
K5多糖の製造方法
本発明は、大腸菌による発酵工程とその後の精製工程とを含むK5多糖の製造方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、大腸菌による発酵工程と精製工程とを順次に行うことによってK5多糖を製造する方法に関する。
【背景技術】
【0002】
K5多糖を得るためのいくつかの方法が公知である。K5多糖は外的腸感染症の原因となる大腸菌株から得ることができる。
【0003】
K5多糖は、等モル量のグルクロン酸とN−アセチルグルコサミンとから成り、これらが基本単位として4−ベータ−グルクロニル−1,4−アルファ−N−アセチルグルコサミンを構成し、基本単位が直線状に反復してK5多糖鎖が形成される。
【0004】
【化1】

【0005】
この基本単位は、完全に脱硫酸化されN−アセチル化されたヘパリンに構造類似性を有しており、従って、動物器官から抽出されるヘパリンの極めて有利な代替材料となり得る、すなわち、発酵によって産生されたK5多糖の化学的修飾によってヘパリンを製造するために使用し得る。
【0006】
生合成によって得られたヘパリンは、以下のようにして製造される。微生物大腸菌を培養してK5多糖を産生させ、次いでこれを化学処理してヘパリンとする。K5多糖からヘパリンを製造する方法は記載されている(Biochem J.,1991,275(1):151−8;国際特許WO92/17507;Semin.Thromb.Hemost.,2001,27(5):437−43;Nat.Biotech.,2003,21(11):1343−46)。一般的に、K5多糖からヘパリンを合成するためには、脱アセチル化およびN−再硫酸化、C−5エピマー化およびO−硫酸化の処理が必要である。
【0007】
ヘパリンの有用性は特にその抗凝血性および抗血栓性にある。しかしながら、ヘパリンは使用条件が限られるという欠点を有している。特にその高い抗凝血活性(aIIa)が出血を引き起こすこともある(Semin.Thromb.Hemost.,5 sup.3,1999)。本発明によれば、酵素的または化学的な解重合によって半合成ヘパリンから低分子量ヘパリン(3000−6500Da)または超低分子量ヘパリン(1500−3000Da)を得ることが可能であり、これらの抗IIa活性は5IU/mg未満であるのに抗Xa活性は特に140−190IU/mgの範囲である。
【0008】
K5多糖の多くの製造方法が従来技術に記載されている(Eur.J.Biochem,1981,116:359−364;EPO489647,WO01/02597、Eur.J.Biochem,1988,117:112−125)。これらの方法は特に、グルコース中、37℃で行う単一増殖段階から成る大腸菌の発酵工程と、本質的にイオン交換カラム処理から成るK5多糖の精製工程とを順次に含む。
【0009】
発酵工程に関しては、本発明方法の発酵工程は、少なくともその1つがフィードを伴う増殖段階である複数の増殖段階と冷却段階とを含み、この発酵工程を精製工程に組合せることによって従来技術に記載された値の10倍にも達する収率が得られる。
【0010】
実際、従来技術に記載された大腸菌の発酵によるK5多糖の生合成方法はいずれも極めて低収率であるという重大な欠点を有している。例えば、従来技術による細胞外形態のK5多糖の製造方法では、単離されたK5多糖の収率は200−850mg/lにすぎない。
【0011】
このような低収率ではK5多糖の工業生産を計画すること、従って、半合成による半合成ヘパリンの工業生産を計画することができない。しかしながら、動物由来でないヘパリンおよび低分子量または超低分子量ヘパリンに対する需要は大きい。
【0012】
従って、本発明の1つの目的は、細胞外形態のK5多糖の新規な製造方法を提供することである。
【0013】
本発明の方法は本質的に2つの工程、すなわち、発酵工程と精製工程とを含む。
【0014】
本発明の方法によれば、従来技術に記載された方法で得られる濃度の10倍に達する濃度のK5多糖を得ることが可能である。従って本発明によれば、10g/lもの濃度のK5多糖を得ることが可能である。このような濃度は、発酵工程と精製工程との組合せによって得られるものである。
【発明の開示】
【0015】
本発明の1つの目的は、
−発酵槽で行うことができ、
−バッチ増殖段階と、
−1回または数回のフェッドバッチ増殖段階と、
−温度を下げ、培地のpHを9までの値に上げる冷却段階と、
−バイオマスを培養培地から分離する段階と、
−バイオマスから混入培養培地を除去する段階と、
を含む、K5多糖を産生する大腸菌株の発酵工程と、
−培養培地および/またはバイオマスを使用する精製工程と、
を含むK5多糖の製造方法である。
【0016】
本発明の目的はより特定的には、上記のようなK5多糖の製造方法であって、培養培地および/またはバイオマスを出発材料とするその精製工程がイオン交換カラムによる処理工程を含むものでなく、
−約7−約11の値へのpHの調整、
−沈殿、
−沈殿物の溶解、
−得られた懸濁液の濾過、および、
−アルコール沈殿、
を記載の順序で含んでいる。
【0017】
発酵
本発明によるK5多糖の製造に使用できる大腸菌株は、パブリックコレクション例えばCNCM(Collection Nationale de Cultures de Microorgansmes,France)またはATCC(American Type Culture Collection,USA)から得られる。あるいは、これらの菌株を臨床的に単離し、次いで文献に記載されたように莢膜の抗原型のキャラクタリゼーションを行ってもよい(Eur.J.Biochem,1988 117:112−125)。
【0018】
本発明の発酵工程は、発酵槽で大腸菌を使用して行うものであり、
−バッチ増殖段階と、
−指数増加量または一定量のフィードを伴う1回または複数のフェッドバッチ増殖サイクルと、
−温度を約10−12℃に下げpHを約7.5−9に上げる冷却段階と、
−バイオマスを培養培地から分離する段階と、
−バイオマスから混入培養培地を除去する段階と、
を含む。
【0019】
本発明によれば“バッチ増殖”という表現は、フィードまたは抜取りを伴うことなく反応装置で微生物を増殖させることを意味する。
【0020】
本発明によれば“フェッドバッチ増殖”という表現は、管理量の基質フィードを伴うが抜取りを伴うことなく反応装置で微生物を増殖させることを意味する。
【0021】
本発明によれば、発酵工程は特に、バッチ増殖段階の後に指数増加量の基質フィードを伴う2回のバッチ増殖段階を含む。
【0022】
本発明によれば、発酵工程は、冷却段階の前に一定量の基質フィードを伴うバッチ増殖段階を含む。
【0023】
本発明の発酵工程では、一回目のフェッドバッチ増殖段階が、バッチ増殖段階の培地中の炭素源の消耗と共に開始される。
【0024】
より特定的には本発明によれば、バッチ増殖段階の培地中の炭素源が消耗したときに一回目のフェッドバッチ増殖段階が、開始され、必要ならば行われるその後のフェッドバッチ増殖段階は酸素移動の極限の結果として溶存酸素の濃度が有意に低下したときに開始される。
【0025】
本発明によれば、バッチ増殖段階の各々は上記の時期に開始され、冷却段階は培養フィード培地の全量が発酵槽に注入されたとき、または、pH変化が検出されたときに開始される。
【0026】
本発明によれば、バッチ増殖段階、(1回または複数の)フェッドバッチ増殖段階および冷却段階の後に、遠心によってバイオマスを培地から分離する。分離後、バイオマスおよび培養培地を除染する。具体的には、これらを約80℃の温度で1−3時間失活させる。次に上清および/またはペレットを使用してK5多糖を精製する。
【0027】
pHはバッチ増殖段階およびフェッドバッチ増殖段階中はNH4OHもしくはNH3の添加によって約6.5に維持するが、最終段階中に約8.5まで上昇する。
【0028】
溶存酸素の濃度は、撹拌速度、空気流量の変更、流入空気の酸素富化または過酸化水素の添加などの操作によって空気飽和量の約15−約80%に維持する。
【0029】
本発明の発酵工程中の増殖温度は約10℃−約40℃である。より特定的には、バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、必要ならば行うその後のフェッドバッチ増殖段階では約25℃である。K5多糖を高収率で得るためには30℃から25℃への移行が特に重要である。
【0030】
使用される培養培地は合成培養培地である。本発明によれば炭素源はグルコースまたはグリセロールである。本発明によれば好ましい炭素源はグリセロールである。
【0031】
より特定的に、本発明の発酵工程の炭素源がグリセロールであり、その濃度はバッチ増殖段階の出発培養培地中では約10−約90g/lの範囲であり、フェッドバッチ増殖段階のフィード培地中では約250−約1200g/lの範囲である。実験実施例1は、K5多糖を高濃度で得るためには、炭素源としてグリセロールを使用すること、および、フィード培地中のグリセロール濃度を増加させることの重要性を示している。
【0032】
本発明によれば、バッチ増殖段階の培地の窒素源が酵母エキス、カザミノ酸、ペプトン、NH4OH、NH3または(NH4)2HPO4でよい。
【0033】
本発明によれば、バッチ増殖段階の出発培養培地中の窒素源は特にNH4OH、NH3または(NH4)2HPO4である。これらは特に約1−約10g/lの範囲の濃度で使用される。
【0034】
本発明によれば、バッチ増殖段階およびフェッドバッチ増殖段階中の窒素源はpHを調整するために、NH4OHまたはNH3によって供給される。その濃度は好ましくは約20%である。
【0035】
より特定的には、本発明の発酵工程で、バッチ増殖段階の培地の窒素源が、出発培養培地では約1−約10g/lの濃度のNH4OH、NH3または(NH4)2HPO4であり、バッチ増殖段階およびフェッドバッチ増殖段階中はpHを調整するために約20%濃度のNH4OHまたはNH3によって供給される。
【0036】
本発明の1つの目的は上記に記載のような発酵工程を含むK5多糖の製造方法であり、該発酵工程では、
−炭素源がグリセロールであり、グリセロールの濃度は、
−バッチ増殖段階の出発培養培地中に約10−約90g/lの範囲であり、
−フェッドバッチ増殖段階のフィード培地中に約250−約1200g/lの範囲であり、
−増殖温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、
−必要ならば行う他のフェッドバッチ増殖段階では約25℃である。
【0037】
本発明の1つの目的はまた、上記に記載のような発酵工程を含むK5多糖の製造方法であり、該発酵工程では、
−炭素源がグリセロールであり、グリセロールの濃度が、
−バッチ増殖段階の出発培養培地中に約10−約90g/lの範囲であり、
−フェッドバッチ増殖段階のフィード培地中に約250−約1200g/lの範囲であり、
−バッチ増殖段階の培地の窒素源が
−出発培養培地では約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4であり、
−バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給される。
【0038】
本発明の1つの目的はまた、上記に記載のような発酵工程を含むK5多糖の製造方法であり、該発酵工程では、
−バッチ増殖段階の培地の窒素源が、
−出発培養培地では約1−約10g/lの範囲のNH4OH、NH3または(NH4)2HPO4であり、
−バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給され、
−増殖温度が、
−バッチ増殖段階およびフェッドバッチ増殖段階では約30℃であり、
−必要ならば行う他のフェッドバッチ増殖段階では約25℃である。
【0039】
最も特定的に本発明の1つの目的はまた、上記に記載のようなK5多糖の製造方法であり、該方法では、
発酵工程が、以下の段階、すなわち、
−バッチ増殖段階、
−先行段階の培地中の炭素源が完全消耗したときに開始する指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する二回目の指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する一定量の基質フィードを伴うバッチ増殖サイクル、
−温度を約12℃に下げpHを約7.5−約9の範囲の値に上げる冷却段階、
−バイオマスから混入培養培地を除去する段階、
を記載の順序で含み、
上記段階を以下のパラメーターで行う、すなわち、
−温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階中は約30℃、
−他の2回のフェッドバッチ増殖段階中は約25℃
であり、
−窒素源は、
−出発培養培地には約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4によって供給され、
−バッチ増殖段階およびフェッドバッチ増殖段階にはpHを調整するために約20%の濃縮NH4OHまたはNH3の添加によって供給され、
−炭素源はグリセロールであり、その濃度は、
−最初のバッチ増殖段階の出発培養培地中で約10−約90g/lの範囲の濃度であり、
−フェッドバッチ増殖段階のフィード培地中で約250−約1200g/lの範囲の濃度である。
【0040】
第二工程:K5多糖の精製
本発明の1つの目的はまた、精製工程を含む上記のようなK5多糖の製造方法である。この精製工程では、発酵ブロスの遠心上清またはこの遠心で得られたペレットの再懸濁液から成るK5多糖含有溶液のpHを約7−約11に調整し、この溶液から多糖を沈殿させる。次に沈殿物を洗浄し、再溶解し、濾過し、次いで沈殿させる。
【0041】
本発明によれば、精製工程の一回目の沈殿は第四級アンモニウム塩を用いて行う。この沈殿はより特定的には塩化ベンゼトニウムを用いて行う。
【0042】
本発明によれば、精製工程の最初の沈殿物を、特に10%濃度で使用する酢酸ナトリウムに再溶解させる。
【0043】
更に、本発明によれば、精製工程で得られた濾液をメタノール沈殿させる。
【0044】
より特定的には、本発明による精製工程の一回目の沈殿が塩化ベンゼトニウムを用いて行われ、この沈殿物を酢酸ナトリウムに再溶解し、次いで濾過し、精製工程で生じた濾液をメタノール沈殿させる。
【0045】
本発明によれば、上記に記載のような精製後に得られた生成物を場合によっては過酸化水素によって1回または数回脱色処理する。より特定的には、精製後の過酸化水素処理を3回行う。
【0046】
必要な場合には、上記のように精製した生成物を過酸化水素で3回脱色処理した後、残留汚染物を除去するために追加の過酸化水素処理を行う。この処理は得られる生成物の純度を改善し得る。従って本発明の1つの目的はまた、精製および3回の過酸化水素処理後に追加の過酸化水素による脱色をさらに含むK5多糖の製造方法である。
【0047】
必要な場合には、上記精製工程のいずれか1つの後でK5多糖をプロテアーゼで処理する。実験実施例8で後述するように、プロテアーゼによる処理は、ほぼ定量的収率で産生されるK5多糖の純度を有意に向上させ得る。従って本発明の目的は特に、精製工程中のプロテアーゼ処理を含むK5多糖の製造方法である。本発明のプロテアーゼは好ましくはアルカラーゼである。
【0048】
最後に、K5多糖を沈殿させ、遠心し、乾燥する。
【0049】
従って本発明の1つの目的は最も特定的には、上記に記載のような精製工程を含むK5多糖の製造方法であって、精製工程で以下の段階:
−バイオマスから培養培地を分離するために培養物を遠心する段階;
−発酵ブロスの遠心上清を使用して
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−同時に、発酵ブロスの遠心によって得られたペレットを使用して
−ペレットを再懸濁化し、
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに再溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−各々の精製工程で得られた2つの調製物を合せる段階;
−K5多糖の沈殿、遠心および乾燥を行う段階;
を行う方法である。
【0050】
従って本発明の1つの目的は好ましくは、発酵工程と精製工程とを含むK5多糖の製造方法であって、発酵工程が、以下の段階:
−バッチ増殖段階、
−先行段階の培地中の炭素源が完全消耗したときに開始する指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する二回目の指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する一定量の基質フィードを伴うバッチ増殖サイクル、
−温度を約12℃に下げpHを約7.5−約9の範囲の値に上げる冷却段階、
−バイオマスから混入培養培地を除去する段階、
を記載の順序で含み、
上記段階が以下のパラメーターで行われる、すなわち:
−温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階中は約30℃、
−他の2回のフェッドバッチ増殖段階中は約25℃
であり、
−窒素源は、
−出発培養培地には約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4によって供給され、
−バッチ増殖段階およびフェッドバッチ増殖段階にはpHを調整するために約20%の濃縮NH4OHまたはNH3の添加によって供給され、
−炭素源はグリセロールであり、その濃度は、
−最初のバッチ増殖段階の出発培養培地中に約10−約90g/lの範囲の濃度であり、
−フェッドバッチ増殖段階のフィード培地中に約250−約1200g/lの範囲の濃度であり、
精製工程が以下の段階:
−バイオマスから培養培地を分離するために培養物を遠心する段階;
−発酵ブロスの遠心上清を使用して
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−同時に、発酵ブロスの遠心によって得られたペレットを使用して
−ペレットを再懸濁化し、
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに再溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−各々の精製工程で得られた2つの調製物を合せる段階;
−K5多糖を沈殿、遠心および乾燥させる段階を含む方法である。
【0051】
このようにして本発明方法では、従来技術に記載された方法で得られる収率の10倍にもなるような収率が得られる。
【0052】
本発明によれば、上記に記載の方法によって製造された多糖が酵素的および/または化学的な脱アセチル化、N−再硫酸化、C−5エピマー化および/またはO−硫酸化の反応基質として使用される。より特定的には本発明によれば、上記に記載の方法によって製造された多糖が半合成ヘパリン(バイオヘパリン)を得るための基質として上記反応の1つで使用される。
【0053】
本発明によれば、得られたバイオヘパリンを次に、約1500−約6500Daの範囲の分子量をもつLMWH(低分子量ヘパリン)を得るためのフラグメンテーション反応で基質として使用する。
【0054】
本発明によればまた、得られたバイオヘパリンを、より特定的に約3500−約5500Daの範囲の分子量をもつLMWH(低分子量ヘパリン)、または、より特定的に約1500−約3000Daの範囲の分子量をもつVLMWH(超低分子量)ヘパリンを得るためのフラグメンテーション反応の基質として使用することが可能である。
【0055】
以下の実施例は本発明の代表例であるが、本発明はこれらの実施例に限定されない。
【0056】
応用例
【実施例1】
【0057】
(1)手順
(a)発酵
発酵は20リットルの発酵槽で大腸菌株ATCC23506を使用して行う。
【0058】
5−60g.l−1のグリセロールと0.5−3g.l−1のKH2PO4と1−10g.l−1の(NH4)2PO4と0.5−5g.l−1のMgSO4と0.2−4g.l−1のクエン酸と微量元素とから成る100mlの培地を収容した500ml容の三角フラスコ中、30℃で一夜凍結させた1.8mlのバイアルを使用して大腸菌株のプレカルチャーを調製する。
【0059】
200rpmで撹拌しながら30℃で増殖させる。生産用発酵槽に接種するためにこのプレカルチャーを使用する。また、グリセロール収容バイアルから直接的に発酵槽に接種してもよい。このプレカルチャーの目的は、発酵槽におけるバッチ増殖段階の持続時間を短縮することだけである。使用した培地は、RFB MIL 11/04培地(J.of Biotechnology,1995,39:59−65)の変種であり、それらの組成を表1に示す。
【0060】
【表1】
【0061】
自己適応形管理アルゴリズムに基づくIRISプログラム(Infors)によって制御されたTECHFORS発酵槽(全容積20l、処理容積12l)で培養物を調製する。培地中の溶存酸素濃度、pH、撹拌速度、温度、通風空気の流速、圧力およびポンプ吐出量(基底調節量、フィード量、抑泡剤)はオンラインで測定し管理する。発酵槽の排出ガスの分析は質量分析計(PRIMA 600S)を使用して行う。
【0062】
以下の特性値のバッチモードで発酵させる。
−処理容量12l、
−初期容量8l、
−追加容量4l
−最終容量は12lプラス基底容量、
−溶存酸素は空気飽和量の15%−80%の範囲(好ましくは30%)に維持、
−温度は20−40℃の範囲(好ましくは30℃)、
−pHは濃NH4OHまたはNH3を加えて6.0−7.5(好ましくは6.5)に調整、
−圧力は0.3バール、0.5バールおよび0.8バール、
−初期通風は4l.分−1(0.5VVM)(4−20l.分−1の範囲で調整)、
−初期撹拌速度は200rpm(200−1500rpmの範囲で調整)。
【0063】
培養は5つの工程、すなわち、バッチ増殖段階、次いで、基質フィードを伴う3回の異なるバッチ増殖段階、および、最終冷却段階を含む。
【0064】
1.バッチ増殖段階
バッチモードの培養段階では、菌株の世代時間を二酸化炭素の発生量の測定値から算定し、管理された基質フィードを伴うバッチ増殖の最初の部分の初期速度を計算する:
Q0=(CO2pmax×VolFer)/(1000×MWCO2×YCO2/グリセロール×G0)
Q0:濃縮培地の初期流速(1.時−1);
CO2pmax:二酸化炭素の最大発生量(mmol.l−1.時−1);
G0:管理された基質フィードを伴うバッチ増殖中に使用された濃縮培地中のグリセロールまたはグルコースの濃度(750g.l−1);
VolFer:発酵槽中の液体容量;
MWCO2:CO2の分子量(g.l−1);
YCO2/グリセロール:グリセロール消費量あたりのCO2の発生量。
【0065】
2.指数増加量の基質フィードを伴う1回目のバッチ増殖段階
炭素源の消耗後、バイオマスの濃度を増加させるために、指数増加量の基質フィードを伴うバッチ増殖サイクルを開始する。新しい培地を供給するポンプのスイッチを入れる。ポンプの吐出量は時間の経過に伴って指数関数的に増加する。
Q(t)=Q0×2[t/(τ×2.4)]
Q0:フィードポンプの初期供給量(l.時−1);
t:指数増加量の基質フィードを伴うバッチ増殖サイクル開始後の経過時間;
τ:バッチ増殖中に測定した菌株世代時間。
【0066】
予め測定した菌株世代時間に1−10(好ましくは2.4)の範囲の補正率を乗算した値をこの供給量に算入する。この供給量は0.05時−1から0.65時−1の範囲の増殖率を微生物に与える。
【0067】
3.指数増加量の基質フィードを伴う2回目のバッチ増殖段階
最初の酸素移動の極限に達すると、(溶存酸素<20%、撹拌速度>1350rpm、空気流速>19,5l.分−1、圧力=0.5バール)、この指数増加量の基質フィードを伴う1回目のバッチ増殖段階を停止する。管理された基質フィードを伴う1回目のバッチ増殖サイクルの開始時の8.4ml.時−1という流速に基づいて次の指数増加量のフィードを開始する。先行の指数増加量の基質フィードを伴うバッチ増殖サイクルと同様に、4−10の範囲(好ましくは6)の新しい補正率を世代時間に乗算する。この供給量は0.05時−1から0.16時−1の範囲の増殖率を微生物に与える。
【0068】
同時に、培地中の酸素溶解度を増加させるために発酵槽の表示温度を20−25℃(好ましくは25℃)に下げる。フィードポンプの新しい制御アルゴリズムは以下の式で表される:
Q’(t)=0.0084×2[t/(τ×6)]
t:指数増加量の基質フィードを伴うバッチ増殖サイクル開始後に経過した時間;
τ:バッチ増殖中に測定した菌株世代時間。
【0069】
酸素移動の極限は約30時間の増殖後(管理された基質フィードを伴う1回目のバッチ増殖サイクル10時間を含む)に出現するが、フィードポンプの実際の初期培地吐出量は約15ml.時−1(t=10時間)である。この低い供給量は、微生物の増殖を維持すると同時に系の溶存酸素を飽和量の30%に調節する機能も果す。2回目の酸素移動の極限が出現するまでこの供給量を継続する。
【0070】
4.一定量の基質フィード段階
2回目の酸素移動の極限(溶存酸素<20%、撹拌速度>1350rpm、空気流速>19.5l.分−1、圧力=0.5バール)が出現すると、指数増加量の培地フィードを1−20ml.l−1.時−1の範囲(好ましくは4.2ml.l−1.時−1)の一定供給量に切換える。4lのフィード培地が注入されるかまたはpHの有意な低下(強力な酸素極限の指標となるNH4OHの完全消費)が検出されるまでこの供給量を継続する。
【0071】
5.冷却段階
次にフィードポンプのスイッチを切り、発酵槽の温度を10℃に下げる。この間に、pHはK5多糖酵素リアーゼの活性に好ましい7.5−9(好ましくは8.5)という値まで自然に上昇するかまたはこのような値に調節できる。これらの条件を0−10時間維持する。
発酵槽が10℃まで冷えた後、バイオハザードレベル2の作業条件下でバイオマスを気密容器に採集する。ブロスを遠心してバイオマスを培地上清から分離する。分離後、80℃で1時間30分の熱処理によって撹拌しながら上清を除染する。バイオマス自体は80℃で2時間30分の処理によって除染する。上清およびバイオマスを−20℃で保存する。
【0072】
これらの試験には以下の条件を使用する:
−COL5K000/001:30℃のバッチ増殖段階/30℃で行う指数増加量の基質フィードを伴う増殖段階/冷却段階;
−COL5K002:30℃のバッチ増殖段階/30℃で行う指数増加量の基質フィードを伴う増殖段階/2回目の30℃の指数増加量の基質フィードを伴う増殖段階/冷却段階;
−COL5K003/004:30℃のバッチ増殖段階/30℃で行う指数増加量の基質フィードを伴う増殖段階/25℃で行う2回目の指数増加量の基質フィードを伴う増殖段階/冷却段階;
−COL5K005/006:30℃のバッチ増殖段階/30℃で行う指数増加量の基質フィードを伴う増殖段階/25℃で行う2回目の指数増加量の基質フィードを伴う増殖段階/25℃で行う一定量の基質フィードを伴う増殖段階/冷却段階。
【0073】
b)多糖の分析
多糖の分析は2工程で行う。
1.最初に多糖をその構成二糖に解重合する。
2.二糖混合物をHPLCによって分析する。
【0074】
1.多糖の解重合
分析すべき約40mgのサンプルを2mlのpH7の酢酸塩バッファ(0.29mlの酢酸、5mgのBSA、15.8mgの酢酸カルシウム、総量を50mlにする量の脱イオン水、2Nの水酸化ナトリウムでpHを7に調整)に再懸濁させ、20mg/mlを含有する溶液を得る。
【0075】
同時に、リン酸塩バッファ(68mgのリン酸二水素カリウム、10mgのBSA、総量を50mlにする量の脱イオン水、1Nの水酸化カリウムでpHを7に調整)中に0.5IU/mlのヘパリナーゼI、IIおよびIIIをそれぞれ含有する溶液を調製する。次にこれらの溶液を等しい割合で混合して最終のヘパリナーゼ123溶液を得る。
【0076】
20mg/mlのサンプル溶液25μl、酢酸塩バッファ25μl、および、ヘパリナーゼ123溶液25μlを300μl容の管で混合する。混合物を十分にホモジナイズし、室温で24時間インキュベートして完全に解重合させる。
【0077】
2.HPLCによる多糖の分析
勾配HPLCを使用し当業者に公知の方法に従って前項の混合物を分析する。以下の分析条件を使用する:
−カラム:Spherisorb SAX 5μm、250×3mm、Waters、
−カラム温度:40℃、
−流速:0.5ml/分、
−注入量:10μl、
−234nmでUV検出、
−移動相:
溶媒A:NaH2PO4、2.5mM pH=2.9
溶媒B:NaClO4、M、NaH2PO4 2.5mM pH=3
勾配:
【0078】
【表2】
【0079】
二糖4−ベータ−グルクロニル−1,4−アルファ−N−アセチル−グルコサミンがHPLCによって検出される。
【0080】
【化2】
2)結果
対照COL5K000に加えて異なる6つの発酵試験(COL5K001−006)を実施した。
【0081】
これらの試験中に得られたK5多糖の濃度を表2に示す。
【0082】
【表3】
【0083】
結果(図1、付記1)は、使用した手順の各々によって対照COL5K000の2−10倍のK5多糖の製造が可能であったことを明らかに示す。COLK006の主要パラメーターのプロフィルは、培地中のK5多糖の濃度増加が2回目のフェッドバッチ増殖段階の付加に関連していることを明らかに示す(図2、付記1)。
【0084】
実施した試験は、培地中に精製した多糖の最大濃度を文献に記載された値に比較して大幅に増加させた。文献の量は900mg.l−1であったが、我々は10g.l−1を上回る濃度を得た。
【0085】
COL5K000の処理プロセスと培養培地中の多糖濃度が増加したCOL5K006の処理プロセスには主として以下の違いが存在した:
−炭素源としてグルコースの代わりにグリセロールを使用する:
−酸素移動の極限が出現したときに、低下させた増殖率の基質フィード段階(2回目および3回目のフィード段階)を付加することによって発酵総時間を延長する:
−フィード溶液のグリセロール濃度を増加する(500g.l−1を750g.l−1にする):
−処理終了時のpHを上げ、これによってK5多糖リアーゼの活性に有利な条件を成立させ、培養培地中に得られる多糖の量を増加させる。
【実施例2】
【0086】
抽出プロセス
以下に記載の抽出工程は、上記で得られた発酵上清からK5多糖を単離するために行った。
【0087】
実施例1で発酵槽COL5K001から6.5リットルの熱失活上清が得られた。これは24.7gのK5多糖を含有している。
【0088】
5Nの水酸化ナトリウムを使用してこの溶液のpHを7.2に調整し、50g/lの塩化ベンゼトニウムを含有している663mlの水溶液を注いだ(すなわち、K5多糖に対して1.2モル当量の塩化ベンゼトニウム)。反応媒体を室温で30分間撹拌し、次いで4000rpmで10分間遠心した。
【0089】
得られたペレットを4リットルの脱イオン水で2回洗浄し、次いで5リットルの10%酢酸ナトリウム溶液に入れて撹拌した。
【0090】
1回目の塩化ベンゼトニウム沈殿で得られた上清を再度処理した。5Nの水酸化ナトリウムを使用してそのpHを7.2に調整し、50g/lの濃度の663mlの塩化ベンゼトニウムを添加し、混合物を30分間撹拌後に遠心した。
【0091】
得られたペレットを4リットルの脱イオン水で2回洗浄し、次いで5リットルの10%酢酸ナトリウム溶液に入れて撹拌した。2種類の再懸濁ペレットを含有している溶液を集め、Clarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0092】
濾液を40リットルのメタノールに注いだ。得られた混合物を25分間撹拌した後、一夜傾瀉した。翌日、4000×gで10分間遠心した。ペレットをメタノールで洗浄し、次いで20Torr下、50℃で36時間乾燥した。
【0093】
このようにして力価54%をもつ31.5gのK5多糖が得られた。収率は68.8%であった。
【実施例3】
【0094】
精製
実施例2で単離した生成物の力価が低かったので(わずか54%)、追加の精製工程を実施した。
【0095】
実施例2で得られた重量31.5gの粗生成物を磁気撹拌付きの三角フラスコで1.2リットルの脱イオン水に再懸濁させた。5Nの水酸化ナトリウムでpHを11に調整した後、15gの塩化ナトリウムと6mlの30%過酸化水素とを溶液に加えた。混合物を10分間撹拌し、次いで撹拌しないで室温で一夜静置した。
【0096】
4倍容のメタノールを溶液に加えることによってK5多糖を沈殿させた。混合物を30分間撹拌し、次いで4000rpmで15分間遠心した。ペレットを20Torr下、45℃で36時間オーブン乾燥した。このようにして、力価が54%から65%に改善された23.5gのK5多糖が得られた。
【実施例4】
【0097】
抽出と精製
実施例1の発酵槽COL5K002から粗K5多糖を抽出して精製した。
【0098】
実施例1で発酵槽COL5K002から5.4リットルの熱失活上清が得られた。これは21.1gのK5多糖を含有している。
【0099】
実施例2に記載したようにCOL5K002上清を塩化ベンゼトニウムで処理し、ペレットを酢酸塩溶液中で撹拌し、次いでClarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0100】
濾液に含まれていたK5多糖を実施例2に記載したようにメタノール沈殿させ、次いで実施例3に詳述したように過酸化水素で脱色し、その後、過酸化水素による2回目の脱色を行った。
【0101】
最終的に、78%の力価をもつ25.3gのK5多糖が90%を上回る収率で得られた。
【実施例5】
【0102】
抽出と精製
実施例4の発酵槽COL5K003から粗K5多糖を抽出して精製した。
【0103】
実施例1で発酵槽COL5K003から6リットルの熱失活上清が得られた。これは45gのK5多糖を含有している。
【0104】
実施例2に記載したようにCOL5K003上清を塩化ベンゼトニウムで処理し、ペレットを酢酸塩溶液中で撹拌し、次いでClarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0105】
濾液に含まれていたK5多糖を実施例2に記載したようにメタノール沈殿させ、次いで実施例4と同様に過酸化水素で2回処理し、その後、3回目の過酸化水素による脱色を行った。
【0106】
最終的に、81%の力価をもつ39.5gのK5多糖が71%の収率で得られた。
【実施例6】
【0107】
ペレットの処理および精製の改善
先行の試験では上清だけを抽出して精製した。遠心沈降物中に存在するK5多糖を回収するために、実施例1の発酵槽COL5K002から得られたペレット(31.9gのK5多糖を含有する5kg)を等容量の脱イオン水に再懸濁させ、5NのNaOHでpHを7.2に調整した。次に混合物を室温で2時間撹拌し、次いで4700×gで90分間遠心した。
【0108】
実施例2に記載したように上清を塩化ベンゼトニウムで処理し、ペレットを酢酸塩溶液中で撹拌し、次いでClarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0109】
濾液に含まれていたK5多糖を実施例2に記載したようにメタノール沈殿させ、次いで実施例5と同様に過酸化水素で3回処理した。
【0110】
力価73%をもつ29.6gのK5多糖が最終収率68%で得られた。
【実施例7】
【0111】
ペレットの処理および精製の改善
実施例1の発酵槽COL5K003から得られたペレット(28.5gのK5多糖を含有する4.75kg)を4.75リットルの脱イオン水に再懸濁させ、5NのNaOHでpHを7.2に調整した。次に混合物を室温で2時間撹拌し、次いで4700×gで2時間遠心した。
【0112】
実施例2に記載したように上清を塩化ベンゼトニウムで処理し、ペレットを酢酸塩溶液中で撹拌し、次いでClarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0113】
実施例2に記載したように濾液に含まれていたK5多糖をメタノール沈殿させ、次いで実施例4と同様に過酸化水素で2回処理した。 力価69%をもつ25.9gのK5多糖が最終収率63%で得られた。
【実施例8】
【0114】
酵素による除タンパク工程を含む精製
最終生成物の純度を改善するために酵素による除タンパク工程を実施した。
【0115】
実施例7で得られた15.9gおよび実施例6で得られた29.6gを含む45.5gのK5多糖を1.8リットルの脱イオン水に再懸濁させた。
【0116】
pHを7.0に調整した後、910μlの液体アルカラーゼを添加し、混合物を撹拌しながら60℃で5時間インキュベートした。
【0117】
次に23gの塩と7.2リットルのメタノールとを添加し、混合物を30分間撹拌し、次いで4000×gで15分間遠心した。
【0118】
ペレットを1.8リットルの脱イオン水に再懸濁させ、実施例3に記載したように過酸化水素で処理した。
【0119】
89%の力価をもつ36.4gのK5多糖が98%の収率で得られた。
【実施例9】
【0120】
酵素による除タンパク処理と脱色処理との有効性の比較
2種類の処理の相対的有効性を評価するために、実施例7で得られた5gのK5多糖を過酸化水素またはアルカリ性アルカラーゼによって処理した。
【0121】
すなわち、実施例7で得られた5gのK5多糖を200mlの脱イオン水に再懸濁させ、溶液を実施例3と同様に過酸化水素で処理した。
【0122】
76.9%の力価をもつ3.75gのK5多糖が収率83%で得られた。
【0123】
同様にして、実施例7で得られた5gのK5多糖を200mlの脱イオン水に再懸濁させた。しかしながら、過酸化水素で処理する代わりに、この溶液を60℃に加熱し、そのpHを7に調整した。次いで100μlのアルカラーゼを溶液に添加し、混合物を60℃で5時間インキュベートした。
【0124】
次にこの混合物を実施例2に記載したようにメタノール沈殿させた。
【0125】
83%の力価をもつ3.62gのK5多糖が収率87%で得られた。これに比べて、過酸化水素処理では力価76.9%および収率83%であった。
【0126】
従って、除タンパク処理では、3回目の過酸化水素処理に比べて収率の改善および純度の向上が得られる。
【0127】
実施例1に記載したような発酵が終了したとき、ペレット減量はCOL5K002で60%、COL5K003で40%であった。実施例6および7に記載したようなこのペレットの再処理プロセスを開発したので、K5多糖の総抽出効率をCOL5K002では37%から78%に向上させ、COL5K003では43%から68%に向上させることが可能になったのである。
【0128】
実施例8に記載したような最後の除タンパク工程は、定量的収率(98%)でK5多糖の純度を89%に向上させ得る。
【図面の簡単な説明】
【0129】
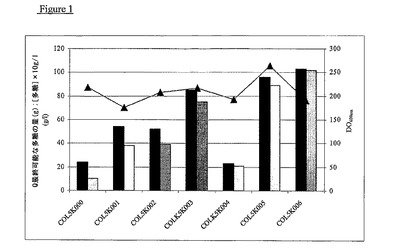
【図1】COL5K000からCOL5K006までの処理で得られた多糖の量。黒い棒グラフは最終可能な多糖の推定量(単位g)を表す;斑点入り棒グラフは上清中の多糖の最終濃度を表す(×20、単位g/l);三角印は培養物が示したOD−600nmを表す。
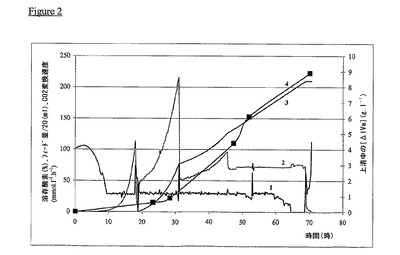
【図2】COL5K006の主要処理パラメーターのプロフィル。曲線1は培地中の溶存酸素のパーセンテージを表す;曲線2は培養物COLK006中のCO2の発生速度を表す(単位nmol.1−1時−1);曲線3はフィード量/20をg.l−1で表す;曲線4は上清中のK5多糖の濃度(単位g.l−1)を表す。
【技術分野】
【0001】
本発明は、大腸菌による発酵工程と精製工程とを順次に行うことによってK5多糖を製造する方法に関する。
【背景技術】
【0002】
K5多糖を得るためのいくつかの方法が公知である。K5多糖は外的腸感染症の原因となる大腸菌株から得ることができる。
【0003】
K5多糖は、等モル量のグルクロン酸とN−アセチルグルコサミンとから成り、これらが基本単位として4−ベータ−グルクロニル−1,4−アルファ−N−アセチルグルコサミンを構成し、基本単位が直線状に反復してK5多糖鎖が形成される。
【0004】
【化1】
【0005】
この基本単位は、完全に脱硫酸化されN−アセチル化されたヘパリンに構造類似性を有しており、従って、動物器官から抽出されるヘパリンの極めて有利な代替材料となり得る、すなわち、発酵によって産生されたK5多糖の化学的修飾によってヘパリンを製造するために使用し得る。
【0006】
生合成によって得られたヘパリンは、以下のようにして製造される。微生物大腸菌を培養してK5多糖を産生させ、次いでこれを化学処理してヘパリンとする。K5多糖からヘパリンを製造する方法は記載されている(Biochem J.,1991,275(1):151−8;国際特許WO92/17507;Semin.Thromb.Hemost.,2001,27(5):437−43;Nat.Biotech.,2003,21(11):1343−46)。一般的に、K5多糖からヘパリンを合成するためには、脱アセチル化およびN−再硫酸化、C−5エピマー化およびO−硫酸化の処理が必要である。
【0007】
ヘパリンの有用性は特にその抗凝血性および抗血栓性にある。しかしながら、ヘパリンは使用条件が限られるという欠点を有している。特にその高い抗凝血活性(aIIa)が出血を引き起こすこともある(Semin.Thromb.Hemost.,5 sup.3,1999)。本発明によれば、酵素的または化学的な解重合によって半合成ヘパリンから低分子量ヘパリン(3000−6500Da)または超低分子量ヘパリン(1500−3000Da)を得ることが可能であり、これらの抗IIa活性は5IU/mg未満であるのに抗Xa活性は特に140−190IU/mgの範囲である。
【0008】
K5多糖の多くの製造方法が従来技術に記載されている(Eur.J.Biochem,1981,116:359−364;EPO489647,WO01/02597、Eur.J.Biochem,1988,117:112−125)。これらの方法は特に、グルコース中、37℃で行う単一増殖段階から成る大腸菌の発酵工程と、本質的にイオン交換カラム処理から成るK5多糖の精製工程とを順次に含む。
【0009】
発酵工程に関しては、本発明方法の発酵工程は、少なくともその1つがフィードを伴う増殖段階である複数の増殖段階と冷却段階とを含み、この発酵工程を精製工程に組合せることによって従来技術に記載された値の10倍にも達する収率が得られる。
【0010】
実際、従来技術に記載された大腸菌の発酵によるK5多糖の生合成方法はいずれも極めて低収率であるという重大な欠点を有している。例えば、従来技術による細胞外形態のK5多糖の製造方法では、単離されたK5多糖の収率は200−850mg/lにすぎない。
【0011】
このような低収率ではK5多糖の工業生産を計画すること、従って、半合成による半合成ヘパリンの工業生産を計画することができない。しかしながら、動物由来でないヘパリンおよび低分子量または超低分子量ヘパリンに対する需要は大きい。
【0012】
従って、本発明の1つの目的は、細胞外形態のK5多糖の新規な製造方法を提供することである。
【0013】
本発明の方法は本質的に2つの工程、すなわち、発酵工程と精製工程とを含む。
【0014】
本発明の方法によれば、従来技術に記載された方法で得られる濃度の10倍に達する濃度のK5多糖を得ることが可能である。従って本発明によれば、10g/lもの濃度のK5多糖を得ることが可能である。このような濃度は、発酵工程と精製工程との組合せによって得られるものである。
【発明の開示】
【0015】
本発明の1つの目的は、
−発酵槽で行うことができ、
−バッチ増殖段階と、
−1回または数回のフェッドバッチ増殖段階と、
−温度を下げ、培地のpHを9までの値に上げる冷却段階と、
−バイオマスを培養培地から分離する段階と、
−バイオマスから混入培養培地を除去する段階と、
を含む、K5多糖を産生する大腸菌株の発酵工程と、
−培養培地および/またはバイオマスを使用する精製工程と、
を含むK5多糖の製造方法である。
【0016】
本発明の目的はより特定的には、上記のようなK5多糖の製造方法であって、培養培地および/またはバイオマスを出発材料とするその精製工程がイオン交換カラムによる処理工程を含むものでなく、
−約7−約11の値へのpHの調整、
−沈殿、
−沈殿物の溶解、
−得られた懸濁液の濾過、および、
−アルコール沈殿、
を記載の順序で含んでいる。
【0017】
発酵
本発明によるK5多糖の製造に使用できる大腸菌株は、パブリックコレクション例えばCNCM(Collection Nationale de Cultures de Microorgansmes,France)またはATCC(American Type Culture Collection,USA)から得られる。あるいは、これらの菌株を臨床的に単離し、次いで文献に記載されたように莢膜の抗原型のキャラクタリゼーションを行ってもよい(Eur.J.Biochem,1988 117:112−125)。
【0018】
本発明の発酵工程は、発酵槽で大腸菌を使用して行うものであり、
−バッチ増殖段階と、
−指数増加量または一定量のフィードを伴う1回または複数のフェッドバッチ増殖サイクルと、
−温度を約10−12℃に下げpHを約7.5−9に上げる冷却段階と、
−バイオマスを培養培地から分離する段階と、
−バイオマスから混入培養培地を除去する段階と、
を含む。
【0019】
本発明によれば“バッチ増殖”という表現は、フィードまたは抜取りを伴うことなく反応装置で微生物を増殖させることを意味する。
【0020】
本発明によれば“フェッドバッチ増殖”という表現は、管理量の基質フィードを伴うが抜取りを伴うことなく反応装置で微生物を増殖させることを意味する。
【0021】
本発明によれば、発酵工程は特に、バッチ増殖段階の後に指数増加量の基質フィードを伴う2回のバッチ増殖段階を含む。
【0022】
本発明によれば、発酵工程は、冷却段階の前に一定量の基質フィードを伴うバッチ増殖段階を含む。
【0023】
本発明の発酵工程では、一回目のフェッドバッチ増殖段階が、バッチ増殖段階の培地中の炭素源の消耗と共に開始される。
【0024】
より特定的には本発明によれば、バッチ増殖段階の培地中の炭素源が消耗したときに一回目のフェッドバッチ増殖段階が、開始され、必要ならば行われるその後のフェッドバッチ増殖段階は酸素移動の極限の結果として溶存酸素の濃度が有意に低下したときに開始される。
【0025】
本発明によれば、バッチ増殖段階の各々は上記の時期に開始され、冷却段階は培養フィード培地の全量が発酵槽に注入されたとき、または、pH変化が検出されたときに開始される。
【0026】
本発明によれば、バッチ増殖段階、(1回または複数の)フェッドバッチ増殖段階および冷却段階の後に、遠心によってバイオマスを培地から分離する。分離後、バイオマスおよび培養培地を除染する。具体的には、これらを約80℃の温度で1−3時間失活させる。次に上清および/またはペレットを使用してK5多糖を精製する。
【0027】
pHはバッチ増殖段階およびフェッドバッチ増殖段階中はNH4OHもしくはNH3の添加によって約6.5に維持するが、最終段階中に約8.5まで上昇する。
【0028】
溶存酸素の濃度は、撹拌速度、空気流量の変更、流入空気の酸素富化または過酸化水素の添加などの操作によって空気飽和量の約15−約80%に維持する。
【0029】
本発明の発酵工程中の増殖温度は約10℃−約40℃である。より特定的には、バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、必要ならば行うその後のフェッドバッチ増殖段階では約25℃である。K5多糖を高収率で得るためには30℃から25℃への移行が特に重要である。
【0030】
使用される培養培地は合成培養培地である。本発明によれば炭素源はグルコースまたはグリセロールである。本発明によれば好ましい炭素源はグリセロールである。
【0031】
より特定的に、本発明の発酵工程の炭素源がグリセロールであり、その濃度はバッチ増殖段階の出発培養培地中では約10−約90g/lの範囲であり、フェッドバッチ増殖段階のフィード培地中では約250−約1200g/lの範囲である。実験実施例1は、K5多糖を高濃度で得るためには、炭素源としてグリセロールを使用すること、および、フィード培地中のグリセロール濃度を増加させることの重要性を示している。
【0032】
本発明によれば、バッチ増殖段階の培地の窒素源が酵母エキス、カザミノ酸、ペプトン、NH4OH、NH3または(NH4)2HPO4でよい。
【0033】
本発明によれば、バッチ増殖段階の出発培養培地中の窒素源は特にNH4OH、NH3または(NH4)2HPO4である。これらは特に約1−約10g/lの範囲の濃度で使用される。
【0034】
本発明によれば、バッチ増殖段階およびフェッドバッチ増殖段階中の窒素源はpHを調整するために、NH4OHまたはNH3によって供給される。その濃度は好ましくは約20%である。
【0035】
より特定的には、本発明の発酵工程で、バッチ増殖段階の培地の窒素源が、出発培養培地では約1−約10g/lの濃度のNH4OH、NH3または(NH4)2HPO4であり、バッチ増殖段階およびフェッドバッチ増殖段階中はpHを調整するために約20%濃度のNH4OHまたはNH3によって供給される。
【0036】
本発明の1つの目的は上記に記載のような発酵工程を含むK5多糖の製造方法であり、該発酵工程では、
−炭素源がグリセロールであり、グリセロールの濃度は、
−バッチ増殖段階の出発培養培地中に約10−約90g/lの範囲であり、
−フェッドバッチ増殖段階のフィード培地中に約250−約1200g/lの範囲であり、
−増殖温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、
−必要ならば行う他のフェッドバッチ増殖段階では約25℃である。
【0037】
本発明の1つの目的はまた、上記に記載のような発酵工程を含むK5多糖の製造方法であり、該発酵工程では、
−炭素源がグリセロールであり、グリセロールの濃度が、
−バッチ増殖段階の出発培養培地中に約10−約90g/lの範囲であり、
−フェッドバッチ増殖段階のフィード培地中に約250−約1200g/lの範囲であり、
−バッチ増殖段階の培地の窒素源が
−出発培養培地では約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4であり、
−バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給される。
【0038】
本発明の1つの目的はまた、上記に記載のような発酵工程を含むK5多糖の製造方法であり、該発酵工程では、
−バッチ増殖段階の培地の窒素源が、
−出発培養培地では約1−約10g/lの範囲のNH4OH、NH3または(NH4)2HPO4であり、
−バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給され、
−増殖温度が、
−バッチ増殖段階およびフェッドバッチ増殖段階では約30℃であり、
−必要ならば行う他のフェッドバッチ増殖段階では約25℃である。
【0039】
最も特定的に本発明の1つの目的はまた、上記に記載のようなK5多糖の製造方法であり、該方法では、
発酵工程が、以下の段階、すなわち、
−バッチ増殖段階、
−先行段階の培地中の炭素源が完全消耗したときに開始する指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する二回目の指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する一定量の基質フィードを伴うバッチ増殖サイクル、
−温度を約12℃に下げpHを約7.5−約9の範囲の値に上げる冷却段階、
−バイオマスから混入培養培地を除去する段階、
を記載の順序で含み、
上記段階を以下のパラメーターで行う、すなわち、
−温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階中は約30℃、
−他の2回のフェッドバッチ増殖段階中は約25℃
であり、
−窒素源は、
−出発培養培地には約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4によって供給され、
−バッチ増殖段階およびフェッドバッチ増殖段階にはpHを調整するために約20%の濃縮NH4OHまたはNH3の添加によって供給され、
−炭素源はグリセロールであり、その濃度は、
−最初のバッチ増殖段階の出発培養培地中で約10−約90g/lの範囲の濃度であり、
−フェッドバッチ増殖段階のフィード培地中で約250−約1200g/lの範囲の濃度である。
【0040】
第二工程:K5多糖の精製
本発明の1つの目的はまた、精製工程を含む上記のようなK5多糖の製造方法である。この精製工程では、発酵ブロスの遠心上清またはこの遠心で得られたペレットの再懸濁液から成るK5多糖含有溶液のpHを約7−約11に調整し、この溶液から多糖を沈殿させる。次に沈殿物を洗浄し、再溶解し、濾過し、次いで沈殿させる。
【0041】
本発明によれば、精製工程の一回目の沈殿は第四級アンモニウム塩を用いて行う。この沈殿はより特定的には塩化ベンゼトニウムを用いて行う。
【0042】
本発明によれば、精製工程の最初の沈殿物を、特に10%濃度で使用する酢酸ナトリウムに再溶解させる。
【0043】
更に、本発明によれば、精製工程で得られた濾液をメタノール沈殿させる。
【0044】
より特定的には、本発明による精製工程の一回目の沈殿が塩化ベンゼトニウムを用いて行われ、この沈殿物を酢酸ナトリウムに再溶解し、次いで濾過し、精製工程で生じた濾液をメタノール沈殿させる。
【0045】
本発明によれば、上記に記載のような精製後に得られた生成物を場合によっては過酸化水素によって1回または数回脱色処理する。より特定的には、精製後の過酸化水素処理を3回行う。
【0046】
必要な場合には、上記のように精製した生成物を過酸化水素で3回脱色処理した後、残留汚染物を除去するために追加の過酸化水素処理を行う。この処理は得られる生成物の純度を改善し得る。従って本発明の1つの目的はまた、精製および3回の過酸化水素処理後に追加の過酸化水素による脱色をさらに含むK5多糖の製造方法である。
【0047】
必要な場合には、上記精製工程のいずれか1つの後でK5多糖をプロテアーゼで処理する。実験実施例8で後述するように、プロテアーゼによる処理は、ほぼ定量的収率で産生されるK5多糖の純度を有意に向上させ得る。従って本発明の目的は特に、精製工程中のプロテアーゼ処理を含むK5多糖の製造方法である。本発明のプロテアーゼは好ましくはアルカラーゼである。
【0048】
最後に、K5多糖を沈殿させ、遠心し、乾燥する。
【0049】
従って本発明の1つの目的は最も特定的には、上記に記載のような精製工程を含むK5多糖の製造方法であって、精製工程で以下の段階:
−バイオマスから培養培地を分離するために培養物を遠心する段階;
−発酵ブロスの遠心上清を使用して
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−同時に、発酵ブロスの遠心によって得られたペレットを使用して
−ペレットを再懸濁化し、
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに再溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−各々の精製工程で得られた2つの調製物を合せる段階;
−K5多糖の沈殿、遠心および乾燥を行う段階;
を行う方法である。
【0050】
従って本発明の1つの目的は好ましくは、発酵工程と精製工程とを含むK5多糖の製造方法であって、発酵工程が、以下の段階:
−バッチ増殖段階、
−先行段階の培地中の炭素源が完全消耗したときに開始する指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する二回目の指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する一定量の基質フィードを伴うバッチ増殖サイクル、
−温度を約12℃に下げpHを約7.5−約9の範囲の値に上げる冷却段階、
−バイオマスから混入培養培地を除去する段階、
を記載の順序で含み、
上記段階が以下のパラメーターで行われる、すなわち:
−温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階中は約30℃、
−他の2回のフェッドバッチ増殖段階中は約25℃
であり、
−窒素源は、
−出発培養培地には約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4によって供給され、
−バッチ増殖段階およびフェッドバッチ増殖段階にはpHを調整するために約20%の濃縮NH4OHまたはNH3の添加によって供給され、
−炭素源はグリセロールであり、その濃度は、
−最初のバッチ増殖段階の出発培養培地中に約10−約90g/lの範囲の濃度であり、
−フェッドバッチ増殖段階のフィード培地中に約250−約1200g/lの範囲の濃度であり、
精製工程が以下の段階:
−バイオマスから培養培地を分離するために培養物を遠心する段階;
−発酵ブロスの遠心上清を使用して
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−同時に、発酵ブロスの遠心によって得られたペレットを使用して
−ペレットを再懸濁化し、
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに再溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−各々の精製工程で得られた2つの調製物を合せる段階;
−K5多糖を沈殿、遠心および乾燥させる段階を含む方法である。
【0051】
このようにして本発明方法では、従来技術に記載された方法で得られる収率の10倍にもなるような収率が得られる。
【0052】
本発明によれば、上記に記載の方法によって製造された多糖が酵素的および/または化学的な脱アセチル化、N−再硫酸化、C−5エピマー化および/またはO−硫酸化の反応基質として使用される。より特定的には本発明によれば、上記に記載の方法によって製造された多糖が半合成ヘパリン(バイオヘパリン)を得るための基質として上記反応の1つで使用される。
【0053】
本発明によれば、得られたバイオヘパリンを次に、約1500−約6500Daの範囲の分子量をもつLMWH(低分子量ヘパリン)を得るためのフラグメンテーション反応で基質として使用する。
【0054】
本発明によればまた、得られたバイオヘパリンを、より特定的に約3500−約5500Daの範囲の分子量をもつLMWH(低分子量ヘパリン)、または、より特定的に約1500−約3000Daの範囲の分子量をもつVLMWH(超低分子量)ヘパリンを得るためのフラグメンテーション反応の基質として使用することが可能である。
【0055】
以下の実施例は本発明の代表例であるが、本発明はこれらの実施例に限定されない。
【0056】
応用例
【実施例1】
【0057】
(1)手順
(a)発酵
発酵は20リットルの発酵槽で大腸菌株ATCC23506を使用して行う。
【0058】
5−60g.l−1のグリセロールと0.5−3g.l−1のKH2PO4と1−10g.l−1の(NH4)2PO4と0.5−5g.l−1のMgSO4と0.2−4g.l−1のクエン酸と微量元素とから成る100mlの培地を収容した500ml容の三角フラスコ中、30℃で一夜凍結させた1.8mlのバイアルを使用して大腸菌株のプレカルチャーを調製する。
【0059】
200rpmで撹拌しながら30℃で増殖させる。生産用発酵槽に接種するためにこのプレカルチャーを使用する。また、グリセロール収容バイアルから直接的に発酵槽に接種してもよい。このプレカルチャーの目的は、発酵槽におけるバッチ増殖段階の持続時間を短縮することだけである。使用した培地は、RFB MIL 11/04培地(J.of Biotechnology,1995,39:59−65)の変種であり、それらの組成を表1に示す。
【0060】
【表1】
【0061】
自己適応形管理アルゴリズムに基づくIRISプログラム(Infors)によって制御されたTECHFORS発酵槽(全容積20l、処理容積12l)で培養物を調製する。培地中の溶存酸素濃度、pH、撹拌速度、温度、通風空気の流速、圧力およびポンプ吐出量(基底調節量、フィード量、抑泡剤)はオンラインで測定し管理する。発酵槽の排出ガスの分析は質量分析計(PRIMA 600S)を使用して行う。
【0062】
以下の特性値のバッチモードで発酵させる。
−処理容量12l、
−初期容量8l、
−追加容量4l
−最終容量は12lプラス基底容量、
−溶存酸素は空気飽和量の15%−80%の範囲(好ましくは30%)に維持、
−温度は20−40℃の範囲(好ましくは30℃)、
−pHは濃NH4OHまたはNH3を加えて6.0−7.5(好ましくは6.5)に調整、
−圧力は0.3バール、0.5バールおよび0.8バール、
−初期通風は4l.分−1(0.5VVM)(4−20l.分−1の範囲で調整)、
−初期撹拌速度は200rpm(200−1500rpmの範囲で調整)。
【0063】
培養は5つの工程、すなわち、バッチ増殖段階、次いで、基質フィードを伴う3回の異なるバッチ増殖段階、および、最終冷却段階を含む。
【0064】
1.バッチ増殖段階
バッチモードの培養段階では、菌株の世代時間を二酸化炭素の発生量の測定値から算定し、管理された基質フィードを伴うバッチ増殖の最初の部分の初期速度を計算する:
Q0=(CO2pmax×VolFer)/(1000×MWCO2×YCO2/グリセロール×G0)
Q0:濃縮培地の初期流速(1.時−1);
CO2pmax:二酸化炭素の最大発生量(mmol.l−1.時−1);
G0:管理された基質フィードを伴うバッチ増殖中に使用された濃縮培地中のグリセロールまたはグルコースの濃度(750g.l−1);
VolFer:発酵槽中の液体容量;
MWCO2:CO2の分子量(g.l−1);
YCO2/グリセロール:グリセロール消費量あたりのCO2の発生量。
【0065】
2.指数増加量の基質フィードを伴う1回目のバッチ増殖段階
炭素源の消耗後、バイオマスの濃度を増加させるために、指数増加量の基質フィードを伴うバッチ増殖サイクルを開始する。新しい培地を供給するポンプのスイッチを入れる。ポンプの吐出量は時間の経過に伴って指数関数的に増加する。
Q(t)=Q0×2[t/(τ×2.4)]
Q0:フィードポンプの初期供給量(l.時−1);
t:指数増加量の基質フィードを伴うバッチ増殖サイクル開始後の経過時間;
τ:バッチ増殖中に測定した菌株世代時間。
【0066】
予め測定した菌株世代時間に1−10(好ましくは2.4)の範囲の補正率を乗算した値をこの供給量に算入する。この供給量は0.05時−1から0.65時−1の範囲の増殖率を微生物に与える。
【0067】
3.指数増加量の基質フィードを伴う2回目のバッチ増殖段階
最初の酸素移動の極限に達すると、(溶存酸素<20%、撹拌速度>1350rpm、空気流速>19,5l.分−1、圧力=0.5バール)、この指数増加量の基質フィードを伴う1回目のバッチ増殖段階を停止する。管理された基質フィードを伴う1回目のバッチ増殖サイクルの開始時の8.4ml.時−1という流速に基づいて次の指数増加量のフィードを開始する。先行の指数増加量の基質フィードを伴うバッチ増殖サイクルと同様に、4−10の範囲(好ましくは6)の新しい補正率を世代時間に乗算する。この供給量は0.05時−1から0.16時−1の範囲の増殖率を微生物に与える。
【0068】
同時に、培地中の酸素溶解度を増加させるために発酵槽の表示温度を20−25℃(好ましくは25℃)に下げる。フィードポンプの新しい制御アルゴリズムは以下の式で表される:
Q’(t)=0.0084×2[t/(τ×6)]
t:指数増加量の基質フィードを伴うバッチ増殖サイクル開始後に経過した時間;
τ:バッチ増殖中に測定した菌株世代時間。
【0069】
酸素移動の極限は約30時間の増殖後(管理された基質フィードを伴う1回目のバッチ増殖サイクル10時間を含む)に出現するが、フィードポンプの実際の初期培地吐出量は約15ml.時−1(t=10時間)である。この低い供給量は、微生物の増殖を維持すると同時に系の溶存酸素を飽和量の30%に調節する機能も果す。2回目の酸素移動の極限が出現するまでこの供給量を継続する。
【0070】
4.一定量の基質フィード段階
2回目の酸素移動の極限(溶存酸素<20%、撹拌速度>1350rpm、空気流速>19.5l.分−1、圧力=0.5バール)が出現すると、指数増加量の培地フィードを1−20ml.l−1.時−1の範囲(好ましくは4.2ml.l−1.時−1)の一定供給量に切換える。4lのフィード培地が注入されるかまたはpHの有意な低下(強力な酸素極限の指標となるNH4OHの完全消費)が検出されるまでこの供給量を継続する。
【0071】
5.冷却段階
次にフィードポンプのスイッチを切り、発酵槽の温度を10℃に下げる。この間に、pHはK5多糖酵素リアーゼの活性に好ましい7.5−9(好ましくは8.5)という値まで自然に上昇するかまたはこのような値に調節できる。これらの条件を0−10時間維持する。
発酵槽が10℃まで冷えた後、バイオハザードレベル2の作業条件下でバイオマスを気密容器に採集する。ブロスを遠心してバイオマスを培地上清から分離する。分離後、80℃で1時間30分の熱処理によって撹拌しながら上清を除染する。バイオマス自体は80℃で2時間30分の処理によって除染する。上清およびバイオマスを−20℃で保存する。
【0072】
これらの試験には以下の条件を使用する:
−COL5K000/001:30℃のバッチ増殖段階/30℃で行う指数増加量の基質フィードを伴う増殖段階/冷却段階;
−COL5K002:30℃のバッチ増殖段階/30℃で行う指数増加量の基質フィードを伴う増殖段階/2回目の30℃の指数増加量の基質フィードを伴う増殖段階/冷却段階;
−COL5K003/004:30℃のバッチ増殖段階/30℃で行う指数増加量の基質フィードを伴う増殖段階/25℃で行う2回目の指数増加量の基質フィードを伴う増殖段階/冷却段階;
−COL5K005/006:30℃のバッチ増殖段階/30℃で行う指数増加量の基質フィードを伴う増殖段階/25℃で行う2回目の指数増加量の基質フィードを伴う増殖段階/25℃で行う一定量の基質フィードを伴う増殖段階/冷却段階。
【0073】
b)多糖の分析
多糖の分析は2工程で行う。
1.最初に多糖をその構成二糖に解重合する。
2.二糖混合物をHPLCによって分析する。
【0074】
1.多糖の解重合
分析すべき約40mgのサンプルを2mlのpH7の酢酸塩バッファ(0.29mlの酢酸、5mgのBSA、15.8mgの酢酸カルシウム、総量を50mlにする量の脱イオン水、2Nの水酸化ナトリウムでpHを7に調整)に再懸濁させ、20mg/mlを含有する溶液を得る。
【0075】
同時に、リン酸塩バッファ(68mgのリン酸二水素カリウム、10mgのBSA、総量を50mlにする量の脱イオン水、1Nの水酸化カリウムでpHを7に調整)中に0.5IU/mlのヘパリナーゼI、IIおよびIIIをそれぞれ含有する溶液を調製する。次にこれらの溶液を等しい割合で混合して最終のヘパリナーゼ123溶液を得る。
【0076】
20mg/mlのサンプル溶液25μl、酢酸塩バッファ25μl、および、ヘパリナーゼ123溶液25μlを300μl容の管で混合する。混合物を十分にホモジナイズし、室温で24時間インキュベートして完全に解重合させる。
【0077】
2.HPLCによる多糖の分析
勾配HPLCを使用し当業者に公知の方法に従って前項の混合物を分析する。以下の分析条件を使用する:
−カラム:Spherisorb SAX 5μm、250×3mm、Waters、
−カラム温度:40℃、
−流速:0.5ml/分、
−注入量:10μl、
−234nmでUV検出、
−移動相:
溶媒A:NaH2PO4、2.5mM pH=2.9
溶媒B:NaClO4、M、NaH2PO4 2.5mM pH=3
勾配:
【0078】
【表2】
【0079】
二糖4−ベータ−グルクロニル−1,4−アルファ−N−アセチル−グルコサミンがHPLCによって検出される。
【0080】
【化2】
2)結果
対照COL5K000に加えて異なる6つの発酵試験(COL5K001−006)を実施した。
【0081】
これらの試験中に得られたK5多糖の濃度を表2に示す。
【0082】
【表3】
【0083】
結果(図1、付記1)は、使用した手順の各々によって対照COL5K000の2−10倍のK5多糖の製造が可能であったことを明らかに示す。COLK006の主要パラメーターのプロフィルは、培地中のK5多糖の濃度増加が2回目のフェッドバッチ増殖段階の付加に関連していることを明らかに示す(図2、付記1)。
【0084】
実施した試験は、培地中に精製した多糖の最大濃度を文献に記載された値に比較して大幅に増加させた。文献の量は900mg.l−1であったが、我々は10g.l−1を上回る濃度を得た。
【0085】
COL5K000の処理プロセスと培養培地中の多糖濃度が増加したCOL5K006の処理プロセスには主として以下の違いが存在した:
−炭素源としてグルコースの代わりにグリセロールを使用する:
−酸素移動の極限が出現したときに、低下させた増殖率の基質フィード段階(2回目および3回目のフィード段階)を付加することによって発酵総時間を延長する:
−フィード溶液のグリセロール濃度を増加する(500g.l−1を750g.l−1にする):
−処理終了時のpHを上げ、これによってK5多糖リアーゼの活性に有利な条件を成立させ、培養培地中に得られる多糖の量を増加させる。
【実施例2】
【0086】
抽出プロセス
以下に記載の抽出工程は、上記で得られた発酵上清からK5多糖を単離するために行った。
【0087】
実施例1で発酵槽COL5K001から6.5リットルの熱失活上清が得られた。これは24.7gのK5多糖を含有している。
【0088】
5Nの水酸化ナトリウムを使用してこの溶液のpHを7.2に調整し、50g/lの塩化ベンゼトニウムを含有している663mlの水溶液を注いだ(すなわち、K5多糖に対して1.2モル当量の塩化ベンゼトニウム)。反応媒体を室温で30分間撹拌し、次いで4000rpmで10分間遠心した。
【0089】
得られたペレットを4リットルの脱イオン水で2回洗浄し、次いで5リットルの10%酢酸ナトリウム溶液に入れて撹拌した。
【0090】
1回目の塩化ベンゼトニウム沈殿で得られた上清を再度処理した。5Nの水酸化ナトリウムを使用してそのpHを7.2に調整し、50g/lの濃度の663mlの塩化ベンゼトニウムを添加し、混合物を30分間撹拌後に遠心した。
【0091】
得られたペレットを4リットルの脱イオン水で2回洗浄し、次いで5リットルの10%酢酸ナトリウム溶液に入れて撹拌した。2種類の再懸濁ペレットを含有している溶液を集め、Clarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0092】
濾液を40リットルのメタノールに注いだ。得られた混合物を25分間撹拌した後、一夜傾瀉した。翌日、4000×gで10分間遠心した。ペレットをメタノールで洗浄し、次いで20Torr下、50℃で36時間乾燥した。
【0093】
このようにして力価54%をもつ31.5gのK5多糖が得られた。収率は68.8%であった。
【実施例3】
【0094】
精製
実施例2で単離した生成物の力価が低かったので(わずか54%)、追加の精製工程を実施した。
【0095】
実施例2で得られた重量31.5gの粗生成物を磁気撹拌付きの三角フラスコで1.2リットルの脱イオン水に再懸濁させた。5Nの水酸化ナトリウムでpHを11に調整した後、15gの塩化ナトリウムと6mlの30%過酸化水素とを溶液に加えた。混合物を10分間撹拌し、次いで撹拌しないで室温で一夜静置した。
【0096】
4倍容のメタノールを溶液に加えることによってK5多糖を沈殿させた。混合物を30分間撹拌し、次いで4000rpmで15分間遠心した。ペレットを20Torr下、45℃で36時間オーブン乾燥した。このようにして、力価が54%から65%に改善された23.5gのK5多糖が得られた。
【実施例4】
【0097】
抽出と精製
実施例1の発酵槽COL5K002から粗K5多糖を抽出して精製した。
【0098】
実施例1で発酵槽COL5K002から5.4リットルの熱失活上清が得られた。これは21.1gのK5多糖を含有している。
【0099】
実施例2に記載したようにCOL5K002上清を塩化ベンゼトニウムで処理し、ペレットを酢酸塩溶液中で撹拌し、次いでClarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0100】
濾液に含まれていたK5多糖を実施例2に記載したようにメタノール沈殿させ、次いで実施例3に詳述したように過酸化水素で脱色し、その後、過酸化水素による2回目の脱色を行った。
【0101】
最終的に、78%の力価をもつ25.3gのK5多糖が90%を上回る収率で得られた。
【実施例5】
【0102】
抽出と精製
実施例4の発酵槽COL5K003から粗K5多糖を抽出して精製した。
【0103】
実施例1で発酵槽COL5K003から6リットルの熱失活上清が得られた。これは45gのK5多糖を含有している。
【0104】
実施例2に記載したようにCOL5K003上清を塩化ベンゼトニウムで処理し、ペレットを酢酸塩溶液中で撹拌し、次いでClarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0105】
濾液に含まれていたK5多糖を実施例2に記載したようにメタノール沈殿させ、次いで実施例4と同様に過酸化水素で2回処理し、その後、3回目の過酸化水素による脱色を行った。
【0106】
最終的に、81%の力価をもつ39.5gのK5多糖が71%の収率で得られた。
【実施例6】
【0107】
ペレットの処理および精製の改善
先行の試験では上清だけを抽出して精製した。遠心沈降物中に存在するK5多糖を回収するために、実施例1の発酵槽COL5K002から得られたペレット(31.9gのK5多糖を含有する5kg)を等容量の脱イオン水に再懸濁させ、5NのNaOHでpHを7.2に調整した。次に混合物を室温で2時間撹拌し、次いで4700×gで90分間遠心した。
【0108】
実施例2に記載したように上清を塩化ベンゼトニウムで処理し、ペレットを酢酸塩溶液中で撹拌し、次いでClarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0109】
濾液に含まれていたK5多糖を実施例2に記載したようにメタノール沈殿させ、次いで実施例5と同様に過酸化水素で3回処理した。
【0110】
力価73%をもつ29.6gのK5多糖が最終収率68%で得られた。
【実施例7】
【0111】
ペレットの処理および精製の改善
実施例1の発酵槽COL5K003から得られたペレット(28.5gのK5多糖を含有する4.75kg)を4.75リットルの脱イオン水に再懸濁させ、5NのNaOHでpHを7.2に調整した。次に混合物を室温で2時間撹拌し、次いで4700×gで2時間遠心した。
【0112】
実施例2に記載したように上清を塩化ベンゼトニウムで処理し、ペレットを酢酸塩溶液中で撹拌し、次いでClarcelプレレイヤーを配備した焼結ガラス漏斗で濾過した。
【0113】
実施例2に記載したように濾液に含まれていたK5多糖をメタノール沈殿させ、次いで実施例4と同様に過酸化水素で2回処理した。 力価69%をもつ25.9gのK5多糖が最終収率63%で得られた。
【実施例8】
【0114】
酵素による除タンパク工程を含む精製
最終生成物の純度を改善するために酵素による除タンパク工程を実施した。
【0115】
実施例7で得られた15.9gおよび実施例6で得られた29.6gを含む45.5gのK5多糖を1.8リットルの脱イオン水に再懸濁させた。
【0116】
pHを7.0に調整した後、910μlの液体アルカラーゼを添加し、混合物を撹拌しながら60℃で5時間インキュベートした。
【0117】
次に23gの塩と7.2リットルのメタノールとを添加し、混合物を30分間撹拌し、次いで4000×gで15分間遠心した。
【0118】
ペレットを1.8リットルの脱イオン水に再懸濁させ、実施例3に記載したように過酸化水素で処理した。
【0119】
89%の力価をもつ36.4gのK5多糖が98%の収率で得られた。
【実施例9】
【0120】
酵素による除タンパク処理と脱色処理との有効性の比較
2種類の処理の相対的有効性を評価するために、実施例7で得られた5gのK5多糖を過酸化水素またはアルカリ性アルカラーゼによって処理した。
【0121】
すなわち、実施例7で得られた5gのK5多糖を200mlの脱イオン水に再懸濁させ、溶液を実施例3と同様に過酸化水素で処理した。
【0122】
76.9%の力価をもつ3.75gのK5多糖が収率83%で得られた。
【0123】
同様にして、実施例7で得られた5gのK5多糖を200mlの脱イオン水に再懸濁させた。しかしながら、過酸化水素で処理する代わりに、この溶液を60℃に加熱し、そのpHを7に調整した。次いで100μlのアルカラーゼを溶液に添加し、混合物を60℃で5時間インキュベートした。
【0124】
次にこの混合物を実施例2に記載したようにメタノール沈殿させた。
【0125】
83%の力価をもつ3.62gのK5多糖が収率87%で得られた。これに比べて、過酸化水素処理では力価76.9%および収率83%であった。
【0126】
従って、除タンパク処理では、3回目の過酸化水素処理に比べて収率の改善および純度の向上が得られる。
【0127】
実施例1に記載したような発酵が終了したとき、ペレット減量はCOL5K002で60%、COL5K003で40%であった。実施例6および7に記載したようなこのペレットの再処理プロセスを開発したので、K5多糖の総抽出効率をCOL5K002では37%から78%に向上させ、COL5K003では43%から68%に向上させることが可能になったのである。
【0128】
実施例8に記載したような最後の除タンパク工程は、定量的収率(98%)でK5多糖の純度を89%に向上させ得る。
【図面の簡単な説明】
【0129】
【図1】COL5K000からCOL5K006までの処理で得られた多糖の量。黒い棒グラフは最終可能な多糖の推定量(単位g)を表す;斑点入り棒グラフは上清中の多糖の最終濃度を表す(×20、単位g/l);三角印は培養物が示したOD−600nmを表す。
【図2】COL5K006の主要処理パラメーターのプロフィル。曲線1は培地中の溶存酸素のパーセンテージを表す;曲線2は培養物COLK006中のCO2の発生速度を表す(単位nmol.1−1時−1);曲線3はフィード量/20をg.l−1で表す;曲線4は上清中のK5多糖の濃度(単位g.l−1)を表す。
【特許請求の範囲】
【請求項1】
a)発酵槽で行うことができ、
−バッチ増殖段階と、
−1回または数回のフェッドバッチ増殖段階と、
−温度を下げ、培地のpHを9までの値に上げる冷却段階と、
−バイオマスを培養培地から分離する段階と、
−バイオマスから混入培養培地を除去する段階と、
を含む、K5多糖を産生する大腸菌株の発酵工程と、
b)培養培地および/またはバイオマスを使用する精製工程と、
を含むK5多糖の製造方法。
【請求項2】
培養培地および/またはバイオマスを出発材料とする精製工程が、
−約7−約11の値へのpHの調整、
−第一回目の沈殿、
−沈殿物の溶解、
−得られた懸濁液の濾過、および、
−アルコール沈殿、
を含む請求項1に記載のK5多糖の製造方法。
【請求項3】
バッチ増殖段階後に、指数増加量の基質フィードを伴う2回のバッチ増殖段階を含む請求項1または2に記載の方法。
【請求項4】
冷却段階の前に、一定量の基質フィードを伴うバッチ増殖段階を含む請求項1から3のいずれか一項に記載の方法。
【請求項5】
バッチ増殖段階で培地中の炭素源が消耗したときに一回目のフェッドバッチ増殖段階を開始することを特徴とする請求項1から4のいずれか一項に記載の方法。
【請求項6】
−バッチ増殖段階で培地中の炭素源が消耗したときに一回目のフェッドバッチ増殖段階を開始すること、
−必要ならば、酸素移動の極限の結果として溶存酸素の濃度が有意に低下したときに、その後のフェッドバッチ増殖段階を開始すること、
を特徴とする請求項1から4のいずれか一項に記載の方法。
【請求項7】
−バッチ増殖段階中の培地中の炭素源が消耗したときに一回目のフェッドバッチ増殖段階を開始すること、
−必要ならば、酸素移動の極限の結果として溶存酸素の濃度が有意に低下したときに、その後のフェッドバッチ増殖段階を開始すること、
−全部の培養フィード培地が発酵槽に注入されたかまたはpHの変化が検出されたときに、冷却段階を開始すること、
を特徴とする請求項1から4のいずれか一項に記載の方法。
【請求項8】
炭素源がグルコースまたはグリセロールであることを特徴とする請求項1から7のいずれか一項に記載の方法。
【請求項9】
炭素源がグリセロールであることを特徴とする請求項1から7のいずれか一項に記載の方法。
【請求項10】
炭素源がグリセロールであり、その濃度がバッチ増殖段階の出発培養培地中で約10−約90g/lの範囲であり、フェッドバッチ増殖段階のフィード培地中では約250−約1200g/lの範囲であることを特徴とする請求項1から7のいずれか一項に記載の方法。
【請求項11】
バッチ増殖段階の培地の窒素源が酵母エキス、カザミノ酸、ペプトン、NH4OH、NH3または(NH4)2HPO4から成ることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項12】
バッチ増殖段階の出発培養培地中の窒素源がNH4OH、NH3または(NH4)2HPO4であることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項13】
バッチ増殖段階の出発培養培地中の窒素源の濃度が約1−約10g/lの範囲であることを特徴とする請求項12に記載の方法。
【請求項14】
バッチ増殖段階およびフェッドバッチ増殖段階のpHを調整するために、窒素源がNH4OHまたはNH3によって供給されることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項15】
バッチ増殖段階およびフェッドバッチ増殖段階のpHを調整するために、窒素源が約20%濃度のNH4OHまたはNH3によって供給されることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項16】
バッチ増殖段階の培地の窒素源が、出発培養培地では約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4であり、バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給されることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項17】
増殖温度が約10℃−約40℃の範囲であることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項18】
増殖温度は、バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、必要ならば行う他のフェッドバッチ増殖段階では約25℃であることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項19】
−炭素源がグリセロールであり、グリセロールの濃度は、
−バッチ増殖段階では出発培養培地中に約10−約90g/lの範囲であり、
−フェッドバッチ増殖段階ではフィード培地中に約250−約1200g/lの範囲であり、
−増殖温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、
−必要ならば行う他のフェッドバッチ増殖段階では約25℃である
ことを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項20】
−炭素源がグリセロールであり、グリセロールの濃度が、
−バッチ増殖段階では出発培養培地中に約10−約90g/lの範囲であり、
−フェッドバッチ増殖段階ではフィード培地中に約250−約1200g/lの範囲であり、
−バッチ増殖段階の培地の窒素源が
−出発培養培地では約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4であり、
−バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給される
ことを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項21】
−バッチ増殖段階の培地の窒素源が、
−出発培養培地では約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4であり、
−バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給され、
−増殖温度が、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、
−必要ならば行う他のフェッドバッチ増殖段階では約25℃である、
ことを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項22】
発酵工程が、以下の段階、すなわち、
−バッチ増殖段階、
−先行段階の培地中の炭素源が完全消耗したときに開始する指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する二回目の指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する一定量の基質フィードを伴うバッチ増殖サイクル、
−温度を約12℃に下げpHを約7.5−約9の範囲の値に上げる冷却段階、
−バイオマスから混入培養培地を除去する段階、
を記載の順序で含むこと、および、
前記工程を以下のパラメーターで行うこと、すなわち、
−温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階中は約30℃、
−他の2回のフェッドバッチ増殖段階中は約25℃
であり、
−窒素源は、
−出発培養培地には約1−約10g/lの範囲の濃度の(NH4)2HPO4、NH4OHまたはNH3によって供給され、
−バッチ増殖段階およびフェッドバッチ増殖段階にはpHを調整するために約20%の濃縮NH4OHまたはNH3の添加によって供給され、
−炭素源はグリセロールであり、その濃度は、
−最初のバッチ増殖段階の出発培養培地中で約10−約90g/lの範囲の濃度であり、
−フェッドバッチ増殖段階のフィード培地中で約250−約1200g/lの範囲の濃度である、
ことを特徴とする請求項1または2に記載の方法。
【請求項23】
バッチ増殖段階、フェッドバッチ増殖段階および冷却段階の後で遠心によってバイオマスを培養培地から分離することを特徴とする請求項1から22のいずれか一項に記載の方法。
【請求項24】
分離後に、バイオマスと培養培地とを約80℃の温度で約1時間−約3時間の範囲の期間維持することによって失活させることを特徴とする請求項1から23のいずれか一項に記載の方法。
【請求項25】
精製工程の一回目の沈殿は第四級アンモニウム塩を用いて行うことを特徴とする請求項2に記載の方法。
【請求項26】
第四級アンモニウム塩が塩化ベンゼトニウムである請求項25に記載の方法。
【請求項27】
精製工程の最初の沈殿物を、特に10%濃度で使用する酢酸ナトリウムに再溶解させることを特徴とする請求項2に記載の方法。
【請求項28】
精製工程で生じた濾液をメタノール沈殿させることを特徴とする請求項2に記載の方法。
【請求項29】
精製工程の一回目の沈殿が塩化ベンゼトニウムを用いて行われること、この沈殿物を酢酸ナトリウムに再溶解させ、濾別し、精製工程で生じた濾液をメタノール沈殿させることを特徴とする請求項2に記載の方法。
【請求項30】
精製後に得られた生成物を過酸化水素によって1回または数回脱色することを特徴とする請求項1から29のいずれか一項に記載の方法。
【請求項31】
精製後に得られた生成物を過酸化水素によって3回脱色することを特徴とする請求項30に記載の方法。
【請求項32】
精製し過酸化水素で3回脱色した後に得られた生成物に過酸化水素による脱色処理をもう一度行うことを特徴とする請求項1から31のいずれか一項に記載の方法。
【請求項33】
プロテアーゼで処理する工程を含むことを特徴とする請求項1から31のいずれか一項に記載の方法。
【請求項34】
使用されるプロテアーゼがアルカラーゼであることを特徴とする請求項33に記載の方法。
【請求項35】
精製工程が以下の段階:
−バイオマスから培養培地を分離するために培養物を遠心する段階;
−発酵ブロスの遠心上清を使用して
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−同時に、発酵ブロスの遠心によって得られたペレットを使用して
−ペレットを再懸濁化し、
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに再溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−各々の精製で得られた2つの調製物を合せる段階;
−K5多糖の沈殿、遠心および乾燥を行う段階;
を含むことを特徴とする請求項1から24のいずれか一項に記載の方法。
【請求項36】
発酵工程が、以下の段階:
−バッチ増殖段階、
−先行段階の培地中の炭素源が完全消耗したときに開始する指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する二回目の指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する一定量の基質フィードを伴うバッチ増殖サイクル、
−温度を約12℃に下げpHを約7.5−約9の範囲の値に上げる冷却段階、
−バイオマスから混入培養培地を除去する段階、
を記載の順序で含むこと、および、
前記工程を以下のパラメーターで行うこと:
−温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階中は約30℃、
−他の2回のフェッドバッチ増殖段階中は約25℃
であり、
−窒素源は、
−出発培養培地には約1−約10g/lの範囲の濃度の(NH4)2HPO4、NH4OHまたはNH3によって供給され、
−バッチ増殖段階およびフェッドバッチ増殖段階にはpHを調整するために約20%の濃縮NH4OHまたはNH3の添加によって供給され、
−炭素源はグリセロールであり、その濃度は、
−最初のバッチ増殖段階の出発培養培地中で約10−約90g/lの範囲の濃度であり、
−フェッドバッチ増殖段階のフィード培地中で約250−約1200g/lの範囲の濃度であること、および、
精製工程が以下の段階:
−バイオマスから培養培地を分離するために培養物を遠心する段階;
−発酵ブロスの遠心上清を使用して
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−同時に、発酵ブロスの遠心によって得られたペレットを使用して
−ペレットを再懸濁化し、
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに再溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−各々の精製で得られた2つの調製物を合せる段階;
−K5多糖の沈殿、遠心および乾燥を行う段階;
を含むことを特徴とする請求項1または2に記載の方法。
【請求項37】
請求項1から36のいずれか一項に記載の方法によって産生された多糖が酵素的および/または化学的な脱アセチル化、N−再硫酸化、C−5エピマー化および/またはO−硫酸化の反応基質として使用されることを特徴とする方法。
【請求項38】
請求項1から36のいずれか一項に記載の方法によって産生された多糖が半合成ヘパリン(バイオヘパリン)を得るための基質として使用されることを特徴とする請求項37に記載の方法。
【請求項39】
得られたバイオヘパリンを次に約1500−約6500Daの範囲の分子量をもつLMWH(低分子量ヘパリン)を得るためのフラグメンテーション反応で基質として使用することを特徴とする請求項38に記載の方法。
【請求項40】
LMWHが、より特定的に約3500−約5500Daの範囲の分子量をもつ低分子量ヘパリンであるか、または、より特定的に約1500−約3000Daの範囲の分子量をもつ超低分子量ヘパリンであることを特徴とする請求項39に記載の方法。
【請求項1】
a)発酵槽で行うことができ、
−バッチ増殖段階と、
−1回または数回のフェッドバッチ増殖段階と、
−温度を下げ、培地のpHを9までの値に上げる冷却段階と、
−バイオマスを培養培地から分離する段階と、
−バイオマスから混入培養培地を除去する段階と、
を含む、K5多糖を産生する大腸菌株の発酵工程と、
b)培養培地および/またはバイオマスを使用する精製工程と、
を含むK5多糖の製造方法。
【請求項2】
培養培地および/またはバイオマスを出発材料とする精製工程が、
−約7−約11の値へのpHの調整、
−第一回目の沈殿、
−沈殿物の溶解、
−得られた懸濁液の濾過、および、
−アルコール沈殿、
を含む請求項1に記載のK5多糖の製造方法。
【請求項3】
バッチ増殖段階後に、指数増加量の基質フィードを伴う2回のバッチ増殖段階を含む請求項1または2に記載の方法。
【請求項4】
冷却段階の前に、一定量の基質フィードを伴うバッチ増殖段階を含む請求項1から3のいずれか一項に記載の方法。
【請求項5】
バッチ増殖段階で培地中の炭素源が消耗したときに一回目のフェッドバッチ増殖段階を開始することを特徴とする請求項1から4のいずれか一項に記載の方法。
【請求項6】
−バッチ増殖段階で培地中の炭素源が消耗したときに一回目のフェッドバッチ増殖段階を開始すること、
−必要ならば、酸素移動の極限の結果として溶存酸素の濃度が有意に低下したときに、その後のフェッドバッチ増殖段階を開始すること、
を特徴とする請求項1から4のいずれか一項に記載の方法。
【請求項7】
−バッチ増殖段階中の培地中の炭素源が消耗したときに一回目のフェッドバッチ増殖段階を開始すること、
−必要ならば、酸素移動の極限の結果として溶存酸素の濃度が有意に低下したときに、その後のフェッドバッチ増殖段階を開始すること、
−全部の培養フィード培地が発酵槽に注入されたかまたはpHの変化が検出されたときに、冷却段階を開始すること、
を特徴とする請求項1から4のいずれか一項に記載の方法。
【請求項8】
炭素源がグルコースまたはグリセロールであることを特徴とする請求項1から7のいずれか一項に記載の方法。
【請求項9】
炭素源がグリセロールであることを特徴とする請求項1から7のいずれか一項に記載の方法。
【請求項10】
炭素源がグリセロールであり、その濃度がバッチ増殖段階の出発培養培地中で約10−約90g/lの範囲であり、フェッドバッチ増殖段階のフィード培地中では約250−約1200g/lの範囲であることを特徴とする請求項1から7のいずれか一項に記載の方法。
【請求項11】
バッチ増殖段階の培地の窒素源が酵母エキス、カザミノ酸、ペプトン、NH4OH、NH3または(NH4)2HPO4から成ることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項12】
バッチ増殖段階の出発培養培地中の窒素源がNH4OH、NH3または(NH4)2HPO4であることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項13】
バッチ増殖段階の出発培養培地中の窒素源の濃度が約1−約10g/lの範囲であることを特徴とする請求項12に記載の方法。
【請求項14】
バッチ増殖段階およびフェッドバッチ増殖段階のpHを調整するために、窒素源がNH4OHまたはNH3によって供給されることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項15】
バッチ増殖段階およびフェッドバッチ増殖段階のpHを調整するために、窒素源が約20%濃度のNH4OHまたはNH3によって供給されることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項16】
バッチ増殖段階の培地の窒素源が、出発培養培地では約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4であり、バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給されることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項17】
増殖温度が約10℃−約40℃の範囲であることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項18】
増殖温度は、バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、必要ならば行う他のフェッドバッチ増殖段階では約25℃であることを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項19】
−炭素源がグリセロールであり、グリセロールの濃度は、
−バッチ増殖段階では出発培養培地中に約10−約90g/lの範囲であり、
−フェッドバッチ増殖段階ではフィード培地中に約250−約1200g/lの範囲であり、
−増殖温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、
−必要ならば行う他のフェッドバッチ増殖段階では約25℃である
ことを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項20】
−炭素源がグリセロールであり、グリセロールの濃度が、
−バッチ増殖段階では出発培養培地中に約10−約90g/lの範囲であり、
−フェッドバッチ増殖段階ではフィード培地中に約250−約1200g/lの範囲であり、
−バッチ増殖段階の培地の窒素源が
−出発培養培地では約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4であり、
−バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給される
ことを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項21】
−バッチ増殖段階の培地の窒素源が、
−出発培養培地では約1−約10g/lの範囲の濃度のNH4OH、NH3または(NH4)2HPO4であり、
−バッチ増殖段階およびフェッドバッチ増殖段階ではpHを調整するために約20%濃度のNH4OHまたはNH3によって供給され、
−増殖温度が、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階では約30℃であり、
−必要ならば行う他のフェッドバッチ増殖段階では約25℃である、
ことを特徴とする請求項1から10のいずれか一項に記載の方法。
【請求項22】
発酵工程が、以下の段階、すなわち、
−バッチ増殖段階、
−先行段階の培地中の炭素源が完全消耗したときに開始する指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する二回目の指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する一定量の基質フィードを伴うバッチ増殖サイクル、
−温度を約12℃に下げpHを約7.5−約9の範囲の値に上げる冷却段階、
−バイオマスから混入培養培地を除去する段階、
を記載の順序で含むこと、および、
前記工程を以下のパラメーターで行うこと、すなわち、
−温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階中は約30℃、
−他の2回のフェッドバッチ増殖段階中は約25℃
であり、
−窒素源は、
−出発培養培地には約1−約10g/lの範囲の濃度の(NH4)2HPO4、NH4OHまたはNH3によって供給され、
−バッチ増殖段階およびフェッドバッチ増殖段階にはpHを調整するために約20%の濃縮NH4OHまたはNH3の添加によって供給され、
−炭素源はグリセロールであり、その濃度は、
−最初のバッチ増殖段階の出発培養培地中で約10−約90g/lの範囲の濃度であり、
−フェッドバッチ増殖段階のフィード培地中で約250−約1200g/lの範囲の濃度である、
ことを特徴とする請求項1または2に記載の方法。
【請求項23】
バッチ増殖段階、フェッドバッチ増殖段階および冷却段階の後で遠心によってバイオマスを培養培地から分離することを特徴とする請求項1から22のいずれか一項に記載の方法。
【請求項24】
分離後に、バイオマスと培養培地とを約80℃の温度で約1時間−約3時間の範囲の期間維持することによって失活させることを特徴とする請求項1から23のいずれか一項に記載の方法。
【請求項25】
精製工程の一回目の沈殿は第四級アンモニウム塩を用いて行うことを特徴とする請求項2に記載の方法。
【請求項26】
第四級アンモニウム塩が塩化ベンゼトニウムである請求項25に記載の方法。
【請求項27】
精製工程の最初の沈殿物を、特に10%濃度で使用する酢酸ナトリウムに再溶解させることを特徴とする請求項2に記載の方法。
【請求項28】
精製工程で生じた濾液をメタノール沈殿させることを特徴とする請求項2に記載の方法。
【請求項29】
精製工程の一回目の沈殿が塩化ベンゼトニウムを用いて行われること、この沈殿物を酢酸ナトリウムに再溶解させ、濾別し、精製工程で生じた濾液をメタノール沈殿させることを特徴とする請求項2に記載の方法。
【請求項30】
精製後に得られた生成物を過酸化水素によって1回または数回脱色することを特徴とする請求項1から29のいずれか一項に記載の方法。
【請求項31】
精製後に得られた生成物を過酸化水素によって3回脱色することを特徴とする請求項30に記載の方法。
【請求項32】
精製し過酸化水素で3回脱色した後に得られた生成物に過酸化水素による脱色処理をもう一度行うことを特徴とする請求項1から31のいずれか一項に記載の方法。
【請求項33】
プロテアーゼで処理する工程を含むことを特徴とする請求項1から31のいずれか一項に記載の方法。
【請求項34】
使用されるプロテアーゼがアルカラーゼであることを特徴とする請求項33に記載の方法。
【請求項35】
精製工程が以下の段階:
−バイオマスから培養培地を分離するために培養物を遠心する段階;
−発酵ブロスの遠心上清を使用して
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−同時に、発酵ブロスの遠心によって得られたペレットを使用して
−ペレットを再懸濁化し、
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに再溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−各々の精製で得られた2つの調製物を合せる段階;
−K5多糖の沈殿、遠心および乾燥を行う段階;
を含むことを特徴とする請求項1から24のいずれか一項に記載の方法。
【請求項36】
発酵工程が、以下の段階:
−バッチ増殖段階、
−先行段階の培地中の炭素源が完全消耗したときに開始する指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する二回目の指数増加量の基質フィードを伴うバッチ増殖段階、
−酸素移動の極限の結果として先行段階の培養培地中の酸素濃度が有意に低下したときに開始する一定量の基質フィードを伴うバッチ増殖サイクル、
−温度を約12℃に下げpHを約7.5−約9の範囲の値に上げる冷却段階、
−バイオマスから混入培養培地を除去する段階、
を記載の順序で含むこと、および、
前記工程を以下のパラメーターで行うこと:
−温度は、
−バッチ増殖段階および一回目のフェッドバッチ増殖段階中は約30℃、
−他の2回のフェッドバッチ増殖段階中は約25℃
であり、
−窒素源は、
−出発培養培地には約1−約10g/lの範囲の濃度の(NH4)2HPO4、NH4OHまたはNH3によって供給され、
−バッチ増殖段階およびフェッドバッチ増殖段階にはpHを調整するために約20%の濃縮NH4OHまたはNH3の添加によって供給され、
−炭素源はグリセロールであり、その濃度は、
−最初のバッチ増殖段階の出発培養培地中で約10−約90g/lの範囲の濃度であり、
−フェッドバッチ増殖段階のフィード培地中で約250−約1200g/lの範囲の濃度であること、および、
精製工程が以下の段階:
−バイオマスから培養培地を分離するために培養物を遠心する段階;
−発酵ブロスの遠心上清を使用して
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−同時に、発酵ブロスの遠心によって得られたペレットを使用して
−ペレットを再懸濁化し、
−pHを約7−約11の値に調整し、
−塩化ベンゼトニウムで沈殿させ、
−沈殿物を洗浄し、
−沈殿物を10%酢酸ナトリウムに再溶解し、
−10%酢酸ナトリウム溶液を濾過し、
−濾液をメタノール沈殿させ、
−濾液を過酸化水素によって3回脱色し、
−アルカラーゼ処理する段階;
−各々の精製で得られた2つの調製物を合せる段階;
−K5多糖の沈殿、遠心および乾燥を行う段階;
を含むことを特徴とする請求項1または2に記載の方法。
【請求項37】
請求項1から36のいずれか一項に記載の方法によって産生された多糖が酵素的および/または化学的な脱アセチル化、N−再硫酸化、C−5エピマー化および/またはO−硫酸化の反応基質として使用されることを特徴とする方法。
【請求項38】
請求項1から36のいずれか一項に記載の方法によって産生された多糖が半合成ヘパリン(バイオヘパリン)を得るための基質として使用されることを特徴とする請求項37に記載の方法。
【請求項39】
得られたバイオヘパリンを次に約1500−約6500Daの範囲の分子量をもつLMWH(低分子量ヘパリン)を得るためのフラグメンテーション反応で基質として使用することを特徴とする請求項38に記載の方法。
【請求項40】
LMWHが、より特定的に約3500−約5500Daの範囲の分子量をもつ低分子量ヘパリンであるか、または、より特定的に約1500−約3000Daの範囲の分子量をもつ超低分子量ヘパリンであることを特徴とする請求項39に記載の方法。
【図1】

【図2】
【図2】
【公表番号】特表2008−512105(P2008−512105A)
【公表日】平成20年4月24日(2008.4.24)
【国際特許分類】
【出願番号】特願2007−530737(P2007−530737)
【出願日】平成17年9月6日(2005.9.6)
【国際出願番号】PCT/FR2005/002210
【国際公開番号】WO2006/030099
【国際公開日】平成18年3月23日(2006.3.23)
【出願人】(500152119)アバンテイス・フアルマ・エス・アー (65)
【Fターム(参考)】
【公表日】平成20年4月24日(2008.4.24)
【国際特許分類】
【出願日】平成17年9月6日(2005.9.6)
【国際出願番号】PCT/FR2005/002210
【国際公開番号】WO2006/030099
【国際公開日】平成18年3月23日(2006.3.23)
【出願人】(500152119)アバンテイス・フアルマ・エス・アー (65)
【Fターム(参考)】
[ Back to top ]