
IL−17A拮抗物質
インターロイキン−17A(IL−17A)抗体拮抗物質、IL−17A抗体拮抗物質又はそのフラグメントをコードするポリヌクレオチド、並びに前述のものを製造及び使用するが開示される。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、インターロイキン−17A(IL−17A)抗体拮抗物質、IL−17A抗体拮抗物質又はそのフラグメントをコードするポリヌクレオチド、並びに上記のものを製造及び使用する方法に関する。
【背景技術】
【0002】
インターロイキン−17A(IL−17A、CTLA−8、IL−17)は、IL−23及びTGF−β等のサイトカインに応答して、活性化Th17細胞、CD8+ T細胞、γδ T細胞、及びNK細胞によって分泌されるサイトカインであり、好中球生態、炎症、臓器破壊、及び宿主細胞防衛に関与する、線維芽細胞及び滑膜細胞等の多数の細胞タイプからの、抗菌性ペプチド(デフェンシン)、炎症促進性サイトカイン、及びケモカイン等のメディエーターの産生を調節する(Weaver et al.,Annu.Rev.Immunol.25:821〜52,2007、Aggarwal et al.,J.Biol.Chem.278:1910〜4,2003、Mangan et al.,Nature 441:231〜4,2006に概説される)。IL−17Aは、TNF−α及びIL−1β等の他のサイトカインと協同して、炎症促進性環境を増強する。
【0003】
IL−17Aサイトカインファミリーは、IL−17A、B、C、D、E、及びFと指定される6つの同族体から成り、各々、相違する特有の生物学的役割を有する(Kawaguchi et al.,J.Allergy Clin.Immunol.114:1265〜73,2004、Kolls and Linden,Immunity 21:467〜76,2004、Moseley et al.,Cytokine Growth Factor Rev.14:155〜74,2003)。このファミリーメンバーのうち、IL−17Fは、IL−17Aと最も相同であり、肺における好中球増加症の誘導及び炎症促進性サイトカインの誘導等の多くの類似した機能特性を共有するが、人間においては、IL−17Fは、IL−17Aよりも約10倍効力が低い(Moseley et al.,Cytokine Growth Factor Rev.14:155〜74,2003、Kolls et al.,Immunity,21:467〜76,2004、McAllister et al.,J.Immunol.175:404〜12,2005)。IL−17A及びIL−17Fはまた、生体外で中間生物活性を有する、ヘテロ二量体を形成することができる(Wright et al.,J.Biol.Chem.282:13447〜55,2007)。
【0004】
IL−17Aは、インターロイキン−17受容体A(IL−17RA)及び受容体C(IL−17RC)と相互作用することによってその効果を媒介する(Moseley et al.,Cytokine Growth Factor Rev.14:155〜74,2003、Toy et al.,J.Immunol.177:36〜9,2006)。IL−17Fは、同じ受容体を通じてシグナル伝達するが、これらの受容体へのIL−17F親和性は、それよりも有意に低い(Kuestner et al.,J.Immunol.179:5462〜73,2007)。ヒトIL−17F及びヒトIL−17F/IL−17RA複合体の結晶構造は、ホモ二量体における推定受容体結合空洞を特定した(Hymowitz et al.,EMBO J.20:5332〜41,2001、Ely et al.,Nat.Immunology 10:1245〜51、2009)。類似した空洞は、中和Fabとの複合体におけるヒトIL−17Aの結晶構造において特定されたが、この空洞は、部分的に占領されていた(Gerhardt et al.,J.Mol.Biol.394:905〜21,2009)。
【0005】
IL−17Aの不適切な又は過剰な産生は、関節リウマチ(Lubberts,Cytokine 41:84〜91,2008)、ぜんそく等のアレルギー性気道疾患を含む気道過敏症(Linden,Curr.Opin.Investig.Drugs.4:1304〜12,2003、Ivanov,Trends Pharmacol.Sci.30:95〜103,2009に概説される)、乾癬(Johansen et al.,Br.J.Dermatol.160:319〜24,2009)、アトピー性皮膚炎を含む皮膚過敏症(Toda et al.,J.Allergy Clin.Immunol.111:875〜81,2003)、全身性硬化症(Fujimoto et al.,J.Dermatolog.Sci.50:240〜42,2008)、潰瘍性大腸炎及びクローン病を含む炎症性腸疾患(Holtta et al.,Inflamm.Bowel Dis.14:1175〜84,2008、Zhang et al.,Inflamm.Bowel Dis.12:382〜88,2006)、並びに慢性閉塞性肺疾患を含む肺疾患(Curtis et al.,Proc.Am.Thorac.Soc 4:512〜21,2007)を含む、種々の疾患及び傷害の病理に関連する。
【発明の概要】
【発明が解決しようとする課題】
【0006】
IL−17Aに対する抗体は、IL−17A媒介型疾患及び傷害の治療における使用のために提案されている(PCT公開第WO08/021156号、第WO07/070750号、第WO07/149032号、第WO06/054059号、第WO06/013107号、第WO08/001063号、第WO10/034443号、米国特許出願第US2008/095775号、第US2009/0175881号)。抗体治療薬の薬物動態学、有効性、及び安全性プロファイルは、具体的な組成に応じるため、IL−17A媒介型疾患及び傷害の治療における使用に好適な、ヒトIL−17Aに対する改善された抗体の必要性が存在する。
【課題を解決するための手段】
【0007】
本発明の一態様は、単離された抗体又はそのフラグメントであり、該抗体は、アミノ酸残基56〜68(配列番号157)及び100〜116(配列番号158)において、又は残基L26、R55、E57、P59、E60、R61、Y62、S64、V65、W67、R101、E102、P103、及びF110において、配列番号105に示される配列を有するヒトIL−17Aに特異的に結合する。
【0008】
本発明の別の態様は、単離された抗体又はそのフラグメントであり、抗体は、ヒトIL−17A上のP2ポケット空洞に特異的に結合し、このP2ポケット空洞は、配列番号105のアミノ酸残基V22、V24、L26、I28、Y62、L99、R101、F110、及びL112を含む。
【0009】
本発明の別の態様は、ヒトIL−17Aに特異的に結合し、ある種の重鎖相補性決定領域(CDR)1、2、及び3(HCDR1、HCDR2、HCDR3)のアミノ酸配列、ある種の軽鎖相補性決定領域(CDR)1、2、及び3(LCDR1、LCDR2、LCDR3)のアミノ酸配列、ある種の重鎖可変領域(VH)のアミノ酸配列又はある種の軽鎖可変領域のアミノ酸配列(VL)を含むモノクローナル抗体と、ヒトIL−17A結合をめぐって競合する、単離された抗体又はフラグメントアミノ酸配列である。
【0010】
本発明の別の態様は、ある種の重鎖可変領域パラトープアミノ酸残基及びある種の軽鎖可変領域パラトープアミノ酸残基を含む、ヒトIL−17Aに特異的に結合し、配列番号105に示されるアミノ酸配列を有するヒトIL−17Aのある種の残基と相互作用する、単離された抗体又はフラグメントである。
【0011】
本発明の別の態様は、重鎖可変領域及び軽鎖可変領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、該抗体が、Chothia残基F56及びY58から選択される重鎖可変領域パラトープ、並びにChothia残基Y91、F93、及びF94から選択される軽鎖可変領域パラトープを含む。
【0012】
本発明の別の態様は、重鎖可変領域(VH)及び軽鎖可変領域(VL)を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、該抗体が、ある種の重鎖相補性決定領域(CDR)1、2、及び3(HCDR1、HCDR2、HCDR3)のアミノ酸配列、ある種の軽鎖相補性決定領域(CDR)1、2、及び3(LCDR1、LCDR2、LCDR3)のアミノ酸配列、ある種の重鎖可変領域(VH)のアミノ酸配列又はある種の軽鎖可変領域(VL)のアミノ酸配列を含む。
【0013】
本発明の別の態様は、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであり、該抗体は、ある種の重鎖のアミノ酸配列及びある種の軽鎖のアミノ酸配列を含む。
【0014】
本発明の別の態様は、本発明の単離された抗体又はフラグメント及び医薬的に許容される担体を含む医薬組成物である。
【0015】
本発明の別の態様は、配列番号67、68、69、81、82、83、84、85、86、92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を含む単離された抗体重鎖である。
【0016】
本発明の別の態様は、配列番号76、77、78、79、80、87、88、89、90、又は91に示されるアミノ酸配列を含む単離された抗体軽鎖である。
【0017】
本発明の別の態様は、配列番号67、68、69、81、82、83、84、85、86、92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を含む抗体重鎖をコードする、単離されたポリヌクレオチドである。
【0018】
本発明の別の態様は、配列番号76、77、78、79、80、87、88、89、90、又は91に示されるアミノ酸配列を含む抗体軽鎖をコードする、単離されたポリヌクレオチドである。
【0019】
本発明の別の態様は、少なくとも1つの本発明のポリヌクレオチドを含むベクターである。
【0020】
本発明の別の態様は、本発明のベクターを有する宿主細胞細胞である。
【0021】
本発明の別の態様は、ヒトIL−17Aの、IL−17RAとの相互作用を阻害する方法であり、ヒトIL−17A及びIL−17RAを提供する工程と、
ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる群から選択される少なくとも1つのアミノ酸残基においてヒトIL−17Aに結合する拮抗物質と接触させる工程と、を含む。
【0022】
本発明の別の態様は、IL−17A生物活性を阻害する方法であり、ヒトIL17−A及びIL−17RAを提供する工程と、ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる群から選択される少なくとも1つのアミノ酸残基においてヒトIL−17Aに結合する拮抗物質と接触させる工程と、を含む。
【0023】
本発明の別の態様は、請求項3又は7に記載の単離された抗体の治療上の有効量を、炎症性状態の治療を必要とする患者に、炎症性状態を治療するうえで充分な時間にわたって投与する工程を含む、炎症性状態を治療する方法である。
【図面の簡単な説明】
【0024】
【図1】A〜Hはファミリー2、6a、6b、19a、及び19b IL−17A抗体拮抗物質のCDR配列。
【図2】例示のA)IGLV3及びIGLJ、並びにB)パラトープ残基をグラグトするための足場としてのIGHV3及びIGHJ生殖細胞系列遺伝子。mAb6785配列は、上に示される。CDR領域は、下線を引かれており、mAb6785軽鎖(Y31、D49、Y90、F92、F93)及び重鎖(S52、T54、F57、Y59、Q99、L100、及びT101)におけるコア接触部位は、アスタリスク「*」によって示される。B)におけるフレームワーク4領域は、二重下線を引かれている。示されている配列は、特に具体的な指示がない限り、*01対立遺伝子。
【図3】選択された抗体A)軽鎖及びB)重鎖のためのKabat及びChotia番号付け。Kabat CDR及びChothia HVの場所は、灰色で強調表示されている。
【図4】ELISA形式での、標識化されたA及びB)mAb1926、C)mAb317、D)mAb3171、E)及びF)mAb7357の、IL−17Aとの競合結合アッセイ。
【図5】A)異なる抗IL−17A mAbと複合体化されたIL−17AのH/D交換マップ。保護ブロックの上の番号付けは、成熟IL−17A(配列番号105)配列の番号付けに相当する。
【図6】A)IL−17A/Fab6468複合体の全体的な分子構造。IL−17Aの二量体は、濃い灰色及び薄い灰色で示される。2つのFab分子は、それぞれ濃い灰色及び薄い灰色で示される。B)IL−17A(薄い灰色)及びIL−17F(濃い灰色)の単量体の比較、C)IL−17Aの二量体(薄い及び濃い灰色)、D)IL−17Fの二量体(薄い及び濃い灰色)。
【図7】Fab6468に対するIL−17A上の2つの結合部位及びコアエピトープ。プロトマー1及び2は、それぞれ濃い及び薄い灰色である。コアエピトープは、黒色の楕円形によって示される。破線は、無秩序残基を表す。
【図8】IL−17A及びIL−17F推定受容体結合ポケットの比較。A)IL−17AのP1及びP2ポケットの正面及び背面図。mAb6468軽鎖CDR3のFFモチーフは、P2ポケットに示される。B)P2ポケットにおけるN末端FFモチーフを有するIL−17F。C)IL−17A及びIL−17Fの配列整列並びにP1及びP2ポケットの保存。
【図9】ELISA形式での、mAb1926の、IL−17Aタンパク質の異なる種への結合特異性。
【発明を実施するための形態】
【0025】
本明細書に引用する、特許及び特許出願を含むがそれらに限定されない刊行物はすべて、それらがあたかも本明細書に完全に記載されているのとまったく同様に本願に援用するものである。
【0026】
本明細書で使用されるとき、用語は、特定の実施形態を記載する目的でのみ使用され、限定を意図するものではないことが理解される。別段の規定がない限り、本明細書で使用されるとき、全ての技術及び科学用語は、本発明が属する技術分野における当業者によって一般的に理解されている意味と同一の意味を有する。
【0027】
本明細書に記載されているものと同様又は同等の任意の方法及び材料を、本発明の試験を実施するために使用できるが、例示となる材料及び方法を本明細書に記載する。本発明の記載及び請求には、以下の用語が使用される。
【0028】
本明細書で使用されるとき、「拮抗物質」という用語は、任意の機序によって、IL−17A活性を部分的に又は完全に阻害する分子を意味する。例示の拮抗物質は、抗体、融合タンパク質、ペプチド、ペプチド模倣体、核酸、オリゴヌクレオチド、及び小分子である。薬剤は、後述されるIL−17A活性についての周知のアッセイを使用して特定することができる。
【0029】
本明細書で使用されるとき、「IL−17A抗体拮抗物質」又は「IL−17Aに反応する抗体」という用語は、直接若しくは間接に、IL−17A生物活性を低減若しくは阻害する、IL−17Aのその受容体への結合を遮断する、又はIL−17A受容体活性化を阻害することができる抗体を指す。例えば、IL−17Aに反応する抗体は、IL−17Aに直接結合し、IL−17A活性を中和する、すなわち、サイトカイン及びケモカイン放出を低減するために、IL−17Aシグナル伝達を遮断することができる。
【0030】
「IL−17A」(CTLA−8、IL−17、インターロイキン−17A)という用語は、GenBank登録番号NP_002181に示されるアミノ酸配列を有する、ヒトIL−17Aポリペプチドを指す。配列番号105は、成熟ヒトIL−17Aのアミノ酸配列を示す。生体内IL−17Aは、2つの単量体のホモ二量体を形成し、それらは単量体A及び単量体B、又はプロトマーA及びプロトマーB、又はプロトマー1及びプロトマー2、又は鎖A及び鎖Bと指定される。IL−17Aはまた、IL−17Fとのヘテロ二量体を形成することができる。「IL−17A」という用語は、単量体、ホモ二量体、及びヘテロ二量体型を含む。「IL−17Amut6」という用語は、A70Q及びA132Q置換を有するIL−17Aの変異型を指す。成熟IL−17Amut6のアミノ酸配列は、配列番号106に、及びcDNA配列は、配列番号112に示される。IL−17A及びIL−17Amut6は、同等の活性を有する(PCT特許出願第WO09/003096号)。
【0031】
本明細書で使用されるとき、「IL−17A受容体」という用語は、は、両方の受容体ポリペプチド、IL−17RA(GenBank登録番号NP_055154、配列番号107)及びIL−17RC(GenBank登録番号NP_703191、配列番号113)、並びに2つのポリペプチドのホモ二量体又はヘテロ二量体を含む。
【0032】
本明細書で使用されるとき、「抗体」という用語は、広義で意図され、ポリクローナル抗体、マウス、ヒト型(human)、ヒト適合性、ヒト化、及びキメラモノクローナル抗体を含むモノクローナル抗体、抗体フラグメント、少なくとも2つの無傷抗体、二量体の、四量体の、又は多量体の抗体から形成される多重特異的抗体を含む、免疫グロブリン分子を含む。
【0033】
本明細書で使用されるとき、「モノクローナル抗体」(mAb)という用語は、実質的に均質な抗体の集団から得られる抗体(又は抗体フラグメント)を意味する。モノクローナル抗体は、高度に特異的であり、典型的に単一のエピトープに対して向けられている。「モノクローナル」という修飾語は、抗体のほぼ均質な性質を指して言うものであり、抗体がいずれかの特定の方法によって作製される必要はない。
【0034】
免疫グロブリンは、重鎖の定常領域のアミノ酸配列に応じて、IgA、IgD、IgE、IgG及びIgMの5つの大きなクラスに分類することができる。IgA及びIgGは、IgA1、IgA2、IgG1、IgG2、IgG3、IgG4というアイソタイプに更に細かく分類される。あらゆる脊椎動物種の抗体の軽鎖は、定常領域のアミノ酸配列に基づいてカッパ(κ)及びラムダ(λ)の2つの明確に異なるタイプのうちの1つに分類することができる。
【0035】
「抗体フラグメント」という用語は、重鎖相補性決定領域(HCDR)、軽鎖相補性決定領域(LCDR)、重鎖可変領域(VH)、軽鎖可変領域(VL)、重鎖定常領域(CH)、軽鎖定常領域(CL)、又は抗体重鎖若しくは軽鎖のいずれかからのフレームワーク領域(FR)等の、免疫グロブリン分子の少なくとも一部分を含む。抗体は、Fab、F(ab’)、F(ab’)2、scFv、dsFv、又は二重特異性抗体であってもよい。上述の抗体フラグメントの構造、並びに抗体及びそのフラグメントの調製及び使用のための技術は、当該技術分野において周知である。
【0036】
「フレームワーク」領域からなる抗体可変領域は、3つの「抗原結合部位」によって妨害される。抗原結合部位は、種々の用語を使用して記載される。(i)3つがVHに(HCDR1、HCDR2、HCDR3)、及び3つがVLに(LCDR1、LCDR2、LCDR3)ある、相補性決定領域(CDR)は、配列可変性に基づく(Wu and Kabat,J.Exp.Med.132:211〜250,1970、Kabat et al.,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.,1991)。(ii)3つがVHに(H1、H2、H3)及び3つがVLに(L1、L2、L3)ある、「高度可変領域」、「HVR」、又は「HV」は、Chothia and Lesk(Chothia and Lesk,Mol.Biol.196:901〜917,1987)によって記載されるように、構造において高度可変である抗体可変ドメインの領域を指す。他の用語には、「IMGT−CDR」(Lefranc et al.,Dev.Comparat.Immunol.27:55〜77,2003)及び「特異性決定残基使用」(SDRU)(Almagro,Mol.Recognit.17:132〜143,2004)が含まれる。International ImMunoGeneTics(IMGT)データベース(http://www_imgt_org)は、抗原結合部位の標準化された番号付け及び記載を提供する。各CDR、HV、及びIMGTの表記間の対応については、Lefranc et al.,Dev.Comparat.Immunol.27:55〜77,2003に記載されている。
【0037】
本明細書で使用されるとき、「Chothia残基」は、Al−Lazikani(Al−Lazikani et al.,J.Mol.Biol.,273,927〜48,1997)に従って番号付けされた抗体VL残基及びVH残基である。ポリペプチド番号付けに関連して2つの最も使用される番号付けシステム、Kabat(Kabat et al.,Sequences of Immunological Interest,5th Ed.Public Health Service,NIH,Bethesda,MD,1991)とChothia(Chothia及びLesk,Mol.Biol.196:901〜17,1987)との間の対応を、本発明の代表的な抗体に対して図3に示す。
【0038】
「フレームワーク」又は「フレームワーク配列」は、抗原結合部位として記載されていない、可変領域の残りの配列である。抗原結合部位は上述のような種々の用語によって記載され得るため、フレームワークの正確なアミノ抗原結合部位がどのように記載されるかによって決まる。
【0039】
本明細書で使用されるとき、「実質的に同一」という用語は、比較されている2つの抗体又は抗体フラグメントアミノ酸配列が、同一であるか、又は「ごく僅かな差異」を有することを意味する。ごく僅かな差異とは、抗体特性に悪影響を及ぼさない、抗体又は抗体フラグメントアミノ酸配列における1、2、3、4、5、6、7、8、9、10、11、12、13、14、又は15個のアミノ酸の置換である。本明細書で開示する配列とほぼ同じアミノ酸配列も本願の一部である。特定の実施形態では、配列の同一性は、約90%、91%、92%、93%、94%、95%、96%、97%、98%、99%又はそれよりも高い。同一性百分率は、例えば、Vector NTI v.9.0.0(Invitrogen,Carslbad,CA)のAlignXモジュールの初期設定を使用する一対整列によって決定することができる。本発明のタンパク質配列を、問い合わせ配列として使用して、例えば、関連配列を特定するために、公的又は特許データベースに対して検索を実行することができる。かかる検索を実行するために使用される例示のプログラムは、初期設定を使用する、XBLAST若しくはBLASTPプログラム(http_//www_ncbi_nlm/nih_gov)、又はGenomeQuest(商標)(GenomeQuest、Westborough、MA)一式である。
【0040】
本願で用いられる「〜と組み合わせて(in combination with)」という用語は、記述の対象である薬剤が、他の薬剤と共に、混合物中で一緒に、又は単独薬剤として同時に、又は単独薬剤として順次に任意の順序で、動物に投与され得ることを意味する。
【0041】
本明細書で使用されるとき、「炎症性状態」という用語は、部分的には、サイトカイン、ケモカイン、又は炎症性細胞(例えば、好中球、単球、リンパ球、マクロファージ)の活性によって媒介される、有害な刺激等(病原体、損傷した細胞、身体的傷害、若しくは刺激物)に対する急性又は慢性の局部的又は全身性反応を指し、ほとんどの例において、疼痛、潮紅、膨張、及び組織機能の障害によって特徴付けられる。
【0042】
本明細書で使用されるとき、「IL−17A媒介型炎症性状態」という用語は、少なくとも部分的にはIL−17A生物活性から結果として生じる、又はIL−17A活性によって引き起こされる、炎症性状態を指す。例示のIL−17A媒介型炎症性状態は、乾癬及び関節リウマチである。
【0043】
本明細書で使用されるとき、「IL−17A媒介型状態」という用語は、IL−17Aが、直接又は間接によらず、疾患又は状態の、因果関係、発症、進行、持続、又は病理を含む、疾患及び病状において役割を果たす、全ての疾患及び病状を包含する。
【0044】
用語「エピトープ」は、本明細書で使用するとき、抗体が特異的に結合する抗原の部分を意味する。エピトープは、通常、アミノ酸又は多糖類側鎖のような化学的に活性な(極性、非極性又は疎水性)部分の表面集団からなり、特定の3次元構造特性並びに特定の帯電特性を有し得る。エピトープは、立体配座空間単位を形成する隣接した又は隣接していないアミノ酸のいずれか又は両方から成り得る。隣接していないエピトープについて、抗原の直鎖配列の異なる部分からのアミノ酸は、タンパク質分子の折り畳みを通じて、3次元空間において近接近する。
【0045】
本明細書で使用されるとき、「パラトープ」という用語は、抗原がそこに特異的に結合する、抗体の一部分を意味する。パラトープは元来線状であるか、又は不連続であることができ、抗体のアミノ酸の線状構造よりも、隣接していないアミノ酸間の空間的な関係により形成される。「軽鎖パラトープ」及び「重鎖パラトープ」又は「軽鎖パラトープアミノ酸残基」及び「重鎖パラトープアミノ酸残基」は、それぞれ、抗原と接触する抗体の軽鎖及び重鎖残基を指す。
【0046】
用語「特異的結合」は、本明細書で使用するとき、抗体が特定の抗原と、他の抗原又はタンパク質に対するよりも高い親和性で結合することを指す。典型的に、抗体は、解離定数(KD)が10-7M以下で結合し、既定の抗原以外の非特異的抗原(例えば、BSA、カゼイン、又はその他任意の特定ポリペプチド)に結合するためのKDよりも、少なくとも10倍低いKDで、既定の抗原に結合する。「抗原を認識する抗体」及び「抗原に特異的な抗体」という語句は、本明細書で、例えば、IL−17A特異的抗体のように「抗原に特異的に結合する抗体」又は「抗原特異的抗体」という用語と、互換可能に使用される。解離定数は標準的な手順を用いて測定することができる。
【0047】
本明細書で使用されるとき、「IL−17A生物活性」又は「IL−17A活性化」という用語は、IL−17A受容体へのIL−17A結合の結果として生じる任意の活性を指す。例示のIL−17A生物活性は、IL−6若しくはIL−8の増加した分泌、NF−κB活性化、又はIL−17A受容体への結合時のERK1、ERK2、及びp38等の下流キナーゼの調節をもたらす。サイトカイン及びケモカインの、細胞、組織からの、若しくは循環中の放出、NF−κB活性化、又はキナーゼリン酸化事象は、周知の方法、例えば、免疫学的アッセイ、免疫ブロット法、又はレポーター遺伝子系を使用して測定することができる(Yao et al.,Immunity 3:811〜21、1995、Awane et al.,J.Immunol.162:5337〜44,1999)。
【0048】
「ベクター」という用語は、生体系内で複製されることができる、又はかかる系の間で移動可能である、ポリヌクレオチドを意味する。ベクターポリヌクレオチドは典型的に、生体系においてこれらのポリヌクレオチドの複製又は維持を促進するように機能する複製起点、ポリアデニル化信号、又は選択マーカー等の因子を含有する。このような生体系の例としては、細胞、ウイルス、動物、植物、及びベクターを複製することのできる生物学的構成成分を利用して再構成された生体系を挙げることができる。ベクターを含むポリヌクレオチドは、DNA若しくはRNA分子又はこれらのハイブリッドであってもよい。
【0049】
「発現ベクター」という用語は、生体系又は再構成された生体系において、その発現ベクター中に存在するポリヌクレオチド配列によってコードされたポリペプチドの翻訳を命令するために利用することができるベクターを意味する。
【0050】
用語「ポリヌクレオチド」は、糖−リン酸骨格鎖又は他の等価な共有結合化学によって共有結合されたヌクレオチド鎖を含む分子を意味する。二本鎖及び一本鎖のDNA及びRNAが、ポリヌクレオチドの典型的な例である。
【0051】
「ポリペプチド」又は「タンパク質」という用語は、ペプチド結合によって連結されてポリペプチドを形成する少なくとも2つのアミノ酸残基を含む分子を意味する。50個のアミノ酸未満の小さなポリペプチドは「ペプチド」と呼ばれる場合がある。
【0052】
本明細書では以下のようにアミノ酸の慣習的な1文字及び3文字の略号を用いる。
【0053】
【表1】
【0054】
物質の組成物
本発明はIL−17Aの生物活性を阻害することが可能なIL−17A抗体拮抗物質及びかかる抗体の使用法を提供する。かかる抗体によってIL−17A活性化が阻害され得る例示の機序としては、IL−17Aのホモ若しくはヘテロ二量化の生体外で、生体内で、若しくはその場での阻害、及びIL−17Aの、IL−17A受容体への結合の遮断、受容体二量化の阻害、下流シグナル伝達経路のキナーゼ活性の阻害、又はIL−17A mRNA転写の阻害が挙げられる。他の機序によってIL−17Aの活性を阻害することが可能な他の抗体拮抗物質もまた、本発明の異なる態様及び実施形態の範囲に含まれるものである。これらの拮抗物質は、研究用試薬、診断用試薬及び治療薬として有用である。
【0055】
本発明は、ヒト免疫グロブリンの遺伝子ライブラリーから誘導される抗原結合部位を提供する。抗原結合部位を担持するための構造は、概して、抗体重鎖若しくは軽鎖又はそれらの部分である。
【0056】
本発明は、重鎖可変領域(VH)及び軽鎖可変領域(VL)を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はそのフラグメントを提供し、該抗体は、表1aに示されるように、重鎖相補性決定領域(CDR)1、2、及び3(HCDR1、HCDR2、及びHCDR3)アミノ酸配列並びに軽鎖相補性決定領域(CDR)1、2、及び3(LCDR1、LCDR2、及びLCDR3)アミノ酸配列を含む。
【0057】
【表2】
【0058】
【表3】
【0059】
ある種の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号23、35、及び52に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含み、配列番号35のHCDR2は更に、式(I):
Xaa1−I−I−P−W−F−G−Xaa2−T−Xaa3−Y−A−Q−K−F−Q−G、
(I)に示すように記載され、式中、
Xaa1は、His、Met、Arg、Ser、又はTyrであり得、
Xaa2は、Trp、Thr、又はTyrであり得、
Xaa3は、Tyr、Phe、Ser、又はAspであり得る。
【0060】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号2、5、及び11に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含み、配列番号11のLCDR3は更に、式(II):
Xaa4−Q−Xaa5−Xaa6−Xaa7−Xaa8−Xaa9−Xaa10、
(II)に示すように記載され、式中、
Xaa4は、His又はGlnであり得、
Xaa5は、Phe又はGlyであり得、
Xaa6は、Thr、Val、又はAsnであり得、
Xaa7は、Ile、Thr、又はTyrであり得、
Xaa8は、Pro又はArgであり得、
Xaa9は、Ser又はProであり得、
Xaa10は、His、Phe、又はLeuであり得る。
【0061】
他の実施形態では、本発明は、VH及びVLを含む、結合ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号2、5、及び17に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含み、配列番号17のLCDR3は更に、式(III):
Xaa11−Q−Xaa12−Xaa13−Xaa14−Xaa15−Xaa16−Xaa17−Xaa18−T、
(III)に示すように記載され、
式中、
Xaa11は、Gln又はThrであり得、
Xaa12は、Ser又はTyrであり得、
Xaa13は、Asn、Arg、Val、又はTyrであり得、
Xaa14は、His又はSerであり得、
Xaa15は、Ile、Thr、Leu、Ala、又はSerであり得、
Xaa16は、Pro、Leu、又はSerであり得、
Xaa17は、Pro、Ser、Phe、又はLeuであり得、
Xaa18は、Ala、Leu、又はAspであり得る。
【0062】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号24、36、及び57に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含み、配列番号57のHCDR3は更に、式(IV):
E−V−D−S−Xaa19−Y−Y−S−Y−F−D−I、
(IV)に示すように記載され、
式中、
Xaa19は、Met、Ile、Leu、又はThrである。
【0063】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号3、6、及び22に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含み、配列番号22のLCDR3は更に、式(V):
G−S−Y−D−F−F−L−G−Xaa20−I−V、
(V)に示すように記載され、
式中、
Xaa20は、Met、Leu、Thr、又はTyrである。
【0064】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号25、46、及び61に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含み、配列番号46のHCDR2は更に、式(VI):
Xaa21−I−Xaa22−Xaa23−Xaa24−Xaa25−Xaa26−Xaa27−Xaa28−Xaa29−Y−A−D−S−V−K−G、
(VI)に示すように記載され、
式中、
Xaa21は、Ala、Gly、Thr、又はValであり得、
Xaa22は、Asn又はSerであり得、
Xaa23は、Gly、Met、Lys、Ile、Leu、又はHisであり得、
Xaa24は、Leu、Asp、Ala、His、Thr、Gly又はSerであり得、
Xaa25は、Gly又はSerであり得、
Xaa26は、Thr、Gly、Tyr、又はAspであり得、
Xaa27は、His、Trp、Tyr、又はPheであり得、
Xaa28は、Lys、Thr、又はIleであり得、
Xaa29は、Tyr、Phe、又はAsnであり得、
【0065】
配列番号61のHCDR3は、式(VII):
Q−L−Xaa30−L−D−V、
、(VII)に示すように記載され、
式中、
Xaa30は、Met、Leu、又はThrであり得る。
【0066】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号25、51、及び58に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含み、配列番号51のHCDR2は更に、式(VIII):
V−T−S−Xaa31−Xaa32−Xaa33−Xaa34−T−Y−Y−A−Xaa35−S−V−K−G、
(VIII)に示すように記載され、
式中、
Xaa31は、Ala、Lys、Met、又はHisであり得、
Xaa32は、Asn、Met、Thr、又はArgであり得、
Xaa33は、Gly又はAspであり得、
Xaa34は、Arg、His、又はAsnであり得、
Xaa35は、Asp又はGlyであり得る。
【0067】
その抗原結合部位アミノ酸配列が表1aに示される配列(配列番号1〜61)と実質的に同一である抗体は、本発明の範囲内に包含される。典型的に、これは、類似した電荷又は疎水性若しくは立体化学的性質を有するアミノ酸との、1つ以上のアミノ酸置換を伴い、抗体特性、例えば、安定性又は親和性を改善するようにされる。例えば、保存的置換は、その位置のアミノ酸残基の極性又は電荷にほとんどあるいは全く影響しないように、天然アミノ酸残基の非天然残基との置換を伴ってもよい。更に、アラニン・スキャニング変異導入法について以前に記載されているように(MacLennan et al.,Acta Physiol.Scand.Suppl.643:55〜67,1998、Sasaki et al.,Adv.Biophys.35:1〜24,1998)、ポリペプチド内の任意の内因性の残基をアラニンで置換することもできる。保存的な置換は、かかる修飾が行われた分子と類似した機能的及び化学的性質を有する分子をもたらす。分子の機能的及び/又は化学的特性における非保存的置換は、(1)置換領域における分子骨格鎖の構造を、例えばシート若しくはヘリックス立体配座として維持する、(2)標的部位における分子の電荷若しくは疎水性を維持する、又は(3)分子のサイズを維持する、効果において、有意に異なるアミノ酸配列中の置換を選択することによって実現することができる。当業者であれば、所望のアミノ酸置換(保存的又は非保存的であるかによらず)を、かかる置換が望ましい時点で決定することができる。例えば、アミノ酸置換を使用して、親和性に影響を及ぼす残基、又は凝集等の望ましくない特性を付与する残基等の、抗体の機能に重要な残基を特定することができる。例示のアミノ酸置換は、表1b、及び図1に示される。
【0068】
抗原結合部位とは対照的に、フレームワーク領域における置換も、それらが抗体の特性に悪影響を及ぼさない限りは行われてもよい。フレームワーク置換は、抗体親和性又は安定性を改善するために、例えば、バーニアゾーン残基において行うことができる(米国特許第6,649,055号)。置換はまた、潜在的な免疫原性を低減するために、相同のヒト生殖細胞系列遺伝子配列と比較したとき配列において異なる、抗体中のフレームワーク位置においても行うことができる。これらの修飾は、例えば、pIXライブラリー等の、de novo抗体ライブラリーに由来する抗体に対して行うことができる。
【0069】
【表4】
【0070】
保存的アミノ酸置換はまた、生物学的系における合成によってよりはむしろ、化学的ペプチド合成によって典型的に組み込まれる、非天然産のアミノ酸残基も包含する。アミノ酸置換は、例えば、PCR突然変異生成(米国特許第4,683,195号)によって行うことができる。変異型のライブラリーは、周知の方法を使用して、例えば、無作為(NNK)又は非無作為コドン、例えば、11個のアミノ酸(ACDEGKNRSYW)をコードするDVKコドンを使用して、及び実施例1に示されるように、所望の特性を有するライブラリー又は変異型をスクリーニングして、生成することができる。図1は、抗体特性を改善するために、LCDR3、HCDR2、及びHCDR3領域内の5つの親IL−17A抗体拮抗物質に対して行われた置換を示す。親和性又は安定性等の、改善された特性は、周知の方法によって測定することができる。
【0071】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、ある種のVH及びVL配列を含み、また表2に示される各々単離されたVH及びVLも提供する。
【0072】
【表5】
【0073】
実施例に例証される実施形態は、1つは重鎖から及び1つは軽鎖からの、可変領域の対、完全長抗体鎖の対、又はCDR1、CDR2、及びCDR3領域の対を含むが、当事者であれば、代替的な実施形態が、単一の重鎖可変領域若しくは単一の軽鎖可変領域、単一の完全長抗体鎖、又は重鎖若しくは軽鎖のいずれかの1つの抗体鎖からのCDR1、CDR2、及びCDR3領域を含んでもよいことを認識するであろう。単一の可変領域、完全長抗体鎖、又は1つの鎖のCDR1、CDR2、及びCDR3領域を使用して、別の鎖中の対応するドメインをスクリーニングすることができ、これらの2つの鎖は、IL−17Aに特異的に結合する抗体を形成することができる。スクリーニングは、例えば、PCT公開第WO92/01047号で開示されている階層的二重コンビナトリアルアプローチを使用する、ファージディスプレイスクリーニング法によって実現されてもよい。このアプローチでは、H又はL鎖のいずれかのクローンを含む個別のコロニーを用いて他方の鎖(L又はH)をコードするクローンの完全なライブラリに感染させ、得られた2鎖特異的抗原結合ドメインを上述のようなファージディスプレイ法に基づいて選択する。
【0074】
他の実施形態では、本発明は、表2に示されるVH及びVLアミノ酸配列と少なくとも90%同一のアミノ酸配列を有するVH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0075】
他の実施形態では、本発明は、表2に示されるVH及びVLアミノ酸配列と少なくとも95%同一のアミノ酸配列を有するVH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0076】
別の態様では、本発明は、表2に示されるある種の重鎖及び軽鎖アミノ酸配列を有する単離された抗体又はフラグメントを提供する。抗体残基を連続して番号付けすることに加えて、ポリペプチドをコードする抗体鎖は、Kabatの又はChothiaの、番号付けに基づいて番号付けすることができる(Kabat et al.,sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.,1991、Chothia and Lesk,Mol.Biol.196:901〜917,1987)。選択された抗体鎖のための連続したKabat及びChotia番号付けの間の対応の例は、図3に示される。灰色で強調表示された位置は、抗体CDR領域を示す。
【0077】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、抗体は、Chothia残基S51、T53、F56、Y58、Q95、L96、及びT97から選択される重鎖可変領域パラトープ、並びにChothia残基Y32、D50、Y91、F93、及びF94から選択される軽鎖可変領域パラトープを含む。重鎖パラトープ及び軽鎖パラトープChothia残基は、配列番号86の重鎖残基S52、T54、F57、Y59、Q99、L100、及びT101、並びに配列番号79の軽鎖残基Y31、D49、Y90、F92、及びF93に相当する。
【0078】
他の実施形態では、本発明は、配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する重鎖可変領域パラトープアミノ酸残基を含み、
ヒトIL−17AのR55又はE57と相互作用する第1のスレオニン残基、
ヒトIL−17AのR55又はE57と相互作用するグルタミン残基、
ヒトIL−17AのE57と相互作用するリジン残基、
ヒトIL−17AのP59、E60、又はR101と相互作用するチロシン残基、
ヒトIL−17AのE60、R101、E102、又はP103と相互作用するフェニルアラニン残基、
ヒトIL−17AのE60と相互作用するセリン残基、及び
ヒトIL−17AのE60と相互作用する第2のスレオニン残基を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0079】
他の実施形態では、本発明は、配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する軽鎖可変領域パラトープアミノ酸残基を含み、
ヒトIL−17AのL26と相互作用する第1のフェニルアラニン残基、
ヒトIL−17AのR55又はW67と相互作用するアスパラギン酸残基、
ヒトIL−17AのP59、S64、又はR101と相互作用する第1のチロシン残基、
ヒトIL−17AのP59、E60、R61、Y62、R101、又はF110と相互作用する第2のフェニルアラニン残基、及び
ヒトIL−17AのV65と相互作用する第2のチロシン残基を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0080】
別の実施形態では、本発明は、配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する重鎖可変領域パラトープアミノ酸残基及び軽鎖可変領域パラトープアミノ酸残基を含み、
ヒトIL−17AのR101と相互作用する重鎖可変領域におけるチロシン残基、
ヒトIL−17AのR101と相互作用する重鎖可変領域におけるフェニルアラニン残基、
ヒトIL−17AのY62及びR101と相互作用する軽鎖可変領域における第1のフェニルアラニン残基、
ヒトIL−17AのL26及びF110と相互作用する軽鎖可変領域における第2のフェニルアラニン残基、及び
ヒトIL−17AのR101と相互作用する軽鎖可変領域におけるチロシン残基を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0081】
別の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、
Chothia残基F56及びY58から選択される重鎖可変領域パラトープ、並びに
Chothia残基Y91、F93、及びF94から選択される軽鎖可変領域パラトープを含む。
【0082】
重鎖パラトープChothia残基F56及びY58並びに軽鎖パラトープChothia残基Y91、F92、及びF94は、IL−17A残基L26、Y62、R101、及びF110と直接接触している残基である。これらのIL−17A残基は、Fab6468エピトープ及びP2ポケット空洞の両方の部分である(下記を参照されたい)。特定の理論に決して束縛されるものではないが、これらの選択された残基におけるFab6468とIL−17Aとの間の相互作用は、抗体がIL−17A活性を遮断するために十分であることが考えられる。
【0083】
いかなる非ヒト配列も欠く完全な型(human)mAbは、例えば、Knappik et al.,J.Mol.Biol.296:57〜86,2000、及びKrebs et al.,J.Immunol.Meth.254:67〜84 2001に参照される技術によって、ファージディスプレイライブラリーから調製及び最適化することができる。例示の方法では、本発明の抗体は、バクテリオファージpIXコートタンパク質を有する融合タンパク質として、抗体重鎖及び軽鎖可変領域を発現するライブラリーから単離される。抗体ライブラリーは、ヒトIL−17mut6(配列番号105)への結合についてスクリーニングされ、得られた陽性クローンは更に特徴付けられ、Fabはクローンライセートから単離され、完全長IgGsとして発現される。例示の抗体ライブラリー及びスクリーニング方法は、Shi et al.,J.Mol.Biol.397:385〜96,2010、PCT特許出願第WO09/085462号,及び米国第12/546850号、米国特許第5,223,409号、第5,969,108号、及び第5,885,793号)に記載されている。
【0084】
結果として得られたmAbは、本明細書に例示されるように、それらのフレームワーク領域において、ある種のフレームワーク残基をマッチングヒト生殖細胞系列に存在する残基に変更するように更に修飾することができる。
【0085】
特異的なIL−17Aエピトープに結合する本発明の抗体は、ヒト免疫グロブリン遺伝子座を発現するヒト化マウス(Lonberg et al.,Nature 368:856〜9,1994、Fishwild et al.,Nature Biotechnology 14:845〜51,1996、Mendez et al.,Nature Genetics 15:146〜56,1997、米国特許第5,770,429号、第7,041,870号、及び第5,939,598号)又はBalb/cマウスに、エピトープをコードするペプチド、例えば、ペプチド56NEDPERYPSVIWE68(配列番号157)又は100RREPPHCPNSFrlEKIL116(配列番号158)で免疫付与することによって、及びKohler et al.,Nature 256:495〜97のハイブリドーマ法を使用して作製することができる。結果として得られた抗体は、標準の方法を使用して、エピトープへのそれらの結合について試験される。特定されたmAbは更に、Queen et al.,Proc.Natl.Acad.Sci.(USA),86:10029〜32,1989及びHodgson et al.,Bio/Technology,9:421,1991で開示されている技術等の技術によって、変化したフレームワーク支持残基を組み込むことによって結合親和性を保存するように修飾することができる。
【0086】
ヒトIL−17Aに特異的に結合する、ある種のパラトープ残基(例えば、表10に記載されるコアパラトープ残基)を有する単離された抗体は、例えば、パラトープ残基を好適な足場にグラグトすること、操作された足場を完全抗体に組み立てること、結果として得られた抗体を発現させること、及び抗体をIL−17Aへの結合について又はIL−17A生物活性に及ぼす効果について試験することによって、作製することができる。例示の足場は、ヒト生殖細胞系列遺伝子によりコードされたヒト抗体可変領域のアミノ酸配列である。足場は、例えば、全体的な配列相同性、パラトープ残基の間の同一性%、又は足場とmAb6785等の例示の抗体との間の正準構造クラス同一性に基づいて選択することができる。ヒト抗体生殖細胞系列遺伝子は、例えば、TomLinson et al.,J.Mol.Biol 227:776〜798、及びInternational ImMunoGeneTics(IMGT)データベース(http_://_www_imgt_org)で開示されている。コンセンサスヒトフレームワーク領域はまた、例えば、米国特許第6,054,297号に記載されているように使用することができる。例えば、PCT公開第WO10/045340号に記載されている方法に従って、好適な足場の選択を行うことができる。
【0087】
パラトープ残基をグラフトする足場として使用可能な例示のヒト生殖細胞系列遺伝子は、Vλ3、Vh3、Jλ、及びJhフレームワークによってコードされた遺伝子である。例示のVκ3遺伝子には、IGLV3−1、IGLV3−9、IGLV3−10、IGLV3−12、IGLV3−16、IGLV3−19、IGLV3−21、IGLV3−22、IGLV3−25、IGLV3−27、及びIGLV3−32(IMGT命名法、*01対立遺伝子)、(それぞれ配列番号117〜127)がある。例示のJλ遺伝子には、IGLJ1、IGLJ2、IGLJ3、IGLJ4、IGLJ5、IGLJ6、及びIGLJ7(それぞれ配列番号128〜134)がある。例示のVh3遺伝子には、IGHV3−7、IGHV3−9、IGHV3−11、IGHV3−16、IGHV3−19、IGHV3−20、IGHV3−21、IGHV3−23、IGHV3−30、IGHV3−30*03、IGHV3−33、IGHV3−45、IGHV3−48、IGHV3−64、及びIGHV3−74(IMGT命名法、異なる対立遺伝子が指定される場合を除いて*01対立遺伝子)(それぞれ配列番号135〜150)がある。例示のJh遺伝子には、IGHJ1、IGHJ2、IGHJ3、IGHJ4、IGHJ5、及びIGHJ6(それぞれ配列番号151〜156)がある。この生殖細胞系列J領域は、FR4配列を選択するために、その全体又は一部を使用される。例えば、mAb6785軽鎖パラトープ残基は、IGLV3−1とIGLJ2配列との間の単一のアミノ酸残基、例えば、メチオニンの挿入により、IGLJ2(配列番号129)によってコードされたJ領域配列に結合される、IGLV3−1(配列番号117)によってコードされたVλ3タンパク質フレームワーク中にグラフトすることができる。IGLV3−1によってコードされたVλ3タンパク質フレームワークは、更なる置換、例えば、配列番号117の33位(「ACW」)におけるシステイン残基の、例えば、アスパラギンとの置換、及び配列番号117の残基1〜3の、IGLV1ファミリーの「QSV」等の他のラムダ鎖ファミリーに共通のアミノ末端配列との置換(「SYE」)を含んでもよい。他の例示の機能的Vλ3及びJλ遺伝子からの配列は、CDR3領域の長さが11アミノ酸になるように、Vλ3遺伝子によってコードされたカルボキシ末端と、Jλ遺伝子によってコードされたアミノ末端と間の0、1、又は2個のアミノ酸残基の挿入により、mAb6785軽鎖パラトープ残基をグラグトするために使用することができる。例えば、メチオニン及びイソロイシンは、IGLV3−22(配列番号124)とIGLJ2(配列番号129)との間に挿入することができる。図2Aは、グラフトに使用可能な例示の軽鎖足場の整列を示す。mAb6785重鎖パラトープ残基は、IGHV3−23(配列番号142)によってコードされた例えば、Vh3フレームワーク上にグラフトすることができ、それはV領域とJ領域との間のHCDR3を構成する、約5〜7つの残基、例えば、6つの残基の挿入により、IGHJ1(配列番号151)の領域FR4配列(11C−末端アミノ酸、例えば「WGQGTLVTVSS」)に結合される。挿入されたHCDR3約5〜7つの残基には、グルタミン、ロイシン、及びスレオニン、例えば、mAb6785Vhからのパラトープ残基のうちの3つの挿入が含まれる(表10)。他の例示の機能的Vh3及びJh遺伝子からの配列は、mAb6785重鎖パラトープ残基をグラグトするために使用することができる。場合によっては、Vh3遺伝子からの1個のC−末端アミノ酸は、FR3配列のみが足場に含まれるように、HCDR3を構成する約5〜7つの残基の挿入の前に欠失され得る。17個の残基(IGHV3−23(配列番号142)の残基50〜66)のCDR2をコードする他のVh3遺伝子からの配列を使用することもでき、他のJh遺伝子のFR4配列は、IGJH1の代わりに置換することができる。
【0088】
ヒトIL−17Aへの特異的結合の及び結果として得られた抗体の生物活性は、標準の方法を使用して評価することができる。mAb6785軽鎖可変領域及び重鎖可変領域の、例示のVh3、Vλ3、Jλ、又はJh遺伝子との整列は、図2A及び2Bに示される。代替的に、表10に記載されるように、拡張されたパラトープの残基mAb6785をコアパラトープ残基の代わりに使用することができる。パラトープグラフトされた操作された抗体は更に、バーニアゾーン残基(米国特許第6,639,055号)又は親和性決定残基(米国特許出願第2010/0261620号、Cobaugh et al.,J Mol Biol.378:622〜33,2008)の置換によって、抗体特性、例えば親和性を改善するように修飾することができる。パラトープグラフトされた抗体がIL−17Aへの結合を保持する限り、パラトープグラフトされた抗体におけるフレームワークアミノ酸配列は、mAb6785フレームワーク配列と70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%同一であり得る。例示の生殖細胞系列遺伝子フレームワークの対立遺伝子の変異型をV及びJ領域タンパク質配列の代わりに使用することができる。対立遺伝子の変異型の配列は、周知であり、International ImMunoGeneTics(IMGT)データベース(http_://_www_imgt_org)で得ることができる。
【0089】
標準的な方法を用いて、パラトープ残基に追加して抗原結合部位由来の配列をグラフトすることもできる。例えば、完全なHCDR3又はLCDR3をグラフトしてもよい。
【0090】
本発明の別の実施形態は、ヒトIL−17Aに特異的に結合し、ある種のHCDR1、HCDR2、及びHCDR3、並びにLCDR1、LCDR2、及びLCDR3アミノ酸配列を含むモノクローナル抗体と、ヒトIL−17A結合をめぐって競合する、単離された抗体又はフラグメントである。本発明の例示のモノクローナル抗体は、配列番号25、43、及び60に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列並びに配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体である。
【0091】
IL−17Aへの特異的結合との間の競合は、周知の方法を使用して生体外でアッセイすることができる。例えば、非標識抗体の存在下でのMSD Sulfo−Tag(登録商標)NHS−エステル標識抗体のIL−17Aへの結合を、ELISAによって評価することができる。
【0092】
本発明の別の実施形態は、単離された抗体又は抗体又はそのフラグメントであり、該抗体は、アミノ酸残基56〜68(配列番号157)及び100〜116(配列番号158)において、又は残基L26、R55、E57、P59、E60、R61、Y62、S64、V65、W67、R101、E102、P103、及びF110において、配列番号105に示される配列を有するヒトIL−17Aに特異的に結合する。
【0093】
幾つかの周知の方法を用いて本発明の抗体の結合エピトープを決定することができる。例えば、両方の個別の構成成分の構造が知られている場合、インシリコでタンパク質−タンパク質のドッキングを行って、適合する相互作用部位を特定することができる。抗原抗体複合体において水素−重水素(H/D)交換を行って、抗体が結合し得る抗原の領域をマッピングすることができる。抗原のセグメント及び点突然変異生成を使用して、抗体の結合に重要なアミノ酸の位置を特定することができる。抗体−抗原複合体の共結晶構造を使用して、エピトープ及びパラトープに寄与する残基を特定することができる。
【0094】
前述の抗IL−17A抗体は、本発明に記載されるFab6468のためのエピトープと異なる、IL−17A上のエピトープに結合する。抗体結合ヒトIL−17A(配列番号105)残基74〜85、46〜53、71〜87、80〜86、11〜18、29〜41、又は54〜62が記載されている(それぞれPCT公開第WO08/021156号、第WO07/106769号、第WO07/149032号、第WO07/070750号、米国出願第US2008/095775号)。立体配座エピトープは、PCT公開第WO09/130459号、及びGerhardt et al.,J.Mol.Biol:394:901〜21,2009に記載されている。
【0095】
本発明の別の実施形態は、単離された抗体又はそのフラグメントであり、ここで抗体は、IL−17A上のP2ポケット空洞に特異的に結合し、このP2ポケット空洞は、配列番号105のアミノ酸残基V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる。
【0096】
抗IL−17A Fab6468とのIL−17Aホモ二量体の共結晶構造は、L−17RA結合に関与する可能性が高い、IL−17Aホモ二量体の表面上の疎水性P2ポケット空洞を特定した(実施例を参照されたい)。本明細書で使用されるとき、「P2ポケット空洞」は、IL−17Aホモ二量体上の第三級疎水性構造的空洞を指し、P2ポケットにおける表面露出残基は、単量体A上のV24、L26、I28、Y62、L99、R101、F110、及びL112、並びに単量体B上のV22、V24、及びL112であり、逆もまた同様である。IL−17A、例えば、Fab6468に反応する、本発明の選択された抗体は、P2ポケット空洞残基L26、Y62、R101、及びF110と直接接触し、これらの残基もまたFab6468エピトープの部分である。特定の理論に決して束縛されるものではないが、選択されたIL−17AP2ポケット空洞残基に結合する本発明の抗体は、IL−17AとIL−17RAとの間の相互作用を遮断する。共結晶構造に基づき、Fab6468の軽鎖(配列番号79)中の残基93及び94におけるフェニルアラニンモチーフ(FF)は、IL−17A/IL−17RA相互作用を遮断し、したがってP2ポケット空洞遮断剤である。新規ペプチド又は小分子等の他のP2ポケット空洞遮断剤拮抗物質もまた、本発明の範囲内である。これらは、IL−17A/Fab6468共構造に基づいてモデル化され、IL−17Aに結合するFab6468を置き換えるそれらの能力についてスクリーニングされ得る。例えば、ペプチド阻害剤は、FFモチーフを組み込んだ無作為ペプチドライブラリー(例えば、XXXXFFXXのライブラリー、Xは任意のアミノ酸を示し、Fはフェニルアラニンである)からスクリーニングされ、例えば、pIII、pVII、又はpIXコートタンパク質との融合として、バクテリオファージ上に表示され得(米国第5,223,409号、Gao et al.,Proc.Natl.Acad.Sci.USA,96:6025〜30,1999,Tornetta et al.,J.Immunol.Methods.360:39〜46,2010、Shi et al.,J.Mol.Biol.397:385〜96,2010)、その後、それらの、IL−17Aに結合するFab6468の阻害及びIL−17A活性の阻害について試験され得る。
【0097】
小分子は、合成若しくは天然の組成物、又はそれらの任意の組み合わせのライブラリーを使用してスクリーニングされ得、結果として得られた第1の陽性ヒットは、薬剤の構造的類似体を産生するように容易に修飾され得る。ペプチドライブラリー及びpIX融合を作製する、並びに結果として得られたライブラリーをスクリーニングする方法は、周知である。
【0098】
ヒトIL−17Aの、IL−17RAとの相互作用を阻害する方法の、本発明の別の実施形態は、
ヒトIL−17A及びIL−17RAを提供する工程と、
ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる群から選択される少なくとも1つのアミノ酸残基において、ヒトIL−17Aに特異的に結合する拮抗物質と接触させる工程と、を含む。
【0099】
本発明の別の実施形態は、ヒトIL−17A生物活性を阻害する方法であり、
ヒトIL17−A及びIL−17RAを提供する工程と、
ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる群から選択される少なくとも1つのアミノ酸残基において、ヒトIL−17Aに特異的に結合する拮抗物質と接触させる工程と、を含む。
【0100】
ヒトIL−17A及びIL−17RAは、単離されたタンパク質又は融合タンパク質として提供され得る。ヒトIL−17Aホモ二量体は、IL−2、IL−23及びIL−1βの存在下での2つの抗CD3/抗CD28刺激によるナイーブCD4T細胞の生体外刺激によって調製された、活性化Th17細胞の培地から精製され得る。IL−17RAは、細胞若しくは細胞膜に関連し得、天然若しくは過剰発現され得るか、又はIL−17RAのフラグメント、例えば、受容体の細胞外ドメインであり得る。IL−17RAは、ヒトIL−17RA、又はマウス、ラット、若しくはサル等の他の種からのIL−17RAであり得る。ヒトIL−17A残基V22、V24、L26、I28、Y62、L99、R101、F110、及びL112に結合する拮抗物質は、突然変異生成研究によって又は共結晶構造によって、拮抗物質の、IL−17Aに結合するFab6468を置き換える能力によって特定することができる。ヒトIL−17A及びIL−17RAの融合タンパク質は、周知の方法によって作製することができる。例示の融合タンパク質は、免疫グロブリンFcドメインに融合された可溶性IL−17RAである。
【0101】
本発明の別の態様は、本発明の抗体重鎖若しくは抗体軽鎖又はそれらのフラグメント、又はそれらの補体うちのいずれかをコードする、単離されたポリヌクレオチドである。ある種のポリヌクレオチドの例が本明細書に開示されるが、遺伝コードの縮重又は所定の発現系におけるコドンの選択性を考慮すると、本発明の抗体拮抗物質をコードする他のポリヌクレオチドも本発明の範囲内に含まれる。例示のポリヌクレオチドは、配列番号101、102、103、及び104に示される。
【0102】
例示の抗体拮抗物質は、IgG、IgD、IgE、IgA、又はIgMアイソタイプの抗体であり得る。更にかかる抗体拮抗物質は、グリコシル化、異性化、脱グリコシル化、又はポリエチレングリコール部分の付加(ペギレーション)及び脂質化等の非天然の共有結合修飾等の反応によって翻訳後修飾され得る。かかる修飾は、生体内あるいは生体外で行われてもよい。例えば、本発明の抗体をポリエチレングリコールと結合させて、薬物動態的なプロファイルを改善することができる。共役は当業者に既知の方法によって行うことができる。治療用抗体のPEGとの共役によって、機能を低下させずに薬力学的動態が向上することが示されている。Deckert et al.,nt.J.Cancer 87:382〜90,2000、Knight et al.,Platelets 15:409〜18,2004、Leong et al.,Cytokine 16:106〜19,2001、及びYang et al.,Protein Eng.16:761〜70,2003を参照されたい。
【0103】
本発明の抗体の薬物動態的な特性は、当業者に既知の技術によるFc修飾を通じて向上することができる。抗体の「Fc」は、抗体の抗原への結合に直接関与しないが、種々のエフェクタ−機能を示す。抗体「Fc」は、周知の用語であり、抗体のパパイン開裂に基づいて記載される。抗体のFcは、補体活性化、C1q結合、及びFc受容体結合に基づいて、ADCC(抗体依存性細胞媒介型細胞傷害性)及びCDC(補体依存性細胞傷害性)に直接関与する。かかる補体及びFc受容体結合部位は、周知であり、例えば、L234、L235、D270、N297、E318、K320、K322、P331、及びP329が含まれる(KabatのEUインデックスに従った番号付け)(Brekke et al.,Eur.J.Immunol.24:2542〜7,1995、米国特許第5,624,821号、第7,597,889号、Canfield and Morrison,J.Exp.Med.173:1483〜91,1991)。例えば、IgG1のヒンジ領域のLeu234/Leu235のL234A/L235Aへの変異、又はIgG4のヒンジ領域のPhe235/Leu236のP235A/L236Aへの変異によって、FcRへの結合性が最小となり、補体依存性細胞傷害活性及びADCCを介在する免疫グロブリンの能力が低減する。イソメラーゼの作用を介して重鎖間又は重鎖内のいずれかの生体内ジスルフィド結合を形成することができる(Aalberse and Schuurman,Immunology 105:9〜19,2002)、IgG4重鎖のヒンジ領域内のCys−Pro−Ser−Cys(CPSC)モチーフにおけるSerからProへの置換は、「IgG1様挙動」をもたらし、すなわち、Pro置換された分子は、重鎖間ジスルフィド結合を形成することができない。CPSCモチーフの位置は典型的に、成熟した重鎖の残基228に見出されるが、CDRの長さに応じて変化し得る。Leu234/Leu235残基を有する例示のIgG1 Fc領域は、配列番号114に示されるアミノ酸配列を有し、残基L117及びL118は、成熟した重鎖におけるLeu234/Leu235残基に相当する。Cys−Pro−Ser−Cys(CPSC)モチーフ及びLeu234/Leu235残基を有する例示のIgG4 Fc領域は、配列番号115に示されるアミノ酸配列を有し、CPSCモチーフは、残基106〜109に位置し、Leu234/Leu235残基は、122及び123位に位置する。
【0104】
安定性、選択性、交差反応性、親和性、免疫原性、又は他の望ましい生物学的若しくは生物物理学的特性を改善するように修飾される、本発明の抗体又はそのフラグメントが発明の範囲内にある。抗体の安定性は、(1)内在的な安定性に影響を及ぼす個々のドメインのコアパッキング、(2)HCとLCとのペアリングに影響を及ぼすタンパク質/タンパク質の界面相互作用、(3)極性及び荷電残基の埋め込み、(4)極性及び荷電残基のためのH結合ネットワーク、並びに(5)他の分子内及び分子間力の間の表面電荷及び極性残基の分布、を含む幾つかの因子によって影響を受ける。(Wornら、J.Mol.Biol.305:989〜1010、2001)。構造を不安定化させる可能性のある残基は、抗体の結晶構造に基づいて、あるいは場合によっては分子モデリングによって特定され得、抗体の安定性に及ぼすこれらの残基の効果を、特定された残基に突然変異を有する変異体を生成し、評価することによって試験することができる。抗体の安定性を高める方法の1つは、示差走査熱量測定法(DSC)によって測定される熱転移中点(Tm)を高くすることである。広くは、タンパク質Tmは、その安定性と相関し、溶液中のほどき及び変性、並びにタンパク質がほどける傾向に応じる分解プロセスに対する、その感受性と逆相関する(Remmele et al.,Biopharm。13:36〜46、2000)。幾つかの研究は、DSCによる熱安定性として測定される製剤の物理的安定性及び他の方法によって測定される物理的安定性の、順位付けの間の相関を見出している(Gupta et al.,AAPS PharmSci.5E8,2003、Zhang et al.,J.Pharm.Sci.93:3076〜89,2004、Maa et al.,Int.J.Pharm.,140:155〜68,1996、Bedu−Addo et al.,Pharm.Res.,21:1353〜61,2004、Remmele et al.,Pharm Res.,15:200〜8,1997)。製剤研究は、FabのTmが対応するmAbの長期の物理的安定性と密接な関係があることを示唆している。フレームワーク又はCDR内のいずれかにおけるアミノ酸の差異は、Fabドメインの熱安定性に有意な影響を及ぼし得る(Yasui、et al.,FEBS Lett.353:143〜6,1994)。
【0105】
本発明の抗体拮抗物質は、約10-7、10-8、10-9、10-10、10-11又は10-12M以下のKdでIL−17Aと結合することができる。抗体等の所定の分子のIL−17Aに対する親和性は、任意の好適な方法を使用して実験的に決定することができる。かかる方法は、当業者に既知のBiacore又はKinExA装置、ELISA又は競合的結合アッセイを利用し得る。
【0106】
所望の親和性でヒトIL−17Aに結合する抗体拮抗物質は、抗体親和性成熟を含む技術によって、変異型又はフラグメントのライブラリーから選択することができる。抗体拮抗物質は、任意の好適な方法を使用してIL−17Aの生物活性の阻害に基づいて特定することができる。かかる方法では、周知の方法を使用した、本出願に記載されるようなレポーター遺伝子アッセイ又はサイトカイン産生を測定するアッセイを利用し得る。
【0107】
本発明の別の実施形態は、少なくとも1つの本発明のポリヌクレオチドを含むベクターである。かかるベクターは、プラスミドベクター、ウイルスベクター、バキュロウイルス発現ベクター、トランスポゾンに基づいたベクター、又は任意の手段によって特定の生物又は遺伝子的バックグラウンドに本発明のポリヌクレオチドを導入するのに適した他の任意のベクターであってもよい。
【0108】
本発明の別の実施形態は、配列番号67〜75及び81〜86に示されるアミノ酸配列を有する免疫グロブリン重鎖可変領域、又は配列番号62〜66及び76〜80に示されるアミノ酸配列を有する免疫グロブリン軽鎖可変領域、又は配列番号92〜100に示されるアミノ酸配列を有する免疫グロブリン重鎖、又は配列番号87〜91に示されるアミノ酸配列を有する免疫グロブリン軽鎖、を含むポリペプチドをコードするポリヌクレオチド等の、本発明のポリヌクレオチドのいずれかを含む宿主細胞細胞である。かかる宿主細胞細胞は、真核細胞、細菌細胞、植物細胞又は古細菌細胞であってもよい。真核細胞の例としては、哺乳動物、昆虫、鳥類又は他の動物由来のものであり得る。哺乳動物真核細胞としては、SP2/0(American Type培養Collection(ATCC),Manassas,VA,CRL−1581)、NS0(European Collection of Cell Cultures(ECACC)、Salisbury,Wiltshire,UK,ECACC No.85110503)、FO(ATCC CRL−1646)、及びAg653(ATCC CRL−1580)マウス細胞株等の、ハイブリドーマ又は骨髄腫細胞株等の不死化細胞株が挙げられる。例示のヒト骨髄腫細胞株には、U266(ATTC CRL−TIB−196)がある。他の有用な細胞株としては、CHO−K1SV(Lonza Biologics,Walkersville,MD)、CHO−K1(ATCCCRL−61)、又はDG44等の、チャイニーズハムスターの卵巣(CHO)細胞に由来するものが挙げられる。
【0109】
本発明の別の実施形態は、本発明の宿主細胞細胞を培養する工程と、宿主細胞細胞によって産生された抗体を回収する工程とを含む、IL−17Aとの反応性を有する抗体を製造する方法である。抗体を製造して精製する方法は、当該技術分野で周知である。発現について、操作されたファミリー2、6a、6b、19a、及び19b重鎖配列は、MAWVWTLLFLMAAAQSIQA(配列番号109)等のN末端リーダー配列を含み得る。リーダー配列を含む候補mAb6785(ファミリー19)及び成熟型(リーダー配列を含まない)の重鎖をコードする例示のヌクレオチド配列は、それぞれ配列番号101及び102に示される。同様に、発現について、本発明のファミリー2、6a、6b抗体の軽鎖配列は、MGVPTQVLGLLLLWLTDARC(配列番号110)等のN末端リーダー配列を含み得、本発明のファミリー19a及び19b抗体の軽鎖配列は、MAWSPLLLTLLAHCTGSWA(配列番号116)等のN末端リーダー配列を含み得る。リーダー配列を有するコドン最適化されたmAb6785及び成熟型(リーダー配列を有さない)の軽鎖をコードする例示のヌクレオチド配列は、それぞれ配列番号103及び104に示される。
【0110】
本発明の別の実施形態は、本発明の抗体を産生するハイブリドーマ細胞株である。
【0111】
治療の方法
本発明のIL−17A拮抗物質、例えば、IL−17A抗体拮抗物質は、動物患者においてIL−17Aの影響を低減することが所望される場合の任意の療法で利用され得る。IL−17Aは、体内で循環している可能性があり、あるいは望ましくない高レベルで体内の特定の部位、例えば、炎症の部位に局在して存在する可能性がある。特定の理論に決して束縛されるものではないが、本発明の拮抗物質は、IL−17Aのその受容体への結合、又はIL−17Aのホモ若しくはヘテロ二量化を予防又は低減することによって、有益な療法を提供する。本発明の方法を用いて任意の分類に属する動物患者を治療することができる。かかる動物の例としては、ヒト、齧歯類、イヌ、ネコ、及び家畜等の哺乳動物が挙げられる。
【0112】
本発明の抗体は、炎症性状態、アレルギー及びアレルギー性状態、過敏症反応、自己免疫疾患、重症感染症、並びに臓器又は組織移植拒絶症等の、IL−17A媒介型の状態の予防及び治療に有用であり得る。本発明の抗体はまた、かかる治療のための薬剤の調製においても有用であり、薬剤は、本明細書に定められる投薬量で投与するために調製される。例示のIL−17A媒介型状態は、関節リウマチ(RA)、強直性脊椎炎、乾癬性関節炎、変形性関節症、骨粗鬆症、ブドウ膜炎、炎症性線維症(例えば、強皮症、肺線維症、及び肝硬変)、炎症性腸障害(例えば、クローン病、潰瘍性大腸炎、及び炎症性腸疾患)、ぜんそく(を含むアレルギー性ぜんそく)、アレルギー、COPD、多発性硬化症、乾癬、全身性紅斑性狼瘡、糖尿病、並びに癌を含む、炎症性状態、免疫並びに増殖性障害である。抗ヒトIL−17A療法で治療された患者におけるプラスの転帰は、関節リウマチ、乾癬、及び非感染性ブドウ膜炎で記載されている(Genovese et al.,Arthritis Rheum.62:929〜39,2010、Hueber et al.,Sci.Transl.Med.2:52ra72.,2010)。
【0113】
炎症性肺状態は、炎症性状態の一例である。例示の炎症性肺状態には、ウイルス性、細菌性、真菌性、寄生生物、又はプリオン感染に関連する状態を含む感染誘発性肺状態、アレルゲン誘発性肺状態、石綿症、珪肺症、又はベリリウム中毒等の汚染物質誘発性肺状態、胃吸引誘発性肺状態、免疫調節異常、嚢胞性線維症等の遺伝性素因による炎症性状態、並びに人工呼吸器傷害等の物理的外傷誘発性肺状態が含まれる。これらの炎症性状態には更に、ぜんそく、気腫、気管支炎、慢性閉塞性肺疾患(COPD)、サルコイドーシス、ヒスチオサイトーシス、リンパ管筋腫症、急性肺傷害、急性呼吸窮迫症候群、慢性肺疾患、気管支肺異形成症、院外感染性肺炎、院内肺炎、人工呼吸器関連肺炎、敗血症、ウイルス性肺炎、インフルエンザ感染、パラインフルエンザ感染、ロタウイルス感染、ヒトメタニューモウイルス感染、呼吸系発疹ウイルス感染、及びアスペルギルス又は他の真菌感染も含まれる。例示の感染関連炎症性疾患としては、重症肺炎、嚢胞性線維症、気管支炎、気道増悪、及び急性呼吸窮迫症候群(ARDS)を含むウイルス又は細菌性肺炎が挙げられ得る。かかる感染に伴う状態は、一次ウイルス感染及び二次ウイルス感染等の複数の感染を伴う場合がある。調節不全のIL−17A産生は、ぜんそく及び慢性閉塞性肺疾患(COPD)等の肺疾患の病理において役割を果たす可能性がある(Alcorn et al.,Annu.Rev.Physiol.72:495〜516,2010に概説される)。IL−17Aは、IL−17Aが、肺常在上皮細胞からの好中球動員、生存、及び活性化において重要な因子を誘発する能力を有するため、重症のぜんそく並びにCOPDの顕著な特徴である肺の好中球性炎症を調節することが示されている(例えば、IL−6、IL−8、GM−CSF、G−CSF)。本発明の抗体は、肺上皮細胞からのIL−6、IL−8、及びGM−CSF分泌を抑制し、したがって、ぜんそく及びCOPD等の肺炎症性状態を有する対象の治療的又は予防的治療において有益であり得る。ぜんそく及び気道炎症のために一般的に使用される動物モデルには、オボアルブミンチャレンジモデル及びメタコリン感作モデルが含まれる(Hessel et al.,Eur.J.Pharmacol.293:401〜12,1995)。培養したヒト気管支上皮細胞、気管支線維芽細胞又は気道平滑筋細胞からのサイトカイン及びケモカインの産生の阻害を生体外モデルとして用いることもできる。本発明の拮抗物質をこれらのモデルのいずれかに投与することによって、ぜんそく、気道炎症、COPD等の症状を改善し、経過を変化させるためのこれらの拮抗物質の使用を評価することができる。
【0114】
乾癬は、炎症性状態の別の例である。乾癬は、角化細胞のT細胞媒介型高度増殖並びに炎症性浸潤によって特徴付けられる。乾癬組織の炎症及び高度増殖は、正常な皮膚とは異なる組織学的、抗原性、及びサイトカインプロファイルに関連する。乾癬に関連するサイトカインの中でもとりわけ、TNFα、IL−19、IL−18、IL−15、IL−12、IL−7、IFNγ、IL−17A、及びIL−23が挙げられる(Gudjonsson et al.,Clin.Exp.Immunol.135:1〜8,2004)。IL−17Aは、乾癬病変で過剰発現されることが見出されており(米国特許第7,776,540号)、抗ヒトIL−17A療法で治療された患者におけるプラスの転帰が記載されている(Hueber et al.,Sci.Transl.Med.2:52ra72.,2010)。
【0115】
変形性関節症、関節リウマチ、傷害の結果として関節炎にかかった関節等を含む関節炎が、一般的な炎症性状態であり、それらは本発明の拮抗物質等の抗炎症性タンパク質の治療的使用から利益を受けるであろう。IL−17Aシグナル伝達の活性化によって、炎症を起こした関節の炎症及び更なる組織損傷が持続し得る。関節リウマチのための複数の動物モデルが既知である。例えばコラーゲン誘導性関節炎(CIA)モデルでは、ヒト関節リウマチとよく似た慢性炎症性関節炎をマウスに発症させる。本発明のIL−17A抗体をCIAモデルマウスに投与することによって、疾患の症状を改善して疾患の経過を変化させるためのこれらの拮抗物質の使用を評価することができる。
【0116】
慢性の消化管炎症性状態の例としては、炎症性腸疾患(IBD)、潰瘍性大腸炎(UC)、及びクローン病(CD)、外界からの刺激によって誘発される大腸炎(例えば、化学療法、放射線療法等の投与等の治療レジメンによって引き起こされるかあるいはこれにともなう(例えば、副作用として)消化管炎症(例えば、大腸炎))、感染性大腸炎、虚血性大腸炎、膠原性又はリンパ性大腸炎、壊死性腸炎、慢性肉芽腫症又はセリアック病などの状態における大腸炎、食品アレルギー、胃炎、感染性胃炎又は小腸結腸炎(例えばヘリコバクター・ピロリの感染による慢性活動性胃炎)、並びに感染性病原体によって引き起こされる消化管炎症の他の形態がある。胃腸炎症性状態のための複数の動物モデルが存在する。最も広く使用されるモデルは、結腸に慢性炎症及び潰瘍形成を誘発する2,4,6−トリニトロベンゼンスルホン酸/エタノール(TNBS)誘発大腸炎モデル又はオキサゾロンモデルである(Neurath et al.,Intern.Rev.Immunol 19:51〜62、2000)。別のモデルでは、血性下痢、体重減少、結腸の短縮、及び好中球の浸潤を伴う粘膜の潰瘍形成として発現する急性大腸炎を誘発する硫酸デキストランナトリウム(DSS)を使用する。別のモデルは、ナイーブなCD45RBhigh CD4 T細胞をRAG又はSCIDマウスに養子移入することを含む。このモデルでは、ドナーのナイーブT細胞がレシピエントの腸を攻撃することにより、ヒトの炎症性腸疾患に似た慢性腸炎症及び症状を引き起こす(Read and Powrie,Curr.Protoc.Immunol.Chapter 15 unit 15.13,2001)。これらのモデルのいずれかにおいて本発明の拮抗物質の投与を用いることによって、これらの拮抗物質が、炎症性腸疾患等の腸内の炎症に伴う症状を改善し、疾患の経過を変化させる潜在的な有効性を評価することができる。
【0117】
腎線維症は、急性刺激(例えば、移植片虚血/再潅流)(Freese et al.,Nephrol.Dial.Transplant.16:2401〜6,2001)又は 慢性状態(例えば、糖尿病)(Ritz et al.,Nephrol.Dial.Transplant.11 Suppl 9:38〜44,1996)のいずれかから発症し得る。その病因は典型的に、腎糸球体濾過器官及び尿細管間質に最初に炎症反応が起こった後、線維化が持続することとして特徴付けられる(Liu,Kidney Int.69:213〜7,2006)。尿細管間質線維症は、末期の腎不全に対する腎損傷の病因において重要な役割を果たしていることが示されており、近位細管細胞が中心的媒介物質であることが明らかにされている(Phillips and Steadman,Histol.Histopathol.17:247〜52,2002、Phillips,Chang Gung Med.J.30:2〜6,2007)。尿細管間質区画における線維化は、部分的に、常在線維芽細胞の活性化により媒介され、この常在線維芽細胞は、炎症促進性のサイトカインを分泌し、これが近位細管上皮を刺激して、局所的な炎症及び線維化の媒介物質を分泌する。更に、走化性サイトカインが線維芽細胞及び上皮細胞によって分泌され、単核白血球/マクロファージ及びT細胞を尿細管間質へと湿潤させるよう導く方向付け勾配を供給する。炎症性湿潤により、更なる線維化及び炎症性のサイトカインが産生され、それらは更に線維芽細胞及び上皮サイトカインの放出を活性化し、同時に上皮も刺激して表現型移行を起こし、これにより細胞が過剰な細胞外マトリックス構成成分を沈積させる(Simonson,Kidney Int.71:846〜54、2007)。IL−17Aは、ヒト腎同種移植片拒絶中に上方調節されることが示されている(Van Kooten et al.,J.Am.Soc.Nephrol.9:1526〜34,1998、Loong et al.,J.Path.197:322〜32,2002)。IL−17Aは、近位尿細管上皮による炎症促進性メディエーターIL−6、IL−8、補体構成要素C3、及びRANTESの産生を刺激する(Van Kooten et al.,J.Am.Soc.Nephrol.9:1526〜34,1998、Woltman et al.,J.Am.Nephrol.11:2044〜55,2000)。これらの因子が次に、他の炎症性細胞タイプの間質への補充を媒介し、これが炎症/免疫反応の維持に寄与し、抑制されない場合には線維症及び慢性同種移植腎症の発症に寄与する(Racusen et al.,Kidney Int.55:713〜23,1999、Mannon,Am.J.Transpl.6:867〜75,2006)。
【0118】
他の例示的な繊維性の病態としては、肝臓線維症(アルコール性肝硬変、ウイルス性肝硬変、自己免疫性肝炎が挙げられるがこれらに限定されない);肺線維症(強皮症、特発性肺線維症が挙げられるがこれらに限定されない);腎臓線維症(強皮症、糖尿病性腎炎、糸球体腎炎、狼瘡腎炎を含むがこれらに限定されない);皮膚線維症(強皮症、肥厚性及びケロイド瘢痕、火傷を含むがこれらに限定されない);骨髄線維症;神経線維腫症;線維腫;腸線維症;及び外科手術の結果としての線維症癒着等が挙げられる。線維症は、臓器特異的な線維症又は全身性線維症であり得る。器官特異性線維症は、肺線維症、肝臓線維症、腎臓線維症、心臓線維症、血管線維症、皮膚線維症、眼線維症、又は骨髄線維症に関連し得る。肺線維症は、特発性肺線維症、薬剤誘発性肺線維症、ぜんそく、サルコイドーシス、又は慢性閉塞性肺疾患に関連し得る。肝臓線維症は、肝硬変、住血吸虫症、又は胆管炎に関連し得る。肝硬変は、アルコール性肝硬変、C型肝炎後肝硬変、原発性胆汁性肝硬変の中から選択され得る。胆管炎は硬化性胆管炎であり得る。腎線維症は、糖尿病性腎障害又はループス糸球体腎炎に伴い得る。心線維症は、心筋梗塞に伴い得る。血管線維症は、血管形成術後動脈再狭窄、又はアテローム性動脈硬化に伴い得る。皮膚線維症は、火傷瘢痕化、肥厚性瘢痕化、ケロイド、又は腎性線維性皮膚症に伴い得る。眼線維症は、後眼窩線維症、白内障手術後又は増殖性硝子体網膜症に伴い得る。骨髄線維症は、特発性骨髄線維症又は薬剤誘発性骨髄線維症に伴い得る。全身性線維症は、全身性硬化症及び移植片体宿主細胞病から選択され得る。
【0119】
本発明の方法によって予防又は治療し得る他の炎症性状態及び神経障害は、自己免疫疾患によって引き起こされるものである。これらの状態及び神経障害としては、多発性硬化症、全身性エリテマトーデス、並びに、アルツハイマー病、パーキンソン病、ハンチントン病、双極性疾患、及び筋萎縮性側索硬化症(ALS)を含む神経変性及び中枢神経系(CNS)疾患、原発性胆汁性硬変、原発性硬化性胆管炎、非アルコール性脂肪性肝疾患/脂肪性肝炎、線維症、C型肝炎ウイルス(HCV)、及びB型肝炎ウイルス(HBV)等の肝臓疾患、糖尿病及びインスリン抵抗性、アテローム性動脈硬化症、脳出血、脳卒中、及び心筋梗塞等の心血管疾患、関節炎、関節リウマチ、乾癬性関節炎及び若年性関節リウマチ(JRA)、骨粗鬆症、変形性関節症、膵炎、線維症、脳炎、乾癬、巨細胞動脈炎、強直性脊椎炎、自己免疫性肝炎、ヒト免疫不全ウイルス(HIV)、炎症性皮膚状態、移植、癌、アレルギー、内分泌疾患、創傷治癒、他の自己免疫性疾患、気道過敏症、並びに、細胞、ウイルス又はプリオンによって介在される感染症又は疾患が挙げられる。
【0120】
投与/医薬組成物
IL−17A活性の抑制が所望される状態の治療に有効な薬剤の「治療上の有効量」は、標準的な研究技術によって決定され得る。例えば、ぜんそく、クローン病、潰瘍性大腸炎、又は関節リウマチ等の炎症性状態の治療において有効となる薬剤の投薬量は、本明細書に記載されるモデル等の、関連する動物モデルに薬剤を投与することによって決定され得る。
【0121】
更に、任意に生体外アッセイを用いて最適な投薬量範囲を特定することができる。特定の有効用量の選択は、当業者であれば幾つかの因子の考慮に基づいて(例えば、臨床試験によって)決定することができる。かかる因子には、治療又は予防しようとする疾患、関与する症状、患者の体重、患者の免疫状態、及び当業者に既知の他の因子が含まれる。製剤に使用される正確な投薬量は、投与経路、及び疾患の重篤度にも依存し、医師の判断及び各患者の状況に基づいて決定されなければならない。有効用量は、生体外又は動物モデル試験系から導出される用量反応曲線から外挿することができる。
【0122】
本発明の薬剤の治療上の使用のための投与方法は、薬剤を宿主細胞に送達する任意の好適な経路でよい。これらの薬剤の医薬組成物は、例えば、皮内、筋肉内、腹腔内、静脈内、皮下、又は鼻腔内等の非経口投与に特に有用である。
【0123】
本発明の薬剤は、有効量の薬剤を製薬上許容され得る担体中の有効成分として含有する医薬組成物として調製することができる。「担体」という用語は、活性化合物が一緒に投与される希釈剤、助剤、賦形剤、又は溶媒のことを指す。かかる医薬用溶媒は、落花生油、大豆油、鉱物油、ゴマ油等の、石油、動物、植物又は合成物由来の水及び油等の液体であってよい。例えば、0.4%生理食塩水及び0.3%グリシンを使用することができる。これらの溶液は滅菌され、粒子状物質を含まない。これらの溶液は、従来の周知の滅菌技術(例えば、濾過)によって滅菌することができる。この組成物は、生理学的条件に近づけるために必要とされるpH調整剤及び緩衝剤等の製薬上許容される補助物質を含むことができる。かかる医薬製剤中の本発明の薬剤の濃度は、約0.5重量%未満、通常は約1重量%又は少なくとも約1重量%か、最大で15又は20重量%までと大きく異なってよく、選択される投与方法に従って、主として必要とされる用量、液体の体積、粘度等に基づいて選択される。
【0124】
したがって、筋肉内注射用の本発明の医薬組成物は、1mLの滅菌緩衝水、及び約1ng〜約100mg、例えば、約50ng〜約30mg、又はより好ましくは約5mg〜約25mgの本発明のIL−17A拮抗物質を含有するように調製することができる。同様に、静脈内注射用の本発明の医薬組成物は、250mLの滅菌リンゲル溶液、及び約1mg〜約30mg、好ましくは5mg〜約25mgの本発明の拮抗物質を含有するように調製することができる。非経口投与可能な組成物を調製する実際の方法は周知であり、例えば、「Remington’s Pharmaceutical Science」,15th ed.,Mack Publishing Company,Easton,PA.に、より詳細に記載されている。
【0125】
本発明の抗体拮抗物質は凍結乾燥して保存することができ、使用前に好適な担体中で再構成することができる。この技術は従来の免疫グロブリン及びタンパクの調製において有効であることが示されており、当該技術分野で既知の凍結乾燥と再構成の技術を用いることができる。
【0126】
ここで、以下の具体的かつ非限定的な実施例を参照して、本発明を説明する。
【実施例】
【0127】
(実施例1)
抗ヒトIL−17A拮抗性mAbの特定
The MorphoSys Human Combinatorial Antibody Library(HuCAL(登録商標))ゴールドファージディスプレイライブラリー(Morphosys AG,Martinsried,Germany)をヒト抗体フラグメントの源として使用し、溶液のサブプール中でパンニングした。第1回目のパンニングでは、サブライブラリーをビオチン化成熟His6タグ付きIL−17A A132Q及びA70Q変異型(IL−17Amut6)(配列番号106)に対して選択した。第2回目では、第1回目の増幅生産物を、IL−17Aに特異的な抗体に反して偏倚した競合物質としての他のIL−17Aファミリーメンバーの存在又は不在下で、ビオチン化His6タグ付きIL−17Amut6に対して選択した。第2回目の増幅生産物を2つのプールに分割した。第1のプールを第1回目と同様にパンニングした。第2のプールのクローンを、最初の選択で使用したサブライブラリーに応じてHCDR2又はLCDR3のいずれかに更に分散させ、次いでIL−17Amut6に対して更に2回のパンニングを行って、スクリーニングのためのクローンの第2の源を得た。クローンライセートからのFabを、ヒツジ抗ヒトFd抗体でコーティングしたELISAプレートウェル中に捕捉し、ビオチン化IL−17Amut6への結合についてスクリーニングした。陽性クローンの粗溶解物を、組み換えヒトIL−17RA受容体(配列番号107)へのIL−17Amut6結合の阻害についてスクリーニングした。
【0128】
選択されたクローンを、全ての配列ファミリーの配列スコアリング、親和性、及び表現に基づいて精製したFabとして選択して更に特徴付け、MOR番号を指定した。MOR7708、MOR7785、MOR7706、MOR7775、及びMOR7700についての更なる変異型を生成して、HCDR2、HCDR3、又はCDR3に常在するTrp又はMetを置き換えた。表3は、生成された変異型を示す。
【0129】
【表6】
【0130】
組み換えヒトIL−17RA受容体へのIL−17Amut6及びcynoIL−17A結合、及びIL−17Amut6へのそれらの結合のFabによる阻害について、Fabを試験した。全ての試験したFabは、IL−17RAへのIL−17Amut6及びcynoIL−17A結合の両方を阻害した。SETアッセイを使用してIL−17Amut6へのFabの親和性を測定した(表4)。特定したFabから、ファミリー2、6a、6b、19a、及び19bからの候補を選択して更に特徴付けた。
【0131】
【表7】
【0132】
(実施例2)
抗IL−17A拮抗性mAbの誘導、操作、及び特徴付け
選択したMOR番号Fabを変換し、ヒトIgG1形式のmAbとして発現させ、対応するMORmAb表記を与えた。生成したMORmAbを、発現及び凝集、ヒトIL−17RAへのヒト及びcyno IL−17A結合を阻害する、並びにNHDF細胞からのIL−8分泌を阻害する、それらの能力について試験した。表5は、MORmAbのために選択したアッセイについてのIC50値を示す。試験したMORmAb(MORmAb番号7702、7708、7785、7786、7706、7775、7700、8095、8096、8097、8098、7768)のいずれも、他のIL−17ファミリーメンバーと交差反応しなかった。
【0133】
【表8】
【0134】
抗IL−17A拮抗性mAbのフレームワーク操作
活性並びに生物物理学的及び生化学的特性に基づき、選択されたMORmAbを、それらの可変領域において、ある種のフレームワーク残基をマッチングヒト生殖細胞系列存在する残基に変更するように、及びコドンを高度に発現した哺乳類タンパク質中で最も頻繁に生じるコドンに変更するように、更に操作した。ファミリー2 VLにおいて、L11V、及びV85T(直鎖配列)置換を行い、フレームワークをVK−1生殖細胞系列Vb−L5(IGKV1−12*01)と厳密に一致するように変換した。V11V及びV85T置換を有する例示の可変領域は、配列番号76に示されるアミノ酸配列を有する可変領域である。ファミリー6a及び6b VLにおいて、D1E、V59I、及びT86V置換を行い(直鎖配列)、フレームワークをVk−3生殖細胞系列Vb−L6(IGKV3−11*01)と厳密に一致するように変換した。D1E、V59I、及びT86V置換を有する例示の可変領域は、配列番号77に示されるアミノ酸配列を有する可変領域である。ファミリー6a及び6b VHにおいて、G44S置換を行って(直鎖配列)、Vh−6生殖細胞系列Vb 6−01(IGHV6−1*01)と一致させた。G44S置換を有する例示の可変領域は、配列番号81に示されるアミノ酸配列を有する可変領域である。ファミリー19a及び19b VLにおいて、アミノ酸1〜3(DIE)をQSVで置換して、人工のカッパN末端をラムダ鎖のN末端に置き換えた。QSV置換を有する例示の可変領域は、配列番号79に示されるアミノ酸配列を有する可変領域である。ファミリー19a及び19b VHにおいて、V5L置換を行って、Vh−3生殖細胞系列Vb3〜23(IGHV3−23*01とほぼ一致させた。また、このプロセスにおいて、重鎖定常領域アミノ酸配列残基353〜357(REEMT)をRDELTで置換した。V5L置換を有する例示の可変領域は、配列番号86に示されるアミノ酸配列を有する可変領域である。定常領域353〜357 REEMT−>RDELT置換を有する例示の重鎖は、配列番号100に示されるアミノ酸配列を有する重鎖である。操作された抗体にmAb番号を与えた。
【0135】
操作された及び元の可変領域並びに完全長抗体の対応する表記及び配列表を表2に列挙する。各ファミリー内のCDRの配列を図1に示す。
【0136】
操作されたmAbを、MORmAbについて上述したように特徴付けた。示されるアッセイを使用して測定したIC50値(pM)を表6に示す。
【0137】
【表9】
【0138】
ビアコアを使用して、選択されたmAbの親和性を査定した。測定の結果を表7に示す。
【0139】
【表10】
*1つの抗IL−17当たりの二量体
【0140】
抗IL−17抗体は、NHBE細胞中のサイトカイン分泌を阻害する。
IL−17Aは、IL−17Aが、好中球動員、生存、及び活性化において重要な因子を誘発する能力を有するため、重症のぜんそく並びにCOPDの顕著な特徴である肺の好中球性炎症を調節することが示されている(例えば、IL−6、IL−8、GM−CSF)。抗IL−17A本発明の抗体が肺常在細胞中のIL−17A誘発性変化を阻害し得るかどうかを決定するために、正常ヒト気管支上皮(NHBE)細胞を、mAb6785の存在下で48時間、ヒトIL−17Aで刺激した。mAb6785は、それぞれ619.0±64.0pM及び564±86pMのIC50で、NHBE細胞によるIL−17A誘発性IL−6及びGM−CSF産生を阻害した。
【0141】
抗IL−17抗体は、IL−17Aの/Fヘテロ二量体の生物活性を阻害する。
正常なヒト皮膚線維芽細胞(NHDF、Lonza)細胞を、48ウェルの平底組織培養プレートの中に、1ウェル当たり10,000細胞でFGM−2培地(Lonza)中に播種し、一晩インキュベートした(37°、5% CO2)。インキュベーション後、rhIL−17A/Fヘテロ二量体(R&D Systems)の50ng/mL最終濃度(1.47nM)を、mAb6785又は対照抗体の希釈系(30μg/mL−0.5ng/mL)を用いて室温で10分間プレインキュベートし、細胞に添加した。細胞を48時間インキュベートし(37°、5% CO2)、培養物の上清を収集し、製造業者の指示に従って、ヒトIL−6 Duo Sets(R&D Systems、Inc.)を使用して、IL−6含量についてELISAによってアッセイした。GraphPad Prismソフトウェア(GraphPad Software,Inc)を使用して、IC50値を非直線回帰によって決定した。mAb6785は、2±2.5nMのEC50で、NHDFによるIL−17A/Fヘテロ二量体誘発型IL−6産生を阻害した。
【0142】
方法
溶液平衡滴定(SET)を使用したピコモル親和性の決定
溶液平衡滴定(SET)によるKD決定のために、Fabタンパク質の単量体分画(少なくとも90%単量体含量、分析用SECによって分析、Superdex75カラム、GE)を使用した。
【0143】
基本的に、前述のように溶液中の電気化学発光(ECL)に基づく親和性決定及びデータ評価を行った(Haenel et al.,Anal Biochem 339:182〜4,2005)。規定の固定濃度の精製したFab(約10〜100pM)を、化学的均衡が達成されるまで、溶液中のIL−17Amut6(5nMの最大濃度)の濃度を増加させながらインキュベートした。溶液中の結合されていないFabを定量化するために、試料を、コーティングされたビオチン化IL−17Amut6と共に、ストレプトアビジンMSDの384ウェルのマイクロタイタープレート(Meso Scale Discovery,Gaithersburg,MD)に移した。検出のために、ルテニウム複合体で標識化された抗ヒトFab/IgG抗体を適用し、プレートをSector(商標)Imager 6000(MSD)で読み取った。後述されるモデルを使用してExcel/XLfitソフトウェアにより、滴定曲線(抗原濃度の関数としての遊離Fabの濃度)をプロットし、当てはめた。
【0144】
Fab分子のKDの決定についてのデータ評価のために、次の当てはめモデルを使用した(Abraham et al.J Mol Recognit.9:456〜461,1996に従って修正):
y=Bmax−(Bmax/(2*cFab)*(x+cFab+KD−sqrt((x+cFab+KD)*(x+cFab+KD)−4*x*cFab)))
他方では:
Bmax:最大結合シグナル(抗原濃度=0で)
cFab:適用されたFab濃度
x:適用された総可溶性抗原濃度(結合部位)
sqrt:平方根
KD:均衡解離定常
【0145】
IL−17RAへのIL−17A結合の阻害(例えば、「IL−17RA阻害」アッセイ
透明のマキシソーププレートを、pH 9.4の0.1M炭酸ナトリウム緩衝液中、100μL/ウェルの2.5μg/mLヒトIL−17RA−Fc(R&D Systems,Minneapolis,MN)でコーティングし、4℃で一晩インキュベートした。ブロッキング及び洗浄後、25ng/mLのビオチン化ヒトIL−17mut6(配列番号106)又はカニクイザルIL−17A(配列番号108)を、試験mAb又は対照mAb(30〜0μg/mL最終濃度)を用いて、100μLの合わせた容量中で5〜10分間プレインキュベートし、次いでプレートに添加した。シグナルを、100μLの1mg/mL SA−HRP(Jackson Immunoresearch,West Grove,PA)の1:10,000希釈を用いて室温(RT)で20分間、続いて100μL/ウェルのOPD基質(Sigma−Aldrich Corp.,St.Louis,MO)で検出した。プレートを492nmで読み取った(Envision,PerkinElmer,Waltham,MA)。IL−17RAへのFab結合を、mAbについて記載されるように試験した。
【0146】
NHDF細胞からのIL−8及びIL−6産生の阻害(例えば「IL−8産生」及び「IL−6産生」アッセイ)
抗IL−17AmAbの、IL−8及びIL−6産生に及ぼす阻害の効果を正常なヒト皮膚線維芽細胞(NHDF)中で査定した。細胞を、48ウェルの平底組織培養プレート中に、FGM−2培地中、1ウェル当たり0.1×105細胞、1ウェル当たり250μLでプレートし、一晩インキュベートした(37°、5% CO2)。インキュベーション後、0.1ng/mLのヒトTNF−αを全てのウェルに添加した。10ng/mLのIL−17mut6又は25ng/mLのカニクイザルIL−17Aを、試験mAb又は対照mAb(30〜0μg/mL最終濃度)を用いて、250μLの合わせた容量中で、室温で10分間プレインキュベートし、次いで250μLの細胞に添加した。本アッセイにおいて、抗体を添加していないIL−17mut6試料を対照試料として含めた一方で、TNF−α又は培養培地のみからなる試料を陰性対照として含めた。細胞を24時間(37°、5% CO2)インキュベートし、調湿した培地を収集し、ヒトIL−6 & IL−8 ELISA Duo Setsを使用して、製造業者(R&D Systems,Minneapolis,MN)の指示に従って、IL−6及びIL−8についてELISAによってアッセイした。Fabを、mAbについて記載されるようにアッセイした。
【0147】
NHBE細胞からのIL−6及びG−CSF産生の阻害
正常なヒト気管支上皮(NHBE、Lonza)細胞を、BEGM培地(Lonza)中、1ウェル当たり20,000細胞で播種し、一晩インキュベートした(37°、5%CO2)。インキュベーション後、細胞をIL−17Amut6を用いて、試験した抗体の存在下で、様々な濃度で48時間刺激した(30μg/mL〜0.5ng.mL)。インキュベーション後、上清を収集し、ヒトIL−6−又はG−CSF特異的ELISA(R&D Systems,Inc.)を使用して、IL−6又はG−CSF含量についてアッセイした。IC50値を、GraphPad Prismソフトウェア(GraphPad Software,Inc)を使用して非直線回帰によって決定した。
【0148】
IL−17Aファミリーメンバーとの交差反応性
透明のマキシソーププレートを、PBS中の5μg/mLのmAb又はアイソタイプ対照mAbの100μL/ウェルでコーティングし、4℃で一晩インキュベートした。プレートを200μL/ウェルで、ELISAブロック緩衝液(1% BSA、0.05% NaN3を含むPBS中5%スクロース)を用いて1時間ブロックし、洗浄緩衝液(PBS、0.01% Tween−20)で3回洗浄した。競合サイトカインをアッセイ希釈緩衝液(PBS中1% BSA)中で、2倍の最終濃度で滴定し、ビオチン化サイトカインを2倍の最終濃度で調製した。2倍の最終濃度の100μLのサイトカインを、アッセイ緩衝液中、2倍の最終濃度の100μLのビオチン化IL−17mut6(25ng/mL最終濃度)と混合した(30〜0μg/mL最終濃度)。組み換えヒトIL−23(R&D Systems,Minneapolis,MN)を陰性対照として、緩衝液のみの試料を背景対照として、及びIL−17mut6を陽性対照として使用した。サイトカイン/ビオチン化IL−17mut6混合物を複製物した、1ウェル当たり100μLを、プレートに添加し、1〜2時間インキュベートした。プレートを洗浄緩衝液で3回洗浄し、100μLの1mg/mL SA−HRPの1:10,000希釈を用いて、室温で20分間インキュベートした(Jackson Immunoresearch,West Grove,PA)。プレートをELISA洗浄緩衝液で3回洗浄した。洗浄後、100μL/ウェルのOPD基質(Sigma−Aldrich Corp.,St.Louis,MO)を各ウェルに添加し、適切な色の変化が検出されるまでインキュベートした。反応を50μLの2N硫酸の添加により停止し、Envision機器を使用してプレートを492nmと読み取った。
【0149】
親和性測定−ビアコアアッセイ
表面プラズモン共鳴(SPR)を用いる親和性測定を、Biacore 3000光バイオセンサ(Biacore)を用いて行った。選択されたFab(約30 RU)又はmAb(約50 RU)を、それぞれFab又はmAb捕捉のためのヒツジ抗Fd抗体又は抗ヒトFc抗体を使用して、センサーチップ表面上に捕捉した。Fab又はmAb捕捉後、溶液中、huIL−17mut6又はcyno IL−17A(0.2から49nM)を注入した。
【0150】
(実施例4)
エピトープマッピング
抗体エピトープを、競合結合、H/D交換分析、及び抗体−IL−17A共構造の組み合わせによって推定した(実施例5を参照されたい)。次の抗体を使用した:mAb1926、MORmAb7700、MORmAb7706、MORmAb7708、mAb7357(ハイブリドーマC1863に由来するマウス抗ヒトIL−17A中和抗体)、mAb2832(ハイブリドーマC1861に由来するマウス/ヒトキメラ抗ヒトIL−17A中和抗体)、mAb317(マウス抗ヒトIL−17A抗体、R&D Systems,Minneapolis,MN)及びmAb3171(マウス抗ヒトIL−17A抗体、R&D Systems,Minneapolis,MN)、並びにmAbeBIO16〜7178(マウス抗ヒトIL−17A抗体、e−Bioscience,San Diego,CA)。この3つの商業用抗体は、異なる度合いの中和活性を示した。
【0151】
競合的エピトープ結合
競合ELISAのために、1ウェル当たり5μL(20μg/mL)IL−17Aのmut6タンパク質を、MSD HighBindプレート(Meso Scale Discovery,Gaithersburg,MD)上に室温で2時間コーティングした。150μLの5% MSD遮断剤A緩衝液(Meso Scale Discovery,Gaithersburg,MD)を各ウェルに添加し、室温で2時間インキュベートした。プレートを0.1M HEPES緩衝液(pH 7.4)で洗浄した。10nMの標識化された抗体(MDS蛍光染色)を、競合物質抗体の濃度を増加させながら(1nM〜2μM)インキュベートし、25μLの混合物を指定されたウェルに添加した。室温で穏やかに振盪しながら2時間のインキュベートした後、プレートを上記のように洗浄し、150μL希釈MSD読み取り緩衝液Tを添加し、プレートをMDSセクターイメージャ6000で読み取った。
【0152】
アッセイを、標識化されたmAb1926、mAb317、mAb3171、又はmAb7357により行った(図4)。競合アッセイに基づき、抗IL−17A抗体を4つの異なるビンに割り当てた。ビンA:mAb1926、MORmAb7706、及びMORmAb7708、ビンB:eBio16〜7178及びmAb7357、ビンC:mAb317、ビンD:mAb3171。
【0153】
H/D交換分析:
H/D交換のために、抗体摂動を分析するために使用した手順は、前述の手順と類似していたが(Hamuro et al.,J.Biomol.Techniques,14:171〜82,2003、Horn et al.,Biochemistry,45:8488〜98,2006)、幾らかの修飾を伴った。組み換えIL−17Amut6(C−末端His−タグによりHEK293E細胞中で発現させた)を、重水素化水溶液中で所定の時間にわたってインキュベートすることにより、交換可能な水素原子における重水素の組み込みを得た。重水素化IL−17Amut6を、固定化した個々の抗IL−17A mAbを含有するカラム上に捕捉し、次いで水性緩衝液で洗浄した。逆交換したIL−17Amut6タンパク質をカラムから溶出し、重水素含有フラグメントの局在性をプロテアーゼ消化及び質量分析によって決定した。抗体に結合した領域は、交換から比較的保護された部位であり、したがって、抗体と複合体化されていないIL−17Amut6と比較してより高い分画の重水素を含有すると推定された。IL−17Amut6のH/D交換摂動マップは、図5に示される。バーの上の番号は、IL−17Amut6残基を指す。
【0154】
MORmAb7700、MORmAb7706、及びMORmAb7708は、IL−17Aの(配列番号105)45NRSTSPWNLH54(配列番号159)、56NEDPERYPSVIWE68(配列番号157)、及び100RREPPHCPNSFRLEKIL116(配列番号158)の3つのセグメントに対して異なる度合いの示差交換を示し、抗体による保護を示唆した。56NEDPERYPSVIWE68(配列番号157)フラグメントは、MORmAb7708によって強力に保護され、MORmAb7700によってわずかに保護され、MORmAb7706によっては保護されなかった。これらの抗体のフラグメント保護パターンにおける重複は、上述の競合アッセイにおけるそれらの交差阻害と一致する。
【0155】
mAb7357及びmAbeBio16〜7178の両方について、上述の競合アッセイにおけるそれらの交差阻害と一致して、71CRHLGCINADGNVDYHM87(配列番号160)に対する強力な保護を観察した。弱い、及びしたがって決定的でない示差交換を、mAb7357、mAb2832、mAb317及びmAb3171を有する他のフラグメントについて観察した。
【0156】
H/D交換研究は、上に記載される4つの競合群のうちの2つについて結合部位の位置を特定した。ビンA抗体(MORmAb7700、MORmAb7706、及びMORmAb7708)は、配列番号105のペプチドセグメント45NRSTSPWNLH54、(配列番号159)、56NEDPERYPSVIWE68(配列番号157)、及び100RREPPHCPNSFRLEKIL116(配列番号158)の領域で結合し、ビンB抗体(mAb7357及びmAbeBio16〜7178)は、ペプチドセグメント71CRHLGCINADGNVDYHM87(配列番号160)の領域で結合した。mAb317及びmAb3171は、互いから、並びにビンA及びビンB抗体から異なる部位に結合した。しかしながら、両方の抗体とのH/D交換研究における弱いシグナルは、IL−17A上のそれらのエピトープの位置を特定するのに十分な証拠を提供しなかった。
【0157】
(実施例5)
IL−17A及び抗IL−17A抗体の共結晶構造
mAb6785の組み換えHis6タグ付きFabである、Fab6468とのIL−17Amut6の共構造をX線結晶学によって決定した。Fab6468の軽鎖のアミノ酸配列は、配列番号90に示され、重鎖アミノ酸配列は、配列番号111に示される。実施例5において、IL−17Aアミノ酸残基は、配列番号105に従った残基を示すように参照され、Fab6468残基は、配列番号79に従った軽鎖可変領域残基を示すように参照され、重鎖可変領域残基は配列番号86に従う。組み換えヒトIL−17Amut6の発現、再折り畳み、及び精製が記載されている(Wu et al.,Cytokine,ePub ahead of print Jul29)。HEK−293F細胞中でFab6468を発現させ、記載される方法と類似した方法を使用して精製した(Zhao et al.,Protein Expr Purif,67:182〜9,2009)。
【0158】
IL−17A/Fab6468複合体の結晶化
IL−17A/Fab6468複合体を、IL−17Amut6及びFab6468を、pH 6.5の20mM MES、0.2M NaCl、及び10%グリセロール中、1:1.1モル比で混合することによって調製し、4℃で一晩インキュベートした。複合体を、pH 6.5の20mM MES、0.2M NaCl、及び10%グリセロール中、Superdex 200 10/300 GLカラム(GE Healthcare,Piscataway,NJ)上でのサイズ排除クロマトグラフィー(SEC)を使用して、過剰の複合体化されていないFabから精製した。複合体に相当する分画をプールし、Amicon Ultra 10000 MWCOデバイスで4.6mg/mLにまで濃縮した。
【0159】
等容量のタンパク質及びリザーバ溶液を、Corningプレート3550(Corning Inc.,Corning,NY)を使用するシッティングドロップ形式で分注するOryx4自動タンパク質結晶化ロボット(Douglas Instruments,East Garston,UK)を使用して、自動化結晶化スクリーニングを行った。最初のスクリーニングを、Hampton結晶スクリーンHT(HR2−130,Hampton Research)を用いて行い、pH 4.5〜4.6の硫酸アンモニウム、PEGを含有する複数の条件から針様の結晶を産生した。これらの小さい結晶を使用して、マイクロシードマトリックススクリーニング(MMS)(D’Arcy et al.,Acta Crystallographica Section D,63:550〜4,2007)のためのシードストックを産生した。回折品質の結晶を、pH 5.5の0.1M酢酸ナトリウム、12%PEG MME 5000、及び0.2M硫酸リチウム中のMMSスクリーンから得た。
【0160】
IL−17A/Fab6468複合体のX線データ収集
X線データ収集のために、結晶を、24%グリセロールを補充した母液に数秒間浸漬し、95°Kの窒素流中で急速冷凍した。X線回折データを収集し、Osmic(商標)VariMax(商標)共焦光学系、Saturn 944 CCD検出器、及びX−stream(商標)2000低温冷却システムを備えたRigaku MicroMax(商標)−007HF微小焦点X線発生装置(Rigaku,Woodlands,TX)を使用して処理した。回折強度を、1/2度の像当たり3分の露出時間から2.2Åの最大分解能で、254°結晶回転にわたって検出した。X線データをprogram D*TREK(Pflugrath,J.,Acta Crystallographica Section D,55:1718〜25,1999)により処理した。結晶は、単斜晶空間群P21に属し、a=73.40Å、b=64.04Å、c=145.61Å、及びβ=95.39°であった。X線データ統計値を表8に示す。
【0161】
【表11】
*最外郭についての値は、()内にある。
【0162】
構造決定
IL−17A/Fab6468の結晶構造を、Phaser(Read,Acta Crystallorg D Biol Crystallorg,57:1373〜82,2001)を使用して分子置換によって決定した。検索モデルは、IL−17F(PDB ID 1JPY)(Hymowitz et al.,EMBO J.,20:5332〜41,2001)及びFv(VH/VL)についての相同性モデルであり、それはModeller(Accelrys,CA)を使用して、VH及びVLの両方について、抗IL−13抗体CNTO607(PDB ID 3G6A)(Teplyakov et al.,J.Mol.Biol.389:115〜23.2009)に基づいて構築された。2つの定常ドメインCL/CH1をPDB ID 8FABから得た(Strong et al.,Biochemistry,30:3739〜48,1991)。構造精密化を、PHENIX(Adams et al.,J.Synchrotron.Radiat.11:53〜5,2004)を用いて行った。二回転非結晶学的対対称を、早期段階の精密化において最初に課したが、Rフリーに基づいて最終段階において緩めた。モデル調整及び手動の再構築を、COOTを使用して行った(Emsley et al.,Acta Crystallogr.D.Biol.Crystallogr.60:2126〜32,2004)。最終のR結晶及びRフリーは、2.2Åに対する全61,706個の独立の反射のそれぞれについて23.4%及び29.7%であった。精密化統計を表9に提供する。
【0163】
【表12】
【0164】
IL−17A/Fab6468複合体構造
複合体の構造を高分解能(約2.2Å)に対して決定した。IL−17Aは、結晶中ほぼ対称のホモ−二量体であり、2つのFab分子に結合していた。抗体−抗原相互作用は、大部分が疎水性であり、ほとんどの抗体とは対照的に、軽鎖CDRは、幾つかの重要な接触を行った。IL−17A/Fab6468複合体の全体的な分子構造を図6Aに示す。IL−17A二量体の単量体は、シスチンノットの全体的なトポロジーを採用した(図6B)。2つの単量体は、77骨格鎖Cα原子についての0.54ÅのCα RMSDと非常に類似していた。IL−17A単量体シスチンノットの全体的な構築は、76 Cα原子についての0.71Åのrmsdを有するIL−17Fの構築と非常に類似していた(図6B)。各IL−17A単量体は、3つのジスルフィド結合によって安定化していた。鎖Bについて、3つの鎖間ジスルフィド結合が観察された一方で(C10〜C106、C71〜C121、C76〜C123)、鎖Aについては、C10〜C106ジスルフィド結合は、単量体のこれらのセグメントにおける無秩序に起因して観察されなかった。後者の2つのジスルフィド結合(C71〜C121、C76〜C123)は、IL−17F及びNGFに類似したシスチンノット構築を安定化した。IL−17Aの鎖Bについての構造的モデルには、全ての残基10〜128(残基1〜9は無秩序であった)が含まれ一方で、鎖A残基については、残基21〜29、41〜104及び109〜127のみが観察され、その他の残基1〜20、30〜40、105〜108、及び128は、構造における無秩序に起因して欠失していた。2つのFabについて、両方の軽鎖の残基1〜2は、無秩序であったか、又は乏しい電子密度を有した。鎖内ジスルフィド結合並びに重鎖上のHisタグ含む、重鎖及び軽鎖の両方のC−末端3残基は、無秩序であった。
【0165】
IL−17A(鎖B)の秩序化N末端セグメントは、短いヘリカル要素を含有した(残基8〜12)。それは、同じ単量体のループ3〜4に向かって再び折り畳み、鎖間ジスルフィド結合(C10〜C106)を形成した。対照的に、IL−17Fの同等のセグメントは、二量体の他方の単量体へと到達し、鎖内ジスルフィド結合を形成し、2つの単量体を共有結合で連結した。2つのIL−17A単量体のセグメント17〜39の秩序化部分を、IL−17Fにおいてと同様にスワップした。このスワッピングは、IL−17A二量体のこれらの部分についての交差をもたらした。分子内ジスルフィド結合(C10〜C106)と組み合わせて、IL−17Aの2つのN末端セグメントは、2つの連鎖単量体を形成し、それはまた、非還元SDS−PAGE上で26kDの見かけの二量体ももたらした。
【0166】
IL−17Aの二量体は、4つの主なβ−ストランド(ストランド1〜4)についてほぼ対称であった(図6C)。76個の残基についてのCα rmsdは、0.71Åである。若干の不斉は、2つの源に由来した。第1に、鎖Aは、主にN末端において、幾つかの無秩序セグメントを含有した。短いβ−ストランド(ストランド0、残基22〜26)のみが見かけ上、秩序化されていた一方で、鎖Bの残基10〜40は、ヘリカルセグメント(残基12〜16)及びβ−ストランド(ストランド0、残基21〜25)で秩序化されていた。第2に、IL−17Aの2つの単量体(40〜128)の、シスチンノットの4つの主なβ−ストランドが、二回転対称によって関連させられた一方で、ストランド0の秩序化部分は、主体が重ねられるとき、完全に重ね合わさらなかった(図示されず)。これがタンパク質再折り畳みのアーチファクトであるか、又はかかる配列が天然に存在するかどうかは、更なる調査なしには明確でない。この種の生物活性は、商業的供給源の参照IL−17A(大腸菌においても産生される)(R&D Systems,Minneapolis,MN)の生物活性に類似しており、C10〜C106についての鎖間又は鎖内ジスルフィド結合がその受容体結合に重要でないことが示唆された。それにもかかわらず、現在の構造は、N末端セグメント(1〜20)及び(30〜39)は、非常に柔軟であり、それらの構造は、折り畳まれたIL−17A二量体の活性に影響を及ぼさない。
【0167】
エピトープ及びパラトープ
IL−17AとFab6468との間の結合に関与する残基を表10に列挙する。IL−17AのプロトマーAにおいて欠失した残基及びIL−17A二量体の若干の不斉性質に起因して、2つの接触部位からの全てのエピトープ残基は、同一でなかった(表10及び図7)。しかしながら、同一であるコアな一組の残基並びにそれらの相互作用が存在した。これらの残基は、IL−17A配列番号105のL26、R55、E57、P59、E60、R61、Y62、S64、V65、W67、R101、E102、P103、及びF110であり(表10に黒色で強調表示される)、それらはFab6468に対するコアエピトープを構成する。
【0168】
表10コアエピトープ残基は、黒色で強調表示される。コアパラトープ残基は、太字にされている。拡張されたパラトープ残基は、括弧内に示される。
【0169】
【表13】
【0170】
同様に、2つの部位における抗体からの接触残基は、完全に同一ではなかった。コアエピトープ残基への同一の接触に関与する残基は、「コアパラトープ」と称され、それは次の残基から成っていた:軽鎖(LC):Y31、D49、Y90、F92、F93(配列番号79);並びに重鎖(HC):S52、T54、F57、Y59、Q99、L100、及びT101(配列番号86)(表10)。コアパラトープ残基は、表10に太字で示される。特異的IL−17A残基に結合する1つの単量体において特定された更なる「拡張されたパラトープ」残基は、括弧内に示される。
【0171】
共結晶構造で特定されたL26を除く全てのコアエピトープ残基は、MORmAb7700についてのH/D交換、56NEDPERYPSVIWE68(配列番号157)及び100RREPPHCPNSFRLEKIL116(配列番号158)によって特定された、保護されたセグメントのうちの2つの境界内又は境界にあったため、MORmAb7700についてのH/D保護データは、共結晶研究に一致していた。MORmAb8302及びmAb1926を含むMORmAb7700抗体誘導体の全ては、それらが多くてもVHのN末端領域における1つの残基(実施例1を参照されたい)、VLのN末端における3つの残基(実施例1を参照されたい)、及び3つのCDR残基(H2、H3、及びL3において1つずつ、表1a)分だけ異なるのみで、それらのうちのいずれも抗体パラトープの一部でないため、Fab6468と同じ結合特異性を有することが想定される。
【0172】
本発明で特徴付けられるIL−17A構造は、以前に公開された構造に非常に類似しているが、欠失したセグメントに起因して、P2ポケット空洞(下記を参照されたい)が以前の研究(構造2VXS、Protein DataBank http_//www_rcsb_org/pdb/home/home_do;Gerhardt et al.,J.Mol.Biol.394:905〜21,2009において入手可能)では特定されなかったことを除く。
【0173】
IL−17RAとの複合体におけるヒトIL−17Fの結晶構造が報告されている(Ely et al.,Nat.Immunology,10:1245〜51,2009)。IL−17AとIL−17Fとの間の配列及び構造的類似性のため、IL−17Aは、IL−17Fと類似した様態でIL−17RAと相互作用する可能性が高い。この研究で得られたFab6468との複合体におけるIL−17A構造を、報告されたIL−17F/IL−17RA複合体におけるIL−17F上に重ねることによる分子モデリングにより、Fab6468のセグメントがIL−17RAとの立体障害を有するであろうことが示された。これらのセグメントのうちの1つは、Fab6468の軽鎖CDR3におけるFFモチーフの周囲に局在化する(配列番号79の残基92及び93)。したがって、特定の理論に決して束縛されるものではないが、Fab6468が、IL−17RA及び類推して、IL−17RCとの、その相互作用を遮断することによって、IL−17A機能を阻害することが示唆されるが、IL−17RCとIL−17Aとの間の相互作用の様式は、分子レベルでは知られていない。
【0174】
IL−17RAに対するIL−17A及びIL−17Fの親和性における有意な差異は、IL−17A及びIL−17RAの相互作用の詳細において有意な差異が存在し得ることを示唆するが、その程度は、IL−17A/IL−17RAの共結晶構造が決定されるとき初めて得られるであろう。これには、この研究におけるP2ポケット空洞の特定が関連するが、それは報告されたIL−17F/IL−17RA結晶構造におけるIL−17F類似領域において部分的に特定されているのみである(Ely et al.,Nat.Immunology,10:1245〜51,2009)。
【0175】
2つの深い、大部分が疎水性のポケットを、二量体境界面に沿ってIL−17Aの表面上で特定した(図8A、5B)。IL−17Fにおいて最初に発見された(Hymowitz et al.,EMBO J,20:5332〜41,2001)ポケットに類似したP1ポケットは、単量体Aの残基Q94、E95、L97、及びK114、並びに単量体BのL53、Y62、P63、V65、I66、W67、I96、V117、及びV119から成り、逆もまた同様である。二量体の一方の側で、P1ポケットは、セグメント30〜40によって部分的にカバーされる一方で、他方の側では、セグメントが無秩序であることに起因してそれは完全に開いている。このセグメントは柔軟であると考えられるため、他の分子がP1ポケットにアクセス可能であろう。P2ポケットもまた、単量体AのV24、L26、I28、Y62、L99、R101、F110、及びL112、並びに単量体BのV22、V24、及びL112の両方の鎖からの残基から成り、逆もまた同様である。
【0176】
P2ポケットの詳細は、上述のようにIL−17A二量体の不斉に起因して若干異なるが、2つのP2ポケットの全体的な形状は、非常に類似している。P1及びP2ポケットを裏打ちする2組の残基は、IL−17AとIL−17Fとの間で非常に良好に保存されている(図8C)。しかしながら、IL−17F構造において、P2ポケットは、残基F10及びF11(F10F11モチーフ)によって占領されている(図8B)(Hymowitz et al.,EMBO J,20:5332〜41,2001)。FFモチーフは、ヒトIL−17Aにおいて不在であり、代わりに、対応するアミノ酸残基は、I4及びP5(配列番号105における残基4及び5)である(図8C)。これらの残基は、それらがフェニルアラニン残基よりもはるかに小さいため、P2ポケット並びにFFモチーフにおいて結合する可能性が低く、P2ポケットに対して十分な親和性を有さない可能性が最も高い。したがって、IL−17FのFFモチーフは恐らく、受容体IL−17RA及びIL−17RCとのヒトIL−17A及びIL−17F相互作用についての構造的判別子である。これらの大部分の疎水性のポケット(P1及びP2)の両方は、IL−17RAへのIL−17A結合に必要とされる可能性が高い。IL−17F/IL−17RA複合体の近年の結晶構造は、FFモチーフがIL−17RAによって置換されていることを示す(Ely et al.,Nat.Immunology 10:1245〜51,2009)。P2からのFFモチーフ排除によるエネルギー損失は、より低い結合親和性をもたらす可能性が高い。これは、IL−17RAが、ヒトにおいて、IL−17Aに高い親和性で結合するが、IL−17Fには低い親和性で結合するという観察と一致し(Kuestner et al.,J Immunol,179:5462〜73,2007)、IL−17A及びIL−17F効力における差異を潜在的に説明し得る。マウスにおいて、FFモチーフは、IL−17A及びIL−17Fの両方で不在であり、それぞれ残基IP及びALによって置き換えられている。AL及びIP残基の対は、小さく、P2ポケットに対して低い親和性を有する可能性が高い。したがって、P2は、マウスIL−17A及びIL−17FにおいてIL−17RA結合のために有効であろう。マウスIL−17A及びIL−17Fの両方は、類似した親和性でマウスIL−17RAに結合し(Kuestner et al.,J Immunol,179:5462〜73,2007)、これは、結合のためのP2ポケットの有効性がリガンドの親和性を増加させるという本発明の示唆と一致する。
【0177】
全体的に、IL−17AとIL−17Fとの間で観察した構造的差異は、それぞれの受容体とのそれらの相互作用を詳細に分析するための基盤を提供する。更に、ペプチド、ペプチド模倣体、及び小分子がいずれか又は両方のポケットにおいて結合して、IL−17A及び/又はIL−17Fがそれらの受容体と相互作用することを遮断するように設計され得ることが考えられる。Fab6468(配列番号79における残基F92及びF93)に存在するFFモチーフは、P2ポケット残基L26、R61、L99、R101、及びR102に結合するため、Fab 6468構造を使用して、ファージディスプレイを使用して無作為化又は設計したペプチドライブラリーからのペプチド等の、更なるIL−17A拮抗物質を選択及び最適化することが可能である。
【0178】
P1及びP2ポケットを裏打ちする残基は、IL−17AとIL−17Fとの間で良好に保存されており、分子モデリングにより、IL−17Aホモ二量体のみと比較したとき、IL−17A/Fヘテロ二量体が、ほぼ同一の全体的構造を採用するであろうとことが示唆される。したがって、P1及びP2ポケットは、類似した全体的なトポロジーによりIL−17A/Fヘテロ二量体中に存在し、その受容体結合部位を構成する可能性が高い。したがって、P2ポケット残基に結合するIL−17A拮抗物質は、IL−17A/Fヘテロ二量体に結合し、拮抗する可能性がある。
【0179】
(実施例6)
交差種結合特異性
mAbtr1926の交差種結合特異性を評価するために、マイクロタイタープレート上にコーティングした異なるIL−17Aタンパク質を用いて結合ELISAを行った。ヒト、マウス、及びラットIL−17Aタンパク質をマイクロタイタープレート上にコーティングした。標識化されたmAb1926の連続希釈液を37℃で2時間インキュベートした。インキュベーション後、マイクロタイタープレートを完全に洗浄し、結合した標識化mAb1926を検出した。mAb1926は、ラット又はマウスIL−17Aタンパク質よりもはるかに強力に、ヒトIL−17Aに結合した(図9)。ラット及びマウスIL−17Aへのこの低減された結合は、タンパク質の両方がFab6468拡張型エピトープの7位においてヒトIL−17Aから異なることと一致する(表10)。更に、Fab6468エピトープの一部に近い位置である、ヒトIL−17Aの残基40と41との間のラット及びマウスIL−17Aにおいて、1つのアミノ酸挿入が存在する。
【図1A】
【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図2A】
【図2B】
【図3A−1】
【図3A−2】
【図3B−1】
【図3B−2】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【技術分野】
【0001】
本発明は、インターロイキン−17A(IL−17A)抗体拮抗物質、IL−17A抗体拮抗物質又はそのフラグメントをコードするポリヌクレオチド、並びに上記のものを製造及び使用する方法に関する。
【背景技術】
【0002】
インターロイキン−17A(IL−17A、CTLA−8、IL−17)は、IL−23及びTGF−β等のサイトカインに応答して、活性化Th17細胞、CD8+ T細胞、γδ T細胞、及びNK細胞によって分泌されるサイトカインであり、好中球生態、炎症、臓器破壊、及び宿主細胞防衛に関与する、線維芽細胞及び滑膜細胞等の多数の細胞タイプからの、抗菌性ペプチド(デフェンシン)、炎症促進性サイトカイン、及びケモカイン等のメディエーターの産生を調節する(Weaver et al.,Annu.Rev.Immunol.25:821〜52,2007、Aggarwal et al.,J.Biol.Chem.278:1910〜4,2003、Mangan et al.,Nature 441:231〜4,2006に概説される)。IL−17Aは、TNF−α及びIL−1β等の他のサイトカインと協同して、炎症促進性環境を増強する。
【0003】
IL−17Aサイトカインファミリーは、IL−17A、B、C、D、E、及びFと指定される6つの同族体から成り、各々、相違する特有の生物学的役割を有する(Kawaguchi et al.,J.Allergy Clin.Immunol.114:1265〜73,2004、Kolls and Linden,Immunity 21:467〜76,2004、Moseley et al.,Cytokine Growth Factor Rev.14:155〜74,2003)。このファミリーメンバーのうち、IL−17Fは、IL−17Aと最も相同であり、肺における好中球増加症の誘導及び炎症促進性サイトカインの誘導等の多くの類似した機能特性を共有するが、人間においては、IL−17Fは、IL−17Aよりも約10倍効力が低い(Moseley et al.,Cytokine Growth Factor Rev.14:155〜74,2003、Kolls et al.,Immunity,21:467〜76,2004、McAllister et al.,J.Immunol.175:404〜12,2005)。IL−17A及びIL−17Fはまた、生体外で中間生物活性を有する、ヘテロ二量体を形成することができる(Wright et al.,J.Biol.Chem.282:13447〜55,2007)。
【0004】
IL−17Aは、インターロイキン−17受容体A(IL−17RA)及び受容体C(IL−17RC)と相互作用することによってその効果を媒介する(Moseley et al.,Cytokine Growth Factor Rev.14:155〜74,2003、Toy et al.,J.Immunol.177:36〜9,2006)。IL−17Fは、同じ受容体を通じてシグナル伝達するが、これらの受容体へのIL−17F親和性は、それよりも有意に低い(Kuestner et al.,J.Immunol.179:5462〜73,2007)。ヒトIL−17F及びヒトIL−17F/IL−17RA複合体の結晶構造は、ホモ二量体における推定受容体結合空洞を特定した(Hymowitz et al.,EMBO J.20:5332〜41,2001、Ely et al.,Nat.Immunology 10:1245〜51、2009)。類似した空洞は、中和Fabとの複合体におけるヒトIL−17Aの結晶構造において特定されたが、この空洞は、部分的に占領されていた(Gerhardt et al.,J.Mol.Biol.394:905〜21,2009)。
【0005】
IL−17Aの不適切な又は過剰な産生は、関節リウマチ(Lubberts,Cytokine 41:84〜91,2008)、ぜんそく等のアレルギー性気道疾患を含む気道過敏症(Linden,Curr.Opin.Investig.Drugs.4:1304〜12,2003、Ivanov,Trends Pharmacol.Sci.30:95〜103,2009に概説される)、乾癬(Johansen et al.,Br.J.Dermatol.160:319〜24,2009)、アトピー性皮膚炎を含む皮膚過敏症(Toda et al.,J.Allergy Clin.Immunol.111:875〜81,2003)、全身性硬化症(Fujimoto et al.,J.Dermatolog.Sci.50:240〜42,2008)、潰瘍性大腸炎及びクローン病を含む炎症性腸疾患(Holtta et al.,Inflamm.Bowel Dis.14:1175〜84,2008、Zhang et al.,Inflamm.Bowel Dis.12:382〜88,2006)、並びに慢性閉塞性肺疾患を含む肺疾患(Curtis et al.,Proc.Am.Thorac.Soc 4:512〜21,2007)を含む、種々の疾患及び傷害の病理に関連する。
【発明の概要】
【発明が解決しようとする課題】
【0006】
IL−17Aに対する抗体は、IL−17A媒介型疾患及び傷害の治療における使用のために提案されている(PCT公開第WO08/021156号、第WO07/070750号、第WO07/149032号、第WO06/054059号、第WO06/013107号、第WO08/001063号、第WO10/034443号、米国特許出願第US2008/095775号、第US2009/0175881号)。抗体治療薬の薬物動態学、有効性、及び安全性プロファイルは、具体的な組成に応じるため、IL−17A媒介型疾患及び傷害の治療における使用に好適な、ヒトIL−17Aに対する改善された抗体の必要性が存在する。
【課題を解決するための手段】
【0007】
本発明の一態様は、単離された抗体又はそのフラグメントであり、該抗体は、アミノ酸残基56〜68(配列番号157)及び100〜116(配列番号158)において、又は残基L26、R55、E57、P59、E60、R61、Y62、S64、V65、W67、R101、E102、P103、及びF110において、配列番号105に示される配列を有するヒトIL−17Aに特異的に結合する。
【0008】
本発明の別の態様は、単離された抗体又はそのフラグメントであり、抗体は、ヒトIL−17A上のP2ポケット空洞に特異的に結合し、このP2ポケット空洞は、配列番号105のアミノ酸残基V22、V24、L26、I28、Y62、L99、R101、F110、及びL112を含む。
【0009】
本発明の別の態様は、ヒトIL−17Aに特異的に結合し、ある種の重鎖相補性決定領域(CDR)1、2、及び3(HCDR1、HCDR2、HCDR3)のアミノ酸配列、ある種の軽鎖相補性決定領域(CDR)1、2、及び3(LCDR1、LCDR2、LCDR3)のアミノ酸配列、ある種の重鎖可変領域(VH)のアミノ酸配列又はある種の軽鎖可変領域のアミノ酸配列(VL)を含むモノクローナル抗体と、ヒトIL−17A結合をめぐって競合する、単離された抗体又はフラグメントアミノ酸配列である。
【0010】
本発明の別の態様は、ある種の重鎖可変領域パラトープアミノ酸残基及びある種の軽鎖可変領域パラトープアミノ酸残基を含む、ヒトIL−17Aに特異的に結合し、配列番号105に示されるアミノ酸配列を有するヒトIL−17Aのある種の残基と相互作用する、単離された抗体又はフラグメントである。
【0011】
本発明の別の態様は、重鎖可変領域及び軽鎖可変領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、該抗体が、Chothia残基F56及びY58から選択される重鎖可変領域パラトープ、並びにChothia残基Y91、F93、及びF94から選択される軽鎖可変領域パラトープを含む。
【0012】
本発明の別の態様は、重鎖可変領域(VH)及び軽鎖可変領域(VL)を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、該抗体が、ある種の重鎖相補性決定領域(CDR)1、2、及び3(HCDR1、HCDR2、HCDR3)のアミノ酸配列、ある種の軽鎖相補性決定領域(CDR)1、2、及び3(LCDR1、LCDR2、LCDR3)のアミノ酸配列、ある種の重鎖可変領域(VH)のアミノ酸配列又はある種の軽鎖可変領域(VL)のアミノ酸配列を含む。
【0013】
本発明の別の態様は、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであり、該抗体は、ある種の重鎖のアミノ酸配列及びある種の軽鎖のアミノ酸配列を含む。
【0014】
本発明の別の態様は、本発明の単離された抗体又はフラグメント及び医薬的に許容される担体を含む医薬組成物である。
【0015】
本発明の別の態様は、配列番号67、68、69、81、82、83、84、85、86、92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を含む単離された抗体重鎖である。
【0016】
本発明の別の態様は、配列番号76、77、78、79、80、87、88、89、90、又は91に示されるアミノ酸配列を含む単離された抗体軽鎖である。
【0017】
本発明の別の態様は、配列番号67、68、69、81、82、83、84、85、86、92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を含む抗体重鎖をコードする、単離されたポリヌクレオチドである。
【0018】
本発明の別の態様は、配列番号76、77、78、79、80、87、88、89、90、又は91に示されるアミノ酸配列を含む抗体軽鎖をコードする、単離されたポリヌクレオチドである。
【0019】
本発明の別の態様は、少なくとも1つの本発明のポリヌクレオチドを含むベクターである。
【0020】
本発明の別の態様は、本発明のベクターを有する宿主細胞細胞である。
【0021】
本発明の別の態様は、ヒトIL−17Aの、IL−17RAとの相互作用を阻害する方法であり、ヒトIL−17A及びIL−17RAを提供する工程と、
ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる群から選択される少なくとも1つのアミノ酸残基においてヒトIL−17Aに結合する拮抗物質と接触させる工程と、を含む。
【0022】
本発明の別の態様は、IL−17A生物活性を阻害する方法であり、ヒトIL17−A及びIL−17RAを提供する工程と、ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる群から選択される少なくとも1つのアミノ酸残基においてヒトIL−17Aに結合する拮抗物質と接触させる工程と、を含む。
【0023】
本発明の別の態様は、請求項3又は7に記載の単離された抗体の治療上の有効量を、炎症性状態の治療を必要とする患者に、炎症性状態を治療するうえで充分な時間にわたって投与する工程を含む、炎症性状態を治療する方法である。
【図面の簡単な説明】
【0024】
【図1】A〜Hはファミリー2、6a、6b、19a、及び19b IL−17A抗体拮抗物質のCDR配列。
【図2】例示のA)IGLV3及びIGLJ、並びにB)パラトープ残基をグラグトするための足場としてのIGHV3及びIGHJ生殖細胞系列遺伝子。mAb6785配列は、上に示される。CDR領域は、下線を引かれており、mAb6785軽鎖(Y31、D49、Y90、F92、F93)及び重鎖(S52、T54、F57、Y59、Q99、L100、及びT101)におけるコア接触部位は、アスタリスク「*」によって示される。B)におけるフレームワーク4領域は、二重下線を引かれている。示されている配列は、特に具体的な指示がない限り、*01対立遺伝子。
【図3】選択された抗体A)軽鎖及びB)重鎖のためのKabat及びChotia番号付け。Kabat CDR及びChothia HVの場所は、灰色で強調表示されている。
【図4】ELISA形式での、標識化されたA及びB)mAb1926、C)mAb317、D)mAb3171、E)及びF)mAb7357の、IL−17Aとの競合結合アッセイ。
【図5】A)異なる抗IL−17A mAbと複合体化されたIL−17AのH/D交換マップ。保護ブロックの上の番号付けは、成熟IL−17A(配列番号105)配列の番号付けに相当する。
【図6】A)IL−17A/Fab6468複合体の全体的な分子構造。IL−17Aの二量体は、濃い灰色及び薄い灰色で示される。2つのFab分子は、それぞれ濃い灰色及び薄い灰色で示される。B)IL−17A(薄い灰色)及びIL−17F(濃い灰色)の単量体の比較、C)IL−17Aの二量体(薄い及び濃い灰色)、D)IL−17Fの二量体(薄い及び濃い灰色)。
【図7】Fab6468に対するIL−17A上の2つの結合部位及びコアエピトープ。プロトマー1及び2は、それぞれ濃い及び薄い灰色である。コアエピトープは、黒色の楕円形によって示される。破線は、無秩序残基を表す。
【図8】IL−17A及びIL−17F推定受容体結合ポケットの比較。A)IL−17AのP1及びP2ポケットの正面及び背面図。mAb6468軽鎖CDR3のFFモチーフは、P2ポケットに示される。B)P2ポケットにおけるN末端FFモチーフを有するIL−17F。C)IL−17A及びIL−17Fの配列整列並びにP1及びP2ポケットの保存。
【図9】ELISA形式での、mAb1926の、IL−17Aタンパク質の異なる種への結合特異性。
【発明を実施するための形態】
【0025】
本明細書に引用する、特許及び特許出願を含むがそれらに限定されない刊行物はすべて、それらがあたかも本明細書に完全に記載されているのとまったく同様に本願に援用するものである。
【0026】
本明細書で使用されるとき、用語は、特定の実施形態を記載する目的でのみ使用され、限定を意図するものではないことが理解される。別段の規定がない限り、本明細書で使用されるとき、全ての技術及び科学用語は、本発明が属する技術分野における当業者によって一般的に理解されている意味と同一の意味を有する。
【0027】
本明細書に記載されているものと同様又は同等の任意の方法及び材料を、本発明の試験を実施するために使用できるが、例示となる材料及び方法を本明細書に記載する。本発明の記載及び請求には、以下の用語が使用される。
【0028】
本明細書で使用されるとき、「拮抗物質」という用語は、任意の機序によって、IL−17A活性を部分的に又は完全に阻害する分子を意味する。例示の拮抗物質は、抗体、融合タンパク質、ペプチド、ペプチド模倣体、核酸、オリゴヌクレオチド、及び小分子である。薬剤は、後述されるIL−17A活性についての周知のアッセイを使用して特定することができる。
【0029】
本明細書で使用されるとき、「IL−17A抗体拮抗物質」又は「IL−17Aに反応する抗体」という用語は、直接若しくは間接に、IL−17A生物活性を低減若しくは阻害する、IL−17Aのその受容体への結合を遮断する、又はIL−17A受容体活性化を阻害することができる抗体を指す。例えば、IL−17Aに反応する抗体は、IL−17Aに直接結合し、IL−17A活性を中和する、すなわち、サイトカイン及びケモカイン放出を低減するために、IL−17Aシグナル伝達を遮断することができる。
【0030】
「IL−17A」(CTLA−8、IL−17、インターロイキン−17A)という用語は、GenBank登録番号NP_002181に示されるアミノ酸配列を有する、ヒトIL−17Aポリペプチドを指す。配列番号105は、成熟ヒトIL−17Aのアミノ酸配列を示す。生体内IL−17Aは、2つの単量体のホモ二量体を形成し、それらは単量体A及び単量体B、又はプロトマーA及びプロトマーB、又はプロトマー1及びプロトマー2、又は鎖A及び鎖Bと指定される。IL−17Aはまた、IL−17Fとのヘテロ二量体を形成することができる。「IL−17A」という用語は、単量体、ホモ二量体、及びヘテロ二量体型を含む。「IL−17Amut6」という用語は、A70Q及びA132Q置換を有するIL−17Aの変異型を指す。成熟IL−17Amut6のアミノ酸配列は、配列番号106に、及びcDNA配列は、配列番号112に示される。IL−17A及びIL−17Amut6は、同等の活性を有する(PCT特許出願第WO09/003096号)。
【0031】
本明細書で使用されるとき、「IL−17A受容体」という用語は、は、両方の受容体ポリペプチド、IL−17RA(GenBank登録番号NP_055154、配列番号107)及びIL−17RC(GenBank登録番号NP_703191、配列番号113)、並びに2つのポリペプチドのホモ二量体又はヘテロ二量体を含む。
【0032】
本明細書で使用されるとき、「抗体」という用語は、広義で意図され、ポリクローナル抗体、マウス、ヒト型(human)、ヒト適合性、ヒト化、及びキメラモノクローナル抗体を含むモノクローナル抗体、抗体フラグメント、少なくとも2つの無傷抗体、二量体の、四量体の、又は多量体の抗体から形成される多重特異的抗体を含む、免疫グロブリン分子を含む。
【0033】
本明細書で使用されるとき、「モノクローナル抗体」(mAb)という用語は、実質的に均質な抗体の集団から得られる抗体(又は抗体フラグメント)を意味する。モノクローナル抗体は、高度に特異的であり、典型的に単一のエピトープに対して向けられている。「モノクローナル」という修飾語は、抗体のほぼ均質な性質を指して言うものであり、抗体がいずれかの特定の方法によって作製される必要はない。
【0034】
免疫グロブリンは、重鎖の定常領域のアミノ酸配列に応じて、IgA、IgD、IgE、IgG及びIgMの5つの大きなクラスに分類することができる。IgA及びIgGは、IgA1、IgA2、IgG1、IgG2、IgG3、IgG4というアイソタイプに更に細かく分類される。あらゆる脊椎動物種の抗体の軽鎖は、定常領域のアミノ酸配列に基づいてカッパ(κ)及びラムダ(λ)の2つの明確に異なるタイプのうちの1つに分類することができる。
【0035】
「抗体フラグメント」という用語は、重鎖相補性決定領域(HCDR)、軽鎖相補性決定領域(LCDR)、重鎖可変領域(VH)、軽鎖可変領域(VL)、重鎖定常領域(CH)、軽鎖定常領域(CL)、又は抗体重鎖若しくは軽鎖のいずれかからのフレームワーク領域(FR)等の、免疫グロブリン分子の少なくとも一部分を含む。抗体は、Fab、F(ab’)、F(ab’)2、scFv、dsFv、又は二重特異性抗体であってもよい。上述の抗体フラグメントの構造、並びに抗体及びそのフラグメントの調製及び使用のための技術は、当該技術分野において周知である。
【0036】
「フレームワーク」領域からなる抗体可変領域は、3つの「抗原結合部位」によって妨害される。抗原結合部位は、種々の用語を使用して記載される。(i)3つがVHに(HCDR1、HCDR2、HCDR3)、及び3つがVLに(LCDR1、LCDR2、LCDR3)ある、相補性決定領域(CDR)は、配列可変性に基づく(Wu and Kabat,J.Exp.Med.132:211〜250,1970、Kabat et al.,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.,1991)。(ii)3つがVHに(H1、H2、H3)及び3つがVLに(L1、L2、L3)ある、「高度可変領域」、「HVR」、又は「HV」は、Chothia and Lesk(Chothia and Lesk,Mol.Biol.196:901〜917,1987)によって記載されるように、構造において高度可変である抗体可変ドメインの領域を指す。他の用語には、「IMGT−CDR」(Lefranc et al.,Dev.Comparat.Immunol.27:55〜77,2003)及び「特異性決定残基使用」(SDRU)(Almagro,Mol.Recognit.17:132〜143,2004)が含まれる。International ImMunoGeneTics(IMGT)データベース(http://www_imgt_org)は、抗原結合部位の標準化された番号付け及び記載を提供する。各CDR、HV、及びIMGTの表記間の対応については、Lefranc et al.,Dev.Comparat.Immunol.27:55〜77,2003に記載されている。
【0037】
本明細書で使用されるとき、「Chothia残基」は、Al−Lazikani(Al−Lazikani et al.,J.Mol.Biol.,273,927〜48,1997)に従って番号付けされた抗体VL残基及びVH残基である。ポリペプチド番号付けに関連して2つの最も使用される番号付けシステム、Kabat(Kabat et al.,Sequences of Immunological Interest,5th Ed.Public Health Service,NIH,Bethesda,MD,1991)とChothia(Chothia及びLesk,Mol.Biol.196:901〜17,1987)との間の対応を、本発明の代表的な抗体に対して図3に示す。
【0038】
「フレームワーク」又は「フレームワーク配列」は、抗原結合部位として記載されていない、可変領域の残りの配列である。抗原結合部位は上述のような種々の用語によって記載され得るため、フレームワークの正確なアミノ抗原結合部位がどのように記載されるかによって決まる。
【0039】
本明細書で使用されるとき、「実質的に同一」という用語は、比較されている2つの抗体又は抗体フラグメントアミノ酸配列が、同一であるか、又は「ごく僅かな差異」を有することを意味する。ごく僅かな差異とは、抗体特性に悪影響を及ぼさない、抗体又は抗体フラグメントアミノ酸配列における1、2、3、4、5、6、7、8、9、10、11、12、13、14、又は15個のアミノ酸の置換である。本明細書で開示する配列とほぼ同じアミノ酸配列も本願の一部である。特定の実施形態では、配列の同一性は、約90%、91%、92%、93%、94%、95%、96%、97%、98%、99%又はそれよりも高い。同一性百分率は、例えば、Vector NTI v.9.0.0(Invitrogen,Carslbad,CA)のAlignXモジュールの初期設定を使用する一対整列によって決定することができる。本発明のタンパク質配列を、問い合わせ配列として使用して、例えば、関連配列を特定するために、公的又は特許データベースに対して検索を実行することができる。かかる検索を実行するために使用される例示のプログラムは、初期設定を使用する、XBLAST若しくはBLASTPプログラム(http_//www_ncbi_nlm/nih_gov)、又はGenomeQuest(商標)(GenomeQuest、Westborough、MA)一式である。
【0040】
本願で用いられる「〜と組み合わせて(in combination with)」という用語は、記述の対象である薬剤が、他の薬剤と共に、混合物中で一緒に、又は単独薬剤として同時に、又は単独薬剤として順次に任意の順序で、動物に投与され得ることを意味する。
【0041】
本明細書で使用されるとき、「炎症性状態」という用語は、部分的には、サイトカイン、ケモカイン、又は炎症性細胞(例えば、好中球、単球、リンパ球、マクロファージ)の活性によって媒介される、有害な刺激等(病原体、損傷した細胞、身体的傷害、若しくは刺激物)に対する急性又は慢性の局部的又は全身性反応を指し、ほとんどの例において、疼痛、潮紅、膨張、及び組織機能の障害によって特徴付けられる。
【0042】
本明細書で使用されるとき、「IL−17A媒介型炎症性状態」という用語は、少なくとも部分的にはIL−17A生物活性から結果として生じる、又はIL−17A活性によって引き起こされる、炎症性状態を指す。例示のIL−17A媒介型炎症性状態は、乾癬及び関節リウマチである。
【0043】
本明細書で使用されるとき、「IL−17A媒介型状態」という用語は、IL−17Aが、直接又は間接によらず、疾患又は状態の、因果関係、発症、進行、持続、又は病理を含む、疾患及び病状において役割を果たす、全ての疾患及び病状を包含する。
【0044】
用語「エピトープ」は、本明細書で使用するとき、抗体が特異的に結合する抗原の部分を意味する。エピトープは、通常、アミノ酸又は多糖類側鎖のような化学的に活性な(極性、非極性又は疎水性)部分の表面集団からなり、特定の3次元構造特性並びに特定の帯電特性を有し得る。エピトープは、立体配座空間単位を形成する隣接した又は隣接していないアミノ酸のいずれか又は両方から成り得る。隣接していないエピトープについて、抗原の直鎖配列の異なる部分からのアミノ酸は、タンパク質分子の折り畳みを通じて、3次元空間において近接近する。
【0045】
本明細書で使用されるとき、「パラトープ」という用語は、抗原がそこに特異的に結合する、抗体の一部分を意味する。パラトープは元来線状であるか、又は不連続であることができ、抗体のアミノ酸の線状構造よりも、隣接していないアミノ酸間の空間的な関係により形成される。「軽鎖パラトープ」及び「重鎖パラトープ」又は「軽鎖パラトープアミノ酸残基」及び「重鎖パラトープアミノ酸残基」は、それぞれ、抗原と接触する抗体の軽鎖及び重鎖残基を指す。
【0046】
用語「特異的結合」は、本明細書で使用するとき、抗体が特定の抗原と、他の抗原又はタンパク質に対するよりも高い親和性で結合することを指す。典型的に、抗体は、解離定数(KD)が10-7M以下で結合し、既定の抗原以外の非特異的抗原(例えば、BSA、カゼイン、又はその他任意の特定ポリペプチド)に結合するためのKDよりも、少なくとも10倍低いKDで、既定の抗原に結合する。「抗原を認識する抗体」及び「抗原に特異的な抗体」という語句は、本明細書で、例えば、IL−17A特異的抗体のように「抗原に特異的に結合する抗体」又は「抗原特異的抗体」という用語と、互換可能に使用される。解離定数は標準的な手順を用いて測定することができる。
【0047】
本明細書で使用されるとき、「IL−17A生物活性」又は「IL−17A活性化」という用語は、IL−17A受容体へのIL−17A結合の結果として生じる任意の活性を指す。例示のIL−17A生物活性は、IL−6若しくはIL−8の増加した分泌、NF−κB活性化、又はIL−17A受容体への結合時のERK1、ERK2、及びp38等の下流キナーゼの調節をもたらす。サイトカイン及びケモカインの、細胞、組織からの、若しくは循環中の放出、NF−κB活性化、又はキナーゼリン酸化事象は、周知の方法、例えば、免疫学的アッセイ、免疫ブロット法、又はレポーター遺伝子系を使用して測定することができる(Yao et al.,Immunity 3:811〜21、1995、Awane et al.,J.Immunol.162:5337〜44,1999)。
【0048】
「ベクター」という用語は、生体系内で複製されることができる、又はかかる系の間で移動可能である、ポリヌクレオチドを意味する。ベクターポリヌクレオチドは典型的に、生体系においてこれらのポリヌクレオチドの複製又は維持を促進するように機能する複製起点、ポリアデニル化信号、又は選択マーカー等の因子を含有する。このような生体系の例としては、細胞、ウイルス、動物、植物、及びベクターを複製することのできる生物学的構成成分を利用して再構成された生体系を挙げることができる。ベクターを含むポリヌクレオチドは、DNA若しくはRNA分子又はこれらのハイブリッドであってもよい。
【0049】
「発現ベクター」という用語は、生体系又は再構成された生体系において、その発現ベクター中に存在するポリヌクレオチド配列によってコードされたポリペプチドの翻訳を命令するために利用することができるベクターを意味する。
【0050】
用語「ポリヌクレオチド」は、糖−リン酸骨格鎖又は他の等価な共有結合化学によって共有結合されたヌクレオチド鎖を含む分子を意味する。二本鎖及び一本鎖のDNA及びRNAが、ポリヌクレオチドの典型的な例である。
【0051】
「ポリペプチド」又は「タンパク質」という用語は、ペプチド結合によって連結されてポリペプチドを形成する少なくとも2つのアミノ酸残基を含む分子を意味する。50個のアミノ酸未満の小さなポリペプチドは「ペプチド」と呼ばれる場合がある。
【0052】
本明細書では以下のようにアミノ酸の慣習的な1文字及び3文字の略号を用いる。
【0053】
【表1】
【0054】
物質の組成物
本発明はIL−17Aの生物活性を阻害することが可能なIL−17A抗体拮抗物質及びかかる抗体の使用法を提供する。かかる抗体によってIL−17A活性化が阻害され得る例示の機序としては、IL−17Aのホモ若しくはヘテロ二量化の生体外で、生体内で、若しくはその場での阻害、及びIL−17Aの、IL−17A受容体への結合の遮断、受容体二量化の阻害、下流シグナル伝達経路のキナーゼ活性の阻害、又はIL−17A mRNA転写の阻害が挙げられる。他の機序によってIL−17Aの活性を阻害することが可能な他の抗体拮抗物質もまた、本発明の異なる態様及び実施形態の範囲に含まれるものである。これらの拮抗物質は、研究用試薬、診断用試薬及び治療薬として有用である。
【0055】
本発明は、ヒト免疫グロブリンの遺伝子ライブラリーから誘導される抗原結合部位を提供する。抗原結合部位を担持するための構造は、概して、抗体重鎖若しくは軽鎖又はそれらの部分である。
【0056】
本発明は、重鎖可変領域(VH)及び軽鎖可変領域(VL)を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はそのフラグメントを提供し、該抗体は、表1aに示されるように、重鎖相補性決定領域(CDR)1、2、及び3(HCDR1、HCDR2、及びHCDR3)アミノ酸配列並びに軽鎖相補性決定領域(CDR)1、2、及び3(LCDR1、LCDR2、及びLCDR3)アミノ酸配列を含む。
【0057】
【表2】
【0058】
【表3】
【0059】
ある種の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号23、35、及び52に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含み、配列番号35のHCDR2は更に、式(I):
Xaa1−I−I−P−W−F−G−Xaa2−T−Xaa3−Y−A−Q−K−F−Q−G、
(I)に示すように記載され、式中、
Xaa1は、His、Met、Arg、Ser、又はTyrであり得、
Xaa2は、Trp、Thr、又はTyrであり得、
Xaa3は、Tyr、Phe、Ser、又はAspであり得る。
【0060】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号2、5、及び11に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含み、配列番号11のLCDR3は更に、式(II):
Xaa4−Q−Xaa5−Xaa6−Xaa7−Xaa8−Xaa9−Xaa10、
(II)に示すように記載され、式中、
Xaa4は、His又はGlnであり得、
Xaa5は、Phe又はGlyであり得、
Xaa6は、Thr、Val、又はAsnであり得、
Xaa7は、Ile、Thr、又はTyrであり得、
Xaa8は、Pro又はArgであり得、
Xaa9は、Ser又はProであり得、
Xaa10は、His、Phe、又はLeuであり得る。
【0061】
他の実施形態では、本発明は、VH及びVLを含む、結合ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号2、5、及び17に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含み、配列番号17のLCDR3は更に、式(III):
Xaa11−Q−Xaa12−Xaa13−Xaa14−Xaa15−Xaa16−Xaa17−Xaa18−T、
(III)に示すように記載され、
式中、
Xaa11は、Gln又はThrであり得、
Xaa12は、Ser又はTyrであり得、
Xaa13は、Asn、Arg、Val、又はTyrであり得、
Xaa14は、His又はSerであり得、
Xaa15は、Ile、Thr、Leu、Ala、又はSerであり得、
Xaa16は、Pro、Leu、又はSerであり得、
Xaa17は、Pro、Ser、Phe、又はLeuであり得、
Xaa18は、Ala、Leu、又はAspであり得る。
【0062】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号24、36、及び57に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含み、配列番号57のHCDR3は更に、式(IV):
E−V−D−S−Xaa19−Y−Y−S−Y−F−D−I、
(IV)に示すように記載され、
式中、
Xaa19は、Met、Ile、Leu、又はThrである。
【0063】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号3、6、及び22に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含み、配列番号22のLCDR3は更に、式(V):
G−S−Y−D−F−F−L−G−Xaa20−I−V、
(V)に示すように記載され、
式中、
Xaa20は、Met、Leu、Thr、又はTyrである。
【0064】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号25、46、及び61に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含み、配列番号46のHCDR2は更に、式(VI):
Xaa21−I−Xaa22−Xaa23−Xaa24−Xaa25−Xaa26−Xaa27−Xaa28−Xaa29−Y−A−D−S−V−K−G、
(VI)に示すように記載され、
式中、
Xaa21は、Ala、Gly、Thr、又はValであり得、
Xaa22は、Asn又はSerであり得、
Xaa23は、Gly、Met、Lys、Ile、Leu、又はHisであり得、
Xaa24は、Leu、Asp、Ala、His、Thr、Gly又はSerであり得、
Xaa25は、Gly又はSerであり得、
Xaa26は、Thr、Gly、Tyr、又はAspであり得、
Xaa27は、His、Trp、Tyr、又はPheであり得、
Xaa28は、Lys、Thr、又はIleであり得、
Xaa29は、Tyr、Phe、又はAsnであり得、
【0065】
配列番号61のHCDR3は、式(VII):
Q−L−Xaa30−L−D−V、
、(VII)に示すように記載され、
式中、
Xaa30は、Met、Leu、又はThrであり得る。
【0066】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、配列番号25、51、及び58に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含み、配列番号51のHCDR2は更に、式(VIII):
V−T−S−Xaa31−Xaa32−Xaa33−Xaa34−T−Y−Y−A−Xaa35−S−V−K−G、
(VIII)に示すように記載され、
式中、
Xaa31は、Ala、Lys、Met、又はHisであり得、
Xaa32は、Asn、Met、Thr、又はArgであり得、
Xaa33は、Gly又はAspであり得、
Xaa34は、Arg、His、又はAsnであり得、
Xaa35は、Asp又はGlyであり得る。
【0067】
その抗原結合部位アミノ酸配列が表1aに示される配列(配列番号1〜61)と実質的に同一である抗体は、本発明の範囲内に包含される。典型的に、これは、類似した電荷又は疎水性若しくは立体化学的性質を有するアミノ酸との、1つ以上のアミノ酸置換を伴い、抗体特性、例えば、安定性又は親和性を改善するようにされる。例えば、保存的置換は、その位置のアミノ酸残基の極性又は電荷にほとんどあるいは全く影響しないように、天然アミノ酸残基の非天然残基との置換を伴ってもよい。更に、アラニン・スキャニング変異導入法について以前に記載されているように(MacLennan et al.,Acta Physiol.Scand.Suppl.643:55〜67,1998、Sasaki et al.,Adv.Biophys.35:1〜24,1998)、ポリペプチド内の任意の内因性の残基をアラニンで置換することもできる。保存的な置換は、かかる修飾が行われた分子と類似した機能的及び化学的性質を有する分子をもたらす。分子の機能的及び/又は化学的特性における非保存的置換は、(1)置換領域における分子骨格鎖の構造を、例えばシート若しくはヘリックス立体配座として維持する、(2)標的部位における分子の電荷若しくは疎水性を維持する、又は(3)分子のサイズを維持する、効果において、有意に異なるアミノ酸配列中の置換を選択することによって実現することができる。当業者であれば、所望のアミノ酸置換(保存的又は非保存的であるかによらず)を、かかる置換が望ましい時点で決定することができる。例えば、アミノ酸置換を使用して、親和性に影響を及ぼす残基、又は凝集等の望ましくない特性を付与する残基等の、抗体の機能に重要な残基を特定することができる。例示のアミノ酸置換は、表1b、及び図1に示される。
【0068】
抗原結合部位とは対照的に、フレームワーク領域における置換も、それらが抗体の特性に悪影響を及ぼさない限りは行われてもよい。フレームワーク置換は、抗体親和性又は安定性を改善するために、例えば、バーニアゾーン残基において行うことができる(米国特許第6,649,055号)。置換はまた、潜在的な免疫原性を低減するために、相同のヒト生殖細胞系列遺伝子配列と比較したとき配列において異なる、抗体中のフレームワーク位置においても行うことができる。これらの修飾は、例えば、pIXライブラリー等の、de novo抗体ライブラリーに由来する抗体に対して行うことができる。
【0069】
【表4】
【0070】
保存的アミノ酸置換はまた、生物学的系における合成によってよりはむしろ、化学的ペプチド合成によって典型的に組み込まれる、非天然産のアミノ酸残基も包含する。アミノ酸置換は、例えば、PCR突然変異生成(米国特許第4,683,195号)によって行うことができる。変異型のライブラリーは、周知の方法を使用して、例えば、無作為(NNK)又は非無作為コドン、例えば、11個のアミノ酸(ACDEGKNRSYW)をコードするDVKコドンを使用して、及び実施例1に示されるように、所望の特性を有するライブラリー又は変異型をスクリーニングして、生成することができる。図1は、抗体特性を改善するために、LCDR3、HCDR2、及びHCDR3領域内の5つの親IL−17A抗体拮抗物質に対して行われた置換を示す。親和性又は安定性等の、改善された特性は、周知の方法によって測定することができる。
【0071】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、ある種のVH及びVL配列を含み、また表2に示される各々単離されたVH及びVLも提供する。
【0072】
【表5】
【0073】
実施例に例証される実施形態は、1つは重鎖から及び1つは軽鎖からの、可変領域の対、完全長抗体鎖の対、又はCDR1、CDR2、及びCDR3領域の対を含むが、当事者であれば、代替的な実施形態が、単一の重鎖可変領域若しくは単一の軽鎖可変領域、単一の完全長抗体鎖、又は重鎖若しくは軽鎖のいずれかの1つの抗体鎖からのCDR1、CDR2、及びCDR3領域を含んでもよいことを認識するであろう。単一の可変領域、完全長抗体鎖、又は1つの鎖のCDR1、CDR2、及びCDR3領域を使用して、別の鎖中の対応するドメインをスクリーニングすることができ、これらの2つの鎖は、IL−17Aに特異的に結合する抗体を形成することができる。スクリーニングは、例えば、PCT公開第WO92/01047号で開示されている階層的二重コンビナトリアルアプローチを使用する、ファージディスプレイスクリーニング法によって実現されてもよい。このアプローチでは、H又はL鎖のいずれかのクローンを含む個別のコロニーを用いて他方の鎖(L又はH)をコードするクローンの完全なライブラリに感染させ、得られた2鎖特異的抗原結合ドメインを上述のようなファージディスプレイ法に基づいて選択する。
【0074】
他の実施形態では、本発明は、表2に示されるVH及びVLアミノ酸配列と少なくとも90%同一のアミノ酸配列を有するVH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0075】
他の実施形態では、本発明は、表2に示されるVH及びVLアミノ酸配列と少なくとも95%同一のアミノ酸配列を有するVH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0076】
別の態様では、本発明は、表2に示されるある種の重鎖及び軽鎖アミノ酸配列を有する単離された抗体又はフラグメントを提供する。抗体残基を連続して番号付けすることに加えて、ポリペプチドをコードする抗体鎖は、Kabatの又はChothiaの、番号付けに基づいて番号付けすることができる(Kabat et al.,sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.,1991、Chothia and Lesk,Mol.Biol.196:901〜917,1987)。選択された抗体鎖のための連続したKabat及びChotia番号付けの間の対応の例は、図3に示される。灰色で強調表示された位置は、抗体CDR領域を示す。
【0077】
他の実施形態では、本発明は、VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、抗体は、Chothia残基S51、T53、F56、Y58、Q95、L96、及びT97から選択される重鎖可変領域パラトープ、並びにChothia残基Y32、D50、Y91、F93、及びF94から選択される軽鎖可変領域パラトープを含む。重鎖パラトープ及び軽鎖パラトープChothia残基は、配列番号86の重鎖残基S52、T54、F57、Y59、Q99、L100、及びT101、並びに配列番号79の軽鎖残基Y31、D49、Y90、F92、及びF93に相当する。
【0078】
他の実施形態では、本発明は、配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する重鎖可変領域パラトープアミノ酸残基を含み、
ヒトIL−17AのR55又はE57と相互作用する第1のスレオニン残基、
ヒトIL−17AのR55又はE57と相互作用するグルタミン残基、
ヒトIL−17AのE57と相互作用するリジン残基、
ヒトIL−17AのP59、E60、又はR101と相互作用するチロシン残基、
ヒトIL−17AのE60、R101、E102、又はP103と相互作用するフェニルアラニン残基、
ヒトIL−17AのE60と相互作用するセリン残基、及び
ヒトIL−17AのE60と相互作用する第2のスレオニン残基を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0079】
他の実施形態では、本発明は、配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する軽鎖可変領域パラトープアミノ酸残基を含み、
ヒトIL−17AのL26と相互作用する第1のフェニルアラニン残基、
ヒトIL−17AのR55又はW67と相互作用するアスパラギン酸残基、
ヒトIL−17AのP59、S64、又はR101と相互作用する第1のチロシン残基、
ヒトIL−17AのP59、E60、R61、Y62、R101、又はF110と相互作用する第2のフェニルアラニン残基、及び
ヒトIL−17AのV65と相互作用する第2のチロシン残基を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0080】
別の実施形態では、本発明は、配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する重鎖可変領域パラトープアミノ酸残基及び軽鎖可変領域パラトープアミノ酸残基を含み、
ヒトIL−17AのR101と相互作用する重鎖可変領域におけるチロシン残基、
ヒトIL−17AのR101と相互作用する重鎖可変領域におけるフェニルアラニン残基、
ヒトIL−17AのY62及びR101と相互作用する軽鎖可変領域における第1のフェニルアラニン残基、
ヒトIL−17AのL26及びF110と相互作用する軽鎖可変領域における第2のフェニルアラニン残基、及び
ヒトIL−17AのR101と相互作用する軽鎖可変領域におけるチロシン残基を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供する。
【0081】
別の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントを提供し、該抗体は、
Chothia残基F56及びY58から選択される重鎖可変領域パラトープ、並びに
Chothia残基Y91、F93、及びF94から選択される軽鎖可変領域パラトープを含む。
【0082】
重鎖パラトープChothia残基F56及びY58並びに軽鎖パラトープChothia残基Y91、F92、及びF94は、IL−17A残基L26、Y62、R101、及びF110と直接接触している残基である。これらのIL−17A残基は、Fab6468エピトープ及びP2ポケット空洞の両方の部分である(下記を参照されたい)。特定の理論に決して束縛されるものではないが、これらの選択された残基におけるFab6468とIL−17Aとの間の相互作用は、抗体がIL−17A活性を遮断するために十分であることが考えられる。
【0083】
いかなる非ヒト配列も欠く完全な型(human)mAbは、例えば、Knappik et al.,J.Mol.Biol.296:57〜86,2000、及びKrebs et al.,J.Immunol.Meth.254:67〜84 2001に参照される技術によって、ファージディスプレイライブラリーから調製及び最適化することができる。例示の方法では、本発明の抗体は、バクテリオファージpIXコートタンパク質を有する融合タンパク質として、抗体重鎖及び軽鎖可変領域を発現するライブラリーから単離される。抗体ライブラリーは、ヒトIL−17mut6(配列番号105)への結合についてスクリーニングされ、得られた陽性クローンは更に特徴付けられ、Fabはクローンライセートから単離され、完全長IgGsとして発現される。例示の抗体ライブラリー及びスクリーニング方法は、Shi et al.,J.Mol.Biol.397:385〜96,2010、PCT特許出願第WO09/085462号,及び米国第12/546850号、米国特許第5,223,409号、第5,969,108号、及び第5,885,793号)に記載されている。
【0084】
結果として得られたmAbは、本明細書に例示されるように、それらのフレームワーク領域において、ある種のフレームワーク残基をマッチングヒト生殖細胞系列に存在する残基に変更するように更に修飾することができる。
【0085】
特異的なIL−17Aエピトープに結合する本発明の抗体は、ヒト免疫グロブリン遺伝子座を発現するヒト化マウス(Lonberg et al.,Nature 368:856〜9,1994、Fishwild et al.,Nature Biotechnology 14:845〜51,1996、Mendez et al.,Nature Genetics 15:146〜56,1997、米国特許第5,770,429号、第7,041,870号、及び第5,939,598号)又はBalb/cマウスに、エピトープをコードするペプチド、例えば、ペプチド56NEDPERYPSVIWE68(配列番号157)又は100RREPPHCPNSFrlEKIL116(配列番号158)で免疫付与することによって、及びKohler et al.,Nature 256:495〜97のハイブリドーマ法を使用して作製することができる。結果として得られた抗体は、標準の方法を使用して、エピトープへのそれらの結合について試験される。特定されたmAbは更に、Queen et al.,Proc.Natl.Acad.Sci.(USA),86:10029〜32,1989及びHodgson et al.,Bio/Technology,9:421,1991で開示されている技術等の技術によって、変化したフレームワーク支持残基を組み込むことによって結合親和性を保存するように修飾することができる。
【0086】
ヒトIL−17Aに特異的に結合する、ある種のパラトープ残基(例えば、表10に記載されるコアパラトープ残基)を有する単離された抗体は、例えば、パラトープ残基を好適な足場にグラグトすること、操作された足場を完全抗体に組み立てること、結果として得られた抗体を発現させること、及び抗体をIL−17Aへの結合について又はIL−17A生物活性に及ぼす効果について試験することによって、作製することができる。例示の足場は、ヒト生殖細胞系列遺伝子によりコードされたヒト抗体可変領域のアミノ酸配列である。足場は、例えば、全体的な配列相同性、パラトープ残基の間の同一性%、又は足場とmAb6785等の例示の抗体との間の正準構造クラス同一性に基づいて選択することができる。ヒト抗体生殖細胞系列遺伝子は、例えば、TomLinson et al.,J.Mol.Biol 227:776〜798、及びInternational ImMunoGeneTics(IMGT)データベース(http_://_www_imgt_org)で開示されている。コンセンサスヒトフレームワーク領域はまた、例えば、米国特許第6,054,297号に記載されているように使用することができる。例えば、PCT公開第WO10/045340号に記載されている方法に従って、好適な足場の選択を行うことができる。
【0087】
パラトープ残基をグラフトする足場として使用可能な例示のヒト生殖細胞系列遺伝子は、Vλ3、Vh3、Jλ、及びJhフレームワークによってコードされた遺伝子である。例示のVκ3遺伝子には、IGLV3−1、IGLV3−9、IGLV3−10、IGLV3−12、IGLV3−16、IGLV3−19、IGLV3−21、IGLV3−22、IGLV3−25、IGLV3−27、及びIGLV3−32(IMGT命名法、*01対立遺伝子)、(それぞれ配列番号117〜127)がある。例示のJλ遺伝子には、IGLJ1、IGLJ2、IGLJ3、IGLJ4、IGLJ5、IGLJ6、及びIGLJ7(それぞれ配列番号128〜134)がある。例示のVh3遺伝子には、IGHV3−7、IGHV3−9、IGHV3−11、IGHV3−16、IGHV3−19、IGHV3−20、IGHV3−21、IGHV3−23、IGHV3−30、IGHV3−30*03、IGHV3−33、IGHV3−45、IGHV3−48、IGHV3−64、及びIGHV3−74(IMGT命名法、異なる対立遺伝子が指定される場合を除いて*01対立遺伝子)(それぞれ配列番号135〜150)がある。例示のJh遺伝子には、IGHJ1、IGHJ2、IGHJ3、IGHJ4、IGHJ5、及びIGHJ6(それぞれ配列番号151〜156)がある。この生殖細胞系列J領域は、FR4配列を選択するために、その全体又は一部を使用される。例えば、mAb6785軽鎖パラトープ残基は、IGLV3−1とIGLJ2配列との間の単一のアミノ酸残基、例えば、メチオニンの挿入により、IGLJ2(配列番号129)によってコードされたJ領域配列に結合される、IGLV3−1(配列番号117)によってコードされたVλ3タンパク質フレームワーク中にグラフトすることができる。IGLV3−1によってコードされたVλ3タンパク質フレームワークは、更なる置換、例えば、配列番号117の33位(「ACW」)におけるシステイン残基の、例えば、アスパラギンとの置換、及び配列番号117の残基1〜3の、IGLV1ファミリーの「QSV」等の他のラムダ鎖ファミリーに共通のアミノ末端配列との置換(「SYE」)を含んでもよい。他の例示の機能的Vλ3及びJλ遺伝子からの配列は、CDR3領域の長さが11アミノ酸になるように、Vλ3遺伝子によってコードされたカルボキシ末端と、Jλ遺伝子によってコードされたアミノ末端と間の0、1、又は2個のアミノ酸残基の挿入により、mAb6785軽鎖パラトープ残基をグラグトするために使用することができる。例えば、メチオニン及びイソロイシンは、IGLV3−22(配列番号124)とIGLJ2(配列番号129)との間に挿入することができる。図2Aは、グラフトに使用可能な例示の軽鎖足場の整列を示す。mAb6785重鎖パラトープ残基は、IGHV3−23(配列番号142)によってコードされた例えば、Vh3フレームワーク上にグラフトすることができ、それはV領域とJ領域との間のHCDR3を構成する、約5〜7つの残基、例えば、6つの残基の挿入により、IGHJ1(配列番号151)の領域FR4配列(11C−末端アミノ酸、例えば「WGQGTLVTVSS」)に結合される。挿入されたHCDR3約5〜7つの残基には、グルタミン、ロイシン、及びスレオニン、例えば、mAb6785Vhからのパラトープ残基のうちの3つの挿入が含まれる(表10)。他の例示の機能的Vh3及びJh遺伝子からの配列は、mAb6785重鎖パラトープ残基をグラグトするために使用することができる。場合によっては、Vh3遺伝子からの1個のC−末端アミノ酸は、FR3配列のみが足場に含まれるように、HCDR3を構成する約5〜7つの残基の挿入の前に欠失され得る。17個の残基(IGHV3−23(配列番号142)の残基50〜66)のCDR2をコードする他のVh3遺伝子からの配列を使用することもでき、他のJh遺伝子のFR4配列は、IGJH1の代わりに置換することができる。
【0088】
ヒトIL−17Aへの特異的結合の及び結果として得られた抗体の生物活性は、標準の方法を使用して評価することができる。mAb6785軽鎖可変領域及び重鎖可変領域の、例示のVh3、Vλ3、Jλ、又はJh遺伝子との整列は、図2A及び2Bに示される。代替的に、表10に記載されるように、拡張されたパラトープの残基mAb6785をコアパラトープ残基の代わりに使用することができる。パラトープグラフトされた操作された抗体は更に、バーニアゾーン残基(米国特許第6,639,055号)又は親和性決定残基(米国特許出願第2010/0261620号、Cobaugh et al.,J Mol Biol.378:622〜33,2008)の置換によって、抗体特性、例えば親和性を改善するように修飾することができる。パラトープグラフトされた抗体がIL−17Aへの結合を保持する限り、パラトープグラフトされた抗体におけるフレームワークアミノ酸配列は、mAb6785フレームワーク配列と70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%同一であり得る。例示の生殖細胞系列遺伝子フレームワークの対立遺伝子の変異型をV及びJ領域タンパク質配列の代わりに使用することができる。対立遺伝子の変異型の配列は、周知であり、International ImMunoGeneTics(IMGT)データベース(http_://_www_imgt_org)で得ることができる。
【0089】
標準的な方法を用いて、パラトープ残基に追加して抗原結合部位由来の配列をグラフトすることもできる。例えば、完全なHCDR3又はLCDR3をグラフトしてもよい。
【0090】
本発明の別の実施形態は、ヒトIL−17Aに特異的に結合し、ある種のHCDR1、HCDR2、及びHCDR3、並びにLCDR1、LCDR2、及びLCDR3アミノ酸配列を含むモノクローナル抗体と、ヒトIL−17A結合をめぐって競合する、単離された抗体又はフラグメントである。本発明の例示のモノクローナル抗体は、配列番号25、43、及び60に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列並びに配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体である。
【0091】
IL−17Aへの特異的結合との間の競合は、周知の方法を使用して生体外でアッセイすることができる。例えば、非標識抗体の存在下でのMSD Sulfo−Tag(登録商標)NHS−エステル標識抗体のIL−17Aへの結合を、ELISAによって評価することができる。
【0092】
本発明の別の実施形態は、単離された抗体又は抗体又はそのフラグメントであり、該抗体は、アミノ酸残基56〜68(配列番号157)及び100〜116(配列番号158)において、又は残基L26、R55、E57、P59、E60、R61、Y62、S64、V65、W67、R101、E102、P103、及びF110において、配列番号105に示される配列を有するヒトIL−17Aに特異的に結合する。
【0093】
幾つかの周知の方法を用いて本発明の抗体の結合エピトープを決定することができる。例えば、両方の個別の構成成分の構造が知られている場合、インシリコでタンパク質−タンパク質のドッキングを行って、適合する相互作用部位を特定することができる。抗原抗体複合体において水素−重水素(H/D)交換を行って、抗体が結合し得る抗原の領域をマッピングすることができる。抗原のセグメント及び点突然変異生成を使用して、抗体の結合に重要なアミノ酸の位置を特定することができる。抗体−抗原複合体の共結晶構造を使用して、エピトープ及びパラトープに寄与する残基を特定することができる。
【0094】
前述の抗IL−17A抗体は、本発明に記載されるFab6468のためのエピトープと異なる、IL−17A上のエピトープに結合する。抗体結合ヒトIL−17A(配列番号105)残基74〜85、46〜53、71〜87、80〜86、11〜18、29〜41、又は54〜62が記載されている(それぞれPCT公開第WO08/021156号、第WO07/106769号、第WO07/149032号、第WO07/070750号、米国出願第US2008/095775号)。立体配座エピトープは、PCT公開第WO09/130459号、及びGerhardt et al.,J.Mol.Biol:394:901〜21,2009に記載されている。
【0095】
本発明の別の実施形態は、単離された抗体又はそのフラグメントであり、ここで抗体は、IL−17A上のP2ポケット空洞に特異的に結合し、このP2ポケット空洞は、配列番号105のアミノ酸残基V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる。
【0096】
抗IL−17A Fab6468とのIL−17Aホモ二量体の共結晶構造は、L−17RA結合に関与する可能性が高い、IL−17Aホモ二量体の表面上の疎水性P2ポケット空洞を特定した(実施例を参照されたい)。本明細書で使用されるとき、「P2ポケット空洞」は、IL−17Aホモ二量体上の第三級疎水性構造的空洞を指し、P2ポケットにおける表面露出残基は、単量体A上のV24、L26、I28、Y62、L99、R101、F110、及びL112、並びに単量体B上のV22、V24、及びL112であり、逆もまた同様である。IL−17A、例えば、Fab6468に反応する、本発明の選択された抗体は、P2ポケット空洞残基L26、Y62、R101、及びF110と直接接触し、これらの残基もまたFab6468エピトープの部分である。特定の理論に決して束縛されるものではないが、選択されたIL−17AP2ポケット空洞残基に結合する本発明の抗体は、IL−17AとIL−17RAとの間の相互作用を遮断する。共結晶構造に基づき、Fab6468の軽鎖(配列番号79)中の残基93及び94におけるフェニルアラニンモチーフ(FF)は、IL−17A/IL−17RA相互作用を遮断し、したがってP2ポケット空洞遮断剤である。新規ペプチド又は小分子等の他のP2ポケット空洞遮断剤拮抗物質もまた、本発明の範囲内である。これらは、IL−17A/Fab6468共構造に基づいてモデル化され、IL−17Aに結合するFab6468を置き換えるそれらの能力についてスクリーニングされ得る。例えば、ペプチド阻害剤は、FFモチーフを組み込んだ無作為ペプチドライブラリー(例えば、XXXXFFXXのライブラリー、Xは任意のアミノ酸を示し、Fはフェニルアラニンである)からスクリーニングされ、例えば、pIII、pVII、又はpIXコートタンパク質との融合として、バクテリオファージ上に表示され得(米国第5,223,409号、Gao et al.,Proc.Natl.Acad.Sci.USA,96:6025〜30,1999,Tornetta et al.,J.Immunol.Methods.360:39〜46,2010、Shi et al.,J.Mol.Biol.397:385〜96,2010)、その後、それらの、IL−17Aに結合するFab6468の阻害及びIL−17A活性の阻害について試験され得る。
【0097】
小分子は、合成若しくは天然の組成物、又はそれらの任意の組み合わせのライブラリーを使用してスクリーニングされ得、結果として得られた第1の陽性ヒットは、薬剤の構造的類似体を産生するように容易に修飾され得る。ペプチドライブラリー及びpIX融合を作製する、並びに結果として得られたライブラリーをスクリーニングする方法は、周知である。
【0098】
ヒトIL−17Aの、IL−17RAとの相互作用を阻害する方法の、本発明の別の実施形態は、
ヒトIL−17A及びIL−17RAを提供する工程と、
ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる群から選択される少なくとも1つのアミノ酸残基において、ヒトIL−17Aに特異的に結合する拮抗物質と接触させる工程と、を含む。
【0099】
本発明の別の実施形態は、ヒトIL−17A生物活性を阻害する方法であり、
ヒトIL17−A及びIL−17RAを提供する工程と、
ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112からなる群から選択される少なくとも1つのアミノ酸残基において、ヒトIL−17Aに特異的に結合する拮抗物質と接触させる工程と、を含む。
【0100】
ヒトIL−17A及びIL−17RAは、単離されたタンパク質又は融合タンパク質として提供され得る。ヒトIL−17Aホモ二量体は、IL−2、IL−23及びIL−1βの存在下での2つの抗CD3/抗CD28刺激によるナイーブCD4T細胞の生体外刺激によって調製された、活性化Th17細胞の培地から精製され得る。IL−17RAは、細胞若しくは細胞膜に関連し得、天然若しくは過剰発現され得るか、又はIL−17RAのフラグメント、例えば、受容体の細胞外ドメインであり得る。IL−17RAは、ヒトIL−17RA、又はマウス、ラット、若しくはサル等の他の種からのIL−17RAであり得る。ヒトIL−17A残基V22、V24、L26、I28、Y62、L99、R101、F110、及びL112に結合する拮抗物質は、突然変異生成研究によって又は共結晶構造によって、拮抗物質の、IL−17Aに結合するFab6468を置き換える能力によって特定することができる。ヒトIL−17A及びIL−17RAの融合タンパク質は、周知の方法によって作製することができる。例示の融合タンパク質は、免疫グロブリンFcドメインに融合された可溶性IL−17RAである。
【0101】
本発明の別の態様は、本発明の抗体重鎖若しくは抗体軽鎖又はそれらのフラグメント、又はそれらの補体うちのいずれかをコードする、単離されたポリヌクレオチドである。ある種のポリヌクレオチドの例が本明細書に開示されるが、遺伝コードの縮重又は所定の発現系におけるコドンの選択性を考慮すると、本発明の抗体拮抗物質をコードする他のポリヌクレオチドも本発明の範囲内に含まれる。例示のポリヌクレオチドは、配列番号101、102、103、及び104に示される。
【0102】
例示の抗体拮抗物質は、IgG、IgD、IgE、IgA、又はIgMアイソタイプの抗体であり得る。更にかかる抗体拮抗物質は、グリコシル化、異性化、脱グリコシル化、又はポリエチレングリコール部分の付加(ペギレーション)及び脂質化等の非天然の共有結合修飾等の反応によって翻訳後修飾され得る。かかる修飾は、生体内あるいは生体外で行われてもよい。例えば、本発明の抗体をポリエチレングリコールと結合させて、薬物動態的なプロファイルを改善することができる。共役は当業者に既知の方法によって行うことができる。治療用抗体のPEGとの共役によって、機能を低下させずに薬力学的動態が向上することが示されている。Deckert et al.,nt.J.Cancer 87:382〜90,2000、Knight et al.,Platelets 15:409〜18,2004、Leong et al.,Cytokine 16:106〜19,2001、及びYang et al.,Protein Eng.16:761〜70,2003を参照されたい。
【0103】
本発明の抗体の薬物動態的な特性は、当業者に既知の技術によるFc修飾を通じて向上することができる。抗体の「Fc」は、抗体の抗原への結合に直接関与しないが、種々のエフェクタ−機能を示す。抗体「Fc」は、周知の用語であり、抗体のパパイン開裂に基づいて記載される。抗体のFcは、補体活性化、C1q結合、及びFc受容体結合に基づいて、ADCC(抗体依存性細胞媒介型細胞傷害性)及びCDC(補体依存性細胞傷害性)に直接関与する。かかる補体及びFc受容体結合部位は、周知であり、例えば、L234、L235、D270、N297、E318、K320、K322、P331、及びP329が含まれる(KabatのEUインデックスに従った番号付け)(Brekke et al.,Eur.J.Immunol.24:2542〜7,1995、米国特許第5,624,821号、第7,597,889号、Canfield and Morrison,J.Exp.Med.173:1483〜91,1991)。例えば、IgG1のヒンジ領域のLeu234/Leu235のL234A/L235Aへの変異、又はIgG4のヒンジ領域のPhe235/Leu236のP235A/L236Aへの変異によって、FcRへの結合性が最小となり、補体依存性細胞傷害活性及びADCCを介在する免疫グロブリンの能力が低減する。イソメラーゼの作用を介して重鎖間又は重鎖内のいずれかの生体内ジスルフィド結合を形成することができる(Aalberse and Schuurman,Immunology 105:9〜19,2002)、IgG4重鎖のヒンジ領域内のCys−Pro−Ser−Cys(CPSC)モチーフにおけるSerからProへの置換は、「IgG1様挙動」をもたらし、すなわち、Pro置換された分子は、重鎖間ジスルフィド結合を形成することができない。CPSCモチーフの位置は典型的に、成熟した重鎖の残基228に見出されるが、CDRの長さに応じて変化し得る。Leu234/Leu235残基を有する例示のIgG1 Fc領域は、配列番号114に示されるアミノ酸配列を有し、残基L117及びL118は、成熟した重鎖におけるLeu234/Leu235残基に相当する。Cys−Pro−Ser−Cys(CPSC)モチーフ及びLeu234/Leu235残基を有する例示のIgG4 Fc領域は、配列番号115に示されるアミノ酸配列を有し、CPSCモチーフは、残基106〜109に位置し、Leu234/Leu235残基は、122及び123位に位置する。
【0104】
安定性、選択性、交差反応性、親和性、免疫原性、又は他の望ましい生物学的若しくは生物物理学的特性を改善するように修飾される、本発明の抗体又はそのフラグメントが発明の範囲内にある。抗体の安定性は、(1)内在的な安定性に影響を及ぼす個々のドメインのコアパッキング、(2)HCとLCとのペアリングに影響を及ぼすタンパク質/タンパク質の界面相互作用、(3)極性及び荷電残基の埋め込み、(4)極性及び荷電残基のためのH結合ネットワーク、並びに(5)他の分子内及び分子間力の間の表面電荷及び極性残基の分布、を含む幾つかの因子によって影響を受ける。(Wornら、J.Mol.Biol.305:989〜1010、2001)。構造を不安定化させる可能性のある残基は、抗体の結晶構造に基づいて、あるいは場合によっては分子モデリングによって特定され得、抗体の安定性に及ぼすこれらの残基の効果を、特定された残基に突然変異を有する変異体を生成し、評価することによって試験することができる。抗体の安定性を高める方法の1つは、示差走査熱量測定法(DSC)によって測定される熱転移中点(Tm)を高くすることである。広くは、タンパク質Tmは、その安定性と相関し、溶液中のほどき及び変性、並びにタンパク質がほどける傾向に応じる分解プロセスに対する、その感受性と逆相関する(Remmele et al.,Biopharm。13:36〜46、2000)。幾つかの研究は、DSCによる熱安定性として測定される製剤の物理的安定性及び他の方法によって測定される物理的安定性の、順位付けの間の相関を見出している(Gupta et al.,AAPS PharmSci.5E8,2003、Zhang et al.,J.Pharm.Sci.93:3076〜89,2004、Maa et al.,Int.J.Pharm.,140:155〜68,1996、Bedu−Addo et al.,Pharm.Res.,21:1353〜61,2004、Remmele et al.,Pharm Res.,15:200〜8,1997)。製剤研究は、FabのTmが対応するmAbの長期の物理的安定性と密接な関係があることを示唆している。フレームワーク又はCDR内のいずれかにおけるアミノ酸の差異は、Fabドメインの熱安定性に有意な影響を及ぼし得る(Yasui、et al.,FEBS Lett.353:143〜6,1994)。
【0105】
本発明の抗体拮抗物質は、約10-7、10-8、10-9、10-10、10-11又は10-12M以下のKdでIL−17Aと結合することができる。抗体等の所定の分子のIL−17Aに対する親和性は、任意の好適な方法を使用して実験的に決定することができる。かかる方法は、当業者に既知のBiacore又はKinExA装置、ELISA又は競合的結合アッセイを利用し得る。
【0106】
所望の親和性でヒトIL−17Aに結合する抗体拮抗物質は、抗体親和性成熟を含む技術によって、変異型又はフラグメントのライブラリーから選択することができる。抗体拮抗物質は、任意の好適な方法を使用してIL−17Aの生物活性の阻害に基づいて特定することができる。かかる方法では、周知の方法を使用した、本出願に記載されるようなレポーター遺伝子アッセイ又はサイトカイン産生を測定するアッセイを利用し得る。
【0107】
本発明の別の実施形態は、少なくとも1つの本発明のポリヌクレオチドを含むベクターである。かかるベクターは、プラスミドベクター、ウイルスベクター、バキュロウイルス発現ベクター、トランスポゾンに基づいたベクター、又は任意の手段によって特定の生物又は遺伝子的バックグラウンドに本発明のポリヌクレオチドを導入するのに適した他の任意のベクターであってもよい。
【0108】
本発明の別の実施形態は、配列番号67〜75及び81〜86に示されるアミノ酸配列を有する免疫グロブリン重鎖可変領域、又は配列番号62〜66及び76〜80に示されるアミノ酸配列を有する免疫グロブリン軽鎖可変領域、又は配列番号92〜100に示されるアミノ酸配列を有する免疫グロブリン重鎖、又は配列番号87〜91に示されるアミノ酸配列を有する免疫グロブリン軽鎖、を含むポリペプチドをコードするポリヌクレオチド等の、本発明のポリヌクレオチドのいずれかを含む宿主細胞細胞である。かかる宿主細胞細胞は、真核細胞、細菌細胞、植物細胞又は古細菌細胞であってもよい。真核細胞の例としては、哺乳動物、昆虫、鳥類又は他の動物由来のものであり得る。哺乳動物真核細胞としては、SP2/0(American Type培養Collection(ATCC),Manassas,VA,CRL−1581)、NS0(European Collection of Cell Cultures(ECACC)、Salisbury,Wiltshire,UK,ECACC No.85110503)、FO(ATCC CRL−1646)、及びAg653(ATCC CRL−1580)マウス細胞株等の、ハイブリドーマ又は骨髄腫細胞株等の不死化細胞株が挙げられる。例示のヒト骨髄腫細胞株には、U266(ATTC CRL−TIB−196)がある。他の有用な細胞株としては、CHO−K1SV(Lonza Biologics,Walkersville,MD)、CHO−K1(ATCCCRL−61)、又はDG44等の、チャイニーズハムスターの卵巣(CHO)細胞に由来するものが挙げられる。
【0109】
本発明の別の実施形態は、本発明の宿主細胞細胞を培養する工程と、宿主細胞細胞によって産生された抗体を回収する工程とを含む、IL−17Aとの反応性を有する抗体を製造する方法である。抗体を製造して精製する方法は、当該技術分野で周知である。発現について、操作されたファミリー2、6a、6b、19a、及び19b重鎖配列は、MAWVWTLLFLMAAAQSIQA(配列番号109)等のN末端リーダー配列を含み得る。リーダー配列を含む候補mAb6785(ファミリー19)及び成熟型(リーダー配列を含まない)の重鎖をコードする例示のヌクレオチド配列は、それぞれ配列番号101及び102に示される。同様に、発現について、本発明のファミリー2、6a、6b抗体の軽鎖配列は、MGVPTQVLGLLLLWLTDARC(配列番号110)等のN末端リーダー配列を含み得、本発明のファミリー19a及び19b抗体の軽鎖配列は、MAWSPLLLTLLAHCTGSWA(配列番号116)等のN末端リーダー配列を含み得る。リーダー配列を有するコドン最適化されたmAb6785及び成熟型(リーダー配列を有さない)の軽鎖をコードする例示のヌクレオチド配列は、それぞれ配列番号103及び104に示される。
【0110】
本発明の別の実施形態は、本発明の抗体を産生するハイブリドーマ細胞株である。
【0111】
治療の方法
本発明のIL−17A拮抗物質、例えば、IL−17A抗体拮抗物質は、動物患者においてIL−17Aの影響を低減することが所望される場合の任意の療法で利用され得る。IL−17Aは、体内で循環している可能性があり、あるいは望ましくない高レベルで体内の特定の部位、例えば、炎症の部位に局在して存在する可能性がある。特定の理論に決して束縛されるものではないが、本発明の拮抗物質は、IL−17Aのその受容体への結合、又はIL−17Aのホモ若しくはヘテロ二量化を予防又は低減することによって、有益な療法を提供する。本発明の方法を用いて任意の分類に属する動物患者を治療することができる。かかる動物の例としては、ヒト、齧歯類、イヌ、ネコ、及び家畜等の哺乳動物が挙げられる。
【0112】
本発明の抗体は、炎症性状態、アレルギー及びアレルギー性状態、過敏症反応、自己免疫疾患、重症感染症、並びに臓器又は組織移植拒絶症等の、IL−17A媒介型の状態の予防及び治療に有用であり得る。本発明の抗体はまた、かかる治療のための薬剤の調製においても有用であり、薬剤は、本明細書に定められる投薬量で投与するために調製される。例示のIL−17A媒介型状態は、関節リウマチ(RA)、強直性脊椎炎、乾癬性関節炎、変形性関節症、骨粗鬆症、ブドウ膜炎、炎症性線維症(例えば、強皮症、肺線維症、及び肝硬変)、炎症性腸障害(例えば、クローン病、潰瘍性大腸炎、及び炎症性腸疾患)、ぜんそく(を含むアレルギー性ぜんそく)、アレルギー、COPD、多発性硬化症、乾癬、全身性紅斑性狼瘡、糖尿病、並びに癌を含む、炎症性状態、免疫並びに増殖性障害である。抗ヒトIL−17A療法で治療された患者におけるプラスの転帰は、関節リウマチ、乾癬、及び非感染性ブドウ膜炎で記載されている(Genovese et al.,Arthritis Rheum.62:929〜39,2010、Hueber et al.,Sci.Transl.Med.2:52ra72.,2010)。
【0113】
炎症性肺状態は、炎症性状態の一例である。例示の炎症性肺状態には、ウイルス性、細菌性、真菌性、寄生生物、又はプリオン感染に関連する状態を含む感染誘発性肺状態、アレルゲン誘発性肺状態、石綿症、珪肺症、又はベリリウム中毒等の汚染物質誘発性肺状態、胃吸引誘発性肺状態、免疫調節異常、嚢胞性線維症等の遺伝性素因による炎症性状態、並びに人工呼吸器傷害等の物理的外傷誘発性肺状態が含まれる。これらの炎症性状態には更に、ぜんそく、気腫、気管支炎、慢性閉塞性肺疾患(COPD)、サルコイドーシス、ヒスチオサイトーシス、リンパ管筋腫症、急性肺傷害、急性呼吸窮迫症候群、慢性肺疾患、気管支肺異形成症、院外感染性肺炎、院内肺炎、人工呼吸器関連肺炎、敗血症、ウイルス性肺炎、インフルエンザ感染、パラインフルエンザ感染、ロタウイルス感染、ヒトメタニューモウイルス感染、呼吸系発疹ウイルス感染、及びアスペルギルス又は他の真菌感染も含まれる。例示の感染関連炎症性疾患としては、重症肺炎、嚢胞性線維症、気管支炎、気道増悪、及び急性呼吸窮迫症候群(ARDS)を含むウイルス又は細菌性肺炎が挙げられ得る。かかる感染に伴う状態は、一次ウイルス感染及び二次ウイルス感染等の複数の感染を伴う場合がある。調節不全のIL−17A産生は、ぜんそく及び慢性閉塞性肺疾患(COPD)等の肺疾患の病理において役割を果たす可能性がある(Alcorn et al.,Annu.Rev.Physiol.72:495〜516,2010に概説される)。IL−17Aは、IL−17Aが、肺常在上皮細胞からの好中球動員、生存、及び活性化において重要な因子を誘発する能力を有するため、重症のぜんそく並びにCOPDの顕著な特徴である肺の好中球性炎症を調節することが示されている(例えば、IL−6、IL−8、GM−CSF、G−CSF)。本発明の抗体は、肺上皮細胞からのIL−6、IL−8、及びGM−CSF分泌を抑制し、したがって、ぜんそく及びCOPD等の肺炎症性状態を有する対象の治療的又は予防的治療において有益であり得る。ぜんそく及び気道炎症のために一般的に使用される動物モデルには、オボアルブミンチャレンジモデル及びメタコリン感作モデルが含まれる(Hessel et al.,Eur.J.Pharmacol.293:401〜12,1995)。培養したヒト気管支上皮細胞、気管支線維芽細胞又は気道平滑筋細胞からのサイトカイン及びケモカインの産生の阻害を生体外モデルとして用いることもできる。本発明の拮抗物質をこれらのモデルのいずれかに投与することによって、ぜんそく、気道炎症、COPD等の症状を改善し、経過を変化させるためのこれらの拮抗物質の使用を評価することができる。
【0114】
乾癬は、炎症性状態の別の例である。乾癬は、角化細胞のT細胞媒介型高度増殖並びに炎症性浸潤によって特徴付けられる。乾癬組織の炎症及び高度増殖は、正常な皮膚とは異なる組織学的、抗原性、及びサイトカインプロファイルに関連する。乾癬に関連するサイトカインの中でもとりわけ、TNFα、IL−19、IL−18、IL−15、IL−12、IL−7、IFNγ、IL−17A、及びIL−23が挙げられる(Gudjonsson et al.,Clin.Exp.Immunol.135:1〜8,2004)。IL−17Aは、乾癬病変で過剰発現されることが見出されており(米国特許第7,776,540号)、抗ヒトIL−17A療法で治療された患者におけるプラスの転帰が記載されている(Hueber et al.,Sci.Transl.Med.2:52ra72.,2010)。
【0115】
変形性関節症、関節リウマチ、傷害の結果として関節炎にかかった関節等を含む関節炎が、一般的な炎症性状態であり、それらは本発明の拮抗物質等の抗炎症性タンパク質の治療的使用から利益を受けるであろう。IL−17Aシグナル伝達の活性化によって、炎症を起こした関節の炎症及び更なる組織損傷が持続し得る。関節リウマチのための複数の動物モデルが既知である。例えばコラーゲン誘導性関節炎(CIA)モデルでは、ヒト関節リウマチとよく似た慢性炎症性関節炎をマウスに発症させる。本発明のIL−17A抗体をCIAモデルマウスに投与することによって、疾患の症状を改善して疾患の経過を変化させるためのこれらの拮抗物質の使用を評価することができる。
【0116】
慢性の消化管炎症性状態の例としては、炎症性腸疾患(IBD)、潰瘍性大腸炎(UC)、及びクローン病(CD)、外界からの刺激によって誘発される大腸炎(例えば、化学療法、放射線療法等の投与等の治療レジメンによって引き起こされるかあるいはこれにともなう(例えば、副作用として)消化管炎症(例えば、大腸炎))、感染性大腸炎、虚血性大腸炎、膠原性又はリンパ性大腸炎、壊死性腸炎、慢性肉芽腫症又はセリアック病などの状態における大腸炎、食品アレルギー、胃炎、感染性胃炎又は小腸結腸炎(例えばヘリコバクター・ピロリの感染による慢性活動性胃炎)、並びに感染性病原体によって引き起こされる消化管炎症の他の形態がある。胃腸炎症性状態のための複数の動物モデルが存在する。最も広く使用されるモデルは、結腸に慢性炎症及び潰瘍形成を誘発する2,4,6−トリニトロベンゼンスルホン酸/エタノール(TNBS)誘発大腸炎モデル又はオキサゾロンモデルである(Neurath et al.,Intern.Rev.Immunol 19:51〜62、2000)。別のモデルでは、血性下痢、体重減少、結腸の短縮、及び好中球の浸潤を伴う粘膜の潰瘍形成として発現する急性大腸炎を誘発する硫酸デキストランナトリウム(DSS)を使用する。別のモデルは、ナイーブなCD45RBhigh CD4 T細胞をRAG又はSCIDマウスに養子移入することを含む。このモデルでは、ドナーのナイーブT細胞がレシピエントの腸を攻撃することにより、ヒトの炎症性腸疾患に似た慢性腸炎症及び症状を引き起こす(Read and Powrie,Curr.Protoc.Immunol.Chapter 15 unit 15.13,2001)。これらのモデルのいずれかにおいて本発明の拮抗物質の投与を用いることによって、これらの拮抗物質が、炎症性腸疾患等の腸内の炎症に伴う症状を改善し、疾患の経過を変化させる潜在的な有効性を評価することができる。
【0117】
腎線維症は、急性刺激(例えば、移植片虚血/再潅流)(Freese et al.,Nephrol.Dial.Transplant.16:2401〜6,2001)又は 慢性状態(例えば、糖尿病)(Ritz et al.,Nephrol.Dial.Transplant.11 Suppl 9:38〜44,1996)のいずれかから発症し得る。その病因は典型的に、腎糸球体濾過器官及び尿細管間質に最初に炎症反応が起こった後、線維化が持続することとして特徴付けられる(Liu,Kidney Int.69:213〜7,2006)。尿細管間質線維症は、末期の腎不全に対する腎損傷の病因において重要な役割を果たしていることが示されており、近位細管細胞が中心的媒介物質であることが明らかにされている(Phillips and Steadman,Histol.Histopathol.17:247〜52,2002、Phillips,Chang Gung Med.J.30:2〜6,2007)。尿細管間質区画における線維化は、部分的に、常在線維芽細胞の活性化により媒介され、この常在線維芽細胞は、炎症促進性のサイトカインを分泌し、これが近位細管上皮を刺激して、局所的な炎症及び線維化の媒介物質を分泌する。更に、走化性サイトカインが線維芽細胞及び上皮細胞によって分泌され、単核白血球/マクロファージ及びT細胞を尿細管間質へと湿潤させるよう導く方向付け勾配を供給する。炎症性湿潤により、更なる線維化及び炎症性のサイトカインが産生され、それらは更に線維芽細胞及び上皮サイトカインの放出を活性化し、同時に上皮も刺激して表現型移行を起こし、これにより細胞が過剰な細胞外マトリックス構成成分を沈積させる(Simonson,Kidney Int.71:846〜54、2007)。IL−17Aは、ヒト腎同種移植片拒絶中に上方調節されることが示されている(Van Kooten et al.,J.Am.Soc.Nephrol.9:1526〜34,1998、Loong et al.,J.Path.197:322〜32,2002)。IL−17Aは、近位尿細管上皮による炎症促進性メディエーターIL−6、IL−8、補体構成要素C3、及びRANTESの産生を刺激する(Van Kooten et al.,J.Am.Soc.Nephrol.9:1526〜34,1998、Woltman et al.,J.Am.Nephrol.11:2044〜55,2000)。これらの因子が次に、他の炎症性細胞タイプの間質への補充を媒介し、これが炎症/免疫反応の維持に寄与し、抑制されない場合には線維症及び慢性同種移植腎症の発症に寄与する(Racusen et al.,Kidney Int.55:713〜23,1999、Mannon,Am.J.Transpl.6:867〜75,2006)。
【0118】
他の例示的な繊維性の病態としては、肝臓線維症(アルコール性肝硬変、ウイルス性肝硬変、自己免疫性肝炎が挙げられるがこれらに限定されない);肺線維症(強皮症、特発性肺線維症が挙げられるがこれらに限定されない);腎臓線維症(強皮症、糖尿病性腎炎、糸球体腎炎、狼瘡腎炎を含むがこれらに限定されない);皮膚線維症(強皮症、肥厚性及びケロイド瘢痕、火傷を含むがこれらに限定されない);骨髄線維症;神経線維腫症;線維腫;腸線維症;及び外科手術の結果としての線維症癒着等が挙げられる。線維症は、臓器特異的な線維症又は全身性線維症であり得る。器官特異性線維症は、肺線維症、肝臓線維症、腎臓線維症、心臓線維症、血管線維症、皮膚線維症、眼線維症、又は骨髄線維症に関連し得る。肺線維症は、特発性肺線維症、薬剤誘発性肺線維症、ぜんそく、サルコイドーシス、又は慢性閉塞性肺疾患に関連し得る。肝臓線維症は、肝硬変、住血吸虫症、又は胆管炎に関連し得る。肝硬変は、アルコール性肝硬変、C型肝炎後肝硬変、原発性胆汁性肝硬変の中から選択され得る。胆管炎は硬化性胆管炎であり得る。腎線維症は、糖尿病性腎障害又はループス糸球体腎炎に伴い得る。心線維症は、心筋梗塞に伴い得る。血管線維症は、血管形成術後動脈再狭窄、又はアテローム性動脈硬化に伴い得る。皮膚線維症は、火傷瘢痕化、肥厚性瘢痕化、ケロイド、又は腎性線維性皮膚症に伴い得る。眼線維症は、後眼窩線維症、白内障手術後又は増殖性硝子体網膜症に伴い得る。骨髄線維症は、特発性骨髄線維症又は薬剤誘発性骨髄線維症に伴い得る。全身性線維症は、全身性硬化症及び移植片体宿主細胞病から選択され得る。
【0119】
本発明の方法によって予防又は治療し得る他の炎症性状態及び神経障害は、自己免疫疾患によって引き起こされるものである。これらの状態及び神経障害としては、多発性硬化症、全身性エリテマトーデス、並びに、アルツハイマー病、パーキンソン病、ハンチントン病、双極性疾患、及び筋萎縮性側索硬化症(ALS)を含む神経変性及び中枢神経系(CNS)疾患、原発性胆汁性硬変、原発性硬化性胆管炎、非アルコール性脂肪性肝疾患/脂肪性肝炎、線維症、C型肝炎ウイルス(HCV)、及びB型肝炎ウイルス(HBV)等の肝臓疾患、糖尿病及びインスリン抵抗性、アテローム性動脈硬化症、脳出血、脳卒中、及び心筋梗塞等の心血管疾患、関節炎、関節リウマチ、乾癬性関節炎及び若年性関節リウマチ(JRA)、骨粗鬆症、変形性関節症、膵炎、線維症、脳炎、乾癬、巨細胞動脈炎、強直性脊椎炎、自己免疫性肝炎、ヒト免疫不全ウイルス(HIV)、炎症性皮膚状態、移植、癌、アレルギー、内分泌疾患、創傷治癒、他の自己免疫性疾患、気道過敏症、並びに、細胞、ウイルス又はプリオンによって介在される感染症又は疾患が挙げられる。
【0120】
投与/医薬組成物
IL−17A活性の抑制が所望される状態の治療に有効な薬剤の「治療上の有効量」は、標準的な研究技術によって決定され得る。例えば、ぜんそく、クローン病、潰瘍性大腸炎、又は関節リウマチ等の炎症性状態の治療において有効となる薬剤の投薬量は、本明細書に記載されるモデル等の、関連する動物モデルに薬剤を投与することによって決定され得る。
【0121】
更に、任意に生体外アッセイを用いて最適な投薬量範囲を特定することができる。特定の有効用量の選択は、当業者であれば幾つかの因子の考慮に基づいて(例えば、臨床試験によって)決定することができる。かかる因子には、治療又は予防しようとする疾患、関与する症状、患者の体重、患者の免疫状態、及び当業者に既知の他の因子が含まれる。製剤に使用される正確な投薬量は、投与経路、及び疾患の重篤度にも依存し、医師の判断及び各患者の状況に基づいて決定されなければならない。有効用量は、生体外又は動物モデル試験系から導出される用量反応曲線から外挿することができる。
【0122】
本発明の薬剤の治療上の使用のための投与方法は、薬剤を宿主細胞に送達する任意の好適な経路でよい。これらの薬剤の医薬組成物は、例えば、皮内、筋肉内、腹腔内、静脈内、皮下、又は鼻腔内等の非経口投与に特に有用である。
【0123】
本発明の薬剤は、有効量の薬剤を製薬上許容され得る担体中の有効成分として含有する医薬組成物として調製することができる。「担体」という用語は、活性化合物が一緒に投与される希釈剤、助剤、賦形剤、又は溶媒のことを指す。かかる医薬用溶媒は、落花生油、大豆油、鉱物油、ゴマ油等の、石油、動物、植物又は合成物由来の水及び油等の液体であってよい。例えば、0.4%生理食塩水及び0.3%グリシンを使用することができる。これらの溶液は滅菌され、粒子状物質を含まない。これらの溶液は、従来の周知の滅菌技術(例えば、濾過)によって滅菌することができる。この組成物は、生理学的条件に近づけるために必要とされるpH調整剤及び緩衝剤等の製薬上許容される補助物質を含むことができる。かかる医薬製剤中の本発明の薬剤の濃度は、約0.5重量%未満、通常は約1重量%又は少なくとも約1重量%か、最大で15又は20重量%までと大きく異なってよく、選択される投与方法に従って、主として必要とされる用量、液体の体積、粘度等に基づいて選択される。
【0124】
したがって、筋肉内注射用の本発明の医薬組成物は、1mLの滅菌緩衝水、及び約1ng〜約100mg、例えば、約50ng〜約30mg、又はより好ましくは約5mg〜約25mgの本発明のIL−17A拮抗物質を含有するように調製することができる。同様に、静脈内注射用の本発明の医薬組成物は、250mLの滅菌リンゲル溶液、及び約1mg〜約30mg、好ましくは5mg〜約25mgの本発明の拮抗物質を含有するように調製することができる。非経口投与可能な組成物を調製する実際の方法は周知であり、例えば、「Remington’s Pharmaceutical Science」,15th ed.,Mack Publishing Company,Easton,PA.に、より詳細に記載されている。
【0125】
本発明の抗体拮抗物質は凍結乾燥して保存することができ、使用前に好適な担体中で再構成することができる。この技術は従来の免疫グロブリン及びタンパクの調製において有効であることが示されており、当該技術分野で既知の凍結乾燥と再構成の技術を用いることができる。
【0126】
ここで、以下の具体的かつ非限定的な実施例を参照して、本発明を説明する。
【実施例】
【0127】
(実施例1)
抗ヒトIL−17A拮抗性mAbの特定
The MorphoSys Human Combinatorial Antibody Library(HuCAL(登録商標))ゴールドファージディスプレイライブラリー(Morphosys AG,Martinsried,Germany)をヒト抗体フラグメントの源として使用し、溶液のサブプール中でパンニングした。第1回目のパンニングでは、サブライブラリーをビオチン化成熟His6タグ付きIL−17A A132Q及びA70Q変異型(IL−17Amut6)(配列番号106)に対して選択した。第2回目では、第1回目の増幅生産物を、IL−17Aに特異的な抗体に反して偏倚した競合物質としての他のIL−17Aファミリーメンバーの存在又は不在下で、ビオチン化His6タグ付きIL−17Amut6に対して選択した。第2回目の増幅生産物を2つのプールに分割した。第1のプールを第1回目と同様にパンニングした。第2のプールのクローンを、最初の選択で使用したサブライブラリーに応じてHCDR2又はLCDR3のいずれかに更に分散させ、次いでIL−17Amut6に対して更に2回のパンニングを行って、スクリーニングのためのクローンの第2の源を得た。クローンライセートからのFabを、ヒツジ抗ヒトFd抗体でコーティングしたELISAプレートウェル中に捕捉し、ビオチン化IL−17Amut6への結合についてスクリーニングした。陽性クローンの粗溶解物を、組み換えヒトIL−17RA受容体(配列番号107)へのIL−17Amut6結合の阻害についてスクリーニングした。
【0128】
選択されたクローンを、全ての配列ファミリーの配列スコアリング、親和性、及び表現に基づいて精製したFabとして選択して更に特徴付け、MOR番号を指定した。MOR7708、MOR7785、MOR7706、MOR7775、及びMOR7700についての更なる変異型を生成して、HCDR2、HCDR3、又はCDR3に常在するTrp又はMetを置き換えた。表3は、生成された変異型を示す。
【0129】
【表6】
【0130】
組み換えヒトIL−17RA受容体へのIL−17Amut6及びcynoIL−17A結合、及びIL−17Amut6へのそれらの結合のFabによる阻害について、Fabを試験した。全ての試験したFabは、IL−17RAへのIL−17Amut6及びcynoIL−17A結合の両方を阻害した。SETアッセイを使用してIL−17Amut6へのFabの親和性を測定した(表4)。特定したFabから、ファミリー2、6a、6b、19a、及び19bからの候補を選択して更に特徴付けた。
【0131】
【表7】
【0132】
(実施例2)
抗IL−17A拮抗性mAbの誘導、操作、及び特徴付け
選択したMOR番号Fabを変換し、ヒトIgG1形式のmAbとして発現させ、対応するMORmAb表記を与えた。生成したMORmAbを、発現及び凝集、ヒトIL−17RAへのヒト及びcyno IL−17A結合を阻害する、並びにNHDF細胞からのIL−8分泌を阻害する、それらの能力について試験した。表5は、MORmAbのために選択したアッセイについてのIC50値を示す。試験したMORmAb(MORmAb番号7702、7708、7785、7786、7706、7775、7700、8095、8096、8097、8098、7768)のいずれも、他のIL−17ファミリーメンバーと交差反応しなかった。
【0133】
【表8】
【0134】
抗IL−17A拮抗性mAbのフレームワーク操作
活性並びに生物物理学的及び生化学的特性に基づき、選択されたMORmAbを、それらの可変領域において、ある種のフレームワーク残基をマッチングヒト生殖細胞系列存在する残基に変更するように、及びコドンを高度に発現した哺乳類タンパク質中で最も頻繁に生じるコドンに変更するように、更に操作した。ファミリー2 VLにおいて、L11V、及びV85T(直鎖配列)置換を行い、フレームワークをVK−1生殖細胞系列Vb−L5(IGKV1−12*01)と厳密に一致するように変換した。V11V及びV85T置換を有する例示の可変領域は、配列番号76に示されるアミノ酸配列を有する可変領域である。ファミリー6a及び6b VLにおいて、D1E、V59I、及びT86V置換を行い(直鎖配列)、フレームワークをVk−3生殖細胞系列Vb−L6(IGKV3−11*01)と厳密に一致するように変換した。D1E、V59I、及びT86V置換を有する例示の可変領域は、配列番号77に示されるアミノ酸配列を有する可変領域である。ファミリー6a及び6b VHにおいて、G44S置換を行って(直鎖配列)、Vh−6生殖細胞系列Vb 6−01(IGHV6−1*01)と一致させた。G44S置換を有する例示の可変領域は、配列番号81に示されるアミノ酸配列を有する可変領域である。ファミリー19a及び19b VLにおいて、アミノ酸1〜3(DIE)をQSVで置換して、人工のカッパN末端をラムダ鎖のN末端に置き換えた。QSV置換を有する例示の可変領域は、配列番号79に示されるアミノ酸配列を有する可変領域である。ファミリー19a及び19b VHにおいて、V5L置換を行って、Vh−3生殖細胞系列Vb3〜23(IGHV3−23*01とほぼ一致させた。また、このプロセスにおいて、重鎖定常領域アミノ酸配列残基353〜357(REEMT)をRDELTで置換した。V5L置換を有する例示の可変領域は、配列番号86に示されるアミノ酸配列を有する可変領域である。定常領域353〜357 REEMT−>RDELT置換を有する例示の重鎖は、配列番号100に示されるアミノ酸配列を有する重鎖である。操作された抗体にmAb番号を与えた。
【0135】
操作された及び元の可変領域並びに完全長抗体の対応する表記及び配列表を表2に列挙する。各ファミリー内のCDRの配列を図1に示す。
【0136】
操作されたmAbを、MORmAbについて上述したように特徴付けた。示されるアッセイを使用して測定したIC50値(pM)を表6に示す。
【0137】
【表9】
【0138】
ビアコアを使用して、選択されたmAbの親和性を査定した。測定の結果を表7に示す。
【0139】
【表10】
*1つの抗IL−17当たりの二量体
【0140】
抗IL−17抗体は、NHBE細胞中のサイトカイン分泌を阻害する。
IL−17Aは、IL−17Aが、好中球動員、生存、及び活性化において重要な因子を誘発する能力を有するため、重症のぜんそく並びにCOPDの顕著な特徴である肺の好中球性炎症を調節することが示されている(例えば、IL−6、IL−8、GM−CSF)。抗IL−17A本発明の抗体が肺常在細胞中のIL−17A誘発性変化を阻害し得るかどうかを決定するために、正常ヒト気管支上皮(NHBE)細胞を、mAb6785の存在下で48時間、ヒトIL−17Aで刺激した。mAb6785は、それぞれ619.0±64.0pM及び564±86pMのIC50で、NHBE細胞によるIL−17A誘発性IL−6及びGM−CSF産生を阻害した。
【0141】
抗IL−17抗体は、IL−17Aの/Fヘテロ二量体の生物活性を阻害する。
正常なヒト皮膚線維芽細胞(NHDF、Lonza)細胞を、48ウェルの平底組織培養プレートの中に、1ウェル当たり10,000細胞でFGM−2培地(Lonza)中に播種し、一晩インキュベートした(37°、5% CO2)。インキュベーション後、rhIL−17A/Fヘテロ二量体(R&D Systems)の50ng/mL最終濃度(1.47nM)を、mAb6785又は対照抗体の希釈系(30μg/mL−0.5ng/mL)を用いて室温で10分間プレインキュベートし、細胞に添加した。細胞を48時間インキュベートし(37°、5% CO2)、培養物の上清を収集し、製造業者の指示に従って、ヒトIL−6 Duo Sets(R&D Systems、Inc.)を使用して、IL−6含量についてELISAによってアッセイした。GraphPad Prismソフトウェア(GraphPad Software,Inc)を使用して、IC50値を非直線回帰によって決定した。mAb6785は、2±2.5nMのEC50で、NHDFによるIL−17A/Fヘテロ二量体誘発型IL−6産生を阻害した。
【0142】
方法
溶液平衡滴定(SET)を使用したピコモル親和性の決定
溶液平衡滴定(SET)によるKD決定のために、Fabタンパク質の単量体分画(少なくとも90%単量体含量、分析用SECによって分析、Superdex75カラム、GE)を使用した。
【0143】
基本的に、前述のように溶液中の電気化学発光(ECL)に基づく親和性決定及びデータ評価を行った(Haenel et al.,Anal Biochem 339:182〜4,2005)。規定の固定濃度の精製したFab(約10〜100pM)を、化学的均衡が達成されるまで、溶液中のIL−17Amut6(5nMの最大濃度)の濃度を増加させながらインキュベートした。溶液中の結合されていないFabを定量化するために、試料を、コーティングされたビオチン化IL−17Amut6と共に、ストレプトアビジンMSDの384ウェルのマイクロタイタープレート(Meso Scale Discovery,Gaithersburg,MD)に移した。検出のために、ルテニウム複合体で標識化された抗ヒトFab/IgG抗体を適用し、プレートをSector(商標)Imager 6000(MSD)で読み取った。後述されるモデルを使用してExcel/XLfitソフトウェアにより、滴定曲線(抗原濃度の関数としての遊離Fabの濃度)をプロットし、当てはめた。
【0144】
Fab分子のKDの決定についてのデータ評価のために、次の当てはめモデルを使用した(Abraham et al.J Mol Recognit.9:456〜461,1996に従って修正):
y=Bmax−(Bmax/(2*cFab)*(x+cFab+KD−sqrt((x+cFab+KD)*(x+cFab+KD)−4*x*cFab)))
他方では:
Bmax:最大結合シグナル(抗原濃度=0で)
cFab:適用されたFab濃度
x:適用された総可溶性抗原濃度(結合部位)
sqrt:平方根
KD:均衡解離定常
【0145】
IL−17RAへのIL−17A結合の阻害(例えば、「IL−17RA阻害」アッセイ
透明のマキシソーププレートを、pH 9.4の0.1M炭酸ナトリウム緩衝液中、100μL/ウェルの2.5μg/mLヒトIL−17RA−Fc(R&D Systems,Minneapolis,MN)でコーティングし、4℃で一晩インキュベートした。ブロッキング及び洗浄後、25ng/mLのビオチン化ヒトIL−17mut6(配列番号106)又はカニクイザルIL−17A(配列番号108)を、試験mAb又は対照mAb(30〜0μg/mL最終濃度)を用いて、100μLの合わせた容量中で5〜10分間プレインキュベートし、次いでプレートに添加した。シグナルを、100μLの1mg/mL SA−HRP(Jackson Immunoresearch,West Grove,PA)の1:10,000希釈を用いて室温(RT)で20分間、続いて100μL/ウェルのOPD基質(Sigma−Aldrich Corp.,St.Louis,MO)で検出した。プレートを492nmで読み取った(Envision,PerkinElmer,Waltham,MA)。IL−17RAへのFab結合を、mAbについて記載されるように試験した。
【0146】
NHDF細胞からのIL−8及びIL−6産生の阻害(例えば「IL−8産生」及び「IL−6産生」アッセイ)
抗IL−17AmAbの、IL−8及びIL−6産生に及ぼす阻害の効果を正常なヒト皮膚線維芽細胞(NHDF)中で査定した。細胞を、48ウェルの平底組織培養プレート中に、FGM−2培地中、1ウェル当たり0.1×105細胞、1ウェル当たり250μLでプレートし、一晩インキュベートした(37°、5% CO2)。インキュベーション後、0.1ng/mLのヒトTNF−αを全てのウェルに添加した。10ng/mLのIL−17mut6又は25ng/mLのカニクイザルIL−17Aを、試験mAb又は対照mAb(30〜0μg/mL最終濃度)を用いて、250μLの合わせた容量中で、室温で10分間プレインキュベートし、次いで250μLの細胞に添加した。本アッセイにおいて、抗体を添加していないIL−17mut6試料を対照試料として含めた一方で、TNF−α又は培養培地のみからなる試料を陰性対照として含めた。細胞を24時間(37°、5% CO2)インキュベートし、調湿した培地を収集し、ヒトIL−6 & IL−8 ELISA Duo Setsを使用して、製造業者(R&D Systems,Minneapolis,MN)の指示に従って、IL−6及びIL−8についてELISAによってアッセイした。Fabを、mAbについて記載されるようにアッセイした。
【0147】
NHBE細胞からのIL−6及びG−CSF産生の阻害
正常なヒト気管支上皮(NHBE、Lonza)細胞を、BEGM培地(Lonza)中、1ウェル当たり20,000細胞で播種し、一晩インキュベートした(37°、5%CO2)。インキュベーション後、細胞をIL−17Amut6を用いて、試験した抗体の存在下で、様々な濃度で48時間刺激した(30μg/mL〜0.5ng.mL)。インキュベーション後、上清を収集し、ヒトIL−6−又はG−CSF特異的ELISA(R&D Systems,Inc.)を使用して、IL−6又はG−CSF含量についてアッセイした。IC50値を、GraphPad Prismソフトウェア(GraphPad Software,Inc)を使用して非直線回帰によって決定した。
【0148】
IL−17Aファミリーメンバーとの交差反応性
透明のマキシソーププレートを、PBS中の5μg/mLのmAb又はアイソタイプ対照mAbの100μL/ウェルでコーティングし、4℃で一晩インキュベートした。プレートを200μL/ウェルで、ELISAブロック緩衝液(1% BSA、0.05% NaN3を含むPBS中5%スクロース)を用いて1時間ブロックし、洗浄緩衝液(PBS、0.01% Tween−20)で3回洗浄した。競合サイトカインをアッセイ希釈緩衝液(PBS中1% BSA)中で、2倍の最終濃度で滴定し、ビオチン化サイトカインを2倍の最終濃度で調製した。2倍の最終濃度の100μLのサイトカインを、アッセイ緩衝液中、2倍の最終濃度の100μLのビオチン化IL−17mut6(25ng/mL最終濃度)と混合した(30〜0μg/mL最終濃度)。組み換えヒトIL−23(R&D Systems,Minneapolis,MN)を陰性対照として、緩衝液のみの試料を背景対照として、及びIL−17mut6を陽性対照として使用した。サイトカイン/ビオチン化IL−17mut6混合物を複製物した、1ウェル当たり100μLを、プレートに添加し、1〜2時間インキュベートした。プレートを洗浄緩衝液で3回洗浄し、100μLの1mg/mL SA−HRPの1:10,000希釈を用いて、室温で20分間インキュベートした(Jackson Immunoresearch,West Grove,PA)。プレートをELISA洗浄緩衝液で3回洗浄した。洗浄後、100μL/ウェルのOPD基質(Sigma−Aldrich Corp.,St.Louis,MO)を各ウェルに添加し、適切な色の変化が検出されるまでインキュベートした。反応を50μLの2N硫酸の添加により停止し、Envision機器を使用してプレートを492nmと読み取った。
【0149】
親和性測定−ビアコアアッセイ
表面プラズモン共鳴(SPR)を用いる親和性測定を、Biacore 3000光バイオセンサ(Biacore)を用いて行った。選択されたFab(約30 RU)又はmAb(約50 RU)を、それぞれFab又はmAb捕捉のためのヒツジ抗Fd抗体又は抗ヒトFc抗体を使用して、センサーチップ表面上に捕捉した。Fab又はmAb捕捉後、溶液中、huIL−17mut6又はcyno IL−17A(0.2から49nM)を注入した。
【0150】
(実施例4)
エピトープマッピング
抗体エピトープを、競合結合、H/D交換分析、及び抗体−IL−17A共構造の組み合わせによって推定した(実施例5を参照されたい)。次の抗体を使用した:mAb1926、MORmAb7700、MORmAb7706、MORmAb7708、mAb7357(ハイブリドーマC1863に由来するマウス抗ヒトIL−17A中和抗体)、mAb2832(ハイブリドーマC1861に由来するマウス/ヒトキメラ抗ヒトIL−17A中和抗体)、mAb317(マウス抗ヒトIL−17A抗体、R&D Systems,Minneapolis,MN)及びmAb3171(マウス抗ヒトIL−17A抗体、R&D Systems,Minneapolis,MN)、並びにmAbeBIO16〜7178(マウス抗ヒトIL−17A抗体、e−Bioscience,San Diego,CA)。この3つの商業用抗体は、異なる度合いの中和活性を示した。
【0151】
競合的エピトープ結合
競合ELISAのために、1ウェル当たり5μL(20μg/mL)IL−17Aのmut6タンパク質を、MSD HighBindプレート(Meso Scale Discovery,Gaithersburg,MD)上に室温で2時間コーティングした。150μLの5% MSD遮断剤A緩衝液(Meso Scale Discovery,Gaithersburg,MD)を各ウェルに添加し、室温で2時間インキュベートした。プレートを0.1M HEPES緩衝液(pH 7.4)で洗浄した。10nMの標識化された抗体(MDS蛍光染色)を、競合物質抗体の濃度を増加させながら(1nM〜2μM)インキュベートし、25μLの混合物を指定されたウェルに添加した。室温で穏やかに振盪しながら2時間のインキュベートした後、プレートを上記のように洗浄し、150μL希釈MSD読み取り緩衝液Tを添加し、プレートをMDSセクターイメージャ6000で読み取った。
【0152】
アッセイを、標識化されたmAb1926、mAb317、mAb3171、又はmAb7357により行った(図4)。競合アッセイに基づき、抗IL−17A抗体を4つの異なるビンに割り当てた。ビンA:mAb1926、MORmAb7706、及びMORmAb7708、ビンB:eBio16〜7178及びmAb7357、ビンC:mAb317、ビンD:mAb3171。
【0153】
H/D交換分析:
H/D交換のために、抗体摂動を分析するために使用した手順は、前述の手順と類似していたが(Hamuro et al.,J.Biomol.Techniques,14:171〜82,2003、Horn et al.,Biochemistry,45:8488〜98,2006)、幾らかの修飾を伴った。組み換えIL−17Amut6(C−末端His−タグによりHEK293E細胞中で発現させた)を、重水素化水溶液中で所定の時間にわたってインキュベートすることにより、交換可能な水素原子における重水素の組み込みを得た。重水素化IL−17Amut6を、固定化した個々の抗IL−17A mAbを含有するカラム上に捕捉し、次いで水性緩衝液で洗浄した。逆交換したIL−17Amut6タンパク質をカラムから溶出し、重水素含有フラグメントの局在性をプロテアーゼ消化及び質量分析によって決定した。抗体に結合した領域は、交換から比較的保護された部位であり、したがって、抗体と複合体化されていないIL−17Amut6と比較してより高い分画の重水素を含有すると推定された。IL−17Amut6のH/D交換摂動マップは、図5に示される。バーの上の番号は、IL−17Amut6残基を指す。
【0154】
MORmAb7700、MORmAb7706、及びMORmAb7708は、IL−17Aの(配列番号105)45NRSTSPWNLH54(配列番号159)、56NEDPERYPSVIWE68(配列番号157)、及び100RREPPHCPNSFRLEKIL116(配列番号158)の3つのセグメントに対して異なる度合いの示差交換を示し、抗体による保護を示唆した。56NEDPERYPSVIWE68(配列番号157)フラグメントは、MORmAb7708によって強力に保護され、MORmAb7700によってわずかに保護され、MORmAb7706によっては保護されなかった。これらの抗体のフラグメント保護パターンにおける重複は、上述の競合アッセイにおけるそれらの交差阻害と一致する。
【0155】
mAb7357及びmAbeBio16〜7178の両方について、上述の競合アッセイにおけるそれらの交差阻害と一致して、71CRHLGCINADGNVDYHM87(配列番号160)に対する強力な保護を観察した。弱い、及びしたがって決定的でない示差交換を、mAb7357、mAb2832、mAb317及びmAb3171を有する他のフラグメントについて観察した。
【0156】
H/D交換研究は、上に記載される4つの競合群のうちの2つについて結合部位の位置を特定した。ビンA抗体(MORmAb7700、MORmAb7706、及びMORmAb7708)は、配列番号105のペプチドセグメント45NRSTSPWNLH54、(配列番号159)、56NEDPERYPSVIWE68(配列番号157)、及び100RREPPHCPNSFRLEKIL116(配列番号158)の領域で結合し、ビンB抗体(mAb7357及びmAbeBio16〜7178)は、ペプチドセグメント71CRHLGCINADGNVDYHM87(配列番号160)の領域で結合した。mAb317及びmAb3171は、互いから、並びにビンA及びビンB抗体から異なる部位に結合した。しかしながら、両方の抗体とのH/D交換研究における弱いシグナルは、IL−17A上のそれらのエピトープの位置を特定するのに十分な証拠を提供しなかった。
【0157】
(実施例5)
IL−17A及び抗IL−17A抗体の共結晶構造
mAb6785の組み換えHis6タグ付きFabである、Fab6468とのIL−17Amut6の共構造をX線結晶学によって決定した。Fab6468の軽鎖のアミノ酸配列は、配列番号90に示され、重鎖アミノ酸配列は、配列番号111に示される。実施例5において、IL−17Aアミノ酸残基は、配列番号105に従った残基を示すように参照され、Fab6468残基は、配列番号79に従った軽鎖可変領域残基を示すように参照され、重鎖可変領域残基は配列番号86に従う。組み換えヒトIL−17Amut6の発現、再折り畳み、及び精製が記載されている(Wu et al.,Cytokine,ePub ahead of print Jul29)。HEK−293F細胞中でFab6468を発現させ、記載される方法と類似した方法を使用して精製した(Zhao et al.,Protein Expr Purif,67:182〜9,2009)。
【0158】
IL−17A/Fab6468複合体の結晶化
IL−17A/Fab6468複合体を、IL−17Amut6及びFab6468を、pH 6.5の20mM MES、0.2M NaCl、及び10%グリセロール中、1:1.1モル比で混合することによって調製し、4℃で一晩インキュベートした。複合体を、pH 6.5の20mM MES、0.2M NaCl、及び10%グリセロール中、Superdex 200 10/300 GLカラム(GE Healthcare,Piscataway,NJ)上でのサイズ排除クロマトグラフィー(SEC)を使用して、過剰の複合体化されていないFabから精製した。複合体に相当する分画をプールし、Amicon Ultra 10000 MWCOデバイスで4.6mg/mLにまで濃縮した。
【0159】
等容量のタンパク質及びリザーバ溶液を、Corningプレート3550(Corning Inc.,Corning,NY)を使用するシッティングドロップ形式で分注するOryx4自動タンパク質結晶化ロボット(Douglas Instruments,East Garston,UK)を使用して、自動化結晶化スクリーニングを行った。最初のスクリーニングを、Hampton結晶スクリーンHT(HR2−130,Hampton Research)を用いて行い、pH 4.5〜4.6の硫酸アンモニウム、PEGを含有する複数の条件から針様の結晶を産生した。これらの小さい結晶を使用して、マイクロシードマトリックススクリーニング(MMS)(D’Arcy et al.,Acta Crystallographica Section D,63:550〜4,2007)のためのシードストックを産生した。回折品質の結晶を、pH 5.5の0.1M酢酸ナトリウム、12%PEG MME 5000、及び0.2M硫酸リチウム中のMMSスクリーンから得た。
【0160】
IL−17A/Fab6468複合体のX線データ収集
X線データ収集のために、結晶を、24%グリセロールを補充した母液に数秒間浸漬し、95°Kの窒素流中で急速冷凍した。X線回折データを収集し、Osmic(商標)VariMax(商標)共焦光学系、Saturn 944 CCD検出器、及びX−stream(商標)2000低温冷却システムを備えたRigaku MicroMax(商標)−007HF微小焦点X線発生装置(Rigaku,Woodlands,TX)を使用して処理した。回折強度を、1/2度の像当たり3分の露出時間から2.2Åの最大分解能で、254°結晶回転にわたって検出した。X線データをprogram D*TREK(Pflugrath,J.,Acta Crystallographica Section D,55:1718〜25,1999)により処理した。結晶は、単斜晶空間群P21に属し、a=73.40Å、b=64.04Å、c=145.61Å、及びβ=95.39°であった。X線データ統計値を表8に示す。
【0161】
【表11】
*最外郭についての値は、()内にある。
【0162】
構造決定
IL−17A/Fab6468の結晶構造を、Phaser(Read,Acta Crystallorg D Biol Crystallorg,57:1373〜82,2001)を使用して分子置換によって決定した。検索モデルは、IL−17F(PDB ID 1JPY)(Hymowitz et al.,EMBO J.,20:5332〜41,2001)及びFv(VH/VL)についての相同性モデルであり、それはModeller(Accelrys,CA)を使用して、VH及びVLの両方について、抗IL−13抗体CNTO607(PDB ID 3G6A)(Teplyakov et al.,J.Mol.Biol.389:115〜23.2009)に基づいて構築された。2つの定常ドメインCL/CH1をPDB ID 8FABから得た(Strong et al.,Biochemistry,30:3739〜48,1991)。構造精密化を、PHENIX(Adams et al.,J.Synchrotron.Radiat.11:53〜5,2004)を用いて行った。二回転非結晶学的対対称を、早期段階の精密化において最初に課したが、Rフリーに基づいて最終段階において緩めた。モデル調整及び手動の再構築を、COOTを使用して行った(Emsley et al.,Acta Crystallogr.D.Biol.Crystallogr.60:2126〜32,2004)。最終のR結晶及びRフリーは、2.2Åに対する全61,706個の独立の反射のそれぞれについて23.4%及び29.7%であった。精密化統計を表9に提供する。
【0163】
【表12】
【0164】
IL−17A/Fab6468複合体構造
複合体の構造を高分解能(約2.2Å)に対して決定した。IL−17Aは、結晶中ほぼ対称のホモ−二量体であり、2つのFab分子に結合していた。抗体−抗原相互作用は、大部分が疎水性であり、ほとんどの抗体とは対照的に、軽鎖CDRは、幾つかの重要な接触を行った。IL−17A/Fab6468複合体の全体的な分子構造を図6Aに示す。IL−17A二量体の単量体は、シスチンノットの全体的なトポロジーを採用した(図6B)。2つの単量体は、77骨格鎖Cα原子についての0.54ÅのCα RMSDと非常に類似していた。IL−17A単量体シスチンノットの全体的な構築は、76 Cα原子についての0.71Åのrmsdを有するIL−17Fの構築と非常に類似していた(図6B)。各IL−17A単量体は、3つのジスルフィド結合によって安定化していた。鎖Bについて、3つの鎖間ジスルフィド結合が観察された一方で(C10〜C106、C71〜C121、C76〜C123)、鎖Aについては、C10〜C106ジスルフィド結合は、単量体のこれらのセグメントにおける無秩序に起因して観察されなかった。後者の2つのジスルフィド結合(C71〜C121、C76〜C123)は、IL−17F及びNGFに類似したシスチンノット構築を安定化した。IL−17Aの鎖Bについての構造的モデルには、全ての残基10〜128(残基1〜9は無秩序であった)が含まれ一方で、鎖A残基については、残基21〜29、41〜104及び109〜127のみが観察され、その他の残基1〜20、30〜40、105〜108、及び128は、構造における無秩序に起因して欠失していた。2つのFabについて、両方の軽鎖の残基1〜2は、無秩序であったか、又は乏しい電子密度を有した。鎖内ジスルフィド結合並びに重鎖上のHisタグ含む、重鎖及び軽鎖の両方のC−末端3残基は、無秩序であった。
【0165】
IL−17A(鎖B)の秩序化N末端セグメントは、短いヘリカル要素を含有した(残基8〜12)。それは、同じ単量体のループ3〜4に向かって再び折り畳み、鎖間ジスルフィド結合(C10〜C106)を形成した。対照的に、IL−17Fの同等のセグメントは、二量体の他方の単量体へと到達し、鎖内ジスルフィド結合を形成し、2つの単量体を共有結合で連結した。2つのIL−17A単量体のセグメント17〜39の秩序化部分を、IL−17Fにおいてと同様にスワップした。このスワッピングは、IL−17A二量体のこれらの部分についての交差をもたらした。分子内ジスルフィド結合(C10〜C106)と組み合わせて、IL−17Aの2つのN末端セグメントは、2つの連鎖単量体を形成し、それはまた、非還元SDS−PAGE上で26kDの見かけの二量体ももたらした。
【0166】
IL−17Aの二量体は、4つの主なβ−ストランド(ストランド1〜4)についてほぼ対称であった(図6C)。76個の残基についてのCα rmsdは、0.71Åである。若干の不斉は、2つの源に由来した。第1に、鎖Aは、主にN末端において、幾つかの無秩序セグメントを含有した。短いβ−ストランド(ストランド0、残基22〜26)のみが見かけ上、秩序化されていた一方で、鎖Bの残基10〜40は、ヘリカルセグメント(残基12〜16)及びβ−ストランド(ストランド0、残基21〜25)で秩序化されていた。第2に、IL−17Aの2つの単量体(40〜128)の、シスチンノットの4つの主なβ−ストランドが、二回転対称によって関連させられた一方で、ストランド0の秩序化部分は、主体が重ねられるとき、完全に重ね合わさらなかった(図示されず)。これがタンパク質再折り畳みのアーチファクトであるか、又はかかる配列が天然に存在するかどうかは、更なる調査なしには明確でない。この種の生物活性は、商業的供給源の参照IL−17A(大腸菌においても産生される)(R&D Systems,Minneapolis,MN)の生物活性に類似しており、C10〜C106についての鎖間又は鎖内ジスルフィド結合がその受容体結合に重要でないことが示唆された。それにもかかわらず、現在の構造は、N末端セグメント(1〜20)及び(30〜39)は、非常に柔軟であり、それらの構造は、折り畳まれたIL−17A二量体の活性に影響を及ぼさない。
【0167】
エピトープ及びパラトープ
IL−17AとFab6468との間の結合に関与する残基を表10に列挙する。IL−17AのプロトマーAにおいて欠失した残基及びIL−17A二量体の若干の不斉性質に起因して、2つの接触部位からの全てのエピトープ残基は、同一でなかった(表10及び図7)。しかしながら、同一であるコアな一組の残基並びにそれらの相互作用が存在した。これらの残基は、IL−17A配列番号105のL26、R55、E57、P59、E60、R61、Y62、S64、V65、W67、R101、E102、P103、及びF110であり(表10に黒色で強調表示される)、それらはFab6468に対するコアエピトープを構成する。
【0168】
表10コアエピトープ残基は、黒色で強調表示される。コアパラトープ残基は、太字にされている。拡張されたパラトープ残基は、括弧内に示される。
【0169】
【表13】
【0170】
同様に、2つの部位における抗体からの接触残基は、完全に同一ではなかった。コアエピトープ残基への同一の接触に関与する残基は、「コアパラトープ」と称され、それは次の残基から成っていた:軽鎖(LC):Y31、D49、Y90、F92、F93(配列番号79);並びに重鎖(HC):S52、T54、F57、Y59、Q99、L100、及びT101(配列番号86)(表10)。コアパラトープ残基は、表10に太字で示される。特異的IL−17A残基に結合する1つの単量体において特定された更なる「拡張されたパラトープ」残基は、括弧内に示される。
【0171】
共結晶構造で特定されたL26を除く全てのコアエピトープ残基は、MORmAb7700についてのH/D交換、56NEDPERYPSVIWE68(配列番号157)及び100RREPPHCPNSFRLEKIL116(配列番号158)によって特定された、保護されたセグメントのうちの2つの境界内又は境界にあったため、MORmAb7700についてのH/D保護データは、共結晶研究に一致していた。MORmAb8302及びmAb1926を含むMORmAb7700抗体誘導体の全ては、それらが多くてもVHのN末端領域における1つの残基(実施例1を参照されたい)、VLのN末端における3つの残基(実施例1を参照されたい)、及び3つのCDR残基(H2、H3、及びL3において1つずつ、表1a)分だけ異なるのみで、それらのうちのいずれも抗体パラトープの一部でないため、Fab6468と同じ結合特異性を有することが想定される。
【0172】
本発明で特徴付けられるIL−17A構造は、以前に公開された構造に非常に類似しているが、欠失したセグメントに起因して、P2ポケット空洞(下記を参照されたい)が以前の研究(構造2VXS、Protein DataBank http_//www_rcsb_org/pdb/home/home_do;Gerhardt et al.,J.Mol.Biol.394:905〜21,2009において入手可能)では特定されなかったことを除く。
【0173】
IL−17RAとの複合体におけるヒトIL−17Fの結晶構造が報告されている(Ely et al.,Nat.Immunology,10:1245〜51,2009)。IL−17AとIL−17Fとの間の配列及び構造的類似性のため、IL−17Aは、IL−17Fと類似した様態でIL−17RAと相互作用する可能性が高い。この研究で得られたFab6468との複合体におけるIL−17A構造を、報告されたIL−17F/IL−17RA複合体におけるIL−17F上に重ねることによる分子モデリングにより、Fab6468のセグメントがIL−17RAとの立体障害を有するであろうことが示された。これらのセグメントのうちの1つは、Fab6468の軽鎖CDR3におけるFFモチーフの周囲に局在化する(配列番号79の残基92及び93)。したがって、特定の理論に決して束縛されるものではないが、Fab6468が、IL−17RA及び類推して、IL−17RCとの、その相互作用を遮断することによって、IL−17A機能を阻害することが示唆されるが、IL−17RCとIL−17Aとの間の相互作用の様式は、分子レベルでは知られていない。
【0174】
IL−17RAに対するIL−17A及びIL−17Fの親和性における有意な差異は、IL−17A及びIL−17RAの相互作用の詳細において有意な差異が存在し得ることを示唆するが、その程度は、IL−17A/IL−17RAの共結晶構造が決定されるとき初めて得られるであろう。これには、この研究におけるP2ポケット空洞の特定が関連するが、それは報告されたIL−17F/IL−17RA結晶構造におけるIL−17F類似領域において部分的に特定されているのみである(Ely et al.,Nat.Immunology,10:1245〜51,2009)。
【0175】
2つの深い、大部分が疎水性のポケットを、二量体境界面に沿ってIL−17Aの表面上で特定した(図8A、5B)。IL−17Fにおいて最初に発見された(Hymowitz et al.,EMBO J,20:5332〜41,2001)ポケットに類似したP1ポケットは、単量体Aの残基Q94、E95、L97、及びK114、並びに単量体BのL53、Y62、P63、V65、I66、W67、I96、V117、及びV119から成り、逆もまた同様である。二量体の一方の側で、P1ポケットは、セグメント30〜40によって部分的にカバーされる一方で、他方の側では、セグメントが無秩序であることに起因してそれは完全に開いている。このセグメントは柔軟であると考えられるため、他の分子がP1ポケットにアクセス可能であろう。P2ポケットもまた、単量体AのV24、L26、I28、Y62、L99、R101、F110、及びL112、並びに単量体BのV22、V24、及びL112の両方の鎖からの残基から成り、逆もまた同様である。
【0176】
P2ポケットの詳細は、上述のようにIL−17A二量体の不斉に起因して若干異なるが、2つのP2ポケットの全体的な形状は、非常に類似している。P1及びP2ポケットを裏打ちする2組の残基は、IL−17AとIL−17Fとの間で非常に良好に保存されている(図8C)。しかしながら、IL−17F構造において、P2ポケットは、残基F10及びF11(F10F11モチーフ)によって占領されている(図8B)(Hymowitz et al.,EMBO J,20:5332〜41,2001)。FFモチーフは、ヒトIL−17Aにおいて不在であり、代わりに、対応するアミノ酸残基は、I4及びP5(配列番号105における残基4及び5)である(図8C)。これらの残基は、それらがフェニルアラニン残基よりもはるかに小さいため、P2ポケット並びにFFモチーフにおいて結合する可能性が低く、P2ポケットに対して十分な親和性を有さない可能性が最も高い。したがって、IL−17FのFFモチーフは恐らく、受容体IL−17RA及びIL−17RCとのヒトIL−17A及びIL−17F相互作用についての構造的判別子である。これらの大部分の疎水性のポケット(P1及びP2)の両方は、IL−17RAへのIL−17A結合に必要とされる可能性が高い。IL−17F/IL−17RA複合体の近年の結晶構造は、FFモチーフがIL−17RAによって置換されていることを示す(Ely et al.,Nat.Immunology 10:1245〜51,2009)。P2からのFFモチーフ排除によるエネルギー損失は、より低い結合親和性をもたらす可能性が高い。これは、IL−17RAが、ヒトにおいて、IL−17Aに高い親和性で結合するが、IL−17Fには低い親和性で結合するという観察と一致し(Kuestner et al.,J Immunol,179:5462〜73,2007)、IL−17A及びIL−17F効力における差異を潜在的に説明し得る。マウスにおいて、FFモチーフは、IL−17A及びIL−17Fの両方で不在であり、それぞれ残基IP及びALによって置き換えられている。AL及びIP残基の対は、小さく、P2ポケットに対して低い親和性を有する可能性が高い。したがって、P2は、マウスIL−17A及びIL−17FにおいてIL−17RA結合のために有効であろう。マウスIL−17A及びIL−17Fの両方は、類似した親和性でマウスIL−17RAに結合し(Kuestner et al.,J Immunol,179:5462〜73,2007)、これは、結合のためのP2ポケットの有効性がリガンドの親和性を増加させるという本発明の示唆と一致する。
【0177】
全体的に、IL−17AとIL−17Fとの間で観察した構造的差異は、それぞれの受容体とのそれらの相互作用を詳細に分析するための基盤を提供する。更に、ペプチド、ペプチド模倣体、及び小分子がいずれか又は両方のポケットにおいて結合して、IL−17A及び/又はIL−17Fがそれらの受容体と相互作用することを遮断するように設計され得ることが考えられる。Fab6468(配列番号79における残基F92及びF93)に存在するFFモチーフは、P2ポケット残基L26、R61、L99、R101、及びR102に結合するため、Fab 6468構造を使用して、ファージディスプレイを使用して無作為化又は設計したペプチドライブラリーからのペプチド等の、更なるIL−17A拮抗物質を選択及び最適化することが可能である。
【0178】
P1及びP2ポケットを裏打ちする残基は、IL−17AとIL−17Fとの間で良好に保存されており、分子モデリングにより、IL−17Aホモ二量体のみと比較したとき、IL−17A/Fヘテロ二量体が、ほぼ同一の全体的構造を採用するであろうとことが示唆される。したがって、P1及びP2ポケットは、類似した全体的なトポロジーによりIL−17A/Fヘテロ二量体中に存在し、その受容体結合部位を構成する可能性が高い。したがって、P2ポケット残基に結合するIL−17A拮抗物質は、IL−17A/Fヘテロ二量体に結合し、拮抗する可能性がある。
【0179】
(実施例6)
交差種結合特異性
mAbtr1926の交差種結合特異性を評価するために、マイクロタイタープレート上にコーティングした異なるIL−17Aタンパク質を用いて結合ELISAを行った。ヒト、マウス、及びラットIL−17Aタンパク質をマイクロタイタープレート上にコーティングした。標識化されたmAb1926の連続希釈液を37℃で2時間インキュベートした。インキュベーション後、マイクロタイタープレートを完全に洗浄し、結合した標識化mAb1926を検出した。mAb1926は、ラット又はマウスIL−17Aタンパク質よりもはるかに強力に、ヒトIL−17Aに結合した(図9)。ラット及びマウスIL−17Aへのこの低減された結合は、タンパク質の両方がFab6468拡張型エピトープの7位においてヒトIL−17Aから異なることと一致する(表10)。更に、Fab6468エピトープの一部に近い位置である、ヒトIL−17Aの残基40と41との間のラット及びマウスIL−17Aにおいて、1つのアミノ酸挿入が存在する。
【図1A】
【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図2A】
【図2B】
【図3A−1】
【図3A−2】
【図3B−1】
【図3B−2】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【特許請求の範囲】
【請求項1】
単離された抗体又はそのフラグメントであって、前記抗体が、アミノ酸残基56〜68(配列番号157)及び100〜116(配列番号158)において、又は残基L26、R55、E57、P59、E60、R61、Y62、S64、V65、W67、R101、E102、P103、及びF110において、配列番号105に示される配列を有するヒトIL−17Aに特異的に結合する、単離された抗体又はそのフラグメント。
【請求項2】
単離された抗体又はそのフラグメントであって、前記抗体が、ヒトIL−17A上のP2ポケット空洞に特異的に結合し、前記P2ポケット空洞が、配列番号105のアミノ酸残基V22、V24、L26、I28、Y62、L99、R101、F110、及びL112を含む、単離された抗体又はそのフラグメント。
【請求項3】
それぞれ配列番号25、43、及び60に示される重鎖CDR1、2、及び3(HCDR1、HCDR2、及びHCDR3)アミノ酸配列、並びにそれぞれ配列番号3、6、及び18に示される軽鎖CDR1、2、及び3(LCDR1、LCDR2、及びLCDR3)アミノ酸配列を含むモノクローナル抗体と、ヒトIL−17A結合をめぐって競合する、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項4】
配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する、重鎖可変領域パラトープアミノ酸残基を含み、
a.ヒトIL−17AのR55又はE57と相互作用する第1のスレオニン残基と、
b.ヒトIL−17AのR55又はE57と相互作用するグルタミン残基と、
c.ヒトIL−17AのE57と相互作用するリジン残基と、
d.ヒトIL−17AのP59、E60、又はR101と相互作用するチロシン残基と、
e.ヒトIL−17AのE60、R101、E102、又はP103と相互作用するフェニルアラニン残基と、
f.ヒトIL−17AのE60と相互作用するセリン残基と、
g.ヒトIL−17AのE60と相互作用する第2のスレオニン残基と、を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項5】
配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する、軽鎖可変領域パラトープアミノ酸残基を含み、
a.ヒトIL−17AのL26と相互作用する第1のフェニルアラニン残基と、
b.ヒトIL−17AのR55又はW67と相互作用するアスパラギン酸残基と、
c.ヒトIL−17AのP59、S64、又はR101と相互作用する第1のチロシン残基と、
d.ヒトIL−17AのP59、E60、R61、Y62、R101、又はF110と相互作用する第2のフェニルアラニン残基と、
e.ヒトIL−17AのV65と相互作用する第2のチロシン残基と、を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項6】
請求項4に記載の重鎖可変領域パラトープ及び請求項5に記載の軽鎖可変領域パラトープを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項7】
重鎖可変領域及び軽鎖可変領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.Chothia残基S51、T53、F56、Y58、Q95、L96、及びT97から選択される重鎖可変領域パラトープか、
b.Chothia残基Y32、D50、Y91、F93、及びF94から選択される軽鎖可変領域パラトープか、
c.Chothia残基S51、T53、F56、Y58、Q95、L96、及びT97から選択される重鎖可変領域パラトープと、
d.Chothia残基Y32、D50、Y91、F93、及びF94から選択される軽鎖可変領域パラトープと、
を含む、単離された抗体又はフラグメント。
【請求項8】
配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する、重鎖可変領域パラトープアミノ酸残基及び軽鎖可変領域パラトープアミノ酸残基を含み、
a.ヒトIL−17AのR101と相互作用する前記重鎖可変領域におけるチロシン残基と、
b.ヒトIL−17AのR101と相互作用する前記重鎖可変領域におけるフェニルアラニン残基と、
c.ヒトIL−17AのY62及びR101と相互作用する前記軽鎖可変領域における第1のフェニルアラニン残基と、
d.ヒトIL−17AのL26及びF110と相互作用する前記軽鎖可変領域における第2のフェニルアラニン残基と、
e.ヒトIL−17AのR101と相互作用する前記軽鎖可変領域におけるチロシン残基と、を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項9】
重鎖可変領域及び軽鎖可変領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.Chothia残基F56及びY58から選択される重鎖可変領域パラトープと、
b.Chothia残基Y91、F93、及びF94から選択される軽鎖可変領域パラトープと、
を含む、単離された抗体又はフラグメント。
【請求項10】
重鎖可変領域(VH)及び軽鎖可変領域(VL)を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はそのフラグメントであって、前記抗体が、
a.それぞれ配列番号23、35、及び52に示される重鎖相補性決定領域(CDR)1、2、及び3(HCDR1、HCDR2、HCDR3)アミノ酸配列であって、配列番号35の前記HCDR2が更に、式(I):
Xaa1−I−I−P−W−F−G−Xaa2−T−Xaa3−Y−A−Q−K−F−Q−G、(I)に示すように記載され、
式中、
Xaa1が、His、Met、Arg、Ser、又はTyrであり得、
Xaa2が、Trp、Thr、又はTyrであり得、
Xaa3が、Tyr、Phe、Ser、又はAspであり得る、アミノ酸配列か、
b.それぞれ配列番号23、35、及び52に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号35の前記HCDR2が更に、式(I):
Xaa1−I−I−P−W−F−G−Xaa2−T−Xaa3−Y−A−Q−K−F−Q−G、(I)に示すように記載され、
式中、
Xaa1が、His、Met、Arg、Ser、又はTyrであり得、
Xaa2が、Trp、Thr、又はTyrであり得、
Xaa3が、Tyr、Phe、Ser、又はAspであり得る、アミノ酸配列、及び
それぞれ配列番号1、4、及び7に示される軽鎖相補性決定領域(CDR)1、2、及び3(LCDR1、LCDR2、LCDR3)アミノ酸配列か、
c.それぞれ配列番号2、5、及び11に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号11の前記LCDR3が更に、式(II):
Xaa4−Q−Xaa5−Xaa6−Xaa7−Xaa8−Xaa9−Xaa10、(II)に示すように記載され、
式中、
Xaa4が、His又はGlnであり得、
Xaa5が、Phe又はGlyであり得、
Xaa6が、Thr、Val、又はAsnであり得、
Xaa7が、Ile、Thr、又はTyrであり得、
Xaa8が、Pro又はArgであり得、
Xaa9が、Ser又はProであり得、
Xaa10が、His、Phe、又はLeuであり得る、アミノ酸配列か、
d.それぞれ配列番号2、5、及び11に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号11の前記LCDR3が更に、式(II):
Xaa4−Q−Xaa5−Xaa6−Xaa7−Xaa8−Xaa9−Xaa10、(II)に示すように記載され、
式中、
Xaa4が、His又はGlnであり得、
Xaa5が、Phe又はGlyであり得、
Xaa6が、Thr、Val、又はAsnであり得、
Xaa7が、Ile、Thr、又はTyrであり得、
Xaa8が、Pro又はArgであり得、
Xaa9が、Ser又はProであり得、
Xaa10が、His、Phe、又はLeuであり得る、アミノ酸配列、及び
それぞれ配列番号24、36、及び53に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列か、
e.それぞれ配列番号24、36、及び57に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号57の前記HCDR3が更に、式(IV):
E−V−D−S−Xaa19−Y−Y−S−Y−F−D−I、(IV)に示すように記載され、
式中、
Xaa19が、Met、Ile、Leu、又はThrである、アミノ酸配列か、
f.それぞれ配列番号2、5、及び17に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号17の前記LCDR3が更に、式(III):
Xaa11−Q−Xaa12−Xaa13−Xaa14−Xaa15−Xaa16−Xaa17−Xaa18−T、(III)に示すように記載され、
式中、
Xaa11が、Gln又はThrであり得、
Xaa12が、Ser又はTyrであり得、
Xaa13が、Asn、Arg、Val、又はTyrであり得、
Xaa14が、His又はSerであり得、
Xaa15が、Ile、Thr、Leu、Ala、又はSerであり得、
Xaa16が、Pro、Leu、又はSerであり得、
Xaa17が、Pro、Ser、Phe、又はLeuであり得、
Xaa18が、Ala、Leu、又はAspであり得る、アミノ酸配列か、
g.それぞれ配列番号24、36、及び57に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号57の前記HCDR3が更に、式(IV):
E−V−D−S−Xaa19−Y−Y−S−Y−F−D−I、(IV)に示すように記載され、
式中、
Xaa19が、Met、Ile、Leu、又はThrである、アミノ酸配列、及び
それぞれ配列番号2、5、及び17に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号17の前記LCDR3が更に、式(III):
Xaa11−Q−Xaa12−Xaa13−Xaa14−Xaa15−Xaa16−Xaa17−Xaa18−T、(III)に示すように記載され、
式中、
Xaa11が、Gln又はThrであり得、
Xaa12が、Ser又はTyrであり得、
Xaa13が、Asn、Arg、Val、又はTyrであり得、
Xaa14が、His又はSerであり得、
Xaa15が、Ile、Thr、Leu、Ala、又はSerであり得、
Xaa16が、Pro、Leu、又はSerであり得、
Xaa17が、Pro、Ser、Phe、又はLeuであり得、
Xaa18が、Ala、Leu、又はAspであり得る、アミノ酸配列か、
h.それぞれ配列番号25、46、及び61に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号46の前記HCDR2が更に、式(VI):
Xaa21−I−Xaa22−Xaa23−Xaa24−Xaa25−Xaa26−Xaa27−Xaa28−Xaa29−Y−A−D−S−V−K−G、(VI)に示すように記載され、
式中、
Xaa21が、Ala、Gly、Thr、又はValであり得、
Xaa22が、Asn又はSerであり得、
Xaa23が、Gly、Met、Lys、Ile、Leu、又はHisであり得、
Xaa24が、Leu、Asp、Ala、His、Thr、Gly又はSerであり得;
Xaa25が、Gly又はSerであり得、
Xaa26が、Thr、Gly、Tyr、又はAspであり得、
Xaa27が、His、Trp、Tyr、又はPheであり得、
Xaa28が、Lys、Thr、又はIleであり得、
Xaa29が、Tyr、Phe、又はAsnであり得、
配列番号61の前記HCDR3が更に、式(VII):
Q−L−Xaa30−L−D−V、(VII)に示すように記載され、
式中、
Xaa30が、Met、Leu、又はThrであり得る、アミノ酸配列か、
i.それぞれ配列番号3、6、及び22に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号22の前記LCDR3が更に、式(V):
G−S−Y−D−F−F−L−G−Xaa20−I−V、(V)に示すように記載され、
式中、
Xaa20が、Met、Leu、Thr、又はTyrである、アミノ酸配列か、
j.それぞれ配列番号25、46、及び61に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号46の前記HCDR2が更に、式(VI):
Xaa21−I−Xaa22−Xaa23−Xaa24−Xaa25−Xaa26−Xaa27−Xaa28−Xaa29−Y−A−D−S−V−K−G、(VI)に示すように記載され、
式中、
Xaa21が、Ala、Gly、Thr、又はValであり得、
Xaa22が、Asn又はSerであり得、
Xaa23が、Gly、Met、Lys、Ile、Leu、又はHisであり得、
Xaa24が、Leu、Asp、Ala、His、Thr、Gly、又はSerであり得、
Xaa25が、Gly又はSerであり得、
Xaa26が、Thr、Gly、Tyr、又はAspであり得、
Xaa27が、His、Trp、Tyr、又はPheであり得、
Xaa28が、Lys、Thr、又はIleであり得、
Xaa29が、Tyr、Phe、又はAsnであり得、
配列番号61のHCDR3が更に、式(VII):
Q−L−Xaa30−L−D−V、(VII)に示すように記載され、
式中、
Xaa30が、Met、Leu、又はThrであり得る、アミノ酸配列、及び
それぞれ配列番号3、6、及び22に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号22の前記LCDR3が更に、式(V):
G−S−Y−D−F−F−L−G−Xaa20−I−V、(V)に示すように記載され、
式中、
Xaa20が、Met、Leu、Thr、又はTyrである、アミノ酸配列か、
k.それぞれ配列番号25、51、及び58に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号51の前記HCDR2が更に、式(VIII):
V−T−S−Xaa31−Xaa32−Xaa33−Xaa34−T−Y−Y−A−Xaa35−S−V−K−G、(VIII)に示すように記載され、
式中、
Xaa31が、Ala、Lys、Met、又はHisであり得、
Xaa32が、Asn、Met、Thr、又はArgであり得、
Xaa33が、Gly又はAspであり得、
Xaa34が、Arg、His、又はAsnであり得、
Xaa35が、Asp又はGlyであり得る、アミノ酸配列か、
l.それぞれ配列番号25、51、及び58に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号51の前記HCDR2が更に、式(VIII):
V−T−S−Xaa31−Xaa32−Xaa33−Xaa34−T−Y−Y−A−Xaa35−S−V−K−G、(VIII)に示すように記載され、
式中、
Xaa31が、Ala、Lys、Met、又はHisであり得、
Xaa32が、Asn、Met、Thr、又はArgであり得、
Xaa33が、Gly又はAspであり得、
Xaa34が、Arg、His、又はAsnであり得、
Xaa35が、Asp又はGlyであり得る、アミノ酸配列、及び
それぞれ配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列、を含む、単離された抗体又はそのフラグメント。
【請求項11】
VH及びVL領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はそのフラグメントであって、前記抗体が、
a.それぞれ配列番号23、26、及び52か、
b.それぞれ配列番号23、27、及び52か、
c.それぞれ配列番号23、28、及び52か、
d.それぞれ配列番号23、29、及び52か、
e.それぞれ配列番号23、30、及び52か、
f.それぞれ配列番号23、31、及び52か、
g.それぞれ配列番号23、32、及び52か、
h.それぞれ配列番号23、33、及び52か
i.それぞれ配列番号23、34、及び52か、に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はそのフラグメント。
【請求項12】
前記抗体が更に、それぞれ配列番号1、4、及び7に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、請求項11に記載の単離された抗体又はフラグメント。
【請求項13】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、それぞれ配列番号1、4、及び7に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項14】
Vh及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号2、5、及び8か、
b.それぞれ配列番号2、5、及び9か、
c.それぞれ配列番号2、5、及び10か、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項15】
請求項14に記載の単離された抗体又はフラグメントであって、前記抗体が更に、それぞれ配列番号24、36、及び53に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項16】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号24、36、及び53か、
b.それぞれ配列番号24、36、及び54か、
c.それぞれ配列番号24、36、及び55か、
d.それぞれ配列番号24、36、及び56か、に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項17】
前記抗体が更に、
a.それぞれ配列番号2、5、及び12か、
b.それぞれ配列番号2、5、及び13か、
c.それぞれ配列番号2、5、及び14か、
d.それぞれ配列番号2、5、及び15か、
e.それぞれ配列番号2、5、及び16か、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、請求項16に記載の単離された抗体又はフラグメント。
【請求項18】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号2、5、及び12、
b.それぞれ配列番号2、5、及び13、
c.それぞれ配列番号2、5、及び14、
d.それぞれ配列番号2、5、及び15、
e.それぞれ配列番号2、5、及び16、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項19】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号25、37、及び58か、
b.それぞれ配列番号25、38、及び58か、
c.それぞれ配列番号25、39、及び58か、
d.それぞれ配列番号25、40、及び58か、
e.それぞれ配列番号25、41、及び58か、
f.それぞれ配列番号25、42、及び58か、
g.それぞれ配列番号25、43、及び58か、
h.それぞれ配列番号25、41、及び59か、
i.それぞれ配列番号25、41、及び60か、
j.それぞれ配列番号25、43、及び60か、
k.それぞれ配列番号25、44、及び58か、
l.それぞれ配列番号25、45、及び58か、に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項20】
前記抗体が更に、
a.それぞれ配列番号3、6、及び18と、
b.それぞれ配列番号3、6、及び19と、
c.それぞれ配列番号3、6、及び20と、
d.それぞれ配列番号3、6、及び21と、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、請求項19に記載の単離された抗体又はフラグメント。
【請求項21】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号3、6、及び18か、
b.それぞれ配列番号3、6、及び19か、
c.それぞれ配列番号3、6、及び20か、
d.それぞれ配列番号3、6、及び21か、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項22】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号25、47、及び58か、
b.それぞれ配列番号25、48、及び58か、
c.それぞれ配列番号25、49、及び58か、
d.それぞれ配列番号25、50、及び58か、に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項23】
前記抗体が更に、それぞれ配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、請求項22に記載の単離された抗体又はフラグメント。
【請求項24】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号25、43、及び60に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列か、
b.それぞれ配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列か、
c.それぞれ配列番号25、43、及び60に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列と、
それぞれ配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列か、
d.配列番号86のVHか、
e.配列番号79のVLか、
f.配列番号86のVHと、
配列番号79のVLと、を含む、単離された抗体又はフラグメント。
【請求項25】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、配列番号67、68、69、81、82、83、84、85、又は86に示されるアミノ酸配列を有する前記VHを含む、単離された抗体又はフラグメント。
【請求項26】
前記抗体が更に、配列番号76、77、78、79、又は80に示されるアミノ酸配列を有する前記VLを含む、請求項25に記載の単離された抗体又はフラグメント。
【請求項27】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、配列番号76、77、78、79、又は80に示されるアミノ酸配列を有する前記VLを含む、単離された抗体又はフラグメント。
【請求項28】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、配列番号67、68、69、81、82、83、84、85、又は86に示されるアミノ酸配列を有するVHと少なくとも90%同一である前記VHを含む、単離された抗体又はフラグメント。
【請求項29】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、配列番号76、77、78、79、又は80に示されるアミノ酸配列を有する可変領域と少なくとも90%同一である前記VLを含む、単離された抗体又はフラグメント。
【請求項30】
配列番号92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を有する抗体重鎖を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項31】
配列番号87、88、89、90、又は91に示されるアミノ酸配列を有する抗体軽鎖を含む、IL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項32】
前記抗体が完全なヒト型(human)である、請求項3又は7に記載の単離された抗体又はフラグメント。
【請求項33】
前記抗体がポリエチレングリコールと結合されている、請求項3又は7に記載の単離された抗体又はフラグメント。
【請求項34】
IgG1又はIgG4アイソタイプを有する、請求項3又は7に記載の単離された抗体又はフラグメント。
【請求項35】
前記Fcドメインが、前記FcドメインにおけるS229P、P235A、又はL236A突然変異を含む、請求項3又は7に記載の単離された抗体又はフラグメント。
【請求項36】
請求項3又は7に記載の単離された抗体又はフラグメント及び製薬上許容され得る担体を含む、医薬組成物。
【請求項37】
配列番号67、68、69、81、82、83、84、85、86、92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を含む、単離された抗体重鎖。
【請求項38】
配列番号76、77、78、79、80、87、88、89、90、又は91に示されるアミノ酸配列を含む、単離された抗体軽鎖。
【請求項39】
配列番号67、68、69、81、82、83、84、85、86、92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を含む抗体重鎖をコードする、単離されたポリヌクレオチド。
【請求項40】
配列番号76、77、78、79、80、87、88、89、90、又は91に示されるアミノ酸配列を含む抗体軽鎖をコードする、単離されたポリヌクレオチド。
【請求項41】
請求項39又は40に記載の少なくとも1つのポリヌクレオチドを含む、ベクター。
【請求項42】
請求項40に記載のベクターを含む、宿主細胞。
【請求項43】
a.ヒトIL−17A及びIL−17RAを提供する工程と、
b.ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112から成る群から選択される少なくとも1つのアミノ酸残基におけるヒトIL−17Aに結合する拮抗物質と接触させる工程と、を含む、ヒトIL−17AのIL−17RAとの相互作用を阻害する方法。
【請求項44】
a.ヒトIL17−A及びIL−17RAを提供する工程と、
b.ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112から成る群から選択される少なくとも1つのアミノ酸残基におけるヒトIL−17Aに結合する拮抗物質と接触させる工程と、を含む、IL−17A生物活性を阻害する方法。
【請求項45】
前記拮抗物質が、抗体、ペプチド、又は小分子である、請求項43又は44に記載の方法。
【請求項46】
請求項3又は7に記載の単離された抗体の治療上の有効量を、炎症性状態の治療を必要とする患者に炎症性状態を治療するうえで充分な時間にわたって投与する工程を含む、炎症性状態を治療する方法。
【請求項47】
前記炎症性状態が、気道、肺、胃腸管、小腸、大腸、結腸、直腸、関節、骨及び滑膜組織、軟骨、上皮、内皮、肝組織、又は皮膚を侵す、請求項46に記載の方法。
【請求項48】
前記炎症性状態が、炎症性肺状態、ぜんそく、慢性閉塞性肺疾患(COPD)、炎症性腸疾患、自己免疫疾患、関節リウマチ、乾癬、又は全身性硬化症である、請求項46に記載の方法。
【請求項1】
単離された抗体又はそのフラグメントであって、前記抗体が、アミノ酸残基56〜68(配列番号157)及び100〜116(配列番号158)において、又は残基L26、R55、E57、P59、E60、R61、Y62、S64、V65、W67、R101、E102、P103、及びF110において、配列番号105に示される配列を有するヒトIL−17Aに特異的に結合する、単離された抗体又はそのフラグメント。
【請求項2】
単離された抗体又はそのフラグメントであって、前記抗体が、ヒトIL−17A上のP2ポケット空洞に特異的に結合し、前記P2ポケット空洞が、配列番号105のアミノ酸残基V22、V24、L26、I28、Y62、L99、R101、F110、及びL112を含む、単離された抗体又はそのフラグメント。
【請求項3】
それぞれ配列番号25、43、及び60に示される重鎖CDR1、2、及び3(HCDR1、HCDR2、及びHCDR3)アミノ酸配列、並びにそれぞれ配列番号3、6、及び18に示される軽鎖CDR1、2、及び3(LCDR1、LCDR2、及びLCDR3)アミノ酸配列を含むモノクローナル抗体と、ヒトIL−17A結合をめぐって競合する、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項4】
配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する、重鎖可変領域パラトープアミノ酸残基を含み、
a.ヒトIL−17AのR55又はE57と相互作用する第1のスレオニン残基と、
b.ヒトIL−17AのR55又はE57と相互作用するグルタミン残基と、
c.ヒトIL−17AのE57と相互作用するリジン残基と、
d.ヒトIL−17AのP59、E60、又はR101と相互作用するチロシン残基と、
e.ヒトIL−17AのE60、R101、E102、又はP103と相互作用するフェニルアラニン残基と、
f.ヒトIL−17AのE60と相互作用するセリン残基と、
g.ヒトIL−17AのE60と相互作用する第2のスレオニン残基と、を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項5】
配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する、軽鎖可変領域パラトープアミノ酸残基を含み、
a.ヒトIL−17AのL26と相互作用する第1のフェニルアラニン残基と、
b.ヒトIL−17AのR55又はW67と相互作用するアスパラギン酸残基と、
c.ヒトIL−17AのP59、S64、又はR101と相互作用する第1のチロシン残基と、
d.ヒトIL−17AのP59、E60、R61、Y62、R101、又はF110と相互作用する第2のフェニルアラニン残基と、
e.ヒトIL−17AのV65と相互作用する第2のチロシン残基と、を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項6】
請求項4に記載の重鎖可変領域パラトープ及び請求項5に記載の軽鎖可変領域パラトープを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項7】
重鎖可変領域及び軽鎖可変領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.Chothia残基S51、T53、F56、Y58、Q95、L96、及びT97から選択される重鎖可変領域パラトープか、
b.Chothia残基Y32、D50、Y91、F93、及びF94から選択される軽鎖可変領域パラトープか、
c.Chothia残基S51、T53、F56、Y58、Q95、L96、及びT97から選択される重鎖可変領域パラトープと、
d.Chothia残基Y32、D50、Y91、F93、及びF94から選択される軽鎖可変領域パラトープと、
を含む、単離された抗体又はフラグメント。
【請求項8】
配列番号105に示されるアミノ酸配列を有するヒトIL−17Aの残基と相互作用する、重鎖可変領域パラトープアミノ酸残基及び軽鎖可変領域パラトープアミノ酸残基を含み、
a.ヒトIL−17AのR101と相互作用する前記重鎖可変領域におけるチロシン残基と、
b.ヒトIL−17AのR101と相互作用する前記重鎖可変領域におけるフェニルアラニン残基と、
c.ヒトIL−17AのY62及びR101と相互作用する前記軽鎖可変領域における第1のフェニルアラニン残基と、
d.ヒトIL−17AのL26及びF110と相互作用する前記軽鎖可変領域における第2のフェニルアラニン残基と、
e.ヒトIL−17AのR101と相互作用する前記軽鎖可変領域におけるチロシン残基と、を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項9】
重鎖可変領域及び軽鎖可変領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.Chothia残基F56及びY58から選択される重鎖可変領域パラトープと、
b.Chothia残基Y91、F93、及びF94から選択される軽鎖可変領域パラトープと、
を含む、単離された抗体又はフラグメント。
【請求項10】
重鎖可変領域(VH)及び軽鎖可変領域(VL)を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はそのフラグメントであって、前記抗体が、
a.それぞれ配列番号23、35、及び52に示される重鎖相補性決定領域(CDR)1、2、及び3(HCDR1、HCDR2、HCDR3)アミノ酸配列であって、配列番号35の前記HCDR2が更に、式(I):
Xaa1−I−I−P−W−F−G−Xaa2−T−Xaa3−Y−A−Q−K−F−Q−G、(I)に示すように記載され、
式中、
Xaa1が、His、Met、Arg、Ser、又はTyrであり得、
Xaa2が、Trp、Thr、又はTyrであり得、
Xaa3が、Tyr、Phe、Ser、又はAspであり得る、アミノ酸配列か、
b.それぞれ配列番号23、35、及び52に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号35の前記HCDR2が更に、式(I):
Xaa1−I−I−P−W−F−G−Xaa2−T−Xaa3−Y−A−Q−K−F−Q−G、(I)に示すように記載され、
式中、
Xaa1が、His、Met、Arg、Ser、又はTyrであり得、
Xaa2が、Trp、Thr、又はTyrであり得、
Xaa3が、Tyr、Phe、Ser、又はAspであり得る、アミノ酸配列、及び
それぞれ配列番号1、4、及び7に示される軽鎖相補性決定領域(CDR)1、2、及び3(LCDR1、LCDR2、LCDR3)アミノ酸配列か、
c.それぞれ配列番号2、5、及び11に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号11の前記LCDR3が更に、式(II):
Xaa4−Q−Xaa5−Xaa6−Xaa7−Xaa8−Xaa9−Xaa10、(II)に示すように記載され、
式中、
Xaa4が、His又はGlnであり得、
Xaa5が、Phe又はGlyであり得、
Xaa6が、Thr、Val、又はAsnであり得、
Xaa7が、Ile、Thr、又はTyrであり得、
Xaa8が、Pro又はArgであり得、
Xaa9が、Ser又はProであり得、
Xaa10が、His、Phe、又はLeuであり得る、アミノ酸配列か、
d.それぞれ配列番号2、5、及び11に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号11の前記LCDR3が更に、式(II):
Xaa4−Q−Xaa5−Xaa6−Xaa7−Xaa8−Xaa9−Xaa10、(II)に示すように記載され、
式中、
Xaa4が、His又はGlnであり得、
Xaa5が、Phe又はGlyであり得、
Xaa6が、Thr、Val、又はAsnであり得、
Xaa7が、Ile、Thr、又はTyrであり得、
Xaa8が、Pro又はArgであり得、
Xaa9が、Ser又はProであり得、
Xaa10が、His、Phe、又はLeuであり得る、アミノ酸配列、及び
それぞれ配列番号24、36、及び53に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列か、
e.それぞれ配列番号24、36、及び57に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号57の前記HCDR3が更に、式(IV):
E−V−D−S−Xaa19−Y−Y−S−Y−F−D−I、(IV)に示すように記載され、
式中、
Xaa19が、Met、Ile、Leu、又はThrである、アミノ酸配列か、
f.それぞれ配列番号2、5、及び17に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号17の前記LCDR3が更に、式(III):
Xaa11−Q−Xaa12−Xaa13−Xaa14−Xaa15−Xaa16−Xaa17−Xaa18−T、(III)に示すように記載され、
式中、
Xaa11が、Gln又はThrであり得、
Xaa12が、Ser又はTyrであり得、
Xaa13が、Asn、Arg、Val、又はTyrであり得、
Xaa14が、His又はSerであり得、
Xaa15が、Ile、Thr、Leu、Ala、又はSerであり得、
Xaa16が、Pro、Leu、又はSerであり得、
Xaa17が、Pro、Ser、Phe、又はLeuであり得、
Xaa18が、Ala、Leu、又はAspであり得る、アミノ酸配列か、
g.それぞれ配列番号24、36、及び57に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号57の前記HCDR3が更に、式(IV):
E−V−D−S−Xaa19−Y−Y−S−Y−F−D−I、(IV)に示すように記載され、
式中、
Xaa19が、Met、Ile、Leu、又はThrである、アミノ酸配列、及び
それぞれ配列番号2、5、及び17に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号17の前記LCDR3が更に、式(III):
Xaa11−Q−Xaa12−Xaa13−Xaa14−Xaa15−Xaa16−Xaa17−Xaa18−T、(III)に示すように記載され、
式中、
Xaa11が、Gln又はThrであり得、
Xaa12が、Ser又はTyrであり得、
Xaa13が、Asn、Arg、Val、又はTyrであり得、
Xaa14が、His又はSerであり得、
Xaa15が、Ile、Thr、Leu、Ala、又はSerであり得、
Xaa16が、Pro、Leu、又はSerであり得、
Xaa17が、Pro、Ser、Phe、又はLeuであり得、
Xaa18が、Ala、Leu、又はAspであり得る、アミノ酸配列か、
h.それぞれ配列番号25、46、及び61に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号46の前記HCDR2が更に、式(VI):
Xaa21−I−Xaa22−Xaa23−Xaa24−Xaa25−Xaa26−Xaa27−Xaa28−Xaa29−Y−A−D−S−V−K−G、(VI)に示すように記載され、
式中、
Xaa21が、Ala、Gly、Thr、又はValであり得、
Xaa22が、Asn又はSerであり得、
Xaa23が、Gly、Met、Lys、Ile、Leu、又はHisであり得、
Xaa24が、Leu、Asp、Ala、His、Thr、Gly又はSerであり得;
Xaa25が、Gly又はSerであり得、
Xaa26が、Thr、Gly、Tyr、又はAspであり得、
Xaa27が、His、Trp、Tyr、又はPheであり得、
Xaa28が、Lys、Thr、又はIleであり得、
Xaa29が、Tyr、Phe、又はAsnであり得、
配列番号61の前記HCDR3が更に、式(VII):
Q−L−Xaa30−L−D−V、(VII)に示すように記載され、
式中、
Xaa30が、Met、Leu、又はThrであり得る、アミノ酸配列か、
i.それぞれ配列番号3、6、及び22に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号22の前記LCDR3が更に、式(V):
G−S−Y−D−F−F−L−G−Xaa20−I−V、(V)に示すように記載され、
式中、
Xaa20が、Met、Leu、Thr、又はTyrである、アミノ酸配列か、
j.それぞれ配列番号25、46、及び61に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号46の前記HCDR2が更に、式(VI):
Xaa21−I−Xaa22−Xaa23−Xaa24−Xaa25−Xaa26−Xaa27−Xaa28−Xaa29−Y−A−D−S−V−K−G、(VI)に示すように記載され、
式中、
Xaa21が、Ala、Gly、Thr、又はValであり得、
Xaa22が、Asn又はSerであり得、
Xaa23が、Gly、Met、Lys、Ile、Leu、又はHisであり得、
Xaa24が、Leu、Asp、Ala、His、Thr、Gly、又はSerであり得、
Xaa25が、Gly又はSerであり得、
Xaa26が、Thr、Gly、Tyr、又はAspであり得、
Xaa27が、His、Trp、Tyr、又はPheであり得、
Xaa28が、Lys、Thr、又はIleであり得、
Xaa29が、Tyr、Phe、又はAsnであり得、
配列番号61のHCDR3が更に、式(VII):
Q−L−Xaa30−L−D−V、(VII)に示すように記載され、
式中、
Xaa30が、Met、Leu、又はThrであり得る、アミノ酸配列、及び
それぞれ配列番号3、6、及び22に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列であって、配列番号22の前記LCDR3が更に、式(V):
G−S−Y−D−F−F−L−G−Xaa20−I−V、(V)に示すように記載され、
式中、
Xaa20が、Met、Leu、Thr、又はTyrである、アミノ酸配列か、
k.それぞれ配列番号25、51、及び58に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号51の前記HCDR2が更に、式(VIII):
V−T−S−Xaa31−Xaa32−Xaa33−Xaa34−T−Y−Y−A−Xaa35−S−V−K−G、(VIII)に示すように記載され、
式中、
Xaa31が、Ala、Lys、Met、又はHisであり得、
Xaa32が、Asn、Met、Thr、又はArgであり得、
Xaa33が、Gly又はAspであり得、
Xaa34が、Arg、His、又はAsnであり得、
Xaa35が、Asp又はGlyであり得る、アミノ酸配列か、
l.それぞれ配列番号25、51、及び58に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列であって、配列番号51の前記HCDR2が更に、式(VIII):
V−T−S−Xaa31−Xaa32−Xaa33−Xaa34−T−Y−Y−A−Xaa35−S−V−K−G、(VIII)に示すように記載され、
式中、
Xaa31が、Ala、Lys、Met、又はHisであり得、
Xaa32が、Asn、Met、Thr、又はArgであり得、
Xaa33が、Gly又はAspであり得、
Xaa34が、Arg、His、又はAsnであり得、
Xaa35が、Asp又はGlyであり得る、アミノ酸配列、及び
それぞれ配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列、を含む、単離された抗体又はそのフラグメント。
【請求項11】
VH及びVL領域を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はそのフラグメントであって、前記抗体が、
a.それぞれ配列番号23、26、及び52か、
b.それぞれ配列番号23、27、及び52か、
c.それぞれ配列番号23、28、及び52か、
d.それぞれ配列番号23、29、及び52か、
e.それぞれ配列番号23、30、及び52か、
f.それぞれ配列番号23、31、及び52か、
g.それぞれ配列番号23、32、及び52か、
h.それぞれ配列番号23、33、及び52か
i.それぞれ配列番号23、34、及び52か、に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はそのフラグメント。
【請求項12】
前記抗体が更に、それぞれ配列番号1、4、及び7に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、請求項11に記載の単離された抗体又はフラグメント。
【請求項13】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、それぞれ配列番号1、4、及び7に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項14】
Vh及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号2、5、及び8か、
b.それぞれ配列番号2、5、及び9か、
c.それぞれ配列番号2、5、及び10か、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項15】
請求項14に記載の単離された抗体又はフラグメントであって、前記抗体が更に、それぞれ配列番号24、36、及び53に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項16】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号24、36、及び53か、
b.それぞれ配列番号24、36、及び54か、
c.それぞれ配列番号24、36、及び55か、
d.それぞれ配列番号24、36、及び56か、に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項17】
前記抗体が更に、
a.それぞれ配列番号2、5、及び12か、
b.それぞれ配列番号2、5、及び13か、
c.それぞれ配列番号2、5、及び14か、
d.それぞれ配列番号2、5、及び15か、
e.それぞれ配列番号2、5、及び16か、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、請求項16に記載の単離された抗体又はフラグメント。
【請求項18】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号2、5、及び12、
b.それぞれ配列番号2、5、及び13、
c.それぞれ配列番号2、5、及び14、
d.それぞれ配列番号2、5、及び15、
e.それぞれ配列番号2、5、及び16、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項19】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号25、37、及び58か、
b.それぞれ配列番号25、38、及び58か、
c.それぞれ配列番号25、39、及び58か、
d.それぞれ配列番号25、40、及び58か、
e.それぞれ配列番号25、41、及び58か、
f.それぞれ配列番号25、42、及び58か、
g.それぞれ配列番号25、43、及び58か、
h.それぞれ配列番号25、41、及び59か、
i.それぞれ配列番号25、41、及び60か、
j.それぞれ配列番号25、43、及び60か、
k.それぞれ配列番号25、44、及び58か、
l.それぞれ配列番号25、45、及び58か、に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項20】
前記抗体が更に、
a.それぞれ配列番号3、6、及び18と、
b.それぞれ配列番号3、6、及び19と、
c.それぞれ配列番号3、6、及び20と、
d.それぞれ配列番号3、6、及び21と、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、請求項19に記載の単離された抗体又はフラグメント。
【請求項21】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号3、6、及び18か、
b.それぞれ配列番号3、6、及び19か、
c.それぞれ配列番号3、6、及び20か、
d.それぞれ配列番号3、6、及び21か、に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項22】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号25、47、及び58か、
b.それぞれ配列番号25、48、及び58か、
c.それぞれ配列番号25、49、及び58か、
d.それぞれ配列番号25、50、及び58か、に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列を含む、単離された抗体又はフラグメント。
【請求項23】
前記抗体が更に、それぞれ配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列を含む、請求項22に記載の単離された抗体又はフラグメント。
【請求項24】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、
a.それぞれ配列番号25、43、及び60に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列か、
b.それぞれ配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列か、
c.それぞれ配列番号25、43、及び60に示されるHCDR1、HCDR2、及びHCDR3アミノ酸配列と、
それぞれ配列番号3、6、及び18に示されるLCDR1、LCDR2、及びLCDR3アミノ酸配列か、
d.配列番号86のVHか、
e.配列番号79のVLか、
f.配列番号86のVHと、
配列番号79のVLと、を含む、単離された抗体又はフラグメント。
【請求項25】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、配列番号67、68、69、81、82、83、84、85、又は86に示されるアミノ酸配列を有する前記VHを含む、単離された抗体又はフラグメント。
【請求項26】
前記抗体が更に、配列番号76、77、78、79、又は80に示されるアミノ酸配列を有する前記VLを含む、請求項25に記載の単離された抗体又はフラグメント。
【請求項27】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、配列番号76、77、78、79、又は80に示されるアミノ酸配列を有する前記VLを含む、単離された抗体又はフラグメント。
【請求項28】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、配列番号67、68、69、81、82、83、84、85、又は86に示されるアミノ酸配列を有するVHと少なくとも90%同一である前記VHを含む、単離された抗体又はフラグメント。
【請求項29】
VH及びVLを含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメントであって、前記抗体が、配列番号76、77、78、79、又は80に示されるアミノ酸配列を有する可変領域と少なくとも90%同一である前記VLを含む、単離された抗体又はフラグメント。
【請求項30】
配列番号92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を有する抗体重鎖を含む、ヒトIL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項31】
配列番号87、88、89、90、又は91に示されるアミノ酸配列を有する抗体軽鎖を含む、IL−17Aに特異的に結合する単離された抗体又はフラグメント。
【請求項32】
前記抗体が完全なヒト型(human)である、請求項3又は7に記載の単離された抗体又はフラグメント。
【請求項33】
前記抗体がポリエチレングリコールと結合されている、請求項3又は7に記載の単離された抗体又はフラグメント。
【請求項34】
IgG1又はIgG4アイソタイプを有する、請求項3又は7に記載の単離された抗体又はフラグメント。
【請求項35】
前記Fcドメインが、前記FcドメインにおけるS229P、P235A、又はL236A突然変異を含む、請求項3又は7に記載の単離された抗体又はフラグメント。
【請求項36】
請求項3又は7に記載の単離された抗体又はフラグメント及び製薬上許容され得る担体を含む、医薬組成物。
【請求項37】
配列番号67、68、69、81、82、83、84、85、86、92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を含む、単離された抗体重鎖。
【請求項38】
配列番号76、77、78、79、80、87、88、89、90、又は91に示されるアミノ酸配列を含む、単離された抗体軽鎖。
【請求項39】
配列番号67、68、69、81、82、83、84、85、86、92、93、94、95、96、97、98、99、又は100に示されるアミノ酸配列を含む抗体重鎖をコードする、単離されたポリヌクレオチド。
【請求項40】
配列番号76、77、78、79、80、87、88、89、90、又は91に示されるアミノ酸配列を含む抗体軽鎖をコードする、単離されたポリヌクレオチド。
【請求項41】
請求項39又は40に記載の少なくとも1つのポリヌクレオチドを含む、ベクター。
【請求項42】
請求項40に記載のベクターを含む、宿主細胞。
【請求項43】
a.ヒトIL−17A及びIL−17RAを提供する工程と、
b.ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112から成る群から選択される少なくとも1つのアミノ酸残基におけるヒトIL−17Aに結合する拮抗物質と接触させる工程と、を含む、ヒトIL−17AのIL−17RAとの相互作用を阻害する方法。
【請求項44】
a.ヒトIL17−A及びIL−17RAを提供する工程と、
b.ヒトIL−17Aを、V22、V24、L26、I28、Y62、L99、R101、F110、及びL112から成る群から選択される少なくとも1つのアミノ酸残基におけるヒトIL−17Aに結合する拮抗物質と接触させる工程と、を含む、IL−17A生物活性を阻害する方法。
【請求項45】
前記拮抗物質が、抗体、ペプチド、又は小分子である、請求項43又は44に記載の方法。
【請求項46】
請求項3又は7に記載の単離された抗体の治療上の有効量を、炎症性状態の治療を必要とする患者に炎症性状態を治療するうえで充分な時間にわたって投与する工程を含む、炎症性状態を治療する方法。
【請求項47】
前記炎症性状態が、気道、肺、胃腸管、小腸、大腸、結腸、直腸、関節、骨及び滑膜組織、軟骨、上皮、内皮、肝組織、又は皮膚を侵す、請求項46に記載の方法。
【請求項48】
前記炎症性状態が、炎症性肺状態、ぜんそく、慢性閉塞性肺疾患(COPD)、炎症性腸疾患、自己免疫疾患、関節リウマチ、乾癬、又は全身性硬化症である、請求項46に記載の方法。
【図5】


【図6】
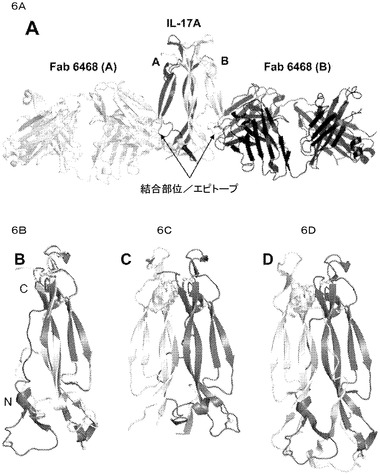
【図7】
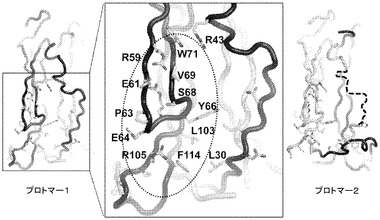
【図8】
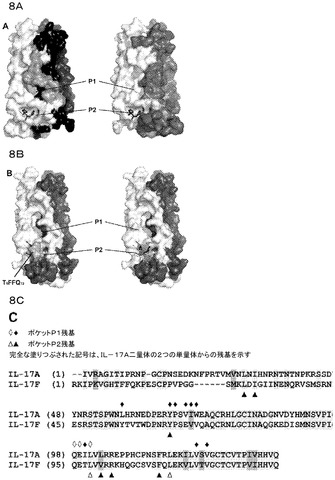

【図9】
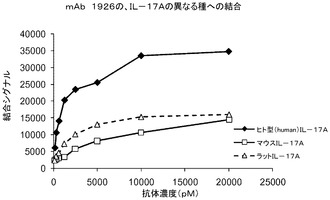
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2013−509193(P2013−509193A)
【公表日】平成25年3月14日(2013.3.14)
【国際特許分類】
【出願番号】特願2012−537102(P2012−537102)
【出願日】平成22年10月29日(2010.10.29)
【国際出願番号】PCT/US2010/054662
【国際公開番号】WO2011/053763
【国際公開日】平成23年5月5日(2011.5.5)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
【公表日】平成25年3月14日(2013.3.14)
【国際特許分類】
【出願日】平成22年10月29日(2010.10.29)
【国際出願番号】PCT/US2010/054662
【国際公開番号】WO2011/053763
【国際公開日】平成23年5月5日(2011.5.5)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
[ Back to top ]