
IL−6オートクライン増殖性ヒト骨髄腫細胞株
【課題】骨髄腫のIL−6依存性増殖機構のモデルとなりえる骨髄腫細胞株を樹立することを目的とする。さらに、該細胞株を移植した実験動物、該細胞株または該実験動物を用いる骨髄腫治療剤のスクリーニング法を提供することも目的とする。
【解決手段】オートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株であって、IL−6タンパク質を分泌し、20%ウシ胎児血清(FCS)および100μg/mlカナマイシンを含むRPMI1640培養液中で増殖可能であり、かつ、抗IL−6抗体により増殖が抑制される、前記ヒト骨髄腫細胞株、該細胞株を移植した実験動物、骨髄腫治療剤を該細胞株に添加して骨髄腫細胞増殖抑制を試験することからなる骨髄腫治療剤のインビトロスクリーニング法、ならびに骨髄腫治療剤を該実験動物に投与して骨髄腫細胞増殖抑制を試験することからなる骨髄腫治療剤のインビボスクリーニング法。
【解決手段】オートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株であって、IL−6タンパク質を分泌し、20%ウシ胎児血清(FCS)および100μg/mlカナマイシンを含むRPMI1640培養液中で増殖可能であり、かつ、抗IL−6抗体により増殖が抑制される、前記ヒト骨髄腫細胞株、該細胞株を移植した実験動物、骨髄腫治療剤を該細胞株に添加して骨髄腫細胞増殖抑制を試験することからなる骨髄腫治療剤のインビトロスクリーニング法、ならびに骨髄腫治療剤を該実験動物に投与して骨髄腫細胞増殖抑制を試験することからなる骨髄腫治療剤のインビボスクリーニング法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はヒト骨髄腫細胞株に関し、さらに詳しくはオートクライン機構によってIL−6依存性で増殖するヒト骨髄腫細胞株、該細胞株を移植した実験動物、ならびに該細胞株または該実験動物を用いる骨髄腫治療剤のスクリーニング法に関する。
【背景技術】
【0002】
B細胞刺激因子2(BSF−2)およびマウスハイブリドーマ/形質細胞腫増殖因子と同一因子であるインターロイキン6(IL−6)は多発性骨髄腫(multiple myeloma:以下MMと記載する場合がある)細胞の主要な増殖因子であると考えられている(Kawano et al., Nature 332:83,1988; Klein et al., Blood 73:517, 1989)。多発性骨髄腫は、形質細胞が悪性化した腫瘍で、骨髄を増殖の場とし、複数の部位に同時に発生する。IL−6はこのような細胞上で2種の膜タンパク質を介してその活性を伝達する。その1つは、IL−6が結合する分子量80kDのリガンド結合性膜タンパク質(IL−6受容体)であり、他の1つは非リガンド結合性のシグナル伝達にかかわる膜タンパク質gp130である(Taga et al., J. Exp. Med. 196:967, 1987)。
【0003】
1988年Kawanoらは新鮮ヒト骨髄腫細胞が構成的にIL−6を産生し、かつIL−6受容体を発現していること、ならびに抗IL−6受容体(R)抗体によりインビトロで増殖が抑制されることから、骨髄腫細胞は成長因子を自ら産生し、自身で受容するというオートクライン機構により増殖する可能性があることを報告し(Kawano et al., Nature 332:83, 1988)、一方Kleinらは、自ら成長因子を産生しないが、周囲からの成長因子を受容するというパラクライン機構により増殖することを提唱した(Klein et al., Blood 73:517, 1989)。また、血清中のIL−6濃度は骨髄腫の病勢と相関していることが知られており(Bataille et al., J. Clin. Invest. 84:2008, 1989)、IL−6が骨髄腫の主要な増殖因子の1つであると考えられている。
【0004】
また、新鮮分離した骨髄腫細胞の場合、骨髄腫細胞以外の細胞の混入が避けられず、正確なアッセイが困難であることから、現在にいたるまで、骨髄腫細胞がオートクライン機構またはパラクライン機構のいずれによって増殖するのかは定かではない。
【0005】
IL−6依存性増殖をするヒト骨髄腫細胞株中にトランスフェクションによってヒトIL−6 cDNAを導入すると、自律的に増殖して腫瘍化することが観察され、これはパラクラインIL−6増殖機構を示唆するものである(Okuno et al., Exp. Hematol. 20:395, 1992)。また、ヒト骨髄腫細胞株U266はIL−6オートクライン機構によって増殖することが報告されている(Jernberg et al., Leukemia 5:255, 1991; Levy et al., J. Clin. Invest. 88:696, 1991)。しかしながら、U266の増殖は外因性IL−6によっても(Jernberg et al., Leukemia 5:255, 1991)、また抗IL−6モノクローナル抗体によっても(Levy et al., J. Clin. Invest. 88:696, 1991)影響を受けなかった、という報告もあり、U266の増殖機構に対するIL−6の関与は不明である。
【0006】
本発明者らは、多発性骨髄腫患者の腹水から得た1例の新鮮骨髄腫細胞において、腫瘍細胞は明らかなIL−6依存性増殖を示し、かつ、自律的増殖とともに抗IL−6受容体抗体で強く抑制されることを報告した(Goto et al., Biotherapy 7:655, 1993)。
【0007】
本発明者らはさらに、新鮮ヒト骨髄腫細胞をヒトIL−6遺伝子導入重症複合免疫不全マウス(IL−6トランスジェニックSCIDマウス)に移植した腫瘍細胞の性質について検討した(Goto et al., 第52回日本癌学会総会講演要旨集、498頁、1993年10月)。その結果、皮下に移植した3匹にはその移植部位に形質細胞腫を認め、腋窩リンパ節転移も認めた。腹腔内移植では腫瘤形成は認めなかった。移植後の腫瘍細胞は表面抗原やインビトロでのIL−6依存性増殖および抗ヒトIL−6受容体抗体による増殖抑制効果などに移植前と比べて変化を認めなかった。
【非特許文献1】Kawano et al., Nature 332:83,1988
【非特許文献2】Klein et al., Blood 73:517, 1989
【非特許文献3】Taga et al., J. Exp. Med. 196:967, 1987
【非特許文献4】Kawano et al., Nature 332:83, 1988
【非特許文献5】Klein et al., Blood 73:517, 1989
【非特許文献6】Bataille et al., J. Clin. Invest. 84:2008, 1989
【非特許文献7】Okuno et al., Exp. Hematol. 20:395, 1992
【非特許文献8】Jernberg et al., Leukemia 5:255, 1991
【非特許文献9】Levy et al., J. Clin. Invest. 88:696, 1991)
【非特許文献10】Jernberg et al., Leukemia 5:255, 1991
【非特許文献11】Levy et al., J. Clin. Invest. 88:696, 1991
【非特許文献12】Goto et al., Biotherapy 7:655, 1993
【非特許文献13】Goto et al., 第52回日本癌学会総会講演要旨集、498頁、1993年10月
【発明の概要】
【発明が解決しようとする課題】
【0008】
上記したように、骨髄腫細胞の増殖機構には未だ不明な点が多く、これを解明することが求められている。
本発明の目的は骨髄腫のIL−6依存性増殖機構のモデルとなりえる骨髄腫細胞株を樹立することにある。該細胞株は抗IL−6抗体、抗IL−6受容体抗体などのIL−6活性阻害剤をはじめとする骨髄腫治療剤による骨髄腫の治療のインビトロモデルとして有用である。
【0009】
本発明は該細胞株を移植した実験動物を提供することも目的とする。上記実験動物は抗IL−6抗体、抗IL−6受容体抗体などのIL−6活性阻害剤をはじめとする骨髄腫治療剤による骨髄腫の治療のインビボモデルとして有用である。
【0010】
本発明は骨髄腫治療剤のスクリーニング法を提供することも目的とする。上記インビトロモデルにおいては、細胞増殖抑制あるいはMタンパク(ミエローマタンパク)分泌抑制を指標とする骨髄腫治療剤のスクリーニング法を使用することができる。また、上記インビボモデルにおいては、細胞増殖抑制、Mタンパク分泌抑制あるいは骨病変の抑制を指標とする骨髄腫治療剤のスクリーニング法を使用することができる。なお、Mタンパクとは、骨髄腫が特異的に産生する免疫グロブリンタンパク質であり、それを産生する骨髄腫によりIgA、IgM、IgG、IgEおよびBence−Jonesタンパクの5種類がある。
【課題を解決するための手段】
【0011】
本発明者らは、上記目的を達成すべく鋭意研究した結果、IL−6依存性増殖機構を有する骨髄腫のモデルとなりうる細胞株を樹立することに成功し、本発明を完成した。
すなわち、本発明はオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株を提供する。
【0012】
また、本発明は該細胞株を移植した実験動物を提供する。
さらに、本発明は骨髄腫治療剤を上記細胞株に添加して骨髄腫細胞増殖抑制あるいはMタンパク分泌抑制を試験することからなる骨髄腫治療剤のインビトロスクリーニング法を提供する。
【0013】
さらに、本発明は骨髄腫治療剤を上記実験動物に投与して骨髄腫細胞増殖抑制、Mタンパク分泌抑制あるいは骨病変の抑制を試験することからなる骨髄腫治療剤のインビボスクリーニング法を提供する。
【図面の簡単な説明】
【0014】
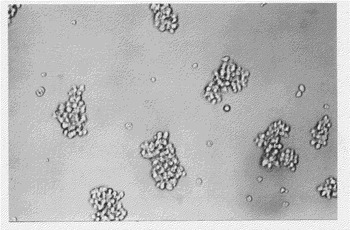
【図1】液体培養におけるKPMM2の自律的凝集を示す図(生物の形態を表す写真)である。
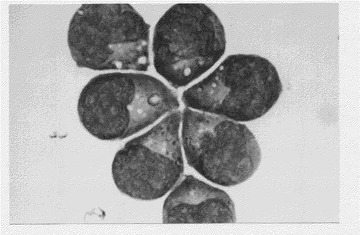
【図2】KPMM2の形態を示す図(生物の形態を表す写真)である。ライト−ギムザ染色では形質細胞の特徴を有する。
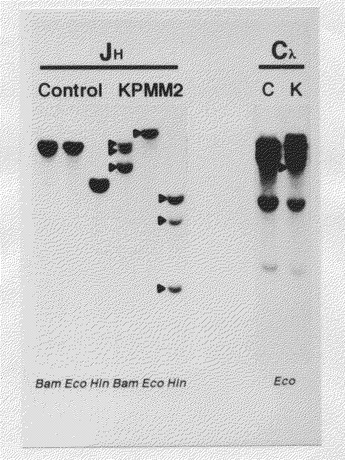
【図3】サザンブロット分析によるKPMM2のJHおよびCλ遺伝子の再構成を示す図(電気泳動の写真)である。KPMM2から得たDNAをBamHI、EcoRIおよびHindIIIで消化し、JHおよびCλ遺伝子プローブを用いてサザンブロット分析を行った。再構成したバンドを(▲)で示す。
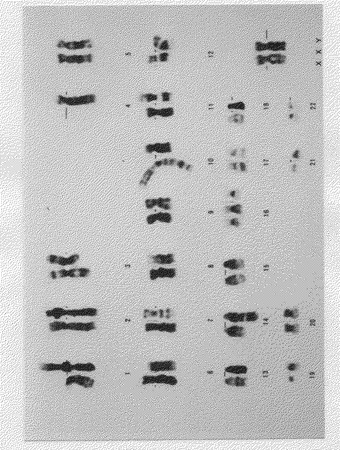
【図4】KPMM2の核型を示す図(生物の形質を表す写真)である。検索した15細胞はすべて46、XX、der(1;19)(q10;q10)、t(3;14)(q21;q32)、−4、t(6;11)(p12;p15)、der(10)add(10)(p13)dic(9;10)(q10;q26)、+16を示した。
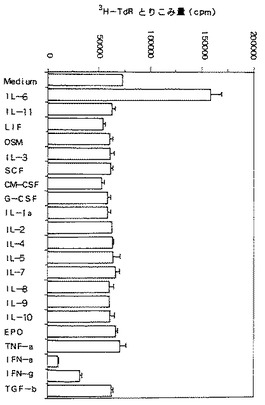
【図5】KPMM2の細胞増殖に対する各種サイトカインの効果を示す図である。使用したサイトカインの濃度は以下の通りである:IL−6、1ng/ml;IFN−αおよびIFN−γ、1000U/ml;その他のサイトカイン、100ng/ml。各数値は3回の試験の平均+標準偏差(SD)を表す。
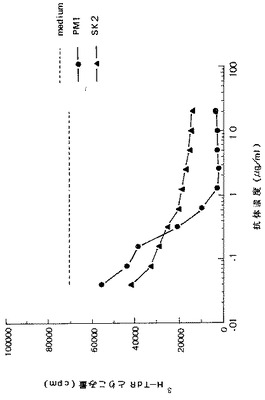
【図6】KPMM2の細胞増殖に対する抗IL−6 mAbおよび抗IL−6R mAbの効果を示す図である。SK2はマウス抗IL−6 mAb(▲);PM1はマウス抗IL−6R mAb(●)。破線は対照を示す。各数値は3回の試験の平均を表す。
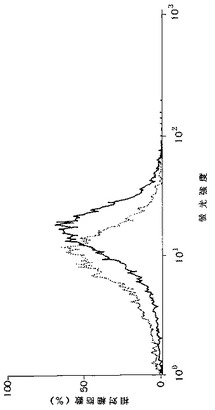
【図7】KPMM2細胞におけるIL−6Rの発現を示す図である。細胞は抗IL−6R mAb(MT18)で染色した。マウスIgG2b抗体を対照として用いた。破線はmIgG2bを、実線はMT18を表す。
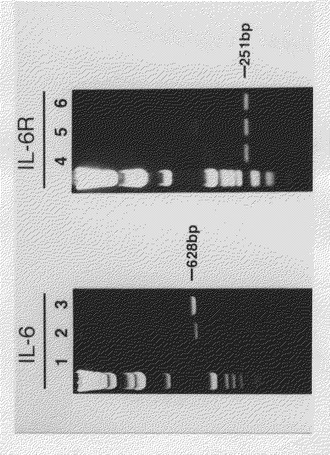
【図8】1.5%アガロースゲル上でのRT−PCR(逆転写PCR)分析により、KPMM2のIL−6およびIL−6R mRNAの発現を示す図(電気泳動の写真)である。レーン1および4、SKW6.4;レーン2および5、KPMM2;レーン3および6、陽性対照。
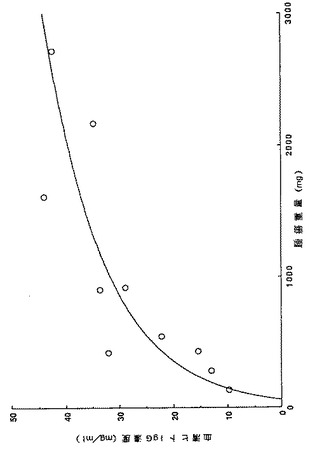
【図9】KPMM2を皮下移植したマウスにおける腫瘍体積と血清ヒトIgG濃度の相関を示す図である。
【図10】(a)はインビトロ培養したKPMM2におけるヒトCD38抗原のFACS解析を示す図である。KPMM2は40から2000までの範囲の蛍光強度を有する。
【0015】
(b)はKPMM2を静脈内移植したSCIDマウスの骨髄細胞におけるヒトCD38抗原のFACS解析を示す図である。71%の細胞が40から2000の範囲の蛍光強度を有する。
【0016】
(c)はKPMM2を皮下移植したSCIDマウスの腫瘍塊から得た細胞におけるヒトCD38抗原のFACS解析を示す図である。全ての細胞が40から2000の範囲の蛍光強度を有する。
【0017】
(d)はKPMM2を移植していないSCIDマウスの骨髄細胞におけるヒトCD38抗原のFACS解析を示す図である。40から2000の範囲の蛍光強度を有する細胞は全くみられない。
【図11】KPMM2に対する抗IL−6 mAbおよび抗IL−6R mAbのインビボにおける腫瘍増殖抑制効果を示す図である。SK2は抗ヒトIL−6 mAb;再構成ヒト型化PM1は抗ヒトIL−6R mAb。
【図12】KPMM2移植ヌードマウス中の血清ヒトIgG濃度に対する抗IL−6 mAbまたは抗IL−6R mAbの効果を示す図である。
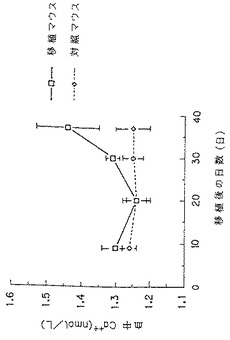
【図13】KPMM2を静脈内移植したSCIDマウスの血中イオン化カルシウム濃度(□)の経時的変化を示す図である。(◇)は対照を示す。各数値はSCIDマウス4匹(37日目のみ5匹)の平均±S.D.を示す。
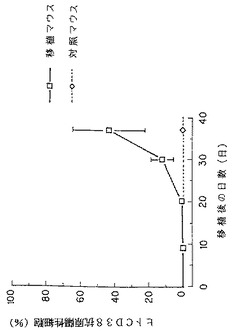
【図14】KPMM2を静脈内移植したSCIDマウスの骨髄中のヒトCD38抗原陽性細胞の割合(□)の経時的変化を示す図である。(◇)は対照を示す。各数値はSCIDマウス4匹(37日目のみ5匹)の平均±S.D.を示す。
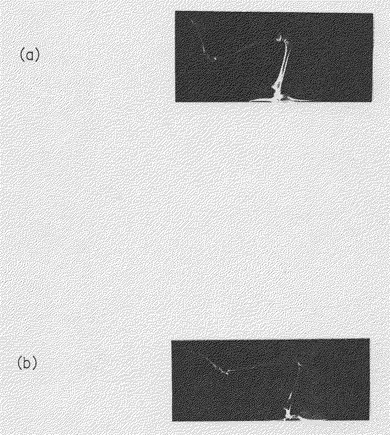
【図15】KPMM2を静脈内移植したSCIDマウスの骨のX線撮影像を示す図(生物の形態を示す写真)である。
【0018】
(a)はKPMM2を移植していない対照群のSCIDマウス
(b)はKPMM2移植後37日目のSCIDマウス
【発明を実施するための形態】
【0019】
本発明の細胞株は、例えば多発性骨髄腫患者の腹水などから採取した骨髄腫細胞を用いて樹立することができる。本発明では特に、IgG、λ型多発性骨髄腫患者の腹水から骨髄腫細胞を採取した。培養開始後1カ月で細胞は安定して増殖し始め、1年以上維持された。このようにして樹立された細胞株はKPMM2と命名され、生命工学工業技術研究所特許微生物寄託センターに受託番号FIRM:P−14170で寄託されている(1994年2月22日寄託)。細胞株KPMM2はインビトロでの継代、ならびに実験動物、例えば重症複合免疫不全(SCID)マウス、IL−6トランスジェニックSCIDマウスおよびヌードマウスなどの免疫不全状態にあるマウス中での安定した継代が可能である。
【0020】
細胞株KPMM2はインビトロでIL−6を産生し、またIL−6受容体(IL−6R)を発現していることが確認された。また、KPMM2の各種サイトカインに対する反応性を3H−チミジンの取り込み実験、MTT(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide)法(J. Immun. Methods 65:55-63, 1983 参照)および生細胞の測定で試験した結果、IL−6とともにインキュベートしたときのみ顕著に刺激され、これはKPMM2がIL−6に特異的に反応して増殖することを示している。さらに、KPMM2の増殖は、抗IL−6 mAb(モノクローナル抗体)および抗IL−6R mAbによって用量依存的に顕著に抑制された。また、RT−PCR(逆転写ポリメレースチェインリアクション)により、KPMM2がIL−6およびIL−6R mRNAを発現していることが確認された。なお、KPMM2は各種の接着分子、すなわちCD44、VLA−β、ICAM−1、NCAM、LFA−3およびVLA−4を発現しており、またインビトロで自律的細胞凝集を示す。
【0021】
このようなKPMM2の各種特徴から、KPMM2がIL−6オートクライン機構により増殖する骨髄腫細胞株であることが示された。本発明の細胞株はその増殖機構がIL−6依存性のオートクライン機構であることが証明された最初の細胞株である。
【0022】
したがって、本発明の細胞株は抗IL−6 mAbあるいは抗IL−6R mAbなどのIL−6活性阻害剤をはじめとする骨髄腫治療剤のスクリーニングに有用である。例えば、抗IL−6抗体または抗IL−6受容体抗体を本発明の細胞株に添加して骨髄腫細胞増殖抑制効果を試験することからなる骨髄腫治療剤のインビトロスクリーニング法が可能である。また、本発明の骨髄腫細胞株は、細胞数の増加に比例してMタンパクの分泌量が増大することにより、Mタンパク分泌抑制を指標として骨髄腫治療剤のインビトロスクリーニングを行うことができる。さらに、本発明の細胞株は、細胞間相互作用やIL−6シグナル伝達を介する骨髄腫細胞の増殖において接着分子の果たす役割を研究するモデルとしても有用である。
【0023】
本発明は、本発明の細胞株を移植した実験動物も提供する。本発明の細胞株を移植する実験動物としては、マウスの他、ラット、ウサギ、モルモット、ハムスター、サルなどが挙げられ、さらには、T細胞あるいはB細胞といった免疫担当細胞の機能に障害が生じ、免疫不全状態にある実験動物に本発明の細胞株を移植するのがよい。既に述べたように、本発明の細胞株はSCIDマウス、IL−6トランスジェニックSCIDマウスおよびヌードマウスなどの免疫不全状態にあるマウス中で安定した継代が可能である。
【0024】
興味深いことに、本発明の上記細胞株をマウスに移植するには、皮下移植、腹腔内移植の外に、静脈内移植によっても行うことができる。皮下移植および腹腔内移植した場合には、それぞれ移植部位皮下および腹腔内に固形腫瘍が観察されるが、静脈内移植した場合には、骨髄への腫瘍細胞の生着が見られ、これを実際の骨髄腫の病態に近いモデルとして使用することができる。例えば、抗IL−6抗体または抗IL−6受容体抗体といったIL−6活性阻害剤をはじめとする骨髄腫治療剤を本発明の実験動物に投与して骨髄腫細胞増殖抑制効果を試験することからなる骨髄腫治療剤のインビボスクリーニング法が可能である。また、本発明の細胞株が生着した実験動物では、腫瘍の増殖に伴って血清中のMタンパク濃度の上昇が観察されるため、Mタンパク濃度の抑制を指標とした骨髄腫治療剤のインビボスクリーニング法も可能である。さらに、本発明の細胞株が実験動物の骨髄へ生着すると骨髄腫細胞の増殖に伴い、血中イオン化カルシウム濃度の上昇、骨破壊、骨融解および骨吸収といった骨病変が観察されることから、これら骨病変の抑制を指標とした骨髄腫治療剤のインビボスクリーニング法が可能である。
【0025】
以下に本発明を実施例によりさらに詳しく説明するが、本発明の範囲はこれに限定されるものではない。
【実施例】
【0026】
実施例1:骨髄腫細胞株の樹立および維持
IgG、λ型多発性骨髄腫患者(76才、女性、ステージIIA)の腹水から骨髄腫細胞を採取した。腹水は多数の骨髄腫細胞を含んでおり、腹水中のIL−6レベルは91.0pg/mlに達していた。採取した腹水をFicoll−Hypaque(ファルマシア社製)を用いた密度勾配遠心法にかけて単核球を分離、プラスチックシャーレにて付着細胞を除去し、さらにヒツジ赤血球にてT細胞を除去し、腫瘍細胞を95%以上に純化した。細胞を20%ウシ胎児血清(FCS:Xavier Investments製、オーストラリア)、組換えヒトIL−6(中外製薬製)4ng/mlおよびカナマイシン(明治製菓社製)100μg/mlを含むRPMI1640(ギブコ社製)培養液中に1×106細胞/mlの濃度で浮遊させた。次いで25mlフラスコ中で10mlの培養液中で培養し、湿潤5%CO2中、37℃でインキュベートした。安定した細胞増殖が観察されるまで3日ごとに培地を部分的に新しいものに取り替えた。
【0027】
培養開始1カ月後で細胞は安定して増殖し始め1年以上維持され、細胞株として樹立された。この細胞株をKPMM2と命名した。IL−6の存在下または非存在下における倍加時間はそれぞれ48時間および72時間であった。
【0028】
KPMM2の形態およびIg分泌は以下の通りである。
KPMM2の形態およびIg分泌
KPMM2細胞は光学顕微鏡下で自律的な細胞凝集を示して増殖することが観察され(図1)、ライト−ギムザ染色では形質細胞様の形態を示した(図2)。KPMM2は酸ホスファターゼが陽性であり、α−ナフチルブチレートエステラーゼがやや陽性であるが、ペルオキシダーゼ、AS−Dクロロアセテートエステラーゼ、パス(過ヨウ素酸シッフ試薬)およびアルカリホスファターゼは陰性であった。細胞質IgGおよびλL鎖が検出されたが、IgA、IgM、およびκL鎖は直接免疫蛍光法で陰性であった。また、細胞(106個/ml)を3日間培養すると、培養上清にはIgGおよびλL鎖の分泌が見られた。
【0029】
実施例2:表面抗原の解析
上記実施例1で樹立した細胞株KPMM2の表面抗原の発現を各種ヒト抗原に対するモノクローナル抗体のパネルを用いて、直接および間接蛍光抗体法(Fried et al., Flow Cytometry, Boca Raton, CRC Press:59-78, 1989)により検討した。
【0030】
実施例1に記載の方法で得られたKPMM2を、106個/チューブとなるように、100μlの蛍光活性化細胞選択装置(FACS)用緩衝液(2% FCSおよび0.1% NaN3を含むリン酸緩衝化生理食塩溶液(PBS)、以下FACS緩衝液という)に浮遊させた。次いで、直接蛍光抗体法においては、飽和量の下記表1に記載の各種ヒト抗原に対するフルオレシンイソチオシアネート(FITC)あるいはフィコエリスリン(PE)標識抗体を添加し、4℃にて30分間インキュベートした。細胞を上記FACS緩衝液で2回洗浄した後、フローサイトメーター(EPICS PROFILE,コールター社製)で分析した。
【0031】
一方、間接蛍光抗体法においては、非標識の下記表1に記載の各種ヒト抗原に対する抗体を添加し、4℃にて30分間インキュベートして、細胞をFACS緩衝液で2回洗浄した後、5μg/mlのFITCあるいはPE標識ヤギ抗マウスIgG抗体F(ab’)2断片(TAGO社製)を加え、4℃にて30分間反応させた。FACS緩衝液で2回洗浄した後、FACS緩衝液に浮遊させ、フローサイトメーター(EPICS PROFILE,コールター社製)で分析した。
【0032】
KPMM2の表面抗原を以下の表1にまとめて示す。
【0033】
【表1】

【0034】
表から明らかなように、KPMM2は形質細胞関連抗原(CD38、PCA−1およびBL3)、接着分子(CD44、VLA−β、ICAM−1、NCAM、LFA−3およびVLA−4)ならびにCD45、CD63、CD71、IgGおよびλなどの抗原が陽性であった。
【0035】
実施例3:免疫グロブリン遺伝子再構成
上記実施例1で樹立した細胞株KPMM2の免疫グロブリン(Ig)遺伝子再構成をサザンブロット法により分析した。
【0036】
実施例1に記載の方法により得られたKPMM2細胞(107個)から、Manual of Clinical Immunology, 3rd edition, American Society for Microbiology, 1986の方法に準じてDNAを調製し、3種類の制限酵素BamHI、EcoRIあるいはHindIII(ベーリンガー・マンハイム社製)で別々に処理し、エタノール沈殿としてDNAを回収して、0.8%アガロースゲル(SEAKEM GTG,FMC社製)により24時間電気泳動を行った。電気泳動したDNAをナイロン膜(ハイボンドN+,アマシャム社製)に移し、これを乾燥させた。次に、タカラランダムプライマーDNAラベリングキット(宝酒造社製)を用いて、32P標識したヒトIg JH、CκおよびCλプローブ(オンコア社製)を用い、添付の処方に従ってサザンブロット分析を行った。なお、対照として健常人の末梢血単核球から得た再構成を生じていない染色体DNAを用いた。その結果、IgHおよびκ鎖遺伝子が再構成していたが、λ鎖遺伝子は再構成していなかった(図3)。以上のことより、KPMM2がモノクローナルな抗体を産生すること、および細胞の単一性が確認された。
【0037】
実施例4:細胞遺伝学的分析
上記実施例1で樹立した細胞株KPMM2の染色体の構造異常を分析した。
実施例1に記載の方法で得られたKPMM2を20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液にて5×105個/mlとなるように培養した。培養48時間後に、KPMM2に0.05μgのコルセミド(ギブコ社製)を15分間処理し、分裂中期で細胞周期が停止したKPMM2細胞を回収した。回収したKPMM2細胞を0.075MのKClで20分間処理し、メタノール−酢酸で固定した。次いで、KPMM2細胞の染色体をトリプシン−ギムザバンド染色法によって分析した。
【0038】
その結果、KPMM2は多くの構造異常をもつ二倍体細胞であることが判明した(図4)。分析した15細胞すべてが46、XX、der(1;19)(q10;q10)、t(3;14)(q21;q32)、−4、t(6;11)(p12;p15)、der(10)add(10)(p13)dic(9;10)(q10;q26)、+16を示した。
【0039】
実施例5:EBVおよびマイコプラズマの検出
実施例1で樹立した細胞株KPMM2のエプスタイン・バールウイルス(EBV)およびマイコプラズマ汚染を試験した。
【0040】
実施例1に記載の方法で得られたKPMM2細胞の染色体からエプスタイン・バールウイルス(EBV)を検出するために、Systemic Genetic Institute社から購入したEBV BamW領域増幅プライマーを用いて、添付の処方に従いPCR(ポリメレース チェイン リアクション)を行った。マイコプラズマ感染の検出はマイコプラズマDNA検出用M.T.C.キット(Gen−Probe Inc.社製)により、添付の処方に従って実施した。
【0041】
その結果、KPMM2はEBVゲノムおよびマイコプラズマゲノムに対して陰性であった。
実施例6:サイトカインに対する反応性
実施例1で樹立した細胞株KPMM2の各種サイトカインへの反応性を試験した。
【0042】
実施例1に記載する方法で得られたKPMM2を20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に浮遊させ、容量が200μlで96穴プレート(ファルコン社製)へ1×104個/穴となるように分注した。
【0043】
96穴プレートの各穴には、下記の濃度となるように各種サイトカインを別々に加えた。
組換えIL−2、同IL−3、同腫瘍壊死因子(TNF)−α、同顆粒球マクロファージコロニー刺激因子(GM−CSF)、同幹細胞成長因子(SCF)(以上、Genzyme社製)、同IL−4、同IL−7、同IL−10、同IL−11、同白血病阻害因子(LIF)、同オンコスタチンM(OSM)(以上、Pepro Tec Inc.社製)、同IL−9、同トランスフォーミング成長因子(TGF)−β(以上、R&D System Inc.社製)、同IL−1α(Boehringer Manheim社製)、同IL−5(Upstate Biotechnology Inc.社製)、同IL−8(Amersham社製)、同エリスロポエチン(EPO)および同顆粒球コロニー刺激因子(G−CSF)(以上、中外製薬株式会社より提供)は100ng/ml。
【0044】
組換えインターフェロン(IFN)−γ(塩野義製薬株式会社より提供)および天然型ヒトIFN−α(住友製薬株式会社より提供)は1000U/ml。
組換えIL−6(中外製薬株式会社より提供)は1ng/ml。
【0045】
なお、対照群は、サイトカインを加えない20%FCSおよび100μg/mlカナマイシンを添加したRPMI1640培養液で培養した。
96穴プレートの各穴に分注したKPMM2を、上記サイトカイン存在下あるいは非存在下で湿潤5%CO2中、37℃で96時間培養した。その培養終了4時間前に、各穴に3H−チミジン(Amersham社製)を1μCi/穴となるように添加した。KPMM2が取り込んだ3H−チミジンの量の測定は、液体シンチレーションカウンター(1205 ベータプレート、ファルマシア社製)を用いた。
【0046】
KPMM2の増殖に対する各種サイトカインの効果を図5に示す。図5から明らかなように、KPMM2細胞はIL−6とともにインキュベートしたときのみに3H−チミジンの取り込みが顕著に刺激された。濃度1ng/mlにおけるIL−6により、3H−チミジンの取り込みが2.2倍に上昇した。一方、IFN−αおよびIFN−γはKPMM2細胞の増殖を顕著に阻害した。
【0047】
また、KPMM2のサイトカインに対する反応性を調べるために、MTT(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide)法(J. Immun. Methods 65:55, 1983 を参照)および生細胞を直接測定する方法を用いたが、同様の結果が得られた。
【0048】
実施例7:抗IL−6 mAbおよび抗IL−6R mAbによる増殖阻害
実施例1で樹立した細胞株KPMM2に対するマウス抗ヒトIL−6R mAb(モノクローナル抗体)(IgG1クラス:PM1)およびマウス抗IL−6 mAb(IgG1クラス:SK2)の効果を調べた。SK2は文献(Y. Ohe et al., Br. J. Cancer 67:939, 1993)に記載されており、またPM1は文献(Hirata et al., J. Immunol. 143:2900, 1989)に記載されている。
【0049】
実施例1に記載する方法で得たKPMM2を20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に浮遊させ、容量が200μlで96穴プレート(ファルコン社製)へ1×104個/穴となるように分注した。
【0050】
96穴プレートの各穴には、各種濃度のIL−6 mAb(モノクローナル抗体)(SK2)および抗IL−6R(受容体)mAb(PM1)を別々に加えた。なお、対照群は、抗体を加えない20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液とした。
【0051】
96穴プレートの各穴に分注したKPMM2を、抗IL−6 mAbおよび抗IL−6R mAb存在下あるいは非存在下で湿潤5%CO2中、37℃で96時間培養した。その培養終了4時間前に、各穴に3H−チミジン(Amersham社製)を1μCi/穴となるように添加した。KPMM2が取り込んだ3H−チミジンの量の測定は、液体シンチレーションカウンター(1205 ベータプレート、ファルマシア社製)を用いた。
【0052】
KPMM2の増殖に対する抗IL−6 mAbおよび抗IL−6R mAbの効果を図6に示す。SK2およびPM1の添加はいずれも用量依存的に細胞増殖を有意に阻害した。特に、PM1は1μg/mlの濃度でKPMM2の増殖を完全に阻害した。
【0053】
また、KPMM2増殖の抗IL−6 mAbおよび抗IL−6R mAbに対する効果を調べるために、MTT(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide)法(J. Immun. Methods 65:55, 1983 を参照)および生細胞を直接測定する方法を用いたが、同様の結果が得られた。
【0054】
実施例8:ELISAによるIL−6産生の測定
上記実施例1で樹立した細胞株KPMM2によるIL−6産生能を試験した。
実施例1に記載する方法で得たKPMM2を、20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に106個/mlとなるように浮遊させ、ヒトIL−6非存在下で湿潤5%CO2中、37℃で培養した。
【0055】
培養72時間後、培養上清中に含まれるKPMM2が産生したIL−6の濃度を、ヒトIL−6用ELISAキット(帝人バイオラボラトリー社製)を用いて添付の処方に従い測定した。なお、陰性対照として、20%FCSおよび100μg/mlカナマイシンを添加したRPMI1640培養液を用いた。培養上清には79.7±19.6(平均値±S.D.)pg/mlのIL−6の産生が検出され、対照培養液において検出限界(4.0pg/ml)以下であったのに比較すると、培養上清中のIL−6濃度が顕著に増加していることが確認された。
【0056】
実施例9:フローサイトメトリーによるIL−6Rの発現の確認
KPMM2細胞上でのIL−6R発現を確認するために、IL−6Rに結合し、IL−6のIL−6Rへの結合を阻害しないマウス抗ヒトIL−6R mAbであるMT18抗体(Hirata et al., J. Immunol. 143:2900, 1989)を用いて、間接蛍光抗体法を実施した。
【0057】
実施例1に記載の方法で得られたKPMM2細胞を106個/チューブとなるように、100μlの実施例2記載のFACS緩衝液に浮遊させ、10μg/mlのMT18抗体を加え、4℃にて3時間反応させた後、FACS緩衝液で2回洗浄し、100μlのFACS緩衝液に浮遊させ、さらに、FITC標識ヤギ抗マウスIgG抗体(TAGO社製)を5μg/ml添加し、4℃にて30分間反応させた。FACS緩衝液で2回洗浄した後、同FACS緩衝液に浮遊させ、フローサイトメーター(EPICS PROFILE、コールター社製)で蛍光を測定した。
【0058】
その結果、図7に示すように、KPMM2細胞上にIL−6Rの発現が示された。
実施例10:ELISAによるヒトIgG(Mタンパク)産生の測定
KPMM2のヒトIgG(Mタンパク)産生能を試験した。
【0059】
実施例1に記載の方法で得られたKPMM2を106個/mlとなるように、20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に浮遊させ、容量2mlで12穴プレート(ファルコン社製)の各穴に分注し、ヒトIL−6非存在下で湿潤5%CO2中、37℃で72時間培養した。なお、実験は3回行った。その後、培養上清中のヒトIgG濃度を、TAGO社製ヤギ抗IgG抗体(NO.4100)およびアルカリフォスファターゼ標識ヤギ抗IgGガンマ鎖特異的抗体(NO.2490)を用いるELISAにて測定した。なお、スタンダードとしてカッペル社製ヒトIgG(NO.0001−860)を用いた。その結果、培養上清中には10.1μg/mlのヒトIgGが検出され、対照培養液では検出限界以下(5ng/ml)であったのに比較して、培養上清中のヒトIgG濃度が顕著に増加していることが確認された。
【0060】
実施例11:RT−PCR(逆転写ポリメレースチェインリアクション)によるIL−6およびIL−6R mRNAの検出
実施例1に記載の方法で得られたKPMM2におけるヒトIL−6およびヒトIL−6Rの発現を確認するために、RT−PCR(逆転写ポリメレースチェインリアクション)法によりヒトIL−6およびヒトIL−6RのメッセンジャーRNA(mRNA)を検出した。
【0061】
グアニジウムセシウムクロライド法(Molecular Cloning, Cold Spring Harbor Laboratory Press)によってKPMM2細胞(108個)から全RNAを調製した。陰性対照としてヒトB細胞リンパ腫細胞株SKW6.4からも同様に全RNAを調製した。1本鎖cDNA合成はcDNA合成キット(Invitrogen社製)を用いて、添付の処方に従い全RNA5μgから直接実施した。
【0062】
陽性対照としてヒトIL−6およびヒトIL−6R検出用PCRプライマーを用いた(Clontech Inc.社製)。PCR溶液各100μlは、10mM Tris−HCl(pH8.3)、50mM 塩化カリウム、Amplitaq(Perkin Elmer Cetus社製)2.5ユニット、1本鎖cDNA合成反応物1μl、各プライマー100pmolを含む。各PCR用チューブに鉱物オイル50μlを上層してPCRに付した。最初に94℃で1分メルトし、60℃で1分と72℃で10分のサイクルを30サイクル繰り返した。最終サイクルの後、最終的に72℃で10分伸長を行った。ヒトIL−6およびヒトIL−6R検出用プライマーを用いた陽性対照群も同様に増幅し、約50pgの陽性対照PCR産物を得た。
【0063】
各反応チューブから10μlを取って1.5%アガロースゲル上で電気泳動を行った。図8に示すように、IL−6(628bp)およびIL−6R(251bp)のPCR産物はいずれもKPMM2のRNAから増幅された。これらの結果は、KPMM2がIL−6のオートクライン機構によって増殖することを遺伝子発現の面から支持する。一方、SKW6.4細胞はIL−6に応答してIgMを分泌する。IL−6RのPCR産物はSKW6.4 mRNAから増幅された。しかし、IL−6のPCR産物はこの実験で検出されず、このことはSKW6.4細胞がIL−6を産生していないことを示唆する。
【0064】
実施例12:細胞株KPMM2のSCIDマウスおよびヌードマウスへの移植
(1)KPMM2の可移植性およびインビボでの継代
上記実施例1で樹立した細胞株KPMM2の可移植性およびインビボでの継代を以下のように検討した。
【0065】
KPMM2を20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に108個/mlで懸濁し、0.2mgのウサギ抗アシアロGM1抗体(Code No.014−09801、和光純薬工業社製)で処理したIL−6トランスジェニック重症免疫不全マウス(以下IL−6−SCID Tmという)(中外製薬製)の腹部皮下に注射針で0.1ml移植した。その結果、1カ月後には3例全例で移植部位に結節型の腫瘍を形成した。
【0066】
この腫瘍を無菌的に摘出し、摘出した腫瘍塊を使い捨て注射器のピストンなどでつぶし、ナイロンメッシュ(70μm、ファルコン社製)を通して細胞を回収した。この細胞を上記と同様にRPMI1640培養液に懸濁し、IL−6−SCID Tm、重症免疫不全マウス(SCIDマウスという)(日本クレア社製)、BALB/c−nu/nuマウス(以下ヌードマウスという)(日本クレア社製)に皮下移植したところ、同じように移植部位に結節型の腫瘍を形成し、インビボでの継代が可能であった。また腫瘍塊を3mm角のブロックにし、移植針を用いてIL−6−SCID Tm、SCIDマウス、ヌードマウスの皮下に移植しても継代可能であった。
(2)KPMM2の異なる移植経路での可移植性
KPMM2の移植経路を皮下(s.c.)、静脈内(i.v.)、腹腔内(i.p.)とした場合の可移植性を以下のように検討した。
【0067】
(1)においてSCIDマウスにて継代したKPMM2腫瘍を無菌的に摘出し、摘出した腫瘍塊を使い捨て注射器のピストンでつぶし、ナイロンメッシュを通して細胞を回収し、108個/mlの細胞懸濁液を作成した。この懸濁液をSCIDマウス、あるいは0.2mgのウサギ抗アシアロGM1抗体および500RのX線で処理したヌードマウスに0.1mlずつ皮下(s.c.)、静脈内(i.v.)、腹腔内(i.p.)の3経路で移植し、40日後に生着の判定を行った(表2)。その結果、約40日後には全例で生着が確認され、s.c.移植では移植部位に、i.p.移植では腹腔内に固形腫瘍を形成した。また、i.v.移植ではKPMM2は骨髄での生着が認められた。
【0068】
【表2】
【0069】
(3)KPMM2移植動物の血清ヒトIgG(Mタンパク:ミエローマタンパク)の濃度
細胞株KPMM2をSCIDマウスへ皮下移植し、形成された腫瘍の体積と血清中ヒトIgG濃度の相関を調べた。上記(2)に記載の方法で細胞株KPMM2をSCIDマウスへ皮下移植し、KPMM2移植前、移植後21日目、42日目の3回、血清サンプルを採取してELISA法にてヒトIgG濃度を測定した。
【0070】
その結果、マウス血清中のヒトIgG濃度はKPMM2を移植した動物ではいずれも経時的に上昇した。また図9に示すように、皮下移植した動物では皮下に形成された腫瘍の体積と血清ヒトIgG濃度に相関が見られ、腫瘍の大きな動物ほど血清ヒトIgG濃度は高値を示した。このことから、血清ヒトIgG濃度を指標としても抗腫瘍効果の判定ができると考えられた。
(4)KPMM2の可移植性および移植細胞数の検討
SCIDマウスとヌードマウスで移植細胞数を変えたときのKPMM2の可移植性を検討した。
【0071】
特別な前処理を行わない雄のSCIDマウスおよびヌードマウスに、上記(2)記載の方法で各々KPMM2細胞を107、3×106、106個腹部皮下(s.c.)に移植し、またSCIDマウスには上記(2)の方法で得たKPMM2細胞を3×106、106個静脈内(i.v.)でも移植した。
【0072】
その結果、SCIDマウスに皮下移植した場合、3×106個以上移植すると、21日までに全例で腫瘍結節を形成し、106個移植した場合でも21日までに2/3、32日までに全例で腫瘍結節を形成した。ヌードマウスに皮下移植すると、107個移植した場合では21日までに2/3、32日までには全例で、また3×106個移植した場合では42日までには全例で腫瘍結節が形成された。しかし、106個の移植では42日までに腫瘍の生着は見られなかった。また、SCIDマウスに静脈内移植した場合、42日までに3×106個移植で全例、106個移植で1/3にKPMM2が生着した。これらの結果を以下の表3にまとめて示す。
【0073】
【表3】
【0074】
(5)KPMM2移植SCIDマウスにおけるヒトCD38抗原の発現の解析
KPMM2を移植したSCIDマウスにおいて、その生着を確認するために、KPMM2の細胞表面上で特徴的に発現している表面抗原であるヒトCD38抗原の発現を試験した。
【0075】
KPMM2を上記(1)に記載する方法でSCIDマウスへ静脈内移植し、移植後37日目にマウスを屠殺した。屠殺したマウスの大腿骨より骨髄を回収して懸濁し、ステンレスメッシュ(100μm)を通し、蛍光活性化細胞選択装置(FACS)用緩衝液(2%FCSおよび0.1%NaN3を含むPBS(−)溶液)にて骨髄細胞の浮遊液を調製した。なお、陰性対照としてKPMM2を移植していないSCIDマウスの骨髄からも同様の方法にて骨髄細胞浮遊液を調製した。
【0076】
また、本実施例に記載する方法によりKPMM2を皮下移植したSCIDマウスからは、移植後37日目に皮下腫瘍塊を外科的に摘出し、摘出した腫瘍塊を2枚のスライドグラスの間にはさんですりつぶし、これをステンレスメッシュ(100μm)を通し、FACS緩衝液にて骨髄細胞の浮遊液を調製した。
【0077】
陽性対照として、実施例1に記載した方法でインビトロにて培養したKPMM2を用いた。インビトロで4日間培養したKPMM2をFACS緩衝液で洗浄した後、同緩衝液中に浮遊させた。
【0078】
次に、このように調製した細胞浮遊液を用いて、FACS解析によるヒトCD38抗原の発現を調べた。
すなわち、100μlのFACS緩衝液中にて、各群の浮遊細胞106個に対し、2.5μg/mlのフィコエリスリン(PE)標識抗ヒトCD38抗体(Leu−17、ベクトン・ディッキンソン社製)を添加し、氷上で30分間反応させた。次いで、1mlのFACS緩衝液で2回洗浄後、500μlのFACS緩衝液で懸濁し、FACScan(ベクトン・ディッキンソン社製)によりFACS解析を行った。
【0079】
陽性対照群であるインビトロ培養KPMM2のFACS解析の結果から、蛍光強度40から2000までの範囲にある細胞をヒトCD38抗原発現細胞とした(図10(a)参照)。KPMM2を静脈内移植したSCIDマウスでは、その骨髄細胞のおよそ71%がヒトCD38陽性細胞で占められていた(図10(b)参照)。このことは、SCIDマウスに静脈内移植したKPMM2がSCIDマウスの骨髄に生着したことを示している。また、KPMM2をSCIDマウスに皮下移植して生じた腫瘍塊から得られた細胞は全てヒトCD38が陽性であり(図10(c)参照)、KPMM2を皮下移植して生ずる腫瘍塊は、全てKPMM2から構成されていることが示された。なお、KPMM2を移植していない陰性対照群のSCIDマウスの骨髄細胞にはヒトCD38陽性細胞は全く検出されなかった(図10(d)参照)。
【0080】
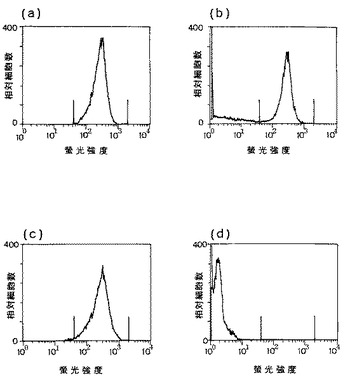
実施例13:抗ヒトIL−6抗体SK2および抗ヒトIL−6R再構成ヒト型化抗体PM1の抗腫瘍効果
KPMM2移植動物における抗ヒトIL−6抗体SK2および抗ヒトIL−6R再構成ヒト型化抗体PM1のインビボ抗腫瘍効果を以下のようにして検討した。
【0081】
上記実施例12(1)に記載の方法でSCIDマウスで継代して得たKPMM2骨髄腫腫瘍塊をウサギ抗アシアロGM1抗体/X線処理ヌードマウス(5週齢、雄)に4mm角ブロックで皮下移植し、翌日に1回だけSK2抗体を1mg/マウスの1用量、再構成ヒト型化PM1抗体(国際公開出願WO92−19759参照)を0.125、0.5および1mg/マウスの3用量で静脈内投与した。各抗体は0.2ml/マウスとなるようにPBS(−)(ニッスイ製)で調製し、陰性対照群には、PBS(−)を0.2ml/マウス投与した。その後腫瘍の大きさを経時的に観察し、対照群の腫瘍が十分大きくなった35日目に全採血を行ってから腫瘍を摘出して重量を測定した。
【0082】
その結果を図11に示す。陰性対照群の平均腫瘍重量が約1gであったのに対し、再構成ヒト型化PM1抗体投与群においては、1mg/マウス投与した場合で腫瘍増殖抑制率(Growth Inhibitory Ratio:GIR)は78%、0.5mg/マウス投与した場合でのGIRは53%、0.125mg/マウス投与した場合でのGIRは66%を示し、強い腫瘍増殖抑制効果が見られた。またSK2においても、1mg/マウス投与群でGIR61%と腫瘍増殖を抑制した。
【0083】
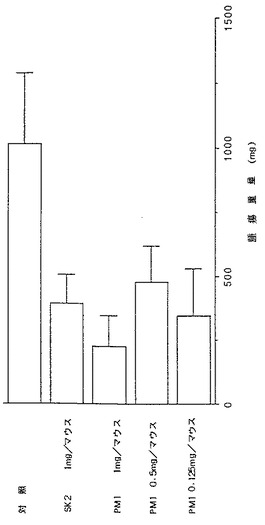
さらに、腫瘍摘出時に採取した血清サンプルのヒトIgG濃度をELISA法にて測定した。その結果を図12に示す。抗体非投与陰性対照群では血清ヒトIgG濃度が平均27.6mg/mlであったものが、再構成ヒト型化PM1抗体を1、0.5、0.125mg/マウス投与することでそれぞれ71%、55%、75%抑制された、SK2 1mg/マウス投与でも43%抑制された。腫瘍重量と血清ヒトIgG濃度は各処理群間でも、個体レベルでもよく相関していた。
【0084】
実施例14:KPMM2静脈内移植SCIDマウスにおけるイオン化カルシウム濃度の上昇および骨吸収の亢進
KPMM2(107個)を上記実施例12(2)に記載する方法でSCIDマウス(日本クレア社製)へ静脈内移植し、移植後経時的に血中イオン化カルシウム濃度および骨吸収の有無を試験した。
【0085】
KPMM2移植後9日目、20日目、30日目および37日目にKPMM2静脈内移植SCIDマウスをエーテル麻酔し、マウス眼窩より60μl容量のキャピラリーカラム(チバ・コーニング社製)で採血し、直ちに血中イオン化カルシウム濃度を634自動Ca++/pHアナライザー(チバ・コーニング社製)にて測定した。なお、対照としてKPMM2を移植していないSCIDマウスからも同様の方法で採血し、血中イオン化カルシウム濃度を測定した。
【0086】
その結果、KPMM2静脈内移植SCIDマウスの血中イオン化カルシウム濃度は、移植後30日目より上昇がみられ、37日目には対照群のマウスに比べ、約20%の血中イオン化カルシウム濃度上昇が観察された(図13)。なお、血中イオン化カルシウム濃度上昇は、KPMM2をSCIDマウスへ静脈内移植したときの、マウス骨髄におけるKPMM2(ヒトCD38抗原陽性細胞)が占める増加の割合の経時的増加とよく相関していた(図14参照)。
【0087】
また、KPMM2静脈内移植37日目のSCIDマウス下肢の骨をX線撮影して形態的に観察したところ、対照群のマウスと比較し、顕著な骨吸収像が確認された(図15参照)。以上の結果は、KPMM2静脈内移植SCIDマウスの骨病変が、実際の骨髄腫の病変とよく一致していることを示している。
【産業上の利用可能性】
【0088】
本発明によってIL−6オートクライン依存性で増殖する骨髄腫が存在することが明らかとなった。本発明のオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株は、骨髄腫のIL−6依存性増殖機構モデルとして有用である。また抗IL−6抗体、抗IL−6受容体抗体などのIL−6活性阻害剤をはじめとする骨髄腫治療剤の治療モデルとしても使用し得るものである。本発明のオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株はインビトロおよびインビボで細胞増殖抑制を指標とした骨髄腫治療剤の評価系を作成するのに有用であるのはもちろん、Mタンパクを産生し、この産生量が骨髄腫の増殖に極めてよく相関することから、Mタンパク産生量の抑制を指標とした骨髄腫治療剤の評価系を作成するのに有用である。
【0089】
さらに、本発明のヒト骨髄腫細胞株を実験動物に静脈内移植して得られる骨髄腫の骨髄生着モデルでは、多発性骨髄腫の増殖に伴い骨髄腫に特徴的な骨病変が観察され、したがってこの骨病変の抑制を指標とした骨髄腫治療剤の評価系を作成することができる。
【0090】
これらの点から本発明のヒト骨髄腫細胞株の利用価値は極めて大きい。
【技術分野】
【0001】
本発明はヒト骨髄腫細胞株に関し、さらに詳しくはオートクライン機構によってIL−6依存性で増殖するヒト骨髄腫細胞株、該細胞株を移植した実験動物、ならびに該細胞株または該実験動物を用いる骨髄腫治療剤のスクリーニング法に関する。
【背景技術】
【0002】
B細胞刺激因子2(BSF−2)およびマウスハイブリドーマ/形質細胞腫増殖因子と同一因子であるインターロイキン6(IL−6)は多発性骨髄腫(multiple myeloma:以下MMと記載する場合がある)細胞の主要な増殖因子であると考えられている(Kawano et al., Nature 332:83,1988; Klein et al., Blood 73:517, 1989)。多発性骨髄腫は、形質細胞が悪性化した腫瘍で、骨髄を増殖の場とし、複数の部位に同時に発生する。IL−6はこのような細胞上で2種の膜タンパク質を介してその活性を伝達する。その1つは、IL−6が結合する分子量80kDのリガンド結合性膜タンパク質(IL−6受容体)であり、他の1つは非リガンド結合性のシグナル伝達にかかわる膜タンパク質gp130である(Taga et al., J. Exp. Med. 196:967, 1987)。
【0003】
1988年Kawanoらは新鮮ヒト骨髄腫細胞が構成的にIL−6を産生し、かつIL−6受容体を発現していること、ならびに抗IL−6受容体(R)抗体によりインビトロで増殖が抑制されることから、骨髄腫細胞は成長因子を自ら産生し、自身で受容するというオートクライン機構により増殖する可能性があることを報告し(Kawano et al., Nature 332:83, 1988)、一方Kleinらは、自ら成長因子を産生しないが、周囲からの成長因子を受容するというパラクライン機構により増殖することを提唱した(Klein et al., Blood 73:517, 1989)。また、血清中のIL−6濃度は骨髄腫の病勢と相関していることが知られており(Bataille et al., J. Clin. Invest. 84:2008, 1989)、IL−6が骨髄腫の主要な増殖因子の1つであると考えられている。
【0004】
また、新鮮分離した骨髄腫細胞の場合、骨髄腫細胞以外の細胞の混入が避けられず、正確なアッセイが困難であることから、現在にいたるまで、骨髄腫細胞がオートクライン機構またはパラクライン機構のいずれによって増殖するのかは定かではない。
【0005】
IL−6依存性増殖をするヒト骨髄腫細胞株中にトランスフェクションによってヒトIL−6 cDNAを導入すると、自律的に増殖して腫瘍化することが観察され、これはパラクラインIL−6増殖機構を示唆するものである(Okuno et al., Exp. Hematol. 20:395, 1992)。また、ヒト骨髄腫細胞株U266はIL−6オートクライン機構によって増殖することが報告されている(Jernberg et al., Leukemia 5:255, 1991; Levy et al., J. Clin. Invest. 88:696, 1991)。しかしながら、U266の増殖は外因性IL−6によっても(Jernberg et al., Leukemia 5:255, 1991)、また抗IL−6モノクローナル抗体によっても(Levy et al., J. Clin. Invest. 88:696, 1991)影響を受けなかった、という報告もあり、U266の増殖機構に対するIL−6の関与は不明である。
【0006】
本発明者らは、多発性骨髄腫患者の腹水から得た1例の新鮮骨髄腫細胞において、腫瘍細胞は明らかなIL−6依存性増殖を示し、かつ、自律的増殖とともに抗IL−6受容体抗体で強く抑制されることを報告した(Goto et al., Biotherapy 7:655, 1993)。
【0007】
本発明者らはさらに、新鮮ヒト骨髄腫細胞をヒトIL−6遺伝子導入重症複合免疫不全マウス(IL−6トランスジェニックSCIDマウス)に移植した腫瘍細胞の性質について検討した(Goto et al., 第52回日本癌学会総会講演要旨集、498頁、1993年10月)。その結果、皮下に移植した3匹にはその移植部位に形質細胞腫を認め、腋窩リンパ節転移も認めた。腹腔内移植では腫瘤形成は認めなかった。移植後の腫瘍細胞は表面抗原やインビトロでのIL−6依存性増殖および抗ヒトIL−6受容体抗体による増殖抑制効果などに移植前と比べて変化を認めなかった。
【非特許文献1】Kawano et al., Nature 332:83,1988
【非特許文献2】Klein et al., Blood 73:517, 1989
【非特許文献3】Taga et al., J. Exp. Med. 196:967, 1987
【非特許文献4】Kawano et al., Nature 332:83, 1988
【非特許文献5】Klein et al., Blood 73:517, 1989
【非特許文献6】Bataille et al., J. Clin. Invest. 84:2008, 1989
【非特許文献7】Okuno et al., Exp. Hematol. 20:395, 1992
【非特許文献8】Jernberg et al., Leukemia 5:255, 1991
【非特許文献9】Levy et al., J. Clin. Invest. 88:696, 1991)
【非特許文献10】Jernberg et al., Leukemia 5:255, 1991
【非特許文献11】Levy et al., J. Clin. Invest. 88:696, 1991
【非特許文献12】Goto et al., Biotherapy 7:655, 1993
【非特許文献13】Goto et al., 第52回日本癌学会総会講演要旨集、498頁、1993年10月
【発明の概要】
【発明が解決しようとする課題】
【0008】
上記したように、骨髄腫細胞の増殖機構には未だ不明な点が多く、これを解明することが求められている。
本発明の目的は骨髄腫のIL−6依存性増殖機構のモデルとなりえる骨髄腫細胞株を樹立することにある。該細胞株は抗IL−6抗体、抗IL−6受容体抗体などのIL−6活性阻害剤をはじめとする骨髄腫治療剤による骨髄腫の治療のインビトロモデルとして有用である。
【0009】
本発明は該細胞株を移植した実験動物を提供することも目的とする。上記実験動物は抗IL−6抗体、抗IL−6受容体抗体などのIL−6活性阻害剤をはじめとする骨髄腫治療剤による骨髄腫の治療のインビボモデルとして有用である。
【0010】
本発明は骨髄腫治療剤のスクリーニング法を提供することも目的とする。上記インビトロモデルにおいては、細胞増殖抑制あるいはMタンパク(ミエローマタンパク)分泌抑制を指標とする骨髄腫治療剤のスクリーニング法を使用することができる。また、上記インビボモデルにおいては、細胞増殖抑制、Mタンパク分泌抑制あるいは骨病変の抑制を指標とする骨髄腫治療剤のスクリーニング法を使用することができる。なお、Mタンパクとは、骨髄腫が特異的に産生する免疫グロブリンタンパク質であり、それを産生する骨髄腫によりIgA、IgM、IgG、IgEおよびBence−Jonesタンパクの5種類がある。
【課題を解決するための手段】
【0011】
本発明者らは、上記目的を達成すべく鋭意研究した結果、IL−6依存性増殖機構を有する骨髄腫のモデルとなりうる細胞株を樹立することに成功し、本発明を完成した。
すなわち、本発明はオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株を提供する。
【0012】
また、本発明は該細胞株を移植した実験動物を提供する。
さらに、本発明は骨髄腫治療剤を上記細胞株に添加して骨髄腫細胞増殖抑制あるいはMタンパク分泌抑制を試験することからなる骨髄腫治療剤のインビトロスクリーニング法を提供する。
【0013】
さらに、本発明は骨髄腫治療剤を上記実験動物に投与して骨髄腫細胞増殖抑制、Mタンパク分泌抑制あるいは骨病変の抑制を試験することからなる骨髄腫治療剤のインビボスクリーニング法を提供する。
【図面の簡単な説明】
【0014】
【図1】液体培養におけるKPMM2の自律的凝集を示す図(生物の形態を表す写真)である。
【図2】KPMM2の形態を示す図(生物の形態を表す写真)である。ライト−ギムザ染色では形質細胞の特徴を有する。
【図3】サザンブロット分析によるKPMM2のJHおよびCλ遺伝子の再構成を示す図(電気泳動の写真)である。KPMM2から得たDNAをBamHI、EcoRIおよびHindIIIで消化し、JHおよびCλ遺伝子プローブを用いてサザンブロット分析を行った。再構成したバンドを(▲)で示す。
【図4】KPMM2の核型を示す図(生物の形質を表す写真)である。検索した15細胞はすべて46、XX、der(1;19)(q10;q10)、t(3;14)(q21;q32)、−4、t(6;11)(p12;p15)、der(10)add(10)(p13)dic(9;10)(q10;q26)、+16を示した。
【図5】KPMM2の細胞増殖に対する各種サイトカインの効果を示す図である。使用したサイトカインの濃度は以下の通りである:IL−6、1ng/ml;IFN−αおよびIFN−γ、1000U/ml;その他のサイトカイン、100ng/ml。各数値は3回の試験の平均+標準偏差(SD)を表す。
【図6】KPMM2の細胞増殖に対する抗IL−6 mAbおよび抗IL−6R mAbの効果を示す図である。SK2はマウス抗IL−6 mAb(▲);PM1はマウス抗IL−6R mAb(●)。破線は対照を示す。各数値は3回の試験の平均を表す。
【図7】KPMM2細胞におけるIL−6Rの発現を示す図である。細胞は抗IL−6R mAb(MT18)で染色した。マウスIgG2b抗体を対照として用いた。破線はmIgG2bを、実線はMT18を表す。
【図8】1.5%アガロースゲル上でのRT−PCR(逆転写PCR)分析により、KPMM2のIL−6およびIL−6R mRNAの発現を示す図(電気泳動の写真)である。レーン1および4、SKW6.4;レーン2および5、KPMM2;レーン3および6、陽性対照。
【図9】KPMM2を皮下移植したマウスにおける腫瘍体積と血清ヒトIgG濃度の相関を示す図である。
【図10】(a)はインビトロ培養したKPMM2におけるヒトCD38抗原のFACS解析を示す図である。KPMM2は40から2000までの範囲の蛍光強度を有する。
【0015】
(b)はKPMM2を静脈内移植したSCIDマウスの骨髄細胞におけるヒトCD38抗原のFACS解析を示す図である。71%の細胞が40から2000の範囲の蛍光強度を有する。
【0016】
(c)はKPMM2を皮下移植したSCIDマウスの腫瘍塊から得た細胞におけるヒトCD38抗原のFACS解析を示す図である。全ての細胞が40から2000の範囲の蛍光強度を有する。
【0017】
(d)はKPMM2を移植していないSCIDマウスの骨髄細胞におけるヒトCD38抗原のFACS解析を示す図である。40から2000の範囲の蛍光強度を有する細胞は全くみられない。
【図11】KPMM2に対する抗IL−6 mAbおよび抗IL−6R mAbのインビボにおける腫瘍増殖抑制効果を示す図である。SK2は抗ヒトIL−6 mAb;再構成ヒト型化PM1は抗ヒトIL−6R mAb。
【図12】KPMM2移植ヌードマウス中の血清ヒトIgG濃度に対する抗IL−6 mAbまたは抗IL−6R mAbの効果を示す図である。
【図13】KPMM2を静脈内移植したSCIDマウスの血中イオン化カルシウム濃度(□)の経時的変化を示す図である。(◇)は対照を示す。各数値はSCIDマウス4匹(37日目のみ5匹)の平均±S.D.を示す。
【図14】KPMM2を静脈内移植したSCIDマウスの骨髄中のヒトCD38抗原陽性細胞の割合(□)の経時的変化を示す図である。(◇)は対照を示す。各数値はSCIDマウス4匹(37日目のみ5匹)の平均±S.D.を示す。
【図15】KPMM2を静脈内移植したSCIDマウスの骨のX線撮影像を示す図(生物の形態を示す写真)である。
【0018】
(a)はKPMM2を移植していない対照群のSCIDマウス
(b)はKPMM2移植後37日目のSCIDマウス
【発明を実施するための形態】
【0019】
本発明の細胞株は、例えば多発性骨髄腫患者の腹水などから採取した骨髄腫細胞を用いて樹立することができる。本発明では特に、IgG、λ型多発性骨髄腫患者の腹水から骨髄腫細胞を採取した。培養開始後1カ月で細胞は安定して増殖し始め、1年以上維持された。このようにして樹立された細胞株はKPMM2と命名され、生命工学工業技術研究所特許微生物寄託センターに受託番号FIRM:P−14170で寄託されている(1994年2月22日寄託)。細胞株KPMM2はインビトロでの継代、ならびに実験動物、例えば重症複合免疫不全(SCID)マウス、IL−6トランスジェニックSCIDマウスおよびヌードマウスなどの免疫不全状態にあるマウス中での安定した継代が可能である。
【0020】
細胞株KPMM2はインビトロでIL−6を産生し、またIL−6受容体(IL−6R)を発現していることが確認された。また、KPMM2の各種サイトカインに対する反応性を3H−チミジンの取り込み実験、MTT(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide)法(J. Immun. Methods 65:55-63, 1983 参照)および生細胞の測定で試験した結果、IL−6とともにインキュベートしたときのみ顕著に刺激され、これはKPMM2がIL−6に特異的に反応して増殖することを示している。さらに、KPMM2の増殖は、抗IL−6 mAb(モノクローナル抗体)および抗IL−6R mAbによって用量依存的に顕著に抑制された。また、RT−PCR(逆転写ポリメレースチェインリアクション)により、KPMM2がIL−6およびIL−6R mRNAを発現していることが確認された。なお、KPMM2は各種の接着分子、すなわちCD44、VLA−β、ICAM−1、NCAM、LFA−3およびVLA−4を発現しており、またインビトロで自律的細胞凝集を示す。
【0021】
このようなKPMM2の各種特徴から、KPMM2がIL−6オートクライン機構により増殖する骨髄腫細胞株であることが示された。本発明の細胞株はその増殖機構がIL−6依存性のオートクライン機構であることが証明された最初の細胞株である。
【0022】
したがって、本発明の細胞株は抗IL−6 mAbあるいは抗IL−6R mAbなどのIL−6活性阻害剤をはじめとする骨髄腫治療剤のスクリーニングに有用である。例えば、抗IL−6抗体または抗IL−6受容体抗体を本発明の細胞株に添加して骨髄腫細胞増殖抑制効果を試験することからなる骨髄腫治療剤のインビトロスクリーニング法が可能である。また、本発明の骨髄腫細胞株は、細胞数の増加に比例してMタンパクの分泌量が増大することにより、Mタンパク分泌抑制を指標として骨髄腫治療剤のインビトロスクリーニングを行うことができる。さらに、本発明の細胞株は、細胞間相互作用やIL−6シグナル伝達を介する骨髄腫細胞の増殖において接着分子の果たす役割を研究するモデルとしても有用である。
【0023】
本発明は、本発明の細胞株を移植した実験動物も提供する。本発明の細胞株を移植する実験動物としては、マウスの他、ラット、ウサギ、モルモット、ハムスター、サルなどが挙げられ、さらには、T細胞あるいはB細胞といった免疫担当細胞の機能に障害が生じ、免疫不全状態にある実験動物に本発明の細胞株を移植するのがよい。既に述べたように、本発明の細胞株はSCIDマウス、IL−6トランスジェニックSCIDマウスおよびヌードマウスなどの免疫不全状態にあるマウス中で安定した継代が可能である。
【0024】
興味深いことに、本発明の上記細胞株をマウスに移植するには、皮下移植、腹腔内移植の外に、静脈内移植によっても行うことができる。皮下移植および腹腔内移植した場合には、それぞれ移植部位皮下および腹腔内に固形腫瘍が観察されるが、静脈内移植した場合には、骨髄への腫瘍細胞の生着が見られ、これを実際の骨髄腫の病態に近いモデルとして使用することができる。例えば、抗IL−6抗体または抗IL−6受容体抗体といったIL−6活性阻害剤をはじめとする骨髄腫治療剤を本発明の実験動物に投与して骨髄腫細胞増殖抑制効果を試験することからなる骨髄腫治療剤のインビボスクリーニング法が可能である。また、本発明の細胞株が生着した実験動物では、腫瘍の増殖に伴って血清中のMタンパク濃度の上昇が観察されるため、Mタンパク濃度の抑制を指標とした骨髄腫治療剤のインビボスクリーニング法も可能である。さらに、本発明の細胞株が実験動物の骨髄へ生着すると骨髄腫細胞の増殖に伴い、血中イオン化カルシウム濃度の上昇、骨破壊、骨融解および骨吸収といった骨病変が観察されることから、これら骨病変の抑制を指標とした骨髄腫治療剤のインビボスクリーニング法が可能である。
【0025】
以下に本発明を実施例によりさらに詳しく説明するが、本発明の範囲はこれに限定されるものではない。
【実施例】
【0026】
実施例1:骨髄腫細胞株の樹立および維持
IgG、λ型多発性骨髄腫患者(76才、女性、ステージIIA)の腹水から骨髄腫細胞を採取した。腹水は多数の骨髄腫細胞を含んでおり、腹水中のIL−6レベルは91.0pg/mlに達していた。採取した腹水をFicoll−Hypaque(ファルマシア社製)を用いた密度勾配遠心法にかけて単核球を分離、プラスチックシャーレにて付着細胞を除去し、さらにヒツジ赤血球にてT細胞を除去し、腫瘍細胞を95%以上に純化した。細胞を20%ウシ胎児血清(FCS:Xavier Investments製、オーストラリア)、組換えヒトIL−6(中外製薬製)4ng/mlおよびカナマイシン(明治製菓社製)100μg/mlを含むRPMI1640(ギブコ社製)培養液中に1×106細胞/mlの濃度で浮遊させた。次いで25mlフラスコ中で10mlの培養液中で培養し、湿潤5%CO2中、37℃でインキュベートした。安定した細胞増殖が観察されるまで3日ごとに培地を部分的に新しいものに取り替えた。
【0027】
培養開始1カ月後で細胞は安定して増殖し始め1年以上維持され、細胞株として樹立された。この細胞株をKPMM2と命名した。IL−6の存在下または非存在下における倍加時間はそれぞれ48時間および72時間であった。
【0028】
KPMM2の形態およびIg分泌は以下の通りである。
KPMM2の形態およびIg分泌
KPMM2細胞は光学顕微鏡下で自律的な細胞凝集を示して増殖することが観察され(図1)、ライト−ギムザ染色では形質細胞様の形態を示した(図2)。KPMM2は酸ホスファターゼが陽性であり、α−ナフチルブチレートエステラーゼがやや陽性であるが、ペルオキシダーゼ、AS−Dクロロアセテートエステラーゼ、パス(過ヨウ素酸シッフ試薬)およびアルカリホスファターゼは陰性であった。細胞質IgGおよびλL鎖が検出されたが、IgA、IgM、およびκL鎖は直接免疫蛍光法で陰性であった。また、細胞(106個/ml)を3日間培養すると、培養上清にはIgGおよびλL鎖の分泌が見られた。
【0029】
実施例2:表面抗原の解析
上記実施例1で樹立した細胞株KPMM2の表面抗原の発現を各種ヒト抗原に対するモノクローナル抗体のパネルを用いて、直接および間接蛍光抗体法(Fried et al., Flow Cytometry, Boca Raton, CRC Press:59-78, 1989)により検討した。
【0030】
実施例1に記載の方法で得られたKPMM2を、106個/チューブとなるように、100μlの蛍光活性化細胞選択装置(FACS)用緩衝液(2% FCSおよび0.1% NaN3を含むリン酸緩衝化生理食塩溶液(PBS)、以下FACS緩衝液という)に浮遊させた。次いで、直接蛍光抗体法においては、飽和量の下記表1に記載の各種ヒト抗原に対するフルオレシンイソチオシアネート(FITC)あるいはフィコエリスリン(PE)標識抗体を添加し、4℃にて30分間インキュベートした。細胞を上記FACS緩衝液で2回洗浄した後、フローサイトメーター(EPICS PROFILE,コールター社製)で分析した。
【0031】
一方、間接蛍光抗体法においては、非標識の下記表1に記載の各種ヒト抗原に対する抗体を添加し、4℃にて30分間インキュベートして、細胞をFACS緩衝液で2回洗浄した後、5μg/mlのFITCあるいはPE標識ヤギ抗マウスIgG抗体F(ab’)2断片(TAGO社製)を加え、4℃にて30分間反応させた。FACS緩衝液で2回洗浄した後、FACS緩衝液に浮遊させ、フローサイトメーター(EPICS PROFILE,コールター社製)で分析した。
【0032】
KPMM2の表面抗原を以下の表1にまとめて示す。
【0033】
【表1】
【0034】
表から明らかなように、KPMM2は形質細胞関連抗原(CD38、PCA−1およびBL3)、接着分子(CD44、VLA−β、ICAM−1、NCAM、LFA−3およびVLA−4)ならびにCD45、CD63、CD71、IgGおよびλなどの抗原が陽性であった。
【0035】
実施例3:免疫グロブリン遺伝子再構成
上記実施例1で樹立した細胞株KPMM2の免疫グロブリン(Ig)遺伝子再構成をサザンブロット法により分析した。
【0036】
実施例1に記載の方法により得られたKPMM2細胞(107個)から、Manual of Clinical Immunology, 3rd edition, American Society for Microbiology, 1986の方法に準じてDNAを調製し、3種類の制限酵素BamHI、EcoRIあるいはHindIII(ベーリンガー・マンハイム社製)で別々に処理し、エタノール沈殿としてDNAを回収して、0.8%アガロースゲル(SEAKEM GTG,FMC社製)により24時間電気泳動を行った。電気泳動したDNAをナイロン膜(ハイボンドN+,アマシャム社製)に移し、これを乾燥させた。次に、タカラランダムプライマーDNAラベリングキット(宝酒造社製)を用いて、32P標識したヒトIg JH、CκおよびCλプローブ(オンコア社製)を用い、添付の処方に従ってサザンブロット分析を行った。なお、対照として健常人の末梢血単核球から得た再構成を生じていない染色体DNAを用いた。その結果、IgHおよびκ鎖遺伝子が再構成していたが、λ鎖遺伝子は再構成していなかった(図3)。以上のことより、KPMM2がモノクローナルな抗体を産生すること、および細胞の単一性が確認された。
【0037】
実施例4:細胞遺伝学的分析
上記実施例1で樹立した細胞株KPMM2の染色体の構造異常を分析した。
実施例1に記載の方法で得られたKPMM2を20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液にて5×105個/mlとなるように培養した。培養48時間後に、KPMM2に0.05μgのコルセミド(ギブコ社製)を15分間処理し、分裂中期で細胞周期が停止したKPMM2細胞を回収した。回収したKPMM2細胞を0.075MのKClで20分間処理し、メタノール−酢酸で固定した。次いで、KPMM2細胞の染色体をトリプシン−ギムザバンド染色法によって分析した。
【0038】
その結果、KPMM2は多くの構造異常をもつ二倍体細胞であることが判明した(図4)。分析した15細胞すべてが46、XX、der(1;19)(q10;q10)、t(3;14)(q21;q32)、−4、t(6;11)(p12;p15)、der(10)add(10)(p13)dic(9;10)(q10;q26)、+16を示した。
【0039】
実施例5:EBVおよびマイコプラズマの検出
実施例1で樹立した細胞株KPMM2のエプスタイン・バールウイルス(EBV)およびマイコプラズマ汚染を試験した。
【0040】
実施例1に記載の方法で得られたKPMM2細胞の染色体からエプスタイン・バールウイルス(EBV)を検出するために、Systemic Genetic Institute社から購入したEBV BamW領域増幅プライマーを用いて、添付の処方に従いPCR(ポリメレース チェイン リアクション)を行った。マイコプラズマ感染の検出はマイコプラズマDNA検出用M.T.C.キット(Gen−Probe Inc.社製)により、添付の処方に従って実施した。
【0041】
その結果、KPMM2はEBVゲノムおよびマイコプラズマゲノムに対して陰性であった。
実施例6:サイトカインに対する反応性
実施例1で樹立した細胞株KPMM2の各種サイトカインへの反応性を試験した。
【0042】
実施例1に記載する方法で得られたKPMM2を20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に浮遊させ、容量が200μlで96穴プレート(ファルコン社製)へ1×104個/穴となるように分注した。
【0043】
96穴プレートの各穴には、下記の濃度となるように各種サイトカインを別々に加えた。
組換えIL−2、同IL−3、同腫瘍壊死因子(TNF)−α、同顆粒球マクロファージコロニー刺激因子(GM−CSF)、同幹細胞成長因子(SCF)(以上、Genzyme社製)、同IL−4、同IL−7、同IL−10、同IL−11、同白血病阻害因子(LIF)、同オンコスタチンM(OSM)(以上、Pepro Tec Inc.社製)、同IL−9、同トランスフォーミング成長因子(TGF)−β(以上、R&D System Inc.社製)、同IL−1α(Boehringer Manheim社製)、同IL−5(Upstate Biotechnology Inc.社製)、同IL−8(Amersham社製)、同エリスロポエチン(EPO)および同顆粒球コロニー刺激因子(G−CSF)(以上、中外製薬株式会社より提供)は100ng/ml。
【0044】
組換えインターフェロン(IFN)−γ(塩野義製薬株式会社より提供)および天然型ヒトIFN−α(住友製薬株式会社より提供)は1000U/ml。
組換えIL−6(中外製薬株式会社より提供)は1ng/ml。
【0045】
なお、対照群は、サイトカインを加えない20%FCSおよび100μg/mlカナマイシンを添加したRPMI1640培養液で培養した。
96穴プレートの各穴に分注したKPMM2を、上記サイトカイン存在下あるいは非存在下で湿潤5%CO2中、37℃で96時間培養した。その培養終了4時間前に、各穴に3H−チミジン(Amersham社製)を1μCi/穴となるように添加した。KPMM2が取り込んだ3H−チミジンの量の測定は、液体シンチレーションカウンター(1205 ベータプレート、ファルマシア社製)を用いた。
【0046】
KPMM2の増殖に対する各種サイトカインの効果を図5に示す。図5から明らかなように、KPMM2細胞はIL−6とともにインキュベートしたときのみに3H−チミジンの取り込みが顕著に刺激された。濃度1ng/mlにおけるIL−6により、3H−チミジンの取り込みが2.2倍に上昇した。一方、IFN−αおよびIFN−γはKPMM2細胞の増殖を顕著に阻害した。
【0047】
また、KPMM2のサイトカインに対する反応性を調べるために、MTT(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide)法(J. Immun. Methods 65:55, 1983 を参照)および生細胞を直接測定する方法を用いたが、同様の結果が得られた。
【0048】
実施例7:抗IL−6 mAbおよび抗IL−6R mAbによる増殖阻害
実施例1で樹立した細胞株KPMM2に対するマウス抗ヒトIL−6R mAb(モノクローナル抗体)(IgG1クラス:PM1)およびマウス抗IL−6 mAb(IgG1クラス:SK2)の効果を調べた。SK2は文献(Y. Ohe et al., Br. J. Cancer 67:939, 1993)に記載されており、またPM1は文献(Hirata et al., J. Immunol. 143:2900, 1989)に記載されている。
【0049】
実施例1に記載する方法で得たKPMM2を20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に浮遊させ、容量が200μlで96穴プレート(ファルコン社製)へ1×104個/穴となるように分注した。
【0050】
96穴プレートの各穴には、各種濃度のIL−6 mAb(モノクローナル抗体)(SK2)および抗IL−6R(受容体)mAb(PM1)を別々に加えた。なお、対照群は、抗体を加えない20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液とした。
【0051】
96穴プレートの各穴に分注したKPMM2を、抗IL−6 mAbおよび抗IL−6R mAb存在下あるいは非存在下で湿潤5%CO2中、37℃で96時間培養した。その培養終了4時間前に、各穴に3H−チミジン(Amersham社製)を1μCi/穴となるように添加した。KPMM2が取り込んだ3H−チミジンの量の測定は、液体シンチレーションカウンター(1205 ベータプレート、ファルマシア社製)を用いた。
【0052】
KPMM2の増殖に対する抗IL−6 mAbおよび抗IL−6R mAbの効果を図6に示す。SK2およびPM1の添加はいずれも用量依存的に細胞増殖を有意に阻害した。特に、PM1は1μg/mlの濃度でKPMM2の増殖を完全に阻害した。
【0053】
また、KPMM2増殖の抗IL−6 mAbおよび抗IL−6R mAbに対する効果を調べるために、MTT(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide)法(J. Immun. Methods 65:55, 1983 を参照)および生細胞を直接測定する方法を用いたが、同様の結果が得られた。
【0054】
実施例8:ELISAによるIL−6産生の測定
上記実施例1で樹立した細胞株KPMM2によるIL−6産生能を試験した。
実施例1に記載する方法で得たKPMM2を、20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に106個/mlとなるように浮遊させ、ヒトIL−6非存在下で湿潤5%CO2中、37℃で培養した。
【0055】
培養72時間後、培養上清中に含まれるKPMM2が産生したIL−6の濃度を、ヒトIL−6用ELISAキット(帝人バイオラボラトリー社製)を用いて添付の処方に従い測定した。なお、陰性対照として、20%FCSおよび100μg/mlカナマイシンを添加したRPMI1640培養液を用いた。培養上清には79.7±19.6(平均値±S.D.)pg/mlのIL−6の産生が検出され、対照培養液において検出限界(4.0pg/ml)以下であったのに比較すると、培養上清中のIL−6濃度が顕著に増加していることが確認された。
【0056】
実施例9:フローサイトメトリーによるIL−6Rの発現の確認
KPMM2細胞上でのIL−6R発現を確認するために、IL−6Rに結合し、IL−6のIL−6Rへの結合を阻害しないマウス抗ヒトIL−6R mAbであるMT18抗体(Hirata et al., J. Immunol. 143:2900, 1989)を用いて、間接蛍光抗体法を実施した。
【0057】
実施例1に記載の方法で得られたKPMM2細胞を106個/チューブとなるように、100μlの実施例2記載のFACS緩衝液に浮遊させ、10μg/mlのMT18抗体を加え、4℃にて3時間反応させた後、FACS緩衝液で2回洗浄し、100μlのFACS緩衝液に浮遊させ、さらに、FITC標識ヤギ抗マウスIgG抗体(TAGO社製)を5μg/ml添加し、4℃にて30分間反応させた。FACS緩衝液で2回洗浄した後、同FACS緩衝液に浮遊させ、フローサイトメーター(EPICS PROFILE、コールター社製)で蛍光を測定した。
【0058】
その結果、図7に示すように、KPMM2細胞上にIL−6Rの発現が示された。
実施例10:ELISAによるヒトIgG(Mタンパク)産生の測定
KPMM2のヒトIgG(Mタンパク)産生能を試験した。
【0059】
実施例1に記載の方法で得られたKPMM2を106個/mlとなるように、20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に浮遊させ、容量2mlで12穴プレート(ファルコン社製)の各穴に分注し、ヒトIL−6非存在下で湿潤5%CO2中、37℃で72時間培養した。なお、実験は3回行った。その後、培養上清中のヒトIgG濃度を、TAGO社製ヤギ抗IgG抗体(NO.4100)およびアルカリフォスファターゼ標識ヤギ抗IgGガンマ鎖特異的抗体(NO.2490)を用いるELISAにて測定した。なお、スタンダードとしてカッペル社製ヒトIgG(NO.0001−860)を用いた。その結果、培養上清中には10.1μg/mlのヒトIgGが検出され、対照培養液では検出限界以下(5ng/ml)であったのに比較して、培養上清中のヒトIgG濃度が顕著に増加していることが確認された。
【0060】
実施例11:RT−PCR(逆転写ポリメレースチェインリアクション)によるIL−6およびIL−6R mRNAの検出
実施例1に記載の方法で得られたKPMM2におけるヒトIL−6およびヒトIL−6Rの発現を確認するために、RT−PCR(逆転写ポリメレースチェインリアクション)法によりヒトIL−6およびヒトIL−6RのメッセンジャーRNA(mRNA)を検出した。
【0061】
グアニジウムセシウムクロライド法(Molecular Cloning, Cold Spring Harbor Laboratory Press)によってKPMM2細胞(108個)から全RNAを調製した。陰性対照としてヒトB細胞リンパ腫細胞株SKW6.4からも同様に全RNAを調製した。1本鎖cDNA合成はcDNA合成キット(Invitrogen社製)を用いて、添付の処方に従い全RNA5μgから直接実施した。
【0062】
陽性対照としてヒトIL−6およびヒトIL−6R検出用PCRプライマーを用いた(Clontech Inc.社製)。PCR溶液各100μlは、10mM Tris−HCl(pH8.3)、50mM 塩化カリウム、Amplitaq(Perkin Elmer Cetus社製)2.5ユニット、1本鎖cDNA合成反応物1μl、各プライマー100pmolを含む。各PCR用チューブに鉱物オイル50μlを上層してPCRに付した。最初に94℃で1分メルトし、60℃で1分と72℃で10分のサイクルを30サイクル繰り返した。最終サイクルの後、最終的に72℃で10分伸長を行った。ヒトIL−6およびヒトIL−6R検出用プライマーを用いた陽性対照群も同様に増幅し、約50pgの陽性対照PCR産物を得た。
【0063】
各反応チューブから10μlを取って1.5%アガロースゲル上で電気泳動を行った。図8に示すように、IL−6(628bp)およびIL−6R(251bp)のPCR産物はいずれもKPMM2のRNAから増幅された。これらの結果は、KPMM2がIL−6のオートクライン機構によって増殖することを遺伝子発現の面から支持する。一方、SKW6.4細胞はIL−6に応答してIgMを分泌する。IL−6RのPCR産物はSKW6.4 mRNAから増幅された。しかし、IL−6のPCR産物はこの実験で検出されず、このことはSKW6.4細胞がIL−6を産生していないことを示唆する。
【0064】
実施例12:細胞株KPMM2のSCIDマウスおよびヌードマウスへの移植
(1)KPMM2の可移植性およびインビボでの継代
上記実施例1で樹立した細胞株KPMM2の可移植性およびインビボでの継代を以下のように検討した。
【0065】
KPMM2を20%FCSおよび100μg/mlカナマイシンを含むRPMI1640培養液に108個/mlで懸濁し、0.2mgのウサギ抗アシアロGM1抗体(Code No.014−09801、和光純薬工業社製)で処理したIL−6トランスジェニック重症免疫不全マウス(以下IL−6−SCID Tmという)(中外製薬製)の腹部皮下に注射針で0.1ml移植した。その結果、1カ月後には3例全例で移植部位に結節型の腫瘍を形成した。
【0066】
この腫瘍を無菌的に摘出し、摘出した腫瘍塊を使い捨て注射器のピストンなどでつぶし、ナイロンメッシュ(70μm、ファルコン社製)を通して細胞を回収した。この細胞を上記と同様にRPMI1640培養液に懸濁し、IL−6−SCID Tm、重症免疫不全マウス(SCIDマウスという)(日本クレア社製)、BALB/c−nu/nuマウス(以下ヌードマウスという)(日本クレア社製)に皮下移植したところ、同じように移植部位に結節型の腫瘍を形成し、インビボでの継代が可能であった。また腫瘍塊を3mm角のブロックにし、移植針を用いてIL−6−SCID Tm、SCIDマウス、ヌードマウスの皮下に移植しても継代可能であった。
(2)KPMM2の異なる移植経路での可移植性
KPMM2の移植経路を皮下(s.c.)、静脈内(i.v.)、腹腔内(i.p.)とした場合の可移植性を以下のように検討した。
【0067】
(1)においてSCIDマウスにて継代したKPMM2腫瘍を無菌的に摘出し、摘出した腫瘍塊を使い捨て注射器のピストンでつぶし、ナイロンメッシュを通して細胞を回収し、108個/mlの細胞懸濁液を作成した。この懸濁液をSCIDマウス、あるいは0.2mgのウサギ抗アシアロGM1抗体および500RのX線で処理したヌードマウスに0.1mlずつ皮下(s.c.)、静脈内(i.v.)、腹腔内(i.p.)の3経路で移植し、40日後に生着の判定を行った(表2)。その結果、約40日後には全例で生着が確認され、s.c.移植では移植部位に、i.p.移植では腹腔内に固形腫瘍を形成した。また、i.v.移植ではKPMM2は骨髄での生着が認められた。
【0068】
【表2】
【0069】
(3)KPMM2移植動物の血清ヒトIgG(Mタンパク:ミエローマタンパク)の濃度
細胞株KPMM2をSCIDマウスへ皮下移植し、形成された腫瘍の体積と血清中ヒトIgG濃度の相関を調べた。上記(2)に記載の方法で細胞株KPMM2をSCIDマウスへ皮下移植し、KPMM2移植前、移植後21日目、42日目の3回、血清サンプルを採取してELISA法にてヒトIgG濃度を測定した。
【0070】
その結果、マウス血清中のヒトIgG濃度はKPMM2を移植した動物ではいずれも経時的に上昇した。また図9に示すように、皮下移植した動物では皮下に形成された腫瘍の体積と血清ヒトIgG濃度に相関が見られ、腫瘍の大きな動物ほど血清ヒトIgG濃度は高値を示した。このことから、血清ヒトIgG濃度を指標としても抗腫瘍効果の判定ができると考えられた。
(4)KPMM2の可移植性および移植細胞数の検討
SCIDマウスとヌードマウスで移植細胞数を変えたときのKPMM2の可移植性を検討した。
【0071】
特別な前処理を行わない雄のSCIDマウスおよびヌードマウスに、上記(2)記載の方法で各々KPMM2細胞を107、3×106、106個腹部皮下(s.c.)に移植し、またSCIDマウスには上記(2)の方法で得たKPMM2細胞を3×106、106個静脈内(i.v.)でも移植した。
【0072】
その結果、SCIDマウスに皮下移植した場合、3×106個以上移植すると、21日までに全例で腫瘍結節を形成し、106個移植した場合でも21日までに2/3、32日までに全例で腫瘍結節を形成した。ヌードマウスに皮下移植すると、107個移植した場合では21日までに2/3、32日までには全例で、また3×106個移植した場合では42日までには全例で腫瘍結節が形成された。しかし、106個の移植では42日までに腫瘍の生着は見られなかった。また、SCIDマウスに静脈内移植した場合、42日までに3×106個移植で全例、106個移植で1/3にKPMM2が生着した。これらの結果を以下の表3にまとめて示す。
【0073】
【表3】
【0074】
(5)KPMM2移植SCIDマウスにおけるヒトCD38抗原の発現の解析
KPMM2を移植したSCIDマウスにおいて、その生着を確認するために、KPMM2の細胞表面上で特徴的に発現している表面抗原であるヒトCD38抗原の発現を試験した。
【0075】
KPMM2を上記(1)に記載する方法でSCIDマウスへ静脈内移植し、移植後37日目にマウスを屠殺した。屠殺したマウスの大腿骨より骨髄を回収して懸濁し、ステンレスメッシュ(100μm)を通し、蛍光活性化細胞選択装置(FACS)用緩衝液(2%FCSおよび0.1%NaN3を含むPBS(−)溶液)にて骨髄細胞の浮遊液を調製した。なお、陰性対照としてKPMM2を移植していないSCIDマウスの骨髄からも同様の方法にて骨髄細胞浮遊液を調製した。
【0076】
また、本実施例に記載する方法によりKPMM2を皮下移植したSCIDマウスからは、移植後37日目に皮下腫瘍塊を外科的に摘出し、摘出した腫瘍塊を2枚のスライドグラスの間にはさんですりつぶし、これをステンレスメッシュ(100μm)を通し、FACS緩衝液にて骨髄細胞の浮遊液を調製した。
【0077】
陽性対照として、実施例1に記載した方法でインビトロにて培養したKPMM2を用いた。インビトロで4日間培養したKPMM2をFACS緩衝液で洗浄した後、同緩衝液中に浮遊させた。
【0078】
次に、このように調製した細胞浮遊液を用いて、FACS解析によるヒトCD38抗原の発現を調べた。
すなわち、100μlのFACS緩衝液中にて、各群の浮遊細胞106個に対し、2.5μg/mlのフィコエリスリン(PE)標識抗ヒトCD38抗体(Leu−17、ベクトン・ディッキンソン社製)を添加し、氷上で30分間反応させた。次いで、1mlのFACS緩衝液で2回洗浄後、500μlのFACS緩衝液で懸濁し、FACScan(ベクトン・ディッキンソン社製)によりFACS解析を行った。
【0079】
陽性対照群であるインビトロ培養KPMM2のFACS解析の結果から、蛍光強度40から2000までの範囲にある細胞をヒトCD38抗原発現細胞とした(図10(a)参照)。KPMM2を静脈内移植したSCIDマウスでは、その骨髄細胞のおよそ71%がヒトCD38陽性細胞で占められていた(図10(b)参照)。このことは、SCIDマウスに静脈内移植したKPMM2がSCIDマウスの骨髄に生着したことを示している。また、KPMM2をSCIDマウスに皮下移植して生じた腫瘍塊から得られた細胞は全てヒトCD38が陽性であり(図10(c)参照)、KPMM2を皮下移植して生ずる腫瘍塊は、全てKPMM2から構成されていることが示された。なお、KPMM2を移植していない陰性対照群のSCIDマウスの骨髄細胞にはヒトCD38陽性細胞は全く検出されなかった(図10(d)参照)。
【0080】
実施例13:抗ヒトIL−6抗体SK2および抗ヒトIL−6R再構成ヒト型化抗体PM1の抗腫瘍効果
KPMM2移植動物における抗ヒトIL−6抗体SK2および抗ヒトIL−6R再構成ヒト型化抗体PM1のインビボ抗腫瘍効果を以下のようにして検討した。
【0081】
上記実施例12(1)に記載の方法でSCIDマウスで継代して得たKPMM2骨髄腫腫瘍塊をウサギ抗アシアロGM1抗体/X線処理ヌードマウス(5週齢、雄)に4mm角ブロックで皮下移植し、翌日に1回だけSK2抗体を1mg/マウスの1用量、再構成ヒト型化PM1抗体(国際公開出願WO92−19759参照)を0.125、0.5および1mg/マウスの3用量で静脈内投与した。各抗体は0.2ml/マウスとなるようにPBS(−)(ニッスイ製)で調製し、陰性対照群には、PBS(−)を0.2ml/マウス投与した。その後腫瘍の大きさを経時的に観察し、対照群の腫瘍が十分大きくなった35日目に全採血を行ってから腫瘍を摘出して重量を測定した。
【0082】
その結果を図11に示す。陰性対照群の平均腫瘍重量が約1gであったのに対し、再構成ヒト型化PM1抗体投与群においては、1mg/マウス投与した場合で腫瘍増殖抑制率(Growth Inhibitory Ratio:GIR)は78%、0.5mg/マウス投与した場合でのGIRは53%、0.125mg/マウス投与した場合でのGIRは66%を示し、強い腫瘍増殖抑制効果が見られた。またSK2においても、1mg/マウス投与群でGIR61%と腫瘍増殖を抑制した。
【0083】
さらに、腫瘍摘出時に採取した血清サンプルのヒトIgG濃度をELISA法にて測定した。その結果を図12に示す。抗体非投与陰性対照群では血清ヒトIgG濃度が平均27.6mg/mlであったものが、再構成ヒト型化PM1抗体を1、0.5、0.125mg/マウス投与することでそれぞれ71%、55%、75%抑制された、SK2 1mg/マウス投与でも43%抑制された。腫瘍重量と血清ヒトIgG濃度は各処理群間でも、個体レベルでもよく相関していた。
【0084】
実施例14:KPMM2静脈内移植SCIDマウスにおけるイオン化カルシウム濃度の上昇および骨吸収の亢進
KPMM2(107個)を上記実施例12(2)に記載する方法でSCIDマウス(日本クレア社製)へ静脈内移植し、移植後経時的に血中イオン化カルシウム濃度および骨吸収の有無を試験した。
【0085】
KPMM2移植後9日目、20日目、30日目および37日目にKPMM2静脈内移植SCIDマウスをエーテル麻酔し、マウス眼窩より60μl容量のキャピラリーカラム(チバ・コーニング社製)で採血し、直ちに血中イオン化カルシウム濃度を634自動Ca++/pHアナライザー(チバ・コーニング社製)にて測定した。なお、対照としてKPMM2を移植していないSCIDマウスからも同様の方法で採血し、血中イオン化カルシウム濃度を測定した。
【0086】
その結果、KPMM2静脈内移植SCIDマウスの血中イオン化カルシウム濃度は、移植後30日目より上昇がみられ、37日目には対照群のマウスに比べ、約20%の血中イオン化カルシウム濃度上昇が観察された(図13)。なお、血中イオン化カルシウム濃度上昇は、KPMM2をSCIDマウスへ静脈内移植したときの、マウス骨髄におけるKPMM2(ヒトCD38抗原陽性細胞)が占める増加の割合の経時的増加とよく相関していた(図14参照)。
【0087】
また、KPMM2静脈内移植37日目のSCIDマウス下肢の骨をX線撮影して形態的に観察したところ、対照群のマウスと比較し、顕著な骨吸収像が確認された(図15参照)。以上の結果は、KPMM2静脈内移植SCIDマウスの骨病変が、実際の骨髄腫の病変とよく一致していることを示している。
【産業上の利用可能性】
【0088】
本発明によってIL−6オートクライン依存性で増殖する骨髄腫が存在することが明らかとなった。本発明のオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株は、骨髄腫のIL−6依存性増殖機構モデルとして有用である。また抗IL−6抗体、抗IL−6受容体抗体などのIL−6活性阻害剤をはじめとする骨髄腫治療剤の治療モデルとしても使用し得るものである。本発明のオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株はインビトロおよびインビボで細胞増殖抑制を指標とした骨髄腫治療剤の評価系を作成するのに有用であるのはもちろん、Mタンパクを産生し、この産生量が骨髄腫の増殖に極めてよく相関することから、Mタンパク産生量の抑制を指標とした骨髄腫治療剤の評価系を作成するのに有用である。
【0089】
さらに、本発明のヒト骨髄腫細胞株を実験動物に静脈内移植して得られる骨髄腫の骨髄生着モデルでは、多発性骨髄腫の増殖に伴い骨髄腫に特徴的な骨病変が観察され、したがってこの骨病変の抑制を指標とした骨髄腫治療剤の評価系を作成することができる。
【0090】
これらの点から本発明のヒト骨髄腫細胞株の利用価値は極めて大きい。
【特許請求の範囲】
【請求項1】
オートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株であって、IL−6タンパク質を分泌し、20%ウシ胎児血清(FCS)および100μg/mlカナマイシンを含むRPMI1640培養液中で増殖可能であり、かつ、抗IL−6抗体により増殖が抑制される、前記ヒト骨髄腫細胞株。
【請求項2】
ヒト骨髄腫がヒト多発性骨髄腫である請求項1に記載の細胞株。
【請求項3】
オートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株であって、重症複合免疫不全マウス及び/又はヌードマウスに静脈内移植した場合に骨髄へ生着する能力を有する請求項1に記載のヒト骨髄腫細胞株。
【請求項4】
請求項1〜3のいずれかに記載の細胞株を移植したヒト以外の実験動物。
【請求項5】
実験動物が免疫不全状態である請求項4に記載のヒト以外の実験動物。
【請求項6】
実験動物がマウスである請求項4に記載の実験動物。
【請求項7】
実験動物にオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株を静脈内移植して得られる請求項4に記載のヒト以外の実験動物。
【請求項8】
実験動物にオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株を皮下移植して得られる請求項4に記載のヒト以外の実験動物。
【請求項9】
実験動物にオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株を腹腔内移植して得られる請求項4に記載のヒト以外の実験動物。
【請求項10】
骨髄腫治療剤を請求項1〜3のいずれかに記載の細胞株に添加して骨髄腫細胞増殖抑制を試験することからなる骨髄腫治療剤のインビトロスクリーニング法。
【請求項11】
骨髄腫細胞増殖抑制試験をMタンパク分泌抑制を指標として行うことからなる請求項10に記載のインビトロスクリーニング法。
【請求項12】
骨髄腫治療剤がIL−6活性阻害剤である請求項10に記載のインビトロスクリーニング法。
【請求項13】
骨髄腫治療剤を請求項4に記載のヒト以外の実験動物に移植して骨髄腫細胞増殖抑制を試験することからなる骨髄腫治療剤のインビボスクリーニング法。
【請求項14】
骨髄腫細胞増殖抑制試験をMタンパク分泌抑制を指標として行うことからなる請求項13に記載のインビボスクリーニング法。
【請求項15】
骨髄腫細胞増殖抑制試験を骨病変の抑制を指標として行うことからなる請求項13に記載のインビボスクリーニング法。
【請求項16】
骨髄腫治療剤がIL−6活性阻害剤である請求項13に記載のインビボスクリーニング法。
【請求項1】
オートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株であって、IL−6タンパク質を分泌し、20%ウシ胎児血清(FCS)および100μg/mlカナマイシンを含むRPMI1640培養液中で増殖可能であり、かつ、抗IL−6抗体により増殖が抑制される、前記ヒト骨髄腫細胞株。
【請求項2】
ヒト骨髄腫がヒト多発性骨髄腫である請求項1に記載の細胞株。
【請求項3】
オートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株であって、重症複合免疫不全マウス及び/又はヌードマウスに静脈内移植した場合に骨髄へ生着する能力を有する請求項1に記載のヒト骨髄腫細胞株。
【請求項4】
請求項1〜3のいずれかに記載の細胞株を移植したヒト以外の実験動物。
【請求項5】
実験動物が免疫不全状態である請求項4に記載のヒト以外の実験動物。
【請求項6】
実験動物がマウスである請求項4に記載の実験動物。
【請求項7】
実験動物にオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株を静脈内移植して得られる請求項4に記載のヒト以外の実験動物。
【請求項8】
実験動物にオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株を皮下移植して得られる請求項4に記載のヒト以外の実験動物。
【請求項9】
実験動物にオートクライン機構によりIL−6依存性で増殖するヒト骨髄腫細胞株を腹腔内移植して得られる請求項4に記載のヒト以外の実験動物。
【請求項10】
骨髄腫治療剤を請求項1〜3のいずれかに記載の細胞株に添加して骨髄腫細胞増殖抑制を試験することからなる骨髄腫治療剤のインビトロスクリーニング法。
【請求項11】
骨髄腫細胞増殖抑制試験をMタンパク分泌抑制を指標として行うことからなる請求項10に記載のインビトロスクリーニング法。
【請求項12】
骨髄腫治療剤がIL−6活性阻害剤である請求項10に記載のインビトロスクリーニング法。
【請求項13】
骨髄腫治療剤を請求項4に記載のヒト以外の実験動物に移植して骨髄腫細胞増殖抑制を試験することからなる骨髄腫治療剤のインビボスクリーニング法。
【請求項14】
骨髄腫細胞増殖抑制試験をMタンパク分泌抑制を指標として行うことからなる請求項13に記載のインビボスクリーニング法。
【請求項15】
骨髄腫細胞増殖抑制試験を骨病変の抑制を指標として行うことからなる請求項13に記載のインビボスクリーニング法。
【請求項16】
骨髄腫治療剤がIL−6活性阻害剤である請求項13に記載のインビボスクリーニング法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2010−91(P2010−91A)
【公開日】平成22年1月7日(2010.1.7)
【国際特許分類】
【出願番号】特願2009−230780(P2009−230780)
【出願日】平成21年10月2日(2009.10.2)
【分割の表示】特願2004−1967(P2004−1967)の分割
【原出願日】平成6年3月3日(1994.3.3)
【出願人】(000003311)中外製薬株式会社 (228)
【Fターム(参考)】
【公開日】平成22年1月7日(2010.1.7)
【国際特許分類】
【出願日】平成21年10月2日(2009.10.2)
【分割の表示】特願2004−1967(P2004−1967)の分割
【原出願日】平成6年3月3日(1994.3.3)
【出願人】(000003311)中外製薬株式会社 (228)
【Fターム(参考)】
[ Back to top ]