
L−システインの製造法

【課題】O−アセチルセリンを利用してL−シスチン、もしくはそれらの誘導体、又はこれらの混合物を製造する新たな方法を提供する。
【解決手段】チオ硫酸とO−アセチル−L−セリンからS−スルホ−L−システインを生成する反応を触媒するO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌を、O−アセチル−L−セリン及びチオ硫酸に、これらを含む反応液中で作用させ、該反応液からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を採取することにより、これらの化合物を製造する。
【解決手段】チオ硫酸とO−アセチル−L−セリンからS−スルホ−L−システインを生成する反応を触媒するO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌を、O−アセチル−L−セリン及びチオ硫酸に、これらを含む反応液中で作用させ、該反応液からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を採取することにより、これらの化合物を製造する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、L−システイン又はその関連物質の製造法に関し、詳しくはL−システイン又はその関連物質の製造に好適な細菌、及びそれらを用いたL−システイン又はその関連物質の製造法に関する。L−システイン及びその関連物質は、医薬品、化粧品及び食品分野で利用されている。
【背景技術】
【0002】
L−システインは、毛髪、角、羽毛等のケラチン含有物質から抽出することにより、あるいはDL−2−アミノチアゾリン−4−カルボン酸を前駆体とする細菌酵素変換により得られている。また、新規な酵素を用いた固定化酵素法によるL−システインの大量生産も計画されている。
【0003】
さらに、細菌を用いた発酵法によるL−システインの生産も試みられている。例えば、本発明者らは、L−システイン分解系が抑制され、かつ、L−システインによるフィードバック阻害が低減されたセリンアセチルトランスフェラーゼ(serine acetyltransferase(EC 2.3.1.30):以下、「SAT」ともいう)を保持するエシェリヒア属細菌を用いたL−システインの製造法を開示している(特許文献1)。また、L−システイン分解系を抑制することによってL−システイン生産能が高められた細菌としては、シスタチオニン−β−リアーゼ(特許文献1)、トリプトフォアナーゼ(特許文献2)、O−アセチルセリン スルフヒドリラーゼB(特許文献3)の活性を低下させたコリネ型細菌又はエシェリヒア属細菌が知られている。また、L−システインによるフィードバック阻害が低減された特定の変異を有するSATをコードするDNA配列により脱制御されたL-システイン物質代謝を有する細菌を用いた、L−システインの製造法が知られている(特許文献4)。
【0004】
さらに、YdeDタンパク質をコードするydeD遺伝子(非特許文献1)、及びYfiKタンパク質をコードするyfiK遺伝子(特許文献5)は、L−システイン経路の代謝産物の排出に関与していることが知られている。また、細胞に毒性の物質を排出するのに適したタンパク質をコードする遺伝子であるmar-遺伝子座、acr-遺伝子座、cmr-遺伝子座、mex-遺伝子座、bmr-遺伝子座、qacA-遺伝子座(特許文献6)、又はemrAB、emrKY、yojIH、acrEF、bcrもしくはcusA遺伝子(特許文献7)の発現を上昇させることによりL−システイン生産能を高める技術が知られている。
【0005】
一方、抗生物質又は細菌に毒性の物質を細胞から直接放出するために好適である蛋白質をコードする遺伝子を過剰発現する細菌を用いたL−システインの製造法が報告されている(特許文献8)。
【0006】
このように、L−システインを排出する因子が知られているが、これらの排出因子はL−システインとその前駆体であるO−アセチル−L−セリン(以下、「OAS」と記載することがある)とを区別することができないため、これらのL−システイン排出因子を増強した微生物では、培地中にL−システインだけでなく副生物としてOASも蓄積することが知られていた(非特許文献1)。また、L−システインは細胞にとって毒性の高いアミノ酸であることから、排出因子を強化したとしても、細胞内で生成したL−システインによって生育が阻害されることが考えられた。
【0007】
オートトランスポーターとは、1987年にナイセリア・ゴノロエ(Neisseria gonorrhoeae)のIgA1プロテアーゼについて初めて発見された(非特許文献2)、グラム陰性菌が持つtype-Vのタンパク質分泌システムである。このシステムでは、N末からシグナル配列、パッセンジャーと呼ばれるペプチド(すなわちIgA1プロテアーゼドメイン)、及びC末にオートトランスポータードメインと呼ばれるリンカー配列及びそれに続くβバレル構造を持ったタンパク質が発現し、このタンパク質はシグナル配列に依存して内膜を通過した後、βバレルが外膜に入り込みパッセンジャーが通過する穴を形成し、パッセンジャー領域が外膜の外に分泌・固定される。パッセンジャーの部分に目的の酵素ペプチドを挿入することで、有用酵素の提示発現システムとして利用されている(非特許文献3)。
【0008】
酵素を微生物の細胞外膜に提示させることにより、細胞外に分泌される酵素と比べて、安定した酵素活性が得られることがある。一方、微生物が生産する酵素を用いて、酵素反応により有用物質の生産を行う場合は、目的酵素を細胞内に蓄積させて、細胞を破砕して酵素を取り出してから反応させる方法や、目的酵素を培地中に分泌させる方法等が考えられる。しかしながら、これらの場合は、酵素を生成するプロセスや、酵素反応後に、生成した反応物と変換酵素を分離するための複雑な精製プロセスが必要となり、高コストとなってしまう。
【0009】
また、他のオートトランスポーターとしては、シュードモナス エスピー.(Pseudomonas sp. ) LP7315株由来の耐熱性モノグリセリドリパーゼのオートトランスポート(AT)システム(非特許文献4)が知られている。今中や山下は、このPseudomonas sp. strain LP7315由来のATシステムを用いて、システイン前駆体であるOASをシステインに変換する酵素(OASS-A)を細胞外膜に提示させて、L−システイン(もしくはS−スルホ−L−システイン)の生産を行うことを示している(非特許文献5、6)。
【0010】
L−システインの前駆体であるOASをL−システインに変換する酵素は、O−アセチルセリンサルフヒドリラーゼ(O-acetylserine sulfhydrylase、以下「OASS」と記載することがある)と呼ばれ、二つのアイソザイムOASS-A、OASS-Bが存在することが知られている。エシェリヒア・コリ(Escherichia coli)では、OASS-A及びOASS-Bは、それぞれcysK、cysMにコードされている(非特許文献7)。OASS-Aは、OASとスルフィド(sulfide)との反応を触媒し、L−システインを産生する。一方、OASS-Bは、OASとスルフィドは又チオサルフェイト(チオ硫酸、thiosulfate)との反応を触媒し、それぞれL−システイン又はS−スルホ−L−システインを産生する。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】特開平11−155571号
【特許文献2】特開2003−169668
【特許文献3】特開2005−245311
【特許文献4】特表2000−504926
【特許文献5】特開2004−49237
【特許文献6】米国特許第5972663号
【特許文献7】特開2005−287333
【特許文献8】特開平11−56381号
【非特許文献】
【0012】
【非特許文献1】Dassler, T. et al., Mol. Microbiol., 36, 1101-1112 (2000)
【非特許文献2】Pohlner J., et al,. Nature 325 (6103):458-62 (1987)
【非特許文献3】Joachin, J., Appl Microbiol. Biotechnol., 69:607-614 (2006)
【非特許文献4】Sakiyama T, et al., J. Biosci. Bioeng., 91(1):27-32 (2001)
【非特許文献5】今中ら、化学工学会第37回秋季大会 (2005)講演要旨
【非特許文献6】山下ら、化学工学会春季大会(2006)講演要旨
【非特許文献7】Kredich N. M. et al., Biosynthesis of Cysteine. In Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed., Vol. 1, pp. 514-527 (1996)
【発明の概要】
【発明が解決しようとする課題】
【0013】
本発明は、OASを利用してL-システイン関連物質を製造する新たな方法を提供することを課題とする。
【課題を解決するための手段】
【0014】
前記したように、OASS-AによるOASとスルフィドとの反応では、直接反応生成物としてシステインができるため有用である反面、S源としてスルフィドを使わなければならない。具体的なスルフィドとしては、例えば硫化ナトリウム(Na2S)が挙げられるが、スルフィドは取り扱い上の安全性や物質自体の安定性の面で問題がある。硫化ナトリウムは、簡単に硫化水素(H2S)になり気中に放出されてしまう。例えば、反応溶液のpHを調整するときにはこの傾向が激しく、反応系に添加したNa2Sのかなりの量がH2Sとして気中に放出される。また、例えば37℃にて変換反応を行うとさらにこの傾向は増大する。工業レベルでの生産を考えるとき、放出されるH2Sの安全性も問題となる。一方、OASS-Bは、チオサルフェイトをS源として用いることができるため安全性が高く使いやすい。また、反応産物であるS−スルホ−L−システインはジチオスレイトール(DTT)等の還元剤により容易にL−システインに変換することができる。
【0015】
本発明者らは、cysMにコードされるOASS-Bをオートトランスポーターと融合することで細胞外膜に提示した菌を用いて、OAS及びチオ硫酸からL-システイン関連物質を生産することができることを見出し、本発明を完成させるに至った。
すなわち本発明は、以下のとおりである。
【0016】
(1)チオ硫酸とO−アセチル−L−セリンからS−スルホ−L−システインを生成する反応を触媒するO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌を、O−アセチル−L−セリン及びチオ硫酸に、これらを含む反応液中で作用させ、該反応液からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を採取することを特徴とする、S−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物の製造法。(2)前記反応液中のS−スルホ−L−システイン、又は前記反応液から採取したS−スルホ−L−システインを還元する工程をさらに含む、前記方法。
(3)前記細菌において、前記O−アセチルセリンサルフヒドリラーゼはオートトランスポーターとの融合タンパク質として発現される、前記方法。
(4)前記融合タンパク質におけるO−アセチルセリンサルフヒドリラーゼ部分が、cysM遺伝子によりコードされる、前記方法。
(5)前記O−アセチルセリンサルフヒドリラーゼ部分が、下記のいずれかのアミノ酸配列を有する、前記方法。
(A)配列番号2のアミノ酸配列。
(B)配列番号2のアミノ酸配列において、1〜10個のアミノ酸の置換、欠失、挿入、または付加を含むアミノ酸配列。
(6)前記オートトランスポーターが、シュードモナス属細菌の耐熱性モノグリセリドリパーゼのオートトランスポーターである、前記方法。
(7)前記反応液に含まれるO−アセチル−L−セリンが、O−アセチル−L−セリン生産能を有する他の細菌により生産されたものである、前記方法。
(8)前記O−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌はO−アセチル−L−セリン生産能を有し、前記反応液に含まれるO−アセチル−L−セリンは該細菌が分泌したものであって、前記反応液は培地である、前記方法。
(9)前記他の細菌又はO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌は、下記の性質の少なくともいずれかを有する、前記方法。
i)L−システイン生合成系酵素活性が増強されるように改変されている。
ii)細胞内のO−アセチルセリン分解酵活性が低下するように改変されている。
iii)O−アセチルセリンを排出する因子の活性が増強されるように改変されている。
(10)前記O−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌がエシェリヒア属細菌である、前記方法。
(11)前記他の細菌がエシェリヒア属細菌である、前記方法。
(12)エシェリヒア属細菌がエシェリヒア・コリである、前記方法。
(13)cysM遺伝子によりコードされるO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌であって、前記O−アセチルセリンサルフヒドリラーゼはオートトランスポーターとの融合タンパク質として発現される、細菌。
(14)前記融合タンパク質におけるO−アセチルセリンサルフヒドリラーゼ部分が、下記のいずれかのアミノ酸配列を有する、前記細菌。
(A)配列番号2のアミノ酸配列。
(B)配列番号2のアミノ酸配列において、1〜10個のアミノ酸の置換、欠失、挿入、または付加を含むアミノ酸配列。
(15)前記オートトランスポーターが、シュードモナス属細菌の耐熱性モノグリセリドリパーゼのオートトランスポーターである、前記細菌。
(16)細胞内のO−アセチルセリンサルフヒドリラーゼ活性が低下するように改変された、前記細菌。
【発明の効果】
【0017】
本発明により、OASからのL-システイン関連物質の生産が可能になる。また、OASを生産する細菌と組み合わせることで、糖を原料としたL-システイン関連物質の効率的な生産が可能になる。本発明の一形態によれば、OASからのL−システイン関連物質の生産におけるイオウ源として、チオ硫酸を用いることができる。
また、本発明により、O−アセチルセリンサルフヒドリラーゼタンパク質をオートトランスポーターの機能を利用することにより細胞外膜に提示した細菌が提供される。
【図面の簡単な説明】
【0018】
【図1】OASS提示菌を用いたOASからのシステイン関連物質の生産を示す図。
【図2】pET-OASSB(X)-AT保持株のSDS-PAGEの結果を示す図(電気泳動写真)。
【図3】pET-OASS(+40)-AT保持株を用いたシステイン関連物質生産の経時変化を示す図。
【図4】L−システイン発酵で副生したOASを利用したOASS提示菌によるシステイン関連物質の生産を示す図。
【発明を実施するための形態】
【0019】
本発明の方法は、チオ硫酸とO−アセチル−L−セリン(OAS)からS−スルホ−L−システインを生成する反応を触媒するO−アセチルセリンサルフヒドリラーゼ(OASS-B)を細胞外膜に提示する細菌を、OAS及びチオ硫酸に、これらを含む反応液中で作用させ、該反応液からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を採取することを特徴とする、S−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物の製造法である。すなわち、本発明では、細菌の細胞外膜に提示されたOASS-Bを用いて、OAS及びチオ硫酸からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を生成させる。本明細書においては、S−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体(OASを除く)、又はこれらの混合物を、「システイン関連物質」と記載することがある。システイン関連物質は、フリー体であってもよく、塩であってもよい。塩としては、例えば硫酸塩、塩酸塩、炭酸塩、アンモニウム塩、ナトリウム塩、カリウム塩が挙げられる。塩であってもよい。システイン関連物質については後述する。また、前記OASS-Bを細胞外膜に提示する細菌を、OASS提示菌(OASS-autodisplaying bacterium)と記載することがある。
【0020】
本発明の第一の形態においては、反応液に含まれるOASは、好ましくはOAS生産能を有する他の細菌(OAS生産菌)により生産されたものである。この形態においては、OAS生産菌が生産したOASを原料として、OASS提示菌によりS−スルホ−L−システインを生成させる。尚、OASは、細菌を用いる方法以外の方法によって生産されたものであってもよい。
【0021】
本発明の第二の形態においては、OASS提示菌は、OAS生産能をも有し、前記反応液に含まれるOASは該細菌が分泌したものであって、前記反応液は培地である。この形態では、OASS提示菌はOAS生産菌でもあり、OASS提示菌自身が培地に分泌したOASを原料として、同細菌の細胞外膜に提示されたOASS-BによりS−スルホ−L−システインを生成させる。
いずれの形態においても、後述するように、S−スルホ−L−システインは、L−システイン、L−シスチン、又はそれらの誘導体に変換され得る。
以下、OAS生産菌及びOASS提示菌について説明する。
【0022】
<1>OAS生産菌
OAS生産能とは、細菌を培地中で培養したときに、培地中または菌体内にOASを生成し、培地中または菌体から回収できる程度に蓄積する能力をいう。また、OAS生産能を有する細菌とは、野生株または親株よりも多い量のOASを生産し培地に蓄積することができる細菌を意味し、好ましくは、0.2g/L以上、より好ましくは0.3g/L以上、特に好ましくは0.4g/L以上の量のL-システインを生産し培地に蓄積することができる細菌が含まれる。
【0023】
OAS生産能を有する細菌としては、本来的にOAS生産能を有するものであってもよいが、下記のような細菌を、変異法や組換えDNA技術を利用して、OAS生産能を有するように改変したものであってもよい。
【0024】
OAS生産菌の育種に用いる細菌としては、OAS生産能を付与又は増強され得るものであれば特に制限されないが、グラム陰性細菌、例えばエシェリヒア属、エンテロバクター属、パントエア属、クレブシエラ属、セラチア属、エルビニア属、サルモネラ属、モルガネラ属など、腸内細菌科に属する細菌、及び、コリネバクテリウム属、バチルス属、シュードモナス属、アグロバクテリウム属、ビブリオ属に属する細菌等が好ましい。具体的にはNCBI(National Center for Biotechnology Information)データベースに記載されている分類により腸内細菌科に属するものが利用できる(http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=91347)。中でも、エシェリヒア属細菌が好ましく、エシェリヒア・コリがより好ましい。
【0025】
エシェリヒア属細菌としては、特に限定されないが、具体的にはNeidhardtらの著書(Backmann, B. J. 1996. Derivations and Genotypes of some mutant derivatives of Escherichia coli K-12, p. 2460-2488. Table 1. In F. D. Neidhardt (ed.), Escherichia
coli and Salmonella Cellular and Molecular Biology/Second Edition, American Society for Microbiology Press, Washington, D.C.)に挙げられるものが利用できる。その中では、例えばエシェリヒア・コリが挙げられる。エシェリヒア・コリとしては具体的には、プロトタイプの野生株K12株由来のエシェリヒア・コリ W3110 (ATCC 27325)、エシェリヒア・コリ MG1655 (ATCC 47076)等が挙げられる。
【0026】
これらを入手するには、例えばアメリカン・タイプ・カルチャー・コレクション(住所
12301 Parklawn Drive, Rockville, Maryland 20852 P.O. Box 1549, Manassas, VA 20108, United States of America)より分譲を受けることが出来る。すなわち各菌株に対応する登録番号が付与されており、この登録番号を利用して分譲を受けることが出来る(http://www.atcc.org/参照)。各菌株に対応する登録番号は、アメリカン・タイプ・カルチャー・コレクションのカタログに記載されている。
【0027】
エンテロバクター属細菌としては、エンテロバクター・アグロメランス(Enterobacter
agglomerans)、エンテロバクター・アエロゲネス(Enterobacter aerogenes)等、パントエア属細菌としてはパントエア・アナナティス(Pantoea ananatis)が挙げられる。尚、近年、エンテロバクター・アグロメランスは、16S rRNAの塩基配列解析などにより、パントエア・アグロメランス(Pantoea agglomerans)又はパントエア・アナナティス(Pantoea ananatis)、パントエア・スチューアルティ(Pantoea stewartii)に再分類されているものがある。本発明においては、腸内細菌科に分類されるものであれば、エンテロバクター属又はパントエア属のいずれに属するものであってもよい。
【0028】
特に、パントエア属細菌、エルビニア属細菌、エンテロバクター属細菌は、γ-プロテオバクテリアに分類される細菌であり、分類学的に非常に近縁である(J Gen Appl Microbiol 1997 Dec;43(6) 355-361, International Journal of Systematic Bacteriology, Oct. 1997,p1061-1067)。近年、DNA-DNAハイブリダイゼーション実験等により、エンテロバクター属に属する細菌には、パントエア・アグロメランス(Pantoea agglomerans)又はパントエア・ディスパーサ(Pantoea dispersa)等に再分類されているものがある(International Journal of Systematic Bacteriology, July 1989;39(3).p.337-345)。また、エルビニア属に属する細菌にはパントエア・アナナス(Pantoea ananas)、パントエア・スチューアルティに再分類されているものがある(International Journal of Systematic Bacteriology, Jan 1993;43(1), p.162-173 参照)。
【0029】
エンテロバクター属細菌としては、エンテロバクター・アグロメランス(Enterobacter
agglomerans)、エンテロバクター・アエロゲネス(Enterobacter aerogenes)等が挙げられる。具体的には、欧州特許出願公開952221号明細書に例示された菌株を使用することが出来る。エンテロバクター属の代表的な株として、エンテロバクター・アグロメランスATCC12287株が挙げられる。
【0030】
パントエア属細菌の代表的な菌株として、パントエア・アナナティス、パントエア・スチューアルティ(Pantoea stewartii)パントエア・アグロメランス、パントエア・シトレア(Pantoea citrea)が挙げられる。パントエア・アナナティスとして具体的には、パントエア・アナナティスAJ13355株、SC17株、及びSC17(0)株が挙げられる。SC17株は、静岡県磐田市の土壌から、低pHでL-グルタミン酸及び炭素源を含む培地で増殖できる株として分離された株AJ13355(FERM BP-6614)から、粘液質低生産変異株として選択された株である(米国特許第6,596,517号)。SC17(0)株は、パントエア・アナナティスにおいて遺伝子破壊を行うために、λ Red遺伝子産物に耐性な菌株として構築された株である(WO2008/075483)。
【0031】
パントエア・アナナティスAJ13355株は、平成10年2月19日に、通産省工業技術院生命工学工業技術研究所(現名称、産業技術総合研究所特許生物寄託センター、住所 郵便番号305-8566 茨城県つくば市東1丁目1番地1中央第6)に、受託番号FERM P-16644として寄託され、平成11年1月11日にブダペスト条約に基づく国際寄託に移管され、受託番号FERM BP-6614が付与されている。尚、同株は、分離された当時はエンテロバクター・アグロメランス(Enterobacter agglomerans)と同定され、エンテロバクター・アグロメランスAJ13355として寄託されたが、近年16S rRNAの塩基配列解析などにより、パントエア・アナナティス(Pantoea ananatis)に再分類されている。また、SC17株は、平成21年2月4日に、産業技術総合研究所特許生物寄託センター(住所 郵便番号305-8566 茨城県つくば市東1丁目1番地1中央第6)に寄託され、受託番号FERM BP-11091が付与されている。また、SC17(0)株は、2005年9月21にロシアン・ナショナル・コレクション・オブ・インダストリアル・マイクロオーガニズム(Russian National Collection of Industrial Microorganisms (VKPM), GNII Genetika)(住所:Russia, 117545 Moscow, 1 Dorozhny proezd. 1)に受託番号VKPM B-9246のもとに寄託されている。
【0032】
エルビニア属細菌としては、エルビニア・アミロボーラ、エルビニア・カロトボーラが挙げられ、クレブシエラ属細菌としては、クレブシエラ・プランティコーラが挙げられる。
【0033】
OAS生産菌、又はOAS生産能を有するOASS提示菌として好ましい細菌は、下記の性質のいずれか1、2又は3種を有するものであるが、これらに限定されない。
i)L−システイン生合成系酵素活性が増強されるように改変されている。
ii)細胞内のO−アセチルセリン分解酵活性が低下するように改変されている。
iii)O−アセチルセリンを排出する因子の活性が増強されるように改変されている。
以下、腸内細菌科に属する細菌にOAS生産能を付与する方法、又はこれらの細菌のOAS生産能を増強する方法について述べる。
【0034】
細菌にOAS生産能を付与するには、栄養要求性変異株、アナログ耐性株又は代謝制御変異株の取得や、L-システインの生合成系酵素の発現が増強された組換え株の創製等、従来、コリネ型細菌又はエシェリヒア属細菌等の育種に採用されてきた方法を適用することができる(アミノ酸発酵、(株)学会出版センター、1986年5月30日初版発行、第77-100頁参照)。ここで、OAS生産菌の育種において、付与される栄養要求性、アナログ耐性、代謝制御変異等の性質は、単独でもよく、2種又は3種以上であってもよい。また、発現が増強されるL-システイン生合成系酵素も、単独であっても、2種又は3種以上であってもよい。さらに、栄養要求性、アナログ耐性、代謝制御変異等の性質の付与と、生合成系酵素の増強が組み合わされてもよい。
【0035】
OAS生産能を有する栄養要求性変異株、L-システインのアナログ耐性株、又は代謝制御変異株を取得するには、親株又は野生株を通常の変異処理、すなわちX線や紫外線の照射、またはN-メチル-N'-ニトロ-N-ニトロソグアニジン(NTG)もしくはエチルメタンスルフォネート(EMS)等の変異剤処理などによって処理し、得られた変異株の中から、栄養要求性、アナログ耐性、又は代謝制御変異を示し、かつOAS生産能を有するものを選択することによって得ることができる。
【0036】
OASはL-システインの前駆体であるため、L-システイン生産菌から容易に構築することができる。L-システイン生産菌として、具体的には、フィードバック阻害耐性のセリンアセチルトランスフェラーゼ(SAT)をコードする複数種のcysEアレルで形質転換されたE. coli JM15(米国特許第6,218,168号)、細胞に毒性の物質を排出するのに適したタンパク質をコードする過剰発現遺伝子を有するE. coli W3110 (米国特許第5,972,663号)、システインデスルフヒドラーゼ活性が低下したE. coli株 (特開平11-155571号公報)、cysB遺伝子によりコードされるシステインレギュロンの正の転写制御因子の活性が上昇したE. coli W3110 (WO01/27307)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。
【0037】
OAS生産菌は、細胞内のOAS分解酵素活性が低下するように改変されていることが好ましい。例えば、L-システイン生産菌からL-システイン分解酵素をコードするcysM及びcysKの遺伝子を同時に欠損させることで、OASを生産するように改変することができる。一方、E. coliでは、前述のとおりL-システインを排出する活性を有する公知のタンパク質としてydeDにコードされるタンパク質(特開2002-233384)が知られているが、このYdeDはL-システインに加えて、OAS等のL-システイン関連化合物も同時に排出することが知られている。したがって、OAS生産菌を得るためには、L-システイン排出因子として知られる因子の活性を上昇させてもよい。
【0038】
〔SATの増強〕
次に、細菌にOAS生産能を付与する方法として、L-システイン生合成酵素活性を増強する方法について説明する。L-システイン生合成酵素活性の増強は、例えば、セリンアセチルトランスフェラーゼ(SAT)活性を増強することによって行うことが出来る。細菌における細胞内のSAT活性の増強は、SATをコードする遺伝子のコピー数を高めることによって達成される。例えば、SATをコードする遺伝子断片を、細菌で機能するベクター、好ましくはマルチコピー型のベクターと連結して組換えDNAを作製し、これで宿主細菌を形質転換すればよい。
【0039】
SAT遺伝子は、エシェリヒア属細菌由来の遺伝子および他の生物由来の遺伝子のいずれも使用することができる。エシェリヒア・コリのSATをコードする遺伝子として、cysEが野生株及びL−システイン分泌変異株よりクローニングされ、塩基配列が明らかになっている(Denk, D. and Boeck, A., J. General Microbiol., 133, 515-525 (1987))。したがって、その塩基配列に基づいて作製したプライマーを用いて、エシェリヒア属細菌の染色体DNAを鋳型とするPCRによって、SAT遺伝子を取得することができる(特開平11-155571号参照)。他の生物のSATをコードする遺伝子も、同様にして取得され得る。このようにして得られるSAT遺伝子は、上記cysE遺伝子と同様にして発現増強を行うことができる。
【0040】
なお、SAT遺伝子の発現に「L−システインによるフィードバック阻害」などの抑制機構が存在する場合には、該抑制機構に非感受性となるように、発現調節配列又は抑制に関与する遺伝子を改変することによっても、SAT遺伝子の発現を増強することができる。
【0041】
〔OASS活性の低下〕
OAS生産菌は、細胞内における、OASをL-システインに変換するOASS活性が低下するように改変されていることが好ましい。ただし、OASS活性を低下させた後にL-システイン生合成経路の強化を行ってもよい。
【0042】
「細胞内のOASS活性が低下する」とは、細胞内のOASS活性が、野生株または親株等の非改変株に対して低下していることを意味し、活性が完全に消失していることを含む。本明細書に記載された他のタンパク質の活性の「低下」についても同様である。
OASSとしては、cysK、cysM遺伝子にコードされるOASS-A、及びOASS-Bが挙げられる。
【0043】
OASS活性は、例えば、実施例4に記載された方法で測定することができる。すなわち、試料をOAS及びイオウ源に作用させ、生成するシステイン関連物質の量を定量することにより、OASS活性を測定することができる。OASからのシステイン関連物質の生成におけるイオウ源は、OASS-Aでは硫化ナトリウム等の硫化物が、OASS-Bではチオ硫酸ナトリウム等のチオ硫酸塩が用いられる。システイン関連物質は、Gaitonde法(Gaitonde M.K.,(1967) Biochem J 133:627-633)で定量することができる。細胞内のOASS活性を測定するには、細胞を破砕し、膜画分を遠心分離等により除去し、可溶性画分についてOASS活性を測定すればよい。
【0044】
OASS活性を低下させるような改変は、例えば、OASS-AをコードするcysK、及びOASS-BをコードするcysM遺伝子の発現を低下させることによって達成される。具体的には例えば、染色体上のcysM、及びcysK遺伝子のそれぞれのコード領域の一部又は全部を欠損させることによって、これらの遺伝子にコードされるOASSタンパク質の細胞内の活性を低下させることができる。
【0045】
またOASSタンパク質活性の低下は、前記遺伝子のプロモーターやシャインダルガノ(SD)配列等の発現調節配列を改変することなどによって、これらの遺伝子の発現を低下させることによっても、達成することができる。また、発現調節配列以外の非翻訳領域の改変によっても、遺伝子の発現量を低下させることができる。さらには、染色体上の遺伝子の前後の配列を含めて、遺伝子全体を欠失させてもよい。また、染色体上のcysM及び/又はcysK遺伝子のコード領域にアミノ酸置換(ミスセンス変異)を導入すること、また終始コドンを導入すること(ナンセンス変異)、あるいは一〜二塩基付加・欠失するフレームシフト変異を導入することによっても、遺伝子の発現を低下させることが出来る(Journal of Biological Chemistry 272:8611-8617(1997) Proceedings of the National Academy of Sciences,USA 95 5511-5515(1998), Journal of Biological Chemistry 266, 20833-20839(1991))。
【0046】
また、OASSタンパク質の活性が低下するような改変であれば、X線もしくは紫外線を照射、またはN-メチル-N'-ニトロ-N-ニトロソグアニジン等の変異剤による通常の変異処理による改変であってもよい。
【0047】
発現調節配列の改変は、好ましくは1塩基以上、より好ましくは2塩基以上、特に好ましくは3塩基以上である。また、コード領域を欠失させる場合は、OASSタンパク質の機能が低下するのであれば、欠失させる領域は、N末端領域、内部領域、C末端領域のいずれの領域であってもよく、コード領域全体であってよい。通常、欠失させる領域は長い方が確実に遺伝子を不活化することができる。また、欠失させる領域の上流と下流のリーディングフレームは一致しないことが好ましい。
【0048】
cysM及び/又はcysK遺伝子のコード領域に他の配列を挿入する場合も、挿入の箇所は遺伝子のいずれの領域であってもよいが、挿入する配列は長い方が、確実に遺伝子を不活化することができる。挿入部位の前後の配列は、リーディングフレームが一致しないことが好ましい。他の配列としては、コードされるCysM及び/又はCysKタンパク質の機能を低下させるものであれば特に制限されないが、例えば、抗生物質耐性遺伝子やL-システイン生産に有用な遺伝子を搭載したトランスポゾン等が挙げられる。
【0049】
染色体上のcysM及び又はcysK遺伝子を上記のように改変するには、例えば、遺伝子の部分配列を欠失し、活性を有するOASSタンパク質を産生しないように改変した欠失型遺伝子を作製し、該遺伝子を含むDNAで細菌を形質転換して、欠失型遺伝子と染色体上の遺伝子とで相同組換えを起こさせることにより、染色体上の遺伝子を欠失型遺伝子に置換することによって達成できる。欠失型遺伝子によってコードされるOASSタンパク質は、生成したとしても、野生型タンパク質とは異なる立体構造を有し、機能が低下する。このような相同組換えを利用した遺伝子置換による遺伝子破壊は既に確立しており、「Redドリブンインテグレーション(Red-driven integration)」と呼ばれる方法(Datsenko, K. A, and Wanner, B. L. Proc. Natl. Acad. Sci. U S A. 97:6640-6645 (2000))、Redドリブンインテグレーション法とλファージ由来の切り出しシステム(Cho, E. H., Gumport, R. I., Gardner, J. F. J. Bacteriol. 184: 5200-5203 (2002))とを組合わせた方法(WO2005/010175号参照)等の直鎖状DNAを用いる方法や、温度感受性複製起点を含むプラスミド、接合伝達可能なプラスミドを用いる方法、宿主内で複製起点を持たないスイサイドベクターを利用する方法などがある(米国特許第6303383号、または特開平05-007491号)。
【0050】
cysM及び/又はcysK遺伝子の転写量が低下したことの確認は、各々の遺伝子から転写されるmRNAの量を野生株、あるいは非改変株と比較することによって行うことが出来る。mRNAの量を評価する方法としては、ノーザンハイブリダイゼーション、RT-PCR等が挙げられる(Molecular cloning(Cold spring Harbor Laboratory Press, Cold spring Harbor (USA), 2001))。
【0051】
OASSタンパク質の量が低下したことの確認は、抗体を用いてウェスタンブロットによって行うことが出来る(Molecular cloning(Cold spring Harbor Laboratory Press, Cold
spring Harbor (USA), 2001))。
【0052】
さらに、OASSタンパク質の量が低下したことの確認は、細胞のOASS活性を測定することによっても行うことができる。
【0053】
OASS-Aをコードする遺伝子として具体的には、配列番号1に示す塩基配列を含む遺伝子を挙げることが出来る。配列番号1には、エシェリヒア・コリの野生型cysK遺伝子の塩基配列を、配列番号2は同遺伝子がコードするOASS-Aのアミノ酸配列を示す。また、OASS-Bをコードする遺伝子として具体的には、配列番号3に示す塩基配列を含む遺伝子を挙げることが出来る。配列番号3には、エシェリヒア・コリの野生型cysM遺伝子の塩基配列を、配列番号4は同遺伝子がコードするOASS-Bのアミノ酸配列をを示す。なお、OASSをコードする遺伝子は上述した遺伝子情報を持つ遺伝子や、公知の配列を有する遺伝子に限られず、それらの遺伝子のバリアント、すなわちOASS活性を有するタンパク質をコードする限りにおいて、それらの遺伝子のホモログや人為的な改変体等、保存的変異を有する遺伝子も使用することができる。すなわち、公知のタンパク質のアミノ酸配列において、1若しくは数個の位置での1又は数個のアミノ酸が置換、欠失、挿入又は付加されたアミノ酸配列を有するタンパク質をコードする遺伝子であっても良い。このようなタンパク質のOASS活性は、1又は数個の置換、欠失、挿入又は付加される前のタンパク質に対して同等の活性、例えば70%以上、好ましくは80%以上、より好ましくは90%以上の活性を維持していることが好ましい。なお上記「1又は数個」とは、アミノ酸残基のタンパク質の立体構造における位置やアミノ酸残基の種類によっても異なるが、具体的には好ましくは1〜20個、より好ましくは1〜10個、さらに好ましくは1〜5個を意味する。保存的変異とは、置換部位が芳香族アミノ酸である場合には、Phe、Trp、Tyr間で、置換部位が疎水性アミノ酸である場合には、Leu、Ile、Val間で、極性アミノ酸である場合には、Gln、Asn間で、塩基性アミノ酸である場合には、Lys、Arg、His間で、酸性アミノ酸である場合には、Asp、Glu間で、ヒドロキシル基を持つアミノ酸である場合には、Ser、Thr間でお互いに置換する変異である。保存的変異の代表的なものは、保存的置換であり、保存的置換とみなされる置換としては、具体的には、AlaからSer又はThrへの置換、ArgからGln、His又はLysへの置換、AsnからGlu、Gln、Lys、His又はAspへの置換、AspからAsn、Glu又はGlnへの置換、CysからSer又はAlaへの置換、GlnからAsn、Glu、Lys、His、Asp又はArgへの置換、GluからGly、Asn、Gln、Lys又はAspへの置換、GlyからProへの置換、HisからAsn、Lys、Gln、Arg又はTyrへの置換、IleからLeu、Met、Val又はPheへの置換、LeuからIle、Met、Val又はPheへの置換、LysからAsn、Glu、Gln、His又はArgへの置換、MetからIle、Leu、Val又はPheへの置換、PheからTrp、Tyr、Met、Ile又はLeuへの置換、SerからThr又はAlaへの置換、ThrからSer又はAlaへの置換、TrpからPhe又はTyrへの置換、TyrからHis、Phe又はTrpへの置換、及び、ValからMet、Ile又はLeuへの置換が挙げられる。また、上記のようなアミノ酸の置換、欠失、挿入、付加、または逆位等には、遺伝子が由来する細菌の個体差、種の違いに基づく場合などの天然に生じる変異(mutant又はvariant)によって生じるものも含まれる。
【0054】
さらに、上記のような保存的変異を有する遺伝子は、コードされるアミノ酸配列全体に対して、80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは97%、特に好ましくは99%以上の相同性を有し、かつ、OASS活性を有するタンパク質であってもよい。このような公知の配列を有するOASSに対して相同性を有すタンパク質をコードする遺伝子は、上記のエシェリヒア・コリ株の野生型cysK又はcysM遺伝子を問い合わせ配列として用いたBLAST検索やFASTA検索によって公開データベースから容易に配列情報を取得することが出来、この公知の遺伝子配列に基づいて作製したオリゴヌクレオチドをプライマーとして、細菌のゲノムDNAを鋳型に用いてPCRにより取得することができる。尚、本明細書において、「相同性」(homology)」は、「同一性」(identity)を指すことがある。
【0055】
また、OASSをコードする遺伝子は、公知の遺伝子配列の相補配列又はその相補配列から調製され得るプローブとストリンジェントな条件でハイブリダイズし、公知の遺伝子産物と同等の機能を有するタンパク質をコードする遺伝子であってもよい。本発明において、ストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。一例を示せば、相同性が高いDNA同士、例えば80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは97%以上の相同性を有するDNA同士がハイブリダイズし、それより相同性が低いDNA同士がハイブリダイズしない条件、あるいは通常のサザンハイブリダイゼーションの洗いの条件である60℃、1XSSC、0.1% SDS、好ましくは0.1×SSC、0.1% SDS、に相当する塩濃度で、1回好ましくは2〜3回洗浄する条件を挙げることができる。
【0056】
上記ハイブリダイゼーションに用いるプローブは、遺伝子の相補配列の一部であってもよい。そのようなプローブは、公知の遺伝子配列に基づいて作製したオリゴヌクレオチドをプライマーとし、これらの塩基配列を含むDNA断片を鋳型とするPCRによって作製することができる。例えば、プローブとして、300 bp程度の長さのDNA断片を用いる場合には、ハイブリダイゼーションの洗いの条件は、50℃、2×SSC、0.1% SDSが挙げられる。
【0057】
<2>OASS提示菌
次に、OASS提示菌について説明する。OASS提示菌は、OASとチオ硫酸からS−スルホ−L−システインを生成する反応を触媒するOASSを細胞外膜に提示する細菌である。尚、OASSは、少なくともチオ硫酸とOASからS−スルホ−L−システインを生成し得る限り、チオ硫酸以外のイオウ源、例えば硫化物とOASからS−スルホ−L−システインを生成するものであってもよい。このようなOASSとしてはOASS-Bが、OASS-Bをコードする遺伝子としては前記のcysM遺伝子及びそのバリアントが挙げられる。
【0058】
OASS提示菌の構築に用いる細菌は、細胞外膜を有するグラム陰性菌であり、細胞外膜にタンパク質を提示することが可能な細菌であれば特に制限されない。具体的には、前述した腸内細菌科に属する細菌が挙げられる。中でも、エシェリヒア属性菌が好ましく、エシェリヒア・コリがより好ましい。
【0059】
「OASSを細胞外膜に提示する細菌」とは、OASSの少なくとも一部又は全部が細菌の細胞外に存在するようにして固定され、OAS及びチオ硫酸に反応液中で作用させたときに、OASからシステイン関連物質が生成することができる細菌である。
【0060】
以下、OASS-Bを細菌の細胞外膜へ提示させる方法について説明する。
OASS-Bを細胞外膜へ提示させるには、例えば、オートトランスポーターを利用する方法が挙げられる。具体的には、遺伝子組換え技術を利用して、OASS-Bをオートトランスポーターとの融合タンパク質として発現させる。オートトランスポーターによるペプチド提示システムでは、提示されるペプチドは、N末から分泌シグナル、パッセンジャー(提示されるペブチドに相当)、及びオートトランスポータードメインからなる前駆体タンパク質として発現し、該ペブチド(パッセンジャー)のみが細胞外膜の外に提示される。本発明において、「オートトランスポーター」とは前記オートトランスポータードメインをいう。パッセンジャー部分をOASSに置換した前駆体をコードする遺伝子を構築して、この遺伝子を細菌に導入することによって、OASSを細胞外膜に提示させることができる。本発明において、「O−アセチルセリンサルフヒドリラーゼ部分(OASS部分; OASS moiety)とは、上記のような前駆体タンパク質中にパッセンジャー部分として含まれるOASSをいう。尚、前駆体タンパク質のデザイン、具体的には前駆体タンパク質を構成するペプチド同士の連結部位のデザインによって、OASSのN末端及び/又はC末端の1又は複数のアミノ酸が変化することがある。そのような場合であっても、OASS部分はOASS活性を有する限り、野生型OASSとは異なる配列を含んでいてもよい。
【0061】
分泌シグナルは、シグナルペプチドに続く配列に内膜を通過させるためのものであり、パッセンジャーより下流の配列がペリブラズムに分泌され、分泌シグナルがシグナルペプチダーゼで切断されることが可能なものであれば特に制限されない。分泌シグナルとしては、例えば、Pseudomonas sp. strain LP7315のモノグリセリドリパーゼの分泌シグナルの他、PelBタンパク質、OmpTタンパク質、ompAタンパク質、β-ラクタマーゼ等の分泌シグナル等、E. coli等の細菌におけるタンパク質の分泌生産に用いられている分泌シグナル(Pines O. et al., Mol. Biotechnol. (1999) 12. 25-34)が挙げられる。
【0062】
オートトランスポーターとしては、Pseudomonas sp. strain LP7315のモノグリセリドリパーゼのオートトランスポータードメインが挙げられる。オートトランスポーターは、αヘリックス構造を有するリンカー部分とβバレル構造を有する部分(以下、「βバレル部分」ともいう。)を有する。Pseudomonas sp. LP7315株のモノグリセリドリパーゼをコードする遺伝子及び同遺伝子がコードするアミノ酸配列を、配列番号5及び6に示す。配列の相同性解析等から、配列番号6において、シグナルペプチドは1〜69位、リパーゼドメインは85〜885位、リンカー部分は937〜981位、βバレル部分は994〜1803位と推定される。リパーゼドメインとリンカー部分との間、リンカー部分とβバレル部分との間、及び、βバレル部分のC末端側には、上記の推定領域に属さない配列(以下、便宜上「スペーサー」という。)が存在するが、これらのスペーサーは、実際はリンカー及び/又はβバレル部分に含まれるか、又は、遷移的な構造を有する部分であると推定される。本発明において「オートトランスポーター」とは、上記の推定上のリンカー部分及びβバレル部分に加えて、スペーサーを含んでいてもよい。また、リパーゼドメインは、相同性解析からは85〜885位と推定されるが、実際のN末端はシグナルペプチドの直後である可能性がある。
【0063】
また、実施例に示すように、オートトランスポーターがリンカー部分と推定される領域を欠失していても、パッセンジャーとオートトランスポーターとの融合タンパク質のうちパッセンジャー部分が細胞外膜に提示されることがある。また、βバレル部分と推定される領域のN末端側を欠失していても、融合タンパク質のパッセンジャー部分が細胞外膜に提示されることがある。これは、パッセンジャー部分のN末端部分及び/又はβバレル部分(又はそのN末端部分を欠失した部分)のN末端部分が、リンカーとして機能することがあるものと推定される。このように、オートトランスポーターが本来のリンカー部分及び/又はβバレル部分のN末端部分を含んでいなくても、βバレル融合タンパク質のパッセンジャー部分のC末端及び/又はβバレル部分のC末端がリンカーとして機能し、パッセンジャー部分が細胞外膜に提示され得る限り、そのようなオートトランスポーターの改変体は「オートトランスポーター」に含まれる。
【0064】
リンカーは、野生型オートトランスポーターのリンカー部分の配列を有するものであってもよく、一部又は全体を任意の配列で置換えたものであってもよい。また、リンカーの長さは、野生型の配列を延長又は短縮したものであってもよい。さらには、前記のとおり、野生型オートトランスポーターのリンカー部分を含まずに、パッセンジャー及び/又はβバレル部分の一部がリンカーとして機能してもよい。また、リンカーの長さは、野生型のリンカー部分の配列と同じであってもよいが、それよりも長くてもよく、短くてもよい。「リンカーの長さ」は、本明細書においては、実施例に記載したpET-OASSA-ATプラスミドにコードされる融合タンパク質におけるOASS-Bとβバレル部分との間の長さを0とし、それに対する相対値で表す。尚、pET-OASSA-ATでは、同プラスミドの構築に用いたEcoRI認識配列によって、OASS-Bとβバレル部分の間に2アミノ酸残基挿入されている。また、βバレル部分は、前記モノグリセリドリパーゼの配列における994〜1803位に対して、N末端の34アミノ酸残基を欠失している。リンカーの長さ(pET-OASSA-ATに対する相対値)は、パッセンジャーの配列によっても異なるが、通常-50〜500アミノ酸、好ましくは20〜300アミノ酸、より好ましくは20〜70アミノ酸が好ましい。
【0065】
このモノグリセリドリパーゼの分泌シグナル及び/又はオートトランスポータードメインを利用する場合、これらをコードするDNAは、配列番号5の塩基配列に基づいて作製したプライマーを用いて、pET-OASSB(+79)-ATプラスミドDNA(参考配列)を鋳型とするPCR法によって取得することができる。他の細菌のオートトランスポータードメインをコードする遺伝子も、公知の配列情報に基づいて作製したプローブを用いたハイブリダイゼーション法によって、細菌の染色体DNA又は染色体DNAライブラリーから取得することができる。pET-OASSB(+79)-ATプラスミドで形質転換したエシェリシア・コリBL21株はAJ110734株と命名され、平成21年3月9日に、産業技術総合研究所特許生物寄託センター(住所 郵便番号305-8566 茨城県つくば市東1丁目1番地1中央第6)に寄託され、受託番号FERM P-21779が付与されている。
【0066】
エシェリヒア・コリのcysM遺伝子は、配列番号3の塩基配列に基づいて作製したプライマーを用いて、エシェリヒア・コリの染色体DNAを鋳型とするPCR法によってcysM遺伝子を取得することができる。他の細菌のcysM遺伝子も、前記配列情報に基づいて作製したプローブを用いたハイブリダイゼーション法によって、細菌の染色体DNA又は染色体DNAライブラリーから取得することができる。
【0067】
分泌シグナル、OASS、及びオートトランスポーターを含む(以下、「OASS-AT」と記載することがある)をコードするDNAは、これらを各々コードするDNA断片を連結することによって取得することができる。また、パッセンジャー及びオートトランスポーターを含む前駆体タンパク質、又はさらに分泌シグナルを含む前駆体タンパク質をコードするDNAの、パッセンジャーをコードする配列を、OASSをコードする配列で置換えることによっても、OASS-ATをコードするDNAを取得することができる。
【0068】
OASS-ATを発現させるためのプロモーターは、オートトランスポーターを含むタンパク質をコードする遺伝子固有のプロモーターであってもよく、他の遺伝子由来のプロモーター又は人工プロモーターであってもよい。プロモーターとしては、例えば、lacプロモーター、trpプロモーター、trcプロモーター、tacプロモーター、araBAプロモーター、ラムダファージのPRプロモーター、PLプロモーター、tetプロモーター、T7プロモーター、φ10プロモーター等が挙げられる。
【0069】
OASS-Bを細胞外膜に提示させるには、上記のようなOASS-ATをコードするDNAを宿主細胞で機能するベクター、好ましくはマルチコピー型ベクターと連結して組み換えDNAを作製し、これで細菌を形質転換すればよい。
【0070】
また、OASS-Bを細胞外膜へ提示させるには、OASS-ATをコードするDNAを細菌の染色体DNA上に多コピー存在させることによっても達成できる。細菌の染色体DNAに上記遺伝子を多コピー導入するには、染色体DNA上に多コピー存在する配列を標的に利用して相同組換えにより行う。染色体DNA上に多コピー存在する配列としては、レペティティブDNA、転移因子の端部に存在するインバーテッド・リピート等が利用できる。あるいは、特開平2-109985号公報に開示されているように、上記融合遺伝子をトランスポゾンに搭載してこれを転移させて染色体DNA上に多コピー導入することも可能である。
【0071】
本発明の第二の形態においては、OASS提示菌は、本発明の第一の形態に用いるOAS生産菌と同様に、細胞内のOASS活性が低下するように改変されていることが好ましい。OASS活性の低下は、OAS生産菌の育種と同様にして行うことができる。
【0072】
細菌の細胞外膜にOASS-Bを提示させるには、オートトランスポーターを用いる方法以外に、何らかのシグナル及びアンカーに連結して細胞表層に固定化する方法(特願2000−202442)を採用することができる。
【0073】
<システイン関連物質の製造法>
上記のようにして得られる細菌を、OAS及びチオ硫酸に、これらを含む反応液中で作用させることにより、S−スルホ−L−システインが生成する。
【0074】
S−スルホ−L−システインは、反応液中で、又は反応液から採取した後に還元することにより、L−システインに変換することができる。還元は、例えばジチオスレイトール(DTT)等の還元剤を用いることにより行うことができる。また、S−スルホ−L−システインは、培地中の成分によってL−システインに変換されることがある。さらに、L−システインは、培地中で、ジスルフィド結合が形成されて一部がL−シスチンに変換することがある。さらに、L−システインは、細胞中や培地中に存在するケトン又はアルデヒド、例えばピルビン酸と縮合し、ヘミチオケタールを中間体としてチアゾリジン誘導体が生成することがある(特許第2992010参照)。これらのチアゾリジン誘導体及びヘミチオケタールは、平衡混合物として存在することがある。したがって、OASから生成される化合物には、S−スルホ−L−システインのみに限られず、L−システイン、L−シスチン、もしくはそれらの誘導体、例えばチアゾリジン誘導体、もしくはヘミチオケタール、又はこれらの混合物も含まれる。
【0075】
本発明の第一の形態においては、OAS生産菌を炭素源を含む培地で培養し、同培地にOASを蓄積させる。得られたOASを含む反応液にOASS提示菌とイオウ源を加え、反応を起させることにより、システイン関連物質が生成する。第一の形態における反応液は、OASが蓄積した培地もしくはその上清、又はその希釈液もしくは濃縮液であってもよく、これらから採取したOASを含む培地又は緩衝液等であってもよい。尚、上記反応に培地を用いる場合、炭素源は不要であるが、含まれていてもよい。また、第一の形態においては、OASS提示菌は、菌体をアクリルアミド、カラギーナン等で固定化した固定化菌体であってもよい。さらに、反応はバッチ式でもカラム式でもよい。さらに、OAS生産菌とOASS提示菌を同時に培養してもよい。
本発明の第二の形態においては、OAS生産能を有するOASS提示菌を、炭素源を含む培地で培養することによって、培地中にシステイン関連物質が蓄積する。この形態においては、OASSの基質としてのイオウ源は、培養当初から培地に加えてもよく、培養途中に培地に加えてもよい。
【0076】
いずれの形態においても、イオウ源としてはチオ硫酸及び硫化物が挙げられるが、チオ硫酸が好ましい。反応液又は培地中のイオウ源の濃度は、OASSに対して通常0.1倍〜10倍、好ましくは0.2倍〜5倍、特に好ましくは0.2倍〜2倍である。また、反応液中のOASS提示菌の量は、適宜設定することができる。例えば、緩衝液又は生産培地にてOASS提示菌を懸濁し、OD660を適当な値、例えば2〜8になるよう調整することによって、菌濃度を設定することができる。
【0077】
培地としては、炭素源、窒素源、イオウ源、無機イオン及び必要に応じその他の有機成分を含有する通常の培地が挙げられる。
【0078】
炭素源としては、グルコース、フラクトース、シュクロース、糖蜜やでんぷんの加水分解物などの糖類、エタノール、グリセロール等のアルコール類、フマール酸、クエン酸、コハク酸等の有機酸類を用いることができる。
【0079】
窒素源としては、硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム等の無機アンモニウム塩、大豆加水分解物などの有機窒素、アンモニアガス、アンモニア水等を用いることができる。
【0080】
イオウ源としては、硫酸塩、亜硫酸塩、硫化物、次亜硫酸塩、チオ硫酸塩等の無機硫黄化合物が挙げられる、尚、OAS生産菌の培養に用いる培地におけるイオウ源は、生育に必要な量含まれていればよい。
【0081】
有機成分としては、ビタミンB1などの要求物質または酵母エキス等を適量含有させることが望ましい。これらの他に、必要に応じてリン酸カリウム、硫酸マグネシウム、鉄イオン、マンガンイオン等が少量添加される。
【0082】
OAS生産培養は、好気的条件下で30〜90時間実施することが好ましく、培養温度は25℃〜37℃に、培養中pHは5〜8に制御することが好ましい。尚、pH調整には無機あるいは有機の酸性あるいはアルカリ性物質、更にアンモニアガス等を使用することができる。OASS提示菌による反応は、好ましくは振盪しながら、30〜90時間実施することが好ましい。
培養物又は反応液からのシステイン関連物質の採取は、通常のイオン交換樹脂法、沈澱法その他の公知の方法を組み合わせることにより実施できる。本発明において採取されるシステイン関連物質は、システイン関連物質以外に微生物菌体、培地成分、水分、及び微生物の代謝副産物を含んでいてもよい。採取されたシステイン関連物質の純度は、50%以上、好ましくは85%以上、特に好ましくは95%以上である (US5,431,933, JP1214636B, US4,956,471, US4,777,051, US4946654, US5,840358, US6,238,714, US2005/0025878)。
【0083】
上記のようにして得られるS−スルホ−L−システインは、L−システインの製造に用いることができる。また、L-システインは、L-システイン誘導体の製造に用いることができる。L−システイン誘導体としては、メチルシステイン、エチルシステイン、カルボシステイン、スルホシステイン、アセチルシステイン等が含まれる。
【実施例】
【0084】
以下、本発明を実施例によりさらに具体的に説明する。
〔実施例1〕オートトランスポータードメインを含む発現ベクターの構築
Pseudomonas sp. strain LP7315のモノグリセリドリパーゼ遺伝子のオートトランスポータードメインをコードする領域をPCRにより増幅した。同株のゲノムDNAをテンプレートとして、5’端にNcoI-EcoRIサイトを付加したプライマー#1(5'-ccatggttgaattcaccctgggcgtgg-3'(配列番号7))と、5'端にHindIIIサイトを付加したプライマー#2(5'-tacaagcttgccggcggcaacaag-3'(配列番号8))を用いて、定法に従い、KOD plus DNA polymerase(TOYOBO)にてPCRを行った。PCR条件は、94℃で5分の後、94℃で15秒、48℃で10秒、68℃で1分、というサイクルを30回繰り返した。こうして増幅した断片をpUC118 HincII/BAP(タカラバイオ社製)と連結した。シークエンシングによりPCRエラーがないことを確認したのち、NcoIとHindIIIによる制限酵素処理を行い、同酵素で切断したpET22b(+)(Novagen社製)ベクターに連結した。こうしてPseudomonas sp. LP7315株由来モノグリセリドリパーゼ中のオートトランスポータードメインをコードするDNAがベクター上のT7プロモーター下流に連結された発現ベクター、pET-ATを作製した。
【0085】
尚、pET22b(+)のマルチクローニングサイト上流には、分泌シグナルPelBをコードする配列が存在し、pET-ATのPelBコード領域とATコード配列の間に、これらのフレームが一致するようにペプチドコード配列を挿入すると、PelBと、前記ペプチドと、オートトランスポータードメイン(AT)との融合タンパク質が発現する。また、前記ペプチドとATとの間の配列によってコードされる領域は、オートトランスポートターによる分泌システムにおけるリンカー配列に相当する。
【0086】
尚、上記のオートトランスポータードメインをコードする領域は、後述のプラスミドpET-OASSB(+79)-ATをテンプレートとし、プライマー#1及びプライマー#2を用いたPCRによっても、取得することができる。
【0087】
〔実施例2〕OASS発現ベクターの構築
(1)まず、OASS-A又はOASS-Bを、PelB及びオートトランスポータードメイン(AT)との融合タンパク質として発現するベクターを構築した。
【0088】
OASS-A遺伝子をクローニングするため、E. coli野生株(MG1655)ゲノムDNAをテンプレートとして、5'端にNcoIサイトを付加したプライマー#3(5'-ccatggaaaagatttttgaagataac-3'(配列番号9))と、5'端にEcoRIサイトを付加したプライマー#4(5'-ctggaattcaatccccgccccctggctaaaatgc-3'(配列番号10))を用いて、PyrobestR DNA polymeraseにてPCRを行った。PCR条件は、94℃で5分の後、94℃で30秒、55℃で30秒、72℃で1分、というサイクルを30回繰り返した。この増幅断片をNcoI、EcoRIで消化後、同酵素で切断した上記pET-ATベクターに挿入した。シークエンシングによりPCRエラーがないことを確認した。こして、OASS-Aをオートトランスポーターにて細胞外提示するベクター、pET-OASSA-ATを構築した。このプラスミドが持つ、PelB、OASS-A、及びATとの融合タンパク質(以下、「PelB-OASSA-AT」と記載することがある)をコードする遺伝子の塩基配列を配列番号11に示し、コードされるアミノ酸配列を配列番号12に示した。配列番号12において、1〜323位はOASS-A部分であり、326位〜570位はオートトランスポータードメインである。324位〜325位は、これらの連結に用いたEcoRI配列によって生じた部分である。
【0089】
同様に、OASS-B遺伝子をクローニングするため、E. coli野生株(MG1655)ゲノムDNAをテンプレートに、プライマー#5(5'-aactgcagccatggaaacattagaacaaacaataggc-3'(配列番号13))と、プライマー#6(5'-ctggaattcaatccccgccccctggctaaaatgc-3'(配列番号14))を用いて、OASS-Aと同じ条件にてDNA断片を増幅した。その断片をNcoI、EcoRIで消化後、同酵素で切断したpET-ATベクターに挿入した。その後、増幅した領域にPCRエラーがないことを確認した。こうして、OASS-Bをオートトランスポーターにて細胞外提示するベクター、pET-OASSB-ATを構築した。このプラスミドが持つ、PelB、OASS-B、及びATとの融合タンパク質(以下、「PelB-OASSB-AT」と記載することがある)をコードする遺伝子の塩基配列を配列番号15に示し、コードされるアミノ酸配列を配列番号16に示した。配列番号16において、1〜303位はOASS-B部分であり、306位〜550位はオートトランスポータードメインのうち、βバレル部分である。304位〜305位は、これらの連結に用いたEcoRI配列によって生じた部分である。尚、PelB-OASSB-ATは、野生型モノグリセリドリパーゼにおける本来のリンカー部分は含んでない。
【0090】
(2)次に、OASS-Bを単独で、又はPelBとの融合タンパク質として発現するベクターを構築した。
OASS-Bを、プライマー#7(5'-ggcatatggccatggaaacattagaacaaacaataggc-3'(配列番号17))と、プライマー#8(5'-ttctcgagaatccccgccccctggctaaaatgc-3'(配列番号18))を用いて上記と同じ条件にてPCR増幅した。プライマー#7にはNdeIとNcoIサイトが付加され、プライマー#8にはXbaIサイトが付加されている。NdeI-XbaIで消化したPCR断片を同酵素で処理したpET22b(+)に連結してpET-OASSB(PelB-)を構築した。また、NcoI-XbaIで消化したPCR断片を同酵素で処理したpET22b(+)に連結してpET-OASSB(PelB+)を構築した。これらの発現ベクターは、OASS-Bのみが単独で、又はPelBとの融合体として発現するようにデザインされ、発現産物はATとの融合体ではない。PelBは、pET22b(+)のマルチクローニングサイト上流に付加されている分泌シグナルであり、NdeI-XbaI部位で挿入したコンストラクトではPelB配列が除かれ、菌体内にてOASS-Bを過剰発現させることができる。一方、pET-OASSB(PelB+)はその分泌シグナルを保持している形であり、OASS-Bを発現し、菌体外へ分泌することができる。
【0091】
〔実施例3〕OASS発現株の培養
pET-AT、pET-OASSA-AT、pET-OASSB-AT、pET-OASSB(PelB-)、pET-OASSB(PelB+)の各ベクターでE. coli BL21株コンピテントセル(Novagen)を形質転換し、形質転換体をアンピシリン100 mg/Lを含むLB寒天培地上で選択した。得られた形質転換体を3 mLのLB培地(Bacto tryptone; 10 g/L、yeast extract; 5g/L、NaCl; 10 g/L、アンピシリン; 100 mg/L)で37℃、14時間、前培養を行ったのち、50 mLのLB培地に前培養液500μLを加え、37℃で本培養を行った。
【0092】
培養液のOD660をBACTOMONITOR BACT-550(JIKCO社製)にて測定し、OD660が0.5前後になった段階で終濃度1mMとなるようIPTGを添加し、さらに37℃で4時間培養した。IPTG添加によりpETベクター上のT7プロモーターが誘導され、その支配下にある遺伝子発現が誘導される。その後、4℃、7,800rpm、5min遠心分離をし、上清を捨て集菌した。その菌体をPBS(NaCl; 8 g/L、KCl; 0.2 g/L、Na2HPO4・12H2O; 2.9 g/L、KH2PO4; 0.2 g/L)に懸濁させ、洗浄を3回行い、以下の各実験に適した菌体濃度に調製した。
【0093】
〔実施例4〕OASSのIn vitro酵素活性測定とシステインの定量
以下の実施例で使用した、OASSのIn vitroでの酵素反応測定法を以下に示す。pH7.5に調整したBuffer A(50 mMリン酸カルシウム、1 mM EDTA、0.5 mM β-メルカプトエタノール)84μLにOD660に調製した菌体懸濁液22μLを添加して室温で2分間放置した。さらに200 mMのOAS溶液13μL(終濃度 20 mM)と、Buffer Aに溶解して調製した20 mMの硫化ナトリウム(NaOH溶液にてpH7.5に調製)もしくはチオ硫酸ナトリウムの溶液13μL(共に終濃度2 mM)を加え、室温にて5分間反応させた後、反応液に20%のトリクロロ酢酸(TCA)67μLを添加して反応を停止させた。15,000 rpm、5min遠心分離を行い、上清のみを回収した。
【0094】
L−システインは、Gaitonde法(Gaitonde M.K.,(1967) Biochem J 133:627-633)により定量した。Gaitonde法では、システインに加えて、L−シスチンやS-スルホシステインのようなシステイン関連物質も合わせて定量される。以下に具体的な方法を示す。培養液上清もしくは適当に希釈した培養液50μLに、等量のBuffer B(200 mM Tris-HCl(pH8.5)、10 mM DTT)を加えてサンプルを作製した。これらを室温で10分間放置した。ニンヒドリン溶液(酢酸:塩酸=3:2の溶液に2.5%のニンヒドリンを加えた溶液)50μLと、酢酸50μLを混合した溶液に、上記サンプル50μLを加え、100℃で10分間加熱した後、4℃に冷却した。この反応液40μLを95%エタノール200 μLと混合し、マイクロプレートリーダー(SPECTRAmax 340PC)で560nmの吸光度を測定した。同様に調製した5 mg/Lから60 mg/Lのシステイン標準液を指標に検量線を描き、サンプル中のシステイン関連物質を定量した。
【0095】
なお、以下の実施例では、OASS-Bとチオ硫酸ナトリウムの反応から生成するS-スルホシステインについても、比較しやすくするため、すべてのシステイン関連物質は同モルのシステインに換算して値を表示した。酵素活性は単位時間、単位容量、単位ODあたりのシステイン生産量とした。つまり1U = システイン生産量(μmol)/OD560/minで示した。以下、システイン関連物質の生成及び定量値に関し、システイン関連物質を単に「システイン」と記載することがある。
【0096】
〔実施例5〕OASS-B発現条件による酵素反応速度の検討
pETベクターを用いた酵素発現系のうち、どのシステムが最も効率よくシステイン関連物質を生産できるかを検討した。pET-OASSB(PelB-)、pET-OASSB(PelB+)、pET-OASSB-AT、pET-ATの各ベクターを導入したE. coli BL21株株を、実施例3に記載したようにして培養した。
【0097】
それぞれの形質転換体でOASS-Bが発現していることをSDS-PAGEにて確認した後、培養液を遠心分離し、培養上清を回収した。さらに、残った菌体をPBSにて2回洗浄後、PBSでOD600=2になるように懸濁した。そこに、PBSに溶解した20mM OASを、菌体懸濁液もしくは培養上清と等量加え(終濃度10mM)、さらに、1Mチオ硫酸ナトリウムを1/50量加えて(終濃度20mM)、37℃で2時間、振とうしながら反応を行った。その後、反応液中のシステイン濃度を実施例4に記載の方法にて測定した。
【0098】
結果を表1に示した。分泌シグナルを付加したOASS-Bを発現するpET-OASSB(PelB-)プラスミドでは、培養上清中にシステイン生成が見出されたが、分泌シグナルを付加しないOASS-Bを発現するpET-OASSB(PelB-)プラスミドとの差は少なかった。一方、菌体懸濁液では、、オートトランスポーターを用いてOASS-Bを細胞外提示した発現系では、効果的、かつ菌体特異的にシステイン関連物質への変換反応が行われることが明らかとなった。
【0099】
【表1】

【0100】
〔実施例6〕オートトランスポーターを用いたOASS細胞表層提示とその活性測定
オートトランスポーターを用いた発現系にて、OASS-Bが細胞外に提示されているかどうか、またその提示された酵素が活性を保持しているかを確認した。実施例5に記載された方法でPelB-OASSB-ATの発現菌を得たのち、in vitro酵素活性法にてOASS-Bの活性を測定した。また、この菌体を用いたシステインへの変換反応が生産培地中にて使用できるかを検討するため、bufferAの代わりに生産培地((NH4)2SO4 15 g/L、KH2PO4 1.5 g/L、MgSO4-7H2O 1 g/L、thiamine HCl 0.1 mg/L、FeSO4・7H2O 1.7 mg/L、Na2MoO4・2H2O 0.15 mg/L、CoCl2・6H2O 0.7 mg/L、MnCl2・4H2O 1.6 mg/L、ZnSO4・7H2O 0.3 mg/L、CuSO4・5H2O 0.25 mg/L、tryptone 0.6 g/L、yeast extract 0.3 g/L、NaCl 0.6 g/L、CaCO3 20 g/L、L-Histidine HCl・H2O 135mg/L、pyridoxine 2 mg/L、glucose 20 g/L)を用いた活性測定も同時に行った。
培地を用いた場合、buffer A中に比べれば活性は落ちるものの、活性を保持していることが明らかとなった(表2)。
【0101】
【表2】
【0102】
オートトランスポーターを用いた発現系にてOASS-Bが活性をすことは確認できたが、さらに、実際にOASS-Bが細胞外膜に提示されているかを確認した。そこで、PelB-OASSB-AT発現菌の菌体をPBSにてOD660=2に調整したのち、4℃で10分間超音波処理を行い、細胞を破砕し、4℃、50,000 rpm、40min超遠心分離を行った。この操作の結果、上清部分を可溶性画分とし、沈殿画分を膜画分に分離することができる。各画分における酵素活性を確認するため、上記in vitroの測定系にてそれぞれの酵素活性を測定したところ、大部分の活性が膜画分に存在していた(表3)。OASS-BとATとの融合タンパク質(OASSB-AT)の細胞質へのミスローカライズは認められず、このATシステムがもくろみ通り働き、OASSを外膜に提示していることが示唆された。
【0103】
【表3】
【0104】
〔実施例7〕OAS生産菌とOASS提示菌を用いたシステイン関連化合物の生産培養
OAS生産菌が生産するOASを、OASS-B提示菌によりL-システイン関連物質に変換するというコンセプトの成立を確認するために、以下の実験を行った。
まず、OAS生産菌として、MG1655ΔcysKΔcysM/pACYC-DESを用いた。
pACYC-DESは、炭素源から効率よくOASを作るために、OASを細胞外に排出する活性を有する排出因子であるYdeDをコードするydeD(Tobias Dassler.,et al,(2000)Molecular Microbiology.,36(5))、L-システイン生合成における鍵酵素の一つであるホスホグリセレートデヒドロゲナーゼのフィードバック阻害解除型変異体をコードするserA*(EP620853B)、および同じくL-システイン生合成における鍵酵素の一つであるセリンアセチルトランスフェラーゼのフィードバック阻害解除型変異体をコードするcysE*(US patent 2005/0124049 A1)の各遺伝子を搭載した多コピープラスミドである同プラスミドの構築は、EP1528108に詳述されている。
【0105】
MG1655ΔcysKΔcysM/pACYC-DESは、上記プラスミドpACYC-DESで、MG1655ΔcysKΔcysM株を形質転換することにより得た。このMG1655ΔcysKΔcysM株は、さらにOASを蓄積させるため、OASからシステイン等への反応を触媒する酵素であるOASSをコードする両遺伝子cysK、cysMを欠損してある。MG1655ΔcysKΔcysMこの菌の作製方法を以下に示す。
【0106】
まずpMIV-5JS(特開2008-99668、EP1942183)を鋳型に、プライマーP1(5'-agtaagatttttgaagataactcgctgactatcggtcgctcaagttagtataaaaaagctgaac-3'(配列番号19))とプライマーP2(5'-ggtaaagctttcatcttcttgtagtttcaacgccgctgaagcctgcttttttatactaagttgg-3'(配列番号20))を用いてクロラムフェニコール耐性遺伝子を含むフラグメントを増幅した。pMIV-5JSは、Muファージ由来のインテグレーションカセットを持つプラスミドpM12、thrLターミネーターフラグメント、pACYC184由来のクロラムフェニコール耐性遺伝子、及びλattL及びλattRフラグメントから構成されるプラスミドである。
【0107】
PCRは、94℃で5分の後、98℃を5秒、55℃にて10秒、72℃にて3分、のプログラムを30サイクル行った。増幅したフラグメント上にはλ依存インテグレーション(「Red-driven integration」と呼ばれる方法(Proc. Natl. Acad. Sci. USA, 2000, vol. 97, No. 12, p6640-6645)によって目的の断片を挿入するための染色体上のターゲットとなる配列(ここではcysKのC末端、N末端の配列)がデザインされている。そのため、取得されたDNA断片を、目的の菌株にこのλ依存インテグレーションによって挿入した場合には、染色体上のcysK遺伝子の中にクロラムフェニコール遺伝子が挿入された構造となる。
【0108】
定法に従い、エレクトロポレーション法にて上記増幅断片をMG1655/pKD46に導入した。プラスミドpKD46は、目的のDNA断片をMG1655株の染色体に組み込むためのヘルパープラスミドである(Datsenko, K.A., Wanner, B.L., Proc.Nat1.Acad.Sci.USA, 97, 6640-6645, (2000))。こうして得られたMG1655ΔcysK株に導入されたクロラムフェニコール耐性マーカーを、ラムダファージ由来の切り出しシステムによって除去した。具体的には、ラムダファージのInt-Xis遺伝子を搭載したpMT-Int-Xis2(WO2005/010175)でMG1655ΔcysK株を形質転換し、得られた形質転換株からクロラムフェニコール感受性を示すMG1655ΔcysK(s)株を取得した。
【0109】
さらにcysM遺伝子を欠損させるために、まずpUC18にcysMをクローニングした。MG16655のゲノムを鋳型として、P3(5'-acgggatccttgcgtttgtcatcagtctcc-3'(配列番号36))とP4(5'-acgaagcttgtcgcatcaggcaacaccacg-3'(配列番号37))をプライマーとして用いたPCRにより、cysM遺伝子を増幅した。PCRは、94℃で5分の後、98℃を5秒、55℃にて10秒、72℃にて1分、のプログラムを30サイクル行った。この増幅したフラグメントをBamHIとHindIIIで切断し、同酵素で消化したpUC18ベクターに挿入し、pUC-cysMを作製した。pUC-cysMのうち、cysM中に唯一存在するEco91I切断配列を利用し、Eco91IにてpUC-CysMを消化した後、pACYC184をEco47IIIを用いて切り出したクロラムフェニコール耐性遺伝子を含む配列を挿入し、pUC-cysM::CmRを作製した。
【0110】
pUC-cysM::CmRでE. coli JC7623(標識遺伝子;thr-1 leu-6 thi-1 lacY1 galK2 ara-14 xyl-5 mtl-1 proA2 his-4 argE3 str-31 tsx-33 supE344 recB21 recC22 sbcB15)(国立遺伝研究所より分与可能)を形質転換した。クロラムフェニコール耐性かつアンピシリン感受性となるプラスミドレスの株、すなわち染色体上のcysM中にクロラムフェニコール耐性遺伝子が挿入された株を得た。その株からP1トランスダクション法にてMG1655ΔcysK(s)株に形質導入し、得られた形質導入株からクロラムフェニコール耐性を示すMG1655ΔcysKΔcysM株を取得した。この株にpACYC-DESをエレクトロポレーションにて導入し、テトラサイクリン耐性株を選択、MG1655ΔcysKΔcysM/pACYC-DES株を取得した。
【0111】
上記MG1655ΔcysKΔcysM/pACYC-DES株を、試験管(内径22mm)に張り込んだ2mLの生産培地中にて、32℃で約19時間振盪培養したのち、4℃、7,800rpm、5min遠心分離を行い、OASを含んだ上清を得た。上記培地は、グルコースが40 g/Lであり、テトラサイクリン 12.5 mg/Lを加えた以外は、実施例6に記載した生産培地と同じ組成である。
【0112】
またOASS提示菌として、BL21/pET-OASSA-AT及びpET-OASSB-ATで形質転換したE. coli BL21を用い、これらの菌を実施例3に記載の方法にて培養した。得られたOASS提示菌を、上記のOASを含有する上清を用いて希釈してOD660=2にそろえた。希釈液2 mLを大試験管に張り込み、そこに1Mチオ硫酸ナトリウムを1/50量加えて(終濃度20mM)、32℃で振盪保温しながら0、30、60、120、180分後にサンプリングを行い、L−システイン関連物質生成の経時変化を検証した。S源にチオ硫酸ナトリウムを用いたが、チオ硫酸ナトリウムは硫化ナトリウムに比べ酸性側でも安定に存在することが知られており、培地中に残存するチオ硫酸ナトリウム量の経時変化を見ることができる。
【0113】
その結果、ネガティブコントロールのATのみ発現した菌(E. coli BL21/pET-AT)、及びチオ硫酸ナトリウムを基質としないOASS-Aを発現した菌では、システイン関連物質の生成はほとんど見られないのに対し、OASS-Bを発現した菌では反応時間に比例してシステイン関連物質の生産量が上がり、最終的に約160 mg/Lのシステイン関連物質を産生した(図1)。尚、ここで生成したシステイン関連物質は概ねS-スルホシステインである。なお、S-スルホシステインはDTT等の還元剤存在下で、容易にシステインに変換することができる。
【0114】
〔実施例8〕リンカーの最適化によるOASS-Bの活性向上
オートトランスポーターを利用したタンパク質発現において、オートトランスポーターと目的タンパク質を繋ぐリンカー部分の長さが活性に大きく影響を与える例が知られている(Jochen Maurer.,et al(1999)Journal Of Bacteriology.,181(22),7014-7020)。そこで、OASS-Bのリンカー最適化を行った。
【0115】
pET-OASSB-ATのリンカー長(モノグリセリドリパーゼにおける本来のリンカーは含まず、βバレル部分のN末端の34アミノ酸を欠失している)を±0とし、それよりも79アミノ酸長い+79型から36アミノ酸短い-36型(βバレル部分をさらに36アミノ酸欠失)の間でいくつかのバリアントを作製してそれぞれの活性を測定することで、リンカー最適化を行った。まず、79アミノ酸長いリンカー型のプラスミドを構築した。プライマー#1の代わりにプライマー#9(5’-ccatggttgaattcaccctgggcgtgg-3’(配列番号21))を用いた以外はpET-ATを作製するのと同様の方法にてpET-(+79)-ATを構築し、これにOASS-Bコード配列を導入してpET-OASSB(+79)-ATを作製した。±0型よりも短いバリアントは、βバレル部分のN末端部分が±0型よりもさらに欠失している。例えば、-9型では、野生型モノグリセリドリパーゼのβバレル部分と推定される領域よりも43(34+9)アミノ酸短い。
【0116】
pET-OASSB(+79)-ATが持つ、PelB、OASS-A、及びATとの融合タンパク質(「PelB-OASSB(+79)-AT」)をコードする遺伝子の塩基配列を配列番号34に示し、コードされるアミノ酸配列を配列番号35に示した。配列番号35において、1〜303位はOASS-B部分であり、306位〜629位はオートトランスポータードメインである。304位〜305位は、これらの連結に用いたEcoRI配列によって生じた部分である。PelB-OASSB(+79)-ATのオートトランスポータードメインは、PelB-OASSB-ATに比べて、モノグリセリドリパーゼ前駆体タンパク質のリパーゼドメインに由来する9アミノ酸、及び、各ドメイン間のスペーサーに由来する21アミノ酸、リンカー部分の15アミノ酸、及びβバレル部分のN末端34アミノ酸の合計79アミノ酸長い。pET-OASSB(+79)-ATから、任意の長さのリンカーを持つ融合タンパク質をコードするプラスミドを構築することができる。尚、配列番号6においては、PelB-OASSB-ATのトランスポータードメインは366〜610位に相当し、PelB-OASSB(+79)-ATのオートトランスポータードメインは287〜610位に相当する。
【0117】
pET-OASSB(+79)-ATを鋳型にLinker(X)-primerとRv-Primerにて、94℃で5分の後、98℃を5秒、55℃にて10秒、72℃にて1分、のプログラムを40サイクルというPCR反応を行い、目的の長さのリンカーフラグメントを得た。Linker(X)-primerにおける「X」は、リンカー長を示す。リンカー長は、pET-OASSB-ATを±0とし、長くしたものを(+)で表した。また、pET-OASSB-ATから、さらにβバレル部分の長さを短くしたものを(-)で表した。作製したリンカーの長さとその作製に用いたプライマーは以下の通りである。
【0118】
【表4】
【0119】
各リンカー側プライマー(上記Linker(X)-primer)にはEcoRIサイトを、オートトランスポーター側プライマー(上記Rv-Primer)にはStuIサイトを付加しており、PCRで増幅後、これらの制限酵素にて処理を行い、同じ酵素で消化してリンカー部をとり除いたpET-OASSB-ATベクターに挿入した。こうして構築したpET-OASSB(X)-ATをE. coli BL21株にエレクトロポレーション法にて導入し、実施例4に記載のin vitro活性測定法にて、それぞれの酵素活性を測定した。
【0120】
まず始めに、リンカーを短くするタイプを中心に作製し、活性を測定した。するとpET-OASSB(+40)-ATが最も活性が高く、pET-OASSB(±0)-ATの約4.7倍であった(表5)。また各コンストラクトにおけるOASS発現をSDS-PAGEにて確認するため、それぞれのコンストラクトの発現菌体をPBSにてOD660=10に揃え、それらを等量のサンプルバッファー(BIO-RAD社製 Laemmli Sample Buffer)に懸濁した。すべてのサンプルを100℃で3分間加熱し、氷上で急冷してからSDS-PAGE(第一化学薬品PAGミニ「第一」4-20%)解析を行った。SDS-PAGEの各レーンには、上記サンプルをそれぞれ10μLをロードし、泳動条件は30 mAで約90minとした。泳動後のゲルをCBB染色をして各タンパク質の発現量を確認したところ、発現されたタンパク質の量はほぼ同等であり(図2)、活性強度は発現量のみに依存するものではないと考えられた。
【0121】
続いて+40型を中心としたリンカータイプを作製し、同様に活性評価を行った。その結果pET-OASSB(+49)-ATが最も活性が高く、pET-OASSB(±0)-ATの約5.9倍もの活性を有していることが判明した(表5)。表5中、リンカー長はpET-OASSB(±0)-ATをゼロとしたときのプラスマイナスで表した。また、OASS-B活性はpET-OASSB(±0)-AT(リンカー長±0)の活性を1.00としたときの相対活性で示した。
【0122】
pET-OASSB(+40)-ATとpET-OASSB(±0)-ATを用いて、実施例5と同様にOAS生産菌との混合によるシステインの生産培養を行ったところ、180分間の反応でpET-OASSB(±0)-ATは163 mg/Lのシステイン関連物質を生産するのに対し、pET-OASSB(+40)-ATは328 mg/Lと約2倍の生産量を得ることに成功した(図3)。このように、リンカー長を変えることで、in vitro酵素活性だけでなく、システイン生産量も向上した。
【0123】
【表5】
【0124】
〔実施例9〕OASS提示菌を利用したシステイン生産菌による副生OASのシステイン関連物質への変換
糖からのシステイン発酵生産においては、システイン排出タンパク質として知られるYdeDを用いることが有効であると考えられているが、同排出因子はシステインだけではなくOASも細胞外へ排出するためOASを著量副生することが知られている(Tobias Dassler.,et al,(2000)Molecular Microbiology.,36(5),1101-1112)。そこでOAS生産菌とOASS提示菌を用い、糖からのシステイン生産の検討を行った。
【0125】
OAS生産菌としてE. coli-MG1655/pACYC-DESを、グルコース濃度を40g/Lとした以外は実施例6に記載の生産培地と同じ組成の培地2mLを入れた大試験管に植菌し、32℃で約20時間振盪培養をした。培養後、グルコースアナライザー(サクラ精機社製 バイオテックアナライザー AS210)を用いて培地中の残糖を測定し、消費糖量を計算したところ21.3 g/Lの糖を消費していた。産生したシステイン関連物質(OASは含まない)はGaitonde法にて定量を行ったところ、3.9mMであった。また、培地中に蓄積したOASの濃度をHPLCにて定量したところ、18.7 mMであった。ただし、サンプルを200 mMのTris-HCl(pH9.0)で希釈することでOASをより安定なNASに変換して検出を行った。OASはアルカリ性条件では用意にNASに変換されることが知られている。HPLCの条件は以下の通りである。
【0126】
カラム;Inertsil ODS-3(疎水性カラム/GLサイエンス社製)
バッファー流速;1.0mL/min
カラム温度;40℃
検出器;UV210nm
サンプルアプライ量;10μL
バッファー;0.1M KH2PO4・H3PO4(pH2.2)、5mM 1-オクタンスルホン酸Na
【0127】
上記培養液を遠心分離し、上清画分のみを回収したのち、この培養上清を用いてOASS提示菌(pET-OASSB(+49)-AT)の菌体量をO.D.660 =8にそろえ、実施例7と同様の方法にてシステイン関連物質生産を行った。OASS-B提示菌では、180分間反応させたところ、反応前のシステイン関連物質量に比べ約70 mg/L生産量が増加した(図4の棒グラフ)。またOASの濃度をHPLCにて確認したところ、システイン関連物質の増加に伴ってOAS濃度が減少していることが分かった(図4の折れ線グラフ)。
以上の結果より、細胞の外膜に提示したOASS-Bを用いて培地中のOASがシステイン関連物質に変換されたことが示された。
【0128】
〔配列表の説明〕
配列番号1:E. coli cysK遺伝子の塩基配列
配列番号2:E. coli cysK遺伝子によりコードされるOASS-Aのアミノ酸配列
配列番号3:E. coli cysM遺伝子の塩基配列
配列番号4:E. coli cysM遺伝子によりコードされるOASS-Bのアミノ酸配列
配列番号5:Pseudomonas sp. LP7315株のモノグリセリドリパーゼ遺伝子の塩基配列
配列番号6:Pseudomonas sp. LP7315株のモノグリセリドリパーゼ遺伝子がコードするアミノ酸配列
配列番号7:プライマー#1
配列番号8:プライマー#2
配列番号9:プライマー#3
配列番号10:プライマー#4
配列番号11:PelB-OASSA-ATをコードする遺伝子の塩基配列
配列番号12:PelB-OASSA-ATのアミノ酸配列
配列番号13:プライマー#5
配列番号14:プライマー#6
配列番号15:PelB-OASSB-ATをコードする塩基配列
配列番号16:PelB-OASSB-ATのアミノ酸配列
配列番号17:プライマー#7
配列番号18:プライマー#8
配列番号19:プライマーP1
配列番号20:プライマーP2
配列番号21:プライマー#9
配列番号22:Linker(+67)プライマー
配列番号23:Linker(+58)プライマー
配列番号24:Linker(+49)プライマー
配列番号25:Linker(+40)プライマー
配列番号26:Linker(+31)プライマー
配列番号27:Linker(+22)プライマー
配列番号28:Linker(+9) プライマー
配列番号29:Linker(-9) プライマー
配列番号30:Linker(-18)プライマー
配列番号31:Linker(-27)プライマー
配列番号32:Linker(-36)プライマー
配列番号33:Rv-primer
配列番号34:PelB-OASSB(+79)-ATをコードする塩基配列
配列番号35:PelB-OASSB(+79)-ATのアミノ酸配列
配列番号36:プライマーP3
配列番号37:プライマーP4
【技術分野】
【0001】
本発明は、L−システイン又はその関連物質の製造法に関し、詳しくはL−システイン又はその関連物質の製造に好適な細菌、及びそれらを用いたL−システイン又はその関連物質の製造法に関する。L−システイン及びその関連物質は、医薬品、化粧品及び食品分野で利用されている。
【背景技術】
【0002】
L−システインは、毛髪、角、羽毛等のケラチン含有物質から抽出することにより、あるいはDL−2−アミノチアゾリン−4−カルボン酸を前駆体とする細菌酵素変換により得られている。また、新規な酵素を用いた固定化酵素法によるL−システインの大量生産も計画されている。
【0003】
さらに、細菌を用いた発酵法によるL−システインの生産も試みられている。例えば、本発明者らは、L−システイン分解系が抑制され、かつ、L−システインによるフィードバック阻害が低減されたセリンアセチルトランスフェラーゼ(serine acetyltransferase(EC 2.3.1.30):以下、「SAT」ともいう)を保持するエシェリヒア属細菌を用いたL−システインの製造法を開示している(特許文献1)。また、L−システイン分解系を抑制することによってL−システイン生産能が高められた細菌としては、シスタチオニン−β−リアーゼ(特許文献1)、トリプトフォアナーゼ(特許文献2)、O−アセチルセリン スルフヒドリラーゼB(特許文献3)の活性を低下させたコリネ型細菌又はエシェリヒア属細菌が知られている。また、L−システインによるフィードバック阻害が低減された特定の変異を有するSATをコードするDNA配列により脱制御されたL-システイン物質代謝を有する細菌を用いた、L−システインの製造法が知られている(特許文献4)。
【0004】
さらに、YdeDタンパク質をコードするydeD遺伝子(非特許文献1)、及びYfiKタンパク質をコードするyfiK遺伝子(特許文献5)は、L−システイン経路の代謝産物の排出に関与していることが知られている。また、細胞に毒性の物質を排出するのに適したタンパク質をコードする遺伝子であるmar-遺伝子座、acr-遺伝子座、cmr-遺伝子座、mex-遺伝子座、bmr-遺伝子座、qacA-遺伝子座(特許文献6)、又はemrAB、emrKY、yojIH、acrEF、bcrもしくはcusA遺伝子(特許文献7)の発現を上昇させることによりL−システイン生産能を高める技術が知られている。
【0005】
一方、抗生物質又は細菌に毒性の物質を細胞から直接放出するために好適である蛋白質をコードする遺伝子を過剰発現する細菌を用いたL−システインの製造法が報告されている(特許文献8)。
【0006】
このように、L−システインを排出する因子が知られているが、これらの排出因子はL−システインとその前駆体であるO−アセチル−L−セリン(以下、「OAS」と記載することがある)とを区別することができないため、これらのL−システイン排出因子を増強した微生物では、培地中にL−システインだけでなく副生物としてOASも蓄積することが知られていた(非特許文献1)。また、L−システインは細胞にとって毒性の高いアミノ酸であることから、排出因子を強化したとしても、細胞内で生成したL−システインによって生育が阻害されることが考えられた。
【0007】
オートトランスポーターとは、1987年にナイセリア・ゴノロエ(Neisseria gonorrhoeae)のIgA1プロテアーゼについて初めて発見された(非特許文献2)、グラム陰性菌が持つtype-Vのタンパク質分泌システムである。このシステムでは、N末からシグナル配列、パッセンジャーと呼ばれるペプチド(すなわちIgA1プロテアーゼドメイン)、及びC末にオートトランスポータードメインと呼ばれるリンカー配列及びそれに続くβバレル構造を持ったタンパク質が発現し、このタンパク質はシグナル配列に依存して内膜を通過した後、βバレルが外膜に入り込みパッセンジャーが通過する穴を形成し、パッセンジャー領域が外膜の外に分泌・固定される。パッセンジャーの部分に目的の酵素ペプチドを挿入することで、有用酵素の提示発現システムとして利用されている(非特許文献3)。
【0008】
酵素を微生物の細胞外膜に提示させることにより、細胞外に分泌される酵素と比べて、安定した酵素活性が得られることがある。一方、微生物が生産する酵素を用いて、酵素反応により有用物質の生産を行う場合は、目的酵素を細胞内に蓄積させて、細胞を破砕して酵素を取り出してから反応させる方法や、目的酵素を培地中に分泌させる方法等が考えられる。しかしながら、これらの場合は、酵素を生成するプロセスや、酵素反応後に、生成した反応物と変換酵素を分離するための複雑な精製プロセスが必要となり、高コストとなってしまう。
【0009】
また、他のオートトランスポーターとしては、シュードモナス エスピー.(Pseudomonas sp. ) LP7315株由来の耐熱性モノグリセリドリパーゼのオートトランスポート(AT)システム(非特許文献4)が知られている。今中や山下は、このPseudomonas sp. strain LP7315由来のATシステムを用いて、システイン前駆体であるOASをシステインに変換する酵素(OASS-A)を細胞外膜に提示させて、L−システイン(もしくはS−スルホ−L−システイン)の生産を行うことを示している(非特許文献5、6)。
【0010】
L−システインの前駆体であるOASをL−システインに変換する酵素は、O−アセチルセリンサルフヒドリラーゼ(O-acetylserine sulfhydrylase、以下「OASS」と記載することがある)と呼ばれ、二つのアイソザイムOASS-A、OASS-Bが存在することが知られている。エシェリヒア・コリ(Escherichia coli)では、OASS-A及びOASS-Bは、それぞれcysK、cysMにコードされている(非特許文献7)。OASS-Aは、OASとスルフィド(sulfide)との反応を触媒し、L−システインを産生する。一方、OASS-Bは、OASとスルフィドは又チオサルフェイト(チオ硫酸、thiosulfate)との反応を触媒し、それぞれL−システイン又はS−スルホ−L−システインを産生する。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】特開平11−155571号
【特許文献2】特開2003−169668
【特許文献3】特開2005−245311
【特許文献4】特表2000−504926
【特許文献5】特開2004−49237
【特許文献6】米国特許第5972663号
【特許文献7】特開2005−287333
【特許文献8】特開平11−56381号
【非特許文献】
【0012】
【非特許文献1】Dassler, T. et al., Mol. Microbiol., 36, 1101-1112 (2000)
【非特許文献2】Pohlner J., et al,. Nature 325 (6103):458-62 (1987)
【非特許文献3】Joachin, J., Appl Microbiol. Biotechnol., 69:607-614 (2006)
【非特許文献4】Sakiyama T, et al., J. Biosci. Bioeng., 91(1):27-32 (2001)
【非特許文献5】今中ら、化学工学会第37回秋季大会 (2005)講演要旨
【非特許文献6】山下ら、化学工学会春季大会(2006)講演要旨
【非特許文献7】Kredich N. M. et al., Biosynthesis of Cysteine. In Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed., Vol. 1, pp. 514-527 (1996)
【発明の概要】
【発明が解決しようとする課題】
【0013】
本発明は、OASを利用してL-システイン関連物質を製造する新たな方法を提供することを課題とする。
【課題を解決するための手段】
【0014】
前記したように、OASS-AによるOASとスルフィドとの反応では、直接反応生成物としてシステインができるため有用である反面、S源としてスルフィドを使わなければならない。具体的なスルフィドとしては、例えば硫化ナトリウム(Na2S)が挙げられるが、スルフィドは取り扱い上の安全性や物質自体の安定性の面で問題がある。硫化ナトリウムは、簡単に硫化水素(H2S)になり気中に放出されてしまう。例えば、反応溶液のpHを調整するときにはこの傾向が激しく、反応系に添加したNa2Sのかなりの量がH2Sとして気中に放出される。また、例えば37℃にて変換反応を行うとさらにこの傾向は増大する。工業レベルでの生産を考えるとき、放出されるH2Sの安全性も問題となる。一方、OASS-Bは、チオサルフェイトをS源として用いることができるため安全性が高く使いやすい。また、反応産物であるS−スルホ−L−システインはジチオスレイトール(DTT)等の還元剤により容易にL−システインに変換することができる。
【0015】
本発明者らは、cysMにコードされるOASS-Bをオートトランスポーターと融合することで細胞外膜に提示した菌を用いて、OAS及びチオ硫酸からL-システイン関連物質を生産することができることを見出し、本発明を完成させるに至った。
すなわち本発明は、以下のとおりである。
【0016】
(1)チオ硫酸とO−アセチル−L−セリンからS−スルホ−L−システインを生成する反応を触媒するO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌を、O−アセチル−L−セリン及びチオ硫酸に、これらを含む反応液中で作用させ、該反応液からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を採取することを特徴とする、S−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物の製造法。(2)前記反応液中のS−スルホ−L−システイン、又は前記反応液から採取したS−スルホ−L−システインを還元する工程をさらに含む、前記方法。
(3)前記細菌において、前記O−アセチルセリンサルフヒドリラーゼはオートトランスポーターとの融合タンパク質として発現される、前記方法。
(4)前記融合タンパク質におけるO−アセチルセリンサルフヒドリラーゼ部分が、cysM遺伝子によりコードされる、前記方法。
(5)前記O−アセチルセリンサルフヒドリラーゼ部分が、下記のいずれかのアミノ酸配列を有する、前記方法。
(A)配列番号2のアミノ酸配列。
(B)配列番号2のアミノ酸配列において、1〜10個のアミノ酸の置換、欠失、挿入、または付加を含むアミノ酸配列。
(6)前記オートトランスポーターが、シュードモナス属細菌の耐熱性モノグリセリドリパーゼのオートトランスポーターである、前記方法。
(7)前記反応液に含まれるO−アセチル−L−セリンが、O−アセチル−L−セリン生産能を有する他の細菌により生産されたものである、前記方法。
(8)前記O−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌はO−アセチル−L−セリン生産能を有し、前記反応液に含まれるO−アセチル−L−セリンは該細菌が分泌したものであって、前記反応液は培地である、前記方法。
(9)前記他の細菌又はO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌は、下記の性質の少なくともいずれかを有する、前記方法。
i)L−システイン生合成系酵素活性が増強されるように改変されている。
ii)細胞内のO−アセチルセリン分解酵活性が低下するように改変されている。
iii)O−アセチルセリンを排出する因子の活性が増強されるように改変されている。
(10)前記O−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌がエシェリヒア属細菌である、前記方法。
(11)前記他の細菌がエシェリヒア属細菌である、前記方法。
(12)エシェリヒア属細菌がエシェリヒア・コリである、前記方法。
(13)cysM遺伝子によりコードされるO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌であって、前記O−アセチルセリンサルフヒドリラーゼはオートトランスポーターとの融合タンパク質として発現される、細菌。
(14)前記融合タンパク質におけるO−アセチルセリンサルフヒドリラーゼ部分が、下記のいずれかのアミノ酸配列を有する、前記細菌。
(A)配列番号2のアミノ酸配列。
(B)配列番号2のアミノ酸配列において、1〜10個のアミノ酸の置換、欠失、挿入、または付加を含むアミノ酸配列。
(15)前記オートトランスポーターが、シュードモナス属細菌の耐熱性モノグリセリドリパーゼのオートトランスポーターである、前記細菌。
(16)細胞内のO−アセチルセリンサルフヒドリラーゼ活性が低下するように改変された、前記細菌。
【発明の効果】
【0017】
本発明により、OASからのL-システイン関連物質の生産が可能になる。また、OASを生産する細菌と組み合わせることで、糖を原料としたL-システイン関連物質の効率的な生産が可能になる。本発明の一形態によれば、OASからのL−システイン関連物質の生産におけるイオウ源として、チオ硫酸を用いることができる。
また、本発明により、O−アセチルセリンサルフヒドリラーゼタンパク質をオートトランスポーターの機能を利用することにより細胞外膜に提示した細菌が提供される。
【図面の簡単な説明】
【0018】
【図1】OASS提示菌を用いたOASからのシステイン関連物質の生産を示す図。
【図2】pET-OASSB(X)-AT保持株のSDS-PAGEの結果を示す図(電気泳動写真)。
【図3】pET-OASS(+40)-AT保持株を用いたシステイン関連物質生産の経時変化を示す図。
【図4】L−システイン発酵で副生したOASを利用したOASS提示菌によるシステイン関連物質の生産を示す図。
【発明を実施するための形態】
【0019】
本発明の方法は、チオ硫酸とO−アセチル−L−セリン(OAS)からS−スルホ−L−システインを生成する反応を触媒するO−アセチルセリンサルフヒドリラーゼ(OASS-B)を細胞外膜に提示する細菌を、OAS及びチオ硫酸に、これらを含む反応液中で作用させ、該反応液からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を採取することを特徴とする、S−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物の製造法である。すなわち、本発明では、細菌の細胞外膜に提示されたOASS-Bを用いて、OAS及びチオ硫酸からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を生成させる。本明細書においては、S−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体(OASを除く)、又はこれらの混合物を、「システイン関連物質」と記載することがある。システイン関連物質は、フリー体であってもよく、塩であってもよい。塩としては、例えば硫酸塩、塩酸塩、炭酸塩、アンモニウム塩、ナトリウム塩、カリウム塩が挙げられる。塩であってもよい。システイン関連物質については後述する。また、前記OASS-Bを細胞外膜に提示する細菌を、OASS提示菌(OASS-autodisplaying bacterium)と記載することがある。
【0020】
本発明の第一の形態においては、反応液に含まれるOASは、好ましくはOAS生産能を有する他の細菌(OAS生産菌)により生産されたものである。この形態においては、OAS生産菌が生産したOASを原料として、OASS提示菌によりS−スルホ−L−システインを生成させる。尚、OASは、細菌を用いる方法以外の方法によって生産されたものであってもよい。
【0021】
本発明の第二の形態においては、OASS提示菌は、OAS生産能をも有し、前記反応液に含まれるOASは該細菌が分泌したものであって、前記反応液は培地である。この形態では、OASS提示菌はOAS生産菌でもあり、OASS提示菌自身が培地に分泌したOASを原料として、同細菌の細胞外膜に提示されたOASS-BによりS−スルホ−L−システインを生成させる。
いずれの形態においても、後述するように、S−スルホ−L−システインは、L−システイン、L−シスチン、又はそれらの誘導体に変換され得る。
以下、OAS生産菌及びOASS提示菌について説明する。
【0022】
<1>OAS生産菌
OAS生産能とは、細菌を培地中で培養したときに、培地中または菌体内にOASを生成し、培地中または菌体から回収できる程度に蓄積する能力をいう。また、OAS生産能を有する細菌とは、野生株または親株よりも多い量のOASを生産し培地に蓄積することができる細菌を意味し、好ましくは、0.2g/L以上、より好ましくは0.3g/L以上、特に好ましくは0.4g/L以上の量のL-システインを生産し培地に蓄積することができる細菌が含まれる。
【0023】
OAS生産能を有する細菌としては、本来的にOAS生産能を有するものであってもよいが、下記のような細菌を、変異法や組換えDNA技術を利用して、OAS生産能を有するように改変したものであってもよい。
【0024】
OAS生産菌の育種に用いる細菌としては、OAS生産能を付与又は増強され得るものであれば特に制限されないが、グラム陰性細菌、例えばエシェリヒア属、エンテロバクター属、パントエア属、クレブシエラ属、セラチア属、エルビニア属、サルモネラ属、モルガネラ属など、腸内細菌科に属する細菌、及び、コリネバクテリウム属、バチルス属、シュードモナス属、アグロバクテリウム属、ビブリオ属に属する細菌等が好ましい。具体的にはNCBI(National Center for Biotechnology Information)データベースに記載されている分類により腸内細菌科に属するものが利用できる(http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=91347)。中でも、エシェリヒア属細菌が好ましく、エシェリヒア・コリがより好ましい。
【0025】
エシェリヒア属細菌としては、特に限定されないが、具体的にはNeidhardtらの著書(Backmann, B. J. 1996. Derivations and Genotypes of some mutant derivatives of Escherichia coli K-12, p. 2460-2488. Table 1. In F. D. Neidhardt (ed.), Escherichia
coli and Salmonella Cellular and Molecular Biology/Second Edition, American Society for Microbiology Press, Washington, D.C.)に挙げられるものが利用できる。その中では、例えばエシェリヒア・コリが挙げられる。エシェリヒア・コリとしては具体的には、プロトタイプの野生株K12株由来のエシェリヒア・コリ W3110 (ATCC 27325)、エシェリヒア・コリ MG1655 (ATCC 47076)等が挙げられる。
【0026】
これらを入手するには、例えばアメリカン・タイプ・カルチャー・コレクション(住所
12301 Parklawn Drive, Rockville, Maryland 20852 P.O. Box 1549, Manassas, VA 20108, United States of America)より分譲を受けることが出来る。すなわち各菌株に対応する登録番号が付与されており、この登録番号を利用して分譲を受けることが出来る(http://www.atcc.org/参照)。各菌株に対応する登録番号は、アメリカン・タイプ・カルチャー・コレクションのカタログに記載されている。
【0027】
エンテロバクター属細菌としては、エンテロバクター・アグロメランス(Enterobacter
agglomerans)、エンテロバクター・アエロゲネス(Enterobacter aerogenes)等、パントエア属細菌としてはパントエア・アナナティス(Pantoea ananatis)が挙げられる。尚、近年、エンテロバクター・アグロメランスは、16S rRNAの塩基配列解析などにより、パントエア・アグロメランス(Pantoea agglomerans)又はパントエア・アナナティス(Pantoea ananatis)、パントエア・スチューアルティ(Pantoea stewartii)に再分類されているものがある。本発明においては、腸内細菌科に分類されるものであれば、エンテロバクター属又はパントエア属のいずれに属するものであってもよい。
【0028】
特に、パントエア属細菌、エルビニア属細菌、エンテロバクター属細菌は、γ-プロテオバクテリアに分類される細菌であり、分類学的に非常に近縁である(J Gen Appl Microbiol 1997 Dec;43(6) 355-361, International Journal of Systematic Bacteriology, Oct. 1997,p1061-1067)。近年、DNA-DNAハイブリダイゼーション実験等により、エンテロバクター属に属する細菌には、パントエア・アグロメランス(Pantoea agglomerans)又はパントエア・ディスパーサ(Pantoea dispersa)等に再分類されているものがある(International Journal of Systematic Bacteriology, July 1989;39(3).p.337-345)。また、エルビニア属に属する細菌にはパントエア・アナナス(Pantoea ananas)、パントエア・スチューアルティに再分類されているものがある(International Journal of Systematic Bacteriology, Jan 1993;43(1), p.162-173 参照)。
【0029】
エンテロバクター属細菌としては、エンテロバクター・アグロメランス(Enterobacter
agglomerans)、エンテロバクター・アエロゲネス(Enterobacter aerogenes)等が挙げられる。具体的には、欧州特許出願公開952221号明細書に例示された菌株を使用することが出来る。エンテロバクター属の代表的な株として、エンテロバクター・アグロメランスATCC12287株が挙げられる。
【0030】
パントエア属細菌の代表的な菌株として、パントエア・アナナティス、パントエア・スチューアルティ(Pantoea stewartii)パントエア・アグロメランス、パントエア・シトレア(Pantoea citrea)が挙げられる。パントエア・アナナティスとして具体的には、パントエア・アナナティスAJ13355株、SC17株、及びSC17(0)株が挙げられる。SC17株は、静岡県磐田市の土壌から、低pHでL-グルタミン酸及び炭素源を含む培地で増殖できる株として分離された株AJ13355(FERM BP-6614)から、粘液質低生産変異株として選択された株である(米国特許第6,596,517号)。SC17(0)株は、パントエア・アナナティスにおいて遺伝子破壊を行うために、λ Red遺伝子産物に耐性な菌株として構築された株である(WO2008/075483)。
【0031】
パントエア・アナナティスAJ13355株は、平成10年2月19日に、通産省工業技術院生命工学工業技術研究所(現名称、産業技術総合研究所特許生物寄託センター、住所 郵便番号305-8566 茨城県つくば市東1丁目1番地1中央第6)に、受託番号FERM P-16644として寄託され、平成11年1月11日にブダペスト条約に基づく国際寄託に移管され、受託番号FERM BP-6614が付与されている。尚、同株は、分離された当時はエンテロバクター・アグロメランス(Enterobacter agglomerans)と同定され、エンテロバクター・アグロメランスAJ13355として寄託されたが、近年16S rRNAの塩基配列解析などにより、パントエア・アナナティス(Pantoea ananatis)に再分類されている。また、SC17株は、平成21年2月4日に、産業技術総合研究所特許生物寄託センター(住所 郵便番号305-8566 茨城県つくば市東1丁目1番地1中央第6)に寄託され、受託番号FERM BP-11091が付与されている。また、SC17(0)株は、2005年9月21にロシアン・ナショナル・コレクション・オブ・インダストリアル・マイクロオーガニズム(Russian National Collection of Industrial Microorganisms (VKPM), GNII Genetika)(住所:Russia, 117545 Moscow, 1 Dorozhny proezd. 1)に受託番号VKPM B-9246のもとに寄託されている。
【0032】
エルビニア属細菌としては、エルビニア・アミロボーラ、エルビニア・カロトボーラが挙げられ、クレブシエラ属細菌としては、クレブシエラ・プランティコーラが挙げられる。
【0033】
OAS生産菌、又はOAS生産能を有するOASS提示菌として好ましい細菌は、下記の性質のいずれか1、2又は3種を有するものであるが、これらに限定されない。
i)L−システイン生合成系酵素活性が増強されるように改変されている。
ii)細胞内のO−アセチルセリン分解酵活性が低下するように改変されている。
iii)O−アセチルセリンを排出する因子の活性が増強されるように改変されている。
以下、腸内細菌科に属する細菌にOAS生産能を付与する方法、又はこれらの細菌のOAS生産能を増強する方法について述べる。
【0034】
細菌にOAS生産能を付与するには、栄養要求性変異株、アナログ耐性株又は代謝制御変異株の取得や、L-システインの生合成系酵素の発現が増強された組換え株の創製等、従来、コリネ型細菌又はエシェリヒア属細菌等の育種に採用されてきた方法を適用することができる(アミノ酸発酵、(株)学会出版センター、1986年5月30日初版発行、第77-100頁参照)。ここで、OAS生産菌の育種において、付与される栄養要求性、アナログ耐性、代謝制御変異等の性質は、単独でもよく、2種又は3種以上であってもよい。また、発現が増強されるL-システイン生合成系酵素も、単独であっても、2種又は3種以上であってもよい。さらに、栄養要求性、アナログ耐性、代謝制御変異等の性質の付与と、生合成系酵素の増強が組み合わされてもよい。
【0035】
OAS生産能を有する栄養要求性変異株、L-システインのアナログ耐性株、又は代謝制御変異株を取得するには、親株又は野生株を通常の変異処理、すなわちX線や紫外線の照射、またはN-メチル-N'-ニトロ-N-ニトロソグアニジン(NTG)もしくはエチルメタンスルフォネート(EMS)等の変異剤処理などによって処理し、得られた変異株の中から、栄養要求性、アナログ耐性、又は代謝制御変異を示し、かつOAS生産能を有するものを選択することによって得ることができる。
【0036】
OASはL-システインの前駆体であるため、L-システイン生産菌から容易に構築することができる。L-システイン生産菌として、具体的には、フィードバック阻害耐性のセリンアセチルトランスフェラーゼ(SAT)をコードする複数種のcysEアレルで形質転換されたE. coli JM15(米国特許第6,218,168号)、細胞に毒性の物質を排出するのに適したタンパク質をコードする過剰発現遺伝子を有するE. coli W3110 (米国特許第5,972,663号)、システインデスルフヒドラーゼ活性が低下したE. coli株 (特開平11-155571号公報)、cysB遺伝子によりコードされるシステインレギュロンの正の転写制御因子の活性が上昇したE. coli W3110 (WO01/27307)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。
【0037】
OAS生産菌は、細胞内のOAS分解酵素活性が低下するように改変されていることが好ましい。例えば、L-システイン生産菌からL-システイン分解酵素をコードするcysM及びcysKの遺伝子を同時に欠損させることで、OASを生産するように改変することができる。一方、E. coliでは、前述のとおりL-システインを排出する活性を有する公知のタンパク質としてydeDにコードされるタンパク質(特開2002-233384)が知られているが、このYdeDはL-システインに加えて、OAS等のL-システイン関連化合物も同時に排出することが知られている。したがって、OAS生産菌を得るためには、L-システイン排出因子として知られる因子の活性を上昇させてもよい。
【0038】
〔SATの増強〕
次に、細菌にOAS生産能を付与する方法として、L-システイン生合成酵素活性を増強する方法について説明する。L-システイン生合成酵素活性の増強は、例えば、セリンアセチルトランスフェラーゼ(SAT)活性を増強することによって行うことが出来る。細菌における細胞内のSAT活性の増強は、SATをコードする遺伝子のコピー数を高めることによって達成される。例えば、SATをコードする遺伝子断片を、細菌で機能するベクター、好ましくはマルチコピー型のベクターと連結して組換えDNAを作製し、これで宿主細菌を形質転換すればよい。
【0039】
SAT遺伝子は、エシェリヒア属細菌由来の遺伝子および他の生物由来の遺伝子のいずれも使用することができる。エシェリヒア・コリのSATをコードする遺伝子として、cysEが野生株及びL−システイン分泌変異株よりクローニングされ、塩基配列が明らかになっている(Denk, D. and Boeck, A., J. General Microbiol., 133, 515-525 (1987))。したがって、その塩基配列に基づいて作製したプライマーを用いて、エシェリヒア属細菌の染色体DNAを鋳型とするPCRによって、SAT遺伝子を取得することができる(特開平11-155571号参照)。他の生物のSATをコードする遺伝子も、同様にして取得され得る。このようにして得られるSAT遺伝子は、上記cysE遺伝子と同様にして発現増強を行うことができる。
【0040】
なお、SAT遺伝子の発現に「L−システインによるフィードバック阻害」などの抑制機構が存在する場合には、該抑制機構に非感受性となるように、発現調節配列又は抑制に関与する遺伝子を改変することによっても、SAT遺伝子の発現を増強することができる。
【0041】
〔OASS活性の低下〕
OAS生産菌は、細胞内における、OASをL-システインに変換するOASS活性が低下するように改変されていることが好ましい。ただし、OASS活性を低下させた後にL-システイン生合成経路の強化を行ってもよい。
【0042】
「細胞内のOASS活性が低下する」とは、細胞内のOASS活性が、野生株または親株等の非改変株に対して低下していることを意味し、活性が完全に消失していることを含む。本明細書に記載された他のタンパク質の活性の「低下」についても同様である。
OASSとしては、cysK、cysM遺伝子にコードされるOASS-A、及びOASS-Bが挙げられる。
【0043】
OASS活性は、例えば、実施例4に記載された方法で測定することができる。すなわち、試料をOAS及びイオウ源に作用させ、生成するシステイン関連物質の量を定量することにより、OASS活性を測定することができる。OASからのシステイン関連物質の生成におけるイオウ源は、OASS-Aでは硫化ナトリウム等の硫化物が、OASS-Bではチオ硫酸ナトリウム等のチオ硫酸塩が用いられる。システイン関連物質は、Gaitonde法(Gaitonde M.K.,(1967) Biochem J 133:627-633)で定量することができる。細胞内のOASS活性を測定するには、細胞を破砕し、膜画分を遠心分離等により除去し、可溶性画分についてOASS活性を測定すればよい。
【0044】
OASS活性を低下させるような改変は、例えば、OASS-AをコードするcysK、及びOASS-BをコードするcysM遺伝子の発現を低下させることによって達成される。具体的には例えば、染色体上のcysM、及びcysK遺伝子のそれぞれのコード領域の一部又は全部を欠損させることによって、これらの遺伝子にコードされるOASSタンパク質の細胞内の活性を低下させることができる。
【0045】
またOASSタンパク質活性の低下は、前記遺伝子のプロモーターやシャインダルガノ(SD)配列等の発現調節配列を改変することなどによって、これらの遺伝子の発現を低下させることによっても、達成することができる。また、発現調節配列以外の非翻訳領域の改変によっても、遺伝子の発現量を低下させることができる。さらには、染色体上の遺伝子の前後の配列を含めて、遺伝子全体を欠失させてもよい。また、染色体上のcysM及び/又はcysK遺伝子のコード領域にアミノ酸置換(ミスセンス変異)を導入すること、また終始コドンを導入すること(ナンセンス変異)、あるいは一〜二塩基付加・欠失するフレームシフト変異を導入することによっても、遺伝子の発現を低下させることが出来る(Journal of Biological Chemistry 272:8611-8617(1997) Proceedings of the National Academy of Sciences,USA 95 5511-5515(1998), Journal of Biological Chemistry 266, 20833-20839(1991))。
【0046】
また、OASSタンパク質の活性が低下するような改変であれば、X線もしくは紫外線を照射、またはN-メチル-N'-ニトロ-N-ニトロソグアニジン等の変異剤による通常の変異処理による改変であってもよい。
【0047】
発現調節配列の改変は、好ましくは1塩基以上、より好ましくは2塩基以上、特に好ましくは3塩基以上である。また、コード領域を欠失させる場合は、OASSタンパク質の機能が低下するのであれば、欠失させる領域は、N末端領域、内部領域、C末端領域のいずれの領域であってもよく、コード領域全体であってよい。通常、欠失させる領域は長い方が確実に遺伝子を不活化することができる。また、欠失させる領域の上流と下流のリーディングフレームは一致しないことが好ましい。
【0048】
cysM及び/又はcysK遺伝子のコード領域に他の配列を挿入する場合も、挿入の箇所は遺伝子のいずれの領域であってもよいが、挿入する配列は長い方が、確実に遺伝子を不活化することができる。挿入部位の前後の配列は、リーディングフレームが一致しないことが好ましい。他の配列としては、コードされるCysM及び/又はCysKタンパク質の機能を低下させるものであれば特に制限されないが、例えば、抗生物質耐性遺伝子やL-システイン生産に有用な遺伝子を搭載したトランスポゾン等が挙げられる。
【0049】
染色体上のcysM及び又はcysK遺伝子を上記のように改変するには、例えば、遺伝子の部分配列を欠失し、活性を有するOASSタンパク質を産生しないように改変した欠失型遺伝子を作製し、該遺伝子を含むDNAで細菌を形質転換して、欠失型遺伝子と染色体上の遺伝子とで相同組換えを起こさせることにより、染色体上の遺伝子を欠失型遺伝子に置換することによって達成できる。欠失型遺伝子によってコードされるOASSタンパク質は、生成したとしても、野生型タンパク質とは異なる立体構造を有し、機能が低下する。このような相同組換えを利用した遺伝子置換による遺伝子破壊は既に確立しており、「Redドリブンインテグレーション(Red-driven integration)」と呼ばれる方法(Datsenko, K. A, and Wanner, B. L. Proc. Natl. Acad. Sci. U S A. 97:6640-6645 (2000))、Redドリブンインテグレーション法とλファージ由来の切り出しシステム(Cho, E. H., Gumport, R. I., Gardner, J. F. J. Bacteriol. 184: 5200-5203 (2002))とを組合わせた方法(WO2005/010175号参照)等の直鎖状DNAを用いる方法や、温度感受性複製起点を含むプラスミド、接合伝達可能なプラスミドを用いる方法、宿主内で複製起点を持たないスイサイドベクターを利用する方法などがある(米国特許第6303383号、または特開平05-007491号)。
【0050】
cysM及び/又はcysK遺伝子の転写量が低下したことの確認は、各々の遺伝子から転写されるmRNAの量を野生株、あるいは非改変株と比較することによって行うことが出来る。mRNAの量を評価する方法としては、ノーザンハイブリダイゼーション、RT-PCR等が挙げられる(Molecular cloning(Cold spring Harbor Laboratory Press, Cold spring Harbor (USA), 2001))。
【0051】
OASSタンパク質の量が低下したことの確認は、抗体を用いてウェスタンブロットによって行うことが出来る(Molecular cloning(Cold spring Harbor Laboratory Press, Cold
spring Harbor (USA), 2001))。
【0052】
さらに、OASSタンパク質の量が低下したことの確認は、細胞のOASS活性を測定することによっても行うことができる。
【0053】
OASS-Aをコードする遺伝子として具体的には、配列番号1に示す塩基配列を含む遺伝子を挙げることが出来る。配列番号1には、エシェリヒア・コリの野生型cysK遺伝子の塩基配列を、配列番号2は同遺伝子がコードするOASS-Aのアミノ酸配列を示す。また、OASS-Bをコードする遺伝子として具体的には、配列番号3に示す塩基配列を含む遺伝子を挙げることが出来る。配列番号3には、エシェリヒア・コリの野生型cysM遺伝子の塩基配列を、配列番号4は同遺伝子がコードするOASS-Bのアミノ酸配列をを示す。なお、OASSをコードする遺伝子は上述した遺伝子情報を持つ遺伝子や、公知の配列を有する遺伝子に限られず、それらの遺伝子のバリアント、すなわちOASS活性を有するタンパク質をコードする限りにおいて、それらの遺伝子のホモログや人為的な改変体等、保存的変異を有する遺伝子も使用することができる。すなわち、公知のタンパク質のアミノ酸配列において、1若しくは数個の位置での1又は数個のアミノ酸が置換、欠失、挿入又は付加されたアミノ酸配列を有するタンパク質をコードする遺伝子であっても良い。このようなタンパク質のOASS活性は、1又は数個の置換、欠失、挿入又は付加される前のタンパク質に対して同等の活性、例えば70%以上、好ましくは80%以上、より好ましくは90%以上の活性を維持していることが好ましい。なお上記「1又は数個」とは、アミノ酸残基のタンパク質の立体構造における位置やアミノ酸残基の種類によっても異なるが、具体的には好ましくは1〜20個、より好ましくは1〜10個、さらに好ましくは1〜5個を意味する。保存的変異とは、置換部位が芳香族アミノ酸である場合には、Phe、Trp、Tyr間で、置換部位が疎水性アミノ酸である場合には、Leu、Ile、Val間で、極性アミノ酸である場合には、Gln、Asn間で、塩基性アミノ酸である場合には、Lys、Arg、His間で、酸性アミノ酸である場合には、Asp、Glu間で、ヒドロキシル基を持つアミノ酸である場合には、Ser、Thr間でお互いに置換する変異である。保存的変異の代表的なものは、保存的置換であり、保存的置換とみなされる置換としては、具体的には、AlaからSer又はThrへの置換、ArgからGln、His又はLysへの置換、AsnからGlu、Gln、Lys、His又はAspへの置換、AspからAsn、Glu又はGlnへの置換、CysからSer又はAlaへの置換、GlnからAsn、Glu、Lys、His、Asp又はArgへの置換、GluからGly、Asn、Gln、Lys又はAspへの置換、GlyからProへの置換、HisからAsn、Lys、Gln、Arg又はTyrへの置換、IleからLeu、Met、Val又はPheへの置換、LeuからIle、Met、Val又はPheへの置換、LysからAsn、Glu、Gln、His又はArgへの置換、MetからIle、Leu、Val又はPheへの置換、PheからTrp、Tyr、Met、Ile又はLeuへの置換、SerからThr又はAlaへの置換、ThrからSer又はAlaへの置換、TrpからPhe又はTyrへの置換、TyrからHis、Phe又はTrpへの置換、及び、ValからMet、Ile又はLeuへの置換が挙げられる。また、上記のようなアミノ酸の置換、欠失、挿入、付加、または逆位等には、遺伝子が由来する細菌の個体差、種の違いに基づく場合などの天然に生じる変異(mutant又はvariant)によって生じるものも含まれる。
【0054】
さらに、上記のような保存的変異を有する遺伝子は、コードされるアミノ酸配列全体に対して、80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは97%、特に好ましくは99%以上の相同性を有し、かつ、OASS活性を有するタンパク質であってもよい。このような公知の配列を有するOASSに対して相同性を有すタンパク質をコードする遺伝子は、上記のエシェリヒア・コリ株の野生型cysK又はcysM遺伝子を問い合わせ配列として用いたBLAST検索やFASTA検索によって公開データベースから容易に配列情報を取得することが出来、この公知の遺伝子配列に基づいて作製したオリゴヌクレオチドをプライマーとして、細菌のゲノムDNAを鋳型に用いてPCRにより取得することができる。尚、本明細書において、「相同性」(homology)」は、「同一性」(identity)を指すことがある。
【0055】
また、OASSをコードする遺伝子は、公知の遺伝子配列の相補配列又はその相補配列から調製され得るプローブとストリンジェントな条件でハイブリダイズし、公知の遺伝子産物と同等の機能を有するタンパク質をコードする遺伝子であってもよい。本発明において、ストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。一例を示せば、相同性が高いDNA同士、例えば80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは97%以上の相同性を有するDNA同士がハイブリダイズし、それより相同性が低いDNA同士がハイブリダイズしない条件、あるいは通常のサザンハイブリダイゼーションの洗いの条件である60℃、1XSSC、0.1% SDS、好ましくは0.1×SSC、0.1% SDS、に相当する塩濃度で、1回好ましくは2〜3回洗浄する条件を挙げることができる。
【0056】
上記ハイブリダイゼーションに用いるプローブは、遺伝子の相補配列の一部であってもよい。そのようなプローブは、公知の遺伝子配列に基づいて作製したオリゴヌクレオチドをプライマーとし、これらの塩基配列を含むDNA断片を鋳型とするPCRによって作製することができる。例えば、プローブとして、300 bp程度の長さのDNA断片を用いる場合には、ハイブリダイゼーションの洗いの条件は、50℃、2×SSC、0.1% SDSが挙げられる。
【0057】
<2>OASS提示菌
次に、OASS提示菌について説明する。OASS提示菌は、OASとチオ硫酸からS−スルホ−L−システインを生成する反応を触媒するOASSを細胞外膜に提示する細菌である。尚、OASSは、少なくともチオ硫酸とOASからS−スルホ−L−システインを生成し得る限り、チオ硫酸以外のイオウ源、例えば硫化物とOASからS−スルホ−L−システインを生成するものであってもよい。このようなOASSとしてはOASS-Bが、OASS-Bをコードする遺伝子としては前記のcysM遺伝子及びそのバリアントが挙げられる。
【0058】
OASS提示菌の構築に用いる細菌は、細胞外膜を有するグラム陰性菌であり、細胞外膜にタンパク質を提示することが可能な細菌であれば特に制限されない。具体的には、前述した腸内細菌科に属する細菌が挙げられる。中でも、エシェリヒア属性菌が好ましく、エシェリヒア・コリがより好ましい。
【0059】
「OASSを細胞外膜に提示する細菌」とは、OASSの少なくとも一部又は全部が細菌の細胞外に存在するようにして固定され、OAS及びチオ硫酸に反応液中で作用させたときに、OASからシステイン関連物質が生成することができる細菌である。
【0060】
以下、OASS-Bを細菌の細胞外膜へ提示させる方法について説明する。
OASS-Bを細胞外膜へ提示させるには、例えば、オートトランスポーターを利用する方法が挙げられる。具体的には、遺伝子組換え技術を利用して、OASS-Bをオートトランスポーターとの融合タンパク質として発現させる。オートトランスポーターによるペプチド提示システムでは、提示されるペプチドは、N末から分泌シグナル、パッセンジャー(提示されるペブチドに相当)、及びオートトランスポータードメインからなる前駆体タンパク質として発現し、該ペブチド(パッセンジャー)のみが細胞外膜の外に提示される。本発明において、「オートトランスポーター」とは前記オートトランスポータードメインをいう。パッセンジャー部分をOASSに置換した前駆体をコードする遺伝子を構築して、この遺伝子を細菌に導入することによって、OASSを細胞外膜に提示させることができる。本発明において、「O−アセチルセリンサルフヒドリラーゼ部分(OASS部分; OASS moiety)とは、上記のような前駆体タンパク質中にパッセンジャー部分として含まれるOASSをいう。尚、前駆体タンパク質のデザイン、具体的には前駆体タンパク質を構成するペプチド同士の連結部位のデザインによって、OASSのN末端及び/又はC末端の1又は複数のアミノ酸が変化することがある。そのような場合であっても、OASS部分はOASS活性を有する限り、野生型OASSとは異なる配列を含んでいてもよい。
【0061】
分泌シグナルは、シグナルペプチドに続く配列に内膜を通過させるためのものであり、パッセンジャーより下流の配列がペリブラズムに分泌され、分泌シグナルがシグナルペプチダーゼで切断されることが可能なものであれば特に制限されない。分泌シグナルとしては、例えば、Pseudomonas sp. strain LP7315のモノグリセリドリパーゼの分泌シグナルの他、PelBタンパク質、OmpTタンパク質、ompAタンパク質、β-ラクタマーゼ等の分泌シグナル等、E. coli等の細菌におけるタンパク質の分泌生産に用いられている分泌シグナル(Pines O. et al., Mol. Biotechnol. (1999) 12. 25-34)が挙げられる。
【0062】
オートトランスポーターとしては、Pseudomonas sp. strain LP7315のモノグリセリドリパーゼのオートトランスポータードメインが挙げられる。オートトランスポーターは、αヘリックス構造を有するリンカー部分とβバレル構造を有する部分(以下、「βバレル部分」ともいう。)を有する。Pseudomonas sp. LP7315株のモノグリセリドリパーゼをコードする遺伝子及び同遺伝子がコードするアミノ酸配列を、配列番号5及び6に示す。配列の相同性解析等から、配列番号6において、シグナルペプチドは1〜69位、リパーゼドメインは85〜885位、リンカー部分は937〜981位、βバレル部分は994〜1803位と推定される。リパーゼドメインとリンカー部分との間、リンカー部分とβバレル部分との間、及び、βバレル部分のC末端側には、上記の推定領域に属さない配列(以下、便宜上「スペーサー」という。)が存在するが、これらのスペーサーは、実際はリンカー及び/又はβバレル部分に含まれるか、又は、遷移的な構造を有する部分であると推定される。本発明において「オートトランスポーター」とは、上記の推定上のリンカー部分及びβバレル部分に加えて、スペーサーを含んでいてもよい。また、リパーゼドメインは、相同性解析からは85〜885位と推定されるが、実際のN末端はシグナルペプチドの直後である可能性がある。
【0063】
また、実施例に示すように、オートトランスポーターがリンカー部分と推定される領域を欠失していても、パッセンジャーとオートトランスポーターとの融合タンパク質のうちパッセンジャー部分が細胞外膜に提示されることがある。また、βバレル部分と推定される領域のN末端側を欠失していても、融合タンパク質のパッセンジャー部分が細胞外膜に提示されることがある。これは、パッセンジャー部分のN末端部分及び/又はβバレル部分(又はそのN末端部分を欠失した部分)のN末端部分が、リンカーとして機能することがあるものと推定される。このように、オートトランスポーターが本来のリンカー部分及び/又はβバレル部分のN末端部分を含んでいなくても、βバレル融合タンパク質のパッセンジャー部分のC末端及び/又はβバレル部分のC末端がリンカーとして機能し、パッセンジャー部分が細胞外膜に提示され得る限り、そのようなオートトランスポーターの改変体は「オートトランスポーター」に含まれる。
【0064】
リンカーは、野生型オートトランスポーターのリンカー部分の配列を有するものであってもよく、一部又は全体を任意の配列で置換えたものであってもよい。また、リンカーの長さは、野生型の配列を延長又は短縮したものであってもよい。さらには、前記のとおり、野生型オートトランスポーターのリンカー部分を含まずに、パッセンジャー及び/又はβバレル部分の一部がリンカーとして機能してもよい。また、リンカーの長さは、野生型のリンカー部分の配列と同じであってもよいが、それよりも長くてもよく、短くてもよい。「リンカーの長さ」は、本明細書においては、実施例に記載したpET-OASSA-ATプラスミドにコードされる融合タンパク質におけるOASS-Bとβバレル部分との間の長さを0とし、それに対する相対値で表す。尚、pET-OASSA-ATでは、同プラスミドの構築に用いたEcoRI認識配列によって、OASS-Bとβバレル部分の間に2アミノ酸残基挿入されている。また、βバレル部分は、前記モノグリセリドリパーゼの配列における994〜1803位に対して、N末端の34アミノ酸残基を欠失している。リンカーの長さ(pET-OASSA-ATに対する相対値)は、パッセンジャーの配列によっても異なるが、通常-50〜500アミノ酸、好ましくは20〜300アミノ酸、より好ましくは20〜70アミノ酸が好ましい。
【0065】
このモノグリセリドリパーゼの分泌シグナル及び/又はオートトランスポータードメインを利用する場合、これらをコードするDNAは、配列番号5の塩基配列に基づいて作製したプライマーを用いて、pET-OASSB(+79)-ATプラスミドDNA(参考配列)を鋳型とするPCR法によって取得することができる。他の細菌のオートトランスポータードメインをコードする遺伝子も、公知の配列情報に基づいて作製したプローブを用いたハイブリダイゼーション法によって、細菌の染色体DNA又は染色体DNAライブラリーから取得することができる。pET-OASSB(+79)-ATプラスミドで形質転換したエシェリシア・コリBL21株はAJ110734株と命名され、平成21年3月9日に、産業技術総合研究所特許生物寄託センター(住所 郵便番号305-8566 茨城県つくば市東1丁目1番地1中央第6)に寄託され、受託番号FERM P-21779が付与されている。
【0066】
エシェリヒア・コリのcysM遺伝子は、配列番号3の塩基配列に基づいて作製したプライマーを用いて、エシェリヒア・コリの染色体DNAを鋳型とするPCR法によってcysM遺伝子を取得することができる。他の細菌のcysM遺伝子も、前記配列情報に基づいて作製したプローブを用いたハイブリダイゼーション法によって、細菌の染色体DNA又は染色体DNAライブラリーから取得することができる。
【0067】
分泌シグナル、OASS、及びオートトランスポーターを含む(以下、「OASS-AT」と記載することがある)をコードするDNAは、これらを各々コードするDNA断片を連結することによって取得することができる。また、パッセンジャー及びオートトランスポーターを含む前駆体タンパク質、又はさらに分泌シグナルを含む前駆体タンパク質をコードするDNAの、パッセンジャーをコードする配列を、OASSをコードする配列で置換えることによっても、OASS-ATをコードするDNAを取得することができる。
【0068】
OASS-ATを発現させるためのプロモーターは、オートトランスポーターを含むタンパク質をコードする遺伝子固有のプロモーターであってもよく、他の遺伝子由来のプロモーター又は人工プロモーターであってもよい。プロモーターとしては、例えば、lacプロモーター、trpプロモーター、trcプロモーター、tacプロモーター、araBAプロモーター、ラムダファージのPRプロモーター、PLプロモーター、tetプロモーター、T7プロモーター、φ10プロモーター等が挙げられる。
【0069】
OASS-Bを細胞外膜に提示させるには、上記のようなOASS-ATをコードするDNAを宿主細胞で機能するベクター、好ましくはマルチコピー型ベクターと連結して組み換えDNAを作製し、これで細菌を形質転換すればよい。
【0070】
また、OASS-Bを細胞外膜へ提示させるには、OASS-ATをコードするDNAを細菌の染色体DNA上に多コピー存在させることによっても達成できる。細菌の染色体DNAに上記遺伝子を多コピー導入するには、染色体DNA上に多コピー存在する配列を標的に利用して相同組換えにより行う。染色体DNA上に多コピー存在する配列としては、レペティティブDNA、転移因子の端部に存在するインバーテッド・リピート等が利用できる。あるいは、特開平2-109985号公報に開示されているように、上記融合遺伝子をトランスポゾンに搭載してこれを転移させて染色体DNA上に多コピー導入することも可能である。
【0071】
本発明の第二の形態においては、OASS提示菌は、本発明の第一の形態に用いるOAS生産菌と同様に、細胞内のOASS活性が低下するように改変されていることが好ましい。OASS活性の低下は、OAS生産菌の育種と同様にして行うことができる。
【0072】
細菌の細胞外膜にOASS-Bを提示させるには、オートトランスポーターを用いる方法以外に、何らかのシグナル及びアンカーに連結して細胞表層に固定化する方法(特願2000−202442)を採用することができる。
【0073】
<システイン関連物質の製造法>
上記のようにして得られる細菌を、OAS及びチオ硫酸に、これらを含む反応液中で作用させることにより、S−スルホ−L−システインが生成する。
【0074】
S−スルホ−L−システインは、反応液中で、又は反応液から採取した後に還元することにより、L−システインに変換することができる。還元は、例えばジチオスレイトール(DTT)等の還元剤を用いることにより行うことができる。また、S−スルホ−L−システインは、培地中の成分によってL−システインに変換されることがある。さらに、L−システインは、培地中で、ジスルフィド結合が形成されて一部がL−シスチンに変換することがある。さらに、L−システインは、細胞中や培地中に存在するケトン又はアルデヒド、例えばピルビン酸と縮合し、ヘミチオケタールを中間体としてチアゾリジン誘導体が生成することがある(特許第2992010参照)。これらのチアゾリジン誘導体及びヘミチオケタールは、平衡混合物として存在することがある。したがって、OASから生成される化合物には、S−スルホ−L−システインのみに限られず、L−システイン、L−シスチン、もしくはそれらの誘導体、例えばチアゾリジン誘導体、もしくはヘミチオケタール、又はこれらの混合物も含まれる。
【0075】
本発明の第一の形態においては、OAS生産菌を炭素源を含む培地で培養し、同培地にOASを蓄積させる。得られたOASを含む反応液にOASS提示菌とイオウ源を加え、反応を起させることにより、システイン関連物質が生成する。第一の形態における反応液は、OASが蓄積した培地もしくはその上清、又はその希釈液もしくは濃縮液であってもよく、これらから採取したOASを含む培地又は緩衝液等であってもよい。尚、上記反応に培地を用いる場合、炭素源は不要であるが、含まれていてもよい。また、第一の形態においては、OASS提示菌は、菌体をアクリルアミド、カラギーナン等で固定化した固定化菌体であってもよい。さらに、反応はバッチ式でもカラム式でもよい。さらに、OAS生産菌とOASS提示菌を同時に培養してもよい。
本発明の第二の形態においては、OAS生産能を有するOASS提示菌を、炭素源を含む培地で培養することによって、培地中にシステイン関連物質が蓄積する。この形態においては、OASSの基質としてのイオウ源は、培養当初から培地に加えてもよく、培養途中に培地に加えてもよい。
【0076】
いずれの形態においても、イオウ源としてはチオ硫酸及び硫化物が挙げられるが、チオ硫酸が好ましい。反応液又は培地中のイオウ源の濃度は、OASSに対して通常0.1倍〜10倍、好ましくは0.2倍〜5倍、特に好ましくは0.2倍〜2倍である。また、反応液中のOASS提示菌の量は、適宜設定することができる。例えば、緩衝液又は生産培地にてOASS提示菌を懸濁し、OD660を適当な値、例えば2〜8になるよう調整することによって、菌濃度を設定することができる。
【0077】
培地としては、炭素源、窒素源、イオウ源、無機イオン及び必要に応じその他の有機成分を含有する通常の培地が挙げられる。
【0078】
炭素源としては、グルコース、フラクトース、シュクロース、糖蜜やでんぷんの加水分解物などの糖類、エタノール、グリセロール等のアルコール類、フマール酸、クエン酸、コハク酸等の有機酸類を用いることができる。
【0079】
窒素源としては、硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム等の無機アンモニウム塩、大豆加水分解物などの有機窒素、アンモニアガス、アンモニア水等を用いることができる。
【0080】
イオウ源としては、硫酸塩、亜硫酸塩、硫化物、次亜硫酸塩、チオ硫酸塩等の無機硫黄化合物が挙げられる、尚、OAS生産菌の培養に用いる培地におけるイオウ源は、生育に必要な量含まれていればよい。
【0081】
有機成分としては、ビタミンB1などの要求物質または酵母エキス等を適量含有させることが望ましい。これらの他に、必要に応じてリン酸カリウム、硫酸マグネシウム、鉄イオン、マンガンイオン等が少量添加される。
【0082】
OAS生産培養は、好気的条件下で30〜90時間実施することが好ましく、培養温度は25℃〜37℃に、培養中pHは5〜8に制御することが好ましい。尚、pH調整には無機あるいは有機の酸性あるいはアルカリ性物質、更にアンモニアガス等を使用することができる。OASS提示菌による反応は、好ましくは振盪しながら、30〜90時間実施することが好ましい。
培養物又は反応液からのシステイン関連物質の採取は、通常のイオン交換樹脂法、沈澱法その他の公知の方法を組み合わせることにより実施できる。本発明において採取されるシステイン関連物質は、システイン関連物質以外に微生物菌体、培地成分、水分、及び微生物の代謝副産物を含んでいてもよい。採取されたシステイン関連物質の純度は、50%以上、好ましくは85%以上、特に好ましくは95%以上である (US5,431,933, JP1214636B, US4,956,471, US4,777,051, US4946654, US5,840358, US6,238,714, US2005/0025878)。
【0083】
上記のようにして得られるS−スルホ−L−システインは、L−システインの製造に用いることができる。また、L-システインは、L-システイン誘導体の製造に用いることができる。L−システイン誘導体としては、メチルシステイン、エチルシステイン、カルボシステイン、スルホシステイン、アセチルシステイン等が含まれる。
【実施例】
【0084】
以下、本発明を実施例によりさらに具体的に説明する。
〔実施例1〕オートトランスポータードメインを含む発現ベクターの構築
Pseudomonas sp. strain LP7315のモノグリセリドリパーゼ遺伝子のオートトランスポータードメインをコードする領域をPCRにより増幅した。同株のゲノムDNAをテンプレートとして、5’端にNcoI-EcoRIサイトを付加したプライマー#1(5'-ccatggttgaattcaccctgggcgtgg-3'(配列番号7))と、5'端にHindIIIサイトを付加したプライマー#2(5'-tacaagcttgccggcggcaacaag-3'(配列番号8))を用いて、定法に従い、KOD plus DNA polymerase(TOYOBO)にてPCRを行った。PCR条件は、94℃で5分の後、94℃で15秒、48℃で10秒、68℃で1分、というサイクルを30回繰り返した。こうして増幅した断片をpUC118 HincII/BAP(タカラバイオ社製)と連結した。シークエンシングによりPCRエラーがないことを確認したのち、NcoIとHindIIIによる制限酵素処理を行い、同酵素で切断したpET22b(+)(Novagen社製)ベクターに連結した。こうしてPseudomonas sp. LP7315株由来モノグリセリドリパーゼ中のオートトランスポータードメインをコードするDNAがベクター上のT7プロモーター下流に連結された発現ベクター、pET-ATを作製した。
【0085】
尚、pET22b(+)のマルチクローニングサイト上流には、分泌シグナルPelBをコードする配列が存在し、pET-ATのPelBコード領域とATコード配列の間に、これらのフレームが一致するようにペプチドコード配列を挿入すると、PelBと、前記ペプチドと、オートトランスポータードメイン(AT)との融合タンパク質が発現する。また、前記ペプチドとATとの間の配列によってコードされる領域は、オートトランスポートターによる分泌システムにおけるリンカー配列に相当する。
【0086】
尚、上記のオートトランスポータードメインをコードする領域は、後述のプラスミドpET-OASSB(+79)-ATをテンプレートとし、プライマー#1及びプライマー#2を用いたPCRによっても、取得することができる。
【0087】
〔実施例2〕OASS発現ベクターの構築
(1)まず、OASS-A又はOASS-Bを、PelB及びオートトランスポータードメイン(AT)との融合タンパク質として発現するベクターを構築した。
【0088】
OASS-A遺伝子をクローニングするため、E. coli野生株(MG1655)ゲノムDNAをテンプレートとして、5'端にNcoIサイトを付加したプライマー#3(5'-ccatggaaaagatttttgaagataac-3'(配列番号9))と、5'端にEcoRIサイトを付加したプライマー#4(5'-ctggaattcaatccccgccccctggctaaaatgc-3'(配列番号10))を用いて、PyrobestR DNA polymeraseにてPCRを行った。PCR条件は、94℃で5分の後、94℃で30秒、55℃で30秒、72℃で1分、というサイクルを30回繰り返した。この増幅断片をNcoI、EcoRIで消化後、同酵素で切断した上記pET-ATベクターに挿入した。シークエンシングによりPCRエラーがないことを確認した。こして、OASS-Aをオートトランスポーターにて細胞外提示するベクター、pET-OASSA-ATを構築した。このプラスミドが持つ、PelB、OASS-A、及びATとの融合タンパク質(以下、「PelB-OASSA-AT」と記載することがある)をコードする遺伝子の塩基配列を配列番号11に示し、コードされるアミノ酸配列を配列番号12に示した。配列番号12において、1〜323位はOASS-A部分であり、326位〜570位はオートトランスポータードメインである。324位〜325位は、これらの連結に用いたEcoRI配列によって生じた部分である。
【0089】
同様に、OASS-B遺伝子をクローニングするため、E. coli野生株(MG1655)ゲノムDNAをテンプレートに、プライマー#5(5'-aactgcagccatggaaacattagaacaaacaataggc-3'(配列番号13))と、プライマー#6(5'-ctggaattcaatccccgccccctggctaaaatgc-3'(配列番号14))を用いて、OASS-Aと同じ条件にてDNA断片を増幅した。その断片をNcoI、EcoRIで消化後、同酵素で切断したpET-ATベクターに挿入した。その後、増幅した領域にPCRエラーがないことを確認した。こうして、OASS-Bをオートトランスポーターにて細胞外提示するベクター、pET-OASSB-ATを構築した。このプラスミドが持つ、PelB、OASS-B、及びATとの融合タンパク質(以下、「PelB-OASSB-AT」と記載することがある)をコードする遺伝子の塩基配列を配列番号15に示し、コードされるアミノ酸配列を配列番号16に示した。配列番号16において、1〜303位はOASS-B部分であり、306位〜550位はオートトランスポータードメインのうち、βバレル部分である。304位〜305位は、これらの連結に用いたEcoRI配列によって生じた部分である。尚、PelB-OASSB-ATは、野生型モノグリセリドリパーゼにおける本来のリンカー部分は含んでない。
【0090】
(2)次に、OASS-Bを単独で、又はPelBとの融合タンパク質として発現するベクターを構築した。
OASS-Bを、プライマー#7(5'-ggcatatggccatggaaacattagaacaaacaataggc-3'(配列番号17))と、プライマー#8(5'-ttctcgagaatccccgccccctggctaaaatgc-3'(配列番号18))を用いて上記と同じ条件にてPCR増幅した。プライマー#7にはNdeIとNcoIサイトが付加され、プライマー#8にはXbaIサイトが付加されている。NdeI-XbaIで消化したPCR断片を同酵素で処理したpET22b(+)に連結してpET-OASSB(PelB-)を構築した。また、NcoI-XbaIで消化したPCR断片を同酵素で処理したpET22b(+)に連結してpET-OASSB(PelB+)を構築した。これらの発現ベクターは、OASS-Bのみが単独で、又はPelBとの融合体として発現するようにデザインされ、発現産物はATとの融合体ではない。PelBは、pET22b(+)のマルチクローニングサイト上流に付加されている分泌シグナルであり、NdeI-XbaI部位で挿入したコンストラクトではPelB配列が除かれ、菌体内にてOASS-Bを過剰発現させることができる。一方、pET-OASSB(PelB+)はその分泌シグナルを保持している形であり、OASS-Bを発現し、菌体外へ分泌することができる。
【0091】
〔実施例3〕OASS発現株の培養
pET-AT、pET-OASSA-AT、pET-OASSB-AT、pET-OASSB(PelB-)、pET-OASSB(PelB+)の各ベクターでE. coli BL21株コンピテントセル(Novagen)を形質転換し、形質転換体をアンピシリン100 mg/Lを含むLB寒天培地上で選択した。得られた形質転換体を3 mLのLB培地(Bacto tryptone; 10 g/L、yeast extract; 5g/L、NaCl; 10 g/L、アンピシリン; 100 mg/L)で37℃、14時間、前培養を行ったのち、50 mLのLB培地に前培養液500μLを加え、37℃で本培養を行った。
【0092】
培養液のOD660をBACTOMONITOR BACT-550(JIKCO社製)にて測定し、OD660が0.5前後になった段階で終濃度1mMとなるようIPTGを添加し、さらに37℃で4時間培養した。IPTG添加によりpETベクター上のT7プロモーターが誘導され、その支配下にある遺伝子発現が誘導される。その後、4℃、7,800rpm、5min遠心分離をし、上清を捨て集菌した。その菌体をPBS(NaCl; 8 g/L、KCl; 0.2 g/L、Na2HPO4・12H2O; 2.9 g/L、KH2PO4; 0.2 g/L)に懸濁させ、洗浄を3回行い、以下の各実験に適した菌体濃度に調製した。
【0093】
〔実施例4〕OASSのIn vitro酵素活性測定とシステインの定量
以下の実施例で使用した、OASSのIn vitroでの酵素反応測定法を以下に示す。pH7.5に調整したBuffer A(50 mMリン酸カルシウム、1 mM EDTA、0.5 mM β-メルカプトエタノール)84μLにOD660に調製した菌体懸濁液22μLを添加して室温で2分間放置した。さらに200 mMのOAS溶液13μL(終濃度 20 mM)と、Buffer Aに溶解して調製した20 mMの硫化ナトリウム(NaOH溶液にてpH7.5に調製)もしくはチオ硫酸ナトリウムの溶液13μL(共に終濃度2 mM)を加え、室温にて5分間反応させた後、反応液に20%のトリクロロ酢酸(TCA)67μLを添加して反応を停止させた。15,000 rpm、5min遠心分離を行い、上清のみを回収した。
【0094】
L−システインは、Gaitonde法(Gaitonde M.K.,(1967) Biochem J 133:627-633)により定量した。Gaitonde法では、システインに加えて、L−シスチンやS-スルホシステインのようなシステイン関連物質も合わせて定量される。以下に具体的な方法を示す。培養液上清もしくは適当に希釈した培養液50μLに、等量のBuffer B(200 mM Tris-HCl(pH8.5)、10 mM DTT)を加えてサンプルを作製した。これらを室温で10分間放置した。ニンヒドリン溶液(酢酸:塩酸=3:2の溶液に2.5%のニンヒドリンを加えた溶液)50μLと、酢酸50μLを混合した溶液に、上記サンプル50μLを加え、100℃で10分間加熱した後、4℃に冷却した。この反応液40μLを95%エタノール200 μLと混合し、マイクロプレートリーダー(SPECTRAmax 340PC)で560nmの吸光度を測定した。同様に調製した5 mg/Lから60 mg/Lのシステイン標準液を指標に検量線を描き、サンプル中のシステイン関連物質を定量した。
【0095】
なお、以下の実施例では、OASS-Bとチオ硫酸ナトリウムの反応から生成するS-スルホシステインについても、比較しやすくするため、すべてのシステイン関連物質は同モルのシステインに換算して値を表示した。酵素活性は単位時間、単位容量、単位ODあたりのシステイン生産量とした。つまり1U = システイン生産量(μmol)/OD560/minで示した。以下、システイン関連物質の生成及び定量値に関し、システイン関連物質を単に「システイン」と記載することがある。
【0096】
〔実施例5〕OASS-B発現条件による酵素反応速度の検討
pETベクターを用いた酵素発現系のうち、どのシステムが最も効率よくシステイン関連物質を生産できるかを検討した。pET-OASSB(PelB-)、pET-OASSB(PelB+)、pET-OASSB-AT、pET-ATの各ベクターを導入したE. coli BL21株株を、実施例3に記載したようにして培養した。
【0097】
それぞれの形質転換体でOASS-Bが発現していることをSDS-PAGEにて確認した後、培養液を遠心分離し、培養上清を回収した。さらに、残った菌体をPBSにて2回洗浄後、PBSでOD600=2になるように懸濁した。そこに、PBSに溶解した20mM OASを、菌体懸濁液もしくは培養上清と等量加え(終濃度10mM)、さらに、1Mチオ硫酸ナトリウムを1/50量加えて(終濃度20mM)、37℃で2時間、振とうしながら反応を行った。その後、反応液中のシステイン濃度を実施例4に記載の方法にて測定した。
【0098】
結果を表1に示した。分泌シグナルを付加したOASS-Bを発現するpET-OASSB(PelB-)プラスミドでは、培養上清中にシステイン生成が見出されたが、分泌シグナルを付加しないOASS-Bを発現するpET-OASSB(PelB-)プラスミドとの差は少なかった。一方、菌体懸濁液では、、オートトランスポーターを用いてOASS-Bを細胞外提示した発現系では、効果的、かつ菌体特異的にシステイン関連物質への変換反応が行われることが明らかとなった。
【0099】
【表1】
【0100】
〔実施例6〕オートトランスポーターを用いたOASS細胞表層提示とその活性測定
オートトランスポーターを用いた発現系にて、OASS-Bが細胞外に提示されているかどうか、またその提示された酵素が活性を保持しているかを確認した。実施例5に記載された方法でPelB-OASSB-ATの発現菌を得たのち、in vitro酵素活性法にてOASS-Bの活性を測定した。また、この菌体を用いたシステインへの変換反応が生産培地中にて使用できるかを検討するため、bufferAの代わりに生産培地((NH4)2SO4 15 g/L、KH2PO4 1.5 g/L、MgSO4-7H2O 1 g/L、thiamine HCl 0.1 mg/L、FeSO4・7H2O 1.7 mg/L、Na2MoO4・2H2O 0.15 mg/L、CoCl2・6H2O 0.7 mg/L、MnCl2・4H2O 1.6 mg/L、ZnSO4・7H2O 0.3 mg/L、CuSO4・5H2O 0.25 mg/L、tryptone 0.6 g/L、yeast extract 0.3 g/L、NaCl 0.6 g/L、CaCO3 20 g/L、L-Histidine HCl・H2O 135mg/L、pyridoxine 2 mg/L、glucose 20 g/L)を用いた活性測定も同時に行った。
培地を用いた場合、buffer A中に比べれば活性は落ちるものの、活性を保持していることが明らかとなった(表2)。
【0101】
【表2】
【0102】
オートトランスポーターを用いた発現系にてOASS-Bが活性をすことは確認できたが、さらに、実際にOASS-Bが細胞外膜に提示されているかを確認した。そこで、PelB-OASSB-AT発現菌の菌体をPBSにてOD660=2に調整したのち、4℃で10分間超音波処理を行い、細胞を破砕し、4℃、50,000 rpm、40min超遠心分離を行った。この操作の結果、上清部分を可溶性画分とし、沈殿画分を膜画分に分離することができる。各画分における酵素活性を確認するため、上記in vitroの測定系にてそれぞれの酵素活性を測定したところ、大部分の活性が膜画分に存在していた(表3)。OASS-BとATとの融合タンパク質(OASSB-AT)の細胞質へのミスローカライズは認められず、このATシステムがもくろみ通り働き、OASSを外膜に提示していることが示唆された。
【0103】
【表3】
【0104】
〔実施例7〕OAS生産菌とOASS提示菌を用いたシステイン関連化合物の生産培養
OAS生産菌が生産するOASを、OASS-B提示菌によりL-システイン関連物質に変換するというコンセプトの成立を確認するために、以下の実験を行った。
まず、OAS生産菌として、MG1655ΔcysKΔcysM/pACYC-DESを用いた。
pACYC-DESは、炭素源から効率よくOASを作るために、OASを細胞外に排出する活性を有する排出因子であるYdeDをコードするydeD(Tobias Dassler.,et al,(2000)Molecular Microbiology.,36(5))、L-システイン生合成における鍵酵素の一つであるホスホグリセレートデヒドロゲナーゼのフィードバック阻害解除型変異体をコードするserA*(EP620853B)、および同じくL-システイン生合成における鍵酵素の一つであるセリンアセチルトランスフェラーゼのフィードバック阻害解除型変異体をコードするcysE*(US patent 2005/0124049 A1)の各遺伝子を搭載した多コピープラスミドである同プラスミドの構築は、EP1528108に詳述されている。
【0105】
MG1655ΔcysKΔcysM/pACYC-DESは、上記プラスミドpACYC-DESで、MG1655ΔcysKΔcysM株を形質転換することにより得た。このMG1655ΔcysKΔcysM株は、さらにOASを蓄積させるため、OASからシステイン等への反応を触媒する酵素であるOASSをコードする両遺伝子cysK、cysMを欠損してある。MG1655ΔcysKΔcysMこの菌の作製方法を以下に示す。
【0106】
まずpMIV-5JS(特開2008-99668、EP1942183)を鋳型に、プライマーP1(5'-agtaagatttttgaagataactcgctgactatcggtcgctcaagttagtataaaaaagctgaac-3'(配列番号19))とプライマーP2(5'-ggtaaagctttcatcttcttgtagtttcaacgccgctgaagcctgcttttttatactaagttgg-3'(配列番号20))を用いてクロラムフェニコール耐性遺伝子を含むフラグメントを増幅した。pMIV-5JSは、Muファージ由来のインテグレーションカセットを持つプラスミドpM12、thrLターミネーターフラグメント、pACYC184由来のクロラムフェニコール耐性遺伝子、及びλattL及びλattRフラグメントから構成されるプラスミドである。
【0107】
PCRは、94℃で5分の後、98℃を5秒、55℃にて10秒、72℃にて3分、のプログラムを30サイクル行った。増幅したフラグメント上にはλ依存インテグレーション(「Red-driven integration」と呼ばれる方法(Proc. Natl. Acad. Sci. USA, 2000, vol. 97, No. 12, p6640-6645)によって目的の断片を挿入するための染色体上のターゲットとなる配列(ここではcysKのC末端、N末端の配列)がデザインされている。そのため、取得されたDNA断片を、目的の菌株にこのλ依存インテグレーションによって挿入した場合には、染色体上のcysK遺伝子の中にクロラムフェニコール遺伝子が挿入された構造となる。
【0108】
定法に従い、エレクトロポレーション法にて上記増幅断片をMG1655/pKD46に導入した。プラスミドpKD46は、目的のDNA断片をMG1655株の染色体に組み込むためのヘルパープラスミドである(Datsenko, K.A., Wanner, B.L., Proc.Nat1.Acad.Sci.USA, 97, 6640-6645, (2000))。こうして得られたMG1655ΔcysK株に導入されたクロラムフェニコール耐性マーカーを、ラムダファージ由来の切り出しシステムによって除去した。具体的には、ラムダファージのInt-Xis遺伝子を搭載したpMT-Int-Xis2(WO2005/010175)でMG1655ΔcysK株を形質転換し、得られた形質転換株からクロラムフェニコール感受性を示すMG1655ΔcysK(s)株を取得した。
【0109】
さらにcysM遺伝子を欠損させるために、まずpUC18にcysMをクローニングした。MG16655のゲノムを鋳型として、P3(5'-acgggatccttgcgtttgtcatcagtctcc-3'(配列番号36))とP4(5'-acgaagcttgtcgcatcaggcaacaccacg-3'(配列番号37))をプライマーとして用いたPCRにより、cysM遺伝子を増幅した。PCRは、94℃で5分の後、98℃を5秒、55℃にて10秒、72℃にて1分、のプログラムを30サイクル行った。この増幅したフラグメントをBamHIとHindIIIで切断し、同酵素で消化したpUC18ベクターに挿入し、pUC-cysMを作製した。pUC-cysMのうち、cysM中に唯一存在するEco91I切断配列を利用し、Eco91IにてpUC-CysMを消化した後、pACYC184をEco47IIIを用いて切り出したクロラムフェニコール耐性遺伝子を含む配列を挿入し、pUC-cysM::CmRを作製した。
【0110】
pUC-cysM::CmRでE. coli JC7623(標識遺伝子;thr-1 leu-6 thi-1 lacY1 galK2 ara-14 xyl-5 mtl-1 proA2 his-4 argE3 str-31 tsx-33 supE344 recB21 recC22 sbcB15)(国立遺伝研究所より分与可能)を形質転換した。クロラムフェニコール耐性かつアンピシリン感受性となるプラスミドレスの株、すなわち染色体上のcysM中にクロラムフェニコール耐性遺伝子が挿入された株を得た。その株からP1トランスダクション法にてMG1655ΔcysK(s)株に形質導入し、得られた形質導入株からクロラムフェニコール耐性を示すMG1655ΔcysKΔcysM株を取得した。この株にpACYC-DESをエレクトロポレーションにて導入し、テトラサイクリン耐性株を選択、MG1655ΔcysKΔcysM/pACYC-DES株を取得した。
【0111】
上記MG1655ΔcysKΔcysM/pACYC-DES株を、試験管(内径22mm)に張り込んだ2mLの生産培地中にて、32℃で約19時間振盪培養したのち、4℃、7,800rpm、5min遠心分離を行い、OASを含んだ上清を得た。上記培地は、グルコースが40 g/Lであり、テトラサイクリン 12.5 mg/Lを加えた以外は、実施例6に記載した生産培地と同じ組成である。
【0112】
またOASS提示菌として、BL21/pET-OASSA-AT及びpET-OASSB-ATで形質転換したE. coli BL21を用い、これらの菌を実施例3に記載の方法にて培養した。得られたOASS提示菌を、上記のOASを含有する上清を用いて希釈してOD660=2にそろえた。希釈液2 mLを大試験管に張り込み、そこに1Mチオ硫酸ナトリウムを1/50量加えて(終濃度20mM)、32℃で振盪保温しながら0、30、60、120、180分後にサンプリングを行い、L−システイン関連物質生成の経時変化を検証した。S源にチオ硫酸ナトリウムを用いたが、チオ硫酸ナトリウムは硫化ナトリウムに比べ酸性側でも安定に存在することが知られており、培地中に残存するチオ硫酸ナトリウム量の経時変化を見ることができる。
【0113】
その結果、ネガティブコントロールのATのみ発現した菌(E. coli BL21/pET-AT)、及びチオ硫酸ナトリウムを基質としないOASS-Aを発現した菌では、システイン関連物質の生成はほとんど見られないのに対し、OASS-Bを発現した菌では反応時間に比例してシステイン関連物質の生産量が上がり、最終的に約160 mg/Lのシステイン関連物質を産生した(図1)。尚、ここで生成したシステイン関連物質は概ねS-スルホシステインである。なお、S-スルホシステインはDTT等の還元剤存在下で、容易にシステインに変換することができる。
【0114】
〔実施例8〕リンカーの最適化によるOASS-Bの活性向上
オートトランスポーターを利用したタンパク質発現において、オートトランスポーターと目的タンパク質を繋ぐリンカー部分の長さが活性に大きく影響を与える例が知られている(Jochen Maurer.,et al(1999)Journal Of Bacteriology.,181(22),7014-7020)。そこで、OASS-Bのリンカー最適化を行った。
【0115】
pET-OASSB-ATのリンカー長(モノグリセリドリパーゼにおける本来のリンカーは含まず、βバレル部分のN末端の34アミノ酸を欠失している)を±0とし、それよりも79アミノ酸長い+79型から36アミノ酸短い-36型(βバレル部分をさらに36アミノ酸欠失)の間でいくつかのバリアントを作製してそれぞれの活性を測定することで、リンカー最適化を行った。まず、79アミノ酸長いリンカー型のプラスミドを構築した。プライマー#1の代わりにプライマー#9(5’-ccatggttgaattcaccctgggcgtgg-3’(配列番号21))を用いた以外はpET-ATを作製するのと同様の方法にてpET-(+79)-ATを構築し、これにOASS-Bコード配列を導入してpET-OASSB(+79)-ATを作製した。±0型よりも短いバリアントは、βバレル部分のN末端部分が±0型よりもさらに欠失している。例えば、-9型では、野生型モノグリセリドリパーゼのβバレル部分と推定される領域よりも43(34+9)アミノ酸短い。
【0116】
pET-OASSB(+79)-ATが持つ、PelB、OASS-A、及びATとの融合タンパク質(「PelB-OASSB(+79)-AT」)をコードする遺伝子の塩基配列を配列番号34に示し、コードされるアミノ酸配列を配列番号35に示した。配列番号35において、1〜303位はOASS-B部分であり、306位〜629位はオートトランスポータードメインである。304位〜305位は、これらの連結に用いたEcoRI配列によって生じた部分である。PelB-OASSB(+79)-ATのオートトランスポータードメインは、PelB-OASSB-ATに比べて、モノグリセリドリパーゼ前駆体タンパク質のリパーゼドメインに由来する9アミノ酸、及び、各ドメイン間のスペーサーに由来する21アミノ酸、リンカー部分の15アミノ酸、及びβバレル部分のN末端34アミノ酸の合計79アミノ酸長い。pET-OASSB(+79)-ATから、任意の長さのリンカーを持つ融合タンパク質をコードするプラスミドを構築することができる。尚、配列番号6においては、PelB-OASSB-ATのトランスポータードメインは366〜610位に相当し、PelB-OASSB(+79)-ATのオートトランスポータードメインは287〜610位に相当する。
【0117】
pET-OASSB(+79)-ATを鋳型にLinker(X)-primerとRv-Primerにて、94℃で5分の後、98℃を5秒、55℃にて10秒、72℃にて1分、のプログラムを40サイクルというPCR反応を行い、目的の長さのリンカーフラグメントを得た。Linker(X)-primerにおける「X」は、リンカー長を示す。リンカー長は、pET-OASSB-ATを±0とし、長くしたものを(+)で表した。また、pET-OASSB-ATから、さらにβバレル部分の長さを短くしたものを(-)で表した。作製したリンカーの長さとその作製に用いたプライマーは以下の通りである。
【0118】
【表4】
【0119】
各リンカー側プライマー(上記Linker(X)-primer)にはEcoRIサイトを、オートトランスポーター側プライマー(上記Rv-Primer)にはStuIサイトを付加しており、PCRで増幅後、これらの制限酵素にて処理を行い、同じ酵素で消化してリンカー部をとり除いたpET-OASSB-ATベクターに挿入した。こうして構築したpET-OASSB(X)-ATをE. coli BL21株にエレクトロポレーション法にて導入し、実施例4に記載のin vitro活性測定法にて、それぞれの酵素活性を測定した。
【0120】
まず始めに、リンカーを短くするタイプを中心に作製し、活性を測定した。するとpET-OASSB(+40)-ATが最も活性が高く、pET-OASSB(±0)-ATの約4.7倍であった(表5)。また各コンストラクトにおけるOASS発現をSDS-PAGEにて確認するため、それぞれのコンストラクトの発現菌体をPBSにてOD660=10に揃え、それらを等量のサンプルバッファー(BIO-RAD社製 Laemmli Sample Buffer)に懸濁した。すべてのサンプルを100℃で3分間加熱し、氷上で急冷してからSDS-PAGE(第一化学薬品PAGミニ「第一」4-20%)解析を行った。SDS-PAGEの各レーンには、上記サンプルをそれぞれ10μLをロードし、泳動条件は30 mAで約90minとした。泳動後のゲルをCBB染色をして各タンパク質の発現量を確認したところ、発現されたタンパク質の量はほぼ同等であり(図2)、活性強度は発現量のみに依存するものではないと考えられた。
【0121】
続いて+40型を中心としたリンカータイプを作製し、同様に活性評価を行った。その結果pET-OASSB(+49)-ATが最も活性が高く、pET-OASSB(±0)-ATの約5.9倍もの活性を有していることが判明した(表5)。表5中、リンカー長はpET-OASSB(±0)-ATをゼロとしたときのプラスマイナスで表した。また、OASS-B活性はpET-OASSB(±0)-AT(リンカー長±0)の活性を1.00としたときの相対活性で示した。
【0122】
pET-OASSB(+40)-ATとpET-OASSB(±0)-ATを用いて、実施例5と同様にOAS生産菌との混合によるシステインの生産培養を行ったところ、180分間の反応でpET-OASSB(±0)-ATは163 mg/Lのシステイン関連物質を生産するのに対し、pET-OASSB(+40)-ATは328 mg/Lと約2倍の生産量を得ることに成功した(図3)。このように、リンカー長を変えることで、in vitro酵素活性だけでなく、システイン生産量も向上した。
【0123】
【表5】
【0124】
〔実施例9〕OASS提示菌を利用したシステイン生産菌による副生OASのシステイン関連物質への変換
糖からのシステイン発酵生産においては、システイン排出タンパク質として知られるYdeDを用いることが有効であると考えられているが、同排出因子はシステインだけではなくOASも細胞外へ排出するためOASを著量副生することが知られている(Tobias Dassler.,et al,(2000)Molecular Microbiology.,36(5),1101-1112)。そこでOAS生産菌とOASS提示菌を用い、糖からのシステイン生産の検討を行った。
【0125】
OAS生産菌としてE. coli-MG1655/pACYC-DESを、グルコース濃度を40g/Lとした以外は実施例6に記載の生産培地と同じ組成の培地2mLを入れた大試験管に植菌し、32℃で約20時間振盪培養をした。培養後、グルコースアナライザー(サクラ精機社製 バイオテックアナライザー AS210)を用いて培地中の残糖を測定し、消費糖量を計算したところ21.3 g/Lの糖を消費していた。産生したシステイン関連物質(OASは含まない)はGaitonde法にて定量を行ったところ、3.9mMであった。また、培地中に蓄積したOASの濃度をHPLCにて定量したところ、18.7 mMであった。ただし、サンプルを200 mMのTris-HCl(pH9.0)で希釈することでOASをより安定なNASに変換して検出を行った。OASはアルカリ性条件では用意にNASに変換されることが知られている。HPLCの条件は以下の通りである。
【0126】
カラム;Inertsil ODS-3(疎水性カラム/GLサイエンス社製)
バッファー流速;1.0mL/min
カラム温度;40℃
検出器;UV210nm
サンプルアプライ量;10μL
バッファー;0.1M KH2PO4・H3PO4(pH2.2)、5mM 1-オクタンスルホン酸Na
【0127】
上記培養液を遠心分離し、上清画分のみを回収したのち、この培養上清を用いてOASS提示菌(pET-OASSB(+49)-AT)の菌体量をO.D.660 =8にそろえ、実施例7と同様の方法にてシステイン関連物質生産を行った。OASS-B提示菌では、180分間反応させたところ、反応前のシステイン関連物質量に比べ約70 mg/L生産量が増加した(図4の棒グラフ)。またOASの濃度をHPLCにて確認したところ、システイン関連物質の増加に伴ってOAS濃度が減少していることが分かった(図4の折れ線グラフ)。
以上の結果より、細胞の外膜に提示したOASS-Bを用いて培地中のOASがシステイン関連物質に変換されたことが示された。
【0128】
〔配列表の説明〕
配列番号1:E. coli cysK遺伝子の塩基配列
配列番号2:E. coli cysK遺伝子によりコードされるOASS-Aのアミノ酸配列
配列番号3:E. coli cysM遺伝子の塩基配列
配列番号4:E. coli cysM遺伝子によりコードされるOASS-Bのアミノ酸配列
配列番号5:Pseudomonas sp. LP7315株のモノグリセリドリパーゼ遺伝子の塩基配列
配列番号6:Pseudomonas sp. LP7315株のモノグリセリドリパーゼ遺伝子がコードするアミノ酸配列
配列番号7:プライマー#1
配列番号8:プライマー#2
配列番号9:プライマー#3
配列番号10:プライマー#4
配列番号11:PelB-OASSA-ATをコードする遺伝子の塩基配列
配列番号12:PelB-OASSA-ATのアミノ酸配列
配列番号13:プライマー#5
配列番号14:プライマー#6
配列番号15:PelB-OASSB-ATをコードする塩基配列
配列番号16:PelB-OASSB-ATのアミノ酸配列
配列番号17:プライマー#7
配列番号18:プライマー#8
配列番号19:プライマーP1
配列番号20:プライマーP2
配列番号21:プライマー#9
配列番号22:Linker(+67)プライマー
配列番号23:Linker(+58)プライマー
配列番号24:Linker(+49)プライマー
配列番号25:Linker(+40)プライマー
配列番号26:Linker(+31)プライマー
配列番号27:Linker(+22)プライマー
配列番号28:Linker(+9) プライマー
配列番号29:Linker(-9) プライマー
配列番号30:Linker(-18)プライマー
配列番号31:Linker(-27)プライマー
配列番号32:Linker(-36)プライマー
配列番号33:Rv-primer
配列番号34:PelB-OASSB(+79)-ATをコードする塩基配列
配列番号35:PelB-OASSB(+79)-ATのアミノ酸配列
配列番号36:プライマーP3
配列番号37:プライマーP4
【特許請求の範囲】
【請求項1】
チオ硫酸とO−アセチル−L−セリンからS−スルホ−L−システインを生成する反応を触媒するO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌を、O−アセチル−L−セリン及びチオ硫酸に、これらを含む反応液中で作用させ、該反応液からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を採取することを特徴とする、S−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物の製造法。
【請求項2】
前記反応液中のS−スルホ−L−システイン、又は前記反応液から採取したS−スルホ−L−システインを還元する工程をさらに含む、請求項1に記載の方法。
【請求項3】
前記細菌において、前記O−アセチルセリンサルフヒドリラーゼはオートトランスポーターとの融合タンパク質として発現される、請求項1又は2に記載の方法。
【請求項4】
前記融合タンパク質におけるO−アセチルセリンサルフヒドリラーゼ部分が、cysM遺伝子によりコードされる、請求項3に記載の方法。
【請求項5】
前記O−アセチルセリンサルフヒドリラーゼ部分が、下記のいずれかのアミノ酸配列を有する、請求項4に記載の方法。
(A)配列番号2のアミノ酸配列。
(B)配列番号2のアミノ酸配列において、1〜10個のアミノ酸の置換、欠失、挿入、または付加を含むアミノ酸配列。
【請求項6】
前記オートトランスポーターが、シュードモナス属細菌の耐熱性モノグリセリドリパーゼのオートトランスポーターである、請求項3〜5のいずれか一項に記載の方法。
【請求項7】
前記反応液に含まれるO−アセチル−L−セリンが、O−アセチル−L−セリン生産能を有する他の細菌により生産されたものである、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
前記O−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌はO−アセチル−L−セリン生産能を有し、前記反応液に含まれるO−アセチル−L−セリンは該細菌がが分泌したものであって、前記反応液は培地である、請求項1〜7のいずれか一項に記載に記載の方法。
【請求項9】
前記他の細菌又はO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌は、下記の性質の少なくともいずれかを有する、請求項7又は8に記載の方法。
i)L−システイン生合成系酵素活性が増強されるように改変されている。
ii)細胞内のO−アセチルセリン分解酵活性が低下するように改変されている。
iii)O−アセチルセリンを排出する因子の活性が増強されるように改変されている。
【請求項10】
前記O−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌がエシェリヒア属細菌である、請求項1〜8のいずれか一項に記載の方法。
【請求項11】
前記他の細菌がエシェリヒア属細菌である、請求項7又は9に記載の方法。
【請求項12】
エシェリヒア属細菌がエシェリヒア・コリである、請求項10又は11に記載の方法。
【請求項13】
cysM遺伝子によりコードされるO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌であって、前記O−アセチルセリンサルフヒドリラーゼはオートトランスポーターとの融合タンパク質として発現される、細菌。
【請求項14】
前記融合タンパク質におけるO−アセチルセリンサルフヒドリラーゼ部分が、下記のいずれかのアミノ酸配列を有する、請求項13に記載の細菌。
(A)配列番号2のアミノ酸配列。
(B)配列番号2のアミノ酸配列において、1〜10個のアミノ酸の置換、欠失、挿入、または付加を含むアミノ酸配列。
【請求項15】
前記オートトランスポーターが、シュードモナス属細菌の耐熱性モノグリセリドリパーゼのオートトランスポーターである、請求項13又は14に記載の細菌。
【請求項16】
細胞内のO−アセチルセリンサルフヒドリラーゼ活性が低下するように改変された、請求項13〜15のいずれか一項に記載の細菌。
【請求項1】
チオ硫酸とO−アセチル−L−セリンからS−スルホ−L−システインを生成する反応を触媒するO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌を、O−アセチル−L−セリン及びチオ硫酸に、これらを含む反応液中で作用させ、該反応液からS−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物を採取することを特徴とする、S−スルホ−L−システイン、L−システイン、L−シスチン、もしくはそれらの誘導体、又はこれらの混合物の製造法。
【請求項2】
前記反応液中のS−スルホ−L−システイン、又は前記反応液から採取したS−スルホ−L−システインを還元する工程をさらに含む、請求項1に記載の方法。
【請求項3】
前記細菌において、前記O−アセチルセリンサルフヒドリラーゼはオートトランスポーターとの融合タンパク質として発現される、請求項1又は2に記載の方法。
【請求項4】
前記融合タンパク質におけるO−アセチルセリンサルフヒドリラーゼ部分が、cysM遺伝子によりコードされる、請求項3に記載の方法。
【請求項5】
前記O−アセチルセリンサルフヒドリラーゼ部分が、下記のいずれかのアミノ酸配列を有する、請求項4に記載の方法。
(A)配列番号2のアミノ酸配列。
(B)配列番号2のアミノ酸配列において、1〜10個のアミノ酸の置換、欠失、挿入、または付加を含むアミノ酸配列。
【請求項6】
前記オートトランスポーターが、シュードモナス属細菌の耐熱性モノグリセリドリパーゼのオートトランスポーターである、請求項3〜5のいずれか一項に記載の方法。
【請求項7】
前記反応液に含まれるO−アセチル−L−セリンが、O−アセチル−L−セリン生産能を有する他の細菌により生産されたものである、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
前記O−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌はO−アセチル−L−セリン生産能を有し、前記反応液に含まれるO−アセチル−L−セリンは該細菌がが分泌したものであって、前記反応液は培地である、請求項1〜7のいずれか一項に記載に記載の方法。
【請求項9】
前記他の細菌又はO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌は、下記の性質の少なくともいずれかを有する、請求項7又は8に記載の方法。
i)L−システイン生合成系酵素活性が増強されるように改変されている。
ii)細胞内のO−アセチルセリン分解酵活性が低下するように改変されている。
iii)O−アセチルセリンを排出する因子の活性が増強されるように改変されている。
【請求項10】
前記O−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌がエシェリヒア属細菌である、請求項1〜8のいずれか一項に記載の方法。
【請求項11】
前記他の細菌がエシェリヒア属細菌である、請求項7又は9に記載の方法。
【請求項12】
エシェリヒア属細菌がエシェリヒア・コリである、請求項10又は11に記載の方法。
【請求項13】
cysM遺伝子によりコードされるO−アセチルセリンサルフヒドリラーゼを細胞外膜に提示する細菌であって、前記O−アセチルセリンサルフヒドリラーゼはオートトランスポーターとの融合タンパク質として発現される、細菌。
【請求項14】
前記融合タンパク質におけるO−アセチルセリンサルフヒドリラーゼ部分が、下記のいずれかのアミノ酸配列を有する、請求項13に記載の細菌。
(A)配列番号2のアミノ酸配列。
(B)配列番号2のアミノ酸配列において、1〜10個のアミノ酸の置換、欠失、挿入、または付加を含むアミノ酸配列。
【請求項15】
前記オートトランスポーターが、シュードモナス属細菌の耐熱性モノグリセリドリパーゼのオートトランスポーターである、請求項13又は14に記載の細菌。
【請求項16】
細胞内のO−アセチルセリンサルフヒドリラーゼ活性が低下するように改変された、請求項13〜15のいずれか一項に記載の細菌。
【図1】

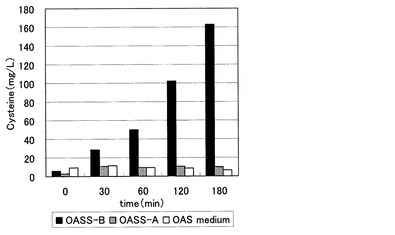
【図2】
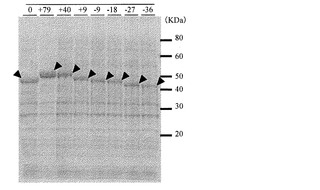
【図3】
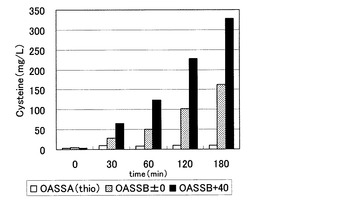
【図4】
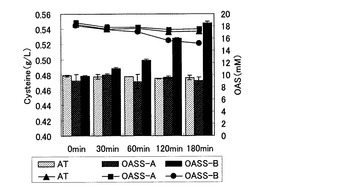
【図2】
【図3】
【図4】
【公開番号】特開2010−252650(P2010−252650A)
【公開日】平成22年11月11日(2010.11.11)
【国際特許分類】
【出願番号】特願2009−104176(P2009−104176)
【出願日】平成21年4月22日(2009.4.22)
【出願人】(000000066)味の素株式会社 (887)
【Fターム(参考)】
【公開日】平成22年11月11日(2010.11.11)
【国際特許分類】
【出願日】平成21年4月22日(2009.4.22)
【出願人】(000000066)味の素株式会社 (887)
【Fターム(参考)】
[ Back to top ]