
HLA−DR特異的抗体、組成物、および方法
本出願において、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能である、HLA−DR特異的モノクローナル抗体が、提供される。特定の例示的なHLA−DR特異的抗体は、減少したレベルの免疫抑制活性を示す。本発明の抗体は、HLA−DRを発現する腫瘍細胞に関連する癌の診断法、ならびにHLA−DRを発現する腫瘍細胞に関連する癌の処置のための組成物およびHLA−DRを発現する腫瘍細胞に関連する癌の処置のための方法において、用途を見出す。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願)
本出願は、2003年12月15日に出願した米国仮特許出願番号60/608,944の優先権を主張する。
【0002】
本発明は、一般的には、免疫学の分野および分子生物学の分野に関する。より具体的には、本発明は、HLA−DR特異的抗体、およびHLA−DR特異的抗体を調製するための方法を提供する。HLA−DR特異的抗体を含む組成物および方法もまた、提供される。本明細書中に提示される抗体は、HLA−DR抗原に特異的に結合し、そしてHLA−DRを発現する腫瘍細胞においてアポトーシスを誘導する。特定の実施形態において、アポトーシス性HLA−DR特異的抗体は、低下した免疫抑制活性レベルを示す。本発明の抗体は、診断方法における使用、ならびにHLA−DR陽性腫瘍細胞に関連する癌の処置のための組成物および方法における使用を見出す。
【背景技術】
【0003】
クラスII主要組織適合遺伝子複合体(MHC)分子(正常な抗原提示細胞(APC)上に構成的に発現される)は、CD4+ヘルパーT(Th)細胞に対する抗原由来ペプチドの提示を担う。非特許文献1および非特許文献2。これらの分子を介するシグナル伝達は、活性化Bリンパ球のプログラムされた細胞死(PCDまたはアポトーシス)をもたらすセカンドメッセンジャーの生成を開始する。抗原提示に加えて、クラスII分子は、細胞増殖を調節し得るシグナルを伝達し、特定のクラスII MHC特異的モノクローナル抗体は、癌細胞のアポトーシスを誘導することが示されている(非特許文献3)。
【0004】
HLAクラスII分子は、ヒトBリンパ球上に構成的に発現され、そして活性化後にヒトTリンパ球上で誘導される。細胞死の60%までが、HLA−DR分子を介するリンパ球の刺激後に観察されている。特定のHLA−DR特異的抗体は、クラスII分子の細胞表面発現における90%までの減少を引き起こす。HLA−DR特異的抗体およびそのフラグメントは、HLA−DP分子およびHLA−DQ分子の発現に影響を与えない(非特許文献4)。
【0005】
癌細胞のアポトーシスを誘導することが示されたクラスII MHC特異的抗体はまた、頻繁に、正常なTh細胞機能を妨害する(非特許文献5)。現在利用可能なアポトーシス誘導性クラスII特異的モノクローナル抗体の大多数は、ペプチド結合部位に見かけ上接近した状態にある、HLA−DRへテロダイマーの第一タンパク質ドメイン上に位置するエピトープを認識する。これらの抗体は、抗原提示を妨害し、Th応答の強力なインビトロおよびインビボでの阻害を引き起こし、さらに、Bリンパ芽球様細胞および少ない割合の正常活性化B細胞に対して細胞傷害性である。そのF(ab’)2フラグメントは、ダウンレギュレーションおよび細胞傷害性の両方を媒介し、一方、その一価Fabフラグメントは、細胞傷害性ではないが、ダウンレギュレーション特性およびT細胞阻害特性を保持する(非特許文献5)。
【非特許文献1】Babbittら、Nature(1985年)317:p.359〜361
【非特許文献2】Trumanら、Blood(1997年)89(6):p.1996〜2007
【非特許文献3】Newellら、Proc.Natl.Acad.Sci.U.S.A.(1993年)90(22):p.10459〜10463
【非特許文献4】Trumanら、Int.Immunol.(1994年)6(6):p.887〜896
【非特許文献5】Vidovicら、Eur.J.Immunol.(1995年)25:p.3349〜3355
【発明の開示】
【発明が解決しようとする課題】
【0006】
従って、当該分野で利用可能なほとんどのHLA−DR特異的アポトーシス性モノクローナル抗体は、HLA−DR陽性細胞を含む癌処置用の組成物および方法におけるそれらの抗体の有用性を限定するのに充分な免疫抑制活性を示す。従って、改善されたHLA−DR特異的抗体(例えば、非免疫抑制アポトーシス性HLA−DR特異的抗体)、ならびにこのような抗体を含む、これらの欠陥を克服する組成物および方法に関して、当該分野において必要性が残っている。
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明は、特に、HLA−DR陽性腫瘍細胞においてアポトーシスを誘発可能なHLA−DR特異的抗体およびその抗原結合フラグメントを提供することによって、これらの必要性および他の関連する必要性に取り組む。本発明は、ヒト主要組織適合遺伝子複合体(MHC)クラスIIと特異的に反応する抗体は、表面上にHLA−DR分子を発現する細胞のアポトーシスを誘導し得るという発見に、部分的に基づく。本発明の抗体は、このHLA−DR特異的抗体が、HLA−DRを発現する非腫瘍細胞/非新形成細胞の生存度にも機能にも影響を与えないという点で、非常に特異的である。特定の実施形態において、本発明のHLA−DR抗体のアポトーシス活性は、クラスII非依存性免疫応答を同時に抑制することなく、達成され得る。
【0008】
本発明の重要な実用上の関連性は、「DN1924」と呼ばれるモノクローナル抗体、および「キメラDN1924」と呼ばれるDN1924のマウス/ヒト改変体が、HLAクラスII陽性新形成(血球新形成、例えば、プラズマ細胞腫/多発性骨髄腫、ホジキンリンパ腫および非ホジキンリンパ腫、ならびにB細胞リンパ腫が挙げられるが、これらに限定されない)の選択的抗体ベース療法のために有効であり得ることである。インビトロ研究によって、DN1924モノクローナル抗体およびキメラDN1924モノクローナル抗体は、正常なTh機能を妨害せず、従って、DN1924および/またはキメラDN1924は、被験体の正常なHLA−DR発現細胞に影響を与えないことが、示される。従って、現在利用可能な治療剤よりも副作用が少ないと予測することが、妥当である。
【0009】
従って、特定の局面において、本発明は、非ヒトモノクローナルHLA−DR特異的アポトーシス性抗体を提供する。本発明の非ヒトモノクローナル抗体は、種々の動物種から単離され得る。この動物種としては、非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、鳥類、ウサギ、マウス、ラット、モルモット、ハムスター、イヌ、およびネコなどの起源が挙げられるが、これらに限定されない。HLA−DR特異的アポトーシス性非ヒトモノクローナル抗体は、本明細書において、(1)配列番号13および配列番号2においてそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(これらは、それぞれ、配列番号46のポリヌクレオチドおよび配列番号45のポリヌクレオチドによってコードされる)を含む、免疫抑制抗体DN1921と、(2)配列番号35および配列番号24においてそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(これらは、それぞれ、配列番号48のポリヌクレオチドおよび配列番号47のポリヌクレオチドによってコードされる)を含む、非免疫抑制抗体DN1924と、によって例証される。
【0010】
本明細書中で例証されるDN1921抗体およびDN1924抗体の各々は、マウスIgG1重鎖定常ドメインおよびマウスIgκ軽鎖定常ドメインをさらに含む。関連する代替的実施形態において、DN1921モノクローナル抗体および/またはDN1924モノクローナル抗体は、上記の重鎖可変ドメインおよび軽鎖可変ドメインと、IgM、IgD、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される抗体アイソタイプ由来のマウス重鎖定常ドメインとを、含み得る。
【0011】
また、DN1921重鎖およびDN1921軽鎖ならびにDN1924重鎖およびDN1924軽鎖の、抗原結合フラグメント、改変体、および誘導体も、提供される。特定の実施形態において、DN1921の改変体は、配列番号13および配列番号2に対してそれぞれ少なくとも70%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1921の改変体は、配列番号13および配列番号2に対してそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1921の改変体は、配列番号13および配列番号2に対してそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0012】
代替的実施形態において、DN1924の改変体は、配列番号35および配列番号24に対してそれぞれ少なくとも70%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1924の改変体は、配列番号35および配列番号24に対してそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1924の改変体は、配列番号35および配列番号24に対してそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0013】
本発明の他の局面は、非ヒト/ヒトキメラHLA−DR特異的モノクローナル抗体を提供する。この抗体において、このキメラ抗体は、ヒト定常ドメインに作動可能に連結された非ヒト可変ドメインを含む。本発明のHLA−DR特異的モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導するが、この抗体が結合する正常細胞においてはアポトーシスを誘導しない。特定の実施形態において、本発明の非ヒト/ヒトキメラHLA−DR特異的アポトーシス性モノクローナル抗体は、被験体にインビボで投与された場合に、非免疫抑制性である。
【0014】
本発明のキメラモノクローナル抗体の構築のために適切な非ヒトモノクローナル抗体可変ドメインは、種々の動物種から単離され得る。この動物種としては、非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、鳥類、ウサギ、マウス、ラット、モルモット、ハムスター、イヌ、およびネコなどの起源が挙げられるが、これらに限定されない。ヒト抗体の重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される抗体アイソタイプから単離され得る。
【0015】
本明細書中に開示される例示的なHLA−DR特異的キメラモノクローナル抗体は、マウスモノクローナル抗体DN1921の重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号13および配列番号2)を、ヒト重鎖定常ドメインおよびヒト軽鎖定常ドメイン(それぞれ、配列番号14および配列番号3)に作動可能に連結された状態で含む。本明細書中に開示されるキメラDN1921モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、そしてインビボで投与された場合に免疫抑制性である。この例示的な全長キメラDN1921モノクローナル抗体の重鎖のアミノ酸配列は、本明細書中では配列番号12として示される。この例示的な全長キメラDN1921モノクローナル抗体の軽鎖のアミノ酸配列は、本明細書中では配列番号1として示される。
【0016】
また、配列番号12のキメラDN1921重鎖および配列番号1のキメラDN1921軽鎖のそれぞれの改変体が、提供される。特定の実施形態において、キメラDN1921重鎖の改変体およびキメラDN1921軽鎖の改変体は、それぞれ、配列番号12および配列番号1に対して少なくとも70%同一である。他の実施形態において、キメラDN1921重鎖の改変体およびキメラDN1921軽鎖の改変体は、それぞれ、配列番号12および配列番号1に対して少なくとも80%、90%、または95%同一である。なおさらなる実施形態において、キメラDN1921重鎖の改変体およびキメラDN1921軽鎖の改変体は、それぞれ、配列番号12および配列番号1に対して少なくとも98%または99%同一である。
【0017】
本明細書中に開示される別の例示的なHLA−DR特異的キメラモノクローナル抗体は、マウスモノクローナル抗体DN1924の重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号35および配列番号24)を、ヒト重鎖定常ドメインおよびヒト軽鎖定常ドメイン(それぞれ、配列番号14および配列番号3)に作動可能に連結された状態で含む。本明細書中に開示されるキメラDN1924モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、そしてインビボで投与された場合に非免疫抑制性である。この例示的な全長キメラDN1924モノクローナル抗体の重鎖のアミノ酸配列は、本明細書中では配列番号34として示される。この例示的な全長キメラDN1924モノクローナル抗体の軽鎖のアミノ酸配列は、本明細書中では配列番号23として示される。
【0018】
代替的実施形態において、配列番号34のキメラDN1924重鎖および配列番号23のキメラDN1924軽鎖のそれぞれの改変体が、提供される。特定の実施形態において、キメラDN1924重鎖の改変体およびキメラDN1924軽鎖の改変体は、それぞれ、配列番号34および配列番号23に対して少なくとも70%同一である。他の実施形態において、キメラDN1924重鎖の改変体およびキメラDN1924軽鎖の改変体は、それぞれ、配列番号34および配列番号23に対して少なくとも80%、90%、または95%同一である。なおさらなる実施形態において、キメラDN1924重鎖の改変体およびキメラDN1924軽鎖の改変体は、それぞれ、配列番号34および配列番号23に対して少なくとも98%または99%同一である。
【0019】
なおさらなる実施形態において、HLA−DR特異的アポトーシス性抗体は、ヒト化モノクローナル抗体であり、このヒト化抗体は、1つ以上の非ヒト相補性決定領域(CDR)を、ヒト可変ドメインフレームワーク領域(FR)に作動可能に連結されて重鎖可変ドメインおよび軽鎖可変ドメインを作製する状態で含み、これらの可変ドメインは、ヒト定常ドメイン重鎖およびヒト定常ドメイン軽鎖(それぞれ、配列番号14および配列番号3において示される重鎖IgG2定常ドメインおよび軽鎖Igκ定常ドメインによって本明細書において例証される)に作動可能に連結された状態である。
【0020】
本発明の例示的なHLA−DR特異的ヒト化モノクローナル抗体は、マウスモノクローナル抗体DN1921重鎖可変ドメイン(配列番号13)の1つ以上の相補性決定領域(CDR)を、ヒト重鎖フレームワーク(FR)ドメインに作動可能に連結されてヒト化HLA−DR特異的重鎖可変ドメインを作製した状態で含む。このヒト化HLA−DR特異的重鎖可変ドメインは、必要に応じて、ヒト重鎖定常ドメインと、配列番号14において示されるヒトIgκ軽鎖定常ドメインとに作動可能に連結される。このヒト重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される。
【0021】
あるいは、またはさらに、本発明の例示的なHLA−DR特異的ヒト化モノクローナル抗体は、マウスモノクローナル抗体DN1921軽鎖可変ドメイン(配列番号2)の1つ以上の相補性決定領域(CDR)を、ヒト軽鎖フレームワーク(FR)ドメイン1、2、3、および/または4に作動可能に連結されてヒト化HLA−DR特異的軽鎖可変ドメインを作製した状態で含む。このヒト化HLA−DR特異的軽鎖可変ドメインは、必要に応じて、ヒト重鎖定常ドメインと、配列番号3において示されるヒトIgκ軽鎖定常ドメインによって本明細書中で例証されるヒト軽鎖定常ドメインとに作動可能に連結される。このヒト重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される。
【0022】
DN1921に基づくヒト化モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、そしてインビボで投与された場合には免疫抑制性である。本発明に従うヒト化抗体を作製するために適切なDN1921 CDRは、配列番号17(DN1921 VH CDR1)、配列番号19(DN1921 VH CDR2)、配列番号21(DN1921 VH CDR3)、配列番号6(DN1921 VL CDR1)、配列番号8(DN1921 VL CDR2)、および配列番号10(DN1921 VL CDR3)として示される。
【0023】
本発明のさらなる例示的なHLA−DR特異的ヒト化モノクローナル抗体は、マウスモノクローナル抗体DN1924重鎖可変ドメイン(配列番号35)の1つ以上の相補性決定領域(CDR)を、ヒトフレームワーク(FR)ドメイン1、2、3、および/または4に作動可能に連結されてヒト化HLA−DR特異的重鎖可変ドメインを作製した状態で含む。このヒト化HLA−DR特異的重鎖可変ドメインは、必要に応じて、ヒト重鎖定常ドメインと、配列番号36において示されるヒトIgκ軽鎖定常ドメインによって本明細書中で例証されるヒト軽鎖定常ドメインとに作動可能に連結される。このヒト重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される。
【0024】
あるいは、またはさらに、本発明のさらなる例示的なHLA−DR特異的ヒト化モノクローナル抗体は、マウスモノクローナル抗体DN1924軽鎖可変ドメイン(配列番号24)の1つ以上の相補性決定領域(CDR)を、ヒトフレームワーク(FR)ドメイン1、2、3、および/または4に作動可能に連結されてヒト化HLA−DR特異的軽鎖可変ドメインを作製した状態で含む。このヒト化HLA−DR特異的軽鎖可変ドメインは、必要に応じて、ヒト重鎖定常ドメインと、配列番号25において示されるヒトIgκ軽鎖定常ドメインによって例証されるヒト軽鎖定常ドメインとに作動可能に連結される。このヒト重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される。
【0025】
DN1924に基づくヒト化モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、そしてインビボで投与された場合には非免疫抑制性である。本発明に従うヒト化抗体を作製するために適切なDN1924 CDRは、配列番号39(DN1924 VH CDR1)、配列番号41(DN1924 VH CDR2)、配列番号43(DN1924 VH CDR3)、配列番号28(DN1924 VL CDR1)、配列番号30(DN1924 VL CDR2)、および配列番号32(DN1924 VL CDR3)として示される。
【0026】
本発明はまた、組成物および方法を提供し、これらは、本明細書中に示される、1つ以上の非ヒトモノクローナルHLA−DR特異的抗体、キメラHLA−DR特異的抗体、および/またはヒト化HLA−DR特異的抗体を含む。そのようなHLA−DR特異的抗体を含む組成物は、インビボ投与のために適切であり、そしてHLA−DRを発現する腫瘍細胞のアポトーシスを誘発する際に有効である。
【0027】
本発明のこれらの局面および他の局面は、以下の詳細な説明および添付の図面を参照すると明らかとなる。本明細書中に開示されるすべての参考文献は、各々が個別に援用されたかのごとく、その全体が本明細書中に参考として援用される。
【0028】
配列番号1は、キメラDN1921全長抗体軽鎖のアミノ酸配列である。
【0029】
配列番号2は、キメラDN1921軽鎖可変領域のアミノ酸配列である。
【0030】
配列番号3は、キメラDN1921軽鎖定常領域(ヒトIgG2κ)のアミノ酸配列である。
【0031】
配列番号4は、キメラDN1921軽鎖リーダー配列のアミノ酸配列である。
【0032】
配列番号5は、キメラDN1921軽鎖フレームワーク領域1(FR1)のアミノ酸配列である。
【0033】
配列番号6は、キメラDN1921軽鎖相補性決定領域1(CDR1)のアミノ酸配列である。
【0034】
配列番号7は、キメラDN1921軽鎖FR2のアミノ酸配列である。
【0035】
配列番号8は、キメラDN1921軽鎖CDR2のアミノ酸配列である。
【0036】
配列番号9は、キメラDN1921軽鎖FR3のアミノ酸配列である。
【0037】
配列番号10は、キメラDN1921軽鎖CDR3のアミノ酸配列である。
【0038】
配列番号11は、キメラDN1921軽鎖FR4のアミノ酸配列である。
【0039】
配列番号12は、キメラDN1921全長抗体重鎖のアミノ酸配列である。
【0040】
配列番号13は、キメラDN1921重鎖可変領域のアミノ酸配列である。
【0041】
配列番号14は、キメラDN1921重鎖定常領域(ヒトIgG2)のアミノ酸配列である。
【0042】
配列番号15は、キメラDN1921重鎖リーダー配列のアミノ酸配列である。
【0043】
配列番号16は、キメラDN1921重鎖FR1のアミノ酸配列である。
【0044】
配列番号17は、キメラDN1921重鎖CDR1のアミノ酸配列である。
【0045】
配列番号18は、キメラDN1921重鎖FR2のアミノ酸配列である。
【0046】
配列番号19は、キメラDN1921重鎖CDR2のアミノ酸配列である。
【0047】
配列番号20は、キメラDN1921重鎖FR3のアミノ酸配列である。
【0048】
配列番号21は、キメラDN1921重鎖CDR3のアミノ酸配列である。
【0049】
配列番号22は、キメラDN1921重鎖FR4のアミノ酸配列である。
【0050】
配列番号23は、キメラDN1924全長抗体軽鎖のアミノ酸配列である。
【0051】
配列番号24は、キメラDN1924軽鎖可変領域のアミノ酸配列である。
【0052】
配列番号25は、キメラDN1924軽鎖定常領域(ヒトIgκ)のアミノ酸配列である。
【0053】
配列番号26は、キメラDN1924軽鎖リーダー配列のアミノ酸配列である。
【0054】
配列番号27は、キメラDN1924軽鎖FR1のアミノ酸配列である。
【0055】
配列番号28は、キメラDN1924軽鎖CDR1のアミノ酸配列である。
【0056】
配列番号29は、キメラDN1924軽鎖FR2のアミノ酸配列である。
【0057】
配列番号30は、キメラDN1924軽鎖CDR2のアミノ酸配列である。
【0058】
配列番号31は、キメラDN1924軽鎖FR3のアミノ酸配列である。
【0059】
配列番号32は、キメラDN1924軽鎖CDR3のアミノ酸配列である。
【0060】
配列番号33は、キメラDN1924軽鎖FR4のアミノ酸配列である。
【0061】
配列番号34は、キメラDN1924全長抗体重鎖のアミノ酸配列である。
【0062】
配列番号35は、キメラDN1924重鎖可変領域のアミノ酸配列である。
【0063】
配列番号36は、キメラDN1924重鎖定常領域(IgG2)のアミノ酸配列である。
【0064】
配列番号37は、キメラDN1924重鎖リーダー配列のアミノ酸配列である。
【0065】
配列番号38は、キメラDN1924重鎖FR1のアミノ酸配列である。
【0066】
配列番号39は、キメラDN1924重鎖CDR1のアミノ酸配列である。
【0067】
配列番号40は、キメラDN1924重鎖FR2のアミノ酸配列である。
【0068】
配列番号41は、キメラDN1924重鎖CDR2のアミノ酸配列である。
【0069】
配列番号42は、キメラDN1924重鎖FR3のアミノ酸配列である。
【0070】
配列番号43は、キメラDN1924重鎖CDR3のアミノ酸配列である。
【0071】
配列番号44は、キメラDN1924重鎖FR4のアミノ酸配列である。
【0072】
配列番号45は、DN1921全長抗体軽鎖をコードするポリヌクレオチドのヌクレオチド配列である。
【0073】
配列番号46は、DN1921全長抗体重鎖をコードするポリヌクレオチドのヌクレオチド配列である。
【0074】
配列番号47は、DN1924全長抗体軽鎖をコードするポリヌクレオチドのヌクレオチド配列である。
【0075】
配列番号48は、DN1924全長抗体重鎖をコードするポリヌクレオチドのヌクレオチド配列である。
【0076】
配列番号49は、ヒトHLA−DRα鎖のアミノ酸配列である。
【0077】
配列番号50は、ヒトHLA−DRβ鎖のアミノ酸配列である。
【0078】
配列番号51は、配列番号49のヒトHLA−DRα鎖をコードするヌクレオチド配列である。
【0079】
配列番号52は、配列番号50のヒトHLA−DRβ鎖をコードするヌクレオチド配列である。
【0080】
配列番号53は、マウスIgG1重鎖定常ドメインのアミノ酸配列である。
【0081】
配列番号54は、配列番号53のマウスIgG1重鎖定常ドメインをコードするヌクレオチド配列である。
【0082】
配列番号55は、マウスIgκ軽鎖定常ドメインのアミノ酸配列である。
【0083】
配列番号56は、配列番号55のマウスIgκ軽鎖定常ドメインをコードするヌクレオチド配列である。
【0084】
配列番号57は、プライマー配列
【0085】
【化1】
である。
【0086】
配列番号58は、プライマー配列
【0087】
【化2】
である。
【0088】
配列番号59は、プライマー配列
【0089】
【化3】
である。
【0090】
配列番号60は、プライマー配列
【0091】
【化4】
である。
【0092】
配列番号61は、プライマー配列
【0093】
【化5】
である。
【0094】
配列番号62は、プライマー配列
【0095】
【化6】
である。
【0096】
配列番号63は、プライマー配列
【0097】
【化7】
である。
【0098】
配列番号64は、プライマー配列
【0099】
【化8】
である。
【0100】
配列番号65は、プライマー配列
【0101】
【化9】
である。
【0102】
配列番号66は、プライマー配列
【0103】
【化10】
である。
【0104】
配列番号67は、プライマー配列
【0105】
【化11】
である。
【0106】
配列番号68は、プライマー配列
【0107】
【化12】
である。
【0108】
配列番号69は、プライマー配列
【0109】
【化13】
である。
【0110】
配列番号70は、プライマー配列
【0111】
【化14】
である。
【発明を実施するための最良の形態】
【0112】
(発明の詳細な説明)
上記のように、本発明は、HLA−DR特異的抗体、ならびにHLA−DR特異的抗体を含む組成物および方法に関する。本明細書に示される抗体は、腫瘍細胞のアポトーシスを誘導することにおいて有効であるが、HLA−DRを発現する正常細胞においては有効でない。特定の実施形態において、本発明のアポトーシス性HLA−DR特異的抗体は、低下した免疫抑制活性をさらに示す。本発明のHLA−DR特異的抗体は、とりわけ、癌(ここでこの癌細胞の少なくとも部分集団は、HLA−DR抗原を発現する)のような疾患の診断および処置における薬剤として有用である。
【0113】
本明細書で使用される場合、用語「HLA−DR」とは、「ヒト白血球抗原」(HLA)DR遺伝子座およびこれらのタンパク質産物をいい、このタンパク質産物は、ヒト白血球で発現される同種異系抗原である。同種異系抗原は、自己を外来組織から見分ける多型遺伝子の産物である。用語「クラスII主要組織適合遺伝子複合体」、「クラスII MHC」、または「クラスII」抗原とは、種々の細胞型で種々のレベルで発現されかつT細胞による全てのタンパク質抗原の認識において必須の役割を果たす抗原をいう。クラスII MHC分子は、代表的には、7〜30またはそれ以上のアミノ酸のペプチドを結合し、抗原特異的CD4+ T細胞によって認識される複合体を形成する。このCD4分子は、クラスII分子の第2のドメインに結合する。
【0114】
用語「アポトーシス性細胞死」、「細胞自己死」または「アポトーシス」とは、本明細書で使用される場合、その死亡をもたらす、正常な一連の細胞中の事象をいう。アポトーシスは、大部分の組織の生理学的維持において極めて重大な役割を果たす。細胞死プログラムの活性化を介して望ましくない細胞の排除を可能にすることは、通常の生物学的プロセスである。アポトーシスは、照射、ウイルス感染または異常な増殖に由来する損傷後に、癌の場合のように、これらの潜在的に危険な細胞を除去することによって、異常な細胞から生物を保護するために、緊急な応答を提供する。アポトーシス性細胞死は、細胞分化の特定の段階において、および特定の刺激に応答して起こる細胞事象の複雑なカスケードから生じ、死細胞の細胞質および核の凝縮によって特徴づけられる。
【0115】
用語「腫瘍」、「癌」または「新生物」とは、本明細書で交換可能に使用され、正常組織から発生するが、構造がなく異常に細胞が増殖している、異常な増殖をいう。腫瘍または癌細胞は、一般に、接触阻害の能力を欠き、侵襲性であり得、そして/または転移する能力を有し得る。本発明の抗体で処置しやすい腫瘍細胞は、クラスII腫瘍組織適合抗原HLA−DRの発現によって特徴づけられる。
【0116】
本明細書および添付の特許請求の範囲で使用される場合、単数形「a」、「an」および「the」は、文脈が明らかに別であることを示さなければ、複数形への参照を包含する。
【0117】
本発明の実施は、反対であることが具体的に示されない限り、当該分野の技術範囲内である、ウイルス学、免疫学、微生物学、分子生物学および組換えDNA技術の従来の方法が採用される。この従来の方法の多くは、例示目的で以下に示される。このような技術は、文献中で十分に説明されている。例えば、Sambrookら,「Molecular Cloning:A Laboratory Manual」(第2版,1989);Maniatisら,「Molecular Cloning:A Laboratory Manual」(1982);「DNA Cloning:A Practical Approach,vol.I & II」(D.Glover編);「Oligonucleotide Synthesis」(N.Gait編,1984);「Nucleic Acid Hybridization」(B.Hames & S.Higgins,編,1985);「Transcription and Translation」(B.Hames & S.Higgins,編,1984);「Animal Cell Culture」(R.Freshney編,1986);Perbal,「A Practical Guide to Molecular Cloning」(1984);Ausubelら,「Current protocols in Molecular Biology」(New York,John Wiley and Sons,1987);Bonifacinoら,「Current Protocols in Cell Biology」(New York,John Wiley & Sons,1999);Coliganら,「Current Protocols in Immunology」(New York,John Wiley & Sons,1999);ならびにHarlow and Lane Antibodies:a Laboratory Manual Cold Spring Harbor Laboratory(1988)を参照のこと。
【0118】
本明細書に引用される全ての刊行物、特許および特許出願は、前出、後出に拘わらず、その全体が本明細書に参考として援用される。本発明は、具体的な実施形態の詳細な説明を通じてよりよく理解される。これらの実施形態の各々は、本明細書で以下に詳細に記載される。
【0119】
(HLA−DR特異的抗体)
本発明は、抗体、より具体的には、モノクローナル抗体、非ヒト/ヒトキメラモノクローナル抗体、ヒト化モノクローナル抗体、および完全ヒトモノクローナル抗体であって、これらの抗体は、細胞表面に発現されたHLA−DRに結合した場合に、HLA−DRに特異的に結合し、かつ腫瘍細胞においてアポトーシスを誘導し得る得が、正常細胞においては誘導しない、抗体に関する。特定の実施形態において、本発明のモノクローナル抗体は、免疫抑制活性の低下したレベルによってさらに特徴づけられる。
【0120】
本明細書中で例示されるのは、マウスIgG重鎖およびIgκ軽鎖定常領域(それぞれ、配列番号53および55)に作動可能に融合された、配列番号13および2(DN1921)ならびに配列番号35および24(DN1924)の重鎖可変ドメインおよび軽鎖可変ドメインを含む、それぞれ、DN1921およびDN1924と称されるマウスモノクローナル抗体である;DN1921および/またはDN1924の可変領域(VHおよびVL)ならびにヒトIgG2重鎖およびIgκ軽鎖定常領域(それぞれ、配列番号14および3(DN1921)ならびに配列番号36および25(DN1924)を含む)を含むキメラ抗体;ならびにヒトフレームワーク領域(FR)およびヒト定常ドメインに作動可能に融合された、DN1921および/またはDN1924重鎖および軽鎖の可変ドメインの1つ以上の相補性決定領域(CDR)を含む、ヒト化抗体である。
【0121】
本明細書で記載される場合、用語「抗体」とは、モノクローナル抗体、キメラ抗体、ヒト化抗体、および完全ヒト抗体、ならびにその生物学的フラグメントもしくは抗原結合フラグメントおよび/またはその一部を包含する。本明細書で「抗体」への言及は、その一部、フラグメント、前駆体形態、誘導体、改変体、および遺伝子操作したかもしくは天然に変異した形態への言及を包含し、かつその得られる誘導体および/または改変体が、HLA−DR結合特異性および/または親和性の少なくとも実質的な量を保持する限りにおいて、アミノ酸置換および化学物質および/もしくは放射性同位体などでの標識を含む。用語「抗体」とは、抗体重鎖および軽鎖の両方、ならびに抗体の全てのアイソタイプ(IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA、およびIgA2が挙げられる)を広く包含し、その抗原結合フラグメント(Fab、F(ab’)2、Fc、およびscFvが挙げられるが、これらに限定されない)もまた包含する。
【0122】
用語「モノクローナル抗体」とは、本明細書で使用される場合、実質的に均一な抗体の集団から得られた抗体(すなわち、その集団を構成する個々の抗体が、抗体結合特異性、親和性、および/または活性に実質的に影響を及ぼさない、天然に存在する変異を除いて、同一である抗体)をいう。
【0123】
HLA−DR特異的抗体、またはその抗原結合フラグメントは、(例えば、ELISAアッセイにおいて)検出可能なレベルでHLA−DRと反応しかつ類似の条件下で関連性のないポリペプチドとは検出可能に反応しない場合、HLA−DRに「特異的に結合する」、「免疫原性であるように(immunogically)結合する」、そして/または「免疫学的に活性である」といわれる。
【0124】
この文脈で使用される場合、免疫学的に結合するとは、一般に、抗体と抗原(この抗原に対してその抗体が特異的である)との間に存在する型の非共有結合的反応をいう。抗体−HLA−DR結合相互作用の強度、すなわち親和性は、相互作用の解離定数(Kd)の観点から表され得、ここでKdが小さいほど、より大きな親和性を表す。HLA−DR特異的抗体の免疫学的結合特性は、当該分野で周知の方法を用いて定量され得る。1つのこのような方法は、HLA−DR特異的抗体/抗原複合体形成および解離の速度を測定する工程を包含し、ここでこの速度は、複合体パートナーの濃度、相互作用に親和性、および両方の方向で速度に等しく影響を与える幾何的パラメーターに依存する。従って、「オン反応速度定数」(Kon)および「オフ反応速度定数」(Koff)は、濃度と、会合および解離の実際の速度の計算によって決定され得る。Koff/Konの比率は、親和性に関しない全てのパラメーターのキャンセルを可能にし、よって、解離定数Kdに等しい。一般には、Daviesら,Annual Rev.Biochem.59:439−473(1990)を参照のこと。本発明の抗HLA−DR抗体は、HLA−DRタンパク質に特異的に結合する。本明細書で「特異的に結合する」とは、その抗体が、少なくとも106〜109M、より一般には、少なくとも107〜109Mの範囲の結合定数でHLA−DRポリペプチドに結合することを意味する。
【0125】
HLA−DR特異的抗体の「抗原結合部位」または「結合部分」とは、HLA−DR結合に関与する抗体分子の一部をいう。この抗原結合部位は、重鎖(「H」)および軽鎖(「L」)のN末端改変(「V」)領域のアミノ酸残基によって形成される。重鎖および軽鎖のV領域内の3つの高度に多様な拡がり(stretch)は、「フレームワーク領域」または「FR」として公知のより保存された隣接する拡がりの間に挟まれている「超可変領域」または「相補性決定領域(CDR)」として言及される。
【0126】
従って、用語「FR」とは、抗体中のCDRの間かつこれに隣接して天然に見出されるアミノ酸配列をいう。HLA−DR特異的抗体分子において、軽鎖の3つのCDRおよび重鎖の3つのCDRは、HLA−DR結合表面を形成するために、三次元空間で互いに対して配置されている。このHLA−DR結合表面は、結合したHLA−DRの三次元表面に相補的である。
【0127】
本明細書の目的に関して「機能的に活性な」または「機能的活性」とは、本明細書に開示されるDN1921抗HLA−DR抗体および/またはDN1924抗HLA−DR抗体の生物学的活性および/または免疫学的活性を保持しているHLA−DR特異的抗体をいう。用語「エピトープ」とは、HLA−DR特異的抗体の相補性決定領域(CDR)と相互作用するHLA−DR抗原の特定の部分をいう。
【0128】
(HLA−DR特異的非ヒトモノクローナル抗体)
本発明は、1つ以上のアポトーシス誘導性ドメインでHLA−DRに結合し、それによって、HLA−DR発現腫瘍細胞においてアポトーシスを誘導するが、HLA−DR発現正常細胞(これに対して抗体が結合する)では誘導しない非ヒトモノクローナル抗体を提供する。本明細書で例示されるのは、DN1921およびDN1924と称される、アポトーシス性HLA−DR特異的マウスモノクローナル抗体である。DN1921は、免疫抑制活性によってさらに特徴づけられる一方で、DN1924は、DN1921と比較して、減少した免疫抑制活性を示す。
【0129】
機能のいかなる理論にも限定されずに、本発明のHLA−DR特異的モノクローナル抗体は、HLA−DRヘテロ二量体の第1のタンパク質ドメイン上の1つ以上のアポトーシス誘導性エピトープに特異的に結合することによって、アポトーシス活性を達成することができると考えられる。免疫抑制性HLA−DR特異的モノクローナル抗体(本明細書ではDN1921によって例示される)が、抗原結合ドメインについてさらに競合し得、従って、ヘルパーT(Th)細胞に抗原を提示し得る一方で、非免疫抑制性HLA−DR特異的モノクローナル抗体(本明細書ではDN1924によって例示される)が、抗原結合について低下した競合能力を有するとさらに考えられる。クラスII MHC分子(HLA−DR)の第1のドメインに特異的に結合する、非ヒトモノクローナル抗体および非ヒト/ヒトキメラ抗体を、本明細書に開示されるように調製した。クラスII分子は、細胞増殖を調節し得るシグナルを伝達し、本明細書で示されるそのクラスII MHC HLA−DR特異的抗体は、癌細胞のアポトーシスを誘導し得る。
【0130】
ヒト主要組織適合遺伝子複合体(MHC)クラスII分子特異的抗体(DN1921、キメラDN1921、DN1924、およびキメラDN1924と称される)は、それらの表面でHLA−DRを発現する腫瘍細胞においてアポトーシスを誘導することが分かった。これらの抗体の抗癌活性は、悪性でないHLA−DR陽性細胞の生存性にも機能にも影響を及ぼさない点で、高度に選択性である。
【0131】
大部分の場合において、HLA−DRタンパク質のエピトープまたはフラグメントに対して行われる抗体は、全長タンパク質に結合することができると同時に、他のタンパク質に対してはほとんどもしくは全く交叉反応性を示さない。特定の実施形態において、本発明の抗体は、αHLA−DR鎖およびβHLA−DR鎖のアミノ酸1〜88およびアミノ酸1〜96(それぞれ、配列番号49および50)に対応するHLA−DR分子の第1のドメインに対して生成される。
【0132】
本明細書で例示される非ヒトモノクローナル抗体は、マウスIgG、重鎖定常領域およびIgκ軽鎖定常領域(それぞれ、配列番号53および55)に作動可能に融合された、配列番号13および2(DN1921)ならびに配列番号35および24(DN1924)の重鎖可変ドメインおよび軽鎖可変ドメインを含む、それぞれ、DN1921およびDN1924と称されるマウスモノクローナル抗体である。
【0133】
従って、特定の局面において、本発明は、非ヒトモノクローナル抗体HLA−DR特異的アポトーシス性抗体を提供する。本発明の非ヒトモノクローナル抗体は、種々の動物種から単離され得る。これらの種としては、非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、鳥類、ウサギ、マウス、ラット、モルモット、ハムスター、イヌおよびネコ起源が挙げられるが、限定されない。HLA−DR特異的アポトーシス性非ヒトモノクローナル抗体は、本明細書で、(1)配列番号13および配列番号2にそれぞれおいて示される重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ配列番号46および45のポリヌクレオチドによってコードされる)を含む免疫抑制性抗体DN1921、ならびに(2)配列番号35および配列番号24においてそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号48および47のポリヌクレオチドによってコードされる)を含む、非免疫抑制性抗体DN1924、によって例示される。
【0134】
本明細書で例示されるこのDN1921抗体およびDN1924抗体の各々は、マウスIgG、重鎖およびIgκ軽鎖定常ドメインをさらに含む。関連した代替実施形態において、DN1921および/またはDN1924モノクローナル抗体は、前述の重鎖可変ドメインおよび軽鎖可変ドメインならびにIgM、IgD、IgG2、IgG3、IgG4、IgE、IgA、およびIgA2からなる群より選択される抗体アイソタイプに由来するマウス重鎖定常ドメインを含み得る。
【0135】
DN1921およびDN1924の重鎖および軽鎖の抗原結合フラグメント、改変体、および誘導体もまた提供される。特定の実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも70%同一である重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0136】
代替的実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも70%同一である重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも80%、90%、または95%同一である重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0137】
本発明のモノクローナル抗体(mAb)の生成のために、従来の方法論が採用され得る。一般に、齧歯類(通常はマウス)が、目的のHLA−DR抗原で繰り返し免疫され、抗体生成B細胞が塗擦した免疫動物から単離され、その抗体生成B細胞が、骨髄腫細胞と融合されてB細胞「ハイブリドーマ」を生じることによって不死化される。ハイブリドーマ細胞のライブラリーは、抗原結合特異性についてスクリーニングされ、適切なクローンが精製および増殖される。モノクローナル抗体は、ハイブリドーマ法(例えば、以前に記載され、当該分野で容易に利用可能なもの)を用いて調製され得る。Kohlerら,Nature 256:495(1975)。
【0138】
不死化細胞株は、例えば、上記のように、HLA−DR抗原で免疫した動物から得た脾細胞より生成され得る。脾細胞は、例えば、骨髄腫細胞融合パートナーと、好ましくは、免疫した動物と同系であるものと融合することにより不死化される。種々の融合技術が、採用され得る。例えば、その脾細胞および骨髄腫細胞は、数分の間非イオン性界面活性剤と合わされ、次いで、低密度で選択培地(この培地は、ハイブリッド細胞の増殖を支持するが、骨髄腫細胞の増殖は支持しない)にプレートされ得る。
【0139】
ハイブリドーマ細胞は、融合していない不死化細胞の増殖または生存を阻害する1種以上の物質を好ましくは含む適切な培養培地中で培養され得る。例示的な不死化細胞株は、are those that 効率的に融合し、その選択された抗体生成細胞による抗体の安定した高レベルの発現を支持し、かつヒポキサンチン、アミノプテリン、およびチミジン(「HAT培地」)のような培地に感受性であるものである。より例示的な不死化細胞株は、マウス骨髄腫細胞株であり、この細胞株は、例えば、the Salk Institute Cell Distribution Center,San Diego,Californiaおよびthe American Type Culture Collection(ATCC),Rockville,Marylandから入手可能である。ヒト骨髄腫細胞株およびマウス−ヒトヘテロ骨髄腫細胞株が、ヒトモノクローナル抗体の生成のために記載された。Kozbor,J.Immunol.133:3001(1984)およびBrodeurら,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.編,New York,1987)。
【0140】
ハイブリドーマ細胞が培養される培養培地は、次いで、HLA−DRポリペプチドに対するモノクローナル抗体の存在についてアッセイされ得る。このハイブリドーマ細胞によって生成されるモノクローナル抗体の結合特異性は、免疫沈降によって、またはインビトロ結合アッセイ(例えば、ラジオイムノアッセイまたは酵素結合イムノソルベントアッセイ(ELISA))によって決定され得る。このような技術およびアッセイは、当該分野で公知である。このモノクローナル抗体の結合親和性は、例えば、Munsonら,Anal.Biocheni.107:220(1980)のスキャッチャード分析によって決定され得る。
【0141】
望ましいハイブリドーマ細胞が同定された後、そのクローンは、限界希釈法によってサブクローニングされ得、標準的な方法によって増殖される。Goding,「Monoclonal Antibodies:Principles and Practice」,59−103(Academic Press,1986)。この目的に関して、適切な培養培地としては、例えば、ダルベッコ改変イーグル培地およびRPMI−1640培地が挙げられる。あるいは、そのハイブリドーマ細胞は、哺乳動物における腹水としてインビボで増殖され得る。
【0142】
HLA−DR特異的モノクローナル抗体は、増殖しつつあるハイブリドーマコロニーの上清から単離され得る。さらに、種々の技術は、収率を高めるために使用され得、例えば、適切な脊椎動物宿主(例えば、マウス)の腹腔にハイブリドーマ細胞株を注射することが挙げられる。モノクローナル抗体は、次いで、腹水または血液から採取され得る。夾雑物は、従来の技術(例えば、クロマトグラフィー、ゲル濾過、沈降および抽出)によって抗体から除去され得る。本発明のHLA−DRポリペプチド(例えば、配列番号49および50に示されるヒトHLA−DR αポリペプチドおよびβポリペプチド)は、例えば、アフィニティークロマトグラフィー工程における精製プロセスで使用され得る。あるいは、サブクローンによって分泌されるモノクローナル抗体は、従来の抗体精製手順(例えば、プロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、または他のアフィニティークロマトグラフィー方法)により、培養培地または腹水から単離または精製され得る。
【0143】
モノクローナル抗体はまた、組換えDNA方法(例えば、米国特許第4,816,567号(本明細書に参考として援用される)に記載されるもの)によって作製され得る。本発明のモノクローナル抗体をコードするDNAは、容易に単離され得、そして従来の手順を用いて(例えば、マウス抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合し得るオリゴヌクレオチドプローブを使用することによって)配列決定され得る。
【0144】
一旦単離されると、そのDNAは、発現ベクターにクローニングされ得る。次いでこのベクターは、宿主細胞(例えば、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、または別の方法で抗体タンパク質を生成しない骨髄腫細胞)にトランスフェクトされて、組換え宿主細胞においてモノクローナル抗体の合成を得る。このDNAはまた、例えば、同種マウス配列の代わりにヒト重鎖定常領域および軽鎖定常領域のコード配列を置換することによって、または非抗体ポリペプチドのコード配列の全てもしくは一部を、抗体コード配列に共有結合することによって、改変され得る。このような非抗体ポリペプチドは、本発明の抗体の定常領域の代わりに用いられ得るか、または本発明の抗体の1つの抗原結合部位の可変ドメインの代わりに用いられて、本明細書で以下により詳細に記載されるキメラ二価抗体が作り出され得る。
【0145】
よって、特定の局面において、本発明は、非ヒトモノクローナルHLA−DR特異的アポトーシス性抗体を提供する。本発明の非ヒトモノクローナル抗体は、種々の動物種(非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、家禽、ウサギ、マウス、ラット、モルモット、ハムスター、イヌ、およびネコ起源が挙げられるが、これらに限定されない)から単離され得る。HLA−DR特異的アポトーシス性非ヒトモノクローナル抗体は、(1)配列番号13および配列番号2にそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(これらは、配列番号46および45のポリヌクレオチドによってそれぞれコードされる)を含む免疫抑制性抗体DN1921、ならびに(2)配列番号35および配列番号24にそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(これは、配列番号48および47のポリヌクレオチドによりそれぞれコードされる)を含む非免疫抑制性抗体DN1924によって、本明細書で例示される。
【0146】
本明細書で例示されるDN1921抗体およびDN1924抗体の各々は、マウスIgG1重鎖およびIgκ軽鎖の定常ドメインをさらに含む。関連する代替的な実施形態において、DN1921モノクローナル抗体および/またはDN1924モノクローナル抗体は、前述の重鎖および軽鎖の可変ドメインならびに抗体アイソタイプ(IgM、IgD、IgG2、IgG3、IgG4、IgE、IgA、およびIgA2からなる群より選択される)由来のマウスの重鎖定常ドメインを含み得る。
【0147】
DN1921およびDN1924の重鎖および軽鎖の抗原結合フラグメント、改変体、および誘導体もまた提供される。特定の実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも70%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも98%または99%同一である重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0148】
代替的実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも70%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0149】
(キメラ抗体)
本発明の他の局面は、非ヒト/ヒトキメラHLA−DR特異的モノクローナル抗体を提供し、ここでこのキメラ抗体は、ヒト定常ドメインに作動可能に融合された非ヒト可変ドメインを含む。本発明の非ヒト/ヒトキメラHLA−DR特異的モノクローナル抗体は、HLA−DR発現腫瘍細胞においてアポトーシスを誘導するが、その抗体が結合される正常細胞においては誘導しない。特定の実施形態において、本発明の非ヒト/ヒトキメラHLA−DR特異的アポトーシス性モノクローナル抗体は、被験体にインビボで投与される場合、非免疫抑制性である。マウス/ヒトキメラDN1921 HLA−DR特異的モノクローナル抗体およびマウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体によって例示される、非ヒト/ヒトキメラHLA−DR特異的抗体が本明細書で示される。これらのキメラ抗体のアミノ酸配列およびそれらの構成的構造領域は、表1(キメラDN1921)および表2(キメラDN1924)において示される。
【0150】
本明細書で使用される場合、用語「キメラ抗体」とは、重鎖および軽鎖を含む抗体分子であって、この抗体分子において、非ヒト抗体の可変ドメインが、ヒト定常ドメインに作動可能に融合されたものをいう。キメラ抗体は、一般に、その親の完全非ヒト抗体に比較して、減少した免疫原性を示す。
【0151】
ヒト定常ドメインに作動可能に融合された非ヒト抗原結合可変ドメインを含むキメラ抗体分子を作製するための方法論は、当該分野で記載されている。例えば、Winterら,Nature 349:293−299(1991);Lobuglioら,Proc.Nat.Acad.Sci.USA 86:4220−4224(1989);Shawら,J.Immunol.138:4534−4538(1987);Brownら,Cancer Res.47:3577−3583(1987);米国特許第4,816,567号(Cabillyによる);Cabillyら,Proc.Natl.Acad.Sci.USA 81:3273−3277(1984);Morrisonら,Proc.Natl.Acad.Sci.USA 81:6851−6855(1984);Boulianneら,Nature 312:643−646(1984);Cabillyら,欧州特許出願第125023(1984年11月14日公開);Neubergerら,Nature 314:268−270(1985);Taniguchiら,欧州特許出願第171496(1985年2月19日公開);Morrisonら,欧州特許出願第173494号(1986年3月5日公開);Neubergerら,PCT出願WO86/01533(1986年3月13日公開);Kudoら,欧州特許出願第184187号(1986年6月11日公開);Morrisonら,欧州特許出願第173494号(1986年3月5日公開);Sahaganら,J.Immunol.137:1066−1074(1986);Robinsonら,国際特許公開番号PCT/US86/02269(1987年公開5月7日);Liuら,Proc.Natl.Acad.Sci.USA 84:3439−3443(1987);Sunら,Proc.Natl.Acad.Sci.USA 84:214−218(1987);Betterら,Science 240:1041−1043(1988);ならびにHarlow and Lane Antibodies:a Laboratory Manual Cold Spring Harbor Laboratory(1988))を参照のこと。これらの参考文献の各々は、本明細書に参考として援用される。
【0152】
キメラ抗体は、一価抗体、二価抗体または多価抗体を包含する。一価キメラ抗体は、キメラL鎖とジスルフィド架橋によって結合したキメラH鎖によって形成される重鎖および軽鎖(HL)の二量体である。二価キメラ抗体は、少なくとも1つのジスルフィド架橋によって結合した2つのHL二量体によって形成される四量体(H2L2)である。例えば、凝集するCH領域(例えば、IgM H鎖、またはμ鎖から)を使用することによって、多価キメラ抗体も生成され得る。
【0153】
本発明のマウス抗体およびキメラ抗体、フラグメントおよび領域は、個々の抗体の重鎖および/または軽鎖を含む。キメラ重鎖は、ヒト重鎖定常領域の少なくとも一部に連結された、HLA−DRに対して特異的な非ヒト抗体の重鎖に由来する抗原結合領域を含む。
【0154】
本発明に従うキメラ軽鎖は、ヒト軽鎖定常領域の少なくとも一部に連結された、HLA−DRに特異的な非ヒト抗体の軽鎖に由来する抗原結合領域を含む。
【0155】
同じまたは異なる可変領域結合特異性のキメラ重鎖およびキメラ軽鎖を有する抗体、フラグメントまたは誘導体はまた、公知の方法に従って(例えば、Ausubel(前出)およびHarlow(前出)に従って)個々のポリペプチド鎖の適切な結合によって調製され得る。
【0156】
特定の実施形態において、キメラ重鎖(またはその誘導体)を発現する宿主細胞は、キメラ軽鎖(またはその誘導体)を発現する宿主とは別個に培養されてもよく、その免疫グロブリン鎖は、別個に回収され、次いで結合されてもよい。あるいは、この宿主細胞は、一緒に培養され得、その鎖は、培養培地中で自発的に会合し、続いて、組み立てられた免疫グロブリン、フラグメントまたは誘導体を回収することが可能になる。
【0157】
本明細書で例示される場合、抗体重鎖および軽鎖は、同じ細胞にトランスフェクトされ、輸送および分泌されるときに細胞によって組み立てられてもよい。
【0158】
ハイブリッド細胞は、非ヒトHLA−DR特異的抗体生成細胞(代表的には、天然もしくは組換えいずれかのヒトHLA−DR、またはヒトHLA−DRタンパク質配列のペプチドフラグメントに対して免疫された動物の脾細胞)の融合によって形成され得る。あるいは、非ヒトHLA−DR特異的抗体生成細胞は、HLA−DRで免疫した動物の血液、脾臓、リンパ節または他の組織から得られたBリンパ球であり得る。
【0159】
第2の融合パートナーは、不死化機能を提供し、リンパ芽球腫細胞またはプラスマ細胞腫細胞または骨髄腫細胞であり得る。これらの細胞は、それら自身抗体生成細胞ではなく、悪性である。例示的な融合パートナー細胞としては、ハイブリドーマSP2/0−Ag14(SP2/0;ATCC CRL1581)および骨髄腫P3X63Ag8(ATCC TIB9)、あまたはその誘導体が挙げられる。例えば、Ausubel(前出)およびHarlow(前出)を参照のこと。
【0160】
ヒトHLA−DRに特異的なモノクローナル抗体を生成するマウスハイブリドーマは、マウス融合パートナー細胞(例えば、SP2/0)と、精製HLA−DR、組換えHLA−DR、天然もしくは合成HLA−DRペプチド(αおよびβ HLA−DR鎖(それぞれ配列番号49および50)のアミノ酸1〜88およびアミノ酸1〜96を含むペプチドが挙げられる)に対して免疫したマウス由来の脾細胞とを融合することによって形成される。
【0161】
マウスを免疫するために、種々の異なる従来のプロトコルが行われ得る。例えば、マウスは、HLA−DRの初回免疫および追加免疫を受容し得る。本発明のキメラ抗体の抗原結合領域をコードするヌクレオチド配列を与えるこの抗体生成細胞はまた、非ヒト(例えば、霊長類)細胞またはヒト細胞の形質転換によって生成され得る。例えば、HLA−DR特異的抗体を生成するBリンパ球は、エプスタイン−バーウイルスのようなウイルスに感染して形質転換されて、不死化HLA−DR特異的抗体生成細胞を生じる。例えば、Kozborら,Immunol.Today 4:72−79(1983)を参照のこと。あるいは、Bリンパ球は、当該分野で周知のように、癌遺伝子または癌遺伝子産物を提供することによって形質転換され得る。例えば、Ausubel(前出)およびHarlow(前出)を参照のこと。
【0162】
本発明のキメラモノクローナル抗体の構築に適した非ヒトモノクローナル抗体可変ドメインは、種々の動物種(非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、家禽、ウサギ、マウス、ラット、モルモット、ハムスター、犬およびネコ起源が挙げられるが、これらに限定されない)から単離され得る。ヒト抗体重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA、およびIgA2からなる群より選択される抗体アイソタイプから単離され得る。
【0163】
本明細書で議論される、例示的なHLA−DR特異的キメラモノクローナル抗体は、ヒト重鎖定常ドメインおよび軽鎖定常ドメイン(そrぞれ、配列番号14および3)に作動可能に融合された、マウスモノクローナル抗体DN1921の重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号13および2)を含む。この本明細書で議論されるキメラDN1921モノクローナル抗体は、インビボで投与される場合に、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導することができ、かつ免疫抑制性である。例示的な全長キメラDN1921モノクローナル抗体の重鎖のアミノ酸配列は、配列番号12として示される。例示的な全長キメラDN1921モノクローナル抗体軽鎖のアミノ酸配列は、配列番号1として示される。
【0164】
キメラDN1921の重鎖および軽鎖(それぞれ配列番号12および1)の改変体もまた提供される。特定の実施形態において、キメラDN1921の重鎖および軽鎖の改変体は、それぞれ配列番号12および1に少なくとも70%同一である。他の実施形態において、キメラDN1921重鎖および軽鎖の改変体は、それぞれ配列番号12および1に少なくとも80%、90%、または95%同一である。なおさらなる実施形態において、キメラDN1921重鎖および軽鎖の改変体は、それぞれ配列番号12および1に少なくとも98%または99%同一である。
【0165】
本明細書で開示される、代替的な例示的HLA−DR特異的キメラモノクローナル抗体は、ヒト重鎖定常ドメインおよび軽鎖定常ドメイン(それぞれ、配列番号35および24)に作動可能に融合された、マウスモノクローナル抗体DN1924の重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号35および24)を含む。本明細書で開示されるキメラDN1924モノクローナル抗体は、インビボで投与された場合に、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導することができ、非免疫抑制性である。例示的な全長キメラDN1924モノクローナル抗体の重鎖のアミノ酸配列は、本明細書で配列番号34として示される。例示的な全長キメラDN1924モノクローナル抗体軽鎖のアミノ酸配列は、本明細書で配列番号23として示される。
【0166】
代替的実施形態において、キメラDN1924重鎖および軽鎖の改変体は、それぞれ、配列番号34および23である。特定の実施形態において、キメラDN1924重鎖および軽鎖の改変体は、それぞれ配列番号34および23に少なくとも70%同一である。他の実施形態において、キメラDN1924重鎖および軽鎖の改変体は、それぞれ配列番号34および23に少なくとも80%、90%、または95%同一である。なおさらなる実施形態において、キメラDN1924重鎖および軽鎖の改変体は、それぞれ配列番号34および23に少なくとも98%または99%同一である。
【0167】
【表1−1】
【0168】
【表1−2】
【0169】
【表1−3】
【0170】
【表2−1】
【0171】
【表2−2】
【0172】
【表2−3】
【0173】
【表2−4】
(ヒト化抗体)
なおさらなる実施形態において、本発明は、ヒト化HLA−DR特異的アポトーシス性モノクローナル抗体を提供し、ここで、このヒト化抗体は、1つ以上の非ヒト相補性決定領域(CDR)、ヒト可変ドメインフレームワーク領域(FR)、およびヒト重鎖定常ドメイン(例えば、IgG2重鎖定常ドメイン)およびヒト軽鎖定常ドメイン(例えば、Igκ軽鎖定常ドメイン)を含む。一般に、「ヒト化」HLA−DR特異的抗体は、そうでなければ、ヒトレシピエントにおけるこれらの部分の治療的適用の持続時間および有効性を制限する、対応する非ヒト(例えば、げっ歯類)抗体またはその非ヒト領域に対する、望ましくない免疫学的応答を最小にするのに有用である。Jonesら,Nature 321:522−525(1986);Reichmannら,Nature 332:323−327(1988);およびVerhoeyernら,Science 239:1534−1536(1988)。
【0174】
本明細書中で使用される場合、用語「ヒト化」は、ヒト抗体(レシピエント抗体)を含むことを意味し、ヒト化抗体においては、レシピエントの相補性決定領域(CDR)に由来する残基が、非ヒト種(ドナー抗体)(例えば、所望の特異性、親和性および能力を有する、マウス、ラットまたはウサギ)のCDRに由来する残基で置き換えられる。いくつかの場合、ヒト抗体の可変ドメインフレームワーク残基は、対応する非ヒト残基で置き換えられる。ヒト化抗体はまた、レシピエント抗体にも、輸入されたCDRまたはフレームワークの配列にも見られない残基を含み得る。
【0175】
一般に、ヒト化抗体は、少なくとも1つ、そして、代表的には2つの、可変ドメインの実質的に全てを含み、ここでは、CDR領域の全てまたは実質的に全ては、非ヒト抗体のCDR領域に対応し、そして、これらの領域の全てまたは実質的に全ては、ヒト抗体のコンセンサス配列の領域である。ヒト化抗体はまた、好適には、抗体の定常領域の少なくとも一部(代表的には、ヒト抗体の一部)を含む。
【0176】
ヒト化モノクローナル抗体は、代表的に、約90〜95%がヒトであり、相補性決定領域(CDR)のみが非ヒトである。一般に、げっ歯類細胞において産生されるヒト化モノクローナル抗体は、それらが由来する種からの対応するキメラモノクローナル抗体よりも免疫原性が低い。Bellら,Lancet 355:858−859(2000)。
【0177】
上述のように、用語「相補性決定領域」(CDR)は、結合抗原の3次元表面と相補的な表面を形成する抗体分子の超可変領域をいう。N末端からC末端に向かって、抗体の重鎖および軽鎖の各々は、3つのCDR(CDR1、CDR2およびCDR3)から構成される。それゆえ、HLA−DR抗原結合部位、は、合計6つのCDRを含み、重鎖V領域および軽鎖V領域の各々からの3つのCDRを含む。CDRのアミノ酸残基は、結合抗原との広範な接触を形成し、最も広範な抗原接触は、重鎖CDR3とである。
【0178】
ヒト化HLA−DR特異的アポトーシス性抗体は、ヒト可変ドメインフレームワーク領域(FR)に作動可能に融合した1つ以上の非ヒト相補性決定領域(CDR)を含んで、重鎖可変ドメインおよび軽鎖可変ドメインを生じ、これらは、ヒト定常ドメインの重鎖および軽鎖(本明細書において、それぞれ、配列番号14および3に示される、重鎖IgG2定常ドメインおよび軽鎖Igκ定常ドメインにより例示される)に作動可能に融合している。
【0179】
本発明の例示的なHLA−DR特異的ヒト化モノクローナル抗体は、ヒト重鎖フレームワーク(FR)ドメインに作動可能に融合された、マウスモノクローナル抗体DN1921重鎖可変ドメイン(配列番号13)の1つ以上の相補性決定領域(CDR)を含んで、ヒト化HLA−DR特異的重鎖可変ドメインを生じ、これは、必要に応じて、定常ドメインが、本明細書において、配列番号14に示されるヒトIgG2重鎖定常ドメインにより例示される、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される、ヒト重鎖定常ドメインに作動可能に融合される。
【0180】
あるいは、またはさらに、本発明の例示的なHLA−DR特異的ヒト化モノクローナル抗体は、ヒト軽鎖フレームワーク(FR)ドメイン1、2、3および/または4に作動可能に融合された、マウスモノクローナル抗体DN1921軽鎖可変ドメイン(配列番号2)の1つ以上の相補性決定領域(CDR)を含んで、ヒト化HLA−DR特異的軽鎖可変ドメインを生じ、これは、必要に応じて、本明細書において配列番号3に示されるヒトIgκ軽鎖定常ドメインにより例示される、ヒト軽鎖定常ドメインに作動可能に融合される。
【0181】
DN1921に基づくヒト化モノクローナル抗体は、HLA−DR発現腫瘍細胞においてアポトーシスを誘導し得、そして、インビボで投与された場合に、免疫抑制的である。本発明に従うヒト化抗体を作製するために適切なDN1921 CDRは、配列番号17(DN1921 VH CDR1)、配列番号19(DN1921 VH CDR2)、配列番号21(DN1921 VH CDR3)、配列番号6(DN1921 VL CDR1)、配列番号8(DN1921 VL CDR2)、および配列番号10(DN1921 VL CDR3)として表される。
【0182】
本発明のさらなる例示的なHLA−DR特異的ヒト化モノクローナル抗体は、ヒトフレームワーク(FR)ドメイン1、2、3および/または4に作動可能に融合された、マウスモノクローナル抗体DN1924重鎖可変ドメイン(配列番号35)の1つ以上の相補性決定領域(CDR)を含んで、ヒト化HLA−DR特異的重鎖可変ドメインを生じ、これは、必要に応じて、定常ドメインが、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択され、そして、本明細書において、配列番号36に示されるヒトIgG2重鎖定常ドメインにより例示される、ヒト重鎖定常ドメインに作動可能に融合される。
【0183】
あるいは、またはさらに、本発明のさらなる例示的なHLA−DR特異的ヒト化モノクローナル抗体は、ヒトフレームワーク(FR)ドメイン1、2、3および/または4に作動可能に融合された、マウスモノクローナル抗体DN1924軽鎖可変ドメイン(配列番号24)の1つ以上の相補性決定領域(CDR)を含んで、ヒト化HLA−DR特異的軽鎖可変ドメインを生じ、これは、必要に応じて、本明細書において、配列番号25に示されるヒトIgκ軽鎖定常ドメインによりヒト軽鎖定常ドメインにより例示される、ヒト軽鎖定常ドメインに作動可能に融合される。
【0184】
DN1924に基づくヒト化モノクローナル抗体は、HLA−DRを発現する細胞においてアポトーシスを誘導し得、そして、インビボで投与された場合に、非免疫抑制的である。本発明に従うヒト化抗体を作製するために適切なDN1924 CDRは、配列番号39(DN1924 VH CDR1)、配列番号41(DN1924 VH CDR2)、配列番号43(DN1924 VH CDR3)、配列番号28(DN1924 VL CDR1)、配列番号30(DN1924 VL CDR2)、および配列番号32(DN1924 VL CDR3)として表される。
【0185】
各CDRに隣接するのは、主にV領域を、抗原結合部位へと折りたたむことを担う、フレームワーク(FR)領域、特に、CDRに直ぐ隣接するFR残基である。いくつかのFR残基は、結合抗原に接触し得る。FR内では、特定のアミノ残基および特定の構造的特徴が、非常に高度に保存されている。この点から、全てのV領域の配列は、約90アミノ酸残基の内部ジスルフィドループを含む。V領域が、結合部位に折りたたまれるとき、CDRは、抗原結合表面を形成する突出したループモチーフとして提示される。CDRループの、特定の正確なCDRアミノ酸配列への折りたたみ形状に影響を与える、FRの保存された構造領域がある。さらに、特定の残基が、抗体の重鎖と軽鎖との相互作用を安定化する、非共有結合性内部ドメイン接点に関与することが知られている。
【0186】
非ヒト抗体をヒト化するための方法は、当該分野で周知である。一般に、ヒト化抗体は、非ヒト供給源から導入された1つ以上のアミノ酸残基を有する。ヒト化は、適切なヒト抗体定常ドメインに融合する前に、ヒト支持FRにCDRを接合することにより達成され得る。Jonesら,Nature 321:522−525(1986);Reichmannら,Nature 332:323−327(1988);およびVerhoeyernら,Science 239:1534−1536(1988)を参照のこと。
【0187】
あるいは、げっ歯類のCDRは、組換え的に操作されたげっ歯類FRにより支持され得る。1992年12月23日に公開された欧州特許第519,596号を参照のこと。Queenに対する、米国特許第5,530,101号、同第5,585,089号、同第5,693,762号、および同第6,180,370号;Adairに対する米国特許第5,859,205号;ならびに、Winterに対する米国特許第5,225,539号(これらの特許の各々は、その全体が、本明細書中に参考として援用される)もまた参照のこと。
【0188】
実際には、ヒト化抗体は、代表的に、いくつかのCDR残基、そして、可能ないくつかのフレームワーク残基が、げっ歯類抗体の類似の部位に由来する残基で置き換えられているヒト抗体である。従って、本発明のヒト化HLA−DR特異的抗体の各々は、それぞれ、重鎖FRと軽鎖FRとの間に挿入された、少なくとも1、2もしくは3の非ヒト重鎖CDR、および/または、少なくとも1、2もしくは3の非ヒト軽鎖CDRを含み、これらは、CDRに対する支持を提供し、そして、互いに関するCDRの空間的関係を規定する。
【0189】
(完全ヒト抗体)
完全ヒト抗体は、ファージディスプレイ技術およびトランスジェニックマウス法により作製され得る。ファージディスプレイ技術により同定された抗体フラグメントをコードするDNAフラグメントは、組換え技術により合わされ、完全な「ヒト」抗体を生成する。トランスジェニックマウスから抗体を作製するために、ハイブリドーマが調製され、そして、従来のモノクローナル抗体法についてスクリーニングされる。
【0190】
ファージディスプレイ技術により完全ヒト抗体を作製するための方法論は、Hoogenboomら,J.Mol.Biol.227:381(1991)およびMarksら,J.Mol.Biol.222:581(1991)に記載される。米国特許第6,248,516号、同第6,291,158号、同第6,291,159号、同第6,291,160号、同第6,291,161号、同第5,969,108号、同第6,172,197号、同第5,885,793号、同第6,265,150号、同第5,223,409号、同第5,403,484号、同第5,571,698号、同第5,837,500号、および同第6,300,064号(この各々は、本明細書中に参考として援用される)もまた参照のこと。
【0191】
完全ヒト抗体を作製するためのトランスジェニック動物系は、米国特許第6,150,584号、同第6,114,598号、同第6,162,963号、同第6,075,181号および同第5,770,429号(この各々は、本明細書中に参考として援用される)に開示される。
【0192】
またヒトモノクローナル抗体の調製に利用可能なさらなる技術は、例えば、「Monoclonal Antibodies and Cancer Therapy」、p.77(Alan R.Liss編,1985)およびBoernerら,J.Immunol.147(1):86−95(1991)に記載される。
【0193】
(HLA−DR抗体の抗原結合フラグメント、改変体および誘導体)
本発明はまた、前述のHLA−DR特異的モノクローナル抗体のいずれかの、HLA−DR抗原結合フラグメント、改変体、および/または誘導体を企図する。本発明のHLA−DR抗原結合フラグメント、改変体、および誘導体の各々は、HLA−DR発現正常細胞ではなく、HLA−DR発現腫瘍細胞において、アポトーシスを誘導し得る。特定の実施形態において、HLA−DR抗原結合フラグメント、改変体および誘導体(例えば、モノクローナル抗体DN1921に基づくもの)は、インビボで投与された場合、免疫抑制的である。他の実施形態において、HLA−DR抗原結合フラグメント、改変体および誘導体(例えば、モノクローナル抗体DN1924に基づくもの)は、インビボで投与された場合、非免疫抑制的である。
【0194】
対応する全長HLA−DR特異的抗体分子の免疫学的結合特性を示し得る、治療的に有用なHLA−DR特異的抗原結合フラグメントを作製するための方法論は、当該分野で容易に利用可能である。本発明の方法は、HLA−DR分子の架橋を促進し、それによりアポトーシスを刺激するために、HLA−DR特異的抗体が二価、三価、または多価であることを必要とする。従って、HLA−DR特異的抗体の1つ以上のフラグメントは、固体支持体に結合され、それにより、HLA−DRの架橋を促進し得る。
【0195】
タンパク質分解酵素であるパパインは、優先的にIgG分子を切断して、いくつかのフラグメントを生じ、このうち2つ(「F(ab)」フラグメント)は、インタクトなこう現結合部位を含む共有結合性のヘテロ二量体を含む。酵素ペプシンは、IgG分子を切断して、いくつかのフラグメントを提供し得、このフラグメントは、両方の抗原結合部位を含む「F(ab’)2」フラグメントを含む。「Fv」フラグメントは、IgM抗体分子の、そして、時として、IgG抗体分子またはIgA抗体分子の優先的なタンパク質分解性切断により生成され得る。Fvフラグメントは、あるいは、当該分野で公知の技術を用いて作製され得る。Fvフラグメントは、ネイティブな抗体分子の抗原認識能および抗原結合能の大半を保持する、抗原結合部位を含む、非共有結合性のVH::VLヘテロ二量体を含む。Inbarら,Proc.Nat.Acad.Sci.USA 69:2659−2662(1972);Hochmanら,Biochem.15:2706−2710(1976);およびEhrlichら,Biochem 19:4091−4096(1980)。
【0196】
単鎖Fv(「sFv」)ポリペプチドは、ペプチドをコードするリンカーにより連結された、VHをコードする遺伝子およびVLをコードする遺伝子を含む遺伝子融合物から発現される、共有結合されたVH::VLヘテロ二量体である。Hustonら,Proc.Nat.Acad.Sci.USA 85(16):5879−5883(1988)。1つの抗体V領域に由来する、自然に凝集しているが、化学的には分かれている、軽ポリペプチド鎖および重ポリペプチド鎖を、sFv分子(抗原結合部位の構造に実質的に類似する3次元構造へと折り畳まれる)へと変換するための化学構造を見分けるための多数の方法が記載されている。例えば、米国特許第5,091,513号、同第5,132,405号および同第4,946,778号を参照のこと。
【0197】
本発明に従う、抗体、およびその抗原結合フラグメントをコードするポリヌクレオチドは、従来の分子生物学および組換えDNA方法論により作製され得る。このような方法論は、文献で完全に説明されている。例えば、Sambrookら,Molecular Cloning:A Laboratory Manual(第2版,1989);Maniatisら,Molecular Cloning:A Laboratory Manual(1982);DNA Cloning:A Practical Approach,第I巻および第II巻(D.Glover編);Oligonucleotide Synthesis(N.Gait編,1984);Nucleic Acid Hybridization(B.HamesおよびS.Higgins編,1985);Transcription and Translation(B.HamesおよびS.Higgins編,1984);Animal Cell Culture(R.Freshney編,1986);Perbal,A Practical Guide to Molecular Cloning(1984)を参照のこと。これらの刊行物の各々は、その全体が参考として援用される。
【0198】
簡単に言うと、例えば、抗体重鎖または抗体軽鎖をコードするDNA配列は、適切な発現ベクター内に連結され、この発現ベクターは、抗体領域をコードするポリペプチドに作動可能に連結された転写プロモーターを含み、そして、この抗体領域をコードするポリヌクレオチドの3’側には転写終止シグナルを含む。適切な発現ベクターはまた、転写開始部位、翻訳開始を命令するためのKozak配列、および翻訳を停止させる終止コドンを提供し得る。さらに、発現ベクターはまた、ポリペプチドをコードする1つ以上のポリヌクレオチド配列(例えば、His−His−His−His−His−HisまたはFLAG(登録商標)配列Asp−Tyr−Lys−Asp−Asp−Asp−Asp−Lys(Sigma−Aldrich,St.Louis,MO))を含み得、この配列は、抗体ペプチドの検出および親和性精製を容易にする。
【0199】
発現は、組換えポリペプチドをコードするDNA分子を含む発現ベクターで形質転換または形質移入された任意の適切な宿主細胞内で達成され得る。適切な宿主細胞としては、原核生物、酵母および高等真核生物細胞が挙げられる。代表的に、使用される宿主細胞は、COSまたはCHOのような哺乳動物細胞株由来である。組換え抗体ポリペプチドを培養培地中に分泌する適切な系からの上清が、まず、市販のフィルタを使用して濃縮され得る。濃縮の後、濃縮物が、適切な精製マトリクス(例えば、親和性マトリクスまたはイオン交換樹脂)に適用され得る。最後に、1つ以上の逆相HPLC工程を用いて、組換えポリペプチドをさらに精製し得る。
【0200】
抗体フラグメント(例えば、Fabフラグメント)は、あるいは、従来のポリペプチド合成方法論により合成され得る。例えば、このような抗体フラグメントは、市販の固相技術(例えば、Merrifield固相合成法)を用いて合成され得、この方法において、アミノ酸が、増大するアミノ酸鎖に連続的に添加される。Merrifield,J.Am.Chem.Soc.85:2149−2146(1963)を参照のこと。ポリペプチドの自動化合成のための設備は、Perkin Elmer/Applied BioSystems Division(Foster City,Calif.)などの供給業者から市販されており、製造業者の説明書に従って操作され得る。
【0201】
従って、本発明に従うHLA−DR特異的抗体およびそのフラグメントは、フラグメント、誘導体および改変体がHLA−DR結合の特異的かつアポトーシス性の非免疫抑制活性の高原結合特性に実質的に影響を与えない限り、これらのフラグメント、誘導体および改変体を包含する。
【0202】
本明細書中で使用される場合、ポリペプチドまたはタンパク質の「フラグメント、誘導体および改変体」は、ネイティブなポリペプチドまたはタンパク質とは、1つ以上の置換、欠失および/または挿入で異なるポリペプチドまたはタンパク質であり、その結果、このポリペプチドまたはタンパク質の抗原結合活性は、実質的に減少されない。換言すると、HLA−DRに特異的に結合する改変体の能力は、親HLA−DR特異的抗体に対して、増強されていても不変であってもよいか、または、得られるHLA−DR特異的抗体もしくはそのフラグメントの効力に影響を与えることなく、親抗体に対して、50%未満、そして代表的には20%未満消失していてもよい。一般に、適切なHLA−DR特異的抗体の誘導体または改変体は、HLA−DRを発現する新生物細胞においてアポトーシスを誘導する、その誘導体または改変体の能力を評価することによって特徴付けられ得る。
【0203】
本明細書中で使用される場合、用語「改変体」とは、具体的に同定された配列と異なるヌクレオチド配列またはアミノ酸配列を包含し、改変体においては、1つ以上のヌクレオチド残基またはアミノ酸残基が、欠失、置換または付加されている。改変体は、天然に存在する対立遺伝子改変体であっても、天然に存在しない改変体であってもよい。改変体の配列(ポリヌクレオチドまたはポリペプチド)は、好ましくは、本発明の配列と、少なくとも70%、より好ましくは、少なくとも80%もしくは少なくとも90%、なおより好ましくは少なくとも95%、そして最も好ましくは、少なくとも98%の同一性を示す。
【0204】
%同一性は、以下に記載されるように、比較されるべき2つの配列を整列し、整列した部分における同一の残基の数を決定し、この数を、本発明の(照会した)配列内の残基の総数で割り、そして、その結果に100をかけることによって決定される。列挙されたレベルの配列類似性を示すことに加え、本発明の改変体の配列は、好ましくは、その改変体が比較される配列の抗原結合に対して実質的に類似する抗原結合性(antigen−bindingity)を示す。
【0205】
改変体は、1つのアミノ酸が、類似の特性を有する別のアミノ酸で置換される、置換として定義される、「保存的アミノ酸置換」を含み得、その結果、ポリペプチドの二次構造および疎水性の性質は、実質的に不変である。アミノ酸置換は、一般に、残基の極性、電荷、可溶性、疎水性、親水性および/または両親媒性の性質における類似性に基づいてなされ得る。例えば、負に帯電したアミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられ;正に帯電したアミノ酸としては、リジンおよびアルギニンが挙げられ;そして、類似の親水性値を有する、無電荷の極性頭基を有するアミノ酸としては、ロイシン、イソロイシンおよびバリン;グリシンおよびアラニン;アスパラギンおよびグルタミン;ならびにセリン、スレオニン、フェニルアラニンおよびチロシンが挙げられる。保存的変化を表し得るアミノ酸の他のグループとしては、以下が挙げられる:(1)ala、pro、gly、glu、asp、gln、asn、ser、thr;(2)cys、ser、tyr、thr;(3)val、ile、leu、met、ala、phe;(4)lys、arg、his;および(5)phe、tyr、trp、his。改変体はまた、あるいは、非保存的な変化を含み得る。
【0206】
ポリペプチドの抗原結合フラグメント、誘導体および改変体は、抗体の化学的もしくは酵素的な消化のいずれかによってか、または、その抗体をコードするポリヌクレオチドおよび結果として生じる変異体のその後の発現の変異分析によって同定され得る。ついで、ポリペプチドのフラグメントまたは変異体ポリペプチドが、例えば、以下に提供される代表的なアッセイを使用して、どの部分が生物活性を保持するかを決定するために試験される。
【0207】
本発明の抗体のフラグメント、誘導体および改変体はまた、合成もしくは組換え手段によって作製され得る。約100より少ないアミノ酸、一般には、約50より少ないアミノ酸を有する合成ポリペプチドは、当業者に周知の技術を使用して作製され得る。例えば、このようなポリペプチドは、上述のMerrifield固相合成法により合成され得る。
【0208】
改変体はまた、オリゴヌクレオチド指向型、部位特異的変異誘発のような標準的な変異誘発技術を用いて調製され得る。Kunkel,Proc.Natl.Acad.Sci.USA 82:488−492(1985)。ポリヌクレオチド配列の部分はまた、短縮型ポリペプチドの調製を可能にする標準的な技術を用いて除去され得る。改変体は、さらに、またはあるいは、例えば、ポリペプチドの二次構造および疎水性の性質に対して最小限の影響を有するアミノ酸の欠失または付加によって修飾され得る。
【0209】
HLA−DR特異的抗体のフラグメント、誘導体および改変体は、好ましくは、ネイティブなHLA−DR特異的抗体に対して、少なくとも約70%、より好ましくは少なくとも約80%もしくは90%、そしてより好ましくは、少なくとも約95%もしくは98%の配列同一性を示す。ポリペプチド配列が整列され得、そして、特定の領域における同一なアミノ酸の百分率が、公的に利用可能なコンピュータアルゴリズムを用いて、別のポリペプチドに対して決定され得る。ポリペプチド配列の整列および同一性は、BLASTPアルゴリズムを用いて試験され得る。BLASTPアルゴリズムは、PearsonおよびLipman,Proc.Natl.Acad.Sci.USA 85:2444−2448(1988);ならびにPearson,Methods in Enzymol.183:63−98(1990)に記載される。
【0210】
BLASTPソフトウェアは、NCBIの匿名のFTPサーバ上で利用可能であり、そして、National Center for Biotechnology Information(NCBI)、National Library of Medicine,Building 38A,Room 8N805,Bethesda,MD 20894から利用可能である。文書に記載され、アルゴリズムと共に配布されているデフォルトパラメータに設定された、BLASTPアルゴリズムバージョン2.0.6[1998年9月10日]およびバージョン2.0.11[2000年1月20日]は、本発明に従う改変体の決定において使用するための例示的なものである。アルゴリズムのBLASTファミリー(BLASTPを含む)の使用は、NCBIのウェブサイトおよびAltschulら,「Gapped BLAST and PSI−BLAST:a new generation of protein database search programs」,Nucleic Acids Res.25:338903402(1997)の刊行物に記載される。
【0211】
BLASTPまたは類似のアルゴリズムにより得られた、照会配列による1つ以上のデータベースの配列に対する「ヒット」は、配列の類似の部分を整列し、そして同定する。このヒットは、配列の重なりの類似性の程度および長さの順に並べられる。データベースの配列に対するヒットは、一般に、照会配列の、配列の長さの一部のみに対する重なりを表す。
【0212】
ポリペプチド配列の%同一性は、デフォルトパラメータに設定されたBLASTPのような適切なアルゴリズムを使用してポリペプチド配列を整列し;整列された部分に対して同一のアミノ酸の数を同定し;同一なアミノ酸の数を、本発明のポリペプチドのアミノ酸の総数で割り;そして、100をかけて、%同一性を決定することによって決定される。
【0213】
BLASTPアルゴリズムはまた、ポリペプチドの整列についての「期待」値を生じる。この期待値(E)は、特定の大きさのデータベースを検索する際に、偶然、特定の数の隣接する配列にわたって見えることが「期待」され得るヒットの数を示す。期待値は、データベースに対するヒットが、真の類似性を示すかどうかを決定するための有意な閾値として使用される。例えば、ポリペプチドヒットに対して割当てられた0.1のE値が、SwissProtデータベースの大きさのデータベースにおける値を意味するものとして解釈される場合、ほんの偶然に、類似のスコアを有する配列の整列された部分にわたって、0.1のマッチを見ることが期待され得る。この基準によれば、配列の整列かつマッチした部分は、90%の関連する確率を有する。整列かつマッチした部分にわたって0.01以下のE値を有する配列について、SwissProtデータベースにおいて偶然にマッチを発見する確率は、BLASTPアルゴリズムを用いた場合の1%以下である。
【0214】
1つの実施形態によれば、「改変体」HLA−DR特異的抗体は、本発明のHLA−DR特異的抗体および抗体フラグメントの各々を参照すれば、好ましくは、本発明のHLA−DR特異的抗体と同じ数であるか、またはこれよりも少ないアミノ酸を有し、かつ、本発明のHLA−DR特異的抗体と比較した場合に、0.01以下のE値を生じる配列を含む。
【0215】
本発明のHLA−DR特異的抗体に対して特定の%同一性を有することに加え、改変体抗体は、好ましくは、本発明のHLA−DR特異的抗体に共通するさらなる構造的および/または抗原結合特徴を有する。本発明のHLA−DR特異的抗体に対して、特定の程度の同一性を有する抗体は、その一次構造において高い程度の類似性を共有し、そして、実質的に類似の抗原結合特性を有する。本発明のHLA−DR特異的抗体に対して、高い程度の一次構造の類似性を共有することに加え、HLA−DRに対して特定の程度の同一性を有するか、または、HLA−DRに特異的に結合し得る抗体は、好ましくは、以下の特徴のうち少なくとも1つを有する:(i)これらは、本発明のHLA−DR特異的抗体と実質的に同じ抗原結合特性を有する;または(ii)例えば、CDRおよび/もしくはフレームワーク領域のような、特定可能なドメインを共通して含む。
【0216】
(結合体)
意図される正確な治療的および/または診断的な適用に依存して、本発明の抗体またはそのフラグメントは、1つ以上の治療剤と結合されることが望ましくあり得る。この点で適切な薬剤としては、放射性核種、分化誘導因子(differentiation inducer)、薬物、毒素およびこれらの誘導体が挙げられる。例示的な適切な放射性核種としては、90Y、123I、125I、131I、186Re、188Re、211Atおよび212Biが挙げられる。例示的な適切な薬物としては、メトトレキサート、ならびにピリミジンアナログおよびプリンアナログが挙げられる。例示的な適切な分化誘導因子としては、エステルおよび酪酸が挙げられる。例示的な適切な毒素としては、リシン、アブリン、ジフテリア毒素、コレラ毒素、ゲロニン、Pseudomonas外毒素、Shigella毒素およびヤマゴボウ抗ウイルスタンパク質が挙げられる。
【0217】
治療剤は、直接または間接(例えば、リンカー基を介して)のいずれかで、適切なモノクローナル抗体に結合(例えば、共有結合)され得る。薬剤と抗体との間の直接的な反応は、各々が、互いと反応し得る置換基を有する場合に可能である。例えば、一方の上にある求核基(例えば、アミノ基またはスルフヒドリル基)は、もう一方の上にある、カルボニル含有基(たとえば、無水物)もしくは、酸ハロゲン化物と、または、良好な脱離基(例えば、ハロゲン化物)を含有するアルキル基と反応し得る。
【0218】
あるいは、治療剤と抗体とをリンカー基を介して結合することが望ましくあり得る。リンカー基は、結合能との干渉を避けるために、薬剤から抗体を遠ざけるためのスペーサーとして機能し得る。リンカー基はまた、薬剤または抗体上の置換基の化学反応性を増加させ、従って、カップリング能を増加させるように機能し得る。化学反応性の増加はまた、そうでなければ、不可能であった、薬剤、または薬剤の官能基の使用を容易にする。
【0219】
ホモ官能性およびヘテロ官能性の両方の、種々の二官能性試薬または多官能性試薬(例えば、Pierce Chemical Co.,Rockford,Illに記載されるもの)が、リンカー基として使用され得ることが、当業者に明らかである。カップリングは、例えば、アミノ基、カルボキシル基、スルフヒドリル基または酸化炭化水素残基を介して達成され得る。このような方法論は、例えば、米国特許第4,671,958号に記載される。
【0220】
治療剤が、本発明の免疫複合体の抗体部分を含まないときにより強力である場合、細胞内への内部移行の間またはその際に切断可能であるリンカー基を使用することが望ましくあり得る。多数の異なる切断可能なリンカー基が記載されている。これらのリンカー基からの薬剤の細胞内放出のための機構としては、ジスルフィド結合の還元による切断(例えば、米国特許第4,489,710号)、感光性結合の照射による切断(例えば、米国特許第4,625,014号)、アミノ酸側鎖の加水分解による切断(例えば、米国特許第4,638,045号)、血清補体が媒介する加水分解による切断(例えば、米国特許第4,671,958号)、および酸が触媒する加水分解による切断(例えば、米国特許第4,569,789号)が挙げられる。
【0221】
1以上の薬剤を1つの抗体に結合させることが望ましくあり得る。1つの実施形態において、薬剤の複数の分子が、1つの抗体分子に結合される。別の実施形態において、1つ以上の型の薬剤が、1つの抗体に結合される。特定の実施形態にかかわらず、1つ以上の薬剤との免疫複合体は、種々の方法で調製され得る。例えば、1つ以上の薬剤は、抗体分子に直接結合され得るか、または、付着のための複数の部位を提供するリンカーが使用され得る。あるいは、キャリアが使用され得る。
【0222】
キャリアは、直接的であるか、もしくはリンカー基を介する共有結合を含む、種々の方法で薬剤を有し得る。適切なキャリアとしては、アルブミンのようなタンパク質(例えば、米国特許第4,507,234号)、アミノデキストランのようなペプチドおよび多糖類(例えば、米国特許第4,699,784号)が挙げられる。キャリアはまた、非共有結合またはリポソーム小胞内(例えば、米国特許第4,429,008号および同第4,873,088号)のようなカプセル化によって薬剤を有し得る。放射性核種剤に固有のキャリアとしては、放射性ハロゲン化低分子およびキレート化合物が挙げられる。例えば、米国特許第4,735,792号は、代表的な放射性ハロゲン化低分子およびその合成を開示する。放射性核種キレートは、キレート化合物から形成され得、これらは、窒素原子および硫黄原子を、金属または金属酸化物、放射性核種と結合するためのドナー原子として含有する化合物を含む。例えば、米国特許第4,673,562号は、代表的なキレート化合物およびその合成を開示する。
【0223】
用語「細胞傷害性薬物」とは、本明細書中で使用される場合、癌細胞の増殖を阻害するか、または癌細胞の死を促進するために使用される薬物を指す。細胞傷害性薬物の例としては、ara−C、ブレオマイシン、シスプラチン、クラドリビン(cladribine)、シクロホスファミド、ドキソルビシン、エトポシドおよび5−フルオロウラシル(5−FU)のような化学療法剤が挙げられる。
【0224】
(抗HLA−A−DR抗体の同定および特徴づけのためのアッセイ系)
以下のアッセイ系は、本発明に従う、HLA−DR特異的なアポトーシス性、非免疫抑制性抗体の慣用的な同定および特徴づけのために適切に使用され得る。
【0225】
(クラスII MHC)
「免疫抑制的であり、かつ細胞傷害性である」抗HLA−DR mAbは、第1のドメインを認識し、ペプチド結合溝の近くに位置するエピトープに結合することによって抗原提示を阻害し得る。本発明の抗HLA−DR mAb(例えば、DN1924)は、「細胞傷害性のみ」である。従って、このような「細胞傷害性のみ」の抗体が結合する、HLA−DRの正確な配列は、「免疫抑制的であり、かつ細胞傷害性である」抗体の配列とは同一でない。Vidovicら,Eur.J.Immunol.25:3349−3355(1995)。
【0226】
(アポトーシスによる細胞死)
上述のように、アポトーシス(プログラムされた細胞死)は、抗MHCクラスII抗体の、クラスIIを発現する細胞への結合と関係している。APC(例えば、Bリンパ球、マクロファージ、単球、樹状細胞など)の表面上に発現されたクラスII MHCをコードする分子は、最終的に、両方の細胞型の活性化および分化をもたらす相互作用である、Tリンパ球に対する抗原提示の制限的要素として機能する。
【0227】
HLA−DR媒介性の細胞死は、非常に迅速で、Fcレセプター非依存性であり、かつ、補完的であり、非壊死性であることが実証されている。Trumanら,Int.Immunol.6(6):887−896(1994)およびTrumanら,Blood 89(6):1996−2007(1997)。
【0228】
本発見は、腫瘍細胞のアポトーシスの誘発における抗HLA−DR抗体の役割に関し、従って、このような細胞においてプログラムされた細胞死を誘導するための方法を提供する。例示的な実施形態において、HLA−DR抗原を架橋し得る抗HLA−DR抗体およびそのフラグメントは、HLA−DR発現腫瘍細胞により媒介される状態を研究または処置する際に有用であり、すなわち、HLA−DR発現腫瘍細胞に関連する障害を処置または予防するのに有用である。従って、本発明の抗体は、種々の疾患を処置するのに有用であり、これらの疾患としては、B細胞起源の癌により特徴付けられる任意の疾患(アポトーシスの増加が望ましくない)、例えば、ホジキンリンパ腫および非ホジキンリンパ腫、慢性リンパ急性白血病、骨髄腫および形質細胞腫が挙げられるがこれらに限定されない。Hollandら,Cancer Med.2:2697−2828(1996)。
【0229】
(細胞死の機構の評価)
この節では、アポトーシスによる細胞死の程度を評価するために有用なインビトロアッセイを記載する。細胞死は、プロピジウムヨーダイド(PI)での細胞染色によってか、または、アポトーシスによる細胞死に特異的なアッセイ(例えば、アネキシンVでの染色)を用いることによって検出され得る。Vermesら,J.Immunol.Meth.184:39−51(1995)。壊死による細胞死は、以下に記載するような細胞の生存度についてのアッセイを、関連のある細胞の形態の顕微鏡での観察との組み合わせの結果を評価することによって、アポトーシスによる細胞死から区別され得る。
【0230】
(壊死性細胞死)
壊死は、内部ホメオスタシスの虚脱が細胞膜の完全性の損失およびその後の膨張を含む細胞の溶解(dissolution)を導き、その後に細胞の溶解(lysis)が続く、受動的プロセスである。Schwartzら、1993。壊死性の細胞死は、細胞膜の完全性の損失、およびヨウ化プロピジウム(PI)のような、一次性の壊死および二次性の壊死を受ける細胞のDNAに結合することが当業者に公知の色素に対する透過性によって、特徴付けられる。Vitaleら、Histochemistry 100:223−229(1993)およびSwatら、J.Immunol.Methods 137:79−87(1991)。壊死は、アポトーシスから区別され得る(アポトーシスの初期段階においては、細胞膜はインタクトなまま残る)。結果として、以下に記載されるようにアポトーシス性細胞死を壊死性細胞死から区別するため、アポトーシスについてのアッセイと並行して、PIを使用した色素排除アッセイ(dye exclusion assay)が使用され得る。PIを用いた蛍光活性化細胞分類機(Fluorescent−activated cell sorter;FACS)ベースのフローサイトメトリーアッセイは、壊死性細胞の迅速な評価および定量化を可能にする。
【0231】
(アポトーシス性細胞死)
プログラムされた細胞死またはアポトーシスの検出は、当業者に理解されるように達成され得る。アポトーシスを受ける細胞のパーセンテージは、抗HLA−DR抗体の投与とともに、またはその投与なしで、アポトーシスの刺激後の種々の時間で測定され得る。アポトーシス性細胞死を受ける細胞の形態は、一般的に、細胞の細胞質および核の萎縮、ならびにクロマチンの凝縮および断片化によって特徴付けられる。Wyllieら、J.Patrol.142:67−77(1984)。
【0232】
アポトーシス性B細胞における部分的DNA分解が以前に報告されている。Trumanら、Int.Immuunol.6:887−896(1994)およびCohenら、Annu.Rev.Immunol.10:267−293(1992)。この観察に一致して、DNA断片化は、以前に記載された腫瘍性B細胞をアポトーシス誘導性(apoptogenic)の抗HLA−DR mAbとともにインキュベーションした後には検出されなかった。しかし、これらの培養物の相対的細胞サイズおよびPI取り込み式フローサイトメトリーのプロフィールは、アポトーシスを受ける細胞について以前に示されたプロフィールと本質的に同じである。Vidovicら、Cancer Lett.128:127−135(1988);Newellら、Proc.Natl.Acad.Sci.U.S.A.90:10459−10463(1993);およびTrumanら(1994)。
【0233】
(有用性)
(抗HLA−DR抗体に関する使用)
本発明のHLA−DR特異的抗体は、診断用および治療用の組成物ならびに診断用および治療用の方法において利用され得る。例えば、HLA−DR特異的抗体は、HLA−DR発現腫瘍細胞についての診断アッセイおよびHLA−DR発現腫瘍細胞に関する治療において(例えば、組織または血清中のこのような細胞の発現を検出すること、およびこのような腫瘍を有する被験体において臨床的結果を改善する治療のための基礎として働くことに)使用され得る。
【0234】
当該分野で公知の種々の診断アッセイ技術(例えば、競合的結合アッセイ、直接もしくは間接サンドイッチアッセイ、および均一もしくは不均一相において実施される免疫沈降アッセイ)が使用され得る。Zola、「Monoclonal Antibodies:A Manual of Techniques」、pp.147−158(CRC Press,Inc.,1987)。この診断アッセイに使用され得る抗体は、検出可能な部分によって標識され得る。この検出可能な部分は、直接的もしくは間接的に、検出可能なシグナルを生成し得る。例えば、この検出可能な部分は、放射性同位体(例えば、3H、14C、32P、35S、または125I)、蛍光化合物または化学発光化合物(例えば、フルオレセインイソシアネート、ローダミン、もしくはルシフェリン)、あるいは酵素(例えば、アルカリホスファターゼ、β−ガラクトシダーゼ、もしくは西洋ワサビペルオキシダーゼ)であり得る。特異的結合を促進する分子としては、対(例えば、ビオチンおよびストレプトアビジン、ジゴキシンおよび抗ジゴキシンなど)が挙げられる。特異的結合を促進する分子の対のメンバーの一方は、標識が、直接的または間接的に検出可能なシグナルを提供する公知の手順による検出を行わせる分子で標識され得る。抗体を検出可能な部分に結合させるための当該分野で公知の任意の方法が利用され得る。この方法としては、Hunterら、Nature 144:945(1962);Davidら、Biochemistry 13:1014(1974);Painら、J.Immunol.Meth.40:219(1981);およびNygren,J.Histochem.and Cytochen.30:407(1982)によって記載される方法が挙げられる。
【0235】
HLA−DR特異的抗体はまた、細胞培養物または天然の供給源からのHLA−DR発現癌細胞のアフィニティー精製のために有用である。このような方法において、HLA−DRに対する抗体は、当該分野で公知の方法論を用いて適切な支持体(例えば、Sephadex樹脂もしくは濾紙)に固定化される。固定された抗体は、精製されるべきHLA−DR発現癌細胞を含むサンプルと接触させられ、そして固体支持体は、固定化された抗体に結合するHLA−DR発現癌細胞を除く、サンプル中の実質的に全ての物質を除去する適切な培地で洗浄される。支持体は、HLA−DR発現癌細胞を固定化された抗体から放出させる別の適切な培地で洗浄される。
【0236】
(抗HLA−DR特異的モノクローナル抗体の生物学的効果)
本発明のHLA−DR特異的モノクローナル抗体は、HLA−DRの第1ドメインを認識する。本発明の抗体または特異的モノクローナル抗体のCDR領域は、検出可能なレベルのHLA−DRを発現する腫瘍細胞と免疫反応し、そしてこの腫瘍細胞におけるアポトーシスを誘導し得る。HLA−DR特異的マウスモノクローナル抗体DN1924、およびマウス/ヒトキメラモノクローナル抗体は、非腫瘍性成細胞と比較して、腫瘍(プラズマ細胞腫 MC/CAR)細胞(両方ともHLA−DRを発現する)におけるアポトーシスの誘導に関するインビトロでの特異性を示した。実施例2および図2〜4、ならびに実施例7および図8A〜Fを参照のこと。さらに、DN1924による正常なTh反応との干渉の欠如がインビトロで示された。実施例2および図5を参照のこと。
【0237】
(アポトーシス性HLA−DR特異的抗体を用いたインビボ癌治療)
本発明の抗体は、治療的に有効であり、HLA−DR抗原を発現する腫瘍細胞のアポトーシス性細胞死を刺激し得る。これらの発見は、HLA−DR陽性の癌(特に血液の癌)に対する選択的な抗体ベースの抗腫瘍治療の可能性を提起する。
【0238】
抗クラスII MHC mAbの殺腫瘍性(tumoricidal)効果は、クラスII依存性免疫応答の同時的な抑制なしで(両特性ともタンパク質の第一ドメインを認識するmAb認識に関連するが)達成され得る。
【0239】
所望の治療的効果を有する抗体は、宿主にとって生理学的に受容可能なキャリア中において投与され得、そして例えば、非経口、皮下(SC)、腹腔内(IP)、静脈内(IV)などの方法で投与され得る。導入の様式に依存して、抗体は、種々の方法で処方され得る。処方物中の治療的に活性な抗体の濃度は、約1mg/mlから約1g/mlまで変動し得る。
【0240】
本発明の目的のための「被験体」は、ヒトおよび他の動物(特に哺乳動物)の両方を包含する。従って、本方法は、ヒト治療および獣医学的適用の両方に対して適用可能である。例示的実施形態において、被験体は哺乳動物であり、最も例示的な実施形態において、被験体はヒトである。
【0241】
本明細書において使用される場合、用語「改善された治療結果」または「腫瘍細胞数の減少」とは、処置前後に患者および/または患者から採取されたサンプルの試験によって決定される場合の、血中および/または罹患する組織もしくは器官における腫瘍サイズまたは検出可能な循環性癌細胞の数のいずれかの50%減少、好ましくは80%減少、より好ましくは90%減少、そしてさらにより好ましくは100%減少を意味する。
【0242】
本明細書において使用される場合、用語「処置する工程」、「処置」および「治療」とは、治癒可能な治療、予防的な(prophylactic)治療、および予防的な(preventative)治療をいう。
【0243】
本明細書において使用される場合、「治療上有効な量」によって、HLA−DR発現腫瘍細胞を、そのアポトーシスを刺激することによって減少または消失させる用量が意味される。正確な用量は、処置の目的に依存し、そして公知の技術を使用して当業者によって確認され得る。
【0244】
好ましくは、抗体は、非経口投与のために、適切な不活性キャリア(例えば、滅菌生理学的食塩水)中に処方される。例えば、キャリア溶液中の抗体の濃度は、代表的に約1〜約100mg/mlの間である。投与される用量は、投与の経路によって決定される。例示的な投与の経路としては、非経口投与またはIV投与が挙げられる。治療的に有効な用量は、HLA−DR発現性新生物細胞のアポトーシス性細胞死の有意な増加を生じるのに有効な用量である。HLA−DR発現性新生物細胞のアポトーシス性細胞死の有意な増加は、検出可能な量のHLA−DRを発現しない細胞と比較して、HLA−DR発現性細胞のアポトーシス性細胞死の2倍の増加、より好ましくは5倍の増加、さらにより好ましくは10倍の増加、最も好ましくは20倍の増加である。
【0245】
発明の重要な特徴に従って、抗HLA−DR抗体は、単独で、または他の抗癌剤(例えば、化学治療剤(例えば、シスプラチン、タキソール、メトトレキサートなど);腫瘍壊死因子−α(TNF−α);FADD、PMA;イオノマイシン;スタウロスポリンまたはRituxan(登録商標))と組み合わせて投与され得る。
【0246】
抗HLA−DR抗体(例えば、DN1924)の治療上有効な量は、本明細書において記載されるように、HLA−DR陽性腫瘍細胞のアポトーシス性細胞死についてのインビトロモデルにおいてDN1924に関して得られたデータと合わせて、公知の抗体についての確立された有効量との比較によって推定され得る。当該分野において公知であるように、抗体の変性、全身送達 対 局所送達、ならびに年齢、体重、健康、性別、食事、投与時間、薬物の相互作用、および状態の重症度に起因して、用量の調節が必要であり得る。当業者によって、慣習的な実験を通じてこのような調節が行われ得、そして適切な用量が決定され得る。
【0247】
特定の実施形態の範囲内において、本発明の方法は、所望の抗原結合特異性および/または抗原結合親和性を有する抗体に関して細胞をスクリーニングする工程をさらに提供し得る。本明細書において使用される場合、用語「抗原」は、抗体が特異的に結合および/または相互作用するこれらの物質、分子、タンパク質、核酸、脂質および/または炭水化物の全てを広範に包含する。
【0248】
抗体は、例示的な抗原結合特異性および/または抗原結合親和性に関して、当該分野で現在利用可能な任意の方法論によってスクリーニングされ得る。例えば、従来の細胞パニング手順、ウエスタンブロッティング手順、およびELISA手順が、特定の特異性を有する抗体をスクリーニングする工程を達成するために利用され得る。米国特許第4,016,043号、同第4,424,279号、および同第4,018,653号(これらの各々は、本命最小において参考として援用される)への参照によって見られ得るように、広範囲の適切なイムノアッセイ技術が利用可能である。
【0249】
1つの型のアッセイにおいて、未標識の抗−抗体が固体支持体上に固定され、そして試験されるべき抗体含有サンプルがその固定された抗−抗体と接触させられる。第一の複合体の形成を可能にするのに十分な適切な時間の後、次に、検出可能なシグナルを生成し得るレポーター分子で標識された標識抗原を添加してインキュベートし、固定された抗−抗体/抗体サンプル/試験抗原の第二の複合体の形成のために十分な時間を与える。複合体化されない物質は、洗い流され、レポーター分子によって生成されたシグナルの観察によって抗原の存在が決定される。この結果は、眼に見えるシグナルの単純な観察による定性的なものであっても、既知の量の抗原または抗体を含むコントロールサンプルとの比較によって定量化されてもいずれでもよい。この型のアッセイの変化は、サンプルおよび標識された抗原の両方が結合抗体に対して同時に添加される同時アッセイを包含する。
【0250】
本明細書中で使用される場合、用語「固体支持体」とは、例えば、マイクロタイタープレート、膜およびビーズなどをいう。例えば、このような固体支持体は、ガラス、プラスチック(例えば、ポリスチレン)、多糖類、ナイロン、ニトロセルロースまたはテフロン(登録商標)などで構成され得る。このような支持体の表面は、固体または多孔質であり得、そして任意の都合よい形状であり得る。
【0251】
第2の型のアッセイにおいて、抗体を探索するための抗原が固体支持体に結合される。この結合プロセスは当該分野において周知であり、そして一般的に、その抗原を固体支持体に対して架橋結合、共有結合、または物理的に吸収することからなる。ポリマー−抗原複合体は、試験サンプルに備えて洗浄される。次いで、試験されるべきサンプルのアリコートが、固相複合体に添加され、そして抗原への抗体の結合を可能にするのに十分な時間(例えば、20〜40分、またはより都合よい場合、一晩)、適切な条件下(例えば、ほぼ室温〜約38℃、例えば25℃)でインキュベートされる。インキュベーション時間に続いて、固体支持体は、洗浄され、乾燥され、そしてレポーター分子が結合し得る抗体とともにインキュベートされ、それによって固定された抗原に複合体化された試験抗体に対する二次抗体の結合の検出を可能にする。
【0252】
適切な固体支持体としては、ガラスまたはポリマーが挙げられ、最も一般的に使用されるポリマーは、セルロース、ポリアクリルアミド、ナイロン、ポリスチレン、ポリ塩化ビニルまたはポリプロピレンである。固体支持体は、チューブ、ビーズ、マイクロプレートのディスク、またはイムノアッセイの実施のために適切な任意の他の表面の形態であり得る。
【0253】
代替的アッセイシステムは、標的抗体を固定化し、そしてその固定化された抗体を抗原(レポーター分子に標的されていても、されていなくてもよい)に暴露する工程を包含する。本明細書において記載されるように、用語「レポーター分子」とは、その化学的、生物学的、および/または物理的性質によって、抗原または二次抗体と複合体化された抗体についてのスクリーニングを可能にする分析的に同定可能なシグナルを提供する分子をいう。検出は、定性的であっても定量的であってもいずれでもよい。本明細書において開示された型のアッセイにおいて利用される最も一般的に使用されるレポーター分子は、酵素、フルオロフォア、放射性同位体、および/または化学発光分子である。1つの特定の有用なアッセイシステムにおいて、B細胞レポーターに関する抗体を細胞表面に発現する細胞ライブラリーの培養物が、標識(例えば、ビオチン)と共役された抗原か、または組換えエピトープ(例えば、FLAGエピトープ)を有する抗原とともにインキュベートされ得る。Castrucciら、J.Virol.66:4647−4653(1992)。適切なインキュベーション時間の後、標識された抗原は、氷冷された培地中の懸濁液から2回または3回細胞をペレット化することによって洗い流される。
【0254】
細胞は、抗原に結合された標識に特異的に結合する試薬(例えば、ビオチン標識に対するストレプトアビジンもしくはアビジン、またはFLAGエピトープに対する抗FLAG抗体)に結合されたビーズの懸濁液を含有する培地中でさらにインキュベートされ得る。目的の抗原を結合したことの間接的な結果としてビーズに結合した細胞は、適切な手段によって残りの細胞から分離され、次いで組織培養物に戻されて増殖する。次第に限られた量の抗原を用いてこのプロセスを繰り返すことは、抗原に特異的におよび/または高い親和性で結合した細胞の濃縮を引き起こす。抗原に対する前結合とは関係なくビーズに直接的に結合する細胞は、抗原なしでビーズとともにインキュベーションすることによって除去され得る。次いで、個々の抗原結合クローンは、適切な蛍光色素(例えば、フルオレセイン)に対して直接的もしくは間接的に結合された抗原で細胞を標識した後に、例えば、蛍光活性化細胞分類法(FACS)などによって精製され得る。
【0255】
酵素イムノアッセイ(EIA)の場合、酵素は、一般的にグルタルアルデヒドまたは過ヨウ素酸塩によって検出抗体に結合される。しかし、容易に認識されるように、種々の異なる結合技術が存在し、これらは当業者に容易に利用され得る。一般的に使用される酵素としては、西洋ワサビペルオキシダーゼ、グルコースオキシダーゼ、β−ガラクトシダーゼおよびアルカリホスファターゼが挙げられる。一般に、酵素標識された抗体は、抗原と抗体との間の潜在的複合体に添加され、結合させられ、そして洗浄されて過剰の試薬が除去される。次いで、適切な基質を含む溶液が抗原/試験抗体/標識抗体の複合体に添加される。この基質は、二次抗体に結合した酵素と反応し、定性的な視覚的シグナルを与える。これはさらに、通常、分光光度法で定量化されて、サンプル中に存在する抗体の量を示し得る。
【0256】
あるいは、蛍光化合物(例えば、フルオレセインおよびローダミン)または蛍光タンパク質(例えば、フィコエリトリン)が、抗体に、その結合能を変えることなく化学結合され得る。特定波長の光による照射によって活性化される場合、蛍光標識された抗体は、その光エネルギーを吸収し、その分子の励起状態を示し、続いて、光学顕微鏡または他の光学装置によって視覚的に検出され得る特徴的な色で光の放射を示す。EIAにおける場合、蛍光標識された抗体は、抗原−抗体複合体に結合させられる。未結合の試薬を除去した後、残りの第3の複合体は、適切な波長の光に曝される。観察された蛍光は、目的の結合抗体の存在を示す。免疫蛍光技術およびEIA技術は、どちらも当該分野において十分確立されている。他のレポーター分子(例えば、放射性同位体および化学発光分子および/もしくは生物発光分子)もまた、本明細書において開示されたスクリーニング法において適切に利用され得ることが理解される。
【0257】
以下の実施例は、上記の発明を使用する様式をより十分に記載するため、ならびに本発明の種々の局面を実施するために企図される最適な様式を示すために役立つ。これらの実施例は、決して、本発明の真の範囲を限定するために役立つのではなく、例示的目的のために提示される。以下の実施例は、例示のために提供されるのであって、限定のために提供されるのではない。
【実施例】
【0258】
(実施例1)
(DN1921マウスモノクローナル抗体およびDN1924マウスモノクローナル抗体の生成)
この実施例は、マウスDN1921HLA−DR特異的モノクローナル抗体およびDN1924 HLA−DR特異的モノクローナル抗体の生成を示す。
【0259】
DN1921およびDN1924と称されるマウスモノクローナルHLA−DR特異的抗体を、当該分野で公知の標準的な技術に従って調製した。HarlowおよびLane,「Antibodies:A Laboratory Manual」(Cold Spring Harbor Laboratory,Cold Spring Harbor,NY,1988)。免疫原で過剰免疫された近交系実験用マウスBALB/c系(Jackson Laboratory,Bar Harbor,ME)がB細胞のドナーであった。BALB/c由来の変異Bリンパ腫株M12.C3を、キメラヒト/マウス クラスII遺伝子でトランスフェクトし、免疫原として使用した。このトランスフェクト体(M12.C3.25と称する)によって発現されるMHCクラスII分子は、HLA−DRの第1の細胞外(α1およびβ1)ドメイン、ならびに対応するマウスMHCクラスII分子H2−Eの第2の細胞外(α2およびβ2)ドメイン、膜貫通ドメインおよび細胞質内ドメインから構成された。Vidovicら、Eur.J.Immunol.25:3349−3355(1995)。マウスを、月1回の間隔で5回のIP注射(各々は、1mLのリン酸緩衝化生理食塩水(PBS)中に再懸濁された、107(−照射された(100Gy)M12.C3.25細胞からなる)によって免疫化した。最後の注射から3日後、免疫脾臓細胞を、HAT感受性Ig陰性マウスミエローマ細胞PAI−0と融合した。Stockerら、Research Disclosure 217 :155−157(1982)。
【0260】
単一のハイブリドーマ培養物の上清流体を、4℃での4時間のインキュベーション後でのEBV−LCL RPMI 1788に対するそれらの毒性、またはヒト末梢血単核細胞(PBMC)のインビトロでの増殖において誘導されたブドウ球菌エンテロトキシンB(SEB)を抑制するそれらの能力についてスクリーニングした。所望の生物活性のモノクローナル抗体を適切な形式で分泌することが同定されたコロニーを、それぞれ「DN1921」および「DN1924」と称し、そして限界希釈法によって2回サブクローン化した。標準的アイソタイプ決定キット(Zymed,South San Francisco,CA)を用いて、DN1921およびDN1924が、両方ともマウスIgG1/κアイソタイプであることを見出した。
【0261】
(実施例2)
(DN1924モノクローナル抗体のインビトロ殺腫瘍性効果の評価)
この実施例は、DN1924と称するマウスHLA−DR特異的モノクローナル抗体が、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導し得るが、HLA−DRを発現する正常(すなわち、非悪性)細胞においてはアポトーシスを誘導しないことを示す。
【0262】
ヒト細胞株MC/CAR(プラズマ細胞腫)およびRPMI 1788(エプスタイン−バーウイルス形質転換リンパ芽球状B細胞株、EBV−LCL)を、ATCC(Rockville,MD)から譲渡された。Rittsら、Int.J.Cancer 31:133−141(1983)。細胞を、10% FCS、2mMのL−グルタミン、0.1mg/mlの硫酸カナマイシン、および3×10−5Mの2−MEを補充したIMDM培地(Gibco,Grand Island,NY)中で、105/mlの密度で、37℃で、5%CO2を含む加湿された雰囲気(組織培養インキュベーター)中で培養した。5×105細胞/mlで培養したHLA−DR−特異的mAb分泌性マウスB細胞ハイブリドーマ細胞株DN1924および10F12の滅菌濾過された上清流体を、最終濃度20%でヒト細胞を添加した。示された共培養期間に続いて、細胞を洗浄し、そしてさらに1μg/mlのヨウ化プロピジウム(PI,Sigma,St.Louis,MO)とともに5分インキュベーションし、その後、CELLQuest 3.1fソフトウェアを用いたFACScan(登録商標)フローサイトメーター(Becton−Dickinson,San Jose,CA)上で、細胞の大きさ 対 赤色PI蛍光の分析した後、その生存度を決定した。Ottenら、「Current Protocols in Immunology」、pp.5.4.1−5.4.19(Coliganら編、Greene & Wiley,New York,1997)。生存細胞が積極的にPIを排除すること、一方死細胞が、それを、細胞のDNAへの接近可能性と直接的に比例して取り込むことが示された。Swatら(1991)。
【0263】
新生物細胞のアポトーシスを誘導するDN1924抗体の能力を、75%を超える細胞死を引き起こした2つの独立なヒトB細胞腫瘍株(EBV−LCL RPMI 1788およびプラズマ細胞腫MC/CAR)の共培養によって示した。図2。
【0264】
ヒト末梢血から得られた正常形成性(すなわち、非新生物性)HLA−DR+リンパ球においては、細胞毒性効果は全くなかった。対照的に、DN1924、10F12(第2のタンパク質ドメイン内に位置する共通エピトープについて特異的な抗DR mAb)は、MC/CARの生存度に影響しなかった。図3。
【0265】
2つの異なるインキュベーション条件下での細胞毒性の時間経過は、図4の上パネルに示される。DN1924誘導性細胞死が温度依存的であることは明らかである;これはヒト体温(37℃)においてより速い速度(細胞毒性効果は30分以内に明らかとなる)であり、そして4℃においては、遅くなるが、まだなお生じる(細胞毒性効果は1〜2時間以内に明らかとなる)。腫瘍細胞との長い(3週間)共培養後でさえも衰えていないDN1924の腫瘍細胞毒性は、この抗体に対して耐性となる能力のないことを示す。図4の下パネル。
【0266】
(実施例3)
(DN1924 HLA−DR特異的マウスモノクローナル抗体は、低下した免疫抑制活性を示す)
この実施例は、DN1924 HLA−DR特異的マウスモノクローナル抗体が、低レベルの免疫抑制活性を示すことを示す。
【0267】
ブドウ球菌エンテロトキシンBに対するヒトTh細胞のHLA−DR依存性インビトロ増殖反応を、先に記載したように生じさせた。Mollickら、J.Immunol.146:463−468(1991)。簡単にいうと、Ficoll分離によってヘパリン処理した血液から得られた2×105個の新鮮ヒトPBMCを、10% FCS、2mMのL−グルタミン、0.1mg/mlの硫酸カナマイシン、および3×10−5Mの2−MEを補充した0.2mlのIMDM培地中で0.1μg/mlのSEB(Toxin Technology,Sarasota,FL)とともに37℃、5%CO2を含む加湿した雰囲気(組織培養インキュベーター)中で、3日間培養した。5×105細胞/mlで培養したHLA−DR−特異的mAb分泌性マウスB細胞ハイブリドーマ細胞株DN1924およびL243(ATCC,Rockville,MD)の滅菌濾過された上清流体を、最終濃度20%でアッセイの開始時に添加した。Lampsonら、J.Imnunol.125:293−299(1980)およびFuら、Human Immunol.39:253−260(1994)。3つ組の培養物のT細胞増殖を、最後の16時間の間の[3H]チミジン取り込みによって測定した。Bradley、「Selected Methods in CellularImmunology」pp.153−174(MishellおよびShiigi(編)、W.H.Freeman & Co.,New York(1980)。L243およびDN1924のmAbの両方とも第1のHLA−DRドメイン上のエピトープを認識するが、mAb L243がSEBで誘発されたT細胞反応の約70%を抑制した一方、DN1924は効果を有さなかった(図5)。
【0268】
(実施例4)
(DN1924 HLA−DR特異的マウスモノクローナル抗体のインビボ抗腫瘍活性の評価)
DN1924のインビボ抗腫瘍活性を、HLA−DR陽性ヒトプラズマ細胞種MC/CARを注射された重症複合型免疫不全近交系(scid)マウスにおいて評価した。20匹の8週齢のマウス(体重に関してランダム化された)に、マウスあたり0.1ml RPMI 1640培地中の107個のMC/CAR細胞を用いてIP注射した。Rittsら、(1983)。その後、動物の半数は、0.1mlのPBS中0.625mgのDN1924抗体の、単回IP用量を受けた。残りのマウスに、0.1ml PBSの対応するIP注入を与えた。マウスを毎日モニタリングし、その生存を記録した。図6に示されるように、DN1924は、顕著な治療活性を示し、腫瘍を有するマウスの生存を延長した。
【0269】
(実施例5)
(マウスDN1921およびDN1924におけるマウス/ヒトキメラHLA−DR特異的モノクローナル抗体の生成)
この実施例は、マウスモノクローナル抗体DN1921およびDN1924の重鎖可変領域および軽鎖可変領域と、ヒトIgG2の重鎖定常領域およびヒトIgκの軽鎖定常領域とを含むマウス/ヒトキメラHLA−DR特異的モノクローナル抗体の構築を記載する。
【0270】
可変領域セグメントを、cDNAを全RNAから「RACE」するプライマーを設計するための定常領域特異的配列を用いて、各抗体(DN1924およびDN1921)のマウス版を発現するハイブリドーマ細胞からクローニングした。可変領域(DN1924−heavy、DN1924−light;DN1921−heavy、DN1922−light)配列を、これらのクローンから決定した。
【0271】
ヒトIgG2重鎖定常領域セグメントおよびヒトIgκ軽鎖定常領域セグメントを、NCBIおよびIMGTのホームページから設計されたプライマーを用いて、ヒト脾臓全RNAからクローン化した。クローンを配列決定して、適切なIgκ定常領域またはヒトIgG2定常領域を確認した。
【0272】
DN1921およびDN1924重鎖および軽鎖ポリヌクレオチド配列を増幅するために使用されたオリゴヌクレオチドプライマーは、表3に提示される。
【0273】
(重鎖構築)
可変領域遺伝子セグメントと定常領域遺伝子セグメントとを組み合わせるため、PCR「縫い付け(sewing)」技術を利用した。簡単にいうと、プライマーを、抗体分子のコード領域の5’末端および3’末端に対応するように設計した(表3、プライマーAおよびB)。プライマーの第2のセットを、別個のプラスミド上に最初に存在する2つの片の間の接合部に重なるように設計した。最初のPCR反応を、プライマーセットA+Dとともに可変領域プラスミドをテンプレートとして用いて、またはプライマーセットB+Cとともに定常領域プラスミドをテンプレートとして用いて、準備した。第2のPCR反応は、プライマーセットA+Dを用いて最初の2つの一次反応の生成物を組み込んだ。最後のPCR生成物をクローニングし、配列を確認し、そして必要な場合、(コード領域の外に)制限酵素を利用して、発現ベクター中にシャトル(shuttle)した。
【0274】
(軽鎖構築)
可変領域遺伝子セグメントと定常領域遺伝子セグメントとを組み合わせるため、重鎖構築に関して上記したように、PCR「縫い付け」技術を利用した。プライマーを、抗体分子のコード領域の5’末端および3’末端に対応するように設計した(表3、プライマーAおよびB)。第3のプライマーを、別個のプラスミド上に最初に存在する2つの片の間の接合部に重なるように設計した。最初のPCR反応を、プライマーセットC+Bとともに定常領域プラスミドをテンプレートとして用いて準備した。第2のPCR反応は、プライマーセットA+Bを用いて、一次反応の生成物、およびプラスミドがコードする軽鎖可変領域を組み込んだ。最後のPCR生成物をクローニングし、配列を確認し、そして必要な場合、(コード領域の外に)制限酵素を利用して、発現ベクター中にシャトルした。
【0275】
(発現系)
DN1924重鎖およびDN1924軽鎖を、pEE14.4発現ベクター(Lonza Biologics,Portsmouth,NH)にサブクローニングし、キメラ免疫グロブリンの両方の鎖が同じベクター骨格上にコードされるようにした。各鎖の発現を、それ自身のCMV主要即時早期(major immediate early)(MIE)プロモーターによって駆動した。重鎖および軽鎖の両方のオープンリーディングフレームを含むプラスミドを、CHO−K1/Sv細胞(Lonza Biologics)中にトランスフェクトし、細胞を、標準的な技術を用いて、50μMのメチオニン−スルホキサミン(sulfoxamine)(MSX)を含む無グルタミン培地中で選択した。細胞株を生成するキメラ抗体をELISAによって同定した。
【0276】
DN1921重鎖および軽鎖の発現に関して、同じプロトコールに従った。
【0277】
(表3)
(キメラDN1921モノクローナル抗体およびキメラDN1924モノクローナル抗体の構築に使用されたオリゴヌクレオチド)
【0278】
【表3】
表3のオリゴ配列において、大文字は、抗体分子のオープンリーディングフレーム中のアミノ酸残基についてのコドンを示し、小文字は、Kozakコンセンサス配列(下線)およびクローニング目的のための制限エンドヌクレアーゼ部位に対して付加されたヌクレオチドを示す。プライマー「C]および「D」中の点線は、マウス可変領域とヒト定常領域との間の接合部を示す。
【0279】
(実施例6)
(HLA−DR細胞に特異的に結合するマウス/ヒトキメラDN1921 HLA−DR特異的モノクローナル抗体およびマウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体)
この実施例は、キメラDN1921 HLA−DR特異的モノクローナル抗体およびキメラDN1924 HLA−DR特異的モノクローナル抗体の両方が、種々の組織培養細胞株において発現されたHLA−DRに特異的に結合することを示す。
【0280】
標準的免疫蛍光アッセイ[Current Protocols in Immunology 1,pp.5.3.1−5.4.13(Coliganら編、New York,Wiley Interscience,1998)]を、ビオチン化ヤギ抗ヒトIgGビオチン結合体(1:200最終希釈;Sigma,St.Louis,MO)、ならびにストレプトアビジン−PE(0.8μg/ml)をそれぞれ第2および第3の試薬として用いて行った。
【0281】
(実施例7)
(マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体は、HLA−DR発現腫瘍細胞株においてアポトーシスを誘導する)
この実施例は、キメラDN1924 HLA−DR特異的モノクローナル抗体が、HLA−DR発現腫瘍細胞株においてアポトーシスを誘導するのに有効であるが、HLA−DR陰性腫瘍細胞または正常HLA−DR発現細胞においては有効でないことを示す。
【0282】
アポトーシスアッセイを、Alexa Flour488結合体をヨウ化プロピジウム(PI)対比染色とともに使用して、Molecular Probe Annexin Vキットを用いて行った。簡単にいうと、5×105個の生存細胞を0.1〜10μgのmAbとともに0.5〜1mlの総容量の培地中で、37℃で10〜30分間共培養し、2mlの氷冷PBSで1回洗浄し、5μlのAnnexin Vおよび1μg/mlでPIを含む100μlの1×緩衝液中に再懸濁し、室温、暗中で15分間インキュベートした。次いで、300μlの1×緩衝液を添加し、そしてサンプルを30分以内にフローサイトメーターによって分析した[Current Protocols in Immunology 1,pp.5.4.1−5.4.13(Coliganら編、New York,Wiley Interscience,1998)]。
【0283】
表4は、マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体が、HLA−DR陽性LcL細胞株の一団の各メンバーに結合し、そしてアポトーシスを誘導することを示す。各LcL細胞株は、異なるDRβ1対立遺伝子を発現する。これらのデータは、DRβ1対立遺伝子の幅広い範囲にわたるHLA−DR発現に関する疾患の処置における、本発明のマウス/ヒトキメラ抗HLA−DR抗体に対する有用性を支持する。
【0284】
(表4)
(マウス/ヒトキメラDN−1924(mut−hIgG2クローンDN16)は、ある範囲のDRβ1対立遺伝子を発現するHLA−DR陽性LcL細胞株の一団に特異的に結合し、アポトーシスを誘導する)
【0285】
【表4】
表5は、マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体が、腫瘍細胞株の一団(3つのHLA−DR陰性腫瘍細胞株および12のHLA−DR陽性細胞株を含む)の各HLA−DR陽性メンバー(KG−1を除く)に結合し、アポトーシスを誘導することを示す。キメラDN1924は、Fcレセプターを発現するもの(すなわちU937およびTHP−1)、またはB細胞腫瘍細胞株(すなわちDB)でさえ、HLA−DR陰性細胞株のアポトーシスを誘導しない。対照的に、キメラDN1924は、12のHLA−DR陽性腫瘍細胞株のうちの11のアポトーシスを誘導する。KG−1(AML腫瘍細胞株)は、キメラDN1924結合によってアポトーシスを受けないHLA−DR陽性細胞株の(特異的キメラDN1924結合活性によって示されるように)唯一の例である。全体で、これらのデータは、キメラDN1924が、広範なHLA−DR陽性腫瘍型の処置において有効であることを示す。
【0286】
(表5)
(マウス/ヒトキメラDN−1924(mut−hIgG2クローンDN16)は、HLA−DR陽性細胞株の一団に特異的に結合し、アポトーシスを誘導する)
【0287】
【表5】
(実施例8)
(マウス/ヒトキメラDN1921 HLA−DR特異的モノクローナル抗体は、免疫抑制性である)
以前に記載されるたように、IL−2分泌アッセイを行った[Current Protocols in Immunology1,pp.3.14.1−3.14.11(Coliganら編、New York,Wiley Interscience,1998)]。簡単にいうと、PAP/HLA−DR1特異的CD4+T細胞ハイブリドーマ株Paperino[Vidovicら、Human Immunol.64:238−244,2003]のハイブリドーマ細胞105個を、抗原提示細胞としての2×105個のヒトHLA−DR1+PBMC、PAP、およびmAb(mDN1921=マウスDN1921;mDN1924=マウスDN1924;DSm89#34=キメラDN1921;DN−GS#16=キメラDN1924)とともに、0.2mlのマイクロウェル中で共培養した。1日後、培養上清を回収し、そしてIL−2依存性細胞株HT−2の104個の細胞の増殖を支持するそれらの能力について、24時間、50%濃度で試験した。最後の6時間の培養期間の間、[3H]チミジンcpm組み込みによって細胞増殖を測定した。
【0288】
(実施例9)
(マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体に誘導されたアポトーシスは、カスパーゼ非依存性である)
この実施例は、マウス/ヒトキメラDN1924のアポトーシス誘導活性がカスパーゼ非依存性であることを示す。
【0289】
外膜に対するホスファチジルセリンの暴露(Annexin V染色で検出される)およびカスパーゼ活性は、両方ともアポトーシスの指標である。しかし、本実施例は、キメラDN1924に誘導されたアポトーシスが、カスパーゼ3、カスパーゼ8、またはカスパーゼ9の活性に関与しないことを示す(図13A−Dおよび表6)。
【0290】
キメラDN1924媒介性アポトーシスを、添加されたZ−VAD−fmk(広範囲のカスパーゼインヒビター)の存在下および非存在下の両方において誘導した。誘導後、アポトーシスを、Annexin V染色によって、ならびにカスパーゼ3、カスパーゼ8、およびカスパーゼ9活性化を測定することによって検出した。Annexin V染色は、アポトーシス誘導を確証し、そしてZ−VAD−fmkがアポトーシス誘導の阻害に不十分であることを確証した。従って、マウス/ヒトDN1924で誘導されたアポトーシスは、カスパーゼ3、カスパーゼ8、またはカスパーゼ9の活性化に非依存性であった。
【0291】
(表6)
(マウス/ヒトキメラDN1924に誘導されたアポトーシスは、カスパーゼ非依存性である)
【0292】
【表6】
(実施例10)
(マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体は、腫瘍増殖の速度を低下させること、およびRAJI異種移植片SCIDマウス動物系において生存の延長を促進することに有効である)
この実施例は、マウス/ヒトキメラDN1924抗HLA−DR抗体のインビボ皮下投与が、腫瘍増殖の速度を低下させること(図14)、およびRaji異種移植片scidマウス動物系において生存の延長を促進すること(図15)に有効であることを示す。
【0293】
0日目、scidマウス(5匹の動物の4つの群)は、0.1ml PBS中の5×106個のRaji腫瘍細胞(バーキットリンパ腫;ATCCカタログ番号CCL−86)で負荷(challenge)された。腫瘍負荷後1日目、4日目、7日目および10日目において、グループ1は、腹腔内注射を介して、300μl PBS中に50μgのマウスDN1924抗HLA−DR抗体を受け、グループ2は、腹腔内注射を介して、300μl PBS中に50μgのマウス/ヒトDN1924抗HLA−DR抗体(クローン番号64)を受け、グループ3は、腹腔内注射を介して、300μl PBS中に50μgのヒトIgG2,κアイソタイプコントロール抗体(Sigma、カタログ番号I5404)を受け、そしてグループ4は、腹腔内注射を介して、300μlのPBSを受けた。
【0294】
図14は、マウスDN1924およびマウス/ヒトキメラDN1924の両方とも、scidマウス動物モデル系において、Raji異種移植片を用いた皮下負荷によって腫瘍増殖の速度を低下させるのに有効であったことを示す。
【0295】
図15は、マウスDN1924およびマウス/ヒトキメラDN1924の両方とも、scidマウス動物モデル系において、Raji異種移植片を用いた皮下負荷後に生存を延長させるのに有効であったことを示す。
【0296】
本発明は、特定の処置方法および組成物に関して記載されたが、本発明から逸脱することなく種々の変更および改変がなされ得ることは、当業者にとって明らかである。
【図面の簡単な説明】
【0297】
【図1】HLA−DR分子の概略的エピトープマップを示す。これは、CY:細胞質テール;TM:膜貫通部分;α1およびβ1:それぞれ、α鎖およびβ鎖の第1(細胞外)ドメイン;ならびにα2およびβ2:それぞれ、α鎖およびβ鎖の第2(細胞外)ドメインを示す。
【図2】図2は、DN1924マウスモノクローナル抗体とともに16時間培養した後の種々の細胞集団の生存度を示す。生細胞が、2次元プロットの右下象限中に位置する。PBMCは、末梢血単核球に対応する。
【図3】図3は、示されるモノクローナル抗体とともに16時間培養した後のヒト細胞株の生存度を示す。
【図4】図4は、示されるような、種々の温度条件下で種々の時間の間DN1924モノクローナル抗体とともにインキュベーションした後のEBV−LCL RPMI 1788の生存度を示す。
【図5】図5は、[3H]チミジン取り込みによって示される、ヒトPBMCのブドウ球菌エンテロトキシンB(SEB)特異的インビトロ増殖応答に対するDN1924モノクローナル抗体の免疫抑制効果が存在しないことを示す。
【図6】図6は、HLA−DR+ヒトプラズマ細胞腫MC/CARを注射し、DN1924(白三角)またはリン酸緩衝化生理食塩水(PBS:黒四角)のいずれかで処理した、scidマウスの生存を示す。
【図7A】図7A〜図7Dは、DN1921と、DN1924(図7A)、キメラDN1924(図7B)、およびキメラDN1921(図7C)とがHLA−DR発現細胞に特異的に結合するのを示す、FACSデータである。
【図7B】図7A〜図7Dは、DN1921と、DN1924(図7A)、キメラDN1924(図7B)、およびキメラDN1921(図7C)とがHLA−DR発現細胞に特異的に結合するのを示す、FACSデータである。
【図7C】図7A〜図7Dは、DN1921と、DN1924(図7A)、キメラDN1924(図7B)、およびキメラDN1921(図7C)とがHLA−DR発現細胞に特異的に結合するのを示す、FACSデータである。
【図7D】図7A〜図7Dは、DN1921と、DN1924(図7A)、キメラDN1924(図7B)、およびキメラDN1921(図7C)とがHLA−DR発現細胞に特異的に結合するのを示す、FACSデータである。
【図8A】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8B】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8C】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8D】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8E】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8F】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図9】図9は、マウスDN1921 HLA−DR特異的モノクローナル抗体およびキメラDN1921 HLA−DR特異的モノクローナル抗体によってはPAP/HLA−DR特異的Paperino応答の阻害を示すが、マウスDN1924 HLA−DR特異的モノクローナル抗体によってもキメラDN1924 HLA−DR特異的モノクローナル抗体によっても、PAP/HLA−DR特異的Paperino応答の阻害は示さない、グラフである。
【図10】図10は、キメラDN1921を発現するプラスミドベクターおよびキメラDN1924を発現するプラスミドベクターの構築の模式図である。
【図11】図11は、Igκ定常領域アライメントである。上の2行は、ヒトIgκについての2つの異なる登録番号(J00241およびM63438)である。下の行は、マウスIgκ(GenBank登録番号L35138)である。
【図12】図12は、IgG定常領域アライメントである。上の3行は、ヒト(h)IgG2、hIgG1、およびhIgG4(それぞれ、GenBank登録番号AJ250170、AF027159、およびAJ294733)である。下の3行は、マウス(m)IgG1、IgG2a、およびIgG2b(それぞれ、GenBank登録番号J00453、J00470、およびJ00461)である。
【図13A】図13A〜図13Dは、マウス/ヒトキメラDN1924抗HLA−DR媒介性アポトーシスが、カスパーゼ非依存性であること(図13A)、およびカスパーゼ3(図13B)も、カスパーゼ8(図13C)も、カスパーゼ9(図13D)も、キメラDN1924処理細胞において活性化されることを示す棒グラフである。
【図13B】図13A〜図13Dは、マウス/ヒトキメラDN1924抗HLA−DR媒介性アポトーシスが、カスパーゼ非依存性であること(図13A)、およびカスパーゼ3(図13B)も、カスパーゼ8(図13C)も、カスパーゼ9(図13D)も、キメラDN1924処理細胞において活性化されることを示す棒グラフである。
【図13C】図13A〜図13Dは、マウス/ヒトキメラDN1924抗HLA−DR媒介性アポトーシスが、カスパーゼ非依存性であること(図13A)、およびカスパーゼ3(図13B)も、カスパーゼ8(図13C)も、カスパーゼ9(図13D)も、キメラDN1924処理細胞において活性化されることを示す棒グラフである。
【図13D】図13A〜図13Dは、マウス/ヒトキメラDN1924抗HLA−DR媒介性アポトーシスが、カスパーゼ非依存性であること(図13A)、およびカスパーゼ3(図13B)も、カスパーゼ8(図13C)も、カスパーゼ9(図13D)も、キメラDN1924処理細胞において活性化されることを示す棒グラフである。
【図14】図14は、マウス/ヒトキメラDN1924抗HLA−DR抗体のインビボ皮下投与が、Raji異種移植scidマウス動物モデル系における腫瘍増殖速度を減少することにおいて有効であることを示すグラフである。
【図15】図15は、マウス/ヒトキメラDN1924抗HLA−DR抗体のインビボ皮下投与が、Raji異種移植scidマウス動物モデル系における生存増加を促進することにおいて有効であることを示すグラフである。
【技術分野】
【0001】
(関連出願)
本出願は、2003年12月15日に出願した米国仮特許出願番号60/608,944の優先権を主張する。
【0002】
本発明は、一般的には、免疫学の分野および分子生物学の分野に関する。より具体的には、本発明は、HLA−DR特異的抗体、およびHLA−DR特異的抗体を調製するための方法を提供する。HLA−DR特異的抗体を含む組成物および方法もまた、提供される。本明細書中に提示される抗体は、HLA−DR抗原に特異的に結合し、そしてHLA−DRを発現する腫瘍細胞においてアポトーシスを誘導する。特定の実施形態において、アポトーシス性HLA−DR特異的抗体は、低下した免疫抑制活性レベルを示す。本発明の抗体は、診断方法における使用、ならびにHLA−DR陽性腫瘍細胞に関連する癌の処置のための組成物および方法における使用を見出す。
【背景技術】
【0003】
クラスII主要組織適合遺伝子複合体(MHC)分子(正常な抗原提示細胞(APC)上に構成的に発現される)は、CD4+ヘルパーT(Th)細胞に対する抗原由来ペプチドの提示を担う。非特許文献1および非特許文献2。これらの分子を介するシグナル伝達は、活性化Bリンパ球のプログラムされた細胞死(PCDまたはアポトーシス)をもたらすセカンドメッセンジャーの生成を開始する。抗原提示に加えて、クラスII分子は、細胞増殖を調節し得るシグナルを伝達し、特定のクラスII MHC特異的モノクローナル抗体は、癌細胞のアポトーシスを誘導することが示されている(非特許文献3)。
【0004】
HLAクラスII分子は、ヒトBリンパ球上に構成的に発現され、そして活性化後にヒトTリンパ球上で誘導される。細胞死の60%までが、HLA−DR分子を介するリンパ球の刺激後に観察されている。特定のHLA−DR特異的抗体は、クラスII分子の細胞表面発現における90%までの減少を引き起こす。HLA−DR特異的抗体およびそのフラグメントは、HLA−DP分子およびHLA−DQ分子の発現に影響を与えない(非特許文献4)。
【0005】
癌細胞のアポトーシスを誘導することが示されたクラスII MHC特異的抗体はまた、頻繁に、正常なTh細胞機能を妨害する(非特許文献5)。現在利用可能なアポトーシス誘導性クラスII特異的モノクローナル抗体の大多数は、ペプチド結合部位に見かけ上接近した状態にある、HLA−DRへテロダイマーの第一タンパク質ドメイン上に位置するエピトープを認識する。これらの抗体は、抗原提示を妨害し、Th応答の強力なインビトロおよびインビボでの阻害を引き起こし、さらに、Bリンパ芽球様細胞および少ない割合の正常活性化B細胞に対して細胞傷害性である。そのF(ab’)2フラグメントは、ダウンレギュレーションおよび細胞傷害性の両方を媒介し、一方、その一価Fabフラグメントは、細胞傷害性ではないが、ダウンレギュレーション特性およびT細胞阻害特性を保持する(非特許文献5)。
【非特許文献1】Babbittら、Nature(1985年)317:p.359〜361
【非特許文献2】Trumanら、Blood(1997年)89(6):p.1996〜2007
【非特許文献3】Newellら、Proc.Natl.Acad.Sci.U.S.A.(1993年)90(22):p.10459〜10463
【非特許文献4】Trumanら、Int.Immunol.(1994年)6(6):p.887〜896
【非特許文献5】Vidovicら、Eur.J.Immunol.(1995年)25:p.3349〜3355
【発明の開示】
【発明が解決しようとする課題】
【0006】
従って、当該分野で利用可能なほとんどのHLA−DR特異的アポトーシス性モノクローナル抗体は、HLA−DR陽性細胞を含む癌処置用の組成物および方法におけるそれらの抗体の有用性を限定するのに充分な免疫抑制活性を示す。従って、改善されたHLA−DR特異的抗体(例えば、非免疫抑制アポトーシス性HLA−DR特異的抗体)、ならびにこのような抗体を含む、これらの欠陥を克服する組成物および方法に関して、当該分野において必要性が残っている。
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明は、特に、HLA−DR陽性腫瘍細胞においてアポトーシスを誘発可能なHLA−DR特異的抗体およびその抗原結合フラグメントを提供することによって、これらの必要性および他の関連する必要性に取り組む。本発明は、ヒト主要組織適合遺伝子複合体(MHC)クラスIIと特異的に反応する抗体は、表面上にHLA−DR分子を発現する細胞のアポトーシスを誘導し得るという発見に、部分的に基づく。本発明の抗体は、このHLA−DR特異的抗体が、HLA−DRを発現する非腫瘍細胞/非新形成細胞の生存度にも機能にも影響を与えないという点で、非常に特異的である。特定の実施形態において、本発明のHLA−DR抗体のアポトーシス活性は、クラスII非依存性免疫応答を同時に抑制することなく、達成され得る。
【0008】
本発明の重要な実用上の関連性は、「DN1924」と呼ばれるモノクローナル抗体、および「キメラDN1924」と呼ばれるDN1924のマウス/ヒト改変体が、HLAクラスII陽性新形成(血球新形成、例えば、プラズマ細胞腫/多発性骨髄腫、ホジキンリンパ腫および非ホジキンリンパ腫、ならびにB細胞リンパ腫が挙げられるが、これらに限定されない)の選択的抗体ベース療法のために有効であり得ることである。インビトロ研究によって、DN1924モノクローナル抗体およびキメラDN1924モノクローナル抗体は、正常なTh機能を妨害せず、従って、DN1924および/またはキメラDN1924は、被験体の正常なHLA−DR発現細胞に影響を与えないことが、示される。従って、現在利用可能な治療剤よりも副作用が少ないと予測することが、妥当である。
【0009】
従って、特定の局面において、本発明は、非ヒトモノクローナルHLA−DR特異的アポトーシス性抗体を提供する。本発明の非ヒトモノクローナル抗体は、種々の動物種から単離され得る。この動物種としては、非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、鳥類、ウサギ、マウス、ラット、モルモット、ハムスター、イヌ、およびネコなどの起源が挙げられるが、これらに限定されない。HLA−DR特異的アポトーシス性非ヒトモノクローナル抗体は、本明細書において、(1)配列番号13および配列番号2においてそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(これらは、それぞれ、配列番号46のポリヌクレオチドおよび配列番号45のポリヌクレオチドによってコードされる)を含む、免疫抑制抗体DN1921と、(2)配列番号35および配列番号24においてそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(これらは、それぞれ、配列番号48のポリヌクレオチドおよび配列番号47のポリヌクレオチドによってコードされる)を含む、非免疫抑制抗体DN1924と、によって例証される。
【0010】
本明細書中で例証されるDN1921抗体およびDN1924抗体の各々は、マウスIgG1重鎖定常ドメインおよびマウスIgκ軽鎖定常ドメインをさらに含む。関連する代替的実施形態において、DN1921モノクローナル抗体および/またはDN1924モノクローナル抗体は、上記の重鎖可変ドメインおよび軽鎖可変ドメインと、IgM、IgD、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される抗体アイソタイプ由来のマウス重鎖定常ドメインとを、含み得る。
【0011】
また、DN1921重鎖およびDN1921軽鎖ならびにDN1924重鎖およびDN1924軽鎖の、抗原結合フラグメント、改変体、および誘導体も、提供される。特定の実施形態において、DN1921の改変体は、配列番号13および配列番号2に対してそれぞれ少なくとも70%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1921の改変体は、配列番号13および配列番号2に対してそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1921の改変体は、配列番号13および配列番号2に対してそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0012】
代替的実施形態において、DN1924の改変体は、配列番号35および配列番号24に対してそれぞれ少なくとも70%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1924の改変体は、配列番号35および配列番号24に対してそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1924の改変体は、配列番号35および配列番号24に対してそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0013】
本発明の他の局面は、非ヒト/ヒトキメラHLA−DR特異的モノクローナル抗体を提供する。この抗体において、このキメラ抗体は、ヒト定常ドメインに作動可能に連結された非ヒト可変ドメインを含む。本発明のHLA−DR特異的モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導するが、この抗体が結合する正常細胞においてはアポトーシスを誘導しない。特定の実施形態において、本発明の非ヒト/ヒトキメラHLA−DR特異的アポトーシス性モノクローナル抗体は、被験体にインビボで投与された場合に、非免疫抑制性である。
【0014】
本発明のキメラモノクローナル抗体の構築のために適切な非ヒトモノクローナル抗体可変ドメインは、種々の動物種から単離され得る。この動物種としては、非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、鳥類、ウサギ、マウス、ラット、モルモット、ハムスター、イヌ、およびネコなどの起源が挙げられるが、これらに限定されない。ヒト抗体の重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される抗体アイソタイプから単離され得る。
【0015】
本明細書中に開示される例示的なHLA−DR特異的キメラモノクローナル抗体は、マウスモノクローナル抗体DN1921の重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号13および配列番号2)を、ヒト重鎖定常ドメインおよびヒト軽鎖定常ドメイン(それぞれ、配列番号14および配列番号3)に作動可能に連結された状態で含む。本明細書中に開示されるキメラDN1921モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、そしてインビボで投与された場合に免疫抑制性である。この例示的な全長キメラDN1921モノクローナル抗体の重鎖のアミノ酸配列は、本明細書中では配列番号12として示される。この例示的な全長キメラDN1921モノクローナル抗体の軽鎖のアミノ酸配列は、本明細書中では配列番号1として示される。
【0016】
また、配列番号12のキメラDN1921重鎖および配列番号1のキメラDN1921軽鎖のそれぞれの改変体が、提供される。特定の実施形態において、キメラDN1921重鎖の改変体およびキメラDN1921軽鎖の改変体は、それぞれ、配列番号12および配列番号1に対して少なくとも70%同一である。他の実施形態において、キメラDN1921重鎖の改変体およびキメラDN1921軽鎖の改変体は、それぞれ、配列番号12および配列番号1に対して少なくとも80%、90%、または95%同一である。なおさらなる実施形態において、キメラDN1921重鎖の改変体およびキメラDN1921軽鎖の改変体は、それぞれ、配列番号12および配列番号1に対して少なくとも98%または99%同一である。
【0017】
本明細書中に開示される別の例示的なHLA−DR特異的キメラモノクローナル抗体は、マウスモノクローナル抗体DN1924の重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号35および配列番号24)を、ヒト重鎖定常ドメインおよびヒト軽鎖定常ドメイン(それぞれ、配列番号14および配列番号3)に作動可能に連結された状態で含む。本明細書中に開示されるキメラDN1924モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、そしてインビボで投与された場合に非免疫抑制性である。この例示的な全長キメラDN1924モノクローナル抗体の重鎖のアミノ酸配列は、本明細書中では配列番号34として示される。この例示的な全長キメラDN1924モノクローナル抗体の軽鎖のアミノ酸配列は、本明細書中では配列番号23として示される。
【0018】
代替的実施形態において、配列番号34のキメラDN1924重鎖および配列番号23のキメラDN1924軽鎖のそれぞれの改変体が、提供される。特定の実施形態において、キメラDN1924重鎖の改変体およびキメラDN1924軽鎖の改変体は、それぞれ、配列番号34および配列番号23に対して少なくとも70%同一である。他の実施形態において、キメラDN1924重鎖の改変体およびキメラDN1924軽鎖の改変体は、それぞれ、配列番号34および配列番号23に対して少なくとも80%、90%、または95%同一である。なおさらなる実施形態において、キメラDN1924重鎖の改変体およびキメラDN1924軽鎖の改変体は、それぞれ、配列番号34および配列番号23に対して少なくとも98%または99%同一である。
【0019】
なおさらなる実施形態において、HLA−DR特異的アポトーシス性抗体は、ヒト化モノクローナル抗体であり、このヒト化抗体は、1つ以上の非ヒト相補性決定領域(CDR)を、ヒト可変ドメインフレームワーク領域(FR)に作動可能に連結されて重鎖可変ドメインおよび軽鎖可変ドメインを作製する状態で含み、これらの可変ドメインは、ヒト定常ドメイン重鎖およびヒト定常ドメイン軽鎖(それぞれ、配列番号14および配列番号3において示される重鎖IgG2定常ドメインおよび軽鎖Igκ定常ドメインによって本明細書において例証される)に作動可能に連結された状態である。
【0020】
本発明の例示的なHLA−DR特異的ヒト化モノクローナル抗体は、マウスモノクローナル抗体DN1921重鎖可変ドメイン(配列番号13)の1つ以上の相補性決定領域(CDR)を、ヒト重鎖フレームワーク(FR)ドメインに作動可能に連結されてヒト化HLA−DR特異的重鎖可変ドメインを作製した状態で含む。このヒト化HLA−DR特異的重鎖可変ドメインは、必要に応じて、ヒト重鎖定常ドメインと、配列番号14において示されるヒトIgκ軽鎖定常ドメインとに作動可能に連結される。このヒト重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される。
【0021】
あるいは、またはさらに、本発明の例示的なHLA−DR特異的ヒト化モノクローナル抗体は、マウスモノクローナル抗体DN1921軽鎖可変ドメイン(配列番号2)の1つ以上の相補性決定領域(CDR)を、ヒト軽鎖フレームワーク(FR)ドメイン1、2、3、および/または4に作動可能に連結されてヒト化HLA−DR特異的軽鎖可変ドメインを作製した状態で含む。このヒト化HLA−DR特異的軽鎖可変ドメインは、必要に応じて、ヒト重鎖定常ドメインと、配列番号3において示されるヒトIgκ軽鎖定常ドメインによって本明細書中で例証されるヒト軽鎖定常ドメインとに作動可能に連結される。このヒト重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される。
【0022】
DN1921に基づくヒト化モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、そしてインビボで投与された場合には免疫抑制性である。本発明に従うヒト化抗体を作製するために適切なDN1921 CDRは、配列番号17(DN1921 VH CDR1)、配列番号19(DN1921 VH CDR2)、配列番号21(DN1921 VH CDR3)、配列番号6(DN1921 VL CDR1)、配列番号8(DN1921 VL CDR2)、および配列番号10(DN1921 VL CDR3)として示される。
【0023】
本発明のさらなる例示的なHLA−DR特異的ヒト化モノクローナル抗体は、マウスモノクローナル抗体DN1924重鎖可変ドメイン(配列番号35)の1つ以上の相補性決定領域(CDR)を、ヒトフレームワーク(FR)ドメイン1、2、3、および/または4に作動可能に連結されてヒト化HLA−DR特異的重鎖可変ドメインを作製した状態で含む。このヒト化HLA−DR特異的重鎖可変ドメインは、必要に応じて、ヒト重鎖定常ドメインと、配列番号36において示されるヒトIgκ軽鎖定常ドメインによって本明細書中で例証されるヒト軽鎖定常ドメインとに作動可能に連結される。このヒト重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される。
【0024】
あるいは、またはさらに、本発明のさらなる例示的なHLA−DR特異的ヒト化モノクローナル抗体は、マウスモノクローナル抗体DN1924軽鎖可変ドメイン(配列番号24)の1つ以上の相補性決定領域(CDR)を、ヒトフレームワーク(FR)ドメイン1、2、3、および/または4に作動可能に連結されてヒト化HLA−DR特異的軽鎖可変ドメインを作製した状態で含む。このヒト化HLA−DR特異的軽鎖可変ドメインは、必要に応じて、ヒト重鎖定常ドメインと、配列番号25において示されるヒトIgκ軽鎖定常ドメインによって例証されるヒト軽鎖定常ドメインとに作動可能に連結される。このヒト重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される。
【0025】
DN1924に基づくヒト化モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、そしてインビボで投与された場合には非免疫抑制性である。本発明に従うヒト化抗体を作製するために適切なDN1924 CDRは、配列番号39(DN1924 VH CDR1)、配列番号41(DN1924 VH CDR2)、配列番号43(DN1924 VH CDR3)、配列番号28(DN1924 VL CDR1)、配列番号30(DN1924 VL CDR2)、および配列番号32(DN1924 VL CDR3)として示される。
【0026】
本発明はまた、組成物および方法を提供し、これらは、本明細書中に示される、1つ以上の非ヒトモノクローナルHLA−DR特異的抗体、キメラHLA−DR特異的抗体、および/またはヒト化HLA−DR特異的抗体を含む。そのようなHLA−DR特異的抗体を含む組成物は、インビボ投与のために適切であり、そしてHLA−DRを発現する腫瘍細胞のアポトーシスを誘発する際に有効である。
【0027】
本発明のこれらの局面および他の局面は、以下の詳細な説明および添付の図面を参照すると明らかとなる。本明細書中に開示されるすべての参考文献は、各々が個別に援用されたかのごとく、その全体が本明細書中に参考として援用される。
【0028】
配列番号1は、キメラDN1921全長抗体軽鎖のアミノ酸配列である。
【0029】
配列番号2は、キメラDN1921軽鎖可変領域のアミノ酸配列である。
【0030】
配列番号3は、キメラDN1921軽鎖定常領域(ヒトIgG2κ)のアミノ酸配列である。
【0031】
配列番号4は、キメラDN1921軽鎖リーダー配列のアミノ酸配列である。
【0032】
配列番号5は、キメラDN1921軽鎖フレームワーク領域1(FR1)のアミノ酸配列である。
【0033】
配列番号6は、キメラDN1921軽鎖相補性決定領域1(CDR1)のアミノ酸配列である。
【0034】
配列番号7は、キメラDN1921軽鎖FR2のアミノ酸配列である。
【0035】
配列番号8は、キメラDN1921軽鎖CDR2のアミノ酸配列である。
【0036】
配列番号9は、キメラDN1921軽鎖FR3のアミノ酸配列である。
【0037】
配列番号10は、キメラDN1921軽鎖CDR3のアミノ酸配列である。
【0038】
配列番号11は、キメラDN1921軽鎖FR4のアミノ酸配列である。
【0039】
配列番号12は、キメラDN1921全長抗体重鎖のアミノ酸配列である。
【0040】
配列番号13は、キメラDN1921重鎖可変領域のアミノ酸配列である。
【0041】
配列番号14は、キメラDN1921重鎖定常領域(ヒトIgG2)のアミノ酸配列である。
【0042】
配列番号15は、キメラDN1921重鎖リーダー配列のアミノ酸配列である。
【0043】
配列番号16は、キメラDN1921重鎖FR1のアミノ酸配列である。
【0044】
配列番号17は、キメラDN1921重鎖CDR1のアミノ酸配列である。
【0045】
配列番号18は、キメラDN1921重鎖FR2のアミノ酸配列である。
【0046】
配列番号19は、キメラDN1921重鎖CDR2のアミノ酸配列である。
【0047】
配列番号20は、キメラDN1921重鎖FR3のアミノ酸配列である。
【0048】
配列番号21は、キメラDN1921重鎖CDR3のアミノ酸配列である。
【0049】
配列番号22は、キメラDN1921重鎖FR4のアミノ酸配列である。
【0050】
配列番号23は、キメラDN1924全長抗体軽鎖のアミノ酸配列である。
【0051】
配列番号24は、キメラDN1924軽鎖可変領域のアミノ酸配列である。
【0052】
配列番号25は、キメラDN1924軽鎖定常領域(ヒトIgκ)のアミノ酸配列である。
【0053】
配列番号26は、キメラDN1924軽鎖リーダー配列のアミノ酸配列である。
【0054】
配列番号27は、キメラDN1924軽鎖FR1のアミノ酸配列である。
【0055】
配列番号28は、キメラDN1924軽鎖CDR1のアミノ酸配列である。
【0056】
配列番号29は、キメラDN1924軽鎖FR2のアミノ酸配列である。
【0057】
配列番号30は、キメラDN1924軽鎖CDR2のアミノ酸配列である。
【0058】
配列番号31は、キメラDN1924軽鎖FR3のアミノ酸配列である。
【0059】
配列番号32は、キメラDN1924軽鎖CDR3のアミノ酸配列である。
【0060】
配列番号33は、キメラDN1924軽鎖FR4のアミノ酸配列である。
【0061】
配列番号34は、キメラDN1924全長抗体重鎖のアミノ酸配列である。
【0062】
配列番号35は、キメラDN1924重鎖可変領域のアミノ酸配列である。
【0063】
配列番号36は、キメラDN1924重鎖定常領域(IgG2)のアミノ酸配列である。
【0064】
配列番号37は、キメラDN1924重鎖リーダー配列のアミノ酸配列である。
【0065】
配列番号38は、キメラDN1924重鎖FR1のアミノ酸配列である。
【0066】
配列番号39は、キメラDN1924重鎖CDR1のアミノ酸配列である。
【0067】
配列番号40は、キメラDN1924重鎖FR2のアミノ酸配列である。
【0068】
配列番号41は、キメラDN1924重鎖CDR2のアミノ酸配列である。
【0069】
配列番号42は、キメラDN1924重鎖FR3のアミノ酸配列である。
【0070】
配列番号43は、キメラDN1924重鎖CDR3のアミノ酸配列である。
【0071】
配列番号44は、キメラDN1924重鎖FR4のアミノ酸配列である。
【0072】
配列番号45は、DN1921全長抗体軽鎖をコードするポリヌクレオチドのヌクレオチド配列である。
【0073】
配列番号46は、DN1921全長抗体重鎖をコードするポリヌクレオチドのヌクレオチド配列である。
【0074】
配列番号47は、DN1924全長抗体軽鎖をコードするポリヌクレオチドのヌクレオチド配列である。
【0075】
配列番号48は、DN1924全長抗体重鎖をコードするポリヌクレオチドのヌクレオチド配列である。
【0076】
配列番号49は、ヒトHLA−DRα鎖のアミノ酸配列である。
【0077】
配列番号50は、ヒトHLA−DRβ鎖のアミノ酸配列である。
【0078】
配列番号51は、配列番号49のヒトHLA−DRα鎖をコードするヌクレオチド配列である。
【0079】
配列番号52は、配列番号50のヒトHLA−DRβ鎖をコードするヌクレオチド配列である。
【0080】
配列番号53は、マウスIgG1重鎖定常ドメインのアミノ酸配列である。
【0081】
配列番号54は、配列番号53のマウスIgG1重鎖定常ドメインをコードするヌクレオチド配列である。
【0082】
配列番号55は、マウスIgκ軽鎖定常ドメインのアミノ酸配列である。
【0083】
配列番号56は、配列番号55のマウスIgκ軽鎖定常ドメインをコードするヌクレオチド配列である。
【0084】
配列番号57は、プライマー配列
【0085】
【化1】
である。
【0086】
配列番号58は、プライマー配列
【0087】
【化2】
である。
【0088】
配列番号59は、プライマー配列
【0089】
【化3】
である。
【0090】
配列番号60は、プライマー配列
【0091】
【化4】
である。
【0092】
配列番号61は、プライマー配列
【0093】
【化5】
である。
【0094】
配列番号62は、プライマー配列
【0095】
【化6】
である。
【0096】
配列番号63は、プライマー配列
【0097】
【化7】
である。
【0098】
配列番号64は、プライマー配列
【0099】
【化8】
である。
【0100】
配列番号65は、プライマー配列
【0101】
【化9】
である。
【0102】
配列番号66は、プライマー配列
【0103】
【化10】
である。
【0104】
配列番号67は、プライマー配列
【0105】
【化11】
である。
【0106】
配列番号68は、プライマー配列
【0107】
【化12】
である。
【0108】
配列番号69は、プライマー配列
【0109】
【化13】
である。
【0110】
配列番号70は、プライマー配列
【0111】
【化14】
である。
【発明を実施するための最良の形態】
【0112】
(発明の詳細な説明)
上記のように、本発明は、HLA−DR特異的抗体、ならびにHLA−DR特異的抗体を含む組成物および方法に関する。本明細書に示される抗体は、腫瘍細胞のアポトーシスを誘導することにおいて有効であるが、HLA−DRを発現する正常細胞においては有効でない。特定の実施形態において、本発明のアポトーシス性HLA−DR特異的抗体は、低下した免疫抑制活性をさらに示す。本発明のHLA−DR特異的抗体は、とりわけ、癌(ここでこの癌細胞の少なくとも部分集団は、HLA−DR抗原を発現する)のような疾患の診断および処置における薬剤として有用である。
【0113】
本明細書で使用される場合、用語「HLA−DR」とは、「ヒト白血球抗原」(HLA)DR遺伝子座およびこれらのタンパク質産物をいい、このタンパク質産物は、ヒト白血球で発現される同種異系抗原である。同種異系抗原は、自己を外来組織から見分ける多型遺伝子の産物である。用語「クラスII主要組織適合遺伝子複合体」、「クラスII MHC」、または「クラスII」抗原とは、種々の細胞型で種々のレベルで発現されかつT細胞による全てのタンパク質抗原の認識において必須の役割を果たす抗原をいう。クラスII MHC分子は、代表的には、7〜30またはそれ以上のアミノ酸のペプチドを結合し、抗原特異的CD4+ T細胞によって認識される複合体を形成する。このCD4分子は、クラスII分子の第2のドメインに結合する。
【0114】
用語「アポトーシス性細胞死」、「細胞自己死」または「アポトーシス」とは、本明細書で使用される場合、その死亡をもたらす、正常な一連の細胞中の事象をいう。アポトーシスは、大部分の組織の生理学的維持において極めて重大な役割を果たす。細胞死プログラムの活性化を介して望ましくない細胞の排除を可能にすることは、通常の生物学的プロセスである。アポトーシスは、照射、ウイルス感染または異常な増殖に由来する損傷後に、癌の場合のように、これらの潜在的に危険な細胞を除去することによって、異常な細胞から生物を保護するために、緊急な応答を提供する。アポトーシス性細胞死は、細胞分化の特定の段階において、および特定の刺激に応答して起こる細胞事象の複雑なカスケードから生じ、死細胞の細胞質および核の凝縮によって特徴づけられる。
【0115】
用語「腫瘍」、「癌」または「新生物」とは、本明細書で交換可能に使用され、正常組織から発生するが、構造がなく異常に細胞が増殖している、異常な増殖をいう。腫瘍または癌細胞は、一般に、接触阻害の能力を欠き、侵襲性であり得、そして/または転移する能力を有し得る。本発明の抗体で処置しやすい腫瘍細胞は、クラスII腫瘍組織適合抗原HLA−DRの発現によって特徴づけられる。
【0116】
本明細書および添付の特許請求の範囲で使用される場合、単数形「a」、「an」および「the」は、文脈が明らかに別であることを示さなければ、複数形への参照を包含する。
【0117】
本発明の実施は、反対であることが具体的に示されない限り、当該分野の技術範囲内である、ウイルス学、免疫学、微生物学、分子生物学および組換えDNA技術の従来の方法が採用される。この従来の方法の多くは、例示目的で以下に示される。このような技術は、文献中で十分に説明されている。例えば、Sambrookら,「Molecular Cloning:A Laboratory Manual」(第2版,1989);Maniatisら,「Molecular Cloning:A Laboratory Manual」(1982);「DNA Cloning:A Practical Approach,vol.I & II」(D.Glover編);「Oligonucleotide Synthesis」(N.Gait編,1984);「Nucleic Acid Hybridization」(B.Hames & S.Higgins,編,1985);「Transcription and Translation」(B.Hames & S.Higgins,編,1984);「Animal Cell Culture」(R.Freshney編,1986);Perbal,「A Practical Guide to Molecular Cloning」(1984);Ausubelら,「Current protocols in Molecular Biology」(New York,John Wiley and Sons,1987);Bonifacinoら,「Current Protocols in Cell Biology」(New York,John Wiley & Sons,1999);Coliganら,「Current Protocols in Immunology」(New York,John Wiley & Sons,1999);ならびにHarlow and Lane Antibodies:a Laboratory Manual Cold Spring Harbor Laboratory(1988)を参照のこと。
【0118】
本明細書に引用される全ての刊行物、特許および特許出願は、前出、後出に拘わらず、その全体が本明細書に参考として援用される。本発明は、具体的な実施形態の詳細な説明を通じてよりよく理解される。これらの実施形態の各々は、本明細書で以下に詳細に記載される。
【0119】
(HLA−DR特異的抗体)
本発明は、抗体、より具体的には、モノクローナル抗体、非ヒト/ヒトキメラモノクローナル抗体、ヒト化モノクローナル抗体、および完全ヒトモノクローナル抗体であって、これらの抗体は、細胞表面に発現されたHLA−DRに結合した場合に、HLA−DRに特異的に結合し、かつ腫瘍細胞においてアポトーシスを誘導し得る得が、正常細胞においては誘導しない、抗体に関する。特定の実施形態において、本発明のモノクローナル抗体は、免疫抑制活性の低下したレベルによってさらに特徴づけられる。
【0120】
本明細書中で例示されるのは、マウスIgG重鎖およびIgκ軽鎖定常領域(それぞれ、配列番号53および55)に作動可能に融合された、配列番号13および2(DN1921)ならびに配列番号35および24(DN1924)の重鎖可変ドメインおよび軽鎖可変ドメインを含む、それぞれ、DN1921およびDN1924と称されるマウスモノクローナル抗体である;DN1921および/またはDN1924の可変領域(VHおよびVL)ならびにヒトIgG2重鎖およびIgκ軽鎖定常領域(それぞれ、配列番号14および3(DN1921)ならびに配列番号36および25(DN1924)を含む)を含むキメラ抗体;ならびにヒトフレームワーク領域(FR)およびヒト定常ドメインに作動可能に融合された、DN1921および/またはDN1924重鎖および軽鎖の可変ドメインの1つ以上の相補性決定領域(CDR)を含む、ヒト化抗体である。
【0121】
本明細書で記載される場合、用語「抗体」とは、モノクローナル抗体、キメラ抗体、ヒト化抗体、および完全ヒト抗体、ならびにその生物学的フラグメントもしくは抗原結合フラグメントおよび/またはその一部を包含する。本明細書で「抗体」への言及は、その一部、フラグメント、前駆体形態、誘導体、改変体、および遺伝子操作したかもしくは天然に変異した形態への言及を包含し、かつその得られる誘導体および/または改変体が、HLA−DR結合特異性および/または親和性の少なくとも実質的な量を保持する限りにおいて、アミノ酸置換および化学物質および/もしくは放射性同位体などでの標識を含む。用語「抗体」とは、抗体重鎖および軽鎖の両方、ならびに抗体の全てのアイソタイプ(IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA、およびIgA2が挙げられる)を広く包含し、その抗原結合フラグメント(Fab、F(ab’)2、Fc、およびscFvが挙げられるが、これらに限定されない)もまた包含する。
【0122】
用語「モノクローナル抗体」とは、本明細書で使用される場合、実質的に均一な抗体の集団から得られた抗体(すなわち、その集団を構成する個々の抗体が、抗体結合特異性、親和性、および/または活性に実質的に影響を及ぼさない、天然に存在する変異を除いて、同一である抗体)をいう。
【0123】
HLA−DR特異的抗体、またはその抗原結合フラグメントは、(例えば、ELISAアッセイにおいて)検出可能なレベルでHLA−DRと反応しかつ類似の条件下で関連性のないポリペプチドとは検出可能に反応しない場合、HLA−DRに「特異的に結合する」、「免疫原性であるように(immunogically)結合する」、そして/または「免疫学的に活性である」といわれる。
【0124】
この文脈で使用される場合、免疫学的に結合するとは、一般に、抗体と抗原(この抗原に対してその抗体が特異的である)との間に存在する型の非共有結合的反応をいう。抗体−HLA−DR結合相互作用の強度、すなわち親和性は、相互作用の解離定数(Kd)の観点から表され得、ここでKdが小さいほど、より大きな親和性を表す。HLA−DR特異的抗体の免疫学的結合特性は、当該分野で周知の方法を用いて定量され得る。1つのこのような方法は、HLA−DR特異的抗体/抗原複合体形成および解離の速度を測定する工程を包含し、ここでこの速度は、複合体パートナーの濃度、相互作用に親和性、および両方の方向で速度に等しく影響を与える幾何的パラメーターに依存する。従って、「オン反応速度定数」(Kon)および「オフ反応速度定数」(Koff)は、濃度と、会合および解離の実際の速度の計算によって決定され得る。Koff/Konの比率は、親和性に関しない全てのパラメーターのキャンセルを可能にし、よって、解離定数Kdに等しい。一般には、Daviesら,Annual Rev.Biochem.59:439−473(1990)を参照のこと。本発明の抗HLA−DR抗体は、HLA−DRタンパク質に特異的に結合する。本明細書で「特異的に結合する」とは、その抗体が、少なくとも106〜109M、より一般には、少なくとも107〜109Mの範囲の結合定数でHLA−DRポリペプチドに結合することを意味する。
【0125】
HLA−DR特異的抗体の「抗原結合部位」または「結合部分」とは、HLA−DR結合に関与する抗体分子の一部をいう。この抗原結合部位は、重鎖(「H」)および軽鎖(「L」)のN末端改変(「V」)領域のアミノ酸残基によって形成される。重鎖および軽鎖のV領域内の3つの高度に多様な拡がり(stretch)は、「フレームワーク領域」または「FR」として公知のより保存された隣接する拡がりの間に挟まれている「超可変領域」または「相補性決定領域(CDR)」として言及される。
【0126】
従って、用語「FR」とは、抗体中のCDRの間かつこれに隣接して天然に見出されるアミノ酸配列をいう。HLA−DR特異的抗体分子において、軽鎖の3つのCDRおよび重鎖の3つのCDRは、HLA−DR結合表面を形成するために、三次元空間で互いに対して配置されている。このHLA−DR結合表面は、結合したHLA−DRの三次元表面に相補的である。
【0127】
本明細書の目的に関して「機能的に活性な」または「機能的活性」とは、本明細書に開示されるDN1921抗HLA−DR抗体および/またはDN1924抗HLA−DR抗体の生物学的活性および/または免疫学的活性を保持しているHLA−DR特異的抗体をいう。用語「エピトープ」とは、HLA−DR特異的抗体の相補性決定領域(CDR)と相互作用するHLA−DR抗原の特定の部分をいう。
【0128】
(HLA−DR特異的非ヒトモノクローナル抗体)
本発明は、1つ以上のアポトーシス誘導性ドメインでHLA−DRに結合し、それによって、HLA−DR発現腫瘍細胞においてアポトーシスを誘導するが、HLA−DR発現正常細胞(これに対して抗体が結合する)では誘導しない非ヒトモノクローナル抗体を提供する。本明細書で例示されるのは、DN1921およびDN1924と称される、アポトーシス性HLA−DR特異的マウスモノクローナル抗体である。DN1921は、免疫抑制活性によってさらに特徴づけられる一方で、DN1924は、DN1921と比較して、減少した免疫抑制活性を示す。
【0129】
機能のいかなる理論にも限定されずに、本発明のHLA−DR特異的モノクローナル抗体は、HLA−DRヘテロ二量体の第1のタンパク質ドメイン上の1つ以上のアポトーシス誘導性エピトープに特異的に結合することによって、アポトーシス活性を達成することができると考えられる。免疫抑制性HLA−DR特異的モノクローナル抗体(本明細書ではDN1921によって例示される)が、抗原結合ドメインについてさらに競合し得、従って、ヘルパーT(Th)細胞に抗原を提示し得る一方で、非免疫抑制性HLA−DR特異的モノクローナル抗体(本明細書ではDN1924によって例示される)が、抗原結合について低下した競合能力を有するとさらに考えられる。クラスII MHC分子(HLA−DR)の第1のドメインに特異的に結合する、非ヒトモノクローナル抗体および非ヒト/ヒトキメラ抗体を、本明細書に開示されるように調製した。クラスII分子は、細胞増殖を調節し得るシグナルを伝達し、本明細書で示されるそのクラスII MHC HLA−DR特異的抗体は、癌細胞のアポトーシスを誘導し得る。
【0130】
ヒト主要組織適合遺伝子複合体(MHC)クラスII分子特異的抗体(DN1921、キメラDN1921、DN1924、およびキメラDN1924と称される)は、それらの表面でHLA−DRを発現する腫瘍細胞においてアポトーシスを誘導することが分かった。これらの抗体の抗癌活性は、悪性でないHLA−DR陽性細胞の生存性にも機能にも影響を及ぼさない点で、高度に選択性である。
【0131】
大部分の場合において、HLA−DRタンパク質のエピトープまたはフラグメントに対して行われる抗体は、全長タンパク質に結合することができると同時に、他のタンパク質に対してはほとんどもしくは全く交叉反応性を示さない。特定の実施形態において、本発明の抗体は、αHLA−DR鎖およびβHLA−DR鎖のアミノ酸1〜88およびアミノ酸1〜96(それぞれ、配列番号49および50)に対応するHLA−DR分子の第1のドメインに対して生成される。
【0132】
本明細書で例示される非ヒトモノクローナル抗体は、マウスIgG、重鎖定常領域およびIgκ軽鎖定常領域(それぞれ、配列番号53および55)に作動可能に融合された、配列番号13および2(DN1921)ならびに配列番号35および24(DN1924)の重鎖可変ドメインおよび軽鎖可変ドメインを含む、それぞれ、DN1921およびDN1924と称されるマウスモノクローナル抗体である。
【0133】
従って、特定の局面において、本発明は、非ヒトモノクローナル抗体HLA−DR特異的アポトーシス性抗体を提供する。本発明の非ヒトモノクローナル抗体は、種々の動物種から単離され得る。これらの種としては、非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、鳥類、ウサギ、マウス、ラット、モルモット、ハムスター、イヌおよびネコ起源が挙げられるが、限定されない。HLA−DR特異的アポトーシス性非ヒトモノクローナル抗体は、本明細書で、(1)配列番号13および配列番号2にそれぞれおいて示される重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ配列番号46および45のポリヌクレオチドによってコードされる)を含む免疫抑制性抗体DN1921、ならびに(2)配列番号35および配列番号24においてそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号48および47のポリヌクレオチドによってコードされる)を含む、非免疫抑制性抗体DN1924、によって例示される。
【0134】
本明細書で例示されるこのDN1921抗体およびDN1924抗体の各々は、マウスIgG、重鎖およびIgκ軽鎖定常ドメインをさらに含む。関連した代替実施形態において、DN1921および/またはDN1924モノクローナル抗体は、前述の重鎖可変ドメインおよび軽鎖可変ドメインならびにIgM、IgD、IgG2、IgG3、IgG4、IgE、IgA、およびIgA2からなる群より選択される抗体アイソタイプに由来するマウス重鎖定常ドメインを含み得る。
【0135】
DN1921およびDN1924の重鎖および軽鎖の抗原結合フラグメント、改変体、および誘導体もまた提供される。特定の実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも70%同一である重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0136】
代替的実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも70%同一である重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも80%、90%、または95%同一である重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0137】
本発明のモノクローナル抗体(mAb)の生成のために、従来の方法論が採用され得る。一般に、齧歯類(通常はマウス)が、目的のHLA−DR抗原で繰り返し免疫され、抗体生成B細胞が塗擦した免疫動物から単離され、その抗体生成B細胞が、骨髄腫細胞と融合されてB細胞「ハイブリドーマ」を生じることによって不死化される。ハイブリドーマ細胞のライブラリーは、抗原結合特異性についてスクリーニングされ、適切なクローンが精製および増殖される。モノクローナル抗体は、ハイブリドーマ法(例えば、以前に記載され、当該分野で容易に利用可能なもの)を用いて調製され得る。Kohlerら,Nature 256:495(1975)。
【0138】
不死化細胞株は、例えば、上記のように、HLA−DR抗原で免疫した動物から得た脾細胞より生成され得る。脾細胞は、例えば、骨髄腫細胞融合パートナーと、好ましくは、免疫した動物と同系であるものと融合することにより不死化される。種々の融合技術が、採用され得る。例えば、その脾細胞および骨髄腫細胞は、数分の間非イオン性界面活性剤と合わされ、次いで、低密度で選択培地(この培地は、ハイブリッド細胞の増殖を支持するが、骨髄腫細胞の増殖は支持しない)にプレートされ得る。
【0139】
ハイブリドーマ細胞は、融合していない不死化細胞の増殖または生存を阻害する1種以上の物質を好ましくは含む適切な培養培地中で培養され得る。例示的な不死化細胞株は、are those that 効率的に融合し、その選択された抗体生成細胞による抗体の安定した高レベルの発現を支持し、かつヒポキサンチン、アミノプテリン、およびチミジン(「HAT培地」)のような培地に感受性であるものである。より例示的な不死化細胞株は、マウス骨髄腫細胞株であり、この細胞株は、例えば、the Salk Institute Cell Distribution Center,San Diego,Californiaおよびthe American Type Culture Collection(ATCC),Rockville,Marylandから入手可能である。ヒト骨髄腫細胞株およびマウス−ヒトヘテロ骨髄腫細胞株が、ヒトモノクローナル抗体の生成のために記載された。Kozbor,J.Immunol.133:3001(1984)およびBrodeurら,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.編,New York,1987)。
【0140】
ハイブリドーマ細胞が培養される培養培地は、次いで、HLA−DRポリペプチドに対するモノクローナル抗体の存在についてアッセイされ得る。このハイブリドーマ細胞によって生成されるモノクローナル抗体の結合特異性は、免疫沈降によって、またはインビトロ結合アッセイ(例えば、ラジオイムノアッセイまたは酵素結合イムノソルベントアッセイ(ELISA))によって決定され得る。このような技術およびアッセイは、当該分野で公知である。このモノクローナル抗体の結合親和性は、例えば、Munsonら,Anal.Biocheni.107:220(1980)のスキャッチャード分析によって決定され得る。
【0141】
望ましいハイブリドーマ細胞が同定された後、そのクローンは、限界希釈法によってサブクローニングされ得、標準的な方法によって増殖される。Goding,「Monoclonal Antibodies:Principles and Practice」,59−103(Academic Press,1986)。この目的に関して、適切な培養培地としては、例えば、ダルベッコ改変イーグル培地およびRPMI−1640培地が挙げられる。あるいは、そのハイブリドーマ細胞は、哺乳動物における腹水としてインビボで増殖され得る。
【0142】
HLA−DR特異的モノクローナル抗体は、増殖しつつあるハイブリドーマコロニーの上清から単離され得る。さらに、種々の技術は、収率を高めるために使用され得、例えば、適切な脊椎動物宿主(例えば、マウス)の腹腔にハイブリドーマ細胞株を注射することが挙げられる。モノクローナル抗体は、次いで、腹水または血液から採取され得る。夾雑物は、従来の技術(例えば、クロマトグラフィー、ゲル濾過、沈降および抽出)によって抗体から除去され得る。本発明のHLA−DRポリペプチド(例えば、配列番号49および50に示されるヒトHLA−DR αポリペプチドおよびβポリペプチド)は、例えば、アフィニティークロマトグラフィー工程における精製プロセスで使用され得る。あるいは、サブクローンによって分泌されるモノクローナル抗体は、従来の抗体精製手順(例えば、プロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、または他のアフィニティークロマトグラフィー方法)により、培養培地または腹水から単離または精製され得る。
【0143】
モノクローナル抗体はまた、組換えDNA方法(例えば、米国特許第4,816,567号(本明細書に参考として援用される)に記載されるもの)によって作製され得る。本発明のモノクローナル抗体をコードするDNAは、容易に単離され得、そして従来の手順を用いて(例えば、マウス抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合し得るオリゴヌクレオチドプローブを使用することによって)配列決定され得る。
【0144】
一旦単離されると、そのDNAは、発現ベクターにクローニングされ得る。次いでこのベクターは、宿主細胞(例えば、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、または別の方法で抗体タンパク質を生成しない骨髄腫細胞)にトランスフェクトされて、組換え宿主細胞においてモノクローナル抗体の合成を得る。このDNAはまた、例えば、同種マウス配列の代わりにヒト重鎖定常領域および軽鎖定常領域のコード配列を置換することによって、または非抗体ポリペプチドのコード配列の全てもしくは一部を、抗体コード配列に共有結合することによって、改変され得る。このような非抗体ポリペプチドは、本発明の抗体の定常領域の代わりに用いられ得るか、または本発明の抗体の1つの抗原結合部位の可変ドメインの代わりに用いられて、本明細書で以下により詳細に記載されるキメラ二価抗体が作り出され得る。
【0145】
よって、特定の局面において、本発明は、非ヒトモノクローナルHLA−DR特異的アポトーシス性抗体を提供する。本発明の非ヒトモノクローナル抗体は、種々の動物種(非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、家禽、ウサギ、マウス、ラット、モルモット、ハムスター、イヌ、およびネコ起源が挙げられるが、これらに限定されない)から単離され得る。HLA−DR特異的アポトーシス性非ヒトモノクローナル抗体は、(1)配列番号13および配列番号2にそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(これらは、配列番号46および45のポリヌクレオチドによってそれぞれコードされる)を含む免疫抑制性抗体DN1921、ならびに(2)配列番号35および配列番号24にそれぞれ示される重鎖可変ドメインおよび軽鎖可変ドメイン(これは、配列番号48および47のポリヌクレオチドによりそれぞれコードされる)を含む非免疫抑制性抗体DN1924によって、本明細書で例示される。
【0146】
本明細書で例示されるDN1921抗体およびDN1924抗体の各々は、マウスIgG1重鎖およびIgκ軽鎖の定常ドメインをさらに含む。関連する代替的な実施形態において、DN1921モノクローナル抗体および/またはDN1924モノクローナル抗体は、前述の重鎖および軽鎖の可変ドメインならびに抗体アイソタイプ(IgM、IgD、IgG2、IgG3、IgG4、IgE、IgA、およびIgA2からなる群より選択される)由来のマウスの重鎖定常ドメインを含み得る。
【0147】
DN1921およびDN1924の重鎖および軽鎖の抗原結合フラグメント、改変体、および誘導体もまた提供される。特定の実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも70%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1921の改変体は、配列番号13および2にそれぞれ少なくとも98%または99%同一である重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0148】
代替的実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも70%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。他の実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも80%、90%、または95%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。なおさらなる実施形態において、DN1924の改変体は、配列番号35および24にそれぞれ少なくとも98%または99%同一である、重鎖可変ドメインおよび軽鎖可変ドメインを含む。
【0149】
(キメラ抗体)
本発明の他の局面は、非ヒト/ヒトキメラHLA−DR特異的モノクローナル抗体を提供し、ここでこのキメラ抗体は、ヒト定常ドメインに作動可能に融合された非ヒト可変ドメインを含む。本発明の非ヒト/ヒトキメラHLA−DR特異的モノクローナル抗体は、HLA−DR発現腫瘍細胞においてアポトーシスを誘導するが、その抗体が結合される正常細胞においては誘導しない。特定の実施形態において、本発明の非ヒト/ヒトキメラHLA−DR特異的アポトーシス性モノクローナル抗体は、被験体にインビボで投与される場合、非免疫抑制性である。マウス/ヒトキメラDN1921 HLA−DR特異的モノクローナル抗体およびマウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体によって例示される、非ヒト/ヒトキメラHLA−DR特異的抗体が本明細書で示される。これらのキメラ抗体のアミノ酸配列およびそれらの構成的構造領域は、表1(キメラDN1921)および表2(キメラDN1924)において示される。
【0150】
本明細書で使用される場合、用語「キメラ抗体」とは、重鎖および軽鎖を含む抗体分子であって、この抗体分子において、非ヒト抗体の可変ドメインが、ヒト定常ドメインに作動可能に融合されたものをいう。キメラ抗体は、一般に、その親の完全非ヒト抗体に比較して、減少した免疫原性を示す。
【0151】
ヒト定常ドメインに作動可能に融合された非ヒト抗原結合可変ドメインを含むキメラ抗体分子を作製するための方法論は、当該分野で記載されている。例えば、Winterら,Nature 349:293−299(1991);Lobuglioら,Proc.Nat.Acad.Sci.USA 86:4220−4224(1989);Shawら,J.Immunol.138:4534−4538(1987);Brownら,Cancer Res.47:3577−3583(1987);米国特許第4,816,567号(Cabillyによる);Cabillyら,Proc.Natl.Acad.Sci.USA 81:3273−3277(1984);Morrisonら,Proc.Natl.Acad.Sci.USA 81:6851−6855(1984);Boulianneら,Nature 312:643−646(1984);Cabillyら,欧州特許出願第125023(1984年11月14日公開);Neubergerら,Nature 314:268−270(1985);Taniguchiら,欧州特許出願第171496(1985年2月19日公開);Morrisonら,欧州特許出願第173494号(1986年3月5日公開);Neubergerら,PCT出願WO86/01533(1986年3月13日公開);Kudoら,欧州特許出願第184187号(1986年6月11日公開);Morrisonら,欧州特許出願第173494号(1986年3月5日公開);Sahaganら,J.Immunol.137:1066−1074(1986);Robinsonら,国際特許公開番号PCT/US86/02269(1987年公開5月7日);Liuら,Proc.Natl.Acad.Sci.USA 84:3439−3443(1987);Sunら,Proc.Natl.Acad.Sci.USA 84:214−218(1987);Betterら,Science 240:1041−1043(1988);ならびにHarlow and Lane Antibodies:a Laboratory Manual Cold Spring Harbor Laboratory(1988))を参照のこと。これらの参考文献の各々は、本明細書に参考として援用される。
【0152】
キメラ抗体は、一価抗体、二価抗体または多価抗体を包含する。一価キメラ抗体は、キメラL鎖とジスルフィド架橋によって結合したキメラH鎖によって形成される重鎖および軽鎖(HL)の二量体である。二価キメラ抗体は、少なくとも1つのジスルフィド架橋によって結合した2つのHL二量体によって形成される四量体(H2L2)である。例えば、凝集するCH領域(例えば、IgM H鎖、またはμ鎖から)を使用することによって、多価キメラ抗体も生成され得る。
【0153】
本発明のマウス抗体およびキメラ抗体、フラグメントおよび領域は、個々の抗体の重鎖および/または軽鎖を含む。キメラ重鎖は、ヒト重鎖定常領域の少なくとも一部に連結された、HLA−DRに対して特異的な非ヒト抗体の重鎖に由来する抗原結合領域を含む。
【0154】
本発明に従うキメラ軽鎖は、ヒト軽鎖定常領域の少なくとも一部に連結された、HLA−DRに特異的な非ヒト抗体の軽鎖に由来する抗原結合領域を含む。
【0155】
同じまたは異なる可変領域結合特異性のキメラ重鎖およびキメラ軽鎖を有する抗体、フラグメントまたは誘導体はまた、公知の方法に従って(例えば、Ausubel(前出)およびHarlow(前出)に従って)個々のポリペプチド鎖の適切な結合によって調製され得る。
【0156】
特定の実施形態において、キメラ重鎖(またはその誘導体)を発現する宿主細胞は、キメラ軽鎖(またはその誘導体)を発現する宿主とは別個に培養されてもよく、その免疫グロブリン鎖は、別個に回収され、次いで結合されてもよい。あるいは、この宿主細胞は、一緒に培養され得、その鎖は、培養培地中で自発的に会合し、続いて、組み立てられた免疫グロブリン、フラグメントまたは誘導体を回収することが可能になる。
【0157】
本明細書で例示される場合、抗体重鎖および軽鎖は、同じ細胞にトランスフェクトされ、輸送および分泌されるときに細胞によって組み立てられてもよい。
【0158】
ハイブリッド細胞は、非ヒトHLA−DR特異的抗体生成細胞(代表的には、天然もしくは組換えいずれかのヒトHLA−DR、またはヒトHLA−DRタンパク質配列のペプチドフラグメントに対して免疫された動物の脾細胞)の融合によって形成され得る。あるいは、非ヒトHLA−DR特異的抗体生成細胞は、HLA−DRで免疫した動物の血液、脾臓、リンパ節または他の組織から得られたBリンパ球であり得る。
【0159】
第2の融合パートナーは、不死化機能を提供し、リンパ芽球腫細胞またはプラスマ細胞腫細胞または骨髄腫細胞であり得る。これらの細胞は、それら自身抗体生成細胞ではなく、悪性である。例示的な融合パートナー細胞としては、ハイブリドーマSP2/0−Ag14(SP2/0;ATCC CRL1581)および骨髄腫P3X63Ag8(ATCC TIB9)、あまたはその誘導体が挙げられる。例えば、Ausubel(前出)およびHarlow(前出)を参照のこと。
【0160】
ヒトHLA−DRに特異的なモノクローナル抗体を生成するマウスハイブリドーマは、マウス融合パートナー細胞(例えば、SP2/0)と、精製HLA−DR、組換えHLA−DR、天然もしくは合成HLA−DRペプチド(αおよびβ HLA−DR鎖(それぞれ配列番号49および50)のアミノ酸1〜88およびアミノ酸1〜96を含むペプチドが挙げられる)に対して免疫したマウス由来の脾細胞とを融合することによって形成される。
【0161】
マウスを免疫するために、種々の異なる従来のプロトコルが行われ得る。例えば、マウスは、HLA−DRの初回免疫および追加免疫を受容し得る。本発明のキメラ抗体の抗原結合領域をコードするヌクレオチド配列を与えるこの抗体生成細胞はまた、非ヒト(例えば、霊長類)細胞またはヒト細胞の形質転換によって生成され得る。例えば、HLA−DR特異的抗体を生成するBリンパ球は、エプスタイン−バーウイルスのようなウイルスに感染して形質転換されて、不死化HLA−DR特異的抗体生成細胞を生じる。例えば、Kozborら,Immunol.Today 4:72−79(1983)を参照のこと。あるいは、Bリンパ球は、当該分野で周知のように、癌遺伝子または癌遺伝子産物を提供することによって形質転換され得る。例えば、Ausubel(前出)およびHarlow(前出)を参照のこと。
【0162】
本発明のキメラモノクローナル抗体の構築に適した非ヒトモノクローナル抗体可変ドメインは、種々の動物種(非ヒト霊長類、ヒツジ、ブタ、ウシ、ウマ、ロバ、家禽、ウサギ、マウス、ラット、モルモット、ハムスター、犬およびネコ起源が挙げられるが、これらに限定されない)から単離され得る。ヒト抗体重鎖定常ドメインは、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA、およびIgA2からなる群より選択される抗体アイソタイプから単離され得る。
【0163】
本明細書で議論される、例示的なHLA−DR特異的キメラモノクローナル抗体は、ヒト重鎖定常ドメインおよび軽鎖定常ドメイン(そrぞれ、配列番号14および3)に作動可能に融合された、マウスモノクローナル抗体DN1921の重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号13および2)を含む。この本明細書で議論されるキメラDN1921モノクローナル抗体は、インビボで投与される場合に、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導することができ、かつ免疫抑制性である。例示的な全長キメラDN1921モノクローナル抗体の重鎖のアミノ酸配列は、配列番号12として示される。例示的な全長キメラDN1921モノクローナル抗体軽鎖のアミノ酸配列は、配列番号1として示される。
【0164】
キメラDN1921の重鎖および軽鎖(それぞれ配列番号12および1)の改変体もまた提供される。特定の実施形態において、キメラDN1921の重鎖および軽鎖の改変体は、それぞれ配列番号12および1に少なくとも70%同一である。他の実施形態において、キメラDN1921重鎖および軽鎖の改変体は、それぞれ配列番号12および1に少なくとも80%、90%、または95%同一である。なおさらなる実施形態において、キメラDN1921重鎖および軽鎖の改変体は、それぞれ配列番号12および1に少なくとも98%または99%同一である。
【0165】
本明細書で開示される、代替的な例示的HLA−DR特異的キメラモノクローナル抗体は、ヒト重鎖定常ドメインおよび軽鎖定常ドメイン(それぞれ、配列番号35および24)に作動可能に融合された、マウスモノクローナル抗体DN1924の重鎖可変ドメインおよび軽鎖可変ドメイン(それぞれ、配列番号35および24)を含む。本明細書で開示されるキメラDN1924モノクローナル抗体は、インビボで投与された場合に、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導することができ、非免疫抑制性である。例示的な全長キメラDN1924モノクローナル抗体の重鎖のアミノ酸配列は、本明細書で配列番号34として示される。例示的な全長キメラDN1924モノクローナル抗体軽鎖のアミノ酸配列は、本明細書で配列番号23として示される。
【0166】
代替的実施形態において、キメラDN1924重鎖および軽鎖の改変体は、それぞれ、配列番号34および23である。特定の実施形態において、キメラDN1924重鎖および軽鎖の改変体は、それぞれ配列番号34および23に少なくとも70%同一である。他の実施形態において、キメラDN1924重鎖および軽鎖の改変体は、それぞれ配列番号34および23に少なくとも80%、90%、または95%同一である。なおさらなる実施形態において、キメラDN1924重鎖および軽鎖の改変体は、それぞれ配列番号34および23に少なくとも98%または99%同一である。
【0167】
【表1−1】
【0168】
【表1−2】
【0169】
【表1−3】
【0170】
【表2−1】
【0171】
【表2−2】
【0172】
【表2−3】
【0173】
【表2−4】
(ヒト化抗体)
なおさらなる実施形態において、本発明は、ヒト化HLA−DR特異的アポトーシス性モノクローナル抗体を提供し、ここで、このヒト化抗体は、1つ以上の非ヒト相補性決定領域(CDR)、ヒト可変ドメインフレームワーク領域(FR)、およびヒト重鎖定常ドメイン(例えば、IgG2重鎖定常ドメイン)およびヒト軽鎖定常ドメイン(例えば、Igκ軽鎖定常ドメイン)を含む。一般に、「ヒト化」HLA−DR特異的抗体は、そうでなければ、ヒトレシピエントにおけるこれらの部分の治療的適用の持続時間および有効性を制限する、対応する非ヒト(例えば、げっ歯類)抗体またはその非ヒト領域に対する、望ましくない免疫学的応答を最小にするのに有用である。Jonesら,Nature 321:522−525(1986);Reichmannら,Nature 332:323−327(1988);およびVerhoeyernら,Science 239:1534−1536(1988)。
【0174】
本明細書中で使用される場合、用語「ヒト化」は、ヒト抗体(レシピエント抗体)を含むことを意味し、ヒト化抗体においては、レシピエントの相補性決定領域(CDR)に由来する残基が、非ヒト種(ドナー抗体)(例えば、所望の特異性、親和性および能力を有する、マウス、ラットまたはウサギ)のCDRに由来する残基で置き換えられる。いくつかの場合、ヒト抗体の可変ドメインフレームワーク残基は、対応する非ヒト残基で置き換えられる。ヒト化抗体はまた、レシピエント抗体にも、輸入されたCDRまたはフレームワークの配列にも見られない残基を含み得る。
【0175】
一般に、ヒト化抗体は、少なくとも1つ、そして、代表的には2つの、可変ドメインの実質的に全てを含み、ここでは、CDR領域の全てまたは実質的に全ては、非ヒト抗体のCDR領域に対応し、そして、これらの領域の全てまたは実質的に全ては、ヒト抗体のコンセンサス配列の領域である。ヒト化抗体はまた、好適には、抗体の定常領域の少なくとも一部(代表的には、ヒト抗体の一部)を含む。
【0176】
ヒト化モノクローナル抗体は、代表的に、約90〜95%がヒトであり、相補性決定領域(CDR)のみが非ヒトである。一般に、げっ歯類細胞において産生されるヒト化モノクローナル抗体は、それらが由来する種からの対応するキメラモノクローナル抗体よりも免疫原性が低い。Bellら,Lancet 355:858−859(2000)。
【0177】
上述のように、用語「相補性決定領域」(CDR)は、結合抗原の3次元表面と相補的な表面を形成する抗体分子の超可変領域をいう。N末端からC末端に向かって、抗体の重鎖および軽鎖の各々は、3つのCDR(CDR1、CDR2およびCDR3)から構成される。それゆえ、HLA−DR抗原結合部位、は、合計6つのCDRを含み、重鎖V領域および軽鎖V領域の各々からの3つのCDRを含む。CDRのアミノ酸残基は、結合抗原との広範な接触を形成し、最も広範な抗原接触は、重鎖CDR3とである。
【0178】
ヒト化HLA−DR特異的アポトーシス性抗体は、ヒト可変ドメインフレームワーク領域(FR)に作動可能に融合した1つ以上の非ヒト相補性決定領域(CDR)を含んで、重鎖可変ドメインおよび軽鎖可変ドメインを生じ、これらは、ヒト定常ドメインの重鎖および軽鎖(本明細書において、それぞれ、配列番号14および3に示される、重鎖IgG2定常ドメインおよび軽鎖Igκ定常ドメインにより例示される)に作動可能に融合している。
【0179】
本発明の例示的なHLA−DR特異的ヒト化モノクローナル抗体は、ヒト重鎖フレームワーク(FR)ドメインに作動可能に融合された、マウスモノクローナル抗体DN1921重鎖可変ドメイン(配列番号13)の1つ以上の相補性決定領域(CDR)を含んで、ヒト化HLA−DR特異的重鎖可変ドメインを生じ、これは、必要に応じて、定常ドメインが、本明細書において、配列番号14に示されるヒトIgG2重鎖定常ドメインにより例示される、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択される、ヒト重鎖定常ドメインに作動可能に融合される。
【0180】
あるいは、またはさらに、本発明の例示的なHLA−DR特異的ヒト化モノクローナル抗体は、ヒト軽鎖フレームワーク(FR)ドメイン1、2、3および/または4に作動可能に融合された、マウスモノクローナル抗体DN1921軽鎖可変ドメイン(配列番号2)の1つ以上の相補性決定領域(CDR)を含んで、ヒト化HLA−DR特異的軽鎖可変ドメインを生じ、これは、必要に応じて、本明細書において配列番号3に示されるヒトIgκ軽鎖定常ドメインにより例示される、ヒト軽鎖定常ドメインに作動可能に融合される。
【0181】
DN1921に基づくヒト化モノクローナル抗体は、HLA−DR発現腫瘍細胞においてアポトーシスを誘導し得、そして、インビボで投与された場合に、免疫抑制的である。本発明に従うヒト化抗体を作製するために適切なDN1921 CDRは、配列番号17(DN1921 VH CDR1)、配列番号19(DN1921 VH CDR2)、配列番号21(DN1921 VH CDR3)、配列番号6(DN1921 VL CDR1)、配列番号8(DN1921 VL CDR2)、および配列番号10(DN1921 VL CDR3)として表される。
【0182】
本発明のさらなる例示的なHLA−DR特異的ヒト化モノクローナル抗体は、ヒトフレームワーク(FR)ドメイン1、2、3および/または4に作動可能に融合された、マウスモノクローナル抗体DN1924重鎖可変ドメイン(配列番号35)の1つ以上の相補性決定領域(CDR)を含んで、ヒト化HLA−DR特異的重鎖可変ドメインを生じ、これは、必要に応じて、定常ドメインが、IgM、IgD、IgG1、IgG2、IgG3、IgG4、IgE、IgA1およびIgA2からなる群より選択され、そして、本明細書において、配列番号36に示されるヒトIgG2重鎖定常ドメインにより例示される、ヒト重鎖定常ドメインに作動可能に融合される。
【0183】
あるいは、またはさらに、本発明のさらなる例示的なHLA−DR特異的ヒト化モノクローナル抗体は、ヒトフレームワーク(FR)ドメイン1、2、3および/または4に作動可能に融合された、マウスモノクローナル抗体DN1924軽鎖可変ドメイン(配列番号24)の1つ以上の相補性決定領域(CDR)を含んで、ヒト化HLA−DR特異的軽鎖可変ドメインを生じ、これは、必要に応じて、本明細書において、配列番号25に示されるヒトIgκ軽鎖定常ドメインによりヒト軽鎖定常ドメインにより例示される、ヒト軽鎖定常ドメインに作動可能に融合される。
【0184】
DN1924に基づくヒト化モノクローナル抗体は、HLA−DRを発現する細胞においてアポトーシスを誘導し得、そして、インビボで投与された場合に、非免疫抑制的である。本発明に従うヒト化抗体を作製するために適切なDN1924 CDRは、配列番号39(DN1924 VH CDR1)、配列番号41(DN1924 VH CDR2)、配列番号43(DN1924 VH CDR3)、配列番号28(DN1924 VL CDR1)、配列番号30(DN1924 VL CDR2)、および配列番号32(DN1924 VL CDR3)として表される。
【0185】
各CDRに隣接するのは、主にV領域を、抗原結合部位へと折りたたむことを担う、フレームワーク(FR)領域、特に、CDRに直ぐ隣接するFR残基である。いくつかのFR残基は、結合抗原に接触し得る。FR内では、特定のアミノ残基および特定の構造的特徴が、非常に高度に保存されている。この点から、全てのV領域の配列は、約90アミノ酸残基の内部ジスルフィドループを含む。V領域が、結合部位に折りたたまれるとき、CDRは、抗原結合表面を形成する突出したループモチーフとして提示される。CDRループの、特定の正確なCDRアミノ酸配列への折りたたみ形状に影響を与える、FRの保存された構造領域がある。さらに、特定の残基が、抗体の重鎖と軽鎖との相互作用を安定化する、非共有結合性内部ドメイン接点に関与することが知られている。
【0186】
非ヒト抗体をヒト化するための方法は、当該分野で周知である。一般に、ヒト化抗体は、非ヒト供給源から導入された1つ以上のアミノ酸残基を有する。ヒト化は、適切なヒト抗体定常ドメインに融合する前に、ヒト支持FRにCDRを接合することにより達成され得る。Jonesら,Nature 321:522−525(1986);Reichmannら,Nature 332:323−327(1988);およびVerhoeyernら,Science 239:1534−1536(1988)を参照のこと。
【0187】
あるいは、げっ歯類のCDRは、組換え的に操作されたげっ歯類FRにより支持され得る。1992年12月23日に公開された欧州特許第519,596号を参照のこと。Queenに対する、米国特許第5,530,101号、同第5,585,089号、同第5,693,762号、および同第6,180,370号;Adairに対する米国特許第5,859,205号;ならびに、Winterに対する米国特許第5,225,539号(これらの特許の各々は、その全体が、本明細書中に参考として援用される)もまた参照のこと。
【0188】
実際には、ヒト化抗体は、代表的に、いくつかのCDR残基、そして、可能ないくつかのフレームワーク残基が、げっ歯類抗体の類似の部位に由来する残基で置き換えられているヒト抗体である。従って、本発明のヒト化HLA−DR特異的抗体の各々は、それぞれ、重鎖FRと軽鎖FRとの間に挿入された、少なくとも1、2もしくは3の非ヒト重鎖CDR、および/または、少なくとも1、2もしくは3の非ヒト軽鎖CDRを含み、これらは、CDRに対する支持を提供し、そして、互いに関するCDRの空間的関係を規定する。
【0189】
(完全ヒト抗体)
完全ヒト抗体は、ファージディスプレイ技術およびトランスジェニックマウス法により作製され得る。ファージディスプレイ技術により同定された抗体フラグメントをコードするDNAフラグメントは、組換え技術により合わされ、完全な「ヒト」抗体を生成する。トランスジェニックマウスから抗体を作製するために、ハイブリドーマが調製され、そして、従来のモノクローナル抗体法についてスクリーニングされる。
【0190】
ファージディスプレイ技術により完全ヒト抗体を作製するための方法論は、Hoogenboomら,J.Mol.Biol.227:381(1991)およびMarksら,J.Mol.Biol.222:581(1991)に記載される。米国特許第6,248,516号、同第6,291,158号、同第6,291,159号、同第6,291,160号、同第6,291,161号、同第5,969,108号、同第6,172,197号、同第5,885,793号、同第6,265,150号、同第5,223,409号、同第5,403,484号、同第5,571,698号、同第5,837,500号、および同第6,300,064号(この各々は、本明細書中に参考として援用される)もまた参照のこと。
【0191】
完全ヒト抗体を作製するためのトランスジェニック動物系は、米国特許第6,150,584号、同第6,114,598号、同第6,162,963号、同第6,075,181号および同第5,770,429号(この各々は、本明細書中に参考として援用される)に開示される。
【0192】
またヒトモノクローナル抗体の調製に利用可能なさらなる技術は、例えば、「Monoclonal Antibodies and Cancer Therapy」、p.77(Alan R.Liss編,1985)およびBoernerら,J.Immunol.147(1):86−95(1991)に記載される。
【0193】
(HLA−DR抗体の抗原結合フラグメント、改変体および誘導体)
本発明はまた、前述のHLA−DR特異的モノクローナル抗体のいずれかの、HLA−DR抗原結合フラグメント、改変体、および/または誘導体を企図する。本発明のHLA−DR抗原結合フラグメント、改変体、および誘導体の各々は、HLA−DR発現正常細胞ではなく、HLA−DR発現腫瘍細胞において、アポトーシスを誘導し得る。特定の実施形態において、HLA−DR抗原結合フラグメント、改変体および誘導体(例えば、モノクローナル抗体DN1921に基づくもの)は、インビボで投与された場合、免疫抑制的である。他の実施形態において、HLA−DR抗原結合フラグメント、改変体および誘導体(例えば、モノクローナル抗体DN1924に基づくもの)は、インビボで投与された場合、非免疫抑制的である。
【0194】
対応する全長HLA−DR特異的抗体分子の免疫学的結合特性を示し得る、治療的に有用なHLA−DR特異的抗原結合フラグメントを作製するための方法論は、当該分野で容易に利用可能である。本発明の方法は、HLA−DR分子の架橋を促進し、それによりアポトーシスを刺激するために、HLA−DR特異的抗体が二価、三価、または多価であることを必要とする。従って、HLA−DR特異的抗体の1つ以上のフラグメントは、固体支持体に結合され、それにより、HLA−DRの架橋を促進し得る。
【0195】
タンパク質分解酵素であるパパインは、優先的にIgG分子を切断して、いくつかのフラグメントを生じ、このうち2つ(「F(ab)」フラグメント)は、インタクトなこう現結合部位を含む共有結合性のヘテロ二量体を含む。酵素ペプシンは、IgG分子を切断して、いくつかのフラグメントを提供し得、このフラグメントは、両方の抗原結合部位を含む「F(ab’)2」フラグメントを含む。「Fv」フラグメントは、IgM抗体分子の、そして、時として、IgG抗体分子またはIgA抗体分子の優先的なタンパク質分解性切断により生成され得る。Fvフラグメントは、あるいは、当該分野で公知の技術を用いて作製され得る。Fvフラグメントは、ネイティブな抗体分子の抗原認識能および抗原結合能の大半を保持する、抗原結合部位を含む、非共有結合性のVH::VLヘテロ二量体を含む。Inbarら,Proc.Nat.Acad.Sci.USA 69:2659−2662(1972);Hochmanら,Biochem.15:2706−2710(1976);およびEhrlichら,Biochem 19:4091−4096(1980)。
【0196】
単鎖Fv(「sFv」)ポリペプチドは、ペプチドをコードするリンカーにより連結された、VHをコードする遺伝子およびVLをコードする遺伝子を含む遺伝子融合物から発現される、共有結合されたVH::VLヘテロ二量体である。Hustonら,Proc.Nat.Acad.Sci.USA 85(16):5879−5883(1988)。1つの抗体V領域に由来する、自然に凝集しているが、化学的には分かれている、軽ポリペプチド鎖および重ポリペプチド鎖を、sFv分子(抗原結合部位の構造に実質的に類似する3次元構造へと折り畳まれる)へと変換するための化学構造を見分けるための多数の方法が記載されている。例えば、米国特許第5,091,513号、同第5,132,405号および同第4,946,778号を参照のこと。
【0197】
本発明に従う、抗体、およびその抗原結合フラグメントをコードするポリヌクレオチドは、従来の分子生物学および組換えDNA方法論により作製され得る。このような方法論は、文献で完全に説明されている。例えば、Sambrookら,Molecular Cloning:A Laboratory Manual(第2版,1989);Maniatisら,Molecular Cloning:A Laboratory Manual(1982);DNA Cloning:A Practical Approach,第I巻および第II巻(D.Glover編);Oligonucleotide Synthesis(N.Gait編,1984);Nucleic Acid Hybridization(B.HamesおよびS.Higgins編,1985);Transcription and Translation(B.HamesおよびS.Higgins編,1984);Animal Cell Culture(R.Freshney編,1986);Perbal,A Practical Guide to Molecular Cloning(1984)を参照のこと。これらの刊行物の各々は、その全体が参考として援用される。
【0198】
簡単に言うと、例えば、抗体重鎖または抗体軽鎖をコードするDNA配列は、適切な発現ベクター内に連結され、この発現ベクターは、抗体領域をコードするポリペプチドに作動可能に連結された転写プロモーターを含み、そして、この抗体領域をコードするポリヌクレオチドの3’側には転写終止シグナルを含む。適切な発現ベクターはまた、転写開始部位、翻訳開始を命令するためのKozak配列、および翻訳を停止させる終止コドンを提供し得る。さらに、発現ベクターはまた、ポリペプチドをコードする1つ以上のポリヌクレオチド配列(例えば、His−His−His−His−His−HisまたはFLAG(登録商標)配列Asp−Tyr−Lys−Asp−Asp−Asp−Asp−Lys(Sigma−Aldrich,St.Louis,MO))を含み得、この配列は、抗体ペプチドの検出および親和性精製を容易にする。
【0199】
発現は、組換えポリペプチドをコードするDNA分子を含む発現ベクターで形質転換または形質移入された任意の適切な宿主細胞内で達成され得る。適切な宿主細胞としては、原核生物、酵母および高等真核生物細胞が挙げられる。代表的に、使用される宿主細胞は、COSまたはCHOのような哺乳動物細胞株由来である。組換え抗体ポリペプチドを培養培地中に分泌する適切な系からの上清が、まず、市販のフィルタを使用して濃縮され得る。濃縮の後、濃縮物が、適切な精製マトリクス(例えば、親和性マトリクスまたはイオン交換樹脂)に適用され得る。最後に、1つ以上の逆相HPLC工程を用いて、組換えポリペプチドをさらに精製し得る。
【0200】
抗体フラグメント(例えば、Fabフラグメント)は、あるいは、従来のポリペプチド合成方法論により合成され得る。例えば、このような抗体フラグメントは、市販の固相技術(例えば、Merrifield固相合成法)を用いて合成され得、この方法において、アミノ酸が、増大するアミノ酸鎖に連続的に添加される。Merrifield,J.Am.Chem.Soc.85:2149−2146(1963)を参照のこと。ポリペプチドの自動化合成のための設備は、Perkin Elmer/Applied BioSystems Division(Foster City,Calif.)などの供給業者から市販されており、製造業者の説明書に従って操作され得る。
【0201】
従って、本発明に従うHLA−DR特異的抗体およびそのフラグメントは、フラグメント、誘導体および改変体がHLA−DR結合の特異的かつアポトーシス性の非免疫抑制活性の高原結合特性に実質的に影響を与えない限り、これらのフラグメント、誘導体および改変体を包含する。
【0202】
本明細書中で使用される場合、ポリペプチドまたはタンパク質の「フラグメント、誘導体および改変体」は、ネイティブなポリペプチドまたはタンパク質とは、1つ以上の置換、欠失および/または挿入で異なるポリペプチドまたはタンパク質であり、その結果、このポリペプチドまたはタンパク質の抗原結合活性は、実質的に減少されない。換言すると、HLA−DRに特異的に結合する改変体の能力は、親HLA−DR特異的抗体に対して、増強されていても不変であってもよいか、または、得られるHLA−DR特異的抗体もしくはそのフラグメントの効力に影響を与えることなく、親抗体に対して、50%未満、そして代表的には20%未満消失していてもよい。一般に、適切なHLA−DR特異的抗体の誘導体または改変体は、HLA−DRを発現する新生物細胞においてアポトーシスを誘導する、その誘導体または改変体の能力を評価することによって特徴付けられ得る。
【0203】
本明細書中で使用される場合、用語「改変体」とは、具体的に同定された配列と異なるヌクレオチド配列またはアミノ酸配列を包含し、改変体においては、1つ以上のヌクレオチド残基またはアミノ酸残基が、欠失、置換または付加されている。改変体は、天然に存在する対立遺伝子改変体であっても、天然に存在しない改変体であってもよい。改変体の配列(ポリヌクレオチドまたはポリペプチド)は、好ましくは、本発明の配列と、少なくとも70%、より好ましくは、少なくとも80%もしくは少なくとも90%、なおより好ましくは少なくとも95%、そして最も好ましくは、少なくとも98%の同一性を示す。
【0204】
%同一性は、以下に記載されるように、比較されるべき2つの配列を整列し、整列した部分における同一の残基の数を決定し、この数を、本発明の(照会した)配列内の残基の総数で割り、そして、その結果に100をかけることによって決定される。列挙されたレベルの配列類似性を示すことに加え、本発明の改変体の配列は、好ましくは、その改変体が比較される配列の抗原結合に対して実質的に類似する抗原結合性(antigen−bindingity)を示す。
【0205】
改変体は、1つのアミノ酸が、類似の特性を有する別のアミノ酸で置換される、置換として定義される、「保存的アミノ酸置換」を含み得、その結果、ポリペプチドの二次構造および疎水性の性質は、実質的に不変である。アミノ酸置換は、一般に、残基の極性、電荷、可溶性、疎水性、親水性および/または両親媒性の性質における類似性に基づいてなされ得る。例えば、負に帯電したアミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられ;正に帯電したアミノ酸としては、リジンおよびアルギニンが挙げられ;そして、類似の親水性値を有する、無電荷の極性頭基を有するアミノ酸としては、ロイシン、イソロイシンおよびバリン;グリシンおよびアラニン;アスパラギンおよびグルタミン;ならびにセリン、スレオニン、フェニルアラニンおよびチロシンが挙げられる。保存的変化を表し得るアミノ酸の他のグループとしては、以下が挙げられる:(1)ala、pro、gly、glu、asp、gln、asn、ser、thr;(2)cys、ser、tyr、thr;(3)val、ile、leu、met、ala、phe;(4)lys、arg、his;および(5)phe、tyr、trp、his。改変体はまた、あるいは、非保存的な変化を含み得る。
【0206】
ポリペプチドの抗原結合フラグメント、誘導体および改変体は、抗体の化学的もしくは酵素的な消化のいずれかによってか、または、その抗体をコードするポリヌクレオチドおよび結果として生じる変異体のその後の発現の変異分析によって同定され得る。ついで、ポリペプチドのフラグメントまたは変異体ポリペプチドが、例えば、以下に提供される代表的なアッセイを使用して、どの部分が生物活性を保持するかを決定するために試験される。
【0207】
本発明の抗体のフラグメント、誘導体および改変体はまた、合成もしくは組換え手段によって作製され得る。約100より少ないアミノ酸、一般には、約50より少ないアミノ酸を有する合成ポリペプチドは、当業者に周知の技術を使用して作製され得る。例えば、このようなポリペプチドは、上述のMerrifield固相合成法により合成され得る。
【0208】
改変体はまた、オリゴヌクレオチド指向型、部位特異的変異誘発のような標準的な変異誘発技術を用いて調製され得る。Kunkel,Proc.Natl.Acad.Sci.USA 82:488−492(1985)。ポリヌクレオチド配列の部分はまた、短縮型ポリペプチドの調製を可能にする標準的な技術を用いて除去され得る。改変体は、さらに、またはあるいは、例えば、ポリペプチドの二次構造および疎水性の性質に対して最小限の影響を有するアミノ酸の欠失または付加によって修飾され得る。
【0209】
HLA−DR特異的抗体のフラグメント、誘導体および改変体は、好ましくは、ネイティブなHLA−DR特異的抗体に対して、少なくとも約70%、より好ましくは少なくとも約80%もしくは90%、そしてより好ましくは、少なくとも約95%もしくは98%の配列同一性を示す。ポリペプチド配列が整列され得、そして、特定の領域における同一なアミノ酸の百分率が、公的に利用可能なコンピュータアルゴリズムを用いて、別のポリペプチドに対して決定され得る。ポリペプチド配列の整列および同一性は、BLASTPアルゴリズムを用いて試験され得る。BLASTPアルゴリズムは、PearsonおよびLipman,Proc.Natl.Acad.Sci.USA 85:2444−2448(1988);ならびにPearson,Methods in Enzymol.183:63−98(1990)に記載される。
【0210】
BLASTPソフトウェアは、NCBIの匿名のFTPサーバ上で利用可能であり、そして、National Center for Biotechnology Information(NCBI)、National Library of Medicine,Building 38A,Room 8N805,Bethesda,MD 20894から利用可能である。文書に記載され、アルゴリズムと共に配布されているデフォルトパラメータに設定された、BLASTPアルゴリズムバージョン2.0.6[1998年9月10日]およびバージョン2.0.11[2000年1月20日]は、本発明に従う改変体の決定において使用するための例示的なものである。アルゴリズムのBLASTファミリー(BLASTPを含む)の使用は、NCBIのウェブサイトおよびAltschulら,「Gapped BLAST and PSI−BLAST:a new generation of protein database search programs」,Nucleic Acids Res.25:338903402(1997)の刊行物に記載される。
【0211】
BLASTPまたは類似のアルゴリズムにより得られた、照会配列による1つ以上のデータベースの配列に対する「ヒット」は、配列の類似の部分を整列し、そして同定する。このヒットは、配列の重なりの類似性の程度および長さの順に並べられる。データベースの配列に対するヒットは、一般に、照会配列の、配列の長さの一部のみに対する重なりを表す。
【0212】
ポリペプチド配列の%同一性は、デフォルトパラメータに設定されたBLASTPのような適切なアルゴリズムを使用してポリペプチド配列を整列し;整列された部分に対して同一のアミノ酸の数を同定し;同一なアミノ酸の数を、本発明のポリペプチドのアミノ酸の総数で割り;そして、100をかけて、%同一性を決定することによって決定される。
【0213】
BLASTPアルゴリズムはまた、ポリペプチドの整列についての「期待」値を生じる。この期待値(E)は、特定の大きさのデータベースを検索する際に、偶然、特定の数の隣接する配列にわたって見えることが「期待」され得るヒットの数を示す。期待値は、データベースに対するヒットが、真の類似性を示すかどうかを決定するための有意な閾値として使用される。例えば、ポリペプチドヒットに対して割当てられた0.1のE値が、SwissProtデータベースの大きさのデータベースにおける値を意味するものとして解釈される場合、ほんの偶然に、類似のスコアを有する配列の整列された部分にわたって、0.1のマッチを見ることが期待され得る。この基準によれば、配列の整列かつマッチした部分は、90%の関連する確率を有する。整列かつマッチした部分にわたって0.01以下のE値を有する配列について、SwissProtデータベースにおいて偶然にマッチを発見する確率は、BLASTPアルゴリズムを用いた場合の1%以下である。
【0214】
1つの実施形態によれば、「改変体」HLA−DR特異的抗体は、本発明のHLA−DR特異的抗体および抗体フラグメントの各々を参照すれば、好ましくは、本発明のHLA−DR特異的抗体と同じ数であるか、またはこれよりも少ないアミノ酸を有し、かつ、本発明のHLA−DR特異的抗体と比較した場合に、0.01以下のE値を生じる配列を含む。
【0215】
本発明のHLA−DR特異的抗体に対して特定の%同一性を有することに加え、改変体抗体は、好ましくは、本発明のHLA−DR特異的抗体に共通するさらなる構造的および/または抗原結合特徴を有する。本発明のHLA−DR特異的抗体に対して、特定の程度の同一性を有する抗体は、その一次構造において高い程度の類似性を共有し、そして、実質的に類似の抗原結合特性を有する。本発明のHLA−DR特異的抗体に対して、高い程度の一次構造の類似性を共有することに加え、HLA−DRに対して特定の程度の同一性を有するか、または、HLA−DRに特異的に結合し得る抗体は、好ましくは、以下の特徴のうち少なくとも1つを有する:(i)これらは、本発明のHLA−DR特異的抗体と実質的に同じ抗原結合特性を有する;または(ii)例えば、CDRおよび/もしくはフレームワーク領域のような、特定可能なドメインを共通して含む。
【0216】
(結合体)
意図される正確な治療的および/または診断的な適用に依存して、本発明の抗体またはそのフラグメントは、1つ以上の治療剤と結合されることが望ましくあり得る。この点で適切な薬剤としては、放射性核種、分化誘導因子(differentiation inducer)、薬物、毒素およびこれらの誘導体が挙げられる。例示的な適切な放射性核種としては、90Y、123I、125I、131I、186Re、188Re、211Atおよび212Biが挙げられる。例示的な適切な薬物としては、メトトレキサート、ならびにピリミジンアナログおよびプリンアナログが挙げられる。例示的な適切な分化誘導因子としては、エステルおよび酪酸が挙げられる。例示的な適切な毒素としては、リシン、アブリン、ジフテリア毒素、コレラ毒素、ゲロニン、Pseudomonas外毒素、Shigella毒素およびヤマゴボウ抗ウイルスタンパク質が挙げられる。
【0217】
治療剤は、直接または間接(例えば、リンカー基を介して)のいずれかで、適切なモノクローナル抗体に結合(例えば、共有結合)され得る。薬剤と抗体との間の直接的な反応は、各々が、互いと反応し得る置換基を有する場合に可能である。例えば、一方の上にある求核基(例えば、アミノ基またはスルフヒドリル基)は、もう一方の上にある、カルボニル含有基(たとえば、無水物)もしくは、酸ハロゲン化物と、または、良好な脱離基(例えば、ハロゲン化物)を含有するアルキル基と反応し得る。
【0218】
あるいは、治療剤と抗体とをリンカー基を介して結合することが望ましくあり得る。リンカー基は、結合能との干渉を避けるために、薬剤から抗体を遠ざけるためのスペーサーとして機能し得る。リンカー基はまた、薬剤または抗体上の置換基の化学反応性を増加させ、従って、カップリング能を増加させるように機能し得る。化学反応性の増加はまた、そうでなければ、不可能であった、薬剤、または薬剤の官能基の使用を容易にする。
【0219】
ホモ官能性およびヘテロ官能性の両方の、種々の二官能性試薬または多官能性試薬(例えば、Pierce Chemical Co.,Rockford,Illに記載されるもの)が、リンカー基として使用され得ることが、当業者に明らかである。カップリングは、例えば、アミノ基、カルボキシル基、スルフヒドリル基または酸化炭化水素残基を介して達成され得る。このような方法論は、例えば、米国特許第4,671,958号に記載される。
【0220】
治療剤が、本発明の免疫複合体の抗体部分を含まないときにより強力である場合、細胞内への内部移行の間またはその際に切断可能であるリンカー基を使用することが望ましくあり得る。多数の異なる切断可能なリンカー基が記載されている。これらのリンカー基からの薬剤の細胞内放出のための機構としては、ジスルフィド結合の還元による切断(例えば、米国特許第4,489,710号)、感光性結合の照射による切断(例えば、米国特許第4,625,014号)、アミノ酸側鎖の加水分解による切断(例えば、米国特許第4,638,045号)、血清補体が媒介する加水分解による切断(例えば、米国特許第4,671,958号)、および酸が触媒する加水分解による切断(例えば、米国特許第4,569,789号)が挙げられる。
【0221】
1以上の薬剤を1つの抗体に結合させることが望ましくあり得る。1つの実施形態において、薬剤の複数の分子が、1つの抗体分子に結合される。別の実施形態において、1つ以上の型の薬剤が、1つの抗体に結合される。特定の実施形態にかかわらず、1つ以上の薬剤との免疫複合体は、種々の方法で調製され得る。例えば、1つ以上の薬剤は、抗体分子に直接結合され得るか、または、付着のための複数の部位を提供するリンカーが使用され得る。あるいは、キャリアが使用され得る。
【0222】
キャリアは、直接的であるか、もしくはリンカー基を介する共有結合を含む、種々の方法で薬剤を有し得る。適切なキャリアとしては、アルブミンのようなタンパク質(例えば、米国特許第4,507,234号)、アミノデキストランのようなペプチドおよび多糖類(例えば、米国特許第4,699,784号)が挙げられる。キャリアはまた、非共有結合またはリポソーム小胞内(例えば、米国特許第4,429,008号および同第4,873,088号)のようなカプセル化によって薬剤を有し得る。放射性核種剤に固有のキャリアとしては、放射性ハロゲン化低分子およびキレート化合物が挙げられる。例えば、米国特許第4,735,792号は、代表的な放射性ハロゲン化低分子およびその合成を開示する。放射性核種キレートは、キレート化合物から形成され得、これらは、窒素原子および硫黄原子を、金属または金属酸化物、放射性核種と結合するためのドナー原子として含有する化合物を含む。例えば、米国特許第4,673,562号は、代表的なキレート化合物およびその合成を開示する。
【0223】
用語「細胞傷害性薬物」とは、本明細書中で使用される場合、癌細胞の増殖を阻害するか、または癌細胞の死を促進するために使用される薬物を指す。細胞傷害性薬物の例としては、ara−C、ブレオマイシン、シスプラチン、クラドリビン(cladribine)、シクロホスファミド、ドキソルビシン、エトポシドおよび5−フルオロウラシル(5−FU)のような化学療法剤が挙げられる。
【0224】
(抗HLA−A−DR抗体の同定および特徴づけのためのアッセイ系)
以下のアッセイ系は、本発明に従う、HLA−DR特異的なアポトーシス性、非免疫抑制性抗体の慣用的な同定および特徴づけのために適切に使用され得る。
【0225】
(クラスII MHC)
「免疫抑制的であり、かつ細胞傷害性である」抗HLA−DR mAbは、第1のドメインを認識し、ペプチド結合溝の近くに位置するエピトープに結合することによって抗原提示を阻害し得る。本発明の抗HLA−DR mAb(例えば、DN1924)は、「細胞傷害性のみ」である。従って、このような「細胞傷害性のみ」の抗体が結合する、HLA−DRの正確な配列は、「免疫抑制的であり、かつ細胞傷害性である」抗体の配列とは同一でない。Vidovicら,Eur.J.Immunol.25:3349−3355(1995)。
【0226】
(アポトーシスによる細胞死)
上述のように、アポトーシス(プログラムされた細胞死)は、抗MHCクラスII抗体の、クラスIIを発現する細胞への結合と関係している。APC(例えば、Bリンパ球、マクロファージ、単球、樹状細胞など)の表面上に発現されたクラスII MHCをコードする分子は、最終的に、両方の細胞型の活性化および分化をもたらす相互作用である、Tリンパ球に対する抗原提示の制限的要素として機能する。
【0227】
HLA−DR媒介性の細胞死は、非常に迅速で、Fcレセプター非依存性であり、かつ、補完的であり、非壊死性であることが実証されている。Trumanら,Int.Immunol.6(6):887−896(1994)およびTrumanら,Blood 89(6):1996−2007(1997)。
【0228】
本発見は、腫瘍細胞のアポトーシスの誘発における抗HLA−DR抗体の役割に関し、従って、このような細胞においてプログラムされた細胞死を誘導するための方法を提供する。例示的な実施形態において、HLA−DR抗原を架橋し得る抗HLA−DR抗体およびそのフラグメントは、HLA−DR発現腫瘍細胞により媒介される状態を研究または処置する際に有用であり、すなわち、HLA−DR発現腫瘍細胞に関連する障害を処置または予防するのに有用である。従って、本発明の抗体は、種々の疾患を処置するのに有用であり、これらの疾患としては、B細胞起源の癌により特徴付けられる任意の疾患(アポトーシスの増加が望ましくない)、例えば、ホジキンリンパ腫および非ホジキンリンパ腫、慢性リンパ急性白血病、骨髄腫および形質細胞腫が挙げられるがこれらに限定されない。Hollandら,Cancer Med.2:2697−2828(1996)。
【0229】
(細胞死の機構の評価)
この節では、アポトーシスによる細胞死の程度を評価するために有用なインビトロアッセイを記載する。細胞死は、プロピジウムヨーダイド(PI)での細胞染色によってか、または、アポトーシスによる細胞死に特異的なアッセイ(例えば、アネキシンVでの染色)を用いることによって検出され得る。Vermesら,J.Immunol.Meth.184:39−51(1995)。壊死による細胞死は、以下に記載するような細胞の生存度についてのアッセイを、関連のある細胞の形態の顕微鏡での観察との組み合わせの結果を評価することによって、アポトーシスによる細胞死から区別され得る。
【0230】
(壊死性細胞死)
壊死は、内部ホメオスタシスの虚脱が細胞膜の完全性の損失およびその後の膨張を含む細胞の溶解(dissolution)を導き、その後に細胞の溶解(lysis)が続く、受動的プロセスである。Schwartzら、1993。壊死性の細胞死は、細胞膜の完全性の損失、およびヨウ化プロピジウム(PI)のような、一次性の壊死および二次性の壊死を受ける細胞のDNAに結合することが当業者に公知の色素に対する透過性によって、特徴付けられる。Vitaleら、Histochemistry 100:223−229(1993)およびSwatら、J.Immunol.Methods 137:79−87(1991)。壊死は、アポトーシスから区別され得る(アポトーシスの初期段階においては、細胞膜はインタクトなまま残る)。結果として、以下に記載されるようにアポトーシス性細胞死を壊死性細胞死から区別するため、アポトーシスについてのアッセイと並行して、PIを使用した色素排除アッセイ(dye exclusion assay)が使用され得る。PIを用いた蛍光活性化細胞分類機(Fluorescent−activated cell sorter;FACS)ベースのフローサイトメトリーアッセイは、壊死性細胞の迅速な評価および定量化を可能にする。
【0231】
(アポトーシス性細胞死)
プログラムされた細胞死またはアポトーシスの検出は、当業者に理解されるように達成され得る。アポトーシスを受ける細胞のパーセンテージは、抗HLA−DR抗体の投与とともに、またはその投与なしで、アポトーシスの刺激後の種々の時間で測定され得る。アポトーシス性細胞死を受ける細胞の形態は、一般的に、細胞の細胞質および核の萎縮、ならびにクロマチンの凝縮および断片化によって特徴付けられる。Wyllieら、J.Patrol.142:67−77(1984)。
【0232】
アポトーシス性B細胞における部分的DNA分解が以前に報告されている。Trumanら、Int.Immuunol.6:887−896(1994)およびCohenら、Annu.Rev.Immunol.10:267−293(1992)。この観察に一致して、DNA断片化は、以前に記載された腫瘍性B細胞をアポトーシス誘導性(apoptogenic)の抗HLA−DR mAbとともにインキュベーションした後には検出されなかった。しかし、これらの培養物の相対的細胞サイズおよびPI取り込み式フローサイトメトリーのプロフィールは、アポトーシスを受ける細胞について以前に示されたプロフィールと本質的に同じである。Vidovicら、Cancer Lett.128:127−135(1988);Newellら、Proc.Natl.Acad.Sci.U.S.A.90:10459−10463(1993);およびTrumanら(1994)。
【0233】
(有用性)
(抗HLA−DR抗体に関する使用)
本発明のHLA−DR特異的抗体は、診断用および治療用の組成物ならびに診断用および治療用の方法において利用され得る。例えば、HLA−DR特異的抗体は、HLA−DR発現腫瘍細胞についての診断アッセイおよびHLA−DR発現腫瘍細胞に関する治療において(例えば、組織または血清中のこのような細胞の発現を検出すること、およびこのような腫瘍を有する被験体において臨床的結果を改善する治療のための基礎として働くことに)使用され得る。
【0234】
当該分野で公知の種々の診断アッセイ技術(例えば、競合的結合アッセイ、直接もしくは間接サンドイッチアッセイ、および均一もしくは不均一相において実施される免疫沈降アッセイ)が使用され得る。Zola、「Monoclonal Antibodies:A Manual of Techniques」、pp.147−158(CRC Press,Inc.,1987)。この診断アッセイに使用され得る抗体は、検出可能な部分によって標識され得る。この検出可能な部分は、直接的もしくは間接的に、検出可能なシグナルを生成し得る。例えば、この検出可能な部分は、放射性同位体(例えば、3H、14C、32P、35S、または125I)、蛍光化合物または化学発光化合物(例えば、フルオレセインイソシアネート、ローダミン、もしくはルシフェリン)、あるいは酵素(例えば、アルカリホスファターゼ、β−ガラクトシダーゼ、もしくは西洋ワサビペルオキシダーゼ)であり得る。特異的結合を促進する分子としては、対(例えば、ビオチンおよびストレプトアビジン、ジゴキシンおよび抗ジゴキシンなど)が挙げられる。特異的結合を促進する分子の対のメンバーの一方は、標識が、直接的または間接的に検出可能なシグナルを提供する公知の手順による検出を行わせる分子で標識され得る。抗体を検出可能な部分に結合させるための当該分野で公知の任意の方法が利用され得る。この方法としては、Hunterら、Nature 144:945(1962);Davidら、Biochemistry 13:1014(1974);Painら、J.Immunol.Meth.40:219(1981);およびNygren,J.Histochem.and Cytochen.30:407(1982)によって記載される方法が挙げられる。
【0235】
HLA−DR特異的抗体はまた、細胞培養物または天然の供給源からのHLA−DR発現癌細胞のアフィニティー精製のために有用である。このような方法において、HLA−DRに対する抗体は、当該分野で公知の方法論を用いて適切な支持体(例えば、Sephadex樹脂もしくは濾紙)に固定化される。固定された抗体は、精製されるべきHLA−DR発現癌細胞を含むサンプルと接触させられ、そして固体支持体は、固定化された抗体に結合するHLA−DR発現癌細胞を除く、サンプル中の実質的に全ての物質を除去する適切な培地で洗浄される。支持体は、HLA−DR発現癌細胞を固定化された抗体から放出させる別の適切な培地で洗浄される。
【0236】
(抗HLA−DR特異的モノクローナル抗体の生物学的効果)
本発明のHLA−DR特異的モノクローナル抗体は、HLA−DRの第1ドメインを認識する。本発明の抗体または特異的モノクローナル抗体のCDR領域は、検出可能なレベルのHLA−DRを発現する腫瘍細胞と免疫反応し、そしてこの腫瘍細胞におけるアポトーシスを誘導し得る。HLA−DR特異的マウスモノクローナル抗体DN1924、およびマウス/ヒトキメラモノクローナル抗体は、非腫瘍性成細胞と比較して、腫瘍(プラズマ細胞腫 MC/CAR)細胞(両方ともHLA−DRを発現する)におけるアポトーシスの誘導に関するインビトロでの特異性を示した。実施例2および図2〜4、ならびに実施例7および図8A〜Fを参照のこと。さらに、DN1924による正常なTh反応との干渉の欠如がインビトロで示された。実施例2および図5を参照のこと。
【0237】
(アポトーシス性HLA−DR特異的抗体を用いたインビボ癌治療)
本発明の抗体は、治療的に有効であり、HLA−DR抗原を発現する腫瘍細胞のアポトーシス性細胞死を刺激し得る。これらの発見は、HLA−DR陽性の癌(特に血液の癌)に対する選択的な抗体ベースの抗腫瘍治療の可能性を提起する。
【0238】
抗クラスII MHC mAbの殺腫瘍性(tumoricidal)効果は、クラスII依存性免疫応答の同時的な抑制なしで(両特性ともタンパク質の第一ドメインを認識するmAb認識に関連するが)達成され得る。
【0239】
所望の治療的効果を有する抗体は、宿主にとって生理学的に受容可能なキャリア中において投与され得、そして例えば、非経口、皮下(SC)、腹腔内(IP)、静脈内(IV)などの方法で投与され得る。導入の様式に依存して、抗体は、種々の方法で処方され得る。処方物中の治療的に活性な抗体の濃度は、約1mg/mlから約1g/mlまで変動し得る。
【0240】
本発明の目的のための「被験体」は、ヒトおよび他の動物(特に哺乳動物)の両方を包含する。従って、本方法は、ヒト治療および獣医学的適用の両方に対して適用可能である。例示的実施形態において、被験体は哺乳動物であり、最も例示的な実施形態において、被験体はヒトである。
【0241】
本明細書において使用される場合、用語「改善された治療結果」または「腫瘍細胞数の減少」とは、処置前後に患者および/または患者から採取されたサンプルの試験によって決定される場合の、血中および/または罹患する組織もしくは器官における腫瘍サイズまたは検出可能な循環性癌細胞の数のいずれかの50%減少、好ましくは80%減少、より好ましくは90%減少、そしてさらにより好ましくは100%減少を意味する。
【0242】
本明細書において使用される場合、用語「処置する工程」、「処置」および「治療」とは、治癒可能な治療、予防的な(prophylactic)治療、および予防的な(preventative)治療をいう。
【0243】
本明細書において使用される場合、「治療上有効な量」によって、HLA−DR発現腫瘍細胞を、そのアポトーシスを刺激することによって減少または消失させる用量が意味される。正確な用量は、処置の目的に依存し、そして公知の技術を使用して当業者によって確認され得る。
【0244】
好ましくは、抗体は、非経口投与のために、適切な不活性キャリア(例えば、滅菌生理学的食塩水)中に処方される。例えば、キャリア溶液中の抗体の濃度は、代表的に約1〜約100mg/mlの間である。投与される用量は、投与の経路によって決定される。例示的な投与の経路としては、非経口投与またはIV投与が挙げられる。治療的に有効な用量は、HLA−DR発現性新生物細胞のアポトーシス性細胞死の有意な増加を生じるのに有効な用量である。HLA−DR発現性新生物細胞のアポトーシス性細胞死の有意な増加は、検出可能な量のHLA−DRを発現しない細胞と比較して、HLA−DR発現性細胞のアポトーシス性細胞死の2倍の増加、より好ましくは5倍の増加、さらにより好ましくは10倍の増加、最も好ましくは20倍の増加である。
【0245】
発明の重要な特徴に従って、抗HLA−DR抗体は、単独で、または他の抗癌剤(例えば、化学治療剤(例えば、シスプラチン、タキソール、メトトレキサートなど);腫瘍壊死因子−α(TNF−α);FADD、PMA;イオノマイシン;スタウロスポリンまたはRituxan(登録商標))と組み合わせて投与され得る。
【0246】
抗HLA−DR抗体(例えば、DN1924)の治療上有効な量は、本明細書において記載されるように、HLA−DR陽性腫瘍細胞のアポトーシス性細胞死についてのインビトロモデルにおいてDN1924に関して得られたデータと合わせて、公知の抗体についての確立された有効量との比較によって推定され得る。当該分野において公知であるように、抗体の変性、全身送達 対 局所送達、ならびに年齢、体重、健康、性別、食事、投与時間、薬物の相互作用、および状態の重症度に起因して、用量の調節が必要であり得る。当業者によって、慣習的な実験を通じてこのような調節が行われ得、そして適切な用量が決定され得る。
【0247】
特定の実施形態の範囲内において、本発明の方法は、所望の抗原結合特異性および/または抗原結合親和性を有する抗体に関して細胞をスクリーニングする工程をさらに提供し得る。本明細書において使用される場合、用語「抗原」は、抗体が特異的に結合および/または相互作用するこれらの物質、分子、タンパク質、核酸、脂質および/または炭水化物の全てを広範に包含する。
【0248】
抗体は、例示的な抗原結合特異性および/または抗原結合親和性に関して、当該分野で現在利用可能な任意の方法論によってスクリーニングされ得る。例えば、従来の細胞パニング手順、ウエスタンブロッティング手順、およびELISA手順が、特定の特異性を有する抗体をスクリーニングする工程を達成するために利用され得る。米国特許第4,016,043号、同第4,424,279号、および同第4,018,653号(これらの各々は、本命最小において参考として援用される)への参照によって見られ得るように、広範囲の適切なイムノアッセイ技術が利用可能である。
【0249】
1つの型のアッセイにおいて、未標識の抗−抗体が固体支持体上に固定され、そして試験されるべき抗体含有サンプルがその固定された抗−抗体と接触させられる。第一の複合体の形成を可能にするのに十分な適切な時間の後、次に、検出可能なシグナルを生成し得るレポーター分子で標識された標識抗原を添加してインキュベートし、固定された抗−抗体/抗体サンプル/試験抗原の第二の複合体の形成のために十分な時間を与える。複合体化されない物質は、洗い流され、レポーター分子によって生成されたシグナルの観察によって抗原の存在が決定される。この結果は、眼に見えるシグナルの単純な観察による定性的なものであっても、既知の量の抗原または抗体を含むコントロールサンプルとの比較によって定量化されてもいずれでもよい。この型のアッセイの変化は、サンプルおよび標識された抗原の両方が結合抗体に対して同時に添加される同時アッセイを包含する。
【0250】
本明細書中で使用される場合、用語「固体支持体」とは、例えば、マイクロタイタープレート、膜およびビーズなどをいう。例えば、このような固体支持体は、ガラス、プラスチック(例えば、ポリスチレン)、多糖類、ナイロン、ニトロセルロースまたはテフロン(登録商標)などで構成され得る。このような支持体の表面は、固体または多孔質であり得、そして任意の都合よい形状であり得る。
【0251】
第2の型のアッセイにおいて、抗体を探索するための抗原が固体支持体に結合される。この結合プロセスは当該分野において周知であり、そして一般的に、その抗原を固体支持体に対して架橋結合、共有結合、または物理的に吸収することからなる。ポリマー−抗原複合体は、試験サンプルに備えて洗浄される。次いで、試験されるべきサンプルのアリコートが、固相複合体に添加され、そして抗原への抗体の結合を可能にするのに十分な時間(例えば、20〜40分、またはより都合よい場合、一晩)、適切な条件下(例えば、ほぼ室温〜約38℃、例えば25℃)でインキュベートされる。インキュベーション時間に続いて、固体支持体は、洗浄され、乾燥され、そしてレポーター分子が結合し得る抗体とともにインキュベートされ、それによって固定された抗原に複合体化された試験抗体に対する二次抗体の結合の検出を可能にする。
【0252】
適切な固体支持体としては、ガラスまたはポリマーが挙げられ、最も一般的に使用されるポリマーは、セルロース、ポリアクリルアミド、ナイロン、ポリスチレン、ポリ塩化ビニルまたはポリプロピレンである。固体支持体は、チューブ、ビーズ、マイクロプレートのディスク、またはイムノアッセイの実施のために適切な任意の他の表面の形態であり得る。
【0253】
代替的アッセイシステムは、標的抗体を固定化し、そしてその固定化された抗体を抗原(レポーター分子に標的されていても、されていなくてもよい)に暴露する工程を包含する。本明細書において記載されるように、用語「レポーター分子」とは、その化学的、生物学的、および/または物理的性質によって、抗原または二次抗体と複合体化された抗体についてのスクリーニングを可能にする分析的に同定可能なシグナルを提供する分子をいう。検出は、定性的であっても定量的であってもいずれでもよい。本明細書において開示された型のアッセイにおいて利用される最も一般的に使用されるレポーター分子は、酵素、フルオロフォア、放射性同位体、および/または化学発光分子である。1つの特定の有用なアッセイシステムにおいて、B細胞レポーターに関する抗体を細胞表面に発現する細胞ライブラリーの培養物が、標識(例えば、ビオチン)と共役された抗原か、または組換えエピトープ(例えば、FLAGエピトープ)を有する抗原とともにインキュベートされ得る。Castrucciら、J.Virol.66:4647−4653(1992)。適切なインキュベーション時間の後、標識された抗原は、氷冷された培地中の懸濁液から2回または3回細胞をペレット化することによって洗い流される。
【0254】
細胞は、抗原に結合された標識に特異的に結合する試薬(例えば、ビオチン標識に対するストレプトアビジンもしくはアビジン、またはFLAGエピトープに対する抗FLAG抗体)に結合されたビーズの懸濁液を含有する培地中でさらにインキュベートされ得る。目的の抗原を結合したことの間接的な結果としてビーズに結合した細胞は、適切な手段によって残りの細胞から分離され、次いで組織培養物に戻されて増殖する。次第に限られた量の抗原を用いてこのプロセスを繰り返すことは、抗原に特異的におよび/または高い親和性で結合した細胞の濃縮を引き起こす。抗原に対する前結合とは関係なくビーズに直接的に結合する細胞は、抗原なしでビーズとともにインキュベーションすることによって除去され得る。次いで、個々の抗原結合クローンは、適切な蛍光色素(例えば、フルオレセイン)に対して直接的もしくは間接的に結合された抗原で細胞を標識した後に、例えば、蛍光活性化細胞分類法(FACS)などによって精製され得る。
【0255】
酵素イムノアッセイ(EIA)の場合、酵素は、一般的にグルタルアルデヒドまたは過ヨウ素酸塩によって検出抗体に結合される。しかし、容易に認識されるように、種々の異なる結合技術が存在し、これらは当業者に容易に利用され得る。一般的に使用される酵素としては、西洋ワサビペルオキシダーゼ、グルコースオキシダーゼ、β−ガラクトシダーゼおよびアルカリホスファターゼが挙げられる。一般に、酵素標識された抗体は、抗原と抗体との間の潜在的複合体に添加され、結合させられ、そして洗浄されて過剰の試薬が除去される。次いで、適切な基質を含む溶液が抗原/試験抗体/標識抗体の複合体に添加される。この基質は、二次抗体に結合した酵素と反応し、定性的な視覚的シグナルを与える。これはさらに、通常、分光光度法で定量化されて、サンプル中に存在する抗体の量を示し得る。
【0256】
あるいは、蛍光化合物(例えば、フルオレセインおよびローダミン)または蛍光タンパク質(例えば、フィコエリトリン)が、抗体に、その結合能を変えることなく化学結合され得る。特定波長の光による照射によって活性化される場合、蛍光標識された抗体は、その光エネルギーを吸収し、その分子の励起状態を示し、続いて、光学顕微鏡または他の光学装置によって視覚的に検出され得る特徴的な色で光の放射を示す。EIAにおける場合、蛍光標識された抗体は、抗原−抗体複合体に結合させられる。未結合の試薬を除去した後、残りの第3の複合体は、適切な波長の光に曝される。観察された蛍光は、目的の結合抗体の存在を示す。免疫蛍光技術およびEIA技術は、どちらも当該分野において十分確立されている。他のレポーター分子(例えば、放射性同位体および化学発光分子および/もしくは生物発光分子)もまた、本明細書において開示されたスクリーニング法において適切に利用され得ることが理解される。
【0257】
以下の実施例は、上記の発明を使用する様式をより十分に記載するため、ならびに本発明の種々の局面を実施するために企図される最適な様式を示すために役立つ。これらの実施例は、決して、本発明の真の範囲を限定するために役立つのではなく、例示的目的のために提示される。以下の実施例は、例示のために提供されるのであって、限定のために提供されるのではない。
【実施例】
【0258】
(実施例1)
(DN1921マウスモノクローナル抗体およびDN1924マウスモノクローナル抗体の生成)
この実施例は、マウスDN1921HLA−DR特異的モノクローナル抗体およびDN1924 HLA−DR特異的モノクローナル抗体の生成を示す。
【0259】
DN1921およびDN1924と称されるマウスモノクローナルHLA−DR特異的抗体を、当該分野で公知の標準的な技術に従って調製した。HarlowおよびLane,「Antibodies:A Laboratory Manual」(Cold Spring Harbor Laboratory,Cold Spring Harbor,NY,1988)。免疫原で過剰免疫された近交系実験用マウスBALB/c系(Jackson Laboratory,Bar Harbor,ME)がB細胞のドナーであった。BALB/c由来の変異Bリンパ腫株M12.C3を、キメラヒト/マウス クラスII遺伝子でトランスフェクトし、免疫原として使用した。このトランスフェクト体(M12.C3.25と称する)によって発現されるMHCクラスII分子は、HLA−DRの第1の細胞外(α1およびβ1)ドメイン、ならびに対応するマウスMHCクラスII分子H2−Eの第2の細胞外(α2およびβ2)ドメイン、膜貫通ドメインおよび細胞質内ドメインから構成された。Vidovicら、Eur.J.Immunol.25:3349−3355(1995)。マウスを、月1回の間隔で5回のIP注射(各々は、1mLのリン酸緩衝化生理食塩水(PBS)中に再懸濁された、107(−照射された(100Gy)M12.C3.25細胞からなる)によって免疫化した。最後の注射から3日後、免疫脾臓細胞を、HAT感受性Ig陰性マウスミエローマ細胞PAI−0と融合した。Stockerら、Research Disclosure 217 :155−157(1982)。
【0260】
単一のハイブリドーマ培養物の上清流体を、4℃での4時間のインキュベーション後でのEBV−LCL RPMI 1788に対するそれらの毒性、またはヒト末梢血単核細胞(PBMC)のインビトロでの増殖において誘導されたブドウ球菌エンテロトキシンB(SEB)を抑制するそれらの能力についてスクリーニングした。所望の生物活性のモノクローナル抗体を適切な形式で分泌することが同定されたコロニーを、それぞれ「DN1921」および「DN1924」と称し、そして限界希釈法によって2回サブクローン化した。標準的アイソタイプ決定キット(Zymed,South San Francisco,CA)を用いて、DN1921およびDN1924が、両方ともマウスIgG1/κアイソタイプであることを見出した。
【0261】
(実施例2)
(DN1924モノクローナル抗体のインビトロ殺腫瘍性効果の評価)
この実施例は、DN1924と称するマウスHLA−DR特異的モノクローナル抗体が、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導し得るが、HLA−DRを発現する正常(すなわち、非悪性)細胞においてはアポトーシスを誘導しないことを示す。
【0262】
ヒト細胞株MC/CAR(プラズマ細胞腫)およびRPMI 1788(エプスタイン−バーウイルス形質転換リンパ芽球状B細胞株、EBV−LCL)を、ATCC(Rockville,MD)から譲渡された。Rittsら、Int.J.Cancer 31:133−141(1983)。細胞を、10% FCS、2mMのL−グルタミン、0.1mg/mlの硫酸カナマイシン、および3×10−5Mの2−MEを補充したIMDM培地(Gibco,Grand Island,NY)中で、105/mlの密度で、37℃で、5%CO2を含む加湿された雰囲気(組織培養インキュベーター)中で培養した。5×105細胞/mlで培養したHLA−DR−特異的mAb分泌性マウスB細胞ハイブリドーマ細胞株DN1924および10F12の滅菌濾過された上清流体を、最終濃度20%でヒト細胞を添加した。示された共培養期間に続いて、細胞を洗浄し、そしてさらに1μg/mlのヨウ化プロピジウム(PI,Sigma,St.Louis,MO)とともに5分インキュベーションし、その後、CELLQuest 3.1fソフトウェアを用いたFACScan(登録商標)フローサイトメーター(Becton−Dickinson,San Jose,CA)上で、細胞の大きさ 対 赤色PI蛍光の分析した後、その生存度を決定した。Ottenら、「Current Protocols in Immunology」、pp.5.4.1−5.4.19(Coliganら編、Greene & Wiley,New York,1997)。生存細胞が積極的にPIを排除すること、一方死細胞が、それを、細胞のDNAへの接近可能性と直接的に比例して取り込むことが示された。Swatら(1991)。
【0263】
新生物細胞のアポトーシスを誘導するDN1924抗体の能力を、75%を超える細胞死を引き起こした2つの独立なヒトB細胞腫瘍株(EBV−LCL RPMI 1788およびプラズマ細胞腫MC/CAR)の共培養によって示した。図2。
【0264】
ヒト末梢血から得られた正常形成性(すなわち、非新生物性)HLA−DR+リンパ球においては、細胞毒性効果は全くなかった。対照的に、DN1924、10F12(第2のタンパク質ドメイン内に位置する共通エピトープについて特異的な抗DR mAb)は、MC/CARの生存度に影響しなかった。図3。
【0265】
2つの異なるインキュベーション条件下での細胞毒性の時間経過は、図4の上パネルに示される。DN1924誘導性細胞死が温度依存的であることは明らかである;これはヒト体温(37℃)においてより速い速度(細胞毒性効果は30分以内に明らかとなる)であり、そして4℃においては、遅くなるが、まだなお生じる(細胞毒性効果は1〜2時間以内に明らかとなる)。腫瘍細胞との長い(3週間)共培養後でさえも衰えていないDN1924の腫瘍細胞毒性は、この抗体に対して耐性となる能力のないことを示す。図4の下パネル。
【0266】
(実施例3)
(DN1924 HLA−DR特異的マウスモノクローナル抗体は、低下した免疫抑制活性を示す)
この実施例は、DN1924 HLA−DR特異的マウスモノクローナル抗体が、低レベルの免疫抑制活性を示すことを示す。
【0267】
ブドウ球菌エンテロトキシンBに対するヒトTh細胞のHLA−DR依存性インビトロ増殖反応を、先に記載したように生じさせた。Mollickら、J.Immunol.146:463−468(1991)。簡単にいうと、Ficoll分離によってヘパリン処理した血液から得られた2×105個の新鮮ヒトPBMCを、10% FCS、2mMのL−グルタミン、0.1mg/mlの硫酸カナマイシン、および3×10−5Mの2−MEを補充した0.2mlのIMDM培地中で0.1μg/mlのSEB(Toxin Technology,Sarasota,FL)とともに37℃、5%CO2を含む加湿した雰囲気(組織培養インキュベーター)中で、3日間培養した。5×105細胞/mlで培養したHLA−DR−特異的mAb分泌性マウスB細胞ハイブリドーマ細胞株DN1924およびL243(ATCC,Rockville,MD)の滅菌濾過された上清流体を、最終濃度20%でアッセイの開始時に添加した。Lampsonら、J.Imnunol.125:293−299(1980)およびFuら、Human Immunol.39:253−260(1994)。3つ組の培養物のT細胞増殖を、最後の16時間の間の[3H]チミジン取り込みによって測定した。Bradley、「Selected Methods in CellularImmunology」pp.153−174(MishellおよびShiigi(編)、W.H.Freeman & Co.,New York(1980)。L243およびDN1924のmAbの両方とも第1のHLA−DRドメイン上のエピトープを認識するが、mAb L243がSEBで誘発されたT細胞反応の約70%を抑制した一方、DN1924は効果を有さなかった(図5)。
【0268】
(実施例4)
(DN1924 HLA−DR特異的マウスモノクローナル抗体のインビボ抗腫瘍活性の評価)
DN1924のインビボ抗腫瘍活性を、HLA−DR陽性ヒトプラズマ細胞種MC/CARを注射された重症複合型免疫不全近交系(scid)マウスにおいて評価した。20匹の8週齢のマウス(体重に関してランダム化された)に、マウスあたり0.1ml RPMI 1640培地中の107個のMC/CAR細胞を用いてIP注射した。Rittsら、(1983)。その後、動物の半数は、0.1mlのPBS中0.625mgのDN1924抗体の、単回IP用量を受けた。残りのマウスに、0.1ml PBSの対応するIP注入を与えた。マウスを毎日モニタリングし、その生存を記録した。図6に示されるように、DN1924は、顕著な治療活性を示し、腫瘍を有するマウスの生存を延長した。
【0269】
(実施例5)
(マウスDN1921およびDN1924におけるマウス/ヒトキメラHLA−DR特異的モノクローナル抗体の生成)
この実施例は、マウスモノクローナル抗体DN1921およびDN1924の重鎖可変領域および軽鎖可変領域と、ヒトIgG2の重鎖定常領域およびヒトIgκの軽鎖定常領域とを含むマウス/ヒトキメラHLA−DR特異的モノクローナル抗体の構築を記載する。
【0270】
可変領域セグメントを、cDNAを全RNAから「RACE」するプライマーを設計するための定常領域特異的配列を用いて、各抗体(DN1924およびDN1921)のマウス版を発現するハイブリドーマ細胞からクローニングした。可変領域(DN1924−heavy、DN1924−light;DN1921−heavy、DN1922−light)配列を、これらのクローンから決定した。
【0271】
ヒトIgG2重鎖定常領域セグメントおよびヒトIgκ軽鎖定常領域セグメントを、NCBIおよびIMGTのホームページから設計されたプライマーを用いて、ヒト脾臓全RNAからクローン化した。クローンを配列決定して、適切なIgκ定常領域またはヒトIgG2定常領域を確認した。
【0272】
DN1921およびDN1924重鎖および軽鎖ポリヌクレオチド配列を増幅するために使用されたオリゴヌクレオチドプライマーは、表3に提示される。
【0273】
(重鎖構築)
可変領域遺伝子セグメントと定常領域遺伝子セグメントとを組み合わせるため、PCR「縫い付け(sewing)」技術を利用した。簡単にいうと、プライマーを、抗体分子のコード領域の5’末端および3’末端に対応するように設計した(表3、プライマーAおよびB)。プライマーの第2のセットを、別個のプラスミド上に最初に存在する2つの片の間の接合部に重なるように設計した。最初のPCR反応を、プライマーセットA+Dとともに可変領域プラスミドをテンプレートとして用いて、またはプライマーセットB+Cとともに定常領域プラスミドをテンプレートとして用いて、準備した。第2のPCR反応は、プライマーセットA+Dを用いて最初の2つの一次反応の生成物を組み込んだ。最後のPCR生成物をクローニングし、配列を確認し、そして必要な場合、(コード領域の外に)制限酵素を利用して、発現ベクター中にシャトル(shuttle)した。
【0274】
(軽鎖構築)
可変領域遺伝子セグメントと定常領域遺伝子セグメントとを組み合わせるため、重鎖構築に関して上記したように、PCR「縫い付け」技術を利用した。プライマーを、抗体分子のコード領域の5’末端および3’末端に対応するように設計した(表3、プライマーAおよびB)。第3のプライマーを、別個のプラスミド上に最初に存在する2つの片の間の接合部に重なるように設計した。最初のPCR反応を、プライマーセットC+Bとともに定常領域プラスミドをテンプレートとして用いて準備した。第2のPCR反応は、プライマーセットA+Bを用いて、一次反応の生成物、およびプラスミドがコードする軽鎖可変領域を組み込んだ。最後のPCR生成物をクローニングし、配列を確認し、そして必要な場合、(コード領域の外に)制限酵素を利用して、発現ベクター中にシャトルした。
【0275】
(発現系)
DN1924重鎖およびDN1924軽鎖を、pEE14.4発現ベクター(Lonza Biologics,Portsmouth,NH)にサブクローニングし、キメラ免疫グロブリンの両方の鎖が同じベクター骨格上にコードされるようにした。各鎖の発現を、それ自身のCMV主要即時早期(major immediate early)(MIE)プロモーターによって駆動した。重鎖および軽鎖の両方のオープンリーディングフレームを含むプラスミドを、CHO−K1/Sv細胞(Lonza Biologics)中にトランスフェクトし、細胞を、標準的な技術を用いて、50μMのメチオニン−スルホキサミン(sulfoxamine)(MSX)を含む無グルタミン培地中で選択した。細胞株を生成するキメラ抗体をELISAによって同定した。
【0276】
DN1921重鎖および軽鎖の発現に関して、同じプロトコールに従った。
【0277】
(表3)
(キメラDN1921モノクローナル抗体およびキメラDN1924モノクローナル抗体の構築に使用されたオリゴヌクレオチド)
【0278】
【表3】
表3のオリゴ配列において、大文字は、抗体分子のオープンリーディングフレーム中のアミノ酸残基についてのコドンを示し、小文字は、Kozakコンセンサス配列(下線)およびクローニング目的のための制限エンドヌクレアーゼ部位に対して付加されたヌクレオチドを示す。プライマー「C]および「D」中の点線は、マウス可変領域とヒト定常領域との間の接合部を示す。
【0279】
(実施例6)
(HLA−DR細胞に特異的に結合するマウス/ヒトキメラDN1921 HLA−DR特異的モノクローナル抗体およびマウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体)
この実施例は、キメラDN1921 HLA−DR特異的モノクローナル抗体およびキメラDN1924 HLA−DR特異的モノクローナル抗体の両方が、種々の組織培養細胞株において発現されたHLA−DRに特異的に結合することを示す。
【0280】
標準的免疫蛍光アッセイ[Current Protocols in Immunology 1,pp.5.3.1−5.4.13(Coliganら編、New York,Wiley Interscience,1998)]を、ビオチン化ヤギ抗ヒトIgGビオチン結合体(1:200最終希釈;Sigma,St.Louis,MO)、ならびにストレプトアビジン−PE(0.8μg/ml)をそれぞれ第2および第3の試薬として用いて行った。
【0281】
(実施例7)
(マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体は、HLA−DR発現腫瘍細胞株においてアポトーシスを誘導する)
この実施例は、キメラDN1924 HLA−DR特異的モノクローナル抗体が、HLA−DR発現腫瘍細胞株においてアポトーシスを誘導するのに有効であるが、HLA−DR陰性腫瘍細胞または正常HLA−DR発現細胞においては有効でないことを示す。
【0282】
アポトーシスアッセイを、Alexa Flour488結合体をヨウ化プロピジウム(PI)対比染色とともに使用して、Molecular Probe Annexin Vキットを用いて行った。簡単にいうと、5×105個の生存細胞を0.1〜10μgのmAbとともに0.5〜1mlの総容量の培地中で、37℃で10〜30分間共培養し、2mlの氷冷PBSで1回洗浄し、5μlのAnnexin Vおよび1μg/mlでPIを含む100μlの1×緩衝液中に再懸濁し、室温、暗中で15分間インキュベートした。次いで、300μlの1×緩衝液を添加し、そしてサンプルを30分以内にフローサイトメーターによって分析した[Current Protocols in Immunology 1,pp.5.4.1−5.4.13(Coliganら編、New York,Wiley Interscience,1998)]。
【0283】
表4は、マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体が、HLA−DR陽性LcL細胞株の一団の各メンバーに結合し、そしてアポトーシスを誘導することを示す。各LcL細胞株は、異なるDRβ1対立遺伝子を発現する。これらのデータは、DRβ1対立遺伝子の幅広い範囲にわたるHLA−DR発現に関する疾患の処置における、本発明のマウス/ヒトキメラ抗HLA−DR抗体に対する有用性を支持する。
【0284】
(表4)
(マウス/ヒトキメラDN−1924(mut−hIgG2クローンDN16)は、ある範囲のDRβ1対立遺伝子を発現するHLA−DR陽性LcL細胞株の一団に特異的に結合し、アポトーシスを誘導する)
【0285】
【表4】
表5は、マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体が、腫瘍細胞株の一団(3つのHLA−DR陰性腫瘍細胞株および12のHLA−DR陽性細胞株を含む)の各HLA−DR陽性メンバー(KG−1を除く)に結合し、アポトーシスを誘導することを示す。キメラDN1924は、Fcレセプターを発現するもの(すなわちU937およびTHP−1)、またはB細胞腫瘍細胞株(すなわちDB)でさえ、HLA−DR陰性細胞株のアポトーシスを誘導しない。対照的に、キメラDN1924は、12のHLA−DR陽性腫瘍細胞株のうちの11のアポトーシスを誘導する。KG−1(AML腫瘍細胞株)は、キメラDN1924結合によってアポトーシスを受けないHLA−DR陽性細胞株の(特異的キメラDN1924結合活性によって示されるように)唯一の例である。全体で、これらのデータは、キメラDN1924が、広範なHLA−DR陽性腫瘍型の処置において有効であることを示す。
【0286】
(表5)
(マウス/ヒトキメラDN−1924(mut−hIgG2クローンDN16)は、HLA−DR陽性細胞株の一団に特異的に結合し、アポトーシスを誘導する)
【0287】
【表5】
(実施例8)
(マウス/ヒトキメラDN1921 HLA−DR特異的モノクローナル抗体は、免疫抑制性である)
以前に記載されるたように、IL−2分泌アッセイを行った[Current Protocols in Immunology1,pp.3.14.1−3.14.11(Coliganら編、New York,Wiley Interscience,1998)]。簡単にいうと、PAP/HLA−DR1特異的CD4+T細胞ハイブリドーマ株Paperino[Vidovicら、Human Immunol.64:238−244,2003]のハイブリドーマ細胞105個を、抗原提示細胞としての2×105個のヒトHLA−DR1+PBMC、PAP、およびmAb(mDN1921=マウスDN1921;mDN1924=マウスDN1924;DSm89#34=キメラDN1921;DN−GS#16=キメラDN1924)とともに、0.2mlのマイクロウェル中で共培養した。1日後、培養上清を回収し、そしてIL−2依存性細胞株HT−2の104個の細胞の増殖を支持するそれらの能力について、24時間、50%濃度で試験した。最後の6時間の培養期間の間、[3H]チミジンcpm組み込みによって細胞増殖を測定した。
【0288】
(実施例9)
(マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体に誘導されたアポトーシスは、カスパーゼ非依存性である)
この実施例は、マウス/ヒトキメラDN1924のアポトーシス誘導活性がカスパーゼ非依存性であることを示す。
【0289】
外膜に対するホスファチジルセリンの暴露(Annexin V染色で検出される)およびカスパーゼ活性は、両方ともアポトーシスの指標である。しかし、本実施例は、キメラDN1924に誘導されたアポトーシスが、カスパーゼ3、カスパーゼ8、またはカスパーゼ9の活性に関与しないことを示す(図13A−Dおよび表6)。
【0290】
キメラDN1924媒介性アポトーシスを、添加されたZ−VAD−fmk(広範囲のカスパーゼインヒビター)の存在下および非存在下の両方において誘導した。誘導後、アポトーシスを、Annexin V染色によって、ならびにカスパーゼ3、カスパーゼ8、およびカスパーゼ9活性化を測定することによって検出した。Annexin V染色は、アポトーシス誘導を確証し、そしてZ−VAD−fmkがアポトーシス誘導の阻害に不十分であることを確証した。従って、マウス/ヒトDN1924で誘導されたアポトーシスは、カスパーゼ3、カスパーゼ8、またはカスパーゼ9の活性化に非依存性であった。
【0291】
(表6)
(マウス/ヒトキメラDN1924に誘導されたアポトーシスは、カスパーゼ非依存性である)
【0292】
【表6】
(実施例10)
(マウス/ヒトキメラDN1924 HLA−DR特異的モノクローナル抗体は、腫瘍増殖の速度を低下させること、およびRAJI異種移植片SCIDマウス動物系において生存の延長を促進することに有効である)
この実施例は、マウス/ヒトキメラDN1924抗HLA−DR抗体のインビボ皮下投与が、腫瘍増殖の速度を低下させること(図14)、およびRaji異種移植片scidマウス動物系において生存の延長を促進すること(図15)に有効であることを示す。
【0293】
0日目、scidマウス(5匹の動物の4つの群)は、0.1ml PBS中の5×106個のRaji腫瘍細胞(バーキットリンパ腫;ATCCカタログ番号CCL−86)で負荷(challenge)された。腫瘍負荷後1日目、4日目、7日目および10日目において、グループ1は、腹腔内注射を介して、300μl PBS中に50μgのマウスDN1924抗HLA−DR抗体を受け、グループ2は、腹腔内注射を介して、300μl PBS中に50μgのマウス/ヒトDN1924抗HLA−DR抗体(クローン番号64)を受け、グループ3は、腹腔内注射を介して、300μl PBS中に50μgのヒトIgG2,κアイソタイプコントロール抗体(Sigma、カタログ番号I5404)を受け、そしてグループ4は、腹腔内注射を介して、300μlのPBSを受けた。
【0294】
図14は、マウスDN1924およびマウス/ヒトキメラDN1924の両方とも、scidマウス動物モデル系において、Raji異種移植片を用いた皮下負荷によって腫瘍増殖の速度を低下させるのに有効であったことを示す。
【0295】
図15は、マウスDN1924およびマウス/ヒトキメラDN1924の両方とも、scidマウス動物モデル系において、Raji異種移植片を用いた皮下負荷後に生存を延長させるのに有効であったことを示す。
【0296】
本発明は、特定の処置方法および組成物に関して記載されたが、本発明から逸脱することなく種々の変更および改変がなされ得ることは、当業者にとって明らかである。
【図面の簡単な説明】
【0297】
【図1】HLA−DR分子の概略的エピトープマップを示す。これは、CY:細胞質テール;TM:膜貫通部分;α1およびβ1:それぞれ、α鎖およびβ鎖の第1(細胞外)ドメイン;ならびにα2およびβ2:それぞれ、α鎖およびβ鎖の第2(細胞外)ドメインを示す。
【図2】図2は、DN1924マウスモノクローナル抗体とともに16時間培養した後の種々の細胞集団の生存度を示す。生細胞が、2次元プロットの右下象限中に位置する。PBMCは、末梢血単核球に対応する。
【図3】図3は、示されるモノクローナル抗体とともに16時間培養した後のヒト細胞株の生存度を示す。
【図4】図4は、示されるような、種々の温度条件下で種々の時間の間DN1924モノクローナル抗体とともにインキュベーションした後のEBV−LCL RPMI 1788の生存度を示す。
【図5】図5は、[3H]チミジン取り込みによって示される、ヒトPBMCのブドウ球菌エンテロトキシンB(SEB)特異的インビトロ増殖応答に対するDN1924モノクローナル抗体の免疫抑制効果が存在しないことを示す。
【図6】図6は、HLA−DR+ヒトプラズマ細胞腫MC/CARを注射し、DN1924(白三角)またはリン酸緩衝化生理食塩水(PBS:黒四角)のいずれかで処理した、scidマウスの生存を示す。
【図7A】図7A〜図7Dは、DN1921と、DN1924(図7A)、キメラDN1924(図7B)、およびキメラDN1921(図7C)とがHLA−DR発現細胞に特異的に結合するのを示す、FACSデータである。
【図7B】図7A〜図7Dは、DN1921と、DN1924(図7A)、キメラDN1924(図7B)、およびキメラDN1921(図7C)とがHLA−DR発現細胞に特異的に結合するのを示す、FACSデータである。
【図7C】図7A〜図7Dは、DN1921と、DN1924(図7A)、キメラDN1924(図7B)、およびキメラDN1921(図7C)とがHLA−DR発現細胞に特異的に結合するのを示す、FACSデータである。
【図7D】図7A〜図7Dは、DN1921と、DN1924(図7A)、キメラDN1924(図7B)、およびキメラDN1921(図7C)とがHLA−DR発現細胞に特異的に結合するのを示す、FACSデータである。
【図8A】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8B】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8C】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8D】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8E】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図8F】図8A〜図8Fは、DN1924が、HLA−DR陽性腫瘍細胞においてアポトーシス活性を誘導すること(図8A);DN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8B);キメラDN1924が、HLA−DR陽性腫瘍細胞においてアポトーシスを誘導すること(図8C);キメラDN1924によるアポトーシスの誘導が、用量依存性であること(図8D);キメラDN1924が、正常なHLA−DR陽性細胞においてアポトーシスを誘導しないこと(図8E);DN1924が、HLA−DR陰性腫瘍細胞においてアポトーシスを誘導しないこと(図8F);を示す、FACSデータである。
【図9】図9は、マウスDN1921 HLA−DR特異的モノクローナル抗体およびキメラDN1921 HLA−DR特異的モノクローナル抗体によってはPAP/HLA−DR特異的Paperino応答の阻害を示すが、マウスDN1924 HLA−DR特異的モノクローナル抗体によってもキメラDN1924 HLA−DR特異的モノクローナル抗体によっても、PAP/HLA−DR特異的Paperino応答の阻害は示さない、グラフである。
【図10】図10は、キメラDN1921を発現するプラスミドベクターおよびキメラDN1924を発現するプラスミドベクターの構築の模式図である。
【図11】図11は、Igκ定常領域アライメントである。上の2行は、ヒトIgκについての2つの異なる登録番号(J00241およびM63438)である。下の行は、マウスIgκ(GenBank登録番号L35138)である。
【図12】図12は、IgG定常領域アライメントである。上の3行は、ヒト(h)IgG2、hIgG1、およびhIgG4(それぞれ、GenBank登録番号AJ250170、AF027159、およびAJ294733)である。下の3行は、マウス(m)IgG1、IgG2a、およびIgG2b(それぞれ、GenBank登録番号J00453、J00470、およびJ00461)である。
【図13A】図13A〜図13Dは、マウス/ヒトキメラDN1924抗HLA−DR媒介性アポトーシスが、カスパーゼ非依存性であること(図13A)、およびカスパーゼ3(図13B)も、カスパーゼ8(図13C)も、カスパーゼ9(図13D)も、キメラDN1924処理細胞において活性化されることを示す棒グラフである。
【図13B】図13A〜図13Dは、マウス/ヒトキメラDN1924抗HLA−DR媒介性アポトーシスが、カスパーゼ非依存性であること(図13A)、およびカスパーゼ3(図13B)も、カスパーゼ8(図13C)も、カスパーゼ9(図13D)も、キメラDN1924処理細胞において活性化されることを示す棒グラフである。
【図13C】図13A〜図13Dは、マウス/ヒトキメラDN1924抗HLA−DR媒介性アポトーシスが、カスパーゼ非依存性であること(図13A)、およびカスパーゼ3(図13B)も、カスパーゼ8(図13C)も、カスパーゼ9(図13D)も、キメラDN1924処理細胞において活性化されることを示す棒グラフである。
【図13D】図13A〜図13Dは、マウス/ヒトキメラDN1924抗HLA−DR媒介性アポトーシスが、カスパーゼ非依存性であること(図13A)、およびカスパーゼ3(図13B)も、カスパーゼ8(図13C)も、カスパーゼ9(図13D)も、キメラDN1924処理細胞において活性化されることを示す棒グラフである。
【図14】図14は、マウス/ヒトキメラDN1924抗HLA−DR抗体のインビボ皮下投与が、Raji異種移植scidマウス動物モデル系における腫瘍増殖速度を減少することにおいて有効であることを示すグラフである。
【図15】図15は、マウス/ヒトキメラDN1924抗HLA−DR抗体のインビボ皮下投与が、Raji異種移植scidマウス動物モデル系における生存増加を促進することにおいて有効であることを示すグラフである。
【特許請求の範囲】
【請求項1】
HLA−DR特異的モノクローナル抗体であって、該抗体は、重鎖可変ドメインおよび軽鎖可変ドメインならびに重鎖定常ドメインおよび軽鎖定常ドメインを含み、該重鎖可変ドメインは、該重鎖定常ドメインに作動可能に融合されており、該軽鎖可変ドメインは、該軽鎖定常ドメインに作動可能に融合されており、該重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも70%同一であり、該軽鎖可変ドメインは、配列番号24のアミノ酸配列と少なくとも70%同一であり、該モノクローナル抗体は、ヒトHLA−DRに特異的に結合する、抗体。
【請求項2】
請求項1に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖定常ドメインは、配列番号36のアミノ酸配列に対して少なくとも70%同一であり、前記軽鎖定常ドメインは、配列番号25のアミノ酸配列に対して少なくとも70%同一である、抗体。
【請求項3】
請求項2に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも80%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも80%同一である、抗体。
【請求項4】
請求項2に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも90%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも90%同一である、抗体。
【請求項5】
請求項2に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも98%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも98%同一である、抗体。
【請求項6】
請求項1に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖定常ドメインおよび前記軽鎖定常ドメインは、それぞれ、ヒト重鎖定常ドメインおよびヒト軽鎖定常ドメインである、抗体。
【請求項7】
請求項6に記載のHLA−DR特異的モノクローナル抗体であって、前記ヒト重鎖定常ドメインおよび前記ヒト軽鎖定常ドメインは、それぞれ、ヒトIgM重鎖定常ドメイン、ヒトIgD重鎖定常ドメイン、ヒトIgG2重鎖定常ドメイン、ヒトIgG3重鎖定常ドメイン、ヒトIgG4重鎖定常ドメイン、ヒトIgE重鎖定常ドメイン、ヒトIgA1重鎖定常ドメイン、およびヒトIgA2重鎖定常ドメイン、ならびにヒトIgM軽鎖定常ドメイン、ヒトIgD軽鎖定常ドメイン、ヒトIgG2軽鎖定常ドメイン、ヒトIgG3軽鎖定常ドメイン、ヒトIgG4軽鎖定常ドメイン、ヒトIgE軽鎖定常ドメイン、ヒトIgA1軽鎖定常ドメイン、およびヒトIgA2軽鎖定常ドメインである、抗体。
【請求項8】
請求項7に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖定常ドメインは、配列番号36において示されるヒトIgG2κ重鎖定常ドメイン配列に対して少なくとも70%同一であり、前記軽鎖定常ドメインは、配列番号25において示されるヒトIgG2κ軽鎖定常ドメイン配列に対して少なくとも70%同一である、抗体。
【請求項9】
請求項8に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも80%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも80%同一である、抗体。
【請求項10】
請求項8に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも90%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも90%同一である、抗体。
【請求項11】
請求項8に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも98%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも98%同一である、抗体。
【請求項12】
請求項8に記載のHLA−DR特異的モノクローナル抗体であって、該モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、該モノクローナル抗体は、HLA−DR依存性免疫応答を抑制しない、抗体。
【請求項13】
請求項1または請求項6に記載の抗体を含む組成物であって、該組成物は、薬学的に受容可能な賦形剤をさらに含む、組成物。
【請求項14】
アポトーシスを誘発する方法であって、該方法は、
HLA−DRを発現する腫瘍細胞を有する被験体に対して、治療上有効な量の請求項1または請求項6に記載の抗体を投与する工程
を包含する、方法。
【請求項15】
請求項14に記載の方法であって、前記HLA−DRを発現する腫瘍細胞は、プラズマ細胞腫/多発性骨髄腫、ホジキンリンパ腫、非ホジキンリンパ腫、およびB細胞白血病からなる群より選択される癌に由来する、方法。
【請求項16】
請求項14に記載の方法であって、前記抗体は、前記被験体に対して静脈内(IV)投与される、方法。
【請求項1】
HLA−DR特異的モノクローナル抗体であって、該抗体は、重鎖可変ドメインおよび軽鎖可変ドメインならびに重鎖定常ドメインおよび軽鎖定常ドメインを含み、該重鎖可変ドメインは、該重鎖定常ドメインに作動可能に融合されており、該軽鎖可変ドメインは、該軽鎖定常ドメインに作動可能に融合されており、該重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも70%同一であり、該軽鎖可変ドメインは、配列番号24のアミノ酸配列と少なくとも70%同一であり、該モノクローナル抗体は、ヒトHLA−DRに特異的に結合する、抗体。
【請求項2】
請求項1に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖定常ドメインは、配列番号36のアミノ酸配列に対して少なくとも70%同一であり、前記軽鎖定常ドメインは、配列番号25のアミノ酸配列に対して少なくとも70%同一である、抗体。
【請求項3】
請求項2に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも80%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも80%同一である、抗体。
【請求項4】
請求項2に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも90%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも90%同一である、抗体。
【請求項5】
請求項2に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも98%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも98%同一である、抗体。
【請求項6】
請求項1に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖定常ドメインおよび前記軽鎖定常ドメインは、それぞれ、ヒト重鎖定常ドメインおよびヒト軽鎖定常ドメインである、抗体。
【請求項7】
請求項6に記載のHLA−DR特異的モノクローナル抗体であって、前記ヒト重鎖定常ドメインおよび前記ヒト軽鎖定常ドメインは、それぞれ、ヒトIgM重鎖定常ドメイン、ヒトIgD重鎖定常ドメイン、ヒトIgG2重鎖定常ドメイン、ヒトIgG3重鎖定常ドメイン、ヒトIgG4重鎖定常ドメイン、ヒトIgE重鎖定常ドメイン、ヒトIgA1重鎖定常ドメイン、およびヒトIgA2重鎖定常ドメイン、ならびにヒトIgM軽鎖定常ドメイン、ヒトIgD軽鎖定常ドメイン、ヒトIgG2軽鎖定常ドメイン、ヒトIgG3軽鎖定常ドメイン、ヒトIgG4軽鎖定常ドメイン、ヒトIgE軽鎖定常ドメイン、ヒトIgA1軽鎖定常ドメイン、およびヒトIgA2軽鎖定常ドメインである、抗体。
【請求項8】
請求項7に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖定常ドメインは、配列番号36において示されるヒトIgG2κ重鎖定常ドメイン配列に対して少なくとも70%同一であり、前記軽鎖定常ドメインは、配列番号25において示されるヒトIgG2κ軽鎖定常ドメイン配列に対して少なくとも70%同一である、抗体。
【請求項9】
請求項8に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも80%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも80%同一である、抗体。
【請求項10】
請求項8に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも90%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも90%同一である、抗体。
【請求項11】
請求項8に記載のHLA−DR特異的モノクローナル抗体であって、前記重鎖可変ドメインは、配列番号35のアミノ酸配列に対して少なくとも98%同一であり、前記軽鎖可変ドメインは、配列番号24のアミノ酸配列に対して少なくとも98%同一である、抗体。
【請求項12】
請求項8に記載のHLA−DR特異的モノクローナル抗体であって、該モノクローナル抗体は、HLA−DRを発現する腫瘍細胞においてアポトーシスを誘導可能であり、該モノクローナル抗体は、HLA−DR依存性免疫応答を抑制しない、抗体。
【請求項13】
請求項1または請求項6に記載の抗体を含む組成物であって、該組成物は、薬学的に受容可能な賦形剤をさらに含む、組成物。
【請求項14】
アポトーシスを誘発する方法であって、該方法は、
HLA−DRを発現する腫瘍細胞を有する被験体に対して、治療上有効な量の請求項1または請求項6に記載の抗体を投与する工程
を包含する、方法。
【請求項15】
請求項14に記載の方法であって、前記HLA−DRを発現する腫瘍細胞は、プラズマ細胞腫/多発性骨髄腫、ホジキンリンパ腫、非ホジキンリンパ腫、およびB細胞白血病からなる群より選択される癌に由来する、方法。
【請求項16】
請求項14に記載の方法であって、前記抗体は、前記被験体に対して静脈内(IV)投与される、方法。
【図1】

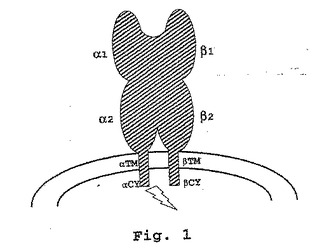
【図2】
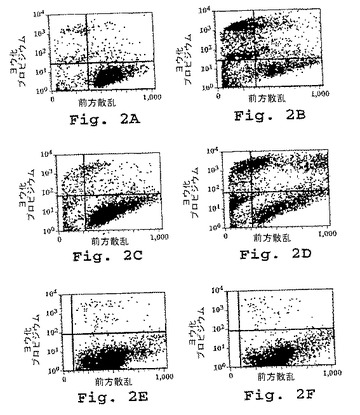
【図3】
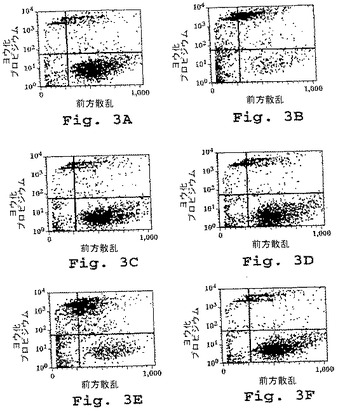
【図4】
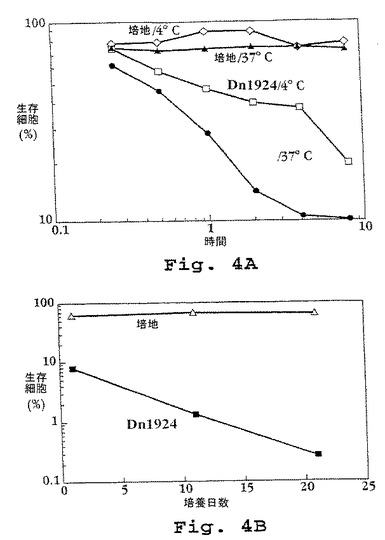
【図5】
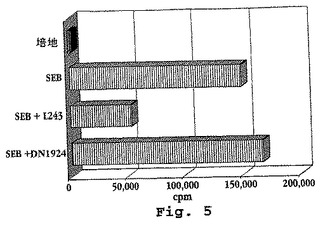
【図6】
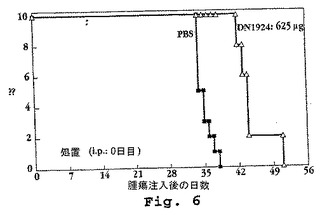
【図7A】
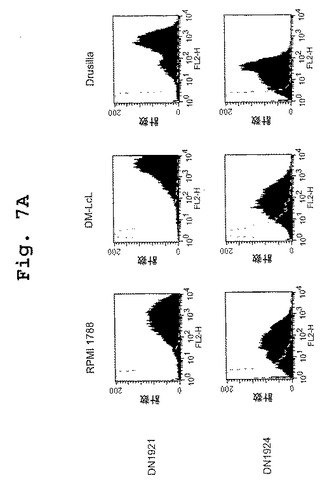
【図7B】
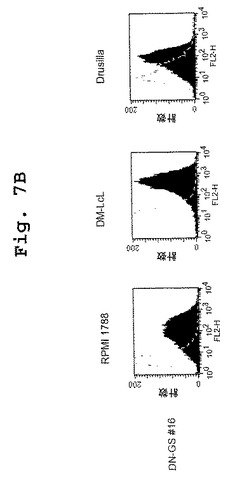
【図7C】
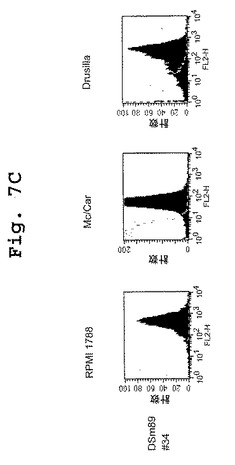
【図7D】
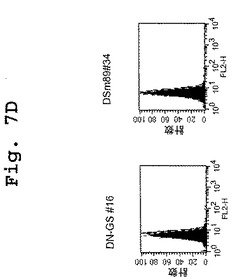
【図8A】
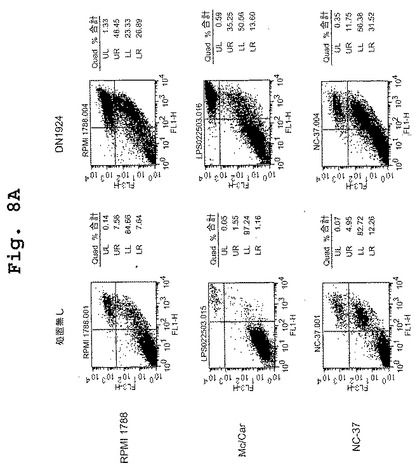
【図8B】
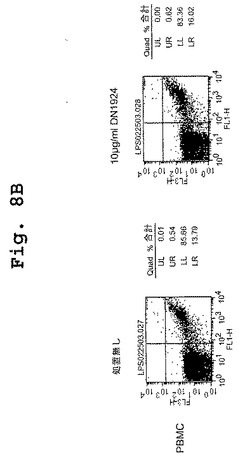
【図8C】
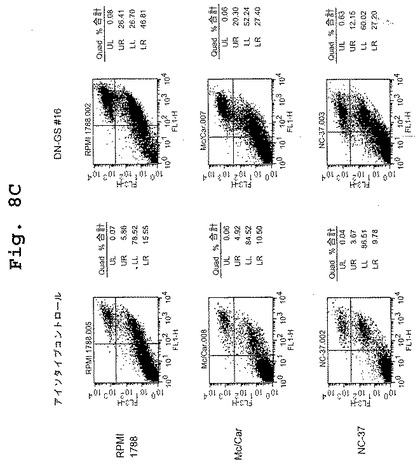
【図8D】
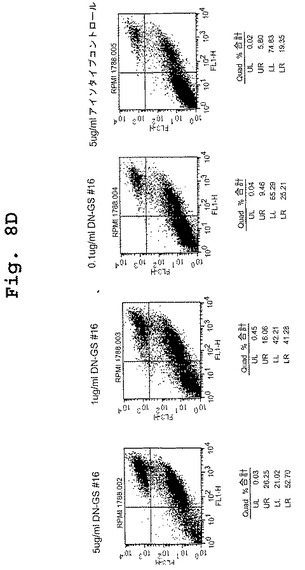
【図8E】
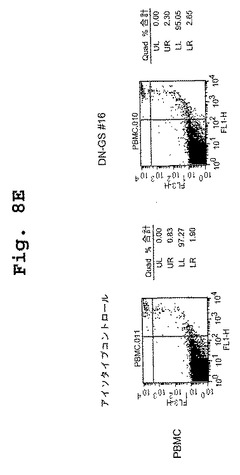
【図8F】
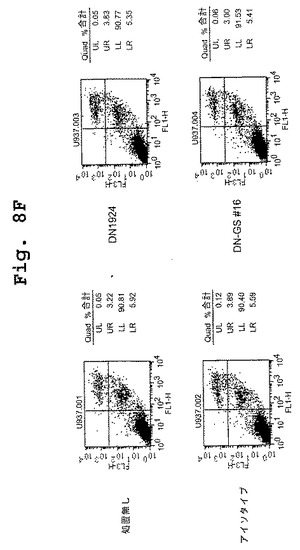
【図9】
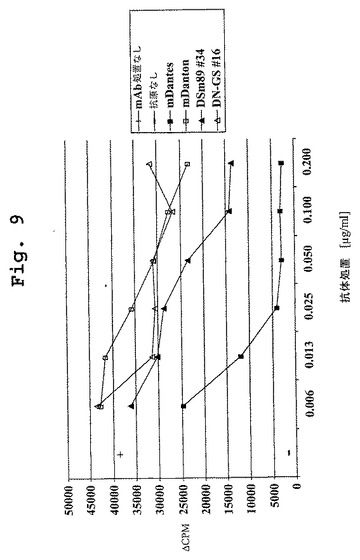
【図10】
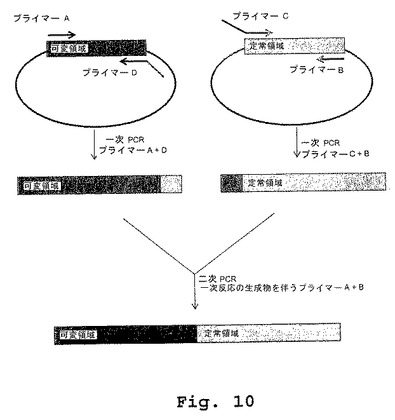
【図11】

【図12】

【図13A】
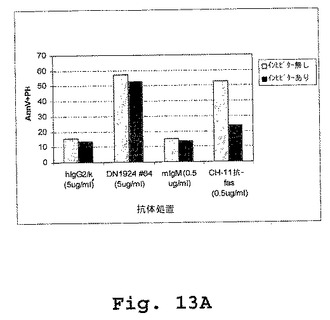
【図13B】
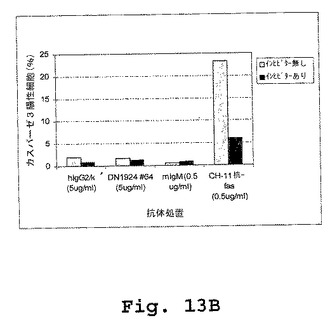
【図13C】
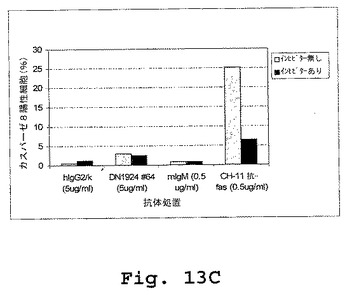
【図13D】
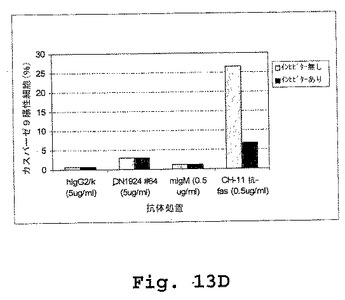
【図14】
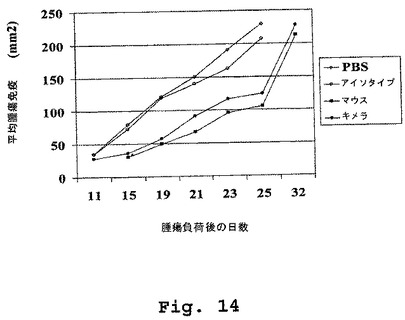
【図15】
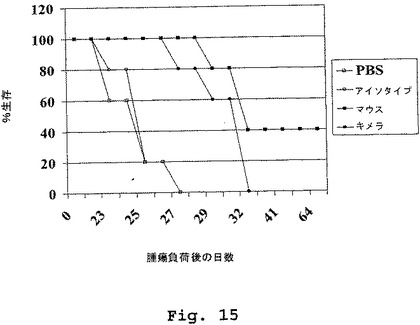
【図2】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図7C】
【図7D】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図9】
【図10】
【図11】
【図12】
【図13A】
【図13B】
【図13C】
【図13D】
【図14】
【図15】
【公表番号】特表2007−513976(P2007−513976A)
【公表日】平成19年5月31日(2007.5.31)
【国際特許分類】
【出願番号】特願2006−544134(P2006−544134)
【出願日】平成16年12月15日(2004.12.15)
【国際出願番号】PCT/US2004/042312
【国際公開番号】WO2005/058251
【国際公開日】平成17年6月30日(2005.6.30)
【出願人】(500072714)デンドレオン コーポレイション (11)
【Fターム(参考)】
【公表日】平成19年5月31日(2007.5.31)
【国際特許分類】
【出願日】平成16年12月15日(2004.12.15)
【国際出願番号】PCT/US2004/042312
【国際公開番号】WO2005/058251
【国際公開日】平成17年6月30日(2005.6.30)
【出願人】(500072714)デンドレオン コーポレイション (11)
【Fターム(参考)】
[ Back to top ]