
KDRペプチド及びこれを含むワクチン
【課題】腫瘍組織内皮細胞に於いて強発現しているVGEF受容体2(KDR)を標的とした癌ワクチン療法の開発を目的に、ワクチンとして有効なペプチドを提供する。
【解決手段】KDRタンパク質の全アミノ酸配列を基に、HLA-A*2402に対し結合親和性の高いノナペプチド、及びデカペプチド。化学合成したこれらのペプチド及びアミノ酸の置換又は付加ペプチドより、高い細胞傷害性T細胞(CLT)誘導能を有するペプチド。更にCLT誘導能の高い抗原提示細胞を誘導する方法、これらペプチドによる腫瘍の治療、予防、及びワクチン。
【解決手段】KDRタンパク質の全アミノ酸配列を基に、HLA-A*2402に対し結合親和性の高いノナペプチド、及びデカペプチド。化学合成したこれらのペプチド及びアミノ酸の置換又は付加ペプチドより、高い細胞傷害性T細胞(CLT)誘導能を有するペプチド。更にCLT誘導能の高い抗原提示細胞を誘導する方法、これらペプチドによる腫瘍の治療、予防、及びワクチン。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、癌ワクチンとして非常に有効な新規ペプチド、及び該ペプチドを含む腫瘍の治療及び予防のための医薬に関する。
【背景技術】
【0002】
腫瘍拒絶抗原遺伝子が悪性黒色腫を中心に同定されるに従って、それを応用した癌免疫療法が開発されている。とりわけ、抗腫瘍免疫応答におけるCD8陽性T細胞の重要性が認識されるに伴い、腫瘍特異的CD8陽性T細胞を生体内で惹起させる癌ワクチン療法が着目され、種々の臨床応用が行われている。また、10個前後のアミノ酸残基からなるペプチドが種々の共刺激分子の補助によりClassI経路を介してT細胞を活性化し、腫瘍特異的細胞傷害性T細胞(CTL)を誘導する機序が解明され、さらに個々のHLA分子に拘束性を示すペプチドの同定も精力的に進んでいる。
【0003】
しかしながら、腫瘍を制御する事は未だ完全に出来ていないのが現状である。その原因として、腫瘍細胞の不均一性、腫瘍細胞におけるMHC-Class Iの発現低下や消失、さらに腫瘍細胞における標的分子の欠落が考えられる。また、現在同定されている腫瘍抗原ペプチドはある種の腫瘍に存在するが、全ての腫瘍を網羅できるものではない。そこで、これらの問題点を解決するため、標的細胞を腫瘍細胞ではなく、腫瘍血管における内皮細胞に着目した。すなわち、内皮細胞はMHC-Class Iの発現低下や消失、また不均一性といった問題がほとんどないので、腫瘍血管を標的にするCTLが誘導できれば、腫瘍の種類に関係なく、またClass Iの消失や標的分子の欠損といった従来の癌ワクチン療法で問題となっていた原因を凌駕でき、治療効果が大いに期待できると考えられる。癌における新生血管の研究は1970年代のFolkmanらの先駆的な仮説の提唱に始まり、様々な角度から研究がなされている。なかでも、VEGF-VEGF受容体(VEGFR)において多くの検討がなされ、腫瘍新生血管におけるその意義が評価されている。特に癌治療においては、target-oriented drugとして新生血管阻害剤の開発が精力的に進められ、すでに臨床治験も行われているが、この考え方を癌ワクチン療法に生かした治療は未だ行われていない。一つの理由としてVEGFRが正常細胞に発現しているため免疫寛容状態になっていると考えられた。しかしながら、1990年代のPlate, Millauer, Risauらの研究により腫瘍組織における内皮細胞においてVEGFRが強発現している事が確認され、また、CEA,HER/neuのように正常細胞でも発現している自己抗原に対する免疫応答が必ずしも免疫寛容状態になっていないことが明らかになった事により、VEGFR が癌ワクチン療法の標的に成り得るのではないかと考えた。
【0004】
最近、VEGFRに対する能動免疫を行うことにより、腫瘍における血管新生及び転移を抑制できることが報告された(非特許文献1;The Journal of experimental medicine 2002年 第195巻 第12号 p.1575-1584)。しかし、この文献では可溶性のVEGFRタンパク質を用いているに過ぎず、有効なペプチドのアミノ酸配列についての検討は何らなされていない。
【非特許文献1】The Journal of experimental medicine 2002年 第195巻 第12号 p.1575-1584
【発明の開示】
【0005】
本発明者等は、VEGFシグナルにおいて内皮細胞の増殖に関与すると考えられ、腫瘍組織内皮細胞において強発現しているVEGFR2 (KDR/flk-1、以下KDRという)を標的とした癌ワクチン療法の可能性に着目し、ワクチンとして有効に用いることができるペプチドの探索を行い、その特異性について検討を行い、本発明を完成するに到った。
【0006】
すなわち、本発明は、以下の(1)〜(22)を提供する。
(1) 配列番号2、3、5、8、11または12に示すアミノ酸配列からなるペプチドから選ばれるノナペプチド。
(2) 配列番号2、3、5、8、11または12に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチド。
(3) N末端から2番目のアミノ酸がフェニルアラニン、チロシン、メチオニンまたはトリプトファンである、上記(2)に記載のペプチド。
(4) C末端のアミノ酸がフェニルアラニン、ロイシン、イソロイシン、トリプトファンまたはメチオニンである、上記(2)または(3)に記載のペプチド。
(5) 配列番号29、30、33、34、40、または46に示すアミノ酸配列からなるペプチドから選ばれるノナペプチドまたはデカペプチド。
(6) 配列番号29、30、33、34、40、または46に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチド。
(7) N末端から2番目のアミノ酸がロイシンまたはメチオニンである、上記(6)に記載のペプチド。
(8) C末端のアミノ酸がバリンまたはロイシンである、上記(6)または(7)に記載のペプチド。
(9) 上記(1)〜(8)のいずれかに記載のペプチドを1種以上含む、腫瘍の治療及び/または予防のための医薬。
(10) 上記(1)〜(8)のいずれかに記載のペプチドを1種以上含む、糖尿病性網膜症、慢性関節リウマチ、乾癬、アテローム性動脈硬化症の治療のための医薬。
(11) 上記(1)〜(8)のいずれかに記載のペプチドとHLA抗原とを含む複合体を表面に提示しているエキソソーム。
(12) HLA抗原がHLA−A24またはHLA−A02である、上記(11)に記載のエキソソーム。
(13) HLA抗原がHLA−A2402またはHLA−0201である、上記(12)に記載のエキソソーム。
(14) 上記(1)〜(8)のいずれかに記載のペプチドを用いてCTL誘導能の高い抗原提示細胞を誘導する方法。
(15) 上記(1)〜(8)のいずれかに記載のペプチドを用いてCTLを誘導する方法。
(16) 上記(1)〜(8)のいずれかに記載のペプチドをコードするポリヌクレオチドを含む遺伝子を抗原提示細胞に導入することを含む、細胞傷害性T細胞誘導能の高い抗原提示細胞を誘導する方法。
(17) 上記(1)〜(8)のいずれかに記載のペプチドを用いて誘導される、単離された細胞傷害性T細胞。
(18) HLA抗原と上記(1)〜(8)のいずれかに記載のペプチドとの複合体を提示してなる抗原提示細胞。
(19) 上記(14)または(15)に記載の方法によって誘導される、上記(18)に記載の抗原提示細胞。
(20) 上記(1)〜(8)のいずれかに記載のペプチドを有効成分とする、病態部位における血管新生阻害のためのワクチン。
(21) HLA抗原がHLA−A24またはHLA−A02である被験者に対して投与するための、上記(20)に記載のワクチン。
(22) 悪性腫瘍の増殖及び/または転移を抑制するために使用される、上記(20)または(21)に記載のワクチン。
本明細書は本願の優先権の基礎である日本国特許出願2002-267285、2003-062003、2003-167042号の明細書および/または図面に記載される内容を包含する。
【0007】
[発明の実施の形態]
本発明者等はまず、in vivoにおいて抗原提示細胞上に提示される場合に、種々のタンパク質が9量体のペプチド(ノナペプチド)に分解されてから提示されることを考慮し、KDRタンパク質の9量体または10量体の部分ペプチドについて、ヒト主要組織適合抗原(MHC抗原)であるHLA抗原との結合親和性を検討した。尚、表1の右側のペプチドにおいて、小文字は5番目のアミノ酸を示す。
ヒトKDRタンパク質のアミノ酸配列は公知であり、例えば米国特許第5,861,301号に記載されており、当業者であれば容易に入手することができる。9量体及び10量体ペプチドは、得られたKDRタンパク質の全アミノ酸配列に基づいて、任意の位置からのペプチドを合成して得ることができる。ペプチドの合成は、通常のペプチド化学において用いられる方法に準じて行うことができる。通常用いられる合成方法は、例えば、Peptide Synthesis, Interscience, New York, 1966; The Proteins, Vol 2, Academic Press Inc., New York, 1976; ペプチド合成、丸善(株)、1975; ペプチド合成の基礎と実験、丸善(株)、1985; 医薬品の開発 続 第14巻・ペプチド合成、広川書店、1991等の文献や、国際公開WO99/67288号等の公報に記載されている。HLA抗原との結合は、細胞表面にHLA抗原を有する細胞、例えば樹状細胞を単離して、細胞へのペプチドの結合を通常行われる手法を用いて測定することができる。
【0008】
あるいはまた、最近インターネット上で利用可能となっているソフトウェア、例えばParker K. C., J. Immunol. 152, 1994に記載されているもの等を用いて、種々のペプチドとHLA抗原との結合親和性をin silicoで計算することもできる。尚、HLA抗原との結合親和性は、例えばParker, K.C., J. Immunol., 152, 1994; Nukaya, I., Int. J. Cancer, 80, 1999等に記載のように測定することができる。
【0009】
HLA抗原としては、例えば日本人において多く発現しているといわれるA−24型またはA−02型等を用いることが有効な結果を得るために好ましく、更に好ましくはA−2402、A−0201等のサブタイプである。しかしながら、臨床においては、治療を必要とする患者のHLA抗原の型を予め調べることにより、これとの結合親和性、あるいは抗原提示による細胞傷害性T細胞(CTL)誘導能の高いペプチドを適宜選択することができる。更に、結合親和性及びCTL誘導能の高いペプチドを得るために、天然に存在するKDR部分ペプチドのアミノ酸配列に基づいて1個、2個または数個のアミノ酸の置換または付加を行うこともできる。ここで、「数個」とは、5個以下、好ましくは3個以下を意味する。更に、天然において提示されるペプチド以外にも、既にHLA抗原に結合して提示されるペプチドの配列の規則性が知られているので(J. Immunol., 152, 3913, 1994; Immunogenetics. 41:178, 1995; J. Immunol. 155:4307, 1994)、得られたペプチドに対してこれらの規則性に基づいた改変を行っても良い。例えば、HLA−24結合親和性の高いものはペプチドのN末端から2番目のアミノ酸をフェニルアラニン、チロシン、メチオニンまたはトリプトファンに置換したり、C末端のアミノ酸をフェニルアラニン、ロイシン、イソロイシン、トリプトファンまたはメチオニンに置換したペプチドも好適に使用することができる。一方、HLA−0201結合親和性の高いものは、ペプチドのN末端から2番目のアミノ酸をロイシンまたはメチオニンに置換したり、C末端のアミノ酸をバリンまたはロイシンに置換したペプチドも好適に使用することができる。更に、ペプチドのN末端及び/またはC末端に1〜2個のアミノ酸が付加していても良い。
【0010】
しかしながら、ペプチドの配列が他の機能を有する内在性または外来性のタンパク質のアミノ酸配列の一部と同一となる場合には、自己免疫疾患等の副作用が生じたり、あるいは特定の物質に対するアレルギー症状を引き起こしたりする可能性があるため、利用可能なデータベースを用いてホモロジー検索を行い、他のタンパク質のアミノ酸配列と一致することを避けるのが好ましい。更に、ホモロジー検索において、アミノ酸が1個または2個異なるペプチドも存在しないことが明らかであれば、HLA抗原との結合親和性及び/またはCTL誘導能を高めるための上記アミノ酸配列の改変もこのような問題を生じるおそれがない。
【0011】
上記のようにしてHLA抗原との結合親和性の高いペプチドは、癌ワクチンとして有効である可能性が高いことが予想されるが、高い結合親和性を有することを指標として選択した候補ペプチドについて、実際にCTL誘導能を有するか否かを検討することが必要である。CTL誘導能の確認は、ヒトMHC抗原を有する抗原提示細胞(例えばB−リンパ球、マクロファージ、樹状細胞)、具体的にはヒト末梢血単核球由来の樹状細胞を誘導し、ペプチドで刺激した後にCD8陽性細胞と混合し、標的細胞に対する細胞傷害活性を測定する。反応系として、ヒトHLA抗原を発現するように作製されたトランスジェニック動物(例えば、Hum. Immunol. 2000 Aug.;61(8):764-79 Related Articles, Books, Linkout Induction of CTL response by a minimal epitope vaccine in HLA A*0201/DR1 transgenic mice: dependence on HLA class II restricted T(H) response., BenMohamed L., Krishnan R., Longmate J., Auge C., Low L., Primus J., Diamond DJ.に記載のもの)を用いることもできる。細胞傷害活性は、例えば標的細胞を51Cr等で放射標識し、標的細胞から遊離した放射活性から計算することができる。あるいはまた、ペプチドを拘束した抗原提示細胞の存在下でCTLが産生・放出したIFN-γ及び抗IFN-γモノクローナル抗体によって培地上に可視化される阻止円を測定することによって観察することもできる。
【0012】
上記のようにしてペプチドのCTL誘導能を検討した結果、HLA抗原との結合親和性が高いものが必ずしも誘導能が高いわけではないことが明らかとなった。そして、VYSSEEAEL(配列番号2)、GYRIYDVVL(配列番号3)、SYMISYAGM(配列番号5)、RFVPDGNRI(配列番号8)、KWEFPRDRL(配列番号11)、またはDFLTLEHLI(配列番号12)に示すアミノ酸配列からなるペプチドから選ばれるノナペプチド、及びAMFFWLLLV(配列番号29)、VIAMFFWLL(配列番号30)、AVIAMFFWL(配列番号33)、KLIEIGVQT(配列番号34)、YMISYAGMV(配列番号40)またはIQSDVWSFGV(配列番号46)に示すアミノ酸配列からなるペプチドから選ばれるノナペプチドまたはデカペプチドが特に高いCTL誘導能を有することが明らかとなった。
【0013】
本発明は更に、配列番号2、3、5、8、11または12に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチドも提供する。配列番号2、3、5、8、11または12に示す9個のアミノ酸からなるアミノ酸配列に基づいて、1個、2個または数個のアミノ酸の置換または付加は、他のタンパク質のアミノ酸配列との一致がない限りにおいて、CTL誘導能を有し得る。特に、アミノ酸の置換として、N末端から2番目のアミノ酸のフェニルアラニン、チロシン、メチオニンまたはトリプトファンへの置換、C末端のアミノ酸のフェニルアラニン、ロイシン、イソロイシン、トリプトファンまたはメチオニンへの置換、アミノ酸の付加として、N末端及び/またはC末端への1〜2個のアミノ酸の付加は好適な例である。
【0014】
本発明はまた、配列番号29、30、33、34、40、または46に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチドも提供する。配列番号29、30、33、34、40、または46に示す9個または10個のアミノ酸からなるアミノ酸配列に基づいて、1個、2個または数個のアミノ酸の置換または付加は、他のタンパク質のアミノ酸配列との一致がない限りにおいて、CTL誘導能を有し得る。特に、アミノ酸の置換として、N末端から2番目のアミノ酸のロイシンまたはメチオニンへの置換、C末端のアミノ酸のバリンまたはロイシンへの置換、アミノ酸の付加として、N末端及び/またはC末端への1〜2個のアミノ酸の付加は好適な例である。こうした改変ペプチドの例として、限定するものではないが、例えば配列番号30のペプチドのN末端から2番目のアミノ酸をロイシンに変換したペプチド(配列番号54)が挙げられる。これらの改変ペプチドによる刺激によって得られるCTLクローンは改変前のペプチドを認識し、傷害することができる。
【0015】
上記の本発明のペプチドは、1種または2種以上の組み合わせとして、生体内でCTLを誘導し得る癌ワクチンとして使用することができる。本発明のペプチドの投与により、抗原提示細胞のHLA抗原に該ペプチドが高密度に提示され、提示されたペプチドとHLA抗原との複合体に対して特異的に反応するCTLが誘導され、標的細胞となるべき腫瘍細胞内の血管内皮細胞に対する攻撃力が高まる。あるいは、被験者から樹状細胞を取り出して本発明のペプチドで刺激することにより、細胞表面に本発明のペプチドを拘束した抗原提示細胞が得られ、これを再度被験者に投与することで被験者においてCTLを誘導し、標的細胞に対する攻撃力を高めることができる。
【0016】
すなわち、本発明は、本発明のペプチドを1種以上含む、腫瘍の治療または腫瘍の増殖・転移等の予防のための医薬を提供するものである。更に、病態部位における血管新生は、腫瘍の他、糖尿病性網膜症、慢性関節リウマチ、乾癬、アテローム性動脈硬化症のような疾患、並びに固形癌の転移と深く結びついている(Folkman, J., Nature Med. 1:27-31 (1995); Bicknell, R., Harris, A.L., Curr. Opin. Oncol. 8:60-65 (1996))。従って、本発明のペプチドは、腫瘍、糖尿病性網膜症、慢性関節リウマチ、乾癬、アテローム性動脈硬化症のような疾患、並びに固形癌の転移の治療に用いることができる。
【0017】
本発明のペプチドは、悪性腫瘍組織において形成され、正常な血管とは形態学的に異なる蛇行性血管の形成を阻害することが確かめられたが、ワクチン接種したマウスにおける創傷治癒及び受精能を解析した結果、正常な生理的血管新生に対しては悪影響を有さないことも確認した。更に、本発明のペプチドを認識するCTLクローンを用いて非増殖性または増殖性内皮細胞に対する細胞傷害性をin vitroで試験したところ、これらのクローンは非増殖性内皮細胞よりも増殖性内皮細胞に対して強い活性を示すことも明らかとなった。すなわち、増殖性内皮細胞が観察される疾患、特に癌において非常に特異的に作用することができる。
尚、in vivo及びin vitroにおいて、本発明のペプチドによる樹状細胞の刺激は、細胞に対して高濃度のペプチドを存在させることによって、該細胞に予め拘束されているペプチドとの交換が生じ、容易に行われる。このため、本発明において使用されるペプチドは、HLA抗原との結合親和性がある程度以上高いことが必要とされる。
【0018】
本発明の医薬は、本発明のペプチドを単独で直接投与しても良いが、通常用いられる製剤学的方法によって製剤化した医薬組成物として投与しても良い。その場合、本発明のペプチドの他に、通常医薬に用いられる担体、賦形剤等を適宜含むことができ、特に限定されるものではない。本発明の医薬の用途としては、胃癌、十二指腸癌、大腸癌、肺癌、乳癌、前立腺癌、脳腫瘍等の種々の腫瘍の治療及び予防に用いることができる。本発明のペプチドは、腫瘍細胞そのものではなく、腫瘍組織内に新たに形成される新生血管の内皮細胞を標的とするものであり、従って、治療対象の腫瘍は広範囲にわたり、特に用途を限定するものではない。
【0019】
本発明のペプチドを有効成分とする腫瘍の治療及び/または予防のための医薬は、細胞性免疫が効果的に成立するようにアジュバントと共に投与したり、他の抗癌剤等の有効成分と共に投与したり、また粒子状の剤型にして投与することができる。アジュバントとしては、文献(C1in. Microbio1.Rev., 7:277-289, 1994)に記載のものなどが応用可能である。また、リポソーム製剤、直径数μmのビーズに結合させた粒子状の製剤、リピッドを結合させた製剤なども考えられる。投与方法としては、経口投与、皮内投与、皮下投与、静脈注射などが利用でき、全身投与、あるいは目的となる腫瘍の近傍に局所投与しても良い。本発明のペプチドの投与量は、治療すべき疾患、患者の年齢、体重、投与方法等により適宜調整することができるが、通常 0.001mg〜1000mg、好ましくは 0.001mg〜1OOOmg、より好ましくはO.1mg〜1Omgであり、数日ないし数月に1回投与するのが好ましい。当業者であれば、適当な投与量を適宜選択することが可能である。
【0020】
あるいはまた、本発明は、本発明のペプチドとHLA抗原との複合体を表面に提示している、エキソソームと呼ばれる細胞内小胞を提供する。エキソソームの調製は、例えば特表平11−510507号、及び特表2000−512161号に詳細に記載されている方法を用いて行うことができるが、好ましくは治療及び/または予防の対象となる被験者から得た抗原提示細胞を用いて調製する。本発明のエキソソームは、上記本発明のペプチドと同様に癌ワクチンとして接種することができる。
HLA抗原としては、治療及び/または予防を必要とする被験者のHLA抗原と同じ型のものであることが必要である。例えば、日本人の場合にはHLA−A24またはHLA−A02、特にHLA−A2402またはHLA−0201とすると好適であることが多い。
【0021】
本発明はまた、本発明のペプチドを用いた抗原提示細胞の誘導方法も提供する。樹状細胞を末梢血単球から誘導した後、in vitroまたはin vivoで本発明のペプチドと接触(刺激)させ、抗原提示細胞を誘導することができる。本発明のペプチドを被験者に投与した場合、被験者の体内で本発明のペプチドを拘束した抗原提示細胞が誘導される。あるいは、抗原提示細胞に本発明のペプチドをin vitroで拘束させた後に被験者にワクチンとして投与することもできる。
【0022】
本発明はまた、上記本発明のペプチドをコードするポリヌクレオチドを含む遺伝子をin vitroで抗原提示細胞に導入することを含む、細胞傷害性T細胞誘導能の高い抗原提示細胞を誘導する方法を提供する。導入する遺伝子はDNAの形態であってもRNAの形態であっても良い。導入の方法は当分野において通常行われるリポフェクション、エレクトロポレーション、リン酸カルシウム法等の種々の方法を用いれば良く、特に限定するものではない。具体的には、例えばCancer Res., 56:5672, 1996; J. Immunol., 161:5607, 1998; J. Exp. Med., 184:465, 1996; 特表2000-509281号に記載のようにして行うことができる。遺伝子を抗原提示細胞に導入することによって、該遺伝子は細胞中で転写、翻訳等の処理の後、得られたタンパク質がMHCクラスIまたはクラスIIのプロセッシング及び提示経路を経て、部分ペプチドが提示される。
【0023】
本発明は更に、上記本発明のペプチドを用いてCTLを誘導する方法を提供する。本発明のペプチドを被験者に投与した場合、被験者の体内でCTLが誘導され、腫瘍組織内の新生血管の内皮細胞を標的とした免疫力が増強される。あるいは、被験者由来の抗原提示細胞及びCD8陽性細胞、または末梢血単核球をin vitroで本発明のペプチドと接触(刺激)させ、CTLを誘導してから被験者にもどすex vivoの治療方法にも用いることができる。
【0024】
本発明は更に、本発明のペプチドを用いて誘導される、単離された細胞傷害性T細胞を提供する。本発明のペプチドを提示した抗原提示細胞による刺激に基づいて誘導された細胞傷害性T細胞は、好ましくは治療及び/または予防の対象である被験者由来のものであり、単独、または、本発明のペプチド、エキソソーム等を含む、他の医薬と共に抗腫瘍効果を目的として投与することができる。得られた細胞傷害性T細胞は、本発明のペプチド、好ましくは誘導に用いられたものと同じペプチドを提示する標的細胞に対して特異的に作用する。標的細胞は、KDRを内在的に発現する細胞であっても、強制的にKDRを発現させた細胞であっても良く、また、本発明のペプチドの刺激によって細胞表面に該ペプチドを提示する細胞であっても攻撃の対象となり得る。
【0025】
本発明は更に、HLA抗原と本発明のペプチドとの複合体を提示してなる抗原提示細胞を提供する。本発明のペプチド、あるいは本発明のペプチドをコードするヌクレオチドとの接触によって得られる該抗原提示細胞は、好ましくは治療及び/または予防の対象である被験者由来のものであり、単独、または、本発明のペプチド、エキソソーム、細胞傷害性T細胞等を含む、他の医薬と共にワクチンとして投与することができる。
【実施例】
【0026】
以下、実施例を挙げて本発明を更に説明するが、本発明はこれら実施例に限定されるものではない。
[実施例1] VEGFR-2(KDR)由来ペプチドの予測−1(HLA-A*2402)
KDR タンパク質の全アミノ酸配列から、BioInformatics & Molecular Analysis Section (BIMAS) HLA Peptide Binding Prediction soft (http://bimas.dcrt.nih.gov/cgi-bin/molbio/ken_parker_comboform)により、HLA-A*2402に対する結合親和性の高いものから順に9量体のペプチドを12種(配列番号1〜12)、10量体のペプチドを12種(配列番号13〜24)予測した(表1)。表1には、9量体及び10量体のそれぞれについて結合親和性の高い順に、そのN末端のKDR タンパク質のアミノ酸配列における位置と共に挙げた。尚、表中のCE652(配列番号25)はNukaya I.によって報告されている腫瘍抗原CEA(癌胎児性抗原)のエピトープの一つである(Int. J. Cancer 80, 1999)。また、陰性対照ペプチドとしてHIVペプチド(ILKEPVHGV(配列番号55)及びRYLRDQQLL(配列番号56))を用いた。
ペプチドは、標準的な固相合成法に従ってCleaved PepSet(Mimotope社、San Diego, LA)で合成し、逆相HPLCで精製した。HPLC及び質量分析によって、ペプチドの純度(>95%)及び種類をそれぞれ決定した。
【0027】
【表1】

【0028】
[実施例2] 予測したペプチドを用いたCTL lineの樹立
Ebstein-Bar virus (EBV)不死化B細胞株であるTISI(HLA-A24/24)とEHM(HLA-A3/3)は宝酒造バイオ研究所より供与された。
健常人の末梢血7mlを採取し、HLAのserotypingを行った。HLA-A24陽性の末梢血から、Ficoll Paque(Pharmacia)を用いた比重遠沈法にて末梢血単核球(PBMC)を分離した。
【0029】
得られたPBMCを培養フラスコ(Corning,43072)にて10時間静置し、浮遊細胞を除去した後にAIM-V(Invitrogen)に2%自己血清を添加した培養液にヒトGM-CSF(キリンビールより供与)1,000U/ml、ヒトIL-4(Genzyme)1,000U/mlを添加しフラスコに付着した細胞を培養した。5日後にOK-432(中外製薬より供与)10μg/mlにて48時間培養し、CTLの誘導のための抗原提示細胞として使用した。樹状細胞(Dendritic cell, DC)はFITCラベルした抗ClassII,CD80,CD86抗体及び、PEラベルした抗ClassI,CD11c,CD40(いずれもBeckton-Dickinson)、CD83(Immunotech)抗体を反応させてCell Quest softwareを使用したFACS-Calibur(Becton-Dickinson)にて表面抗原の解析を行い、DCであることを確認した。
【0030】
あらかじめ誘導しておいたDCに、実施例1で結合親和性が高いとして予測された9量体のペプチド12種類(配列番号1〜12)(20μg/ml)を3μg/mlのβ2-ミクログロブリンの存在下で20℃、4時間パルスし、その後DCをPBMCから磁気ビーズ(Dynabeads M-450とDetachabeds)にて選択されたCD8陽性細胞に対して1:20または1:2の比で、10ng/mlのヒトIL-7(Genzyme)存在下で48 well plate(Corning)にて混合培養を行った。3日後に最終10U/mlのIL-2(SIGMA)を加えた。7日後、14日後に同様のDCにて刺激を加えてCTL誘導を行い、20日後にペプチドをパルスしたTISI細胞を標的として各wellの細胞傷害活性を測定し、陽性wellのみをアロのPBMC、EBV不死化B細胞株(EHM)及び抗CD3抗体30ng/mlにて刺激する方法で培養し、その14日後にCTLの機能解析を行った。
【0031】
細胞傷害活性は4時間51Cr遊離法にて測定した。ペプチドをパルスする標的細胞は20μg/mlの濃度にて一晩パルスを行った。標的細胞を100μCiのNa251CrO4にて37℃、1時間ラベルした後RPMI1640にて3回洗浄し、96穴U型マイクロプレート(Corning)に標的細胞(1×104/100μl)と種々の濃度の効果細胞を100μl加え、総量200μlとして4時間、37℃のCO2インキュベーターにて培養した。培養終了後、各wellより上清を100μl採取し、γカウンターにて測定した。自然解離は標的細胞と培地で、最大解離は標的細胞と1M HCLで得られる放射活性とした。細胞傷害活性(特異的溶解のパーセンテージ)は次式により算出した。
細胞傷害活性(% Cytotoxicity)
=(実験解離-自然解離)/(最大解離-自然解離)×100
【0032】
その結果、表2に示すように6種類のCTL lineが樹立でき、配列番号2(アミノ酸開始位置KDR1318)、配列番号3(KDR220)、配列番号5(KDR189)、配列番号8(KDR169)、配列番号11(KDR826)、及び配列番号12(KDR998)に示すノナペプチドがエピトープペプチドとして特に有効であることが示された。
【0033】
【表2】
【0034】
[実施例3] 予測したペプチドから誘導されるCTL クローンの樹立
実施例2において樹立した6種類のCTL lineを96穴U型プレートに0.3、1、または3 cells/wellになるように希釈した。そこにアロ のPBMC 7×104 cells/well, EHM 1×104 cells/well, 抗CD3抗体 30ng/ml,IL-2 125U/ml となるようにして、AIM-Vに5%自己血清を添加して合計 150μl/wellになるようにした。10日後にIL-2が最終濃度125U/mlになるように調節したIL-2添加mediumを50μl/wellにて加えた。14日目にCTLの機能解析を行い、活性のあるものを大量培養した。
【0035】
その結果、配列番号8(アミノ酸開始位置KDR169)、配列番号5(KDR189)、配列番号3(KDR220)、配列番号2(KDR1318)、及び配列番号11(KDR826)の5種のノナペプチドよりそれぞれ8種類(C13-169、C19-169、C29-169、C53-169、C72-169、C61-169、K5-169及びK25-169)、2種類(C12-189及びC18-189)、11種類(KWC3、23、25、26、36、42、46、58、61、70及び77)、1種類(C7-1318)、1種類(C65-826)のCTL クローンを樹立した(表3)。
【表3】
【0036】
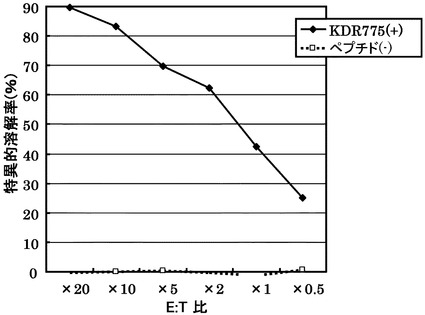
[実施例4] 樹立したCTLクローンの細胞傷害活性の測定
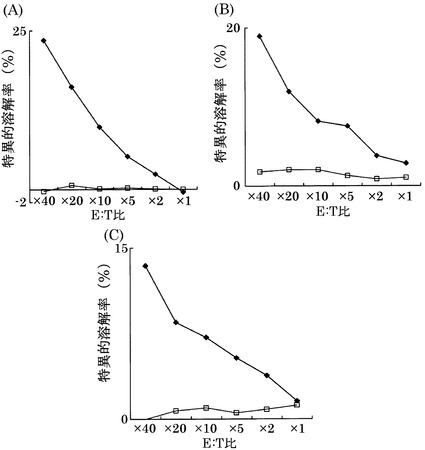
実施例3において樹立した10種類のCTL クローンの細胞傷害活性の強度を、標的細胞との比率(E:T比)を変化させて検討した(図1)。
その結果、配列番号8のノナペプチド(アミノ酸開始位置KDR169)から誘導されたC53-169が最も強力な細胞傷害活性を示した。
【0037】
[実施例5] 樹立したCTLクローンのHLA-テトラマー解析
HLA-A*2402と配列番号8のペプチド(アミノ酸開始位置KDR169)でHLA-テトラマーを合成した。すなわち、H鎖(約35kDa)とL鎖(約11kDa)を発現するプラスミドベクターをそれぞれ作製し、大腸菌を用いてこれらの組換えタンパク質を発現させた。 H鎖のプラスミドベクターは細胞外ドメインのみで構成され、カルボキシル末端側はビオチン化酵素認識配列に置換してある。H鎖およびL鎖は抗原ペプチドと共にリフォールディングすることにより、HLAペプチド複合体を形成する。HLAペプチド複合体をビオチン化酵素によりビオチン化し,ゲルろ過クロマトグラフィー、イオン交換クロマトグラフィーにより過剰のビオチンを除去し,ビオチン化HLAペプチド複合体を分離精製した。得られたビオチン化HLAペプチド複合体をストレプトアビジン(streptavidin; SA)と4:1のモル比で反応させて四量体化し、HLA-テトラマーを合成した。試料を解析する場合には,フィコエリスリン(phycoerythrin; PE)で蛍光標識したSA(SA-PE)を使用し四量体化した。CD8分子はHLA-A,B,Cの一部を除き,α3ドメインの245番目の一アミノ酸であるアラニンに結合することが解っており、そこでアラニンをバリンへ置換した変異導入HLA-テトラマーを作製することで,CD8分子とα3ドメインの結合に依存しないHLAペプチド複合体に対し高いavidityをもつCTLを選択的に検出することが可能である変異導入HLA-テトラマーを合成した。
【0038】
合成した変異型HLA-A24 KDR169-テトラマーにより、樹立したCTL クローンを検出した。その結果、図2に示すように、樹立したCTLクローンはテトラマー陽性かつCD8陽性のフラクションに強く染色するCTLであり、本発明のペプチド及びHLA抗原の複合体を特異的に認識するものであることが明らかになった。尚、テトラマー陰性かつCD8陰性のフラクションのシグナルはフィーダー細胞によるものである。
【0039】
[実施例6] VEGFR-2(KDR)由来ペプチドの予測−2(HLA-A*0201)
実施例1と同様にして、KDR タンパク質の全アミノ酸配列から、HLA Peptide Binding Prediction soft(http://bimas.dcrt.nih.gov/cgi-bin/molbio/ken_parker_comboform)によりHLA-A*0201に対する結合親和性の高いものから順に、9量体のペプチドを15種(配列番号26〜40)、10量体のペプチドを12種(配列番号41〜52)予測した(表4)。表4には、9量体及び10量体のそれぞれについて結合親和性の高い順に、そのN末端のKDR タンパク質のアミノ酸配列における位置と共に挙げた。尚、表中のCEA588(配列番号53)はTanaka, H.らによって報告されている腫瘍抗原CEA(癌胎児性抗原)のエピトープの一つである(ポスター発表、AACR #3669, vol.42, p681-682, March 2001)。
【0040】
【表4】
【0041】
[実施例7] HLA-A*0201拘束性エピトープペプチドの決定
実施例6において予測された9量体または10量体のペプチド16種類(配列番号26〜41)を、ヒトHLA(A0201)を発現するトランスジェニックマウス(Hum. Immunol. 2000 Aug.;61(8):764-79 Related Articles, Books, Linkout Induction of CTL response by a minimal epitope vaccine in HLA A*0201/DR1 transgenic mice: dependence on HLA class II restricted T(H) response., BenMohamed L., Krishnan R., Longmate J., Auge C., Low L., Primus J., Diamond DJ.に記載)を用い、vaccination法にてCTLの誘導を行った。
【0042】
6-8週令BALB/C(H-2d)マウスは日本クレア株式会社から手に入れた。A2/Kb トランスジェニックマウス (TGM) はF Jim Primus Ph.D. Vanderbilt-Ingram Cancer Centerより供与された。またT2細胞(TAP欠損細胞、HLA-A*0201陽性)は三重大学第三内科珠玖洋教授より供与された。
【0043】
マウス大腿骨、脛骨より骨髄を採取し、抗CD4,CD8,Gr-1(Beckton-Dickinson)、B220(バイオサイエンス)抗体、及びウサギ補体(PeL-Freez)にてリンパ球及び顆粒球を除去した。細胞は6well plateにて培養し、翌日浮遊細胞を回収して別の6well plateにRPMI1640(Invitrogen)に10%FBSを加えた培溶液にマウスGM-CSF(キリンビールより供与)1,000U/ml,マウスIL-4(PEPRO TECH)1,000U/mlを添加し培養した。その3日後に上記培養液にてhalf medium changeを行い、その2日後にOK-432 10μg/mlにて20時間培養し、浮遊細胞をDCとして使用した。DCはFITCラベルした抗ClassII, CD40抗体及び、PEラベルした抗ClassI,CD11c,CD80,CD86(いずれもBeckton-Dickinson)抗体を反応させてCell Quest softwareを使用したFACS-Calibur (Becton-Dickinson)にて表面抗原の解析を行い、DCであることを確認した。
【0044】
次いで、各ペプチド100μg、HbcAg120-140 helper peptide 140μg、IFA 100μlを混和(計 200μl)し、day 0に右側腹部にday11に左側腹部に皮下摂取した。Day 21 にワクチン接種したマウスの脾臓を採取し、赤血球をred lysis buffer (sigma)にて溶血除去し、その細胞の一部をIFN-γ ELISPOT assayのresponder cellとして使用した。また、残りの細胞は24 well plate (Corning)に6×106 cells/wellにてフィーダー細胞と3:1の比率にて再刺激を加え、5日後に細胞傷害活性を測定した。フィーダー細胞は同種別マウスの赤血球を除去した脾細胞をlipopolysaccharide (LPS) 25μg/ml存在下で3日間培養したものを使用した。
【0045】
本実施例においては、CTLの誘導能をIFN-γを産生する細胞が作るSPOTで評価するIFN-γ ELISPOT法を用いた。96穴平坦底マルチスクリーンプレートMAHA S45(ミリポア)を4℃、一晩抗マウスIFN-γ単クローン抗体(Pharmingen)にて処理し、翌日0.05% Tween 20を含んだPBSにて洗浄し、室温で2時間blocking bufferと反応させた。その後、ペプチドをパルスしたT2細胞とパルスしていないT2細胞を各wellに105 cell/100μlずつ散布した。そこに、ワクチンしたマウスの碑細胞を最大4×106 cell/100μl散布して、全200μlとして一晩37℃で培養した。翌日、各wellを洗浄後、ビオチン化したラット抗マウスIFN-γ抗体(Pharmingen)と2時間反応させ、十分に洗浄し、Extravidinを加えて2時間室温で反応させた。プレートを洗浄後、Alkaline Phosphatase Conjugate Substrate(BIO-RAD)を加えて5分間室温で放置し、IFN-γの産生による青色班をKS ELISPOT compact release(Carl Zeiss)にて計測した。そして
特異的青色班=(TISI(+)SPOT-TISI(-)SPOT)
で決定し、
TISI(+)SPOT/TISI(-)SPOT≧2を満たしたものを有効とした。
その結果、IFN-γELISPOT アッセイにて5種類のエピトープペプチド、配列番号29(アミノ酸開始位置KDR775)、配列番号30(KDR773)、配列番号33(KDR772)、配列番号34(KDR1328)及び配列番号40(KDR190)を得た(表5)。
【0046】
【表5】
【0047】
[実施例8] in vivoにおける抗腫瘍効果の確認1
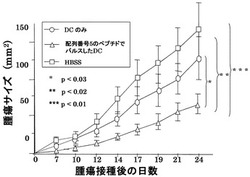
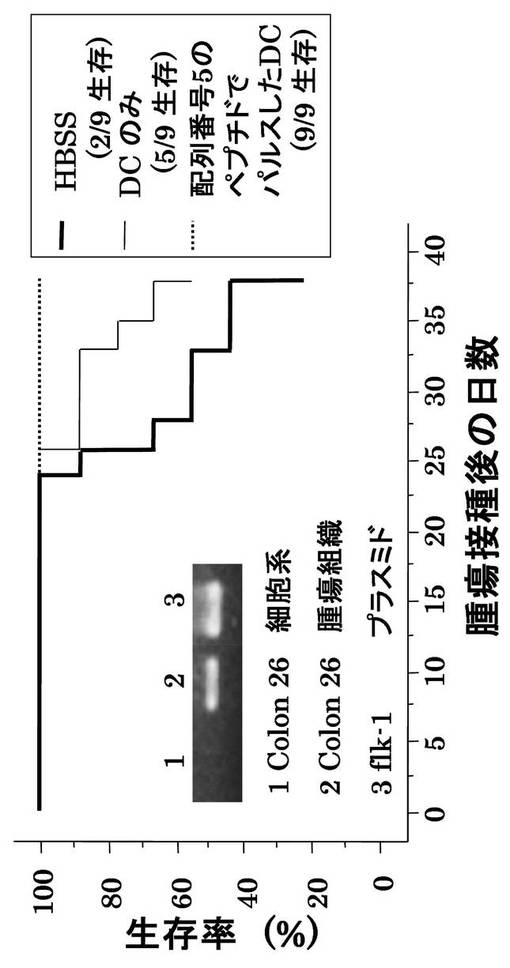
ヒトHLA−A2402とアンカーモチーフが同じである6〜8週齢の雄BALB/cマウス(CLEA Japan Inc.)の骨髄を採取し、実施例7と同様にGM-CSF(キリンビール)(1000U/ml)及びIL-4(Genzyme)(1000U/ml)を添加して培養して、樹状細胞を調製した。この樹状細胞に配列番号5のペプチド(アミノ酸開始位置KDR189)をパルスしたもの(1×106/マウス)をBALB/cマウス9匹に7日間隔で2回右下腹部に投与し、その7日後に結腸癌細胞株Colon26(大鵬薬品株式会社)(5×105個)を右腹部に接種し、マウスの生存と腫瘍増殖を検討した。
【0048】
まず、KDRの発現をRT-PCRで確認したところ、図3に示すように、Colon26細胞系単独ではKDRを発現していないが、Colon26を接種したマウスの腫瘍組織からKDRの発現が確認できた。すなわち、腫瘍血管が腫瘍組織に出現し、腫瘍を形成していることが確かめられた。
【0049】
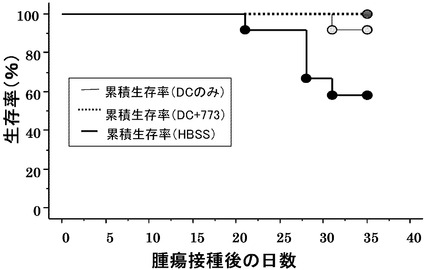
図3にColon26を接種されたマウスの生存曲線を示す。対照であるHBSS(Hank's Balanced Salt Solution)のみ(100μl)の投与の場合、腫瘍接種から35日後において、9匹中2匹の生存、ペプチドをパルスしない樹状細胞の接種では9匹中5匹の生存であったが、配列番号5のペプチドでパルスした樹状細胞の接種の場合には全9匹が生存していた。
【0050】
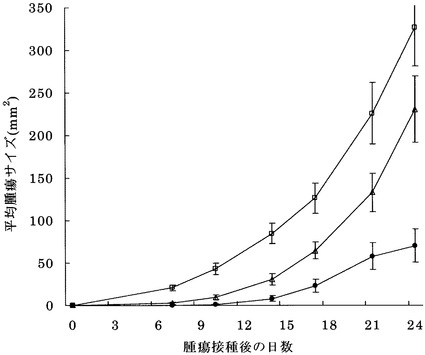
図4にマウスにおけるColon26由来の腫瘍の増殖に対する効果を示す。対照と比較して、樹状細胞を接種した場合に腫瘍増殖に対する抑制効果が認められ、配列番号5のペプチドでパルスした樹状細胞の場合に顕著な抑制効果が認められた。
【0051】
[実施例9] 本発明のペプチドのホモロジー検索
本発明のペプチド(配列番号2(アミノ酸開始位置KDR1318)、配列番号3(KDR220)、配列番号5(KDR189)、配列番号8(KDR169)、配列番号11(KDR826)、配列番号30(KDR773)、及び配列番号40(KDR190))のホモロジー検索をBLAST(http://www.ncbi.nlm.nih.gov/blast/blast.cgi)にて行った結果、いずれのペプチドにおいても配列が完全に一致したものは認められなかった(表6)。実施例4において強いCTL活性を誘導できた配列番号8の9量体ペプチド(KDR169)に関しては、2個のミスマッチ配列が1つ(77.8% homology)、3個のミスマッチ配列が2つ(66.7% homology)あるのみであった。実施例8においてin vivoにおける顕著な抗腫瘍効果が認められたペプチド配列番号5のペプチド(KDR 189)に関しては、3個のミスマッチ配列が1つ(66.7% homology)あるのみであった。
【0052】
【表6】
【0053】
[実施例10] in vivoにおける抗腫瘍効果の確認2
ヒトHLA−A0201を発現する6〜8週齢の雄A2/Kbトランスジェニックマウス(TGM)の骨髄を採取し、実施例8と同様にGM-CSF(キリンビール)(1000U/ml)及びIL-4(Genzyme)(1000U/ml)を添加して培養して、樹状細胞を調製した。この樹状細胞に配列番号30(アミノ酸開始位置KDR773)のペプチドをパルスしたもの(1×106/マウス)をA2/Kbトランスジェニックマウス12匹に7日間隔で2回左下腹部に投与し、その7日後にB16メラノーマ細胞(ATCC)(1×106個)を右腹部に接種し、マウスの生存と腫瘍増殖を検討した。
【0054】
図5にB16を接種されたマウスの生存率を示す。対照であるHBSSのみ(100μl)の投与の場合、腫瘍接種から35日後において、12匹中8匹の生存、ペプチドをパルスしない樹状細胞の接種では12匹中11匹の生存であったが、配列番号30のペプチドでパルスした樹状細胞の接種の場合には全12匹が生存していた。
【0055】
図6にマウスにおけるB16マウス由来の腫瘍の増殖に対する効果を示す。対照と比較して、樹状細胞を接種した場合に腫瘍増殖に対する抑制効果が認められ、配列番号30のペプチドでパルスした樹状細胞の場合に顕著な抑制効果が認められた。
【0056】
[実施例11] KDRを強制発現させた細胞に対する細胞障害活性
Miyake, S.ら、Proc. Natl. Acad. Sci. USA. 93, 1320-1324 (1996)に記載の方法に従い、HLA-A24陽性であるヒト大腸癌HT29株(ATCC)にアデノウイルスを用いてKDR(J. Biol. Chem. 1994 Oct 28;269(43):26988-95参照)を強制発現した。具体的には、ヒト293細胞のプラークアッセイによって決定した力価に基いて、細胞を特定の多重度で感染させた。HT29細胞を60mmのディッシュ中に105個の細胞密度で接種し、24時間インキュベートした。次の日、培地を捨て、新しい培地で連続希釈したウイルス200μlと交換した。37℃で1時間インキュベーションした後、増殖培地を添加し、細胞を48時間培養して標的細胞として使用した。対照としてEGFP(Leukemia 1999 Apr; 13(4):605-13参照)をアデノウイルスにて強制発現したHLA-A24陽性HT29株を用いた。KDRとEGFPを強制発現したHT29株は、細胞表面に各々その部分ペプチドを提示している可能性がある。
【0057】
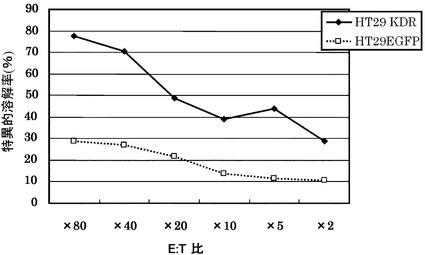
HLA-A24に結合する配列番号8のペプチド(アミノ酸開始位置KDR169)から誘導された実施例3に記載のCTLクローンC29-169を用い、KDR強制発現HLA-A24陽性HT29細胞に対する殺細胞効果をクロムリリースアッセイにて測定した。結果を図7に示す。
【0058】
細胞障害活性は4時間51Cr遊離法にて測定した。標的細胞は6 well plate に1×106cells/wellの濃度にて24時間培養し、KDR又はEGFPを導入したアデノウイルス(Ad-KDR又はAd-EGFP)をMOI 50にて感染させ、48時間後に使用した。標的細胞を100μCiのNa251CrO4にて37℃、1時間ラベルした後、RPMI1640にて3回洗浄し、96穴U型マイクロプレート(Corning)に標的細胞(1×104 cells/100μl)と種々の濃度の効果細胞を100μl加え、総量200μlとして4時間、37℃のCO2インキュベーターにて培養した。培養終了後、各wellより上清を100μl採取し、γカウンターにて測定し、実施例2と同様にして細胞障害活性を算出した。
【0059】
その結果、図7に示すように、CTLクローンC29-169は、KDRを強制発現したHLA-A24陽性HT29に対し、EGFPを強制発現した細胞と比較して顕著に高い殺細胞効果を示した。
【0060】
[実施例12] 内因的にKDRを発現している細胞に対する細胞障害活性
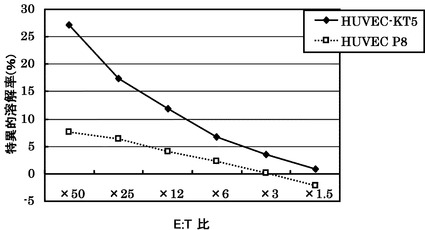
KDRを内因的に発現しているHLA-A24陽性ヒト臍帯血管内皮細胞由来HUVEC KT5株に対する実施例3に記載のCTLクローンC29-169の殺細胞効果について、KDR陽性、HLA-A24陰性HUVEC株 P8を対照として検討した。これらの細胞はKDRの部分ペプチドを細胞表面に提示する。
【0061】
細胞障害活性は4時間51Cr遊離法にて測定した。標的細胞を100μCiのNa251CrO4にて37℃、1時間ラベルした後RPMI1640にて3回洗浄し、96穴U型マイクロプレート(Corning)に標的細胞(1×104 cells/100μl)と種々の濃度の効果細胞を100μl加え、総量200μlとして4時間、37℃のCO2インキュベーターにて培養した。培養終了後、各wellより上清を100μl採取し、γカウンターにて測定し、実施例2と同様にして細胞障害活性を算出した。
【0062】
図8の結果から明らかなように、CTLクローンC29-169は、内在的にKDRを発現しているHLA-A24陽性HUVECに対して殺細胞効果を示し、HLA-A24陰性HUVECに対しては低い殺細胞効果を示した。
【0063】
[実施例13] HLA-A0201結合性ペプチドを用いたCTLクローンの樹立
実施例7のELISPOT解析において有効とされたHLA-A0201に結合する配列番号40のペプチド(アミノ酸開始位置KDR190)、配列番号30のペプチド(KDR773)のN末端から2番目のアミノ酸をロイシンに変換したペプチドKDR773-2L(配列番号54)、配列番号29のペプチド(KDR775)、配列番号34のペプチド(KDR1328)、及び配列番号33(KDR772)を用いて、実施例3と同様にして、それぞれ2種類(KWC14-190、KWC65-190)、4種類(KWC44-773-2L、KWC76-773-2L、KWC129-773-2L、KWC134-773-2L)、2種類(KWC81-775、KWC85-775)、12種類(KWC16、KWC21、KWC22、KWC47、KWC51、KWC108、KWC117、KWC132、KWC151、KWC153、KWC156、KWC159)、1種類(KWC72-772)のCTLクローンを得た。それぞれの細胞傷害活性を表7に示す。
【0064】
【表7】
【0065】
表7の結果から明らかなように、配列番号29、33、34、または40に示すペプチド、及び配列番号30に示すペプチドのN末端から2番目のアミノ酸がロイシンである配列番号54に示すペプチドのいずれも、顕著な細胞傷害活性を有するCTLを誘導することができ、エピトープペプチドとして有効であることが示された。
【0066】
[実施例14]
HLA-A0201陽性T2株に配列番号29のペプチド(アミノ酸開始位置KDR775)を加えて標的細胞とし、実施例13で配列番号29のペプチドから得られたCTLクローン(KDR C85-775(KWC85-775))の殺細胞効果をクロムリリースアッセイを用いて検討した。
その結果、図9に示すように、CTLクローンC85-775(KWC85-775)は、配列番号29のペプチドを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、配列番号29のペプチドを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0067】
[実施例15]
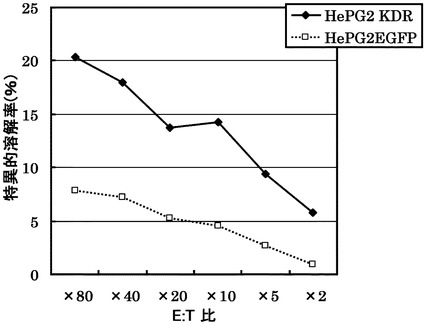
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対するCTLクローン(KDR C85-775(KWC85-775))の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A0201陽性HePG2を用いた。
その結果、図10に示すように、CTLクローンC85-775(KWC85-775)は、KDRペプチドを表面に提示したHLA-A0201陽性細胞に対し、EGFPペプチドを提示した細胞と比較して顕著に高い殺細胞効果を示した。
【0068】
[実施例16]
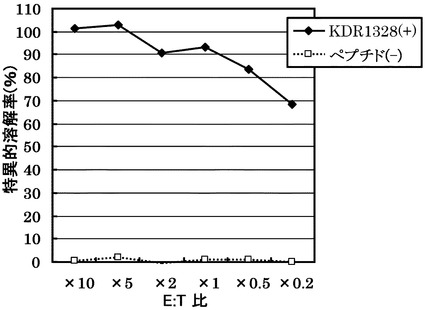
HLA-A0201陽性T2株に配列番号34のペプチド(アミノ酸開始位置KDR1328)を加えて標的細胞とし、実施例13で配列番号34のペプチドから得られたCTLクローン(KDR C51-1328(KWC51))の殺細胞効果をクロムリリースアッセイを用いて検討した。
その結果、図11に示すように、CTLクローンKDR C51-1328(KWC51)は、配列番号34のペプチドを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、配列番号34のペプチドを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0069】
[実施例17]
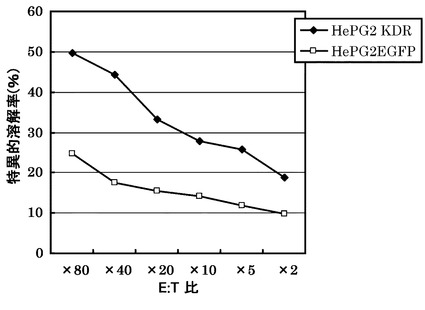
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対するCTLクローン(KDR C51-1328(KWC51))の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A0201陽性HePG2を用いた。
その結果、図12に示すように、CTLクローンKDR C51-1328(KWC51)は、KDRペプチドを表面に提示したHLA-A0201陽性細胞に対し、EGFPペプチドを提示した細胞と比較して顕著に高い殺細胞効果を示した。
【0070】
[実施例18]
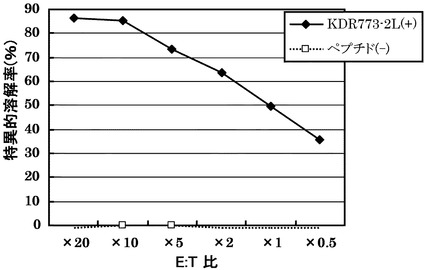
HLA-A0201陽性T2株に配列番号30の2番目のペプチドをロイシンに変換した配列番号54のペプチド(KDR773-2L)を加えて標的細胞とし、実施例13で配列番号34のペプチドから得られたCTLクローン(KDR C44-773-2L(KWC44-773-2L))の殺細胞効果をクロムリリースアッセイを用いて検討した。結果を図13に示す。
その結果、図13に示すように、CTLクローンKDR C44-773-2L(KWC44-773-2L)は、KDR773-2Lを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、KDR773-2Lを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0071】
[実施例19]
HLA-A0201陽性T2株に配列番号30の2番目のペプチドをロイシンに変換した配列番号54に示すペプチド(KDR773-2L)より得られたCTLクローン(KDR C44-773-2L(KWC44-773-2L))のクロムリリースアッセイでの殺細胞効果を、標的細胞としてT2に改変していない配列番号30に示すペプチド(アミノ酸開始位置KDR773)をパルスしたもので検討した。
その結果、図14に示すように、改変したペプチドで刺激し、得られたCTLクローンは改変前のペプチドを認識し、傷害することができた。
【0072】
[実施例20]
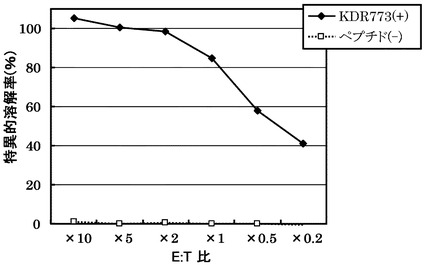
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対するCTLクローン(KDR C44-773-2L(KWC44-773-2L))の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A0201陽性HePG2を用いた。
その結果、図15に示すように、CTLクローンKDR C44-773-2L(KWC44-773-2L)は、KDRペプチドを表面に提示したHLA-A0201陽性細胞に対し、EGFPペプチドを提示した細胞と比較して顕著に高い殺細胞効果を示した。
【0073】
[実施例21]
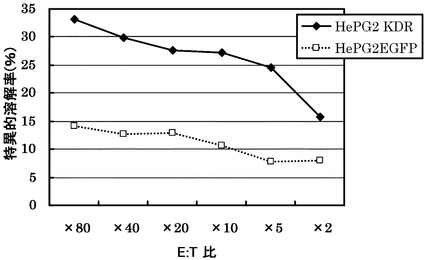
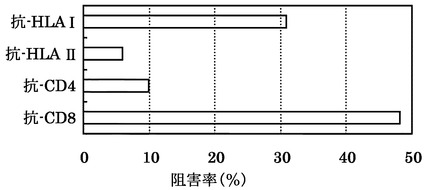
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対する配列番号34のペプチドから得られたCTLクローン(KDR C51-1328(KWC51))の殺細胞効果が各種抗体でブロックされるかどうかについて検討した。
その結果、図16に示すように、標的細胞が提示するHLA抗原に対する抗HLA ClassI抗体と、細胞傷害性T細胞のマーカーとなるCD8に対する抗CD8抗体がその作用を阻害した。
【0074】
[実施例22] Dorsal Air Sac assay(DAS)による血管新生阻害活性の検討
ペプチドエピトープの実際の血管新生阻害活性をマウスでDASアッセイ(Clinical Cancer Research Vol.5, 2185-2191, August 1999; Cancer Research 62, 6116-6123, November 1, 2002参照)にて評価した。
【0075】
チャンバーリングの両面にメンブレンフィルターを固定し、エチレンオキサイトガスで滅菌した。B16メラノーマ細胞を5×105 cells/0.15mlに調整し、チャンバー内に注入した。マウス(A2/kbトランスジェニックマウス(B16由来))を腹臥位として固定し、マウス背部皮下にシリンジにて5〜10mlの空気を注入しair sacを作製した。Sac下部を約1cm切開し、細胞浮遊液で満たされたチャンバーリングを背部皮下に移植し、皮膚をスキンステープラで閉じた。移植日より6日目にマウス皮膚をチャンバーごと剥皮し、伸展させた状態で固定した。黒色ゴムリングでマーキングし、実態顕微鏡を用いて新生血管を計測した。悪性腫瘍細胞から放出される血管新生因子によって新たに形成される血管は、ジグザグに進行する特徴があり、本来存在するバックグラウンドの血管とは形態学的に異なっている。従って、新生血管の判定はゴムリング内の蛇行走行を示す血管で、長さが3mm以上のものとし、血管新生指数(Angiogenesis Index(AI))としてカウントした。AIは蛇行血管数に応じて0〜5までの6段階で評価し、蛇行血管が5本以上のものはすべて5とした。
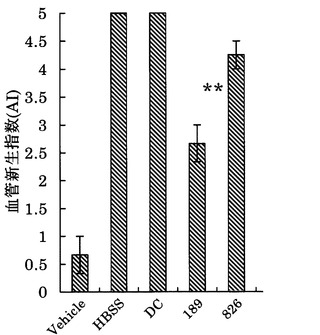
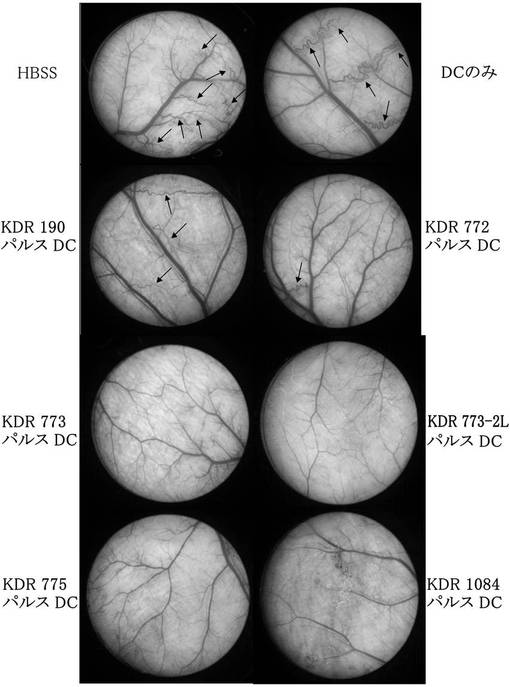
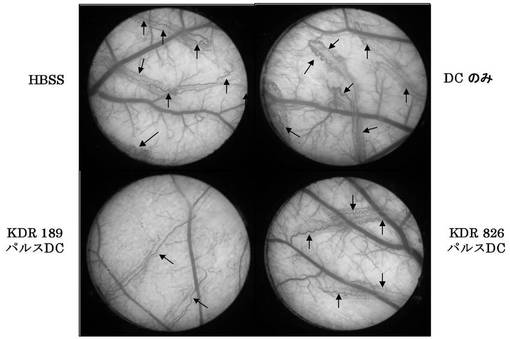
ワクチン接種はチャンバー移植の14日前にHBSS、DC単独、DCにペプチドパルスしたものをそれぞれ5×104cells尾静脈に注入し、それを1週間後に再度行い、計2回接種した。結果を図17及び以下の表8に示す。
【0076】
【表8】
【0077】
図17及び表8の結果から明らかなように、DCに配列番号40に示すペプチド(アミノ酸開始位置KDR190)、配列番号33に示すペプチド(KDR772)、配列番号30に示すペプチド(KDR773)、配列番号54に示すペプチド(KDR773-2L)、配列番号29に示すペプチド(KDR775)、及び配列番号46に示すペプチド(KDR1084)をワクチン接種した群において、DC単独投与群と比較して、蛇行血管の形成が明らかに抑制され、統計学的有意さをもって血管新生阻害効果があった。
【0078】
尚、これらのエピトープペプチドを用いたワクチン接種による正常な生理的血管新生に対する悪影響を観察するために、ワクチン接種したマウスにおける創傷治癒及び受精能を解析した。しかしながら、ワクチン接種したマウスにおいて有意な悪影響はなかった。更に、KDRペプチドを認識するCTLクローンを用いて非増殖性または増殖性内皮細胞に対する細胞傷害性をin vitroで試験し、ヒトにおける悪影響を検討したところ、これらのクローンは非増殖性内皮細胞よりも増殖性内皮細胞に対して強い活性を示した。
【0079】
[実施例23]
ヒトHLA-A0201を発現するA2/Kbトランスジェニックマウスを用いて大腸癌株MC38移植担癌マウスに対する配列番号30(アミノ酸開始位置KDR773)と配列番号34(KDR1084)ペプチドを不完全フロイントアジュバントと混合させてそれぞれday3,13に投与した場合の腫瘍体積の変化を示した。
【0080】
実施例22と同様のワクチン接種方法でHLA-A*0201拘束性エピトープペプチド(KDR773)による治療モデルにおけるin vivo抗腫瘍効果を腫瘍面積により評価した。その結果、図18から明らかなように、配列番号30のペプチド及び配列番号34のペプチドのワクチン接種により、有意な腫瘍増殖抑制効果が確認できた。
【0081】
[実施例24]
HLA-A0201陽性T2株に配列番号40のペプチド(アミノ酸開始位置KDR190)を加えて標的細胞とし、実施例13で配列番号40のペプチドから得られたCTLクローン(KWC65-190)の殺細胞効果をクロムリリースアッセイを用いて検討した。
【0082】
その結果、図19に示すように、CTLクローンKWC65-190は、配列番号40のペプチドを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、配列番号40のペプチドを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0083】
[実施例25]
HLA-A0201陽性T2株に配列番号33のペプチド(アミノ酸開始位置KDR772)を加えて標的細胞とし、実施例13で配列番号33のペプチドから得られたCTLクローン(KWC72-772)の殺細胞効果をクロムリリースアッセイを用いて検討した。
その結果、図20に示すように、CTLクローンKWC72-772は、配列番号33のペプチドを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、配列番号33のペプチドを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0084】
[実施例26]
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対するCTLクローン(KWC72-772)の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A0201陽性HePG2を用いた。
その結果、図21に示すように、CTLクローンKWC72-772は、KDRペプチドを表面に提示したHLA-A0201陽性細胞に対し、EGFPペプチドを提示した細胞と比較して顕著に高い殺細胞効果を示した。
【0085】
[実施例27]
HLA-A24陽性であるヒトB-リンパ芽球細胞系A24-LCL(HLA-A24/24、宝酒造製)に配列番号2(アミノ酸開始位置KDR1318)、3(KDR220)、5(KDR189)、及び11(アミノ酸開始位置KDR826)のペプチドを加えて標的細胞とし、実施例3に記載のCTLクローンC7-1318、KWC46、C18-189、及びC65-826の殺細胞効果をクロムリリースアッセイを用いて検討した。
その結果、図22〜25に示すように、CTLクローンC7-1318、KWC46、C18-189、及びC65-826は、それぞれ配列番号2、3、5、及び11のペプチドを提示したHLA-A24陽性細胞に対して明らかに殺細胞効果を示した。これに対して、各ペプチドを提示しない対照細胞に対しては、全く、またはほとんど殺細胞効果が見られなかった。
【0086】
[実施例28]
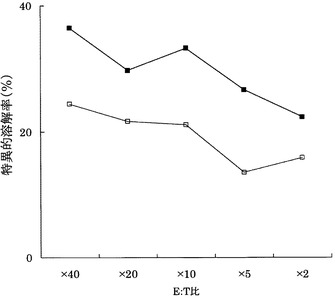
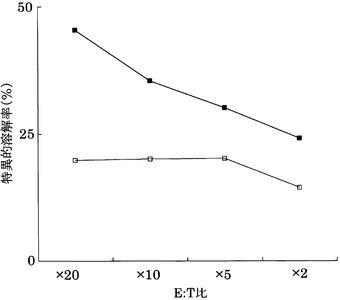
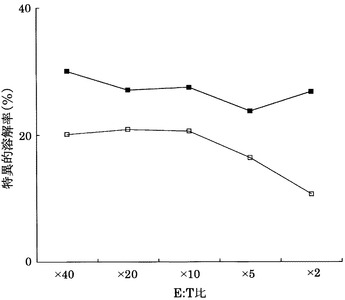
実施例11と同様にして、KDRをアデノウイルスで強制発現させたヒト大腸癌HT29株(ATCC)に対する実施例3に記載のCTLクローンC7-1318、KWC46、C18-189、及びC65-826の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A24陽性HT29株を用いた。
その結果、図26〜29に示すように、CTLクローンC7-1318、KWC46、C18-189、及びC65-826は、いずれもKDRを強制発現したHLA-A24陽性HT29に対し、EGFPを強制発現した細胞と比較して顕著に高い殺細胞効果を示した。
【0087】
[実施例29]
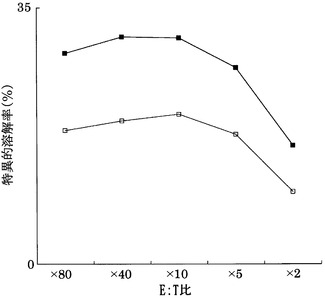
実施例22と同様にして、BALB/cマウスにおいてcolon26細胞によって誘導される血管新生応答に対するペプチドエピトープの血管新生阻害活性をDASアッセイにて評価した。
図30及び31の結果から明らかなように、DCに配列番号5に示すペプチド(アミノ酸開始位置KDR189)及び配列番号11に示すペプチド(KDR826)をワクチン接種した群において、DC単独投与群と比較して、蛇行血管の形成が明らかに抑制され、統計学的有意さをもって血管新生阻害効果があった。
【0088】
[実施例30]
癌患者のPBMCを用いて、各種エピトープペプチド(HLA-24結合親和性の高いペプチドとして配列番号8(アミノ酸開始位置KDR169)、配列番号5(KDR189)、配列番号3(KDR220)、配列番号11(KDR826)、及び配列番号17(KDR1318)、並びにHLA-A0201結合親和性の高いペプチドとして配列番号40(KDR190)、配列番号33(KDR772)、配列番号30(KDR773)、配列番号29(KDR775)、及び配列番号34(KDR1328))で刺激した後の各ペプチドに対するCTL応答を調べた。
【0089】
Maeda, Y.ら、Br. J. Cancer 87, 796-804 (2002)に報告されている方法を用いてPBMC中のペプチド特異的CTLを検出した。まず、患者から採取したPBMC(1×105個)を200μlの培地を入れたu底型の96ウェルプレートのウェルで各ペプチド10μMと共にインキュベートした。培地は45%のRPMI-1640培地、45%のAIM-V培地、10%のFBS、100U/mlのインターロイキン-2(IL-2)、及び0.1μM MEM非必須アミノ酸溶液から構成される。3日ごとに培地の半分を除去し、対応するペプチド(20μM)を含有する新しい培地と交換した。12日間のインキュベーション後、細胞を回収して各ペプチドに応答したIFN-γ産生能を試験した。対応するペプチドに応答してペプチドで刺激したPBMCによって産生されるIFN-γの量が対照であるHIVペプチドに応答して産生される量の2倍以上多い場合には陽性と判定した。
【0090】
その結果、HLA-A2及び-A24の癌患者においても、健康なドナーと同様にCTL前駆体が産生されることが示された(表9)。表中、患者A、B及びCはHLA-A24型の患者であり、患者DはHLA-A02型の患者である。また患者A及び患者Dは結腸癌患者、患者B及び患者Cはメラノーマ患者である。尚、CMVは陽性対照として用いたサイトメガロウイルス由来ペプチドであり、該ペプチドについてはA−24結合親和性の高いものは、例えばKuzushima, K.ら, Blood 2001, 98(6):p1872-1881に、A−2結合親和性の高いものは、例えばSolache A.ら, J. of Immunol. 1999, 163(10);p5512-5518にそれぞれ記載されている。
【0091】
【表9】
【0092】
更に、実施例14等と同様にして標準的な4時間の51Cr遊離法を行い、これらのCTL前駆体からCTL系を確立した(図32)。
【産業上の利用可能性】
【0093】
本発明により、広範囲にわたる腫瘍組織に形成される内皮細胞を標的として細胞傷害性T細胞を誘導し、癌ワクチンとして非常に有効な新規ペプチド、及びこれを含有する腫瘍の治療及び予防のための医薬が提供される。
本明細書で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書にとり入れるものとする。
【図面の簡単な説明】
【0094】
【図1】図1は、標的細胞に対するCTLクローンによる細胞傷害活性を示す。
【図2】図2は、樹立したCTLクローンのHLA-テトラマー解析の結果を示す。(a)本発明のペプチドによって誘導されたCTLクローン及びフィーダー細胞(b)フィーダー細胞(対照)
【図3】図3は、colon26を接種したBALB/cマウスの生存に対する本発明のペプチドを用いたワクチン接種の効果を示す。
【図4】図4は、colon26由来の腫瘍の増殖に対する本発明のペプチドを用いたワクチン接種の効果を示す。
【図5】図5は、B16を接種したマウスの生存に対する本発明のペプチドを用いたワクチン接種の効果を示す。
【図6】図6は、B16メラノーマ由来の腫瘍の増殖に対する本発明のペプチドを用いたワクチン接種の効果を示す。A2/Kb TGMに対してB16腫瘍細胞を皮下注射でチャレンジする前に、KDR773でパルスしたDC(●)、DC単独(△)、またはHBSS(□)で1週間の間隔をおいて2回ワクチン接種した。平均の腫瘍増殖を図示した。P<0.01。
【図7】図7は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC29-169の殺細胞効果を示す。
【図8】図8は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC29-169の殺細胞効果を示す。
【図9】図9は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C85-775(KWC85-775)の殺細胞効果を示す。
【図10】図10は、KDRペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C85-775(KWC85-775)の殺細胞効果を示す。
【図11】図11は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C51-1328(KWC51)の殺細胞効果を示す。
【図12】図12は、KDRペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C51-1328(KWC51)の殺細胞効果を示す。
【図13】図13は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C44-773-2L(KWC44-773-2L)の殺細胞効果を示す。
【図14】図14は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C44-773-2L(KWC44-773-2L)の殺細胞効果を示す。
【図15】図15は、KDRペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C44-773-2L(KWC44-773-2L)の殺細胞効果を示す。
【図16】図16は、CTLクローンの殺細胞効果に対する各種抗体の阻害作用を示す。
【図17】図17は、A2/Kb TGMにおいてB16細胞によって誘導される血管新生応答に対する本発明のペプチドの阻害活性を示す。マウスをHBSS、パルスしていないDC、または本発明のエピトープペプチド(KDR190、772、773、773-2L、775、または1084)でパルスしたDCで2回ワクチン接種した。矢印は特徴的なジグザグに進む新たに形成された血管を示す。
【図18】図18は、本発明のペプチドのトランスジェニックマウスにおけるヒト腫瘍体積の増加に対する抑制効果を示す。
【図19】図19は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKWC65-190の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図20】図20は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKWC72-772の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図21】図21は、KDRペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKWC72-772の殺細胞効果を示す。黒四角:HePG2-VEGFR2、□:HePG2-EGFP
【図22】図22は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC7-1318の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図23】図23は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンKWC46の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図24】図24は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC18-189の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図25】図25は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC65-826の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図26】図26は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC7-1318の殺細胞効果を示す。黒四角:HT29-VEGFR2、□:HT29-EGFP
【図27】図27は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンKWC46の殺細胞効果を示す。黒四角:HT29-VEGFR2、□:HT29-EGFP
【図28】図28は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC18-189の殺細胞効果を示す。黒四角:HT29-VEGFR2、□:HT29-EGFP
【図29】図29は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC65-826の殺細胞効果を示す。黒四角:HT29-VEGFR2、□:HT29-EGFP
【図30】図30は、BALB/cにおいてcolon26細胞によって誘導される血管新生応答に対する本発明のペプチドの阻害活性を示す。マウスをHBSS、パルスしていないDC、または本発明のエピトープペプチド(KDR189または826)でパルスしたDCで2回ワクチン接種した。矢印は特徴的なジグザグに進む新たに形成された血管を示す。
【図31】図31は、BALB/cにおいてcolon26細胞によって誘導される血管新生応答に対する本発明のペプチドの阻害を示す。柱は平均を、縦棒は標準誤差を示す。
【図32】図32は、本発明のペプチドで刺激した患者由来のCTLによるHLA-A24陽性細胞に対する殺細胞効果を示す。黒ひし形:ペプチド存在下、□:ペプチド不存在下(A)配列番号8(アミノ酸開始位置KDR169)で刺激して得られた結腸癌患者由来のCTLの殺細胞効果(B)配列番号5(アミノ酸開始位置KDR189)で刺激して得られた結腸癌患者由来のCTLの殺細胞効果(C)配列番号3(アミノ酸開始位置KDR220)で刺激して得られた結腸癌患者由来のCTLの殺細胞効果
【技術分野】
【0001】
本発明は、癌ワクチンとして非常に有効な新規ペプチド、及び該ペプチドを含む腫瘍の治療及び予防のための医薬に関する。
【背景技術】
【0002】
腫瘍拒絶抗原遺伝子が悪性黒色腫を中心に同定されるに従って、それを応用した癌免疫療法が開発されている。とりわけ、抗腫瘍免疫応答におけるCD8陽性T細胞の重要性が認識されるに伴い、腫瘍特異的CD8陽性T細胞を生体内で惹起させる癌ワクチン療法が着目され、種々の臨床応用が行われている。また、10個前後のアミノ酸残基からなるペプチドが種々の共刺激分子の補助によりClassI経路を介してT細胞を活性化し、腫瘍特異的細胞傷害性T細胞(CTL)を誘導する機序が解明され、さらに個々のHLA分子に拘束性を示すペプチドの同定も精力的に進んでいる。
【0003】
しかしながら、腫瘍を制御する事は未だ完全に出来ていないのが現状である。その原因として、腫瘍細胞の不均一性、腫瘍細胞におけるMHC-Class Iの発現低下や消失、さらに腫瘍細胞における標的分子の欠落が考えられる。また、現在同定されている腫瘍抗原ペプチドはある種の腫瘍に存在するが、全ての腫瘍を網羅できるものではない。そこで、これらの問題点を解決するため、標的細胞を腫瘍細胞ではなく、腫瘍血管における内皮細胞に着目した。すなわち、内皮細胞はMHC-Class Iの発現低下や消失、また不均一性といった問題がほとんどないので、腫瘍血管を標的にするCTLが誘導できれば、腫瘍の種類に関係なく、またClass Iの消失や標的分子の欠損といった従来の癌ワクチン療法で問題となっていた原因を凌駕でき、治療効果が大いに期待できると考えられる。癌における新生血管の研究は1970年代のFolkmanらの先駆的な仮説の提唱に始まり、様々な角度から研究がなされている。なかでも、VEGF-VEGF受容体(VEGFR)において多くの検討がなされ、腫瘍新生血管におけるその意義が評価されている。特に癌治療においては、target-oriented drugとして新生血管阻害剤の開発が精力的に進められ、すでに臨床治験も行われているが、この考え方を癌ワクチン療法に生かした治療は未だ行われていない。一つの理由としてVEGFRが正常細胞に発現しているため免疫寛容状態になっていると考えられた。しかしながら、1990年代のPlate, Millauer, Risauらの研究により腫瘍組織における内皮細胞においてVEGFRが強発現している事が確認され、また、CEA,HER/neuのように正常細胞でも発現している自己抗原に対する免疫応答が必ずしも免疫寛容状態になっていないことが明らかになった事により、VEGFR が癌ワクチン療法の標的に成り得るのではないかと考えた。
【0004】
最近、VEGFRに対する能動免疫を行うことにより、腫瘍における血管新生及び転移を抑制できることが報告された(非特許文献1;The Journal of experimental medicine 2002年 第195巻 第12号 p.1575-1584)。しかし、この文献では可溶性のVEGFRタンパク質を用いているに過ぎず、有効なペプチドのアミノ酸配列についての検討は何らなされていない。
【非特許文献1】The Journal of experimental medicine 2002年 第195巻 第12号 p.1575-1584
【発明の開示】
【0005】
本発明者等は、VEGFシグナルにおいて内皮細胞の増殖に関与すると考えられ、腫瘍組織内皮細胞において強発現しているVEGFR2 (KDR/flk-1、以下KDRという)を標的とした癌ワクチン療法の可能性に着目し、ワクチンとして有効に用いることができるペプチドの探索を行い、その特異性について検討を行い、本発明を完成するに到った。
【0006】
すなわち、本発明は、以下の(1)〜(22)を提供する。
(1) 配列番号2、3、5、8、11または12に示すアミノ酸配列からなるペプチドから選ばれるノナペプチド。
(2) 配列番号2、3、5、8、11または12に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチド。
(3) N末端から2番目のアミノ酸がフェニルアラニン、チロシン、メチオニンまたはトリプトファンである、上記(2)に記載のペプチド。
(4) C末端のアミノ酸がフェニルアラニン、ロイシン、イソロイシン、トリプトファンまたはメチオニンである、上記(2)または(3)に記載のペプチド。
(5) 配列番号29、30、33、34、40、または46に示すアミノ酸配列からなるペプチドから選ばれるノナペプチドまたはデカペプチド。
(6) 配列番号29、30、33、34、40、または46に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチド。
(7) N末端から2番目のアミノ酸がロイシンまたはメチオニンである、上記(6)に記載のペプチド。
(8) C末端のアミノ酸がバリンまたはロイシンである、上記(6)または(7)に記載のペプチド。
(9) 上記(1)〜(8)のいずれかに記載のペプチドを1種以上含む、腫瘍の治療及び/または予防のための医薬。
(10) 上記(1)〜(8)のいずれかに記載のペプチドを1種以上含む、糖尿病性網膜症、慢性関節リウマチ、乾癬、アテローム性動脈硬化症の治療のための医薬。
(11) 上記(1)〜(8)のいずれかに記載のペプチドとHLA抗原とを含む複合体を表面に提示しているエキソソーム。
(12) HLA抗原がHLA−A24またはHLA−A02である、上記(11)に記載のエキソソーム。
(13) HLA抗原がHLA−A2402またはHLA−0201である、上記(12)に記載のエキソソーム。
(14) 上記(1)〜(8)のいずれかに記載のペプチドを用いてCTL誘導能の高い抗原提示細胞を誘導する方法。
(15) 上記(1)〜(8)のいずれかに記載のペプチドを用いてCTLを誘導する方法。
(16) 上記(1)〜(8)のいずれかに記載のペプチドをコードするポリヌクレオチドを含む遺伝子を抗原提示細胞に導入することを含む、細胞傷害性T細胞誘導能の高い抗原提示細胞を誘導する方法。
(17) 上記(1)〜(8)のいずれかに記載のペプチドを用いて誘導される、単離された細胞傷害性T細胞。
(18) HLA抗原と上記(1)〜(8)のいずれかに記載のペプチドとの複合体を提示してなる抗原提示細胞。
(19) 上記(14)または(15)に記載の方法によって誘導される、上記(18)に記載の抗原提示細胞。
(20) 上記(1)〜(8)のいずれかに記載のペプチドを有効成分とする、病態部位における血管新生阻害のためのワクチン。
(21) HLA抗原がHLA−A24またはHLA−A02である被験者に対して投与するための、上記(20)に記載のワクチン。
(22) 悪性腫瘍の増殖及び/または転移を抑制するために使用される、上記(20)または(21)に記載のワクチン。
本明細書は本願の優先権の基礎である日本国特許出願2002-267285、2003-062003、2003-167042号の明細書および/または図面に記載される内容を包含する。
【0007】
[発明の実施の形態]
本発明者等はまず、in vivoにおいて抗原提示細胞上に提示される場合に、種々のタンパク質が9量体のペプチド(ノナペプチド)に分解されてから提示されることを考慮し、KDRタンパク質の9量体または10量体の部分ペプチドについて、ヒト主要組織適合抗原(MHC抗原)であるHLA抗原との結合親和性を検討した。尚、表1の右側のペプチドにおいて、小文字は5番目のアミノ酸を示す。
ヒトKDRタンパク質のアミノ酸配列は公知であり、例えば米国特許第5,861,301号に記載されており、当業者であれば容易に入手することができる。9量体及び10量体ペプチドは、得られたKDRタンパク質の全アミノ酸配列に基づいて、任意の位置からのペプチドを合成して得ることができる。ペプチドの合成は、通常のペプチド化学において用いられる方法に準じて行うことができる。通常用いられる合成方法は、例えば、Peptide Synthesis, Interscience, New York, 1966; The Proteins, Vol 2, Academic Press Inc., New York, 1976; ペプチド合成、丸善(株)、1975; ペプチド合成の基礎と実験、丸善(株)、1985; 医薬品の開発 続 第14巻・ペプチド合成、広川書店、1991等の文献や、国際公開WO99/67288号等の公報に記載されている。HLA抗原との結合は、細胞表面にHLA抗原を有する細胞、例えば樹状細胞を単離して、細胞へのペプチドの結合を通常行われる手法を用いて測定することができる。
【0008】
あるいはまた、最近インターネット上で利用可能となっているソフトウェア、例えばParker K. C., J. Immunol. 152, 1994に記載されているもの等を用いて、種々のペプチドとHLA抗原との結合親和性をin silicoで計算することもできる。尚、HLA抗原との結合親和性は、例えばParker, K.C., J. Immunol., 152, 1994; Nukaya, I., Int. J. Cancer, 80, 1999等に記載のように測定することができる。
【0009】
HLA抗原としては、例えば日本人において多く発現しているといわれるA−24型またはA−02型等を用いることが有効な結果を得るために好ましく、更に好ましくはA−2402、A−0201等のサブタイプである。しかしながら、臨床においては、治療を必要とする患者のHLA抗原の型を予め調べることにより、これとの結合親和性、あるいは抗原提示による細胞傷害性T細胞(CTL)誘導能の高いペプチドを適宜選択することができる。更に、結合親和性及びCTL誘導能の高いペプチドを得るために、天然に存在するKDR部分ペプチドのアミノ酸配列に基づいて1個、2個または数個のアミノ酸の置換または付加を行うこともできる。ここで、「数個」とは、5個以下、好ましくは3個以下を意味する。更に、天然において提示されるペプチド以外にも、既にHLA抗原に結合して提示されるペプチドの配列の規則性が知られているので(J. Immunol., 152, 3913, 1994; Immunogenetics. 41:178, 1995; J. Immunol. 155:4307, 1994)、得られたペプチドに対してこれらの規則性に基づいた改変を行っても良い。例えば、HLA−24結合親和性の高いものはペプチドのN末端から2番目のアミノ酸をフェニルアラニン、チロシン、メチオニンまたはトリプトファンに置換したり、C末端のアミノ酸をフェニルアラニン、ロイシン、イソロイシン、トリプトファンまたはメチオニンに置換したペプチドも好適に使用することができる。一方、HLA−0201結合親和性の高いものは、ペプチドのN末端から2番目のアミノ酸をロイシンまたはメチオニンに置換したり、C末端のアミノ酸をバリンまたはロイシンに置換したペプチドも好適に使用することができる。更に、ペプチドのN末端及び/またはC末端に1〜2個のアミノ酸が付加していても良い。
【0010】
しかしながら、ペプチドの配列が他の機能を有する内在性または外来性のタンパク質のアミノ酸配列の一部と同一となる場合には、自己免疫疾患等の副作用が生じたり、あるいは特定の物質に対するアレルギー症状を引き起こしたりする可能性があるため、利用可能なデータベースを用いてホモロジー検索を行い、他のタンパク質のアミノ酸配列と一致することを避けるのが好ましい。更に、ホモロジー検索において、アミノ酸が1個または2個異なるペプチドも存在しないことが明らかであれば、HLA抗原との結合親和性及び/またはCTL誘導能を高めるための上記アミノ酸配列の改変もこのような問題を生じるおそれがない。
【0011】
上記のようにしてHLA抗原との結合親和性の高いペプチドは、癌ワクチンとして有効である可能性が高いことが予想されるが、高い結合親和性を有することを指標として選択した候補ペプチドについて、実際にCTL誘導能を有するか否かを検討することが必要である。CTL誘導能の確認は、ヒトMHC抗原を有する抗原提示細胞(例えばB−リンパ球、マクロファージ、樹状細胞)、具体的にはヒト末梢血単核球由来の樹状細胞を誘導し、ペプチドで刺激した後にCD8陽性細胞と混合し、標的細胞に対する細胞傷害活性を測定する。反応系として、ヒトHLA抗原を発現するように作製されたトランスジェニック動物(例えば、Hum. Immunol. 2000 Aug.;61(8):764-79 Related Articles, Books, Linkout Induction of CTL response by a minimal epitope vaccine in HLA A*0201/DR1 transgenic mice: dependence on HLA class II restricted T(H) response., BenMohamed L., Krishnan R., Longmate J., Auge C., Low L., Primus J., Diamond DJ.に記載のもの)を用いることもできる。細胞傷害活性は、例えば標的細胞を51Cr等で放射標識し、標的細胞から遊離した放射活性から計算することができる。あるいはまた、ペプチドを拘束した抗原提示細胞の存在下でCTLが産生・放出したIFN-γ及び抗IFN-γモノクローナル抗体によって培地上に可視化される阻止円を測定することによって観察することもできる。
【0012】
上記のようにしてペプチドのCTL誘導能を検討した結果、HLA抗原との結合親和性が高いものが必ずしも誘導能が高いわけではないことが明らかとなった。そして、VYSSEEAEL(配列番号2)、GYRIYDVVL(配列番号3)、SYMISYAGM(配列番号5)、RFVPDGNRI(配列番号8)、KWEFPRDRL(配列番号11)、またはDFLTLEHLI(配列番号12)に示すアミノ酸配列からなるペプチドから選ばれるノナペプチド、及びAMFFWLLLV(配列番号29)、VIAMFFWLL(配列番号30)、AVIAMFFWL(配列番号33)、KLIEIGVQT(配列番号34)、YMISYAGMV(配列番号40)またはIQSDVWSFGV(配列番号46)に示すアミノ酸配列からなるペプチドから選ばれるノナペプチドまたはデカペプチドが特に高いCTL誘導能を有することが明らかとなった。
【0013】
本発明は更に、配列番号2、3、5、8、11または12に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチドも提供する。配列番号2、3、5、8、11または12に示す9個のアミノ酸からなるアミノ酸配列に基づいて、1個、2個または数個のアミノ酸の置換または付加は、他のタンパク質のアミノ酸配列との一致がない限りにおいて、CTL誘導能を有し得る。特に、アミノ酸の置換として、N末端から2番目のアミノ酸のフェニルアラニン、チロシン、メチオニンまたはトリプトファンへの置換、C末端のアミノ酸のフェニルアラニン、ロイシン、イソロイシン、トリプトファンまたはメチオニンへの置換、アミノ酸の付加として、N末端及び/またはC末端への1〜2個のアミノ酸の付加は好適な例である。
【0014】
本発明はまた、配列番号29、30、33、34、40、または46に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチドも提供する。配列番号29、30、33、34、40、または46に示す9個または10個のアミノ酸からなるアミノ酸配列に基づいて、1個、2個または数個のアミノ酸の置換または付加は、他のタンパク質のアミノ酸配列との一致がない限りにおいて、CTL誘導能を有し得る。特に、アミノ酸の置換として、N末端から2番目のアミノ酸のロイシンまたはメチオニンへの置換、C末端のアミノ酸のバリンまたはロイシンへの置換、アミノ酸の付加として、N末端及び/またはC末端への1〜2個のアミノ酸の付加は好適な例である。こうした改変ペプチドの例として、限定するものではないが、例えば配列番号30のペプチドのN末端から2番目のアミノ酸をロイシンに変換したペプチド(配列番号54)が挙げられる。これらの改変ペプチドによる刺激によって得られるCTLクローンは改変前のペプチドを認識し、傷害することができる。
【0015】
上記の本発明のペプチドは、1種または2種以上の組み合わせとして、生体内でCTLを誘導し得る癌ワクチンとして使用することができる。本発明のペプチドの投与により、抗原提示細胞のHLA抗原に該ペプチドが高密度に提示され、提示されたペプチドとHLA抗原との複合体に対して特異的に反応するCTLが誘導され、標的細胞となるべき腫瘍細胞内の血管内皮細胞に対する攻撃力が高まる。あるいは、被験者から樹状細胞を取り出して本発明のペプチドで刺激することにより、細胞表面に本発明のペプチドを拘束した抗原提示細胞が得られ、これを再度被験者に投与することで被験者においてCTLを誘導し、標的細胞に対する攻撃力を高めることができる。
【0016】
すなわち、本発明は、本発明のペプチドを1種以上含む、腫瘍の治療または腫瘍の増殖・転移等の予防のための医薬を提供するものである。更に、病態部位における血管新生は、腫瘍の他、糖尿病性網膜症、慢性関節リウマチ、乾癬、アテローム性動脈硬化症のような疾患、並びに固形癌の転移と深く結びついている(Folkman, J., Nature Med. 1:27-31 (1995); Bicknell, R., Harris, A.L., Curr. Opin. Oncol. 8:60-65 (1996))。従って、本発明のペプチドは、腫瘍、糖尿病性網膜症、慢性関節リウマチ、乾癬、アテローム性動脈硬化症のような疾患、並びに固形癌の転移の治療に用いることができる。
【0017】
本発明のペプチドは、悪性腫瘍組織において形成され、正常な血管とは形態学的に異なる蛇行性血管の形成を阻害することが確かめられたが、ワクチン接種したマウスにおける創傷治癒及び受精能を解析した結果、正常な生理的血管新生に対しては悪影響を有さないことも確認した。更に、本発明のペプチドを認識するCTLクローンを用いて非増殖性または増殖性内皮細胞に対する細胞傷害性をin vitroで試験したところ、これらのクローンは非増殖性内皮細胞よりも増殖性内皮細胞に対して強い活性を示すことも明らかとなった。すなわち、増殖性内皮細胞が観察される疾患、特に癌において非常に特異的に作用することができる。
尚、in vivo及びin vitroにおいて、本発明のペプチドによる樹状細胞の刺激は、細胞に対して高濃度のペプチドを存在させることによって、該細胞に予め拘束されているペプチドとの交換が生じ、容易に行われる。このため、本発明において使用されるペプチドは、HLA抗原との結合親和性がある程度以上高いことが必要とされる。
【0018】
本発明の医薬は、本発明のペプチドを単独で直接投与しても良いが、通常用いられる製剤学的方法によって製剤化した医薬組成物として投与しても良い。その場合、本発明のペプチドの他に、通常医薬に用いられる担体、賦形剤等を適宜含むことができ、特に限定されるものではない。本発明の医薬の用途としては、胃癌、十二指腸癌、大腸癌、肺癌、乳癌、前立腺癌、脳腫瘍等の種々の腫瘍の治療及び予防に用いることができる。本発明のペプチドは、腫瘍細胞そのものではなく、腫瘍組織内に新たに形成される新生血管の内皮細胞を標的とするものであり、従って、治療対象の腫瘍は広範囲にわたり、特に用途を限定するものではない。
【0019】
本発明のペプチドを有効成分とする腫瘍の治療及び/または予防のための医薬は、細胞性免疫が効果的に成立するようにアジュバントと共に投与したり、他の抗癌剤等の有効成分と共に投与したり、また粒子状の剤型にして投与することができる。アジュバントとしては、文献(C1in. Microbio1.Rev., 7:277-289, 1994)に記載のものなどが応用可能である。また、リポソーム製剤、直径数μmのビーズに結合させた粒子状の製剤、リピッドを結合させた製剤なども考えられる。投与方法としては、経口投与、皮内投与、皮下投与、静脈注射などが利用でき、全身投与、あるいは目的となる腫瘍の近傍に局所投与しても良い。本発明のペプチドの投与量は、治療すべき疾患、患者の年齢、体重、投与方法等により適宜調整することができるが、通常 0.001mg〜1000mg、好ましくは 0.001mg〜1OOOmg、より好ましくはO.1mg〜1Omgであり、数日ないし数月に1回投与するのが好ましい。当業者であれば、適当な投与量を適宜選択することが可能である。
【0020】
あるいはまた、本発明は、本発明のペプチドとHLA抗原との複合体を表面に提示している、エキソソームと呼ばれる細胞内小胞を提供する。エキソソームの調製は、例えば特表平11−510507号、及び特表2000−512161号に詳細に記載されている方法を用いて行うことができるが、好ましくは治療及び/または予防の対象となる被験者から得た抗原提示細胞を用いて調製する。本発明のエキソソームは、上記本発明のペプチドと同様に癌ワクチンとして接種することができる。
HLA抗原としては、治療及び/または予防を必要とする被験者のHLA抗原と同じ型のものであることが必要である。例えば、日本人の場合にはHLA−A24またはHLA−A02、特にHLA−A2402またはHLA−0201とすると好適であることが多い。
【0021】
本発明はまた、本発明のペプチドを用いた抗原提示細胞の誘導方法も提供する。樹状細胞を末梢血単球から誘導した後、in vitroまたはin vivoで本発明のペプチドと接触(刺激)させ、抗原提示細胞を誘導することができる。本発明のペプチドを被験者に投与した場合、被験者の体内で本発明のペプチドを拘束した抗原提示細胞が誘導される。あるいは、抗原提示細胞に本発明のペプチドをin vitroで拘束させた後に被験者にワクチンとして投与することもできる。
【0022】
本発明はまた、上記本発明のペプチドをコードするポリヌクレオチドを含む遺伝子をin vitroで抗原提示細胞に導入することを含む、細胞傷害性T細胞誘導能の高い抗原提示細胞を誘導する方法を提供する。導入する遺伝子はDNAの形態であってもRNAの形態であっても良い。導入の方法は当分野において通常行われるリポフェクション、エレクトロポレーション、リン酸カルシウム法等の種々の方法を用いれば良く、特に限定するものではない。具体的には、例えばCancer Res., 56:5672, 1996; J. Immunol., 161:5607, 1998; J. Exp. Med., 184:465, 1996; 特表2000-509281号に記載のようにして行うことができる。遺伝子を抗原提示細胞に導入することによって、該遺伝子は細胞中で転写、翻訳等の処理の後、得られたタンパク質がMHCクラスIまたはクラスIIのプロセッシング及び提示経路を経て、部分ペプチドが提示される。
【0023】
本発明は更に、上記本発明のペプチドを用いてCTLを誘導する方法を提供する。本発明のペプチドを被験者に投与した場合、被験者の体内でCTLが誘導され、腫瘍組織内の新生血管の内皮細胞を標的とした免疫力が増強される。あるいは、被験者由来の抗原提示細胞及びCD8陽性細胞、または末梢血単核球をin vitroで本発明のペプチドと接触(刺激)させ、CTLを誘導してから被験者にもどすex vivoの治療方法にも用いることができる。
【0024】
本発明は更に、本発明のペプチドを用いて誘導される、単離された細胞傷害性T細胞を提供する。本発明のペプチドを提示した抗原提示細胞による刺激に基づいて誘導された細胞傷害性T細胞は、好ましくは治療及び/または予防の対象である被験者由来のものであり、単独、または、本発明のペプチド、エキソソーム等を含む、他の医薬と共に抗腫瘍効果を目的として投与することができる。得られた細胞傷害性T細胞は、本発明のペプチド、好ましくは誘導に用いられたものと同じペプチドを提示する標的細胞に対して特異的に作用する。標的細胞は、KDRを内在的に発現する細胞であっても、強制的にKDRを発現させた細胞であっても良く、また、本発明のペプチドの刺激によって細胞表面に該ペプチドを提示する細胞であっても攻撃の対象となり得る。
【0025】
本発明は更に、HLA抗原と本発明のペプチドとの複合体を提示してなる抗原提示細胞を提供する。本発明のペプチド、あるいは本発明のペプチドをコードするヌクレオチドとの接触によって得られる該抗原提示細胞は、好ましくは治療及び/または予防の対象である被験者由来のものであり、単独、または、本発明のペプチド、エキソソーム、細胞傷害性T細胞等を含む、他の医薬と共にワクチンとして投与することができる。
【実施例】
【0026】
以下、実施例を挙げて本発明を更に説明するが、本発明はこれら実施例に限定されるものではない。
[実施例1] VEGFR-2(KDR)由来ペプチドの予測−1(HLA-A*2402)
KDR タンパク質の全アミノ酸配列から、BioInformatics & Molecular Analysis Section (BIMAS) HLA Peptide Binding Prediction soft (http://bimas.dcrt.nih.gov/cgi-bin/molbio/ken_parker_comboform)により、HLA-A*2402に対する結合親和性の高いものから順に9量体のペプチドを12種(配列番号1〜12)、10量体のペプチドを12種(配列番号13〜24)予測した(表1)。表1には、9量体及び10量体のそれぞれについて結合親和性の高い順に、そのN末端のKDR タンパク質のアミノ酸配列における位置と共に挙げた。尚、表中のCE652(配列番号25)はNukaya I.によって報告されている腫瘍抗原CEA(癌胎児性抗原)のエピトープの一つである(Int. J. Cancer 80, 1999)。また、陰性対照ペプチドとしてHIVペプチド(ILKEPVHGV(配列番号55)及びRYLRDQQLL(配列番号56))を用いた。
ペプチドは、標準的な固相合成法に従ってCleaved PepSet(Mimotope社、San Diego, LA)で合成し、逆相HPLCで精製した。HPLC及び質量分析によって、ペプチドの純度(>95%)及び種類をそれぞれ決定した。
【0027】
【表1】
【0028】
[実施例2] 予測したペプチドを用いたCTL lineの樹立
Ebstein-Bar virus (EBV)不死化B細胞株であるTISI(HLA-A24/24)とEHM(HLA-A3/3)は宝酒造バイオ研究所より供与された。
健常人の末梢血7mlを採取し、HLAのserotypingを行った。HLA-A24陽性の末梢血から、Ficoll Paque(Pharmacia)を用いた比重遠沈法にて末梢血単核球(PBMC)を分離した。
【0029】
得られたPBMCを培養フラスコ(Corning,43072)にて10時間静置し、浮遊細胞を除去した後にAIM-V(Invitrogen)に2%自己血清を添加した培養液にヒトGM-CSF(キリンビールより供与)1,000U/ml、ヒトIL-4(Genzyme)1,000U/mlを添加しフラスコに付着した細胞を培養した。5日後にOK-432(中外製薬より供与)10μg/mlにて48時間培養し、CTLの誘導のための抗原提示細胞として使用した。樹状細胞(Dendritic cell, DC)はFITCラベルした抗ClassII,CD80,CD86抗体及び、PEラベルした抗ClassI,CD11c,CD40(いずれもBeckton-Dickinson)、CD83(Immunotech)抗体を反応させてCell Quest softwareを使用したFACS-Calibur(Becton-Dickinson)にて表面抗原の解析を行い、DCであることを確認した。
【0030】
あらかじめ誘導しておいたDCに、実施例1で結合親和性が高いとして予測された9量体のペプチド12種類(配列番号1〜12)(20μg/ml)を3μg/mlのβ2-ミクログロブリンの存在下で20℃、4時間パルスし、その後DCをPBMCから磁気ビーズ(Dynabeads M-450とDetachabeds)にて選択されたCD8陽性細胞に対して1:20または1:2の比で、10ng/mlのヒトIL-7(Genzyme)存在下で48 well plate(Corning)にて混合培養を行った。3日後に最終10U/mlのIL-2(SIGMA)を加えた。7日後、14日後に同様のDCにて刺激を加えてCTL誘導を行い、20日後にペプチドをパルスしたTISI細胞を標的として各wellの細胞傷害活性を測定し、陽性wellのみをアロのPBMC、EBV不死化B細胞株(EHM)及び抗CD3抗体30ng/mlにて刺激する方法で培養し、その14日後にCTLの機能解析を行った。
【0031】
細胞傷害活性は4時間51Cr遊離法にて測定した。ペプチドをパルスする標的細胞は20μg/mlの濃度にて一晩パルスを行った。標的細胞を100μCiのNa251CrO4にて37℃、1時間ラベルした後RPMI1640にて3回洗浄し、96穴U型マイクロプレート(Corning)に標的細胞(1×104/100μl)と種々の濃度の効果細胞を100μl加え、総量200μlとして4時間、37℃のCO2インキュベーターにて培養した。培養終了後、各wellより上清を100μl採取し、γカウンターにて測定した。自然解離は標的細胞と培地で、最大解離は標的細胞と1M HCLで得られる放射活性とした。細胞傷害活性(特異的溶解のパーセンテージ)は次式により算出した。
細胞傷害活性(% Cytotoxicity)
=(実験解離-自然解離)/(最大解離-自然解離)×100
【0032】
その結果、表2に示すように6種類のCTL lineが樹立でき、配列番号2(アミノ酸開始位置KDR1318)、配列番号3(KDR220)、配列番号5(KDR189)、配列番号8(KDR169)、配列番号11(KDR826)、及び配列番号12(KDR998)に示すノナペプチドがエピトープペプチドとして特に有効であることが示された。
【0033】
【表2】
【0034】
[実施例3] 予測したペプチドから誘導されるCTL クローンの樹立
実施例2において樹立した6種類のCTL lineを96穴U型プレートに0.3、1、または3 cells/wellになるように希釈した。そこにアロ のPBMC 7×104 cells/well, EHM 1×104 cells/well, 抗CD3抗体 30ng/ml,IL-2 125U/ml となるようにして、AIM-Vに5%自己血清を添加して合計 150μl/wellになるようにした。10日後にIL-2が最終濃度125U/mlになるように調節したIL-2添加mediumを50μl/wellにて加えた。14日目にCTLの機能解析を行い、活性のあるものを大量培養した。
【0035】
その結果、配列番号8(アミノ酸開始位置KDR169)、配列番号5(KDR189)、配列番号3(KDR220)、配列番号2(KDR1318)、及び配列番号11(KDR826)の5種のノナペプチドよりそれぞれ8種類(C13-169、C19-169、C29-169、C53-169、C72-169、C61-169、K5-169及びK25-169)、2種類(C12-189及びC18-189)、11種類(KWC3、23、25、26、36、42、46、58、61、70及び77)、1種類(C7-1318)、1種類(C65-826)のCTL クローンを樹立した(表3)。
【表3】
【0036】
[実施例4] 樹立したCTLクローンの細胞傷害活性の測定
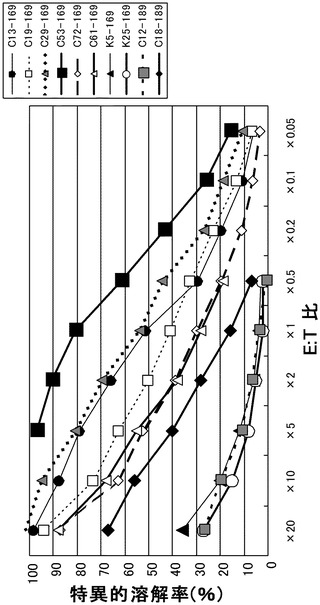
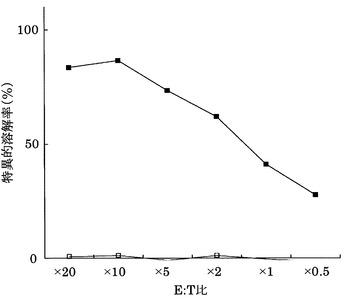
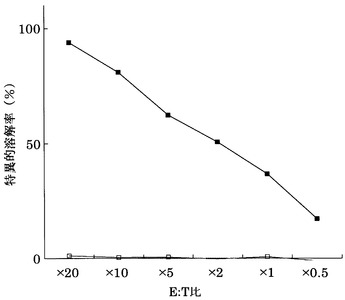
実施例3において樹立した10種類のCTL クローンの細胞傷害活性の強度を、標的細胞との比率(E:T比)を変化させて検討した(図1)。
その結果、配列番号8のノナペプチド(アミノ酸開始位置KDR169)から誘導されたC53-169が最も強力な細胞傷害活性を示した。
【0037】
[実施例5] 樹立したCTLクローンのHLA-テトラマー解析
HLA-A*2402と配列番号8のペプチド(アミノ酸開始位置KDR169)でHLA-テトラマーを合成した。すなわち、H鎖(約35kDa)とL鎖(約11kDa)を発現するプラスミドベクターをそれぞれ作製し、大腸菌を用いてこれらの組換えタンパク質を発現させた。 H鎖のプラスミドベクターは細胞外ドメインのみで構成され、カルボキシル末端側はビオチン化酵素認識配列に置換してある。H鎖およびL鎖は抗原ペプチドと共にリフォールディングすることにより、HLAペプチド複合体を形成する。HLAペプチド複合体をビオチン化酵素によりビオチン化し,ゲルろ過クロマトグラフィー、イオン交換クロマトグラフィーにより過剰のビオチンを除去し,ビオチン化HLAペプチド複合体を分離精製した。得られたビオチン化HLAペプチド複合体をストレプトアビジン(streptavidin; SA)と4:1のモル比で反応させて四量体化し、HLA-テトラマーを合成した。試料を解析する場合には,フィコエリスリン(phycoerythrin; PE)で蛍光標識したSA(SA-PE)を使用し四量体化した。CD8分子はHLA-A,B,Cの一部を除き,α3ドメインの245番目の一アミノ酸であるアラニンに結合することが解っており、そこでアラニンをバリンへ置換した変異導入HLA-テトラマーを作製することで,CD8分子とα3ドメインの結合に依存しないHLAペプチド複合体に対し高いavidityをもつCTLを選択的に検出することが可能である変異導入HLA-テトラマーを合成した。
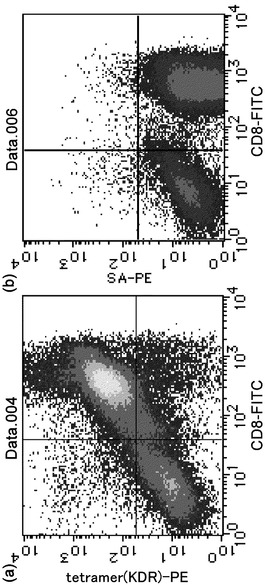
【0038】
合成した変異型HLA-A24 KDR169-テトラマーにより、樹立したCTL クローンを検出した。その結果、図2に示すように、樹立したCTLクローンはテトラマー陽性かつCD8陽性のフラクションに強く染色するCTLであり、本発明のペプチド及びHLA抗原の複合体を特異的に認識するものであることが明らかになった。尚、テトラマー陰性かつCD8陰性のフラクションのシグナルはフィーダー細胞によるものである。
【0039】
[実施例6] VEGFR-2(KDR)由来ペプチドの予測−2(HLA-A*0201)
実施例1と同様にして、KDR タンパク質の全アミノ酸配列から、HLA Peptide Binding Prediction soft(http://bimas.dcrt.nih.gov/cgi-bin/molbio/ken_parker_comboform)によりHLA-A*0201に対する結合親和性の高いものから順に、9量体のペプチドを15種(配列番号26〜40)、10量体のペプチドを12種(配列番号41〜52)予測した(表4)。表4には、9量体及び10量体のそれぞれについて結合親和性の高い順に、そのN末端のKDR タンパク質のアミノ酸配列における位置と共に挙げた。尚、表中のCEA588(配列番号53)はTanaka, H.らによって報告されている腫瘍抗原CEA(癌胎児性抗原)のエピトープの一つである(ポスター発表、AACR #3669, vol.42, p681-682, March 2001)。
【0040】
【表4】
【0041】
[実施例7] HLA-A*0201拘束性エピトープペプチドの決定
実施例6において予測された9量体または10量体のペプチド16種類(配列番号26〜41)を、ヒトHLA(A0201)を発現するトランスジェニックマウス(Hum. Immunol. 2000 Aug.;61(8):764-79 Related Articles, Books, Linkout Induction of CTL response by a minimal epitope vaccine in HLA A*0201/DR1 transgenic mice: dependence on HLA class II restricted T(H) response., BenMohamed L., Krishnan R., Longmate J., Auge C., Low L., Primus J., Diamond DJ.に記載)を用い、vaccination法にてCTLの誘導を行った。
【0042】
6-8週令BALB/C(H-2d)マウスは日本クレア株式会社から手に入れた。A2/Kb トランスジェニックマウス (TGM) はF Jim Primus Ph.D. Vanderbilt-Ingram Cancer Centerより供与された。またT2細胞(TAP欠損細胞、HLA-A*0201陽性)は三重大学第三内科珠玖洋教授より供与された。
【0043】
マウス大腿骨、脛骨より骨髄を採取し、抗CD4,CD8,Gr-1(Beckton-Dickinson)、B220(バイオサイエンス)抗体、及びウサギ補体(PeL-Freez)にてリンパ球及び顆粒球を除去した。細胞は6well plateにて培養し、翌日浮遊細胞を回収して別の6well plateにRPMI1640(Invitrogen)に10%FBSを加えた培溶液にマウスGM-CSF(キリンビールより供与)1,000U/ml,マウスIL-4(PEPRO TECH)1,000U/mlを添加し培養した。その3日後に上記培養液にてhalf medium changeを行い、その2日後にOK-432 10μg/mlにて20時間培養し、浮遊細胞をDCとして使用した。DCはFITCラベルした抗ClassII, CD40抗体及び、PEラベルした抗ClassI,CD11c,CD80,CD86(いずれもBeckton-Dickinson)抗体を反応させてCell Quest softwareを使用したFACS-Calibur (Becton-Dickinson)にて表面抗原の解析を行い、DCであることを確認した。
【0044】
次いで、各ペプチド100μg、HbcAg120-140 helper peptide 140μg、IFA 100μlを混和(計 200μl)し、day 0に右側腹部にday11に左側腹部に皮下摂取した。Day 21 にワクチン接種したマウスの脾臓を採取し、赤血球をred lysis buffer (sigma)にて溶血除去し、その細胞の一部をIFN-γ ELISPOT assayのresponder cellとして使用した。また、残りの細胞は24 well plate (Corning)に6×106 cells/wellにてフィーダー細胞と3:1の比率にて再刺激を加え、5日後に細胞傷害活性を測定した。フィーダー細胞は同種別マウスの赤血球を除去した脾細胞をlipopolysaccharide (LPS) 25μg/ml存在下で3日間培養したものを使用した。
【0045】
本実施例においては、CTLの誘導能をIFN-γを産生する細胞が作るSPOTで評価するIFN-γ ELISPOT法を用いた。96穴平坦底マルチスクリーンプレートMAHA S45(ミリポア)を4℃、一晩抗マウスIFN-γ単クローン抗体(Pharmingen)にて処理し、翌日0.05% Tween 20を含んだPBSにて洗浄し、室温で2時間blocking bufferと反応させた。その後、ペプチドをパルスしたT2細胞とパルスしていないT2細胞を各wellに105 cell/100μlずつ散布した。そこに、ワクチンしたマウスの碑細胞を最大4×106 cell/100μl散布して、全200μlとして一晩37℃で培養した。翌日、各wellを洗浄後、ビオチン化したラット抗マウスIFN-γ抗体(Pharmingen)と2時間反応させ、十分に洗浄し、Extravidinを加えて2時間室温で反応させた。プレートを洗浄後、Alkaline Phosphatase Conjugate Substrate(BIO-RAD)を加えて5分間室温で放置し、IFN-γの産生による青色班をKS ELISPOT compact release(Carl Zeiss)にて計測した。そして
特異的青色班=(TISI(+)SPOT-TISI(-)SPOT)
で決定し、
TISI(+)SPOT/TISI(-)SPOT≧2を満たしたものを有効とした。
その結果、IFN-γELISPOT アッセイにて5種類のエピトープペプチド、配列番号29(アミノ酸開始位置KDR775)、配列番号30(KDR773)、配列番号33(KDR772)、配列番号34(KDR1328)及び配列番号40(KDR190)を得た(表5)。
【0046】
【表5】
【0047】
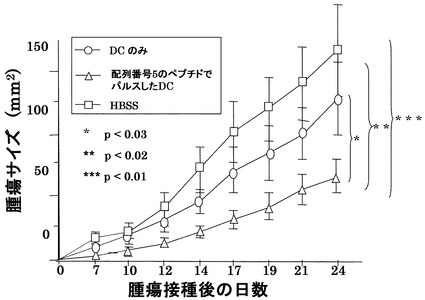
[実施例8] in vivoにおける抗腫瘍効果の確認1
ヒトHLA−A2402とアンカーモチーフが同じである6〜8週齢の雄BALB/cマウス(CLEA Japan Inc.)の骨髄を採取し、実施例7と同様にGM-CSF(キリンビール)(1000U/ml)及びIL-4(Genzyme)(1000U/ml)を添加して培養して、樹状細胞を調製した。この樹状細胞に配列番号5のペプチド(アミノ酸開始位置KDR189)をパルスしたもの(1×106/マウス)をBALB/cマウス9匹に7日間隔で2回右下腹部に投与し、その7日後に結腸癌細胞株Colon26(大鵬薬品株式会社)(5×105個)を右腹部に接種し、マウスの生存と腫瘍増殖を検討した。
【0048】
まず、KDRの発現をRT-PCRで確認したところ、図3に示すように、Colon26細胞系単独ではKDRを発現していないが、Colon26を接種したマウスの腫瘍組織からKDRの発現が確認できた。すなわち、腫瘍血管が腫瘍組織に出現し、腫瘍を形成していることが確かめられた。
【0049】
図3にColon26を接種されたマウスの生存曲線を示す。対照であるHBSS(Hank's Balanced Salt Solution)のみ(100μl)の投与の場合、腫瘍接種から35日後において、9匹中2匹の生存、ペプチドをパルスしない樹状細胞の接種では9匹中5匹の生存であったが、配列番号5のペプチドでパルスした樹状細胞の接種の場合には全9匹が生存していた。
【0050】
図4にマウスにおけるColon26由来の腫瘍の増殖に対する効果を示す。対照と比較して、樹状細胞を接種した場合に腫瘍増殖に対する抑制効果が認められ、配列番号5のペプチドでパルスした樹状細胞の場合に顕著な抑制効果が認められた。
【0051】
[実施例9] 本発明のペプチドのホモロジー検索
本発明のペプチド(配列番号2(アミノ酸開始位置KDR1318)、配列番号3(KDR220)、配列番号5(KDR189)、配列番号8(KDR169)、配列番号11(KDR826)、配列番号30(KDR773)、及び配列番号40(KDR190))のホモロジー検索をBLAST(http://www.ncbi.nlm.nih.gov/blast/blast.cgi)にて行った結果、いずれのペプチドにおいても配列が完全に一致したものは認められなかった(表6)。実施例4において強いCTL活性を誘導できた配列番号8の9量体ペプチド(KDR169)に関しては、2個のミスマッチ配列が1つ(77.8% homology)、3個のミスマッチ配列が2つ(66.7% homology)あるのみであった。実施例8においてin vivoにおける顕著な抗腫瘍効果が認められたペプチド配列番号5のペプチド(KDR 189)に関しては、3個のミスマッチ配列が1つ(66.7% homology)あるのみであった。
【0052】
【表6】
【0053】
[実施例10] in vivoにおける抗腫瘍効果の確認2
ヒトHLA−A0201を発現する6〜8週齢の雄A2/Kbトランスジェニックマウス(TGM)の骨髄を採取し、実施例8と同様にGM-CSF(キリンビール)(1000U/ml)及びIL-4(Genzyme)(1000U/ml)を添加して培養して、樹状細胞を調製した。この樹状細胞に配列番号30(アミノ酸開始位置KDR773)のペプチドをパルスしたもの(1×106/マウス)をA2/Kbトランスジェニックマウス12匹に7日間隔で2回左下腹部に投与し、その7日後にB16メラノーマ細胞(ATCC)(1×106個)を右腹部に接種し、マウスの生存と腫瘍増殖を検討した。
【0054】
図5にB16を接種されたマウスの生存率を示す。対照であるHBSSのみ(100μl)の投与の場合、腫瘍接種から35日後において、12匹中8匹の生存、ペプチドをパルスしない樹状細胞の接種では12匹中11匹の生存であったが、配列番号30のペプチドでパルスした樹状細胞の接種の場合には全12匹が生存していた。
【0055】
図6にマウスにおけるB16マウス由来の腫瘍の増殖に対する効果を示す。対照と比較して、樹状細胞を接種した場合に腫瘍増殖に対する抑制効果が認められ、配列番号30のペプチドでパルスした樹状細胞の場合に顕著な抑制効果が認められた。
【0056】
[実施例11] KDRを強制発現させた細胞に対する細胞障害活性
Miyake, S.ら、Proc. Natl. Acad. Sci. USA. 93, 1320-1324 (1996)に記載の方法に従い、HLA-A24陽性であるヒト大腸癌HT29株(ATCC)にアデノウイルスを用いてKDR(J. Biol. Chem. 1994 Oct 28;269(43):26988-95参照)を強制発現した。具体的には、ヒト293細胞のプラークアッセイによって決定した力価に基いて、細胞を特定の多重度で感染させた。HT29細胞を60mmのディッシュ中に105個の細胞密度で接種し、24時間インキュベートした。次の日、培地を捨て、新しい培地で連続希釈したウイルス200μlと交換した。37℃で1時間インキュベーションした後、増殖培地を添加し、細胞を48時間培養して標的細胞として使用した。対照としてEGFP(Leukemia 1999 Apr; 13(4):605-13参照)をアデノウイルスにて強制発現したHLA-A24陽性HT29株を用いた。KDRとEGFPを強制発現したHT29株は、細胞表面に各々その部分ペプチドを提示している可能性がある。
【0057】
HLA-A24に結合する配列番号8のペプチド(アミノ酸開始位置KDR169)から誘導された実施例3に記載のCTLクローンC29-169を用い、KDR強制発現HLA-A24陽性HT29細胞に対する殺細胞効果をクロムリリースアッセイにて測定した。結果を図7に示す。
【0058】
細胞障害活性は4時間51Cr遊離法にて測定した。標的細胞は6 well plate に1×106cells/wellの濃度にて24時間培養し、KDR又はEGFPを導入したアデノウイルス(Ad-KDR又はAd-EGFP)をMOI 50にて感染させ、48時間後に使用した。標的細胞を100μCiのNa251CrO4にて37℃、1時間ラベルした後、RPMI1640にて3回洗浄し、96穴U型マイクロプレート(Corning)に標的細胞(1×104 cells/100μl)と種々の濃度の効果細胞を100μl加え、総量200μlとして4時間、37℃のCO2インキュベーターにて培養した。培養終了後、各wellより上清を100μl採取し、γカウンターにて測定し、実施例2と同様にして細胞障害活性を算出した。
【0059】
その結果、図7に示すように、CTLクローンC29-169は、KDRを強制発現したHLA-A24陽性HT29に対し、EGFPを強制発現した細胞と比較して顕著に高い殺細胞効果を示した。
【0060】
[実施例12] 内因的にKDRを発現している細胞に対する細胞障害活性
KDRを内因的に発現しているHLA-A24陽性ヒト臍帯血管内皮細胞由来HUVEC KT5株に対する実施例3に記載のCTLクローンC29-169の殺細胞効果について、KDR陽性、HLA-A24陰性HUVEC株 P8を対照として検討した。これらの細胞はKDRの部分ペプチドを細胞表面に提示する。
【0061】
細胞障害活性は4時間51Cr遊離法にて測定した。標的細胞を100μCiのNa251CrO4にて37℃、1時間ラベルした後RPMI1640にて3回洗浄し、96穴U型マイクロプレート(Corning)に標的細胞(1×104 cells/100μl)と種々の濃度の効果細胞を100μl加え、総量200μlとして4時間、37℃のCO2インキュベーターにて培養した。培養終了後、各wellより上清を100μl採取し、γカウンターにて測定し、実施例2と同様にして細胞障害活性を算出した。
【0062】
図8の結果から明らかなように、CTLクローンC29-169は、内在的にKDRを発現しているHLA-A24陽性HUVECに対して殺細胞効果を示し、HLA-A24陰性HUVECに対しては低い殺細胞効果を示した。
【0063】
[実施例13] HLA-A0201結合性ペプチドを用いたCTLクローンの樹立
実施例7のELISPOT解析において有効とされたHLA-A0201に結合する配列番号40のペプチド(アミノ酸開始位置KDR190)、配列番号30のペプチド(KDR773)のN末端から2番目のアミノ酸をロイシンに変換したペプチドKDR773-2L(配列番号54)、配列番号29のペプチド(KDR775)、配列番号34のペプチド(KDR1328)、及び配列番号33(KDR772)を用いて、実施例3と同様にして、それぞれ2種類(KWC14-190、KWC65-190)、4種類(KWC44-773-2L、KWC76-773-2L、KWC129-773-2L、KWC134-773-2L)、2種類(KWC81-775、KWC85-775)、12種類(KWC16、KWC21、KWC22、KWC47、KWC51、KWC108、KWC117、KWC132、KWC151、KWC153、KWC156、KWC159)、1種類(KWC72-772)のCTLクローンを得た。それぞれの細胞傷害活性を表7に示す。
【0064】
【表7】
【0065】
表7の結果から明らかなように、配列番号29、33、34、または40に示すペプチド、及び配列番号30に示すペプチドのN末端から2番目のアミノ酸がロイシンである配列番号54に示すペプチドのいずれも、顕著な細胞傷害活性を有するCTLを誘導することができ、エピトープペプチドとして有効であることが示された。
【0066】
[実施例14]
HLA-A0201陽性T2株に配列番号29のペプチド(アミノ酸開始位置KDR775)を加えて標的細胞とし、実施例13で配列番号29のペプチドから得られたCTLクローン(KDR C85-775(KWC85-775))の殺細胞効果をクロムリリースアッセイを用いて検討した。
その結果、図9に示すように、CTLクローンC85-775(KWC85-775)は、配列番号29のペプチドを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、配列番号29のペプチドを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0067】
[実施例15]
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対するCTLクローン(KDR C85-775(KWC85-775))の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A0201陽性HePG2を用いた。
その結果、図10に示すように、CTLクローンC85-775(KWC85-775)は、KDRペプチドを表面に提示したHLA-A0201陽性細胞に対し、EGFPペプチドを提示した細胞と比較して顕著に高い殺細胞効果を示した。
【0068】
[実施例16]
HLA-A0201陽性T2株に配列番号34のペプチド(アミノ酸開始位置KDR1328)を加えて標的細胞とし、実施例13で配列番号34のペプチドから得られたCTLクローン(KDR C51-1328(KWC51))の殺細胞効果をクロムリリースアッセイを用いて検討した。
その結果、図11に示すように、CTLクローンKDR C51-1328(KWC51)は、配列番号34のペプチドを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、配列番号34のペプチドを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0069】
[実施例17]
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対するCTLクローン(KDR C51-1328(KWC51))の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A0201陽性HePG2を用いた。
その結果、図12に示すように、CTLクローンKDR C51-1328(KWC51)は、KDRペプチドを表面に提示したHLA-A0201陽性細胞に対し、EGFPペプチドを提示した細胞と比較して顕著に高い殺細胞効果を示した。
【0070】
[実施例18]
HLA-A0201陽性T2株に配列番号30の2番目のペプチドをロイシンに変換した配列番号54のペプチド(KDR773-2L)を加えて標的細胞とし、実施例13で配列番号34のペプチドから得られたCTLクローン(KDR C44-773-2L(KWC44-773-2L))の殺細胞効果をクロムリリースアッセイを用いて検討した。結果を図13に示す。
その結果、図13に示すように、CTLクローンKDR C44-773-2L(KWC44-773-2L)は、KDR773-2Lを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、KDR773-2Lを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0071】
[実施例19]
HLA-A0201陽性T2株に配列番号30の2番目のペプチドをロイシンに変換した配列番号54に示すペプチド(KDR773-2L)より得られたCTLクローン(KDR C44-773-2L(KWC44-773-2L))のクロムリリースアッセイでの殺細胞効果を、標的細胞としてT2に改変していない配列番号30に示すペプチド(アミノ酸開始位置KDR773)をパルスしたもので検討した。
その結果、図14に示すように、改変したペプチドで刺激し、得られたCTLクローンは改変前のペプチドを認識し、傷害することができた。
【0072】
[実施例20]
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対するCTLクローン(KDR C44-773-2L(KWC44-773-2L))の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A0201陽性HePG2を用いた。
その結果、図15に示すように、CTLクローンKDR C44-773-2L(KWC44-773-2L)は、KDRペプチドを表面に提示したHLA-A0201陽性細胞に対し、EGFPペプチドを提示した細胞と比較して顕著に高い殺細胞効果を示した。
【0073】
[実施例21]
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対する配列番号34のペプチドから得られたCTLクローン(KDR C51-1328(KWC51))の殺細胞効果が各種抗体でブロックされるかどうかについて検討した。
その結果、図16に示すように、標的細胞が提示するHLA抗原に対する抗HLA ClassI抗体と、細胞傷害性T細胞のマーカーとなるCD8に対する抗CD8抗体がその作用を阻害した。
【0074】
[実施例22] Dorsal Air Sac assay(DAS)による血管新生阻害活性の検討
ペプチドエピトープの実際の血管新生阻害活性をマウスでDASアッセイ(Clinical Cancer Research Vol.5, 2185-2191, August 1999; Cancer Research 62, 6116-6123, November 1, 2002参照)にて評価した。
【0075】
チャンバーリングの両面にメンブレンフィルターを固定し、エチレンオキサイトガスで滅菌した。B16メラノーマ細胞を5×105 cells/0.15mlに調整し、チャンバー内に注入した。マウス(A2/kbトランスジェニックマウス(B16由来))を腹臥位として固定し、マウス背部皮下にシリンジにて5〜10mlの空気を注入しair sacを作製した。Sac下部を約1cm切開し、細胞浮遊液で満たされたチャンバーリングを背部皮下に移植し、皮膚をスキンステープラで閉じた。移植日より6日目にマウス皮膚をチャンバーごと剥皮し、伸展させた状態で固定した。黒色ゴムリングでマーキングし、実態顕微鏡を用いて新生血管を計測した。悪性腫瘍細胞から放出される血管新生因子によって新たに形成される血管は、ジグザグに進行する特徴があり、本来存在するバックグラウンドの血管とは形態学的に異なっている。従って、新生血管の判定はゴムリング内の蛇行走行を示す血管で、長さが3mm以上のものとし、血管新生指数(Angiogenesis Index(AI))としてカウントした。AIは蛇行血管数に応じて0〜5までの6段階で評価し、蛇行血管が5本以上のものはすべて5とした。
ワクチン接種はチャンバー移植の14日前にHBSS、DC単独、DCにペプチドパルスしたものをそれぞれ5×104cells尾静脈に注入し、それを1週間後に再度行い、計2回接種した。結果を図17及び以下の表8に示す。
【0076】
【表8】
【0077】
図17及び表8の結果から明らかなように、DCに配列番号40に示すペプチド(アミノ酸開始位置KDR190)、配列番号33に示すペプチド(KDR772)、配列番号30に示すペプチド(KDR773)、配列番号54に示すペプチド(KDR773-2L)、配列番号29に示すペプチド(KDR775)、及び配列番号46に示すペプチド(KDR1084)をワクチン接種した群において、DC単独投与群と比較して、蛇行血管の形成が明らかに抑制され、統計学的有意さをもって血管新生阻害効果があった。
【0078】
尚、これらのエピトープペプチドを用いたワクチン接種による正常な生理的血管新生に対する悪影響を観察するために、ワクチン接種したマウスにおける創傷治癒及び受精能を解析した。しかしながら、ワクチン接種したマウスにおいて有意な悪影響はなかった。更に、KDRペプチドを認識するCTLクローンを用いて非増殖性または増殖性内皮細胞に対する細胞傷害性をin vitroで試験し、ヒトにおける悪影響を検討したところ、これらのクローンは非増殖性内皮細胞よりも増殖性内皮細胞に対して強い活性を示した。
【0079】
[実施例23]
ヒトHLA-A0201を発現するA2/Kbトランスジェニックマウスを用いて大腸癌株MC38移植担癌マウスに対する配列番号30(アミノ酸開始位置KDR773)と配列番号34(KDR1084)ペプチドを不完全フロイントアジュバントと混合させてそれぞれday3,13に投与した場合の腫瘍体積の変化を示した。
【0080】
実施例22と同様のワクチン接種方法でHLA-A*0201拘束性エピトープペプチド(KDR773)による治療モデルにおけるin vivo抗腫瘍効果を腫瘍面積により評価した。その結果、図18から明らかなように、配列番号30のペプチド及び配列番号34のペプチドのワクチン接種により、有意な腫瘍増殖抑制効果が確認できた。
【0081】
[実施例24]
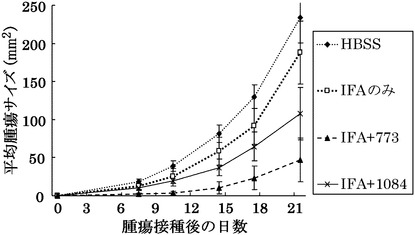
HLA-A0201陽性T2株に配列番号40のペプチド(アミノ酸開始位置KDR190)を加えて標的細胞とし、実施例13で配列番号40のペプチドから得られたCTLクローン(KWC65-190)の殺細胞効果をクロムリリースアッセイを用いて検討した。
【0082】
その結果、図19に示すように、CTLクローンKWC65-190は、配列番号40のペプチドを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、配列番号40のペプチドを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0083】
[実施例25]
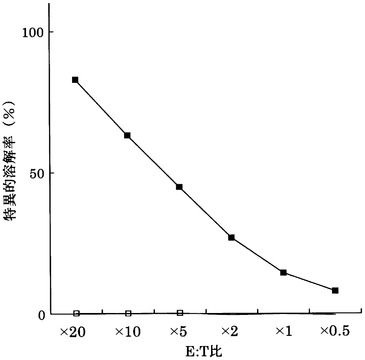
HLA-A0201陽性T2株に配列番号33のペプチド(アミノ酸開始位置KDR772)を加えて標的細胞とし、実施例13で配列番号33のペプチドから得られたCTLクローン(KWC72-772)の殺細胞効果をクロムリリースアッセイを用いて検討した。
その結果、図20に示すように、CTLクローンKWC72-772は、配列番号33のペプチドを提示したHLA-A0201陽性細胞に対して明らかに殺細胞効果を示した。これに対して、配列番号33のペプチドを提示しない対照細胞に対しては、全く殺細胞効果が見られなかった。
【0084】
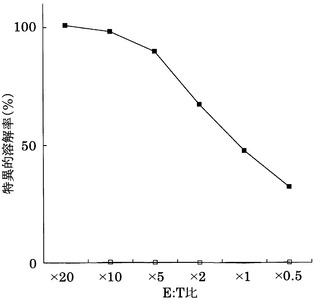
[実施例26]
KDRをアデノウイルスで強制発現させた肝細胞癌株HLA-A0201陽性HePG2に対するCTLクローン(KWC72-772)の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A0201陽性HePG2を用いた。
その結果、図21に示すように、CTLクローンKWC72-772は、KDRペプチドを表面に提示したHLA-A0201陽性細胞に対し、EGFPペプチドを提示した細胞と比較して顕著に高い殺細胞効果を示した。
【0085】
[実施例27]
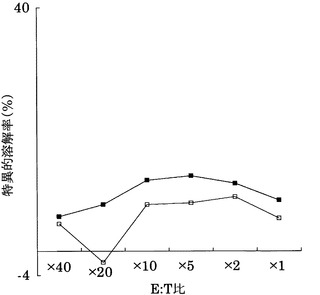
HLA-A24陽性であるヒトB-リンパ芽球細胞系A24-LCL(HLA-A24/24、宝酒造製)に配列番号2(アミノ酸開始位置KDR1318)、3(KDR220)、5(KDR189)、及び11(アミノ酸開始位置KDR826)のペプチドを加えて標的細胞とし、実施例3に記載のCTLクローンC7-1318、KWC46、C18-189、及びC65-826の殺細胞効果をクロムリリースアッセイを用いて検討した。
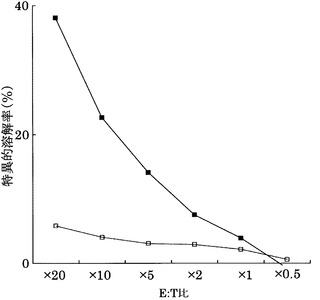
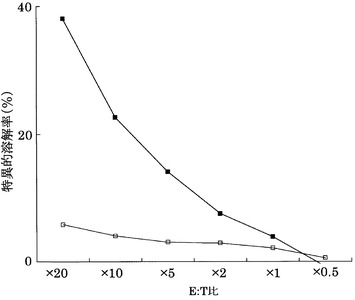
その結果、図22〜25に示すように、CTLクローンC7-1318、KWC46、C18-189、及びC65-826は、それぞれ配列番号2、3、5、及び11のペプチドを提示したHLA-A24陽性細胞に対して明らかに殺細胞効果を示した。これに対して、各ペプチドを提示しない対照細胞に対しては、全く、またはほとんど殺細胞効果が見られなかった。
【0086】
[実施例28]
実施例11と同様にして、KDRをアデノウイルスで強制発現させたヒト大腸癌HT29株(ATCC)に対する実施例3に記載のCTLクローンC7-1318、KWC46、C18-189、及びC65-826の殺細胞効果をクロムリリースアッセイを用いて検討した。対照としてEGFPを強制発現させたHLA-A24陽性HT29株を用いた。
その結果、図26〜29に示すように、CTLクローンC7-1318、KWC46、C18-189、及びC65-826は、いずれもKDRを強制発現したHLA-A24陽性HT29に対し、EGFPを強制発現した細胞と比較して顕著に高い殺細胞効果を示した。
【0087】
[実施例29]
実施例22と同様にして、BALB/cマウスにおいてcolon26細胞によって誘導される血管新生応答に対するペプチドエピトープの血管新生阻害活性をDASアッセイにて評価した。
図30及び31の結果から明らかなように、DCに配列番号5に示すペプチド(アミノ酸開始位置KDR189)及び配列番号11に示すペプチド(KDR826)をワクチン接種した群において、DC単独投与群と比較して、蛇行血管の形成が明らかに抑制され、統計学的有意さをもって血管新生阻害効果があった。
【0088】
[実施例30]
癌患者のPBMCを用いて、各種エピトープペプチド(HLA-24結合親和性の高いペプチドとして配列番号8(アミノ酸開始位置KDR169)、配列番号5(KDR189)、配列番号3(KDR220)、配列番号11(KDR826)、及び配列番号17(KDR1318)、並びにHLA-A0201結合親和性の高いペプチドとして配列番号40(KDR190)、配列番号33(KDR772)、配列番号30(KDR773)、配列番号29(KDR775)、及び配列番号34(KDR1328))で刺激した後の各ペプチドに対するCTL応答を調べた。
【0089】
Maeda, Y.ら、Br. J. Cancer 87, 796-804 (2002)に報告されている方法を用いてPBMC中のペプチド特異的CTLを検出した。まず、患者から採取したPBMC(1×105個)を200μlの培地を入れたu底型の96ウェルプレートのウェルで各ペプチド10μMと共にインキュベートした。培地は45%のRPMI-1640培地、45%のAIM-V培地、10%のFBS、100U/mlのインターロイキン-2(IL-2)、及び0.1μM MEM非必須アミノ酸溶液から構成される。3日ごとに培地の半分を除去し、対応するペプチド(20μM)を含有する新しい培地と交換した。12日間のインキュベーション後、細胞を回収して各ペプチドに応答したIFN-γ産生能を試験した。対応するペプチドに応答してペプチドで刺激したPBMCによって産生されるIFN-γの量が対照であるHIVペプチドに応答して産生される量の2倍以上多い場合には陽性と判定した。
【0090】
その結果、HLA-A2及び-A24の癌患者においても、健康なドナーと同様にCTL前駆体が産生されることが示された(表9)。表中、患者A、B及びCはHLA-A24型の患者であり、患者DはHLA-A02型の患者である。また患者A及び患者Dは結腸癌患者、患者B及び患者Cはメラノーマ患者である。尚、CMVは陽性対照として用いたサイトメガロウイルス由来ペプチドであり、該ペプチドについてはA−24結合親和性の高いものは、例えばKuzushima, K.ら, Blood 2001, 98(6):p1872-1881に、A−2結合親和性の高いものは、例えばSolache A.ら, J. of Immunol. 1999, 163(10);p5512-5518にそれぞれ記載されている。
【0091】
【表9】
【0092】
更に、実施例14等と同様にして標準的な4時間の51Cr遊離法を行い、これらのCTL前駆体からCTL系を確立した(図32)。
【産業上の利用可能性】
【0093】
本発明により、広範囲にわたる腫瘍組織に形成される内皮細胞を標的として細胞傷害性T細胞を誘導し、癌ワクチンとして非常に有効な新規ペプチド、及びこれを含有する腫瘍の治療及び予防のための医薬が提供される。
本明細書で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書にとり入れるものとする。
【図面の簡単な説明】
【0094】
【図1】図1は、標的細胞に対するCTLクローンによる細胞傷害活性を示す。
【図2】図2は、樹立したCTLクローンのHLA-テトラマー解析の結果を示す。(a)本発明のペプチドによって誘導されたCTLクローン及びフィーダー細胞(b)フィーダー細胞(対照)
【図3】図3は、colon26を接種したBALB/cマウスの生存に対する本発明のペプチドを用いたワクチン接種の効果を示す。
【図4】図4は、colon26由来の腫瘍の増殖に対する本発明のペプチドを用いたワクチン接種の効果を示す。
【図5】図5は、B16を接種したマウスの生存に対する本発明のペプチドを用いたワクチン接種の効果を示す。
【図6】図6は、B16メラノーマ由来の腫瘍の増殖に対する本発明のペプチドを用いたワクチン接種の効果を示す。A2/Kb TGMに対してB16腫瘍細胞を皮下注射でチャレンジする前に、KDR773でパルスしたDC(●)、DC単独(△)、またはHBSS(□)で1週間の間隔をおいて2回ワクチン接種した。平均の腫瘍増殖を図示した。P<0.01。
【図7】図7は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC29-169の殺細胞効果を示す。
【図8】図8は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC29-169の殺細胞効果を示す。
【図9】図9は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C85-775(KWC85-775)の殺細胞効果を示す。
【図10】図10は、KDRペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C85-775(KWC85-775)の殺細胞効果を示す。
【図11】図11は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C51-1328(KWC51)の殺細胞効果を示す。
【図12】図12は、KDRペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C51-1328(KWC51)の殺細胞効果を示す。
【図13】図13は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C44-773-2L(KWC44-773-2L)の殺細胞効果を示す。
【図14】図14は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C44-773-2L(KWC44-773-2L)の殺細胞効果を示す。
【図15】図15は、KDRペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKDR C44-773-2L(KWC44-773-2L)の殺細胞効果を示す。
【図16】図16は、CTLクローンの殺細胞効果に対する各種抗体の阻害作用を示す。
【図17】図17は、A2/Kb TGMにおいてB16細胞によって誘導される血管新生応答に対する本発明のペプチドの阻害活性を示す。マウスをHBSS、パルスしていないDC、または本発明のエピトープペプチド(KDR190、772、773、773-2L、775、または1084)でパルスしたDCで2回ワクチン接種した。矢印は特徴的なジグザグに進む新たに形成された血管を示す。
【図18】図18は、本発明のペプチドのトランスジェニックマウスにおけるヒト腫瘍体積の増加に対する抑制効果を示す。
【図19】図19は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKWC65-190の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図20】図20は、本発明のペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKWC72-772の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図21】図21は、KDRペプチドを提示したHLA-A0201陽性細胞に対するCTLクローンKWC72-772の殺細胞効果を示す。黒四角:HePG2-VEGFR2、□:HePG2-EGFP
【図22】図22は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC7-1318の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図23】図23は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンKWC46の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図24】図24は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC18-189の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図25】図25は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC65-826の殺細胞効果を示す。黒四角:ペプチド存在下、□:ペプチド不存在下
【図26】図26は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC7-1318の殺細胞効果を示す。黒四角:HT29-VEGFR2、□:HT29-EGFP
【図27】図27は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンKWC46の殺細胞効果を示す。黒四角:HT29-VEGFR2、□:HT29-EGFP
【図28】図28は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC18-189の殺細胞効果を示す。黒四角:HT29-VEGFR2、□:HT29-EGFP
【図29】図29は、本発明のペプチドを提示したHLA-A24陽性細胞に対するCTLクローンC65-826の殺細胞効果を示す。黒四角:HT29-VEGFR2、□:HT29-EGFP
【図30】図30は、BALB/cにおいてcolon26細胞によって誘導される血管新生応答に対する本発明のペプチドの阻害活性を示す。マウスをHBSS、パルスしていないDC、または本発明のエピトープペプチド(KDR189または826)でパルスしたDCで2回ワクチン接種した。矢印は特徴的なジグザグに進む新たに形成された血管を示す。
【図31】図31は、BALB/cにおいてcolon26細胞によって誘導される血管新生応答に対する本発明のペプチドの阻害を示す。柱は平均を、縦棒は標準誤差を示す。
【図32】図32は、本発明のペプチドで刺激した患者由来のCTLによるHLA-A24陽性細胞に対する殺細胞効果を示す。黒ひし形:ペプチド存在下、□:ペプチド不存在下(A)配列番号8(アミノ酸開始位置KDR169)で刺激して得られた結腸癌患者由来のCTLの殺細胞効果(B)配列番号5(アミノ酸開始位置KDR189)で刺激して得られた結腸癌患者由来のCTLの殺細胞効果(C)配列番号3(アミノ酸開始位置KDR220)で刺激して得られた結腸癌患者由来のCTLの殺細胞効果
【特許請求の範囲】
【請求項1】
配列番号2、3、5、8、11または12に示すアミノ酸配列からなるペプチドから選ばれるノナペプチド。
【請求項2】
配列番号2、3、5、8、11または12に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチド。
【請求項3】
N末端から2番目のアミノ酸がフェニルアラニン、チロシン、メチオニンまたはトリプトファンである、請求項2に記載のペプチド。
【請求項4】
C末端のアミノ酸がフェニルアラニン、ロイシン、イソロイシン、トリプトファンまたはメチオニンである、請求項2または3に記載のペプチド。
【請求項5】
配列番号29、30、33、34、40、または46に示すアミノ酸配列からなるペプチドから選ばれるノナペプチドまたはデカペプチド。
【請求項6】
配列番号29、30、33、34、40、または46に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチド。
【請求項7】
N末端から2番目のアミノ酸がロイシンまたはメチオニンである、請求項6に記載のペプチド。
【請求項8】
C末端のアミノ酸がバリンまたはロイシンである、請求項6または7に記載のペプチド。
【請求項9】
請求項1〜8のいずれか1項に記載のペプチドを1種以上含む、腫瘍の治療及び/または予防のための医薬。
【請求項10】
請求項1〜8のいずれか1項に記載のペプチドを1種以上含む、糖尿病性網膜症、慢性関節リウマチ、乾癬、アテローム性動脈硬化症の治療のための医薬。
【請求項11】
請求項1〜8のいずれか1項に記載のペプチドとHLA抗原とを含む複合体を表面に提示しているエキソソーム。
【請求項12】
HLA抗原がHLA−A24またはHLA−A02である、請求項11に記載のエキソソーム。
【請求項13】
HLA抗原がHLA−A2402またはHLA−0201である、請求項12に記載のエキソソーム。
【請求項14】
請求項1〜8のいずれか1項に記載のペプチドを用いて細胞傷害性T細胞誘導能の高い抗原提示細胞を誘導する方法。
【請求項15】
請求項1〜8のいずれか1項に記載のペプチドを用いて細胞傷害性T細胞を誘導する方法。
【請求項16】
請求項1〜8のいずれか1項に記載のペプチドをコードするポリヌクレオチドを含む遺伝子を抗原提示細胞に導入することを含む、細胞傷害性T細胞誘導能の高い抗原提示細胞を誘導する方法。
【請求項17】
請求項1〜8のいずれか1項に記載のペプチドを用いて誘導される、単離された細胞傷害性T細胞。
【請求項18】
HLA抗原と請求項1〜8のいずれか1項に記載のペプチドとの複合体を提示してなる抗原提示細胞。
【請求項19】
請求項14または15に記載の方法によって誘導される、請求項18に記載の抗原提示細胞。
【請求項20】
請求項1〜8のいずれか1項に記載のペプチドを有効成分とする、病態部位における血管新生阻害のためのワクチン。
【請求項21】
HLA抗原がHLA−A24またはHLA−A02である被験者に対して投与するための、請求項20に記載のワクチン。
【請求項22】
悪性腫瘍の増殖及び/または転移を抑制するために使用される、請求項20または21に記載のワクチン。
【請求項1】
配列番号2、3、5、8、11または12に示すアミノ酸配列からなるペプチドから選ばれるノナペプチド。
【請求項2】
配列番号2、3、5、8、11または12に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチド。
【請求項3】
N末端から2番目のアミノ酸がフェニルアラニン、チロシン、メチオニンまたはトリプトファンである、請求項2に記載のペプチド。
【請求項4】
C末端のアミノ酸がフェニルアラニン、ロイシン、イソロイシン、トリプトファンまたはメチオニンである、請求項2または3に記載のペプチド。
【請求項5】
配列番号29、30、33、34、40、または46に示すアミノ酸配列からなるペプチドから選ばれるノナペプチドまたはデカペプチド。
【請求項6】
配列番号29、30、33、34、40、または46に示すアミノ酸配列において1個、2個または数個のアミノ酸が置換または付加されており、細胞傷害性T細胞の誘導能を有するペプチド。
【請求項7】
N末端から2番目のアミノ酸がロイシンまたはメチオニンである、請求項6に記載のペプチド。
【請求項8】
C末端のアミノ酸がバリンまたはロイシンである、請求項6または7に記載のペプチド。
【請求項9】
請求項1〜8のいずれか1項に記載のペプチドを1種以上含む、腫瘍の治療及び/または予防のための医薬。
【請求項10】
請求項1〜8のいずれか1項に記載のペプチドを1種以上含む、糖尿病性網膜症、慢性関節リウマチ、乾癬、アテローム性動脈硬化症の治療のための医薬。
【請求項11】
請求項1〜8のいずれか1項に記載のペプチドとHLA抗原とを含む複合体を表面に提示しているエキソソーム。
【請求項12】
HLA抗原がHLA−A24またはHLA−A02である、請求項11に記載のエキソソーム。
【請求項13】
HLA抗原がHLA−A2402またはHLA−0201である、請求項12に記載のエキソソーム。
【請求項14】
請求項1〜8のいずれか1項に記載のペプチドを用いて細胞傷害性T細胞誘導能の高い抗原提示細胞を誘導する方法。
【請求項15】
請求項1〜8のいずれか1項に記載のペプチドを用いて細胞傷害性T細胞を誘導する方法。
【請求項16】
請求項1〜8のいずれか1項に記載のペプチドをコードするポリヌクレオチドを含む遺伝子を抗原提示細胞に導入することを含む、細胞傷害性T細胞誘導能の高い抗原提示細胞を誘導する方法。
【請求項17】
請求項1〜8のいずれか1項に記載のペプチドを用いて誘導される、単離された細胞傷害性T細胞。
【請求項18】
HLA抗原と請求項1〜8のいずれか1項に記載のペプチドとの複合体を提示してなる抗原提示細胞。
【請求項19】
請求項14または15に記載の方法によって誘導される、請求項18に記載の抗原提示細胞。
【請求項20】
請求項1〜8のいずれか1項に記載のペプチドを有効成分とする、病態部位における血管新生阻害のためのワクチン。
【請求項21】
HLA抗原がHLA−A24またはHLA−A02である被験者に対して投与するための、請求項20に記載のワクチン。
【請求項22】
悪性腫瘍の増殖及び/または転移を抑制するために使用される、請求項20または21に記載のワクチン。
【図1】

【図2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図31】
【図32】
【図3】
【図17】
【図30】
【図2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図31】
【図32】
【図3】
【図17】
【図30】
【公開番号】特開2007−277251(P2007−277251A)
【公開日】平成19年10月25日(2007.10.25)
【国際特許分類】
【出願番号】特願2007−123850(P2007−123850)
【出願日】平成19年5月8日(2007.5.8)
【分割の表示】特願2004−571938(P2004−571938)の分割
【原出願日】平成15年9月12日(2003.9.12)
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【Fターム(参考)】
【公開日】平成19年10月25日(2007.10.25)
【国際特許分類】
【出願日】平成19年5月8日(2007.5.8)
【分割の表示】特願2004−571938(P2004−571938)の分割
【原出願日】平成15年9月12日(2003.9.12)
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【Fターム(参考)】
[ Back to top ]