
IL−23に結合する、フィブロネクチンをベースとする足場ドメインタンパク質
本発明はインターロイキン−23(IL−23)、特にIL−23のp19サブユニットに結合する、フィブロネクチンをベースとする足場ドメインタンパク質に関する。本発明はまた、自己免疫疾患を治療するための治療的適応における革新的なタンパク質の使用に関する。本発明はさらに、このようなタンパク質、このようなタンパク質またはその断片をコードするポリヌクレオチドを含む細胞、およびこの革新的なタンパク質をコードするポリヌクレオチドを含むベクターに関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明はインターロイキン−23(IL−23)、特にIL−23のp19サブユニットに結合する、フィブロネクチンをベースとする足場ドメインタンパク質に関する。本発明はまた、自己免疫疾患を治療するための治療的適応における革新的なタンパク質の使用に関する。本発明はさらに、このようなタンパク質、このようなタンパク質またはその断片をコードするポリヌクレオチドを含む細胞、およびこの革新的なタンパク質をコードするポリヌクレオチドを含むベクターに関する。
【背景技術】
【0002】
序論
IL−23はIL−12ヘテロ二量体サイトカインファミリーのメンバーである。それはIL−12に共通しているp40サブユニットおよび固有のp19サブユニットを含有する。IL−23はIL−12Rβ1およびIL−23からなるヘテロ二量体受容体複合体を通ってシグナルを送る(Aggarwal, Sら, “Interleukin−23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin−17”, J. Biol. Chem., 278:1910−1914 (2003))。IL−23は、多発性硬化症、関節リウマチ、乾癬およびクローン病などの慢性炎症性疾患の標的候補である。
【0003】
フィブロネクチンをベースとする足場は、対象とする任意の化合物と結合するよう進化させることができるタンパク質のファミリーである。通常、フィブロネクチンIII型(Fn3)またはFn3様ドメインに由来する足場を使用するこれらのタンパク質は、天然の抗体または操作された抗体(すなわち、ポリクローナル、モノクローナルまたは一本鎖抗体)に特徴的な形で機能し、さらに、構造的利点を有する。具体的には、これらの抗体模倣物の構造は、普通は抗体における構造および機能の喪失につながる条件下でも、フォールディング、安定性および溶解性が最適となるように設計されている。フィブロネクチンをベースとする足場タンパク質の一例として、アドネクチン(Adnexus,a Bristol−Myers Squibb R&D Company)がある。
【0004】
フィブロネクチンIII型(Fn3)ドメインは、N末端からC末端の順に、ベータまたはベータ様鎖、A;ループ、AB;ベータまたはベータ様鎖、B;ループ、BC;ベータまたはベータ様鎖C;ループCD;ベータまたはベータ様鎖D;ループDE;ベータまたはベータ様鎖、E;ループ、EF;ベータまたはベータ様鎖F;ループFG;およびベータまたはベータ様鎖Gを含む。ループAB、BC、CD、DE、EFおよびFGのいずれかまたはすべてが、標的結合に関与し得る。BC、DEおよびFGループは、免疫グロブリン由来の相補性決定領域(CDR)と構造上および機能上の両面で類似している。米国特許第7,115,396号には、BC、DEおよびFGループの変化が高親和性TNFαバインダーをもたらすFn3ドメインタンパク質が記載されている。米国公開第2007/0148126号には、BC、DEおよびFGループの変化が高親和性VEGFR2バインダーをもたらすFn3ドメインタンパク質が記載されている。
【発明の概要】
【発明が解決しようとする課題】
【0005】
自己免疫疾患の治療法のための改善されたフィブロネクチンドメイン足場タンパク質を得ることは有利であろう。インターロイキン−17を産生するエフェクターT細胞のサブセット(IL−17;「Th17細胞」)は非常に炎症性であり、激しい自己免疫を誘導する。Th17細胞は、Th1およびTh2細胞に比べて、IL−6、腫瘍壊死因子(TNF)、IL−22、IL−17AおよびIL−17Fと同様にケモカイン受容体CCR6を含む、異なるサイトカインおよびケモカインのサブセットを発現する。IL−23は活性化T細胞によるIL−17の産生を促進させ(Aggarwal, Sら, “Interleukin−23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin−17”, J. Biol. Chem., 278:1910−1914 (2003))、かつ、IL−17産生CD4+T細胞の増殖を誘導するための重要なサイトカインである。IL−23への暴露は、Th17細胞の病原性を決定する重要な特徴であるように思われる。
【課題を解決するための手段】
【0006】
本出願は、ヒトIL−23特異的p19サブユニットに対するアドネクチン商標を提供する。本発明の一態様は、一以上の溶媒接触可能ループが無作為化されまたは変異した、Fn3ドメインを含むポリペプチドを提供する。いくつかの実施形態では、前記Fn3ドメインは、ヒトフィブロネクチンIII型ドメインの野生型第10モジュール(10Fn3)に由来するFn3ドメインである。いくつかの実施形態では、本発明の10Fn3ポリペプチドは、ヒト10Fn3ドメインと、少なくとも40%、50%、60%、65%、70%、75%、80%、85%、または90%同一である。
【0007】
いくつかの実施形態では、BC、DEおよびFGから選択される一以上のループが、対応するヒトフィブロネクチンループと比較して伸長または短縮されてもよい。
【0008】
いくつかの実施形態では、本発明のポリペプチドは第10フィブロネクチンIII型(10Fn3)ドメインを含み、ここに10Fn3ドメインは、ループ、AB;ループ、BC;ループ、CD;ループ、DE;ループEF;およびループFGを含み;かつ、ヒト10Fn3ドメインの対応するループの配列と比較して変化したアミノ酸配列を備える、ループBC、DEおよびFGから選択される少なくとも一つのループを有するものである。
【0009】
いくつかの実施形態では、本発明のポリペプチドは、非ループ領域と少なくとも80、85、90、95、98、99または100%同一であるアミノ酸配列を含むFn3ドメインを含む。
【0010】
いくつかの実施形態では、本発明のタンパク質のBCループは、配列番号2〜6からなる群から選択されるアミノ酸配列を含む。
【0011】
いくつかの実施形態では、本発明のタンパク質のDEループは、配列番号7〜48からなる群から選択されるアミノ酸配列を含む。
【0012】
いくつかの実施形態では、本発明のタンパク質のFGループは、配列番号49〜59からなる群から選択されるアミノ酸配列を含む。
【0013】
いくつかの実施形態では、10Fn3ドメインは、アミノ酸置換、挿入、もしくは欠失で開始および/または終止してもよい。
【0014】
いくつかの実施形態では、本発明のタンパク質は、配列番号2〜6に示されるBCループ配列からの一つのループ配列、配列番号7〜48に示される一つのDEループ配列、および配列番号49〜59に示される一つのFGループ配列を含む。
【0015】
いくつかの実施形態では、本発明のタンパク質は、配列番号2〜59のいずれか一つと少なくとも70、75、80、85、90、95、98、99または100%同一であるBC、DEおよびFGループアミノ酸配列を含む。
【0016】
いくつかの実施形態では、抗IL−23アドネクチンは配列番号60〜100のいずれか一つのアミノ酸配列を含む。
【0017】
いくつかの実施形態では、抗IL−23アドネクチンは配列番号60〜100のいずれか一つの3〜96位由来のFn3ドメインアミノ酸配列を含む。
【0018】
いくつかの実施形態では、抗IL−23アドネクチンは、配列番号60〜100のいずれか一つと少なくとも70、75、80、85、90、95、98、99または100%同一であるアミノ酸配列を含む。
【0019】
一態様では、抗IL−23アドネクチンはさらに、薬物動態(PK)部分を含む。いくつかの実施形態では、PK部分はポリエチレングリコール(PEG)を含む。
【0020】
一態様では、本出願は自己免疫疾患の治療において有益な抗IL−23アドネクチンを提供する。
【0021】
一態様では、本発明はフィブロネクチンIII型第10(10Fn3)ドメインおよび抗IL−23アドネクチンを含む融合ポリペプチドを提供し、ここに前記10Fn3ドメインは1uM以下のKdでHSAに結合するものである。特定の実施形態では、10Fn3ドメインは配列番号103と少なくとも70%同一であるアミノ酸配列を含む。一の実施形態では、10Fn3ドメインは、配列番号104に説明されるアミノ酸配列を有するBCループ、配列番号105に説明されるアミノ酸配列を有するDEループ、および配列番号106に説明されるアミノ酸配列を有するFGループを含む。別の実施態様では、10Fn3ドメインは、一以上の、配列番号104に説明されるアミノ酸配列を有するBCループ、配列番号105に説明されるアミノ酸配列を有するDEループ、および配列番号106に説明されるアミノ酸配列を有するFGループを含む。
【0022】
一の実施形態では、融合ポリペプチドの10Fn3ドメインはまた、一以上のアカゲザル血清アルブミン(RhSA)、カニクイザル血清アルブミン(CySA)、またはマウス血清アルブミン(MuSA)に結合する。他の実施形態では、10Fn3ドメインは一以上のRhSA、CySA、またはMuSAと交差反応しない。
【0023】
特定の実施形態では、融合ポリペプチドの10Fn3ドメインは1uM以下のKdでHSAに結合する。いくつかの実施形態では、10Fn3ドメインは500nM以下のKdでHSAに結合する。他の実施形態では、10Fn3ドメインは少なくとも200nM、100nM、50nM、20nM、10nM、または5nMのKdでHSAに結合する。
【0024】
他の実施形態では、融合ポリペプチドの10Fn3ドメインはHSAのドメインIまたはIIに結合する。一の実施形態では、10Fn3ドメインはHSAのドメインIおよびIIの両方に結合する。いくつかの実施形態では、10Fn3ドメインは5.5から7.4のpH範囲でHSAに結合する。他の実施形態では、10Fn3ドメインはpH5.5で200nM以下のKdでHSAに結合する。別の実施形態では、5.5から7.4のpH範囲で少なくとも500nM、200nM、100nM、50nM、20nM、10nM、または5nMのKdでHSAに結合する。一の実施形態では、10Fn3ドメインはpH5.5で少なくとも500nM、200nM、100nM、50nM、20nM、10nM、または5nMのKdでHSAに結合する。
【0025】
いくつかの実施形態では、血清アルブミン存在下での融合ポリペプチドの血清半減期は、血清アルブミン非存在下での前記ポリペプチドの血清半減期より少なくとも5倍長い。特定の実施形態では、血清アルブミン存在下での融合ポリペプチドの血清半減期は、血清アルブミン非存在下での前記ポリペプチドの血清半減期より少なくとも2倍、5倍、7倍、10倍、12倍、15倍、20倍、22倍、25倍、27倍、または30倍長い。いくつかの実施形態では、血清アルブミンはHSA、RhSA、CySA、またはMuSAのいずれか一つである。
【0026】
特定の実施形態では、血清アルブミン存在下での融合ポリペプチドの血清半減期は少なくとも20時間である。特定の実施形態では、血清アルブミン存在下での融合ポリペプチドの血清半減期は少なくとも10時間、12時間、15時間、20時間、25時間、30時間、40時間、50時間、75時間、90時間、100時間、110時間、120時間、130時間、150時間、170時間、または200時間である。いくつかの実施形態では、融合ポリペプチドの血清半減期は霊長類(例えば、ヒトもしくはサル)またはマウスにおいて観察される。
【0027】
前述の態様および実施形態のいずれにおいても、10Fn3ドメインは配列番号107、111、115、119、および123〜143から選択される配列を含む。
【図面の簡単な説明】
【0028】
【図1】図1は、本発明の抗IL−23アドネクチンの全長DNA配列アラインメントを示す。
【0029】
【図2】図2は、実施例2において記載されるように、pBMS2008/ATI001044タンパク質発現ベクターを示す。
【0030】
【図3】図3は、本発明の抗IL−23アドネクチンの全長アミノ酸配列アラインメントを示す。
【0031】
【図4】図4は、実施例4において記載されるように、抗IL−23アドネクチンによるPBMC pSTAT3阻害由来の代表的なIC50曲線を示す。
【0032】
【図5】図5は、実施例4において記載されるように、抗IL−23アドネクチンおよび抗p40モノクローナル抗体(MAB1510)によるIL−23依存性IL−17Aの阻害に対する代表的なIC50曲線を示す。
【0033】
【図6】図6は、ドナー228(実施例4において記載されるように試験された4ドナーの一つ)のPBMCによるIL−23誘導性IL−17産生のATI001045阻害を示す。
【0034】
【図7】図7は、抗IL−23アドネクチンに対する代表的な選択性データを示す。実施例4において記載されるように、回収されたATI001016に結合する10nM IL−23および1uM IL−12の結合相および解離相を示すバッファー除去センサーグラムが示される。
【0035】
【図8】図8は、実施例4において記載されるように、抗IL−23アドネクチンがNK−92細胞におけるIL−12誘導性IFN−γ産生を阻害しないことを示す。
【0036】
【図9A】図9Aは、実施例4において記載されるように、ATI001045がマウス薬力学的モデルにおいて血清IL−17レベルを阻害することを示す。
【0037】
【図9B】図9Bは、実施例4において記載されるように、マウス薬力学的モデルにおける抗IL−23アドネクチンの阻害活性の比較を示す。
【0038】
【図10A】図10Aは、実施例4において記載されるように、ヒトIL−23誘導表皮肥厚におけるATI000934用量反応を示す。
【0039】
【図10B】図10Bは、実施例4において記載されるように、ヒトIL−23誘導表皮肥厚におけるATI001045用量反応を示す。
【0040】
【図11】図11は、マウスにおけるインビボ(in vivo)のHSA半減期を示す。HSAは20mg/kg(図11A)または50mg/kg(図11B)でマウスへ注射された。
【0041】
【図12】図12A−Dは、マウスにおけるSABA1−SABA4の半減期決定を示す。
【0042】
【図13】図13Aは、HSAと同時注射した際のSABA1−4のマウスにおける半減期増強のグラフの概要を示す。図13bは、カニクイザルおよびマウス由来のデータを比較する。
【0043】
【図14】図14A−Bは、カニクイザルにおけるSABA1.1およびSABA5.1に対する半減期決定を示す。
【0044】
【図15】図15は、直接結合ELISAアッセイによるヒト、マウス、およびラット由来のアルブミンに結合するSABA1.2を示す。
【0045】
【図16】図16は、SABA1.1およびHSAの化学量(stoichiometry)の決定を示す。
【0046】
【図17】図17は、HSAの組換えドメイン断片に結合するSABA1.2のビアコア解析を示す。
【0047】
【図18】図18は、1mpkおよび10mpkで投与されたサルにおけるSABA1.2に対する薬物動態プロファイルを示す。
【0048】
【図19】図19は、1mpkで静脈内投与または皮下投与されたサルにおけるSABA1.2に対する薬物動態プロファイルを示す。
【発明を実施するための形態】
【0049】
発明の詳細な説明
定義
「ポリペプチド」によって、長さ、翻訳後修飾または機能にかかわらず、二以上のアミノ酸の任意の配列が意味される。「ポリペプチド」、「ペプチド」および「タンパク質」は、本明細書において同義的に使用される。ポリペプチドは、出典明示により本明細書に組み込まれる米国特許第6,559,126号に記載されるものなどの天然アミノ酸および非天然アミノ酸を含み得る。ポリペプチドはまた、種々の標準的な化学法のいずれかで修飾されていてもよい(例えば、アミノ酸は保護基で修飾されていてもよい;カルボキシ末端アミノ酸は、末端アミド基にされてもよい;アミノ末端残基は、例えば、親油性を増強するための基で修飾されてもよい;またはポリペプチドは、化学的にグリコシル化されていてもよく、あるいは、安定性もしくはインビボ(in vivo)半減期を増大するよう修飾されていてもよい)。ポリペプチド修飾は、ポリペプチドへの、環状化合物または他の分子などの別の構造の結合(attachment)を含み得、また一以上のアミノ酸を変化した立体配置(すなわち、RもしくはS;または、LもしくはD)で含有するポリペプチドを含み得る。本発明のポリペプチドは、IL−23のp19サブユニットに特異的に結合するよう修飾された、第10フィブロネクチンIII型ドメインに由来するタンパク質であり、本明細書において「アドネクチン」または「抗IL−23アドネクチン」と呼ばれる。
【0050】
用語「PK」は、「薬物動態(pharmokinetic)」の頭字語であり、例として、被験体による吸収、分布、代謝および排出を含む化合物の特性を包含する。「PK調節タンパク質」または「PK部分」とは、生物学的に活性な分子と融合されるか、一緒に投与される場合に、生物学的に活性な分子の薬物動態特性に影響を及ぼす任意のタンパク質、ペプチドまたは部分を指す。PK調節タンパク質またはPK部分の例として、PEG、ヒト血清アルブミン(HSA)バインダー(米国公開第2005/0287153号および同2007/0003549号に開示されるような)、ヒト血清アルブミン、FcまたはFc断片、および糖(例えば、シアル酸)が挙げられる。
【0051】
本明細書において、「アミノ酸配列同一性パーセント(%)」とは、配列同一性の一部として保存的置換は全く考慮することなく、最大配列同一性パーセントを達成するために、配列をアラインし、必要に応じてギャップを導入した後の、選択された配列中のアミノ酸残基と同一である、候補配列中のアミノ酸残基のパーセンテージとして定義される。アミノ酸配列同一性パーセントを決定する目的上、アラインメントは、当技術分野における技術の範囲内の種々の方法で、例えば、BLAST、BLAST−2、ALIGN、ALIGN−2またはMegalign(DNASTAR登録商標)ソフトウェアなどの公的に入手可能なコンピュータソフトウェアを使用して達成できる。当業者ならば、比較されている配列の全長にわたって最大アラインメントを達成するのに必要な任意のアルゴリズムを含む、アラインメントを測定するための適当なパラメータを決定できる。
【0052】
「単離された」ポリペプチドとは、同定および分離および/またはその天然環境の成分から回収されているものである。その天然環境の夾雑成分とは、ポリペプチドの診断または治療的使用を干渉する物質であり、酵素、ホルモンおよび他のタンパク質性または非タンパク質性溶質を挙げることができる。好ましい実施形態では、ポリペプチドは、(1)ローリー法によって決定される、95重量%を超える、最も好ましくは、99重量%を超えるポリペプチドに、(2)スピニングカップシークエネーターの使用によってN末端もしくは内部アミノ酸配列の少なくとも残基(複数)を得るのに十分な程度に、または(3)クーマシーブルーもしくは、好ましくは、銀染色を使用する還元もしくは非還元条件下でのSDS−PAGEによる均一性に精製される。単離されたポリペプチドは、ポリペプチドの天然環境の少なくとも1種の成分が存在しないので、組換え細胞内でインサイチュ(in situ)でのポリペプチドを含む。しかし、通常、単離されたポリペプチドは、少なくとも1つの精製工程によって調製される。
【0053】
アミノ酸配列または化合物の「半減期」は、通常、インビボ(in vivo)でポリペプチドの血清濃度が、例えば、天然の機序による、配列もしくは化合物の分解および/または配列もしくは化合物のクリアランスもしくは隔離によって50%低減されるのにかかる時間として定義され得る。半減期は、薬物動態分析によってなど、それ自体公知の任意の方法で決定できる。適した技術は、当業者には明らかであり、例えば、通常、本発明のアミノ酸配列または化合物の適した用量を霊長類に適宜投与する工程;前記霊長類から定期的に血液サンプルまたは他のサンプルを採取する工程;前記血液サンプル中の本発明のアミノ酸配列または化合物のレベルまたは濃度を決定する工程;およびこのように得られたデータ(のプロット)から、本発明のアミノ酸配列または化合物のレベルまたは濃度が、投与時の初期レベルと比較して50%低減されるまでの時間を算出する工程を含み得る。例えば、Kenneth, A ら, Chemical Stability of Pharmaceuticals: A. Handbook for Pharmacists および Lee, P. I. D. ら, Pharmacokinetic analysis: A Practical Approach (1996)などの標準的なハンドブックが参照される。Gibaldi M. ら,Pharmacokinetics, 2nd Rev. edition, Marcel Dekker, 刊行(1982)もまた参照される。
【0054】
半減期は、t1/2−アルファ、t1/2−ベータおよび曲線下面積(AUC)などのパラメータを使用して表現され得る。本明細書において、「半減期の増大」とは、これらのパラメータのうちいずれか1つ、これらのパラメータのうちいずれか2つ、またはこれらのパラメータのうち3つ全ての増大を指す。「半減期の増大」とは、特に、t1/2−アルファおよび/またはAUCまたは両方の増大を伴うか、または伴わない、t1/2−ベータの増大を指す。
【0055】
概説
本出願は、ヒトIL−23特異的p19サブユニットに対するアドネクチンを提供する。IL−23特異的アンタゴニストを同定するために、抗p40モノクローナル抗体を使用して、IL−23をアドネクチンの大規模合成ライブラリーへ提示した。IL−23 p19サブユニットに結合するアドネクチンは、ヒトIL−23との結合、IL−23/IL−23R相互作用の競合、およびT細胞株におけるIL−23誘導性シグナル伝達の阻害に関してスクリーニングされた。抗IL−23アドネクチンは、標的濃度を低下させ、遅い解離速度(slow off−rate)で抗IL−23アドネクチンを選択することによる、さらなる選択圧に供された。この最適化工程から、アドネクチンのファミリーが、好ましい生化学的および生物物理学的特性を備えるIL−23特異的阻害剤として同定された。
【0056】
フィブロネクチンベースの足場
本出願の一態様は、一以上の溶媒接触可能ループが無作為化または変異した、Fn3ドメインを含むポリペプチドを提供する。いくつかの実施形態では、前記Fn3ドメインは、ヒトフィブロネクチンIII型ドメインの野生型第10モジュール(10Fn3)に由来するFn3ドメイン:VSDVPRDLEVVAATPTSLLISWDAPAVTVRYYRITYGETGGNSPVQEFTVPGSKSTATISGLKPGVDYTITVYAVTGRGDSPASSKPISINYRT(配列番号1)である。上記の10Fn3配列において、BC、DEおよびFGループには下線が引かれている。
【0057】
種々の変異10Fn3足場が報告されている。一態様では、Asp7、Glu9およびAsp23の一以上が、例えば、負の電荷を有さないアミノ酸残基(例えば、Asn、Lys等)などの別のアミノ酸によって置換されている。これらの突然変異は、野生型形態と比較して、中性pHでの変異10Fn3のより大きな安定性を促進する効果を有すると報告されている(PCT公開番号WO02/04523参照)。有益または中立のいずれかである10Fn3足場における種々のさらなる変化が開示されている。例えば、Batori ら, Protein Eng., 15(12):1015−1020 (Dec. 2002); Koide ら, Biochemistry, 40(34):10326−10333 (Aug. 28, 2001)参照。
【0058】
変種および野生型10Fn3タンパク質の両方とも、同一構造、すなわち、AからGと表される7つのβ鎖ドメイン配列および7つのβ鎖ドメイン配列を接続する6つのループ領域(ABループ、BCループ、CDループ、DEループ、EFループおよびFGルー
プ)を特徴とする。N末端およびC末端の最も近くに位置するβ鎖は、溶液中でβ様コンホメーションをとり得る。配列番号1中、ABループは、残基15〜16に対応し、BCループは、残基21〜30に対応し、CDループは、残基39〜45に対応し、DEループは、残基51〜56に対応し、EFループは、残基60〜66に対応し、FGループは、残基76〜87に対応する(Xu ら, Chemistry & Biology, 9:933−942 (2002))。
【0059】
いくつかの実施形態では、10Fn3ポリペプチドは、配列番号1に示されるヒト10Fn3ドメインと、少なくとも40%、50%、60%、65%、70%、75%、80%、85%または90%同一であり得る。変異性の多くは、通常、一以上のループ中で起こる。10Fn3ポリペプチドのβ鎖またはβ様鎖の各々は、このような変動が、生理学的条件におけるポリペプチドの安定性を乱さないという条件で、本質的に、配列番号1の対応するβ鎖またはβ様鎖の配列と、少なくとも80%、85%、90%、95%または100%同一であるアミノ酸配列からなり得る。
【0060】
いくつかの実施形態では、本開示内容は、第10フィブロネクチンIII型(10Fn3)ドメインを含むポリペプチドであって、10Fn3ドメインが、ループ、AB;ループ、BC;ループ、CD;ループ、DE;ループEF;およびループFGを含むものであり、かつ、ヒト10Fn3ドメインの対応するループの配列と比較してアミノ酸配列が変化したループBC、DEおよびFGから選択される少なくとも1つのループを有するポリペプチドを提供する。いくつかの実施形態では、BCおよびFGループが変化し、いくつかの実施形態では、BC、DEおよびFGループが変化している、すなわち、Fn3ドメインが、天然に存在しないループを含む。「変化した」とは、(ヒトフィブロネクチンドメインに対応する)鋳型配列に対する一以上のアミノ酸配列の変化を意味し、アミノ酸付加、欠失および置換を含む。アミノ酸配列の変化は、通常、核酸コード配列の意図的な、盲検的な(blind)または自発的な配列変動によって達成され得、任意の技術、例えば、PCR、エラー−プローンPCRまたは化学的DNA合成によって起こり得る。
【0061】
いくつかの実施形態では、BC、DEおよびFGから選択される一以上のループは、対応するヒトフィブロネクチンループと比較して、長さが伸長または短縮され得る。いくつかの実施形態では、ループの長さは、2〜25アミノ酸によって伸長され得る。いくつかの実施形態では、ループの長さは、1〜11アミノ酸によって減少され得る。したがって、抗原結合を最適化するために、抗原結合において最大のあり得る可動性および親和性を得るよう、10Fn3のFGループの長さを、長さにおいて、ならびに、配列において変化させてもよい。
【0062】
いくつかの実施形態では、ポリペプチドは、配列番号1の非ループ領域と少なくとも80、85、90、95、98、99または100%同一であるアミノ酸配列を含むFn3ドメインを含み、BC、DEおよびFGから選択される少なくとも1つのループが変化している。いくつかの実施形態では、変化したBCループは、最大10のアミノ酸置換、最大4のアミノ酸欠失、最大10のアミノ酸挿入またはそれらの組合せを有する。いくつかの実施形態では、変化したDEループは、最大6のアミノ酸置換、最大4のアミノ酸欠失、最大13のアミノ酸挿入またはそれらの組合せを有する。いくつかの実施形態では、FGループは、最大12のアミノ酸置換、最大11のアミノ酸欠失、最大25のアミノ酸挿入またはそれらの組合せを有する。
【0063】
いくつかの実施形態では、本発明のタンパク質のBCループは、GHYPMHV(配列番号2)、GHYPLHV(配列番号3)、GHYPMHI(配列番号4)、GHYPLHI(配列番号5)およびGHYPLHL(配列番号6)からなる群から選択されるアミノ酸配列を含む。
【0064】
いくつかの実施形態では、本発明のタンパク質のDEループは、HRTH(配列番号7)、YYHY(配列番号8)、SKQH(配列番号9)、SNVH(配列番号10)、NRAH(配列番号11)、RKTY(配列番号12)、RSRY(配列番号13)、SRYY(配列番号14)、PHRY(配列番号15)、RSTH(配列番号16)、SRIY(配列番号17)、HQRY(配列番号18)、KQVY(配列番号19)、AHRY(配列番号20)、RSRH(配列番号21)、ARQY(配列番号22)、RTQY(配列番号23)、PRYH(配列番号24)、MRQH(配列番号25)、SRKY(配列番号26)、RQKY(配列番号27)、HAKY(配列番号28)、SNRY(配列番号29)、NTSH(配列番号30)、SQVY(配列番号31)、NRVY(配列番号32)、PRSH(配列番号33)、RTKY(配列番号34)、SRYH(配列番号35)、PRRY(配列番号36)、RQKY(配列番号37)、RYKY(配列番号38)、VPRH(配列番号39)、TPKH(配列番号40)、RSKY(配列番号41)、SRKY(配列番号42)、VPRY(配列番号43)、PRRY(配列番号44)、RMRH(配列番号45)、PPRH(配列番号46)、RQIY(配列番号47)、およびMRQH(配列番号48)からなる群から選択されるアミノ酸配列を含む。
【0065】
いくつかの実施形態では、本発明のタンパク質のFGループは、YYNEADYSQI(配列番号49)、YYQEYEYRYI(配列番号50)、YYMEEKYAVI(配列番号51)、YYAQENYKEI(配列番号52)、YYKEANYREI(配列番号53)、YYAQEEYHII(配列番号54)、YYKEADYSQI(配列番号55)、YYEQVEYREI(配列番号56)、YYEQPIYATI(配列番号57)、YYEQVEYREI(配列番号58)およびYYSEELYKYI(配列番号59)からなる群から選択されるアミノ酸配列を含む。
【0066】
10Fn3ドメインは、アミノ酸変異で始まってもよい。例えば、Fn3ドメインのN末端に、さらなるMG配列が配置されてもよい。Mは、通常、切断除去され、N末端にGが残る。いくつかの実施形態では、10Fn3ドメインのC末端に、配列が配置されていてもよい。例えば、GSGC(配列番号101)などのシステイン含有リンカーがC末端へ付加される、部位特異的PEG化において。あるいは、システイン含有リンカーEIDKPCQ(配列番号102)のため、セリンをシステインに変更することによって変異させられた、天然に存在するC末端テールのPEG化。GSGCリンカーを含む本発明の抗IL−23アドネクチンの例として、ATI001014、ATI001015、ATI001016、ATI001044、ATI001045およびATI001047が挙げられる。ATI000934は、EIDKPCQリンカーを含む本発明の抗IL−23アドネクチンの例である。
【0067】
いくつかの実施形態では、本発明のタンパク質は、配列番号2〜6に示されるBCループ配列からの一のループ配列、配列番号7〜48に示される一のDEループ配列、および配列番号49〜59に示される一のFGループ配列を含む。いくつかの実施形態では、本発明のタンパク質は、配列番号2〜59のいずれか一つと少なくとも70、75、80、85、90、95、98、99または100%同一であるBC、DEおよびFGループアミノ酸配列を含む。
【0068】
さらに、当業者であれば、配列番号2〜6に示されるBCループ配列が、X1がMまたはLのいずれかであり、X2がIまたはVのいずれかである共通の配列モチーフGHYPX1HX2(配列番号257)を共有し、かつ、配列番号49〜59に示されるFGループが、X3が任意のアミノ酸である共通の配列モチーフYYX3X3X3X3YX3X3I(配列番号258)を共有することを認識するであろう。それゆえ、コンセンサス配列GHYPX1HX2に適合するBCループを備え、かつ/または、配列番号49〜59に明確に示されるものの域を超えて、パターンYYX3X3X3X3YX3X3Iに適合する他のFGループを備える、IL−23に結合するさらなるアドネクチンを生成することが可能であろう。
【0069】
いくつかの実施形態では、抗IL−23アドネクチンは配列番号60〜100のいずれか一つのアミノ酸配列を含む。いくつかの実施形態では、抗IL−23アドネクチンは配列番号60〜100のいずれか一つの3〜96位由来のFn3ドメインアミノ酸配列を含む。いくつかの実施形態では、抗IL−23アドネクチンは、配列番号60〜100のいずれか一つと少なくとも70、75、80、85、90、95、98、99または100%同一であるアミノ酸配列を含む。いくつかの実施形態では、抗IL−23アドネクチンは、配列番号60〜100のいずれか一つの3〜96位由来のアミノ酸配列と少なくとも70、75、80、85、90、95、98、99または100%同一であるアミノ酸配列を含む。
【0070】
いくつかの実施形態では、抗IL−23アドネクチンはPEG化され、かつ/またはhisタグを含有してもよい。本明細書において用いる場合、ATI000934は、ループ配列が構築体1571G06(配列番号87)のものと同一であるタンパク質を指し、前記タンパク質はC末端でEIDKPCQ残基を含み、PEG化され、かつhisタグを含有する。ATI001014は、ループ配列が構築体1571G04(配列番号86)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化され、かつhisタグを含有する。ATI001015は、ループ配列が構築体1572G06(配列番号91)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化され、かつhisタグを含有する。ATI001016は、ループ配列が構築体1490B03(配列番号79)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化され、かつhisタグを含有する。ATI001044は、ループ配列が構築体1490B03(配列番号79)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含むが、前記タンパク質はPEG化されておらず、かつhisタグが存在しない。ATI001045は、ループ配列が構築体1490B03(配列番号79)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化されているが、hisタグが存在しない。ATI001047は、ループ配列が構築体1571G04(配列番号86)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化されているが、hisタグが存在しない。
【0071】
フィブロネクチンは、そのインテグリン結合モチーフ、「アルギニン−グリシン−アスパラギン酸」(RGD)を介して特定の種類のインテグリンと天然に結合する。いくつかの実施形態では、ポリペプチドは、(RGD)インテグリン結合モチーフを欠く10Fn3ドメインを含む。
【0072】
薬物動態部分
一態様では、本出願は、薬物動態(PK)部分をさらに含む抗IL−23アドネクチンを提供する。改善される薬物動態は、認識される治療的必要性に従って評価できる。おそらくは、タンパク質が、投与後の血清において利用可能なままである時間を増大することによって、生物学的利用能(bioavailability)を増大すること、および/または用量間の時間を増大することが望ましいことが多い。場合によっては、経時的なタンパク質の血清濃度の連続性を改善すること(例えば、投与の直後および次の投与の直前のタンパク質の血清濃度の相違の減少)が望ましい。抗IL−23アドネクチンは、哺乳類(例えば、マウス、ラットまたはヒト)においてポリペプチドのクリアランス速度を、修飾されていないアドネクチンと比較して3倍超低減する部分に結合させてもよい。改善される薬物動態の他の尺度として、血清半減期を挙げることができ、これはアルファ相およびベータ相に分けられることが多い。いずれかの相または両方の相が、適当な部分の付加によって大幅に改善され得る。
【0073】
本明細書において「PK部分」と呼ばれる、血液からのタンパク質のクリアランスを遅延させる傾向がある部分として、ポリオキシアルキレン部分、例えば、ポリエチレングリコール、糖(例えば、シアル酸)および耐容性良好であるタンパク質部分(例えば、Fc、Fc断片、トランスフェリンまたは血清アルブミン)が挙げられる。アドネクチンは、米国公開第2007/0048282号に記載される、アルブミンまたはアルブミンの断片(部分)もしくは変種と融合させてもよい。
【0074】
いくつかの実施形態では、PK部分は、例えば米国公開第2007/0178082号および同2007/0269422号に記載されるものなどの血清アルブミン結合タンパク質である。
【0075】
いくつかの実施形態では、PK部分は、例えば米国公開第2007/0178082号に記載されるものなどの血清免疫グロブリン結合タンパク質である。
【0076】
いくつかの実施形態では、アドネクチンは、ポリエチレングリコール(PEG)を含む。一以上のPEG分子を、タンパク質上の異なる位置に結合してもよく、このような結合は、アミン、チオールまたは他の適した反応性基を用いる反応によって達成できる。アミン部分は、例えば、ポリペプチドのN末端に見られる第一級アミンまたはリシンもしくはアルギニンなどのアミノ酸中に存在するアミン基であり得る。いくつかの実施形態では、PEG部分は:a)N末端;b)N末端と最もN末端側のβ鎖またはβ様鎖との間;c)標的結合部位と反対側のポリペプチドの面上に位置するループ;d)C末端と最もC末端側のβ鎖またはβ様鎖との間;およびe)C末端、からなる群から選択されるポリペプチド上の位置に結合している。
【0077】
PEG化は、部位特異的PEG化によって達成でき、この方法においては、PEG化が優先的に起こる部位を作り出すために、適した反応性基がタンパク質に導入される。いくつかの実施形態では、タンパク質は、所望の位置にシステイン残基を導入するよう修飾され、システイン上で部位特異的PEG化が可能となる。PEGは、分子量が大きく変わってもよく、分岐または直鎖であってもよい。
【0078】
いくつかの実施形態では、アドネクチンは、Fn3ドメインおよびPK部分を含む。いくつかの実施形態では、Fn3ドメインは、10Fn3ドメインである。いくつかの実施形態では、PK部分は、ポリペプチドの血清半減期を、Fn3ドメイン単独と比較して、5、10、20、30、40、50、60、70、80、90、100、120、150、200、400、600、800、1000%以上を超えて増大する。
【0079】
いくつかの実施形態では、PK部分はポリマー糖である。いくつかの実施形態では、PK部分はポリエチレングリコール部分である。いくつかの実施形態では、PK部分は血清アルブミン結合タンパク質である。いくつかの実施形態では、PK部分はヒト血清アルブミンである。いくつかの実施形態では、PK部分は血清免疫グロブリン結合タンパク質である。いくつかの実施形態では、PK部分はトランスフェリンである。いくつかの実施形態では、PK部分は血清タンパク質に対して特異的な別のアドネクチンである。
【0080】
生化学的および生物物理学的特性
本出願は、IL−23のp19サブユニットに結合するFn3ドメインを含むアドネクチンを提供する。表1および実施例4に示されるように、標的分子に結合するポリペプチドは、平衡定数(例えば、解離、KD)に関して、および速度定数(例えば、結合速度定数(on−rate constant)、Konおよび解離速度定数(off−rate constant)、Koff)の点で評価できる。アドネクチンは通常、500nM、100nM、10nM、1nM、500pM、200pM、100pM未満のKDで標的分子と結合するが、Koffが十分に低い、またはKonが十分に高い場合には、より高いKD値も許容され得る。
【0081】
本発明の抗IL−23アドネクチンのファミリーのBC、DEおよびFGループ配列は、対応する全長配列番号と共に下記の表1に提示される。
【表1−1】
【表1−2】
【表1−3】
【0082】
抗IL−23アドネクチンのさらなる特性は表2に記載される。
【表2】
(n.d. 検出不可) (詳細な方法は実施例4に記載される)。
【0083】
核酸−タンパク質融合技術
一態様では、本出願は、IL−23のp19サブユニットと結合するフィブロネクチンIII型ドメインを含むアドネクチンを提供する。特異的結合特性を有するFn3ドメインを、迅速に生成し、試験する一つの方法は、Adnexus,Bristol−Myers Squibb R&D Companyの核酸−タンパク質融合技術である。この開示内容は、インビトロ(in vitro)発現および核酸−タンパク質融合物(RNA−およびDNA−タンパク質融合物)を利用してタンパク質との結合にとって重要である、新規ポリペプチドおよびアミノ酸モチーフを同定する、PROfusionと呼ばれるタグ技術を利用する。核酸−タンパク質融合技術は、タンパク質をそのコーディング遺伝情報と共有結合によってカップリングする技術である。RNA−タンパク質融合技術およびフィブロネクチンベースの足場タンパク質ライブラリースクリーニング法の詳細な説明については、出典明示により本明細書に組み込まれる、Szostak ら,米国特許第6,258,558号、同6,261,804号、同6,214,553号、同6,281,344号、同6,207,446号、同6,518,018号、同6,818,418号;およびRoberts ら, Proc Natl. Acad. Sci. 94:12297−12302(1997), 参照。
【0084】
ベクターおよびポリヌクレオチド実施形態
本明細書に開示される種々のタンパク質またはポリペプチドのいずれかをコードする核酸は、化学的に合成されてもよい。コドン使用頻度(codon usage)は、細胞における発現を改善するよう選択されてもよい。このようなコドン使用頻度は、選択される細胞種に依存する。大腸菌および他の細菌、ならびに哺乳類細胞、植物細胞、酵母細胞および昆虫細胞のため、特定化されたコドン使用パターンが開発されている。例えば、 Mayfield ら, Proc. Natl. Acad. Sci. USA, 100(2):438−442 (Jan. 21, 2003); Sinclair ら, Protein Expr. Purif., 26(I):96−105 (Oct. 2002); Connell, N.D., Curr. Opin. Biotechnol., 12(5):446−449 (Oct. 2001); Makrides ら, Microbiol. Rev., 60(3):512−538 (Sept. 1996); および Sharp ら, Yeast, 7(7):657−678 (Oct. 1991) 参照。
【0085】
核酸操作の一般的な技術は、例えば、出典明示により本明細書に組み込まれる、Sambrook ら, Molecular Cloning: A Laboratory Manual, 2nd Edition, Vols. 1−3, Cold Spring Harbor Laboratory Press, publ. (1989), またはAusubel, F. ら, Current Protocols in Molecular Biology, Green Publishing and Wiley−Interscience, New York, publ. (1987)および定期的更新に記載されている。通常、ポリペプチドをコードするDNAは、哺乳類、ウイルスまたは昆虫遺伝子に由来する適した転写または翻訳調節エレメントと作動可能なように連結されている。このような調節エレメントとして、転写プロモーター、転写を制御するための所望によるオペレーター配列、適したmRNAリボソーム結合部位をコードする配列ならびに転写および翻訳の終結を制御する配列が挙げられる。宿主において複製する能力は、通常、複製起点によって付与され、形質転換体の認識を容易にする選択遺伝子がさらに組み込まれる。
【0086】
本明細書に記載されるタンパク質は、直接的にだけではなく、好ましくは、シグナル配列または成熟タンパク質またはポリペプチドのN末端に特異的切断部位を有する他のポリペプチドである異種ポリペプチドとの融合ポリペプチドとしても組換えによって生成してもよい。選択される異種シグナル配列は、宿主細胞によって認識され、プロセシングされる(すなわち、シグナルペプチダーゼによって切断される)ものであることが好ましい。
【0087】
天然シグナル配列を認識およびプロセシングしない原核生物宿主細胞のために、シグナル配列は、例えば、アルカリホスファターゼ、ペニシリナーゼ、lppまたは熱安定性エンテロトキシンIIリーダーの群から選択される原核生物シグナル配列で置換される。
【0088】
酵母分泌のためには、天然シグナル配列を、例えば、酵母インベルターゼリーダー、因子リーダー(サッカロミセス(Saccharomyces)およびクルイベロマイセス(Kluyveromyces)アルファ因子リーダーを含む)、または酸性ホスファターゼリーダー、C.アルビカンス(albicans)グルコアミラーゼリーダー、または米国特許第5,631,144号に記載されるシグナルによって置換してもよい。哺乳類細胞発現では、哺乳類シグナル配列ならびにウイルスの分泌リーダー、例えば、単純ヘルペスgDシグナルが利用可能である。このような前駆体領域のDNAを、リーディングフレームでタンパク質をコードするDNAとライゲートしてもよい。
【0089】
発現ベクターおよびクローニングベクターは、両者とも、ベクターが一以上の選択された宿主細胞において複製するのを可能にする核酸配列を含有する。通常、クローニングベクターでは、この配列は、ベクターが宿主染色体DNAとは無関係に複製するのを可能にするものであり、複製起点または自立複製配列を含む。このような配列は、種々の細菌、酵母およびウイルスについてよく知られている。プラスミドpBR322に由来する複製起点は、ほとんどのグラム陰性菌にとって適しており、2ミクロンのプラスミド起点が酵母に適しており、種々のウイルス起点(SV40、ポリオーマ、アデノウイルス、VSVまたはBPV)が、哺乳類細胞中のクローニングベクターにとって有用である。一般に、複製起点成分は、哺乳類発現ベクターにとって必要ではない(SV40起点は、通常、単に、それが初期プロモーターを含むために使用され得る)。
【0090】
発現ベクターおよびクローニングベクターは、選択マーカーとも呼ばれる選択遺伝子を含有し得る。通常の選択遺伝子は、(a)抗生物質または他の毒素、例えば、アンピシリン、ネオマイシン、メトトレキセートもしくはテトラサイクリンに対する耐性を付与する、(b)栄養要求欠損を補完する、または(c)複合培地からは利用できない決定的な栄養素を供給するタンパク質をコードするものであり、例えば、バチルス(Bacilli)のD−アラニンラセマーゼをコードする遺伝子が挙げられる。
【0091】
発現ベクターおよびクローニングベクターは、通常、宿主生物によって認識され、本発明のタンパク質、例えば、フィブロネクチンベースの足場タンパク質をコードする核酸と作動可能なように連結しているプロモーターを含有する。原核生物宿主とともに使用するのに適したプロモーターとして、phoAプロモーター、β−ラクタマーゼおよびラクトースプロモーター系、アルカリホスファターゼ、トリプトファン(trp)プロモーター系およびtacプロモーターなどのハイブリッドプロモーターが挙げられる。しかしながら、他の既知細菌プロモーターも適している。細菌系において使用するためのプロモーターはまた、本発明のタンパク質をコードするDNAと作動可能なように連結しているシャイン−ダルガーノ(S.D.)配列を含む。真核生物のためのプロモーター配列は公知である。実質的にすべての真核細胞遺伝子は、転写が開始される部位からおよそ25〜30塩基上流に位置するATリッチ領域を有する。多数の遺伝子の転写の開始から70〜80塩基上流に見られる別の配列は、CNCAAT領域である(ここで、Nは、任意のヌクレオチドであり得る)。ほとんどの真核細胞遺伝子の3’末端には、コード配列の3’末端へのポリAテールの付加のためのシグナルであり得るAATAAA配列がある。これらの配列のすべてが、真核細胞発現ベクターに適宜挿入される。
【0092】
酵母宿主とともに使用するのに適したプロモーター配列の例として、3−ホスホグリセレートキナーゼまたはエノラーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース−6−リン酸イソメラーゼ、3−ホスホグリセレートムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼおよびグルコキナーゼなどの他の解糖酵素のプロモーターが挙げられる。
【0093】
哺乳類宿主細胞におけるベクターからの転写は、例えば、ポリオーマウイルス、鶏痘ウイルス、アデノウイルス(アデノウイルス2など)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよび最も好ましくは、サルウイルス40(SV40)などのウイルスのゲノムから、異種哺乳類プロモーター、例えば、アクチンプロモーターまたは免疫グロブリンプロモーターから、ヒートショックプロモーターから得られたプロモーターによって、このようなプロモーターが宿主細胞系と適合するならば、制御され得る。
【0094】
高等真核生物による本発明のタンパク質をコードするDNAの転写は、ベクター中にエンハンサー配列を挿入することによって増大されることが多い。哺乳類遺伝子(グロビン、エラスターゼ、アルブミン、アルファ−フェトプロテインおよびインスリン)に由来する多数のエンハンサー配列が現在知られている。しかしながら、通常、真核細胞ウイルス由来のエンハンサーを使用する。例として、複製起点の後ろ側のSV40エンハンサー(bp100〜270)、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後ろ側のポリオーマエンハンサーおよびアデノウイルスエンハンサーが挙げられる。真核細胞プロモーターの活性化のためのエンハンサーエレメントに関しては、Yaniv, Nature 297:17−18(1982)も参照。エンハンサーは、ペプチドをコードする配列の5’または3’側の位置でベクター中にスプライシングされ得るが、プロモーターの5’側に位置することが好ましい。
【0095】
真核宿主細胞(例えば、酵母、真菌、昆虫、植物、動物、ヒトまたは他の多細胞生物に由来する有核細胞)において使用される発現ベクターはまた、転写の終結のために、またmRNAを安定化するために必要な配列も含有する。このような配列は通常、真核細胞またはウイルスのDNAまたはcDNAの5’非翻訳領域、および場合により、3’非翻訳領域から得ることができる。これらの領域は、本発明のタンパク質をコードするmRNAの非翻訳部分中のポリアデニル化断片として転写されるヌクレオチドセグメントを含有する。一つの有用な転写終結成分として、ウシ成長ホルモンポリアデニル化領域がある。WO94/11026およびそこで開示される発現ベクターを参照。
【0096】
組換えDNAはまた、タンパク質を精製するのに有用であり得る任意の種類のタンパクン質タグ配列を含み得る。タンパク質タグの例として、それだけには限らないが、ヒスチジンタグ、FLAGタグ、mycタグ、HAタグまたはGSTタグが挙げられる。細菌、真菌、酵母および哺乳類細胞宿主とともに使用するのに適当なクローニングおよび発現ベクターは、Cloning Vector: A Laboratory Manual, (Elsevier, New York, 1985))中に見ることができ、その関連の開示内容は、出典明示により本明細書に組み込まれる。
【0097】
発現コンストラクトは、当業者には明らかであるように、宿主細胞にとって適当な方法を使用して宿主細胞に導入される。核酸を宿主細胞に導入するための種々の方法が当技術分野で公知であり、これらには限らないが、エレクトロポレーション;塩化カルシウム、塩化ルビジウム、リン酸カルシウム、DEAE−デキストランまたは他の物質を使用するトランスフェクション;微粒子銃;リポフェクション;および感染(ベクターが感染性物質である)が挙げられる。
【0098】
適した宿主細胞として、原核生物、酵母、哺乳類細胞または細菌細胞が挙げられる。適した細菌として、グラム陰性またはグラム陽性菌、例えば、大腸菌またはバチルス種が挙げられる。好ましくは、サッカロミセス種に由来する酵母、例えば、S.セレビシエ(cerevisiae)も、ポリペプチドの生成のために使用してよい。種々の哺乳類または昆虫細胞培養系を使用して、組換えタンパク質を発現することもできる。昆虫細胞において異種タンパク質を生成するためのバキュロウイルス系は、Luckowら(Bio/Technology, 6:47(1988))に総説されている。適した哺乳類宿主細胞株の例として、内皮細胞、COS−7サル腎臓細胞、CV−1、L細胞、C127、3T3、チャイニーズハムスター卵巣(CHO)、ヒト胚性腎細胞、HeLa、293、293TおよびBHK細胞株が挙げられる。精製ポリペプチドは、組換えタンパク質を発現するのに適した宿主/ベクター系を培養することによって調製される。多数の適用のために、本明細書に開示される小さいサイズの多数のポリペプチドは、発現のための好ましい方法として大腸菌において発現を行う。次いで、タンパク質は、培養培地または細胞抽出物から精製される。
【0099】
タンパク質生成
宿主細胞は、タンパク質生成のために本明細書に記載される発現またはクローニングベクターを用いて形質転換され、プロモーターを誘導し、形質転換体を選択し、または所望の配列をコードする遺伝子を増幅するために適当に改変された従来の栄養培地で培養される。本明細書に示される実施例では、ハイスループットタンパク質生成(HTPP)および中規模生成のために使用される宿主細胞は、BL21 DE3 plyS−細菌株であった。本発明のタンパク質を生成するために使用される宿主細胞は、Hamら, Meth. Enzymol. 58:44 (1979)、Baritesら, Anal. Biochem.102:255 (1980)、米国特許第4,767,704号、同4,657,866号、同4,927,762号、同4,560,655号、同5,122,469号、同6,048,728号、同5,672,502号または米国特許第RE30,985号に記載されるものなどの、種々の培地で培養してもよい。任意の他の必要な栄養補助剤を、当業者に公知である適当な濃度で含めてもよい。温度、pHなどといった培養条件は、発現のために選択された宿主細胞とともにこれまでに使用されたものであり、当業者には明らかであろう。
【0100】
本明細書に開示されるタンパク質はまた、細胞−翻訳系を使用して生成できる。このような目的上、ポリペプチドをコードする核酸は、インビトロ(in vitro)転写を可能にしてmRNAを生成し、使用されている特定の無細胞系(哺乳類または酵母などの真核細胞を含まない翻訳系または細菌などの原核細胞を含まない翻訳系)におけるmRNAの無細胞翻訳を可能にするよう改変されなければならない。
【0101】
本発明のタンパク質はまた、化学合成によって(例えば、Solid Phase Peptide Synthesis, 2nd Edition, The Pierce Chemical Co., Rockford, IL, publ. (1984)に記載される方法によって)生成できる。タンパク質への修飾も化学合成によって生成できる。
【0102】
本発明のタンパク質は、タンパク質化学の分野で一般に知られている、タンパク質の単離/精製法によって精製できる。非限定的な例として、抽出、再結晶化、塩析(例えば、硫酸アンモニウムまたは硫酸ナトリウムを用いる)、遠心分離、透析、限外濾過、吸着クロマトグラフィー、イオン交換クロマトグラフィー、疎水性クロマトグラフィー、順相クロマトグラフィー、逆相クロマトグラフィー、ゲル濾過、ゲル浸透クロマトグラフィー、アフィニティークロマトグラフィー、電気泳動、向流分配またはこれらの任意の組合せが挙げられる。精製後、ポリペプチドを、これらには限らないが、濾過および透析を含む、当技術分野で公知の種々の方法のいずれかによって、異なるバッファーに交換してもよく、かつ/または、濃縮してもよい。
【0103】
精製されたポリペプチドは、好ましくは少なくとも85%純粋、より好ましくは少なくとも95%純粋、最も好ましくは少なくとも98%純粋である。純度の正確な数値にかかわらず、ポリペプチドは医薬製剤として使用するのに十分に純粋である。
【0104】
プラットフォーム製造工程(platform manufacturing process)を使用して、抗IL−23アドネクチンを調製した。実施例1には製造工程の一例が記載されている。アドネクチンは大腸菌(E.coli)内で生成された。大腸菌MG1655細胞を、封入体として不溶型でタンパク質を生成する発現ベクター(pBMS2008/ATI001044)で形質転換した。組換え株を撹拌式発酵タンク内で生育させる。発酵終了時に、精製に向けて、封入体を回収し、可溶化し、かつリフォールドさせた。精製したアドネクチンを、マレイミドリンカーを用いて40kDaの分岐メトキシPEGへコンジュゲートした。次いで、コンジュゲート化物質を再精製し、遊離PEG、遊離アドネクチン、および製品に関連する不純物を除去した。品質管理テストをバルク薬剤原料に対して行った。
【0105】
治療的インビボ(in vivo)使用
一態様では、本出願は、狼蒼(たとえば、紅斑性狼蒼、腎炎性狼蒼)、橋本甲状腺炎、原発性粘液水腫、グレーブス病、悪性貧血、自己免疫萎縮性胃炎、アジソン病、糖尿病(たとえば、インスリン依存型糖尿病、I型糖尿病)、グッドパスチャー症候群、重症筋無力症、天疱瘡、クローン病、交感神経性眼炎、自己免疫ブドウ膜炎、多発性硬化症、自己免疫性溶血性貧血、突発性血小板減少症、原発性胆汁性肝硬変、慢性活動性肝炎、潰瘍性大腸炎、シェーグレン症候群、リウマチ性疾患(たとえば、関節リウマチ)、多発性筋炎、強皮症および混合性結合組織病などの、自己免疫疾患の治療において有用である抗IL−23アドネクチンを提供する。
【0106】
本出願はまた、被験体に抗IL−23アドネクチンを投与する方法を提供する。いくつかの実施形態では、被験体はヒトである。いくつかの実施形態では、抗IL−23アドネクチンは、哺乳類、特に、ヒトにとって医薬上許容される。「医薬上許容される」ポリペプチドとは、本質的にエンドトキシンを含まない、または極めて低いエンドトキシンレベルを有するなどの、著しく有害な医学的結果を伴わずに動物に投与されるポリペプチドを指す。
【0107】
製剤および投与
本出願はさらに、本明細書に記載される抗IL−23アドネクチンを含む医薬上許容される組成物を提供し、該組成物は本質的にエンドトキシンを含まない。抗IL−23アドネクチンを含む治療用製剤は、所望の程度の純度を有する記載されるアドネクチンを、任意の生理学的に許容される担体、賦形剤または安定剤と混合することによって、水溶液、凍結乾燥された、または他の乾燥された製剤の形態で、保存のために調製される(Osol, A., ed., Remington’s Pharmaceutical Sciences, 16th Edition (1980))。許容される担体、賦形剤または安定剤は、使用される用量および濃度でレシピエントにとって非毒性であり、これらの例として、リン酸、クエン酸および他の有機酸などのバッファー;アスコルビン酸およびメチオニンを含む抗酸化物質;保存料(オクタデシルジメチルベンジル塩化アンモニウム;塩化ヘキサメトニウム;塩化ベンザルコニウム、塩化ベンゼトニウム;フェノール、ブチルもしくはベンジルアルコール;メチルもしくはプロピルパラベンなどのアルキルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3−ペンタノール;およびm−クレゾールなど);低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチンまたは免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニンまたはリシンなどのアミノ酸;単糖、二糖およびグルコース、マンノースまたはデキストランを含む他の炭水化物;EDTAなどのキレート化剤;スクロース、マンニトール、トレハロースまたはソルビトールなどの糖;ナトリウムなどの塩形成性対イオン;金属錯体(例えば、Zn−タンパク質複合体);ならびに/あるいはTWEEN、PLURONIC登録商標またはポリエチレングリコール(PEG)などの非イオン性界面活性剤が挙げられる。
【0108】
また、本明細書において製剤とは、治療されている特定の適応症にとって必要な二以上の活性化合物、好ましくは、互いに有害な影響を及ぼさない相補的な活性を有するものを含み得る。このような分子は、意図される目的上効果的である量の組合せで適宜存在する。
【0109】
インビボ(in vivo)投与に使用される製剤は、無菌でなければならない。これは、滅菌濾過メンブレンを通した濾過によって容易に達成される。
【0110】
当業者ならば、各治療薬の用量が、薬剤の独自性によって決まることを理解するであろう。
【0111】
治療適用のためには、抗IL−23アドネクチンは、医薬上許容される剤形で被験体に投与される。それは、ボーラス(bolus)として、または一定時間にわたる連続注入によって静脈内に、または皮下経路によって投与され得る。適した医薬上許容される担体、希釈剤および賦形剤は周知であり、臨床状況が必要とする際に、当業者によって決定され得る。適した担体、希釈剤および/または賦形剤の例として、(1)ダルベッコのリン酸緩衝生理食塩水、(2)0.9%生理食塩水(0.9% w/v NaCl)、および(3)5%(w/v)デキストロースが挙げられる。
【0112】
本発明の方法は、インビトロ(in vitro)、インビボ(in vivo)またはエキソビボ(ex vivo)で実施できる。
【0113】
抗IL−23アドネクチンおよび一以上のさらなる治療薬の投与は、同時投与されるか逐次投与されるかにかかわらず、治療適用のために、上記のように起こり得る。同時投与に適した医薬上許容される担体、希釈剤および賦形剤は、投与されている特定の治療薬の独自性によって決まる、と当業者に理解されるであろう。
【0114】
凍結乾燥されているのではなく水性の剤形で存在する場合には、タンパク質は、通常、約0.1mg/ml〜100mg/mlの濃度で製剤化されるが、これらの範囲の外側の広い変動も容認される。疾患の治療のための、抗IL−23アドネクチンの適当な用量は、治療される疾患の種類、疾患の重篤度および経過、アドネクチンが予防目的で投与されるのか治療目的で投与されるのか、これまでの治療の経過、患者の病歴およびアドネクチンに対する反応、ならびに主治医の裁量によって決まるであろう。タンパク質は、1回で、または一連の治療にわたって、患者に適宜投与される。
【0115】
血清アルブミン結合アドネクチン(SABA)の融合
特定の態様では、本出願は、ヒト血清アルブミンに結合する10Fn3ドメインに融合した抗IL−23アドネクチンを含む融合タンパク質(血清アルブミン結合アドネクチン Serum Albumin Binding Adnectin(10Fn3ドメイン)またはSABA)を提供する。このような融合タンパク質は、抗IL−23アドネクチン単独に比べて、アルブミン存在下で延長した半減期を有する。
【0116】
特定の態様では、本出願は、融合タンパク質のt1/2を延長するため、血清アルブミン、例えばヒト血清アルブミン(HSA)に特異的に結合する10Fn3ドメインを含む融合タンパク質を提供する。
【0117】
特定の実施形態では、SABAに融合された抗IL−23アドネクチンの血清半減期は、SABAへコンジュゲートされなかった場合の抗IL−23アドネクチンの血清半減期と比べて増大する。特定の実施形態では、SABA融合体の血清半減期は、SABAに融合されなかった場合の抗IL−23アドネクチンの血清半減期と比べて、少なくとも20、40、60、80、100、120、150、180、200、400、600、800、1000、1200、1500、1800、1900、2000、2500、または3000%長い。他の実施形態では、SABA融合体の血清半減期は、SABAに融合されなかった場合の抗IL−23アドネクチンの血清半減期より、少なくとも1.5倍、2倍、2.5倍、3倍、3.5倍、4倍、4.5倍、5倍、6倍、7倍、8倍、10倍、12倍、13倍、15倍、17倍、20倍、22倍、25倍、27倍、30倍、35倍、40倍、または50倍大きい。いくつかの実施態様では、SABA融合体の血清半減期は、少なくとも10時間、15時間、20時間、25時間、30時間、35時間、40時間、50時間、60時間、70時間、80時間、90時間、100時間、110時間、120時間、130時間、135時間、140時間、150時間、160時間、または200時間である。
【0118】
従って、本明細書に記載されるSABA融合分子は、抗IL−23アドネクチンおよびSABA間の融合体の構築によって抗IL−23アドネクチンの半減期を増大させるために有用である。このような融合分子を使用して、IL−23の生物活性へ応答する症状を治療してもよい。本発明は、IL−23の異常調節によって引き起こされる疾患におけるSABA融合分子の使用を企図する。
【0119】
融合体は、抗IL−23アドネクチンをSABA分子の末端のいずれかへ付着させることこと、すなわち、SABA−抗IL−23アドネクチン配置または抗IL−23アドネクチン−SABA配置で付着させることによって、形成される。
【0120】
一態様では、本開示は血清アルブミン結合10Fn3ドメインを含む抗IL−23アドネクチンを含む融合タンパク質を提供する。例示的な実施形態では、本明細書に記載される血清アルブミン結合10Fn3タンパク質は、3uM、2.5uM、2uM、1.5uM、1uM、500nM、100nM、50nM、10nM、1nM、500pM、200pM、100pM、50pMまたは10pM未満のKdでHSAに結合する。特定の実施形態では、本明細書に記載される血清アルブミン結合10Fn3タンパク質は、25℃または37℃で、5.5から7.4のpH範囲で、3uM、2.5uM、2uM、1.5uM、1uM、500nM、100nM、50nM、10nM、1nM、500pM、200pM、100pM、50pMまたは10pM未満のKdでHSAに結合する。いくつかの実施形態では、本明細書に記載される血清アルブミン結合10Fn3タンパク質は、7.4以上のpHでのHSAに対する結合親和性と比較して、pH7.4未満でより強くHSAに結合する。
【0121】
特定の実施形態では、本明細書に記載されるHSA結合10Fn3ドメインを含む融合タンパク質はまた、サル、ラット、またはマウスの一以上に由来する血清アルブミンにも結合する。特定の実施形態では、本明細書に記載される血清アルブミン結合10Fn3タンパク質は、3uM、2.5uM、2uM、1.5uM、1uM、500nM、100nM、50nM、10nM、1nM、500pMまたは100pM未満のKdで、アカゲザル血清アルブミン(RhSA)またはカニクイザル血清アルブミン(CySA)に結合する。
【0122】
特定の実施形態では、本明細書に記載される血清アルブミン結合10Fn3ドメインを含む融合タンパク質は、HSAのドメインIおよび/またはドメインIIに結合する。一の実施形態では、本明細書に記載される血清アルブミン結合10Fn3ドメインを含む融合タンパク質は、HSAのドメインIIIに結合しない。
【0123】
特定の実施形態では、融合タンパク質の血清アルブミン結合10Fn3部分(SABA)は、野生型10Fn3ドメイン(配列番号1)と少なくとも40%、50%、60%、70%、75%、80%または85%同一性を有する配列を含む。一の実施形態では、野生型10Fn3ドメインと比べて、BC、DEまたはFGループのうち少なくとも1つが修飾されている。別の実施形態では、野生型10Fn3ドメインと比べて、BC、DEまたはFGループのうち少なくとも2つが修飾されている。別の実施形態では、野生型10Fn3ドメインと比べて、BC、DEおよびFGループのうち3つ全てが修飾されている。他の実施形態では、SABAは、表3に示される26のコア(core)SABA配列(すなわち、配列番号103、107、111、115、119、および123〜143)のいずれか1つ、または、表3に示される伸長したSABA配列(すなわち、配列番号188〜215、6×HISタグを引く)のいずれか1つと、少なくとも40%、50%、60%、70%、75%、80%、85%、90%、または95%同一性を有する配列を含む。
【0124】
特定の実施形態では、コアアミノ酸残基は固定されており、任意の置換、保存的置換、欠失または付加は、コアアミノ酸残基以外の残基で生じる。例示的な実施態様では、BC、DEおよびFGループは、以下表3に示されるHSAバインダーのいずれかに由来するBC、DEおよびFGループ配列(すなわち、表3の配列番号103、107、111、115、119、および123〜143)を含むポリペプチドで置換される。
【0125】
特定の実施形態では、SABA(例えば、SABAコア配列または上述のそれらに由来する配列)は、N末端伸長配列および/またはC末端伸長配列を含むように改変されてもよい。例示的な伸長配列は、表3に示される。例えば、SABA1.1として表される配列番号188は、N末端配列MGVSDVPRDLE(配列番号144,AdNT1と表す)およびC末端配列EIDKPSQ(配列番号153)を備える、コアSABA1配列(配列番号103)を含む。SABA1.1はさらにHis6タグをC末端で含む、しかしながら、His6タグは完全に任意であり、N−またはC−末端伸長配列内のどこにでも設置されうる。さらに、表3に提供される例示的なN−またはC−末端伸長配列(配列番号144〜163)およびそれらの変種のいずれかを使用して、表3に提供される任意の与えられるSABAコア配列を修飾することができる。
【0126】
他の実施形態では、SABA融合分子を設計する場合に必要であれば、テール配列は他の知られたリンカー配列(例えば、表3、配列番号164〜187)と結合させてもよい。
【0127】
コンジュゲーションリンカー
SABA融合体は共有結合的に、または非共有結合的に連結されてもよい。いくつかの実施形態では、血清アルブミン結合10Fn3は、直接的にまたは間接的に、ポリペプチドリンカーを介して抗IL−23アドネクチンに連結された。Fn3の連結のための適したリンカーは、分離されたドメインが、標的分子に結合する高い親和性を可能にするような三次元構造の形成をお互い独立してフォールドできるようにするものである。
【0128】
本開示はこれらの必要条件を満たす多数の適したリンカーを提供し、グリシン−セリンベースのリンカー、グリシン−プロリンベースのリンカー、ならびにアミノ酸配列PSTSTST(配列番号184)を有するリンカーが挙げられる。本明細書に記載される実施例では、ポリペプチドリンカーを介して連結されたFn3ドメインが、その標的結合機能を保持することを実証する。いくつかの実施形態では、リンカーは、グリシン−セリンベースのリンカーである。これらのリンカーはグリシンおよびセリン残基を含み、8から50、10から30、および10から20個の間のアミノ酸の長さであり得る。例として、アミノ酸配列(GS)7(配列番号171)、G(GS)6(配列番号166)、およびG(GS)7G(配列番号168)を有するリンカーが挙げられる。他のリンカーはグルタミン酸を含有し、例えば、(GSE)5(配列番号173)およびGGSE GGSE(配列番号177)が挙げられる。他の例示的なグリシン−セリンリンカーとして、(GS)4(配列番号170)、(GGGGS)7(配列番号179)、(GGGGS)5(配列番号180)、および(GGGGS)3G(配列番号181)が挙げられる。いくつかの実施形態では、リンカーは、グリシン−プロリンベースのリンカーである。これらのリンカーは、グリシンおよびプロリン残基を含み、3から30、10から30、および3から20個の間のアミノ酸の長さであり得る。例として、(GP)3G(配列番号182)および(GP)5G(配列番号183)を有するリンカーが挙げられる。他の実施形態では、リンカーは3から30、10から30、および3から20アミノ酸の間の長さを有するプロリン−アラニンベースのリンカーであってもよい。プロリン−アラニンベースのリンカーの例として、例えば、(PA)3(配列番号185)、(PA)6(配列番号186)および(PA)9(配列番号187)が挙げられる。最適リンカー長およびアミノ酸組成は、当技術分野で周知の方法によって日常的な実験によって決定され得ると考えられる。
【0129】
いくつかの実施形態では、本明細書に記載される融合体は、血液または標的組織においてプロテアーゼによって切断可能であるプロテアーゼ部位を有するポリペプチドリンカーを介して連結される。このような実施形態を使用して、より良好なデリバリーまたは治療的特性、またはより効率的な生成のために、治療用タンパク質を放出できる。
【0130】
さらなるリンカーまたはスペーサーは、Fn3ドメインのC末端において、Fn3ドメインとポリペプチドリンカーの間に導入され得る。さらなるリンカーまたはスペーサーは、Fn3ドメインのN末端において、Fn3ドメインとポリペプチドリンカーの間に導入され得る。
【0131】
いくつかの実施形態では、治療的部分(therapeutic moiety)は、ポリマーリンカーを介して直接的または間接的にSABAに連結されてもよい。ポリマーリンカーを使用して、以下の特徴のうち一以上を有するタンパク質融合体を作り出すために、融合体の各構成要素(component)間の距離を最適に変えることができる:1)対象とするタンパク質との結合時の、一以上のタンパク質ドメインの結合の立体障害の低減または増大、2)タンパク質安定性または溶解度の増大、3)タンパク質凝集の減少、および4)タンパク質の全結合力または親和性の増大。
【0132】
いくつかの実施形態では、治療的部分は、重合糖などの生体適合性ポリマーを介してSABAに連結している。重合糖は、血液または標的組織において酵素によって切断可能である酵素切断部位を含み得る。このような実施形態を使用して、より良好なデリバリーまたは治療的特性、またはより効率的な生成のために、治療用タンパク質を放出できる。
【0133】
血清アルブミン結合アドネクチン(SABA)配列の概要
本願で参照されるSABA配列の多くは、以下の表3に要約される。特別の定めのない限り、全てのN末端伸長は下線で示され、全てのC末端テール/伸長は二重下線で示され、リンカー配列は囲まれている。ループ領域BC、DEおよびFGは、各コアSABA配列に対して網掛けされている。
【表3−1】
【表3−2】
【表3−3】
【表3−4】
【表3−5】
【表3−6】
【表3−7】
【表3−8】
【表3−9】
【表3−10】
【表3−11】
実施例
【実施例1】
【0134】
製造過程
発酵および集菌
生産発酵(production fermentation)は無菌の基礎培地で調製される。バイアルを融解して、成長培地を含む移動容器に接種するために使用する。接種材料は直ちに生産発酵へ移される。培養は撹拌して34℃の温度で維持され、OD600が5〜10(1OD単位は約1x109細胞/mL)に到達するまで生育させる。流加培地の添加はこのODで開始する。発酵は、培養がイソプロピルβ−D−1−チオガラクトピラノシド(IPTG)の添加によりアドネクチンを生成するために誘導される点である、OD600=25まで継続する。容器の温度は誘導の際、34℃から39℃へ上昇される。サンプルは毎時間、無菌的に採取され、細胞密度を試験する。
【0135】
9〜12時間の誘導発酵の後、25℃へ温度を低下させ、終濃度10mMでエチレンジアミン四酢酸(EDTA)を添加し、水酸化ナトリウムの添加によりpHを上昇させ、かつ、撹拌を弱めることにより、容器を回収(harvest)のために準備する。一時間の保持期間(hold period)の後、発酵容器内容物を集菌容器へ流し入れる。
【0136】
封入体の調製
回収プールの細胞破砕は、細胞を破砕してその内容物を放出させるMICROFLUIDIZER登録商標に通すことによって行う。細胞破砕に続いて、非常に強い遠心力により、連続工程で、固体および液相を分離するための、ディスクスタック遠心分離機を使用して、封入体が回収される。次いで封入体を、バッファー(20〜25℃)で2回、水(20〜25℃)で2回、洗浄する。封入体を洗浄するたび、遠心分離によって回収する。洗浄された封入体をスラリーとして回収する。
【0137】
封入体の可溶化およびタンパク質リフォールディング
可溶化バッファーを封入体スラリーへ添加し、続いて室温で1時間撹拌する。OD280=20(総タンパク質)がこの工程の間の目標とされる。
【0138】
タンパク質リフォールディングは二段階希釈工程を使用して実行する。希釈バッファーを、可溶化封入体一部に対して希釈バッファー二分の一部の比率(v/v)で、可溶化封入体へ添加する。第二の希釈は、OD280=0.7(総タンパク質)を目標として、可溶化封入体をリフォールドバッファーへ添加することにより実行する。前記の希釈は室温で撹拌しながら実行する。一時間の混合を実行し、撹拌を止め、タンパク質溶液を室温で一晩保つ。可溶化されリフォールディングされたアドネクチンは、0.8μm〜22μmのフィルターに通され、A280およびRP−HPLCによりタンパク質含有量に関して試験される。
【0139】
精製およびPEGへのコンジュゲーション
リフォールドされフィルター処理されたアドネクチンは、初期回収(initial capture)のため、陽イオン交換(CEX1)カラムへ直接ロードされる。結合した物質を洗浄バッファーで洗浄し、50mM酢酸ナトリウム、500mM塩化ナトリウム、1.5%プロピレングリコール、pH5.5で溶出する。溶出プールを、純度、同一性、濃度、およびエンドトキシンに関して測定する。
【0140】
回収クロマトグラフィー(capture chromatography)からの溶出物をさらに、疎水性相互作用クロマトグラフィー(HIC)を使用して精製する。CEX1溶出物をHICカラムへ直接ロードし、洗浄し、続いて50mM酢酸ナトリウム、30%プロピレングリコール、pH5.5で溶出する。溶出プールを、純度、同一性、および濃度に関して測定する。
【0141】
精製されたアドネクチンはその後、40kDaの分岐PEGのマレイミド誘導体(mPEG2−MAL)で直接整形(format)される。HIC溶出物は室温で撹拌され、mPEG2−MALが添加される。室温で1時間混合後、反応混合液を同じ温度で一晩インキュベートしてもよい。次いで、PEG化溶液を最終CEXカラム(CEX2)へと進める。サンプルはタンパク質含量、純度、およびエンドトキシンのために採取される。
【0142】
PEG化溶液のpHおよび伝導度は、再精製のため、最終陽イオン交換カラム(CEX2)へロードするより前に、75mM酢酸でそれぞれ4.0および1.0mS/cmに調節される。一度ロードしたら、結合した物質を洗浄バッファーで洗浄し、続いて50mM酢酸ナトリウム、25mM塩化ナトリウム、pH5.0で溶出する。サンプルはタンパク質含量、純度、およびエンドトキシンのために採取される。
【0143】
CEX2溶出物は、V−スクリーンを有する30kDaの名目分子量カットオフメンブレンを備えるタンジェンシャルフロー濾過ユニットで、15mg/mLへ濃縮される。50mM酢酸ナトリウム、25mM塩化ナトリウム、pH5.0中のバルク薬剤原料を0.22μmのフィルターに通し、−80℃で凍結した。
【実施例2】
【0144】
遺伝子、ベクターおよび宿主細胞
T7プロモーターの制御下でタンパク質をコードするプラスミドを、株構築(strain construction)での使用のために生成した。このプラスミドDNAを使用して、大腸菌K−12 MG1655コンピテントセル(F−ラムダ−、ilvG−rfb−50 rph−1)を形質転換させた。宿主株はIPTGの添加で遺伝子からの発現の誘導ができるように設計された。形質転換されたMG1655株はカナマイシンに対して耐性である。タンパク質発現ベクターは図2に示される。プレートからのシングルコロニー選択を使用して、発酵培養物を接種し、その後それを分注し、研究細胞バンクとして使用するため凍結させた。
【実施例3】
【0145】
生物物理学的特性評価および生化学的特性評価
本発明のタンパク質の構造および品質は、数種の総合的解析方法により試験した。
【0146】
MALDI−MS
質量スペクトルプロファイルはMALDIによって解析した。サンプル上のMALDI解析の正確さを評価するため、各サンプルに対してスチールプレート上に20の個別のスポットを設置し、連続して解析した。合計20スペクトルが生成された。
【0147】
ペプチドマッピングを使用して、本発明のタンパク質並びに対応する非PEG化タンパク質に関して、cDNA配列から予測されるアミノ酸(第一構造)の正確な発現を確認した。完全な配列カバー率を得るために、トリプシン(Lys残基およびArg残基のC末端側への切断)およびエンドプロテイナーゼGlu−C(Glu残基のC末端側への切断)を採用して、ペプチド断片の2つのオーバーラップするセットを得た。ペプチドマッピングを使用して、共有結合的な翻訳後修飾として挙げられる残存N末端メチオニン、ジスルフィド結合、アスパラギンの脱アミノ化、メチオニン酸化(等)も決定した。ペプチドは、分子量による液体クロマトグラフィー質量分析(LC−MS)、および衝突誘起解離(CID)によって部分的な配列情報を提供するタンデム質量分析(MSMS)により、同定され、かつ特徴づけられた。
【0148】
SDS−PAGE
ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)を使用して、非PEG化およびPEG化の抗IL−23アドネクチンの分子量バンドパターンを可視化した。サンプルは、還元剤有りまたは無しのサンプルバッファー中に調製された。SDS中で加熱の後、サンプルおよび分子量マーカーを、プレ−キャストのグラジエント(4−20%)ポリアクリルアミドゲル上で、電気泳動的に解析した。電気泳動後、ゲルを固定し、クーマシーブルーを使用して染色した。サンプルのバンドパターンの同等性を視覚的に評価した。
【0149】
サイズ排除クロマトグラフィー/マルチアングル光散乱(SECMALS)
サイズ排除クロマトグラフィー(SEC)を、高分子量(HMW)種および低分子量(LMW)種のモノマーの定量的解析のために使用した。続くSEC分離で、分離された種の分子量を、示差屈折率計を並んで備えるマルチアングル光散乱により決定した。
【実施例4】
【0150】
インビトロ(in vitro)非臨床薬理
SPRによるKD
結合特性は、表面プラズモン共鳴(SPR)により特徴づけられる。ヒトIL−23を、ProteOn XRP(Bio−Rad)チップ表面の一次元に2から4レベルで固定化し、同じSPRチップ表面の他の次元で6種の異なる濃度の抗IL−23アドネクチンへと暴露させた。これにより再生無しで速度論的定量が可能となった。二重の(duplicate)チップを25℃および37℃での速度論的定量のために使用した。速度論的パラメータの評価は、ラングミュア相互作用モデル(Langmuir interaction model)およびProteOnマネージャーソフトウェアに適合する(fit with)定数パラメータを用いて行った。
【0151】
以下表4に示されるように、これらの抗IL−23アドネクチンに対する解離速度(off−rate)は、25℃では遅い(約10−5s−1)。37℃でさえ、その解離速度(off−rate)はSPR技術の検出限界に近かかったので、報告された解離定数測定は過小評価であるかもしれない。
【表4】
【0152】
溶液相親和性
ヒトIL−23に対するATI001045の溶液相親和性を、カイネティックエクスクルージョンアッセイ(Kinetic Exclusion Assay)(KinExA)を用いて測定した。一形式において、hIL−23の二重滴定は3つの濃度各々に対して行った。相対非結合ATI001045濃度をヒトIL−23固体マトリックス上への回収によって測定し、続いてアドネクチン足場を認識する蛍光標識抗体で検出した。技術的な制限により、試験できる最小の濃度は0.75nMであった。それゆえ、表5に示す包括的なKD解析はKDに対して51pMの見積もりを与えるものの、親和性は95%信頼区間で、1桁pMまで低く、あるいは、150pMまで高くなり得る。
【表5】
【0153】
ヒトIL−23に対するATI001045およびATI001047の溶液親和性は、KinExAにおいて代替形式を使用して測定された。アドネクチンの二重滴定は、3つの濃度(ATI001045)または単一の濃度(ATI001047)のヒトIL−23の各々に対して行われた(最も低い濃度に対しては4重測定)。相対非結合ヒトIL−23濃度を非PEG化ATI001045固体マトリックス上への回収によって測定し、続いてhIL−23のp40サブユニットを認識する蛍光標識抗体で検出した。表6に示す包括的なKD解析は、ATI001045に関して95%信頼区間で22〜2.4pMの、9.4pMのKDを与え、95%信頼区間で60.1から19.4pMの、36.3pMのKDを与える。
【表6】
【0154】
Kit225細胞でのSTAT3リン酸化
Parhamら (“A receptor for the heterodimeric cytokine IL−23 is composed of IL−12Rbeta1 and a novel cytokine receptor subunit, IL−23R”, J. Immunol., 168(11):5699−5708 (Jun. 1, 2002))は、ヒトIL−2依存性T細胞株、Kit225からIL−23Rをクローン化した。これらの細胞は、IL−12RB1ならびにIL−23Rの両方の発現に対してFACS解析により特徴づけられ、かつ、pSTAT3の刺激によるIL−23ならびにpSTAT4の刺激によるIL−12に応答した。Kit225細胞を96ウェルプレートへ播種し、37℃で3時間、FBSおよびIL−2の非存在下で静置した(quiesced)。このインキュベートに続き、ヒト組換えIL−23(または、1時間アンタゴニストとプレインキュベートしたIL−23)を添加し、STAT3のリン酸化(p−STAT3として略される)を刺激するために、細胞を37℃で15分間、インキュベーターへ戻した。各条件を96ウェルプレートにて二重測定で測定した。刺激は細胞の氷上静置および氷冷PBSの添加により停止させた。最後に、標準的なプロトコールに従って細胞をペレット化および溶解し、ELISAによってpSTAT3生成を検出した。
【0155】
刺激のためのIL−23の最適濃度は35pMであった。IL−23誘導性pSTAT3の阻害は、抗p40モノクローナル抗体(mAb1510)ならびに抗p19ポリクローナル抗体(AF1716)の滴定によって実証した。ATI001045、ATI001047、ATI001014およびATI001016は、抗p19ポリクローナル抗体よりも約150倍強力な、〜300pMのIC50を備える同等の活性を有した一方、ATI001015は、抗p19ポリクローナル抗体よりも約40倍強力な、〜1.2nMのIC50を有した。アドネクチンATI000934は、1nMのIC50を備え、ATI001045の1/3の効力である(表7)。
【表7】
【0156】
ヒトPBMCでのSTAT3リン酸化
二代細胞をベースとする確認測定は、初代ヒト細胞における作用メカニズムとして、STAT3のリン酸化を評価する目的で開発された。健常ドナー由来の末梢血単核細胞(PBMC)は、低レベルのIL−23Rを名目上発現し、外因性IL−23で刺激された場合ほとんど応答しない、ナイーブで休眠状態のT細胞から主として成る。しかしながら、IL−2でのナイーブPBMCのポリクローナルな活性化は、その後IL−23Rの発現を増大させる、ナイーブT細胞の活性化および分化を引き起こす。これらの活性化した細胞はその後、STAT3のリン酸化を引き起こすSTAT経路を活性化する、外因性IL−23での刺激の影響を受けやすい。
【0157】
市販の抗体(AF1716、抗p19 pAbおよびmAb1510、抗p40 mAb、両者ともR&D Systemsから)を、IL−23誘導性STAT3リン酸化の阻害のための陽性対照として使用した。6種のアドネクチンの阻害活性を、多数のドナー由来の血液を使用する10の別実験において比較した(表8で要約する)。サブセットに関する例示的データを図4に示す。抗IL−23アドネクチンは、STAT3リン酸化において抗p19よりも著しく強力(>150倍)であったが、抗p40モノクローナル抗体より5倍弱い効力と同様であった。
【表8】
【0158】
マウス脾細胞によるIL−23誘導性サイトカイン産生
初代細胞を用いる初代細胞アッセイ(initial cellular assay)を設計し、抗IL−23アドネクチンの、マウスTh17細胞からのIL−23依存性サイトカイン分泌を阻害する能力を評価した。解析のためのマウスTh17細胞を分化させるために、CD4+T細胞を磁気ビーズで濃縮し、照射した脾細胞(irradiated splenocytes)と共培養し、TGF−βおよびIL−6ならびにIL−4およびIFN−γに対する中和抗体の存在下、抗CD3で活性化させた。6日間の培養後、極性化(polarized)Th17細胞を回収し、96ウェルプレートへ再播種し、100ng/ml ヒトIL−23および5ng/ml マウスIL−2で刺激した。IL−2の添加は、細胞生存を維持するために必要とされ、IL−23に応答して強力なサイトカイン産生を可能にするが、単独でIL−17AあるいはIL−22産生を強く誘導はしなかった。IL−2は低レベルのサイトカイン分泌を誘導するので、各サンプルセットには、IL−23の非存在下で産生されたサイトカインのベースラインレベルに関して調整するために、IL−2単独で刺激された細胞を含めた。IL−23依存性応答は、IL−2およびIL−23の組合せにより誘導されたサイトカインのレベルと、IL−2単独により誘導されたベースラインレベルの違いを計算することによって評価した。アドネクチンの用量範囲は、その抑制可能性を試験するために、IL−2およびIL−23でのTh17細胞の再刺激の間、追加された。ヒト抗p40抗体(R&D Systems MAB1510)の用量範囲は、IL−23阻害を評価するための陽性対照として、同時に実施された。各条件を96ウェルプレートにて三重測定で試験した。4日後、三重測定からの馴化培地をプールし、細胞残渣を除去し、ELISAによってIL−17A濃度およびIL−22濃度の両方を測定した。
【0159】
IL−2およびIL−23でのTh17細胞の刺激は、IL−2単独により誘導されたレベルと比較して、2から3倍のIL−17Aの増加、および、少なくとも5倍のIL−22の増強を誘導した。ATI000934、ATI001014、ATI001015、ATI001016、ATI001045および陽性対照の抗p40モノクローナル抗体は、IL−23依存性IL−17AおよびIL−22分泌において、用量依存的減少を調節した。IL−17AおよびIL−22分泌の両方の阻害に対するIC50値は、各アドネクチンならびに抗p40対照に対して計算され、これらのデータは表9に要約される。試験された全てのアドネクチンは、IL−23依存性IL−17A分泌の阻害に対して抗p40対照の2倍以内で強力であり、IL−23依存性IL−22産生の阻害に対して2から3倍以内で強力であった。
【表9】
【0160】
ヒトT細胞によるIL−23誘導性サイトカイン産生
PBMCは、健常ドナー由来のEDTA処理全血の密度勾配分離によって獲得した。T細胞は、ヒツジ赤血球(SRBC)と共にロゼット形成したPBMCのE+画分から調製した。このT細胞を96ウェル平底プレートへ1ウェルあたり100,000細胞で播種し、37℃で1時間、抗CD3(10μg/mlでのOKT)で覆い、PBSで洗浄した。抗CD28(1μg/mlでの9.3)およびIL−1β(10ng/ml)またはIL−1β+IL−23(1ng/ml)を含有するRPMI−FCS培地の混合物を調製した。このサイトカインの組合せは、ヒトT細胞のIL−17分泌T細胞への分化を促進するということが示されている。開始濃度(starting concentration)1μg/mlのATI001045をIL−1β+IL−23を含有する混合物へ添加した。IL−17はDUOSET登録商標 ELISA development kits(R&D Systems)を使用して、上清中に検出した。ATI001045は、バックグラウンドとしてIL−1β単独を使用して、2.0±1.6nMのEC50でIL−17産生を阻害した(n=4人の異なるドナー)。市販の抗p40抗体(MAB1510)が内部標準として使用され、2.2±1.4nMのEC50でIL−17産生を阻害した(n=3)。ドナー228からの例示的データを図6に示す。
【0161】
IL−12を超えるIL−23に対する抗IL−23アドネクチンの選択性
表2に示されるアドネクチンならびにATI001016を使用して、IL−23/IL−12に対する生化学的選択性を試験した。結合解析には、固定化抗His抗体での抗IL−23アドネクチンの回収、続いてアドネクチン上へのIL−23またはIL−12の流入が含まれた。IL−23に対するアドネクチンの選択性は、IL−23よりも100倍高い濃度のIL−12に対する結合シグナルを比較することによって評価した。表7に示す例示的データは、ATI001016は10nM ヒトIL−23に対して強力な結合(〜40RU)を示すが、一方で、1μM ヒトIL−12に対しては検出可能な結合は観察されなかったことを示す。
【0162】
NK−92細胞は、IFN−γを分泌することによりIL−2依存的にIL−12に応答することが知られているヒトナチュラルキラー細胞株である。細胞を典型的に洗浄してIL−2を除去し、次いで96ウェルプレートへ播種し、その後25pMの組換えヒトIL−12(または、アンタゴニストとプレインキュベートしたIL−12)で処理し、さらに20時間インキュベートした。清澄化された上清をELISAによりIFN−γに関して測定した。
【0163】
4点の5μMで開始する5倍希釈系列を、表2に示されるアドネクチンクローン各々について調製し、NK−92細胞への添加の前に、37℃で30分間25pM IL−12とインキュベートした。12点の5μMのATI001045およびATI001016で開始する5倍希釈系列を、NK−92細胞への添加の前に、37℃で30分間25pM IL−12とインキュベートした。表2に示されるクローンもATI001045あるいはATI001016も、これらの抗IL−23アドネクチンは、NK−92細胞表面上の受容体とのIL−12の相互作用を阻害しないことを示し、試験されたいずれの濃度でも、IFN−γ分泌を検出可能に阻害しなかった。それらは、陰性対照および100nM 抗p19ポリクローナル抗体と同等であるようである。陽性対照として、抗p40モノクローナル抗体(mAb1510)はIL−12誘導性IFN−γ分泌を0.07nMのIC50で阻害した(図8)。
【0164】
抗IL−23アドネクチンは薬力学的モデルにおいてIL−23誘導性IL−17を阻害(block)する
以下のスケジュールに従って、メスC57Bl/6マウスに組換えマウスIL−2およびヒトIL−23を腹腔内(IP)注射した。
【表10】
【0165】
全てのマウスを、IL−2およびIL−23の最終投与の7−8時間後、時間=30hで安楽死させた。血清を回収し、ELISAによりIL−17およびIL−23に関して測定した。
【0166】
ヒトIL−23はマウス受容体に結合し、IL−17およびIL−22などのサイトカインの産生を誘導する。IL−2およびヒトIL−23を腹腔内(IP)投与された動物由来の脾細胞は、エキソビボ(ex vivo)培養において抗マウスCD3eで刺激された時、IL−17を分泌する。IL−17の著しいレベルが表10に記載される処置計画に供された動物の血清中に検出でき、ここで、C57Bl/6マウスは、24時間のさらなる期間にわたるIL−2+IL−23の3回の二重注射(dual injection)の前に、IL−2で24時間、前処理した。おそらく、IL−2はポリクローナルに活性化し、インサイチュ(in situ)でTh集団を拡大させ、IL−23受容体の発現を上方制御する。これは、インビボ(in vivo)環境における、薬物作用のメカニズムおよび薬物濃度と効果の間の関係を調査することができる方法を提供する。このモデルは、抗p40モノクローナル抗体、mAb1510で確認された(データは示さず)。8つの別実験において、8つの別の実験でのIL−2+IL−23の初回投与(initial dose)の2時間前に、0.5、0.15、0.05および0.015mg/kgで、皮下(SC)投与した場合に、5種の抗IL−23アドネクチンを、マウスIL−17の産生を阻害する能力に関して試験した。ATI001045に対する例示的な用量反応データを図9aに示す(0.03mg/kgの計算上の平均ED50)。抑制範囲はアドネクチン全体で可変的であるが、試験された全ての抗IL−23アドネクチンは、血清中のヒトIL−23 マウスIL−17産生の用量依存的阻害を示した。
【0167】
IL−23誘導性皮膚表皮肥厚モデルにおける抗IL−23アドネクチンの活性
マウスの背中の皮膚または外耳介(external ear pinna)へのIL−23の皮内注射は、皮膚炎症および表皮の過形成(表皮肥厚)を誘発する(Zheng, Y., “Interleukin−22, a TH17 cytokine, mediates IL−23−induced dermal inflammation and acanthosis”, Nature, Vol. 445/8 (Feb. 2007))。これらの研究では、組換えヒトIL−23(rHuIL−23)がマウスの耳へ注射され、異常な皮膚IL−23暴露の下流の結果が調査された。
【0168】
6から8週齢のC57BL/6メスマウスに対し、5μgの二重鎖、組換え、ヒトIL−23を一日おきに12日目まで、右耳へ注射した。PBSを対照として反対側の耳に注射した。一試験では、ATI001045での処置は、最初のIL−23注射の約2時間前に開始し、1週間あたり3回、12日目まで継続した。ATI001045は、0.1、0.3、1、3mg/kgの用量で皮下(SC)投与された。第二の試験では、媒体またはATI000934−123 (1753E02)を、IL−23投与の1時間前に、1、3または10mg/kgで腹腔内(IP)投与し、その後1週間あたり3回、10日目まで投与した。抗HuIL−12/IL−23 p40抗体(R&D mAb1510)を10mg/kgで、陽性対照として0および4日目にIPで与えた。耳の厚さを(千分の1インチ単位で)、次の投与の前に、MITUTOYO登録商標(#2412F)ダイヤルキャリパーを使用して、一日おきに測定した。耳の厚さは、各動物に対し、IL−23注射された耳の測定結果から対照の耳の値を引くことによって計算した。試験の最後(ATI001045に対して14日目、およびATI000934に対して12日目)に、CO2ガスでの安楽死に続いて、耳を生え際で切除し、ホルマリン固定/パラフィン包埋組織をH&E染色したスライドで組織学的に観察した。
【0169】
全体としては、この試験において、1、3および10mg/kgの用量のATI000934は、同様のレベルのIL−23誘導性の耳肥厚の阻害を提供した(図10)。抗p40群を含む、全ての処置群における耳の厚さは、5日目から試験の最後の12日目にかけて、媒体よりも著しく薄かった(p<0.01 ANOVA/Dunnett’s)。12日目、最終的な血漿サンプルを最終投与48時間後に獲得し、ATI000934の血中濃度(circulating level)について解析し、それぞれ11、18、36μg/mlであると決定された。
【0170】
12日目の最終測定に続いて、1群あたり10動物から、所定の組織学的検査のために剖検で耳を回収した。ATI000934を投与された動物の大部分は、表皮肥厚および浸潤を有したが、組織学的重症度スコアは、媒体処置動物で観察されたものからは減少した。明らかな用量反応は見られなかった。抗p40を投与された全ての動物もまた、表皮肥厚および浸潤を有したが、組織学的重症度スコアは、媒体処置動物で観察されたものからは減少した。
【0171】
ATI001045(1mg/kgおよび3mg/kg)は、5日目から14日目にかけて、媒体(PBS)処置動物と比較して、用量依存的に耳の厚さを減少させた(p<0.01 対媒体 ANOVA/Dunnett’s、図10)。対照的に、0.1mg/kgの用量レベルでは、どの試験日でも、媒体処置と統計的に違いはなかった(p>0.05)。0.3mg/kgでの処置は、5日目、7日目、9日目において媒体よりも統計的に弱い、中間的な減少(intermediate reduction)を提供した。最終投与48時間後に回収された血漿サンプルを、ATI001045の血中濃度(circulating levels)について評価し、0.1、0.3、1、3mg/kgの用量に対し、それぞれ0.698、2.72、8、22.5μg/mlであると決定された。組織学的解析から、ATI001045の投与が、IL−23誘導性細胞性浸潤および耳の厚さのスコアに相関する表皮肥厚の用量依存的減少をもたらした、ということが明らかとなった。
【実施例5】
【0172】
本明細書で使用される材料および方法
ハイスループットタンパク質生成(HTPP)
選択されたバインダーをpET9dベクターにクローニングし、大腸菌BL21 DE3 plysS株へ形質転換し、50μg/mLカナマイシンを含有する5mlのLB培地に24ウェルフォーマットで播種し、37℃で一晩増殖させた。一晩培養物から200μlを吸引することおよび適当なウェルに分配することにより、誘導性発現のための新鮮な5mLのLB培地(50μg/ml カナマイシン)培養物を調製した。培養物を37℃でA600 0.6〜0.9まで増殖させた。1mM イソプロピル−β−D−1−チオガラクトシド(IPTG)を用いて誘導した後、培養物を、30℃で6時間発現させ、4℃で2750gでの10分間の遠心分離によって回収した。
【0173】
細胞ペレット(24ウェルフォーマット中)を、450μlの溶解バッファー(50mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、1mM PMSF、10mM CHAPS、40mM イミダゾール、1mg/ml リゾチーム、30μg/ml DNAse、2μg/ml アプロトニン(aprotonin)、pH8.0)に再懸濁し、室温で1〜3時間振盪することによって溶解した。溶解物を清澄化し、96ウェル、1.2mlキャッチプレートが取り付けられた96ウェルワットマンGF/Dユニフィルタ登録商標に移すことによって96ウェルフォーマットに再び入れ、陽圧によって濾過した。清澄化した溶解物を、平衡バッファー(50mM NaH2PO4、0.5M NaCl、40mM イミダゾール、pH8.0)で平衡化された96ウェルのNi−キレートプレートへ移し、5分間インキュベートした。非結合物質を陽圧により除去した。樹脂を、洗浄バッファー#1(50mM NaH2PO4、0.5M NaCl、5mM CHAPS、40mM イミダゾール、pH8.0)を用いて、2x 0.3ml/ウェルで洗浄し、各洗浄液を陽圧により除去した。溶出に先立って、各ウェルを50μl 溶出バッファー(PBS + 20mM EDTA)で洗浄し、5分間インキュベートし、洗浄液を陽圧により廃棄した。各ウェルにさらなる100μlの溶出バッファーを注ぐことによってタンパク質を溶出した。室温で30分インキュベートした後、プレート(単数または複数)を、200gで5分間遠心分離にかけ、溶出されたタンパク質を、溶出に先立って溶出キャッチプレートの底に加えた5μlの0.5M MgCl2を含有する96ウェルキャッチプレート中に回収する。溶出したタンパク質を、SGEをタンパク質標準として用いる総タンパク質アッセイ(BCA)を使用して定量した。
【0174】
不溶性のフィブロネクチンをベースとする足場タンパク質バインダーの中規模発現および精製
発現のために、HIS6タグが続く、選択されたクローン(単数または複数)を、pET9dベクターにクローニングし、大腸菌BL21 DE3 plysS株中で発現させた。20mlの種菌培養物(単一プレーティングされたコロニーから作製した)を使用して、50μg/ml カナマイシンおよび34μg/ml クロラムフェニコールを含有する1リットルのLB培地またはTB−オーバーナイト発現培地(自己誘導)に播種した。LB培地での培養物は、37℃でA600 0.6〜1.0までインキュベートし、その時点でイソプロピル−β−D−1−チオガラクトシド(IPTG)で誘導し、30℃で4時間増殖させた。TB−オーバーナイト発現培地で増殖させた培養物は、37℃で5時間インキュベートし、その時点で温度を19時間、18℃へ下げた。培養物を4℃で≧10,000gでの30分間の遠心分離によって回収した。細胞ペレットを−80℃で凍結させた。細胞ペレットを、氷上でULTRA−TURRAX登録商標ホモジナイザー(IKA works)を使用して、25mlの溶解バッファー(20mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、pH7.4)中に再懸濁した。細胞溶解は、モデルM−110S MICROFLUIDIZER登録商標(Microfluidics)を使用して高圧ホモジナイゼーション(≧18,000 psi)によって達成した。不溶性画分を、4℃で≧23,300gでの30分間の遠心分離によって分離した。溶解物の遠心分離から回収された不溶性ペレットを、20mM リン酸ナトリウム/500mM NaCl、pH7.4で洗浄した。ペレットを、超音波処理を用いて20mM リン酸ナトリウム/500mM NaCl pH7.4中、6.0M 塩酸グアニジンに再可溶化し、続いて37度で1〜2時間インキュベートした。再可溶化されたペレットを0.45μmで濾過し、20mM リン酸ナトリウム/500mM NaCl/6.0M グアニジン pH7.4バッファーで平衡化したHISTRAP登録商標カラムにロードした。ローディング後、カラムを同一のバッファーを用いて、さらなる25カラム容量(CV)に対して洗浄した。結合したタンパク質を、20mM リン酸ナトリウム/500mM NaCl/6.0M Guan−HCl、pH7.4中、50mM イミダゾールを用いて溶出した。精製されたタンパク質は、50mM 酢酸ナトリウム/150mM NaCl pH4.5またはPBS、pH7.2に対する透析によって、リフォールディングされた。
【0175】
可溶性のフィブロネクチンをベースとする足場タンパク質バインダーの中規模発現および精製
不溶性バインダーの精製の代替として、可溶性バインダーの精製を使用してもよい。発現のために、HIS6タグが続く、選択されたクローン(単数または複数)を、pET9dベクターにクローニングし、大腸菌BL21 DE3 plysS株中で発現させた。20mlの種菌培養物(単一プレーティングされたコロニーから作製した)を使用して、50μg/ml カナマイシンおよび34μg/ml クロラムフェニコールを含有する1リットルのLB培地またはTB−オーバーナイト発現培地(自己誘導)に播種した。LB培地での培養物は、37℃でA600 0.6〜1.0までインキュベートし、その時点でイソプロピル−β−D−1−チオガラクトシド(IPTG)で誘導し、30℃で4時間増殖させた。TB−オーバーナイト発現培地で増殖させた培養物は、37℃で5時間インキュベートし、その時点で温度を19時間、18℃へ下げた。培養物を4℃で≧10,000gでの30分間の遠心分離によって回収した。細胞ペレットを−80℃で凍結させた。細胞ペレットを、氷上でULTRA−TURRAX登録商標ホモジナイザー(IKA works)を使用して、25mlの溶解バッファー(20mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、pH7.4)中に再懸濁する。細胞溶解は、モデルM−110S MICROFLUIDIZER登録商標(Microfluidics)を使用して高圧ホモジナイゼーション(≧18,000 psi)によって達成される。可溶性画分を、4℃で≧23,300gで30分間の遠心分離によって分離する。上清を0.45μmフィルターへ通して清澄化する。清澄化された溶解物を、20mM リン酸ナトリウム/500mM NaCl pH7.4で事前に平衡化されたHISTRAP登録商標カラム(GE)にロードする。次いでカラムを、25カラム容量の同一のバッファーで洗浄し、続いて20カラム容量の20mM リン酸ナトリウム/500mM NaCl/25mM イミダゾール、pH7.4、その後35カラム容量の20mM リン酸ナトリウム/500mM NaCl/40mM イミダゾール、pH7.4を用いて洗浄する。タンパク質を、15カラム容量の20mM リン酸ナトリウム/500mM NaCl/500mM イミダゾール、pH7.4で溶出し、A280での吸光度に基づいて画分をプールし、1x PBS、50mM トリス、150mM NaCl、pH8.5または50mM NaOAc;150mM NaCl;pH4.5に対して透析する。いかなる沈殿物も0.22μmでの濾過によって除去される。
【0176】
フィブロネクチンをベースとする足場タンパク質(アドネクチン)は、種々の大きさおよび種類のPEGを用いてPEG化させることができる。PEG化を可能にするために、10FN3タンパク質のC末端で見られる天然に存在する残基EIDKPSQが、アミノ酸の、典型的にはセリンのシステインへの、単一点突然変異によって、修飾される。単一のシステイン残基におけるタンパク質のPEG化は、種々のマレイミド誘導体化されたPEGの形をコンジュゲートすること、PEG試薬をタンパク質溶液と組み合わせること、およびインキュベートすることによって達成される。代替方法は、EIDKPSQテールをGSGCリンカーで置換し、同様にシステイン残基をPEG化のために使用することである。操作されたシステイン残基を含有するアドネクチンを、マイケル付加反応を介して、システイン上のチオール基とPEG試薬のマレイミド官能基の間でPEGとコンジュゲートした。簡単に説明すると、40kDaのPEGを、弱酸性から中性条件下で、タンパク質溶液へ、過剰のモル濃度で添加する。反応は、2時間から一晩、室温で続行することができる。次いで反応をイオン交換カラムへ供し、PEG化アドネクチンを未反応のPEG−マレイミドおよび非PEG化アドネクチンから分離する。SE/HPLC方法が使用されてもよい。精製されたPEG化アドネクチンは典型的にSDS−PAGEおよびサイズ排除クロマトグラフィーによって解析される。
【実施例6】
【0177】
候補血清アルブミン結合アドネクチン(SABA)のスクリーニングおよび選択
PROfusionとして知られる選択技術(例えば、Roberts ら, Proc. Natl. Acad. Sci. USA, 94(23):12297−12302 (1997)および WO 2008/066752参照)が、10Fn3のBC、DEおよびFGループに設計された可変領域を有するDNAライブラリーへ適用された。1013分子を越えるランダムライブラリーがこの設計から作り出され、所望の結合特性を備える候補血清アルブミン結合アドネクチン(SABA)を単離するために、HSAのビオチン化形態に対して、選択圧が適用された。
【0178】
ハイスループットタンパク質生成(HTTP)工程
様々なHSA結合アドネクチンがハイスループットタンパク質生成(HTPP)を使用して精製された。選択されたバインダーを、HIS6タグを含有するpET9dベクターにクローニングし、大腸菌BL21(DE3)pLysS株へ形質転換した。形質転換された細胞を、50μg/mLカナマイシンを含有する5mlのLB培地に24ウェルフォーマットで播種し、37℃で一晩増殖させた。一晩培養物から200μlを吸引することおよび適当なウェルに分配することによって、誘導性発現のために、新鮮な5mlのLB培地(50μg/ml カナマイシン)培養物を調製した。培養物は37℃でA600 0.6〜0.9まで増殖させた。1mM イソプロピル−β−D−1−チオガラクトシド(IPTG)を用いて誘導した後、培養物を、30℃でさらに4時間発現させ、4℃で3220x gでの10分間の遠心分離によって回収した。細胞ペレットを−80度で凍結した。
【0179】
細胞ペレット(24ウェルフォーマット中)を、450μlの溶解バッファー(50mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、1mM PMSF、10mM CHAPS、40mM イミダゾール、1mg/ml リゾチーム、30μg/ml DNAse、2μg/ml アプロトニン(aprotonin)、pH8.0)に再懸濁し、室温で1時間振盪することによって溶解した。溶解物を清澄化し、96ウェル、650μlキャッチプレートが取り付けられた96ウェルワットマンGF/Dユニフィルタ登録商標へ移すことによって96ウェルフォーマットに再び入れ、200x gで5分間遠心分離した。清澄化した溶解物を、平衡バッファー(50mM NaH2PO4、0.5M NaCl、10mM CHAPS、40mM イミダゾール、pH8.0)で平衡化された96ウェルNi−キレートプレートに移し、5分間インキュベートした。非結合物質を除去した。樹脂を、洗浄バッファー#1(50mM NaH2PO4、0.5M NaCl、5mM CHAPS、40mM イミダゾール、pH8.0)を用いて、2x 0.3ml/ウェルで洗浄した。次いで、樹脂をPBSを用いて3x 0.3ml/ウェルで洗浄した。溶出に先立って、各ウェルを50μl 溶出バッファー(PBS + 20mM EDTA)で洗浄し、5分間インキュベートし、洗浄液を真空により廃棄した。各ウェルにさらなる100μlの溶出バッファーを注ぐことによってタンパク質を溶出した。室温で30分インキュベートした後、プレート(単数または複数)を、200x gで5分間遠心分離にかけ、溶出されたタンパク質を、溶出キャッチプレートの底に加えた5μlの0.5M MgCl2を含有する96ウェルキャッチプレート中に回収した。溶出されたタンパク質を、SGE(対照アドネクチン)をタンパク質標準として用いるBCAタンパク質アッセイを使用して定量した。SGEアドネクチンは、インテグリン結合ドメイン(78〜80位のアミノ酸RGD)がSGEで置換された、野生型10Fn3ドメイン(配列番号1)である。
【0180】
HSA、RhSAおよびMuSA直接結合ELISA
HSAへの直接的なバインダーをアッセイするために、MaxiSorpプレート(Nunc International, Rochester, NY)を4℃で一晩、PBS中10μg/mL HSA(Sigma, St. Louis, MO)で覆い、続いてカゼインブロックバッファー(Thermo Scientific, Rockford, IL)中で1〜3時間、室温でブロッキングを行った。シングルポイントスクリーニングアッセイのため、精製されたHTPPアドネクチンをカゼインブロックバッファー中に1:20で希釈し、室温で1時間、各ウェル内でHSAに結合させた。用量反応アッセイのため、0.1nMから1μMの範囲の濃度が使用された。非結合アドネクチンを除去するためのPBSTでの洗浄の後、カゼインブロックバッファー中に1:2500で希釈された抗His mAb−HRPコンジュゲート(R&D Systems, MN)を、室温で1時間、結合されたHisタグ化アドネクチンへ添加した。過剰のコンジュゲートはPBSTを用いる洗浄によって除去され、製造業者の使用説明書に従い、TMB検出試薬(BD Biosciences)を使用して、結合アドネクチンを検出した。
【0181】
候補血清アルブミン結合アドネクチン(SABA)の同定
HSA/RhSA/MuSA結合基準および生物物理学的基準に関するスクリーニングの結果を受けて、4つの特有の血清アルブミン結合アドネクチン(SABA)を同定し、それらの半減期をマウスで評価させるために選択した。インビトロ(in vitro)およびインビボ(in vivo)での特性評価を行うために、4つのSABAについて中規模生成が行われた。表3は、SABA1〜26と表される、PROfusionから同定された26の特有のSABAコア配列の配列を提供する。SABA4は中規模生成に先立って固定の足場変異を有した。SABA4の足場完全型はSABA5である。SABA4およびSABA5はBC、DEおよびFGループにおいて同一配列を有する。
【実施例7】
【0182】
候補SABAの生成および製剤処方(Formulation)
SABAの中規模タンパク質生成
His6タグが続く選択されたSABAを、pET9dベクターにクローニングし、大腸菌BL21(DE3)pLysS株中で発現させた (SABA1.1、SABA2.1、SABA3.1およびSABA5.1と表される、各Hisタグ化SABA配列に関する表3を参照)。20mlの種菌培養物(単一プレーティングされたコロニーから作製した)を使用して、50μg/ml カナマイシンを含有する1リットルのLB培地に播種した。培養物を37℃でA600 0.6〜1.0まで増殖させた。1mM イソプロピル−β−D−1−チオガラクトシド(IPTG)を用いて誘導した後、培養物を、30℃でさらに4時間増殖させ、4℃で≧10,000x gでの30分間の遠心分離によって回収した。細胞ペレットを−80℃で凍結させた。細胞ペレットを、氷上でULTRA−TURRAX登録商標ホモジナイザー(IKA works)を使用して、25mlの溶解バッファー(20mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、pH7.4)中に再懸濁した。細胞溶解は、モデルM−110S MICROFLUIDIZER登録商標(Microfluidics)を使用して高圧ホモジナイゼーション(≧18,000 psi)によって達成した。可溶性画分を、4℃で23,300x gでの30分間の遠心分離によって分離した。上清を0.45μmフィルターへ通して清澄化した。清澄化された溶解物を、20mM NaH2PO4、0.5M NaCl、pH7.4で事前に平衡化されたHISTRAP登録商標カラム(GE)にロードした。次いでカラムを、25カラム容量の20mM NaH2PO4、0.5M NaCl、pH7.4で、続いて、20カラム容量の20mM NaH2PO4、0.5M NaCl、25mM イミダゾール pH7.4、次いで、35カラム容量の20mM NaH2PO4、0.5M NaCl、40mM イミダゾール pH7.4を用いて洗浄した。タンパク質を、15カラム容量の20mM NaH2PO4、0.5M NaCl、500mM イミダゾール pH7.4で溶出し、A280での吸光度に基づいて画分をプールし、1x PBS、50mM トリス、150mM NaCl pH8.5または50mM NaOAc;150mM NaCl;pH4.5に対して透析した。いかなる沈殿物も0.22μmでの濾過によって除去された。
【0183】
中規模発現および精製は、可溶型で発現され、細菌性サイトゾルの可溶性画分から精製された、高純度かつ活性なアドネクチンを生じた。100mM NaPO4、100mM NaSO4、150mM NaCl、pH6.8(GE Healthcare)の移動相でのSUPERDEX登録商標 200またはSUPERDEX登録商標 75 10/30GLでのSEC解析は、大部分、単量体アドネクチンを示した。
【0184】
SABA1.2の製剤処方(Formulation)
一つの具体的なSABA、SABA1.2(配列番号180)、が、初期製剤処方(preliminary formulation)スクリーニングのために選択された。SABA1.2は、10Fn3の「コア1」配列上に(ED)5伸長を含む。SABA1.2に対して、pH6.0で10mM コハク酸、8% ソルビトール、5% グリセリン、かつ製品濃度5mg/mLでの、安定な製剤処方が同定された。この製剤処方において、1.25mg/mLのタンパク質濃度を使用して、示差走査熱量測定(DSC)により決定されたタンパク質の融点は75℃であった。この製剤処方は、4℃および25℃で、1.2%の初期凝集濃度(initial aggregate level)を有し、十分な物理的安定性および化学的安定性をもたらした。一ヶ月の静置の後、凝集のレベルは非常に低かった(4℃で1.6%かつ25℃で3.8%)。該タンパク質はまた、この製剤処方において、−80℃および−20℃から外気温へ推移させる5回の凍結融解後も安定であった。加えて、このSABA1.2製剤処方は、4℃および外気温で、沈殿もしくは凝集の増加も無く、少なくとも20mg/mLのタンパク質濃度で可溶性であった。
【実施例8】
【0185】
候補SABAの生物物理学的特性評価
サイズ排除クロマトグラフィー
標準的なサイズ排除クロマトグラフィー(SEC)を中規模工程によって生じる候補SABAに対して行った。中規模物質のSECは、A214nmならびにA280nmでのUV検出器付き、および蛍光検出器(励起=280nm、蛍光=350nm)付きのAgilent 1100もしくは1200 HPLCシステム上で、SUPERDEX登録商標 200 10/30またはSUPERDEX登録商標 75 10/30カラム(GE Healthcare)を使用して行った。SECカラムの適した流量で、100mM 硫酸ナトリウム、100mM リン酸ナトリウム、150mM 塩化ナトリウム、pH6.8のバッファーが採用された。ゲル濾過標準(Bio−Rad Laboratories, Hercules, CA)が分子量校正のために使用された。
【0186】
中規模精製SABAでのSECの結果は、大部分、単量体アドネクチンを、および、示されるように、球状ゲル濾過標準(BioRad)に対しておよそ10kDaの範囲での溶出を示した。
【0187】
耐熱性
中規模SABAの示差走査熱量測定(DSC)解析を行って、それらのそれぞれのTmを決定した。1mg/mlの溶液を、3atm圧力下、1分あたり1℃の速度で、5℃から95℃へ温度を上昇させる(ramping)ことにより、N−DSC II熱量計(Calorimetry Sciences Corp)でスキャンした。データを、Orgin Software(OrginLab Corp)を使用する適合度(best fit)を用いて、適切なバッファーの対照測定(control run)に対して解析した。SEC解析およびDSC解析の結果を表11に要約する。
【表11】
【実施例9】
【0188】
血清アルブミンに結合する候補SABA1の特性評価
HTPPおよび/または中規模物質から精製された選択されたSABAクローンの動態(kinetics)を、Biasensor CM5チップの表面上にそれぞれの血清アルブミン(HSA/RhSA/MuSA)を捕捉すること、および参照フローセルおよび捕捉されたアルブミンの両方にわたってSABAの濃度系列を流すことにより決定した。加えて、アルブミンとの結合を、pH5.5からpH7.4にわたる様々なpH条件下で実行した。HSA結合アドネクチン、SABA2.1、SABA3.1、SABA4.1(SABA5.1)&SABA1.1は、RhSAへ公差反応したが、MuSAへは公差反応しなかった。SABA2およびSABA4結合はpH感受性である一方、クローンSABA3はpH6.0に至るまでHSAとのpH耐性結合を明示した。SABA1.1はpH5.5に至るまでpH耐性および親和性/動態(kinetics)に関する生化学的基準に適合する。
【0189】
ドメインマッピングはビアコアにより決定した。HTPPおよび/または中規模物質から精製された選択されたSABAクローンは、Biasensor CM5チップの表面上にHSA、またはHSA−ドメインI&IIあるいはHSA−ドメインIIIのみから成る構築物を捕捉すること、および参照フローセルおよび捕捉されたアルブミンの両方にわたってSABAの濃度系列を流すことにより決定した。クローンSABA2&SABA1はHSAおよびHSA−ドメインI−II構築物と結合し、HSA−ドメインIII構築物とは結合しなかった。クローンSABA3&SABA4はHSAと結合したが、HSA−ドメインI−II構築物またはHSA−ドメインIII構築物のいずれとも結合しなかった。結果を表12に要約する。
【表12】
【実施例10】
【0190】
候補SABAのインビボ(in vivo)t1/2の調査
HSA結合アドネクチンはMuSAと交差反応しないため、マウスでのHSA結合アドネクチンの評価を可能にするために、マウスにおけるHSAの半減期を決定した。HSAを、20mg/kg用量(図11A)および50mg/kg用量(図11B)で、約6週齢のNcrヌードメスマウスの尾静脈へ注射し、注射後周期的に採取した血液サンプル中のHSA濃度をELISAにより決定した。20mg/kgおよび50mg/kgでマウスに注射されたHSAのt1/2は、それぞれ、〜24時間および〜20時間であると決定された。
【0191】
マウスでのSABA1〜4の半減期決定
1リットルのHSA結合クローンSABA1.1、SABA2.1、SABA3.1およびSABA4.1の大腸菌増殖液を調製し、精製し、エンドトキシンを除去した。各SABA変種をマウスの尾静脈へ注射し、注射後周期的に採取した血液サンプル中の濃度をELISAにより決定した。
【0192】
各SABAの薬物動態プロファイルを、約6週齢のNcrヌードメスマウスで、HSAの存在下または非存在下において比較した。結合クローンはHSAおよびRhSAに対して選択性であり、マウス血清アルブミンとは結合しなかったため、HSAを同時注射されたマウスは、HSAを各SABAと予め混合させた(3〜4モル濃度過剰のHSA)。マウス血漿中のSABA1.1の半減期は0.56時間であるのに対し、HSAと同時注射されたSABA1.1の半減期は5.6時間、半減期において〜10倍増加していた(図12A)。マウス血漿中のSABA2.1の半減期0.24時間であるのに対し、HSAと同時注射されたSABA2.1の半減期は2.8時間、半減期において〜12倍増加していた(図12B)。マウス血漿中のSABA3.1の半減期は0.28時間であるのに対し、HSAと同時注射されたSABA3.1の半減期は0.53時間、半減期において〜2倍増加していた(図12C)。マウス血漿中のSABA4.1の半減期は0.66時間であるのに対し、HSAと同時注射されたSABA4.1の半減期は4.6時間、半減期において〜7倍増加していた(図12D)。本実施例の概要を図13Aに示す。
【0193】
カニクイザルでのSABA1.1およびSABA5.1の半減期決定
SABA1.1およびSABA5.1の三週間単回投与概念実証試験(proof of concept study)をカニクイザルで実施し、2匹のカニクイザルにおける1mg毎kg(mpk)用量、IVでの薬物動態を評価した。薬物動態は、血漿サンプル中のアドネクチンを検出するために開発された定量的ELISAをベースとするアッセイを使用して評価した。SABA1.1は96〜137時間の範囲の半減期を有する。SABA5.1は約12時間の半減期を有し、唯一120時間までELISAで測定可能であった。図14AおよびBは、これらのクローンに対するデータを要約し、カニクイザルからのデータを比較するものである。
【実施例11】
【0194】
血清アルブミンと結合するSABA1の特性評価
SABA1.1およびSABA1.2はHSAおよびRhSAと結合する
SABA1.2、(ED)5伸長(配列番号190)を含む「コア1」10Fn3、は、ヒト血清アルブミン(HSA)と、中性pHかつ25℃で、8.21E+03 M−1s−1の平均結合速度定数(ka)、かつ4.43E−04 s−1の平均解離速度定数(kd)、55.3 nMの計算平均Kdで結合した(表13)。アカゲザル血清アルブミン(RhSA)に対しては、測定された平均結合速度定数は6.6E+03 M−1s−1であり、解離速度定数は3.78E−03 s−1であり、580 nMの計算平均Kdを提供した。1μMに至るまで、SABA1.2とマウスまたはラット血清アルブミンの間に測定可能な相互作用は観察されなかった(表13および図15)。37℃で、kaおよびkdは2から5倍増加し、HSAに対する親和性において〜2倍の増加およびRhSAに対して1/2の親和性をもたらした(表13)。
【表13】
【0195】
さらに、熱量滴定を行い、SABA1およびHSAの間の化学量論組成(stoichiometry)を決定した。この試験では、SABA1.1、His6伸長(配列番号189)を含む「コア1」10Fn3、が使用された。HSA(115μMのタンパク質溶液の注射あたり10μl)を、8.1μMの濃度でSABA1.1を含有する熱量測定セル(calorimetric cell)へ注入した。実験はPBSバッファーpH7.4中、37℃にて行った。図16はSABA1.1がHSAと1:1の化学量論組成で結合することを示す。
【0196】
SABA1.2は低pHでHSAと強く結合する
アルブミンの長い半減期(例えば、HSAのt1/2は19日である)は、主に、それらがエンドソームの内側に存在する低pH条件下で、新生児型Fc受容体、FcRn、に結合することにより、エンドサイトーシス経路からリサイクルされる、という事実に起因している。表14に示されるように、エンドソームpH5.5でHSAと強く結合したSABA1.2は、SABA1のt1/2を示唆し、一度HSAに結合すると、FcRnリサイクル機構の利益を享受する。
【表14】
【0197】
SABA1.2はHSAのドメインIおよびIIと結合するが、ドメインIIIとは結合しない
アルブミン上のSABA1.2結合部位は、組換えHSA断片を使用して、N末端ドメインIまたはIIへマッピングされ、ドメインIIIとの検出可能な結合は無かった(図17)。ドメインIIIは、最初にFcRnと相互作用するHSAのドメインであるため、SABA1.2は、FcRnへのHSA結合と競合する可能性が低く、半減期を増強するためのリサイクル機構を十分に活用する可能性をかさねて増大させる。
【実施例12】
【0198】
SABA1.2のインビボ(in vivo)薬理
SABA1.2の四週間単回投与前毒性試験(pre−toxicology study)をカニクイザルで実施し、二つの異なる用量レベルで薬物動態および免疫原性を評価した。薬物動態および免疫原性はまた、静脈投与群および皮下投与群の両者を含む、三週間単回投与前毒性試験においても評価した。さらに、SABA1.2の薬物動態を、血漿サンプル中のSABA1.2を検出するために開発された定量的ELISAをベースとするアッセイを使用して、カニクイザルで、二つの別個の単回投与前毒性試験においても評価した。
【0199】
SABA1.2を1mpkおよび10mpk、IVでサルへ投与した。図18および下記に記載されるパラメータに示されるように、CmxおよびAUCは、用量とともにほぼ直線的に増加した。WINNONLIN登録商標ソフトウェアを使用して、ノンコンパートメント解析を行い、薬物動態パラメータを評価した。10mpkでのSABA1.2に対するクリアランス(CL)は、0.15ml/時/kg、ベータ相半減期(t1/2)は143時間、分布容積(Vz)は30mL/kg、そして、総薬物暴露量(total drug exposure)(AUCall)は5,609,457時*nmol/Lであった(表15)。1mpkでのSABA1.2に対するクリアランス(CL)は、0.4ml/時/kg、半減期(t1/2)は124時間、分布容積(Vz)は72mL/kg、そして、総薬物暴露量(AUCall)は214,636時*nmol/Lであった(表15)。
【0200】
SABA1.2のSCまたはIV投与の後、ベータ相薬物動態プロファイルは類似していた(図19)。WINNONLIN登録商標ソフトウェアを使用して、ノンコンパートメント解析を行い、薬物動態パラメータを評価した。1mpk IVでのSABA1.2に対するクリアランス(CL)は、0.22ml/時/kg、ベータ相半減期(t1/2)は125時間、分布容積(Vz)は40mL/kg、そして、総薬物暴露量(AUCall)は357,993時*nmol/Lであった(表15)。1mpk SCでのSABA1.2に対するクリアランス(CL)は、0.32ml/時/kg、ベータ相半減期(t1/2)は134時間、分布容積(Vz)は62mL/kg、そして、総薬物暴露量(AUCall)は251,339時*nmol/Lであった(表15)。IVに比べてSCの相対的生物学的利用能(F)は0.7であった。
【表15】
【図1−1】
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図1−6】
【技術分野】
【0001】
本発明はインターロイキン−23(IL−23)、特にIL−23のp19サブユニットに結合する、フィブロネクチンをベースとする足場ドメインタンパク質に関する。本発明はまた、自己免疫疾患を治療するための治療的適応における革新的なタンパク質の使用に関する。本発明はさらに、このようなタンパク質、このようなタンパク質またはその断片をコードするポリヌクレオチドを含む細胞、およびこの革新的なタンパク質をコードするポリヌクレオチドを含むベクターに関する。
【背景技術】
【0002】
序論
IL−23はIL−12ヘテロ二量体サイトカインファミリーのメンバーである。それはIL−12に共通しているp40サブユニットおよび固有のp19サブユニットを含有する。IL−23はIL−12Rβ1およびIL−23からなるヘテロ二量体受容体複合体を通ってシグナルを送る(Aggarwal, Sら, “Interleukin−23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin−17”, J. Biol. Chem., 278:1910−1914 (2003))。IL−23は、多発性硬化症、関節リウマチ、乾癬およびクローン病などの慢性炎症性疾患の標的候補である。
【0003】
フィブロネクチンをベースとする足場は、対象とする任意の化合物と結合するよう進化させることができるタンパク質のファミリーである。通常、フィブロネクチンIII型(Fn3)またはFn3様ドメインに由来する足場を使用するこれらのタンパク質は、天然の抗体または操作された抗体(すなわち、ポリクローナル、モノクローナルまたは一本鎖抗体)に特徴的な形で機能し、さらに、構造的利点を有する。具体的には、これらの抗体模倣物の構造は、普通は抗体における構造および機能の喪失につながる条件下でも、フォールディング、安定性および溶解性が最適となるように設計されている。フィブロネクチンをベースとする足場タンパク質の一例として、アドネクチン(Adnexus,a Bristol−Myers Squibb R&D Company)がある。
【0004】
フィブロネクチンIII型(Fn3)ドメインは、N末端からC末端の順に、ベータまたはベータ様鎖、A;ループ、AB;ベータまたはベータ様鎖、B;ループ、BC;ベータまたはベータ様鎖C;ループCD;ベータまたはベータ様鎖D;ループDE;ベータまたはベータ様鎖、E;ループ、EF;ベータまたはベータ様鎖F;ループFG;およびベータまたはベータ様鎖Gを含む。ループAB、BC、CD、DE、EFおよびFGのいずれかまたはすべてが、標的結合に関与し得る。BC、DEおよびFGループは、免疫グロブリン由来の相補性決定領域(CDR)と構造上および機能上の両面で類似している。米国特許第7,115,396号には、BC、DEおよびFGループの変化が高親和性TNFαバインダーをもたらすFn3ドメインタンパク質が記載されている。米国公開第2007/0148126号には、BC、DEおよびFGループの変化が高親和性VEGFR2バインダーをもたらすFn3ドメインタンパク質が記載されている。
【発明の概要】
【発明が解決しようとする課題】
【0005】
自己免疫疾患の治療法のための改善されたフィブロネクチンドメイン足場タンパク質を得ることは有利であろう。インターロイキン−17を産生するエフェクターT細胞のサブセット(IL−17;「Th17細胞」)は非常に炎症性であり、激しい自己免疫を誘導する。Th17細胞は、Th1およびTh2細胞に比べて、IL−6、腫瘍壊死因子(TNF)、IL−22、IL−17AおよびIL−17Fと同様にケモカイン受容体CCR6を含む、異なるサイトカインおよびケモカインのサブセットを発現する。IL−23は活性化T細胞によるIL−17の産生を促進させ(Aggarwal, Sら, “Interleukin−23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin−17”, J. Biol. Chem., 278:1910−1914 (2003))、かつ、IL−17産生CD4+T細胞の増殖を誘導するための重要なサイトカインである。IL−23への暴露は、Th17細胞の病原性を決定する重要な特徴であるように思われる。
【課題を解決するための手段】
【0006】
本出願は、ヒトIL−23特異的p19サブユニットに対するアドネクチン商標を提供する。本発明の一態様は、一以上の溶媒接触可能ループが無作為化されまたは変異した、Fn3ドメインを含むポリペプチドを提供する。いくつかの実施形態では、前記Fn3ドメインは、ヒトフィブロネクチンIII型ドメインの野生型第10モジュール(10Fn3)に由来するFn3ドメインである。いくつかの実施形態では、本発明の10Fn3ポリペプチドは、ヒト10Fn3ドメインと、少なくとも40%、50%、60%、65%、70%、75%、80%、85%、または90%同一である。
【0007】
いくつかの実施形態では、BC、DEおよびFGから選択される一以上のループが、対応するヒトフィブロネクチンループと比較して伸長または短縮されてもよい。
【0008】
いくつかの実施形態では、本発明のポリペプチドは第10フィブロネクチンIII型(10Fn3)ドメインを含み、ここに10Fn3ドメインは、ループ、AB;ループ、BC;ループ、CD;ループ、DE;ループEF;およびループFGを含み;かつ、ヒト10Fn3ドメインの対応するループの配列と比較して変化したアミノ酸配列を備える、ループBC、DEおよびFGから選択される少なくとも一つのループを有するものである。
【0009】
いくつかの実施形態では、本発明のポリペプチドは、非ループ領域と少なくとも80、85、90、95、98、99または100%同一であるアミノ酸配列を含むFn3ドメインを含む。
【0010】
いくつかの実施形態では、本発明のタンパク質のBCループは、配列番号2〜6からなる群から選択されるアミノ酸配列を含む。
【0011】
いくつかの実施形態では、本発明のタンパク質のDEループは、配列番号7〜48からなる群から選択されるアミノ酸配列を含む。
【0012】
いくつかの実施形態では、本発明のタンパク質のFGループは、配列番号49〜59からなる群から選択されるアミノ酸配列を含む。
【0013】
いくつかの実施形態では、10Fn3ドメインは、アミノ酸置換、挿入、もしくは欠失で開始および/または終止してもよい。
【0014】
いくつかの実施形態では、本発明のタンパク質は、配列番号2〜6に示されるBCループ配列からの一つのループ配列、配列番号7〜48に示される一つのDEループ配列、および配列番号49〜59に示される一つのFGループ配列を含む。
【0015】
いくつかの実施形態では、本発明のタンパク質は、配列番号2〜59のいずれか一つと少なくとも70、75、80、85、90、95、98、99または100%同一であるBC、DEおよびFGループアミノ酸配列を含む。
【0016】
いくつかの実施形態では、抗IL−23アドネクチンは配列番号60〜100のいずれか一つのアミノ酸配列を含む。
【0017】
いくつかの実施形態では、抗IL−23アドネクチンは配列番号60〜100のいずれか一つの3〜96位由来のFn3ドメインアミノ酸配列を含む。
【0018】
いくつかの実施形態では、抗IL−23アドネクチンは、配列番号60〜100のいずれか一つと少なくとも70、75、80、85、90、95、98、99または100%同一であるアミノ酸配列を含む。
【0019】
一態様では、抗IL−23アドネクチンはさらに、薬物動態(PK)部分を含む。いくつかの実施形態では、PK部分はポリエチレングリコール(PEG)を含む。
【0020】
一態様では、本出願は自己免疫疾患の治療において有益な抗IL−23アドネクチンを提供する。
【0021】
一態様では、本発明はフィブロネクチンIII型第10(10Fn3)ドメインおよび抗IL−23アドネクチンを含む融合ポリペプチドを提供し、ここに前記10Fn3ドメインは1uM以下のKdでHSAに結合するものである。特定の実施形態では、10Fn3ドメインは配列番号103と少なくとも70%同一であるアミノ酸配列を含む。一の実施形態では、10Fn3ドメインは、配列番号104に説明されるアミノ酸配列を有するBCループ、配列番号105に説明されるアミノ酸配列を有するDEループ、および配列番号106に説明されるアミノ酸配列を有するFGループを含む。別の実施態様では、10Fn3ドメインは、一以上の、配列番号104に説明されるアミノ酸配列を有するBCループ、配列番号105に説明されるアミノ酸配列を有するDEループ、および配列番号106に説明されるアミノ酸配列を有するFGループを含む。
【0022】
一の実施形態では、融合ポリペプチドの10Fn3ドメインはまた、一以上のアカゲザル血清アルブミン(RhSA)、カニクイザル血清アルブミン(CySA)、またはマウス血清アルブミン(MuSA)に結合する。他の実施形態では、10Fn3ドメインは一以上のRhSA、CySA、またはMuSAと交差反応しない。
【0023】
特定の実施形態では、融合ポリペプチドの10Fn3ドメインは1uM以下のKdでHSAに結合する。いくつかの実施形態では、10Fn3ドメインは500nM以下のKdでHSAに結合する。他の実施形態では、10Fn3ドメインは少なくとも200nM、100nM、50nM、20nM、10nM、または5nMのKdでHSAに結合する。
【0024】
他の実施形態では、融合ポリペプチドの10Fn3ドメインはHSAのドメインIまたはIIに結合する。一の実施形態では、10Fn3ドメインはHSAのドメインIおよびIIの両方に結合する。いくつかの実施形態では、10Fn3ドメインは5.5から7.4のpH範囲でHSAに結合する。他の実施形態では、10Fn3ドメインはpH5.5で200nM以下のKdでHSAに結合する。別の実施形態では、5.5から7.4のpH範囲で少なくとも500nM、200nM、100nM、50nM、20nM、10nM、または5nMのKdでHSAに結合する。一の実施形態では、10Fn3ドメインはpH5.5で少なくとも500nM、200nM、100nM、50nM、20nM、10nM、または5nMのKdでHSAに結合する。
【0025】
いくつかの実施形態では、血清アルブミン存在下での融合ポリペプチドの血清半減期は、血清アルブミン非存在下での前記ポリペプチドの血清半減期より少なくとも5倍長い。特定の実施形態では、血清アルブミン存在下での融合ポリペプチドの血清半減期は、血清アルブミン非存在下での前記ポリペプチドの血清半減期より少なくとも2倍、5倍、7倍、10倍、12倍、15倍、20倍、22倍、25倍、27倍、または30倍長い。いくつかの実施形態では、血清アルブミンはHSA、RhSA、CySA、またはMuSAのいずれか一つである。
【0026】
特定の実施形態では、血清アルブミン存在下での融合ポリペプチドの血清半減期は少なくとも20時間である。特定の実施形態では、血清アルブミン存在下での融合ポリペプチドの血清半減期は少なくとも10時間、12時間、15時間、20時間、25時間、30時間、40時間、50時間、75時間、90時間、100時間、110時間、120時間、130時間、150時間、170時間、または200時間である。いくつかの実施形態では、融合ポリペプチドの血清半減期は霊長類(例えば、ヒトもしくはサル)またはマウスにおいて観察される。
【0027】
前述の態様および実施形態のいずれにおいても、10Fn3ドメインは配列番号107、111、115、119、および123〜143から選択される配列を含む。
【図面の簡単な説明】
【0028】
【図1】図1は、本発明の抗IL−23アドネクチンの全長DNA配列アラインメントを示す。
【0029】
【図2】図2は、実施例2において記載されるように、pBMS2008/ATI001044タンパク質発現ベクターを示す。
【0030】
【図3】図3は、本発明の抗IL−23アドネクチンの全長アミノ酸配列アラインメントを示す。
【0031】
【図4】図4は、実施例4において記載されるように、抗IL−23アドネクチンによるPBMC pSTAT3阻害由来の代表的なIC50曲線を示す。
【0032】
【図5】図5は、実施例4において記載されるように、抗IL−23アドネクチンおよび抗p40モノクローナル抗体(MAB1510)によるIL−23依存性IL−17Aの阻害に対する代表的なIC50曲線を示す。
【0033】
【図6】図6は、ドナー228(実施例4において記載されるように試験された4ドナーの一つ)のPBMCによるIL−23誘導性IL−17産生のATI001045阻害を示す。
【0034】
【図7】図7は、抗IL−23アドネクチンに対する代表的な選択性データを示す。実施例4において記載されるように、回収されたATI001016に結合する10nM IL−23および1uM IL−12の結合相および解離相を示すバッファー除去センサーグラムが示される。
【0035】
【図8】図8は、実施例4において記載されるように、抗IL−23アドネクチンがNK−92細胞におけるIL−12誘導性IFN−γ産生を阻害しないことを示す。
【0036】
【図9A】図9Aは、実施例4において記載されるように、ATI001045がマウス薬力学的モデルにおいて血清IL−17レベルを阻害することを示す。
【0037】
【図9B】図9Bは、実施例4において記載されるように、マウス薬力学的モデルにおける抗IL−23アドネクチンの阻害活性の比較を示す。
【0038】
【図10A】図10Aは、実施例4において記載されるように、ヒトIL−23誘導表皮肥厚におけるATI000934用量反応を示す。
【0039】
【図10B】図10Bは、実施例4において記載されるように、ヒトIL−23誘導表皮肥厚におけるATI001045用量反応を示す。
【0040】
【図11】図11は、マウスにおけるインビボ(in vivo)のHSA半減期を示す。HSAは20mg/kg(図11A)または50mg/kg(図11B)でマウスへ注射された。
【0041】
【図12】図12A−Dは、マウスにおけるSABA1−SABA4の半減期決定を示す。
【0042】
【図13】図13Aは、HSAと同時注射した際のSABA1−4のマウスにおける半減期増強のグラフの概要を示す。図13bは、カニクイザルおよびマウス由来のデータを比較する。
【0043】
【図14】図14A−Bは、カニクイザルにおけるSABA1.1およびSABA5.1に対する半減期決定を示す。
【0044】
【図15】図15は、直接結合ELISAアッセイによるヒト、マウス、およびラット由来のアルブミンに結合するSABA1.2を示す。
【0045】
【図16】図16は、SABA1.1およびHSAの化学量(stoichiometry)の決定を示す。
【0046】
【図17】図17は、HSAの組換えドメイン断片に結合するSABA1.2のビアコア解析を示す。
【0047】
【図18】図18は、1mpkおよび10mpkで投与されたサルにおけるSABA1.2に対する薬物動態プロファイルを示す。
【0048】
【図19】図19は、1mpkで静脈内投与または皮下投与されたサルにおけるSABA1.2に対する薬物動態プロファイルを示す。
【発明を実施するための形態】
【0049】
発明の詳細な説明
定義
「ポリペプチド」によって、長さ、翻訳後修飾または機能にかかわらず、二以上のアミノ酸の任意の配列が意味される。「ポリペプチド」、「ペプチド」および「タンパク質」は、本明細書において同義的に使用される。ポリペプチドは、出典明示により本明細書に組み込まれる米国特許第6,559,126号に記載されるものなどの天然アミノ酸および非天然アミノ酸を含み得る。ポリペプチドはまた、種々の標準的な化学法のいずれかで修飾されていてもよい(例えば、アミノ酸は保護基で修飾されていてもよい;カルボキシ末端アミノ酸は、末端アミド基にされてもよい;アミノ末端残基は、例えば、親油性を増強するための基で修飾されてもよい;またはポリペプチドは、化学的にグリコシル化されていてもよく、あるいは、安定性もしくはインビボ(in vivo)半減期を増大するよう修飾されていてもよい)。ポリペプチド修飾は、ポリペプチドへの、環状化合物または他の分子などの別の構造の結合(attachment)を含み得、また一以上のアミノ酸を変化した立体配置(すなわち、RもしくはS;または、LもしくはD)で含有するポリペプチドを含み得る。本発明のポリペプチドは、IL−23のp19サブユニットに特異的に結合するよう修飾された、第10フィブロネクチンIII型ドメインに由来するタンパク質であり、本明細書において「アドネクチン」または「抗IL−23アドネクチン」と呼ばれる。
【0050】
用語「PK」は、「薬物動態(pharmokinetic)」の頭字語であり、例として、被験体による吸収、分布、代謝および排出を含む化合物の特性を包含する。「PK調節タンパク質」または「PK部分」とは、生物学的に活性な分子と融合されるか、一緒に投与される場合に、生物学的に活性な分子の薬物動態特性に影響を及ぼす任意のタンパク質、ペプチドまたは部分を指す。PK調節タンパク質またはPK部分の例として、PEG、ヒト血清アルブミン(HSA)バインダー(米国公開第2005/0287153号および同2007/0003549号に開示されるような)、ヒト血清アルブミン、FcまたはFc断片、および糖(例えば、シアル酸)が挙げられる。
【0051】
本明細書において、「アミノ酸配列同一性パーセント(%)」とは、配列同一性の一部として保存的置換は全く考慮することなく、最大配列同一性パーセントを達成するために、配列をアラインし、必要に応じてギャップを導入した後の、選択された配列中のアミノ酸残基と同一である、候補配列中のアミノ酸残基のパーセンテージとして定義される。アミノ酸配列同一性パーセントを決定する目的上、アラインメントは、当技術分野における技術の範囲内の種々の方法で、例えば、BLAST、BLAST−2、ALIGN、ALIGN−2またはMegalign(DNASTAR登録商標)ソフトウェアなどの公的に入手可能なコンピュータソフトウェアを使用して達成できる。当業者ならば、比較されている配列の全長にわたって最大アラインメントを達成するのに必要な任意のアルゴリズムを含む、アラインメントを測定するための適当なパラメータを決定できる。
【0052】
「単離された」ポリペプチドとは、同定および分離および/またはその天然環境の成分から回収されているものである。その天然環境の夾雑成分とは、ポリペプチドの診断または治療的使用を干渉する物質であり、酵素、ホルモンおよび他のタンパク質性または非タンパク質性溶質を挙げることができる。好ましい実施形態では、ポリペプチドは、(1)ローリー法によって決定される、95重量%を超える、最も好ましくは、99重量%を超えるポリペプチドに、(2)スピニングカップシークエネーターの使用によってN末端もしくは内部アミノ酸配列の少なくとも残基(複数)を得るのに十分な程度に、または(3)クーマシーブルーもしくは、好ましくは、銀染色を使用する還元もしくは非還元条件下でのSDS−PAGEによる均一性に精製される。単離されたポリペプチドは、ポリペプチドの天然環境の少なくとも1種の成分が存在しないので、組換え細胞内でインサイチュ(in situ)でのポリペプチドを含む。しかし、通常、単離されたポリペプチドは、少なくとも1つの精製工程によって調製される。
【0053】
アミノ酸配列または化合物の「半減期」は、通常、インビボ(in vivo)でポリペプチドの血清濃度が、例えば、天然の機序による、配列もしくは化合物の分解および/または配列もしくは化合物のクリアランスもしくは隔離によって50%低減されるのにかかる時間として定義され得る。半減期は、薬物動態分析によってなど、それ自体公知の任意の方法で決定できる。適した技術は、当業者には明らかであり、例えば、通常、本発明のアミノ酸配列または化合物の適した用量を霊長類に適宜投与する工程;前記霊長類から定期的に血液サンプルまたは他のサンプルを採取する工程;前記血液サンプル中の本発明のアミノ酸配列または化合物のレベルまたは濃度を決定する工程;およびこのように得られたデータ(のプロット)から、本発明のアミノ酸配列または化合物のレベルまたは濃度が、投与時の初期レベルと比較して50%低減されるまでの時間を算出する工程を含み得る。例えば、Kenneth, A ら, Chemical Stability of Pharmaceuticals: A. Handbook for Pharmacists および Lee, P. I. D. ら, Pharmacokinetic analysis: A Practical Approach (1996)などの標準的なハンドブックが参照される。Gibaldi M. ら,Pharmacokinetics, 2nd Rev. edition, Marcel Dekker, 刊行(1982)もまた参照される。
【0054】
半減期は、t1/2−アルファ、t1/2−ベータおよび曲線下面積(AUC)などのパラメータを使用して表現され得る。本明細書において、「半減期の増大」とは、これらのパラメータのうちいずれか1つ、これらのパラメータのうちいずれか2つ、またはこれらのパラメータのうち3つ全ての増大を指す。「半減期の増大」とは、特に、t1/2−アルファおよび/またはAUCまたは両方の増大を伴うか、または伴わない、t1/2−ベータの増大を指す。
【0055】
概説
本出願は、ヒトIL−23特異的p19サブユニットに対するアドネクチンを提供する。IL−23特異的アンタゴニストを同定するために、抗p40モノクローナル抗体を使用して、IL−23をアドネクチンの大規模合成ライブラリーへ提示した。IL−23 p19サブユニットに結合するアドネクチンは、ヒトIL−23との結合、IL−23/IL−23R相互作用の競合、およびT細胞株におけるIL−23誘導性シグナル伝達の阻害に関してスクリーニングされた。抗IL−23アドネクチンは、標的濃度を低下させ、遅い解離速度(slow off−rate)で抗IL−23アドネクチンを選択することによる、さらなる選択圧に供された。この最適化工程から、アドネクチンのファミリーが、好ましい生化学的および生物物理学的特性を備えるIL−23特異的阻害剤として同定された。
【0056】
フィブロネクチンベースの足場
本出願の一態様は、一以上の溶媒接触可能ループが無作為化または変異した、Fn3ドメインを含むポリペプチドを提供する。いくつかの実施形態では、前記Fn3ドメインは、ヒトフィブロネクチンIII型ドメインの野生型第10モジュール(10Fn3)に由来するFn3ドメイン:VSDVPRDLEVVAATPTSLLISWDAPAVTVRYYRITYGETGGNSPVQEFTVPGSKSTATISGLKPGVDYTITVYAVTGRGDSPASSKPISINYRT(配列番号1)である。上記の10Fn3配列において、BC、DEおよびFGループには下線が引かれている。
【0057】
種々の変異10Fn3足場が報告されている。一態様では、Asp7、Glu9およびAsp23の一以上が、例えば、負の電荷を有さないアミノ酸残基(例えば、Asn、Lys等)などの別のアミノ酸によって置換されている。これらの突然変異は、野生型形態と比較して、中性pHでの変異10Fn3のより大きな安定性を促進する効果を有すると報告されている(PCT公開番号WO02/04523参照)。有益または中立のいずれかである10Fn3足場における種々のさらなる変化が開示されている。例えば、Batori ら, Protein Eng., 15(12):1015−1020 (Dec. 2002); Koide ら, Biochemistry, 40(34):10326−10333 (Aug. 28, 2001)参照。
【0058】
変種および野生型10Fn3タンパク質の両方とも、同一構造、すなわち、AからGと表される7つのβ鎖ドメイン配列および7つのβ鎖ドメイン配列を接続する6つのループ領域(ABループ、BCループ、CDループ、DEループ、EFループおよびFGルー
プ)を特徴とする。N末端およびC末端の最も近くに位置するβ鎖は、溶液中でβ様コンホメーションをとり得る。配列番号1中、ABループは、残基15〜16に対応し、BCループは、残基21〜30に対応し、CDループは、残基39〜45に対応し、DEループは、残基51〜56に対応し、EFループは、残基60〜66に対応し、FGループは、残基76〜87に対応する(Xu ら, Chemistry & Biology, 9:933−942 (2002))。
【0059】
いくつかの実施形態では、10Fn3ポリペプチドは、配列番号1に示されるヒト10Fn3ドメインと、少なくとも40%、50%、60%、65%、70%、75%、80%、85%または90%同一であり得る。変異性の多くは、通常、一以上のループ中で起こる。10Fn3ポリペプチドのβ鎖またはβ様鎖の各々は、このような変動が、生理学的条件におけるポリペプチドの安定性を乱さないという条件で、本質的に、配列番号1の対応するβ鎖またはβ様鎖の配列と、少なくとも80%、85%、90%、95%または100%同一であるアミノ酸配列からなり得る。
【0060】
いくつかの実施形態では、本開示内容は、第10フィブロネクチンIII型(10Fn3)ドメインを含むポリペプチドであって、10Fn3ドメインが、ループ、AB;ループ、BC;ループ、CD;ループ、DE;ループEF;およびループFGを含むものであり、かつ、ヒト10Fn3ドメインの対応するループの配列と比較してアミノ酸配列が変化したループBC、DEおよびFGから選択される少なくとも1つのループを有するポリペプチドを提供する。いくつかの実施形態では、BCおよびFGループが変化し、いくつかの実施形態では、BC、DEおよびFGループが変化している、すなわち、Fn3ドメインが、天然に存在しないループを含む。「変化した」とは、(ヒトフィブロネクチンドメインに対応する)鋳型配列に対する一以上のアミノ酸配列の変化を意味し、アミノ酸付加、欠失および置換を含む。アミノ酸配列の変化は、通常、核酸コード配列の意図的な、盲検的な(blind)または自発的な配列変動によって達成され得、任意の技術、例えば、PCR、エラー−プローンPCRまたは化学的DNA合成によって起こり得る。
【0061】
いくつかの実施形態では、BC、DEおよびFGから選択される一以上のループは、対応するヒトフィブロネクチンループと比較して、長さが伸長または短縮され得る。いくつかの実施形態では、ループの長さは、2〜25アミノ酸によって伸長され得る。いくつかの実施形態では、ループの長さは、1〜11アミノ酸によって減少され得る。したがって、抗原結合を最適化するために、抗原結合において最大のあり得る可動性および親和性を得るよう、10Fn3のFGループの長さを、長さにおいて、ならびに、配列において変化させてもよい。
【0062】
いくつかの実施形態では、ポリペプチドは、配列番号1の非ループ領域と少なくとも80、85、90、95、98、99または100%同一であるアミノ酸配列を含むFn3ドメインを含み、BC、DEおよびFGから選択される少なくとも1つのループが変化している。いくつかの実施形態では、変化したBCループは、最大10のアミノ酸置換、最大4のアミノ酸欠失、最大10のアミノ酸挿入またはそれらの組合せを有する。いくつかの実施形態では、変化したDEループは、最大6のアミノ酸置換、最大4のアミノ酸欠失、最大13のアミノ酸挿入またはそれらの組合せを有する。いくつかの実施形態では、FGループは、最大12のアミノ酸置換、最大11のアミノ酸欠失、最大25のアミノ酸挿入またはそれらの組合せを有する。
【0063】
いくつかの実施形態では、本発明のタンパク質のBCループは、GHYPMHV(配列番号2)、GHYPLHV(配列番号3)、GHYPMHI(配列番号4)、GHYPLHI(配列番号5)およびGHYPLHL(配列番号6)からなる群から選択されるアミノ酸配列を含む。
【0064】
いくつかの実施形態では、本発明のタンパク質のDEループは、HRTH(配列番号7)、YYHY(配列番号8)、SKQH(配列番号9)、SNVH(配列番号10)、NRAH(配列番号11)、RKTY(配列番号12)、RSRY(配列番号13)、SRYY(配列番号14)、PHRY(配列番号15)、RSTH(配列番号16)、SRIY(配列番号17)、HQRY(配列番号18)、KQVY(配列番号19)、AHRY(配列番号20)、RSRH(配列番号21)、ARQY(配列番号22)、RTQY(配列番号23)、PRYH(配列番号24)、MRQH(配列番号25)、SRKY(配列番号26)、RQKY(配列番号27)、HAKY(配列番号28)、SNRY(配列番号29)、NTSH(配列番号30)、SQVY(配列番号31)、NRVY(配列番号32)、PRSH(配列番号33)、RTKY(配列番号34)、SRYH(配列番号35)、PRRY(配列番号36)、RQKY(配列番号37)、RYKY(配列番号38)、VPRH(配列番号39)、TPKH(配列番号40)、RSKY(配列番号41)、SRKY(配列番号42)、VPRY(配列番号43)、PRRY(配列番号44)、RMRH(配列番号45)、PPRH(配列番号46)、RQIY(配列番号47)、およびMRQH(配列番号48)からなる群から選択されるアミノ酸配列を含む。
【0065】
いくつかの実施形態では、本発明のタンパク質のFGループは、YYNEADYSQI(配列番号49)、YYQEYEYRYI(配列番号50)、YYMEEKYAVI(配列番号51)、YYAQENYKEI(配列番号52)、YYKEANYREI(配列番号53)、YYAQEEYHII(配列番号54)、YYKEADYSQI(配列番号55)、YYEQVEYREI(配列番号56)、YYEQPIYATI(配列番号57)、YYEQVEYREI(配列番号58)およびYYSEELYKYI(配列番号59)からなる群から選択されるアミノ酸配列を含む。
【0066】
10Fn3ドメインは、アミノ酸変異で始まってもよい。例えば、Fn3ドメインのN末端に、さらなるMG配列が配置されてもよい。Mは、通常、切断除去され、N末端にGが残る。いくつかの実施形態では、10Fn3ドメインのC末端に、配列が配置されていてもよい。例えば、GSGC(配列番号101)などのシステイン含有リンカーがC末端へ付加される、部位特異的PEG化において。あるいは、システイン含有リンカーEIDKPCQ(配列番号102)のため、セリンをシステインに変更することによって変異させられた、天然に存在するC末端テールのPEG化。GSGCリンカーを含む本発明の抗IL−23アドネクチンの例として、ATI001014、ATI001015、ATI001016、ATI001044、ATI001045およびATI001047が挙げられる。ATI000934は、EIDKPCQリンカーを含む本発明の抗IL−23アドネクチンの例である。
【0067】
いくつかの実施形態では、本発明のタンパク質は、配列番号2〜6に示されるBCループ配列からの一のループ配列、配列番号7〜48に示される一のDEループ配列、および配列番号49〜59に示される一のFGループ配列を含む。いくつかの実施形態では、本発明のタンパク質は、配列番号2〜59のいずれか一つと少なくとも70、75、80、85、90、95、98、99または100%同一であるBC、DEおよびFGループアミノ酸配列を含む。
【0068】
さらに、当業者であれば、配列番号2〜6に示されるBCループ配列が、X1がMまたはLのいずれかであり、X2がIまたはVのいずれかである共通の配列モチーフGHYPX1HX2(配列番号257)を共有し、かつ、配列番号49〜59に示されるFGループが、X3が任意のアミノ酸である共通の配列モチーフYYX3X3X3X3YX3X3I(配列番号258)を共有することを認識するであろう。それゆえ、コンセンサス配列GHYPX1HX2に適合するBCループを備え、かつ/または、配列番号49〜59に明確に示されるものの域を超えて、パターンYYX3X3X3X3YX3X3Iに適合する他のFGループを備える、IL−23に結合するさらなるアドネクチンを生成することが可能であろう。
【0069】
いくつかの実施形態では、抗IL−23アドネクチンは配列番号60〜100のいずれか一つのアミノ酸配列を含む。いくつかの実施形態では、抗IL−23アドネクチンは配列番号60〜100のいずれか一つの3〜96位由来のFn3ドメインアミノ酸配列を含む。いくつかの実施形態では、抗IL−23アドネクチンは、配列番号60〜100のいずれか一つと少なくとも70、75、80、85、90、95、98、99または100%同一であるアミノ酸配列を含む。いくつかの実施形態では、抗IL−23アドネクチンは、配列番号60〜100のいずれか一つの3〜96位由来のアミノ酸配列と少なくとも70、75、80、85、90、95、98、99または100%同一であるアミノ酸配列を含む。
【0070】
いくつかの実施形態では、抗IL−23アドネクチンはPEG化され、かつ/またはhisタグを含有してもよい。本明細書において用いる場合、ATI000934は、ループ配列が構築体1571G06(配列番号87)のものと同一であるタンパク質を指し、前記タンパク質はC末端でEIDKPCQ残基を含み、PEG化され、かつhisタグを含有する。ATI001014は、ループ配列が構築体1571G04(配列番号86)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化され、かつhisタグを含有する。ATI001015は、ループ配列が構築体1572G06(配列番号91)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化され、かつhisタグを含有する。ATI001016は、ループ配列が構築体1490B03(配列番号79)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化され、かつhisタグを含有する。ATI001044は、ループ配列が構築体1490B03(配列番号79)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含むが、前記タンパク質はPEG化されておらず、かつhisタグが存在しない。ATI001045は、ループ配列が構築体1490B03(配列番号79)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化されているが、hisタグが存在しない。ATI001047は、ループ配列が構築体1571G04(配列番号86)のものと同一であるタンパク質を指し、前記タンパク質はC末端でGSGCリンカーを含み、PEG化されているが、hisタグが存在しない。
【0071】
フィブロネクチンは、そのインテグリン結合モチーフ、「アルギニン−グリシン−アスパラギン酸」(RGD)を介して特定の種類のインテグリンと天然に結合する。いくつかの実施形態では、ポリペプチドは、(RGD)インテグリン結合モチーフを欠く10Fn3ドメインを含む。
【0072】
薬物動態部分
一態様では、本出願は、薬物動態(PK)部分をさらに含む抗IL−23アドネクチンを提供する。改善される薬物動態は、認識される治療的必要性に従って評価できる。おそらくは、タンパク質が、投与後の血清において利用可能なままである時間を増大することによって、生物学的利用能(bioavailability)を増大すること、および/または用量間の時間を増大することが望ましいことが多い。場合によっては、経時的なタンパク質の血清濃度の連続性を改善すること(例えば、投与の直後および次の投与の直前のタンパク質の血清濃度の相違の減少)が望ましい。抗IL−23アドネクチンは、哺乳類(例えば、マウス、ラットまたはヒト)においてポリペプチドのクリアランス速度を、修飾されていないアドネクチンと比較して3倍超低減する部分に結合させてもよい。改善される薬物動態の他の尺度として、血清半減期を挙げることができ、これはアルファ相およびベータ相に分けられることが多い。いずれかの相または両方の相が、適当な部分の付加によって大幅に改善され得る。
【0073】
本明細書において「PK部分」と呼ばれる、血液からのタンパク質のクリアランスを遅延させる傾向がある部分として、ポリオキシアルキレン部分、例えば、ポリエチレングリコール、糖(例えば、シアル酸)および耐容性良好であるタンパク質部分(例えば、Fc、Fc断片、トランスフェリンまたは血清アルブミン)が挙げられる。アドネクチンは、米国公開第2007/0048282号に記載される、アルブミンまたはアルブミンの断片(部分)もしくは変種と融合させてもよい。
【0074】
いくつかの実施形態では、PK部分は、例えば米国公開第2007/0178082号および同2007/0269422号に記載されるものなどの血清アルブミン結合タンパク質である。
【0075】
いくつかの実施形態では、PK部分は、例えば米国公開第2007/0178082号に記載されるものなどの血清免疫グロブリン結合タンパク質である。
【0076】
いくつかの実施形態では、アドネクチンは、ポリエチレングリコール(PEG)を含む。一以上のPEG分子を、タンパク質上の異なる位置に結合してもよく、このような結合は、アミン、チオールまたは他の適した反応性基を用いる反応によって達成できる。アミン部分は、例えば、ポリペプチドのN末端に見られる第一級アミンまたはリシンもしくはアルギニンなどのアミノ酸中に存在するアミン基であり得る。いくつかの実施形態では、PEG部分は:a)N末端;b)N末端と最もN末端側のβ鎖またはβ様鎖との間;c)標的結合部位と反対側のポリペプチドの面上に位置するループ;d)C末端と最もC末端側のβ鎖またはβ様鎖との間;およびe)C末端、からなる群から選択されるポリペプチド上の位置に結合している。
【0077】
PEG化は、部位特異的PEG化によって達成でき、この方法においては、PEG化が優先的に起こる部位を作り出すために、適した反応性基がタンパク質に導入される。いくつかの実施形態では、タンパク質は、所望の位置にシステイン残基を導入するよう修飾され、システイン上で部位特異的PEG化が可能となる。PEGは、分子量が大きく変わってもよく、分岐または直鎖であってもよい。
【0078】
いくつかの実施形態では、アドネクチンは、Fn3ドメインおよびPK部分を含む。いくつかの実施形態では、Fn3ドメインは、10Fn3ドメインである。いくつかの実施形態では、PK部分は、ポリペプチドの血清半減期を、Fn3ドメイン単独と比較して、5、10、20、30、40、50、60、70、80、90、100、120、150、200、400、600、800、1000%以上を超えて増大する。
【0079】
いくつかの実施形態では、PK部分はポリマー糖である。いくつかの実施形態では、PK部分はポリエチレングリコール部分である。いくつかの実施形態では、PK部分は血清アルブミン結合タンパク質である。いくつかの実施形態では、PK部分はヒト血清アルブミンである。いくつかの実施形態では、PK部分は血清免疫グロブリン結合タンパク質である。いくつかの実施形態では、PK部分はトランスフェリンである。いくつかの実施形態では、PK部分は血清タンパク質に対して特異的な別のアドネクチンである。
【0080】
生化学的および生物物理学的特性
本出願は、IL−23のp19サブユニットに結合するFn3ドメインを含むアドネクチンを提供する。表1および実施例4に示されるように、標的分子に結合するポリペプチドは、平衡定数(例えば、解離、KD)に関して、および速度定数(例えば、結合速度定数(on−rate constant)、Konおよび解離速度定数(off−rate constant)、Koff)の点で評価できる。アドネクチンは通常、500nM、100nM、10nM、1nM、500pM、200pM、100pM未満のKDで標的分子と結合するが、Koffが十分に低い、またはKonが十分に高い場合には、より高いKD値も許容され得る。
【0081】
本発明の抗IL−23アドネクチンのファミリーのBC、DEおよびFGループ配列は、対応する全長配列番号と共に下記の表1に提示される。
【表1−1】
【表1−2】
【表1−3】
【0082】
抗IL−23アドネクチンのさらなる特性は表2に記載される。
【表2】
(n.d. 検出不可) (詳細な方法は実施例4に記載される)。
【0083】
核酸−タンパク質融合技術
一態様では、本出願は、IL−23のp19サブユニットと結合するフィブロネクチンIII型ドメインを含むアドネクチンを提供する。特異的結合特性を有するFn3ドメインを、迅速に生成し、試験する一つの方法は、Adnexus,Bristol−Myers Squibb R&D Companyの核酸−タンパク質融合技術である。この開示内容は、インビトロ(in vitro)発現および核酸−タンパク質融合物(RNA−およびDNA−タンパク質融合物)を利用してタンパク質との結合にとって重要である、新規ポリペプチドおよびアミノ酸モチーフを同定する、PROfusionと呼ばれるタグ技術を利用する。核酸−タンパク質融合技術は、タンパク質をそのコーディング遺伝情報と共有結合によってカップリングする技術である。RNA−タンパク質融合技術およびフィブロネクチンベースの足場タンパク質ライブラリースクリーニング法の詳細な説明については、出典明示により本明細書に組み込まれる、Szostak ら,米国特許第6,258,558号、同6,261,804号、同6,214,553号、同6,281,344号、同6,207,446号、同6,518,018号、同6,818,418号;およびRoberts ら, Proc Natl. Acad. Sci. 94:12297−12302(1997), 参照。
【0084】
ベクターおよびポリヌクレオチド実施形態
本明細書に開示される種々のタンパク質またはポリペプチドのいずれかをコードする核酸は、化学的に合成されてもよい。コドン使用頻度(codon usage)は、細胞における発現を改善するよう選択されてもよい。このようなコドン使用頻度は、選択される細胞種に依存する。大腸菌および他の細菌、ならびに哺乳類細胞、植物細胞、酵母細胞および昆虫細胞のため、特定化されたコドン使用パターンが開発されている。例えば、 Mayfield ら, Proc. Natl. Acad. Sci. USA, 100(2):438−442 (Jan. 21, 2003); Sinclair ら, Protein Expr. Purif., 26(I):96−105 (Oct. 2002); Connell, N.D., Curr. Opin. Biotechnol., 12(5):446−449 (Oct. 2001); Makrides ら, Microbiol. Rev., 60(3):512−538 (Sept. 1996); および Sharp ら, Yeast, 7(7):657−678 (Oct. 1991) 参照。
【0085】
核酸操作の一般的な技術は、例えば、出典明示により本明細書に組み込まれる、Sambrook ら, Molecular Cloning: A Laboratory Manual, 2nd Edition, Vols. 1−3, Cold Spring Harbor Laboratory Press, publ. (1989), またはAusubel, F. ら, Current Protocols in Molecular Biology, Green Publishing and Wiley−Interscience, New York, publ. (1987)および定期的更新に記載されている。通常、ポリペプチドをコードするDNAは、哺乳類、ウイルスまたは昆虫遺伝子に由来する適した転写または翻訳調節エレメントと作動可能なように連結されている。このような調節エレメントとして、転写プロモーター、転写を制御するための所望によるオペレーター配列、適したmRNAリボソーム結合部位をコードする配列ならびに転写および翻訳の終結を制御する配列が挙げられる。宿主において複製する能力は、通常、複製起点によって付与され、形質転換体の認識を容易にする選択遺伝子がさらに組み込まれる。
【0086】
本明細書に記載されるタンパク質は、直接的にだけではなく、好ましくは、シグナル配列または成熟タンパク質またはポリペプチドのN末端に特異的切断部位を有する他のポリペプチドである異種ポリペプチドとの融合ポリペプチドとしても組換えによって生成してもよい。選択される異種シグナル配列は、宿主細胞によって認識され、プロセシングされる(すなわち、シグナルペプチダーゼによって切断される)ものであることが好ましい。
【0087】
天然シグナル配列を認識およびプロセシングしない原核生物宿主細胞のために、シグナル配列は、例えば、アルカリホスファターゼ、ペニシリナーゼ、lppまたは熱安定性エンテロトキシンIIリーダーの群から選択される原核生物シグナル配列で置換される。
【0088】
酵母分泌のためには、天然シグナル配列を、例えば、酵母インベルターゼリーダー、因子リーダー(サッカロミセス(Saccharomyces)およびクルイベロマイセス(Kluyveromyces)アルファ因子リーダーを含む)、または酸性ホスファターゼリーダー、C.アルビカンス(albicans)グルコアミラーゼリーダー、または米国特許第5,631,144号に記載されるシグナルによって置換してもよい。哺乳類細胞発現では、哺乳類シグナル配列ならびにウイルスの分泌リーダー、例えば、単純ヘルペスgDシグナルが利用可能である。このような前駆体領域のDNAを、リーディングフレームでタンパク質をコードするDNAとライゲートしてもよい。
【0089】
発現ベクターおよびクローニングベクターは、両者とも、ベクターが一以上の選択された宿主細胞において複製するのを可能にする核酸配列を含有する。通常、クローニングベクターでは、この配列は、ベクターが宿主染色体DNAとは無関係に複製するのを可能にするものであり、複製起点または自立複製配列を含む。このような配列は、種々の細菌、酵母およびウイルスについてよく知られている。プラスミドpBR322に由来する複製起点は、ほとんどのグラム陰性菌にとって適しており、2ミクロンのプラスミド起点が酵母に適しており、種々のウイルス起点(SV40、ポリオーマ、アデノウイルス、VSVまたはBPV)が、哺乳類細胞中のクローニングベクターにとって有用である。一般に、複製起点成分は、哺乳類発現ベクターにとって必要ではない(SV40起点は、通常、単に、それが初期プロモーターを含むために使用され得る)。
【0090】
発現ベクターおよびクローニングベクターは、選択マーカーとも呼ばれる選択遺伝子を含有し得る。通常の選択遺伝子は、(a)抗生物質または他の毒素、例えば、アンピシリン、ネオマイシン、メトトレキセートもしくはテトラサイクリンに対する耐性を付与する、(b)栄養要求欠損を補完する、または(c)複合培地からは利用できない決定的な栄養素を供給するタンパク質をコードするものであり、例えば、バチルス(Bacilli)のD−アラニンラセマーゼをコードする遺伝子が挙げられる。
【0091】
発現ベクターおよびクローニングベクターは、通常、宿主生物によって認識され、本発明のタンパク質、例えば、フィブロネクチンベースの足場タンパク質をコードする核酸と作動可能なように連結しているプロモーターを含有する。原核生物宿主とともに使用するのに適したプロモーターとして、phoAプロモーター、β−ラクタマーゼおよびラクトースプロモーター系、アルカリホスファターゼ、トリプトファン(trp)プロモーター系およびtacプロモーターなどのハイブリッドプロモーターが挙げられる。しかしながら、他の既知細菌プロモーターも適している。細菌系において使用するためのプロモーターはまた、本発明のタンパク質をコードするDNAと作動可能なように連結しているシャイン−ダルガーノ(S.D.)配列を含む。真核生物のためのプロモーター配列は公知である。実質的にすべての真核細胞遺伝子は、転写が開始される部位からおよそ25〜30塩基上流に位置するATリッチ領域を有する。多数の遺伝子の転写の開始から70〜80塩基上流に見られる別の配列は、CNCAAT領域である(ここで、Nは、任意のヌクレオチドであり得る)。ほとんどの真核細胞遺伝子の3’末端には、コード配列の3’末端へのポリAテールの付加のためのシグナルであり得るAATAAA配列がある。これらの配列のすべてが、真核細胞発現ベクターに適宜挿入される。
【0092】
酵母宿主とともに使用するのに適したプロモーター配列の例として、3−ホスホグリセレートキナーゼまたはエノラーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース−6−リン酸イソメラーゼ、3−ホスホグリセレートムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼおよびグルコキナーゼなどの他の解糖酵素のプロモーターが挙げられる。
【0093】
哺乳類宿主細胞におけるベクターからの転写は、例えば、ポリオーマウイルス、鶏痘ウイルス、アデノウイルス(アデノウイルス2など)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよび最も好ましくは、サルウイルス40(SV40)などのウイルスのゲノムから、異種哺乳類プロモーター、例えば、アクチンプロモーターまたは免疫グロブリンプロモーターから、ヒートショックプロモーターから得られたプロモーターによって、このようなプロモーターが宿主細胞系と適合するならば、制御され得る。
【0094】
高等真核生物による本発明のタンパク質をコードするDNAの転写は、ベクター中にエンハンサー配列を挿入することによって増大されることが多い。哺乳類遺伝子(グロビン、エラスターゼ、アルブミン、アルファ−フェトプロテインおよびインスリン)に由来する多数のエンハンサー配列が現在知られている。しかしながら、通常、真核細胞ウイルス由来のエンハンサーを使用する。例として、複製起点の後ろ側のSV40エンハンサー(bp100〜270)、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後ろ側のポリオーマエンハンサーおよびアデノウイルスエンハンサーが挙げられる。真核細胞プロモーターの活性化のためのエンハンサーエレメントに関しては、Yaniv, Nature 297:17−18(1982)も参照。エンハンサーは、ペプチドをコードする配列の5’または3’側の位置でベクター中にスプライシングされ得るが、プロモーターの5’側に位置することが好ましい。
【0095】
真核宿主細胞(例えば、酵母、真菌、昆虫、植物、動物、ヒトまたは他の多細胞生物に由来する有核細胞)において使用される発現ベクターはまた、転写の終結のために、またmRNAを安定化するために必要な配列も含有する。このような配列は通常、真核細胞またはウイルスのDNAまたはcDNAの5’非翻訳領域、および場合により、3’非翻訳領域から得ることができる。これらの領域は、本発明のタンパク質をコードするmRNAの非翻訳部分中のポリアデニル化断片として転写されるヌクレオチドセグメントを含有する。一つの有用な転写終結成分として、ウシ成長ホルモンポリアデニル化領域がある。WO94/11026およびそこで開示される発現ベクターを参照。
【0096】
組換えDNAはまた、タンパク質を精製するのに有用であり得る任意の種類のタンパクン質タグ配列を含み得る。タンパク質タグの例として、それだけには限らないが、ヒスチジンタグ、FLAGタグ、mycタグ、HAタグまたはGSTタグが挙げられる。細菌、真菌、酵母および哺乳類細胞宿主とともに使用するのに適当なクローニングおよび発現ベクターは、Cloning Vector: A Laboratory Manual, (Elsevier, New York, 1985))中に見ることができ、その関連の開示内容は、出典明示により本明細書に組み込まれる。
【0097】
発現コンストラクトは、当業者には明らかであるように、宿主細胞にとって適当な方法を使用して宿主細胞に導入される。核酸を宿主細胞に導入するための種々の方法が当技術分野で公知であり、これらには限らないが、エレクトロポレーション;塩化カルシウム、塩化ルビジウム、リン酸カルシウム、DEAE−デキストランまたは他の物質を使用するトランスフェクション;微粒子銃;リポフェクション;および感染(ベクターが感染性物質である)が挙げられる。
【0098】
適した宿主細胞として、原核生物、酵母、哺乳類細胞または細菌細胞が挙げられる。適した細菌として、グラム陰性またはグラム陽性菌、例えば、大腸菌またはバチルス種が挙げられる。好ましくは、サッカロミセス種に由来する酵母、例えば、S.セレビシエ(cerevisiae)も、ポリペプチドの生成のために使用してよい。種々の哺乳類または昆虫細胞培養系を使用して、組換えタンパク質を発現することもできる。昆虫細胞において異種タンパク質を生成するためのバキュロウイルス系は、Luckowら(Bio/Technology, 6:47(1988))に総説されている。適した哺乳類宿主細胞株の例として、内皮細胞、COS−7サル腎臓細胞、CV−1、L細胞、C127、3T3、チャイニーズハムスター卵巣(CHO)、ヒト胚性腎細胞、HeLa、293、293TおよびBHK細胞株が挙げられる。精製ポリペプチドは、組換えタンパク質を発現するのに適した宿主/ベクター系を培養することによって調製される。多数の適用のために、本明細書に開示される小さいサイズの多数のポリペプチドは、発現のための好ましい方法として大腸菌において発現を行う。次いで、タンパク質は、培養培地または細胞抽出物から精製される。
【0099】
タンパク質生成
宿主細胞は、タンパク質生成のために本明細書に記載される発現またはクローニングベクターを用いて形質転換され、プロモーターを誘導し、形質転換体を選択し、または所望の配列をコードする遺伝子を増幅するために適当に改変された従来の栄養培地で培養される。本明細書に示される実施例では、ハイスループットタンパク質生成(HTPP)および中規模生成のために使用される宿主細胞は、BL21 DE3 plyS−細菌株であった。本発明のタンパク質を生成するために使用される宿主細胞は、Hamら, Meth. Enzymol. 58:44 (1979)、Baritesら, Anal. Biochem.102:255 (1980)、米国特許第4,767,704号、同4,657,866号、同4,927,762号、同4,560,655号、同5,122,469号、同6,048,728号、同5,672,502号または米国特許第RE30,985号に記載されるものなどの、種々の培地で培養してもよい。任意の他の必要な栄養補助剤を、当業者に公知である適当な濃度で含めてもよい。温度、pHなどといった培養条件は、発現のために選択された宿主細胞とともにこれまでに使用されたものであり、当業者には明らかであろう。
【0100】
本明細書に開示されるタンパク質はまた、細胞−翻訳系を使用して生成できる。このような目的上、ポリペプチドをコードする核酸は、インビトロ(in vitro)転写を可能にしてmRNAを生成し、使用されている特定の無細胞系(哺乳類または酵母などの真核細胞を含まない翻訳系または細菌などの原核細胞を含まない翻訳系)におけるmRNAの無細胞翻訳を可能にするよう改変されなければならない。
【0101】
本発明のタンパク質はまた、化学合成によって(例えば、Solid Phase Peptide Synthesis, 2nd Edition, The Pierce Chemical Co., Rockford, IL, publ. (1984)に記載される方法によって)生成できる。タンパク質への修飾も化学合成によって生成できる。
【0102】
本発明のタンパク質は、タンパク質化学の分野で一般に知られている、タンパク質の単離/精製法によって精製できる。非限定的な例として、抽出、再結晶化、塩析(例えば、硫酸アンモニウムまたは硫酸ナトリウムを用いる)、遠心分離、透析、限外濾過、吸着クロマトグラフィー、イオン交換クロマトグラフィー、疎水性クロマトグラフィー、順相クロマトグラフィー、逆相クロマトグラフィー、ゲル濾過、ゲル浸透クロマトグラフィー、アフィニティークロマトグラフィー、電気泳動、向流分配またはこれらの任意の組合せが挙げられる。精製後、ポリペプチドを、これらには限らないが、濾過および透析を含む、当技術分野で公知の種々の方法のいずれかによって、異なるバッファーに交換してもよく、かつ/または、濃縮してもよい。
【0103】
精製されたポリペプチドは、好ましくは少なくとも85%純粋、より好ましくは少なくとも95%純粋、最も好ましくは少なくとも98%純粋である。純度の正確な数値にかかわらず、ポリペプチドは医薬製剤として使用するのに十分に純粋である。
【0104】
プラットフォーム製造工程(platform manufacturing process)を使用して、抗IL−23アドネクチンを調製した。実施例1には製造工程の一例が記載されている。アドネクチンは大腸菌(E.coli)内で生成された。大腸菌MG1655細胞を、封入体として不溶型でタンパク質を生成する発現ベクター(pBMS2008/ATI001044)で形質転換した。組換え株を撹拌式発酵タンク内で生育させる。発酵終了時に、精製に向けて、封入体を回収し、可溶化し、かつリフォールドさせた。精製したアドネクチンを、マレイミドリンカーを用いて40kDaの分岐メトキシPEGへコンジュゲートした。次いで、コンジュゲート化物質を再精製し、遊離PEG、遊離アドネクチン、および製品に関連する不純物を除去した。品質管理テストをバルク薬剤原料に対して行った。
【0105】
治療的インビボ(in vivo)使用
一態様では、本出願は、狼蒼(たとえば、紅斑性狼蒼、腎炎性狼蒼)、橋本甲状腺炎、原発性粘液水腫、グレーブス病、悪性貧血、自己免疫萎縮性胃炎、アジソン病、糖尿病(たとえば、インスリン依存型糖尿病、I型糖尿病)、グッドパスチャー症候群、重症筋無力症、天疱瘡、クローン病、交感神経性眼炎、自己免疫ブドウ膜炎、多発性硬化症、自己免疫性溶血性貧血、突発性血小板減少症、原発性胆汁性肝硬変、慢性活動性肝炎、潰瘍性大腸炎、シェーグレン症候群、リウマチ性疾患(たとえば、関節リウマチ)、多発性筋炎、強皮症および混合性結合組織病などの、自己免疫疾患の治療において有用である抗IL−23アドネクチンを提供する。
【0106】
本出願はまた、被験体に抗IL−23アドネクチンを投与する方法を提供する。いくつかの実施形態では、被験体はヒトである。いくつかの実施形態では、抗IL−23アドネクチンは、哺乳類、特に、ヒトにとって医薬上許容される。「医薬上許容される」ポリペプチドとは、本質的にエンドトキシンを含まない、または極めて低いエンドトキシンレベルを有するなどの、著しく有害な医学的結果を伴わずに動物に投与されるポリペプチドを指す。
【0107】
製剤および投与
本出願はさらに、本明細書に記載される抗IL−23アドネクチンを含む医薬上許容される組成物を提供し、該組成物は本質的にエンドトキシンを含まない。抗IL−23アドネクチンを含む治療用製剤は、所望の程度の純度を有する記載されるアドネクチンを、任意の生理学的に許容される担体、賦形剤または安定剤と混合することによって、水溶液、凍結乾燥された、または他の乾燥された製剤の形態で、保存のために調製される(Osol, A., ed., Remington’s Pharmaceutical Sciences, 16th Edition (1980))。許容される担体、賦形剤または安定剤は、使用される用量および濃度でレシピエントにとって非毒性であり、これらの例として、リン酸、クエン酸および他の有機酸などのバッファー;アスコルビン酸およびメチオニンを含む抗酸化物質;保存料(オクタデシルジメチルベンジル塩化アンモニウム;塩化ヘキサメトニウム;塩化ベンザルコニウム、塩化ベンゼトニウム;フェノール、ブチルもしくはベンジルアルコール;メチルもしくはプロピルパラベンなどのアルキルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3−ペンタノール;およびm−クレゾールなど);低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチンまたは免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニンまたはリシンなどのアミノ酸;単糖、二糖およびグルコース、マンノースまたはデキストランを含む他の炭水化物;EDTAなどのキレート化剤;スクロース、マンニトール、トレハロースまたはソルビトールなどの糖;ナトリウムなどの塩形成性対イオン;金属錯体(例えば、Zn−タンパク質複合体);ならびに/あるいはTWEEN、PLURONIC登録商標またはポリエチレングリコール(PEG)などの非イオン性界面活性剤が挙げられる。
【0108】
また、本明細書において製剤とは、治療されている特定の適応症にとって必要な二以上の活性化合物、好ましくは、互いに有害な影響を及ぼさない相補的な活性を有するものを含み得る。このような分子は、意図される目的上効果的である量の組合せで適宜存在する。
【0109】
インビボ(in vivo)投与に使用される製剤は、無菌でなければならない。これは、滅菌濾過メンブレンを通した濾過によって容易に達成される。
【0110】
当業者ならば、各治療薬の用量が、薬剤の独自性によって決まることを理解するであろう。
【0111】
治療適用のためには、抗IL−23アドネクチンは、医薬上許容される剤形で被験体に投与される。それは、ボーラス(bolus)として、または一定時間にわたる連続注入によって静脈内に、または皮下経路によって投与され得る。適した医薬上許容される担体、希釈剤および賦形剤は周知であり、臨床状況が必要とする際に、当業者によって決定され得る。適した担体、希釈剤および/または賦形剤の例として、(1)ダルベッコのリン酸緩衝生理食塩水、(2)0.9%生理食塩水(0.9% w/v NaCl)、および(3)5%(w/v)デキストロースが挙げられる。
【0112】
本発明の方法は、インビトロ(in vitro)、インビボ(in vivo)またはエキソビボ(ex vivo)で実施できる。
【0113】
抗IL−23アドネクチンおよび一以上のさらなる治療薬の投与は、同時投与されるか逐次投与されるかにかかわらず、治療適用のために、上記のように起こり得る。同時投与に適した医薬上許容される担体、希釈剤および賦形剤は、投与されている特定の治療薬の独自性によって決まる、と当業者に理解されるであろう。
【0114】
凍結乾燥されているのではなく水性の剤形で存在する場合には、タンパク質は、通常、約0.1mg/ml〜100mg/mlの濃度で製剤化されるが、これらの範囲の外側の広い変動も容認される。疾患の治療のための、抗IL−23アドネクチンの適当な用量は、治療される疾患の種類、疾患の重篤度および経過、アドネクチンが予防目的で投与されるのか治療目的で投与されるのか、これまでの治療の経過、患者の病歴およびアドネクチンに対する反応、ならびに主治医の裁量によって決まるであろう。タンパク質は、1回で、または一連の治療にわたって、患者に適宜投与される。
【0115】
血清アルブミン結合アドネクチン(SABA)の融合
特定の態様では、本出願は、ヒト血清アルブミンに結合する10Fn3ドメインに融合した抗IL−23アドネクチンを含む融合タンパク質(血清アルブミン結合アドネクチン Serum Albumin Binding Adnectin(10Fn3ドメイン)またはSABA)を提供する。このような融合タンパク質は、抗IL−23アドネクチン単独に比べて、アルブミン存在下で延長した半減期を有する。
【0116】
特定の態様では、本出願は、融合タンパク質のt1/2を延長するため、血清アルブミン、例えばヒト血清アルブミン(HSA)に特異的に結合する10Fn3ドメインを含む融合タンパク質を提供する。
【0117】
特定の実施形態では、SABAに融合された抗IL−23アドネクチンの血清半減期は、SABAへコンジュゲートされなかった場合の抗IL−23アドネクチンの血清半減期と比べて増大する。特定の実施形態では、SABA融合体の血清半減期は、SABAに融合されなかった場合の抗IL−23アドネクチンの血清半減期と比べて、少なくとも20、40、60、80、100、120、150、180、200、400、600、800、1000、1200、1500、1800、1900、2000、2500、または3000%長い。他の実施形態では、SABA融合体の血清半減期は、SABAに融合されなかった場合の抗IL−23アドネクチンの血清半減期より、少なくとも1.5倍、2倍、2.5倍、3倍、3.5倍、4倍、4.5倍、5倍、6倍、7倍、8倍、10倍、12倍、13倍、15倍、17倍、20倍、22倍、25倍、27倍、30倍、35倍、40倍、または50倍大きい。いくつかの実施態様では、SABA融合体の血清半減期は、少なくとも10時間、15時間、20時間、25時間、30時間、35時間、40時間、50時間、60時間、70時間、80時間、90時間、100時間、110時間、120時間、130時間、135時間、140時間、150時間、160時間、または200時間である。
【0118】
従って、本明細書に記載されるSABA融合分子は、抗IL−23アドネクチンおよびSABA間の融合体の構築によって抗IL−23アドネクチンの半減期を増大させるために有用である。このような融合分子を使用して、IL−23の生物活性へ応答する症状を治療してもよい。本発明は、IL−23の異常調節によって引き起こされる疾患におけるSABA融合分子の使用を企図する。
【0119】
融合体は、抗IL−23アドネクチンをSABA分子の末端のいずれかへ付着させることこと、すなわち、SABA−抗IL−23アドネクチン配置または抗IL−23アドネクチン−SABA配置で付着させることによって、形成される。
【0120】
一態様では、本開示は血清アルブミン結合10Fn3ドメインを含む抗IL−23アドネクチンを含む融合タンパク質を提供する。例示的な実施形態では、本明細書に記載される血清アルブミン結合10Fn3タンパク質は、3uM、2.5uM、2uM、1.5uM、1uM、500nM、100nM、50nM、10nM、1nM、500pM、200pM、100pM、50pMまたは10pM未満のKdでHSAに結合する。特定の実施形態では、本明細書に記載される血清アルブミン結合10Fn3タンパク質は、25℃または37℃で、5.5から7.4のpH範囲で、3uM、2.5uM、2uM、1.5uM、1uM、500nM、100nM、50nM、10nM、1nM、500pM、200pM、100pM、50pMまたは10pM未満のKdでHSAに結合する。いくつかの実施形態では、本明細書に記載される血清アルブミン結合10Fn3タンパク質は、7.4以上のpHでのHSAに対する結合親和性と比較して、pH7.4未満でより強くHSAに結合する。
【0121】
特定の実施形態では、本明細書に記載されるHSA結合10Fn3ドメインを含む融合タンパク質はまた、サル、ラット、またはマウスの一以上に由来する血清アルブミンにも結合する。特定の実施形態では、本明細書に記載される血清アルブミン結合10Fn3タンパク質は、3uM、2.5uM、2uM、1.5uM、1uM、500nM、100nM、50nM、10nM、1nM、500pMまたは100pM未満のKdで、アカゲザル血清アルブミン(RhSA)またはカニクイザル血清アルブミン(CySA)に結合する。
【0122】
特定の実施形態では、本明細書に記載される血清アルブミン結合10Fn3ドメインを含む融合タンパク質は、HSAのドメインIおよび/またはドメインIIに結合する。一の実施形態では、本明細書に記載される血清アルブミン結合10Fn3ドメインを含む融合タンパク質は、HSAのドメインIIIに結合しない。
【0123】
特定の実施形態では、融合タンパク質の血清アルブミン結合10Fn3部分(SABA)は、野生型10Fn3ドメイン(配列番号1)と少なくとも40%、50%、60%、70%、75%、80%または85%同一性を有する配列を含む。一の実施形態では、野生型10Fn3ドメインと比べて、BC、DEまたはFGループのうち少なくとも1つが修飾されている。別の実施形態では、野生型10Fn3ドメインと比べて、BC、DEまたはFGループのうち少なくとも2つが修飾されている。別の実施形態では、野生型10Fn3ドメインと比べて、BC、DEおよびFGループのうち3つ全てが修飾されている。他の実施形態では、SABAは、表3に示される26のコア(core)SABA配列(すなわち、配列番号103、107、111、115、119、および123〜143)のいずれか1つ、または、表3に示される伸長したSABA配列(すなわち、配列番号188〜215、6×HISタグを引く)のいずれか1つと、少なくとも40%、50%、60%、70%、75%、80%、85%、90%、または95%同一性を有する配列を含む。
【0124】
特定の実施形態では、コアアミノ酸残基は固定されており、任意の置換、保存的置換、欠失または付加は、コアアミノ酸残基以外の残基で生じる。例示的な実施態様では、BC、DEおよびFGループは、以下表3に示されるHSAバインダーのいずれかに由来するBC、DEおよびFGループ配列(すなわち、表3の配列番号103、107、111、115、119、および123〜143)を含むポリペプチドで置換される。
【0125】
特定の実施形態では、SABA(例えば、SABAコア配列または上述のそれらに由来する配列)は、N末端伸長配列および/またはC末端伸長配列を含むように改変されてもよい。例示的な伸長配列は、表3に示される。例えば、SABA1.1として表される配列番号188は、N末端配列MGVSDVPRDLE(配列番号144,AdNT1と表す)およびC末端配列EIDKPSQ(配列番号153)を備える、コアSABA1配列(配列番号103)を含む。SABA1.1はさらにHis6タグをC末端で含む、しかしながら、His6タグは完全に任意であり、N−またはC−末端伸長配列内のどこにでも設置されうる。さらに、表3に提供される例示的なN−またはC−末端伸長配列(配列番号144〜163)およびそれらの変種のいずれかを使用して、表3に提供される任意の与えられるSABAコア配列を修飾することができる。
【0126】
他の実施形態では、SABA融合分子を設計する場合に必要であれば、テール配列は他の知られたリンカー配列(例えば、表3、配列番号164〜187)と結合させてもよい。
【0127】
コンジュゲーションリンカー
SABA融合体は共有結合的に、または非共有結合的に連結されてもよい。いくつかの実施形態では、血清アルブミン結合10Fn3は、直接的にまたは間接的に、ポリペプチドリンカーを介して抗IL−23アドネクチンに連結された。Fn3の連結のための適したリンカーは、分離されたドメインが、標的分子に結合する高い親和性を可能にするような三次元構造の形成をお互い独立してフォールドできるようにするものである。
【0128】
本開示はこれらの必要条件を満たす多数の適したリンカーを提供し、グリシン−セリンベースのリンカー、グリシン−プロリンベースのリンカー、ならびにアミノ酸配列PSTSTST(配列番号184)を有するリンカーが挙げられる。本明細書に記載される実施例では、ポリペプチドリンカーを介して連結されたFn3ドメインが、その標的結合機能を保持することを実証する。いくつかの実施形態では、リンカーは、グリシン−セリンベースのリンカーである。これらのリンカーはグリシンおよびセリン残基を含み、8から50、10から30、および10から20個の間のアミノ酸の長さであり得る。例として、アミノ酸配列(GS)7(配列番号171)、G(GS)6(配列番号166)、およびG(GS)7G(配列番号168)を有するリンカーが挙げられる。他のリンカーはグルタミン酸を含有し、例えば、(GSE)5(配列番号173)およびGGSE GGSE(配列番号177)が挙げられる。他の例示的なグリシン−セリンリンカーとして、(GS)4(配列番号170)、(GGGGS)7(配列番号179)、(GGGGS)5(配列番号180)、および(GGGGS)3G(配列番号181)が挙げられる。いくつかの実施形態では、リンカーは、グリシン−プロリンベースのリンカーである。これらのリンカーは、グリシンおよびプロリン残基を含み、3から30、10から30、および3から20個の間のアミノ酸の長さであり得る。例として、(GP)3G(配列番号182)および(GP)5G(配列番号183)を有するリンカーが挙げられる。他の実施形態では、リンカーは3から30、10から30、および3から20アミノ酸の間の長さを有するプロリン−アラニンベースのリンカーであってもよい。プロリン−アラニンベースのリンカーの例として、例えば、(PA)3(配列番号185)、(PA)6(配列番号186)および(PA)9(配列番号187)が挙げられる。最適リンカー長およびアミノ酸組成は、当技術分野で周知の方法によって日常的な実験によって決定され得ると考えられる。
【0129】
いくつかの実施形態では、本明細書に記載される融合体は、血液または標的組織においてプロテアーゼによって切断可能であるプロテアーゼ部位を有するポリペプチドリンカーを介して連結される。このような実施形態を使用して、より良好なデリバリーまたは治療的特性、またはより効率的な生成のために、治療用タンパク質を放出できる。
【0130】
さらなるリンカーまたはスペーサーは、Fn3ドメインのC末端において、Fn3ドメインとポリペプチドリンカーの間に導入され得る。さらなるリンカーまたはスペーサーは、Fn3ドメインのN末端において、Fn3ドメインとポリペプチドリンカーの間に導入され得る。
【0131】
いくつかの実施形態では、治療的部分(therapeutic moiety)は、ポリマーリンカーを介して直接的または間接的にSABAに連結されてもよい。ポリマーリンカーを使用して、以下の特徴のうち一以上を有するタンパク質融合体を作り出すために、融合体の各構成要素(component)間の距離を最適に変えることができる:1)対象とするタンパク質との結合時の、一以上のタンパク質ドメインの結合の立体障害の低減または増大、2)タンパク質安定性または溶解度の増大、3)タンパク質凝集の減少、および4)タンパク質の全結合力または親和性の増大。
【0132】
いくつかの実施形態では、治療的部分は、重合糖などの生体適合性ポリマーを介してSABAに連結している。重合糖は、血液または標的組織において酵素によって切断可能である酵素切断部位を含み得る。このような実施形態を使用して、より良好なデリバリーまたは治療的特性、またはより効率的な生成のために、治療用タンパク質を放出できる。
【0133】
血清アルブミン結合アドネクチン(SABA)配列の概要
本願で参照されるSABA配列の多くは、以下の表3に要約される。特別の定めのない限り、全てのN末端伸長は下線で示され、全てのC末端テール/伸長は二重下線で示され、リンカー配列は囲まれている。ループ領域BC、DEおよびFGは、各コアSABA配列に対して網掛けされている。
【表3−1】
【表3−2】
【表3−3】
【表3−4】
【表3−5】
【表3−6】
【表3−7】
【表3−8】
【表3−9】
【表3−10】
【表3−11】
実施例
【実施例1】
【0134】
製造過程
発酵および集菌
生産発酵(production fermentation)は無菌の基礎培地で調製される。バイアルを融解して、成長培地を含む移動容器に接種するために使用する。接種材料は直ちに生産発酵へ移される。培養は撹拌して34℃の温度で維持され、OD600が5〜10(1OD単位は約1x109細胞/mL)に到達するまで生育させる。流加培地の添加はこのODで開始する。発酵は、培養がイソプロピルβ−D−1−チオガラクトピラノシド(IPTG)の添加によりアドネクチンを生成するために誘導される点である、OD600=25まで継続する。容器の温度は誘導の際、34℃から39℃へ上昇される。サンプルは毎時間、無菌的に採取され、細胞密度を試験する。
【0135】
9〜12時間の誘導発酵の後、25℃へ温度を低下させ、終濃度10mMでエチレンジアミン四酢酸(EDTA)を添加し、水酸化ナトリウムの添加によりpHを上昇させ、かつ、撹拌を弱めることにより、容器を回収(harvest)のために準備する。一時間の保持期間(hold period)の後、発酵容器内容物を集菌容器へ流し入れる。
【0136】
封入体の調製
回収プールの細胞破砕は、細胞を破砕してその内容物を放出させるMICROFLUIDIZER登録商標に通すことによって行う。細胞破砕に続いて、非常に強い遠心力により、連続工程で、固体および液相を分離するための、ディスクスタック遠心分離機を使用して、封入体が回収される。次いで封入体を、バッファー(20〜25℃)で2回、水(20〜25℃)で2回、洗浄する。封入体を洗浄するたび、遠心分離によって回収する。洗浄された封入体をスラリーとして回収する。
【0137】
封入体の可溶化およびタンパク質リフォールディング
可溶化バッファーを封入体スラリーへ添加し、続いて室温で1時間撹拌する。OD280=20(総タンパク質)がこの工程の間の目標とされる。
【0138】
タンパク質リフォールディングは二段階希釈工程を使用して実行する。希釈バッファーを、可溶化封入体一部に対して希釈バッファー二分の一部の比率(v/v)で、可溶化封入体へ添加する。第二の希釈は、OD280=0.7(総タンパク質)を目標として、可溶化封入体をリフォールドバッファーへ添加することにより実行する。前記の希釈は室温で撹拌しながら実行する。一時間の混合を実行し、撹拌を止め、タンパク質溶液を室温で一晩保つ。可溶化されリフォールディングされたアドネクチンは、0.8μm〜22μmのフィルターに通され、A280およびRP−HPLCによりタンパク質含有量に関して試験される。
【0139】
精製およびPEGへのコンジュゲーション
リフォールドされフィルター処理されたアドネクチンは、初期回収(initial capture)のため、陽イオン交換(CEX1)カラムへ直接ロードされる。結合した物質を洗浄バッファーで洗浄し、50mM酢酸ナトリウム、500mM塩化ナトリウム、1.5%プロピレングリコール、pH5.5で溶出する。溶出プールを、純度、同一性、濃度、およびエンドトキシンに関して測定する。
【0140】
回収クロマトグラフィー(capture chromatography)からの溶出物をさらに、疎水性相互作用クロマトグラフィー(HIC)を使用して精製する。CEX1溶出物をHICカラムへ直接ロードし、洗浄し、続いて50mM酢酸ナトリウム、30%プロピレングリコール、pH5.5で溶出する。溶出プールを、純度、同一性、および濃度に関して測定する。
【0141】
精製されたアドネクチンはその後、40kDaの分岐PEGのマレイミド誘導体(mPEG2−MAL)で直接整形(format)される。HIC溶出物は室温で撹拌され、mPEG2−MALが添加される。室温で1時間混合後、反応混合液を同じ温度で一晩インキュベートしてもよい。次いで、PEG化溶液を最終CEXカラム(CEX2)へと進める。サンプルはタンパク質含量、純度、およびエンドトキシンのために採取される。
【0142】
PEG化溶液のpHおよび伝導度は、再精製のため、最終陽イオン交換カラム(CEX2)へロードするより前に、75mM酢酸でそれぞれ4.0および1.0mS/cmに調節される。一度ロードしたら、結合した物質を洗浄バッファーで洗浄し、続いて50mM酢酸ナトリウム、25mM塩化ナトリウム、pH5.0で溶出する。サンプルはタンパク質含量、純度、およびエンドトキシンのために採取される。
【0143】
CEX2溶出物は、V−スクリーンを有する30kDaの名目分子量カットオフメンブレンを備えるタンジェンシャルフロー濾過ユニットで、15mg/mLへ濃縮される。50mM酢酸ナトリウム、25mM塩化ナトリウム、pH5.0中のバルク薬剤原料を0.22μmのフィルターに通し、−80℃で凍結した。
【実施例2】
【0144】
遺伝子、ベクターおよび宿主細胞
T7プロモーターの制御下でタンパク質をコードするプラスミドを、株構築(strain construction)での使用のために生成した。このプラスミドDNAを使用して、大腸菌K−12 MG1655コンピテントセル(F−ラムダ−、ilvG−rfb−50 rph−1)を形質転換させた。宿主株はIPTGの添加で遺伝子からの発現の誘導ができるように設計された。形質転換されたMG1655株はカナマイシンに対して耐性である。タンパク質発現ベクターは図2に示される。プレートからのシングルコロニー選択を使用して、発酵培養物を接種し、その後それを分注し、研究細胞バンクとして使用するため凍結させた。
【実施例3】
【0145】
生物物理学的特性評価および生化学的特性評価
本発明のタンパク質の構造および品質は、数種の総合的解析方法により試験した。
【0146】
MALDI−MS
質量スペクトルプロファイルはMALDIによって解析した。サンプル上のMALDI解析の正確さを評価するため、各サンプルに対してスチールプレート上に20の個別のスポットを設置し、連続して解析した。合計20スペクトルが生成された。
【0147】
ペプチドマッピングを使用して、本発明のタンパク質並びに対応する非PEG化タンパク質に関して、cDNA配列から予測されるアミノ酸(第一構造)の正確な発現を確認した。完全な配列カバー率を得るために、トリプシン(Lys残基およびArg残基のC末端側への切断)およびエンドプロテイナーゼGlu−C(Glu残基のC末端側への切断)を採用して、ペプチド断片の2つのオーバーラップするセットを得た。ペプチドマッピングを使用して、共有結合的な翻訳後修飾として挙げられる残存N末端メチオニン、ジスルフィド結合、アスパラギンの脱アミノ化、メチオニン酸化(等)も決定した。ペプチドは、分子量による液体クロマトグラフィー質量分析(LC−MS)、および衝突誘起解離(CID)によって部分的な配列情報を提供するタンデム質量分析(MSMS)により、同定され、かつ特徴づけられた。
【0148】
SDS−PAGE
ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)を使用して、非PEG化およびPEG化の抗IL−23アドネクチンの分子量バンドパターンを可視化した。サンプルは、還元剤有りまたは無しのサンプルバッファー中に調製された。SDS中で加熱の後、サンプルおよび分子量マーカーを、プレ−キャストのグラジエント(4−20%)ポリアクリルアミドゲル上で、電気泳動的に解析した。電気泳動後、ゲルを固定し、クーマシーブルーを使用して染色した。サンプルのバンドパターンの同等性を視覚的に評価した。
【0149】
サイズ排除クロマトグラフィー/マルチアングル光散乱(SECMALS)
サイズ排除クロマトグラフィー(SEC)を、高分子量(HMW)種および低分子量(LMW)種のモノマーの定量的解析のために使用した。続くSEC分離で、分離された種の分子量を、示差屈折率計を並んで備えるマルチアングル光散乱により決定した。
【実施例4】
【0150】
インビトロ(in vitro)非臨床薬理
SPRによるKD
結合特性は、表面プラズモン共鳴(SPR)により特徴づけられる。ヒトIL−23を、ProteOn XRP(Bio−Rad)チップ表面の一次元に2から4レベルで固定化し、同じSPRチップ表面の他の次元で6種の異なる濃度の抗IL−23アドネクチンへと暴露させた。これにより再生無しで速度論的定量が可能となった。二重の(duplicate)チップを25℃および37℃での速度論的定量のために使用した。速度論的パラメータの評価は、ラングミュア相互作用モデル(Langmuir interaction model)およびProteOnマネージャーソフトウェアに適合する(fit with)定数パラメータを用いて行った。
【0151】
以下表4に示されるように、これらの抗IL−23アドネクチンに対する解離速度(off−rate)は、25℃では遅い(約10−5s−1)。37℃でさえ、その解離速度(off−rate)はSPR技術の検出限界に近かかったので、報告された解離定数測定は過小評価であるかもしれない。
【表4】
【0152】
溶液相親和性
ヒトIL−23に対するATI001045の溶液相親和性を、カイネティックエクスクルージョンアッセイ(Kinetic Exclusion Assay)(KinExA)を用いて測定した。一形式において、hIL−23の二重滴定は3つの濃度各々に対して行った。相対非結合ATI001045濃度をヒトIL−23固体マトリックス上への回収によって測定し、続いてアドネクチン足場を認識する蛍光標識抗体で検出した。技術的な制限により、試験できる最小の濃度は0.75nMであった。それゆえ、表5に示す包括的なKD解析はKDに対して51pMの見積もりを与えるものの、親和性は95%信頼区間で、1桁pMまで低く、あるいは、150pMまで高くなり得る。
【表5】
【0153】
ヒトIL−23に対するATI001045およびATI001047の溶液親和性は、KinExAにおいて代替形式を使用して測定された。アドネクチンの二重滴定は、3つの濃度(ATI001045)または単一の濃度(ATI001047)のヒトIL−23の各々に対して行われた(最も低い濃度に対しては4重測定)。相対非結合ヒトIL−23濃度を非PEG化ATI001045固体マトリックス上への回収によって測定し、続いてhIL−23のp40サブユニットを認識する蛍光標識抗体で検出した。表6に示す包括的なKD解析は、ATI001045に関して95%信頼区間で22〜2.4pMの、9.4pMのKDを与え、95%信頼区間で60.1から19.4pMの、36.3pMのKDを与える。
【表6】
【0154】
Kit225細胞でのSTAT3リン酸化
Parhamら (“A receptor for the heterodimeric cytokine IL−23 is composed of IL−12Rbeta1 and a novel cytokine receptor subunit, IL−23R”, J. Immunol., 168(11):5699−5708 (Jun. 1, 2002))は、ヒトIL−2依存性T細胞株、Kit225からIL−23Rをクローン化した。これらの細胞は、IL−12RB1ならびにIL−23Rの両方の発現に対してFACS解析により特徴づけられ、かつ、pSTAT3の刺激によるIL−23ならびにpSTAT4の刺激によるIL−12に応答した。Kit225細胞を96ウェルプレートへ播種し、37℃で3時間、FBSおよびIL−2の非存在下で静置した(quiesced)。このインキュベートに続き、ヒト組換えIL−23(または、1時間アンタゴニストとプレインキュベートしたIL−23)を添加し、STAT3のリン酸化(p−STAT3として略される)を刺激するために、細胞を37℃で15分間、インキュベーターへ戻した。各条件を96ウェルプレートにて二重測定で測定した。刺激は細胞の氷上静置および氷冷PBSの添加により停止させた。最後に、標準的なプロトコールに従って細胞をペレット化および溶解し、ELISAによってpSTAT3生成を検出した。
【0155】
刺激のためのIL−23の最適濃度は35pMであった。IL−23誘導性pSTAT3の阻害は、抗p40モノクローナル抗体(mAb1510)ならびに抗p19ポリクローナル抗体(AF1716)の滴定によって実証した。ATI001045、ATI001047、ATI001014およびATI001016は、抗p19ポリクローナル抗体よりも約150倍強力な、〜300pMのIC50を備える同等の活性を有した一方、ATI001015は、抗p19ポリクローナル抗体よりも約40倍強力な、〜1.2nMのIC50を有した。アドネクチンATI000934は、1nMのIC50を備え、ATI001045の1/3の効力である(表7)。
【表7】
【0156】
ヒトPBMCでのSTAT3リン酸化
二代細胞をベースとする確認測定は、初代ヒト細胞における作用メカニズムとして、STAT3のリン酸化を評価する目的で開発された。健常ドナー由来の末梢血単核細胞(PBMC)は、低レベルのIL−23Rを名目上発現し、外因性IL−23で刺激された場合ほとんど応答しない、ナイーブで休眠状態のT細胞から主として成る。しかしながら、IL−2でのナイーブPBMCのポリクローナルな活性化は、その後IL−23Rの発現を増大させる、ナイーブT細胞の活性化および分化を引き起こす。これらの活性化した細胞はその後、STAT3のリン酸化を引き起こすSTAT経路を活性化する、外因性IL−23での刺激の影響を受けやすい。
【0157】
市販の抗体(AF1716、抗p19 pAbおよびmAb1510、抗p40 mAb、両者ともR&D Systemsから)を、IL−23誘導性STAT3リン酸化の阻害のための陽性対照として使用した。6種のアドネクチンの阻害活性を、多数のドナー由来の血液を使用する10の別実験において比較した(表8で要約する)。サブセットに関する例示的データを図4に示す。抗IL−23アドネクチンは、STAT3リン酸化において抗p19よりも著しく強力(>150倍)であったが、抗p40モノクローナル抗体より5倍弱い効力と同様であった。
【表8】
【0158】
マウス脾細胞によるIL−23誘導性サイトカイン産生
初代細胞を用いる初代細胞アッセイ(initial cellular assay)を設計し、抗IL−23アドネクチンの、マウスTh17細胞からのIL−23依存性サイトカイン分泌を阻害する能力を評価した。解析のためのマウスTh17細胞を分化させるために、CD4+T細胞を磁気ビーズで濃縮し、照射した脾細胞(irradiated splenocytes)と共培養し、TGF−βおよびIL−6ならびにIL−4およびIFN−γに対する中和抗体の存在下、抗CD3で活性化させた。6日間の培養後、極性化(polarized)Th17細胞を回収し、96ウェルプレートへ再播種し、100ng/ml ヒトIL−23および5ng/ml マウスIL−2で刺激した。IL−2の添加は、細胞生存を維持するために必要とされ、IL−23に応答して強力なサイトカイン産生を可能にするが、単独でIL−17AあるいはIL−22産生を強く誘導はしなかった。IL−2は低レベルのサイトカイン分泌を誘導するので、各サンプルセットには、IL−23の非存在下で産生されたサイトカインのベースラインレベルに関して調整するために、IL−2単独で刺激された細胞を含めた。IL−23依存性応答は、IL−2およびIL−23の組合せにより誘導されたサイトカインのレベルと、IL−2単独により誘導されたベースラインレベルの違いを計算することによって評価した。アドネクチンの用量範囲は、その抑制可能性を試験するために、IL−2およびIL−23でのTh17細胞の再刺激の間、追加された。ヒト抗p40抗体(R&D Systems MAB1510)の用量範囲は、IL−23阻害を評価するための陽性対照として、同時に実施された。各条件を96ウェルプレートにて三重測定で試験した。4日後、三重測定からの馴化培地をプールし、細胞残渣を除去し、ELISAによってIL−17A濃度およびIL−22濃度の両方を測定した。
【0159】
IL−2およびIL−23でのTh17細胞の刺激は、IL−2単独により誘導されたレベルと比較して、2から3倍のIL−17Aの増加、および、少なくとも5倍のIL−22の増強を誘導した。ATI000934、ATI001014、ATI001015、ATI001016、ATI001045および陽性対照の抗p40モノクローナル抗体は、IL−23依存性IL−17AおよびIL−22分泌において、用量依存的減少を調節した。IL−17AおよびIL−22分泌の両方の阻害に対するIC50値は、各アドネクチンならびに抗p40対照に対して計算され、これらのデータは表9に要約される。試験された全てのアドネクチンは、IL−23依存性IL−17A分泌の阻害に対して抗p40対照の2倍以内で強力であり、IL−23依存性IL−22産生の阻害に対して2から3倍以内で強力であった。
【表9】
【0160】
ヒトT細胞によるIL−23誘導性サイトカイン産生
PBMCは、健常ドナー由来のEDTA処理全血の密度勾配分離によって獲得した。T細胞は、ヒツジ赤血球(SRBC)と共にロゼット形成したPBMCのE+画分から調製した。このT細胞を96ウェル平底プレートへ1ウェルあたり100,000細胞で播種し、37℃で1時間、抗CD3(10μg/mlでのOKT)で覆い、PBSで洗浄した。抗CD28(1μg/mlでの9.3)およびIL−1β(10ng/ml)またはIL−1β+IL−23(1ng/ml)を含有するRPMI−FCS培地の混合物を調製した。このサイトカインの組合せは、ヒトT細胞のIL−17分泌T細胞への分化を促進するということが示されている。開始濃度(starting concentration)1μg/mlのATI001045をIL−1β+IL−23を含有する混合物へ添加した。IL−17はDUOSET登録商標 ELISA development kits(R&D Systems)を使用して、上清中に検出した。ATI001045は、バックグラウンドとしてIL−1β単独を使用して、2.0±1.6nMのEC50でIL−17産生を阻害した(n=4人の異なるドナー)。市販の抗p40抗体(MAB1510)が内部標準として使用され、2.2±1.4nMのEC50でIL−17産生を阻害した(n=3)。ドナー228からの例示的データを図6に示す。
【0161】
IL−12を超えるIL−23に対する抗IL−23アドネクチンの選択性
表2に示されるアドネクチンならびにATI001016を使用して、IL−23/IL−12に対する生化学的選択性を試験した。結合解析には、固定化抗His抗体での抗IL−23アドネクチンの回収、続いてアドネクチン上へのIL−23またはIL−12の流入が含まれた。IL−23に対するアドネクチンの選択性は、IL−23よりも100倍高い濃度のIL−12に対する結合シグナルを比較することによって評価した。表7に示す例示的データは、ATI001016は10nM ヒトIL−23に対して強力な結合(〜40RU)を示すが、一方で、1μM ヒトIL−12に対しては検出可能な結合は観察されなかったことを示す。
【0162】
NK−92細胞は、IFN−γを分泌することによりIL−2依存的にIL−12に応答することが知られているヒトナチュラルキラー細胞株である。細胞を典型的に洗浄してIL−2を除去し、次いで96ウェルプレートへ播種し、その後25pMの組換えヒトIL−12(または、アンタゴニストとプレインキュベートしたIL−12)で処理し、さらに20時間インキュベートした。清澄化された上清をELISAによりIFN−γに関して測定した。
【0163】
4点の5μMで開始する5倍希釈系列を、表2に示されるアドネクチンクローン各々について調製し、NK−92細胞への添加の前に、37℃で30分間25pM IL−12とインキュベートした。12点の5μMのATI001045およびATI001016で開始する5倍希釈系列を、NK−92細胞への添加の前に、37℃で30分間25pM IL−12とインキュベートした。表2に示されるクローンもATI001045あるいはATI001016も、これらの抗IL−23アドネクチンは、NK−92細胞表面上の受容体とのIL−12の相互作用を阻害しないことを示し、試験されたいずれの濃度でも、IFN−γ分泌を検出可能に阻害しなかった。それらは、陰性対照および100nM 抗p19ポリクローナル抗体と同等であるようである。陽性対照として、抗p40モノクローナル抗体(mAb1510)はIL−12誘導性IFN−γ分泌を0.07nMのIC50で阻害した(図8)。
【0164】
抗IL−23アドネクチンは薬力学的モデルにおいてIL−23誘導性IL−17を阻害(block)する
以下のスケジュールに従って、メスC57Bl/6マウスに組換えマウスIL−2およびヒトIL−23を腹腔内(IP)注射した。
【表10】
【0165】
全てのマウスを、IL−2およびIL−23の最終投与の7−8時間後、時間=30hで安楽死させた。血清を回収し、ELISAによりIL−17およびIL−23に関して測定した。
【0166】
ヒトIL−23はマウス受容体に結合し、IL−17およびIL−22などのサイトカインの産生を誘導する。IL−2およびヒトIL−23を腹腔内(IP)投与された動物由来の脾細胞は、エキソビボ(ex vivo)培養において抗マウスCD3eで刺激された時、IL−17を分泌する。IL−17の著しいレベルが表10に記載される処置計画に供された動物の血清中に検出でき、ここで、C57Bl/6マウスは、24時間のさらなる期間にわたるIL−2+IL−23の3回の二重注射(dual injection)の前に、IL−2で24時間、前処理した。おそらく、IL−2はポリクローナルに活性化し、インサイチュ(in situ)でTh集団を拡大させ、IL−23受容体の発現を上方制御する。これは、インビボ(in vivo)環境における、薬物作用のメカニズムおよび薬物濃度と効果の間の関係を調査することができる方法を提供する。このモデルは、抗p40モノクローナル抗体、mAb1510で確認された(データは示さず)。8つの別実験において、8つの別の実験でのIL−2+IL−23の初回投与(initial dose)の2時間前に、0.5、0.15、0.05および0.015mg/kgで、皮下(SC)投与した場合に、5種の抗IL−23アドネクチンを、マウスIL−17の産生を阻害する能力に関して試験した。ATI001045に対する例示的な用量反応データを図9aに示す(0.03mg/kgの計算上の平均ED50)。抑制範囲はアドネクチン全体で可変的であるが、試験された全ての抗IL−23アドネクチンは、血清中のヒトIL−23 マウスIL−17産生の用量依存的阻害を示した。
【0167】
IL−23誘導性皮膚表皮肥厚モデルにおける抗IL−23アドネクチンの活性
マウスの背中の皮膚または外耳介(external ear pinna)へのIL−23の皮内注射は、皮膚炎症および表皮の過形成(表皮肥厚)を誘発する(Zheng, Y., “Interleukin−22, a TH17 cytokine, mediates IL−23−induced dermal inflammation and acanthosis”, Nature, Vol. 445/8 (Feb. 2007))。これらの研究では、組換えヒトIL−23(rHuIL−23)がマウスの耳へ注射され、異常な皮膚IL−23暴露の下流の結果が調査された。
【0168】
6から8週齢のC57BL/6メスマウスに対し、5μgの二重鎖、組換え、ヒトIL−23を一日おきに12日目まで、右耳へ注射した。PBSを対照として反対側の耳に注射した。一試験では、ATI001045での処置は、最初のIL−23注射の約2時間前に開始し、1週間あたり3回、12日目まで継続した。ATI001045は、0.1、0.3、1、3mg/kgの用量で皮下(SC)投与された。第二の試験では、媒体またはATI000934−123 (1753E02)を、IL−23投与の1時間前に、1、3または10mg/kgで腹腔内(IP)投与し、その後1週間あたり3回、10日目まで投与した。抗HuIL−12/IL−23 p40抗体(R&D mAb1510)を10mg/kgで、陽性対照として0および4日目にIPで与えた。耳の厚さを(千分の1インチ単位で)、次の投与の前に、MITUTOYO登録商標(#2412F)ダイヤルキャリパーを使用して、一日おきに測定した。耳の厚さは、各動物に対し、IL−23注射された耳の測定結果から対照の耳の値を引くことによって計算した。試験の最後(ATI001045に対して14日目、およびATI000934に対して12日目)に、CO2ガスでの安楽死に続いて、耳を生え際で切除し、ホルマリン固定/パラフィン包埋組織をH&E染色したスライドで組織学的に観察した。
【0169】
全体としては、この試験において、1、3および10mg/kgの用量のATI000934は、同様のレベルのIL−23誘導性の耳肥厚の阻害を提供した(図10)。抗p40群を含む、全ての処置群における耳の厚さは、5日目から試験の最後の12日目にかけて、媒体よりも著しく薄かった(p<0.01 ANOVA/Dunnett’s)。12日目、最終的な血漿サンプルを最終投与48時間後に獲得し、ATI000934の血中濃度(circulating level)について解析し、それぞれ11、18、36μg/mlであると決定された。
【0170】
12日目の最終測定に続いて、1群あたり10動物から、所定の組織学的検査のために剖検で耳を回収した。ATI000934を投与された動物の大部分は、表皮肥厚および浸潤を有したが、組織学的重症度スコアは、媒体処置動物で観察されたものからは減少した。明らかな用量反応は見られなかった。抗p40を投与された全ての動物もまた、表皮肥厚および浸潤を有したが、組織学的重症度スコアは、媒体処置動物で観察されたものからは減少した。
【0171】
ATI001045(1mg/kgおよび3mg/kg)は、5日目から14日目にかけて、媒体(PBS)処置動物と比較して、用量依存的に耳の厚さを減少させた(p<0.01 対媒体 ANOVA/Dunnett’s、図10)。対照的に、0.1mg/kgの用量レベルでは、どの試験日でも、媒体処置と統計的に違いはなかった(p>0.05)。0.3mg/kgでの処置は、5日目、7日目、9日目において媒体よりも統計的に弱い、中間的な減少(intermediate reduction)を提供した。最終投与48時間後に回収された血漿サンプルを、ATI001045の血中濃度(circulating levels)について評価し、0.1、0.3、1、3mg/kgの用量に対し、それぞれ0.698、2.72、8、22.5μg/mlであると決定された。組織学的解析から、ATI001045の投与が、IL−23誘導性細胞性浸潤および耳の厚さのスコアに相関する表皮肥厚の用量依存的減少をもたらした、ということが明らかとなった。
【実施例5】
【0172】
本明細書で使用される材料および方法
ハイスループットタンパク質生成(HTPP)
選択されたバインダーをpET9dベクターにクローニングし、大腸菌BL21 DE3 plysS株へ形質転換し、50μg/mLカナマイシンを含有する5mlのLB培地に24ウェルフォーマットで播種し、37℃で一晩増殖させた。一晩培養物から200μlを吸引することおよび適当なウェルに分配することにより、誘導性発現のための新鮮な5mLのLB培地(50μg/ml カナマイシン)培養物を調製した。培養物を37℃でA600 0.6〜0.9まで増殖させた。1mM イソプロピル−β−D−1−チオガラクトシド(IPTG)を用いて誘導した後、培養物を、30℃で6時間発現させ、4℃で2750gでの10分間の遠心分離によって回収した。
【0173】
細胞ペレット(24ウェルフォーマット中)を、450μlの溶解バッファー(50mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、1mM PMSF、10mM CHAPS、40mM イミダゾール、1mg/ml リゾチーム、30μg/ml DNAse、2μg/ml アプロトニン(aprotonin)、pH8.0)に再懸濁し、室温で1〜3時間振盪することによって溶解した。溶解物を清澄化し、96ウェル、1.2mlキャッチプレートが取り付けられた96ウェルワットマンGF/Dユニフィルタ登録商標に移すことによって96ウェルフォーマットに再び入れ、陽圧によって濾過した。清澄化した溶解物を、平衡バッファー(50mM NaH2PO4、0.5M NaCl、40mM イミダゾール、pH8.0)で平衡化された96ウェルのNi−キレートプレートへ移し、5分間インキュベートした。非結合物質を陽圧により除去した。樹脂を、洗浄バッファー#1(50mM NaH2PO4、0.5M NaCl、5mM CHAPS、40mM イミダゾール、pH8.0)を用いて、2x 0.3ml/ウェルで洗浄し、各洗浄液を陽圧により除去した。溶出に先立って、各ウェルを50μl 溶出バッファー(PBS + 20mM EDTA)で洗浄し、5分間インキュベートし、洗浄液を陽圧により廃棄した。各ウェルにさらなる100μlの溶出バッファーを注ぐことによってタンパク質を溶出した。室温で30分インキュベートした後、プレート(単数または複数)を、200gで5分間遠心分離にかけ、溶出されたタンパク質を、溶出に先立って溶出キャッチプレートの底に加えた5μlの0.5M MgCl2を含有する96ウェルキャッチプレート中に回収する。溶出したタンパク質を、SGEをタンパク質標準として用いる総タンパク質アッセイ(BCA)を使用して定量した。
【0174】
不溶性のフィブロネクチンをベースとする足場タンパク質バインダーの中規模発現および精製
発現のために、HIS6タグが続く、選択されたクローン(単数または複数)を、pET9dベクターにクローニングし、大腸菌BL21 DE3 plysS株中で発現させた。20mlの種菌培養物(単一プレーティングされたコロニーから作製した)を使用して、50μg/ml カナマイシンおよび34μg/ml クロラムフェニコールを含有する1リットルのLB培地またはTB−オーバーナイト発現培地(自己誘導)に播種した。LB培地での培養物は、37℃でA600 0.6〜1.0までインキュベートし、その時点でイソプロピル−β−D−1−チオガラクトシド(IPTG)で誘導し、30℃で4時間増殖させた。TB−オーバーナイト発現培地で増殖させた培養物は、37℃で5時間インキュベートし、その時点で温度を19時間、18℃へ下げた。培養物を4℃で≧10,000gでの30分間の遠心分離によって回収した。細胞ペレットを−80℃で凍結させた。細胞ペレットを、氷上でULTRA−TURRAX登録商標ホモジナイザー(IKA works)を使用して、25mlの溶解バッファー(20mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、pH7.4)中に再懸濁した。細胞溶解は、モデルM−110S MICROFLUIDIZER登録商標(Microfluidics)を使用して高圧ホモジナイゼーション(≧18,000 psi)によって達成した。不溶性画分を、4℃で≧23,300gでの30分間の遠心分離によって分離した。溶解物の遠心分離から回収された不溶性ペレットを、20mM リン酸ナトリウム/500mM NaCl、pH7.4で洗浄した。ペレットを、超音波処理を用いて20mM リン酸ナトリウム/500mM NaCl pH7.4中、6.0M 塩酸グアニジンに再可溶化し、続いて37度で1〜2時間インキュベートした。再可溶化されたペレットを0.45μmで濾過し、20mM リン酸ナトリウム/500mM NaCl/6.0M グアニジン pH7.4バッファーで平衡化したHISTRAP登録商標カラムにロードした。ローディング後、カラムを同一のバッファーを用いて、さらなる25カラム容量(CV)に対して洗浄した。結合したタンパク質を、20mM リン酸ナトリウム/500mM NaCl/6.0M Guan−HCl、pH7.4中、50mM イミダゾールを用いて溶出した。精製されたタンパク質は、50mM 酢酸ナトリウム/150mM NaCl pH4.5またはPBS、pH7.2に対する透析によって、リフォールディングされた。
【0175】
可溶性のフィブロネクチンをベースとする足場タンパク質バインダーの中規模発現および精製
不溶性バインダーの精製の代替として、可溶性バインダーの精製を使用してもよい。発現のために、HIS6タグが続く、選択されたクローン(単数または複数)を、pET9dベクターにクローニングし、大腸菌BL21 DE3 plysS株中で発現させた。20mlの種菌培養物(単一プレーティングされたコロニーから作製した)を使用して、50μg/ml カナマイシンおよび34μg/ml クロラムフェニコールを含有する1リットルのLB培地またはTB−オーバーナイト発現培地(自己誘導)に播種した。LB培地での培養物は、37℃でA600 0.6〜1.0までインキュベートし、その時点でイソプロピル−β−D−1−チオガラクトシド(IPTG)で誘導し、30℃で4時間増殖させた。TB−オーバーナイト発現培地で増殖させた培養物は、37℃で5時間インキュベートし、その時点で温度を19時間、18℃へ下げた。培養物を4℃で≧10,000gでの30分間の遠心分離によって回収した。細胞ペレットを−80℃で凍結させた。細胞ペレットを、氷上でULTRA−TURRAX登録商標ホモジナイザー(IKA works)を使用して、25mlの溶解バッファー(20mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、pH7.4)中に再懸濁する。細胞溶解は、モデルM−110S MICROFLUIDIZER登録商標(Microfluidics)を使用して高圧ホモジナイゼーション(≧18,000 psi)によって達成される。可溶性画分を、4℃で≧23,300gで30分間の遠心分離によって分離する。上清を0.45μmフィルターへ通して清澄化する。清澄化された溶解物を、20mM リン酸ナトリウム/500mM NaCl pH7.4で事前に平衡化されたHISTRAP登録商標カラム(GE)にロードする。次いでカラムを、25カラム容量の同一のバッファーで洗浄し、続いて20カラム容量の20mM リン酸ナトリウム/500mM NaCl/25mM イミダゾール、pH7.4、その後35カラム容量の20mM リン酸ナトリウム/500mM NaCl/40mM イミダゾール、pH7.4を用いて洗浄する。タンパク質を、15カラム容量の20mM リン酸ナトリウム/500mM NaCl/500mM イミダゾール、pH7.4で溶出し、A280での吸光度に基づいて画分をプールし、1x PBS、50mM トリス、150mM NaCl、pH8.5または50mM NaOAc;150mM NaCl;pH4.5に対して透析する。いかなる沈殿物も0.22μmでの濾過によって除去される。
【0176】
フィブロネクチンをベースとする足場タンパク質(アドネクチン)は、種々の大きさおよび種類のPEGを用いてPEG化させることができる。PEG化を可能にするために、10FN3タンパク質のC末端で見られる天然に存在する残基EIDKPSQが、アミノ酸の、典型的にはセリンのシステインへの、単一点突然変異によって、修飾される。単一のシステイン残基におけるタンパク質のPEG化は、種々のマレイミド誘導体化されたPEGの形をコンジュゲートすること、PEG試薬をタンパク質溶液と組み合わせること、およびインキュベートすることによって達成される。代替方法は、EIDKPSQテールをGSGCリンカーで置換し、同様にシステイン残基をPEG化のために使用することである。操作されたシステイン残基を含有するアドネクチンを、マイケル付加反応を介して、システイン上のチオール基とPEG試薬のマレイミド官能基の間でPEGとコンジュゲートした。簡単に説明すると、40kDaのPEGを、弱酸性から中性条件下で、タンパク質溶液へ、過剰のモル濃度で添加する。反応は、2時間から一晩、室温で続行することができる。次いで反応をイオン交換カラムへ供し、PEG化アドネクチンを未反応のPEG−マレイミドおよび非PEG化アドネクチンから分離する。SE/HPLC方法が使用されてもよい。精製されたPEG化アドネクチンは典型的にSDS−PAGEおよびサイズ排除クロマトグラフィーによって解析される。
【実施例6】
【0177】
候補血清アルブミン結合アドネクチン(SABA)のスクリーニングおよび選択
PROfusionとして知られる選択技術(例えば、Roberts ら, Proc. Natl. Acad. Sci. USA, 94(23):12297−12302 (1997)および WO 2008/066752参照)が、10Fn3のBC、DEおよびFGループに設計された可変領域を有するDNAライブラリーへ適用された。1013分子を越えるランダムライブラリーがこの設計から作り出され、所望の結合特性を備える候補血清アルブミン結合アドネクチン(SABA)を単離するために、HSAのビオチン化形態に対して、選択圧が適用された。
【0178】
ハイスループットタンパク質生成(HTTP)工程
様々なHSA結合アドネクチンがハイスループットタンパク質生成(HTPP)を使用して精製された。選択されたバインダーを、HIS6タグを含有するpET9dベクターにクローニングし、大腸菌BL21(DE3)pLysS株へ形質転換した。形質転換された細胞を、50μg/mLカナマイシンを含有する5mlのLB培地に24ウェルフォーマットで播種し、37℃で一晩増殖させた。一晩培養物から200μlを吸引することおよび適当なウェルに分配することによって、誘導性発現のために、新鮮な5mlのLB培地(50μg/ml カナマイシン)培養物を調製した。培養物は37℃でA600 0.6〜0.9まで増殖させた。1mM イソプロピル−β−D−1−チオガラクトシド(IPTG)を用いて誘導した後、培養物を、30℃でさらに4時間発現させ、4℃で3220x gでの10分間の遠心分離によって回収した。細胞ペレットを−80度で凍結した。
【0179】
細胞ペレット(24ウェルフォーマット中)を、450μlの溶解バッファー(50mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、1mM PMSF、10mM CHAPS、40mM イミダゾール、1mg/ml リゾチーム、30μg/ml DNAse、2μg/ml アプロトニン(aprotonin)、pH8.0)に再懸濁し、室温で1時間振盪することによって溶解した。溶解物を清澄化し、96ウェル、650μlキャッチプレートが取り付けられた96ウェルワットマンGF/Dユニフィルタ登録商標へ移すことによって96ウェルフォーマットに再び入れ、200x gで5分間遠心分離した。清澄化した溶解物を、平衡バッファー(50mM NaH2PO4、0.5M NaCl、10mM CHAPS、40mM イミダゾール、pH8.0)で平衡化された96ウェルNi−キレートプレートに移し、5分間インキュベートした。非結合物質を除去した。樹脂を、洗浄バッファー#1(50mM NaH2PO4、0.5M NaCl、5mM CHAPS、40mM イミダゾール、pH8.0)を用いて、2x 0.3ml/ウェルで洗浄した。次いで、樹脂をPBSを用いて3x 0.3ml/ウェルで洗浄した。溶出に先立って、各ウェルを50μl 溶出バッファー(PBS + 20mM EDTA)で洗浄し、5分間インキュベートし、洗浄液を真空により廃棄した。各ウェルにさらなる100μlの溶出バッファーを注ぐことによってタンパク質を溶出した。室温で30分インキュベートした後、プレート(単数または複数)を、200x gで5分間遠心分離にかけ、溶出されたタンパク質を、溶出キャッチプレートの底に加えた5μlの0.5M MgCl2を含有する96ウェルキャッチプレート中に回収した。溶出されたタンパク質を、SGE(対照アドネクチン)をタンパク質標準として用いるBCAタンパク質アッセイを使用して定量した。SGEアドネクチンは、インテグリン結合ドメイン(78〜80位のアミノ酸RGD)がSGEで置換された、野生型10Fn3ドメイン(配列番号1)である。
【0180】
HSA、RhSAおよびMuSA直接結合ELISA
HSAへの直接的なバインダーをアッセイするために、MaxiSorpプレート(Nunc International, Rochester, NY)を4℃で一晩、PBS中10μg/mL HSA(Sigma, St. Louis, MO)で覆い、続いてカゼインブロックバッファー(Thermo Scientific, Rockford, IL)中で1〜3時間、室温でブロッキングを行った。シングルポイントスクリーニングアッセイのため、精製されたHTPPアドネクチンをカゼインブロックバッファー中に1:20で希釈し、室温で1時間、各ウェル内でHSAに結合させた。用量反応アッセイのため、0.1nMから1μMの範囲の濃度が使用された。非結合アドネクチンを除去するためのPBSTでの洗浄の後、カゼインブロックバッファー中に1:2500で希釈された抗His mAb−HRPコンジュゲート(R&D Systems, MN)を、室温で1時間、結合されたHisタグ化アドネクチンへ添加した。過剰のコンジュゲートはPBSTを用いる洗浄によって除去され、製造業者の使用説明書に従い、TMB検出試薬(BD Biosciences)を使用して、結合アドネクチンを検出した。
【0181】
候補血清アルブミン結合アドネクチン(SABA)の同定
HSA/RhSA/MuSA結合基準および生物物理学的基準に関するスクリーニングの結果を受けて、4つの特有の血清アルブミン結合アドネクチン(SABA)を同定し、それらの半減期をマウスで評価させるために選択した。インビトロ(in vitro)およびインビボ(in vivo)での特性評価を行うために、4つのSABAについて中規模生成が行われた。表3は、SABA1〜26と表される、PROfusionから同定された26の特有のSABAコア配列の配列を提供する。SABA4は中規模生成に先立って固定の足場変異を有した。SABA4の足場完全型はSABA5である。SABA4およびSABA5はBC、DEおよびFGループにおいて同一配列を有する。
【実施例7】
【0182】
候補SABAの生成および製剤処方(Formulation)
SABAの中規模タンパク質生成
His6タグが続く選択されたSABAを、pET9dベクターにクローニングし、大腸菌BL21(DE3)pLysS株中で発現させた (SABA1.1、SABA2.1、SABA3.1およびSABA5.1と表される、各Hisタグ化SABA配列に関する表3を参照)。20mlの種菌培養物(単一プレーティングされたコロニーから作製した)を使用して、50μg/ml カナマイシンを含有する1リットルのLB培地に播種した。培養物を37℃でA600 0.6〜1.0まで増殖させた。1mM イソプロピル−β−D−1−チオガラクトシド(IPTG)を用いて誘導した後、培養物を、30℃でさらに4時間増殖させ、4℃で≧10,000x gでの30分間の遠心分離によって回収した。細胞ペレットを−80℃で凍結させた。細胞ペレットを、氷上でULTRA−TURRAX登録商標ホモジナイザー(IKA works)を使用して、25mlの溶解バッファー(20mM NaH2PO4、0.5M NaCl、1xコンプリートプロテアーゼインヒビターカクテル−EDTAフリー(Roche)、pH7.4)中に再懸濁した。細胞溶解は、モデルM−110S MICROFLUIDIZER登録商標(Microfluidics)を使用して高圧ホモジナイゼーション(≧18,000 psi)によって達成した。可溶性画分を、4℃で23,300x gでの30分間の遠心分離によって分離した。上清を0.45μmフィルターへ通して清澄化した。清澄化された溶解物を、20mM NaH2PO4、0.5M NaCl、pH7.4で事前に平衡化されたHISTRAP登録商標カラム(GE)にロードした。次いでカラムを、25カラム容量の20mM NaH2PO4、0.5M NaCl、pH7.4で、続いて、20カラム容量の20mM NaH2PO4、0.5M NaCl、25mM イミダゾール pH7.4、次いで、35カラム容量の20mM NaH2PO4、0.5M NaCl、40mM イミダゾール pH7.4を用いて洗浄した。タンパク質を、15カラム容量の20mM NaH2PO4、0.5M NaCl、500mM イミダゾール pH7.4で溶出し、A280での吸光度に基づいて画分をプールし、1x PBS、50mM トリス、150mM NaCl pH8.5または50mM NaOAc;150mM NaCl;pH4.5に対して透析した。いかなる沈殿物も0.22μmでの濾過によって除去された。
【0183】
中規模発現および精製は、可溶型で発現され、細菌性サイトゾルの可溶性画分から精製された、高純度かつ活性なアドネクチンを生じた。100mM NaPO4、100mM NaSO4、150mM NaCl、pH6.8(GE Healthcare)の移動相でのSUPERDEX登録商標 200またはSUPERDEX登録商標 75 10/30GLでのSEC解析は、大部分、単量体アドネクチンを示した。
【0184】
SABA1.2の製剤処方(Formulation)
一つの具体的なSABA、SABA1.2(配列番号180)、が、初期製剤処方(preliminary formulation)スクリーニングのために選択された。SABA1.2は、10Fn3の「コア1」配列上に(ED)5伸長を含む。SABA1.2に対して、pH6.0で10mM コハク酸、8% ソルビトール、5% グリセリン、かつ製品濃度5mg/mLでの、安定な製剤処方が同定された。この製剤処方において、1.25mg/mLのタンパク質濃度を使用して、示差走査熱量測定(DSC)により決定されたタンパク質の融点は75℃であった。この製剤処方は、4℃および25℃で、1.2%の初期凝集濃度(initial aggregate level)を有し、十分な物理的安定性および化学的安定性をもたらした。一ヶ月の静置の後、凝集のレベルは非常に低かった(4℃で1.6%かつ25℃で3.8%)。該タンパク質はまた、この製剤処方において、−80℃および−20℃から外気温へ推移させる5回の凍結融解後も安定であった。加えて、このSABA1.2製剤処方は、4℃および外気温で、沈殿もしくは凝集の増加も無く、少なくとも20mg/mLのタンパク質濃度で可溶性であった。
【実施例8】
【0185】
候補SABAの生物物理学的特性評価
サイズ排除クロマトグラフィー
標準的なサイズ排除クロマトグラフィー(SEC)を中規模工程によって生じる候補SABAに対して行った。中規模物質のSECは、A214nmならびにA280nmでのUV検出器付き、および蛍光検出器(励起=280nm、蛍光=350nm)付きのAgilent 1100もしくは1200 HPLCシステム上で、SUPERDEX登録商標 200 10/30またはSUPERDEX登録商標 75 10/30カラム(GE Healthcare)を使用して行った。SECカラムの適した流量で、100mM 硫酸ナトリウム、100mM リン酸ナトリウム、150mM 塩化ナトリウム、pH6.8のバッファーが採用された。ゲル濾過標準(Bio−Rad Laboratories, Hercules, CA)が分子量校正のために使用された。
【0186】
中規模精製SABAでのSECの結果は、大部分、単量体アドネクチンを、および、示されるように、球状ゲル濾過標準(BioRad)に対しておよそ10kDaの範囲での溶出を示した。
【0187】
耐熱性
中規模SABAの示差走査熱量測定(DSC)解析を行って、それらのそれぞれのTmを決定した。1mg/mlの溶液を、3atm圧力下、1分あたり1℃の速度で、5℃から95℃へ温度を上昇させる(ramping)ことにより、N−DSC II熱量計(Calorimetry Sciences Corp)でスキャンした。データを、Orgin Software(OrginLab Corp)を使用する適合度(best fit)を用いて、適切なバッファーの対照測定(control run)に対して解析した。SEC解析およびDSC解析の結果を表11に要約する。
【表11】
【実施例9】
【0188】
血清アルブミンに結合する候補SABA1の特性評価
HTPPおよび/または中規模物質から精製された選択されたSABAクローンの動態(kinetics)を、Biasensor CM5チップの表面上にそれぞれの血清アルブミン(HSA/RhSA/MuSA)を捕捉すること、および参照フローセルおよび捕捉されたアルブミンの両方にわたってSABAの濃度系列を流すことにより決定した。加えて、アルブミンとの結合を、pH5.5からpH7.4にわたる様々なpH条件下で実行した。HSA結合アドネクチン、SABA2.1、SABA3.1、SABA4.1(SABA5.1)&SABA1.1は、RhSAへ公差反応したが、MuSAへは公差反応しなかった。SABA2およびSABA4結合はpH感受性である一方、クローンSABA3はpH6.0に至るまでHSAとのpH耐性結合を明示した。SABA1.1はpH5.5に至るまでpH耐性および親和性/動態(kinetics)に関する生化学的基準に適合する。
【0189】
ドメインマッピングはビアコアにより決定した。HTPPおよび/または中規模物質から精製された選択されたSABAクローンは、Biasensor CM5チップの表面上にHSA、またはHSA−ドメインI&IIあるいはHSA−ドメインIIIのみから成る構築物を捕捉すること、および参照フローセルおよび捕捉されたアルブミンの両方にわたってSABAの濃度系列を流すことにより決定した。クローンSABA2&SABA1はHSAおよびHSA−ドメインI−II構築物と結合し、HSA−ドメインIII構築物とは結合しなかった。クローンSABA3&SABA4はHSAと結合したが、HSA−ドメインI−II構築物またはHSA−ドメインIII構築物のいずれとも結合しなかった。結果を表12に要約する。
【表12】
【実施例10】
【0190】
候補SABAのインビボ(in vivo)t1/2の調査
HSA結合アドネクチンはMuSAと交差反応しないため、マウスでのHSA結合アドネクチンの評価を可能にするために、マウスにおけるHSAの半減期を決定した。HSAを、20mg/kg用量(図11A)および50mg/kg用量(図11B)で、約6週齢のNcrヌードメスマウスの尾静脈へ注射し、注射後周期的に採取した血液サンプル中のHSA濃度をELISAにより決定した。20mg/kgおよび50mg/kgでマウスに注射されたHSAのt1/2は、それぞれ、〜24時間および〜20時間であると決定された。
【0191】
マウスでのSABA1〜4の半減期決定
1リットルのHSA結合クローンSABA1.1、SABA2.1、SABA3.1およびSABA4.1の大腸菌増殖液を調製し、精製し、エンドトキシンを除去した。各SABA変種をマウスの尾静脈へ注射し、注射後周期的に採取した血液サンプル中の濃度をELISAにより決定した。
【0192】
各SABAの薬物動態プロファイルを、約6週齢のNcrヌードメスマウスで、HSAの存在下または非存在下において比較した。結合クローンはHSAおよびRhSAに対して選択性であり、マウス血清アルブミンとは結合しなかったため、HSAを同時注射されたマウスは、HSAを各SABAと予め混合させた(3〜4モル濃度過剰のHSA)。マウス血漿中のSABA1.1の半減期は0.56時間であるのに対し、HSAと同時注射されたSABA1.1の半減期は5.6時間、半減期において〜10倍増加していた(図12A)。マウス血漿中のSABA2.1の半減期0.24時間であるのに対し、HSAと同時注射されたSABA2.1の半減期は2.8時間、半減期において〜12倍増加していた(図12B)。マウス血漿中のSABA3.1の半減期は0.28時間であるのに対し、HSAと同時注射されたSABA3.1の半減期は0.53時間、半減期において〜2倍増加していた(図12C)。マウス血漿中のSABA4.1の半減期は0.66時間であるのに対し、HSAと同時注射されたSABA4.1の半減期は4.6時間、半減期において〜7倍増加していた(図12D)。本実施例の概要を図13Aに示す。
【0193】
カニクイザルでのSABA1.1およびSABA5.1の半減期決定
SABA1.1およびSABA5.1の三週間単回投与概念実証試験(proof of concept study)をカニクイザルで実施し、2匹のカニクイザルにおける1mg毎kg(mpk)用量、IVでの薬物動態を評価した。薬物動態は、血漿サンプル中のアドネクチンを検出するために開発された定量的ELISAをベースとするアッセイを使用して評価した。SABA1.1は96〜137時間の範囲の半減期を有する。SABA5.1は約12時間の半減期を有し、唯一120時間までELISAで測定可能であった。図14AおよびBは、これらのクローンに対するデータを要約し、カニクイザルからのデータを比較するものである。
【実施例11】
【0194】
血清アルブミンと結合するSABA1の特性評価
SABA1.1およびSABA1.2はHSAおよびRhSAと結合する
SABA1.2、(ED)5伸長(配列番号190)を含む「コア1」10Fn3、は、ヒト血清アルブミン(HSA)と、中性pHかつ25℃で、8.21E+03 M−1s−1の平均結合速度定数(ka)、かつ4.43E−04 s−1の平均解離速度定数(kd)、55.3 nMの計算平均Kdで結合した(表13)。アカゲザル血清アルブミン(RhSA)に対しては、測定された平均結合速度定数は6.6E+03 M−1s−1であり、解離速度定数は3.78E−03 s−1であり、580 nMの計算平均Kdを提供した。1μMに至るまで、SABA1.2とマウスまたはラット血清アルブミンの間に測定可能な相互作用は観察されなかった(表13および図15)。37℃で、kaおよびkdは2から5倍増加し、HSAに対する親和性において〜2倍の増加およびRhSAに対して1/2の親和性をもたらした(表13)。
【表13】
【0195】
さらに、熱量滴定を行い、SABA1およびHSAの間の化学量論組成(stoichiometry)を決定した。この試験では、SABA1.1、His6伸長(配列番号189)を含む「コア1」10Fn3、が使用された。HSA(115μMのタンパク質溶液の注射あたり10μl)を、8.1μMの濃度でSABA1.1を含有する熱量測定セル(calorimetric cell)へ注入した。実験はPBSバッファーpH7.4中、37℃にて行った。図16はSABA1.1がHSAと1:1の化学量論組成で結合することを示す。
【0196】
SABA1.2は低pHでHSAと強く結合する
アルブミンの長い半減期(例えば、HSAのt1/2は19日である)は、主に、それらがエンドソームの内側に存在する低pH条件下で、新生児型Fc受容体、FcRn、に結合することにより、エンドサイトーシス経路からリサイクルされる、という事実に起因している。表14に示されるように、エンドソームpH5.5でHSAと強く結合したSABA1.2は、SABA1のt1/2を示唆し、一度HSAに結合すると、FcRnリサイクル機構の利益を享受する。
【表14】
【0197】
SABA1.2はHSAのドメインIおよびIIと結合するが、ドメインIIIとは結合しない
アルブミン上のSABA1.2結合部位は、組換えHSA断片を使用して、N末端ドメインIまたはIIへマッピングされ、ドメインIIIとの検出可能な結合は無かった(図17)。ドメインIIIは、最初にFcRnと相互作用するHSAのドメインであるため、SABA1.2は、FcRnへのHSA結合と競合する可能性が低く、半減期を増強するためのリサイクル機構を十分に活用する可能性をかさねて増大させる。
【実施例12】
【0198】
SABA1.2のインビボ(in vivo)薬理
SABA1.2の四週間単回投与前毒性試験(pre−toxicology study)をカニクイザルで実施し、二つの異なる用量レベルで薬物動態および免疫原性を評価した。薬物動態および免疫原性はまた、静脈投与群および皮下投与群の両者を含む、三週間単回投与前毒性試験においても評価した。さらに、SABA1.2の薬物動態を、血漿サンプル中のSABA1.2を検出するために開発された定量的ELISAをベースとするアッセイを使用して、カニクイザルで、二つの別個の単回投与前毒性試験においても評価した。
【0199】
SABA1.2を1mpkおよび10mpk、IVでサルへ投与した。図18および下記に記載されるパラメータに示されるように、CmxおよびAUCは、用量とともにほぼ直線的に増加した。WINNONLIN登録商標ソフトウェアを使用して、ノンコンパートメント解析を行い、薬物動態パラメータを評価した。10mpkでのSABA1.2に対するクリアランス(CL)は、0.15ml/時/kg、ベータ相半減期(t1/2)は143時間、分布容積(Vz)は30mL/kg、そして、総薬物暴露量(total drug exposure)(AUCall)は5,609,457時*nmol/Lであった(表15)。1mpkでのSABA1.2に対するクリアランス(CL)は、0.4ml/時/kg、半減期(t1/2)は124時間、分布容積(Vz)は72mL/kg、そして、総薬物暴露量(AUCall)は214,636時*nmol/Lであった(表15)。
【0200】
SABA1.2のSCまたはIV投与の後、ベータ相薬物動態プロファイルは類似していた(図19)。WINNONLIN登録商標ソフトウェアを使用して、ノンコンパートメント解析を行い、薬物動態パラメータを評価した。1mpk IVでのSABA1.2に対するクリアランス(CL)は、0.22ml/時/kg、ベータ相半減期(t1/2)は125時間、分布容積(Vz)は40mL/kg、そして、総薬物暴露量(AUCall)は357,993時*nmol/Lであった(表15)。1mpk SCでのSABA1.2に対するクリアランス(CL)は、0.32ml/時/kg、ベータ相半減期(t1/2)は134時間、分布容積(Vz)は62mL/kg、そして、総薬物暴露量(AUCall)は251,339時*nmol/Lであった(表15)。IVに比べてSCの相対的生物学的利用能(F)は0.7であった。
【表15】
【図1−1】
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図1−6】
【特許請求の範囲】
【請求項1】
フィブロネクチンIII型第10(10Fn3)ドメインを含むポリペプチドであって、ここに該10Fn3ドメインは、ヒト10Fn3ドメインの対応するループの配列と比較してアミノ酸配列が変化したループBC、DEおよびFGから選択される少なくとも1つのループを有し、該ポリペプチドは500nM未満のKdでIL−23のp19サブユニットに結合する、ポリペプチド。
【請求項2】
フィブロネクチンIII型第10(10Fn3)ドメインを含むポリペプチドであって、ここに該10Fn3ドメインは、ヒト10Fn3ドメインの対応するループの配列と比較してアミノ酸配列が変化したループBC、DEおよびFGから選択される少なくとも1つのループを有し、該ポリペプチドはIL−23のp19サブユニットの構造的エピトープに結合する、ポリペプチド。
【請求項3】
BCループが配列番号2〜6から選択される、請求項1または2に記載のポリペプチド。
【請求項4】
DEループが配列番号7〜48から選択される、請求項1または2に記載のポリペプチド。
【請求項5】
FGループが配列番号49〜59から選択される、請求項1または2に記載のポリペプチド。
【請求項6】
BC、DEまたはFGループのアミノ酸配列が、配列番号2〜59のいずれか一つと少なくとも80%同一である、請求項1または2に記載のポリペプチド。
【請求項7】
ポリペプチドのアミノ酸配列が、配列番号60〜100のいずれか一つと少なくとも90%同一である、請求項1または2に記載のポリペプチド。
【請求項8】
ポリペプチドのアミノ酸配列が、配列番号60〜100のアミノ酸3〜96と少なくとも90%同一である、請求項1または2に記載のポリペプチド。
【請求項9】
BCループの配列モチーフが、X1がメチオニンまたはロイシンのいずれかであり、X2がイソロイシンまたはバリンのいずれかである、配列番号257で示されるGHYPX1HX2である、請求項1または2に記載のポリペプチド。
【請求項10】
FGループの配列モチーフが、X3が任意のアミノ酸であり得る、配列番号258で示されるYYX3X3X3X3YX3X3Iである、請求項1または2に記載のポリペプチド。
【請求項11】
ポリエチレングリコール、シアル酸、Fc、Fc断片、トランスフェリン、血清アルブミン、血清アルブミン結合タンパク質および血清免疫グロブリン結合タンパク質からなる群から選択される一以上の薬物動態(PK)部分をさらに含む、請求項1または2に記載のポリペプチド。
【請求項12】
PK部分がポリエチレングリコールである、請求項11に記載のポリペプチド。
【請求項13】
システインリンカーをさらに含む、請求項1または2に記載のポリペプチド。
【請求項14】
システインリンカーが、配列番号101に示されるアミノ酸GSGCおよび配列番号102に示されるアミノ酸EIDKPCQからなる群から選択される、請求項13に記載のポリペプチド。
【請求項15】
組成物が本質的にエンドトキシンを含まない、請求項1〜14のいずれか一項に記載のポリペプチドを含む医薬上許容される組成物。
【請求項16】
内因性IL−23のTh17細胞との反応を干渉するために効果的な量で、IL−23と請求項1〜14のいずれか一項に記載のポリペプチドを接触させることを含む、Th17細胞の病原性を調節する方法。
【請求項1】
フィブロネクチンIII型第10(10Fn3)ドメインを含むポリペプチドであって、ここに該10Fn3ドメインは、ヒト10Fn3ドメインの対応するループの配列と比較してアミノ酸配列が変化したループBC、DEおよびFGから選択される少なくとも1つのループを有し、該ポリペプチドは500nM未満のKdでIL−23のp19サブユニットに結合する、ポリペプチド。
【請求項2】
フィブロネクチンIII型第10(10Fn3)ドメインを含むポリペプチドであって、ここに該10Fn3ドメインは、ヒト10Fn3ドメインの対応するループの配列と比較してアミノ酸配列が変化したループBC、DEおよびFGから選択される少なくとも1つのループを有し、該ポリペプチドはIL−23のp19サブユニットの構造的エピトープに結合する、ポリペプチド。
【請求項3】
BCループが配列番号2〜6から選択される、請求項1または2に記載のポリペプチド。
【請求項4】
DEループが配列番号7〜48から選択される、請求項1または2に記載のポリペプチド。
【請求項5】
FGループが配列番号49〜59から選択される、請求項1または2に記載のポリペプチド。
【請求項6】
BC、DEまたはFGループのアミノ酸配列が、配列番号2〜59のいずれか一つと少なくとも80%同一である、請求項1または2に記載のポリペプチド。
【請求項7】
ポリペプチドのアミノ酸配列が、配列番号60〜100のいずれか一つと少なくとも90%同一である、請求項1または2に記載のポリペプチド。
【請求項8】
ポリペプチドのアミノ酸配列が、配列番号60〜100のアミノ酸3〜96と少なくとも90%同一である、請求項1または2に記載のポリペプチド。
【請求項9】
BCループの配列モチーフが、X1がメチオニンまたはロイシンのいずれかであり、X2がイソロイシンまたはバリンのいずれかである、配列番号257で示されるGHYPX1HX2である、請求項1または2に記載のポリペプチド。
【請求項10】
FGループの配列モチーフが、X3が任意のアミノ酸であり得る、配列番号258で示されるYYX3X3X3X3YX3X3Iである、請求項1または2に記載のポリペプチド。
【請求項11】
ポリエチレングリコール、シアル酸、Fc、Fc断片、トランスフェリン、血清アルブミン、血清アルブミン結合タンパク質および血清免疫グロブリン結合タンパク質からなる群から選択される一以上の薬物動態(PK)部分をさらに含む、請求項1または2に記載のポリペプチド。
【請求項12】
PK部分がポリエチレングリコールである、請求項11に記載のポリペプチド。
【請求項13】
システインリンカーをさらに含む、請求項1または2に記載のポリペプチド。
【請求項14】
システインリンカーが、配列番号101に示されるアミノ酸GSGCおよび配列番号102に示されるアミノ酸EIDKPCQからなる群から選択される、請求項13に記載のポリペプチド。
【請求項15】
組成物が本質的にエンドトキシンを含まない、請求項1〜14のいずれか一項に記載のポリペプチドを含む医薬上許容される組成物。
【請求項16】
内因性IL−23のTh17細胞との反応を干渉するために効果的な量で、IL−23と請求項1〜14のいずれか一項に記載のポリペプチドを接触させることを含む、Th17細胞の病原性を調節する方法。
【図2】

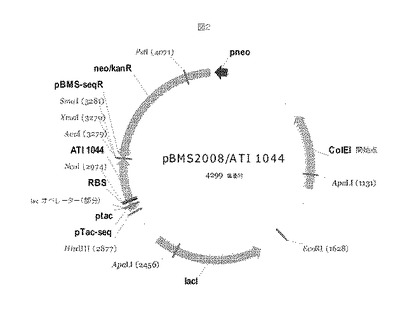
【図3−1】

【図3−2】

【図4】
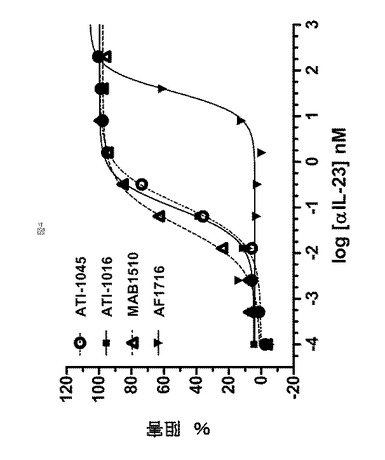
【図5】
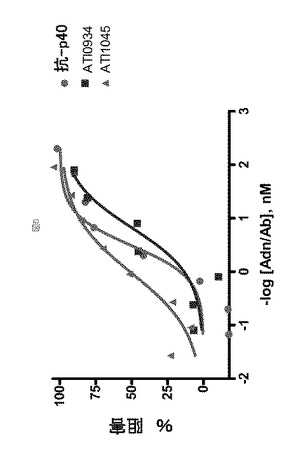
【図6】
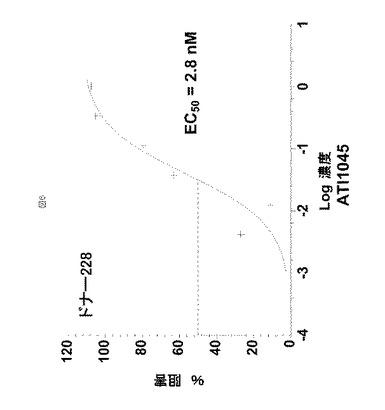
【図7】
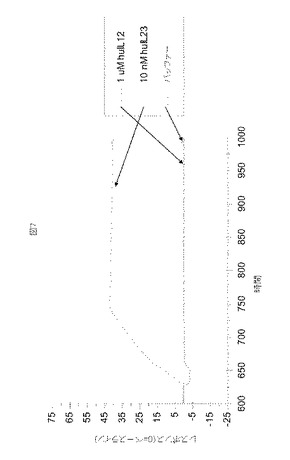
【図8】
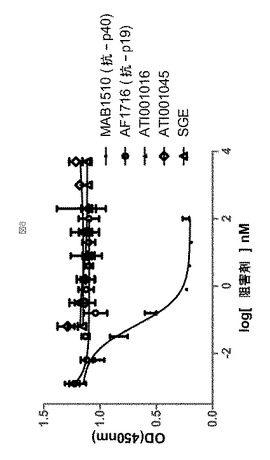
【図9A】
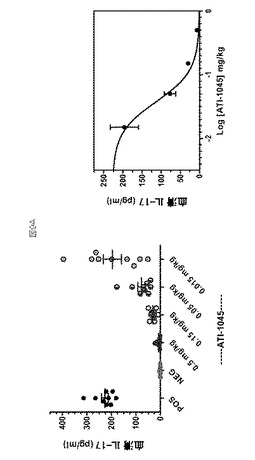
【図9B】
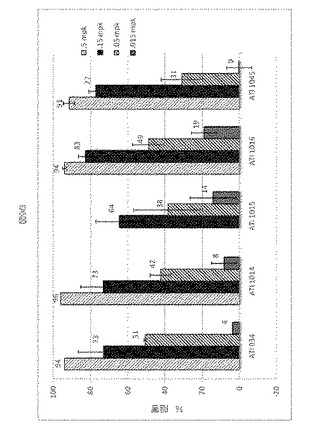
【図10A】
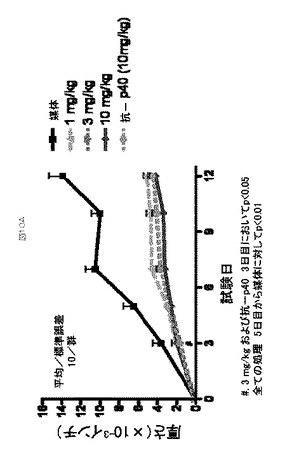
【図10B】
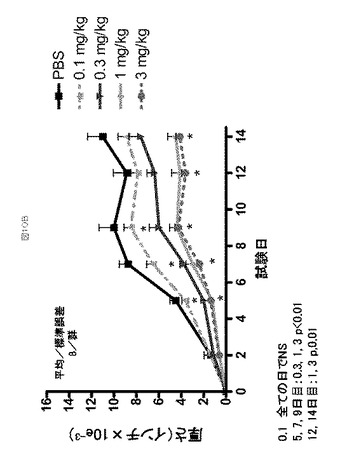
【図11A】
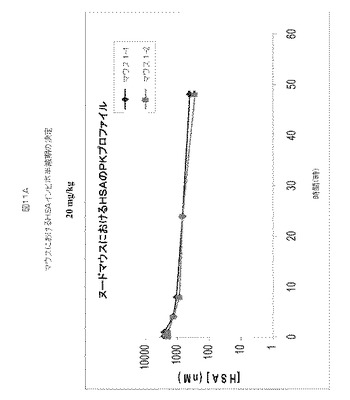
【図11B】
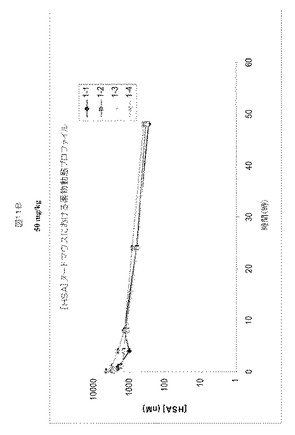
【図12A】
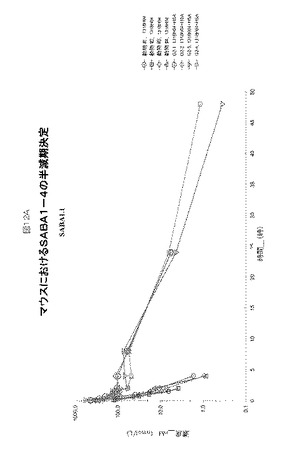
【図12B】
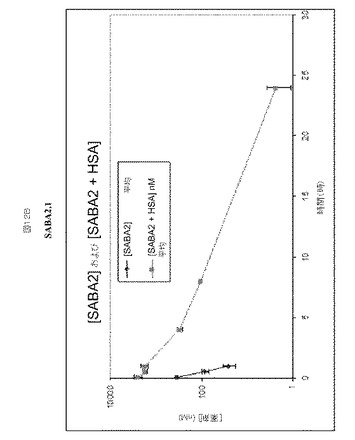
【図12C】
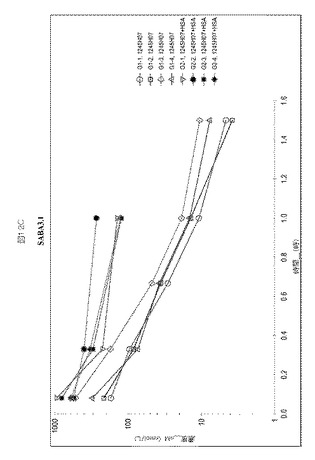
【図12D】
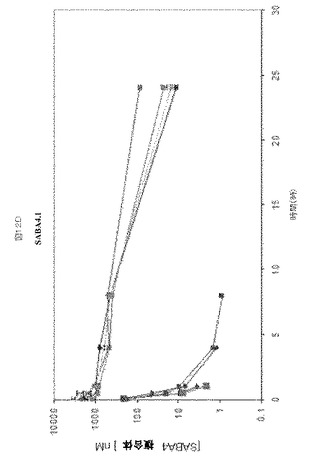
【図13A】
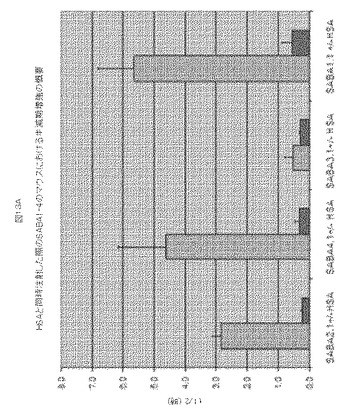
【図13B】
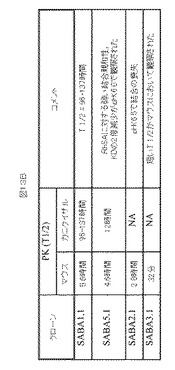
【図14A】
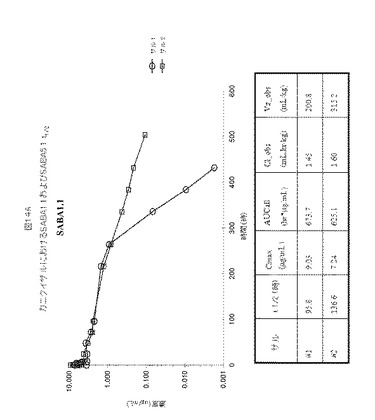
【図14B】
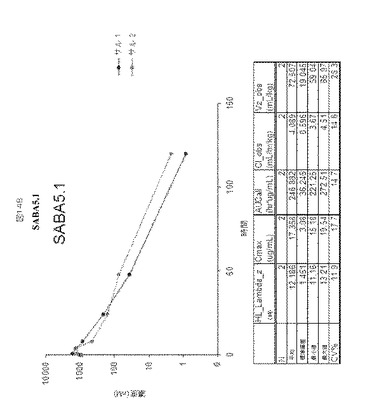
【図15】
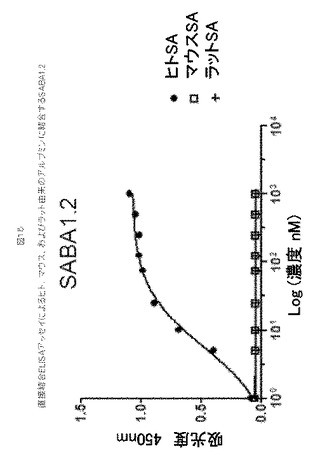
【図16】
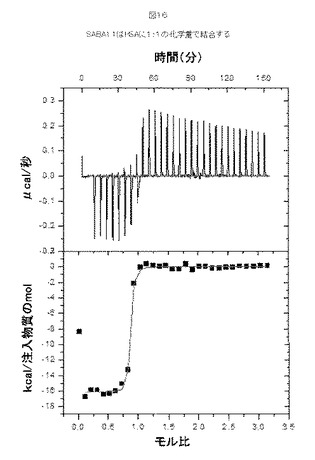
【図17】
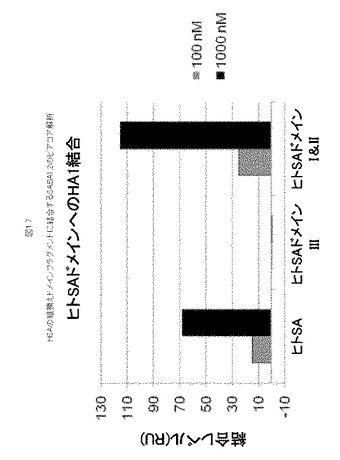
【図18】
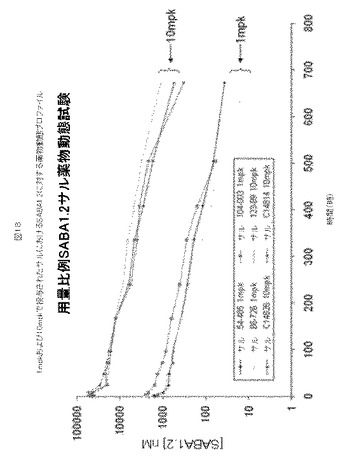
【図19】
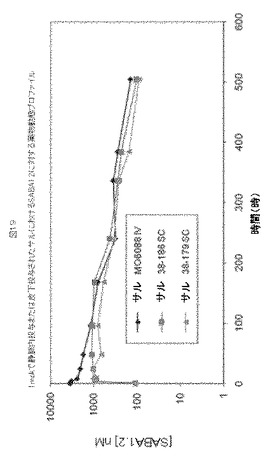
【図3−1】
【図3−2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図10A】
【図10B】
【図11A】
【図11B】
【図12A】
【図12B】
【図12C】
【図12D】
【図13A】
【図13B】
【図14A】
【図14B】
【図15】
【図16】
【図17】
【図18】
【図19】
【公表番号】特表2013−520426(P2013−520426A)
【公表日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願番号】特願2012−553974(P2012−553974)
【出願日】平成23年2月16日(2011.2.16)
【国際出願番号】PCT/US2011/024959
【国際公開番号】WO2011/103105
【国際公開日】平成23年8月25日(2011.8.25)
【出願人】(391015708)ブリストル−マイヤーズ スクイブ カンパニー (494)
【氏名又は名称原語表記】BRISTOL−MYERS SQUIBB COMPANY
【Fターム(参考)】
【公表日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願日】平成23年2月16日(2011.2.16)
【国際出願番号】PCT/US2011/024959
【国際公開番号】WO2011/103105
【国際公開日】平成23年8月25日(2011.8.25)
【出願人】(391015708)ブリストル−マイヤーズ スクイブ カンパニー (494)
【氏名又は名称原語表記】BRISTOL−MYERS SQUIBB COMPANY
【Fターム(参考)】
[ Back to top ]