
HSF−1の改変方法
本発明は、HSF1のDNA結合ドメインのアセチル化を修飾する工程を含む、HSF1活性の調節方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本願は、2008年12月8日に出願された米国特許仮出願第61/120,542号および2009年6月29日に出願された米国特許仮出願第61/221,315号の恩典を主張する。上記出願の全教示は、参照により本明細書に援用される。
【0002】
政府支援
本発明は、国立衛生研究所により授与された補助金番号AG026647号およびGM038109号のもと、政府支援によってなされたものである。本発明において、政府は一定の権利を有する。
【背景技術】
【0003】
発明の背景
細胞は、センサーおよび経路のネットワークを利用してタンパク質ホメオスタシスと称されるタンパク質の合成、フォールディング、輸送、凝集、および分解のバランスを正常に維持している[Sitia et al., Nature 426: 891-894, 2003; Ron et al., Nat Rev Mol Cell Biol 8: 519-529, 2007]。細胞におけるタンパク質ホメオスタシスの維持、つまりプロテオスタシスとは、プロテオームを形成する個々のタンパク質のコンホメーション、結合相互作用、局在および濃度の制御のことである。インビボでのタンパク質フォールディングは、多くの主類のシャペロンおよびフォールディング酵素を含むフォールディングポリペプチド鎖と巨大分子細胞成分の間の相互作用により達成され、凝集を最小化する[Wiseman et al., Cell 131: 809-821, 2007]。また、所定の区画で生成された有機および無機溶質が、フォールディングの物理的化学性質に影響を及ぼす疎水性作用などの非共有結合力によってポリペプチド鎖の救出(salvation)に影響するので、代謝酵素も細胞タンパク質フォールディング効率に影響を及ぼす。代謝経路で、特定のタンパク質の折りたたまれた状態に結合して安定化する小分子リガンドが生成され、フォールディング平衡を移動させることによりフォールディングが増強される[Fan et al., Nature Med., 5, 112 (Jan 1999); Hammarstrom et al., Science 299, 713 (2003)]。所定のタンパク質がある細胞型で折りたたまれるかどうかは、シャペロン、フォールディング酵素、代謝産物等の分布、濃度および細胞内局在に依存している[Wiseman et al.]。ヒトの機能消失疾患は典型的に、効果的な分解をもたらす所定のタンパク質の細胞フォールディングを含む所定のタンパク質中の突然変異により引き起こされる正常なタンパク質ホメオスタシスの乱れの結果であることがしばしばある[Cohen et al., Nature 426: 905-909, 2003]。ヒトの機能獲得疾患も同様に、タンパク質凝集をもたらすタンパク質ホメオスタシスの乱れの結果であることが良くある[Balch et al. (2008), Science 319: 916-919]。
【0004】
細胞レベルで、熱ショック応答は、種々の急性および慢性のストレス条件から細胞を保護する[Westerheide et al., J Biol. Chem. 280(39): 33097 (2005)]。熱ショック応答(HSR)は、環境的および生理学的ストレス因子に対する遺伝子応答であり、正常細胞の代謝の抑制、ならびにタンパク質ミスフォールディングおよび凝集の結果である細胞の損傷からの保護および回復に有用な分子シャペロン、プロテアーゼおよび他のタンパク質を発現する熱ショックタンパク質(HSP)遺伝子の迅速な誘導をもたらす[Westerheide et al.]。熱ショック応答は、転写因子である熱ショック因子-1(HSF-1)により媒介される。HSRは種々のストレス因子により生じる損傷から細胞を保護するが、大量のHSPの蓄積は、細胞の成長および分裂に悪影響を及ぼすことがある[Morimoto et al. (1998), Genes Dev. 12, 3788]。
【0005】
プロテオスタシスおよび熱ショック応答の両方における機能障害は、例えば、癌、神経変性疾患、代謝疾患、炎症性疾患および心臓血管疾患を含む種々の疾患に関連している。当該技術分野において、プロテオスタシス機能障害および/または熱ショックタンパク質の改変された誘導に関連する状態を治療するための治療アプローチの必要性が依然として存在している。
【発明の概要】
【0006】
発明の概要
本発明は、細胞HSF1活性がそのDNA結合ドメインのアセチル化状態により調節されるという発見に基づくものである。例えば、実施例1には、SIRT1(HSF1アセチル化を阻害する)の活性化は、HSF1を脱アセチル化状態に維持することでhsp70プロモーターへのHSF1 DNA結合を長引かせるが、SIRT1のダウンレギュレーションはHSF1活性の減衰を加速させることが示されている。
【0007】
一態様において、本発明は、HSF1のDNA結合ドメイン中のリジン残基のアセチル化を調節する工程を含む、細胞中の熱ショック転写因子1(HSF1)の活性を調節する方法である。別の態様において、HSF1の活性は、リジン残基のアセチル化を抑制することにより増強される。さらに別の態様において、HSF1の活性は、リジン残基のアセチル化を促進することにより低減される。リジン残基は、ヒトHSF1のリジン80(HSF1 K80)または非ヒトHSF1中の対応する保存されたアミノ酸であり得る。
【0008】
別の態様において、HSF1のアセチル化は、細胞に、HSF1のDNA結合ドメイン中のリジン残基のアセチル化を阻害する薬剤を投与することにより阻害される。いくつかの局面において、リジン残基のアセチル化を阻害する薬剤は、単離サーチュイン、サーチュイン活性化剤またはヒストンアセチルトランスフェラーゼ(HAT)阻害剤である。
【0009】
いくつかの局面において、HSF1のアセチル化は、細胞に、有効量の単離サーチュインまたはサーチュイン活性化剤を投与することにより阻害される。単離SIRT1またはSIRT1活性化剤が投与され得る。
【0010】
さらに別の態様において、HSF1のアセチル化は、細胞にHAT阻害剤を投与することにより阻害される。
【0011】
別の態様において、HSF1のアセチル化は、細胞にSIRT1阻害剤またはHAT活性化剤などのサーチュイン阻害剤を投与することにより促進される。
【0012】
さらなる態様において、本発明は、被験体においてHSF1のアセチル化を阻害する工程を含む、HSF1の活性の増加を必要とする被験体におけるHSF1の活性を増加する方法である。一態様において、被験体はヒトである。
【0013】
別の態様において、本発明は、被験体においてHSF1のアセチル化を促進する工程を含む、HSF1の活性の低減を必要とする被験体におけるHSF1の活性を低減する方法である。一態様において、被験体はヒトである。
【0014】
さらなる態様において、本発明は、タンパク質のホメオスタシスの機能障害に関連する状態に苦しむ患者に、治療有効量のHSF1のアセチル化を阻害する薬剤を投与する工程を含む、該患者を治療する方法である。一態様において、該状態は機能消失障害である。別の態様において、該状態は機能獲得障害である。別の態様において、HSF1のアセチル化を阻害する薬剤は、単離サーチュイン、サーチュイン活性化剤またはHAT阻害剤である。さらなる態様において、該薬剤は、単離SIRT1またはSIRT1活性化剤である。さらなる態様において、該薬剤は、HAT阻害剤である。
【0015】
別の態様において、本発明は、熱ショックタンパク質の発現の増加に関連する状態に苦しむ患者に、治療有効量のHSF1のアセチル化を促進する薬剤を投与する工程を含む、該患者を治療する方法である。一態様において、該状態は癌または腫瘍である。別の態様において、該状態はウイルス感染である、別の態様において、HSF1の活性を促進する薬剤は、サーチュイン阻害剤またはHAT活性化剤である。さらなる態様において、該薬剤はSIRT1阻害剤である。
【0016】
本発明はさらに、熱ショック応答の増加およびプロテオスタシス機能障害の疾患の治療を必要とする患者に、有効量の熱ショックアクチベーターおよび有効量のHSF1のアセチル化を阻害する薬剤を投与する工程を含む、該患者において熱ショック応答を増加する方法およびプロテオスタシス機能障害の疾患を治療する方法を包含する。別の態様において、本発明は、熱ショックアクチベーターおよびHSF1のアセチル化を阻害する薬剤を含む医薬組成物である。
【0017】
本発明はまた、細胞に試験薬剤を投与する工程;およびHSF1のDNA結合ドメイン中のリジン残基のアセチル化をモニタリングする工程を含む、細胞においてHSF1活性を調節する薬剤を同定する方法を包含し、試験薬剤の非存在下のDNA結合ドメインのアセチル化と比較したDNA結合ドメインのアセチル化の変化は、細胞中で該試験薬剤がHSF1活性を調節することを示す。さらなる態様において、HSF1 K80または対応する保存されたアミノ酸のアセチル化がモニタリングされる。
【0018】
別の態様において、本発明は、細胞または細胞溶解物に試験薬剤を投与する工程、およびサーチュイン基質のアセチル化を測定する工程を含む、細胞においてHSF1活性を調節する薬剤を同定する方法であり、該試験薬剤の非存在下のアセチル化と比較した基質のアセチル化の変化は、細胞中で該試験薬剤がHSF1活性を調節することを示す。一態様において、サーチュインはSIRT1である。
【0019】
さらなる態様において、本発明は、細胞または細胞溶解物に試験薬剤を投与する工程、およびヒストンアセチルトランスフェラーゼ(HAT)基質のアセチル化を測定する工程を含む、細胞においてHSF1活性を調節する薬剤を同定する方法であり、該試験薬剤の非存在下でのアセチル化と比較した基質のアセチル化の変化は、細胞中で該試験薬剤がHSF1活性を調節することを示す。
【図面の簡単な説明】
【0020】
本発明の前述および他の記載、特徴および利点は、添付の図面に示されるように、本発明の好ましい態様の以下のより具体的な記載から明白であろう。図面において、同様の参照記号は、異なる図面を通じて同じ部分を参照する。図面は必ずしも同じ縮尺ではなく、発明の原理の説明が強調される。
【0021】
【図1】図1A:シャペロン遺伝子発現に対するサーチュイン阻害剤ニコチンアミドの効果。熱ショック(HS)、セラストール、CdCl2またはMG 132に曝露する前に、HeLa細胞をニコチンアミド(NAM)で処理し、示されるプライマーを用いてRT-PCR解析を行なった。図1B:SIRT1 siRNAはhsp70の転写を阻害する。SIRT1に対するsiRNAまたは対照siRNAで形質転換されたHeLa細胞を、指定の時間、HSで処理した。RT-PCR解析を行ない、密度測定によりhsp70 mRNAアバンダンスの増加倍数を計算し、18S rRNAに対して標準化した。図1C:SIRT1 siRNAはhsp70プロモーターに対するHSF1結合を阻害する。上述のようにHeLa細胞を処理した(2B)。HSF1抗体およびqPCRを使用してChIP解析を行ない、1%の入力で行なった反応に対して結果を標準化した。実験は3回行なった。エラーバーは±SDを示す。
【図2】図2A:HSR誘導因子に応答するHSF1のアセチル化。Flag-HSF1およびp300で形質転換された293T細胞を、HS、セラストール、CdCl2またはMG 132で処理した。免疫沈降およびウェスタンブロットを用いたアセチル化アッセイにより細胞溶解物を解析した。リン酸化の増加によりゆっくり遊走するHSF1は、アスタリスクで示される。図2B:HSF1アセチル化におけるニコチンアミドおよびトリコスタチンAの効果。Flag-HSF1およびp300で形質転換した293T細胞を、トリコスタチンA(TSA)、ニコチンアミド(NAM)または両方で処理してHSに曝露し、アセチル化アッセイにより細胞溶解物を解析した。図2C:触媒変異体ではなく野生型SIRT1はHSF1アセチル化を阻害する。293T細胞をFlag-HSF1、p300、およびSIRT1 WTまたはSIRT1 H363Y変異体のいずれかで形質転換した後に、HSで処理し、アセチル化アッセイにより解析した。図2D:HSに応答したHSF1アセチル化。Myc-HSF1で形質転換したCos7細胞を、HSで指定時間処理し、アセチル化アッセイにより解析した。SIRT1に対する抗体またはMyc抗体を用いてSIRT1およびHSF1を検出した。Hsc70を負荷対照とした。図2E:レスベラトロールの効果。HeLa細胞を溶媒(EtOH)またはレスベラトロールで処理した後、指定時間、HS処理を行なった。オリゴヌクレオチドプルダウンアッセイおよびウェスタンブロットにより細胞抽出物を解析した。HSF1のリン酸化の増加はアスタリスクで示す。図2F:SIRT1過剰発現は、HSF1 DNA結合に影響する。293T細胞をSIRT1 WTで形質転換し、次いで指定時間HSに供した。HSF1抗体およびqPCRを用いてChIP解析を行なった。A〜Gの実験は3回行なった。エラーバーは±SDを示す。
【図3】図3A:K80でのHSF1の変異は、DNA結合活性を乱す。指定のHSF1構築物で形質転換したhsf1-/-細胞(HS有または無し)由来の抽出物を用いてEMSA反応を行なった(上パネル)。EMSAプローブは、ヒトhsp70プロモーター由来の近位HSEを含む。同じ試料でウェスタンブロット解析を行って、HSF1およびHsc70のレベルを示した。図3B:K80での組み換えHSF1の突然変異は、DNA結合能を乱す。組み換えWT HSF1またはHSF1 K80Qの量を増加させ(5、20、40、80または120ng)、プローブがHSEを含むEMSA反応を示す(上パネル)。HSF1タンパク質無し(-)の試料を対照とした。同じ試料でウェスタンブロットを行なってHSF1発現レベルを示した。図3C: K80でのHSF1変異の、hsf1-/-細胞におけるHSRレスキューの失敗。hsf1-/-細胞を指定の形態のヒトHSF1で形質転換して、HS有または無しで処理した。指定の遺伝子についてのプライマーによるqPCRを用いてRNAを定量した。グリセルアルデヒド3-リン酸デヒドロゲナーゼについて得られた値に対してデータを標準化し、HSなしで処理したWT HSF1細胞中の各mRNAのアバンダンスと比較する(値セット1)。A〜Cの実験は3回行なった。エラーバーは±SDを示す。
【図4】図4A:SIRT1過剰発現による厳しいストレスからの細胞の保護。293T細胞を空ベクター(モック)またはSIRT1で形質転換して45℃ HSで指定時間処理し、その後37℃で回復させた。24時間後、トリプシンブルー取り込みにより細胞死を決定した。実験は3回行なった。エラーバーは±SDを示す。図4B:年齢依存的なHSRの減退とSIRT1のアバンダンスの低下の相関。早期継代(継代21)および老齢(継代44)WI-38線維芽細胞由来の細胞抽出物をHS、またはHS後に3時間の37℃での回復(R)で処理し、HSEを含むプローブを用いたEMSAにより解析した(上パネル)。同じ試料でウェスタンブロットを行ない、Hsp70、HSF1およびSIRT1発現レベルを示した(下パネル)。
【図5】図5A:pCAFではなくp300およびCBPは、HSF1アセチル化を誘導する。293T細胞をFlag-HSF1およびp300、CBP、またはpCAFで形質転換し、HSで処理してアセチル化アッセイで解析した。図5B:p300は、インビボでhsp70プロモーターに結合する。指定の42℃熱ショック時点ならびに37℃の指定の回復有およびなしの時点で行なったクロマチン免疫沈降実験は、p300のhsp70プロモーターへの結合を示す。クロマチンを架橋して、回収し、p300または HSF1に特異的な抗体で免疫沈降した。次いで試料をhsp70.1プロモーターに特異的なプライマーを用いたPCRで解析した。1%の投入量、抗体無しでも反応を行なった。熱ショックの際に、HSF1およびp300は両方1分以内にhsp70プロモーターに結合し、結合は回復中徐々に弱まる。図5C:SIRT1はインビボでhsp70プロモーターに結合する。指定の42℃熱ショック時点で行なったクロマチン免疫沈降実験は、SIRT1がhsp70プロモーターに結合することを示す。クロマチンを架橋して、回収し、HSF1またはSIRT1に特異的な抗体で免疫沈降した。次いで、試料は、hsp70プロモーターに特異的なプライマーを用いてPCRにより解析した。抗体なしで対照反応も行なった。SIRT1は基底および熱ショック条件の両方でhsp70プロモーターに結合する。
【図6】図6:アセチル化の前提条件ではないバルクHSF1リン酸化。293T細胞をMyc-HSF1 WTまたはHSF1の10個のセリンのリン変異体およびp300で共形質転換した。形質転換の24時間後、細胞を1時間で42℃の熱ショックで処理した。細胞溶解物を免疫沈降に供してアセチル化HSF1を、ウェスタンブロットを用いて検出した、全HSF1を検出するためにMyc抗体を使用した。
【図7−1】図7A:HSF1は9個のリジンでアセチル化され得る。p300で共形質転換した293T細胞で発現させて精製したFlag-HSF1を熱ショック(42℃、1時間)またはセラストール処理(5μM、1時間)のいずれかに供し、LC-MS/MSで解析した。マウスHSF1中で同定されたアセチル化リジン残基の位置を示す。DBD、DNA-結合ドメイン;L、リンカー領域;HR-A/Bおよび-C、7回繰り返しドメイン;RD、制御ドメイン;N、核局在配列;AD1およびAD2、活性化ドメイン。図7B:アセチル化リジンは保存される。T-COFFEEバージョン5.13を使用して、多種類のアセチル化リジンの配列アライメントを行なった。アセチル化リジンを赤で示し、アミノ酸番号を右に示す。Hs、ヒト(NP_005517);Mm、マウス(NP_032322);Dr、ゼブラフィッシュ(NP 571675);Xl、アフリカツメガエル(NP 001084036.1);Dm、キイロショウジョウバエ(P22813);Ce、線虫(NM_060630);Sc、酵母(P10961)。
【図7−2】図7C;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−3】図7D;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−4】図7E;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−5】図7F;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−6】図7G;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−7】図7H;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−8】図7I;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−9】図7J;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−10】図7K;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図8】図8:HSEと複合体化したK.ラクチスHSF DNA-結合ドメインの結晶構造を補充した座標(13)を使用して、発明者らは、整列したヒトHSF1の配列を、Swiss Modelを使用してつなげた。K80の側鎖およびDNAリン酸主鎖までの距離を緑で示す。上パネル中の四角で囲んだ領域は、下パネルをより拡大して示す。
【図9】図9:HSF1 K80Q変異体は、三量体化能力を欠損していない。HSFのDNAへの安定した結合の条件は、三量体HSF複合体の形成である。従って、K80の変異が適切なHSF1フォールディングまたはHSF三量体のストレス誘導形成を乱したと考えられる。この可能性を調べるために、交差結合アッセイを使用して、熱ショック時の高次複合体の形成を測定した。K562細胞を、構成的な三量体であるHSF1変異体であると以前に特徴付けられたWT Myc-HSF1、Myc-HSF1 K80QまたはMyc-HSF1 M391K、L395Pで形質転換した(14)。細胞を熱ショック(HS、42℃、20分)で処理するかまたは処理せず、その後EGS架橋剤で処理した。タンパク質抽出物をSDS-PAGEで分離し、指定の抗体でウェスタンブロット解析を行なった。矢印は単量体HSF1を示し、矢印の先端は三量体HSF1を示す。WTとK80Q HSF1の両方は、熱ショックに応答して単量体から三量体へ移行し、変異がHSF1三量体化に影響を及ぼさないことが示される。
【図10】図10:HSF1 DNA結合には、残基80にリジンを必要とする。指定のHSF1構築物で形質転換され、熱ショック(HS、42℃、20分)有また無しで処理されたhsf1-/-細胞におけるEMSA反応を示す。プローブとして使用したオリゴヌクレオチドは、ヒトhsp70プロモーター由来の近位HSEを含む。同じ試料でウェスタンブロット解析を行ない、HSF1とHsc70の発現レベルを示した。等量負荷についてHsc70を示す。K80のA、H、NおよびTへの変異は全て、熱ショック時のHSF1のDNAへの結合能を障害した。
【図11】図11:核ストレス体の形成にはHSF1の残基80にリジンを必要とする。指定のMyc-HSF1型で形質転換して、1時間、42℃の熱処理(HS)有または無しで処理したHeLa細胞の共焦点画像を示す。α-Myc抗体(赤)を用いて外因性HSF1を染色し、DAPI(青)を使用してDNAを染色した。バー=5μm。HSF1 K80のR、Q、A、H、NおよびTへの変異は全て、熱ショック時の特徴的な核ストレス体へのHSF1の再局在能を阻害した。
【図12】図12:ドミナントネガティブ様式のHSF1 K80Q機能。HSEへのHSF1の安定な結合には、3つのDNA結合ドメインと3つのnGAAnリピートの同時接触が必要である。1つまたは2つの単量体のアセチル化が関連のある非修飾単量体の結合を破壊するので、アセチル化HSF1 DNA結合ドメインの小集団は、ドミナントネガティブ効果を有すると思われる。この問題を調べるために、発明者らはHeLa細胞をMyc-HSF1のWTまたはK80Q変異体で形質転換し、HSF1またはMycに対する抗体で該細胞を二重染色して、内在性HSF1がnSBを形成する能力を解析した。指定の型のMyc-HSF1で形質転換して、熱ショック(HS、42℃、1時間)有またはなしで処理したHeLa細胞の共焦点画像を示す。HSF1タンパク質は、HSF1に対する抗体(緑)およびMycに対する抗体(赤)で染色した。DAPI(青)を使用してDNAを染色した。バー=5μm。内因性および外因性のWT HSF1は、熱ショック時にnSBに局在したが、HSF1のK80Q変異体を発現する細胞においてはいずれの抗体を用いてもnSBは検出されなかった。従って、K80アセチル化の生物学的な意味は、K80アセチル化HSF1分子の実際のレベルよりも高いと思われる。
【図13】図13:HSF1活性化サイクルのモデル。工程1:休止状態のHSF1は細胞質性または核性の不活性単量体である。工程2:ストレス時、HSF1はHSEに結合するDNA結合ホモ三量体を形成する。工程3:HSF1は転写活性を獲得する。工程4:HSF1転写活性は、減衰時にはなくなる。減衰は、DNAに結合したHSF1のトランス活性化機能(示さず)およびHSF1のDNA結合ドメインのアセチル化を介したDNA結合の阻害を抑制するhsp発現からの負のフィードバックを含む2つの調節された工程を有する。SIRT1は、HSF1アセチル化の阻害によるHSRの減衰段階を調節する。
【図14】図14:HSF1-SIRT1制御ネットワークのモデル。加齢および細胞代謝状態によるSIRT1の調節は、HSF1などの転写因子のネットワークの活性に影響を及ぼし、長生およびストレス耐性の増大をもたらす。
【図15】図15は、hsp70プロモーターレポーターを示す図である;hsp70.1pr-luc HeLa安定細胞株は-188から+150にhsp70.1プロモーター配列を含み、近位熱ショックエレメント(HSE)を含む。
【図16】図16は、ルシフェラーゼ解析前に、表示したとおりのレスベラトロールおよび41℃熱ショックもしくは42℃熱ショックいずれかで処理したHeLa hsp70.1pr-luc細胞の倍数活性を示す棒グラフである。
【図17】図17は、ルシフェラーゼ解析前に、表示したとおりのセラストールおよび/またはレスベラトロールで処理したHeLa hsp70.1pr-luc細胞の倍数活性を示す棒グラフである。
【発明を実施するための形態】
【0022】
発明の詳細な説明
本発明の好ましい態様の記載は以下の通りである。
本明細書で使用する場合、単語「a」および「an」は、特定されることがなければ1つ以上を意味する。例えば用語「細胞(a cell)」は、単一の細胞および2つ以上の細胞の組合せの両方を包含する。
【0023】
用語「熱ショック因子」、「熱ショック転写因子」および「HSF」は、ストレス誘導性遺伝子発現に関連する転写因子のファミリーのことをいう。少なくとも4種類のHSFが、脊椎動物および植物で記載されている。ショウジョウバエおよび線虫は、1つのみのHSFを発現する。(Wu, Ann. Rev. Cell Dev. Biol. 1995; 11:441-469)。ヒト細胞では、3種類のHSF(HSF1、HSF2およびHSF4)が特徴付けられている(Morimoto, et al, Genes Dev. 1998; 12:3788-3796)。HSF1は偏在的に発現し、HSPのストレス誘導発現に主要な役割を担っている。
【0024】
HSF1の配列は種間で高度に保存されている。HSF1のDNA結合ドメインは100アミノ酸超よりも長く、N末端の近位に位置している。ヒトHSF1とショウジョウバエHSFのDNA結合ドメイン間には約70%の相同性があり、ヒトHSF1と線虫HSF間には約55%の相同性がある。ヒトHSF1では、DNA結合ドメインはアミノ酸残基13からアミノ酸残基121まで伸びている。本明細書で使用する場合、用語「HSF1」は、任意の種または細胞型由来のHSF1を包含し、ショウジョウバエおよび線虫由来のHSFも包含する。かかるHSFおよびHSF1タンパク質は、HSF1 K80に対応するDNA結合ドメイン中のアセチル化のためのリジンを有する。用語「非ヒトHSF1」は、非ヒト種由来のHSF1を示す。用語HSF1は、GenBank受託番号NP_005517、NP_032322、NP_571675、NP_001084036.1、P22813、NM_060630およびP10961のアミノ酸配列を有するペプチドを含むがこれに限定されない。
【0025】
一態様において、HSF1は哺乳動物HSF1である。別の態様において、HSF1はヒトHSF1である。一態様において、HSF1はGenBank受託番号NP_005517(配列番号:1)と少なくとも約85%、90%、95%、98%および99%の配列同一性を有するアミノ酸配列を有する。別の態様において、HSF1は、GenBank受託番号NP_005517のアミノ酸配列を有する。
【0026】
用語「配列同一性」または配列に関する「同一性」は、2種類のアミノ酸配列間または2種類のヌクレオチド配列間の配列同一性のことをいう。同一性はそれぞれ、比較目的で整列されたそれぞれの配列の位置を比較することにより決定され得る。用語「配列相同性」または配列に関する「相同性」は、2種類のアミノ酸配列間または2種類のヌクレオチド配列間の配列相同性のことをいう。比較した配列中の同等の位置が同じ塩基またはアミノ酸で占められる場合、該分子はその位置で同一であり;同等の部位が同じまたは同様のアミノ酸残基(例えば、立体的および/または電気的性質が類似)で占められる場合、該分子はその位置で相同(同様)であるという。相同性、類似性または同一性のパーセントでの表示は、比較された配列で占められる位置での同一また同様なアミノ酸の数の関数を示す。相同性、類似性または同一性のパーセントでの表示は、比較された配列で占められる位置での同一または同様なアミノ酸の数の関数を示す。FASTA、BLASTまたはENTREZなどの種々の整列アルゴリズムおよび/またはプログラムが使用され得る。FASTAおよびBLASTは、GCG配列解析パッケージ(University of Wisconsin, Madison, Wis.)の一部として利用可能であり、例えばデフォルトを設定して使用することができる。ENTREZは、国立衛生研究所、国立医学図書館、全米バイオテクノロジー情報センター、Bethesda、Mdを通じて利用可能である。一態様において、2種類の配列のパーセント同一性は、ギャップウェイト1、例えば、2種類の配列間に一アミノ酸またはヌクレオチドミスマットがある場合、各アミノ酸ギャップを埋めてGCGプログラムにより決定され得る。
【0027】
本発明は、HSF1活性が、DNA結合ドメイン中のアミノ酸残基のアセチル化および脱アセチル化により媒介されるという発見に基づくものである。HSF1のアセチル化によりDNAへの結合が低下し、HSF1の脱アセチル化状態の維持によりDNAへの結合が延長されることが見出された。一態様において、細胞中のHSF1活性は、HSF1のDNA結合ドメイン中の1つ以上の塩基性アミノ酸のアセチル化を阻害することで増加する。HSF1のDNA結合ドメインのアセチル化は、DNA結合ドメイン中の1つ以上の塩基性アミノ酸残基のアセチル化を阻害する薬剤を投与することにより阻害され得る。アセチル化が減少または脱アセチル化が増加される場合にアミノ酸のアセチル化が阻害されることは理解されよう。別の態様において、HSF1のDNA結合ドメイン中のリジン残基のアセチル化を阻害することによりHSF1活性が増加される。さらなる態様において、細胞のHSF1活性は、ヒトHSF1のリジン残基80(HSF1 K80)のアセチル化を阻害することまたは非ヒトHSF1中の対応する保存されたアミノ酸のアセチル化を阻害することにより増加される。
【0028】
用語「HSF1 K80」は、ヒトHSF1のリジン80を示す。「対応する保存されたアミノ酸」は、非ヒト種のHSF1中の、保存され、ヒトHSF1 K80に対応するリジン残基を示す。本明細書で使用する場合、「保存されたアミノ酸」または「保存された残基」は、2種類のタンパク質間に共通することが分かっているおよび/または異なる種由来の相同タンパク質中などのペプチドモチーフ中の特定の位置を占める残基のことをいう。本明細書で使用する場合、「対応する保存されたアミノ酸」またはHSF1 K80に関する「対応する保存された残基」は、相同HSF1配列または非ヒトHSF1配列中の保存されたアミノ酸についてのアミノ酸位置のことをいう。HSF1の他のホモログ中のアミノ酸のナンバリングは、ヒトHSF1中のナンバリングとは異なり得ることは、当業者には容易に明らかであろう。HSF1の他のホモログ中の対応する保存されたアミノ酸は、例えば、市販の相同性モデリングソフトウェアパッケージまたは従来の配列アライメントパッケージを使用したアミノ酸配列の比較により同定され得る。
【0029】
別の態様において、細胞中のHSF1活性は、HSF1のDNA結合ドメイン中の1つ以上の塩基性アミノ酸のアセチル化を促進することにより低減される。一態様において、DNA結合ドメインのアセチル化を促進する薬剤が細胞に投与される。アミノ酸のアセチル化は、アミノ酸のアセチル化が増加される場合または脱アセチル化が低減される場合に促進されることが理解されよう。別の態様において、HSF1活性は、HSF1のDNA結合ドメイン中のリジン残基のアセチル化を促進することで低減される。さらなる態様において、ヒト細胞中のHSF1活性は、ヒトHSF1 K80のアセチル化を促進することまたは非ヒトHSF1中の対応する保存されたアミノ酸のアセチル化を促進することで低減される。
【0030】
本明細書で使用する場合、「阻害」または「低減」は、直接または間接的な手段により、正味の低減を引き起こすことを包含する。用語「増加」は、直接または間接的な手段により、正味の増大を引き起こすことを意味する。
【0031】
HSF1活性は、熱ショック応答エレメント(HSE)配列を有する遺伝子の転写調節因子としてのHSF1の活性のことをいう。HSE配列を有する遺伝子から転写されたタンパク質は、熱ショック(HS)タンパク質と称される。熱ショックは、特定の遺伝子の転写の増大を引き起こす熱ショック応答のトリガーとして最初に発見された[Snoecx et al. (2001), Physiol. Rev. 81: 1461-1497]。この転写活性の産物は、熱ショックタンパク質と称される[Snoeckx et al.]。ほとんどの熱ショックタンパク質(Hsp)は、分子質量の表示を参照して、例えばHsp27と命名されている。ファミリー中の種々のHspの分類は、その関連のある機能およびサイズに基づく。熱ショックタンパク質のサイズは、10〜170kDaである。ファミリーネームは慣例的に大文字で書かれる。例えば「HSP70」は、HSP70ファミリーのことをいう。HSP70ファミリーは、重量が70〜78kDaである。HSP70ファミリーの一員の一例は、Hsp72(一般的にHsp70と示される)である。熱ショック応答は、熱ショックタンパク質をコードする遺伝子の誘導を包含する。本発明の方法に従って誘導され得る熱ショックタンパク質遺伝子としては、限定されないが、HSP10ファミリー、HSP40ファミリー、HSP60ファミリー、HSP70ファミリー、HSP90ファミリー、HSP100ファミリー、HSP27ファミリー、αA-クリスタリンファミリーおよびαB-クリスタリンファミリーのタンパク質から選択されるファミリー由来のタンパク質をコードする遺伝子が挙げられる。
【0032】
HSF1のアセチル化は、HSF1のアセチル化の増大もしくは脱アセチル化の低減をもたらすか、またはHSF1をアセチル化形態で維持する(直接または間接的な)任意の手段により増大され得る。いくつかの態様において、細胞中のHSF1のアセチル化は、HSF1のアセチル化を増大する薬剤を投与することで増大される。同様に、HSF1のアセチル化は、アセチル化の低減または脱アセチル化の増大をもたらすか、またはHSF1を脱アセチル化形態で維持する任意の手段により低減または阻害され得る。用語「薬剤」は、本明細書において、化合物、化合物の混合物、生物学的巨大分子(例えば、核酸、抗体、タンパク質もしくはその一部、例えばペプチドなど)または細菌、植物、真菌もしくは動物(特に哺乳動物)の細胞または組織などの生物学的物質から作製される抽出物を示すように使用される。一態様において、HSF1のDNA結合ドメインのアセチル化を調節する薬剤は、HSF1を含む細胞と試験化合物を接触させて、試験化合物の存在下または非存在下でDNA結合ドメインのアセチル化の程度を測定することにより同定され得、アセチル化の増大は、該薬剤がHSF1のアセチル化を増大させることを示し、アセチル化の低減は該薬剤がHSF1のアセチル化を阻害することを示す。
【0033】
HSF1のアセチル化を阻害する例示的な方法としては、限定されないが、単離サーチュインを投与すること、細胞中のサーチュインの活性を上げること、およびヒストンアセチルトランスフェラーゼ(HAT)の活性を阻害することが挙げられる。ヒストンアセチルトランスフェラーゼ(HAT)は、ヒストン、Hsp40/Dna J熱ショックタンパク質および種々の転写因子などの特定のタンパク質中のリジン残基のアセチル化を触媒する酵素である[Saha et al. (2006). HATs and HDACs in neurodegeneration: a tale of disconcerted acetylation homeostasis. Cell Death and Differentiation 13:539-550; WO 2009/134131 A1]。アセチルトランスフェラーゼのいくつかのファミリーが同定されており、例えば、prGcn5およびPCAFなどのGNATスーパーファミリー(Gcn5-関連N-アセチルトランスフェラーゼ)、例えばMOZ、Ybf2/Sas3、Sas2およびTip60などのMYSTファミリー、例えばp300およびCBPタンパク質などのp300/CBP HATファミリーが挙げられる(Lau et al. (2000), Cell 5(3): 589-95; 米国特許出願公開公報第20080227752号。以下の実施例1に示されるように、p300およびCBPはHSF1アセチル化を誘導する。
【0034】
ヒストンデアセチルトランスフェラーゼ(HDAC)は基質中のリジン残基の脱アセチル化を触媒する酵素である。脱アセチル化酵素は3つのファミリーに分類される;クラスIおよびII HDACファミリーは、トリコスタチンAにより阻害され、NAD+依存型クラスIIIサーチュインファミリーは、ニコチンアミドにより阻害される。
【0035】
一態様において、HSF1のアセチル化は、細胞に、単離サーチュインを投与すること、または細胞中のサーチュインのレベルもしくは活性を上げることにより阻害され得る。サーチュインは、サーチュイン脱アセチル化酵素タンパク質ファミリーに属するタンパク質である。一態様において、サーチュインはSir2ファミリーに属し、これは、限定されないが酵母Sir2(例えば、GenBank受託番号P53685)、線虫Sir-2.1(例えば、GenBank受託番号NP_501912)、ヒトSIRT1(例えば、GenBank受託番号NM_012238およびNP_036370(またはAF083106))ならびにヒトSIRT2(例えば、GenBank受託番号NM_012237、NM_030593、NP_036369、NP_085096およびAF083107)のタンパク質が挙げられる。本明細書で使用する場合、用語「Sir2タンパク質」は、Sir2ファミリーに属するサーチュインのことをいう。一態様において、Sir2タンパク質はSIRT1である。別の態様において、SIRT1はヒトSIRT1である。
【0036】
上述のように、本発明の一局面は、細胞に単離サーチュインまたはその生物学的に活性な断片もしくはバリアントをHSF1のDNA結合ドメイン中のアミノ酸のアセチル化を阻害するのに充分な量で投与する工程を含む、細胞中のHSF1の活性を増加する方法である。
【0037】
本明細書で使用する場合、細胞または被験体(下記のように)に投与される単離サーチュインまたはその断片もしくはバリアントは、実質的に純粋であるか、またはそれに天然に付随する成分から単離された実質的に純粋な単離ポリペプチドである。用語「単離サーチュイン」または「単離SIRT1」は、薬学的に許容され得る担体または賦形剤を含むことを除外せず、単にサーチュインが細胞およびサーチュインに天然に付随する他の成分から単離されていることを示すことが理解されよう。単離サーチュイン、そのバリアントの断片は、上述の天然に存在するサーチュインと同じまたは実質的に同一のアミノ酸配列を有する組み換えにより生成されたポリペプチドであり得る。
【0038】
サーチュインの生物学的に活性な断片は、脱アセチラーゼ活性または完全長サーチュインの別の生物活性を有する断片である。サーチュインのバリアントは、例えば、サーチュインの配列中に保存的アミノ酸置換を作製することで作製され得る。生物学的に活性なバリアントは、脱アセチラーゼ活性または天然に存在するサーチュインの別の生物学的活性を有する。保存的アミノ酸置換は、同様の側鎖を有する残基と交換可能であることをいう。保存的置換アミノ酸は、側鎖の化学的性質によって分類され得る。例えば、アミノ酸の一グループは、中性で疎水性の側鎖(a、v、l、i、p、w、fおよびm)を有するアミノ酸を含み;別のグループは、中性で極性の側鎖を有するアミノ酸(g、s、t、y、c、nおよびq)であり;別のグループは塩基性側鎖を有するアミノ酸(k、rおよびh)であり;別のグループは酸性側鎖を有するアミノ酸(dおよびe)であり;別のグループは、脂肪族側鎖を有するアミノ酸(g、a、v、lおよびi)であり;別のグループは、脂肪族水酸基側鎖を有するアミノ酸(sおよびt)であり;別のグループは、アミン含有側鎖を有するアミノ酸(n、q、k、rおよびh)であり;別のグループは、芳香族側鎖を有するアミノ酸(f、yおよびw)であり;別のグループは、硫黄含有側鎖を有するアミノ酸(cおよびm)である。例示的な保存的アミノ酸置換基は:r-k;e-d、y-f、l-m;v-iおよびq-hである。アミノ酸は、その一般的な一文字表記を用いて上記され:アラニンはa;システインはc;アスパラギン酸はd;グルタミン酸はe;フェニルアラニンはf;グリシンはg;ヒスチジンはh;イソロイシンはi;リジンはk;ロイシンはl;メチオニンはm;アスパラギンはn;プロリンはp;グルタミンはq;アルギニンはr;セリンはs;トレオニンはt;バリンはv;トリプトファンはw;およびチロシンはyである。
【0039】
一態様において、サーチュインはSir2タンパク質またはその生物学的に活性な断片もしくはバリアントである。別の態様において、サーチュインは、天然に存在するSir2タンパク質またはその断片と少なくとも約80%、85%、90%、95%、98%および99%の配列相同性を有するペプチドである。別の態様において、サーチュインは、天然に存在するSir2タンパク質またはその断片と少なくとも約80%、85%、90%、95%、98%および99%の配列同一性を有するペプチドである。別の態様において、Sir2タンパク質は、哺乳動物SIRT1と少なくとも約80%、85%、90%、95%、98%および99%の配列同一性を有するペプチドである。別の態様において、Sir2タンパク質は、ヒトSIRT1と少なくとも約80%、85%、90%、95%、98%および99%の配列同一性を有するペプチドである。さらなる態様において、Sir2タンパク質は配列番号:2と少なくとも80%、85%、90%、95%、98%および99%の配列同一性を有するペプチドである。
【0040】
別の態様において、Sir2タンパク質は、配列番号:2の250番アミノ酸が保存されたSIRT1触媒ドメイン、アミノ酸残基258〜451と少なくとも約80%の同一性を有する。配列番号:2はヒトSIRT1のアミノ酸配列を示す。好ましい態様において、Sir2タンパク質は、配列番号:2のアミノ酸残基258〜451の間のアミノ酸配列に少なくとも約80、85、90、95、99%の配列同一性を有する配列を含む。他の態様において、Sir2タンパク質は、完全長Sir2タンパク質の生物学的に活性な断片、例えば脱アセチル化が可能なSIRT1の断片である。Sir2タンパク質はまた、天然に存在するSir2ファミリーメンバーに対して1つ以上の置換、例えば1〜10個の置換を含む配列またはバリアントを含む。
【0041】
サーチュインのレベルまたは活性は、細胞中でサーチュインのレベルまたは活性を増加する薬剤(「サーチュイン活性化剤」)を投与することによっても増加し得る。サーチュインの活性とは、脱アセチラーゼ活性またはタンパク質が基質を脱アセチル化する活性またはサーチュインの別の生物活性のことをいう。
【0042】
サーチュイン活性化剤は、サーチュインの活性を増加させる薬剤である。SIRT1活性化剤は、細胞中でSIRT1の活性を増加させる薬剤である。サーチュインおよびSIRT1活性化化合物は、例えば米国特許第7,345,178号、米国特許出願公開公報第20080249103号、20080194803号、200070037865号、20070014833号、20060229265号および20060025337号ならびに国際出願番号WO 2005/002672、WO 2005/002555、WO 2009015180、WO 2009015179、WO 2006094248、WO 2006004722、WO 2006094235、WO 2006094233、WO 2006094209、WO 2006105440、WO 2006078941、WO 2006094236、WO 2006/094210ならびにMilne et al. (2007), Nature 450: 712-716およびSzczepankiewicz et al. (2008), Current Topics in Medical Chemistry 8: 1533-1544に記載されており、各文献の内容は参照により本明細書に援用される。具体的なSIRT1活性化化合物としては、限定されないが、フラボン、スチルベン、フラバノン、イソフラバノン、カテキン、カルコン、タンニンおよびアントシアジン(anthocyadins)、キノキサリン(例えば、ピロロキノキサリン)、SRT-1460、SRT-2183、SRT-1720、SRT-2104、オキサロ酢酸、レスベラトロール、オキサゾロ[4,5-b]ピリジン、アザベンゾイミダゾール、ベンゾイミダゾール、およびイミダゾチアゾール(例えば、SRT1460、SRT2183およびSRT1720)、(Szczepankiewicz et al.)が挙げられる。例示的なスチルベンとしては、限定されないが、トリヒドロキシスチルなどのヒドロキシスチルベン、例えば3,5,4'-トリヒドロキシスチルベン(「レスベラトロール」)が挙げられる。レスベラトロールはまた、3,4',5-スチルベントリオールとしても公知である。特定のレスベラトロールエステルアナログもサーチュインアクチベーターとして記載されている(例えば、WO 2005069998参照)。テトラヒドロキシ-スチルベン、例えばピセタノールも包含される。イソリキリチゲニンなどのトリヒドロキシケイロンを含むヒドロキシケイロン、およびブテイン(butein)などのテトラヒドロキシケイロンもSIRT1活性化化合物として記載されている。フィセチンなどのテトラヒドロキシフラボンを含むヒドロキシフラボン、およびケルセチンなどのペンタヒドロキシフラボンも使用することができる。SIRT1アクチベーターである例示的なヒドロキシフラボンとしては、限定されないが、イソリキリチゲニン、フィセチン、ブテインおよびケルセチンが挙げられる。
【0043】
別の態様において、HSF1のアセチル化は、細胞にヒストンアセチルトランスフェラーゼ(HAT)の活性を阻害する薬剤またはヒストンアセチルトランスフェラーゼ(HAT)の阻害剤を投与することにより阻害することができる。例示的なHAT阻害剤としては、米国特許出願公開公報第20090076155号に記載されるアナカルジン酸(6-ペンタデシルサリチル酸)および例えばその誘導体が挙げられ、その内容は、参照により本明細書中に明確に援用される。クルクマノイド(例えばクルクミンなど)、エピガラクトガテキン-3-没食子酸(epigallogatechin-3-gallate)およびガルシノール(garcinol)もHAT阻害剤として記載されている(例えば、米国特許出願公開公報第20060020027号, Cancer Research 69, 583 (2009), およびJBC 279: 33716-33726参照、それぞれその内容は参照により本明細書に明確に援用される)。HATを阻害するための他の化合物および方法は、米国特許出願公開公報第20090076155、WO 02070675、米国特許出願公開公報第2008227752および米国特許第6,369,030号および6,747,005号に記載されており、それぞれの内容は、参照により本明細書に明確に援用される。p300阻害剤は、例えば、米国特許出願公開公報第20050069986号およびLau et al. (2000), Cell 5(3): 589-95, Mai et al. (2009), Bioorganic and Medicinal Chemical Letters 19(4): 1132-35に記載されており、それぞれの内容は参照により本明細書に明確に援用される。
【0044】
本発明はまた、サーチュインの活性を低減させる工程を含む、HSF1のDNA結合ドメインのアセチル化を促進することにより細胞中のHSF1の活性を低減させる方法に関する。一態様において、サーチュインはSir2タンパク質である。別の態様において、サーチュインはSIRT1である。一態様において、サーチュインの活性を阻害する薬剤(「サーチュイン阻害剤」)は、細胞に投与され得る。一態様において、SIRT1阻害剤が投与される。
【0045】
当業者が理解するように、サーチュイン阻害剤は、小分子、タンパク質、ペプチド、ペプチド模倣物、抗体または核酸であり得る。核酸としては限定されないが、DNA、RNA、RNA干渉剤およびPNAが挙げられる。サーチュイン阻害剤は米国特許7,345,178号、米国特許出願公開公報第20080287653号、20060229265号20060084135号、20050136537号および20070197459号、国際特許出願WO 2008155390、WO 2008011476、WO 2008082646、WO 2008156866、WO 2008086400、WO 2007047604、WO 2007084162、WO 2006099245、WO 2006094209、WO 2006/094210、WO 2005026112、およびWO 2005060711に記載され、それぞれの内容は参照により本明細書に援用される。例示的な小分子サーチュイン阻害剤は、ニコチンアミド、サーチノール、パラサーチノール、HR-73、2,3,4,9-テトラヒドロ-1H-カルバゾール-1-カルボキシアミド、スプリトマイシン、SEN-196、2-フェニルアミノベンズアミド、スラミン、NSC-112546、カンビノール、テノビン-1(tenovin-1)、ガッチフェロン(guttiferone)、ハイパーフォリン、アーチソホリン(artisoforin)およびタンパク質キナーゼC阻害剤Ro31-8220、ロットレリン、ZM449829およびインジルビン-3'-モノオキシム(Szczepankiewicz et al.)である。
【0046】
別の態様において、サーチュイン阻害剤は、抗サーチュイン抗体である。用語抗体は、モノクローナル抗体、ポリクローナル抗体、ヒト化抗体、キメラ抗体、単鎖Fv(scFv)、Fab断片、F(ab')断片、イントラボディおよび合成抗体である。一態様において、サーチュイン阻害剤は、抗Sir2タンパク質抗体である。さらなる態様において、サーチュイン阻害剤は、抗SIRT1抗体である。抗サーチュイン抗体は、サーチュインタンパク質またはそのポリペプチドもしくは断片などの適切な免疫原(合成ペプチド等の合成分子など)に対して生じ得る。免疫抗原の調製、ならびにポリクローナル抗体およびモノクローナル抗体の産生は、例えばファージディスプレイなどの任意の適切な技術を使用して行われ得る。種々の方法が記載されている(例えば、Kohler et al., Nature, 256:495-497 (1975)) およびEur. J. Immunol. 6:511-519 (1976)); Milstein et al., Nature 266:550-552 (1977)); 米国特許第4,172,124号; Harlow, E. and D. Lane, 1988, Antibodies: A Laboratory Manual, (Cold SpringHarbor Laboratory: Cold Spring Harbor, N. Y.); ならびにCurrent Protocols In Molecular Biology, Vol. 2 (Supplement 27, Summer '94), Ausubel, F. M. et al., Eds., (John Wiley & Sons: New York, N.Y.), Chapter 11, 1991)を参照、それぞれの教示は参照により本明細書に援用される)。本発明に従い抗体の断片を使用することもできる。抗体は、従来技術を使用して断片化することができ、該断片は、完全抗体と同様の方法で有用性についてスクリーニングされる。免疫グロブリン分子のFab断片は、共有結合により共に連結された免疫グロブリン重鎖および免疫グロブリン軽鎖の免疫学的に活性な部分を含む免疫グロブリン分子の一部からなる多量体タンパク質であり、抗原と特異的に結合することができる。Fab断片は、当該技術分野において周知の方法を使用して、実質的に完全な免疫グロブリン分子のパパインによるタンパク質溶解性消化により調製し得る。しかしながら、Fab断片はまた、適切な宿主細胞中で、免疫グロブリン重鎖および免疫グロブリン軽鎖の所望の一部を、当該技術分野で公知の任意の方法を用いて発現させることによっても調製し得る。
【0047】
抗サーチュイン抗体はまた、イントラボディでもあり得る。イントラボディは、細胞内発現抗体であり、特異的に細胞内の標的分子に結合して、不活性化するように設計された単鎖抗体分子である。イントラボディは、細胞アッセイおよび完全生物体において使用されている[Chen et al., Hum. Gen. Ther. (1994) 5:595-601; Hassanzadeh et al., Febs Lett. (1998) 16(1, 2):75-80 and 81-86]。サーチュインと特異的に反応するイントラボディを用いて誘導性発現ベクターを構築することができる。
【0048】
さらなる態様において、サーチュイン阻害剤は、核酸である。一態様において、核酸は、アンチセンス核酸である。アンチセンス核酸は、RNA、DNA、PNAまたは任意の他の適切な核酸であり得る。アンチセンス配列とその相補的センス配列の間で二本鎖が形成され得、遺伝子の不活性化が生じる。アンチセンス核酸は、遺伝子から転写されたRNAと二本鎖を形成する、二本鎖DNAと三本鎖を形成することなどにより遺伝子発現を阻害し得る。アンチセンス核酸は、例えばコード配列が公知である任意の遺伝子に対して作製することができるか、またはいくつかの充分に確立された技術(例えば、アンチセンスRNAまたはオリゴヌクレオチドの化学合成(任意に、分解に対して抵抗性を増加させるかもしくは細胞取り込みを向上させる修飾ヌクレオチドおよび/または結合を含む)またはインビトロ転写)により決定することができる。アンチセンス核酸およびその使用は、例えば、米国特許第6,242,258号、6,500,615号、6,498,035号、6,395,544号および5,563,050号に記載され、それぞれの内容は参照により本明細書に援用される。
【0049】
別の態様において、サーチュイン阻害剤はRNA干渉剤である。「RNA干渉剤」は、本明細書で使用する場合、標的遺伝子またはゲノム配列の発現を、RNA干渉(RNAi)により干渉または阻害する任意の薬剤として規定される。かかるRNA干渉剤としては、限定されないが、標的遺伝子もしくはゲノム配列またはその断片に相同なRNA分子を含む核酸分子、短鎖干渉RNA(siRNA)、短鎖ヘアピンまたは低分子ヘアピンRNA(shRNA)、およびRNA干渉(RNAi)により標的遺伝子の発現を干渉または阻害する小分子が挙げられる。本発明の標的遺伝子は、サーチュインをコードする遺伝子である。一態様において、該遺伝子は、Sir2タンパク質をコードする。別の態様において、該遺伝子は、SIRT1をコードする。さらに別の態様において、該遺伝子はヒトSIRT1をコードする。本明細書で使用する場合、「標的遺伝子発現の阻害」は、標的遺伝子もしくは該標的遺伝子にコードされるタンパク質の発現またはタンパク質活性またはレベルの任意の低減を含む。
【0050】
一態様において、RNA干渉剤はsiRNAである。siRNAは、化学的に合成、インビトロ転写により産生、または宿主細胞中で産生され得る。一態様において、siRNAは、約15〜約40ヌクレオチド長または約15〜約28ヌクレオチドまたは約19〜約25ヌクレオチド長または約19、20、21または22ヌクレオチド長の二本鎖RNA(dsRNA)分子であり、各鎖において長さ約0、1、2、3、4、5または6ヌクレオチドの3'および/または5'突出を含み得る。突出の長さは、二本の鎖で独立であり、すなわち一方の鎖の突出の長さは第二鎖の突出の長さに依存しない。
【0051】
RNAiはまた、低分子ヘアピン(ステムループとも称される)RNA(shRNA)を含む。一態様において、これらのshRNAは、短(例えば、約19〜約25ヌクレオチド)アンチセンス鎖、続いて約5〜約9ヌクレオチドのヌクレオチドループ、および類似のセンス鎖からなる。代替的に、センス鎖は、ヌクレオチドループ構造の前にあり得、アンチセンス鎖が続いてもよい。これらのshRNAは、プラスミド、レトロウイルス、およびレンチウイルスに含まれ得、例えばpol III U6プロモーターまたは別のプロモーターから発現され得る(例えば、Stewart, et al. (2003) RNA Apr;9(4):493-501参照、参照により本明細書に援用される)。
【0052】
RNAに加えて、RNA干渉剤はまた、化学的に修飾されたヌクレオチドおよび非ヌクレオチドを含み得、また、リボース糖分子が別の糖分子または同様の機能を果たす分子に置換された分子を含み得る。また、ヌクレオチド残基間に、ホスホロチオエート結合などの非天然結合が使用されてもよい。該RNA鎖は、フルオロフォアなどのレポーター基の反応性官能基により誘導体化され得る。例示的な誘導体は、RNA鎖の両末端または一方の末端、典型的にはセンス鎖の3'末端で修飾される。他の例示的な誘導体は、2'O-アルキル化残基または2'-O-メチルリボシル誘導体および2'-O-フルオロリボシル誘導体などの修飾炭水化物部分を有するヌクレオチドを取り込む。RNA塩基が修飾されていてもよく、例えばアルキル化またはハロゲン化され得る。例えば、ハロゲン化塩基、例えば5-ブロモウラシルおよび5-ヨードウラシルが取り込まれ得る。該塩基がアルキル化されてもよく、例えば7-メチルグアノシンがグアノシン残基の代わりに取り込まれ得る。成功裡の阻害を生じる非天然塩基がさらに取り込まれ得る。
【0053】
別の態様において、核酸はリボザイムまたはデオキシリボザイムである。リボザイムおよびデオキシリボザイムは、核酸分子の配列特異的切断を触媒することが示されている。切断部位は、RNAまたはDNA酵素中のヌクレオチドと標的核酸中のヌクレオチドの相補対形成により決定される。そのため、RNAおよびDNAの酵素は、核酸分子を切断するように設計されて、分解の速度が増大され得る[Cotten et al, EMBOJ. 8: 3861-3866, 1989; Usman et al., Nucl. Acids Mol. Biol. 10: 243, 1996; Usman, et al., Curr. Opin. Struct. Biol. 1: 527, 1996; Sun, et al., Pharmacol. Rev., 52: 325, 2000]。
【0054】
サーチュイン阻害化合物はまた、ペプチドアプタマーを含む。ペプチドアプタマーは、タンパク質機能のドミナント阻害剤として作用するペプチドまたは小ポリペプチドである。ペプチドアプタマーは、標的タンパク質に特異的に結合して、その機能をブロックする(Kolonin and Finley, PNAS (1998) 95:14266-14271)。高い親和性および特異性でサーチュインに結合するペプチドアプタマーは、当該技術分野に公知の種々の技術により単離することができる。ペプチドアプタマーは、酵母ツーハイブリッドスクリーンによりランダムペプチドライブラリーから単離し得る(Xu et al, PNAS (1997) 94:12473-12478)。ペプチドアプタマーはまた、ファージライブラリー(Hoogenboom et al., Immunotechnology (1998) 4:1-20)、または化学的に合成したペプチド/ライブラリーから単離し得る。
【0055】
特定の局面において、HSF1の活性は、HATの活性を増大し得る薬剤(本明細書においてはHATアクチベーターともいう)を投与することにより阻害され得る。HATを活性化し得る小分子化合物の例は、N-(4-クロロ-3-トリフルオロメチル-フェニル)-2-エトキシ-6-ペンタデシル-ベンズアミド(CTPB)である(Balasubrmanyam et al. (2003). Small molecule modulators of histone acetyltransferase p300. The Journal of Biological Chemistry, 278, 19134-19140)。
【0056】
本発明の方法によると、HSF1の活性は、HSF1のDNA結合ドメイン中の塩基性アミノ酸残基のアセチル化を改変することにより細胞内で調節することができる。細胞としては、原核細胞、真核細胞、脊椎動物細胞および無脊椎動物細胞が挙げられる。一態様において、細胞は哺乳動物細胞である。別の態様において、細胞はヒト細胞である。例示的な細胞としては、限定されないが、筋肉細胞(例えば、骨格筋細胞、ならびにペースメーカー細胞、心房細胞、心房心室結節細胞、左心室細胞、右心室細胞、乳頭筋細胞、プルキンエ線維細胞および平滑筋細胞などの心筋細胞)、血液細胞、腎臓細胞、上皮細胞、腸細胞、リンパ節細胞、脾臓細胞、肝細胞、胸腺細胞、唾液腺細胞、下垂体細胞、膀胱細胞、骨細胞、乳房細胞、頚部細胞、結腸直腸細胞、腎臓細胞、咽頭細胞、肺細胞、リンパ細胞、皮膚細胞および造血性細胞(例えばT細胞、B細胞、マクロファージ、樹状細胞およびそれらの前駆細胞)が挙げられる。
【0057】
特定の局面において、本発明は、HSF1の活性の調節を必要とする被験体に、HSF1のDNA結合ドメイン中の塩基性アミノ酸残基のアセチル化を調節する薬剤の有効量を投与する工程を含む、前記被験体においてHSF1の活性を調節する方法である。一態様において、本発明は、HSF1の活性の増大を必要とする被験体に、HSF1のDNA結合ドメインの塩基性アミノ酸のアセチル化を阻害する薬剤の有効量を投与する工程を含む、前記被験体においてHSF1の活性を増大する方法である。さらなる態様において、HSF1のDNA結合ドメインのアセチル化を阻害する薬剤は、単離サーチュイン、サーチュイン活性化剤またはHAT阻害剤である。別の態様において、本発明は、HSF1の活性の低減を必要とする被験体に、HSF1のDNA結合ドメイン中の塩基性アミノ酸のアセチル化を促進する薬剤の有効量を投与する工程を含む、前記被験体においてHSF1の活性を低減する方法である。さらなる態様において、HSF1のDNA結合ドメインのアセチル化を促進する薬剤は、サーチュイン阻害剤またはHATアクチベーターである。
【0058】
また、本発明は、タンパク質ホメオスタシスの機能障害に関連のある状態の治療を必要とする被験体に、HS1のDNA結合ドメイン中の塩基性アミノ酸のアセチル化を阻害する薬剤の治療有効量を投与する工程を含む、前記被験体において、タンパク質ホメオスタシスの機能障害に関連のある状態を治療する方法を包含する。別の態様において、該薬剤は、DNA結合ドメイン中のリジン残基のアセチル化を阻害する。さらなる態様において、該薬剤は、HSF1 K80のアセチル化を阻害する。さらに別の態様において、該方法は、前記被験体に、HAT阻害剤、単離サーチュインおよびサーチュイン活性化剤からなる群より選択される薬剤の治療有効量を投与する工程を含む。一態様において、該薬剤は、HAT阻害剤である。別の態様において、該薬剤は、単離サーチュインである。別の態様において、サーチュインはSir2タンパク質である。さらなる態様において、サーチュインはSIRT1である。さらなる態様において、サーチュインはヒトSIRT1である。さらに別の態様において、該薬剤は、サーチュイン活性化剤またはSIRT1活性化剤である。サーチュイン活性化剤は、先に規定され記載されている。上述のように、サーチュイン活性化剤およびSIRT1活性化剤としては、例えば、フラボン、スチルベン、フラバノン、イソフラバノン、カテキン、カルコン、タンニンおよびアントシアジン、ピロロキノキサリン、SRT-1460、SRT-2183、SRT-1720、オキサロ酢酸、レスベラトロールおよびその他のポリフェノールが挙げられる。単離サーチュインまたはサーチュイン活性化剤は、薬学的な担体または賦形剤を含む医薬組成物中で投与され得る。
【0059】
「治療する」または「治療」は、症状、合併症、または疾患の生化学的徴候の開始を予防または遅延すること、症状を改善または緩和すること、あるいは疾患、状態または障害のさらなる発症を停止または阻害することを含む。「患者」は、治療を必要とするヒト被験体である。「治療有効量」は、障害の1つ以上の症状を改善および/または障害の進行を予防、障害の後退を引き起こすのに充分な治療剤の量のことをいう。
【0060】
本発明は、タンパク質のホメオスタシスにおける機能障害に関連する状態の治療を包含する。例示的なタンパク質としては、グルコセレブロシダーゼ、ヘキソサミンA、嚢胞線維症膜貫通伝導性調節因子、アスパルチルグルコサミニダーゼ、α-ガラクトシダーゼA、システイントランスポーター、酸性セラミダーゼ、酸性α-L-フコシダーゼ、保護タンパク質、カテプシンA、酸性β-グルコシダーゼ、酸性β-ガラクトシダーゼ、イズロン酸2-スルファターゼ、α-L-イズロニダーゼ、ガラクトセレブロシダーゼ、酸性α-マンノシダーゼ、酸性β-マンノシダーゼ、アリルスルファターゼB、アリルスルファターゼA、N-アセチルガラクトサミン-6-硫酸スルファターゼ、酸性β-ガラクトシダーゼ、N-アセチルグルコサミン-1-ホスホトランスフェラーゼ、酸性スフィンゴミエリナーゼ、NPC-I、酸性α-グルコシダーゼ、β-ヘキソサミンB、ヘパリンN-スルファターゼ、α-N-アセチルグルコサミニダーゼ、α-グルコサミニドN-アセチルトランスフェラーゼ、N-アセチルグルコサミン-6-硫酸スルファターゼ、α1抗トリプシン、α-N-アセチルガラクトサミニダーゼ、α-ノイラミニダーゼ、β-グルクロニダーゼ、β-ヘキソサミンAおよび酸性リパーゼ、ポリグルタミン、α-シヌクレイン、Abペプチド、タウタンパク質、hERGカリウムチャネル、島アミロイドポリペプチド、およびトランスチレチンが挙げられる。
【0061】
一態様において、プロテオスタシスの機能障害に関連する疾患は、機能獲得障害である。用語「機能獲得障害、「機能獲得疾患」、「毒性機能獲得障害」および「毒性機能獲得疾患」は交換可能に使用される。機能獲得障害は、凝集関連タンパク質毒性の増加を特徴とする疾患である。これらの疾患において、凝集は、細胞の内側および/または外側のクリアランスを越える。機能獲得疾患としては、限定されないが、ポリグルタミンの凝集に関連する神経変性疾患、レヴィー小体疾患、筋萎縮性側索硬化症、トランスチレチン関連凝集疾患、アルツハイマー病、II型糖尿病、肝臓疾患およびプリオン疾患が挙げられる。ポリグルタミンの凝集に関連する神経変性疾患としては限定されないが、ハンチントン病、室頂核脳幹および淡蒼球ルイ体萎縮症、いくつかの種類の脊椎小脳性運動失調、ならびに脊椎および延髄筋萎縮症が挙げられる。アルツハイマー病は2種類の凝集物:Aβペプチドの細胞外凝集物とタウタンパク質に関連する微小管の細胞内凝集物の形成を特徴とする。トランスチレチン関連凝集疾患としては、例えば、老年性全身アミロイドーシスおよび家族性アミロイド神経障害が挙げられる。レヴィー小体疾患は、α-シヌクレインタンパク質の凝集を特徴として、例えばパーキンソン病が挙げられる。プリオン疾患(感染性海綿状脳症またはTSEとしても知られる)は、プリオンタンパク質の凝集と特徴とする。例示的なヒトプリオン疾患は、クロイツフェルト・ヤコブ病(CJD)、変異型クロイツフェルト・ヤコブ病、ゲルストマン-Straussler-Scheinker症候群、致死性家族性不眠症およびクールーである。
【0062】
さらなる態様において、タンパク質ホメオスタシスの機能障害に関連のある疾患は、機能消失障害である。用語「機能消失疾患」および「機能消失障害」は交換可能に使用される。機能消失疾患は、タンパク質の過剰な分解を引き起こす、タンパク質の高率の悪いフォールディングを特徴とする疾患のグループである。機能消失疾患としては、例えば、嚢胞線維症、気腫、およびリソソーム貯蔵疾患が挙げられる。嚢胞線維症では、変異または欠損酵素が嚢胞線維症膜貫通伝導制御因子(CFTR)である。このタンパク質の最も一般的な変異の一つは、3ヌクレオチド欠失(Δ)であるΔF508であり、タンパク質の508番目(508)の位置でアミノ酸フェニルアラニン(F)の欠失を生じる。リソソーム貯蔵疾患は、特異的なリソソーム酵素の欠失を特徴とする疾患の群であり、これは種々の組織で起こり、欠損酵素により、正常に分解されるタンパク質の蓄積が生じる。リソソーム酵素の欠失は、リソソームヒドロラーゼまたはリソソーム輸送に関連のあるタンパク質に生じ得る。リソソーム貯蔵疾患としては、限定されないが、アスパルチルグルコサミン尿症、ファブリー病、バッテン病、シスチン症、ファーバー、フコイドーシス、ガラクトシドシアリドーシス(Galactasidosialidosis)、ゴーシェ病(1型、2型および3型を含む)、Gm1ガングリオシドーシス、ハンター病、フルラー-シャイエ病、クラッベ病、a-マンノシドーシス、B-マンノシドーシス、マロトー-ラミー病、異染性白質萎縮症、モルキオA症候群、モルキオB症候群、ムコリピドーシスII、ムコリピドーシスIII、ニーマン-ピック病(A型、B型およびC型を含む)、ポンペ病、サンドホフ病、サンフィリポ病(A型、B型、C型およびD型を含む)、シンドラー病、シンドラー-カンザキ病、シアリドーシス、Sly症候群、テイ-サックス病およびウォルマン病が挙げられる。
【0063】
別の態様において、プロテオスタシスおよび/または熱ショックタンパク質の機能障害に関連のある疾患は、心臓血管疾患である。心臓血管疾患としては限定されないが、冠状動脈疾患、心筋梗塞、脳卒中、再狭窄および動脈硬化症が挙げられる。また、プロテオスタシスの機能障害に関連する状態としては、虚血/再かん流傷害、心筋虚血、安定性狭心症、不安定性狭心症、脳卒中、虚血性心臓疾患および脳虚血などの虚血状態が挙げられる。
【0064】
また、本発明は、薬理学的シャペロンの投与と併用して、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤の治療有効量を投与する工程を含む、プロテオスタシスの機能障害と関連している状態に苦しむ患者の治療方法を包含する。薬理学的シャペロンまたは速度論的安定剤は、既存の定常状態レベルのフォールディングされた変異タンパク質に結合し、フォールディングを安定化させることによりフォールディングの平衡を化学的に促進させる化合物をいう[Bouvier、Chem Biol 14:241-242、2007;Fan et al、Nat Med 5:112-115、1999;Sawkar et al、Proc Natl Acad Sci USA 99:15428-15433、2002;Johnson and Kelly、Accounts of Chemical Research 38:911-921、2005]。薬理学的シャペロンは、サーチュイン活性化化合物と併用してプロテオスタシスの機能障害と関連している状態に苦しむ患者を治療するのに充分な量で投与される。例示的な薬理学的シャペロンは、米国特許出願公開第20080056994号、同第20080009516号、同第20070281975号、同第20050130972号、同第20050137223号、同第20050203019号、同第20060264467号および同第20060287358号に記載されており、その内容は参照により本明細書に援用される。
【0065】
別の態様において、本発明は、機構的が異なる(mechanistically distinct)プロテオスタシス調節因子の投与と併用したHSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤により、プロテオスタシスの機能障害と関連している状態に苦しむ患者の治療方法である。用語「プロテオスタシス調節因子」は、細胞内タンパク質のホメオスタシスを増強する小分子、siRNAおよび生物製剤(例えば、タンパク質など)をいう。例えば、プロテオスタシス調節因子は、タンパク質の合成、フォールディング、輸送および分解の経路に影響を及ぼす薬剤であり得る。プロテオスタシス調節因子は、HSRシグナル伝達活性を刺激する薬理学的薬剤を包含する。プロテオスタシス調節因子は、シグナル伝達経路、例えば限定されないが、熱ショック応答またはアンフォールディングタンパク質の応答または両方を操作し、プロテオスタシスネットワーク成分の転写および翻訳をもたらすことにより機能を果たす。プロテオスタシス調節因子は、タンパク質(例えば、変異タンパク質)のフォールディング、輸送および機能を増強し得る。また、プロテオスタシス調節因子は、タンパク質シャペロンの転写または翻訳を上方調節すること、またはタンパク質シャペロンの分解を阻害することによりタンパク質シャペロンを調節し得る。プロテオスタシス調節因子は、しばしば、シャペロンレベルとフォールディング酵素レベルと、一部フォールディングされたコンホメーションアンサンブルに結合する巨大分子との協調的増大により、フォールディングの生物学に影響し得、したがって、より天然の構造を有する中間体へと進行させ、最終的に、輸送のためにフォールディングされた変異タンパク質の濃度を増大させる。一局面において、プロテオスタシス調節因子は、変異タンパク質のホメオスタシスを増強し得るが、変異タンパク質には結合しない点でプロテオスタシス調節因子はシャペロンと異なる。また、プロテオスタシス調節因子は、凝集経路または脱凝集酵素(disaggregase)活性を上方調節し得る。機構的が異なるプロテオスタシス調節因子は、HSF1のアセチル化の調節以外の機構によって細胞内プロテオスタシスを増強するプロテオスタシス調節因子である。例示的なプロテオスタシス調節因子はセラストロール、MG-132およびL型Ca2+チャネル遮断薬(例えば、ジリチアゼムおよびベラパミル)である。用語「セラストロール」は、セラストロールおよびその誘導体またはアナログをいい、限定されないが、その内容が参照により本明細書に明白に援用されるWesterheide et al.,J Biol Chem、2004. 279(53):p. 56053-60に記載されたセラストロール誘導体が挙げられる。セラストロール誘導体としては、例えば、セラストロールメチルエステル、二酢酸ジヒドロセラストロール、セラストロールブチルエーテル、ジヒドロセラストロール、セラストロール ベンジルエステル、プリメステロール、二酢酸プリメステロールおよびセラストロール三酢酸塩が挙げられる。
【0066】
特定の局面において、本発明は、機構的が異なるプロテオスタシス調節因子の投与と併用して、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤を投与する工程を含み、プロテオスタシス調節因子が熱ショック応答活性化因子である、プロテオスタシスの機能障害と関連している状態に苦しむ患者の治療方法である。熱ショック応答活性化因子は、例えば、熱ショック転写因子1(HSF1)を直接もしくは間接的に活性化させること、Hsp70の転写もしくはタンパク質発現を増大させること、Hsp90を阻害すること、および/またはシャペロンmRNAもしくはタンパク質発現を活性化させる、もしくは増大させることにより、熱ショック応答を間接的または直接活性化するか、または増大させる薬剤である(Westerheide et al.,J Biol Chem、2004. 279(53):pp.56053-60、その内容は、参照により本明細書に明白に援用される)。熱ショック応答活性化因子は、基底熱ショック応答を間接的または直接増大させる薬剤を包含することを理解されたい。用語「熱ショック応答活性化因子」、「熱ショック活性化因子」、「熱ショック応答誘導薬」および「熱ショック誘導薬」は本明細書において互換的に使用される。熱ショック応答活性化因子の非限定的な例は、セラストロール、非ステロイド系抗炎症薬、アンサマイシン、ゲルデナマイシン、ラジシオール、グルクロン酸、およびトリブチリン(tributylin)である。また、熱ショック応答活性化因子は、例えば、米国特許公開公報第20070259820号、同第20070207992号、同第20070179087号、同第20060148767号に記載されており、これらのそれぞれの内容は、参照により本明細書に明白に援用される。いくつかの態様において、熱ショック応答活性化因子は、小分子熱ショック応答活性化因子である。HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤および熱ショック応答活性化因子は、それぞれ、熱ショック応答活性化因子とアセチル化を阻害する薬剤を合わせた用量が一緒になって熱ショック応答を増大させるのに充分な用量で投与され得る。熱ショック活性化因子とHSF1のアセチル化を阻害する薬剤の併用により、該薬剤のいずれかを単独で投与する場合よりも大きい熱ショック応答の増大がもたらされる。いくつかの態様において、投与される熱ショック活性化因子の用量は、単独では熱ショック応答を誘導するに充分でないが、該用量は、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤と組み合わせて熱ショック応答を活性化するには充分である。本発明のいくつかの局面において、熱ショック活性化因子とHSF1のDNA結合ドメインのアセチル化を阻害する薬剤の組合せは、熱ショック応答の誘導に対して相加効果より大きい効果を有する。他の局面において、熱ショック応答の増大は、Hsp70の転写またはタンパク質発現の増大を測定することにより測定される。熱ショック応答の増大の例示的な測定方法は、Westerheide et al.(2004)Celastrols as inducers of 熱 shock reipose and cytoprotection. J Biol Chem、2004. 279(53):pp. 56053-60に記載されており、その内容は、参照により本明細書に援用される。さらなる局面において、熱ショック応答活性化因子とHSF1のアセチル化を阻害する薬剤の組合せの投与後の熱ショック応答の増大は、該薬剤の組合せの非存在下での熱ショック応答の少なくとも約2倍、少なくとも約3 倍、少なくとも約5倍、少なくとも約10倍、少なくとも約15倍、または少なくとも約20倍大きいHsp70の転写またはタンパク質発現の増大を包含する。
【0067】
実施例2に示すように、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤も投与する場合、熱ショック応答を誘導するのに、熱ショック活性化因子は、より少ない用量で投与され得ることがわかった。熱ショック応答活性化因子の用量またはEC50が低減されることは、熱ショック応答を誘導するために必要とされる熱ショック応答活性化因子の用量が毒性をもたらす状況があるため、有利であり得る。したがって、いくつかの態様において、本発明は、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤と併用して、熱ショック応答活性化因子を投与する工程を含み、熱ショック応答活性化因子とHSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤を合わせた用量が一緒になって、熱ショック応答を活性化または増大するのに充分である、熱ショック応答を活性化または増大する方法に関する。他の態様において、本発明は、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤と、熱ショック応答活性化因子を投与する工程を含む、熱ショック応答の誘導を必要とする患者において熱ショック応答を誘導するのに有効な熱ショック応答活性化因子の用量を低減する方法である。HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤は、両方の薬剤が同じ組成物の一部である場合、および/または両方の薬剤を同時に投与する場合、および/または両方の薬剤を異なる時点もしくは逐次投与する場合、別の薬剤と併用して投与してもよいことは理解されよう。一態様において、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤は、熱ショック応答活性化因子の前に投与される。
【0068】
特定の他の態様において、本発明は、熱ショック応答活性化因子とHSF1のアセチル化を阻害する薬剤を含む医薬組成物に関する。他の態様において、医薬組成物は、熱ショック活性化因子と、HAT阻害薬、単離サーチュインおよびサーチュイン活性化剤からなる群より選択されるHSF1のアセチル化を阻害する薬剤とを含む。他の態様において、本発明は、熱ショック応答活性化因子とSIRT1活性化剤を含む医薬組成物である。さらなる態様において、本発明は、熱ショック応答活性化因子とHAT阻害薬を含む医薬組成物である。
【0069】
別の局面において、本発明は、治療を必要とする患者に、HSF1のDNA結合ドメインのアセチル化を促進させる薬剤の治療有効量を投与する工程を含む、治療を必要とする患者において熱ショックタンパク質の発現の増大と関連している状態の治療方法である。一態様において、HSF1のアセチル化を促進する薬剤はサーチュイン阻害剤である。別の態様において、HSF1のアセチル化を促進する薬剤はHAT活性化因子である。サーチュイン阻害剤は上記で説明した。別の態様において、熱ショックタンパク質の発現の増大と関連している状態は癌または腫瘍である。また別の態様において、熱ショックタンパク質の発現の増大と関連している状態はウイルス感染である。HSF1のアセチル化を促進させる薬剤は、医薬用担体または賦形剤を含む医薬組成物にて投与され得る。
【0070】
熱ショックタンパク質の発現の増大と関連している状態は癌または腫瘍であり得る。本発明の方法に従って治療され得る癌としては、限定されないが、乳癌、結腸癌、膵臓癌、前立腺癌、肺癌、卵巣癌、頚部癌、多発性骨髄腫、基底細胞癌、神経芽腫、血液の癌、横紋筋肉腫、肝臓癌、皮膚癌、白血病、基底細胞癌、膀胱癌、子宮内膜癌、グリオーマ、リンパ種、および胃腸の癌が挙げられる。
【0071】
別の態様において、本発明は、化学療法剤の投与と併用して、HSF1のアセチル化を促進させる薬剤を投与する工程を含む、癌または腫瘍の治療方法である。使用され得る化学療法剤としては、限定されないが、シクロホスファミド(Cytoxan(登録商標))などのアルキル化剤;ブスルファン、インプロスルファンおよびピポスルファンなどのアルキルスルホン酸塩;ベンゾドパ、カルボクオン、メツレドパ、およびウレドパなどのアジリジン;エチレンイミンおよびメチラメラミン、例えば、アルトレタンミン、トリエチレンメラミン、トリエチレン(etylene)ホスホラミド、トリエチレンチオホスファオ(phosphao)ラミドおよびトリメチロロメラミン;クロラムブシル、クロルナファジン、コロホスファミド、エストラムスチン、イホスファミド、メクロレタミン、塩酸メクロレタミンオキシド、メルファラン、ノベンビシン、フェンステリン、プレドニムスチン、トロホスファミド、ウラシルマスタードなどのナイトロジェンマスタード;カルムスチン、クロロゾトシン、フォテムスチン、ロムスチン、ニムスチン、ラニムスチンなどのニトロソ尿素;アクラシノマイシン、アクチノマイシン、アウトラマイシン、アザセリン、ブレオマイシン、カクチノマイシン、カリケアマイシン、カラビシン、カミノマイシン、カルジノフィリン、クロモマイシン、ダクチノマイシン、ダウノルビシン、デトルビシン、6-ジアゾ-5-オキソ-L-ノルロイシン、ドキソルビシン、エピルビシン、エソルビシン、イダルビシン、マルセロマイシン、マイトマイシン、ミコフェノール酸、ノガラマイシン、オリボマイシン、ペプロマイシン、ポトフィロマイシン、ピューロマイシン、ケラマイシン、ロドルビシン、ストレプトニグリン、ストレプトゾシン、ツベルシジン、ウベニメックス、ジノスタチン、ゾルビシンなどの抗生物質;メトトレキサートおよび5-フルオロウラシル(5-FU)などの代謝拮抗物質;デノプテリン、メトトレキサート、プテロプテリン、トリメトレキサートなどの葉酸アナログ;フルダラビン、6-メルカプトプリン、チアミプリン、チオグアニンなどのプリンアナログ;アンシタビン、アザシチジン、6-アザウリジン、カルモフル、シタラビン、ジデオキシウリジン、ドキシフルウリジン、エノシタビン、フロクスウリジンなどのピリミジンアナログ;カルステロン、プロピオン酸ドロモスタノロン、エピチオスタノール、メピチオスタン、テストラクトンなどのアンドロゲン;アミノグルテチミド、マイトタン、トリロスタンなどの抗アドレナリン薬;フロリン酸などの葉酸補給薬;アセグラトン;アルドホスファミドグリコシド;アミノレブリン酸;アムサクリン;ベストラブシル;ビサントレン;エダトラキサート;デホファミン;デメコルチン;ジアジクオン;エルフオルニチン;酢酸エリプチニウム;エトグルシド;硝酸ガリウム;ヒドロキシ尿素;レンチナン;ロニダミン;ミトグアゾン;ミトザントロン;モピダモール;ニトラクリン;ペントスタチン;フェナメト;ピラルビシン;ポドフィリン酸;2-エチルヒドラジド;プロカルバジン;PSK(登録商標);ラゾキサン;シゾフィラン;スピロゲルマニウム;テヌアゾン酸;トリアジクオン;2,2’,2”-トリクロロトリエチルアミン;ウレタン;ビンデシン;ダカルバジン;マンノムスチン;ミトブロニトール;ミトラクトール;ピポブロマン;ガシトシン;アラビノシド(「Ara-C」);シクロホスファミド;チオテパ;タキサン、例えば、パクリタキセル(Taxol(登録商標)、Bristol-Myers Squibb Oncology、Princeton、N.J.)およびドセタキセル(Taxotere(登録商標);Aventis Antony、France);クロラムブシル;ゲムシタビン;6-チオグアニン;メルカプトプリン;メトトレキサート;シスプラチンおよびカルボプラチンなどの白金アナログ;ビンブラスチン;白金;エトポシド(VP-16);イホスファミド;マイトマイシンC;ミトザントロン;ビンクリスチン;ビノレルビン;ナベルビン;ノバントロン;テニポシド;ダウノマイシン;アミノプテリン;キセロダ;イバンドロネート;CPT-11;トポイソメラーゼ阻害薬RFS 2000;ジフルオロメチルオルニチン(DMFO);レチノイン酸;エスペラマイシン;カペシタビン;ならびに上記のもののいずれかの薬学的に許容され得る塩、酸または誘導体が挙げられる。また、この定義には、腫瘍に対するホルモン作用を調節または阻害する作用をする抗ホルモン薬、例えば、タモキシフェン、ラロキシフェン、アロマターゼ阻害薬4(5)-イミダゾール、4-ヒドロキシタモキシフェン、トリオキシフェン、ケオキシフェン、LY 117018、オナプリストン、およびトレミフェン(ファレストン)などの抗エストロゲン;ならびにフルタミド、ニルタミド、ビカルタミド、ロイプロリド、およびゴセレリンなどの抗アンドロゲン;ならびに上記のもののいずれかの薬学的に許容され得る塩、酸または誘導体も含まれる。
【0072】
さらなる態様において、本発明は、放射線療法と併用してHSF1のアセチル化を促進する工程を含む、癌または腫瘍の治療方法である。
【0073】
別の態様において、熱ショックタンパク質の発現の増大と関連している状態はウイルス感染である。さらなる態様において、ウイルス感染は、腫瘍ウイルスおよびRNAウイルスから選択されるウイルスによって引き起こされるものである。例示的な腫瘍ウイルスは、ヘルペスウイルス、パピローマウイルス、ポリオーマウイルスおよびHTLV-1である[McCance et al. Human Tumor Viruses、1998、American Society for Microbiology]。ヘルペスウイルスとしては、限定されないが、EBV(HHV-4)、HHV-6およびHHV-8が挙げられる。パピローマウイルス としては、限定されないが、HPV-1、-2、-4、-5、-6、-8、-6、-11、-16、-18、-31、-33、-35、-45、-51、-52、-58および-58が挙げられる。
【0074】
RNAウイルスとしては、例えば、アレナウイルス科、ブンヤウイルス科、カルシウイルス科、コロナウイルス科、フィロウイルス科、フラウイルス科、オルトミクソウイルス科、パラミクソウイルス科、ピコルナウイルス科、レオウイルス科、ラブドウイルス科、レトロウイルス科、またはトガウイルス科が挙げられる。例示的なRNAウイルスとしては、限定されないが、SARS関連コロナウイルス、腸疾患および呼吸器疾患に関連しているヒトトロウイルスなどのヒトコロナウイルス;ノーウォークウイルス、黄熱病ウイルス、西ナイルウイルス、C型肝炎ウイルス、デング熱ウイルス、ポリオウイルス、感冒ウイルス、A型肝炎ウイルス、E型肝炎、ロタウイルス、ボルナ病ウイルス;ハンタウイルス、カリフォルニア脳炎ウイルス、日本脳炎ウイルス、ラクロスウイルス、リフトバレー熱ウイルス、ブンヤウイルス、アルボウイルス、エボラウイルスおよびマルブルグウイルスなどのブンヤウイルス;インフルエンザウイルスA型、インフルエンザウイルスB型、インフルエンザウイルスC型、流行性耳下腺炎ウイルス、はしかウイルス、亜急性硬化性汎脳炎(SSPE)ウイルスならびに呼吸器合胞体ウイルス(RSV)が挙げられる。
【0075】
さらなる態様において、本発明は、抗ウイルス薬の投与と併用して、HSF1のアセチル化を促進させる薬剤を投与する工程を含む、ウイルス感染に苦しむ患者の治療方法である。
【0076】
本発明のまた別の局面において、熱ショックタンパク質の発現の増大と関連している状態は、炎症状態および/または自己免疫疾患である。
【0077】
本発明の治療方法に従って使用される薬理学的薬剤または医薬組成物の形態は、意図される投与様式および治療適用に依存する。また、組成物は、所望される製剤に応じて、動物またはヒトへの投与のための医薬組成物を製剤化するために一般に使用されるビヒクルと定義される薬学的に許容され得る無毒性の担体または希釈剤を含み得る。希釈剤は、薬理学的薬剤または組成物の生物学的活性に影響を及ぼさないように選択される。かかる希釈剤の例は、蒸留水、リン酸緩衝生理食塩水、リンゲル溶液、デキストロース溶液、およびハンクス溶液である。また、医薬組成物または製剤は、他の担体、補助剤または無毒性、非治療性、非免疫原性安定剤なども含んでいてもよい。また、医薬組成物は、タンパク質、キトサンなどの多糖、ポリ乳酸、ポリグリコール酸およびコポリマー(例えば、ラテックス官能化SepharoseTM、アガロース、セルロースなど)、ポリマーアミノ酸、アミノ酸コポリマー、ならびに脂質凝集物(油滴またはリポソームなど)などの大型のゆっくり代謝される巨大分子を含んでいてもよい。
【0078】
非経口投与では、医薬組成物または薬理学的薬剤は、水油、生理食塩水、グリセロール、またはエタノールなどの滅菌液であり得る医薬用担体を有する生理学的に許容され得る希釈剤中の該物質の注射用投薬量の溶液または懸濁液として投与され得る。さらに、湿潤剤または乳化剤、界面活性剤、pH緩衝物質などの補助物質が組成物中に存在していてもよい。医薬組成物の他の成分は、石油、動物、植物または合成起源のもの、例えば、ピーナッツ油、ダイズ油、および鉱物油である。一般に、プロピレングリコールまたはポリエチレングリコールなどのグリコールは、特に注射用溶液に好ましい液体担体である。
【0079】
組成物は、液状溶液または懸濁液のいずれかとしての注射用製剤として調製され得る;注射前の液体ビヒクル中での溶液もしくは懸濁液に適した固形形態も調製され得る。また、該調製物を乳化してもよく、リポソームもしくはポリラクチド、ポリグリコリドなどの微粒子、または上記のようなアジュバント効果の増強のためのコポリマー内にカプセル封入してもよい。Langer、Science 249:1527、1990およびHanes、Advanced Drug Delivery Reviews 28:97-119、1997。本明細書に記載の組成物および薬理学的薬剤は、デポー注射または活性成分の持続的もしくはパルス的放出を可能にするような様式で製剤化され得る埋め込み調製物の形態で投与され得る。
【0080】
他の投与様式に適したさらなる製剤としては、経口、鼻腔内および肺経由製剤、坐剤、または経皮適用が挙げられる。
【0081】
坐剤用では、結合剤および担体として、例えば、ポリアルキレングリコールまたはトリグリセリドが挙げられる;かかる坐剤は、0.5%〜10%、好ましくは1%〜2%の範囲の活性成分を含む混合物から形成され得る。経口製剤は、医薬等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、および炭酸マグネシウムなどの賦形剤を含む。局所適用により、経皮または皮内送達がもたらされ得る。経皮送達は、皮膚パッチを用いて、またはトランスフェロソーム(transferosome)を用いて達成され得る[Paul et al.,Eur. J. Immunol. 25:3521-24、1995;Cevc et al.,Biochem. Biophys. Acta 1368:201-15、1998]。
【0082】
別の態様において、本発明は、
試験薬剤を細胞に投与する工程;および
HSF1のDNA結合ドメイン内のリジン残基のアセチル化をモニタリングする工程
を含み、
試験薬剤の非存在下のものと比べたときのDNA結合ドメインのアセチル化の変化は、試験薬剤が細胞においてHSF1活性を調節することを示す、
細胞においてHSF1活性を調節する薬剤を同定する方法である。
【0083】
試験薬剤は、広範な種々の供給源、例えば、合成または天然化合物のライブラリーから得られ得る。例えば、広範な種々の有機化合物および他の薬剤のランダムおよび指向的合成のための数多くの手段が利用可能である。細菌、真菌、植物および動物の抽出物の形態の天然化合物のライブラリーもまた利用可能であり、あるいは容易に作製できる。さらに、天然または合成により作製されたライブラリーおよび化合物は、従来の化学的、物理的および生化学的手段によって容易に修飾され、コンビナトリアルライブラリーを作製するために使用され得る。また、薬理学的薬剤が、構造的アナログを作製するために、アシル化、アルキル化、エステル化、アミド化などの指向的またはランダムな化学的修飾に供され得る。試験薬剤は、数多くの化学的種類、典型的には合成、半合成または天然の無機または有機分子を包含する。試験薬剤としては、合成または天然化合物の大きなライブラリーに見られるものが挙げられる。
【0084】
一態様において、試験薬剤はDNA結合ドメインのアセチル化を阻害する。別の態様において、試験薬剤はDNA結合ドメインのアセチル化を増大させる。さらなる態様において、DNA結合ドメイン内のリジンのアセチル化がモニタリングされる。例えば、非ヒトHSF1内のHSF1 K80または対応する保存アミノ酸がモニタリングされ得る。さらなる態様において、細胞はヒト細胞である。
【0085】
また別の態様において、本発明は、
a. 試験薬剤を細胞または細胞溶解物に投与する工程;
b. サーチュイン基質のアセチル化を測定する工程;
c. 工程(b)のサーチュイン基質のアセチル化を、試験薬剤の非存在下でのサーチュイン基質のアセチル化と比較する工程
を含み、
試験薬剤の投与後のサーチュイン基質のアセチル化の増大は、試験薬剤がHSF1活性を阻害することを示し、試験薬剤の投与後のサーチュイン基質のアセチル化の減少は、試験薬剤がHSF1活性を増大させることを示す、
HSF1活性を調節する薬剤を同定する方法である。サーチュイン基質は、サーチュインによって脱アセチル化され得る因子(タンパク質またはペプチドなど)である。一態様において、サーチュイン基質はHSF1である。別の態様において、サーチュイン基質はHSF1以外の基質である。サーチュイン基質のアセチル化は、例えば、細胞ホモジネートにおいて測定され得る。一態様において、試験薬剤はサーチュイン活性を阻害する。別の態様において、試験薬剤はサーチュイン活性を活性化する。さらなる態様において、サーチュインはSIRT1である。また別の態様において、サーチュインはヒトSIRT1である。別の態様において、細胞はヒト細胞である。サーチュインデアセチラーゼ活性を測定するため、およびサーチュイン基質のアセチル化を測定するためのアッセイは、例えば米国特許公開公報第20080249103号に記載されており、その内容は参照により本明細書に援用される。
【0086】
また別の態様において、本発明は、
a. 試験薬剤を細胞または細胞溶解物に投与する工程;
b. HAT基質のアセチル化を測定する工程;
c. 工程(b)のHAT基質のアセチル化を、試験薬剤の非存在下でのHAT基質のアセチル化と比較する工程
を含み、
試験薬剤の投与後のHAT基質のアセチル化の増大は、試験薬剤がHSF1活性を阻害することを示し、試験薬剤の投与後のHAT基質のアセチル化の減少は、試験薬剤がHSF1活性を増大させることを示す、
HSF1活性を調節する薬剤を同定する方法である。HAT基質は、ヒストンアセチルトランスフェラーゼによってアセチル化され得る因子(タンパク質またはペプチドなど)である。いくつかの局面において、HAT基質はHSF1ではない。
【0087】
別の局面において、本発明は、本明細書に記載の方法によって同定されるHSF1活性を調節する薬剤である。
【0088】
本発明を、なんら限定を意図しない以下の実施例によって説明する。
【実施例】
【0089】
実施例
実施例1:デアセチラーゼSIRT1による熱ショック因子1のストレス誘導性調節
熱ショック因子1(HSF1)は、タンパク質のミスフォールディングと関連するタンパク質損傷性ストレスから細胞を保護するのに必須であり、インスリンシグナル伝達経路および加齢を調節する。ここに、本発明者らは、ヒトHSF1が、DNA結合活性を負に調節する重要な残基において誘導的にアセチル化されることを示す。デアセチラーゼおよび長寿因子SIRT1の活性化により、HSF1が、脱アセチル化DNA結合コンピテント状態に維持されることによって、Hsp70プロモーターへのHSF1結合が長期化された。逆に、SIRT1の下方調節によって、熱ショック応答(HSR)の減衰およびその対応プロモーターエレメントからのHSF1の放出が加速された。この結果は、寿命の調節におけるHSF1の要件の機構的根拠を示し、タンパク質のホメオスタシスおよびHSRにおけるSIRT1の役割を確立する。
【0090】
多様な環境的および生理学的ストレスによるHSF1の一過的活性化は、不活性HSF1単量体の構成的発現、該単量体のDNA結合コンピテント三量体への変換、セリン残基でのHSF1のリン酸化の増大、転写の増強、ならびにHSF1 DNA結合および転写活性の減衰を伴う多工程プロセスである(1)。HSF1は、分子シャペロンHsp70およびHsp90などのタンパク質のホメオスタシスを調節する多数の遺伝子の転写を活性化する。これらのシャペロンはHSF1と結合して、負のフィードバックループを開始させ、HSF1転写活性を阻害する(2)。しかしながら、HSF1は、その標的プロモーター部位から放出されず(3)、これは、HSF1サイクルを完結させるさらなる機構が存在するはずであることを示す。
【0091】
ストレス抵抗性および代謝状態は、タンパク質のホメオスタシスと寿命の増大に密接に関連している。C. elegansでは、インスリンシグナル伝達の低減の保護効果には、タンパク質のミスフォールディング損傷を防ぎ、長寿を促進するために、HSF1とFOXO転写因子DAF-16が必要とされる(4、5)。低カロリー摂取の有益な効果は、代謝制御下にあるNAD依存性デアセチラーゼであるサーチュインファミリー構成員Sir2によって媒介される(6)。哺乳動物Sir2ホモログSIRT1は、数ある細胞内保護経路のなかでも転写因子FOXO3を調節する(7)。したがって、本発明者らは、サーチュイン、具体的にはSIRT1がHSF1活性を調節し、それによりこれらの3つの長寿因子間に直接的な関連性を提供するかどうかを試験した。
【0092】
本発明者らは、HeLa細胞をサーチュイン阻害薬ニコチンアミドで処理し(8)、次いで、細胞を、HSRを誘導することが知られた種々のストレスに曝露した(9)。ニコチンアミド治療により、すべての主要な種類の熱ショック遺伝子(hsp70、hsp90、hsp40およびhsp27、図1A)由来のストレス誘導性mRNAのアバンダンスが減少し、これは、HSRの充分な誘導にサーチュインが必要とされることを示す。3つの核サーチュインのうち、SIRT1は、充分特徴付けられた標的を有する(10)。したがって、本発明者らは、SIRT1をHSRの調節のための候補として調査した。siRNAによりSIRT1が枯渇されると、6時間の熱ショック(HS)中に生成されるhsp70 mRNAの量は、対照siRNAでトランスフェクトされた細胞のものの4分の1であった(図1B)。
【0093】
SIRT1がhsp70プロモーターへのHSF1の漸増に影響を及ぼすかどうかを調べるため、本発明者らは、HSの前に対照またはSIRT1 siRNAでトランスフェクトされた細胞を用いてクロマチン免疫沈降(ChIP)アッセイを行なった(図1C)。対照siRNA処理細胞では、hsp70プロモーターへのHSF1の結合は、急速に起こるが、熱ショックの30分の時点で減衰し始め(11)、6時間にわたって徐々に減少する。しかしながら、SIRT1 siRNAトランスフェクト細胞では、この時間中にHSF1の約4分の1程度がプロモーターと結合した。この結果は、HSF1 DNA結合活性およびhsp70発現のインビボ調節因子としてのSIRT1の役割を裏付ける。
【0094】
HSF1がSIRT1の直接的な標的であるかどうかを調べるため、本発明者らは、HSF1のアセチル化状態を調べた。本発明者らは、293T細胞を、Flag-HSF1融合タンパク質およびp300をコードするベクターでトランスフェクトし、いくつかのHSR誘導薬に曝露した。免疫沈降させたHSF1をアセチル化リジンに結合する抗体を用いてウエスタンブロッティングによって解析した。アセチル化HSF1は、未処理細胞では検出されなかったが、種々のストレス状態に曝露した細胞には存在した(図2A)。p300またはCBPのいずれかの過剰発現では、HSF1のアセチル化がもたらされたが、pCAFではもたらされず(図5A)HS後、p300はhsp70プロモーターに漸増したため、HSF1アセチル化を調節する内因性アセチルトランスフェラーゼはp300/CBPであり得る(図5B)。また、SIRT1は、基底状態およびストレス状態の両方でhsp70プロモーターに結合する(図5C)。
【0095】
デアセチラーゼは、トリコスタチンAによって阻害されるI類およびII類HDACファミリー(12)ならびにニコチンアミドによって阻害されるNAD+依存性のIII類サーチュインファミリー(8)の3つのファミリーに分類される。トリコスタチンAは、HSF1の脱アセチル化に対して効果を有しなかったが、ニコチンアミドは、単独またはトリコスタチンAの存在下で脱アセチル化を阻害した(図2B)。SIRT1 WTの過剰発現は、HSF1アセチル化を阻害したが、障害されたNAD依存性デアセチラーゼ活性を有する点変異[SIRT1 H363Y(13)]はアセチル化を阻害せず(図2C)、これは、HSF1機能におけるSIRT1の役割を裏付ける。HSF1アセチル化は293TおよびCos7細胞において検出されたため、HSF1アセチル化は細胞型特異的ではなかった。HSF1アセチル化はp300の過剰発現によって増強されたが、p300過剰発現を必要としなかった(図2、AおよびD)。HSF1アセチル化の速度論はHSF1 活性化の速度論と適合しない。アセチル化は遅延し、HSF1活性およびDNA結合が減衰している期間中、持続する(11)。さらに、潜在的にリン酸化された10個のセリンがアラニンで置換されたHSF1は、アセチル化に対してコンピテントのままであり、HSF1のリン酸化はアセチル化のための必要条件でないことを示す(図6)。
【0096】
HSR後の時点でのHSF1アセチル化の持続およびSIRT1とHSF1の一緒の共免疫沈降(図2D)は、本発明者らに、SIRT1がHSF1活性の減衰において役割を有するかどうかを調べることを導いた。本発明者らは、HeLa細胞を、SIRT1活性の小分子誘導薬であるレスベラトロールで処理し(14)、HSF1 DNA結合活性をオリゴヌクレオチド系プルダウンアッセイにおいてアッセイした(15)(図2E)。HSおよびビヒクル単独で処理した細胞では、HSF1 DNA結合が2時間以内に誘導され、6時間後に減衰された。HSF1の一過性の活性化は、HSF1のストレス誘導性リン酸化状態を検出するSDS-PAGE上の移動度の変化によって反映された(16)。対照的に、レスベラトロール処理細胞におけるHSF1は、8時間の連続HSの後であっても、DNA結合コンピテントおよびリン酸化状態の状態が持続した。SIRT1を過剰発現する細胞では、ChIP実験によって測定したとき、HSF1 DNA結合はが増強され、減衰が抑制された(図2F)。この結果は、SIRT1のアバンダンスおよび活性の変化によりHSRの減衰が調節されることを示唆する。
【0097】
アセチル化がHSF1 DNA結合を調節する機構を解明するため、本発明者らは、293T細胞から精製したFlag-HSF1由来のペプチドの質量分析によってHSF1上のアセチル化部位を特定した。HSF1内の少なくとも9つのリジンが、ストレスに応答してアセチル化され(図7)、DNA結合ドメイン内に位置するそのK80は、酵母HSFの対応するリジンの変異は機能低下表現型を引き起こすため(17、18)、特に興味をそそられた。さらに、K. lactis HSFの結晶構造の解析により、
ヒトHSF1 K80に対応するリジンは、主要DNA結合ヘリックスと、柔軟で溶媒曝露ループと連結し、DNAのリン酸主鎖と水素結合を形成する短いドメイン内に位置することが示された(19)。HSF-HSE結晶構造の比較タンパク質モデリングでは、ヒトHSF1 K80はDNA主鎖と密に接触していることが示され(図8)、アセチル化によるリジンの正電荷の中和によりDNA結合が妨げられるはずであることを示す。
【0098】
したがって、本発明者らは、構成的アセチル化を模倣するためにK80をグルタミンで置換した。HSF1 WTおよびHSF1 K80Q発現構築物でトランスフェクトされたhsf1-/-線維芽細胞(20)由来の抽出物では、変異タンパク質は、電気泳動移動度シフトアッセイ(EMSA)において、DNAに結合しなかった(図3A)。しかしながら、K80Q変異体は、なお、DNA結合状態のホールマークであるHS誘導性三量体に合成された(図9)。また、K80での他のアミノ酸の置換(K80R、-A、-H、-N、および-T)により、欠陥性DNA結合がもたらされた(図3A、図10)。インビトロでは、組換え非アセチル化WT HSF1は、合成HSEに容易に結合したが、K80Q 変異タンパク質は結合しなかった(図3B)。本発明者らは、WTおよびK80変異体をhsf1-/-線維芽細胞に導入し、HSF1標的遺伝子のHS誘導性発現をqPCRによって解析した。WT HSF1はhsp mRNAの発現を誘導したが、HSF1 K80変異体は非機能性であった(図3C)。変異体は、HS時、核に局在したが、熱ショックヒト細胞において起こる核ストレス体内への再局在において障害された(図11〜12)(21)。残基80の非修飾リジン側鎖は、HSF1 HSE結合能力、核ストレス体内への再局在、および標的遺伝子の発現に必要と思われる。したがって、本発明者らは、HSF1 K80のアセチル化が、DNAからのHSF1三量体の放出の調節を引き起こし、したがって、HSRの減衰における調節工程を表すことを提案する(図13)。
【0099】
SIRT1によるHSRの調節の生物学的重要性を検証するため、本発明者らは、シャペロンの発現によって耐熱性の増大がもたらされるストレス抵抗性のアッセイを使用した(22)。293T細胞をSIRT1でトランスフェクトし、またはトランスフェクトせず、45℃のHSに20または30分間曝露し、24時間回復させ、細胞死について解析した。予測どおり、45℃のHSにより細胞死がもたらされ、これは、処理時間とともに増大した(図4A)。両方の時間点で、SIRT1を過剰発現する細胞は、約3分の1の細胞が細胞死を受けた(図4A)。SIRT1における加齢調節の変化がHSF1活性およびHSRに影響を及ぼすかどうかを調べるため、本発明者らは、加齢プロセスにおける分子的変化に関する研究に広く使用されているヒトWI-38線維芽細胞を使用した。初期および後期の継代数を比較すると、本発明者らは、加齢により、SIRT1のアバンダンスの低下と相関しているHSRの減少およびHSF1 DNA結合活性の活性化の低下がもたらされることを見い出した(図4B)。
【0100】
SIRT1がHSF1を調節するという所見は、寿命の調節におけるHSF1の役割に関する先の観察(4、5)を補完する。HSF1は、細胞の栄養、ストレスおよび寿命が関連する調節ネットワークの中心であると思われる。多くのSIRT1調節転写因子、例えば、FOXO3、p53およびNF-κBは、細胞ストレス応答において重要な役割を有する(7、23、24)。このストレス調節ネットワークへのHSF1の添加は、SIRT1媒介性細胞保護におけるタンパク質のホメオスタシスの中心的役割を強調し(図14)、HSRの分子応答を代謝的要求と関連付け得る。細胞系および動物試験における観察の一致はHSRの加齢関連減少であり(22)、これは、少なくとも一部は、HSF1活性のSIRT1制御に起因し得る。生物レベルでは、本発明者らは、HSF1標的遺伝子の調節は食事と栄養に影響され得ると予測する。
【0101】
材料および方法
構築物、抗体および試薬
mHSF1-FlagおよびhHSF1-Mycは以前に記載された(25,26)。HSF1点変異は、
QuikChange部位特異的変異誘発(Stratagene)によって作出し、配列決定によって確認した。Myc-HSF1の10個のセリン変異体について、リン酸化セリン残基 S230、S303、S307、S314、S319、S320、S338、S363、S368およびS369(参考文献27および本発明者らの未公表データ)はすべて、アラニンに変異した。SIRT1 WTおよびH363Y変異体発現構築物は、Tony Kouzarides博士(ケンブリッジ大学)から、p300発現構築物はDavid Livingston博士(ハーバード大学)からご提供いただいた。この試験で使用した抗体は、α-Flag M2(Sigma)、α-AcK(Cell Signaling 9441)、α-HSF1(4)、α-SIR2(Upstate Biotech #07-131)、α-Myc(Clontech)およびα-p300(Santa Cruz)である。使用した化合物は、ニコチンアミド(Sigma)、トリコスタチンA(Upstate)、EGS(Sigma)、セラストロール(GAIA Chemical Corporation)、CdCl2(Sigma)およびMG132(Calbiochem)である。
【0102】
細胞培養物、トランスフェクションおよび処理条件
すべての細胞は、5%CO2加湿雰囲気中、37℃に維持した。K562細胞は、10%FCSおよび抗生物質(ペニシリンおよびストレプトマイシン)を添加したRPMI 1640培地中で培養した。293T、HeLa、Cos7およびWI-38細胞は、10%FCSおよび抗生物質を添加したDMEM中で培養した。hsf1-/-マウス胚線維芽細胞(29)は、10%FCS、10mM非必須アミノ酸、0.96μlの2-メルカプトエタノール/100 ml、および抗生物質を有するDMEM中で培養した。293T細胞をPolyfect(Qiagen)を用いて、製造業者の使用説明書に従ってトランスフェクトした。K562、Cos7およびhsf1-/-細胞は、Gene Pulserエレクトロポレーション装置(Bio-Rad)を用いてエレクトロポレーションによってトランスフェクトした。細胞を予備加温循環水浴中に42℃で1時間浸漬することによって熱ショックを誘導した。セラストロール処理は、5μMの濃度で1時間行ない、CdCl2処理は、50μMで6時間、MG132は20μMで6時間行なった。細胞に、トリコスタチンAは1mMで一晩、ニコチンアミドは5mMで一晩、レスベラトロールは50μMで40時間添加した。
【0103】
siRNAトランスフェクション
HeLa細胞を、Oligofectamine(Invitrogen)で、製造業者のプロトコルに従って、200nMのDharmacon SmartPool SIRT1 siRNAを用いてトランスフェクトした。24時間後、細胞を分割し、再度トランスフェクトした。2回目のトランスフェクションの48時間後、Trizolを用いてRNAを単離した。SIRT1のノックダウンを確認するため、プライマーを使用した。
【0104】
アセチル化アッセイ
293TまたはCos7細胞を、Flag-HSF1またはMyc-HSF1およびp300でトランスフェクトし、次いで、種々のストレスでの処理前に、ニコチンアミドで処理するか、または非処理とした。細胞溶解物を、Flag抗体またはMyc抗体を用いた免疫沈降に供し、アセチル化HSF1を、アセチル化リジンを認識する抗体を使用するウエスタンブロッティングによって検出した。
【0105】
クロマチン免疫沈降(ChIP)
ChIP反応は、本質的に先に記載のとおりに行なった(30)。HeLa細胞(3x107)から作製した試料を、10μlのα-HSF1(28)を用いて4℃で一晩免疫沈降させた。hsp70.1プロモーター(GenBank Acc# M11717)および近位HSEのためのプライマーを使用した。結果を1%投入量を用いて行なった反応に対して標準化した。
【0106】
293T細胞からのHSF1の精製
293T細胞を、CMV-p300とともに、またはなしで、mHSF1-Flagで製造業者のプロトコルに従ってPolyfect(Qiagen)に示されたとおりにトランスフェクトした。細胞をトリコスタチンAまたはニコチンアミドで表示のとおりに4時間処理した後、ストレス処理を行なった。次いで、細胞ペレットを収集し、RIPAバッファー中で溶解させた。mHSF1-Flagをα-Flag M2親和性ゲルビーズ(Sigma F2426)で免疫沈降させ、Flagペプチドで溶出させた。試料をSDS-PAGEによって分離した後、ウエスタンブロット解析を行なった。
【0107】
質量分析
精製mHSF1-Flagを、mHSF1-FlagおよびCMV-p300でトランスフェクトした293T細胞から上記のとおりに入手し、1μMトリコスタチンAおよび5mMニコチンアミドで18時間処理した後、HSまたはセラストロール処理した。免疫沈降したタンパク質をSDS-PAGEによって分離し、ゲルから切り出し、トリプシンで消化し、ナノエレクトロスプレー源を備えたハイブリッド四重極飛行時間型装置(QSTAR、Applied Biosystems、Foster City、CA)によるタンデム質量分析に供した。Mascot(Matrix Science、Boston、MA;version 1.9.05)およびX! Tandem(www.thegpm.org;version 2006.04.01.2)データベース検索アルゴリズムを使用し、IPIマウス配列 データベース(68,222エントリー;バージョン3.15)に対してMS/MSスペクトルを検索した。MascotおよびX! Tandemは、消化酵素トリプシン切断を1回失敗すると仮定した断片および前駆体イオン質量許容度0.3Daを用いて検索した。システインのカルバミドメチル化は固定修飾として含まれたが、メチオニン酸化、N末端タンパク質およびリジンアセチル化は、データベース検索における可変修飾として含まれた。ペプチド同定は、Peptide Prophetアルゴリズム(31)で測定し、MS/MSスペクトルの手作業検査によって確認さた場合、95.0%より高い確率で許容された。
【0108】
組換えタンパク質
マウスHis-HSF1を大腸菌株BL-21において先に記載のようにして(32)作製した。収集した細菌を冷溶解バッファー [50mM Tris-HCl(pH7.4)、140mM NaCl、10%グリセロール]中に再懸濁し、室温で15分間1mg/mlリゾチームで溶解した。1mM PMSF(フッ化フェニルメチルスルホニル)、1mM DTT(ジチオトレイトール)および無EDTAプロテアーゼ阻害薬カクテル(Roche)の添加後、溶解物を超音波処理し、Talon金属親和性樹脂(Clontech)とともに4℃で2時間インキュベートした。樹脂を0.5%Triton X-100を含む溶解バッファーで充分に洗浄し、50mM NaAc(pH5.0)、300mM NaCl、200mM イミダゾール中で溶出させた。溶出されたタンパク質をPBSに対して透析し、等量に希釈した。
【0109】
電気泳動の移動度シフトアッセイ(EMSA)
WI-38細胞およびトランスフェクトhsf1-/-細胞からのバッファーC抽出物(15μg)または組換えHSF1タンパク質を、ヒトhsp70プロモーター由来の近位HSEを提示する32P標識オリゴヌクレオチドとともにインキュベートした(33)。タンパク質-DNA複合体を4%非変性ポリアクリルアミドゲル上で解析した。
【0110】
オリゴヌクレオチドプルダウンアッセイ
アッセイは、わずかに修正をともなって、本質的に先に記載のとおりに行なった(32)。HeLa細胞をプロテアーゼ阻害薬を含む冷溶解バッファー [25mM HEPES(pH7.4)、100mM NaCl、5mM EDTA、20mM p-グリセロホスフェート、20mM p-ニトロ-フェニル-ホスフェート、0.5%Triton X-100、20mM 100μM オルトバナジウム酸ナトリウム]中に溶解した。細胞抽出物を、1μgのビオチン化オリゴヌクレオチド(Oligomer、Helsinki、Finland)とともにインキュベートし、室温で30分間タンパク質をオリゴヌクレオチドに結合させた。オリゴヌクレオチドをUltraLinkストレプトアビジンビーズ(Pierce)で4℃で1時間沈殿させた。結合した画分を洗浄バッファー [20mM Tris-HCl(pH7.5)、1mM EDTA、10%グリセロール、0.1%Triton X-100)で3回洗浄し、変性バッファーで溶出させ、ウエスタンブロッティングによって解析した。
【0111】
RT-PCR解析
HeLa細胞を収集し、製造業者の使用説明書に従ってTrizol試薬(Gibco-BRL、Gaithersburg、Maryland)を用いてRNAを生成させた。逆転写反応およびPCRを用いて、以下のタンパク質:hsp90、hsp70、hsp40、hsp 25、hsp27、18S、およびgapdhを作製した。
【0112】
定量的リアルタイムRT-PCR解析
トランスフェクトhsf1-/-細胞におけるhsp70およびhsp25遺伝子発現の解析を、本質的に記載のとおりに行なった(34)。簡単には、RNeasyキット(Qiagen)を用いてRNAを単離した。各試料について、1μgのRNAをRQ1 DNase(Promega)で処理し、Moloneyマウス白血病ウイルスRNase H(-)(Promega)を使用することによって逆転写した。ABsolute QPCR ROX Mix(Advanced Biotechnologies)を用いて、反応混合物を調製した。hsp mRNAの相対量を、gapdhに対して標準化した。すべての反応は、3つの生物学的リピート由来の試料を用いて三重に行なった。
【0113】
共焦点顕微鏡検査
カバースリップ上で培養中のHeLa細胞を制御温度に維持するか、または42℃で1時間熱ショックを与えた。細胞をPBSで洗浄し、同時に、0.5%Triton X-100を含有する3.7%パラホルムアルデヒド中に固定および透過させた。細胞をPBSで2回洗浄し、ブロック溶液(PBS-0.05%Tween 20中20%正常ヤギ血清)中で1時間インキュベートした。一次抗体[ウサギα-HSF1(11)、マウスα-Myc(Sigma)] を、5%BSA含有PBS-0.05%Tween 20中、1:500希釈で4℃で一晩使用した。PBS-0.05%Tween 20で3回洗浄後、二次ヤギα-マウスIgG(Rhodamine Red-X and Alexa Fluor 488、Molecular Probes)またはヤギα-ウサギIgG(Alexa Fluor 546、Molecular 488、Molecular Probes)を用いて一次抗体を検出した。すべての二次抗体は、5%BSA含有PBS-0.05%Tween 20中1:500希釈で、室温で1時間使用した。3回の洗浄後、DAPI(4’,6’-ジアミジノ-2-フェニルインドール;Vector Laboratories)を含むVectashieldスライドに細胞を載せ、Zeiss LSM510 META共焦点顕微鏡を用いて解析した。Adobe Photoshopソフトウェアを用いて画像をさらに処理した。
【0114】
架橋
細胞を37℃で維持するか、または42℃ で15分間熱ショックを与え、冷溶解バッファー [25mM HEPES(pH7.4)、100mM NaCl、5mM EDTA、0.5%Triton X-100、20mM β-グリセロホスフェート、20mM p-ニトロ-フェニル-ホスフェート、100μM オルトバナジウム酸ナトリウム、0.5mM フッ化フェニルメチルスルホニル、1mM ジチオトレイトール、プロテアーゼ阻害薬カクテル]中に溶解し、4℃で10分間、15,000×gで遠心分離した。100μgのタンパク質を2mM EGS [エチレングリコールビス(スクシンイミジルスクシネート)]と室温で15分間架橋させた。100mM グリシンの添加による架橋のクエンチ後、試料を変性バッファー中で煮沸し、5%SDS-PAGEゲル上、続いてウエスタンブロッティングによって解析した。
【0115】
熱ストレス抵抗性アッセイ
ベクター対照(Mock)またはSIRT1でトランスフェクトされた293T細胞を、45℃の熱ショックに対する抵抗性について比較した。細胞を45℃の水浴中に20または30分間浸漬し、次いで、37℃で回復させた。細胞を、24時間後のトリパンブルー取込みによって細胞死の割合についてアッセイした。
【0116】

【0117】
実施例2:SIRT1の活性化は、熱ショック応答の小分子活性化因子であるセラストロールと相乗作用する
熱ショック応答のいくつかの活性化因子が同定された[2]。小分子熱ショック活性化因子のいくつかは、タンパク質ミスフォールディング疾患における治療有用性について試験中である。熱ショック応答を活性化させるために、可能な限り低い用量で小分子を使用することは、治療用量と毒性用量の境界はしばしば狭いため有益であり得る。本発明の実験の組は、SIRT1の活性化が熱ショック応答の誘導薬と相乗作用し、それにより、熱または熱ショック応答の小分子活性化因子であるセラストロールが低レベルで応答の誘導を可能にすることを示す[3]。
【0118】
熱ショック応答の活性化によってタンパク質ミスフォールディング疾患を治療するための現行のストラテジーは、生物を高レベルの熱またはこの応答が誘導されるのに充分な量の小分子で治療することを伴う。いくつかの場合において、熱ショック応答を誘導するために必要とされる熱または化合物のレベルはまた、細胞損傷または毒性をもたらし得る。熱ショック応答を誘導するのに必要とされる熱ショックまたは小分子化合物のこの量は、SIRT1を同時に活性化することにより低下され得る。実施例1に記載のように、SIRT1は、熱ショックと組み合わせて、HSF1活性サイクル[I]の減衰工程を妨げることにより、熱ショック応答を活性化することがわかった。本発明の試験は、SIRT1の活性化が、熱ショックおよび熱ショック応答の小分子活性化因子と相乗作用して、応答の強固な活性化に必要とされる化合物の熱ショック温度またはEC50を低下させることを示す。
【0119】
図16および17に示されるように、SIRT1活性化因子レスベラトロール[4]は、レポーターアッセイによって測定されるように、熱ショックおよびセラストロールの両方と相乗作用して熱ショック応答を誘導する。熱ショック応答の活性化を検出するツールとして、ルシフェラーゼに融合したヒトhsp70.1プロモーターの-188から+150までの配列(図15、[4])を含むHeLa安定細胞株を作製した。この細胞株を使用し、SIRT1の活性化がレスベラトロールとともに、熱ショック応答の活性化の温度閾値を低下させ得るかどうかを調べた。図16に示されるように、42℃の熱ショックによりレポーターは15倍誘導されたが、41℃の熱ショックではレポーター5倍しか誘導されなかった。レスベラトロールでの細胞の前処理により、レポーター誘導は両方の温度で増大し、SIRT1の活性化が熱と相乗作用して熱ショック応答を活性化し、より低温での応答の誘導が可能となり得ることを示す。同様に、図17に示されるように、レスベラトロールは、5μMのEC50を有する公知の熱ショック応答誘導薬セラストロールと相乗作用し得ることがわかった[4]。1μMのセラストロール単独では、レポーターを2倍しか活性化しなかったが、レスベラトロールと1μM セラストロールの組合せでは、40倍を超える強固なレポーター活性化を誘導した。
【0120】
hsp70.1pr-luc HeLa安定細胞株は、hsp70.1プロモーター配列の-188から+150までを含み、近位熱ショックエレメント(HSE)を含む。この細胞株の作製は、Westerheide et al.(2004). Celastrols as inducers of the heat shock response and cytoprotection. J Biol Chem、2004. 279(53):p. 56053-60に記載されており、その内容は参照により本明細書に援用される。
【0121】
HeLa hsp70.1pr-luc細胞を白色96ウェルプレートにおいて、ウェル1つあたり15 X 103細胞の密度で100ulのDMEM培地+10%ウシ胎仔血清中にプレーティングした。細胞を37℃、5%CO2で20時間インキュベートした後、レスベラトロールで処理するか、または処理しなかった。レスベラトロールはEtOH中で希釈し、細胞に0.5uMで添加した。2時間後、次いで、水浴中に1時間浸漬することにより、細胞に、表示した温度の熱ショックを与えた。熱ショック処理後、37℃、5%CO2でプレートをインキュベータ内に戻し、18時間インキュベートした後、ルシフェラーゼアッセイを行なった。
【0122】
HeLa hsp70.1pr-luc細胞を白色96ウェルプレートにおいて、ウェル1つあたり15 X 103細胞の密度で100ulのDMEM培地+10%ウシ胎仔血清中にプレーティングした。細胞を37℃、5%CO2で20時間インキュベートした後、レスベラトロールで処理するか、または処理しなかった。レスベラトロールはEtOH中で希釈し、細胞に1.0uMで添加した。2時間後、次いで、細胞を、表示したとおりに1mM セラストロールで処理するか(セラストロールはDMSO中に溶解した)、または処理しなかった。セラストロール添加の18時間後、細胞をルシフェラーゼアッセイによってアッセイした。
【0123】
図の説明:
図16. SIRT1活性化因子レスベラトロールが熱ショックと相乗作用して、hsp70.1pr-lucレポーターを活性化させる。
図17. SIRT1活性化因子レスベラトロールは、セラストロールと相乗作用してhsp70.1pr-lucレポーターを活性化させる。
【0124】
【0125】
本発明を、その好ましい態様を参照して具体的に示し、記載したが、形態および詳細における種々の変形が、添付の特許請求の範囲によって包含される本発明の範囲から逸脱せずになされ得ることは、当業者によって理解されよう。
【0126】
【技術分野】
【0001】
関連出願
本願は、2008年12月8日に出願された米国特許仮出願第61/120,542号および2009年6月29日に出願された米国特許仮出願第61/221,315号の恩典を主張する。上記出願の全教示は、参照により本明細書に援用される。
【0002】
政府支援
本発明は、国立衛生研究所により授与された補助金番号AG026647号およびGM038109号のもと、政府支援によってなされたものである。本発明において、政府は一定の権利を有する。
【背景技術】
【0003】
発明の背景
細胞は、センサーおよび経路のネットワークを利用してタンパク質ホメオスタシスと称されるタンパク質の合成、フォールディング、輸送、凝集、および分解のバランスを正常に維持している[Sitia et al., Nature 426: 891-894, 2003; Ron et al., Nat Rev Mol Cell Biol 8: 519-529, 2007]。細胞におけるタンパク質ホメオスタシスの維持、つまりプロテオスタシスとは、プロテオームを形成する個々のタンパク質のコンホメーション、結合相互作用、局在および濃度の制御のことである。インビボでのタンパク質フォールディングは、多くの主類のシャペロンおよびフォールディング酵素を含むフォールディングポリペプチド鎖と巨大分子細胞成分の間の相互作用により達成され、凝集を最小化する[Wiseman et al., Cell 131: 809-821, 2007]。また、所定の区画で生成された有機および無機溶質が、フォールディングの物理的化学性質に影響を及ぼす疎水性作用などの非共有結合力によってポリペプチド鎖の救出(salvation)に影響するので、代謝酵素も細胞タンパク質フォールディング効率に影響を及ぼす。代謝経路で、特定のタンパク質の折りたたまれた状態に結合して安定化する小分子リガンドが生成され、フォールディング平衡を移動させることによりフォールディングが増強される[Fan et al., Nature Med., 5, 112 (Jan 1999); Hammarstrom et al., Science 299, 713 (2003)]。所定のタンパク質がある細胞型で折りたたまれるかどうかは、シャペロン、フォールディング酵素、代謝産物等の分布、濃度および細胞内局在に依存している[Wiseman et al.]。ヒトの機能消失疾患は典型的に、効果的な分解をもたらす所定のタンパク質の細胞フォールディングを含む所定のタンパク質中の突然変異により引き起こされる正常なタンパク質ホメオスタシスの乱れの結果であることがしばしばある[Cohen et al., Nature 426: 905-909, 2003]。ヒトの機能獲得疾患も同様に、タンパク質凝集をもたらすタンパク質ホメオスタシスの乱れの結果であることが良くある[Balch et al. (2008), Science 319: 916-919]。
【0004】
細胞レベルで、熱ショック応答は、種々の急性および慢性のストレス条件から細胞を保護する[Westerheide et al., J Biol. Chem. 280(39): 33097 (2005)]。熱ショック応答(HSR)は、環境的および生理学的ストレス因子に対する遺伝子応答であり、正常細胞の代謝の抑制、ならびにタンパク質ミスフォールディングおよび凝集の結果である細胞の損傷からの保護および回復に有用な分子シャペロン、プロテアーゼおよび他のタンパク質を発現する熱ショックタンパク質(HSP)遺伝子の迅速な誘導をもたらす[Westerheide et al.]。熱ショック応答は、転写因子である熱ショック因子-1(HSF-1)により媒介される。HSRは種々のストレス因子により生じる損傷から細胞を保護するが、大量のHSPの蓄積は、細胞の成長および分裂に悪影響を及ぼすことがある[Morimoto et al. (1998), Genes Dev. 12, 3788]。
【0005】
プロテオスタシスおよび熱ショック応答の両方における機能障害は、例えば、癌、神経変性疾患、代謝疾患、炎症性疾患および心臓血管疾患を含む種々の疾患に関連している。当該技術分野において、プロテオスタシス機能障害および/または熱ショックタンパク質の改変された誘導に関連する状態を治療するための治療アプローチの必要性が依然として存在している。
【発明の概要】
【0006】
発明の概要
本発明は、細胞HSF1活性がそのDNA結合ドメインのアセチル化状態により調節されるという発見に基づくものである。例えば、実施例1には、SIRT1(HSF1アセチル化を阻害する)の活性化は、HSF1を脱アセチル化状態に維持することでhsp70プロモーターへのHSF1 DNA結合を長引かせるが、SIRT1のダウンレギュレーションはHSF1活性の減衰を加速させることが示されている。
【0007】
一態様において、本発明は、HSF1のDNA結合ドメイン中のリジン残基のアセチル化を調節する工程を含む、細胞中の熱ショック転写因子1(HSF1)の活性を調節する方法である。別の態様において、HSF1の活性は、リジン残基のアセチル化を抑制することにより増強される。さらに別の態様において、HSF1の活性は、リジン残基のアセチル化を促進することにより低減される。リジン残基は、ヒトHSF1のリジン80(HSF1 K80)または非ヒトHSF1中の対応する保存されたアミノ酸であり得る。
【0008】
別の態様において、HSF1のアセチル化は、細胞に、HSF1のDNA結合ドメイン中のリジン残基のアセチル化を阻害する薬剤を投与することにより阻害される。いくつかの局面において、リジン残基のアセチル化を阻害する薬剤は、単離サーチュイン、サーチュイン活性化剤またはヒストンアセチルトランスフェラーゼ(HAT)阻害剤である。
【0009】
いくつかの局面において、HSF1のアセチル化は、細胞に、有効量の単離サーチュインまたはサーチュイン活性化剤を投与することにより阻害される。単離SIRT1またはSIRT1活性化剤が投与され得る。
【0010】
さらに別の態様において、HSF1のアセチル化は、細胞にHAT阻害剤を投与することにより阻害される。
【0011】
別の態様において、HSF1のアセチル化は、細胞にSIRT1阻害剤またはHAT活性化剤などのサーチュイン阻害剤を投与することにより促進される。
【0012】
さらなる態様において、本発明は、被験体においてHSF1のアセチル化を阻害する工程を含む、HSF1の活性の増加を必要とする被験体におけるHSF1の活性を増加する方法である。一態様において、被験体はヒトである。
【0013】
別の態様において、本発明は、被験体においてHSF1のアセチル化を促進する工程を含む、HSF1の活性の低減を必要とする被験体におけるHSF1の活性を低減する方法である。一態様において、被験体はヒトである。
【0014】
さらなる態様において、本発明は、タンパク質のホメオスタシスの機能障害に関連する状態に苦しむ患者に、治療有効量のHSF1のアセチル化を阻害する薬剤を投与する工程を含む、該患者を治療する方法である。一態様において、該状態は機能消失障害である。別の態様において、該状態は機能獲得障害である。別の態様において、HSF1のアセチル化を阻害する薬剤は、単離サーチュイン、サーチュイン活性化剤またはHAT阻害剤である。さらなる態様において、該薬剤は、単離SIRT1またはSIRT1活性化剤である。さらなる態様において、該薬剤は、HAT阻害剤である。
【0015】
別の態様において、本発明は、熱ショックタンパク質の発現の増加に関連する状態に苦しむ患者に、治療有効量のHSF1のアセチル化を促進する薬剤を投与する工程を含む、該患者を治療する方法である。一態様において、該状態は癌または腫瘍である。別の態様において、該状態はウイルス感染である、別の態様において、HSF1の活性を促進する薬剤は、サーチュイン阻害剤またはHAT活性化剤である。さらなる態様において、該薬剤はSIRT1阻害剤である。
【0016】
本発明はさらに、熱ショック応答の増加およびプロテオスタシス機能障害の疾患の治療を必要とする患者に、有効量の熱ショックアクチベーターおよび有効量のHSF1のアセチル化を阻害する薬剤を投与する工程を含む、該患者において熱ショック応答を増加する方法およびプロテオスタシス機能障害の疾患を治療する方法を包含する。別の態様において、本発明は、熱ショックアクチベーターおよびHSF1のアセチル化を阻害する薬剤を含む医薬組成物である。
【0017】
本発明はまた、細胞に試験薬剤を投与する工程;およびHSF1のDNA結合ドメイン中のリジン残基のアセチル化をモニタリングする工程を含む、細胞においてHSF1活性を調節する薬剤を同定する方法を包含し、試験薬剤の非存在下のDNA結合ドメインのアセチル化と比較したDNA結合ドメインのアセチル化の変化は、細胞中で該試験薬剤がHSF1活性を調節することを示す。さらなる態様において、HSF1 K80または対応する保存されたアミノ酸のアセチル化がモニタリングされる。
【0018】
別の態様において、本発明は、細胞または細胞溶解物に試験薬剤を投与する工程、およびサーチュイン基質のアセチル化を測定する工程を含む、細胞においてHSF1活性を調節する薬剤を同定する方法であり、該試験薬剤の非存在下のアセチル化と比較した基質のアセチル化の変化は、細胞中で該試験薬剤がHSF1活性を調節することを示す。一態様において、サーチュインはSIRT1である。
【0019】
さらなる態様において、本発明は、細胞または細胞溶解物に試験薬剤を投与する工程、およびヒストンアセチルトランスフェラーゼ(HAT)基質のアセチル化を測定する工程を含む、細胞においてHSF1活性を調節する薬剤を同定する方法であり、該試験薬剤の非存在下でのアセチル化と比較した基質のアセチル化の変化は、細胞中で該試験薬剤がHSF1活性を調節することを示す。
【図面の簡単な説明】
【0020】
本発明の前述および他の記載、特徴および利点は、添付の図面に示されるように、本発明の好ましい態様の以下のより具体的な記載から明白であろう。図面において、同様の参照記号は、異なる図面を通じて同じ部分を参照する。図面は必ずしも同じ縮尺ではなく、発明の原理の説明が強調される。
【0021】
【図1】図1A:シャペロン遺伝子発現に対するサーチュイン阻害剤ニコチンアミドの効果。熱ショック(HS)、セラストール、CdCl2またはMG 132に曝露する前に、HeLa細胞をニコチンアミド(NAM)で処理し、示されるプライマーを用いてRT-PCR解析を行なった。図1B:SIRT1 siRNAはhsp70の転写を阻害する。SIRT1に対するsiRNAまたは対照siRNAで形質転換されたHeLa細胞を、指定の時間、HSで処理した。RT-PCR解析を行ない、密度測定によりhsp70 mRNAアバンダンスの増加倍数を計算し、18S rRNAに対して標準化した。図1C:SIRT1 siRNAはhsp70プロモーターに対するHSF1結合を阻害する。上述のようにHeLa細胞を処理した(2B)。HSF1抗体およびqPCRを使用してChIP解析を行ない、1%の入力で行なった反応に対して結果を標準化した。実験は3回行なった。エラーバーは±SDを示す。
【図2】図2A:HSR誘導因子に応答するHSF1のアセチル化。Flag-HSF1およびp300で形質転換された293T細胞を、HS、セラストール、CdCl2またはMG 132で処理した。免疫沈降およびウェスタンブロットを用いたアセチル化アッセイにより細胞溶解物を解析した。リン酸化の増加によりゆっくり遊走するHSF1は、アスタリスクで示される。図2B:HSF1アセチル化におけるニコチンアミドおよびトリコスタチンAの効果。Flag-HSF1およびp300で形質転換した293T細胞を、トリコスタチンA(TSA)、ニコチンアミド(NAM)または両方で処理してHSに曝露し、アセチル化アッセイにより細胞溶解物を解析した。図2C:触媒変異体ではなく野生型SIRT1はHSF1アセチル化を阻害する。293T細胞をFlag-HSF1、p300、およびSIRT1 WTまたはSIRT1 H363Y変異体のいずれかで形質転換した後に、HSで処理し、アセチル化アッセイにより解析した。図2D:HSに応答したHSF1アセチル化。Myc-HSF1で形質転換したCos7細胞を、HSで指定時間処理し、アセチル化アッセイにより解析した。SIRT1に対する抗体またはMyc抗体を用いてSIRT1およびHSF1を検出した。Hsc70を負荷対照とした。図2E:レスベラトロールの効果。HeLa細胞を溶媒(EtOH)またはレスベラトロールで処理した後、指定時間、HS処理を行なった。オリゴヌクレオチドプルダウンアッセイおよびウェスタンブロットにより細胞抽出物を解析した。HSF1のリン酸化の増加はアスタリスクで示す。図2F:SIRT1過剰発現は、HSF1 DNA結合に影響する。293T細胞をSIRT1 WTで形質転換し、次いで指定時間HSに供した。HSF1抗体およびqPCRを用いてChIP解析を行なった。A〜Gの実験は3回行なった。エラーバーは±SDを示す。
【図3】図3A:K80でのHSF1の変異は、DNA結合活性を乱す。指定のHSF1構築物で形質転換したhsf1-/-細胞(HS有または無し)由来の抽出物を用いてEMSA反応を行なった(上パネル)。EMSAプローブは、ヒトhsp70プロモーター由来の近位HSEを含む。同じ試料でウェスタンブロット解析を行って、HSF1およびHsc70のレベルを示した。図3B:K80での組み換えHSF1の突然変異は、DNA結合能を乱す。組み換えWT HSF1またはHSF1 K80Qの量を増加させ(5、20、40、80または120ng)、プローブがHSEを含むEMSA反応を示す(上パネル)。HSF1タンパク質無し(-)の試料を対照とした。同じ試料でウェスタンブロットを行なってHSF1発現レベルを示した。図3C: K80でのHSF1変異の、hsf1-/-細胞におけるHSRレスキューの失敗。hsf1-/-細胞を指定の形態のヒトHSF1で形質転換して、HS有または無しで処理した。指定の遺伝子についてのプライマーによるqPCRを用いてRNAを定量した。グリセルアルデヒド3-リン酸デヒドロゲナーゼについて得られた値に対してデータを標準化し、HSなしで処理したWT HSF1細胞中の各mRNAのアバンダンスと比較する(値セット1)。A〜Cの実験は3回行なった。エラーバーは±SDを示す。
【図4】図4A:SIRT1過剰発現による厳しいストレスからの細胞の保護。293T細胞を空ベクター(モック)またはSIRT1で形質転換して45℃ HSで指定時間処理し、その後37℃で回復させた。24時間後、トリプシンブルー取り込みにより細胞死を決定した。実験は3回行なった。エラーバーは±SDを示す。図4B:年齢依存的なHSRの減退とSIRT1のアバンダンスの低下の相関。早期継代(継代21)および老齢(継代44)WI-38線維芽細胞由来の細胞抽出物をHS、またはHS後に3時間の37℃での回復(R)で処理し、HSEを含むプローブを用いたEMSAにより解析した(上パネル)。同じ試料でウェスタンブロットを行ない、Hsp70、HSF1およびSIRT1発現レベルを示した(下パネル)。
【図5】図5A:pCAFではなくp300およびCBPは、HSF1アセチル化を誘導する。293T細胞をFlag-HSF1およびp300、CBP、またはpCAFで形質転換し、HSで処理してアセチル化アッセイで解析した。図5B:p300は、インビボでhsp70プロモーターに結合する。指定の42℃熱ショック時点ならびに37℃の指定の回復有およびなしの時点で行なったクロマチン免疫沈降実験は、p300のhsp70プロモーターへの結合を示す。クロマチンを架橋して、回収し、p300または HSF1に特異的な抗体で免疫沈降した。次いで試料をhsp70.1プロモーターに特異的なプライマーを用いたPCRで解析した。1%の投入量、抗体無しでも反応を行なった。熱ショックの際に、HSF1およびp300は両方1分以内にhsp70プロモーターに結合し、結合は回復中徐々に弱まる。図5C:SIRT1はインビボでhsp70プロモーターに結合する。指定の42℃熱ショック時点で行なったクロマチン免疫沈降実験は、SIRT1がhsp70プロモーターに結合することを示す。クロマチンを架橋して、回収し、HSF1またはSIRT1に特異的な抗体で免疫沈降した。次いで、試料は、hsp70プロモーターに特異的なプライマーを用いてPCRにより解析した。抗体なしで対照反応も行なった。SIRT1は基底および熱ショック条件の両方でhsp70プロモーターに結合する。
【図6】図6:アセチル化の前提条件ではないバルクHSF1リン酸化。293T細胞をMyc-HSF1 WTまたはHSF1の10個のセリンのリン変異体およびp300で共形質転換した。形質転換の24時間後、細胞を1時間で42℃の熱ショックで処理した。細胞溶解物を免疫沈降に供してアセチル化HSF1を、ウェスタンブロットを用いて検出した、全HSF1を検出するためにMyc抗体を使用した。
【図7−1】図7A:HSF1は9個のリジンでアセチル化され得る。p300で共形質転換した293T細胞で発現させて精製したFlag-HSF1を熱ショック(42℃、1時間)またはセラストール処理(5μM、1時間)のいずれかに供し、LC-MS/MSで解析した。マウスHSF1中で同定されたアセチル化リジン残基の位置を示す。DBD、DNA-結合ドメイン;L、リンカー領域;HR-A/Bおよび-C、7回繰り返しドメイン;RD、制御ドメイン;N、核局在配列;AD1およびAD2、活性化ドメイン。図7B:アセチル化リジンは保存される。T-COFFEEバージョン5.13を使用して、多種類のアセチル化リジンの配列アライメントを行なった。アセチル化リジンを赤で示し、アミノ酸番号を右に示す。Hs、ヒト(NP_005517);Mm、マウス(NP_032322);Dr、ゼブラフィッシュ(NP 571675);Xl、アフリカツメガエル(NP 001084036.1);Dm、キイロショウジョウバエ(P22813);Ce、線虫(NM_060630);Sc、酵母(P10961)。
【図7−2】図7C;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−3】図7D;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−4】図7E;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−5】図7F;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−6】図7G;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−7】図7H;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−8】図7I;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−9】図7J;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図7−10】図7K;HSF1中の9個のアセチル化部位の質量分析データ。MS/MSスペクトルを抽出して、トリプシンによる消化を推定するMascot and Scaffoldプログラムを使用して解析した。同定されたそれぞれのアセチル化トリプシンペプチドについて、HSF1由来前駆体イオンの断片化から生じた質量スペクトルを上パネルに示し、対応断片イオン適用範囲を下パネルに示す。42ダルトンの付加質量でアセチル化を同定する。アセチル化リジンimmoniumイオンマイナスNH3を表すm/z126での低質量イオンを最大MS/MSスペクトル中で同定して(アスタリスクで標識)アセチル化のさらなる検証として使用した(12)。
【図8】図8:HSEと複合体化したK.ラクチスHSF DNA-結合ドメインの結晶構造を補充した座標(13)を使用して、発明者らは、整列したヒトHSF1の配列を、Swiss Modelを使用してつなげた。K80の側鎖およびDNAリン酸主鎖までの距離を緑で示す。上パネル中の四角で囲んだ領域は、下パネルをより拡大して示す。
【図9】図9:HSF1 K80Q変異体は、三量体化能力を欠損していない。HSFのDNAへの安定した結合の条件は、三量体HSF複合体の形成である。従って、K80の変異が適切なHSF1フォールディングまたはHSF三量体のストレス誘導形成を乱したと考えられる。この可能性を調べるために、交差結合アッセイを使用して、熱ショック時の高次複合体の形成を測定した。K562細胞を、構成的な三量体であるHSF1変異体であると以前に特徴付けられたWT Myc-HSF1、Myc-HSF1 K80QまたはMyc-HSF1 M391K、L395Pで形質転換した(14)。細胞を熱ショック(HS、42℃、20分)で処理するかまたは処理せず、その後EGS架橋剤で処理した。タンパク質抽出物をSDS-PAGEで分離し、指定の抗体でウェスタンブロット解析を行なった。矢印は単量体HSF1を示し、矢印の先端は三量体HSF1を示す。WTとK80Q HSF1の両方は、熱ショックに応答して単量体から三量体へ移行し、変異がHSF1三量体化に影響を及ぼさないことが示される。
【図10】図10:HSF1 DNA結合には、残基80にリジンを必要とする。指定のHSF1構築物で形質転換され、熱ショック(HS、42℃、20分)有また無しで処理されたhsf1-/-細胞におけるEMSA反応を示す。プローブとして使用したオリゴヌクレオチドは、ヒトhsp70プロモーター由来の近位HSEを含む。同じ試料でウェスタンブロット解析を行ない、HSF1とHsc70の発現レベルを示した。等量負荷についてHsc70を示す。K80のA、H、NおよびTへの変異は全て、熱ショック時のHSF1のDNAへの結合能を障害した。
【図11】図11:核ストレス体の形成にはHSF1の残基80にリジンを必要とする。指定のMyc-HSF1型で形質転換して、1時間、42℃の熱処理(HS)有または無しで処理したHeLa細胞の共焦点画像を示す。α-Myc抗体(赤)を用いて外因性HSF1を染色し、DAPI(青)を使用してDNAを染色した。バー=5μm。HSF1 K80のR、Q、A、H、NおよびTへの変異は全て、熱ショック時の特徴的な核ストレス体へのHSF1の再局在能を阻害した。
【図12】図12:ドミナントネガティブ様式のHSF1 K80Q機能。HSEへのHSF1の安定な結合には、3つのDNA結合ドメインと3つのnGAAnリピートの同時接触が必要である。1つまたは2つの単量体のアセチル化が関連のある非修飾単量体の結合を破壊するので、アセチル化HSF1 DNA結合ドメインの小集団は、ドミナントネガティブ効果を有すると思われる。この問題を調べるために、発明者らはHeLa細胞をMyc-HSF1のWTまたはK80Q変異体で形質転換し、HSF1またはMycに対する抗体で該細胞を二重染色して、内在性HSF1がnSBを形成する能力を解析した。指定の型のMyc-HSF1で形質転換して、熱ショック(HS、42℃、1時間)有またはなしで処理したHeLa細胞の共焦点画像を示す。HSF1タンパク質は、HSF1に対する抗体(緑)およびMycに対する抗体(赤)で染色した。DAPI(青)を使用してDNAを染色した。バー=5μm。内因性および外因性のWT HSF1は、熱ショック時にnSBに局在したが、HSF1のK80Q変異体を発現する細胞においてはいずれの抗体を用いてもnSBは検出されなかった。従って、K80アセチル化の生物学的な意味は、K80アセチル化HSF1分子の実際のレベルよりも高いと思われる。
【図13】図13:HSF1活性化サイクルのモデル。工程1:休止状態のHSF1は細胞質性または核性の不活性単量体である。工程2:ストレス時、HSF1はHSEに結合するDNA結合ホモ三量体を形成する。工程3:HSF1は転写活性を獲得する。工程4:HSF1転写活性は、減衰時にはなくなる。減衰は、DNAに結合したHSF1のトランス活性化機能(示さず)およびHSF1のDNA結合ドメインのアセチル化を介したDNA結合の阻害を抑制するhsp発現からの負のフィードバックを含む2つの調節された工程を有する。SIRT1は、HSF1アセチル化の阻害によるHSRの減衰段階を調節する。
【図14】図14:HSF1-SIRT1制御ネットワークのモデル。加齢および細胞代謝状態によるSIRT1の調節は、HSF1などの転写因子のネットワークの活性に影響を及ぼし、長生およびストレス耐性の増大をもたらす。
【図15】図15は、hsp70プロモーターレポーターを示す図である;hsp70.1pr-luc HeLa安定細胞株は-188から+150にhsp70.1プロモーター配列を含み、近位熱ショックエレメント(HSE)を含む。
【図16】図16は、ルシフェラーゼ解析前に、表示したとおりのレスベラトロールおよび41℃熱ショックもしくは42℃熱ショックいずれかで処理したHeLa hsp70.1pr-luc細胞の倍数活性を示す棒グラフである。
【図17】図17は、ルシフェラーゼ解析前に、表示したとおりのセラストールおよび/またはレスベラトロールで処理したHeLa hsp70.1pr-luc細胞の倍数活性を示す棒グラフである。
【発明を実施するための形態】
【0022】
発明の詳細な説明
本発明の好ましい態様の記載は以下の通りである。
本明細書で使用する場合、単語「a」および「an」は、特定されることがなければ1つ以上を意味する。例えば用語「細胞(a cell)」は、単一の細胞および2つ以上の細胞の組合せの両方を包含する。
【0023】
用語「熱ショック因子」、「熱ショック転写因子」および「HSF」は、ストレス誘導性遺伝子発現に関連する転写因子のファミリーのことをいう。少なくとも4種類のHSFが、脊椎動物および植物で記載されている。ショウジョウバエおよび線虫は、1つのみのHSFを発現する。(Wu, Ann. Rev. Cell Dev. Biol. 1995; 11:441-469)。ヒト細胞では、3種類のHSF(HSF1、HSF2およびHSF4)が特徴付けられている(Morimoto, et al, Genes Dev. 1998; 12:3788-3796)。HSF1は偏在的に発現し、HSPのストレス誘導発現に主要な役割を担っている。
【0024】
HSF1の配列は種間で高度に保存されている。HSF1のDNA結合ドメインは100アミノ酸超よりも長く、N末端の近位に位置している。ヒトHSF1とショウジョウバエHSFのDNA結合ドメイン間には約70%の相同性があり、ヒトHSF1と線虫HSF間には約55%の相同性がある。ヒトHSF1では、DNA結合ドメインはアミノ酸残基13からアミノ酸残基121まで伸びている。本明細書で使用する場合、用語「HSF1」は、任意の種または細胞型由来のHSF1を包含し、ショウジョウバエおよび線虫由来のHSFも包含する。かかるHSFおよびHSF1タンパク質は、HSF1 K80に対応するDNA結合ドメイン中のアセチル化のためのリジンを有する。用語「非ヒトHSF1」は、非ヒト種由来のHSF1を示す。用語HSF1は、GenBank受託番号NP_005517、NP_032322、NP_571675、NP_001084036.1、P22813、NM_060630およびP10961のアミノ酸配列を有するペプチドを含むがこれに限定されない。
【0025】
一態様において、HSF1は哺乳動物HSF1である。別の態様において、HSF1はヒトHSF1である。一態様において、HSF1はGenBank受託番号NP_005517(配列番号:1)と少なくとも約85%、90%、95%、98%および99%の配列同一性を有するアミノ酸配列を有する。別の態様において、HSF1は、GenBank受託番号NP_005517のアミノ酸配列を有する。
【0026】
用語「配列同一性」または配列に関する「同一性」は、2種類のアミノ酸配列間または2種類のヌクレオチド配列間の配列同一性のことをいう。同一性はそれぞれ、比較目的で整列されたそれぞれの配列の位置を比較することにより決定され得る。用語「配列相同性」または配列に関する「相同性」は、2種類のアミノ酸配列間または2種類のヌクレオチド配列間の配列相同性のことをいう。比較した配列中の同等の位置が同じ塩基またはアミノ酸で占められる場合、該分子はその位置で同一であり;同等の部位が同じまたは同様のアミノ酸残基(例えば、立体的および/または電気的性質が類似)で占められる場合、該分子はその位置で相同(同様)であるという。相同性、類似性または同一性のパーセントでの表示は、比較された配列で占められる位置での同一また同様なアミノ酸の数の関数を示す。相同性、類似性または同一性のパーセントでの表示は、比較された配列で占められる位置での同一または同様なアミノ酸の数の関数を示す。FASTA、BLASTまたはENTREZなどの種々の整列アルゴリズムおよび/またはプログラムが使用され得る。FASTAおよびBLASTは、GCG配列解析パッケージ(University of Wisconsin, Madison, Wis.)の一部として利用可能であり、例えばデフォルトを設定して使用することができる。ENTREZは、国立衛生研究所、国立医学図書館、全米バイオテクノロジー情報センター、Bethesda、Mdを通じて利用可能である。一態様において、2種類の配列のパーセント同一性は、ギャップウェイト1、例えば、2種類の配列間に一アミノ酸またはヌクレオチドミスマットがある場合、各アミノ酸ギャップを埋めてGCGプログラムにより決定され得る。
【0027】
本発明は、HSF1活性が、DNA結合ドメイン中のアミノ酸残基のアセチル化および脱アセチル化により媒介されるという発見に基づくものである。HSF1のアセチル化によりDNAへの結合が低下し、HSF1の脱アセチル化状態の維持によりDNAへの結合が延長されることが見出された。一態様において、細胞中のHSF1活性は、HSF1のDNA結合ドメイン中の1つ以上の塩基性アミノ酸のアセチル化を阻害することで増加する。HSF1のDNA結合ドメインのアセチル化は、DNA結合ドメイン中の1つ以上の塩基性アミノ酸残基のアセチル化を阻害する薬剤を投与することにより阻害され得る。アセチル化が減少または脱アセチル化が増加される場合にアミノ酸のアセチル化が阻害されることは理解されよう。別の態様において、HSF1のDNA結合ドメイン中のリジン残基のアセチル化を阻害することによりHSF1活性が増加される。さらなる態様において、細胞のHSF1活性は、ヒトHSF1のリジン残基80(HSF1 K80)のアセチル化を阻害することまたは非ヒトHSF1中の対応する保存されたアミノ酸のアセチル化を阻害することにより増加される。
【0028】
用語「HSF1 K80」は、ヒトHSF1のリジン80を示す。「対応する保存されたアミノ酸」は、非ヒト種のHSF1中の、保存され、ヒトHSF1 K80に対応するリジン残基を示す。本明細書で使用する場合、「保存されたアミノ酸」または「保存された残基」は、2種類のタンパク質間に共通することが分かっているおよび/または異なる種由来の相同タンパク質中などのペプチドモチーフ中の特定の位置を占める残基のことをいう。本明細書で使用する場合、「対応する保存されたアミノ酸」またはHSF1 K80に関する「対応する保存された残基」は、相同HSF1配列または非ヒトHSF1配列中の保存されたアミノ酸についてのアミノ酸位置のことをいう。HSF1の他のホモログ中のアミノ酸のナンバリングは、ヒトHSF1中のナンバリングとは異なり得ることは、当業者には容易に明らかであろう。HSF1の他のホモログ中の対応する保存されたアミノ酸は、例えば、市販の相同性モデリングソフトウェアパッケージまたは従来の配列アライメントパッケージを使用したアミノ酸配列の比較により同定され得る。
【0029】
別の態様において、細胞中のHSF1活性は、HSF1のDNA結合ドメイン中の1つ以上の塩基性アミノ酸のアセチル化を促進することにより低減される。一態様において、DNA結合ドメインのアセチル化を促進する薬剤が細胞に投与される。アミノ酸のアセチル化は、アミノ酸のアセチル化が増加される場合または脱アセチル化が低減される場合に促進されることが理解されよう。別の態様において、HSF1活性は、HSF1のDNA結合ドメイン中のリジン残基のアセチル化を促進することで低減される。さらなる態様において、ヒト細胞中のHSF1活性は、ヒトHSF1 K80のアセチル化を促進することまたは非ヒトHSF1中の対応する保存されたアミノ酸のアセチル化を促進することで低減される。
【0030】
本明細書で使用する場合、「阻害」または「低減」は、直接または間接的な手段により、正味の低減を引き起こすことを包含する。用語「増加」は、直接または間接的な手段により、正味の増大を引き起こすことを意味する。
【0031】
HSF1活性は、熱ショック応答エレメント(HSE)配列を有する遺伝子の転写調節因子としてのHSF1の活性のことをいう。HSE配列を有する遺伝子から転写されたタンパク質は、熱ショック(HS)タンパク質と称される。熱ショックは、特定の遺伝子の転写の増大を引き起こす熱ショック応答のトリガーとして最初に発見された[Snoecx et al. (2001), Physiol. Rev. 81: 1461-1497]。この転写活性の産物は、熱ショックタンパク質と称される[Snoeckx et al.]。ほとんどの熱ショックタンパク質(Hsp)は、分子質量の表示を参照して、例えばHsp27と命名されている。ファミリー中の種々のHspの分類は、その関連のある機能およびサイズに基づく。熱ショックタンパク質のサイズは、10〜170kDaである。ファミリーネームは慣例的に大文字で書かれる。例えば「HSP70」は、HSP70ファミリーのことをいう。HSP70ファミリーは、重量が70〜78kDaである。HSP70ファミリーの一員の一例は、Hsp72(一般的にHsp70と示される)である。熱ショック応答は、熱ショックタンパク質をコードする遺伝子の誘導を包含する。本発明の方法に従って誘導され得る熱ショックタンパク質遺伝子としては、限定されないが、HSP10ファミリー、HSP40ファミリー、HSP60ファミリー、HSP70ファミリー、HSP90ファミリー、HSP100ファミリー、HSP27ファミリー、αA-クリスタリンファミリーおよびαB-クリスタリンファミリーのタンパク質から選択されるファミリー由来のタンパク質をコードする遺伝子が挙げられる。
【0032】
HSF1のアセチル化は、HSF1のアセチル化の増大もしくは脱アセチル化の低減をもたらすか、またはHSF1をアセチル化形態で維持する(直接または間接的な)任意の手段により増大され得る。いくつかの態様において、細胞中のHSF1のアセチル化は、HSF1のアセチル化を増大する薬剤を投与することで増大される。同様に、HSF1のアセチル化は、アセチル化の低減または脱アセチル化の増大をもたらすか、またはHSF1を脱アセチル化形態で維持する任意の手段により低減または阻害され得る。用語「薬剤」は、本明細書において、化合物、化合物の混合物、生物学的巨大分子(例えば、核酸、抗体、タンパク質もしくはその一部、例えばペプチドなど)または細菌、植物、真菌もしくは動物(特に哺乳動物)の細胞または組織などの生物学的物質から作製される抽出物を示すように使用される。一態様において、HSF1のDNA結合ドメインのアセチル化を調節する薬剤は、HSF1を含む細胞と試験化合物を接触させて、試験化合物の存在下または非存在下でDNA結合ドメインのアセチル化の程度を測定することにより同定され得、アセチル化の増大は、該薬剤がHSF1のアセチル化を増大させることを示し、アセチル化の低減は該薬剤がHSF1のアセチル化を阻害することを示す。
【0033】
HSF1のアセチル化を阻害する例示的な方法としては、限定されないが、単離サーチュインを投与すること、細胞中のサーチュインの活性を上げること、およびヒストンアセチルトランスフェラーゼ(HAT)の活性を阻害することが挙げられる。ヒストンアセチルトランスフェラーゼ(HAT)は、ヒストン、Hsp40/Dna J熱ショックタンパク質および種々の転写因子などの特定のタンパク質中のリジン残基のアセチル化を触媒する酵素である[Saha et al. (2006). HATs and HDACs in neurodegeneration: a tale of disconcerted acetylation homeostasis. Cell Death and Differentiation 13:539-550; WO 2009/134131 A1]。アセチルトランスフェラーゼのいくつかのファミリーが同定されており、例えば、prGcn5およびPCAFなどのGNATスーパーファミリー(Gcn5-関連N-アセチルトランスフェラーゼ)、例えばMOZ、Ybf2/Sas3、Sas2およびTip60などのMYSTファミリー、例えばp300およびCBPタンパク質などのp300/CBP HATファミリーが挙げられる(Lau et al. (2000), Cell 5(3): 589-95; 米国特許出願公開公報第20080227752号。以下の実施例1に示されるように、p300およびCBPはHSF1アセチル化を誘導する。
【0034】
ヒストンデアセチルトランスフェラーゼ(HDAC)は基質中のリジン残基の脱アセチル化を触媒する酵素である。脱アセチル化酵素は3つのファミリーに分類される;クラスIおよびII HDACファミリーは、トリコスタチンAにより阻害され、NAD+依存型クラスIIIサーチュインファミリーは、ニコチンアミドにより阻害される。
【0035】
一態様において、HSF1のアセチル化は、細胞に、単離サーチュインを投与すること、または細胞中のサーチュインのレベルもしくは活性を上げることにより阻害され得る。サーチュインは、サーチュイン脱アセチル化酵素タンパク質ファミリーに属するタンパク質である。一態様において、サーチュインはSir2ファミリーに属し、これは、限定されないが酵母Sir2(例えば、GenBank受託番号P53685)、線虫Sir-2.1(例えば、GenBank受託番号NP_501912)、ヒトSIRT1(例えば、GenBank受託番号NM_012238およびNP_036370(またはAF083106))ならびにヒトSIRT2(例えば、GenBank受託番号NM_012237、NM_030593、NP_036369、NP_085096およびAF083107)のタンパク質が挙げられる。本明細書で使用する場合、用語「Sir2タンパク質」は、Sir2ファミリーに属するサーチュインのことをいう。一態様において、Sir2タンパク質はSIRT1である。別の態様において、SIRT1はヒトSIRT1である。
【0036】
上述のように、本発明の一局面は、細胞に単離サーチュインまたはその生物学的に活性な断片もしくはバリアントをHSF1のDNA結合ドメイン中のアミノ酸のアセチル化を阻害するのに充分な量で投与する工程を含む、細胞中のHSF1の活性を増加する方法である。
【0037】
本明細書で使用する場合、細胞または被験体(下記のように)に投与される単離サーチュインまたはその断片もしくはバリアントは、実質的に純粋であるか、またはそれに天然に付随する成分から単離された実質的に純粋な単離ポリペプチドである。用語「単離サーチュイン」または「単離SIRT1」は、薬学的に許容され得る担体または賦形剤を含むことを除外せず、単にサーチュインが細胞およびサーチュインに天然に付随する他の成分から単離されていることを示すことが理解されよう。単離サーチュイン、そのバリアントの断片は、上述の天然に存在するサーチュインと同じまたは実質的に同一のアミノ酸配列を有する組み換えにより生成されたポリペプチドであり得る。
【0038】
サーチュインの生物学的に活性な断片は、脱アセチラーゼ活性または完全長サーチュインの別の生物活性を有する断片である。サーチュインのバリアントは、例えば、サーチュインの配列中に保存的アミノ酸置換を作製することで作製され得る。生物学的に活性なバリアントは、脱アセチラーゼ活性または天然に存在するサーチュインの別の生物学的活性を有する。保存的アミノ酸置換は、同様の側鎖を有する残基と交換可能であることをいう。保存的置換アミノ酸は、側鎖の化学的性質によって分類され得る。例えば、アミノ酸の一グループは、中性で疎水性の側鎖(a、v、l、i、p、w、fおよびm)を有するアミノ酸を含み;別のグループは、中性で極性の側鎖を有するアミノ酸(g、s、t、y、c、nおよびq)であり;別のグループは塩基性側鎖を有するアミノ酸(k、rおよびh)であり;別のグループは酸性側鎖を有するアミノ酸(dおよびe)であり;別のグループは、脂肪族側鎖を有するアミノ酸(g、a、v、lおよびi)であり;別のグループは、脂肪族水酸基側鎖を有するアミノ酸(sおよびt)であり;別のグループは、アミン含有側鎖を有するアミノ酸(n、q、k、rおよびh)であり;別のグループは、芳香族側鎖を有するアミノ酸(f、yおよびw)であり;別のグループは、硫黄含有側鎖を有するアミノ酸(cおよびm)である。例示的な保存的アミノ酸置換基は:r-k;e-d、y-f、l-m;v-iおよびq-hである。アミノ酸は、その一般的な一文字表記を用いて上記され:アラニンはa;システインはc;アスパラギン酸はd;グルタミン酸はe;フェニルアラニンはf;グリシンはg;ヒスチジンはh;イソロイシンはi;リジンはk;ロイシンはl;メチオニンはm;アスパラギンはn;プロリンはp;グルタミンはq;アルギニンはr;セリンはs;トレオニンはt;バリンはv;トリプトファンはw;およびチロシンはyである。
【0039】
一態様において、サーチュインはSir2タンパク質またはその生物学的に活性な断片もしくはバリアントである。別の態様において、サーチュインは、天然に存在するSir2タンパク質またはその断片と少なくとも約80%、85%、90%、95%、98%および99%の配列相同性を有するペプチドである。別の態様において、サーチュインは、天然に存在するSir2タンパク質またはその断片と少なくとも約80%、85%、90%、95%、98%および99%の配列同一性を有するペプチドである。別の態様において、Sir2タンパク質は、哺乳動物SIRT1と少なくとも約80%、85%、90%、95%、98%および99%の配列同一性を有するペプチドである。別の態様において、Sir2タンパク質は、ヒトSIRT1と少なくとも約80%、85%、90%、95%、98%および99%の配列同一性を有するペプチドである。さらなる態様において、Sir2タンパク質は配列番号:2と少なくとも80%、85%、90%、95%、98%および99%の配列同一性を有するペプチドである。
【0040】
別の態様において、Sir2タンパク質は、配列番号:2の250番アミノ酸が保存されたSIRT1触媒ドメイン、アミノ酸残基258〜451と少なくとも約80%の同一性を有する。配列番号:2はヒトSIRT1のアミノ酸配列を示す。好ましい態様において、Sir2タンパク質は、配列番号:2のアミノ酸残基258〜451の間のアミノ酸配列に少なくとも約80、85、90、95、99%の配列同一性を有する配列を含む。他の態様において、Sir2タンパク質は、完全長Sir2タンパク質の生物学的に活性な断片、例えば脱アセチル化が可能なSIRT1の断片である。Sir2タンパク質はまた、天然に存在するSir2ファミリーメンバーに対して1つ以上の置換、例えば1〜10個の置換を含む配列またはバリアントを含む。
【0041】
サーチュインのレベルまたは活性は、細胞中でサーチュインのレベルまたは活性を増加する薬剤(「サーチュイン活性化剤」)を投与することによっても増加し得る。サーチュインの活性とは、脱アセチラーゼ活性またはタンパク質が基質を脱アセチル化する活性またはサーチュインの別の生物活性のことをいう。
【0042】
サーチュイン活性化剤は、サーチュインの活性を増加させる薬剤である。SIRT1活性化剤は、細胞中でSIRT1の活性を増加させる薬剤である。サーチュインおよびSIRT1活性化化合物は、例えば米国特許第7,345,178号、米国特許出願公開公報第20080249103号、20080194803号、200070037865号、20070014833号、20060229265号および20060025337号ならびに国際出願番号WO 2005/002672、WO 2005/002555、WO 2009015180、WO 2009015179、WO 2006094248、WO 2006004722、WO 2006094235、WO 2006094233、WO 2006094209、WO 2006105440、WO 2006078941、WO 2006094236、WO 2006/094210ならびにMilne et al. (2007), Nature 450: 712-716およびSzczepankiewicz et al. (2008), Current Topics in Medical Chemistry 8: 1533-1544に記載されており、各文献の内容は参照により本明細書に援用される。具体的なSIRT1活性化化合物としては、限定されないが、フラボン、スチルベン、フラバノン、イソフラバノン、カテキン、カルコン、タンニンおよびアントシアジン(anthocyadins)、キノキサリン(例えば、ピロロキノキサリン)、SRT-1460、SRT-2183、SRT-1720、SRT-2104、オキサロ酢酸、レスベラトロール、オキサゾロ[4,5-b]ピリジン、アザベンゾイミダゾール、ベンゾイミダゾール、およびイミダゾチアゾール(例えば、SRT1460、SRT2183およびSRT1720)、(Szczepankiewicz et al.)が挙げられる。例示的なスチルベンとしては、限定されないが、トリヒドロキシスチルなどのヒドロキシスチルベン、例えば3,5,4'-トリヒドロキシスチルベン(「レスベラトロール」)が挙げられる。レスベラトロールはまた、3,4',5-スチルベントリオールとしても公知である。特定のレスベラトロールエステルアナログもサーチュインアクチベーターとして記載されている(例えば、WO 2005069998参照)。テトラヒドロキシ-スチルベン、例えばピセタノールも包含される。イソリキリチゲニンなどのトリヒドロキシケイロンを含むヒドロキシケイロン、およびブテイン(butein)などのテトラヒドロキシケイロンもSIRT1活性化化合物として記載されている。フィセチンなどのテトラヒドロキシフラボンを含むヒドロキシフラボン、およびケルセチンなどのペンタヒドロキシフラボンも使用することができる。SIRT1アクチベーターである例示的なヒドロキシフラボンとしては、限定されないが、イソリキリチゲニン、フィセチン、ブテインおよびケルセチンが挙げられる。
【0043】
別の態様において、HSF1のアセチル化は、細胞にヒストンアセチルトランスフェラーゼ(HAT)の活性を阻害する薬剤またはヒストンアセチルトランスフェラーゼ(HAT)の阻害剤を投与することにより阻害することができる。例示的なHAT阻害剤としては、米国特許出願公開公報第20090076155号に記載されるアナカルジン酸(6-ペンタデシルサリチル酸)および例えばその誘導体が挙げられ、その内容は、参照により本明細書中に明確に援用される。クルクマノイド(例えばクルクミンなど)、エピガラクトガテキン-3-没食子酸(epigallogatechin-3-gallate)およびガルシノール(garcinol)もHAT阻害剤として記載されている(例えば、米国特許出願公開公報第20060020027号, Cancer Research 69, 583 (2009), およびJBC 279: 33716-33726参照、それぞれその内容は参照により本明細書に明確に援用される)。HATを阻害するための他の化合物および方法は、米国特許出願公開公報第20090076155、WO 02070675、米国特許出願公開公報第2008227752および米国特許第6,369,030号および6,747,005号に記載されており、それぞれの内容は、参照により本明細書に明確に援用される。p300阻害剤は、例えば、米国特許出願公開公報第20050069986号およびLau et al. (2000), Cell 5(3): 589-95, Mai et al. (2009), Bioorganic and Medicinal Chemical Letters 19(4): 1132-35に記載されており、それぞれの内容は参照により本明細書に明確に援用される。
【0044】
本発明はまた、サーチュインの活性を低減させる工程を含む、HSF1のDNA結合ドメインのアセチル化を促進することにより細胞中のHSF1の活性を低減させる方法に関する。一態様において、サーチュインはSir2タンパク質である。別の態様において、サーチュインはSIRT1である。一態様において、サーチュインの活性を阻害する薬剤(「サーチュイン阻害剤」)は、細胞に投与され得る。一態様において、SIRT1阻害剤が投与される。
【0045】
当業者が理解するように、サーチュイン阻害剤は、小分子、タンパク質、ペプチド、ペプチド模倣物、抗体または核酸であり得る。核酸としては限定されないが、DNA、RNA、RNA干渉剤およびPNAが挙げられる。サーチュイン阻害剤は米国特許7,345,178号、米国特許出願公開公報第20080287653号、20060229265号20060084135号、20050136537号および20070197459号、国際特許出願WO 2008155390、WO 2008011476、WO 2008082646、WO 2008156866、WO 2008086400、WO 2007047604、WO 2007084162、WO 2006099245、WO 2006094209、WO 2006/094210、WO 2005026112、およびWO 2005060711に記載され、それぞれの内容は参照により本明細書に援用される。例示的な小分子サーチュイン阻害剤は、ニコチンアミド、サーチノール、パラサーチノール、HR-73、2,3,4,9-テトラヒドロ-1H-カルバゾール-1-カルボキシアミド、スプリトマイシン、SEN-196、2-フェニルアミノベンズアミド、スラミン、NSC-112546、カンビノール、テノビン-1(tenovin-1)、ガッチフェロン(guttiferone)、ハイパーフォリン、アーチソホリン(artisoforin)およびタンパク質キナーゼC阻害剤Ro31-8220、ロットレリン、ZM449829およびインジルビン-3'-モノオキシム(Szczepankiewicz et al.)である。
【0046】
別の態様において、サーチュイン阻害剤は、抗サーチュイン抗体である。用語抗体は、モノクローナル抗体、ポリクローナル抗体、ヒト化抗体、キメラ抗体、単鎖Fv(scFv)、Fab断片、F(ab')断片、イントラボディおよび合成抗体である。一態様において、サーチュイン阻害剤は、抗Sir2タンパク質抗体である。さらなる態様において、サーチュイン阻害剤は、抗SIRT1抗体である。抗サーチュイン抗体は、サーチュインタンパク質またはそのポリペプチドもしくは断片などの適切な免疫原(合成ペプチド等の合成分子など)に対して生じ得る。免疫抗原の調製、ならびにポリクローナル抗体およびモノクローナル抗体の産生は、例えばファージディスプレイなどの任意の適切な技術を使用して行われ得る。種々の方法が記載されている(例えば、Kohler et al., Nature, 256:495-497 (1975)) およびEur. J. Immunol. 6:511-519 (1976)); Milstein et al., Nature 266:550-552 (1977)); 米国特許第4,172,124号; Harlow, E. and D. Lane, 1988, Antibodies: A Laboratory Manual, (Cold SpringHarbor Laboratory: Cold Spring Harbor, N. Y.); ならびにCurrent Protocols In Molecular Biology, Vol. 2 (Supplement 27, Summer '94), Ausubel, F. M. et al., Eds., (John Wiley & Sons: New York, N.Y.), Chapter 11, 1991)を参照、それぞれの教示は参照により本明細書に援用される)。本発明に従い抗体の断片を使用することもできる。抗体は、従来技術を使用して断片化することができ、該断片は、完全抗体と同様の方法で有用性についてスクリーニングされる。免疫グロブリン分子のFab断片は、共有結合により共に連結された免疫グロブリン重鎖および免疫グロブリン軽鎖の免疫学的に活性な部分を含む免疫グロブリン分子の一部からなる多量体タンパク質であり、抗原と特異的に結合することができる。Fab断片は、当該技術分野において周知の方法を使用して、実質的に完全な免疫グロブリン分子のパパインによるタンパク質溶解性消化により調製し得る。しかしながら、Fab断片はまた、適切な宿主細胞中で、免疫グロブリン重鎖および免疫グロブリン軽鎖の所望の一部を、当該技術分野で公知の任意の方法を用いて発現させることによっても調製し得る。
【0047】
抗サーチュイン抗体はまた、イントラボディでもあり得る。イントラボディは、細胞内発現抗体であり、特異的に細胞内の標的分子に結合して、不活性化するように設計された単鎖抗体分子である。イントラボディは、細胞アッセイおよび完全生物体において使用されている[Chen et al., Hum. Gen. Ther. (1994) 5:595-601; Hassanzadeh et al., Febs Lett. (1998) 16(1, 2):75-80 and 81-86]。サーチュインと特異的に反応するイントラボディを用いて誘導性発現ベクターを構築することができる。
【0048】
さらなる態様において、サーチュイン阻害剤は、核酸である。一態様において、核酸は、アンチセンス核酸である。アンチセンス核酸は、RNA、DNA、PNAまたは任意の他の適切な核酸であり得る。アンチセンス配列とその相補的センス配列の間で二本鎖が形成され得、遺伝子の不活性化が生じる。アンチセンス核酸は、遺伝子から転写されたRNAと二本鎖を形成する、二本鎖DNAと三本鎖を形成することなどにより遺伝子発現を阻害し得る。アンチセンス核酸は、例えばコード配列が公知である任意の遺伝子に対して作製することができるか、またはいくつかの充分に確立された技術(例えば、アンチセンスRNAまたはオリゴヌクレオチドの化学合成(任意に、分解に対して抵抗性を増加させるかもしくは細胞取り込みを向上させる修飾ヌクレオチドおよび/または結合を含む)またはインビトロ転写)により決定することができる。アンチセンス核酸およびその使用は、例えば、米国特許第6,242,258号、6,500,615号、6,498,035号、6,395,544号および5,563,050号に記載され、それぞれの内容は参照により本明細書に援用される。
【0049】
別の態様において、サーチュイン阻害剤はRNA干渉剤である。「RNA干渉剤」は、本明細書で使用する場合、標的遺伝子またはゲノム配列の発現を、RNA干渉(RNAi)により干渉または阻害する任意の薬剤として規定される。かかるRNA干渉剤としては、限定されないが、標的遺伝子もしくはゲノム配列またはその断片に相同なRNA分子を含む核酸分子、短鎖干渉RNA(siRNA)、短鎖ヘアピンまたは低分子ヘアピンRNA(shRNA)、およびRNA干渉(RNAi)により標的遺伝子の発現を干渉または阻害する小分子が挙げられる。本発明の標的遺伝子は、サーチュインをコードする遺伝子である。一態様において、該遺伝子は、Sir2タンパク質をコードする。別の態様において、該遺伝子は、SIRT1をコードする。さらに別の態様において、該遺伝子はヒトSIRT1をコードする。本明細書で使用する場合、「標的遺伝子発現の阻害」は、標的遺伝子もしくは該標的遺伝子にコードされるタンパク質の発現またはタンパク質活性またはレベルの任意の低減を含む。
【0050】
一態様において、RNA干渉剤はsiRNAである。siRNAは、化学的に合成、インビトロ転写により産生、または宿主細胞中で産生され得る。一態様において、siRNAは、約15〜約40ヌクレオチド長または約15〜約28ヌクレオチドまたは約19〜約25ヌクレオチド長または約19、20、21または22ヌクレオチド長の二本鎖RNA(dsRNA)分子であり、各鎖において長さ約0、1、2、3、4、5または6ヌクレオチドの3'および/または5'突出を含み得る。突出の長さは、二本の鎖で独立であり、すなわち一方の鎖の突出の長さは第二鎖の突出の長さに依存しない。
【0051】
RNAiはまた、低分子ヘアピン(ステムループとも称される)RNA(shRNA)を含む。一態様において、これらのshRNAは、短(例えば、約19〜約25ヌクレオチド)アンチセンス鎖、続いて約5〜約9ヌクレオチドのヌクレオチドループ、および類似のセンス鎖からなる。代替的に、センス鎖は、ヌクレオチドループ構造の前にあり得、アンチセンス鎖が続いてもよい。これらのshRNAは、プラスミド、レトロウイルス、およびレンチウイルスに含まれ得、例えばpol III U6プロモーターまたは別のプロモーターから発現され得る(例えば、Stewart, et al. (2003) RNA Apr;9(4):493-501参照、参照により本明細書に援用される)。
【0052】
RNAに加えて、RNA干渉剤はまた、化学的に修飾されたヌクレオチドおよび非ヌクレオチドを含み得、また、リボース糖分子が別の糖分子または同様の機能を果たす分子に置換された分子を含み得る。また、ヌクレオチド残基間に、ホスホロチオエート結合などの非天然結合が使用されてもよい。該RNA鎖は、フルオロフォアなどのレポーター基の反応性官能基により誘導体化され得る。例示的な誘導体は、RNA鎖の両末端または一方の末端、典型的にはセンス鎖の3'末端で修飾される。他の例示的な誘導体は、2'O-アルキル化残基または2'-O-メチルリボシル誘導体および2'-O-フルオロリボシル誘導体などの修飾炭水化物部分を有するヌクレオチドを取り込む。RNA塩基が修飾されていてもよく、例えばアルキル化またはハロゲン化され得る。例えば、ハロゲン化塩基、例えば5-ブロモウラシルおよび5-ヨードウラシルが取り込まれ得る。該塩基がアルキル化されてもよく、例えば7-メチルグアノシンがグアノシン残基の代わりに取り込まれ得る。成功裡の阻害を生じる非天然塩基がさらに取り込まれ得る。
【0053】
別の態様において、核酸はリボザイムまたはデオキシリボザイムである。リボザイムおよびデオキシリボザイムは、核酸分子の配列特異的切断を触媒することが示されている。切断部位は、RNAまたはDNA酵素中のヌクレオチドと標的核酸中のヌクレオチドの相補対形成により決定される。そのため、RNAおよびDNAの酵素は、核酸分子を切断するように設計されて、分解の速度が増大され得る[Cotten et al, EMBOJ. 8: 3861-3866, 1989; Usman et al., Nucl. Acids Mol. Biol. 10: 243, 1996; Usman, et al., Curr. Opin. Struct. Biol. 1: 527, 1996; Sun, et al., Pharmacol. Rev., 52: 325, 2000]。
【0054】
サーチュイン阻害化合物はまた、ペプチドアプタマーを含む。ペプチドアプタマーは、タンパク質機能のドミナント阻害剤として作用するペプチドまたは小ポリペプチドである。ペプチドアプタマーは、標的タンパク質に特異的に結合して、その機能をブロックする(Kolonin and Finley, PNAS (1998) 95:14266-14271)。高い親和性および特異性でサーチュインに結合するペプチドアプタマーは、当該技術分野に公知の種々の技術により単離することができる。ペプチドアプタマーは、酵母ツーハイブリッドスクリーンによりランダムペプチドライブラリーから単離し得る(Xu et al, PNAS (1997) 94:12473-12478)。ペプチドアプタマーはまた、ファージライブラリー(Hoogenboom et al., Immunotechnology (1998) 4:1-20)、または化学的に合成したペプチド/ライブラリーから単離し得る。
【0055】
特定の局面において、HSF1の活性は、HATの活性を増大し得る薬剤(本明細書においてはHATアクチベーターともいう)を投与することにより阻害され得る。HATを活性化し得る小分子化合物の例は、N-(4-クロロ-3-トリフルオロメチル-フェニル)-2-エトキシ-6-ペンタデシル-ベンズアミド(CTPB)である(Balasubrmanyam et al. (2003). Small molecule modulators of histone acetyltransferase p300. The Journal of Biological Chemistry, 278, 19134-19140)。
【0056】
本発明の方法によると、HSF1の活性は、HSF1のDNA結合ドメイン中の塩基性アミノ酸残基のアセチル化を改変することにより細胞内で調節することができる。細胞としては、原核細胞、真核細胞、脊椎動物細胞および無脊椎動物細胞が挙げられる。一態様において、細胞は哺乳動物細胞である。別の態様において、細胞はヒト細胞である。例示的な細胞としては、限定されないが、筋肉細胞(例えば、骨格筋細胞、ならびにペースメーカー細胞、心房細胞、心房心室結節細胞、左心室細胞、右心室細胞、乳頭筋細胞、プルキンエ線維細胞および平滑筋細胞などの心筋細胞)、血液細胞、腎臓細胞、上皮細胞、腸細胞、リンパ節細胞、脾臓細胞、肝細胞、胸腺細胞、唾液腺細胞、下垂体細胞、膀胱細胞、骨細胞、乳房細胞、頚部細胞、結腸直腸細胞、腎臓細胞、咽頭細胞、肺細胞、リンパ細胞、皮膚細胞および造血性細胞(例えばT細胞、B細胞、マクロファージ、樹状細胞およびそれらの前駆細胞)が挙げられる。
【0057】
特定の局面において、本発明は、HSF1の活性の調節を必要とする被験体に、HSF1のDNA結合ドメイン中の塩基性アミノ酸残基のアセチル化を調節する薬剤の有効量を投与する工程を含む、前記被験体においてHSF1の活性を調節する方法である。一態様において、本発明は、HSF1の活性の増大を必要とする被験体に、HSF1のDNA結合ドメインの塩基性アミノ酸のアセチル化を阻害する薬剤の有効量を投与する工程を含む、前記被験体においてHSF1の活性を増大する方法である。さらなる態様において、HSF1のDNA結合ドメインのアセチル化を阻害する薬剤は、単離サーチュイン、サーチュイン活性化剤またはHAT阻害剤である。別の態様において、本発明は、HSF1の活性の低減を必要とする被験体に、HSF1のDNA結合ドメイン中の塩基性アミノ酸のアセチル化を促進する薬剤の有効量を投与する工程を含む、前記被験体においてHSF1の活性を低減する方法である。さらなる態様において、HSF1のDNA結合ドメインのアセチル化を促進する薬剤は、サーチュイン阻害剤またはHATアクチベーターである。
【0058】
また、本発明は、タンパク質ホメオスタシスの機能障害に関連のある状態の治療を必要とする被験体に、HS1のDNA結合ドメイン中の塩基性アミノ酸のアセチル化を阻害する薬剤の治療有効量を投与する工程を含む、前記被験体において、タンパク質ホメオスタシスの機能障害に関連のある状態を治療する方法を包含する。別の態様において、該薬剤は、DNA結合ドメイン中のリジン残基のアセチル化を阻害する。さらなる態様において、該薬剤は、HSF1 K80のアセチル化を阻害する。さらに別の態様において、該方法は、前記被験体に、HAT阻害剤、単離サーチュインおよびサーチュイン活性化剤からなる群より選択される薬剤の治療有効量を投与する工程を含む。一態様において、該薬剤は、HAT阻害剤である。別の態様において、該薬剤は、単離サーチュインである。別の態様において、サーチュインはSir2タンパク質である。さらなる態様において、サーチュインはSIRT1である。さらなる態様において、サーチュインはヒトSIRT1である。さらに別の態様において、該薬剤は、サーチュイン活性化剤またはSIRT1活性化剤である。サーチュイン活性化剤は、先に規定され記載されている。上述のように、サーチュイン活性化剤およびSIRT1活性化剤としては、例えば、フラボン、スチルベン、フラバノン、イソフラバノン、カテキン、カルコン、タンニンおよびアントシアジン、ピロロキノキサリン、SRT-1460、SRT-2183、SRT-1720、オキサロ酢酸、レスベラトロールおよびその他のポリフェノールが挙げられる。単離サーチュインまたはサーチュイン活性化剤は、薬学的な担体または賦形剤を含む医薬組成物中で投与され得る。
【0059】
「治療する」または「治療」は、症状、合併症、または疾患の生化学的徴候の開始を予防または遅延すること、症状を改善または緩和すること、あるいは疾患、状態または障害のさらなる発症を停止または阻害することを含む。「患者」は、治療を必要とするヒト被験体である。「治療有効量」は、障害の1つ以上の症状を改善および/または障害の進行を予防、障害の後退を引き起こすのに充分な治療剤の量のことをいう。
【0060】
本発明は、タンパク質のホメオスタシスにおける機能障害に関連する状態の治療を包含する。例示的なタンパク質としては、グルコセレブロシダーゼ、ヘキソサミンA、嚢胞線維症膜貫通伝導性調節因子、アスパルチルグルコサミニダーゼ、α-ガラクトシダーゼA、システイントランスポーター、酸性セラミダーゼ、酸性α-L-フコシダーゼ、保護タンパク質、カテプシンA、酸性β-グルコシダーゼ、酸性β-ガラクトシダーゼ、イズロン酸2-スルファターゼ、α-L-イズロニダーゼ、ガラクトセレブロシダーゼ、酸性α-マンノシダーゼ、酸性β-マンノシダーゼ、アリルスルファターゼB、アリルスルファターゼA、N-アセチルガラクトサミン-6-硫酸スルファターゼ、酸性β-ガラクトシダーゼ、N-アセチルグルコサミン-1-ホスホトランスフェラーゼ、酸性スフィンゴミエリナーゼ、NPC-I、酸性α-グルコシダーゼ、β-ヘキソサミンB、ヘパリンN-スルファターゼ、α-N-アセチルグルコサミニダーゼ、α-グルコサミニドN-アセチルトランスフェラーゼ、N-アセチルグルコサミン-6-硫酸スルファターゼ、α1抗トリプシン、α-N-アセチルガラクトサミニダーゼ、α-ノイラミニダーゼ、β-グルクロニダーゼ、β-ヘキソサミンAおよび酸性リパーゼ、ポリグルタミン、α-シヌクレイン、Abペプチド、タウタンパク質、hERGカリウムチャネル、島アミロイドポリペプチド、およびトランスチレチンが挙げられる。
【0061】
一態様において、プロテオスタシスの機能障害に関連する疾患は、機能獲得障害である。用語「機能獲得障害、「機能獲得疾患」、「毒性機能獲得障害」および「毒性機能獲得疾患」は交換可能に使用される。機能獲得障害は、凝集関連タンパク質毒性の増加を特徴とする疾患である。これらの疾患において、凝集は、細胞の内側および/または外側のクリアランスを越える。機能獲得疾患としては、限定されないが、ポリグルタミンの凝集に関連する神経変性疾患、レヴィー小体疾患、筋萎縮性側索硬化症、トランスチレチン関連凝集疾患、アルツハイマー病、II型糖尿病、肝臓疾患およびプリオン疾患が挙げられる。ポリグルタミンの凝集に関連する神経変性疾患としては限定されないが、ハンチントン病、室頂核脳幹および淡蒼球ルイ体萎縮症、いくつかの種類の脊椎小脳性運動失調、ならびに脊椎および延髄筋萎縮症が挙げられる。アルツハイマー病は2種類の凝集物:Aβペプチドの細胞外凝集物とタウタンパク質に関連する微小管の細胞内凝集物の形成を特徴とする。トランスチレチン関連凝集疾患としては、例えば、老年性全身アミロイドーシスおよび家族性アミロイド神経障害が挙げられる。レヴィー小体疾患は、α-シヌクレインタンパク質の凝集を特徴として、例えばパーキンソン病が挙げられる。プリオン疾患(感染性海綿状脳症またはTSEとしても知られる)は、プリオンタンパク質の凝集と特徴とする。例示的なヒトプリオン疾患は、クロイツフェルト・ヤコブ病(CJD)、変異型クロイツフェルト・ヤコブ病、ゲルストマン-Straussler-Scheinker症候群、致死性家族性不眠症およびクールーである。
【0062】
さらなる態様において、タンパク質ホメオスタシスの機能障害に関連のある疾患は、機能消失障害である。用語「機能消失疾患」および「機能消失障害」は交換可能に使用される。機能消失疾患は、タンパク質の過剰な分解を引き起こす、タンパク質の高率の悪いフォールディングを特徴とする疾患のグループである。機能消失疾患としては、例えば、嚢胞線維症、気腫、およびリソソーム貯蔵疾患が挙げられる。嚢胞線維症では、変異または欠損酵素が嚢胞線維症膜貫通伝導制御因子(CFTR)である。このタンパク質の最も一般的な変異の一つは、3ヌクレオチド欠失(Δ)であるΔF508であり、タンパク質の508番目(508)の位置でアミノ酸フェニルアラニン(F)の欠失を生じる。リソソーム貯蔵疾患は、特異的なリソソーム酵素の欠失を特徴とする疾患の群であり、これは種々の組織で起こり、欠損酵素により、正常に分解されるタンパク質の蓄積が生じる。リソソーム酵素の欠失は、リソソームヒドロラーゼまたはリソソーム輸送に関連のあるタンパク質に生じ得る。リソソーム貯蔵疾患としては、限定されないが、アスパルチルグルコサミン尿症、ファブリー病、バッテン病、シスチン症、ファーバー、フコイドーシス、ガラクトシドシアリドーシス(Galactasidosialidosis)、ゴーシェ病(1型、2型および3型を含む)、Gm1ガングリオシドーシス、ハンター病、フルラー-シャイエ病、クラッベ病、a-マンノシドーシス、B-マンノシドーシス、マロトー-ラミー病、異染性白質萎縮症、モルキオA症候群、モルキオB症候群、ムコリピドーシスII、ムコリピドーシスIII、ニーマン-ピック病(A型、B型およびC型を含む)、ポンペ病、サンドホフ病、サンフィリポ病(A型、B型、C型およびD型を含む)、シンドラー病、シンドラー-カンザキ病、シアリドーシス、Sly症候群、テイ-サックス病およびウォルマン病が挙げられる。
【0063】
別の態様において、プロテオスタシスおよび/または熱ショックタンパク質の機能障害に関連のある疾患は、心臓血管疾患である。心臓血管疾患としては限定されないが、冠状動脈疾患、心筋梗塞、脳卒中、再狭窄および動脈硬化症が挙げられる。また、プロテオスタシスの機能障害に関連する状態としては、虚血/再かん流傷害、心筋虚血、安定性狭心症、不安定性狭心症、脳卒中、虚血性心臓疾患および脳虚血などの虚血状態が挙げられる。
【0064】
また、本発明は、薬理学的シャペロンの投与と併用して、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤の治療有効量を投与する工程を含む、プロテオスタシスの機能障害と関連している状態に苦しむ患者の治療方法を包含する。薬理学的シャペロンまたは速度論的安定剤は、既存の定常状態レベルのフォールディングされた変異タンパク質に結合し、フォールディングを安定化させることによりフォールディングの平衡を化学的に促進させる化合物をいう[Bouvier、Chem Biol 14:241-242、2007;Fan et al、Nat Med 5:112-115、1999;Sawkar et al、Proc Natl Acad Sci USA 99:15428-15433、2002;Johnson and Kelly、Accounts of Chemical Research 38:911-921、2005]。薬理学的シャペロンは、サーチュイン活性化化合物と併用してプロテオスタシスの機能障害と関連している状態に苦しむ患者を治療するのに充分な量で投与される。例示的な薬理学的シャペロンは、米国特許出願公開第20080056994号、同第20080009516号、同第20070281975号、同第20050130972号、同第20050137223号、同第20050203019号、同第20060264467号および同第20060287358号に記載されており、その内容は参照により本明細書に援用される。
【0065】
別の態様において、本発明は、機構的が異なる(mechanistically distinct)プロテオスタシス調節因子の投与と併用したHSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤により、プロテオスタシスの機能障害と関連している状態に苦しむ患者の治療方法である。用語「プロテオスタシス調節因子」は、細胞内タンパク質のホメオスタシスを増強する小分子、siRNAおよび生物製剤(例えば、タンパク質など)をいう。例えば、プロテオスタシス調節因子は、タンパク質の合成、フォールディング、輸送および分解の経路に影響を及ぼす薬剤であり得る。プロテオスタシス調節因子は、HSRシグナル伝達活性を刺激する薬理学的薬剤を包含する。プロテオスタシス調節因子は、シグナル伝達経路、例えば限定されないが、熱ショック応答またはアンフォールディングタンパク質の応答または両方を操作し、プロテオスタシスネットワーク成分の転写および翻訳をもたらすことにより機能を果たす。プロテオスタシス調節因子は、タンパク質(例えば、変異タンパク質)のフォールディング、輸送および機能を増強し得る。また、プロテオスタシス調節因子は、タンパク質シャペロンの転写または翻訳を上方調節すること、またはタンパク質シャペロンの分解を阻害することによりタンパク質シャペロンを調節し得る。プロテオスタシス調節因子は、しばしば、シャペロンレベルとフォールディング酵素レベルと、一部フォールディングされたコンホメーションアンサンブルに結合する巨大分子との協調的増大により、フォールディングの生物学に影響し得、したがって、より天然の構造を有する中間体へと進行させ、最終的に、輸送のためにフォールディングされた変異タンパク質の濃度を増大させる。一局面において、プロテオスタシス調節因子は、変異タンパク質のホメオスタシスを増強し得るが、変異タンパク質には結合しない点でプロテオスタシス調節因子はシャペロンと異なる。また、プロテオスタシス調節因子は、凝集経路または脱凝集酵素(disaggregase)活性を上方調節し得る。機構的が異なるプロテオスタシス調節因子は、HSF1のアセチル化の調節以外の機構によって細胞内プロテオスタシスを増強するプロテオスタシス調節因子である。例示的なプロテオスタシス調節因子はセラストロール、MG-132およびL型Ca2+チャネル遮断薬(例えば、ジリチアゼムおよびベラパミル)である。用語「セラストロール」は、セラストロールおよびその誘導体またはアナログをいい、限定されないが、その内容が参照により本明細書に明白に援用されるWesterheide et al.,J Biol Chem、2004. 279(53):p. 56053-60に記載されたセラストロール誘導体が挙げられる。セラストロール誘導体としては、例えば、セラストロールメチルエステル、二酢酸ジヒドロセラストロール、セラストロールブチルエーテル、ジヒドロセラストロール、セラストロール ベンジルエステル、プリメステロール、二酢酸プリメステロールおよびセラストロール三酢酸塩が挙げられる。
【0066】
特定の局面において、本発明は、機構的が異なるプロテオスタシス調節因子の投与と併用して、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤を投与する工程を含み、プロテオスタシス調節因子が熱ショック応答活性化因子である、プロテオスタシスの機能障害と関連している状態に苦しむ患者の治療方法である。熱ショック応答活性化因子は、例えば、熱ショック転写因子1(HSF1)を直接もしくは間接的に活性化させること、Hsp70の転写もしくはタンパク質発現を増大させること、Hsp90を阻害すること、および/またはシャペロンmRNAもしくはタンパク質発現を活性化させる、もしくは増大させることにより、熱ショック応答を間接的または直接活性化するか、または増大させる薬剤である(Westerheide et al.,J Biol Chem、2004. 279(53):pp.56053-60、その内容は、参照により本明細書に明白に援用される)。熱ショック応答活性化因子は、基底熱ショック応答を間接的または直接増大させる薬剤を包含することを理解されたい。用語「熱ショック応答活性化因子」、「熱ショック活性化因子」、「熱ショック応答誘導薬」および「熱ショック誘導薬」は本明細書において互換的に使用される。熱ショック応答活性化因子の非限定的な例は、セラストロール、非ステロイド系抗炎症薬、アンサマイシン、ゲルデナマイシン、ラジシオール、グルクロン酸、およびトリブチリン(tributylin)である。また、熱ショック応答活性化因子は、例えば、米国特許公開公報第20070259820号、同第20070207992号、同第20070179087号、同第20060148767号に記載されており、これらのそれぞれの内容は、参照により本明細書に明白に援用される。いくつかの態様において、熱ショック応答活性化因子は、小分子熱ショック応答活性化因子である。HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤および熱ショック応答活性化因子は、それぞれ、熱ショック応答活性化因子とアセチル化を阻害する薬剤を合わせた用量が一緒になって熱ショック応答を増大させるのに充分な用量で投与され得る。熱ショック活性化因子とHSF1のアセチル化を阻害する薬剤の併用により、該薬剤のいずれかを単独で投与する場合よりも大きい熱ショック応答の増大がもたらされる。いくつかの態様において、投与される熱ショック活性化因子の用量は、単独では熱ショック応答を誘導するに充分でないが、該用量は、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤と組み合わせて熱ショック応答を活性化するには充分である。本発明のいくつかの局面において、熱ショック活性化因子とHSF1のDNA結合ドメインのアセチル化を阻害する薬剤の組合せは、熱ショック応答の誘導に対して相加効果より大きい効果を有する。他の局面において、熱ショック応答の増大は、Hsp70の転写またはタンパク質発現の増大を測定することにより測定される。熱ショック応答の増大の例示的な測定方法は、Westerheide et al.(2004)Celastrols as inducers of 熱 shock reipose and cytoprotection. J Biol Chem、2004. 279(53):pp. 56053-60に記載されており、その内容は、参照により本明細書に援用される。さらなる局面において、熱ショック応答活性化因子とHSF1のアセチル化を阻害する薬剤の組合せの投与後の熱ショック応答の増大は、該薬剤の組合せの非存在下での熱ショック応答の少なくとも約2倍、少なくとも約3 倍、少なくとも約5倍、少なくとも約10倍、少なくとも約15倍、または少なくとも約20倍大きいHsp70の転写またはタンパク質発現の増大を包含する。
【0067】
実施例2に示すように、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤も投与する場合、熱ショック応答を誘導するのに、熱ショック活性化因子は、より少ない用量で投与され得ることがわかった。熱ショック応答活性化因子の用量またはEC50が低減されることは、熱ショック応答を誘導するために必要とされる熱ショック応答活性化因子の用量が毒性をもたらす状況があるため、有利であり得る。したがって、いくつかの態様において、本発明は、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤と併用して、熱ショック応答活性化因子を投与する工程を含み、熱ショック応答活性化因子とHSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤を合わせた用量が一緒になって、熱ショック応答を活性化または増大するのに充分である、熱ショック応答を活性化または増大する方法に関する。他の態様において、本発明は、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤と、熱ショック応答活性化因子を投与する工程を含む、熱ショック応答の誘導を必要とする患者において熱ショック応答を誘導するのに有効な熱ショック応答活性化因子の用量を低減する方法である。HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤は、両方の薬剤が同じ組成物の一部である場合、および/または両方の薬剤を同時に投与する場合、および/または両方の薬剤を異なる時点もしくは逐次投与する場合、別の薬剤と併用して投与してもよいことは理解されよう。一態様において、HSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する薬剤は、熱ショック応答活性化因子の前に投与される。
【0068】
特定の他の態様において、本発明は、熱ショック応答活性化因子とHSF1のアセチル化を阻害する薬剤を含む医薬組成物に関する。他の態様において、医薬組成物は、熱ショック活性化因子と、HAT阻害薬、単離サーチュインおよびサーチュイン活性化剤からなる群より選択されるHSF1のアセチル化を阻害する薬剤とを含む。他の態様において、本発明は、熱ショック応答活性化因子とSIRT1活性化剤を含む医薬組成物である。さらなる態様において、本発明は、熱ショック応答活性化因子とHAT阻害薬を含む医薬組成物である。
【0069】
別の局面において、本発明は、治療を必要とする患者に、HSF1のDNA結合ドメインのアセチル化を促進させる薬剤の治療有効量を投与する工程を含む、治療を必要とする患者において熱ショックタンパク質の発現の増大と関連している状態の治療方法である。一態様において、HSF1のアセチル化を促進する薬剤はサーチュイン阻害剤である。別の態様において、HSF1のアセチル化を促進する薬剤はHAT活性化因子である。サーチュイン阻害剤は上記で説明した。別の態様において、熱ショックタンパク質の発現の増大と関連している状態は癌または腫瘍である。また別の態様において、熱ショックタンパク質の発現の増大と関連している状態はウイルス感染である。HSF1のアセチル化を促進させる薬剤は、医薬用担体または賦形剤を含む医薬組成物にて投与され得る。
【0070】
熱ショックタンパク質の発現の増大と関連している状態は癌または腫瘍であり得る。本発明の方法に従って治療され得る癌としては、限定されないが、乳癌、結腸癌、膵臓癌、前立腺癌、肺癌、卵巣癌、頚部癌、多発性骨髄腫、基底細胞癌、神経芽腫、血液の癌、横紋筋肉腫、肝臓癌、皮膚癌、白血病、基底細胞癌、膀胱癌、子宮内膜癌、グリオーマ、リンパ種、および胃腸の癌が挙げられる。
【0071】
別の態様において、本発明は、化学療法剤の投与と併用して、HSF1のアセチル化を促進させる薬剤を投与する工程を含む、癌または腫瘍の治療方法である。使用され得る化学療法剤としては、限定されないが、シクロホスファミド(Cytoxan(登録商標))などのアルキル化剤;ブスルファン、インプロスルファンおよびピポスルファンなどのアルキルスルホン酸塩;ベンゾドパ、カルボクオン、メツレドパ、およびウレドパなどのアジリジン;エチレンイミンおよびメチラメラミン、例えば、アルトレタンミン、トリエチレンメラミン、トリエチレン(etylene)ホスホラミド、トリエチレンチオホスファオ(phosphao)ラミドおよびトリメチロロメラミン;クロラムブシル、クロルナファジン、コロホスファミド、エストラムスチン、イホスファミド、メクロレタミン、塩酸メクロレタミンオキシド、メルファラン、ノベンビシン、フェンステリン、プレドニムスチン、トロホスファミド、ウラシルマスタードなどのナイトロジェンマスタード;カルムスチン、クロロゾトシン、フォテムスチン、ロムスチン、ニムスチン、ラニムスチンなどのニトロソ尿素;アクラシノマイシン、アクチノマイシン、アウトラマイシン、アザセリン、ブレオマイシン、カクチノマイシン、カリケアマイシン、カラビシン、カミノマイシン、カルジノフィリン、クロモマイシン、ダクチノマイシン、ダウノルビシン、デトルビシン、6-ジアゾ-5-オキソ-L-ノルロイシン、ドキソルビシン、エピルビシン、エソルビシン、イダルビシン、マルセロマイシン、マイトマイシン、ミコフェノール酸、ノガラマイシン、オリボマイシン、ペプロマイシン、ポトフィロマイシン、ピューロマイシン、ケラマイシン、ロドルビシン、ストレプトニグリン、ストレプトゾシン、ツベルシジン、ウベニメックス、ジノスタチン、ゾルビシンなどの抗生物質;メトトレキサートおよび5-フルオロウラシル(5-FU)などの代謝拮抗物質;デノプテリン、メトトレキサート、プテロプテリン、トリメトレキサートなどの葉酸アナログ;フルダラビン、6-メルカプトプリン、チアミプリン、チオグアニンなどのプリンアナログ;アンシタビン、アザシチジン、6-アザウリジン、カルモフル、シタラビン、ジデオキシウリジン、ドキシフルウリジン、エノシタビン、フロクスウリジンなどのピリミジンアナログ;カルステロン、プロピオン酸ドロモスタノロン、エピチオスタノール、メピチオスタン、テストラクトンなどのアンドロゲン;アミノグルテチミド、マイトタン、トリロスタンなどの抗アドレナリン薬;フロリン酸などの葉酸補給薬;アセグラトン;アルドホスファミドグリコシド;アミノレブリン酸;アムサクリン;ベストラブシル;ビサントレン;エダトラキサート;デホファミン;デメコルチン;ジアジクオン;エルフオルニチン;酢酸エリプチニウム;エトグルシド;硝酸ガリウム;ヒドロキシ尿素;レンチナン;ロニダミン;ミトグアゾン;ミトザントロン;モピダモール;ニトラクリン;ペントスタチン;フェナメト;ピラルビシン;ポドフィリン酸;2-エチルヒドラジド;プロカルバジン;PSK(登録商標);ラゾキサン;シゾフィラン;スピロゲルマニウム;テヌアゾン酸;トリアジクオン;2,2’,2”-トリクロロトリエチルアミン;ウレタン;ビンデシン;ダカルバジン;マンノムスチン;ミトブロニトール;ミトラクトール;ピポブロマン;ガシトシン;アラビノシド(「Ara-C」);シクロホスファミド;チオテパ;タキサン、例えば、パクリタキセル(Taxol(登録商標)、Bristol-Myers Squibb Oncology、Princeton、N.J.)およびドセタキセル(Taxotere(登録商標);Aventis Antony、France);クロラムブシル;ゲムシタビン;6-チオグアニン;メルカプトプリン;メトトレキサート;シスプラチンおよびカルボプラチンなどの白金アナログ;ビンブラスチン;白金;エトポシド(VP-16);イホスファミド;マイトマイシンC;ミトザントロン;ビンクリスチン;ビノレルビン;ナベルビン;ノバントロン;テニポシド;ダウノマイシン;アミノプテリン;キセロダ;イバンドロネート;CPT-11;トポイソメラーゼ阻害薬RFS 2000;ジフルオロメチルオルニチン(DMFO);レチノイン酸;エスペラマイシン;カペシタビン;ならびに上記のもののいずれかの薬学的に許容され得る塩、酸または誘導体が挙げられる。また、この定義には、腫瘍に対するホルモン作用を調節または阻害する作用をする抗ホルモン薬、例えば、タモキシフェン、ラロキシフェン、アロマターゼ阻害薬4(5)-イミダゾール、4-ヒドロキシタモキシフェン、トリオキシフェン、ケオキシフェン、LY 117018、オナプリストン、およびトレミフェン(ファレストン)などの抗エストロゲン;ならびにフルタミド、ニルタミド、ビカルタミド、ロイプロリド、およびゴセレリンなどの抗アンドロゲン;ならびに上記のもののいずれかの薬学的に許容され得る塩、酸または誘導体も含まれる。
【0072】
さらなる態様において、本発明は、放射線療法と併用してHSF1のアセチル化を促進する工程を含む、癌または腫瘍の治療方法である。
【0073】
別の態様において、熱ショックタンパク質の発現の増大と関連している状態はウイルス感染である。さらなる態様において、ウイルス感染は、腫瘍ウイルスおよびRNAウイルスから選択されるウイルスによって引き起こされるものである。例示的な腫瘍ウイルスは、ヘルペスウイルス、パピローマウイルス、ポリオーマウイルスおよびHTLV-1である[McCance et al. Human Tumor Viruses、1998、American Society for Microbiology]。ヘルペスウイルスとしては、限定されないが、EBV(HHV-4)、HHV-6およびHHV-8が挙げられる。パピローマウイルス としては、限定されないが、HPV-1、-2、-4、-5、-6、-8、-6、-11、-16、-18、-31、-33、-35、-45、-51、-52、-58および-58が挙げられる。
【0074】
RNAウイルスとしては、例えば、アレナウイルス科、ブンヤウイルス科、カルシウイルス科、コロナウイルス科、フィロウイルス科、フラウイルス科、オルトミクソウイルス科、パラミクソウイルス科、ピコルナウイルス科、レオウイルス科、ラブドウイルス科、レトロウイルス科、またはトガウイルス科が挙げられる。例示的なRNAウイルスとしては、限定されないが、SARS関連コロナウイルス、腸疾患および呼吸器疾患に関連しているヒトトロウイルスなどのヒトコロナウイルス;ノーウォークウイルス、黄熱病ウイルス、西ナイルウイルス、C型肝炎ウイルス、デング熱ウイルス、ポリオウイルス、感冒ウイルス、A型肝炎ウイルス、E型肝炎、ロタウイルス、ボルナ病ウイルス;ハンタウイルス、カリフォルニア脳炎ウイルス、日本脳炎ウイルス、ラクロスウイルス、リフトバレー熱ウイルス、ブンヤウイルス、アルボウイルス、エボラウイルスおよびマルブルグウイルスなどのブンヤウイルス;インフルエンザウイルスA型、インフルエンザウイルスB型、インフルエンザウイルスC型、流行性耳下腺炎ウイルス、はしかウイルス、亜急性硬化性汎脳炎(SSPE)ウイルスならびに呼吸器合胞体ウイルス(RSV)が挙げられる。
【0075】
さらなる態様において、本発明は、抗ウイルス薬の投与と併用して、HSF1のアセチル化を促進させる薬剤を投与する工程を含む、ウイルス感染に苦しむ患者の治療方法である。
【0076】
本発明のまた別の局面において、熱ショックタンパク質の発現の増大と関連している状態は、炎症状態および/または自己免疫疾患である。
【0077】
本発明の治療方法に従って使用される薬理学的薬剤または医薬組成物の形態は、意図される投与様式および治療適用に依存する。また、組成物は、所望される製剤に応じて、動物またはヒトへの投与のための医薬組成物を製剤化するために一般に使用されるビヒクルと定義される薬学的に許容され得る無毒性の担体または希釈剤を含み得る。希釈剤は、薬理学的薬剤または組成物の生物学的活性に影響を及ぼさないように選択される。かかる希釈剤の例は、蒸留水、リン酸緩衝生理食塩水、リンゲル溶液、デキストロース溶液、およびハンクス溶液である。また、医薬組成物または製剤は、他の担体、補助剤または無毒性、非治療性、非免疫原性安定剤なども含んでいてもよい。また、医薬組成物は、タンパク質、キトサンなどの多糖、ポリ乳酸、ポリグリコール酸およびコポリマー(例えば、ラテックス官能化SepharoseTM、アガロース、セルロースなど)、ポリマーアミノ酸、アミノ酸コポリマー、ならびに脂質凝集物(油滴またはリポソームなど)などの大型のゆっくり代謝される巨大分子を含んでいてもよい。
【0078】
非経口投与では、医薬組成物または薬理学的薬剤は、水油、生理食塩水、グリセロール、またはエタノールなどの滅菌液であり得る医薬用担体を有する生理学的に許容され得る希釈剤中の該物質の注射用投薬量の溶液または懸濁液として投与され得る。さらに、湿潤剤または乳化剤、界面活性剤、pH緩衝物質などの補助物質が組成物中に存在していてもよい。医薬組成物の他の成分は、石油、動物、植物または合成起源のもの、例えば、ピーナッツ油、ダイズ油、および鉱物油である。一般に、プロピレングリコールまたはポリエチレングリコールなどのグリコールは、特に注射用溶液に好ましい液体担体である。
【0079】
組成物は、液状溶液または懸濁液のいずれかとしての注射用製剤として調製され得る;注射前の液体ビヒクル中での溶液もしくは懸濁液に適した固形形態も調製され得る。また、該調製物を乳化してもよく、リポソームもしくはポリラクチド、ポリグリコリドなどの微粒子、または上記のようなアジュバント効果の増強のためのコポリマー内にカプセル封入してもよい。Langer、Science 249:1527、1990およびHanes、Advanced Drug Delivery Reviews 28:97-119、1997。本明細書に記載の組成物および薬理学的薬剤は、デポー注射または活性成分の持続的もしくはパルス的放出を可能にするような様式で製剤化され得る埋め込み調製物の形態で投与され得る。
【0080】
他の投与様式に適したさらなる製剤としては、経口、鼻腔内および肺経由製剤、坐剤、または経皮適用が挙げられる。
【0081】
坐剤用では、結合剤および担体として、例えば、ポリアルキレングリコールまたはトリグリセリドが挙げられる;かかる坐剤は、0.5%〜10%、好ましくは1%〜2%の範囲の活性成分を含む混合物から形成され得る。経口製剤は、医薬等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、および炭酸マグネシウムなどの賦形剤を含む。局所適用により、経皮または皮内送達がもたらされ得る。経皮送達は、皮膚パッチを用いて、またはトランスフェロソーム(transferosome)を用いて達成され得る[Paul et al.,Eur. J. Immunol. 25:3521-24、1995;Cevc et al.,Biochem. Biophys. Acta 1368:201-15、1998]。
【0082】
別の態様において、本発明は、
試験薬剤を細胞に投与する工程;および
HSF1のDNA結合ドメイン内のリジン残基のアセチル化をモニタリングする工程
を含み、
試験薬剤の非存在下のものと比べたときのDNA結合ドメインのアセチル化の変化は、試験薬剤が細胞においてHSF1活性を調節することを示す、
細胞においてHSF1活性を調節する薬剤を同定する方法である。
【0083】
試験薬剤は、広範な種々の供給源、例えば、合成または天然化合物のライブラリーから得られ得る。例えば、広範な種々の有機化合物および他の薬剤のランダムおよび指向的合成のための数多くの手段が利用可能である。細菌、真菌、植物および動物の抽出物の形態の天然化合物のライブラリーもまた利用可能であり、あるいは容易に作製できる。さらに、天然または合成により作製されたライブラリーおよび化合物は、従来の化学的、物理的および生化学的手段によって容易に修飾され、コンビナトリアルライブラリーを作製するために使用され得る。また、薬理学的薬剤が、構造的アナログを作製するために、アシル化、アルキル化、エステル化、アミド化などの指向的またはランダムな化学的修飾に供され得る。試験薬剤は、数多くの化学的種類、典型的には合成、半合成または天然の無機または有機分子を包含する。試験薬剤としては、合成または天然化合物の大きなライブラリーに見られるものが挙げられる。
【0084】
一態様において、試験薬剤はDNA結合ドメインのアセチル化を阻害する。別の態様において、試験薬剤はDNA結合ドメインのアセチル化を増大させる。さらなる態様において、DNA結合ドメイン内のリジンのアセチル化がモニタリングされる。例えば、非ヒトHSF1内のHSF1 K80または対応する保存アミノ酸がモニタリングされ得る。さらなる態様において、細胞はヒト細胞である。
【0085】
また別の態様において、本発明は、
a. 試験薬剤を細胞または細胞溶解物に投与する工程;
b. サーチュイン基質のアセチル化を測定する工程;
c. 工程(b)のサーチュイン基質のアセチル化を、試験薬剤の非存在下でのサーチュイン基質のアセチル化と比較する工程
を含み、
試験薬剤の投与後のサーチュイン基質のアセチル化の増大は、試験薬剤がHSF1活性を阻害することを示し、試験薬剤の投与後のサーチュイン基質のアセチル化の減少は、試験薬剤がHSF1活性を増大させることを示す、
HSF1活性を調節する薬剤を同定する方法である。サーチュイン基質は、サーチュインによって脱アセチル化され得る因子(タンパク質またはペプチドなど)である。一態様において、サーチュイン基質はHSF1である。別の態様において、サーチュイン基質はHSF1以外の基質である。サーチュイン基質のアセチル化は、例えば、細胞ホモジネートにおいて測定され得る。一態様において、試験薬剤はサーチュイン活性を阻害する。別の態様において、試験薬剤はサーチュイン活性を活性化する。さらなる態様において、サーチュインはSIRT1である。また別の態様において、サーチュインはヒトSIRT1である。別の態様において、細胞はヒト細胞である。サーチュインデアセチラーゼ活性を測定するため、およびサーチュイン基質のアセチル化を測定するためのアッセイは、例えば米国特許公開公報第20080249103号に記載されており、その内容は参照により本明細書に援用される。
【0086】
また別の態様において、本発明は、
a. 試験薬剤を細胞または細胞溶解物に投与する工程;
b. HAT基質のアセチル化を測定する工程;
c. 工程(b)のHAT基質のアセチル化を、試験薬剤の非存在下でのHAT基質のアセチル化と比較する工程
を含み、
試験薬剤の投与後のHAT基質のアセチル化の増大は、試験薬剤がHSF1活性を阻害することを示し、試験薬剤の投与後のHAT基質のアセチル化の減少は、試験薬剤がHSF1活性を増大させることを示す、
HSF1活性を調節する薬剤を同定する方法である。HAT基質は、ヒストンアセチルトランスフェラーゼによってアセチル化され得る因子(タンパク質またはペプチドなど)である。いくつかの局面において、HAT基質はHSF1ではない。
【0087】
別の局面において、本発明は、本明細書に記載の方法によって同定されるHSF1活性を調節する薬剤である。
【0088】
本発明を、なんら限定を意図しない以下の実施例によって説明する。
【実施例】
【0089】
実施例
実施例1:デアセチラーゼSIRT1による熱ショック因子1のストレス誘導性調節
熱ショック因子1(HSF1)は、タンパク質のミスフォールディングと関連するタンパク質損傷性ストレスから細胞を保護するのに必須であり、インスリンシグナル伝達経路および加齢を調節する。ここに、本発明者らは、ヒトHSF1が、DNA結合活性を負に調節する重要な残基において誘導的にアセチル化されることを示す。デアセチラーゼおよび長寿因子SIRT1の活性化により、HSF1が、脱アセチル化DNA結合コンピテント状態に維持されることによって、Hsp70プロモーターへのHSF1結合が長期化された。逆に、SIRT1の下方調節によって、熱ショック応答(HSR)の減衰およびその対応プロモーターエレメントからのHSF1の放出が加速された。この結果は、寿命の調節におけるHSF1の要件の機構的根拠を示し、タンパク質のホメオスタシスおよびHSRにおけるSIRT1の役割を確立する。
【0090】
多様な環境的および生理学的ストレスによるHSF1の一過的活性化は、不活性HSF1単量体の構成的発現、該単量体のDNA結合コンピテント三量体への変換、セリン残基でのHSF1のリン酸化の増大、転写の増強、ならびにHSF1 DNA結合および転写活性の減衰を伴う多工程プロセスである(1)。HSF1は、分子シャペロンHsp70およびHsp90などのタンパク質のホメオスタシスを調節する多数の遺伝子の転写を活性化する。これらのシャペロンはHSF1と結合して、負のフィードバックループを開始させ、HSF1転写活性を阻害する(2)。しかしながら、HSF1は、その標的プロモーター部位から放出されず(3)、これは、HSF1サイクルを完結させるさらなる機構が存在するはずであることを示す。
【0091】
ストレス抵抗性および代謝状態は、タンパク質のホメオスタシスと寿命の増大に密接に関連している。C. elegansでは、インスリンシグナル伝達の低減の保護効果には、タンパク質のミスフォールディング損傷を防ぎ、長寿を促進するために、HSF1とFOXO転写因子DAF-16が必要とされる(4、5)。低カロリー摂取の有益な効果は、代謝制御下にあるNAD依存性デアセチラーゼであるサーチュインファミリー構成員Sir2によって媒介される(6)。哺乳動物Sir2ホモログSIRT1は、数ある細胞内保護経路のなかでも転写因子FOXO3を調節する(7)。したがって、本発明者らは、サーチュイン、具体的にはSIRT1がHSF1活性を調節し、それによりこれらの3つの長寿因子間に直接的な関連性を提供するかどうかを試験した。
【0092】
本発明者らは、HeLa細胞をサーチュイン阻害薬ニコチンアミドで処理し(8)、次いで、細胞を、HSRを誘導することが知られた種々のストレスに曝露した(9)。ニコチンアミド治療により、すべての主要な種類の熱ショック遺伝子(hsp70、hsp90、hsp40およびhsp27、図1A)由来のストレス誘導性mRNAのアバンダンスが減少し、これは、HSRの充分な誘導にサーチュインが必要とされることを示す。3つの核サーチュインのうち、SIRT1は、充分特徴付けられた標的を有する(10)。したがって、本発明者らは、SIRT1をHSRの調節のための候補として調査した。siRNAによりSIRT1が枯渇されると、6時間の熱ショック(HS)中に生成されるhsp70 mRNAの量は、対照siRNAでトランスフェクトされた細胞のものの4分の1であった(図1B)。
【0093】
SIRT1がhsp70プロモーターへのHSF1の漸増に影響を及ぼすかどうかを調べるため、本発明者らは、HSの前に対照またはSIRT1 siRNAでトランスフェクトされた細胞を用いてクロマチン免疫沈降(ChIP)アッセイを行なった(図1C)。対照siRNA処理細胞では、hsp70プロモーターへのHSF1の結合は、急速に起こるが、熱ショックの30分の時点で減衰し始め(11)、6時間にわたって徐々に減少する。しかしながら、SIRT1 siRNAトランスフェクト細胞では、この時間中にHSF1の約4分の1程度がプロモーターと結合した。この結果は、HSF1 DNA結合活性およびhsp70発現のインビボ調節因子としてのSIRT1の役割を裏付ける。
【0094】
HSF1がSIRT1の直接的な標的であるかどうかを調べるため、本発明者らは、HSF1のアセチル化状態を調べた。本発明者らは、293T細胞を、Flag-HSF1融合タンパク質およびp300をコードするベクターでトランスフェクトし、いくつかのHSR誘導薬に曝露した。免疫沈降させたHSF1をアセチル化リジンに結合する抗体を用いてウエスタンブロッティングによって解析した。アセチル化HSF1は、未処理細胞では検出されなかったが、種々のストレス状態に曝露した細胞には存在した(図2A)。p300またはCBPのいずれかの過剰発現では、HSF1のアセチル化がもたらされたが、pCAFではもたらされず(図5A)HS後、p300はhsp70プロモーターに漸増したため、HSF1アセチル化を調節する内因性アセチルトランスフェラーゼはp300/CBPであり得る(図5B)。また、SIRT1は、基底状態およびストレス状態の両方でhsp70プロモーターに結合する(図5C)。
【0095】
デアセチラーゼは、トリコスタチンAによって阻害されるI類およびII類HDACファミリー(12)ならびにニコチンアミドによって阻害されるNAD+依存性のIII類サーチュインファミリー(8)の3つのファミリーに分類される。トリコスタチンAは、HSF1の脱アセチル化に対して効果を有しなかったが、ニコチンアミドは、単独またはトリコスタチンAの存在下で脱アセチル化を阻害した(図2B)。SIRT1 WTの過剰発現は、HSF1アセチル化を阻害したが、障害されたNAD依存性デアセチラーゼ活性を有する点変異[SIRT1 H363Y(13)]はアセチル化を阻害せず(図2C)、これは、HSF1機能におけるSIRT1の役割を裏付ける。HSF1アセチル化は293TおよびCos7細胞において検出されたため、HSF1アセチル化は細胞型特異的ではなかった。HSF1アセチル化はp300の過剰発現によって増強されたが、p300過剰発現を必要としなかった(図2、AおよびD)。HSF1アセチル化の速度論はHSF1 活性化の速度論と適合しない。アセチル化は遅延し、HSF1活性およびDNA結合が減衰している期間中、持続する(11)。さらに、潜在的にリン酸化された10個のセリンがアラニンで置換されたHSF1は、アセチル化に対してコンピテントのままであり、HSF1のリン酸化はアセチル化のための必要条件でないことを示す(図6)。
【0096】
HSR後の時点でのHSF1アセチル化の持続およびSIRT1とHSF1の一緒の共免疫沈降(図2D)は、本発明者らに、SIRT1がHSF1活性の減衰において役割を有するかどうかを調べることを導いた。本発明者らは、HeLa細胞を、SIRT1活性の小分子誘導薬であるレスベラトロールで処理し(14)、HSF1 DNA結合活性をオリゴヌクレオチド系プルダウンアッセイにおいてアッセイした(15)(図2E)。HSおよびビヒクル単独で処理した細胞では、HSF1 DNA結合が2時間以内に誘導され、6時間後に減衰された。HSF1の一過性の活性化は、HSF1のストレス誘導性リン酸化状態を検出するSDS-PAGE上の移動度の変化によって反映された(16)。対照的に、レスベラトロール処理細胞におけるHSF1は、8時間の連続HSの後であっても、DNA結合コンピテントおよびリン酸化状態の状態が持続した。SIRT1を過剰発現する細胞では、ChIP実験によって測定したとき、HSF1 DNA結合はが増強され、減衰が抑制された(図2F)。この結果は、SIRT1のアバンダンスおよび活性の変化によりHSRの減衰が調節されることを示唆する。
【0097】
アセチル化がHSF1 DNA結合を調節する機構を解明するため、本発明者らは、293T細胞から精製したFlag-HSF1由来のペプチドの質量分析によってHSF1上のアセチル化部位を特定した。HSF1内の少なくとも9つのリジンが、ストレスに応答してアセチル化され(図7)、DNA結合ドメイン内に位置するそのK80は、酵母HSFの対応するリジンの変異は機能低下表現型を引き起こすため(17、18)、特に興味をそそられた。さらに、K. lactis HSFの結晶構造の解析により、
ヒトHSF1 K80に対応するリジンは、主要DNA結合ヘリックスと、柔軟で溶媒曝露ループと連結し、DNAのリン酸主鎖と水素結合を形成する短いドメイン内に位置することが示された(19)。HSF-HSE結晶構造の比較タンパク質モデリングでは、ヒトHSF1 K80はDNA主鎖と密に接触していることが示され(図8)、アセチル化によるリジンの正電荷の中和によりDNA結合が妨げられるはずであることを示す。
【0098】
したがって、本発明者らは、構成的アセチル化を模倣するためにK80をグルタミンで置換した。HSF1 WTおよびHSF1 K80Q発現構築物でトランスフェクトされたhsf1-/-線維芽細胞(20)由来の抽出物では、変異タンパク質は、電気泳動移動度シフトアッセイ(EMSA)において、DNAに結合しなかった(図3A)。しかしながら、K80Q変異体は、なお、DNA結合状態のホールマークであるHS誘導性三量体に合成された(図9)。また、K80での他のアミノ酸の置換(K80R、-A、-H、-N、および-T)により、欠陥性DNA結合がもたらされた(図3A、図10)。インビトロでは、組換え非アセチル化WT HSF1は、合成HSEに容易に結合したが、K80Q 変異タンパク質は結合しなかった(図3B)。本発明者らは、WTおよびK80変異体をhsf1-/-線維芽細胞に導入し、HSF1標的遺伝子のHS誘導性発現をqPCRによって解析した。WT HSF1はhsp mRNAの発現を誘導したが、HSF1 K80変異体は非機能性であった(図3C)。変異体は、HS時、核に局在したが、熱ショックヒト細胞において起こる核ストレス体内への再局在において障害された(図11〜12)(21)。残基80の非修飾リジン側鎖は、HSF1 HSE結合能力、核ストレス体内への再局在、および標的遺伝子の発現に必要と思われる。したがって、本発明者らは、HSF1 K80のアセチル化が、DNAからのHSF1三量体の放出の調節を引き起こし、したがって、HSRの減衰における調節工程を表すことを提案する(図13)。
【0099】
SIRT1によるHSRの調節の生物学的重要性を検証するため、本発明者らは、シャペロンの発現によって耐熱性の増大がもたらされるストレス抵抗性のアッセイを使用した(22)。293T細胞をSIRT1でトランスフェクトし、またはトランスフェクトせず、45℃のHSに20または30分間曝露し、24時間回復させ、細胞死について解析した。予測どおり、45℃のHSにより細胞死がもたらされ、これは、処理時間とともに増大した(図4A)。両方の時間点で、SIRT1を過剰発現する細胞は、約3分の1の細胞が細胞死を受けた(図4A)。SIRT1における加齢調節の変化がHSF1活性およびHSRに影響を及ぼすかどうかを調べるため、本発明者らは、加齢プロセスにおける分子的変化に関する研究に広く使用されているヒトWI-38線維芽細胞を使用した。初期および後期の継代数を比較すると、本発明者らは、加齢により、SIRT1のアバンダンスの低下と相関しているHSRの減少およびHSF1 DNA結合活性の活性化の低下がもたらされることを見い出した(図4B)。
【0100】
SIRT1がHSF1を調節するという所見は、寿命の調節におけるHSF1の役割に関する先の観察(4、5)を補完する。HSF1は、細胞の栄養、ストレスおよび寿命が関連する調節ネットワークの中心であると思われる。多くのSIRT1調節転写因子、例えば、FOXO3、p53およびNF-κBは、細胞ストレス応答において重要な役割を有する(7、23、24)。このストレス調節ネットワークへのHSF1の添加は、SIRT1媒介性細胞保護におけるタンパク質のホメオスタシスの中心的役割を強調し(図14)、HSRの分子応答を代謝的要求と関連付け得る。細胞系および動物試験における観察の一致はHSRの加齢関連減少であり(22)、これは、少なくとも一部は、HSF1活性のSIRT1制御に起因し得る。生物レベルでは、本発明者らは、HSF1標的遺伝子の調節は食事と栄養に影響され得ると予測する。
【0101】
材料および方法
構築物、抗体および試薬
mHSF1-FlagおよびhHSF1-Mycは以前に記載された(25,26)。HSF1点変異は、
QuikChange部位特異的変異誘発(Stratagene)によって作出し、配列決定によって確認した。Myc-HSF1の10個のセリン変異体について、リン酸化セリン残基 S230、S303、S307、S314、S319、S320、S338、S363、S368およびS369(参考文献27および本発明者らの未公表データ)はすべて、アラニンに変異した。SIRT1 WTおよびH363Y変異体発現構築物は、Tony Kouzarides博士(ケンブリッジ大学)から、p300発現構築物はDavid Livingston博士(ハーバード大学)からご提供いただいた。この試験で使用した抗体は、α-Flag M2(Sigma)、α-AcK(Cell Signaling 9441)、α-HSF1(4)、α-SIR2(Upstate Biotech #07-131)、α-Myc(Clontech)およびα-p300(Santa Cruz)である。使用した化合物は、ニコチンアミド(Sigma)、トリコスタチンA(Upstate)、EGS(Sigma)、セラストロール(GAIA Chemical Corporation)、CdCl2(Sigma)およびMG132(Calbiochem)である。
【0102】
細胞培養物、トランスフェクションおよび処理条件
すべての細胞は、5%CO2加湿雰囲気中、37℃に維持した。K562細胞は、10%FCSおよび抗生物質(ペニシリンおよびストレプトマイシン)を添加したRPMI 1640培地中で培養した。293T、HeLa、Cos7およびWI-38細胞は、10%FCSおよび抗生物質を添加したDMEM中で培養した。hsf1-/-マウス胚線維芽細胞(29)は、10%FCS、10mM非必須アミノ酸、0.96μlの2-メルカプトエタノール/100 ml、および抗生物質を有するDMEM中で培養した。293T細胞をPolyfect(Qiagen)を用いて、製造業者の使用説明書に従ってトランスフェクトした。K562、Cos7およびhsf1-/-細胞は、Gene Pulserエレクトロポレーション装置(Bio-Rad)を用いてエレクトロポレーションによってトランスフェクトした。細胞を予備加温循環水浴中に42℃で1時間浸漬することによって熱ショックを誘導した。セラストロール処理は、5μMの濃度で1時間行ない、CdCl2処理は、50μMで6時間、MG132は20μMで6時間行なった。細胞に、トリコスタチンAは1mMで一晩、ニコチンアミドは5mMで一晩、レスベラトロールは50μMで40時間添加した。
【0103】
siRNAトランスフェクション
HeLa細胞を、Oligofectamine(Invitrogen)で、製造業者のプロトコルに従って、200nMのDharmacon SmartPool SIRT1 siRNAを用いてトランスフェクトした。24時間後、細胞を分割し、再度トランスフェクトした。2回目のトランスフェクションの48時間後、Trizolを用いてRNAを単離した。SIRT1のノックダウンを確認するため、プライマーを使用した。
【0104】
アセチル化アッセイ
293TまたはCos7細胞を、Flag-HSF1またはMyc-HSF1およびp300でトランスフェクトし、次いで、種々のストレスでの処理前に、ニコチンアミドで処理するか、または非処理とした。細胞溶解物を、Flag抗体またはMyc抗体を用いた免疫沈降に供し、アセチル化HSF1を、アセチル化リジンを認識する抗体を使用するウエスタンブロッティングによって検出した。
【0105】
クロマチン免疫沈降(ChIP)
ChIP反応は、本質的に先に記載のとおりに行なった(30)。HeLa細胞(3x107)から作製した試料を、10μlのα-HSF1(28)を用いて4℃で一晩免疫沈降させた。hsp70.1プロモーター(GenBank Acc# M11717)および近位HSEのためのプライマーを使用した。結果を1%投入量を用いて行なった反応に対して標準化した。
【0106】
293T細胞からのHSF1の精製
293T細胞を、CMV-p300とともに、またはなしで、mHSF1-Flagで製造業者のプロトコルに従ってPolyfect(Qiagen)に示されたとおりにトランスフェクトした。細胞をトリコスタチンAまたはニコチンアミドで表示のとおりに4時間処理した後、ストレス処理を行なった。次いで、細胞ペレットを収集し、RIPAバッファー中で溶解させた。mHSF1-Flagをα-Flag M2親和性ゲルビーズ(Sigma F2426)で免疫沈降させ、Flagペプチドで溶出させた。試料をSDS-PAGEによって分離した後、ウエスタンブロット解析を行なった。
【0107】
質量分析
精製mHSF1-Flagを、mHSF1-FlagおよびCMV-p300でトランスフェクトした293T細胞から上記のとおりに入手し、1μMトリコスタチンAおよび5mMニコチンアミドで18時間処理した後、HSまたはセラストロール処理した。免疫沈降したタンパク質をSDS-PAGEによって分離し、ゲルから切り出し、トリプシンで消化し、ナノエレクトロスプレー源を備えたハイブリッド四重極飛行時間型装置(QSTAR、Applied Biosystems、Foster City、CA)によるタンデム質量分析に供した。Mascot(Matrix Science、Boston、MA;version 1.9.05)およびX! Tandem(www.thegpm.org;version 2006.04.01.2)データベース検索アルゴリズムを使用し、IPIマウス配列 データベース(68,222エントリー;バージョン3.15)に対してMS/MSスペクトルを検索した。MascotおよびX! Tandemは、消化酵素トリプシン切断を1回失敗すると仮定した断片および前駆体イオン質量許容度0.3Daを用いて検索した。システインのカルバミドメチル化は固定修飾として含まれたが、メチオニン酸化、N末端タンパク質およびリジンアセチル化は、データベース検索における可変修飾として含まれた。ペプチド同定は、Peptide Prophetアルゴリズム(31)で測定し、MS/MSスペクトルの手作業検査によって確認さた場合、95.0%より高い確率で許容された。
【0108】
組換えタンパク質
マウスHis-HSF1を大腸菌株BL-21において先に記載のようにして(32)作製した。収集した細菌を冷溶解バッファー [50mM Tris-HCl(pH7.4)、140mM NaCl、10%グリセロール]中に再懸濁し、室温で15分間1mg/mlリゾチームで溶解した。1mM PMSF(フッ化フェニルメチルスルホニル)、1mM DTT(ジチオトレイトール)および無EDTAプロテアーゼ阻害薬カクテル(Roche)の添加後、溶解物を超音波処理し、Talon金属親和性樹脂(Clontech)とともに4℃で2時間インキュベートした。樹脂を0.5%Triton X-100を含む溶解バッファーで充分に洗浄し、50mM NaAc(pH5.0)、300mM NaCl、200mM イミダゾール中で溶出させた。溶出されたタンパク質をPBSに対して透析し、等量に希釈した。
【0109】
電気泳動の移動度シフトアッセイ(EMSA)
WI-38細胞およびトランスフェクトhsf1-/-細胞からのバッファーC抽出物(15μg)または組換えHSF1タンパク質を、ヒトhsp70プロモーター由来の近位HSEを提示する32P標識オリゴヌクレオチドとともにインキュベートした(33)。タンパク質-DNA複合体を4%非変性ポリアクリルアミドゲル上で解析した。
【0110】
オリゴヌクレオチドプルダウンアッセイ
アッセイは、わずかに修正をともなって、本質的に先に記載のとおりに行なった(32)。HeLa細胞をプロテアーゼ阻害薬を含む冷溶解バッファー [25mM HEPES(pH7.4)、100mM NaCl、5mM EDTA、20mM p-グリセロホスフェート、20mM p-ニトロ-フェニル-ホスフェート、0.5%Triton X-100、20mM 100μM オルトバナジウム酸ナトリウム]中に溶解した。細胞抽出物を、1μgのビオチン化オリゴヌクレオチド(Oligomer、Helsinki、Finland)とともにインキュベートし、室温で30分間タンパク質をオリゴヌクレオチドに結合させた。オリゴヌクレオチドをUltraLinkストレプトアビジンビーズ(Pierce)で4℃で1時間沈殿させた。結合した画分を洗浄バッファー [20mM Tris-HCl(pH7.5)、1mM EDTA、10%グリセロール、0.1%Triton X-100)で3回洗浄し、変性バッファーで溶出させ、ウエスタンブロッティングによって解析した。
【0111】
RT-PCR解析
HeLa細胞を収集し、製造業者の使用説明書に従ってTrizol試薬(Gibco-BRL、Gaithersburg、Maryland)を用いてRNAを生成させた。逆転写反応およびPCRを用いて、以下のタンパク質:hsp90、hsp70、hsp40、hsp 25、hsp27、18S、およびgapdhを作製した。
【0112】
定量的リアルタイムRT-PCR解析
トランスフェクトhsf1-/-細胞におけるhsp70およびhsp25遺伝子発現の解析を、本質的に記載のとおりに行なった(34)。簡単には、RNeasyキット(Qiagen)を用いてRNAを単離した。各試料について、1μgのRNAをRQ1 DNase(Promega)で処理し、Moloneyマウス白血病ウイルスRNase H(-)(Promega)を使用することによって逆転写した。ABsolute QPCR ROX Mix(Advanced Biotechnologies)を用いて、反応混合物を調製した。hsp mRNAの相対量を、gapdhに対して標準化した。すべての反応は、3つの生物学的リピート由来の試料を用いて三重に行なった。
【0113】
共焦点顕微鏡検査
カバースリップ上で培養中のHeLa細胞を制御温度に維持するか、または42℃で1時間熱ショックを与えた。細胞をPBSで洗浄し、同時に、0.5%Triton X-100を含有する3.7%パラホルムアルデヒド中に固定および透過させた。細胞をPBSで2回洗浄し、ブロック溶液(PBS-0.05%Tween 20中20%正常ヤギ血清)中で1時間インキュベートした。一次抗体[ウサギα-HSF1(11)、マウスα-Myc(Sigma)] を、5%BSA含有PBS-0.05%Tween 20中、1:500希釈で4℃で一晩使用した。PBS-0.05%Tween 20で3回洗浄後、二次ヤギα-マウスIgG(Rhodamine Red-X and Alexa Fluor 488、Molecular Probes)またはヤギα-ウサギIgG(Alexa Fluor 546、Molecular 488、Molecular Probes)を用いて一次抗体を検出した。すべての二次抗体は、5%BSA含有PBS-0.05%Tween 20中1:500希釈で、室温で1時間使用した。3回の洗浄後、DAPI(4’,6’-ジアミジノ-2-フェニルインドール;Vector Laboratories)を含むVectashieldスライドに細胞を載せ、Zeiss LSM510 META共焦点顕微鏡を用いて解析した。Adobe Photoshopソフトウェアを用いて画像をさらに処理した。
【0114】
架橋
細胞を37℃で維持するか、または42℃ で15分間熱ショックを与え、冷溶解バッファー [25mM HEPES(pH7.4)、100mM NaCl、5mM EDTA、0.5%Triton X-100、20mM β-グリセロホスフェート、20mM p-ニトロ-フェニル-ホスフェート、100μM オルトバナジウム酸ナトリウム、0.5mM フッ化フェニルメチルスルホニル、1mM ジチオトレイトール、プロテアーゼ阻害薬カクテル]中に溶解し、4℃で10分間、15,000×gで遠心分離した。100μgのタンパク質を2mM EGS [エチレングリコールビス(スクシンイミジルスクシネート)]と室温で15分間架橋させた。100mM グリシンの添加による架橋のクエンチ後、試料を変性バッファー中で煮沸し、5%SDS-PAGEゲル上、続いてウエスタンブロッティングによって解析した。
【0115】
熱ストレス抵抗性アッセイ
ベクター対照(Mock)またはSIRT1でトランスフェクトされた293T細胞を、45℃の熱ショックに対する抵抗性について比較した。細胞を45℃の水浴中に20または30分間浸漬し、次いで、37℃で回復させた。細胞を、24時間後のトリパンブルー取込みによって細胞死の割合についてアッセイした。
【0116】
【0117】
実施例2:SIRT1の活性化は、熱ショック応答の小分子活性化因子であるセラストロールと相乗作用する
熱ショック応答のいくつかの活性化因子が同定された[2]。小分子熱ショック活性化因子のいくつかは、タンパク質ミスフォールディング疾患における治療有用性について試験中である。熱ショック応答を活性化させるために、可能な限り低い用量で小分子を使用することは、治療用量と毒性用量の境界はしばしば狭いため有益であり得る。本発明の実験の組は、SIRT1の活性化が熱ショック応答の誘導薬と相乗作用し、それにより、熱または熱ショック応答の小分子活性化因子であるセラストロールが低レベルで応答の誘導を可能にすることを示す[3]。
【0118】
熱ショック応答の活性化によってタンパク質ミスフォールディング疾患を治療するための現行のストラテジーは、生物を高レベルの熱またはこの応答が誘導されるのに充分な量の小分子で治療することを伴う。いくつかの場合において、熱ショック応答を誘導するために必要とされる熱または化合物のレベルはまた、細胞損傷または毒性をもたらし得る。熱ショック応答を誘導するのに必要とされる熱ショックまたは小分子化合物のこの量は、SIRT1を同時に活性化することにより低下され得る。実施例1に記載のように、SIRT1は、熱ショックと組み合わせて、HSF1活性サイクル[I]の減衰工程を妨げることにより、熱ショック応答を活性化することがわかった。本発明の試験は、SIRT1の活性化が、熱ショックおよび熱ショック応答の小分子活性化因子と相乗作用して、応答の強固な活性化に必要とされる化合物の熱ショック温度またはEC50を低下させることを示す。
【0119】
図16および17に示されるように、SIRT1活性化因子レスベラトロール[4]は、レポーターアッセイによって測定されるように、熱ショックおよびセラストロールの両方と相乗作用して熱ショック応答を誘導する。熱ショック応答の活性化を検出するツールとして、ルシフェラーゼに融合したヒトhsp70.1プロモーターの-188から+150までの配列(図15、[4])を含むHeLa安定細胞株を作製した。この細胞株を使用し、SIRT1の活性化がレスベラトロールとともに、熱ショック応答の活性化の温度閾値を低下させ得るかどうかを調べた。図16に示されるように、42℃の熱ショックによりレポーターは15倍誘導されたが、41℃の熱ショックではレポーター5倍しか誘導されなかった。レスベラトロールでの細胞の前処理により、レポーター誘導は両方の温度で増大し、SIRT1の活性化が熱と相乗作用して熱ショック応答を活性化し、より低温での応答の誘導が可能となり得ることを示す。同様に、図17に示されるように、レスベラトロールは、5μMのEC50を有する公知の熱ショック応答誘導薬セラストロールと相乗作用し得ることがわかった[4]。1μMのセラストロール単独では、レポーターを2倍しか活性化しなかったが、レスベラトロールと1μM セラストロールの組合せでは、40倍を超える強固なレポーター活性化を誘導した。
【0120】
hsp70.1pr-luc HeLa安定細胞株は、hsp70.1プロモーター配列の-188から+150までを含み、近位熱ショックエレメント(HSE)を含む。この細胞株の作製は、Westerheide et al.(2004). Celastrols as inducers of the heat shock response and cytoprotection. J Biol Chem、2004. 279(53):p. 56053-60に記載されており、その内容は参照により本明細書に援用される。
【0121】
HeLa hsp70.1pr-luc細胞を白色96ウェルプレートにおいて、ウェル1つあたり15 X 103細胞の密度で100ulのDMEM培地+10%ウシ胎仔血清中にプレーティングした。細胞を37℃、5%CO2で20時間インキュベートした後、レスベラトロールで処理するか、または処理しなかった。レスベラトロールはEtOH中で希釈し、細胞に0.5uMで添加した。2時間後、次いで、水浴中に1時間浸漬することにより、細胞に、表示した温度の熱ショックを与えた。熱ショック処理後、37℃、5%CO2でプレートをインキュベータ内に戻し、18時間インキュベートした後、ルシフェラーゼアッセイを行なった。
【0122】
HeLa hsp70.1pr-luc細胞を白色96ウェルプレートにおいて、ウェル1つあたり15 X 103細胞の密度で100ulのDMEM培地+10%ウシ胎仔血清中にプレーティングした。細胞を37℃、5%CO2で20時間インキュベートした後、レスベラトロールで処理するか、または処理しなかった。レスベラトロールはEtOH中で希釈し、細胞に1.0uMで添加した。2時間後、次いで、細胞を、表示したとおりに1mM セラストロールで処理するか(セラストロールはDMSO中に溶解した)、または処理しなかった。セラストロール添加の18時間後、細胞をルシフェラーゼアッセイによってアッセイした。
【0123】
図の説明:
図16. SIRT1活性化因子レスベラトロールが熱ショックと相乗作用して、hsp70.1pr-lucレポーターを活性化させる。
図17. SIRT1活性化因子レスベラトロールは、セラストロールと相乗作用してhsp70.1pr-lucレポーターを活性化させる。
【0124】
【0125】
本発明を、その好ましい態様を参照して具体的に示し、記載したが、形態および詳細における種々の変形が、添付の特許請求の範囲によって包含される本発明の範囲から逸脱せずになされ得ることは、当業者によって理解されよう。
【0126】
【特許請求の範囲】
【請求項1】
熱ショック転写因子1(HSF1)のDNA結合ドメイン内のリジン残基のアセチル化を修飾する工程を含む、細胞内のHSF1の活性の調節方法。
【請求項2】
該リジン残基のアセチル化を阻害することによりHSF1の活性を増大させる、請求項1記載の方法。
【請求項3】
該リジン残基のアセチル化を促進させることによりHSF1の活性を減少させる、請求項1記載の方法。
【請求項4】
細胞がヒト細胞である、請求項1記載の方法。
【請求項5】
該残基がリジン80(HSF1 K80)である、請求項4記載の方法。
【請求項6】
HSF1 K80のアセチル化を阻害することによりHSF1の活性を増大させる、請求項1記載の方法。
【請求項7】
HSF1 K80のアセチル化を促進させることによりHSF1の活性を減少させる、請求項1記載の方法。
【請求項8】
細胞にHSF1のアセチル化を阻害する薬剤の有効量を投与することによりHSF1のアセチル化を阻害し、該薬剤が、単離サーチュイン、サーチュイン活性化剤およびヒストンアセチルトランスフェラーゼ(HAT)阻害剤からなる群より選択される、請求項2記載の方法。
【請求項9】
細胞にHSF1のアセチル化を阻害する薬剤の有効量を投与することによりHSF1のアセチル化を阻害し、該薬剤が、単離サーチュイン、サーチュイン活性化剤およびヒストンアセチルトランスフェラーゼ(HAT)阻害剤からなる群より選択される、請求項6記載の方法。
【請求項10】
単離サーチュインがサーチュイン1(SIRT1)である、請求項8記載の方法。
【請求項11】
SIRT1活性化剤が投与される、請求項8記載の方法。
【請求項12】
HAT阻害剤が投与される、請求項8記載の方法。
【請求項13】
細胞にHSF1のアセチル化を促進させる薬剤を投与することによりHSF1のアセチル化を促進させ、該薬剤が、サーチュイン阻害剤およびHAT活性化剤からなる群より選択される、請求項3記載の方法。
【請求項14】
SIRT1阻害剤が投与される、請求項13記載の方法。
【請求項15】
SIRT1阻害剤が小分子、核酸または抗体である、請求項13記載の方法。
【請求項16】
HSF1の活性の増大を必要とする被検体におけるその増大方法であって、前記被検体の細胞においてHSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する工程を含む方法。
【請求項17】
該リジン残基がHSF1 K80または対応する保存リジン残基である、請求項16記載の方法。
【請求項18】
HSF1のアセチル化を阻害する薬剤が前記被検体に有効量で投与される、請求項16記載の方法。
【請求項19】
HSF1 K80のアセチル化を阻害する薬剤が単離サーチュイン、サーチュイン活性化剤またはHAT阻害剤である、請求項18記載の方法。
【請求項20】
被検体がヒトである、請求項16記載の方法。
【請求項21】
SIRT1活性化剤が投与される、請求項20記載の方法。
【請求項22】
HAT阻害剤が投与される、請求項20記載の方法。
【請求項23】
HSF1の活性の減少を必要とする被検体におけるその減少方法であって、前記被検体の細胞においてHSF1のDNA結合ドメイン内のリジン残基のアセチル化を促進させる工程を含む方法。
【請求項24】
該リジン残基がHSF1 K80である、請求項23記載の方法。
【請求項25】
HSF1 K80または対応する保存リジン残基のアセチル化を促進させる薬剤が前記被検体に投与される、請求項23記載の方法。
【請求項26】
前記被検体にサーチュイン阻害剤またはHAT活性化剤を投与することによりHSF1 K80のアセチル化が促進される、請求項25記載の方法。
【請求項27】
サーチュイン阻害剤がSIRT1阻害剤である、請求項26記載の方法。
【請求項28】
被検体がヒトである、請求項23記載の方法。
【請求項29】
患者にHSF1のアセチル化を阻害する薬剤の治療有効量を投与する工程を含む、タンパク質のホメオスタシスの機能障害と関連している状態に苦しむ患者の治療方法。
【請求項30】
該薬剤がHSF1 K80のアセチル化を阻害する、請求項29記載の方法。
【請求項31】
HSF1 K80のアセチル化を阻害する薬剤が単離サーチュイン、サーチュイン活性化剤またはHAT阻害剤である、請求項29記載の方法。
【請求項32】
タンパク質のホメオスタシスにおける機能障害と関連している状態が機能消失障害である、請求項29記載の方法。
【請求項33】
タンパク質のホメオスタシスにおける機能障害と関連している状態が機能獲得障害である、請求項29記載の方法。
【請求項34】
HSF1のアセチル化を阻害する薬剤がHAT阻害剤である、請求項33記載の方法。
【請求項35】
機能消失障害が、嚢胞性線維症およびリソソーム蓄積症からなる群より選択される、請求項32記載の方法。
【請求項36】
熱ショックタンパク質の発現の増大と関連している状態に苦しむ患者の治療方法であって、前記患者に、HSF1のアセチル化を促進させる薬剤の治療有効量を投与する工程を含む方法。
【請求項37】
該薬剤がHSF1 K80のアセチル化を促進させる、請求項36記載の方法。
【請求項38】
HSF1 K80のアセチル化を促進させる薬剤がサーチュイン阻害剤またはHAT活性化剤である、請求項36記載の方法。
【請求項39】
熱ショックタンパク質の発現の増大と関連している状態が癌である、請求項36記載の方法。
【請求項40】
さらに、化学療法薬の投与を含む、請求項39記載の方法。
【請求項41】
さらに、放射線療法での治療を含む、請求項39記載の方法。
【請求項42】
熱ショックタンパク質の発現の増大と関連している状態がウイルス感染である、請求項39記載の方法。
【請求項43】
ウイルス感染が、腫瘍ウイルスおよびRNA ウイルスからなる群より選択されるウイルスによって引き起こされる、請求項42記載の方法。
【請求項44】
試験薬剤を細胞に投与する工程;および
HSF1のDNA結合ドメイン内のリジン残基のアセチル化をモニタリングする工程
を含み、
ここで、該試験薬剤の非存在下と比べたときのDNA結合ドメインのアセチル化の変化は、細胞において試験薬剤がHSF1活性を調節することを示す、
細胞においてHSF1活性を調節する薬剤の同定方法。
【請求項45】
試験薬剤がDNA結合ドメインのアセチル化を阻害する、請求項44記載の方法。
【請求項46】
試験薬剤がDNA結合ドメインのアセチル化を増大させる、請求項44記載の方法。
【請求項47】
HSF1 K80のアセチル化または対応する保存アミノ酸がモニタリングされる、請求項44記載の方法。
【請求項48】
プロテオスタシスの機能障害と関連している状態に苦しむ患者の治療方法であって、前記患者に、HSF1のアセチル化を阻害する薬剤の治療有効量を、機構的が異なるプロテオスタシス調節因子と組み合わせて投与することを含む方法。
【請求項49】
機構的が異なるプロテオスタシス調節因子が熱ショック応答活性化因子である、請求項48記載の方法。
【請求項50】
熱ショック応答活性化因子およびHSF1のアセチル化を阻害する薬剤が、それぞれ、これらの併合用量で熱ショック応答を増大させるのに充分な用量で投与される、請求項48記載の方法。
【請求項51】
熱ショック応答活性化因子が小分子である、請求項48記載の方法。
【請求項52】
HSF1のアセチル化を阻害する薬剤がサーチュインであるか、またはサーチュイン(sirutin)活性化剤が単離サーチュイン、サーチュイン活性化剤もしくはHAT阻害薬である、請求項48記載の方法。
【請求項53】
細胞に熱ショック応答活性化因子およびHSF1のアセチル化を阻害する薬剤を投与する工程を含む、細胞における熱ショック応答の活性化方法であって、HSF1のアセチル化を阻害する該薬剤と該熱ショック応答活性化因子の併合用量が熱ショック応答を増大させるのに充分である方法。
【請求項54】
熱ショック応答の活性化を必要とする患者におけるその活性化方法であって、前記患者に、HSF1のアセチル化を阻害する薬剤および熱ショック応答活性化因子を投与する工程を含み、HSF1のアセチル化を阻害する該薬剤と該熱ショック応答活性化因子の併合用量が熱ショック応答を増大させるのに充分である方法。
【請求項55】
薬学的に許容され得る担体、熱ショック応答活性化因子およびHSF1のアセチル化を阻害する薬剤を含む医薬組成物であって、HSF1のアセチル化を阻害する該薬剤と該熱ショック応答活性化因子の併合用量が熱ショック応答を増大させるのに充分である医薬組成物。
【請求項56】
HSF1のアセチル化を阻害する薬剤がHAT阻害薬である、請求項55記載の組成物。
【請求項57】
HSF1のアセチル化を阻害する薬剤がサーチュイン活性化剤である、請求項55記載の組成物。
【請求項58】
HSF1のアセチル化を阻害する薬剤がレスベラトロールである、請求項55記載の組成物。
【請求項59】
熱ショック応答活性化因子がセラストロールである、請求項55記載の組成物。
【請求項1】
熱ショック転写因子1(HSF1)のDNA結合ドメイン内のリジン残基のアセチル化を修飾する工程を含む、細胞内のHSF1の活性の調節方法。
【請求項2】
該リジン残基のアセチル化を阻害することによりHSF1の活性を増大させる、請求項1記載の方法。
【請求項3】
該リジン残基のアセチル化を促進させることによりHSF1の活性を減少させる、請求項1記載の方法。
【請求項4】
細胞がヒト細胞である、請求項1記載の方法。
【請求項5】
該残基がリジン80(HSF1 K80)である、請求項4記載の方法。
【請求項6】
HSF1 K80のアセチル化を阻害することによりHSF1の活性を増大させる、請求項1記載の方法。
【請求項7】
HSF1 K80のアセチル化を促進させることによりHSF1の活性を減少させる、請求項1記載の方法。
【請求項8】
細胞にHSF1のアセチル化を阻害する薬剤の有効量を投与することによりHSF1のアセチル化を阻害し、該薬剤が、単離サーチュイン、サーチュイン活性化剤およびヒストンアセチルトランスフェラーゼ(HAT)阻害剤からなる群より選択される、請求項2記載の方法。
【請求項9】
細胞にHSF1のアセチル化を阻害する薬剤の有効量を投与することによりHSF1のアセチル化を阻害し、該薬剤が、単離サーチュイン、サーチュイン活性化剤およびヒストンアセチルトランスフェラーゼ(HAT)阻害剤からなる群より選択される、請求項6記載の方法。
【請求項10】
単離サーチュインがサーチュイン1(SIRT1)である、請求項8記載の方法。
【請求項11】
SIRT1活性化剤が投与される、請求項8記載の方法。
【請求項12】
HAT阻害剤が投与される、請求項8記載の方法。
【請求項13】
細胞にHSF1のアセチル化を促進させる薬剤を投与することによりHSF1のアセチル化を促進させ、該薬剤が、サーチュイン阻害剤およびHAT活性化剤からなる群より選択される、請求項3記載の方法。
【請求項14】
SIRT1阻害剤が投与される、請求項13記載の方法。
【請求項15】
SIRT1阻害剤が小分子、核酸または抗体である、請求項13記載の方法。
【請求項16】
HSF1の活性の増大を必要とする被検体におけるその増大方法であって、前記被検体の細胞においてHSF1のDNA結合ドメイン内のリジン残基のアセチル化を阻害する工程を含む方法。
【請求項17】
該リジン残基がHSF1 K80または対応する保存リジン残基である、請求項16記載の方法。
【請求項18】
HSF1のアセチル化を阻害する薬剤が前記被検体に有効量で投与される、請求項16記載の方法。
【請求項19】
HSF1 K80のアセチル化を阻害する薬剤が単離サーチュイン、サーチュイン活性化剤またはHAT阻害剤である、請求項18記載の方法。
【請求項20】
被検体がヒトである、請求項16記載の方法。
【請求項21】
SIRT1活性化剤が投与される、請求項20記載の方法。
【請求項22】
HAT阻害剤が投与される、請求項20記載の方法。
【請求項23】
HSF1の活性の減少を必要とする被検体におけるその減少方法であって、前記被検体の細胞においてHSF1のDNA結合ドメイン内のリジン残基のアセチル化を促進させる工程を含む方法。
【請求項24】
該リジン残基がHSF1 K80である、請求項23記載の方法。
【請求項25】
HSF1 K80または対応する保存リジン残基のアセチル化を促進させる薬剤が前記被検体に投与される、請求項23記載の方法。
【請求項26】
前記被検体にサーチュイン阻害剤またはHAT活性化剤を投与することによりHSF1 K80のアセチル化が促進される、請求項25記載の方法。
【請求項27】
サーチュイン阻害剤がSIRT1阻害剤である、請求項26記載の方法。
【請求項28】
被検体がヒトである、請求項23記載の方法。
【請求項29】
患者にHSF1のアセチル化を阻害する薬剤の治療有効量を投与する工程を含む、タンパク質のホメオスタシスの機能障害と関連している状態に苦しむ患者の治療方法。
【請求項30】
該薬剤がHSF1 K80のアセチル化を阻害する、請求項29記載の方法。
【請求項31】
HSF1 K80のアセチル化を阻害する薬剤が単離サーチュイン、サーチュイン活性化剤またはHAT阻害剤である、請求項29記載の方法。
【請求項32】
タンパク質のホメオスタシスにおける機能障害と関連している状態が機能消失障害である、請求項29記載の方法。
【請求項33】
タンパク質のホメオスタシスにおける機能障害と関連している状態が機能獲得障害である、請求項29記載の方法。
【請求項34】
HSF1のアセチル化を阻害する薬剤がHAT阻害剤である、請求項33記載の方法。
【請求項35】
機能消失障害が、嚢胞性線維症およびリソソーム蓄積症からなる群より選択される、請求項32記載の方法。
【請求項36】
熱ショックタンパク質の発現の増大と関連している状態に苦しむ患者の治療方法であって、前記患者に、HSF1のアセチル化を促進させる薬剤の治療有効量を投与する工程を含む方法。
【請求項37】
該薬剤がHSF1 K80のアセチル化を促進させる、請求項36記載の方法。
【請求項38】
HSF1 K80のアセチル化を促進させる薬剤がサーチュイン阻害剤またはHAT活性化剤である、請求項36記載の方法。
【請求項39】
熱ショックタンパク質の発現の増大と関連している状態が癌である、請求項36記載の方法。
【請求項40】
さらに、化学療法薬の投与を含む、請求項39記載の方法。
【請求項41】
さらに、放射線療法での治療を含む、請求項39記載の方法。
【請求項42】
熱ショックタンパク質の発現の増大と関連している状態がウイルス感染である、請求項39記載の方法。
【請求項43】
ウイルス感染が、腫瘍ウイルスおよびRNA ウイルスからなる群より選択されるウイルスによって引き起こされる、請求項42記載の方法。
【請求項44】
試験薬剤を細胞に投与する工程;および
HSF1のDNA結合ドメイン内のリジン残基のアセチル化をモニタリングする工程
を含み、
ここで、該試験薬剤の非存在下と比べたときのDNA結合ドメインのアセチル化の変化は、細胞において試験薬剤がHSF1活性を調節することを示す、
細胞においてHSF1活性を調節する薬剤の同定方法。
【請求項45】
試験薬剤がDNA結合ドメインのアセチル化を阻害する、請求項44記載の方法。
【請求項46】
試験薬剤がDNA結合ドメインのアセチル化を増大させる、請求項44記載の方法。
【請求項47】
HSF1 K80のアセチル化または対応する保存アミノ酸がモニタリングされる、請求項44記載の方法。
【請求項48】
プロテオスタシスの機能障害と関連している状態に苦しむ患者の治療方法であって、前記患者に、HSF1のアセチル化を阻害する薬剤の治療有効量を、機構的が異なるプロテオスタシス調節因子と組み合わせて投与することを含む方法。
【請求項49】
機構的が異なるプロテオスタシス調節因子が熱ショック応答活性化因子である、請求項48記載の方法。
【請求項50】
熱ショック応答活性化因子およびHSF1のアセチル化を阻害する薬剤が、それぞれ、これらの併合用量で熱ショック応答を増大させるのに充分な用量で投与される、請求項48記載の方法。
【請求項51】
熱ショック応答活性化因子が小分子である、請求項48記載の方法。
【請求項52】
HSF1のアセチル化を阻害する薬剤がサーチュインであるか、またはサーチュイン(sirutin)活性化剤が単離サーチュイン、サーチュイン活性化剤もしくはHAT阻害薬である、請求項48記載の方法。
【請求項53】
細胞に熱ショック応答活性化因子およびHSF1のアセチル化を阻害する薬剤を投与する工程を含む、細胞における熱ショック応答の活性化方法であって、HSF1のアセチル化を阻害する該薬剤と該熱ショック応答活性化因子の併合用量が熱ショック応答を増大させるのに充分である方法。
【請求項54】
熱ショック応答の活性化を必要とする患者におけるその活性化方法であって、前記患者に、HSF1のアセチル化を阻害する薬剤および熱ショック応答活性化因子を投与する工程を含み、HSF1のアセチル化を阻害する該薬剤と該熱ショック応答活性化因子の併合用量が熱ショック応答を増大させるのに充分である方法。
【請求項55】
薬学的に許容され得る担体、熱ショック応答活性化因子およびHSF1のアセチル化を阻害する薬剤を含む医薬組成物であって、HSF1のアセチル化を阻害する該薬剤と該熱ショック応答活性化因子の併合用量が熱ショック応答を増大させるのに充分である医薬組成物。
【請求項56】
HSF1のアセチル化を阻害する薬剤がHAT阻害薬である、請求項55記載の組成物。
【請求項57】
HSF1のアセチル化を阻害する薬剤がサーチュイン活性化剤である、請求項55記載の組成物。
【請求項58】
HSF1のアセチル化を阻害する薬剤がレスベラトロールである、請求項55記載の組成物。
【請求項59】
熱ショック応答活性化因子がセラストロールである、請求項55記載の組成物。
【図1】

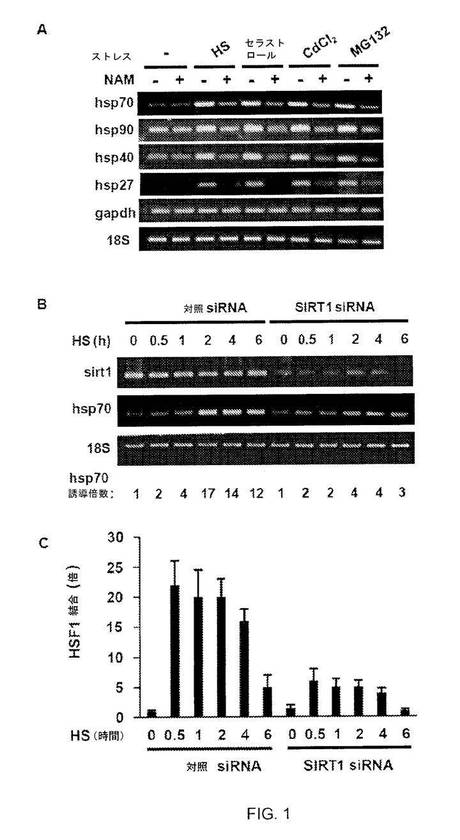
【図2】
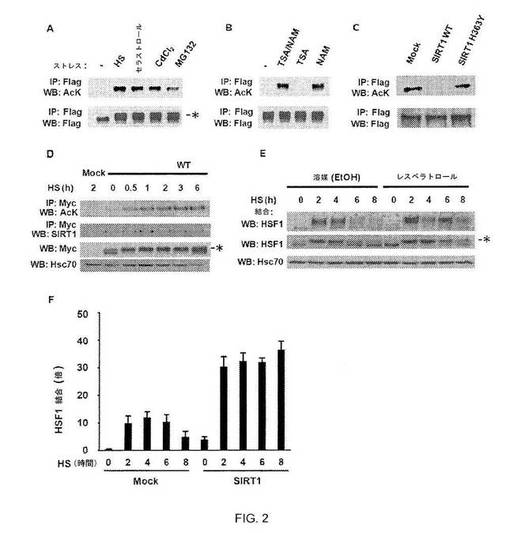
【図3】
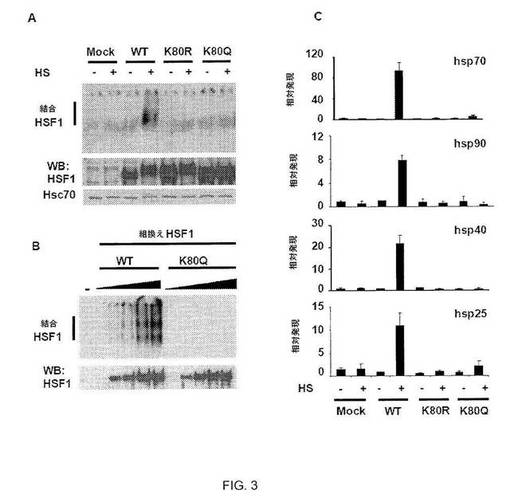
【図4】
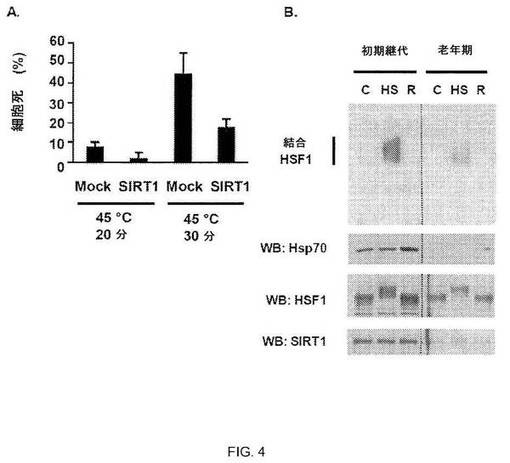
【図5】
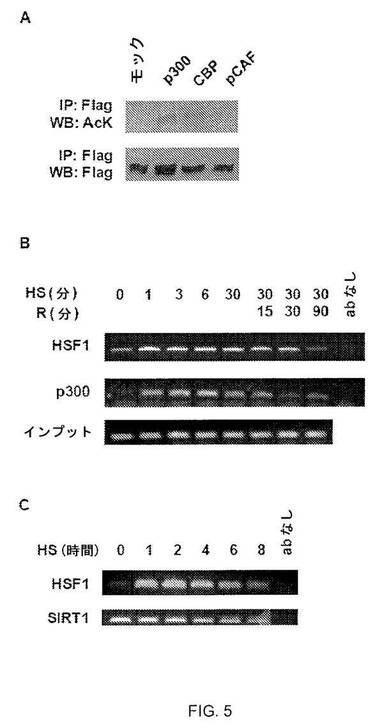
【図6】
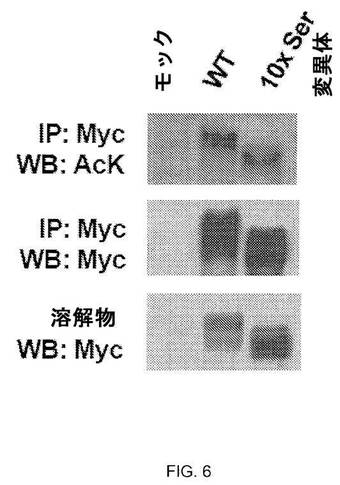
【図7−1】
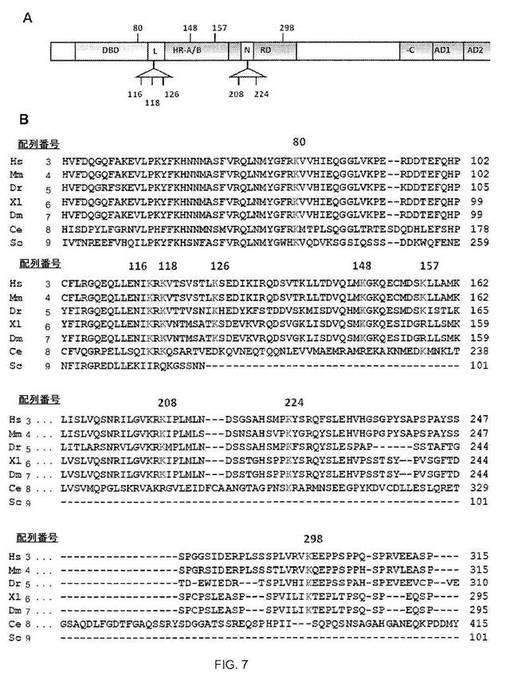
【図7−2】
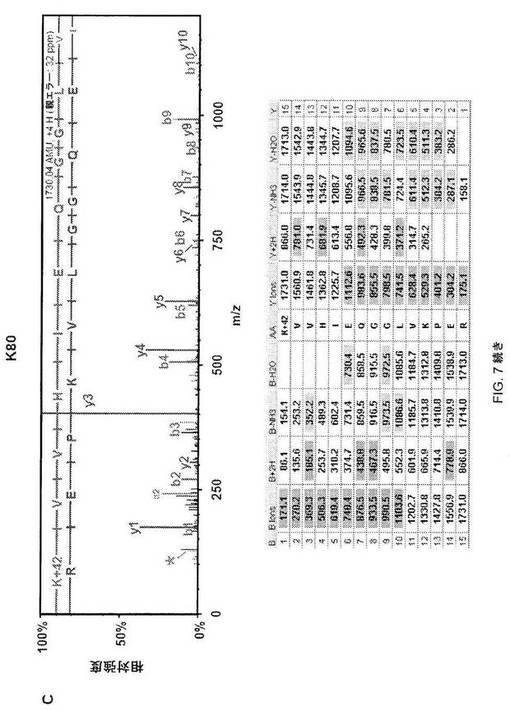
【図7−3】

【図7−4】

【図7−5】
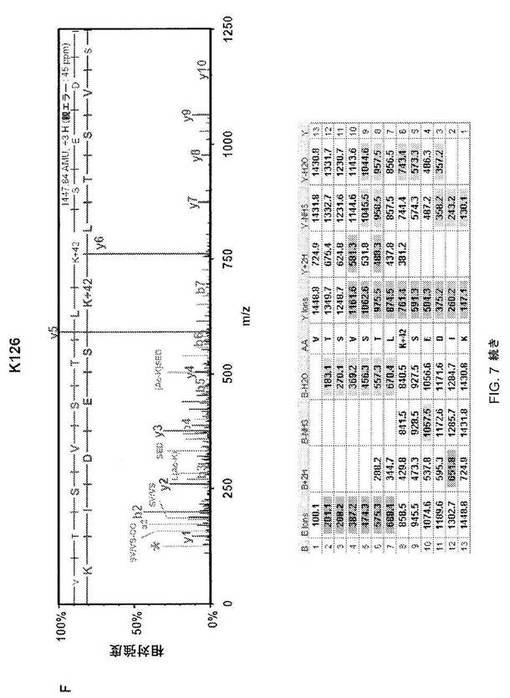
【図7−6】
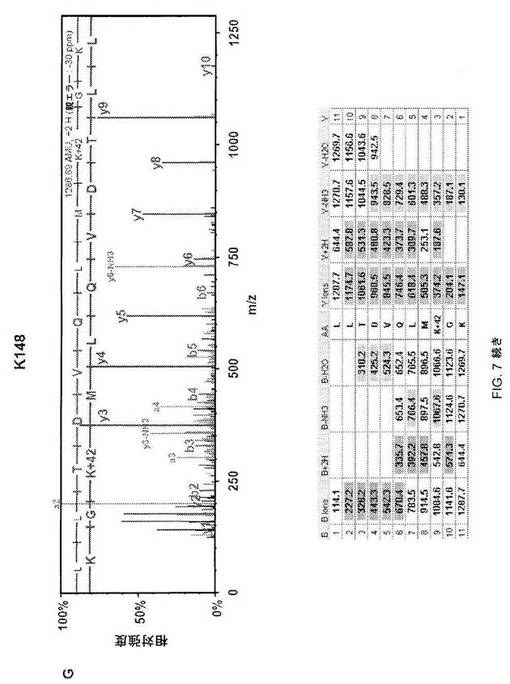
【図7−7】
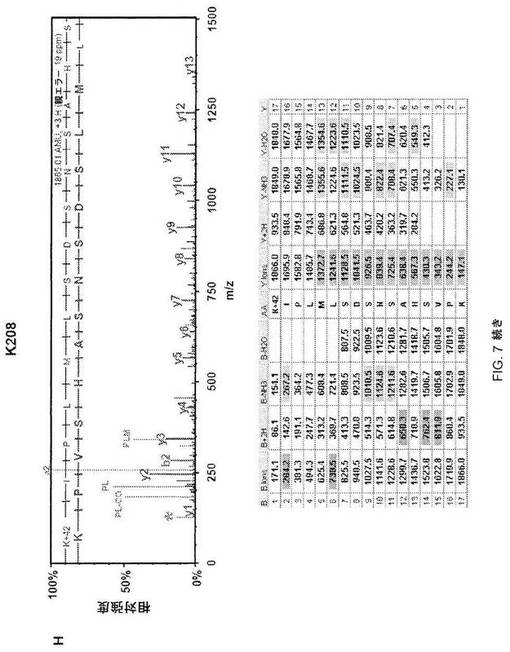
【図7−8】
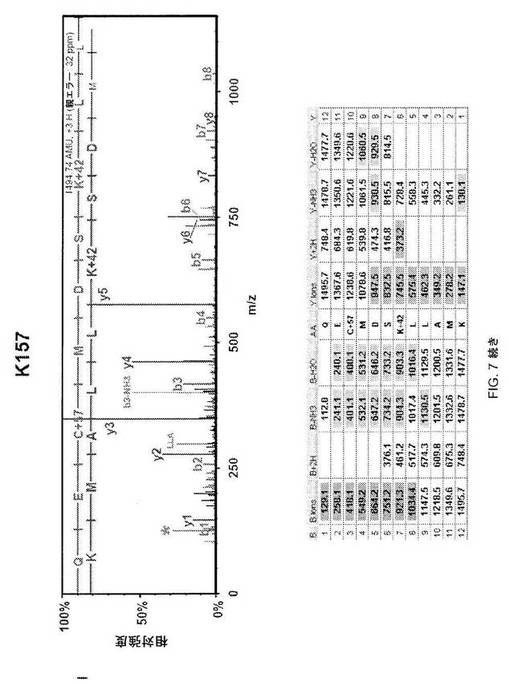
【図7−9】

【図7−10】
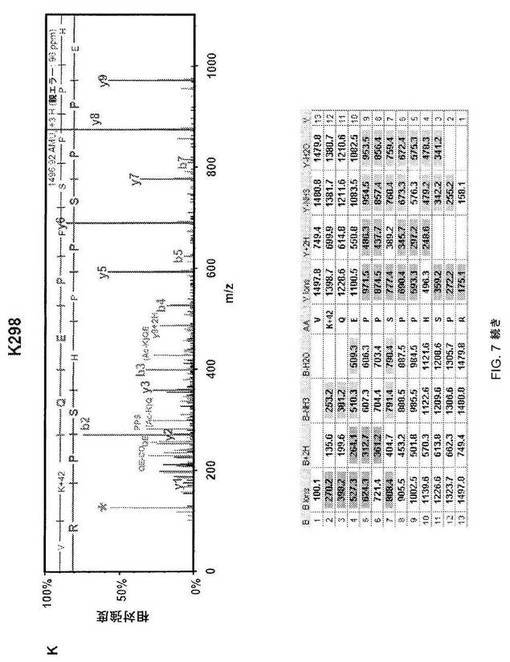
【図8】
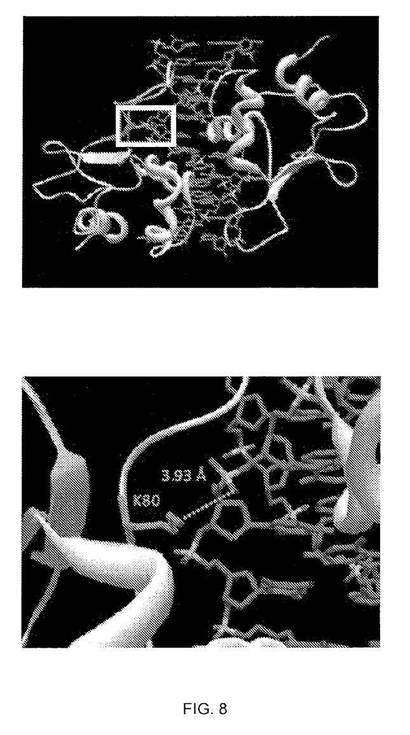
【図9】
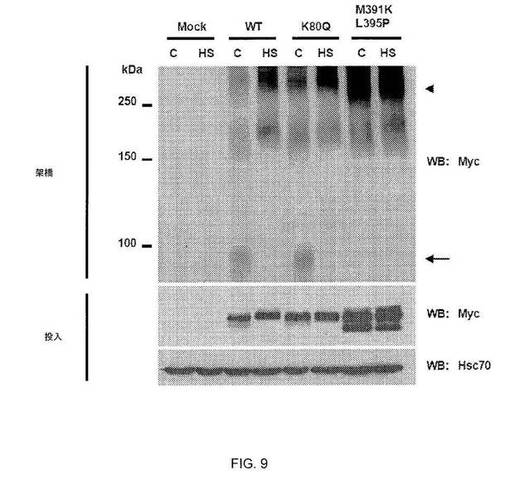
【図10】
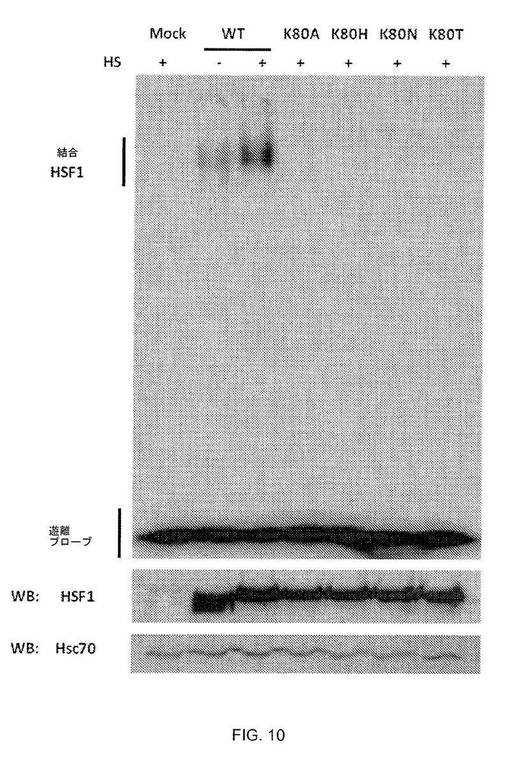
【図11】
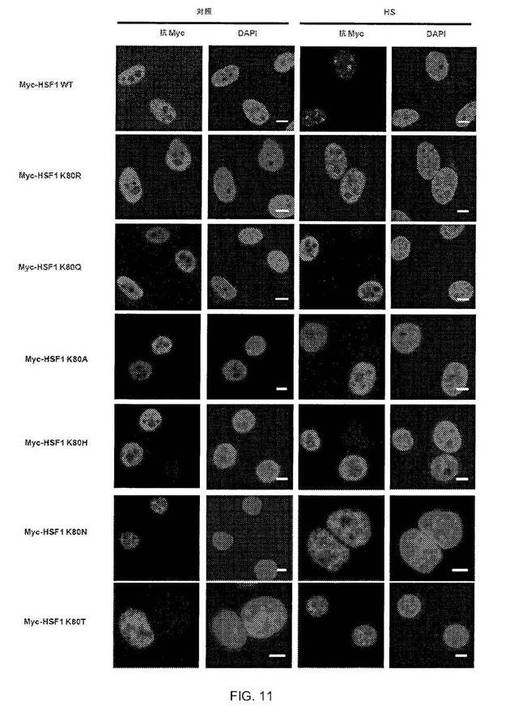
【図12】
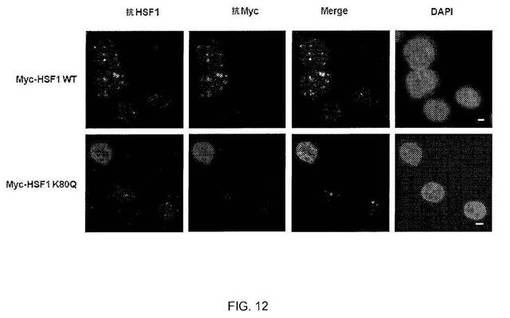
【図13】
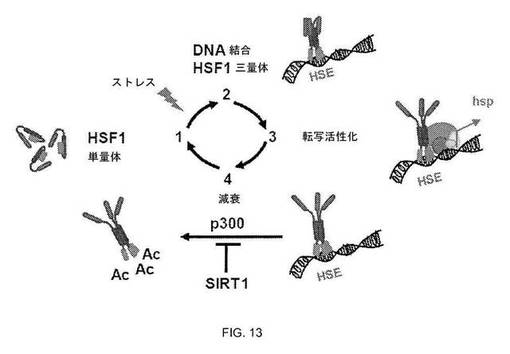
【図14】
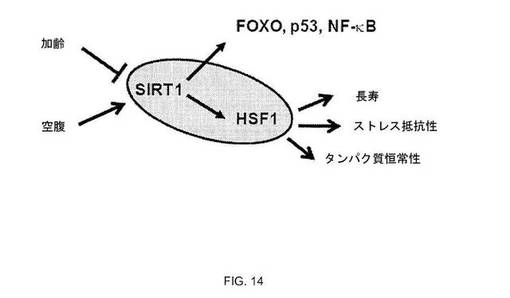
【図15】
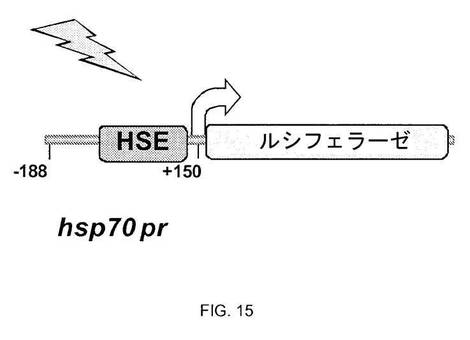
【図16】
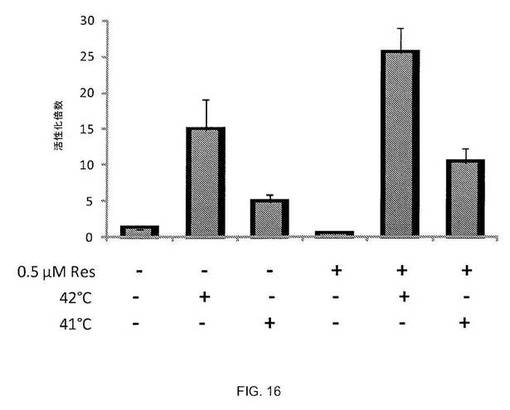
【図17】
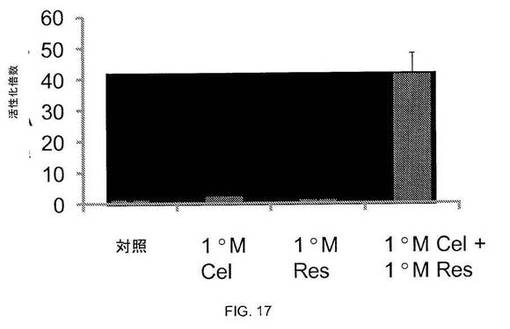
【図2】
【図3】
【図4】
【図5】
【図6】
【図7−1】
【図7−2】
【図7−3】
【図7−4】
【図7−5】
【図7−6】
【図7−7】
【図7−8】
【図7−9】
【図7−10】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2012−511048(P2012−511048A)
【公表日】平成24年5月17日(2012.5.17)
【国際特許分類】
【出願番号】特願2011−540821(P2011−540821)
【出願日】平成21年12月8日(2009.12.8)
【国際出願番号】PCT/US2009/067118
【国際公開番号】WO2010/077642
【国際公開日】平成22年7月8日(2010.7.8)
【出願人】(511124183)ノースウェスタン ユニバーシティ (9)
【Fターム(参考)】
【公表日】平成24年5月17日(2012.5.17)
【国際特許分類】
【出願日】平成21年12月8日(2009.12.8)
【国際出願番号】PCT/US2009/067118
【国際公開番号】WO2010/077642
【国際公開日】平成22年7月8日(2010.7.8)
【出願人】(511124183)ノースウェスタン ユニバーシティ (9)
【Fターム(参考)】
[ Back to top ]