
HIV治療のための抗CXCR4抗体
本発明は、CXCR4と結合することができるだけでなく、CXCR4ホモ二量体のコンフォメーション変化を誘導することができ、かつ、PBMCにおけるHIV−1一次単離物の複製を阻害することができる新規な単離された抗体、またはその誘導化合物もしくは機能的フラグメントに関する。より詳細には、本発明は、CXCR4タンパク質に特異的な515H7および301aE5モノクローナル抗体、ならびにHIV感染の処置のためのそれらの使用に関する。このような抗体から構成される医薬組成物およびこのような抗体の選択のための方法も包含される。
【発明の詳細な説明】
【発明の背景】
【0001】
発明の分野
本発明は、ケモカイン受容体(CXCR)と特異的に結合することができる新規な抗体、特に、ネズミモノクローナル抗体、キメラ抗体およびヒト化抗体、ならびにこのような抗体をコードするアミノ酸配列および核酸配列に関する。一つの態様から、本発明は、CXCR4と特異的に結合することができ、かつ、ヒト免疫不全ウイルス(HIV)感染に対して強い活性を有する新規な抗体、機能的フラグメントまたは誘導体に関する。本発明はまた、このような抗体、機能的フラグメントまたは誘導体の、HIV感染の予防的治療および/または治療的治療のための薬剤としての使用も含んでなる。
【0002】
背景技術
ケモカインは、特に免疫反応中に、ケモカイン勾配として知られるリガンドの化学的勾配によって白血球の移動を制御する小型の分泌型ペプチドである(Zlotnick A. et al., 2000)。ケモカインは、それらのNH2末端システイン残基の位置に基づいてCCおよびCXCという二つの主要なサブファミリーに分かれ、Gタンパク質共役型受容体と結合し、その受容体の二つの主要なサブファミリーはCCRおよびCXCRと呼ばれている。これまでに50を超えるヒトケモカインと18のケモカイン受容体が見出されている。
【0003】
ケモカイン受容体ファミリーのいくつかのメンバーは、主要受容体CD4とともに共受容体として働き、種々の1型HIV株を細胞へ侵入させるが、この主要な共受容体がCCR5およびCXCR4である。T細胞指向性(T-cell tropic)X4 HIV−1は細胞への侵入にCD4とCXCR4を用いるが、マクロファージ指向性(macrophage-tropic)R5 HIV−1はCD4とCCR5を用いる。二重指向性株は、CXCR4とCCR5の双方を共受容体として使用することができる。他のケモカイン受容体の中でも、CCR3、CCR2、CCR8、CXCR6、CXCR7、CX3CR1は、HIV株のより限られたサブセットにより、共受容体として機能する。
【0004】
CXCR4の天然リガンドであるSDF−1、ならびにCCR5の天然リガンドであるCCL3、CCL4、CCL4−L1およびCCL5リガンドは、種々のHIV−1株による細胞融合および感染を阻害することができる。これらの知見は、ケモカイン受容体を標的とする抗HIV治療薬の開発を促し、CCR5指向性HIV−1に感染した患者において、CCR5の小分子アンタゴニストであるマラビロク(maraviroc)(CELSENTRI(商標))と、他の抗HIV−1薬との併用の認可に至らせた。しかしながらやはり、二重指向性HIV−1に感染した患者にも、CXCR4指向性HIV−1に感染した患者にもマラビロクは使用されない(VIDAL 2009)。従って、X4指向性HIV複製を阻害することができるCXCR4アンタゴニストを同定することにより、X4指向性感染患者および二重指向性HIV感染患者の双方にこの種の療法を拡張するという明確な医学的必要がある。
【0005】
ケモカイン受容体4(フーシン、CD184、LESTR、またはHUMSTRとしても知られる)は、352または360個のアミノ酸を含んでなる二つのイソ型として存在する。残基Asn11はグリコシル化され、残基Tyr21は硫酸基の付加によって修飾され、Cys109および186は、受容体の細胞外部分でジスルフィド架橋によって結合されている(Juarez J.ら、2004)。
【0006】
この受容体は、様々な種の正常組織、ナイーブ、非メモリーT細胞、調節T細胞、B細胞、好中球、内皮細胞、一次単球、樹状細胞、ナチュラルキラー細胞、CD34+造血幹細胞、また、低レベルではあるが、心臓、結腸、肝臓、腎臓、および脳によって発現される。CXCR4は、白血球輸送、B細胞のリンパ球生成、および骨髄造血に重要な役割を果たす。
【0007】
これまでに記載されているCXCR4受容体のリガンドの独特なリガンドは、ストロマ細胞由来因子(SDF−1)またはCXCL12である。SDF−1は、リンパ節、骨髄、肝臓、肺において多量に、また、程度は少ないが、腎臓、脳、および皮膚によっても分泌される。CXCR4はまた、III型ヒトヘルペスウイルスによってコードされているウイルスマクロファージ炎症性タンパク質II(vMIP−II)であるアンタゴニストケモカインによっても認識される。
【0008】
従前に記載されているように、CXCR4受容体は、T細胞指向性HIV−1単離物(X4ウイルス)に対する主要な共受容体である。この受容体に干渉すれば、極めて効率的にX4ウイルスの複製を阻害できるはずである。
【発明の概要】
【0009】
本発明の発明態様の一つは、HIV複製を阻害するマウスモノクローナル抗体(Mab)を作製することである。本発明は、CXCR4ホモ二量体と結合することができ、かつ、HIV感染に対して強い活性を有するCXCR4 Mab 515H7(またはそのフラグメント)を包含する。本発明はまた、CXCR4ホモ二量体と結合することができ、かつ、HIV感染に対して強い活性を有するCXCR4 Mab 301aE5(またはそのフラグメント)を包含する。
【0010】
驚くべきことに、本発明者らは、CXCR4と結合することができるだけでなく、CXCR4ホモ二量体のコンフォメーション変化を誘導することができ、かつ、PBMCにおけるX4−HIV−1一次単離物の複製を阻害することができるモノクローナル抗体の作製を成し遂げた。より詳細には、本発明の抗体は、PBMCにおけるX4/R5−HIV−1一次単離物の複製を阻害することができる。
【図面の簡単な説明】
【0011】
【図1】図1Aおよび1Bは、単球およびリンパ球におけるCXCR4発現のゲーティング戦略を示す図である。図1A:CD3−PE抗体によるT細胞の染色、図1B:CD14−PE抗体による単球の染色
【図2】図2Aおよび2Bは、単球およびTリンパ球に対する抗CXCR4 Mab 515H7および301aE5の結合を示す図である。
【図3】図3Aおよび3Bは、HEK293細胞における生物発光共鳴エネルギー移動(BRET)アプローチによる、それぞれSDF−1ならびに抗CXCR4 Mab 515H7および301aE5によるCXCR4受容体二量体の調節を示す図である。
【図4】図4Aおよび4Bは、抗CXCR4 Mab 515H7および301aE5の、ヒトPBMCにおけるHIV−1単離物KON(X4ウイルス)の複製を阻害する能力を示す図である。
【図5】図5は、抗CXCR4 Mab 515H7および301aE5の、ヒトPBMCにおけるHIV−1単離物KON(X4ウイルス)の複製を阻害する能力を示す図である。
【図6】図6A、6Bおよび6Cは、CHO−CXCR4細胞におけるSDF−1により誘発されたカルシウム放出の、Mab 515H7(図6A)、301aE5(図6B)およびc515H7(図6C)による阻害を示す図である。
【図7】図7は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、ヒトPBMCにおけるHIV−1 X4ウイルス一次単離物MN(図7)および92UG024(図8)の複製を阻害する能力を示す図である。
【図8】図8は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、ヒトPBMCにおけるHIV−1 X4ウイルス一次単離物MN(図7)および92UG024(図8)の複製を阻害する能力を示す図である。
【図9】図9は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、ヒトPBMCにおけるHIV−1 X4ウイルス一次単離物KON、MN、および92UG024の複製を阻害する能力を示す図である。
【図10】図10は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、ヒトPBMCにおけるHIV−1 X4/R5二重ウイルス一次単離物89.6の複製を阻害する能力を示す図である。
【図11】図11は、抗CXCR4 Mab 515H7、c515H7および301aE5の、ヒトPBMCにおけるHIV−1 X4/R5二重ウイルス一次単離物89.6の複製を阻害する能力を示す図である。
【図12】図12は、Mab c515H7とマラビロクの併用の、ヒトPBMCにおけるHIV−1一次単離物89.6(X4/R5二重ウイルス)の複製を阻害する有益な効果を示す図である。
【図13】図13は、Mab c515H7とマラビロクの併用の、ヒトPBMCにおけるHIV−1一次単離物UG93067(X4/R5二重ウイルス)の複製を阻害する有益な効果を示す図である。
【図14】図14は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、PBMCにおけるHIV−1 X4ウイルス一次単離物KONの複製を阻害する能力を示す図である。
【図15】図15は、FACS分析によるc515H7 Mabの結合特異性を示す図である。
【図16】図16は、生物発光共鳴エネルギー移動(BRET)アプローチによる、CXCR4ホモ二量体に対するc515H7 Mabの効果を示す図である。
【図17】図17は、515H7重鎖可変ドメインとヒト生殖細胞系IGHV3−49*04およびIGHJ4*01とのアミノ酸配列アライメント。515H7 VHアミノ酸配列が、選択されたヒトアクセプターフレームワーク配列とアラインされている。VH Var1 (VH1)配列は、515H7 VHドメインのヒト化変異体に相当する。76番の位置の一つの逆突然変異が太字で示されている。
【図18】図18は、515H7軽鎖と、ヒト生殖細胞系IGKV4−l*01およびIGKJ1*01とのアミノ酸配列アライメント。515H7 VLアミノ酸配列が、選択されたヒトアクセプターフレームワーク配列とアラインされている。VL Var2.1、Var2.2、およびVar2.3配列は、ヒト化515H7 VL Var2の実施されたヒト化変異体に相当する(変異残基を太字で示す)。Var2.1およびVar2.2はさらに四つのヒト化残基を持ち、Var2.3はさらに五つのヒト残基を含む。
【図19】図19は、キメラ515H7およびヒト化515H7の種々の変異体による、ビオチン化ネズミ抗体515H7の交差遮断。515H7のヒト化変異体(hz515H7)の、親ネズミ抗体515H7を交差遮断する活性を、CXCR4トランスフェクトNIH3T3細胞を用いたフローサイトメトリーによって評価した。ヒト化変異体の活性をキメラ515H7と比較した。変異体VH1とキメラVL(cVL)併用の交差遮断活性は、キメラ(A)の場合と極めてよく似ていた。VLの変異体2と併用した場合(B)には、VH変異体1(VH1、逆突然変異の無い変異体)の活性の低下は見られなかった。
【図20】図20は、キメラ515H7およびヒト化515H7の種々の変異体による、ビオチン化SDF−1結合の阻害。515H7のヒト化変異体(hz515H7)の、SDF−1結合を阻害する能力を、細胞系統RAMOSを用いたフローサイトメトリーによって評価した。ヒト化変異体の阻害能をキメラ515H7と比較した。ヒト化変異体hz515H7 VH1 D76N VL2は、キメラ抗体と同等のSDF−1結合阻害能を有する。ヒト化抗体フラグメントhz515 VH1 VL2は、RAMOS細胞に対するSDF−1の結合阻害に完全な活性を示した。
【図21】図21は、NIH3T3−CXCR4においてCXCR4と特異的に結合するヒト化515H7 Mab(hz515H7 VH1 D76N VL2、hz515H7 VH1 D76N VL2.1、hz515H7 VH1 D76N VL2.2、およびhz515H7 VH1 D76N VL2.3)を示す図である。
【図22】図22は、生物発光共鳴エネルギー移動(BRET)アプローチによる、CXCR4ホモ二量体に対するヒト化515H7 Mab(hz515H7 VH1 D76N VL2、hz515H7 VH1 D76N VL2.1、hz515H7 VH1 D76N VL2.2およびhz515H7 VH1 D76N VL2.3)の効果を示す図である。
【発明の具体的説明】
【0012】
好ましくは、CXCR4化合物は、
Genbank受託番号NP_003458配列番号27:
MEGISIYTSDNYTEEMGSGDYDSMKEPCFREENANFNKIFLPTIYSIIFLTGIVGNGLVILVMGYQKKLRSMTDKYRLHLSVADLLFVITLPFWAVDAVANWYFGNFLCKAVHVIYTVNLYSSVLILAFISLDRYLAIVHATNSQRPRKLLAEKVVYVGVWIPALLLTIPDFIFANVSEADDRYICDRFYPNDLWVVVFQFQHIMVGLILPGIVILSCYCIIISKLSHSKGHQKRKALKTTVILILAFFACWLPYYIGISIDSFILLEIIKQGCEFENTVHKWISITEALAFFHCCLNPILYAFLGAKFKTSAQHALTSVSRGSSLKILSKGKRGGHSSVSTESESSSFHSS、
として表される配列を有するケモカイン(C−X−Cモチーフ)受容体4イソ型b[ホモ・サピエンス]、
Genbank受託番号NP_001008540配列番号28:
MSIPLPLLQIYTSDNYTEEMGSGDYDSMKEPCFREENANFNKIFLPTIYSIIFLTGIVGNGLVILVMGYQKKLRSMTDKYRLHLSVADLLFVITLPFWAVDAVANWYFGNFLCKAVHVIYTVNLYSSVLILAFISLDRYLAIVHATNSQRPRKLLAEKVVYVGVWIPALLLTIPDFIFANVSEADDRYICDRFYPNDLWVVVFQFQHIMVGLILPGIVILSCYCIIISKLSHSKGHQKRKALKTTVILILAFFACWLPYYIGISIDSFILLEIIKQGCEFENTVHKWISITEALAFFHCCLNPILYAFLGAKFKTSAQHALTSVSRGSSLKILS KGKRGGHSSVSTESESSSFHSS、
として表される配列を有するケモカイン(C−X−Cモチーフ)受容体4イソ型a[ホモ・サピエンス]、
配列番号27または28を有するこれらのbまたはaイソ型の一つと少なくとも95%の同一性を有する選択的転写スプライス変異体またはその天然変異体、および
その天然リガンドであるストロマ細胞由来因子−1(SDF−1)によって特異的に認識されることができ、かつ好ましくは、少なくとも100、150、および200のアミノ酸長を有するそのフラグメント
からなる群から選択される、二つのヒトCXCR4イソ型のうちの一つである。
【0013】
CXCR2は、
Genbank受託番号NP_001548配列番号29:
MEDFNMESDSFEDFWKGEDLSNYSYSSTLPPFLLDAAPCEPESLEINKYFVVIIYALVFLLSLLGNSLVMLVILYSRVGRSVTDVYLLNLALADLLFALTLPIWAASKVNGWIFGTFLCKVVSLLKEVNFYSGILLLACISVDRYLAIVHATRTLTQKRYLVKFICLSIWGLSLLLALPVLLFRRTVYSSNVSPACYEDMGNNTANWRMLLRILPQSFGFIVPLLIMLFCYGFTLRTLFKAHMGQKHRAMRVIFAVVLIFLLCWLPYNLVLLADTLMRTQVIQETCERRNHIDRALDATEILGILHSCLNPLIYAFIGQKFRHGLLKILAIHGLISKDSLPKDSRPSFVGSSSGHTSTTL、
として表される配列を有するインターロイキン8受容体β[ホモ・サピエンス]、
配列番号29を有するこのインターロイキン8受容体βと少なくとも95%の同一性を有する、選択的転写スプライス変異体またはその天然変異体、および
IL−8によって特異的に認識されることができ、かつ、好ましくは、少なくとも100、150、および200のアミノ酸長を有するそのフラグメント
からなる群から選択される。
【0014】
本発明はまた、抗HIV活性を有するか、またはHIV感染の治療のための組成物の製造のために使用することができる化合物を選択する方法であって、以下の工程を含むことを特徴とする方法も含む。
【0015】
第1の態様において、本発明の主題は、本発明による抗体を作製および選択する方法である。
【0016】
より詳細には、本発明は、HIV複製を阻害することができる抗CXCR4抗体、またはその一つの機能的フラグメントもしくは誘導体を選択する方法であって、
i)作製された抗体をスクリーニングし、CXCR4と特異的に結合することができる抗体を選択する工程、
ii)工程i)の選択抗体を検定し、末梢血単核細胞(PBMC)と結合できる抗体を選択する工程、
iii)工程ii)の選択抗体を検定し、CXCR4ホモ二量体と結合することができる抗体を選択する工程、および次いで
iv)工程iii)の選択抗体を検定し、PBMCにおけるX4指向性HIV−1一次単離物の複製を阻害することができる抗体を選択する工程
を含んでなる方法に関する。
【0017】
別の実施形態では、本発明は、HIV複製を阻害することができる抗CXCR4抗体、またはその一つの機能的フラグメントもしくは誘導体を選択する方法であって、
i)作製された抗体をスクリーニングし、CXCR4と特異的に結合することができる抗体を選択する工程、
ii)工程i)の選択抗体を検定し、末梢血単核細胞(PBMC)と結合できる抗体を選択する工程、
iii)工程ii)の選択抗体を検定し、CXCR4ホモ二量体と結合することができる抗体を選択する工程、および次いで
iv)工程iii)の選択抗体を検定し、PBMCにおけるX4指向性HIV−1一次単離物の複製を阻害することができ、および/またはPBMCにおいてX4/R5指向性HIV−1一次単離物の複製を阻害することができる抗体を選択する工程
を含んでなる方法に関する。
【0018】
抗体の作製は、例えば、骨髄腫細胞と免疫マウスまたは選択された骨髄腫細胞と適合する他の種に由来する脾臓細胞との融合などの、当業者に公知の方法[Kohler & Milstein, 1975, Nature, 256:495-497]によって実現することができる。この免疫動物は、ヒト免疫グロブリン遺伝子座を有し、その後、直接ヒト抗体を産生するトランスジェニックマウスを含み得る。もう一つの可能性のある実施形態は、ライブラリーをスクリーニングするためのファージディスプレー技術の使用からなるものであり得る。
【0019】
スクリーニング工程i)およびii)は、当業者に公知のいずれの方法または工程によっても実現可能である。限定されない例として、ELISA、BIAcore、免疫組織化学、CXCR4発現細胞膜抽出物、または精製CXCR4を用いたウエスタンブロット解析、FACS分析、および機能的スクリーニングが挙げられる。好ましい方法は、産生された抗体が、標的細胞表面において天然CXCR4受容体のコンフォメーションも認識できることを確認するための、CXCR4形質転換体(工程1)および少なくともPBMC(工程2)に対するFACS分析によるスクリーニングからなる。この方法を以下の例にさらに厳密に記載する。
【0020】
スクリーニング工程iii)は、当業者に公知のいずれの方法または方法によっても実現可能である。限定されるものではないが、好ましい例として、CXCR4トランスフェクト細胞またはPBMC由来の膜抽出物に対して着目する抗体を用いるウエスタンブロット法および/または免疫沈降法が挙げられる。
【0021】
スクリーニング工程iv)は、当業者に公知のいずれの方法または方法によっても実現可能である。限定されるものではないが、好ましい例として、Hollら(J. Immunol. 2004, 173, 6274-83)において記載されているプロトコールを用いることにより、PBMCにおけるX4一次HIV−1単離物および/またはX4/R5一次HIV−1単離物の複製を阻害することができるか否かに関し、抗体をスクリーニングすることからなる方法が挙げられる。
【0022】
本発明の方法の選択の工程iii)の好ましい実施形態では、この工程iii)は、CXCR4−RLuc/CXCR4−YFPを発現する細胞に対するBRET分析によって抗体を評価すること、およびBRETシグナルの少なくとも40%、好ましくは45%、50%、55%、最も好ましくは60%を阻害することができる抗体を選択することからなる。
【0023】
BRET法は、タンパク質二量体形成の代表例として知られている技術である[Angers et al., PNAS, 2000, 97:3684-89]。
【0024】
本方法の工程iii)に用いられるBRET法は当業者によく知られており、以下の例に記載する。より詳細には、BRET(生物発光共鳴エネルギー移動)は、生物発光ドナー(ウミシイタケ・ルシフェラーゼ(Rluc))と、蛍光アクセプターであるGFP(緑色蛍光タンパク質)またはYFP(黄色蛍光タンパク質)突然変異体との間で起こる非放射性のエネルギー移動である。本ケースでは、EYFP(増強黄色蛍光タンパク質)を用いた。移動の有効性は、ドナーとアクセプターの配向および距離に依存する。そして、エネルギー移動は、この二つの分子が近接している(1〜10nm)場合にのみ起こり得る。この特性を用いて、タンパク質−タンパク質相互作用アッセイを作り出す。実際に、二つのパートナー間の相互作用を検討するために、第一のものをウミシイタケ・ルシフェラーゼと、そして、第二のものをGFPの黄色突然変異体と遺伝子融合する。融合タンパク質は、必須ではないが一般に、哺乳類細胞で発現される。その膜透過基質(セレンテラジン)の存在下で、Rlucは青色の光を放出する。GFP突然変異体がRlucから10nmより近ければエネルギー移動が起こり、さらに黄色シグナルが検出できる。BRETシグナルは、アクセプターによって放出された光とドナーによって放出された光の間の比として測定される。従って、BRETシグナルは二つの融合タンパク質が近接するほど、またはコンフォメーション変化がRlucと、GFP突然変異体とを近接させるほど増大する。
【0025】
BRET分析が好ましい実施形態からなる場合、CXCR4二量体のコンフォメーション変化を測定するために、当業者に公知のいずれの方法も使用可能である。限定されるものではないが、以下の技術が挙げられる:FRET(蛍光共鳴エネルギー移動)、HTRF(均一時間分解蛍光)、FLIM(蛍光寿命画像測定法)またはSW−FCCS(単波長蛍光相関分光法)。
【0026】
共免疫沈降法、アルファースクリーン、化学架橋、二重ハイブリッド、アフィニティークロマトグラフィー、ELISA、またはファーウエスタンブロットなどの他の従来法も使用可能である。
【0027】
本発明による方法の特定の態様では、工程iii)は、CXCR4−RLuc/CXCR4−YFPの双方を発現する細胞に対するBRET分析によって抗体を評価すること、およびBRETシグナルの少なくとも40%を阻害することができる抗体を選択することからなる。
【0028】
第2の態様において、本発明の主題は、該方法によって得られる単離された抗体、またはそれらの一つの機能的フラグメントもしくは誘導体である。該抗体またはそれらの一つのフラグメントもしくは誘導体は、ヒトCXCR4と特異的に結合することができ、該抗体はまた、CXCR4ホモ二量体のコンフォメーション変化を誘導することもできる。
【0029】
例えばクローンA120のようなCXCR4 MabがHIV−1実験株(X4HIV−1NL4−3)のPBMCへの侵入を阻害することができることが文献から知られている(Tanaka R. et al, J. Virol. 2001, 75, 11534-11543)。さらに、CXCR4 Mabが、HIV−1 X4一次単離物の、CXCR4を発現する細胞系統への侵入を阻害することができることも知られている。これに対し、このようなウイルスをそれらの天然環境において、すなわち、実験的ウイルスまたは細胞系統に対してだけでなく、阻害することができる抗体は開示されていない。しかしながらやはり、CXCR4 MabがPBMCへのHIV−1 X4一次単離物の侵入を阻害することができるということは新規であり、本発明の自明の態様ではない。
【0030】
「機能的フラグメントおよび誘導体」という表現は、本明細書の後段で詳細に定義される。
ここで、本発明は天然型の抗体に関するものではなく、すなわち、それらはそれらの天然環境になく、単離可能であったか、もしくは天然源から精製によって得られたものであるか、またはさらには遺伝子組換えもしくは化学合成によって得られたものであり、また、それらは、さらに記載されるように、非天然アミノ酸を含むことができることを理解しなければならない。
【0031】
より詳細には、本発明の別の態様によれば、単離された抗体、またはそれらの一つの機能的フラグメントもしくは誘導体が記載され、該抗体は、IMGTナンバリングシステムによって定義される配列番号1〜6および30〜33のアミノ酸配列を含んでなるCDRから選択される少なくとも一つの相補性決定領域(complementary determining region)CDRを含んでなることを特徴とする。
【0032】
第一の態様によれば、本発明は、IMGTナンバリングシステムに従って定義される配列番号1〜6の配列のCDRの中から選択される少なくとも一つのCDR、または配列が最適なアライメントの後に配列番号1〜6の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つのCDRを含んでなる、単離された抗体、またはその機能的フラグメント、もしくは誘導体に関する。
【0033】
第二の態様によれば、本発明は、IMGTナンバリングシステムに従って定義される配列番号1、2および30〜33の配列のCDRの中から選択される少なくとも一つのCDR、または配列が最適なアライメントの後に配列番号1、2、および30〜33の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つのCDRを含んでなる、単離された抗体、またはその機能的フラグメントまたは誘導体に関する。
【0034】
抗体の「機能的フラグメント」とは、特に、フラグメントFv、scFv(sc=一本鎖)、Fab、F(ab’)2、Fab’、scFv−Fc、もしくはダイアボディー(diabody)、または半減期が延長された任意のフラグメントなどの抗体フラグメントを意味する。このような機能的フラグメントは本明細書の後段で詳細に記載される。
【0035】
抗体の「誘導化合物」または「誘導体」とは、特に、ペプチド骨格(peptide scaffold)と、CXCR4を認識する能力を保持するための原抗体の少なくとも一つのCDRとから構成される結合タンパク質を意味する。当業者に周知のこのような誘導化合物は、本明細書の後段でさらに詳細に記載される。
【0036】
より好ましくは、本発明は、本発明による抗体、それらの誘導化合物またはそれらの機能的フラグメント、特に、キメラまたはヒト化抗体、遺伝子組換え、または化学合成によって得られる抗体を含んでなる。
【0037】
好ましい実施形態によれば、本発明による抗体、またはその誘導化合物もしくは機能的フラグメントは、モノクローナル抗体からなることを特徴とする。
【0038】
「モノクローナル抗体」とは、ほぼ均質な抗体集団から生じる抗体を意味すると理解される。より詳細には、集団の個々の抗体は、最小の割合で見られる少数の潜在的に自然に生じる突然変異以外は同一である。言い換えれば、モノクローナル抗体は、単細胞クローン(例えば、ハイブリドーマ、その均質な抗体をコードするDNA分子でトランスフェクトされた真核生物宿主細胞、その均質な抗体をコードするDNA分子でトランスフェクトされた原核生物宿主細胞など)の増殖から生じる均質な抗体からなり、一般に、一つおよびただ一つのクラスおよびサブクラスの重鎖と、ただ一つの種類の軽鎖を特徴とする。モノクローナル抗体は特異性が高く、単一の抗原に対するものである。加えて、種々の決定基またはエピトープに対する種々の抗体を一般に含むポリクローナル抗体の作製とは対照的に、各モノクローナル抗体は抗原の単一のエピトープに対するものである。
【0039】
より詳細には、本発明の第一の好ましい実施形態によれば、抗体、またはその誘導化合物、もしくは機能的フラグメントは、CDR−L1、CDR−L2、およびCDR−L3から選択される少なくとも一つのCDRを含んでなる軽鎖を含んでなることを特徴とし、ここで、
CDR−L1は配列番号1のアミノ酸配列を含んでなり、
CDR−L2は配列番号2のアミノ酸配列を含んでなり、
CDR−L3は配列番号3のアミノ酸配列を含んでなる。
【0040】
別の実施形態によれば、本発明の抗体、またはそれらの誘導化合物もしくは機能的フラグメントの一つは、配列番号1、2、もしくは3の配列の三つのCDRのうち少なくとも一つ、または最適なアライメントの後に配列番号1、2、または3の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる軽鎖を含んでなることを特徴とする。
【0041】
本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体はまた、CDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖を含んでなることを特徴とし、ここで、CDR−L1は配列番号1のアミノ酸配列を含んでなり、CDR−L2は配列番号2のアミノ酸配列を含んでなり、CDR−L3は配列番号3のアミノ酸配列を含んでなる。
【0042】
別の実施形態では、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号7のアミノ酸配列、または最適なアライメントの後に配列番号7の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる配列の軽鎖を含んでなることを特徴とする。
【0043】
本発明の第二の好ましい実施形態によれば、抗体、またはその誘導化合物もしくは機能的フラグメントは、CDR−L1、CDR−L2、およびCDR−L3から選択される少なくとも一つのCDRを含んでなる軽鎖を含んでなることを特徴とし、ここで、
CDR−L1は配列番号1のアミノ酸配列を含んでなり、
CDR−L2は配列番号2のアミノ酸配列を含んでなり、
CDR−L3は配列番号30のアミノ酸配列を含んでなる。
【0044】
別の実施形態によれば、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、配列番号1、2、もしくは30の配列の三つのCDRのうち少なくとも一つ、または最適なアライメントの後に配列番号1、2、もしくは30の配列と少なくとも80%、好ましくは85%、90%、95%および98%の同一性を有する少なくとも一つの配列を含んでなる軽鎖を含んでなることを特徴とする。
【0045】
本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、CDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖を含んでなることを特徴とし、ここで、CDR−L1は配列番号1のアミノ酸配列を含んでなり、CDR−L2は配列番号2のアミノ酸配列を含んでなり、CDR−L3は配列番号30のアミノ酸配列を含んでなる。
【0046】
別の実施形態では、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号34のアミノ酸配列、または最適なアライメントの後に配列番号34の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる配列の軽鎖を含んでなることを特徴とする。
【0047】
より詳細には、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、CDR−H1、CDR−H2、およびCDR−H3から選択される少なくとも一つのCDRを含んでなる重鎖を含んでなることを特徴とし、ここで、
CDR−H1は配列番号4のアミノ酸配列を含んでなり、
CDR−H2は配列番号5のアミノ酸配列を含んでなり、
CDR−H3は配列番号6のアミノ酸配列を含んでなる。
【0048】
別の実施形態によれば、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、配列番号4、5、もしくは6の配列の三つのCDRのうち少なくとも一つ、または最適なアライメントの後に配列番号4、5、もしくは6の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる重鎖を含んでなることを特徴とする。
【0049】
別の特定の実施形態によれば、抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、CDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖を含んでなることを特徴とし、ここで、CDR−H1は配列番号4のアミノ酸配列を含んでなり、CDR−H2は配列番号5のアミノ酸配列を含んでなり、CDR−H3は配列番号6のアミノ酸配列を含んでなる。
【0050】
別の実施形態では、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号8のアミノ酸配列、または最適なアライメントの後に配列番号8の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる配列の重鎖を含んでなることを特徴とする。
【0051】
より詳細には、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、CDR−H1、CDR−H2、およびCDR−H3から選択される少なくとも一つのCDRを含んでなる重鎖を含んでなることを特徴とし、ここで、
CDR−H1は配列番号31のアミノ酸配列を含んでなり、
CDR−H2は配列番号32のアミノ酸配列を含んでなり、
CDR−H3は配列番号33のアミノ酸配列を含んでなる。
【0052】
別の実施形態によれば、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、配列番号31、32、もしくは33の配列の三つのCDRのうち少なくとも一つ、または最適なアライメントの後に配列番号31、32、もしくは33の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる重鎖を含んでなることを特徴とする。
【0053】
別の特定の実施形態によれば、抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、CDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖を含んでなることを特徴とし、ここで、CDR−H1は配列番号31のアミノ酸配列を含んでなり、CDR−H2は配列番号32のアミノ酸配列を含んでなり、CDR−H3は配列番号33のアミノ酸配列を含んでなる。
【0054】
別の実施形態では、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号35のアミノ酸配列、または最適なアライメントの後に配列番号35の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる配列の重鎖を含んでなることを特徴とする。
【0055】
本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号1、2、および3のアミノ酸配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖と、配列番4、5、および6のアミノ酸配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖を含んでなることを特徴とする。
【0056】
最後に、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体はまた、配列番号7のアミノ酸配列を含んでなる軽鎖と、配列番号8のアミノ酸配列を含んでなる重鎖を含んでなることを特徴としうる。
【0057】
本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号1、2、および30のアミノ酸配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖と、配列番号31、32、および33のアミノ酸配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖を含んでなることを特徴とする。
【0058】
最後に、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体はまた、配列番号34のアミノ酸配列を含んでなる軽鎖と、配列番号35のアミノ酸配列とを含んでなる重鎖を含んでなることを特徴とし得る。
【0059】
本明細書において「ポリペプチド」、「ポリペプチド配列」、「ペプチド」、および「抗体化合物またはそれらの配列と結合しているタンパク質」は互換的である。ここで、本発明は天然型の抗体に関するものではなく、すなわち、それらはそれらの天然環境から採取されたものではなく、単離されているか、または天然源から精製によって得られたものであるか、または遺伝子組換えもしくは化学合成によって得られたものであり、従って、それらは、以下に記載されるように、非天然アミノ酸を含むことができることを理解しなければならない。
【0060】
第一の実施形態において、相補性決定領域、またはCDRは、Kabatら(Kabat et al., Sequences of proteins of immunological interest, 第5版, U.S. Department of Health and Human Services, NIH, 1991および後続版)によって定義される免疫グロブリンの重鎖および軽鎖の超可変領域を意味する。三つの重鎖CDRと、三つの軽鎖CDRとが存在する。ここで、「CDR」とは、抗体が認識する抗原またはエピトープに対する抗体の結合親和性を担うアミノ酸残基の大部分を含む領域のうち1以上、またはさらには総てを示すのに用いられる。
【0061】
第二の実施形態において、CDR領域またはCDRにより、IMGTによって定義される免疫グロブリンの重鎖および軽鎖の超可変領域を示すものとする。
【0062】
IMGTユニークナンバリングは、どんな抗原受容体、鎖の種類、または種であれ、可変ドメインを比較するために定義されたものである[Lefranc M. -P., Immunology Today 18, 509 (1997) / Lefranc M.-P., The Immunologist, 7, 132-136 (1999) / Lefranc, M.-P., Pommie, C, Ruiz, M., Giudicelli, V., Foulquier, E., Truong, L., Thouvenin-Contet, V. and Lefranc, Dev. Comp. Immunol, 27, 55-77 (2003)]。IMGTユニークナンバリングでは、例えば、システイン23(1st−CYS)、トリプトファン41(CONSERVED−TRP)、疎水性アミノ酸89、システイン104(2nd−CYS)、フェニルアラニン、またはトリプトファン118(J−PHEまたはJ−TRP)など、保存されているアミノ酸は常に同じ位置を持つ。IMGTユニークナンバリングでは、フレームワーク領域(FR1−IMGT:1〜26番、FR2−IMGT:39〜55番、FR3−IMGT:66〜104番およびFR4−IMGT:118〜128番)および相補性決定領域:CDR1−IMGT:27〜38番、CDR2−IMGT:56〜65番およびCDR3−IMGT:105〜117番の標準化された画定が得られる。ギャップは占有されていない位置を表すので、CDR−IMGT長(例えば、[8.8.13]などと括弧の中に点で区切って示される)は重要な情報となる。IMGTユニークナンバリングは、IMGT Colliers de Perles [Ruiz, M. and Lefranc, M.-P., Immunogenetics, 53, 857-883 (2002) / Kaas, Q. and Lefranc, M.-P., Current Bioinformatics, 2, 21-30 (2007)]として表される2Dグラフ、およびIMGT/3Dstructure-DB [Kaas, Q., Ruiz, M. and Lefranc, M.-P., T cell receptor and MHC structural data. Nucl. Acids. Res., 32, D208-D210 (2004)]に用いられる。
【0063】
三つの重鎖CDRと、三つの軽鎖CDRとが存在する。ここで、CDRは、場合によって、抗体が認識する抗原またはエピトープに対する抗体の親和性による結合を担うアミノ酸残基の大部分を含む、これらの領域の一つまたはいくつか、またはさらにはこれらの領域の全部を示すために用いられる。
【0064】
より明瞭にするために、以下の説明およびより詳細には表2および3では、CDRはIMGTナンバリングシステムおよびKabatナンバリングシステムによって定義されていることを理解しなければならない。
【0065】
IMGTナンバリングシステムは、上記で定義されたIMGTシステムに従ってCDRを定義し、Kabatナンバリングシステムは、上記で定義されたKabatシステムに従ってCDRを定義する。
【0066】
より詳細には、515H7と呼ばれる抗体に関しては、CDR−L1はIMGTナンバリングシステムでは配列番号1からなり、Kabatナンバリングシステムでは配列番号9からなる。CDR−L2に関しては、IMGTナンバリングシステムでは配列番号2からなり、Kabatナンバリングシステムでは配列番号10からなる。CDR−L3は、この二つのナンバリングシステムのそれぞれで配列番号3からなる。重鎖について、CDR−H1は、IMGTナンバリングシステムでは配列番号4からなり、Kabatナンバリングシステムでは配列番号11からなる。CDR−H2は、IMGTナンバリングシステムでは配列番号5からなり、Kabatナンバリングシステムでは配列番号12からなる。最後に、CDR−H3は、IMGTナンバリングシステムでは配列番号6からなり、Kabatナンバリングシステムでは配列番号13からなる。
【0067】
そして、301aE5と呼ばれる抗体に関しては、CDR−L1は、IMGTナンバリングシステムでは配列番号1からなり、Kabatナンバリングシステムでは配列番号9からなる。CDR−L2に関しては、IMGTナンバリングシステムでは配列番号2からなり、Kabatナンバリングシステムでは配列番号36からなる。CDR−L3は、IMGTナンバリングシステムでは配列番号30からなり、Kabatナンバリングシステムでは配列番号37からなる。重鎖について、CDR−H1は、IMGTナンバリングシステムでは配列番号31からなり、Kabatナンバリングシステムでは配列番号38からなる。CDR−H2は、IMGTナンバリングシステムでは配列番号32からなり、Kabatナンバリングシステムでは配列番号39からなる。最後に、CDR−H3は、IMGTナンバリングシステムでは配列番号33からなり、Kabatナンバリングシステムでは配列番号40からなる。
【0068】
本発明の意味において、二つの核酸配列またはアミノ酸配列間の「同一性割合」とは、最適なアライメントの後に得られる、比較する二つの配列間で同一のヌクレオチドまたはアミノ酸残基の割合を意味し、この割合は純粋に統計学的なものであり、その2配列間の違いはそれらの長さに沿ってランダムに分布している。二つの核酸配列またはアミノ酸配列の比較は従来から、それらを最適にアラインした後に配列を比較することによって行われ、この比較はセグメントによって、または「アライメントウインドウ」の使用によって行うことができる。比較のための配列の最適なアライメントは、手による比較の他、Smith and Waterman (1981) [Ad. App. Math. 2:482]のローカル・ホモロジー・アルゴリズム、Neddleman and Wunsch (1970) [J. Mol. Biol. 48:443]のローカル・ホモロジー・アルゴリズム、Pearson and Lipman (1988) [Proc. Natl. Acad. Sci. USA 85:2444]またはこれらのアルゴリズムを用いたコンピューターソフトウエア(Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WIのGAP、BESTFIT、FASTAおよびTFASTA、または匹敵するソフトウエアBLAST NRもしくはBLAST Pによる)によって行うことができる。
【0069】
二つの核酸配列またはアミノ酸配列間の同一性割合は、二つの最適にアラインされた配列を比較することにより決定され、ここで、比較する核酸配列またはアミノ酸配列はその二つの配列間の最適なアライメントのために参照配列に対して付加または欠失を持ちうる。同一性割合は、アミノ酸ヌクレオチドまたは残基がその二つの配列間、好ましくは二つの全配列間で同一である位置の数を求め、その同一の位置の数をアライメントウインドウの位置の総数で割り、その商に100をかけて、その二つの配列間の同一性割合を得ることによって計算される。
【0070】
例えば、http://www.ncbi.nlm.nih.gov/gorf/bl2.htmlのサイトで利用可能なBLASTプログラム、「BLAST2 sequences」(Tatusova et al., "Blast 2 sequences - a new tool for comparing protein and nucleotide sequences", FEMS Microbiol, 1999, Lett. 174:247-250)をデフォルトパラメーター(特に、パラメーターとして、「オープンギャップペナルティー」:5、および「エクステンションギャップペナルティー」:2、選択マトリックスは、例えば、このプログラムによって提案される「BLOSUM 62」である)とともに使用することができ、この2配列間の同一性割合は、このプログラムによって直接計算される。
【0071】
参照アミノ酸配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を示すアミノ酸配列として、好ましい例には、参照配列、ある特定の修飾、特に、少なくとも一つのアミノ酸の欠失、付加、もしくは置換、末端切断または延長を含むものが含まれる。1以上の保存的または非保存的アミノ酸の置換の場合、置換されるアミノ酸が「等価な」アミノ酸に置き換わる置換が好ましい。ここで、「等価なアミノ酸」とは、対応する抗体および以下に定義される具体例の生物活性にいかなる修飾も与えずに構造アミノ酸の一つの取って代わり得るアミノ酸を示すものとする。
【0072】
等価なアミノ酸は、置換されるアミノ酸との構造的相同性または生成され得る種々の抗体間の生物活性の比較試験の結果に基づいて決定される。限定されない例として、下記表1に、対応する修飾抗体の生物活性の有意な修飾をもたらさずに行い得る置換をまとめる。なお、逆の置換も同じ条件下で本来可能である。
【0073】
【表1】

【0074】
特定の実施形態において、本発明は、ネズミ抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0075】
上記に見られるように、本発明はまた、本明細書に記載される抗体に由来する化合物に関する。
【0076】
より詳細には、本発明の抗体、またはその誘導化合物もしくは機能的フラグメントは、該誘導化合物が、最初の抗体のパラトープ認識特性の総てまたは一部を保存するように、少なくとも一つのCDRがグラフトされているペプチド骨格を含んでなる結合タンパク質からなることを特徴とする。
【0077】
また、本発明に記載されるCDR配列のうち1以上の配列が種々の免疫グロブリンタンパク質骨格(scaffolding)に存在してもよい。この場合、このタンパク質配列によって、グラフトされたCDRの折りたたみに有利なペプチド骨格を再び作り出して、それらのパラトープ抗原認識特性を保存可能とすることができる。
【0078】
一般に、当業者は、元の抗体に由来するCDRの少なくとも一つをグラフトするためのタンパク質骨格(scaffolding)の種類をどのように決めればよいかを知っている。より詳細には、選択されたこのような骨格は次のような基準の最大数を満たさなければならないことが知られている(Skerra A., J. Mol. Recogn., 2000, 13:167-187):
−系統発生的保存が良好であること、
−三次元構造が既知であること(例えば、結晶学、NMR分光法または当業者に公知の他のいずれかの方法による)、
−小型であること、
−転写後修飾が少ないか、または全く無いこと、および/または
−産生、発現、および精製が容易なこと。
【0079】
このようなタンパク質骨格の起源は、限定されるものではないが、フィブロネクチンおよび優先的にはフィブロネクチンIII型ドメイン10、リポカリン、アンチカリン(Skerra A., J. Biotechnol., 2001, 74(4):257-75)、黄色ブドウ球菌(Staphylococcus aureus)のタンパク質AのドメインBに由来するタンパク質Z、チオレドキシンAまたは「アンキリンリピート」(Kohl et al., PNAS, 2003, vol. 100, No. 4, 1700-1705)、「アルマジロリピート」、「ロイシンリッチリピート」および「テトラトリコペプチドリピート」などの繰り返しモチーフを有するタンパク質の中から選択される構造物であり得る。
【0080】
例えば、サソリ、昆虫、植物、軟体動物などの毒素、およびニューロンNOシンターゼ(PIN)のタンパク質阻害剤といった毒素に由来する骨格も挙げておくべきであろう。
限定されるものではないが、このようなハイブリッド構築物の例として、PINのループの一つにおける抗CD4抗体、すなわち13B8.2のCDR−H1(重鎖)挿入があり、このようにして得られた新たな結合タンパク質は元の抗体と同じ結合特性を保存している(Bes et al., Biochem. Biophys. Res. Commun., 2006, 343(1), 334-344)。単に例示であるが、ネオカルチノスタチンのループの一つに抗リゾチームVHH抗体のCDR−H3(重鎖)をグラフトすることも挙げられる。
【0081】
最後に、上記のように、このようなペプチド骨格は、元の抗体に由来するCDRの少なくとも一つを含んでなり得る。必ずしも必要というわけではないが、好ましくは、当業者は重鎖由来の少なくとも一つのCDRを選択することができ、これは抗体の特異性を主として担うことが知られている。1以上の適切なCDRを選択することは当業者には自明であり、当業者ならば、次に好適な既知技術を選択することができる(Bes et al., FEBS letters 508, 2001, 67-74)。
【0082】
本発明の特定の態様は、本発明による抗体に由来する化合物を選択する方法に関し、該誘導化合物はin vitroおよび/またはin vivoにおいてHIVの細胞侵入を阻害することができ、該誘導化合物は、少なくとも一つの抗体CDRがグラフトされているペプチド骨格を含んでなり、該方法は、
a)少なくとも一つの抗体CDRがグラフトされているペプチド骨格ペプチド骨格から構成される化合物を、HIV 1型とPBMCを含有する生体サンプルと接触させる工程、および
b)該化合物がHIV−1複製を阻害することができる場合、その化合物を選択する工程
を含んでなり、
少なくとも一つのグラフトされるCDRが、配列番号1〜6および30〜33の配列、または最適なアライメントの後に配列番号1〜6および30〜33の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する配列のCDRの中から選択されることを特徴とする。
【0083】
好ましい様式によれば、該方法は、工程a)において、少なくとも二つまたは三つの抗体CDRがグラフトされているペプチド骨格を含んでなる化合物をin vitroで接触させることを含み得る。
【0084】
この方法のさらに好ましい様式によれば、ペプチド骨格は、構造が上記された骨格または結合タンパク質の中から選択される。
【0085】
明らかに、これらの例は何ら限定されず、当業者に既知または自明の他のいずれの構造も、本特許出願によって付与される保護に包含されるものと考えるべきである。
【0086】
よって、本発明は、ペプチド骨格が、a)系統発生的によく保存されているタンパク質、b)ロバストな(robust)構造のタンパク質、c)周知の3D分子構成を有するタンパク質、d)小型のタンパク質、および/またはe)安定性を変化させずに欠失および/または挿入によって改変することができる領域を含んでなるタンパク質の中から選択されることを特徴とする抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0087】
好ましい実施形態によれば、本発明の抗体、またはその誘導化合物もしくは機能的フラグメントは、該ペプチド骨格が、i)フィブロネクチンに由来する骨格、優先的にはフィブロネクチン3型ドメイン10、リポカリン、アンチカリン、黄色ブドウ球菌(Staphylococcus aureus)のタンパク質AのドメインBに由来するタンパク質Z、チオレドキシンAまたは「アンキリンリピート」(Kohl et al., PNAS, 2003, vol. 100, No. 4, 1700-1705)、「アルマジロリピート」、「ロイシンリッチリピート」および「テトラトリコペプチドリピート」などの繰り返しモチーフを有するタンパク質、またはiii)ニューロンNOシンターゼ(PIN)タンパク質阻害剤の中から選択されることを特徴とする。
【0088】
本発明の別の態様は、上記抗体の機能的フラグメントに関する。
【0089】
より詳細には、本発明は、機能的フラグメントがフラグメントFv、Fab、(Fab’)2、Fab’、scFv、scFv−Fc、およびダイアボディー、またはPEG化フラグメントなどの半減期が延長された任意のフラグメントの中から選択されることを特徴とする抗体、またはその誘導化合物もしくは機能的フラグメントを対象とする。
【0090】
本発明による抗体のこのような機能的フラグメントは、例えば、フラグメントFv、scFv(sc=一本鎖)、Fab、F(ab’)2、Fab’、scFv−Fcまたはダイアボディー、またはポリエチレングリコール(ペグ化)などのポリアルキレングリコールの付加といった化学修飾によって(PEG化フラグメントはFv−PEG、scFv−PEG、Fab−PEG、F(ab’)2−PEGおよびFab’−PEGと呼ばれる)、またはリポソーム、ミロスフェア、またはPLGAに組み込むことによって半減期が延長された任意のフラグメントからなり、該フラグメントは、特に、それが由来する抗体の全般的活性、さらには部分的活性であっても示し得る本発明の特徴的なCDRの少なくとも一つを有する。
【0091】
好ましくは、該機能的フラグメントは、それらが由来する抗体の可変重鎖または軽鎖の部分的配列を含んでなるか、または含み、該部分的配列は、それが由来する抗体と同様の結合特異性および十分な親和性、好ましくはそれが由来する抗体の親和性の少なくとも1/100、より好ましくは少なくとも1/10の親和性を保持するのに十分なものである。
【0092】
このような機能的フラグメントは、それが由来する抗体の配列の少なくとも5つのアミノ酸、好ましくは、6、7、8、10、15、25、50、または100個の連続するアミノ酸を含む。
好ましくは、これらの機能的フラグメントは、それらが由来する抗体と一般に同様の結合特異性を有する、Fv、scFv、Fab、F(ab’)2、F(ab’)、scFv−Fc、またはダイアボディータイプのものである。本発明によれば、本発明の抗体のフラグメントは、上記の抗体からペプシンもしくはパパインを含む酵素消化、および/または化学的還元によるジスルフィド結合の切断などの方法によって得ることができる。該抗体フラグメントはまた、これもまた当業者に公知の組換え遺伝子技術によって、または例えば、Applied BioSystemsなどによって販売されているものなどの自動ペプチド合成装置の手段によるペプチド合成によって得ることができる。
さらに明確にするために、下表2に、本発明の抗体に相当する種々のアミノ酸配列をまとめる。
【0093】
【表2】
【0094】
本発明の抗体対象の特に重要な付加的態様は、それらが抗体依存性細胞傷害性(ADCC)および/または補体依存性細胞傷害性(CDC)などのエフェクター機能を示さないことを特徴とする。
【0095】
より詳細には、一つの例として、本発明の抗体、またはそれらの一つの機能的フラグメントもしくは誘導体は、FcγR(I、II、もしくはIII)に対して、またはC1qに対して、またはその双方に対して親和性を持たない。
【0096】
構造的に、これは、当業者にとって、本発明の抗体、またはそれらの一つの機能的フラグメントもしくは誘導体が、Fc部分を欠いていること、またはそれらのFc部分がエフェクター機能を付与し得る適切なグリコシル化を与えないことを意味する。
【0097】
この結果が、本発明の抗体は好ましくはIgG4またはIgG2イソ型、最も好ましくはIgG4イソ型から選択されるということである。
【0098】
同様に、好ましいフラグメントは、Fv、scFv(scは一本鎖)、Fab、F(ab’)2、Fab’、scFv−Fcフラグメントもしくはダイアボディー、またはポリ(エチレン)グリコール(「ペグ化」)(ペグ化フラグメントはFv−PEG、scFv−PEG、Fab−PEG、F(ab’)2−PEG、またはFab’−PEGと呼ばれる)(「PEG」はポリ(エチレン)グリコール)などのポリ(アルキレン)グリコールの付加のような化学的修飾によって、またはリポソームへの組み込みによって半減期が延長された任意のフラグメントといった、ADCCを欠いているフラグメントである。
【0099】
より詳細には、抗体515H7由来の本発明の好ましい機能的フラグメントはscFvであり、以下、515H7 scFv−Ckフラグメントと呼ばれ、配列番号54のアミノ酸配列を含んでなる。
【0100】
該scFvに相当するヌクレオチド配列は、配列番号55の配列を含んでなる。
【0101】
本発明の別の特定の態様は、抗体がまたマウスとは異種の、特にヒトの抗体に由来する軽鎖および重鎖定常領域も含んでなることを特徴とする、キメラ抗体、またはそれらの誘導化合物もしくは機能的フラグメントに関する。
【0102】
本発明のさらに別の態様は、ヒト抗体に由来する軽鎖および重鎖の定常領域がそれぞれλまたはκ領域およびγ−2または好ましくはγ−4領域であることを特徴とする、ヒト化抗体、またはそれらの誘導化合物もしくは機能的フラグメントに関する。
【0103】
本発明の抗体はまた、キメラ抗体またはヒト化抗体も含んでなる。
【0104】
キメラ抗体は、所与の種に由来する天然可変領域(軽鎖および重鎖)を所与の種とは異種の抗体の軽鎖および重鎖の定常領域と組み合わせて含むものである。
【0105】
抗体、またはそのキメラフラグメントは、組換え遺伝子技術を用いて作製することができる。例えば、該キメラ抗体は、プロモーターと、本発明の非ヒト、特にネズミモノクローナル抗体の可変領域をコードする配列、およびヒト抗体定常領域をコードする配列を含む組換えDNAをクローニングすることによって作製することができる。このような組換え遺伝子の一つによってコードされる本発明によるキメラ抗体は、例えば、マウス−ヒトキメラであってよく、この抗体の特異性はネズミDNAに由来する可変領域によって決定され、そのイソ型はヒトDNAに由来する定常領域によって決定される。キメラ抗体の作製方法については、Verhoeynら(BioEssays, 8:74, 1988)を参照。
【0106】
以下の表3に、本発明によるキメラ抗体515H7(c515H7またはC515H7と呼ばれる)の種々の重鎖および軽鎖のアミノ酸配列をまとめる。
【0107】
【表3】
【0108】
この抗体c515H7重鎖配列番号56〜58および軽鎖配列番号59に相当するヌクレオチド配列はそれぞれ、配列番号60〜63(重鎖)および配列番号64(軽鎖)の配列に相当する。
【0109】
好ましい実施形態では、これらの重鎖配列はそれらのC末端からリシン残基(LonzaからのpConPlusベクター系統原株:pConPlusγ4ΔK、pConPlusγ4PROΔK、およびpConPlusγ2ΔKに見られる)が欠失されている。
【0110】
さらに、G4PRO重鎖は、半抗体の形成を避けるためにヒンジ領域に突然変異を有するヒトIgG4イソ型に相当する。この突然変異は、Lonzaからの親株pConPlusγ4PROΔKに見られる[Angal S, King DJ, Bodmer MW, Turner A, Lawson AD, Roberts G, Pedley B, Adair JR. A single amino acid substitution abolishes the heterogeneity of chimeric mouse/human (IgG4) antibody. Mol Immunol. (1993);30(1):105-108]。
【0111】
より詳細には、本発明は、種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRを含んでなることを特徴とするキメラ抗体重鎖に関し、ここで、該CDRは、IMGTによれば、配列番号4、5、および6の配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3からなる。
【0112】
より詳細には、本発明は、種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRを含んでなることを特徴とするキメラ抗体軽鎖に関し、ここで、該CDRは、IMGTによれば、配列番号1、2、および3の配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3からなる。
【0113】
より詳細には、本発明は、種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRをそれぞれ有する重鎖および軽鎖を含んでなることを特徴とするキメラ抗体、またはその誘導化合物もしくは機能的フラグメントに関し、ここで、該CDRは、IMGTによれば、配列番号4、5、および6の配列をそれぞれ含んでなる重鎖のCDR−H1、CDR−H2、およびCDR−H3と、配列番号1、2、および3の配列をそれぞれ含んでなる軽鎖のCDR−L1、CDR−L2、およびCDR−L3からなる。
【0114】
別の実施形態では、本発明は、配列番号8からなる配列の重鎖可変領域と、配列番号7の配列の軽鎖可変領域とを含んでなるキメラ抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0115】
さらに別の実施形態では、本発明は、配列番号56、57、または58からなる群から選択される配列の重鎖と、配列番号59の配列の軽鎖とを含んでなるキメラ抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0116】
好ましい実施形態では、本発明によるキメラ抗体c515H7 VH(G4wt)/VL−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号56の配列の重鎖可変領域と、配列番号59の配列の軽鎖可変領域とを含んでなる。
【0117】
好ましい実施形態では、本発明によるキメラ抗体c515H7 VH(G4PRO)/VL−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号57の配列の重鎖可変領域と、配列番号59の配列の軽鎖可変領域とを含んでなる。
【0118】
好ましい実施形態では、本発明によるキメラ抗体c515H7 VH(G2wt)/VL−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号58の配列の重鎖可変領域と、配列番号59の配列の軽鎖可変領域とを含んでなる。
【0119】
「ヒト化抗体」とは、非ヒト起源の抗体に由来するCDR領域を含み、その抗体分子の他の部分が一つの(または複数の)ヒト抗体に由来する抗体を意味する。さらに、骨格セグメント残基(FRと呼ばれる)のいくつかは、結合親和性を保存するように改変することができる(Jones et al., Nature, 321:522-525, 1986、 Verhoeyen et al., Science, 239:1534-1536, 1988、 Riechmann et al., Nature, 332:323-327, 1988)。
【0120】
本発明のヒト化抗体またはそのフラグメントは、当業者に公知の技術(例えば、Singer et al., J. Immun., 150:2844-2857, 1992、 Mountain et al., Biotechnol. Genet. Eng. Rev., 10:1-142, 1992、およびBebbington et al., Bio/Technology, 10:169-175, 1992の文献に記載されているもの)によって作製することができる。このようなヒト化抗体は、in vitro診断を含む方法またはin vivoにおける予防的および/もしくは治療的処置における使用に好ましい。他のヒト化技術も当業者に公知であり、例えば、欧州特許第0451261号公報、同第0682040号公報、同第0939127号公報、同第0566647号公報、または米国特許第5,530,101号公報、同第6,18,370号公報、同第5,585,089号公報、および同第5,693,761号公報においてPDLが記載している「CDRグラフト」法がある。米国特許第5,639,641号公報または同第6,054,297号公報、同第5,886,152号公報、および同第5,877,293号公報も引用することができる。
【0121】
さらに、本発明はまた、上記のネズミ抗体に由来するヒト化抗体に関する。
好ましい様式では、ヒト抗体に由来する軽鎖および重鎖の定常領域はそれぞれ、λまたはκおよびγ−2または好ましくはγ−4領域である。
【0122】
より詳細には、本発明は、i)対応するヒト抗体重鎖のフレームワーク領域に相同なフレームワーク領域と、ii)種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRとを含んでなることを特徴とするヒト化抗体重鎖に関し、ここで、該CDRは、IMGTによれば、配列番号4、5、および6の配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3からなる。
【0123】
別の実施形態では、本発明は、配列番号64からなる配列の可変領域を含んでなるヒト化抗体重鎖に関する。
さらに別の実施形態では、本発明は、配列番号67、68、および69からなる群から選択される完全な配列を含んでなるヒト化抗体重鎖に関する。より詳細には、本発明は、ヒト抗体軽鎖の対応するフレームワーク領域に相同なフレームワーク領域と、ii)種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRとを含んでなることを特徴とするヒト化抗体軽鎖に関し、ここで、該CDRは、IMGTによれば、配列番号1、2、および3の配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3からなる。
【0124】
別の実施形態では、本発明は、配列番号65、66、82、または83からなる群から選択される配列の可変領域を含んでなるヒト化抗体軽鎖に関する。
【0125】
さらに別の実施形態では、本発明は、配列番号70、71、84、または85のからなる群から選択される完全な配列を含んでなるヒト化抗体軽鎖に関する。
【0126】
より詳細には、本発明は、i)ヒト抗体の対応するフレームワーク領域に相同なフレームワーク領域と、ii)種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRをそれぞれ有する重鎖および軽鎖を含んでなることを特徴とするヒト化抗体、またはその誘導化合物もしくは機能的フラグメントに関し、ここで、該CDRは、IMGTによれば、配列番号4、5、および6の配列をそれぞれ含んでなる重鎖のCDR−H1、CDR−H2、およびCDR−H3と、配列番号1、2、および3の配列をそれぞれ含んでなる軽鎖のCDR−L1、CDR−L2、およびCDR−L3からなる。
【0127】
別の実施形態では、本発明は、配列番号64からなる配列の重鎖可変領域と、配列番号65、66、82、または83からなる群から選択される配列の軽鎖可変領域とを含んでなるヒト化抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0128】
さらに別の実施形態では、本発明は、配列番号67、68、または69からなる群から選択される配列の重鎖と、配列番号70、71、84または85からなる群から選択される配列の軽鎖とを含んでなるヒト化抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4wt)/VL2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号67の配列の重鎖と、配列番号70の配列の軽鎖とを含んでなる。
【0129】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4PRO)/VL2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号68の配列の重鎖と、配列番号70の配列の軽鎖とを含んでなる。
【0130】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G2wt)/VL2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号69の配列の重鎖と、配列番号70の配列の軽鎖とを含んでなる。
【0131】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4wt)/VL2.1−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号67の配列の重鎖と配列番号71の配列の軽鎖とを含んでなる。
【0132】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4PRO)/VL2.1−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号68の配列の重鎖と配列番号71の配列の軽鎖とを含んでなる。
【0133】
別の好ましい実施形態では、本発明によるヒト化抗体 Hz515H7 VH1 D76N(G2wt)/VL2.1−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号69の配列の重鎖と、配列番号71の配列の軽鎖とを含んでなる。
【0134】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4wt)/VL2.2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号67の配列の重鎖と、配列番号84の配列の軽鎖とを含んでなる。
【0135】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4PRO)/VL2.2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号68の配列の重鎖と、配列番号84の配列の軽鎖とを含んでなる。
【0136】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G2wt)/VL2.2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号69の配列の重鎖と、配列番号84の配列の軽鎖とを含んでなる。
【0137】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4wt)/VL2.3−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号67の配列重鎖と、配列番号85の配列の軽鎖とを含んでなる。
【0138】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4PRO)/VL2.3−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号68の配列の重鎖と、配列番号85の配列の軽鎖とを含んでなる。
【0139】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G2wt)/VL2.3−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号69の配列の重鎖と、配列番号85の配列の軽鎖とを含んでなる。
【0140】
以下の表4に、本発明によるヒト化抗体515H7の、それぞれ種々の重鎖および軽鎖可変ドメインおよび全長(または完全)アミノ酸配列をまとめる。
【0141】
【表4】
【0142】
好ましい実施形態では、これらの重鎖配列はそれらのC末端からリシン残基(LonzaからのpConPlusベクター系統原株:pConPlusγ4ΔK、pConPlusγ4PROΔK、およびpConPlusγ2ΔKに見られる)が欠失されている。
【0143】
さらに、G4PRO重鎖は、半抗体の形成を避けるためにヒンジ領域に突然変異を有するヒトIgG4イソ型に相当する。この突然変異は、Lonzaからの親株pConPlusγ4PROΔKに見られる[Angal S, King DJ, Bodmer MW, Turner A, Lawson AD, Roberts G, Pedley B, Adair JR. A single amino acid substitution abolishes the heterogeneity of chimeric mouse/human (IgG4) antibody. Mol Immunol. (1993);30(1):105-108]。
【0144】
一例として、疑念を避けるため、「VH1」は、「VH Variant 1」、「VH variant 1」、「VH Var 1」、または「VH var 1」と同様である。
【0145】
上記で例示されたVH/VLの組合せに限定されないと理解すべきである。当業者は、当然のことながら、過度な負担無く、また、発明性のある技術を用いずに、本明細書で開示されたVHおよびVLを総て再配列することができるであろう。
【0146】
本発明の新規な態様は、以下の核酸(縮重遺伝コードを含む):
a)本発明による抗体、またはその一つの機能的フラグメントもしくは誘導体をコードする核酸、DNA、またはRNA、
b)配列番号14〜19および41〜45からなる配列群から選択されるDNA配列を含んでなる核酸、
c)配列番号20、21、46、および47からなる配列群から選択されるDNA配列を含んでなる核酸、
d)b)またはc)に定義される核酸の、対応するRNA核酸、
e)a)、b)およびc)に定義される核酸の相補的核酸、および
f)配列番号14〜19および41〜45の配列のCDRの少なくとも一つと高ストリンジェンシー条件下でハイブリダイズすることができる少なくとも18ヌクレオチドの核酸
から選択されることを特徴とする単離された核酸に関する。
【0147】
下表5に、本発明の抗体に関する種々のヌクレオチド配列をまとめる。
【0148】
【表5】
【0149】
本明細書において互換的に用いられる「核酸」、「核配列」、「核酸配列」、「ポリヌクレオチド」、「オリゴヌクレオチド」、「ポリヌクレオチド配列」、および「ヌクレオチド配列」は、非天然ヌクレオチドを含む、または含まない、また二本鎖DNA、一本鎖DNA、またはこれらのDNAの転写産物のいずれかである核酸のフラグメントまたは領域を規定する、修飾または非修飾のヌクレオチドの正確な配列を意味する。
【0150】
ここでまた、本発明はそれらの天然の染色体環境の、すなわち天然状態のヌクレオチド配列には関しないということを含めておくべきである。本発明の配列は単離および/または精製されており、すなわち、それらは直接、または例えばコピーによって間接的にサンプリングされており、それらの環境は少なくとも部分的に改変されている。例えば宿主細胞の手段による組換え遺伝子技術によって得られた、または化学合成によって得られた単離された核酸もここに挙げておくべきであろう。
【0151】
「最適なアライメントの後に好ましい配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性割合を示す核配列」とは、参照核配列に比べて、特に、欠失、末端切断、延長、キメラ融合、および/または置換、特に規則的なものなどの、ある種の改変を示す核配列を意味する。好ましくは、これらは参照配列と同じアミノ酸配列をコードする配列であり、これは遺伝コードの縮重、または好ましくは高ストリンジェント条件(特に以下に定義されるもの)下で参照配列と特異的にハイブリダイズし得る相補的配列に関連づけられる。
【0152】
高ストリンジェント条件下でのハイブリダイゼーションとは、温度およびイオン強度に関する条件が、相補的DNAフラグメントの間でハイブリダイゼーションの維持を可能となるように選択される。単に例として、上記のポリヌクレオチドフラグメントを定義する目的でのハイブリダイゼーション工程の高ストリンジェント条件は有利には次の通りである。DNA−DNAまたはDNA−RNAハイブリダイゼーションは、(1)5×SSC(1×SSCは、0.15M NaCl+0.015Mクエン酸ナトリウムの溶液に相当する)、50%ホルムアミド、7%ドデシル硫酸ナトリウム(SDS)、10×デンハート溶液、5%デキストラン硫酸、および1%サケ精子DNAを含むリン酸バッファー(20mM、pH7.5)中、42℃で3時間のプレハイブリダイゼーション、(2)プローブの長さに応じた温度(すなわち、プローブ>100ヌクレオチド長では42℃)で20時間の主ハイブリダイゼーションと、その後の2×SSC+2%SDS中、20℃での20分の洗浄2回、0.1×SSC+0.1%SDS中、20℃での20分の洗浄1回という2段階で行う。最後の洗浄は、プローブ>100ヌクレオチド長では60℃で、0.1×SSC+0.1%SDS中、30分間行う。大きさの定義されたポリヌクレオチドに対する上記の高ストリンジェントハイブリダイゼーション条件は、それより長いまたは短いオリゴヌクレオチドに関しては、Sambrookら(Molecular cloning: a laboratory manual, Cold Spring Harbor Laboratory、第3版, 2001)に記載されている手順に従い、当業者によって適合させることができる。
【0153】
本発明はまた、以下の核酸:
a)本発明によるヒト化抗体重鎖、またはその誘導化合物もしくは機能的フラグメントをコードする核酸、DNA、またはRNA、
b)本発明によるヒト化抗体軽鎖、またはその誘導化合物もしくは機能的フラグメントをコードする核酸、DNA、またはRNA、
c)本発明によるヒト化抗体、またはその誘導化合物もしくは機能的フラグメンをコードする核酸、DNA、またはRNA、
d)a)、b)、またはc)に定義される核酸と相補的な核酸、
e)高ストリンジェント条件下で配列番号72または75〜77の核酸配列を含んでなる少なくとも重鎖とハイブリダイズすることができる少なくとも18ヌクレオチドの核酸、
f)高ストリンジェント条件下で配列番号73、74、86、87、または78、79、88、89の核酸配列を含んでなる少なくとも軽鎖とハイブリダイズすることができる少なくとも18ヌクレオチドの核酸、
の中から選択されることを特徴とする、単離された核酸分子を包含する。
【0154】
以下の表6に、本発明によるヒト化抗体515H7のそれぞれ種々の重鎖および軽鎖可変ドメインおよび全長(または完全)ヌクレオチド配列をまとめる。
【0155】
【表6】
【0156】
以下の表7に、本発明によるキメラ抗体515H7の種々の重鎖および軽鎖のヌクレオチド配列をまとめる。
【0157】
【表7】
【0158】
言い換えれば、本発明は、以下の核酸:
a)本発明による抗体、またはその一つの機能的フラグメントもしくは誘導体をコードする核酸、DNA、またはRNA、
b)配列番号14〜19および41〜45からなるCDR配列群から選択されるDNA配列を含んでなる核酸、
c)配列番号20、21、46、47、72、73、74、86、および87からなる重鎖および軽鎖可変ドメイン配列群から選択されるDNA配列を含んでなる核酸、
d)配列番号60〜63、75〜79、88、および89からなる重鎖および軽鎖配列群から選択されるDNA配列を含んでなる核酸、
e)配列番号55からなるDNA配列を含んでなる核酸、
f)b)、c)、d)、またはe)に定義される核酸の、対応するRNA核酸、
g)a)、b)、c)、d)、およびe)に定義される核酸の相補的核酸、および
h)高ストリンジェンシー条件下で配列番号14〜19および41〜45の配列のCDRの少なくとも一つとハイブリダイズすることができる少なくとも18のヌクレオチドの核酸
から選択されることを特徴とする、単離された核酸を取り扱う。
【0159】
本発明はまた、本明細書に記載される核酸を含んでなるベクターに関する。
本発明は特に、このようなヌクレオチド配列を含むクローニングおよび/または発現ベクターを対象とする。
【0160】
本発明のベクターは、好ましくは、所与の宿主細胞においてヌクレオチド配列の発現および/または分泌を可能とするエレメントを含む。よって、このベクターは、プロモーター、翻訳開始、および終結シグナル、ならびに好適な転写調節領域を含まなければならない。ベクターは宿主細胞において安定な様式で維持可能でなければならず、所望により、翻訳されたタンパク質の分泌を条件として指定する特異的シグナルを有してもよい。これらの種々のエレメントは、使用する宿主細胞に応じて、当業者により選択され、至適化される。この目的で、これらのヌクレオチド配列は、選択された宿主内で自己複製するベクターに挿入することもできるし、または選択された宿主の組込型ベクターであってもよい。
【0161】
このようなベクターは、当業者により一般に用いられる方法によって作製され、得られたクローンは、リポフェクション、エレクトロポレーション、熱ショック、また化学法などの標準的な方法によって好適な宿主に導入することができる。これらのベクターは、例えば、プラスミドまたはウイルス起源のベクターである。それらを用いて、本発明のヌクレオチド配列をクローニングする、または発現させるために宿主細胞を形質転換することができる。
【0162】
本発明はまた、本発明に記載されているベクターにより形質転換された、または本発明に記載されているベクターを含んでなる宿主細胞も含んでなる。
【0163】
宿主細胞は、細菌細胞、例えば、酵母細胞または動物細胞、特に、哺乳類細胞などの原核生物系、または真核生物系の中から選択することができる。また、昆虫または植物細胞も使用可能である。
【0164】
本発明はまた、本発明による形質転換細胞を有するヒト以外の動物に関する。
【0165】
本発明の別の態様は、本発明による抗体、またはその一つの機能的フラグメントの作製方法であって、以下の工程を含んでなることを特徴とする方法に関する:
a)本発明による宿主細胞の培地および好適な培養条件での培養工程、および
b)このようにして生産された該抗体、またはその一つの機能的フラグメントの、該培養培地からの、または該培養細胞からの回収工程。
【0166】
本発明による形質転換細胞は、本発明による組換えポリペプチドの作製方法に用いられる。また、本発明によるベクターおよび/またはベクターによって形質転換された細胞を使用することを特徴とする、本発明によるポリペプチドの組換え型での作製方法も本発明に含まれる。好ましくは、本発明によるベクターによって形質転換された細胞は、上記のポリペプチドの発現および該組換えペプチドの回収を可能とする条件下で培養される。
【0167】
すでに述べたように、宿主細胞は原核生物系または真核生物系の中から選択することができる。特に、このような原核生物系または真核生物系において分泌を助ける本発明のヌクレオチド配列を同定することが可能である。よって、このような配列を有する本発明によるベクターは、有利には分泌させる組換えタンパク質の生産のために使用することができる。実際、これらの着目する組換えタンパク質の精製は、宿主細胞内ではなく細胞培養の上清に存在するということによって容易となる。
【0168】
本発明のポリペプチドはまた、化学合成によっても製造することができる。このような製造方法の一つも本発明の目的である。当業者は、固相技術(特に、Steward et al., 1984, Solid phase peptides synthesis, Pierce Chem. Company, Rockford, 111,第2版参照)または部分固相技術、フラグメントの縮合または溶液中での従来の合成などの化学合成法を知る。化学合成によって得られ、対応する非天然アミノ酸を含み得るポリペプチドもまた、本発明に含まれる。
【0169】
本発明の方法によって得られる可能性のある抗体、またはその誘導化合物もしくは機能的フラグメントも本発明に含まれる。
【0170】
さらに別の態様によれば、本発明は、ヒトケモカインファミリー受容体と特異的に結合することができ、および/またはX4指向性HIV複製を特異的に阻害することができることをさらに特徴とする、上記のような抗体に関する。
【0171】
さらに別の態様によれば、本発明は、ヒトケモカインファミリー受容体と特異的に結合することができ、および/またはX4/R5指向性HIV複製を特異的に阻害することができることを特徴とする、上記のような抗体に関する。
【0172】
新規な実施形態によれば、本発明は、例えば、CCR5、CD4、CXCR4(本発明の抗体以外の、すなわち、別のエピトープを標的とする)またはCCR3、CCR2、CCR8、CXCR6、CXCR7、CX3CR1などのHIVの細胞侵入に関与する受容体と相互作用することができる第二のモチーフを含んでなるという意味で二重特異性である抗体からなる抗体、またはそれらの誘導化合物もしくは機能的フラグメントに関する。
【0173】
二重特異性または二官能性抗体は、二つの異なる可変領域が同じ分子において組み合わされている第二世代のモノクローナル抗体を構成する(Hollinger and Bohlen, 1999, Cancer and metastasis, rev. 18:411-419)。それらの有用性は、細胞表面のいくつかの分子を標的とするそれらの能力に関して診断用ドメインおよび治療用ドメインの双方で実証されており、このような抗体は化学法(Glennie MJ et al., 1987, J. Immunol. 139, 2367-2375、 Repp R. et al., 1995, J. Hemat., 377-382)または体細胞法(Staerz U.D. and Bevan M.J., 1986, PNAS 83, 1453-1457、 Suresh M.R. et al., 1986, Method Enzymol, 121:210-228)によるだけでなく、優先的には、求める抗体にヘテロ二量体形成を課し、従ってその抗体の精製を容易にすることを可能とする遺伝子操作技術(Merchand et al., 1998, Nature Biotech., 16:677-681)によって得ることができる。
【0174】
これらの二重特異性抗体は、完全なIgG、二重特異性Fab’2、Fab’PEG、ダイアボディー、または二重特異性scFvとして構築できるだけではなく、標的とされる各抗原に対して二つの結合部位が存在する四価二重特異性抗体(Park et al, 2000, Mol. Immunol, 37(18):1123-30)または上記のようなそのフラグメントとしても構築できる。
【0175】
一つの二重特異性抗体の製造および投与が、二つの特異的抗体の製造よりも安くつくことを考えた場合の経済的利点に加え、このような二重特異性抗体の使用は処置の毒性を軽減するという利点も持つ。実際、二重特異性抗体を使用すると、循環する抗体の総量を減らすことができ、結果として毒性が軽減される。
【0176】
本発明の好ましい実施形態においては、二重特異性抗体は二価または四価抗体である。
【0177】
最後に、本発明は、薬剤としての上記の抗体、またはそれらの一つの機能的フラグメントもしくは誘導体に関する。
【0178】
本発明はまた、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体からなる化合物を有効成分として含んでなる医薬組成物に関する。好ましくは、該抗体は賦形剤および/または薬学上許容される担体を追加する。
【0179】
本発明はまた、薬剤としての上記のような組成物に関する。
本発明の特定の態様では、抗体、またはその一つの機能的フラグメントもしくは誘導体、PBMCにおけるHIV−1 KON一次単離物の複製を、少なくとも5μg/ml、好ましくは少なくとも10μg/mlのIC90で阻害する。
【0180】
本発明はまた、HIV感染の予防または治療のための薬物および/または薬剤の製造のための本発明による抗体または組成物の使用を含んでなる。
【0181】
より詳細には、限定されるものではないが、該HIV感染はX4指向性HIV感染である。
【0182】
別の実施形態では、限定されるものではないが、該HIV感染はX4/R5指向性HIV感染である。
【0183】
本発明はまた、HIV複製を阻害するための薬物の製造のための、本発明による抗体、またはその機能的フラグメントもしくは誘導体、好ましくは、ヒト化されたもの、および/または組成物の使用に関する。一般に、本発明は、HIV疾患の予防または治療のための薬物の製造のための、抗体、またはその機能的フラグメントもしくは誘導体、好ましくは、ヒト化されたもの、および/または組成物の使用に関する。
【0184】
本明細書において、「医薬ビヒクル」とは、二次反応を引き起こさず、かつ、例えば、有効化合物の投与を容易にするか、生物内でのその寿命を延長し、および/または有効性を高めるか、溶液中でのその溶解度を高めるか、またはその貯蔵性を高める化合物、または化合物の組合せ(医薬組成物を含む)を意味する。このような医薬担体は周知であり、当業者により、選択される有効化合物の性質および投与経路に応じて適合される。
【0185】
好ましくは、このような化合物は、全身経路、特に、静脈内経路、筋肉内経路、皮内経路、腹腔内経路、皮下経路、腟内経路、または経口経路によって投与される。より好ましくは、本発明による抗体から構成される組成物は、一定時間あけた数回の投与で投与される。
【0186】
それらの投与経路、投与計画および最適な剤形は、例えば、患者の年齢または体重、患者の健康状態の重篤度、患者の処置に対する耐性および受ける副作用など、患者に適合する処置を確立する際に一般に考慮される基準に従って決定することができる。
【0187】
本発明はまた、同時、個別または延長様式で用いるための組合せ製剤として、CXCR4に対する抗体以外の抗HIV抗体または抗HIV細胞侵入抗体または抗HIV複製抗体をさらに含んでなる組成物に関する。
【0188】
さらに別の実施形態によれば、本発明はまた、本発明に記載されているもの以外の抗CCR5、抗CD4化合物、および抗CXCR4化合物などの、HIVの侵入および/または複製を特異的に阻害することができる化合物、または当業者に知られている他のいずれかの抗HIV化合物の中から選択される少なくとも一つの第二の抗HIV化合物を含んでなる上記のような医薬組成物に関する。
【0189】
本発明の補足的な別の実施形態は、同時、個別、または延長使用のための組合せまたは複合製剤として抗HIV化合物をさらに含んでなる上記のような組成物からなる。
【0190】
「同時使用」とは、単一の投与形に含まれる組成物に含まれる両化合物を投与することを意味する。
【0191】
「個別使用」とは、異なる投与形に含まれる組成物の両化合物を同時に投与することを意味する。
【0192】
「延長使用」とは、異なる投与形にそれぞれ含まれる組成物の両化合物の連続的投与を意味する。
【0193】
一般に、本発明による組成物は、HIV処置の有効性を著しく高める。言い換えれば、本発明の抗体の治療効果は、抗HIV剤の投与によって予期されないほど増強される。本発明の組成物によってもたらされる別の大きな二次的利点は、有効成分のより低量の有効用量を使用できる可能性に関し、従って、特に抗HIV剤の作用である副作用の出現の危険を回避または軽減することが可能となる。さらに、この組成物は、予想される治療効果をより迅速に達成することを可能とする。
【0194】
「治療用抗HIV剤」とは、患者に投与した際に、患者におけるHIVの複製を治療または予防する物質を意味する。このような薬剤の限定されない例としては、HIVプロテアーゼ阻害剤(PI)、ヌクレオシド/ヌクレオチドHIV逆転写酵素阻害剤(NRTI/NtRTI)、非ヌクレオシドHIV逆転写酵素阻害剤(NNRTI)、HIV侵入阻害剤、HIVインテグラーゼ阻害剤等の「抗レトロウイルス剤」が挙げられる。
【0195】
このような薬剤は、例えば、VIDALの、「抗HIV化合物」に関する化合物のために費やされた頁に挙げられており、この文献の参照文献によって挙げられている抗HIV化合物は、本明細書において、限定的されない好ましい抗HIV剤として挙げられる。
【0196】
HIVプロテアーゼ阻害剤は、HIVプロテアーゼ活性を阻害することができるいずれの物質も表す。このようなHIVプロテアーゼ阻害剤の例としては、限定されるものではないが、メシル酸サキナビルまたはSQV(Invirase(商標))、インジナビルまたはIDV(Crixivan(商標))、リトナビルまたはRTV(Norvir(商標))、ネルフィナビルまたはNFV(Viracept(商標))、アンプレナビル(Agenerase(商標)、Prozei(商標))、ロピナビル/リトナビルまたはLPV/r(Kaletra(商標)、Aluvia(商標))、アタザナビルまたはATV(Reyataz(商標)、Zrivada(商標))、ホスアンプレナビルまたはFPV(Lexiva(商標)、Telzir(商標))、チプラナビルまたはTPV(Aptivus(商標))、ダルナビルまたはDRV(Prezista(商標))が挙げられる。
【0197】
HIVヌクレオシドまたはヌクレオチド逆転写酵素阻害剤(NRTI)は、HIV RNAの逆転写を遮断するヌクレオシドまたはヌクレオチド類似体である物質を表す。NRTIの例としては、限定されるものではないが、ジドブジンまたはAZT,ZDV(Retrovir/combivir/trixivir(商標))、ジダノシンまたはddi(Videx(商標))、ザルシタビン(HIVID(商標))、スタブジンまたはd4T(Zerit(商標))、ラミブジンまたは3TC(Epivir/combivir/epzicom/trixivir(商標))、アバカビルまたはABC(Ziagen/trixivir/epzicom(商標))、フマル酸テノホビルジソプロキシルまたはTDF(Viread/atripla/truvada(商標))、エムトリシタビンまたはFTC(Emtriva/atripla/truvada(商標))が挙げられる。
【0198】
非ヌクレオシドHIV逆転写酵素阻害剤(NNRTI)は、HIV RNAの逆転写を遮断するヌクレオシドまたはヌクレオチド類似体ではない物質を表す。NNRTIの例としては、限定されるものではないが、ネビラピンまたはNVP(Viramune(商標))、エファビレンツまたはEFV(Sustiva/atripla(商標)、Stocrin(商標))、デラビルジンまたはDLV(Rescriptor(商標))、およびエトラビリンまたはETR(Intelence(商標))が挙げられる。
【0199】
HIV侵入阻害剤は、HIVの細胞侵入を遮断する物質を表す。HIV侵入阻害剤の例としては、限定されるものではないが、エンフビルチドまたはT20(Fuzeon(商標))、マラビロクまたはMVC(Celsentri(商標)、Celzentry(商標))が挙げられる。
【0200】
HIVインテグラーゼ阻害剤は、HIVインテグラーゼ活性を阻害する物質を表す。インテグラーゼ阻害剤の例としては、限定されるものではないが、ラルテグラビルまたはRAL(Isentress(商標))が挙げられる。
【0201】
このような薬剤はまた、例えば、限定されるものではないが、ビクリビロク、PRO140、TNX−355、AMD070、ラシビル、アプリシタビン、エルブシタビン、フロサルブジン(Flosalvudine)、リルピビリン、エルビテグラビルなど、現在臨床試験中の、VIDALに記載されている同種の薬物に属する化合物である。
【0202】
このような薬剤はまた、例えば、限定されるものではないが、成熟阻害剤(ベビリマット)、β−ガラクトシル−セラミドのグリコシド類似体、炭水化物結合剤、RNアーゼH阻害剤、HIV遺伝子発現阻害剤、潜在型T細胞からのHIV放出の刺激剤(バルプロ酸など)など、他の可能性のある種の薬物に属する化合物である。
【0203】
特に好ましい実施形態においては、組合せ製剤としての本発明の組成物は、前記抗HIV剤が同時使用のための前記抗体に化学的に結合されていることを特徴とする。
【0204】
前記抗HIV剤と本発明による抗体の間の結合を容易にするために、結合する二つの化合物の間にポリ(アルキレン)グリコール、ポリエチレングリコール、またはアミノ酸などのスペーサー分子を導入することができ、あるいは、別の実施形態においては、前記抗体と反応することができる官能基を導入されている前記抗HIV剤の活性誘導体を使用することもできる。これらの結合技術は当業者によく知られており、本明細書ではこれ以上詳細には述べない。
【0205】
また、好ましくは、前記の複合体を形成する本発明の抗体は、その機能的フラグメント、特に、それらのFc成分が欠失されたフラグメント、例えばscFvフラグメントの中から選択される。
【0206】
本発明はまた、薬物として用いられる、本発明による、組合せ製剤としての組成物、または抗CXCR4 Mab/抗HIV薬物複合体に関する。
【0207】
好ましくは、組合せ製剤としての前記組成物または前記複合体には、賦形剤および/または医薬ビヒクルが添加される。
【0208】
よって、本発明は、HIV複製に対して生物的に活性な化合物の特異的標的化のための薬物の製造のための、抗体、またはその一つの機能的フラグメントもしくは誘導体の使用に関する。
【0209】
従前に示したように、CXCR4 Mab 515H7および301aE5は、PBMCにおけるHIV−1複製に対して強い活性を有し、従って、このような抗体は、HIV−1感染を処置するためのCXCR4アンタゴニスト抗ウイルス薬の同定のためのスクリーニングアッセイに使用することができる。これらのアッセイの第一の工程では、CXCR4を発現する細胞をMab 515H7および/または301aE5とともに培養し、その後、Mab 515H7および/または301aE5との結合の潜在的な阻害のための分子を評価することができる。この種のアッセイで用いられる細胞は、CHO−CXCR4、NIH3T3−CXCR4またはCXCR4トランスフェクトヒト細胞系統(例えば、U373−MAGI−CXCR4)などのトランスフェクト細胞系統、NALM6などのCXCR4を発現するヒト細胞系統またはPBMCなどの一次細胞であり得る。CXCR4発現細胞に対するMab 515H7および/または301aE5の結合を阻害するCXCR4のアンタゴニストをスクリーニングするために使用される方法は、Zhao Q.ら(AIDS Research And Human Retroviruses, 2003, 19, pp947-955)によって記載されている細胞に基づく競合的酵素結合免疫吸着アッセイ(ELISA)またはJuarez J.ら(Leukemia 2003, 17, ppl294-1300)によって記載されている蛍光活性化細胞選別法(FACS)を用いるプロトコールであり得る。
【0210】
よって、本発明の特定の態様では、CXCR4アンタゴニスト抗ウイルス薬としての分子をスクリーニングおよび/または同定する方法であって、以下の工程を含んでなる方法が考えられる:
a)CXCR4を発現する細胞を選択する工程、
b)該細胞を、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体とともに培養する工程、
c)抗体、またはその一つの機能的フラグメントもしくは誘導体と、CXCR4との間の結合の潜在的な阻害のための供試分子を評価する工程、および
d)該阻害が可能な分子を選択する工程。
【0211】
別の特定の実施形態では、
e)これらの分子をHIV−1複製アッセイで試験する工程
を追加することができる。
【0212】
本発明の他の特徴および利点は、実施例および図面(凡例は下記に示される)とともに説明を読めばさらに明らかになる。
【実施例】
【0213】
実施例1:ヒトCXCR4に対するモノクローナル抗体(Mab)の作製
CXCR4に対するモノクローナル抗体を作製するため、Balb/cマウスに組換えNIH3T3−CXCR4細胞および/またはCXCR4細胞外N末端およびループに相当するペプチドを感作させた。6〜16週齢のマウスに、完全フロイントアジュバントにて抗原を1回、皮下(s.c.)感作させた後、フロイントの不完全アジュバントにて抗原を2〜6回s.c.感作させた。免疫応答を眼窩後方採血によって測定した。血清をELISAによりスクリーニングし(下記の通り)、抗CXCR4抗体力価が高いマウスを融合に用いた。屠殺および脾臓摘出の2日前にマウスの静脈内に抗原を追加投与した。
【0214】
ELISA
抗CXCR4抗体を産生するマウスを選択するため、免疫マウスからの血清をELISAによって調べた。要するに、マイクロタイタープレートを、BSAと結合された精製[1−41]N末端ペプチドで5μg相当量のペプチド/mL、100μL/ウェルにてコーティングし、4℃での一晩培養した後、PBS中0.5%ゼラチン250μL/ウェルでブロックした。CXCR4免疫マウスからの血漿希釈液を各ウェルに加え、37℃で2時間培養した。これらのプレートをPBSで洗浄した後、HRP(Jackson Laboratories)と結合されたヤギ抗マウスIgG抗体とともに37℃で1時間培養した。洗浄後、プレートをTMB基質で現像し、5分後に100μL/ウェルの1M H2SO4を加えることによって反応を停止させた。最高力価の抗CXCR4抗体を生じたマウスを抗体作製に用いた。
【0215】
CXCR4に対するMabを産生するハイブリドーマの作製
最高力価の抗CXCR4抗体を生じたBalb/cマウスから単離したマウス脾細胞を、PEGを用いてマウス骨髄腫細胞系統Sp2/Oと融合させた。細胞をマイクロタイタープレートにおよそ1×105/ウェルで播種した後、ウルトラカルチャー培地+2mM L−グルタミン+1mMピルビン酸ナトリウム+1×HATを含有する選択培地で2週間インキュべートした。次に、ウェルをELISAによって、抗CXCR4モノクローナルIgG抗体に関してスクリーニングした。その後、抗体を分泌するハイブリドーマを制限希釈により少なくとも2回サブクローニングし、in vitroで培養し、さらなる分析のための抗体を作製した。
【0216】
実施例2:FACS分析による抗CXCR4 Mab 515H7および301aE5結合特異性(NIH3T3−CXCR4形質転換体)の特性決定
この実験では、抗CXCR4 Mab 515H7および301aE5のヒトCXCR4(hCXCR4)への特異的結合をFACS分析によって調べた。NIH3T3およびNIH3T3−hCXCR4トランスフェクト細胞を、10μg/mlのモノクローナル抗体515H7および301aE5とともに培養した。その後、これらの細胞を1%BSA/PBS/0.01%NaN3で洗浄した。次に、Alexa標識された二次抗体をこれらの細胞に加え、4℃で20分間培養した。その後、細胞を再び2回洗浄した。2回目の洗浄後、FACS分析を行った。これらの結合試験の結果を、[FACSによって得られた平均蛍光強度(MFI)]を示す下表8に示す。抗CXCR4 Mab 515H7および301aE5はヒトCXCR4−NIH3T3トランスフェクト細胞系統と特異的に結合するが、親NIH3T3細胞に対する認識は見られなかった。
【0217】
【表8】
【0218】
実施例3:FACS分析による末梢血単核細胞(PBMC)に対する抗CXCR4 Mab 515H7および301aE5の結合の特性決定
健康なドナーから血液をバフィーコートとして採取した。100μlの全血を示された濃度の抗ヒトCXCR4抗体(クローン515H7および301aE5)とともに4℃で20分間培養した。血液をPBS−BSA 1%−NaN3 0.01%中で3回洗浄し、1:500希釈のヤギ抗ヒトAlexa 488 IgG(Invitrogen)とともに4℃で20分間培養した。その後、細胞を洗浄し、CD14−PE(Caltag)またはCD3−PE(Caltag)とともに4℃で10分間培養し、3回洗浄した。赤血球をHigh-Yield溶解液(Caltag)で室温にて10分間溶解させた。すぐにFacscalibur (Becton-Dickinson)を用いて細胞を分析した。単球でのCXCR4発現はCD14陽性細胞で行い、T細胞でのCXCR4発現はCD3陽性細胞で行った(図1)。結果は抗原結合能(ABC)で表す。
【0219】
図2Aおよび2Bに示されるように、抗ヒトCXCR4クローン515H7および301aE5はTリンパ球(図2A)および単球(図2B)の双方を染色したが、このことは、515H7および301aE5 Mabが単球およびTリンパ球の細胞表面で発現された天然型のCXCR4を認識可能であることを示している。
【0220】
実施例4:生物発光共鳴エネルギー移動(BRET)アプローチによるCXCR4ホモ二量体に対する515H7および301aE5 Mabの効果
この機能的アッセイは、SDF−1および/または515H7 MabがCXCR4受容体に結合した際に誘発されるコンフォメーション変化をCXCR4ホモ二量体のレベルで評価することができる。
【0221】
検討する相互作用相手の発現ベクターを、従来の分子生物学的技術を適用することにより、対応する色素(ウミシイタケ(Renilla reniformis)ルシフェラーゼRlucおよび黄色蛍光タンパク質YFP)との融合タンパク質として構築した。BRET試験を行う2日前に、HEK293細胞を、対応するBRET相手:[CXCR4/Rluc+CXCR4/YFP]をコードする発現ベクターで一時的にトランスフェクトし、CXCR4ホモ二量体形成を調べた。翌日、細胞を、ポリリシンをプレコーティングした白色96MWプレートの完全培養培地[10%FBSを添加したDMEM]に分注した。まず、細胞をプレートに接着させるために、CO2 5%、37℃で培養した。その後、200μl DMEM/ウェルで一晩、細胞を飢餓状態にした。BRET試験の直前に、DMEMを除去し、細胞を手早くPBSで洗浄した。その後、細胞をPBS中、抗体の存在下または非存在下、37℃にて10分間培養した後、最終量50μl中、SDF−1 300nMとともに、または伴わずにセレンテラジンH 5μMを添加した。37℃でさらに10分間培養した後、Mithras LB940マルチラベルリーダー(Berthold)(1s/波長/ウェル、室温で15反復)を用い、485nmおよび530nmでの発光獲得を誘導した。
BRET比の計算は従前に記載されているように行った(Angersら、2000):[(発光530nm)−(発光485nm)×Cf]/(発光485nm)、式中、Cf=同じ試験条件下でRluc融合タンパク質単独を発現する細胞の(発光530nm)/(発光485nm)。この式を簡単にすると、BRET比が、二つのBRET相手が存在する場合に得られる530/485nm比を、同じ試験条件下で、Rlucと融合された相手だけがアッセイ中に存在する場合に得られる530/485nm比により補正したものに相当することが示される。読み取りやすいように、結果はミリBRET単位(mBU)で表すが、mBUはBRET比に1000を掛けたものである。
【0222】
SDF1(300nM)は、CXCR4受容体と融合されたアダプタータンパク質とアクセプタータンパク質が空間的に近接していることから生じるBRETシグナルを約20%増強したが、それはおそらくCXCR4/CXCR4ホモ二量体形成または既存の二量体のコンフォメーション変化を示唆する(図3AおよびB)。515H7および301aE5 Mabは、CXCR4ホモ二量体に関してSDF−1により誘発されるコンフォメーション変化を調節することができた(515H7および301aE5に関して、SDF−1により誘発されるBRET増強の69%阻害、図3AおよびB)。515H7および301aE5 Mabはまた、それ自体、CXCR4/CXCR4の空間的近接を調節することもできたが、このことはCXCR4/CXCR4ホモ二量体コンフォメーションに対する515H7および301aE5 Mabの影響を示唆する(図3Aおよび3B)。
【0223】
実施例5:抗CXCR4 Mab 515H7および301aE5による、ヒトPBMCにおけるHIV−1一次単離物KON(X4ウイルス)複製の阻害
HIV−1に対して血清反応陰性の健康なドナーからのPBMCをバフィーコートから、またはフィコール−ハイパーク勾配遠心分離による血球成分分離法(cytapheresis)から単離した。PBMCを、25mM HEPES、5mlペニシリン(10000U/ml)−ストレプトマイシン(10000μg/ml)2mM L−グルタミンを含有し、10%熱不活性化FCSを添加したRPMI 1640細胞培養培地中、PHAの存在下で活性化し、1サイクルの中和アッセイで細胞標的として用いた。一次ヒトPBMCにおけるHIV−1複製は、ウイルスp24抗原の細胞内染色をFACS分析によって分析することにより行った。要するに、25μl/ウェルの、種々の希釈率のMab 515H7、301aE5および12G5(R&D Systems)または対照として培養培地(RPMI 1640、10%FCS、0.1%IL−2)を、25μl/ウェルのHIV−1 KON X4一次単離物希釈液とともに37℃で1時間、二反復で培養した。PHA活性化ヒトPBMC(25μl/ウェル)を96ウェルプレート(U底、Costar 3599)中のMab/ウイルス混合物に20×106細胞/mlで加え、RPMI 1640、10%FCSおよび0.1%IL−2中、37℃で24〜36時間培養した。Mabを含まない培地中、非感染PBMCからなる対照を導入した。HIV感染PBMCを検出するために、ウイルスp24抗原の細胞内染色を行い、フローサイトメトリーによって分析した。細胞を固定し、Cytofix/Cytopermキット(Becton Dickinson)を製造者のプロトコールに従って用いて透過処理を施し、1/160希釈で用い、暗所にて4℃で10分間培養した蛍光抗p24 Mab(クローンKC57−Coulter Beckman)で染色した。PBS−3%FCS培地で洗浄した後、フローサイトメトリー分析前にPBMCをPBSで希釈した。種々のサンプルにおけるp24陽性細胞の割合を、生細胞集団に対する20,000事象をゲートすることにより決定した。この生細胞のサブセットのp24発現を、非感染細胞のバックグラウンド染色と比べて分析した。p24抗原陽性値は、mock感染細胞におけるバックグラウンド事象を差し引いた後に得た。中和の割合は、Mabを含まない対照感染ウェルと比べてのp24陽性細胞の減少と定義した。中和力価は、感染細胞の割合を90%低下させるMabの希釈率と定義した。抗CXCR4 Mab 515H7および301aE5を、HIV適用の参照の抗CXCR4 Mabとして知られる12G5 Mabと比較した。図4AおよびBおよび5に示されるように、抗CXCR4 Mab 515H7および301aE5は、それぞれIC90 10μg/ml(66nM)および150μg/ml(1μM)で、PBMCにおけるHIV−1 KON一次単離物の複製を阻害することができるが、12G5 MabはPBMCにおけるHIV−1 KON一次単離物の複製を阻害することができなかった(図4A)。
【0224】
実施例6: CXCR4受容体により媒介される細胞内カルシウム貯蔵の動員
この機能的アッセイは、小胞体の細胞内貯蔵からのカルシウム遊離を誘導するホスホリパーゼC経路の刺激を介したCXCR4受容体シグナル伝達を測定するように設計された。
【0225】
ヒトCXCR4受容体を安定的かつ構成的に発現するCHO−K1細胞を、ナイーブCHO−K1細胞(ATCC CCL−61)は、ヒトCXCR4受容体の全コード配列を含む哺乳類発現ベクター(RefSeq NM_003467)でトランスフェクトした際に得られたものである。細胞を完全培養培地[5%ウシ胎仔血清(FCS)および500μg/mlのゲネチシンを添加したDMEM−ハムF12培地]で増殖させた。細胞を黒色96MWプレートに適当な培養培地中100,000細胞/ウェルで播種した。試験前に細胞を一晩飢餓状態にした。細胞にローディングバッファー[HBSS 1×、HEPES 20mM、プロベネシド酸(Probenicid acid)25mM]中、蛍光カルシウム色素(Fluo-4 No Wash, Invitrogen US)を、37℃で30分間、その後、25℃で30分間負荷した。SDF−1による刺激は、各ウェルに直接注入することにより行った。拮抗作用試験では、10μlのMab溶液をSDF−1の少なくとも10分前にローディングバッファーに直接添加する。動的蛍光測定は、マルチモード蛍光マイクロプレートリーダーMithras LB940(Berthold)にて、以下の設定で行う:485nmでの励起、535nmでの発光、10000任意単位での励起エネルギー。各ウェルの蛍光をSDF−1注入前に毎秒0.1秒、20秒間記録する(基底シグナル)。その後、20μlのSDF−1を注入し、データ記録を2分間続ける。各試験条件を2反復で行う。各ウェルの値をまず、基底蛍光と細胞を含まない対照ウェルによって発せられた蛍光を差し引くことによって補正する。相対的データをSDF−1(100nM)によって得られた最大刺激の割合として表す。
【0226】
SDF1(100nM)は、組換えCHO/CXCR4において細胞内カルシウムの迅速かつ強い放出を誘導したが、ナイーブCHO−K1細胞では蛍光シグナルは検出されなかった。最大強度は基底蛍光の140%超に達し、SDF−1による刺激時に約40秒で見られた(図6A、6B、および6C)。Mab 515H7(133nM)(図6A)およびc515H7(133nM)(図6C)は、SDF−1(100nM)により誘発されたカルシウムシグナルの強い阻害をもたらした。Mab 301aE5(133nM)(図6B)は、SDF−1(100nM)により誘発されたカルシウムシグナルの部分的阻害をもたらした。
【0227】
実施例7:抗CXCR4 Mab 515H7、c515H7および301aE5による、ヒトPBMCにおけるHIV−1一次単離物KON、MN、および92UG024(X4ウイルス)の複製の阻害
1サイクル中和アッセイ
このアッセイは、一次単離物KON、MN、および92UG024を用いて36時間行い、感染2日後に2%の感染CD4 Tリンパ球の検出を可能とするように相応に濃縮および希釈する。
【0228】
種々の希釈率のMab 515H7、c515H7および301aE5 25マイクロリットルを、25μlのウイルスとともに37℃で1時間培養した。20×106細胞/mlのヒトPBMC(25μl)を96ウェルプレート(U底、Costar 3599)中のMab/ウイルス混合物に加え、RPMI 1640 10%FCSおよび20U/ml IL−2(R&D Systems, Minneapolis, MN)中で36時間培養した。
【0229】
2日間の培養後、HIV感染リンパ球を、ウイルスp24 Agの細胞内染色により検出した。細胞を固定し、製造者に従いCytofix/CytopermおよびPerm/Washキット(BD Biosciences)の双方を用いて透過処理を施し、Perm/Wash溶液中1/160希釈で用い、4℃で15分間加える蛍光抗p24 Mab(FITC−またはPE−抗p24、クローンKC57、Beckman Coulter/Immunotech, Hialeah, FL)で染色した。3%FBSを含むPBSで洗浄した後、PBMCを300μlのPBSに希釈した後、DIVAソフトウエア(BD Biosciences)を用いてフローサイトメトリー分析(LSRII、 BD Biosciences)を行った。種々のサンプル中のp24陽性細胞の割合を、前方および側方散乱パラメーターによって同定された生細胞集団に対する20,000事象をゲートすることにより決定した。これらの生細胞のサブセットは、live/dead solution kit (Invitrogen)を用いて分析した。p24 Ag陽性値は、mock感染細胞におけるバックグラウンド事象を差し引いた後に得た。
【0230】
中和パーセントは、Mabを含まない対照感染ウェルと比べてのp24陽性細胞の減少と定義した。中和力価は、感染細胞の割合を90%低下させる抗体の濃度(3反復で行った連続希釈率の間に挿入)と定義した。
【0231】
図7、8および9に示されるように、抗CXCR4 Mab 515H7、c515H7、および301aE5は、PBMCにおけるHIV−1 X4 MN、KON、および92UG024一次単離物の複製を阻害することができる。ICs(μg/mlにおいて)の結果を表9にまとめる。
【0232】
【表9】
【0233】
実施例8:抗CXCR4 Mab 515H7、c515H7および301aE5による、ヒトPBMCにおけるHIV−1一次単離物89.6(X4/R5二重ウイルス)の複製の阻害
1サイクル中和アッセイ
このアッセイは、一次単離物89.6を用いて36時間行い、感染2日後に2%の感染CD4 Tリンパ球の検出を可能とするように相応に濃縮および希釈する。
【0234】
種々の希釈率のMab 515H7、c515H7および301aE5 25マイクロリットルを、25μlのウイルスとともに37℃で1時間培養した。20×106細胞/mlのヒトPBMC(25μl)を96ウェルプレート(U底、Costar 3599)中のMab/ウイルス混合物に加え、RPMI 1640 10%FCSおよび20U/ml IL−2(R&D Systems, Minneapolis, MN)中で36時間培養した。
【0235】
2日間の培養後、HIV感染リンパ球を、ウイルスp24 Agの細胞内染色により検出した。細胞を固定し、製造者に従いCytofix/CytopermおよびPerm/Washキット(BD Biosciences)の双方を用いて透過処理を施し、Perm/Wash溶液中1/160希釈で用い、4℃で15分間加える蛍光抗p24 Mab(FITC−またはPE−抗p24、クローンKC57、Beckman Coulter/Immunotech, Hialeah, FL)で染色した。3%FBSを含むPBSで洗浄した後、PBMCを300μlのPBSに希釈した後、DIVAソフトウエア(BD Biosciences)を用いてフローサイトメトリー分析(LSRII、 BD Biosciences)を行った。種々のサンプル中のp24陽性細胞の割合を、前方および側方散乱パラメーターによって同定された生細胞集団に対する20,000事象をゲートすることにより決定した。これらの生細胞のサブセットは、live/dead solution kit (Invitrogen)を用いて分析した。p24 Ag陽性値は、mock感染細胞におけるバックグラウンド事象を差し引いた後に得た。
【0236】
中和パーセントは、Mabを含まない対照感染ウェルと比べてのp24陽性細胞の減少と定義した。中和力価は、感染細胞の割合を90%低下させる抗体の濃度(3反復で行った連続希釈率の間に挿入)と定義した。
【0237】
図10および11に示されるように、抗CXCR4 Mab 515H7、c515H7、および301aE5は、PBMCにおけるHIV−1 89.6一次単離物の複製を阻害することができる。ICs(μg/ml)の結果を表10にまとめる。
【0238】
【表10】
【0239】
実施例9:抗CXCR4 Mab c515H7と抗CCR5分子マラビロクの併用による、ヒトPBMCにおけるHIV−1一次単離物89.6およびUG93067(X4/R5二重ウイルス)の複製の阻害
中和アッセイ、一次PBMCにおける複数回のHIV一次単離物複製の分析:
c515H7 Mabまたはマラビロクの連続希釈液を併用する、または両者とウイルスの連続希釈液を併用するこのアッセイは、PBMC(末梢血単核細胞)に対する複数回の感染を分析する。要するに、c515H7 Mabまたはマラビロクまたは両者の組合せの連続希釈液(2倍)の25μlアリコート四つをそれぞれ、予め含水させた96ウェルフィルタープレート(孔径1.25μm、Durapor Dv、 Millipore, Molsheim, France)にて、ウイルスの連続希釈液25μlとともに培養した。ウイルスの対照滴定(希釈c515H7 Mabまたはマラビロクの代わりに25μlのRPMI)を、c515H7 Mabまたはマラビロクまたは両者の組合せの希釈液の存在下での滴定と同じプレートで行った。37℃で1時間後、4×106細胞/mlの濃度のPHA刺激PBMC 25μl(5人の健康なドナーからのPHA活性化PBMCのプール)を、RPMIの最終培養容量75μl、10%ウシ胎仔血清(FCS)および20IUのインターロイキン−2(IL−2)/ml(R&D System)となるように加えた。37℃で24時間後、100μlの同じ培養培地を加えた。4日目に濾過により2回の洗浄(各200μlのRPMI)を行ってc515H7 Mabおよびマラビロクを除去し、200μlの新鮮培養培地を加えた。7日目に、培養上清におけるp24の存在をELISAにより測定し、陰性対照(ウイルス希釈液を感染させ、10−6Mジドブジン[AZT]の存在下で維持した培養物)の場合と比べ、陽性ウェルを判定した。4反復のウェルを用い、各希釈率のc515H7 Mabまたはマラビロクまたは両者の組合せの不在下(F0)および存在下(Vn)でウイルス力価(50%組織培養感染量[TCID50])を決定した。中和力価は、ウイルス力価の90%低下をもたらす(Vn/V0=0.1)c515H7 Mabまたはマラビロクまたは両者の組合せの希釈率として定義した。
【0240】
図12に示されるように、二重指向性X4R5ウイルス89.6の複製は、c515H7 Mabにより、IC90 2μg/mlで阻害された(図12)。マラビロク50μg/mlでは、IC90阻害活性に達しなかった(図12)。さらに、2μg/mlのマラビロクを抗体515H7に添加したところ、IC90 0.2μg/mlでc515H7 Mabの阻害活性を増強した(図12)。
【0241】
c515H7 Mabとおよびマラビロクの組合せの有益な効果を、種々の希釈率のこれらの2分子および別の二重指向性ウイルスUG93067を用いて評価した。図13に示されるように、Mab c515H7およびマラビロクの阻害活性は同等であった。これらの結果は、ウイルスUG93067の、CCR5受容体またはCXCR4受容体のいずれかを用いる能力が匹敵していたことを示唆する。UG93067ウイルスを用いると、より良い活性を証明することができ、これらのX4(c515H7 Mab)とR5(マラビロク)阻害剤(各10μg/ml)の組合せだけが、ウイルス力価の90%低下を可能とした(図13)。
【0242】
実施例10:抗CXCR4キメラMab c515H7の生産
ネズミ515H7 Mabのキメラ形式を設計した。これは、着目するネズミ抗体の軽鎖および重鎖可変ドメインを、ヒトCκおよびIgG1/IgG2/IgG4定常ドメインと遺伝的に融合したものに相当する。この組換えMabは、HEK293/EBNA系をpCEP4発現ベクター(InVitrogen, US)とともに使用することによって、一時的トランスフェクション時に作製されたものである。
【0243】
個々のアミノ酸配列およびヌクレオチド配列は本明細書に上記した。さらに、上記の表3にIgG2および4イソ型(好ましいイソ型)の配列が開示されているため、ここでも、IgG1イソ型の重鎖、すなわち、配列番号80のアミノ酸配列および配列番号81のヌクレオチド配列に相当するc515H7 VH(G1wt)の配列を記載することができる。
【0244】
515H7 Mab軽鎖および重鎖の可変ドメインに相当する全ヌクレオチド配列は、グローバル遺伝子合成(Genecust, Luxembourg)によって合成されたものである。それらを、ヒトIgG1/IgG2/IgG4免疫グロブリンの軽鎖[Cκ]または重鎖[CH1−Hinge−CH2−CH3]のいずれかの定常ドメインの全コード配列を有するpCEP4ベクター(InVitrogen, US)にサブクローニングした。全クローニング工程は、the Laboratory manual (Sambrook and Russel, 2001)に記載されている従来の分子生物学的技術に従って、または供給者の説明書に従って行った。各遺伝子構築物を、Big Dyeターミネーター・サイクル・シーケンシング・キット(Applied Biosystems, US)を用いたヌクレオチド配列決定によって完全にバリデートし、3100ジェネティック・アナライザー(Applied Biosystems, US)を用いて分析した。
【0245】
懸濁適合HEK293 EBNA細胞(InVitrogen, US)を通常、オービタルシェーカー(回転速度110rpm)上、250mlフラスコにて、6mMグルタミンを添加した血清不含培地Excell 293(SAFC Biosciences)50ml中で増殖させた。一時的トランスフェクションは、水中終濃度1mg/mlで調製した直鎖25kDaポリエチレンイミン(PEI)(Polysciences)、混合およびプラスミドDNA(重鎖:軽鎖プラスミド比1:1で、終濃度1.25μg/ml)を用い、2.106細胞/mlで行った。トランスフェクション4時間後に、培養物を1容量の新鮮培養培地で希釈して、最終細胞密度を106細胞/mlとした。培養方法を細胞の生存率およびMab生産に関して測定した。一般に、培養は4〜5日間測定した。Mabは、プロテインA樹脂(GE Healthcare, US)での従来のクロマトグラフィーアプローチを用いて精製した。Mabは機能的評価に好適なレベルで生産された。生産性レベルは一般に、精製Mab 6〜15mg/lの範囲である。
【0246】
実施例11:FACS分析による抗CXCR4キメラMab c515H7結合特異性の特性決定
この試験では、抗CXCR4キメラMab c515H7のヒトCXCR4に対する特異的結合をFACS分析によって調べた。
【0247】
NIH3T3−hCXCR4トランスフェクト細胞を0μg/ml〜10μg/mlの用量範囲のモノクローナル抗体c515H7とともに培養した。その後、これらの細胞を1%BSA/PBS/0.01%NaN3で洗浄した。次に、Alexaで標識した二次抗体をこれらの細胞に加え、4℃で20分間インキュベートした。その後、これらの細胞を再び2回洗浄した。2回目の洗浄後に、FACS分析を行った。この結合試験の結果を図15に示すが、これは、抗CXCR4キメラMab C515H7がヒトCXCR4−NIH3T3トランスフェクト細胞系統と特異的に結合することを示す。NIH3T3wt細胞との結合は検出されなかった(データは示されていない)。
【0248】
実施例12:生物発光共鳴エネルギー移動(BRET)アプローチによるCXCR4ホモ二量体に対するc515H7 Mabの効果
この機能的アッセイは、SDF−1および/またはc515H7 MabがCXCR4受容体に結合した際に誘発されるコンフォメーション変化をCXCR4ホモ二量体のレベルで評価することができる。
【0249】
検討する相互作用相手の発現ベクターを、従来の分子生物学的技術を適用することにより、対応する色素(ウミシイタケ(Renilla reniformis)ルシフェラーゼRlucおよび黄色蛍光タンパク質YFP)との融合タンパク質として構築した。BRET試験を行う2日前に、HEK293細胞を、対応するBRET相手:[CXCR4/Rluc+CXCR4/YFP]をコードする発現ベクターで一時的にトランスフェクトし、CXCR4ホモ二量体形成を調べた。翌日、細胞を、ポリリシンをプレコーティングした白色96MWプレートの完全培養培地[10%FBSを添加したDMEM]に分注した。まず、細胞をプレートに接着させるために、CO2 5%、37℃で培養した。その後、200μl DMEM/ウェルで一晩、細胞を飢餓状態にした。BRET試験の直前に、DMEMを除去し、細胞を手早くPBSで洗浄した。その後、細胞をPBS中、抗体の存在下または非存在下、37℃にて10分間インキュベートした後、最終量50μl中、SDF−1 100nMとともに、または伴わずにセレンテラジンH 5μMを添加した。37℃でさらに10分間インキュベートした後、Mithras LB940マルチラベルリーダー(Berthold)(1s/波長/ウェル、室温で15反復)を用い、485nmおよび530nmでの発光獲得を誘導した。
【0250】
BRET比の計算は従前に記載されているように行った(Angersら 2000):[(発光530nm)−(発光485nm)×Cf]/(発光485nm)、式中、Cf=同じ試験条件下でRluc融合タンパク質単独を発現する細胞の(発光530nm)/(発光485nm)。この式を簡単にすると、BRET比が、二つのBRET相手が存在する場合に得られる530/485nm比を、同じ試験条件下で、Rlucと融合された相手だけがアッセイ中に存在する場合に得られる530/485nm比により補正したものに相当することが示される。読み取りやすいように、結果はミリBRET単位(mBU)で表すが、mBUはBRET比に1000を掛けたものである。
【0251】
SDF1(100nM)は、CXCR4受容体と融合されたドナータンパク質と、アクセプタータンパク質とが空間的に近接していることから生じるBRETシグナルを約10%増強したが、それはおそらくCXCR4/CXCR4ホモ二量体形成または既存の二量体のコンフォメーション変化を示唆する(図16)。Mab c515H7は、CXCR4ホモ二量体に関して、SDF−1により誘発されるコンフォメーション変化を調節することができた(SDF−1により誘発されるBRET増強の96%阻害、図16)。Mab c515H7はまた、それ自体、CXCR4/CXCR4の空間的近接を調節することもできたが、このことはCXCR4/CXCR4ホモ二量体コンフォメーションに対するこのMabの影響を示唆する(図16)。
【0252】
実施例13:CD4およびCXCR4またはCCR5を発現するGFP形質導入ヒト骨肉腫(GHOST)細胞を用いたMab 515H7抗HIV−1活性のin vitro評価
CXCR4 515H7 Mabの特異性を判定するために、本発明者らは、CD4およびCXCR4またはCCR5を発現するGHOSTを用い、このMabの抗HIV−1活性を評価した。
【0253】
このアッセイは、X4 HIV−1 LAIウイルス(CXCR4を発現するGhost細胞を持つ)またはR5 HIV−1 BaLウイルス(CCR5を発現するGhost細胞を持つ)のいずれかを用いて48時間行う。500μlのGhost細胞を、10%FCSを添加したダルベッコの培養培地で24時間平板培養した(2.5 105細胞/ml)。種々の希釈率のMab 515H7を37℃で1時間インキュベートした後、これらの細胞に希釈したHIV−1 LAIウイルス(1/10)およびHIV−1 BaLウイルス(1/7)を48時間加えた。細胞をトリプシン処理し、PBS 1×で洗浄した。細胞を固定し、また、ウイルスを不活性化するために、この細胞ペレットに300μLの1.5%パラホルムアルデヒドを暗所、+4℃で2時間加えた。GFP陽性細胞をフローサイトメトリーにより分析し、HIV−1感染の阻害を算出した。
感染細胞の阻害パーセントは、Mabを含まない対照感染ウェルと比較して定義した。ICs(μg/ml)の結果を表11にまとめる。抗CXCR4 Mab 515H7は、Ghost細胞を発現するCXCR4においてHIV−1 X4 Laiウイルスの感染を阻害することができたが、Ghost細胞を発現するCCR5においてHIV−1 R5 BaLウイルスの感染阻害には全く不活性であった。
【0254】
【表11】
【0255】
実施例14:515H7抗CXCR4ネズミ抗体のヒト化および該h515H7のフラグメントの作製
一般手順
515H7抗CXCR4抗体のヒト化は、CDRグラフト法のグローバルルールを適用することで行った。免疫原性分析ならびにCDRおよびフレームワーク(FR)領域の定義は、IMGTユニークナンバリングスキームならびにIMGTライブラリーおよびツール(Lefranc, 1997 - www.imgt.org)を適用することで行った。
【0256】
515H7のヒト化変異体の結合は、ヒトCXCR4で安定的にトランスフェクトされたNIH3T3細胞系統にて判定した。結合活性は、ビオチン化マウス抗体を用いた競合アッセイによって評価した。第二の試みにおいて、ヒト化抗体を、ビオチン化SDF−1のRAMOS細胞との結合を阻害する能力に関して評価した。RAMOS細胞は、CXCR4の発現が高く、CXCR7およびSDF−1の発現が低いために選択された。
【0257】
これらのアッセイを用いて、抗CXCR4抗体の組換えヒト化型を特性決定した。可変ドメインをヒトIgG1/k定常ドメインでフォーマットし、哺乳類発現ベクターpCEPにクローニングした。組換えIgG1/κ由来抗体をHEK293細胞で一時的に発現させた。発現培養上清を濾過し、プロテインAセファロースを用いて抗体を精製した。精製抗体に再びPBSバッファーを加え、抗体濃度をELISAにより測定した。
【0258】
組換え抗体フラグメントを、ヒト化抗体の可変ドメインに特異的なオリゴヌクレオチドを用いたPCRによって作製し、これらを大腸菌系にサブクローニングした。抗体フラグメントの精製は、固定化金属イオンアフィニティークロマトグラフィー(IMAC)によって行った。
【0259】
515H7可変ドメインのヒト化
重鎖および軽鎖可変ドメインの種々の配列アライメントを図17および18に示す。
【0260】
最初の一連の試験では、三つの第一のヒト化変異体の抗CXCR4結合活性を分析した。VH変異体1(VH1)をネズミVLと組合せ、これらの構築物の、ビオチン化ネズミ515H7親抗体の結合を阻害する能力を評価した。VH1の可変ドメインのアミノ酸配列は配列番号90を含んでなり、ヌクレオチド配列は配列番号91を含んでなる。全長VH1のアミノ酸配列は配列番号92を含んでなり、ヌクレオチド配列は配列番号93を含んでなる。この構築物は、キメラ抗体と同等の、ネズミ抗体との競合能を示した(図19A)。このことは、ほとんどのヒトVH変異体がキメラと同等の結合能を有することを示唆する。よって、VH1をVLの変異体2と組み合わせた(図19B)。
【0261】
さらなる試験では、抗体515H7のヒト化変異体がSDF−1のCXCR4発現細胞への結合を阻害するか否かを判定した(図20)。このhz515H7のヒト化変異体の阻害能は、フローサイトメトリーにおいてビオチン化SDF−1を検出することによって評価した。ヒト化抗体hz515H7 VH1 D76N VL2は、キメラc515H7と同等のSDF−1結合阻害能を有する。
【0262】
また、ヒト化変異体hz515H7 VH1 VL2の抗体フラグメントも試験したところ、この抗体フラグメントがSDF−1の結合を完全に阻害することができることが分かった(図20)。
【0263】
実施例15:FACS分析による抗CXCR4ヒト化Mab 515H7結合特異性の特性決定
この試験では、抗CXCR4ヒト化Mab 515H7の、ヒトCXCR4に対する特異的結合をFACS分析により調べた。
【0264】
NIH3T3、NIH3T3−hCXCR4トランスフェクト体を、100μlのFacsバッファー中、暗所、4℃にて20分間、0〜10μg/mLのヒト化 Mab 515H7(hz515H7 VH1 D76N VL2、hz515H7 VH1 D76N VL2.1、hz515H7 VH1 D76N VL2.2、hz515H7 VH1 D76N VL2.3)とともにインキュベートした。Facsバッファー中で3回洗浄した後、細胞を、暗所、4℃にて20分間、二次抗体であるヤギ抗ヒトAlexa 488(1/500希釈)とともにインキュベートした。Facsバッファー中で3回洗浄した後、各ウェルにヨウ化プロピジウムを加え、生存細胞のみをFacsにより分析した。少なくとも5000の生存細胞を評価し、各条件について蛍光強度の平均値を見積もった。
【0265】
これらの結合試験の結果を、[FACSによって得られた平均蛍光強度(MFI)]を示す図21に示す。抗CXCR4ヒト化Mab hz515H7は、ヒトCXCR4−NIH3T3トランスフェクト細胞系統と特異的に結合した(NIH3T3親細胞ではMFI=2.2)。
【0266】
実施例16:生物発光共鳴エネルギー移動(BRET)アプローチによるCXCR4ホモ二量体に対するhz515H7 Mabの効果
この機能的アッセイは、SDF−1および/またはhz515H7 VH1 D76N VL2、hz515H7 VH1 D76N VL2.1、hz515H7 VH1 D76N VL2.2、hz515H7 VH1 D76N VL2.3がCXCR4受容体に結合した際に誘発されるコンフォメーション変化をCXCR4ホモ二量体のレベルで評価することができる。
【0267】
検討する相互作用相手の発現ベクターを、従来の分子生物学的技術を適用することにより、対応する色素(ウミシイタケ(Renilla reniformis)ルシフェラーゼRlucおよび黄色蛍光タンパク質YFP)との融合タンパク質として構築した。BRET試験を行う2日前に、HEK293細胞を、対応するBRET相手:[CXCR4/Rluc+CXCR4/YFP]をコードする発現ベクターで一時的にトランスフェクトし、CXCR4ホモ二量体形成を調べた。翌日、細胞を、ポリリシンをプレコーティングした白色96MWプレートの完全培養培地[10%FBSを添加したDMEM]に分注した。まず、細胞をプレートに接着させるために、CO2 5%、37℃で培養した。その後、200μl DMEM/ウェルで一晩、細胞を飢餓状態にした。BRET試験の直前に、DMEMを除去し、細胞を手早くPBSで洗浄した。その後、細胞をPBS中、抗体の存在下または非存在下、37℃にて10分間インキュベートした後、最終量50μl中、SDF−1 100nMとともに、または伴わずにセレンテラジンH 5μMを添加した。37℃でさらに10分間インキュベートした後、Mithras LB940マルチラベルリーダー(Berthold)(1s/波長/ウェル、室温で15反復)を用い、485nmおよび530nmでの発光獲得を誘導した。
【0268】
BRET比の計算は従前に記載されているように行った(Angersら、2000):[(発光530nm)−(発光485nm)×Cf]/(発光485nm)、式中、Cf=同じ試験条件下でRluc融合タンパク質単独を発現する細胞の(発光530nm)/(発光485nm)。この式を簡単にすると、BRET比が、二つのBRET相手が存在する場合に得られる530/485nm比を、同じ試験条件下で、Rlucと融合された相手だけがアッセイ中に存在する場合に得られる530/485nm比により補正したものに相当することが示される。読み取りやすいように、結果はミリBRET単位(mBU)で表すが、mBUはBRET比に1000を掛けたものである。
【0269】
SDF1(100nM)は、CXCR4受容体と融合されたドナータンパク質と、アクセプタータンパク質とが、空間的に近接していることから生じるBRETシグナルを約12%増強したが、それはおそらくCXCR4/CXCR4ホモ二量体形成または既存の二量体のコンフォメーション変化を示唆する(図22)。
【0270】
515H7ヒト化Mabは、CXCR4ホモ二量体に関してSDF−1により誘発されるコンフォメーション変化を調節することができ、SDF−1により誘発されるBRET増強の阻害割合は、hz515H7 VH1D76N−VL2 Mabで約88%、hz515H7 VH1D76N−VL2.1 Mabで65%、hz515H7 VH1D76N−VL2.2 Mabで33%およびhz515H7 VH1D76N−VL2.3 Mabで21%であった(図22)。
【発明の背景】
【0001】
発明の分野
本発明は、ケモカイン受容体(CXCR)と特異的に結合することができる新規な抗体、特に、ネズミモノクローナル抗体、キメラ抗体およびヒト化抗体、ならびにこのような抗体をコードするアミノ酸配列および核酸配列に関する。一つの態様から、本発明は、CXCR4と特異的に結合することができ、かつ、ヒト免疫不全ウイルス(HIV)感染に対して強い活性を有する新規な抗体、機能的フラグメントまたは誘導体に関する。本発明はまた、このような抗体、機能的フラグメントまたは誘導体の、HIV感染の予防的治療および/または治療的治療のための薬剤としての使用も含んでなる。
【0002】
背景技術
ケモカインは、特に免疫反応中に、ケモカイン勾配として知られるリガンドの化学的勾配によって白血球の移動を制御する小型の分泌型ペプチドである(Zlotnick A. et al., 2000)。ケモカインは、それらのNH2末端システイン残基の位置に基づいてCCおよびCXCという二つの主要なサブファミリーに分かれ、Gタンパク質共役型受容体と結合し、その受容体の二つの主要なサブファミリーはCCRおよびCXCRと呼ばれている。これまでに50を超えるヒトケモカインと18のケモカイン受容体が見出されている。
【0003】
ケモカイン受容体ファミリーのいくつかのメンバーは、主要受容体CD4とともに共受容体として働き、種々の1型HIV株を細胞へ侵入させるが、この主要な共受容体がCCR5およびCXCR4である。T細胞指向性(T-cell tropic)X4 HIV−1は細胞への侵入にCD4とCXCR4を用いるが、マクロファージ指向性(macrophage-tropic)R5 HIV−1はCD4とCCR5を用いる。二重指向性株は、CXCR4とCCR5の双方を共受容体として使用することができる。他のケモカイン受容体の中でも、CCR3、CCR2、CCR8、CXCR6、CXCR7、CX3CR1は、HIV株のより限られたサブセットにより、共受容体として機能する。
【0004】
CXCR4の天然リガンドであるSDF−1、ならびにCCR5の天然リガンドであるCCL3、CCL4、CCL4−L1およびCCL5リガンドは、種々のHIV−1株による細胞融合および感染を阻害することができる。これらの知見は、ケモカイン受容体を標的とする抗HIV治療薬の開発を促し、CCR5指向性HIV−1に感染した患者において、CCR5の小分子アンタゴニストであるマラビロク(maraviroc)(CELSENTRI(商標))と、他の抗HIV−1薬との併用の認可に至らせた。しかしながらやはり、二重指向性HIV−1に感染した患者にも、CXCR4指向性HIV−1に感染した患者にもマラビロクは使用されない(VIDAL 2009)。従って、X4指向性HIV複製を阻害することができるCXCR4アンタゴニストを同定することにより、X4指向性感染患者および二重指向性HIV感染患者の双方にこの種の療法を拡張するという明確な医学的必要がある。
【0005】
ケモカイン受容体4(フーシン、CD184、LESTR、またはHUMSTRとしても知られる)は、352または360個のアミノ酸を含んでなる二つのイソ型として存在する。残基Asn11はグリコシル化され、残基Tyr21は硫酸基の付加によって修飾され、Cys109および186は、受容体の細胞外部分でジスルフィド架橋によって結合されている(Juarez J.ら、2004)。
【0006】
この受容体は、様々な種の正常組織、ナイーブ、非メモリーT細胞、調節T細胞、B細胞、好中球、内皮細胞、一次単球、樹状細胞、ナチュラルキラー細胞、CD34+造血幹細胞、また、低レベルではあるが、心臓、結腸、肝臓、腎臓、および脳によって発現される。CXCR4は、白血球輸送、B細胞のリンパ球生成、および骨髄造血に重要な役割を果たす。
【0007】
これまでに記載されているCXCR4受容体のリガンドの独特なリガンドは、ストロマ細胞由来因子(SDF−1)またはCXCL12である。SDF−1は、リンパ節、骨髄、肝臓、肺において多量に、また、程度は少ないが、腎臓、脳、および皮膚によっても分泌される。CXCR4はまた、III型ヒトヘルペスウイルスによってコードされているウイルスマクロファージ炎症性タンパク質II(vMIP−II)であるアンタゴニストケモカインによっても認識される。
【0008】
従前に記載されているように、CXCR4受容体は、T細胞指向性HIV−1単離物(X4ウイルス)に対する主要な共受容体である。この受容体に干渉すれば、極めて効率的にX4ウイルスの複製を阻害できるはずである。
【発明の概要】
【0009】
本発明の発明態様の一つは、HIV複製を阻害するマウスモノクローナル抗体(Mab)を作製することである。本発明は、CXCR4ホモ二量体と結合することができ、かつ、HIV感染に対して強い活性を有するCXCR4 Mab 515H7(またはそのフラグメント)を包含する。本発明はまた、CXCR4ホモ二量体と結合することができ、かつ、HIV感染に対して強い活性を有するCXCR4 Mab 301aE5(またはそのフラグメント)を包含する。
【0010】
驚くべきことに、本発明者らは、CXCR4と結合することができるだけでなく、CXCR4ホモ二量体のコンフォメーション変化を誘導することができ、かつ、PBMCにおけるX4−HIV−1一次単離物の複製を阻害することができるモノクローナル抗体の作製を成し遂げた。より詳細には、本発明の抗体は、PBMCにおけるX4/R5−HIV−1一次単離物の複製を阻害することができる。
【図面の簡単な説明】
【0011】
【図1】図1Aおよび1Bは、単球およびリンパ球におけるCXCR4発現のゲーティング戦略を示す図である。図1A:CD3−PE抗体によるT細胞の染色、図1B:CD14−PE抗体による単球の染色
【図2】図2Aおよび2Bは、単球およびTリンパ球に対する抗CXCR4 Mab 515H7および301aE5の結合を示す図である。
【図3】図3Aおよび3Bは、HEK293細胞における生物発光共鳴エネルギー移動(BRET)アプローチによる、それぞれSDF−1ならびに抗CXCR4 Mab 515H7および301aE5によるCXCR4受容体二量体の調節を示す図である。
【図4】図4Aおよび4Bは、抗CXCR4 Mab 515H7および301aE5の、ヒトPBMCにおけるHIV−1単離物KON(X4ウイルス)の複製を阻害する能力を示す図である。
【図5】図5は、抗CXCR4 Mab 515H7および301aE5の、ヒトPBMCにおけるHIV−1単離物KON(X4ウイルス)の複製を阻害する能力を示す図である。
【図6】図6A、6Bおよび6Cは、CHO−CXCR4細胞におけるSDF−1により誘発されたカルシウム放出の、Mab 515H7(図6A)、301aE5(図6B)およびc515H7(図6C)による阻害を示す図である。
【図7】図7は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、ヒトPBMCにおけるHIV−1 X4ウイルス一次単離物MN(図7)および92UG024(図8)の複製を阻害する能力を示す図である。
【図8】図8は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、ヒトPBMCにおけるHIV−1 X4ウイルス一次単離物MN(図7)および92UG024(図8)の複製を阻害する能力を示す図である。
【図9】図9は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、ヒトPBMCにおけるHIV−1 X4ウイルス一次単離物KON、MN、および92UG024の複製を阻害する能力を示す図である。
【図10】図10は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、ヒトPBMCにおけるHIV−1 X4/R5二重ウイルス一次単離物89.6の複製を阻害する能力を示す図である。
【図11】図11は、抗CXCR4 Mab 515H7、c515H7および301aE5の、ヒトPBMCにおけるHIV−1 X4/R5二重ウイルス一次単離物89.6の複製を阻害する能力を示す図である。
【図12】図12は、Mab c515H7とマラビロクの併用の、ヒトPBMCにおけるHIV−1一次単離物89.6(X4/R5二重ウイルス)の複製を阻害する有益な効果を示す図である。
【図13】図13は、Mab c515H7とマラビロクの併用の、ヒトPBMCにおけるHIV−1一次単離物UG93067(X4/R5二重ウイルス)の複製を阻害する有益な効果を示す図である。
【図14】図14は、抗CXCR4 Mab 515H7、c515H7、および301aE5の、PBMCにおけるHIV−1 X4ウイルス一次単離物KONの複製を阻害する能力を示す図である。
【図15】図15は、FACS分析によるc515H7 Mabの結合特異性を示す図である。
【図16】図16は、生物発光共鳴エネルギー移動(BRET)アプローチによる、CXCR4ホモ二量体に対するc515H7 Mabの効果を示す図である。
【図17】図17は、515H7重鎖可変ドメインとヒト生殖細胞系IGHV3−49*04およびIGHJ4*01とのアミノ酸配列アライメント。515H7 VHアミノ酸配列が、選択されたヒトアクセプターフレームワーク配列とアラインされている。VH Var1 (VH1)配列は、515H7 VHドメインのヒト化変異体に相当する。76番の位置の一つの逆突然変異が太字で示されている。
【図18】図18は、515H7軽鎖と、ヒト生殖細胞系IGKV4−l*01およびIGKJ1*01とのアミノ酸配列アライメント。515H7 VLアミノ酸配列が、選択されたヒトアクセプターフレームワーク配列とアラインされている。VL Var2.1、Var2.2、およびVar2.3配列は、ヒト化515H7 VL Var2の実施されたヒト化変異体に相当する(変異残基を太字で示す)。Var2.1およびVar2.2はさらに四つのヒト化残基を持ち、Var2.3はさらに五つのヒト残基を含む。
【図19】図19は、キメラ515H7およびヒト化515H7の種々の変異体による、ビオチン化ネズミ抗体515H7の交差遮断。515H7のヒト化変異体(hz515H7)の、親ネズミ抗体515H7を交差遮断する活性を、CXCR4トランスフェクトNIH3T3細胞を用いたフローサイトメトリーによって評価した。ヒト化変異体の活性をキメラ515H7と比較した。変異体VH1とキメラVL(cVL)併用の交差遮断活性は、キメラ(A)の場合と極めてよく似ていた。VLの変異体2と併用した場合(B)には、VH変異体1(VH1、逆突然変異の無い変異体)の活性の低下は見られなかった。
【図20】図20は、キメラ515H7およびヒト化515H7の種々の変異体による、ビオチン化SDF−1結合の阻害。515H7のヒト化変異体(hz515H7)の、SDF−1結合を阻害する能力を、細胞系統RAMOSを用いたフローサイトメトリーによって評価した。ヒト化変異体の阻害能をキメラ515H7と比較した。ヒト化変異体hz515H7 VH1 D76N VL2は、キメラ抗体と同等のSDF−1結合阻害能を有する。ヒト化抗体フラグメントhz515 VH1 VL2は、RAMOS細胞に対するSDF−1の結合阻害に完全な活性を示した。
【図21】図21は、NIH3T3−CXCR4においてCXCR4と特異的に結合するヒト化515H7 Mab(hz515H7 VH1 D76N VL2、hz515H7 VH1 D76N VL2.1、hz515H7 VH1 D76N VL2.2、およびhz515H7 VH1 D76N VL2.3)を示す図である。
【図22】図22は、生物発光共鳴エネルギー移動(BRET)アプローチによる、CXCR4ホモ二量体に対するヒト化515H7 Mab(hz515H7 VH1 D76N VL2、hz515H7 VH1 D76N VL2.1、hz515H7 VH1 D76N VL2.2およびhz515H7 VH1 D76N VL2.3)の効果を示す図である。
【発明の具体的説明】
【0012】
好ましくは、CXCR4化合物は、
Genbank受託番号NP_003458配列番号27:
MEGISIYTSDNYTEEMGSGDYDSMKEPCFREENANFNKIFLPTIYSIIFLTGIVGNGLVILVMGYQKKLRSMTDKYRLHLSVADLLFVITLPFWAVDAVANWYFGNFLCKAVHVIYTVNLYSSVLILAFISLDRYLAIVHATNSQRPRKLLAEKVVYVGVWIPALLLTIPDFIFANVSEADDRYICDRFYPNDLWVVVFQFQHIMVGLILPGIVILSCYCIIISKLSHSKGHQKRKALKTTVILILAFFACWLPYYIGISIDSFILLEIIKQGCEFENTVHKWISITEALAFFHCCLNPILYAFLGAKFKTSAQHALTSVSRGSSLKILSKGKRGGHSSVSTESESSSFHSS、
として表される配列を有するケモカイン(C−X−Cモチーフ)受容体4イソ型b[ホモ・サピエンス]、
Genbank受託番号NP_001008540配列番号28:
MSIPLPLLQIYTSDNYTEEMGSGDYDSMKEPCFREENANFNKIFLPTIYSIIFLTGIVGNGLVILVMGYQKKLRSMTDKYRLHLSVADLLFVITLPFWAVDAVANWYFGNFLCKAVHVIYTVNLYSSVLILAFISLDRYLAIVHATNSQRPRKLLAEKVVYVGVWIPALLLTIPDFIFANVSEADDRYICDRFYPNDLWVVVFQFQHIMVGLILPGIVILSCYCIIISKLSHSKGHQKRKALKTTVILILAFFACWLPYYIGISIDSFILLEIIKQGCEFENTVHKWISITEALAFFHCCLNPILYAFLGAKFKTSAQHALTSVSRGSSLKILS KGKRGGHSSVSTESESSSFHSS、
として表される配列を有するケモカイン(C−X−Cモチーフ)受容体4イソ型a[ホモ・サピエンス]、
配列番号27または28を有するこれらのbまたはaイソ型の一つと少なくとも95%の同一性を有する選択的転写スプライス変異体またはその天然変異体、および
その天然リガンドであるストロマ細胞由来因子−1(SDF−1)によって特異的に認識されることができ、かつ好ましくは、少なくとも100、150、および200のアミノ酸長を有するそのフラグメント
からなる群から選択される、二つのヒトCXCR4イソ型のうちの一つである。
【0013】
CXCR2は、
Genbank受託番号NP_001548配列番号29:
MEDFNMESDSFEDFWKGEDLSNYSYSSTLPPFLLDAAPCEPESLEINKYFVVIIYALVFLLSLLGNSLVMLVILYSRVGRSVTDVYLLNLALADLLFALTLPIWAASKVNGWIFGTFLCKVVSLLKEVNFYSGILLLACISVDRYLAIVHATRTLTQKRYLVKFICLSIWGLSLLLALPVLLFRRTVYSSNVSPACYEDMGNNTANWRMLLRILPQSFGFIVPLLIMLFCYGFTLRTLFKAHMGQKHRAMRVIFAVVLIFLLCWLPYNLVLLADTLMRTQVIQETCERRNHIDRALDATEILGILHSCLNPLIYAFIGQKFRHGLLKILAIHGLISKDSLPKDSRPSFVGSSSGHTSTTL、
として表される配列を有するインターロイキン8受容体β[ホモ・サピエンス]、
配列番号29を有するこのインターロイキン8受容体βと少なくとも95%の同一性を有する、選択的転写スプライス変異体またはその天然変異体、および
IL−8によって特異的に認識されることができ、かつ、好ましくは、少なくとも100、150、および200のアミノ酸長を有するそのフラグメント
からなる群から選択される。
【0014】
本発明はまた、抗HIV活性を有するか、またはHIV感染の治療のための組成物の製造のために使用することができる化合物を選択する方法であって、以下の工程を含むことを特徴とする方法も含む。
【0015】
第1の態様において、本発明の主題は、本発明による抗体を作製および選択する方法である。
【0016】
より詳細には、本発明は、HIV複製を阻害することができる抗CXCR4抗体、またはその一つの機能的フラグメントもしくは誘導体を選択する方法であって、
i)作製された抗体をスクリーニングし、CXCR4と特異的に結合することができる抗体を選択する工程、
ii)工程i)の選択抗体を検定し、末梢血単核細胞(PBMC)と結合できる抗体を選択する工程、
iii)工程ii)の選択抗体を検定し、CXCR4ホモ二量体と結合することができる抗体を選択する工程、および次いで
iv)工程iii)の選択抗体を検定し、PBMCにおけるX4指向性HIV−1一次単離物の複製を阻害することができる抗体を選択する工程
を含んでなる方法に関する。
【0017】
別の実施形態では、本発明は、HIV複製を阻害することができる抗CXCR4抗体、またはその一つの機能的フラグメントもしくは誘導体を選択する方法であって、
i)作製された抗体をスクリーニングし、CXCR4と特異的に結合することができる抗体を選択する工程、
ii)工程i)の選択抗体を検定し、末梢血単核細胞(PBMC)と結合できる抗体を選択する工程、
iii)工程ii)の選択抗体を検定し、CXCR4ホモ二量体と結合することができる抗体を選択する工程、および次いで
iv)工程iii)の選択抗体を検定し、PBMCにおけるX4指向性HIV−1一次単離物の複製を阻害することができ、および/またはPBMCにおいてX4/R5指向性HIV−1一次単離物の複製を阻害することができる抗体を選択する工程
を含んでなる方法に関する。
【0018】
抗体の作製は、例えば、骨髄腫細胞と免疫マウスまたは選択された骨髄腫細胞と適合する他の種に由来する脾臓細胞との融合などの、当業者に公知の方法[Kohler & Milstein, 1975, Nature, 256:495-497]によって実現することができる。この免疫動物は、ヒト免疫グロブリン遺伝子座を有し、その後、直接ヒト抗体を産生するトランスジェニックマウスを含み得る。もう一つの可能性のある実施形態は、ライブラリーをスクリーニングするためのファージディスプレー技術の使用からなるものであり得る。
【0019】
スクリーニング工程i)およびii)は、当業者に公知のいずれの方法または工程によっても実現可能である。限定されない例として、ELISA、BIAcore、免疫組織化学、CXCR4発現細胞膜抽出物、または精製CXCR4を用いたウエスタンブロット解析、FACS分析、および機能的スクリーニングが挙げられる。好ましい方法は、産生された抗体が、標的細胞表面において天然CXCR4受容体のコンフォメーションも認識できることを確認するための、CXCR4形質転換体(工程1)および少なくともPBMC(工程2)に対するFACS分析によるスクリーニングからなる。この方法を以下の例にさらに厳密に記載する。
【0020】
スクリーニング工程iii)は、当業者に公知のいずれの方法または方法によっても実現可能である。限定されるものではないが、好ましい例として、CXCR4トランスフェクト細胞またはPBMC由来の膜抽出物に対して着目する抗体を用いるウエスタンブロット法および/または免疫沈降法が挙げられる。
【0021】
スクリーニング工程iv)は、当業者に公知のいずれの方法または方法によっても実現可能である。限定されるものではないが、好ましい例として、Hollら(J. Immunol. 2004, 173, 6274-83)において記載されているプロトコールを用いることにより、PBMCにおけるX4一次HIV−1単離物および/またはX4/R5一次HIV−1単離物の複製を阻害することができるか否かに関し、抗体をスクリーニングすることからなる方法が挙げられる。
【0022】
本発明の方法の選択の工程iii)の好ましい実施形態では、この工程iii)は、CXCR4−RLuc/CXCR4−YFPを発現する細胞に対するBRET分析によって抗体を評価すること、およびBRETシグナルの少なくとも40%、好ましくは45%、50%、55%、最も好ましくは60%を阻害することができる抗体を選択することからなる。
【0023】
BRET法は、タンパク質二量体形成の代表例として知られている技術である[Angers et al., PNAS, 2000, 97:3684-89]。
【0024】
本方法の工程iii)に用いられるBRET法は当業者によく知られており、以下の例に記載する。より詳細には、BRET(生物発光共鳴エネルギー移動)は、生物発光ドナー(ウミシイタケ・ルシフェラーゼ(Rluc))と、蛍光アクセプターであるGFP(緑色蛍光タンパク質)またはYFP(黄色蛍光タンパク質)突然変異体との間で起こる非放射性のエネルギー移動である。本ケースでは、EYFP(増強黄色蛍光タンパク質)を用いた。移動の有効性は、ドナーとアクセプターの配向および距離に依存する。そして、エネルギー移動は、この二つの分子が近接している(1〜10nm)場合にのみ起こり得る。この特性を用いて、タンパク質−タンパク質相互作用アッセイを作り出す。実際に、二つのパートナー間の相互作用を検討するために、第一のものをウミシイタケ・ルシフェラーゼと、そして、第二のものをGFPの黄色突然変異体と遺伝子融合する。融合タンパク質は、必須ではないが一般に、哺乳類細胞で発現される。その膜透過基質(セレンテラジン)の存在下で、Rlucは青色の光を放出する。GFP突然変異体がRlucから10nmより近ければエネルギー移動が起こり、さらに黄色シグナルが検出できる。BRETシグナルは、アクセプターによって放出された光とドナーによって放出された光の間の比として測定される。従って、BRETシグナルは二つの融合タンパク質が近接するほど、またはコンフォメーション変化がRlucと、GFP突然変異体とを近接させるほど増大する。
【0025】
BRET分析が好ましい実施形態からなる場合、CXCR4二量体のコンフォメーション変化を測定するために、当業者に公知のいずれの方法も使用可能である。限定されるものではないが、以下の技術が挙げられる:FRET(蛍光共鳴エネルギー移動)、HTRF(均一時間分解蛍光)、FLIM(蛍光寿命画像測定法)またはSW−FCCS(単波長蛍光相関分光法)。
【0026】
共免疫沈降法、アルファースクリーン、化学架橋、二重ハイブリッド、アフィニティークロマトグラフィー、ELISA、またはファーウエスタンブロットなどの他の従来法も使用可能である。
【0027】
本発明による方法の特定の態様では、工程iii)は、CXCR4−RLuc/CXCR4−YFPの双方を発現する細胞に対するBRET分析によって抗体を評価すること、およびBRETシグナルの少なくとも40%を阻害することができる抗体を選択することからなる。
【0028】
第2の態様において、本発明の主題は、該方法によって得られる単離された抗体、またはそれらの一つの機能的フラグメントもしくは誘導体である。該抗体またはそれらの一つのフラグメントもしくは誘導体は、ヒトCXCR4と特異的に結合することができ、該抗体はまた、CXCR4ホモ二量体のコンフォメーション変化を誘導することもできる。
【0029】
例えばクローンA120のようなCXCR4 MabがHIV−1実験株(X4HIV−1NL4−3)のPBMCへの侵入を阻害することができることが文献から知られている(Tanaka R. et al, J. Virol. 2001, 75, 11534-11543)。さらに、CXCR4 Mabが、HIV−1 X4一次単離物の、CXCR4を発現する細胞系統への侵入を阻害することができることも知られている。これに対し、このようなウイルスをそれらの天然環境において、すなわち、実験的ウイルスまたは細胞系統に対してだけでなく、阻害することができる抗体は開示されていない。しかしながらやはり、CXCR4 MabがPBMCへのHIV−1 X4一次単離物の侵入を阻害することができるということは新規であり、本発明の自明の態様ではない。
【0030】
「機能的フラグメントおよび誘導体」という表現は、本明細書の後段で詳細に定義される。
ここで、本発明は天然型の抗体に関するものではなく、すなわち、それらはそれらの天然環境になく、単離可能であったか、もしくは天然源から精製によって得られたものであるか、またはさらには遺伝子組換えもしくは化学合成によって得られたものであり、また、それらは、さらに記載されるように、非天然アミノ酸を含むことができることを理解しなければならない。
【0031】
より詳細には、本発明の別の態様によれば、単離された抗体、またはそれらの一つの機能的フラグメントもしくは誘導体が記載され、該抗体は、IMGTナンバリングシステムによって定義される配列番号1〜6および30〜33のアミノ酸配列を含んでなるCDRから選択される少なくとも一つの相補性決定領域(complementary determining region)CDRを含んでなることを特徴とする。
【0032】
第一の態様によれば、本発明は、IMGTナンバリングシステムに従って定義される配列番号1〜6の配列のCDRの中から選択される少なくとも一つのCDR、または配列が最適なアライメントの後に配列番号1〜6の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つのCDRを含んでなる、単離された抗体、またはその機能的フラグメント、もしくは誘導体に関する。
【0033】
第二の態様によれば、本発明は、IMGTナンバリングシステムに従って定義される配列番号1、2および30〜33の配列のCDRの中から選択される少なくとも一つのCDR、または配列が最適なアライメントの後に配列番号1、2、および30〜33の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つのCDRを含んでなる、単離された抗体、またはその機能的フラグメントまたは誘導体に関する。
【0034】
抗体の「機能的フラグメント」とは、特に、フラグメントFv、scFv(sc=一本鎖)、Fab、F(ab’)2、Fab’、scFv−Fc、もしくはダイアボディー(diabody)、または半減期が延長された任意のフラグメントなどの抗体フラグメントを意味する。このような機能的フラグメントは本明細書の後段で詳細に記載される。
【0035】
抗体の「誘導化合物」または「誘導体」とは、特に、ペプチド骨格(peptide scaffold)と、CXCR4を認識する能力を保持するための原抗体の少なくとも一つのCDRとから構成される結合タンパク質を意味する。当業者に周知のこのような誘導化合物は、本明細書の後段でさらに詳細に記載される。
【0036】
より好ましくは、本発明は、本発明による抗体、それらの誘導化合物またはそれらの機能的フラグメント、特に、キメラまたはヒト化抗体、遺伝子組換え、または化学合成によって得られる抗体を含んでなる。
【0037】
好ましい実施形態によれば、本発明による抗体、またはその誘導化合物もしくは機能的フラグメントは、モノクローナル抗体からなることを特徴とする。
【0038】
「モノクローナル抗体」とは、ほぼ均質な抗体集団から生じる抗体を意味すると理解される。より詳細には、集団の個々の抗体は、最小の割合で見られる少数の潜在的に自然に生じる突然変異以外は同一である。言い換えれば、モノクローナル抗体は、単細胞クローン(例えば、ハイブリドーマ、その均質な抗体をコードするDNA分子でトランスフェクトされた真核生物宿主細胞、その均質な抗体をコードするDNA分子でトランスフェクトされた原核生物宿主細胞など)の増殖から生じる均質な抗体からなり、一般に、一つおよびただ一つのクラスおよびサブクラスの重鎖と、ただ一つの種類の軽鎖を特徴とする。モノクローナル抗体は特異性が高く、単一の抗原に対するものである。加えて、種々の決定基またはエピトープに対する種々の抗体を一般に含むポリクローナル抗体の作製とは対照的に、各モノクローナル抗体は抗原の単一のエピトープに対するものである。
【0039】
より詳細には、本発明の第一の好ましい実施形態によれば、抗体、またはその誘導化合物、もしくは機能的フラグメントは、CDR−L1、CDR−L2、およびCDR−L3から選択される少なくとも一つのCDRを含んでなる軽鎖を含んでなることを特徴とし、ここで、
CDR−L1は配列番号1のアミノ酸配列を含んでなり、
CDR−L2は配列番号2のアミノ酸配列を含んでなり、
CDR−L3は配列番号3のアミノ酸配列を含んでなる。
【0040】
別の実施形態によれば、本発明の抗体、またはそれらの誘導化合物もしくは機能的フラグメントの一つは、配列番号1、2、もしくは3の配列の三つのCDRのうち少なくとも一つ、または最適なアライメントの後に配列番号1、2、または3の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる軽鎖を含んでなることを特徴とする。
【0041】
本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体はまた、CDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖を含んでなることを特徴とし、ここで、CDR−L1は配列番号1のアミノ酸配列を含んでなり、CDR−L2は配列番号2のアミノ酸配列を含んでなり、CDR−L3は配列番号3のアミノ酸配列を含んでなる。
【0042】
別の実施形態では、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号7のアミノ酸配列、または最適なアライメントの後に配列番号7の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる配列の軽鎖を含んでなることを特徴とする。
【0043】
本発明の第二の好ましい実施形態によれば、抗体、またはその誘導化合物もしくは機能的フラグメントは、CDR−L1、CDR−L2、およびCDR−L3から選択される少なくとも一つのCDRを含んでなる軽鎖を含んでなることを特徴とし、ここで、
CDR−L1は配列番号1のアミノ酸配列を含んでなり、
CDR−L2は配列番号2のアミノ酸配列を含んでなり、
CDR−L3は配列番号30のアミノ酸配列を含んでなる。
【0044】
別の実施形態によれば、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、配列番号1、2、もしくは30の配列の三つのCDRのうち少なくとも一つ、または最適なアライメントの後に配列番号1、2、もしくは30の配列と少なくとも80%、好ましくは85%、90%、95%および98%の同一性を有する少なくとも一つの配列を含んでなる軽鎖を含んでなることを特徴とする。
【0045】
本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、CDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖を含んでなることを特徴とし、ここで、CDR−L1は配列番号1のアミノ酸配列を含んでなり、CDR−L2は配列番号2のアミノ酸配列を含んでなり、CDR−L3は配列番号30のアミノ酸配列を含んでなる。
【0046】
別の実施形態では、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号34のアミノ酸配列、または最適なアライメントの後に配列番号34の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる配列の軽鎖を含んでなることを特徴とする。
【0047】
より詳細には、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、CDR−H1、CDR−H2、およびCDR−H3から選択される少なくとも一つのCDRを含んでなる重鎖を含んでなることを特徴とし、ここで、
CDR−H1は配列番号4のアミノ酸配列を含んでなり、
CDR−H2は配列番号5のアミノ酸配列を含んでなり、
CDR−H3は配列番号6のアミノ酸配列を含んでなる。
【0048】
別の実施形態によれば、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、配列番号4、5、もしくは6の配列の三つのCDRのうち少なくとも一つ、または最適なアライメントの後に配列番号4、5、もしくは6の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる重鎖を含んでなることを特徴とする。
【0049】
別の特定の実施形態によれば、抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、CDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖を含んでなることを特徴とし、ここで、CDR−H1は配列番号4のアミノ酸配列を含んでなり、CDR−H2は配列番号5のアミノ酸配列を含んでなり、CDR−H3は配列番号6のアミノ酸配列を含んでなる。
【0050】
別の実施形態では、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号8のアミノ酸配列、または最適なアライメントの後に配列番号8の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる配列の重鎖を含んでなることを特徴とする。
【0051】
より詳細には、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、CDR−H1、CDR−H2、およびCDR−H3から選択される少なくとも一つのCDRを含んでなる重鎖を含んでなることを特徴とし、ここで、
CDR−H1は配列番号31のアミノ酸配列を含んでなり、
CDR−H2は配列番号32のアミノ酸配列を含んでなり、
CDR−H3は配列番号33のアミノ酸配列を含んでなる。
【0052】
別の実施形態によれば、本発明の抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、配列番号31、32、もしくは33の配列の三つのCDRのうち少なくとも一つ、または最適なアライメントの後に配列番号31、32、もしくは33の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる重鎖を含んでなることを特徴とする。
【0053】
別の特定の実施形態によれば、抗体、またはそれらの一つの誘導化合物もしくは機能的フラグメントは、CDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖を含んでなることを特徴とし、ここで、CDR−H1は配列番号31のアミノ酸配列を含んでなり、CDR−H2は配列番号32のアミノ酸配列を含んでなり、CDR−H3は配列番号33のアミノ酸配列を含んでなる。
【0054】
別の実施形態では、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号35のアミノ酸配列、または最適なアライメントの後に配列番号35の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する少なくとも一つの配列を含んでなる配列の重鎖を含んでなることを特徴とする。
【0055】
本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号1、2、および3のアミノ酸配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖と、配列番4、5、および6のアミノ酸配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖を含んでなることを特徴とする。
【0056】
最後に、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体はまた、配列番号7のアミノ酸配列を含んでなる軽鎖と、配列番号8のアミノ酸配列を含んでなる重鎖を含んでなることを特徴としうる。
【0057】
本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体は、配列番号1、2、および30のアミノ酸配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖と、配列番号31、32、および33のアミノ酸配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖を含んでなることを特徴とする。
【0058】
最後に、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体はまた、配列番号34のアミノ酸配列を含んでなる軽鎖と、配列番号35のアミノ酸配列とを含んでなる重鎖を含んでなることを特徴とし得る。
【0059】
本明細書において「ポリペプチド」、「ポリペプチド配列」、「ペプチド」、および「抗体化合物またはそれらの配列と結合しているタンパク質」は互換的である。ここで、本発明は天然型の抗体に関するものではなく、すなわち、それらはそれらの天然環境から採取されたものではなく、単離されているか、または天然源から精製によって得られたものであるか、または遺伝子組換えもしくは化学合成によって得られたものであり、従って、それらは、以下に記載されるように、非天然アミノ酸を含むことができることを理解しなければならない。
【0060】
第一の実施形態において、相補性決定領域、またはCDRは、Kabatら(Kabat et al., Sequences of proteins of immunological interest, 第5版, U.S. Department of Health and Human Services, NIH, 1991および後続版)によって定義される免疫グロブリンの重鎖および軽鎖の超可変領域を意味する。三つの重鎖CDRと、三つの軽鎖CDRとが存在する。ここで、「CDR」とは、抗体が認識する抗原またはエピトープに対する抗体の結合親和性を担うアミノ酸残基の大部分を含む領域のうち1以上、またはさらには総てを示すのに用いられる。
【0061】
第二の実施形態において、CDR領域またはCDRにより、IMGTによって定義される免疫グロブリンの重鎖および軽鎖の超可変領域を示すものとする。
【0062】
IMGTユニークナンバリングは、どんな抗原受容体、鎖の種類、または種であれ、可変ドメインを比較するために定義されたものである[Lefranc M. -P., Immunology Today 18, 509 (1997) / Lefranc M.-P., The Immunologist, 7, 132-136 (1999) / Lefranc, M.-P., Pommie, C, Ruiz, M., Giudicelli, V., Foulquier, E., Truong, L., Thouvenin-Contet, V. and Lefranc, Dev. Comp. Immunol, 27, 55-77 (2003)]。IMGTユニークナンバリングでは、例えば、システイン23(1st−CYS)、トリプトファン41(CONSERVED−TRP)、疎水性アミノ酸89、システイン104(2nd−CYS)、フェニルアラニン、またはトリプトファン118(J−PHEまたはJ−TRP)など、保存されているアミノ酸は常に同じ位置を持つ。IMGTユニークナンバリングでは、フレームワーク領域(FR1−IMGT:1〜26番、FR2−IMGT:39〜55番、FR3−IMGT:66〜104番およびFR4−IMGT:118〜128番)および相補性決定領域:CDR1−IMGT:27〜38番、CDR2−IMGT:56〜65番およびCDR3−IMGT:105〜117番の標準化された画定が得られる。ギャップは占有されていない位置を表すので、CDR−IMGT長(例えば、[8.8.13]などと括弧の中に点で区切って示される)は重要な情報となる。IMGTユニークナンバリングは、IMGT Colliers de Perles [Ruiz, M. and Lefranc, M.-P., Immunogenetics, 53, 857-883 (2002) / Kaas, Q. and Lefranc, M.-P., Current Bioinformatics, 2, 21-30 (2007)]として表される2Dグラフ、およびIMGT/3Dstructure-DB [Kaas, Q., Ruiz, M. and Lefranc, M.-P., T cell receptor and MHC structural data. Nucl. Acids. Res., 32, D208-D210 (2004)]に用いられる。
【0063】
三つの重鎖CDRと、三つの軽鎖CDRとが存在する。ここで、CDRは、場合によって、抗体が認識する抗原またはエピトープに対する抗体の親和性による結合を担うアミノ酸残基の大部分を含む、これらの領域の一つまたはいくつか、またはさらにはこれらの領域の全部を示すために用いられる。
【0064】
より明瞭にするために、以下の説明およびより詳細には表2および3では、CDRはIMGTナンバリングシステムおよびKabatナンバリングシステムによって定義されていることを理解しなければならない。
【0065】
IMGTナンバリングシステムは、上記で定義されたIMGTシステムに従ってCDRを定義し、Kabatナンバリングシステムは、上記で定義されたKabatシステムに従ってCDRを定義する。
【0066】
より詳細には、515H7と呼ばれる抗体に関しては、CDR−L1はIMGTナンバリングシステムでは配列番号1からなり、Kabatナンバリングシステムでは配列番号9からなる。CDR−L2に関しては、IMGTナンバリングシステムでは配列番号2からなり、Kabatナンバリングシステムでは配列番号10からなる。CDR−L3は、この二つのナンバリングシステムのそれぞれで配列番号3からなる。重鎖について、CDR−H1は、IMGTナンバリングシステムでは配列番号4からなり、Kabatナンバリングシステムでは配列番号11からなる。CDR−H2は、IMGTナンバリングシステムでは配列番号5からなり、Kabatナンバリングシステムでは配列番号12からなる。最後に、CDR−H3は、IMGTナンバリングシステムでは配列番号6からなり、Kabatナンバリングシステムでは配列番号13からなる。
【0067】
そして、301aE5と呼ばれる抗体に関しては、CDR−L1は、IMGTナンバリングシステムでは配列番号1からなり、Kabatナンバリングシステムでは配列番号9からなる。CDR−L2に関しては、IMGTナンバリングシステムでは配列番号2からなり、Kabatナンバリングシステムでは配列番号36からなる。CDR−L3は、IMGTナンバリングシステムでは配列番号30からなり、Kabatナンバリングシステムでは配列番号37からなる。重鎖について、CDR−H1は、IMGTナンバリングシステムでは配列番号31からなり、Kabatナンバリングシステムでは配列番号38からなる。CDR−H2は、IMGTナンバリングシステムでは配列番号32からなり、Kabatナンバリングシステムでは配列番号39からなる。最後に、CDR−H3は、IMGTナンバリングシステムでは配列番号33からなり、Kabatナンバリングシステムでは配列番号40からなる。
【0068】
本発明の意味において、二つの核酸配列またはアミノ酸配列間の「同一性割合」とは、最適なアライメントの後に得られる、比較する二つの配列間で同一のヌクレオチドまたはアミノ酸残基の割合を意味し、この割合は純粋に統計学的なものであり、その2配列間の違いはそれらの長さに沿ってランダムに分布している。二つの核酸配列またはアミノ酸配列の比較は従来から、それらを最適にアラインした後に配列を比較することによって行われ、この比較はセグメントによって、または「アライメントウインドウ」の使用によって行うことができる。比較のための配列の最適なアライメントは、手による比較の他、Smith and Waterman (1981) [Ad. App. Math. 2:482]のローカル・ホモロジー・アルゴリズム、Neddleman and Wunsch (1970) [J. Mol. Biol. 48:443]のローカル・ホモロジー・アルゴリズム、Pearson and Lipman (1988) [Proc. Natl. Acad. Sci. USA 85:2444]またはこれらのアルゴリズムを用いたコンピューターソフトウエア(Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WIのGAP、BESTFIT、FASTAおよびTFASTA、または匹敵するソフトウエアBLAST NRもしくはBLAST Pによる)によって行うことができる。
【0069】
二つの核酸配列またはアミノ酸配列間の同一性割合は、二つの最適にアラインされた配列を比較することにより決定され、ここで、比較する核酸配列またはアミノ酸配列はその二つの配列間の最適なアライメントのために参照配列に対して付加または欠失を持ちうる。同一性割合は、アミノ酸ヌクレオチドまたは残基がその二つの配列間、好ましくは二つの全配列間で同一である位置の数を求め、その同一の位置の数をアライメントウインドウの位置の総数で割り、その商に100をかけて、その二つの配列間の同一性割合を得ることによって計算される。
【0070】
例えば、http://www.ncbi.nlm.nih.gov/gorf/bl2.htmlのサイトで利用可能なBLASTプログラム、「BLAST2 sequences」(Tatusova et al., "Blast 2 sequences - a new tool for comparing protein and nucleotide sequences", FEMS Microbiol, 1999, Lett. 174:247-250)をデフォルトパラメーター(特に、パラメーターとして、「オープンギャップペナルティー」:5、および「エクステンションギャップペナルティー」:2、選択マトリックスは、例えば、このプログラムによって提案される「BLOSUM 62」である)とともに使用することができ、この2配列間の同一性割合は、このプログラムによって直接計算される。
【0071】
参照アミノ酸配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を示すアミノ酸配列として、好ましい例には、参照配列、ある特定の修飾、特に、少なくとも一つのアミノ酸の欠失、付加、もしくは置換、末端切断または延長を含むものが含まれる。1以上の保存的または非保存的アミノ酸の置換の場合、置換されるアミノ酸が「等価な」アミノ酸に置き換わる置換が好ましい。ここで、「等価なアミノ酸」とは、対応する抗体および以下に定義される具体例の生物活性にいかなる修飾も与えずに構造アミノ酸の一つの取って代わり得るアミノ酸を示すものとする。
【0072】
等価なアミノ酸は、置換されるアミノ酸との構造的相同性または生成され得る種々の抗体間の生物活性の比較試験の結果に基づいて決定される。限定されない例として、下記表1に、対応する修飾抗体の生物活性の有意な修飾をもたらさずに行い得る置換をまとめる。なお、逆の置換も同じ条件下で本来可能である。
【0073】
【表1】
【0074】
特定の実施形態において、本発明は、ネズミ抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0075】
上記に見られるように、本発明はまた、本明細書に記載される抗体に由来する化合物に関する。
【0076】
より詳細には、本発明の抗体、またはその誘導化合物もしくは機能的フラグメントは、該誘導化合物が、最初の抗体のパラトープ認識特性の総てまたは一部を保存するように、少なくとも一つのCDRがグラフトされているペプチド骨格を含んでなる結合タンパク質からなることを特徴とする。
【0077】
また、本発明に記載されるCDR配列のうち1以上の配列が種々の免疫グロブリンタンパク質骨格(scaffolding)に存在してもよい。この場合、このタンパク質配列によって、グラフトされたCDRの折りたたみに有利なペプチド骨格を再び作り出して、それらのパラトープ抗原認識特性を保存可能とすることができる。
【0078】
一般に、当業者は、元の抗体に由来するCDRの少なくとも一つをグラフトするためのタンパク質骨格(scaffolding)の種類をどのように決めればよいかを知っている。より詳細には、選択されたこのような骨格は次のような基準の最大数を満たさなければならないことが知られている(Skerra A., J. Mol. Recogn., 2000, 13:167-187):
−系統発生的保存が良好であること、
−三次元構造が既知であること(例えば、結晶学、NMR分光法または当業者に公知の他のいずれかの方法による)、
−小型であること、
−転写後修飾が少ないか、または全く無いこと、および/または
−産生、発現、および精製が容易なこと。
【0079】
このようなタンパク質骨格の起源は、限定されるものではないが、フィブロネクチンおよび優先的にはフィブロネクチンIII型ドメイン10、リポカリン、アンチカリン(Skerra A., J. Biotechnol., 2001, 74(4):257-75)、黄色ブドウ球菌(Staphylococcus aureus)のタンパク質AのドメインBに由来するタンパク質Z、チオレドキシンAまたは「アンキリンリピート」(Kohl et al., PNAS, 2003, vol. 100, No. 4, 1700-1705)、「アルマジロリピート」、「ロイシンリッチリピート」および「テトラトリコペプチドリピート」などの繰り返しモチーフを有するタンパク質の中から選択される構造物であり得る。
【0080】
例えば、サソリ、昆虫、植物、軟体動物などの毒素、およびニューロンNOシンターゼ(PIN)のタンパク質阻害剤といった毒素に由来する骨格も挙げておくべきであろう。
限定されるものではないが、このようなハイブリッド構築物の例として、PINのループの一つにおける抗CD4抗体、すなわち13B8.2のCDR−H1(重鎖)挿入があり、このようにして得られた新たな結合タンパク質は元の抗体と同じ結合特性を保存している(Bes et al., Biochem. Biophys. Res. Commun., 2006, 343(1), 334-344)。単に例示であるが、ネオカルチノスタチンのループの一つに抗リゾチームVHH抗体のCDR−H3(重鎖)をグラフトすることも挙げられる。
【0081】
最後に、上記のように、このようなペプチド骨格は、元の抗体に由来するCDRの少なくとも一つを含んでなり得る。必ずしも必要というわけではないが、好ましくは、当業者は重鎖由来の少なくとも一つのCDRを選択することができ、これは抗体の特異性を主として担うことが知られている。1以上の適切なCDRを選択することは当業者には自明であり、当業者ならば、次に好適な既知技術を選択することができる(Bes et al., FEBS letters 508, 2001, 67-74)。
【0082】
本発明の特定の態様は、本発明による抗体に由来する化合物を選択する方法に関し、該誘導化合物はin vitroおよび/またはin vivoにおいてHIVの細胞侵入を阻害することができ、該誘導化合物は、少なくとも一つの抗体CDRがグラフトされているペプチド骨格を含んでなり、該方法は、
a)少なくとも一つの抗体CDRがグラフトされているペプチド骨格ペプチド骨格から構成される化合物を、HIV 1型とPBMCを含有する生体サンプルと接触させる工程、および
b)該化合物がHIV−1複製を阻害することができる場合、その化合物を選択する工程
を含んでなり、
少なくとも一つのグラフトされるCDRが、配列番号1〜6および30〜33の配列、または最適なアライメントの後に配列番号1〜6および30〜33の配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性を有する配列のCDRの中から選択されることを特徴とする。
【0083】
好ましい様式によれば、該方法は、工程a)において、少なくとも二つまたは三つの抗体CDRがグラフトされているペプチド骨格を含んでなる化合物をin vitroで接触させることを含み得る。
【0084】
この方法のさらに好ましい様式によれば、ペプチド骨格は、構造が上記された骨格または結合タンパク質の中から選択される。
【0085】
明らかに、これらの例は何ら限定されず、当業者に既知または自明の他のいずれの構造も、本特許出願によって付与される保護に包含されるものと考えるべきである。
【0086】
よって、本発明は、ペプチド骨格が、a)系統発生的によく保存されているタンパク質、b)ロバストな(robust)構造のタンパク質、c)周知の3D分子構成を有するタンパク質、d)小型のタンパク質、および/またはe)安定性を変化させずに欠失および/または挿入によって改変することができる領域を含んでなるタンパク質の中から選択されることを特徴とする抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0087】
好ましい実施形態によれば、本発明の抗体、またはその誘導化合物もしくは機能的フラグメントは、該ペプチド骨格が、i)フィブロネクチンに由来する骨格、優先的にはフィブロネクチン3型ドメイン10、リポカリン、アンチカリン、黄色ブドウ球菌(Staphylococcus aureus)のタンパク質AのドメインBに由来するタンパク質Z、チオレドキシンAまたは「アンキリンリピート」(Kohl et al., PNAS, 2003, vol. 100, No. 4, 1700-1705)、「アルマジロリピート」、「ロイシンリッチリピート」および「テトラトリコペプチドリピート」などの繰り返しモチーフを有するタンパク質、またはiii)ニューロンNOシンターゼ(PIN)タンパク質阻害剤の中から選択されることを特徴とする。
【0088】
本発明の別の態様は、上記抗体の機能的フラグメントに関する。
【0089】
より詳細には、本発明は、機能的フラグメントがフラグメントFv、Fab、(Fab’)2、Fab’、scFv、scFv−Fc、およびダイアボディー、またはPEG化フラグメントなどの半減期が延長された任意のフラグメントの中から選択されることを特徴とする抗体、またはその誘導化合物もしくは機能的フラグメントを対象とする。
【0090】
本発明による抗体のこのような機能的フラグメントは、例えば、フラグメントFv、scFv(sc=一本鎖)、Fab、F(ab’)2、Fab’、scFv−Fcまたはダイアボディー、またはポリエチレングリコール(ペグ化)などのポリアルキレングリコールの付加といった化学修飾によって(PEG化フラグメントはFv−PEG、scFv−PEG、Fab−PEG、F(ab’)2−PEGおよびFab’−PEGと呼ばれる)、またはリポソーム、ミロスフェア、またはPLGAに組み込むことによって半減期が延長された任意のフラグメントからなり、該フラグメントは、特に、それが由来する抗体の全般的活性、さらには部分的活性であっても示し得る本発明の特徴的なCDRの少なくとも一つを有する。
【0091】
好ましくは、該機能的フラグメントは、それらが由来する抗体の可変重鎖または軽鎖の部分的配列を含んでなるか、または含み、該部分的配列は、それが由来する抗体と同様の結合特異性および十分な親和性、好ましくはそれが由来する抗体の親和性の少なくとも1/100、より好ましくは少なくとも1/10の親和性を保持するのに十分なものである。
【0092】
このような機能的フラグメントは、それが由来する抗体の配列の少なくとも5つのアミノ酸、好ましくは、6、7、8、10、15、25、50、または100個の連続するアミノ酸を含む。
好ましくは、これらの機能的フラグメントは、それらが由来する抗体と一般に同様の結合特異性を有する、Fv、scFv、Fab、F(ab’)2、F(ab’)、scFv−Fc、またはダイアボディータイプのものである。本発明によれば、本発明の抗体のフラグメントは、上記の抗体からペプシンもしくはパパインを含む酵素消化、および/または化学的還元によるジスルフィド結合の切断などの方法によって得ることができる。該抗体フラグメントはまた、これもまた当業者に公知の組換え遺伝子技術によって、または例えば、Applied BioSystemsなどによって販売されているものなどの自動ペプチド合成装置の手段によるペプチド合成によって得ることができる。
さらに明確にするために、下表2に、本発明の抗体に相当する種々のアミノ酸配列をまとめる。
【0093】
【表2】
【0094】
本発明の抗体対象の特に重要な付加的態様は、それらが抗体依存性細胞傷害性(ADCC)および/または補体依存性細胞傷害性(CDC)などのエフェクター機能を示さないことを特徴とする。
【0095】
より詳細には、一つの例として、本発明の抗体、またはそれらの一つの機能的フラグメントもしくは誘導体は、FcγR(I、II、もしくはIII)に対して、またはC1qに対して、またはその双方に対して親和性を持たない。
【0096】
構造的に、これは、当業者にとって、本発明の抗体、またはそれらの一つの機能的フラグメントもしくは誘導体が、Fc部分を欠いていること、またはそれらのFc部分がエフェクター機能を付与し得る適切なグリコシル化を与えないことを意味する。
【0097】
この結果が、本発明の抗体は好ましくはIgG4またはIgG2イソ型、最も好ましくはIgG4イソ型から選択されるということである。
【0098】
同様に、好ましいフラグメントは、Fv、scFv(scは一本鎖)、Fab、F(ab’)2、Fab’、scFv−Fcフラグメントもしくはダイアボディー、またはポリ(エチレン)グリコール(「ペグ化」)(ペグ化フラグメントはFv−PEG、scFv−PEG、Fab−PEG、F(ab’)2−PEG、またはFab’−PEGと呼ばれる)(「PEG」はポリ(エチレン)グリコール)などのポリ(アルキレン)グリコールの付加のような化学的修飾によって、またはリポソームへの組み込みによって半減期が延長された任意のフラグメントといった、ADCCを欠いているフラグメントである。
【0099】
より詳細には、抗体515H7由来の本発明の好ましい機能的フラグメントはscFvであり、以下、515H7 scFv−Ckフラグメントと呼ばれ、配列番号54のアミノ酸配列を含んでなる。
【0100】
該scFvに相当するヌクレオチド配列は、配列番号55の配列を含んでなる。
【0101】
本発明の別の特定の態様は、抗体がまたマウスとは異種の、特にヒトの抗体に由来する軽鎖および重鎖定常領域も含んでなることを特徴とする、キメラ抗体、またはそれらの誘導化合物もしくは機能的フラグメントに関する。
【0102】
本発明のさらに別の態様は、ヒト抗体に由来する軽鎖および重鎖の定常領域がそれぞれλまたはκ領域およびγ−2または好ましくはγ−4領域であることを特徴とする、ヒト化抗体、またはそれらの誘導化合物もしくは機能的フラグメントに関する。
【0103】
本発明の抗体はまた、キメラ抗体またはヒト化抗体も含んでなる。
【0104】
キメラ抗体は、所与の種に由来する天然可変領域(軽鎖および重鎖)を所与の種とは異種の抗体の軽鎖および重鎖の定常領域と組み合わせて含むものである。
【0105】
抗体、またはそのキメラフラグメントは、組換え遺伝子技術を用いて作製することができる。例えば、該キメラ抗体は、プロモーターと、本発明の非ヒト、特にネズミモノクローナル抗体の可変領域をコードする配列、およびヒト抗体定常領域をコードする配列を含む組換えDNAをクローニングすることによって作製することができる。このような組換え遺伝子の一つによってコードされる本発明によるキメラ抗体は、例えば、マウス−ヒトキメラであってよく、この抗体の特異性はネズミDNAに由来する可変領域によって決定され、そのイソ型はヒトDNAに由来する定常領域によって決定される。キメラ抗体の作製方法については、Verhoeynら(BioEssays, 8:74, 1988)を参照。
【0106】
以下の表3に、本発明によるキメラ抗体515H7(c515H7またはC515H7と呼ばれる)の種々の重鎖および軽鎖のアミノ酸配列をまとめる。
【0107】
【表3】
【0108】
この抗体c515H7重鎖配列番号56〜58および軽鎖配列番号59に相当するヌクレオチド配列はそれぞれ、配列番号60〜63(重鎖)および配列番号64(軽鎖)の配列に相当する。
【0109】
好ましい実施形態では、これらの重鎖配列はそれらのC末端からリシン残基(LonzaからのpConPlusベクター系統原株:pConPlusγ4ΔK、pConPlusγ4PROΔK、およびpConPlusγ2ΔKに見られる)が欠失されている。
【0110】
さらに、G4PRO重鎖は、半抗体の形成を避けるためにヒンジ領域に突然変異を有するヒトIgG4イソ型に相当する。この突然変異は、Lonzaからの親株pConPlusγ4PROΔKに見られる[Angal S, King DJ, Bodmer MW, Turner A, Lawson AD, Roberts G, Pedley B, Adair JR. A single amino acid substitution abolishes the heterogeneity of chimeric mouse/human (IgG4) antibody. Mol Immunol. (1993);30(1):105-108]。
【0111】
より詳細には、本発明は、種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRを含んでなることを特徴とするキメラ抗体重鎖に関し、ここで、該CDRは、IMGTによれば、配列番号4、5、および6の配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3からなる。
【0112】
より詳細には、本発明は、種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRを含んでなることを特徴とするキメラ抗体軽鎖に関し、ここで、該CDRは、IMGTによれば、配列番号1、2、および3の配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3からなる。
【0113】
より詳細には、本発明は、種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRをそれぞれ有する重鎖および軽鎖を含んでなることを特徴とするキメラ抗体、またはその誘導化合物もしくは機能的フラグメントに関し、ここで、該CDRは、IMGTによれば、配列番号4、5、および6の配列をそれぞれ含んでなる重鎖のCDR−H1、CDR−H2、およびCDR−H3と、配列番号1、2、および3の配列をそれぞれ含んでなる軽鎖のCDR−L1、CDR−L2、およびCDR−L3からなる。
【0114】
別の実施形態では、本発明は、配列番号8からなる配列の重鎖可変領域と、配列番号7の配列の軽鎖可変領域とを含んでなるキメラ抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0115】
さらに別の実施形態では、本発明は、配列番号56、57、または58からなる群から選択される配列の重鎖と、配列番号59の配列の軽鎖とを含んでなるキメラ抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0116】
好ましい実施形態では、本発明によるキメラ抗体c515H7 VH(G4wt)/VL−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号56の配列の重鎖可変領域と、配列番号59の配列の軽鎖可変領域とを含んでなる。
【0117】
好ましい実施形態では、本発明によるキメラ抗体c515H7 VH(G4PRO)/VL−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号57の配列の重鎖可変領域と、配列番号59の配列の軽鎖可変領域とを含んでなる。
【0118】
好ましい実施形態では、本発明によるキメラ抗体c515H7 VH(G2wt)/VL−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号58の配列の重鎖可変領域と、配列番号59の配列の軽鎖可変領域とを含んでなる。
【0119】
「ヒト化抗体」とは、非ヒト起源の抗体に由来するCDR領域を含み、その抗体分子の他の部分が一つの(または複数の)ヒト抗体に由来する抗体を意味する。さらに、骨格セグメント残基(FRと呼ばれる)のいくつかは、結合親和性を保存するように改変することができる(Jones et al., Nature, 321:522-525, 1986、 Verhoeyen et al., Science, 239:1534-1536, 1988、 Riechmann et al., Nature, 332:323-327, 1988)。
【0120】
本発明のヒト化抗体またはそのフラグメントは、当業者に公知の技術(例えば、Singer et al., J. Immun., 150:2844-2857, 1992、 Mountain et al., Biotechnol. Genet. Eng. Rev., 10:1-142, 1992、およびBebbington et al., Bio/Technology, 10:169-175, 1992の文献に記載されているもの)によって作製することができる。このようなヒト化抗体は、in vitro診断を含む方法またはin vivoにおける予防的および/もしくは治療的処置における使用に好ましい。他のヒト化技術も当業者に公知であり、例えば、欧州特許第0451261号公報、同第0682040号公報、同第0939127号公報、同第0566647号公報、または米国特許第5,530,101号公報、同第6,18,370号公報、同第5,585,089号公報、および同第5,693,761号公報においてPDLが記載している「CDRグラフト」法がある。米国特許第5,639,641号公報または同第6,054,297号公報、同第5,886,152号公報、および同第5,877,293号公報も引用することができる。
【0121】
さらに、本発明はまた、上記のネズミ抗体に由来するヒト化抗体に関する。
好ましい様式では、ヒト抗体に由来する軽鎖および重鎖の定常領域はそれぞれ、λまたはκおよびγ−2または好ましくはγ−4領域である。
【0122】
より詳細には、本発明は、i)対応するヒト抗体重鎖のフレームワーク領域に相同なフレームワーク領域と、ii)種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRとを含んでなることを特徴とするヒト化抗体重鎖に関し、ここで、該CDRは、IMGTによれば、配列番号4、5、および6の配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3からなる。
【0123】
別の実施形態では、本発明は、配列番号64からなる配列の可変領域を含んでなるヒト化抗体重鎖に関する。
さらに別の実施形態では、本発明は、配列番号67、68、および69からなる群から選択される完全な配列を含んでなるヒト化抗体重鎖に関する。より詳細には、本発明は、ヒト抗体軽鎖の対応するフレームワーク領域に相同なフレームワーク領域と、ii)種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRとを含んでなることを特徴とするヒト化抗体軽鎖に関し、ここで、該CDRは、IMGTによれば、配列番号1、2、および3の配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3からなる。
【0124】
別の実施形態では、本発明は、配列番号65、66、82、または83からなる群から選択される配列の可変領域を含んでなるヒト化抗体軽鎖に関する。
【0125】
さらに別の実施形態では、本発明は、配列番号70、71、84、または85のからなる群から選択される完全な配列を含んでなるヒト化抗体軽鎖に関する。
【0126】
より詳細には、本発明は、i)ヒト抗体の対応するフレームワーク領域に相同なフレームワーク領域と、ii)種々の哺乳類種に由来する抗体の対応するCDRに相同なCDRをそれぞれ有する重鎖および軽鎖を含んでなることを特徴とするヒト化抗体、またはその誘導化合物もしくは機能的フラグメントに関し、ここで、該CDRは、IMGTによれば、配列番号4、5、および6の配列をそれぞれ含んでなる重鎖のCDR−H1、CDR−H2、およびCDR−H3と、配列番号1、2、および3の配列をそれぞれ含んでなる軽鎖のCDR−L1、CDR−L2、およびCDR−L3からなる。
【0127】
別の実施形態では、本発明は、配列番号64からなる配列の重鎖可変領域と、配列番号65、66、82、または83からなる群から選択される配列の軽鎖可変領域とを含んでなるヒト化抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
【0128】
さらに別の実施形態では、本発明は、配列番号67、68、または69からなる群から選択される配列の重鎖と、配列番号70、71、84または85からなる群から選択される配列の軽鎖とを含んでなるヒト化抗体、またはその誘導化合物もしくは機能的フラグメントに関する。
好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4wt)/VL2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号67の配列の重鎖と、配列番号70の配列の軽鎖とを含んでなる。
【0129】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4PRO)/VL2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号68の配列の重鎖と、配列番号70の配列の軽鎖とを含んでなる。
【0130】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G2wt)/VL2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号69の配列の重鎖と、配列番号70の配列の軽鎖とを含んでなる。
【0131】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4wt)/VL2.1−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号67の配列の重鎖と配列番号71の配列の軽鎖とを含んでなる。
【0132】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4PRO)/VL2.1−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号68の配列の重鎖と配列番号71の配列の軽鎖とを含んでなる。
【0133】
別の好ましい実施形態では、本発明によるヒト化抗体 Hz515H7 VH1 D76N(G2wt)/VL2.1−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号69の配列の重鎖と、配列番号71の配列の軽鎖とを含んでなる。
【0134】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4wt)/VL2.2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号67の配列の重鎖と、配列番号84の配列の軽鎖とを含んでなる。
【0135】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4PRO)/VL2.2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号68の配列の重鎖と、配列番号84の配列の軽鎖とを含んでなる。
【0136】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G2wt)/VL2.2−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号69の配列の重鎖と、配列番号84の配列の軽鎖とを含んでなる。
【0137】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4wt)/VL2.3−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号67の配列重鎖と、配列番号85の配列の軽鎖とを含んでなる。
【0138】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G4PRO)/VL2.3−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号68の配列の重鎖と、配列番号85の配列の軽鎖とを含んでなる。
【0139】
別の好ましい実施形態では、本発明によるヒト化抗体Hz515H7 VH1 D76N(G2wt)/VL2.3−Ck、またはその誘導化合物もしくは機能的フラグメントは、配列番号69の配列の重鎖と、配列番号85の配列の軽鎖とを含んでなる。
【0140】
以下の表4に、本発明によるヒト化抗体515H7の、それぞれ種々の重鎖および軽鎖可変ドメインおよび全長(または完全)アミノ酸配列をまとめる。
【0141】
【表4】
【0142】
好ましい実施形態では、これらの重鎖配列はそれらのC末端からリシン残基(LonzaからのpConPlusベクター系統原株:pConPlusγ4ΔK、pConPlusγ4PROΔK、およびpConPlusγ2ΔKに見られる)が欠失されている。
【0143】
さらに、G4PRO重鎖は、半抗体の形成を避けるためにヒンジ領域に突然変異を有するヒトIgG4イソ型に相当する。この突然変異は、Lonzaからの親株pConPlusγ4PROΔKに見られる[Angal S, King DJ, Bodmer MW, Turner A, Lawson AD, Roberts G, Pedley B, Adair JR. A single amino acid substitution abolishes the heterogeneity of chimeric mouse/human (IgG4) antibody. Mol Immunol. (1993);30(1):105-108]。
【0144】
一例として、疑念を避けるため、「VH1」は、「VH Variant 1」、「VH variant 1」、「VH Var 1」、または「VH var 1」と同様である。
【0145】
上記で例示されたVH/VLの組合せに限定されないと理解すべきである。当業者は、当然のことながら、過度な負担無く、また、発明性のある技術を用いずに、本明細書で開示されたVHおよびVLを総て再配列することができるであろう。
【0146】
本発明の新規な態様は、以下の核酸(縮重遺伝コードを含む):
a)本発明による抗体、またはその一つの機能的フラグメントもしくは誘導体をコードする核酸、DNA、またはRNA、
b)配列番号14〜19および41〜45からなる配列群から選択されるDNA配列を含んでなる核酸、
c)配列番号20、21、46、および47からなる配列群から選択されるDNA配列を含んでなる核酸、
d)b)またはc)に定義される核酸の、対応するRNA核酸、
e)a)、b)およびc)に定義される核酸の相補的核酸、および
f)配列番号14〜19および41〜45の配列のCDRの少なくとも一つと高ストリンジェンシー条件下でハイブリダイズすることができる少なくとも18ヌクレオチドの核酸
から選択されることを特徴とする単離された核酸に関する。
【0147】
下表5に、本発明の抗体に関する種々のヌクレオチド配列をまとめる。
【0148】
【表5】
【0149】
本明細書において互換的に用いられる「核酸」、「核配列」、「核酸配列」、「ポリヌクレオチド」、「オリゴヌクレオチド」、「ポリヌクレオチド配列」、および「ヌクレオチド配列」は、非天然ヌクレオチドを含む、または含まない、また二本鎖DNA、一本鎖DNA、またはこれらのDNAの転写産物のいずれかである核酸のフラグメントまたは領域を規定する、修飾または非修飾のヌクレオチドの正確な配列を意味する。
【0150】
ここでまた、本発明はそれらの天然の染色体環境の、すなわち天然状態のヌクレオチド配列には関しないということを含めておくべきである。本発明の配列は単離および/または精製されており、すなわち、それらは直接、または例えばコピーによって間接的にサンプリングされており、それらの環境は少なくとも部分的に改変されている。例えば宿主細胞の手段による組換え遺伝子技術によって得られた、または化学合成によって得られた単離された核酸もここに挙げておくべきであろう。
【0151】
「最適なアライメントの後に好ましい配列と少なくとも80%、好ましくは85%、90%、95%、および98%の同一性割合を示す核配列」とは、参照核配列に比べて、特に、欠失、末端切断、延長、キメラ融合、および/または置換、特に規則的なものなどの、ある種の改変を示す核配列を意味する。好ましくは、これらは参照配列と同じアミノ酸配列をコードする配列であり、これは遺伝コードの縮重、または好ましくは高ストリンジェント条件(特に以下に定義されるもの)下で参照配列と特異的にハイブリダイズし得る相補的配列に関連づけられる。
【0152】
高ストリンジェント条件下でのハイブリダイゼーションとは、温度およびイオン強度に関する条件が、相補的DNAフラグメントの間でハイブリダイゼーションの維持を可能となるように選択される。単に例として、上記のポリヌクレオチドフラグメントを定義する目的でのハイブリダイゼーション工程の高ストリンジェント条件は有利には次の通りである。DNA−DNAまたはDNA−RNAハイブリダイゼーションは、(1)5×SSC(1×SSCは、0.15M NaCl+0.015Mクエン酸ナトリウムの溶液に相当する)、50%ホルムアミド、7%ドデシル硫酸ナトリウム(SDS)、10×デンハート溶液、5%デキストラン硫酸、および1%サケ精子DNAを含むリン酸バッファー(20mM、pH7.5)中、42℃で3時間のプレハイブリダイゼーション、(2)プローブの長さに応じた温度(すなわち、プローブ>100ヌクレオチド長では42℃)で20時間の主ハイブリダイゼーションと、その後の2×SSC+2%SDS中、20℃での20分の洗浄2回、0.1×SSC+0.1%SDS中、20℃での20分の洗浄1回という2段階で行う。最後の洗浄は、プローブ>100ヌクレオチド長では60℃で、0.1×SSC+0.1%SDS中、30分間行う。大きさの定義されたポリヌクレオチドに対する上記の高ストリンジェントハイブリダイゼーション条件は、それより長いまたは短いオリゴヌクレオチドに関しては、Sambrookら(Molecular cloning: a laboratory manual, Cold Spring Harbor Laboratory、第3版, 2001)に記載されている手順に従い、当業者によって適合させることができる。
【0153】
本発明はまた、以下の核酸:
a)本発明によるヒト化抗体重鎖、またはその誘導化合物もしくは機能的フラグメントをコードする核酸、DNA、またはRNA、
b)本発明によるヒト化抗体軽鎖、またはその誘導化合物もしくは機能的フラグメントをコードする核酸、DNA、またはRNA、
c)本発明によるヒト化抗体、またはその誘導化合物もしくは機能的フラグメンをコードする核酸、DNA、またはRNA、
d)a)、b)、またはc)に定義される核酸と相補的な核酸、
e)高ストリンジェント条件下で配列番号72または75〜77の核酸配列を含んでなる少なくとも重鎖とハイブリダイズすることができる少なくとも18ヌクレオチドの核酸、
f)高ストリンジェント条件下で配列番号73、74、86、87、または78、79、88、89の核酸配列を含んでなる少なくとも軽鎖とハイブリダイズすることができる少なくとも18ヌクレオチドの核酸、
の中から選択されることを特徴とする、単離された核酸分子を包含する。
【0154】
以下の表6に、本発明によるヒト化抗体515H7のそれぞれ種々の重鎖および軽鎖可変ドメインおよび全長(または完全)ヌクレオチド配列をまとめる。
【0155】
【表6】
【0156】
以下の表7に、本発明によるキメラ抗体515H7の種々の重鎖および軽鎖のヌクレオチド配列をまとめる。
【0157】
【表7】
【0158】
言い換えれば、本発明は、以下の核酸:
a)本発明による抗体、またはその一つの機能的フラグメントもしくは誘導体をコードする核酸、DNA、またはRNA、
b)配列番号14〜19および41〜45からなるCDR配列群から選択されるDNA配列を含んでなる核酸、
c)配列番号20、21、46、47、72、73、74、86、および87からなる重鎖および軽鎖可変ドメイン配列群から選択されるDNA配列を含んでなる核酸、
d)配列番号60〜63、75〜79、88、および89からなる重鎖および軽鎖配列群から選択されるDNA配列を含んでなる核酸、
e)配列番号55からなるDNA配列を含んでなる核酸、
f)b)、c)、d)、またはe)に定義される核酸の、対応するRNA核酸、
g)a)、b)、c)、d)、およびe)に定義される核酸の相補的核酸、および
h)高ストリンジェンシー条件下で配列番号14〜19および41〜45の配列のCDRの少なくとも一つとハイブリダイズすることができる少なくとも18のヌクレオチドの核酸
から選択されることを特徴とする、単離された核酸を取り扱う。
【0159】
本発明はまた、本明細書に記載される核酸を含んでなるベクターに関する。
本発明は特に、このようなヌクレオチド配列を含むクローニングおよび/または発現ベクターを対象とする。
【0160】
本発明のベクターは、好ましくは、所与の宿主細胞においてヌクレオチド配列の発現および/または分泌を可能とするエレメントを含む。よって、このベクターは、プロモーター、翻訳開始、および終結シグナル、ならびに好適な転写調節領域を含まなければならない。ベクターは宿主細胞において安定な様式で維持可能でなければならず、所望により、翻訳されたタンパク質の分泌を条件として指定する特異的シグナルを有してもよい。これらの種々のエレメントは、使用する宿主細胞に応じて、当業者により選択され、至適化される。この目的で、これらのヌクレオチド配列は、選択された宿主内で自己複製するベクターに挿入することもできるし、または選択された宿主の組込型ベクターであってもよい。
【0161】
このようなベクターは、当業者により一般に用いられる方法によって作製され、得られたクローンは、リポフェクション、エレクトロポレーション、熱ショック、また化学法などの標準的な方法によって好適な宿主に導入することができる。これらのベクターは、例えば、プラスミドまたはウイルス起源のベクターである。それらを用いて、本発明のヌクレオチド配列をクローニングする、または発現させるために宿主細胞を形質転換することができる。
【0162】
本発明はまた、本発明に記載されているベクターにより形質転換された、または本発明に記載されているベクターを含んでなる宿主細胞も含んでなる。
【0163】
宿主細胞は、細菌細胞、例えば、酵母細胞または動物細胞、特に、哺乳類細胞などの原核生物系、または真核生物系の中から選択することができる。また、昆虫または植物細胞も使用可能である。
【0164】
本発明はまた、本発明による形質転換細胞を有するヒト以外の動物に関する。
【0165】
本発明の別の態様は、本発明による抗体、またはその一つの機能的フラグメントの作製方法であって、以下の工程を含んでなることを特徴とする方法に関する:
a)本発明による宿主細胞の培地および好適な培養条件での培養工程、および
b)このようにして生産された該抗体、またはその一つの機能的フラグメントの、該培養培地からの、または該培養細胞からの回収工程。
【0166】
本発明による形質転換細胞は、本発明による組換えポリペプチドの作製方法に用いられる。また、本発明によるベクターおよび/またはベクターによって形質転換された細胞を使用することを特徴とする、本発明によるポリペプチドの組換え型での作製方法も本発明に含まれる。好ましくは、本発明によるベクターによって形質転換された細胞は、上記のポリペプチドの発現および該組換えペプチドの回収を可能とする条件下で培養される。
【0167】
すでに述べたように、宿主細胞は原核生物系または真核生物系の中から選択することができる。特に、このような原核生物系または真核生物系において分泌を助ける本発明のヌクレオチド配列を同定することが可能である。よって、このような配列を有する本発明によるベクターは、有利には分泌させる組換えタンパク質の生産のために使用することができる。実際、これらの着目する組換えタンパク質の精製は、宿主細胞内ではなく細胞培養の上清に存在するということによって容易となる。
【0168】
本発明のポリペプチドはまた、化学合成によっても製造することができる。このような製造方法の一つも本発明の目的である。当業者は、固相技術(特に、Steward et al., 1984, Solid phase peptides synthesis, Pierce Chem. Company, Rockford, 111,第2版参照)または部分固相技術、フラグメントの縮合または溶液中での従来の合成などの化学合成法を知る。化学合成によって得られ、対応する非天然アミノ酸を含み得るポリペプチドもまた、本発明に含まれる。
【0169】
本発明の方法によって得られる可能性のある抗体、またはその誘導化合物もしくは機能的フラグメントも本発明に含まれる。
【0170】
さらに別の態様によれば、本発明は、ヒトケモカインファミリー受容体と特異的に結合することができ、および/またはX4指向性HIV複製を特異的に阻害することができることをさらに特徴とする、上記のような抗体に関する。
【0171】
さらに別の態様によれば、本発明は、ヒトケモカインファミリー受容体と特異的に結合することができ、および/またはX4/R5指向性HIV複製を特異的に阻害することができることを特徴とする、上記のような抗体に関する。
【0172】
新規な実施形態によれば、本発明は、例えば、CCR5、CD4、CXCR4(本発明の抗体以外の、すなわち、別のエピトープを標的とする)またはCCR3、CCR2、CCR8、CXCR6、CXCR7、CX3CR1などのHIVの細胞侵入に関与する受容体と相互作用することができる第二のモチーフを含んでなるという意味で二重特異性である抗体からなる抗体、またはそれらの誘導化合物もしくは機能的フラグメントに関する。
【0173】
二重特異性または二官能性抗体は、二つの異なる可変領域が同じ分子において組み合わされている第二世代のモノクローナル抗体を構成する(Hollinger and Bohlen, 1999, Cancer and metastasis, rev. 18:411-419)。それらの有用性は、細胞表面のいくつかの分子を標的とするそれらの能力に関して診断用ドメインおよび治療用ドメインの双方で実証されており、このような抗体は化学法(Glennie MJ et al., 1987, J. Immunol. 139, 2367-2375、 Repp R. et al., 1995, J. Hemat., 377-382)または体細胞法(Staerz U.D. and Bevan M.J., 1986, PNAS 83, 1453-1457、 Suresh M.R. et al., 1986, Method Enzymol, 121:210-228)によるだけでなく、優先的には、求める抗体にヘテロ二量体形成を課し、従ってその抗体の精製を容易にすることを可能とする遺伝子操作技術(Merchand et al., 1998, Nature Biotech., 16:677-681)によって得ることができる。
【0174】
これらの二重特異性抗体は、完全なIgG、二重特異性Fab’2、Fab’PEG、ダイアボディー、または二重特異性scFvとして構築できるだけではなく、標的とされる各抗原に対して二つの結合部位が存在する四価二重特異性抗体(Park et al, 2000, Mol. Immunol, 37(18):1123-30)または上記のようなそのフラグメントとしても構築できる。
【0175】
一つの二重特異性抗体の製造および投与が、二つの特異的抗体の製造よりも安くつくことを考えた場合の経済的利点に加え、このような二重特異性抗体の使用は処置の毒性を軽減するという利点も持つ。実際、二重特異性抗体を使用すると、循環する抗体の総量を減らすことができ、結果として毒性が軽減される。
【0176】
本発明の好ましい実施形態においては、二重特異性抗体は二価または四価抗体である。
【0177】
最後に、本発明は、薬剤としての上記の抗体、またはそれらの一つの機能的フラグメントもしくは誘導体に関する。
【0178】
本発明はまた、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体からなる化合物を有効成分として含んでなる医薬組成物に関する。好ましくは、該抗体は賦形剤および/または薬学上許容される担体を追加する。
【0179】
本発明はまた、薬剤としての上記のような組成物に関する。
本発明の特定の態様では、抗体、またはその一つの機能的フラグメントもしくは誘導体、PBMCにおけるHIV−1 KON一次単離物の複製を、少なくとも5μg/ml、好ましくは少なくとも10μg/mlのIC90で阻害する。
【0180】
本発明はまた、HIV感染の予防または治療のための薬物および/または薬剤の製造のための本発明による抗体または組成物の使用を含んでなる。
【0181】
より詳細には、限定されるものではないが、該HIV感染はX4指向性HIV感染である。
【0182】
別の実施形態では、限定されるものではないが、該HIV感染はX4/R5指向性HIV感染である。
【0183】
本発明はまた、HIV複製を阻害するための薬物の製造のための、本発明による抗体、またはその機能的フラグメントもしくは誘導体、好ましくは、ヒト化されたもの、および/または組成物の使用に関する。一般に、本発明は、HIV疾患の予防または治療のための薬物の製造のための、抗体、またはその機能的フラグメントもしくは誘導体、好ましくは、ヒト化されたもの、および/または組成物の使用に関する。
【0184】
本明細書において、「医薬ビヒクル」とは、二次反応を引き起こさず、かつ、例えば、有効化合物の投与を容易にするか、生物内でのその寿命を延長し、および/または有効性を高めるか、溶液中でのその溶解度を高めるか、またはその貯蔵性を高める化合物、または化合物の組合せ(医薬組成物を含む)を意味する。このような医薬担体は周知であり、当業者により、選択される有効化合物の性質および投与経路に応じて適合される。
【0185】
好ましくは、このような化合物は、全身経路、特に、静脈内経路、筋肉内経路、皮内経路、腹腔内経路、皮下経路、腟内経路、または経口経路によって投与される。より好ましくは、本発明による抗体から構成される組成物は、一定時間あけた数回の投与で投与される。
【0186】
それらの投与経路、投与計画および最適な剤形は、例えば、患者の年齢または体重、患者の健康状態の重篤度、患者の処置に対する耐性および受ける副作用など、患者に適合する処置を確立する際に一般に考慮される基準に従って決定することができる。
【0187】
本発明はまた、同時、個別または延長様式で用いるための組合せ製剤として、CXCR4に対する抗体以外の抗HIV抗体または抗HIV細胞侵入抗体または抗HIV複製抗体をさらに含んでなる組成物に関する。
【0188】
さらに別の実施形態によれば、本発明はまた、本発明に記載されているもの以外の抗CCR5、抗CD4化合物、および抗CXCR4化合物などの、HIVの侵入および/または複製を特異的に阻害することができる化合物、または当業者に知られている他のいずれかの抗HIV化合物の中から選択される少なくとも一つの第二の抗HIV化合物を含んでなる上記のような医薬組成物に関する。
【0189】
本発明の補足的な別の実施形態は、同時、個別、または延長使用のための組合せまたは複合製剤として抗HIV化合物をさらに含んでなる上記のような組成物からなる。
【0190】
「同時使用」とは、単一の投与形に含まれる組成物に含まれる両化合物を投与することを意味する。
【0191】
「個別使用」とは、異なる投与形に含まれる組成物の両化合物を同時に投与することを意味する。
【0192】
「延長使用」とは、異なる投与形にそれぞれ含まれる組成物の両化合物の連続的投与を意味する。
【0193】
一般に、本発明による組成物は、HIV処置の有効性を著しく高める。言い換えれば、本発明の抗体の治療効果は、抗HIV剤の投与によって予期されないほど増強される。本発明の組成物によってもたらされる別の大きな二次的利点は、有効成分のより低量の有効用量を使用できる可能性に関し、従って、特に抗HIV剤の作用である副作用の出現の危険を回避または軽減することが可能となる。さらに、この組成物は、予想される治療効果をより迅速に達成することを可能とする。
【0194】
「治療用抗HIV剤」とは、患者に投与した際に、患者におけるHIVの複製を治療または予防する物質を意味する。このような薬剤の限定されない例としては、HIVプロテアーゼ阻害剤(PI)、ヌクレオシド/ヌクレオチドHIV逆転写酵素阻害剤(NRTI/NtRTI)、非ヌクレオシドHIV逆転写酵素阻害剤(NNRTI)、HIV侵入阻害剤、HIVインテグラーゼ阻害剤等の「抗レトロウイルス剤」が挙げられる。
【0195】
このような薬剤は、例えば、VIDALの、「抗HIV化合物」に関する化合物のために費やされた頁に挙げられており、この文献の参照文献によって挙げられている抗HIV化合物は、本明細書において、限定的されない好ましい抗HIV剤として挙げられる。
【0196】
HIVプロテアーゼ阻害剤は、HIVプロテアーゼ活性を阻害することができるいずれの物質も表す。このようなHIVプロテアーゼ阻害剤の例としては、限定されるものではないが、メシル酸サキナビルまたはSQV(Invirase(商標))、インジナビルまたはIDV(Crixivan(商標))、リトナビルまたはRTV(Norvir(商標))、ネルフィナビルまたはNFV(Viracept(商標))、アンプレナビル(Agenerase(商標)、Prozei(商標))、ロピナビル/リトナビルまたはLPV/r(Kaletra(商標)、Aluvia(商標))、アタザナビルまたはATV(Reyataz(商標)、Zrivada(商標))、ホスアンプレナビルまたはFPV(Lexiva(商標)、Telzir(商標))、チプラナビルまたはTPV(Aptivus(商標))、ダルナビルまたはDRV(Prezista(商標))が挙げられる。
【0197】
HIVヌクレオシドまたはヌクレオチド逆転写酵素阻害剤(NRTI)は、HIV RNAの逆転写を遮断するヌクレオシドまたはヌクレオチド類似体である物質を表す。NRTIの例としては、限定されるものではないが、ジドブジンまたはAZT,ZDV(Retrovir/combivir/trixivir(商標))、ジダノシンまたはddi(Videx(商標))、ザルシタビン(HIVID(商標))、スタブジンまたはd4T(Zerit(商標))、ラミブジンまたは3TC(Epivir/combivir/epzicom/trixivir(商標))、アバカビルまたはABC(Ziagen/trixivir/epzicom(商標))、フマル酸テノホビルジソプロキシルまたはTDF(Viread/atripla/truvada(商標))、エムトリシタビンまたはFTC(Emtriva/atripla/truvada(商標))が挙げられる。
【0198】
非ヌクレオシドHIV逆転写酵素阻害剤(NNRTI)は、HIV RNAの逆転写を遮断するヌクレオシドまたはヌクレオチド類似体ではない物質を表す。NNRTIの例としては、限定されるものではないが、ネビラピンまたはNVP(Viramune(商標))、エファビレンツまたはEFV(Sustiva/atripla(商標)、Stocrin(商標))、デラビルジンまたはDLV(Rescriptor(商標))、およびエトラビリンまたはETR(Intelence(商標))が挙げられる。
【0199】
HIV侵入阻害剤は、HIVの細胞侵入を遮断する物質を表す。HIV侵入阻害剤の例としては、限定されるものではないが、エンフビルチドまたはT20(Fuzeon(商標))、マラビロクまたはMVC(Celsentri(商標)、Celzentry(商標))が挙げられる。
【0200】
HIVインテグラーゼ阻害剤は、HIVインテグラーゼ活性を阻害する物質を表す。インテグラーゼ阻害剤の例としては、限定されるものではないが、ラルテグラビルまたはRAL(Isentress(商標))が挙げられる。
【0201】
このような薬剤はまた、例えば、限定されるものではないが、ビクリビロク、PRO140、TNX−355、AMD070、ラシビル、アプリシタビン、エルブシタビン、フロサルブジン(Flosalvudine)、リルピビリン、エルビテグラビルなど、現在臨床試験中の、VIDALに記載されている同種の薬物に属する化合物である。
【0202】
このような薬剤はまた、例えば、限定されるものではないが、成熟阻害剤(ベビリマット)、β−ガラクトシル−セラミドのグリコシド類似体、炭水化物結合剤、RNアーゼH阻害剤、HIV遺伝子発現阻害剤、潜在型T細胞からのHIV放出の刺激剤(バルプロ酸など)など、他の可能性のある種の薬物に属する化合物である。
【0203】
特に好ましい実施形態においては、組合せ製剤としての本発明の組成物は、前記抗HIV剤が同時使用のための前記抗体に化学的に結合されていることを特徴とする。
【0204】
前記抗HIV剤と本発明による抗体の間の結合を容易にするために、結合する二つの化合物の間にポリ(アルキレン)グリコール、ポリエチレングリコール、またはアミノ酸などのスペーサー分子を導入することができ、あるいは、別の実施形態においては、前記抗体と反応することができる官能基を導入されている前記抗HIV剤の活性誘導体を使用することもできる。これらの結合技術は当業者によく知られており、本明細書ではこれ以上詳細には述べない。
【0205】
また、好ましくは、前記の複合体を形成する本発明の抗体は、その機能的フラグメント、特に、それらのFc成分が欠失されたフラグメント、例えばscFvフラグメントの中から選択される。
【0206】
本発明はまた、薬物として用いられる、本発明による、組合せ製剤としての組成物、または抗CXCR4 Mab/抗HIV薬物複合体に関する。
【0207】
好ましくは、組合せ製剤としての前記組成物または前記複合体には、賦形剤および/または医薬ビヒクルが添加される。
【0208】
よって、本発明は、HIV複製に対して生物的に活性な化合物の特異的標的化のための薬物の製造のための、抗体、またはその一つの機能的フラグメントもしくは誘導体の使用に関する。
【0209】
従前に示したように、CXCR4 Mab 515H7および301aE5は、PBMCにおけるHIV−1複製に対して強い活性を有し、従って、このような抗体は、HIV−1感染を処置するためのCXCR4アンタゴニスト抗ウイルス薬の同定のためのスクリーニングアッセイに使用することができる。これらのアッセイの第一の工程では、CXCR4を発現する細胞をMab 515H7および/または301aE5とともに培養し、その後、Mab 515H7および/または301aE5との結合の潜在的な阻害のための分子を評価することができる。この種のアッセイで用いられる細胞は、CHO−CXCR4、NIH3T3−CXCR4またはCXCR4トランスフェクトヒト細胞系統(例えば、U373−MAGI−CXCR4)などのトランスフェクト細胞系統、NALM6などのCXCR4を発現するヒト細胞系統またはPBMCなどの一次細胞であり得る。CXCR4発現細胞に対するMab 515H7および/または301aE5の結合を阻害するCXCR4のアンタゴニストをスクリーニングするために使用される方法は、Zhao Q.ら(AIDS Research And Human Retroviruses, 2003, 19, pp947-955)によって記載されている細胞に基づく競合的酵素結合免疫吸着アッセイ(ELISA)またはJuarez J.ら(Leukemia 2003, 17, ppl294-1300)によって記載されている蛍光活性化細胞選別法(FACS)を用いるプロトコールであり得る。
【0210】
よって、本発明の特定の態様では、CXCR4アンタゴニスト抗ウイルス薬としての分子をスクリーニングおよび/または同定する方法であって、以下の工程を含んでなる方法が考えられる:
a)CXCR4を発現する細胞を選択する工程、
b)該細胞を、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体とともに培養する工程、
c)抗体、またはその一つの機能的フラグメントもしくは誘導体と、CXCR4との間の結合の潜在的な阻害のための供試分子を評価する工程、および
d)該阻害が可能な分子を選択する工程。
【0211】
別の特定の実施形態では、
e)これらの分子をHIV−1複製アッセイで試験する工程
を追加することができる。
【0212】
本発明の他の特徴および利点は、実施例および図面(凡例は下記に示される)とともに説明を読めばさらに明らかになる。
【実施例】
【0213】
実施例1:ヒトCXCR4に対するモノクローナル抗体(Mab)の作製
CXCR4に対するモノクローナル抗体を作製するため、Balb/cマウスに組換えNIH3T3−CXCR4細胞および/またはCXCR4細胞外N末端およびループに相当するペプチドを感作させた。6〜16週齢のマウスに、完全フロイントアジュバントにて抗原を1回、皮下(s.c.)感作させた後、フロイントの不完全アジュバントにて抗原を2〜6回s.c.感作させた。免疫応答を眼窩後方採血によって測定した。血清をELISAによりスクリーニングし(下記の通り)、抗CXCR4抗体力価が高いマウスを融合に用いた。屠殺および脾臓摘出の2日前にマウスの静脈内に抗原を追加投与した。
【0214】
ELISA
抗CXCR4抗体を産生するマウスを選択するため、免疫マウスからの血清をELISAによって調べた。要するに、マイクロタイタープレートを、BSAと結合された精製[1−41]N末端ペプチドで5μg相当量のペプチド/mL、100μL/ウェルにてコーティングし、4℃での一晩培養した後、PBS中0.5%ゼラチン250μL/ウェルでブロックした。CXCR4免疫マウスからの血漿希釈液を各ウェルに加え、37℃で2時間培養した。これらのプレートをPBSで洗浄した後、HRP(Jackson Laboratories)と結合されたヤギ抗マウスIgG抗体とともに37℃で1時間培養した。洗浄後、プレートをTMB基質で現像し、5分後に100μL/ウェルの1M H2SO4を加えることによって反応を停止させた。最高力価の抗CXCR4抗体を生じたマウスを抗体作製に用いた。
【0215】
CXCR4に対するMabを産生するハイブリドーマの作製
最高力価の抗CXCR4抗体を生じたBalb/cマウスから単離したマウス脾細胞を、PEGを用いてマウス骨髄腫細胞系統Sp2/Oと融合させた。細胞をマイクロタイタープレートにおよそ1×105/ウェルで播種した後、ウルトラカルチャー培地+2mM L−グルタミン+1mMピルビン酸ナトリウム+1×HATを含有する選択培地で2週間インキュべートした。次に、ウェルをELISAによって、抗CXCR4モノクローナルIgG抗体に関してスクリーニングした。その後、抗体を分泌するハイブリドーマを制限希釈により少なくとも2回サブクローニングし、in vitroで培養し、さらなる分析のための抗体を作製した。
【0216】
実施例2:FACS分析による抗CXCR4 Mab 515H7および301aE5結合特異性(NIH3T3−CXCR4形質転換体)の特性決定
この実験では、抗CXCR4 Mab 515H7および301aE5のヒトCXCR4(hCXCR4)への特異的結合をFACS分析によって調べた。NIH3T3およびNIH3T3−hCXCR4トランスフェクト細胞を、10μg/mlのモノクローナル抗体515H7および301aE5とともに培養した。その後、これらの細胞を1%BSA/PBS/0.01%NaN3で洗浄した。次に、Alexa標識された二次抗体をこれらの細胞に加え、4℃で20分間培養した。その後、細胞を再び2回洗浄した。2回目の洗浄後、FACS分析を行った。これらの結合試験の結果を、[FACSによって得られた平均蛍光強度(MFI)]を示す下表8に示す。抗CXCR4 Mab 515H7および301aE5はヒトCXCR4−NIH3T3トランスフェクト細胞系統と特異的に結合するが、親NIH3T3細胞に対する認識は見られなかった。
【0217】
【表8】
【0218】
実施例3:FACS分析による末梢血単核細胞(PBMC)に対する抗CXCR4 Mab 515H7および301aE5の結合の特性決定
健康なドナーから血液をバフィーコートとして採取した。100μlの全血を示された濃度の抗ヒトCXCR4抗体(クローン515H7および301aE5)とともに4℃で20分間培養した。血液をPBS−BSA 1%−NaN3 0.01%中で3回洗浄し、1:500希釈のヤギ抗ヒトAlexa 488 IgG(Invitrogen)とともに4℃で20分間培養した。その後、細胞を洗浄し、CD14−PE(Caltag)またはCD3−PE(Caltag)とともに4℃で10分間培養し、3回洗浄した。赤血球をHigh-Yield溶解液(Caltag)で室温にて10分間溶解させた。すぐにFacscalibur (Becton-Dickinson)を用いて細胞を分析した。単球でのCXCR4発現はCD14陽性細胞で行い、T細胞でのCXCR4発現はCD3陽性細胞で行った(図1)。結果は抗原結合能(ABC)で表す。
【0219】
図2Aおよび2Bに示されるように、抗ヒトCXCR4クローン515H7および301aE5はTリンパ球(図2A)および単球(図2B)の双方を染色したが、このことは、515H7および301aE5 Mabが単球およびTリンパ球の細胞表面で発現された天然型のCXCR4を認識可能であることを示している。
【0220】
実施例4:生物発光共鳴エネルギー移動(BRET)アプローチによるCXCR4ホモ二量体に対する515H7および301aE5 Mabの効果
この機能的アッセイは、SDF−1および/または515H7 MabがCXCR4受容体に結合した際に誘発されるコンフォメーション変化をCXCR4ホモ二量体のレベルで評価することができる。
【0221】
検討する相互作用相手の発現ベクターを、従来の分子生物学的技術を適用することにより、対応する色素(ウミシイタケ(Renilla reniformis)ルシフェラーゼRlucおよび黄色蛍光タンパク質YFP)との融合タンパク質として構築した。BRET試験を行う2日前に、HEK293細胞を、対応するBRET相手:[CXCR4/Rluc+CXCR4/YFP]をコードする発現ベクターで一時的にトランスフェクトし、CXCR4ホモ二量体形成を調べた。翌日、細胞を、ポリリシンをプレコーティングした白色96MWプレートの完全培養培地[10%FBSを添加したDMEM]に分注した。まず、細胞をプレートに接着させるために、CO2 5%、37℃で培養した。その後、200μl DMEM/ウェルで一晩、細胞を飢餓状態にした。BRET試験の直前に、DMEMを除去し、細胞を手早くPBSで洗浄した。その後、細胞をPBS中、抗体の存在下または非存在下、37℃にて10分間培養した後、最終量50μl中、SDF−1 300nMとともに、または伴わずにセレンテラジンH 5μMを添加した。37℃でさらに10分間培養した後、Mithras LB940マルチラベルリーダー(Berthold)(1s/波長/ウェル、室温で15反復)を用い、485nmおよび530nmでの発光獲得を誘導した。
BRET比の計算は従前に記載されているように行った(Angersら、2000):[(発光530nm)−(発光485nm)×Cf]/(発光485nm)、式中、Cf=同じ試験条件下でRluc融合タンパク質単独を発現する細胞の(発光530nm)/(発光485nm)。この式を簡単にすると、BRET比が、二つのBRET相手が存在する場合に得られる530/485nm比を、同じ試験条件下で、Rlucと融合された相手だけがアッセイ中に存在する場合に得られる530/485nm比により補正したものに相当することが示される。読み取りやすいように、結果はミリBRET単位(mBU)で表すが、mBUはBRET比に1000を掛けたものである。
【0222】
SDF1(300nM)は、CXCR4受容体と融合されたアダプタータンパク質とアクセプタータンパク質が空間的に近接していることから生じるBRETシグナルを約20%増強したが、それはおそらくCXCR4/CXCR4ホモ二量体形成または既存の二量体のコンフォメーション変化を示唆する(図3AおよびB)。515H7および301aE5 Mabは、CXCR4ホモ二量体に関してSDF−1により誘発されるコンフォメーション変化を調節することができた(515H7および301aE5に関して、SDF−1により誘発されるBRET増強の69%阻害、図3AおよびB)。515H7および301aE5 Mabはまた、それ自体、CXCR4/CXCR4の空間的近接を調節することもできたが、このことはCXCR4/CXCR4ホモ二量体コンフォメーションに対する515H7および301aE5 Mabの影響を示唆する(図3Aおよび3B)。
【0223】
実施例5:抗CXCR4 Mab 515H7および301aE5による、ヒトPBMCにおけるHIV−1一次単離物KON(X4ウイルス)複製の阻害
HIV−1に対して血清反応陰性の健康なドナーからのPBMCをバフィーコートから、またはフィコール−ハイパーク勾配遠心分離による血球成分分離法(cytapheresis)から単離した。PBMCを、25mM HEPES、5mlペニシリン(10000U/ml)−ストレプトマイシン(10000μg/ml)2mM L−グルタミンを含有し、10%熱不活性化FCSを添加したRPMI 1640細胞培養培地中、PHAの存在下で活性化し、1サイクルの中和アッセイで細胞標的として用いた。一次ヒトPBMCにおけるHIV−1複製は、ウイルスp24抗原の細胞内染色をFACS分析によって分析することにより行った。要するに、25μl/ウェルの、種々の希釈率のMab 515H7、301aE5および12G5(R&D Systems)または対照として培養培地(RPMI 1640、10%FCS、0.1%IL−2)を、25μl/ウェルのHIV−1 KON X4一次単離物希釈液とともに37℃で1時間、二反復で培養した。PHA活性化ヒトPBMC(25μl/ウェル)を96ウェルプレート(U底、Costar 3599)中のMab/ウイルス混合物に20×106細胞/mlで加え、RPMI 1640、10%FCSおよび0.1%IL−2中、37℃で24〜36時間培養した。Mabを含まない培地中、非感染PBMCからなる対照を導入した。HIV感染PBMCを検出するために、ウイルスp24抗原の細胞内染色を行い、フローサイトメトリーによって分析した。細胞を固定し、Cytofix/Cytopermキット(Becton Dickinson)を製造者のプロトコールに従って用いて透過処理を施し、1/160希釈で用い、暗所にて4℃で10分間培養した蛍光抗p24 Mab(クローンKC57−Coulter Beckman)で染色した。PBS−3%FCS培地で洗浄した後、フローサイトメトリー分析前にPBMCをPBSで希釈した。種々のサンプルにおけるp24陽性細胞の割合を、生細胞集団に対する20,000事象をゲートすることにより決定した。この生細胞のサブセットのp24発現を、非感染細胞のバックグラウンド染色と比べて分析した。p24抗原陽性値は、mock感染細胞におけるバックグラウンド事象を差し引いた後に得た。中和の割合は、Mabを含まない対照感染ウェルと比べてのp24陽性細胞の減少と定義した。中和力価は、感染細胞の割合を90%低下させるMabの希釈率と定義した。抗CXCR4 Mab 515H7および301aE5を、HIV適用の参照の抗CXCR4 Mabとして知られる12G5 Mabと比較した。図4AおよびBおよび5に示されるように、抗CXCR4 Mab 515H7および301aE5は、それぞれIC90 10μg/ml(66nM)および150μg/ml(1μM)で、PBMCにおけるHIV−1 KON一次単離物の複製を阻害することができるが、12G5 MabはPBMCにおけるHIV−1 KON一次単離物の複製を阻害することができなかった(図4A)。
【0224】
実施例6: CXCR4受容体により媒介される細胞内カルシウム貯蔵の動員
この機能的アッセイは、小胞体の細胞内貯蔵からのカルシウム遊離を誘導するホスホリパーゼC経路の刺激を介したCXCR4受容体シグナル伝達を測定するように設計された。
【0225】
ヒトCXCR4受容体を安定的かつ構成的に発現するCHO−K1細胞を、ナイーブCHO−K1細胞(ATCC CCL−61)は、ヒトCXCR4受容体の全コード配列を含む哺乳類発現ベクター(RefSeq NM_003467)でトランスフェクトした際に得られたものである。細胞を完全培養培地[5%ウシ胎仔血清(FCS)および500μg/mlのゲネチシンを添加したDMEM−ハムF12培地]で増殖させた。細胞を黒色96MWプレートに適当な培養培地中100,000細胞/ウェルで播種した。試験前に細胞を一晩飢餓状態にした。細胞にローディングバッファー[HBSS 1×、HEPES 20mM、プロベネシド酸(Probenicid acid)25mM]中、蛍光カルシウム色素(Fluo-4 No Wash, Invitrogen US)を、37℃で30分間、その後、25℃で30分間負荷した。SDF−1による刺激は、各ウェルに直接注入することにより行った。拮抗作用試験では、10μlのMab溶液をSDF−1の少なくとも10分前にローディングバッファーに直接添加する。動的蛍光測定は、マルチモード蛍光マイクロプレートリーダーMithras LB940(Berthold)にて、以下の設定で行う:485nmでの励起、535nmでの発光、10000任意単位での励起エネルギー。各ウェルの蛍光をSDF−1注入前に毎秒0.1秒、20秒間記録する(基底シグナル)。その後、20μlのSDF−1を注入し、データ記録を2分間続ける。各試験条件を2反復で行う。各ウェルの値をまず、基底蛍光と細胞を含まない対照ウェルによって発せられた蛍光を差し引くことによって補正する。相対的データをSDF−1(100nM)によって得られた最大刺激の割合として表す。
【0226】
SDF1(100nM)は、組換えCHO/CXCR4において細胞内カルシウムの迅速かつ強い放出を誘導したが、ナイーブCHO−K1細胞では蛍光シグナルは検出されなかった。最大強度は基底蛍光の140%超に達し、SDF−1による刺激時に約40秒で見られた(図6A、6B、および6C)。Mab 515H7(133nM)(図6A)およびc515H7(133nM)(図6C)は、SDF−1(100nM)により誘発されたカルシウムシグナルの強い阻害をもたらした。Mab 301aE5(133nM)(図6B)は、SDF−1(100nM)により誘発されたカルシウムシグナルの部分的阻害をもたらした。
【0227】
実施例7:抗CXCR4 Mab 515H7、c515H7および301aE5による、ヒトPBMCにおけるHIV−1一次単離物KON、MN、および92UG024(X4ウイルス)の複製の阻害
1サイクル中和アッセイ
このアッセイは、一次単離物KON、MN、および92UG024を用いて36時間行い、感染2日後に2%の感染CD4 Tリンパ球の検出を可能とするように相応に濃縮および希釈する。
【0228】
種々の希釈率のMab 515H7、c515H7および301aE5 25マイクロリットルを、25μlのウイルスとともに37℃で1時間培養した。20×106細胞/mlのヒトPBMC(25μl)を96ウェルプレート(U底、Costar 3599)中のMab/ウイルス混合物に加え、RPMI 1640 10%FCSおよび20U/ml IL−2(R&D Systems, Minneapolis, MN)中で36時間培養した。
【0229】
2日間の培養後、HIV感染リンパ球を、ウイルスp24 Agの細胞内染色により検出した。細胞を固定し、製造者に従いCytofix/CytopermおよびPerm/Washキット(BD Biosciences)の双方を用いて透過処理を施し、Perm/Wash溶液中1/160希釈で用い、4℃で15分間加える蛍光抗p24 Mab(FITC−またはPE−抗p24、クローンKC57、Beckman Coulter/Immunotech, Hialeah, FL)で染色した。3%FBSを含むPBSで洗浄した後、PBMCを300μlのPBSに希釈した後、DIVAソフトウエア(BD Biosciences)を用いてフローサイトメトリー分析(LSRII、 BD Biosciences)を行った。種々のサンプル中のp24陽性細胞の割合を、前方および側方散乱パラメーターによって同定された生細胞集団に対する20,000事象をゲートすることにより決定した。これらの生細胞のサブセットは、live/dead solution kit (Invitrogen)を用いて分析した。p24 Ag陽性値は、mock感染細胞におけるバックグラウンド事象を差し引いた後に得た。
【0230】
中和パーセントは、Mabを含まない対照感染ウェルと比べてのp24陽性細胞の減少と定義した。中和力価は、感染細胞の割合を90%低下させる抗体の濃度(3反復で行った連続希釈率の間に挿入)と定義した。
【0231】
図7、8および9に示されるように、抗CXCR4 Mab 515H7、c515H7、および301aE5は、PBMCにおけるHIV−1 X4 MN、KON、および92UG024一次単離物の複製を阻害することができる。ICs(μg/mlにおいて)の結果を表9にまとめる。
【0232】
【表9】
【0233】
実施例8:抗CXCR4 Mab 515H7、c515H7および301aE5による、ヒトPBMCにおけるHIV−1一次単離物89.6(X4/R5二重ウイルス)の複製の阻害
1サイクル中和アッセイ
このアッセイは、一次単離物89.6を用いて36時間行い、感染2日後に2%の感染CD4 Tリンパ球の検出を可能とするように相応に濃縮および希釈する。
【0234】
種々の希釈率のMab 515H7、c515H7および301aE5 25マイクロリットルを、25μlのウイルスとともに37℃で1時間培養した。20×106細胞/mlのヒトPBMC(25μl)を96ウェルプレート(U底、Costar 3599)中のMab/ウイルス混合物に加え、RPMI 1640 10%FCSおよび20U/ml IL−2(R&D Systems, Minneapolis, MN)中で36時間培養した。
【0235】
2日間の培養後、HIV感染リンパ球を、ウイルスp24 Agの細胞内染色により検出した。細胞を固定し、製造者に従いCytofix/CytopermおよびPerm/Washキット(BD Biosciences)の双方を用いて透過処理を施し、Perm/Wash溶液中1/160希釈で用い、4℃で15分間加える蛍光抗p24 Mab(FITC−またはPE−抗p24、クローンKC57、Beckman Coulter/Immunotech, Hialeah, FL)で染色した。3%FBSを含むPBSで洗浄した後、PBMCを300μlのPBSに希釈した後、DIVAソフトウエア(BD Biosciences)を用いてフローサイトメトリー分析(LSRII、 BD Biosciences)を行った。種々のサンプル中のp24陽性細胞の割合を、前方および側方散乱パラメーターによって同定された生細胞集団に対する20,000事象をゲートすることにより決定した。これらの生細胞のサブセットは、live/dead solution kit (Invitrogen)を用いて分析した。p24 Ag陽性値は、mock感染細胞におけるバックグラウンド事象を差し引いた後に得た。
【0236】
中和パーセントは、Mabを含まない対照感染ウェルと比べてのp24陽性細胞の減少と定義した。中和力価は、感染細胞の割合を90%低下させる抗体の濃度(3反復で行った連続希釈率の間に挿入)と定義した。
【0237】
図10および11に示されるように、抗CXCR4 Mab 515H7、c515H7、および301aE5は、PBMCにおけるHIV−1 89.6一次単離物の複製を阻害することができる。ICs(μg/ml)の結果を表10にまとめる。
【0238】
【表10】
【0239】
実施例9:抗CXCR4 Mab c515H7と抗CCR5分子マラビロクの併用による、ヒトPBMCにおけるHIV−1一次単離物89.6およびUG93067(X4/R5二重ウイルス)の複製の阻害
中和アッセイ、一次PBMCにおける複数回のHIV一次単離物複製の分析:
c515H7 Mabまたはマラビロクの連続希釈液を併用する、または両者とウイルスの連続希釈液を併用するこのアッセイは、PBMC(末梢血単核細胞)に対する複数回の感染を分析する。要するに、c515H7 Mabまたはマラビロクまたは両者の組合せの連続希釈液(2倍)の25μlアリコート四つをそれぞれ、予め含水させた96ウェルフィルタープレート(孔径1.25μm、Durapor Dv、 Millipore, Molsheim, France)にて、ウイルスの連続希釈液25μlとともに培養した。ウイルスの対照滴定(希釈c515H7 Mabまたはマラビロクの代わりに25μlのRPMI)を、c515H7 Mabまたはマラビロクまたは両者の組合せの希釈液の存在下での滴定と同じプレートで行った。37℃で1時間後、4×106細胞/mlの濃度のPHA刺激PBMC 25μl(5人の健康なドナーからのPHA活性化PBMCのプール)を、RPMIの最終培養容量75μl、10%ウシ胎仔血清(FCS)および20IUのインターロイキン−2(IL−2)/ml(R&D System)となるように加えた。37℃で24時間後、100μlの同じ培養培地を加えた。4日目に濾過により2回の洗浄(各200μlのRPMI)を行ってc515H7 Mabおよびマラビロクを除去し、200μlの新鮮培養培地を加えた。7日目に、培養上清におけるp24の存在をELISAにより測定し、陰性対照(ウイルス希釈液を感染させ、10−6Mジドブジン[AZT]の存在下で維持した培養物)の場合と比べ、陽性ウェルを判定した。4反復のウェルを用い、各希釈率のc515H7 Mabまたはマラビロクまたは両者の組合せの不在下(F0)および存在下(Vn)でウイルス力価(50%組織培養感染量[TCID50])を決定した。中和力価は、ウイルス力価の90%低下をもたらす(Vn/V0=0.1)c515H7 Mabまたはマラビロクまたは両者の組合せの希釈率として定義した。
【0240】
図12に示されるように、二重指向性X4R5ウイルス89.6の複製は、c515H7 Mabにより、IC90 2μg/mlで阻害された(図12)。マラビロク50μg/mlでは、IC90阻害活性に達しなかった(図12)。さらに、2μg/mlのマラビロクを抗体515H7に添加したところ、IC90 0.2μg/mlでc515H7 Mabの阻害活性を増強した(図12)。
【0241】
c515H7 Mabとおよびマラビロクの組合せの有益な効果を、種々の希釈率のこれらの2分子および別の二重指向性ウイルスUG93067を用いて評価した。図13に示されるように、Mab c515H7およびマラビロクの阻害活性は同等であった。これらの結果は、ウイルスUG93067の、CCR5受容体またはCXCR4受容体のいずれかを用いる能力が匹敵していたことを示唆する。UG93067ウイルスを用いると、より良い活性を証明することができ、これらのX4(c515H7 Mab)とR5(マラビロク)阻害剤(各10μg/ml)の組合せだけが、ウイルス力価の90%低下を可能とした(図13)。
【0242】
実施例10:抗CXCR4キメラMab c515H7の生産
ネズミ515H7 Mabのキメラ形式を設計した。これは、着目するネズミ抗体の軽鎖および重鎖可変ドメインを、ヒトCκおよびIgG1/IgG2/IgG4定常ドメインと遺伝的に融合したものに相当する。この組換えMabは、HEK293/EBNA系をpCEP4発現ベクター(InVitrogen, US)とともに使用することによって、一時的トランスフェクション時に作製されたものである。
【0243】
個々のアミノ酸配列およびヌクレオチド配列は本明細書に上記した。さらに、上記の表3にIgG2および4イソ型(好ましいイソ型)の配列が開示されているため、ここでも、IgG1イソ型の重鎖、すなわち、配列番号80のアミノ酸配列および配列番号81のヌクレオチド配列に相当するc515H7 VH(G1wt)の配列を記載することができる。
【0244】
515H7 Mab軽鎖および重鎖の可変ドメインに相当する全ヌクレオチド配列は、グローバル遺伝子合成(Genecust, Luxembourg)によって合成されたものである。それらを、ヒトIgG1/IgG2/IgG4免疫グロブリンの軽鎖[Cκ]または重鎖[CH1−Hinge−CH2−CH3]のいずれかの定常ドメインの全コード配列を有するpCEP4ベクター(InVitrogen, US)にサブクローニングした。全クローニング工程は、the Laboratory manual (Sambrook and Russel, 2001)に記載されている従来の分子生物学的技術に従って、または供給者の説明書に従って行った。各遺伝子構築物を、Big Dyeターミネーター・サイクル・シーケンシング・キット(Applied Biosystems, US)を用いたヌクレオチド配列決定によって完全にバリデートし、3100ジェネティック・アナライザー(Applied Biosystems, US)を用いて分析した。
【0245】
懸濁適合HEK293 EBNA細胞(InVitrogen, US)を通常、オービタルシェーカー(回転速度110rpm)上、250mlフラスコにて、6mMグルタミンを添加した血清不含培地Excell 293(SAFC Biosciences)50ml中で増殖させた。一時的トランスフェクションは、水中終濃度1mg/mlで調製した直鎖25kDaポリエチレンイミン(PEI)(Polysciences)、混合およびプラスミドDNA(重鎖:軽鎖プラスミド比1:1で、終濃度1.25μg/ml)を用い、2.106細胞/mlで行った。トランスフェクション4時間後に、培養物を1容量の新鮮培養培地で希釈して、最終細胞密度を106細胞/mlとした。培養方法を細胞の生存率およびMab生産に関して測定した。一般に、培養は4〜5日間測定した。Mabは、プロテインA樹脂(GE Healthcare, US)での従来のクロマトグラフィーアプローチを用いて精製した。Mabは機能的評価に好適なレベルで生産された。生産性レベルは一般に、精製Mab 6〜15mg/lの範囲である。
【0246】
実施例11:FACS分析による抗CXCR4キメラMab c515H7結合特異性の特性決定
この試験では、抗CXCR4キメラMab c515H7のヒトCXCR4に対する特異的結合をFACS分析によって調べた。
【0247】
NIH3T3−hCXCR4トランスフェクト細胞を0μg/ml〜10μg/mlの用量範囲のモノクローナル抗体c515H7とともに培養した。その後、これらの細胞を1%BSA/PBS/0.01%NaN3で洗浄した。次に、Alexaで標識した二次抗体をこれらの細胞に加え、4℃で20分間インキュベートした。その後、これらの細胞を再び2回洗浄した。2回目の洗浄後に、FACS分析を行った。この結合試験の結果を図15に示すが、これは、抗CXCR4キメラMab C515H7がヒトCXCR4−NIH3T3トランスフェクト細胞系統と特異的に結合することを示す。NIH3T3wt細胞との結合は検出されなかった(データは示されていない)。
【0248】
実施例12:生物発光共鳴エネルギー移動(BRET)アプローチによるCXCR4ホモ二量体に対するc515H7 Mabの効果
この機能的アッセイは、SDF−1および/またはc515H7 MabがCXCR4受容体に結合した際に誘発されるコンフォメーション変化をCXCR4ホモ二量体のレベルで評価することができる。
【0249】
検討する相互作用相手の発現ベクターを、従来の分子生物学的技術を適用することにより、対応する色素(ウミシイタケ(Renilla reniformis)ルシフェラーゼRlucおよび黄色蛍光タンパク質YFP)との融合タンパク質として構築した。BRET試験を行う2日前に、HEK293細胞を、対応するBRET相手:[CXCR4/Rluc+CXCR4/YFP]をコードする発現ベクターで一時的にトランスフェクトし、CXCR4ホモ二量体形成を調べた。翌日、細胞を、ポリリシンをプレコーティングした白色96MWプレートの完全培養培地[10%FBSを添加したDMEM]に分注した。まず、細胞をプレートに接着させるために、CO2 5%、37℃で培養した。その後、200μl DMEM/ウェルで一晩、細胞を飢餓状態にした。BRET試験の直前に、DMEMを除去し、細胞を手早くPBSで洗浄した。その後、細胞をPBS中、抗体の存在下または非存在下、37℃にて10分間インキュベートした後、最終量50μl中、SDF−1 100nMとともに、または伴わずにセレンテラジンH 5μMを添加した。37℃でさらに10分間インキュベートした後、Mithras LB940マルチラベルリーダー(Berthold)(1s/波長/ウェル、室温で15反復)を用い、485nmおよび530nmでの発光獲得を誘導した。
【0250】
BRET比の計算は従前に記載されているように行った(Angersら 2000):[(発光530nm)−(発光485nm)×Cf]/(発光485nm)、式中、Cf=同じ試験条件下でRluc融合タンパク質単独を発現する細胞の(発光530nm)/(発光485nm)。この式を簡単にすると、BRET比が、二つのBRET相手が存在する場合に得られる530/485nm比を、同じ試験条件下で、Rlucと融合された相手だけがアッセイ中に存在する場合に得られる530/485nm比により補正したものに相当することが示される。読み取りやすいように、結果はミリBRET単位(mBU)で表すが、mBUはBRET比に1000を掛けたものである。
【0251】
SDF1(100nM)は、CXCR4受容体と融合されたドナータンパク質と、アクセプタータンパク質とが空間的に近接していることから生じるBRETシグナルを約10%増強したが、それはおそらくCXCR4/CXCR4ホモ二量体形成または既存の二量体のコンフォメーション変化を示唆する(図16)。Mab c515H7は、CXCR4ホモ二量体に関して、SDF−1により誘発されるコンフォメーション変化を調節することができた(SDF−1により誘発されるBRET増強の96%阻害、図16)。Mab c515H7はまた、それ自体、CXCR4/CXCR4の空間的近接を調節することもできたが、このことはCXCR4/CXCR4ホモ二量体コンフォメーションに対するこのMabの影響を示唆する(図16)。
【0252】
実施例13:CD4およびCXCR4またはCCR5を発現するGFP形質導入ヒト骨肉腫(GHOST)細胞を用いたMab 515H7抗HIV−1活性のin vitro評価
CXCR4 515H7 Mabの特異性を判定するために、本発明者らは、CD4およびCXCR4またはCCR5を発現するGHOSTを用い、このMabの抗HIV−1活性を評価した。
【0253】
このアッセイは、X4 HIV−1 LAIウイルス(CXCR4を発現するGhost細胞を持つ)またはR5 HIV−1 BaLウイルス(CCR5を発現するGhost細胞を持つ)のいずれかを用いて48時間行う。500μlのGhost細胞を、10%FCSを添加したダルベッコの培養培地で24時間平板培養した(2.5 105細胞/ml)。種々の希釈率のMab 515H7を37℃で1時間インキュベートした後、これらの細胞に希釈したHIV−1 LAIウイルス(1/10)およびHIV−1 BaLウイルス(1/7)を48時間加えた。細胞をトリプシン処理し、PBS 1×で洗浄した。細胞を固定し、また、ウイルスを不活性化するために、この細胞ペレットに300μLの1.5%パラホルムアルデヒドを暗所、+4℃で2時間加えた。GFP陽性細胞をフローサイトメトリーにより分析し、HIV−1感染の阻害を算出した。
感染細胞の阻害パーセントは、Mabを含まない対照感染ウェルと比較して定義した。ICs(μg/ml)の結果を表11にまとめる。抗CXCR4 Mab 515H7は、Ghost細胞を発現するCXCR4においてHIV−1 X4 Laiウイルスの感染を阻害することができたが、Ghost細胞を発現するCCR5においてHIV−1 R5 BaLウイルスの感染阻害には全く不活性であった。
【0254】
【表11】
【0255】
実施例14:515H7抗CXCR4ネズミ抗体のヒト化および該h515H7のフラグメントの作製
一般手順
515H7抗CXCR4抗体のヒト化は、CDRグラフト法のグローバルルールを適用することで行った。免疫原性分析ならびにCDRおよびフレームワーク(FR)領域の定義は、IMGTユニークナンバリングスキームならびにIMGTライブラリーおよびツール(Lefranc, 1997 - www.imgt.org)を適用することで行った。
【0256】
515H7のヒト化変異体の結合は、ヒトCXCR4で安定的にトランスフェクトされたNIH3T3細胞系統にて判定した。結合活性は、ビオチン化マウス抗体を用いた競合アッセイによって評価した。第二の試みにおいて、ヒト化抗体を、ビオチン化SDF−1のRAMOS細胞との結合を阻害する能力に関して評価した。RAMOS細胞は、CXCR4の発現が高く、CXCR7およびSDF−1の発現が低いために選択された。
【0257】
これらのアッセイを用いて、抗CXCR4抗体の組換えヒト化型を特性決定した。可変ドメインをヒトIgG1/k定常ドメインでフォーマットし、哺乳類発現ベクターpCEPにクローニングした。組換えIgG1/κ由来抗体をHEK293細胞で一時的に発現させた。発現培養上清を濾過し、プロテインAセファロースを用いて抗体を精製した。精製抗体に再びPBSバッファーを加え、抗体濃度をELISAにより測定した。
【0258】
組換え抗体フラグメントを、ヒト化抗体の可変ドメインに特異的なオリゴヌクレオチドを用いたPCRによって作製し、これらを大腸菌系にサブクローニングした。抗体フラグメントの精製は、固定化金属イオンアフィニティークロマトグラフィー(IMAC)によって行った。
【0259】
515H7可変ドメインのヒト化
重鎖および軽鎖可変ドメインの種々の配列アライメントを図17および18に示す。
【0260】
最初の一連の試験では、三つの第一のヒト化変異体の抗CXCR4結合活性を分析した。VH変異体1(VH1)をネズミVLと組合せ、これらの構築物の、ビオチン化ネズミ515H7親抗体の結合を阻害する能力を評価した。VH1の可変ドメインのアミノ酸配列は配列番号90を含んでなり、ヌクレオチド配列は配列番号91を含んでなる。全長VH1のアミノ酸配列は配列番号92を含んでなり、ヌクレオチド配列は配列番号93を含んでなる。この構築物は、キメラ抗体と同等の、ネズミ抗体との競合能を示した(図19A)。このことは、ほとんどのヒトVH変異体がキメラと同等の結合能を有することを示唆する。よって、VH1をVLの変異体2と組み合わせた(図19B)。
【0261】
さらなる試験では、抗体515H7のヒト化変異体がSDF−1のCXCR4発現細胞への結合を阻害するか否かを判定した(図20)。このhz515H7のヒト化変異体の阻害能は、フローサイトメトリーにおいてビオチン化SDF−1を検出することによって評価した。ヒト化抗体hz515H7 VH1 D76N VL2は、キメラc515H7と同等のSDF−1結合阻害能を有する。
【0262】
また、ヒト化変異体hz515H7 VH1 VL2の抗体フラグメントも試験したところ、この抗体フラグメントがSDF−1の結合を完全に阻害することができることが分かった(図20)。
【0263】
実施例15:FACS分析による抗CXCR4ヒト化Mab 515H7結合特異性の特性決定
この試験では、抗CXCR4ヒト化Mab 515H7の、ヒトCXCR4に対する特異的結合をFACS分析により調べた。
【0264】
NIH3T3、NIH3T3−hCXCR4トランスフェクト体を、100μlのFacsバッファー中、暗所、4℃にて20分間、0〜10μg/mLのヒト化 Mab 515H7(hz515H7 VH1 D76N VL2、hz515H7 VH1 D76N VL2.1、hz515H7 VH1 D76N VL2.2、hz515H7 VH1 D76N VL2.3)とともにインキュベートした。Facsバッファー中で3回洗浄した後、細胞を、暗所、4℃にて20分間、二次抗体であるヤギ抗ヒトAlexa 488(1/500希釈)とともにインキュベートした。Facsバッファー中で3回洗浄した後、各ウェルにヨウ化プロピジウムを加え、生存細胞のみをFacsにより分析した。少なくとも5000の生存細胞を評価し、各条件について蛍光強度の平均値を見積もった。
【0265】
これらの結合試験の結果を、[FACSによって得られた平均蛍光強度(MFI)]を示す図21に示す。抗CXCR4ヒト化Mab hz515H7は、ヒトCXCR4−NIH3T3トランスフェクト細胞系統と特異的に結合した(NIH3T3親細胞ではMFI=2.2)。
【0266】
実施例16:生物発光共鳴エネルギー移動(BRET)アプローチによるCXCR4ホモ二量体に対するhz515H7 Mabの効果
この機能的アッセイは、SDF−1および/またはhz515H7 VH1 D76N VL2、hz515H7 VH1 D76N VL2.1、hz515H7 VH1 D76N VL2.2、hz515H7 VH1 D76N VL2.3がCXCR4受容体に結合した際に誘発されるコンフォメーション変化をCXCR4ホモ二量体のレベルで評価することができる。
【0267】
検討する相互作用相手の発現ベクターを、従来の分子生物学的技術を適用することにより、対応する色素(ウミシイタケ(Renilla reniformis)ルシフェラーゼRlucおよび黄色蛍光タンパク質YFP)との融合タンパク質として構築した。BRET試験を行う2日前に、HEK293細胞を、対応するBRET相手:[CXCR4/Rluc+CXCR4/YFP]をコードする発現ベクターで一時的にトランスフェクトし、CXCR4ホモ二量体形成を調べた。翌日、細胞を、ポリリシンをプレコーティングした白色96MWプレートの完全培養培地[10%FBSを添加したDMEM]に分注した。まず、細胞をプレートに接着させるために、CO2 5%、37℃で培養した。その後、200μl DMEM/ウェルで一晩、細胞を飢餓状態にした。BRET試験の直前に、DMEMを除去し、細胞を手早くPBSで洗浄した。その後、細胞をPBS中、抗体の存在下または非存在下、37℃にて10分間インキュベートした後、最終量50μl中、SDF−1 100nMとともに、または伴わずにセレンテラジンH 5μMを添加した。37℃でさらに10分間インキュベートした後、Mithras LB940マルチラベルリーダー(Berthold)(1s/波長/ウェル、室温で15反復)を用い、485nmおよび530nmでの発光獲得を誘導した。
【0268】
BRET比の計算は従前に記載されているように行った(Angersら、2000):[(発光530nm)−(発光485nm)×Cf]/(発光485nm)、式中、Cf=同じ試験条件下でRluc融合タンパク質単独を発現する細胞の(発光530nm)/(発光485nm)。この式を簡単にすると、BRET比が、二つのBRET相手が存在する場合に得られる530/485nm比を、同じ試験条件下で、Rlucと融合された相手だけがアッセイ中に存在する場合に得られる530/485nm比により補正したものに相当することが示される。読み取りやすいように、結果はミリBRET単位(mBU)で表すが、mBUはBRET比に1000を掛けたものである。
【0269】
SDF1(100nM)は、CXCR4受容体と融合されたドナータンパク質と、アクセプタータンパク質とが、空間的に近接していることから生じるBRETシグナルを約12%増強したが、それはおそらくCXCR4/CXCR4ホモ二量体形成または既存の二量体のコンフォメーション変化を示唆する(図22)。
【0270】
515H7ヒト化Mabは、CXCR4ホモ二量体に関してSDF−1により誘発されるコンフォメーション変化を調節することができ、SDF−1により誘発されるBRET増強の阻害割合は、hz515H7 VH1D76N−VL2 Mabで約88%、hz515H7 VH1D76N−VL2.1 Mabで65%、hz515H7 VH1D76N−VL2.2 Mabで33%およびhz515H7 VH1D76N−VL2.3 Mabで21%であった(図22)。
【特許請求の範囲】
【請求項1】
IMGTナンバリングシステムによって定義される配列番号1〜6および30〜33のアミノ酸配列を含んでなるCDRから選択される少なくとも一つの相補性決定領域CDRを含んでなる、単離された抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項2】
配列番号1、2、および3のアミノ酸配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖と、配列番4、5、および6のアミノ酸配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖とを含んでなる、請求項1に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項3】
配列番号7のアミノ酸配列を含んでなる軽鎖および配列番号8のアミノ酸配列を含んでなる重鎖を含んでなる、請求項2に記載の抗体、またはその誘導化合物もしくは機能的フラグメント。
【請求項4】
キメラ抗体であり、かつ配列番号56、57、または58からなる群から選択される配列の重鎖および配列番号59の配列の軽鎖を含んでなる、請求項2に記載の抗体、またはその誘導化合物もしくは機能的フラグメント。
【請求項5】
ヒト化抗体であり、かつ配列番号64からなる配列の重鎖可変領域および配列番号65、66、82、または83からなる群から選択される配列の軽鎖可変領域を含んでなる、請求項2に記載の抗体。
【請求項6】
ヒト化抗体であり、かつ配列番号67、68、または69からなる群から選択される配列の重鎖および配列番号70、71、84、または85からなる群から選択される配列の軽鎖を含んでなるヒト化抗体、またはその誘導化合物もしくは機能的フラグメントを含んでなる、請求項2に記載の抗体。
【請求項7】
機能的フラグメントがフラグメントFv、scFv、Fab、F(ab’)2、Fab’、scFv−Fc、ダイアボディー、または半減期が化学修飾によって延長された任意のフラグメントからなる、請求項2に記載の抗体。
【請求項8】
配列番号54のアミノ酸配列を含んでなるscFvである、請求項7に記載の抗体。
【請求項9】
配列番号1、2、および30のアミノ酸配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖と、配列番31、32、および33のアミノ酸配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖とを含んでなる、請求項1に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項10】
配列番号34のアミノ酸配列を含んでなる軽鎖と、配列番35のアミノ酸配列を含んでなる重鎖とを含んでなる、請求項9に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項11】
以下の核酸から選択された、単離核酸:
a)請求項1〜10のいずれか一項に記載の抗体、またはその機能的フラグメントもしくは誘導体の一つをコードする核酸、DNA、またはRNA、
b)配列番号14〜19および41〜45からなるCDR配列群から選択されるDNA配列を含んでなる核酸、
c)配列番号20、21、46、47、72、73、74、86、および87からなる重鎖および軽鎖可変ドメイン配列群から選択されるDNA配列を含んでなる核酸、
d)配列番号60〜63、75〜79、88、および89からなる重鎖および軽鎖配列群から選択されるDNA配列を含んでなる核酸、
e)配列番号55からなるDNA配列を含んでなる核酸、
f)b)、c)、d)、またはe)に定義される核酸の、対応するRNA核酸、
g)a)、b)、c)、d)、およびe)に定義される核酸の相補的核酸、および
h)高ストリンジェンシー条件下で配列番号14〜19および41〜45の配列のCDRの少なくとも一つとハイブリダイズすることができる少なくとも18のヌクレオチドの核酸。
【請求項12】
請求項11に記載の核酸を含んでなる、ベクター。
【請求項13】
請求項12に記載のベクターを含んでなる、宿主細胞。
【請求項14】
請求項12に記載のベクターによって形質転換された少なくとも一つの細胞を含んでなる、ヒト以外のトランスジェニック動物。
【請求項15】
請求項1〜10のいずれか一項に記載の抗体、またはその機能的フラグメントもしくは誘導体の一つを作製する方法であって、以下の工程を含んでなる、方法:
a)請求項13に記載の細胞の、培地および適当な培養条件での培養工程、および
b)このようにして生産された該抗体、またはそれらの機能的フラグメントもしくは誘導体の一つの、該培養培地または該培養細胞からの回収工程。
【請求項16】
請求項15に記載の方法によって得ることができる、または得られた抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項17】
薬剤としての、請求項1〜10および16に記載の抗体。
【請求項18】
PBMCにおけるHIV−1 KON一次単離物の複製を、IC90少なくとも5μg/ml、好ましくは少なくとも10μg/mlで阻害する、請求項1〜10または16〜17に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項19】
請求項1〜10および16〜18のいずれか一項に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体からなる化合物を有効成分として含んでなる、組成物。
【請求項20】
薬剤としての、請求項19に記載の組成物。
【請求項21】
HIV感染の予防または治療のための、請求項19および20に記載の組成物。
【請求項22】
前記HIV感染がX4指向性HIV感染である、請求項21に記載の組成物。
【請求項23】
前記HIV感染がX4/R5指向性HIV感染である、請求項21に記載の組成物。
【請求項24】
HIVの侵入および/または複製を特異的に阻害することができる化合物の中から選択される少なくとも一つの第二の抗HIV化合物を含んでなる、請求項19〜23のいずれか一項に記載の組成物。
【請求項25】
前記の少なくとも第二の抗HIV化合物が、HIVプロテアーゼ阻害剤(PI)、ヌクレオシド/ヌクレオチドHIV逆転写酵素阻害剤(NRTI/NtRTI)、非ヌクレオシドHIV逆転写酵素阻害剤(NNRTI)、HIV侵入阻害剤、HIVインテグラーゼ阻害剤等の抗レトロウイルス剤からなる群から選択される、請求項24に記載の組成物。
【請求項26】
前記の少なくとも第二の抗HIV化合物がマラビロクである、請求項24または25に記載の組成物。
【請求項27】
CXCR4アンタゴニスト抗ウイルス薬としての分子をスクリーニングおよび/または同定する方法であって、以下の工程を含んでなる方法:
a)CXCR4を発現する細胞を選択する工程、
b)前記細胞を、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体とともに培養する工程、
c)抗体、またはその一つの機能的フラグメントもしくは誘導体と、CXCR4との間の結合の潜在的な阻害のための供試分子を評価する工程、および
d)前記阻害が可能な分子を選択する工程。
【請求項1】
IMGTナンバリングシステムによって定義される配列番号1〜6および30〜33のアミノ酸配列を含んでなるCDRから選択される少なくとも一つの相補性決定領域CDRを含んでなる、単離された抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項2】
配列番号1、2、および3のアミノ酸配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖と、配列番4、5、および6のアミノ酸配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖とを含んでなる、請求項1に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項3】
配列番号7のアミノ酸配列を含んでなる軽鎖および配列番号8のアミノ酸配列を含んでなる重鎖を含んでなる、請求項2に記載の抗体、またはその誘導化合物もしくは機能的フラグメント。
【請求項4】
キメラ抗体であり、かつ配列番号56、57、または58からなる群から選択される配列の重鎖および配列番号59の配列の軽鎖を含んでなる、請求項2に記載の抗体、またはその誘導化合物もしくは機能的フラグメント。
【請求項5】
ヒト化抗体であり、かつ配列番号64からなる配列の重鎖可変領域および配列番号65、66、82、または83からなる群から選択される配列の軽鎖可変領域を含んでなる、請求項2に記載の抗体。
【請求項6】
ヒト化抗体であり、かつ配列番号67、68、または69からなる群から選択される配列の重鎖および配列番号70、71、84、または85からなる群から選択される配列の軽鎖を含んでなるヒト化抗体、またはその誘導化合物もしくは機能的フラグメントを含んでなる、請求項2に記載の抗体。
【請求項7】
機能的フラグメントがフラグメントFv、scFv、Fab、F(ab’)2、Fab’、scFv−Fc、ダイアボディー、または半減期が化学修飾によって延長された任意のフラグメントからなる、請求項2に記載の抗体。
【請求項8】
配列番号54のアミノ酸配列を含んでなるscFvである、請求項7に記載の抗体。
【請求項9】
配列番号1、2、および30のアミノ酸配列をそれぞれ含んでなるCDR−L1、CDR−L2、およびCDR−L3を含んでなる軽鎖と、配列番31、32、および33のアミノ酸配列をそれぞれ含んでなるCDR−H1、CDR−H2、およびCDR−H3を含んでなる重鎖とを含んでなる、請求項1に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項10】
配列番号34のアミノ酸配列を含んでなる軽鎖と、配列番35のアミノ酸配列を含んでなる重鎖とを含んでなる、請求項9に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項11】
以下の核酸から選択された、単離核酸:
a)請求項1〜10のいずれか一項に記載の抗体、またはその機能的フラグメントもしくは誘導体の一つをコードする核酸、DNA、またはRNA、
b)配列番号14〜19および41〜45からなるCDR配列群から選択されるDNA配列を含んでなる核酸、
c)配列番号20、21、46、47、72、73、74、86、および87からなる重鎖および軽鎖可変ドメイン配列群から選択されるDNA配列を含んでなる核酸、
d)配列番号60〜63、75〜79、88、および89からなる重鎖および軽鎖配列群から選択されるDNA配列を含んでなる核酸、
e)配列番号55からなるDNA配列を含んでなる核酸、
f)b)、c)、d)、またはe)に定義される核酸の、対応するRNA核酸、
g)a)、b)、c)、d)、およびe)に定義される核酸の相補的核酸、および
h)高ストリンジェンシー条件下で配列番号14〜19および41〜45の配列のCDRの少なくとも一つとハイブリダイズすることができる少なくとも18のヌクレオチドの核酸。
【請求項12】
請求項11に記載の核酸を含んでなる、ベクター。
【請求項13】
請求項12に記載のベクターを含んでなる、宿主細胞。
【請求項14】
請求項12に記載のベクターによって形質転換された少なくとも一つの細胞を含んでなる、ヒト以外のトランスジェニック動物。
【請求項15】
請求項1〜10のいずれか一項に記載の抗体、またはその機能的フラグメントもしくは誘導体の一つを作製する方法であって、以下の工程を含んでなる、方法:
a)請求項13に記載の細胞の、培地および適当な培養条件での培養工程、および
b)このようにして生産された該抗体、またはそれらの機能的フラグメントもしくは誘導体の一つの、該培養培地または該培養細胞からの回収工程。
【請求項16】
請求項15に記載の方法によって得ることができる、または得られた抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項17】
薬剤としての、請求項1〜10および16に記載の抗体。
【請求項18】
PBMCにおけるHIV−1 KON一次単離物の複製を、IC90少なくとも5μg/ml、好ましくは少なくとも10μg/mlで阻害する、請求項1〜10または16〜17に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体。
【請求項19】
請求項1〜10および16〜18のいずれか一項に記載の抗体、またはその一つの機能的フラグメントもしくは誘導体からなる化合物を有効成分として含んでなる、組成物。
【請求項20】
薬剤としての、請求項19に記載の組成物。
【請求項21】
HIV感染の予防または治療のための、請求項19および20に記載の組成物。
【請求項22】
前記HIV感染がX4指向性HIV感染である、請求項21に記載の組成物。
【請求項23】
前記HIV感染がX4/R5指向性HIV感染である、請求項21に記載の組成物。
【請求項24】
HIVの侵入および/または複製を特異的に阻害することができる化合物の中から選択される少なくとも一つの第二の抗HIV化合物を含んでなる、請求項19〜23のいずれか一項に記載の組成物。
【請求項25】
前記の少なくとも第二の抗HIV化合物が、HIVプロテアーゼ阻害剤(PI)、ヌクレオシド/ヌクレオチドHIV逆転写酵素阻害剤(NRTI/NtRTI)、非ヌクレオシドHIV逆転写酵素阻害剤(NNRTI)、HIV侵入阻害剤、HIVインテグラーゼ阻害剤等の抗レトロウイルス剤からなる群から選択される、請求項24に記載の組成物。
【請求項26】
前記の少なくとも第二の抗HIV化合物がマラビロクである、請求項24または25に記載の組成物。
【請求項27】
CXCR4アンタゴニスト抗ウイルス薬としての分子をスクリーニングおよび/または同定する方法であって、以下の工程を含んでなる方法:
a)CXCR4を発現する細胞を選択する工程、
b)前記細胞を、本発明の抗体、またはその一つの機能的フラグメントもしくは誘導体とともに培養する工程、
c)抗体、またはその一つの機能的フラグメントもしくは誘導体と、CXCR4との間の結合の潜在的な阻害のための供試分子を評価する工程、および
d)前記阻害が可能な分子を選択する工程。
【図1】

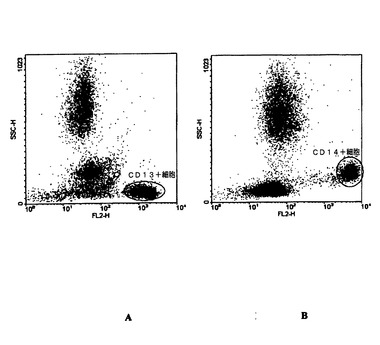
【図2】
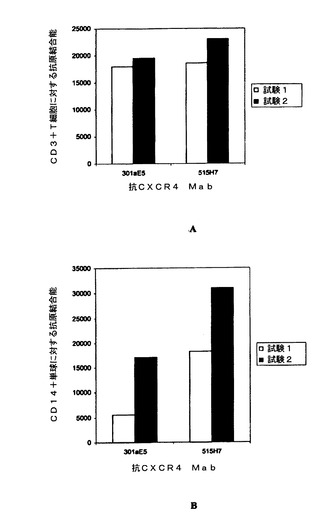
【図3】
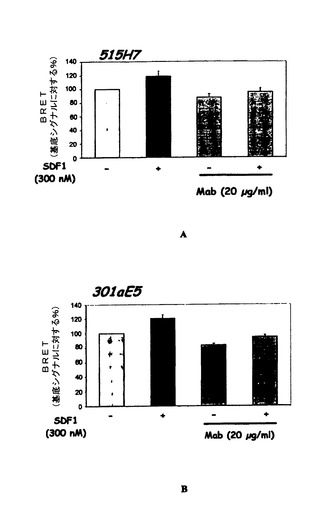
【図4】
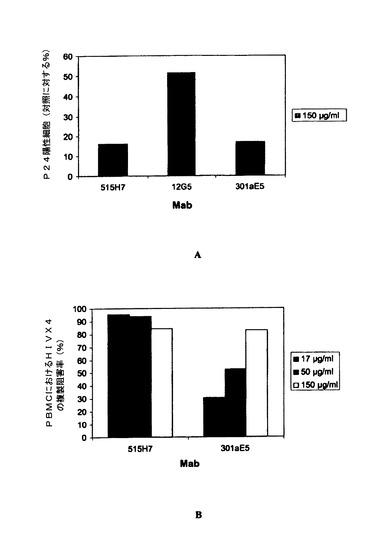
【図5】
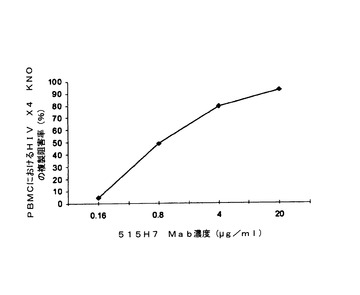
【図6A】
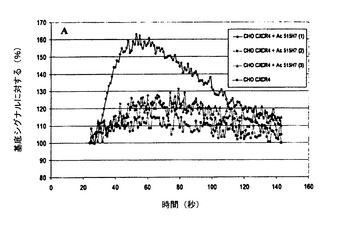
【図6B】
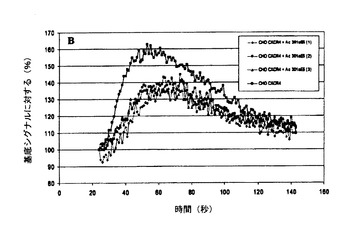
【図6C】
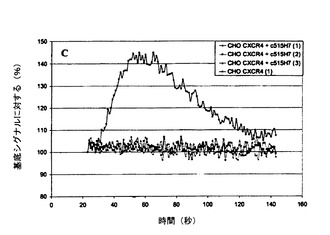
【図7】
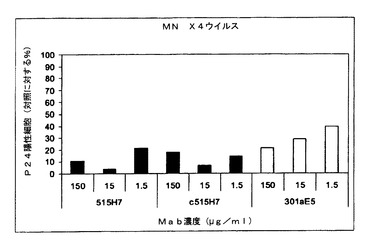
【図8】
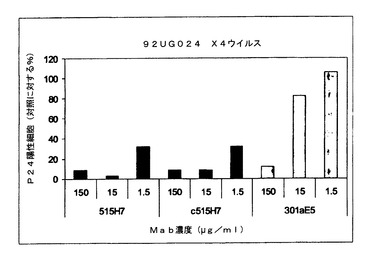
【図9】
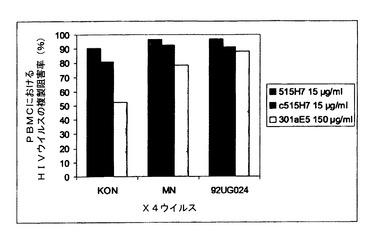
【図10】
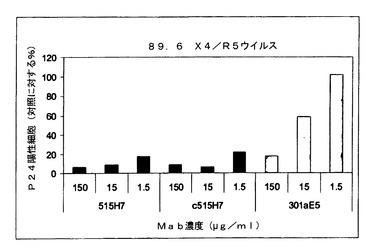
【図11】
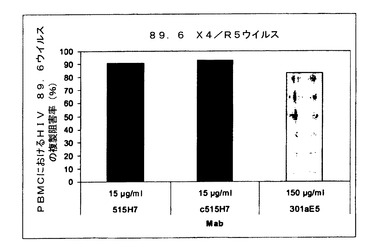
【図12】
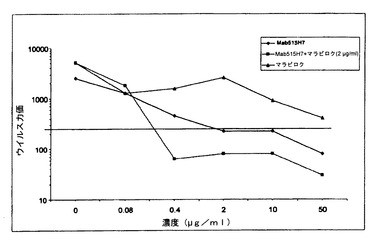
【図13】
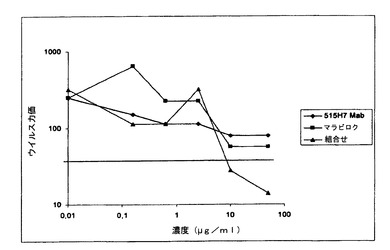
【図14】
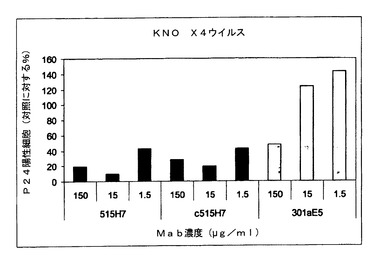
【図15】
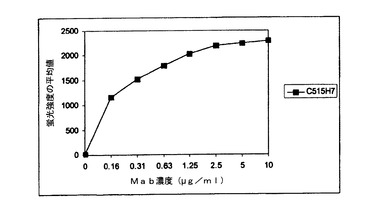
【図16】
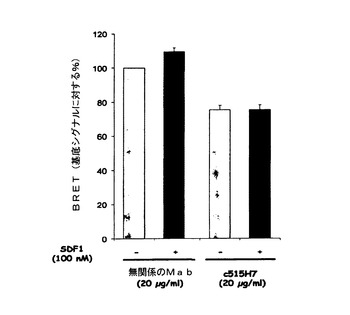
【図17】
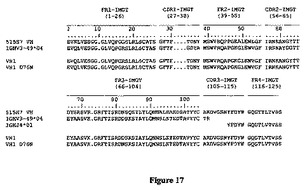
【図18】
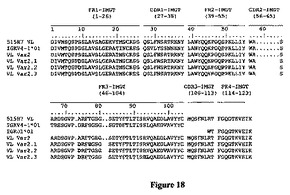
【図19】
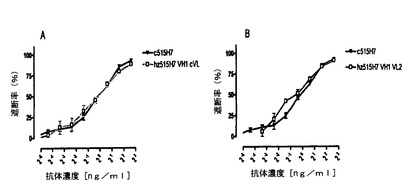
【図20】
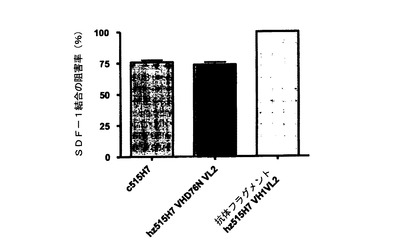
【図21】
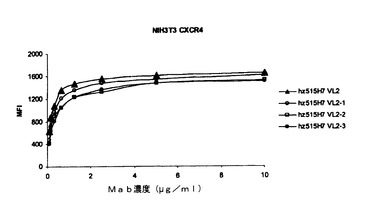
【図22】
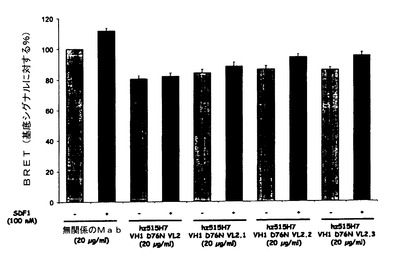
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図6C】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【公表番号】特表2012−525129(P2012−525129A)
【公表日】平成24年10月22日(2012.10.22)
【国際特許分類】
【出願番号】特願2012−507772(P2012−507772)
【出願日】平成22年4月29日(2010.4.29)
【国際出願番号】PCT/EP2010/055863
【国際公開番号】WO2010/125162
【国際公開日】平成22年11月4日(2010.11.4)
【出願人】(500033483)ピエール、ファーブル、メディカマン (73)
【Fターム(参考)】
【公表日】平成24年10月22日(2012.10.22)
【国際特許分類】
【出願日】平成22年4月29日(2010.4.29)
【国際出願番号】PCT/EP2010/055863
【国際公開番号】WO2010/125162
【国際公開日】平成22年11月4日(2010.11.4)
【出願人】(500033483)ピエール、ファーブル、メディカマン (73)
【Fターム(参考)】
[ Back to top ]