
IP3レセプター欠損マウス
【課題】外分泌不全または外分泌過多を伴う障害の治療または予防薬の開発および発症機序の解明等に有用な、唾液分泌および膵液分泌不全モデル動物の提供。
【解決手段】機能的なタイプ3 イノシトール1,4,5-三リン酸レセプター(IP3R3)および/またはタイプ2 イノシトール1,4,5-三リン酸レセプター(IP3R2)を有していない非ヒト脊椎動物の提供。
【解決手段】機能的なタイプ3 イノシトール1,4,5-三リン酸レセプター(IP3R3)および/またはタイプ2 イノシトール1,4,5-三リン酸レセプター(IP3R2)を有していない非ヒト脊椎動物の提供。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、唾液分泌および膵液分泌不全モデル動物に関する。また本発明は、外分泌不全を改善する薬剤をスクリーニングする方法に関する。
【背景技術】
【0002】
顎下腺および膵臓からの外分泌が、食物消化のために重要であり、食物からの養分の吸収に不可欠であり(Solomon, 1984; Kutchai, 1998)、これらの外分泌機能の障害は、動物の成長と生存に重大な影響を及ぼすことが判っている。したがって、ヒトのみならず、家畜動物、愛玩動物における外分泌機能障害を治療または予防するための方法および薬剤の開発が必要である。しかしながら、これらの開発に必要不可欠である外分泌機能障害の適切なモデル動物が存在しない。
【0003】
顎下腺および膵臓などの外分泌組織では、消化酵素等のタンパク質および水を分泌するが、この際、細胞内のカルシウム濃度増加が重要であると考えられている。小胞体(ER)などにおける細胞内カルシウムストアからの細胞質へのカルシウム(Ca2+)動員は、イノシトール1,4,5-三リン酸レセプター(IP3R)とそのリガンドであるイノシトール1,4,5-三リン酸(IP3)との結合により誘導されることは公知の事実である。
【0004】
IP3Rに、セカンドメッセンジャーであるイノシトール1,4,5-三リン酸(IP3)が結合すると、小胞体(ER)などの細胞内ストアから細胞質へのカルシウム動員が起こり、このIP3によって誘発されるカルシウム放出(IICR)(これがその下流の細胞内の情報伝達カスケードを誘発する)は、分泌、増殖、分化、細胞死、遊走、シナプス可塑性の調節等のさまざまな細胞内プロセスに関与すると考えられている。
【0005】
哺乳動物には、それぞれ異なる遺伝子に由来する3つのサブタイプ(タイプ1〜3)のIP3Rがあり(非特許文献1)、興味深いことに、分泌機能を有する数種の細胞では、タイプ3 IP3R(IP3R3)の発現が高いという報告があるが、IP3Rと分泌機能との関連性については直接的な証明はなされていない。タイプ3 IP3R(IP3R3)cDNAは、ラット膵島細胞腺腫細胞系(非特許文献2)から最初にクローニングされたが、種々の組織で発現されている。タイプ2 IP3R(IP3R2)もまた顎下腺、膵臓をはじめ、心臓、免疫系細胞、造血細胞等の種々の組織ならびに細胞系において低レベルの発現が認められる(非特許文献3〜7)。
【0006】
哺乳動物の、3つのサブタイプのIP3Rのうち、タイプ1 IP3R(IP3R1)は、現在最も機能についての解析が進んでおり、これは脳と平滑筋に豊富に存在し、IP3R1欠損マウスは、胃平滑筋の徐波生成(非特許文献8および9)、重篤な運動失調、てんかん発作(非特許文献10)を示し、シナプス可塑性に影響を与えることが判っているが(非特許文献11〜15)、外分泌機能との関連性は報告されていない。しかしながら、IP3Rのサブタイプに特異的なIP3Rブロッカーが未だ開発されていないため、特にタイプ2とタイプ3のIP3Rの生理学的機能に関してはあまり解明されていない。
【0007】
【非特許文献1】Furuichiら、 Intracellular channels. Curr. Opin. Neurobiol. 4, 294-303, 1994
【非特許文献2】Blondelら、J. Biol. Chem. 268, 11356-11363, 1993
【非特許文献3】Fujinoら、Cell Tissue Res. 280, 201-210, 1995
【非特許文献4】Yamamoto-Hinoら、 Receptors and Cnannels. 2, 9-22, 1998
【非特許文献5】MoschellaおよびMarks、J. Cell Biol. 120, 1137-1146, 1993
【非特許文献6】Kijimaら、J. Biol. Chem. 268, 3499-3506, 1993
【非特許文献7】Sugiyamaら、FEBS Lett. 349, 191-196, 1994
【非特許文献8】Suzukiら、J. Physiol. 525, 105-111, 2000
【非特許文献9】Takanoら、Auton. Neurosci. 87, 249-257, 2001
【非特許文献10】Matsumotoら、Nature 379, 168-171, 1996
【非特許文献11】Inoueら、Neurosci. 117, 821-830, 1998
【非特許文献12】Nishiyamaら、Nature 408, 584-588, 2000
【非特許文献13】Itohら、Brain Res. 901, 237-246, 2001
【非特許文献14】Fujiiら、Learning and Memory 7, 312-320, 2000
【非特許文献15】Nagaseら、Neurosci. 117, 821-830, 2003
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明は、外分泌不全または外分泌過多を伴う障害の治療または予防薬の開発および発症機序の解明等に有用な、唾液分泌および膵液分泌不全モデル動物を提供することを課題とする。
【課題を解決するための手段】
【0009】
本発明者らは、外分泌組織との関連性が考えられるタイプ2 IP3レセプター(IP3R2)およびタイプ3 IP3レセプター(IP3R3)を共に欠失するダブルノックアウトマウスが、唾液分泌および膵液分泌不全を呈することを見出し、本発明を完成するに至った。また、これらの一方を欠失したマウスでもアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇がある程度低減され、これらの両方を欠失したマウスではアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇がさらに劇的に低減されることを見出した。これらの知見から、本発明は、以下の態様に関する:
1.機能的なタイプ3 イノシトール1,4,5-三リン酸レセプター(IP3R3)およびタイプ2 イノシトール1,4,5-三リン酸レセプター(IP3R2)のいずれか一方を有していない非ヒト脊椎動物;
2.機能的なIP3R2と機能的なIP3R3の両方を有していない非ヒト脊椎動物;
3.機能的な上記イノシトール1,4,5-三リン酸レセプター(IP3R)を有していないことが、該レセプターをコードしている染色体上の遺伝子が破壊されていることに起因する、上記1または2に記載の非ヒト脊椎動物;
4.IP3R3をコードしている染色体上の遺伝子がノックアウトされているために機能的なIP3R3を遺伝学的に有していない非ヒト脊椎動物と、IP3R2をコードしている染色体上の遺伝子がノックアウトされているために機能的なIP3R2を遺伝学的に有していない非ヒト脊椎動物とを交配して得られる、機能的なIP3R2と機能的なIP3R3の両方を遺伝学的に有していない非ヒト脊椎動物;
5.唾液の分泌が、対応する野生型動物に比較して低減されていることを特徴とする、上記1〜4のいずれかに記載の非ヒト脊椎動物;
6.膵臓細胞からの消化酵素の分泌が、対応する野生型動物に比較して低減されていることを特徴とする、上記1〜5のいずれかに記載の非ヒト脊椎動物;
7.顎下腺細胞および/または膵臓細胞におけるアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇が、対応する野生型の動物に比べて低減されていることを特徴とする、上記1〜6のいずれかに記載の非ヒト脊椎動物;
8.上記非ヒト脊椎動物が哺乳動物である、上記1〜7のいずれかに記載の非ヒト脊椎動物;
9.上記非ヒト脊椎動物が囓歯動物である、上記1〜7のいずれかに記載の非ヒト脊椎動物;
10.上記非ヒト脊椎動物がマウスである、上記1〜7のいずれかに記載の非ヒト脊椎動物;
11.上記1〜10のいずれかに記載の非ヒト脊椎動物に由来する細胞;
12.上記11に記載の細胞であって、野生型の同種の非ヒト脊椎動物に由来する同種の細胞は有している機能的なIP3R3およびIP3R2のいずれか一方または両方を有していないことを特徴とする、上記細胞;
13.不死化されている、上記11または12記載の細胞;
14.上記1〜10のいずれかに記載の非ヒト脊椎動物に薬剤を投与し、該動物の唾液の分泌量、膵臓細胞からの消化酵素の分泌量、および顎下腺細胞および/または膵臓細胞におけるアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇量からなる群より選択される少なくとも1つを測定することを含み、該測定した量が、薬剤を投与していない上記動物個体に比べて、該薬剤を投与した非ヒト脊椎動物個体で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法;
15.上記11〜13のいずれかに記載の細胞でアセチルコリンレセプターを発現している細胞に、薬剤を接触させ、該細胞のアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇量を測定し、該測定した量が、薬剤と接触させていない上記細胞に比べて、該薬剤を接触させた細胞で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法;
16.上記2〜10のいずれかに記載の非ヒト脊椎動物に由来する顎下腺細胞または膵臓細胞に、薬剤を接触させ、該細胞からの外分泌液量または消化酵素分泌量を測定し、該測定した量が、薬剤と接触させていない上記細胞に比べて、該薬剤を接触させた細胞で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法;
17.上記外分泌不全を改善し得る薬剤が、シェーグレン症候群、唾液または涙腺における自己免疫疾患、口内乾燥、乾燥性角結膜炎からなる群から選択される障害を治療または予防するための薬剤である、上記14〜16のいずれかに記載の方法;ならびに、
18.上記外分泌不全が唾液分泌不全または膵液分泌不全である、上記14〜16のいずれかに記載の方法。
【発明の効果】
【0010】
本発明により、外分泌不全または外分泌過多を伴う障害の治療または予防薬の開発および発症機序の解明等に有用な、唾液分泌および膵液分泌不全モデル動物となり得るIP3R2およびIP3R3の双方を欠失するマウスが提供される。該マウスは、唾液分泌と膵液分泌に重篤な不全、およびそれによる成長障害が認められた。さらに、該マウスからIP3R2およびIP3R3の双方を欠失する各種組織細胞を得ることが可能である。
【発明を実施するための最良の形態】
【0011】
以下、本発明を詳細に説明する。
本明細書中、非ヒト脊椎動物とは、好ましくはヒト以外の哺乳動物であり、さらに好ましくは、囓歯動物(ウサギ、モルモット、ラット、マウス等)であり、特に好ましくはマウスである。
【0012】
本明細書中「機能的なIP3R2」または「機能的なIP3R3」とは、イノシトール1,4,5-三リン酸と結合することによりCa2+を細胞内ストアから放出させるというIP3Rの機能を有しているIP3R2またはIP3R3を指す。したがって、「機能的なIP3R2(またはIP3R3)を有していない」とは、IP3R2(またはIP3R3)の発現が実質的に欠失しているか、IP3R2またはIP3R3分子が発現している場合でも、それがIP3Rとしての機能を消失した変異体であるか、あるいは機能的なIP3R2(またはIP3R3)を発現している場合であっても遺伝子操作によりドミナントネガティブ変異体が発現しているためにIP3R2(またはIP3R3)を介した、イノシトール三リン酸に応答して細胞内Ca2+のストアからの放出が実質的に低減されていることを指す。また、「機能的なIP3R2(またはIP3R3)を遺伝学的に有していない」とは、機能的なIP3R2(またはIP3R3)をコードする遺伝子が例えば、ノックアウトなどにより欠損していることを指す。
【0013】
「発現が実質的に欠失している」とは、IP3R2(またはIP3R3)の発現量が正常(野生型)の対応する動物個体(または細胞)に対して30%以下、好ましくは20%以下、より好ましくは10%以下、さらに好ましくは5%以下、最も好ましくはその発現が分子生物学的手法(即ち、mRNAまたはタンパク質の検出)によって検出されない程度であることを指す。機能的なIP3R2(またはIP3R3)を有していない動物は、例えばIP3R2(またはIP3R3)をコードする染色体上の遺伝子を破壊することによりノックアウト動物として得ることができる。ノックアウト動物の作製方法は当業者には公知である。IP3R2およびIP3R3をコードする遺伝子配列は、データベースから入手可能である。この配列に基づいて、ゲノムDNAをクローニングし、常法に従って、遺伝子ターゲティングベクターに組込み、細胞に導入して、該細胞から個体を発生させるとよい。遺伝子破壊によるノックアウト動物の作製方法は種々の公知文献が多数存在しており、当業者は容易に作製することができる。本発明に用いる場合、ホモノックアウト動物が好ましい。
【0014】
機能的なIP3R2と機能的なIP3R3の両方を有していない非ヒト動物は、これらを有していなくとも、生殖能力には実質的な影響はなく、繁殖可能であるため、機能的なIP3R2を遺伝学的に有していない動物と機能的なIP3R3を遺伝学的に有していない動物とを交配させて、その子孫から両方を有していない個体を選択することにより、ダブルノックアウト動物として得ることができる。
【0015】
したがって、機能的なIP3R3またはIP3R2を遺伝学的に有していない非ヒト動物は、機能的なIP3R2と機能的なIP3R3の両方を有していない非ヒト動物の作製に利用することができ、本発明の範囲に包含される。
【0016】
IP3R3またはIP3R2を遺伝学的に欠失している動物は、ダブルノックアウト動物を得るためにも有用である。本発明に用いるダブルノックアウト動物は、ホモダブルノックアウト動物が好ましい。
【0017】
機能的なIP3R2およびIP3R3のいずれか一方または両方を有していない脊椎動物は、顎下腺および/または膵臓細胞において、イノシトール1,4,5-三リン酸介在性細胞内Ca2+動員が、野生型のものに比べて低減される。例えば、唾液腺細胞においてカルバコールでアセチルコリンレセプターを刺激した場合、IP3レセプターを介して細胞内Ca2+濃度([Ca2+]i)が上昇するが、IP3R2およびIP3R3のいずれか一方を欠失している場合(すなわち機能的なIP3R2およびIP3R3のいずれか一方を有していない場合)、野生型に比べてその上昇が約70%以下に低減され、両方を欠失している場合では、野生型に比べて約30%以下に低減され得る。本明細書中、「対応する野生型の動物に比べて低減されている」とは、対象とする、機能的なIP3R2および/またはIP3R3を有していない動物の膵臓細胞からの消化酵素の分泌、唾液の分泌または細胞内カルシウム濃度の上昇が、同様の条件下にある機能的なIP3R2およびIP3R3を有している野生型の同系の動物に比べた場合に、約80%以下、好ましくは50%以下、より好ましくは30%以下、さらに好ましくは20%以下に低減されていることをいう。上記の機能的なIP3R2およびIP3R3の両方を有していない動物では、唾液腺細胞および膵臓細胞の外分泌を誘導するためのシグナルが十分に伝達できず、唾液および膵液の外分泌が低減され得る。例えば該ダブルノックアウトマウスでは、唾液分泌促進剤であるピロカルピンで刺激した場合の唾液の分泌量が、野生型のそれに対して約30%以下に低減され得る。しかし、ピロカルピン刺激により誘導される唾液分泌のみならず、通常の唾液分泌も低減され、これにより、離乳後に固形飼料を摂食できず、致死性の発育不良を呈する。
【0018】
さらにまた、機能的なIP3R2およびIP3R3の両方を有していない動物は、膵液分泌不全を呈する。すなわち、本来、アミラーゼ、トリプシンおよびリパーゼ等の消化酵素を含む膵液が膵臓細胞から小腸内に分泌されることにより、食餌を消化して栄養素を吸収するが、消化酵素の分泌が不十分であるため、食餌摂取をしたとしても、消化吸収効率が悪く、発育不良を起こす。例えば、IP3R2/IP3R3ダブルノックアウトマウスでは、上述のように離乳後に固形飼料を摂食できないが、水分を十分に含むペースト状の飼料を与えた場合は、充分量摂食することができるにもかかわらず、消化不良により、糞便中にタンパク質および脂質を多量に排泄し、発育不良を呈する。
【0019】
上記のとおり、機能的なIP3R2およびIP3R3の両方を有していない動物では、重篤な外分泌不全を起こす。いずれか一方を欠失した動物で、外分泌不全の症状が認められないのは、おそらく、欠失していないIP3R2またはIP3R3が機能することで、欠失した一方の機能が補填されているためと考えられる。
【0020】
機能的なIP3R2およびIP3R3の両方を欠失した動物は、シェーグレン症候群、唾液または涙腺における自己免疫疾患、口内乾燥(ドライマウス)、乾燥性角結膜炎等の外分泌不全を伴う障害のモデル動物として有用であり得る。
【0021】
すなわち、上記障害を含む、外分泌不全を呈する障害の治療または予防のための薬剤をスクリーニングするために、上記動物を用いることができる。上記動物に、候補薬剤を投与し、該動物の唾液の分泌量または膵臓細胞からの消化酵素の分泌量を測定する。ここで、候補薬剤の投与は、薬剤の性質に応じて、経口投与、皮下、静脈または筋肉注射であってもよく、または種々の局所投与であってもよく、当業者であれば適宜、適切な投与方法を選択し得る。唾液の分泌量の測定は、候補薬剤の投与後適切な時間を経て、麻酔下で一定時間内に分泌された唾液を回収してその容量を測定してもよく、また唾液分泌促進作用を有する薬剤(ピロカルピンおよびカルバコール等)で唾液分泌を刺激して、その分泌量を測定してもよい。膵臓細胞からの消化酵素の分泌量を測定するためには、候補薬剤投与後、適切な時間を経て、動物から膵臓を摘出し、生理的な条件下(各種生理的緩衝液中など)で、場合によっては膵臓細胞を分散させて、膵液分泌促進作用を有する薬剤(カルバコール等)で膵液分泌を刺激して、細胞外液の消化酵素(例えば、アミラーゼ、リパーゼまたはトリプシン)の活性を測定するか、またはウェスタンブロット等により細胞外液中の消化酵素量を評価する。さらにまた候補薬剤の投与後、適切な時間を経て、動物から顎下腺または膵臓を摘出し、これらをアセチルコリンレセプター刺激作用を有する薬剤に応答する細胞内Ca2+濃度の上昇の程度を、fura-2を用いる方法等、公知の細胞内Ca2+濃度測定法により測定してもよい。ネガティブコントロールとして候補薬剤を投与していない動物を用いて、同様の処理および測定を行い、ネガティブコントロールについての測定値に比べて、候補薬剤を投与した動物についての測定値の方が大きければ、該候補薬剤は外分泌不全の予防または治療薬として有望であると判定することができる。
【0022】
さらに、上記動物に由来する細胞もまた本発明の範囲に包含される。IP3R2およびIP3R3は、あらゆる組織に広く分布していることから、機能的なIP3R2およびIP3R3のいずれか一方または両方を有していない動物に由来する任意の組織の細胞は、その形質を保持している。したがって、各組織または組織内の各細胞における、IP3R2およびIP3R3の機能を解明する上で、これらの細胞は有用である。したがって、上記動物に由来する細胞は、野生型の同種の細胞は有しているはずの機能的なIP3R2およびIP3R3のいずれか一方または両方を有していないという特徴を有する。かかる特徴により、該細胞を外分泌不全を改善し得る薬剤をスクリーニングするのに用いることができる。
【0023】
これらの細胞は、該動物の個体および胎生個体から単離可能であり、初代培養系細胞として用いることも可能である。さらにまた、これらの細胞を不死化することも、当業者であれば容易に実施可能である。不死化の方法は種々のものが公知であり、任意の方法を適宜用いることができる。不死化された細胞株もまた、それが由来する動物個体または胎生個体と同じように機能的なIP3R2およびIP3R3のいずれか一方または両方を有しておらず、IP3R2およびIP3R3の機能を解明する上で有用であり、不死化細胞であることから、保存、維持および使用が簡便である点において有利である。
【0024】
これらの細胞を用いて、外分泌不全を改善し得る薬剤をスクリーニングする方法は、それぞれの細胞を適切な生理的条件下に維持して、候補薬剤と接触させて、これらをアセチルコリンレセプター刺激作用を有する薬剤に応答する細胞内Ca2+濃度の上昇の程度を評価することにより達成できる。候補薬剤と接触させていない場合の細胞内Ca2+濃度の上昇の程度に比して、上昇の程度が大きい場合、該候補薬剤は外分泌不全の予防または治療薬として有望であると判定することができる。さらに、細胞が顎下腺細胞またはこれに由来する細胞の場合は、候補薬剤と接触させた場合の唾液分泌量を測定するか、または細胞外液に分泌されたアミラーゼの量を測定してもよい。細胞が膵臓細胞またはこれに由来する細胞の場合は、候補薬剤と接触させた場合の細胞外液中の膵液消化酵素の量を測定してもよい。
【0025】
かかるスクリーニングの方法については、当業者が日常的に行い得る改良または変更により、適切な方法および条件を当業者であれば実施し得る。
【0026】
以下、実施例によって本発明を更に具体的に説明するが、これらの実施例は本発明を限定するものではない。
【実施例1】
【0027】
IP3R2ノックアウトマウス、IP3R3ノックアウトマウスとIP3R2/IP3R3ダブルノックアウトマウスの作製
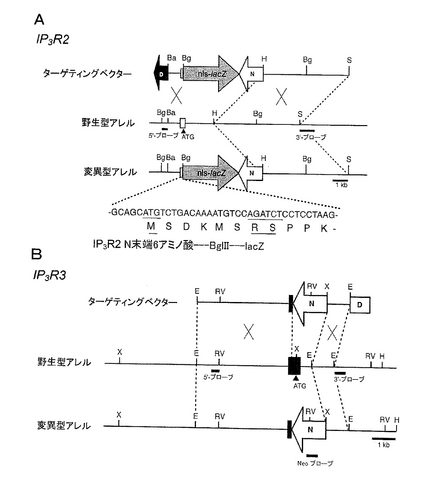
マウスゲノムのIP3R2とIP3R3の第1コーディングエキソンとその両側のイントロンとをそれぞれ含むマウスゲノムクローンは、129マウスゲノムライブラリー(熊本大学・山村研一博士より分与)から単離した。IP3R2またはIP3R3が欠損するマウスを作製するために、当該遺伝子それぞれの第1コーディング・エキソンを破壊した(図1AおよびB参照のこと)。
【0028】
PCRで得たIP3R2 cDNA断片(Morikawaら、Gene 196, 181-185, 1997)および600bp長のヒトIP3R3 cDNA断片(第1エキソンを含む)をプローブとして用いた。上記ゲノムクローンから得られた断片、陽性選択マーカーとしてpKJ1(McBurneyら、Nucleic Acids Res. 19, 5755-57, 1991)から得たneo遺伝子カセットpGK-neo(1.8kb)、および陰性選択マーカーとしてpMC1DT-A(Yagiら、Proc. Natl. Acad. Sci. USA 87, 9918-9922, 1990)から得たXhoI-SalIジフテリア毒素A断片(dT-A)遺伝子カセットを用いて常法に従い、ターゲッティングベクターを構築した。IP3R2ターゲッティングベクターでは、最初の6つのアミノ酸をコードしている第1のエキソンの領域は、LacZ染色(Sanesら、EMBO J. 5, 133-142, 1986)によってIP3R2プロモーター活性をモニターできるよう、lacZ遺伝子と核移行シグナルとをインフレームで連結した。
【0029】
IP3R2とIP3R3のターゲッティングベクターDNAを、直鎖状にして、ES細胞(それぞれJ1とR1)に常法による電気穿孔法にて形質転換した。G418(150μg/ml)に抵抗性を示すクローンを、サザンブロット法により解析して所望の通り相同的組換えを起こしたクローンを得た。IP3R2破壊クローンは、ゲノムDNAをBglIIで切断し、5’末端側の外部プローブとしてBglII-BamHI断片を用いてこれとハイブリダイズさせることにより確認した。野生型アレルからは7kb断片が、変異型アレルからは1.5kb断片が得られた。IP3R3破壊クローンは、ゲノムDNAをEcoRVで切断し、3’末端側の外部プローブとしてEcoRI-SacII断片を用いてこれとハイブリダイズさせることにより確認した。野生型アレルからは8.5kb断片が、変異型アレルからは4.5kb断片が得られた(図1AおよびB)。所望の相同組換えは、neoプローブと他方の外部プローブとによっても確認した(データ示さず)。IP3R2とIP3R3について、所望の相同組換えを起こしたそれぞれのESクローンが、6および12クローン得られた。公知の方法(Bradley Production and analysis of chimeric mice. In Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E.J. Robertson , ed. (Oxford: IRL press), pp. 113-151, 1987; Woodら, Proc. Natl. Acad. Sci. USA 90, 4582-4585, 1993)にしたがって、各々のキメラマウスを得た。C57BL/6Jマウスとキメラマウスを交配させることによって、ヘテロ接合体マウスを得、これをさらに3〜6回、C57BL/6Jマウスと交配させた。
【0030】
得られた各々の遺伝子のヘテロ接合変異体を交配させて、IP3R2とIP3R3とのダブルノックアウトヘテロ接合変異体を得、さらに、本発明のダブルノックアウトホモ接合体を得るために、ダブルノックアウトヘテロ接合体どうしを交配させた。マウスの遺伝型は、尾部組織から得たゲノムDNAのサザンブロット分析またはPCRで決定した。
【0031】
IP3R2およびIP3R3のいずれか一方のノックアウトマウスでは、胎生期および生後数ヶ月間においては、致死性障害および外見上の異常は認められなかった。IP3R3ノックアウトマウスにおいては、脱毛症状を示し、生後数ヶ月以降に体重の増加が認められない傾向にあったが、生殖能も寿命(2年以上)も正常であった。
【0032】
イムノブロット分析により、野生型マウスの顎下腺と膵臓においては、IP3Rの全てのサブタイプが発現されていたが、3つのサブタイプの中でIP3R2とIP3R3が他の組織と比較して豊富に発現されたのに対して、IP3R1発現レベルは他の組織における発現より低かった。作製した変異体マウスにおいて、ターゲットとしたIP3R2および/またはIP3R3の発現は欠損しており、ターゲット以外のサブタイプの発現は認められ、意図したとおりの発現パターンが得られた。
【実施例2】
【0033】
IP3R2/IP3R3ダブルノックアウト変異体マウスの特徴
(1)外観と生存率
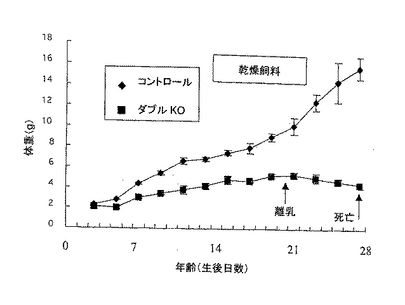
出生時、IP3R2/IP3R3ダブルノックアウトホモ接合体の体重および外見は他の同腹の仔と差はなかったが、出生後に、ダブルノックアウトホモ接合体だけ体重増加が遅れた。通常、マウスは自由に標準の乾燥ペレットCA-1(CLEA)と水を摂取可能な環境で飼育した。離乳(概して出産後約20日:P20)後、ダブルノックアウトホモ接合体は、体重が減少し始め、4週齢に達する前に死亡した(図2A)。これらの現象は性別に関係なく観察され、ダブルノックアウトホモ接合体は通常の食餌(すなわち固形乾燥飼料)では、4週齢以上生存できなかった。
【0034】
(2)摂食不能
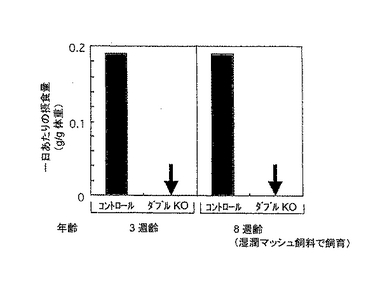
IP3R2/IP3R3ダブルノックアウト変異体は、乾燥食物ペレットを摂取できず、IP3R2-/-/IP3R3-/-ダブルノックアウト変異体は離乳期間の直後から体重が減り始めた。これは、離乳の後、乾燥食物ダイエットへ切り替えることにより、摂食不能が起こるのではないかと考えられた。
【0035】
そこで、出生3週間後の離乳の後、2日の期間にわたって1日の摂食量(標準的乾燥ペレット飼料)をモニターしたところ、ダブルノックアウトマウスは、ケージ内に飼料があることを認識しながらも、全く食べることができなかった(図2B左のパネル)。
【0036】
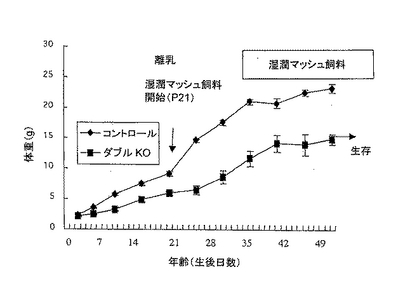
そこで、20日齢の時点で、乾燥ペレットCA-1(CLEA)を2倍(重量比)の滅菌水中で粉砕・混合した湿潤マッシュ飼料を与えた。該マッシュ飼料は、毎晩、新たに調製して与えた。すると、湿潤マッシュ飼料を与えた場合は、ダブルノックアウト変異体は摂食可能であり、生存し続けた。しかしながら、同様に湿潤マッシュ飼料で飼育した同腹のマウス間で比較した場合、ダブルノックアウト変異体の体重増加は少なかった(図2C)。
【0037】
さらに、離乳後から8週齢まで湿潤マッシュ飼料で飼育したマウスを、8週齢の時点で乾燥飼料を与えたところ、やはり、ダブルノックアウトマウスは、全くこれを食べなかった(図2B右のパネル)。このことから、ダブルノックアウトマウスが、乾燥飼料を摂食できないのは、離乳前の発達および離乳の遅れ等が原因ではないと考えられる。
【0038】
(3)唾液生産の不足
上記ダブルノックアウトマウスの歯は正常であった。したがって、唾液生産が不足している可能性が考えられた。そこで、該ダブルノックアウトマウスの唾液分泌を調べた。
3週齢のマウスまたは成熟マウスを37℃の保温プレート上に置き、ネンブタール(50mg/kg、腹腔投与)麻酔下で、ピロカルピン-HCl(1または10mg/kg(Tocris))またはイソプロテレノール(0.3mg/kg(Sigma))を皮下投与した。口腔内で分泌される唾液は手動ピペッタ(ギルソン(P100))を使用してピロカルピンまたはイソプロテレノールで投与後各30分間に分泌された唾液を、遠心管に回収し、その回収された唾液の重量を求めた。
【0039】
母乳から乾燥飼料に切り替える離乳期(生後21日齢:P21)のマウスで調べたところ、ピロカルピンによって促進される唾液分泌は、ダブルノックアウトで顕著に消失していた(図3Aの4のグラフ)。1.0mg/kgのピロカルピンを投与した場合、IP3R2ノックアウト(IP3R2-/-/IP3R3+/+)とIP3R3ノックアウト(IP3R2+/+/IP3R3-/-)マウスと野生型(IP3R2+/+/IP3R3+/+)マウスとの間では唾液分泌に大きな差は認められなかったが、ダブルノックアウトマウスでは無唾液症が観察された。ピロカルピンを10mg/kgで投与した場合は、ダブルノックアウトマウスでも唾液分泌が認められたが、分泌量はコントロールに比べて3分の1程度であった。かかる機能障害は、IP3R2ノックアウトマウスおよびIP3R3ノックアウトマウスでは観察されなかった。
【0040】
次に、離乳後湿潤マッシュ飼料で飼育した成熟マウス(2〜3ヵ月齢)における唾液分泌を測定した。結果は、P21マウスでの結果と基本的に類似していた。ピロカルピンによって促進される唾液分泌は、ダブルノックアウトマウスだけで、顕著に抑制されていた(図3Bの4のグラフ)。
【0041】
常法に従いイムノブロット解析を行って、唾液腺分泌に必須の水量浸透性チャネルであるアクアポリン5(AQP5)の発現量を調べたところ、AQP5はダブルノックアウトマウスでも正常に発現していることが確認された(図3C)。したがって、ダブルノックアウトの唾液分泌の機能障害は、水量浸透性チャネルの減少に依存しているわけではない。
【実施例3】
【0042】
顎下腺と膵臓におけるIP3Rの発現
上記のとおり、IP3R2-/-/IP3R3-/-マウスでは唾液分泌が劇的に損なわれることから、顎下腺、およびこれと並ぶ主要な外分泌組織である膵臓におけるIP3R2とIP3R3の発現を調べた。ウェスタンブロット法は、顎下腺と膵臓を摘出し、0.32M スクロース、5mM Tris-HCl(pH 7.4)、1mM EDTA、0.1M PMSF、10μM ロイペプチン、10μM ペプスタチンAおよび1mM 2-メルカプトエタノールを含む溶液(ホモジナイズバッファー)中でホモジナイズし、50μgを5% SDS-PAGEゲルにアプライして泳動後、ウェスタンブロット解析に供した。IP3R1、IP3R2およびIP3R3はそれぞれ、抗体KM1112、KM1083とKM1082にて検出した。
【0043】
ダブルノックアウトマウスの顎下腺細胞(腺房で管の細胞)と膵臓腺細胞のウェスタンブロットの結果、IP3R2およびIP3R3とも野生型に比べて有意に発現が低減していた(データは示さず)。
【実施例4】
【0044】
膵臓腺細胞のコリン作動性アゴニストにより誘発される消化酵素分泌は、IP3R2-/-/IP3R3-/-ダブルノックアウト変異体では欠損している
次に、ダブルノックアウトマウスの膵臓機能障害について調べた。
【0045】
麻酔下で心臓穿刺により屠殺したマウスの膵臓を直ちに摘出し、冷却したカルシウム不含平衡塩類溶液(BSS:115mM NaCl、5.4mM KCl、2mM Ca2+、1mMマグネシウムイオン、20mM Hepes、10mMブドウ糖、0.125%のウシ血清アルブミン(BSA)、pH 7.4)中に入れ、速やかに砕片し、2mg/mlのコラゲナーゼ(和光034-10533)で消化した、消化後の細胞含有液を細胞濾過器(100μmナイロン・メッシュ)に通して、再懸濁、遠心による洗浄を3回繰り返した。このようにして得た細胞懸濁液を、1〜10μMのカルバコールと、BSA、Ca2+(2mM)を含むBSS 400μl中で、室温で30分間静置してカルバコールに暴露させた。0.22μmフィルタ(ミリポア)にてろ過して得た300μlの上清を分泌タンパク質サンプル液として、膵臓から分泌される消化酵素の分析に用いた。
【0046】
カルバコールが細胞のムスカリン性アセチルコリン受容体に作用することを確認するために、1μMのアトロピン(ムスカリン性アセチルコリン受容体アンタゴニスト)を、カルバコール投与の1分前に添加しておいた。イオノマイシン(3μM)を添加したコントロール実験は、Ca2+に依存する分泌機序がノックアウトマウスでも完全に機能していたかどうか調べるために行った。
【0047】
アミラーゼ、リパーゼおよびトリプシン(トリプシノーゲン)の放出は、分泌タンパク質サンプル液中の各酵素活性を測定し、分泌量は細胞中に含まれる量に対するパーセンテージとして表した。
【0048】
アミラーゼ活性は、AMY(SP)キット(関東Chemicals社)を、リパーゼ活性は、カラー自動試験キット(和光)を使用して測定した。トリプシン活性は、サンプル中のトリプシノーゲンをEKマックス・エンテロキナーゼ(Invitrogen)によって活性化させて、その活性を基質としてN-CBZ-Gly-Pro-Arg-p-ニトロアニリドを用いて検定した(Szmolaら、J. Biol. Chem. 278, 48580-48589, 2003)。
【0049】
非選択的なコリン作動性アゴニストであるカルバコール(CCh)を細胞懸濁液中に添加し、30分間膵臓細胞を刺激した後、細胞外間隙に分泌されたタンパク質を分析し、α-アミラーゼの分泌を調べた結果を図4Aに示す。図4Aにおいて、野生型、IP3R2ノックアウトおよびIP3R3ノックアウトマウスのいずれの膵臓細胞でも、膵臓細胞中の全アミラーゼ活性に対して約1〜2%の活性が分泌物中で検出されたが、IP3R2-/-/IP3R3-/-ダブルノックアウトマウス由来の膵臓細胞ではその分泌物中の活性は検出されなかった(図4Aの4のグラフ)。
【0050】
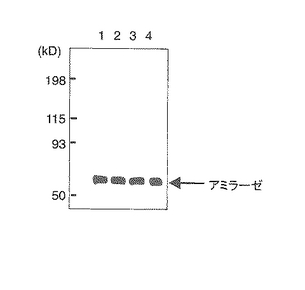
ダブルノックアウトマウス由来の膵臓細胞のアミラーゼタンパク質の分泌量が低減されていることを、イムノブロット分析によって確認した(データ示さず)。しかしながら、膵臓組織のウェスタンブロット解析では、ダブルノックアウトマウスにおいても、膵臓アミラーゼは検出されたことから(図4Bの4のカラム)、アミラーゼの産生は正常であると考えられる。
【0051】
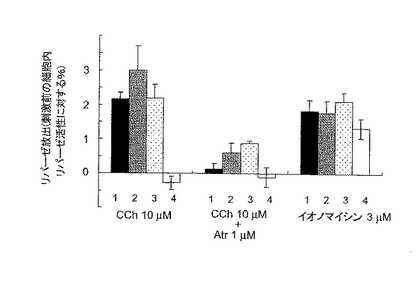
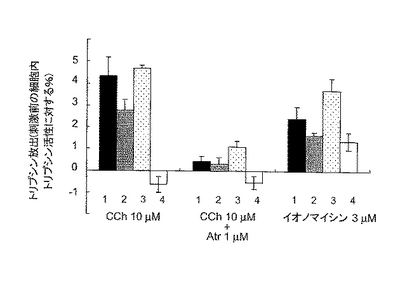
次いで、同様に膵臓細胞からのリパーゼとトリプシノーゲンの分泌を調べた。その結果をそれぞれ図4Cおよび4Dに示す。膵臓細胞からのリパーゼとトリプシノーゲンの分泌は、ダブルノックアウトマウスのみ顕著に消失していた(図4Cおよび4Dの4のグラフ)。
【0052】
すなわち、ダブルノックアウトマウスにおいてのみ、膵臓からの外分泌が顕著に阻害されており、糖、脂質、タンパク質の全ての消化能が著しく低下していると考えられ、これにより、栄養不良が起こり体重増加の抑制または体重の減少が引き起こされると考えられる。
【実施例5】
【0053】
SMG腺房細胞におけるコリン作動性ニューロン刺激による細胞内Ca2+の変化
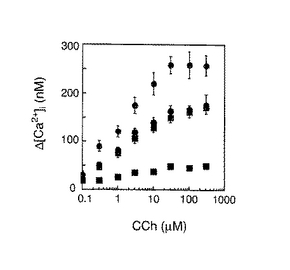
本発明のノックアウトマウスについて、コリン作動性ニューロン刺激によるCa2+シグナル伝達能を調べるために、野生型および各ノックアウトマウスから摘出したSMG細胞を酵素的に分散させて、カルバコール刺激応答性細胞内Ca2+の変動をfura-2を用いて観察した。カルバコール濃度依存的な細胞内Ca2+の変動を図5に示す。IP3R2ノックアウトおよびIP3R3ノックアウトマウスは、野生型に比して約70%程度の細胞内Ca2+の上昇が認められるに止まっており、IP3R2-/-/IP3R3-/-ダブルノックアウトマウスにおいては、野生型の約20%以下の上昇しか認められなかった。すなわち、IP3R2およびIP3R3のいずれかを欠失したノックアウトマウスにおいても、コリン作動性ニューロン刺激によるCa2+シグナル伝達能は低減されていたが、IP3R2およびIP3R3の両方を欠失したマウスでは、コリン作動性ニューロン刺激によるCa2+シグナル伝達能はほとんど喪失されていた。IP3R2およびIP3R3のいずれかを欠失したノックアウトマウスでは、唾液分泌に顕著な影響が認められなかったことは、野生型の70%程度のCa2+シグナル伝達能が保持されていることにより、唾液分泌能はさほど顕著に抑制されなかったものと考えられる。一方、IP3R2およびIP3R3の両方を欠失したマウスでは、野生型の約20%以下しかCa2+シグナル伝達能を有していないため、唾液分泌に顕著に影響を受けたものと考えられる。
【実施例6】
【0054】
本実施例においては、本発明のノックアウトマウスについて、味覚障害、聴覚障害および毛の異常を試験した。
(1)味覚障害
野生型および各ノックアウトマウス(IP3R2ノックアウトマウス、IP3R3ノックアウトマウス)の味覚は、二瓶法に従って評価した。
【0055】
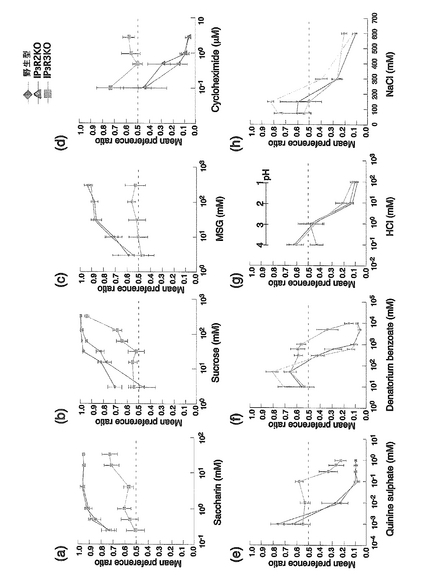
その結果を図6に示す。野生型マウス(菱形)およびIP3R2ノックアウトマウス(三角)に対して、IP3R3ノックアウトマウス(四角)は、甘味(サッカリン、スクロース)、苦味(シクロヘキシミド、硫酸キニーネ、安息香酸デナトリウム)、旨味(グルタミン酸ナトリウム(MSG))に対する味覚が劣っていることがわかる。一方、酸(HCl)および塩(NaCl)については、IP3R3ノックアウトマウスは野生型およびIP3R2ノックアウトマウスと同等の味覚認識を示した。従って、IP3R3ノックアウトマウスは味覚障害のモデルとなりうる。
【0056】
(2)聴覚障害
野生型(n=14)、ならびにIP3R3ヘテロノックアウトマウス(n=7)およびIP3R3ホモノックアウトマウス(n=19)の聴覚(dB)を、Auditory Startle Response and Prepulse inhibitionの方法を用いて評価した。
【0057】
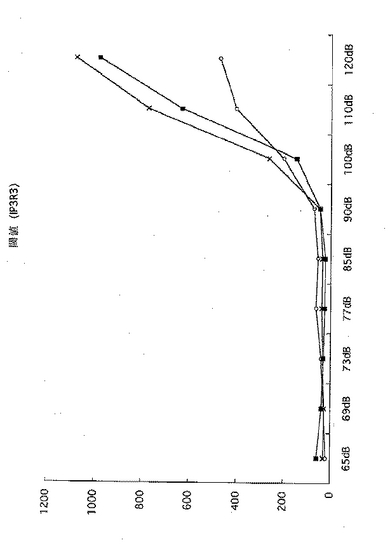
その結果を図7に示す。野生型マウス(黒四角)およびIP3R3ヘテロノックアウトマウス(×印)に対して、IP3R3ホモノックアウトマウス(白丸)は、聴覚障害を有することが判明した。従って、IP3R3ノックアウトマウスは聴覚障害のモデルとなり得、またこのマウスを用いて聴覚障害治療のためのモデルマウスを作出し得る。
【0058】
(3)毛の異常
野生型およびIP3R3ノックアウトマウスの毛を評価した。その結果を図8に示す。野生型マウス(図8A)と比べてIP3R3ノックアウトマウス(図8B)は毛の生育に異常があり、体表に毛のない部分が存在することが明らかとなった。従って、IP3R3ノックアウトマウスは毛発育疾患のモデルとなり、育毛剤の開発に貢献すると考えられる。
【産業上の利用可能性】
【0059】
本発明のIP3R2/IP3R3ダブルノックアウトマウスは、上記特徴により、外分泌組織(例えば唾液腺と膵臓)の機能低下症の疾患モデル動物として利用することができる。例えば、本発明のマウスの唾液腺の機能低下症は、シェーグレン症候群の症状、唾液または涙腺における自己免疫疾患(これらは口渇(口内乾燥)または乾燥性角結膜炎を引き起こす)等の外分泌機能不全を伴う疾患と、機序が共通する可能性が考えられる。
【0060】
膵臓の外分泌においては、IP3R2/IP3R3の欠損は、痩せ症および栄養不良をもたらす機能不全につながるため、膵臓外分泌細胞のIP3R活性は、栄養の利用性および体重を制御することができる主要因のうちの1つである可能性がある。また、急性膵炎では、膵臓外分泌細胞から放出されるトリプシノーゲンが活性型トリプシンに変換されて膵臓細胞を攻撃することが知られている。膵臓腺房細胞からのトリプシノーゲンの放出は、本発明のIP3R2/IP3R3ダブルノックアウトマウスでは阻害されたことから、IP3Rは急性膵炎の治療薬のターゲットとなり得る。すなわち、IP3R2/IP3R3の活性またはIP3生産に至っている情報伝達カスケードを増強または低減することが可能な薬剤は、ヒトの外分泌不全を伴うまたはこれに起因する疾患の有効な治療および/または予防薬となり得る。
【0061】
したがって、本発明のノックアウトマウスは、これらの疾患の発症機序の解明ならびに予防・治療方法および薬剤の開発において有用である。
【図面の簡単な説明】
【0062】
【図1】IP3R2(A)またはIP3R3(B)のノックアウトマウス作製のための、ターゲッティングベクターと遺伝子組換えの概略図である。IP3R2およびIP3R3の遺伝子のそれぞれ第1コーディング・エキソン内(それぞれ白い四角および黒い四角)で破壊した。遺伝子組換を検出するためのプローブは、黒いバーで示している。IP3R2変異遺伝子を核局在化信号を有するlacZ遺伝子(nls-lacZ)で置き換えた。ターゲッティングベクターは、それぞれ、正および負の選択標識として、PGKプロモーターの制御下にあるネオマイシン耐性遺伝子(N)、MC1プロモーターの制御下にあるジフテリア毒素A断片遺伝子(D)をそれぞれ含む。
【図2A】標準の乾燥ペレット飼料を与えた場合の雄のマウスの成長曲線である。各群のマウス(n=6)の平均体重(±SEM)をプロットした。
【図2B】3週齢と8週齢のマウスの1日の乾燥飼料摂取量を示す。8週齢のダブルノックアウトマウスは、実験時まで湿潤マッシュ飼料で飼育した。摂食量は、2日間にわたってモニターした。ダブルノックアウトマウスは乾燥ペレット飼料を全く食べられなかった。
【図2C】湿潤マッシュ飼料を与えた雄マウスの成長曲線を示す。各群のマウス(n=6)の平均体重(±SEM)をプロットした。
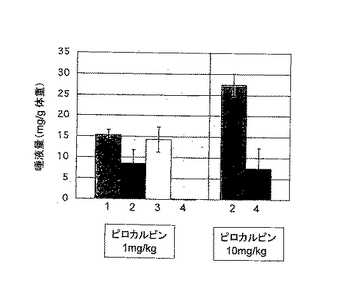
【図3A】1mg/kgまたは10mg/kgのムスカリン性作用薬(ピロカルピン)で刺激した場合のP21マウス(1群あたりn=4または5)の唾液分泌量を示す。唾液分泌は30分間測定した。図3A中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
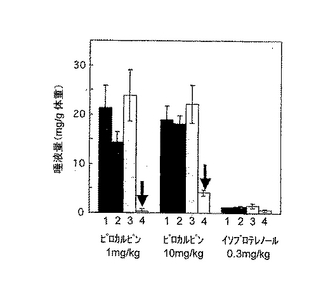
【図3B】ピロカルピン(1mg/kgまたは10mg/kg)またはβアドレナリン作用薬(イソプロテレノール:0.3mg/kg)で刺激した場合の成熟マウス(2〜3ヵ月齢、1群あたりn=4または5)の唾液分泌量を示す。唾液分泌は30分間測定した。図3B中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
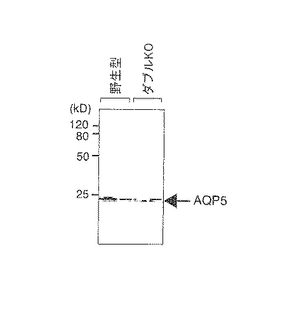
【図3C】野生型とダブルノックアウトマウスの顎下腺のAQP5発現のウェスタンブロット解析の結果である。
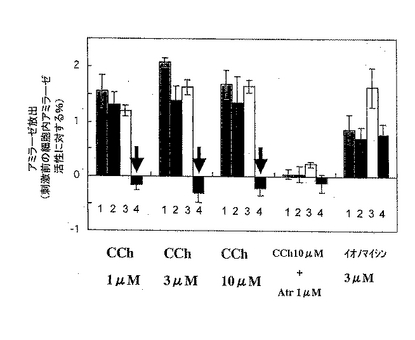
【図4A】膵臓腺細胞からのカルバコール(CCh)刺激によるアミラーゼの放出量を示す。細胞の培養上清中のアミラーゼ活性のカルバコール刺激による増加を、刺激前の細胞内アミラーゼ活性に対するパーセンテージとして表す。各値は、各々の群(n=6〜8)の平均±SEMを示す。図4A中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図4B】膵臓のアミラーゼ発現のウェスタンブロット分析結果である。タンパク質10μgを、抗アミラーゼ抗体によるウェスタンブロットに供した。図4B中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図4C】膵臓腺細胞からのカルバコール刺激によるリパーゼの放出量を示す。細胞の培養上清中のリパーゼ活性のカルバコール刺激による増加を、刺激前の細胞内リパーゼ活性に対するパーセンテージとして表す。各値は、各々の群(n=6〜8)の平均±SEMを示す。図4C中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図4D】膵臓腺細胞からのカルバコール刺激によるトリプシノーゲンの放出量を示す。放出後、上清中のトリプシノーゲンをエンテロキナーゼによって活性化してトリプシンに変換し、トリプシン活性を調べた。各々の群(n=6〜8)の平均±SEMを示す。図4D中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図5】野生型、IP3R2ノックアウトマウス(ホモ:IP3R2-/-)、IP3R3ノックアウトマウス(ホモ:IP3R3-/-)またはIP3R2およびIP3R3のダブルノックアウトマウス(ホモ:IP3R2-/-/ IP3R3-/-)について、カルバコール濃度依存的なアセチルコリンレセプター誘導性細胞内Ca2+上昇(すなわち[Ca2+]i)を示す。fura-2をロードしたSMG細胞の懸濁液を、各濃度のカルバコールで刺激した。各シンボルは以下の通りである:野生型;灰色の丸、IP3R2ノックアウトマウス;灰色の四角、IP3R3ノックアウトマウス;黒色の丸、およびIP3R2およびIP3R3のダブルノックアウトマウス;黒色の四角。各群はn=4とし、平均値±SEMで示す。
【図6】野生型(菱形)、IP3R2ノックアウトマウス(三角)、またはIP3R3ノックアウトマウス(四角)の、サッカリン(a)、スクロース(b)、MSG(c)、シクロヘキシミド(d)、硫酸キニーネ(e)、安息香酸デナトリウム(f)、HCl(g)およびNaCl(h)に対する反応(味覚)を示す。
【図7】野生型(黒四角)、IP3R3ヘテロノックアウトマウス(×印)、またはIP3R3ホモノックアウトマウス(白丸)の聴覚(dB)を示す。
【図8】野生型(A)及びIP3R3ノックアウトマウス(B)の毛の状態を示す。
【技術分野】
【0001】
本発明は、唾液分泌および膵液分泌不全モデル動物に関する。また本発明は、外分泌不全を改善する薬剤をスクリーニングする方法に関する。
【背景技術】
【0002】
顎下腺および膵臓からの外分泌が、食物消化のために重要であり、食物からの養分の吸収に不可欠であり(Solomon, 1984; Kutchai, 1998)、これらの外分泌機能の障害は、動物の成長と生存に重大な影響を及ぼすことが判っている。したがって、ヒトのみならず、家畜動物、愛玩動物における外分泌機能障害を治療または予防するための方法および薬剤の開発が必要である。しかしながら、これらの開発に必要不可欠である外分泌機能障害の適切なモデル動物が存在しない。
【0003】
顎下腺および膵臓などの外分泌組織では、消化酵素等のタンパク質および水を分泌するが、この際、細胞内のカルシウム濃度増加が重要であると考えられている。小胞体(ER)などにおける細胞内カルシウムストアからの細胞質へのカルシウム(Ca2+)動員は、イノシトール1,4,5-三リン酸レセプター(IP3R)とそのリガンドであるイノシトール1,4,5-三リン酸(IP3)との結合により誘導されることは公知の事実である。
【0004】
IP3Rに、セカンドメッセンジャーであるイノシトール1,4,5-三リン酸(IP3)が結合すると、小胞体(ER)などの細胞内ストアから細胞質へのカルシウム動員が起こり、このIP3によって誘発されるカルシウム放出(IICR)(これがその下流の細胞内の情報伝達カスケードを誘発する)は、分泌、増殖、分化、細胞死、遊走、シナプス可塑性の調節等のさまざまな細胞内プロセスに関与すると考えられている。
【0005】
哺乳動物には、それぞれ異なる遺伝子に由来する3つのサブタイプ(タイプ1〜3)のIP3Rがあり(非特許文献1)、興味深いことに、分泌機能を有する数種の細胞では、タイプ3 IP3R(IP3R3)の発現が高いという報告があるが、IP3Rと分泌機能との関連性については直接的な証明はなされていない。タイプ3 IP3R(IP3R3)cDNAは、ラット膵島細胞腺腫細胞系(非特許文献2)から最初にクローニングされたが、種々の組織で発現されている。タイプ2 IP3R(IP3R2)もまた顎下腺、膵臓をはじめ、心臓、免疫系細胞、造血細胞等の種々の組織ならびに細胞系において低レベルの発現が認められる(非特許文献3〜7)。
【0006】
哺乳動物の、3つのサブタイプのIP3Rのうち、タイプ1 IP3R(IP3R1)は、現在最も機能についての解析が進んでおり、これは脳と平滑筋に豊富に存在し、IP3R1欠損マウスは、胃平滑筋の徐波生成(非特許文献8および9)、重篤な運動失調、てんかん発作(非特許文献10)を示し、シナプス可塑性に影響を与えることが判っているが(非特許文献11〜15)、外分泌機能との関連性は報告されていない。しかしながら、IP3Rのサブタイプに特異的なIP3Rブロッカーが未だ開発されていないため、特にタイプ2とタイプ3のIP3Rの生理学的機能に関してはあまり解明されていない。
【0007】
【非特許文献1】Furuichiら、 Intracellular channels. Curr. Opin. Neurobiol. 4, 294-303, 1994
【非特許文献2】Blondelら、J. Biol. Chem. 268, 11356-11363, 1993
【非特許文献3】Fujinoら、Cell Tissue Res. 280, 201-210, 1995
【非特許文献4】Yamamoto-Hinoら、 Receptors and Cnannels. 2, 9-22, 1998
【非特許文献5】MoschellaおよびMarks、J. Cell Biol. 120, 1137-1146, 1993
【非特許文献6】Kijimaら、J. Biol. Chem. 268, 3499-3506, 1993
【非特許文献7】Sugiyamaら、FEBS Lett. 349, 191-196, 1994
【非特許文献8】Suzukiら、J. Physiol. 525, 105-111, 2000
【非特許文献9】Takanoら、Auton. Neurosci. 87, 249-257, 2001
【非特許文献10】Matsumotoら、Nature 379, 168-171, 1996
【非特許文献11】Inoueら、Neurosci. 117, 821-830, 1998
【非特許文献12】Nishiyamaら、Nature 408, 584-588, 2000
【非特許文献13】Itohら、Brain Res. 901, 237-246, 2001
【非特許文献14】Fujiiら、Learning and Memory 7, 312-320, 2000
【非特許文献15】Nagaseら、Neurosci. 117, 821-830, 2003
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明は、外分泌不全または外分泌過多を伴う障害の治療または予防薬の開発および発症機序の解明等に有用な、唾液分泌および膵液分泌不全モデル動物を提供することを課題とする。
【課題を解決するための手段】
【0009】
本発明者らは、外分泌組織との関連性が考えられるタイプ2 IP3レセプター(IP3R2)およびタイプ3 IP3レセプター(IP3R3)を共に欠失するダブルノックアウトマウスが、唾液分泌および膵液分泌不全を呈することを見出し、本発明を完成するに至った。また、これらの一方を欠失したマウスでもアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇がある程度低減され、これらの両方を欠失したマウスではアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇がさらに劇的に低減されることを見出した。これらの知見から、本発明は、以下の態様に関する:
1.機能的なタイプ3 イノシトール1,4,5-三リン酸レセプター(IP3R3)およびタイプ2 イノシトール1,4,5-三リン酸レセプター(IP3R2)のいずれか一方を有していない非ヒト脊椎動物;
2.機能的なIP3R2と機能的なIP3R3の両方を有していない非ヒト脊椎動物;
3.機能的な上記イノシトール1,4,5-三リン酸レセプター(IP3R)を有していないことが、該レセプターをコードしている染色体上の遺伝子が破壊されていることに起因する、上記1または2に記載の非ヒト脊椎動物;
4.IP3R3をコードしている染色体上の遺伝子がノックアウトされているために機能的なIP3R3を遺伝学的に有していない非ヒト脊椎動物と、IP3R2をコードしている染色体上の遺伝子がノックアウトされているために機能的なIP3R2を遺伝学的に有していない非ヒト脊椎動物とを交配して得られる、機能的なIP3R2と機能的なIP3R3の両方を遺伝学的に有していない非ヒト脊椎動物;
5.唾液の分泌が、対応する野生型動物に比較して低減されていることを特徴とする、上記1〜4のいずれかに記載の非ヒト脊椎動物;
6.膵臓細胞からの消化酵素の分泌が、対応する野生型動物に比較して低減されていることを特徴とする、上記1〜5のいずれかに記載の非ヒト脊椎動物;
7.顎下腺細胞および/または膵臓細胞におけるアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇が、対応する野生型の動物に比べて低減されていることを特徴とする、上記1〜6のいずれかに記載の非ヒト脊椎動物;
8.上記非ヒト脊椎動物が哺乳動物である、上記1〜7のいずれかに記載の非ヒト脊椎動物;
9.上記非ヒト脊椎動物が囓歯動物である、上記1〜7のいずれかに記載の非ヒト脊椎動物;
10.上記非ヒト脊椎動物がマウスである、上記1〜7のいずれかに記載の非ヒト脊椎動物;
11.上記1〜10のいずれかに記載の非ヒト脊椎動物に由来する細胞;
12.上記11に記載の細胞であって、野生型の同種の非ヒト脊椎動物に由来する同種の細胞は有している機能的なIP3R3およびIP3R2のいずれか一方または両方を有していないことを特徴とする、上記細胞;
13.不死化されている、上記11または12記載の細胞;
14.上記1〜10のいずれかに記載の非ヒト脊椎動物に薬剤を投与し、該動物の唾液の分泌量、膵臓細胞からの消化酵素の分泌量、および顎下腺細胞および/または膵臓細胞におけるアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇量からなる群より選択される少なくとも1つを測定することを含み、該測定した量が、薬剤を投与していない上記動物個体に比べて、該薬剤を投与した非ヒト脊椎動物個体で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法;
15.上記11〜13のいずれかに記載の細胞でアセチルコリンレセプターを発現している細胞に、薬剤を接触させ、該細胞のアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇量を測定し、該測定した量が、薬剤と接触させていない上記細胞に比べて、該薬剤を接触させた細胞で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法;
16.上記2〜10のいずれかに記載の非ヒト脊椎動物に由来する顎下腺細胞または膵臓細胞に、薬剤を接触させ、該細胞からの外分泌液量または消化酵素分泌量を測定し、該測定した量が、薬剤と接触させていない上記細胞に比べて、該薬剤を接触させた細胞で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法;
17.上記外分泌不全を改善し得る薬剤が、シェーグレン症候群、唾液または涙腺における自己免疫疾患、口内乾燥、乾燥性角結膜炎からなる群から選択される障害を治療または予防するための薬剤である、上記14〜16のいずれかに記載の方法;ならびに、
18.上記外分泌不全が唾液分泌不全または膵液分泌不全である、上記14〜16のいずれかに記載の方法。
【発明の効果】
【0010】
本発明により、外分泌不全または外分泌過多を伴う障害の治療または予防薬の開発および発症機序の解明等に有用な、唾液分泌および膵液分泌不全モデル動物となり得るIP3R2およびIP3R3の双方を欠失するマウスが提供される。該マウスは、唾液分泌と膵液分泌に重篤な不全、およびそれによる成長障害が認められた。さらに、該マウスからIP3R2およびIP3R3の双方を欠失する各種組織細胞を得ることが可能である。
【発明を実施するための最良の形態】
【0011】
以下、本発明を詳細に説明する。
本明細書中、非ヒト脊椎動物とは、好ましくはヒト以外の哺乳動物であり、さらに好ましくは、囓歯動物(ウサギ、モルモット、ラット、マウス等)であり、特に好ましくはマウスである。
【0012】
本明細書中「機能的なIP3R2」または「機能的なIP3R3」とは、イノシトール1,4,5-三リン酸と結合することによりCa2+を細胞内ストアから放出させるというIP3Rの機能を有しているIP3R2またはIP3R3を指す。したがって、「機能的なIP3R2(またはIP3R3)を有していない」とは、IP3R2(またはIP3R3)の発現が実質的に欠失しているか、IP3R2またはIP3R3分子が発現している場合でも、それがIP3Rとしての機能を消失した変異体であるか、あるいは機能的なIP3R2(またはIP3R3)を発現している場合であっても遺伝子操作によりドミナントネガティブ変異体が発現しているためにIP3R2(またはIP3R3)を介した、イノシトール三リン酸に応答して細胞内Ca2+のストアからの放出が実質的に低減されていることを指す。また、「機能的なIP3R2(またはIP3R3)を遺伝学的に有していない」とは、機能的なIP3R2(またはIP3R3)をコードする遺伝子が例えば、ノックアウトなどにより欠損していることを指す。
【0013】
「発現が実質的に欠失している」とは、IP3R2(またはIP3R3)の発現量が正常(野生型)の対応する動物個体(または細胞)に対して30%以下、好ましくは20%以下、より好ましくは10%以下、さらに好ましくは5%以下、最も好ましくはその発現が分子生物学的手法(即ち、mRNAまたはタンパク質の検出)によって検出されない程度であることを指す。機能的なIP3R2(またはIP3R3)を有していない動物は、例えばIP3R2(またはIP3R3)をコードする染色体上の遺伝子を破壊することによりノックアウト動物として得ることができる。ノックアウト動物の作製方法は当業者には公知である。IP3R2およびIP3R3をコードする遺伝子配列は、データベースから入手可能である。この配列に基づいて、ゲノムDNAをクローニングし、常法に従って、遺伝子ターゲティングベクターに組込み、細胞に導入して、該細胞から個体を発生させるとよい。遺伝子破壊によるノックアウト動物の作製方法は種々の公知文献が多数存在しており、当業者は容易に作製することができる。本発明に用いる場合、ホモノックアウト動物が好ましい。
【0014】
機能的なIP3R2と機能的なIP3R3の両方を有していない非ヒト動物は、これらを有していなくとも、生殖能力には実質的な影響はなく、繁殖可能であるため、機能的なIP3R2を遺伝学的に有していない動物と機能的なIP3R3を遺伝学的に有していない動物とを交配させて、その子孫から両方を有していない個体を選択することにより、ダブルノックアウト動物として得ることができる。
【0015】
したがって、機能的なIP3R3またはIP3R2を遺伝学的に有していない非ヒト動物は、機能的なIP3R2と機能的なIP3R3の両方を有していない非ヒト動物の作製に利用することができ、本発明の範囲に包含される。
【0016】
IP3R3またはIP3R2を遺伝学的に欠失している動物は、ダブルノックアウト動物を得るためにも有用である。本発明に用いるダブルノックアウト動物は、ホモダブルノックアウト動物が好ましい。
【0017】
機能的なIP3R2およびIP3R3のいずれか一方または両方を有していない脊椎動物は、顎下腺および/または膵臓細胞において、イノシトール1,4,5-三リン酸介在性細胞内Ca2+動員が、野生型のものに比べて低減される。例えば、唾液腺細胞においてカルバコールでアセチルコリンレセプターを刺激した場合、IP3レセプターを介して細胞内Ca2+濃度([Ca2+]i)が上昇するが、IP3R2およびIP3R3のいずれか一方を欠失している場合(すなわち機能的なIP3R2およびIP3R3のいずれか一方を有していない場合)、野生型に比べてその上昇が約70%以下に低減され、両方を欠失している場合では、野生型に比べて約30%以下に低減され得る。本明細書中、「対応する野生型の動物に比べて低減されている」とは、対象とする、機能的なIP3R2および/またはIP3R3を有していない動物の膵臓細胞からの消化酵素の分泌、唾液の分泌または細胞内カルシウム濃度の上昇が、同様の条件下にある機能的なIP3R2およびIP3R3を有している野生型の同系の動物に比べた場合に、約80%以下、好ましくは50%以下、より好ましくは30%以下、さらに好ましくは20%以下に低減されていることをいう。上記の機能的なIP3R2およびIP3R3の両方を有していない動物では、唾液腺細胞および膵臓細胞の外分泌を誘導するためのシグナルが十分に伝達できず、唾液および膵液の外分泌が低減され得る。例えば該ダブルノックアウトマウスでは、唾液分泌促進剤であるピロカルピンで刺激した場合の唾液の分泌量が、野生型のそれに対して約30%以下に低減され得る。しかし、ピロカルピン刺激により誘導される唾液分泌のみならず、通常の唾液分泌も低減され、これにより、離乳後に固形飼料を摂食できず、致死性の発育不良を呈する。
【0018】
さらにまた、機能的なIP3R2およびIP3R3の両方を有していない動物は、膵液分泌不全を呈する。すなわち、本来、アミラーゼ、トリプシンおよびリパーゼ等の消化酵素を含む膵液が膵臓細胞から小腸内に分泌されることにより、食餌を消化して栄養素を吸収するが、消化酵素の分泌が不十分であるため、食餌摂取をしたとしても、消化吸収効率が悪く、発育不良を起こす。例えば、IP3R2/IP3R3ダブルノックアウトマウスでは、上述のように離乳後に固形飼料を摂食できないが、水分を十分に含むペースト状の飼料を与えた場合は、充分量摂食することができるにもかかわらず、消化不良により、糞便中にタンパク質および脂質を多量に排泄し、発育不良を呈する。
【0019】
上記のとおり、機能的なIP3R2およびIP3R3の両方を有していない動物では、重篤な外分泌不全を起こす。いずれか一方を欠失した動物で、外分泌不全の症状が認められないのは、おそらく、欠失していないIP3R2またはIP3R3が機能することで、欠失した一方の機能が補填されているためと考えられる。
【0020】
機能的なIP3R2およびIP3R3の両方を欠失した動物は、シェーグレン症候群、唾液または涙腺における自己免疫疾患、口内乾燥(ドライマウス)、乾燥性角結膜炎等の外分泌不全を伴う障害のモデル動物として有用であり得る。
【0021】
すなわち、上記障害を含む、外分泌不全を呈する障害の治療または予防のための薬剤をスクリーニングするために、上記動物を用いることができる。上記動物に、候補薬剤を投与し、該動物の唾液の分泌量または膵臓細胞からの消化酵素の分泌量を測定する。ここで、候補薬剤の投与は、薬剤の性質に応じて、経口投与、皮下、静脈または筋肉注射であってもよく、または種々の局所投与であってもよく、当業者であれば適宜、適切な投与方法を選択し得る。唾液の分泌量の測定は、候補薬剤の投与後適切な時間を経て、麻酔下で一定時間内に分泌された唾液を回収してその容量を測定してもよく、また唾液分泌促進作用を有する薬剤(ピロカルピンおよびカルバコール等)で唾液分泌を刺激して、その分泌量を測定してもよい。膵臓細胞からの消化酵素の分泌量を測定するためには、候補薬剤投与後、適切な時間を経て、動物から膵臓を摘出し、生理的な条件下(各種生理的緩衝液中など)で、場合によっては膵臓細胞を分散させて、膵液分泌促進作用を有する薬剤(カルバコール等)で膵液分泌を刺激して、細胞外液の消化酵素(例えば、アミラーゼ、リパーゼまたはトリプシン)の活性を測定するか、またはウェスタンブロット等により細胞外液中の消化酵素量を評価する。さらにまた候補薬剤の投与後、適切な時間を経て、動物から顎下腺または膵臓を摘出し、これらをアセチルコリンレセプター刺激作用を有する薬剤に応答する細胞内Ca2+濃度の上昇の程度を、fura-2を用いる方法等、公知の細胞内Ca2+濃度測定法により測定してもよい。ネガティブコントロールとして候補薬剤を投与していない動物を用いて、同様の処理および測定を行い、ネガティブコントロールについての測定値に比べて、候補薬剤を投与した動物についての測定値の方が大きければ、該候補薬剤は外分泌不全の予防または治療薬として有望であると判定することができる。
【0022】
さらに、上記動物に由来する細胞もまた本発明の範囲に包含される。IP3R2およびIP3R3は、あらゆる組織に広く分布していることから、機能的なIP3R2およびIP3R3のいずれか一方または両方を有していない動物に由来する任意の組織の細胞は、その形質を保持している。したがって、各組織または組織内の各細胞における、IP3R2およびIP3R3の機能を解明する上で、これらの細胞は有用である。したがって、上記動物に由来する細胞は、野生型の同種の細胞は有しているはずの機能的なIP3R2およびIP3R3のいずれか一方または両方を有していないという特徴を有する。かかる特徴により、該細胞を外分泌不全を改善し得る薬剤をスクリーニングするのに用いることができる。
【0023】
これらの細胞は、該動物の個体および胎生個体から単離可能であり、初代培養系細胞として用いることも可能である。さらにまた、これらの細胞を不死化することも、当業者であれば容易に実施可能である。不死化の方法は種々のものが公知であり、任意の方法を適宜用いることができる。不死化された細胞株もまた、それが由来する動物個体または胎生個体と同じように機能的なIP3R2およびIP3R3のいずれか一方または両方を有しておらず、IP3R2およびIP3R3の機能を解明する上で有用であり、不死化細胞であることから、保存、維持および使用が簡便である点において有利である。
【0024】
これらの細胞を用いて、外分泌不全を改善し得る薬剤をスクリーニングする方法は、それぞれの細胞を適切な生理的条件下に維持して、候補薬剤と接触させて、これらをアセチルコリンレセプター刺激作用を有する薬剤に応答する細胞内Ca2+濃度の上昇の程度を評価することにより達成できる。候補薬剤と接触させていない場合の細胞内Ca2+濃度の上昇の程度に比して、上昇の程度が大きい場合、該候補薬剤は外分泌不全の予防または治療薬として有望であると判定することができる。さらに、細胞が顎下腺細胞またはこれに由来する細胞の場合は、候補薬剤と接触させた場合の唾液分泌量を測定するか、または細胞外液に分泌されたアミラーゼの量を測定してもよい。細胞が膵臓細胞またはこれに由来する細胞の場合は、候補薬剤と接触させた場合の細胞外液中の膵液消化酵素の量を測定してもよい。
【0025】
かかるスクリーニングの方法については、当業者が日常的に行い得る改良または変更により、適切な方法および条件を当業者であれば実施し得る。
【0026】
以下、実施例によって本発明を更に具体的に説明するが、これらの実施例は本発明を限定するものではない。
【実施例1】
【0027】
IP3R2ノックアウトマウス、IP3R3ノックアウトマウスとIP3R2/IP3R3ダブルノックアウトマウスの作製
マウスゲノムのIP3R2とIP3R3の第1コーディングエキソンとその両側のイントロンとをそれぞれ含むマウスゲノムクローンは、129マウスゲノムライブラリー(熊本大学・山村研一博士より分与)から単離した。IP3R2またはIP3R3が欠損するマウスを作製するために、当該遺伝子それぞれの第1コーディング・エキソンを破壊した(図1AおよびB参照のこと)。
【0028】
PCRで得たIP3R2 cDNA断片(Morikawaら、Gene 196, 181-185, 1997)および600bp長のヒトIP3R3 cDNA断片(第1エキソンを含む)をプローブとして用いた。上記ゲノムクローンから得られた断片、陽性選択マーカーとしてpKJ1(McBurneyら、Nucleic Acids Res. 19, 5755-57, 1991)から得たneo遺伝子カセットpGK-neo(1.8kb)、および陰性選択マーカーとしてpMC1DT-A(Yagiら、Proc. Natl. Acad. Sci. USA 87, 9918-9922, 1990)から得たXhoI-SalIジフテリア毒素A断片(dT-A)遺伝子カセットを用いて常法に従い、ターゲッティングベクターを構築した。IP3R2ターゲッティングベクターでは、最初の6つのアミノ酸をコードしている第1のエキソンの領域は、LacZ染色(Sanesら、EMBO J. 5, 133-142, 1986)によってIP3R2プロモーター活性をモニターできるよう、lacZ遺伝子と核移行シグナルとをインフレームで連結した。
【0029】
IP3R2とIP3R3のターゲッティングベクターDNAを、直鎖状にして、ES細胞(それぞれJ1とR1)に常法による電気穿孔法にて形質転換した。G418(150μg/ml)に抵抗性を示すクローンを、サザンブロット法により解析して所望の通り相同的組換えを起こしたクローンを得た。IP3R2破壊クローンは、ゲノムDNAをBglIIで切断し、5’末端側の外部プローブとしてBglII-BamHI断片を用いてこれとハイブリダイズさせることにより確認した。野生型アレルからは7kb断片が、変異型アレルからは1.5kb断片が得られた。IP3R3破壊クローンは、ゲノムDNAをEcoRVで切断し、3’末端側の外部プローブとしてEcoRI-SacII断片を用いてこれとハイブリダイズさせることにより確認した。野生型アレルからは8.5kb断片が、変異型アレルからは4.5kb断片が得られた(図1AおよびB)。所望の相同組換えは、neoプローブと他方の外部プローブとによっても確認した(データ示さず)。IP3R2とIP3R3について、所望の相同組換えを起こしたそれぞれのESクローンが、6および12クローン得られた。公知の方法(Bradley Production and analysis of chimeric mice. In Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E.J. Robertson , ed. (Oxford: IRL press), pp. 113-151, 1987; Woodら, Proc. Natl. Acad. Sci. USA 90, 4582-4585, 1993)にしたがって、各々のキメラマウスを得た。C57BL/6Jマウスとキメラマウスを交配させることによって、ヘテロ接合体マウスを得、これをさらに3〜6回、C57BL/6Jマウスと交配させた。
【0030】
得られた各々の遺伝子のヘテロ接合変異体を交配させて、IP3R2とIP3R3とのダブルノックアウトヘテロ接合変異体を得、さらに、本発明のダブルノックアウトホモ接合体を得るために、ダブルノックアウトヘテロ接合体どうしを交配させた。マウスの遺伝型は、尾部組織から得たゲノムDNAのサザンブロット分析またはPCRで決定した。
【0031】
IP3R2およびIP3R3のいずれか一方のノックアウトマウスでは、胎生期および生後数ヶ月間においては、致死性障害および外見上の異常は認められなかった。IP3R3ノックアウトマウスにおいては、脱毛症状を示し、生後数ヶ月以降に体重の増加が認められない傾向にあったが、生殖能も寿命(2年以上)も正常であった。
【0032】
イムノブロット分析により、野生型マウスの顎下腺と膵臓においては、IP3Rの全てのサブタイプが発現されていたが、3つのサブタイプの中でIP3R2とIP3R3が他の組織と比較して豊富に発現されたのに対して、IP3R1発現レベルは他の組織における発現より低かった。作製した変異体マウスにおいて、ターゲットとしたIP3R2および/またはIP3R3の発現は欠損しており、ターゲット以外のサブタイプの発現は認められ、意図したとおりの発現パターンが得られた。
【実施例2】
【0033】
IP3R2/IP3R3ダブルノックアウト変異体マウスの特徴
(1)外観と生存率
出生時、IP3R2/IP3R3ダブルノックアウトホモ接合体の体重および外見は他の同腹の仔と差はなかったが、出生後に、ダブルノックアウトホモ接合体だけ体重増加が遅れた。通常、マウスは自由に標準の乾燥ペレットCA-1(CLEA)と水を摂取可能な環境で飼育した。離乳(概して出産後約20日:P20)後、ダブルノックアウトホモ接合体は、体重が減少し始め、4週齢に達する前に死亡した(図2A)。これらの現象は性別に関係なく観察され、ダブルノックアウトホモ接合体は通常の食餌(すなわち固形乾燥飼料)では、4週齢以上生存できなかった。
【0034】
(2)摂食不能
IP3R2/IP3R3ダブルノックアウト変異体は、乾燥食物ペレットを摂取できず、IP3R2-/-/IP3R3-/-ダブルノックアウト変異体は離乳期間の直後から体重が減り始めた。これは、離乳の後、乾燥食物ダイエットへ切り替えることにより、摂食不能が起こるのではないかと考えられた。
【0035】
そこで、出生3週間後の離乳の後、2日の期間にわたって1日の摂食量(標準的乾燥ペレット飼料)をモニターしたところ、ダブルノックアウトマウスは、ケージ内に飼料があることを認識しながらも、全く食べることができなかった(図2B左のパネル)。
【0036】
そこで、20日齢の時点で、乾燥ペレットCA-1(CLEA)を2倍(重量比)の滅菌水中で粉砕・混合した湿潤マッシュ飼料を与えた。該マッシュ飼料は、毎晩、新たに調製して与えた。すると、湿潤マッシュ飼料を与えた場合は、ダブルノックアウト変異体は摂食可能であり、生存し続けた。しかしながら、同様に湿潤マッシュ飼料で飼育した同腹のマウス間で比較した場合、ダブルノックアウト変異体の体重増加は少なかった(図2C)。
【0037】
さらに、離乳後から8週齢まで湿潤マッシュ飼料で飼育したマウスを、8週齢の時点で乾燥飼料を与えたところ、やはり、ダブルノックアウトマウスは、全くこれを食べなかった(図2B右のパネル)。このことから、ダブルノックアウトマウスが、乾燥飼料を摂食できないのは、離乳前の発達および離乳の遅れ等が原因ではないと考えられる。
【0038】
(3)唾液生産の不足
上記ダブルノックアウトマウスの歯は正常であった。したがって、唾液生産が不足している可能性が考えられた。そこで、該ダブルノックアウトマウスの唾液分泌を調べた。
3週齢のマウスまたは成熟マウスを37℃の保温プレート上に置き、ネンブタール(50mg/kg、腹腔投与)麻酔下で、ピロカルピン-HCl(1または10mg/kg(Tocris))またはイソプロテレノール(0.3mg/kg(Sigma))を皮下投与した。口腔内で分泌される唾液は手動ピペッタ(ギルソン(P100))を使用してピロカルピンまたはイソプロテレノールで投与後各30分間に分泌された唾液を、遠心管に回収し、その回収された唾液の重量を求めた。
【0039】
母乳から乾燥飼料に切り替える離乳期(生後21日齢:P21)のマウスで調べたところ、ピロカルピンによって促進される唾液分泌は、ダブルノックアウトで顕著に消失していた(図3Aの4のグラフ)。1.0mg/kgのピロカルピンを投与した場合、IP3R2ノックアウト(IP3R2-/-/IP3R3+/+)とIP3R3ノックアウト(IP3R2+/+/IP3R3-/-)マウスと野生型(IP3R2+/+/IP3R3+/+)マウスとの間では唾液分泌に大きな差は認められなかったが、ダブルノックアウトマウスでは無唾液症が観察された。ピロカルピンを10mg/kgで投与した場合は、ダブルノックアウトマウスでも唾液分泌が認められたが、分泌量はコントロールに比べて3分の1程度であった。かかる機能障害は、IP3R2ノックアウトマウスおよびIP3R3ノックアウトマウスでは観察されなかった。
【0040】
次に、離乳後湿潤マッシュ飼料で飼育した成熟マウス(2〜3ヵ月齢)における唾液分泌を測定した。結果は、P21マウスでの結果と基本的に類似していた。ピロカルピンによって促進される唾液分泌は、ダブルノックアウトマウスだけで、顕著に抑制されていた(図3Bの4のグラフ)。
【0041】
常法に従いイムノブロット解析を行って、唾液腺分泌に必須の水量浸透性チャネルであるアクアポリン5(AQP5)の発現量を調べたところ、AQP5はダブルノックアウトマウスでも正常に発現していることが確認された(図3C)。したがって、ダブルノックアウトの唾液分泌の機能障害は、水量浸透性チャネルの減少に依存しているわけではない。
【実施例3】
【0042】
顎下腺と膵臓におけるIP3Rの発現
上記のとおり、IP3R2-/-/IP3R3-/-マウスでは唾液分泌が劇的に損なわれることから、顎下腺、およびこれと並ぶ主要な外分泌組織である膵臓におけるIP3R2とIP3R3の発現を調べた。ウェスタンブロット法は、顎下腺と膵臓を摘出し、0.32M スクロース、5mM Tris-HCl(pH 7.4)、1mM EDTA、0.1M PMSF、10μM ロイペプチン、10μM ペプスタチンAおよび1mM 2-メルカプトエタノールを含む溶液(ホモジナイズバッファー)中でホモジナイズし、50μgを5% SDS-PAGEゲルにアプライして泳動後、ウェスタンブロット解析に供した。IP3R1、IP3R2およびIP3R3はそれぞれ、抗体KM1112、KM1083とKM1082にて検出した。
【0043】
ダブルノックアウトマウスの顎下腺細胞(腺房で管の細胞)と膵臓腺細胞のウェスタンブロットの結果、IP3R2およびIP3R3とも野生型に比べて有意に発現が低減していた(データは示さず)。
【実施例4】
【0044】
膵臓腺細胞のコリン作動性アゴニストにより誘発される消化酵素分泌は、IP3R2-/-/IP3R3-/-ダブルノックアウト変異体では欠損している
次に、ダブルノックアウトマウスの膵臓機能障害について調べた。
【0045】
麻酔下で心臓穿刺により屠殺したマウスの膵臓を直ちに摘出し、冷却したカルシウム不含平衡塩類溶液(BSS:115mM NaCl、5.4mM KCl、2mM Ca2+、1mMマグネシウムイオン、20mM Hepes、10mMブドウ糖、0.125%のウシ血清アルブミン(BSA)、pH 7.4)中に入れ、速やかに砕片し、2mg/mlのコラゲナーゼ(和光034-10533)で消化した、消化後の細胞含有液を細胞濾過器(100μmナイロン・メッシュ)に通して、再懸濁、遠心による洗浄を3回繰り返した。このようにして得た細胞懸濁液を、1〜10μMのカルバコールと、BSA、Ca2+(2mM)を含むBSS 400μl中で、室温で30分間静置してカルバコールに暴露させた。0.22μmフィルタ(ミリポア)にてろ過して得た300μlの上清を分泌タンパク質サンプル液として、膵臓から分泌される消化酵素の分析に用いた。
【0046】
カルバコールが細胞のムスカリン性アセチルコリン受容体に作用することを確認するために、1μMのアトロピン(ムスカリン性アセチルコリン受容体アンタゴニスト)を、カルバコール投与の1分前に添加しておいた。イオノマイシン(3μM)を添加したコントロール実験は、Ca2+に依存する分泌機序がノックアウトマウスでも完全に機能していたかどうか調べるために行った。
【0047】
アミラーゼ、リパーゼおよびトリプシン(トリプシノーゲン)の放出は、分泌タンパク質サンプル液中の各酵素活性を測定し、分泌量は細胞中に含まれる量に対するパーセンテージとして表した。
【0048】
アミラーゼ活性は、AMY(SP)キット(関東Chemicals社)を、リパーゼ活性は、カラー自動試験キット(和光)を使用して測定した。トリプシン活性は、サンプル中のトリプシノーゲンをEKマックス・エンテロキナーゼ(Invitrogen)によって活性化させて、その活性を基質としてN-CBZ-Gly-Pro-Arg-p-ニトロアニリドを用いて検定した(Szmolaら、J. Biol. Chem. 278, 48580-48589, 2003)。
【0049】
非選択的なコリン作動性アゴニストであるカルバコール(CCh)を細胞懸濁液中に添加し、30分間膵臓細胞を刺激した後、細胞外間隙に分泌されたタンパク質を分析し、α-アミラーゼの分泌を調べた結果を図4Aに示す。図4Aにおいて、野生型、IP3R2ノックアウトおよびIP3R3ノックアウトマウスのいずれの膵臓細胞でも、膵臓細胞中の全アミラーゼ活性に対して約1〜2%の活性が分泌物中で検出されたが、IP3R2-/-/IP3R3-/-ダブルノックアウトマウス由来の膵臓細胞ではその分泌物中の活性は検出されなかった(図4Aの4のグラフ)。
【0050】
ダブルノックアウトマウス由来の膵臓細胞のアミラーゼタンパク質の分泌量が低減されていることを、イムノブロット分析によって確認した(データ示さず)。しかしながら、膵臓組織のウェスタンブロット解析では、ダブルノックアウトマウスにおいても、膵臓アミラーゼは検出されたことから(図4Bの4のカラム)、アミラーゼの産生は正常であると考えられる。
【0051】
次いで、同様に膵臓細胞からのリパーゼとトリプシノーゲンの分泌を調べた。その結果をそれぞれ図4Cおよび4Dに示す。膵臓細胞からのリパーゼとトリプシノーゲンの分泌は、ダブルノックアウトマウスのみ顕著に消失していた(図4Cおよび4Dの4のグラフ)。
【0052】
すなわち、ダブルノックアウトマウスにおいてのみ、膵臓からの外分泌が顕著に阻害されており、糖、脂質、タンパク質の全ての消化能が著しく低下していると考えられ、これにより、栄養不良が起こり体重増加の抑制または体重の減少が引き起こされると考えられる。
【実施例5】
【0053】
SMG腺房細胞におけるコリン作動性ニューロン刺激による細胞内Ca2+の変化
本発明のノックアウトマウスについて、コリン作動性ニューロン刺激によるCa2+シグナル伝達能を調べるために、野生型および各ノックアウトマウスから摘出したSMG細胞を酵素的に分散させて、カルバコール刺激応答性細胞内Ca2+の変動をfura-2を用いて観察した。カルバコール濃度依存的な細胞内Ca2+の変動を図5に示す。IP3R2ノックアウトおよびIP3R3ノックアウトマウスは、野生型に比して約70%程度の細胞内Ca2+の上昇が認められるに止まっており、IP3R2-/-/IP3R3-/-ダブルノックアウトマウスにおいては、野生型の約20%以下の上昇しか認められなかった。すなわち、IP3R2およびIP3R3のいずれかを欠失したノックアウトマウスにおいても、コリン作動性ニューロン刺激によるCa2+シグナル伝達能は低減されていたが、IP3R2およびIP3R3の両方を欠失したマウスでは、コリン作動性ニューロン刺激によるCa2+シグナル伝達能はほとんど喪失されていた。IP3R2およびIP3R3のいずれかを欠失したノックアウトマウスでは、唾液分泌に顕著な影響が認められなかったことは、野生型の70%程度のCa2+シグナル伝達能が保持されていることにより、唾液分泌能はさほど顕著に抑制されなかったものと考えられる。一方、IP3R2およびIP3R3の両方を欠失したマウスでは、野生型の約20%以下しかCa2+シグナル伝達能を有していないため、唾液分泌に顕著に影響を受けたものと考えられる。
【実施例6】
【0054】
本実施例においては、本発明のノックアウトマウスについて、味覚障害、聴覚障害および毛の異常を試験した。
(1)味覚障害
野生型および各ノックアウトマウス(IP3R2ノックアウトマウス、IP3R3ノックアウトマウス)の味覚は、二瓶法に従って評価した。
【0055】
その結果を図6に示す。野生型マウス(菱形)およびIP3R2ノックアウトマウス(三角)に対して、IP3R3ノックアウトマウス(四角)は、甘味(サッカリン、スクロース)、苦味(シクロヘキシミド、硫酸キニーネ、安息香酸デナトリウム)、旨味(グルタミン酸ナトリウム(MSG))に対する味覚が劣っていることがわかる。一方、酸(HCl)および塩(NaCl)については、IP3R3ノックアウトマウスは野生型およびIP3R2ノックアウトマウスと同等の味覚認識を示した。従って、IP3R3ノックアウトマウスは味覚障害のモデルとなりうる。
【0056】
(2)聴覚障害
野生型(n=14)、ならびにIP3R3ヘテロノックアウトマウス(n=7)およびIP3R3ホモノックアウトマウス(n=19)の聴覚(dB)を、Auditory Startle Response and Prepulse inhibitionの方法を用いて評価した。
【0057】
その結果を図7に示す。野生型マウス(黒四角)およびIP3R3ヘテロノックアウトマウス(×印)に対して、IP3R3ホモノックアウトマウス(白丸)は、聴覚障害を有することが判明した。従って、IP3R3ノックアウトマウスは聴覚障害のモデルとなり得、またこのマウスを用いて聴覚障害治療のためのモデルマウスを作出し得る。
【0058】
(3)毛の異常
野生型およびIP3R3ノックアウトマウスの毛を評価した。その結果を図8に示す。野生型マウス(図8A)と比べてIP3R3ノックアウトマウス(図8B)は毛の生育に異常があり、体表に毛のない部分が存在することが明らかとなった。従って、IP3R3ノックアウトマウスは毛発育疾患のモデルとなり、育毛剤の開発に貢献すると考えられる。
【産業上の利用可能性】
【0059】
本発明のIP3R2/IP3R3ダブルノックアウトマウスは、上記特徴により、外分泌組織(例えば唾液腺と膵臓)の機能低下症の疾患モデル動物として利用することができる。例えば、本発明のマウスの唾液腺の機能低下症は、シェーグレン症候群の症状、唾液または涙腺における自己免疫疾患(これらは口渇(口内乾燥)または乾燥性角結膜炎を引き起こす)等の外分泌機能不全を伴う疾患と、機序が共通する可能性が考えられる。
【0060】
膵臓の外分泌においては、IP3R2/IP3R3の欠損は、痩せ症および栄養不良をもたらす機能不全につながるため、膵臓外分泌細胞のIP3R活性は、栄養の利用性および体重を制御することができる主要因のうちの1つである可能性がある。また、急性膵炎では、膵臓外分泌細胞から放出されるトリプシノーゲンが活性型トリプシンに変換されて膵臓細胞を攻撃することが知られている。膵臓腺房細胞からのトリプシノーゲンの放出は、本発明のIP3R2/IP3R3ダブルノックアウトマウスでは阻害されたことから、IP3Rは急性膵炎の治療薬のターゲットとなり得る。すなわち、IP3R2/IP3R3の活性またはIP3生産に至っている情報伝達カスケードを増強または低減することが可能な薬剤は、ヒトの外分泌不全を伴うまたはこれに起因する疾患の有効な治療および/または予防薬となり得る。
【0061】
したがって、本発明のノックアウトマウスは、これらの疾患の発症機序の解明ならびに予防・治療方法および薬剤の開発において有用である。
【図面の簡単な説明】
【0062】
【図1】IP3R2(A)またはIP3R3(B)のノックアウトマウス作製のための、ターゲッティングベクターと遺伝子組換えの概略図である。IP3R2およびIP3R3の遺伝子のそれぞれ第1コーディング・エキソン内(それぞれ白い四角および黒い四角)で破壊した。遺伝子組換を検出するためのプローブは、黒いバーで示している。IP3R2変異遺伝子を核局在化信号を有するlacZ遺伝子(nls-lacZ)で置き換えた。ターゲッティングベクターは、それぞれ、正および負の選択標識として、PGKプロモーターの制御下にあるネオマイシン耐性遺伝子(N)、MC1プロモーターの制御下にあるジフテリア毒素A断片遺伝子(D)をそれぞれ含む。
【図2A】標準の乾燥ペレット飼料を与えた場合の雄のマウスの成長曲線である。各群のマウス(n=6)の平均体重(±SEM)をプロットした。
【図2B】3週齢と8週齢のマウスの1日の乾燥飼料摂取量を示す。8週齢のダブルノックアウトマウスは、実験時まで湿潤マッシュ飼料で飼育した。摂食量は、2日間にわたってモニターした。ダブルノックアウトマウスは乾燥ペレット飼料を全く食べられなかった。
【図2C】湿潤マッシュ飼料を与えた雄マウスの成長曲線を示す。各群のマウス(n=6)の平均体重(±SEM)をプロットした。
【図3A】1mg/kgまたは10mg/kgのムスカリン性作用薬(ピロカルピン)で刺激した場合のP21マウス(1群あたりn=4または5)の唾液分泌量を示す。唾液分泌は30分間測定した。図3A中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図3B】ピロカルピン(1mg/kgまたは10mg/kg)またはβアドレナリン作用薬(イソプロテレノール:0.3mg/kg)で刺激した場合の成熟マウス(2〜3ヵ月齢、1群あたりn=4または5)の唾液分泌量を示す。唾液分泌は30分間測定した。図3B中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図3C】野生型とダブルノックアウトマウスの顎下腺のAQP5発現のウェスタンブロット解析の結果である。
【図4A】膵臓腺細胞からのカルバコール(CCh)刺激によるアミラーゼの放出量を示す。細胞の培養上清中のアミラーゼ活性のカルバコール刺激による増加を、刺激前の細胞内アミラーゼ活性に対するパーセンテージとして表す。各値は、各々の群(n=6〜8)の平均±SEMを示す。図4A中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図4B】膵臓のアミラーゼ発現のウェスタンブロット分析結果である。タンパク質10μgを、抗アミラーゼ抗体によるウェスタンブロットに供した。図4B中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図4C】膵臓腺細胞からのカルバコール刺激によるリパーゼの放出量を示す。細胞の培養上清中のリパーゼ活性のカルバコール刺激による増加を、刺激前の細胞内リパーゼ活性に対するパーセンテージとして表す。各値は、各々の群(n=6〜8)の平均±SEMを示す。図4C中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図4D】膵臓腺細胞からのカルバコール刺激によるトリプシノーゲンの放出量を示す。放出後、上清中のトリプシノーゲンをエンテロキナーゼによって活性化してトリプシンに変換し、トリプシン活性を調べた。各々の群(n=6〜8)の平均±SEMを示す。図4D中、1は野生型、2はIP3R2のみのノックアウト、3はIP3R3のみのノックアウト、4はIP3R2とIP3R3とのダブルノックアウトマウスの結果である。
【図5】野生型、IP3R2ノックアウトマウス(ホモ:IP3R2-/-)、IP3R3ノックアウトマウス(ホモ:IP3R3-/-)またはIP3R2およびIP3R3のダブルノックアウトマウス(ホモ:IP3R2-/-/ IP3R3-/-)について、カルバコール濃度依存的なアセチルコリンレセプター誘導性細胞内Ca2+上昇(すなわち[Ca2+]i)を示す。fura-2をロードしたSMG細胞の懸濁液を、各濃度のカルバコールで刺激した。各シンボルは以下の通りである:野生型;灰色の丸、IP3R2ノックアウトマウス;灰色の四角、IP3R3ノックアウトマウス;黒色の丸、およびIP3R2およびIP3R3のダブルノックアウトマウス;黒色の四角。各群はn=4とし、平均値±SEMで示す。
【図6】野生型(菱形)、IP3R2ノックアウトマウス(三角)、またはIP3R3ノックアウトマウス(四角)の、サッカリン(a)、スクロース(b)、MSG(c)、シクロヘキシミド(d)、硫酸キニーネ(e)、安息香酸デナトリウム(f)、HCl(g)およびNaCl(h)に対する反応(味覚)を示す。
【図7】野生型(黒四角)、IP3R3ヘテロノックアウトマウス(×印)、またはIP3R3ホモノックアウトマウス(白丸)の聴覚(dB)を示す。
【図8】野生型(A)及びIP3R3ノックアウトマウス(B)の毛の状態を示す。
【特許請求の範囲】
【請求項1】
機能的なタイプ3 イノシトール1,4,5-三リン酸レセプター(IP3R3)およびタイプ2 イノシトール1,4,5-三リン酸レセプター(IP3R2)のいずれか一方を有していない非ヒト脊椎動物。
【請求項2】
機能的なIP3R2と機能的なIP3R3の両方を有していない非ヒト脊椎動物。
【請求項3】
機能的な上記イノシトール1,4,5-三リン酸レセプター(IP3R)を有していないことが、該レセプターをコードしている染色体上の遺伝子が破壊されていることに起因する、請求項1または2に記載の非ヒト脊椎動物。
【請求項4】
IP3R3をコードしている染色体上の遺伝子がノックアウトされているために機能的なIP3R3を遺伝学的に有していない非ヒト脊椎動物と、IP3R2をコードしている染色体上の遺伝子がノックアウトされているために機能的なIP3R2を遺伝学的に有していない非ヒト脊椎動物とを交配して得られる、機能的なIP3R2と機能的なIP3R3の両方を遺伝学的に有していない非ヒト脊椎動物。
【請求項5】
唾液の分泌が、対応する野生型動物に比較して低減されていることを特徴とする、請求項1〜4のいずれか1項に記載の非ヒト脊椎動物。
【請求項6】
膵臓細胞からの消化酵素の分泌が、対応する野生型動物に比較して低減されていることを特徴とする、請求項1〜5のいずれか1項に記載の非ヒト脊椎動物。
【請求項7】
顎下腺細胞および/または膵臓細胞におけるアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇が、対応する野生型の動物に比べて低減されていることを特徴とする、請求項1〜6のいずれか1項に記載の非ヒト脊椎動物。
【請求項8】
上記非ヒト脊椎動物が哺乳動物である、請求項1〜7のいずれか1項記載の非ヒト脊椎動物。
【請求項9】
上記非ヒト脊椎動物が囓歯動物である、請求項1〜7のいずれか1項記載の非ヒト脊椎動物。
【請求項10】
上記非ヒト脊椎動物がマウスである、請求項1〜7のいずれか1項記載の非ヒト脊椎動物。
【請求項11】
請求項1〜10のいずれか1項に記載の非ヒト脊椎動物に由来する細胞。
【請求項12】
請求項11に記載の細胞であって、野生型の同種の非ヒト脊椎動物に由来する同種の細胞は有している機能的なIP3R3およびIP3R2のいずれか一方または両方を有していないことを特徴とする、上記細胞。
【請求項13】
不死化されている、請求項11または12記載の細胞。
【請求項14】
請求項1〜10のいずれか1項に記載の非ヒト脊椎動物に薬剤を投与し、該動物の唾液の分泌量、膵臓細胞からの消化酵素の分泌量、および顎下腺細胞および/または膵臓細胞におけるアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇量からなる群より選択される少なくとも1つを測定することを含み、該測定した量が、薬剤を投与していない上記動物個体に比べて、該薬剤を投与した非ヒト脊椎動物個体で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法。
【請求項15】
請求項11〜13のいずれか1項に記載の細胞でアセチルコリンレセプターを発現している細胞に、薬剤を接触させ、該細胞のアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇量を測定し、該測定した量が、薬剤と接触させていない上記細胞に比べて、該薬剤を接触させた細胞で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法。
【請求項16】
請求項2〜10のいずれか1項に記載の非ヒト脊椎動物に由来する顎下腺細胞または膵臓細胞に、薬剤を接触させ、該細胞からの外分泌液量または消化酵素分泌量を測定し、該測定した量が、薬剤と接触させていない上記細胞に比べて、該薬剤を接触させた細胞で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法。
【請求項17】
上記外分泌不全を改善し得る薬剤が、シェーグレン症候群、唾液または涙腺における自己免疫疾患、口内乾燥、乾燥性角結膜炎からなる群から選択される障害を治療または予防するための薬剤である、請求項14〜16のいずれか1項に記載の方法。
【請求項18】
上記外分泌不全が唾液分泌不全または膵液分泌不全である、請求項14〜16のいずれか1項に記載の方法。
【請求項1】
機能的なタイプ3 イノシトール1,4,5-三リン酸レセプター(IP3R3)およびタイプ2 イノシトール1,4,5-三リン酸レセプター(IP3R2)のいずれか一方を有していない非ヒト脊椎動物。
【請求項2】
機能的なIP3R2と機能的なIP3R3の両方を有していない非ヒト脊椎動物。
【請求項3】
機能的な上記イノシトール1,4,5-三リン酸レセプター(IP3R)を有していないことが、該レセプターをコードしている染色体上の遺伝子が破壊されていることに起因する、請求項1または2に記載の非ヒト脊椎動物。
【請求項4】
IP3R3をコードしている染色体上の遺伝子がノックアウトされているために機能的なIP3R3を遺伝学的に有していない非ヒト脊椎動物と、IP3R2をコードしている染色体上の遺伝子がノックアウトされているために機能的なIP3R2を遺伝学的に有していない非ヒト脊椎動物とを交配して得られる、機能的なIP3R2と機能的なIP3R3の両方を遺伝学的に有していない非ヒト脊椎動物。
【請求項5】
唾液の分泌が、対応する野生型動物に比較して低減されていることを特徴とする、請求項1〜4のいずれか1項に記載の非ヒト脊椎動物。
【請求項6】
膵臓細胞からの消化酵素の分泌が、対応する野生型動物に比較して低減されていることを特徴とする、請求項1〜5のいずれか1項に記載の非ヒト脊椎動物。
【請求項7】
顎下腺細胞および/または膵臓細胞におけるアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇が、対応する野生型の動物に比べて低減されていることを特徴とする、請求項1〜6のいずれか1項に記載の非ヒト脊椎動物。
【請求項8】
上記非ヒト脊椎動物が哺乳動物である、請求項1〜7のいずれか1項記載の非ヒト脊椎動物。
【請求項9】
上記非ヒト脊椎動物が囓歯動物である、請求項1〜7のいずれか1項記載の非ヒト脊椎動物。
【請求項10】
上記非ヒト脊椎動物がマウスである、請求項1〜7のいずれか1項記載の非ヒト脊椎動物。
【請求項11】
請求項1〜10のいずれか1項に記載の非ヒト脊椎動物に由来する細胞。
【請求項12】
請求項11に記載の細胞であって、野生型の同種の非ヒト脊椎動物に由来する同種の細胞は有している機能的なIP3R3およびIP3R2のいずれか一方または両方を有していないことを特徴とする、上記細胞。
【請求項13】
不死化されている、請求項11または12記載の細胞。
【請求項14】
請求項1〜10のいずれか1項に記載の非ヒト脊椎動物に薬剤を投与し、該動物の唾液の分泌量、膵臓細胞からの消化酵素の分泌量、および顎下腺細胞および/または膵臓細胞におけるアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇量からなる群より選択される少なくとも1つを測定することを含み、該測定した量が、薬剤を投与していない上記動物個体に比べて、該薬剤を投与した非ヒト脊椎動物個体で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法。
【請求項15】
請求項11〜13のいずれか1項に記載の細胞でアセチルコリンレセプターを発現している細胞に、薬剤を接触させ、該細胞のアセチルコリンレセプター刺激による細胞内カルシウム濃度の上昇量を測定し、該測定した量が、薬剤と接触させていない上記細胞に比べて、該薬剤を接触させた細胞で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法。
【請求項16】
請求項2〜10のいずれか1項に記載の非ヒト脊椎動物に由来する顎下腺細胞または膵臓細胞に、薬剤を接触させ、該細胞からの外分泌液量または消化酵素分泌量を測定し、該測定した量が、薬剤と接触させていない上記細胞に比べて、該薬剤を接触させた細胞で増加していることを指標とする、外分泌不全を改善し得る薬剤をスクリーニングする方法。
【請求項17】
上記外分泌不全を改善し得る薬剤が、シェーグレン症候群、唾液または涙腺における自己免疫疾患、口内乾燥、乾燥性角結膜炎からなる群から選択される障害を治療または予防するための薬剤である、請求項14〜16のいずれか1項に記載の方法。
【請求項18】
上記外分泌不全が唾液分泌不全または膵液分泌不全である、請求項14〜16のいずれか1項に記載の方法。
【図1】


【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7】
【図8】

【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2006−187284(P2006−187284A)
【公開日】平成18年7月20日(2006.7.20)
【国際特許分類】
【出願番号】特願2005−353560(P2005−353560)
【出願日】平成17年12月7日(2005.12.7)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
【公開日】平成18年7月20日(2006.7.20)
【国際特許分類】
【出願日】平成17年12月7日(2005.12.7)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
[ Back to top ]