
IgGデコイ受容体融合タンパク質の血液脳関門送達のための組成物および方法
IgGデコイ受容体のCNSへの送達のための組成物および関連する方法が提供される。該方法は、血液脳関門の表面に発現された受容体(BBB受容体)に対する抗体に共有結合した受容体細胞外ドメイン(ECD)を含む、二官能性デコイ受容体−BBB受容体抗体融合抗体の全身投与を包含する。いくつかの具体例において、本明細書に記載の組成物は、CNS疾患に罹患している対象を治療するために投与される。
【発明の詳細な説明】
【技術分野】
【0001】
相互参照
本願は、2009年3月18日出願の米国仮出願第61/161,320号(出典明示により全体として本明細書の一部とする)の利益を請求する。
【背景技術】
【0002】
発明の背景
腫瘍壊死因子受容体(TNFR)などの標的受容体の可溶性細胞外ドメイン(ECD)は、ヒトの疾患において治療作用を有する。受容体ECDは、内因性リガンド、例えば、腫瘍壊死因子(TNF)−αを封鎖し、それにより、内因性リガンドの内因性標的受容体へのアクセスを遮断する外因性デコイ(decoy)受容体として作用する。デコイ受容体は、脳疾患の強力な新規治療となる可能性があった。しかしながら、デコイ受容体は、他の巨大分子薬物のように、血液脳関門(BBB)を横断しない。かくして、今日までに、組み換えデコイ受容体の全身投与によって脳障害患者を治療することはできなかった。
【発明の概要】
【課題を解決するための手段】
【0003】
発明の概要
本明細書には、BBBを横断してIgG−受容体ECD(デコイ受容体)融合タンパク質を対象のCNSへ送達するための組成物および関連する方法が記載される。特に、該方法は、受容体ECDおよびBBBの表面に発現される受容体の細胞外ドメインに結合する抗体を含む二官能性デコイ受容体融合抗体の治療上有効量を全身投与することによって、デコイ受容体のCNSへの送達を可能にする。
【0004】
したがって、一の態様において、受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体であって、BBB上に発現した受容体および受容体細胞外ドメインのリガンドに結合する融合抗体が提供される。いくつかの具体例において、BBB上に発現される受容体は、インスリン受容体、トランスフェリン受容体、またはリポタンパク質受容体である。いくつかの具体例において、BBB上に発現される受容体は、ヒトインスリン受容体である。いくつかの具体例において、該二官能性デコイ受容体融合抗体は、ヒトインスリン受容体との結合に関して、配列番号4および6または配列番号6および7のアミノ酸配列を含む二官能性デコイ受容体融合抗体と競合する。いくつかの具体例において、受容体細胞外ドメインは、サイトカイン受容体、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン(ephrin)受容体由来である。いくつかの具体例において、受容体細胞外ドメインは、TNF−α受容体細胞外ドメインを含む。いくつかの場合、TNF−α受容体細胞外ドメインは、ヒト、マウス、ラットまたはブタTNF−α受容体細胞外ドメインのアミノ酸配列に対して少なくとも85%(例えば、90%、95%または100%)同一のアミノ酸配列を含む。ある特定の具体例において、TNF−α受容体細胞外ドメインのアミノ酸配列は、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に融合される。ある特定の具体例において、TNF−α受容体細胞外ドメインのアミノ酸配列は、重鎖免疫グロブリンのカルボキシル末端に融合される。
【0005】
関連する態様において、脳への取り込みがタンパク質100グラムあたり1%、2%、3%、5%、7%または10%以上のIDである二官能性デコイ受容体融合抗体(例えば、HIRMAb−TNFR融合タンパク質)が提供される。いくつかの具体例において、該二官能性デコイ受容体融合抗体(例えば、HIRMAb−TNFR融合タンパク質)は、デコイ受容体およびヒトIgGのFcフラグメントの融合タンパク質、例えば、TNFR:Fcの脳への取り込み量よりも1−、2−、5−、10−、13−、15−、17−、20−、25−、30−、35−、40−、45−または50−倍大きい脳内取り込み量を示す。いくつかの具体例において、本明細書に記載の二官能性デコイ受容体融合抗体(例えば、HIRMAb−TNFR融合タンパク質)は、他の器官と比べて、選択的に脳に豊富にある。いくつかの具体例において、TNFR:Fc融合タンパク質に関する器官PS積(organ PS product)に対する本明細書に記載のデコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)に関する器官PS積の比率を複数の器官について決定する場合、脳の比率は、脂肪、筋肉、心臓、肺、肝臓および/または脾臓の比率よりも1、2、5、10、13、15、17、20、25、30、35、40、45、または50倍大きい。
【0006】
関連する態様において、(i)重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列、(ii)軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列、または(iii)(i)または(ii)の相補性配列を含む核酸であって、ここに、該重鎖および軽鎖免疫グロブリンがBBB受容体に対する抗体由来である核酸が提供される。いくつかの具体例において、該コードされた受容体細胞外ドメインは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン(ephrin)受容体由来である。いくつかの具体例において、該コードされた受容体細胞外ドメインは、TNF−α受容体(例えば、ヒトTNF−α受容体)由来である。他の具体例において、該TNF−α受容体由来のコードされた受容体細胞外ドメインは、ヒト、マウス、ラットまたはブタTNF−α受容体細胞外ドメインのアミノ酸配列に対して少なくとも85%同一のアミノ酸配列を含む。いくつかの具体例において、該コードされた免疫グロブリン重鎖または軽鎖は、ヒトインスリン受容体、トランスフェリン受容体またはリポタンパク質受容体に対する抗体由来である。ある特定の具体例において、上記の第1の配列は、配列番号4または7に対応するアミノ酸配列に対して少なくとも85%(例えば、90%、95%または100%)同一のアミノ酸配列をコードし、または上記の第2の配列は、配列番号6に対応するアミノ酸配列と少なくとも85%(例えば、90%、95%または100%)同一のアミノ酸配列をコードする。他の具体例において、該核酸は、高ストリンジェンシー条件下で、配列番号4、6または7をコードする核酸(例えば、配列番号3または5のヌクレオチド配列を含む核酸)にハイブリダイズする。いくつかの具体例において、該TNF−α受容体由来のコードされた細胞外ドメインは、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に共有結合される。いくつかの具体例において、該TNF−α受容体由来のコードされた細胞外ドメインは、重鎖免疫グロブリンのカルボキシル末端に共有結合される。
【0007】
いくつかの具体例において、核酸は、核酸ベクターとして提供される。いくつかの具体例において、核酸ベクターは、(i)重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列、(ii)軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列、または(iii)(i)または(ii)の相補性配列を含み、ここに、該重鎖および軽鎖免疫グロブリンは、BBB受容体に対する抗体由来である。いくつかの具体例において、該核酸ベクターは、(i)を含み、さらに、BBB受容体に対する抗体由来の軽鎖免疫グロブリンをコードする核酸配列を含むか、または(ii)を含み、さらに、BBB受容体に対する抗体由来の重鎖免疫グロブリンをコードする核酸を含む。関連する態様において、上記核酸のいずれかを含む細胞(例えば、哺乳動物細胞)が提供される。
【0008】
さらなる態様において、受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体を含む医薬組成物を対象に全身投与することを含む、血液脳関門を横切ってデコイ受容体を送達する方法であって、該融合抗体がBBB上に発現された受容体および受容体細胞外ドメインのリガンドに結合するところの方法が提供される。いくつかの具体例において、該BBB上に発現された受容体は、インスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である。いくつかの具体例において、該受容体細胞外ドメインは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である。いくつかの具体例において、該TNF−α受容体由来の細胞外ドメインは、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に共有結合している。いくつかの具体例において、該TNF−α受容体由来の細胞外ドメインは、重鎖免疫グロブリンのカルボキシル末端に共有結合している。
【0009】
別の態様において、受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体を含む医薬組成物の治療上有効量を対象に全身投与することを含むCNS疾患の治療方法であって、該融合抗体がBBB上に発現された受容体および受容体細胞外ドメインのリガンドに結合するところの方法が提供される。いくつかの具体例において、該BBB上に発現された受容体は、インスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である。いくつかの具体例において、該受容体細胞外ドメインは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮長因子受容体、またはエフリン受容体由来である。いくつかの具体例において、治療されるべきCNS疾患は、急性CNS疾患、例えば、全脳虚血、局所脳虚血、外傷性脳損傷、または脊髄損傷である。他の具体例において、治療されるべきCNS疾患は、慢性CNS疾患、例えば、神経変性疾患、例えば、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症、ハンチントン病、多発性硬化症、横断性脊髄炎、運動ニューロン疾患、ピック病、結節硬化症、カナバン病、レット症候群、脊髄小脳失調症、フリードライヒ失調症、視神経萎縮、または網膜変性である。
【0010】
また別の態様において、(i)受容体細胞外ドメインに融合した免疫グロブリン重鎖、および免疫グロブリン軽鎖の両方、または
(ii)受容体細胞外ドメインに融合した免疫グロブリン軽鎖、および免疫グロブリン重鎖の両方
をコードする単一タンデム発現ベクターを真核細胞中に安定に組み込むことを含む、二官能性デコイ受容体融合抗体を製造する方法であって、該コードされた免疫グロブリン重鎖および免疫グロブリン軽鎖がBBB上に発現された受容体に対する抗体由来であるところの方法が提供される。いくつかの具体例において、該BBB上に発現された受容体は、インスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である。いくつかの具体例において、該コードされた受容体細胞外ドメインは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である。いくつかの具体例において、該TNF−α受容体由来の受容体細胞外ドメインは、免疫グロブリン重鎖または免疫グロブリン軽鎖のカルボキシル末端に共有結合される。いくつかの具体例において、該TNF−α受容体由来のコードされた受容体細胞外ドメインは、免疫グロブリン重鎖のカルボキシル末端に共有結合される。
【0011】
出典明示による組み込み
本明細書に挙げられた全ての出版物、特許、および特許出願は、出典明示により、あたかも個々の出版物、特許または特許出願が詳細かつ個々に出典明示により本明細書の一部とされることが示されたかの如く、同程度まで本明細書の一部とされる。
【0012】
図面の簡単な説明
本発明の新規な特徴は、特に、添付の特許請求の範囲で示される。本発明の特徴および利益のより良い理解は、発明の本質を利用する実例となる具体例を示す下記の詳細な説明、および添付の図面を参照して得られるであろう。
【図面の簡単な説明】
【0013】
【図1】HIRMAb−TNFR融合タンパク質は、TNFR ECDのアミノ末端の、キメラHIRMAbの重鎖のCH3領域のカルボキシル末端への融合によって形成される。該融合タンパク質は、二官能性分子である。すなわち、該融合タンパク質は、BBBにて、HIRを結合して脳中への輸送を媒介し、かつ、TNF−αを結合して該サイトカインの炎症性を抑制する。
【0014】
【図2】(A)ヒトU87グリア細胞由来のRNAの逆転写によって生産されたcDNAからPCRによって生産されたヒトTNFR ECD cDNAのアガロースゲルのエチジウムブロマイド染色(レーン1)、およびTNFR−特異的ODNプライマー(表2)。レーン2および3:DNAサイズ標準。(B)pHIRMAb−TNFR、TNFR ECDおよびキメラHIRMAbの重鎖(HC)の融合タンパク質をコードする真核細胞発現プラスミドの遺伝子操作。融合遺伝子の5’側には、サイトメガロウイルス(CMV)プローモーターが配置され、3’側には、ウシ成長ホルモンポリA(pA)配列が配置される。
【0015】
【図3】タンパク質Aアフィニティー精製キメラHIRMAbおよびHIRMAb−TNFR融合タンパク質の還元SDS−PAGEおよびクーマシーブルー染色。どちらも均質に精製され、重鎖および軽鎖から構成される。
【0016】
【図4】抗ヒト(h)IgG一次抗体(左パネル)または抗ヒトTNFR−II一次抗血清(右パネル)のいずれかでのウェスタンブロット。HIRMAb−TNFR融合タンパク質の免疫反応性をキメラHIRMAbと比較する。HIRMAb−TNFR融合タンパク質およびHIRMAbの両方は、抗−hIgGウェスタンにおいて同一の軽鎖を有する。HIRMAb−TNFR融合重鎖は、抗−hIgGおよび抗−ヒトTNFR抗体の両方と反応するが、HIRMAb重鎖は、抗−hIgG抗体とのみ反応する。HIRMAb−TNFR融合重鎖のサイズは、30kDa TNFR ECDの55kDa HIRMAb重鎖への融合のために、HIRMAbの重鎖のサイズよりも約30kDa大きい。
【0017】
【図5】キメラHIRMAbまたはHIRMAb−TNFR融合タンパク質のいずれかのHIR細胞外ドメイン(ECD)に対する結合は飽和可能である。HIR ECDに結合するHIRMAb−TNFRのED50は、キメラHIRMAbの結合のED50に匹敵する。
【0018】
【図6】TNFR:Fc融合タンパク質(A)またはHIRMAb−TNFR融合タンパク質(B)のTNFαに対する結合は飽和可能である。パネルAに示されるように、ヒトIgG1のTNFαに対する結合はない。線形回帰分析の傾きは、KD/Amax比を生じ(ここに、KDは、TNFαの結合定数であり、Amaxは、最大吸光度である)、TNFαに対する結合のKDの相対的指標である。TNFR:Fc融合タンパク質およびHIRMAb−TNFR融合タンパク質のどちらも、比較可能なアフィニティーで、TNFαに結合する。
【0019】
【図7】(A)TNFαのHIRMAb−TNFR融合タンパク質に対する結合に関する放射性受容体アッセイのアウトライン。マウス抗−ヒト(MAH)IgG1 Fcを播種し、それは、HIRMAb−TNFR融合タンパク質のFc領域に結合した。次いで、該融合タンパク質のTNFR細胞外ドメイン(ECD)領域が[125I]−TNFαに結合し、それを非標識TNFαの添加によって置換した。(B)飽和可能な結合を非線形回帰分析によって分析して、HIRMAb−TNFR融合タンパク質に結合しているTNFαの50%阻害を与えた濃度KDを得た。
【0020】
【図8】TNFαは、アクチノマイシンDで処理したヒトWEHI−13VAR細胞において、ED50 約10pg/mLで細胞毒性を引き起こす。しかしながら、1.4nM TNFR:Fcまたは1.4nM HIRMAb−TNFRのいずれかの存在下、高濃度のTNFαによって細胞毒性は引き起こされない。
【0021】
【図9】HIRMAb−TNFR融合タンパク質の重鎖および軽鎖をコードする分離したタンデムな発現カセット(各遺伝子は、CMVプロモーターを有する分離したタンデムなイントロンによって駆動し、各々、ウシ成長ホルモン(BGH)ポリアデニル化(pA)配列を末端に有する)を含有するタンデムベクター(TV)であるpTV−HIRMAb−TNFRの遺伝子操作。pTV−HIRMAb−TNFRは、HIRMAbをコードするユニバーサルTV(pTV−HIRMAbと称する)内で、TNFR ECD cDNAをHIRMAb HCカセットの3’末端にて、独特なHpaI部位にサブクローニングすることによって生じる。TNFR ECD cDNAは、pCD−HIRMAb−TNFRプラスミド(図2B)を鋳型として用いるPCRによって生産された。
【0022】
【図10】HIRMAb−TNFR融合タンパク質の重鎖のドメイン構造。19アミノ酸IgGシグナルペプチドの次にHIRMAb重鎖のVHが配置され、それは、3つのCDR(CDR1、CDR2、CDR3)および4つのFR(FR1、FR2、FR3、FR4)からなり、次に、ヒトIgG1重鎖C領域のドメイン(CH1、ヒンジ、CH2、CH3)が配置され、次に、3−アミノ酸リンカー(Ser−Ser−Ser)が配置され、次に、ヒトTNFR−II ECDの235−アミノ酸配列が配置される。3つのN−結合型グリコシル化部位に下線を付し、CH2領域内に1部位およびTNFR領域内に2部位を含む。
【0023】
【図11】HIRMAb−TNFR融合タンパク質の軽鎖ドメイン構造。20アミノ酸IgGシグナルペプチドの次に、HIRMAb軽鎖のVLが配置され、それは、3つのCDR(CDR1、CDR2、CDR3)および4つのFR(FR1、FR2、FR3、FR4)からなり、その次に、ヒトカッパ軽鎖C領域が配置される。
【0024】
【図12】(A)[125I]−TNFR:Fc融合タンパク質および[3H]−HIRMAb−TNFR融合タンパク質の血漿濃度を、成体アカゲザルにおける該タンパク質の単回静脈内注射後の時間に対してプロットする。データは、%注射用量(I.D.)/mLとして表される。(B)10%トリクロロ酢酸(TCA)によって沈澱可能な血漿放射活性の%を両タンパク質の注射後の時間に対してプロットする。データは、平均±SE(n=1時点につき3回繰り返す)である。
【0025】
【図13】濃度曲線下の血漿面積またはAUC(A)、脳100グラムあたりの脳取り込み量または注射用量(I.D.)%(B)、およびBBBの透過係数表面積(permeability surface area: PS)の積(C)をTNFR:Fc融合タンパク質、HIRMAb−TNFR融合タンパク質、および脳血漿量マーカー、ヒトIgG1(hIgG1)についてプロットする。全ての測定値は、該タンパク質の静脈内投与後2時間で得られた。データは、平均±SE(n=1時点につき3回繰り返す)である。
【0026】
【図14】TNFR:Fc融合タンパク質に関する器官PS積に対する、HIRMAb−TNFR融合タンパク質に関する器官PS積の比率を各器官についてプロットする。データは、平均±SE(n=1時点につき3回繰り返す)である。脳に関する比率は、前頭灰白質、前頭白質、小脳灰白質、および小脳白質についての値の平均であり、22〜37で変動した。
【発明を実施するための形態】
【0027】
発明の詳細な説明
I.序文
II.いくつかの定義
III.血液脳関門
IV.BBBを横断する輸送のためのデコイ受容体融合抗体
V.組成物
VI.核酸、ベクター、細胞および製造
VII.方法
VIII.実施例
IX.配列
【0028】
略語
AA アミノ酸
BBB 血液脳関門
BCA ビシンコニン酸
BGH ウシ成長ホルモン
CDR 相補性決定領域
CHO チャイニーズハムスター卵巣
CMV サイトメガロウイルス
DC 希釈クローニング
DHFR ジヒドロ葉酸還元酵素
ECD 細胞外ドメイン
ED50 50%飽和をもたらす有効用量
FR フレームワーク領域
FS フランキング領域
FWD 順方向
HC 重鎖
HIR ヒトインスリン受容体
HIRMAb HIRに対するMAb
HIRMAb HC HIRMAbの重鎖
HIRMAb LC HIRMAbの軽鎖
HIRMAb−TNFR HIRMAbおよびTNFR ECDの融合タンパク質(ここに、TNFRがHCカルボキシル末端に融合する)
HT ヒポキサンチン−チミジン
IgG 免疫グロブリンG
IGF インスリン様成長因子
LC 軽鎖
MAb モノクローナル抗体
MAH マウス抗−ヒトIgG
MTX メトトレキサート
MW 分子量
N アスパラギン
nt ヌクレオチド
ODN オリゴデオキシヌクレオチド
orf オープンリーディングフレーム
pA ポリ−アデニル化
PAGE ポリアクリルアミドゲル電気泳動
PBS リン酸緩衝化セーライン
PBST PBSおよびTween−20
PCR ポリメラーゼ連鎖反応
pI 等電点
R 受容体
REV 逆方向
RNase A リボヌクレアーゼA
RT 逆転写
RT 室温
SDM 部位特異的変異誘発
SDS ドデシル硫酸ナトリウム
SEC サイズ排除クロマトグラフィー
Ser セリン
SFM 無血清培地
TH トロイの木馬
TNF 腫瘍壊死因子
TNFR TNF受容体
TV タンデムベクター
UTV ユニバーサルTV
VH 重鎖の可変領域
VL 軽鎖の可変領域
【0029】
I.序文
遺伝子操作されたデコイ受容体は、特に受容体ECDがヒト免疫グロブリンG、例えばIgG1のFcフラグメントのアミノ末端に融合される場合、強力な新規治療剤である。Fcフラグメントは、2つの定常領域鎖からなる二量体タンパク質である。ほとんどの受容体は膜内で二量体を形成するので、受容体ECDのFcフラグメントへの融合により、受容体タンパク質の天然二量体コンフィギュレーションが可能となる。デコイ受容体のよく記載される一例は、TNFαデコイ受容体−Fc融合タンパク質である。TNFαデコイ受容体−Fc融合タンパク質は、プロ炎症性サイトカイン、TNFαを封鎖し、それにより、その内因性リガンドによる内因性TNFRの活性化を阻害する能力のために、末梢性炎症疾患の強力な生物学的治療剤である。しかしながら、TNFR:Fc融合タンパク質は、CNS疾患のために処方することはできない。
【0030】
血液脳関門は、全身投与された可溶性デコイ受容体の中枢神経系への送達に対する厳しい障害であり、ここに、それらは、いくつかの神経病理、例えば、神経炎症において役割を果たすある種のリガンド(例えば、TNF−α)を中和するように作用する。本明細書に記載の組成物および方法は、BBBを横断してデコイ受容体をCNSへ送達することにおける重要な3つの因子、すなわち、1)BBBを横断できるようにするための受容体細胞外ドメイン(ECD)の修飾(本明細書中で、「デコイ受容体」という)、2)全身投与された修飾されたIgG−受容体融合抗体のCNS中への取り込み量および率、および3)一旦BBBを横断したデコイ受容体活性の保持に向けられる。本明細書に記載の方法および組成物の種々の態様は、介在するリンカー配列を用いて、またはリンカー配列を用いずに、BBB上に発現された受容体(例えば、ヒトインスリン受容体)の細胞外ドメインに対する免疫グロブリン(重鎖または軽鎖)に融合したデコイ受容体を含む、全身投与することができる融合抗体を提供することによって、これらの因子に向けられる。
【0031】
したがって、本発明は、その必要のある対象に、BBB上に発現された受容体(例えば、hIR)に対する抗体および受容体ECD(例えば、ヒトTNF−α受容体ECD)を含む二官能性デコイ受容体融合抗体の治療上有効量を全身投与することによる、必要とする対象、例えば、CNS炎症に罹患した対象またはその危険性のある対象の中枢神経系にデコイ受容体(例えば、可溶性TNF−α受容体ECD)を送達するための組成物および方法を提供する。
【0032】
II.いくつかの定義
「抗体」なる語は、天然または部分合成もしくは全合成免疫グロブリンをいう。該用語は、また、抗原結合ドメインである結合ドメインまたは抗原結合ドメインに相同性の結合ドメインを有するいずれかのポリペプチドまたはタンパク質を包含する。CDR移植(grafted)抗体もまた、該用語によって意図される。
【0033】
「天然抗体」および「天然免疫グロブリン」は、通常、2つの同一の軽(L)鎖および2つの同一の重(H)鎖からなる約150,000ダルトンのヘテロ四量体糖タンパク質である。各軽鎖は、典型的に、1つの共有ジスルフィド結合によって重鎖に連結され、一方、異なる免疫グロブリンアイソタイプの重鎖によってジスルフィド結合の数は変化する。各重鎖および軽鎖は、また、規則的に間隔をあけた鎖内ジスルフィド架橋を有する。各重鎖は、一方の端に可変ドメイン(「VH」)を有し、続いて、いくつかの定常ドメイン(「CH」)を配置する。各軽鎖は、一方の端に可変ドメイン(「VL」)、もう一方の端に定常ドメイン(「CL」)を有し、軽鎖の定常ドメインは、重鎖の第1の定常ドメインと並んでおり、軽鎖可変ドメインは、重鎖の可変ドメインと並んでいる。特定のアミノ酸残基は、軽鎖および重鎖可変ドメイン間にインターフェースを形成すると考えられる。
【0034】
「可変ドメイン」なる語は、ファミリーメンバーのなかで(すなわち、異なるアイソ形態のなかで、または異なる種において)広範囲に配列が異なるタンパク質ドメインをいう。抗体に関して、「可変ドメイン」なる語は、特定の抗原に対する各特定の抗体の結合および特異性において使用される抗体の可変ドメインをいう。しかしながら、変化性(variability)は、抗体の可変ドメインを通して均等に分布されているわけではない。それは、軽鎖および重鎖可変ドメインともに、超可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存された部分を「フレームワーク領域」または「FR」と呼ぶ。非修飾重鎖および軽鎖の各々の可変ドメインは、4つのFR(各々、FR1、FR2、FR3およびFR4)を含み、それは、主として、場合によっては、β−シート構造の一部を形成するβ−シート構造を連結するループを形成する3つの超可変領域によって連結されたβ−シートコンフィギュレーションを採用している。各鎖の超可変領域は、FRによってごく接近して結びついており、他の鎖由来の超可変領域と共に、抗体の抗原結合部位の形成に寄与する(Kabatら、(1991),Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md.,第647−669頁参照)。定常ドメインは、抗原に対する抗体の結合に直接関与しないが、抗体依存性細胞毒における抗体の関与などの種々のエフェクター機能を示す。
【0035】
「超可変領域」なる語は、本明細書中で使用される場合、抗原−結合に関与する抗体のアミノ酸残基をいう。超可変領域は、相補的に抗原に直接結合し、各々、CDR1、CDR2およびCDR3として知られる3つの「相補性決定領域」または「CDR」由来のアミノ酸残基を含む。軽鎖可変ドメインにおいて、CDRは、典型的に、およその残基24−34(CDRL1)、50−56(CDRL2)および89−97(CDRL3)に相当し、重鎖可変ドメインにおいて、CDRは、典型的に、およその残基31−35(CDRH1)、50−65(CDRH2)および95−102(CDRH3)に相当し(Kabatら、(1991), Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md.参照)、および/または「超可変ループ」由来の残基(すなわち、軽鎖可変ドメインにおいて、残基26−32(L1)、50−52(L2)および91−96(L3)、および重鎖可変ドメインにおいて、26−32(H1)、53−55(H2)および96−101(H3))に相当する(ChothiaおよびLesk,(1987), J. Mol. Biol., 196:901−917参照)。
【0036】
本明細書中で使用される場合、「可変フレームワーク領域」または「VFR」なる語は、抗原結合ポケットまたは溝の一部を形成し、および/または抗原に接触しうるフレームワーク残基をいう。いくつかの具体例において、フレームワーク残基は、抗原結合ポケットまたは溝の一部であるループを形成する。該ループにおけるアミノ酸残基は、抗原に接触しても、しなくてもよい。ある具体例において、VFRのループアミノ酸は、抗体の三次元構造、抗体重鎖または抗体軽鎖の検査により決定される。溶媒にアクセス可能なアミノ酸位置はおそらく、ループを形成し、および/または抗体可変ドメインに接触する抗原を提供するので、該三次元構造は、溶媒にアクセス可能なアミノ酸位置について分析することができる。溶媒にアクセス可能な位置のいくつかは、アミノ酸配列多様性を容認でき、その他は(例えば、構造的位置)あまり多様化されることがない。抗体可変ドメインの三次元構造は、結晶構造またはタンパク質モデリングから由来することができる。いくつかの具体例において、VFRは、重鎖可変ドメインのアミノ酸位置71〜78(該位置は、Kabatら、1991にしたがって規定される)に対応するアミノ酸位置を含む、該アミノ酸位置から実質的になる、または該アミノ酸位置からなる。いくつかの具体例において、VFRは、CDRH2とCDRH3の間に配置されたフレームワーク領域3の一部を形成する。VFRは、標的抗原と接触するように、または抗原結合ポケットの一部を形成するように配置されたループを形成することができる。
【0037】
重鎖の定常ドメインのアミノ酸配列に依存して、免疫グロブリンは種々のクラスに割り当てることができる。免疫グロブリンの5つの主要なクラス、すなわち、IgA、IgD、IgE、IgGおよびIgMがあり、このうちいくつかは、さらにサブクラス(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgAおよびIgA2に分けることができる。免疫グロブリンの異なるクラスに対応する重鎖定常ドメイン(Fc)は、各々、α、δ、ε、γおよびμと呼ばれる。免疫グロブリンの異なるクラスのサブユニット構造および三次元コンフィギュレーションは、よく知られている。
【0038】
いずれかの脊椎動物種由来の抗体(免疫グロブリン)の「軽鎖」は、定常ドメインのアミノ酸配列に基づいて、カッパまたは(「κ」)およびラムダまたは(「λ」)と呼ばれる2つの明らかに別個の型のうちの1つに割り当てることができる。
【0039】
本明細書に記載の抗体または融合抗体への言及において、「選択的に結合する」、「選択的に結合」、「特異的に結合する」または「特異的に結合」なる語は、解離定数(Kd)が約10−6M以下、すなわち、10−7、10−8、10−9、10−10、10−11または10−12Mである、抗体または融合抗体のその標的抗原に対する結合をいう。
【0040】
本明細書中で使用される場合、「抗体」なる語は、また、抗原に特異的に結合する能力を保持する抗体の1以上のフラグメントを意味すると理解される(概して、Holligerら、(2005), Nature Biotech., 23(9):1126−1129参照)。かかる抗体の非限定的な例は、(i)Fabフラグメント、VL、VH、CLおよびCH1ドメインからなる1価のフラグメント、(ii)F(ab’)2フラグメント、ヒンジ領域でジスルフィド架橋によって連結された2個のFabフラグメントを含む二価のフラグメント、(iii)VHおよびCH1ドメインからなるFdフラグメント、(iv)抗体の単一アームのVLおよびVHドメインからなるFvフラグメント、(v)VHドメインからなるdAbフラグメント(Wardら、(1989) Nature, 341:544 546)、および(vi)単離された相補性決定領域(CDR)を包含する。さらに、Fvフラグメントの2つのドメイン、VLおよびVHは、別々の遺伝子によってコードされるが、組み換え法を用いて、これらのドメインを、VLおよびVH領域が対になって一価の分子を形成する単一タンパク質鎖とすることが可能な合成リンカーによって連結することができる(一本鎖Fv(scFv)として知られる;例えば、Birdら、(1988) Science, 242:423 426; and Huston et al., (1988) Proc. Natl. Acad. Sci. USA, 85:5879 5883;およびOsbournら、(1998), Nat. Biotechnol., 16:778参照)。かかる一本鎖抗体もまた、「抗体」なる語に包含されることが意図される。特異的scFvのいずれのVHおよびVL配列も、完全IgG分子または他のアイソタイプをコードする発現ベクターを生成するために、ヒト免疫グロブリン定常領域cDNAまたはゲノム配列に連結することができる。VHおよびVLは、また、タンパク質化学または組み換えDNA技術を用いて、Fab、Fvまたは免疫グロブリンの他のフラグメントの生成に使用することができる。一本鎖抗体の他の形態、例えば、ダイアボディもまた包含される。
【0041】
「F(ab’)2」および「Fab’」部分は、免疫グロブリン(モノクローナル抗体)をペプシンおよびパパインなどのプロテアーゼで処理することによって製造でき、2つのH鎖の各々において、ヒンジ領域間に存在するジスルフィド結合付近で免疫グロブリンを消化することによって生成される抗体を包含する。例えば、パパインは、2つのH鎖の各々において、ヒンジ領域間に存在するジスルフィド結合の上流でIgGを切断して、VL(L鎖可変領域)およびCL(L鎖定常領域)からなるL鎖、およびVH(H鎖可変領域)およびCHγ1(H鎖の定常領域におけるγ1領域)からなるH鎖フラグメントがジスルフィド結合を介してそれらのC末端領域で連結した2つの相同性抗体フラグメントを生成する。これらの2つの相同性抗体フラグメントの各々は、Fab’と呼ばれる。また、ペプシンは、2つのH鎖の各々において、ヒンジ領域間に存在するジスルフィド結合の下流でIgGを切断して、2つの上記Fab’がヒンジ領域で連結したフラグメントよりも僅かに大きい抗体フラグメントを生成する。該抗体フラグメントをF(ab’)2と呼ぶ。
【0042】
Fabフラグメントは、また、軽鎖の定常ドメインおよび重鎖の第1の定常ドメイン(CH1)を含有する。Fab’フラグメントは、抗体ヒンジ領域由来の1以上のシステインを含む重鎖CH1ドメインのカルボキシル末端にて2、3個の残基の付加によってFabフラグメントと相違する。Fab’−SHは、定常ドメインのシステイン残基が遊離チオール基を有するFab’のための本明細書中での名称である。F(ab’)2抗体フラグメントは、元々、Fab’フラグメントの対として生産されたフラグメントであり、該フラグメント間にヒンジシステインを有する。抗体フラグメントの他の化学的カップリングもまた、知られている。
【0043】
「Fv」なる語は、完全な抗原認識および抗原結合部位を含有する最小抗体フラグメントである。該領域は、密接な非共有結合において、1つの重鎖および1つの軽鎖可変ドメインの二量体からなる。該コンフィギュレーションにおいて、各可変ドメインの3つの超可変領域が相互作用して、VH−VL二量体の表面における抗原−結合部位を明確に規定する。集合的に、6つの超可変領域が抗原−結合特異性を抗体に付与する。しかしながら、単一の可変ドメイン(または抗原に特異的な3つの超可変領域のみを含むFvの半分)であっても、抗原を認識し、結合する能力を有するが、完全な結合部位よりもアフィニティーは低い。
【0044】
「一本鎖Fv」または「sFv」抗体フラグメントは、抗体のVH、VL、またはVHおよびVLの両方のドメインを含み、ここに、両ドメインは、単一のポリペプチド鎖に存在する。いくつかの具体例において、Fvポリペプチドは、さらに、VHおよびVLドメイン間にポリペプチドリンカーを含み、それにより、該sFvが抗原結合のための所望の構造を形成できるようにする。sFvの概要に関し、例えば、Pluckthun、The Pharmacology of Monoclonal Antibodies, Vol. 113, Rosenburg and Moore eds. Springer−Verlag, New York, pp. 269 315 (1994)を参照のこと。
【0045】
「キメラ」抗体は、異なる哺乳動物起源の組合せから得られる抗体を包含する。該哺乳動物は、例えば、ウサギ、マウス、ラット、ヤギ、またはヒトであってもよい。異なる哺乳動物の組合せは、ヒトおよびマウス起源のフラグメントの組合せを包含する。
【0046】
いくつかの具体例において、本発明の抗体は、モノクローナル抗体(MAb)、典型的にはマウスモノクローナル抗体のヒト化によって誘導される、キメラヒト−マウス抗体である。かかる抗体は、例えば、抗原投与に応答して特異的なヒト抗体を産生するように「操作」されたトランスジェニックマウスから得られる。該技術において、ヒト重鎖および軽鎖上の座にある要素が、内在性重鎖および軽鎖上の座の標的とされた破壊を含有する胚性幹細胞系列から由来するマウス系統中に導入される。該トランスジェニックマウスは、ヒト抗原に特異的なヒト抗体を合成することができ、該マウスは、ヒト抗体を分泌するハイブリドーマを生産するために使用することができる。
【0047】
本明細書中で使用される場合、「治療」または「治療する」なる語は、治療的利益および/または予防的利益を達成することを包含する。治療的利益によって、治療されている原因となる障害または疾患の根絶または改善が意味される。治療の予防的利益には、疾患の危険性を減少すること、疾患の進行を遅らせること、または疾患の発症の可能性を減少することが包含される。本明細書中で使用される場合、「治療する」または「治療」には、予防が包含される。
【0048】
本明細書中で使用される場合、「有効量」は、全身投与される場合、CNSにおいて有益な結果または所望の結果をもたらすのに十分な量であることができる。有効量は、また、予防的効果を生じる量、例えば、急性の病理学的状態または望ましくない状態の出現を遅延させる、減少させる、または排除する量である。有効量は、1以上の投与において投与することができる。治療に関して、本発明の組成物の「有効量」は、障害、例えば、神経学的障害の進行を緩和、改善、安定化、逆転または遅延させるのに十分な量である。「有効量」は、単独または疾患もしくは障害を治療するために使用される1以上の薬剤と共に使用される本発明の組成物のいずれかであってもよい。本発明の意味内で治療剤の「有効量」は、患者の担当医師または獣医師によって決定されるであろう。かかる量は、当業者によって容易に確かめられ、本発明にしたがって投与される場合、治療効果をもたらすであろう。
【0049】
本明細書中で使用される場合、「分子のトロイの木馬」なる語は、BBBを横断して輸送される分子であって、それ自体でBBBを横断しない別の分子に共有または非共有結合した場合、CNS中へのBBB横断輸送(trans-BBB transport)のための渡し船として作用することができる分子をいう。分子のトロイの木馬の例は、限定するものではないが、BBB上に発現された受容体、例えば、インスリン受容体、トランスフェリン受容体、IGF受容体、リポタンパク質受容体、またはレプチン受容体のECDに結合するポリペプチド(例えば、抗体)を包含する。
【0050】
本明細書中で使用される場合、「対象」または「個体」は、動物、例えば、哺乳動物である。いくつかの具体例において、「対象」または「個体」は、ヒトである。いくつかの非限定的な具体例において、対象は、慢性または急性CNS疾患に罹患している。
【0051】
いくつかの具体例において、デコイ受容体ECD−BBB受容体Ab融合抗体を含む薬理学的組成物は、「末梢に投与される」または「末梢投与される」。本明細書中で使用される場合、これらの用語は、CNSに対する直接的投与ではない、すなわち、薬剤を血液脳関門の脳ではない側と接触させる、個体に対する薬剤、例えば、治療剤の投与のいずれかの形態をいう。本明細書中で使用される場合、「末梢投与」は、静脈内、動脈内、皮下、筋内、腹腔内、経皮、吸入によるもの、経頬、鼻腔内、経腸、経口、非経口、舌下、または経鼻を包含する。
【0052】
本明細書において、「医薬上許容される担体」または「医薬上許容される賦形剤」は、それ自体が組成物を受け取る個体に有害な抗体産生を誘導しないいずれかの担体をいう。かかる担体は、当業者に周知である。医薬上許容される担体/賦形剤のディスカッション全体は、Remington’s Pharmaceutical Sciences, Gennaro, AR, ed., 20th edition, 2000: Williams and Wilkins PA, USAに見ることができる。例示的な医薬上許容される担体は、塩、例えば、無機酸塩、例えば、塩酸塩、臭化水素酸塩、リン酸塩、硫酸塩など、および有機酸の塩、例えば、酢酸塩、プロピオン酸塩、マロン酸塩、安息香酸塩等を包含することができる。例えば、本発明の組成物は、液体形態で提供されてもよく、0.01−1%のポリソルベート−80などの界面活性剤、または炭水化物添加剤、例えば、マンニトール、ソルビトールまたはトレハロースを用いて、または用いずに、種々のpH(pH5〜8)のセーラインを基礎とする水性溶液中で処方されてもよい。一般的に使用されるバッファーは、ヒスチジン、酢酸塩、リン酸塩、またはクエン酸塩を包含する。
【0053】
「組み換え宿主細胞」または「宿主細胞」は、挿入方法、例えば、直接的取り込み、形質導入、f−交配、または組み換え宿主細胞を作出するための当該分野で既知の他の方法に関係なく、外因性ポリヌクレオチドを含む細胞をいう。該外因性ポリヌクレオチドは、非組み込みベクター、例えば、プラスミドとして維持されてもよく、または別法では、宿主ゲノム中に組み込まれてもよい。
【0054】
「ポリペプチド」、「ペプチド」および「タンパク質」なる語は、本明細書中で交換可能に使用され、アミノ酸残基のポリマーを示す。すなわち、ポリペプチドに向けられた記載は、等しく、ペプチドの記載およびタンパク質の記載に適用され、その逆も同様である。該用語は、天然のアミノ酸ポリマーならびに1以上のアミノ酸残基が非天然アミノ酸、例えば、アミノ酸類似体であるアミノ酸ポリマーに適用される。本明細書中で使用される場合、該用語は、全長タンパク質(すなわち、抗原)を包含するいずれの長さのアミノ酸鎖も包含し、ここに、該アミノ酸残基は共有ペプチド結合によって連結される。
【0055】
「アミノ酸」なる語は、天然および非天然アミノ酸、ならびに天然アミノ酸と同様に機能するアミノ酸類似体およびアミノ酸模倣物をいう。天然にコードされるアミノ酸は、20個の一般的なアミノ酸(アラニン、アルギニン、アスパラギン、アスパラギン酸、システイン、グルタミン、グルタミン酸、グリシン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、プロリン、セリン、スレオニン、トリプトファン、チロシン、およびバリン)およびピロリジンおよびセレノシステインである。アミノ酸類似体は、天然アミノ酸と同じ基本化学構造、すなわち、水素に結合したα炭素、カルボキシル基、アミノ基、およびR基を有する化合物、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムをいう。かかる類似体は、修飾されたR基(例えば、ノルロイシン)または修飾されたペプチド骨格を有するが、天然アミノ酸と同じ基本化学構造を保持する。
【0056】
アミノ酸は、本明細書において、一般的に知られた3文字記号によって、またはIUPAC−IUB Biochemical Nomenclature Commissionによって推奨される1文字記号によって言及されうる。同様に、ヌクレオチドは、一般的に許容される1文字コードによって言及されうる。
【0057】
「核酸」なる語は、デオキシリボヌクレオチド、デオキシリボヌクレオシド、リボヌクレオシド、またはリボヌクレオチドおよび一本鎖または二本鎖形態のいずれかのそのポリマーをいう。特に限定しない限り、該用語は、参照核酸と類似の結合特性を有し、天然ヌクレオチドと同様に代謝される天然ヌクレオチドの既知の類似体を含有する核酸を包含する。別段に特に限定しない限り、該用語は、また、PNA(ペプチド核酸)、アンチセンス技術において使用されるDNAの類似体(ホスホロチオエート、ホスホロアミデートなど)を包含するオリゴヌクレオチド類似体をいう。別記しない限り、特定の核酸配列は、また、その保存的に修飾された変種(限定するものではないが、縮重コドン置換を包含する)および相補性配列ならびに明示された配列を暗示的に包含する。詳細には、縮重コドン置換は、1以上の選択された(または全ての)コドンの3番目の位置を混合塩基および/またはデオキシイノシン残基で置換された配列を生成することによって達成されうる(Batzerら(1991), Nucleic Acid Res., 19:5081; Ohtsukaら(1985), J. Biol. Chem., 260:2605−2608;およびRossoliniら(1994), Mol. Cell. Probes 8:91−98)。
【0058】
「単離」および「精製」なる語は、その天然環境から実質的または本質的に除去された、またはその天然環境において濃縮された物質をいう。例えば、単離された核酸は、通常それと隣接する核酸から分離させたもの、または試料中の他の核酸または成分(タンパク質、脂質など)から分離させたものであってもよい。別の例では、ポリペプチドがその天然環境から実質的に除去された、またはその天然環境において濃縮された場合、それは精製されている。核酸およびタンパク質の精製および単離方法は、当該分野で周知である。
【0059】
「BBB受容体Ab」なる語は、血液脳関門上に発現された受容体の細胞外ドメインに対する抗体をいう。BBB受容体の非限定的な例は、インスリン受容体(例えば、ヒトインスリン受容体)、トランスフェリン受容体、リポタンパク質受容体、およびレプチン受容体を包含する。
【0060】
III.血液脳関門
BBBは、脳および脊髄の毛細管を形成する内皮細胞を一緒に固める密着な結合によって形成される(Pardridge, 2008, Bioconj Chem, 19: 1327−1338)。ヒトの脳において、400マイルの脳毛細管が存在する。BBBをイン・ビボで形成する脳毛細管内皮原形質膜を横断する電気抵抗は、いずれの生物膜と同様に高い。血液と器官との間の遊離溶質交換のための通常の細胞間隙および細胞間経路は、CNSには存在しない。結果的に、血液中の薬物は、2つのメカニズム、すなわち、(i)小型分子の高い脂溶性のための自由拡散、および(ii)内因性BBBトランスポーターを介する輸送のうちの1つだけを介して脳にアクセスすることができる。デコイ受容体組成物のためのBBB薬物送達問題の解決に対する1のアプローチは、BBB内のある特定の内因性輸送システムにアクセスできるように、デコイ受容体を再操作することである。本明細書に記載の方法は、本明細書に記載のデコイ受容体融合抗体組成物の全身投与後、機能的デコイ受容体が末梢血からCNS中へBBBを横断できるようにする。本明細書中に記載の方法は、BBB上の表面に発現された発現受容体(例えば、ヒトインスリン受容体)を利用して、所望の二官能性受容体ECD−BBB受容体融合抗体を末梢血からCNS中へ輸送する。
【0061】
IV.BBBを横断する輸送のためのデコイ受容体融合抗体
一の態様において、本発明は、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列に共有結合した受容体ECD(例えば、ヒトTNF−α受容体ECD)のアミノ酸配列を含有する二官能性デコイ受容体融合抗体を利用する組成物および方法を提供する。かかる二官能性デコイ受容体融合抗体は、BBB上に発現された受容体および該受容体ECDのリガンドに結合することができる。該組成物および方法は、BBBを横断して末梢血からCNS中へデコイ受容体を輸送するのに有用である。
【0062】
BBBは、いくつかの巨大分子の血液から脳への輸送を可能にする特定の受容体を有することが示された(これらのトランスポーターは、本発明の組成物のトランスポーターとして適当である)。本発明に有用な内因性BBB受容体介在性輸送システムは、限定するものではないが、インスリン、トランスフェリン、インスリン様生長因子1および2(IGF1およびIGF2)、レプチン、およびリポタンパク質を輸送するものを包含する。いくつかの具体例において、本発明は、内因性インスリンBBB受容体介在性輸送システム、例えば、ヒト内因性インスリンBBB受容体介在性輸送システムを介してBBBを横断することができる抗体を利用する。いくつかの具体例において、該二官能性デコイ受容体融合抗体は、HIR抗体を含む。本明細書に記載のデコイ受容体−HIRAb融合抗体は、ヒトインスリン受容体のECDに結合する。いくつかの具体例において、該デコイ受容体ECDは、重鎖免疫グロブリン(例えば、HIRAb重鎖免疫グロブリン)のカルボキシル末端に融合する。他の具体例において、該デコイ受容体ECDは、軽鎖免疫グロブリン(例えば、HIRAb軽鎖免疫グロブリン)のC末端に融合する。いくつかの場合、上記の受容体ECDは、ヒト、マウス、ラットまたはブタサイトカイン受容体、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエピフリン受容体由来のECDに対して、少なくとも80%(例えば、85%、90%、95%、97%、99%、または少なくとも80%〜100%のうちの別のパーセント)同一のアミノ酸配列を含有する。いくつかの具体例において、上記の受容体ECDは、サイトカイン受容体由来である。いくつかの具体例において、受容体ECDは、TNF−α受容体ドメイン(例えば、ヒトTNF−α受容体ECD)を含有する。上記の受容体のアミノ酸配列についてGenBankアクセッション番号を下記の表1に提供する。
【0063】
【表1】

【0064】
インスリン受容体およびその細胞外インスリン結合ドメイン(ECD)は、構造的および機能的に当該分野において広範囲に特徴付けられている。例えば、Yipら(2003), J. Biol. Chem., 278(30):27329−27332;およびWhittakerら(2005), J. Biol. Chem., 280(22):20932−20936参照のこと。ヒトインスリン受容体のアミノ酸およびヌクレオチド配列は、GenBankアクセッション番号NM 000208の下に見出すことができる。
【0065】
BBB上に発現されたインスリン受容体は、それにより、BBBを横断するデコイ受容体、例えば、TNF−α受容体ECDの輸送のためのベクターとして作用することができる。ある特定のインスリン受容体ECD特異的抗体は、内因性リガンドを模倣していてもよく、それにより、特異的受容体システム上の輸送を介して、原形質膜障壁を横断しうる。ある特定の具体例において、受容体デコイ−HIRAb融合抗体は、ヒトBBB HIR上の外表面エピトープを結合し、該結合により、ヒトBBBインスリン受容体が介在する輸送反応を介して、該融合抗体のBBBの横断を可能にする。
【0066】
ヒトにおける使用のために、キメラHIRAbは、ヒトへの投与時に有意に免疫原性ではない十分なヒト配列、例えば、約80%ヒトおよび約20%マウス、または約85%ヒトおよび約15%マウス、または約90%ヒトおよび約10%マウス、または約95%ヒトおよび約5%マウス、または約95%より多くのヒトおよび約5%未満のマウス配列を含有することが好ましい。本発明における使用のために、十分なヒト配列を有するヒトBBBインスリン受容体に対するキメラ抗体が、例えば、Boadoら(2007), Biotechnol Bioeng, 96(2):381−391に記載される。また、HIRMAbのより高度にヒト化した形態を操作することもでき、該ヒト化HIR Abは、ネズミHIR Abに匹敵する活性を有し、本発明の具体例において使用することができる。例えば、米国特許出願公開第20040101904号(2002年11月27日出願)および第20050142141号(2005年2月17日出願)を参照のこと。
【0067】
例示的具体例において、二官能性デコイ受容体融合抗体は、ヒトインスリン受容体に対する結合について、配列番号4および6、または配列番号6および7のアミノ酸配列を含有する二官能性デコイ受容体融合抗体と競合する。いくつかの場合、該競合する二官能性デコイ受容体は、配列番号4および6、または配列番号6および7のアミノ酸配列を含有する機能的デコイ受容体融合抗体に対して、少なくとも約20%、例えば、25%、30%、35%、40%、45%、50%、60%、65%、70%、75%、80%、85%、90%、または少なくとも約20%〜100%のうちの別のパーセント同一のヒトインスリン受容体に関するアフィニティーを有する。他の具体例において、該二官能性デコイ受容体融合抗体は、配列番号4および6、または配列番号6および7に対して、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一のアミノ酸配列を含有する。
【0068】
本発明で使用されるBBB受容体抗体は、グリコシル化されていてもよく、またはグリコシル化されていなくてもよい。該抗体がグリコシル化されている場合、該抗体の機能に有意に影響を及ぼさないいずれかのグリコシル化パターンが用いられうる。グリコシル化は、抗体が産生される細胞の典型的なパターンで生じることができ、細胞型によって変化しうる。例えば、マウスミエローマ細胞によって産生されたモノクローナル抗体のグリコシル化パターンは、トランスフェクトされたチャイニーズハムスター卵巣(CHO)細胞によって産生されたモノクローナル抗体のグリコシル化パターンとは相違することができる。いくつかの具体例において、該抗体は、トランスフェクトされたチャイニーズハムスター卵巣(CHO)細胞によって産生されるパターンでグリコシル化される。
【0069】
現行の技術により、BBB受容体Absまたはデコイ受容体ECDの大量の配列変種を容易に生成し(例えば、イン・ビトロで)、ヒトインスリン受容体のECDなどの標的抗原への結合について、またはデコイ受容体ECDのリガンドへの結合についてスクリーンすることが可能になることは、当業者に明らかであろう。例えば、抗体配列変種のウルトラハイスループットスクリーニングの一例として、Fukudaら(2006), Nuc. Acid Res., 34(19)(オンラインで公開)を参照のこと。また、Chenら、(1999), Prot Eng, 12(4): 349−356を参照のこと。配列変種を単離するために、全配列または特定のドメインに対応する特定のサブ配列の無作為変異誘発を行ってもよい。別法では、BBB受容体またはデコイ受容体リガンド結合に不可欠であることが知られている残基の変異を回避しつつ、部位特異的変異誘発を繰り返し実施することができる。例えば、Mukaiら(2009), J Mol Biol, 385(4):1221−1229に記載されているように、例えば、TNF−α受容体の構造機能が当該分野で知られている。さらに、デコイ受容体ECD配列の複数の変種、例えば、ヒトTNF−α受容体の変種の生成において、変異認容予測(mutation tolerance prediction)プログラムを用いて、厳密な無作為変異誘発によって生じるであろう非機能的配列変種の数を大幅に減らすことができる。タンパク質配列におけるアミノ酸置換のタンパク質機能に対する影響を予測するための種々のプラグラム(例えば、SIFT、PolyPhen、PANTHER PSEC、PMUT、およびTopoSNP)が例えば、Henikoffら(2006), Annu. Rev. Genomics Hum. Genet., 7:61−80に記載されている。したがって、上記のように、当該分野でルーチンな方法によって、デコイ受容体(例えば、ヒトTNF−α受容体ECD配列)配列変種の非常に多様な「ライブラリー」を作製し、スクリーニングすることによって、非常に大量の操作可能なデコイ受容体ECD配列変種を得ることができることは、当業者に明らかであろう。
【0070】
パーセント配列同一性は、常法によって決定される。例えば、Altschulら(1986), Bull. Math. Bio., 48:603、およびHenikoffおよびHenikoff,(1992), Proc. Natl. Acad. Sci. USA, 89:10915を参照のこと。簡単に言えば、ギャップオープニングペナルティ10、ギャップエクステンションペナルティ1、およびHenikoffおよびHenikoff(上掲)の「BLOSUM62」スコアリングマトリックスを用いて、2つのアミノ酸配列をアラインメントスコアが最適になるようにアラインする。次いで、パーセント同一性は、{(完全に一致した全数)/[(より長い配列の長さ)+(2つの配列をアラインするためにより長い配列中に導入したギャップの数)]}(100)として計算される。
【0071】
2つのアミノ酸配列をアラインするために利用可能な多くの確立されたアルゴリズムが存在することは、当業者に明らかである。PearsonおよびLipmanの「FASTA」類似性サーチアルゴリズムは、本明細書中に開示されるアミノ酸配列および別のペプチドのアミノ酸配列によって共有される同一性のレベルを調べるための適当なタンパク質アラインメント法である。FASTAアルゴリズムは、Pearsonら(1988), Proc. Nat’l Acad. Sci. USA, 85:2444、およびPearson(1990), Meth. Enzymol. 183:63によって記載される。簡単に言えば、FASTAは、まず、保存的アミノ酸置換、挿入または欠失を考慮せずに、クエリー配列(例えば、配列番号4または配列番号6)および最大密度の同一性(ktup変数が1の場合)または同一性対(ktup=2の場合)のいずれかを有する試験配列によって共有される領域を同定することによって、配列類似性を特徴付ける。次いで、最大密度の同一性を有する10個の領域を、アミノ酸置換マトリックスを用いて、全ての対になったアミノ酸の類似性を比較することによって再スコア化し、該領域の末端は、最大スコアに寄与する残基のみを含むように「トリミング」する。「カットオフ」値(配列の長さおよびktup値に基づく所定の式によって計算される)よりも大きいスコアを有するいくつかの領域が存在する場合、トリミングされた最初の領域を、ギャップを有する近似のアラインメントを形成するように該領域を結合できるか否か決定するために試験する。最終的に、2つのアミノ酸配列の最大スコア領域を、アミノ酸挿入および欠失を可能にするNeedleman−Wunsch−Sellersアルゴリズム(Needlemanら(1970), J. Mol. Biol. 48:444; Sellers (1974), SIAM J. Appl. Math., 26:787)の修飾を用いてアラインする。FASTA分析の実例的パラメーターは、ktup=1、ギャップオープニングペナルティ=10、ギャップエクステンションペナルティ=1、および置換マトリックス=BLOSUM62である。これらのパラメーターは、Pearson, (1990), Meth. Enzymol., 183:63のAppendix 2において説明されるように、スコアリングマトリックスファイル(「SMATRIX」)を修飾することによって、FASTAプログラム中に導入することができる。
【0072】
本発明は、また、本明細書中に開示されるアミノ酸配列と比較して、保存的アミノ酸変化を有するタンパク質を包含する。一般的なアミノ酸のなかでも、例えば、「保存的アミノ酸置換」は、下記の各群内でのアミノ酸間の置換によって説明される。(1)グリシン、アラニン、バリン、ロイシンおよびイソロイシン、(2)フェニルアラニン、チロシン、およびトリプトファン、(3)セリンおよびスレオニン、(4)アスパラギン酸およびグルタミン酸、(5)グルタミンおよびアスパラギン、および(6)リジン、アルギニンおよびヒスチジン。BLOSUM62表は、タンパク質配列セグメントの約2,000の局所的な複数のアラインメントに由来するアミノ酸置換マトリックスであり、500以上のグループの関連タンパク質の非常に保存された領域を示す(Henikoffら(1992), Proc. Nat’l Acad. Sci., USA, 89:10915)。したがって、BLOSUM62置換頻度を用いて、本発明のアミノ酸配列中に導入されうる保存的アミノ酸置換を規定することができる。化学的特性のみに基づいて(上記のように)アミノ酸置換を設計することが可能であるが、「保存的アミノ酸置換」なる語は、好ましくは、−1よりも大きいBLOSUM62値によって表される置換をいう。例えば、アミノ酸置換は、置換が0、1、2または3のBLOSUM62値によって特徴付けられるが場合、保存的である。該システムによると、好ましい保存的アミノ酸置換は、少なくとも1(例えば、1、2または3)のBLOSUM62値によって特徴付けられるが、より好ましい保存的アミノ酸置換は、少なくとも2(例えば、2または3)のBLOSUM62値によって特徴付けられる。
【0073】
また、アミノ酸配列は、該配列が本発明の組成物および方法において機能的であるのに十分な生物学的タンパク質活性を保持する限り、付加的な残基、例えば、付加的なN−またはC−末端アミノ酸を含んでいてもよく、依然として実質的に、本明細書中に開示される配列の1つにおいて示されるとおりであることは、理解されよう。
【0074】
V.組成物
顕著なことには、本明細書に記載の二官能性デコイ受容体融合抗体は、高い割合で、それらの分離した構成タンパク質の活性、すなわち、BBB受容体AbのBBB受容体、例えば、ヒトインスリン受容体ECDに対する結合、およびデコイ受容体ECD(例えば、TNF−α受容体)の同族(cognate)リガンドに対する結合を保持する。
【0075】
本明細書には、デコイ受容体ECDに融合したBBBを横断することができるBBB受容体Abを含有する二官能性デコイ受容体−BBB受容体融合抗体であって、ここに、該血液脳関門を横断できるBBB受容体Abおよび該受容体ECDは各々、別々に存在した場合のそれらの活性と比較して、それらの活性の少なくとも約5、10、15、18、20、25、30、35、40、40、45または50%を保持する該融合抗体が記載される。BBB受容体Abの例は、インスリン受容体(例えば、ヒトインスリン受容体)、トランスフェリン受容体、およびリポタンパク質受容体に対するAbs(例えば、モノクローナルAbs)を包含する。適当な受容体ECDの例は、限定するものではないが、ヒト、マウス、ラット、またはブタサイトカイン受容体、TNF−α受容体、TNF−関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体に対して、少なくとも80%(例えば、85%、90%、95%、97%、99%または少なくとも80%〜100%のうちの別のパーセント)同一のアミノ酸配列を含有するものを包含する。
【0076】
いくつかの具体例において、BBB受容体Abは、ヒトインスリン受容体(HIR)のECDに対する抗体である。本明細書に記載の二官能性デコイ受容体−BBB受容体Ab融合抗体において、BBB受容体抗体および受容体ECD間の共有結合は、該結合が該デコイ受容体ECD−BBB受容体Ab融合抗体の該BBB受容体のECDに対する結合および血液脳関門の横断を可能にし、かつ、該デコイ受容体ECDの活性の治療上有用な部分の保持、例えば、該活性の少なくとも10%、20%、30%、40%、50%、60%、70%、80%、または約10%〜100%のうちの別のパーセントの保持を可能にする限り、BBB受容体抗体重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシまたはアミノ末端にあってもよい。ある特定の具体例において、共有結合は、該抗体のHCおよび該デコイ受容体ECD間にある。他の具体例において、共有結合は、該抗体のLCおよび該デコイ受容体ECD間にある。いずれの適当な結合を用いてもよく、例えば、デコイ受容体ECDのアミノ末端に対して軽鎖のカルボキシル末端、デコイ受容体ECDのアミノ末端に対して重鎖のカルボキシル末端、またはデコイ受容体ECDのカルボキシル末端に対して軽鎖のアミノ末端を用いてもよい。好ましい具体例において、該結合は、HCのカルボキシル末端からデコイ受容体ECDのアミノ末端へのものである。いくつかの具体例において、該融合抗体組成物は、そのN末端を介してヒトインスリン受容体抗体の重鎖のC末端に共有結合したヒトTNF−α受容体ECDを含む。
【0077】
末端アミノ酸間の結合が融合アミノ酸配列の一部を形成する介在性ペプチドリンカー配列によって達成できることは、明らかであろう。該ペプチド配列リンカーは、1、2、3、4、5、6、7、8、9、10または10以上のアミノ酸長であってもよい。いくつかの具体例において、2−アミノ酸リンカーを用いる。いくつかの具体例において、該リンカーは、配列ser−serを有する。該ペプチドリンカー配列は、プロテアーゼ切断部位を含みうるが、これは、デコイ受容体ECDの活性の必要条件ではない。実際、本発明のこれらの具体例の利点は、該二官能性デコイ受容体−BBB受容体抗体融合抗体が切断を伴わずに、一旦BBBを横断すれば、輸送および活性の両方に関して部分的または完全に活性であることである。図10は、デコイ受容体ECD−BBB受容体抗体融合抗体のアミノ酸配列の例示的具体例を示し、それは、HCがそのカルボキシル末端によって、3アミノ酸「ser−ser−ser」リンカーを介して、該TNF−α受容体ECDのアミノ末端に融合しているヒトTNF−α受容体ECD−HIR抗体融合抗体(配列番号4)である。
【0078】
いくつかの具体例において、デコイ受容体−BBB受容体Ab融合抗体は、HCおよびLCの両方を含む。いくつかの具体例において、該デコイ受容体−BBB受容体Ab融合個体は一価抗体である。他の具体例において、該デコイ受容体−BBB受容体Ab融合抗体は、実施例セクションにいおいて本明細書中で記載されるような二価抗体である。
【0079】
該デコイ−BBB受容体Ab融合抗体の一部として用いられるBBB受容体Abは、グリコシル化または非グリコシル化することができる。いくつかの具体例において、該抗体は、例えば、CHO細胞におけるその合成によって生じるグリコシル化パターンにおいて、グリコシル化される。
【0080】
本明細書中で使用される場合、「活性」は、生理学的活性(例えば、BBB横断能および/または治療活性)、BBB受容体Abのその標的BBB受容体に関する結合アフィニティー、またはデコイ受容体の同族リガンド(例えば、TNF−αデコイ受容体の場合、TNF)に関するアフィニティーを包含する。
デコイ受容体−BBB受容体Ab融合抗体のBBBを横断する輸送は、標準的な方法によってBBB受容体Abを単独でBBBを横断して輸送する場合と比較してもよい。例えば、動物モデル、例えば、非ヒト霊長類のような哺乳動物によるデコイ受容体−BBB受容体Ab融合抗体の薬物動力学および脳への取り込みを用いてもよい。同様に、デコイ受容体リガンド結合を決定するための標準的なモデル(例えば、ELISA)を用いて、デコイ受容体ECD単独の機能およびデコイ受容体−BBB受容体Ab融合抗体の一部としての機能を比較してもよい。例えば、TNFαのHIRMAb−TNF−デコイ受容体融合タンパク質に対する結合 対 TNF受容体(TNFR)−II:ヒトIgG1 Fc融合タンパク質(TNFR:Fcと称する)に対する結合を明らかにする実施例4を参照のこと。BBB受容体に関する結合アフィニティーをまた、デコイ受容体−BBB受容体Ab融合抗体 対 BBB受容体Ab単独に関して比較することができる。例えば、実施例4を参照のこと。
【0081】
いくつかの場合、デコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)の脳への取り込みは、タンパク質100グラムあたり1%、2%、3%、5%、7%または10%ID以上である。デコイ受容体−BBB受容体Ab融合抗体のBBBを横断する輸送は、また、デコイ受容体およびヒトIgGのFcフラグメントの融合タンパク質、例えば、TNFR:FcのBBBを横断する輸送と比較してもよい。デコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)の脳への取り込みは、デコイ受容体およびヒトIgGのFcフラグメントの融合タンパク質、例えば、TNFR:Fcの脳への取り込みより1、2、5、10、13、15、17、20、25、30、35、40、45、または50倍以上大きくてもよい。脳および他の器官について、器官クリアランス定数(本明細書中では、透過係数表面積(PS)積という)は、本明細書に記載の融合タンパク質について計算されうる。本明細書に記載のデコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)は、他の器官と比較した場合、脳において選択的に豊富に存在しうる。例えば、TNFR:Fc融合タンパク質に関する器官PS積に対する、本明細書に記載のデコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)に関する器官PS積の比率を複数の器官について測定した場合(例えば、図14参照)、脳での比率は、他の器官、例えば、脂肪、筋肉、心臓、肺、肝臓または脾臓の比率より1、2、5、10、13、15、17、20、25、30、35、40、45、または50倍以上大きくてもよい。
【0082】
また、本明細書中には、本明細書に記載の1以上のデコイ受容体−BBB受容体Ab融合抗体、および医薬上許容される賦形剤を含有する医薬組成物が包含される。医薬上許容される担体/賦形剤の全体的な議論は、Remington’s Pharmaceutical Sciences, Gennaro, AR, ed., 20th edition, 2000: Williams and Wilkins PA, USAに見出すことができる。本発明の医薬組成物は、静脈内、皮下、筋内、腹腔内注射、経口、経腸、経頬、経皮、鼻腔内、またはいずれか他の末梢投与に適当な経路を包含するいずれかの末梢経路を介する投与に適当な組成物を包含する。
【0083】
本発明の組成物は、例えば、静脈内、皮下、筋内、または腹腔内投与のための医薬組成物として、特に注射に適する。本発明の水性組成物は、本発明の組成物の有効量を含み、医薬上許容される担体または水性媒体中に溶解または分散させてもよい。「医薬上」または「薬理学上許容される」なる語は、適宜、動物、例えば、ヒトに投与時に、有害な反応、アレルギー性反応または他の都合の悪い反応を生じない分子および組成物をいう。本明細書中で使用される場合、「医薬上許容される担体」は、いずれかおよび全ての溶媒、分散媒体、コーティング、抗菌および抗真菌剤、等張剤および吸収遅延剤などを包含する。医薬上活性な物質のためのかかる媒体および剤の使用は、当該分野で周知である。いずれかの通常の媒体または剤が活性成分と混合できない場合を除き、治療組成物におけるその使用が意図される。補足的活性成分もまた、該組成物中に配合することができる。
【0084】
注射組成物のための例示的医薬上許容される担体は、カルシウム塩、例えば、塩化カルシウム、臭化カルシウム、硫化カルシウムなど、および有機酸の塩、例えば、酢酸塩、プロピオン酸塩、マロン酸塩、安息香酸塩などを包含することができる。例えば、本発明の組成物は、液体形態で提供されてもよく、0.01−1%のポリソルベート−80などの界面活性剤、または炭水化物添加剤、例えば、マンニトール、ソルビトールまたはトレハロースを用いて、または用いずに、種々のpH(pH5〜8)のセーラインを基礎とする水性溶液中で処方されてもよい。一般的に使用されるバッファーは、ヒスチジン、酢酸塩、リン酸塩、またはクエン酸塩を包含する。通常の保管および使用条件下、これらの調製物は、微生物の増殖を防止するための保存剤を含有することができる。微生物の作用の防止は、種々の抗菌剤および抗真菌剤、例えば、パラベン、クロロブタノール、フェノール、ソルビン酸、チメロサールなどによってもたらされることができる。多くの場合、等張剤、例えば、糖類または塩化ナトリウムを含むことが好ましい。注射組成物の吸収延長は、該組成物における吸収を遅延させる剤、例えば、モノステアリン酸アルミニウム、およびゼラチンの使用によってもたらされることができる。
【0085】
ヒト投与の場合、調製物は、FDAおよび他の規制当局標準によって要求される滅菌性、発熱性、一般的な安全性および純度標準を満たす。活性化合物は、一般に、非経口投与のために処方され、例えば、静脈内、筋内、皮下、病巣内または腹腔内経路を介する注射のために処方される。活性成分を含有する水性組成物の調製は、本明細書の開示を考慮して、当業者に既知であろう。典型的には、かかる組成物は、液体溶液または懸濁液のいずれかの注射剤として調製でき、注射前に、液体の添加による溶液または懸濁液の調製において使用するのに適当な固体形態も調製することができ、該調製物は乳化されることもできる。
【0086】
滅菌注射溶液は、必要量の活性化合物を上記した種々の他の成分と共に、適当な溶媒中に配合し、次いで、フィルター滅菌することによって調製される。一般に、分散液は、種々の滅菌活性成分を、基礎分散媒体および上記した必要な他の成分を含有する滅菌ビヒクル中に配合することによって調製される。滅菌注射溶液の調製のための滅菌粉末の場合、調製方法は、予めフィルター滅菌した溶液から活性成分およびいずれかの付加的な所望の成分の粉末を生じる真空乾燥および凍結乾燥技術を包含する。
【0087】
処方において、溶液は、投与処方に適合する方法で、かつ、本明細書に記載の基準に基づいて治療上有効量で、全身投与される。該処方は、種々の投与形態で、例えば、上記の注射溶液の形で容易に投与されるが、薬物放出カプセルなども使用できる。
【0088】
投与されるべき医薬組成物の適当な量、治療の回数、および単位投与量は、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体のCNS取り込み特徴によって、および治療されるべき対象および対象の病態によって変化する。いずれの事象においても、投与責任者が個々の対象の適当な投与量を決定する。
【0089】
静脈内または筋内注射などの非経口投与のために処方された化合物の他に、限定するものではないが、皮内投与(米国特許第5,997,501号、第5,848,991号、および第5,527,288号参照)、肺投与(米国特許第6,361,760号、第6,060,069号、および第6,041,775号参照)、頬投与(米国特許第6,375,975号および第6,284,262号参照)、経皮投与(米国特許第6,348,210号および第6,322,808号参照)および経粘膜投与(米国特許第5,656,284参照)を包含する本発明の他の投与方法を用いてもよい。かかる投与方法は、当該分野で周知である。また、例えば、点鼻溶液またはスプレー、エーロゾルまたは吸入薬と共に、本発明の鼻腔内投与を使用してもよい。点鼻溶液は、通常、滴またはスプレーで鼻道に投与されるように設計された水性溶液である。点鼻溶液は、多くの点で、鼻汁に類似するように調製される。かくして、水性点鼻溶液は、通常、等張性であり、pH5.5〜6.5を維持するようにわずかに緩衝化される。さらに、必要に応じて、眼科用調製物において使用されるものと同様な抗微生物保存剤および適当な安定化剤を処方中に含ませてもよい。種々の市販の点鼻調製物が知られており、例えば、抗生物質および抗ヒスタミン剤を包含し、喘息予防に使用される。
【0090】
他の投与様式に適当なさらなる処方は、座剤およびペッサリーを包含する。直腸ペッサリーまたは座剤もまた、使用してもよい。座剤は、通常、薬物を加えられた、直腸または尿道に挿入するための種々の重量および形状の固体投与形態である。挿入後、腔液中で座剤が軟化、溶融または溶解する。座剤の場合、伝統的な結合剤および担体は、例えば、ポリアルキレングリコールまたはトリグリセリドを包含し、かかる座剤は、活性成分を適当な範囲、例えば、0.5%〜10%、好ましくは1%−2%で含有する混合物から形成されうる。
【0091】
経口処方は、例えば、製薬等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、ナトリウムサッカリン、セルロース、炭酸マグネシウムなどのような通常使用される賦形剤を含む。これらの組成物は、溶液、懸濁液、錠剤、丸薬、カプセル、徐放性処方、または粉末の形態を取る。ある規定された具体例において、経口医薬組成物は、不活性希釈剤、または同化可能な食用担体を含み、または硬もしくは軟外皮ゼラチンカプセル中に被包されてもよく、または打錠されてもよく、または食事の食物と直接混合してもよい。経口治療投与の場合、活性化合物は賦形剤と混合してもよく、摂取可能な錠剤、バッカル錠、トローチ、カプセル、エリキシル、懸濁、シロップ、ウウェハーなどの形態で使用されうる。かかる組成物および調製物は、少なくとも0.1%の活性化合物を含有することができる。組成物および調製物のパーセンテージは、もちろん、変動してもよく、通常、単位重量の約2〜75%または約25−60%でありうる。かかる治療上有用な組成物中における活性化合物の量は、適当な投与量が得られるような量である。
【0092】
錠剤、トローチ、丸薬、カプセルなどは、また、結合剤、例えば、トラガカントゴム、アラビアゴム、トウモロコシデンプン、またはゼラチン、賦形剤、例えば、リン酸二カルシウム、崩壊剤、例えば、トウモロコシデンプン、ジャガイモデンプン、アルギン酸など、滑沢剤、例えば、ステアリン酸マグネシウム、および甘味剤、例えば、シュークロース、ラクトースまたはサッカリン、またはフレーバー剤、例えば、ペパーミント、冬緑油、またはチェリーフレーバーを含有していてもよい。投与単位形態がカプセルである場合、上記のタイプの材料のほかに、液体担体を含有していてもよい。種々の他の材料がコーティングとして、または投与単位の物理的形態を修飾するために存在していてもよい。例えば、錠剤、丸薬またはカプセルは、シェラック、糖またはその両方で被覆されていてもよい。エリキシルのシロップは、活性化合物、甘味剤としてシュークロース、保存剤としてメチレンおよびプロピルパラベン、色素およびフレーバー、例えば、チェリーまたはオレンジフレーバーを含有していてもよい。いくつかの具体例において、経口医薬組成物は、活性成分を胃環境から保護するために、腸溶性被覆されていてもよく、腸溶性被覆方法および処方は当該分野で周知である。
【0093】
VI.核酸、ベクター、細胞および製造
本発明は、また、核酸、ベクター、細胞、および製造方法を提供する。いくつかの具体例において、本発明は、本明細書に記載のポリペプチドをコードする核酸、例えば、(i)重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列、(ii)軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列、または(iii)(i)または(ii)の相補性配列を含む核酸を提供する。該上記の核酸によってコードされる重鎖および軽鎖免疫グロブリンは、BBB受容体、例えば、ヒトインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体に対する抗体由来である。いくつかの場合、該コードされる受容体ECDは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である。いくつかの場合、該コードされる受容体のアミノ酸配列は、外来受容体の一つ由来のECDのアミノ酸配列に対し、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一である。いくつかの場合、該核酸は、ヒト、マウス、ラット、またはブタTNF−α受容体の細胞外ドメインに対し、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一のアミノ酸配列を含むECDをコードする。いくつかの具体例において、第1の配列は、配列番号4または7に相当するアミノ酸配列に対し、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一のアミノ酸配列をコードする。他の具体例において、第2の配列は、配列番号6に相当するアミノ酸配列に対し、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一のアミノ酸配列をコードする。
【0094】
本明細書中に提供される核酸は、いくつかの場合、さらに、MAbの重鎖および受容体ECD間のペプチドリンカーをコードする核酸配列を含有することができる。いくつかの具体例において、該リンカーは、S−S−Mである。他の具体例において、該リンカーは、S−Sである。別の具体例において、該リンカーは、S−S−Sである。該核酸は、さらに、シグナルペプチドをコードする核酸配列を含有していてもよく、ここに、該シグナルペプチドは、該重鎖に連結される。当該分野で既知の、または続いて開発されたいずれかの適当なシグナル配列が使用されうる。いくつかの具体例において、重鎖免疫グロブリンに結合したシグナルペプチドは、配列番号4のアミノ酸1−19と約60、70、80、90、95、99、または100%同一の配列を含む。いくつかの具体例において、該核酸は、他のシグナルペプチドをコードする核酸配列を含有し、ここに、該他のシグナルペプチドは該軽鎖に連結される。該軽鎖に連結したシグナルペプチドは、配列番号6のアミノ酸1−20と約60、70、80、90、95、99、または100%同一の配列を含むことができる。
【0095】
本発明は、また、核酸ベクターを提供する。該ベクターは、本明細書に記載の核酸配列のいずれかを含有することができる。いくつかの具体例において、該ベクターは、(i)BBB受容体に対する抗体由来の重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列(またはその相補体)、または(ii)BBB受容体に対する抗体由来の軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列(またはその相補体)を含む。いくつかの具体例において、本発明は、核酸配列を提供し、いくつかの具体例において、本発明は、特定のヌクレオチド配列と少なくとも約60、70、80、90、95、99または100%同一の核酸配列を提供する。例えば、いくつかの具体例において、本発明は、配列番号3または5に対し、少なくとも約60、70、80、90、95、99または100%同一の配列を含有する核酸を提供する。
【0096】
いくつかの具体例において、本発明は、(i)BBB受容体に対する抗体由来の重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列(またはその相補体)、および(ii)BBB受容体に対する抗体由来の軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列(またはその相補体)の両方を含有する単一タンデム発現ベクター(全てが一片の核酸、例えば、一片のDNA中に組み込まれている)(本明細書中では、「タンデムベクター」という)を提供する。該単一タンデムベクターは、また、一以上の選択および/または増幅遺伝子、例えば、DHFR、ネオマイシンホスホトランスフェラーゼ、ハイブロマイシンホスホトランスフェラーゼ、またはピューロマイシンN−アセチルトランスフェラーゼを含むことができる。いくつかの具体例において、該コードされる選択マーカーは、DHFRである。いくつかの具体例において、該タンデムベクターは、DHFRおよび第2の選択/増幅マーカー(例えば、ネオマイシンホスホトランスフェラーゼ)をコードする。本発明の例示的なタンデムベクターの作製方法は、実施例に提供される。しかしながら、該ベクターを構築するために、当該分野で既知のいずれの適当な技術を用いてもよい。
【0097】
単一タンデムベクターの使用は、いくつかの利益を有する。免疫グロブリンG(IgG)遺伝子での真核細胞系統のトランスフェクションは、一般的に、該細胞系統と、IgGを含む重鎖(HC)および軽鎖(LC)をコードする別のプラスミドとの共トランスフェクションを含む。IgG融合タンパク質の場合、該組み換え治療タンパク質をコードする遺伝子は、HCまたはLC遺伝子のいずれかに融合していてもよい。しかしながら、該共トランスフェクションアプローチは、HCおよびLC−融合遺伝子、またはHC−融合およびLC遺伝子を等しく高い割合で組み込んだ細胞系統の選択を困難にする。本発明のある特定の具体例において利用される融合タンパク質の製造に対するアプローチは、DNAの一本鎖上に、HC−融合タンパク質遺伝子、LC遺伝子、選択遺伝子、例えば、neo、および増幅遺伝子、例えば、ジヒドロ葉酸還元酵素遺伝子を包含する必要とする遺伝子の全てを含有する単一プラスミドDNAで永続的にトランスフェクトされた細胞系統の生産である。図9における融合タンパク質タンデムベクターのダイアグラムに示されるように、HC−デコイ受容体融合遺伝子、LC遺伝子、neo遺伝子、およびDHFR遺伝子は全て、別々であるがタンデムなプロモーター、および別々であるがタンデムな転写終止配列の調節下にある。したがって、治療タンパク質とHCまたはLC IgG遺伝子のいずれかとの融合遺伝子を包含する全ての遺伝子は、宿主細胞ゲノム中に等しく組み込まれる。
【0098】
本発明は、さらに、本発明の1以上のベクターを組み込んだ細胞を提供する。該細胞は、原核細胞または真核細胞であってもよい。いくつかの具体例において、該細胞は真核細胞である。いくつかの具体例において、該細胞は、マウスミエローマハイブリドーマ細胞である。いくつかの具体例において、該細胞は、チャイニーズハムスター卵巣(CHO)細胞である。ベクターを細胞中に組み込むための例示的方法は、実施例に提供する。しかしながら、ベクターを細胞中に組み込むために、当該分野で既知のいずれの適当な技術を用いてもよい。いくつかの具体例において、本発明は、免疫グロブリン融合タンパク質を発現することができる細胞を提供し、ここに、該細胞は、免疫グロブリン軽鎖遺伝子および治療剤に融合した免疫グロブリン重鎖の遺伝子の両方が一片の核酸、例えば、DNA中に組み込まれている単一タンデム発現ベクターが永続的に組み込まれた細胞である。いくつかの具体例において、本発明は、免疫グロブリン融合タンパク質を発現することができる細胞を提供し、ここに、該細胞は、免疫グロブリン重鎖遺伝子およびデコイ受容体ECDに融合した免疫グロブリン軽鎖の遺伝子の両方が一片の核酸、例えば、DNA中に組み込まれている単一タンデム発現ベクターが安定に組み込まれた細胞である。該タンデムベクターの細胞中への導入は、例えば、染色体核酸中への導入、またはエピソーム遺伝子エレメントの導入によるものであってもよい。
【0099】
2つの核酸の配列比較のために、典型的には1つの配列が、試験配列を比較するための参照配列として作用する。配列比較アルゴリズムを用いる場合、試験および参照配列をコンピューターに入力し、必要に応じてサブ配列座標を指定し、配列アルゴリズムプログラムパラメーターを指定する。デフォルトプログラムパラメーターを用いることができ、または別のパラメーターを指定することができる。次いで、配列比較アルゴリズムは、プログラムパラメーターに基づいて、参照配列に対する試験配列のパーセント配列同一性を計算する。
【0100】
本明細書中で使用される場合、「比較ウィンドウ」は、20〜600、通常、約50〜約200、より普通には、約100〜約150からなる群から選択される隣接位置の数のいずれか1つのセグメントに対するリファレンスを含み、ここに、配列は、2つの配列を最適にアラインした後に、同じナンバーの隣接位置の参照配列と比較されうる。比較のための配列をアラインメントする方法は、当該分野で周知である。比較のための配列の最適アラインメントは、限定するものではないが、SmithおよびWaterman(1970), Adv. Appl. Math., 2:482cの局所ホモロジーアルゴリズムによって、NeedlemanおよびWunsch,(1970), J. Mol. Biol., 48:443のホモロジーアラインメントアルゴリズムによって、PearsonおよびLipman,(1988), Proc. Nat’l. Acad. Sci. USA, 85:2444の類似性検索方法によって、これらのアルゴリズムのコンピューターによる手段によって(GAP、BESTFIT、FASTA、およびTFASTA,Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI)、または手動アラインメントおよび目視によって(例えば、Ausubelら(1995 supplement), Current Protocols in Molecular Biology参照)実施することができる。
【0101】
パーセント配列同一性および配列類似性を決定するために適当なアルゴリズムの一例は、BLASTおよびBLAST 2.0アルゴリズムであり、各々、Altschulら(1977), Nuc. Acids Res., 25:3389−3402、およびAltschulら(1990), J. Mol. Biol., 215:403−410に記載されている。BLAST分析を実施するためのソフトウェアは、National Center for Biotechnology Informationから公的に入手可能である。BLASTアルゴリズムパラメーターW、TおよびXは、アラインメントの感度および速度を決定する。BLASTNプログラム(ヌクレオチド配列用)は、デフォルトとして、11のワードレングス(W)、10のエクスペクテーション(E)、M=5、N=−4および両鎖の比較を用いる。BLASTアルゴリズムは、典型的に、「低複雑性」フィルターを切って(turned off)実施される。BLASTアルゴリズムはまた、2配列間の類似性の統計学的分析を行う(例えば、KarlinおよびAltschul, (1993), Proc. Natl. Acad. Sci. USA, 90:5873−5787参照)。BLASTアルゴリズムによって提供される類似性の1測定値は、最小合計確率(P(N))であり、それは、2つのヌクレオチドまたはアミノ酸配列間の合致が偶然に起こる確率を示す。例えば、参照核酸に対する試験核酸の比較において、最小合計確率が約0.2未満、より好ましくは、約0.01未満、最も好ましくは約0.001未満である場合、核酸は参照配列に類似するとみなされる。
【0102】
当該分野で周知のように、遺伝子コードの縮重のために、適当なコドンのいずれかの組合せを用いて所望の融合タンパク質をコードしうる。さらに、組み換え技術に有用な他のエレメント、例えば、プロモーター、終止シグナルなどもまた、核酸配列中に含まれうる。かかるエレメントは、当該分野で周知である。さらに、本明細書中に記載され、請求される全ての核酸配列は、該配列の相補体を包含する。
【0103】
いくつかの具体例において、本発明の核酸は、低、中程度または高ストリンジェンシー条件下で、配列番号4、6または7のアミノ酸配列をコードする核酸に特異的にハイブリダイズする。例えば、かかる核酸は、低、中程度または高ストリンジェンシー条件下で、配列番号3、5、その両方のヌクレオチド配列、またはその相補配列を含む核酸配列にハイブリダイズしうる。低ストリンジェンシーハイブリダイゼーション条件は、例えば、2XSSCおよび0.1%SDS中42℃にて、約40%〜約70%GC含量の100ヌクレオチドプローブを用いるハイブリダイゼーションを包含する。中程度ストリンジェンシーハイブリダイゼーション条件は、例えば、0.5X SSCおよび0.1%SDS中50℃でのハイブリダイゼーションを包含する。高ストリンジェンシーハイブリダイゼーション条件は、例えば、0.2X SSCおよび0.1%SDS中65℃にて、上記のプローブを用いるハイブリダイゼーションを包含する。これらの条件下では、ハイブリダイゼーション温度が高いので、より高いホモロジーを有する核酸を得ることができる。
【0104】
さらに、本発明は、二官能性デコイ受容体融合抗体を製造する方法を提供する。いくつかの具体例において、本発明は、二官能性デコイ受容体融合抗体を製造する方法であって、真核細胞中に、(i)受容体細胞外ドメインに融合した免疫グロブリン重鎖、および免疫グロブリン軽鎖の両方、または(ii)受容体細胞外ドメインに融合した免疫グロブリン軽鎖、および免疫グロブリン重鎖の両方(ここに、コードされる免疫グロブリン重鎖および免疫グロブリン軽鎖は、BBB上に発現された受容体に対する抗体由来である)をコードする単一タンデム発現ベクターを安定に組み込むことを含む方法を提供する。BBB受容体に対する適当な抗体、および受容体ECDは、上記の抗体および受容体ECDのいずれかを包含する。
【0105】
本発明は、免疫グロブリン融合タンパク質を発現すること、および/または免疫グロブリン融合タンパク質を精製することを包含する。発現および精製を包含する例示的な製造方法は、実施例に提供される。
【0106】
しかしながら、該タンパク質を製造、所望により発現、および精製するために、当該分野で既知のいずれの適当な技術を用いてもよい。これらは、タンパク質合成の非組み換え技術、例えば、Merrifieldによって最初に開発され、Stewartら(1984),Solid Phase Peptide Synthesisに記載されるような手動または自動の固相合成を包含する。化学合成は、アミノ酸をC末端で開始する所定の配列中に加える。基本的な固相方法は、C末端を保護したα−アミノ酸を適当な不溶性樹脂支持体に結合させることを必要とする。合成のためのアミノ酸は、その前の残基(または樹脂支持体)との適当なペプチド結合形成を保証するために、α−アミノ基上の保護を必要とする。カルボキシル末端での縮合反応の完了後、α−アミノ保護基を除去して、次の残基の付加を可能にする。いくつかのクラスのα−保護基が記載されており(Stewartら(1984),Solid Phase Peptide Synthesis参照)、酸に不安定なウレタンを基礎とするターシャリー−ブチルオキシカルボニル(Boc)が歴史的に好ましい。他の保護基、および関連する化学手法を用いてもよく、塩基に不安定な9−フルオレニルメチルオキシカルボニル(FMOC)を包含する。また、合成が完了するまで、反応性アミノ酸側鎖官能基は、ブロックする必要がある。官能性ブロッキング基の複雑なアレイは、それらの使用に対する手法および制限とともに、Bodansky(1976),Peptide SynthesisおよびStewartら(1984),Solid Phase Peptide Synthesisに概説されている。
【0107】
固相合成は、記載されるC末端α−保護アミノ酸残基のカップリングによって開始する。カップリングは、活性化剤、例えば、1−ヒドロキシベンゾ−トリアゾール(HOBT)を伴う、または伴わないジシクロヘキシカルボジイミド(DCC)、ジイソプロピルカルボジイミド(DIIPC)、またはエチルジメチルアミノプロピルカルボジイミド(EDC)を必要とする。C末端残基をカップリング後、α−アミノ保護基は、酸に不安定なターシャリー−ブチルオキシカルボニル(Boc)基の場合、ジクロロメタン中のトリフルオロ酢酸(25%以上)によって除去される。ジクロロメタン中のトリエチルアミン(10%)での中和工程は、遊離アミン(対 塩)を回収する。C末端残基を樹脂に付加した後、脱保護、中和およびカップリングサイクルを中間の洗浄工程と共に繰り返して、保護されたペプチド鎖を伸長する。各保護されたアミノ酸は、適当な溶媒中の等モル量のカップリング試薬を用いて、過剰に(3〜5倍)導入される。最終的に、完全にブロックしたペプチドを樹脂支持体上に構築した後、試薬をアプライして該樹脂から該ペプチドを切断し、側鎖ブロッキング基を除去する。無水フッ化水素(HF)は、酸に不安定なターシャリー−ブチルオキシカルボニル(Boc)化学基を切断する。特に側鎖官能基上の副反応を回避するために、いくつかの求核性スカベンジャー、例えば、ジメチルスルフィドおよびアニソールが含まれる。
【0108】
VII.方法
本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体のいずれかを全身投与することによって、デコイ受容体をBBBを横断して送達するための方法が本明細書中に記載される。いくつかの具体例において、受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体であって、BBB上に発現された受容体および該受容体細胞外ドメインのリガンドに結合する該融合抗体を含む医薬組成物を対象に全身投与することによって、CNS疾患を治療するために、本明細書に記載の組成物を投与する。いくつかの場合、治療されるべきCNS疾患は、急性CNS疾患、例えば、局所脳虚血、全脳虚血、外傷性脳損傷、または脊髄損傷である。いくつかの場合、治療されるべきCNS疾患は、慢性CNS疾患である。いくつかの具体例において、慢性CNS疾患は、神経変性CNS疾患、例えば、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症、ハンチントン病、多発性硬化症、横断性脊髄炎、運動ニューロン疾患、ピック病、結節硬化症、カナバン病、レット症候群、脊髄小脳失調症、フリードライヒ失調症、視神経萎縮、または網膜変性である。
【0109】
いくつかの具体例において、卒中、頭部損傷、脊髄損傷、または神経変性疾患に罹患している対象を治療するために、TNF−α受容体ECD−BBB受容体融合抗体が全身投与される。他の具体例において、血管内皮成長因子受容体(VEGFR)−ECD−BBB受容体Ab融合抗体を対象に全身投与して、脳腫瘍における血管新生を阻害する。いくつかの場合、後天性免疫不全症候群(AIDS)由来の認知症を治療するために、TNF関連アポトーシス誘導性リガンド受容体(TRAIL−R)ECD−BBB受容体Ab融合抗体を投与する。いくつかの具体例において、多発性硬化症(MS)を治療するために、インターロイキン(IL)−6デコイ受容体ECD−BBB受容体Ab融合抗体を全身投与する。さらなる具体例において、卒中を治療するために、アポトーシスのTNF様弱インデューサー(TWEAK)受容体デコイECD−BBB受容体Ab融合抗体を全身投与する。また別の具体例において、卒中または脳損傷後の神経修復を促進するために、エフリン受容体、EphA ECD−BBB受容体Ab融合抗体を全身投与する。
【0110】
デコイ受容体−BBB受容体Ab融合抗体の送達に適当な全身投与量は、投与されるべき特定のデコイ受容体−BBB受容体Ab融合抗体、そのCNS取り込み特徴およびデコイ受容体リガンドに対するそのアフィニティーに基づいて、変化するであろう。
【0111】
いくつかの具体例において、投与されるべきデコイ受容体−BBB受容体Ab融合抗体は、インスリン受容体(例えば、ヒトインスリン受容体)、トランスフェリン受容体、
インスリン様成長お因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体に対する抗体を含有し、該デコイ受容体は、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来のECDを含有する。いくつかの具体例において、投与されるべき該デコイ受容体−BBB受容体Ab融合抗体は、TNF−α受容体−ヒトインスリン受容体Ab融合抗体である。
【0112】
本明細書中で使用される場合、「全身投与」または「末梢投与」なる語は、CNS中への直接的投与ではない、すなわち、BBBの物理的貫通または破壊を含まないいずれかの投与方法を包含する。「全身投与」には、限定するものではないが、静脈内、動脈内、筋内、皮下、腹腔内、鼻腔内、経頬、経皮、直腸、経肺胞(吸入)、または経口投与が包含される。本明細書中に記載されるように、いずれの適当なデコイ受容体−BBB受容体Ab融合抗体を使用してもよい。
【0113】
本発明の組成物は、組合せ療法の一部として投与されうる。組合せ療法は、上記のCNS疾患のいずれかに罹患している患者において典型的に見出される症状の治療または軽減のための別の療法と組み合わせた本発明の組成物の投与を含む。本発明の組成物を別のCNS障害方法または組成物と組み合わせて用いる場合、本発明の組成物および付加的な方法または組成物のいずれの組合せを用いてもよい。かくして、例えば、本発明の組成物の使用が別のCNS障害治療剤と組み合わせられている場合、該2つを同時に、連続的に、持続時間が重なるように、類似した頻度で、同じ頻度で、または異なる頻度などで投与してもよい。いくつかの場合、本発明の組成物を1以上の他のCNS障害治療剤と組み合わせて含有する組成物が用いられるであろう。
【0114】
いくつかの具体例において、該組成物、例えば、二官能性TNF−α受容体ECD−BBB受容体Ab融合抗体を患者に、同じ処方内で、別の薬物療法と共に、または別々の組成物として共投与する。例えば、該二官能性TNF−α受容体−ヒトインスリン受容体Ab融合抗体は、別の二官能性デコイ受容体−BBB受容体Ab融合抗体と共に処方することができる。さらに、TNF−α受容体ECD−BBB受容体Ab融合抗体は、他の大きいまたは小さい分子と組み合わせて処方してもよい。CNSまたはCNS関連疾患を治療するために、デコイ受容体−BBB受容体Ab融合抗体と組み合わせて使用するための例示的薬剤は、下記に提供される。
【0115】
CNS炎症疾患を治療するために共投与するための例示的薬剤
対象が自己免疫、炎症疾患、または神経系に影響を及ぼすアレルギー疾患(例えば、Allanら(2003), Philos Trans R Soc Lond B Biol Sci, 358(1438):1669−1677参照)に罹患しているか、または罹患する危険性がある場合、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体(例えば、TNF−α受容体ECD−BBB受容体Ab融合抗体)をいずれかの組合せにおいて1以上の下記の治療剤、すなわち、免疫抑制剤(例えば、タクロリマス(tacrolimus)、シクロスポリン(cyclosporin)、ラパマイシン(rapamicin)、メトトレキサート(methotrexate)、シクロホスファミド(cyclophosphamide)、アザチオプリン(azathioprine)、メルカプトウリン(mercaptopurine)、マイコフェノラート(mycophenolate)、またはFTY720)、グルココルチコイド(例えば、プレドニソン、コルチゾンアセテート、プレドニソロン、メチルプレドニソロン、デクサメタゾン、ベータメタゾン、トリアムシノロン(triamcinolone)、ベクロメタゾン、フルドロコルチゾンアセテート、デオキシコルチコステロンアセテート、アルドステロン)、非スイテロイド性抗炎症薬(例えば、サリチル酸塩、アリールアルカン酸、2−アリールプロピオン酸、N−アリールアントラニル酸、オキシカム、コキシブ(coxibs)、またはスルホンアニリド)、Cox−2−特異的阻害剤(例えば、バルデコキシブ(valdecoxib)、セレコキシブ(celecoxib)、またはロフェコキシブ(rofecoxib))、レフルノミド(leflunomide)、金チオグルコース、金チオマレート、オーロフィン(aurofin)、スルファザラジン(sulfasalazine)、ヒドロキシクロロキニン、ミノサイクリン、TNF−α結合タンパク質(例えば、インフリキシマブ(infliximab)、エタネルセプト(etanercept)、またはアダリムマブ(adalimumab))、アバタセプト(abatacept)、アナキンラ(anakinra)、インターフェロン−β、インターフェロン−γ、インターロイキン−2、アレルギーワクチン、抗ヒスタミン、抗ロイコトリエン、ベータ−アゴニスト、テオフィリン、または抗コリン作用剤と一緒に使用することができる。
【0116】
多発性硬化症を治療するために共投与することができる例示的薬剤
対象が多発性硬化症に罹患しているか、または罹患する危険性がある場合、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体(例えば、インターロイキン(IL)−6デコイ受容体ECD−BBB受容体Ab融合抗体)をいずれかの組合せにおいて1以上の下記の例示的多発性硬化症治療剤、すなわち、インターフェロンβ−1a、インターフェロンβ−1b、グラチラマーアセテート(CopaxoneR(登録商標))、ミトキサントロン(NovantroneR)、低用量ナルトレキソン、ナタリズマブ(Natalizumab)(TysabriR)、SativexR、エイムスプロ(Aimspro)(ヤギ血清)、トリメスタ(Trimesta)(経口エストリオール)、ラキニモド(Laquinimod)、FTY720(フィンゴリモド(Fingolimod))、MBP8298、NeuroVaxR、TovaxinR、レビムン(Revimmune)、CHR−1103、BHT−3009、BG−12、クラドリビン(Cladribine)、ダクリズマブ(daclizumab)(Zenapax)リツキシマブ(Rituximab)(Rituxan)、シクロホスファミド、キャンパス(Campath)、ファムプリジン(Fampridine)−SR、MN−166、テムシロリマス(Temsirolimus)、またはRPI−78Mと一緒に使用することができる。
【0117】
例えば、アルツハイマー病またはAIDS関連認知症の治療のために共投与することができる例示的薬剤
対象が認知症に罹患しているか、または罹患する危険性がある場合、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体(例えば、TNF関連アポトーシス誘導性リガンド受容体(TRAIL−R)ECD−BBB受容体Ab融合抗体)をいずれかの組合せにおいて認知症を治療するための1以上の薬剤または方法と一緒に使用することができる。認知症を治療するための治療剤/治療の例は、限定するものではないが、FlurizanTM(登録商標)(MPC−7869,r フルルビプロフェン(r flurbiprofen))、メマンチン(memantine)、ガラタミン(galantamine)、リバスチグミン(rivastigmine)、ドネジピル(donezipil)、タクリン(tacrine)、Aβ1−42免疫療法、レスベラトロール(resveratrol)、(−)−エピガロカテキン−3−ガラート、スタチン、ビタミンC、またはビタミンEを包含する。
【0118】
血栓塞栓性障害を治療するための例示的薬剤
対象が血栓塞栓性障害(例えば、卒中)に罹患しているか、または罹患する危険性がある場合、1以上の他の抗血栓塞栓剤とのいずれかの組合せにおいて、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体(例えば、アポトーシスのTNF様弱インデューサー(TWEAK)受容体デコイECD−BBB受容体Ab融合抗体)を用いて該対象を治療することができる。抗血栓塞栓剤の例は、限定するものではないが、血栓溶解剤(例えば、アルテプラーゼ、アニストレプラーゼ、ストレプトキナーゼ、ウロキナーゼ、または組織プラスミノーゲンアクチベーター)、ヘパリン、チンザパリン(tinzaparin)、ワーファリン(warfarin)、ダビガトラン(dabigatran)(例えば、ダビガトランエテキレート(dabigatran etexilate))、因子Xa阻害剤(例えば、フォンダパリヌクス(fondaparinux)、ドラパリヌクス(draparinux)、リバロキサバン(rivaroxaban)、DX−9065a、オタミキサバン(otamixaban)、LY517717、またはYM150)、チクロピジン(ticlopidine)、クロピドグレル(clopidogrel)、CS−747(プラスグレル(prasugrel),LY640315)、キシメラガトラン(ximelagatran)、またはBIBR 1048を包含する。
【実施例】
【0119】
下記の具体例は、単なる例示として解釈されるべきものであり、如何なる方法においても、該開示の残りの部分を限定するものとして解釈されるべきではない。当業者は、さらに工夫することなく、本発明の記載に基づいて、本発明をその完全な範囲まで利用することができると確信する。本明細書中に引用された全ての出版物は、出典明示により全体として本明細書の一部とされる。URLまたは他のかかる識別子またはアドレスに言及する場合、かかる識別子は変化することができ、インターネット上の特定の情報は移り変わることができるが、インターネットを検索することによって、等しい情報を見出すことができると理解される。その言及は、かかる情報の入手可能性および公的な流布を証明するものである。
【0120】
実施例1 ヒトTNFR ECD cDNAのクローニングおよび発現
NP 001057のアミノ酸23−257に対応するヒトTNFR−II細胞外ドメイン(ECD)を、表2に記載のオリゴデオキシヌクレオチド(ODNs)を用いるポリメラーゼ連鎖反応(PCR)によってクローン化し、ヒトU87グリア細胞から単離したポリA+RNAの逆転写からcDNAを得た。該TNFR cDNAを、25ng ポリA+RNA−由来のcDNA、0.2μM 順方向および逆方向ODNプライマー(表Iの各々、配列番号1および配列番号2)、0.2mM デオキシヌクレオシドトリホスフェート、および50μl Pfuバッファー中の2.5U PfuUltra DNAポリメラーゼを用いるPCRによってクローン化した。Mastercycler温度サイクラー中、95℃で2分間の開始変性工程、次いで、95℃で30秒間の変性を30サイクル、55℃で30秒間のアニーリング工程、および72℃で1分間の増幅工程を用いて増幅を行い、次いで、72℃で10分間、最終インキュベーションを行った。PCR産物を0.8%アガロースゲル電気泳動中で分析し、ヒトTNFR cDNAに対応する〜0.6kbの予想される主要単一バンドを生じた(図2A)。TNFR ECDのアミノ酸配列は、クローン化したTFR ECD cDNAのヌクレオチド配列から推定し、Leu23−Asp257を含んでいた(NP 001057)。
【0121】
【表2】
【0122】
実施例2 TNFRがHIRMAb重鎖のカルボキシル末端に融合した重鎖−TNFR融合タンパク質をコードする発現プラスミドの遺伝子操作
重鎖(HC)およびTNFR ECDの融合タンパク質を発現する発現プラスミドをpCD−HIRMAb−TNFRと称する。ヒトTNFR−II ECD(NP 001057)のアミノ酸Leu23−Asp257に対応する成熟ヒトTNFR cDNAを該pCD−HIRMAb−HC発現プラスミド(図2B)のHpaI部位に挿入することによって該プラスミドを操作して、pCD−HIRMAb−TNFRを製造した(図2B)。該pCD−HIRMAb−HCプラスミドは、キメラHIRMAbのHCをコードし、該プラスミドおよび軽鎖(LC)発現プラスミドpHIRMAb−LCを用いるCOS細胞のデュアルトランスフェクションにより、キメラHIRMAbの一過性発現を可能にする。TNFR順方向(FWD)PCRプライマー(表I)は、「CA」ヌクレオチドを導入して、オープンリーディングフレームを維持し、かつ、HIRMAb HCのCH3領域のカルボキシル末端とそのシグナルペプチドを差し引いたTNFR ECDのアミノ末端との間に、Ser−Ser−Serリンカーを導入する。該TNFRモノマーの各HCのカルボキシル末端への融合は、図1に示される。該設計は、該TNFRを、TNFR ECDの好ましいコンフォメーションである二量体コンフィギュレーションに立体的に限定し、受容体ダイマーとして結晶化する(Chanら(2000), Immunity, 13:419−422)。該TNFR逆方向(REV)PCRプライマー(表I)は、TNFR ECDタンパク質の末端のアスパラギン酸の直後に、終止コドン「TGA」を導入する。該操作されたpCD−HIRMAb−TNFR発現ベクターは、DNA配列決定によって確認された。
【0123】
該HIRMAb HCおよびLC cDNA発現カセットは、サイトメガロウイルス(CMV)プロモーターによって駆動し、ウシ成長ホルモン(BGH)ポリアデニル化(pA)配列を含有する(図2B)。該ユニバーサルpCD−HIRMAb−HCベクターの操作は、以前に記載されたように(Boadoら(2007b) Biotechnol. Bioeng., 97:1376−1386)、部位特異的変異誘発(SDM)によって、HIRMAb HC CH3オープンリーディングフレーム(orf)の末端に単一のHpaI部位を挿入することによって実施された。
【0124】
表2のODNsを用いるPCRによって、235アミノ酸TNFR−II ECDに対応するcDNAを増幅し、図2Bに概説されるように、該cDNAをpCD−HIRMAb−HCプラスミドのHpaI部位にサブクローン化した。DNA配列決定は、該pCD−HIRMAb−TNFRプラスミドの発現カセットが、714ヌクレオチド(nt)のCMVプロモーター、9ntの全Kozak部位(GCCGCCACC)、2,100ntのHIRMAb HC−TNFR融合タンパク質オープンリーディングフレーム、および370ntのBGH配列(配列番号3)を包含する3,193個のヌクレオチド(nt)を含むことを示した。該2,100ntオープンリーディングフレームは、配列番号3のnt724〜2,823に相当する。該プラスミドは、19アミノ酸IgGシグナルペプチド(AA1〜19、配列番号4)、442アミノ酸HIRMAb HC(AA20〜461、配列番号4)、3アミノ酸リンカー(Ser−Ser−Ser)、およびそのシグナルペプチドを差し引いた235アミノ酸ヒトTNFR−II ECD(AA465〜699、配列番号4)からなる、699アミノ酸(AA)タンパク質をコードした。グリコシル化を差し引いた該重鎖融合タンパク質の予測分子量は、73,900Daであり、予測等電点(pI)は8.45である。該融合タンパク質のTNFR ECD部分の推定アミノ酸配列は、22個のシステイン残基および2個のN−連結コンセンサスグリコシル化部位を該TNFR ECD内のAsn−149およびAsn−171に含んだ(配列番号4)。
【0125】
該HIRMAb−TNFR融合タンパク質は、2個の融合重鎖(AA20−699、配列番号4)および2個の軽鎖(LC)からなる。該LCは、pCD−HIRMAb−LC発現プラスミドによって発現され、該LC発現カセットのnt配列は、配列番号5に示す。該pCD−HIRMAb−LCプラスミドの発現カセットのDNA配列決定は、731ntのCMVプロモーター、9ntの全Kozak部位(GCCGCCACC)、705ntのHIRMAb LC融合タンパク質オープンリーディングフレーム、および370ntのBGH配列を包含する1,809ntを含んでいた(配列番号5)。該705ntオープンリーディングフレームは、配列番号5のnt741〜1,445に相当する。該プラスミドは、20アミノ酸IgGシグナルペプチド(AA1〜20、配列番号6)、および214アミノ酸HIRMAb LC(AA21〜234、配列番号6)からなる234AAタンパク質をコードする。
【0126】
実施例3 トランスフェクトしたCOS細胞によるHIRMAb−TNFR融合タンパク質の分泌
COS細胞は、μg DNA:uL リポフェクタミンの比率1:2.5でリポフェクタミン2000を用いて、pCD−HIRMAb−LCおよびpCD−HIRMAb−TNFR(ここに、pCD−HIRMAb−LCは、HIRMAb−TNFR融合タンパク質中に組み込まれた同じ軽鎖であるキメラHIRMAbの軽鎖をコードする発現プラスミドである)でデュアルトランスフェクトした。トランスフェクション後、該細胞を無血清培地中で培養した。COS細胞を最初、ヒトIgG特異的ELISAで発現をスクリーニングするために、6ウェルクラスター皿に播種した。次いで、10xT500フラスコ中、トランスフェクトしたCOS細胞の培養のために、該トランスフェクションをスケールアップした。馴化した無血清培地を3日目および7日目に収集した。該融合タンパク質をタンパク質Aアフィニティークロマトグラフィーによって精製した。
【0127】
ヒトIgG ELISAは、Immulon 2高結合プレート中で、COS細胞馴化培地を用いて行った。ヤギ抗−ヒトIgG一次抗体を0.1M NaHCO3(100μl、2μg/ml)中で播種し、4℃で一晩インキュベートした。プレートを0.01M Na2HPO4/0.15 M NaCl/pH=7.4/0.05% Tween−20(PBST)で洗浄し、PBST中における1%ゼラチンで22℃にて30分間ブロックした。プレートを100μl/ウェルのヒトIgG1標準または融合タンパク質のいずれかと共に、室温(RT)でて60分間インキュベートした。PBSTで洗浄後、アルカリホスファターゼにコンジュゲートしたヤギ抗−ヒトカッパLC抗体を37℃で60分間培養した。暗所中、pH=10.4でp−ニトロフェニルホスフェートを用いて、発色を行った。NaOHを用いて該反応を停止し、405nmの吸光度をELISAプレートリーダーで測定した。
【0128】
COS細胞によって産生されるタンパク質A精製融合タンパク質の均質性を還元性12%ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)、次いで、クーマシーブルー染色を用いて評価した。ウェスタンブロッティングの場合、ヒトIgG免疫反応性をヒトIgG(H+L)に対する一次ヤギ抗体を用いて試験し、ヒトTNFR免疫反応性をヒトTNFR−II ECDに対するマウスモノクローナル抗体を用いて評価した。SDS−PAGEおよびクーマシーブルー染色後、軽鎖(LC)のサイズは、HIRMAbおよびHIRMAb−TNFR融合タンパク質の両方について同じである(図3)。該融合タンパク質の重鎖(HC)のサイズは、HIRMAbのHCよりも約30kDa大きい(図3)。ウェスタンブロッティングにおいて、HIRMAbまたはHIRMAb−TNFR融合タンパク質のいずれかのLCは、図4(左パネル)に示されるように、該ウェスタン上で、ヒトIgG(H+L)に向けられた一次抗体と等しく反応する。該融合タンパク質のHCのサイズは、抗−ヒトIgG一次抗体(図4、左パネル)または抗−ヒトTNFR一次抗体(図4、右パネル)のいずれかを用いる両方のウェスタンブロットにおいて、HIRMAbのHCのサイズよりも約30kDa大きい。これらの結果は、該HIRMAb−TNFR融合タンパク質が宿主細胞によって首尾よく翻訳され、分泌されることを示す。
【0129】
実施例4 HIRMAb−TNFR融合タンパク質はHIRおよびTNFαの両方について高アフィニティー結合を保持する
本発明の作業において、デコイ受容体のアミノ末端は、IgG鎖のカルボキシル末端に融合される。予想外にも、該コンフィギュレーションは、二官能性デコイ受容体−BBB受容体Ab融合抗体を生じた。
【0130】
HIR細胞外ドメイン(ECD)に対する該融合タンパク質のアフィニティーは、レクチンアフィニティー精製HIR ECDを用いるELISAで決定した。該HIR ECDで永続的にトランスフェクトされたCHO細胞を無血清培地(SFM)中で成育させ、該HIR ECDをコムギ胚芽凝集素アフィニティーカラムで精製した。該HIR ECD(0.2μg/ウェル)をImmulon 2高結合96ウェルプレート上に播種し、キメラHIRMAb、HIRMAb−TNFR融合タンパク質、またはヒトIgG1の該HIR ECDに対する結合を、ビオチン化ヤギ抗−ヒトIgG(H+L)抗体(0.3μg/ウェル)、およびアビジン/ビオチン化ペルオキシダーゼ検出システムを用いて検出した。HIR ECDに対する50%の結合をもたらした濃度、ED50を非線形回帰分析によって決定した。各々、0.44±0.19nMおよび0.62±0.06nMのED50を伴うキメラHIRMAbおよびHIRMAb−TNFR融合タンパク質のHIR ECDに対する比較可能な結合が存在する(図5)。これは、TNFRのHIRMAbへの融合にもかかわらず、HIRMAbによるHIRへの高いアフィニティー結合の保持が存在することを示す。
【0131】
HIRMAb−TNFR融合タンパク質のTNFαに対する結合は、ELISAを用いて決定された。捕獲試薬は、市販のヒトTNFαであった。該アッセイにおける陽性対照は、市販の組み換えヒトTNFR−II:ヒトIgG1 Fc融合タンパク質(TNFR:Fcと称する)であり、陰性対照は、ヒトIgG1/κであった。TNFαは、0.1M NaHCO3/pH=9.0中に溶解し、100μl/ウェル(0.2μg/ウェル)中4℃で一晩培養した。0.01M Tris/0.15M NaCl/pH=7.4(TBS)での洗浄後、該ウェルをTBS中における1%ウシ血清アルブミン(BSA)で30分間ブロックした。100μl/ウェルの容量のHIRMAb−TNFR、TNFR:Fc、またはヒトIgG1kを室温で60分間培養した。TBSおよび0.05%Tween−20(TBST)で洗浄後、ヤギ抗−ヒトIgG−アルカリホスファターゼコンジュゲートを60分間インキュベートした(0.2μg/ウェル)。TBSTで洗浄した後、パラ−ニトロフェニルホスフェートでの発色および1.2M NaOHでの反応停止後、ELISAプレートリーダーを用いて405nmでの色検出を行った。該ヒトIgG1/カッパ(hIgG1/k)は、図6Aに示されるように、TNFαに結合しなかった。該TNFR:Fc、または該HIRMAb−TNFRは、0−600ng/mLの試験濃度範囲で飽和しない直線関係で、培養したTNFαに結合した。したがって、該データを線形回帰分析にフィットさせて、結合曲線の傾きを計算した。該傾きの逆数は、KD/Amax比に等しく、ここに、KDは、培養したTNFαに結合しているTNFRの結合定数であり、Amaxは、最大吸光度である。該分析において、結合曲線の傾きは、該TNFR融合タンパク質のTNFαに対するアフィニティーに正比例する。組み換えTNFR:Fcまたは該HIRMAb−TNFR融合タンパク質のヒトTNFαに対するアフィニティーをELISAで測定した。該TNFR:Fcは、0.88±0.02nMのKD/Amax比でTNFαに結合した(図6A)。該HIRMAb−TNFR融合タンパク質は、1.40±0.08nMの比較可能なKD/Amax比でTNFαに結合した(図6B)。これは、HIRMAbのカルボキシル末端へのTNFRの融合にもかかわらず、TNFR ECDによるTNFαへの高アフィニティー結合の保持が存在することを示す。
【0132】
ヒトTNFαの該HIRMAb−TNFR融合タンパク質への飽和可能な結合は、放射性受容体アッセイ(RRA)を用いて決定された。マウス抗−ヒトIgG1 Fc抗体を96ウェルプレート(0.4μg/ウェル)に播種し、0.1M NaHCO3/pH=8.3中で一晩インキュベーションし、次いで、洗浄し、0.01M Na2HPO4/0.15M NaCl/pH=7.4(PBS)中における1%ウシ血清アルブミン(BSA)でブロックした。次いで、下記の溶液、すなわち(a)PBS中1%BSA、(b)100ng/ウェルのヒトIgG1/カッパ、または(c)100ng/ウェルの該HIRMAb−TNFR融合タンパク質の1つを100μl/ウェルにて加え、次いで、室温で1時間インキュベーションした。次いで、該ウェルをPBSで洗浄し、次いで、0.01uCi/ウェル(0.2ng/ウェル)濃度の[125I]−TNFαおよび種々の濃度の標識していないヒトTNFαの共混合物200μl/ウェルを加え、次いで、室温で3時間インキュベーションした。吸引によりウェルを空にし、冷PBSで洗浄し、250μl/ウェルの1N NaOHを加え、次いで、60℃で30分間加熱した。Perkin Elmer液体シンチレーションカウンターで放射能をカウントし、ウェルあたりの部分(fractional)結合をコンピューターで算出した。該HIRMAb−TNFR融合タンパク質に結合しているTNFの半飽和定数、KDは、統計学的ソフトウェアパッケージを用いる非線形回帰分析によって決定した。該HIRMAb−TNFR融合タンパク質に対するTNFαの飽和可能な結合は、放射性受容体アッセイを用いて検出可能であり、図7Aに概説される。該結合データ(方法)の非線形回帰分析は、結合のKDが0.34±0.17nMであることを示した(図7B)。これらの結果は、該TNFR ECDが、該HIRMAb重鎖のカルボキシル末端へのそのアミノ末端での融合にかかわらず、まだ、非常に高いアフィニティーでその同族リガンドを結合することを示す。
【0133】
実施例5 ヒト細胞におけるHIRMAb−TNFR融合タンパク質の生物学的活性
ヒトWEHI−13VAR細胞(CRL−2148)をAmerican Type Culture Collection(Manassas,VA)から得、TNFα細胞毒性のバイオアッセイとして用いた(Espevik and Nissen−Meyer, (1986), J. Immunol. Methods, 95:99−105)。該細胞を24ウェルクラスター皿に、10%胎仔ウシ血清(FBS)を含有するRPMI−1640培地中300,000細胞/ウェルで播種した。一晩培養後、該培地の半分を吸引除去し、200μlの新鮮なRPMI−1640培地、50μl/ウェルの10μg/mLのアクチノマイシンD(最終濃度=1.0μg/mL)、および最終濃度が1〜100pg/mLのヒト組み換えTNFαで置き換えた。いくつかのウェルにおいて、TNFαをウェルに加える前の30分間、TNFαを組み換えTNFR:Fc、またはHIRMAb−TNFR融合タンパク質に複合体化させた。該TNFR:FcまたはHIRMAb−TNFR融合タンパク質の最終濃度は、1.4nMであった。加湿インキュべーター中、37℃で一晩(20時間)インキュベーション後、該培地にチアゾイルブルーテトラゾリウムブロミド(MTT)を最終濃度0.5mg/mLまで補足した。37℃で3時間インキュベーション後、可溶化溶液(48%イソプロパノール、2% 1N HCl)の添加によって、該反応を停止した。570nmおよび650nmでの吸光度を測定し、A570−A650差をコンピューターで計算した。MTTは、健康細胞中のミトコンドリアによって、ホルマザン結晶に酸化され、該反応は、細胞の生存に反比例する。アクチノマイシンDの不在下で、該TNFαは、1−100pg/mL濃度にて、細胞に対して毒性ではなかった。しかしながら、アクチノマイシンDの存在下で、TNFαのこれらの濃度は、ED50値 5−10pg/mLヒトTNFαを伴う用量依存性細胞毒を生じた(図8)。しかしながら、該TNFαの細胞毒効果は、1.4nM濃度のTNFR:Fc融合タンパク質またはHIRMAb−TNFR融合タンパク質と共に該サイトカインを共インキュベーションすることによって遮断された(図8)。これは、該TNFRデコイ受容体がHIRMAbのカルボキシル末端への融合にかかわらず、生物学的に活性であることを示す。
【0134】
実施例6 TNFRのアミノ酸配列の部位特異的変異誘発
ヒトU87グリア細胞からクローン化された該デコイ受容体の235AA TNFR配列(AA465〜699,配列番号4)内に、174および210位置に多形が存在する。該HIRMAb−TNFR重鎖(配列番号4)の638位置のアルギニン(Arg)は、235AA TNFR ECDのArg−174に対応する。メチオニン(Met)が該ヒトTNFR配列(GenBank AAA36755)の174位置に見出されるので、該Arg−174は天然のR174M多形である。R638M部位特異的変異誘発(SDM)を周知の方法によって実施でき、配列番号7に示されるように、Arg−638をMet−638に変換する。該HIRMAb−TNFR重鎖(配列番号4)の674位置のリジン(Lys)は、235AA TNFR ECDのLys−210に対応する。該ヒトTNFR配列(GenBank AAA36755)のグルタミン酸(Glu)が210位置に見出されるので、該Lys−210は天然のK210E多形である。K674E部位特異的変異誘発(SDM)を周知の方法によって実施でき、配列番号7に示されるように、Lys−674をGlu−674に変換する。pCD−HIRMAb−TNFR発現ベクターのTNFR−R638M多形(配列番号7)での操作は、周知の部位特異的変異誘発法を用いて、ヒトTNFR−R638M FWD(表3、配列番号8)およびヒトTNFR−R638M REV(表3、配列番号9)ODNプライマーを用いるSDMによって実施される。同様に、pCD−HIRMAb−TNFR発現ベクターのTNFR−K674E多形での操作は、ヒトTNFR−K674E FWD(表3、配列番号10)およびヒトTNFR−K674E REV(表3、配列番号11)ODNプライマーを用いるSDMによって実施される。
【0135】
実施例7 HIRMAb−TNFR融合タンパク質をコードするタンデムベクターの遺伝子操作
HIRMAb−TNFR融合タンパク質は、図1に示されるように、2つの重鎖(HC)および2つの軽鎖(LC)からなる。したがって、宿主細胞は、HC遺伝子およびLC遺伝子の両方で永続的にトランスフェクトされなければならない。さらに、宿主細胞は、トランス遺伝子挿入部位の周囲の増幅を有する細胞系統の単離を可能にする遺伝子で永続的にトランスフェクトされなければならない。これは、ジヒドロ葉酸還元酵素(DHFR)をコードしている遺伝子で宿主細胞をトランスフェクション後の、メトトレキサート(MTX)での細胞系統の選択を用いて達成される。したがって、最終的には製造のためのマスター・セル・バンク(Master Cell Bank)をもたらす単一細胞における3つの遺伝子全ての高産生を得る必要がある。3つの遺伝子全ての高い発現を保証するために、タンデムベクター(TV)と呼ばれる一片のDNAを図9に概説されるように操作した。pTV−HIRMAb−TNFRと称するHIRMAb−TNFR融合タンパク質のためのTVの遺伝子操作は、pTV−HIRMAbのHpaI部位へのTNFR ECD cDNAの挿入によって完了した。該TNFR ECD cDNAは、カスタムODNsおよびpCD−HIRMAb−TNFR(図2B)を鋳型として用いるPCRによって生成された。pTV−HIRMAbは、HIRMAb HCオープンリーディングフレームの3’末端に独特のHpaI制限酵部位を有するキメラHIRMAbをコードするタンデムベクターである。pTV−HIRMAbは、HIRMAb LCおよびHC遺伝子のタンデム発現カセットを含有し、各カセットは、イントロンを有するCMVプロモーターによって駆動し、各カセットは、ウシ成長ホルモン(BGH)ポリアデニル化(pA)配列によって終了する。pTV−HIRMAbは、また、ネズミジヒドロ葉酸還元酵素(DHFR)のための第3の発現カセットを含有し、それは、SV40プロモーターによって駆動し、B型肝炎ウイルス(HBV)pA配列によって終了する。HIRMAb−TNFR HC発現カセットは、イントロンを有するCMVプロモーター、9ntの全Kozak部位(GCCGCCACC)、2,100ntのHIRMAb HC−TNFR融合タンパク質オープンリーディングフレーム、およびBGH pA配列からなる。図9に示されるタンデムベクターによってコードされる699 AA HIRMAb−TNFR HCタンパク質は、配列番号4におけるものと同一であり、19アミノ酸IgGシグナルペプチド(AA1〜19、配列番号4)、442アミノ酸HIRMAb HC(AA20〜461、配列番号4)、3アミノ酸リンカー(Ser−Ser−Ser)、およびそのシグナルペプチドを差し引いた235アミノ酸ヒトTNFR−II ECD(AA465〜699、配列番号4)からなる。該重鎖融合タンパク質の予測分子量は、グリコシル化を差し引いて、73,900Daであり、予測等電点(pI)は8.45である。該融合タンパク質のTNFR ECD部分の推定アミノ酸配列は、22個のシステイン残基を含み、2個のN−結合コンセンサスグリコシル化部位をTNFR ECD内のAsn−149およびAsn−171に含んだ(配列番号4)。図9に記載のタンデムベクターによって産生されるHIRMAb−TNFR融合タンパク質は、2つの融合重鎖(AA20−699、配列番号4)および2つの軽鎖(LC)からなる。LC発現カセットは、イントロンを有するCMVプロモーター、9ntの全Kozak部位(GCCGCCACC)、705ntのHIRMAb LC融合タンパク質オープンリーディングフレーム、およびBGH pA配列を含む。図9に示されるタンデムベクターによってコードされる214AA HIRMAb−LCタンパク質は、配列番号6におけるものと同一であり、20アミノ酸IgGシグナルペプチド(AA1〜20、配列番号6)、および214アミノ酸HIRMAb LC(AA21〜234、配列番号6)からなる。
【0136】
実施例8 CHO細胞の安定なトランスフェクションおよび希釈クローニング
無血清培地(SFM)適応性DG44チャイニーズハムスター卵巣(CHO)細胞を、エレクトロポレーターを用いて、PvuIで線形化後の5μgのpTV−HIRMAb−TNFRでエレクトロポレーションした。5 x 106細胞を200μLのリン酸緩衝化セーライン(PBS)および0.2cmキュベット中、方形波および160ボルトを用いて、該DNAでエレクトロポレーションした。細胞をCHO無血清培地(SFM)中に懸濁し、4x96ウェルプレート中で培養した。安定なトランスフェクタントの選択は、エレクトロポレーションの2日後、0.54mg/ml G418を用いて開始した。トランスフェクトタントのコロニーが明らかになったとき、すなわち、21目に、ヒトIgG ELISAのために、上清のアリコートを採取した。陽性コロニーを単離し、さらなる特徴付のために個々に培養した。DG44細胞は、内因性DHFRを欠き、内因性葉酸合成に関して、栄養分、ヒポキサンチンおよびチミジン(HT)に依存する。該TVを有するトランスフェクト細胞は、外因性DHFRを発現する。トランスフェクト細胞系統は、さらに、HT−欠損培地中に置くことによって選択された。トランス遺伝子挿入部位周囲の増幅を有する系統は、細胞を、20nM MTXで開始する増加濃度のMTXに付すことによって選択された。80−160nM MTXにて細胞系統の安定化後、1ウェルあたり1細胞の限界希釈クローニング(DC)によって、高生産性クローンを単離し、DCの各ラウンドにて全部で4000個のウェルに播種し、高容量マイクロプレートディスペンサーおよびマイクロプレートウォッシャーを用いるヒトIgG ELISAを用いて、培地IgGを測定した。クローン化した細胞を、軌道振盪機上の125mLのプラスチック角瓶中、1−2百万細胞/mLの生存細胞密度にて増殖させ、IgG ELISAによる測定の場合、無血清培地中約10mg/LのヒトIgGレベルを生産した。
【0137】
実施例9 イン・ビボにおけるデコイ受容体の脳に対する選択的標的化
HIRMAbは、旧世界霊長類のインスリン受容体と交差反応するが、低級種のインスリン受容体を認識しないので、HIRMAb−TNFR融合タンパク質の脳浸透を成体アカゲザル(Rhesus monkey)において測定した。さらに、ヒトIgG1 Fc領域に融合した市販のTNFRデコイ受容体(TNFR:Fcと称する)をR&D Systemsから購入した。該TNFR:Fc融合タンパク質は、デコイ受容体融合タンパク質に関する当該分野の状況を示す。該TNFR:Fcは、いずれのBBB受容体に対しても特異性を示さず、BBBに浸透しないと予測される。しかしながら、該TNFR:Fc融合タンパク質のBBB輸送は、測定されたことがない。さらに、IgG分子がBBBを横断するという文献における証拠があり[Zlokovicら、A saturable mechanism for transport of immunoglobulin G across the blood−brain barrier of the guinea pig(モルモットの血液脳関門を横断する免疫グロブリンGの輸送のための飽和可能なメカニズム)、Exp. Neurol, 107, 263−270, 1990]、TNFR:Fc融合タンパク質は、ヒトIgG1の定常領域の大部分を含有するので、該タンパク質もまた、BBBを横断することを推測できた。脳への取り込み研究について、CHO−由来HIRMAb−TNFR融合タンパク質は、該TNFR:Fc融合タンパク質の非酸化的放射性ヨウ素化と平行して、トリチウム化された。該[3H]−HIRMAb−TNFRおよび[125I]−TNFR:Fc融合タンパク質を共に、アカゲザルに注射した。結果は、TNFR:Fc融合タンパク質はBBBを横断せず、BBB分子のトロイの木馬に融合後、TNFRの脳浸透における顕著な増加があることを実証する。また、霊長類の非脳器官における該融合タンパク質の取り込みを測定し、それは、該デコイ受容体のHIRMAbへの融合が該医薬のCNSに対する選択的標的化をもたらすことを示す。
【0138】
R&D Systems(Minneapolis,MN)から購入し、SDS−PAGEによって均質であることが示されたTNFR:Fc融合タンパク質(#726−R2)を放射性標識するために、[125I]−Bolton−Hunter試薬を用いた。該TNFR:Fcは、1.0x28cmカラムのSephadex G−25での精製および0.01M NaH2PO4/0.15M NaCl/pH=7.4/0.05% Tween−20(PBST)での溶出後、新鮮なBolton−Hunter試薬で11.5uCi/ugの比活性および>99%のトリクロロ酢酸(TCA)沈澱性まで放射性標識した。標識化TNFR:Fc融合タンパク質のTCA沈澱は、ヨウ素化の24時間後で>99%のままであり、放射性標識後24時間以内に、該TNFR:Fc融合タンパク質を該霊長類に投与した。[3H]−N−スクシンイミジルプロピオネート(NSP)を用いて、HIRMAb−TNFR融合タンパク質を放射性標識し、それは、1.0x28cmカラムのSephadex G−25での精製および0.02M Tris/0.15M NaCl/pH=6.5(TBS)での溶出後、3.0uCi/ugの比活性および95%のTCA沈澱性をもたらした。該溶液は、TBSおよびUltra−15マイクロコンセントレーター(Millipore, Bedford, MA)とバッファー交換し、TCA沈澱性が99%に増加した。該3H−標識HIRMAb−TNFR融合タンパク質は、霊長類研究の前に標識され、−70℃で保存された。
【0139】
4.1kgのアカゲザル雌成体の左大腿静脈に、1806uCiの[3H]−HIRMAb−TNFR融合タンパク質、428uCiの[125I]−TNFR:Fc融合タンパク質を3.1mLのTBS中、ボーラス注射によって30秒かけて静脈内(IV)注射した。HIRMAb−TNFR融合タンパク質の投与量は、0.15mg/kgであった。該動物を最初、筋内ケタミンで麻酔し、吸入による1%イソフランによって、麻酔を維持した。全ての手順は、米国国立衛生研究所(U.S. National Institutes of Health)によって採用され、公表された研究動物のケアおよび使用のためのガイド(The Guide for the Care and Use of Laboratory Animals)にしたがって行われた。静脈内薬物投与後、3Hおよび125I放射能測定のために、大腿静脈血漿を1、2.5、5、15、30、60および120分に得た。該動物を安楽死させ、主要器官(心臓、肝臓、脾臓、肺、骨格筋、および大網脂肪)の試料を採取し、重量を量り、放射能測定のために処理した。頭蓋を開け、脳を取り出した。放射能測定のために、前頭皮質灰白質、前頭皮質白質、小脳灰白質、および小脳白質の試料を採取した。
【0140】
前頭皮質の試料(〜2グラム)を毛細管除去(capillary depletion)分析のために採取した。脳を組織グラインダー中、8mLの冷PBS中でホモジナイズした。該ホモジネートに9.4mLの冷40%デキストラン(70kDa, Sigma Chemical Co.)を補足し、放射能測定のために、該ホモジネートのアリコートを得た。該ホモジネートを固定角ローター中、3200gにて、4℃で10分間遠心分離した。脳微小血管系が高分子量デキストランの該密度にて、ペレットとして定量的に沈澱し、血管後(post-vascular)上清は、毛細管が除去された(depleted)脳実質の測定値である。血管ペレットおよび上清を3Hおよび125I放射能について、ホモジネートと平行して、カウントした。分布容量(VD)は、120分末梢血漿(terminal plasma)における全125Iまたは3H放射能によって割られた分数における全125Iまたは3H放射能の比率から、3つのフラクションの各々について決定した。
【0141】
血漿および組織試料は、ガンマカウンターを用いて125I放射能について分析し、液体シンチレーションカウンターを用いて3H放射能について分析した。125I同位体元素は、液体シンチレーションカウンター(LSC)の3Hチャンネル(0−12keV)において検出される放射を放つ。したがって、クエンチ剤としてクロロホルムを用いてクエンチ曲線を作製して、3Hウィンドウにおける125Iの計数効率をコンピューターで計算した。3H計数について全ての試料をSoluene−350(Perkin Elmer)中に可溶化し、Opti−Fluor O(Perkin Elmer)中におけるLSCにおいてカウントした。
【0142】
血漿中の3Hまたは125I放射能(DPM/mL)を%注入用量(ID)/mLに変換し、%ID/mLを双指数(bi-exponential)方程式 %ID/mL=A1e−k1t+A2e−k2tにフィットさせた。インターセプト(A1、A2)および傾斜(k1、k2)を用いて、中央滞留時間(MRT)、分布の中心体積(Vc)、分布の定常体積(Vss)、血漿濃度曲線下の面積(AUC)、および全身性クリアランス(CL)をコンピューター計算した。非線形回帰分析を用いて、PKパラメーターをコンピューター計算し、データに1/(%ID/mL)2のウェイトをかけた。
【0143】
透過性−表面積(PS)積(permeability-surface area product)とも呼ばれる器官クリアランス(uL/min/g)を末端器官取り込み(%ID/g)および120分血漿AUC(%IDmin/mL)から次のようにコンピューター計算する。
器官PS積=[(%ID/g)/AUC]*1000
HIRMAb−TNFR融合タンパク質は、[3H]で放射能標識され、TNFR:Fc融合タンパク質は、[125I]で放射能標識され、これらのタンパク質を成体アカゲザルの静脈内に共注射した。血漿放射能のクリアランスを図12Aに示し、TCAにより沈澱性であった血漿放射能を図12Bに示す。PKパラメーターの概算のために、血漿クリアランスプロフィール(図12A)を双指数関数(Methods)にフィットさせ、それを各融合タンパク質につき、表4に示す。脳および末梢器官による該融合タンパク質の取り込みは、% I.D./100グラム組織として測定され、これらの値を表5に示す。該融合タンパク質の分布の脳体積(VD)は、毛細管除去(capillary depletion)法で測定され、ホモジネートについてのVD値、血管ペレット、および血管後上清を表6に示す。
【0144】
HIRMAb−TNFRおよびTNFR:Fc融合タンパク質のBBB PS積は、2時間血漿AUC(図13A)および脳への取り込みまたは%ID/100g(図13B)からコンピューター計算され、該PS積を図13Cに示す。比較のために、図13のデータは、血管性間隙マーカー、ヒトIgG1のAUC、%ID/100g、およびBBB PS積も示す。該PS積は、同様に、末梢器官におけるHIRMAb−TNFRおよびTNFR:Fc融合タンパク質についてコンピューター計算され、これらのデータを表7に示す。各器官におけるTNFR:Fc融合タンパク質についてのPS積に対する、HIRMAb−TNFR融合タンパク質についてのPS積の比率を図14にプロットする。
【0145】
【表3】
図12の血漿クリアランスデータから概算される。
【0146】
【表4】
データは、% I.D./100グラム;平均±SE(n=3)
【0147】
【表5】
平均±SE(n=3). VD=分布の体積(uL/g); TCA=トリクロロ酢酸
【0148】
【表6】
データは、平均±SE(n=3)
【0149】
TNFR:Fc融合タンパク質と比較したHIRMAb−TNFR融合タンパク質の霊長類BBBをイン・ビボで横断する選択的輸送を表5に示し、それにより、%ID/100グラムとして表される該タンパク質の脳への取り込みを与える。アカゲザルの脳の重量が100グラムなので、該取り込みデータは、組織100グラムとして表される。しかしながら、表5の脳取り込みパラメーターは、HIRMAb−TNFRおよびTNFR:Fc融合タンパク質の相対的な脳浸透の直接的な測定値ではなく、以下を考慮すべきである。まず、脳取り込みまたは%ID/gは、該タンパク質の血漿AUCの関数である。%ID/gを表4の血漿AUC値で標準化した結果、器官PS積の算定数値が得られ、PS積は、脳について図13に、末梢器官について表7に示される。第2に、脳取り込みまたは%ID/gは、器官血液容量について補正されなければならない。所定のタンパク質に関する器官取り込み値は、該器官の血液腔における該タンパク質の隔離を単純に反映することができ、それは、組織間で幅広く変化することができる。器官の血液量因子は、HIRMAbのアイソタイプ対照であるヒトIgG1のような血液量マーカーについてのPS積の算定数値によって標準化される。ヒトIgG1についてのBBB PS積を図13に示す。ヒトIgG1およびTNFR:Fc融合タンパク質についてのBBB PS積が等価であることは、TNFR:Fc融合タンパク質がBBBを横断しないことを示す定量的証拠である。脳血液量マーカー、例えば、IgG1またはTNFR:Fc融合タンパク質についてのPS積の算定数値は、どちらのタンパク質も実際のPS積が、脳血液量について補正するとゼロになるので、近似値である。第3に、HIRMAb−TNFR融合タンパク質についての高いBBB PS積(図13)は、脳微小血管系による該融合タンパク質の隔離を反映することができ、BBBを横断する実際のトランスサイトーシスおよび脳実質中への浸透を反映しなかった。このため、毛細管除去分析を行った。血管後上清中のHIRMAb−TNFR融合タンパク質の高いVD、および血管ペレット中の低いVD(表6)は、脳によって取り込まれたHIRMAb−TNFR融合タンパク質の>90%が血管後実質(post-vascular parenchyma)中に浸透したことを明らかにする。TNFR:Fc融合タンパク質のホモジネートVD 13±3uL/g(表6)は、脳血液量に等しく、それは、TNFR:Fc融合タンパク質が血液中、脳の方向へBBBを横断しないことを示すさらなる証拠である。
【0150】
HIRMAb−TNFRおよびTNFR:Fc融合タンパク質のPS積は、また、末梢器官についてコンピューター計算された(表7)。脳および末梢器官に関し、TNFR:Fc融合タンパク質についてのPS積に対する、HIRMAb−TNFR融合タンパク質についてのPS積の比を図14でプロットする。これらのデータは、TNFR:Fc融合タンパク質についてのPS積に対する、HIRMAb−TNFR融合タンパク質についてのPS積の比が、心臓、肺、骨格筋および脂肪などの末梢器官についてほぼ均一であり、肝臓または脾臓などの器官について2〜5倍にやや増加し、脳について、選択的かつ顕著に増加することを示す(図14)。脳におけるPS積比30は、TNFR:Fc融合タンパク質に関する実際のBBB PS積がゼロなので、過小評価である。
【0151】
霊長類の薬物動態(PK)および脳取り込みデータは、HIRMAb−TNFR融合タンパク質を用いる治療的介入の最初の投薬についての検討を可能にする。0.2mg/kgの注射用量での脳取り込み 3.0%ID/100グラム(表5)は、HIRMAb−TNFR融合タンパク質の脳濃度 1.1pmol/グラム融合タンパク質をもたらし、この値は、個々の融合タンパク質あたり2つのTNFR部分が存在するので(図1)、2.2pmol/グラムに等しい。正常な脳における免疫活性TNFαの濃度は、検出不可能であるが、外傷性脳損傷において、0.4pmol/グラムに増加する[E. Shohami, M. Novikov, R. Bass, A. YaminおよびR. Gallily:閉鎖性頭部外傷は、脳組織によるTNFアルファおよびIL−6の早期産生を引き起こす(Closed head injury triggers early production of TNF alpha and IL-6 by brain tissue)、J. Cereb. Blood Flow Metab. 14: 615−9 (1994)]。HIRMAb−TNFR融合タンパク質のTNFαに関するアフィニティーは高いので(図4B)、低用量のHIRMAb−TNFR融合タンパク質 0.2mg/kgは、外傷性脳損傷において、脳における大脳TNFαのほとんどを隔離するであろう。より高用量の該融合タンパク質は、病態において、脳におけるTNFαの本質的に100%を隔離するであろう。
【0152】
まとめると、該実験は、モデルデコイ受容体医薬 TNFRをBBB分子のトロイの木馬 HIRMAbとの融合タンパク質として再操作することにより、イン・ビボでBBBに迅速に浸透する新規な化学物質が生じることを明らかにする。対照的に、当該分野の現状を代表するTNFR:Fc融合タンパク質は、BBBを横断しない。比較的低い全身用量の投与後に生じるHIRMAb−TNFR融合タンパク質の大脳濃度は、病態において、脳における標的サイトカインのほぼ全てを隔離するのに十分である。
【0153】
実施例10 ヒト定常領域の変化
該キメラHIRMAb HCの相補性決定領域(CDRs)およびフレームワーク領域(FR)を包含する該融合タンパク質のHCのドメイン構造を図10に示す。定常領域もまた、図10に示し、ヒトIgG1に由来する。CH1、ヒンジ、CH2、およびCH3ドメインを含むC領域のアミノ酸配列を図10に示す。キメラHIRMAb LCのCDRsおよびFRsを包含するLCのドメイン構造は、図11に示す。定常領域は、ヒトカッパLCに由来し、ヒトカッパ定常領域を含むアミノ酸配列を図11に示す。
【0154】
HIRMAb HC−TNFR融合タンパク質の定常(C)−領域は、配列番号4のアミノ酸133〜461からなり、ヒトIgG1アイソタイプ由来である。さらに、重鎖C−領域は、ヒトIgG2、IgG3、およびIgG4を包含する他のヒトIgGアイソタイプのC−領域に由来することができた。異なるC−領域アイソタイプは、各々、ヒンジ領域周辺の柔軟性、プロテアーゼ感受性、相補体の活性化またはFc受容体への結合に関係する周知の利益または不利益を提供する。HIRMAb LCのC領域は、配列番号6のアミノ酸129〜234からなり、ヒトカッパアイソタイプ由来である。さらに、軽鎖C−領域は、ヒトラムダ鎖アイソタイプ由来であることができた。
【0155】
実施例11 HIRMAb−TNFR融合タンパク質を用いる脳疾患の治療
腫瘍壊死因子(TNF)−αは、脳の急性および慢性障害において病原的役割を果たすプロ炎症性サイトカインである。TNF−αおよびTNFRのどちらも、脳虚血においてアップレギュレートされる(Lambertsenら(2007) Neurosci., 144:934−949)。TNFR細胞外ドメイン(ECD)の経頭蓋投与は、中大脳動脈閉塞(MCAO)モデルにおいて、梗塞のサイズを減少させる(Nawashiroら(1997), Brain Res., 778:265−271)。TNFR ECDは、血液脳関門(BBB)を横断しない大型分子なので、該可溶性デコイ受容体は、脳に直接注射しなければならない。脊髄損傷(SCI)において、ヒトIgG1 FcフラグメントおよびヒトTNFRタイプIIのECDの融合タンパク質のクモ膜下投与は、SCIに関連ずるニューロパシー疼痛を減少させる(Marchandら(2008), Eur. J. Pain, 1−12)。同様に、外傷性脳損傷(TBI)モデルにおけるTNFR−II:Fc融合タンパク質の経頭蓋投与は、治療的である(Knoblachら(1999), J. Neuroimmunol., 95:115−125)。しかしながら、TNFR:Fc融合タンパク質はBBBを横断しないので、TBIにおけるTNFR:Fc融合タンパク質の静脈内投与は、治療的ではない(Knoblachら(1999), J. Neuroimmunol., 95:115−125)。虚血または脳もしくは脊髄損傷などの急性脳障害のほかに、BBB輸送可能TNFR:Fc融合タンパク質の使用はまた、慢性神経変性においても治療的でありうる(Tweedieら(2007), Curr. Alzheimer Res., 4:375−378)。
【0156】
実施例12 MAb−VEGFRデコイ受容体融合タンパク質を用いる脳癌の治療
デコイ血管内皮成長因子(VEGF)受容体(VEGF):Fc融合タンパク質は、操作し、発現させることができる(Holashら(2002), Proc. Natl. Acad. Sci. USA, 99:11393−11398)。VEGF:Fc融合タンパク質は、新規な血管成長のための成長因子である内因性VEGFを隔離すると予想される。かかる医薬は、脳癌を包含する癌における抗血管形成因子としての特定の使用を有しうる。しかしながら、VEGF:Fc融合タンパク質は、BBBを横断することが予測されない大型分子薬物である(Pardridge, (2008), Bioconj. Chem., (19:1327−1338)。IgG融合タンパク質、例えば、TNFRに関して図1に示されるような融合タンパク質として、VEGFRデコイ受容体を再操作することが必要である。VEGFのかかる形態は、内因性インスリン受容体上の輸送を介してヒトBBBに浸透するであろう。いったんBBBを横断すれば、次いで、MAb−VEGR融合タンパク質は、BBBの後ろに、脳内にVEGFを隔離し、脳癌における新規な血管成長を減少させるであろう。
【0157】
実施例13 MAb−Fn14デコイ受容体融合タンパク質を用いる脳虚血の治療
アポトーシスの腫瘍壊死因子様弱インデューサー(TWEAK)は、TNF遺伝子ファミリーのメンバーである。TWEAK受容体は、Fn14と呼ばれる膜タンパク質である。可溶性Fn14デコイ受容体は、Fn14のECDがヒトIgG1 Fcフラグメントのアミノ末端に融合したFc融合タンパク質として操作されている。TWEAK:Fc融合タンパク質は、中大脳動脈閉塞モデルにおいて卒中のサイズを減少させ、TWEAK:Fc融合タンパク質は、該大型分子医薬はBBBを包含しないので、経頭蓋注射によって脳に投与されなければならない(Yepesら(2005), Am. J. Pathol., 166:511−520; Zhangら(2007), J. Cereb. Blood Flow Metab., 27:534−544)。しかしながら、ヒトにおける急性卒中治療を施すために、頭部に穴を開けることは実際的ではない。したがって、Fn14 ECDがBBB浸透性IgGのカルボキシル末端に融合した図1に示されるようなIgG−Fn14融合タンパク質として、Fn14デコイ受容体を再操作することが必要である。かかるタンパク質は、静脈内注射によって非侵襲性で与えることができ、次いで、BBBを横断して虚血脳中への受容体を介して輸送される。
【0158】
実施例14 MAb−LtαRデコイ受容体融合タンパク質を用いる多発性硬化症の治療
リンホトキシンα(LTα)は、また、TNFαとしても知られ、リンホトキシンβ(Ltβ)と共に膜において、ヘテロ−三量体複合体を形成することができる。Ltβ複合体は、Ltβ受容体(LtβR)を活性化させて、細胞内シグナル変換現象および炎症カスケードを開始する。LtβR ECDは、デコイ受容体として作用し、内因性Ltβを隔離する場合があり、それは、脳疾患において治療的であることができる。LtβR:Fc融合タンパク質は、実験的脱髄において治療的であり(Plantら、2007)、ヒト脱髄、例えば、多発性硬化症において治療的でありうる。しかしながら、LtβR:Fc融合タンパク質は、ヒトBBBを横断することが予測されない。LtβRデコイ受容体は、図1に示されるようなMAb−LtβR融合タンパク質として再操作されることができる。かかる融合タンパク質は、BBBを横断し、BBBの後ろに、脳内で内因性Ltβを隔離するであろう。
【0159】
実施例15 MAb−TRAIL−Rデコイ受容体融合タンパク質を用いるニューロAIDS(neuro-AIDS)の治療
TNF関連アポトーシス誘導性リガンド(TRAIL)は、炎症性サイトカインであり、TRAIL受容体(TRAIL−R)への結合を介して作用する。TRAILは、ヒト免疫不全ウイルス(HIV)−1による脳におけるニューロンの感染後、後天性免疫不全症候群(AIDS)の認知症において病原的役割を果たす(Ryanら(2004), J. Neuroimmunol., 148:127−139)。AIDSまたはニューロAIDSの神経学的発現の治療に対する新規なアプローチは、可溶性TRAIL−Rデコイ受容体での、脳におけるTRAILの隔離でありうる。しかしながら、TRAIL−RのECDまたはそのFc融合タンパク質は、BBBを横断する輸送を欠くため、脳に浸透しないであろう。該問題は、TRAIL−Rデコイ受容体を図1に示されるようなMAb−TRAIL−R融合タンパク質として再操作することによって、解決することができた。MAb−TRAIL−R融合タンパク質は、BBBを横断する受容体介在性輸送を経験し、脳に侵入し、そこで、該融合タンパク質は、TRAILを脳内に隔離するであろう。
【0160】
実施例16 MAb−IL6−Rデコイ受容体融合タンパク質を用いる多発性硬化症の治療
インターロイキン(IL)−6のような炎症性サイトカインは、脱髄疾患、例えば、実験的自己免疫脳脊髄炎(EAE)、または多発性硬化症において役割を果たしうる。IL−6受容体(IL−6R)は、可溶性IL−6Rデコイ受容体タンパク質の投与後、脳内で内因性IL−6の作用を抑制することができた。IL−6RのECDがヒトIgG1 Fcフラグメントのアミノ末端に融合したIL−6R:Fc融合タンパク質の投与は、EAEにおいて治療的である(Linkerら(2008), J. Neuroimmunol., 205:64−72)。しかしながら、多発性硬化症におけるIL−6R:Fc融合タンパク質の脳中への浸透は、イン・ビボでのBBBを介する輸送を欠くので、制限されうる。対照的に、図1に示されるようなMAb−IL−6R融合タンパク質は、受容体−媒介性輸送を介してBBBに浸透し、それにより、BBBの後ろに、脳においてIL−6を隔離することができた。
【0161】
本発明の好ましい具体例を本明細書中に示し、記載するが、かかる具体例が例示目的のためだけに提供されることは、当業者に明らかであろう。現在、当業者は、本発明から逸脱することなく、多くの変更、変化および置換を思い付くであろう。本明細書に記載の本発明の具体例の種々の代替物が本発明の実施に用いられうることは、理解されるべきである。添付の特許請求の範囲は、本発明の範囲を明確にすることを目的とし、それにより、特許請求の範囲内の方法および構造およびその等価物が包含される。
【0162】
配列表
表3
【技術分野】
【0001】
相互参照
本願は、2009年3月18日出願の米国仮出願第61/161,320号(出典明示により全体として本明細書の一部とする)の利益を請求する。
【背景技術】
【0002】
発明の背景
腫瘍壊死因子受容体(TNFR)などの標的受容体の可溶性細胞外ドメイン(ECD)は、ヒトの疾患において治療作用を有する。受容体ECDは、内因性リガンド、例えば、腫瘍壊死因子(TNF)−αを封鎖し、それにより、内因性リガンドの内因性標的受容体へのアクセスを遮断する外因性デコイ(decoy)受容体として作用する。デコイ受容体は、脳疾患の強力な新規治療となる可能性があった。しかしながら、デコイ受容体は、他の巨大分子薬物のように、血液脳関門(BBB)を横断しない。かくして、今日までに、組み換えデコイ受容体の全身投与によって脳障害患者を治療することはできなかった。
【発明の概要】
【課題を解決するための手段】
【0003】
発明の概要
本明細書には、BBBを横断してIgG−受容体ECD(デコイ受容体)融合タンパク質を対象のCNSへ送達するための組成物および関連する方法が記載される。特に、該方法は、受容体ECDおよびBBBの表面に発現される受容体の細胞外ドメインに結合する抗体を含む二官能性デコイ受容体融合抗体の治療上有効量を全身投与することによって、デコイ受容体のCNSへの送達を可能にする。
【0004】
したがって、一の態様において、受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体であって、BBB上に発現した受容体および受容体細胞外ドメインのリガンドに結合する融合抗体が提供される。いくつかの具体例において、BBB上に発現される受容体は、インスリン受容体、トランスフェリン受容体、またはリポタンパク質受容体である。いくつかの具体例において、BBB上に発現される受容体は、ヒトインスリン受容体である。いくつかの具体例において、該二官能性デコイ受容体融合抗体は、ヒトインスリン受容体との結合に関して、配列番号4および6または配列番号6および7のアミノ酸配列を含む二官能性デコイ受容体融合抗体と競合する。いくつかの具体例において、受容体細胞外ドメインは、サイトカイン受容体、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン(ephrin)受容体由来である。いくつかの具体例において、受容体細胞外ドメインは、TNF−α受容体細胞外ドメインを含む。いくつかの場合、TNF−α受容体細胞外ドメインは、ヒト、マウス、ラットまたはブタTNF−α受容体細胞外ドメインのアミノ酸配列に対して少なくとも85%(例えば、90%、95%または100%)同一のアミノ酸配列を含む。ある特定の具体例において、TNF−α受容体細胞外ドメインのアミノ酸配列は、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に融合される。ある特定の具体例において、TNF−α受容体細胞外ドメインのアミノ酸配列は、重鎖免疫グロブリンのカルボキシル末端に融合される。
【0005】
関連する態様において、脳への取り込みがタンパク質100グラムあたり1%、2%、3%、5%、7%または10%以上のIDである二官能性デコイ受容体融合抗体(例えば、HIRMAb−TNFR融合タンパク質)が提供される。いくつかの具体例において、該二官能性デコイ受容体融合抗体(例えば、HIRMAb−TNFR融合タンパク質)は、デコイ受容体およびヒトIgGのFcフラグメントの融合タンパク質、例えば、TNFR:Fcの脳への取り込み量よりも1−、2−、5−、10−、13−、15−、17−、20−、25−、30−、35−、40−、45−または50−倍大きい脳内取り込み量を示す。いくつかの具体例において、本明細書に記載の二官能性デコイ受容体融合抗体(例えば、HIRMAb−TNFR融合タンパク質)は、他の器官と比べて、選択的に脳に豊富にある。いくつかの具体例において、TNFR:Fc融合タンパク質に関する器官PS積(organ PS product)に対する本明細書に記載のデコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)に関する器官PS積の比率を複数の器官について決定する場合、脳の比率は、脂肪、筋肉、心臓、肺、肝臓および/または脾臓の比率よりも1、2、5、10、13、15、17、20、25、30、35、40、45、または50倍大きい。
【0006】
関連する態様において、(i)重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列、(ii)軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列、または(iii)(i)または(ii)の相補性配列を含む核酸であって、ここに、該重鎖および軽鎖免疫グロブリンがBBB受容体に対する抗体由来である核酸が提供される。いくつかの具体例において、該コードされた受容体細胞外ドメインは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン(ephrin)受容体由来である。いくつかの具体例において、該コードされた受容体細胞外ドメインは、TNF−α受容体(例えば、ヒトTNF−α受容体)由来である。他の具体例において、該TNF−α受容体由来のコードされた受容体細胞外ドメインは、ヒト、マウス、ラットまたはブタTNF−α受容体細胞外ドメインのアミノ酸配列に対して少なくとも85%同一のアミノ酸配列を含む。いくつかの具体例において、該コードされた免疫グロブリン重鎖または軽鎖は、ヒトインスリン受容体、トランスフェリン受容体またはリポタンパク質受容体に対する抗体由来である。ある特定の具体例において、上記の第1の配列は、配列番号4または7に対応するアミノ酸配列に対して少なくとも85%(例えば、90%、95%または100%)同一のアミノ酸配列をコードし、または上記の第2の配列は、配列番号6に対応するアミノ酸配列と少なくとも85%(例えば、90%、95%または100%)同一のアミノ酸配列をコードする。他の具体例において、該核酸は、高ストリンジェンシー条件下で、配列番号4、6または7をコードする核酸(例えば、配列番号3または5のヌクレオチド配列を含む核酸)にハイブリダイズする。いくつかの具体例において、該TNF−α受容体由来のコードされた細胞外ドメインは、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に共有結合される。いくつかの具体例において、該TNF−α受容体由来のコードされた細胞外ドメインは、重鎖免疫グロブリンのカルボキシル末端に共有結合される。
【0007】
いくつかの具体例において、核酸は、核酸ベクターとして提供される。いくつかの具体例において、核酸ベクターは、(i)重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列、(ii)軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列、または(iii)(i)または(ii)の相補性配列を含み、ここに、該重鎖および軽鎖免疫グロブリンは、BBB受容体に対する抗体由来である。いくつかの具体例において、該核酸ベクターは、(i)を含み、さらに、BBB受容体に対する抗体由来の軽鎖免疫グロブリンをコードする核酸配列を含むか、または(ii)を含み、さらに、BBB受容体に対する抗体由来の重鎖免疫グロブリンをコードする核酸を含む。関連する態様において、上記核酸のいずれかを含む細胞(例えば、哺乳動物細胞)が提供される。
【0008】
さらなる態様において、受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体を含む医薬組成物を対象に全身投与することを含む、血液脳関門を横切ってデコイ受容体を送達する方法であって、該融合抗体がBBB上に発現された受容体および受容体細胞外ドメインのリガンドに結合するところの方法が提供される。いくつかの具体例において、該BBB上に発現された受容体は、インスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である。いくつかの具体例において、該受容体細胞外ドメインは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である。いくつかの具体例において、該TNF−α受容体由来の細胞外ドメインは、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に共有結合している。いくつかの具体例において、該TNF−α受容体由来の細胞外ドメインは、重鎖免疫グロブリンのカルボキシル末端に共有結合している。
【0009】
別の態様において、受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体を含む医薬組成物の治療上有効量を対象に全身投与することを含むCNS疾患の治療方法であって、該融合抗体がBBB上に発現された受容体および受容体細胞外ドメインのリガンドに結合するところの方法が提供される。いくつかの具体例において、該BBB上に発現された受容体は、インスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である。いくつかの具体例において、該受容体細胞外ドメインは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮長因子受容体、またはエフリン受容体由来である。いくつかの具体例において、治療されるべきCNS疾患は、急性CNS疾患、例えば、全脳虚血、局所脳虚血、外傷性脳損傷、または脊髄損傷である。他の具体例において、治療されるべきCNS疾患は、慢性CNS疾患、例えば、神経変性疾患、例えば、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症、ハンチントン病、多発性硬化症、横断性脊髄炎、運動ニューロン疾患、ピック病、結節硬化症、カナバン病、レット症候群、脊髄小脳失調症、フリードライヒ失調症、視神経萎縮、または網膜変性である。
【0010】
また別の態様において、(i)受容体細胞外ドメインに融合した免疫グロブリン重鎖、および免疫グロブリン軽鎖の両方、または
(ii)受容体細胞外ドメインに融合した免疫グロブリン軽鎖、および免疫グロブリン重鎖の両方
をコードする単一タンデム発現ベクターを真核細胞中に安定に組み込むことを含む、二官能性デコイ受容体融合抗体を製造する方法であって、該コードされた免疫グロブリン重鎖および免疫グロブリン軽鎖がBBB上に発現された受容体に対する抗体由来であるところの方法が提供される。いくつかの具体例において、該BBB上に発現された受容体は、インスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である。いくつかの具体例において、該コードされた受容体細胞外ドメインは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である。いくつかの具体例において、該TNF−α受容体由来の受容体細胞外ドメインは、免疫グロブリン重鎖または免疫グロブリン軽鎖のカルボキシル末端に共有結合される。いくつかの具体例において、該TNF−α受容体由来のコードされた受容体細胞外ドメインは、免疫グロブリン重鎖のカルボキシル末端に共有結合される。
【0011】
出典明示による組み込み
本明細書に挙げられた全ての出版物、特許、および特許出願は、出典明示により、あたかも個々の出版物、特許または特許出願が詳細かつ個々に出典明示により本明細書の一部とされることが示されたかの如く、同程度まで本明細書の一部とされる。
【0012】
図面の簡単な説明
本発明の新規な特徴は、特に、添付の特許請求の範囲で示される。本発明の特徴および利益のより良い理解は、発明の本質を利用する実例となる具体例を示す下記の詳細な説明、および添付の図面を参照して得られるであろう。
【図面の簡単な説明】
【0013】
【図1】HIRMAb−TNFR融合タンパク質は、TNFR ECDのアミノ末端の、キメラHIRMAbの重鎖のCH3領域のカルボキシル末端への融合によって形成される。該融合タンパク質は、二官能性分子である。すなわち、該融合タンパク質は、BBBにて、HIRを結合して脳中への輸送を媒介し、かつ、TNF−αを結合して該サイトカインの炎症性を抑制する。
【0014】
【図2】(A)ヒトU87グリア細胞由来のRNAの逆転写によって生産されたcDNAからPCRによって生産されたヒトTNFR ECD cDNAのアガロースゲルのエチジウムブロマイド染色(レーン1)、およびTNFR−特異的ODNプライマー(表2)。レーン2および3:DNAサイズ標準。(B)pHIRMAb−TNFR、TNFR ECDおよびキメラHIRMAbの重鎖(HC)の融合タンパク質をコードする真核細胞発現プラスミドの遺伝子操作。融合遺伝子の5’側には、サイトメガロウイルス(CMV)プローモーターが配置され、3’側には、ウシ成長ホルモンポリA(pA)配列が配置される。
【0015】
【図3】タンパク質Aアフィニティー精製キメラHIRMAbおよびHIRMAb−TNFR融合タンパク質の還元SDS−PAGEおよびクーマシーブルー染色。どちらも均質に精製され、重鎖および軽鎖から構成される。
【0016】
【図4】抗ヒト(h)IgG一次抗体(左パネル)または抗ヒトTNFR−II一次抗血清(右パネル)のいずれかでのウェスタンブロット。HIRMAb−TNFR融合タンパク質の免疫反応性をキメラHIRMAbと比較する。HIRMAb−TNFR融合タンパク質およびHIRMAbの両方は、抗−hIgGウェスタンにおいて同一の軽鎖を有する。HIRMAb−TNFR融合重鎖は、抗−hIgGおよび抗−ヒトTNFR抗体の両方と反応するが、HIRMAb重鎖は、抗−hIgG抗体とのみ反応する。HIRMAb−TNFR融合重鎖のサイズは、30kDa TNFR ECDの55kDa HIRMAb重鎖への融合のために、HIRMAbの重鎖のサイズよりも約30kDa大きい。
【0017】
【図5】キメラHIRMAbまたはHIRMAb−TNFR融合タンパク質のいずれかのHIR細胞外ドメイン(ECD)に対する結合は飽和可能である。HIR ECDに結合するHIRMAb−TNFRのED50は、キメラHIRMAbの結合のED50に匹敵する。
【0018】
【図6】TNFR:Fc融合タンパク質(A)またはHIRMAb−TNFR融合タンパク質(B)のTNFαに対する結合は飽和可能である。パネルAに示されるように、ヒトIgG1のTNFαに対する結合はない。線形回帰分析の傾きは、KD/Amax比を生じ(ここに、KDは、TNFαの結合定数であり、Amaxは、最大吸光度である)、TNFαに対する結合のKDの相対的指標である。TNFR:Fc融合タンパク質およびHIRMAb−TNFR融合タンパク質のどちらも、比較可能なアフィニティーで、TNFαに結合する。
【0019】
【図7】(A)TNFαのHIRMAb−TNFR融合タンパク質に対する結合に関する放射性受容体アッセイのアウトライン。マウス抗−ヒト(MAH)IgG1 Fcを播種し、それは、HIRMAb−TNFR融合タンパク質のFc領域に結合した。次いで、該融合タンパク質のTNFR細胞外ドメイン(ECD)領域が[125I]−TNFαに結合し、それを非標識TNFαの添加によって置換した。(B)飽和可能な結合を非線形回帰分析によって分析して、HIRMAb−TNFR融合タンパク質に結合しているTNFαの50%阻害を与えた濃度KDを得た。
【0020】
【図8】TNFαは、アクチノマイシンDで処理したヒトWEHI−13VAR細胞において、ED50 約10pg/mLで細胞毒性を引き起こす。しかしながら、1.4nM TNFR:Fcまたは1.4nM HIRMAb−TNFRのいずれかの存在下、高濃度のTNFαによって細胞毒性は引き起こされない。
【0021】
【図9】HIRMAb−TNFR融合タンパク質の重鎖および軽鎖をコードする分離したタンデムな発現カセット(各遺伝子は、CMVプロモーターを有する分離したタンデムなイントロンによって駆動し、各々、ウシ成長ホルモン(BGH)ポリアデニル化(pA)配列を末端に有する)を含有するタンデムベクター(TV)であるpTV−HIRMAb−TNFRの遺伝子操作。pTV−HIRMAb−TNFRは、HIRMAbをコードするユニバーサルTV(pTV−HIRMAbと称する)内で、TNFR ECD cDNAをHIRMAb HCカセットの3’末端にて、独特なHpaI部位にサブクローニングすることによって生じる。TNFR ECD cDNAは、pCD−HIRMAb−TNFRプラスミド(図2B)を鋳型として用いるPCRによって生産された。
【0022】
【図10】HIRMAb−TNFR融合タンパク質の重鎖のドメイン構造。19アミノ酸IgGシグナルペプチドの次にHIRMAb重鎖のVHが配置され、それは、3つのCDR(CDR1、CDR2、CDR3)および4つのFR(FR1、FR2、FR3、FR4)からなり、次に、ヒトIgG1重鎖C領域のドメイン(CH1、ヒンジ、CH2、CH3)が配置され、次に、3−アミノ酸リンカー(Ser−Ser−Ser)が配置され、次に、ヒトTNFR−II ECDの235−アミノ酸配列が配置される。3つのN−結合型グリコシル化部位に下線を付し、CH2領域内に1部位およびTNFR領域内に2部位を含む。
【0023】
【図11】HIRMAb−TNFR融合タンパク質の軽鎖ドメイン構造。20アミノ酸IgGシグナルペプチドの次に、HIRMAb軽鎖のVLが配置され、それは、3つのCDR(CDR1、CDR2、CDR3)および4つのFR(FR1、FR2、FR3、FR4)からなり、その次に、ヒトカッパ軽鎖C領域が配置される。
【0024】
【図12】(A)[125I]−TNFR:Fc融合タンパク質および[3H]−HIRMAb−TNFR融合タンパク質の血漿濃度を、成体アカゲザルにおける該タンパク質の単回静脈内注射後の時間に対してプロットする。データは、%注射用量(I.D.)/mLとして表される。(B)10%トリクロロ酢酸(TCA)によって沈澱可能な血漿放射活性の%を両タンパク質の注射後の時間に対してプロットする。データは、平均±SE(n=1時点につき3回繰り返す)である。
【0025】
【図13】濃度曲線下の血漿面積またはAUC(A)、脳100グラムあたりの脳取り込み量または注射用量(I.D.)%(B)、およびBBBの透過係数表面積(permeability surface area: PS)の積(C)をTNFR:Fc融合タンパク質、HIRMAb−TNFR融合タンパク質、および脳血漿量マーカー、ヒトIgG1(hIgG1)についてプロットする。全ての測定値は、該タンパク質の静脈内投与後2時間で得られた。データは、平均±SE(n=1時点につき3回繰り返す)である。
【0026】
【図14】TNFR:Fc融合タンパク質に関する器官PS積に対する、HIRMAb−TNFR融合タンパク質に関する器官PS積の比率を各器官についてプロットする。データは、平均±SE(n=1時点につき3回繰り返す)である。脳に関する比率は、前頭灰白質、前頭白質、小脳灰白質、および小脳白質についての値の平均であり、22〜37で変動した。
【発明を実施するための形態】
【0027】
発明の詳細な説明
I.序文
II.いくつかの定義
III.血液脳関門
IV.BBBを横断する輸送のためのデコイ受容体融合抗体
V.組成物
VI.核酸、ベクター、細胞および製造
VII.方法
VIII.実施例
IX.配列
【0028】
略語
AA アミノ酸
BBB 血液脳関門
BCA ビシンコニン酸
BGH ウシ成長ホルモン
CDR 相補性決定領域
CHO チャイニーズハムスター卵巣
CMV サイトメガロウイルス
DC 希釈クローニング
DHFR ジヒドロ葉酸還元酵素
ECD 細胞外ドメイン
ED50 50%飽和をもたらす有効用量
FR フレームワーク領域
FS フランキング領域
FWD 順方向
HC 重鎖
HIR ヒトインスリン受容体
HIRMAb HIRに対するMAb
HIRMAb HC HIRMAbの重鎖
HIRMAb LC HIRMAbの軽鎖
HIRMAb−TNFR HIRMAbおよびTNFR ECDの融合タンパク質(ここに、TNFRがHCカルボキシル末端に融合する)
HT ヒポキサンチン−チミジン
IgG 免疫グロブリンG
IGF インスリン様成長因子
LC 軽鎖
MAb モノクローナル抗体
MAH マウス抗−ヒトIgG
MTX メトトレキサート
MW 分子量
N アスパラギン
nt ヌクレオチド
ODN オリゴデオキシヌクレオチド
orf オープンリーディングフレーム
pA ポリ−アデニル化
PAGE ポリアクリルアミドゲル電気泳動
PBS リン酸緩衝化セーライン
PBST PBSおよびTween−20
PCR ポリメラーゼ連鎖反応
pI 等電点
R 受容体
REV 逆方向
RNase A リボヌクレアーゼA
RT 逆転写
RT 室温
SDM 部位特異的変異誘発
SDS ドデシル硫酸ナトリウム
SEC サイズ排除クロマトグラフィー
Ser セリン
SFM 無血清培地
TH トロイの木馬
TNF 腫瘍壊死因子
TNFR TNF受容体
TV タンデムベクター
UTV ユニバーサルTV
VH 重鎖の可変領域
VL 軽鎖の可変領域
【0029】
I.序文
遺伝子操作されたデコイ受容体は、特に受容体ECDがヒト免疫グロブリンG、例えばIgG1のFcフラグメントのアミノ末端に融合される場合、強力な新規治療剤である。Fcフラグメントは、2つの定常領域鎖からなる二量体タンパク質である。ほとんどの受容体は膜内で二量体を形成するので、受容体ECDのFcフラグメントへの融合により、受容体タンパク質の天然二量体コンフィギュレーションが可能となる。デコイ受容体のよく記載される一例は、TNFαデコイ受容体−Fc融合タンパク質である。TNFαデコイ受容体−Fc融合タンパク質は、プロ炎症性サイトカイン、TNFαを封鎖し、それにより、その内因性リガンドによる内因性TNFRの活性化を阻害する能力のために、末梢性炎症疾患の強力な生物学的治療剤である。しかしながら、TNFR:Fc融合タンパク質は、CNS疾患のために処方することはできない。
【0030】
血液脳関門は、全身投与された可溶性デコイ受容体の中枢神経系への送達に対する厳しい障害であり、ここに、それらは、いくつかの神経病理、例えば、神経炎症において役割を果たすある種のリガンド(例えば、TNF−α)を中和するように作用する。本明細書に記載の組成物および方法は、BBBを横断してデコイ受容体をCNSへ送達することにおける重要な3つの因子、すなわち、1)BBBを横断できるようにするための受容体細胞外ドメイン(ECD)の修飾(本明細書中で、「デコイ受容体」という)、2)全身投与された修飾されたIgG−受容体融合抗体のCNS中への取り込み量および率、および3)一旦BBBを横断したデコイ受容体活性の保持に向けられる。本明細書に記載の方法および組成物の種々の態様は、介在するリンカー配列を用いて、またはリンカー配列を用いずに、BBB上に発現された受容体(例えば、ヒトインスリン受容体)の細胞外ドメインに対する免疫グロブリン(重鎖または軽鎖)に融合したデコイ受容体を含む、全身投与することができる融合抗体を提供することによって、これらの因子に向けられる。
【0031】
したがって、本発明は、その必要のある対象に、BBB上に発現された受容体(例えば、hIR)に対する抗体および受容体ECD(例えば、ヒトTNF−α受容体ECD)を含む二官能性デコイ受容体融合抗体の治療上有効量を全身投与することによる、必要とする対象、例えば、CNS炎症に罹患した対象またはその危険性のある対象の中枢神経系にデコイ受容体(例えば、可溶性TNF−α受容体ECD)を送達するための組成物および方法を提供する。
【0032】
II.いくつかの定義
「抗体」なる語は、天然または部分合成もしくは全合成免疫グロブリンをいう。該用語は、また、抗原結合ドメインである結合ドメインまたは抗原結合ドメインに相同性の結合ドメインを有するいずれかのポリペプチドまたはタンパク質を包含する。CDR移植(grafted)抗体もまた、該用語によって意図される。
【0033】
「天然抗体」および「天然免疫グロブリン」は、通常、2つの同一の軽(L)鎖および2つの同一の重(H)鎖からなる約150,000ダルトンのヘテロ四量体糖タンパク質である。各軽鎖は、典型的に、1つの共有ジスルフィド結合によって重鎖に連結され、一方、異なる免疫グロブリンアイソタイプの重鎖によってジスルフィド結合の数は変化する。各重鎖および軽鎖は、また、規則的に間隔をあけた鎖内ジスルフィド架橋を有する。各重鎖は、一方の端に可変ドメイン(「VH」)を有し、続いて、いくつかの定常ドメイン(「CH」)を配置する。各軽鎖は、一方の端に可変ドメイン(「VL」)、もう一方の端に定常ドメイン(「CL」)を有し、軽鎖の定常ドメインは、重鎖の第1の定常ドメインと並んでおり、軽鎖可変ドメインは、重鎖の可変ドメインと並んでいる。特定のアミノ酸残基は、軽鎖および重鎖可変ドメイン間にインターフェースを形成すると考えられる。
【0034】
「可変ドメイン」なる語は、ファミリーメンバーのなかで(すなわち、異なるアイソ形態のなかで、または異なる種において)広範囲に配列が異なるタンパク質ドメインをいう。抗体に関して、「可変ドメイン」なる語は、特定の抗原に対する各特定の抗体の結合および特異性において使用される抗体の可変ドメインをいう。しかしながら、変化性(variability)は、抗体の可変ドメインを通して均等に分布されているわけではない。それは、軽鎖および重鎖可変ドメインともに、超可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存された部分を「フレームワーク領域」または「FR」と呼ぶ。非修飾重鎖および軽鎖の各々の可変ドメインは、4つのFR(各々、FR1、FR2、FR3およびFR4)を含み、それは、主として、場合によっては、β−シート構造の一部を形成するβ−シート構造を連結するループを形成する3つの超可変領域によって連結されたβ−シートコンフィギュレーションを採用している。各鎖の超可変領域は、FRによってごく接近して結びついており、他の鎖由来の超可変領域と共に、抗体の抗原結合部位の形成に寄与する(Kabatら、(1991),Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md.,第647−669頁参照)。定常ドメインは、抗原に対する抗体の結合に直接関与しないが、抗体依存性細胞毒における抗体の関与などの種々のエフェクター機能を示す。
【0035】
「超可変領域」なる語は、本明細書中で使用される場合、抗原−結合に関与する抗体のアミノ酸残基をいう。超可変領域は、相補的に抗原に直接結合し、各々、CDR1、CDR2およびCDR3として知られる3つの「相補性決定領域」または「CDR」由来のアミノ酸残基を含む。軽鎖可変ドメインにおいて、CDRは、典型的に、およその残基24−34(CDRL1)、50−56(CDRL2)および89−97(CDRL3)に相当し、重鎖可変ドメインにおいて、CDRは、典型的に、およその残基31−35(CDRH1)、50−65(CDRH2)および95−102(CDRH3)に相当し(Kabatら、(1991), Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md.参照)、および/または「超可変ループ」由来の残基(すなわち、軽鎖可変ドメインにおいて、残基26−32(L1)、50−52(L2)および91−96(L3)、および重鎖可変ドメインにおいて、26−32(H1)、53−55(H2)および96−101(H3))に相当する(ChothiaおよびLesk,(1987), J. Mol. Biol., 196:901−917参照)。
【0036】
本明細書中で使用される場合、「可変フレームワーク領域」または「VFR」なる語は、抗原結合ポケットまたは溝の一部を形成し、および/または抗原に接触しうるフレームワーク残基をいう。いくつかの具体例において、フレームワーク残基は、抗原結合ポケットまたは溝の一部であるループを形成する。該ループにおけるアミノ酸残基は、抗原に接触しても、しなくてもよい。ある具体例において、VFRのループアミノ酸は、抗体の三次元構造、抗体重鎖または抗体軽鎖の検査により決定される。溶媒にアクセス可能なアミノ酸位置はおそらく、ループを形成し、および/または抗体可変ドメインに接触する抗原を提供するので、該三次元構造は、溶媒にアクセス可能なアミノ酸位置について分析することができる。溶媒にアクセス可能な位置のいくつかは、アミノ酸配列多様性を容認でき、その他は(例えば、構造的位置)あまり多様化されることがない。抗体可変ドメインの三次元構造は、結晶構造またはタンパク質モデリングから由来することができる。いくつかの具体例において、VFRは、重鎖可変ドメインのアミノ酸位置71〜78(該位置は、Kabatら、1991にしたがって規定される)に対応するアミノ酸位置を含む、該アミノ酸位置から実質的になる、または該アミノ酸位置からなる。いくつかの具体例において、VFRは、CDRH2とCDRH3の間に配置されたフレームワーク領域3の一部を形成する。VFRは、標的抗原と接触するように、または抗原結合ポケットの一部を形成するように配置されたループを形成することができる。
【0037】
重鎖の定常ドメインのアミノ酸配列に依存して、免疫グロブリンは種々のクラスに割り当てることができる。免疫グロブリンの5つの主要なクラス、すなわち、IgA、IgD、IgE、IgGおよびIgMがあり、このうちいくつかは、さらにサブクラス(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgAおよびIgA2に分けることができる。免疫グロブリンの異なるクラスに対応する重鎖定常ドメイン(Fc)は、各々、α、δ、ε、γおよびμと呼ばれる。免疫グロブリンの異なるクラスのサブユニット構造および三次元コンフィギュレーションは、よく知られている。
【0038】
いずれかの脊椎動物種由来の抗体(免疫グロブリン)の「軽鎖」は、定常ドメインのアミノ酸配列に基づいて、カッパまたは(「κ」)およびラムダまたは(「λ」)と呼ばれる2つの明らかに別個の型のうちの1つに割り当てることができる。
【0039】
本明細書に記載の抗体または融合抗体への言及において、「選択的に結合する」、「選択的に結合」、「特異的に結合する」または「特異的に結合」なる語は、解離定数(Kd)が約10−6M以下、すなわち、10−7、10−8、10−9、10−10、10−11または10−12Mである、抗体または融合抗体のその標的抗原に対する結合をいう。
【0040】
本明細書中で使用される場合、「抗体」なる語は、また、抗原に特異的に結合する能力を保持する抗体の1以上のフラグメントを意味すると理解される(概して、Holligerら、(2005), Nature Biotech., 23(9):1126−1129参照)。かかる抗体の非限定的な例は、(i)Fabフラグメント、VL、VH、CLおよびCH1ドメインからなる1価のフラグメント、(ii)F(ab’)2フラグメント、ヒンジ領域でジスルフィド架橋によって連結された2個のFabフラグメントを含む二価のフラグメント、(iii)VHおよびCH1ドメインからなるFdフラグメント、(iv)抗体の単一アームのVLおよびVHドメインからなるFvフラグメント、(v)VHドメインからなるdAbフラグメント(Wardら、(1989) Nature, 341:544 546)、および(vi)単離された相補性決定領域(CDR)を包含する。さらに、Fvフラグメントの2つのドメイン、VLおよびVHは、別々の遺伝子によってコードされるが、組み換え法を用いて、これらのドメインを、VLおよびVH領域が対になって一価の分子を形成する単一タンパク質鎖とすることが可能な合成リンカーによって連結することができる(一本鎖Fv(scFv)として知られる;例えば、Birdら、(1988) Science, 242:423 426; and Huston et al., (1988) Proc. Natl. Acad. Sci. USA, 85:5879 5883;およびOsbournら、(1998), Nat. Biotechnol., 16:778参照)。かかる一本鎖抗体もまた、「抗体」なる語に包含されることが意図される。特異的scFvのいずれのVHおよびVL配列も、完全IgG分子または他のアイソタイプをコードする発現ベクターを生成するために、ヒト免疫グロブリン定常領域cDNAまたはゲノム配列に連結することができる。VHおよびVLは、また、タンパク質化学または組み換えDNA技術を用いて、Fab、Fvまたは免疫グロブリンの他のフラグメントの生成に使用することができる。一本鎖抗体の他の形態、例えば、ダイアボディもまた包含される。
【0041】
「F(ab’)2」および「Fab’」部分は、免疫グロブリン(モノクローナル抗体)をペプシンおよびパパインなどのプロテアーゼで処理することによって製造でき、2つのH鎖の各々において、ヒンジ領域間に存在するジスルフィド結合付近で免疫グロブリンを消化することによって生成される抗体を包含する。例えば、パパインは、2つのH鎖の各々において、ヒンジ領域間に存在するジスルフィド結合の上流でIgGを切断して、VL(L鎖可変領域)およびCL(L鎖定常領域)からなるL鎖、およびVH(H鎖可変領域)およびCHγ1(H鎖の定常領域におけるγ1領域)からなるH鎖フラグメントがジスルフィド結合を介してそれらのC末端領域で連結した2つの相同性抗体フラグメントを生成する。これらの2つの相同性抗体フラグメントの各々は、Fab’と呼ばれる。また、ペプシンは、2つのH鎖の各々において、ヒンジ領域間に存在するジスルフィド結合の下流でIgGを切断して、2つの上記Fab’がヒンジ領域で連結したフラグメントよりも僅かに大きい抗体フラグメントを生成する。該抗体フラグメントをF(ab’)2と呼ぶ。
【0042】
Fabフラグメントは、また、軽鎖の定常ドメインおよび重鎖の第1の定常ドメイン(CH1)を含有する。Fab’フラグメントは、抗体ヒンジ領域由来の1以上のシステインを含む重鎖CH1ドメインのカルボキシル末端にて2、3個の残基の付加によってFabフラグメントと相違する。Fab’−SHは、定常ドメインのシステイン残基が遊離チオール基を有するFab’のための本明細書中での名称である。F(ab’)2抗体フラグメントは、元々、Fab’フラグメントの対として生産されたフラグメントであり、該フラグメント間にヒンジシステインを有する。抗体フラグメントの他の化学的カップリングもまた、知られている。
【0043】
「Fv」なる語は、完全な抗原認識および抗原結合部位を含有する最小抗体フラグメントである。該領域は、密接な非共有結合において、1つの重鎖および1つの軽鎖可変ドメインの二量体からなる。該コンフィギュレーションにおいて、各可変ドメインの3つの超可変領域が相互作用して、VH−VL二量体の表面における抗原−結合部位を明確に規定する。集合的に、6つの超可変領域が抗原−結合特異性を抗体に付与する。しかしながら、単一の可変ドメイン(または抗原に特異的な3つの超可変領域のみを含むFvの半分)であっても、抗原を認識し、結合する能力を有するが、完全な結合部位よりもアフィニティーは低い。
【0044】
「一本鎖Fv」または「sFv」抗体フラグメントは、抗体のVH、VL、またはVHおよびVLの両方のドメインを含み、ここに、両ドメインは、単一のポリペプチド鎖に存在する。いくつかの具体例において、Fvポリペプチドは、さらに、VHおよびVLドメイン間にポリペプチドリンカーを含み、それにより、該sFvが抗原結合のための所望の構造を形成できるようにする。sFvの概要に関し、例えば、Pluckthun、The Pharmacology of Monoclonal Antibodies, Vol. 113, Rosenburg and Moore eds. Springer−Verlag, New York, pp. 269 315 (1994)を参照のこと。
【0045】
「キメラ」抗体は、異なる哺乳動物起源の組合せから得られる抗体を包含する。該哺乳動物は、例えば、ウサギ、マウス、ラット、ヤギ、またはヒトであってもよい。異なる哺乳動物の組合せは、ヒトおよびマウス起源のフラグメントの組合せを包含する。
【0046】
いくつかの具体例において、本発明の抗体は、モノクローナル抗体(MAb)、典型的にはマウスモノクローナル抗体のヒト化によって誘導される、キメラヒト−マウス抗体である。かかる抗体は、例えば、抗原投与に応答して特異的なヒト抗体を産生するように「操作」されたトランスジェニックマウスから得られる。該技術において、ヒト重鎖および軽鎖上の座にある要素が、内在性重鎖および軽鎖上の座の標的とされた破壊を含有する胚性幹細胞系列から由来するマウス系統中に導入される。該トランスジェニックマウスは、ヒト抗原に特異的なヒト抗体を合成することができ、該マウスは、ヒト抗体を分泌するハイブリドーマを生産するために使用することができる。
【0047】
本明細書中で使用される場合、「治療」または「治療する」なる語は、治療的利益および/または予防的利益を達成することを包含する。治療的利益によって、治療されている原因となる障害または疾患の根絶または改善が意味される。治療の予防的利益には、疾患の危険性を減少すること、疾患の進行を遅らせること、または疾患の発症の可能性を減少することが包含される。本明細書中で使用される場合、「治療する」または「治療」には、予防が包含される。
【0048】
本明細書中で使用される場合、「有効量」は、全身投与される場合、CNSにおいて有益な結果または所望の結果をもたらすのに十分な量であることができる。有効量は、また、予防的効果を生じる量、例えば、急性の病理学的状態または望ましくない状態の出現を遅延させる、減少させる、または排除する量である。有効量は、1以上の投与において投与することができる。治療に関して、本発明の組成物の「有効量」は、障害、例えば、神経学的障害の進行を緩和、改善、安定化、逆転または遅延させるのに十分な量である。「有効量」は、単独または疾患もしくは障害を治療するために使用される1以上の薬剤と共に使用される本発明の組成物のいずれかであってもよい。本発明の意味内で治療剤の「有効量」は、患者の担当医師または獣医師によって決定されるであろう。かかる量は、当業者によって容易に確かめられ、本発明にしたがって投与される場合、治療効果をもたらすであろう。
【0049】
本明細書中で使用される場合、「分子のトロイの木馬」なる語は、BBBを横断して輸送される分子であって、それ自体でBBBを横断しない別の分子に共有または非共有結合した場合、CNS中へのBBB横断輸送(trans-BBB transport)のための渡し船として作用することができる分子をいう。分子のトロイの木馬の例は、限定するものではないが、BBB上に発現された受容体、例えば、インスリン受容体、トランスフェリン受容体、IGF受容体、リポタンパク質受容体、またはレプチン受容体のECDに結合するポリペプチド(例えば、抗体)を包含する。
【0050】
本明細書中で使用される場合、「対象」または「個体」は、動物、例えば、哺乳動物である。いくつかの具体例において、「対象」または「個体」は、ヒトである。いくつかの非限定的な具体例において、対象は、慢性または急性CNS疾患に罹患している。
【0051】
いくつかの具体例において、デコイ受容体ECD−BBB受容体Ab融合抗体を含む薬理学的組成物は、「末梢に投与される」または「末梢投与される」。本明細書中で使用される場合、これらの用語は、CNSに対する直接的投与ではない、すなわち、薬剤を血液脳関門の脳ではない側と接触させる、個体に対する薬剤、例えば、治療剤の投与のいずれかの形態をいう。本明細書中で使用される場合、「末梢投与」は、静脈内、動脈内、皮下、筋内、腹腔内、経皮、吸入によるもの、経頬、鼻腔内、経腸、経口、非経口、舌下、または経鼻を包含する。
【0052】
本明細書において、「医薬上許容される担体」または「医薬上許容される賦形剤」は、それ自体が組成物を受け取る個体に有害な抗体産生を誘導しないいずれかの担体をいう。かかる担体は、当業者に周知である。医薬上許容される担体/賦形剤のディスカッション全体は、Remington’s Pharmaceutical Sciences, Gennaro, AR, ed., 20th edition, 2000: Williams and Wilkins PA, USAに見ることができる。例示的な医薬上許容される担体は、塩、例えば、無機酸塩、例えば、塩酸塩、臭化水素酸塩、リン酸塩、硫酸塩など、および有機酸の塩、例えば、酢酸塩、プロピオン酸塩、マロン酸塩、安息香酸塩等を包含することができる。例えば、本発明の組成物は、液体形態で提供されてもよく、0.01−1%のポリソルベート−80などの界面活性剤、または炭水化物添加剤、例えば、マンニトール、ソルビトールまたはトレハロースを用いて、または用いずに、種々のpH(pH5〜8)のセーラインを基礎とする水性溶液中で処方されてもよい。一般的に使用されるバッファーは、ヒスチジン、酢酸塩、リン酸塩、またはクエン酸塩を包含する。
【0053】
「組み換え宿主細胞」または「宿主細胞」は、挿入方法、例えば、直接的取り込み、形質導入、f−交配、または組み換え宿主細胞を作出するための当該分野で既知の他の方法に関係なく、外因性ポリヌクレオチドを含む細胞をいう。該外因性ポリヌクレオチドは、非組み込みベクター、例えば、プラスミドとして維持されてもよく、または別法では、宿主ゲノム中に組み込まれてもよい。
【0054】
「ポリペプチド」、「ペプチド」および「タンパク質」なる語は、本明細書中で交換可能に使用され、アミノ酸残基のポリマーを示す。すなわち、ポリペプチドに向けられた記載は、等しく、ペプチドの記載およびタンパク質の記載に適用され、その逆も同様である。該用語は、天然のアミノ酸ポリマーならびに1以上のアミノ酸残基が非天然アミノ酸、例えば、アミノ酸類似体であるアミノ酸ポリマーに適用される。本明細書中で使用される場合、該用語は、全長タンパク質(すなわち、抗原)を包含するいずれの長さのアミノ酸鎖も包含し、ここに、該アミノ酸残基は共有ペプチド結合によって連結される。
【0055】
「アミノ酸」なる語は、天然および非天然アミノ酸、ならびに天然アミノ酸と同様に機能するアミノ酸類似体およびアミノ酸模倣物をいう。天然にコードされるアミノ酸は、20個の一般的なアミノ酸(アラニン、アルギニン、アスパラギン、アスパラギン酸、システイン、グルタミン、グルタミン酸、グリシン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、プロリン、セリン、スレオニン、トリプトファン、チロシン、およびバリン)およびピロリジンおよびセレノシステインである。アミノ酸類似体は、天然アミノ酸と同じ基本化学構造、すなわち、水素に結合したα炭素、カルボキシル基、アミノ基、およびR基を有する化合物、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムをいう。かかる類似体は、修飾されたR基(例えば、ノルロイシン)または修飾されたペプチド骨格を有するが、天然アミノ酸と同じ基本化学構造を保持する。
【0056】
アミノ酸は、本明細書において、一般的に知られた3文字記号によって、またはIUPAC−IUB Biochemical Nomenclature Commissionによって推奨される1文字記号によって言及されうる。同様に、ヌクレオチドは、一般的に許容される1文字コードによって言及されうる。
【0057】
「核酸」なる語は、デオキシリボヌクレオチド、デオキシリボヌクレオシド、リボヌクレオシド、またはリボヌクレオチドおよび一本鎖または二本鎖形態のいずれかのそのポリマーをいう。特に限定しない限り、該用語は、参照核酸と類似の結合特性を有し、天然ヌクレオチドと同様に代謝される天然ヌクレオチドの既知の類似体を含有する核酸を包含する。別段に特に限定しない限り、該用語は、また、PNA(ペプチド核酸)、アンチセンス技術において使用されるDNAの類似体(ホスホロチオエート、ホスホロアミデートなど)を包含するオリゴヌクレオチド類似体をいう。別記しない限り、特定の核酸配列は、また、その保存的に修飾された変種(限定するものではないが、縮重コドン置換を包含する)および相補性配列ならびに明示された配列を暗示的に包含する。詳細には、縮重コドン置換は、1以上の選択された(または全ての)コドンの3番目の位置を混合塩基および/またはデオキシイノシン残基で置換された配列を生成することによって達成されうる(Batzerら(1991), Nucleic Acid Res., 19:5081; Ohtsukaら(1985), J. Biol. Chem., 260:2605−2608;およびRossoliniら(1994), Mol. Cell. Probes 8:91−98)。
【0058】
「単離」および「精製」なる語は、その天然環境から実質的または本質的に除去された、またはその天然環境において濃縮された物質をいう。例えば、単離された核酸は、通常それと隣接する核酸から分離させたもの、または試料中の他の核酸または成分(タンパク質、脂質など)から分離させたものであってもよい。別の例では、ポリペプチドがその天然環境から実質的に除去された、またはその天然環境において濃縮された場合、それは精製されている。核酸およびタンパク質の精製および単離方法は、当該分野で周知である。
【0059】
「BBB受容体Ab」なる語は、血液脳関門上に発現された受容体の細胞外ドメインに対する抗体をいう。BBB受容体の非限定的な例は、インスリン受容体(例えば、ヒトインスリン受容体)、トランスフェリン受容体、リポタンパク質受容体、およびレプチン受容体を包含する。
【0060】
III.血液脳関門
BBBは、脳および脊髄の毛細管を形成する内皮細胞を一緒に固める密着な結合によって形成される(Pardridge, 2008, Bioconj Chem, 19: 1327−1338)。ヒトの脳において、400マイルの脳毛細管が存在する。BBBをイン・ビボで形成する脳毛細管内皮原形質膜を横断する電気抵抗は、いずれの生物膜と同様に高い。血液と器官との間の遊離溶質交換のための通常の細胞間隙および細胞間経路は、CNSには存在しない。結果的に、血液中の薬物は、2つのメカニズム、すなわち、(i)小型分子の高い脂溶性のための自由拡散、および(ii)内因性BBBトランスポーターを介する輸送のうちの1つだけを介して脳にアクセスすることができる。デコイ受容体組成物のためのBBB薬物送達問題の解決に対する1のアプローチは、BBB内のある特定の内因性輸送システムにアクセスできるように、デコイ受容体を再操作することである。本明細書に記載の方法は、本明細書に記載のデコイ受容体融合抗体組成物の全身投与後、機能的デコイ受容体が末梢血からCNS中へBBBを横断できるようにする。本明細書中に記載の方法は、BBB上の表面に発現された発現受容体(例えば、ヒトインスリン受容体)を利用して、所望の二官能性受容体ECD−BBB受容体融合抗体を末梢血からCNS中へ輸送する。
【0061】
IV.BBBを横断する輸送のためのデコイ受容体融合抗体
一の態様において、本発明は、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列に共有結合した受容体ECD(例えば、ヒトTNF−α受容体ECD)のアミノ酸配列を含有する二官能性デコイ受容体融合抗体を利用する組成物および方法を提供する。かかる二官能性デコイ受容体融合抗体は、BBB上に発現された受容体および該受容体ECDのリガンドに結合することができる。該組成物および方法は、BBBを横断して末梢血からCNS中へデコイ受容体を輸送するのに有用である。
【0062】
BBBは、いくつかの巨大分子の血液から脳への輸送を可能にする特定の受容体を有することが示された(これらのトランスポーターは、本発明の組成物のトランスポーターとして適当である)。本発明に有用な内因性BBB受容体介在性輸送システムは、限定するものではないが、インスリン、トランスフェリン、インスリン様生長因子1および2(IGF1およびIGF2)、レプチン、およびリポタンパク質を輸送するものを包含する。いくつかの具体例において、本発明は、内因性インスリンBBB受容体介在性輸送システム、例えば、ヒト内因性インスリンBBB受容体介在性輸送システムを介してBBBを横断することができる抗体を利用する。いくつかの具体例において、該二官能性デコイ受容体融合抗体は、HIR抗体を含む。本明細書に記載のデコイ受容体−HIRAb融合抗体は、ヒトインスリン受容体のECDに結合する。いくつかの具体例において、該デコイ受容体ECDは、重鎖免疫グロブリン(例えば、HIRAb重鎖免疫グロブリン)のカルボキシル末端に融合する。他の具体例において、該デコイ受容体ECDは、軽鎖免疫グロブリン(例えば、HIRAb軽鎖免疫グロブリン)のC末端に融合する。いくつかの場合、上記の受容体ECDは、ヒト、マウス、ラットまたはブタサイトカイン受容体、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエピフリン受容体由来のECDに対して、少なくとも80%(例えば、85%、90%、95%、97%、99%、または少なくとも80%〜100%のうちの別のパーセント)同一のアミノ酸配列を含有する。いくつかの具体例において、上記の受容体ECDは、サイトカイン受容体由来である。いくつかの具体例において、受容体ECDは、TNF−α受容体ドメイン(例えば、ヒトTNF−α受容体ECD)を含有する。上記の受容体のアミノ酸配列についてGenBankアクセッション番号を下記の表1に提供する。
【0063】
【表1】
【0064】
インスリン受容体およびその細胞外インスリン結合ドメイン(ECD)は、構造的および機能的に当該分野において広範囲に特徴付けられている。例えば、Yipら(2003), J. Biol. Chem., 278(30):27329−27332;およびWhittakerら(2005), J. Biol. Chem., 280(22):20932−20936参照のこと。ヒトインスリン受容体のアミノ酸およびヌクレオチド配列は、GenBankアクセッション番号NM 000208の下に見出すことができる。
【0065】
BBB上に発現されたインスリン受容体は、それにより、BBBを横断するデコイ受容体、例えば、TNF−α受容体ECDの輸送のためのベクターとして作用することができる。ある特定のインスリン受容体ECD特異的抗体は、内因性リガンドを模倣していてもよく、それにより、特異的受容体システム上の輸送を介して、原形質膜障壁を横断しうる。ある特定の具体例において、受容体デコイ−HIRAb融合抗体は、ヒトBBB HIR上の外表面エピトープを結合し、該結合により、ヒトBBBインスリン受容体が介在する輸送反応を介して、該融合抗体のBBBの横断を可能にする。
【0066】
ヒトにおける使用のために、キメラHIRAbは、ヒトへの投与時に有意に免疫原性ではない十分なヒト配列、例えば、約80%ヒトおよび約20%マウス、または約85%ヒトおよび約15%マウス、または約90%ヒトおよび約10%マウス、または約95%ヒトおよび約5%マウス、または約95%より多くのヒトおよび約5%未満のマウス配列を含有することが好ましい。本発明における使用のために、十分なヒト配列を有するヒトBBBインスリン受容体に対するキメラ抗体が、例えば、Boadoら(2007), Biotechnol Bioeng, 96(2):381−391に記載される。また、HIRMAbのより高度にヒト化した形態を操作することもでき、該ヒト化HIR Abは、ネズミHIR Abに匹敵する活性を有し、本発明の具体例において使用することができる。例えば、米国特許出願公開第20040101904号(2002年11月27日出願)および第20050142141号(2005年2月17日出願)を参照のこと。
【0067】
例示的具体例において、二官能性デコイ受容体融合抗体は、ヒトインスリン受容体に対する結合について、配列番号4および6、または配列番号6および7のアミノ酸配列を含有する二官能性デコイ受容体融合抗体と競合する。いくつかの場合、該競合する二官能性デコイ受容体は、配列番号4および6、または配列番号6および7のアミノ酸配列を含有する機能的デコイ受容体融合抗体に対して、少なくとも約20%、例えば、25%、30%、35%、40%、45%、50%、60%、65%、70%、75%、80%、85%、90%、または少なくとも約20%〜100%のうちの別のパーセント同一のヒトインスリン受容体に関するアフィニティーを有する。他の具体例において、該二官能性デコイ受容体融合抗体は、配列番号4および6、または配列番号6および7に対して、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一のアミノ酸配列を含有する。
【0068】
本発明で使用されるBBB受容体抗体は、グリコシル化されていてもよく、またはグリコシル化されていなくてもよい。該抗体がグリコシル化されている場合、該抗体の機能に有意に影響を及ぼさないいずれかのグリコシル化パターンが用いられうる。グリコシル化は、抗体が産生される細胞の典型的なパターンで生じることができ、細胞型によって変化しうる。例えば、マウスミエローマ細胞によって産生されたモノクローナル抗体のグリコシル化パターンは、トランスフェクトされたチャイニーズハムスター卵巣(CHO)細胞によって産生されたモノクローナル抗体のグリコシル化パターンとは相違することができる。いくつかの具体例において、該抗体は、トランスフェクトされたチャイニーズハムスター卵巣(CHO)細胞によって産生されるパターンでグリコシル化される。
【0069】
現行の技術により、BBB受容体Absまたはデコイ受容体ECDの大量の配列変種を容易に生成し(例えば、イン・ビトロで)、ヒトインスリン受容体のECDなどの標的抗原への結合について、またはデコイ受容体ECDのリガンドへの結合についてスクリーンすることが可能になることは、当業者に明らかであろう。例えば、抗体配列変種のウルトラハイスループットスクリーニングの一例として、Fukudaら(2006), Nuc. Acid Res., 34(19)(オンラインで公開)を参照のこと。また、Chenら、(1999), Prot Eng, 12(4): 349−356を参照のこと。配列変種を単離するために、全配列または特定のドメインに対応する特定のサブ配列の無作為変異誘発を行ってもよい。別法では、BBB受容体またはデコイ受容体リガンド結合に不可欠であることが知られている残基の変異を回避しつつ、部位特異的変異誘発を繰り返し実施することができる。例えば、Mukaiら(2009), J Mol Biol, 385(4):1221−1229に記載されているように、例えば、TNF−α受容体の構造機能が当該分野で知られている。さらに、デコイ受容体ECD配列の複数の変種、例えば、ヒトTNF−α受容体の変種の生成において、変異認容予測(mutation tolerance prediction)プログラムを用いて、厳密な無作為変異誘発によって生じるであろう非機能的配列変種の数を大幅に減らすことができる。タンパク質配列におけるアミノ酸置換のタンパク質機能に対する影響を予測するための種々のプラグラム(例えば、SIFT、PolyPhen、PANTHER PSEC、PMUT、およびTopoSNP)が例えば、Henikoffら(2006), Annu. Rev. Genomics Hum. Genet., 7:61−80に記載されている。したがって、上記のように、当該分野でルーチンな方法によって、デコイ受容体(例えば、ヒトTNF−α受容体ECD配列)配列変種の非常に多様な「ライブラリー」を作製し、スクリーニングすることによって、非常に大量の操作可能なデコイ受容体ECD配列変種を得ることができることは、当業者に明らかであろう。
【0070】
パーセント配列同一性は、常法によって決定される。例えば、Altschulら(1986), Bull. Math. Bio., 48:603、およびHenikoffおよびHenikoff,(1992), Proc. Natl. Acad. Sci. USA, 89:10915を参照のこと。簡単に言えば、ギャップオープニングペナルティ10、ギャップエクステンションペナルティ1、およびHenikoffおよびHenikoff(上掲)の「BLOSUM62」スコアリングマトリックスを用いて、2つのアミノ酸配列をアラインメントスコアが最適になるようにアラインする。次いで、パーセント同一性は、{(完全に一致した全数)/[(より長い配列の長さ)+(2つの配列をアラインするためにより長い配列中に導入したギャップの数)]}(100)として計算される。
【0071】
2つのアミノ酸配列をアラインするために利用可能な多くの確立されたアルゴリズムが存在することは、当業者に明らかである。PearsonおよびLipmanの「FASTA」類似性サーチアルゴリズムは、本明細書中に開示されるアミノ酸配列および別のペプチドのアミノ酸配列によって共有される同一性のレベルを調べるための適当なタンパク質アラインメント法である。FASTAアルゴリズムは、Pearsonら(1988), Proc. Nat’l Acad. Sci. USA, 85:2444、およびPearson(1990), Meth. Enzymol. 183:63によって記載される。簡単に言えば、FASTAは、まず、保存的アミノ酸置換、挿入または欠失を考慮せずに、クエリー配列(例えば、配列番号4または配列番号6)および最大密度の同一性(ktup変数が1の場合)または同一性対(ktup=2の場合)のいずれかを有する試験配列によって共有される領域を同定することによって、配列類似性を特徴付ける。次いで、最大密度の同一性を有する10個の領域を、アミノ酸置換マトリックスを用いて、全ての対になったアミノ酸の類似性を比較することによって再スコア化し、該領域の末端は、最大スコアに寄与する残基のみを含むように「トリミング」する。「カットオフ」値(配列の長さおよびktup値に基づく所定の式によって計算される)よりも大きいスコアを有するいくつかの領域が存在する場合、トリミングされた最初の領域を、ギャップを有する近似のアラインメントを形成するように該領域を結合できるか否か決定するために試験する。最終的に、2つのアミノ酸配列の最大スコア領域を、アミノ酸挿入および欠失を可能にするNeedleman−Wunsch−Sellersアルゴリズム(Needlemanら(1970), J. Mol. Biol. 48:444; Sellers (1974), SIAM J. Appl. Math., 26:787)の修飾を用いてアラインする。FASTA分析の実例的パラメーターは、ktup=1、ギャップオープニングペナルティ=10、ギャップエクステンションペナルティ=1、および置換マトリックス=BLOSUM62である。これらのパラメーターは、Pearson, (1990), Meth. Enzymol., 183:63のAppendix 2において説明されるように、スコアリングマトリックスファイル(「SMATRIX」)を修飾することによって、FASTAプログラム中に導入することができる。
【0072】
本発明は、また、本明細書中に開示されるアミノ酸配列と比較して、保存的アミノ酸変化を有するタンパク質を包含する。一般的なアミノ酸のなかでも、例えば、「保存的アミノ酸置換」は、下記の各群内でのアミノ酸間の置換によって説明される。(1)グリシン、アラニン、バリン、ロイシンおよびイソロイシン、(2)フェニルアラニン、チロシン、およびトリプトファン、(3)セリンおよびスレオニン、(4)アスパラギン酸およびグルタミン酸、(5)グルタミンおよびアスパラギン、および(6)リジン、アルギニンおよびヒスチジン。BLOSUM62表は、タンパク質配列セグメントの約2,000の局所的な複数のアラインメントに由来するアミノ酸置換マトリックスであり、500以上のグループの関連タンパク質の非常に保存された領域を示す(Henikoffら(1992), Proc. Nat’l Acad. Sci., USA, 89:10915)。したがって、BLOSUM62置換頻度を用いて、本発明のアミノ酸配列中に導入されうる保存的アミノ酸置換を規定することができる。化学的特性のみに基づいて(上記のように)アミノ酸置換を設計することが可能であるが、「保存的アミノ酸置換」なる語は、好ましくは、−1よりも大きいBLOSUM62値によって表される置換をいう。例えば、アミノ酸置換は、置換が0、1、2または3のBLOSUM62値によって特徴付けられるが場合、保存的である。該システムによると、好ましい保存的アミノ酸置換は、少なくとも1(例えば、1、2または3)のBLOSUM62値によって特徴付けられるが、より好ましい保存的アミノ酸置換は、少なくとも2(例えば、2または3)のBLOSUM62値によって特徴付けられる。
【0073】
また、アミノ酸配列は、該配列が本発明の組成物および方法において機能的であるのに十分な生物学的タンパク質活性を保持する限り、付加的な残基、例えば、付加的なN−またはC−末端アミノ酸を含んでいてもよく、依然として実質的に、本明細書中に開示される配列の1つにおいて示されるとおりであることは、理解されよう。
【0074】
V.組成物
顕著なことには、本明細書に記載の二官能性デコイ受容体融合抗体は、高い割合で、それらの分離した構成タンパク質の活性、すなわち、BBB受容体AbのBBB受容体、例えば、ヒトインスリン受容体ECDに対する結合、およびデコイ受容体ECD(例えば、TNF−α受容体)の同族(cognate)リガンドに対する結合を保持する。
【0075】
本明細書には、デコイ受容体ECDに融合したBBBを横断することができるBBB受容体Abを含有する二官能性デコイ受容体−BBB受容体融合抗体であって、ここに、該血液脳関門を横断できるBBB受容体Abおよび該受容体ECDは各々、別々に存在した場合のそれらの活性と比較して、それらの活性の少なくとも約5、10、15、18、20、25、30、35、40、40、45または50%を保持する該融合抗体が記載される。BBB受容体Abの例は、インスリン受容体(例えば、ヒトインスリン受容体)、トランスフェリン受容体、およびリポタンパク質受容体に対するAbs(例えば、モノクローナルAbs)を包含する。適当な受容体ECDの例は、限定するものではないが、ヒト、マウス、ラット、またはブタサイトカイン受容体、TNF−α受容体、TNF−関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体に対して、少なくとも80%(例えば、85%、90%、95%、97%、99%または少なくとも80%〜100%のうちの別のパーセント)同一のアミノ酸配列を含有するものを包含する。
【0076】
いくつかの具体例において、BBB受容体Abは、ヒトインスリン受容体(HIR)のECDに対する抗体である。本明細書に記載の二官能性デコイ受容体−BBB受容体Ab融合抗体において、BBB受容体抗体および受容体ECD間の共有結合は、該結合が該デコイ受容体ECD−BBB受容体Ab融合抗体の該BBB受容体のECDに対する結合および血液脳関門の横断を可能にし、かつ、該デコイ受容体ECDの活性の治療上有用な部分の保持、例えば、該活性の少なくとも10%、20%、30%、40%、50%、60%、70%、80%、または約10%〜100%のうちの別のパーセントの保持を可能にする限り、BBB受容体抗体重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシまたはアミノ末端にあってもよい。ある特定の具体例において、共有結合は、該抗体のHCおよび該デコイ受容体ECD間にある。他の具体例において、共有結合は、該抗体のLCおよび該デコイ受容体ECD間にある。いずれの適当な結合を用いてもよく、例えば、デコイ受容体ECDのアミノ末端に対して軽鎖のカルボキシル末端、デコイ受容体ECDのアミノ末端に対して重鎖のカルボキシル末端、またはデコイ受容体ECDのカルボキシル末端に対して軽鎖のアミノ末端を用いてもよい。好ましい具体例において、該結合は、HCのカルボキシル末端からデコイ受容体ECDのアミノ末端へのものである。いくつかの具体例において、該融合抗体組成物は、そのN末端を介してヒトインスリン受容体抗体の重鎖のC末端に共有結合したヒトTNF−α受容体ECDを含む。
【0077】
末端アミノ酸間の結合が融合アミノ酸配列の一部を形成する介在性ペプチドリンカー配列によって達成できることは、明らかであろう。該ペプチド配列リンカーは、1、2、3、4、5、6、7、8、9、10または10以上のアミノ酸長であってもよい。いくつかの具体例において、2−アミノ酸リンカーを用いる。いくつかの具体例において、該リンカーは、配列ser−serを有する。該ペプチドリンカー配列は、プロテアーゼ切断部位を含みうるが、これは、デコイ受容体ECDの活性の必要条件ではない。実際、本発明のこれらの具体例の利点は、該二官能性デコイ受容体−BBB受容体抗体融合抗体が切断を伴わずに、一旦BBBを横断すれば、輸送および活性の両方に関して部分的または完全に活性であることである。図10は、デコイ受容体ECD−BBB受容体抗体融合抗体のアミノ酸配列の例示的具体例を示し、それは、HCがそのカルボキシル末端によって、3アミノ酸「ser−ser−ser」リンカーを介して、該TNF−α受容体ECDのアミノ末端に融合しているヒトTNF−α受容体ECD−HIR抗体融合抗体(配列番号4)である。
【0078】
いくつかの具体例において、デコイ受容体−BBB受容体Ab融合抗体は、HCおよびLCの両方を含む。いくつかの具体例において、該デコイ受容体−BBB受容体Ab融合個体は一価抗体である。他の具体例において、該デコイ受容体−BBB受容体Ab融合抗体は、実施例セクションにいおいて本明細書中で記載されるような二価抗体である。
【0079】
該デコイ−BBB受容体Ab融合抗体の一部として用いられるBBB受容体Abは、グリコシル化または非グリコシル化することができる。いくつかの具体例において、該抗体は、例えば、CHO細胞におけるその合成によって生じるグリコシル化パターンにおいて、グリコシル化される。
【0080】
本明細書中で使用される場合、「活性」は、生理学的活性(例えば、BBB横断能および/または治療活性)、BBB受容体Abのその標的BBB受容体に関する結合アフィニティー、またはデコイ受容体の同族リガンド(例えば、TNF−αデコイ受容体の場合、TNF)に関するアフィニティーを包含する。
デコイ受容体−BBB受容体Ab融合抗体のBBBを横断する輸送は、標準的な方法によってBBB受容体Abを単独でBBBを横断して輸送する場合と比較してもよい。例えば、動物モデル、例えば、非ヒト霊長類のような哺乳動物によるデコイ受容体−BBB受容体Ab融合抗体の薬物動力学および脳への取り込みを用いてもよい。同様に、デコイ受容体リガンド結合を決定するための標準的なモデル(例えば、ELISA)を用いて、デコイ受容体ECD単独の機能およびデコイ受容体−BBB受容体Ab融合抗体の一部としての機能を比較してもよい。例えば、TNFαのHIRMAb−TNF−デコイ受容体融合タンパク質に対する結合 対 TNF受容体(TNFR)−II:ヒトIgG1 Fc融合タンパク質(TNFR:Fcと称する)に対する結合を明らかにする実施例4を参照のこと。BBB受容体に関する結合アフィニティーをまた、デコイ受容体−BBB受容体Ab融合抗体 対 BBB受容体Ab単独に関して比較することができる。例えば、実施例4を参照のこと。
【0081】
いくつかの場合、デコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)の脳への取り込みは、タンパク質100グラムあたり1%、2%、3%、5%、7%または10%ID以上である。デコイ受容体−BBB受容体Ab融合抗体のBBBを横断する輸送は、また、デコイ受容体およびヒトIgGのFcフラグメントの融合タンパク質、例えば、TNFR:FcのBBBを横断する輸送と比較してもよい。デコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)の脳への取り込みは、デコイ受容体およびヒトIgGのFcフラグメントの融合タンパク質、例えば、TNFR:Fcの脳への取り込みより1、2、5、10、13、15、17、20、25、30、35、40、45、または50倍以上大きくてもよい。脳および他の器官について、器官クリアランス定数(本明細書中では、透過係数表面積(PS)積という)は、本明細書に記載の融合タンパク質について計算されうる。本明細書に記載のデコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)は、他の器官と比較した場合、脳において選択的に豊富に存在しうる。例えば、TNFR:Fc融合タンパク質に関する器官PS積に対する、本明細書に記載のデコイ受容体−BBB受容体AB融合抗体(例えば、HIRMAb−TNFR融合タンパク質)に関する器官PS積の比率を複数の器官について測定した場合(例えば、図14参照)、脳での比率は、他の器官、例えば、脂肪、筋肉、心臓、肺、肝臓または脾臓の比率より1、2、5、10、13、15、17、20、25、30、35、40、45、または50倍以上大きくてもよい。
【0082】
また、本明細書中には、本明細書に記載の1以上のデコイ受容体−BBB受容体Ab融合抗体、および医薬上許容される賦形剤を含有する医薬組成物が包含される。医薬上許容される担体/賦形剤の全体的な議論は、Remington’s Pharmaceutical Sciences, Gennaro, AR, ed., 20th edition, 2000: Williams and Wilkins PA, USAに見出すことができる。本発明の医薬組成物は、静脈内、皮下、筋内、腹腔内注射、経口、経腸、経頬、経皮、鼻腔内、またはいずれか他の末梢投与に適当な経路を包含するいずれかの末梢経路を介する投与に適当な組成物を包含する。
【0083】
本発明の組成物は、例えば、静脈内、皮下、筋内、または腹腔内投与のための医薬組成物として、特に注射に適する。本発明の水性組成物は、本発明の組成物の有効量を含み、医薬上許容される担体または水性媒体中に溶解または分散させてもよい。「医薬上」または「薬理学上許容される」なる語は、適宜、動物、例えば、ヒトに投与時に、有害な反応、アレルギー性反応または他の都合の悪い反応を生じない分子および組成物をいう。本明細書中で使用される場合、「医薬上許容される担体」は、いずれかおよび全ての溶媒、分散媒体、コーティング、抗菌および抗真菌剤、等張剤および吸収遅延剤などを包含する。医薬上活性な物質のためのかかる媒体および剤の使用は、当該分野で周知である。いずれかの通常の媒体または剤が活性成分と混合できない場合を除き、治療組成物におけるその使用が意図される。補足的活性成分もまた、該組成物中に配合することができる。
【0084】
注射組成物のための例示的医薬上許容される担体は、カルシウム塩、例えば、塩化カルシウム、臭化カルシウム、硫化カルシウムなど、および有機酸の塩、例えば、酢酸塩、プロピオン酸塩、マロン酸塩、安息香酸塩などを包含することができる。例えば、本発明の組成物は、液体形態で提供されてもよく、0.01−1%のポリソルベート−80などの界面活性剤、または炭水化物添加剤、例えば、マンニトール、ソルビトールまたはトレハロースを用いて、または用いずに、種々のpH(pH5〜8)のセーラインを基礎とする水性溶液中で処方されてもよい。一般的に使用されるバッファーは、ヒスチジン、酢酸塩、リン酸塩、またはクエン酸塩を包含する。通常の保管および使用条件下、これらの調製物は、微生物の増殖を防止するための保存剤を含有することができる。微生物の作用の防止は、種々の抗菌剤および抗真菌剤、例えば、パラベン、クロロブタノール、フェノール、ソルビン酸、チメロサールなどによってもたらされることができる。多くの場合、等張剤、例えば、糖類または塩化ナトリウムを含むことが好ましい。注射組成物の吸収延長は、該組成物における吸収を遅延させる剤、例えば、モノステアリン酸アルミニウム、およびゼラチンの使用によってもたらされることができる。
【0085】
ヒト投与の場合、調製物は、FDAおよび他の規制当局標準によって要求される滅菌性、発熱性、一般的な安全性および純度標準を満たす。活性化合物は、一般に、非経口投与のために処方され、例えば、静脈内、筋内、皮下、病巣内または腹腔内経路を介する注射のために処方される。活性成分を含有する水性組成物の調製は、本明細書の開示を考慮して、当業者に既知であろう。典型的には、かかる組成物は、液体溶液または懸濁液のいずれかの注射剤として調製でき、注射前に、液体の添加による溶液または懸濁液の調製において使用するのに適当な固体形態も調製することができ、該調製物は乳化されることもできる。
【0086】
滅菌注射溶液は、必要量の活性化合物を上記した種々の他の成分と共に、適当な溶媒中に配合し、次いで、フィルター滅菌することによって調製される。一般に、分散液は、種々の滅菌活性成分を、基礎分散媒体および上記した必要な他の成分を含有する滅菌ビヒクル中に配合することによって調製される。滅菌注射溶液の調製のための滅菌粉末の場合、調製方法は、予めフィルター滅菌した溶液から活性成分およびいずれかの付加的な所望の成分の粉末を生じる真空乾燥および凍結乾燥技術を包含する。
【0087】
処方において、溶液は、投与処方に適合する方法で、かつ、本明細書に記載の基準に基づいて治療上有効量で、全身投与される。該処方は、種々の投与形態で、例えば、上記の注射溶液の形で容易に投与されるが、薬物放出カプセルなども使用できる。
【0088】
投与されるべき医薬組成物の適当な量、治療の回数、および単位投与量は、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体のCNS取り込み特徴によって、および治療されるべき対象および対象の病態によって変化する。いずれの事象においても、投与責任者が個々の対象の適当な投与量を決定する。
【0089】
静脈内または筋内注射などの非経口投与のために処方された化合物の他に、限定するものではないが、皮内投与(米国特許第5,997,501号、第5,848,991号、および第5,527,288号参照)、肺投与(米国特許第6,361,760号、第6,060,069号、および第6,041,775号参照)、頬投与(米国特許第6,375,975号および第6,284,262号参照)、経皮投与(米国特許第6,348,210号および第6,322,808号参照)および経粘膜投与(米国特許第5,656,284参照)を包含する本発明の他の投与方法を用いてもよい。かかる投与方法は、当該分野で周知である。また、例えば、点鼻溶液またはスプレー、エーロゾルまたは吸入薬と共に、本発明の鼻腔内投与を使用してもよい。点鼻溶液は、通常、滴またはスプレーで鼻道に投与されるように設計された水性溶液である。点鼻溶液は、多くの点で、鼻汁に類似するように調製される。かくして、水性点鼻溶液は、通常、等張性であり、pH5.5〜6.5を維持するようにわずかに緩衝化される。さらに、必要に応じて、眼科用調製物において使用されるものと同様な抗微生物保存剤および適当な安定化剤を処方中に含ませてもよい。種々の市販の点鼻調製物が知られており、例えば、抗生物質および抗ヒスタミン剤を包含し、喘息予防に使用される。
【0090】
他の投与様式に適当なさらなる処方は、座剤およびペッサリーを包含する。直腸ペッサリーまたは座剤もまた、使用してもよい。座剤は、通常、薬物を加えられた、直腸または尿道に挿入するための種々の重量および形状の固体投与形態である。挿入後、腔液中で座剤が軟化、溶融または溶解する。座剤の場合、伝統的な結合剤および担体は、例えば、ポリアルキレングリコールまたはトリグリセリドを包含し、かかる座剤は、活性成分を適当な範囲、例えば、0.5%〜10%、好ましくは1%−2%で含有する混合物から形成されうる。
【0091】
経口処方は、例えば、製薬等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、ナトリウムサッカリン、セルロース、炭酸マグネシウムなどのような通常使用される賦形剤を含む。これらの組成物は、溶液、懸濁液、錠剤、丸薬、カプセル、徐放性処方、または粉末の形態を取る。ある規定された具体例において、経口医薬組成物は、不活性希釈剤、または同化可能な食用担体を含み、または硬もしくは軟外皮ゼラチンカプセル中に被包されてもよく、または打錠されてもよく、または食事の食物と直接混合してもよい。経口治療投与の場合、活性化合物は賦形剤と混合してもよく、摂取可能な錠剤、バッカル錠、トローチ、カプセル、エリキシル、懸濁、シロップ、ウウェハーなどの形態で使用されうる。かかる組成物および調製物は、少なくとも0.1%の活性化合物を含有することができる。組成物および調製物のパーセンテージは、もちろん、変動してもよく、通常、単位重量の約2〜75%または約25−60%でありうる。かかる治療上有用な組成物中における活性化合物の量は、適当な投与量が得られるような量である。
【0092】
錠剤、トローチ、丸薬、カプセルなどは、また、結合剤、例えば、トラガカントゴム、アラビアゴム、トウモロコシデンプン、またはゼラチン、賦形剤、例えば、リン酸二カルシウム、崩壊剤、例えば、トウモロコシデンプン、ジャガイモデンプン、アルギン酸など、滑沢剤、例えば、ステアリン酸マグネシウム、および甘味剤、例えば、シュークロース、ラクトースまたはサッカリン、またはフレーバー剤、例えば、ペパーミント、冬緑油、またはチェリーフレーバーを含有していてもよい。投与単位形態がカプセルである場合、上記のタイプの材料のほかに、液体担体を含有していてもよい。種々の他の材料がコーティングとして、または投与単位の物理的形態を修飾するために存在していてもよい。例えば、錠剤、丸薬またはカプセルは、シェラック、糖またはその両方で被覆されていてもよい。エリキシルのシロップは、活性化合物、甘味剤としてシュークロース、保存剤としてメチレンおよびプロピルパラベン、色素およびフレーバー、例えば、チェリーまたはオレンジフレーバーを含有していてもよい。いくつかの具体例において、経口医薬組成物は、活性成分を胃環境から保護するために、腸溶性被覆されていてもよく、腸溶性被覆方法および処方は当該分野で周知である。
【0093】
VI.核酸、ベクター、細胞および製造
本発明は、また、核酸、ベクター、細胞、および製造方法を提供する。いくつかの具体例において、本発明は、本明細書に記載のポリペプチドをコードする核酸、例えば、(i)重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列、(ii)軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列、または(iii)(i)または(ii)の相補性配列を含む核酸を提供する。該上記の核酸によってコードされる重鎖および軽鎖免疫グロブリンは、BBB受容体、例えば、ヒトインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体に対する抗体由来である。いくつかの場合、該コードされる受容体ECDは、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である。いくつかの場合、該コードされる受容体のアミノ酸配列は、外来受容体の一つ由来のECDのアミノ酸配列に対し、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一である。いくつかの場合、該核酸は、ヒト、マウス、ラット、またはブタTNF−α受容体の細胞外ドメインに対し、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一のアミノ酸配列を含むECDをコードする。いくつかの具体例において、第1の配列は、配列番号4または7に相当するアミノ酸配列に対し、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一のアミノ酸配列をコードする。他の具体例において、第2の配列は、配列番号6に相当するアミノ酸配列に対し、少なくとも約80%、85%、90%、95%、または少なくとも約80%〜約100%のうちの他のパーセント同一のアミノ酸配列をコードする。
【0094】
本明細書中に提供される核酸は、いくつかの場合、さらに、MAbの重鎖および受容体ECD間のペプチドリンカーをコードする核酸配列を含有することができる。いくつかの具体例において、該リンカーは、S−S−Mである。他の具体例において、該リンカーは、S−Sである。別の具体例において、該リンカーは、S−S−Sである。該核酸は、さらに、シグナルペプチドをコードする核酸配列を含有していてもよく、ここに、該シグナルペプチドは、該重鎖に連結される。当該分野で既知の、または続いて開発されたいずれかの適当なシグナル配列が使用されうる。いくつかの具体例において、重鎖免疫グロブリンに結合したシグナルペプチドは、配列番号4のアミノ酸1−19と約60、70、80、90、95、99、または100%同一の配列を含む。いくつかの具体例において、該核酸は、他のシグナルペプチドをコードする核酸配列を含有し、ここに、該他のシグナルペプチドは該軽鎖に連結される。該軽鎖に連結したシグナルペプチドは、配列番号6のアミノ酸1−20と約60、70、80、90、95、99、または100%同一の配列を含むことができる。
【0095】
本発明は、また、核酸ベクターを提供する。該ベクターは、本明細書に記載の核酸配列のいずれかを含有することができる。いくつかの具体例において、該ベクターは、(i)BBB受容体に対する抗体由来の重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列(またはその相補体)、または(ii)BBB受容体に対する抗体由来の軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列(またはその相補体)を含む。いくつかの具体例において、本発明は、核酸配列を提供し、いくつかの具体例において、本発明は、特定のヌクレオチド配列と少なくとも約60、70、80、90、95、99または100%同一の核酸配列を提供する。例えば、いくつかの具体例において、本発明は、配列番号3または5に対し、少なくとも約60、70、80、90、95、99または100%同一の配列を含有する核酸を提供する。
【0096】
いくつかの具体例において、本発明は、(i)BBB受容体に対する抗体由来の重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列(またはその相補体)、および(ii)BBB受容体に対する抗体由来の軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列(またはその相補体)の両方を含有する単一タンデム発現ベクター(全てが一片の核酸、例えば、一片のDNA中に組み込まれている)(本明細書中では、「タンデムベクター」という)を提供する。該単一タンデムベクターは、また、一以上の選択および/または増幅遺伝子、例えば、DHFR、ネオマイシンホスホトランスフェラーゼ、ハイブロマイシンホスホトランスフェラーゼ、またはピューロマイシンN−アセチルトランスフェラーゼを含むことができる。いくつかの具体例において、該コードされる選択マーカーは、DHFRである。いくつかの具体例において、該タンデムベクターは、DHFRおよび第2の選択/増幅マーカー(例えば、ネオマイシンホスホトランスフェラーゼ)をコードする。本発明の例示的なタンデムベクターの作製方法は、実施例に提供される。しかしながら、該ベクターを構築するために、当該分野で既知のいずれの適当な技術を用いてもよい。
【0097】
単一タンデムベクターの使用は、いくつかの利益を有する。免疫グロブリンG(IgG)遺伝子での真核細胞系統のトランスフェクションは、一般的に、該細胞系統と、IgGを含む重鎖(HC)および軽鎖(LC)をコードする別のプラスミドとの共トランスフェクションを含む。IgG融合タンパク質の場合、該組み換え治療タンパク質をコードする遺伝子は、HCまたはLC遺伝子のいずれかに融合していてもよい。しかしながら、該共トランスフェクションアプローチは、HCおよびLC−融合遺伝子、またはHC−融合およびLC遺伝子を等しく高い割合で組み込んだ細胞系統の選択を困難にする。本発明のある特定の具体例において利用される融合タンパク質の製造に対するアプローチは、DNAの一本鎖上に、HC−融合タンパク質遺伝子、LC遺伝子、選択遺伝子、例えば、neo、および増幅遺伝子、例えば、ジヒドロ葉酸還元酵素遺伝子を包含する必要とする遺伝子の全てを含有する単一プラスミドDNAで永続的にトランスフェクトされた細胞系統の生産である。図9における融合タンパク質タンデムベクターのダイアグラムに示されるように、HC−デコイ受容体融合遺伝子、LC遺伝子、neo遺伝子、およびDHFR遺伝子は全て、別々であるがタンデムなプロモーター、および別々であるがタンデムな転写終止配列の調節下にある。したがって、治療タンパク質とHCまたはLC IgG遺伝子のいずれかとの融合遺伝子を包含する全ての遺伝子は、宿主細胞ゲノム中に等しく組み込まれる。
【0098】
本発明は、さらに、本発明の1以上のベクターを組み込んだ細胞を提供する。該細胞は、原核細胞または真核細胞であってもよい。いくつかの具体例において、該細胞は真核細胞である。いくつかの具体例において、該細胞は、マウスミエローマハイブリドーマ細胞である。いくつかの具体例において、該細胞は、チャイニーズハムスター卵巣(CHO)細胞である。ベクターを細胞中に組み込むための例示的方法は、実施例に提供する。しかしながら、ベクターを細胞中に組み込むために、当該分野で既知のいずれの適当な技術を用いてもよい。いくつかの具体例において、本発明は、免疫グロブリン融合タンパク質を発現することができる細胞を提供し、ここに、該細胞は、免疫グロブリン軽鎖遺伝子および治療剤に融合した免疫グロブリン重鎖の遺伝子の両方が一片の核酸、例えば、DNA中に組み込まれている単一タンデム発現ベクターが永続的に組み込まれた細胞である。いくつかの具体例において、本発明は、免疫グロブリン融合タンパク質を発現することができる細胞を提供し、ここに、該細胞は、免疫グロブリン重鎖遺伝子およびデコイ受容体ECDに融合した免疫グロブリン軽鎖の遺伝子の両方が一片の核酸、例えば、DNA中に組み込まれている単一タンデム発現ベクターが安定に組み込まれた細胞である。該タンデムベクターの細胞中への導入は、例えば、染色体核酸中への導入、またはエピソーム遺伝子エレメントの導入によるものであってもよい。
【0099】
2つの核酸の配列比較のために、典型的には1つの配列が、試験配列を比較するための参照配列として作用する。配列比較アルゴリズムを用いる場合、試験および参照配列をコンピューターに入力し、必要に応じてサブ配列座標を指定し、配列アルゴリズムプログラムパラメーターを指定する。デフォルトプログラムパラメーターを用いることができ、または別のパラメーターを指定することができる。次いで、配列比較アルゴリズムは、プログラムパラメーターに基づいて、参照配列に対する試験配列のパーセント配列同一性を計算する。
【0100】
本明細書中で使用される場合、「比較ウィンドウ」は、20〜600、通常、約50〜約200、より普通には、約100〜約150からなる群から選択される隣接位置の数のいずれか1つのセグメントに対するリファレンスを含み、ここに、配列は、2つの配列を最適にアラインした後に、同じナンバーの隣接位置の参照配列と比較されうる。比較のための配列をアラインメントする方法は、当該分野で周知である。比較のための配列の最適アラインメントは、限定するものではないが、SmithおよびWaterman(1970), Adv. Appl. Math., 2:482cの局所ホモロジーアルゴリズムによって、NeedlemanおよびWunsch,(1970), J. Mol. Biol., 48:443のホモロジーアラインメントアルゴリズムによって、PearsonおよびLipman,(1988), Proc. Nat’l. Acad. Sci. USA, 85:2444の類似性検索方法によって、これらのアルゴリズムのコンピューターによる手段によって(GAP、BESTFIT、FASTA、およびTFASTA,Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI)、または手動アラインメントおよび目視によって(例えば、Ausubelら(1995 supplement), Current Protocols in Molecular Biology参照)実施することができる。
【0101】
パーセント配列同一性および配列類似性を決定するために適当なアルゴリズムの一例は、BLASTおよびBLAST 2.0アルゴリズムであり、各々、Altschulら(1977), Nuc. Acids Res., 25:3389−3402、およびAltschulら(1990), J. Mol. Biol., 215:403−410に記載されている。BLAST分析を実施するためのソフトウェアは、National Center for Biotechnology Informationから公的に入手可能である。BLASTアルゴリズムパラメーターW、TおよびXは、アラインメントの感度および速度を決定する。BLASTNプログラム(ヌクレオチド配列用)は、デフォルトとして、11のワードレングス(W)、10のエクスペクテーション(E)、M=5、N=−4および両鎖の比較を用いる。BLASTアルゴリズムは、典型的に、「低複雑性」フィルターを切って(turned off)実施される。BLASTアルゴリズムはまた、2配列間の類似性の統計学的分析を行う(例えば、KarlinおよびAltschul, (1993), Proc. Natl. Acad. Sci. USA, 90:5873−5787参照)。BLASTアルゴリズムによって提供される類似性の1測定値は、最小合計確率(P(N))であり、それは、2つのヌクレオチドまたはアミノ酸配列間の合致が偶然に起こる確率を示す。例えば、参照核酸に対する試験核酸の比較において、最小合計確率が約0.2未満、より好ましくは、約0.01未満、最も好ましくは約0.001未満である場合、核酸は参照配列に類似するとみなされる。
【0102】
当該分野で周知のように、遺伝子コードの縮重のために、適当なコドンのいずれかの組合せを用いて所望の融合タンパク質をコードしうる。さらに、組み換え技術に有用な他のエレメント、例えば、プロモーター、終止シグナルなどもまた、核酸配列中に含まれうる。かかるエレメントは、当該分野で周知である。さらに、本明細書中に記載され、請求される全ての核酸配列は、該配列の相補体を包含する。
【0103】
いくつかの具体例において、本発明の核酸は、低、中程度または高ストリンジェンシー条件下で、配列番号4、6または7のアミノ酸配列をコードする核酸に特異的にハイブリダイズする。例えば、かかる核酸は、低、中程度または高ストリンジェンシー条件下で、配列番号3、5、その両方のヌクレオチド配列、またはその相補配列を含む核酸配列にハイブリダイズしうる。低ストリンジェンシーハイブリダイゼーション条件は、例えば、2XSSCおよび0.1%SDS中42℃にて、約40%〜約70%GC含量の100ヌクレオチドプローブを用いるハイブリダイゼーションを包含する。中程度ストリンジェンシーハイブリダイゼーション条件は、例えば、0.5X SSCおよび0.1%SDS中50℃でのハイブリダイゼーションを包含する。高ストリンジェンシーハイブリダイゼーション条件は、例えば、0.2X SSCおよび0.1%SDS中65℃にて、上記のプローブを用いるハイブリダイゼーションを包含する。これらの条件下では、ハイブリダイゼーション温度が高いので、より高いホモロジーを有する核酸を得ることができる。
【0104】
さらに、本発明は、二官能性デコイ受容体融合抗体を製造する方法を提供する。いくつかの具体例において、本発明は、二官能性デコイ受容体融合抗体を製造する方法であって、真核細胞中に、(i)受容体細胞外ドメインに融合した免疫グロブリン重鎖、および免疫グロブリン軽鎖の両方、または(ii)受容体細胞外ドメインに融合した免疫グロブリン軽鎖、および免疫グロブリン重鎖の両方(ここに、コードされる免疫グロブリン重鎖および免疫グロブリン軽鎖は、BBB上に発現された受容体に対する抗体由来である)をコードする単一タンデム発現ベクターを安定に組み込むことを含む方法を提供する。BBB受容体に対する適当な抗体、および受容体ECDは、上記の抗体および受容体ECDのいずれかを包含する。
【0105】
本発明は、免疫グロブリン融合タンパク質を発現すること、および/または免疫グロブリン融合タンパク質を精製することを包含する。発現および精製を包含する例示的な製造方法は、実施例に提供される。
【0106】
しかしながら、該タンパク質を製造、所望により発現、および精製するために、当該分野で既知のいずれの適当な技術を用いてもよい。これらは、タンパク質合成の非組み換え技術、例えば、Merrifieldによって最初に開発され、Stewartら(1984),Solid Phase Peptide Synthesisに記載されるような手動または自動の固相合成を包含する。化学合成は、アミノ酸をC末端で開始する所定の配列中に加える。基本的な固相方法は、C末端を保護したα−アミノ酸を適当な不溶性樹脂支持体に結合させることを必要とする。合成のためのアミノ酸は、その前の残基(または樹脂支持体)との適当なペプチド結合形成を保証するために、α−アミノ基上の保護を必要とする。カルボキシル末端での縮合反応の完了後、α−アミノ保護基を除去して、次の残基の付加を可能にする。いくつかのクラスのα−保護基が記載されており(Stewartら(1984),Solid Phase Peptide Synthesis参照)、酸に不安定なウレタンを基礎とするターシャリー−ブチルオキシカルボニル(Boc)が歴史的に好ましい。他の保護基、および関連する化学手法を用いてもよく、塩基に不安定な9−フルオレニルメチルオキシカルボニル(FMOC)を包含する。また、合成が完了するまで、反応性アミノ酸側鎖官能基は、ブロックする必要がある。官能性ブロッキング基の複雑なアレイは、それらの使用に対する手法および制限とともに、Bodansky(1976),Peptide SynthesisおよびStewartら(1984),Solid Phase Peptide Synthesisに概説されている。
【0107】
固相合成は、記載されるC末端α−保護アミノ酸残基のカップリングによって開始する。カップリングは、活性化剤、例えば、1−ヒドロキシベンゾ−トリアゾール(HOBT)を伴う、または伴わないジシクロヘキシカルボジイミド(DCC)、ジイソプロピルカルボジイミド(DIIPC)、またはエチルジメチルアミノプロピルカルボジイミド(EDC)を必要とする。C末端残基をカップリング後、α−アミノ保護基は、酸に不安定なターシャリー−ブチルオキシカルボニル(Boc)基の場合、ジクロロメタン中のトリフルオロ酢酸(25%以上)によって除去される。ジクロロメタン中のトリエチルアミン(10%)での中和工程は、遊離アミン(対 塩)を回収する。C末端残基を樹脂に付加した後、脱保護、中和およびカップリングサイクルを中間の洗浄工程と共に繰り返して、保護されたペプチド鎖を伸長する。各保護されたアミノ酸は、適当な溶媒中の等モル量のカップリング試薬を用いて、過剰に(3〜5倍)導入される。最終的に、完全にブロックしたペプチドを樹脂支持体上に構築した後、試薬をアプライして該樹脂から該ペプチドを切断し、側鎖ブロッキング基を除去する。無水フッ化水素(HF)は、酸に不安定なターシャリー−ブチルオキシカルボニル(Boc)化学基を切断する。特に側鎖官能基上の副反応を回避するために、いくつかの求核性スカベンジャー、例えば、ジメチルスルフィドおよびアニソールが含まれる。
【0108】
VII.方法
本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体のいずれかを全身投与することによって、デコイ受容体をBBBを横断して送達するための方法が本明細書中に記載される。いくつかの具体例において、受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体であって、BBB上に発現された受容体および該受容体細胞外ドメインのリガンドに結合する該融合抗体を含む医薬組成物を対象に全身投与することによって、CNS疾患を治療するために、本明細書に記載の組成物を投与する。いくつかの場合、治療されるべきCNS疾患は、急性CNS疾患、例えば、局所脳虚血、全脳虚血、外傷性脳損傷、または脊髄損傷である。いくつかの場合、治療されるべきCNS疾患は、慢性CNS疾患である。いくつかの具体例において、慢性CNS疾患は、神経変性CNS疾患、例えば、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症、ハンチントン病、多発性硬化症、横断性脊髄炎、運動ニューロン疾患、ピック病、結節硬化症、カナバン病、レット症候群、脊髄小脳失調症、フリードライヒ失調症、視神経萎縮、または網膜変性である。
【0109】
いくつかの具体例において、卒中、頭部損傷、脊髄損傷、または神経変性疾患に罹患している対象を治療するために、TNF−α受容体ECD−BBB受容体融合抗体が全身投与される。他の具体例において、血管内皮成長因子受容体(VEGFR)−ECD−BBB受容体Ab融合抗体を対象に全身投与して、脳腫瘍における血管新生を阻害する。いくつかの場合、後天性免疫不全症候群(AIDS)由来の認知症を治療するために、TNF関連アポトーシス誘導性リガンド受容体(TRAIL−R)ECD−BBB受容体Ab融合抗体を投与する。いくつかの具体例において、多発性硬化症(MS)を治療するために、インターロイキン(IL)−6デコイ受容体ECD−BBB受容体Ab融合抗体を全身投与する。さらなる具体例において、卒中を治療するために、アポトーシスのTNF様弱インデューサー(TWEAK)受容体デコイECD−BBB受容体Ab融合抗体を全身投与する。また別の具体例において、卒中または脳損傷後の神経修復を促進するために、エフリン受容体、EphA ECD−BBB受容体Ab融合抗体を全身投与する。
【0110】
デコイ受容体−BBB受容体Ab融合抗体の送達に適当な全身投与量は、投与されるべき特定のデコイ受容体−BBB受容体Ab融合抗体、そのCNS取り込み特徴およびデコイ受容体リガンドに対するそのアフィニティーに基づいて、変化するであろう。
【0111】
いくつかの具体例において、投与されるべきデコイ受容体−BBB受容体Ab融合抗体は、インスリン受容体(例えば、ヒトインスリン受容体)、トランスフェリン受容体、
インスリン様成長お因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体に対する抗体を含有し、該デコイ受容体は、TNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来のECDを含有する。いくつかの具体例において、投与されるべき該デコイ受容体−BBB受容体Ab融合抗体は、TNF−α受容体−ヒトインスリン受容体Ab融合抗体である。
【0112】
本明細書中で使用される場合、「全身投与」または「末梢投与」なる語は、CNS中への直接的投与ではない、すなわち、BBBの物理的貫通または破壊を含まないいずれかの投与方法を包含する。「全身投与」には、限定するものではないが、静脈内、動脈内、筋内、皮下、腹腔内、鼻腔内、経頬、経皮、直腸、経肺胞(吸入)、または経口投与が包含される。本明細書中に記載されるように、いずれの適当なデコイ受容体−BBB受容体Ab融合抗体を使用してもよい。
【0113】
本発明の組成物は、組合せ療法の一部として投与されうる。組合せ療法は、上記のCNS疾患のいずれかに罹患している患者において典型的に見出される症状の治療または軽減のための別の療法と組み合わせた本発明の組成物の投与を含む。本発明の組成物を別のCNS障害方法または組成物と組み合わせて用いる場合、本発明の組成物および付加的な方法または組成物のいずれの組合せを用いてもよい。かくして、例えば、本発明の組成物の使用が別のCNS障害治療剤と組み合わせられている場合、該2つを同時に、連続的に、持続時間が重なるように、類似した頻度で、同じ頻度で、または異なる頻度などで投与してもよい。いくつかの場合、本発明の組成物を1以上の他のCNS障害治療剤と組み合わせて含有する組成物が用いられるであろう。
【0114】
いくつかの具体例において、該組成物、例えば、二官能性TNF−α受容体ECD−BBB受容体Ab融合抗体を患者に、同じ処方内で、別の薬物療法と共に、または別々の組成物として共投与する。例えば、該二官能性TNF−α受容体−ヒトインスリン受容体Ab融合抗体は、別の二官能性デコイ受容体−BBB受容体Ab融合抗体と共に処方することができる。さらに、TNF−α受容体ECD−BBB受容体Ab融合抗体は、他の大きいまたは小さい分子と組み合わせて処方してもよい。CNSまたはCNS関連疾患を治療するために、デコイ受容体−BBB受容体Ab融合抗体と組み合わせて使用するための例示的薬剤は、下記に提供される。
【0115】
CNS炎症疾患を治療するために共投与するための例示的薬剤
対象が自己免疫、炎症疾患、または神経系に影響を及ぼすアレルギー疾患(例えば、Allanら(2003), Philos Trans R Soc Lond B Biol Sci, 358(1438):1669−1677参照)に罹患しているか、または罹患する危険性がある場合、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体(例えば、TNF−α受容体ECD−BBB受容体Ab融合抗体)をいずれかの組合せにおいて1以上の下記の治療剤、すなわち、免疫抑制剤(例えば、タクロリマス(tacrolimus)、シクロスポリン(cyclosporin)、ラパマイシン(rapamicin)、メトトレキサート(methotrexate)、シクロホスファミド(cyclophosphamide)、アザチオプリン(azathioprine)、メルカプトウリン(mercaptopurine)、マイコフェノラート(mycophenolate)、またはFTY720)、グルココルチコイド(例えば、プレドニソン、コルチゾンアセテート、プレドニソロン、メチルプレドニソロン、デクサメタゾン、ベータメタゾン、トリアムシノロン(triamcinolone)、ベクロメタゾン、フルドロコルチゾンアセテート、デオキシコルチコステロンアセテート、アルドステロン)、非スイテロイド性抗炎症薬(例えば、サリチル酸塩、アリールアルカン酸、2−アリールプロピオン酸、N−アリールアントラニル酸、オキシカム、コキシブ(coxibs)、またはスルホンアニリド)、Cox−2−特異的阻害剤(例えば、バルデコキシブ(valdecoxib)、セレコキシブ(celecoxib)、またはロフェコキシブ(rofecoxib))、レフルノミド(leflunomide)、金チオグルコース、金チオマレート、オーロフィン(aurofin)、スルファザラジン(sulfasalazine)、ヒドロキシクロロキニン、ミノサイクリン、TNF−α結合タンパク質(例えば、インフリキシマブ(infliximab)、エタネルセプト(etanercept)、またはアダリムマブ(adalimumab))、アバタセプト(abatacept)、アナキンラ(anakinra)、インターフェロン−β、インターフェロン−γ、インターロイキン−2、アレルギーワクチン、抗ヒスタミン、抗ロイコトリエン、ベータ−アゴニスト、テオフィリン、または抗コリン作用剤と一緒に使用することができる。
【0116】
多発性硬化症を治療するために共投与することができる例示的薬剤
対象が多発性硬化症に罹患しているか、または罹患する危険性がある場合、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体(例えば、インターロイキン(IL)−6デコイ受容体ECD−BBB受容体Ab融合抗体)をいずれかの組合せにおいて1以上の下記の例示的多発性硬化症治療剤、すなわち、インターフェロンβ−1a、インターフェロンβ−1b、グラチラマーアセテート(CopaxoneR(登録商標))、ミトキサントロン(NovantroneR)、低用量ナルトレキソン、ナタリズマブ(Natalizumab)(TysabriR)、SativexR、エイムスプロ(Aimspro)(ヤギ血清)、トリメスタ(Trimesta)(経口エストリオール)、ラキニモド(Laquinimod)、FTY720(フィンゴリモド(Fingolimod))、MBP8298、NeuroVaxR、TovaxinR、レビムン(Revimmune)、CHR−1103、BHT−3009、BG−12、クラドリビン(Cladribine)、ダクリズマブ(daclizumab)(Zenapax)リツキシマブ(Rituximab)(Rituxan)、シクロホスファミド、キャンパス(Campath)、ファムプリジン(Fampridine)−SR、MN−166、テムシロリマス(Temsirolimus)、またはRPI−78Mと一緒に使用することができる。
【0117】
例えば、アルツハイマー病またはAIDS関連認知症の治療のために共投与することができる例示的薬剤
対象が認知症に罹患しているか、または罹患する危険性がある場合、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体(例えば、TNF関連アポトーシス誘導性リガンド受容体(TRAIL−R)ECD−BBB受容体Ab融合抗体)をいずれかの組合せにおいて認知症を治療するための1以上の薬剤または方法と一緒に使用することができる。認知症を治療するための治療剤/治療の例は、限定するものではないが、FlurizanTM(登録商標)(MPC−7869,r フルルビプロフェン(r flurbiprofen))、メマンチン(memantine)、ガラタミン(galantamine)、リバスチグミン(rivastigmine)、ドネジピル(donezipil)、タクリン(tacrine)、Aβ1−42免疫療法、レスベラトロール(resveratrol)、(−)−エピガロカテキン−3−ガラート、スタチン、ビタミンC、またはビタミンEを包含する。
【0118】
血栓塞栓性障害を治療するための例示的薬剤
対象が血栓塞栓性障害(例えば、卒中)に罹患しているか、または罹患する危険性がある場合、1以上の他の抗血栓塞栓剤とのいずれかの組合せにおいて、本明細書に記載のデコイ受容体−BBB受容体Ab融合抗体(例えば、アポトーシスのTNF様弱インデューサー(TWEAK)受容体デコイECD−BBB受容体Ab融合抗体)を用いて該対象を治療することができる。抗血栓塞栓剤の例は、限定するものではないが、血栓溶解剤(例えば、アルテプラーゼ、アニストレプラーゼ、ストレプトキナーゼ、ウロキナーゼ、または組織プラスミノーゲンアクチベーター)、ヘパリン、チンザパリン(tinzaparin)、ワーファリン(warfarin)、ダビガトラン(dabigatran)(例えば、ダビガトランエテキレート(dabigatran etexilate))、因子Xa阻害剤(例えば、フォンダパリヌクス(fondaparinux)、ドラパリヌクス(draparinux)、リバロキサバン(rivaroxaban)、DX−9065a、オタミキサバン(otamixaban)、LY517717、またはYM150)、チクロピジン(ticlopidine)、クロピドグレル(clopidogrel)、CS−747(プラスグレル(prasugrel),LY640315)、キシメラガトラン(ximelagatran)、またはBIBR 1048を包含する。
【実施例】
【0119】
下記の具体例は、単なる例示として解釈されるべきものであり、如何なる方法においても、該開示の残りの部分を限定するものとして解釈されるべきではない。当業者は、さらに工夫することなく、本発明の記載に基づいて、本発明をその完全な範囲まで利用することができると確信する。本明細書中に引用された全ての出版物は、出典明示により全体として本明細書の一部とされる。URLまたは他のかかる識別子またはアドレスに言及する場合、かかる識別子は変化することができ、インターネット上の特定の情報は移り変わることができるが、インターネットを検索することによって、等しい情報を見出すことができると理解される。その言及は、かかる情報の入手可能性および公的な流布を証明するものである。
【0120】
実施例1 ヒトTNFR ECD cDNAのクローニングおよび発現
NP 001057のアミノ酸23−257に対応するヒトTNFR−II細胞外ドメイン(ECD)を、表2に記載のオリゴデオキシヌクレオチド(ODNs)を用いるポリメラーゼ連鎖反応(PCR)によってクローン化し、ヒトU87グリア細胞から単離したポリA+RNAの逆転写からcDNAを得た。該TNFR cDNAを、25ng ポリA+RNA−由来のcDNA、0.2μM 順方向および逆方向ODNプライマー(表Iの各々、配列番号1および配列番号2)、0.2mM デオキシヌクレオシドトリホスフェート、および50μl Pfuバッファー中の2.5U PfuUltra DNAポリメラーゼを用いるPCRによってクローン化した。Mastercycler温度サイクラー中、95℃で2分間の開始変性工程、次いで、95℃で30秒間の変性を30サイクル、55℃で30秒間のアニーリング工程、および72℃で1分間の増幅工程を用いて増幅を行い、次いで、72℃で10分間、最終インキュベーションを行った。PCR産物を0.8%アガロースゲル電気泳動中で分析し、ヒトTNFR cDNAに対応する〜0.6kbの予想される主要単一バンドを生じた(図2A)。TNFR ECDのアミノ酸配列は、クローン化したTFR ECD cDNAのヌクレオチド配列から推定し、Leu23−Asp257を含んでいた(NP 001057)。
【0121】
【表2】
【0122】
実施例2 TNFRがHIRMAb重鎖のカルボキシル末端に融合した重鎖−TNFR融合タンパク質をコードする発現プラスミドの遺伝子操作
重鎖(HC)およびTNFR ECDの融合タンパク質を発現する発現プラスミドをpCD−HIRMAb−TNFRと称する。ヒトTNFR−II ECD(NP 001057)のアミノ酸Leu23−Asp257に対応する成熟ヒトTNFR cDNAを該pCD−HIRMAb−HC発現プラスミド(図2B)のHpaI部位に挿入することによって該プラスミドを操作して、pCD−HIRMAb−TNFRを製造した(図2B)。該pCD−HIRMAb−HCプラスミドは、キメラHIRMAbのHCをコードし、該プラスミドおよび軽鎖(LC)発現プラスミドpHIRMAb−LCを用いるCOS細胞のデュアルトランスフェクションにより、キメラHIRMAbの一過性発現を可能にする。TNFR順方向(FWD)PCRプライマー(表I)は、「CA」ヌクレオチドを導入して、オープンリーディングフレームを維持し、かつ、HIRMAb HCのCH3領域のカルボキシル末端とそのシグナルペプチドを差し引いたTNFR ECDのアミノ末端との間に、Ser−Ser−Serリンカーを導入する。該TNFRモノマーの各HCのカルボキシル末端への融合は、図1に示される。該設計は、該TNFRを、TNFR ECDの好ましいコンフォメーションである二量体コンフィギュレーションに立体的に限定し、受容体ダイマーとして結晶化する(Chanら(2000), Immunity, 13:419−422)。該TNFR逆方向(REV)PCRプライマー(表I)は、TNFR ECDタンパク質の末端のアスパラギン酸の直後に、終止コドン「TGA」を導入する。該操作されたpCD−HIRMAb−TNFR発現ベクターは、DNA配列決定によって確認された。
【0123】
該HIRMAb HCおよびLC cDNA発現カセットは、サイトメガロウイルス(CMV)プロモーターによって駆動し、ウシ成長ホルモン(BGH)ポリアデニル化(pA)配列を含有する(図2B)。該ユニバーサルpCD−HIRMAb−HCベクターの操作は、以前に記載されたように(Boadoら(2007b) Biotechnol. Bioeng., 97:1376−1386)、部位特異的変異誘発(SDM)によって、HIRMAb HC CH3オープンリーディングフレーム(orf)の末端に単一のHpaI部位を挿入することによって実施された。
【0124】
表2のODNsを用いるPCRによって、235アミノ酸TNFR−II ECDに対応するcDNAを増幅し、図2Bに概説されるように、該cDNAをpCD−HIRMAb−HCプラスミドのHpaI部位にサブクローン化した。DNA配列決定は、該pCD−HIRMAb−TNFRプラスミドの発現カセットが、714ヌクレオチド(nt)のCMVプロモーター、9ntの全Kozak部位(GCCGCCACC)、2,100ntのHIRMAb HC−TNFR融合タンパク質オープンリーディングフレーム、および370ntのBGH配列(配列番号3)を包含する3,193個のヌクレオチド(nt)を含むことを示した。該2,100ntオープンリーディングフレームは、配列番号3のnt724〜2,823に相当する。該プラスミドは、19アミノ酸IgGシグナルペプチド(AA1〜19、配列番号4)、442アミノ酸HIRMAb HC(AA20〜461、配列番号4)、3アミノ酸リンカー(Ser−Ser−Ser)、およびそのシグナルペプチドを差し引いた235アミノ酸ヒトTNFR−II ECD(AA465〜699、配列番号4)からなる、699アミノ酸(AA)タンパク質をコードした。グリコシル化を差し引いた該重鎖融合タンパク質の予測分子量は、73,900Daであり、予測等電点(pI)は8.45である。該融合タンパク質のTNFR ECD部分の推定アミノ酸配列は、22個のシステイン残基および2個のN−連結コンセンサスグリコシル化部位を該TNFR ECD内のAsn−149およびAsn−171に含んだ(配列番号4)。
【0125】
該HIRMAb−TNFR融合タンパク質は、2個の融合重鎖(AA20−699、配列番号4)および2個の軽鎖(LC)からなる。該LCは、pCD−HIRMAb−LC発現プラスミドによって発現され、該LC発現カセットのnt配列は、配列番号5に示す。該pCD−HIRMAb−LCプラスミドの発現カセットのDNA配列決定は、731ntのCMVプロモーター、9ntの全Kozak部位(GCCGCCACC)、705ntのHIRMAb LC融合タンパク質オープンリーディングフレーム、および370ntのBGH配列を包含する1,809ntを含んでいた(配列番号5)。該705ntオープンリーディングフレームは、配列番号5のnt741〜1,445に相当する。該プラスミドは、20アミノ酸IgGシグナルペプチド(AA1〜20、配列番号6)、および214アミノ酸HIRMAb LC(AA21〜234、配列番号6)からなる234AAタンパク質をコードする。
【0126】
実施例3 トランスフェクトしたCOS細胞によるHIRMAb−TNFR融合タンパク質の分泌
COS細胞は、μg DNA:uL リポフェクタミンの比率1:2.5でリポフェクタミン2000を用いて、pCD−HIRMAb−LCおよびpCD−HIRMAb−TNFR(ここに、pCD−HIRMAb−LCは、HIRMAb−TNFR融合タンパク質中に組み込まれた同じ軽鎖であるキメラHIRMAbの軽鎖をコードする発現プラスミドである)でデュアルトランスフェクトした。トランスフェクション後、該細胞を無血清培地中で培養した。COS細胞を最初、ヒトIgG特異的ELISAで発現をスクリーニングするために、6ウェルクラスター皿に播種した。次いで、10xT500フラスコ中、トランスフェクトしたCOS細胞の培養のために、該トランスフェクションをスケールアップした。馴化した無血清培地を3日目および7日目に収集した。該融合タンパク質をタンパク質Aアフィニティークロマトグラフィーによって精製した。
【0127】
ヒトIgG ELISAは、Immulon 2高結合プレート中で、COS細胞馴化培地を用いて行った。ヤギ抗−ヒトIgG一次抗体を0.1M NaHCO3(100μl、2μg/ml)中で播種し、4℃で一晩インキュベートした。プレートを0.01M Na2HPO4/0.15 M NaCl/pH=7.4/0.05% Tween−20(PBST)で洗浄し、PBST中における1%ゼラチンで22℃にて30分間ブロックした。プレートを100μl/ウェルのヒトIgG1標準または融合タンパク質のいずれかと共に、室温(RT)でて60分間インキュベートした。PBSTで洗浄後、アルカリホスファターゼにコンジュゲートしたヤギ抗−ヒトカッパLC抗体を37℃で60分間培養した。暗所中、pH=10.4でp−ニトロフェニルホスフェートを用いて、発色を行った。NaOHを用いて該反応を停止し、405nmの吸光度をELISAプレートリーダーで測定した。
【0128】
COS細胞によって産生されるタンパク質A精製融合タンパク質の均質性を還元性12%ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)、次いで、クーマシーブルー染色を用いて評価した。ウェスタンブロッティングの場合、ヒトIgG免疫反応性をヒトIgG(H+L)に対する一次ヤギ抗体を用いて試験し、ヒトTNFR免疫反応性をヒトTNFR−II ECDに対するマウスモノクローナル抗体を用いて評価した。SDS−PAGEおよびクーマシーブルー染色後、軽鎖(LC)のサイズは、HIRMAbおよびHIRMAb−TNFR融合タンパク質の両方について同じである(図3)。該融合タンパク質の重鎖(HC)のサイズは、HIRMAbのHCよりも約30kDa大きい(図3)。ウェスタンブロッティングにおいて、HIRMAbまたはHIRMAb−TNFR融合タンパク質のいずれかのLCは、図4(左パネル)に示されるように、該ウェスタン上で、ヒトIgG(H+L)に向けられた一次抗体と等しく反応する。該融合タンパク質のHCのサイズは、抗−ヒトIgG一次抗体(図4、左パネル)または抗−ヒトTNFR一次抗体(図4、右パネル)のいずれかを用いる両方のウェスタンブロットにおいて、HIRMAbのHCのサイズよりも約30kDa大きい。これらの結果は、該HIRMAb−TNFR融合タンパク質が宿主細胞によって首尾よく翻訳され、分泌されることを示す。
【0129】
実施例4 HIRMAb−TNFR融合タンパク質はHIRおよびTNFαの両方について高アフィニティー結合を保持する
本発明の作業において、デコイ受容体のアミノ末端は、IgG鎖のカルボキシル末端に融合される。予想外にも、該コンフィギュレーションは、二官能性デコイ受容体−BBB受容体Ab融合抗体を生じた。
【0130】
HIR細胞外ドメイン(ECD)に対する該融合タンパク質のアフィニティーは、レクチンアフィニティー精製HIR ECDを用いるELISAで決定した。該HIR ECDで永続的にトランスフェクトされたCHO細胞を無血清培地(SFM)中で成育させ、該HIR ECDをコムギ胚芽凝集素アフィニティーカラムで精製した。該HIR ECD(0.2μg/ウェル)をImmulon 2高結合96ウェルプレート上に播種し、キメラHIRMAb、HIRMAb−TNFR融合タンパク質、またはヒトIgG1の該HIR ECDに対する結合を、ビオチン化ヤギ抗−ヒトIgG(H+L)抗体(0.3μg/ウェル)、およびアビジン/ビオチン化ペルオキシダーゼ検出システムを用いて検出した。HIR ECDに対する50%の結合をもたらした濃度、ED50を非線形回帰分析によって決定した。各々、0.44±0.19nMおよび0.62±0.06nMのED50を伴うキメラHIRMAbおよびHIRMAb−TNFR融合タンパク質のHIR ECDに対する比較可能な結合が存在する(図5)。これは、TNFRのHIRMAbへの融合にもかかわらず、HIRMAbによるHIRへの高いアフィニティー結合の保持が存在することを示す。
【0131】
HIRMAb−TNFR融合タンパク質のTNFαに対する結合は、ELISAを用いて決定された。捕獲試薬は、市販のヒトTNFαであった。該アッセイにおける陽性対照は、市販の組み換えヒトTNFR−II:ヒトIgG1 Fc融合タンパク質(TNFR:Fcと称する)であり、陰性対照は、ヒトIgG1/κであった。TNFαは、0.1M NaHCO3/pH=9.0中に溶解し、100μl/ウェル(0.2μg/ウェル)中4℃で一晩培養した。0.01M Tris/0.15M NaCl/pH=7.4(TBS)での洗浄後、該ウェルをTBS中における1%ウシ血清アルブミン(BSA)で30分間ブロックした。100μl/ウェルの容量のHIRMAb−TNFR、TNFR:Fc、またはヒトIgG1kを室温で60分間培養した。TBSおよび0.05%Tween−20(TBST)で洗浄後、ヤギ抗−ヒトIgG−アルカリホスファターゼコンジュゲートを60分間インキュベートした(0.2μg/ウェル)。TBSTで洗浄した後、パラ−ニトロフェニルホスフェートでの発色および1.2M NaOHでの反応停止後、ELISAプレートリーダーを用いて405nmでの色検出を行った。該ヒトIgG1/カッパ(hIgG1/k)は、図6Aに示されるように、TNFαに結合しなかった。該TNFR:Fc、または該HIRMAb−TNFRは、0−600ng/mLの試験濃度範囲で飽和しない直線関係で、培養したTNFαに結合した。したがって、該データを線形回帰分析にフィットさせて、結合曲線の傾きを計算した。該傾きの逆数は、KD/Amax比に等しく、ここに、KDは、培養したTNFαに結合しているTNFRの結合定数であり、Amaxは、最大吸光度である。該分析において、結合曲線の傾きは、該TNFR融合タンパク質のTNFαに対するアフィニティーに正比例する。組み換えTNFR:Fcまたは該HIRMAb−TNFR融合タンパク質のヒトTNFαに対するアフィニティーをELISAで測定した。該TNFR:Fcは、0.88±0.02nMのKD/Amax比でTNFαに結合した(図6A)。該HIRMAb−TNFR融合タンパク質は、1.40±0.08nMの比較可能なKD/Amax比でTNFαに結合した(図6B)。これは、HIRMAbのカルボキシル末端へのTNFRの融合にもかかわらず、TNFR ECDによるTNFαへの高アフィニティー結合の保持が存在することを示す。
【0132】
ヒトTNFαの該HIRMAb−TNFR融合タンパク質への飽和可能な結合は、放射性受容体アッセイ(RRA)を用いて決定された。マウス抗−ヒトIgG1 Fc抗体を96ウェルプレート(0.4μg/ウェル)に播種し、0.1M NaHCO3/pH=8.3中で一晩インキュベーションし、次いで、洗浄し、0.01M Na2HPO4/0.15M NaCl/pH=7.4(PBS)中における1%ウシ血清アルブミン(BSA)でブロックした。次いで、下記の溶液、すなわち(a)PBS中1%BSA、(b)100ng/ウェルのヒトIgG1/カッパ、または(c)100ng/ウェルの該HIRMAb−TNFR融合タンパク質の1つを100μl/ウェルにて加え、次いで、室温で1時間インキュベーションした。次いで、該ウェルをPBSで洗浄し、次いで、0.01uCi/ウェル(0.2ng/ウェル)濃度の[125I]−TNFαおよび種々の濃度の標識していないヒトTNFαの共混合物200μl/ウェルを加え、次いで、室温で3時間インキュベーションした。吸引によりウェルを空にし、冷PBSで洗浄し、250μl/ウェルの1N NaOHを加え、次いで、60℃で30分間加熱した。Perkin Elmer液体シンチレーションカウンターで放射能をカウントし、ウェルあたりの部分(fractional)結合をコンピューターで算出した。該HIRMAb−TNFR融合タンパク質に結合しているTNFの半飽和定数、KDは、統計学的ソフトウェアパッケージを用いる非線形回帰分析によって決定した。該HIRMAb−TNFR融合タンパク質に対するTNFαの飽和可能な結合は、放射性受容体アッセイを用いて検出可能であり、図7Aに概説される。該結合データ(方法)の非線形回帰分析は、結合のKDが0.34±0.17nMであることを示した(図7B)。これらの結果は、該TNFR ECDが、該HIRMAb重鎖のカルボキシル末端へのそのアミノ末端での融合にかかわらず、まだ、非常に高いアフィニティーでその同族リガンドを結合することを示す。
【0133】
実施例5 ヒト細胞におけるHIRMAb−TNFR融合タンパク質の生物学的活性
ヒトWEHI−13VAR細胞(CRL−2148)をAmerican Type Culture Collection(Manassas,VA)から得、TNFα細胞毒性のバイオアッセイとして用いた(Espevik and Nissen−Meyer, (1986), J. Immunol. Methods, 95:99−105)。該細胞を24ウェルクラスター皿に、10%胎仔ウシ血清(FBS)を含有するRPMI−1640培地中300,000細胞/ウェルで播種した。一晩培養後、該培地の半分を吸引除去し、200μlの新鮮なRPMI−1640培地、50μl/ウェルの10μg/mLのアクチノマイシンD(最終濃度=1.0μg/mL)、および最終濃度が1〜100pg/mLのヒト組み換えTNFαで置き換えた。いくつかのウェルにおいて、TNFαをウェルに加える前の30分間、TNFαを組み換えTNFR:Fc、またはHIRMAb−TNFR融合タンパク質に複合体化させた。該TNFR:FcまたはHIRMAb−TNFR融合タンパク質の最終濃度は、1.4nMであった。加湿インキュべーター中、37℃で一晩(20時間)インキュベーション後、該培地にチアゾイルブルーテトラゾリウムブロミド(MTT)を最終濃度0.5mg/mLまで補足した。37℃で3時間インキュベーション後、可溶化溶液(48%イソプロパノール、2% 1N HCl)の添加によって、該反応を停止した。570nmおよび650nmでの吸光度を測定し、A570−A650差をコンピューターで計算した。MTTは、健康細胞中のミトコンドリアによって、ホルマザン結晶に酸化され、該反応は、細胞の生存に反比例する。アクチノマイシンDの不在下で、該TNFαは、1−100pg/mL濃度にて、細胞に対して毒性ではなかった。しかしながら、アクチノマイシンDの存在下で、TNFαのこれらの濃度は、ED50値 5−10pg/mLヒトTNFαを伴う用量依存性細胞毒を生じた(図8)。しかしながら、該TNFαの細胞毒効果は、1.4nM濃度のTNFR:Fc融合タンパク質またはHIRMAb−TNFR融合タンパク質と共に該サイトカインを共インキュベーションすることによって遮断された(図8)。これは、該TNFRデコイ受容体がHIRMAbのカルボキシル末端への融合にかかわらず、生物学的に活性であることを示す。
【0134】
実施例6 TNFRのアミノ酸配列の部位特異的変異誘発
ヒトU87グリア細胞からクローン化された該デコイ受容体の235AA TNFR配列(AA465〜699,配列番号4)内に、174および210位置に多形が存在する。該HIRMAb−TNFR重鎖(配列番号4)の638位置のアルギニン(Arg)は、235AA TNFR ECDのArg−174に対応する。メチオニン(Met)が該ヒトTNFR配列(GenBank AAA36755)の174位置に見出されるので、該Arg−174は天然のR174M多形である。R638M部位特異的変異誘発(SDM)を周知の方法によって実施でき、配列番号7に示されるように、Arg−638をMet−638に変換する。該HIRMAb−TNFR重鎖(配列番号4)の674位置のリジン(Lys)は、235AA TNFR ECDのLys−210に対応する。該ヒトTNFR配列(GenBank AAA36755)のグルタミン酸(Glu)が210位置に見出されるので、該Lys−210は天然のK210E多形である。K674E部位特異的変異誘発(SDM)を周知の方法によって実施でき、配列番号7に示されるように、Lys−674をGlu−674に変換する。pCD−HIRMAb−TNFR発現ベクターのTNFR−R638M多形(配列番号7)での操作は、周知の部位特異的変異誘発法を用いて、ヒトTNFR−R638M FWD(表3、配列番号8)およびヒトTNFR−R638M REV(表3、配列番号9)ODNプライマーを用いるSDMによって実施される。同様に、pCD−HIRMAb−TNFR発現ベクターのTNFR−K674E多形での操作は、ヒトTNFR−K674E FWD(表3、配列番号10)およびヒトTNFR−K674E REV(表3、配列番号11)ODNプライマーを用いるSDMによって実施される。
【0135】
実施例7 HIRMAb−TNFR融合タンパク質をコードするタンデムベクターの遺伝子操作
HIRMAb−TNFR融合タンパク質は、図1に示されるように、2つの重鎖(HC)および2つの軽鎖(LC)からなる。したがって、宿主細胞は、HC遺伝子およびLC遺伝子の両方で永続的にトランスフェクトされなければならない。さらに、宿主細胞は、トランス遺伝子挿入部位の周囲の増幅を有する細胞系統の単離を可能にする遺伝子で永続的にトランスフェクトされなければならない。これは、ジヒドロ葉酸還元酵素(DHFR)をコードしている遺伝子で宿主細胞をトランスフェクション後の、メトトレキサート(MTX)での細胞系統の選択を用いて達成される。したがって、最終的には製造のためのマスター・セル・バンク(Master Cell Bank)をもたらす単一細胞における3つの遺伝子全ての高産生を得る必要がある。3つの遺伝子全ての高い発現を保証するために、タンデムベクター(TV)と呼ばれる一片のDNAを図9に概説されるように操作した。pTV−HIRMAb−TNFRと称するHIRMAb−TNFR融合タンパク質のためのTVの遺伝子操作は、pTV−HIRMAbのHpaI部位へのTNFR ECD cDNAの挿入によって完了した。該TNFR ECD cDNAは、カスタムODNsおよびpCD−HIRMAb−TNFR(図2B)を鋳型として用いるPCRによって生成された。pTV−HIRMAbは、HIRMAb HCオープンリーディングフレームの3’末端に独特のHpaI制限酵部位を有するキメラHIRMAbをコードするタンデムベクターである。pTV−HIRMAbは、HIRMAb LCおよびHC遺伝子のタンデム発現カセットを含有し、各カセットは、イントロンを有するCMVプロモーターによって駆動し、各カセットは、ウシ成長ホルモン(BGH)ポリアデニル化(pA)配列によって終了する。pTV−HIRMAbは、また、ネズミジヒドロ葉酸還元酵素(DHFR)のための第3の発現カセットを含有し、それは、SV40プロモーターによって駆動し、B型肝炎ウイルス(HBV)pA配列によって終了する。HIRMAb−TNFR HC発現カセットは、イントロンを有するCMVプロモーター、9ntの全Kozak部位(GCCGCCACC)、2,100ntのHIRMAb HC−TNFR融合タンパク質オープンリーディングフレーム、およびBGH pA配列からなる。図9に示されるタンデムベクターによってコードされる699 AA HIRMAb−TNFR HCタンパク質は、配列番号4におけるものと同一であり、19アミノ酸IgGシグナルペプチド(AA1〜19、配列番号4)、442アミノ酸HIRMAb HC(AA20〜461、配列番号4)、3アミノ酸リンカー(Ser−Ser−Ser)、およびそのシグナルペプチドを差し引いた235アミノ酸ヒトTNFR−II ECD(AA465〜699、配列番号4)からなる。該重鎖融合タンパク質の予測分子量は、グリコシル化を差し引いて、73,900Daであり、予測等電点(pI)は8.45である。該融合タンパク質のTNFR ECD部分の推定アミノ酸配列は、22個のシステイン残基を含み、2個のN−結合コンセンサスグリコシル化部位をTNFR ECD内のAsn−149およびAsn−171に含んだ(配列番号4)。図9に記載のタンデムベクターによって産生されるHIRMAb−TNFR融合タンパク質は、2つの融合重鎖(AA20−699、配列番号4)および2つの軽鎖(LC)からなる。LC発現カセットは、イントロンを有するCMVプロモーター、9ntの全Kozak部位(GCCGCCACC)、705ntのHIRMAb LC融合タンパク質オープンリーディングフレーム、およびBGH pA配列を含む。図9に示されるタンデムベクターによってコードされる214AA HIRMAb−LCタンパク質は、配列番号6におけるものと同一であり、20アミノ酸IgGシグナルペプチド(AA1〜20、配列番号6)、および214アミノ酸HIRMAb LC(AA21〜234、配列番号6)からなる。
【0136】
実施例8 CHO細胞の安定なトランスフェクションおよび希釈クローニング
無血清培地(SFM)適応性DG44チャイニーズハムスター卵巣(CHO)細胞を、エレクトロポレーターを用いて、PvuIで線形化後の5μgのpTV−HIRMAb−TNFRでエレクトロポレーションした。5 x 106細胞を200μLのリン酸緩衝化セーライン(PBS)および0.2cmキュベット中、方形波および160ボルトを用いて、該DNAでエレクトロポレーションした。細胞をCHO無血清培地(SFM)中に懸濁し、4x96ウェルプレート中で培養した。安定なトランスフェクタントの選択は、エレクトロポレーションの2日後、0.54mg/ml G418を用いて開始した。トランスフェクトタントのコロニーが明らかになったとき、すなわち、21目に、ヒトIgG ELISAのために、上清のアリコートを採取した。陽性コロニーを単離し、さらなる特徴付のために個々に培養した。DG44細胞は、内因性DHFRを欠き、内因性葉酸合成に関して、栄養分、ヒポキサンチンおよびチミジン(HT)に依存する。該TVを有するトランスフェクト細胞は、外因性DHFRを発現する。トランスフェクト細胞系統は、さらに、HT−欠損培地中に置くことによって選択された。トランス遺伝子挿入部位周囲の増幅を有する系統は、細胞を、20nM MTXで開始する増加濃度のMTXに付すことによって選択された。80−160nM MTXにて細胞系統の安定化後、1ウェルあたり1細胞の限界希釈クローニング(DC)によって、高生産性クローンを単離し、DCの各ラウンドにて全部で4000個のウェルに播種し、高容量マイクロプレートディスペンサーおよびマイクロプレートウォッシャーを用いるヒトIgG ELISAを用いて、培地IgGを測定した。クローン化した細胞を、軌道振盪機上の125mLのプラスチック角瓶中、1−2百万細胞/mLの生存細胞密度にて増殖させ、IgG ELISAによる測定の場合、無血清培地中約10mg/LのヒトIgGレベルを生産した。
【0137】
実施例9 イン・ビボにおけるデコイ受容体の脳に対する選択的標的化
HIRMAbは、旧世界霊長類のインスリン受容体と交差反応するが、低級種のインスリン受容体を認識しないので、HIRMAb−TNFR融合タンパク質の脳浸透を成体アカゲザル(Rhesus monkey)において測定した。さらに、ヒトIgG1 Fc領域に融合した市販のTNFRデコイ受容体(TNFR:Fcと称する)をR&D Systemsから購入した。該TNFR:Fc融合タンパク質は、デコイ受容体融合タンパク質に関する当該分野の状況を示す。該TNFR:Fcは、いずれのBBB受容体に対しても特異性を示さず、BBBに浸透しないと予測される。しかしながら、該TNFR:Fc融合タンパク質のBBB輸送は、測定されたことがない。さらに、IgG分子がBBBを横断するという文献における証拠があり[Zlokovicら、A saturable mechanism for transport of immunoglobulin G across the blood−brain barrier of the guinea pig(モルモットの血液脳関門を横断する免疫グロブリンGの輸送のための飽和可能なメカニズム)、Exp. Neurol, 107, 263−270, 1990]、TNFR:Fc融合タンパク質は、ヒトIgG1の定常領域の大部分を含有するので、該タンパク質もまた、BBBを横断することを推測できた。脳への取り込み研究について、CHO−由来HIRMAb−TNFR融合タンパク質は、該TNFR:Fc融合タンパク質の非酸化的放射性ヨウ素化と平行して、トリチウム化された。該[3H]−HIRMAb−TNFRおよび[125I]−TNFR:Fc融合タンパク質を共に、アカゲザルに注射した。結果は、TNFR:Fc融合タンパク質はBBBを横断せず、BBB分子のトロイの木馬に融合後、TNFRの脳浸透における顕著な増加があることを実証する。また、霊長類の非脳器官における該融合タンパク質の取り込みを測定し、それは、該デコイ受容体のHIRMAbへの融合が該医薬のCNSに対する選択的標的化をもたらすことを示す。
【0138】
R&D Systems(Minneapolis,MN)から購入し、SDS−PAGEによって均質であることが示されたTNFR:Fc融合タンパク質(#726−R2)を放射性標識するために、[125I]−Bolton−Hunter試薬を用いた。該TNFR:Fcは、1.0x28cmカラムのSephadex G−25での精製および0.01M NaH2PO4/0.15M NaCl/pH=7.4/0.05% Tween−20(PBST)での溶出後、新鮮なBolton−Hunter試薬で11.5uCi/ugの比活性および>99%のトリクロロ酢酸(TCA)沈澱性まで放射性標識した。標識化TNFR:Fc融合タンパク質のTCA沈澱は、ヨウ素化の24時間後で>99%のままであり、放射性標識後24時間以内に、該TNFR:Fc融合タンパク質を該霊長類に投与した。[3H]−N−スクシンイミジルプロピオネート(NSP)を用いて、HIRMAb−TNFR融合タンパク質を放射性標識し、それは、1.0x28cmカラムのSephadex G−25での精製および0.02M Tris/0.15M NaCl/pH=6.5(TBS)での溶出後、3.0uCi/ugの比活性および95%のTCA沈澱性をもたらした。該溶液は、TBSおよびUltra−15マイクロコンセントレーター(Millipore, Bedford, MA)とバッファー交換し、TCA沈澱性が99%に増加した。該3H−標識HIRMAb−TNFR融合タンパク質は、霊長類研究の前に標識され、−70℃で保存された。
【0139】
4.1kgのアカゲザル雌成体の左大腿静脈に、1806uCiの[3H]−HIRMAb−TNFR融合タンパク質、428uCiの[125I]−TNFR:Fc融合タンパク質を3.1mLのTBS中、ボーラス注射によって30秒かけて静脈内(IV)注射した。HIRMAb−TNFR融合タンパク質の投与量は、0.15mg/kgであった。該動物を最初、筋内ケタミンで麻酔し、吸入による1%イソフランによって、麻酔を維持した。全ての手順は、米国国立衛生研究所(U.S. National Institutes of Health)によって採用され、公表された研究動物のケアおよび使用のためのガイド(The Guide for the Care and Use of Laboratory Animals)にしたがって行われた。静脈内薬物投与後、3Hおよび125I放射能測定のために、大腿静脈血漿を1、2.5、5、15、30、60および120分に得た。該動物を安楽死させ、主要器官(心臓、肝臓、脾臓、肺、骨格筋、および大網脂肪)の試料を採取し、重量を量り、放射能測定のために処理した。頭蓋を開け、脳を取り出した。放射能測定のために、前頭皮質灰白質、前頭皮質白質、小脳灰白質、および小脳白質の試料を採取した。
【0140】
前頭皮質の試料(〜2グラム)を毛細管除去(capillary depletion)分析のために採取した。脳を組織グラインダー中、8mLの冷PBS中でホモジナイズした。該ホモジネートに9.4mLの冷40%デキストラン(70kDa, Sigma Chemical Co.)を補足し、放射能測定のために、該ホモジネートのアリコートを得た。該ホモジネートを固定角ローター中、3200gにて、4℃で10分間遠心分離した。脳微小血管系が高分子量デキストランの該密度にて、ペレットとして定量的に沈澱し、血管後(post-vascular)上清は、毛細管が除去された(depleted)脳実質の測定値である。血管ペレットおよび上清を3Hおよび125I放射能について、ホモジネートと平行して、カウントした。分布容量(VD)は、120分末梢血漿(terminal plasma)における全125Iまたは3H放射能によって割られた分数における全125Iまたは3H放射能の比率から、3つのフラクションの各々について決定した。
【0141】
血漿および組織試料は、ガンマカウンターを用いて125I放射能について分析し、液体シンチレーションカウンターを用いて3H放射能について分析した。125I同位体元素は、液体シンチレーションカウンター(LSC)の3Hチャンネル(0−12keV)において検出される放射を放つ。したがって、クエンチ剤としてクロロホルムを用いてクエンチ曲線を作製して、3Hウィンドウにおける125Iの計数効率をコンピューターで計算した。3H計数について全ての試料をSoluene−350(Perkin Elmer)中に可溶化し、Opti−Fluor O(Perkin Elmer)中におけるLSCにおいてカウントした。
【0142】
血漿中の3Hまたは125I放射能(DPM/mL)を%注入用量(ID)/mLに変換し、%ID/mLを双指数(bi-exponential)方程式 %ID/mL=A1e−k1t+A2e−k2tにフィットさせた。インターセプト(A1、A2)および傾斜(k1、k2)を用いて、中央滞留時間(MRT)、分布の中心体積(Vc)、分布の定常体積(Vss)、血漿濃度曲線下の面積(AUC)、および全身性クリアランス(CL)をコンピューター計算した。非線形回帰分析を用いて、PKパラメーターをコンピューター計算し、データに1/(%ID/mL)2のウェイトをかけた。
【0143】
透過性−表面積(PS)積(permeability-surface area product)とも呼ばれる器官クリアランス(uL/min/g)を末端器官取り込み(%ID/g)および120分血漿AUC(%IDmin/mL)から次のようにコンピューター計算する。
器官PS積=[(%ID/g)/AUC]*1000
HIRMAb−TNFR融合タンパク質は、[3H]で放射能標識され、TNFR:Fc融合タンパク質は、[125I]で放射能標識され、これらのタンパク質を成体アカゲザルの静脈内に共注射した。血漿放射能のクリアランスを図12Aに示し、TCAにより沈澱性であった血漿放射能を図12Bに示す。PKパラメーターの概算のために、血漿クリアランスプロフィール(図12A)を双指数関数(Methods)にフィットさせ、それを各融合タンパク質につき、表4に示す。脳および末梢器官による該融合タンパク質の取り込みは、% I.D./100グラム組織として測定され、これらの値を表5に示す。該融合タンパク質の分布の脳体積(VD)は、毛細管除去(capillary depletion)法で測定され、ホモジネートについてのVD値、血管ペレット、および血管後上清を表6に示す。
【0144】
HIRMAb−TNFRおよびTNFR:Fc融合タンパク質のBBB PS積は、2時間血漿AUC(図13A)および脳への取り込みまたは%ID/100g(図13B)からコンピューター計算され、該PS積を図13Cに示す。比較のために、図13のデータは、血管性間隙マーカー、ヒトIgG1のAUC、%ID/100g、およびBBB PS積も示す。該PS積は、同様に、末梢器官におけるHIRMAb−TNFRおよびTNFR:Fc融合タンパク質についてコンピューター計算され、これらのデータを表7に示す。各器官におけるTNFR:Fc融合タンパク質についてのPS積に対する、HIRMAb−TNFR融合タンパク質についてのPS積の比率を図14にプロットする。
【0145】
【表3】
図12の血漿クリアランスデータから概算される。
【0146】
【表4】
データは、% I.D./100グラム;平均±SE(n=3)
【0147】
【表5】
平均±SE(n=3). VD=分布の体積(uL/g); TCA=トリクロロ酢酸
【0148】
【表6】
データは、平均±SE(n=3)
【0149】
TNFR:Fc融合タンパク質と比較したHIRMAb−TNFR融合タンパク質の霊長類BBBをイン・ビボで横断する選択的輸送を表5に示し、それにより、%ID/100グラムとして表される該タンパク質の脳への取り込みを与える。アカゲザルの脳の重量が100グラムなので、該取り込みデータは、組織100グラムとして表される。しかしながら、表5の脳取り込みパラメーターは、HIRMAb−TNFRおよびTNFR:Fc融合タンパク質の相対的な脳浸透の直接的な測定値ではなく、以下を考慮すべきである。まず、脳取り込みまたは%ID/gは、該タンパク質の血漿AUCの関数である。%ID/gを表4の血漿AUC値で標準化した結果、器官PS積の算定数値が得られ、PS積は、脳について図13に、末梢器官について表7に示される。第2に、脳取り込みまたは%ID/gは、器官血液容量について補正されなければならない。所定のタンパク質に関する器官取り込み値は、該器官の血液腔における該タンパク質の隔離を単純に反映することができ、それは、組織間で幅広く変化することができる。器官の血液量因子は、HIRMAbのアイソタイプ対照であるヒトIgG1のような血液量マーカーについてのPS積の算定数値によって標準化される。ヒトIgG1についてのBBB PS積を図13に示す。ヒトIgG1およびTNFR:Fc融合タンパク質についてのBBB PS積が等価であることは、TNFR:Fc融合タンパク質がBBBを横断しないことを示す定量的証拠である。脳血液量マーカー、例えば、IgG1またはTNFR:Fc融合タンパク質についてのPS積の算定数値は、どちらのタンパク質も実際のPS積が、脳血液量について補正するとゼロになるので、近似値である。第3に、HIRMAb−TNFR融合タンパク質についての高いBBB PS積(図13)は、脳微小血管系による該融合タンパク質の隔離を反映することができ、BBBを横断する実際のトランスサイトーシスおよび脳実質中への浸透を反映しなかった。このため、毛細管除去分析を行った。血管後上清中のHIRMAb−TNFR融合タンパク質の高いVD、および血管ペレット中の低いVD(表6)は、脳によって取り込まれたHIRMAb−TNFR融合タンパク質の>90%が血管後実質(post-vascular parenchyma)中に浸透したことを明らかにする。TNFR:Fc融合タンパク質のホモジネートVD 13±3uL/g(表6)は、脳血液量に等しく、それは、TNFR:Fc融合タンパク質が血液中、脳の方向へBBBを横断しないことを示すさらなる証拠である。
【0150】
HIRMAb−TNFRおよびTNFR:Fc融合タンパク質のPS積は、また、末梢器官についてコンピューター計算された(表7)。脳および末梢器官に関し、TNFR:Fc融合タンパク質についてのPS積に対する、HIRMAb−TNFR融合タンパク質についてのPS積の比を図14でプロットする。これらのデータは、TNFR:Fc融合タンパク質についてのPS積に対する、HIRMAb−TNFR融合タンパク質についてのPS積の比が、心臓、肺、骨格筋および脂肪などの末梢器官についてほぼ均一であり、肝臓または脾臓などの器官について2〜5倍にやや増加し、脳について、選択的かつ顕著に増加することを示す(図14)。脳におけるPS積比30は、TNFR:Fc融合タンパク質に関する実際のBBB PS積がゼロなので、過小評価である。
【0151】
霊長類の薬物動態(PK)および脳取り込みデータは、HIRMAb−TNFR融合タンパク質を用いる治療的介入の最初の投薬についての検討を可能にする。0.2mg/kgの注射用量での脳取り込み 3.0%ID/100グラム(表5)は、HIRMAb−TNFR融合タンパク質の脳濃度 1.1pmol/グラム融合タンパク質をもたらし、この値は、個々の融合タンパク質あたり2つのTNFR部分が存在するので(図1)、2.2pmol/グラムに等しい。正常な脳における免疫活性TNFαの濃度は、検出不可能であるが、外傷性脳損傷において、0.4pmol/グラムに増加する[E. Shohami, M. Novikov, R. Bass, A. YaminおよびR. Gallily:閉鎖性頭部外傷は、脳組織によるTNFアルファおよびIL−6の早期産生を引き起こす(Closed head injury triggers early production of TNF alpha and IL-6 by brain tissue)、J. Cereb. Blood Flow Metab. 14: 615−9 (1994)]。HIRMAb−TNFR融合タンパク質のTNFαに関するアフィニティーは高いので(図4B)、低用量のHIRMAb−TNFR融合タンパク質 0.2mg/kgは、外傷性脳損傷において、脳における大脳TNFαのほとんどを隔離するであろう。より高用量の該融合タンパク質は、病態において、脳におけるTNFαの本質的に100%を隔離するであろう。
【0152】
まとめると、該実験は、モデルデコイ受容体医薬 TNFRをBBB分子のトロイの木馬 HIRMAbとの融合タンパク質として再操作することにより、イン・ビボでBBBに迅速に浸透する新規な化学物質が生じることを明らかにする。対照的に、当該分野の現状を代表するTNFR:Fc融合タンパク質は、BBBを横断しない。比較的低い全身用量の投与後に生じるHIRMAb−TNFR融合タンパク質の大脳濃度は、病態において、脳における標的サイトカインのほぼ全てを隔離するのに十分である。
【0153】
実施例10 ヒト定常領域の変化
該キメラHIRMAb HCの相補性決定領域(CDRs)およびフレームワーク領域(FR)を包含する該融合タンパク質のHCのドメイン構造を図10に示す。定常領域もまた、図10に示し、ヒトIgG1に由来する。CH1、ヒンジ、CH2、およびCH3ドメインを含むC領域のアミノ酸配列を図10に示す。キメラHIRMAb LCのCDRsおよびFRsを包含するLCのドメイン構造は、図11に示す。定常領域は、ヒトカッパLCに由来し、ヒトカッパ定常領域を含むアミノ酸配列を図11に示す。
【0154】
HIRMAb HC−TNFR融合タンパク質の定常(C)−領域は、配列番号4のアミノ酸133〜461からなり、ヒトIgG1アイソタイプ由来である。さらに、重鎖C−領域は、ヒトIgG2、IgG3、およびIgG4を包含する他のヒトIgGアイソタイプのC−領域に由来することができた。異なるC−領域アイソタイプは、各々、ヒンジ領域周辺の柔軟性、プロテアーゼ感受性、相補体の活性化またはFc受容体への結合に関係する周知の利益または不利益を提供する。HIRMAb LCのC領域は、配列番号6のアミノ酸129〜234からなり、ヒトカッパアイソタイプ由来である。さらに、軽鎖C−領域は、ヒトラムダ鎖アイソタイプ由来であることができた。
【0155】
実施例11 HIRMAb−TNFR融合タンパク質を用いる脳疾患の治療
腫瘍壊死因子(TNF)−αは、脳の急性および慢性障害において病原的役割を果たすプロ炎症性サイトカインである。TNF−αおよびTNFRのどちらも、脳虚血においてアップレギュレートされる(Lambertsenら(2007) Neurosci., 144:934−949)。TNFR細胞外ドメイン(ECD)の経頭蓋投与は、中大脳動脈閉塞(MCAO)モデルにおいて、梗塞のサイズを減少させる(Nawashiroら(1997), Brain Res., 778:265−271)。TNFR ECDは、血液脳関門(BBB)を横断しない大型分子なので、該可溶性デコイ受容体は、脳に直接注射しなければならない。脊髄損傷(SCI)において、ヒトIgG1 FcフラグメントおよびヒトTNFRタイプIIのECDの融合タンパク質のクモ膜下投与は、SCIに関連ずるニューロパシー疼痛を減少させる(Marchandら(2008), Eur. J. Pain, 1−12)。同様に、外傷性脳損傷(TBI)モデルにおけるTNFR−II:Fc融合タンパク質の経頭蓋投与は、治療的である(Knoblachら(1999), J. Neuroimmunol., 95:115−125)。しかしながら、TNFR:Fc融合タンパク質はBBBを横断しないので、TBIにおけるTNFR:Fc融合タンパク質の静脈内投与は、治療的ではない(Knoblachら(1999), J. Neuroimmunol., 95:115−125)。虚血または脳もしくは脊髄損傷などの急性脳障害のほかに、BBB輸送可能TNFR:Fc融合タンパク質の使用はまた、慢性神経変性においても治療的でありうる(Tweedieら(2007), Curr. Alzheimer Res., 4:375−378)。
【0156】
実施例12 MAb−VEGFRデコイ受容体融合タンパク質を用いる脳癌の治療
デコイ血管内皮成長因子(VEGF)受容体(VEGF):Fc融合タンパク質は、操作し、発現させることができる(Holashら(2002), Proc. Natl. Acad. Sci. USA, 99:11393−11398)。VEGF:Fc融合タンパク質は、新規な血管成長のための成長因子である内因性VEGFを隔離すると予想される。かかる医薬は、脳癌を包含する癌における抗血管形成因子としての特定の使用を有しうる。しかしながら、VEGF:Fc融合タンパク質は、BBBを横断することが予測されない大型分子薬物である(Pardridge, (2008), Bioconj. Chem., (19:1327−1338)。IgG融合タンパク質、例えば、TNFRに関して図1に示されるような融合タンパク質として、VEGFRデコイ受容体を再操作することが必要である。VEGFのかかる形態は、内因性インスリン受容体上の輸送を介してヒトBBBに浸透するであろう。いったんBBBを横断すれば、次いで、MAb−VEGR融合タンパク質は、BBBの後ろに、脳内にVEGFを隔離し、脳癌における新規な血管成長を減少させるであろう。
【0157】
実施例13 MAb−Fn14デコイ受容体融合タンパク質を用いる脳虚血の治療
アポトーシスの腫瘍壊死因子様弱インデューサー(TWEAK)は、TNF遺伝子ファミリーのメンバーである。TWEAK受容体は、Fn14と呼ばれる膜タンパク質である。可溶性Fn14デコイ受容体は、Fn14のECDがヒトIgG1 Fcフラグメントのアミノ末端に融合したFc融合タンパク質として操作されている。TWEAK:Fc融合タンパク質は、中大脳動脈閉塞モデルにおいて卒中のサイズを減少させ、TWEAK:Fc融合タンパク質は、該大型分子医薬はBBBを包含しないので、経頭蓋注射によって脳に投与されなければならない(Yepesら(2005), Am. J. Pathol., 166:511−520; Zhangら(2007), J. Cereb. Blood Flow Metab., 27:534−544)。しかしながら、ヒトにおける急性卒中治療を施すために、頭部に穴を開けることは実際的ではない。したがって、Fn14 ECDがBBB浸透性IgGのカルボキシル末端に融合した図1に示されるようなIgG−Fn14融合タンパク質として、Fn14デコイ受容体を再操作することが必要である。かかるタンパク質は、静脈内注射によって非侵襲性で与えることができ、次いで、BBBを横断して虚血脳中への受容体を介して輸送される。
【0158】
実施例14 MAb−LtαRデコイ受容体融合タンパク質を用いる多発性硬化症の治療
リンホトキシンα(LTα)は、また、TNFαとしても知られ、リンホトキシンβ(Ltβ)と共に膜において、ヘテロ−三量体複合体を形成することができる。Ltβ複合体は、Ltβ受容体(LtβR)を活性化させて、細胞内シグナル変換現象および炎症カスケードを開始する。LtβR ECDは、デコイ受容体として作用し、内因性Ltβを隔離する場合があり、それは、脳疾患において治療的であることができる。LtβR:Fc融合タンパク質は、実験的脱髄において治療的であり(Plantら、2007)、ヒト脱髄、例えば、多発性硬化症において治療的でありうる。しかしながら、LtβR:Fc融合タンパク質は、ヒトBBBを横断することが予測されない。LtβRデコイ受容体は、図1に示されるようなMAb−LtβR融合タンパク質として再操作されることができる。かかる融合タンパク質は、BBBを横断し、BBBの後ろに、脳内で内因性Ltβを隔離するであろう。
【0159】
実施例15 MAb−TRAIL−Rデコイ受容体融合タンパク質を用いるニューロAIDS(neuro-AIDS)の治療
TNF関連アポトーシス誘導性リガンド(TRAIL)は、炎症性サイトカインであり、TRAIL受容体(TRAIL−R)への結合を介して作用する。TRAILは、ヒト免疫不全ウイルス(HIV)−1による脳におけるニューロンの感染後、後天性免疫不全症候群(AIDS)の認知症において病原的役割を果たす(Ryanら(2004), J. Neuroimmunol., 148:127−139)。AIDSまたはニューロAIDSの神経学的発現の治療に対する新規なアプローチは、可溶性TRAIL−Rデコイ受容体での、脳におけるTRAILの隔離でありうる。しかしながら、TRAIL−RのECDまたはそのFc融合タンパク質は、BBBを横断する輸送を欠くため、脳に浸透しないであろう。該問題は、TRAIL−Rデコイ受容体を図1に示されるようなMAb−TRAIL−R融合タンパク質として再操作することによって、解決することができた。MAb−TRAIL−R融合タンパク質は、BBBを横断する受容体介在性輸送を経験し、脳に侵入し、そこで、該融合タンパク質は、TRAILを脳内に隔離するであろう。
【0160】
実施例16 MAb−IL6−Rデコイ受容体融合タンパク質を用いる多発性硬化症の治療
インターロイキン(IL)−6のような炎症性サイトカインは、脱髄疾患、例えば、実験的自己免疫脳脊髄炎(EAE)、または多発性硬化症において役割を果たしうる。IL−6受容体(IL−6R)は、可溶性IL−6Rデコイ受容体タンパク質の投与後、脳内で内因性IL−6の作用を抑制することができた。IL−6RのECDがヒトIgG1 Fcフラグメントのアミノ末端に融合したIL−6R:Fc融合タンパク質の投与は、EAEにおいて治療的である(Linkerら(2008), J. Neuroimmunol., 205:64−72)。しかしながら、多発性硬化症におけるIL−6R:Fc融合タンパク質の脳中への浸透は、イン・ビボでのBBBを介する輸送を欠くので、制限されうる。対照的に、図1に示されるようなMAb−IL−6R融合タンパク質は、受容体−媒介性輸送を介してBBBに浸透し、それにより、BBBの後ろに、脳においてIL−6を隔離することができた。
【0161】
本発明の好ましい具体例を本明細書中に示し、記載するが、かかる具体例が例示目的のためだけに提供されることは、当業者に明らかであろう。現在、当業者は、本発明から逸脱することなく、多くの変更、変化および置換を思い付くであろう。本明細書に記載の本発明の具体例の種々の代替物が本発明の実施に用いられうることは、理解されるべきである。添付の特許請求の範囲は、本発明の範囲を明確にすることを目的とし、それにより、特許請求の範囲内の方法および構造およびその等価物が包含される。
【0162】
配列表
表3
【特許請求の範囲】
【請求項1】
受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体であって、BBB上に発現された受容体および該受容体細胞外ドメインのリガンドに結合する融合抗体。
【請求項2】
該BBB上に発現された受容体がインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である、請求項1記載の二官能性デコイ受容体融合抗体。
【請求項3】
該BBB上に発現された受容体がインスリン受容体である、請求項2記載の二官能性デコイ受容体融合抗体。
【請求項4】
該インスリン受容体がヒトインスリン受容体である、請求項3記載の二官能性デコイ受容体融合抗体。
【請求項5】
該二官能性デコイ受容体融合抗体が、該ヒトインスリン受容体との結合に関して、配列番号4および6または配列番号6および7のアミノ酸配列を含む二官能性デコイ受容体融合抗体と競合する、請求項4記載の二官能性デコイ受容体融合抗体。
【請求項6】
該受容体細胞外ドメインがサイトカイン受容体由来である、請求項1記載の二官能性デコイ受容体融合抗体。
【請求項7】
該受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン(ephrin)受容体由来である、請求項1記載の二官能性デコイ受容体融合抗体。
【請求項8】
該受容体細胞外ドメインがTNF−α受容体細胞外ドメインを含む、請求項7記載の二官能性デコイ受容体融合抗体。
【請求項9】
該TNF−α受容体細胞外ドメインが、ヒト、マウス、ラットまたはブタTNF−α受容体細胞外ドメインのアミノ酸配列に対して少なくとも85%同一のアミノ酸配列を含む、請求項8記載の二官能性デコイ受容体融合抗体。
【請求項10】
TNF−α受容体細胞外ドメインが、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に融合している、請求項8記載の二官能性デコイ受容体融合抗体。
【請求項11】
TNF−α受容体細胞外ドメインが重鎖免疫グロブリンのカルボキシル末端に融合している、請求項8記載の二官能性デコイ受容体融合抗体。
【請求項12】
(i)重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列、
(ii)軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列、または
(iii)(i)または(ii)の相補性配列
を含む核酸であって、該重鎖および軽鎖免疫グロブリンがBBB受容体に対する抗体に由来する、核酸。
【請求項13】
該コードされた受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である、請求項12記載の核酸。
【請求項14】
該コードされた受容体細胞外ドメインがTNF−α受容体由来である、請求項13記載の核酸。
【請求項15】
該TNF−α受容体由来のコードされた細胞外ドメインが、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に融合している、請求項14記載の核酸。
【請求項16】
該TNF−α受容体由来のコードされた細胞外ドメインが、重鎖免疫グロブリンのカルボキシル末端に融合している、請求項14記載の核酸。
【請求項17】
該TNF−α受容体由来のコードされた細胞外ドメインが、ヒト、マウス、ラットまたはブタTNF−α受容体細胞外ドメインのアミノ酸配列に対して少なくとも85%同一のアミノ酸配列を含む、請求項14記載の核酸。
【請求項18】
該BBB受容体に対する抗体が、ヒトインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体に対する抗体である、請求項12記載の核酸。
【請求項19】
(i)またはその相補体を含む、請求項12記載の核酸。
【請求項20】
さらに、免疫グロブリン軽鎖をコードする核酸配列またはその相補配列を含み、ここに、該コードされた免疫グロブリン軽鎖がBBB受容体に対する抗体由来である、請求項19記載の核酸。
【請求項21】
(ii)またはその相補体を含む、請求項12記載の核酸。
【請求項22】
さらに、免疫グロブリン重鎖をコードする核酸配列またはその相補配列を含み、ここに、該コードされた免疫グロブリン重鎖がBBB受容体に対する抗体由来である、請求項21記載の核酸。
【請求項23】
該第1の配列が配列番号4または7に対応するアミノ酸配列に対して少なくとも85%同一のアミノ酸配列をコードし、または該第2の配列が配列番号6に対応するアミノ酸配列に対して少なくとも85%同一のアミノ酸配列をコードする、請求項12記載の核酸。
【請求項24】
高ストリンジェンシー条件下で、配列番号3または5のアミノ酸配列をコードする核酸にハイブリダイズする、請求項12記載の核酸。
【請求項25】
請求項12記載の核酸を含む、核酸ベクター。
【請求項26】
(i)を含み、さらに、BBB受容体に対する抗体由来の軽鎖免疫グロブリンをコードする核酸配列を含むか、または
(ii)を含み、さらに、BBB受容体に対する抗体由来の重鎖免疫グロブリンをコードする核酸を含む、請求項25記載の核酸ベクター。
【請求項27】
請求項25記載の核酸ベクターを含む細胞。
【請求項28】
請求項26記載の核酸ベクターを含む細胞。
【請求項29】
哺乳動物細胞である、請求項27記載の細胞。
【請求項30】
受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体を含む医薬組成物を対象に全身投与することを含む、血液脳関門を横切ってデコイ受容体を送達する方法であって、該融合抗体がBBB上に発現された受容体および受容体細胞外ドメインのリガンドに結合するところの、方法。
【請求項31】
該BBB上に発現された受容体がインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である、請求項30記載の方法。
【請求項32】
該BBB上に発現された受容体がインスリン受容体である、請求項31記載の方法。
【請求項33】
該受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である、請求項30記載の方法。
【請求項34】
該受容体細胞外ドメインがTNF−α受容体由来である、請求項30記載の方法。
【請求項35】
該TNF−α受容体由来の細胞外ドメインが、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に共有結合している、請求項34記載の方法。
【請求項36】
該TNF−α受容体由来の細胞外ドメインが重鎖免疫グロブリンのカルボキシル末端に共有結合している、請求項34記載の方法。
【請求項37】
受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体を含む医薬組成物の治療上有効量を対象に全身投与することを含むCNS疾患の治療方法であって、該融合抗体がBBB上に発現された受容体および受容体細胞外ドメインのリガンドに結合するところの、方法。
【請求項38】
該BBB上に発現された受容体がインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である、請求項37記載の方法。
【請求項39】
該BBB上に発現された受容体がインスリン受容体である、請求項38記載の方法。
【請求項40】
該受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である、請求項37記載の方法。
【請求項41】
該受容体細胞外ドメインがTNF−α受容体由来である、請求項40記載の方法。
【請求項42】
該CNS疾患が急性CNS疾患である、請求項37記載の方法。
【請求項43】
該急性CNS疾患が全脳虚血、局所脳虚血、外傷性脳損傷、または脊髄損傷である、請求項42記載の方法。
【請求項44】
該CNS疾患が慢性CNS疾患である、請求項37記載の方法。
【請求項45】
該慢性CNS疾患が神経変性CNS疾患である、請求項44記載の方法。
【請求項46】
該神経変性CNS疾患がアルツハイマー病、パーキンソン病、筋萎縮性側索硬化症、ハンチントン病、多発性硬化症、横断性脊髄炎、運動ニューロン疾患、ピック病、結節硬化症、カナバン病、レット症候群、脊髄小脳失調症、フリードライヒ失調症、視神経萎縮、または網膜変性である、請求項45記載の方法。
【請求項47】
(i)受容体細胞外ドメインに融合した免疫グロブリン重鎖、および免疫グロブリン軽鎖の両方、または
(ii)受容体細胞外ドメインに融合した免疫グロブリン軽鎖、および免疫グロブリン重鎖の両方
をコードする単一タンデム発現ベクターを真核細胞中に安定に組み込むことを含む、二官能性デコイ受容体融合抗体を製造する方法であって、該コードされた免疫グロブリン重鎖および免疫グロブリン軽鎖がBBB上に発現された受容体に対する抗体由来であるところの、方法。
【請求項48】
該単一タンデム発現ベクターが(i)をコードする、請求項47記載の方法。
【請求項49】
該単一タンデム発現ベクターが(ii)をコードする、請求項47記載の方法。
【請求項50】
該BBB上に発現された受容体がインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である、請求項47記載の方法。
【請求項51】
該BBB上に発現された受容体がインスリン受容体である、請求項50記載の方法。
【請求項52】
該コードされた受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である、請求項47記載の方法。
【請求項53】
該受容体細胞外ドメインがTNF−α受容体由来である、請求項52記載の方法。
【請求項54】
該TNF−α受容体由来の受容体細胞外ドメインが免疫グロブリン重鎖または免疫グロブリン軽鎖のカルボキシル末端に融合している、請求項53記載の方法。
【請求項55】
該TNF−α受容体由来の受容体細胞外ドメインが免疫グロブリン重鎖のカルボキシル末端に融合している、請求項53記載の方法。
【請求項1】
受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体であって、BBB上に発現された受容体および該受容体細胞外ドメインのリガンドに結合する融合抗体。
【請求項2】
該BBB上に発現された受容体がインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である、請求項1記載の二官能性デコイ受容体融合抗体。
【請求項3】
該BBB上に発現された受容体がインスリン受容体である、請求項2記載の二官能性デコイ受容体融合抗体。
【請求項4】
該インスリン受容体がヒトインスリン受容体である、請求項3記載の二官能性デコイ受容体融合抗体。
【請求項5】
該二官能性デコイ受容体融合抗体が、該ヒトインスリン受容体との結合に関して、配列番号4および6または配列番号6および7のアミノ酸配列を含む二官能性デコイ受容体融合抗体と競合する、請求項4記載の二官能性デコイ受容体融合抗体。
【請求項6】
該受容体細胞外ドメインがサイトカイン受容体由来である、請求項1記載の二官能性デコイ受容体融合抗体。
【請求項7】
該受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン(ephrin)受容体由来である、請求項1記載の二官能性デコイ受容体融合抗体。
【請求項8】
該受容体細胞外ドメインがTNF−α受容体細胞外ドメインを含む、請求項7記載の二官能性デコイ受容体融合抗体。
【請求項9】
該TNF−α受容体細胞外ドメインが、ヒト、マウス、ラットまたはブタTNF−α受容体細胞外ドメインのアミノ酸配列に対して少なくとも85%同一のアミノ酸配列を含む、請求項8記載の二官能性デコイ受容体融合抗体。
【請求項10】
TNF−α受容体細胞外ドメインが、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に融合している、請求項8記載の二官能性デコイ受容体融合抗体。
【請求項11】
TNF−α受容体細胞外ドメインが重鎖免疫グロブリンのカルボキシル末端に融合している、請求項8記載の二官能性デコイ受容体融合抗体。
【請求項12】
(i)重鎖免疫グロブリンおよび該重鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第1の配列、
(ii)軽鎖免疫グロブリンおよび該軽鎖免疫グロブリンとフレーム内の受容体細胞外ドメインをコードする第2の配列、または
(iii)(i)または(ii)の相補性配列
を含む核酸であって、該重鎖および軽鎖免疫グロブリンがBBB受容体に対する抗体に由来する、核酸。
【請求項13】
該コードされた受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である、請求項12記載の核酸。
【請求項14】
該コードされた受容体細胞外ドメインがTNF−α受容体由来である、請求項13記載の核酸。
【請求項15】
該TNF−α受容体由来のコードされた細胞外ドメインが、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に融合している、請求項14記載の核酸。
【請求項16】
該TNF−α受容体由来のコードされた細胞外ドメインが、重鎖免疫グロブリンのカルボキシル末端に融合している、請求項14記載の核酸。
【請求項17】
該TNF−α受容体由来のコードされた細胞外ドメインが、ヒト、マウス、ラットまたはブタTNF−α受容体細胞外ドメインのアミノ酸配列に対して少なくとも85%同一のアミノ酸配列を含む、請求項14記載の核酸。
【請求項18】
該BBB受容体に対する抗体が、ヒトインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体に対する抗体である、請求項12記載の核酸。
【請求項19】
(i)またはその相補体を含む、請求項12記載の核酸。
【請求項20】
さらに、免疫グロブリン軽鎖をコードする核酸配列またはその相補配列を含み、ここに、該コードされた免疫グロブリン軽鎖がBBB受容体に対する抗体由来である、請求項19記載の核酸。
【請求項21】
(ii)またはその相補体を含む、請求項12記載の核酸。
【請求項22】
さらに、免疫グロブリン重鎖をコードする核酸配列またはその相補配列を含み、ここに、該コードされた免疫グロブリン重鎖がBBB受容体に対する抗体由来である、請求項21記載の核酸。
【請求項23】
該第1の配列が配列番号4または7に対応するアミノ酸配列に対して少なくとも85%同一のアミノ酸配列をコードし、または該第2の配列が配列番号6に対応するアミノ酸配列に対して少なくとも85%同一のアミノ酸配列をコードする、請求項12記載の核酸。
【請求項24】
高ストリンジェンシー条件下で、配列番号3または5のアミノ酸配列をコードする核酸にハイブリダイズする、請求項12記載の核酸。
【請求項25】
請求項12記載の核酸を含む、核酸ベクター。
【請求項26】
(i)を含み、さらに、BBB受容体に対する抗体由来の軽鎖免疫グロブリンをコードする核酸配列を含むか、または
(ii)を含み、さらに、BBB受容体に対する抗体由来の重鎖免疫グロブリンをコードする核酸を含む、請求項25記載の核酸ベクター。
【請求項27】
請求項25記載の核酸ベクターを含む細胞。
【請求項28】
請求項26記載の核酸ベクターを含む細胞。
【請求項29】
哺乳動物細胞である、請求項27記載の細胞。
【請求項30】
受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体を含む医薬組成物を対象に全身投与することを含む、血液脳関門を横切ってデコイ受容体を送達する方法であって、該融合抗体がBBB上に発現された受容体および受容体細胞外ドメインのリガンドに結合するところの、方法。
【請求項31】
該BBB上に発現された受容体がインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である、請求項30記載の方法。
【請求項32】
該BBB上に発現された受容体がインスリン受容体である、請求項31記載の方法。
【請求項33】
該受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である、請求項30記載の方法。
【請求項34】
該受容体細胞外ドメインがTNF−α受容体由来である、請求項30記載の方法。
【請求項35】
該TNF−α受容体由来の細胞外ドメインが、重鎖免疫グロブリンまたは軽鎖免疫グロブリンのカルボキシル末端に共有結合している、請求項34記載の方法。
【請求項36】
該TNF−α受容体由来の細胞外ドメインが重鎖免疫グロブリンのカルボキシル末端に共有結合している、請求項34記載の方法。
【請求項37】
受容体細胞外ドメインのアミノ酸配列に共有結合した重鎖免疫グロブリンまたは軽鎖免疫グロブリンのアミノ酸配列を含む二官能性デコイ受容体融合抗体を含む医薬組成物の治療上有効量を対象に全身投与することを含むCNS疾患の治療方法であって、該融合抗体がBBB上に発現された受容体および受容体細胞外ドメインのリガンドに結合するところの、方法。
【請求項38】
該BBB上に発現された受容体がインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である、請求項37記載の方法。
【請求項39】
該BBB上に発現された受容体がインスリン受容体である、請求項38記載の方法。
【請求項40】
該受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である、請求項37記載の方法。
【請求項41】
該受容体細胞外ドメインがTNF−α受容体由来である、請求項40記載の方法。
【請求項42】
該CNS疾患が急性CNS疾患である、請求項37記載の方法。
【請求項43】
該急性CNS疾患が全脳虚血、局所脳虚血、外傷性脳損傷、または脊髄損傷である、請求項42記載の方法。
【請求項44】
該CNS疾患が慢性CNS疾患である、請求項37記載の方法。
【請求項45】
該慢性CNS疾患が神経変性CNS疾患である、請求項44記載の方法。
【請求項46】
該神経変性CNS疾患がアルツハイマー病、パーキンソン病、筋萎縮性側索硬化症、ハンチントン病、多発性硬化症、横断性脊髄炎、運動ニューロン疾患、ピック病、結節硬化症、カナバン病、レット症候群、脊髄小脳失調症、フリードライヒ失調症、視神経萎縮、または網膜変性である、請求項45記載の方法。
【請求項47】
(i)受容体細胞外ドメインに融合した免疫グロブリン重鎖、および免疫グロブリン軽鎖の両方、または
(ii)受容体細胞外ドメインに融合した免疫グロブリン軽鎖、および免疫グロブリン重鎖の両方
をコードする単一タンデム発現ベクターを真核細胞中に安定に組み込むことを含む、二官能性デコイ受容体融合抗体を製造する方法であって、該コードされた免疫グロブリン重鎖および免疫グロブリン軽鎖がBBB上に発現された受容体に対する抗体由来であるところの、方法。
【請求項48】
該単一タンデム発現ベクターが(i)をコードする、請求項47記載の方法。
【請求項49】
該単一タンデム発現ベクターが(ii)をコードする、請求項47記載の方法。
【請求項50】
該BBB上に発現された受容体がインスリン受容体、トランスフェリン受容体、インスリン様成長因子(IGF)受容体、レプチン受容体、またはリポタンパク質受容体である、請求項47記載の方法。
【請求項51】
該BBB上に発現された受容体がインスリン受容体である、請求項50記載の方法。
【請求項52】
該コードされた受容体細胞外ドメインがTNF−α受容体、TNF関連アポトーシス誘導性リガンド(TRAIL)受容体、アポトーシスのTNF様弱インデューサー(TWEAK)受容体、IL−6受容体、血管内皮成長因子受容体、またはエフリン受容体由来である、請求項47記載の方法。
【請求項53】
該受容体細胞外ドメインがTNF−α受容体由来である、請求項52記載の方法。
【請求項54】
該TNF−α受容体由来の受容体細胞外ドメインが免疫グロブリン重鎖または免疫グロブリン軽鎖のカルボキシル末端に融合している、請求項53記載の方法。
【請求項55】
該TNF−α受容体由来の受容体細胞外ドメインが免疫グロブリン重鎖のカルボキシル末端に融合している、請求項53記載の方法。
【図1】

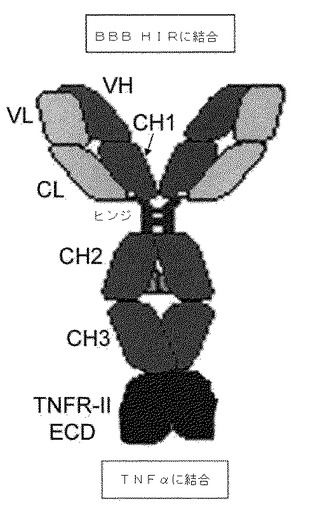
【図2】

【図3】

【図4】

【図5】
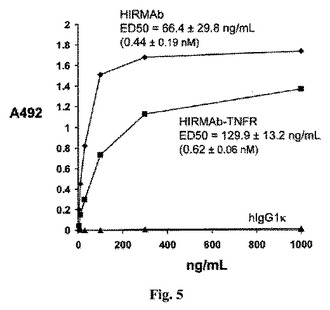
【図6】
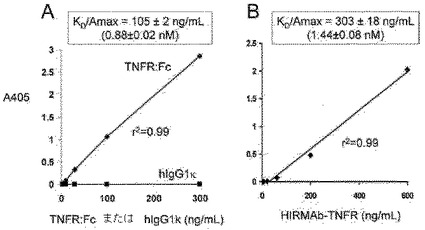
【図7】
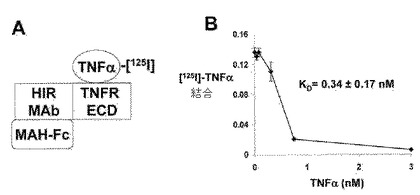
【図8】
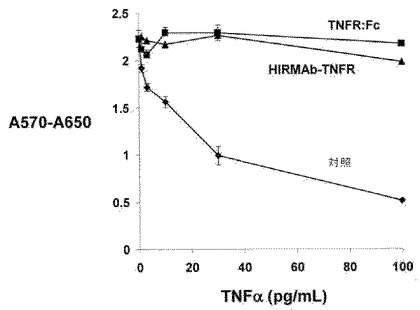
【図9】
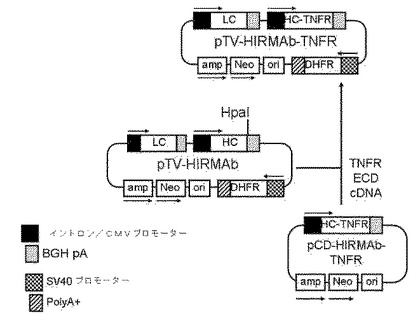
【図10】

【図11】
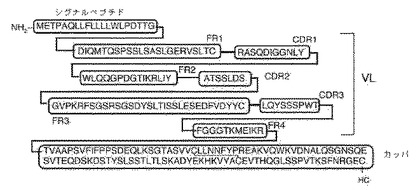
【図12】
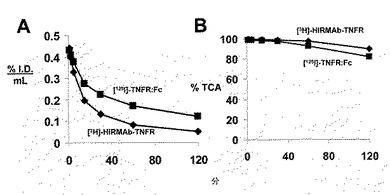
【図13】
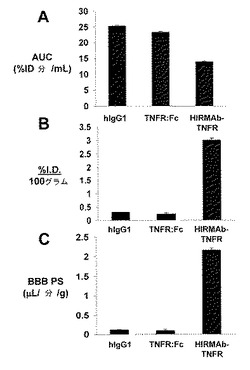
【図14】
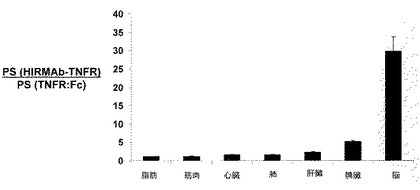
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公表番号】特表2012−521194(P2012−521194A)
【公表日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願番号】特願2012−500979(P2012−500979)
【出願日】平成22年3月18日(2010.3.18)
【国際出願番号】PCT/US2010/027882
【国際公開番号】WO2010/108048
【国際公開日】平成22年9月23日(2010.9.23)
【出願人】(508105751)アーメイゲン・テクノロジーズ・インコーポレイテッド (5)
【氏名又は名称原語表記】ARMAGEN TECHNOLOGIES, INC.
【Fターム(参考)】
【公表日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願日】平成22年3月18日(2010.3.18)
【国際出願番号】PCT/US2010/027882
【国際公開番号】WO2010/108048
【国際公開日】平成22年9月23日(2010.9.23)
【出願人】(508105751)アーメイゲン・テクノロジーズ・インコーポレイテッド (5)
【氏名又は名称原語表記】ARMAGEN TECHNOLOGIES, INC.
【Fターム(参考)】
[ Back to top ]