
IL−13結合タンパク質
本発明は、抗IL−13抗体および抗IL−3/抗IL−4 mAbdAbを含むヒトIL−13に対する抗原結合タンパク質、それらを含む医薬品ならびに炎症性疾患(喘息またはIPFなど)の治療および/もしくは予防におけるかかる抗原結合タンパク質の使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、インターロイキン13(IL−13)と結合し、その活性を中和する抗原結合タンパク質(特に抗体)、かかる抗原結合タンパク質をコードするポリヌクレオチド、前記抗原結合タンパク質を含む医薬品ならびに炎症(喘息など)関連疾患の治療および/もしくは予防におけるかかる抗原結合タンパク質の使用に関する。本発明の他の態様、目的および利益は、以下の本明細書から明らかとなるであろう。
【背景技術】
【0002】
インターロイキン−13(IL−13)
IL−13は、炎症サイトカイン産生を阻害するT細胞由来サイトカインとして元々記載されている12kDa分泌サイトカインである。構造的試験により、IL−13は2つのジスルフィド結合により保持されている4つのヘリックスバンドルの配置を有することが示される。IL−13は4つの潜在的グリコシル化部位を有するが、ラット肺由来の天然IL−13は、非グリコシル化分子として産生されることが分析により示されている。NSOおよびCOS−7細胞におけるヒトIL−13発現により、この観察所見が確認されている(Eisenmesser et al, J. Mol. Biol. 2001 310(1):231−241; Moy et al, J. Mol. Biol 2001 310(1):219−230; Cannon−Carlson et al, Protein Expression and Purification 1998 12(2):239−248)。
【0003】
IL−13は、喘息、慢性閉塞性肺疾患(COPD)、アトピー性皮膚炎およびアレルギー性鼻炎を含むアレルギー疾患、食道好酸球増加症、腫瘍学適応症、例えばB細胞慢性リンパ性白血病(B−CLL)およびホジキン病、炎症腸疾患、例えば潰瘍性大腸炎、クローン病および不確定性大腸炎、乾癬および乾癬性関節炎、急性移植片対宿主病、糖尿病性腎症、線維状態(肺線維症、例えば特発性肺線維症(IPF)など)に関係している。
【発明の概要】
【0004】
本発明は、IL−13(例えばIL−13抗体)、およびかかるIL−13抗体とIL−4アンタゴニストおよび/またはIL−5アンタゴニストの組み合わせと結合する抗原結合タンパク質を提供する。本発明のIL−13抗体は、CDRH3が変異しているマウスmAb 6A1に関連または由来する。6A1マウス重鎖可変領域アミノ酸配列は、配列番号58として提供される。6A1マウス軽鎖可変領域アミノ酸配列は、配列番号59として提供される。
【0005】
本発明の重鎖可変領域(VH)は、(Kabat(Kabat et al; Sequences of proteins of Immunological Interest NIH, 1987)により定義される)以下のCDRを含む:
本発明の重鎖可変領域のCDRは、以下のCDRを含み得る:

【0006】
本発明の軽鎖可変領域は、(Kabatにより定義される)以下のCDRを含む:
【0007】
抗体のCDR配列は、上表に説明するようにKabat番号付け系(Kabat et al; Sequences of proteins of Immunological Interest NIH, 1987)により決定することもでき、あるいはChothia番号付け系(Al−Lazikani et al., (1997) JMB 273,927−948)、接触点の定義方法(MacCallum R.M., and Martin A.C.R. and Thornton J.M, (1996), Journal of Molecular Biology, 262 (5), 732−745)または当業者にとって既知である抗体中残基の番号付けおよびCDR決定のため他の任意の確立された方法を用いて決定することもできる。本明細書に記載の本発明のCDRは、これらのいずれかの方法によって、またはChothia番号付けとKabat番号付けの組み合わせを用いて定義し得る。例えばCDRH1は、FYIKDTYMH(配列番号60)またはGFYIKDTYMH(配列番号61)を含むものとして定義し得る。
【0008】
本発明は、1つまたは複数のエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、例えばIL−4と結合できるエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、またはIL−5と結合できるエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、またはIL−4と結合できる第1のエピトープ結合ドメインおよびIL−5と結合できる第2のエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質も提供する。
【0009】
本発明は、残基89(kabat番号付け)の、「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)、「A」(アラニン)、「T」(トレオニン)および「E」(グルタミン酸)から選択される残基への変異による、免疫グロブリン単一可変ドメイン(例えばヒトdAb、例えばヒトVKドメイン抗体)の凝集傾向の低減方法も提供する。一実施形態では、この方法は、残基89(kabat番号付け)を「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)および「E」(グルタミン酸)から選択される残基に変異させることを含む。さらなる実施形態では、この方法は、残基89(kabat番号付け)を「Q」(グルタミン)に変異させることを含む。
【0010】
一実施形態では、この方法は配列番号80の抗IL−4ドメイン抗体に適用して、変異dAb配列(例えば配列番号94)をもたらすことができる。
【0011】
かかる変異dAbは、単独またはより大きな配列の一部(例えばmAbdAb配列の一部)であってよく、例えば、配列番号117〜134から選択される配列を含むdAbをもたらし得る。
【0012】
本発明は、凝集プロファイルが改善されているヒトVK dAb、例えばVK dAbの残基89(kabat番号付け)が「Q」(グルタミン)であるIGKV1−17、IGKV1D−17、IGKV1/OR2−108、IGKV1−6、IGKV5−2、IGKV1D−42、IGKV2−24、IGKV2−28、IGKV2−30、IGKV2−40、IGKV2D−29、IGKV2D−30、IGKV2D−24およびIGKV6−21から選択される生殖系列フレームワークに由来するヒトVK dAbも提供する。1つのかかる実施形態では、VK dAbは、VK dAbの残基89(kabat番号付け)が「Q」(グルタミン)である生殖系列フレームワークIGKV1−17、IGKV1D−17、IGKV1/OR2−108、IGKV1−6、IGKV5−2、IGKV1D−42、IGKV2−24、IGKV2−28、IGKV2−30、IGKV2−40、IGKV2D−29、IGKV2D−30、IGKV2D−24およびIGKV6−21から選択される生殖系列フレームワーク領域を含む。
【0013】
一実施形態では、本発明は、89位(kabat番号付け)に変異を含み、89位が「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)、「A」(アラニン)、「T」(トレオニン)および「E」(グルタミン酸)から選択されるものに変異している配列番号80の配列を含むヒトdAbを提供する。例えば本発明は、89位(kabat番号付け)が、「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)および「E」(グルタミン酸)から選択される残基に変異している配列番号80の配列を含むヒトdAbを提供する。
【0014】
一実施形態では、本発明は、配列番号94の配列を含むヒトdAbを提供する。
【0015】
本発明は、本明細書に記載の任意の抗原結合タンパク質重鎖をコードするポリヌクレオチド配列、および本明細書に記載の任意の抗原結合タンパク質軽鎖をコードするポリヌクレオチドも提供する。かかるポリヌクレオチドは等価物のポリペプチド配列に相当するコード配列を表すが、かかるポリヌクレオチド配列は開始コドン、適切なシグナル配列および停止コドンと共に発現ベクター中にクローン化し得ることが理解されるであろう。
【0016】
本発明は、本明細書に記載の任意の抗原結合タンパク質の重鎖および軽鎖をコードする1つまたは複数のポリヌクレオチドを含む組換え形質転換またはトランスフェクトされた宿主細胞も提供する。
【0017】
本発明はさらに、本明細書に記載の任意の抗原結合タンパク質の産生方法を提供し、この方法は、第1および第2ベクターを含む宿主細胞の培養工程を含み、前記第1ベクターは本明細書に記載の任意の抗原結合タンパク質重鎖をコードするポリヌクレオチドを含み、前記第2ベクターは、適切な培地(例えば無血清培地)中、本明細書に記載の任意の抗原結合タンパク質軽鎖をコードするポリヌクレオチドを含む。
【0018】
本発明はさらに、本明細書に記載の抗原結合タンパク質および医薬上許容可能な担体を含む医薬組成物を提供する。
【0019】
さらなる態様では、本発明は、本発明の抗原結合タンパク質の投与によりアトピー性疾患/障害および慢性炎症疾患/障害と関連した疾患または障害の治療または予防の方法を提供する。特定の興味は、喘息、例えばアレルギー性喘息、特に重症喘息(すなわちコルチコステロイドの全身投与を含む現在の治療が奏効しない喘息;Busse WW et al, J Allergy Clin. Immunol 2000, 106: 1033−1042を参照されたい)、「困難性」喘息(difficult asthma)(最大推奨用量の吸入ステロイド処方にもかかわらず制御不成功を特徴とする喘息性形質として定義、Barnes PJ (1998), Eur Respir J 12:1208−1218を参照されたい)、「ブリットル」喘息(Brittle asthma)(高用量の吸入ステロイドにもかかわらず大きく変動する最大呼気流量(PEF)を維持する重度の不安定喘息患者のサブグループを定義、Ayres JG et al (1998) Thorax 58:315−321を参照されたい)、夜間性喘息、月経前喘息、ステロイド抵抗性喘息(Woodcock AJ (1993) Eur Respir J 6:743−747を参照されたい)、ステロイド依存性喘息(高用量の経口ステロイドでのみ制御できる喘息として定義)、アスピリン誘発性喘息、成人発症型喘息、小児喘息の治療における本発明の抗原結合タンパク質の使用である。本発明の抗体は、急性喘息エピソード(喘息発作重積状態)を予防する、頻度を減らす、または作用を緩和するために使用してよい。本発明の抗体は、喘息の治療に使用される他の薬の必要な投薬を(投薬の投与量または投与頻度のいずれかに関して)減らすためにも使用してよい。例えば、本発明の抗体は、喘息のステロイド治療(コルチコステロイド治療など)に必要な投薬を減らす(「ステロイド使用を控える」)ために使用してよい。本発明の抗体で治療し得る他の疾患または障害としては、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、慢性閉塞性肺疾患(COPD)、好酸球性食道炎、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症(強皮症)、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、および細胞周期調節疾患、例えばホジキン病、B細胞慢性リンパ球性白血病が挙げられる。
【0020】
別の態様では、本発明は、アトピー性疾患/障害および慢性炎症疾患/障害の治療または予防のための薬の調製における本発明の抗原結合タンパク質の使用を提供する。特定の興味は、喘息、例えばアレルギー性喘息、特に重症喘息(すなわちコルチコステロイドの全身投与を含む現在の治療が奏効しない喘息;Busse WW et al, J Allergy Clin. Immunol 2000, 106: 1033−1042を参照されたい)、「困難性」喘息(最大推奨用量の吸入ステロイド処方にもかかわらず制御不成功を特徴とする喘息性形質として定義、Barnes PJ (1998), Eur Respir J 12:1208−1218を参照されたい)、「ブリットル」喘息(高用量の吸入ステロイドにもかかわらず大きく変動する最大呼気流量(PEF)を維持する重度の不安定喘息患者のサブグループを定義、Ayres JG et al (1998) Thorax 58:315−321を参照されたい)、夜間性喘息、月経前喘息、ステロイド抵抗性喘息(Woodcock AJ (1993) Eur Respir J 6:743−747を参照されたい)、ステロイド依存性喘息(高用量の経口ステロイドでのみ制御できる喘息として定義)、アスピリン誘発性喘息、成人発症型喘息、小児喘息の治療における本発明の抗原結合タンパク質の使用である。本発明の抗体は、急性喘息エピソード(喘息発作重積状態)を予防する、頻度を減らす、または作用を緩和するために使用してよい。本発明の抗体は、喘息の治療に使用される他の薬の必要な投薬を(投薬の投与量または投与頻度のいずれかに関して)減らすためにも使用してよい。例えば、本発明の抗体は、喘息のステロイド治療(コルチコステロイド治療など)に必要な投薬を減らす(「ステロイド使用を控える」)ために使用してよい。本発明の抗体で治療し得る他の疾患または障害としては、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、慢性閉塞性肺疾患(COPD)、好酸球性食道炎、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症(強皮症)、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、および細胞周期調節疾患、例えばホジキン病、B細胞慢性リンパ球性白血病が挙げられる。
【0021】
本発明の他の態様および利益を本明細書およびその実施形態に詳細に記載する。
【0022】
定義
本発明のその抗原結合タンパク質に関して本明細書を通して使用される「ヒトIL−13と結合する」という用語は、抗原結合タンパク質が他のヒトタンパク質(IL−4など)と結合しないかわずかしか結合せずに、ヒトIL−13(以下、hIL−13と呼ぶ)と結合することを意味する。特に、本発明の抗原結合タンパク質は、Biacoreアッセイ(例えば実施例3に記載のBiacoreアッセイ)にてヒトIL−13と結合することを見ることができる点から、ヒトIL−13と結合する。しかしながら、この用語は、本発明のある抗原結合タンパク質は他種由来のIL−13(例えばカニクイザルIL−13)と交差反応もし得るという事実を除外しない。
【0023】
本明細書で使用する「抗原結合タンパク質」という用語は、ヒトIL−13と結合でき、中和できる抗体、抗体断片および他のタンパク質構築物を指す。
【0024】
Fv、Fc、Fd、Fab、またはF(ab)2という用語は、標準的な意味で使用される(例えば、Harlow et al., Antibodies A Laboratory Manual, Cold Spring Harbor Laboratory, (1988)を参照されたい)。
【0025】
「キメラ抗体」とは、アクセプター抗体に由来する軽鎖および重鎖定常領域に関連したドナー抗体由来の天然可変領域(軽鎖および重鎖)を含む加工された抗体の一種を指す。
【0026】
「ヒト化抗体」とは、非ヒトドナー免疫グロブリンに由来するCDRを有し、分子の残りの免疫グロブリン派生部分は1つ(以上)のヒト免疫グロブリンに由来する加工された抗体の一種を指す。加えて、フレームワーク支持残基は、結合親和性を保持するために改変し得る(例えば、Queen et al., Proc. Natl Acad Sci USA, 86:10029−10032 (1989), Hodgson et al., Bio/Technology, 9:421 (1991)を参照されたい)。適切なヒトアクセプター抗体は、ドナー抗体のヌクレオチド配列およびアミノ酸配列への相同により、従来のデータベース、例えば、KABAT(登録商標)データベース、Los Alamosデータベース、およびSwissタンパク質データベースから選択し得る。ドナー抗体のフレームワーク領域との(アミノ酸に基づく)相同を特徴とするヒト抗体は、ドナーCDRを挿入するための重鎖定常領域および/または重鎖可変フレームワーク領域の提供に適し得る。軽鎖の定常または可変フレームワーク領域を供与できる適切なアクセプター抗体を類似の形態にて選択し得る。アクセプター抗体の重鎖と軽鎖は同じアクセプター抗体に由来している必要がないことに留意されたい。先行技術にて、かかるヒト化抗体の産生方法がいくつか記載されている−例えば欧州特許第A−0239400号および欧州特許第A−054951号を参照されたい。
【0027】
「ドナー抗体」という用語は、領域をコードする改変された免疫グロブリンならびにその結果発現したドナー抗体の抗原特異性および中和活性特徴を有する改変抗体を提供するようにその可変領域、CDR、または他の、その機能的断片もしくは第1免疫グロブリンパートナーアナログのアミノ酸配列に寄与する(モノクローナル、および/または組換え)抗体を指す。
【0028】
「アクセプター抗体」という用語は、第1免疫グロブリンパートナーに対するその重鎖および/もしくは軽鎖フレームワーク領域ならびに/またはその重鎖および/もしくは軽鎖定常領域をコードするアミノ酸配列のすべて(または任意の一部だが、いくつかの実施形態では、すべて)に寄与する、ドナー抗体と異種の(モノクローナルおよび/または組換え)抗体を指す。ある実施形態では、ヒト抗体はアクセプター抗体である。
【0029】
「CDR」は、免疫グロブリン重鎖および軽鎖の超可変領域である抗体の相補性決定領域アミノ酸配列として定義する。例えば、Kabat et al., Sequences of Proteins of Immunological Interest, 4th Ed., U.S. Department of Health and Human Services, National Institutes of Health (1987)を参照されたい。免疫グロブリンの可変部分には3つの重鎖および3つの軽鎖CDR(またはCDR領域)がある。したがって、本明細書で使用する「CDR」とは、全3つの重鎖CDRまたは全3つの軽鎖CDR(または適切な場合、全重鎖CDRと全軽鎖CDRの両方)を指す。抗体の構造およびタンパク質の折り畳み部分は、他の残基が抗原結合領域の一部とみなされ、当業者によりそのように理解されることを意味し得る。例えばChothia et al., (1989) Conformations of immunoglobulin hypervariable regions; Nature 342, p877−883を参照されたい。
【0030】
本明細書で使用する「ドメイン」という用語は、残りのタンパク質とは独立した三次構造を有する折り畳まれたタンパク質構造を指す。一般に、ドメインは、タンパク質の別々の機能特性を担い、多くの場合、残るタンパク質および/またはドメインの機能を失わずに他のタンパク質に添加、除去または移動し得る。「単一抗体可変ドメイン」とは、抗体可変ドメインに特徴的な配列を含む折り畳まれたポリペプチドドメインである。したがって、完全な抗体可変ドメインおよび修飾可変ドメイン(例えば、1つまたは複数のループが抗体可変ドメインに特徴的でない配列に置換されている)、またはNもしくはC末端伸長部分が切断されているか含まれる抗体可変ドメイン、ならびに完全長ドメインの少なくとも結合活性および特異性を保持する可変ドメインの折り畳まれた断片を含む。
【0031】
「免疫グロブリン単一可変ドメイン」という語句は、異なるV領域またはVドメインとは独立して抗原またはエピトープと特異的結合する抗体可変ドメイン(VH、VHH、VL)を指す。免疫グロブリン単一可変ドメインは、単一免疫グロブリン可変ドメインによる抗原結合のための他の領域またはドメインが不要である他の異なる可変領域または可変ドメインと1つの型式(例えば、ホモまたはヘテロ多量体)で存在できる(すなわち免疫グロブリン単一可変ドメインは、追加の可変ドメインとは独立して抗原と結合する)。「ドメイン抗体」または「dAb」とは、本明細書で使用する、抗原と結合できる「免疫グロブリン単一可変ドメイン」という用語と同義である。免疫グロブリン単一可変ドメインは、ヒト抗体可変ドメインであり得るが、齧歯類(例えば、WO00/29004号に開示のもの)、テンジクザメおよびラクダVHH dAb(ナノボディ)などの他種に由来する単一抗体可変ドメインも含む。ラクダVHHは、天然で軽鎖を欠いている重鎖抗体を産生するラクダ、ラマ、アルパカ、ヒトコブラクダ、およびグアナコを含む種に由来する免疫グロブリン単一可変ドメインポリペプチドである。かかるVHHドメインは、当技術分野で利用可能な標準的な技術に従いヒト化し得、かかるドメインは、依然として本発明による「ドメイン抗体」とみなされる。本明細書で使用する「VH」は、ラクダVHHドメインを含む。NARVは、テンジクザメを含む軟骨魚において同定された別の種類の免疫グロブリン単一可変ドメインである。これらのドメインは、新規の抗原受容体可変領域(一般にV(NAR)またはNARVと略される)としても知られている。詳細については、Mol. Immunol. 44, 656−665 (2006)および米国特許第20050043519A1号を参照されたい。
【0032】
「エピトープ結合ドメイン」という用語は、異なるV領域またはVドメインとは独立して抗原またはエピトープと特異的結合するドメインを指し、これはドメイン抗体(dAb)、例えばヒト、ラクダまたはサメ免疫グロブリン単一可変ドメインであってもよく、天然リガンド以外のリガンドへの結合を達成するためにタンパク質工学にかけられた非免疫グロブリン足場、例えばCTLA−4(Evibody);リポカリン;プロテインA派生分子(プロテインAのZドメイン(Affibody、SpA)、Aドメイン(Avimer/Maxibody)など);熱ショックタンパク質(GroElおよびGroESなど);トランスフェリン(trans−body);アンキリン反復タンパク質(DARPin);ペプチドアプタマー;C型レクチンドメイン(Tetranectin);ヒトγクリスタリンおよびヒトユビキチン(affilin);PDZドメイン;ヒトプロテアーゼ阻害剤のサソリ毒クニッツ型ドメイン;ならびにフィブロネクチン(adnectin)からなる群から選択される非免疫グロブリン足場の誘導体であるドメインであってもよい。
【0033】
CTLA−4(細胞傷害性Tリンパ球結合抗原4)は主にCD4+T細胞上で発現するCD28ファミリー受容体である。その細胞外ドメインは、可変ドメイン様Ig折り畳み部分を有する。抗体のCDRに対応するループは、異なる結合特性を与えるために異種配列と置換することができる。異なる結合特異性を有するように加工されたCTLA−4分子は、Evibodyとしても知られている。詳細については、Journal of Immunological Methods 248 (1−2), 31−45 (2001)を参照されたい。
【0034】
リポカリンは、小さな疎水性分子(ステロイド、ビリン、レチノイドおよび脂質など)を輸送する細胞外タンパク質ファミリーである。それらは、異なる標的抗原と結合するように加工することができる、円錐構造の開端にループをいくつか有する硬βシート状二次構造を有する。Anticalinは、サイズが160〜180個のアミノ酸であり、リポカリンに由来する。詳細については、Biochim Biophys Acta 1482: 337−350 (2000)、米国特許第7250297B1号および米国特許第20070224633号を参照されたい。
【0035】
Affibody(アフィボディ)は、抗原と結合するように加工することができる黄色ブドウ球菌のプロテインAに由来する足場である。ドメインは、約58個のアミノ酸の3つのヘリックスバンドルからなる。ライブラリは、表面残基の無作為化により生成されている。詳細については、Protein Eng. Des. Sel. 17, 455−462 (2004)および欧州特許第1641818A1号を参照されたい。
【0036】
Avimer(アビマー)は、Aドメイン足場ファミリーに由来する多ドメインタンパク質である。約35個のアミノ酸の天然ドメインは、定義されたジスルフィド結合構造をとる。多様性は、Aドメインファミリーによって示される天然変異をシャッフルすることによって生成される。詳細については、Nature Biotechnology 23(12), 1556 − 1561 (2005) and Expert Opinion on Investigational Drugs 16(6), 909−917 (June 2007)を参照されたい。
【0037】
トランスフェリンは、単量体血清輸送糖タンパク質である。トランスフェリンは、許容状態の表面ループへのペプチド配列の挿入によって、異なる標的抗原と結合するように加工することができる。加工されたトランスフェリン足場の例としては、Trans−bodyが挙げられる。詳細については、J. Biol. Chem 274, 24066−24073 (1999)を参照されたい。
【0038】
設計されたアンキリン反復タンパク質(DARPin)は、内在性膜タンパク質の細胞骨格への結合を媒介するタンパク質ファミリーであるアンキリンに由来する。単一アンキリン反復は、2つのα−ヘリックスおよびβ−ターンからなる33残基のモチーフである。それらは、各反復の第1α−ヘリックスおよびβ−ターンにおける残基を無作為化することによって、異なる標的抗原と結合するように加工することができる。それらの結合界面は、モジュール数を増大することによって増大できる(親和性成熟の方法)。詳細については、J. Mol. Biol. 332, 489−503 (2003), PNAS 100(4), 1700−1705 (2003)およびJ. Mol. Biol. 369, 1015−1028 (2007)ならびに米国特許第20040132028A1号を参照されたい。
【0039】
フィブロネクチンは、抗原と結合するように加工することができる足場である。Adnectinは、ヒトフィブロネクチンIII型(FN3)15反復単位の10番目ドメイン天然アミノ酸配列の主鎖からなる。β−サンドイッチの片端における3つのループは、Adnectinが興味の治療標的を特異的に認識することを可能にするように加工することができる。詳細については、Protein Eng. Des. Sel. 18, 435−444 (2005)、米国特許第20080139791号、WO2005056764号および米国特許第6818418B1号を参照されたい。
【0040】
ペプチドアプタマーは、普遍的な足場タンパク質、典型的には、活性部位に挿入された制限された可変ペプチドループを含むチオレドキシン(TrxA)からなるコンビナトリアル認識分子である。詳細については、Expert Opin. Biol. Ther.5, 783−797 (2005)を参照されたい。
【0041】
微小体(Microbody)は、3〜4個のシステイン架橋を含む25〜50個のアミノ酸の長さの天然マイクロタンパク質に由来し、マイクロタンパク質の例としては、KalataB1およびコノトキシンおよびノッチンが挙げられる。マイクロタンパク質は、マイクロタンパク質の全体的な折り畳み部分に影響を与えずに最大25個のアミノ酸を含むように加工することができるループを有する。加工されたノッチンドメインの詳細については、WO2008098796号を参照されたい。
【0042】
他のエピトープ結合ドメインとしては、異なる標的抗原結合特性を加工するために足場として使用されてきたタンパク質が挙げられ、ヒトγクリスタリンおよびヒトユビキチン(affilin)、ヒトプロテアーゼ阻害剤のクニッツ型ドメイン、Ras結合プロテインAF−6のPDZドメイン、サソリ毒(カリブドトキシン)、C型レクチンドメイン(tetranectin)が挙げられ、Chapter 7 − Non−Antibody Scaffolds from Handbook of Therapeutic Antibodies (2007, edited by Stefan Dubel)およびProtein Science 15:14−27 (2006)に概説されている。本発明のエピトープ結合ドメインは、これらの任意の代替タンパク質ドメインに由来し得る。
【0043】
本明細書で使用する「抗原結合部位」という用語は、抗原と特異的結合できるタンパク質上の部位を指し、これは単一ドメイン、例えばエピトープ結合ドメインであってもよく、または標準的な抗体上に見つけることができるような対になったVH/VLドメインであってもよい。本発明のいくつかの態様では、単鎖Fv(ScFv)ドメインは、抗原結合部位を提供できる。
【0044】
本明細書で使用する「mAbdAb」および「dAbmAb」という用語は、本発明の抗原結合タンパク質を指す。2つの用語は同義的に使用でき、本明細書で使用する場合に同じ意味を有することが意図される。
【0045】
本明細書で使用する「抗原結合タンパク質」という用語は、抗体、抗体断片、例えばドメイン抗体(dAb)、ScFv、FAb、FAb2、およびIL−13と結合できる他のタンパク質構築物を指す。抗原結合分子は、Ig可変ドメイン(例えば抗体、ドメイン抗体、Fab、Fab’、F(ab’)2、Fv、ScFv、二重特異性抗体、mAbdAb、Affibody、ヘテロ抱合体抗体もしくは二重特異性抗体)を少なくとも1つ含み得る。一実施形態では、抗原結合分子は抗体である。別の実施形態では、抗原結合分子はdAb、すなわち異なるV領域またはVドメインとは独立して抗原またはエピトープと特異的結合するVH、VHHまたはVLなどの免疫グロブリン単一可変ドメインである。抗原結合分子は、2つの標的と結合可能であり得る、すなわち二重標的タンパク質であり得る。抗原結合分子は、抗体および抗原結合断片(例えば、1つもしくは複数のドメイン抗体ならびに/またはモノクローナル抗体と結合した1つもしくは複数のScFvなど)の組み合わせであり得る。抗原結合分子は、IL−13への結合を達成するためにタンパク質工学にかけられた非Igドメイン、例えばCTLA−4(Evibody);リポカリン;プロテインA派生分子(プロテインAのZドメイン(Affibody、SpA)、Aドメイン(Avimer/Maxibody)など);熱ショックタンパク質(GroElおよびGroESなど);トランスフェリン(trans−body);アンキリン反復タンパク質(DARPin);ペプチドアプタマー;C型レクチンドメイン(Tetranectin);ヒトγクリスタリンおよびヒトユビキチン(affilin);PDZドメイン;ヒトプロテアーゼ阻害剤のサソリ毒クニッツ型ドメイン;ならびにフィブロネクチン(adnectin)からなる群から選択される足場の誘導体であるドメインも含み得る。本明細書で使用する「抗原結合タンパク質」はヒトIL−13を拮抗および/または中和できる。加えて、抗原結合タンパク質はIL−13と結合することによってIL−13活性をブロックし得、天然リガンドによる受容体への結合および/または受容体の活性化を防止し得る。
【0046】
本明細書で使用する「IL−13アンタゴニスト」は、IL−13の活性を少なくとも1つ低減およびまたは除去できる任意の化合物を含む。例として、IL−13アンタゴニストはIL−13と結合し、この結合はIL−13活性を直接低減または除去する場合もあり、リガンドの受容体結合を少なくとも1つブロックすることによって間接的に低減または除去する場合もある。
【0047】
本明細書で使用する「IL−4アンタゴニスト」は、IL−4の活性を少なくとも1つ低減およびまたは除去できる任意の化合物を含む。例として、IL−4アンタゴニストはIL−4と結合し、この結合はIL−4活性を直接低減または除去する場合もあり、リガンドの受容体結合を少なくとも1つブロックすることによって間接的に低減または除去する場合もある。
【0048】
本明細書で使用する「IL−5アンタゴニスト」は、IL−5の活性を少なくとも1つ低減およびまたは除去できる任意の化合物を含む。例として、IL−5アンタゴニストはIL−5と結合し、この結合はIL−5活性を直接低減または除去する場合もあり、リガンドの受容体結合を少なくとも1つブロックすることによって間接的に低減または除去する場合もある。
【図面の簡単な説明】
【0049】
【図1】TF−1細胞増殖アッセイにおいてY100B変異体を含むmAbdAbのヒトIL−13を中和能力を示すグラフ
【図2】TF−1細胞増殖アッセイにおいてY100B変異体を含むmAbdAbのヒトIL−13を中和能力を示すグラフ
【図3】BPC2222のSECトレース
【図4】BPC2223のSECトレース
【図5】BPC2230のSECトレース
【図6】BPC2231のSECトレース
【図7】BPC1085のSECトレース
【図8】BPC1086のSECトレース
【図9】BPC1087のSECトレース
【図10】ELISAにより決定した、精製されたmAbdAb(BPC1085、BPC1086およびBPC1087)のヒトIL−4への結合を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図11】TF−1細胞バイオアッセイにおいて、ヒトIL−4に対し精製されたmAbdAb(BPC1085、BPC1086およびBPC1087)によるヒトIL−4中和を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図12】TF−1細胞バイオアッセイにおいて、ヒトIL−13に対し精製されたmAbdAb(BPC1085、BPC1086およびBPC1087)によるヒトIL−13中和を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図13】BPC3214のSECプロファイル。
【図14】BPC3215のSECプロファイル。
【図15】ELISAにより決定した、精製されたmAbdAb BPC3214、BPC3215、BPC1085および対照mAb A1Y100BVAL1および抗IL−4 mAbのヒトIL−13への結合を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図16】ELISAにより決定した、精製されたmAbdAb BPC3214、BPC3215、BPC1085および対照mAb A1Y100BVAL1および抗IL−4 mAbのヒトIL−4への結合を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図17】ELISAにより決定した、一過性発現したmAbdAbの組換え大腸菌発現ヒトIL−4への結合を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【発明を実施するための形態】
【0050】
一実施形態では、本発明の抗原結合タンパク質は、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8および配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14および配列番号15、配列番号16、配列番号17および配列番号18からなる一覧から選択されるCDRH3ならびに適切なCDRH1およびCDRH2を含む重鎖可変領域を含み、適切なCDRL1、CDRL2およびCDRL3を含む軽鎖可変領域と対をなして、ヒトIL−13と結合する抗原結合Fv単位を形成する。一実施形態では、本発明の抗原結合タンパク質は、ヒトIL−13活性を中和する。
【0051】
この実施形態の一態様では、配列番号1もしくは配列番号60、もしくは配列番号61に説明されるCDRH1ならびに配列番号2に説明されるCDRH2も重鎖可変領域に存在する。この実施形態のさらなる態様では、配列番号19に説明されるCDRHL1、配列番号20に説明されるCDRL2および配列番号21に説明されるCDRL3も軽鎖可変領域に存在する。
【0052】
別の態様では、抗原結合タンパク質は、Biacoreにより10nM以下、より特に2nM以下、例えば約0.8nM〜2nM、1nM以下、または100pM以下、例えば約20pM〜約100pMまたは約20pM〜約80pM、または約20pM〜約60pMと測定された高親和性でヒトIL−13と結合する。1つのかかる実施形態では、これは例えば実施例3に説明するように、バイオセンサーチップ上に捕捉された抗原結合タンパク質を用いてBiacoreにより測定される。
【0053】
本発明の重鎖可変領域はヒトIL−13と結合させる軽鎖可変領域と共に、従来の免疫グロブリン形態で(例えば、ヒトIgG、IgA、IgMなど)またはヒトIL−13との結合を可能にする他の任意の「抗体様」型式(例えば、単一鎖Fv、二重特異性抗体、Tandab(商標)など)において構成し得る(代替「抗体」型式の概要については、Holliger and Hudson, Nature Biotechnology, 2005, Vol 23, No. 9, 1126−1136を参照されたい)。
【0054】
本発明の抗原結合タンパク質は、配列番号58および配列番号59に記載の可変領域を有するマウス抗体、または非マウスのその等価物(ラット、ヒト、キメラまたはそのヒト化変異体など)に由来し、例えば配列番号22および配列番号24に記載の重鎖および軽鎖を有するヒト化抗体に由来する。
【0055】
本発明の一態様では、ヒトIL−13と結合し、CDRH3 SIYDDYHYDDYYAMDY(配列番号3)の変異体を含み、CDRH3が1つもしくは複数の以下の位置(Kabat番号付けを使用)で以下に説明する代替アミノ酸に置換されている抗原結合タンパク質(例えば抗体)を提供する:
a)1位のS95は、トリプトファン(W)と置換されている
b)2位のI96は、バリン(V)と置換されている
c)3位のY97は、フェニルアラニン(F)と置換されている
d)4位のD98は、グルタミン(E)と置換されている
e)7位のH100Aは、アラニン(A)、グルタミン酸(E)、グルタミン(Q)、アルギニン(R)、セリン(S)、トレオニン(T)もしくはバリン(V)と置換されている、ならびに
f)8位のY100Bは、アラニン(A)、イソロイシン、(I)、トリプトファン(W)もしくはバリン(V)と置換されている。
【0056】
本発明の別の態様では、ヒトIL−13と結合し、配列番号3に説明されるCDRH3を含む抗原結合タンパク質(例えば抗体)を提供し、このCDRH3は、S95W、I96V、Y97F、D98E、H100A_A、H100A_E、H100A_Q、H100A_R、H100A_S、H100A_T、H100A_V、Y100B_A、Y100B_I、Y100B_W、およびY100B_Vの置換を1つまたは複数含む。
【0057】
一態様では、本発明の抗原結合タンパク質(例えば本発明の抗体)は、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8および配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14および配列番号15、配列番号16、配列番号17および配列番号18に説明されるものから選択されるCDRH3配列を含む。かかる抗原結合タンパク質はさらに以下のCDR配列
CDRH1:配列番号1、60および61から選択される、
CDRH2:配列番号2、
CDRL1:配列番号19、
CDRL2:配列番号20、ならびに
CDRL3:配列番号21
を含み得る。
【0058】
一実施形態では、抗原結合タンパク質は、以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号18、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21
を含む。
【0059】
別の実施形態では、抗原結合タンパク質は、以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号17、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21
を含む。
【0060】
別の実施形態では、抗原結合タンパク質は、以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号16、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21
を含む。
【0061】
別の実施形態では、抗原結合タンパク質は、以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号15、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21
を含む。
【0062】
本明細書を通して、抗体配列中のアミノ酸残基は、Kabatスキームに従い番号付けされる。同様に、「CDR」、「CDRL1」、「CDRL2」、「CDRL3」、「CDRH1」、「CDRH2」、「CDRH3」(例えば配列番号60および61に説明されるCDRH3)という用語は、別に示されない限り、Kabat et al; “Sequences of proteins of Immunological Interest” NIH, 1987に説明されるようにKabat番号付け系に従う。
【0063】
本発明の別の態様では、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号88、配列番号89、配列番号90および配列番号91から選択される配列を有する重鎖;ならびに配列番号24の軽鎖を含む抗原結合タンパク質(ヒト化抗体またはその抗原結合断片など)を提供する。
【0064】
本発明は、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号88、配列番号89、配列番号90から選択される重鎖を含む抗原結合タンパク質(ヒト化抗体またはその抗原結合断片など)を提供する。
【0065】
本発明は、配列番号108、配列番号110、配列番号112および配列番号114から選択される軽鎖を含む抗原結合タンパク質(ヒト化抗体またはその抗原結合断片など)も提供する。
【0066】
本発明はさらに、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号88、配列番号89、配列番号90および配列番号91から選択される配列を有する重鎖;ならびに配列番号108、配列番号110、配列番号112および配列番号114の軽鎖を含む抗原結合タンパク質(ヒト化抗体またはその抗原結合断片など)を提供する。
【0067】
一実施形態では、本発明の抗原結合タンパク質は、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号88、配列番号89、配列番号90および配列番号91から選択される重鎖、ならびに配列番号24、配列番号108、配列番号110、配列番号112および配列番号114から選択される軽鎖を含む抗体を含む。
【0068】
一実施形態では、本発明の抗原結合タンパク質は、配列番号48、配列番号50、配列番号52および配列番号54から選択される重鎖;ならびに配列番号24、配列番号108、配列番号110、配列番号112および配列番号114から選択される軽鎖を含み、例えば抗原結合タンパク質は配列番号48の重鎖および配列番号24の軽鎖、または配列番号50の重鎖および配列番号24の軽鎖、または配列番号52の重鎖および配列番号24の軽鎖、または配列番号54の重鎖および配列番号24の軽鎖、または配列番号88の重鎖および配列番号24の軽鎖または配列番号89の重鎖および配列番号24の軽鎖、または配列番号90の重鎖および配列番号24の軽鎖、または配列番号91の重鎖および配列番号24の軽鎖、または配列番号48の重鎖および配列番号108の軽鎖、または配列番号50の重鎖および配列番号108の軽鎖、または配列番号52の重鎖および配列番号108の軽鎖、または配列番号54の重鎖および配列番号108の軽鎖、または配列番号88の重鎖および配列番号108の軽鎖または配列番号89の重鎖および配列番号108の軽鎖、または配列番号90の重鎖および配列番号108の軽鎖、または配列番号91の重鎖および配列番号108の軽鎖、または配列番号48の重鎖および配列番号110の軽鎖、または配列番号50の重鎖および配列番号110の軽鎖、または配列番号52の重鎖および配列番号110の軽鎖、または配列番号54の重鎖および配列番号110の軽鎖、または配列番号88の重鎖および配列番号110の軽鎖または配列番号89の重鎖および配列番号110の軽鎖、または配列番号90の重鎖および配列番号110の軽鎖、または配列番号91の重鎖および配列番号110の軽鎖を含む。
【0069】
1つのかかる実施形態では、本発明の抗原結合タンパク質は、配列番号48、配列番号50、配列番号52および配列番号54から選択される重鎖;ならびに配列番号108および配列番号110から選択される軽鎖(例えば配列番号48の重鎖および配列番号108の軽鎖を含む抗原結合タンパク質、または配列番号48の重鎖および配列番号110の軽鎖を含む抗原結合タンパク質、または配列番号50の重鎖および配列番号108の軽鎖を含む抗原結合タンパク質、または配列番号50の重鎖および配列番号110の軽鎖を含む抗原結合タンパク質、または配列番号52の重鎖および配列番号108の軽鎖を含む抗原結合タンパク質、または配列番号52の重鎖および配列番号110の軽鎖を含む抗原結合タンパク質、または配列番号54の重鎖および配列番号108の軽鎖を含む抗原結合タンパク質、または配列番号54の重鎖および配列番号110の軽鎖を含む抗原結合タンパク質)を含む。
【0070】
本発明のIL−13抗体はIL−4および/もしくはIL−5アンタゴニスト、例えばIL−4抗体もしくはエピトープ結合ドメイン、ならびに/またはIL−5抗体もしくはエピトープ結合ドメインとの組み合わせであり得る。これらは、同時に投与する、すなわち同時投与するか、または互いに24時間以内(例えば互いに20時間以内、または15時間以内または12時間以内、または10時間以内、または8時間以内、または6時間以内、または4時間以内、または2時間以内、または1時間以内、または30分以内)に投与する分離する分子の混合物として投与し得る。
【0071】
さらなる実施形態では、アンタゴニストは、2つ以上の抗原と結合できる1分子として存在し、例えば本発明は、IL−13と結合でき、IL−4へも結合できるまたはIL−5へも結合できる、またはIL−4とIL−5へも結合できる本発明のIL−13抗体を含む抗原結合タンパク質を提供する。
【0072】
一実施形態では、本発明の抗原結合タンパク質は、本発明の少なくともCDRH3、ならびに任意選択的に1つもしくは複数のCDRH1、CDRH2、CDRL1、CDRL2、およびCDRL3を含み、IL−13と結合でき、1つもしくは複数のIL−4もしくはIL−5へも結合できる多特異的な抗体であり得る。1つのかかる実施形態では、CDRH3、すなわち本明細書において定義される抗原結合タンパク質を含み、IL−4、またはIL−5と結合できる抗原結合部位をさらに含む多特異的な抗体を提供する。
【0073】
本発明の抗原結合タンパク質の1例は、IL−4もしくはIL−5に特異性の1つもしくは複数のエピトープ結合ドメインと結合している、本明細書において定義されるCDRH1、CDRH2、CDRH3、CDRL1、CDRL2、およびCDRL3を含むIL−13に特異的な抗体、例えばIL−13とIL−4もしくはIL−13とIL−5と結合できる二重特異性抗原結合タンパク質、またはIL−13、IL−4およびIL−5と結合できる三重特異性抗原結合タンパク質である。
【0074】
本明細書に記載の任意の抗原結合タンパク質は、例えば、実施例8に記載したものなどの適切なアッセイを用いて化学量論分析により決定されるように2つ以上の抗原に同時と結合可能であり得ることが理解されるであろう。
【0075】
本発明は、1つまたは複数のエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、例えばIL−4と結合できるエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、またはIL−5と結合できるエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、またはIL−4と結合できる第1のエピトープ結合ドメインおよびIL−5と結合できる第2のエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質も提供する。
【0076】
エピトープ結合ドメインは、IL−13抗体重鎖のC末端もしくはN末端またはIL−13抗体軽鎖のC末端もしくはN末端と結合し得る。
【0077】
本発明の抗体はリンカーを用いることによりエピトープ結合ドメインと結合し得る。適切なリンカーの例としては、1個のアミノ酸〜150個のアミノ酸の長さ、または1個のアミノ酸〜140個のアミノ酸、例えば、1個のアミノ酸〜130個のアミノ酸、または1〜120個のアミノ酸、または1〜80個のアミノ酸、または1〜50個のアミノ酸、または1〜20個のアミノ酸、または1〜10個のアミノ酸、または5〜18個のアミノ酸であり得るアミノ酸配列が挙げられる。かかる配列はそれら自体の三次構造を有し得る、例えば、本発明のリンカーは単一可変ドメインを含み得る。リンカーのサイズは一実施形態では、単一可変ドメインと等価物である。適切なリンカーのサイズは、1〜20オングストローム、例えば15オングストローム未満、または10オングストローム未満、または5オングストローム未満であり得る。
【0078】
本発明の一実施形態では、少なくとも1つのエピトープ結合ドメインは、1〜150個のアミノ酸、例えば1〜20個のアミノ酸、例えば1〜10個のアミノ酸を含むリンカーでIL−13抗体と直接結合している。
【0079】
かかるリンカーは、配列番号82〜87、92〜93に説明されるものの任意の1つまたはかかるリンカーの複数から選択され得る。
【0080】
本発明の抗原結合タンパク質において使用するリンカーは、単独または他のリンカーに加えて、1つまたは複数のGS残基セット、例えば「GSTVAAPS」(配列番号92)または「TVAAPSGS」(配列番号87)または「GSTVAAPSGS」(配列番号93)を含み得る。一実施形態では、リンカーは、配列番号83を含む。
【0081】
一実施形態では、エピトープ結合ドメインはリンカー「(PAS)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(GGGGS)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(TVAAPS)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(GS)m(TVAAPSGS)n」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(PAVPPP)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(TVSDVP)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(TGLDSP)n(GS)m」によりIL−13抗体と結合している。かかる実施形態のすべてにおいて、n=1〜10、およびm=0〜4である。
【0082】
かかるリンカーの例としては、(PAS)n(GS)m(式中n=1およびm=1)、(PAS)n(GS)m(式中n=2およびm=1)、(PAS)n(GS)m(式中n=3およびm=1)、(PAS)n(GS)m(式中n=4およびm=1)、(PAS)n(GS)m(式中n=2およびm=0)、(PAS)n(GS)m(式中n=3およびm=0)、(PAS)n(GS)m式中n=4およびm=0が挙げられる。
【0083】
かかるリンカーの例としては、(GGGGS)n(GS)m(式中n=1およびm=1)、(GGGGS)n(GS)m(式中n=2およびm=1)、(GGGGS)n(GS)m(式中n=3およびm=1)、(GGGGS)n(GS)m(式中n=4およびm=1)、(GGGGS)n(GS)m(式中n=2およびm=0)、(GGGGS)n(GS)m(式中n=3およびm=0)、(GGGGS)n(GS)m式中n=4およびm=0が挙げられる。
【0084】
かかるリンカーの例としては、(TVAAPS)n(GS)m式中n=1およびm=1(配列番号87)、(TVAAPS)n(GS)m式中n=2およびm=1(配列番号145)、(TVAAPS)n(GS)m式中n=3およびm=1(配列番号146)、(TVAAPS)n(GS)m(式中n=4およびm=1)、(TVAAPS)n(GS)m(式中n=2およびm=0)、(TVAAPS)n(GS)m(式中n=3およびm=0)、(TVAAPS)n(GS)m式中n=4およびm=0が挙げられる。
【0085】
かかるリンカーの例としては、(GS)m(TVAAPSGS)n式中n=1およびm=1(配列番号139)、(GS)m(TVAAPSGS)n式中n=2およびm=1(配列番号140)、(GS)m(TVAAPSGS)n式中n=3およびm=1(配列番号141)、または(GS)m(TVAAPSGS)n式中n=4およびm=1(配列番号142)、(GS)m(TVAAPSGS)n式中n=5およびm=1(配列番号143)、(GS)m(TVAAPSGS)n式中n=6およびm=1(配列番号144)、(GS)m(TVAAPSGS)n式中n=1およびm=0(配列番号87)、(GS)m(TVAAPSGS)n(式中n=2およびm=10)、(GS)m(TVAAPSGS)n(式中n=3およびm=0)、または(GS)m(TVAAPSGS)n式中n=0が挙げられる。
【0086】
かかるリンカーの例としては、(PAVPPP)n(GS)m(式中n=1およびm=1)、(PAVPPP)n(GS)m式中n=2およびm=1(配列番号65)、(PAVPPP)n(GS)m(式中n=3およびm=1)、(PAVPPP)n(GS)m(式中n=4およびm=1)、(PAVPPP)n(GS)m(式中n=2およびm=0)、(PAVPPP)n(GS)m(式中n=3およびm=0)、(PAVPPP)n(GS)m式中n=4およびm=0が挙げられる。
【0087】
かかるリンカーの例としては、(TVSDVP)n(GS)m式中n=1およびm=1(配列番号67)、(TVSDVP)n(GS)m(式中n=2およびm=1)、(TVSDVP)n(GS)m(式中n=3およびm=1)、(TVSDVP)n(GS)m(式中n=4およびm=1)、(TVSDVP)n(GS)m(式中n=2およびm=0)、(TVSDVP)n(GS)m(式中n=3およびm=0)、(TVSDVP)n(GS)m式中n=4およびm=0が挙げられる。
【0088】
かかるリンカーの例としては、(TGLDSP)n(GS)m(式中n=1およびm=1)、(TGLDSP)n(GS)m(式中n=2およびm=1)、(TGLDSP)n(GS)m(式中n=3およびm=1)、(TGLDSP)n(GS)m(式中n=4およびm=1)、(TGLDSP)n(GS)m(式中n=2およびm=0)、(TGLDSP)n(GS)m(式中n=3およびm=0)、(TGLDSP)n(GS)m式中n=4およびm=0が挙げられる。
【0089】
別の実施形態では、エピトープ結合ドメインとIL−13抗体の間にリンカーはない。別の実施形態では、エピトープ結合ドメインはリンカー「TVAAPS」(配列番号83)によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「TVAAPSGS」(配列番号87)によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「GS」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「ASTKGPT」(配列番号84)によりIL−13抗体と結合している。
【0090】
本発明において使用するエピトープ結合ドメインは、異なるV領域またはVドメインとは独立して抗原またはエピトープと特異的結合するドメインであり、これはドメイン抗体であってもよいし、天然リガンド以外のリガンドへの結合を達成するためにタンパク質工学にかけられた非免疫グロブリンドメイン、例えばCTLA−4(Evibody);リポカリン;プロテインA派生分子(プロテインAのZドメイン(Affibody、SpA)、Aドメイン(Avimer/Maxibody)など);熱ショックタンパク質(GroElおよびGroESなど);トランスフェリン(trans−body);アンキリン反復タンパク質(DARPin);ペプチドアプタマー;C型レクチンドメイン(Tetranectin);ヒトγクリスタリンおよびヒトユビキチン(affilin);PDZドメイン;ヒトプロテアーゼ阻害剤のサソリ毒クニッツ型ドメイン;ならびにフィブロネクチン(adnectin)からなる群から選択される非免疫グロブリン足場の誘導体であるドメインであってもよい。一実施形態では、これはドメイン抗体または他の適切なドメイン(CTLA−4、リポカリン、SpA、Affibody、avimer、GroEl、トランスフェリン、GroESおよびフィブロネクチンからなる群から選択されるドメインなど)であり得る。一実施形態では、これは免疫グロブリン単一可変ドメイン、Affibody、アンキリン反復タンパク質(DARPin)およびadnectinから選択し得る。別の実施形態では、これはAffibody、アンキリン反復タンパク質(DARPin)およびadnectinから選択し得る。別の実施形態では、これはドメイン抗体、例えばヒト、ラクダ(ナノボディ)、またはサメ(NARV)ドメイン抗体から選択されるドメイン抗体であり得る。
【0091】
かかる抗原結合タンパク質の例としては、重鎖のC末端もしくはN末端またはC末端と結合するIL−4アンタゴニストであるエピトープ結合ドメインを有する本発明のIL−13抗体が挙げられる。例としては、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号98、配列番号100、配列番号102、配列番号104、配列番号106または配列番号117〜138に説明される重鎖配列、ならびに配列番号24、配列番号108、配列番号110、配列番号112もしくは配列番号114に説明される軽鎖配列を含む抗原結合タンパク質が挙げられ、重鎖および軽鎖の片方または両方はさらにIL−4と拮抗できる1つまたは複数のエピトープ結合ドメイン、例えばIL−4と結合できる単一可変ドメインを含む。かかるエピトープ結合ドメインは、配列番号78〜81および94に説明されるものから選択できる。
【0092】
一実施形態では、本発明の抗原結合構築物は、配列番号62の重鎖配列および配列番号24の軽鎖配列、または配列番号64の重鎖配列および配列番号24の軽鎖配列、または配列番号66の重鎖配列および配列番号24の軽鎖配列、または配列番号68の重鎖配列および配列番号24の軽鎖配列、または配列番号70の重鎖配列および配列番号24の軽鎖配列、または配列番号72の重鎖配列および配列番号24の軽鎖配列、または配列番号74の重鎖配列および配列番号24の軽鎖配列、または配列番号76の重鎖配列および配列番号24の軽鎖配列を含む。
【0093】
一実施形態では、本発明の抗原結合構築物は、配列番号94の重鎖配列および配列番号24の軽鎖配列、または配列番号96の重鎖配列および配列番号24の軽鎖配列、または配列番号98の重鎖配列および配列番号24の軽鎖配列、または配列番号100の重鎖配列および配列番号24の軽鎖配列、または配列番号102の重鎖配列および配列番号24の軽鎖配列、または配列番号104の重鎖配列および配列番号24の軽鎖配列、または配列番号106の重鎖配列および配列番号24の軽鎖配列を含む。
【0094】
一実施形態では、本発明の抗原結合構築物は、配列番号62の重鎖配列および配列番号108の軽鎖配列、または配列番号64の重鎖配列および配列番号108の軽鎖配列、または配列番号66の重鎖配列および配列番号108の軽鎖配列、または配列番号68の重鎖配列および配列番号108の軽鎖配列、または配列番号70の重鎖配列および配列番号108の軽鎖配列、または配列番号72の重鎖配列および配列番号108の軽鎖配列、または配列番号74の重鎖配列および配列番号108の軽鎖配列、または配列番号76の重鎖配列および配列番号108の軽鎖配列、または配列番号62の重鎖配列および配列番号110の軽鎖配列、または配列番号64の重鎖配列および配列番号110の軽鎖配列、または配列番号66の重鎖配列および配列番号110の軽鎖配列、または配列番号68の重鎖配列および配列番号110の軽鎖配列、または配列番号70の重鎖配列および配列番号110の軽鎖配列、または配列番号72の重鎖配列および配列番号110の軽鎖配列、または配列番号74の重鎖配列および配列番号110の軽鎖配列、または配列番号76の重鎖配列および配列番号110の軽鎖配列を含む。
【0095】
一実施形態では、本発明の抗原結合構築物は、配列番号96の重鎖配列および配列番号108の軽鎖配列、または配列番号98の重鎖配列および配列番号108の軽鎖配列、または配列番号100の重鎖配列および配列番号108の軽鎖配列、または配列番号102の重鎖配列および配列番号108の軽鎖配列、または配列番号104の重鎖配列および配列番号108の軽鎖配列、または配列番号106の重鎖配列および配列番号108の軽鎖配列、または配列番号96の重鎖配列および配列番号110の軽鎖配列、または配列番号98の重鎖配列および配列番号110の軽鎖配列、または配列番号100の重鎖配列および配列番号110の軽鎖配列、または配列番号102の重鎖配列および配列番号110の軽鎖配列、または配列番号104の重鎖配列および配列番号110の軽鎖配列、または配列番号106の重鎖配列および配列番号110の軽鎖配列を含む。
【0096】
一実施形態では、IL−13抗体の重鎖は、配列番号48、配列番号50、配列番号52および配列番号54に説明されるものから選択される。別の実施形態では、重鎖は配列番号88〜91、配列番号96〜106、および配列番号117〜138に説明されるものから選択される。1つのかかる実施形態では、重鎖は配列番号96、配列番号98、配列番号100、配列番号102、配列番号104および配列番号106から選択される。
【0097】
一実施形態では、抗原結合タンパク質は、IL−5アンタゴニストであるエピトープ結合ドメインと結合した抗IL−13抗体を含み、この抗IL−13抗体は配列番号3〜18、例えば配列番号15〜18に説明されるものおよび配列番号24の軽鎖配列から選択されるCDRH3を含む。
【0098】
例としては、配列番号26、28、30、32、34、36、38、40、42、44、46、48、50、52もしくは54に説明される重鎖配列ならびに配列番号24に説明される軽鎖配列を含み、重鎖および軽鎖の片方または両方がさらにIL−5と拮抗できる1つまたは複数のエピトープ結合ドメイン、例えばIL−5と結合できる免疫グロブリン単一可変ドメインを含む抗原結合タンパク質が挙げられる。
【0099】
さらなる実施形態では、抗原結合タンパク質は、配列番号26、28、30、32、34、36、38、40、42、44、46、48、50、52もしくは54に説明される重鎖配列ならびに配列番号24に説明される軽鎖配列を含み、この重鎖および軽鎖の片方または両方はさらにIL−4と拮抗できる1つまたは複数のエピトープ結合ドメイン(例えばIL−4と結合できる免疫グロブリン単一可変ドメイン)、およびIL−5と拮抗できる1つまたは複数のエピトープ結合ドメイン(例えばIL−5と結合できる免疫グロブリン単一可変ドメイン)を含む。
【0100】
一実施形態では、本発明の抗原結合タンパク質は、ヒト血清アルブミンと結合できるエピトープ結合ドメインを少なくとも1つ含む。
【0101】
一実施形態では、少なくとも3つの抗原結合部位がある、例えば4、または5または6または8または10個の抗原結合部位があり、抗原結合タンパク質は少なくとも3または4または5または6または8または10個の抗原と結合できる、例えば抗原結合タンパク質は、3または4または5または6または8または10個の抗原と同時に結合できる。
【0102】
一実施形態では、第1のエピトープ結合ドメインはタンパク質足場と結合しており、第2のエピトープ結合ドメインは第1のエピトープ結合ドメインと結合している。例えばタンパク質足場がIgG足場である場合、第1のエピトープ結合ドメインはIgG足場の重鎖のC末端と結合していてもよく、そのエピトープ結合ドメインはそのC末端で第2のエピトープ結合ドメインと結合でき、または例えば第1のエピトープ結合ドメインはIgG足場の軽鎖のC末端と結合していてもよく、第1のエピトープ結合ドメインはさらにそのC末端で第2のエピトープ結合ドメインと結合していてもよいし、または例えば第1のエピトープ結合ドメインはIgG足場の軽鎖のN末端と結合していてもよく、第1のエピトープ結合ドメインはさらにそのN末端で第2のエピトープ結合ドメインと結合していてもよいし、または例えば第1のエピトープ結合ドメインは、IgG足場の重鎖のN末端と結合していてもよく、第1のエピトープ結合ドメインはさらにそのN末端で第2のエピトープ結合ドメインと結合していてもよい。
【0103】
エピトープ結合ドメインがドメイン抗体である場合、一部のドメイン抗体は足場内の特定の位置に適合し得る。
【0104】
本発明において使用する免疫グロブリン単一可変ドメインは、IL−13 mAbの重鎖および/または軽鎖のC末端と結合できる。加えて一部の免疫グロブリン単一可変ドメインは、従来の抗体の重鎖および軽鎖の両C末端と結合できる。
【0105】
免疫グロブリン単一可変ドメインのN末端が抗体定常ドメイン(CH3またはCLのいずれか)に融合している構築物において、ペプチドリンカーは免疫グロブリン単一可変ドメインが抗原と結合するのを補助し得る。実際、dAbのN末端は、抗原結合活性に関与するCDRに近接して位置する。したがって短いペプチドリンカーは、エピトープ結合ドメインと抗体の定常ドメイン間のスペーサーとして働き、これはdAb CDRがより容易に抗原に達することを可能にし得るため、高親和性で結合し得る。
【0106】
免疫グロブリン単一可変ドメインがIgGと結合する環境は、それらがどの抗体鎖へ融合しているかによって異なる:
抗体軽鎖のC末端で融合している場合、各免疫グロブリン単一可変ドメインは、抗体ヒンジおよびFc部分に近接して位置することが予期される。かかる免疫グロブリン単一可変ドメインは互いに遠く離れて位置している可能性が高い。従来の抗体では、Fab断片間の角度および各Fab断片とFc部分間の角度は非常に著しく変わり得る。mAbdAbと、Fab断片間の角度は著しく異なることはないであろうが、各Fab断片とFc部分間の角度にはいくらか制限が観察され得る。
【0107】
抗体重鎖のC末端で融合している場合、各免疫グロブリン単一可変ドメインは、Fc部分のCH3ドメインに近接して位置することが予期される。これはFc受容体(例えばFcγRI、II、III、FcRn)に対するFc結合特性に影響を与えることは予期されない。なぜなら、これらの受容体はCH2ドメイン(FcγRI、IIおよびIIIクラスの受容体について)またはCH2ドメインとCH3ドメイン間のヒンジ(例えばFcRn受容体)と結合するためである。かかる抗原結合タンパク質はさらに、両免疫グロブリン単一可変ドメインが空間的に互いに近いことを特徴とし、適切なリンカーの供給によって柔軟性が得られる場合、これらの免疫グロブリン単一可変ドメインはホモ二量体種さえも形成し得るため、Fc部分の「ジッパー型」四次構造を増やして構築物の安定性を高め得ることが予期される。
【0108】
かかる構造に関する考察は、抗体にエピトープ結合ドメイン、例えば免疫グロブリン単一可変ドメインを結合させるために最も適した位置の選択に役立つことができる。
【0109】
抗原のサイズ、その局在性(血中または細胞表面上)、その四次構造(単量体または多量体)は変わり得る。従来の抗体は、ヒンジ領域の存在のためにアダプター構築物として機能するように当然設計され、Fab断片の先端の2つの抗原結合部位の配向は広範囲に変わり得るため、抗原の分子特性およびその環境に適応し得る。一方、ヒンジ領域のない抗体と結合した免疫グロブリン単一可変ドメインは、直接的にも間接的にも構造的な柔軟性に欠け得る。
【0110】
溶液状態および免疫グロブリン単一可変ドメインへの結合形態の理解も役立つ。インビトロのヒトdAbは大部分が溶液中に単量体、ホモ二量体または多量体形態で存在し得るという証拠が増えている(Reiter et al. (1999) J Mol Biol 290 p685−698; Ewert et al (2003) J Mol Biol 325, p531−553, Jespers et al (2004) J Mol Biol 337 p893−903; Jespers et al (2004) Nat Biotechnol 22 p1161−1165; Martin et al (1997) Protein Eng. 10 p607−614; Sepulvada et al (2003) J Mol Biol 333 p355−365)。これは、免疫グロブリン軽鎖の二量体であるベンス−ジョーンズタンパク質(Epp et al (1975) Biochemistry 14 p4943−4952; Huan et al (1994) Biochemistry 33 p14848−14857; Huang et al (1997) Mol immunol 34 p1291−1301)およびアミロイド線維(James et al. (2007) J Mol Biol. 367:603−8)などのIgドメインでインビボにおいて観察される多量体化事象を十分に暗示する。
【0111】
例えば、溶液中で二量体化する傾向があるdAbは、軽鎖C末端よりもFc部分のC末端と結合させることが本発明の抗原結合タンパク質との関連で所望され得る。なぜなら、FcのC末端への結合はそれらのdAbの二量体化を可能にさせるためである。
【0112】
本発明の抗原結合タンパク質は、単一抗原に特異的な抗原結合部位を含んでいてもよいし、2つ以上の抗原もしくは単一抗原上の2つ以上のエピトープに特異的な抗原結合部位を有してもよく、またはそれぞれの抗原結合部位が同じもしくは異なる抗原上の異なるエピトープに特異的である抗原結合部位があってもよい。
【0113】
本発明の抗原結合タンパク質は、天然抗体または機能的断片またはその等価物の構造内に構成し得る本発明の重鎖可変領域および軽鎖可変領域を含み得る。したがって本発明の抗原結合タンパク質は、適切な軽鎖と対をなす場合、完全長抗体、(Fab」)2断片、Fab断片、またはその等価物(scFV、二量体、三量体、または四量体、Tandabなど)内に形成された本発明のVH領域を含み得る。抗体は、IgG1、IgG2、IgG3、もしくはIgG4;またはIgM;IgA、IgEもしくはIgDまたはその修飾変異型であり得る。それに応じて抗体重鎖の定常ドメインを選択し得る。軽鎖定常ドメインは、κ定常ドメインであってもλ定常ドメインであってもよい。さらに、抗原結合タンパク質は、すべてのクラスの修飾、例えばIgG二量体、Fc受容体と結合しなくなったまたはC1q結合を媒介しなくなったFc変異体を含み得る。抗原結合タンパク質は、抗原結合領域および非免疫グロブリン領域を含むWO86/01533号に記載されている種類のキメラ抗体でもあり得る。
【0114】
定常領域は、必要な任意の機能性に応じて選択される。IgG1は、補体結合を介して溶解能を示し得るおよび/またはADCC(抗体依存性細胞傷害)を媒介し得る。非細胞傷害性ブロック抗体が必要である場合、IgG4を使用できる。しかしながら、IgG4抗体は産生において不安定性を示し得るため、代替的に、一般にさらに安定したIgG1に修飾する。推奨される修飾は、欧州特許第0307434号に記載されている(例えば235位および237位の変異)。したがって本発明は、本発明による抗原結合タンパク質(例えば抗体)の溶解または非溶解形態を提供する。
【0115】
ある形態では、本発明の抗体は本明細書に記載の任意の重鎖可変領域を有する完全長(例えばH2L2四量体)溶解または非溶解IgG1抗体である。さらなる態様では、本発明は、本明細書に記載の軽鎖および重鎖可変領域をコードするポリヌクレオチドを提供する。
【0116】
本発明の一実施形態では、抗原結合部位はBiacore(商標)の測定に基づき少なくとも約1mMのKdで抗原と結合する、例えば少なくとも約10nM、少なくとも約1nM、少なくとも約500pM、少なくとも約200pM、少なくとも約100pM、または少なくとも約50pMのKdで各抗原と結合する。
【0117】
本発明の一実施形態では、抗原結合部位はBiacore(商標)の測定に基づき少なくとも約1mMのKdで抗原と結合する、例えば少なくとも約10nM、少なくとも約1nM、少なくとも約500pM、少なくとも約200pM、少なくとも約100pM、または少なくとも約50pMのKdで各抗原と結合する。
【0118】
一実施形態では、本発明は、Biacoreの測定に基づき、ヒトIL−13に対して抗IL=13抗体よりも少なくとも2倍高い親和性、または少なくとも4倍高い親和性、または少なくとも6倍高い親和性、または少なくとも8倍高い親和性、または少なくとも10倍高い親和性を有する、配列番号22に説明される重鎖配列および配列番号24に説明される軽鎖配列を含む抗原結合タンパク質を提供する。
【0119】
本明細書を通して使用される、本発明の抗原結合タンパク質に関する「中和」という用語およびその文法的変形は、本発明の抗原結合タンパク質の存在下においてIL−13の生体活性が、かかる抗原結合タンパク質の不在下のIL−13の活性よりも完全にまたは部分的に低下することを意味する。中和は、これらに限定されないが、1つまたは複数のリガンド結合をブロックし、リガンドによる受容体活性化を防止し、IL−13受容体をダウンレギュレートし、またはエフェクター機能に影響を与えたために起こり得る。中和レベルはいくつかの方法において、例えば以下の実施例に説明されるアッセイ、例えばTF1アッセイ(例えば実施例4に記載のように実行し得る)を用いて測定できる。このアッセイにおけるIL−13、IL−4またはこれら両サイトカインの中和は、中和する抗原結合タンパク質の存在下、TF1細胞増殖の阻害を評価することによって測定する。
【0120】
例えば、中和する抗原結合タンパク質の存在下でIL−13とその受容体間の低減した結合を評価することによる他の中和の評価方法は当該分野において知られており、例えば、Biacoreアッセイなどが挙げられる。
【0121】
本発明の代替態様では、本明細書において例示した抗体、例えば実施例4に説明するように実行できるTF1細胞増殖アッセイにおけるA1Y100BAlaL1、A1Y100BIleL1、A1 Y100BTrpL1またはA1Y100BValL1の中性活性を保持する抗原結合タンパク質と少なくとも実質的に等しい中性活性を有する抗原結合タンパク質を提供する。
【0122】
抗原結合タンパク質(例えば本発明の抗体)は、本発明の抗原結合タンパク質コード配列を含む発現ベクターを用いた宿主細胞のトランスフェクションにより産生し得る。発現ベクターまたは組換えプラスミドは、宿主細胞における複製および発現ならびに/または宿主細胞からの分泌を制御できる従来の調節制御配列との操作可能に結合して抗原結合タンパク質に対するこれらのコード配列を配置することにより産生される。調節配列としては、プロモーター配列、例えば、CMVプロモーター、および他の既知の抗体に由来し得るシグナル配列が挙げられる。同様に、相補的な抗原結合タンパク質軽鎖または重鎖をコードするDNA配列を有する第2の発現ベクターを産生できる。ある実施形態では、この第2の発現ベクターは、各ポリペプチド鎖が機能的に発現することを可能な限り確実にするためにコード配列および選択マーカーが考慮される点を除き第1の発現ベクターと同一である。あるいは、抗原結合タンパク質の重鎖および軽鎖コード配列は単一ベクター上に存在し得る。選択された宿主細胞を第1ベクターと第2ベクターの両方を従来技術により共トランスフェクトして(または単一ベクターにより単にトランスフェクトして)、組換えまたは合成の軽鎖および重鎖の両方を含むトランスフェクトされた本発明の宿主細胞を作製する。次いでトランスフェクトされた細胞を従来技術により培養し、本発明の加工された抗原結合タンパク質を産生する。組換え重鎖および/または軽鎖の両結合を含む抗原結合タンパク質は、適切なアッセイ(ELISAまたはRIAなど)により培養からスクリーニングされる。他の抗原結合タンパク質を構築するために類似の従来技術を適用し得る。
【0123】
本発明の組成物の方法および構築において使用されるクローニング工程およびサブクローニング工程に適したベクターは、当業者によって選択され得る。例えば、クローニングベクターの従来のpUCシリーズを使用してよい。1つのベクターであるpUC19は、Amersham(Buckinghamshire, United Kingdom)またはPharmacia (Uppsala, Sweden)などの供給源から市販されている。さらに、容易に複製でき、豊富なクローニング部位および選択可能な遺伝子(例えば、抗生剤耐性)を有し、容易に操作される任意のベクターをクローニングに使用してよい。したがって、クローニングベクターの選択は、本発明における限定要素ではない。
【0124】
発現ベクターは、異種DNA配列の発現を増幅するのに適した遺伝子、例えば、哺乳動物ジヒドロ葉酸還元酵素遺伝子(DHFR)も特徴とし得る。他のベクター配列としては、ウシ成長ホルモン(BGH)由来などのポリAシグナル配列およびベータグロビンプロモーター配列(betaglopro)が挙げられる。本明細書において有用な発現ベクターは、当業者にとって周知の技術により合成され得る。
【0125】
かかるベクターの成分、例えばレプリコン、選択遺伝子、エンハンサー、プロモーター、シグナル配列などは、市販または天然の供給源から得てもよいし、選択された宿主において組換えDNA生成物の発現および/または分泌の対象における使用のための既知の手段によって合成してもよい。当該分野において知られている、哺乳動物、細菌、昆虫、酵母、および真菌の発現に適した他の多くの種類の発現ベクターもこの目的のために選択してよい。
【0126】
本発明は、本発明の抗原結合タンパク質のコード配列を含む組換えプラスミドを用いてトランスフェクトされた細胞系も包含する。これらのクローニングベクターのクローニングおよび他の操作に有用な宿主細胞も従来のものである。しかしながら、様々な大腸菌株由来の細胞を、クローニングベクターの複製および本発明の抗原結合タンパク質の構築における他の工程に使用してよい。
【0127】
本発明の抗原結合タンパク質の発現に適した宿主細胞または細胞系としては、NS0、Sp2/0、CHO(例えばDG44)、COS、HEK、線維芽細胞(例えば、3T3)、および骨髄腫細胞などの哺乳動物細胞が挙げられ、例えばCHOまたは骨髄腫細胞において発現し得る。分子がヒトグリコシル化パターンで修飾されることを可能にするようにヒト細胞を使用してよい。あるいは、他の真核細胞系を適用し得る。形質転換、培養、増幅、スクリーニングならびに生成物の産生および精製に適した哺乳動物宿主細胞の選択および方法は当該分野において知られている。例えば、上に引用したSambrook et al.を参照されたい。
【0128】
細菌細胞は、本発明の組換えFab、または他の実施形態の発現に適した宿主細胞として有用であることが分かるかもしれない(例えば、Pluckthun, A., Immunol. Rev., 130:151−188 (1992)を参照されたい)。しかしながら、細菌細胞内で発現するタンパク質は折り畳まれていないか不適切に折り畳まれた形態または非グリコシル化形態となる傾向があるために、細菌細胞内で産生した組換えFabはいずれも抗原結合能力の保持についてスクリーニングする必要があるであろう。細菌細胞によって発現している分子が適切に折り畳まれた形態で産生した場合、その細菌細胞は所望される宿主であり得、または代替的な実施形態では、分子は細菌宿主内で発現し、続いて再び折り畳まれ得る。例えば、発現のために使用される様々な大腸菌株が生物工学分野において宿主細胞として周知である。様々な枯草菌、ストレプトミセス属、他の桿菌株などもこの方法に使用してよい。
【0129】
所望の場合、当業者に知られている酵母細胞の株、ならびに昆虫細胞(例えばショウジョウバエおよび鱗翅類)およびウイルス発現系も宿主細胞として利用可能である。例えばMiller et al., Genetic Engineering, 8:277−298, Plenum Press (1986)およびそこで引用されている参考文献を参照されたい。
【0130】
ベクターを構築し得る一般的な方法、本発明の宿主細胞を産生するために必要とされるトランスフェクション方法、およびかかる宿主細胞から本発明の抗原結合タンパク質を産生するために必要な培養方法はすべて従来技術であってよい。典型的には、本発明の培養方法は、無血清培養方法、通常、懸濁液中での無血清細胞培養である。同様に、産生後は、本発明の抗原結合タンパク質は、硫酸アンモニウム沈殿法、親和性カラム、カラムクロマトグラフィー、ゲル電気泳動などを含む当該技術分野の標準的な手段に従い細胞培養の内容物から精製し得る。かかる技術は、当該技術分野の技術範囲内であり、本発明を限定しない。例えば、改変抗体の調製については、WO99/58679号およびWO96/16990号に記載されている。
【0131】
抗原結合タンパク質のさらに別の発現方法は、米国特許第4,873,316号に記載されるように、遺伝子導入動物における発現の利用である。これは、哺乳動物に遺伝子導入で組み込まれた場合、雌の母乳において所望の組換えタンパク質の産生を可能にする動物のカゼインプロモーターを用いる発現系に関する。
【0132】
本発明のさらなる態様では、本発明の抗体の産生方法を提供し、この方法は、本発明の抗体の軽鎖および/もしくは重鎖をコードするベクターを用いて形質転換またはトランスフェクトされた宿主細胞を培養する工程、およびその結果産生した抗体を回収する工程を含む。
【0133】
本発明に従い、ヒトIL−13と結合してその活性を中和する本発明の抗IL−13抗体の産生方法を提供し、この方法は以下;
(a)抗体重鎖をコードする第1ベクターを提供する工程;
(b)抗体軽鎖をコードする第2ベクターを提供する工程;
(c)前記第1および第2ベクターを用いて哺乳動物宿主細胞(例えばCHO)を形質転換する工程;
(d)前記培地内へ前記宿主細胞から抗体の分泌を促す条件下で工程(c)の宿主細胞を培養する工程;
(e)工程(d)の分泌した抗体を回収する工程
を含む。
【0134】
所望の方法によって発現後は、適切なアッセイを用いて抗体のインビトロ活性を検討する。現在、抗体のIL−13に対する定性および定量的結合を評価するために従来のELISAアッセイ形態が使用されている。さらに、通常のクリアランス機序にもかかわらず体内における抗体の持続性を評価するために実施するヒト臨床研究の前に、中和効力を検証するために他のインビトロアッセイを使用してよい。
【0135】
治療の用量および期間は、ヒト循環における本発明の分子の相対的な持続時間に関し、治療する状態および患者の一般的な健康状態に応じて、当業者によって調整されることができる。最大の治療効果に達するために、長期(例えば4〜6ヶ月)にわたる反復投薬(例えば週1回または隔週)が必要な場合があることが想定される。
【0136】
本発明の治療薬の投与方法は、その作用物質を宿主へ送達する上で適した任意の経路であってよい。本発明の抗原結合タンパク質および医薬組成物は、非経口投与、すなわち、皮下(s.c.)、髄腔内、腹腔内、筋肉内(i.m.)、静脈内(i.v.)、または鼻腔内投与に特に有用である。
【0137】
本発明の治療薬は、医薬上許容可能な担体中に有効成分として有効量の本発明の抗原結合タンパク質を含む医薬組成物として調製し得る。一実施形態では、本発明の予防薬は、抗原結合タンパク質を注射形態で含む水性懸濁液または溶液である。一実施形態では、懸濁液または溶液は生理的pHに緩衝されており、一実施形態では、非経口投与用の組成物は、本発明の抗原結合タンパク質の溶液または医薬上許容可能な担体に溶解したその混合物を含む。一実施形態では、担体は水性担体である。様々な水性担体、例えば、0.9%食塩水、0.3%グリシンなどを適用し得る。これらの溶液は、滅菌で作製し得、一般に粒子物質を含まない。これらの溶液は、従来の周知の滅菌技術(例えば、濾過)により滅菌し得る。組成物には、生理的状態に近づけるために必要に応じて医薬上許容可能な補助物質(pH調節剤および緩衝剤など)を含み得る。かかる医薬製剤中の本発明の抗原結合タンパク質の濃度は広範囲、すなわち、約0.5重量%未満、通常、約1重量%以上から約15または20重量%程度に変わり得、選択された特定の投与方法に従い、主に体液の容量および粘性などに基づき選択されるであろう。
【0138】
したがって、筋肉内注射用の本発明の医薬組成物は、滅菌緩衝水約1mL、ならびに本発明の抗原結合タンパク質、例えば抗体を約1ng〜約100mg、例えば約50ng〜約30mgもしくは約5mg〜約25mg含むように調製できる。同様に、静脈内注射用の本発明の医薬組成物は、滅菌リンガー溶液を最大約250ml、ならびに本発明の抗原結合タンパク質を約1〜約30もしくは5mg〜約25mg/mlリンガー溶液含むように作製し得る。非経口投与可能な組成物を調製するための実際の方法は当業者に周知または明白であり、例えば、Remington’s Pharmaceutical Science, 15th ed., Mack Publishing Company, Easton, Pennsylvaniaに詳述されている。静脈内に投与可能な本発明の抗原結合タンパク質製剤の調製については、Lasmar U and Parkins D “The formulation of Biopharmaceutical products”, Pharma. Sci.Tech.today, page 129−137, Vol.3 (3rd April 2000); Wang, W “Instability, stabilisation and formulation of liquid protein pharmaceuticals”, Int. J. Pharm 185 (1999) 129−188; Stability of Protein Pharmaceuticals Part A and B ed Ahern T.J., Manning M.C., New York, NY: Plenum Press (1992); Akers,M.J. “Excipient−Drug interactions in Parenteral Formulations”, J.Pharm Sci 91 (2002) 2283−2300; Imamura, K et al “Effects of types of sugar on stabilization of Protein in the dried state”, J Pharm Sci 92 (2003) 266−274; Izutsu, Kkojima, S. “Excipient crystalinity and its protein−structure−stabilizing effect during freeze−drying”, J Pharm. Pharmacol, 54 (2002) 1033−1039; Johnson, R, “Mannitol−sucrose mixtures−versatile formulations for protein lyophilization”, J. Pharm. Sci, 91 (2002) 914−922;およびHa,E Wang W, Wang Y.j. “Peroxide formation in polysorbate 80 and protein stability”, J. Pharm Sci, 91, 2252−2264,(2002)を参照されたく、その全体の内容は参照によって本明細書に組み込まれ、読者は、この全内容を特に参照する。
【0139】
一実施形態では、本発明の治療薬は、医薬調製物中にある場合、単位用量形態で存在する。適切な治療有効量は当業者によって容易に決定されるであろう。患者に適した用量は体重に応じて算出し得、例えば適切な用量は、約0.1〜約20mg/kg、例えば約1〜約20mg/kg、例えば約10〜約20mg/kgまたは例えば約1〜約15mg/kg、例えば約10〜約15mg/kgの範囲であり得る。本発明の抗原結合タンパク質の、ヒトにおける喘息またはIPFなどの状態を効果的に治療するために適した用量は、約0.1〜約1000mg、例えば約0.1〜約500mg、例えば約500mg、例えば約0.1〜約100mg、または約0.1〜約80mg、または約0.1〜約60mg、または約0.1〜約40mg、または例えば約1〜約100mg、または約1〜約50mgの範囲内であり得、これは非経口(例えば皮下、静脈内または筋肉内)投与し得る。かかる用量は、必要な場合、医師によって適切に選択された適切な時間間隔で反復し得る。
【0140】
本明細書に記載の抗原結合タンパク質は、保存用に凍結乾燥し、使用前に適切な担体中で再構成することができる。この技術は従来の免疫グロブリンで有効であることが示されており、当技術分野において知られている凍結乾燥および再構成技術を使用できる。
【0141】
別の態様では、本発明は、アトピー性疾患/障害および慢性炎症疾患/障害、例えば、喘息、例えばアレルギー性喘息、特に重症喘息(すなわちコルチコステロイドの全身投与を含む現在の治療が奏効しない喘息;Busse WW et al, J Allergy Clin. Immunol 2000, 106: 1033−1042を参照されたい)、「困難性」喘息(最大推奨用量の吸入ステロイド処方にもかかわらず制御不成功を特徴とする喘息性形質として定義、Barnes PJ (1998), Eur Respir J 12:1208−1218を参照されたい)、「ブリットル」喘息(高用量の吸入ステロイドにもかかわらず大きく変動する最大呼気流量(PEF)を維持する重度の不安定喘息患者のサブグループを定義、Ayres JG et al (1998) Thorax 58:315−321を参照されたい)、夜間性喘息、月経前喘息、ステロイド抵抗性喘息(Woodcock AJ (1993) Eur Respir J 6:743−747を参照されたい)、ステロイド依存性喘息(高用量の経口ステロイドでのみ制御できる喘息として定義)、アスピリン誘発性喘息、成人発症型喘息、小児喘息の治療または予防のための本発明の抗原結合タンパク質もしくはその機能的断片ならびに医薬上許容可能な担体を含む医薬組成物を提供する。本発明の抗体は、急性喘息エピソード(喘息発作重積状態)を予防する、頻度を減らす、または作用を緩和するために使用してよい。本発明の抗体は、喘息の治療に使用される他の薬の必要な投薬を(投薬の投与量または投与頻度のいずれかに関して)減らすためにも使用してよい。例えば、本発明の抗体は、喘息のステロイド治療(コルチコステロイド治療など)に必要な投薬を減らす(「ステロイド使用を控える」)ために使用してよい。本発明の抗体で治療し得る他の疾患または障害としては、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、慢性閉塞性肺疾患(COPD)、好酸球性食道炎、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症(強皮症)、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、および細胞周期調節疾患、例えばホジキン病、B細胞慢性リンパ球性白血病が挙げられる。一実施形態では、障害は重症喘息である。さらなる実施形態では、障害は線維性障害(IPFなど)である。
【0142】
よりさらなる態様では、本発明は、本発明の抗原結合タンパク質および医薬上許容可能な担体を含む医薬組成物を、アトピー性疾患/障害および慢性炎症疾患/障害、例えば、喘息、例えばアレルギー性喘息、特に重症喘息(すなわちコルチコステロイドの全身投与を含む現在の治療が奏効しない喘息;Busse WW et al, J Allergy Clin. Immunol 2000, 106: 1033−1042を参照されたい)、「困難性」喘息(最大推奨用量の吸入ステロイド処方にもかかわらず制御不成功を特徴とする喘息性形質として定義、Barnes PJ (1998), Eur Respir J 12:1208−1218を参照されたい)、「ブリットル」喘息(高用量の吸入ステロイドにもかかわらず大きく変動する最大呼気流量(PEF)を維持する重度の不安定喘息患者のサブグループを定義、Ayres JG et al (1998) Thorax 58:315−321を参照されたい)、夜間性喘息、月経前喘息、ステロイド抵抗性喘息(Woodcock AJ (1993) Eur Respir J 6:743−747を参照されたい)、ステロイド依存性喘息(高用量の経口ステロイドでのみ制御できる喘息として定義)、アスピリン誘発性喘息、成人発症型喘息、小児喘息の治療のために提供する。本発明の抗体は、急性喘息エピソード(喘息発作重積状態)を予防する、頻度を減らす、または作用を緩和するために使用してよい。本発明の抗体は、喘息の治療に使用される他の薬の必要な投薬を(投薬の投与量または投与頻度のいずれかに関して)減らすためにも使用してよい。例えば、本発明の抗体は、喘息のステロイド治療(コルチコステロイド治療など)に必要な投薬を減らす(「ステロイド使用を控える」)ために使用してよい。本発明の抗体で治療し得る他の疾患または障害としては、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、慢性閉塞性肺疾患(COPD)、好酸球性食道炎、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症(強皮症)、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、および細胞周期調節疾患、例えばホジキン病、B細胞慢性リンパ球性白血病が挙げられる。一実施形態では、障害は重症喘息である。さらなる実施形態では、障害は線維性障害(IPFなど)である。
【0143】
本明細書に記載の配列(配列番号26〜配列番号55および配列番号62〜配列番号146)は、本明細書に記載の配列と実質的に同一である、例えば、少なくとも90%同一である、例えば、少なくとも91%、または少なくとも92%、または少なくとも93%、または少なくとも94%または少なくとも95%、または少なくとも96%、または少なくとも97%または少なくとも98%、または少なくとも99%同一である配列を含むことが理解されるであろう。
【0144】
核酸において、「実質的な同一性」という用語は、適切にヌクレオチドを挿入してまたは欠失させて至適整列および比較時に、少なくとも約80%のヌクレオチド、少なくとも約90%〜約95%、または少なくとも約98%〜約99.5%のヌクレオチドが同一である2つの核酸、またはその設計された配列を示す。あるいは、実質的な同一性は、選択的ハイブリダイゼーション条件下でセグメントが鎖の補体にハイブリダイズする場合に存在する。
【0145】
ヌクレオチド配列およびアミノ酸配列において、「同一」という用語は、2つの核酸またはアミノ酸配列を適切に挿入してまたは欠失させて至適整列および比較時の同一度を示す。あるいは、実質的な同一性は、選択的ハイブリダイゼーション条件下でDNAセグメントが鎖の補体にハイブリダイズする場合に存在する。
【0146】
2配列間の同一性%は、2配列の至適整列のために導入する必要があるギャップ数、および各ギャップの長さを考慮した、配列の共有する同一の位置数の関数(すなわち、同一性%=同一の位置数/総位置数×100)である。配列の比較および2配列間の同一性%の決定は、以下の非制限的な実施例に記載のように数学アルゴリズムを用いて成し遂げることができる。
【0147】
2つのヌクレオチド配列間の同一性%は、GCGソフトウェアパッケージ内のGAPプログラムを用いて、NWSgapdna.CMP行列、ならびに40、50、60、70、もしくは80のギャップウェイト、ならびに1、2、3、4、5、もしくは6のレングスウェイトを用いて決定することができる。2つのヌクレオチド配列またはアミノ酸配列間の同一性%は、ALIGNプログラム(バージョン2.0)に組み込まれているE. Meyers and W. Miller (Comput. Appl. Biosci., 4:11−17 (1988))のアルゴリズムを用いて、PAM120ウェイト残基表、ギャップレングスペナルティ12、およびギャップペナルティ4を用いて決定することもできる。加えて、2つのアミノ酸配列間の同一性%は、GCGソフトウェアパッケージ内のGAPプログラムに組み込まれているNeedleman and Wunsch (J. Mol. Biol. 48:444−453 (1970))アルゴリズムを用いて、Blossum62行列もしくはPAM250行列のいずれか、ならびに16、14、12、10、8、6、もしくは4のギャップウェイト、ならびに1、2、3、4、5、もしくは6のレングスウェイトを用いて決定することができる。
【0148】
例として、本発明のポリヌクレオチド配列は、配列番号25の基準配列と同一、すなわち100%同一であってもよく、または、基準配列と比較してヌクレオチド改変を特定の整数の数まで含んでもよい。かかる改変は、少なくとも1つのヌクレオチド欠失、置換(転位および塩基転換を含む)、または挿入からなる群から選択され、前記改変は基準ヌクレオチド配列の5’もしくは3’末端位またはそれら末端位の間のどこかで、基準配列中のヌクレオチド間に個々に散在してまたは基準配列中の1つもしくは複数の連続基としてのいずれかで生じ得る。ヌクレオチド改変数は、配列番号25の総ヌクレオチド数にそれぞれの同一性%の%数値を乗じて(100で割って)、その積を前記配列番号23の総ヌクレオチド数から減算するか、または:
nn≦xn−(xn × y)、
式中nnはヌクレオチド改変数であり、xnは配列番号25の総ヌクレオチド数であり、yは、50%の場合は0.50、60%の場合は0.60、70%の場合は0.70、80%の場合は0.80、85%の場合は0.85、90%の場合は0.90、95%の場合は0.95、97%の場合は0.97または100%の場合は1.00であり、xnとyの積が整数ではない場合はいずれも端数を切り捨てて最も近い整数にした後でxnから減算することによって決定する。配列番号25のポリヌクレオチド配列改変はこのコード配列中にナンセンス、ミスセンスまたはフレームシフト変異を生じ得、その結果かかる改変に続いてポリヌクレオチドによりコードされるポリペプチドを改変する。
【0149】
同様に、別の例では、本発明のポリペプチド配列は、配列番号24によりコードされた基準配列と同一、すなわち100%同一であってもよく、または、基準配列と比較して同一性%が100%未満であるようにアミノ酸改変を特定の整数の数まで含んでもよい。かかる改変は、少なくとも1つのアミノ酸欠失、置換(保存的および非保存的置換を含む)、または挿入からなる群から選択され、前記改変は基準ポリペプチド配列のアミノもしくはカルボキシ末端位またはそれら末端位の間のどこかで、基準配列中のアミノ酸間に個々に散在してまたは基準配列中の1つもしくは複数の連続基としてのいずれかで生じ得る。所定の同一性%のアミノ酸改変数は、配列番号24によりコードされるポリペプチド配列の総アミノ酸数にそれぞれの同一性%の%数値を乗じて(100で割って)、次いでその積を前記配列番号24によりコードされるポリペプチド配列の総アミノ酸数から減算するか、または:
na≦xa−(xa × y)、
式中naは、アミノ酸改変数であり、xaは配列番号24によりコードされるポリペプチド配列の総アミノ酸数であり、yは、例えば70%の場合は0.70、80%の場合は0.80、85%の場合は0.85、などであり、xaとyの積が整数ではない場合はいずれも端数を切り捨てて最も近い整数にした後でxaから減算することによって決定する。
【0150】
以下の実施例により例証するが、これらは本発明を限定しない。
実施例
【実施例1】
【0151】
組換え抗IL−13抗体の構築
オリジナルのマウスmAbを組換えヒトIL−13を有するマウスの免疫化により産生した。反応した動物由来の脾臓を回収して、骨髄腫細胞に融合させてハイブリドーマを生成した。ハイブリドーマ上清物質の結合をスクリーニングした。標準的な技術を用いて興味のハイブリドーマをモノクローン化した。生成したマウス抗体(6A1)は配列番号58および配列番号59に示す可変領域を含む。このマウス抗体およびこの抗体A1L1(配列番号22および24)のヒト化バージョンの詳細については、WO2006/003407号に記載されており、これは参照によって本明細書に組み込まれる。抗IL−13 mAb抗体A1L1を比較抗体として以下の実施例のいくつかに使用した。
【0152】
配列番号22に説明した重鎖を含むヒト化抗体の変異体をいくつか産生した。これらはすべて抗体のCDRH3領域が異なる(配列番号3)。
【0153】
RldおよびRln哺乳動物発現ベクター内へクローニングのための制限部位を含む重複オリゴヌクレオチドならびにヒトシグナル配列を形成して本発明の抗体のための塩基性DNA発現構築物、配列番号23(重鎖)および配列番号25(軽鎖)をde novoで調製した。Hind IIIおよびSpe I制限部位を導入して、ヒトγ1定常領域を含むRld内へのクローニングのためにシグナル配列(配列番号56)を含むVHドメインを枠組みした。Hind IIIおよびBsiWI制限部位を導入して、ヒトκ定常領域を含むRln内へのクローニングのためにシグナル配列(配列番号56)を含むVLドメインを枠組みした。代替構築物は同様にヒト定常領域を含むpTTベクターを用いて産生した。適切な場合、部位指向変異(SDM)を用いて異なるヒト化構築物を生成した。
【実施例2】
【0154】
HEK 293 6E細胞における抗体発現
重鎖と軽鎖をそれぞれコードするpTTプラスミドをHEK 293 6E細胞内に一過性共トランスフェクトし、少量発現させて抗体を産生した。抗体を組織培養上清から直接評価した。固定化プロテインAカラムを用いて他の抗体を精製して、吸光度を280nmで読み取ることにより定量化し、指定の場合、精製された抗体物質を以下に説明する実施例に記載のアッセイにおいて評価した。
【0155】
我々がコード(すなわちA1Y100BTrpL1)により抗体を指す場合、我々は記載の第1プラスミドと第2プラスミドの共トランスフェクションおよび発現により生成したmAbを指している。例えば「A1Y100BTrpL1」とは、適切な細胞系においてA1Y100BTrp配列を含むプラスミドとL1配列を含むプラスミドの共トランスフェクションにより生成したmAbに関する。
【実施例3】
【0156】
抗IL−13ヒト化mAbのBiacore分析
動力学分析
CDRH3変異体の初回スクリーニングをProteOn XPR36(Biorad)上で実施した。方法は以下のとおりであった。抗ヒトIgG(Biacore BR−1008〜39)を第1級アミンカップリングによりGLMチップ上に固定化してから、CDRH3変異体抗体をこの表面上に捕捉して、IL13を256、64、16、4、1nMで通過させ、0nM注入(すなわち緩衝液単独)を二重照合に使用した。3M MgCl2を用いて捕捉表面を再生し、結合CDRH3変異体抗体を除去して、別のサイクルの捕捉およびアナライト注入を準備した。データは、機械に固有のソフトウェアを用いて1:1モデルに適合させた。精製された物質であった親抗体を除きすべての作業を組織培養上清から直接抗体を用いて実施した。
【0157】
スクリーニングにより、親分子より良好な動力学プロファイルを有するように思われた抗体をいくつか同定してから、これらの同一試料をBiacore T100上で分析して類似の方法を用いて結果を確認し、同じ抗ヒトIgG捕捉抗体を第1級アミンカップリングによりCM5チップ上に固定化して、IL13を表面上に256、64、16、4、1および0.25nMで通過させ、0nMを二重照合に使用して、3M MgCl2で再生し、T100に固有の1:1モデルにデータを適合させた。ProteOnスクリーニングおよびT100実行から選択された構築物の全体の親和性(平衡解離定数KD)を表1に詳述する。
【表1】
【0158】
データにより、Y100B残基のいくつかの変異により全体の親和性が改善されると思われることが強調された。この観点から、先に記載のものと同じ方法でProteOnを用いて、組織培養上清から再び抗体を直接用いて、初回スクリーニング時に存在しなかったこの残基の変異を試験した。試験した変異のうち、A1Y100B ValL1において、親の0.390〜0.460nM値と比較して0.166nM値が得られ、全体の親和性(平衡解離定数KD)が改善されると思われた。先に記載のものと同じ方法を用いてBiacore T100上でY100B Valを試験時、平衡解離定数KDは親の0.346nM値と比較して0.025nMで測定された。
【0159】
抗体変異体を組織培養上清から直接用いてこれまで実行した作業の点から、精製された抗体をA1Y100BL1変異体Ala、IleおよびTrpのために産生した。これらは、この段階で精製しなかったA1Y100B ValL1変異体を対象として、前と同じ方法を用いてBiacore T100上で実行した。この実験から得られたデータを表2に示す。
【表2】
【0160】
実験により、変異体はIL13に対する結合親和性を親分子と比較して改善しなかったことを確認した。
【0161】
精製されたA1Y100BL1変異体では組織培養上清から得られるよりも良好な結合親和性が得られることから、A1Y100B ValL1を精製し、同様に親和性を改善した他の精製されたA1Y100BL1変異体と共に、Biacore T100機械で先に記載の方法を用いて実行した。この実験から得られたデータを表3に示す。この実験は表2のデータと十分一致し、Y100B変異の親和性の改善が確認された。
【表3】
【実施例4】
【0162】
TF−1細胞増殖バイオアッセイにおける大腸菌発現組換えヒトIL−13の中和
ヒトIL−13を含むいくつかの異なるサイトカインに対する反応において、TF−1細胞は増殖する。したがってIL−13のためのこれらの細胞の増殖反応を用いてIL−13の生体活性を測定し、続いてアッセイを開発してmAbのIL−13中和効能(IL−13生体活性の阻害)を決定することができる。
【0163】
アッセイは滅菌96ウェル組織培養プレートにおいて滅菌条件下で実施して、すべての試験ウェルは三連で実施した。14ng/mlの組換え大腸菌発現ヒトIL−13を様々な希釈のmAb(通常3倍希釈液で滴定した93.4nM〜0.014nM)と37℃で1時間プレインキュベートした。無関係な特異性抗体を陰性対照として同様に滴定した。次いでこれらの試料を、滅菌96ウェル組織培養プレート中TF−1細胞50μl(濃度2×105細胞/ml)に添加した。したがって最終100μlアッセイ体積には、様々な希釈のmAb(3倍希釈液で滴定した最終濃度46.7nM〜0.007nM)、組換え大腸菌発現ヒトIL−13(最終濃度7ng/ml)およびTF−1細胞(最終濃度1×105細胞/ml)を含有した。アッセイプレートは加湿CO2インキュベーター内で37℃で約3日間インキュベートした。次いでPromega製「Cell Titre96(登録商標)Non−Radioactive Cell Proliferation Assay」(カタログ番号G4100)を製造業者の説明書に記載どおりに用いて、細胞増殖量を決定した。96ウェルプレート中の試料の吸光度はプレートリーダーで570nmで読み取った。
【0164】
組換え大腸菌発現ヒトIL−13の生体活性を中和するmAbの能力は定義した量のヒトIL−13(7ng/ml)の生体活性を50%中和するために必要なmAb濃度(=ND50)として表した。必要なmAb濃度が低いほど、中和能力は強力である。本明細書に提供するND50データ(表4)はマイクロソフトエクセルのRobosageを用いて算出した。データの図解は図1に見ることができる。
【表4】
【実施例5】
【0165】
CDRH3変異型抗IL−13 mAbを含むmAb−dAbの構築および発現
標準的な分子生物学技術を用いて、A1抗体のCDRH3変異体の可変重鎖領域の各配列をコードする遺伝子を既存の構築物から、hIgG1定常領域のC末端にてTVAAPSまたはASTKGPSリンカーを介して抗ヒトIL−4ドメイン抗体(DOM9−112−210)に融合したhIgG1定常領域を含む発現ベクターに移した。構築した重鎖の詳細を表5に列挙する。
【表5】
【0166】
BPC1624、BPC1625、BPC1626およびBPC1627をHEK293細胞内で発現した。簡単に述べると、250mlのHEK293細胞1.5×106細胞/mlを、293fectin試薬(Invitrogen#51〜0031)で先にインキュベートした重鎖および軽鎖発現プラスミドと共トランスフェクトした。これらを37℃、5%CO2、および95%相対湿度で攪拌インキュベーター内に入れた。24時間後にトリプトン供給培養を添加して、細胞をさらに5日間増殖させた。上清を遠心分離により収穫してフィルター滅菌した。固定化プロテインAカラムを用いて発現した分子を親和性クロマトグラフィーにより精製して、吸光度を280nmで測定することによって濃度を決定した。精製された試料中のタンパク質凝集レベルをサイズ排除クロマトグラフィーにより決定した。精製されたタンパク質の収率および凝集レベルを表5bに示す。
【0167】
【実施例6】
【0168】
CDRH3変異型抗IL−13 mAbを含むmAb−dAbの中和活性データ
TF−1細胞増殖バイオアッセイにおいてCDRH3変異型抗IL−13 mAbを含むmAb−dAbによる大腸菌発現組換えヒトIL−13の中和を試験した。
【0169】
アッセイは滅菌96ウェル組織培養プレートにおいて滅菌条件下で実施して、すべての試験ウェルは三連で実施した。約20ng/mlの組換え大腸菌発現ヒトIL−13を様々な希釈のmAbdAb(通常3倍希釈液で滴定した、50nM〜0.02nM)(HEK細胞内で作製し、実施例5に記載のように精製されたmAbdAb)の総体積50μlと37℃で1時間プレインキュベートした。無関係な特異性抗体を陰性対照として同様に滴定した(データは示さず)。次いでこれらの試料を、滅菌96ウェル組織培養プレート中TF−1細胞50μl(濃度2×105細胞/ml)に添加した。したがって最終100μlアッセイ体積には、様々な希釈のmAbdAb(3倍希釈液で滴定した最終濃度25nM〜0.01nM)、組換え大腸菌発現ヒトIL−13(最終濃度10ng/ml)およびTF−1細胞(最終濃度1×105細胞/ml)を含有した。アッセイプレートは加湿CO2インキュベーター内で37℃で約3日間インキュベートした。次いでPromega製「Cell Titre96(登録商標)Non−Radioactive Cell Proliferation Assay」(カタログ番号G4100)を製造業者の説明書に記載どおりに用いて、細胞増殖量を決定した。96ウェルプレート中の試料の吸光度はプレートリーダーで570nmで読み取った。
【0170】
ヒトIL−13の生体活性を中和するmAbdAbの能力は定義した量のヒトIL−13(10ng/ml)の生体活性を50%中和するために必要なmAb−dAb濃度(=ND50)として表した。必要なmAbdAb濃度が低いほど、中和能力は強力である。本明細書に提供するND50データ(表6)はGraphPad Prismを用いて算出した。これらのデータを図2に図式的に表す。
【表6】
【実施例7】
【0171】
CHOE1a発現系におけるCDRH3変異型抗IL−13 mAbを含むmAb−dAbの発現
表5に示す分子BPC1624〜1631もCHOE1a細胞内で発現した。重鎖および軽鎖をコードするDNAベクターを懸濁CHO細胞内に共電気穿孔した。攪拌フラスコ内のMR1基底選択的培地で、37℃、5%CO2、130rpmで細胞生存能および細胞数が改善されるまで細胞を継代させた。次いでCHO細胞をMR1基底×2選択的培地内で播種し、34℃、5%CO2、130rpmで8〜12日間インキュベートした。細胞を遠心分離によりペレットして、上清を滅菌濾過した。
【0172】
固定化プロテインAカラムを用いて発現物質を親和性クロマトグラフィーにより精製して、吸光度を280nmで測定することによって収率を決定した。凝集レベルをサイズ排除クロマトグラフィーにより決定した。凝集を調製用サイズ排除クロマトグラフィーにより除去し、収率を再評価した。この発現系から得られる収率および凝集レベルを表7に列挙する。
【表7】
【実施例8】
【0173】
抗原結合タンパク質の化学量論評価(Biacore(商標)を使用)
この実施例は予測である。本発明の抗原結合タンパク質を試験できる追加アッセイを実行するための指針を提供する。
【0174】
抗ヒトIgGは、第1級アミンカップリングによりCM5バイオセンサーチップ上に固定化する。抗原結合タンパク質をこの表面上に捕捉後、単一濃度のIL−13またはIL−4またはIL−5を通過させる。この濃度は結合表面を飽和するのに十分であり、観察される結合シグナルは完全なRmaxに達する。次いで以下の式:
化学量論値=Rmax × Mw(リガンド) / Mw(アナライト) × R(固定化または捕捉されたリガンド)
を用いて化学量論値を算出する。
【0175】
複数のアナライト結合の化学量論値を同時に算出する場合、異なる抗原を飽和抗原濃度および上で算出した化学量論で連続通過させる。この作業は、Biacore 3000上、HBS−EPランニング緩衝液を用いて25℃で実行できる。
【実施例9】
【0176】
改善されたヒト化変異型mAbの用量予測
異なるモノクローナル抗体(mAb)候補を結合親和性に基づき順位付けるために抗体リガンド結合PK−PDモデルを開発して、ヒトにおける潜在的治療用量を予測した。
【0177】
ヒトにおける予測した潜在的治療用量をこの目的のために、mAbの月1回静脈内投与1時間後に肺(作用部位)中標的IL−13を定常状態で90%阻害する用量として定義した。各分子の分子量は同じであり、mAbの標準分子量、すなわち150kDaと等しいと推定した。加えて、異なる候補の動物またはヒト薬力学データの不在下、A1L1抗体のヒト薬力学をすべての候補と推測した。
【0178】
同一の抗体リガンド結合PK−PDモデルならびに標的濃度、標的ターンオーバー、標的組織:血漿比率およびmAb組織浸透に関する同じ仮定を各mAbにおいて使用する。したがって、モデルにより提供される順位付けは、もっぱら分子の結合親和性に基づき、変数のみ異なる。かかる状態において、ヒトにおける4つの候補A1Y100BlleL1、A1Y100BValL1、A1Y100BAlaL1およびA1Y100BTrpL1の潜在的治療用量は、ヒトにおけるA1L1の予測される潜在的治療用量を超える実質的な改善を提供すると予測される。
【実施例10】
【0179】
変異型IL−4dAbを伴う抗IL13/IL4 mAbdAb
10.1構築および発現
凝集予測アルゴリズムを用いて、抗IL−4dAb(DOM9−155−154、配列番号80)の凝集傾向がある残基を調査した。Kabat89位のロイシン残基を凝集促進のための主要残基として同定した。
【0180】
このdAbを含むmAbdAbの潜在的凝集を減らすために、このアミノ酸残基を他のアミノ酸と置換していくつかのmAb−dAb変異体を生成した。既存のmAbdAb構築物の重鎖をコードするDNA発現ベクターを用いて、部位指向変異により発現構築物を生成した。変異dAb配列を含む生成した新規mAbdAb重鎖のタンパク質配列は、配列番号117〜134において提示される。
【0181】
89位に別の変異を挿入された他の重鎖配列は、配列番号96〜106である。これらを実施例11に詳述する。
【0182】
発現した分子の一覧を表8に提供する。
【表8】
【0183】
【0184】
10.2 HEK 293 6E細胞における分子発現
重鎖と軽鎖をそれぞれコードするプラスミドをHEK 293 6E細胞内に一過性共トランスフェクトして少量発現させて、抗体分子を産生した。トランスフェクションから最大24時間後にトリプトン供給物を各細胞培養に添加して、3日後に細胞を回収した。
【0185】
抗体分子を組織培養上清から直接評価して、Gyrolabワークステーションを用いて定量化した。
【0186】
細胞上清中の抗体分子の定量化のためのGyrolabワークステーション方法
Gyrolab Bioaffyワークステーション(Gyros)を用いた定量免疫アッセイにより、少量の一過性HEK 2936Eトランスフェクション(0.75〜2.0ml)から産生した抗体を組織培養上清から定量化した。コンパクトディスク(CD)マイクロ実験室(Gyros)上にストレプトアビジンコーティングした粒子に固定化したビオチン化抗IgGAffibody分子(Abcam)を用いたFc領域を介して抗体を捕捉された。Affibody試薬を簡単にボルテックスして、PBS−Tween 20(0.01%v/v)で最終作業濃度0.1mg/mlに希釈した。次いで、レーザ誘発蛍光を用いてALEXA 647標識化Fab2抗ヒトIgGκ軽鎖分子により抗体を検出した。ALEXA 647標識化検出試薬は、簡単にボルテックスすることによって、および13000rpmで4分間遠心分離することによって調製した。標識化Fab2検出試薬を非標識化Fab2に添加して、これをRexcip F検出試薬希釈液(Gyros)を用いて最終濃度75nMおよび1.5μMにそれぞれ希釈した。抗体定量化範囲は抗CD23モノクローナル抗体標準曲線と比較して0.244〜250μg/mlであった。組織培地(Freestyle 293発現培地、Pluronic F68およびGeneticin、Invitrogen)で抗体を連続希釈して抗CD23(1mg/ml)標準曲線を作成した。
【0187】
場合により、固定化プロテインAカラムを用いて抗体分子を精製して、吸光度を280nmで読み取ることにより定量化し、指定の場合、精製された抗体分子を以下に説明する実施例に記載のアッセイにおいて評価した。
【0188】
10.3 IL−4結合ELISA
以下の方法を用いた直接結合ELISAにおいて、これらのmAbdAbのIL−4への結合を試験した。
【0189】
5μg/mlヒトIL−4(GSK製)のNaHCO3溶液を96ウェル高結合プレートにコーティングして、4℃で一晩保存した。プレートを0.05%のTween−20(TBST)含有トリス緩衝食塩水で2回洗浄した。100μLのブロック溶液(1%BSAのTBST緩衝液)を各ウェルに添加して、プレートを室温で少なくとも1時間インキュベートした。引き続いてmAbdAbをプレート全体でブロック溶液で希釈した。1時間インキュベーション後、プレートを3回洗浄した。ヤギ抗ヒトκ軽鎖特異的ペルオキシダーゼ抱合抗体(Sigma A7164)をブロック溶液で1μg/mLに希釈して、50μLを各ウェルに添加した。プレートを1時間インキュベートした。さらに3つの洗浄工程後、50μlのOPD(o−フェニレンジアミン二塩酸塩)SigmaFast基質溶液を各ウェルに添加して、3M硫酸25μL添加により約5分後に反応は停止した。基本終点プロトコルを用いて、VersaMax Tunableマイクロプレートリーダー(Molecular Devices)を用いて、吸光度を490nmで読み取った。
【0190】
精製された物質であった陽性対照(抗IL−4 mAb)および抗IL13陰性対照mAbを除き、gyrolabプラットフォームを用いて定量化されている組織培養上清から直接mAbdAbを用いて実験を実施した。これらのデータを図17に示す。
【0191】
ELISA結果は、これらの一過性発現抗IL13 mAb−抗IL4dAb結合IL−4のほとんど(ただし、IL−4結合活性において一部変形)が観察されたことを示す。精製された陽性対照の抗IL−4 mAbもIL−4への結合を示したのに対し、精製された陰性対照mAbはヒトIL−4と結合しないことが示された。
【0192】
10.4 mAbdAb発現における凝集レベルの分析
293fectin(Invitrogen、12347019)を用いて、BPC1090、BPC1091、BPC1092、BPC1093、BPC1094およびBPC1095の重鎖および軽鎖をコードするpTTプラスミドをHEK 293 6E細胞内に一過性共トランスフェクトして、実施例10.2に上述したmAbdAbよりもわずかに多く(200〜600ml)発現させた。同じ方法論を用いてBPC1111およびBPC1085を独立してHEK293 6E細胞内で一過性発現させた。上のトランスフェクションに使用したプラスミドは、EndoFree Plasmid Maxi Kit(Qiagen,12362)を用いて生成した。
【0193】
24時間後にトリプトン供給物を各細胞培養に添加して、72時間後に細胞を回収した。抗体をプロテインAカラムを用いて精製し、280nmでの吸光度の読み取りにより定量化し、サイズ排除クロマトグラフィー(SEC)により分析した。
【0194】
これらのmAbdAbを、独立して発現しているBPC2223、抗IL13 mAb(829)および抗Il−13 mAb(586)と比較した。
【0195】
変異dAbを含むmAbdAb(BPC1090、BPC1091、BPC1093、BPC1094およびBPC1095)と同様に、両抗体(オリジナルのCDRH3を有する586および変異CDRH3を有する829)は低い凝集レベルを示した。オリジナルのdAbを含む、すなわち89位が変異していなかったBPC2223はL89H変異を有するBPC1092よりも凝集レベルが高かった。代表的な凝集データを表8bに示す。
【表8b】
【0196】
10.5 BIAcore分析
BIAcoreアッセイにおいて、精製されたmAbおよびmAbdAb構築物について、89位の変異がdAbのIL−4への結合に対して何らかの影響を与えるかどうかを決定するために試験した。
【0197】
プロテインAを第1級アミンカップリングによりCM5チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。組換え大腸菌発現ヒトIL4をアナライトとして256、64、16、4および1、0.25および0.0625で使用し、0nM(すなわち緩衝液単独)を結合曲線の二重照合に使用した。抗プロテインA表面を50mM NaOHで再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【表8c】
【実施例11】
【0198】
CDRH3変異型抗IL−13 mAbおよび変異dAbを含む抗原結合タンパク質(BPC1085、BPC1086およびBPC1087)の構築および試験
11.1構築および発現
代替プラスミド構築物を生成するための塩基構築物として抗IL−13 mAbおよび抗IL−4dAbからなる重鎖をコードするプラスミドを使用した。2工程のクローニング戦略を要した。工程1では、HindIIIおよびSpeIを用いた制限クローニングにより、H鎖の抗IL13 mAb成分のVHをコードするDNA配列を別のヒト化抗IL13抗体のVH(配列番号54)をコードするDNA配列と置換した。工程2では、mAbdAbの抗IL4dAb(DOM9−155−154、配列番号80)成分のKabat89位でロイシンコードコドンを部位指向変異によりグルタミンに変異した。生成した重鎖DNA配列はすべて配列番号96、98および100に提示される。構築および発現した分子の一覧を表9に提供する。
【表9】
【0199】
293fectin(Invitrogen、12347019)を用いて、BPC1085、BPC1086およびBPC1087 mAbdAbをコードする重鎖および軽鎖発現プラスミドをHEK 2936E細胞内に共トランスフェクトした。24時間後にトリプトン供給物を各細胞培養に添加して、72時間後に細胞を回収した。プロテインAカラムを用いて抗体を精製後、結合アッセイにおいて試験した。
【0200】
プロテインA親和性を用いてBPC1085、BPC1086およびBPC1087 mAbdAbを精製した。AKTA Xpress系上で1mlプロテインAカラム(GE Healthcare)を用いて、カラムをPBS(Gibco/Invitrogen)中で平衡させて、Pierce IgG溶出液を用いて抗体を溶出した。1Mトリス(ヒドロキシメチル)アミノメタン緩衝液(通常5〜10%v/v)を用いて溶出した留分を中和した。溶出した抗体留分をプールして、サイズ排除クロマトグラフィーにより凝集分析し、分光光度計を用いてOD280nmでの読み取りにより定量化した。
【0201】
これらを、表10に記載する等価物mAbdAb(2222、2223、2230および2231)と比較した。これらは、以下を含む:
i)BPC2222、BPC2223、BPC2230およびBPC2231において「L」であり、BPC1085、BPC1086およびBPC1087において「Q」である89位を除きBPC1085、BPC1086およびBPC1087に使用されるものと同一である1つのdAb。
【0202】
ii)同一リンカー
iii)BPC2222、BPC2223、BPC2230およびBPC2231において「Y」であり、BPC1085、BPC1086およびBPC1087において「V」である100B位を除きBPC1085、BPC1086およびBPC1087に使用されるものと同一である1つのIL−13 mAb配列。
【表10】
【0203】
プロテインA親和性を用いてBPC2222、2223、2230および2231 mAbdAbを精製した。AKTA Xpress系上で1mlプロテインAカラム(GE Healthcare)を用いて、カラムをPBS(Gibco/Invitrogen)中で平衡させて、Pierce IgG溶出液を用いて抗体を溶出した。1Mトリス(ヒドロキシメチル)アミノメタン緩衝液(通常5〜10%v/v)を用いて溶出した留分を中和した。溶出した抗体留分をプールして、サイズ排除クロマトグラフィーにより凝集分析し、分光光度計を用いてOD280nmでの読み取りにより定量化した。
【0204】
BPC2222、2223、2230および2231は30〜40%の凝集を示し、凝集した物質は10分前に溶出した。
【0205】
サイズ排除クロマトグラフィーにより評価したところ、構築物BPC1085、1086および1087はBPC2222、2223、2230および2231より低い凝集レベルを示した。これらの分子のSECプロファイルを図3〜9に示す。
【0206】
11.2 IL−4結合ELISA
実施例10.3に記載の方法に従った直接結合ELISAにおいて、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbのIL−4への結合を試験した。
【0207】
これらのデータを図10に示す。ELISA結果により、これらの精製されたmAbdAbはIL−4と結合したことを確認した。陽性対照の抗IL−4 mAbおよびBPC2231もIL−4への結合を示したのに対し、陰性対照mAb(抗IL−13 mAb)はIL−4と結合しないことが示された。これは、このELISAにおいて、リンカーの長さをGS(TVAAPSGS)からGS(TVAAPSGS)2〜4に伸長時にdAb効能が向上したことを示した。
【0208】
11.3 TF−1細胞増殖バイオアッセイにおけるIL−4中和
TF−1細胞バイオアッセイにおいて、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbのヒトIL−4中和を試験した。
【0209】
ヒトIL−4を含むいくつかの異なるサイトカインに対する反応においてTF−1細胞は増殖する。したがってIL−4のためのこれらの細胞の増殖反応を用いてIL−4の生体活性を測定し、続いてアッセイを開発してmAbdAbのIL−4中和効能(IL−4生体活性の阻害)を決定することができる。
【0210】
アッセイは滅菌96ウェル組織培養プレートにおいて滅菌条件下で実施して、すべての試験ウェルは二連で実施した。約2.2ng/mlの組換え大腸菌発現ヒトIL−4を様々な希釈のmAbdAb(通常3倍希釈液で滴定した、560nM〜0.009nM)の総体積120μlと37℃で1時間プレインキュベートした。無関係な特異性抗体を陰性対照(抗IL13 mAb)として同様に滴定した。次いでこれらの試料50μlを、滅菌96ウェル組織培養プレート中TF−1細胞50μl(濃度2×105細胞/ml)に添加した。したがって最終100μlアッセイ体積には、様々な希釈のmAbdAb(3倍希釈液で滴定した最終濃度270nM〜0.005nM)、組換え大腸菌発現ヒトIL−4(最終濃度1.1ng/ml)およびTF−1細胞(最終濃度1×105細胞/ml)を含有した。アッセイプレートは加湿CO2インキュベーター内で37℃で約4日間インキュベートした。次いでPromega製「Cell Titre96(登録商標)Non−Radioactive Cell Proliferation Assay」(カタログ番号G4100)を製造業者の説明書に記載どおりに用いて、細胞増殖量を決定した。96ウェルプレート中の試料の吸光度はプレートリーダーで570nmで読み取った。これらのデータをエクセルシートに入力し、2回の試験ウェル値を平均化して、平均背景値(mAb−dAbなしおよびIL−4試験ウェルなし)を差し引いた。
【0211】
組換え大腸菌発現ヒトIL−4の生体活性を中和するmAbdAbの能力は定義した量のヒトIL−4(1.1ng/ml)の生体活性を50%中和するために必要なmAb−dAb濃度(=ND50)として表した。必要なmAbdAb濃度が低いほど、中和能力は強力である。本明細書に提供するND50データ(表11)はエクセルのRobosage関数を用いて算出した。これらのデータを図11に図式的に表す。
【0212】
TF−1細胞バイオアッセイにおいて、抗IL−4 mAbおよびDOM9−155−154(配列番号80)をヒトおよびカニクイザルIL−4中和の陽性対照として含んだ。さらに、TF−1細胞バイオアッセイにおいて、無関係な抗原(ダミーdAb)に特異的なdAbもヒトまたはカニクイザルIL−4中和の陰性対照として含んだ。
【0213】
これらを数回反復し、これらの実験の1つの結果を図11に示す。ND50値をデータセットから算出した。ND50値は、IL−4の生体活性を50%中和できるmAbdAbまたはmAbまたはdAb濃度である。平均ND50値および試験回数(n)を表11に示す。
【表11】
【0214】
これらのデータにより、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbは、ヒトおよびカニクイザルIL−4の生体活性を中和したことを確認した。抗IL−4 mAbおよびDOM9−155−154もヒトおよびカニクイザルIL−4の生体活性を中和し、陰性dAb(ダミーdAb)は同じバイオアッセイにおいて中和を示さなかった。
【0215】
全3つのmAbdAbは良好な効能を示し、リンカーの長さが伸びるにつれてdAb効能の明らかな増大傾向があることが中和アッセイから明らかであった(ただし、より粗いELISAではこの効能差が見出されなかった)。陰性対照の抗IL−4 mAbは同じバイオアッセイにおいて中和を示さなかった。
【0216】
11.4 TF−1細胞増殖バイオアッセイにおけるヒトIL−13の中和
TF−1細胞バイオアッセイにおいて、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbのヒトIL−13の中和を下記のとおり試験した。
【0217】
TF−1細胞はいくつかの異なるサイトカイン(ヒトIL−13を含む)に対する反応において増殖する。したがってIL−13のためのこれらの細胞の増殖反応を用いてIL−13の生体活性を測定し、続いてアッセイを開発してmAbdAbのIL−13中和効能(IL−13生体活性の阻害)を決定することができる。
【0218】
アッセイは滅菌96ウェル組織培養プレートにおいて滅菌条件下で実施して、すべての試験ウェルは二連で実施した。約14ng/mlの組換え大腸菌発現ヒトIL−13を様々な希釈のmAbdAb(通常3倍希釈液で滴定した560nM〜0.009nM)の総体積120μlと37℃で1時間プレインキュベートした。無関係な特異性の抗体およびdAbを陰性対照として同様に滴定した(それぞれ抗IL−4 mAbおよびDOM9−155−154)。次いでこれらの試料50μlを、滅菌96ウェル組織培養プレート中TF−1細胞50μl(濃度2×105細胞/ml)に添加した。したがって最終100μlアッセイ体積には様々な希釈のmAbdAb(3倍希釈液で滴定した最終濃度270nM〜0.005nM)、組換え大腸菌発現ヒトIL−13(最終濃度7ng/ml)およびTF−1細胞(最終濃度1×105細胞/ml)を含有した。アッセイプレートは加湿CO2インキュベーター内で37℃で約4日間インキュベートした。次いでPromega製「Cell Titre96(登録商標)Non−Radioactive Cell Proliferation Assay」(カタログ番号G4100)を製造業者の説明書に記載どおりに用いて、細胞増殖量を決定した。96ウェルプレート中の試料の吸光度はプレートリーダーで570nmで読み取った。これらのデータをエクセルシートに入力し、2回の試験ウェル値を平均化して、平均背景値(mAb−dAbなしおよびIL−13試験ウェルなし)を差し引いた。
【0219】
組換え大腸菌発現ヒトIL−13の生体活性を中和するmAbdAbの能力は定義した量のヒトIL−13(7ng/ml)の生体活性を50%中和するために必要なmAb−dAb濃度(=ND50)として表した。必要なmAbdAb濃度が低いほど、中和能力は強力である。本明細書に提供するND50データ(表12)はエクセルのRobosage関数を用いて算出した。これらのデータを図12に図式的に表す。
【0220】
TF−1細胞バイオアッセイにおいて、ヒトIL−13中和の陽性対照として抗IL−13 mAb(配列番号22および24)を含んだ。さらに、抗IL−4 mAbも陰性対照として含んだ。
【0221】
図12は、TF−1細胞中和アッセイ結果を示す。ND50値をデータセットから算出した。ND50値は、IL−13の生体活性を50%中和できるmAbdAbまたはmAbまたはdAb濃度である。
【0222】
平均ND50値および試験回数を表12に示す。
【表12】
【0223】
これらのデータにより、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbは組換えヒトおよびカニクイザルIL−13の生体活性を中和したことが確認される。陰性対照の抗IL−4 mAbは同じバイオアッセイにおいて中和を示さなかった。
【実施例12】
【0224】
抗IL−13 mAb軽鎖の再ヒト化
12.1再ヒト化
一部の抗IL−13 mAb−抗IL−4dAb分子(BPC1085)の発現を改善するために、マウス抗体6A1の軽鎖CDR(配列番号59に説明される軽鎖)を新規フレームワーク上に再グラフト化した。PCRベース戦略およびオリゴヌクレオチド重複を用いてコドン至適化軽鎖可変領域配列(表13に要約する)をde novoで構築した。PCRプライマーはシグナル配列(配列番号56)を組み込み、VLドメインを枠組みするように設計されたHindIIIおよびBsiWI制限部位を含むように設計し、ヒトκC領域を含むpTTおよびRln哺乳動物発現ベクター内へのクローニングを可能にした。構築されている再ヒト化軽鎖を表13に要約する。
【表13】
【0225】
12.2 HEK 293 6E細胞における分子発現
再ヒト化軽鎖の発現特性を初めにmAb型式において検討した。293fectin(Invitrogen、12347019)を用いて、A1Y100BVAL1(配列番号54)重鎖をコードするプラスミド、既存の軽鎖(配列番号24)および再ヒト化軽鎖をHEK 293 6E細胞内に一過性共トランスフェクトした。プラスミドは少量(2×0.75ml培養体積)発現して抗体を産生した。24時間後にトリプトン供給物を細胞培養に添加して、さらに72時間後に細胞を回収した。構築して発現したmAbのすべてを表14に要約する。
【表14】
【0226】
Gyrolabワークステーションを用いた定量免疫アッセイにより、抗体発現を組織培養上清から直接評価した。再ヒト化軽鎖を含む抗体BPC3208およびBPC3211(それぞれP0およびP1で示す)は、A1Y100BVAL1 mAbより改善された発現収率を示した。Q0およびQ1軽鎖(BPC3219およびBPC3220)は抗IL−13 mAbの発現を改善しなかった。発現データを表15に提示する。
【表15】
【0227】
12.3 HEK 293 6E細胞内のmAb−dAb発現
BPC3208およびBPC3211の再ヒト化軽鎖は抗IL−13 mAbの改善された発現を示したため、それらを抗IL−13mAb−抗IL−4dAbとの関連で検討した。293fectin(Invitrogen、12347019)を用いて、再ヒト化軽鎖P0およびP1ならびに586(L1)軽鎖を829H−GS(TVAAPSGS)2−256重鎖(配列番号96、詳細は表16に要約する)とHEK 293 6E細胞内に共トランスフェクトした。プラスミドを50〜500ml発現させて、抗体分子を産生した。24時間後にトリプトン供給物を細胞培養に添加して、さらに48時間後に細胞を回収した。固定化プロテインAカラムを用いて抗体を精製して、吸光度を280nmで読み取ることにより定量化し、指定の場合、精製された抗体分子を以下に説明する実施例に記載のアッセイにおいて評価した。図13および14に例証するとおりBPC3214およびBPC3215をサイズ排除クロマトグラフィー(SEC)により分析した。
【表16】
【0228】
mAb型式における観察と一致して、再ヒト化軽鎖を含むmAbdAb(BPC3214およびBPC3215)は、BPC1085より改善された発現を示した。代表的な発現データを表17 5に要約する。
【表17】
【0229】
12.4 ヒトIL−13結合ELISA
直接結合ELISAを介して、精製されたBPC3214およびBPC3215のヒトIL−13への結合についてBPC1085(実施例10に記載)と比較して試験した。抗IL−13 mAb A1Y100BVAL1および抗IL−4 mAbもそれぞれ陽性対照および陰性対照として検討した。組換え大腸菌発現ヒトIL−13(バッチ番号GRITS31061)5μg/mlを96ウェル高結合プレート50μl/ウェルにコーティングして、+4℃で一晩インキュベートした。続く工程をすべて室温で行った。プレートを0.05%のTween−20含有リン酸緩衝食塩水で3回洗浄した。100μLブロック溶液(0.05%のTween−20含有1%BSAのリン酸緩衝食塩水)を各ウェルに添加して、プレートを室温で少なくとも1時間インキュベートした。次いで別の洗浄工程を実施した。引き続いて精製された抗体をプレート全体でブロック溶液で希釈した。1時間インキュベーション後、プレートを洗浄した。ヤギ抗ヒトκ軽鎖特異的ペルオキシダーゼ抱合抗体をブロック溶液で0.75μg/mlに希釈して、50μlを各ウェルに添加した。プレートを1時間インキュベートした。さらに2つの洗浄工程後、50μlのOPD(o−フェニレンジアミン二塩酸塩)SigmaFast基質溶液を各ウェルに添加して、3M硫酸50μLを添加することによって反応を停止させた。基本終点プロトコルを用いて、VersaMaxマイクロプレートリーダー(Molecular Devices)を用いて吸光度を490nmで読み取った。これらのデータを図15に示す。直接結合ELISAにより、BPC3214およびBPC3215はヒトIL−13と結合することを確認した。BPC3214およびBPC3215はBPC1085と類似のIL−13結合効能を示す。陽性対照の抗IL−13 mAb A1Y100BVAL1も組換えIL−13への結合を示したのに対し、陰性対照の抗IL−4 mAbはヒトIL−13と結合しないことが示された。
【0230】
12.5 ヒトIL−4結合ELISA
直接結合ELISAにおいて、精製されたBPC3214およびBPC3215の組換え大腸菌発現ヒトIL−4への結合も試験した。ELISAを実施例4に記載のように実施し、組換え大腸菌発現ヒトIL−4 5μg/mlを96ウェル高結合プレート50μl/ウェルにコーティングして、+4℃で一晩インキュベートした。これらのデータを図16に示す。直接結合ELISAにより、BPC3214およびBPC3215はヒトIL−4と結合することが確認される。BPC1085も検討した。BPC3214はBPC1085と類似のIL−4結合効能を示す。陽性対照の抗IL−4 mAbも組換えIL−4への結合を示したのに対し、陰性対照の抗IL−13 mAb A1Y100BVAL1はヒトIL−4に対して結合しないことが示された。
【実施例13】
【0231】
BIAcore(商標)分析により評価したオリジナルのIL−13 mAb CDRH3を含むmAbdAb(BPC2222、BPC2223およびBPC2230〜2231)のIL−13およびIL−4結合親和性
方法
プロテインAを第1級アミンカップリングによりC1チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。組換え大腸菌発現ヒトIL13を256、64、16、4、および1nMで使用し、組換え大腸菌発現ヒトIL4を64、16、4、1および0.25nMで使用し、0nM(すなわち緩衝液単独)をIL4結合とIL13結合の両方の結合曲線の二重照合に使用した。プロテインA表面を100mMリン酸で再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【0232】
ヒトIL13への結合結果を表18に示し、ヒトIL4への結合結果を表19に示す。
【表18】
【0233】
【表19】
【実施例14】
【0234】
BIAcore(商標)分析により評価したオリジナルのIL−13 mAb CDRH3を含むmAbdAb(BPC2222、BPC2231)および変異型抗IL−13 mAb CDRH3(BPC1085〜1087)のIL−13およびIL−4結合親和性
方法
プロテインAを第1級アミンカップリングによりCM5チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。組換え大腸菌発現ヒトIL13を256nMのみで使用し、組換え大腸菌発現ヒトIL4を64、16、4および1nMで使用し、0nM(すなわち緩衝液単独)をIL4結合とIL13結合の両方の結合曲線の二重照合に使用した。プロテインA表面を50mM NaOHで再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【0235】
ヒトIL13への結合結果を表20に示し、ヒトIL4への結合結果を表21に示す。
【表20】
【0236】
BPC1086およびBPC1087の結合速度はBiacoreの感度外であるが、我々がこのデータを精密に分析できないという事実から、IL4との相互作用は速い結合速度での高親和性のものである可能性が高いことが示される。
【表21】
【実施例15】
【0237】
BIAcore(商標)分析により評価したいくつかの変異型抗IL−13 mAb CDRH3を含むmAbdAb(BPC1085、BPC1090−BPC1095、およびBPC1108−BPC1119)のIL−4結合親和性
方法(ヒトIL−4結合親和性)
プロテインAを第1級アミンカップリングによりCM5チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。
【0238】
組換え大腸菌発現ヒトIL4を64、16、4、1および0.25nMで使用した。すべての結合曲線を0nM注入(すなわち緩衝液単独)で二重照合した。
【0239】
プロテインA表面を50mM NaOHで再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【0240】
ヒトIL4への結合結果を表22に示す。
【表22】
【実施例16】
【0241】
BIAcore(商標)分析により評価して、オリジナルの軽鎖(BPC1085)と比較した再ヒト化軽鎖を含むmAbdAb(BPC3214およびBPC3215)のIL−13結合親和性
方法(ヒトおよびカニクイザルIL−13結合親和性)
プロテインAを第1級アミンカップリングによりCM5チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。
【0242】
組換え大腸菌発現ヒトIL13およびカニクイザルIL13を64、16、4、1および0.25nM使用した。すべての結合曲線を0nM注入(すなわち緩衝液単独)で二重照合した。
【0243】
プロテインA表面を50mM NaOHで再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【0244】
ヒトおよびカニクイザルIL13への結合結果を表23に示す。
【表23】
【実施例17】
【0245】
変異dAbを伴うおよび伴わないmAbdAbのストレッサー試験
いくつかのmAbdAbをPBSまたは50mM酢酸緩衝液に入れて、37℃で最大14日間インキュベートした。次いで、視認可能な沈殿物の存在、溶解性凝集および濃度安定性アドヒアランスについて分析した。
【0246】
結果により、変異dAbを含むmAbdAb(BPC2222、2223、2230、2231)は、非変異dAb(BPC1085、1086、1087)と同様に動作することが示され、両種類のmAbdAbはPBSと酢酸緩衝液の両方において37℃、2週間のインキュベーション期間に溶液中のタンパク質濃度に変化がなかったことから、安定していると思われた。加えて溶液中の凝集レベルには変化はないかほとんど認められず、沈殿は観察されなかった。
【実施例18】
【0247】
PK評価
ラットへのIV投与後の分離した試験において、BPC1085、BPC1086、およびBPC1087の薬力学を調査した。BPC1085のPKもカニクイザルへのIV投与後に調査した。
【0248】
ラットにおける全3分子およびサルにおけるBPC1085のPKが標準的mAbのPKと一致することを見出した。
【表24】
【0249】
【0250】
【0251】
【0252】
【0253】
【0254】
【0255】
【0256】
【0257】
【0258】
【0259】
【0260】
【0261】
【0262】
【0263】
【0264】
【0265】
【0266】
【0267】
【0268】
【0269】
【0270】
【0271】
【0272】
【0273】
【0274】
【0275】
【0276】
【0277】
【0278】
【0279】
【0280】
【0281】
【0282】
【0283】
【0284】
【0285】
【0286】
【0287】
【0288】
【技術分野】
【0001】
本発明は、インターロイキン13(IL−13)と結合し、その活性を中和する抗原結合タンパク質(特に抗体)、かかる抗原結合タンパク質をコードするポリヌクレオチド、前記抗原結合タンパク質を含む医薬品ならびに炎症(喘息など)関連疾患の治療および/もしくは予防におけるかかる抗原結合タンパク質の使用に関する。本発明の他の態様、目的および利益は、以下の本明細書から明らかとなるであろう。
【背景技術】
【0002】
インターロイキン−13(IL−13)
IL−13は、炎症サイトカイン産生を阻害するT細胞由来サイトカインとして元々記載されている12kDa分泌サイトカインである。構造的試験により、IL−13は2つのジスルフィド結合により保持されている4つのヘリックスバンドルの配置を有することが示される。IL−13は4つの潜在的グリコシル化部位を有するが、ラット肺由来の天然IL−13は、非グリコシル化分子として産生されることが分析により示されている。NSOおよびCOS−7細胞におけるヒトIL−13発現により、この観察所見が確認されている(Eisenmesser et al, J. Mol. Biol. 2001 310(1):231−241; Moy et al, J. Mol. Biol 2001 310(1):219−230; Cannon−Carlson et al, Protein Expression and Purification 1998 12(2):239−248)。
【0003】
IL−13は、喘息、慢性閉塞性肺疾患(COPD)、アトピー性皮膚炎およびアレルギー性鼻炎を含むアレルギー疾患、食道好酸球増加症、腫瘍学適応症、例えばB細胞慢性リンパ性白血病(B−CLL)およびホジキン病、炎症腸疾患、例えば潰瘍性大腸炎、クローン病および不確定性大腸炎、乾癬および乾癬性関節炎、急性移植片対宿主病、糖尿病性腎症、線維状態(肺線維症、例えば特発性肺線維症(IPF)など)に関係している。
【発明の概要】
【0004】
本発明は、IL−13(例えばIL−13抗体)、およびかかるIL−13抗体とIL−4アンタゴニストおよび/またはIL−5アンタゴニストの組み合わせと結合する抗原結合タンパク質を提供する。本発明のIL−13抗体は、CDRH3が変異しているマウスmAb 6A1に関連または由来する。6A1マウス重鎖可変領域アミノ酸配列は、配列番号58として提供される。6A1マウス軽鎖可変領域アミノ酸配列は、配列番号59として提供される。
【0005】
本発明の重鎖可変領域(VH)は、(Kabat(Kabat et al; Sequences of proteins of Immunological Interest NIH, 1987)により定義される)以下のCDRを含む:
本発明の重鎖可変領域のCDRは、以下のCDRを含み得る:
【0006】
本発明の軽鎖可変領域は、(Kabatにより定義される)以下のCDRを含む:
【0007】
抗体のCDR配列は、上表に説明するようにKabat番号付け系(Kabat et al; Sequences of proteins of Immunological Interest NIH, 1987)により決定することもでき、あるいはChothia番号付け系(Al−Lazikani et al., (1997) JMB 273,927−948)、接触点の定義方法(MacCallum R.M., and Martin A.C.R. and Thornton J.M, (1996), Journal of Molecular Biology, 262 (5), 732−745)または当業者にとって既知である抗体中残基の番号付けおよびCDR決定のため他の任意の確立された方法を用いて決定することもできる。本明細書に記載の本発明のCDRは、これらのいずれかの方法によって、またはChothia番号付けとKabat番号付けの組み合わせを用いて定義し得る。例えばCDRH1は、FYIKDTYMH(配列番号60)またはGFYIKDTYMH(配列番号61)を含むものとして定義し得る。
【0008】
本発明は、1つまたは複数のエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、例えばIL−4と結合できるエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、またはIL−5と結合できるエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、またはIL−4と結合できる第1のエピトープ結合ドメインおよびIL−5と結合できる第2のエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質も提供する。
【0009】
本発明は、残基89(kabat番号付け)の、「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)、「A」(アラニン)、「T」(トレオニン)および「E」(グルタミン酸)から選択される残基への変異による、免疫グロブリン単一可変ドメイン(例えばヒトdAb、例えばヒトVKドメイン抗体)の凝集傾向の低減方法も提供する。一実施形態では、この方法は、残基89(kabat番号付け)を「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)および「E」(グルタミン酸)から選択される残基に変異させることを含む。さらなる実施形態では、この方法は、残基89(kabat番号付け)を「Q」(グルタミン)に変異させることを含む。
【0010】
一実施形態では、この方法は配列番号80の抗IL−4ドメイン抗体に適用して、変異dAb配列(例えば配列番号94)をもたらすことができる。
【0011】
かかる変異dAbは、単独またはより大きな配列の一部(例えばmAbdAb配列の一部)であってよく、例えば、配列番号117〜134から選択される配列を含むdAbをもたらし得る。
【0012】
本発明は、凝集プロファイルが改善されているヒトVK dAb、例えばVK dAbの残基89(kabat番号付け)が「Q」(グルタミン)であるIGKV1−17、IGKV1D−17、IGKV1/OR2−108、IGKV1−6、IGKV5−2、IGKV1D−42、IGKV2−24、IGKV2−28、IGKV2−30、IGKV2−40、IGKV2D−29、IGKV2D−30、IGKV2D−24およびIGKV6−21から選択される生殖系列フレームワークに由来するヒトVK dAbも提供する。1つのかかる実施形態では、VK dAbは、VK dAbの残基89(kabat番号付け)が「Q」(グルタミン)である生殖系列フレームワークIGKV1−17、IGKV1D−17、IGKV1/OR2−108、IGKV1−6、IGKV5−2、IGKV1D−42、IGKV2−24、IGKV2−28、IGKV2−30、IGKV2−40、IGKV2D−29、IGKV2D−30、IGKV2D−24およびIGKV6−21から選択される生殖系列フレームワーク領域を含む。
【0013】
一実施形態では、本発明は、89位(kabat番号付け)に変異を含み、89位が「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)、「A」(アラニン)、「T」(トレオニン)および「E」(グルタミン酸)から選択されるものに変異している配列番号80の配列を含むヒトdAbを提供する。例えば本発明は、89位(kabat番号付け)が、「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)および「E」(グルタミン酸)から選択される残基に変異している配列番号80の配列を含むヒトdAbを提供する。
【0014】
一実施形態では、本発明は、配列番号94の配列を含むヒトdAbを提供する。
【0015】
本発明は、本明細書に記載の任意の抗原結合タンパク質重鎖をコードするポリヌクレオチド配列、および本明細書に記載の任意の抗原結合タンパク質軽鎖をコードするポリヌクレオチドも提供する。かかるポリヌクレオチドは等価物のポリペプチド配列に相当するコード配列を表すが、かかるポリヌクレオチド配列は開始コドン、適切なシグナル配列および停止コドンと共に発現ベクター中にクローン化し得ることが理解されるであろう。
【0016】
本発明は、本明細書に記載の任意の抗原結合タンパク質の重鎖および軽鎖をコードする1つまたは複数のポリヌクレオチドを含む組換え形質転換またはトランスフェクトされた宿主細胞も提供する。
【0017】
本発明はさらに、本明細書に記載の任意の抗原結合タンパク質の産生方法を提供し、この方法は、第1および第2ベクターを含む宿主細胞の培養工程を含み、前記第1ベクターは本明細書に記載の任意の抗原結合タンパク質重鎖をコードするポリヌクレオチドを含み、前記第2ベクターは、適切な培地(例えば無血清培地)中、本明細書に記載の任意の抗原結合タンパク質軽鎖をコードするポリヌクレオチドを含む。
【0018】
本発明はさらに、本明細書に記載の抗原結合タンパク質および医薬上許容可能な担体を含む医薬組成物を提供する。
【0019】
さらなる態様では、本発明は、本発明の抗原結合タンパク質の投与によりアトピー性疾患/障害および慢性炎症疾患/障害と関連した疾患または障害の治療または予防の方法を提供する。特定の興味は、喘息、例えばアレルギー性喘息、特に重症喘息(すなわちコルチコステロイドの全身投与を含む現在の治療が奏効しない喘息;Busse WW et al, J Allergy Clin. Immunol 2000, 106: 1033−1042を参照されたい)、「困難性」喘息(difficult asthma)(最大推奨用量の吸入ステロイド処方にもかかわらず制御不成功を特徴とする喘息性形質として定義、Barnes PJ (1998), Eur Respir J 12:1208−1218を参照されたい)、「ブリットル」喘息(Brittle asthma)(高用量の吸入ステロイドにもかかわらず大きく変動する最大呼気流量(PEF)を維持する重度の不安定喘息患者のサブグループを定義、Ayres JG et al (1998) Thorax 58:315−321を参照されたい)、夜間性喘息、月経前喘息、ステロイド抵抗性喘息(Woodcock AJ (1993) Eur Respir J 6:743−747を参照されたい)、ステロイド依存性喘息(高用量の経口ステロイドでのみ制御できる喘息として定義)、アスピリン誘発性喘息、成人発症型喘息、小児喘息の治療における本発明の抗原結合タンパク質の使用である。本発明の抗体は、急性喘息エピソード(喘息発作重積状態)を予防する、頻度を減らす、または作用を緩和するために使用してよい。本発明の抗体は、喘息の治療に使用される他の薬の必要な投薬を(投薬の投与量または投与頻度のいずれかに関して)減らすためにも使用してよい。例えば、本発明の抗体は、喘息のステロイド治療(コルチコステロイド治療など)に必要な投薬を減らす(「ステロイド使用を控える」)ために使用してよい。本発明の抗体で治療し得る他の疾患または障害としては、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、慢性閉塞性肺疾患(COPD)、好酸球性食道炎、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症(強皮症)、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、および細胞周期調節疾患、例えばホジキン病、B細胞慢性リンパ球性白血病が挙げられる。
【0020】
別の態様では、本発明は、アトピー性疾患/障害および慢性炎症疾患/障害の治療または予防のための薬の調製における本発明の抗原結合タンパク質の使用を提供する。特定の興味は、喘息、例えばアレルギー性喘息、特に重症喘息(すなわちコルチコステロイドの全身投与を含む現在の治療が奏効しない喘息;Busse WW et al, J Allergy Clin. Immunol 2000, 106: 1033−1042を参照されたい)、「困難性」喘息(最大推奨用量の吸入ステロイド処方にもかかわらず制御不成功を特徴とする喘息性形質として定義、Barnes PJ (1998), Eur Respir J 12:1208−1218を参照されたい)、「ブリットル」喘息(高用量の吸入ステロイドにもかかわらず大きく変動する最大呼気流量(PEF)を維持する重度の不安定喘息患者のサブグループを定義、Ayres JG et al (1998) Thorax 58:315−321を参照されたい)、夜間性喘息、月経前喘息、ステロイド抵抗性喘息(Woodcock AJ (1993) Eur Respir J 6:743−747を参照されたい)、ステロイド依存性喘息(高用量の経口ステロイドでのみ制御できる喘息として定義)、アスピリン誘発性喘息、成人発症型喘息、小児喘息の治療における本発明の抗原結合タンパク質の使用である。本発明の抗体は、急性喘息エピソード(喘息発作重積状態)を予防する、頻度を減らす、または作用を緩和するために使用してよい。本発明の抗体は、喘息の治療に使用される他の薬の必要な投薬を(投薬の投与量または投与頻度のいずれかに関して)減らすためにも使用してよい。例えば、本発明の抗体は、喘息のステロイド治療(コルチコステロイド治療など)に必要な投薬を減らす(「ステロイド使用を控える」)ために使用してよい。本発明の抗体で治療し得る他の疾患または障害としては、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、慢性閉塞性肺疾患(COPD)、好酸球性食道炎、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症(強皮症)、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、および細胞周期調節疾患、例えばホジキン病、B細胞慢性リンパ球性白血病が挙げられる。
【0021】
本発明の他の態様および利益を本明細書およびその実施形態に詳細に記載する。
【0022】
定義
本発明のその抗原結合タンパク質に関して本明細書を通して使用される「ヒトIL−13と結合する」という用語は、抗原結合タンパク質が他のヒトタンパク質(IL−4など)と結合しないかわずかしか結合せずに、ヒトIL−13(以下、hIL−13と呼ぶ)と結合することを意味する。特に、本発明の抗原結合タンパク質は、Biacoreアッセイ(例えば実施例3に記載のBiacoreアッセイ)にてヒトIL−13と結合することを見ることができる点から、ヒトIL−13と結合する。しかしながら、この用語は、本発明のある抗原結合タンパク質は他種由来のIL−13(例えばカニクイザルIL−13)と交差反応もし得るという事実を除外しない。
【0023】
本明細書で使用する「抗原結合タンパク質」という用語は、ヒトIL−13と結合でき、中和できる抗体、抗体断片および他のタンパク質構築物を指す。
【0024】
Fv、Fc、Fd、Fab、またはF(ab)2という用語は、標準的な意味で使用される(例えば、Harlow et al., Antibodies A Laboratory Manual, Cold Spring Harbor Laboratory, (1988)を参照されたい)。
【0025】
「キメラ抗体」とは、アクセプター抗体に由来する軽鎖および重鎖定常領域に関連したドナー抗体由来の天然可変領域(軽鎖および重鎖)を含む加工された抗体の一種を指す。
【0026】
「ヒト化抗体」とは、非ヒトドナー免疫グロブリンに由来するCDRを有し、分子の残りの免疫グロブリン派生部分は1つ(以上)のヒト免疫グロブリンに由来する加工された抗体の一種を指す。加えて、フレームワーク支持残基は、結合親和性を保持するために改変し得る(例えば、Queen et al., Proc. Natl Acad Sci USA, 86:10029−10032 (1989), Hodgson et al., Bio/Technology, 9:421 (1991)を参照されたい)。適切なヒトアクセプター抗体は、ドナー抗体のヌクレオチド配列およびアミノ酸配列への相同により、従来のデータベース、例えば、KABAT(登録商標)データベース、Los Alamosデータベース、およびSwissタンパク質データベースから選択し得る。ドナー抗体のフレームワーク領域との(アミノ酸に基づく)相同を特徴とするヒト抗体は、ドナーCDRを挿入するための重鎖定常領域および/または重鎖可変フレームワーク領域の提供に適し得る。軽鎖の定常または可変フレームワーク領域を供与できる適切なアクセプター抗体を類似の形態にて選択し得る。アクセプター抗体の重鎖と軽鎖は同じアクセプター抗体に由来している必要がないことに留意されたい。先行技術にて、かかるヒト化抗体の産生方法がいくつか記載されている−例えば欧州特許第A−0239400号および欧州特許第A−054951号を参照されたい。
【0027】
「ドナー抗体」という用語は、領域をコードする改変された免疫グロブリンならびにその結果発現したドナー抗体の抗原特異性および中和活性特徴を有する改変抗体を提供するようにその可変領域、CDR、または他の、その機能的断片もしくは第1免疫グロブリンパートナーアナログのアミノ酸配列に寄与する(モノクローナル、および/または組換え)抗体を指す。
【0028】
「アクセプター抗体」という用語は、第1免疫グロブリンパートナーに対するその重鎖および/もしくは軽鎖フレームワーク領域ならびに/またはその重鎖および/もしくは軽鎖定常領域をコードするアミノ酸配列のすべて(または任意の一部だが、いくつかの実施形態では、すべて)に寄与する、ドナー抗体と異種の(モノクローナルおよび/または組換え)抗体を指す。ある実施形態では、ヒト抗体はアクセプター抗体である。
【0029】
「CDR」は、免疫グロブリン重鎖および軽鎖の超可変領域である抗体の相補性決定領域アミノ酸配列として定義する。例えば、Kabat et al., Sequences of Proteins of Immunological Interest, 4th Ed., U.S. Department of Health and Human Services, National Institutes of Health (1987)を参照されたい。免疫グロブリンの可変部分には3つの重鎖および3つの軽鎖CDR(またはCDR領域)がある。したがって、本明細書で使用する「CDR」とは、全3つの重鎖CDRまたは全3つの軽鎖CDR(または適切な場合、全重鎖CDRと全軽鎖CDRの両方)を指す。抗体の構造およびタンパク質の折り畳み部分は、他の残基が抗原結合領域の一部とみなされ、当業者によりそのように理解されることを意味し得る。例えばChothia et al., (1989) Conformations of immunoglobulin hypervariable regions; Nature 342, p877−883を参照されたい。
【0030】
本明細書で使用する「ドメイン」という用語は、残りのタンパク質とは独立した三次構造を有する折り畳まれたタンパク質構造を指す。一般に、ドメインは、タンパク質の別々の機能特性を担い、多くの場合、残るタンパク質および/またはドメインの機能を失わずに他のタンパク質に添加、除去または移動し得る。「単一抗体可変ドメイン」とは、抗体可変ドメインに特徴的な配列を含む折り畳まれたポリペプチドドメインである。したがって、完全な抗体可変ドメインおよび修飾可変ドメイン(例えば、1つまたは複数のループが抗体可変ドメインに特徴的でない配列に置換されている)、またはNもしくはC末端伸長部分が切断されているか含まれる抗体可変ドメイン、ならびに完全長ドメインの少なくとも結合活性および特異性を保持する可変ドメインの折り畳まれた断片を含む。
【0031】
「免疫グロブリン単一可変ドメイン」という語句は、異なるV領域またはVドメインとは独立して抗原またはエピトープと特異的結合する抗体可変ドメイン(VH、VHH、VL)を指す。免疫グロブリン単一可変ドメインは、単一免疫グロブリン可変ドメインによる抗原結合のための他の領域またはドメインが不要である他の異なる可変領域または可変ドメインと1つの型式(例えば、ホモまたはヘテロ多量体)で存在できる(すなわち免疫グロブリン単一可変ドメインは、追加の可変ドメインとは独立して抗原と結合する)。「ドメイン抗体」または「dAb」とは、本明細書で使用する、抗原と結合できる「免疫グロブリン単一可変ドメイン」という用語と同義である。免疫グロブリン単一可変ドメインは、ヒト抗体可変ドメインであり得るが、齧歯類(例えば、WO00/29004号に開示のもの)、テンジクザメおよびラクダVHH dAb(ナノボディ)などの他種に由来する単一抗体可変ドメインも含む。ラクダVHHは、天然で軽鎖を欠いている重鎖抗体を産生するラクダ、ラマ、アルパカ、ヒトコブラクダ、およびグアナコを含む種に由来する免疫グロブリン単一可変ドメインポリペプチドである。かかるVHHドメインは、当技術分野で利用可能な標準的な技術に従いヒト化し得、かかるドメインは、依然として本発明による「ドメイン抗体」とみなされる。本明細書で使用する「VH」は、ラクダVHHドメインを含む。NARVは、テンジクザメを含む軟骨魚において同定された別の種類の免疫グロブリン単一可変ドメインである。これらのドメインは、新規の抗原受容体可変領域(一般にV(NAR)またはNARVと略される)としても知られている。詳細については、Mol. Immunol. 44, 656−665 (2006)および米国特許第20050043519A1号を参照されたい。
【0032】
「エピトープ結合ドメイン」という用語は、異なるV領域またはVドメインとは独立して抗原またはエピトープと特異的結合するドメインを指し、これはドメイン抗体(dAb)、例えばヒト、ラクダまたはサメ免疫グロブリン単一可変ドメインであってもよく、天然リガンド以外のリガンドへの結合を達成するためにタンパク質工学にかけられた非免疫グロブリン足場、例えばCTLA−4(Evibody);リポカリン;プロテインA派生分子(プロテインAのZドメイン(Affibody、SpA)、Aドメイン(Avimer/Maxibody)など);熱ショックタンパク質(GroElおよびGroESなど);トランスフェリン(trans−body);アンキリン反復タンパク質(DARPin);ペプチドアプタマー;C型レクチンドメイン(Tetranectin);ヒトγクリスタリンおよびヒトユビキチン(affilin);PDZドメイン;ヒトプロテアーゼ阻害剤のサソリ毒クニッツ型ドメイン;ならびにフィブロネクチン(adnectin)からなる群から選択される非免疫グロブリン足場の誘導体であるドメインであってもよい。
【0033】
CTLA−4(細胞傷害性Tリンパ球結合抗原4)は主にCD4+T細胞上で発現するCD28ファミリー受容体である。その細胞外ドメインは、可変ドメイン様Ig折り畳み部分を有する。抗体のCDRに対応するループは、異なる結合特性を与えるために異種配列と置換することができる。異なる結合特異性を有するように加工されたCTLA−4分子は、Evibodyとしても知られている。詳細については、Journal of Immunological Methods 248 (1−2), 31−45 (2001)を参照されたい。
【0034】
リポカリンは、小さな疎水性分子(ステロイド、ビリン、レチノイドおよび脂質など)を輸送する細胞外タンパク質ファミリーである。それらは、異なる標的抗原と結合するように加工することができる、円錐構造の開端にループをいくつか有する硬βシート状二次構造を有する。Anticalinは、サイズが160〜180個のアミノ酸であり、リポカリンに由来する。詳細については、Biochim Biophys Acta 1482: 337−350 (2000)、米国特許第7250297B1号および米国特許第20070224633号を参照されたい。
【0035】
Affibody(アフィボディ)は、抗原と結合するように加工することができる黄色ブドウ球菌のプロテインAに由来する足場である。ドメインは、約58個のアミノ酸の3つのヘリックスバンドルからなる。ライブラリは、表面残基の無作為化により生成されている。詳細については、Protein Eng. Des. Sel. 17, 455−462 (2004)および欧州特許第1641818A1号を参照されたい。
【0036】
Avimer(アビマー)は、Aドメイン足場ファミリーに由来する多ドメインタンパク質である。約35個のアミノ酸の天然ドメインは、定義されたジスルフィド結合構造をとる。多様性は、Aドメインファミリーによって示される天然変異をシャッフルすることによって生成される。詳細については、Nature Biotechnology 23(12), 1556 − 1561 (2005) and Expert Opinion on Investigational Drugs 16(6), 909−917 (June 2007)を参照されたい。
【0037】
トランスフェリンは、単量体血清輸送糖タンパク質である。トランスフェリンは、許容状態の表面ループへのペプチド配列の挿入によって、異なる標的抗原と結合するように加工することができる。加工されたトランスフェリン足場の例としては、Trans−bodyが挙げられる。詳細については、J. Biol. Chem 274, 24066−24073 (1999)を参照されたい。
【0038】
設計されたアンキリン反復タンパク質(DARPin)は、内在性膜タンパク質の細胞骨格への結合を媒介するタンパク質ファミリーであるアンキリンに由来する。単一アンキリン反復は、2つのα−ヘリックスおよびβ−ターンからなる33残基のモチーフである。それらは、各反復の第1α−ヘリックスおよびβ−ターンにおける残基を無作為化することによって、異なる標的抗原と結合するように加工することができる。それらの結合界面は、モジュール数を増大することによって増大できる(親和性成熟の方法)。詳細については、J. Mol. Biol. 332, 489−503 (2003), PNAS 100(4), 1700−1705 (2003)およびJ. Mol. Biol. 369, 1015−1028 (2007)ならびに米国特許第20040132028A1号を参照されたい。
【0039】
フィブロネクチンは、抗原と結合するように加工することができる足場である。Adnectinは、ヒトフィブロネクチンIII型(FN3)15反復単位の10番目ドメイン天然アミノ酸配列の主鎖からなる。β−サンドイッチの片端における3つのループは、Adnectinが興味の治療標的を特異的に認識することを可能にするように加工することができる。詳細については、Protein Eng. Des. Sel. 18, 435−444 (2005)、米国特許第20080139791号、WO2005056764号および米国特許第6818418B1号を参照されたい。
【0040】
ペプチドアプタマーは、普遍的な足場タンパク質、典型的には、活性部位に挿入された制限された可変ペプチドループを含むチオレドキシン(TrxA)からなるコンビナトリアル認識分子である。詳細については、Expert Opin. Biol. Ther.5, 783−797 (2005)を参照されたい。
【0041】
微小体(Microbody)は、3〜4個のシステイン架橋を含む25〜50個のアミノ酸の長さの天然マイクロタンパク質に由来し、マイクロタンパク質の例としては、KalataB1およびコノトキシンおよびノッチンが挙げられる。マイクロタンパク質は、マイクロタンパク質の全体的な折り畳み部分に影響を与えずに最大25個のアミノ酸を含むように加工することができるループを有する。加工されたノッチンドメインの詳細については、WO2008098796号を参照されたい。
【0042】
他のエピトープ結合ドメインとしては、異なる標的抗原結合特性を加工するために足場として使用されてきたタンパク質が挙げられ、ヒトγクリスタリンおよびヒトユビキチン(affilin)、ヒトプロテアーゼ阻害剤のクニッツ型ドメイン、Ras結合プロテインAF−6のPDZドメイン、サソリ毒(カリブドトキシン)、C型レクチンドメイン(tetranectin)が挙げられ、Chapter 7 − Non−Antibody Scaffolds from Handbook of Therapeutic Antibodies (2007, edited by Stefan Dubel)およびProtein Science 15:14−27 (2006)に概説されている。本発明のエピトープ結合ドメインは、これらの任意の代替タンパク質ドメインに由来し得る。
【0043】
本明細書で使用する「抗原結合部位」という用語は、抗原と特異的結合できるタンパク質上の部位を指し、これは単一ドメイン、例えばエピトープ結合ドメインであってもよく、または標準的な抗体上に見つけることができるような対になったVH/VLドメインであってもよい。本発明のいくつかの態様では、単鎖Fv(ScFv)ドメインは、抗原結合部位を提供できる。
【0044】
本明細書で使用する「mAbdAb」および「dAbmAb」という用語は、本発明の抗原結合タンパク質を指す。2つの用語は同義的に使用でき、本明細書で使用する場合に同じ意味を有することが意図される。
【0045】
本明細書で使用する「抗原結合タンパク質」という用語は、抗体、抗体断片、例えばドメイン抗体(dAb)、ScFv、FAb、FAb2、およびIL−13と結合できる他のタンパク質構築物を指す。抗原結合分子は、Ig可変ドメイン(例えば抗体、ドメイン抗体、Fab、Fab’、F(ab’)2、Fv、ScFv、二重特異性抗体、mAbdAb、Affibody、ヘテロ抱合体抗体もしくは二重特異性抗体)を少なくとも1つ含み得る。一実施形態では、抗原結合分子は抗体である。別の実施形態では、抗原結合分子はdAb、すなわち異なるV領域またはVドメインとは独立して抗原またはエピトープと特異的結合するVH、VHHまたはVLなどの免疫グロブリン単一可変ドメインである。抗原結合分子は、2つの標的と結合可能であり得る、すなわち二重標的タンパク質であり得る。抗原結合分子は、抗体および抗原結合断片(例えば、1つもしくは複数のドメイン抗体ならびに/またはモノクローナル抗体と結合した1つもしくは複数のScFvなど)の組み合わせであり得る。抗原結合分子は、IL−13への結合を達成するためにタンパク質工学にかけられた非Igドメイン、例えばCTLA−4(Evibody);リポカリン;プロテインA派生分子(プロテインAのZドメイン(Affibody、SpA)、Aドメイン(Avimer/Maxibody)など);熱ショックタンパク質(GroElおよびGroESなど);トランスフェリン(trans−body);アンキリン反復タンパク質(DARPin);ペプチドアプタマー;C型レクチンドメイン(Tetranectin);ヒトγクリスタリンおよびヒトユビキチン(affilin);PDZドメイン;ヒトプロテアーゼ阻害剤のサソリ毒クニッツ型ドメイン;ならびにフィブロネクチン(adnectin)からなる群から選択される足場の誘導体であるドメインも含み得る。本明細書で使用する「抗原結合タンパク質」はヒトIL−13を拮抗および/または中和できる。加えて、抗原結合タンパク質はIL−13と結合することによってIL−13活性をブロックし得、天然リガンドによる受容体への結合および/または受容体の活性化を防止し得る。
【0046】
本明細書で使用する「IL−13アンタゴニスト」は、IL−13の活性を少なくとも1つ低減およびまたは除去できる任意の化合物を含む。例として、IL−13アンタゴニストはIL−13と結合し、この結合はIL−13活性を直接低減または除去する場合もあり、リガンドの受容体結合を少なくとも1つブロックすることによって間接的に低減または除去する場合もある。
【0047】
本明細書で使用する「IL−4アンタゴニスト」は、IL−4の活性を少なくとも1つ低減およびまたは除去できる任意の化合物を含む。例として、IL−4アンタゴニストはIL−4と結合し、この結合はIL−4活性を直接低減または除去する場合もあり、リガンドの受容体結合を少なくとも1つブロックすることによって間接的に低減または除去する場合もある。
【0048】
本明細書で使用する「IL−5アンタゴニスト」は、IL−5の活性を少なくとも1つ低減およびまたは除去できる任意の化合物を含む。例として、IL−5アンタゴニストはIL−5と結合し、この結合はIL−5活性を直接低減または除去する場合もあり、リガンドの受容体結合を少なくとも1つブロックすることによって間接的に低減または除去する場合もある。
【図面の簡単な説明】
【0049】
【図1】TF−1細胞増殖アッセイにおいてY100B変異体を含むmAbdAbのヒトIL−13を中和能力を示すグラフ
【図2】TF−1細胞増殖アッセイにおいてY100B変異体を含むmAbdAbのヒトIL−13を中和能力を示すグラフ
【図3】BPC2222のSECトレース
【図4】BPC2223のSECトレース
【図5】BPC2230のSECトレース
【図6】BPC2231のSECトレース
【図7】BPC1085のSECトレース
【図8】BPC1086のSECトレース
【図9】BPC1087のSECトレース
【図10】ELISAにより決定した、精製されたmAbdAb(BPC1085、BPC1086およびBPC1087)のヒトIL−4への結合を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図11】TF−1細胞バイオアッセイにおいて、ヒトIL−4に対し精製されたmAbdAb(BPC1085、BPC1086およびBPC1087)によるヒトIL−4中和を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図12】TF−1細胞バイオアッセイにおいて、ヒトIL−13に対し精製されたmAbdAb(BPC1085、BPC1086およびBPC1087)によるヒトIL−13中和を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図13】BPC3214のSECプロファイル。
【図14】BPC3215のSECプロファイル。
【図15】ELISAにより決定した、精製されたmAbdAb BPC3214、BPC3215、BPC1085および対照mAb A1Y100BVAL1および抗IL−4 mAbのヒトIL−13への結合を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図16】ELISAにより決定した、精製されたmAbdAb BPC3214、BPC3215、BPC1085および対照mAb A1Y100BVAL1および抗IL−4 mAbのヒトIL−4への結合を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【図17】ELISAにより決定した、一過性発現したmAbdAbの組換え大腸菌発現ヒトIL−4への結合を示すグラフ。IL−4対照mAbは「パスコリズマブ」と表示する。
【発明を実施するための形態】
【0050】
一実施形態では、本発明の抗原結合タンパク質は、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8および配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14および配列番号15、配列番号16、配列番号17および配列番号18からなる一覧から選択されるCDRH3ならびに適切なCDRH1およびCDRH2を含む重鎖可変領域を含み、適切なCDRL1、CDRL2およびCDRL3を含む軽鎖可変領域と対をなして、ヒトIL−13と結合する抗原結合Fv単位を形成する。一実施形態では、本発明の抗原結合タンパク質は、ヒトIL−13活性を中和する。
【0051】
この実施形態の一態様では、配列番号1もしくは配列番号60、もしくは配列番号61に説明されるCDRH1ならびに配列番号2に説明されるCDRH2も重鎖可変領域に存在する。この実施形態のさらなる態様では、配列番号19に説明されるCDRHL1、配列番号20に説明されるCDRL2および配列番号21に説明されるCDRL3も軽鎖可変領域に存在する。
【0052】
別の態様では、抗原結合タンパク質は、Biacoreにより10nM以下、より特に2nM以下、例えば約0.8nM〜2nM、1nM以下、または100pM以下、例えば約20pM〜約100pMまたは約20pM〜約80pM、または約20pM〜約60pMと測定された高親和性でヒトIL−13と結合する。1つのかかる実施形態では、これは例えば実施例3に説明するように、バイオセンサーチップ上に捕捉された抗原結合タンパク質を用いてBiacoreにより測定される。
【0053】
本発明の重鎖可変領域はヒトIL−13と結合させる軽鎖可変領域と共に、従来の免疫グロブリン形態で(例えば、ヒトIgG、IgA、IgMなど)またはヒトIL−13との結合を可能にする他の任意の「抗体様」型式(例えば、単一鎖Fv、二重特異性抗体、Tandab(商標)など)において構成し得る(代替「抗体」型式の概要については、Holliger and Hudson, Nature Biotechnology, 2005, Vol 23, No. 9, 1126−1136を参照されたい)。
【0054】
本発明の抗原結合タンパク質は、配列番号58および配列番号59に記載の可変領域を有するマウス抗体、または非マウスのその等価物(ラット、ヒト、キメラまたはそのヒト化変異体など)に由来し、例えば配列番号22および配列番号24に記載の重鎖および軽鎖を有するヒト化抗体に由来する。
【0055】
本発明の一態様では、ヒトIL−13と結合し、CDRH3 SIYDDYHYDDYYAMDY(配列番号3)の変異体を含み、CDRH3が1つもしくは複数の以下の位置(Kabat番号付けを使用)で以下に説明する代替アミノ酸に置換されている抗原結合タンパク質(例えば抗体)を提供する:
a)1位のS95は、トリプトファン(W)と置換されている
b)2位のI96は、バリン(V)と置換されている
c)3位のY97は、フェニルアラニン(F)と置換されている
d)4位のD98は、グルタミン(E)と置換されている
e)7位のH100Aは、アラニン(A)、グルタミン酸(E)、グルタミン(Q)、アルギニン(R)、セリン(S)、トレオニン(T)もしくはバリン(V)と置換されている、ならびに
f)8位のY100Bは、アラニン(A)、イソロイシン、(I)、トリプトファン(W)もしくはバリン(V)と置換されている。
【0056】
本発明の別の態様では、ヒトIL−13と結合し、配列番号3に説明されるCDRH3を含む抗原結合タンパク質(例えば抗体)を提供し、このCDRH3は、S95W、I96V、Y97F、D98E、H100A_A、H100A_E、H100A_Q、H100A_R、H100A_S、H100A_T、H100A_V、Y100B_A、Y100B_I、Y100B_W、およびY100B_Vの置換を1つまたは複数含む。
【0057】
一態様では、本発明の抗原結合タンパク質(例えば本発明の抗体)は、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8および配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14および配列番号15、配列番号16、配列番号17および配列番号18に説明されるものから選択されるCDRH3配列を含む。かかる抗原結合タンパク質はさらに以下のCDR配列
CDRH1:配列番号1、60および61から選択される、
CDRH2:配列番号2、
CDRL1:配列番号19、
CDRL2:配列番号20、ならびに
CDRL3:配列番号21
を含み得る。
【0058】
一実施形態では、抗原結合タンパク質は、以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号18、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21
を含む。
【0059】
別の実施形態では、抗原結合タンパク質は、以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号17、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21
を含む。
【0060】
別の実施形態では、抗原結合タンパク質は、以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号16、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21
を含む。
【0061】
別の実施形態では、抗原結合タンパク質は、以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号15、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21
を含む。
【0062】
本明細書を通して、抗体配列中のアミノ酸残基は、Kabatスキームに従い番号付けされる。同様に、「CDR」、「CDRL1」、「CDRL2」、「CDRL3」、「CDRH1」、「CDRH2」、「CDRH3」(例えば配列番号60および61に説明されるCDRH3)という用語は、別に示されない限り、Kabat et al; “Sequences of proteins of Immunological Interest” NIH, 1987に説明されるようにKabat番号付け系に従う。
【0063】
本発明の別の態様では、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号88、配列番号89、配列番号90および配列番号91から選択される配列を有する重鎖;ならびに配列番号24の軽鎖を含む抗原結合タンパク質(ヒト化抗体またはその抗原結合断片など)を提供する。
【0064】
本発明は、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号88、配列番号89、配列番号90から選択される重鎖を含む抗原結合タンパク質(ヒト化抗体またはその抗原結合断片など)を提供する。
【0065】
本発明は、配列番号108、配列番号110、配列番号112および配列番号114から選択される軽鎖を含む抗原結合タンパク質(ヒト化抗体またはその抗原結合断片など)も提供する。
【0066】
本発明はさらに、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号88、配列番号89、配列番号90および配列番号91から選択される配列を有する重鎖;ならびに配列番号108、配列番号110、配列番号112および配列番号114の軽鎖を含む抗原結合タンパク質(ヒト化抗体またはその抗原結合断片など)を提供する。
【0067】
一実施形態では、本発明の抗原結合タンパク質は、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号88、配列番号89、配列番号90および配列番号91から選択される重鎖、ならびに配列番号24、配列番号108、配列番号110、配列番号112および配列番号114から選択される軽鎖を含む抗体を含む。
【0068】
一実施形態では、本発明の抗原結合タンパク質は、配列番号48、配列番号50、配列番号52および配列番号54から選択される重鎖;ならびに配列番号24、配列番号108、配列番号110、配列番号112および配列番号114から選択される軽鎖を含み、例えば抗原結合タンパク質は配列番号48の重鎖および配列番号24の軽鎖、または配列番号50の重鎖および配列番号24の軽鎖、または配列番号52の重鎖および配列番号24の軽鎖、または配列番号54の重鎖および配列番号24の軽鎖、または配列番号88の重鎖および配列番号24の軽鎖または配列番号89の重鎖および配列番号24の軽鎖、または配列番号90の重鎖および配列番号24の軽鎖、または配列番号91の重鎖および配列番号24の軽鎖、または配列番号48の重鎖および配列番号108の軽鎖、または配列番号50の重鎖および配列番号108の軽鎖、または配列番号52の重鎖および配列番号108の軽鎖、または配列番号54の重鎖および配列番号108の軽鎖、または配列番号88の重鎖および配列番号108の軽鎖または配列番号89の重鎖および配列番号108の軽鎖、または配列番号90の重鎖および配列番号108の軽鎖、または配列番号91の重鎖および配列番号108の軽鎖、または配列番号48の重鎖および配列番号110の軽鎖、または配列番号50の重鎖および配列番号110の軽鎖、または配列番号52の重鎖および配列番号110の軽鎖、または配列番号54の重鎖および配列番号110の軽鎖、または配列番号88の重鎖および配列番号110の軽鎖または配列番号89の重鎖および配列番号110の軽鎖、または配列番号90の重鎖および配列番号110の軽鎖、または配列番号91の重鎖および配列番号110の軽鎖を含む。
【0069】
1つのかかる実施形態では、本発明の抗原結合タンパク質は、配列番号48、配列番号50、配列番号52および配列番号54から選択される重鎖;ならびに配列番号108および配列番号110から選択される軽鎖(例えば配列番号48の重鎖および配列番号108の軽鎖を含む抗原結合タンパク質、または配列番号48の重鎖および配列番号110の軽鎖を含む抗原結合タンパク質、または配列番号50の重鎖および配列番号108の軽鎖を含む抗原結合タンパク質、または配列番号50の重鎖および配列番号110の軽鎖を含む抗原結合タンパク質、または配列番号52の重鎖および配列番号108の軽鎖を含む抗原結合タンパク質、または配列番号52の重鎖および配列番号110の軽鎖を含む抗原結合タンパク質、または配列番号54の重鎖および配列番号108の軽鎖を含む抗原結合タンパク質、または配列番号54の重鎖および配列番号110の軽鎖を含む抗原結合タンパク質)を含む。
【0070】
本発明のIL−13抗体はIL−4および/もしくはIL−5アンタゴニスト、例えばIL−4抗体もしくはエピトープ結合ドメイン、ならびに/またはIL−5抗体もしくはエピトープ結合ドメインとの組み合わせであり得る。これらは、同時に投与する、すなわち同時投与するか、または互いに24時間以内(例えば互いに20時間以内、または15時間以内または12時間以内、または10時間以内、または8時間以内、または6時間以内、または4時間以内、または2時間以内、または1時間以内、または30分以内)に投与する分離する分子の混合物として投与し得る。
【0071】
さらなる実施形態では、アンタゴニストは、2つ以上の抗原と結合できる1分子として存在し、例えば本発明は、IL−13と結合でき、IL−4へも結合できるまたはIL−5へも結合できる、またはIL−4とIL−5へも結合できる本発明のIL−13抗体を含む抗原結合タンパク質を提供する。
【0072】
一実施形態では、本発明の抗原結合タンパク質は、本発明の少なくともCDRH3、ならびに任意選択的に1つもしくは複数のCDRH1、CDRH2、CDRL1、CDRL2、およびCDRL3を含み、IL−13と結合でき、1つもしくは複数のIL−4もしくはIL−5へも結合できる多特異的な抗体であり得る。1つのかかる実施形態では、CDRH3、すなわち本明細書において定義される抗原結合タンパク質を含み、IL−4、またはIL−5と結合できる抗原結合部位をさらに含む多特異的な抗体を提供する。
【0073】
本発明の抗原結合タンパク質の1例は、IL−4もしくはIL−5に特異性の1つもしくは複数のエピトープ結合ドメインと結合している、本明細書において定義されるCDRH1、CDRH2、CDRH3、CDRL1、CDRL2、およびCDRL3を含むIL−13に特異的な抗体、例えばIL−13とIL−4もしくはIL−13とIL−5と結合できる二重特異性抗原結合タンパク質、またはIL−13、IL−4およびIL−5と結合できる三重特異性抗原結合タンパク質である。
【0074】
本明細書に記載の任意の抗原結合タンパク質は、例えば、実施例8に記載したものなどの適切なアッセイを用いて化学量論分析により決定されるように2つ以上の抗原に同時と結合可能であり得ることが理解されるであろう。
【0075】
本発明は、1つまたは複数のエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、例えばIL−4と結合できるエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、またはIL−5と結合できるエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質、またはIL−4と結合できる第1のエピトープ結合ドメインおよびIL−5と結合できる第2のエピトープ結合ドメインと結合している本発明のIL−13抗体を含む抗原結合タンパク質も提供する。
【0076】
エピトープ結合ドメインは、IL−13抗体重鎖のC末端もしくはN末端またはIL−13抗体軽鎖のC末端もしくはN末端と結合し得る。
【0077】
本発明の抗体はリンカーを用いることによりエピトープ結合ドメインと結合し得る。適切なリンカーの例としては、1個のアミノ酸〜150個のアミノ酸の長さ、または1個のアミノ酸〜140個のアミノ酸、例えば、1個のアミノ酸〜130個のアミノ酸、または1〜120個のアミノ酸、または1〜80個のアミノ酸、または1〜50個のアミノ酸、または1〜20個のアミノ酸、または1〜10個のアミノ酸、または5〜18個のアミノ酸であり得るアミノ酸配列が挙げられる。かかる配列はそれら自体の三次構造を有し得る、例えば、本発明のリンカーは単一可変ドメインを含み得る。リンカーのサイズは一実施形態では、単一可変ドメインと等価物である。適切なリンカーのサイズは、1〜20オングストローム、例えば15オングストローム未満、または10オングストローム未満、または5オングストローム未満であり得る。
【0078】
本発明の一実施形態では、少なくとも1つのエピトープ結合ドメインは、1〜150個のアミノ酸、例えば1〜20個のアミノ酸、例えば1〜10個のアミノ酸を含むリンカーでIL−13抗体と直接結合している。
【0079】
かかるリンカーは、配列番号82〜87、92〜93に説明されるものの任意の1つまたはかかるリンカーの複数から選択され得る。
【0080】
本発明の抗原結合タンパク質において使用するリンカーは、単独または他のリンカーに加えて、1つまたは複数のGS残基セット、例えば「GSTVAAPS」(配列番号92)または「TVAAPSGS」(配列番号87)または「GSTVAAPSGS」(配列番号93)を含み得る。一実施形態では、リンカーは、配列番号83を含む。
【0081】
一実施形態では、エピトープ結合ドメインはリンカー「(PAS)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(GGGGS)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(TVAAPS)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(GS)m(TVAAPSGS)n」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(PAVPPP)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(TVSDVP)n(GS)m」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「(TGLDSP)n(GS)m」によりIL−13抗体と結合している。かかる実施形態のすべてにおいて、n=1〜10、およびm=0〜4である。
【0082】
かかるリンカーの例としては、(PAS)n(GS)m(式中n=1およびm=1)、(PAS)n(GS)m(式中n=2およびm=1)、(PAS)n(GS)m(式中n=3およびm=1)、(PAS)n(GS)m(式中n=4およびm=1)、(PAS)n(GS)m(式中n=2およびm=0)、(PAS)n(GS)m(式中n=3およびm=0)、(PAS)n(GS)m式中n=4およびm=0が挙げられる。
【0083】
かかるリンカーの例としては、(GGGGS)n(GS)m(式中n=1およびm=1)、(GGGGS)n(GS)m(式中n=2およびm=1)、(GGGGS)n(GS)m(式中n=3およびm=1)、(GGGGS)n(GS)m(式中n=4およびm=1)、(GGGGS)n(GS)m(式中n=2およびm=0)、(GGGGS)n(GS)m(式中n=3およびm=0)、(GGGGS)n(GS)m式中n=4およびm=0が挙げられる。
【0084】
かかるリンカーの例としては、(TVAAPS)n(GS)m式中n=1およびm=1(配列番号87)、(TVAAPS)n(GS)m式中n=2およびm=1(配列番号145)、(TVAAPS)n(GS)m式中n=3およびm=1(配列番号146)、(TVAAPS)n(GS)m(式中n=4およびm=1)、(TVAAPS)n(GS)m(式中n=2およびm=0)、(TVAAPS)n(GS)m(式中n=3およびm=0)、(TVAAPS)n(GS)m式中n=4およびm=0が挙げられる。
【0085】
かかるリンカーの例としては、(GS)m(TVAAPSGS)n式中n=1およびm=1(配列番号139)、(GS)m(TVAAPSGS)n式中n=2およびm=1(配列番号140)、(GS)m(TVAAPSGS)n式中n=3およびm=1(配列番号141)、または(GS)m(TVAAPSGS)n式中n=4およびm=1(配列番号142)、(GS)m(TVAAPSGS)n式中n=5およびm=1(配列番号143)、(GS)m(TVAAPSGS)n式中n=6およびm=1(配列番号144)、(GS)m(TVAAPSGS)n式中n=1およびm=0(配列番号87)、(GS)m(TVAAPSGS)n(式中n=2およびm=10)、(GS)m(TVAAPSGS)n(式中n=3およびm=0)、または(GS)m(TVAAPSGS)n式中n=0が挙げられる。
【0086】
かかるリンカーの例としては、(PAVPPP)n(GS)m(式中n=1およびm=1)、(PAVPPP)n(GS)m式中n=2およびm=1(配列番号65)、(PAVPPP)n(GS)m(式中n=3およびm=1)、(PAVPPP)n(GS)m(式中n=4およびm=1)、(PAVPPP)n(GS)m(式中n=2およびm=0)、(PAVPPP)n(GS)m(式中n=3およびm=0)、(PAVPPP)n(GS)m式中n=4およびm=0が挙げられる。
【0087】
かかるリンカーの例としては、(TVSDVP)n(GS)m式中n=1およびm=1(配列番号67)、(TVSDVP)n(GS)m(式中n=2およびm=1)、(TVSDVP)n(GS)m(式中n=3およびm=1)、(TVSDVP)n(GS)m(式中n=4およびm=1)、(TVSDVP)n(GS)m(式中n=2およびm=0)、(TVSDVP)n(GS)m(式中n=3およびm=0)、(TVSDVP)n(GS)m式中n=4およびm=0が挙げられる。
【0088】
かかるリンカーの例としては、(TGLDSP)n(GS)m(式中n=1およびm=1)、(TGLDSP)n(GS)m(式中n=2およびm=1)、(TGLDSP)n(GS)m(式中n=3およびm=1)、(TGLDSP)n(GS)m(式中n=4およびm=1)、(TGLDSP)n(GS)m(式中n=2およびm=0)、(TGLDSP)n(GS)m(式中n=3およびm=0)、(TGLDSP)n(GS)m式中n=4およびm=0が挙げられる。
【0089】
別の実施形態では、エピトープ結合ドメインとIL−13抗体の間にリンカーはない。別の実施形態では、エピトープ結合ドメインはリンカー「TVAAPS」(配列番号83)によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「TVAAPSGS」(配列番号87)によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「GS」によりIL−13抗体と結合している。別の実施形態では、エピトープ結合ドメインはリンカー「ASTKGPT」(配列番号84)によりIL−13抗体と結合している。
【0090】
本発明において使用するエピトープ結合ドメインは、異なるV領域またはVドメインとは独立して抗原またはエピトープと特異的結合するドメインであり、これはドメイン抗体であってもよいし、天然リガンド以外のリガンドへの結合を達成するためにタンパク質工学にかけられた非免疫グロブリンドメイン、例えばCTLA−4(Evibody);リポカリン;プロテインA派生分子(プロテインAのZドメイン(Affibody、SpA)、Aドメイン(Avimer/Maxibody)など);熱ショックタンパク質(GroElおよびGroESなど);トランスフェリン(trans−body);アンキリン反復タンパク質(DARPin);ペプチドアプタマー;C型レクチンドメイン(Tetranectin);ヒトγクリスタリンおよびヒトユビキチン(affilin);PDZドメイン;ヒトプロテアーゼ阻害剤のサソリ毒クニッツ型ドメイン;ならびにフィブロネクチン(adnectin)からなる群から選択される非免疫グロブリン足場の誘導体であるドメインであってもよい。一実施形態では、これはドメイン抗体または他の適切なドメイン(CTLA−4、リポカリン、SpA、Affibody、avimer、GroEl、トランスフェリン、GroESおよびフィブロネクチンからなる群から選択されるドメインなど)であり得る。一実施形態では、これは免疫グロブリン単一可変ドメイン、Affibody、アンキリン反復タンパク質(DARPin)およびadnectinから選択し得る。別の実施形態では、これはAffibody、アンキリン反復タンパク質(DARPin)およびadnectinから選択し得る。別の実施形態では、これはドメイン抗体、例えばヒト、ラクダ(ナノボディ)、またはサメ(NARV)ドメイン抗体から選択されるドメイン抗体であり得る。
【0091】
かかる抗原結合タンパク質の例としては、重鎖のC末端もしくはN末端またはC末端と結合するIL−4アンタゴニストであるエピトープ結合ドメインを有する本発明のIL−13抗体が挙げられる。例としては、配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号98、配列番号100、配列番号102、配列番号104、配列番号106または配列番号117〜138に説明される重鎖配列、ならびに配列番号24、配列番号108、配列番号110、配列番号112もしくは配列番号114に説明される軽鎖配列を含む抗原結合タンパク質が挙げられ、重鎖および軽鎖の片方または両方はさらにIL−4と拮抗できる1つまたは複数のエピトープ結合ドメイン、例えばIL−4と結合できる単一可変ドメインを含む。かかるエピトープ結合ドメインは、配列番号78〜81および94に説明されるものから選択できる。
【0092】
一実施形態では、本発明の抗原結合構築物は、配列番号62の重鎖配列および配列番号24の軽鎖配列、または配列番号64の重鎖配列および配列番号24の軽鎖配列、または配列番号66の重鎖配列および配列番号24の軽鎖配列、または配列番号68の重鎖配列および配列番号24の軽鎖配列、または配列番号70の重鎖配列および配列番号24の軽鎖配列、または配列番号72の重鎖配列および配列番号24の軽鎖配列、または配列番号74の重鎖配列および配列番号24の軽鎖配列、または配列番号76の重鎖配列および配列番号24の軽鎖配列を含む。
【0093】
一実施形態では、本発明の抗原結合構築物は、配列番号94の重鎖配列および配列番号24の軽鎖配列、または配列番号96の重鎖配列および配列番号24の軽鎖配列、または配列番号98の重鎖配列および配列番号24の軽鎖配列、または配列番号100の重鎖配列および配列番号24の軽鎖配列、または配列番号102の重鎖配列および配列番号24の軽鎖配列、または配列番号104の重鎖配列および配列番号24の軽鎖配列、または配列番号106の重鎖配列および配列番号24の軽鎖配列を含む。
【0094】
一実施形態では、本発明の抗原結合構築物は、配列番号62の重鎖配列および配列番号108の軽鎖配列、または配列番号64の重鎖配列および配列番号108の軽鎖配列、または配列番号66の重鎖配列および配列番号108の軽鎖配列、または配列番号68の重鎖配列および配列番号108の軽鎖配列、または配列番号70の重鎖配列および配列番号108の軽鎖配列、または配列番号72の重鎖配列および配列番号108の軽鎖配列、または配列番号74の重鎖配列および配列番号108の軽鎖配列、または配列番号76の重鎖配列および配列番号108の軽鎖配列、または配列番号62の重鎖配列および配列番号110の軽鎖配列、または配列番号64の重鎖配列および配列番号110の軽鎖配列、または配列番号66の重鎖配列および配列番号110の軽鎖配列、または配列番号68の重鎖配列および配列番号110の軽鎖配列、または配列番号70の重鎖配列および配列番号110の軽鎖配列、または配列番号72の重鎖配列および配列番号110の軽鎖配列、または配列番号74の重鎖配列および配列番号110の軽鎖配列、または配列番号76の重鎖配列および配列番号110の軽鎖配列を含む。
【0095】
一実施形態では、本発明の抗原結合構築物は、配列番号96の重鎖配列および配列番号108の軽鎖配列、または配列番号98の重鎖配列および配列番号108の軽鎖配列、または配列番号100の重鎖配列および配列番号108の軽鎖配列、または配列番号102の重鎖配列および配列番号108の軽鎖配列、または配列番号104の重鎖配列および配列番号108の軽鎖配列、または配列番号106の重鎖配列および配列番号108の軽鎖配列、または配列番号96の重鎖配列および配列番号110の軽鎖配列、または配列番号98の重鎖配列および配列番号110の軽鎖配列、または配列番号100の重鎖配列および配列番号110の軽鎖配列、または配列番号102の重鎖配列および配列番号110の軽鎖配列、または配列番号104の重鎖配列および配列番号110の軽鎖配列、または配列番号106の重鎖配列および配列番号110の軽鎖配列を含む。
【0096】
一実施形態では、IL−13抗体の重鎖は、配列番号48、配列番号50、配列番号52および配列番号54に説明されるものから選択される。別の実施形態では、重鎖は配列番号88〜91、配列番号96〜106、および配列番号117〜138に説明されるものから選択される。1つのかかる実施形態では、重鎖は配列番号96、配列番号98、配列番号100、配列番号102、配列番号104および配列番号106から選択される。
【0097】
一実施形態では、抗原結合タンパク質は、IL−5アンタゴニストであるエピトープ結合ドメインと結合した抗IL−13抗体を含み、この抗IL−13抗体は配列番号3〜18、例えば配列番号15〜18に説明されるものおよび配列番号24の軽鎖配列から選択されるCDRH3を含む。
【0098】
例としては、配列番号26、28、30、32、34、36、38、40、42、44、46、48、50、52もしくは54に説明される重鎖配列ならびに配列番号24に説明される軽鎖配列を含み、重鎖および軽鎖の片方または両方がさらにIL−5と拮抗できる1つまたは複数のエピトープ結合ドメイン、例えばIL−5と結合できる免疫グロブリン単一可変ドメインを含む抗原結合タンパク質が挙げられる。
【0099】
さらなる実施形態では、抗原結合タンパク質は、配列番号26、28、30、32、34、36、38、40、42、44、46、48、50、52もしくは54に説明される重鎖配列ならびに配列番号24に説明される軽鎖配列を含み、この重鎖および軽鎖の片方または両方はさらにIL−4と拮抗できる1つまたは複数のエピトープ結合ドメイン(例えばIL−4と結合できる免疫グロブリン単一可変ドメイン)、およびIL−5と拮抗できる1つまたは複数のエピトープ結合ドメイン(例えばIL−5と結合できる免疫グロブリン単一可変ドメイン)を含む。
【0100】
一実施形態では、本発明の抗原結合タンパク質は、ヒト血清アルブミンと結合できるエピトープ結合ドメインを少なくとも1つ含む。
【0101】
一実施形態では、少なくとも3つの抗原結合部位がある、例えば4、または5または6または8または10個の抗原結合部位があり、抗原結合タンパク質は少なくとも3または4または5または6または8または10個の抗原と結合できる、例えば抗原結合タンパク質は、3または4または5または6または8または10個の抗原と同時に結合できる。
【0102】
一実施形態では、第1のエピトープ結合ドメインはタンパク質足場と結合しており、第2のエピトープ結合ドメインは第1のエピトープ結合ドメインと結合している。例えばタンパク質足場がIgG足場である場合、第1のエピトープ結合ドメインはIgG足場の重鎖のC末端と結合していてもよく、そのエピトープ結合ドメインはそのC末端で第2のエピトープ結合ドメインと結合でき、または例えば第1のエピトープ結合ドメインはIgG足場の軽鎖のC末端と結合していてもよく、第1のエピトープ結合ドメインはさらにそのC末端で第2のエピトープ結合ドメインと結合していてもよいし、または例えば第1のエピトープ結合ドメインはIgG足場の軽鎖のN末端と結合していてもよく、第1のエピトープ結合ドメインはさらにそのN末端で第2のエピトープ結合ドメインと結合していてもよいし、または例えば第1のエピトープ結合ドメインは、IgG足場の重鎖のN末端と結合していてもよく、第1のエピトープ結合ドメインはさらにそのN末端で第2のエピトープ結合ドメインと結合していてもよい。
【0103】
エピトープ結合ドメインがドメイン抗体である場合、一部のドメイン抗体は足場内の特定の位置に適合し得る。
【0104】
本発明において使用する免疫グロブリン単一可変ドメインは、IL−13 mAbの重鎖および/または軽鎖のC末端と結合できる。加えて一部の免疫グロブリン単一可変ドメインは、従来の抗体の重鎖および軽鎖の両C末端と結合できる。
【0105】
免疫グロブリン単一可変ドメインのN末端が抗体定常ドメイン(CH3またはCLのいずれか)に融合している構築物において、ペプチドリンカーは免疫グロブリン単一可変ドメインが抗原と結合するのを補助し得る。実際、dAbのN末端は、抗原結合活性に関与するCDRに近接して位置する。したがって短いペプチドリンカーは、エピトープ結合ドメインと抗体の定常ドメイン間のスペーサーとして働き、これはdAb CDRがより容易に抗原に達することを可能にし得るため、高親和性で結合し得る。
【0106】
免疫グロブリン単一可変ドメインがIgGと結合する環境は、それらがどの抗体鎖へ融合しているかによって異なる:
抗体軽鎖のC末端で融合している場合、各免疫グロブリン単一可変ドメインは、抗体ヒンジおよびFc部分に近接して位置することが予期される。かかる免疫グロブリン単一可変ドメインは互いに遠く離れて位置している可能性が高い。従来の抗体では、Fab断片間の角度および各Fab断片とFc部分間の角度は非常に著しく変わり得る。mAbdAbと、Fab断片間の角度は著しく異なることはないであろうが、各Fab断片とFc部分間の角度にはいくらか制限が観察され得る。
【0107】
抗体重鎖のC末端で融合している場合、各免疫グロブリン単一可変ドメインは、Fc部分のCH3ドメインに近接して位置することが予期される。これはFc受容体(例えばFcγRI、II、III、FcRn)に対するFc結合特性に影響を与えることは予期されない。なぜなら、これらの受容体はCH2ドメイン(FcγRI、IIおよびIIIクラスの受容体について)またはCH2ドメインとCH3ドメイン間のヒンジ(例えばFcRn受容体)と結合するためである。かかる抗原結合タンパク質はさらに、両免疫グロブリン単一可変ドメインが空間的に互いに近いことを特徴とし、適切なリンカーの供給によって柔軟性が得られる場合、これらの免疫グロブリン単一可変ドメインはホモ二量体種さえも形成し得るため、Fc部分の「ジッパー型」四次構造を増やして構築物の安定性を高め得ることが予期される。
【0108】
かかる構造に関する考察は、抗体にエピトープ結合ドメイン、例えば免疫グロブリン単一可変ドメインを結合させるために最も適した位置の選択に役立つことができる。
【0109】
抗原のサイズ、その局在性(血中または細胞表面上)、その四次構造(単量体または多量体)は変わり得る。従来の抗体は、ヒンジ領域の存在のためにアダプター構築物として機能するように当然設計され、Fab断片の先端の2つの抗原結合部位の配向は広範囲に変わり得るため、抗原の分子特性およびその環境に適応し得る。一方、ヒンジ領域のない抗体と結合した免疫グロブリン単一可変ドメインは、直接的にも間接的にも構造的な柔軟性に欠け得る。
【0110】
溶液状態および免疫グロブリン単一可変ドメインへの結合形態の理解も役立つ。インビトロのヒトdAbは大部分が溶液中に単量体、ホモ二量体または多量体形態で存在し得るという証拠が増えている(Reiter et al. (1999) J Mol Biol 290 p685−698; Ewert et al (2003) J Mol Biol 325, p531−553, Jespers et al (2004) J Mol Biol 337 p893−903; Jespers et al (2004) Nat Biotechnol 22 p1161−1165; Martin et al (1997) Protein Eng. 10 p607−614; Sepulvada et al (2003) J Mol Biol 333 p355−365)。これは、免疫グロブリン軽鎖の二量体であるベンス−ジョーンズタンパク質(Epp et al (1975) Biochemistry 14 p4943−4952; Huan et al (1994) Biochemistry 33 p14848−14857; Huang et al (1997) Mol immunol 34 p1291−1301)およびアミロイド線維(James et al. (2007) J Mol Biol. 367:603−8)などのIgドメインでインビボにおいて観察される多量体化事象を十分に暗示する。
【0111】
例えば、溶液中で二量体化する傾向があるdAbは、軽鎖C末端よりもFc部分のC末端と結合させることが本発明の抗原結合タンパク質との関連で所望され得る。なぜなら、FcのC末端への結合はそれらのdAbの二量体化を可能にさせるためである。
【0112】
本発明の抗原結合タンパク質は、単一抗原に特異的な抗原結合部位を含んでいてもよいし、2つ以上の抗原もしくは単一抗原上の2つ以上のエピトープに特異的な抗原結合部位を有してもよく、またはそれぞれの抗原結合部位が同じもしくは異なる抗原上の異なるエピトープに特異的である抗原結合部位があってもよい。
【0113】
本発明の抗原結合タンパク質は、天然抗体または機能的断片またはその等価物の構造内に構成し得る本発明の重鎖可変領域および軽鎖可変領域を含み得る。したがって本発明の抗原結合タンパク質は、適切な軽鎖と対をなす場合、完全長抗体、(Fab」)2断片、Fab断片、またはその等価物(scFV、二量体、三量体、または四量体、Tandabなど)内に形成された本発明のVH領域を含み得る。抗体は、IgG1、IgG2、IgG3、もしくはIgG4;またはIgM;IgA、IgEもしくはIgDまたはその修飾変異型であり得る。それに応じて抗体重鎖の定常ドメインを選択し得る。軽鎖定常ドメインは、κ定常ドメインであってもλ定常ドメインであってもよい。さらに、抗原結合タンパク質は、すべてのクラスの修飾、例えばIgG二量体、Fc受容体と結合しなくなったまたはC1q結合を媒介しなくなったFc変異体を含み得る。抗原結合タンパク質は、抗原結合領域および非免疫グロブリン領域を含むWO86/01533号に記載されている種類のキメラ抗体でもあり得る。
【0114】
定常領域は、必要な任意の機能性に応じて選択される。IgG1は、補体結合を介して溶解能を示し得るおよび/またはADCC(抗体依存性細胞傷害)を媒介し得る。非細胞傷害性ブロック抗体が必要である場合、IgG4を使用できる。しかしながら、IgG4抗体は産生において不安定性を示し得るため、代替的に、一般にさらに安定したIgG1に修飾する。推奨される修飾は、欧州特許第0307434号に記載されている(例えば235位および237位の変異)。したがって本発明は、本発明による抗原結合タンパク質(例えば抗体)の溶解または非溶解形態を提供する。
【0115】
ある形態では、本発明の抗体は本明細書に記載の任意の重鎖可変領域を有する完全長(例えばH2L2四量体)溶解または非溶解IgG1抗体である。さらなる態様では、本発明は、本明細書に記載の軽鎖および重鎖可変領域をコードするポリヌクレオチドを提供する。
【0116】
本発明の一実施形態では、抗原結合部位はBiacore(商標)の測定に基づき少なくとも約1mMのKdで抗原と結合する、例えば少なくとも約10nM、少なくとも約1nM、少なくとも約500pM、少なくとも約200pM、少なくとも約100pM、または少なくとも約50pMのKdで各抗原と結合する。
【0117】
本発明の一実施形態では、抗原結合部位はBiacore(商標)の測定に基づき少なくとも約1mMのKdで抗原と結合する、例えば少なくとも約10nM、少なくとも約1nM、少なくとも約500pM、少なくとも約200pM、少なくとも約100pM、または少なくとも約50pMのKdで各抗原と結合する。
【0118】
一実施形態では、本発明は、Biacoreの測定に基づき、ヒトIL−13に対して抗IL=13抗体よりも少なくとも2倍高い親和性、または少なくとも4倍高い親和性、または少なくとも6倍高い親和性、または少なくとも8倍高い親和性、または少なくとも10倍高い親和性を有する、配列番号22に説明される重鎖配列および配列番号24に説明される軽鎖配列を含む抗原結合タンパク質を提供する。
【0119】
本明細書を通して使用される、本発明の抗原結合タンパク質に関する「中和」という用語およびその文法的変形は、本発明の抗原結合タンパク質の存在下においてIL−13の生体活性が、かかる抗原結合タンパク質の不在下のIL−13の活性よりも完全にまたは部分的に低下することを意味する。中和は、これらに限定されないが、1つまたは複数のリガンド結合をブロックし、リガンドによる受容体活性化を防止し、IL−13受容体をダウンレギュレートし、またはエフェクター機能に影響を与えたために起こり得る。中和レベルはいくつかの方法において、例えば以下の実施例に説明されるアッセイ、例えばTF1アッセイ(例えば実施例4に記載のように実行し得る)を用いて測定できる。このアッセイにおけるIL−13、IL−4またはこれら両サイトカインの中和は、中和する抗原結合タンパク質の存在下、TF1細胞増殖の阻害を評価することによって測定する。
【0120】
例えば、中和する抗原結合タンパク質の存在下でIL−13とその受容体間の低減した結合を評価することによる他の中和の評価方法は当該分野において知られており、例えば、Biacoreアッセイなどが挙げられる。
【0121】
本発明の代替態様では、本明細書において例示した抗体、例えば実施例4に説明するように実行できるTF1細胞増殖アッセイにおけるA1Y100BAlaL1、A1Y100BIleL1、A1 Y100BTrpL1またはA1Y100BValL1の中性活性を保持する抗原結合タンパク質と少なくとも実質的に等しい中性活性を有する抗原結合タンパク質を提供する。
【0122】
抗原結合タンパク質(例えば本発明の抗体)は、本発明の抗原結合タンパク質コード配列を含む発現ベクターを用いた宿主細胞のトランスフェクションにより産生し得る。発現ベクターまたは組換えプラスミドは、宿主細胞における複製および発現ならびに/または宿主細胞からの分泌を制御できる従来の調節制御配列との操作可能に結合して抗原結合タンパク質に対するこれらのコード配列を配置することにより産生される。調節配列としては、プロモーター配列、例えば、CMVプロモーター、および他の既知の抗体に由来し得るシグナル配列が挙げられる。同様に、相補的な抗原結合タンパク質軽鎖または重鎖をコードするDNA配列を有する第2の発現ベクターを産生できる。ある実施形態では、この第2の発現ベクターは、各ポリペプチド鎖が機能的に発現することを可能な限り確実にするためにコード配列および選択マーカーが考慮される点を除き第1の発現ベクターと同一である。あるいは、抗原結合タンパク質の重鎖および軽鎖コード配列は単一ベクター上に存在し得る。選択された宿主細胞を第1ベクターと第2ベクターの両方を従来技術により共トランスフェクトして(または単一ベクターにより単にトランスフェクトして)、組換えまたは合成の軽鎖および重鎖の両方を含むトランスフェクトされた本発明の宿主細胞を作製する。次いでトランスフェクトされた細胞を従来技術により培養し、本発明の加工された抗原結合タンパク質を産生する。組換え重鎖および/または軽鎖の両結合を含む抗原結合タンパク質は、適切なアッセイ(ELISAまたはRIAなど)により培養からスクリーニングされる。他の抗原結合タンパク質を構築するために類似の従来技術を適用し得る。
【0123】
本発明の組成物の方法および構築において使用されるクローニング工程およびサブクローニング工程に適したベクターは、当業者によって選択され得る。例えば、クローニングベクターの従来のpUCシリーズを使用してよい。1つのベクターであるpUC19は、Amersham(Buckinghamshire, United Kingdom)またはPharmacia (Uppsala, Sweden)などの供給源から市販されている。さらに、容易に複製でき、豊富なクローニング部位および選択可能な遺伝子(例えば、抗生剤耐性)を有し、容易に操作される任意のベクターをクローニングに使用してよい。したがって、クローニングベクターの選択は、本発明における限定要素ではない。
【0124】
発現ベクターは、異種DNA配列の発現を増幅するのに適した遺伝子、例えば、哺乳動物ジヒドロ葉酸還元酵素遺伝子(DHFR)も特徴とし得る。他のベクター配列としては、ウシ成長ホルモン(BGH)由来などのポリAシグナル配列およびベータグロビンプロモーター配列(betaglopro)が挙げられる。本明細書において有用な発現ベクターは、当業者にとって周知の技術により合成され得る。
【0125】
かかるベクターの成分、例えばレプリコン、選択遺伝子、エンハンサー、プロモーター、シグナル配列などは、市販または天然の供給源から得てもよいし、選択された宿主において組換えDNA生成物の発現および/または分泌の対象における使用のための既知の手段によって合成してもよい。当該分野において知られている、哺乳動物、細菌、昆虫、酵母、および真菌の発現に適した他の多くの種類の発現ベクターもこの目的のために選択してよい。
【0126】
本発明は、本発明の抗原結合タンパク質のコード配列を含む組換えプラスミドを用いてトランスフェクトされた細胞系も包含する。これらのクローニングベクターのクローニングおよび他の操作に有用な宿主細胞も従来のものである。しかしながら、様々な大腸菌株由来の細胞を、クローニングベクターの複製および本発明の抗原結合タンパク質の構築における他の工程に使用してよい。
【0127】
本発明の抗原結合タンパク質の発現に適した宿主細胞または細胞系としては、NS0、Sp2/0、CHO(例えばDG44)、COS、HEK、線維芽細胞(例えば、3T3)、および骨髄腫細胞などの哺乳動物細胞が挙げられ、例えばCHOまたは骨髄腫細胞において発現し得る。分子がヒトグリコシル化パターンで修飾されることを可能にするようにヒト細胞を使用してよい。あるいは、他の真核細胞系を適用し得る。形質転換、培養、増幅、スクリーニングならびに生成物の産生および精製に適した哺乳動物宿主細胞の選択および方法は当該分野において知られている。例えば、上に引用したSambrook et al.を参照されたい。
【0128】
細菌細胞は、本発明の組換えFab、または他の実施形態の発現に適した宿主細胞として有用であることが分かるかもしれない(例えば、Pluckthun, A., Immunol. Rev., 130:151−188 (1992)を参照されたい)。しかしながら、細菌細胞内で発現するタンパク質は折り畳まれていないか不適切に折り畳まれた形態または非グリコシル化形態となる傾向があるために、細菌細胞内で産生した組換えFabはいずれも抗原結合能力の保持についてスクリーニングする必要があるであろう。細菌細胞によって発現している分子が適切に折り畳まれた形態で産生した場合、その細菌細胞は所望される宿主であり得、または代替的な実施形態では、分子は細菌宿主内で発現し、続いて再び折り畳まれ得る。例えば、発現のために使用される様々な大腸菌株が生物工学分野において宿主細胞として周知である。様々な枯草菌、ストレプトミセス属、他の桿菌株などもこの方法に使用してよい。
【0129】
所望の場合、当業者に知られている酵母細胞の株、ならびに昆虫細胞(例えばショウジョウバエおよび鱗翅類)およびウイルス発現系も宿主細胞として利用可能である。例えばMiller et al., Genetic Engineering, 8:277−298, Plenum Press (1986)およびそこで引用されている参考文献を参照されたい。
【0130】
ベクターを構築し得る一般的な方法、本発明の宿主細胞を産生するために必要とされるトランスフェクション方法、およびかかる宿主細胞から本発明の抗原結合タンパク質を産生するために必要な培養方法はすべて従来技術であってよい。典型的には、本発明の培養方法は、無血清培養方法、通常、懸濁液中での無血清細胞培養である。同様に、産生後は、本発明の抗原結合タンパク質は、硫酸アンモニウム沈殿法、親和性カラム、カラムクロマトグラフィー、ゲル電気泳動などを含む当該技術分野の標準的な手段に従い細胞培養の内容物から精製し得る。かかる技術は、当該技術分野の技術範囲内であり、本発明を限定しない。例えば、改変抗体の調製については、WO99/58679号およびWO96/16990号に記載されている。
【0131】
抗原結合タンパク質のさらに別の発現方法は、米国特許第4,873,316号に記載されるように、遺伝子導入動物における発現の利用である。これは、哺乳動物に遺伝子導入で組み込まれた場合、雌の母乳において所望の組換えタンパク質の産生を可能にする動物のカゼインプロモーターを用いる発現系に関する。
【0132】
本発明のさらなる態様では、本発明の抗体の産生方法を提供し、この方法は、本発明の抗体の軽鎖および/もしくは重鎖をコードするベクターを用いて形質転換またはトランスフェクトされた宿主細胞を培養する工程、およびその結果産生した抗体を回収する工程を含む。
【0133】
本発明に従い、ヒトIL−13と結合してその活性を中和する本発明の抗IL−13抗体の産生方法を提供し、この方法は以下;
(a)抗体重鎖をコードする第1ベクターを提供する工程;
(b)抗体軽鎖をコードする第2ベクターを提供する工程;
(c)前記第1および第2ベクターを用いて哺乳動物宿主細胞(例えばCHO)を形質転換する工程;
(d)前記培地内へ前記宿主細胞から抗体の分泌を促す条件下で工程(c)の宿主細胞を培養する工程;
(e)工程(d)の分泌した抗体を回収する工程
を含む。
【0134】
所望の方法によって発現後は、適切なアッセイを用いて抗体のインビトロ活性を検討する。現在、抗体のIL−13に対する定性および定量的結合を評価するために従来のELISAアッセイ形態が使用されている。さらに、通常のクリアランス機序にもかかわらず体内における抗体の持続性を評価するために実施するヒト臨床研究の前に、中和効力を検証するために他のインビトロアッセイを使用してよい。
【0135】
治療の用量および期間は、ヒト循環における本発明の分子の相対的な持続時間に関し、治療する状態および患者の一般的な健康状態に応じて、当業者によって調整されることができる。最大の治療効果に達するために、長期(例えば4〜6ヶ月)にわたる反復投薬(例えば週1回または隔週)が必要な場合があることが想定される。
【0136】
本発明の治療薬の投与方法は、その作用物質を宿主へ送達する上で適した任意の経路であってよい。本発明の抗原結合タンパク質および医薬組成物は、非経口投与、すなわち、皮下(s.c.)、髄腔内、腹腔内、筋肉内(i.m.)、静脈内(i.v.)、または鼻腔内投与に特に有用である。
【0137】
本発明の治療薬は、医薬上許容可能な担体中に有効成分として有効量の本発明の抗原結合タンパク質を含む医薬組成物として調製し得る。一実施形態では、本発明の予防薬は、抗原結合タンパク質を注射形態で含む水性懸濁液または溶液である。一実施形態では、懸濁液または溶液は生理的pHに緩衝されており、一実施形態では、非経口投与用の組成物は、本発明の抗原結合タンパク質の溶液または医薬上許容可能な担体に溶解したその混合物を含む。一実施形態では、担体は水性担体である。様々な水性担体、例えば、0.9%食塩水、0.3%グリシンなどを適用し得る。これらの溶液は、滅菌で作製し得、一般に粒子物質を含まない。これらの溶液は、従来の周知の滅菌技術(例えば、濾過)により滅菌し得る。組成物には、生理的状態に近づけるために必要に応じて医薬上許容可能な補助物質(pH調節剤および緩衝剤など)を含み得る。かかる医薬製剤中の本発明の抗原結合タンパク質の濃度は広範囲、すなわち、約0.5重量%未満、通常、約1重量%以上から約15または20重量%程度に変わり得、選択された特定の投与方法に従い、主に体液の容量および粘性などに基づき選択されるであろう。
【0138】
したがって、筋肉内注射用の本発明の医薬組成物は、滅菌緩衝水約1mL、ならびに本発明の抗原結合タンパク質、例えば抗体を約1ng〜約100mg、例えば約50ng〜約30mgもしくは約5mg〜約25mg含むように調製できる。同様に、静脈内注射用の本発明の医薬組成物は、滅菌リンガー溶液を最大約250ml、ならびに本発明の抗原結合タンパク質を約1〜約30もしくは5mg〜約25mg/mlリンガー溶液含むように作製し得る。非経口投与可能な組成物を調製するための実際の方法は当業者に周知または明白であり、例えば、Remington’s Pharmaceutical Science, 15th ed., Mack Publishing Company, Easton, Pennsylvaniaに詳述されている。静脈内に投与可能な本発明の抗原結合タンパク質製剤の調製については、Lasmar U and Parkins D “The formulation of Biopharmaceutical products”, Pharma. Sci.Tech.today, page 129−137, Vol.3 (3rd April 2000); Wang, W “Instability, stabilisation and formulation of liquid protein pharmaceuticals”, Int. J. Pharm 185 (1999) 129−188; Stability of Protein Pharmaceuticals Part A and B ed Ahern T.J., Manning M.C., New York, NY: Plenum Press (1992); Akers,M.J. “Excipient−Drug interactions in Parenteral Formulations”, J.Pharm Sci 91 (2002) 2283−2300; Imamura, K et al “Effects of types of sugar on stabilization of Protein in the dried state”, J Pharm Sci 92 (2003) 266−274; Izutsu, Kkojima, S. “Excipient crystalinity and its protein−structure−stabilizing effect during freeze−drying”, J Pharm. Pharmacol, 54 (2002) 1033−1039; Johnson, R, “Mannitol−sucrose mixtures−versatile formulations for protein lyophilization”, J. Pharm. Sci, 91 (2002) 914−922;およびHa,E Wang W, Wang Y.j. “Peroxide formation in polysorbate 80 and protein stability”, J. Pharm Sci, 91, 2252−2264,(2002)を参照されたく、その全体の内容は参照によって本明細書に組み込まれ、読者は、この全内容を特に参照する。
【0139】
一実施形態では、本発明の治療薬は、医薬調製物中にある場合、単位用量形態で存在する。適切な治療有効量は当業者によって容易に決定されるであろう。患者に適した用量は体重に応じて算出し得、例えば適切な用量は、約0.1〜約20mg/kg、例えば約1〜約20mg/kg、例えば約10〜約20mg/kgまたは例えば約1〜約15mg/kg、例えば約10〜約15mg/kgの範囲であり得る。本発明の抗原結合タンパク質の、ヒトにおける喘息またはIPFなどの状態を効果的に治療するために適した用量は、約0.1〜約1000mg、例えば約0.1〜約500mg、例えば約500mg、例えば約0.1〜約100mg、または約0.1〜約80mg、または約0.1〜約60mg、または約0.1〜約40mg、または例えば約1〜約100mg、または約1〜約50mgの範囲内であり得、これは非経口(例えば皮下、静脈内または筋肉内)投与し得る。かかる用量は、必要な場合、医師によって適切に選択された適切な時間間隔で反復し得る。
【0140】
本明細書に記載の抗原結合タンパク質は、保存用に凍結乾燥し、使用前に適切な担体中で再構成することができる。この技術は従来の免疫グロブリンで有効であることが示されており、当技術分野において知られている凍結乾燥および再構成技術を使用できる。
【0141】
別の態様では、本発明は、アトピー性疾患/障害および慢性炎症疾患/障害、例えば、喘息、例えばアレルギー性喘息、特に重症喘息(すなわちコルチコステロイドの全身投与を含む現在の治療が奏効しない喘息;Busse WW et al, J Allergy Clin. Immunol 2000, 106: 1033−1042を参照されたい)、「困難性」喘息(最大推奨用量の吸入ステロイド処方にもかかわらず制御不成功を特徴とする喘息性形質として定義、Barnes PJ (1998), Eur Respir J 12:1208−1218を参照されたい)、「ブリットル」喘息(高用量の吸入ステロイドにもかかわらず大きく変動する最大呼気流量(PEF)を維持する重度の不安定喘息患者のサブグループを定義、Ayres JG et al (1998) Thorax 58:315−321を参照されたい)、夜間性喘息、月経前喘息、ステロイド抵抗性喘息(Woodcock AJ (1993) Eur Respir J 6:743−747を参照されたい)、ステロイド依存性喘息(高用量の経口ステロイドでのみ制御できる喘息として定義)、アスピリン誘発性喘息、成人発症型喘息、小児喘息の治療または予防のための本発明の抗原結合タンパク質もしくはその機能的断片ならびに医薬上許容可能な担体を含む医薬組成物を提供する。本発明の抗体は、急性喘息エピソード(喘息発作重積状態)を予防する、頻度を減らす、または作用を緩和するために使用してよい。本発明の抗体は、喘息の治療に使用される他の薬の必要な投薬を(投薬の投与量または投与頻度のいずれかに関して)減らすためにも使用してよい。例えば、本発明の抗体は、喘息のステロイド治療(コルチコステロイド治療など)に必要な投薬を減らす(「ステロイド使用を控える」)ために使用してよい。本発明の抗体で治療し得る他の疾患または障害としては、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、慢性閉塞性肺疾患(COPD)、好酸球性食道炎、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症(強皮症)、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、および細胞周期調節疾患、例えばホジキン病、B細胞慢性リンパ球性白血病が挙げられる。一実施形態では、障害は重症喘息である。さらなる実施形態では、障害は線維性障害(IPFなど)である。
【0142】
よりさらなる態様では、本発明は、本発明の抗原結合タンパク質および医薬上許容可能な担体を含む医薬組成物を、アトピー性疾患/障害および慢性炎症疾患/障害、例えば、喘息、例えばアレルギー性喘息、特に重症喘息(すなわちコルチコステロイドの全身投与を含む現在の治療が奏効しない喘息;Busse WW et al, J Allergy Clin. Immunol 2000, 106: 1033−1042を参照されたい)、「困難性」喘息(最大推奨用量の吸入ステロイド処方にもかかわらず制御不成功を特徴とする喘息性形質として定義、Barnes PJ (1998), Eur Respir J 12:1208−1218を参照されたい)、「ブリットル」喘息(高用量の吸入ステロイドにもかかわらず大きく変動する最大呼気流量(PEF)を維持する重度の不安定喘息患者のサブグループを定義、Ayres JG et al (1998) Thorax 58:315−321を参照されたい)、夜間性喘息、月経前喘息、ステロイド抵抗性喘息(Woodcock AJ (1993) Eur Respir J 6:743−747を参照されたい)、ステロイド依存性喘息(高用量の経口ステロイドでのみ制御できる喘息として定義)、アスピリン誘発性喘息、成人発症型喘息、小児喘息の治療のために提供する。本発明の抗体は、急性喘息エピソード(喘息発作重積状態)を予防する、頻度を減らす、または作用を緩和するために使用してよい。本発明の抗体は、喘息の治療に使用される他の薬の必要な投薬を(投薬の投与量または投与頻度のいずれかに関して)減らすためにも使用してよい。例えば、本発明の抗体は、喘息のステロイド治療(コルチコステロイド治療など)に必要な投薬を減らす(「ステロイド使用を控える」)ために使用してよい。本発明の抗体で治療し得る他の疾患または障害としては、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、慢性閉塞性肺疾患(COPD)、好酸球性食道炎、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症(強皮症)、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、および細胞周期調節疾患、例えばホジキン病、B細胞慢性リンパ球性白血病が挙げられる。一実施形態では、障害は重症喘息である。さらなる実施形態では、障害は線維性障害(IPFなど)である。
【0143】
本明細書に記載の配列(配列番号26〜配列番号55および配列番号62〜配列番号146)は、本明細書に記載の配列と実質的に同一である、例えば、少なくとも90%同一である、例えば、少なくとも91%、または少なくとも92%、または少なくとも93%、または少なくとも94%または少なくとも95%、または少なくとも96%、または少なくとも97%または少なくとも98%、または少なくとも99%同一である配列を含むことが理解されるであろう。
【0144】
核酸において、「実質的な同一性」という用語は、適切にヌクレオチドを挿入してまたは欠失させて至適整列および比較時に、少なくとも約80%のヌクレオチド、少なくとも約90%〜約95%、または少なくとも約98%〜約99.5%のヌクレオチドが同一である2つの核酸、またはその設計された配列を示す。あるいは、実質的な同一性は、選択的ハイブリダイゼーション条件下でセグメントが鎖の補体にハイブリダイズする場合に存在する。
【0145】
ヌクレオチド配列およびアミノ酸配列において、「同一」という用語は、2つの核酸またはアミノ酸配列を適切に挿入してまたは欠失させて至適整列および比較時の同一度を示す。あるいは、実質的な同一性は、選択的ハイブリダイゼーション条件下でDNAセグメントが鎖の補体にハイブリダイズする場合に存在する。
【0146】
2配列間の同一性%は、2配列の至適整列のために導入する必要があるギャップ数、および各ギャップの長さを考慮した、配列の共有する同一の位置数の関数(すなわち、同一性%=同一の位置数/総位置数×100)である。配列の比較および2配列間の同一性%の決定は、以下の非制限的な実施例に記載のように数学アルゴリズムを用いて成し遂げることができる。
【0147】
2つのヌクレオチド配列間の同一性%は、GCGソフトウェアパッケージ内のGAPプログラムを用いて、NWSgapdna.CMP行列、ならびに40、50、60、70、もしくは80のギャップウェイト、ならびに1、2、3、4、5、もしくは6のレングスウェイトを用いて決定することができる。2つのヌクレオチド配列またはアミノ酸配列間の同一性%は、ALIGNプログラム(バージョン2.0)に組み込まれているE. Meyers and W. Miller (Comput. Appl. Biosci., 4:11−17 (1988))のアルゴリズムを用いて、PAM120ウェイト残基表、ギャップレングスペナルティ12、およびギャップペナルティ4を用いて決定することもできる。加えて、2つのアミノ酸配列間の同一性%は、GCGソフトウェアパッケージ内のGAPプログラムに組み込まれているNeedleman and Wunsch (J. Mol. Biol. 48:444−453 (1970))アルゴリズムを用いて、Blossum62行列もしくはPAM250行列のいずれか、ならびに16、14、12、10、8、6、もしくは4のギャップウェイト、ならびに1、2、3、4、5、もしくは6のレングスウェイトを用いて決定することができる。
【0148】
例として、本発明のポリヌクレオチド配列は、配列番号25の基準配列と同一、すなわち100%同一であってもよく、または、基準配列と比較してヌクレオチド改変を特定の整数の数まで含んでもよい。かかる改変は、少なくとも1つのヌクレオチド欠失、置換(転位および塩基転換を含む)、または挿入からなる群から選択され、前記改変は基準ヌクレオチド配列の5’もしくは3’末端位またはそれら末端位の間のどこかで、基準配列中のヌクレオチド間に個々に散在してまたは基準配列中の1つもしくは複数の連続基としてのいずれかで生じ得る。ヌクレオチド改変数は、配列番号25の総ヌクレオチド数にそれぞれの同一性%の%数値を乗じて(100で割って)、その積を前記配列番号23の総ヌクレオチド数から減算するか、または:
nn≦xn−(xn × y)、
式中nnはヌクレオチド改変数であり、xnは配列番号25の総ヌクレオチド数であり、yは、50%の場合は0.50、60%の場合は0.60、70%の場合は0.70、80%の場合は0.80、85%の場合は0.85、90%の場合は0.90、95%の場合は0.95、97%の場合は0.97または100%の場合は1.00であり、xnとyの積が整数ではない場合はいずれも端数を切り捨てて最も近い整数にした後でxnから減算することによって決定する。配列番号25のポリヌクレオチド配列改変はこのコード配列中にナンセンス、ミスセンスまたはフレームシフト変異を生じ得、その結果かかる改変に続いてポリヌクレオチドによりコードされるポリペプチドを改変する。
【0149】
同様に、別の例では、本発明のポリペプチド配列は、配列番号24によりコードされた基準配列と同一、すなわち100%同一であってもよく、または、基準配列と比較して同一性%が100%未満であるようにアミノ酸改変を特定の整数の数まで含んでもよい。かかる改変は、少なくとも1つのアミノ酸欠失、置換(保存的および非保存的置換を含む)、または挿入からなる群から選択され、前記改変は基準ポリペプチド配列のアミノもしくはカルボキシ末端位またはそれら末端位の間のどこかで、基準配列中のアミノ酸間に個々に散在してまたは基準配列中の1つもしくは複数の連続基としてのいずれかで生じ得る。所定の同一性%のアミノ酸改変数は、配列番号24によりコードされるポリペプチド配列の総アミノ酸数にそれぞれの同一性%の%数値を乗じて(100で割って)、次いでその積を前記配列番号24によりコードされるポリペプチド配列の総アミノ酸数から減算するか、または:
na≦xa−(xa × y)、
式中naは、アミノ酸改変数であり、xaは配列番号24によりコードされるポリペプチド配列の総アミノ酸数であり、yは、例えば70%の場合は0.70、80%の場合は0.80、85%の場合は0.85、などであり、xaとyの積が整数ではない場合はいずれも端数を切り捨てて最も近い整数にした後でxaから減算することによって決定する。
【0150】
以下の実施例により例証するが、これらは本発明を限定しない。
実施例
【実施例1】
【0151】
組換え抗IL−13抗体の構築
オリジナルのマウスmAbを組換えヒトIL−13を有するマウスの免疫化により産生した。反応した動物由来の脾臓を回収して、骨髄腫細胞に融合させてハイブリドーマを生成した。ハイブリドーマ上清物質の結合をスクリーニングした。標準的な技術を用いて興味のハイブリドーマをモノクローン化した。生成したマウス抗体(6A1)は配列番号58および配列番号59に示す可変領域を含む。このマウス抗体およびこの抗体A1L1(配列番号22および24)のヒト化バージョンの詳細については、WO2006/003407号に記載されており、これは参照によって本明細書に組み込まれる。抗IL−13 mAb抗体A1L1を比較抗体として以下の実施例のいくつかに使用した。
【0152】
配列番号22に説明した重鎖を含むヒト化抗体の変異体をいくつか産生した。これらはすべて抗体のCDRH3領域が異なる(配列番号3)。
【0153】
RldおよびRln哺乳動物発現ベクター内へクローニングのための制限部位を含む重複オリゴヌクレオチドならびにヒトシグナル配列を形成して本発明の抗体のための塩基性DNA発現構築物、配列番号23(重鎖)および配列番号25(軽鎖)をde novoで調製した。Hind IIIおよびSpe I制限部位を導入して、ヒトγ1定常領域を含むRld内へのクローニングのためにシグナル配列(配列番号56)を含むVHドメインを枠組みした。Hind IIIおよびBsiWI制限部位を導入して、ヒトκ定常領域を含むRln内へのクローニングのためにシグナル配列(配列番号56)を含むVLドメインを枠組みした。代替構築物は同様にヒト定常領域を含むpTTベクターを用いて産生した。適切な場合、部位指向変異(SDM)を用いて異なるヒト化構築物を生成した。
【実施例2】
【0154】
HEK 293 6E細胞における抗体発現
重鎖と軽鎖をそれぞれコードするpTTプラスミドをHEK 293 6E細胞内に一過性共トランスフェクトし、少量発現させて抗体を産生した。抗体を組織培養上清から直接評価した。固定化プロテインAカラムを用いて他の抗体を精製して、吸光度を280nmで読み取ることにより定量化し、指定の場合、精製された抗体物質を以下に説明する実施例に記載のアッセイにおいて評価した。
【0155】
我々がコード(すなわちA1Y100BTrpL1)により抗体を指す場合、我々は記載の第1プラスミドと第2プラスミドの共トランスフェクションおよび発現により生成したmAbを指している。例えば「A1Y100BTrpL1」とは、適切な細胞系においてA1Y100BTrp配列を含むプラスミドとL1配列を含むプラスミドの共トランスフェクションにより生成したmAbに関する。
【実施例3】
【0156】
抗IL−13ヒト化mAbのBiacore分析
動力学分析
CDRH3変異体の初回スクリーニングをProteOn XPR36(Biorad)上で実施した。方法は以下のとおりであった。抗ヒトIgG(Biacore BR−1008〜39)を第1級アミンカップリングによりGLMチップ上に固定化してから、CDRH3変異体抗体をこの表面上に捕捉して、IL13を256、64、16、4、1nMで通過させ、0nM注入(すなわち緩衝液単独)を二重照合に使用した。3M MgCl2を用いて捕捉表面を再生し、結合CDRH3変異体抗体を除去して、別のサイクルの捕捉およびアナライト注入を準備した。データは、機械に固有のソフトウェアを用いて1:1モデルに適合させた。精製された物質であった親抗体を除きすべての作業を組織培養上清から直接抗体を用いて実施した。
【0157】
スクリーニングにより、親分子より良好な動力学プロファイルを有するように思われた抗体をいくつか同定してから、これらの同一試料をBiacore T100上で分析して類似の方法を用いて結果を確認し、同じ抗ヒトIgG捕捉抗体を第1級アミンカップリングによりCM5チップ上に固定化して、IL13を表面上に256、64、16、4、1および0.25nMで通過させ、0nMを二重照合に使用して、3M MgCl2で再生し、T100に固有の1:1モデルにデータを適合させた。ProteOnスクリーニングおよびT100実行から選択された構築物の全体の親和性(平衡解離定数KD)を表1に詳述する。
【表1】
【0158】
データにより、Y100B残基のいくつかの変異により全体の親和性が改善されると思われることが強調された。この観点から、先に記載のものと同じ方法でProteOnを用いて、組織培養上清から再び抗体を直接用いて、初回スクリーニング時に存在しなかったこの残基の変異を試験した。試験した変異のうち、A1Y100B ValL1において、親の0.390〜0.460nM値と比較して0.166nM値が得られ、全体の親和性(平衡解離定数KD)が改善されると思われた。先に記載のものと同じ方法を用いてBiacore T100上でY100B Valを試験時、平衡解離定数KDは親の0.346nM値と比較して0.025nMで測定された。
【0159】
抗体変異体を組織培養上清から直接用いてこれまで実行した作業の点から、精製された抗体をA1Y100BL1変異体Ala、IleおよびTrpのために産生した。これらは、この段階で精製しなかったA1Y100B ValL1変異体を対象として、前と同じ方法を用いてBiacore T100上で実行した。この実験から得られたデータを表2に示す。
【表2】
【0160】
実験により、変異体はIL13に対する結合親和性を親分子と比較して改善しなかったことを確認した。
【0161】
精製されたA1Y100BL1変異体では組織培養上清から得られるよりも良好な結合親和性が得られることから、A1Y100B ValL1を精製し、同様に親和性を改善した他の精製されたA1Y100BL1変異体と共に、Biacore T100機械で先に記載の方法を用いて実行した。この実験から得られたデータを表3に示す。この実験は表2のデータと十分一致し、Y100B変異の親和性の改善が確認された。
【表3】
【実施例4】
【0162】
TF−1細胞増殖バイオアッセイにおける大腸菌発現組換えヒトIL−13の中和
ヒトIL−13を含むいくつかの異なるサイトカインに対する反応において、TF−1細胞は増殖する。したがってIL−13のためのこれらの細胞の増殖反応を用いてIL−13の生体活性を測定し、続いてアッセイを開発してmAbのIL−13中和効能(IL−13生体活性の阻害)を決定することができる。
【0163】
アッセイは滅菌96ウェル組織培養プレートにおいて滅菌条件下で実施して、すべての試験ウェルは三連で実施した。14ng/mlの組換え大腸菌発現ヒトIL−13を様々な希釈のmAb(通常3倍希釈液で滴定した93.4nM〜0.014nM)と37℃で1時間プレインキュベートした。無関係な特異性抗体を陰性対照として同様に滴定した。次いでこれらの試料を、滅菌96ウェル組織培養プレート中TF−1細胞50μl(濃度2×105細胞/ml)に添加した。したがって最終100μlアッセイ体積には、様々な希釈のmAb(3倍希釈液で滴定した最終濃度46.7nM〜0.007nM)、組換え大腸菌発現ヒトIL−13(最終濃度7ng/ml)およびTF−1細胞(最終濃度1×105細胞/ml)を含有した。アッセイプレートは加湿CO2インキュベーター内で37℃で約3日間インキュベートした。次いでPromega製「Cell Titre96(登録商標)Non−Radioactive Cell Proliferation Assay」(カタログ番号G4100)を製造業者の説明書に記載どおりに用いて、細胞増殖量を決定した。96ウェルプレート中の試料の吸光度はプレートリーダーで570nmで読み取った。
【0164】
組換え大腸菌発現ヒトIL−13の生体活性を中和するmAbの能力は定義した量のヒトIL−13(7ng/ml)の生体活性を50%中和するために必要なmAb濃度(=ND50)として表した。必要なmAb濃度が低いほど、中和能力は強力である。本明細書に提供するND50データ(表4)はマイクロソフトエクセルのRobosageを用いて算出した。データの図解は図1に見ることができる。
【表4】
【実施例5】
【0165】
CDRH3変異型抗IL−13 mAbを含むmAb−dAbの構築および発現
標準的な分子生物学技術を用いて、A1抗体のCDRH3変異体の可変重鎖領域の各配列をコードする遺伝子を既存の構築物から、hIgG1定常領域のC末端にてTVAAPSまたはASTKGPSリンカーを介して抗ヒトIL−4ドメイン抗体(DOM9−112−210)に融合したhIgG1定常領域を含む発現ベクターに移した。構築した重鎖の詳細を表5に列挙する。
【表5】
【0166】
BPC1624、BPC1625、BPC1626およびBPC1627をHEK293細胞内で発現した。簡単に述べると、250mlのHEK293細胞1.5×106細胞/mlを、293fectin試薬(Invitrogen#51〜0031)で先にインキュベートした重鎖および軽鎖発現プラスミドと共トランスフェクトした。これらを37℃、5%CO2、および95%相対湿度で攪拌インキュベーター内に入れた。24時間後にトリプトン供給培養を添加して、細胞をさらに5日間増殖させた。上清を遠心分離により収穫してフィルター滅菌した。固定化プロテインAカラムを用いて発現した分子を親和性クロマトグラフィーにより精製して、吸光度を280nmで測定することによって濃度を決定した。精製された試料中のタンパク質凝集レベルをサイズ排除クロマトグラフィーにより決定した。精製されたタンパク質の収率および凝集レベルを表5bに示す。
【0167】
【実施例6】
【0168】
CDRH3変異型抗IL−13 mAbを含むmAb−dAbの中和活性データ
TF−1細胞増殖バイオアッセイにおいてCDRH3変異型抗IL−13 mAbを含むmAb−dAbによる大腸菌発現組換えヒトIL−13の中和を試験した。
【0169】
アッセイは滅菌96ウェル組織培養プレートにおいて滅菌条件下で実施して、すべての試験ウェルは三連で実施した。約20ng/mlの組換え大腸菌発現ヒトIL−13を様々な希釈のmAbdAb(通常3倍希釈液で滴定した、50nM〜0.02nM)(HEK細胞内で作製し、実施例5に記載のように精製されたmAbdAb)の総体積50μlと37℃で1時間プレインキュベートした。無関係な特異性抗体を陰性対照として同様に滴定した(データは示さず)。次いでこれらの試料を、滅菌96ウェル組織培養プレート中TF−1細胞50μl(濃度2×105細胞/ml)に添加した。したがって最終100μlアッセイ体積には、様々な希釈のmAbdAb(3倍希釈液で滴定した最終濃度25nM〜0.01nM)、組換え大腸菌発現ヒトIL−13(最終濃度10ng/ml)およびTF−1細胞(最終濃度1×105細胞/ml)を含有した。アッセイプレートは加湿CO2インキュベーター内で37℃で約3日間インキュベートした。次いでPromega製「Cell Titre96(登録商標)Non−Radioactive Cell Proliferation Assay」(カタログ番号G4100)を製造業者の説明書に記載どおりに用いて、細胞増殖量を決定した。96ウェルプレート中の試料の吸光度はプレートリーダーで570nmで読み取った。
【0170】
ヒトIL−13の生体活性を中和するmAbdAbの能力は定義した量のヒトIL−13(10ng/ml)の生体活性を50%中和するために必要なmAb−dAb濃度(=ND50)として表した。必要なmAbdAb濃度が低いほど、中和能力は強力である。本明細書に提供するND50データ(表6)はGraphPad Prismを用いて算出した。これらのデータを図2に図式的に表す。
【表6】
【実施例7】
【0171】
CHOE1a発現系におけるCDRH3変異型抗IL−13 mAbを含むmAb−dAbの発現
表5に示す分子BPC1624〜1631もCHOE1a細胞内で発現した。重鎖および軽鎖をコードするDNAベクターを懸濁CHO細胞内に共電気穿孔した。攪拌フラスコ内のMR1基底選択的培地で、37℃、5%CO2、130rpmで細胞生存能および細胞数が改善されるまで細胞を継代させた。次いでCHO細胞をMR1基底×2選択的培地内で播種し、34℃、5%CO2、130rpmで8〜12日間インキュベートした。細胞を遠心分離によりペレットして、上清を滅菌濾過した。
【0172】
固定化プロテインAカラムを用いて発現物質を親和性クロマトグラフィーにより精製して、吸光度を280nmで測定することによって収率を決定した。凝集レベルをサイズ排除クロマトグラフィーにより決定した。凝集を調製用サイズ排除クロマトグラフィーにより除去し、収率を再評価した。この発現系から得られる収率および凝集レベルを表7に列挙する。
【表7】
【実施例8】
【0173】
抗原結合タンパク質の化学量論評価(Biacore(商標)を使用)
この実施例は予測である。本発明の抗原結合タンパク質を試験できる追加アッセイを実行するための指針を提供する。
【0174】
抗ヒトIgGは、第1級アミンカップリングによりCM5バイオセンサーチップ上に固定化する。抗原結合タンパク質をこの表面上に捕捉後、単一濃度のIL−13またはIL−4またはIL−5を通過させる。この濃度は結合表面を飽和するのに十分であり、観察される結合シグナルは完全なRmaxに達する。次いで以下の式:
化学量論値=Rmax × Mw(リガンド) / Mw(アナライト) × R(固定化または捕捉されたリガンド)
を用いて化学量論値を算出する。
【0175】
複数のアナライト結合の化学量論値を同時に算出する場合、異なる抗原を飽和抗原濃度および上で算出した化学量論で連続通過させる。この作業は、Biacore 3000上、HBS−EPランニング緩衝液を用いて25℃で実行できる。
【実施例9】
【0176】
改善されたヒト化変異型mAbの用量予測
異なるモノクローナル抗体(mAb)候補を結合親和性に基づき順位付けるために抗体リガンド結合PK−PDモデルを開発して、ヒトにおける潜在的治療用量を予測した。
【0177】
ヒトにおける予測した潜在的治療用量をこの目的のために、mAbの月1回静脈内投与1時間後に肺(作用部位)中標的IL−13を定常状態で90%阻害する用量として定義した。各分子の分子量は同じであり、mAbの標準分子量、すなわち150kDaと等しいと推定した。加えて、異なる候補の動物またはヒト薬力学データの不在下、A1L1抗体のヒト薬力学をすべての候補と推測した。
【0178】
同一の抗体リガンド結合PK−PDモデルならびに標的濃度、標的ターンオーバー、標的組織:血漿比率およびmAb組織浸透に関する同じ仮定を各mAbにおいて使用する。したがって、モデルにより提供される順位付けは、もっぱら分子の結合親和性に基づき、変数のみ異なる。かかる状態において、ヒトにおける4つの候補A1Y100BlleL1、A1Y100BValL1、A1Y100BAlaL1およびA1Y100BTrpL1の潜在的治療用量は、ヒトにおけるA1L1の予測される潜在的治療用量を超える実質的な改善を提供すると予測される。
【実施例10】
【0179】
変異型IL−4dAbを伴う抗IL13/IL4 mAbdAb
10.1構築および発現
凝集予測アルゴリズムを用いて、抗IL−4dAb(DOM9−155−154、配列番号80)の凝集傾向がある残基を調査した。Kabat89位のロイシン残基を凝集促進のための主要残基として同定した。
【0180】
このdAbを含むmAbdAbの潜在的凝集を減らすために、このアミノ酸残基を他のアミノ酸と置換していくつかのmAb−dAb変異体を生成した。既存のmAbdAb構築物の重鎖をコードするDNA発現ベクターを用いて、部位指向変異により発現構築物を生成した。変異dAb配列を含む生成した新規mAbdAb重鎖のタンパク質配列は、配列番号117〜134において提示される。
【0181】
89位に別の変異を挿入された他の重鎖配列は、配列番号96〜106である。これらを実施例11に詳述する。
【0182】
発現した分子の一覧を表8に提供する。
【表8】
【0183】
【0184】
10.2 HEK 293 6E細胞における分子発現
重鎖と軽鎖をそれぞれコードするプラスミドをHEK 293 6E細胞内に一過性共トランスフェクトして少量発現させて、抗体分子を産生した。トランスフェクションから最大24時間後にトリプトン供給物を各細胞培養に添加して、3日後に細胞を回収した。
【0185】
抗体分子を組織培養上清から直接評価して、Gyrolabワークステーションを用いて定量化した。
【0186】
細胞上清中の抗体分子の定量化のためのGyrolabワークステーション方法
Gyrolab Bioaffyワークステーション(Gyros)を用いた定量免疫アッセイにより、少量の一過性HEK 2936Eトランスフェクション(0.75〜2.0ml)から産生した抗体を組織培養上清から定量化した。コンパクトディスク(CD)マイクロ実験室(Gyros)上にストレプトアビジンコーティングした粒子に固定化したビオチン化抗IgGAffibody分子(Abcam)を用いたFc領域を介して抗体を捕捉された。Affibody試薬を簡単にボルテックスして、PBS−Tween 20(0.01%v/v)で最終作業濃度0.1mg/mlに希釈した。次いで、レーザ誘発蛍光を用いてALEXA 647標識化Fab2抗ヒトIgGκ軽鎖分子により抗体を検出した。ALEXA 647標識化検出試薬は、簡単にボルテックスすることによって、および13000rpmで4分間遠心分離することによって調製した。標識化Fab2検出試薬を非標識化Fab2に添加して、これをRexcip F検出試薬希釈液(Gyros)を用いて最終濃度75nMおよび1.5μMにそれぞれ希釈した。抗体定量化範囲は抗CD23モノクローナル抗体標準曲線と比較して0.244〜250μg/mlであった。組織培地(Freestyle 293発現培地、Pluronic F68およびGeneticin、Invitrogen)で抗体を連続希釈して抗CD23(1mg/ml)標準曲線を作成した。
【0187】
場合により、固定化プロテインAカラムを用いて抗体分子を精製して、吸光度を280nmで読み取ることにより定量化し、指定の場合、精製された抗体分子を以下に説明する実施例に記載のアッセイにおいて評価した。
【0188】
10.3 IL−4結合ELISA
以下の方法を用いた直接結合ELISAにおいて、これらのmAbdAbのIL−4への結合を試験した。
【0189】
5μg/mlヒトIL−4(GSK製)のNaHCO3溶液を96ウェル高結合プレートにコーティングして、4℃で一晩保存した。プレートを0.05%のTween−20(TBST)含有トリス緩衝食塩水で2回洗浄した。100μLのブロック溶液(1%BSAのTBST緩衝液)を各ウェルに添加して、プレートを室温で少なくとも1時間インキュベートした。引き続いてmAbdAbをプレート全体でブロック溶液で希釈した。1時間インキュベーション後、プレートを3回洗浄した。ヤギ抗ヒトκ軽鎖特異的ペルオキシダーゼ抱合抗体(Sigma A7164)をブロック溶液で1μg/mLに希釈して、50μLを各ウェルに添加した。プレートを1時間インキュベートした。さらに3つの洗浄工程後、50μlのOPD(o−フェニレンジアミン二塩酸塩)SigmaFast基質溶液を各ウェルに添加して、3M硫酸25μL添加により約5分後に反応は停止した。基本終点プロトコルを用いて、VersaMax Tunableマイクロプレートリーダー(Molecular Devices)を用いて、吸光度を490nmで読み取った。
【0190】
精製された物質であった陽性対照(抗IL−4 mAb)および抗IL13陰性対照mAbを除き、gyrolabプラットフォームを用いて定量化されている組織培養上清から直接mAbdAbを用いて実験を実施した。これらのデータを図17に示す。
【0191】
ELISA結果は、これらの一過性発現抗IL13 mAb−抗IL4dAb結合IL−4のほとんど(ただし、IL−4結合活性において一部変形)が観察されたことを示す。精製された陽性対照の抗IL−4 mAbもIL−4への結合を示したのに対し、精製された陰性対照mAbはヒトIL−4と結合しないことが示された。
【0192】
10.4 mAbdAb発現における凝集レベルの分析
293fectin(Invitrogen、12347019)を用いて、BPC1090、BPC1091、BPC1092、BPC1093、BPC1094およびBPC1095の重鎖および軽鎖をコードするpTTプラスミドをHEK 293 6E細胞内に一過性共トランスフェクトして、実施例10.2に上述したmAbdAbよりもわずかに多く(200〜600ml)発現させた。同じ方法論を用いてBPC1111およびBPC1085を独立してHEK293 6E細胞内で一過性発現させた。上のトランスフェクションに使用したプラスミドは、EndoFree Plasmid Maxi Kit(Qiagen,12362)を用いて生成した。
【0193】
24時間後にトリプトン供給物を各細胞培養に添加して、72時間後に細胞を回収した。抗体をプロテインAカラムを用いて精製し、280nmでの吸光度の読み取りにより定量化し、サイズ排除クロマトグラフィー(SEC)により分析した。
【0194】
これらのmAbdAbを、独立して発現しているBPC2223、抗IL13 mAb(829)および抗Il−13 mAb(586)と比較した。
【0195】
変異dAbを含むmAbdAb(BPC1090、BPC1091、BPC1093、BPC1094およびBPC1095)と同様に、両抗体(オリジナルのCDRH3を有する586および変異CDRH3を有する829)は低い凝集レベルを示した。オリジナルのdAbを含む、すなわち89位が変異していなかったBPC2223はL89H変異を有するBPC1092よりも凝集レベルが高かった。代表的な凝集データを表8bに示す。
【表8b】
【0196】
10.5 BIAcore分析
BIAcoreアッセイにおいて、精製されたmAbおよびmAbdAb構築物について、89位の変異がdAbのIL−4への結合に対して何らかの影響を与えるかどうかを決定するために試験した。
【0197】
プロテインAを第1級アミンカップリングによりCM5チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。組換え大腸菌発現ヒトIL4をアナライトとして256、64、16、4および1、0.25および0.0625で使用し、0nM(すなわち緩衝液単独)を結合曲線の二重照合に使用した。抗プロテインA表面を50mM NaOHで再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【表8c】
【実施例11】
【0198】
CDRH3変異型抗IL−13 mAbおよび変異dAbを含む抗原結合タンパク質(BPC1085、BPC1086およびBPC1087)の構築および試験
11.1構築および発現
代替プラスミド構築物を生成するための塩基構築物として抗IL−13 mAbおよび抗IL−4dAbからなる重鎖をコードするプラスミドを使用した。2工程のクローニング戦略を要した。工程1では、HindIIIおよびSpeIを用いた制限クローニングにより、H鎖の抗IL13 mAb成分のVHをコードするDNA配列を別のヒト化抗IL13抗体のVH(配列番号54)をコードするDNA配列と置換した。工程2では、mAbdAbの抗IL4dAb(DOM9−155−154、配列番号80)成分のKabat89位でロイシンコードコドンを部位指向変異によりグルタミンに変異した。生成した重鎖DNA配列はすべて配列番号96、98および100に提示される。構築および発現した分子の一覧を表9に提供する。
【表9】
【0199】
293fectin(Invitrogen、12347019)を用いて、BPC1085、BPC1086およびBPC1087 mAbdAbをコードする重鎖および軽鎖発現プラスミドをHEK 2936E細胞内に共トランスフェクトした。24時間後にトリプトン供給物を各細胞培養に添加して、72時間後に細胞を回収した。プロテインAカラムを用いて抗体を精製後、結合アッセイにおいて試験した。
【0200】
プロテインA親和性を用いてBPC1085、BPC1086およびBPC1087 mAbdAbを精製した。AKTA Xpress系上で1mlプロテインAカラム(GE Healthcare)を用いて、カラムをPBS(Gibco/Invitrogen)中で平衡させて、Pierce IgG溶出液を用いて抗体を溶出した。1Mトリス(ヒドロキシメチル)アミノメタン緩衝液(通常5〜10%v/v)を用いて溶出した留分を中和した。溶出した抗体留分をプールして、サイズ排除クロマトグラフィーにより凝集分析し、分光光度計を用いてOD280nmでの読み取りにより定量化した。
【0201】
これらを、表10に記載する等価物mAbdAb(2222、2223、2230および2231)と比較した。これらは、以下を含む:
i)BPC2222、BPC2223、BPC2230およびBPC2231において「L」であり、BPC1085、BPC1086およびBPC1087において「Q」である89位を除きBPC1085、BPC1086およびBPC1087に使用されるものと同一である1つのdAb。
【0202】
ii)同一リンカー
iii)BPC2222、BPC2223、BPC2230およびBPC2231において「Y」であり、BPC1085、BPC1086およびBPC1087において「V」である100B位を除きBPC1085、BPC1086およびBPC1087に使用されるものと同一である1つのIL−13 mAb配列。
【表10】
【0203】
プロテインA親和性を用いてBPC2222、2223、2230および2231 mAbdAbを精製した。AKTA Xpress系上で1mlプロテインAカラム(GE Healthcare)を用いて、カラムをPBS(Gibco/Invitrogen)中で平衡させて、Pierce IgG溶出液を用いて抗体を溶出した。1Mトリス(ヒドロキシメチル)アミノメタン緩衝液(通常5〜10%v/v)を用いて溶出した留分を中和した。溶出した抗体留分をプールして、サイズ排除クロマトグラフィーにより凝集分析し、分光光度計を用いてOD280nmでの読み取りにより定量化した。
【0204】
BPC2222、2223、2230および2231は30〜40%の凝集を示し、凝集した物質は10分前に溶出した。
【0205】
サイズ排除クロマトグラフィーにより評価したところ、構築物BPC1085、1086および1087はBPC2222、2223、2230および2231より低い凝集レベルを示した。これらの分子のSECプロファイルを図3〜9に示す。
【0206】
11.2 IL−4結合ELISA
実施例10.3に記載の方法に従った直接結合ELISAにおいて、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbのIL−4への結合を試験した。
【0207】
これらのデータを図10に示す。ELISA結果により、これらの精製されたmAbdAbはIL−4と結合したことを確認した。陽性対照の抗IL−4 mAbおよびBPC2231もIL−4への結合を示したのに対し、陰性対照mAb(抗IL−13 mAb)はIL−4と結合しないことが示された。これは、このELISAにおいて、リンカーの長さをGS(TVAAPSGS)からGS(TVAAPSGS)2〜4に伸長時にdAb効能が向上したことを示した。
【0208】
11.3 TF−1細胞増殖バイオアッセイにおけるIL−4中和
TF−1細胞バイオアッセイにおいて、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbのヒトIL−4中和を試験した。
【0209】
ヒトIL−4を含むいくつかの異なるサイトカインに対する反応においてTF−1細胞は増殖する。したがってIL−4のためのこれらの細胞の増殖反応を用いてIL−4の生体活性を測定し、続いてアッセイを開発してmAbdAbのIL−4中和効能(IL−4生体活性の阻害)を決定することができる。
【0210】
アッセイは滅菌96ウェル組織培養プレートにおいて滅菌条件下で実施して、すべての試験ウェルは二連で実施した。約2.2ng/mlの組換え大腸菌発現ヒトIL−4を様々な希釈のmAbdAb(通常3倍希釈液で滴定した、560nM〜0.009nM)の総体積120μlと37℃で1時間プレインキュベートした。無関係な特異性抗体を陰性対照(抗IL13 mAb)として同様に滴定した。次いでこれらの試料50μlを、滅菌96ウェル組織培養プレート中TF−1細胞50μl(濃度2×105細胞/ml)に添加した。したがって最終100μlアッセイ体積には、様々な希釈のmAbdAb(3倍希釈液で滴定した最終濃度270nM〜0.005nM)、組換え大腸菌発現ヒトIL−4(最終濃度1.1ng/ml)およびTF−1細胞(最終濃度1×105細胞/ml)を含有した。アッセイプレートは加湿CO2インキュベーター内で37℃で約4日間インキュベートした。次いでPromega製「Cell Titre96(登録商標)Non−Radioactive Cell Proliferation Assay」(カタログ番号G4100)を製造業者の説明書に記載どおりに用いて、細胞増殖量を決定した。96ウェルプレート中の試料の吸光度はプレートリーダーで570nmで読み取った。これらのデータをエクセルシートに入力し、2回の試験ウェル値を平均化して、平均背景値(mAb−dAbなしおよびIL−4試験ウェルなし)を差し引いた。
【0211】
組換え大腸菌発現ヒトIL−4の生体活性を中和するmAbdAbの能力は定義した量のヒトIL−4(1.1ng/ml)の生体活性を50%中和するために必要なmAb−dAb濃度(=ND50)として表した。必要なmAbdAb濃度が低いほど、中和能力は強力である。本明細書に提供するND50データ(表11)はエクセルのRobosage関数を用いて算出した。これらのデータを図11に図式的に表す。
【0212】
TF−1細胞バイオアッセイにおいて、抗IL−4 mAbおよびDOM9−155−154(配列番号80)をヒトおよびカニクイザルIL−4中和の陽性対照として含んだ。さらに、TF−1細胞バイオアッセイにおいて、無関係な抗原(ダミーdAb)に特異的なdAbもヒトまたはカニクイザルIL−4中和の陰性対照として含んだ。
【0213】
これらを数回反復し、これらの実験の1つの結果を図11に示す。ND50値をデータセットから算出した。ND50値は、IL−4の生体活性を50%中和できるmAbdAbまたはmAbまたはdAb濃度である。平均ND50値および試験回数(n)を表11に示す。
【表11】
【0214】
これらのデータにより、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbは、ヒトおよびカニクイザルIL−4の生体活性を中和したことを確認した。抗IL−4 mAbおよびDOM9−155−154もヒトおよびカニクイザルIL−4の生体活性を中和し、陰性dAb(ダミーdAb)は同じバイオアッセイにおいて中和を示さなかった。
【0215】
全3つのmAbdAbは良好な効能を示し、リンカーの長さが伸びるにつれてdAb効能の明らかな増大傾向があることが中和アッセイから明らかであった(ただし、より粗いELISAではこの効能差が見出されなかった)。陰性対照の抗IL−4 mAbは同じバイオアッセイにおいて中和を示さなかった。
【0216】
11.4 TF−1細胞増殖バイオアッセイにおけるヒトIL−13の中和
TF−1細胞バイオアッセイにおいて、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbのヒトIL−13の中和を下記のとおり試験した。
【0217】
TF−1細胞はいくつかの異なるサイトカイン(ヒトIL−13を含む)に対する反応において増殖する。したがってIL−13のためのこれらの細胞の増殖反応を用いてIL−13の生体活性を測定し、続いてアッセイを開発してmAbdAbのIL−13中和効能(IL−13生体活性の阻害)を決定することができる。
【0218】
アッセイは滅菌96ウェル組織培養プレートにおいて滅菌条件下で実施して、すべての試験ウェルは二連で実施した。約14ng/mlの組換え大腸菌発現ヒトIL−13を様々な希釈のmAbdAb(通常3倍希釈液で滴定した560nM〜0.009nM)の総体積120μlと37℃で1時間プレインキュベートした。無関係な特異性の抗体およびdAbを陰性対照として同様に滴定した(それぞれ抗IL−4 mAbおよびDOM9−155−154)。次いでこれらの試料50μlを、滅菌96ウェル組織培養プレート中TF−1細胞50μl(濃度2×105細胞/ml)に添加した。したがって最終100μlアッセイ体積には様々な希釈のmAbdAb(3倍希釈液で滴定した最終濃度270nM〜0.005nM)、組換え大腸菌発現ヒトIL−13(最終濃度7ng/ml)およびTF−1細胞(最終濃度1×105細胞/ml)を含有した。アッセイプレートは加湿CO2インキュベーター内で37℃で約4日間インキュベートした。次いでPromega製「Cell Titre96(登録商標)Non−Radioactive Cell Proliferation Assay」(カタログ番号G4100)を製造業者の説明書に記載どおりに用いて、細胞増殖量を決定した。96ウェルプレート中の試料の吸光度はプレートリーダーで570nmで読み取った。これらのデータをエクセルシートに入力し、2回の試験ウェル値を平均化して、平均背景値(mAb−dAbなしおよびIL−13試験ウェルなし)を差し引いた。
【0219】
組換え大腸菌発現ヒトIL−13の生体活性を中和するmAbdAbの能力は定義した量のヒトIL−13(7ng/ml)の生体活性を50%中和するために必要なmAb−dAb濃度(=ND50)として表した。必要なmAbdAb濃度が低いほど、中和能力は強力である。本明細書に提供するND50データ(表12)はエクセルのRobosage関数を用いて算出した。これらのデータを図12に図式的に表す。
【0220】
TF−1細胞バイオアッセイにおいて、ヒトIL−13中和の陽性対照として抗IL−13 mAb(配列番号22および24)を含んだ。さらに、抗IL−4 mAbも陰性対照として含んだ。
【0221】
図12は、TF−1細胞中和アッセイ結果を示す。ND50値をデータセットから算出した。ND50値は、IL−13の生体活性を50%中和できるmAbdAbまたはmAbまたはdAb濃度である。
【0222】
平均ND50値および試験回数を表12に示す。
【表12】
【0223】
これらのデータにより、精製されたBPC1085、BPC1086およびBPC1087 mAbdAbは組換えヒトおよびカニクイザルIL−13の生体活性を中和したことが確認される。陰性対照の抗IL−4 mAbは同じバイオアッセイにおいて中和を示さなかった。
【実施例12】
【0224】
抗IL−13 mAb軽鎖の再ヒト化
12.1再ヒト化
一部の抗IL−13 mAb−抗IL−4dAb分子(BPC1085)の発現を改善するために、マウス抗体6A1の軽鎖CDR(配列番号59に説明される軽鎖)を新規フレームワーク上に再グラフト化した。PCRベース戦略およびオリゴヌクレオチド重複を用いてコドン至適化軽鎖可変領域配列(表13に要約する)をde novoで構築した。PCRプライマーはシグナル配列(配列番号56)を組み込み、VLドメインを枠組みするように設計されたHindIIIおよびBsiWI制限部位を含むように設計し、ヒトκC領域を含むpTTおよびRln哺乳動物発現ベクター内へのクローニングを可能にした。構築されている再ヒト化軽鎖を表13に要約する。
【表13】
【0225】
12.2 HEK 293 6E細胞における分子発現
再ヒト化軽鎖の発現特性を初めにmAb型式において検討した。293fectin(Invitrogen、12347019)を用いて、A1Y100BVAL1(配列番号54)重鎖をコードするプラスミド、既存の軽鎖(配列番号24)および再ヒト化軽鎖をHEK 293 6E細胞内に一過性共トランスフェクトした。プラスミドは少量(2×0.75ml培養体積)発現して抗体を産生した。24時間後にトリプトン供給物を細胞培養に添加して、さらに72時間後に細胞を回収した。構築して発現したmAbのすべてを表14に要約する。
【表14】
【0226】
Gyrolabワークステーションを用いた定量免疫アッセイにより、抗体発現を組織培養上清から直接評価した。再ヒト化軽鎖を含む抗体BPC3208およびBPC3211(それぞれP0およびP1で示す)は、A1Y100BVAL1 mAbより改善された発現収率を示した。Q0およびQ1軽鎖(BPC3219およびBPC3220)は抗IL−13 mAbの発現を改善しなかった。発現データを表15に提示する。
【表15】
【0227】
12.3 HEK 293 6E細胞内のmAb−dAb発現
BPC3208およびBPC3211の再ヒト化軽鎖は抗IL−13 mAbの改善された発現を示したため、それらを抗IL−13mAb−抗IL−4dAbとの関連で検討した。293fectin(Invitrogen、12347019)を用いて、再ヒト化軽鎖P0およびP1ならびに586(L1)軽鎖を829H−GS(TVAAPSGS)2−256重鎖(配列番号96、詳細は表16に要約する)とHEK 293 6E細胞内に共トランスフェクトした。プラスミドを50〜500ml発現させて、抗体分子を産生した。24時間後にトリプトン供給物を細胞培養に添加して、さらに48時間後に細胞を回収した。固定化プロテインAカラムを用いて抗体を精製して、吸光度を280nmで読み取ることにより定量化し、指定の場合、精製された抗体分子を以下に説明する実施例に記載のアッセイにおいて評価した。図13および14に例証するとおりBPC3214およびBPC3215をサイズ排除クロマトグラフィー(SEC)により分析した。
【表16】
【0228】
mAb型式における観察と一致して、再ヒト化軽鎖を含むmAbdAb(BPC3214およびBPC3215)は、BPC1085より改善された発現を示した。代表的な発現データを表17 5に要約する。
【表17】
【0229】
12.4 ヒトIL−13結合ELISA
直接結合ELISAを介して、精製されたBPC3214およびBPC3215のヒトIL−13への結合についてBPC1085(実施例10に記載)と比較して試験した。抗IL−13 mAb A1Y100BVAL1および抗IL−4 mAbもそれぞれ陽性対照および陰性対照として検討した。組換え大腸菌発現ヒトIL−13(バッチ番号GRITS31061)5μg/mlを96ウェル高結合プレート50μl/ウェルにコーティングして、+4℃で一晩インキュベートした。続く工程をすべて室温で行った。プレートを0.05%のTween−20含有リン酸緩衝食塩水で3回洗浄した。100μLブロック溶液(0.05%のTween−20含有1%BSAのリン酸緩衝食塩水)を各ウェルに添加して、プレートを室温で少なくとも1時間インキュベートした。次いで別の洗浄工程を実施した。引き続いて精製された抗体をプレート全体でブロック溶液で希釈した。1時間インキュベーション後、プレートを洗浄した。ヤギ抗ヒトκ軽鎖特異的ペルオキシダーゼ抱合抗体をブロック溶液で0.75μg/mlに希釈して、50μlを各ウェルに添加した。プレートを1時間インキュベートした。さらに2つの洗浄工程後、50μlのOPD(o−フェニレンジアミン二塩酸塩)SigmaFast基質溶液を各ウェルに添加して、3M硫酸50μLを添加することによって反応を停止させた。基本終点プロトコルを用いて、VersaMaxマイクロプレートリーダー(Molecular Devices)を用いて吸光度を490nmで読み取った。これらのデータを図15に示す。直接結合ELISAにより、BPC3214およびBPC3215はヒトIL−13と結合することを確認した。BPC3214およびBPC3215はBPC1085と類似のIL−13結合効能を示す。陽性対照の抗IL−13 mAb A1Y100BVAL1も組換えIL−13への結合を示したのに対し、陰性対照の抗IL−4 mAbはヒトIL−13と結合しないことが示された。
【0230】
12.5 ヒトIL−4結合ELISA
直接結合ELISAにおいて、精製されたBPC3214およびBPC3215の組換え大腸菌発現ヒトIL−4への結合も試験した。ELISAを実施例4に記載のように実施し、組換え大腸菌発現ヒトIL−4 5μg/mlを96ウェル高結合プレート50μl/ウェルにコーティングして、+4℃で一晩インキュベートした。これらのデータを図16に示す。直接結合ELISAにより、BPC3214およびBPC3215はヒトIL−4と結合することが確認される。BPC1085も検討した。BPC3214はBPC1085と類似のIL−4結合効能を示す。陽性対照の抗IL−4 mAbも組換えIL−4への結合を示したのに対し、陰性対照の抗IL−13 mAb A1Y100BVAL1はヒトIL−4に対して結合しないことが示された。
【実施例13】
【0231】
BIAcore(商標)分析により評価したオリジナルのIL−13 mAb CDRH3を含むmAbdAb(BPC2222、BPC2223およびBPC2230〜2231)のIL−13およびIL−4結合親和性
方法
プロテインAを第1級アミンカップリングによりC1チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。組換え大腸菌発現ヒトIL13を256、64、16、4、および1nMで使用し、組換え大腸菌発現ヒトIL4を64、16、4、1および0.25nMで使用し、0nM(すなわち緩衝液単独)をIL4結合とIL13結合の両方の結合曲線の二重照合に使用した。プロテインA表面を100mMリン酸で再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【0232】
ヒトIL13への結合結果を表18に示し、ヒトIL4への結合結果を表19に示す。
【表18】
【0233】
【表19】
【実施例14】
【0234】
BIAcore(商標)分析により評価したオリジナルのIL−13 mAb CDRH3を含むmAbdAb(BPC2222、BPC2231)および変異型抗IL−13 mAb CDRH3(BPC1085〜1087)のIL−13およびIL−4結合親和性
方法
プロテインAを第1級アミンカップリングによりCM5チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。組換え大腸菌発現ヒトIL13を256nMのみで使用し、組換え大腸菌発現ヒトIL4を64、16、4および1nMで使用し、0nM(すなわち緩衝液単独)をIL4結合とIL13結合の両方の結合曲線の二重照合に使用した。プロテインA表面を50mM NaOHで再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【0235】
ヒトIL13への結合結果を表20に示し、ヒトIL4への結合結果を表21に示す。
【表20】
【0236】
BPC1086およびBPC1087の結合速度はBiacoreの感度外であるが、我々がこのデータを精密に分析できないという事実から、IL4との相互作用は速い結合速度での高親和性のものである可能性が高いことが示される。
【表21】
【実施例15】
【0237】
BIAcore(商標)分析により評価したいくつかの変異型抗IL−13 mAb CDRH3を含むmAbdAb(BPC1085、BPC1090−BPC1095、およびBPC1108−BPC1119)のIL−4結合親和性
方法(ヒトIL−4結合親和性)
プロテインAを第1級アミンカップリングによりCM5チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。
【0238】
組換え大腸菌発現ヒトIL4を64、16、4、1および0.25nMで使用した。すべての結合曲線を0nM注入(すなわち緩衝液単独)で二重照合した。
【0239】
プロテインA表面を50mM NaOHで再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【0240】
ヒトIL4への結合結果を表22に示す。
【表22】
【実施例16】
【0241】
BIAcore(商標)分析により評価して、オリジナルの軽鎖(BPC1085)と比較した再ヒト化軽鎖を含むmAbdAb(BPC3214およびBPC3215)のIL−13結合親和性
方法(ヒトおよびカニクイザルIL−13結合親和性)
プロテインAを第1級アミンカップリングによりCM5チップ上に固定化した;この表面を、試験する抗体分子の捕捉表面として使用した。
【0242】
組換え大腸菌発現ヒトIL13およびカニクイザルIL13を64、16、4、1および0.25nM使用した。すべての結合曲線を0nM注入(すなわち緩衝液単独)で二重照合した。
【0243】
プロテインA表面を50mM NaOHで再生した。HBS−EPをランニング緩衝液として用いてアッセイを25℃で実行した。データは、Biacore T100分析ソフトウェアに固有の1:1モデルに適合させた。
【0244】
ヒトおよびカニクイザルIL13への結合結果を表23に示す。
【表23】
【実施例17】
【0245】
変異dAbを伴うおよび伴わないmAbdAbのストレッサー試験
いくつかのmAbdAbをPBSまたは50mM酢酸緩衝液に入れて、37℃で最大14日間インキュベートした。次いで、視認可能な沈殿物の存在、溶解性凝集および濃度安定性アドヒアランスについて分析した。
【0246】
結果により、変異dAbを含むmAbdAb(BPC2222、2223、2230、2231)は、非変異dAb(BPC1085、1086、1087)と同様に動作することが示され、両種類のmAbdAbはPBSと酢酸緩衝液の両方において37℃、2週間のインキュベーション期間に溶液中のタンパク質濃度に変化がなかったことから、安定していると思われた。加えて溶液中の凝集レベルには変化はないかほとんど認められず、沈殿は観察されなかった。
【実施例18】
【0247】
PK評価
ラットへのIV投与後の分離した試験において、BPC1085、BPC1086、およびBPC1087の薬力学を調査した。BPC1085のPKもカニクイザルへのIV投与後に調査した。
【0248】
ラットにおける全3分子およびサルにおけるBPC1085のPKが標準的mAbのPKと一致することを見出した。
【表24】
【0249】
【0250】
【0251】
【0252】
【0253】
【0254】
【0255】
【0256】
【0257】
【0258】
【0259】
【0260】
【0261】
【0262】
【0263】
【0264】
【0265】
【0266】
【0267】
【0268】
【0269】
【0270】
【0271】
【0272】
【0273】
【0274】
【0275】
【0276】
【0277】
【0278】
【0279】
【0280】
【0281】
【0282】
【0283】
【0284】
【0285】
【0286】
【0287】
【0288】
【特許請求の範囲】
【請求項1】
ヒトIL−13と結合し、CDRH3 SIYDDYHYDDYYAMDY(配列番号3)を含む抗原結合タンパク質であって、前記CDRH3が以下の置換:
g)1位のS95がトリプトファン(W)に置換されている
h)2位のI96がバリン(V)に置換されている
i)3位のY97がフェニルアラニン(F)に置換されている
j)4位のD98がグルタミン(E)に置換されている
k)7位のH100Aがアラニン(A)、グルタミン酸(E)、グルタミン(Q)、アルギニン(R)、セリン(S)、トレオニン(T)もしくはバリン(V)に置換されている、ならびに
l)8位のY100Bがアラニン(A)、イソロイシン、(I)、トリプトファン(W)もしくはバリン(V)に置換されている
のうち1つ以上を含む、抗原結合タンパク質。
【請求項2】
CDRH3の8位のY100Bがアラニン(A)、イソロイシン、(I)、トリプトファン(W)またはバリン(V)から選択されるアミノ酸に置換されている、請求項1に記載の抗原結合タンパク質。
【請求項3】
ヒトIL−13と結合し、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8および配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14および配列番号15、配列番号16、配列番号17および配列番号18に記載のものから選択されるCDRH3配列を含む、請求項1または2に記載の抗原結合タンパク質。
【請求項4】
前記抗原結合タンパク質がさらに以下のCDR配列:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項5】
前記抗原結合タンパク質が以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号18、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項6】
前記抗原結合タンパク質が以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号17、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項7】
前記抗原結合タンパク質が以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号16、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項8】
前記抗原結合タンパク質が以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号15、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項9】
前記抗原結合タンパク質が抗体を含む、請求項1〜8のいずれか一項に記載の抗原結合タンパク質。
【請求項10】
前記抗体がヒト化抗体である請求項9に記載の抗体。
【請求項11】
前記抗体がIgGアイソタイプである請求項10の抗原結合タンパク質。
【請求項12】
配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52および配列番号54から選択される重鎖;ならびに配列番号24、108、110、112および114の軽鎖を含む、請求項1〜11のいずれか一項に記載の抗原結合タンパク質。
【請求項13】
配列番号48、配列番号50、配列番号52および配列番号54から選択される重鎖;ならびに配列番号108および110の軽鎖を含む、請求項13に記載の抗原結合タンパク質。
【請求項14】
配列番号54の重鎖および配列番号108の軽鎖を含む、請求項13に記載の抗原結合タンパク質。
【請求項15】
配列番号54の重鎖および配列番号110の軽鎖を含む、請求項13に記載の抗原結合タンパク質。
【請求項16】
前記抗体が低減したADCCおよび/または補体活性化を示すように変異したFc領域を含む、請求項10〜請求項15のいずれか一項に記載の抗体。
【請求項17】
IL−13と結合でき、IL−4およびIL−5のうち少なくとも1つとも結合できる、請求項1〜16のいずれか一項に記載の抗原結合タンパク質。
【請求項18】
配列番号78、79、80、81および94から選択される少なくとも1つのdAbを含む、請求項17に記載の抗原結合タンパク質。
【請求項19】
配列番号62、64、66、68、70、72、74、76、96、98、100、102、104、106、および117〜138のうちいずれか1つから選択される重鎖ならびに配列番号24、108、110、112および114の軽鎖を含む、請求項18に記載の抗原結合タンパク質。
【請求項20】
配列番号96、98、100、102、104、106のいずれか1つから選択される重鎖ならびに配列番号24、108および110の軽鎖を含む、請求項19に記載の抗原結合タンパク質。
【請求項21】
配列番号96の重鎖および配列番号24の軽鎖、または配列番号96の重鎖および配列番号108の軽鎖、または配列番号96の重鎖および配列番号110の軽鎖、または配列番号98の重鎖および配列番号24の軽鎖、または配列番号98の重鎖および配列番号108の軽鎖、または配列番号98の重鎖および配列番号110の軽鎖、または配列番号100の重鎖および配列番号24の軽鎖、または配列番号100の重鎖および配列番号108の軽鎖、または配列番号100の重鎖および配列番号110の軽鎖を含む、請求項20に記載の抗原結合タンパク質。
【請求項22】
第1および第2ベクターを含む形質転換またはトランスフェクトされた組換え宿主細胞であって、前記第1ベクターは請求項1〜21のいずれか一項に記載の抗体の重鎖をコードするポリヌクレオチドを含み、前記第2ベクターは請求項1〜21のいずれか一項に記載の軽鎖をコードするポリヌクレオチドを含む、細胞。
【請求項23】
請求項1〜請求項21のいずれか一項に記載の抗原結合タンパク質および医薬上許容可能な担体を含む医薬組成物。
【請求項24】
アレルギー性喘息、重症喘息、困難性喘息、ブリットル喘息、夜間性喘息、月経前喘息、ステロイド抵抗性喘息、ステロイド依存性喘息、アスピリン誘発性喘息、成人発症型喘息、小児喘息、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、COPD、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、細胞周期調節疾患(ホジキン病など)、B細胞慢性リンパ球性白血病に罹患しているヒト患者の治療方法であって、治療有効量の請求項1〜21の抗原結合タンパク質を投与する工程を含む、方法。
【請求項25】
アレルギー性喘息、重症喘息、困難性喘息、ブリットル喘息、夜間性喘息、月経前喘息、ステロイド抵抗性喘息、ステロイド依存性喘息、アスピリン誘発性喘息、成人発症型喘息、小児喘息、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、COPD、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、細胞周期調節疾患(ホジキン病など)、B細胞慢性リンパ球性白血病の治療または予防のための医薬の調製における、請求項1〜請求項21のいずれか一項に記載の抗原結合タンパク質の使用。
【請求項26】
アレルギー性喘息、重症喘息、困難性喘息、ブリットル喘息、夜間性喘息、月経前喘息、ステロイド抵抗性喘息、ステロイド依存性喘息、アスピリン誘発性喘息、成人発症型喘息、小児喘息、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、COPD、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、細胞周期調節疾患(ホジキン病など)、B細胞慢性リンパ球性白血病の治療または予防における使用のための、請求項1〜請求項21のいずれか一項に記載の抗原結合タンパク質。
【請求項27】
免疫グロブリン単一可変ドメイン凝集の低減方法であって、
a)免疫グロブリン単一可変ドメインの凝集レベルを測定する工程と、
b)kabat89位を「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)、「A」(アラニン)、「T」(トレオニン)および「E」(グルタミン酸)から選択される残基に変異させる工程と、
c)変異免疫グロブリン単一可変ドメインの凝集レベルを測定する工程と
を含み、
その結合活性は保持されているが、前記変異免疫グロブリン単一可変ドメインの凝集レベルは非変異免疫グロブリン単一可変ドメインの凝集レベルより低い、方法。
【請求項28】
前記kabat残基89が「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)および「E」(グルタミン酸)から選択される残基に変異する、請求項27に記載の方法。
【請求項29】
前記kabat残基89が「Q」(グルタミン)に変異する、請求項28に記載の方法。
【請求項30】
ヒトVK dAbであって、IGKV1−17、IGKV1D−17、IGKV1/OR2−108、IGKV1−6、IGKV5−2、IGKV1D−42、IGKV2−24、IGKV2−28、IGKV2−30、IGKV2−40、IGKV2D−29、IGKV2D−30、IGKV2D−24およびIGKV6−21から選択される生殖系列フレームワークに由来し、かつその残基89(kabat番号付け)がグルタミンである、ヒトVK dAb。
【請求項31】
配列番号94の配列を含む、免疫グロブリン単一可変ドメイン。
【請求項1】
ヒトIL−13と結合し、CDRH3 SIYDDYHYDDYYAMDY(配列番号3)を含む抗原結合タンパク質であって、前記CDRH3が以下の置換:
g)1位のS95がトリプトファン(W)に置換されている
h)2位のI96がバリン(V)に置換されている
i)3位のY97がフェニルアラニン(F)に置換されている
j)4位のD98がグルタミン(E)に置換されている
k)7位のH100Aがアラニン(A)、グルタミン酸(E)、グルタミン(Q)、アルギニン(R)、セリン(S)、トレオニン(T)もしくはバリン(V)に置換されている、ならびに
l)8位のY100Bがアラニン(A)、イソロイシン、(I)、トリプトファン(W)もしくはバリン(V)に置換されている
のうち1つ以上を含む、抗原結合タンパク質。
【請求項2】
CDRH3の8位のY100Bがアラニン(A)、イソロイシン、(I)、トリプトファン(W)またはバリン(V)から選択されるアミノ酸に置換されている、請求項1に記載の抗原結合タンパク質。
【請求項3】
ヒトIL−13と結合し、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8および配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14および配列番号15、配列番号16、配列番号17および配列番号18に記載のものから選択されるCDRH3配列を含む、請求項1または2に記載の抗原結合タンパク質。
【請求項4】
前記抗原結合タンパク質がさらに以下のCDR配列:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項5】
前記抗原結合タンパク質が以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号18、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項6】
前記抗原結合タンパク質が以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号17、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項7】
前記抗原結合タンパク質が以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号16、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項8】
前記抗原結合タンパク質が以下のCDR:
CDRH1:配列番号1、
CDRH2:配列番号2、
CDRH3:配列番号15、
CDRL1:配列番号19、
CDRL2:配列番号20、および
CDRL3:配列番号21、
を含む、請求項1〜請求項3のいずれか一項に記載の抗原結合タンパク質。
【請求項9】
前記抗原結合タンパク質が抗体を含む、請求項1〜8のいずれか一項に記載の抗原結合タンパク質。
【請求項10】
前記抗体がヒト化抗体である請求項9に記載の抗体。
【請求項11】
前記抗体がIgGアイソタイプである請求項10の抗原結合タンパク質。
【請求項12】
配列番号26、配列番号28、配列番号30、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52および配列番号54から選択される重鎖;ならびに配列番号24、108、110、112および114の軽鎖を含む、請求項1〜11のいずれか一項に記載の抗原結合タンパク質。
【請求項13】
配列番号48、配列番号50、配列番号52および配列番号54から選択される重鎖;ならびに配列番号108および110の軽鎖を含む、請求項13に記載の抗原結合タンパク質。
【請求項14】
配列番号54の重鎖および配列番号108の軽鎖を含む、請求項13に記載の抗原結合タンパク質。
【請求項15】
配列番号54の重鎖および配列番号110の軽鎖を含む、請求項13に記載の抗原結合タンパク質。
【請求項16】
前記抗体が低減したADCCおよび/または補体活性化を示すように変異したFc領域を含む、請求項10〜請求項15のいずれか一項に記載の抗体。
【請求項17】
IL−13と結合でき、IL−4およびIL−5のうち少なくとも1つとも結合できる、請求項1〜16のいずれか一項に記載の抗原結合タンパク質。
【請求項18】
配列番号78、79、80、81および94から選択される少なくとも1つのdAbを含む、請求項17に記載の抗原結合タンパク質。
【請求項19】
配列番号62、64、66、68、70、72、74、76、96、98、100、102、104、106、および117〜138のうちいずれか1つから選択される重鎖ならびに配列番号24、108、110、112および114の軽鎖を含む、請求項18に記載の抗原結合タンパク質。
【請求項20】
配列番号96、98、100、102、104、106のいずれか1つから選択される重鎖ならびに配列番号24、108および110の軽鎖を含む、請求項19に記載の抗原結合タンパク質。
【請求項21】
配列番号96の重鎖および配列番号24の軽鎖、または配列番号96の重鎖および配列番号108の軽鎖、または配列番号96の重鎖および配列番号110の軽鎖、または配列番号98の重鎖および配列番号24の軽鎖、または配列番号98の重鎖および配列番号108の軽鎖、または配列番号98の重鎖および配列番号110の軽鎖、または配列番号100の重鎖および配列番号24の軽鎖、または配列番号100の重鎖および配列番号108の軽鎖、または配列番号100の重鎖および配列番号110の軽鎖を含む、請求項20に記載の抗原結合タンパク質。
【請求項22】
第1および第2ベクターを含む形質転換またはトランスフェクトされた組換え宿主細胞であって、前記第1ベクターは請求項1〜21のいずれか一項に記載の抗体の重鎖をコードするポリヌクレオチドを含み、前記第2ベクターは請求項1〜21のいずれか一項に記載の軽鎖をコードするポリヌクレオチドを含む、細胞。
【請求項23】
請求項1〜請求項21のいずれか一項に記載の抗原結合タンパク質および医薬上許容可能な担体を含む医薬組成物。
【請求項24】
アレルギー性喘息、重症喘息、困難性喘息、ブリットル喘息、夜間性喘息、月経前喘息、ステロイド抵抗性喘息、ステロイド依存性喘息、アスピリン誘発性喘息、成人発症型喘息、小児喘息、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、COPD、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、細胞周期調節疾患(ホジキン病など)、B細胞慢性リンパ球性白血病に罹患しているヒト患者の治療方法であって、治療有効量の請求項1〜21の抗原結合タンパク質を投与する工程を含む、方法。
【請求項25】
アレルギー性喘息、重症喘息、困難性喘息、ブリットル喘息、夜間性喘息、月経前喘息、ステロイド抵抗性喘息、ステロイド依存性喘息、アスピリン誘発性喘息、成人発症型喘息、小児喘息、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、COPD、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、細胞周期調節疾患(ホジキン病など)、B細胞慢性リンパ球性白血病の治療または予防のための医薬の調製における、請求項1〜請求項21のいずれか一項に記載の抗原結合タンパク質の使用。
【請求項26】
アレルギー性喘息、重症喘息、困難性喘息、ブリットル喘息、夜間性喘息、月経前喘息、ステロイド抵抗性喘息、ステロイド依存性喘息、アスピリン誘発性喘息、成人発症型喘息、小児喘息、アトピー性皮膚炎、アレルギー性鼻炎、クローン病、COPD、線維性疾患もしくは障害(特発性肺線維症など)、進行性全身性硬化症、肝線維症、肝肉芽腫、住血吸虫症、リーシュマニア症、細胞周期調節疾患(ホジキン病など)、B細胞慢性リンパ球性白血病の治療または予防における使用のための、請求項1〜請求項21のいずれか一項に記載の抗原結合タンパク質。
【請求項27】
免疫グロブリン単一可変ドメイン凝集の低減方法であって、
a)免疫グロブリン単一可変ドメインの凝集レベルを測定する工程と、
b)kabat89位を「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)、「A」(アラニン)、「T」(トレオニン)および「E」(グルタミン酸)から選択される残基に変異させる工程と、
c)変異免疫グロブリン単一可変ドメインの凝集レベルを測定する工程と
を含み、
その結合活性は保持されているが、前記変異免疫グロブリン単一可変ドメインの凝集レベルは非変異免疫グロブリン単一可変ドメインの凝集レベルより低い、方法。
【請求項28】
前記kabat残基89が「Q」(グルタミン)、「G」(グリシン)、「S」(セリン)、「M」(メチオニン)および「E」(グルタミン酸)から選択される残基に変異する、請求項27に記載の方法。
【請求項29】
前記kabat残基89が「Q」(グルタミン)に変異する、請求項28に記載の方法。
【請求項30】
ヒトVK dAbであって、IGKV1−17、IGKV1D−17、IGKV1/OR2−108、IGKV1−6、IGKV5−2、IGKV1D−42、IGKV2−24、IGKV2−28、IGKV2−30、IGKV2−40、IGKV2D−29、IGKV2D−30、IGKV2D−24およびIGKV6−21から選択される生殖系列フレームワークに由来し、かつその残基89(kabat番号付け)がグルタミンである、ヒトVK dAb。
【請求項31】
配列番号94の配列を含む、免疫グロブリン単一可変ドメイン。
【図1】

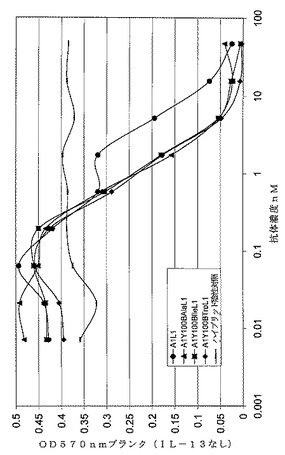
【図2】
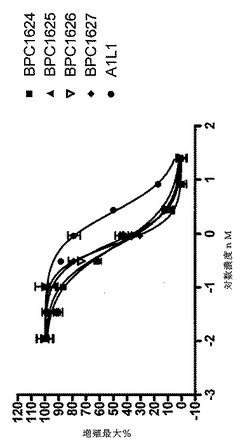
【図3】
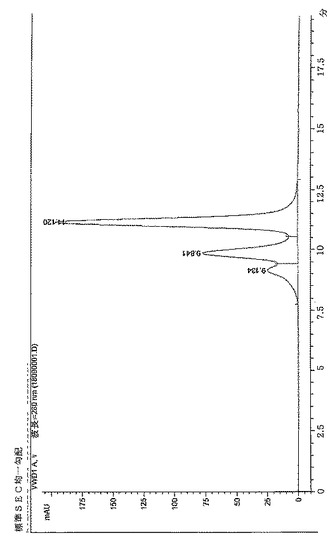
【図4】
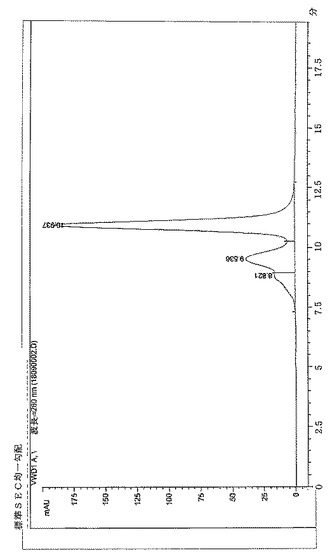
【図5】
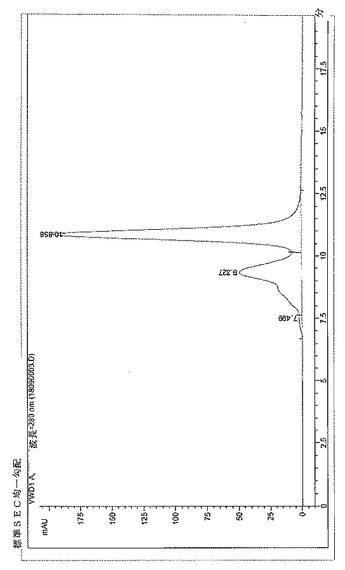
【図6】
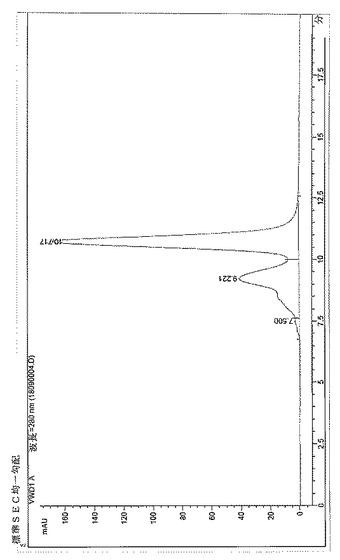
【図7】
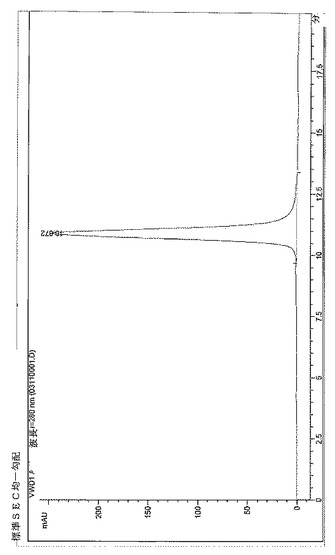
【図8】
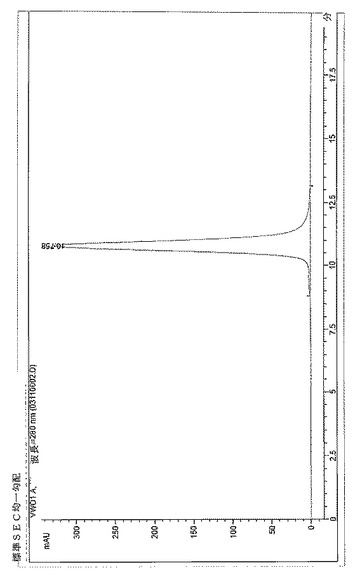
【図9】
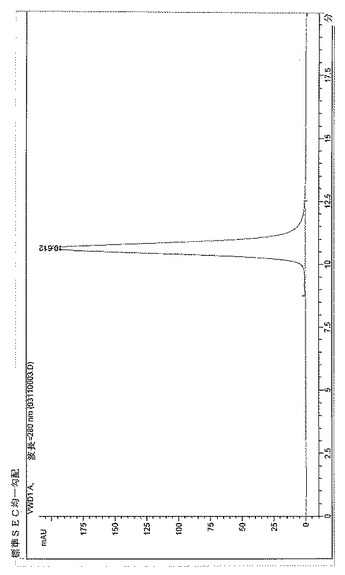
【図10】
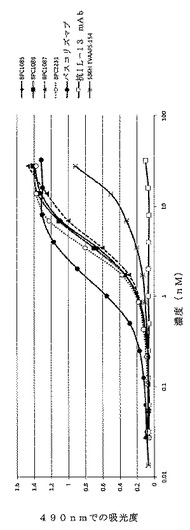
【図11】
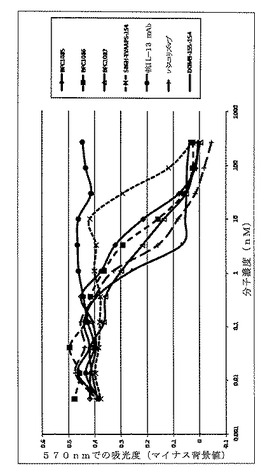
【図12】
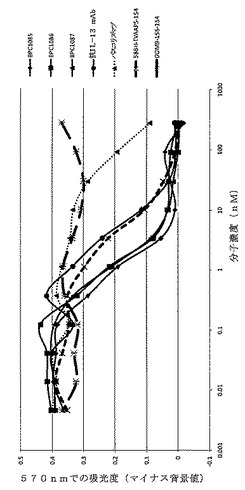
【図13】
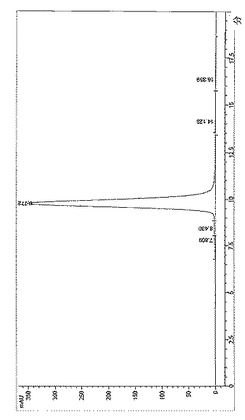
【図14】
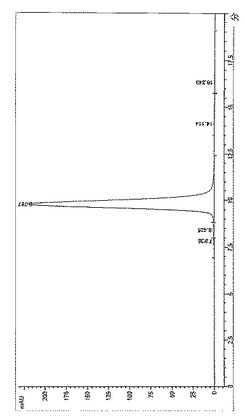
【図15】
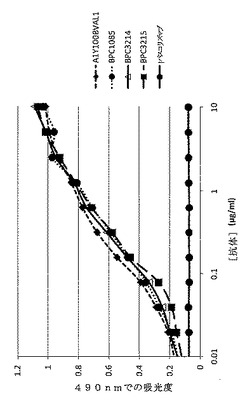
【図16】
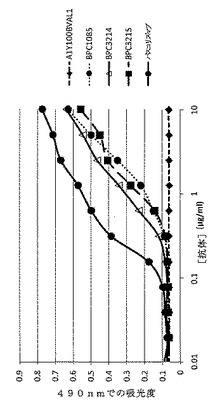
【図17】
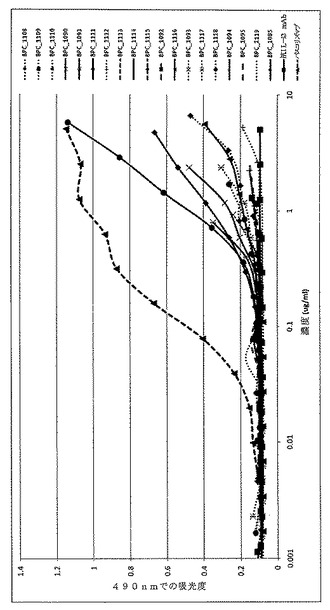
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2012−528110(P2012−528110A)
【公表日】平成24年11月12日(2012.11.12)
【国際特許分類】
【出願番号】特願2012−512352(P2012−512352)
【出願日】平成22年5月26日(2010.5.26)
【国際出願番号】PCT/EP2010/057228
【国際公開番号】WO2010/136481
【国際公開日】平成22年12月2日(2010.12.2)
【出願人】(397009934)グラクソ グループ リミテッド (832)
【氏名又は名称原語表記】GLAXO GROUP LIMITED
【住所又は居所原語表記】Glaxo Wellcome House,Berkeley Avenue Greenford,Middlesex UB6 0NN,Great Britain
【Fターム(参考)】
【公表日】平成24年11月12日(2012.11.12)
【国際特許分類】
【出願日】平成22年5月26日(2010.5.26)
【国際出願番号】PCT/EP2010/057228
【国際公開番号】WO2010/136481
【国際公開日】平成22年12月2日(2010.12.2)
【出願人】(397009934)グラクソ グループ リミテッド (832)
【氏名又は名称原語表記】GLAXO GROUP LIMITED
【住所又は居所原語表記】Glaxo Wellcome House,Berkeley Avenue Greenford,Middlesex UB6 0NN,Great Britain
【Fターム(参考)】
[ Back to top ]