での処置に陽性に応答しているかどうかを決定するために使用することができる。例えば、本発明のIsoT調節化合物での処置後、対象のCD4+ T細胞数、あるいは対象のウイルス量をモニターすることができる。CD4+ T細胞の増加は対象が抗ウイルス治療の投与によって恩恵を受けていることを示す。これ、そして当業者に既知の他の方法を、本発明の化合物が対象におけるHIV感染およびAIDSの処置に有効である程度を測定するために使用することができる。
【0062】
本発明のIsoTモジュレーターを処置する対象に滅菌条件下で直接投与することができる。モジュレーターは単独で、または医薬組成物の有効成分として投与され得る。本発明の治療組成物はまた、他の治療剤と組み合わせて、あるいは関連して使用することができる。ある適用では、第1のIsoTモジュレーターを第2のIsoTモジュレーターと組み合わせて、1種のIsoTモジュレーターを個別に使用したときには達成できないようなより高度にHIV感染を阻害するために、使用される。他の適用において、本発明のIsoT調節化合物をAZTのような既知の抗HIV薬剤と組み合わせて使用することができる。
【0063】
本発明の医薬組成物は典型的には、少なくとも1種の有効成分と1種以上の許容される担体を含む。薬学的に許容される担体は組成物を改良または安定化し、あるいは組成物の製造を容易にする。薬学的に許容される担体は投与される具体的な組成物(例えば核酸、タンパク質または調節化合物)によって、そして組成物を投与するために使用する具体的な方法によって、ある程度決定される。それらはまた、他の成分と適合し、対象に有害でないという意味において、薬学的および生理学的に許容されるべきである。この担体は投与が望まれる製剤の形態、例えば経口、舌下、直腸、鼻、静脈内または非経腸に依存して広い範囲の形態を取り得る。例えば安定性または薬理学的特性を向上するためにIsoT調節化合物を投与の前に担体タンパク質、例えばオブアルブミンまたは血清アルブミンと組み合わせることができる。
【0064】
医薬組成物を様々な形態、例えば顆粒、錠剤、ピル、座薬、カプセル剤などに製剤することができる。製剤における治療活性化合物の濃度は、約0.1〜100重量%まで変化し得る。治療製剤を薬学の分野で周知の方法によって製造する。治療製剤を処置に使用することができるあらゆる有効な手段によって送達することができる。例えばGoodman & Gilman’s The Pharmacological Bases of Therapeutics, Hardman et al., eds., McGraw-Hill Professional (10th ed., 2001); Remington: The Science and Practice of Pharmacy, Gennaro (ed.), Lippincott Williams & Wilkins (20th ed., 2003);およびPharmaceutical Dosage Forms and Drug Delivery Systems, Ansel et al. (eds.), Lippincott Williams & Wilkins (7th ed., 1999)参照。
【0065】
治療製剤は便宜には単位投与形態で存在し、適当な治療用量で投与することができる。適当な治療用量はあらゆる周知の方法、例えば最大耐用用量を決定するための哺乳類での、そして安全用量を決定するための正常ヒト対象での臨床試験によって決定することができる。高用量が必要とされ得る特的の状況を除き、IsoTモジュレーターの好ましい用量は通常、1日当たり約0.001〜約1000mg、より通常は約0.01〜約500mgの範囲である。
【0066】
IsoTモジュレーターの好ましい投与量および投与形態は、処置している医師によって個別に見出される要素、例えば状態または処置される状態、具体的なIsoTモジュレーターを含む投与する組成物の選択、個々の対象の年齢、体重および応答、対象の状態の重症度、ならびに選択した投与経路に依存して、異なる対象について変化し得る。一般的な法則として、投与されるIsoTモジュレーターの量は対象の状態を効果的かつ確実に予防または縮小させる最小用量である。したがって、上記用量範囲は一般的なガイドラインを提供し、本明細書の記載を補助することを意図しており、本発明の範囲を限定することを意図していない。
【実施例】
【0067】
下記実施例は説明のために提示しており、本発明を限定しない。
【0068】
実施例1 VprはIsoTのC末端領域と相互作用する
Vpr−IsoT相互作用をヒト白血球ライブラリーを使用して酵母2ハイブリッドスクリーニングによって同定した。酵母におけるタンパク質−タンパク質のアッセイをGAL4とLexA融合タンパク質で行った。GAL4発現ベクター(Clontech)にクローン化したヒト白血球cDNAライブラリーを、HIV HXB2 Vprをおとりとして使用してスクリーニングした。Vpr cDNAをVpr特異的プライマーを使用してPCRによって増幅した。cDNAをLexA DBD発現ベクターpSLANSに挿入した。pSLANSはpBTM116(Bartel et al Biotechniques 14: 920-924, 1993)の改変版である。NotI挿入を受け入れ、gly4−ser−gly4−serをLexAとおとりの間に設置するため、改変した。LexA DBD−Vpr融合タンパク質をコード化するcDNAをL40 MATa酵母株に形質転換した。Gal4 AD−白血球cDNA融合タンパク質をコード化するcDNAを酵母株540 MATαに形質転換した。2種の酵母株を交配し、両プラスミドを含む形質転換体をTHUKL欠乏合成培地で選択し、タンパク質相互作用をβ−ガラクトシダーゼフィルターアッセイによって分析した。
【0069】
HIV−1 HXB2 Vprを、ヒト白血球cDNAライブラリーにおいてコード化される新規な相互作用する宿主細胞因子を同定するための酵母2ハイブリッドスクリーニングにおいておとりとして使用した。約200万の2倍形質転換体をスクリーニングし、多くの陽性候補を単離した。陽性コロニーをβ−ガラクトシダーゼ活性について、タンパク質−タンパク質相互作用の更なる確認のためのフィルターリフトアッセイを使用してアッセイした。独立の候補cDNAクローンの配列決定およびGenBankデータベースとの比較によって、IsoTである相互作用パートナー、26Sプロテアソームの成分を同定した。IsoTは遊離ポリユビキチン鎖中に配列を有するユビキチンの再循環に主に関与している。プレイ長(PL)IsoTの配列分析によってC末端227アミノ酸およびさらに635bpの3’−UTRを含むクローンを確認した(図1A)。興味深いことに、PLクローンはUBA(2)ドメインを通じてVprと相互作用することが既に同定されているhHR23A DNAダメージ修復酵素と類似の2つのUBAドメインを有する。β−ガラクトシダーゼ活性によってIsoTとVprの酵母中での相互作用を確認したが、Vprと陰性対照ベクターVP16はβ−ガラクトシダーゼ活性を示さなかった(図1B)。HXB2に由来するVprタンパク質をシングルTヌクレオチドの挿入によってフレームシフトして、74〜78位で5個のフレームシフトしたアミノ酸に導く。したがって、VprのC末端領域(アミノ酸位置74〜96)はIsoTとの結合を媒介するのに必要でない。これらの残基(84〜94)はウイルス粒子取り込みに重要であると思われる(Paxton et al., 1993)。
【0070】
本発明者らはまた、動物細胞におけるVpr−IsoT相互作用を分析するために共免疫沈降実験を行った。IsoT哺乳類発現ベクターをFLAGエピトープを全長IsoT遺伝子の5’末端と融合することによって構成した。VIP3ベクターを修飾してMycタグ(Myc:Vpr融合タンパク質)またはFLAGタグ(FLAG:IsoT融合タンパク質)のいずれかを発現させた。IsoTの全長クローンを、IsoT特異的プライマーを使用した膵臓由来のcDNAライブラリーのPCR増幅によって得て、IsoTであることを配列分析によって確認した後、FLAGタグ化VIP3ベクターにサブクローニングした。約3×106個の293T細胞(10cmプレート)を20μgのFLAG−IsoTおよび/またはMyc−Vprと、CaP(Clontech)を用いてトランスフェクトした。トランスフェクション後24時間で、細胞を氷上で、緩い低浸透圧バッファー(0.5%のトリトン−X、20mMのTris−HCl、50mMのNaCl、10%のグリセロールおよびプロテアーゼ阻害剤カクテル) [Complete, Roche Molecular Biochemicals]中で30分間溶解させた。細胞溶解物を16,000×gで5分間(チェック)遠心分離することによって澄まし、上清を抗FLAG抗体M2(Sigma−チェック)で免疫沈降(2時間、4℃)し、その後タンパク質A結合ゲルビーズとインキュベートした。結合タンパク質をSDSサンプルバッファー中で沸騰させて溶出し、アリコートをSDS−PAGEおよびヤギ抗Myc抗体でウェスタンブロッティングによって分析した。
【0071】
結果は、エピトープ−タグ化IsoTとVprのHEK293T細胞における過剰発現によって、VprとIsoTのヒト細胞における相互作用を示した(データは示さず)。
【0072】
実施例2 IsoT cDNA過剰発現はHIV生産を増加させる
HIVライフサイクルにおけるIsoTの役割を明らかにするため、IsoTを偽ウイルスプラスミドで平行してトランスフェクトし、標的293T細胞で形質導入し、ルシフェラーゼ受容体活性についてアッセイしてウイルス上清を分析した。293T HEK細胞の偽ウイルス生産のための一時的トランスフェクションを、6ウェルマイクロタイタープレート(3×105細胞/ウェル)中で、カルシウムホスファターゼ法(Clontech)を使用して行った。ルシフェラーゼベクタープラスミド(ゲノムRNAをコード化する)、デルタプサイパッケージング構造物(全構造タンパク質ならびに制御および修飾タンパク質Tat、Rev、VprおよびVifをコード化する)およびVSV−gエンベロープ発現プラスミドを2.0:1.5:0.5(それぞれ2、1.5、0.5μgのDNA/ウェル)の比でトランスフェクトした。ルシフェラーゼ受容体活性を、ウイルス上清の等量アリコートで293T標的細胞を、96ウェルまたは384ウェルフォーマットで形質導入し、Bright−Gloルシフェラーゼ基質および適当な蛍光フィルターを有するAcquestプレートリーダーを使用して、測定した。値を相対光単位(RLU)で表す。
【0073】
製造した偽ウイルスは標的細胞ゲノムへの逆転写および集積によって発現するルシフェラーゼ遺伝子のみをコード化する。第4プラスミドのコトランスフェクションにるウイルス出芽への効果の上昇または減少は、偽ウイルスプラスミド単独のときと比較して標的細胞のルシフェラーゼ活性の変化に反映される。過剰発現したIsoTの存在下でトランスフェクション後24時間で回収したウイルス上清はウイルス生産で2倍(50%)の増加を導いた(10,000rlu対20,000rlu)。トランスフェクション後48時間で、ウイルス生産はIsoTの存在下で3倍増加した(図2A)。偽ウイルス感染性における24および48時間後のウイルス生産の観察された増加に伴って、本発明者らはウェスタン分析によって外因性IsoT発現が平行して増加することを見出した。
【0074】
IsoTの役割をさらに評価するために、残基335の活性部位システインをアラニンで置換して触媒的に不活性の変異体を製造した。IsoT−C335A変異体を、PCRを利用した部位特異的突然変異誘発法によって製造した部位特異的ヌクレオチド置換体を使用して製造した。PCRを天然Taq DNAポリメラーゼで、製造者(Perkin−Elmer)が特定した条件下で行った。PCR産物をサブクローニング of the C335AフラグメントのIsoT cDNA:VIP3プラスミドへのサブクローニングに利用する適当な制限部位を有するように加工した。カスタムオリゴヌクレオチドプライマーをNNNから商業的に得た。PCRのテンプレートはIsoT:VIP3構造物であった。
【0075】
IsoT−C335A変異体の発現が、不活性のときHIV偽ウイルス生産を増加させるとは予期されなかった。これは図2Bに示す。全長野生型(FL−WT)IsoTの過剰発現はC335A変異体と比較して約2.5倍力価が増加した。プレイ長クローンは全長クローンで観察された表現型と置換することができず、正しいコンフォメーションにおいて機能的に活性なタンパク質がウイルス力価の増加を媒介するのに重要であることを示唆した。FL−WT IsoT発現のHIV力価の増加に対する用量依存効果を生産細胞へHIV−1ベクタープラスミドで平行して力価判定した。逆に、C335Aの発現は内因的に発現したIsoTのみで観察されたレベルと比較して、約50%HIV力価が減少したトランスドミナント陰性方法において作用すると思われる(図2C、0μg)。FLおよびC335A変異体タンパク質の発現レベルは、ウェスタン分析によって測定されたものと同等であった(データは示さず)。
【0076】
実施例3 ブロッキングVpr取り込みがウイルス生産レベルを減少させる
VprのHIVビリオン取り込み変異体を、デルタプサイパッケージングプラスミドにおけるVprのコドン30で、アラニンをリジンで置換することによって製造した。案の定、Vpr−A30Lビリオン取り込み変異体はWTウイルス生産と比較して、標的293T細胞におけるレポーターウイルスのルシフェラーゼ活性を測定すると、ウイルス生産において40%の減少を導いた(図3A)。対照的に、核局在化に欠陥がある変異体Vpr(W54R)はウイルス生産にほとんど効果がなかった。p24レベルもウイルス上清のアリコートから定量したが、これは図3Aの感染性の減少が生産細胞から放出されたウイルス粒子の数が40%減少したことと関連した(データは示さず)。製造したウイルスの感染性はVprに導入し、p24レベルに正規化した変異体には影響しなかった(図3B)。
【0077】
実施例4 IsoT減少によるHIV粒子生産の阻害
IsoTの過剰発現が上昇したウイルスベクター生産を導き、Vprのブロッキング取り込みがウイルスベクター生産の減少を導くという観察から、本発明者らは次に、IsoTおよびVprのHIV−1ベクター生産における減少効果を試験した。IsoTおよびVpr標的mRNAのsiRNA効果を確認するため、前日にIsoTおよびVpr RNAそれぞれを標的とする低分子干渉siRNAを導入した後、FLAGタグ化IsoTおよびMycタグ化Vprを293Tにおいて過剰発現させた。2つのsiRNAをIsoT、VprおよびTatに対して各々設計し、QiagenにHPPグレードを発注した。具体的には、IsoT特異的siRNAの配列は:IsoT 1: 5’-AGUUCACCUUCGGCUUAGAUU-3’ (SEQ ID NO:1);およびIsoT 2: 5’-GGCAGAUGGGUGAUCUACA-dTdT-3’ (SEQ ID NO:2)である。Vpr特異的siRNAの配列は: Vpr 1: 5’-GGAGUGGAAGCCAUAAUAA-UU-3’ (SEQ ID NO:3);およびVpr 2: 5’-ACAACUGCUGUUUAUCCAUUU-3’ (SEQ ID NO:4)である。Tat特異的siRNAの配列は:Tat 1: 5’-CUGCUUGUACCAAUUGCUA-dTdT-3’ (SEQ ID NO:5);およびTat 2: 5’-GCCUUAGGCAUCUCCUAUG-dTdT-3’ (SEQ ID NO:6)である。2つのsiRNAを1:1の比で組合せ、他に記載がない限りプールとして使用した。実験で使用した他のsiRNAはDharmacon Researchに発注した。
【0078】
試験した全てのsiRNAがIsoTまたはVprのレベルを効果的に減少させた(データは示さず)ため、選択したsiRNAは有効である。別注のIsoT抗体の生産に際して、内因性IsoTの減少を確認した。内因性レベルの高度な減少を、48時間と比較して72時間で観察した。アラマーブルー(ミトコンドリア活性)および細胞力価glo(ATP放出)細胞生存率キット試薬によって評価した293T細胞生存率に効果はなかった(データは示さず)。
【0079】
IsoT減少のウイルスベクター生産に対する影響を評価するため、293T細胞をsiRNAで処理し、翌日偽ウイルスプラスミドでトランスフェクションし、そしてウイルスを2日後に収穫し、293T標的細胞における感染性を測定した。RNA干渉によるIsoTおよびVprタンパク質の減少は、293T標的細胞において評価すると力価の減少を導いた(図4)。IsoT減少は60%の力価(感染性)の減少を導いた(図4A)。これは非飽和状態に依存し;ウイルス生産量は生産細胞へのデルタプサイ(構造遺伝子)のトランスフェクト量、および/またはウイルス収穫までの時間の長さ(48時間後、ウイルス生産量は典型的には飽和した)によって規制されるように線形の範囲であった。驚くべきことに、Vprレベルの減少はTatレベルの減少と比較して重度にウイルス生産を減少させた。デルタプサイパッケージングプラスミドから非飽和状態下で生産されるVprのレベルの減少は、明らかにこのアッセイにおいて感染性ウイルス粒子形成に重要である。使用したsiRNAはいずれも、293Tにおいて全く毒性を示さなかった(図4B)。
【0080】
VprおよびTatの減少が効果的にウイルス生産を減少させるが、IsoT減少は感染性ウイルス生産を60%のみ減少させるという観察は、大部分が、TatおよびVprのデノボ合成の前に低分子干渉RNAを添加したためであろう。これは外因性対内因性IsoTのIsoT減少で観察された結果と類似している。RNA干渉の前に合成されたIsoTタンパク質がVprによってなおも使用され、出芽および放出を促進していると思われる。IsoTを標的とする5つの独立した、個性的なsiRNAがタンパク質発現およびHIV偽ウイルス力価を効果的に減少させた(データは示さず)。
【0081】
実施例5 IsoTおよびVprレベルの減少はHIV−1 IIIb複製を減少させる
本発明者らは次に、HIV複製可能な細胞におけるIsoTおよびVpr減少の効果を試験しようとした。この目的のために、HeLa−T4−βGal HIV受容体細胞をHIV IIIbで感染させ、その後siRNA処理を行った。HeLa−CD4−βgal細胞(Kimpton et al., J Virol 66:2232-2239, 1992)はAIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIHを通じてDr. Michael Emermanから得た。結果は、感染の72時間後、IsoTまたはVpr減少によって、全く細胞毒性が観察されることなく、HIV複製がおよそ40%減少した(図5)。Tatレベルの減少によって約85%複製が減少した。宿主細胞因子の減少による細胞毒性の対照として、RNA干渉を使用してRNA Pol IIレベルを減少させた。減少したRNA Pol IIレベルの存在下での細胞生存率の顕著な減少がウイルス複製の観察された阻害を十分に説明した。これらのデータは、許容細胞におけるHIV−1複製を効果的に補助し、機構が細胞毒性に貢献しないというIsoTおよびVprの役割と一致する。
【0082】
本発明者らはまた、正常上皮細胞におけるIsoTレベルの減少の影響を評価した。この目的のため、IsoTレベルを低分子干渉RNAを使用してIOSE−80細胞において減少させた。結果は、IsoTレベルの減少が正常上皮細胞の生存能力には影響しないが、RNA Pol IIレベルの減少は生存率を65%減少させることを示した(データは示さず)。これらの結果は、IsoT機能の下方制御が正常の、非感染細胞の生存能力に影響しないことを示す。
【0083】
実施例6 本発明のスクリーニング法によるHIV阻害化合物の同定
この実施例は上記方法を使用したHIV阻害剤の同定について記載する。具体的には、低分子有機化合物のライブラリーをハイスループット形式でスクリーニングした。試験化合物を、IsoTの阻害能についてスクリーニングした。1次スクリーニングによって同定したヒットした化合物を、さらにHIV感染の阻害効果について試験した。簡潔に述べると、ユビキチン−7−アミド−4−メチルクマリン(Ub−AMC)を基質として使用したIsoT酵素活性の生化学アッセイを開発した。IMCのIsoTによる分裂および遊離が380nmでの励起および460nmでの放射蛍光シグナルを発生する。化合物の存在下での切断活性の阻害はスクリーニングヒットを構成する。内部での低分子化合物回収(1.7×106化合物)を、1536ウェルHTS形式で、内部で作成したIsoT酵素を使用してスクリーニングした。スクリーニングヒット率は0.4%であった。オリジナルヒット(1,761の化合物の87.3%が入手可能と再確認)を、第1に細胞毒性化合物の再確認、除去に供し、そして精製した化合物のアッセイを行った。同定された化合物はHIV複製の阻害活性について試験し、他の優先順位の特徴(例えば有効性、選択窓、化学的な取り扱いやすさ)を試験した。潜在的なHIV阻害剤として同定した6個の化合物が存在した。
【0084】
かくして同定したHIV阻害化合物の代表は、化合物Aである。この化合物のIsoTプロテアーゼ阻害についてのIC50は129nMである。化合物のHeLa−T4−βGal感染モデルにおけるHIV IIIbに対する抗ウイルス活性についてのEC50は6.14μMであり、細胞毒性は観察されなかった。脱ユビキチン化(DUB)酵素に対してアッセイを行ったとき、当該化合物はIsoT、ならびにミクロレベルのUCH−L1およびUSP7よりも低い程度のUCH−L3およびUSP2iso2の阻害が示された。阻害はイソペプチダーゼ活性に特異的であり、システインプロテアーゼのパネルには阻害が観察されなかった(データは示さず)。IsoTプロテアーゼ活性の直接阻害についての更なる支持を、テトラユビキチン切断アッセイ、次いで抗Ub抗体を使用したウェスタン分析を使用して得た。結果は、化合物を力価評価すると、テトラユビキチンの阻害に良好な用量依存性が存在することを示した。化合物Aのモル濃度が増加すると、製造された遊離(切断)ユビキチンレベルの減少が存在する。見かけのIC50は約100nmであり、これは生化学アッセイにおいてIsoT阻害について観察されたIC50と良好に相関する。これらのデータはさらに、化合物によるHIV複製の阻害がIsoT活性の阻害によることを支持する。ハイスループットスクリーニングデザインならびにスクリーニングに使用した物質および方法は、以下に詳述する。
【0085】
IsoTハイスループットスクリーニングデザイン:アッセイデザインはDang et al. (Biochemistry. 37:1868-79, 1998)の方法を基礎とし、さらに1536ウェル形式におけるウルトラHTSについて最適化した。簡潔に述べると、イソペプチダーゼ活性をユビキチンAMC基質からAMCの遊離を通じて検出し、蛍光シグナルを測定する。阻害剤化合物またはユビキチンアルデヒドの存在下では、プロテアーゼ活性を阻害する:完全反応バッファー(20mM HEPES、0.5mM EDTA、1mg/ml BSA、10mM DTT、pH7.8)中、5μL/ウェルの360pMのIsoTを黒色個体非TC1536ウェルプレート(Greiner Bio-one, Cat# 789176)に、カスタムBottleValve液体分配機によって分注し、50nL 1mMの化合物および対照(Ubアルデヒド)を本発明者らの社内取り付けPinToolを使用して移し(最終化合物濃度10μM)、そして相互作用を3時間、37℃で進行させる。1μL 300nMのUb−AMCを分注し、反応物を20分間、30℃でインキュベートする。蛍光強度をViewLuxプレートリーダー(PerkinElmer)を使用して測定する。化合物の用量依存性およびIC50測定についての滴定を、Minitrak装置(Packard BioScience)を使用して、DMSOでの半対数(half-log)連続希釈について行う。
【0086】
IsoT酵素の製造:IsoTをpRSETaへと、標準的なクローン化方法を使用してクローン化し、株BL21(DE3)へと形質転換する。タンパク質の発現を、タンパク質発現を1mMのIPTGで誘導したことを除き既出のとおり(Lesley et al., 2002)に行った。タンパク質の精製を以下のとおりに行った:4グラムの細胞ペレットを50mM Tris−HCl、50mM NaCl、10mM イミダゾール、pH7.9からなる溶解バッファー40ml中に再懸濁し、4×1分間超音波を加えた。15,000×gで20分間スピンさせた後、澄明な溶解物を、ニッケルを入れた1.5mlのベッド体積のChelating-sepharose FastFlow (Amersham)を含むカラムに注いだ。カラムを50mM Tris−HCl、300mM NaCl、40mM イミダゾールおよび10% グリセロール、pH7.9からなる洗浄バッファー7.5mlで洗浄した。タンパク質を20mM Tris−HCl、300mM イミダゾール、10% グリセロール、pH8.0からなる溶出バッファー4.5mlで溶出した。
【0087】
IsoT阻害剤のHIV IIIbに対する試験:IsoTスクリーニングからDMSOで希釈した化合物を、1mMから出発して、384ウェルポリプロピレン化合物プレート(Greiner)に並べ、次いでDMSOでの半対数連続希釈で、合計8個、Minitrak装置(Packard Bioscience)を使用して製造した。HeLaCD4βgal細胞(1500/50μL/ウェル)を384ウェル白色プレート(Greiner)に播種し、一晩密着の後、500nLの化合物を3つの複製細胞プレートに、Minitrak(Packard Bioscience)を使用して、感染性および細胞毒性試験のために移し、10μMから出発して最終化合物連続希釈を得た。30分後、HIV−IIIb(16ng/10μL/ウェル)を感染性試験細胞プレートに加えた。化合物およびウィルス添加の3日後、50μLのGal Screen(Applied Biosystems)を感染細胞に加え、1時間後にCLIPR (Molecular Devices)で蛍光を読み取って、感染を評価した。50μLの1:4希釈Cell Titer Glo (Promega)を感染細胞に加え、1時間後にCLIPRで読み取って、細胞毒性を評価した。用量応答測定を標準のシグモイド型用量応答式を使用して解釈した。社内の曲線適合コンピュータプログラムをIC50の計算に使用した。
【0088】
IsoT阻害剤によるテトラユビキチン(Ub4)切断の阻害:IsoTを反応バッファー(20mM HEPES、0.5mM EDTA、1mg/mL BSA、10mM DTT、pH=7.8)で最終濃度760pMとなるように希釈し、18μLを9個のエッペンドルフチューブそれぞれに加えた。化合物2096をDMSOで、200μMから出発して合計8個、半分対数連続希釈した。1μLの各希釈物をIsoTのチューブに加え、アッセイチューブ中10μMから出発する連続希釈を得た。最大切断についての対照として、1個のチューブに1μLのDMSOのみを加えた。37℃で30分間インキュベーションした後、1μLの反応バッファー中テトラユビキチン(Ub4;Biomol)の250μg/mL溶液を各チューブに加え、当該チューブを37℃で18時間さらにインキュベートした。変性タンパク質サンプルバッファー(Invitrogen)を加え、全サンプルを沸騰させ、4〜12%NuPage bis-Tris ゲル(Invitrogen)にロードし、製造者の指示通りにゲル電気泳動を行った。ニトロセルロースへと移した後、ブロットをPBST(0.05% Tween20を含むリン酸緩衝化食塩水)中5%無脂肪ミルクでブロックし、ユビキチンに対するウサギポリクローナル抗体(上記PBST中5%無脂肪ミルクで1:1000に希釈)で、次いでHRP結合ヤギ抗ウサギ2次抗体(PBST中5%無脂肪ミルクで1:3000、Southern Biotechnologies)でのイムノブロッティングに供した。バンドをECLプラス検出試薬(Amersham)を使用して可視化した。
【0089】
本明細書に記載の例および態様は例示のみを目的としており、その範囲内で様々な修飾または改変が当業者には提案され、そして本願および添付の特許請求の範囲の技術的範囲に含まれることが理解される。本明細書に記載のものと類似または同様のあらゆる方法および物質を本発明の実施または試験に使用することができるが、好ましい方法および物質は記載されている。
【0090】
本明細書に記載した全ての文献、GenBank配列、特許および特許公報は、各々そのように記載されているが如く、その全体について、あらゆる目的のために本明細書の一部とする。
【図面の簡単な説明】
【0091】
【図1】図1A−1BはVprのIsoTへの結合を示す:(A)概略図およびIsoTプレイクローンの位置を酵母−2−ハイブリッドを全長IsoT cDNAと比較して同定した。プレイクローンは2個のUBAドメインを有するC末端227アミノ酸を含む。本明細書で使用する全長IsoT cDNAクローンを膵臓由来のcDNAライブラリから増幅した。C335の相対位置(活性部位システイン)を示す。(B)VprとIsoTの特異的相互作用を示すために、β−ガラクトシダーゼフィルターリフトアッセイを使用して酵母−2−ハイブリッドをアッセイする。VP16は空ベクターである。
【0092】
【図2】図2A−2Cは野生型IsoT過剰発現がHIV−1ウイルスの力価を増加させるが、活性部位変異はそうではないことを示す:(A)全長(FL)野生型IsoTの過剰発現がトランスフェクション後24および48時間で偽ウイルス過剰発現を増加させる。293T細胞をIsoTをコード化する第4cDNAの存在下(+)または非存在下(−)において3個の偽ウイルスプラスミドとコトランスフェクトした。レポーターウイルス上清をトランスフェクション後24または48時間で回収し、293T標的細胞におけるレポーター活性についてアッセイした。細胞溶解物を平行してウェスタン分析で、IsoT特異的抗体を使用して内因性IsoT発現ならびにIsoT過剰発現について分析した。1列目はプラスミドでトランスフェクトしていない、トランスフェクション試薬のみのものである。(B)293T細胞を(A)のとおりに、第4プラスミドとしてVIP3(プラスミド骨格)、FL−野生型、PL−C末端227、またはFL−C335Aでコトランスフェクトした。トランスフェクション後30時間に回収したウイルス上清由来のルシフェラーゼレポーター活性を293T標的細胞においてアッセイした。(C)HIV力価に対する用量依存関数効果。FL−WTおよびFL−C335Aを力価判定し、平行して第4プラスミドとして偽プラスミドとトランスフェクトした。
【0093】
【図3】図3A−3BはVpr(A30L)ウイルスの減少した出芽が感染性ではなく減少した粒子生産と関連することを示す。293T細胞を偽ウイルスDNA発現WTおよびVprの変異型とトランスフェクトした。(A)WTまたは核局在化変異体W54Rではなく、A30L Vpr取り込み変異体は、293T標的細胞における感染性によって測定するとウイルス力価を減少した。(B)p24レベルを標的293T細胞を感染するために使用した偽ウイルスインプットについて、(A)から標準化(9.38ng)した。
【0094】
【図4】図4A−4BはRNA干渉によるIsoTまたはVprの欠乏が293T細胞における偽ウイルス生産を阻害することを示す。(A)293T細胞をHIV Tat、IsoT、VprまたはルシフェラーゼGL3(陰性対照)を標的とする示したsiRNAでトランスフェクトした。細胞を一晩培養し、ウェルあたり2:0.05:0.5または100ng:2.5ng:25ngのベクターRNA:デルタプサイ:envの比で、3プラスミド偽ウイルス系のcDNAトランスフェクションに供した。3つの複写ウェルを条件毎に行った。ウイルス上清を48時間後に回収し、標的293T細胞に感染させるために使用した。これらの形質導入細胞におけるルシフェラーゼ活性を48時間後に測定し、Aに示す。(B)293T細胞をHIV Tat、IsoT、VprまたはルシフェラーゼGL3に対するsiRNAとトランスフェクトした。72時間後細胞生存率をCellTiterGloを使用し、蛍光を測定して決定した。
【0095】
【図5】図5はIsoTまたはVprの欠乏がHeLa−CD4−β−gal細胞におけるHIV−1 IIIb複製を阻害することを示す。細胞をTat、IsoT、Vpr、ルシフェラーゼGL2(陰性対照)、またはPolR2A(細胞毒性についての陽性対照)を標的とする示したsiRNAとトランスフェクトした。24時間培養後、細胞をHIV−1 IIIbに感染させる。β−ガラクトシダーゼ活性を、感染の測定として感染後72時間で測定した。細胞毒性をsiRNAトランスフェクト非感染細胞について平行して測定した。データを陰性対照GL2 siRNAトランスフェクト細胞によって発生したシグナルの割合として表現する。
【技術分野】
【0001】
関連出願の相互参照
本願は米国仮出願第60/673,623(出願日:2005年4月21日)に基づき合衆国法典第35巻、119条(e)の優先権を主張する。先の出願の開示はその全体をあらゆる目的のために引用し、本明細書の一部とする。
【0002】
発明分野
本発明は、一般的にHIV感染を阻害する化合物を同定するための方法と、当該化合物の治療的に関する。より具体的には、本発明はイソペプチダーゼT(IsoT)の阻害剤をスクリーニングすること、そしてかかる阻害剤をHIVの処置または予防のために使用することに関する。
【背景技術】
【0003】
発明の背景
ヒト免疫不全ウイルス(HIV)はレトロウイルス科に属するレンチウイルスである。性的接触および妊娠期間中のHIVの感染は世界中のAIDS症例のほぼ90%を占めると概算された。この感染は、感染個体の体液、例えば精液、膣分泌物または血液に曝露されたとき、性器の粘膜または胎盤を通じてHIVが通過して引き起こされる。のこりのAIDS症例はHIVに汚染された血液の輸血、静脈内薬剤使用者間の針の共有、侵襲処置におけるHIV汚染体液への事故的曝露、そして感染ウイルスが受容ヒト組織へ直接接触することができる他の状況のためである。
【発明の開示】
【発明が解決しようとする課題】
【0004】
現在利用可能なHIV感染およびAIDSの処置用薬剤は満足のいくものではない。HIVを処置するための一般薬剤、例えばAZTまたはHIVプロテアーゼ阻害剤の毒性または望ましくない副作用は、有効医薬濃度で使用したときそれらの抗ウイルス活性と相容れない。したがって、AIDSおよびHIV感染を予防および処置するためのよりよい他の化合物に対するこの分野における要求は未だに存在する。本発明は、このおよび他の要求に対処する。
【課題を解決するための手段】
【0005】
発明の要約
ある態様において、本発明はHIV感染を阻害する薬剤を同定するための方法を提供する。当該方法は、イソペプチダーゼT(IsoT)分子の生物学的活性を試験化合物の存在下でアッセイして、IsoT分子の生物学的活性を阻害する化合物を同定することを必要とする。これらの方法は、同定した化合物のHIV感染阻害能をさらに試験することを含むことができる。ある方法では、アッセイする生物学的活性はIsoT分子のイソペプチダーゼ活性または他の分子(例えばユビキタンまたはVpr)とのその結合である。また、IsoT分子をコード化する遺伝子の発現であってもよい。典型的には、スクリーニングに使用するIsoT分子は哺乳類細胞に由来する。好ましい態様において、ヒトIsoT分子を使用する。
【0006】
これらのスクリーニング法のいくつかにおいて、同定した化合物によるHIV−1感染の阻害能を、当該化合物で処理した人工HIV許容細胞におけるHIV複製と、当該化合物で処理しなかった対照細胞におけるHIV複製を比較することによって試験する。これらの方法のいくつかにおいて、使用するHIV許容細胞はHeLa−T4−βGal HIV細胞である。ある方法において、HIV複製をp24抗原ELISAアッセイまたは逆転写酵素活性アッセイによってモニターする。
【0007】
本発明の他の方法において、同定されたIsoT調節化合物によるHIV感染の阻害能を、当該化合物で処理した宿主細胞における偽ウイルス生産と、当該化合物で処理しなかった対照宿主細胞における偽ウイルス生産を比較することによって試験する。使用される宿主細胞は293T HEK細胞である。いくつかの方法では、宿主細胞を、細胞においてHIV偽ウイルスを生産する偽ウイルスプラスミドでトランスフェクトする。
【0008】
関連する局面において、本発明はHIV感染を阻害する薬剤を同定する方法を提供する。これらの方法には、第1に試験化合物をスクリーニングしてIsoTイソペプチダーゼ活性またはVprとのその結合を下方制御するIsoT調節化合物を同定すること、次に同定したIsoT調節化合物のHIV感染阻害能について試験することを含む。典型的には、スクリーニングに使用するIsoT分子は哺乳類細胞に由来する。好ましい態様において、スクリーニングに使用するIsoT分子はヒトIsoTである。
【0009】
これらの方法のいくつかにおいて、IsoT調節化合物によるHIV−1感染の阻害能を、当該化合物で処理したHIV許容細胞におけるHIV複製と、当該化合物で処理しなかった対照細胞におけるHIV複製を比較することによって試験する。いくつかの方法において、使用されるHIV許容細胞はHeLa−T4−βGal HIV細胞である。いくつかのスクリーニング法において、HIV複製をp24抗原ELISAアッセイまたは逆転写酵素活性アッセイによってモニターする。
【0010】
他の方法において、IsoT調節化合物によるHIV感染の阻害能を、当該化合物で処理した宿主細胞における偽ウイルス生産と、当該化合物で処理しなかった対照宿主細胞における偽ウイルス生産を比較することによって試験する。使用される宿主細胞は293T HEK細胞である。いくつかの方法では、宿主細胞を、当該細胞においてHIV偽ウイルスを生産する偽ウイルスプラスミドでトランスフェクトする。
【0011】
他の態様において、本発明は対象のHIV感染を処置する方法を提供する。かかる方法は、HIV感染を有する対象に、有効量のIsoT調節化合物を含む医薬組成物を投与することを必要とする。IsoT調節化合物はIsoTイソペプチダーゼ活性、VprとのIsoT結合またはIsoTをコード化する遺伝子の発現を下方制御することができる。いくつかの方法において、使用するIsoT調節化合物はヒトIsoTを下方制御することができる。いくつかの方法において、使用するIsoT調節化合物はインビトロでIsoT発現細胞におけるHIV複製を阻害することができる。これらの治療法において、IsoT調節化合物を既知の抗HIV薬剤と共に投与することがある。いくつかの方法において、IsoT阻害薬剤はIsoT活性を特異的に阻害する本発明によって同定された化合物である。他の方法において、IsoT阻害薬剤はIsoTの発現または細胞レベルを特異的に下方制御する核酸分子、例えばsiRNAまたはアンチセンスDNAである。
【0012】
本発明の性質および利点の更なる理解を、明細書および特許請求の範囲の残りの部分を参照することによって理解することができる。
【0013】
図面の簡単な説明
図1A−1BはVprのIsoTへの結合を示す:(A)概略図およびIsoTプレイクローンの位置を酵母−2−ハイブリッドを全長IsoT cDNAと比較して同定した。プレイクローンは2個のUBAドメインを有するC末端227アミノ酸を含む。本明細書で使用する全長IsoT cDNAクローンを膵臓由来のcDNAライブラリから増幅した。C335の相対位置(活性部位システイン)を示す。(B)VprとIsoTの特異的相互作用を示すために、β−ガラクトシダーゼフィルターリフトアッセイを使用して酵母−2−ハイブリッドをアッセイする。VP16は空ベクターである。
【0014】
図2A−2Cは野生型IsoT過剰発現がHIV−1ウイルスの力価を増加させるが、活性部位変異はそうではないことを示す:(A)全長(FL)野生型IsoTの過剰発現がトランスフェクション後24および48時間で偽ウイルス過剰発現を増加させる。293T細胞をIsoTをコード化する第4cDNAの存在下(+)または非存在下(−)において3個の偽ウイルスプラスミドとコトランスフェクトした。レポーターウイルス上清をトランスフェクション後24または48時間で回収し、293T標的細胞におけるレポーター活性についてアッセイした。細胞溶解物を平行してウェスタン分析で、IsoT特異的抗体を使用して内因性IsoT発現ならびにIsoT過剰発現について分析した。1列目はプラスミドでトランスフェクトしていない、トランスフェクション試薬のみのものである。(B)293T細胞を(A)のとおりに、第4プラスミドとしてVIP3(プラスミド骨格)、FL−野生型、PL−C末端227、またはFL−C335Aでコトランスフェクトした。トランスフェクション後30時間に回収したウイルス上清由来のルシフェラーゼレポーター活性を293T標的細胞においてアッセイした。(C)HIV力価に対する用量依存関数効果。FL−WTおよびFL−C335Aを力価判定し、平行して第4プラスミドとして偽プラスミドとトランスフェクトした。
【0015】
図3A−3BはVpr(A30L)ウイルスの減少した出芽が感染性ではなく減少した粒子生産と関連することを示す。293T細胞を偽ウイルスDNA発現WTおよびVprの変異型とトランスフェクトした。(A)WTまたは核局在化変異体W54Rではなく、A30L Vpr取り込み変異体は、293T標的細胞における感染性によって測定するとウイルス力価を減少した。(B)p24レベルを標的293T細胞を感染するために使用した偽ウイルスインプットについて、(A)から標準化(9.38ng)した。
【0016】
図4A−4BはRNA干渉によるIsoTまたはVprの欠乏が293T細胞における偽ウイルス生産を阻害することを示す。(A)293T細胞をHIV Tat、IsoT、VprまたはルシフェラーゼGL3(陰性対照)を標的とする示したsiRNAでトランスフェクトした。細胞を一晩培養し、ウェルあたり2:0.05:0.5または100ng:2.5ng:25ngのベクターRNA:デルタプサイ:envの比で、3プラスミド偽ウイルス系のcDNAトランスフェクションに供した。3つの複写ウェルを条件毎に行った。ウイルス上清を48時間後に回収し、標的293T細胞に感染させるために使用した。これらの形質導入細胞におけるルシフェラーゼ活性を48時間後に測定し、Aに示す。(B)293T細胞をHIV Tat、IsoT、VprまたはルシフェラーゼGL3に対するsiRNAとトランスフェクトした。72時間後細胞生存率をCellTiterGloを使用し、蛍光を測定して決定した。
【0017】
図5はIsoTまたはVprの欠乏がHeLa−CD4−β−gal細胞におけるHIV−1 IIIb複製を阻害することを示す。細胞をTat、IsoT、Vpr、ルシフェラーゼGL2(陰性対照)、またはPolR2A(細胞毒性についての陽性対照)を標的とする示したsiRNAとトランスフェクトした。24時間培養後、細胞をHIV−1 IIIbに感染させる。β−ガラクトシダーゼ活性を、感染の測定として感染後72時間で測定した。細胞毒性をsiRNAトランスフェクト非感染細胞について平行して測定した。データを陰性対照GL2 siRNAトランスフェクト細胞によって発生したシグナルの割合として表現する。
【発明を実施するための最良の形態】
【0018】
詳細な説明
I.概説
本発明は一部において、IsoTがHIV感染に関与するという本発明者らの発見に基づく。ヒトIsoTは、遊離K48結合ポリユビキチン鎖のモノマーへの分解に関与する93kDaの亜鉛結合脱ユビキチン化酵素である。本発明者らはIsoTがHIV−1ウイルスのウイルス性タンパク質R(Vpr)の結合パートナーであることを見出した。Vprは、単球およびマクロファージにおけるHIVウイルス複製に必須であり、T細胞およびT細胞株におけるウイルス複製を増加させる96アミノ酸、14kDa付属タンパク質である(例えば、Cohen et al., J Virol 64:30973099, 1990;Paxton et al., J Virol 67:72297237, 1993;およびAndersen et al., Curr HIV Res. 3:43-51, 2005参照)。本発明者らはまた、活性部位変異体ではなく野生型IsoTの過剰発現が生産細胞由来のウイルス力価を上昇させることを見出した。さらに、低分子干渉RNAを用いたIsoTレベルの欠乏がHEK293T生産細胞におけるウイルス力価を減少させ、そして実験室株HIV−1(IIIb)およびHeLa−T4−βGalにおけるHIV−1 BaL、ならびにU373 HIV−1インジケーター細胞株それぞれの感染性を減少させることを発見した。これらのデータはIsoTが宿主細胞からのHIV−1放出の効果を上昇させるのに重要な役割を果たしていることを示す。
【0019】
これらの発見により、本発明はHIV感染を阻害する新規薬剤をスクリーニングする方法を提供する。試験化合物を、第1にIsoT分子の生物学的活性、例えばその発現、そのVprとの相互作用、またはそのイソペプチダーゼ活性を調節する能力についてスクリーニングする。したがって同定された薬剤を、典型的にはさらにHIV感染またはHIV感染を示す活性を調節する能力について試験する。様々なIsoT分子をスクリーニングアッセイにおいて使用することができる。例えば、ヒト、ウサギ、ラットまたはマウス由来のIsoTを調節剤のスクリーニングのために使用することができる。いくつかの好ましい態様において、ヒトIsoT分子を使用する。本発明に開示したスクリーニング法の有用性を例示するために、本発明者らはヒトIsoTの阻害剤のために低分子化合物のハイスループットスクリーニングを行った。下記実施例に詳述するとおり、IsoTを阻害することによる抗ウイルス活性を有する化合物がスクリーニングによって同定された。
【0020】
本発明はまた、治療使用を含む。IsoT活性の薬理学的阻害はHIV感染に関連する状態の処置または予防の新規アプローチを提供する。典型的には、アプローチには本発明によって同定することができるIsoT阻害剤(例えばsiRNAまたは低分子有機化合物)を対象に投与することを含む。
【0021】
以下の項は本発明の組成物を製造し、使用するための、そして本発明の方法を実施するための指標を提供する。
【0022】
II.定義
他に定義がない限り、本明細書で使用されている全ての技術および科学用語は本発明が属する分野における当業者が通常理解するとおりの意味を有する。下記文献は当業者に本発明において使用される多くの用語の一般的な定義を提供する: Oxford Dictionary of Biochemistry and Molecular Biology, Smith et al. (eds.), Oxford University Press (revised ed., 2000); Dictionary of Microbiology and Molecular Biology, Singleton et al. (Eds.), John Wiley & Sons (3rd ed., 2002);および A Dictionary of Biology (Oxford Paperback Reference), Martin and Hine (Eds.), Oxford University Press (4th ed., 2000)。さらに、下記定義を本発明の実施において読者を補助するために提供する。
【0023】
「試験薬剤」または「試験化合物」なる用語には、あらゆる物質、分子、要素、化合物、実体、またはそれらの組合せを含む。限定されないが、例えばタンパク質、ポリペプチド、小有機分子、ポリサッカライド、ポリヌクレオチド等が含まれる。天然物、合成化合物または化学化合物または2種以上の物質の組合せであり得る。特に記載がない限り、「薬剤」、「物質」および「化合物」なる用語は相互に交換可能に使用することができる。
【0024】
本明細書において「アナログ」なる用語は、参照分子と構造的に類似しているが参照分子の特定の置換基を別の置換基で置換することによって標的化および制御方法が修正されている分子を意味するものとして使用する。参照分子と比較して、アナログは同一の、類似のまたは改善された有用性を示すものと、当業者には予期される。アナログの合成およびスクリーニングは改善された特徴(例えば標的分子に対する高い結合親和性)を有する既知の化合物の変異体を同定するための薬学化学において周知の方法である。
【0025】
HIVはレトロウイルスのファミリーであるヒト免疫不全ウイルス(HIV)を意味する。限定されないが、これらのウイルスにはHIV−I、HIV−II、HIV−III(HTLV−II、LAV−1、LAV−2としても知られる)等が含まれる。本明細書において使用するとき、HIVはHIVファミリーにおけるあらゆる株、形態、サブタイプおよび変異体であり得る。
【0026】
2個の核酸配列またはアミノ酸配列の文脈中での「同一」または「配列同一」なる用語は、特定の比較窓にわたって最大対応を並べたとき、2個の配列における残基が同じであることを意味する。比較のための配列のアラインメント法は、当業者に既知である。比較のための配列の最適なアラインメントを、Smith and Waterman (1981) Adv. Appl. Math. 2:482の局所ホモロジーアルゴリズムによって; Needleman and Wunsch (1970) J. Mol. Biol. 48:443のアラインメントアルゴリズムによって; Pearson and Lipman (1988) Proc. Nat. Acad. Sci U.S.A. 85:2444の同一性の調査法によって;by computerized implementations of these algorithms これらのアルゴリズムのコンピュータによる実行(限定されないが、Intelligentics, Mountain View, CAによるPC/GeneプログラムのCLUSTAL;およびWisconsin Genetics Software Package, Genetics Computer Group (GCG), 575 Science Dr., Madison, Wis., U.S.A.におけるGAP、BESTFIT、BLAST、FASTA、またはTFASTAを含む)によって、行うことができる。アラインメントは、検査およびマニュアルアラインメントによってしばしば行うこともできる。
【0027】
「実質的に同一の」核酸またはアミノ酸配列なる用語は、上記プログラム(例えばBLAST)を標準的なパラメータを用いて使用して標準配列と比較したとき、少なくとも90%、好ましくは少なくとも95%、より好ましくは少なくとも98%、そしてもっとも好ましくは少なくとも99%以上の配列同一性を有する配列を含む核酸またはアミノ酸を意味する。好ましくは、実質的同一性は少なくとも約50残基長である配列領域にわたって、より好ましくは少なくとも約100残基の領域にわたって存在し、そしてもっとも好ましくは配列は少なくとも約150残基にわたって実質的に同一である。もっとも好ましい態様において、配列はコーディング領域の全長にわたって実質的に同一である。
【0028】
IsoT分子の生物学的活性に関する「調節」なる用語は、IsoTの細胞レベル、細胞内局在性または他の生物学的活性(例えばそのイソペプチダーゼ活性)における変化を意味する。IsoT活性の調節は上方制御(すなわち、活性化または刺激)あるいは下方制御(すなわち阻害または抑制)であってよい。例えば、調節は細胞レベル、あるいはIsoTの酵素的修飾(例えばリン酸化)、結合特性(例えばVprまたは基質への結合)、またはIsoTタンパク質の他の生物学的、機能的または免疫学的特徴における変化を引き起こし得る。活性の変化は例えば、IsoTコード化遺伝子の発現、IsoTタンパク質をコード化するmRNAの安定性、翻訳効率の増加もしくは減少、またはIsoT酵素の他の生物学的活性(例えばそのイソペプチダーゼ活性)の変化によって引き起こされ得る。IsoTモジュレーターの作用の形態は、直接、例えばIsoTタンパク質またはIsoTタンパク質をコード化する遺伝子に結合することによっていても良い。変化はまた、関節、例えばIsoTを調節する他の分子と結合および/または修飾(例えば酵素的に)することによっていても良い。
【0029】
「対象」なる用語は、哺乳類、とりわけヒトを意味する。これは他の非ヒト動物、例えばウシ、ウマ、ヒツジ、ブタ、ネコ、イヌ、マウス、ラット、ウサギ、モルモット、サルを含む。
【0030】
IsoTのような分子の「変異体」は、分子全体またはそのフラグメントと構造および生物学的活性が実質的に同一の分子を意味する。したがって、2個の分子が類似の活性を有するならば、一方の分子の構成または2次、3次もしくは4次構造が他方のものと同一でなくとも、あるいはアミノ酸残基の配列が同一でなくとも、それらは本明細書で使用する用語としての変異体であると考える。
【0031】
III.スクリーニングスキーム
本発明によると、HIV感染の新規な阻害剤を、第1に試験化合物をIsoTの生物学的活性を調節(例えば阻害)する能力についてスクリーニングすることによって同定する。スクリーニングアッセイにおいてモニターするIsoTの生物学的活性は、そのイソペプチダーゼ活性であってもよい。それは他の分子、例えばVprまたはユビキチンへのその結合であってもよい。生物学的活性はまた、IsoT発現またはその細胞レベルであり得る。IsoTの生物学的活性を調節する試験化合物を同定した後、それらを典型的にはさらに、HIV感染を調節する、あるいはHIV感染もしくはHIV複製の指標である活性を調節する能力について試験する。この工程は、IsoTの生物学的活性を調節することによって第1の工程で同定された化合物が実際にHIV感染を制御(例えば阻害)することができることを確認するために行われる。
【0032】
当業者に周知の様々な生化学および分子生物学の技術またはアッセイをIsoTモジュレーターをスクリーニングするために使用することができる。かかる技術は、例えばHandbook of Drug Screening, Seethala et al. (eds.), Marcel Dekker (1st ed., 2001);High Throughput Screening: Methods and Protocols (Methods in Molecular Biology, 190), Janzen (ed.), Humana Press (1st ed., 2002); Current Protocols in Immunology, Coligan et al. (Ed.), John Wiley & Sons Inc (2002); Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press (3rd ed., 2001); および Brent et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (ringbou ed., 2003)に記載されている。
【0033】
様々な種由来のIsoTを、本発明のIsoTモジュレーターのスクリーニングにおいて使用することができる。好ましくは、哺乳類細胞由来のIsoT分子を使用する。例えば、ヒトIsoT(受託番号 NM_003481 および NP_003472)がクローン化され、文献、例えば Falquet et al., FEBS Lett. 376: 233-237; 1995; Wilkinson et al., Biochemistry 34:14535-14546, 1995; and Lacombe et al., FEBS Lett. 531:469-474, 2002に記載された。他の例には、チンパンジーIsoT(受託番号 XM_526393 および XP_526393)およびマウスIsoT(受託番号 BC066993 および AAH66993)が含まれる。あらゆるこれらのIsoT配列または実質的にその配列と同一の配列を、本発明のIsoTモジュレーターを同定するためのスクリーニングアッセイに使用することができる。下記実施例に例示したとおり、IsoTポリヌクレオチドのクローニングおよび発現、ならびにIsoTポリペプチドの精製を、通常行われている方法および技術で行うことができる。かかる方法および技術は例えば、Sambrook et al., supra;および Brent et al., supraに記載されている。IsoTを得る具体的な方法は、文献、例えばGabriel et al., Biochemistry 41:13755 -13766, 2002にも提供されている。
【0034】
無傷のIsoT分子または無傷のIsoT分子をコード化する核酸に加えて、IsoTフラグメント(例えば触媒ドメインまたはVpr結合ドメイン)、アナログまたは機能的誘導体も使用することができる。これらのアッセイにおいて使用することができるIsoTフラグメントは通常、IsoT分子の生物学的活性(例えばそのイソペプチダーゼ活性またはVprとの結合)の1種以上を保持している。上記のとおり、異なる種由来のIsoTは既に配列決定され、十分に特徴付けられている。したがって、それらのフラグメント、アナログ、誘導体または融合タンパク質を当業者に周知の方法を使用して容易に得ることができる。例えば、IsoTの機能的誘導体をタンパク質分解的切断によって天然に発生した、あるいは組み換えによって発現したタンパク質から、当業者に既知の常套の精製方法によって製造することができる。あるいは、機能的誘導体をそのイソペプチダーゼ活性を保持しているIsoTのフラグメントのみを発現することによって、組み換えDNA技術により製造することができる。
【0035】
IV.試験化合物
本発明の方法でスクリーニングすることができる試験化合物には、ポリペプチド、βターン模倣薬、ポリサッカライド、リン脂質、ホルモン、プロスタグランジン、ステロイド、芳香族化合物、ヘテロ環式化合物、ベンゾジアゼピン、オリゴマーN−置換グリシン、オリゴカルバメート、ポリペプチド、サッカライド、脂肪酸、ステロイド、プリン類、ピリミジン類、誘導体、構造アナログまたはそれらの組合せが含まれる。試験化合物は合成分子であることもあるし、天然分子であることもある。
【0036】
試験化合物を、合成または天然化合物のライブラリーを含む広い種類のソースから得る。コンビナトリアルライブラリーを段階的方法で合成することができる多くのタイプの化合物のために製造することができる。化合物の大きなコンビナトリアルライブラリーをWO 95/12608、WO 93/06121、WO 94/08051、WO 95/35503およびWO 95/30642に記載のコード化合成ライブラリー(ESL)法によって構成することができる。ペプチドライブラリーをファージディスプレイ法(例えばDevlin, WO 91/18980参照)によって製造することもできる。バクテリア、菌、植物および動物抽出物の形態の天然化合物のライブラリーを商業的ソースから得るか、あるいは実際に収集することができる。既知の薬理学的薬剤は、構造アナログを製造するためのアシル化、アルキル化、エステル化、アミド化のような指向性またはランダム化学修飾に供することができる。
【0037】
他の化合物またはペプチドのコンビナトリアルライブラリーを任意の位置で配列優先または固定なしで十分にランダム化することができる。あるいは、ライブラリーを偏向させることができる、すなわち配列中のいくつかの位置が一定であるか、あるいは限定された数の可能性から選択される。例えば、いくつかの場合において、ヌクレオチドまたはアミノ酸残基を、例えば疎水性アミノ酸、親水性残基、立体的に偏向した(小または大)残基、システインの製造の方向、架橋、SH−3ドメインに関するプロリン、セリン、スレオニン、チロシンまたはリン酸化部位に関するヒスチジン、またはプリンのような明らかなクラス内でランダム化する。
【0038】
試験化合物は、天然に発生するタンパク質またはそれらのフラグメントであり得る。かかる試験化合物は天然ソース、例えば細胞または組織溶解物から得ることができる。ポリペプチド薬剤のライブラリーを、例えば商業的に入手可能なcDNAライブラリーから製造するか、あるいは常套の方法で製造することができる。試験化合物はまた、約5〜約30個、好ましくは約5〜約20個、とりわけ好ましくは約7〜約15個のアミノ酸のペプチドのようなペプチドであり得る。ペプチドは天然に発生したタンパク質の消化物、ランダムペプチドまたは「偏向した」ランダムペプチドであってもよい。いくつかの方法において、試験化合物はポリペプチドまたはタンパク質である。
【0039】
試験化合物は核酸でもあり得る。核酸試験化合物は天然に発生した核酸、ランダム核酸または「偏向した」ランダム核酸であってもよい。例えば、原核または真核ゲノムの消化物、を上記タンパク質と同様に使用することができる。
【0040】
好ましい態様において、試験化合物は低分子、例えば約500または1000未満の分子量の分子である。好ましくはハイスループットアッセイを適用し、かかる低分子をスクリーニングするために使用することができる。いくつかの方法において、上記低分子試験化合物のコンビナトリアルライブラリーを容易に使用してIsoTの低分子モジュレーターをスクリーニングすることができる。かかるスクリーニングには多くのアッセイ、例えばSchultz et al., Bioorg Med Chem Lett 8:2409-2414, 1998; Weller et al., Mol Divers. 3:61-70, 1997; Fernandes et al., Curr Opin Chem Biol 2:597-603, 1998;およびSittampalam et al., Curr Opin Chem Biol 1:384-91, 1997に記載のものを使用することができる。
【0041】
本発明の方法でスクリーニングする試験化合物のライブラリーはまた、IsoTポリペプチド、それらのフラグメントまたはアナログの構造的研究に基づき製造することができる。かかる構造的研究はIsoポリペプチドとより結合しやすい試験化合物を同定することができる。IsoTポリペプチドの3次元構造を多くの方法、例えば結晶構造および分子モデリングで研究することができる。X線結晶解析を使用したタンパク質の構造研究法は文献に周知である。Physical Bio-chemistry, Van Holde, K. E. (Prentice-Hall, New Jersey 1971), pp. 221-239, および Physical Chemistry with Applications to the Life Sciences, D. Eisenberg & D. C. Crothers (Benjamin Cummings, Menlo Park 1979)参照。IsoTポリペプチド構造のコンピューターモデリングは、IsoTモジュレーターをスクリーニングするための試験化合物を設計する他の方法を提供する。分子モデリング法は文献、例えばU.S. Patent No. 5,612,894、表題“System and method for molecular modeling utilizing a sensitivity factor”およびU.S. Patent No. 5,583,973、表題“Molecular modeling method and system”に記載されている。さらに、タンパク質構造も中性子回折および核磁気共鳴(NMR)によって決定することができる。例えばPhysical Chemistry, 4th Ed. Moore, W. J. (Prentice-Hall, New Jersey 1972), およびNMR of Proteins and Nucleic Acids, K. Wuthrich (Wiley-Interscience, New York 1986)参照。
【0042】
本発明のモジュレーターにはIsoTポリペプチドに特異的に結合する抗体も含むことができる。かかる抗体はモノクローナルまたはポリクローナルであり得る。かかる抗体は当業者に周知の方法を使用して製造することができる。例えば、非ヒト、例えばマウスもしくはラットモノクローナル抗体の製造を、例えばIsoT ポリペプチドまたはそのフラグメントで動物を免疫することによって行うことができる(Harlow & Lane, Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York, 1988参照)。かかる免疫源は天然ソースから、ペプチド合成または組換発現によって得ることができる。
【0043】
マウス抗体のヒト化形態を、非ヒト抗体のCDR領域をヒト定常領域と組み換えDNA技術によって結合することによって製造することができる。Queen et al., Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989) および WO 90/07861参照。ヒト抗体をファージディスプレー法を使用して得ることができる。例えばDower et al., WO 91/17271; McCafferty et al., WO 92/01047参照。これらの方法において、ファージのライブラリーが形成され、そこではメンバーは外部表面に異なる抗体を提示する。抗体はFvまたはFabフラグメントとして通常提示される。所望の特異性を有するファージ提示抗体を本発明のIsoTポリペプチドに対する親和性濃縮によって選択する。
【0044】
IsoTポリペプチドに対するヒト抗体を、少なくともヒト免疫グロブリン遺伝子座および不活性内因性免疫グロブリン遺伝子座のセグメントをコード化する導入遺伝子を有する非ヒトトランスジェニック哺乳類から製造することもできる。例えばLonberg et al., WO93/12227 (1993); Kucherlapati, WO 91/10741 (1991)参照。ヒト抗体を競合的結合実験またはその他によって選択して、特定のマウス抗体と同じエピトープ特異性を有することができる。かかる抗体はとりわけ、マウス抗体の有用な機能的特徴を共有する可能性がある。ヒトポリクローナル抗体はまた、免疫源で免疫したヒト由来の血清の形態で提供され得る。所望により、かかるポリクローナル抗体はIsoTポリペプチドまたはそのフラグメントを使用した親和性精製によって濃縮することができる。
【0045】
V.IsoTモジュレーターのための試験化合物スクリーニング
HIV感染を阻害する新規化合物を同定するために、本明細書に記載のとおり、試験化合物を第1にIsoTの生物学的活性を調節する能力についてスクリーニングする。好ましい態様において、試験化合物はIsoTのイソペプチダーゼ活性を調節(例えば阻害)する能力について試験する。IsoTの酵素活性を調節(例えば阻害)する化合物を多くのアッセイ形式で同定することができる。例えば、下記実施例に示すとおり、スクリーニングは精製IsoT酵素およびユビキチン−7−アミド−4−メチルクマリン(methlcoumarin)(Ub−AMC)を基質として使用して生化学アッセイを行うことができる。試験化合物のIsoTのイソペプチダーゼ活性に対する効果を当業者に既知の、例えばLacombe et al., FEBS Lett. 20:531:469-74, 2002;および Gabriel et al., Biochemistry. 19:41:13755-66, 2002のIsoT活性アッセイを使用してモニターすることができる。例えば、IsoTの酵素活性をユビキチンダイマー基質でアッセイすることができる。IsoTによる基質の切断は、Lacombe et al., FEBS Lett. 20:531:469-74, 2002に記載のとおり、電気泳動およびクーマシー染色によって分析することができる。分枝および直線ユビキチンダイマー基質を文献、例えばGabriel et al., Biochemistry 41:13755 -13766, 2002に記載のとおり製造することができる。ユビキチンダイマーの他に、他の基質もIsoT酵素活性のモジュレーターをスクリーニングするために使用することができる。例えば多くのアミノメチルクマリン(AMC)ベースのペプチド基質を、IsoT活性をモニターするために使用することができる(Stein et al., Biochemistry 34:12616-12623, 1995)。これらの基質(例えばZ-Leu-Arg-Gly-Gly-AMC または ユビキチン−アルデヒド)は、IsoTイソペプチダーゼの反応速度論およびIsoT活性のユビキチンアルデヒドによる阻害を研究するために文献において使用されている(Stein et al., Biochemistry 34:12616-12623, 1995;および Melandri et al., Biochemistry 35: 12893 -12900, 1996)。
【0046】
他の態様において、試験化合物を第1にIsoTのVprまたはユビキチンのような他の分子との結合を調節する活性についてスクリーニングすることができる。Vprはクローン化され、下記実施例および文献、例えばAmini et al., J Biol Chem. 279:46046-56, 2004; Iijima et al., Virol. 327:249-61, 2004; Yao et al., Retrovirology. 1:21-31, 2004; および Muthumani et al., Int Immunol. 17:103-16, 2005に既知の方法を使用して発現される。多くのアッセイおよび方法をIsoTポリペプチドのVprまたはユビキチンとの結合に対する試験化合物の効果を試験するために使用することができる。これらにはタンパク質と他の化合物(他のタンパク質を含む)の相互作用を研究するために当業者に一般的に知られている方法、例えば標識インビトロタンパク質−タンパク質結合アッセイ、免疫沈降法、GSTプルダウンアッセイ、酵母または哺乳類2種ハイブリッドスクリーニングおよび蛍光消光または蛍光偏光が含まれる。さらに、IsoTの他の分子(例えばユビキチン)との結合を試験するための具体的な方法は、下記実施例および文献(Amini et al., J. Biol. Chem., 279:46046-46056, 2004;および Yao et al., Retrovirology. 1:21-31, 2004参照)に提供される。
【0047】
ある態様において、IsoTとVpr結合の物理的相互作用を下記実施例に例示のような共免疫沈降法によって分析することができる。標識または分子タグIsoT(例えばFLAG−IsoT)およびVpr(例えばMyc−Vpr)を宿主細胞、例えば293T細胞において共発現させることができる。IsoTのVprとの結合を細胞溶解物を抗FLAG抗体で免疫沈降し、その後ウェスタンブロット分析することによって試験することができる。同様に、IsoTのユビキチンとの結合も当業者に既知の方法で分析することができる。例えば、IsoT−ユビキチン結合をタンデムKClおよびウレア直線グラジエントを用いるユビキチン−セファロースカラムを使用して試験することができる。この方法はLacombe et al. (FEBS Lett. 20:531:469-74, 2002) およびGabriel et al. (Biochemistry 41:13755 -13766, 2002)に記載されている。
【0048】
他の方法において、試験化合物をIsoTの発現または細胞レベル(転写、翻訳または翻訳後修飾)を調節する活性についてアッセイする。周知の様々な生化学および分子生物学技術をIsoT遺伝子の発現またはIsoTポリペプチドの細胞レベルを試験するために使用することができる。かかる技術は例えば、Sambrook et al., supra;および Brent et al., supraに記載されている。ある態様において、IsoT分子の内因性レベルを細胞が通常発現しているIsoTにおいて直接モニターすることができる。ある態様において、IsoT分子の発言または細胞レベルをクローンcDNAまたはIsoTをコード化するゲノム配列を使用して発現系において試験することができる。
【0049】
あるいは、IsoT遺伝子の発現の調節を細胞ベースの系において、発現ベクターの培養細胞株への短期または安定トランスフェクションによって試験することができる。レポーター遺伝子と作動可能なように結合しているIsoT遺伝子の転写制御配列(例えばプロモーター)を有するアッセイベクターを、プロモーター活性のアッセイのためにあらゆる哺乳類宿主細胞株にトランスフェクトすることができる。レポーター遺伝子と作動可能なように結合しているIsoT遺伝子(またはIsoT遺伝子の転写制御因子)を含む構成物を、分子生物学において通常実施されている技術および方法のみを使用して製造することができる(例えばSambrook et al. および Brent et al., supra参照)。細胞培養、トランスフェクションおよびレポーター遺伝子アッセイの一般的な方法は、文献、例えばBrent et al., supra; および Transfection Guide, Promega Corporation, Madison, WI (1998)に記載されている。あらゆる容易にトランスフェクト可能な哺乳類細胞株をIsoTプロモーター機能をアッセイするために、またはIsoTを発現するために使用することができる。例えばCHO、COS、HCT116、HEK293、MCF−7、およびHepG2は全て好適な細胞株である。
【0050】
適当な宿主細胞に挿入したとき、発現ベクターにおける転写制御因子は宿主RNAポリメラーゼによるレポーター遺伝子の転写を誘導する。レポーター遺伝子は典型的には、宿主細胞には本来存在しない容易にアッセイされる酵素活性を有するポリペプチドをコード化する。真核細胞プロモーターの典型的なレポーターポリペプチドには例えば、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、ホタルもしくはRenillaルシフェラーゼ、β−ガラクトシダーゼ、β−グルクロニダーゼ、アルカリホスファターゼおよび緑蛍光タンパク質(GFP)が含まれる。
【0051】
一般的に、スクリーニングで同定されたIsoT−調節化合物はIsoTの生物学的活性(例えばそのイソペプチダーゼ活性またはVprとの結合)を顕著に改変することができる。調節の程度を同じ化合物の対照タンパク質(例えばルシフェラーゼまたはタンパク質キナーゼ)に対する活性と、あるいは対照化合物の同じIsoT分子に対する活性と比較して測定する。例えば、IsoT発現またはIsoTイソペプチダーゼ活性を阻害するIsoT調節化合物は典型的には、対照酵素についてのIC50よりも少なくとも2、5、10、25、50、100、500または1000倍低いIsoT分子(例えばヒトIsoT)についてのIC50(最大阻害の50%を引き起こす有効濃度)を有する。IsoT分子の同じ活性を阻害する陰性対照化合物のIC50よりもIsoTのIC50は少なくとも5、10、25、50、100、500または1000倍低いと定義することもできる。同様に、調節をIsoTのVprとの結合で測定するならば、同じ程度の阻害を達成するための対照化合物の濃度よりも5、10、25、50または100倍低い濃度でのIsoT調節化合物が存在することで、通常IsoT−Vpr結合活性を予防または阻害することができる。本明細書において使用するとき、IsoTの他の分子への結合の阻害(例えばVprまたはユビキチン多量体)はIsoTのその分子への結合親和性を少なくとも2倍、好ましくは少なくとも5倍、より好ましくは少なくとも10、20、50または100倍減少させることを意味する。
【0052】
VI.HIV阻害化合物のスクリーニング
HIVの新規阻害剤を同定するために、上記IsoTモジュレーターを典型的にはさらに、それらのHIV感染に対する阻害作用を確認するために試験する。典型的には、化合物をHIV感染またはHIV複製の指標である活性を調節する能力についてスクリーニングする。多くのアッセイおよび方法をIsoT調節化合物のHIV阻害活性を試験するために使用することができる。ある方法では、IsoT調節化合物のHIV感染に対する潜在的な阻害活性を、当業者が通常実施している方法を使用して、インビトロでの培養細胞のHIV感染に対するそれらの効果を試験することによって試験することができる。例えば下記実施例に記載のように、化合物の抗ウイルス活性をHIVウイルスに感染する前に化合物と接触させたHeLaCD4βgal細胞においてウイルス複製をモニターすることによって評価することができる。化合物を、Seddiki et al., AIDS Res Hum Retroviruses. 15:381-90, 1999に記載のように第1マクロファージ培養物のHIV感染について試験することもできる。それらを、Fujii et al., J Vet Med Sci. 66:115-21, 2004に記載のように他のT細胞および単球細胞株のHIV感染について試験することもできる。HIV感染をモニターするためのさらなるインビトロ系は文献に記載されている。例えばLi et al., Pediatr Res. 54:282-8, 2003; Steinberg et al., Virol. 193:524-7, 1993; Hansen et al., Antiviral Res. 16:233-42, 1991;および Piedimonte et al., AIDS Res Hum Retroviruses. 6:251-60, 1990参照。
【0053】
これらのアッセイにおいて、細胞のHIV感染を形態学的に、例えば顕微鏡細胞変異効果アッセイ(例えばFujii et al., J Vet Med Sci. 66:115-21, 2004参照)によってモニターすることができる。酵素的に、例えば細胞培養物の上清におけるHIV逆転写(RT)活性をアッセイすることによって評価することもできる。かかるアッセイは文献、例えばReynolds et al., Proc Natl Acad Sci U S A. 100:1615-20, 2003;および Li et al., Pediatr Res. 54:282-8, 2003に記載されている。他のアッセイではHIV感染をウイルス核酸またはウイルス抗原の定量的集積によってモニターする。例えば、Winters et al. (PCR Methods Appl. 1:257-62, 1992)はHIV感染細胞培養物からHIV gag RNAおよびDNAをアッセイする方法を記載した。Vanitharani et al.はウイルスp24抗原の生産を測定するHIV感染アッセイを記載した(Virology 289:334-42, 2001)。ウイルス複製はまた、例えばChargelegue et al., J Virol Methods. 38(3):323-32, 1992;およびKlein et al., J Virol Methods. 107(2):169-75, 2003に記載のとおり、p24抗原ELISAアッセイによってインビトロでモニターすることができる。全てのこれらのアッセイを、本発明のIsoT調節化合物の抗HIV活性を評価するために使用し、そして改変することができる。
【0054】
ある方法において、IsoT調節化合物のHIV感染に対する潜在的な阻害効果をHIV複製が許容状態である人工レポーター細胞において試験することができる。これらの細胞において、HIV転写制御因子、例えばHIV−LTRの制御下でのレポーター遺伝子の発現を試験することによって、HIV感染および複製をモニターする。かかる細胞の一つの例がHeLa−T4−βGal HIVレポーター細胞である。下記実施例に説明のとおり、IsoT調節化合物で処理した後HeLa−T4−βGalレポーター細胞をHIV−IIIbで感染させるることができる。β−ガラクトシダーゼ活性を測定することによってモニターすると、化合物処理細胞のウイルス感染を化合物で処理しなかった対照細胞のものと比較することができる。IsoT調節化合物で処理した細胞のウイルス力価が減少しており、あるいは感染が減少していることで、当該化合物が実際にHIV感染またはウイルス複製を阻害することができると確認する。
【0055】
本明細書で試験したHela−T4−βGal細胞に加えて、多くの類似のレポーターアッセイも文献に記載されている。例えばGervaix et al. (Proc Natl Acad Sci USA. 94:4653-8, 1997)はヒト化緑蛍光タンパク質(GFP)をコード化するプラスミドをHIV−I LTRプロモーターの制御下で発現する安定なT細胞株を開発した。HIV−Iの感染によって、非感染細胞と比較して100倍〜1000倍の感染細胞の蛍光の増加が観察される。これらのアッセイ系のいずれかを本発明においてIsoT調節化合物のHIV感染に対する効果をリアルタイムで観察するために使用することができる。これらのインビトロ系は感染細胞の時間にわたる定量と、化合物の感受性の測定を行うことができる。
【0056】
他の方法において、IsoT調節化合物のHIV複製に対する効果を当該化合物で処理した細胞におけるHIV−1偽ウイルスの生産を試験することによって試験することができる。細胞はIsoTを内因的にまたは外因的に発現し得る。IsoT−コード化構造を宿主細胞に、とりわけIsoTを内因的に発現していない宿主細胞にトランスフェクトすることができる。下記実施例に記載のとおり、HIV−1偽ウイルスの生産を、レポーター遺伝子(例えばルシフェラーゼ遺伝子)をコード化するプサイ−陽性RNA、全構造タンパク質をコード化するデルタプサイパッケージング構造、ならびに制御または修飾タンパク質、例えばTat、Rev、VprおよびVif、ならびにVSV−gエンベロープ発現プラスミドで生産細胞(例えば293T HEK細胞)をトランスフェクトすることによって得ることができる。生産細胞で生産される偽ウイルスはレポーター遺伝子のみをコード化する。標的細胞を生産細胞由来の上清中の偽ウイルスでトランスフェクトした後、レポーター遺伝子を下記逆転写および標的細胞ゲノムへの集積によって発現する。
【0057】
HIV複製の阻害剤をスクリーニングするために、生産宿主細胞を偽ウイルスプラスミドのトランスフェクションの前、同時、または後にIsoT調節化合物で処理することができる。好ましくは、化合物を偽ウイルスプラスミドのトランスフェクション前に宿主細胞に投与し、アッセイプロセス全体で存在させる。生産された偽ウイルスの力価を生産細胞由来の上清における偽ウイルスで標的細胞を感染させること、そして標的細胞におけるレポーターの活性(例えばルシフェラーゼ活性)を評価することによってモニターすることができる。対照として化合物で処理しなかった生産細胞由来の上清で感染させた標的細胞におけるレポーター活性も測定する。IsoT調節化合物がウイルス出芽に対する阻害効果を有するならば、化合物で処理した生産細胞由来の上清と接触させた標的細胞は対照細胞と比較して減少したレポーター活性を有する。
【0058】
VII.治療適用
(i)IsoTイソペプチダーゼ活性;(ii)IsoTのVprとの結合、または(iii)IsoTコード化遺伝子の発現を下方制御することによって、上記IsoT調節化合物は本発明の有用な治療適用を提供する。それらはHIVに感染した細胞におけるHIV複製を阻害するために容易に使用することができる。それらはまた、HIV感染ならびにHIV感染に関連する疾患または状態(例えばAIDS)を有する対象を予防的または治療的に処置するために有用である。本発明のIsoT調節化合物での処置に適しているHIV感染には、あらゆるレトロウイルスのHIVファミリー(例えばHIV−1またはHIV−2)による対象、とりわけヒト対象の感染を含む。IsoT調節化合物は、レトロウイルスのHIVファミリーのあらゆるメンバーのキャリアーである対象の処置に有用である。それらは活性AIDSで診断される対象の処置に有用であり得る。化合物はまた、かかる対象におけるAIDS関連状態の処置または予防に有用である。HIV感染を有するとは診断されていないがHIVによる感染の危険があると考えられる対象も、本発明のIsoT調節化合物での処置に適している。
【0059】
本発明の治療適用に使用することができるIsoT阻害化合物には、下記実施例に記載のような本発明の方法によって同定され得る低分子有機化合物が含まれる。一度同定されると、かかる化合物を有機化学の通常使用される方法を用いて容易に合成することができ、そして/または商業的供給者から容易に入手することができる。本発明の治療法を実施するために好適なIsoTモジュレーターには、IsoT細胞レベルまたはその生物学的活性(例えばその酵素活性)を特異的に調節することができる他の化合物も含む。かかる化合物には、IsoTポリペプチドを特異的に認識するアンタゴニストまたはアゴニスト抗体が含まれる。それらにはまた、低分子干渉RNA(siRNA)、マイクロRNA(miRNA)および合成ヘアピンRNA(shRNA)、アンチセンス核酸、ならびに相補DNA(cDNA)が含まれる。例えば、下記実施例に記載のIsoT特異的またはVpr特異的siRNAを使用することができる。さらに、治療的適用は、上記スクリーニング方法によって同定することができる他のIsoT阻害剤を使用することができる。
【0060】
いずれかのAIDS関連状態を有する対象は本発明のIsoT調節化合物で処置するのに適している。かかる状態にはAIDS関連複合(ARC)、進行性全身性リンパ節腫(PGL)、抗HIV抗体陽性状態およびHIV陽性状態、AIDS関連神経学的状態(例えば認知症または熱帯性麻痺)、カポジ肉腫、血小板減少症紫斑病および関連する日和見感染、例えばニューモシスティス・カリニ肺炎、マイコバクテリア結核、食道カンジダ症、脳トキソプラズマ症、CMV網膜炎、HIV関連脳障害、HIV関連消耗症候群等が含まれる。
【0061】
インビボでHIV感染およびAIDSの進行を測定するための標準的な方法を、対象が本発明のIsoT調節化合物での処置に陽性に応答しているかどうかを決定するために使用することができる。例えば、本発明のIsoT調節化合物での処置後、対象のCD4+ T細胞数、あるいは対象のウイルス量をモニターすることができる。CD4+ T細胞の増加は対象が抗ウイルス治療の投与によって恩恵を受けていることを示す。これ、そして当業者に既知の他の方法を、本発明の化合物が対象におけるHIV感染およびAIDSの処置に有効である程度を測定するために使用することができる。
【0062】
本発明のIsoTモジュレーターを処置する対象に滅菌条件下で直接投与することができる。モジュレーターは単独で、または医薬組成物の有効成分として投与され得る。本発明の治療組成物はまた、他の治療剤と組み合わせて、あるいは関連して使用することができる。ある適用では、第1のIsoTモジュレーターを第2のIsoTモジュレーターと組み合わせて、1種のIsoTモジュレーターを個別に使用したときには達成できないようなより高度にHIV感染を阻害するために、使用される。他の適用において、本発明のIsoT調節化合物をAZTのような既知の抗HIV薬剤と組み合わせて使用することができる。
【0063】
本発明の医薬組成物は典型的には、少なくとも1種の有効成分と1種以上の許容される担体を含む。薬学的に許容される担体は組成物を改良または安定化し、あるいは組成物の製造を容易にする。薬学的に許容される担体は投与される具体的な組成物(例えば核酸、タンパク質または調節化合物)によって、そして組成物を投与するために使用する具体的な方法によって、ある程度決定される。それらはまた、他の成分と適合し、対象に有害でないという意味において、薬学的および生理学的に許容されるべきである。この担体は投与が望まれる製剤の形態、例えば経口、舌下、直腸、鼻、静脈内または非経腸に依存して広い範囲の形態を取り得る。例えば安定性または薬理学的特性を向上するためにIsoT調節化合物を投与の前に担体タンパク質、例えばオブアルブミンまたは血清アルブミンと組み合わせることができる。
【0064】
医薬組成物を様々な形態、例えば顆粒、錠剤、ピル、座薬、カプセル剤などに製剤することができる。製剤における治療活性化合物の濃度は、約0.1〜100重量%まで変化し得る。治療製剤を薬学の分野で周知の方法によって製造する。治療製剤を処置に使用することができるあらゆる有効な手段によって送達することができる。例えばGoodman & Gilman’s The Pharmacological Bases of Therapeutics, Hardman et al., eds., McGraw-Hill Professional (10th ed., 2001); Remington: The Science and Practice of Pharmacy, Gennaro (ed.), Lippincott Williams & Wilkins (20th ed., 2003);およびPharmaceutical Dosage Forms and Drug Delivery Systems, Ansel et al. (eds.), Lippincott Williams & Wilkins (7th ed., 1999)参照。
【0065】
治療製剤は便宜には単位投与形態で存在し、適当な治療用量で投与することができる。適当な治療用量はあらゆる周知の方法、例えば最大耐用用量を決定するための哺乳類での、そして安全用量を決定するための正常ヒト対象での臨床試験によって決定することができる。高用量が必要とされ得る特的の状況を除き、IsoTモジュレーターの好ましい用量は通常、1日当たり約0.001〜約1000mg、より通常は約0.01〜約500mgの範囲である。
【0066】
IsoTモジュレーターの好ましい投与量および投与形態は、処置している医師によって個別に見出される要素、例えば状態または処置される状態、具体的なIsoTモジュレーターを含む投与する組成物の選択、個々の対象の年齢、体重および応答、対象の状態の重症度、ならびに選択した投与経路に依存して、異なる対象について変化し得る。一般的な法則として、投与されるIsoTモジュレーターの量は対象の状態を効果的かつ確実に予防または縮小させる最小用量である。したがって、上記用量範囲は一般的なガイドラインを提供し、本明細書の記載を補助することを意図しており、本発明の範囲を限定することを意図していない。
【実施例】
【0067】
下記実施例は説明のために提示しており、本発明を限定しない。
【0068】
実施例1 VprはIsoTのC末端領域と相互作用する
Vpr−IsoT相互作用をヒト白血球ライブラリーを使用して酵母2ハイブリッドスクリーニングによって同定した。酵母におけるタンパク質−タンパク質のアッセイをGAL4とLexA融合タンパク質で行った。GAL4発現ベクター(Clontech)にクローン化したヒト白血球cDNAライブラリーを、HIV HXB2 Vprをおとりとして使用してスクリーニングした。Vpr cDNAをVpr特異的プライマーを使用してPCRによって増幅した。cDNAをLexA DBD発現ベクターpSLANSに挿入した。pSLANSはpBTM116(Bartel et al Biotechniques 14: 920-924, 1993)の改変版である。NotI挿入を受け入れ、gly4−ser−gly4−serをLexAとおとりの間に設置するため、改変した。LexA DBD−Vpr融合タンパク質をコード化するcDNAをL40 MATa酵母株に形質転換した。Gal4 AD−白血球cDNA融合タンパク質をコード化するcDNAを酵母株540 MATαに形質転換した。2種の酵母株を交配し、両プラスミドを含む形質転換体をTHUKL欠乏合成培地で選択し、タンパク質相互作用をβ−ガラクトシダーゼフィルターアッセイによって分析した。
【0069】
HIV−1 HXB2 Vprを、ヒト白血球cDNAライブラリーにおいてコード化される新規な相互作用する宿主細胞因子を同定するための酵母2ハイブリッドスクリーニングにおいておとりとして使用した。約200万の2倍形質転換体をスクリーニングし、多くの陽性候補を単離した。陽性コロニーをβ−ガラクトシダーゼ活性について、タンパク質−タンパク質相互作用の更なる確認のためのフィルターリフトアッセイを使用してアッセイした。独立の候補cDNAクローンの配列決定およびGenBankデータベースとの比較によって、IsoTである相互作用パートナー、26Sプロテアソームの成分を同定した。IsoTは遊離ポリユビキチン鎖中に配列を有するユビキチンの再循環に主に関与している。プレイ長(PL)IsoTの配列分析によってC末端227アミノ酸およびさらに635bpの3’−UTRを含むクローンを確認した(図1A)。興味深いことに、PLクローンはUBA(2)ドメインを通じてVprと相互作用することが既に同定されているhHR23A DNAダメージ修復酵素と類似の2つのUBAドメインを有する。β−ガラクトシダーゼ活性によってIsoTとVprの酵母中での相互作用を確認したが、Vprと陰性対照ベクターVP16はβ−ガラクトシダーゼ活性を示さなかった(図1B)。HXB2に由来するVprタンパク質をシングルTヌクレオチドの挿入によってフレームシフトして、74〜78位で5個のフレームシフトしたアミノ酸に導く。したがって、VprのC末端領域(アミノ酸位置74〜96)はIsoTとの結合を媒介するのに必要でない。これらの残基(84〜94)はウイルス粒子取り込みに重要であると思われる(Paxton et al., 1993)。
【0070】
本発明者らはまた、動物細胞におけるVpr−IsoT相互作用を分析するために共免疫沈降実験を行った。IsoT哺乳類発現ベクターをFLAGエピトープを全長IsoT遺伝子の5’末端と融合することによって構成した。VIP3ベクターを修飾してMycタグ(Myc:Vpr融合タンパク質)またはFLAGタグ(FLAG:IsoT融合タンパク質)のいずれかを発現させた。IsoTの全長クローンを、IsoT特異的プライマーを使用した膵臓由来のcDNAライブラリーのPCR増幅によって得て、IsoTであることを配列分析によって確認した後、FLAGタグ化VIP3ベクターにサブクローニングした。約3×106個の293T細胞(10cmプレート)を20μgのFLAG−IsoTおよび/またはMyc−Vprと、CaP(Clontech)を用いてトランスフェクトした。トランスフェクション後24時間で、細胞を氷上で、緩い低浸透圧バッファー(0.5%のトリトン−X、20mMのTris−HCl、50mMのNaCl、10%のグリセロールおよびプロテアーゼ阻害剤カクテル) [Complete, Roche Molecular Biochemicals]中で30分間溶解させた。細胞溶解物を16,000×gで5分間(チェック)遠心分離することによって澄まし、上清を抗FLAG抗体M2(Sigma−チェック)で免疫沈降(2時間、4℃)し、その後タンパク質A結合ゲルビーズとインキュベートした。結合タンパク質をSDSサンプルバッファー中で沸騰させて溶出し、アリコートをSDS−PAGEおよびヤギ抗Myc抗体でウェスタンブロッティングによって分析した。
【0071】
結果は、エピトープ−タグ化IsoTとVprのHEK293T細胞における過剰発現によって、VprとIsoTのヒト細胞における相互作用を示した(データは示さず)。
【0072】
実施例2 IsoT cDNA過剰発現はHIV生産を増加させる
HIVライフサイクルにおけるIsoTの役割を明らかにするため、IsoTを偽ウイルスプラスミドで平行してトランスフェクトし、標的293T細胞で形質導入し、ルシフェラーゼ受容体活性についてアッセイしてウイルス上清を分析した。293T HEK細胞の偽ウイルス生産のための一時的トランスフェクションを、6ウェルマイクロタイタープレート(3×105細胞/ウェル)中で、カルシウムホスファターゼ法(Clontech)を使用して行った。ルシフェラーゼベクタープラスミド(ゲノムRNAをコード化する)、デルタプサイパッケージング構造物(全構造タンパク質ならびに制御および修飾タンパク質Tat、Rev、VprおよびVifをコード化する)およびVSV−gエンベロープ発現プラスミドを2.0:1.5:0.5(それぞれ2、1.5、0.5μgのDNA/ウェル)の比でトランスフェクトした。ルシフェラーゼ受容体活性を、ウイルス上清の等量アリコートで293T標的細胞を、96ウェルまたは384ウェルフォーマットで形質導入し、Bright−Gloルシフェラーゼ基質および適当な蛍光フィルターを有するAcquestプレートリーダーを使用して、測定した。値を相対光単位(RLU)で表す。
【0073】
製造した偽ウイルスは標的細胞ゲノムへの逆転写および集積によって発現するルシフェラーゼ遺伝子のみをコード化する。第4プラスミドのコトランスフェクションにるウイルス出芽への効果の上昇または減少は、偽ウイルスプラスミド単独のときと比較して標的細胞のルシフェラーゼ活性の変化に反映される。過剰発現したIsoTの存在下でトランスフェクション後24時間で回収したウイルス上清はウイルス生産で2倍(50%)の増加を導いた(10,000rlu対20,000rlu)。トランスフェクション後48時間で、ウイルス生産はIsoTの存在下で3倍増加した(図2A)。偽ウイルス感染性における24および48時間後のウイルス生産の観察された増加に伴って、本発明者らはウェスタン分析によって外因性IsoT発現が平行して増加することを見出した。
【0074】
IsoTの役割をさらに評価するために、残基335の活性部位システインをアラニンで置換して触媒的に不活性の変異体を製造した。IsoT−C335A変異体を、PCRを利用した部位特異的突然変異誘発法によって製造した部位特異的ヌクレオチド置換体を使用して製造した。PCRを天然Taq DNAポリメラーゼで、製造者(Perkin−Elmer)が特定した条件下で行った。PCR産物をサブクローニング of the C335AフラグメントのIsoT cDNA:VIP3プラスミドへのサブクローニングに利用する適当な制限部位を有するように加工した。カスタムオリゴヌクレオチドプライマーをNNNから商業的に得た。PCRのテンプレートはIsoT:VIP3構造物であった。
【0075】
IsoT−C335A変異体の発現が、不活性のときHIV偽ウイルス生産を増加させるとは予期されなかった。これは図2Bに示す。全長野生型(FL−WT)IsoTの過剰発現はC335A変異体と比較して約2.5倍力価が増加した。プレイ長クローンは全長クローンで観察された表現型と置換することができず、正しいコンフォメーションにおいて機能的に活性なタンパク質がウイルス力価の増加を媒介するのに重要であることを示唆した。FL−WT IsoT発現のHIV力価の増加に対する用量依存効果を生産細胞へHIV−1ベクタープラスミドで平行して力価判定した。逆に、C335Aの発現は内因的に発現したIsoTのみで観察されたレベルと比較して、約50%HIV力価が減少したトランスドミナント陰性方法において作用すると思われる(図2C、0μg)。FLおよびC335A変異体タンパク質の発現レベルは、ウェスタン分析によって測定されたものと同等であった(データは示さず)。
【0076】
実施例3 ブロッキングVpr取り込みがウイルス生産レベルを減少させる
VprのHIVビリオン取り込み変異体を、デルタプサイパッケージングプラスミドにおけるVprのコドン30で、アラニンをリジンで置換することによって製造した。案の定、Vpr−A30Lビリオン取り込み変異体はWTウイルス生産と比較して、標的293T細胞におけるレポーターウイルスのルシフェラーゼ活性を測定すると、ウイルス生産において40%の減少を導いた(図3A)。対照的に、核局在化に欠陥がある変異体Vpr(W54R)はウイルス生産にほとんど効果がなかった。p24レベルもウイルス上清のアリコートから定量したが、これは図3Aの感染性の減少が生産細胞から放出されたウイルス粒子の数が40%減少したことと関連した(データは示さず)。製造したウイルスの感染性はVprに導入し、p24レベルに正規化した変異体には影響しなかった(図3B)。
【0077】
実施例4 IsoT減少によるHIV粒子生産の阻害
IsoTの過剰発現が上昇したウイルスベクター生産を導き、Vprのブロッキング取り込みがウイルスベクター生産の減少を導くという観察から、本発明者らは次に、IsoTおよびVprのHIV−1ベクター生産における減少効果を試験した。IsoTおよびVpr標的mRNAのsiRNA効果を確認するため、前日にIsoTおよびVpr RNAそれぞれを標的とする低分子干渉siRNAを導入した後、FLAGタグ化IsoTおよびMycタグ化Vprを293Tにおいて過剰発現させた。2つのsiRNAをIsoT、VprおよびTatに対して各々設計し、QiagenにHPPグレードを発注した。具体的には、IsoT特異的siRNAの配列は:IsoT 1: 5’-AGUUCACCUUCGGCUUAGAUU-3’ (SEQ ID NO:1);およびIsoT 2: 5’-GGCAGAUGGGUGAUCUACA-dTdT-3’ (SEQ ID NO:2)である。Vpr特異的siRNAの配列は: Vpr 1: 5’-GGAGUGGAAGCCAUAAUAA-UU-3’ (SEQ ID NO:3);およびVpr 2: 5’-ACAACUGCUGUUUAUCCAUUU-3’ (SEQ ID NO:4)である。Tat特異的siRNAの配列は:Tat 1: 5’-CUGCUUGUACCAAUUGCUA-dTdT-3’ (SEQ ID NO:5);およびTat 2: 5’-GCCUUAGGCAUCUCCUAUG-dTdT-3’ (SEQ ID NO:6)である。2つのsiRNAを1:1の比で組合せ、他に記載がない限りプールとして使用した。実験で使用した他のsiRNAはDharmacon Researchに発注した。
【0078】
試験した全てのsiRNAがIsoTまたはVprのレベルを効果的に減少させた(データは示さず)ため、選択したsiRNAは有効である。別注のIsoT抗体の生産に際して、内因性IsoTの減少を確認した。内因性レベルの高度な減少を、48時間と比較して72時間で観察した。アラマーブルー(ミトコンドリア活性)および細胞力価glo(ATP放出)細胞生存率キット試薬によって評価した293T細胞生存率に効果はなかった(データは示さず)。
【0079】
IsoT減少のウイルスベクター生産に対する影響を評価するため、293T細胞をsiRNAで処理し、翌日偽ウイルスプラスミドでトランスフェクションし、そしてウイルスを2日後に収穫し、293T標的細胞における感染性を測定した。RNA干渉によるIsoTおよびVprタンパク質の減少は、293T標的細胞において評価すると力価の減少を導いた(図4)。IsoT減少は60%の力価(感染性)の減少を導いた(図4A)。これは非飽和状態に依存し;ウイルス生産量は生産細胞へのデルタプサイ(構造遺伝子)のトランスフェクト量、および/またはウイルス収穫までの時間の長さ(48時間後、ウイルス生産量は典型的には飽和した)によって規制されるように線形の範囲であった。驚くべきことに、Vprレベルの減少はTatレベルの減少と比較して重度にウイルス生産を減少させた。デルタプサイパッケージングプラスミドから非飽和状態下で生産されるVprのレベルの減少は、明らかにこのアッセイにおいて感染性ウイルス粒子形成に重要である。使用したsiRNAはいずれも、293Tにおいて全く毒性を示さなかった(図4B)。
【0080】
VprおよびTatの減少が効果的にウイルス生産を減少させるが、IsoT減少は感染性ウイルス生産を60%のみ減少させるという観察は、大部分が、TatおよびVprのデノボ合成の前に低分子干渉RNAを添加したためであろう。これは外因性対内因性IsoTのIsoT減少で観察された結果と類似している。RNA干渉の前に合成されたIsoTタンパク質がVprによってなおも使用され、出芽および放出を促進していると思われる。IsoTを標的とする5つの独立した、個性的なsiRNAがタンパク質発現およびHIV偽ウイルス力価を効果的に減少させた(データは示さず)。
【0081】
実施例5 IsoTおよびVprレベルの減少はHIV−1 IIIb複製を減少させる
本発明者らは次に、HIV複製可能な細胞におけるIsoTおよびVpr減少の効果を試験しようとした。この目的のために、HeLa−T4−βGal HIV受容体細胞をHIV IIIbで感染させ、その後siRNA処理を行った。HeLa−CD4−βgal細胞(Kimpton et al., J Virol 66:2232-2239, 1992)はAIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIHを通じてDr. Michael Emermanから得た。結果は、感染の72時間後、IsoTまたはVpr減少によって、全く細胞毒性が観察されることなく、HIV複製がおよそ40%減少した(図5)。Tatレベルの減少によって約85%複製が減少した。宿主細胞因子の減少による細胞毒性の対照として、RNA干渉を使用してRNA Pol IIレベルを減少させた。減少したRNA Pol IIレベルの存在下での細胞生存率の顕著な減少がウイルス複製の観察された阻害を十分に説明した。これらのデータは、許容細胞におけるHIV−1複製を効果的に補助し、機構が細胞毒性に貢献しないというIsoTおよびVprの役割と一致する。
【0082】
本発明者らはまた、正常上皮細胞におけるIsoTレベルの減少の影響を評価した。この目的のため、IsoTレベルを低分子干渉RNAを使用してIOSE−80細胞において減少させた。結果は、IsoTレベルの減少が正常上皮細胞の生存能力には影響しないが、RNA Pol IIレベルの減少は生存率を65%減少させることを示した(データは示さず)。これらの結果は、IsoT機能の下方制御が正常の、非感染細胞の生存能力に影響しないことを示す。
【0083】
実施例6 本発明のスクリーニング法によるHIV阻害化合物の同定
この実施例は上記方法を使用したHIV阻害剤の同定について記載する。具体的には、低分子有機化合物のライブラリーをハイスループット形式でスクリーニングした。試験化合物を、IsoTの阻害能についてスクリーニングした。1次スクリーニングによって同定したヒットした化合物を、さらにHIV感染の阻害効果について試験した。簡潔に述べると、ユビキチン−7−アミド−4−メチルクマリン(Ub−AMC)を基質として使用したIsoT酵素活性の生化学アッセイを開発した。IMCのIsoTによる分裂および遊離が380nmでの励起および460nmでの放射蛍光シグナルを発生する。化合物の存在下での切断活性の阻害はスクリーニングヒットを構成する。内部での低分子化合物回収(1.7×106化合物)を、1536ウェルHTS形式で、内部で作成したIsoT酵素を使用してスクリーニングした。スクリーニングヒット率は0.4%であった。オリジナルヒット(1,761の化合物の87.3%が入手可能と再確認)を、第1に細胞毒性化合物の再確認、除去に供し、そして精製した化合物のアッセイを行った。同定された化合物はHIV複製の阻害活性について試験し、他の優先順位の特徴(例えば有効性、選択窓、化学的な取り扱いやすさ)を試験した。潜在的なHIV阻害剤として同定した6個の化合物が存在した。
【0084】
かくして同定したHIV阻害化合物の代表は、化合物Aである。この化合物のIsoTプロテアーゼ阻害についてのIC50は129nMである。化合物のHeLa−T4−βGal感染モデルにおけるHIV IIIbに対する抗ウイルス活性についてのEC50は6.14μMであり、細胞毒性は観察されなかった。脱ユビキチン化(DUB)酵素に対してアッセイを行ったとき、当該化合物はIsoT、ならびにミクロレベルのUCH−L1およびUSP7よりも低い程度のUCH−L3およびUSP2iso2の阻害が示された。阻害はイソペプチダーゼ活性に特異的であり、システインプロテアーゼのパネルには阻害が観察されなかった(データは示さず)。IsoTプロテアーゼ活性の直接阻害についての更なる支持を、テトラユビキチン切断アッセイ、次いで抗Ub抗体を使用したウェスタン分析を使用して得た。結果は、化合物を力価評価すると、テトラユビキチンの阻害に良好な用量依存性が存在することを示した。化合物Aのモル濃度が増加すると、製造された遊離(切断)ユビキチンレベルの減少が存在する。見かけのIC50は約100nmであり、これは生化学アッセイにおいてIsoT阻害について観察されたIC50と良好に相関する。これらのデータはさらに、化合物によるHIV複製の阻害がIsoT活性の阻害によることを支持する。ハイスループットスクリーニングデザインならびにスクリーニングに使用した物質および方法は、以下に詳述する。
【0085】
IsoTハイスループットスクリーニングデザイン:アッセイデザインはDang et al. (Biochemistry. 37:1868-79, 1998)の方法を基礎とし、さらに1536ウェル形式におけるウルトラHTSについて最適化した。簡潔に述べると、イソペプチダーゼ活性をユビキチンAMC基質からAMCの遊離を通じて検出し、蛍光シグナルを測定する。阻害剤化合物またはユビキチンアルデヒドの存在下では、プロテアーゼ活性を阻害する:完全反応バッファー(20mM HEPES、0.5mM EDTA、1mg/ml BSA、10mM DTT、pH7.8)中、5μL/ウェルの360pMのIsoTを黒色個体非TC1536ウェルプレート(Greiner Bio-one, Cat# 789176)に、カスタムBottleValve液体分配機によって分注し、50nL 1mMの化合物および対照(Ubアルデヒド)を本発明者らの社内取り付けPinToolを使用して移し(最終化合物濃度10μM)、そして相互作用を3時間、37℃で進行させる。1μL 300nMのUb−AMCを分注し、反応物を20分間、30℃でインキュベートする。蛍光強度をViewLuxプレートリーダー(PerkinElmer)を使用して測定する。化合物の用量依存性およびIC50測定についての滴定を、Minitrak装置(Packard BioScience)を使用して、DMSOでの半対数(half-log)連続希釈について行う。
【0086】
IsoT酵素の製造:IsoTをpRSETaへと、標準的なクローン化方法を使用してクローン化し、株BL21(DE3)へと形質転換する。タンパク質の発現を、タンパク質発現を1mMのIPTGで誘導したことを除き既出のとおり(Lesley et al., 2002)に行った。タンパク質の精製を以下のとおりに行った:4グラムの細胞ペレットを50mM Tris−HCl、50mM NaCl、10mM イミダゾール、pH7.9からなる溶解バッファー40ml中に再懸濁し、4×1分間超音波を加えた。15,000×gで20分間スピンさせた後、澄明な溶解物を、ニッケルを入れた1.5mlのベッド体積のChelating-sepharose FastFlow (Amersham)を含むカラムに注いだ。カラムを50mM Tris−HCl、300mM NaCl、40mM イミダゾールおよび10% グリセロール、pH7.9からなる洗浄バッファー7.5mlで洗浄した。タンパク質を20mM Tris−HCl、300mM イミダゾール、10% グリセロール、pH8.0からなる溶出バッファー4.5mlで溶出した。
【0087】
IsoT阻害剤のHIV IIIbに対する試験:IsoTスクリーニングからDMSOで希釈した化合物を、1mMから出発して、384ウェルポリプロピレン化合物プレート(Greiner)に並べ、次いでDMSOでの半対数連続希釈で、合計8個、Minitrak装置(Packard Bioscience)を使用して製造した。HeLaCD4βgal細胞(1500/50μL/ウェル)を384ウェル白色プレート(Greiner)に播種し、一晩密着の後、500nLの化合物を3つの複製細胞プレートに、Minitrak(Packard Bioscience)を使用して、感染性および細胞毒性試験のために移し、10μMから出発して最終化合物連続希釈を得た。30分後、HIV−IIIb(16ng/10μL/ウェル)を感染性試験細胞プレートに加えた。化合物およびウィルス添加の3日後、50μLのGal Screen(Applied Biosystems)を感染細胞に加え、1時間後にCLIPR (Molecular Devices)で蛍光を読み取って、感染を評価した。50μLの1:4希釈Cell Titer Glo (Promega)を感染細胞に加え、1時間後にCLIPRで読み取って、細胞毒性を評価した。用量応答測定を標準のシグモイド型用量応答式を使用して解釈した。社内の曲線適合コンピュータプログラムをIC50の計算に使用した。
【0088】
IsoT阻害剤によるテトラユビキチン(Ub4)切断の阻害:IsoTを反応バッファー(20mM HEPES、0.5mM EDTA、1mg/mL BSA、10mM DTT、pH=7.8)で最終濃度760pMとなるように希釈し、18μLを9個のエッペンドルフチューブそれぞれに加えた。化合物2096をDMSOで、200μMから出発して合計8個、半分対数連続希釈した。1μLの各希釈物をIsoTのチューブに加え、アッセイチューブ中10μMから出発する連続希釈を得た。最大切断についての対照として、1個のチューブに1μLのDMSOのみを加えた。37℃で30分間インキュベーションした後、1μLの反応バッファー中テトラユビキチン(Ub4;Biomol)の250μg/mL溶液を各チューブに加え、当該チューブを37℃で18時間さらにインキュベートした。変性タンパク質サンプルバッファー(Invitrogen)を加え、全サンプルを沸騰させ、4〜12%NuPage bis-Tris ゲル(Invitrogen)にロードし、製造者の指示通りにゲル電気泳動を行った。ニトロセルロースへと移した後、ブロットをPBST(0.05% Tween20を含むリン酸緩衝化食塩水)中5%無脂肪ミルクでブロックし、ユビキチンに対するウサギポリクローナル抗体(上記PBST中5%無脂肪ミルクで1:1000に希釈)で、次いでHRP結合ヤギ抗ウサギ2次抗体(PBST中5%無脂肪ミルクで1:3000、Southern Biotechnologies)でのイムノブロッティングに供した。バンドをECLプラス検出試薬(Amersham)を使用して可視化した。
【0089】
本明細書に記載の例および態様は例示のみを目的としており、その範囲内で様々な修飾または改変が当業者には提案され、そして本願および添付の特許請求の範囲の技術的範囲に含まれることが理解される。本明細書に記載のものと類似または同様のあらゆる方法および物質を本発明の実施または試験に使用することができるが、好ましい方法および物質は記載されている。
【0090】
本明細書に記載した全ての文献、GenBank配列、特許および特許公報は、各々そのように記載されているが如く、その全体について、あらゆる目的のために本明細書の一部とする。
【図面の簡単な説明】
【0091】
【図1】図1A−1BはVprのIsoTへの結合を示す:(A)概略図およびIsoTプレイクローンの位置を酵母−2−ハイブリッドを全長IsoT cDNAと比較して同定した。プレイクローンは2個のUBAドメインを有するC末端227アミノ酸を含む。本明細書で使用する全長IsoT cDNAクローンを膵臓由来のcDNAライブラリから増幅した。C335の相対位置(活性部位システイン)を示す。(B)VprとIsoTの特異的相互作用を示すために、β−ガラクトシダーゼフィルターリフトアッセイを使用して酵母−2−ハイブリッドをアッセイする。VP16は空ベクターである。
【0092】
【図2】図2A−2Cは野生型IsoT過剰発現がHIV−1ウイルスの力価を増加させるが、活性部位変異はそうではないことを示す:(A)全長(FL)野生型IsoTの過剰発現がトランスフェクション後24および48時間で偽ウイルス過剰発現を増加させる。293T細胞をIsoTをコード化する第4cDNAの存在下(+)または非存在下(−)において3個の偽ウイルスプラスミドとコトランスフェクトした。レポーターウイルス上清をトランスフェクション後24または48時間で回収し、293T標的細胞におけるレポーター活性についてアッセイした。細胞溶解物を平行してウェスタン分析で、IsoT特異的抗体を使用して内因性IsoT発現ならびにIsoT過剰発現について分析した。1列目はプラスミドでトランスフェクトしていない、トランスフェクション試薬のみのものである。(B)293T細胞を(A)のとおりに、第4プラスミドとしてVIP3(プラスミド骨格)、FL−野生型、PL−C末端227、またはFL−C335Aでコトランスフェクトした。トランスフェクション後30時間に回収したウイルス上清由来のルシフェラーゼレポーター活性を293T標的細胞においてアッセイした。(C)HIV力価に対する用量依存関数効果。FL−WTおよびFL−C335Aを力価判定し、平行して第4プラスミドとして偽プラスミドとトランスフェクトした。
【0093】
【図3】図3A−3BはVpr(A30L)ウイルスの減少した出芽が感染性ではなく減少した粒子生産と関連することを示す。293T細胞を偽ウイルスDNA発現WTおよびVprの変異型とトランスフェクトした。(A)WTまたは核局在化変異体W54Rではなく、A30L Vpr取り込み変異体は、293T標的細胞における感染性によって測定するとウイルス力価を減少した。(B)p24レベルを標的293T細胞を感染するために使用した偽ウイルスインプットについて、(A)から標準化(9.38ng)した。
【0094】
【図4】図4A−4BはRNA干渉によるIsoTまたはVprの欠乏が293T細胞における偽ウイルス生産を阻害することを示す。(A)293T細胞をHIV Tat、IsoT、VprまたはルシフェラーゼGL3(陰性対照)を標的とする示したsiRNAでトランスフェクトした。細胞を一晩培養し、ウェルあたり2:0.05:0.5または100ng:2.5ng:25ngのベクターRNA:デルタプサイ:envの比で、3プラスミド偽ウイルス系のcDNAトランスフェクションに供した。3つの複写ウェルを条件毎に行った。ウイルス上清を48時間後に回収し、標的293T細胞に感染させるために使用した。これらの形質導入細胞におけるルシフェラーゼ活性を48時間後に測定し、Aに示す。(B)293T細胞をHIV Tat、IsoT、VprまたはルシフェラーゼGL3に対するsiRNAとトランスフェクトした。72時間後細胞生存率をCellTiterGloを使用し、蛍光を測定して決定した。
【0095】
【図5】図5はIsoTまたはVprの欠乏がHeLa−CD4−β−gal細胞におけるHIV−1 IIIb複製を阻害することを示す。細胞をTat、IsoT、Vpr、ルシフェラーゼGL2(陰性対照)、またはPolR2A(細胞毒性についての陽性対照)を標的とする示したsiRNAとトランスフェクトした。24時間培養後、細胞をHIV−1 IIIbに感染させる。β−ガラクトシダーゼ活性を、感染の測定として感染後72時間で測定した。細胞毒性をsiRNAトランスフェクト非感染細胞について平行して測定した。データを陰性対照GL2 siRNAトランスフェクト細胞によって発生したシグナルの割合として表現する。
【特許請求の範囲】
【請求項1】
HIV感染を阻害する薬剤を同定する方法であって、イソペプチダーゼT(IsoT)分子またはそのフラグメントの生物学的活性を、試験化合物の存在下でアッセイして、IsoT分子の生物学的活性を阻害する化合物を同定し;それによってHIV感染を阻害する化合物を同定することを含んでなる方法。
【請求項2】
さらに、同定された化合物をHIV感染阻害能について試験することを含む、請求項1に記載の方法。
【請求項3】
生物学的活性が、IsoTのVprまたはユビキチンへの結合である請求項1に記載の方法。
【請求項4】
生物学的活性がIsoT分子のイソペプチダーゼ活性またはIsoT分子をコード化する遺伝子の発現である、請求項1に記載の方法。
【請求項5】
IsoT分子が哺乳類細胞由来である、請求項1に記載の方法。
【請求項6】
IsoT分子がヒトIsoTである、請求項5に記載の方法。
【請求項7】
化合物によるHIV−1感染の阻害能を、当該化合物で処理した人工HIV許容細胞におけるHIV複製と、当該化合物で処理しなかった対照細胞におけるHIV複製を比較することによって試験する、請求項2に記載の方法。
【請求項8】
HIV許容細胞がHeLa−T4−βGal HIV細胞である、請求項7に記載の方法。
【請求項9】
HIV複製をp24抗原ELISAアッセイまたは逆転写酵素活性アッセイによってモニターする、請求項7に記載の方法。
【請求項10】
化合物によるHIV感染の阻害能を、当該化合物で処理した宿主細胞における偽ウイルス生産と、当該化合物で処理しなかった対照宿主細胞における偽ウイルス生産を比較することによって試験する、請求項2に記載の方法。
【請求項11】
宿主細胞が293T HEK細胞である、請求項10に記載の方法。
【請求項12】
宿主細胞が、当該細胞においてHIV偽ウイルスを生産する偽ウイルスプラスミドでトランスフェクトされている、請求項10に記載の方法。
【請求項13】
HIV感染を阻害する薬剤を同定する方法であって、
(a)試験化合物をスクリーニングして、IsoTイソペプチダーゼ活性またはそのVprとの結合を下方制御するIsoT調節化合物を同定する工程、そして
(b)同定されたIsoT調節化合物をHIV感染阻害能について試験する工程
を含んでなる方法。
【請求項14】
IsoT分子が哺乳類細胞に由来する、請求項13に記載の方法。
【請求項15】
IsoT分子がヒトIsoTである、請求項14に記載の方法。
【請求項16】
化合物によるHIV−1感染の阻害能を、当該化合物で処理したHIV許容細胞におけるHIV複製と、当該化合物で処理しなかった対照細胞におけるHIV複製を比較することによって試験する、請求項13に記載の方法。
【請求項17】
HIV許容細胞がHeLa−T4−βGal HIV細胞である、請求項16に記載の方法。
【請求項18】
HIV複製をp24抗原ELISAアッセイまたは逆転写酵素活性アッセイによってモニターする、請求項16に記載の方法。
【請求項19】
化合物によるHIV感染の阻害能を、当該化合物で処理した宿主細胞における偽ウイルス生産と、当該化合物で処理しなかった対照宿主細胞における偽ウイルス生産を比較することによって試験する、請求項13に記載の方法。
【請求項20】
宿主細胞が293T HEK細胞である、請求項19に記載の方法。
【請求項21】
宿主細胞が、当該細胞においてHIV偽ウイルスを生産する偽ウイルスプラスミドでトランスフェクトされている、請求項19に記載の方法。
【請求項22】
HIV感染細胞におけるHIV複製を阻害する方法であって、(i)IsoTイソペプチダーゼ活性;(ii)VprとのIsoTの結合;または(iii)IsoTコード化遺伝子の発現;を下方制御する化合物を当該細胞に接触させ;それによって対象におけるHIV感染を処置することを含んでなる方法。
【請求項23】
HIV感染細胞がHIV感染を有する対象に存在し、そして当該対象に有効量の化合物を含む医薬組成物を投与することを含む、請求項22に記載の方法。
【請求項24】
IsoTがヒトIsoTである、請求項22に記載の方法。
【請求項25】
化合物がインビトロでIsoT発現細胞におけるHIV複製を阻害する、請求項22に記載の方法。
【請求項26】
化合物が低分子干渉RNA(siRNA)、マイクロRNA(miRNA)および合成ヘアピンRNA(shRNA)、アンチセンス核酸、ならびに相補DNA(cDNA)からなる群から選択される、請求項25に記載の方法。
【請求項27】
化合物がIsoTまたはVprに対するsiRNAである、請求項26に記載の方法。
【請求項28】
化合物を既知の抗HIV薬と共に対象に投与する、請求項22に記載の方法。
【請求項1】
HIV感染を阻害する薬剤を同定する方法であって、イソペプチダーゼT(IsoT)分子またはそのフラグメントの生物学的活性を、試験化合物の存在下でアッセイして、IsoT分子の生物学的活性を阻害する化合物を同定し;それによってHIV感染を阻害する化合物を同定することを含んでなる方法。
【請求項2】
さらに、同定された化合物をHIV感染阻害能について試験することを含む、請求項1に記載の方法。
【請求項3】
生物学的活性が、IsoTのVprまたはユビキチンへの結合である請求項1に記載の方法。
【請求項4】
生物学的活性がIsoT分子のイソペプチダーゼ活性またはIsoT分子をコード化する遺伝子の発現である、請求項1に記載の方法。
【請求項5】
IsoT分子が哺乳類細胞由来である、請求項1に記載の方法。
【請求項6】
IsoT分子がヒトIsoTである、請求項5に記載の方法。
【請求項7】
化合物によるHIV−1感染の阻害能を、当該化合物で処理した人工HIV許容細胞におけるHIV複製と、当該化合物で処理しなかった対照細胞におけるHIV複製を比較することによって試験する、請求項2に記載の方法。
【請求項8】
HIV許容細胞がHeLa−T4−βGal HIV細胞である、請求項7に記載の方法。
【請求項9】
HIV複製をp24抗原ELISAアッセイまたは逆転写酵素活性アッセイによってモニターする、請求項7に記載の方法。
【請求項10】
化合物によるHIV感染の阻害能を、当該化合物で処理した宿主細胞における偽ウイルス生産と、当該化合物で処理しなかった対照宿主細胞における偽ウイルス生産を比較することによって試験する、請求項2に記載の方法。
【請求項11】
宿主細胞が293T HEK細胞である、請求項10に記載の方法。
【請求項12】
宿主細胞が、当該細胞においてHIV偽ウイルスを生産する偽ウイルスプラスミドでトランスフェクトされている、請求項10に記載の方法。
【請求項13】
HIV感染を阻害する薬剤を同定する方法であって、
(a)試験化合物をスクリーニングして、IsoTイソペプチダーゼ活性またはそのVprとの結合を下方制御するIsoT調節化合物を同定する工程、そして
(b)同定されたIsoT調節化合物をHIV感染阻害能について試験する工程
を含んでなる方法。
【請求項14】
IsoT分子が哺乳類細胞に由来する、請求項13に記載の方法。
【請求項15】
IsoT分子がヒトIsoTである、請求項14に記載の方法。
【請求項16】
化合物によるHIV−1感染の阻害能を、当該化合物で処理したHIV許容細胞におけるHIV複製と、当該化合物で処理しなかった対照細胞におけるHIV複製を比較することによって試験する、請求項13に記載の方法。
【請求項17】
HIV許容細胞がHeLa−T4−βGal HIV細胞である、請求項16に記載の方法。
【請求項18】
HIV複製をp24抗原ELISAアッセイまたは逆転写酵素活性アッセイによってモニターする、請求項16に記載の方法。
【請求項19】
化合物によるHIV感染の阻害能を、当該化合物で処理した宿主細胞における偽ウイルス生産と、当該化合物で処理しなかった対照宿主細胞における偽ウイルス生産を比較することによって試験する、請求項13に記載の方法。
【請求項20】
宿主細胞が293T HEK細胞である、請求項19に記載の方法。
【請求項21】
宿主細胞が、当該細胞においてHIV偽ウイルスを生産する偽ウイルスプラスミドでトランスフェクトされている、請求項19に記載の方法。
【請求項22】
HIV感染細胞におけるHIV複製を阻害する方法であって、(i)IsoTイソペプチダーゼ活性;(ii)VprとのIsoTの結合;または(iii)IsoTコード化遺伝子の発現;を下方制御する化合物を当該細胞に接触させ;それによって対象におけるHIV感染を処置することを含んでなる方法。
【請求項23】
HIV感染細胞がHIV感染を有する対象に存在し、そして当該対象に有効量の化合物を含む医薬組成物を投与することを含む、請求項22に記載の方法。
【請求項24】
IsoTがヒトIsoTである、請求項22に記載の方法。
【請求項25】
化合物がインビトロでIsoT発現細胞におけるHIV複製を阻害する、請求項22に記載の方法。
【請求項26】
化合物が低分子干渉RNA(siRNA)、マイクロRNA(miRNA)および合成ヘアピンRNA(shRNA)、アンチセンス核酸、ならびに相補DNA(cDNA)からなる群から選択される、請求項25に記載の方法。
【請求項27】
化合物がIsoTまたはVprに対するsiRNAである、請求項26に記載の方法。
【請求項28】
化合物を既知の抗HIV薬と共に対象に投与する、請求項22に記載の方法。
【図1】

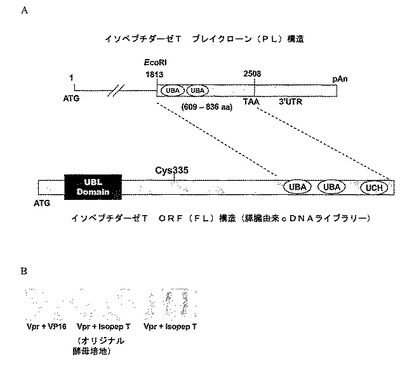
【図2】
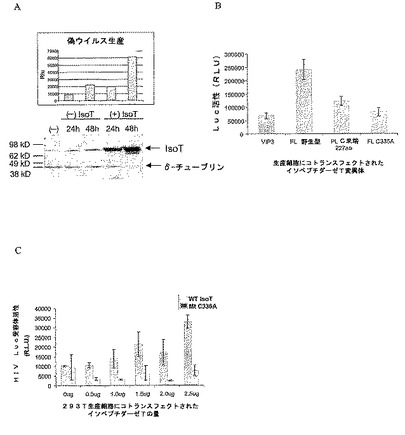
【図3】
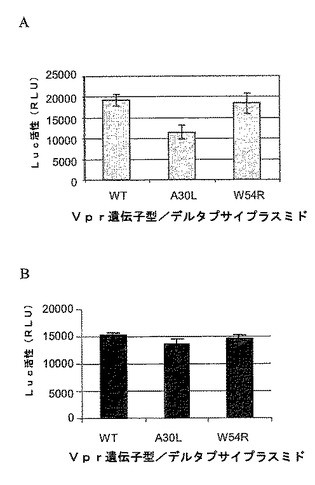
【図4】
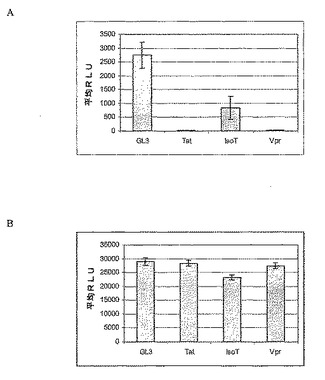
【図5】
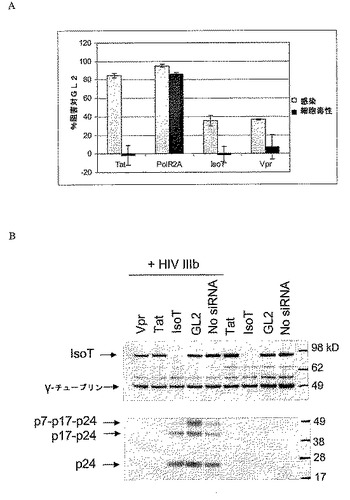
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2008−538504(P2008−538504A)
【公表日】平成20年10月30日(2008.10.30)
【国際特許分類】
【出願番号】特願2008−507915(P2008−507915)
【出願日】平成18年4月21日(2006.4.21)
【国際出願番号】PCT/US2006/015081
【国際公開番号】WO2006/116075
【国際公開日】平成18年11月2日(2006.11.2)
【出願人】(503261524)アイアールエム・リミテッド・ライアビリティ・カンパニー (158)
【氏名又は名称原語表記】IRM,LLC
【Fターム(参考)】
【公表日】平成20年10月30日(2008.10.30)
【国際特許分類】
【出願日】平成18年4月21日(2006.4.21)
【国際出願番号】PCT/US2006/015081
【国際公開番号】WO2006/116075
【国際公開日】平成18年11月2日(2006.11.2)
【出願人】(503261524)アイアールエム・リミテッド・ライアビリティ・カンパニー (158)
【氏名又は名称原語表記】IRM,LLC
【Fターム(参考)】
[ Back to top ]