
HIV調節/アクセサリータンパク質の融合タンパク質
本発明は、Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4つのHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない融合タンパク質に関する。さらに本発明は、前記タンパク質をコードする核酸、前記核酸を含むベクター、および前記タンパク質を製造する方法に関する。本融合タンパク質、核酸およびベクターは、少なくとも部分的なHIV感染予防のためのワクチンとして使用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4つのHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない融合タンパク質に関する。さらに本発明は、前記タンパク質をコードする核酸、前記核酸を含むベクター、および前記タンパク質を製造する方法に関する。本融合タンパク質、核酸およびベクターは、少なくとも部分的なHIV感染予防のためのワクチンとして使用することができる。
【背景技術】
【0002】
ヒト免疫不全ウイルス(HIV)は後天性免疫不全症候群(AIDS)の原因因子である。すべてのレトロウイルスと同様に、このウイルスのゲノムも、Gag、PolおよびEnvタンパク質をコードしている。また、このウイルスゲノムは調節タンパク質、すなわちTatおよびRevと、アクセサリータンパク質、すなわちVpr、Vpx、Vpu、VifおよびNefもコードしている。
【0003】
AIDSの流行が蔓延するのを抑制しようとする公衆衛生的努力にもかかわらず、新規感染数は依然として増加している。世界保健機関は、この世界的伝染病を、2000年末の時点で感染者数3610万人と推定したが、これは10年前のデータに基づいて予測された数値よりも50%高い値であった(WHOおよびUNAIDS,UNAIDS,2000)。2000年の新規HIV−1感染者数は、全世界では、530万人と推定される。
【0004】
この伝染病が着実に蔓延していることを考えると、有効なワクチンを臨床施設に提供する必要は依然として存在する。新規なベクターやアジュバント系など、多種多様なHIV−1ワクチン送達戦略が、現在既に開発され、さまざまな前臨床設定および臨床試験で評価されている。第III相臨床試験に入った最初のワクチン候補は、アルミニウムアジュバント中のエンベロープgp120タンパク質に基づいている(非特許文献1)。第III相試験は開始されているが、このワクチンは先の第II相試験ではあまりはかばかしい結果を残していない。
【0005】
エンベロープ抗原に基づく予防ワクチンが長年にわたって試みられていたが、最近では、Tat、NefおよびRevなどの調節タンパク質を候補ワクチン抗原として使用することに努力が集中している。これらの調節抗原は、治療的には、数年前から使用されている(非特許文献2)。さらに最近になって、予防ワクチンの研究でも、小規模な前臨床試験で、これらの抗原の使用は有望であることが明らかになった。TatおよびRevまたはTatのみを予防ワクチン候補として使用するとSIVmacは抑制されることが実証された(非特許文献3)。また、感染細胞が成熟ウイルス粒子を高レベルに産生し始める前に感染細胞を排除するには、ウイルス初期調節タンパク質に対するCTLが重要であることも指摘されている(非特許文献4)。
【0006】
HIVの調節/アクセサリータンパク質は有効な免疫応答を誘発するが、それらは、全部ではないにしても、その大半が重大な副作用を持ち、今までは、その副作用によって、ワクチンとしてのそれらの使用が制限されてきた。Nef、TatおよびVpuはCD4+および(または)MHCクラスI発現のダウンレギュレーションに関与することが示されている(非特許文献5)。Tatは生体内で急性免疫抑制を媒介することが知られている(非特許文献6)。Vprについても免疫抑制効果が記載されている(非特許文献7)。酵母細胞ではVprとVpxが異なる細胞分裂抑制効果および細胞毒性を持つことが記載されている(非特許文献8)。したがって、HIVのアクセサリー/調節タンパク質は、全部ではないにしても、その大半が、ワクチン製剤には望ましくない機能的特性を持つようである。
【非特許文献1】Francisら、AIDS Res.Hum.Retroviruses 1998;14(Suppl 3)(5):S325−31
【非特許文献2】Millerら、Nature Medicine 1997,3,389−94、Calarotaら、Lancet 1998,351,1320−5、Ayyavooら、AIDS,2000,14,1−9
【非特許文献3】Osterhausら、Vaccine 1999,17,2713−4
【非特許文献4】van Baalenら、J.Gen.Virol 1997,78,1913−8;Addoら、PNAS,2001,98,1781−6
【非特許文献5】Howcroftら、Science,1993,260,1320−2;Schwartzら、Nature Med.1996,2,338−42;Swannら、Virology,2001,282,267−77;Janvierら、J.Virol.,2001,78,3971−6;Weissmannら、PNAS 1998,95,11601−6
【非特許文献6】Cohenら、PNAS,1999,96,10842−10847
【非特許文献7】Ayyavooら、Nature Med.,1997,3:1117−1123
【非特許文献8】Zhangら、Virology,1997,230,103−12
【発明の開示】
【発明が解決しようとする課題】
【0007】
HIVタンパク質の有害作用を軽減する試みが、WO 02/06303に開示されている。特に、WO 02/06303には、HIV Vif、VpuおよびNefのアミノ酸配列を含む融合タンパク質が開示されている。この融合タンパク質では、成分タンパク質がもう1つの成分タンパク質と連続しているか、またはタンパク質分解切断部位を構成するアミノ酸配列などの非成分タンパク質で隔てられている。成分タンパク質間にタンパク質分解切断部位を含む融合タンパク質を使用することが好ましいと記載されている。成分タンパク質はタンパク質分解切断部位によって隔てられているので、有害であることがわかっている天然のHIVタンパク質が生成される。融合タンパク質の切断によって生じるHIVタンパク質の有害作用を軽減するために、WO 02/06303では、弱毒化タンパク質を使用することが提案されている。したがって、WO 02/06303では、HIV Vif、VprおよびNefタンパク質を含む融合タンパク質であって、HIVタンパク質間に切断部位が挿入されており、HIVタンパク質が弱毒化タンパク質である融合タンパク質が教示されている。しかし、弱毒化タンパク質の欠点は、弱毒化タンパク質のアミノ酸配列が天然タンパク質のアミノ酸配列とは異なるので、弱毒化タンパク質による免疫化で生成する免疫応答は、天然タンパク質を弱く認識するに過ぎないか、天然タンパク質を全く認識しない場合さえありうるということである。
【課題を解決するための手段】
【0008】
HIVの調節/アクセサリータンパク質のいくつかまたは全部に対する有効な免疫応答の発生、特に、有効な細胞傷害性T細胞応答の発生を可能にするワクチンであって、ワクチン中の調節/アクセサリーHIVタンパク質またはワクチンが産生する調節/アクセサリーHIVタンパク質の機能が天然の個々の調節/アクセサリータンパク質よりも低下しているために、ワクチン中のアクセサリー/調節タンパク質が望ましくない副作用を発揮する危険性が減少していると共に、それら低活性HIVタンパク質が天然HIVタンパク質と同様の免疫応答を誘発するようなワクチンを提供することが、本発明の目的だった。
【発明を実施するための最良の形態】
【0009】
この目的は、Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4種類のHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない融合タンパク質を提供することによって達成された。特に、本発明の目的は、前記融合タンパク質をコードする核酸およびベクターによって達成された。
【0010】
本融合タンパク質をヒト細胞などの動物細胞中で産生させた場合、本融合タンパク質は、天然のN末端およびC末端を持つアクセサリー/調節タンパク質を与えるような形では、細胞性プロテアーゼによって切断されない。
【0011】
融合タンパク質の一部であるHIVタンパク質が持つ二次/三次構造は、個々のHIVタンパク質と比較すると変化しているという事実により、融合タンパク質中のHIVタンパク質は、全く機能しないわけではないとしても、個々のタンパク質よりも機能が低下している。機能が低下しているか、または全く機能しない調節/アクセサリータンパク質は、天然のコンフォメーションをとっているHIVタンパク質の望ましくない副作用を持たない。免疫原性に関する限り、融合タンパク質の免疫原性と、その融合タンパク質を構成している個々のHIV調節/アクセサリータンパク質の免疫原性とを比較しても、実質的な違いはない。特に、免疫系に対して提示されるエピトープは同じであるから、細胞傷害性T細胞(CTL)応答に関して、実質的な違いはない。融合タンパク質を患者に投与する場合にも、同じことが言える。
【0012】
本発明に関して「HIV」という用語は、当業者に知られている任意のHIVグループ、サブタイプ(クレード)、株または分離株を表す。具体的には、HIVは、HIV−1でもHIV−2でもよい。HIV−1は9つのサブタイプ(クレードA〜I)に分類されており、HIV−2は5つのサブタイプ(A〜E)に分類されているが、これらはすべて本発明の範囲に包含される。本発明の場合、最も好ましいHIVクレードは、HIV−1クレードA、BおよびCである。しかし本発明は最も好ましいこれらのクレードに限定されるわけではない。
【0013】
HIV調節タンパク質Vif、Vpr、Vpu、Rev、Tat、VpxおよびNefのタンパク質配列は、当業者に知られている。一例として、GenBankデータベースに開示されているさまざまな配列、特にGenBankアクセッション番号KO3455を持つHIV−1分離株HXB2Rの配列が挙げられるが、本発明はこの態様に限定されるわけではない。このGenBankエントリーでは、さまざまなHIV1遺伝子の配列とそれらの遺伝子がコードするタンパク質が特定されている。
【0014】
本融合タンパク質を構成するHIVタンパク質は、好ましくは、同じクレードに由来する。もう1つの態様では、本融合タンパク質を構成するHIVタンパク質が、2つ以上のクレードに由来する。また、本融合タンパク質を構成するHIVタンパク質の1つ以上がHIV−1タンパク質であり、本融合タンパク質を構成するHIVタンパク質の1つ以上がHIV−2タンパク質であることも可能である。
【0015】
本融合タンパク質を構成するHIVタンパク質のアミノ酸配列は、好ましくは、既知のHIV分離株がコードする配列である。すなわち、本融合タンパク質中のHIVタンパク質のアミノ酸配列は、天然のHIV分離株がコードする対応タンパク質のアミノ酸配列と同一である。もう1つの選択肢として、本融合タンパク質中の1種以上のHIVタンパク質のアミノ酸配列は、コンセンサス配列、すなわち、既知HIV分離株中にそのままの形で見いだすことはできないが、既知HIV分離株のいくつかまたはすべてに対して(特にCTLエピトープに関して)最適な相同性を示す配列であってもよい。コンセンサス配列を割り出すためのコンピュータアルゴリズムは当業者に知られている。
【0016】
もう1つの態様として、本融合タンパク質は、融合タンパク質の一部である1種以上のHIVタンパク質のアミノ酸配列の誘導体を含んでもよい。本明細書で使用する「HIVタンパク質のアミノ酸配列の誘導体」という用語は、対応する天然HIVタンパク質と比較して変化したアミノ酸配列を持つHIVタンパク質を表す。変化したアミノ酸配列として、HIVタンパク質の配列の1種以上のアミノ酸が置換もしくは挿入されているか、または欠失している配列が挙げられる。より具体的に述べると、「HIVタンパク質のアミノ酸配列の誘導体」は、本融合タンパク質中のアミノ酸配列の対応する部分を、既知HIV分離株の各HIVタンパク質のアミノ酸配列と比較した場合に、少なくとも50%、より好ましくは少なくとも70%、さらに好ましくは少なくとも80%、最も好ましくは少なくとも90%の相同性を示すアミノ酸配列である。あるアミノ酸配列は、その相同性がたった1つのHIV分離株の対応するタンパク質について見いだされるに過ぎない場合でも、他の分離株の対応するタンパク質にはそれより低い相同性を示すものがあるかもしれないという事実とは関わりなく、上記の配列相同性を持つものと見なされる。例えば、本融合タンパク質中のVpr誘導体が、あるHIV分離株のVpr配列には95%の相同性を示すが、他のHIV分離株(のすべて)には50〜70%の相同性しか示さない場合、当該Vpr誘導体の相同性は少なくとも90%であると見なされる。
【0017】
本融合タンパク質中のタンパク質のコンフォメーションは生物学的に活性なタンパク質の天然のコンフォメーションとは異なるので、本融合タンパク質中のHIVタンパク質が個々のタンパク質と比較して低下した活性を持つこと、または全く活性を持たないことは、既に指摘したとおりである。しかし、融合タンパク質中のHIVタンパク質が生物学的に活性である危険性は、さらに減少させることが望ましいかもしれない。この目的にとって、融合タンパク質の一部である個々のHIVタンパク質の特に好ましい「誘導体」は、低下した活性を持つHIVタンパク質または活性を全く持たないHIVタンパク質が得られるように、いくつかのアミノ酸、より好ましくは10個を超えないアミノ酸、最も好ましくは5個を超えないアミノ酸が、欠失しているか、または挿入もしくは置換されているアミノ酸配列誘導体である。あるHIVタンパク質の生物活性が低下しているかどうかを判定するための試験は、当業者に知られている。
【0018】
生体内でのウイルス複製に不可欠なVifタンパク質の分子機序はまだ分かっていないが、Vifは強い自己会合傾向を持っている。この多量体化は、ウイルスの生活環におけるVifの機能にとって重要であることが示されている(Yang S.ら、J Biol Chem 2001;276:4889−4893)。また、vifはウイルス核タンパク質複合体と特異的に会合することも示されており、これが機能的に重要であるのかもしれない(Khan M.A.ら、J Virol.2001;75(16):7252−65)。したがって、低下した活性を持つvifタンパク質は、多量体化および(または)核タンパク質複合体への会合の減少を示す。
【0019】
Vprタンパク質はウイルスの生活環に重要な役割を果たす。Vprはウイルスのプレインテグレーション複合体の核内移行を調節し、マクロファージなどの非分裂細胞の感染を促進する(Agostiniら、AIDS Res Hum Retroviruses 2002;18(4):283−8)。また、それはLTRとの相互作用によって媒介されるトランス活性化活性も持っている(Vanitharani R.ら、Virology 2001;289(2):334−42)。したがって、低下した活性を持つvprは、トランス活性化および(または)ウイルスプレインテグレーション複合体との相互作用の減少を示すか、またはトランス活性化および(または)ウイルスプレインテグレーション複合体との相互作用を全く示さない。
【0020】
Vprに対して高い相同性を持つVpxも、非分裂細胞における効率のよいウイルスの複製にとって重要である。Vpxは、gag前駆体ポリタンパク質のp6ドメインとの相互作用によって、ウイルス粒子にパッケージングされる。Vprと同様に、Vpxも核内へのプレインテグレーション複合体の輸送に関与する(Mahalingamら、J.Virol 2001;75(1):362−74)。したがって、低下した活性を持つVpxは、gag前駆体を介してプレインテグレーション複合体に会合する能力が減少している。
【0021】
Vpuタンパク質はCD4の細胞質尾部と相互作用することが知られており、CD4分解を引き起こす(Bourら、Virology 1995;69(3):1510−20)。したがって、低下した活性を持つVpuは、CD4分解を誘発する能力が低下している。
【0022】
詳しく特徴づけられているTatタンパク質の関連する生物活性は、トランス活性化応答要素(TAR)との相互作用を介した転写のトランス活性化である。TatはTAR以外のHIV配列を欠く異種プロモーターをトランス活性化できることが実証されている(Han P.ら、Nucleic Acid Res 1991;19(25):7225−9)。したがって、低下した活性を持つtatタンパク質は、TAR要素を介したプロモーターのトランス活性化の減少を示す。
【0023】
Nefタンパク質は、CD4の細胞表面ダウンレギュレーションを誘発することから、疾患進行の原因となるウイルス複製にとって不可欠である(Lou Tら、J Biomed Sci 1997;4(4):132)。このダウンレギュレーションは、CD4とNefの間の直接的相互作用によって開始される(Preusser A.ら、Biochem Biophys Res Commun 2002;292(3):734−40)。したがって、低下した機能を持つNefタンパク質は、CD4との相互作用の減少を示す。
【0024】
Revの関連する機能は、ウイルスRNAのRev応答要素(RRE)との相互作用によって開始される転写後トランス活性化である(Iwaiら、1992;Nuceic Acids Res 1992;20(24):6465−72)。したがって、低下した活性を持つRevは、RREとの相互作用の減少を示す。
【0025】
本発明の融合タンパク質は、Vif、Vpr、Vpu、Rev、Vpx、TatおよびNefから選択される少なくとも4種類のHIVタンパク質のアミノ酸配列を含む。本融合タンパク質は、好ましくは、前記HIVタンパク質のうち、5つ、6つまたはすべてを含みうる。本融合タンパク質におけるHIVタンパク質の順序は決定的な問題ではない。
【0026】
上記少なくとも4種類のHIVタンパク質のうちの1つ以上が、本融合タンパク質中に2コピー以上含まれていてもよい。したがって例えば、本発明の融合タンパク質は、Vif、Vpr、Vpuおよび2コピーのRevを含みうる。それら2コピー以上のHIVタンパク質のアミノ酸配列は同一でありうる。もう1つの選択肢として、それらのコピーのアミノ酸配列は異なっていてもよく、特に、異なるHIV株またはHIVクレードに由来するタンパク質配列(例えば1コピーのHIV−1 Revと1コピーのHIV−2 Rev)を使用する場合には異なっていてもよい。
【0027】
融合タンパク質中の隣り合うHIVタンパク質は、アミノ酸を追加せずに融合するか、または融合タンパク質中の隣り合う2つのHIVタンパク質が少なくとも1つの追加アミノ酸で隔てられるような形で融合することができる。両者の組合せも本発明の範囲に包含される。例えば、4つのHIVタンパク質のアミノ酸配列を含む本発明の融合タンパク質では、隣り合う2つのHIVタンパク質を互いに直接連結し、3番目と4番目のHIVタンパク質は追加アミノ酸を介して連結することができる。この態様に関して「追加アミノ酸」という用語は、天然HIVタンパク質のその位置には見いだされないアミノ酸を表す。
【0028】
したがって本発明の融合タンパク質は、好ましくは、以下の一般式:
+P1−−−P2−−−P3−−−P4−−−P5*−−−P6*−−−P7*+
[式中、P1〜P7は、Vif、Vpr、Vpx、Vpu、Tat、RevおよびNefから選択される異なるHIVタンパク質を表す]
を持ち、本融合タンパク質は、少なくとも4種類の前記HIVタンパク質、すなわちP1〜P4と、要すればさらに1つ(P5*)、2つ(P5*−−−P6*)または3つ(P5*−−−P6*−−−P7*)追加の前記HIVタンパク質を含む。略号「−−−」は、独立して、0個またはn個の追加アミノ酸を意味する。「−−−」が0個のアミノ酸を意味する場合、隣り合うHIVタンパク質は、アミノ酸を追加せずに互いに直接融合される。「−−−」が1〜n個のアミノ酸を意味する場合、隣り合うHIVタンパク質は1〜n個のアミノ酸によって隔てられる。追加アミノ酸数の上限、すなわち整数nは、細胞中で産生または発現されうる融合タンパク質の最大サイズに依存する。
【0029】
ある態様では、すべての「−−−」が独立して0〜20個、より好ましくは0〜10個、さらに好ましくは0〜5個の追加アミノ酸を意味する。
【0030】
もう1つの態様では、「−−−」の少なくとも1つが、Vif、Vpr、Vpx、Vpu、Rev、TatおよびNefから選択されるHIVタンパク質ではない追加タンパク質またはその一部のアミノ酸配列を意味する。したがって、このもう1つの態様では、追加タンパク質に、調節/アクセサリーHIVタンパク質が隣接している。追加タンパク質はどのタンパク質であってもよい。追加タンパク質は、より好ましくは、HIVに対してより良い免疫応答を誘発するのに役立ちうる追加エピトープを含む。したがって、追加タンパク質は、HIV Env、Gagおよび(または)Polタンパク質またはその一部でありうる。この場合、Env、GagおよびPolの「一部」という用語は、前記タンパク質の1つに由来するアミノ酸ストレッチであって、少なくとも1つのエピトープを含むものを表す。一部という用語は、より好ましくは、前記タンパク質の1つに由来する少なくとも10個、さらに好ましくは少なくとも20個、最も好ましくは少なくとも50個のアミノ酸を表す。これに関連する態様では、「−−−」のうち少なくとも1つが、本融合タンパク質の一部であるタンパク質P1〜P7の1種以上のアミノ酸配列を意味する。したがってこの場合、本融合タンパク質は、本融合タンパク質の一部であるタンパク質のうちの1つ以上について、1コピー以上を含みうる。上述のように、それらのタンパク質コピーは、同じアミノ酸配列を持っていてもよいし、同じアミノ酸配列を持っていなくてもよい。
【0031】
上記の式において、略号「+」は、独立して、0〜n個の追加末端アミノ酸を意味する。したがって、本発明の融合タンパク質は、そのタンパク質のC末端および(または)N末端に追加アミノ酸を含んでもよいし、含まなくてもよい。ある態様では、「+」の少なくとも一方が、Vif、Vpr、Vpx、Vpu、Rev、TatおよびNefから選択されるHIVタンパク質ではない追加タンパク質のアミノ酸配列またはその一部を意味する。したがって、この態様では、本融合タンパク質は、そのC末端および(または)N末端に、Vif、Vpr、Vpx、Vpu、Rev、TatまたはNefではない追加タンパク質を含む。追加タンパク質はどのタンパク質であってもよい。追加タンパク質は、より好ましくは、HIVに対してより良い免疫応答を誘発するのに役立ちうる追加エピトープを含む。例えば、追加タンパク質は、HIV Env、Gagおよび(または)Polタンパク質またはその一部でありうる。この場合、Env、GagおよびPolの「一部」という用語は、前記タンパク質の1つに由来するアミノ酸ストレッチであって、少なくとも1つのエピトープを含むものを表す。一部という用語は、より好ましくは、前記タンパク質の1つに由来する少なくとも10個、さらに好ましくは少なくとも20個、最も好ましくは少なくとも50個のアミノ酸を表す。
【0032】
もう1つの態様では、「+」の少なくとも一方が、融合タンパク質の検出または精製を容易にするアミノ酸配列を意味する。したがって、「+」の少なくとも一方は、例えばHisタグなどのタグであるかもしれない。
【0033】
本発明によれば、本融合タンパク質は、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない。より具体的に述べると、本発明の融合タンパク質は、ヒト細胞中で発現させた場合に、天然のN末端およびC末端を持つ個々のHIVタンパク質にプロセシングされない。ヒト細胞中で発現させた場合に、ある融合タンパク質が天然のN末端およびC末端を持つ個々のHIVタンパク質にプロセシングされるかどうかを調べる方法は、当業者に知られている。これについては、Ayyavooら、AIDS 2000,14,1−9、特に同文献の図2に記載の実験を参照されたい。簡単に述べると、当業者は各融合タンパク質をHeLa細胞などのヒト細胞中で容易に発現させることができるだろう。次に、細胞を溶解し、その細胞溶解物を、集合して各HIV融合タンパク質を形成している個々のHIVタンパク質に特異的な抗体によるウェスタンブロット実験または免疫沈降アッセイにかける。本発明の融合タンパク質の場合、個々のHIV調節/アクセサリータンパク質のサイズに一致するサイズを持つ有意な量のHIVタンパク質は検出されない。
【0034】
本発明の融合タンパク質が天然のN末端およびC末端を持つ個々のHIVタンパク質にプロセシングされないことを保証するために、融合タンパク質は、天然のN末端およびC末端を持つHIVタンパク質の生成を誘発するかもしれない細胞性プロテアーゼの特異的切断配列を、融合タンパク質を構成するHIVタンパク質のアミノ酸配列間に含んではならない。したがって、上記の一般式で略記したアミノ酸配列「−−−」は、天然のN末端およびC末端を持つHIVタンパク質の生成を誘発するかもしれない細胞性プロテアーゼの特異的切断配列を含有しない。特に、本融合タンパク質は、融合タンパク質を構成する異なるHIVタンパク質のアミノ酸配列間に切断配列REKRAVVG(アミノ酸の一文字表記)を含有しない。当業者には細胞性プロテアーゼの切断配列が他にも知られている。したがって、当業者は、天然のN末端およびC末端を持つ個々のHIVタンパク質をもたらすかもしれない(細胞性)プロテアーゼの切断部位が含まれることを、容易に防ぐことができる。システインプロテアーゼの切断配列の一例は、Ile/leu−X−Thr−X−Glyである。
【0035】
本発明のタンパク質は、天然N末端と天然C末端の両方を持つHIVタンパク質をもたらす特異的切断配列を含まない。しかしこれは、当該切断部位が天然N末端と天然C末端の両方を持つHIVタンパク質の生成を媒介するのでない限り、融合タンパク質中のタンパク質間に細胞性プロテアーゼの切断部位が存在することを、広く排除しているわけではない。特に、上記の一般式で略記したアミノ酸配列「−−−」は、MHCIまたはMHCII上に提示される短いペプチドの生成に関与するプロテアーゼの切断部位を含みうる。この態様では、切断反応の結果は、好ましくは20アミノ酸未満の短いペプチドストレッチであり、そのN末端またはC末端は、HIVアクセサリー/調節タンパク質の1つのN末端またはC末端と一致してもよい。ただし、これらの短いペプチドが抗原提示の過程で生成する場合、これらの短いペプチドは、それらが由来するHIVタンパク質の活性を、もはや持っていない。
【0036】
また、本発明は、上に定義した本発明の融合タンパク質をコードする核酸に関する。この核酸はDNAでもRNAでもよい。プラスミドまたはDNAウイルスに基づくベクターなどのDNAベクターを使って核酸をヒト細胞中に挿入しようとする場合、核酸はDNAであることが好ましい。
【0037】
本発明の融合タンパク質をコードする核酸を構築する方法は、当業者に知られている。以下の方法に限定されるわけではないが、当業者は、ゲノムHIVクローンから、サブゲノムHIVクローンから、または調節/アクセサリーHIVタンパク質の1種以上のコード配列を含む任意の出発物(例えばプラスミド)から、出発することができる。調節/アクセサリータンパク質のコード配列が連続した読み枠の形をとっている場合は、適当な制限酵素で当該コード配列を含む核酸を切断することにより、そのコード配列を単離することができる。このようにして得たDNA断片は、さらなるクローニングに使用することができる。もう1つの選択肢として、アクセサリー/調節タンパク質のコード配列は、適当なプライマーとポリメラーゼ連鎖反応(PCR)法とを使って得ることもできる。調節/アクセサリータンパク質が、例えばTatおよびRevがそうであるように、2つ以上のエクソンによってコードされている場合は、異なるエクソンを別個にクローニングし、それらを融合して、調節/アクセサリータンパク質の連続した読み枠を生成させるか、またはRT−PCRなどの逆転写技術を用いる必要がありうる。
【0038】
コード配列は遺伝子合成によって、すなわち一組の相補的なかつ/またはオーバーラップしたオリゴヌクレオチドを使って遺伝子を生成させることによって得ることもできる。
【0039】
融合タンパク質を得るために、当該融合タンパク質をコードする核酸は連続した読み枠を持つことが好ましい。したがって、HIVタンパク質または追加タンパク質をコードする配列の停止コドンは、最後の配列を除くすべての配列について、アミノ酸をコードするコドンに突然変異させるか、完全に欠失させることが好ましい。これは、好ましくは、停止コドンを含まないコード配列を増幅させる特異的プライマーをPCRに使用すると、容易に達成することができる。言い換えると、この選択肢では、下流プライマーが停止コドンに相補的であってはならない。したがって、増幅されるDNA断片は停止コドンを含まないことになり、これをクローニングベクターにクローニングすることができる。もう1つの選択肢として、停止コドンを持つコード配列をクローニングベクターにクローニングすることもできる。この停止コドンは後に、例えば特異的エンドヌクレアーゼを使って、または突然変異誘発によって除去することができる。
【0040】
これらのクローニング工程により、本発明の融合タンパク質をコードする連続した読み枠がもたらされるはずである。
【0041】
融合タンパク質を発現させるのに必要な調節要素は、所望する発現系での発現を駆動する調節要素であれば何でもよい。大腸菌などの原核細胞中で融合タンパク質を生成させようとする場合は、細菌プロモーターまたはファージプロモーターを使用することが好ましい。真核細胞中で融合タンパク質を発現させようとする場合は、真核またはウイルスプロモーター/エンハンサーを使用することが好ましい。ポックスウイルスプロモーターを使って融合タンパク質を発現させようとする場合(下記参照)は、7.5プロモーターまたはATIプロモーターなどのポックスウイルスプロモーターを使用することが好ましい。
【0042】
上述のように、本融合タンパク質は、Vif、Vpr、Vpx、Vpu、Tat、RevおよびNefから選択されるHIVタンパク質ではない融合パートナーを含みうる。したがって、本融合タンパク質は、他のタンパク質のアミノ酸配列またはその一部を含みうる。他のタンパク質の例は、HIV Gag、PolおよびEnvタンパク質である。したがって、本発明の核酸は、少なくとも4つの調節/アクセサリーHIVタンパク質またはその誘導体をコードするオープンリーディングフレーム中に、1種以上の追加タンパク質のコード配列またはその一部も含みうる。
【0043】
本発明のさらにもう1つの態様では、本核酸はさらに、HIVに対する免疫応答をより一層改善するのに役立ちうる追加タンパク質をコードする独立した発現カセットも含みうる。好ましい一態様として、本核酸はさらに、Gag、PolおよびEnvから選択される少なくとも1つの追加HIVタンパク質のコード配列またはその一部を含む発現カセットを含みうる。さらに好ましくは、本核酸は、融合タンパク質のコード配列に加えて、HIVタンパク質Gag、PolおよびEnvのすべてのコード配列を含みうる。本核酸は、好ましくは、ベクターの一部である。本核酸は、ウイルスベクター(好ましくはMVAなどのポックスウイルスベクター)のウイルスゲノムまたはその一部であることもできる。したがって、ポックスウイルスベクターから本融合タンパク質および追加HIVタンパク質(例えばGag、PolおよびEnvから選択される少なくとも1つの追加HIVタンパク質)を発現させることができる。
【0044】
さらに本発明は本発明の核酸を含むベクターに関する。「ベクター」という用語は、当業者に知られる任意のベクターを表す。ベクターは、pBR322またはpUC系ベクターなどのプラスミドベクターであることができる。ベクターは、より好ましくは、ウイルスベクターである。本発明に関して「ウイルスベクター」という用語は、ウイルスゲノムを含む感染性および(または)弱毒化ウイルスを表す。この場合、本発明の核酸は、各ウイルスベクターのウイルスゲノムの一部であり、かつ/またはウイルスゲノムを構成している。本組換えベクターは、細胞および細胞株の感染、特にヒトを含む生きている動物の感染に使用することができる。典型的な本発明のウイルスベクターはアデノウイルスベクター、レトロウイルスベクターまたはアデノ関連ウイルス2(AAV2)に基づくベクターである。ポックスウイルスベクターが最も好ましい。ポックスウイルスとして、好ましくは、カナリア痘瘡ウイルス、鶏痘ウイルスまたはワクシニアウイルスを挙げることができる。変異ワクシニアウイルスアンカラ(modified vaccinia virus Ankara:MVA)(Sutter,G.ら[1994]Vaccine 12:1032−40)が、さらに好ましい。典型的なMVA株は、ヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(European Collection of Animal Cell Cultures)に受託番号ECACC V00120707として寄託されているMVA575である。2001年11月22日に「Modified Vaccinia Ankara Virus Variant」という名称で欧州特許庁に出願されたPCT出願PCT/EP01/13628に記載されているMVA−BNまたはその誘導体が、最も好ましい。MVA−BNはヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズに受託番号ECACC V00083008として寄託されている。MVA−BNウイルスベクターは変異ワクシニアアンカラに由来する著しく弱毒化されたウイルスであり、ヒト細胞で増殖的に複製する能力を持たないことを特徴とするので、MVA−BNまたはその誘導体を用いることにより、HIVに対して特定の安全なウイルスワクチンを提供するというもう1つの技術的課題が解決された。MVA−BNはヒトでは複製しないので他のどの既知ワクシニアウイルス株よりも安全である。好ましい一態様では、本発明は、本発明のDNAを含むウイルスベクターとして、MVA−BNおよびMVA−BNの誘導体に関する。MVA−BNの特徴、あるMVAがMVA−BNまたはその誘導体であるかどうかを調べることを可能にする生物学的アッセイの説明、およびMVA−BNまたはその誘導体の取得を可能にする方法は、上記のPCT出願PCT/EP01/13628に開示されており、この文献は参照により本明細書に組み込まれる。
【0045】
したがってこれらの態様では、本発明は、好ましくは、本発明の融合タンパク質をコードする発現カセットをウイルスゲノム中に含む組換えMVA(例えばMVA−BN)に関する。
【0046】
本発明の核酸をウイルスゲノムに挿入する方法、および組換えウイルスを取得する方法は、当業者に知られている。
【0047】
組換えワクシニアウイルスにおいて、本発明DNAの発現は、好ましくはポックスウイルスプロモーターの転写制御、より好ましくはワクシニアウイルスプロモーターの転写制御を受けるが、これに限るわけではない。本発明のDNAは、好ましくは、ウイルスゲノムの非必須領域に挿入される。本発明のもう1つの好ましい態様では、異種核酸配列を、ポックスウイルスゲノムの天然の欠失部位(PCT/EP96/02926に記載されているもの)に挿入する。しかし、挿入部位の性質は、組換えワクシニアウイルスが得られる限り、本発明にとって決定的な問題ではない。したがって当業者は他の適切な挿入部位を容易に思いつくことができる。
【0048】
ウイルスベクター、特にポックスウイルスベクターは、好ましくは、本発明の融合タンパク質のコード配列に加えて、HIV Gag、PolおよびEnv遺伝子から選択される追加のレトロウイルス遺伝子を、ウイルスゲノム中に含みうる。ウイルスベクター、特にポックスウイルスベクターは、より好ましくは、本発明の融合タンパク質のコード配列に加えて、Gag、PolおよびEnvをコードするHIV遺伝子をすべて含みうる。本発明では同じ核酸を使ってこれらの追加遺伝子を挿入することができる。この態様では、すべてのHIV遺伝子が、ウイルスゲノムの同じ挿入部位に位置することになる。もう1つの態様では、追加遺伝子はウイルスゲノムの異なる位置に挿入される。
【0049】
好ましい一態様として、本発明は、HIV感染およびAIDSを少なくとも部分的に予防するためのワクチンとしての、本発明の核酸、ベクターまたは融合タンパク質に関する。「ワクチン」は、特異的免疫応答を誘導する化合物、すなわち核酸、融合タンパク質、ベクターまたはウイルスである。
【0050】
この態様の一選択肢として、本発明の「ワクチン」は、本発明の融合タンパク質に基づく。
【0051】
好ましい一態様では、本発明の核酸、特にDNAを、ワクチンとして使用する。本発明の場合と同様に真核発現カセットを含んでいる裸のDNAを投与すると(特にDNAを筋肉内投与すると)、その発現カセットによってコードされているタンパク質の発現が起こることが、当業者に知られている。そのタンパク質は免疫系にばく露され、特異的免疫応答を生じさせる。もう1つの態様では、本発明のベクター、特にウイルスベクター、より好ましくはポックスウイルスベクター、最も好ましくはMVAベクターなどのワクシニアウイルスベクターを投与することによって、予防接種を行う。
【0052】
ワクシニアウイルス系ワクチンを製造するには、本発明のウイルスを生理学的に許容できる形態に変換する。これは、天然痘の予防接種に用いられるポックスウイルスワクチン(Stickl,H.ら[1974]Dtsch.med.Wschr.99,2386−2392に記載されているもの)を製造する際の経験に基づいて行うことができる。例えば、精製ウイルスは、5×108 TCID50/mlの力価で約10mMトリスおよび140mM NaCl(pH7.4)中に製剤化して、−80℃で保存する。ワクチン注射剤を製造するには、例えば、2%ペプトンおよび1%ヒトアルブミンの存在下で、リン酸緩衝食塩水(PBS)100mL中のウイルス粒子102〜108個を、アンプル(好ましくはガラスアンプル)中で凍結乾燥する。もう1つの選択肢として、ワクチン注射剤は、製剤中のウイルスの段階的凍結乾燥によって製造することもできる。この製剤は、さらに、例えばマンニトール、デキストラン、糖、グリシン、ラクトースもしくはポリビニルピロリドンなどの添加剤、または、生体内投与に適した酸化防止剤もしくは不活性ガス、安定剤または組換えタンパク質(例えばヒト血清アルブミン)などの他の添加剤も含むことができる。その場合、このガラスアンプルを封印して、4℃〜室温で数ヶ月間保存することができる。しかし、必要がない限り、アンプルは−20℃未満の温度で保存することが好ましい。予防接種を行うには、上記の凍結乾燥物を0.1〜0.5mlの水性溶液、好ましくは生理食塩水またはトリス緩衝液に溶解し、それを全身的または局所的に、すなわち非経口投与、筋肉内投与、または当業者に知られる他の任意の投与経路によって、投与することができる。当業者であれば、投与様式、投与量および投与回数を、既知の方法で最適化することができる。ポックスウイルスベクターの場合は、皮下投与または筋肉内投与が最も好ましい。
【0053】
ワクチンが本発明のDNAを含むMVA−BNベクターまたはその誘導体である場合、本発明の一特定態様は、第1接種(「初回免疫接種」)および第2接種(「追加免疫接種」)で、治療有効量の当該ワクチンを投与することに関する。
【0054】
ワクチンが本発明のDNAを含むMVA−BNベクターまたはその誘導体である場合、本発明の一特定態様は、第1バイアル/容器に入った第1予防接種(「初回免疫」)用の本発明MVA−BNウイルスベクターと、第2バイアル/容器に入った第2予防接種(「追加免疫」)用の本発明MVA−BNウイルスベクターとを含む予防接種用キットに関する。したがって本発明は、ワクチン態様として、本発明の核酸、ベクターまたは融合タンパク質を含むワクチン、およびワクチンの製造を目的とする前記核酸、ベクターまたはタンパク質の使用に関する。
【0055】
さらにもう1つの態様として、本発明は、本発明の融合タンパク質、本発明の核酸、または本発明のベクターを保護を必要とする、ヒトを含む動物に投与することによって、ヒトを含む動物をHIV感染から保護する方法に関する。
【0056】
さらに本発明は、本発明のタンパク質を製造する方法であって、(i)本発明の核酸またはベクターを宿主細胞にトランスフェクトする工程、または(ii)宿主細胞に本発明のウイルスベクターを感染させる工程、(iii)工程(i)のトランスフェクト宿主細胞または工程(ii)の感染宿主細胞中で融合タンパク質を発現させる工程、および(iv)融合タンパク質を回収する工程を含む方法に関する。
【0057】
さらに本発明は、本発明の核酸またはベクターをトランスフェクトした宿主細胞、または本発明のウイルスベクターに感染した宿主細胞に関する。
【0058】
もう1つの態様として、本融合タンパク質は、Vif、Vpr、Vpu、Rev、VpxおよびTatから選択される少なくとも3種類のHIVタンパク質を含みうる。本融合タンパク質は、好ましくは、前記HIVタンパク質の4個、5個またはすべてを含みうる。この態様の融合タンパク質の典型例は、HIVタンパク質Vpr、Vif、Vpu、RevおよびTatのアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む。上述のように、融合タンパク質中のHIVタンパク質の順序は決定的な問題ではない。上述した好ましい態様はすべてこの代替態様にも当てはまる。
【0059】
図面の簡単な説明
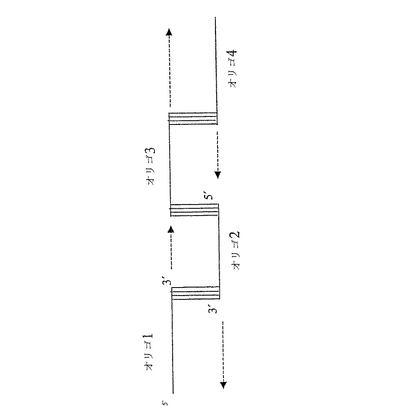
図1:図1は、オリゴヌクレオチドのアニーリングを表す略図である。この図は4つのオリゴヌクレオチドのアニーリングを示している。各オリゴヌクレオチドは一本鎖であり、相補的末端によってアニールすることができる。プルーフリーディング活性を示すポリメラーゼ(例えばPfxポリメラーゼ)を使ってギャップを埋める。
【0060】
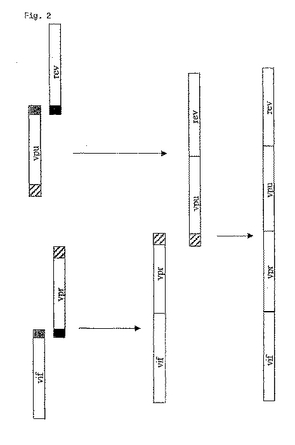
図2:図2は、ブロブ(blob)の4つの遺伝子のアニーリングを表す略図である。vif遺伝子はvpr断片とのオーバーラップ配列を示し、vpuコード断片はrev遺伝子とのオーバーラップ配列を示す(灰色の部分)。PCR断片を変性し、オーバーラップする相補的末端をハイブリダイズさせる。その結果生じたギャップをPfxポリメラーゼで埋める。vif−vpr断片をvpu−rev断片のオーバーラップ配列に融合し、それを再び融合に使用する。
【0061】
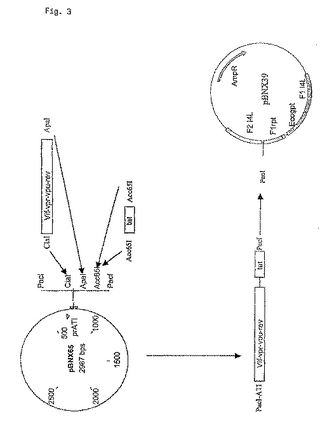
図3:図3は、MVAゲノムに外来遺伝子を挿入するための組換えベクターに、本発明の融合タンパク質をコードする配列をクローニングするための戦略である。融合したvif、vpr、vpuおよびrevポリタンパク質コード領域を、ClaIおよびApaI制限酵素部位を含むプライマーを使って増幅した。このpCR産物を、ポックスウイルスATIプロモーターを持つベクターpBNX65のClaI/ApaI切断物にクローニングした。Acc65I制限酵素部位を含むプライマーを使ったPCRによってtatコード領域を増幅し、Acc65Iで線状化したpBNX65+vif−revにライゲートした。得られた発現カセット(ATIプロモーター+本発明の融合タンパク質をコードする配列)をPacI制限酵素消化によって単離し、MVAゲノムI4L遺伝子間領域に外来遺伝子を挿入するための組換えベクター(pBNX39)に挿入した。pBNX39は、MVAゲノムの挿入部位の隣接配列に相同な配列(F1 I4LおよびF2 I4L)を持っている。MVAゲノムとpBNX39との相同組換え後に組換えウイルスを選択することができるように、このベクターは大腸菌gpt遺伝子(ホスホリボシルトランスフェラーゼ遺伝子)も持っている。この選択カセットは、組換えウイルスの精製後に、Flank1とflank1の反復配列(F1rpt)との間で起こる相同組換えによって欠失する。
【0062】
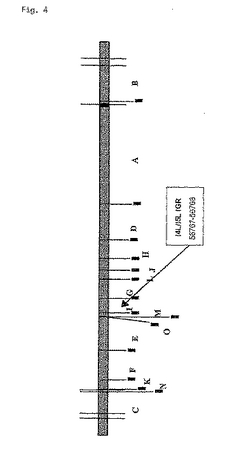
図4:図4は、MVAゲノムの概略図である。MVAは、HindIIIによる制限酵素消化後に特徴的な断片(A〜O)を示す線状ゲノムを持っている。I4L遺伝子とI5L遺伝子の間の非機能領域はI断片中に位置している。pBNX39を使った外来遺伝子の挿入は56767〜56768位で起こる。
【実施例】
【0063】
(HIV Vif−Vpr−Vpu−Rev−Tat融合タンパク質をコードするDNAの作製)
HIVゲノムの単一遺伝子は、標準的PCRプロトコールを用いるPCRにより、ゲノムDNAから調製するか、またはオーバーラップ配列を介したオリゴヌクレオチドのアニーリングと、その結果生じた一本鎖ギャップの充填とに基づく技術によって合成的に調製した。
【0064】
本発明の融合タンパク質をコードする核酸に挿入しようとする遺伝子のコード領域をオリゴヌクレオチドに基づいて作製するために、15bpのオーバーラップを持つ40マーのオリゴヌクレオチドを設計した。これらのオリゴヌクレオチドの配列は、IIIB株に由来するHIV1分離株HXB2Rのゲノム地図に基づいている。アニーリング反応用のオリゴヌクレオチド、または必要な配列を単離するためのPCR用のオリゴヌクレオチドは、結果として得られるコード領域から翻訳終結用の停止コドンが欠失するように設計した。tat遺伝子は停止コドンを含むオリゴを使って合成した。というのも、この遺伝子は本発明の融合タンパク質をコードする核酸の最後に挿入されるものなので、ポリタンパク質の翻訳が正しく終結するように停止トリプレットを含むべきだからである。
【0065】
オリゴアニーリング反応については、10サイクルの2段階Pfxポリメラーゼ(Gibco−BRL)反応(95℃で変性および68℃でアニーニング/伸長)を行った。この反応中に、オリゴのオーバーラップ配列はアニールした状態になり、ギャップがPfxプルーフリーディングポリメラーゼによって充填される(図1)。融合タンパク質をコードするヌクレオチド配列中の第1コード遺伝子であるvifコード領域を合成するために、ゲノムHIV cDNAを使ったPCRを行った。このPCRは、続いて行うvifとvprのアニーリングに備えて、vifコード領域が次のvpr遺伝子の最初の15bpに融合されるように行った。HIV HXB2Rゲノムのbp5559−5847に広がるVprコード領域は、10個のヌクレオチドのアニーリングによって調製した。生じたギャップを充填し、増幅のためのPCRを行った後、vif用の隣接領域と、vprコード領域の後ろに挿入する予定であるvpu用の隣接領域とに融合したvprコード領域を含む生成物を得た。
【0066】
vifの合成に使用したものと同じcDNAから、PCRによってVpuコード領域を増幅することにより、vprおよびrevと融合させるための隣接領域を含む生成物を得た。
【0067】
revコード領域は、HIV HXB2Rゲノムのbp5970−6045および8379−8650の領域を覆うと共に、vpuおよびtatとのアニーリング用の15bpオーバーラップを持つ14個のオリゴヌクレオチドのアニーリングによって合成した。
【0068】
tatコード領域は、HIV HXB2Rゲノムのbp5831−6045および8379−8466を覆う10個のオリゴヌクレオチドを使って作製した。
【0069】
vifおよびvprコード領域ならびにvpuおよびrevコード領域は、それら2つの断片をそれぞれのオーバーラップを使ってアニールさせることにより、2段階Pfxポリメラーゼ反応で融合させた(図2)。その融合生成物をさらにPCR増幅した後、断片を精製し、vprおよびvpuのオーバーラップを使って互いにライゲートした(図2)。得られた生成物(vif−vpr−vpu−revのコード配列)をPCR増幅した後、ポックスウイルスATIプロモーターを含むpBluescriptKS+ベクター(図3、pBNX65)中の隣り合うクローニング部位に、vif−vpr−vpu−rev断片とtatとをクローニングすることにより、tatコード領域を融合した。次に、PacI制限酵素消化によって発現カセット全体を単離し、pBNX39(図3)に挿入した。pBNX39は、MVAゲノムに相同な配列を含んでおり、それが相同組換えによるゲノム(図4)の非コード領域(I4L)への挿入を可能にする。
【図面の簡単な説明】
【0070】
【図1】オリゴヌクレオチドのアニーリングを表す略図である。
【図2】ブロブ(blob)の4つの遺伝子のアニーリングを表す略図である。
【図3】MVAゲノムに外来遺伝子を挿入するための組換えベクターに、本発明の融合タンパク質をコードする配列をクローニングするための戦略図である。
【図4】MVAゲノムの概略図である。
【技術分野】
【0001】
本発明は、Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4つのHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない融合タンパク質に関する。さらに本発明は、前記タンパク質をコードする核酸、前記核酸を含むベクター、および前記タンパク質を製造する方法に関する。本融合タンパク質、核酸およびベクターは、少なくとも部分的なHIV感染予防のためのワクチンとして使用することができる。
【背景技術】
【0002】
ヒト免疫不全ウイルス(HIV)は後天性免疫不全症候群(AIDS)の原因因子である。すべてのレトロウイルスと同様に、このウイルスのゲノムも、Gag、PolおよびEnvタンパク質をコードしている。また、このウイルスゲノムは調節タンパク質、すなわちTatおよびRevと、アクセサリータンパク質、すなわちVpr、Vpx、Vpu、VifおよびNefもコードしている。
【0003】
AIDSの流行が蔓延するのを抑制しようとする公衆衛生的努力にもかかわらず、新規感染数は依然として増加している。世界保健機関は、この世界的伝染病を、2000年末の時点で感染者数3610万人と推定したが、これは10年前のデータに基づいて予測された数値よりも50%高い値であった(WHOおよびUNAIDS,UNAIDS,2000)。2000年の新規HIV−1感染者数は、全世界では、530万人と推定される。
【0004】
この伝染病が着実に蔓延していることを考えると、有効なワクチンを臨床施設に提供する必要は依然として存在する。新規なベクターやアジュバント系など、多種多様なHIV−1ワクチン送達戦略が、現在既に開発され、さまざまな前臨床設定および臨床試験で評価されている。第III相臨床試験に入った最初のワクチン候補は、アルミニウムアジュバント中のエンベロープgp120タンパク質に基づいている(非特許文献1)。第III相試験は開始されているが、このワクチンは先の第II相試験ではあまりはかばかしい結果を残していない。
【0005】
エンベロープ抗原に基づく予防ワクチンが長年にわたって試みられていたが、最近では、Tat、NefおよびRevなどの調節タンパク質を候補ワクチン抗原として使用することに努力が集中している。これらの調節抗原は、治療的には、数年前から使用されている(非特許文献2)。さらに最近になって、予防ワクチンの研究でも、小規模な前臨床試験で、これらの抗原の使用は有望であることが明らかになった。TatおよびRevまたはTatのみを予防ワクチン候補として使用するとSIVmacは抑制されることが実証された(非特許文献3)。また、感染細胞が成熟ウイルス粒子を高レベルに産生し始める前に感染細胞を排除するには、ウイルス初期調節タンパク質に対するCTLが重要であることも指摘されている(非特許文献4)。
【0006】
HIVの調節/アクセサリータンパク質は有効な免疫応答を誘発するが、それらは、全部ではないにしても、その大半が重大な副作用を持ち、今までは、その副作用によって、ワクチンとしてのそれらの使用が制限されてきた。Nef、TatおよびVpuはCD4+および(または)MHCクラスI発現のダウンレギュレーションに関与することが示されている(非特許文献5)。Tatは生体内で急性免疫抑制を媒介することが知られている(非特許文献6)。Vprについても免疫抑制効果が記載されている(非特許文献7)。酵母細胞ではVprとVpxが異なる細胞分裂抑制効果および細胞毒性を持つことが記載されている(非特許文献8)。したがって、HIVのアクセサリー/調節タンパク質は、全部ではないにしても、その大半が、ワクチン製剤には望ましくない機能的特性を持つようである。
【非特許文献1】Francisら、AIDS Res.Hum.Retroviruses 1998;14(Suppl 3)(5):S325−31
【非特許文献2】Millerら、Nature Medicine 1997,3,389−94、Calarotaら、Lancet 1998,351,1320−5、Ayyavooら、AIDS,2000,14,1−9
【非特許文献3】Osterhausら、Vaccine 1999,17,2713−4
【非特許文献4】van Baalenら、J.Gen.Virol 1997,78,1913−8;Addoら、PNAS,2001,98,1781−6
【非特許文献5】Howcroftら、Science,1993,260,1320−2;Schwartzら、Nature Med.1996,2,338−42;Swannら、Virology,2001,282,267−77;Janvierら、J.Virol.,2001,78,3971−6;Weissmannら、PNAS 1998,95,11601−6
【非特許文献6】Cohenら、PNAS,1999,96,10842−10847
【非特許文献7】Ayyavooら、Nature Med.,1997,3:1117−1123
【非特許文献8】Zhangら、Virology,1997,230,103−12
【発明の開示】
【発明が解決しようとする課題】
【0007】
HIVタンパク質の有害作用を軽減する試みが、WO 02/06303に開示されている。特に、WO 02/06303には、HIV Vif、VpuおよびNefのアミノ酸配列を含む融合タンパク質が開示されている。この融合タンパク質では、成分タンパク質がもう1つの成分タンパク質と連続しているか、またはタンパク質分解切断部位を構成するアミノ酸配列などの非成分タンパク質で隔てられている。成分タンパク質間にタンパク質分解切断部位を含む融合タンパク質を使用することが好ましいと記載されている。成分タンパク質はタンパク質分解切断部位によって隔てられているので、有害であることがわかっている天然のHIVタンパク質が生成される。融合タンパク質の切断によって生じるHIVタンパク質の有害作用を軽減するために、WO 02/06303では、弱毒化タンパク質を使用することが提案されている。したがって、WO 02/06303では、HIV Vif、VprおよびNefタンパク質を含む融合タンパク質であって、HIVタンパク質間に切断部位が挿入されており、HIVタンパク質が弱毒化タンパク質である融合タンパク質が教示されている。しかし、弱毒化タンパク質の欠点は、弱毒化タンパク質のアミノ酸配列が天然タンパク質のアミノ酸配列とは異なるので、弱毒化タンパク質による免疫化で生成する免疫応答は、天然タンパク質を弱く認識するに過ぎないか、天然タンパク質を全く認識しない場合さえありうるということである。
【課題を解決するための手段】
【0008】
HIVの調節/アクセサリータンパク質のいくつかまたは全部に対する有効な免疫応答の発生、特に、有効な細胞傷害性T細胞応答の発生を可能にするワクチンであって、ワクチン中の調節/アクセサリーHIVタンパク質またはワクチンが産生する調節/アクセサリーHIVタンパク質の機能が天然の個々の調節/アクセサリータンパク質よりも低下しているために、ワクチン中のアクセサリー/調節タンパク質が望ましくない副作用を発揮する危険性が減少していると共に、それら低活性HIVタンパク質が天然HIVタンパク質と同様の免疫応答を誘発するようなワクチンを提供することが、本発明の目的だった。
【発明を実施するための最良の形態】
【0009】
この目的は、Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4種類のHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない融合タンパク質を提供することによって達成された。特に、本発明の目的は、前記融合タンパク質をコードする核酸およびベクターによって達成された。
【0010】
本融合タンパク質をヒト細胞などの動物細胞中で産生させた場合、本融合タンパク質は、天然のN末端およびC末端を持つアクセサリー/調節タンパク質を与えるような形では、細胞性プロテアーゼによって切断されない。
【0011】
融合タンパク質の一部であるHIVタンパク質が持つ二次/三次構造は、個々のHIVタンパク質と比較すると変化しているという事実により、融合タンパク質中のHIVタンパク質は、全く機能しないわけではないとしても、個々のタンパク質よりも機能が低下している。機能が低下しているか、または全く機能しない調節/アクセサリータンパク質は、天然のコンフォメーションをとっているHIVタンパク質の望ましくない副作用を持たない。免疫原性に関する限り、融合タンパク質の免疫原性と、その融合タンパク質を構成している個々のHIV調節/アクセサリータンパク質の免疫原性とを比較しても、実質的な違いはない。特に、免疫系に対して提示されるエピトープは同じであるから、細胞傷害性T細胞(CTL)応答に関して、実質的な違いはない。融合タンパク質を患者に投与する場合にも、同じことが言える。
【0012】
本発明に関して「HIV」という用語は、当業者に知られている任意のHIVグループ、サブタイプ(クレード)、株または分離株を表す。具体的には、HIVは、HIV−1でもHIV−2でもよい。HIV−1は9つのサブタイプ(クレードA〜I)に分類されており、HIV−2は5つのサブタイプ(A〜E)に分類されているが、これらはすべて本発明の範囲に包含される。本発明の場合、最も好ましいHIVクレードは、HIV−1クレードA、BおよびCである。しかし本発明は最も好ましいこれらのクレードに限定されるわけではない。
【0013】
HIV調節タンパク質Vif、Vpr、Vpu、Rev、Tat、VpxおよびNefのタンパク質配列は、当業者に知られている。一例として、GenBankデータベースに開示されているさまざまな配列、特にGenBankアクセッション番号KO3455を持つHIV−1分離株HXB2Rの配列が挙げられるが、本発明はこの態様に限定されるわけではない。このGenBankエントリーでは、さまざまなHIV1遺伝子の配列とそれらの遺伝子がコードするタンパク質が特定されている。
【0014】
本融合タンパク質を構成するHIVタンパク質は、好ましくは、同じクレードに由来する。もう1つの態様では、本融合タンパク質を構成するHIVタンパク質が、2つ以上のクレードに由来する。また、本融合タンパク質を構成するHIVタンパク質の1つ以上がHIV−1タンパク質であり、本融合タンパク質を構成するHIVタンパク質の1つ以上がHIV−2タンパク質であることも可能である。
【0015】
本融合タンパク質を構成するHIVタンパク質のアミノ酸配列は、好ましくは、既知のHIV分離株がコードする配列である。すなわち、本融合タンパク質中のHIVタンパク質のアミノ酸配列は、天然のHIV分離株がコードする対応タンパク質のアミノ酸配列と同一である。もう1つの選択肢として、本融合タンパク質中の1種以上のHIVタンパク質のアミノ酸配列は、コンセンサス配列、すなわち、既知HIV分離株中にそのままの形で見いだすことはできないが、既知HIV分離株のいくつかまたはすべてに対して(特にCTLエピトープに関して)最適な相同性を示す配列であってもよい。コンセンサス配列を割り出すためのコンピュータアルゴリズムは当業者に知られている。
【0016】
もう1つの態様として、本融合タンパク質は、融合タンパク質の一部である1種以上のHIVタンパク質のアミノ酸配列の誘導体を含んでもよい。本明細書で使用する「HIVタンパク質のアミノ酸配列の誘導体」という用語は、対応する天然HIVタンパク質と比較して変化したアミノ酸配列を持つHIVタンパク質を表す。変化したアミノ酸配列として、HIVタンパク質の配列の1種以上のアミノ酸が置換もしくは挿入されているか、または欠失している配列が挙げられる。より具体的に述べると、「HIVタンパク質のアミノ酸配列の誘導体」は、本融合タンパク質中のアミノ酸配列の対応する部分を、既知HIV分離株の各HIVタンパク質のアミノ酸配列と比較した場合に、少なくとも50%、より好ましくは少なくとも70%、さらに好ましくは少なくとも80%、最も好ましくは少なくとも90%の相同性を示すアミノ酸配列である。あるアミノ酸配列は、その相同性がたった1つのHIV分離株の対応するタンパク質について見いだされるに過ぎない場合でも、他の分離株の対応するタンパク質にはそれより低い相同性を示すものがあるかもしれないという事実とは関わりなく、上記の配列相同性を持つものと見なされる。例えば、本融合タンパク質中のVpr誘導体が、あるHIV分離株のVpr配列には95%の相同性を示すが、他のHIV分離株(のすべて)には50〜70%の相同性しか示さない場合、当該Vpr誘導体の相同性は少なくとも90%であると見なされる。
【0017】
本融合タンパク質中のタンパク質のコンフォメーションは生物学的に活性なタンパク質の天然のコンフォメーションとは異なるので、本融合タンパク質中のHIVタンパク質が個々のタンパク質と比較して低下した活性を持つこと、または全く活性を持たないことは、既に指摘したとおりである。しかし、融合タンパク質中のHIVタンパク質が生物学的に活性である危険性は、さらに減少させることが望ましいかもしれない。この目的にとって、融合タンパク質の一部である個々のHIVタンパク質の特に好ましい「誘導体」は、低下した活性を持つHIVタンパク質または活性を全く持たないHIVタンパク質が得られるように、いくつかのアミノ酸、より好ましくは10個を超えないアミノ酸、最も好ましくは5個を超えないアミノ酸が、欠失しているか、または挿入もしくは置換されているアミノ酸配列誘導体である。あるHIVタンパク質の生物活性が低下しているかどうかを判定するための試験は、当業者に知られている。
【0018】
生体内でのウイルス複製に不可欠なVifタンパク質の分子機序はまだ分かっていないが、Vifは強い自己会合傾向を持っている。この多量体化は、ウイルスの生活環におけるVifの機能にとって重要であることが示されている(Yang S.ら、J Biol Chem 2001;276:4889−4893)。また、vifはウイルス核タンパク質複合体と特異的に会合することも示されており、これが機能的に重要であるのかもしれない(Khan M.A.ら、J Virol.2001;75(16):7252−65)。したがって、低下した活性を持つvifタンパク質は、多量体化および(または)核タンパク質複合体への会合の減少を示す。
【0019】
Vprタンパク質はウイルスの生活環に重要な役割を果たす。Vprはウイルスのプレインテグレーション複合体の核内移行を調節し、マクロファージなどの非分裂細胞の感染を促進する(Agostiniら、AIDS Res Hum Retroviruses 2002;18(4):283−8)。また、それはLTRとの相互作用によって媒介されるトランス活性化活性も持っている(Vanitharani R.ら、Virology 2001;289(2):334−42)。したがって、低下した活性を持つvprは、トランス活性化および(または)ウイルスプレインテグレーション複合体との相互作用の減少を示すか、またはトランス活性化および(または)ウイルスプレインテグレーション複合体との相互作用を全く示さない。
【0020】
Vprに対して高い相同性を持つVpxも、非分裂細胞における効率のよいウイルスの複製にとって重要である。Vpxは、gag前駆体ポリタンパク質のp6ドメインとの相互作用によって、ウイルス粒子にパッケージングされる。Vprと同様に、Vpxも核内へのプレインテグレーション複合体の輸送に関与する(Mahalingamら、J.Virol 2001;75(1):362−74)。したがって、低下した活性を持つVpxは、gag前駆体を介してプレインテグレーション複合体に会合する能力が減少している。
【0021】
Vpuタンパク質はCD4の細胞質尾部と相互作用することが知られており、CD4分解を引き起こす(Bourら、Virology 1995;69(3):1510−20)。したがって、低下した活性を持つVpuは、CD4分解を誘発する能力が低下している。
【0022】
詳しく特徴づけられているTatタンパク質の関連する生物活性は、トランス活性化応答要素(TAR)との相互作用を介した転写のトランス活性化である。TatはTAR以外のHIV配列を欠く異種プロモーターをトランス活性化できることが実証されている(Han P.ら、Nucleic Acid Res 1991;19(25):7225−9)。したがって、低下した活性を持つtatタンパク質は、TAR要素を介したプロモーターのトランス活性化の減少を示す。
【0023】
Nefタンパク質は、CD4の細胞表面ダウンレギュレーションを誘発することから、疾患進行の原因となるウイルス複製にとって不可欠である(Lou Tら、J Biomed Sci 1997;4(4):132)。このダウンレギュレーションは、CD4とNefの間の直接的相互作用によって開始される(Preusser A.ら、Biochem Biophys Res Commun 2002;292(3):734−40)。したがって、低下した機能を持つNefタンパク質は、CD4との相互作用の減少を示す。
【0024】
Revの関連する機能は、ウイルスRNAのRev応答要素(RRE)との相互作用によって開始される転写後トランス活性化である(Iwaiら、1992;Nuceic Acids Res 1992;20(24):6465−72)。したがって、低下した活性を持つRevは、RREとの相互作用の減少を示す。
【0025】
本発明の融合タンパク質は、Vif、Vpr、Vpu、Rev、Vpx、TatおよびNefから選択される少なくとも4種類のHIVタンパク質のアミノ酸配列を含む。本融合タンパク質は、好ましくは、前記HIVタンパク質のうち、5つ、6つまたはすべてを含みうる。本融合タンパク質におけるHIVタンパク質の順序は決定的な問題ではない。
【0026】
上記少なくとも4種類のHIVタンパク質のうちの1つ以上が、本融合タンパク質中に2コピー以上含まれていてもよい。したがって例えば、本発明の融合タンパク質は、Vif、Vpr、Vpuおよび2コピーのRevを含みうる。それら2コピー以上のHIVタンパク質のアミノ酸配列は同一でありうる。もう1つの選択肢として、それらのコピーのアミノ酸配列は異なっていてもよく、特に、異なるHIV株またはHIVクレードに由来するタンパク質配列(例えば1コピーのHIV−1 Revと1コピーのHIV−2 Rev)を使用する場合には異なっていてもよい。
【0027】
融合タンパク質中の隣り合うHIVタンパク質は、アミノ酸を追加せずに融合するか、または融合タンパク質中の隣り合う2つのHIVタンパク質が少なくとも1つの追加アミノ酸で隔てられるような形で融合することができる。両者の組合せも本発明の範囲に包含される。例えば、4つのHIVタンパク質のアミノ酸配列を含む本発明の融合タンパク質では、隣り合う2つのHIVタンパク質を互いに直接連結し、3番目と4番目のHIVタンパク質は追加アミノ酸を介して連結することができる。この態様に関して「追加アミノ酸」という用語は、天然HIVタンパク質のその位置には見いだされないアミノ酸を表す。
【0028】
したがって本発明の融合タンパク質は、好ましくは、以下の一般式:
+P1−−−P2−−−P3−−−P4−−−P5*−−−P6*−−−P7*+
[式中、P1〜P7は、Vif、Vpr、Vpx、Vpu、Tat、RevおよびNefから選択される異なるHIVタンパク質を表す]
を持ち、本融合タンパク質は、少なくとも4種類の前記HIVタンパク質、すなわちP1〜P4と、要すればさらに1つ(P5*)、2つ(P5*−−−P6*)または3つ(P5*−−−P6*−−−P7*)追加の前記HIVタンパク質を含む。略号「−−−」は、独立して、0個またはn個の追加アミノ酸を意味する。「−−−」が0個のアミノ酸を意味する場合、隣り合うHIVタンパク質は、アミノ酸を追加せずに互いに直接融合される。「−−−」が1〜n個のアミノ酸を意味する場合、隣り合うHIVタンパク質は1〜n個のアミノ酸によって隔てられる。追加アミノ酸数の上限、すなわち整数nは、細胞中で産生または発現されうる融合タンパク質の最大サイズに依存する。
【0029】
ある態様では、すべての「−−−」が独立して0〜20個、より好ましくは0〜10個、さらに好ましくは0〜5個の追加アミノ酸を意味する。
【0030】
もう1つの態様では、「−−−」の少なくとも1つが、Vif、Vpr、Vpx、Vpu、Rev、TatおよびNefから選択されるHIVタンパク質ではない追加タンパク質またはその一部のアミノ酸配列を意味する。したがって、このもう1つの態様では、追加タンパク質に、調節/アクセサリーHIVタンパク質が隣接している。追加タンパク質はどのタンパク質であってもよい。追加タンパク質は、より好ましくは、HIVに対してより良い免疫応答を誘発するのに役立ちうる追加エピトープを含む。したがって、追加タンパク質は、HIV Env、Gagおよび(または)Polタンパク質またはその一部でありうる。この場合、Env、GagおよびPolの「一部」という用語は、前記タンパク質の1つに由来するアミノ酸ストレッチであって、少なくとも1つのエピトープを含むものを表す。一部という用語は、より好ましくは、前記タンパク質の1つに由来する少なくとも10個、さらに好ましくは少なくとも20個、最も好ましくは少なくとも50個のアミノ酸を表す。これに関連する態様では、「−−−」のうち少なくとも1つが、本融合タンパク質の一部であるタンパク質P1〜P7の1種以上のアミノ酸配列を意味する。したがってこの場合、本融合タンパク質は、本融合タンパク質の一部であるタンパク質のうちの1つ以上について、1コピー以上を含みうる。上述のように、それらのタンパク質コピーは、同じアミノ酸配列を持っていてもよいし、同じアミノ酸配列を持っていなくてもよい。
【0031】
上記の式において、略号「+」は、独立して、0〜n個の追加末端アミノ酸を意味する。したがって、本発明の融合タンパク質は、そのタンパク質のC末端および(または)N末端に追加アミノ酸を含んでもよいし、含まなくてもよい。ある態様では、「+」の少なくとも一方が、Vif、Vpr、Vpx、Vpu、Rev、TatおよびNefから選択されるHIVタンパク質ではない追加タンパク質のアミノ酸配列またはその一部を意味する。したがって、この態様では、本融合タンパク質は、そのC末端および(または)N末端に、Vif、Vpr、Vpx、Vpu、Rev、TatまたはNefではない追加タンパク質を含む。追加タンパク質はどのタンパク質であってもよい。追加タンパク質は、より好ましくは、HIVに対してより良い免疫応答を誘発するのに役立ちうる追加エピトープを含む。例えば、追加タンパク質は、HIV Env、Gagおよび(または)Polタンパク質またはその一部でありうる。この場合、Env、GagおよびPolの「一部」という用語は、前記タンパク質の1つに由来するアミノ酸ストレッチであって、少なくとも1つのエピトープを含むものを表す。一部という用語は、より好ましくは、前記タンパク質の1つに由来する少なくとも10個、さらに好ましくは少なくとも20個、最も好ましくは少なくとも50個のアミノ酸を表す。
【0032】
もう1つの態様では、「+」の少なくとも一方が、融合タンパク質の検出または精製を容易にするアミノ酸配列を意味する。したがって、「+」の少なくとも一方は、例えばHisタグなどのタグであるかもしれない。
【0033】
本発明によれば、本融合タンパク質は、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない。より具体的に述べると、本発明の融合タンパク質は、ヒト細胞中で発現させた場合に、天然のN末端およびC末端を持つ個々のHIVタンパク質にプロセシングされない。ヒト細胞中で発現させた場合に、ある融合タンパク質が天然のN末端およびC末端を持つ個々のHIVタンパク質にプロセシングされるかどうかを調べる方法は、当業者に知られている。これについては、Ayyavooら、AIDS 2000,14,1−9、特に同文献の図2に記載の実験を参照されたい。簡単に述べると、当業者は各融合タンパク質をHeLa細胞などのヒト細胞中で容易に発現させることができるだろう。次に、細胞を溶解し、その細胞溶解物を、集合して各HIV融合タンパク質を形成している個々のHIVタンパク質に特異的な抗体によるウェスタンブロット実験または免疫沈降アッセイにかける。本発明の融合タンパク質の場合、個々のHIV調節/アクセサリータンパク質のサイズに一致するサイズを持つ有意な量のHIVタンパク質は検出されない。
【0034】
本発明の融合タンパク質が天然のN末端およびC末端を持つ個々のHIVタンパク質にプロセシングされないことを保証するために、融合タンパク質は、天然のN末端およびC末端を持つHIVタンパク質の生成を誘発するかもしれない細胞性プロテアーゼの特異的切断配列を、融合タンパク質を構成するHIVタンパク質のアミノ酸配列間に含んではならない。したがって、上記の一般式で略記したアミノ酸配列「−−−」は、天然のN末端およびC末端を持つHIVタンパク質の生成を誘発するかもしれない細胞性プロテアーゼの特異的切断配列を含有しない。特に、本融合タンパク質は、融合タンパク質を構成する異なるHIVタンパク質のアミノ酸配列間に切断配列REKRAVVG(アミノ酸の一文字表記)を含有しない。当業者には細胞性プロテアーゼの切断配列が他にも知られている。したがって、当業者は、天然のN末端およびC末端を持つ個々のHIVタンパク質をもたらすかもしれない(細胞性)プロテアーゼの切断部位が含まれることを、容易に防ぐことができる。システインプロテアーゼの切断配列の一例は、Ile/leu−X−Thr−X−Glyである。
【0035】
本発明のタンパク質は、天然N末端と天然C末端の両方を持つHIVタンパク質をもたらす特異的切断配列を含まない。しかしこれは、当該切断部位が天然N末端と天然C末端の両方を持つHIVタンパク質の生成を媒介するのでない限り、融合タンパク質中のタンパク質間に細胞性プロテアーゼの切断部位が存在することを、広く排除しているわけではない。特に、上記の一般式で略記したアミノ酸配列「−−−」は、MHCIまたはMHCII上に提示される短いペプチドの生成に関与するプロテアーゼの切断部位を含みうる。この態様では、切断反応の結果は、好ましくは20アミノ酸未満の短いペプチドストレッチであり、そのN末端またはC末端は、HIVアクセサリー/調節タンパク質の1つのN末端またはC末端と一致してもよい。ただし、これらの短いペプチドが抗原提示の過程で生成する場合、これらの短いペプチドは、それらが由来するHIVタンパク質の活性を、もはや持っていない。
【0036】
また、本発明は、上に定義した本発明の融合タンパク質をコードする核酸に関する。この核酸はDNAでもRNAでもよい。プラスミドまたはDNAウイルスに基づくベクターなどのDNAベクターを使って核酸をヒト細胞中に挿入しようとする場合、核酸はDNAであることが好ましい。
【0037】
本発明の融合タンパク質をコードする核酸を構築する方法は、当業者に知られている。以下の方法に限定されるわけではないが、当業者は、ゲノムHIVクローンから、サブゲノムHIVクローンから、または調節/アクセサリーHIVタンパク質の1種以上のコード配列を含む任意の出発物(例えばプラスミド)から、出発することができる。調節/アクセサリータンパク質のコード配列が連続した読み枠の形をとっている場合は、適当な制限酵素で当該コード配列を含む核酸を切断することにより、そのコード配列を単離することができる。このようにして得たDNA断片は、さらなるクローニングに使用することができる。もう1つの選択肢として、アクセサリー/調節タンパク質のコード配列は、適当なプライマーとポリメラーゼ連鎖反応(PCR)法とを使って得ることもできる。調節/アクセサリータンパク質が、例えばTatおよびRevがそうであるように、2つ以上のエクソンによってコードされている場合は、異なるエクソンを別個にクローニングし、それらを融合して、調節/アクセサリータンパク質の連続した読み枠を生成させるか、またはRT−PCRなどの逆転写技術を用いる必要がありうる。
【0038】
コード配列は遺伝子合成によって、すなわち一組の相補的なかつ/またはオーバーラップしたオリゴヌクレオチドを使って遺伝子を生成させることによって得ることもできる。
【0039】
融合タンパク質を得るために、当該融合タンパク質をコードする核酸は連続した読み枠を持つことが好ましい。したがって、HIVタンパク質または追加タンパク質をコードする配列の停止コドンは、最後の配列を除くすべての配列について、アミノ酸をコードするコドンに突然変異させるか、完全に欠失させることが好ましい。これは、好ましくは、停止コドンを含まないコード配列を増幅させる特異的プライマーをPCRに使用すると、容易に達成することができる。言い換えると、この選択肢では、下流プライマーが停止コドンに相補的であってはならない。したがって、増幅されるDNA断片は停止コドンを含まないことになり、これをクローニングベクターにクローニングすることができる。もう1つの選択肢として、停止コドンを持つコード配列をクローニングベクターにクローニングすることもできる。この停止コドンは後に、例えば特異的エンドヌクレアーゼを使って、または突然変異誘発によって除去することができる。
【0040】
これらのクローニング工程により、本発明の融合タンパク質をコードする連続した読み枠がもたらされるはずである。
【0041】
融合タンパク質を発現させるのに必要な調節要素は、所望する発現系での発現を駆動する調節要素であれば何でもよい。大腸菌などの原核細胞中で融合タンパク質を生成させようとする場合は、細菌プロモーターまたはファージプロモーターを使用することが好ましい。真核細胞中で融合タンパク質を発現させようとする場合は、真核またはウイルスプロモーター/エンハンサーを使用することが好ましい。ポックスウイルスプロモーターを使って融合タンパク質を発現させようとする場合(下記参照)は、7.5プロモーターまたはATIプロモーターなどのポックスウイルスプロモーターを使用することが好ましい。
【0042】
上述のように、本融合タンパク質は、Vif、Vpr、Vpx、Vpu、Tat、RevおよびNefから選択されるHIVタンパク質ではない融合パートナーを含みうる。したがって、本融合タンパク質は、他のタンパク質のアミノ酸配列またはその一部を含みうる。他のタンパク質の例は、HIV Gag、PolおよびEnvタンパク質である。したがって、本発明の核酸は、少なくとも4つの調節/アクセサリーHIVタンパク質またはその誘導体をコードするオープンリーディングフレーム中に、1種以上の追加タンパク質のコード配列またはその一部も含みうる。
【0043】
本発明のさらにもう1つの態様では、本核酸はさらに、HIVに対する免疫応答をより一層改善するのに役立ちうる追加タンパク質をコードする独立した発現カセットも含みうる。好ましい一態様として、本核酸はさらに、Gag、PolおよびEnvから選択される少なくとも1つの追加HIVタンパク質のコード配列またはその一部を含む発現カセットを含みうる。さらに好ましくは、本核酸は、融合タンパク質のコード配列に加えて、HIVタンパク質Gag、PolおよびEnvのすべてのコード配列を含みうる。本核酸は、好ましくは、ベクターの一部である。本核酸は、ウイルスベクター(好ましくはMVAなどのポックスウイルスベクター)のウイルスゲノムまたはその一部であることもできる。したがって、ポックスウイルスベクターから本融合タンパク質および追加HIVタンパク質(例えばGag、PolおよびEnvから選択される少なくとも1つの追加HIVタンパク質)を発現させることができる。
【0044】
さらに本発明は本発明の核酸を含むベクターに関する。「ベクター」という用語は、当業者に知られる任意のベクターを表す。ベクターは、pBR322またはpUC系ベクターなどのプラスミドベクターであることができる。ベクターは、より好ましくは、ウイルスベクターである。本発明に関して「ウイルスベクター」という用語は、ウイルスゲノムを含む感染性および(または)弱毒化ウイルスを表す。この場合、本発明の核酸は、各ウイルスベクターのウイルスゲノムの一部であり、かつ/またはウイルスゲノムを構成している。本組換えベクターは、細胞および細胞株の感染、特にヒトを含む生きている動物の感染に使用することができる。典型的な本発明のウイルスベクターはアデノウイルスベクター、レトロウイルスベクターまたはアデノ関連ウイルス2(AAV2)に基づくベクターである。ポックスウイルスベクターが最も好ましい。ポックスウイルスとして、好ましくは、カナリア痘瘡ウイルス、鶏痘ウイルスまたはワクシニアウイルスを挙げることができる。変異ワクシニアウイルスアンカラ(modified vaccinia virus Ankara:MVA)(Sutter,G.ら[1994]Vaccine 12:1032−40)が、さらに好ましい。典型的なMVA株は、ヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(European Collection of Animal Cell Cultures)に受託番号ECACC V00120707として寄託されているMVA575である。2001年11月22日に「Modified Vaccinia Ankara Virus Variant」という名称で欧州特許庁に出願されたPCT出願PCT/EP01/13628に記載されているMVA−BNまたはその誘導体が、最も好ましい。MVA−BNはヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズに受託番号ECACC V00083008として寄託されている。MVA−BNウイルスベクターは変異ワクシニアアンカラに由来する著しく弱毒化されたウイルスであり、ヒト細胞で増殖的に複製する能力を持たないことを特徴とするので、MVA−BNまたはその誘導体を用いることにより、HIVに対して特定の安全なウイルスワクチンを提供するというもう1つの技術的課題が解決された。MVA−BNはヒトでは複製しないので他のどの既知ワクシニアウイルス株よりも安全である。好ましい一態様では、本発明は、本発明のDNAを含むウイルスベクターとして、MVA−BNおよびMVA−BNの誘導体に関する。MVA−BNの特徴、あるMVAがMVA−BNまたはその誘導体であるかどうかを調べることを可能にする生物学的アッセイの説明、およびMVA−BNまたはその誘導体の取得を可能にする方法は、上記のPCT出願PCT/EP01/13628に開示されており、この文献は参照により本明細書に組み込まれる。
【0045】
したがってこれらの態様では、本発明は、好ましくは、本発明の融合タンパク質をコードする発現カセットをウイルスゲノム中に含む組換えMVA(例えばMVA−BN)に関する。
【0046】
本発明の核酸をウイルスゲノムに挿入する方法、および組換えウイルスを取得する方法は、当業者に知られている。
【0047】
組換えワクシニアウイルスにおいて、本発明DNAの発現は、好ましくはポックスウイルスプロモーターの転写制御、より好ましくはワクシニアウイルスプロモーターの転写制御を受けるが、これに限るわけではない。本発明のDNAは、好ましくは、ウイルスゲノムの非必須領域に挿入される。本発明のもう1つの好ましい態様では、異種核酸配列を、ポックスウイルスゲノムの天然の欠失部位(PCT/EP96/02926に記載されているもの)に挿入する。しかし、挿入部位の性質は、組換えワクシニアウイルスが得られる限り、本発明にとって決定的な問題ではない。したがって当業者は他の適切な挿入部位を容易に思いつくことができる。
【0048】
ウイルスベクター、特にポックスウイルスベクターは、好ましくは、本発明の融合タンパク質のコード配列に加えて、HIV Gag、PolおよびEnv遺伝子から選択される追加のレトロウイルス遺伝子を、ウイルスゲノム中に含みうる。ウイルスベクター、特にポックスウイルスベクターは、より好ましくは、本発明の融合タンパク質のコード配列に加えて、Gag、PolおよびEnvをコードするHIV遺伝子をすべて含みうる。本発明では同じ核酸を使ってこれらの追加遺伝子を挿入することができる。この態様では、すべてのHIV遺伝子が、ウイルスゲノムの同じ挿入部位に位置することになる。もう1つの態様では、追加遺伝子はウイルスゲノムの異なる位置に挿入される。
【0049】
好ましい一態様として、本発明は、HIV感染およびAIDSを少なくとも部分的に予防するためのワクチンとしての、本発明の核酸、ベクターまたは融合タンパク質に関する。「ワクチン」は、特異的免疫応答を誘導する化合物、すなわち核酸、融合タンパク質、ベクターまたはウイルスである。
【0050】
この態様の一選択肢として、本発明の「ワクチン」は、本発明の融合タンパク質に基づく。
【0051】
好ましい一態様では、本発明の核酸、特にDNAを、ワクチンとして使用する。本発明の場合と同様に真核発現カセットを含んでいる裸のDNAを投与すると(特にDNAを筋肉内投与すると)、その発現カセットによってコードされているタンパク質の発現が起こることが、当業者に知られている。そのタンパク質は免疫系にばく露され、特異的免疫応答を生じさせる。もう1つの態様では、本発明のベクター、特にウイルスベクター、より好ましくはポックスウイルスベクター、最も好ましくはMVAベクターなどのワクシニアウイルスベクターを投与することによって、予防接種を行う。
【0052】
ワクシニアウイルス系ワクチンを製造するには、本発明のウイルスを生理学的に許容できる形態に変換する。これは、天然痘の予防接種に用いられるポックスウイルスワクチン(Stickl,H.ら[1974]Dtsch.med.Wschr.99,2386−2392に記載されているもの)を製造する際の経験に基づいて行うことができる。例えば、精製ウイルスは、5×108 TCID50/mlの力価で約10mMトリスおよび140mM NaCl(pH7.4)中に製剤化して、−80℃で保存する。ワクチン注射剤を製造するには、例えば、2%ペプトンおよび1%ヒトアルブミンの存在下で、リン酸緩衝食塩水(PBS)100mL中のウイルス粒子102〜108個を、アンプル(好ましくはガラスアンプル)中で凍結乾燥する。もう1つの選択肢として、ワクチン注射剤は、製剤中のウイルスの段階的凍結乾燥によって製造することもできる。この製剤は、さらに、例えばマンニトール、デキストラン、糖、グリシン、ラクトースもしくはポリビニルピロリドンなどの添加剤、または、生体内投与に適した酸化防止剤もしくは不活性ガス、安定剤または組換えタンパク質(例えばヒト血清アルブミン)などの他の添加剤も含むことができる。その場合、このガラスアンプルを封印して、4℃〜室温で数ヶ月間保存することができる。しかし、必要がない限り、アンプルは−20℃未満の温度で保存することが好ましい。予防接種を行うには、上記の凍結乾燥物を0.1〜0.5mlの水性溶液、好ましくは生理食塩水またはトリス緩衝液に溶解し、それを全身的または局所的に、すなわち非経口投与、筋肉内投与、または当業者に知られる他の任意の投与経路によって、投与することができる。当業者であれば、投与様式、投与量および投与回数を、既知の方法で最適化することができる。ポックスウイルスベクターの場合は、皮下投与または筋肉内投与が最も好ましい。
【0053】
ワクチンが本発明のDNAを含むMVA−BNベクターまたはその誘導体である場合、本発明の一特定態様は、第1接種(「初回免疫接種」)および第2接種(「追加免疫接種」)で、治療有効量の当該ワクチンを投与することに関する。
【0054】
ワクチンが本発明のDNAを含むMVA−BNベクターまたはその誘導体である場合、本発明の一特定態様は、第1バイアル/容器に入った第1予防接種(「初回免疫」)用の本発明MVA−BNウイルスベクターと、第2バイアル/容器に入った第2予防接種(「追加免疫」)用の本発明MVA−BNウイルスベクターとを含む予防接種用キットに関する。したがって本発明は、ワクチン態様として、本発明の核酸、ベクターまたは融合タンパク質を含むワクチン、およびワクチンの製造を目的とする前記核酸、ベクターまたはタンパク質の使用に関する。
【0055】
さらにもう1つの態様として、本発明は、本発明の融合タンパク質、本発明の核酸、または本発明のベクターを保護を必要とする、ヒトを含む動物に投与することによって、ヒトを含む動物をHIV感染から保護する方法に関する。
【0056】
さらに本発明は、本発明のタンパク質を製造する方法であって、(i)本発明の核酸またはベクターを宿主細胞にトランスフェクトする工程、または(ii)宿主細胞に本発明のウイルスベクターを感染させる工程、(iii)工程(i)のトランスフェクト宿主細胞または工程(ii)の感染宿主細胞中で融合タンパク質を発現させる工程、および(iv)融合タンパク質を回収する工程を含む方法に関する。
【0057】
さらに本発明は、本発明の核酸またはベクターをトランスフェクトした宿主細胞、または本発明のウイルスベクターに感染した宿主細胞に関する。
【0058】
もう1つの態様として、本融合タンパク質は、Vif、Vpr、Vpu、Rev、VpxおよびTatから選択される少なくとも3種類のHIVタンパク質を含みうる。本融合タンパク質は、好ましくは、前記HIVタンパク質の4個、5個またはすべてを含みうる。この態様の融合タンパク質の典型例は、HIVタンパク質Vpr、Vif、Vpu、RevおよびTatのアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む。上述のように、融合タンパク質中のHIVタンパク質の順序は決定的な問題ではない。上述した好ましい態様はすべてこの代替態様にも当てはまる。
【0059】
図面の簡単な説明
図1:図1は、オリゴヌクレオチドのアニーリングを表す略図である。この図は4つのオリゴヌクレオチドのアニーリングを示している。各オリゴヌクレオチドは一本鎖であり、相補的末端によってアニールすることができる。プルーフリーディング活性を示すポリメラーゼ(例えばPfxポリメラーゼ)を使ってギャップを埋める。
【0060】
図2:図2は、ブロブ(blob)の4つの遺伝子のアニーリングを表す略図である。vif遺伝子はvpr断片とのオーバーラップ配列を示し、vpuコード断片はrev遺伝子とのオーバーラップ配列を示す(灰色の部分)。PCR断片を変性し、オーバーラップする相補的末端をハイブリダイズさせる。その結果生じたギャップをPfxポリメラーゼで埋める。vif−vpr断片をvpu−rev断片のオーバーラップ配列に融合し、それを再び融合に使用する。
【0061】
図3:図3は、MVAゲノムに外来遺伝子を挿入するための組換えベクターに、本発明の融合タンパク質をコードする配列をクローニングするための戦略である。融合したvif、vpr、vpuおよびrevポリタンパク質コード領域を、ClaIおよびApaI制限酵素部位を含むプライマーを使って増幅した。このpCR産物を、ポックスウイルスATIプロモーターを持つベクターpBNX65のClaI/ApaI切断物にクローニングした。Acc65I制限酵素部位を含むプライマーを使ったPCRによってtatコード領域を増幅し、Acc65Iで線状化したpBNX65+vif−revにライゲートした。得られた発現カセット(ATIプロモーター+本発明の融合タンパク質をコードする配列)をPacI制限酵素消化によって単離し、MVAゲノムI4L遺伝子間領域に外来遺伝子を挿入するための組換えベクター(pBNX39)に挿入した。pBNX39は、MVAゲノムの挿入部位の隣接配列に相同な配列(F1 I4LおよびF2 I4L)を持っている。MVAゲノムとpBNX39との相同組換え後に組換えウイルスを選択することができるように、このベクターは大腸菌gpt遺伝子(ホスホリボシルトランスフェラーゼ遺伝子)も持っている。この選択カセットは、組換えウイルスの精製後に、Flank1とflank1の反復配列(F1rpt)との間で起こる相同組換えによって欠失する。
【0062】
図4:図4は、MVAゲノムの概略図である。MVAは、HindIIIによる制限酵素消化後に特徴的な断片(A〜O)を示す線状ゲノムを持っている。I4L遺伝子とI5L遺伝子の間の非機能領域はI断片中に位置している。pBNX39を使った外来遺伝子の挿入は56767〜56768位で起こる。
【実施例】
【0063】
(HIV Vif−Vpr−Vpu−Rev−Tat融合タンパク質をコードするDNAの作製)
HIVゲノムの単一遺伝子は、標準的PCRプロトコールを用いるPCRにより、ゲノムDNAから調製するか、またはオーバーラップ配列を介したオリゴヌクレオチドのアニーリングと、その結果生じた一本鎖ギャップの充填とに基づく技術によって合成的に調製した。
【0064】
本発明の融合タンパク質をコードする核酸に挿入しようとする遺伝子のコード領域をオリゴヌクレオチドに基づいて作製するために、15bpのオーバーラップを持つ40マーのオリゴヌクレオチドを設計した。これらのオリゴヌクレオチドの配列は、IIIB株に由来するHIV1分離株HXB2Rのゲノム地図に基づいている。アニーリング反応用のオリゴヌクレオチド、または必要な配列を単離するためのPCR用のオリゴヌクレオチドは、結果として得られるコード領域から翻訳終結用の停止コドンが欠失するように設計した。tat遺伝子は停止コドンを含むオリゴを使って合成した。というのも、この遺伝子は本発明の融合タンパク質をコードする核酸の最後に挿入されるものなので、ポリタンパク質の翻訳が正しく終結するように停止トリプレットを含むべきだからである。
【0065】
オリゴアニーリング反応については、10サイクルの2段階Pfxポリメラーゼ(Gibco−BRL)反応(95℃で変性および68℃でアニーニング/伸長)を行った。この反応中に、オリゴのオーバーラップ配列はアニールした状態になり、ギャップがPfxプルーフリーディングポリメラーゼによって充填される(図1)。融合タンパク質をコードするヌクレオチド配列中の第1コード遺伝子であるvifコード領域を合成するために、ゲノムHIV cDNAを使ったPCRを行った。このPCRは、続いて行うvifとvprのアニーリングに備えて、vifコード領域が次のvpr遺伝子の最初の15bpに融合されるように行った。HIV HXB2Rゲノムのbp5559−5847に広がるVprコード領域は、10個のヌクレオチドのアニーリングによって調製した。生じたギャップを充填し、増幅のためのPCRを行った後、vif用の隣接領域と、vprコード領域の後ろに挿入する予定であるvpu用の隣接領域とに融合したvprコード領域を含む生成物を得た。
【0066】
vifの合成に使用したものと同じcDNAから、PCRによってVpuコード領域を増幅することにより、vprおよびrevと融合させるための隣接領域を含む生成物を得た。
【0067】
revコード領域は、HIV HXB2Rゲノムのbp5970−6045および8379−8650の領域を覆うと共に、vpuおよびtatとのアニーリング用の15bpオーバーラップを持つ14個のオリゴヌクレオチドのアニーリングによって合成した。
【0068】
tatコード領域は、HIV HXB2Rゲノムのbp5831−6045および8379−8466を覆う10個のオリゴヌクレオチドを使って作製した。
【0069】
vifおよびvprコード領域ならびにvpuおよびrevコード領域は、それら2つの断片をそれぞれのオーバーラップを使ってアニールさせることにより、2段階Pfxポリメラーゼ反応で融合させた(図2)。その融合生成物をさらにPCR増幅した後、断片を精製し、vprおよびvpuのオーバーラップを使って互いにライゲートした(図2)。得られた生成物(vif−vpr−vpu−revのコード配列)をPCR増幅した後、ポックスウイルスATIプロモーターを含むpBluescriptKS+ベクター(図3、pBNX65)中の隣り合うクローニング部位に、vif−vpr−vpu−rev断片とtatとをクローニングすることにより、tatコード領域を融合した。次に、PacI制限酵素消化によって発現カセット全体を単離し、pBNX39(図3)に挿入した。pBNX39は、MVAゲノムに相同な配列を含んでおり、それが相同組換えによるゲノム(図4)の非コード領域(I4L)への挿入を可能にする。
【図面の簡単な説明】
【0070】
【図1】オリゴヌクレオチドのアニーリングを表す略図である。
【図2】ブロブ(blob)の4つの遺伝子のアニーリングを表す略図である。
【図3】MVAゲノムに外来遺伝子を挿入するための組換えベクターに、本発明の融合タンパク質をコードする配列をクローニングするための戦略図である。
【図4】MVAゲノムの概略図である。
【特許請求の範囲】
【請求項1】
Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4つのHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない融合タンパク質。
【請求項2】
HIVタンパク質がVif、Vpr、Vpx、Vpu、RevおよびTatから選択される、請求項1記載の融合タンパク質。
【請求項3】
HIVタンパク質Vif、Vpr、Vpu、RevおよびTatのアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む、請求項1又は2記載の融合タンパク質。
【請求項4】
少なくとも2つのHIVタンパク質のアミノ酸配列が追加アミノ酸なしで互いに融合される、請求項1〜3のいずれか1つに記載の融合タンパク質。
【請求項5】
少なくとも2つのHIVタンパク質のアミノ酸配列が少なくとも1つの追加アミノ酸によって隔てられる、請求項1〜4のいずれか1つに記載の融合タンパク質。
【請求項6】
少なくとも1つのHIVタンパク質のアミノ酸配列が、Vif、Vpr、Vpx、Vpu、Rev、TatおよびNefから選択されるHIVタンパク質ではない融合パートナーに融合される、請求項1〜5のいずれか1つに記載の融合タンパク質。
【請求項7】
請求項1〜6のいずれか1つに記載の融合タンパク質をコードする核酸。
【請求項8】
核酸がDNAである、請求項7記載の核酸。
【請求項9】
DNAからの融合タンパク質の発現が、真核プロモーター、原核プロモーターおよびウイルスプロモーターから選択される調節要素によって制御される、請求項8記載の核酸。
【請求項10】
ウイルスプロモーターがポックスウイルスプロモーターである、請求項9記載の核酸。
【請求項11】
Gag、PolおよびEnvから選択される少なくとも1つの追加HIVタンパク質のコード配列をさらに含む、請求項7〜10のいずれか1つに記載の核酸。
【請求項12】
HIV Gag、PolおよびEnvタンパク質のコード配列を含む、請求項11記載の核酸。
【請求項13】
請求項7〜12のいずれか1つに記載の核酸を含むベクター。
【請求項14】
ベクターがウイルスベクターである、請求項13記載のベクター。
【請求項15】
ウイルスベクターがポックスウイルスベクター、特にワクシニアウイルスベクターである、請求項14記載のベクター。
【請求項16】
ワクシニアウイルスベクターが変異ワクシニアウイルスアンカラ(MVA)である、請求項15記載のベクター。
【請求項17】
MVAがヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(ECACC)に受託番号V00120707として寄託されているMVA−575、およびECACCに受託番号V00083008として寄託されているMVA−BNから選択される、請求項16記載のベクター。
【請求項18】
請求項1〜6のいずれか1つに記載のタンパク質を製造する方法であって、
請求項7〜12のいずれか1つに記載の核酸もしくは請求項13記載のベクターを宿主細胞にトランスフェクトする工程、または
宿主細胞に請求項14〜17のいずれか1つに記載のウイルスベクターを感染させる工程と、
トランスフェクトした宿主細胞または感染させた宿主細胞中で、融合タンパク質を発現させる工程と、
融合タンパク質を回収する工程と、
を含む、上記方法。
【請求項19】
請求項7〜12のいずれか1つに記載の核酸もしくは請求項13に記載のベクターをトランスフェクトするか、または請求項14〜17のいずれか1つに記載のウイルスベクターに感染させた宿主細胞。
【請求項20】
医薬としての、請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクター。
【請求項21】
ワクチンとしての、請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクター。
【請求項22】
請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクターを含むワクチン。
【請求項23】
ワクチンの製造を目的とする、請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクターの使用方法。
【請求項24】
ヒトを含む動物をHIV感染から保護する方法であって、この保護を必要とする、ヒトを含む動物に、請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクターを投与することによってHIV感染から保護する上記方法。
【特許請求の範囲】
【請求項1】
Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4つのHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、当該融合タンパク質は天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされず、かつHIVタンパク質のアミノ酸配列の誘導体は、融合タンパク質中のアミノ酸配列の対応部分を既知HIV単離株の各HIVタンパク質のアミノ酸配列と比較した場合に、少なくとも50%の相同性を示すアミノ酸配列である、融合タンパク質。
【請求項2】
前記相同性が少なくとも80%である、請求項1記載の融合タンパク質。
【請求項3】
アミノ酸配列の誘導体では、低下した活性を持つかまたは活性を全く持たないHIVタンパク質が得られるように、既知HIV単離株の各HIVタンパク質のアミノ酸配列と比較して、10個を超えないアミノ酸配列が欠失しているか、または挿入もしくは置換されている、請求項1に記載の融合タンパク質。
【請求項4】
HIVタンパク質がVif、Vpr、Vpx、Vpu、RevおよびTatから選択される、請求項1〜3のいずれか1つに記載の融合タンパク質。
【請求項5】
HIVタンパク質Vif、Vpr、Vpu、RevおよびTatのアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む、請求項1〜4のいずれか1つに記載の融合タンパク質。
【請求項6】
少なくとも2つのHIVタンパク質のアミノ酸配列が追加アミノ酸なしで互いに融合される、請求項1〜5のいずれか1つに記載の融合タンパク質。
【請求項7】
少なくとも2つのHIVタンパク質のアミノ酸配列が少なくとも1つの追加アミノ酸によって隔てられる、請求項1〜6のいずれか1つに記載の融合タンパク質。
【請求項8】
少なくとも1つのHIVタンパク質のアミノ酸配列が、Vif、Vpr、Vpx、Vpu、Rev、TatおよびNefから選択されるHIVタンパク質ではない融合パートナーに融合される、請求項1〜7のいずれか1つに記載の融合タンパク質。
【請求項9】
請求項1〜8のいずれか1つに記載の融合タンパク質をコードする核酸。
【請求項10】
核酸がDNAである、請求項9記載の核酸。
【請求項11】
DNAからの融合タンパク質の発現が、真核プロモーター、原核プロモーターおよびウイルスプロモーターから選択される調節要素によって制御される、請求項10記載の核酸。
【請求項12】
ウイルスプロモーターがポックスウイルスプロモーターである、請求項11記載の核酸。
【請求項13】
Gag、PolおよびEnvから選択される少なくとも1つの追加HIVタンパク質のコード配列をさらに含む、請求項9〜12のいずれか1つに記載の核酸。
【請求項14】
HIV Gag、PolおよびEnvタンパク質のコード配列を含む、請求項13記載の核酸。
【請求項15】
請求項9〜14のいずれか1つに記載の核酸を含むベクター。
【請求項16】
ベクターがウイルスベクターである、請求項15記載のベクター。
【請求項17】
ウイルスベクターがポックスウイルスベクター、特にワクシニアウイルスベクターである、請求項16記載のベクター。
【請求項18】
ワクシニアウイルスベクターが変異ワクシニアウイルスアンカラ(MVA)である、請求項17に記載のベクター。
【請求項19】
MVAがヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(ECACC)に受託番号V00120707として寄託されているMVA−575、およびECACCに受託番号V00083008として寄託されているMVA−BNから選択される、請求項18記載のベクター。
【請求項20】
請求項1〜8のいずれか1つに記載のタンパク質を製造する方法であって、
請求項9〜14のいずれか1つに記載の核酸もしくは請求項15記載のベクターを宿主細胞にトランスフェクトする工程、または
宿主細胞に請求項16〜19のいずれか1つに記載のウイルスベクターを感染させる工程と、
トランスフェクトした宿主細胞または感染させた宿主細胞中で、融合タンパク質を発現させる工程と、
融合タンパク質を回収する工程と、
を含む、上記方法。
【請求項21】
請求項9〜14のいずれか1つに記載の核酸もしくは請求項15記載のベクターをトランスフェクトするか、または請求項16〜19のいずれか1つに記載のウイルスベクターに感染させた宿主細胞。
【請求項22】
医薬としての、請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクター。
【請求項23】
ワクチンとしての、請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクター。
【請求項24】
請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクターを含むワクチン。
【請求項25】
ワクチンの製造を目的とする、請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクターの使用。
【請求項26】
ヒトを含む動物をHIV感染から保護する方法であって、保護を必要とする、ヒトを含む動物に、請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクターを投与することによってHIV感染から保護する方法。
【請求項1】
Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4つのHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされない融合タンパク質。
【請求項2】
HIVタンパク質がVif、Vpr、Vpx、Vpu、RevおよびTatから選択される、請求項1記載の融合タンパク質。
【請求項3】
HIVタンパク質Vif、Vpr、Vpu、RevおよびTatのアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む、請求項1又は2記載の融合タンパク質。
【請求項4】
少なくとも2つのHIVタンパク質のアミノ酸配列が追加アミノ酸なしで互いに融合される、請求項1〜3のいずれか1つに記載の融合タンパク質。
【請求項5】
少なくとも2つのHIVタンパク質のアミノ酸配列が少なくとも1つの追加アミノ酸によって隔てられる、請求項1〜4のいずれか1つに記載の融合タンパク質。
【請求項6】
少なくとも1つのHIVタンパク質のアミノ酸配列が、Vif、Vpr、Vpx、Vpu、Rev、TatおよびNefから選択されるHIVタンパク質ではない融合パートナーに融合される、請求項1〜5のいずれか1つに記載の融合タンパク質。
【請求項7】
請求項1〜6のいずれか1つに記載の融合タンパク質をコードする核酸。
【請求項8】
核酸がDNAである、請求項7記載の核酸。
【請求項9】
DNAからの融合タンパク質の発現が、真核プロモーター、原核プロモーターおよびウイルスプロモーターから選択される調節要素によって制御される、請求項8記載の核酸。
【請求項10】
ウイルスプロモーターがポックスウイルスプロモーターである、請求項9記載の核酸。
【請求項11】
Gag、PolおよびEnvから選択される少なくとも1つの追加HIVタンパク質のコード配列をさらに含む、請求項7〜10のいずれか1つに記載の核酸。
【請求項12】
HIV Gag、PolおよびEnvタンパク質のコード配列を含む、請求項11記載の核酸。
【請求項13】
請求項7〜12のいずれか1つに記載の核酸を含むベクター。
【請求項14】
ベクターがウイルスベクターである、請求項13記載のベクター。
【請求項15】
ウイルスベクターがポックスウイルスベクター、特にワクシニアウイルスベクターである、請求項14記載のベクター。
【請求項16】
ワクシニアウイルスベクターが変異ワクシニアウイルスアンカラ(MVA)である、請求項15記載のベクター。
【請求項17】
MVAがヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(ECACC)に受託番号V00120707として寄託されているMVA−575、およびECACCに受託番号V00083008として寄託されているMVA−BNから選択される、請求項16記載のベクター。
【請求項18】
請求項1〜6のいずれか1つに記載のタンパク質を製造する方法であって、
請求項7〜12のいずれか1つに記載の核酸もしくは請求項13記載のベクターを宿主細胞にトランスフェクトする工程、または
宿主細胞に請求項14〜17のいずれか1つに記載のウイルスベクターを感染させる工程と、
トランスフェクトした宿主細胞または感染させた宿主細胞中で、融合タンパク質を発現させる工程と、
融合タンパク質を回収する工程と、
を含む、上記方法。
【請求項19】
請求項7〜12のいずれか1つに記載の核酸もしくは請求項13に記載のベクターをトランスフェクトするか、または請求項14〜17のいずれか1つに記載のウイルスベクターに感染させた宿主細胞。
【請求項20】
医薬としての、請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクター。
【請求項21】
ワクチンとしての、請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクター。
【請求項22】
請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクターを含むワクチン。
【請求項23】
ワクチンの製造を目的とする、請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクターの使用方法。
【請求項24】
ヒトを含む動物をHIV感染から保護する方法であって、この保護を必要とする、ヒトを含む動物に、請求項1〜6のいずれか1つに記載の融合タンパク質、請求項7〜12のいずれか1つに記載の核酸、または請求項13〜17のいずれか1つに記載のベクターを投与することによってHIV感染から保護する上記方法。
【特許請求の範囲】
【請求項1】
Vif、Vpr、Vpu、Vpx、Rev、TatおよびNefから選択される少なくとも4つのHIVタンパク質のアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む融合タンパク質であって、当該融合タンパク質は天然のN末端およびC末端を持つ個々のHIVタンパク質にはプロセシングされず、かつHIVタンパク質のアミノ酸配列の誘導体は、融合タンパク質中のアミノ酸配列の対応部分を既知HIV単離株の各HIVタンパク質のアミノ酸配列と比較した場合に、少なくとも50%の相同性を示すアミノ酸配列である、融合タンパク質。
【請求項2】
前記相同性が少なくとも80%である、請求項1記載の融合タンパク質。
【請求項3】
アミノ酸配列の誘導体では、低下した活性を持つかまたは活性を全く持たないHIVタンパク質が得られるように、既知HIV単離株の各HIVタンパク質のアミノ酸配列と比較して、10個を超えないアミノ酸配列が欠失しているか、または挿入もしくは置換されている、請求項1に記載の融合タンパク質。
【請求項4】
HIVタンパク質がVif、Vpr、Vpx、Vpu、RevおよびTatから選択される、請求項1〜3のいずれか1つに記載の融合タンパク質。
【請求項5】
HIVタンパク質Vif、Vpr、Vpu、RevおよびTatのアミノ酸配列、または1種以上の前記タンパク質のアミノ酸配列の誘導体を含む、請求項1〜4のいずれか1つに記載の融合タンパク質。
【請求項6】
少なくとも2つのHIVタンパク質のアミノ酸配列が追加アミノ酸なしで互いに融合される、請求項1〜5のいずれか1つに記載の融合タンパク質。
【請求項7】
少なくとも2つのHIVタンパク質のアミノ酸配列が少なくとも1つの追加アミノ酸によって隔てられる、請求項1〜6のいずれか1つに記載の融合タンパク質。
【請求項8】
少なくとも1つのHIVタンパク質のアミノ酸配列が、Vif、Vpr、Vpx、Vpu、Rev、TatおよびNefから選択されるHIVタンパク質ではない融合パートナーに融合される、請求項1〜7のいずれか1つに記載の融合タンパク質。
【請求項9】
請求項1〜8のいずれか1つに記載の融合タンパク質をコードする核酸。
【請求項10】
核酸がDNAである、請求項9記載の核酸。
【請求項11】
DNAからの融合タンパク質の発現が、真核プロモーター、原核プロモーターおよびウイルスプロモーターから選択される調節要素によって制御される、請求項10記載の核酸。
【請求項12】
ウイルスプロモーターがポックスウイルスプロモーターである、請求項11記載の核酸。
【請求項13】
Gag、PolおよびEnvから選択される少なくとも1つの追加HIVタンパク質のコード配列をさらに含む、請求項9〜12のいずれか1つに記載の核酸。
【請求項14】
HIV Gag、PolおよびEnvタンパク質のコード配列を含む、請求項13記載の核酸。
【請求項15】
請求項9〜14のいずれか1つに記載の核酸を含むベクター。
【請求項16】
ベクターがウイルスベクターである、請求項15記載のベクター。
【請求項17】
ウイルスベクターがポックスウイルスベクター、特にワクシニアウイルスベクターである、請求項16記載のベクター。
【請求項18】
ワクシニアウイルスベクターが変異ワクシニアウイルスアンカラ(MVA)である、請求項17に記載のベクター。
【請求項19】
MVAがヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(ECACC)に受託番号V00120707として寄託されているMVA−575、およびECACCに受託番号V00083008として寄託されているMVA−BNから選択される、請求項18記載のベクター。
【請求項20】
請求項1〜8のいずれか1つに記載のタンパク質を製造する方法であって、
請求項9〜14のいずれか1つに記載の核酸もしくは請求項15記載のベクターを宿主細胞にトランスフェクトする工程、または
宿主細胞に請求項16〜19のいずれか1つに記載のウイルスベクターを感染させる工程と、
トランスフェクトした宿主細胞または感染させた宿主細胞中で、融合タンパク質を発現させる工程と、
融合タンパク質を回収する工程と、
を含む、上記方法。
【請求項21】
請求項9〜14のいずれか1つに記載の核酸もしくは請求項15記載のベクターをトランスフェクトするか、または請求項16〜19のいずれか1つに記載のウイルスベクターに感染させた宿主細胞。
【請求項22】
医薬としての、請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクター。
【請求項23】
ワクチンとしての、請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクター。
【請求項24】
請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクターを含むワクチン。
【請求項25】
ワクチンの製造を目的とする、請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクターの使用。
【請求項26】
ヒトを含む動物をHIV感染から保護する方法であって、保護を必要とする、ヒトを含む動物に、請求項1〜8のいずれか1つに記載の融合タンパク質、請求項9〜14のいずれか1つに記載の核酸、または請求項15〜19のいずれか1つに記載のベクターを投与することによってHIV感染から保護する方法。
【図1】


【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公表番号】特表2006−502704(P2006−502704A)
【公表日】平成18年1月26日(2006.1.26)
【国際特許分類】
【出願番号】特願2004−506348(P2004−506348)
【出願日】平成15年5月14日(2003.5.14)
【国際出願番号】PCT/EP2003/005039
【国際公開番号】WO2003/097675
【国際公開日】平成15年11月27日(2003.11.27)
【出願人】(502240076)バヴァリアン・ノルディック・アクティーゼルスカブ (18)
【Fターム(参考)】
【公表日】平成18年1月26日(2006.1.26)
【国際特許分類】
【出願日】平成15年5月14日(2003.5.14)
【国際出願番号】PCT/EP2003/005039
【国際公開番号】WO2003/097675
【国際公開日】平成15年11月27日(2003.11.27)
【出願人】(502240076)バヴァリアン・ノルディック・アクティーゼルスカブ (18)
【Fターム(参考)】
[ Back to top ]