
IL−5により媒介される疾病の治療に有用な組み換えIL−5アンタゴニスト
【課題】ヒト・インターロイキン5に対する中和モノクローナル抗体のごとき高アフィニティーIL−5アンタゴニストであって、好酸球の分化および増殖(すなわち、好酸球の蓄積)を抑制し、好酸球による炎症を抑制する高アフィニティーIL−5アンタゴニストを提供する。
【解決手段】重鎖および軽鎖を含んでなるヒト・インターロイキン−5に特異的なヒト化抗体。
【解決手段】重鎖および軽鎖を含んでなるヒト・インターロイキン−5に特異的なヒト化抗体。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本願は、1994年12月23日出願のU.S.第08/363,131号の一部継続出願であるU.S.第08/470,110号および08/467,420号(ともに1995年6月6日出願)の一部継続出願である。
【0002】
発明の分野
一般的には、本発明は、IL−5および過剰な好酸球産生により媒介される症状の治療および診断に有用な抗体および変化した抗体、そしてより詳細には、mAbs、Fab、キメラおよびヒト化抗体の分野に関する。
【背景技術】
【0003】
発明の背景
好酸球は、肺組織の過敏反応に関連したアレルギー性疾患を包含する広範囲の炎症性疾病状態の発生に関与している(非特許文献1)。注目すべき例は喘息であり、喘息は、非特異的気管支過敏症を引き起こす気道の可逆的閉塞により特徴づけられる。さらに喘息は、気管支粘膜のレベルの慢性的炎症反応ならびにマクロファージ、リンパ球および好酸球による特徴的な炎症の発生と因果関係がある。好酸球は、疾病の典型的な粘膜ダメージの開始において中心的役割を果たしているようである(非特許文献2)。数が増加した活性化された好酸球は、慢性喘息の患者の循環系、気管支分泌物および肺の実質において報告されており、種々の肺機能試験により測定される疾病の重さは血中好酸球数と相関関係がある(非特許文献3)。また、しばしば、脱顆粒反応の過程における増加した好酸球が、後期喘息反応中の患者気管支肺胞洗浄液(BAL)中に回収され、通常には、ステロイド療法の結果としての好酸球の減少は臨床的徴候の改善に関連している(非特許文献4)。
【0004】
インターロイキン5(IL−5)は、活性化されたCD4およびTリンパ球により産生されるホモダイマー糖蛋白である。ヒトにおいて、IL−5は好酸球の増殖および分化の調節に大いに関係している。IL−5の上昇したレベルは喘息性気管支肺胞洗浄液中に検出される(非特許文献5)。IL−5に関してトランスジェニックなマウスは、抗原刺激の不存在下の末梢血および組織における著しい好酸球増加を示し(非特許文献6)、抗−ネズミ・IL−5モノクローナル抗体は、マウスの血液および組織における好酸球増加(非特許文献7)ならびに実験動物における寄生虫感染およびアレルゲン攻撃に関連した好酸球増加(非特許文献8〜10)の抑制における効果が示されている。
【0005】
コルチコステロイドは、好酸球数および喘息の他の炎症性成分の減少にきわめて効果的であるが、重い喘息およびより最近になってからは軽い喘息ないしは中程度の喘息のいずれにおいても副作用の心配がある。唯一の他の主要抗−炎症薬剤療法−クロモグリケート類(クロモリンナトリウムおよびネドクロミル)−はコルチコステロイドよりもかなり効果が少なく、それらの正確な作用機構は不明のままである。
【0006】
より最近の開発は、新たな吸入ステロイド、長時間作用する気管支拡張剤、および新規生化学的もしくは薬理学的標的(例えば、カリウムチャンネル活性化剤、ロイコトリエンアンタゴニスト、5−リポオキシゲナーゼ(5−LO)阻害剤等)に作用する薬剤に焦点を絞っている。理想的な薬剤は、ステロイドの有効性をクロモリンナトリウムの安全性と結び付けるものであり、選択性が増加していてより迅速に作用を発揮するものである。中和IL−5抗体は潜在的にヒトにおける好酸球増加関連徴候の軽減に有用でありうる。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Butterfieldら,Immunopharmacology of Eosinophils(H.SmithおよびR.Cook編,)中,p151-192,Academic Press, London(1993)
【非特許文献2】Corrigan et al., Immunol.Today,13:501-507(1992)
【非特許文献3】Griffen et al.,J.Aller.Clin.Immunol.67,548-557(1991)
【非特許文献4】Bousquet et al.,N.Eng.J.Med.,323:1033-1039(1990)
【非特許文献5】Motojima et al.,Allergy,48:98(1993)
【非特許文献6】Dent et al.,J.Exp.Med.,172, 1425(1990)
【非特許文献7】Hitoshi et al.,Int.Immunol.,315(1991)
【非特許文献8】Coffman et al.,Science,245,308-310(1989)
【非特許文献9】Sher et al.,Proc.Natl.Acad.Sci.,83:61-65(1990)
【非特許文献10】Chand et al.,Eur.J.Pharmacol.,211:121-123(1992)
【発明の概要】
【発明が解決しようとする課題】
【0008】
ゆえに、ヒト・インターロイキン5に対する中和モノクローナル抗体のごとき高アフィニティーIL−5アンタゴニストであって、好酸球の分化および増殖(すなわち、好酸球の蓄積)を抑制し、かくして好酸球により炎症を抑制する高アフィニティーIL−5アンタゴニストに対する必要性が当該分野において存在する。
【課題を解決するための手段】
【0009】
発明の概要
第1の態様において、本発明は、ヒト・インターロイキン5に特異的で、詳細な説明の欄において説明するように、約3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる結合アフィニティーを有する齧歯類(例えば、ラットおよびマウス)の中和モノクローナル抗体を提供する。かかるモノクローナル抗体の典型例はマウスのモノクローナル抗体2B6、2E6および2F2ならびに4A6のごときラットのモノクローナル抗体である。本発明のもう1つの態様は、SK119−2B6.206.75(1)、SK119−2E3.39.40.2、SK119−2F2.37.80.12、4A6(1)G1F7および5D3(1)F5D6のごときハイブリドーマである。
【0010】
関連態様において、本発明は、本発明の齧歯類中和モノクローナル抗体のFc領域を欠失させることにより製造される、ヒト・インターロイキン−5に特異的な中和FabフラグメントまたはそのF(ab')2フラグメントを提供する。
【0011】
さらにもう1つの関連態様において、本発明は、チェイン・シャフリング(chain shuffling)法により製造される、ヒト・インターロイキン−5に特異的な中和FabフラグメントまたはそのF(ab')2フラグメントを提供する。該方法により、線維状ファージFabディスプレイライブラリーにおいてインターロイキン5で免疫された齧歯類から単離された軽鎖(または重鎖)免疫グロブリンライブラリーとともに本発明齧歯類中和モノクローナル抗体から単離された重鎖(または軽鎖)免疫グロブリンが発現される。
【0012】
さらにもう1つの関連態様において、本発明は、ヒト・インターロイキン5に対して約3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる非ヒト・中和モノクローナル抗体(mAb(s))由来の相補性決定領域(CDR(s))を含んでなる、ヒト・インターロイキン5に特異的な変化した抗体ならびにそれをコードしている核酸分子を提供する。変化した抗体がヒト化抗体である場合には、非ヒト・免疫グロブリン由来の相補性決定領域(CDRs)をコードしている配列が、少なくとも1つ、好ましくは全部の相補性決定領域(CDRs)が非ヒト・モノクローナル抗体由来のCDRsにより置換されている第1の免疫グロブリンパートナー中に挿入される。好ましくは、第1の免疫グロブリンパートナーは、同様に免疫グロブリンの不変鎖全部またはその一部を含んでなる第2の免疫グロブリンパートナーに作動可能に連結される。
【0013】
関連態様において、本発明は、ヒト・インターロイキン5に対して約3.5x10−11Mに等しいかまたはそれ未満の解離定数を有すことにより特徴づけられる非ヒト・中和モノクローナル抗体(mAbs)由来のCDRs、ならびにかかるCDRsをコードしている核酸分子を提供する。
【0014】
さらにもう1つの態様において、ヒト・重鎖および軽鎖の不変領域ならびにヒト・インターロイキン5に対して約3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる非ヒト・中和モノクローナル抗体由来の重鎖および軽鎖の可変領域を含むキメラ抗体を提供する。
【0015】
さらにもう1つの態様において、本発明は、上記の変化した抗体のうちの1種(またはそれ以上)および医薬上許容される担体を含む医薬組成物を提供する。
【0016】
さらなる態様において、本発明は、ヒトに有効量の本発明医薬組成物を投与することによる、過剰の好酸球産生に関連したヒトにおける症状の治療方法を提供する。
【0017】
さらなる態様において、本発明は、IL−5に対して約3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる非ヒト・中和モノクローナル抗体(mAbs)由来の変化した抗体(例えば、組み換え抗体、CDRs、FabまたはそのF(ab')2フラグメントもしくはアナログ)の組み換え製造方法および該組み換え製造において有用な成分を提供する。これらの成分は、上記のものをコードしている単離核酸配列、ならびに選択された調節配列の制御下の当該核酸配列を含む組み換えプラスミドを包含し、該調節配列は、該組み換えプラスミドでトランスフェクションされた宿主細胞(好ましくは哺乳動物細胞)における該核酸配列の発現を指令することができるものである。該製造方法は、変化した抗体、好ましくはヒト化抗体がトランスフェクションされた細胞中で発現されるような条件下で本発明のトランスフェクションされた宿主細胞系を培養し、次いで、そこから発現産物を単離することを包含する。
【0018】
本発明のさらなる態様において、患者から生物学的液体試料を得て、IL−5/(モノクローナルまたはポリクローナル)抗体複合体が形成されるような条件下で本発明抗体および変化した抗体をかかる試料と接触させ、次いで、該IL−5/抗体複合体の存在または不存在を検出することを特徴とする、ヒトにおける過剰な好酸球産生に関連した症状の診断方法が提供される。
【0019】
本発明の他の態様および利点を、詳細な説明および好ましい具体例の欄においてさらに説明する。
【発明の効果】
【0020】
本発明により、高い結合アフィニティーを有するヒト・インターロイキン5に対する中和モノクローナル抗体などが提供される。
【図面の簡単な説明】
【0021】
【図1】図1[配列番号:1および15]は、マウス・抗体2B6およびマウス/ヒト・2B6キメラ抗体に関する重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図2】図2[配列番号:2および16]は、マウス・抗体2B6およびマウス/ヒト・2B6キメラ抗体に関する軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【図3】図3[配列番号:3]は、マウス・抗体2F2に関する重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図4】図4[配列番号:4]は、マウス・抗体2F2に関する軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【図5】図5[配列番号:5]は、マウス・抗体2E3に関する重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図6】図6[配列番号:6]は、マウス・抗体2E3に関する軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【図7】図7[配列番号:7−14]は、マウス抗体・2B6、2F2および2E3由来の重鎖および軽鎖のCDRsを示す。
【図8】図8[配列番号:18、19]は、ヒト化抗体2B6に関する重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図9】図9[配列番号:20、21]は、ヒト化抗体2B6に関する軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【図10】図10は、哺乳動物細胞におけるヒト化重鎖遺伝子の発現に用いるプラスミドpCDIL5HZHC1.0のスキーム図である。該プラスミドはベータラクタマーゼ遺伝子(BETA LAC)、SV−40複製開始点(SV40)、サイトメガロウイルスプロモーター配列(CMV)、シグナル配列、ヒト化重鎖、ウシ・成長ホルモン(BGH)由来のポリAシグナル、ベータグロビンプロモーター(ベータグロプロ)、ジヒドロ葉酸レダクターゼ遺伝子(DHFR)、およびもう1つのBGH配列のポリAシグナルをpUC19骨格中に含んでいる。
【図11】図11は、哺乳動物細胞におけるヒト化軽鎖遺伝子の発現に用いるプラスミドpCNIL5HZLC1.0のスキーム図である。
【図12】図12[配列番号:61、62]は、ヒト化抗体2B6に関するNewM重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図13】図13[配列番号:69、70]は、ヒト化抗体2B6に関するREI軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【発明を実施するための形態】
【0022】
発明の詳細な説明
本発明は、マウス・モノクローナル抗体2B6のおいて例示されるヒト・IL−5特異性、中和活性、およびヒト・IL−5に対する高アフィニティーにより特徴づけられる種々の抗体、変化した抗体およびそれらのフラグメントを提供する。新規中和抗体を得るための慣用的なハイブリドーマ法、ファージディスプレイ組み合わせライブラリー、免疫グロブリンのチェイン・シャフリング、およびヒト化法により本発明抗体は製造された。これらの製造物は、例えば喘息のごときIL−5により媒介される疾病の治療のための治療および医薬組成物において有用である。また、これらの製造物は、ヒトにおける内在性IL−5レベルまたは活性化細胞からex vivoで放出されるIL−5レベルの測定(例えば、酵素結合免疫吸着アッセイ(ELISA))による、IL−5により媒介される症状の診断において有用である。
【0023】
I.定義
「変化した抗体」は、変化した免疫グロブリンコーディング領域によりコードされている蛋白をいい、選択された宿主細胞における発現により得ることができる。かかる変化した抗体は遺伝子組み換えによる抗体(例えば、キメラ抗体またはヒト化抗体)または免疫グロブリン不変領域の全部もしくは一部を欠く抗体フラグメント、例えば、Fv、Fab、またはF(ab')2等である。
【0024】
「変化した免疫グロブリンコーディング領域」は、本発明の変化した抗体をコードしている核酸配列をいう。変化した抗体がCDRを継ぎ足した抗体またはヒト化抗体である場合、非ヒト・免疫グロブリン由来の相補性決定領域(CDRs)をコードしている配列が、ヒト・可変枠組み配列を含んでなる第1の免疫グロブリンパートナー中に挿入される。第1の免疫グロブリンパートナーが第2の免疫グロブリンパートナーに作動可能に連結されていてもよい。
【0025】
「第1の免疫グロブリンパートナー」は、無傷(または天然に存在する)のCDRコーディング領域がドナー抗体のCDRコーディング領域により置換されているヒト・枠組みまたはヒト・免疫グロブリン可変領域をコードしている核酸配列をいう。ヒト・可変領域は免疫グロブリン重鎖、軽鎖(または両方)、それらの機能的フラグメントのアナログであってよい。抗体(免疫グロブリン)の可変領域中に存在するかかるCDR領域を、当該分野で知られた方法により決定することができる。例えば、Kabatら(Sequences of Proteins of Immunological Interest,4th Ed.,U.S.Department of Health and Human Services,National Institutes of Health(1987))は、CDRsの存在位置に関する規則を開示している。さらに、CDR領域/構造の同定に有用なコンピュータープログラムが知られている。
【0026】
「中和」は、ヒト・IL−5のその特異的受容体への結合を妨げることにより、あるいはその受容体を通じてのIL−5のシグナリングを阻害することによりIL−5活性を阻害する抗体をいう。B13細胞バイオアッセイ(IL−5中和アッセイ、実施例2C参照)において測定されるIL−5活性の阻害において90%有効、好ましくは95%有効、最も好ましくは100%有効な場合、mAbは中和抗体である。
【0027】
用語「高アフィニティー」は、光学的バイオセンサー分析(実施例2参照)により測定されるヒト・IL−5に対する3.5x10−11Mに等しいかまたはそれ未満のKdにより特徴づけられる結合アフィニティーを有する抗体をいう。
【0028】
「ヒト・IL−5に対する結合特異性」は、マウスではなくヒトのIL−5に対する高アフィニティーを意味する。
【0029】
「第2の免疫グロブリンパートナー」は、イン・フレームで、あるいは所望により慣用的リンカー配列により第1の免疫グロブリンパートナーが融合(すなわち、作動可能に連結)している蛋白またはペプチドをコードしているもう1つのヌクレオチド配列をいう。好ましくは、それは免疫グロブリン遺伝子である。第2の免疫グロブリンパートナーは、その不変領域全体(すなわち、同種のもの−第1および第2の変化した抗体は同じ起源由来である)または対象とするさらなる抗体(すなわち、異種のもの)をコードしている核酸配列を包含しうる。それは免疫グロブリン重鎖または軽鎖(または1本のポリペプチドの一部としての両鎖)であってもよい。第2の免疫グロブリンパートナーは特定の免疫グロブリンクラスまたはイソタイプに限らない。さらに、第2の免疫グロブリンパートナーは、FabまたはF(ab')2のごとき免疫グロブリン不変領域の一部(すなわち、適当なヒト・不変領域または枠組み領域の別個の部分)を含んでなっていてもよい。また、かかる第2の免疫グロブリンパートナーは、例えばファージディスプレイライブラリーとして宿主細胞の外表面上に露出した内在性膜蛋白をコードしている配列、あるいは分析または診断的検出のための蛋白、例えばセイヨウワサビペルオキシダーゼ、β−ガラクトシダーゼ等をコードしている配列を含んでいてもよい。
【0030】
用語Fv、Fc、Fd、Fab、またはF(ab')2を、その標準的な意味で用いる(例えば、Harlow et al.,Antibodies A Laboratory Manual,Cold SpringHarbor Laboratory,(1988)参照)。
【0031】
本明細書の用語「組み換え法による抗体」は、変化した抗体の1のタイプ、すなわち、選択されたアクセプター抗体の軽鎖および/または重鎖可変ドメインの一部分が、選択されたエピトープに特異性を有する1種またはそれ以上のドナー抗体由来の類似部分により置換されている全長の合成抗体(抗体フラグメントに対立するものとしてのキメラ抗体またはヒト化抗体)をいう。例えば、かかる分子は、未修飾軽鎖(またはキメラ軽鎖)に結合したヒト化重鎖(あるいはその逆)によって特徴づけられる抗体を包含しうる。組み換え法による抗体もまた、ドナー抗体の結合特異性を保持するようなアクセプター抗体の軽鎖および/または重鎖の可変ドメイン枠組み領域をコードしている核酸配列の変化によって特徴づけられる。これらの抗体は、アクセプター抗体由来の1種またはそれ以上のCDRs(好ましくは全部のCDRs)の本明細書記載のドナー抗体由来のCDRsでの置換を含んでいてもよい。
【0032】
「キメラ抗体」は、アクセプター抗体由来の軽鎖および重鎖不変領域に結合したドナー抗体由来の天然に存在する可変領域(軽鎖および重鎖)を含んでいる組み換え法による抗体の1のタイプをいう。
【0033】
「ヒト化抗体」は、非ヒト・ドナー免疫グロブリン由来のCDRsを有する組み換え法による抗体の1のタイプであって、その残りの免疫グロブリン由来の部分は1種(またはそれ以上)のヒト由来であるものをいう。さらに、結合アフィニティーを保持するように枠組み支持残基を変化させてもよい(例えば、Queen et al.,Proc.Natl.Acad.Sci.USA,86:10029-10032(1989)、Hodgson et al.,Bio/Technology,9:421(1991)参照)。
【0034】
用語「ドナー抗体」は、その可変領域、CDRs、またはその他のフラグメントもしくはアナログの核酸配列が第1の免疫グロブリンパートナーのものになっている抗体をいい、その結果、変化した免疫グロブリンコーディング領域を提供し、発現した変化した抗体がドナー抗体の抗原特異性および中和活性を有する。本発明における使用に適した1のドナー抗体は、2B6と命名された非ヒト・中和モノクローナル抗体(すなわち、マウスの)である。抗体2B6は、イソタイプIgG1に属する高アフィニティーヒト・IL−5特異性(すなわち、マウス・IL−5を認識しない)中和抗体であると定義され、適当なマウス・IgG不変領域上に配列番号:2および16の可変軽鎖DNAおよびアミノ酸配列、ならびに配列番号:1および15の可変重鎖DNAおよびアミノ酸配列を有している。
【0035】
用語「アクセプター抗体」は、ドナー抗体とは異種の抗体(モノクローナル、または組み換え型)をいい、その重鎖および/または軽鎖枠組み領域ならびに/あるいはその重鎖および/または軽鎖不変領域のすべて(またはいずれかの部分、しかし、好ましくはすべての部分)が第1の免疫グロブリンパートナーのものとなっている。好ましくは、ヒト・抗体がアクセプター抗体である。
【0036】
「CDRs」は、免疫グロブリン重鎖および軽鎖の超可変領域である抗体の相補性決定領域のアミノ酸配列をいう。例えば、Kabat et al.,Sequences of Proteins of Immunological Interest,4th Ed,U.S.Department of Health and Human Services,National Institutes of Health(1987)参照。免疫グロブリン可変領域には3本の重鎖CDRsおよび3本の軽鎖CDRsがある。よって、本明細書の用語「CDRs」は、3本の重鎖CDRsすべて、あるいは3本の軽鎖CDRsすべて(あるいは適当な場合には、すべての重鎖および軽鎖CDRsの両方)をいう。
【0037】
CDRsは、抗原またはエピトープへの抗体結合のための接触残基の大部分を提供する。本発明の対象CDRsはドナー抗体可変重鎖および軽鎖配列由来であり、天然に存在するCDRsのアナログを包含し、該アナログはまた、それらが由来するドナー抗体と同じ抗原結合特異性および/または中和能を共有している。
【0038】
「抗原結合特異性または中和能を共有」とは、例えば、mAb 2B6は一定レベルの抗原アフィニティーにより特徴づけられるが、適当な構造的環境においては2B6の核酸配列によりコードされているCDRは低いまたは高いアフィニティーを有するかもしれないということを意味する。それにもかかわらず、かかる環境において、2B6のCDRsは2B6と同じエピトープを認識するであろうと予想される。典型的な2B6の重鎖CDRsは配列番号:7、配列番号:8および配列番号:9を包含し、典型的な2B6の軽鎖CDRは配列番号:10、配列番号:11および配列番号:12を包含する。
【0039】
「機能的フラグメント」は、そのフラグメントが由来している抗体と同じ抗原結合特異性および/または中和能を保持している部分的な重鎖または軽鎖可変配列(例えば、免疫グロブリン可変領域のアミノ末端またはカルボキシ末端における小規模な欠失を有する)である。
【0040】
「アナログ」は、少なくとも1個のアミノ酸によって修飾されているアミノ酸配列であり、該修飾は化学的なものあるいは少数(すなわち、10個未満)のアミノ酸の置換または転移であってもよく、修飾されているとしてもアミノ酸配列が未修飾配列の生物学的特性、例えば、抗原特異性および高アフィニティーを保持しているものである。例えば、ある種のエンドヌクレアーゼ制限部位をCDRコーディング配列中またはその周辺に作成する場合には、置換により(サイレント)変異を構築することができる。
【0041】
アナログは対立遺伝子変化を生じるものであってもよい。「対立遺伝子変化または修飾」とは、本発明アミノ酸またはペプチド配列をコードしている核酸配列中の変化である。かかる変化または修飾は遺伝コードの縮重によるものであってもよく、あるいは所望特性を得るように慎重に組み換え法を用いて作成されたものであってもよい。これらの変化または修飾は、コードされているアミノ酸配列の変化を生じるものであってもよく、生じないものであってもよい。
【0042】
用語「エフェクター剤」は、変化した抗体および/またはドナー抗体もしくはその他のフラグメントの天然または合成の軽鎖または重鎖が慣用的手段により結合されていてもよい非蛋白担体分子をいう。かかる非蛋白担体は、診断分野において用いられる慣用的担体、例えば、ポリスチレンまたは他のプラスチックビーズ、多糖類、例えば、BIAcore[Pharmacia]システムに用いるもの、または医学分野で使用され、ヒトおよび動物への投与が安全である他の非蛋白物質を包含する。他のエフェクター剤は、重金属原子またはラジオアイソトープをキレートするためのマクロサイトを包含する。かかるエフェクター剤は、変化した抗体の半減期を延長することにも有用であり、例えば、ポリエチレングリコールが挙げられる。
【0043】
II.高アフィニティーIL−5モノクローナル抗体
本発明抗体、変化した抗体およびフラグメントの構築に使用するために、無傷のヒト・IL−5またはそれに由来するペプチドエピトープを与えることにより、非ヒト種(例えば、ウシ、ヒツジ、サル、ニワトリ、齧歯類(例えばマウスおよびラット)等)を用いて所望免疫グロブリンを得てもよい。慣用的なハイブリドーマ法を用いて、IL−5に対する非ヒト・mAbを分泌するハイブリドーマ細胞系を得る。次いで、実施例セクションに記載のごとくIL−5で被覆した96ウェルプレートを用いて、あるいは別法としてストレプトアビジンで被覆したプレートに結合したビオチン化IL−5を用いてかかるハイブリドーマを結合に関してスクリーニングする。
【0044】
本発明の1の典型的な高アフィニティー中和mAbはマウス・抗体であるmAb 2B6であり、実施例1においてより詳細に説明されるキメラ抗体またはヒト化抗体の開発にそれを用いることができる。2B6 mAbは、ヒト・IL−5に対する抗原結合特異性により特徴づけられ、IL−5に対して3.5x10−11M未満(約2.2x10−11M)のKdを有する。光学的バイオセンサーによる測定により2B6由来のFabフラグメントのIL−5に対するKdを評価したところ(実施例3H参照)、約9x10−11Mである。mAb 2B6は、ヒト・IL−5とヒト・IL−5受容体のα−鎖との間の結合相互作用をブロックするように思われる。
【0045】
もう1つの望ましいドナー抗体はマウス・mAb 2E3である。このmAbは、イソタイプIgG2aであり、3.5x10−11M未満(約2.0x10−11M)のヒト・IL−5に対する解離定数を有することにより特徴づけられる。
【0046】
さらにもう1つの望ましいドナー抗体はラット・mAb 4A6である。このmAbは、3.5x10−11M未満(約1.8x10−11M)のヒト・IL−5に対する解離定数を有することにより特徴づけられる。さらに、mAb 4A6は、ヒト・IL−5とヒト・IL−5受容体のβ−鎖との間の結合相互作用をブロックするように思われる。
【0047】
本発明は2B6 mAb、2E3 mAb、またはそれらの超可変(すなわち、CDR)配列の使用に限定されない。ヒト・IL−5に対する3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる他の適当な高アフィニティーIL−5抗体および対応する抗−IL−5 CDRsをそれらに代えて用いてもよい。以下の説明のいずれの箇所においても、ドナー抗体は2B6または2E3と同定されるが、この命名は例示および単なる説明のためのものである。
【0048】
III.抗体フラグメント
本発明は、ヒト・IL−5に指向されたmAb由来のFabフラグメントまたはF(ab')2フラグメントの使用も包含する。これらのフラグメントは、in vivoにおけるIL−5および好酸球により媒介される症状に対する防御剤として、あるいはin vitroでのIL−5診断薬の一部として有用である。Fabフラグメントは軽鎖全体および重鎖のアミノ末端部分を含んでおり;F(ab')2フラグメントはジスルフィド結合により結合している2個のFabフラグメントにより形成されるフラグメントである。mAbs 2B6、2E3および他の類似高アフィニティーIL−5結合抗体は、FabフラグメントおよびF(ab')2フラグメントのソースを提供するものであり、それらのフラグメントは慣用的手段、例えば、適当な蛋白分解酵素パパインおよび/またはペプシンでのmAbの開裂により、あるいは組み換え法により得ることができる。本明細書記載のごとく、これらのFabおよびF(ab')2フラグメントはそれら自体、治療、予防または診断薬、ならびに組み換え型抗体またはヒト化抗体の形成において有用な可変領域およびCDR配列を含む配列のドナーとして有用である。
【0049】
組み合わせファージライブラリー(例えば、Winter et al.,Ann.Rev.Immunol.,12:433-455(1994)参照)または免疫グロブリンのチェイン・シャフリング(例えば、Marks et al.,Bio/Technology,10:779-783(1992)参照)によりFabおよびF(ab')2フラグメントを構築することができ(両文献を参照により本明細書に記載されているものとみなす)、それらの方法において、選択された抗体(例えば、2B6)由来のFdまたはVH免疫グロブリンを軽鎖免疫グロブリンVL(VK)のレパートリーと結合させて新規Fabsを得る。逆に、選択された抗体由来の軽鎖免疫グロブリンを重鎖免疫グロブリンVH(またはFd)のレパートリーと結合させて新規Fabを得てもよい。実施例セクションにおいてより詳細に説明するように、mAb 2B6のFdを軽鎖免疫グロブリンのレパートリーと結合させた場合に中和IL−5 Fabsが得られた。よって、チェイン・シャフリング法により、ユニークな配列(ヌクレオチドおよびアミノ酸)を有する中和Fabsを得ることができる。
【0050】
IV.対象とする抗−IL−5アミノ酸およびヌクレオチド配列
上記mAb 2B6または他の抗体は、ドナー抗体の抗原結合特異性により特徴づけられる種々の変化した抗体の設計および取得に有用な配列、例えば、可変重鎖および/または軽鎖ペプチド配列、枠組み配列、CDR配列、機能的フラグメント、およびそれらのアナログ、およびそれらをコードしている核酸配列を提供する。
【0051】
よって、一例として、本発明は、IL−5マウス抗体2B6由来の可変軽鎖および可変重鎖配列ならびにそれら由来の配列を提供する。2B6の重鎖可変領域を図1に示す。CDRをコードしている領域はボックス領域により示され、それらは配列番号:7;配列番号:8;および配列番号:9に示される。2B6の軽鎖クローン可変領域を図2に示す。CDRをコードしている領域を配列番号:10;配列番号:11;および配列番号:12に示す。
【0052】
ヒト化重鎖可変領域を図8[配列番号:18および19]に示す。シグナル配列も配列番号:17に示す。当業者に知られた他の適当なシグナル配列を、本明細書に例示したシグナル配列に代えて使用してもよい。この構築物のCDRアミノ酸配列は無傷のマウスのものおよびキメラ重鎖CDRsと同一であり、それらを配列番号:7、配列番号:8、および配列番号:9に示す。典型的な(合成)ヒト化軽鎖可変配列を図9[配列番号:20および21]に示す。
【0053】
可変軽鎖および重鎖ペプチド配列をコードしている本発明核酸配列またはそのフラグメントは、CDRsまたは枠組み領域をコードしている核酸配列中の特異的変化の突然変異による導入、ならびに生じた修飾または融合核酸配列の発現用プラスミド中への導入にも有用である。例えば、枠組みおよびCDRをコードしている領域におけるサイレント置換を用いて突然変異されたCDR(および/または枠組み)領域の挿入を容易にする制限酵素部位を作成した。これらのCDRをコードしている領域を、本発明ヒト化抗体の構築に使用した。
【0054】
遺伝コードの縮重を考慮して、本発明の重鎖および軽鎖アミノ酸配列およびCDR配列ならびにドナー抗体の抗原特異性を共有しているそれらの機能的フラグメントおよびアナログをコードしている種々のコーディング配列を構築してもよい。第2の免疫グロブリンパートナーに作動可能に結合する場合には、可変鎖ペプチド配列またはCDRsをコードしている本発明の単離核酸配列またはそのフラグメントを用いて、本発明の変化した抗体、例えば、キメラまたはヒト化抗体、あるいは他の組み換え法による抗体を得ることができる。
【0055】
本明細書記載の変化した抗体および抗体の一部をコードしている単離核酸配列のほかに、他のかかる核酸配列、例えば、無傷のCDRをコードしている配列の相補的な配列またはCDRをコードしている領域周辺の修飾されたヒト・枠組み領域に相補的な配列も本発明に包含されることに注意すべきである。有用なDNA配列は、厳密なハイブリダイゼーション条件下[T.Maniatis et al.,Molecular Cloning(A Laboratory Manual),Cold Spring Harbor Laboratory(1982),387−389頁参照]でDNA配列にハイブリダイゼーションする配列を包含する。1のかかる厳密なハイブリダイゼーション条件の例は、4XSSC、65℃、次いで、65℃の0.1XSSCでの1時間の洗浄である。あるいはまた、厳密なハイブリダイゼーション条件の典型例は、50%ホルムアミド、4XSSC中、42℃である。好ましくは、これらのハイブリダイゼーションするDNA配列は少なくとも18ヌクレオチドの長さ、すなわち、ほぼCDRのサイズである。
【0056】
V.変化した免疫グロブリン分子および変化した抗体
変化した免疫グロブリン分子は、キメラ抗体およびヒト化抗体のごとき組み換え法により抗体を包含する変化した抗体をコードしていてもよい。望ましい変化した免疫グロブリンコーディング領域はCDRをコードしている領域を含み、該領域は、IL−5抗体の抗原特異性、好ましくは本発明により提供されるような高アフィニティー抗体の抗原特異性を有するペプチドをコードしており、第1の免疫グロブリンパートナー(ヒト・枠組みまたはヒト・免疫グロブリン可変領域)中に挿入されている。
【0057】
好ましくは、第1の免疫グロブリンパートナーは第2の免疫グロブリンパートナーに作動可能に連結される。第2の免疫グロブリンパートナーは上で定義されており、興味ある第2の抗体領域、例えばFc領域をコードしている配列を含んでもよい。第2の免疫グロブリンパートナーは、軽鎖または重鎖不変領域がイン・フレームまたはリンカー配列手段により融合しているもう1つの免疫グロブリンをコードしている配列を含んでもよい。機能的フラグメントまたはIL−5のアナログに対して結合が促進されるように、それらに指向された組み換え法による抗体を設計してもよい。
【0058】
第2の免疫グロブリンパートナーを非蛋白担体分子を包含する上記定義のエフェクター剤と結合させてもよく、慣用的手段により第2の免疫グロブリンパートナーを作動可能に結合させる。
【0059】
第2の免疫グロブリンパートナー、例えば抗体配列とエフェクター剤との間の融合または結合は、例えば慣用的な共有結合またはイオン結合、蛋白融合、あるいは異種二官能基架橋(例、カルボジイミド、グルタルアルデヒド等)のような適当な手段によるものであってよい。かかる方法は当該分野において知られており、普通の化学および生化学の教科書に記載されている。
【0060】
さらに、第2の免疫グロブリンパートナーとエフェクター剤との間に所望量のスペースを単に提供するだけの慣用的リンカー配列を、変化した免疫グロブリンコーディング領域中に構築してもよい。かかるリンカーの設計は当業者によく知られている。
【0061】
さらに、本発明分子のシグナル配列を修飾して発現を促進してもよい。一例として、シグナル配列およびマウス・重鎖配列由来のCDRsを有する2B6ヒト化抗体はもとのシグナルペプチドを別のシグナル配列[配列番号:17]に置き換えられている。
【0062】
典型的な変化した抗体は、mAb 2B6の抗原特異性を有する可変重鎖および/または軽鎖ペプチドまたは蛋白配列、例えば、VHおよびVL鎖を含んでいる。さらにもう1つの望ましい本発明の変化した抗体は、少なくとも1つ、マウス・抗体分子2B6の重鎖および/または軽鎖の可変領域の好ましくは全部のCDRsを含んでおり、残りの配列はヒト起源のもの、またはその機能的フラグメントもしくはアナログである。例えば、ヒト化VHおよびVL領域(図8および9)参照。
【0063】
さらなる具体例において、組み換え法による本発明抗体にさらなる剤が結合していてもよい。例えば、組み換えDNA法の手順を用いて、完全な抗体分子のFcフラグメントまたはCH2 CH3ドメインが酵素または他の検出可能な分子(すなわち、ポリペプチドエフェクターまたはレポーター分子)に置き換えられている本発明の組み換え法による抗体を得てもよい。
【0064】
第2の免疫グロブリンパートナーを、マウス2B6の抗原特異性を有するCDR含有配列とは異種の非免疫グロブリンペプチド、蛋白またはそのフラグメントに作動可能に結合させてもよい。得られる蛋白は、発現すると、IL−5抗原特異性および非免疫グロブリン特性を示すことができる。その融合パートナーの特性は、例えば、別の結合ドメインまたは受容体ドメインのごとき機能的特性、あるいは融合パートナー自体が治療蛋白である場合には治療特性またはさらなる抗原特性であってもよい。
【0065】
もう1つの望ましい本発明蛋白は、全長の重鎖および軽鎖を有する完全な抗体分子、あるいはFabまたはF(ab')2フラグメントのごときそれらの別々のフラグメント、重鎖ダイマー、またはFVまたは1本鎖抗体(SCA)のごときそれらの最小の組み換えフラグメント、または例えばmAb 2B6または2E3のような選択されたドナーmAbと同じ特異性を有する他の分子を含んでなっていてもよい。かかる蛋白を変化した抗体の形態で用いてもよく、あるいはその未融合形態で使用してもよい。
【0066】
第2の免疫グロブリンパートナーがドナー抗体とは異なる抗体、例えば、異なるイソタイプまたはクラスの免疫グロブリン枠組みまたは不変領域に由来する場合はいつでも、組み換え法による抗体が得られる。組み換え法による抗体は、1の起源、例えばアクセプター抗体由来の免疫グロブリン(Ig)不変領域および可変枠組み領域、ならびにドナー抗体、例えば本明細書記載の抗−IL−5抗体由来の1つまたはそれ以上(好ましくは全部)のCDRsを含んでなっていてもよい。さらに、核酸またはアミノ酸レベルでのアクセプターmAb軽鎖および/または重鎖可変ドメイン枠組み領域の変化、例えば、欠失、置換、または付加、あるいはドナーCDRの変化を行ってドナー抗体の抗原結合特異性を保持するようにしてもよい。
【0067】
IL−5 mAbの可変重鎖および/または軽鎖の一方(または両方)(所望により説明したように修飾されていてもよい)あるいは1つまたはそれ以上の下で定義する重鎖または軽鎖CDRs(図7も参照)を用いるようにかかる組み換え法による抗体を設計する。組み換え法による本発明抗体は中和抗体、すなわち、望ましくはIL−5受容体への結合をブロックし、さらにまたIL−5依存性細胞の増殖をブロックまたは防止するものである。
【0068】
かかる組み換え法による抗体は、選択されたヒト・免疫グロブリンもしくはサブタイプの枠組み領域を含むヒト化抗体、またはIL−5抗体の機能的フラグメントに融合したヒト・重鎖および軽鎖不変領域を含むキメラ抗体を包含しうる。適当なヒト(または他の動物)のアクセプター抗体は、ドナー抗体の核酸およびアミノ酸配列に対する相同性により、慣用的データベース、例えば、KABATRデータベース、Los Alamosデータベース、およびSwiss Proteinデータベースから選択されるものであってもよい。ドナー抗体の枠組み領域に対する相同性(アミノ酸による)により特徴づけられるヒト・抗体は、ドナーCDRsの挿入用の重鎖不変領域および/または重鎖可変領域枠組み領域の提供に適するかもしれない。軽鎖不変または可変枠組み領域を提供しうる適当なアクセプター抗体を同様の方法で選択してもよい。アクセプター抗体の重鎖および軽鎖は同じアクセプター抗体に由来する必要がないことに注意すべきである。
【0069】
望ましくは、異種枠組みおよび不変領域をヒト・免疫グロブリンのクラスおよびイソタイプ、例えば、IgG(サブタイプ1から4)、IgM、IgAおよびIgEから選択する。しかしながら、受容体抗体はヒト・免疫グロブリン蛋白配列のみを含んでなることを必要としない。例えば、ヒト・免疫グロブリン鎖の一部をコードしているDNA配列が、ポリペプチドエフェクターまたはレポーター分子のごとき非免疫グロブリンアミノ酸配列をコードしているDNA配列に融合している遺伝子を構築してもよい。
【0070】
特に望ましいヒト化抗体の一例は、選択されたヒト・抗体配列の枠組み領域上に挿入された2B6のCDRsを含む。中和ヒト化抗体とするには、IL−5抗体の重鎖および/または軽鎖可変領域由来の1個、2個または好ましくは3個のCDRsを、選択されたヒト・抗体配列の枠組み領域中に挿入し、ヒト・抗体の無傷のCDRsと入れ替える。
【0071】
好ましくは、ヒト化抗体において、ヒト・重鎖および軽鎖双方中の可変ドメインは組み換え法により1個またはそれ以上のCDRが置換されたものである。6個すべてのCDRsまたは6個未満のCDRsの種々の組み合わせを用いることが可能である。好ましくは、6個全部のCDRを置換する。ヒト・アクセプター抗体由来の未修飾軽鎖を軽鎖として用いて、ヒト・重鎖中でのみCDRsを置換することが可能である。さらに別法として、慣用的な抗体のデータベースを用いて、適合する軽鎖を別のヒト・抗体から選択してもよい。組み換え法による抗体の残りの部分はいずれかの適当なアクセプターヒト・免疫グロブリン由来であってもよい。
【0072】
よって、好ましくは、組み換え法によるヒト化抗体は、天然のヒト・抗体またはそのフラグメントの構造を有し、効果的な治療的使用、例えば、ヒトにおけるIL−5により媒介される炎症性疾患の治療、または診断用途に必要な特性の組み合わせを有する。
【0073】
もう1つの例として、組み換え法による抗体は、2E3の可変軽鎖領域の3つのCDRs[配列番号:10、11および13]および2B6の可変重鎖領域の3つのCDRs[配列番号:7、8および9]を含む。得られるヒト化抗体はmAb 2B6と同じ抗原結合特異性および高アフィニティーにより特徴づけられる。
【0074】
必ずしもドナー抗体の特異性および高アフィニティーに影響することなく、可変ドメインのアミノ酸を変化させることにより組み換え法による抗体をさらに修飾(すなわち、アナログを得る)してもよいことが当業者により理解されるであろう。重鎖および軽鎖のアミノ酸を、可変ドメイン枠組みまたはCDRsまたはそれらの両方のいずれかに存在する他のアミノ酸により置換してもよいと予想される。
【0075】
さらに、不変領域を変化させて本発明分子の選択特性を増加または減少させてもよい。例えば、ダイマー化、Fc受容体への結合、または補体結合および活性化能が挙げられる(例えば、Angal et al.,Mol.Immunol,30:105-108(1993)、Xu et al.,J.Biol.Chem,269,3469-3474(1994)、Winter et al.,EP307,434-B参照)。
【0076】
キメラ抗体である変化した抗体は、非ヒト・ドナー抗体の重鎖および軽鎖可変領域全体を提供し、両鎖についてヒト・免疫グロブリン不変領域に結合した枠組み領域を包含するという点で上記ヒト化抗体とは異なる。本発明ヒト化抗体に関連したさらなる非ヒト配列を保有しているキメラ抗体はヒトにおいて有意な免疫応答を誘導しうると予想される。
【0077】
かかる抗体は、上述のIL−5により媒介される疾病の予防および治療に有用である。
【0078】
VI.変化した抗体および組み換え法による抗体の製造
好ましくは、2B6または他の適当なドナーmAbs(例えば、2E3、2F2、4A6等)の可変軽鎖および/または重鎖配列およびCDRs、およびそれらをコードしている核酸配列を、変化した抗体、好ましくはヒト化抗体の構築に用いる。同一または類似の方法を用いて本発明の他の具体例を得てもよい。 選択されたドナーmAb、例えばマウス抗体2B6を産生するハイブリドーマを慣用的にクローンし、当業者に知られた方法、例えば、Sambrook et al., molecular Cloning (A Laboratory Manual),2nd Edition,Cold Spring Harbor Laboratory(1989)に記載の方法によりその重鎖および軽鎖可変領域のDNAを得る。ドナーmAb結合特異性を保持するのに必要な少なくともCDRをコードしている領域およびアクセプターmAb軽鎖および/または重鎖可変ドメイン枠組み領域含んでいる2B6の可変重鎖および軽鎖、ならびにヒト・免疫グロブリン由来の抗体鎖の残りの部分を、ポリヌクレオチドプライマーおよび逆転写酵素を用いて得る。既知データベースを用い、他の抗体と比較することによりCDRをコードしている領域を同定する。
【0079】
次いで、マウス/ヒト・キメラ抗体を調製し、結合能についてアッセイすることができる。かかるキメラ抗体は、ヒト・Ig不変領域に結合した非ヒト・ドナー抗体のVHおよびVL領域全体を含んでいる。
【0080】
コンピューターデータベース、例えばKABATRを用いてヒト抗体由来の重鎖可変領域の同種枠組み領域を同定し、2B6に対して相同性を有するヒト・抗体をアクセプター抗体として選択した。ヒト・抗体枠組み中の2B6のCDRをコードしている領域を含む合成重鎖可変領域の配列を、制限部位を含むように枠組み中のヌクレオチド置換を所望により行って設計した。次いで、この設計された配列を、長い合成オリゴマーを用いて合成した。別法として、ポリメラーゼ連鎖反応(PCR)により増幅され、エラーを修正されたオリゴヌクレオチドを重複させることにより、設計配列を合成することもできる。
適当な軽鎖可変枠組み領域を同様の方法で設計した。
【0081】
ヒト化抗体はキメラ抗体由来であってもよく、あるいは好ましくは、重鎖および軽鎖由来のドナーmAbのCDRをコードしている領域を選択された重鎖および軽鎖枠組み中に適当に挿入することにより合成的に得てもよい。別法として、本発明ヒト化抗体を標準的な突然変異法を用いて調製してもよい。かくして得られたヒト化抗体はヒト・枠組み領域およびドナーmAbのCDRをコードしている領域を含む。枠組み残基について引き続き操作を行ってよい。得られるヒト化抗体は組み換え宿主細胞、例えば、COS、CHOまたはミエローマ細胞において発現されうる。他の適当なIL−5特異的中和高アフィニティー非ヒト・抗体に対してこの方法を用いて他のヒト化抗体を調製してもよい。
【0082】
宿主細胞における複製および発現および/または宿主細胞からの分泌を制御しうる慣用的な調節的制御配列に作動可能に結合した変化した抗体の配列にもとのプラスミドのコーディング配列を置き換えることにより、慣用的な発現ベクターまたは組み換えプラスミドを製造する。調節配列はプロモーター配列、例えば、CMVプロモーター、およびシグナル配列を包含し、それらは他の既知抗体由来であってもよい。同様にして、相補的な抗体の軽鎖または重鎖をコードしているDNA配列を有する第2の発現ベクターを作成することもできる。好ましくは、この第2の発現ベクターは、コーディング配列および選択可能マーカーを除き、各ポリペプチド鎖が機能的に発現されることが可能なかぎり、第1のものと同じである。
【0083】
第1および第2の両方のベクターを用いる慣用的方法によって選択された宿主細胞を同時トランスフェクションして(あるいは単一ベクターによりトランスフェクションして)、組み換えまたは合成の軽鎖および重鎖両方を含んでなる本発明トランスフェクション宿主細胞を作成する。次いで、トランスフェクション細胞を慣用的方法により培養して本発明の組み換え法による抗体を得る。組み換え重鎖および/または軽鎖との結合を含むヒト化抗体を、ELISAまたはRIAのごとき適当なアッセイにより培養物からスクリーニングする。同様の慣用的方法を用いて本発明の他の変化した抗体および分子を構築してもよい。
【0084】
本発明方法および組成物の構築に用いるクローニングおよびサブクローニング工程に適したベクターが当業者により選択される。例えば、慣用的なpUCシリーズのクローニングベクターを用いてもよい。使用される1のベクターはpUC19であり、Amersham(英国Buckinghamshire)またはPharmacia(スウェーデンUppsala)のごとき業者から市販されている。さらに、容易に複製しうるいずれかのベクターは多くのクローニング部位および選択可能遺伝子(例えば、抗生物質耐性)を有しており、クローニングのための容易に取り扱いすることができる。よって、クローニングベクターの選択は本発明における制限因子ではない。
【0085】
同様に、本発明による組み換え法により得られた抗体の発現に用いるベクターは、いずれかの慣用的ベクターから当業者により選択されうる。またベクターは、選択された細胞において異種DNA配列の複製および発現を指令する選択された調節配列(CMVプロモーターのごとき)を含んでいる。これらのベクターは、組み換え法による抗体または変化した免疫グロブリンのコーディング領域をコードしている上記DNA配列を含んでいる。さらに、ベクターは、取り扱いが容易なように所望制限部位を挿入することにより修飾された選択免疫グロブリン配列を含んでいてもよい。
【0086】
異種DNA配列、例えば、哺乳動物ジヒドロ葉酸レダクターゼ(DHFR)の発現の増幅に適した遺伝子により発現ベクターを特徴づけてもよい。他の好ましいベクター配列は、ウシ・成長ホルモン(BGH)およびベータグロブリンプロモーター配列(betaglopro)のごときポリAシグナル配列を包含する。当業者によく知られた方法により本発明において有用な発現ベクターを合成してもよい。
【0087】
選択宿主における組み換えDNAからの生成物の発現および/または分泌の指令に有用なかかるベクターの成分、例えば、レプリコン、選択遺伝子、エンハンサー、プロモーター、シグナル配列等を、市販または天然ソースから得てもよく、あるいは既知方法により合成してもよい。
【0088】
また本発明は、組み換え法による抗体またはその変化した免疫グロブリン分子のコーディング配列を含む組み換えプラスミドでトランスフェクションした細胞系に関する。これらのクローニングベクターのクローニングおよび他の操作に有用な宿主細胞も慣用的である。しかしながら、最も望ましくは、種々のE.coli株の細胞をクローニングベクターの複製および本発明の変化した抗体の構築における他のステップに使用する。
【0089】
本発明の組み換え法による抗体または変化した抗体の発現に適する宿主または細胞系は、好ましくは、CHO、COS、線維芽細胞(例えば、3T3)、および骨髄細胞のごとき哺乳動物細胞、より好ましくは、骨髄細胞またはCHOである。ヒト・細胞を用いてもよく、かくして、分子をヒトのグリコシレーションパターンで修飾することが可能となる。別法として、他の真核細胞系を用いてもよい。形質転換、培養、増幅、スクリーニングならびに生成物の生産および精製に適した哺乳動物宿主細胞および方法の選択は当該分野において知られている。例えば、上で引用したSambrookらの文献参照。
【0090】
細菌細胞は本発明組み換えFabの発現に適した宿主細胞であることがわかるだろう(例えば、Pluckthun,A.,Immunol.Rev.,130:151-188(1992)参照)。しかしながら、細菌細胞において発現された蛋白の、変性または正しく折り畳まれていない形態あるいはグリコシレーションされていない形態となる傾向のため、細菌細胞において生産された組み換えFabを抗原結合能の保持についてスクリーニングしなければならない。細菌細胞により発現された分子が正しく折り畳まれた形態で生産された場合には、当該細菌細胞は望ましい宿主であろう。例えば、発現に使用される種々のE.coli株は、バイオテクノロジーの分野において宿主としてよく知られている。種々のB.subtilis、Streptomyces、他の枯草菌等の株も本発明方法に使用できる。
【0091】
所望ならば、当業者に知られた酵母細胞株ならびに昆虫細胞、例えば、DrosophilaおよびLepidoptera、およびウイルス発現系も宿主細胞として用いられる。例えば、Miller et al.,Genetic Engineering,8:277-298,Plenum Press(1986)およびその中の引用文献参照。
【0092】
本発明ベクターを構築することのできる一般的方法、本発明宿主細胞を製造するのに必要なトランスフェクション法、およびかかる宿主細胞から本発明の変化した抗体を製造するのに必要な培養法はすべて慣用的方法である。同様に、生産されたならば、硫酸アンモニウム沈殿、アフィニティーカラム、カラムクロマトグラフィー、ゲル電気泳動等を包含する当該分野の標準的方法により本発明の変化した抗体を細胞培養物から精製することができる。かかる方法は当業者の範囲内であり、本発明を制限するものでない。
【0093】
ヒト化抗体のさらなる発現方法は、米国特許第4,873,316号記載のごとく、トランスジェニック動物における発現を用いるものであってもよい。これは、動物のカゼインプロモーター(トランスジェニック的に哺乳動物に導入された場合、メスの哺乳動物はその乳中の所望組み換え蛋白を生産する)を用いる発現系に関連している。所望方法により発現されたならば、適当なアッセイを用いて、組み換え法による抗体をインビトロ活性について試験する。現在慣用的となっているELISAアッセイフォーマットを用いて、組み換え法により抗体のIL−5への結合を定量的かつ定性的に評価する。さらに、他のインビトロアッセイを用いて、通常のクリアランス機構存在下の体内での組み換え抗体の維持を評価するために引き続き行われるヒトの臨床研究を行う前に中和効率を確認しておいてもよい。
【0094】
2B6から調製されたヒト化抗体について説明された方法に従って、当業者は、本明細書記載の他のドナーIL−5抗体、可変領域配列およびCDRペプチドからヒト化抗体を構築することができる。レシピエントにより「自己」であると潜在的に認識される可変領域枠組みを用いて組み換え法による抗体を製造することができる。可変領域枠組みに対する小さな修飾を行って、レシピエントに対する免疫原性を有意に上昇させずに抗原結合を促進することができる。かかる組み換え法による抗体は、ヒトのIL−5により媒介される症状を効果的に治療する可能性がある。
【0095】
VII. 治療的/予防的使用
また本発明は、有効量の本明細書記載の1種またはそれ以上の組み換え法による抗体または変化した抗体を包含する抗体、あるいはそれらのフラグメントを投与することを特徴とする、喘息のごとき好酸球増加症関連症候群にかかっているヒトの治療方法に関する。
【0096】
ヒト・IL−5への結合、次いで、好酸球刺激のブロックにより、本発明分子の使用による治療的応答を得る。よって、本発明分子は、治療用途に適した調合品および処方に含まれた場合、アレルギー性鼻炎、喘息、慢性好酸球性肺炎、アレルギー性気管支肺アスペルギルス症、セリアック病、好酸球性胃腸炎、Churg-Strauss症候群(結節性動脈周囲炎およびアトピー)、好酸球性筋肉痛症候群、過好酸球増加症候群、血管皮膚炎を包含する水腫性反応、ぜん虫感染(ここで、好酸球は防御的役割をしている可能性がある)、オンコセルカ皮膚炎およびアトピー性皮膚炎のごときアレルギー性および/またはアトピー性応答、あるいは好酸球増加症に関連した応答をしているヒトに対して非常に望ましい。
【0097】
他の抗体、詳細には本発明組み換え法による抗体が指向する症状の原因となりうる他のマーカー(エピトープ)と反応するヒト・mAbと組み合わせて本発明の変化した抗体、抗体およびそれらのフラグメントを用いてもよい。
【0098】
本発明治療剤は、約2日ないし約3週間、あるいは必要な期間のアレルギー症状の治療に望ましいと考えられる。例えば、季節性鼻炎等を治療する場合には、より長期の治療が望ましい。このことは、IL−5により媒介される疾病に対する現用の先行技術である輸液プロトコールよりもかなり有利であることを示す。治療のための用量および期間は、ヒトの循環系における本発明分子の相対的持続時間に関係し、治療すべき症状および患者の一般的健康状態に応じて当業者により調整されうる。
【0099】
本発明治療剤の投与モードは、宿主に対して剤を送達するいずれの適切な経路であってもよい。変化した抗体、抗体、組み換え法による抗体、およびそれらのフラグメント、ならびに本発明医薬成分は、詳細には、非経口投与、すなわち、皮下、筋肉内、静脈または鼻腔内投与において有用である。
【0100】
本発明治療剤を、医薬上許容される担体中の有効成分としての有効量の本発明の組み換え法による(例えば、ヒト化)抗体を含有する医薬組成物として調製してもよい。本発明の予防剤において、組み換え法による抗体を含有する、好ましくは生理学的pHに調節された即座に注射可能な形態の水性懸濁液または溶液が好ましい。通常には、非経口投与用組成物は本発明の組み換え法による抗体の溶液または医薬上許容される担体(好ましくは水性担体)に溶解されたそのカクテルを含んでなる。種々の水性担体を用いることができ、例えば、0.4%食塩水、0.3%グリシン等を用いることができる。これらの溶液は滅菌されており、一般的には微粒子不含である。よく知られた滅菌法(例えば、濾過)によりこれらの溶液を滅菌することができる。組成物は、pH調節および緩衝化剤等のごとき、所望により医薬上許容される添加物を含んでいてもよい。かかる医薬処方中の本発明抗体の濃度を広範囲に、すなわち、重量で約0.5%未満から、通常には約1%またはそれ未満ないし15%または20%まで変化させることができ、主として液体の体積、粘度等に基づいて選択され、選択された個々の投与モードに応じて選択されるであろう。
【0101】
よって、筋肉内注射用の本発明医薬組成物を、1mlの滅菌緩衝水および約1ngないし約100mg、例えば約50ngないし約30mg、またはより好ましくは約5mgなし約25mgの本発明の組み換え法による抗体を含有するように調製することができる。同様に、静脈輸液用の本発明医薬組成物を、約250mlの滅菌リンゲル溶液、および約1ないし約30mg、好ましくは5mgないし約25mgの本発明の組み換え法による抗体を含有するように調製することができる。非経口的投与可能組成物の調製のための実際的な方法は当業者によく知られているかまたは明らかであり、例えばRemington's Pharmaceutical Science,15th ed.,Mark Publishing Company,Easton,Pennsylvaniaにおいてより詳細に記載されている。
【0102】
本発明治療剤は、医薬組成物中に入れる場合、1回分の剤型として提供するのが好ましい。適当な治療上有効量は当業者により容易に決定されうる。ヒトまたは他の動物における炎症性疾患を有効に治療するためには、1回分の用量は、体重70kgにつき約0.1mgはいし約20mgの本発明抗体を非経口的に、好ましくは静脈または筋肉内投与すべきである。必要ならば、炎症応答中に医師がかかる用量を適当時間間隔をおいて調製してもよい。
【0103】
本発明の変化した抗体および組み換え法による抗体を、IL−5により媒介される疾病の決定またはかかる疾病の治療進行の追跡のごとき診断に用いてもよい。診断には、これらの変化した抗体を、慣用的には、血清、血漿または他の適当な組織中のIL−5レベルの測定あるいは培養中のヒト細胞による放出の測定のためにELISAまたは他の慣用的アッセイフォーマット用に標識することができる。変化した抗体が用いられるアッセイの性質は慣用的なものであり、本開示を限定するものでない。
【0104】
よって、本発明の1の具体例は、患者における過剰な好酸球産生に関連したアレルギーおよび他の症状の診断を援助する方法であって、患者から得た試料(血漿または組織)中のヒト・IL−5の量を決定し、次いで、決定した量を正常集団におけるヒト・IL−5平均量と比較することを特徴とし、そのことにより患者試料中の有意に上昇したIL−5量が過剰な好酸球産生に関連したアレルギーおよび他の症状を示すものとなる方法に関する。
【0105】
本明細書記載の抗体、変化した抗体またはそれらのフラグメントを保存のための凍結乾燥し、使用のために適当な担体中で復元することができる。この方法は、慣用的免疫グロブリンに関して効果的であることが示されており、当該分野で知られている凍結乾燥および復元方法を用いることができる。
【0106】
下記実施例は、典型的な組み換え法による抗体の構築および適当なベクターおよび宿主細胞中でのその発現を包含する本発明の種々の態様を説明するが、本発明の範囲を限定するものと解してはならない。すべてのアミノ酸は慣用的な3文字または1文字法により表される。特記しないかぎり、すべての必要な制限酵素、プラスミド、ならびに他の試薬および材料は市販のものである。すべての一般的クローニング、連結および他のDNA組み換え法は、T.Maniatisら(すでに引用)、または同じ出版社からのその第2版(1989)(Sambrookら編)(「Sambrookら」という)において行われたものである。
【実施例1】
【0107】
実施例1 hIL−5(ヒト・IL−5)に対するMabs(モノクローナル抗体)の製造
ヒト・IL−5をDrosophila Schneider 2(S2)細胞において発現させ、均一に精製した。Spodoptera frugiperda 21(Sf21)細胞を用いてマウス・IL−5をバキュロウイルス中で発現させ、均一に精製した。モノクローナル抗体TRFK−5(中和ラット・抗−マウスIL−5抗体)をGenzyme Corp.(Cambridge,MA)から得た。
【0108】
A.免疫手順
組み換えヒト・IL−5(IL−5)を、7匹のCAF1マウス(メス)(Charls River,Wilmington,MA)のパネルに対する免疫原として用いた。4カ月の期間にわたり、1:1の割合のTiterMAX(登録商標)(CytoRx Corp.,Norcross,GA)を用いて乳化したリン酸緩衝化食塩水(PBS)中のIL−5を動物に3回皮下注射した。最初の抗原量は50μgであり、追加免疫は25μgおよび10μgであった。追加免疫後、血清試料を集め、受容体結合阻害アッセイおよびB13増殖アッセイ(またはIL−5中和アッセイ(実施例2C))により、IL−5への結合および中和活性についてアッセイした。すべてのマウスは、IL−5に結合する血清試料を産生した。脾臓ドナーとして選択された動物を安楽死させる3日前に10μgの組み換えヒト・IL−5を静脈から追加免疫した。
【0109】
B.ハイブリドーマ取得
Kohler et al.,Nature,256:495(1975)により初めて報告された融合法を変更して用いて、細胞単層(Kennet et al.,Eds.,"Hybridomas:A new dimension in biological analysis",pp.368-377,Plenum Press,New York)を用いる方法を行った。2匹のドナーマウスからの脾臓細胞をプールし、1千万個のSP2/0/Ag14骨髄腫細胞に対して5千万個の脾臓細胞の割合で融合を行った。融合が確認された細胞の上清を、IL−5に対する結合に関してELISAによりアッセイした。IL−5に対する抗体を産生する細胞を含むウェルを増殖させ、上清をIL−5受容体結合阻害アッセイおよびB13(中和)増殖アッセイにおいてスクリーニングした(下記)。
【0110】
IL−5と反応するmAbsを分泌する16個のハイブリドーマを単離した。ハイブリドーマ上清をヨウ素化IL−5と混合し、IL−5受容体(IL−5R)のα−鎖を発現するDrosophila細胞から調製した抽出物に添加し、受容体結合の阻害に関してアッセイした。11個のハイブリドーマ上清は、IL−5受容体α−鎖へのヨウ素化IL−5の結合を60%よりも多く阻害した。3個のmAbs(2B6、2E3および2F2)も、ヒト・IL−5に応答してマウス・B13細胞の増殖を70%よいも多く阻害したが、マウス・IL−5には応答しなかった。5個のハイブリドーマ(そのうち4個(1C6、2B6、2E3および2F2)は結合および/または増殖をブロックし、1個(24G9)は中和的でなかった)を軟寒天で繰り返しサブクローンして安定なクローン細胞系を得た。クローン細胞系からの上清を交差反応性についてELISAによりスクリーニングしたが、ヒト・IL−1α、IL−1β、IL−4、IL−8、M−CSFまたはTGFαには結合しなかった。mABsを精製し、光学的バイオセンサー(BIAcore)分析により結合アフィニティーを評価したところ、10ないし100pMの範囲であった。マウス・イソタイプ分類試薬(PhaMingen,San Diego,CA)を用いて細胞系からの上清をイソタイプに分類した。MAbsの中和活性に関するアフィニティーおよびIC50のまとめを表1(実施例2)に示す。
【0111】
同様の方法により、対応する免疫プロトコールおよびラット・骨髄腫をマウスについて上記した融合に用いてラット・ハイブリドーマを免疫ラットから得た。2つのラット・ハイブリドーマ4A6および5D3は、IL−5に結合するmAbsを産生することが同定された。2B6、2E3および2F2と同様に、mAbs4A6および5D3は、下記B13アッセイにおいて中和的であることがわかった。
【0112】
C.ハイブリドーマの寄託
モノクローナル抗体2B6を産生するハイブリドーマ細胞系SK119−2B6.206.75(1)は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11783として寄託され、特許寄託物として受託された。
【0113】
モノクローナル抗体2E3を産生するハイブリドーマ細胞系SK119−2E3.39.40.2は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11782として寄託され、特許寄託物として受託された。
【0114】
モノクローナル抗体2F2を産生するハイブリドーマ細胞系SK119−2F2.37.80.12は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11781として寄託され、特許寄託物として受託された。
【0115】
モノクローナル抗体24G9を産生するハイブリドーマ細胞系SK119−24G9.8.20.5は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11780として寄託され、特許寄託物として受託された。
【0116】
モノクローナル抗体4A6を産生するハイブリドーマ細胞系4A6(1)G1F7は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11943として寄託され、特許寄託物として受託された。
【0117】
モノクローナル抗体5D3を産生するハイブリドーマ細胞系5D3(1)F5D6は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11942として寄託され、特許寄託物として受託された。
【実施例2】
【0118】
実施例2 アッセイ
A. ELISA
MaxiSorb(登録商標)イムノプレート(Nunc,Naperville,IL)の個々のウェルを、0.05M炭酸バッファーpH9.6中0.2μgのIL−5で被覆した。4℃で一晩インキュベーション後、0.025% TweenR20含有PBSでプレートをすすぎ、0.025% TweenR20含有PBS中1% BSAにて室温で2時間ブロックした。未希釈ハイブリッド上清をIL−5被覆ウェルに添加し、室温で2時間インキュベーションした。プレートをすすいだ後、ヤギ・抗−マウスIgGおよびIgM標識ペルオキシダーゼ(Boehrionger Mannheim,Indianapolis,IN)を、1% BSAおよび0.025%TweenR20含有PBS中1/7500希釈して添加した。15分後、VMax(登録商標)マイクロプレートリーダー(Molecular Devices, Menlo Park,CA)により450nmにおいてプレートを読んだ。
【0119】
B.受容体結合阻害アッセイ
ヒト・IL−5受容体(IL−5R)のα−鎖を発現するDrosophila S2細胞の膜抽出物を用いて受容体へのIL−5の結合に対する抗体の効果を測定した。膜を調製するために、109個の細胞を1000xg、4℃、10分間ペレット化した。細胞ペレットをドライアイス/エタノール浴で15分凍結した。ペレットを融解し、4℃の10mlのPBSに再懸濁し、1000xgで10分間ペレット化した。細胞ペレットをPBSで2回洗浄し、13.5mlの低張バッファー(10mM Tris pH7.5、3mM MgCl2、1mM ジチオスレイトール、1mM フェニルメチルスルホニルフルオリド、1μM ロイペプチン、1μM ペプスタチンA)に再懸濁し、氷上で5分間インキュベーションした。細胞懸濁液を15mlのDounceホモジナイザーでホモジナイズし、2.5M蔗糖溶液を用いて蔗糖最終濃度0.25Mとした。1000xgで15分の遠心分離により細胞残渣を除去した。細胞膜を100000xg、4℃で90分間ペレット化し、10mM Tris pH7.5、3mM MgCl2、250mM 蔗糖中に再懸濁し、−70℃で保存した。
【0120】
受容体を含むDrosophila膜を用いるアッセイを、25mM HEPESバッファー pH7.2および0.1% BSA(結合バッファー)を含有するDrosophila組織培養培地M3(Lindquist et al.,Drosophila Inf.Serv.,58:163(1982))を用いて、MultiscreenGV(登録商標)プレート(Millipore Corp.,Bedford,MA)中で行った。0.1mlの結合バッファーを用いてウェルをプレブロック(preblock)した。50μlの試験試料(3系とした)をウェルに添加し、次いで、25μlのヨウ素化(125I)IL−5を添加した。20分室温でインキュベーションした後、ヒト・IL−5Rのα−鎖を発現しているDrosophila S2細胞の膜抽出物25μlをウェルに添加した。さらに1時間インキュベーションした後、減圧濾過により膜を集め、結合バッファーで3回洗浄した。フィルターを乾燥し、カウントした。
【0121】
C. IL−5中和アッセイ
親切にも、R.Palacios(Basel Institute of Immunology,Switzerland)からマウス・IL−5/IL−3依存性細胞系LyH7.B13(B13)を得た。L−グルタミン、非必須アミノ酸、ピルビン酸ナトリウム、ペニシリン−ストレプトマイシン(すべてGibcoBRL)ならびに2−メルカプトエタノール(5x10−5M,Sigma)、10% ウシ胎児血清(Globepharm,Surrey,UK)および1〜10ユニットのマウス・IL−5を補足したRPMI 1640培地(GibcoBRL, Renfrewshire,UK)中で1週間に2回細胞をサブクローンした。アッセイのために、96ウェル丸底プレート中、適当に希釈した試験試料の存在下、3系で48時間培養し、次いで、0.5μCiの3H−チミジン(Amersham,Bucks,UK)を添加して最後の4時間のインキュベーションを行った。それらを1205 Beteplate(LKB Wallac,Beds,UK)におけるシンチレーションカウンティングのために処理した。
【0122】
D.光学的バイオセンサー
固定化hIL−5と抗体に関する速度論的平衡結合特性を、BIAcore光学的バイオセンサー(Pharmacia Biosensor,Uppsala,Sweden)を用いて行った。以前に記載されているようにして(Karlsson et al.,J.Immunol.Meth.,145:229-240(1991)(その全体を参照により本明細書に記載されているものと見なす))速度論的データを評価した。
【0123】
3つの中和mAbs、すなわち、2B6、2E3および2F2は、膜受容体への125I−IL−5の結合に対する阻害およびB細胞増殖の中和について類似した能力を有しおり、またIL−5に対して非常に類似したアフィニティーを有していた(表1参照)。これら3つのmAbs、すなわち2つのIgG1および1つのIgG2aからのVHおよびVLのヌクレオチド配列をそれぞれ決定した。得られた配列は非常に類似しており、ほんの少数の残基のみが異なっていた。
【0124】
【表1】

【実施例3】
【0125】
実施例3 組み合わせライブラリーからのIL−5 Fabsの単離および特徴づけ
A. PCRおよび組み合わせライブラリーの構築
Huse et al.Science,246:1275(1989)およびKang,S.A.Methods:Comparison Methods Enzymol.,2:111(1991)(それら全体を参照により本明細書に記載されているものと見なす)により記載されたようにして、3匹のマウスの脾臓から精製したRNAを、cDNAキット(Boehringer Mannheim,Indianapolis,IN)により逆転写した(キット付属のプライマー(dT)15または3’Fd(IgG1、IgG2aおよびIgG3)およびカッパ軽鎖プライマーを用いる)。記載されたプライマーおよび温度サイクリング条件(上記Huseら)を用いてPCRにより免疫グロブリンcDNAを増幅した。AmpliWaxTMPCR Gem 100(Perkin Elmer Cetus,Norwalk,CT)ビーズおよび製造者のプロトコールを用いるホット・スタート(Hot Start)法をすべての反応に使用した。PCR生成物をゲル精製し、消化し、pMKFabGene3ベクター(Ames et al.,J.Immunol.,512:4572(1994))中に連結した。Fd cDNAを用いる連結後のライブラリー力価は5.1X107CFUであり、カッパcDNAを用いるライゲーション後の力価は1.5X106CFUであった。ファージミドライブラリーを用いて形質転換されたXL1-Blue細胞(Stratagene,La Jolla,CA)を、ヘルパーファージVCSM13(Stratagene)に感染させ、Barbas and Lerner,Methosa:Comparison Methods Enzymol.,2:119(1991)により記載されたようにしてファージを調製した。
【0126】
B.バイオパンニング(Biopanning)
4つのマイクロタイターウェル(Immulon II Removawell Strips,Dynatech Laboratories Inc.,Chantilly,VA)を、0.1M重炭酸,pH8.6中IL−5(1μg/ウェル)で4℃において被覆した。ウェルを水洗し、3% BSA含有PBSを用いて37℃で1時間ブロックした。ブロッキング溶液を除去し、ライブラリーをマイクロタイターウェルに添加し(50μl/ウェル)、37℃で2時間インキュベーションした。ウェルをTBS/TweenR(50mM Tris−HCl,pH7.5、150mM NaCl、0.5% TweenR20)で10回洗浄し、水で1回洗浄し、次いで、0.1M HCl(1mg/ml BSA含有グリシンでpH2.2に調節)で付着ファージを溶離した。
【0127】
C.コロニー釣菌
バイオパンニングの第3ラウンドおよび第4ラウンドから単離されたコロニーからのコロニー釣菌を記載されたようにして(Barbas and Lerner、上記)行った。0.5〜1.0μCiの125I−IL−5とともにフィルターを室温で1時間インキュベーションし、製造者の推奨手順に従ってBolton-Hunter試薬(NEN,Billerica,MA)を用いて1% BSA含有PBS中でヨウ素化し、PBS 0.25% Tweenで洗浄し、Kodak XARフィルムにさらした。IL−5反応性Fabsを発現しているコロニーをオートラジオグラフィーにより検出した。
【0128】
D.可溶性FABsの調製
ファージミドDNAをNheIおよびSpeIで消化して遺伝子IIIを除去し、自己連結させた。XL1−Blue細胞を形質転換し、単離されたコロニーを、1%グルコースおよび50μg/mlカルベニシリンを含有する5.0mlのスーパーブロス(SB)培地(30gトリプトン、20g酵母エキス、10gの3−[N−モルホリノ]プロパンスルホン酸(MOPS)、pHを7に調節)中で37℃で一晩増殖させた。この培養1mlからの細胞をBeckman GS-6R遠心分離機にて3500rpmで10分間ペレット化し、50μg/mlのカルベニシリンを含有する5mlのSBに植菌した。培養物を37℃で1時間振盪し、イソプロピル−b−D−チオガラクトピラノシド(IPTG;1mM)を添加し、培養物を28℃に移し、一晩おいた。30mM Tris pH8.0に懸濁した20%蔗糖中、4℃で20分間細胞を溶解し、次いで、微量遠心機で10分間遠心分離することによりペリプラスム抽出物から可溶性Fabを調製した。ウェルタンブロットにより、既知量のマウス・Fabを含有する試料と比較することによってFab濃度を評価した。別の細菌のペリプラスム抽出物は同様の濃度のFabを含んでおり、ウェスタンブロット分析による分析では1ないし20μg/mlの範囲であった。
【0129】
E. FABsの精製
重鎖のカルボキシ末端上にキレートするペプチドを加工作成して蛋白精製の助けとした。NheIおよびSpeIでの消化によるM13遺伝子IIIコーディング領域の除去後、6個のヒスチジン残基をコードしている1対の重複オリゴヌクレオチド:[配列番号:43]5’−CTAGCCACCACCACCACCACCACTAA−3’;[配列番号:44]3’−GGTGGTGGTGGTGGTGGTGATTGATC−5’をFab発現ベクター中にサブクローンした。上記のごとくFab発現の誘導を行った。28℃で一晩インキュベーションした後、20%蔗糖、30mM Tris pH8.0中4℃で30分インキュベーションすることにより細胞ペレットのペリプラスム溶解物を調製した。尿素およびBrij−35界面活性剤を清澄化した上清に添加して、最終濃度をそれぞれ、2Mおよび1%とした。室温で1時間撹拌後、処理され清澄化された上清を、5mlのニッケル−NTA金属キレーティングカラム(1.5x3cm)(バッファーA(100mM Na−リン酸、10mM Tris、0.3M NaCl、2M尿素,pH8.0)で平衡化されている)に、流速0.5ml/分で直接負荷した。4カラム体積分(20ml)の洗浄後、上と同じバッファー中pHを8から4にする6カラム体積分(30ml)の逆pHグラジエントで結合物質を溶離した。精製されたFabsは、pH5.5において対称形の鋭いピークとなってカラムから溶出した。精製Fabsは>90%の純度であり、DNA不含であった。
【0130】
F. FAB ELISA
0.1M重炭酸バッファー,pH8.6に懸濁された(1mg/ml;ウェル1個あたり50ml)蛋白でImmulon IIプレート(Dynatech)を4℃で一晩被覆した。0.05% Tween(登録商標)20含有PBSにて希釈および洗浄を行った。プレートを洗浄し、1%BSA含有PBSにて室温で1時間ブロッキングした。可溶性Fabを含有する細菌懸濁液の種々の希釈物、または精製Fabsをプレートに添加した。1時間インキュベーション後、プレートを洗浄し、ビオチン化ヤギ・抗−マウスカッパ(Southern Biotechnology Associates,Inc.,Birmingham,AL)を添加(1:2000希釈;50μl/ウェル)して1時間おいた。プレートを洗浄し、ストレプトアビジン標識セイヨウワサビペルオキシダーゼを添加し(1:2000希釈;50μl/ウェル)1時間おいた。プレートを洗浄し、ABTSペルオキシダーゼ基質を添加し(100μl/ウェル;Kirkegaard & Perry Laboratories,Gaithersberg,MD)、405nmにおける光学密度をUVmax(登録商標)(Molecular Devices)マイクロプレートリーダーで読んだ。
【0131】
G.組み合わせライブラリーからのFabsの単離および特徴付け
IL−5で被覆したマイクロタイターウェルに対する複数ラウンドのバイオパンニングにより、IL−5に対するFabsを有するファージをライブラリーから選択した。4ラウンドの選択後、125I−IL−5を用いるコロニー釣菌アッセイによりIL−5反応性Fabsが同定された。第3ラウンドから34個のコロニーが、第4ラウンドから4個のコロニーが同定され、それらは標識IL−5に結合した。Fab−遺伝子III融合蛋白を発現している培養上清を用いる直接結合ELISAによりIL−5への結合を確認した。これらのコロニーからDNAを単離し、M13遺伝子IIIのコーディング領域を除去後、可溶性Fabの発現を誘導した。ペリプラスムフラクションを調製し、IL−5への結合についてELISAによりアッセイした。Fabは、別の蛋白rC5aへの結合を示すことなくIL−5に特異的に結合した。
【0132】
未希釈ペリプラスム抽出物(1ないし20μg/mlのFabを含有)を、IL−5R結合阻害アッセイ(実施例2)においてアッセイした。L−5Rαへのヨウ素化IL−5の結合を35%よりも多く阻害するFabはなかった。
【0133】
H.中和mAbのFABへの変換
FdおよびmAb(2B6)のκcDNAを、上記条件を用いるPCRにより単離した。ゲル精製したフラグメントをpMKFabGene3ベクター中にサブクローンし、これを修飾して遺伝子IIIのcDNAの3’末端にヘキサ−His配列を含むようにして、プラスミドpMKFabGene3Hを得た。2B6重鎖および軽鎖を有する機能的なIL−5結合Fabクローンをコロニー釣菌アッセイにより同定した。NheI/SpeI消化による遺伝子IIIの除去および自己連結を行って、重鎖をイン−フレームでヘキサ−Hisに融合させ、上記のごとく精製を可能にした。このFabは用量依存的に受容体結合を阻害し、IC50は約7.5μg/mlであり、親mAbであるマウス・2B6のIC50と同様であった。
【0134】
I.チェイン−シャッフル(chain-shuffle)されたライブラリーの構築およびスクリーニング
中和mAb 2B6のFdをコードしているcDNAを、XhoI/SpeIフラグメントとしてpMKFabGene3H中にサブクローンして、軽鎖cDNAのかわりにSstI/XbaIフラグメントを含ませた。このファージミドをSstIおよびXbaIで消化し、IL−5免疫マウス由来(上記)のSstI/XbaI消化された軽鎖のPCR生成物と連結した。連結後のライブラリー力価は4x105CFUであった。バイオパンニングおよびコロニー釣菌アッセイを、組み合わせライブラリーに関して上で説明したように行った。
【0135】
中和mAb 2B6のFdをコードしているcDNAを、IL−5免疫マウスから回収された同じ軽鎖レパートリーと対合させることによりライブラリーを構築した。このチェインシャッフルされたライブラリーを、固定化IL−5に対する4ラウンドのバイオパンニングに供し、得られたコロニーをコロニー釣菌アッセイを用いてIL−5反応性に関してアッセイした。ヨウ素化IL−5に結合する陽性コロニーを、ELISAおよびIL−5Rα結合アッセイによりさらにアッセイした。チェインシャッフルされたライブラリーから回収されたFabのうち2個(2および15)がIL−5RαへのIL−5の結合をブロックし、B13アッセイにおいてIL−5依存的増殖を抑制した。これら2個のVkの配列は2B6のVk(2B6のVHに対するもとの軽鎖パートナー)の配列と類似であった。Fab2および15に関する軽鎖配列は、それぞれ配列番号:45および46である。Fab2については、CDR1〜3はそれぞれ配列番号:10、11および47である。Fab15については、CDR1〜3はそれぞれ配列番号:10、11および48である。
【0136】
下記実施例4および5に記載したすべての抗体のアミノ酸配列はKABATナンバーリングシステムを用い、このシステムにおいてはCDRおよび枠組み長が変化してもかまわない。すなわち、鍵アミノ酸は、それに先行するアミノ酸数にかかわらず常に同じ番号を与えられる。例えば、すべての軽鎖のCDR1に先行するシステインは、常にKABAT位置23であり、たとえCDR1が17個までのアミノ酸を含む場合であってもCDR1の後に続くトリプトファン残基は常にKABAT位置35である。
【実施例4】
【0137】
実施例4 ヒト化抗体
マウス・CDRsをヒト化抗体枠組み中に含むように1のヒト化抗体を設計した。下記操作を行うことにより、IL−5特異的マウス・抗体2B6のこのヒト化バージョンを調製した。
【0138】
A.遺伝子クローニング
Boehringer Mannheim(Indianapolis,IN)から得たキットを用いて2B6、2F2および2E3ハイブリドーマ細胞系(実施例1)のそれぞれからmRNAを単離し、次いで、cDNAキット(Boehringer Mannheim)付属のプライマー(dT)15を用いて逆転写してcDNAを作成した。マウス・免疫グロブリンに特異的なPCRプライマーを用いて、重鎖可変領域のアミノ酸9(KABATナンバーリングシステム)からヒンジ領域まで伸長しているドメインならびに軽鎖可変領域のアミノ酸9(KABATナンバーリングシステム)から不変領域の末端まで伸長しているドメインをコードしているDNAを増幅した。独立別個のPCR反応により各抗体鎖の数個のクローンを得た。
【0139】
使用したマウス・ガンマ1ヒンジ領域プライマーは[配列番号:22]:5’GTACATATGCAAGGCTTACAACCACAATC3’であった。
【0140】
使用したマウス・ガンマ2aヒンジ領域プライマーは[配列番号:23]:5’GGACAGGGCTTACTAGTGGGCCCTCTGGGCTC3’であった。
【0141】
使用したマウス・重鎖可変領域プライマーは[配列番号:24]:5’AGGT(CまたはG)(CまたはA)A(GまたはA)CT(GまたはT)TCTCGAGTC(TまたはA)GG3’であった。
【0142】
使用したマウス・カッパ鎖不変領域プライマーは[配列番号:25]:5’CTAACACTCATTCCTGTTGAAGCTCTTGACAATGGG3’であった。
マウス・軽鎖可変領域プライマーは[配列番号:26]:5’CCAGATGTGAGCTCGTGATGACCCAGACTCCA3’であった。
【0143】
PCRフラグメントをプラスミドpGEM7f+(Promega)中にクローンし、次いで、それをE.coli DH5a(Bethesda Reaearch Labs)中に形質転換した。
【0144】
B. DNA配列決定
上記Aで得た重鎖および軽鎖マウスcDNAクローンを配列決定した。これらのクローンの可変領域の配列決定結果を配列番号:1〜6(図1〜6)に示す。各クローンは、マウス・重鎖可変領域または軽鎖可変領域間で保存されていることが知られているアミノ酸を含んでいた。CDRアミノ酸配列を以下に記載する。 2B6重鎖に関するCDR領域は配列番号:7、8および9である。図7参照。これらの配列は配列番号:1によりコードされている。軽鎖に関するCDR領域は配列番号:10、11および12である。図7参照。これらの配列は配列番号:2によりコードされている。
【0145】
2F2重鎖に関するCDR領域は配列番号:7、8および9である。図7参照。これらの配列は配列番号:3によりコードされている。軽鎖に関するCDR領域は配列番号:10、11および13である。図7参照。これらの配列は配列番号:4によりコードされている。
【0146】
2E3重鎖に関するCDR領域は配列番号:7、8および14である。図7参照。これらの配列は配列番号:5によりコードされている。軽鎖に関するCDR領域は配列番号:10、11および13である。図7参照。これらの配列は配列番号:6によりコードされている。
【0147】
C.ヒト・枠組みの選択
2B6のクローニング後、KABATおよびSWISS−PROT(Nuc. Acids Res.,20:2019-2022(1992))蛋白配列データベースにおいて可変領域重鎖および軽鎖のアミノ酸配列(図1および2)(それぞれ配列番号:15および16)を既知マウス・免疫グロブリン配列と比較してN末端残基に割り当てた。次いで、2B6重鎖および軽鎖可変領域の推定アミノ酸配列をヒト・免疫グロブリン蛋白配列データベースと比較して、マウス配列に最もぴったりと合致する重鎖および軽鎖両方のヒト・枠組みを同定した。さらに、CDR表現に影響しうるアミノ酸による潜在的矛盾を評価するためにFabドメインの構造的モデルから得られた位置のデータベースを用いて重鎖および軽鎖を評価した。矛盾位置における対応マウス・アミノ酸の置換により、ヒト化可変領域枠組みの合成中に矛盾は解決された。
【0148】
ヒト骨髄腫免疫グロブリン(COR)から得られた抗体の重鎖枠組み領域を用いた(E.M.Press and N.M.Hogg,Biochem.J.,117:641-660(1970))。ヒト・重鎖枠組みアミノ酸配列は2B6枠組みに対して約66%の相同性があることが見いだされた。
【0149】
適当な軽鎖可変領域枠組みについては、Bence-Jones蛋白(LEN)(Schneider et al.,Hoppe-Seyler's Z.Physiol.Chem.,356:507-557(1975))の軽鎖可変枠組み配列を用いた。ヒト・軽鎖枠組み領域は、マウス・2B6軽鎖枠組み領域に対して、アミノ酸レベルで約82%の相同性があった。
【0150】
選択されたヒト・枠組みを戻し翻訳(back translate)してDNA配列を得た。
【0151】
D.ヒト化MAB遺伝子の構築
2B6重鎖CDRs[図7および配列番号:1〜2]およびヒト・抗体の枠組み配列を得て、合成重鎖可変領域[配列番号:18]を作成した。結合した場合にアミノ酸21〜106(KABAT番号付け)をコードすることとなる4つの合成オリゴヌクレオチド[配列番号:27および28][配列番号:29および30]を用いてこれを作成した。次いで、COR枠組み(上記)に基づく別のヒト化重鎖由来の配列を含むpUC18をベースにしたプラスミドのHpaI−KpnI制限部位中にオリゴヌクレオチドを連結した。このプラスミドはシグナル配列[配列番号:17]および残りの可変領域配列を提供する。変異原性プライマーを用いるPCRまたは存在する制限部位中への合成リンカーの付加により、マッピングされた配列中のエラーを修正した。
【0152】
シグナル配列およびヒト化重鎖可変領域をpUCをベースとしたプラスミドからEcoRI−ApaIフラグメントとして切り出し、IgG1ヒト・不変領域を含んでいる発現ベクターpCD中に連結した。合成重鎖可変領域ヌクレオチドおよびアミノ酸配列を図8[配列番号:18および19]に示す。ヒト・枠組み残基は配列番号:19のアミノ酸1〜30、36〜49、66〜97および109〜119である。CDRsのアミノ酸配列はマウス・2B6 CDRsと同じである。得られた発現ベクターpCDIL5HZHC1.0を図10に示す。
【0153】
2B6軽鎖CDRs[図7および配列番号:10、11および12]およびヒト・抗体の枠組み配列を得てから、合成軽鎖可変領域[配列番号:20]を作成した。それぞれSacI−KpnIおよびPstI−HindIII末端を有するヒト化VLのアミノ酸27〜58(KABAT番号付け)[配列番号:31および32]およびアミノ酸80〜109[配列番号:33および34]をコードしている4つの合成オリゴヌクレオチドを、LEN枠組みに対して高度な相同性を有する別のヒト・軽鎖枠組み(B17)(Marsh et al,Nuc.Acids Res.,13:6531-6544(1985))由来の配列を含むpUC18をベースにしたプラスミド中に挿入した。このプラスミドは残りの可変領域配列を提供する。マッピングされた配列中のエラーおよびLENとB17との間の1個のアミノ酸の相違を、変異原性プライマーを用いるPCTまたは存在する制限部位中への合成リンカーの付加により修正した。
【0154】
ヒト化軽鎖領域をpUCプラスミドからEcoRV−NarIフラグメントとして単離し、カッパヒト不変領域ならびにシグナル配列[配列番号:17]を含む発現ベクターpCN中に連結した。合成軽鎖可変領域ヌクレオチドおよびアミノ酸配列を図9[配列番号:20および21]に示す。ヒト・枠組み残基は配列番号:21のアミノ酸1〜23、41〜55、63〜94および104〜113である。CDRsのアミノ酸配列はマウス・2B6 CDRsと同じである。しかしながら、これらのCDRsに関するコーディング配列はマウス・2B6コーディング配列とは異なっており、制限酵素部位の作成が可能である。得られた発現ベクターの1つであるpCNIL5HZLC1.0を11に示す。これらの合成可変軽鎖および/または重鎖配列をヒト化抗体の構築に使用する。
【0155】
E.ヒト化MAbの発現
IgG1イソタイプ由来のヒト化重鎖には配列番号:19に示す合成重鎖可変領域を用いる。2B6重鎖CDRsを含んでいるこの合成VHを上記のごとく設計し、合成した。
【0156】
ヒト化軽鎖、ヒト・カッパ鎖には配列番号:21に示す合成軽鎖可変領域を用いる。2B6軽鎖CDRsを含んでいるこの合成VLを上記のごとく設計し、合成した。ヒト化可変領域をコードしているDNAフラグメントをpUC19をベースにした哺乳動物細胞発現プラスミド中に挿入してプラスミドpCDIL5HZHC1.0(重鎖)[配列番号:49、図10も参照]およびpCNIL5HLC1.0(軽鎖)[配列番号:50、図11も参照]。該発現プラスミドはシグナル配列を使用するものであり、CMVプロモーターおよび下記実施例5にて慣用的方法(すでに引用したManiatisらの文献)により製造されるキメラのヒト・重鎖またはヒト・軽鎖不変領域を含んでいる。プラスミドをCOS細胞中に同時トランスフェクションし、実施例5記載のELISAにより3日後および4日後に上清をヒト・抗体の存在についてアッセイした。
【0157】
上記実施例は典型的な組み換え法による抗体の調製を説明する。慣用的手段により得られた他の抗−IL−5抗体(例えば、2F2、2E3、4A6、5D3、24G9等)を用いて、他の組み換え抗体の取得に関して同様の手順を用いることができる。
【0158】
F.精製
CHOにより発現されたキメラおよびヒト化2B6の精製を、慣用的なプロテインA(またはG)アフィニティークロマトグラフィー、次いで、イオン交換および分子ふるいクロマトグラフィーにより行うことができる。同様のプロセスをうまく用いて他のmAbs(例えば、呼吸器合胞体ウイルス、インターロイキン−4およびマラリアサーカムスポロゾイトに対するもの)の純度95%以上の精製が行われている。
【0159】
G.さらなるヒト化mAbsおよび発現プラスミド
プラスミドpCDIL5HZHC1.0[配列番号:49]を得てから、枠組み位置73のスレオニンをアスパラギンに置換した発現プラスミドpCDIL5HZHC1.1を作成した。EcoRVおよびXhoI末端[配列番号:51および配列番号:52]を有する合成リンカーを、同様にして消化されたpCDIL5HZHC1.0中に連結することにより、これを作成した。同様にして、発現プラスミドpCDIL5HZH1.2は、枠組み位置37においてバリンのかわりにイソロイシンを有する。HpaIおよびXbaI末端[配列番号:53および配列番号:54]を有する合成リンカーを、同様にして消化されたpCDIL5HZHC1.0中に連結することにより、これを作成した。さらに、HpaIおよびXbaI末端[配列番号:53および配列番号:54]を有する合成リンカーを、同様にして消化されたpCDIL5HZHC1.1中に連結することにより、pCDIL5HZHC1.3を作成した。
【0160】
4つの合成オリゴヌクレオチド[配列番号:31、32、33および34]のDNA配列を含むすでに記載されているpUC18をベースにしたプラスミドを得て、枠組み位置15がロイシンからアラニンに変更されているヒト化軽鎖可変領域を作成した。このプラスミドをNheIおよびSacI制限エンドヌクレアーゼで消化し、合成リンカー[配列番号:55および56]を挿入した。次いで、EcoRV−NarIフラグメントを単離し、同様に消化した発現ベクターpCNIL5HZLC1.0中に連結してpCNIL5HZLC1.1を得た。
【0161】
免疫グロブリン(NEW)(Saul et al,J.Biol.Chem.253:585-597(1978))から得た重鎖枠組み領域および2B6重鎖CDRs[図7および配列番号:1〜2]を用いて合成可変領域を作成した。CDR表現に影響しうる枠組みアミノ酸を同定し、ずでに記載されている方法を用いて置換を行った。4つの重複合成オリゴヌクレオチド[配列番号:57、58、59および60]を得て、それらはアニーリングされ伸長された場合に、シグナル配列[配列番号:17]および重鎖可変領域を表わすアミノ酸をコードする。次いで、この合成遺伝子を、PCRプライマー[配列番号:63および64]を用いて増幅し、COR枠組みに基づく別のヒト化重鎖由来の配列を含むpUC18をベースにしたプラスミド中にBstXI−HindIII制限フラグメントとして連結した。合成オリゴヌクレオチドリンカー[配列番号:75および76]をSacIIおよびKpnI制限部位に挿入することにより、枠組みのフェニルアラニンからチロシンへの置換をアミノ酸位置91(KABAT番号付けシステム)(図12の位置94と同じ)において行った。得られた重鎖可変領域[図12および配列番号:61、62]をNEWMヒト化重鎖という。変異原性プライマーを用いるPCRまたは存在する制限部位中への合成リンカーの付加により、マッピングされた配列中のエラーを修正した。シグナル配列およびヒト化重鎖可変領域をpUCをベースにしたプラスミドからEcoRI−ApaIフラグメントとして切り出し、ヒト・IgG1不変領域を含む発現ベクターpCD中に連結してプラスミドpCDIL5NEWMを得た。CDRsのアミノ酸配列はマウス・2B6重鎖CDRsと同じである。
【0162】
免疫グロブリン(REI)(Palm et al,Hoppe-Seyler's Z.Physiol.Chem.356:167-191(1975))から得た軽鎖枠組み領域および2B6軽鎖CDRs[図7および配列番号:10、11および12]を用いて合成可変領域を作成した。CDRの表現に影響しうる枠組みアミノ酸を同定し、すでに記載された方法を用いて置換を行った。4つの重複合成オリゴヌクレオチド[配列番号:65、66、67および68]を得て、それらはアニーリンされ伸長された場合、REIヒト化軽鎖と呼ばれる軽鎖可変領域[図13および配列番号:69、70]を表すアミノ酸をコードする。次いで、この合成遺伝子をPCRプライマー[配列番号:71および72]を用いて増幅し、EcoRI−HindIII制限フラグメントとしてpGEM−7Zf(+)(Promega Corporation,Madison,WI)中に連結した。
【0163】
変異原性プライマーを用いるPCRまたは存在する制限部位中への合成リンカーの付加によりマッピングされた配列中のエラーを修正した。ヒト化軽鎖可変領域をpGEM−7Zf(+)をベースにしたプラスミドからEcoRV−NarIフラグメントとして切り出し、ヒト・カッパ不変領域ならびにシグナル配列[配列番号:17]を含む発現ベクターpCN中に連結した。CDRsのアミノ酸配列はマウス・2B6軽鎖CDRsと同じである。しかしながら、これらのCDRsのコーディング配列はマウス・2B6のコーディング配列とは異なり、制限酵素部位の作成を可能にする。これらの合成可変軽鎖および/または重鎖配列をヒト化抗体の構築に使用する。
【0164】
すでに記載されたpGEM−7Zf(+)をベースにしたプラスミドを得ると、枠組み位置15がバリンからアラニンに変化しているヒト化軽鎖可変領域を作成することができる。このプラスミドをNheIおよびSacI制限エンドヌクレアーゼで消化することができ、合成リンカー[配列番号:73および74]を挿入する。次いで、EcoRV−NarIフラグメントを単離し、同様に消化された発現ベクターpCNIL5HZREI中に連結してプラスミドpCNIL5REIV15Aを得ることができる。
【実施例5】
【0165】
実施例5 キメラ抗体の構築
マウス・mAb 2B6重鎖可変領域のアミノ酸9〜104(KABAT番号付け)をコードしているDNAを、2B6ハイブリドーマ細胞系から得たcDNA(実施例4参照)のpGEM7Zf(+)をベースにしたPCRクローンpGEM7Zf(+)からAvaII−StyI制限フラグメントとして単離した。このフラグメントを4つの小型合成オリゴマーリンカー[配列番号:35および36][配列番号:37および38]とともにpUC18をベースにしたプラスミド(BstXI−HindIIIで消化)中で一緒にすることにより、隣接重鎖可変領域配列およびシグナル配列[配列番号:17]を提供した。密接に関連したマウス・重鎖から推定されたN末端アミノ酸のコンセンサスを、最初の8個のVH残基に関して帰属し、それらは配列番号:35および36中にコードされている。2B6重鎖の最初の15個のN末端アミノ酸を配列決定することにより重鎖の推定アミノ酸配列を確かめた。
【0166】
シグナルおよびVH領域に関する配列を含むEcoRI−ApaIフラグメントを単離し、すでにヒト・IgG1不変領域をコードしているプラスミドpCD中に連結した。
【0167】
マウス・mAb 2B6軽鎖可変領域のアミノ酸12〜99(KABAT番号付け)をコードしているDNAを、2B6ハイブリドーマ細胞系から得たcDNA(実施例4参照)のpGEM7Zf(+)に基づくPCRクローンからDdeII−AvaI制限フラグメントとして単離した。pUC18をベースにしたプラスミド(EcoRV−HindIIIで消化)中においてこのフラグメントを4つの小型合成オリゴマーリンカー[配列番号:39および40][配列番号:41および42]と一緒にすることにより、隣接軽鎖可変領域配列を提供した。密接に関連したマウス・軽鎖から推定されたN末端アミノ酸のコンセンサスを、最初の8個のVL残基に関して帰属し、それらは配列番号:39および
40中にコードされている。2B6軽鎖の最初の15個のN末端アミノ酸を配列決定することにより軽鎖の推定アミノ酸配列を確かめた。次いで、この可変領域をEcoRV−NarIフラグメントとして単離し、すでにヒト・カッパ領域およびシグナル配列を含んでいる発現ベクターpCN中に連結した。
【0168】
pCDおよびpCNをベースにしたプラスミドをCOS細胞中に同時トランスフェクションすることによりキメラ抗体の発現を行った。3日後および5日後に培養上清を集め、下記のごとくELISAにより免疫グロブリン発現に関してアッセイした。最終工程以外の各工程の後にPBS洗浄を行う。ヒト・抗体のRc領域に特異的なヤギ・抗体100ng/50μl/ウェルを用いてマイクロタイタープレートを一晩被覆した。培養上清を添加し、1時間インキュベーションした。次いで、セイヨウワサビ・ペルオキシダーゼ抱合ヤギ・抗−ヒトIgG抗体を添加し、1時間インキュベーションした。次いで、ABTSペルオキシダーゼ基質(Kirkegaard & Perry Laboratories Inc.,Gaithersberg,MD)を添加した。1時間インキュベーションした後、マイクロタイタープレートリーダー(Molecular Devices Corporation,Menlo Park,CA)により405nmの吸光度を読んだ。キメラ抗体の発現を検出した。同様のELISAにおいて、キメラ抗体を含むCOS細胞上清は、ヒト・IL−5蛋白で被覆したマイクロタイターウェルに特異的に結合した。この結果は、IL−5に対する抗体をコードしている遺伝子が合成され、発現されたことを示すものである。
【0169】
上の実施例は典型的な組み換え法による抗体の調製を説明するものである。慣用的手段により得られた他の抗−IL−5抗体(例えば、2F2、2E3、4A6、5D3、24G9等)を用いて、他の組み換え抗体の取得に関して同様の手順を用いることができる。
【産業上の利用可能性】
【0170】
本発明により、高い結合アフィニティーを有するヒト・インターロイキン5に対する中和モノクローナル抗体などが提供されるので、IL−5により媒介されるえ疾病の治療または予防用医薬組成物などの分野において利用可能である。
【配列表フリーテキスト】
【0171】
SEQ ID NO: 1: First base corresponds to Kabat position 24
SEQ ID NO: 2: First base corresponds to Kabat position 25
SEQ ID NO: 3: First base corresponds to Kabat position 24
SEQ ID NO: 4: First base corresponds to Kabat 25
SEQ ID NO: 5: First base corresponds to Kabat position 24
SEQ ID NO: 6: First base corresponds to Kabat position 25
SEQ ID NO: 17: signal sequence
SEQ ID NO: 18: heavy chain variable region
SEQ ID NO: 19: heavy chain variable region
SEQ ID NO: 20: light chain variable region
SEQ ID NO: 21: light chain variable region
SEQ ID NO: 22: mouse gamma 1 hinge region primer
SEQ ID NO: 23: mouse gamma 2a hinge region primer
SEQ ID NO: 24: mouse heavy chain variable region primer
SEQ ID NO: 25: mouse kappa chain constant region primer
SEQ ID NO: 26: mouse light chain variable region primer
SEQ ID NO: 27: oligonucleotide encodes a part of heavy chain variable region
SEQ ID NO: 28: oligonucleotide encodes a part of heavy chain variable region
SEQ ID NO: 29: oligonucleotide encodes a part of heavy chain variable region
SEQ ID NO: 30: oligonucleotide encodes a part of heavy chain variable region
SEQ ID NO: 31: oligonucleotide encodes a part of light chain variable region
SEQ ID NO: 32: oligonucleotide encodes a part of light chain variable region
SEQ ID NO: 33: oligonucleotide encodes a part of light chain variable region
SEQ ID NO: 34: oligonucleotide encodes a part of light chain variable region
SEQ ID NO: 35: small synthetic oligomer linker
SEQ ID NO: 36: small synthetic oligomer linker
SEQ ID NO: 37: small synthetic oligomer linker
SEQ ID NO: 38: small synthetic oligomer linker
SEQ ID NO: 39: small synthetic oligomer linker
SEQ ID NO: 40: small synthetic oligomer linker
SEQ ID NO: 41: small synthetic oligomer linker
SEQ ID NO: 42: small synthetic oligomer linker
SEQ ID NO: 43: overlapping oligonucleotide
SEQ ID NO: 44: overlapping oligonucleotide
SEQ ID NO: 45: light chain
SEQ ID NO: 46: light chain
SEQ ID NO: 47: CDR3
SEQ ID NO: 48: CDR3
SEQ ID NO: 49: heavy chain
SEQ ID NO: 50: light chain
SEQ ID NO: 51: linker
SEQ ID NO: 52: linker
SEQ ID NO: 53: linker
SEQ ID NO: 54: linker
SEQ ID NO: 55: linker
SEQ ID NO: 56: linker
SEQ ID NO: 57: overlapping synthetic oligonucleotide
SEQ ID NO: 58: overlapping synthetic oligonucleotide
SEQ ID NO: 59: overlapping synthetic oligonucleotide
SEQ ID NO: 60: overlapping synthetic oligonucleotide
SEQ ID NO: 61: heavy chain variable region
SEQ ID NO: 62: heavy chain variable region
SEQ ID NO: 63: PCR primer
SEQ ID NO: 64: PCR primer
SEQ ID NO: 65: overlapping synthetic oligonucleotide
SEQ ID NO: 66: overlapping synthetic oligonucleotide
SEQ ID NO: 67: overlapping synthetic oligonucleotide
SEQ ID NO: 68: overlapping synthetic oligonucleotide
SEQ ID NO: 69: light chain variable region
SEQ ID NO: 70: light chain variable region
SEQ ID NO: 71: PCR primer
SEQ ID NO: 72: PCR primer
SEQ ID NO: 73: synthetic linker
SEQ ID NO: 74: synthetic linker
SEQ ID NO: 75: synthetic oligonucleotide linker
SEQ ID NO: 76: synthetic oligonucleotide linker
【技術分野】
【0001】
関連出願の相互参照
本願は、1994年12月23日出願のU.S.第08/363,131号の一部継続出願であるU.S.第08/470,110号および08/467,420号(ともに1995年6月6日出願)の一部継続出願である。
【0002】
発明の分野
一般的には、本発明は、IL−5および過剰な好酸球産生により媒介される症状の治療および診断に有用な抗体および変化した抗体、そしてより詳細には、mAbs、Fab、キメラおよびヒト化抗体の分野に関する。
【背景技術】
【0003】
発明の背景
好酸球は、肺組織の過敏反応に関連したアレルギー性疾患を包含する広範囲の炎症性疾病状態の発生に関与している(非特許文献1)。注目すべき例は喘息であり、喘息は、非特異的気管支過敏症を引き起こす気道の可逆的閉塞により特徴づけられる。さらに喘息は、気管支粘膜のレベルの慢性的炎症反応ならびにマクロファージ、リンパ球および好酸球による特徴的な炎症の発生と因果関係がある。好酸球は、疾病の典型的な粘膜ダメージの開始において中心的役割を果たしているようである(非特許文献2)。数が増加した活性化された好酸球は、慢性喘息の患者の循環系、気管支分泌物および肺の実質において報告されており、種々の肺機能試験により測定される疾病の重さは血中好酸球数と相関関係がある(非特許文献3)。また、しばしば、脱顆粒反応の過程における増加した好酸球が、後期喘息反応中の患者気管支肺胞洗浄液(BAL)中に回収され、通常には、ステロイド療法の結果としての好酸球の減少は臨床的徴候の改善に関連している(非特許文献4)。
【0004】
インターロイキン5(IL−5)は、活性化されたCD4およびTリンパ球により産生されるホモダイマー糖蛋白である。ヒトにおいて、IL−5は好酸球の増殖および分化の調節に大いに関係している。IL−5の上昇したレベルは喘息性気管支肺胞洗浄液中に検出される(非特許文献5)。IL−5に関してトランスジェニックなマウスは、抗原刺激の不存在下の末梢血および組織における著しい好酸球増加を示し(非特許文献6)、抗−ネズミ・IL−5モノクローナル抗体は、マウスの血液および組織における好酸球増加(非特許文献7)ならびに実験動物における寄生虫感染およびアレルゲン攻撃に関連した好酸球増加(非特許文献8〜10)の抑制における効果が示されている。
【0005】
コルチコステロイドは、好酸球数および喘息の他の炎症性成分の減少にきわめて効果的であるが、重い喘息およびより最近になってからは軽い喘息ないしは中程度の喘息のいずれにおいても副作用の心配がある。唯一の他の主要抗−炎症薬剤療法−クロモグリケート類(クロモリンナトリウムおよびネドクロミル)−はコルチコステロイドよりもかなり効果が少なく、それらの正確な作用機構は不明のままである。
【0006】
より最近の開発は、新たな吸入ステロイド、長時間作用する気管支拡張剤、および新規生化学的もしくは薬理学的標的(例えば、カリウムチャンネル活性化剤、ロイコトリエンアンタゴニスト、5−リポオキシゲナーゼ(5−LO)阻害剤等)に作用する薬剤に焦点を絞っている。理想的な薬剤は、ステロイドの有効性をクロモリンナトリウムの安全性と結び付けるものであり、選択性が増加していてより迅速に作用を発揮するものである。中和IL−5抗体は潜在的にヒトにおける好酸球増加関連徴候の軽減に有用でありうる。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Butterfieldら,Immunopharmacology of Eosinophils(H.SmithおよびR.Cook編,)中,p151-192,Academic Press, London(1993)
【非特許文献2】Corrigan et al., Immunol.Today,13:501-507(1992)
【非特許文献3】Griffen et al.,J.Aller.Clin.Immunol.67,548-557(1991)
【非特許文献4】Bousquet et al.,N.Eng.J.Med.,323:1033-1039(1990)
【非特許文献5】Motojima et al.,Allergy,48:98(1993)
【非特許文献6】Dent et al.,J.Exp.Med.,172, 1425(1990)
【非特許文献7】Hitoshi et al.,Int.Immunol.,315(1991)
【非特許文献8】Coffman et al.,Science,245,308-310(1989)
【非特許文献9】Sher et al.,Proc.Natl.Acad.Sci.,83:61-65(1990)
【非特許文献10】Chand et al.,Eur.J.Pharmacol.,211:121-123(1992)
【発明の概要】
【発明が解決しようとする課題】
【0008】
ゆえに、ヒト・インターロイキン5に対する中和モノクローナル抗体のごとき高アフィニティーIL−5アンタゴニストであって、好酸球の分化および増殖(すなわち、好酸球の蓄積)を抑制し、かくして好酸球により炎症を抑制する高アフィニティーIL−5アンタゴニストに対する必要性が当該分野において存在する。
【課題を解決するための手段】
【0009】
発明の概要
第1の態様において、本発明は、ヒト・インターロイキン5に特異的で、詳細な説明の欄において説明するように、約3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる結合アフィニティーを有する齧歯類(例えば、ラットおよびマウス)の中和モノクローナル抗体を提供する。かかるモノクローナル抗体の典型例はマウスのモノクローナル抗体2B6、2E6および2F2ならびに4A6のごときラットのモノクローナル抗体である。本発明のもう1つの態様は、SK119−2B6.206.75(1)、SK119−2E3.39.40.2、SK119−2F2.37.80.12、4A6(1)G1F7および5D3(1)F5D6のごときハイブリドーマである。
【0010】
関連態様において、本発明は、本発明の齧歯類中和モノクローナル抗体のFc領域を欠失させることにより製造される、ヒト・インターロイキン−5に特異的な中和FabフラグメントまたはそのF(ab')2フラグメントを提供する。
【0011】
さらにもう1つの関連態様において、本発明は、チェイン・シャフリング(chain shuffling)法により製造される、ヒト・インターロイキン−5に特異的な中和FabフラグメントまたはそのF(ab')2フラグメントを提供する。該方法により、線維状ファージFabディスプレイライブラリーにおいてインターロイキン5で免疫された齧歯類から単離された軽鎖(または重鎖)免疫グロブリンライブラリーとともに本発明齧歯類中和モノクローナル抗体から単離された重鎖(または軽鎖)免疫グロブリンが発現される。
【0012】
さらにもう1つの関連態様において、本発明は、ヒト・インターロイキン5に対して約3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる非ヒト・中和モノクローナル抗体(mAb(s))由来の相補性決定領域(CDR(s))を含んでなる、ヒト・インターロイキン5に特異的な変化した抗体ならびにそれをコードしている核酸分子を提供する。変化した抗体がヒト化抗体である場合には、非ヒト・免疫グロブリン由来の相補性決定領域(CDRs)をコードしている配列が、少なくとも1つ、好ましくは全部の相補性決定領域(CDRs)が非ヒト・モノクローナル抗体由来のCDRsにより置換されている第1の免疫グロブリンパートナー中に挿入される。好ましくは、第1の免疫グロブリンパートナーは、同様に免疫グロブリンの不変鎖全部またはその一部を含んでなる第2の免疫グロブリンパートナーに作動可能に連結される。
【0013】
関連態様において、本発明は、ヒト・インターロイキン5に対して約3.5x10−11Mに等しいかまたはそれ未満の解離定数を有すことにより特徴づけられる非ヒト・中和モノクローナル抗体(mAbs)由来のCDRs、ならびにかかるCDRsをコードしている核酸分子を提供する。
【0014】
さらにもう1つの態様において、ヒト・重鎖および軽鎖の不変領域ならびにヒト・インターロイキン5に対して約3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる非ヒト・中和モノクローナル抗体由来の重鎖および軽鎖の可変領域を含むキメラ抗体を提供する。
【0015】
さらにもう1つの態様において、本発明は、上記の変化した抗体のうちの1種(またはそれ以上)および医薬上許容される担体を含む医薬組成物を提供する。
【0016】
さらなる態様において、本発明は、ヒトに有効量の本発明医薬組成物を投与することによる、過剰の好酸球産生に関連したヒトにおける症状の治療方法を提供する。
【0017】
さらなる態様において、本発明は、IL−5に対して約3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる非ヒト・中和モノクローナル抗体(mAbs)由来の変化した抗体(例えば、組み換え抗体、CDRs、FabまたはそのF(ab')2フラグメントもしくはアナログ)の組み換え製造方法および該組み換え製造において有用な成分を提供する。これらの成分は、上記のものをコードしている単離核酸配列、ならびに選択された調節配列の制御下の当該核酸配列を含む組み換えプラスミドを包含し、該調節配列は、該組み換えプラスミドでトランスフェクションされた宿主細胞(好ましくは哺乳動物細胞)における該核酸配列の発現を指令することができるものである。該製造方法は、変化した抗体、好ましくはヒト化抗体がトランスフェクションされた細胞中で発現されるような条件下で本発明のトランスフェクションされた宿主細胞系を培養し、次いで、そこから発現産物を単離することを包含する。
【0018】
本発明のさらなる態様において、患者から生物学的液体試料を得て、IL−5/(モノクローナルまたはポリクローナル)抗体複合体が形成されるような条件下で本発明抗体および変化した抗体をかかる試料と接触させ、次いで、該IL−5/抗体複合体の存在または不存在を検出することを特徴とする、ヒトにおける過剰な好酸球産生に関連した症状の診断方法が提供される。
【0019】
本発明の他の態様および利点を、詳細な説明および好ましい具体例の欄においてさらに説明する。
【発明の効果】
【0020】
本発明により、高い結合アフィニティーを有するヒト・インターロイキン5に対する中和モノクローナル抗体などが提供される。
【図面の簡単な説明】
【0021】
【図1】図1[配列番号:1および15]は、マウス・抗体2B6およびマウス/ヒト・2B6キメラ抗体に関する重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図2】図2[配列番号:2および16]は、マウス・抗体2B6およびマウス/ヒト・2B6キメラ抗体に関する軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【図3】図3[配列番号:3]は、マウス・抗体2F2に関する重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図4】図4[配列番号:4]は、マウス・抗体2F2に関する軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【図5】図5[配列番号:5]は、マウス・抗体2E3に関する重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図6】図6[配列番号:6]は、マウス・抗体2E3に関する軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【図7】図7[配列番号:7−14]は、マウス抗体・2B6、2F2および2E3由来の重鎖および軽鎖のCDRsを示す。
【図8】図8[配列番号:18、19]は、ヒト化抗体2B6に関する重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図9】図9[配列番号:20、21]は、ヒト化抗体2B6に関する軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【図10】図10は、哺乳動物細胞におけるヒト化重鎖遺伝子の発現に用いるプラスミドpCDIL5HZHC1.0のスキーム図である。該プラスミドはベータラクタマーゼ遺伝子(BETA LAC)、SV−40複製開始点(SV40)、サイトメガロウイルスプロモーター配列(CMV)、シグナル配列、ヒト化重鎖、ウシ・成長ホルモン(BGH)由来のポリAシグナル、ベータグロビンプロモーター(ベータグロプロ)、ジヒドロ葉酸レダクターゼ遺伝子(DHFR)、およびもう1つのBGH配列のポリAシグナルをpUC19骨格中に含んでいる。
【図11】図11は、哺乳動物細胞におけるヒト化軽鎖遺伝子の発現に用いるプラスミドpCNIL5HZLC1.0のスキーム図である。
【図12】図12[配列番号:61、62]は、ヒト化抗体2B6に関するNewM重鎖可変領域を示す。ボックス領域はCDRsを示す。
【図13】図13[配列番号:69、70]は、ヒト化抗体2B6に関するREI軽鎖可変領域を示す。ボックス領域はCDRsを示す。
【発明を実施するための形態】
【0022】
発明の詳細な説明
本発明は、マウス・モノクローナル抗体2B6のおいて例示されるヒト・IL−5特異性、中和活性、およびヒト・IL−5に対する高アフィニティーにより特徴づけられる種々の抗体、変化した抗体およびそれらのフラグメントを提供する。新規中和抗体を得るための慣用的なハイブリドーマ法、ファージディスプレイ組み合わせライブラリー、免疫グロブリンのチェイン・シャフリング、およびヒト化法により本発明抗体は製造された。これらの製造物は、例えば喘息のごときIL−5により媒介される疾病の治療のための治療および医薬組成物において有用である。また、これらの製造物は、ヒトにおける内在性IL−5レベルまたは活性化細胞からex vivoで放出されるIL−5レベルの測定(例えば、酵素結合免疫吸着アッセイ(ELISA))による、IL−5により媒介される症状の診断において有用である。
【0023】
I.定義
「変化した抗体」は、変化した免疫グロブリンコーディング領域によりコードされている蛋白をいい、選択された宿主細胞における発現により得ることができる。かかる変化した抗体は遺伝子組み換えによる抗体(例えば、キメラ抗体またはヒト化抗体)または免疫グロブリン不変領域の全部もしくは一部を欠く抗体フラグメント、例えば、Fv、Fab、またはF(ab')2等である。
【0024】
「変化した免疫グロブリンコーディング領域」は、本発明の変化した抗体をコードしている核酸配列をいう。変化した抗体がCDRを継ぎ足した抗体またはヒト化抗体である場合、非ヒト・免疫グロブリン由来の相補性決定領域(CDRs)をコードしている配列が、ヒト・可変枠組み配列を含んでなる第1の免疫グロブリンパートナー中に挿入される。第1の免疫グロブリンパートナーが第2の免疫グロブリンパートナーに作動可能に連結されていてもよい。
【0025】
「第1の免疫グロブリンパートナー」は、無傷(または天然に存在する)のCDRコーディング領域がドナー抗体のCDRコーディング領域により置換されているヒト・枠組みまたはヒト・免疫グロブリン可変領域をコードしている核酸配列をいう。ヒト・可変領域は免疫グロブリン重鎖、軽鎖(または両方)、それらの機能的フラグメントのアナログであってよい。抗体(免疫グロブリン)の可変領域中に存在するかかるCDR領域を、当該分野で知られた方法により決定することができる。例えば、Kabatら(Sequences of Proteins of Immunological Interest,4th Ed.,U.S.Department of Health and Human Services,National Institutes of Health(1987))は、CDRsの存在位置に関する規則を開示している。さらに、CDR領域/構造の同定に有用なコンピュータープログラムが知られている。
【0026】
「中和」は、ヒト・IL−5のその特異的受容体への結合を妨げることにより、あるいはその受容体を通じてのIL−5のシグナリングを阻害することによりIL−5活性を阻害する抗体をいう。B13細胞バイオアッセイ(IL−5中和アッセイ、実施例2C参照)において測定されるIL−5活性の阻害において90%有効、好ましくは95%有効、最も好ましくは100%有効な場合、mAbは中和抗体である。
【0027】
用語「高アフィニティー」は、光学的バイオセンサー分析(実施例2参照)により測定されるヒト・IL−5に対する3.5x10−11Mに等しいかまたはそれ未満のKdにより特徴づけられる結合アフィニティーを有する抗体をいう。
【0028】
「ヒト・IL−5に対する結合特異性」は、マウスではなくヒトのIL−5に対する高アフィニティーを意味する。
【0029】
「第2の免疫グロブリンパートナー」は、イン・フレームで、あるいは所望により慣用的リンカー配列により第1の免疫グロブリンパートナーが融合(すなわち、作動可能に連結)している蛋白またはペプチドをコードしているもう1つのヌクレオチド配列をいう。好ましくは、それは免疫グロブリン遺伝子である。第2の免疫グロブリンパートナーは、その不変領域全体(すなわち、同種のもの−第1および第2の変化した抗体は同じ起源由来である)または対象とするさらなる抗体(すなわち、異種のもの)をコードしている核酸配列を包含しうる。それは免疫グロブリン重鎖または軽鎖(または1本のポリペプチドの一部としての両鎖)であってもよい。第2の免疫グロブリンパートナーは特定の免疫グロブリンクラスまたはイソタイプに限らない。さらに、第2の免疫グロブリンパートナーは、FabまたはF(ab')2のごとき免疫グロブリン不変領域の一部(すなわち、適当なヒト・不変領域または枠組み領域の別個の部分)を含んでなっていてもよい。また、かかる第2の免疫グロブリンパートナーは、例えばファージディスプレイライブラリーとして宿主細胞の外表面上に露出した内在性膜蛋白をコードしている配列、あるいは分析または診断的検出のための蛋白、例えばセイヨウワサビペルオキシダーゼ、β−ガラクトシダーゼ等をコードしている配列を含んでいてもよい。
【0030】
用語Fv、Fc、Fd、Fab、またはF(ab')2を、その標準的な意味で用いる(例えば、Harlow et al.,Antibodies A Laboratory Manual,Cold SpringHarbor Laboratory,(1988)参照)。
【0031】
本明細書の用語「組み換え法による抗体」は、変化した抗体の1のタイプ、すなわち、選択されたアクセプター抗体の軽鎖および/または重鎖可変ドメインの一部分が、選択されたエピトープに特異性を有する1種またはそれ以上のドナー抗体由来の類似部分により置換されている全長の合成抗体(抗体フラグメントに対立するものとしてのキメラ抗体またはヒト化抗体)をいう。例えば、かかる分子は、未修飾軽鎖(またはキメラ軽鎖)に結合したヒト化重鎖(あるいはその逆)によって特徴づけられる抗体を包含しうる。組み換え法による抗体もまた、ドナー抗体の結合特異性を保持するようなアクセプター抗体の軽鎖および/または重鎖の可変ドメイン枠組み領域をコードしている核酸配列の変化によって特徴づけられる。これらの抗体は、アクセプター抗体由来の1種またはそれ以上のCDRs(好ましくは全部のCDRs)の本明細書記載のドナー抗体由来のCDRsでの置換を含んでいてもよい。
【0032】
「キメラ抗体」は、アクセプター抗体由来の軽鎖および重鎖不変領域に結合したドナー抗体由来の天然に存在する可変領域(軽鎖および重鎖)を含んでいる組み換え法による抗体の1のタイプをいう。
【0033】
「ヒト化抗体」は、非ヒト・ドナー免疫グロブリン由来のCDRsを有する組み換え法による抗体の1のタイプであって、その残りの免疫グロブリン由来の部分は1種(またはそれ以上)のヒト由来であるものをいう。さらに、結合アフィニティーを保持するように枠組み支持残基を変化させてもよい(例えば、Queen et al.,Proc.Natl.Acad.Sci.USA,86:10029-10032(1989)、Hodgson et al.,Bio/Technology,9:421(1991)参照)。
【0034】
用語「ドナー抗体」は、その可変領域、CDRs、またはその他のフラグメントもしくはアナログの核酸配列が第1の免疫グロブリンパートナーのものになっている抗体をいい、その結果、変化した免疫グロブリンコーディング領域を提供し、発現した変化した抗体がドナー抗体の抗原特異性および中和活性を有する。本発明における使用に適した1のドナー抗体は、2B6と命名された非ヒト・中和モノクローナル抗体(すなわち、マウスの)である。抗体2B6は、イソタイプIgG1に属する高アフィニティーヒト・IL−5特異性(すなわち、マウス・IL−5を認識しない)中和抗体であると定義され、適当なマウス・IgG不変領域上に配列番号:2および16の可変軽鎖DNAおよびアミノ酸配列、ならびに配列番号:1および15の可変重鎖DNAおよびアミノ酸配列を有している。
【0035】
用語「アクセプター抗体」は、ドナー抗体とは異種の抗体(モノクローナル、または組み換え型)をいい、その重鎖および/または軽鎖枠組み領域ならびに/あるいはその重鎖および/または軽鎖不変領域のすべて(またはいずれかの部分、しかし、好ましくはすべての部分)が第1の免疫グロブリンパートナーのものとなっている。好ましくは、ヒト・抗体がアクセプター抗体である。
【0036】
「CDRs」は、免疫グロブリン重鎖および軽鎖の超可変領域である抗体の相補性決定領域のアミノ酸配列をいう。例えば、Kabat et al.,Sequences of Proteins of Immunological Interest,4th Ed,U.S.Department of Health and Human Services,National Institutes of Health(1987)参照。免疫グロブリン可変領域には3本の重鎖CDRsおよび3本の軽鎖CDRsがある。よって、本明細書の用語「CDRs」は、3本の重鎖CDRsすべて、あるいは3本の軽鎖CDRsすべて(あるいは適当な場合には、すべての重鎖および軽鎖CDRsの両方)をいう。
【0037】
CDRsは、抗原またはエピトープへの抗体結合のための接触残基の大部分を提供する。本発明の対象CDRsはドナー抗体可変重鎖および軽鎖配列由来であり、天然に存在するCDRsのアナログを包含し、該アナログはまた、それらが由来するドナー抗体と同じ抗原結合特異性および/または中和能を共有している。
【0038】
「抗原結合特異性または中和能を共有」とは、例えば、mAb 2B6は一定レベルの抗原アフィニティーにより特徴づけられるが、適当な構造的環境においては2B6の核酸配列によりコードされているCDRは低いまたは高いアフィニティーを有するかもしれないということを意味する。それにもかかわらず、かかる環境において、2B6のCDRsは2B6と同じエピトープを認識するであろうと予想される。典型的な2B6の重鎖CDRsは配列番号:7、配列番号:8および配列番号:9を包含し、典型的な2B6の軽鎖CDRは配列番号:10、配列番号:11および配列番号:12を包含する。
【0039】
「機能的フラグメント」は、そのフラグメントが由来している抗体と同じ抗原結合特異性および/または中和能を保持している部分的な重鎖または軽鎖可変配列(例えば、免疫グロブリン可変領域のアミノ末端またはカルボキシ末端における小規模な欠失を有する)である。
【0040】
「アナログ」は、少なくとも1個のアミノ酸によって修飾されているアミノ酸配列であり、該修飾は化学的なものあるいは少数(すなわち、10個未満)のアミノ酸の置換または転移であってもよく、修飾されているとしてもアミノ酸配列が未修飾配列の生物学的特性、例えば、抗原特異性および高アフィニティーを保持しているものである。例えば、ある種のエンドヌクレアーゼ制限部位をCDRコーディング配列中またはその周辺に作成する場合には、置換により(サイレント)変異を構築することができる。
【0041】
アナログは対立遺伝子変化を生じるものであってもよい。「対立遺伝子変化または修飾」とは、本発明アミノ酸またはペプチド配列をコードしている核酸配列中の変化である。かかる変化または修飾は遺伝コードの縮重によるものであってもよく、あるいは所望特性を得るように慎重に組み換え法を用いて作成されたものであってもよい。これらの変化または修飾は、コードされているアミノ酸配列の変化を生じるものであってもよく、生じないものであってもよい。
【0042】
用語「エフェクター剤」は、変化した抗体および/またはドナー抗体もしくはその他のフラグメントの天然または合成の軽鎖または重鎖が慣用的手段により結合されていてもよい非蛋白担体分子をいう。かかる非蛋白担体は、診断分野において用いられる慣用的担体、例えば、ポリスチレンまたは他のプラスチックビーズ、多糖類、例えば、BIAcore[Pharmacia]システムに用いるもの、または医学分野で使用され、ヒトおよび動物への投与が安全である他の非蛋白物質を包含する。他のエフェクター剤は、重金属原子またはラジオアイソトープをキレートするためのマクロサイトを包含する。かかるエフェクター剤は、変化した抗体の半減期を延長することにも有用であり、例えば、ポリエチレングリコールが挙げられる。
【0043】
II.高アフィニティーIL−5モノクローナル抗体
本発明抗体、変化した抗体およびフラグメントの構築に使用するために、無傷のヒト・IL−5またはそれに由来するペプチドエピトープを与えることにより、非ヒト種(例えば、ウシ、ヒツジ、サル、ニワトリ、齧歯類(例えばマウスおよびラット)等)を用いて所望免疫グロブリンを得てもよい。慣用的なハイブリドーマ法を用いて、IL−5に対する非ヒト・mAbを分泌するハイブリドーマ細胞系を得る。次いで、実施例セクションに記載のごとくIL−5で被覆した96ウェルプレートを用いて、あるいは別法としてストレプトアビジンで被覆したプレートに結合したビオチン化IL−5を用いてかかるハイブリドーマを結合に関してスクリーニングする。
【0044】
本発明の1の典型的な高アフィニティー中和mAbはマウス・抗体であるmAb 2B6であり、実施例1においてより詳細に説明されるキメラ抗体またはヒト化抗体の開発にそれを用いることができる。2B6 mAbは、ヒト・IL−5に対する抗原結合特異性により特徴づけられ、IL−5に対して3.5x10−11M未満(約2.2x10−11M)のKdを有する。光学的バイオセンサーによる測定により2B6由来のFabフラグメントのIL−5に対するKdを評価したところ(実施例3H参照)、約9x10−11Mである。mAb 2B6は、ヒト・IL−5とヒト・IL−5受容体のα−鎖との間の結合相互作用をブロックするように思われる。
【0045】
もう1つの望ましいドナー抗体はマウス・mAb 2E3である。このmAbは、イソタイプIgG2aであり、3.5x10−11M未満(約2.0x10−11M)のヒト・IL−5に対する解離定数を有することにより特徴づけられる。
【0046】
さらにもう1つの望ましいドナー抗体はラット・mAb 4A6である。このmAbは、3.5x10−11M未満(約1.8x10−11M)のヒト・IL−5に対する解離定数を有することにより特徴づけられる。さらに、mAb 4A6は、ヒト・IL−5とヒト・IL−5受容体のβ−鎖との間の結合相互作用をブロックするように思われる。
【0047】
本発明は2B6 mAb、2E3 mAb、またはそれらの超可変(すなわち、CDR)配列の使用に限定されない。ヒト・IL−5に対する3.5x10−11Mに等しいかまたはそれ未満の解離定数により特徴づけられる他の適当な高アフィニティーIL−5抗体および対応する抗−IL−5 CDRsをそれらに代えて用いてもよい。以下の説明のいずれの箇所においても、ドナー抗体は2B6または2E3と同定されるが、この命名は例示および単なる説明のためのものである。
【0048】
III.抗体フラグメント
本発明は、ヒト・IL−5に指向されたmAb由来のFabフラグメントまたはF(ab')2フラグメントの使用も包含する。これらのフラグメントは、in vivoにおけるIL−5および好酸球により媒介される症状に対する防御剤として、あるいはin vitroでのIL−5診断薬の一部として有用である。Fabフラグメントは軽鎖全体および重鎖のアミノ末端部分を含んでおり;F(ab')2フラグメントはジスルフィド結合により結合している2個のFabフラグメントにより形成されるフラグメントである。mAbs 2B6、2E3および他の類似高アフィニティーIL−5結合抗体は、FabフラグメントおよびF(ab')2フラグメントのソースを提供するものであり、それらのフラグメントは慣用的手段、例えば、適当な蛋白分解酵素パパインおよび/またはペプシンでのmAbの開裂により、あるいは組み換え法により得ることができる。本明細書記載のごとく、これらのFabおよびF(ab')2フラグメントはそれら自体、治療、予防または診断薬、ならびに組み換え型抗体またはヒト化抗体の形成において有用な可変領域およびCDR配列を含む配列のドナーとして有用である。
【0049】
組み合わせファージライブラリー(例えば、Winter et al.,Ann.Rev.Immunol.,12:433-455(1994)参照)または免疫グロブリンのチェイン・シャフリング(例えば、Marks et al.,Bio/Technology,10:779-783(1992)参照)によりFabおよびF(ab')2フラグメントを構築することができ(両文献を参照により本明細書に記載されているものとみなす)、それらの方法において、選択された抗体(例えば、2B6)由来のFdまたはVH免疫グロブリンを軽鎖免疫グロブリンVL(VK)のレパートリーと結合させて新規Fabsを得る。逆に、選択された抗体由来の軽鎖免疫グロブリンを重鎖免疫グロブリンVH(またはFd)のレパートリーと結合させて新規Fabを得てもよい。実施例セクションにおいてより詳細に説明するように、mAb 2B6のFdを軽鎖免疫グロブリンのレパートリーと結合させた場合に中和IL−5 Fabsが得られた。よって、チェイン・シャフリング法により、ユニークな配列(ヌクレオチドおよびアミノ酸)を有する中和Fabsを得ることができる。
【0050】
IV.対象とする抗−IL−5アミノ酸およびヌクレオチド配列
上記mAb 2B6または他の抗体は、ドナー抗体の抗原結合特異性により特徴づけられる種々の変化した抗体の設計および取得に有用な配列、例えば、可変重鎖および/または軽鎖ペプチド配列、枠組み配列、CDR配列、機能的フラグメント、およびそれらのアナログ、およびそれらをコードしている核酸配列を提供する。
【0051】
よって、一例として、本発明は、IL−5マウス抗体2B6由来の可変軽鎖および可変重鎖配列ならびにそれら由来の配列を提供する。2B6の重鎖可変領域を図1に示す。CDRをコードしている領域はボックス領域により示され、それらは配列番号:7;配列番号:8;および配列番号:9に示される。2B6の軽鎖クローン可変領域を図2に示す。CDRをコードしている領域を配列番号:10;配列番号:11;および配列番号:12に示す。
【0052】
ヒト化重鎖可変領域を図8[配列番号:18および19]に示す。シグナル配列も配列番号:17に示す。当業者に知られた他の適当なシグナル配列を、本明細書に例示したシグナル配列に代えて使用してもよい。この構築物のCDRアミノ酸配列は無傷のマウスのものおよびキメラ重鎖CDRsと同一であり、それらを配列番号:7、配列番号:8、および配列番号:9に示す。典型的な(合成)ヒト化軽鎖可変配列を図9[配列番号:20および21]に示す。
【0053】
可変軽鎖および重鎖ペプチド配列をコードしている本発明核酸配列またはそのフラグメントは、CDRsまたは枠組み領域をコードしている核酸配列中の特異的変化の突然変異による導入、ならびに生じた修飾または融合核酸配列の発現用プラスミド中への導入にも有用である。例えば、枠組みおよびCDRをコードしている領域におけるサイレント置換を用いて突然変異されたCDR(および/または枠組み)領域の挿入を容易にする制限酵素部位を作成した。これらのCDRをコードしている領域を、本発明ヒト化抗体の構築に使用した。
【0054】
遺伝コードの縮重を考慮して、本発明の重鎖および軽鎖アミノ酸配列およびCDR配列ならびにドナー抗体の抗原特異性を共有しているそれらの機能的フラグメントおよびアナログをコードしている種々のコーディング配列を構築してもよい。第2の免疫グロブリンパートナーに作動可能に結合する場合には、可変鎖ペプチド配列またはCDRsをコードしている本発明の単離核酸配列またはそのフラグメントを用いて、本発明の変化した抗体、例えば、キメラまたはヒト化抗体、あるいは他の組み換え法による抗体を得ることができる。
【0055】
本明細書記載の変化した抗体および抗体の一部をコードしている単離核酸配列のほかに、他のかかる核酸配列、例えば、無傷のCDRをコードしている配列の相補的な配列またはCDRをコードしている領域周辺の修飾されたヒト・枠組み領域に相補的な配列も本発明に包含されることに注意すべきである。有用なDNA配列は、厳密なハイブリダイゼーション条件下[T.Maniatis et al.,Molecular Cloning(A Laboratory Manual),Cold Spring Harbor Laboratory(1982),387−389頁参照]でDNA配列にハイブリダイゼーションする配列を包含する。1のかかる厳密なハイブリダイゼーション条件の例は、4XSSC、65℃、次いで、65℃の0.1XSSCでの1時間の洗浄である。あるいはまた、厳密なハイブリダイゼーション条件の典型例は、50%ホルムアミド、4XSSC中、42℃である。好ましくは、これらのハイブリダイゼーションするDNA配列は少なくとも18ヌクレオチドの長さ、すなわち、ほぼCDRのサイズである。
【0056】
V.変化した免疫グロブリン分子および変化した抗体
変化した免疫グロブリン分子は、キメラ抗体およびヒト化抗体のごとき組み換え法により抗体を包含する変化した抗体をコードしていてもよい。望ましい変化した免疫グロブリンコーディング領域はCDRをコードしている領域を含み、該領域は、IL−5抗体の抗原特異性、好ましくは本発明により提供されるような高アフィニティー抗体の抗原特異性を有するペプチドをコードしており、第1の免疫グロブリンパートナー(ヒト・枠組みまたはヒト・免疫グロブリン可変領域)中に挿入されている。
【0057】
好ましくは、第1の免疫グロブリンパートナーは第2の免疫グロブリンパートナーに作動可能に連結される。第2の免疫グロブリンパートナーは上で定義されており、興味ある第2の抗体領域、例えばFc領域をコードしている配列を含んでもよい。第2の免疫グロブリンパートナーは、軽鎖または重鎖不変領域がイン・フレームまたはリンカー配列手段により融合しているもう1つの免疫グロブリンをコードしている配列を含んでもよい。機能的フラグメントまたはIL−5のアナログに対して結合が促進されるように、それらに指向された組み換え法による抗体を設計してもよい。
【0058】
第2の免疫グロブリンパートナーを非蛋白担体分子を包含する上記定義のエフェクター剤と結合させてもよく、慣用的手段により第2の免疫グロブリンパートナーを作動可能に結合させる。
【0059】
第2の免疫グロブリンパートナー、例えば抗体配列とエフェクター剤との間の融合または結合は、例えば慣用的な共有結合またはイオン結合、蛋白融合、あるいは異種二官能基架橋(例、カルボジイミド、グルタルアルデヒド等)のような適当な手段によるものであってよい。かかる方法は当該分野において知られており、普通の化学および生化学の教科書に記載されている。
【0060】
さらに、第2の免疫グロブリンパートナーとエフェクター剤との間に所望量のスペースを単に提供するだけの慣用的リンカー配列を、変化した免疫グロブリンコーディング領域中に構築してもよい。かかるリンカーの設計は当業者によく知られている。
【0061】
さらに、本発明分子のシグナル配列を修飾して発現を促進してもよい。一例として、シグナル配列およびマウス・重鎖配列由来のCDRsを有する2B6ヒト化抗体はもとのシグナルペプチドを別のシグナル配列[配列番号:17]に置き換えられている。
【0062】
典型的な変化した抗体は、mAb 2B6の抗原特異性を有する可変重鎖および/または軽鎖ペプチドまたは蛋白配列、例えば、VHおよびVL鎖を含んでいる。さらにもう1つの望ましい本発明の変化した抗体は、少なくとも1つ、マウス・抗体分子2B6の重鎖および/または軽鎖の可変領域の好ましくは全部のCDRsを含んでおり、残りの配列はヒト起源のもの、またはその機能的フラグメントもしくはアナログである。例えば、ヒト化VHおよびVL領域(図8および9)参照。
【0063】
さらなる具体例において、組み換え法による本発明抗体にさらなる剤が結合していてもよい。例えば、組み換えDNA法の手順を用いて、完全な抗体分子のFcフラグメントまたはCH2 CH3ドメインが酵素または他の検出可能な分子(すなわち、ポリペプチドエフェクターまたはレポーター分子)に置き換えられている本発明の組み換え法による抗体を得てもよい。
【0064】
第2の免疫グロブリンパートナーを、マウス2B6の抗原特異性を有するCDR含有配列とは異種の非免疫グロブリンペプチド、蛋白またはそのフラグメントに作動可能に結合させてもよい。得られる蛋白は、発現すると、IL−5抗原特異性および非免疫グロブリン特性を示すことができる。その融合パートナーの特性は、例えば、別の結合ドメインまたは受容体ドメインのごとき機能的特性、あるいは融合パートナー自体が治療蛋白である場合には治療特性またはさらなる抗原特性であってもよい。
【0065】
もう1つの望ましい本発明蛋白は、全長の重鎖および軽鎖を有する完全な抗体分子、あるいはFabまたはF(ab')2フラグメントのごときそれらの別々のフラグメント、重鎖ダイマー、またはFVまたは1本鎖抗体(SCA)のごときそれらの最小の組み換えフラグメント、または例えばmAb 2B6または2E3のような選択されたドナーmAbと同じ特異性を有する他の分子を含んでなっていてもよい。かかる蛋白を変化した抗体の形態で用いてもよく、あるいはその未融合形態で使用してもよい。
【0066】
第2の免疫グロブリンパートナーがドナー抗体とは異なる抗体、例えば、異なるイソタイプまたはクラスの免疫グロブリン枠組みまたは不変領域に由来する場合はいつでも、組み換え法による抗体が得られる。組み換え法による抗体は、1の起源、例えばアクセプター抗体由来の免疫グロブリン(Ig)不変領域および可変枠組み領域、ならびにドナー抗体、例えば本明細書記載の抗−IL−5抗体由来の1つまたはそれ以上(好ましくは全部)のCDRsを含んでなっていてもよい。さらに、核酸またはアミノ酸レベルでのアクセプターmAb軽鎖および/または重鎖可変ドメイン枠組み領域の変化、例えば、欠失、置換、または付加、あるいはドナーCDRの変化を行ってドナー抗体の抗原結合特異性を保持するようにしてもよい。
【0067】
IL−5 mAbの可変重鎖および/または軽鎖の一方(または両方)(所望により説明したように修飾されていてもよい)あるいは1つまたはそれ以上の下で定義する重鎖または軽鎖CDRs(図7も参照)を用いるようにかかる組み換え法による抗体を設計する。組み換え法による本発明抗体は中和抗体、すなわち、望ましくはIL−5受容体への結合をブロックし、さらにまたIL−5依存性細胞の増殖をブロックまたは防止するものである。
【0068】
かかる組み換え法による抗体は、選択されたヒト・免疫グロブリンもしくはサブタイプの枠組み領域を含むヒト化抗体、またはIL−5抗体の機能的フラグメントに融合したヒト・重鎖および軽鎖不変領域を含むキメラ抗体を包含しうる。適当なヒト(または他の動物)のアクセプター抗体は、ドナー抗体の核酸およびアミノ酸配列に対する相同性により、慣用的データベース、例えば、KABATRデータベース、Los Alamosデータベース、およびSwiss Proteinデータベースから選択されるものであってもよい。ドナー抗体の枠組み領域に対する相同性(アミノ酸による)により特徴づけられるヒト・抗体は、ドナーCDRsの挿入用の重鎖不変領域および/または重鎖可変領域枠組み領域の提供に適するかもしれない。軽鎖不変または可変枠組み領域を提供しうる適当なアクセプター抗体を同様の方法で選択してもよい。アクセプター抗体の重鎖および軽鎖は同じアクセプター抗体に由来する必要がないことに注意すべきである。
【0069】
望ましくは、異種枠組みおよび不変領域をヒト・免疫グロブリンのクラスおよびイソタイプ、例えば、IgG(サブタイプ1から4)、IgM、IgAおよびIgEから選択する。しかしながら、受容体抗体はヒト・免疫グロブリン蛋白配列のみを含んでなることを必要としない。例えば、ヒト・免疫グロブリン鎖の一部をコードしているDNA配列が、ポリペプチドエフェクターまたはレポーター分子のごとき非免疫グロブリンアミノ酸配列をコードしているDNA配列に融合している遺伝子を構築してもよい。
【0070】
特に望ましいヒト化抗体の一例は、選択されたヒト・抗体配列の枠組み領域上に挿入された2B6のCDRsを含む。中和ヒト化抗体とするには、IL−5抗体の重鎖および/または軽鎖可変領域由来の1個、2個または好ましくは3個のCDRsを、選択されたヒト・抗体配列の枠組み領域中に挿入し、ヒト・抗体の無傷のCDRsと入れ替える。
【0071】
好ましくは、ヒト化抗体において、ヒト・重鎖および軽鎖双方中の可変ドメインは組み換え法により1個またはそれ以上のCDRが置換されたものである。6個すべてのCDRsまたは6個未満のCDRsの種々の組み合わせを用いることが可能である。好ましくは、6個全部のCDRを置換する。ヒト・アクセプター抗体由来の未修飾軽鎖を軽鎖として用いて、ヒト・重鎖中でのみCDRsを置換することが可能である。さらに別法として、慣用的な抗体のデータベースを用いて、適合する軽鎖を別のヒト・抗体から選択してもよい。組み換え法による抗体の残りの部分はいずれかの適当なアクセプターヒト・免疫グロブリン由来であってもよい。
【0072】
よって、好ましくは、組み換え法によるヒト化抗体は、天然のヒト・抗体またはそのフラグメントの構造を有し、効果的な治療的使用、例えば、ヒトにおけるIL−5により媒介される炎症性疾患の治療、または診断用途に必要な特性の組み合わせを有する。
【0073】
もう1つの例として、組み換え法による抗体は、2E3の可変軽鎖領域の3つのCDRs[配列番号:10、11および13]および2B6の可変重鎖領域の3つのCDRs[配列番号:7、8および9]を含む。得られるヒト化抗体はmAb 2B6と同じ抗原結合特異性および高アフィニティーにより特徴づけられる。
【0074】
必ずしもドナー抗体の特異性および高アフィニティーに影響することなく、可変ドメインのアミノ酸を変化させることにより組み換え法による抗体をさらに修飾(すなわち、アナログを得る)してもよいことが当業者により理解されるであろう。重鎖および軽鎖のアミノ酸を、可変ドメイン枠組みまたはCDRsまたはそれらの両方のいずれかに存在する他のアミノ酸により置換してもよいと予想される。
【0075】
さらに、不変領域を変化させて本発明分子の選択特性を増加または減少させてもよい。例えば、ダイマー化、Fc受容体への結合、または補体結合および活性化能が挙げられる(例えば、Angal et al.,Mol.Immunol,30:105-108(1993)、Xu et al.,J.Biol.Chem,269,3469-3474(1994)、Winter et al.,EP307,434-B参照)。
【0076】
キメラ抗体である変化した抗体は、非ヒト・ドナー抗体の重鎖および軽鎖可変領域全体を提供し、両鎖についてヒト・免疫グロブリン不変領域に結合した枠組み領域を包含するという点で上記ヒト化抗体とは異なる。本発明ヒト化抗体に関連したさらなる非ヒト配列を保有しているキメラ抗体はヒトにおいて有意な免疫応答を誘導しうると予想される。
【0077】
かかる抗体は、上述のIL−5により媒介される疾病の予防および治療に有用である。
【0078】
VI.変化した抗体および組み換え法による抗体の製造
好ましくは、2B6または他の適当なドナーmAbs(例えば、2E3、2F2、4A6等)の可変軽鎖および/または重鎖配列およびCDRs、およびそれらをコードしている核酸配列を、変化した抗体、好ましくはヒト化抗体の構築に用いる。同一または類似の方法を用いて本発明の他の具体例を得てもよい。 選択されたドナーmAb、例えばマウス抗体2B6を産生するハイブリドーマを慣用的にクローンし、当業者に知られた方法、例えば、Sambrook et al., molecular Cloning (A Laboratory Manual),2nd Edition,Cold Spring Harbor Laboratory(1989)に記載の方法によりその重鎖および軽鎖可変領域のDNAを得る。ドナーmAb結合特異性を保持するのに必要な少なくともCDRをコードしている領域およびアクセプターmAb軽鎖および/または重鎖可変ドメイン枠組み領域含んでいる2B6の可変重鎖および軽鎖、ならびにヒト・免疫グロブリン由来の抗体鎖の残りの部分を、ポリヌクレオチドプライマーおよび逆転写酵素を用いて得る。既知データベースを用い、他の抗体と比較することによりCDRをコードしている領域を同定する。
【0079】
次いで、マウス/ヒト・キメラ抗体を調製し、結合能についてアッセイすることができる。かかるキメラ抗体は、ヒト・Ig不変領域に結合した非ヒト・ドナー抗体のVHおよびVL領域全体を含んでいる。
【0080】
コンピューターデータベース、例えばKABATRを用いてヒト抗体由来の重鎖可変領域の同種枠組み領域を同定し、2B6に対して相同性を有するヒト・抗体をアクセプター抗体として選択した。ヒト・抗体枠組み中の2B6のCDRをコードしている領域を含む合成重鎖可変領域の配列を、制限部位を含むように枠組み中のヌクレオチド置換を所望により行って設計した。次いで、この設計された配列を、長い合成オリゴマーを用いて合成した。別法として、ポリメラーゼ連鎖反応(PCR)により増幅され、エラーを修正されたオリゴヌクレオチドを重複させることにより、設計配列を合成することもできる。
適当な軽鎖可変枠組み領域を同様の方法で設計した。
【0081】
ヒト化抗体はキメラ抗体由来であってもよく、あるいは好ましくは、重鎖および軽鎖由来のドナーmAbのCDRをコードしている領域を選択された重鎖および軽鎖枠組み中に適当に挿入することにより合成的に得てもよい。別法として、本発明ヒト化抗体を標準的な突然変異法を用いて調製してもよい。かくして得られたヒト化抗体はヒト・枠組み領域およびドナーmAbのCDRをコードしている領域を含む。枠組み残基について引き続き操作を行ってよい。得られるヒト化抗体は組み換え宿主細胞、例えば、COS、CHOまたはミエローマ細胞において発現されうる。他の適当なIL−5特異的中和高アフィニティー非ヒト・抗体に対してこの方法を用いて他のヒト化抗体を調製してもよい。
【0082】
宿主細胞における複製および発現および/または宿主細胞からの分泌を制御しうる慣用的な調節的制御配列に作動可能に結合した変化した抗体の配列にもとのプラスミドのコーディング配列を置き換えることにより、慣用的な発現ベクターまたは組み換えプラスミドを製造する。調節配列はプロモーター配列、例えば、CMVプロモーター、およびシグナル配列を包含し、それらは他の既知抗体由来であってもよい。同様にして、相補的な抗体の軽鎖または重鎖をコードしているDNA配列を有する第2の発現ベクターを作成することもできる。好ましくは、この第2の発現ベクターは、コーディング配列および選択可能マーカーを除き、各ポリペプチド鎖が機能的に発現されることが可能なかぎり、第1のものと同じである。
【0083】
第1および第2の両方のベクターを用いる慣用的方法によって選択された宿主細胞を同時トランスフェクションして(あるいは単一ベクターによりトランスフェクションして)、組み換えまたは合成の軽鎖および重鎖両方を含んでなる本発明トランスフェクション宿主細胞を作成する。次いで、トランスフェクション細胞を慣用的方法により培養して本発明の組み換え法による抗体を得る。組み換え重鎖および/または軽鎖との結合を含むヒト化抗体を、ELISAまたはRIAのごとき適当なアッセイにより培養物からスクリーニングする。同様の慣用的方法を用いて本発明の他の変化した抗体および分子を構築してもよい。
【0084】
本発明方法および組成物の構築に用いるクローニングおよびサブクローニング工程に適したベクターが当業者により選択される。例えば、慣用的なpUCシリーズのクローニングベクターを用いてもよい。使用される1のベクターはpUC19であり、Amersham(英国Buckinghamshire)またはPharmacia(スウェーデンUppsala)のごとき業者から市販されている。さらに、容易に複製しうるいずれかのベクターは多くのクローニング部位および選択可能遺伝子(例えば、抗生物質耐性)を有しており、クローニングのための容易に取り扱いすることができる。よって、クローニングベクターの選択は本発明における制限因子ではない。
【0085】
同様に、本発明による組み換え法により得られた抗体の発現に用いるベクターは、いずれかの慣用的ベクターから当業者により選択されうる。またベクターは、選択された細胞において異種DNA配列の複製および発現を指令する選択された調節配列(CMVプロモーターのごとき)を含んでいる。これらのベクターは、組み換え法による抗体または変化した免疫グロブリンのコーディング領域をコードしている上記DNA配列を含んでいる。さらに、ベクターは、取り扱いが容易なように所望制限部位を挿入することにより修飾された選択免疫グロブリン配列を含んでいてもよい。
【0086】
異種DNA配列、例えば、哺乳動物ジヒドロ葉酸レダクターゼ(DHFR)の発現の増幅に適した遺伝子により発現ベクターを特徴づけてもよい。他の好ましいベクター配列は、ウシ・成長ホルモン(BGH)およびベータグロブリンプロモーター配列(betaglopro)のごときポリAシグナル配列を包含する。当業者によく知られた方法により本発明において有用な発現ベクターを合成してもよい。
【0087】
選択宿主における組み換えDNAからの生成物の発現および/または分泌の指令に有用なかかるベクターの成分、例えば、レプリコン、選択遺伝子、エンハンサー、プロモーター、シグナル配列等を、市販または天然ソースから得てもよく、あるいは既知方法により合成してもよい。
【0088】
また本発明は、組み換え法による抗体またはその変化した免疫グロブリン分子のコーディング配列を含む組み換えプラスミドでトランスフェクションした細胞系に関する。これらのクローニングベクターのクローニングおよび他の操作に有用な宿主細胞も慣用的である。しかしながら、最も望ましくは、種々のE.coli株の細胞をクローニングベクターの複製および本発明の変化した抗体の構築における他のステップに使用する。
【0089】
本発明の組み換え法による抗体または変化した抗体の発現に適する宿主または細胞系は、好ましくは、CHO、COS、線維芽細胞(例えば、3T3)、および骨髄細胞のごとき哺乳動物細胞、より好ましくは、骨髄細胞またはCHOである。ヒト・細胞を用いてもよく、かくして、分子をヒトのグリコシレーションパターンで修飾することが可能となる。別法として、他の真核細胞系を用いてもよい。形質転換、培養、増幅、スクリーニングならびに生成物の生産および精製に適した哺乳動物宿主細胞および方法の選択は当該分野において知られている。例えば、上で引用したSambrookらの文献参照。
【0090】
細菌細胞は本発明組み換えFabの発現に適した宿主細胞であることがわかるだろう(例えば、Pluckthun,A.,Immunol.Rev.,130:151-188(1992)参照)。しかしながら、細菌細胞において発現された蛋白の、変性または正しく折り畳まれていない形態あるいはグリコシレーションされていない形態となる傾向のため、細菌細胞において生産された組み換えFabを抗原結合能の保持についてスクリーニングしなければならない。細菌細胞により発現された分子が正しく折り畳まれた形態で生産された場合には、当該細菌細胞は望ましい宿主であろう。例えば、発現に使用される種々のE.coli株は、バイオテクノロジーの分野において宿主としてよく知られている。種々のB.subtilis、Streptomyces、他の枯草菌等の株も本発明方法に使用できる。
【0091】
所望ならば、当業者に知られた酵母細胞株ならびに昆虫細胞、例えば、DrosophilaおよびLepidoptera、およびウイルス発現系も宿主細胞として用いられる。例えば、Miller et al.,Genetic Engineering,8:277-298,Plenum Press(1986)およびその中の引用文献参照。
【0092】
本発明ベクターを構築することのできる一般的方法、本発明宿主細胞を製造するのに必要なトランスフェクション法、およびかかる宿主細胞から本発明の変化した抗体を製造するのに必要な培養法はすべて慣用的方法である。同様に、生産されたならば、硫酸アンモニウム沈殿、アフィニティーカラム、カラムクロマトグラフィー、ゲル電気泳動等を包含する当該分野の標準的方法により本発明の変化した抗体を細胞培養物から精製することができる。かかる方法は当業者の範囲内であり、本発明を制限するものでない。
【0093】
ヒト化抗体のさらなる発現方法は、米国特許第4,873,316号記載のごとく、トランスジェニック動物における発現を用いるものであってもよい。これは、動物のカゼインプロモーター(トランスジェニック的に哺乳動物に導入された場合、メスの哺乳動物はその乳中の所望組み換え蛋白を生産する)を用いる発現系に関連している。所望方法により発現されたならば、適当なアッセイを用いて、組み換え法による抗体をインビトロ活性について試験する。現在慣用的となっているELISAアッセイフォーマットを用いて、組み換え法により抗体のIL−5への結合を定量的かつ定性的に評価する。さらに、他のインビトロアッセイを用いて、通常のクリアランス機構存在下の体内での組み換え抗体の維持を評価するために引き続き行われるヒトの臨床研究を行う前に中和効率を確認しておいてもよい。
【0094】
2B6から調製されたヒト化抗体について説明された方法に従って、当業者は、本明細書記載の他のドナーIL−5抗体、可変領域配列およびCDRペプチドからヒト化抗体を構築することができる。レシピエントにより「自己」であると潜在的に認識される可変領域枠組みを用いて組み換え法による抗体を製造することができる。可変領域枠組みに対する小さな修飾を行って、レシピエントに対する免疫原性を有意に上昇させずに抗原結合を促進することができる。かかる組み換え法による抗体は、ヒトのIL−5により媒介される症状を効果的に治療する可能性がある。
【0095】
VII. 治療的/予防的使用
また本発明は、有効量の本明細書記載の1種またはそれ以上の組み換え法による抗体または変化した抗体を包含する抗体、あるいはそれらのフラグメントを投与することを特徴とする、喘息のごとき好酸球増加症関連症候群にかかっているヒトの治療方法に関する。
【0096】
ヒト・IL−5への結合、次いで、好酸球刺激のブロックにより、本発明分子の使用による治療的応答を得る。よって、本発明分子は、治療用途に適した調合品および処方に含まれた場合、アレルギー性鼻炎、喘息、慢性好酸球性肺炎、アレルギー性気管支肺アスペルギルス症、セリアック病、好酸球性胃腸炎、Churg-Strauss症候群(結節性動脈周囲炎およびアトピー)、好酸球性筋肉痛症候群、過好酸球増加症候群、血管皮膚炎を包含する水腫性反応、ぜん虫感染(ここで、好酸球は防御的役割をしている可能性がある)、オンコセルカ皮膚炎およびアトピー性皮膚炎のごときアレルギー性および/またはアトピー性応答、あるいは好酸球増加症に関連した応答をしているヒトに対して非常に望ましい。
【0097】
他の抗体、詳細には本発明組み換え法による抗体が指向する症状の原因となりうる他のマーカー(エピトープ)と反応するヒト・mAbと組み合わせて本発明の変化した抗体、抗体およびそれらのフラグメントを用いてもよい。
【0098】
本発明治療剤は、約2日ないし約3週間、あるいは必要な期間のアレルギー症状の治療に望ましいと考えられる。例えば、季節性鼻炎等を治療する場合には、より長期の治療が望ましい。このことは、IL−5により媒介される疾病に対する現用の先行技術である輸液プロトコールよりもかなり有利であることを示す。治療のための用量および期間は、ヒトの循環系における本発明分子の相対的持続時間に関係し、治療すべき症状および患者の一般的健康状態に応じて当業者により調整されうる。
【0099】
本発明治療剤の投与モードは、宿主に対して剤を送達するいずれの適切な経路であってもよい。変化した抗体、抗体、組み換え法による抗体、およびそれらのフラグメント、ならびに本発明医薬成分は、詳細には、非経口投与、すなわち、皮下、筋肉内、静脈または鼻腔内投与において有用である。
【0100】
本発明治療剤を、医薬上許容される担体中の有効成分としての有効量の本発明の組み換え法による(例えば、ヒト化)抗体を含有する医薬組成物として調製してもよい。本発明の予防剤において、組み換え法による抗体を含有する、好ましくは生理学的pHに調節された即座に注射可能な形態の水性懸濁液または溶液が好ましい。通常には、非経口投与用組成物は本発明の組み換え法による抗体の溶液または医薬上許容される担体(好ましくは水性担体)に溶解されたそのカクテルを含んでなる。種々の水性担体を用いることができ、例えば、0.4%食塩水、0.3%グリシン等を用いることができる。これらの溶液は滅菌されており、一般的には微粒子不含である。よく知られた滅菌法(例えば、濾過)によりこれらの溶液を滅菌することができる。組成物は、pH調節および緩衝化剤等のごとき、所望により医薬上許容される添加物を含んでいてもよい。かかる医薬処方中の本発明抗体の濃度を広範囲に、すなわち、重量で約0.5%未満から、通常には約1%またはそれ未満ないし15%または20%まで変化させることができ、主として液体の体積、粘度等に基づいて選択され、選択された個々の投与モードに応じて選択されるであろう。
【0101】
よって、筋肉内注射用の本発明医薬組成物を、1mlの滅菌緩衝水および約1ngないし約100mg、例えば約50ngないし約30mg、またはより好ましくは約5mgなし約25mgの本発明の組み換え法による抗体を含有するように調製することができる。同様に、静脈輸液用の本発明医薬組成物を、約250mlの滅菌リンゲル溶液、および約1ないし約30mg、好ましくは5mgないし約25mgの本発明の組み換え法による抗体を含有するように調製することができる。非経口的投与可能組成物の調製のための実際的な方法は当業者によく知られているかまたは明らかであり、例えばRemington's Pharmaceutical Science,15th ed.,Mark Publishing Company,Easton,Pennsylvaniaにおいてより詳細に記載されている。
【0102】
本発明治療剤は、医薬組成物中に入れる場合、1回分の剤型として提供するのが好ましい。適当な治療上有効量は当業者により容易に決定されうる。ヒトまたは他の動物における炎症性疾患を有効に治療するためには、1回分の用量は、体重70kgにつき約0.1mgはいし約20mgの本発明抗体を非経口的に、好ましくは静脈または筋肉内投与すべきである。必要ならば、炎症応答中に医師がかかる用量を適当時間間隔をおいて調製してもよい。
【0103】
本発明の変化した抗体および組み換え法による抗体を、IL−5により媒介される疾病の決定またはかかる疾病の治療進行の追跡のごとき診断に用いてもよい。診断には、これらの変化した抗体を、慣用的には、血清、血漿または他の適当な組織中のIL−5レベルの測定あるいは培養中のヒト細胞による放出の測定のためにELISAまたは他の慣用的アッセイフォーマット用に標識することができる。変化した抗体が用いられるアッセイの性質は慣用的なものであり、本開示を限定するものでない。
【0104】
よって、本発明の1の具体例は、患者における過剰な好酸球産生に関連したアレルギーおよび他の症状の診断を援助する方法であって、患者から得た試料(血漿または組織)中のヒト・IL−5の量を決定し、次いで、決定した量を正常集団におけるヒト・IL−5平均量と比較することを特徴とし、そのことにより患者試料中の有意に上昇したIL−5量が過剰な好酸球産生に関連したアレルギーおよび他の症状を示すものとなる方法に関する。
【0105】
本明細書記載の抗体、変化した抗体またはそれらのフラグメントを保存のための凍結乾燥し、使用のために適当な担体中で復元することができる。この方法は、慣用的免疫グロブリンに関して効果的であることが示されており、当該分野で知られている凍結乾燥および復元方法を用いることができる。
【0106】
下記実施例は、典型的な組み換え法による抗体の構築および適当なベクターおよび宿主細胞中でのその発現を包含する本発明の種々の態様を説明するが、本発明の範囲を限定するものと解してはならない。すべてのアミノ酸は慣用的な3文字または1文字法により表される。特記しないかぎり、すべての必要な制限酵素、プラスミド、ならびに他の試薬および材料は市販のものである。すべての一般的クローニング、連結および他のDNA組み換え法は、T.Maniatisら(すでに引用)、または同じ出版社からのその第2版(1989)(Sambrookら編)(「Sambrookら」という)において行われたものである。
【実施例1】
【0107】
実施例1 hIL−5(ヒト・IL−5)に対するMabs(モノクローナル抗体)の製造
ヒト・IL−5をDrosophila Schneider 2(S2)細胞において発現させ、均一に精製した。Spodoptera frugiperda 21(Sf21)細胞を用いてマウス・IL−5をバキュロウイルス中で発現させ、均一に精製した。モノクローナル抗体TRFK−5(中和ラット・抗−マウスIL−5抗体)をGenzyme Corp.(Cambridge,MA)から得た。
【0108】
A.免疫手順
組み換えヒト・IL−5(IL−5)を、7匹のCAF1マウス(メス)(Charls River,Wilmington,MA)のパネルに対する免疫原として用いた。4カ月の期間にわたり、1:1の割合のTiterMAX(登録商標)(CytoRx Corp.,Norcross,GA)を用いて乳化したリン酸緩衝化食塩水(PBS)中のIL−5を動物に3回皮下注射した。最初の抗原量は50μgであり、追加免疫は25μgおよび10μgであった。追加免疫後、血清試料を集め、受容体結合阻害アッセイおよびB13増殖アッセイ(またはIL−5中和アッセイ(実施例2C))により、IL−5への結合および中和活性についてアッセイした。すべてのマウスは、IL−5に結合する血清試料を産生した。脾臓ドナーとして選択された動物を安楽死させる3日前に10μgの組み換えヒト・IL−5を静脈から追加免疫した。
【0109】
B.ハイブリドーマ取得
Kohler et al.,Nature,256:495(1975)により初めて報告された融合法を変更して用いて、細胞単層(Kennet et al.,Eds.,"Hybridomas:A new dimension in biological analysis",pp.368-377,Plenum Press,New York)を用いる方法を行った。2匹のドナーマウスからの脾臓細胞をプールし、1千万個のSP2/0/Ag14骨髄腫細胞に対して5千万個の脾臓細胞の割合で融合を行った。融合が確認された細胞の上清を、IL−5に対する結合に関してELISAによりアッセイした。IL−5に対する抗体を産生する細胞を含むウェルを増殖させ、上清をIL−5受容体結合阻害アッセイおよびB13(中和)増殖アッセイにおいてスクリーニングした(下記)。
【0110】
IL−5と反応するmAbsを分泌する16個のハイブリドーマを単離した。ハイブリドーマ上清をヨウ素化IL−5と混合し、IL−5受容体(IL−5R)のα−鎖を発現するDrosophila細胞から調製した抽出物に添加し、受容体結合の阻害に関してアッセイした。11個のハイブリドーマ上清は、IL−5受容体α−鎖へのヨウ素化IL−5の結合を60%よりも多く阻害した。3個のmAbs(2B6、2E3および2F2)も、ヒト・IL−5に応答してマウス・B13細胞の増殖を70%よいも多く阻害したが、マウス・IL−5には応答しなかった。5個のハイブリドーマ(そのうち4個(1C6、2B6、2E3および2F2)は結合および/または増殖をブロックし、1個(24G9)は中和的でなかった)を軟寒天で繰り返しサブクローンして安定なクローン細胞系を得た。クローン細胞系からの上清を交差反応性についてELISAによりスクリーニングしたが、ヒト・IL−1α、IL−1β、IL−4、IL−8、M−CSFまたはTGFαには結合しなかった。mABsを精製し、光学的バイオセンサー(BIAcore)分析により結合アフィニティーを評価したところ、10ないし100pMの範囲であった。マウス・イソタイプ分類試薬(PhaMingen,San Diego,CA)を用いて細胞系からの上清をイソタイプに分類した。MAbsの中和活性に関するアフィニティーおよびIC50のまとめを表1(実施例2)に示す。
【0111】
同様の方法により、対応する免疫プロトコールおよびラット・骨髄腫をマウスについて上記した融合に用いてラット・ハイブリドーマを免疫ラットから得た。2つのラット・ハイブリドーマ4A6および5D3は、IL−5に結合するmAbsを産生することが同定された。2B6、2E3および2F2と同様に、mAbs4A6および5D3は、下記B13アッセイにおいて中和的であることがわかった。
【0112】
C.ハイブリドーマの寄託
モノクローナル抗体2B6を産生するハイブリドーマ細胞系SK119−2B6.206.75(1)は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11783として寄託され、特許寄託物として受託された。
【0113】
モノクローナル抗体2E3を産生するハイブリドーマ細胞系SK119−2E3.39.40.2は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11782として寄託され、特許寄託物として受託された。
【0114】
モノクローナル抗体2F2を産生するハイブリドーマ細胞系SK119−2F2.37.80.12は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11781として寄託され、特許寄託物として受託された。
【0115】
モノクローナル抗体24G9を産生するハイブリドーマ細胞系SK119−24G9.8.20.5は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11780として寄託され、特許寄託物として受託された。
【0116】
モノクローナル抗体4A6を産生するハイブリドーマ細胞系4A6(1)G1F7は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11943として寄託され、特許寄託物として受託された。
【0117】
モノクローナル抗体5D3を産生するハイブリドーマ細胞系5D3(1)F5D6は、特許手続上の微生物の寄託に関するブダペスト条約に従って、American Type Culture Collection(ATCC),Rockville,MD,USAに受託番号HB 11942として寄託され、特許寄託物として受託された。
【実施例2】
【0118】
実施例2 アッセイ
A. ELISA
MaxiSorb(登録商標)イムノプレート(Nunc,Naperville,IL)の個々のウェルを、0.05M炭酸バッファーpH9.6中0.2μgのIL−5で被覆した。4℃で一晩インキュベーション後、0.025% TweenR20含有PBSでプレートをすすぎ、0.025% TweenR20含有PBS中1% BSAにて室温で2時間ブロックした。未希釈ハイブリッド上清をIL−5被覆ウェルに添加し、室温で2時間インキュベーションした。プレートをすすいだ後、ヤギ・抗−マウスIgGおよびIgM標識ペルオキシダーゼ(Boehrionger Mannheim,Indianapolis,IN)を、1% BSAおよび0.025%TweenR20含有PBS中1/7500希釈して添加した。15分後、VMax(登録商標)マイクロプレートリーダー(Molecular Devices, Menlo Park,CA)により450nmにおいてプレートを読んだ。
【0119】
B.受容体結合阻害アッセイ
ヒト・IL−5受容体(IL−5R)のα−鎖を発現するDrosophila S2細胞の膜抽出物を用いて受容体へのIL−5の結合に対する抗体の効果を測定した。膜を調製するために、109個の細胞を1000xg、4℃、10分間ペレット化した。細胞ペレットをドライアイス/エタノール浴で15分凍結した。ペレットを融解し、4℃の10mlのPBSに再懸濁し、1000xgで10分間ペレット化した。細胞ペレットをPBSで2回洗浄し、13.5mlの低張バッファー(10mM Tris pH7.5、3mM MgCl2、1mM ジチオスレイトール、1mM フェニルメチルスルホニルフルオリド、1μM ロイペプチン、1μM ペプスタチンA)に再懸濁し、氷上で5分間インキュベーションした。細胞懸濁液を15mlのDounceホモジナイザーでホモジナイズし、2.5M蔗糖溶液を用いて蔗糖最終濃度0.25Mとした。1000xgで15分の遠心分離により細胞残渣を除去した。細胞膜を100000xg、4℃で90分間ペレット化し、10mM Tris pH7.5、3mM MgCl2、250mM 蔗糖中に再懸濁し、−70℃で保存した。
【0120】
受容体を含むDrosophila膜を用いるアッセイを、25mM HEPESバッファー pH7.2および0.1% BSA(結合バッファー)を含有するDrosophila組織培養培地M3(Lindquist et al.,Drosophila Inf.Serv.,58:163(1982))を用いて、MultiscreenGV(登録商標)プレート(Millipore Corp.,Bedford,MA)中で行った。0.1mlの結合バッファーを用いてウェルをプレブロック(preblock)した。50μlの試験試料(3系とした)をウェルに添加し、次いで、25μlのヨウ素化(125I)IL−5を添加した。20分室温でインキュベーションした後、ヒト・IL−5Rのα−鎖を発現しているDrosophila S2細胞の膜抽出物25μlをウェルに添加した。さらに1時間インキュベーションした後、減圧濾過により膜を集め、結合バッファーで3回洗浄した。フィルターを乾燥し、カウントした。
【0121】
C. IL−5中和アッセイ
親切にも、R.Palacios(Basel Institute of Immunology,Switzerland)からマウス・IL−5/IL−3依存性細胞系LyH7.B13(B13)を得た。L−グルタミン、非必須アミノ酸、ピルビン酸ナトリウム、ペニシリン−ストレプトマイシン(すべてGibcoBRL)ならびに2−メルカプトエタノール(5x10−5M,Sigma)、10% ウシ胎児血清(Globepharm,Surrey,UK)および1〜10ユニットのマウス・IL−5を補足したRPMI 1640培地(GibcoBRL, Renfrewshire,UK)中で1週間に2回細胞をサブクローンした。アッセイのために、96ウェル丸底プレート中、適当に希釈した試験試料の存在下、3系で48時間培養し、次いで、0.5μCiの3H−チミジン(Amersham,Bucks,UK)を添加して最後の4時間のインキュベーションを行った。それらを1205 Beteplate(LKB Wallac,Beds,UK)におけるシンチレーションカウンティングのために処理した。
【0122】
D.光学的バイオセンサー
固定化hIL−5と抗体に関する速度論的平衡結合特性を、BIAcore光学的バイオセンサー(Pharmacia Biosensor,Uppsala,Sweden)を用いて行った。以前に記載されているようにして(Karlsson et al.,J.Immunol.Meth.,145:229-240(1991)(その全体を参照により本明細書に記載されているものと見なす))速度論的データを評価した。
【0123】
3つの中和mAbs、すなわち、2B6、2E3および2F2は、膜受容体への125I−IL−5の結合に対する阻害およびB細胞増殖の中和について類似した能力を有しおり、またIL−5に対して非常に類似したアフィニティーを有していた(表1参照)。これら3つのmAbs、すなわち2つのIgG1および1つのIgG2aからのVHおよびVLのヌクレオチド配列をそれぞれ決定した。得られた配列は非常に類似しており、ほんの少数の残基のみが異なっていた。
【0124】
【表1】
【実施例3】
【0125】
実施例3 組み合わせライブラリーからのIL−5 Fabsの単離および特徴づけ
A. PCRおよび組み合わせライブラリーの構築
Huse et al.Science,246:1275(1989)およびKang,S.A.Methods:Comparison Methods Enzymol.,2:111(1991)(それら全体を参照により本明細書に記載されているものと見なす)により記載されたようにして、3匹のマウスの脾臓から精製したRNAを、cDNAキット(Boehringer Mannheim,Indianapolis,IN)により逆転写した(キット付属のプライマー(dT)15または3’Fd(IgG1、IgG2aおよびIgG3)およびカッパ軽鎖プライマーを用いる)。記載されたプライマーおよび温度サイクリング条件(上記Huseら)を用いてPCRにより免疫グロブリンcDNAを増幅した。AmpliWaxTMPCR Gem 100(Perkin Elmer Cetus,Norwalk,CT)ビーズおよび製造者のプロトコールを用いるホット・スタート(Hot Start)法をすべての反応に使用した。PCR生成物をゲル精製し、消化し、pMKFabGene3ベクター(Ames et al.,J.Immunol.,512:4572(1994))中に連結した。Fd cDNAを用いる連結後のライブラリー力価は5.1X107CFUであり、カッパcDNAを用いるライゲーション後の力価は1.5X106CFUであった。ファージミドライブラリーを用いて形質転換されたXL1-Blue細胞(Stratagene,La Jolla,CA)を、ヘルパーファージVCSM13(Stratagene)に感染させ、Barbas and Lerner,Methosa:Comparison Methods Enzymol.,2:119(1991)により記載されたようにしてファージを調製した。
【0126】
B.バイオパンニング(Biopanning)
4つのマイクロタイターウェル(Immulon II Removawell Strips,Dynatech Laboratories Inc.,Chantilly,VA)を、0.1M重炭酸,pH8.6中IL−5(1μg/ウェル)で4℃において被覆した。ウェルを水洗し、3% BSA含有PBSを用いて37℃で1時間ブロックした。ブロッキング溶液を除去し、ライブラリーをマイクロタイターウェルに添加し(50μl/ウェル)、37℃で2時間インキュベーションした。ウェルをTBS/TweenR(50mM Tris−HCl,pH7.5、150mM NaCl、0.5% TweenR20)で10回洗浄し、水で1回洗浄し、次いで、0.1M HCl(1mg/ml BSA含有グリシンでpH2.2に調節)で付着ファージを溶離した。
【0127】
C.コロニー釣菌
バイオパンニングの第3ラウンドおよび第4ラウンドから単離されたコロニーからのコロニー釣菌を記載されたようにして(Barbas and Lerner、上記)行った。0.5〜1.0μCiの125I−IL−5とともにフィルターを室温で1時間インキュベーションし、製造者の推奨手順に従ってBolton-Hunter試薬(NEN,Billerica,MA)を用いて1% BSA含有PBS中でヨウ素化し、PBS 0.25% Tweenで洗浄し、Kodak XARフィルムにさらした。IL−5反応性Fabsを発現しているコロニーをオートラジオグラフィーにより検出した。
【0128】
D.可溶性FABsの調製
ファージミドDNAをNheIおよびSpeIで消化して遺伝子IIIを除去し、自己連結させた。XL1−Blue細胞を形質転換し、単離されたコロニーを、1%グルコースおよび50μg/mlカルベニシリンを含有する5.0mlのスーパーブロス(SB)培地(30gトリプトン、20g酵母エキス、10gの3−[N−モルホリノ]プロパンスルホン酸(MOPS)、pHを7に調節)中で37℃で一晩増殖させた。この培養1mlからの細胞をBeckman GS-6R遠心分離機にて3500rpmで10分間ペレット化し、50μg/mlのカルベニシリンを含有する5mlのSBに植菌した。培養物を37℃で1時間振盪し、イソプロピル−b−D−チオガラクトピラノシド(IPTG;1mM)を添加し、培養物を28℃に移し、一晩おいた。30mM Tris pH8.0に懸濁した20%蔗糖中、4℃で20分間細胞を溶解し、次いで、微量遠心機で10分間遠心分離することによりペリプラスム抽出物から可溶性Fabを調製した。ウェルタンブロットにより、既知量のマウス・Fabを含有する試料と比較することによってFab濃度を評価した。別の細菌のペリプラスム抽出物は同様の濃度のFabを含んでおり、ウェスタンブロット分析による分析では1ないし20μg/mlの範囲であった。
【0129】
E. FABsの精製
重鎖のカルボキシ末端上にキレートするペプチドを加工作成して蛋白精製の助けとした。NheIおよびSpeIでの消化によるM13遺伝子IIIコーディング領域の除去後、6個のヒスチジン残基をコードしている1対の重複オリゴヌクレオチド:[配列番号:43]5’−CTAGCCACCACCACCACCACCACTAA−3’;[配列番号:44]3’−GGTGGTGGTGGTGGTGGTGATTGATC−5’をFab発現ベクター中にサブクローンした。上記のごとくFab発現の誘導を行った。28℃で一晩インキュベーションした後、20%蔗糖、30mM Tris pH8.0中4℃で30分インキュベーションすることにより細胞ペレットのペリプラスム溶解物を調製した。尿素およびBrij−35界面活性剤を清澄化した上清に添加して、最終濃度をそれぞれ、2Mおよび1%とした。室温で1時間撹拌後、処理され清澄化された上清を、5mlのニッケル−NTA金属キレーティングカラム(1.5x3cm)(バッファーA(100mM Na−リン酸、10mM Tris、0.3M NaCl、2M尿素,pH8.0)で平衡化されている)に、流速0.5ml/分で直接負荷した。4カラム体積分(20ml)の洗浄後、上と同じバッファー中pHを8から4にする6カラム体積分(30ml)の逆pHグラジエントで結合物質を溶離した。精製されたFabsは、pH5.5において対称形の鋭いピークとなってカラムから溶出した。精製Fabsは>90%の純度であり、DNA不含であった。
【0130】
F. FAB ELISA
0.1M重炭酸バッファー,pH8.6に懸濁された(1mg/ml;ウェル1個あたり50ml)蛋白でImmulon IIプレート(Dynatech)を4℃で一晩被覆した。0.05% Tween(登録商標)20含有PBSにて希釈および洗浄を行った。プレートを洗浄し、1%BSA含有PBSにて室温で1時間ブロッキングした。可溶性Fabを含有する細菌懸濁液の種々の希釈物、または精製Fabsをプレートに添加した。1時間インキュベーション後、プレートを洗浄し、ビオチン化ヤギ・抗−マウスカッパ(Southern Biotechnology Associates,Inc.,Birmingham,AL)を添加(1:2000希釈;50μl/ウェル)して1時間おいた。プレートを洗浄し、ストレプトアビジン標識セイヨウワサビペルオキシダーゼを添加し(1:2000希釈;50μl/ウェル)1時間おいた。プレートを洗浄し、ABTSペルオキシダーゼ基質を添加し(100μl/ウェル;Kirkegaard & Perry Laboratories,Gaithersberg,MD)、405nmにおける光学密度をUVmax(登録商標)(Molecular Devices)マイクロプレートリーダーで読んだ。
【0131】
G.組み合わせライブラリーからのFabsの単離および特徴付け
IL−5で被覆したマイクロタイターウェルに対する複数ラウンドのバイオパンニングにより、IL−5に対するFabsを有するファージをライブラリーから選択した。4ラウンドの選択後、125I−IL−5を用いるコロニー釣菌アッセイによりIL−5反応性Fabsが同定された。第3ラウンドから34個のコロニーが、第4ラウンドから4個のコロニーが同定され、それらは標識IL−5に結合した。Fab−遺伝子III融合蛋白を発現している培養上清を用いる直接結合ELISAによりIL−5への結合を確認した。これらのコロニーからDNAを単離し、M13遺伝子IIIのコーディング領域を除去後、可溶性Fabの発現を誘導した。ペリプラスムフラクションを調製し、IL−5への結合についてELISAによりアッセイした。Fabは、別の蛋白rC5aへの結合を示すことなくIL−5に特異的に結合した。
【0132】
未希釈ペリプラスム抽出物(1ないし20μg/mlのFabを含有)を、IL−5R結合阻害アッセイ(実施例2)においてアッセイした。L−5Rαへのヨウ素化IL−5の結合を35%よりも多く阻害するFabはなかった。
【0133】
H.中和mAbのFABへの変換
FdおよびmAb(2B6)のκcDNAを、上記条件を用いるPCRにより単離した。ゲル精製したフラグメントをpMKFabGene3ベクター中にサブクローンし、これを修飾して遺伝子IIIのcDNAの3’末端にヘキサ−His配列を含むようにして、プラスミドpMKFabGene3Hを得た。2B6重鎖および軽鎖を有する機能的なIL−5結合Fabクローンをコロニー釣菌アッセイにより同定した。NheI/SpeI消化による遺伝子IIIの除去および自己連結を行って、重鎖をイン−フレームでヘキサ−Hisに融合させ、上記のごとく精製を可能にした。このFabは用量依存的に受容体結合を阻害し、IC50は約7.5μg/mlであり、親mAbであるマウス・2B6のIC50と同様であった。
【0134】
I.チェイン−シャッフル(chain-shuffle)されたライブラリーの構築およびスクリーニング
中和mAb 2B6のFdをコードしているcDNAを、XhoI/SpeIフラグメントとしてpMKFabGene3H中にサブクローンして、軽鎖cDNAのかわりにSstI/XbaIフラグメントを含ませた。このファージミドをSstIおよびXbaIで消化し、IL−5免疫マウス由来(上記)のSstI/XbaI消化された軽鎖のPCR生成物と連結した。連結後のライブラリー力価は4x105CFUであった。バイオパンニングおよびコロニー釣菌アッセイを、組み合わせライブラリーに関して上で説明したように行った。
【0135】
中和mAb 2B6のFdをコードしているcDNAを、IL−5免疫マウスから回収された同じ軽鎖レパートリーと対合させることによりライブラリーを構築した。このチェインシャッフルされたライブラリーを、固定化IL−5に対する4ラウンドのバイオパンニングに供し、得られたコロニーをコロニー釣菌アッセイを用いてIL−5反応性に関してアッセイした。ヨウ素化IL−5に結合する陽性コロニーを、ELISAおよびIL−5Rα結合アッセイによりさらにアッセイした。チェインシャッフルされたライブラリーから回収されたFabのうち2個(2および15)がIL−5RαへのIL−5の結合をブロックし、B13アッセイにおいてIL−5依存的増殖を抑制した。これら2個のVkの配列は2B6のVk(2B6のVHに対するもとの軽鎖パートナー)の配列と類似であった。Fab2および15に関する軽鎖配列は、それぞれ配列番号:45および46である。Fab2については、CDR1〜3はそれぞれ配列番号:10、11および47である。Fab15については、CDR1〜3はそれぞれ配列番号:10、11および48である。
【0136】
下記実施例4および5に記載したすべての抗体のアミノ酸配列はKABATナンバーリングシステムを用い、このシステムにおいてはCDRおよび枠組み長が変化してもかまわない。すなわち、鍵アミノ酸は、それに先行するアミノ酸数にかかわらず常に同じ番号を与えられる。例えば、すべての軽鎖のCDR1に先行するシステインは、常にKABAT位置23であり、たとえCDR1が17個までのアミノ酸を含む場合であってもCDR1の後に続くトリプトファン残基は常にKABAT位置35である。
【実施例4】
【0137】
実施例4 ヒト化抗体
マウス・CDRsをヒト化抗体枠組み中に含むように1のヒト化抗体を設計した。下記操作を行うことにより、IL−5特異的マウス・抗体2B6のこのヒト化バージョンを調製した。
【0138】
A.遺伝子クローニング
Boehringer Mannheim(Indianapolis,IN)から得たキットを用いて2B6、2F2および2E3ハイブリドーマ細胞系(実施例1)のそれぞれからmRNAを単離し、次いで、cDNAキット(Boehringer Mannheim)付属のプライマー(dT)15を用いて逆転写してcDNAを作成した。マウス・免疫グロブリンに特異的なPCRプライマーを用いて、重鎖可変領域のアミノ酸9(KABATナンバーリングシステム)からヒンジ領域まで伸長しているドメインならびに軽鎖可変領域のアミノ酸9(KABATナンバーリングシステム)から不変領域の末端まで伸長しているドメインをコードしているDNAを増幅した。独立別個のPCR反応により各抗体鎖の数個のクローンを得た。
【0139】
使用したマウス・ガンマ1ヒンジ領域プライマーは[配列番号:22]:5’GTACATATGCAAGGCTTACAACCACAATC3’であった。
【0140】
使用したマウス・ガンマ2aヒンジ領域プライマーは[配列番号:23]:5’GGACAGGGCTTACTAGTGGGCCCTCTGGGCTC3’であった。
【0141】
使用したマウス・重鎖可変領域プライマーは[配列番号:24]:5’AGGT(CまたはG)(CまたはA)A(GまたはA)CT(GまたはT)TCTCGAGTC(TまたはA)GG3’であった。
【0142】
使用したマウス・カッパ鎖不変領域プライマーは[配列番号:25]:5’CTAACACTCATTCCTGTTGAAGCTCTTGACAATGGG3’であった。
マウス・軽鎖可変領域プライマーは[配列番号:26]:5’CCAGATGTGAGCTCGTGATGACCCAGACTCCA3’であった。
【0143】
PCRフラグメントをプラスミドpGEM7f+(Promega)中にクローンし、次いで、それをE.coli DH5a(Bethesda Reaearch Labs)中に形質転換した。
【0144】
B. DNA配列決定
上記Aで得た重鎖および軽鎖マウスcDNAクローンを配列決定した。これらのクローンの可変領域の配列決定結果を配列番号:1〜6(図1〜6)に示す。各クローンは、マウス・重鎖可変領域または軽鎖可変領域間で保存されていることが知られているアミノ酸を含んでいた。CDRアミノ酸配列を以下に記載する。 2B6重鎖に関するCDR領域は配列番号:7、8および9である。図7参照。これらの配列は配列番号:1によりコードされている。軽鎖に関するCDR領域は配列番号:10、11および12である。図7参照。これらの配列は配列番号:2によりコードされている。
【0145】
2F2重鎖に関するCDR領域は配列番号:7、8および9である。図7参照。これらの配列は配列番号:3によりコードされている。軽鎖に関するCDR領域は配列番号:10、11および13である。図7参照。これらの配列は配列番号:4によりコードされている。
【0146】
2E3重鎖に関するCDR領域は配列番号:7、8および14である。図7参照。これらの配列は配列番号:5によりコードされている。軽鎖に関するCDR領域は配列番号:10、11および13である。図7参照。これらの配列は配列番号:6によりコードされている。
【0147】
C.ヒト・枠組みの選択
2B6のクローニング後、KABATおよびSWISS−PROT(Nuc. Acids Res.,20:2019-2022(1992))蛋白配列データベースにおいて可変領域重鎖および軽鎖のアミノ酸配列(図1および2)(それぞれ配列番号:15および16)を既知マウス・免疫グロブリン配列と比較してN末端残基に割り当てた。次いで、2B6重鎖および軽鎖可変領域の推定アミノ酸配列をヒト・免疫グロブリン蛋白配列データベースと比較して、マウス配列に最もぴったりと合致する重鎖および軽鎖両方のヒト・枠組みを同定した。さらに、CDR表現に影響しうるアミノ酸による潜在的矛盾を評価するためにFabドメインの構造的モデルから得られた位置のデータベースを用いて重鎖および軽鎖を評価した。矛盾位置における対応マウス・アミノ酸の置換により、ヒト化可変領域枠組みの合成中に矛盾は解決された。
【0148】
ヒト骨髄腫免疫グロブリン(COR)から得られた抗体の重鎖枠組み領域を用いた(E.M.Press and N.M.Hogg,Biochem.J.,117:641-660(1970))。ヒト・重鎖枠組みアミノ酸配列は2B6枠組みに対して約66%の相同性があることが見いだされた。
【0149】
適当な軽鎖可変領域枠組みについては、Bence-Jones蛋白(LEN)(Schneider et al.,Hoppe-Seyler's Z.Physiol.Chem.,356:507-557(1975))の軽鎖可変枠組み配列を用いた。ヒト・軽鎖枠組み領域は、マウス・2B6軽鎖枠組み領域に対して、アミノ酸レベルで約82%の相同性があった。
【0150】
選択されたヒト・枠組みを戻し翻訳(back translate)してDNA配列を得た。
【0151】
D.ヒト化MAB遺伝子の構築
2B6重鎖CDRs[図7および配列番号:1〜2]およびヒト・抗体の枠組み配列を得て、合成重鎖可変領域[配列番号:18]を作成した。結合した場合にアミノ酸21〜106(KABAT番号付け)をコードすることとなる4つの合成オリゴヌクレオチド[配列番号:27および28][配列番号:29および30]を用いてこれを作成した。次いで、COR枠組み(上記)に基づく別のヒト化重鎖由来の配列を含むpUC18をベースにしたプラスミドのHpaI−KpnI制限部位中にオリゴヌクレオチドを連結した。このプラスミドはシグナル配列[配列番号:17]および残りの可変領域配列を提供する。変異原性プライマーを用いるPCRまたは存在する制限部位中への合成リンカーの付加により、マッピングされた配列中のエラーを修正した。
【0152】
シグナル配列およびヒト化重鎖可変領域をpUCをベースとしたプラスミドからEcoRI−ApaIフラグメントとして切り出し、IgG1ヒト・不変領域を含んでいる発現ベクターpCD中に連結した。合成重鎖可変領域ヌクレオチドおよびアミノ酸配列を図8[配列番号:18および19]に示す。ヒト・枠組み残基は配列番号:19のアミノ酸1〜30、36〜49、66〜97および109〜119である。CDRsのアミノ酸配列はマウス・2B6 CDRsと同じである。得られた発現ベクターpCDIL5HZHC1.0を図10に示す。
【0153】
2B6軽鎖CDRs[図7および配列番号:10、11および12]およびヒト・抗体の枠組み配列を得てから、合成軽鎖可変領域[配列番号:20]を作成した。それぞれSacI−KpnIおよびPstI−HindIII末端を有するヒト化VLのアミノ酸27〜58(KABAT番号付け)[配列番号:31および32]およびアミノ酸80〜109[配列番号:33および34]をコードしている4つの合成オリゴヌクレオチドを、LEN枠組みに対して高度な相同性を有する別のヒト・軽鎖枠組み(B17)(Marsh et al,Nuc.Acids Res.,13:6531-6544(1985))由来の配列を含むpUC18をベースにしたプラスミド中に挿入した。このプラスミドは残りの可変領域配列を提供する。マッピングされた配列中のエラーおよびLENとB17との間の1個のアミノ酸の相違を、変異原性プライマーを用いるPCTまたは存在する制限部位中への合成リンカーの付加により修正した。
【0154】
ヒト化軽鎖領域をpUCプラスミドからEcoRV−NarIフラグメントとして単離し、カッパヒト不変領域ならびにシグナル配列[配列番号:17]を含む発現ベクターpCN中に連結した。合成軽鎖可変領域ヌクレオチドおよびアミノ酸配列を図9[配列番号:20および21]に示す。ヒト・枠組み残基は配列番号:21のアミノ酸1〜23、41〜55、63〜94および104〜113である。CDRsのアミノ酸配列はマウス・2B6 CDRsと同じである。しかしながら、これらのCDRsに関するコーディング配列はマウス・2B6コーディング配列とは異なっており、制限酵素部位の作成が可能である。得られた発現ベクターの1つであるpCNIL5HZLC1.0を11に示す。これらの合成可変軽鎖および/または重鎖配列をヒト化抗体の構築に使用する。
【0155】
E.ヒト化MAbの発現
IgG1イソタイプ由来のヒト化重鎖には配列番号:19に示す合成重鎖可変領域を用いる。2B6重鎖CDRsを含んでいるこの合成VHを上記のごとく設計し、合成した。
【0156】
ヒト化軽鎖、ヒト・カッパ鎖には配列番号:21に示す合成軽鎖可変領域を用いる。2B6軽鎖CDRsを含んでいるこの合成VLを上記のごとく設計し、合成した。ヒト化可変領域をコードしているDNAフラグメントをpUC19をベースにした哺乳動物細胞発現プラスミド中に挿入してプラスミドpCDIL5HZHC1.0(重鎖)[配列番号:49、図10も参照]およびpCNIL5HLC1.0(軽鎖)[配列番号:50、図11も参照]。該発現プラスミドはシグナル配列を使用するものであり、CMVプロモーターおよび下記実施例5にて慣用的方法(すでに引用したManiatisらの文献)により製造されるキメラのヒト・重鎖またはヒト・軽鎖不変領域を含んでいる。プラスミドをCOS細胞中に同時トランスフェクションし、実施例5記載のELISAにより3日後および4日後に上清をヒト・抗体の存在についてアッセイした。
【0157】
上記実施例は典型的な組み換え法による抗体の調製を説明する。慣用的手段により得られた他の抗−IL−5抗体(例えば、2F2、2E3、4A6、5D3、24G9等)を用いて、他の組み換え抗体の取得に関して同様の手順を用いることができる。
【0158】
F.精製
CHOにより発現されたキメラおよびヒト化2B6の精製を、慣用的なプロテインA(またはG)アフィニティークロマトグラフィー、次いで、イオン交換および分子ふるいクロマトグラフィーにより行うことができる。同様のプロセスをうまく用いて他のmAbs(例えば、呼吸器合胞体ウイルス、インターロイキン−4およびマラリアサーカムスポロゾイトに対するもの)の純度95%以上の精製が行われている。
【0159】
G.さらなるヒト化mAbsおよび発現プラスミド
プラスミドpCDIL5HZHC1.0[配列番号:49]を得てから、枠組み位置73のスレオニンをアスパラギンに置換した発現プラスミドpCDIL5HZHC1.1を作成した。EcoRVおよびXhoI末端[配列番号:51および配列番号:52]を有する合成リンカーを、同様にして消化されたpCDIL5HZHC1.0中に連結することにより、これを作成した。同様にして、発現プラスミドpCDIL5HZH1.2は、枠組み位置37においてバリンのかわりにイソロイシンを有する。HpaIおよびXbaI末端[配列番号:53および配列番号:54]を有する合成リンカーを、同様にして消化されたpCDIL5HZHC1.0中に連結することにより、これを作成した。さらに、HpaIおよびXbaI末端[配列番号:53および配列番号:54]を有する合成リンカーを、同様にして消化されたpCDIL5HZHC1.1中に連結することにより、pCDIL5HZHC1.3を作成した。
【0160】
4つの合成オリゴヌクレオチド[配列番号:31、32、33および34]のDNA配列を含むすでに記載されているpUC18をベースにしたプラスミドを得て、枠組み位置15がロイシンからアラニンに変更されているヒト化軽鎖可変領域を作成した。このプラスミドをNheIおよびSacI制限エンドヌクレアーゼで消化し、合成リンカー[配列番号:55および56]を挿入した。次いで、EcoRV−NarIフラグメントを単離し、同様に消化した発現ベクターpCNIL5HZLC1.0中に連結してpCNIL5HZLC1.1を得た。
【0161】
免疫グロブリン(NEW)(Saul et al,J.Biol.Chem.253:585-597(1978))から得た重鎖枠組み領域および2B6重鎖CDRs[図7および配列番号:1〜2]を用いて合成可変領域を作成した。CDR表現に影響しうる枠組みアミノ酸を同定し、ずでに記載されている方法を用いて置換を行った。4つの重複合成オリゴヌクレオチド[配列番号:57、58、59および60]を得て、それらはアニーリングされ伸長された場合に、シグナル配列[配列番号:17]および重鎖可変領域を表わすアミノ酸をコードする。次いで、この合成遺伝子を、PCRプライマー[配列番号:63および64]を用いて増幅し、COR枠組みに基づく別のヒト化重鎖由来の配列を含むpUC18をベースにしたプラスミド中にBstXI−HindIII制限フラグメントとして連結した。合成オリゴヌクレオチドリンカー[配列番号:75および76]をSacIIおよびKpnI制限部位に挿入することにより、枠組みのフェニルアラニンからチロシンへの置換をアミノ酸位置91(KABAT番号付けシステム)(図12の位置94と同じ)において行った。得られた重鎖可変領域[図12および配列番号:61、62]をNEWMヒト化重鎖という。変異原性プライマーを用いるPCRまたは存在する制限部位中への合成リンカーの付加により、マッピングされた配列中のエラーを修正した。シグナル配列およびヒト化重鎖可変領域をpUCをベースにしたプラスミドからEcoRI−ApaIフラグメントとして切り出し、ヒト・IgG1不変領域を含む発現ベクターpCD中に連結してプラスミドpCDIL5NEWMを得た。CDRsのアミノ酸配列はマウス・2B6重鎖CDRsと同じである。
【0162】
免疫グロブリン(REI)(Palm et al,Hoppe-Seyler's Z.Physiol.Chem.356:167-191(1975))から得た軽鎖枠組み領域および2B6軽鎖CDRs[図7および配列番号:10、11および12]を用いて合成可変領域を作成した。CDRの表現に影響しうる枠組みアミノ酸を同定し、すでに記載された方法を用いて置換を行った。4つの重複合成オリゴヌクレオチド[配列番号:65、66、67および68]を得て、それらはアニーリンされ伸長された場合、REIヒト化軽鎖と呼ばれる軽鎖可変領域[図13および配列番号:69、70]を表すアミノ酸をコードする。次いで、この合成遺伝子をPCRプライマー[配列番号:71および72]を用いて増幅し、EcoRI−HindIII制限フラグメントとしてpGEM−7Zf(+)(Promega Corporation,Madison,WI)中に連結した。
【0163】
変異原性プライマーを用いるPCRまたは存在する制限部位中への合成リンカーの付加によりマッピングされた配列中のエラーを修正した。ヒト化軽鎖可変領域をpGEM−7Zf(+)をベースにしたプラスミドからEcoRV−NarIフラグメントとして切り出し、ヒト・カッパ不変領域ならびにシグナル配列[配列番号:17]を含む発現ベクターpCN中に連結した。CDRsのアミノ酸配列はマウス・2B6軽鎖CDRsと同じである。しかしながら、これらのCDRsのコーディング配列はマウス・2B6のコーディング配列とは異なり、制限酵素部位の作成を可能にする。これらの合成可変軽鎖および/または重鎖配列をヒト化抗体の構築に使用する。
【0164】
すでに記載されたpGEM−7Zf(+)をベースにしたプラスミドを得ると、枠組み位置15がバリンからアラニンに変化しているヒト化軽鎖可変領域を作成することができる。このプラスミドをNheIおよびSacI制限エンドヌクレアーゼで消化することができ、合成リンカー[配列番号:73および74]を挿入する。次いで、EcoRV−NarIフラグメントを単離し、同様に消化された発現ベクターpCNIL5HZREI中に連結してプラスミドpCNIL5REIV15Aを得ることができる。
【実施例5】
【0165】
実施例5 キメラ抗体の構築
マウス・mAb 2B6重鎖可変領域のアミノ酸9〜104(KABAT番号付け)をコードしているDNAを、2B6ハイブリドーマ細胞系から得たcDNA(実施例4参照)のpGEM7Zf(+)をベースにしたPCRクローンpGEM7Zf(+)からAvaII−StyI制限フラグメントとして単離した。このフラグメントを4つの小型合成オリゴマーリンカー[配列番号:35および36][配列番号:37および38]とともにpUC18をベースにしたプラスミド(BstXI−HindIIIで消化)中で一緒にすることにより、隣接重鎖可変領域配列およびシグナル配列[配列番号:17]を提供した。密接に関連したマウス・重鎖から推定されたN末端アミノ酸のコンセンサスを、最初の8個のVH残基に関して帰属し、それらは配列番号:35および36中にコードされている。2B6重鎖の最初の15個のN末端アミノ酸を配列決定することにより重鎖の推定アミノ酸配列を確かめた。
【0166】
シグナルおよびVH領域に関する配列を含むEcoRI−ApaIフラグメントを単離し、すでにヒト・IgG1不変領域をコードしているプラスミドpCD中に連結した。
【0167】
マウス・mAb 2B6軽鎖可変領域のアミノ酸12〜99(KABAT番号付け)をコードしているDNAを、2B6ハイブリドーマ細胞系から得たcDNA(実施例4参照)のpGEM7Zf(+)に基づくPCRクローンからDdeII−AvaI制限フラグメントとして単離した。pUC18をベースにしたプラスミド(EcoRV−HindIIIで消化)中においてこのフラグメントを4つの小型合成オリゴマーリンカー[配列番号:39および40][配列番号:41および42]と一緒にすることにより、隣接軽鎖可変領域配列を提供した。密接に関連したマウス・軽鎖から推定されたN末端アミノ酸のコンセンサスを、最初の8個のVL残基に関して帰属し、それらは配列番号:39および
40中にコードされている。2B6軽鎖の最初の15個のN末端アミノ酸を配列決定することにより軽鎖の推定アミノ酸配列を確かめた。次いで、この可変領域をEcoRV−NarIフラグメントとして単離し、すでにヒト・カッパ領域およびシグナル配列を含んでいる発現ベクターpCN中に連結した。
【0168】
pCDおよびpCNをベースにしたプラスミドをCOS細胞中に同時トランスフェクションすることによりキメラ抗体の発現を行った。3日後および5日後に培養上清を集め、下記のごとくELISAにより免疫グロブリン発現に関してアッセイした。最終工程以外の各工程の後にPBS洗浄を行う。ヒト・抗体のRc領域に特異的なヤギ・抗体100ng/50μl/ウェルを用いてマイクロタイタープレートを一晩被覆した。培養上清を添加し、1時間インキュベーションした。次いで、セイヨウワサビ・ペルオキシダーゼ抱合ヤギ・抗−ヒトIgG抗体を添加し、1時間インキュベーションした。次いで、ABTSペルオキシダーゼ基質(Kirkegaard & Perry Laboratories Inc.,Gaithersberg,MD)を添加した。1時間インキュベーションした後、マイクロタイタープレートリーダー(Molecular Devices Corporation,Menlo Park,CA)により405nmの吸光度を読んだ。キメラ抗体の発現を検出した。同様のELISAにおいて、キメラ抗体を含むCOS細胞上清は、ヒト・IL−5蛋白で被覆したマイクロタイターウェルに特異的に結合した。この結果は、IL−5に対する抗体をコードしている遺伝子が合成され、発現されたことを示すものである。
【0169】
上の実施例は典型的な組み換え法による抗体の調製を説明するものである。慣用的手段により得られた他の抗−IL−5抗体(例えば、2F2、2E3、4A6、5D3、24G9等)を用いて、他の組み換え抗体の取得に関して同様の手順を用いることができる。
【産業上の利用可能性】
【0170】
本発明により、高い結合アフィニティーを有するヒト・インターロイキン5に対する中和モノクローナル抗体などが提供されるので、IL−5により媒介されるえ疾病の治療または予防用医薬組成物などの分野において利用可能である。
【配列表フリーテキスト】
【0171】
SEQ ID NO: 1: First base corresponds to Kabat position 24
SEQ ID NO: 2: First base corresponds to Kabat position 25
SEQ ID NO: 3: First base corresponds to Kabat position 24
SEQ ID NO: 4: First base corresponds to Kabat 25
SEQ ID NO: 5: First base corresponds to Kabat position 24
SEQ ID NO: 6: First base corresponds to Kabat position 25
SEQ ID NO: 17: signal sequence
SEQ ID NO: 18: heavy chain variable region
SEQ ID NO: 19: heavy chain variable region
SEQ ID NO: 20: light chain variable region
SEQ ID NO: 21: light chain variable region
SEQ ID NO: 22: mouse gamma 1 hinge region primer
SEQ ID NO: 23: mouse gamma 2a hinge region primer
SEQ ID NO: 24: mouse heavy chain variable region primer
SEQ ID NO: 25: mouse kappa chain constant region primer
SEQ ID NO: 26: mouse light chain variable region primer
SEQ ID NO: 27: oligonucleotide encodes a part of heavy chain variable region
SEQ ID NO: 28: oligonucleotide encodes a part of heavy chain variable region
SEQ ID NO: 29: oligonucleotide encodes a part of heavy chain variable region
SEQ ID NO: 30: oligonucleotide encodes a part of heavy chain variable region
SEQ ID NO: 31: oligonucleotide encodes a part of light chain variable region
SEQ ID NO: 32: oligonucleotide encodes a part of light chain variable region
SEQ ID NO: 33: oligonucleotide encodes a part of light chain variable region
SEQ ID NO: 34: oligonucleotide encodes a part of light chain variable region
SEQ ID NO: 35: small synthetic oligomer linker
SEQ ID NO: 36: small synthetic oligomer linker
SEQ ID NO: 37: small synthetic oligomer linker
SEQ ID NO: 38: small synthetic oligomer linker
SEQ ID NO: 39: small synthetic oligomer linker
SEQ ID NO: 40: small synthetic oligomer linker
SEQ ID NO: 41: small synthetic oligomer linker
SEQ ID NO: 42: small synthetic oligomer linker
SEQ ID NO: 43: overlapping oligonucleotide
SEQ ID NO: 44: overlapping oligonucleotide
SEQ ID NO: 45: light chain
SEQ ID NO: 46: light chain
SEQ ID NO: 47: CDR3
SEQ ID NO: 48: CDR3
SEQ ID NO: 49: heavy chain
SEQ ID NO: 50: light chain
SEQ ID NO: 51: linker
SEQ ID NO: 52: linker
SEQ ID NO: 53: linker
SEQ ID NO: 54: linker
SEQ ID NO: 55: linker
SEQ ID NO: 56: linker
SEQ ID NO: 57: overlapping synthetic oligonucleotide
SEQ ID NO: 58: overlapping synthetic oligonucleotide
SEQ ID NO: 59: overlapping synthetic oligonucleotide
SEQ ID NO: 60: overlapping synthetic oligonucleotide
SEQ ID NO: 61: heavy chain variable region
SEQ ID NO: 62: heavy chain variable region
SEQ ID NO: 63: PCR primer
SEQ ID NO: 64: PCR primer
SEQ ID NO: 65: overlapping synthetic oligonucleotide
SEQ ID NO: 66: overlapping synthetic oligonucleotide
SEQ ID NO: 67: overlapping synthetic oligonucleotide
SEQ ID NO: 68: overlapping synthetic oligonucleotide
SEQ ID NO: 69: light chain variable region
SEQ ID NO: 70: light chain variable region
SEQ ID NO: 71: PCR primer
SEQ ID NO: 72: PCR primer
SEQ ID NO: 73: synthetic linker
SEQ ID NO: 74: synthetic linker
SEQ ID NO: 75: synthetic oligonucleotide linker
SEQ ID NO: 76: synthetic oligonucleotide linker
【特許請求の範囲】
【請求項1】
重鎖および軽鎖を含んでなるヒト・インターロイキン−5に特異的なヒト化抗体であって、
(a)該重鎖の可変領域が、配列番号:7のアミノ酸配列からなるCDR1領域、配列番号:8のアミノ酸配列からなるCDR2領域、および配列番号:9のアミノ酸配列からなるCDR3領域を含み、
(b)該軽鎖の可変領域が、配列番号:10のアミノ酸配列からなるCDR1領域、配列番号:11のアミノ酸配列からなるCDR2領域、および配列番号:12のアミノ酸配列からなるCDR3領域を含む、
ヒト化抗体。
【請求項2】
重鎖および軽鎖を含んでなるヒト・インターロイキン−5に特異的なヒト化抗体であって、
(a)該重鎖の可変領域が、配列番号:15のアミノ酸残基31〜35からなるCDR1領域、配列番号:15のアミノ酸残基50〜65からなるCDR2領域、および配列番号:15のアミノ酸残基98〜108からなるCDR3領域を含み、
(b)該軽鎖の可変領域が、配列番号:16のアミノ酸残基24〜40からなるCDR1領域、配列番号:16のアミノ酸残基56〜62からなるCDR2領域、および配列番号:16のアミノ酸残基95〜103からなるCDR3領域を含む、
ヒト化抗体。
【請求項3】
ヒト・インターロイキン−5のその特異的受容体への結合を妨げる、あるいは、その受容体を通じてのインターロイキン−5のシグナリングを阻害する、請求項1または2に記載のヒト化抗体。
【請求項4】
3.5×10−11Mまたはそれ未満のKdでヒト・インターロイキン−5に結合する、請求項1〜3のいずれか一項に記載のヒト化抗体。
【請求項5】
該重鎖および軽鎖の枠組み領域が少なくとも1の選択されたヒト抗体に由来する、請求項1〜4のいずれか一項に記載のヒト化抗体。
【請求項6】
少なくとも1の選択されたヒト抗体が、ATCC受託番号HB 11783を有するハイブリドーマ細胞系により産生されるヒト・インターロイキン−5に特異的なマウスの中和モノクローナル抗体の重鎖および軽鎖可変領域の推定アミノ酸配列を、ヒト・免疫グロブリン蛋白配列データベースとそれぞれ比較することにより、該中和モノクローナル抗体の重鎖および軽鎖可変領域の推定アミノ酸配列のそれぞれと最も合致すると同定された、重鎖および軽鎖可変領域のヒト枠組み領域を含む、請求項5に記載のヒト化抗体。
【請求項7】
該重鎖の枠組み領域のアミノ酸配列が、ATCC受託番号HB 11783を有するハイブリドーマ細胞系により産生される、ヒト・インターロイキン−5に特異的なマウスの中和モノクローナル抗体の重鎖枠組み領域のアミノ酸配列に対して66%の相同性がある、請求項5に記載のヒト化抗体。
【請求項8】
該軽鎖の枠組み領域のアミノ酸配列が、ATCC受託番号HB 11783を有するハイブリドーマ細胞系により産生される、ヒト・インターロイキン−5に特異的なマウスの中和モノクローナル抗体の軽鎖枠組み領域のアミノ酸配列に対して82%の相同性がある、請求項5に記載のヒト化抗体。
【請求項9】
配列番号:62の重鎖可変領域および配列番号:70の軽鎖可変領域を含む、ヒト化抗体。
【請求項10】
少なくとも1個のアミノ酸によって修飾されているアミノ酸配列を有する、請求項1〜9のいずれか一項に記載のヒト化抗体。
【請求項11】
前記修飾が10個未満のアミノ酸の置換または転位であり、未修飾配列の抗原特異性および高アフィニティーを保持している、請求項10に記載のヒト化抗体。
【請求項12】
ATCC受託番号HB 11783を有するハイブリドーマ細胞系により産生される、ヒト・インターロイキン−5に特異的なマウスの中和モノクローナル抗体と同じ特異性および高アフィニティー性を有し、重鎖および軽鎖のアミノ酸が可変ドメインの枠組み領域またはCDRまたはそれらの両方のいずれかに存在する他のアミノ酸により置換されている、請求項1〜11のいずれか一項に記載のヒト化抗体。
【請求項13】
請求項1〜12のいずれか一項に記載のヒト化抗体および医薬上許容される担体を含んでなる、インターロイキン−5を中和するための組成物であって、喘息および過好酸球増加症候群からなる群より選択される症状の処置のための組成物。
【請求項14】
請求項1〜12のいずれか一項に記載のヒト化抗体および医薬上許容される担体を含んでなる、インターロイキン−5により媒介される疾病の診断のための組成物。
【請求項1】
重鎖および軽鎖を含んでなるヒト・インターロイキン−5に特異的なヒト化抗体であって、
(a)該重鎖の可変領域が、配列番号:7のアミノ酸配列からなるCDR1領域、配列番号:8のアミノ酸配列からなるCDR2領域、および配列番号:9のアミノ酸配列からなるCDR3領域を含み、
(b)該軽鎖の可変領域が、配列番号:10のアミノ酸配列からなるCDR1領域、配列番号:11のアミノ酸配列からなるCDR2領域、および配列番号:12のアミノ酸配列からなるCDR3領域を含む、
ヒト化抗体。
【請求項2】
重鎖および軽鎖を含んでなるヒト・インターロイキン−5に特異的なヒト化抗体であって、
(a)該重鎖の可変領域が、配列番号:15のアミノ酸残基31〜35からなるCDR1領域、配列番号:15のアミノ酸残基50〜65からなるCDR2領域、および配列番号:15のアミノ酸残基98〜108からなるCDR3領域を含み、
(b)該軽鎖の可変領域が、配列番号:16のアミノ酸残基24〜40からなるCDR1領域、配列番号:16のアミノ酸残基56〜62からなるCDR2領域、および配列番号:16のアミノ酸残基95〜103からなるCDR3領域を含む、
ヒト化抗体。
【請求項3】
ヒト・インターロイキン−5のその特異的受容体への結合を妨げる、あるいは、その受容体を通じてのインターロイキン−5のシグナリングを阻害する、請求項1または2に記載のヒト化抗体。
【請求項4】
3.5×10−11Mまたはそれ未満のKdでヒト・インターロイキン−5に結合する、請求項1〜3のいずれか一項に記載のヒト化抗体。
【請求項5】
該重鎖および軽鎖の枠組み領域が少なくとも1の選択されたヒト抗体に由来する、請求項1〜4のいずれか一項に記載のヒト化抗体。
【請求項6】
少なくとも1の選択されたヒト抗体が、ATCC受託番号HB 11783を有するハイブリドーマ細胞系により産生されるヒト・インターロイキン−5に特異的なマウスの中和モノクローナル抗体の重鎖および軽鎖可変領域の推定アミノ酸配列を、ヒト・免疫グロブリン蛋白配列データベースとそれぞれ比較することにより、該中和モノクローナル抗体の重鎖および軽鎖可変領域の推定アミノ酸配列のそれぞれと最も合致すると同定された、重鎖および軽鎖可変領域のヒト枠組み領域を含む、請求項5に記載のヒト化抗体。
【請求項7】
該重鎖の枠組み領域のアミノ酸配列が、ATCC受託番号HB 11783を有するハイブリドーマ細胞系により産生される、ヒト・インターロイキン−5に特異的なマウスの中和モノクローナル抗体の重鎖枠組み領域のアミノ酸配列に対して66%の相同性がある、請求項5に記載のヒト化抗体。
【請求項8】
該軽鎖の枠組み領域のアミノ酸配列が、ATCC受託番号HB 11783を有するハイブリドーマ細胞系により産生される、ヒト・インターロイキン−5に特異的なマウスの中和モノクローナル抗体の軽鎖枠組み領域のアミノ酸配列に対して82%の相同性がある、請求項5に記載のヒト化抗体。
【請求項9】
配列番号:62の重鎖可変領域および配列番号:70の軽鎖可変領域を含む、ヒト化抗体。
【請求項10】
少なくとも1個のアミノ酸によって修飾されているアミノ酸配列を有する、請求項1〜9のいずれか一項に記載のヒト化抗体。
【請求項11】
前記修飾が10個未満のアミノ酸の置換または転位であり、未修飾配列の抗原特異性および高アフィニティーを保持している、請求項10に記載のヒト化抗体。
【請求項12】
ATCC受託番号HB 11783を有するハイブリドーマ細胞系により産生される、ヒト・インターロイキン−5に特異的なマウスの中和モノクローナル抗体と同じ特異性および高アフィニティー性を有し、重鎖および軽鎖のアミノ酸が可変ドメインの枠組み領域またはCDRまたはそれらの両方のいずれかに存在する他のアミノ酸により置換されている、請求項1〜11のいずれか一項に記載のヒト化抗体。
【請求項13】
請求項1〜12のいずれか一項に記載のヒト化抗体および医薬上許容される担体を含んでなる、インターロイキン−5を中和するための組成物であって、喘息および過好酸球増加症候群からなる群より選択される症状の処置のための組成物。
【請求項14】
請求項1〜12のいずれか一項に記載のヒト化抗体および医薬上許容される担体を含んでなる、インターロイキン−5により媒介される疾病の診断のための組成物。
【図1】


【図2】

【図3】

【図4】

【図5】

【図6】

【図7】

【図8】

【図9】
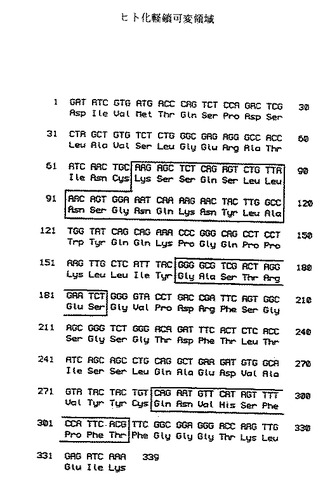
【図10】
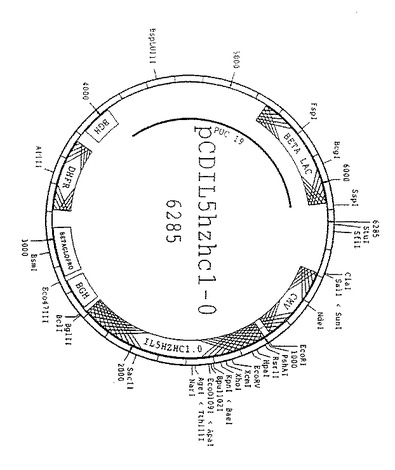
【図11】
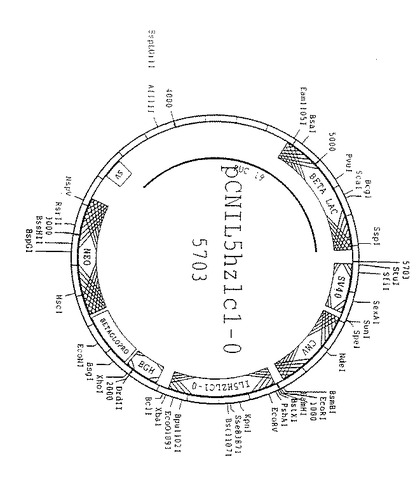
【図12】

【図13】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公開番号】特開2013−91636(P2013−91636A)
【公開日】平成25年5月16日(2013.5.16)
【国際特許分類】
【出願番号】特願2012−220614(P2012−220614)
【出願日】平成24年10月2日(2012.10.2)
【分割の表示】特願2009−244448(P2009−244448)の分割
【原出願日】平成7年12月22日(1995.12.22)
【出願人】(591002957)グラクソスミスクライン・リミテッド・ライアビリティ・カンパニー (341)
【氏名又は名称原語表記】GlaxoSmithKline LLC
【出願人】(595047190)スミスクライン ビーチャム リミテッド (34)
【氏名又は名称原語表記】SmithKline Beecham Limited
【Fターム(参考)】
【公開日】平成25年5月16日(2013.5.16)
【国際特許分類】
【出願日】平成24年10月2日(2012.10.2)
【分割の表示】特願2009−244448(P2009−244448)の分割
【原出願日】平成7年12月22日(1995.12.22)
【出願人】(591002957)グラクソスミスクライン・リミテッド・ライアビリティ・カンパニー (341)
【氏名又は名称原語表記】GlaxoSmithKline LLC
【出願人】(595047190)スミスクライン ビーチャム リミテッド (34)
【氏名又は名称原語表記】SmithKline Beecham Limited
【Fターム(参考)】
[ Back to top ]