
L−メチオニン前駆体産生菌株
【課題】L−メチオニン前駆体産生菌株を使用して作製されたL−メチオニン前駆体から酵素転換反応を用いてL−メチオニンおよび有機酸を高収率で産生する方法の提供。
【解決手段】L−メチオニン前駆体を産生することができる菌株を作製し、この菌株を培養して発酵によりL−メチオニン前駆体を産生する工程および前記L−メチオニン前駆体を基質とし転換酵素によって酵素反応を起すことによりL−メチオニンおよび有機酸を産生する工程とを含むL−メチオニンおよび有機酸を産生する方法。
【解決手段】L−メチオニン前駆体を産生することができる菌株を作製し、この菌株を培養して発酵によりL−メチオニン前駆体を産生する工程および前記L−メチオニン前駆体を基質とし転換酵素によって酵素反応を起すことによりL−メチオニンおよび有機酸を産生する工程とを含むL−メチオニンおよび有機酸を産生する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はL−メチオニン前駆体産生菌株に関するものであって、より詳しくは本発明によって作製されたL−メチオニン前駆体産生菌株を使用して作製されたL−メチオニン前駆体から酵素転換反応を用いてL−メチオニンおよび有機酸を高収率で産生することによって、既存の方法に比べて環境にやさしい方法でL−メチオニンを産生し、L−メチオニンを選択的に産生することによって飼料および食品添加剤、医薬用および医薬品の原料などの多様な分野に広く使用するためのものである。
【背景技術】
【0002】
メチオニンは生体内の必須アミノ酸の一種類で飼料および食品添加剤として広く使用され、医薬用として輸液剤、医薬品の合成原料としても使用される。メチオニンはコリン(レシチン)とクレアチンのような化合物の前駆体で作用し、システインとタウリンの合成原料でも使われる。また、硫黄を提供する役割をする。S−アデノシル−メチオニンはL−メチオニンから由来して生体内でメチル基を提供する役割をして、脳の多様な神経伝達物質(neurotransmitter)合成に関連されている。メチオニンおよび/またはS−アデノシル−L−メチオニン(SAM)は生体内で肝と動脈において脂肪蓄積を抑制し、鬱、炎症、肝疾患、筋肉痛を緩和するなどの多様な役割をする。
【0003】
今まで知られたメチオニンおよび/またはS−アデノシル−L−メチオニンの生体内の役割は次の通りである。
【0004】
1)脂肪代謝を促進する肝と動脈において脂肪沈着を抑制し、脳、心臓、腎臓の血流を増進させる役割をする(JHepatol. Jeon BR et al., 2001 Mar; 34(3):395-401(非特許文献1))。
【0005】
2)消化を促進し、毒性物質の解毒と排泄を促進、鉛のような重金属の排泄を促進する。
【0006】
3)メチオニンは毎日1日に800〜1600mgの投与で優れた抗鬱剤になると知られている(Am J Clin Nutr. Mischoulon D. et al., 2002 Nov; 76(5): 1158S-61S(非特許文献2))。
【0007】
4)肝疾患において肝機能の向上作用をする(FASEB J Mato JM., 2002 Jan;16(1): 15-26)。特にアルコールによる肝疾患に効果的である(Cochrane DatabaseSyst Rev., Rambaldi A., 2001; (4): CD002235(非特許文献3))。
【0008】
5)骨関節疾患に抗炎症効果が良く、関節回復を促進する(ACP J Club. Sander O., 2003Jan-Feb; 138(1): 21、J Fam Pract., Soeken KL et al.,2002 May; 51(5): 425-30(非特許文献4))。
【0009】
6)メチオニンは毛髪の必須栄養素である。毛髪に栄養を供給し脱毛を防止する(AudiolNeurootol., Lockwood DS et al., 2000 Sep-Oct; 5(5): 263-266(非特許文献5))。
【0010】
メチオニンは前記のように動物飼料を含めた食品と医薬品で用いるために化学合成と生物学的合成を通じて産生することができる。
【0011】
化学合成は主に5−(β−メチルメルカプトエチル)ヒダントインを加水分解させる反応を通じてL−メチオニンを産生する。しかし、このような化学合成を通じて産生されたメチオニンはL−型とD−型が混合された形態でのみ産生されるという短所がある。
【0012】
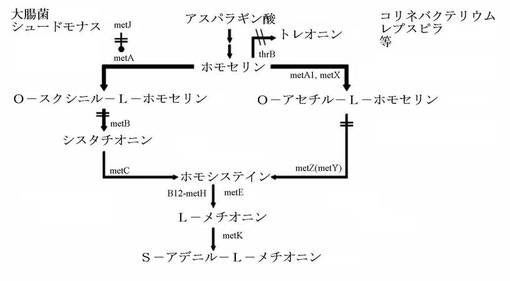
生物学的合成はメチオニン生成に関与する蛋白質を用いる方法である。L−メチオニンはmetA、metB、metC、metE、metH遺伝子から発現される酵素蛋白質の作用によってホモセリンからメチオニンに生合成される。具体的に、metAはメチオニン生合成の一番目の酵素であるホモセリンO−スクシニルトランスフェラーゼ(homoserineO-succinyltransterase)をコードする遺伝子でホモセリンをO−スクシニル−L−ホモセリンに転換する機能をする。metBがコードするO−スクシニルホモセリンリアーゼ(O-succinylhomoserinelyase)、またはシスタチオニンγ合成酵素酵素はO−スクシニル−L−ホモセリンをシスタチオニン(cystathionine)に転換する機能をする。metCがコードするシスタチオニンベタリアーゼ(cystathionine beta lyase)酵素はシスタチオニンをL−ホモシステインに転換する機能をする。metEがコードするコバラミン−非依存的メチオニンシンターゼとmetHがコードするコバラミン−依存的メチオニンシンターゼがL−ホモシステインをL−メチオニンに転換させる。この際、metFがコードする5,10−メチレンテトラヒドロフォレートリダクターゼ(5,10-methylenetetrahydrofolatereductase)とglyAがコードするセリンヒドロキシメチルトラスフェラーゼ(serin hydroxymethytransferase)は共に作用してL−メチオニン合成に必要なメチル基を提供するN(5)−メチルテトラヒドロフォレート(N(5)-methyltetrahydrofolate)を合成する機能をする。
【0013】
L−メチオニンは前記酵素による一連の有機的な反応を通じて合成される。前記蛋白質または前記蛋白質に影響を与えるその他蛋白質の遺伝的な操作は、L−メチオニン合成の調節を生じさせる可能性がある。例えば、特開2000−139471号(特許文献1)にはエシェリキア属に属するゲノム遺伝子上のthrBC、metJを除去し、metBLを過剰発現させ、metKを漏出変異体として作製し、L−メチオニンを産生する方法が記述されている。また、米国特許出願公開第2003/0092026号(特許文献2)にはコリネバクテリウム属に属し、L−メチオニン合成の阻害因子であるmetD遺伝子を除去した微生物を用いる方法が記述されている。米国特許出願公開第2002/0049305号(特許文献3)には5,10−メチレンテトラヒドロフォレートリダクターゼ(metF)の発現を増加させてL−メチオニンの産生を増加させる方法が記載されている。
【0014】
米国特許出願公開第2005/0054060号(特許文献4)では、L−メチオニンを産生する微生物を作製するためにシスタチオニンシンターゼに変異を加えてシステインを利用せず、直接H2SまたはCH3SHを用いてホモシステインまたはメチオニンを合成することが記載されている。この方法では変異されたシスタチオニンシンターゼを直接細胞に導入して細胞内のメチオニン合成過程に沿ってメチオニンを合成した。しかし、この方法では細胞内のメチオニン代謝経路を用いるから酵素の反応過程が非効率的である。また、H2SまたはCH3SHが細胞に毒性を有するという点から見て実効性に劣る。さらに、メチルメルカプタン(CH3SH)に対する基質特異性がシュードモナスまたはクロモバクテリウム属由来のスクシニルホモセリンリアーゼに比べて非常に低いことが我々の実験において確認された。
【0015】
シスタチオニンシンターゼの場合、文献によると、互いに異なる基質と反応して互いに異なる物質を産生する傾向が大きい。シスタチオニンシンターゼはホモシステインとO−スクシニルホモセリンを結合して非常に高い効率でホモランチオニンという物質を産生するという報告がある(J.Bacteriol (2006) vol 188:p609-618(非特許文献6))。前記のようにシスタチオニンシンターゼが細胞内に存在する場合、互いに異なるメチオニン前駆体と結合することができ、この結合によって副産物を作る効率性も非常に高い。特に、シスタチオニンシンターゼを過剰発現する場合はより多くの副産物を生成する可能性があり、細胞内の反応の効率は非常に劣る。
【0016】
なお、既存の生物学的合成で産生されるメチオニンはL型という長所があるが、その量が非常に微量である。これはメチオニンの合成経路がとてもよく調節されるフィードバック調節システムを有しているからである。メチオニンが一定水準以上に合成されると、最終産物であるメチオニンがメチオニン合成を開始する初期蛋白質metAの遺伝子転写をフィードバックで抑制する。metA遺伝子は転写段階においてメチオニンによって阻害を受け、翻訳段階において細胞内の蛋白質分解酵素によって分解されるメカニズムにより、メチオニンの量が調節されるため、metA遺伝子の高発現だけではメチオニンの量を一定水準以上に増加させることができない(DvoraBiran, Eyal Gur, Leora Gollan and Eliora Z. Ron: Control of methioninebiosynthesis in Escherichia coli by proteolysis: Molecular Microbiology (2000)37(6), 1436-1443(非特許文献7))。従って、既存の多数の特許はmetA遺伝子のフィードバックを解除する研究に重点をおいていた(国際公開公報第2005/108561号(特許文献5)、国際公開公報第1403813号(特許文献6))。
【0017】
また、生物学的システムでメチオニンを産生する場合、メチオニンの分解代謝の経路であるS−アデノシルメチオニンシンターゼによって生成されたメチオニンがS−アデノシルメチオニンに変換される。S−アデノシルメチオニンは細胞に必須的な物質であってS−アデノシルメチオニンシンターゼを欠損させることができないため、既存にはこの遺伝子を弱化させる方向でメチオニンの産生を増加しようとする特許が出願されたことがある(国際公開公報第2005/108561号(特許文献7))。
【先行技術文献】
【特許文献】
【0018】
【特許文献1】特開2000−139471号公報
【特許文献2】米国特許出願公開第2003/0092026号明細書
【特許文献3】米国特許出願公開第2002/0049305号明細書
【特許文献4】米国特許出願公開第2005/0054060号明細書
【特許文献5】国際公開公報第2005/108561号パンフレット
【特許文献6】国際公開公報第1403813号パンフレット
【特許文献7】国際公開公報第2005/108561号パンフレット
【非特許文献】
【0019】
【非特許文献1】J Hepatol. Jeon BR et al., 2001 Mar;34(3):395-401
【非特許文献2】Am J Clin Nutr. Mischoulon D. et al., 2002Nov; 76(5): 1158S-61S
【非特許文献3】Cochrane Database Syst Rev., Rambaldi A.,2001; (4): CD002235
【非特許文献4】ACP J Club. Sander O., 2003 Jan-Feb; 138(1):21、J Fam Pract., Soeken KL et al., 2002 May; 51(5):425-30
【非特許文献5】Audiol Neurootol., Lockwood DS et al., 2000Sep-Oct; 5(5): 263-266
【非特許文献6】J.Bacteriol (2006) vol 188:p609-618
【非特許文献7】Dvora Biran, Eyal Gur, Leora Gollan andEliora Z. Ron: Control of methionine biosynthesis in Escherichia coli byproteolysis: Molecular Microbiology (2000) 37(6), 1436-1443
【発明の概要】
【発明が解決しようとする課題】
【0020】
前記の従来技術は細胞内のメチオニン代謝経路の中、シスタチオニンシンターゼ代謝経路を用いることから酵素の反応過程が非効率的であり、硫化物特有の基質毒性問題および副産物の生成の問題があり、またメチオニン合成経路のフィードバック調節によって十分な量のメチオニンを合成することができないという問題がある。
【0021】
従って、本発明は既存の生物学的メチオニンの産生工程を代替して発酵によるL−メチオニン前駆体の産生および前記L−メチオニン前駆体の酵素転換工程の2つの工程を使用することによって、従来の前記問題を克服したL−メチオニンの産生方法を提供することを目的とする。
【0022】
また、本発明はL−メチオニンを選択的に産生する方法を提供することを目的とする。
【0023】
また、本発明は別途の産生工程無しで副産物として有機酸を同時に産生する方法を提供することを目的とする。
【課題を解決するための手段】
【0024】
本発明について以下に詳細に説明する。
本発明は1)L−メチオニン前駆体を産生することができる菌株を作製し、この菌株を培養して発酵によりL−メチオニン前駆体を産生する工程、2)産生された前記L−メチオニン前駆体との酵素反応によりL−メチオニンおよび有機酸を産生する工程で構成されるL−メチオニンおよび有機酸を産生する方法を提供する。
【0025】
より詳しくは、前記第1工程はL−メチオニン前駆体産生菌株を作製し、この菌株を用いた発酵法でL−メチオニン前駆体を培養液に蓄積する工程である。この際、L−メチオニン前駆体産生菌株は本発明者の意図した方法によって作製される。従って、本発明はこのような菌株および菌株の作製方法も含まれる。
【0026】
前記L−メチオニン前駆体は下記の化学式から構成されたO−アシルホモセリン(O-acylhomoserine)基のうちの一つで表され;
【化1】

ここでRは、C、H、O、Nおよびその他化合物を含む最大15個の炭素分子である物質である。例えば、前記O−アシルホモセリン基はO−アセチルホモセリン(O-acetyl homoserine)、O−スクシニルホモセリン(O-succinylhomoserine)、プロピオニルホモセリン(propionyl homoserine)、アセトアセチルホモセリン(acetoacetyl homoserine)、クマロイルホモセリン(coumaroylhomoserine)、マロニルホモセリン(malonyl homoserine)、ヒドロキシメチルグルタリルホモセリン(hydroxymethylglutaryl homoserine)およびピメリルホモセリン(pimelyl homoserine)を含むが、これに限られるものではない。
【0027】
本発明において、前記L−メチオニン前駆体は好ましくはO−アセチルホモセリンまたはO−スクシニルホモセリンである。
【0028】
本発明において、「L−メチオニン前駆体産生菌株」とはL−メチオニン前駆体を生物体内で産生することができる原核および真核の微生物菌株であって、本発明による操作によってL−メチオニン前駆体を蓄積することができる微生物菌株をいう。例えば、L−メチオニンを産生するエシェリキア属、エルウィニア属、セラチア属、プロビデンシア属、コリネバクテリウム属、シュードモナス属、レプトスピラ属、サルモネラ属、ブレビバクテリア属、ヒポモナス属、クロモバクテリウム属およびノルカジア属、または菌類または酵母類に属する微生物菌株が含まれる。好ましくは、O−スクシニルホモセリンを産生するためにシュードモナス属、ノルカジア属、エシェリキア属の微生物が使用され、O−アセチルホモセリンを産生するためにエシェリキア属、コリネバクテリウム属、レプトスピラ属、酵母類などが使用される。より好ましくは、エシェリキア属に属する微生物菌株、より好ましくは大腸菌(Escherichia coli; 以下E.coliと称する)が使用される。また、エシェリキア属菌株に外来の遺伝子を導入することによってO−スクシニルホモセリンおよびO−アセチルホモセリンを選択的に産生することができる。
【0029】
本発明は前記の菌株からO−スクシニルホモセリンまたはO−アセチルホモセリンの分解に関与する遺伝子を欠損または弱化させ、O−スクシニルホモセリンまたはO−アセチルホモセリンの合成に関与する遺伝子を導入または強化させる菌株をL−メチオニン前駆体産生菌株で提供する。なお、選択的にはO−スクシニルホモセリンまたはO−アセチルホモセリンの産生強化のためにトレオニン生合成の経路を遮断または弱化した菌株を提供する。また、選択的にはO−スクシニルホモセリンまたはO−アセチルホモセリンの合成に関与する蛋白質のフィードバック調節が解除された遺伝子を導入し、過剰発現または活性を強化させた菌株を提供する。
【0030】
より詳しくは、本発明はL−メチオニン前駆体の分解に関与するmetB遺伝子、トレオニン生合成の経路に関与するthrB遺伝子およびL−メチオニン前駆体の産生遺伝子の転写を抑制するmetJ遺伝子を欠損させた菌株ならびにL−メチオニン前駆体の生合成に関与するmetAもしくはmetX遺伝子の発現を強化させ、またはフィードバック制御が解除されたmetAもしくはmetX遺伝子を導入し、またはmetA遺伝子を欠損させる代わりにmetX遺伝子を導入し、またはmetX遺伝子を欠損させる代わりにmetA遺伝子を導入したL−メチオニン前駆体産生菌株を提供する。
【0031】
本発明において、前記遺伝子の欠損は遺伝体上で前記遺伝子の部位を切除したり特定遺伝子の配列を挿入して蛋白質の配列を変形させることによって行われ、遺伝子の弱化は前記遺伝子のプロモータ部位または5’−UTR部位の塩基配列を変化させることによって蛋白質の発現を弱化させたりまたは当該遺伝子のORF部位に変異を導入することによって蛋白質の活性を弱化させることができる。
【0032】
本発明において、前記遺伝子の発現強化は前記遺伝子のプロモータ部位または5’−UTR部位の塩基配列を変化させることによって蛋白質の発現を強化させることができ、当該遺伝子のORF部位に変異を導入することによって蛋白質の活性を強化させることができ、当該遺伝子を染色体上に追加導入することによって発現を強化させることができ、当該遺伝子をベクター上に自己プロモータまたは強化された別個のプロモータと共に導入して菌株に形質転換することによって蛋白質の発現量を強化させることができる。
【0033】
本発明の具体的実施様態において、前記L−メチオニン前駆体産生菌株を作製する方法は次のとおりである。
【0034】
第1工程として、L−メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンを蓄積できるようにするため、菌株からシスタチオニンγ合成酵素またはO−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼなどの蛋白質をコードする遺伝子を欠損または弱化させる。
【0035】
シスタチオニンγ合成酵素をコードする遺伝子をmetB、O−スクシニルホモセリンサルフヒドリラーゼをコードする遺伝子をmetZ、O−アセチルホモセリンサルフヒドリラーゼをコードする遺伝子をmetYと表記する。上述のような活性を有する蛋白質をコードする遺伝子は例えば、大腸菌(E.coli)から公知のmetBがある。これは文献(Blattner et. Al., Science 277: 1453-1462 (1997))によって既に報告されている大腸菌のゲノム配列から得ることができる(Accession no. AAC75876)。また、前記遺伝子配列はNCBI(米国国立バイオテクノロジー情報センター)およびDDBJ(日本DNAデータバンク)のようなデータベースからも得られる。この他にも同じ活性を有する遺伝子としてコリネバクテリウムのmetBおよびmetY、シュードモナス由来のmetZなどが報告されている。
【0036】
シスタチオニンγ合成酵素、O−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼは下記の反応式に示されるようにO−スクシニルホモセリンまたはO−アセチルホモセリンをシスタチオニンまたはホモシステインに合成する活性を有している。従って、このような活性を有する遺伝子を欠損または弱化させる場合、O−スクシニルホモセリンまたはO−アセチルホモセリンが培養液中に過剰に蓄積される。
【0037】
L−システイン+O−スクシニル−L−ホモセリン <=> コハク酸+シスタチオニン
L−システイン+O−アセチル−L−ホモセリン <=> 酢酸+シスタチオニン
HS−+O−スクシニル−L−ホモセリン <=> コハク酸+ホモシステイン
HS−+O−アセチル−L−ホモセリン <=> 酢酸+ホモシステイン
【0038】
第2工程として、第1工程から作製された菌株でホモセリンキナーゼをコードするthrB遺伝子を弱化または欠損させる。thrB遺伝子はホモセリンからO−ホスホホモセリン(O-phosphohomoserine)を合成した後、thrC遺伝子によってトレオニンに転換される。合成されたホモセリンが全量メチオニンの前駆体合成に用いられるようにthrB遺伝子を欠損または弱化させる。
【0039】
第3工程として、metA遺伝子の転写を調節する転写調節因子であるmetJの遺伝子を欠損または弱化させる。メチオニン前駆体の合成に関与するmetA遺伝子はメチオニンによってフィードバック調節を受けるものであり、metJ遺伝子はmetA遺伝子の転写に関与するリプレッサーである。恒常的にmetA遺伝子を過剰発現することおよびメチオニン前駆体の合成を活性化するためには、metA遺伝子の転写リプレッサーの除去が有益である。従って、E.coliからmetJ遺伝子が除去されると、metA遺伝子の発現は増加し、L−メチオニン前駆体を大量合成することができる。
【0040】
上記第2、第3工程は前駆体産生菌株によって改変可能であり、前駆体産生菌株を作製するために必須ではない。より好ましくは、エシェリキア属の微生物で前駆体の産生経路を強化するために実施することができる。
【0041】
第4工程として、メチオニン前駆体の合成を増加させるために、メチオニン生合成の経路の初期段階を媒介する酵素のホモセリンO−スクシニルトランスフェラーゼまたはホモセリンO−アセチルトランスフェラーゼをコードする遺伝子であるmetA遺伝子またはmetX遺伝子の発現を強化させる。metA遺伝子はホモセリンO−スクシニルトランスフェラーゼをコードする遺伝子の通称であり、metX遺伝子はホモセリンO−アセチルトランスフェラーゼをコードする遺伝子の通称である。前記metAまたはmetX遺伝子の発現を強化するために遺伝子の追加導入または5’−UTRまたはプロモータを変化させたり各遺伝子ORFに変異を導入する。この遺伝子の発現を強化すると、L−メチオニン前駆体の合成を大きく増加することができる。
【0042】
菌株の生育にメチオニンが必要な場合、フィードバック制御が解除されたmetAまたはmetX遺伝子を導入することができる。この場合、培地中に添加されたメチオニンの濃度に関わらずL−メチオニン前駆体を合成することができ、培地中にメチオニンを添加することによってL−メチオニン前駆体を合成するでき、細胞の成長を促す。
【0043】
また、O−スクシニルホモセリンを産生する菌株からO−アセチルホモセリンの産生を増加させるために、染色体内に存在するホモセリンO−スクシニルトランスフェラーゼをコードする遺伝子のmetA遺伝子を欠損させることができる。metA遺伝子を欠損させてO−スクシニルホモセリンの産生を止め、さらにmetX遺伝子を導入してO−アセチルホモセリンを産生する場合、metA遺伝子が存在時にmetX遺伝子を導入する場合に比べて高い収率のO−アセチルホモセリンを産生することができる。
【0044】
なお、O−アセチルホモセリンを産生する菌株からO−スクシニルホモセリンの産生を増加させるために、染色体内に存在するホモセリンO−アセチルトランスフェラーゼをコードする遺伝子のmetX遺伝子を欠損させることができる。metX遺伝子を欠損させてO−アセチルホモセリンの産生を止め、さらにmetA遺伝子を導入してO−スクシニルホモセリンを産生する場合、最も高い収率のO−スクシニルホモセリンを産生することができる。
【0045】
上記1〜4工程のうち、一部または全体を導入した菌株を作製してL−メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンを蓄積することができる。
【0046】
L−メチオニン前駆体産生菌株はL−リシン、L−トレオニンまたはL−イソロイシン産生菌株を用いて作製することもできる。好ましくは、L−トレオニン産生菌株を用いて作製することができる。この場合には既にホモセリンまでの合成が円滑になされる菌株であって、最も多い量のメチオニン前駆体を合成することができる。従って、L−トレオニン産生菌株を用いてトレオニンの合成経路に関与する遺伝子を欠損または弱化させ、metBまたはmetY、またはmetZ遺伝子を欠損または弱化させてメチオニン前駆体を蓄積することができる。より好ましくは、thrB遺伝子を欠損または弱化させた後、metBまたはmetYまたはmetZ遺伝子を欠損または弱化させてメチオニン前駆体を合成することができる。また、metAまたはmetX遺伝子の発現を強化して最も多量のメチオニン前駆体を合成することができる。
【0047】
本発明において、「L−トレオニン産生菌株」とはL−トレオニンを生体内で産生することができる原核または真核の微生物菌株をいう。例えば、L−トレオニンを産生するエシェリキア属、エルウィニア属、セラチア属、プロビデンシア属、コリネバクテリウム属およびブレビバクテリウム属に属する微生物菌株が含まれる。好ましくは、エシェリキア属に属する微生物菌株、より好ましくは大腸菌が使用される。
【0048】
また、L−トレオニン産生菌株は天然微生物菌株のみならず、変異微生物も含む。このような変異微生物は例えば、イソロイシン漏出形要求性、L−リシン類似体に対する耐性およびα−アミノ酪酸の耐性を有する微生物;内在的なホスホエノールピルベートカルボキシラーゼ(ppc)遺伝子がゲノム含有された遺伝子の他に追加で1コピー以上の染色体DNA中に挿入された変異微生物;L−メチオニン生合成の中間体であるオキサロ酢酸(oxaloacetate, OAA)をホスホエノールピルビン酸(PEP)に転換するに関与するpckA遺伝子が不活性化された微生物;L−メチオニンの生合成に関与するtyrB遺伝子の発現を抑制するtyrR遺伝子が不活性化された微生物;および糖伝達に関連されたgalP遺伝子の発現を抑制するgalR遺伝子が不活性化された微生物が含まれる。前記L−リシン類似体はS−(2−アミノエチル)−L−システインおよびδ−メチル−L−リシンからなる群から選択される一つ以上の化合物であってよい。
【0049】
本発明の好ましい実施態様ではL−トレオニンを産生するE.coli変異菌株TF4076(KFCC10718、韓国特許公告第92-8365号)を変異させたL−トレオニン産生およびL−メチオニン−非要求性CJM002を使用した。TF4076はメチオニン要求性、メチオニン類似体(例、α-アミノ-β-ヒドロキシ吉草酸、AHV)に対する耐性、リシン類似体(例、S-(2-アミノエチル)-L-システイン、AEC)に対する耐性、イソロイシン類似体(例、α-アミノ酪酸)に対する耐性、メチオニンの類似体(例、エチオニン)に対する耐性などの特性を有している。上記韓国特許の全体内容は請求項によって本明細書に含まれる。このようなTF4076はメチオニン要求性菌株でインビボにおいてメチオニンを合成できない。本発明者らはこのようなメチオニン要求性を解除してメチオニン産生菌株として用いるためにNTGを用いた人工的な突然変異法を使用してメチオニン要求性が解除されたトレオニン産生菌株のE.coli CJM002を作製した。E.coli CJM002は大腸菌 MF001としてKCCM(Korean Culture Center of Microorganism、大韓民国ソウル西大門区弘済1洞ユリムビル、361-221)に2004年4月9日付で寄託した(寄託番号:KCCM−10568)。また、前記の方法を通じて作製したO−スクシニルホモセリン産生菌株の大腸菌CJM−BTJ(pMetA-CL)菌株は2006年7月21日に寄託し(寄託番号:KCCM−10767)、大腸菌CJM−BTJ(pCJ-MetA-CL)菌株は2007年7月5日に寄託した(寄託番号:KCCM−10872)。また、上記方法によって作製したO−アセチルホモセリン産生菌株の大腸菌CJM−BTJA(pCJ-MetX-CL)菌株は2007年7月5日に寄託した(寄託番号:KCCM−10873)。
【0050】
上述のとおり作製されたL−メチオニン前駆体産生菌株の培養過程は当業者に知られた適当な培地と培養条件に応じてなされる。このような培養過程は当業者であれば選ばれる菌株によって容易に調整して使用することができる。前記培養方法の例にはバッチ式、連続式および流加式培養が含まれるが、これに限定されるものではない。このような多様な培養方法は、例えば文献("Biochemical Engineering" by James M. Lee, Prentice-HallInternational Editions, pp138-176)に開示されている。
【0051】
培養に使用される培地は特別な菌株の要求条件を適切に満たさなければならない。多様な微生物の培地は例えば文献("Manualof Methods for General Bacteriology" by the American Society forBacteriology, Washington D.C., USA, 1981)に開示されている。前記培地は多様な炭素源、窒素源および微量元素成分を含む。使用できる炭素源の例にはブドウ糖、ショ糖、乳糖、果糖(fructose)、マルトース、でんぶん、セルロースのような炭水化物、大豆油、ヒマワリ油、ヒマシ油、ココナッツ油のような脂肪、パルミチン酸、ステアリン酸、リノール酸のような脂肪酸、グリセロールおよびエタノールのようなアルコール、アセト酸のような有機酸が含まれる。これらの化合物のうちの一つまたはその混合物は、炭素源として使用され得る。使用できる窒素源の例には、ペプトン、酵母抽出物、肉汁、麦芽抽出物、トウモロコシ浸漬液(CSL)および大豆粉のような有機窒素源および尿素、硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム、炭酸アンモニウムおよび硝酸アンモニウムのような有機窒素源が含まれる。これらの化合物のうちの一つまたはその混合物は、窒素源として使用され得る。ここで、培地は付加的にリン源としてリン酸二水素カリウム、リン酸水素二カリウムおよび対応するナトリウム−含有塩を含むことができる。また、硫酸マグネシウムや硫酸鉄のような金属塩も含むことができる。さらに、アミノ酸、ビタミンおよび適切な前駆体なども加えられ得る。これらの培地または前駆体はバッチ式または連続式で培養物に添加され得る。
【0052】
また、培養中に水酸化アンモニウム、水酸化カリウム、アンモニア、リン酸および硫酸のような化合物を培養物に適切な方式で添加して培養物のpHを調整することができる。また、培養中に脂肪酸ポリグリコールエステルのような消泡剤を使用して気泡の生成を抑制することができる。また、培養物の好気状態を保持するために、培養物内に酸素または酸素含有気体(例、空気)を注入する。培養物の温度は通常20℃〜45℃であり、好ましくは25℃〜40℃である。培養期間は望むL−メチオニン前駆体の生成量が得られるまで続けられ、好ましくは10〜160時間である。
【0053】
第2工程は、前記L−メチオニン前駆体産生菌株によって産生されたO−スクシニルホモセリンまたはO−アセチルホモセリンとメチルメルカプタンに基質を用いてシスタチオニンシンターゼ、O−スクシニルホモセリンサルフヒドリラーゼもしくはO−アセチルホモセリンサルフヒドリラーゼ活性を有する酵素または前記酵素を含む菌株を用いた酵素反応を通じてL−メチオニンおよび有機酸を産生する工程を含む。
【0054】
より詳細には、本発明は上記方法で蓄積されたホモセリン、O−ホスホホモセリン、O−スクシニルホモセリン、またはO−アセチルホモセリンを基質として用いてシスタチオニンγ合成酵素、O−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼなどの酵素反応をによりL−メチオニンを産生する方法を提供する。好ましくは、本発明はO−スクシニルホモセリンまたはO−アセチルホモセリンを基質で用いる。
【0055】
本発明において、前記シスタチオニンγ合成酵素またはO−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼはエシェリキア属(Escherichia sp.)、シュードモナス属(Pseudomonas sp.)、レプトスピラ属(Leptospira sp.)、コリネバクテリウム属(Corynebacteriumsp.)、サッカロミセス属(Saccharomyces sp.)、クロモバクテリウム属(Chromobacterium sp.)、ノルカジア属(Norcardia sp.)、ブラジリゾビウム属(Bradyrhizobium sp.)、ヒホモナス属、メチロコックス属(Methylococcussp.)、メチロバシルス属(Methylobacillus sp.)、ニトロソモナス属(Nitrosomonas sp.)、クレシエラ属(Klesiella sp.)、バシルス属(Bacillus sp.)、シゲラ属(Shigella sp.)、コルウェリア属(Colwellia sp.)、サルモネラ属(Salmonella sp.)、酵母(yeast)、菌類(fungi)に属する微生物菌株に由来するものであり得る。
【0056】
第2工程において、O−スクシニルホモセリンをL−メチオニン前駆体で使用する場合、好ましくはシュードモナス属、ノルカジア属、クロモバクテリウム属に属する微生物菌株で、より好ましくは、シュードモナスアウロゲノサ(Pseudomonas aurogenosa)、ノルカジアファルシニカ(NorcardiaFarcinica)、シュードモナスプチダ(Pseudomonas putida)、クロモバクテリウムビオラシウム(Chromobacterium Violaceum)に属する微生物菌株に由来するシスタチオニンγ合成酵素またはO−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼが使用される。
【0057】
第2工程において、O−アセチルホモセリンをL−メチオニン前駆体として使用する場合、好ましくはレプトスピラ属、クロモバクテリウム属、ヒポモナス属に属する微生物菌株であって、より好ましくは、レプトスピラアウロゲノサ(Leptospira aurogenosa)、ヒポモナスネプチュニウム(HyphomonasNeptunium)、クロモバクテリウムビオラシウム(Chromobacterium Violaceum)に属する微生物菌株に由来するシスタチオニンγ合成酵素またはO−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼが使用される。
【0058】
前記の反応は下記示すとおりであり、その構造式は図2に示される。
CH3SH+O−スクシニル−L−ホモセリン <=> コハク酸+メチオニン
CH3SH+O−アセチル−L−ホモセリン <=> 酢酸+メチオニン
【0059】
上記反応では図2に表記された化学式のようにメチルメルカプタンのCH3S-残基がO−スクシニルホモセリンまたはO−アセチルホモセリンのコハク酸または酢酸の残基と置換されてメチオニンを生成するようになる。反応時にメチルメルカプタン(CH3SH)は様々な形態で添加が可能である。
【0060】
前記活性を有する酵素をコードする遺伝子の配列はNCBI(米国国立バイオテクノロジー情報センター)および日本のDNAデータバンク(KEGG)のようなデータベースからも得られる。
【0061】
生物学的転換反応のために得られた遺伝子配列から遺伝子をクローニングして発現ベクターに挿入した後、組換え菌株由来の酵素を活性型で発現させる。酵素発現菌株および発現された酵素の両方を前記反応に直接用いることができる。
【0062】
上記遺伝子を発現させた酵素および酵素発現菌株はL−メチオニン前駆体を蓄積した発酵上澄液または発酵液と直接混合または部分的な混合反応を用いて反応を導く。本発明の好ましい実施態様において、発酵液に蓄積されたO−スクシニルホモセリンまたはO−アセチルホモセリンはシュードモナス属、クロモバクテリウム属、レプトスピラ属、ヒポモナス属由来のシスタチオニンγ合成酵素またはO−アセチルホモセリンサルフヒドリラーゼまたはO−スクシニルホモセリンサルフヒドリラーゼによってメチオニンに転換することができる。
【0063】
より好ましくは、発酵液に蓄積されたO−スクシニルホモセリンはシュードモナスアウロゲノサまたはシュードモナスプチダまたはクロモバクテリウムビオラシウム由来のシスタチオニンγ合成酵素およびO−アセチルホモセリンサルフヒドリラーゼおよびO−スクシニルホモセリンサルフヒドリラーゼによってメチオニンに転換することができる。
【0064】
または発酵液に蓄積されたO−アセチルホモセリンはレプトスピラメイエリ(Leptospira meyeri)、ヒポモナスネプチュニウム(Hyphomonas Neptunium)、クロモバクテリウムビオラシウム(ChromobacteriumViolaceum)由来のシスタチオニンγ合成酵素およびO−アセチルホモセリンサルフヒドリラーゼおよびO−スクシニルホモセリンサルフヒドリラーゼによってメチオニンに転換することができる。
【0065】
各遺伝子を大腸菌発現ベクターであるpCL−CJ1ベクター(韓国CJ社)で発現させ、超音波破砕を行い、細胞溶解液から酵素液を調製し、発現蛋白を得た。得られた酵素液をO−スクシニルホモセリンおよびO−アセチルホモセリンが蓄積された発酵液に添加し、これと共にメチルメルカプタン溶液を共に添加して反応を誘導した。反応はDTNB(5,5-dithiobis(2-nitro-benzoic acid, 米国sigma社)を用いて確認し、反応産物はHPLCを用いて分析した。
【0066】
本発明はO−スクシニルホモセリン、O−アセチルホモセリンとCH3SHとの反応を通じてL−メチオニンの他にもO−スクシニルホモセリンの場合はコハク酸を、O−アセチルホモセリンの場合は酢酸を別途の産生工程無しに副産物として得ることができる。
【図面の簡単な説明】
【0067】
【図1】メチオニン前駆体産生菌株の遺伝子操作図である。
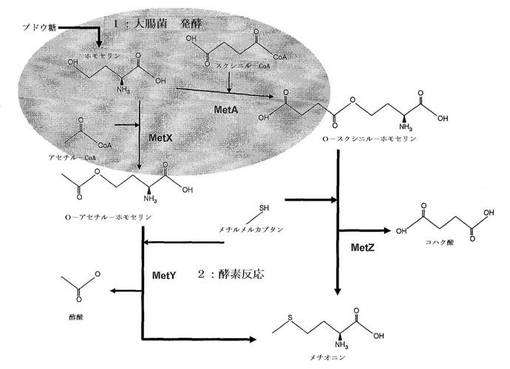
【図2】第2工程でメチオニンを産生するための化学構造図である。
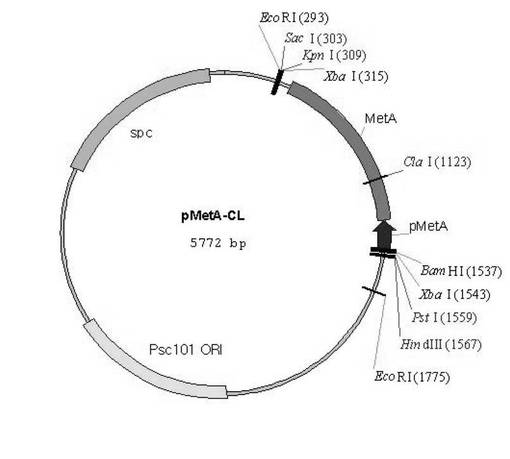
【図3】metA遺伝子の発現のためのベクターpMetA-CLの模式図である。
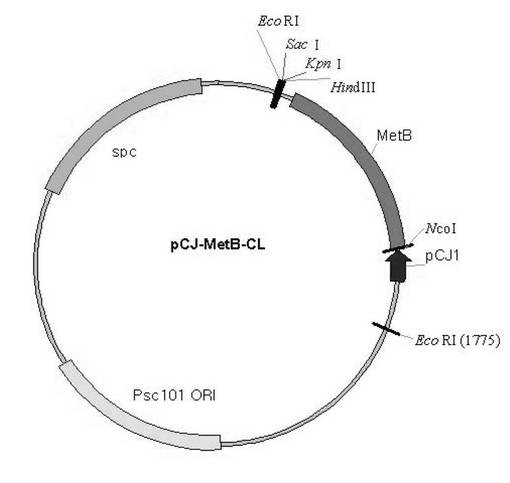
【図4】metB遺伝子の発現のためのベクターpCJ-MetB-CLの模式図である。
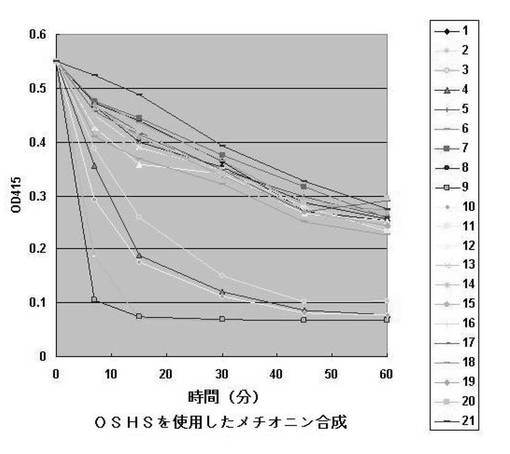
【図5】多様な酵素のO−スクシニルホモセリン消費に対する反応曲線である。各番号に使用された酵素液の由来は次のとおりである。酵素液21番は特定遺伝子を含まない細胞抽出液を用いた。
【表1】
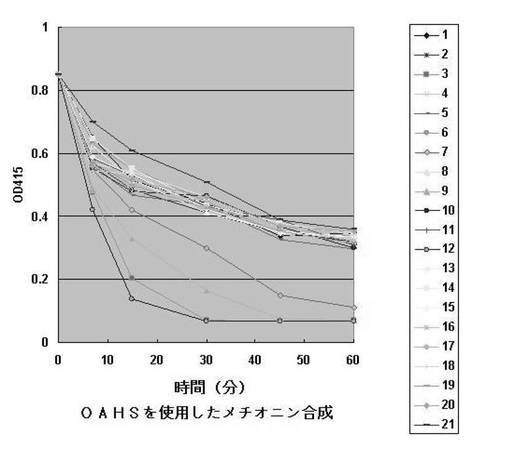
【図6】多様な酵素のO−アセチルホモセリン消費に対する反応曲線である。各番号は図5と同じである。
【図7】転換反応に使用された各酵素のアミノ酸配列をDNAstarのmegalign方法を用いて整列した図である。
【図8】転換反応に使用された各酵素のアミノ酸配列をDNAstarのmegalign方法を用いて整列した図である。
【発明を実施するための形態】
【0068】
以下、本発明の現時点での現実的で好ましい実施態様を以下の実施例に示すとおり説明する。しかし、下記実施例は本発明を例示して説明するためのものであって、本発明の権利範囲が下記実施例によって限定されるものではない。
【実施例1】
【0069】
実施例1:メチオニン前駆体産生菌株の作製
【0070】
<1−1> metB遺伝子の欠損
大腸菌株においてシスタチオニンシンターゼをコードする遺伝子であるMetB遺伝子を欠損させるために、FRT-one-step PCR deletion方法を使用した(PNAS (2000)vol97:P6640-6645)。配列番号1、2番のプライマーを用いてpKD3ベクター(PNAS (2000) vol97:P6640-6645)を鋳型にしてPCR反応で欠損カセットを作製した。変性(denaturation)工程は94℃で30秒、アニーリング(annealing)工程は55℃で30秒、伸長(extension)工程は72℃で1分間施し、これを30サイクルでPCR行った。
【0071】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.2kbpサイズのバンドからDNAを精製した。回収されたDNA断片はpKD46ベクター(PNAS (2000) vol97:P6640-6645)を予め形質転換させたE.coli(K12)W3110菌株にエレクトロポレーションを行った。エレクトロポレーションの前に、pKD46で形質転換を行ったW3110菌株は100μg/Lアンピシリンと5mMの1−アラビノースが含まれたLB培地を用いて30℃でOD600が0.6になるまで培養した。その後に、培養した菌株を滅菌蒸留水で2回、10%グリセロールで1回洗浄した。エレクトロポレーションは2500Vで行った。回収した菌株は25μg/Lクロラムフェニコールを含むLBプレート培地に塗布して37℃で一夜培養した後、耐性を示す菌株を選別した。
【0072】
上記と同様の条件で同じプライマーを鋳型として選別した菌株に対してPCRを行った。1.0%アガロースゲル上で遺伝子のサイズが1.2Kbに観察されるものを確認することによってmetB遺伝子の欠損を同定した。次に、確認された菌株はpCP20ベクター(PNAS (2000) vol97:P6640-6645)で形質転換を行い、LB培地で培養した。最終的に、同じ条件でPCRを行い、1.0%アガロースゲル上で遺伝子のサイズが150bpまで小さくなったmetB遺伝子の欠損菌株を作製し、クロラムフェニコールマーカーが除去されたことを確認した。作製された菌株をW3−Bと命名した。
【0073】
<1−2> thrB遺伝子の欠損
ホモセリンキナーゼをコードするthrB遺伝子を欠損させることによって、ホモセリンからO−スクシニルホモセリンの合成量を増加させようとした。特にトレオニン産生菌株を用いる場合、ホモセリンを利用する活性が強力なため、この遺伝子の欠損は必ず必要である。上記のように作製されたW3−B菌株でthrB遺伝子を欠損させるために、上記metB遺伝子の欠損と同様にFRT-one-step PCR deletionを行った。
【0074】
thrB欠損カセットを作製するために、pKD4ベクター(PNAS (2000) vol97:P6640-6645)を鋳型にして、配列番号3および4のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で1分間施し、これを30サイクル行うPCRを行った。その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.6kbpサイズのバンドからDNAを精製した。回収したDNA断片をpKD46ベクターで予め形質転換させたW3−B菌株にエレクトロポレーションによって導入した。回収された菌株を50μg/Lカナマイシンを含むLBプレート培地に塗布して37℃で一夜培養した後、耐性を示す菌株を選別した。
【0075】
上記と同一の条件で、選別された菌株を鋳型にして配列番号3、4のプライマーを用いてPCRを行った後、1.0%アガロースゲル上で遺伝子のサイズが1.6Kbに確認される菌株を選別することによってthrB遺伝子の欠損を確認した。次に、確認された菌株をpCP20ベクターで形質転換してLB培地で培養した。最終的に、同一条件下のPCRを行い1.0%アガロースゲル上で遺伝子のサイズが150bpに小さくなったthrB遺伝子の欠損菌株を作製し、カナマイシンマーカーが除去されたことを確認した。作製された菌株はW3−BT菌株と命名した。
【0076】
<1−3> metJ遺伝子の欠損
メチオニン前駆体の合成に関与するmetA遺伝子の調節遺伝子であるmetJを欠損させるために、metB遺伝子の欠損と同様にFRT-one-step PCR deletion法を行った。
【0077】
metJ遺伝子欠損カセットを作製するために配列番号5、6のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で1分間施し、これを30サイクルでPCRを行った。
【0078】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.2kbpサイズのバンドからDNAを精製した。回収されたDNA断片をpKD46ベクターで予め形質転換させたW3−BT菌株にエレクトロポレーションによって導入した。回収された菌株はクロラムフェニコールを含むLBプレート培地に塗布して37℃で一夜培養した後、耐性を示す菌株を選別した。
【0079】
選別された菌株を鋳型にして配列番号7および8のプライマーを用いて同じ条件でPCRした後、1.0%アガロースゲル上で遺伝子のサイズが1.6Kbに変化することを確認することによってmetJ遺伝子の欠損を確認した。確認された菌株は再びpCP20ベクターで形質転換してLB培地で培養し、再び同一な条件のPCRを通じて1.0%アガロースゲル上で遺伝子のサイズが600bpに小さくなった最終metJ遺伝子欠損菌株を作製し、クロラムフェニコールマーカーが除去されたことを確認した。作製された菌株をW3−BTJと命名した。
【0080】
<1−4−1> metA遺伝子の過剰発現
より多いメチオニン前駆体の合成のために、メチオニン前駆体のO−スクシニルホモセリン合成に関与するホモセリンO−スクシニルトランスフェラーゼ酵素をコードするmetA遺伝子を過剰発現を行った。
【0081】
これのためにE.coli W3110の染色体を鋳型にして、配列番号9および10番のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを25サイクル行うPCR反応を施した。
【0082】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.2kbpサイズのバンドからDNAを精製した。回収されたDNA断片はpCL1920ベクターをSmaIで切断して得られたDNA断片と連結させた。連結されたベクターをE.coliに形質転換させて50μg/Lスペクチノマイシンが含まれたLB培地で培養した後に選別した。このように作製されたベクターをpMetA-CLと命名した。pMetA-CLベクターの模式図は図3のとおりである。上記ベクターをW3-BTJ菌株に形質転換させて作製された菌株をW3-BTJ/pMetA-CLと命名し、O−スクシニルホモセリンの増加を観察した。
【0083】
metA遺伝子の発現をさらに増加させるための他の方法として、前記のmetA遺伝子をpCL1920ベクターにCJ1プロモータ(韓国、CJ社)とEcoRV制限酵素を用いて連結させた。連結されたベクターをE.coli菌株に形質転換させ50μg/Lスペクチノマイシンが含まれたLB培地において培養した後に選別した。このように作製されたベクターをpCJ-MetA-CLと命名した。前記ベクターをW3-BTJ菌株に形質転換させて作製された菌株をW3-BTJ/pCJ-MetA-CLと命名し、O−スクシニルホモセリンの増加を観察した。
【0084】
<1−4−2> metX遺伝子の過剰発現
O−アセチルホモセリンの合成のために、メチオニン前駆体のO−アセチルホモセリン合成遺伝子であるホモセリンO−アセチルトランスフェラーゼをコードするmetX遺伝子の過剰発現を行った。
【0085】
これのためにレプトスピラメイエリの染色体を鋳型にして配列番号11および12番のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを25サイクル行うPCR反応を施した。
【0086】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.1kbpサイズのバンドからDNAを精製した。回収されたDNA断片をpCL1920ベクターにCJ1プロモータとEcoRV制限酵素を用いて連結させた。連結されたベクターをE.coliに形質転換させて50μg/Lスペクチノマイシンが含まれたLB培地で培養した後に選別した。このように作製されたベクターをpCJ1-MetXlme-CLと命名した。前記ベクターをW3-BTJ菌株に形質転換させて作製された菌株をW3-BTJ/pCJ-MetXlme-CLと命名し、O−アセチルホモセリンの増加を観察した。
【0087】
metX遺伝子の過剰発現のための他の方法として、コリネバクテリウム染色体を鋳型にして配列番号68、69番のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを25サイクル行うPCR反応を施した。
【0088】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、DNAを精製した。回収されたDNA断片をpCL1920ベクターにCJ1プロモータとEcoRV制限酵素を用いて連結させた。連結されたベクターをE.coliに形質転換させて50μg/Lスペクチノマイシンが含まれたLB培地で培養した後に選別した。このように作製されたベクターをpCJ-MetXcgl-CLと命名した。前記ベクターをW3-BTJ菌株に形質転換させて作製された菌株をW3-BTJ/pCJ-MetXcgl-CLと命名し、O−アセチルホモセリンの増加を観察した。
【0089】
<1−4−3> metA遺伝子の欠損
O−アセチルホモセリンの産生量を増加させるために、W3-BTJ菌株でホモセリンO−スクシニルトランスフェラーゼをコードする遺伝子であるmetAを欠損させた。metA遺伝子のみを導入した場合、一定量のO−スクシニルホモセリンが蓄積されることが観察されたから、metA遺伝子を欠損させる場合最も多い量のO−アセチルホモセリンを蓄積することができると推定した(表4参照)。metA遺伝子を欠損させるためにFRT one step PCR deletion方法を使用した。
【0090】
metA遺伝子欠損カセットを作製するために配列番号70および71のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で1分間施し、これを30サイクルでPCR行った。
【0091】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.2kbpサイズのバンドからDNAを精製した。回収されたDNA断片をpKD46ベクターで予め形質転換させたE.coli W3-BTJ菌株にエレクトロポレーションによって導入した。回収された菌株をクロラムフェニコールを含むLBプレート培地に塗布して37℃で一夜培養した後、耐性を示す菌株を選別した。
【0092】
選別された菌株を鋳型にして配列番号70および71のプライマーを用いて同じ条件でPCRした後、1.0%アガロースゲル上で遺伝子のサイズが1.1Kbに変化されることを確認することによってmetA遺伝子の欠損を確認した。確認された菌株はpCP20ベクターで形質転換させてLB培地で培養し、同一な条件のPCRを通じて1.0%アガロースゲル上で遺伝子のサイズが100bpに小さくなった最終metA遺伝子欠損菌株を作製し、クロラムフェニコールマーカーが除去されたことを確認した。作製された菌株をW3−BTJAと命名した。W3−BTJA菌株を前記pCJ-MetXlme-CLベクターで形質転換させて作製された菌株をW3-BTJA/pCJ-MetX-CLと命名した。前記菌株を同一な方法で培養した結果、O−スクシニルホモセリンの蓄積は観察されず、O−アセチルホモセリンの産生量がW3-BTJに比べて約20%有意に増加することを確認した。
【0093】
<1−5> L−トレオニン産生菌株の転換
メチオニン要求性が解除されたL−トレオニン産生菌株の大腸菌CJM002(KCCM-10568号)を用いて前記1−1〜1−3と同一な方法でメチオニン前駆体産生菌株を作製した。作製された菌株はそれぞれCJM-BTJ、CJM-BTJ/pMetA-CLおよびCJM-BTJ/pCJ-MetA-CLと命名した。また、このCJM-BTJ菌株を用いて前記<1−4−3>の方法でmetA遺伝子を欠損させた菌株を作製し、作製された菌株をCJM-BTJAと命名した。
【実施例2】
【0094】
実施例2:L−メチオニン前駆体の産生のための発酵
【0095】
<2−1> フラスコ培養実験
実施例1から作製された菌株のメチオニン前駆体の産生量を実験するために三角フラスコ培養を施した。抗生剤スペクチノマイシンが含有されたLBプレート培地にW3-BTJとCJM-BTJおよびmetA、metX発現ベクターで形質転換させたW3-BTJ菌株を播種して31℃で一夜培養した後、単一コロニーをスペクチノマイシンが含まれた3mlのLB培地に播種した後、31℃で5時間培養し、再び25mlメチオニン前駆体の産生培地を含む250ml三角フラスコに200倍希釈して31℃200rpmで64時間培養してHPLC分析を通じてメチオニン前駆体の産生量を比較した(表3および表4)。その結果、メチオニン要求性が解除されたトレオニン産生菌株を使用して作製されたメチオニン前駆体産生菌株の場合、メチオニン前駆体の産生量が著しく増加したことがわかった。
【0096】
【表2】
【0097】
【表3】
【0098】
【表4】
【0099】
<2−2> 大型発酵槽実験
実施例1で最も優れたメチオニン前駆体の産生能を見せた菌株を用いてメチオニン前駆体を大量生産するために、5L発酵槽培養を施した。抗生剤スペクチノマイシンが含有されたLBプレート培地にCJM-BTJ/pCJ-metA-CL菌株またはCJM-BTJA/pCJ-metXlme-CL菌株を播種して31℃で一夜培養した。その後、単一コロニーをスペクチノマイシンが含まれた10mlLB培地に播種した後、31℃で5時間培養し、再び200mlメチオニン前駆体のシード培地を含む1000ml三角フラスコに100倍希釈して31℃、200rpmで3〜10時間培養した後、5L発酵槽に播種して流加式培養(Fed batch)発酵法で50〜100時間培養した。このように培養した発酵液においてメチオニン前駆体の濃度をHPLCで分析した結果は表6の通りである。
【0100】
【表5】
【0101】
【表6】
【実施例3】
【0102】
実施例3:メチオニン転換酵素の産生
【0103】
<3−1> E.coli由来のシスタチオニンγ合成酵素
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるE.coli由来のシスタチオニンγ合成酵素をコードしているmetB遺伝子をクローニングした。
【0104】
E.coliの染色体を鋳型にしてプライマー配列番号13および14を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを25サイクル行うPCR反応を通じて獲得した。
【0105】
獲得したDNA断片をNcol/HindIIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。最終作製されたベクターをpCL-MetB-CLと命名した。模式図は図4のとおりである。クローニングされたベクターをE.coli W3110細胞に形質転換させ、50μg/Lのスペクチノマイシンが含まれたLBプレート培地で培養した後、コロニーを選別した。選別されたコロニーを50μg/Lのスペクチノマイシンが含まれたLB培地3mlに播種して一夜の間37℃で培養した。培養された細胞は再び回収して0.1Mリン酸カリウム緩衝液(Potassium phosphate, pH7.5)で洗浄して再び200μlのリン酸カリウム緩衝液に懸濁し、30秒おきに5回超音波処理をして細胞を破砕した。破砕された細胞抽出液を12000rpmで10分間遠心分離した後、上澄液を取ってBio-Rad蛋白質定量液(米国BIO-Rad社)を用いて蛋白質の総量を定量した。また、SDS-PAGE法を用いて蛋白質の発現を確認した。回収された細胞抽出液の上澄液を酵素転換反応に使用した。
【0106】
<3−2> シュードモナス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるシュードモナス属(Pseudomonas Sp.)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。シュードモナスはシュードモナスアルギノサおよびシュードモナスプチダの2種を使用した。
【0107】
それぞれの菌株の染色体を鋳型にしてシュードモナスアルギノサの場合はプライマー配列番号15および16を、シュードモナスプチダの場合は配列番号17および18を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0108】
獲得したDNA断片をNdeI/PacIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0109】
<3−3> シュードモナス属由来のO−アセチルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるシュードモナス属由来のO−アセチルホモセリンサルフヒドリラーゼをコードするmetY遺伝子をクローニングした。
【0110】
シュードモナスアルギノサ菌株の染色体を鋳型にし、配列番号19および20を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0111】
獲得したDNA断片をNdeI/PacIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0112】
<3−4> コリネバクテリウムグルタミクム由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるコリネバクテリウムグルタミクム(Corynebacterium glutamicum)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0113】
コリネバクテリウムグルタミクムの染色体を鋳型にしてプライマー配列番号21および22を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0114】
獲得したDNA断片をNcoI/Hind3で切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0115】
<3−5> コリネバクテリウムグルタミクム由来のO−アセチルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるコリネバクテリウムグルタミクム由来のO−アセチルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0116】
コリネバクテリウムグルタミクムの染色体を鋳型にしてプライマー配列番号23および24を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0117】
獲得したDNA断片をNdeI/Avr2で切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0118】
<3−6> レプトスピラ属由来のO−アセチルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるレプトスピラメイエリ(Leptospira meyeri)由来のO−アセチルホモセリンサルフヒドリラーゼをコードするmetY遺伝子をクローニングした。
【0119】
レプトスピラメイエリの染色体を鋳型にしてプライマー配列番号25および26を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0120】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0121】
<3−7> サッカロミセス属由来のO−アセチルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用される出芽酵母(Saccharomyces cerevisiae)由来のO−アセチルホモセリンサルフヒドリラーゼをコードするmet25遺伝子をクローニングした。
【0122】
出芽酵母の染色体を鋳型にしてプライマー配列番号27および28を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0123】
獲得したDNA断片をNdeI/PacIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0124】
<3−8> クロモバクテリウム属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるクロモバクテリウムビオラシウム(Chromobacterium Violaceum)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0125】
クロモバクテリウムビオラシウムの染色体を鋳型にしてプライマー配列番号29および30を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0126】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)のにクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0127】
<3−9> ノルカジア属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるノルカジアファルシニカ(Norcardia Farcinica)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0128】
ノルカジアファルシニカの染色体を鋳型にしてプライマー配列番号31および32を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応通じて獲得した。
【0129】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0130】
<3−10> ブラジリゾビウム属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるブラジリゾビウムジャポニクム(Bradyrhizobium Japonicum)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0131】
ブラジリゾビウムジャポニクムの染色体を鋳型にしてプライマー配列33および34を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0132】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0133】
<3−11> ヒポモナス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるヒポモナスネプチュニウム(Hyphomonas Neptunium)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0134】
ヒポモナスネプチュニウムの染色体を鋳型にしてプライマー配列番号35および36を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0135】
獲得したDNA断片をBamHII/HindIIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0136】
<3−12> メチロコックス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるメチロコックスカプスラツス(Methylococcus Capsulatus)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0137】
メチロコックスカプスラツスの染色体を鋳型にしてプライマー配列番号37および38を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0138】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0139】
<3−13> メチロバシルス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるメチロバシルスフラゲラツス(Methylobacillus Flagellatus)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0140】
メチロバシルスフラゲラツスの染色体を鋳型にしてプライマー配列番号39および40を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0141】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0142】
<3−14> ニトロソモナス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるニトロソモナスヨーロパエア由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0143】
ニトロソモナスヨーロパエアの染色体を鋳型にしてプライマー配列番号41および42を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0144】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0145】
<3−15> クレシエラ属由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるクレシエラニュモニアエ(Klesiella Pneumoniae)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0146】
クレシエラニュモニアエの染色体を鋳型にしてプライマー配列番号43および44を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0147】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0148】
<3−16> バシルス属由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用される枯草菌(Bacillus Subtilis)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0149】
枯草菌の染色体を鋳型にしてプライマー配列番号45および46を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0150】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0151】
<3−17> シゲラ属由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるシゲラフレキシネリ2457T(Shigella Flexneri 2457T)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0152】
シゲラフレキシネリ2457Tの染色体を鋳型にしてプライマー配列番号47および48を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0153】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0154】
<3−18> コルウェリア属由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるコルウェリアサイクレリトラエア(Colwellia Psychrerythraea)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0155】
コルウェリアサイクレリトラエアの染色体を鋳型にしてプライマー配列番号49および50を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0156】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0157】
<3−19> サルモネラ由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるサルモネラエンタリカ血清型パラチフス(Salmonella enterica serovar Paratyphi A)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0158】
サルモネラエンタリカ血清型パラチフスの染色体を鋳型にしてプライマー配列番号51および52を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0159】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0160】
<3−20> OSHSを基質にした転換酵素の活性比較
前記の<3−1>から<3−19>の方法で獲得したそれぞれの酵素液を活性比較して最適のメチオニン転換酵素を選別した。
【0161】
このために、O−スクシニルホモセリン(Sigma社)を3mMの濃度で0.1Mリン酸カリウム緩衝液(pH7.5)に溶解した。また、この反応液に助酵素で使用されるピリドキサールホスフェート(Pyridoxal5'-phosphate、米国Sigma社)を最終濃度10μMになるように添加した。反応液にもう一つの基質として使用されるメチルメルカプタン(Methylmercaptan、日本東京化成工業株式会社)を最終濃度2mMで添加した。この反応液1mlを37℃に位置した後、各酵素抽出液を蛋白質濃度5mg/mlに合わせて10μl添加した。反応の進行如何は毎5〜10分おきに100μlの反応液を回収して4mg/mlのDTNB(米国Sigma社)溶液900μlに添加した後、OD415における吸光度を測定することによって確認した。
【0162】
DTNBは反応液中に残存するメチルメルカプタンのSH基と反応して黄色いの物質を合成する。従って、反応液状のメチルメルカプタンが反応によってメチオニンに転換されることで反応液の黄色が消えるものであって、反応の進行が確認できる。
【0163】
実験結果、図5のようにクロモバクテリウム由来のO−スクシニルホモセリンサルフヒドリラーゼ、ノルカジア由来のO−スクシニルホモセリンサルフヒドリラーゼ、シュードモナス属由来のO−スクシニルホモセリンサルフヒドリラーゼおよびO−アセチルホモセリンサルフヒドリラーゼが非常に高い活性を見せた。他の酵素も全てある程度の活性は見せているが、反応進行速度が相対的に遅いことが確認された。各酵素の基質に対する反応性を要約した表は表7のとおりである。また、1時間の反応が終了された後に選別された反応液からHPLC分析を通じて最終生成されたメチオニンおよびコハク酸の生成を確認した結果は表8に示した。
【0164】
<3−21> OAHSを基質にした転換酵素の活性比較。
前記<3−20>と同一の反応をO−アセチルホモセリンを用いて反応させた。O−アセチルホモセリンは発酵上澄液から精製した。O−スクシニルホモセリンと同一な反応液および同一な酵素液を用いて反応させた結果、図6のようにヒポモナス由来のO−スクシニルホモセリンサルフヒドリラーゼ、シュードモナス属由来のO−アセチルホモセリンサルフヒドリラーゼ、クロモバクテリウム属由来のO−スクシニルホモセリンサルフヒドリラーゼおよびレプトスピラ属由来のO−アセチルホモセリンサルフヒドリラーゼが高い活性を見せた。他の酵素も全てある程度の活性は見せているが、反応進行速度が相対的に遅いことが確認された。各酵素の基質に対する反応性を要約して表7に示した。また、1時間の反応が終了された後に選別された反応液からHPLC分析を通じて最終生成されたメチオニンおよび酢酸の量を確認した結果は表9に示した。
【0165】
【表7】
【0166】
【表8】
【0167】
【表9】
【0168】
<3−22> 転換酵素のフィードバック阻害確認
前記3−20および21と同一の方法を使用してメチオニンの存在有無によるフィードバックの阻害反応を確認した。前記と同一な反応液を作製し、各反応液にそれぞれ5g/Lのメチオニンを添加または非添加して同一な反応進行を行った。メチオニンを添加しない場合の反応進行速度を100%にしたとき、メチオニンの存在時の残存活性を%で表記して表10に示した。
【0169】
その結果、シュードモナス由来のO−アセチルホモセリンサルフヒドリラーゼ、ノルカジア由来のO−スクシニルホモセリンサルフヒドリラーゼおよびレプトスピラ由来のO−アセチルホモセリンサルフヒドリラーゼがメチオニンによる活性阻害を見せたことを確認した。従って、これらの酵素はメチオニンによるフィードバック阻害を有しているものと推定され、後ほど反応にはフィードバック阻害がない酵素を使用した。前記の実施例においてフィードバック阻害を受ける酵素もフィードバック阻害が解除される変異株を使用する場合、同一な反応に使用できると推定された。
【0170】
【表10】
【0171】
<3−23> 転換酵素の相同性比較
転換反応に使用された各酵素間の相同性を比較してO−スクシニルホモセリンおよびO−アセチルホモセリンに対する反応性およびフィードバック調節阻害との相関関係を確認した。
【0172】
使用された酵素の相同性を比較した結果、O−スクシニルホモセリンサルフヒドリラーゼで命名されるmetZ間の相同性およびO−アセチルホモセリンサルフヒドリラーゼで命名されるmetY間の相同性が相互間の相同性に比べて相対的に高いことがわかった。前記の実施例と連関してみてO−スクシニルホモセリンサルフヒドリラーゼで命名されたmetZの場合にはフィードバック調節阻害を見せていない場合が最も多いのに比べて、O−アセチルホモセリンサルフヒドリラーゼで命名されたmetYの場合には実施例に使用された酵素の全てがフィードバック調節阻害を見せて相対的に高いフィードバック調節阻害を有していることと確認された。O−スクシニルホモセリンおよびO−アセチルホモセリンに対する選択性について、metZ遺伝子群はO−スクシニルホモセリンに対する選択性が高く、metY遺伝子群はO−アセチルホモセリンに対する選択性は高かったが、シュードモナスプチダ由来のmetYおよびクロモバクテリウムビオラシウム由来のmetZは2つの両基質に対して特異的に高い反応性を見せた。
【0173】
実施例に使用された全ての酵素をアミノ酸配列に基づいて配列整列プログラムのクラスタルWプログラム(ClustalW program; DNAstar)で整列(align)した結果、次のような配列で明記されるドメインを共通的に有していることが確認された。
【0174】
ドメイン1:Y-(S, I, T, V)-R-X-X-(N,S)
ドメイン2:
(V,A,I)-(V,L,I)-D-N-X-(F,V,M,I)-X-(T,S)-(P,A)-X-(L,I)-(Q,C,V)-X-(P,G)-(L,F)
-X-(L,M,H)-G-(A,V)-(D,H)
ドメイン3:
(S,A,G,P)-(P,A,V)-F-(N,D)-(A,S)-(W,F,Y)-X-X-X-(K,Q,R,S)-G-(L,M,V,I,M)-(
E,K,D,R)-T-(L,M)
ドメイン5:(H,Y)-(P,A)-(A,S)-(T,S)-(T,M,Q)-(T,S)-H-
ドメイン6:(V,I,L)-R-(V,I,L,F)-(S,A)-(V,I,T)-G-(L,I)-E-
【実施例4】
【0175】
実施例4:メチオニン転換酵素によるメチオニン転換反応
【0176】
<4−1> 転換酵素の大量産生
実施例2(2−2および2−8)から作製されたメチオニン転換酵素の産生菌株を大量産生するために1L発酵槽培養を施した。抗生剤スペクチノマイシンが含有されたLBプレート培地にW3110をシュードモナス由来metZ発現ベクターで形質転換またはヒポモナス由来metZ発現ベクターで形質転換させた菌株をそれぞれ播種して30〜40℃で一夜培養した後、単一コロニーをスペクチノマイシンが含まれた40mlLB培地に播種した後、30〜40℃で5時間培養した。このように培養されたシュードモナス由来metZ発現菌株およびヒポモナス由来metZ発現菌株を1L発酵槽で600〜900rpm、30〜40℃、15〜30時間培養した。培養に使用された培地組成は表11に示した。
【0177】
培養が終わったメチオニン転換酵素発酵液を超音波電動を用いて細胞を破砕してメチオニン転換酵素液を作製した。
【0178】
【表11】
【0179】
<4−2> メチオニン転換反応
前記の実施例2(2−2)から産生されたO−スクシニルホモセリン発酵液とO−アセチルホモセリン発酵液とにそれぞれ実施例4(4−1)から作製されたシュードモナス由来O−スクシニルホモセリン転換酵素液およびヒホモナス由来のO−アセチルホモセリン転換酵素液を使用してメチオニン転換反応を行った。
【0180】
細胞を除去しないメチオニン前駆体発酵液2.0Lに破砕した酵素培養液0.1Lを添加した後、15%Na-メチルメルカプタン0.3Lを添加して反応を開始し、2時間経過後に発酵液を回収して細胞を除去した後、HPLCを通じてメチオニン生成を確認した。その結果を表12に示した。
【0181】
【表12】
【0182】
前記の結果からわかるように、既存の発明では10g/L以下の低い濃度のL−メチオニンを産生することに比べて前記の発明では30g/L以上のL−メチオニンを生成してL−メチオニンを大量に産生可能であることが確認できた。
【産業上の利用可能性】
【0183】
L−メチオニンを選択的に産生することによって飼料および食品添加剤、医薬用および医薬品の原料などの多様な分野で広く使用することができる。上述のとおり本発明の具体例をついて詳細に説明したが、本発明の範疇および技術思想範囲内で当業者にとって多様な変形および修正が可能であることは明らかであり、このような変形および修正が添付された本発明の特許請求範囲に属することも当然である。
【技術分野】
【0001】
本発明はL−メチオニン前駆体産生菌株に関するものであって、より詳しくは本発明によって作製されたL−メチオニン前駆体産生菌株を使用して作製されたL−メチオニン前駆体から酵素転換反応を用いてL−メチオニンおよび有機酸を高収率で産生することによって、既存の方法に比べて環境にやさしい方法でL−メチオニンを産生し、L−メチオニンを選択的に産生することによって飼料および食品添加剤、医薬用および医薬品の原料などの多様な分野に広く使用するためのものである。
【背景技術】
【0002】
メチオニンは生体内の必須アミノ酸の一種類で飼料および食品添加剤として広く使用され、医薬用として輸液剤、医薬品の合成原料としても使用される。メチオニンはコリン(レシチン)とクレアチンのような化合物の前駆体で作用し、システインとタウリンの合成原料でも使われる。また、硫黄を提供する役割をする。S−アデノシル−メチオニンはL−メチオニンから由来して生体内でメチル基を提供する役割をして、脳の多様な神経伝達物質(neurotransmitter)合成に関連されている。メチオニンおよび/またはS−アデノシル−L−メチオニン(SAM)は生体内で肝と動脈において脂肪蓄積を抑制し、鬱、炎症、肝疾患、筋肉痛を緩和するなどの多様な役割をする。
【0003】
今まで知られたメチオニンおよび/またはS−アデノシル−L−メチオニンの生体内の役割は次の通りである。
【0004】
1)脂肪代謝を促進する肝と動脈において脂肪沈着を抑制し、脳、心臓、腎臓の血流を増進させる役割をする(JHepatol. Jeon BR et al., 2001 Mar; 34(3):395-401(非特許文献1))。
【0005】
2)消化を促進し、毒性物質の解毒と排泄を促進、鉛のような重金属の排泄を促進する。
【0006】
3)メチオニンは毎日1日に800〜1600mgの投与で優れた抗鬱剤になると知られている(Am J Clin Nutr. Mischoulon D. et al., 2002 Nov; 76(5): 1158S-61S(非特許文献2))。
【0007】
4)肝疾患において肝機能の向上作用をする(FASEB J Mato JM., 2002 Jan;16(1): 15-26)。特にアルコールによる肝疾患に効果的である(Cochrane DatabaseSyst Rev., Rambaldi A., 2001; (4): CD002235(非特許文献3))。
【0008】
5)骨関節疾患に抗炎症効果が良く、関節回復を促進する(ACP J Club. Sander O., 2003Jan-Feb; 138(1): 21、J Fam Pract., Soeken KL et al.,2002 May; 51(5): 425-30(非特許文献4))。
【0009】
6)メチオニンは毛髪の必須栄養素である。毛髪に栄養を供給し脱毛を防止する(AudiolNeurootol., Lockwood DS et al., 2000 Sep-Oct; 5(5): 263-266(非特許文献5))。
【0010】
メチオニンは前記のように動物飼料を含めた食品と医薬品で用いるために化学合成と生物学的合成を通じて産生することができる。
【0011】
化学合成は主に5−(β−メチルメルカプトエチル)ヒダントインを加水分解させる反応を通じてL−メチオニンを産生する。しかし、このような化学合成を通じて産生されたメチオニンはL−型とD−型が混合された形態でのみ産生されるという短所がある。
【0012】
生物学的合成はメチオニン生成に関与する蛋白質を用いる方法である。L−メチオニンはmetA、metB、metC、metE、metH遺伝子から発現される酵素蛋白質の作用によってホモセリンからメチオニンに生合成される。具体的に、metAはメチオニン生合成の一番目の酵素であるホモセリンO−スクシニルトランスフェラーゼ(homoserineO-succinyltransterase)をコードする遺伝子でホモセリンをO−スクシニル−L−ホモセリンに転換する機能をする。metBがコードするO−スクシニルホモセリンリアーゼ(O-succinylhomoserinelyase)、またはシスタチオニンγ合成酵素酵素はO−スクシニル−L−ホモセリンをシスタチオニン(cystathionine)に転換する機能をする。metCがコードするシスタチオニンベタリアーゼ(cystathionine beta lyase)酵素はシスタチオニンをL−ホモシステインに転換する機能をする。metEがコードするコバラミン−非依存的メチオニンシンターゼとmetHがコードするコバラミン−依存的メチオニンシンターゼがL−ホモシステインをL−メチオニンに転換させる。この際、metFがコードする5,10−メチレンテトラヒドロフォレートリダクターゼ(5,10-methylenetetrahydrofolatereductase)とglyAがコードするセリンヒドロキシメチルトラスフェラーゼ(serin hydroxymethytransferase)は共に作用してL−メチオニン合成に必要なメチル基を提供するN(5)−メチルテトラヒドロフォレート(N(5)-methyltetrahydrofolate)を合成する機能をする。
【0013】
L−メチオニンは前記酵素による一連の有機的な反応を通じて合成される。前記蛋白質または前記蛋白質に影響を与えるその他蛋白質の遺伝的な操作は、L−メチオニン合成の調節を生じさせる可能性がある。例えば、特開2000−139471号(特許文献1)にはエシェリキア属に属するゲノム遺伝子上のthrBC、metJを除去し、metBLを過剰発現させ、metKを漏出変異体として作製し、L−メチオニンを産生する方法が記述されている。また、米国特許出願公開第2003/0092026号(特許文献2)にはコリネバクテリウム属に属し、L−メチオニン合成の阻害因子であるmetD遺伝子を除去した微生物を用いる方法が記述されている。米国特許出願公開第2002/0049305号(特許文献3)には5,10−メチレンテトラヒドロフォレートリダクターゼ(metF)の発現を増加させてL−メチオニンの産生を増加させる方法が記載されている。
【0014】
米国特許出願公開第2005/0054060号(特許文献4)では、L−メチオニンを産生する微生物を作製するためにシスタチオニンシンターゼに変異を加えてシステインを利用せず、直接H2SまたはCH3SHを用いてホモシステインまたはメチオニンを合成することが記載されている。この方法では変異されたシスタチオニンシンターゼを直接細胞に導入して細胞内のメチオニン合成過程に沿ってメチオニンを合成した。しかし、この方法では細胞内のメチオニン代謝経路を用いるから酵素の反応過程が非効率的である。また、H2SまたはCH3SHが細胞に毒性を有するという点から見て実効性に劣る。さらに、メチルメルカプタン(CH3SH)に対する基質特異性がシュードモナスまたはクロモバクテリウム属由来のスクシニルホモセリンリアーゼに比べて非常に低いことが我々の実験において確認された。
【0015】
シスタチオニンシンターゼの場合、文献によると、互いに異なる基質と反応して互いに異なる物質を産生する傾向が大きい。シスタチオニンシンターゼはホモシステインとO−スクシニルホモセリンを結合して非常に高い効率でホモランチオニンという物質を産生するという報告がある(J.Bacteriol (2006) vol 188:p609-618(非特許文献6))。前記のようにシスタチオニンシンターゼが細胞内に存在する場合、互いに異なるメチオニン前駆体と結合することができ、この結合によって副産物を作る効率性も非常に高い。特に、シスタチオニンシンターゼを過剰発現する場合はより多くの副産物を生成する可能性があり、細胞内の反応の効率は非常に劣る。
【0016】
なお、既存の生物学的合成で産生されるメチオニンはL型という長所があるが、その量が非常に微量である。これはメチオニンの合成経路がとてもよく調節されるフィードバック調節システムを有しているからである。メチオニンが一定水準以上に合成されると、最終産物であるメチオニンがメチオニン合成を開始する初期蛋白質metAの遺伝子転写をフィードバックで抑制する。metA遺伝子は転写段階においてメチオニンによって阻害を受け、翻訳段階において細胞内の蛋白質分解酵素によって分解されるメカニズムにより、メチオニンの量が調節されるため、metA遺伝子の高発現だけではメチオニンの量を一定水準以上に増加させることができない(DvoraBiran, Eyal Gur, Leora Gollan and Eliora Z. Ron: Control of methioninebiosynthesis in Escherichia coli by proteolysis: Molecular Microbiology (2000)37(6), 1436-1443(非特許文献7))。従って、既存の多数の特許はmetA遺伝子のフィードバックを解除する研究に重点をおいていた(国際公開公報第2005/108561号(特許文献5)、国際公開公報第1403813号(特許文献6))。
【0017】
また、生物学的システムでメチオニンを産生する場合、メチオニンの分解代謝の経路であるS−アデノシルメチオニンシンターゼによって生成されたメチオニンがS−アデノシルメチオニンに変換される。S−アデノシルメチオニンは細胞に必須的な物質であってS−アデノシルメチオニンシンターゼを欠損させることができないため、既存にはこの遺伝子を弱化させる方向でメチオニンの産生を増加しようとする特許が出願されたことがある(国際公開公報第2005/108561号(特許文献7))。
【先行技術文献】
【特許文献】
【0018】
【特許文献1】特開2000−139471号公報
【特許文献2】米国特許出願公開第2003/0092026号明細書
【特許文献3】米国特許出願公開第2002/0049305号明細書
【特許文献4】米国特許出願公開第2005/0054060号明細書
【特許文献5】国際公開公報第2005/108561号パンフレット
【特許文献6】国際公開公報第1403813号パンフレット
【特許文献7】国際公開公報第2005/108561号パンフレット
【非特許文献】
【0019】
【非特許文献1】J Hepatol. Jeon BR et al., 2001 Mar;34(3):395-401
【非特許文献2】Am J Clin Nutr. Mischoulon D. et al., 2002Nov; 76(5): 1158S-61S
【非特許文献3】Cochrane Database Syst Rev., Rambaldi A.,2001; (4): CD002235
【非特許文献4】ACP J Club. Sander O., 2003 Jan-Feb; 138(1):21、J Fam Pract., Soeken KL et al., 2002 May; 51(5):425-30
【非特許文献5】Audiol Neurootol., Lockwood DS et al., 2000Sep-Oct; 5(5): 263-266
【非特許文献6】J.Bacteriol (2006) vol 188:p609-618
【非特許文献7】Dvora Biran, Eyal Gur, Leora Gollan andEliora Z. Ron: Control of methionine biosynthesis in Escherichia coli byproteolysis: Molecular Microbiology (2000) 37(6), 1436-1443
【発明の概要】
【発明が解決しようとする課題】
【0020】
前記の従来技術は細胞内のメチオニン代謝経路の中、シスタチオニンシンターゼ代謝経路を用いることから酵素の反応過程が非効率的であり、硫化物特有の基質毒性問題および副産物の生成の問題があり、またメチオニン合成経路のフィードバック調節によって十分な量のメチオニンを合成することができないという問題がある。
【0021】
従って、本発明は既存の生物学的メチオニンの産生工程を代替して発酵によるL−メチオニン前駆体の産生および前記L−メチオニン前駆体の酵素転換工程の2つの工程を使用することによって、従来の前記問題を克服したL−メチオニンの産生方法を提供することを目的とする。
【0022】
また、本発明はL−メチオニンを選択的に産生する方法を提供することを目的とする。
【0023】
また、本発明は別途の産生工程無しで副産物として有機酸を同時に産生する方法を提供することを目的とする。
【課題を解決するための手段】
【0024】
本発明について以下に詳細に説明する。
本発明は1)L−メチオニン前駆体を産生することができる菌株を作製し、この菌株を培養して発酵によりL−メチオニン前駆体を産生する工程、2)産生された前記L−メチオニン前駆体との酵素反応によりL−メチオニンおよび有機酸を産生する工程で構成されるL−メチオニンおよび有機酸を産生する方法を提供する。
【0025】
より詳しくは、前記第1工程はL−メチオニン前駆体産生菌株を作製し、この菌株を用いた発酵法でL−メチオニン前駆体を培養液に蓄積する工程である。この際、L−メチオニン前駆体産生菌株は本発明者の意図した方法によって作製される。従って、本発明はこのような菌株および菌株の作製方法も含まれる。
【0026】
前記L−メチオニン前駆体は下記の化学式から構成されたO−アシルホモセリン(O-acylhomoserine)基のうちの一つで表され;
【化1】
ここでRは、C、H、O、Nおよびその他化合物を含む最大15個の炭素分子である物質である。例えば、前記O−アシルホモセリン基はO−アセチルホモセリン(O-acetyl homoserine)、O−スクシニルホモセリン(O-succinylhomoserine)、プロピオニルホモセリン(propionyl homoserine)、アセトアセチルホモセリン(acetoacetyl homoserine)、クマロイルホモセリン(coumaroylhomoserine)、マロニルホモセリン(malonyl homoserine)、ヒドロキシメチルグルタリルホモセリン(hydroxymethylglutaryl homoserine)およびピメリルホモセリン(pimelyl homoserine)を含むが、これに限られるものではない。
【0027】
本発明において、前記L−メチオニン前駆体は好ましくはO−アセチルホモセリンまたはO−スクシニルホモセリンである。
【0028】
本発明において、「L−メチオニン前駆体産生菌株」とはL−メチオニン前駆体を生物体内で産生することができる原核および真核の微生物菌株であって、本発明による操作によってL−メチオニン前駆体を蓄積することができる微生物菌株をいう。例えば、L−メチオニンを産生するエシェリキア属、エルウィニア属、セラチア属、プロビデンシア属、コリネバクテリウム属、シュードモナス属、レプトスピラ属、サルモネラ属、ブレビバクテリア属、ヒポモナス属、クロモバクテリウム属およびノルカジア属、または菌類または酵母類に属する微生物菌株が含まれる。好ましくは、O−スクシニルホモセリンを産生するためにシュードモナス属、ノルカジア属、エシェリキア属の微生物が使用され、O−アセチルホモセリンを産生するためにエシェリキア属、コリネバクテリウム属、レプトスピラ属、酵母類などが使用される。より好ましくは、エシェリキア属に属する微生物菌株、より好ましくは大腸菌(Escherichia coli; 以下E.coliと称する)が使用される。また、エシェリキア属菌株に外来の遺伝子を導入することによってO−スクシニルホモセリンおよびO−アセチルホモセリンを選択的に産生することができる。
【0029】
本発明は前記の菌株からO−スクシニルホモセリンまたはO−アセチルホモセリンの分解に関与する遺伝子を欠損または弱化させ、O−スクシニルホモセリンまたはO−アセチルホモセリンの合成に関与する遺伝子を導入または強化させる菌株をL−メチオニン前駆体産生菌株で提供する。なお、選択的にはO−スクシニルホモセリンまたはO−アセチルホモセリンの産生強化のためにトレオニン生合成の経路を遮断または弱化した菌株を提供する。また、選択的にはO−スクシニルホモセリンまたはO−アセチルホモセリンの合成に関与する蛋白質のフィードバック調節が解除された遺伝子を導入し、過剰発現または活性を強化させた菌株を提供する。
【0030】
より詳しくは、本発明はL−メチオニン前駆体の分解に関与するmetB遺伝子、トレオニン生合成の経路に関与するthrB遺伝子およびL−メチオニン前駆体の産生遺伝子の転写を抑制するmetJ遺伝子を欠損させた菌株ならびにL−メチオニン前駆体の生合成に関与するmetAもしくはmetX遺伝子の発現を強化させ、またはフィードバック制御が解除されたmetAもしくはmetX遺伝子を導入し、またはmetA遺伝子を欠損させる代わりにmetX遺伝子を導入し、またはmetX遺伝子を欠損させる代わりにmetA遺伝子を導入したL−メチオニン前駆体産生菌株を提供する。
【0031】
本発明において、前記遺伝子の欠損は遺伝体上で前記遺伝子の部位を切除したり特定遺伝子の配列を挿入して蛋白質の配列を変形させることによって行われ、遺伝子の弱化は前記遺伝子のプロモータ部位または5’−UTR部位の塩基配列を変化させることによって蛋白質の発現を弱化させたりまたは当該遺伝子のORF部位に変異を導入することによって蛋白質の活性を弱化させることができる。
【0032】
本発明において、前記遺伝子の発現強化は前記遺伝子のプロモータ部位または5’−UTR部位の塩基配列を変化させることによって蛋白質の発現を強化させることができ、当該遺伝子のORF部位に変異を導入することによって蛋白質の活性を強化させることができ、当該遺伝子を染色体上に追加導入することによって発現を強化させることができ、当該遺伝子をベクター上に自己プロモータまたは強化された別個のプロモータと共に導入して菌株に形質転換することによって蛋白質の発現量を強化させることができる。
【0033】
本発明の具体的実施様態において、前記L−メチオニン前駆体産生菌株を作製する方法は次のとおりである。
【0034】
第1工程として、L−メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンを蓄積できるようにするため、菌株からシスタチオニンγ合成酵素またはO−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼなどの蛋白質をコードする遺伝子を欠損または弱化させる。
【0035】
シスタチオニンγ合成酵素をコードする遺伝子をmetB、O−スクシニルホモセリンサルフヒドリラーゼをコードする遺伝子をmetZ、O−アセチルホモセリンサルフヒドリラーゼをコードする遺伝子をmetYと表記する。上述のような活性を有する蛋白質をコードする遺伝子は例えば、大腸菌(E.coli)から公知のmetBがある。これは文献(Blattner et. Al., Science 277: 1453-1462 (1997))によって既に報告されている大腸菌のゲノム配列から得ることができる(Accession no. AAC75876)。また、前記遺伝子配列はNCBI(米国国立バイオテクノロジー情報センター)およびDDBJ(日本DNAデータバンク)のようなデータベースからも得られる。この他にも同じ活性を有する遺伝子としてコリネバクテリウムのmetBおよびmetY、シュードモナス由来のmetZなどが報告されている。
【0036】
シスタチオニンγ合成酵素、O−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼは下記の反応式に示されるようにO−スクシニルホモセリンまたはO−アセチルホモセリンをシスタチオニンまたはホモシステインに合成する活性を有している。従って、このような活性を有する遺伝子を欠損または弱化させる場合、O−スクシニルホモセリンまたはO−アセチルホモセリンが培養液中に過剰に蓄積される。
【0037】
L−システイン+O−スクシニル−L−ホモセリン <=> コハク酸+シスタチオニン
L−システイン+O−アセチル−L−ホモセリン <=> 酢酸+シスタチオニン
HS−+O−スクシニル−L−ホモセリン <=> コハク酸+ホモシステイン
HS−+O−アセチル−L−ホモセリン <=> 酢酸+ホモシステイン
【0038】
第2工程として、第1工程から作製された菌株でホモセリンキナーゼをコードするthrB遺伝子を弱化または欠損させる。thrB遺伝子はホモセリンからO−ホスホホモセリン(O-phosphohomoserine)を合成した後、thrC遺伝子によってトレオニンに転換される。合成されたホモセリンが全量メチオニンの前駆体合成に用いられるようにthrB遺伝子を欠損または弱化させる。
【0039】
第3工程として、metA遺伝子の転写を調節する転写調節因子であるmetJの遺伝子を欠損または弱化させる。メチオニン前駆体の合成に関与するmetA遺伝子はメチオニンによってフィードバック調節を受けるものであり、metJ遺伝子はmetA遺伝子の転写に関与するリプレッサーである。恒常的にmetA遺伝子を過剰発現することおよびメチオニン前駆体の合成を活性化するためには、metA遺伝子の転写リプレッサーの除去が有益である。従って、E.coliからmetJ遺伝子が除去されると、metA遺伝子の発現は増加し、L−メチオニン前駆体を大量合成することができる。
【0040】
上記第2、第3工程は前駆体産生菌株によって改変可能であり、前駆体産生菌株を作製するために必須ではない。より好ましくは、エシェリキア属の微生物で前駆体の産生経路を強化するために実施することができる。
【0041】
第4工程として、メチオニン前駆体の合成を増加させるために、メチオニン生合成の経路の初期段階を媒介する酵素のホモセリンO−スクシニルトランスフェラーゼまたはホモセリンO−アセチルトランスフェラーゼをコードする遺伝子であるmetA遺伝子またはmetX遺伝子の発現を強化させる。metA遺伝子はホモセリンO−スクシニルトランスフェラーゼをコードする遺伝子の通称であり、metX遺伝子はホモセリンO−アセチルトランスフェラーゼをコードする遺伝子の通称である。前記metAまたはmetX遺伝子の発現を強化するために遺伝子の追加導入または5’−UTRまたはプロモータを変化させたり各遺伝子ORFに変異を導入する。この遺伝子の発現を強化すると、L−メチオニン前駆体の合成を大きく増加することができる。
【0042】
菌株の生育にメチオニンが必要な場合、フィードバック制御が解除されたmetAまたはmetX遺伝子を導入することができる。この場合、培地中に添加されたメチオニンの濃度に関わらずL−メチオニン前駆体を合成することができ、培地中にメチオニンを添加することによってL−メチオニン前駆体を合成するでき、細胞の成長を促す。
【0043】
また、O−スクシニルホモセリンを産生する菌株からO−アセチルホモセリンの産生を増加させるために、染色体内に存在するホモセリンO−スクシニルトランスフェラーゼをコードする遺伝子のmetA遺伝子を欠損させることができる。metA遺伝子を欠損させてO−スクシニルホモセリンの産生を止め、さらにmetX遺伝子を導入してO−アセチルホモセリンを産生する場合、metA遺伝子が存在時にmetX遺伝子を導入する場合に比べて高い収率のO−アセチルホモセリンを産生することができる。
【0044】
なお、O−アセチルホモセリンを産生する菌株からO−スクシニルホモセリンの産生を増加させるために、染色体内に存在するホモセリンO−アセチルトランスフェラーゼをコードする遺伝子のmetX遺伝子を欠損させることができる。metX遺伝子を欠損させてO−アセチルホモセリンの産生を止め、さらにmetA遺伝子を導入してO−スクシニルホモセリンを産生する場合、最も高い収率のO−スクシニルホモセリンを産生することができる。
【0045】
上記1〜4工程のうち、一部または全体を導入した菌株を作製してL−メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンを蓄積することができる。
【0046】
L−メチオニン前駆体産生菌株はL−リシン、L−トレオニンまたはL−イソロイシン産生菌株を用いて作製することもできる。好ましくは、L−トレオニン産生菌株を用いて作製することができる。この場合には既にホモセリンまでの合成が円滑になされる菌株であって、最も多い量のメチオニン前駆体を合成することができる。従って、L−トレオニン産生菌株を用いてトレオニンの合成経路に関与する遺伝子を欠損または弱化させ、metBまたはmetY、またはmetZ遺伝子を欠損または弱化させてメチオニン前駆体を蓄積することができる。より好ましくは、thrB遺伝子を欠損または弱化させた後、metBまたはmetYまたはmetZ遺伝子を欠損または弱化させてメチオニン前駆体を合成することができる。また、metAまたはmetX遺伝子の発現を強化して最も多量のメチオニン前駆体を合成することができる。
【0047】
本発明において、「L−トレオニン産生菌株」とはL−トレオニンを生体内で産生することができる原核または真核の微生物菌株をいう。例えば、L−トレオニンを産生するエシェリキア属、エルウィニア属、セラチア属、プロビデンシア属、コリネバクテリウム属およびブレビバクテリウム属に属する微生物菌株が含まれる。好ましくは、エシェリキア属に属する微生物菌株、より好ましくは大腸菌が使用される。
【0048】
また、L−トレオニン産生菌株は天然微生物菌株のみならず、変異微生物も含む。このような変異微生物は例えば、イソロイシン漏出形要求性、L−リシン類似体に対する耐性およびα−アミノ酪酸の耐性を有する微生物;内在的なホスホエノールピルベートカルボキシラーゼ(ppc)遺伝子がゲノム含有された遺伝子の他に追加で1コピー以上の染色体DNA中に挿入された変異微生物;L−メチオニン生合成の中間体であるオキサロ酢酸(oxaloacetate, OAA)をホスホエノールピルビン酸(PEP)に転換するに関与するpckA遺伝子が不活性化された微生物;L−メチオニンの生合成に関与するtyrB遺伝子の発現を抑制するtyrR遺伝子が不活性化された微生物;および糖伝達に関連されたgalP遺伝子の発現を抑制するgalR遺伝子が不活性化された微生物が含まれる。前記L−リシン類似体はS−(2−アミノエチル)−L−システインおよびδ−メチル−L−リシンからなる群から選択される一つ以上の化合物であってよい。
【0049】
本発明の好ましい実施態様ではL−トレオニンを産生するE.coli変異菌株TF4076(KFCC10718、韓国特許公告第92-8365号)を変異させたL−トレオニン産生およびL−メチオニン−非要求性CJM002を使用した。TF4076はメチオニン要求性、メチオニン類似体(例、α-アミノ-β-ヒドロキシ吉草酸、AHV)に対する耐性、リシン類似体(例、S-(2-アミノエチル)-L-システイン、AEC)に対する耐性、イソロイシン類似体(例、α-アミノ酪酸)に対する耐性、メチオニンの類似体(例、エチオニン)に対する耐性などの特性を有している。上記韓国特許の全体内容は請求項によって本明細書に含まれる。このようなTF4076はメチオニン要求性菌株でインビボにおいてメチオニンを合成できない。本発明者らはこのようなメチオニン要求性を解除してメチオニン産生菌株として用いるためにNTGを用いた人工的な突然変異法を使用してメチオニン要求性が解除されたトレオニン産生菌株のE.coli CJM002を作製した。E.coli CJM002は大腸菌 MF001としてKCCM(Korean Culture Center of Microorganism、大韓民国ソウル西大門区弘済1洞ユリムビル、361-221)に2004年4月9日付で寄託した(寄託番号:KCCM−10568)。また、前記の方法を通じて作製したO−スクシニルホモセリン産生菌株の大腸菌CJM−BTJ(pMetA-CL)菌株は2006年7月21日に寄託し(寄託番号:KCCM−10767)、大腸菌CJM−BTJ(pCJ-MetA-CL)菌株は2007年7月5日に寄託した(寄託番号:KCCM−10872)。また、上記方法によって作製したO−アセチルホモセリン産生菌株の大腸菌CJM−BTJA(pCJ-MetX-CL)菌株は2007年7月5日に寄託した(寄託番号:KCCM−10873)。
【0050】
上述のとおり作製されたL−メチオニン前駆体産生菌株の培養過程は当業者に知られた適当な培地と培養条件に応じてなされる。このような培養過程は当業者であれば選ばれる菌株によって容易に調整して使用することができる。前記培養方法の例にはバッチ式、連続式および流加式培養が含まれるが、これに限定されるものではない。このような多様な培養方法は、例えば文献("Biochemical Engineering" by James M. Lee, Prentice-HallInternational Editions, pp138-176)に開示されている。
【0051】
培養に使用される培地は特別な菌株の要求条件を適切に満たさなければならない。多様な微生物の培地は例えば文献("Manualof Methods for General Bacteriology" by the American Society forBacteriology, Washington D.C., USA, 1981)に開示されている。前記培地は多様な炭素源、窒素源および微量元素成分を含む。使用できる炭素源の例にはブドウ糖、ショ糖、乳糖、果糖(fructose)、マルトース、でんぶん、セルロースのような炭水化物、大豆油、ヒマワリ油、ヒマシ油、ココナッツ油のような脂肪、パルミチン酸、ステアリン酸、リノール酸のような脂肪酸、グリセロールおよびエタノールのようなアルコール、アセト酸のような有機酸が含まれる。これらの化合物のうちの一つまたはその混合物は、炭素源として使用され得る。使用できる窒素源の例には、ペプトン、酵母抽出物、肉汁、麦芽抽出物、トウモロコシ浸漬液(CSL)および大豆粉のような有機窒素源および尿素、硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム、炭酸アンモニウムおよび硝酸アンモニウムのような有機窒素源が含まれる。これらの化合物のうちの一つまたはその混合物は、窒素源として使用され得る。ここで、培地は付加的にリン源としてリン酸二水素カリウム、リン酸水素二カリウムおよび対応するナトリウム−含有塩を含むことができる。また、硫酸マグネシウムや硫酸鉄のような金属塩も含むことができる。さらに、アミノ酸、ビタミンおよび適切な前駆体なども加えられ得る。これらの培地または前駆体はバッチ式または連続式で培養物に添加され得る。
【0052】
また、培養中に水酸化アンモニウム、水酸化カリウム、アンモニア、リン酸および硫酸のような化合物を培養物に適切な方式で添加して培養物のpHを調整することができる。また、培養中に脂肪酸ポリグリコールエステルのような消泡剤を使用して気泡の生成を抑制することができる。また、培養物の好気状態を保持するために、培養物内に酸素または酸素含有気体(例、空気)を注入する。培養物の温度は通常20℃〜45℃であり、好ましくは25℃〜40℃である。培養期間は望むL−メチオニン前駆体の生成量が得られるまで続けられ、好ましくは10〜160時間である。
【0053】
第2工程は、前記L−メチオニン前駆体産生菌株によって産生されたO−スクシニルホモセリンまたはO−アセチルホモセリンとメチルメルカプタンに基質を用いてシスタチオニンシンターゼ、O−スクシニルホモセリンサルフヒドリラーゼもしくはO−アセチルホモセリンサルフヒドリラーゼ活性を有する酵素または前記酵素を含む菌株を用いた酵素反応を通じてL−メチオニンおよび有機酸を産生する工程を含む。
【0054】
より詳細には、本発明は上記方法で蓄積されたホモセリン、O−ホスホホモセリン、O−スクシニルホモセリン、またはO−アセチルホモセリンを基質として用いてシスタチオニンγ合成酵素、O−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼなどの酵素反応をによりL−メチオニンを産生する方法を提供する。好ましくは、本発明はO−スクシニルホモセリンまたはO−アセチルホモセリンを基質で用いる。
【0055】
本発明において、前記シスタチオニンγ合成酵素またはO−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼはエシェリキア属(Escherichia sp.)、シュードモナス属(Pseudomonas sp.)、レプトスピラ属(Leptospira sp.)、コリネバクテリウム属(Corynebacteriumsp.)、サッカロミセス属(Saccharomyces sp.)、クロモバクテリウム属(Chromobacterium sp.)、ノルカジア属(Norcardia sp.)、ブラジリゾビウム属(Bradyrhizobium sp.)、ヒホモナス属、メチロコックス属(Methylococcussp.)、メチロバシルス属(Methylobacillus sp.)、ニトロソモナス属(Nitrosomonas sp.)、クレシエラ属(Klesiella sp.)、バシルス属(Bacillus sp.)、シゲラ属(Shigella sp.)、コルウェリア属(Colwellia sp.)、サルモネラ属(Salmonella sp.)、酵母(yeast)、菌類(fungi)に属する微生物菌株に由来するものであり得る。
【0056】
第2工程において、O−スクシニルホモセリンをL−メチオニン前駆体で使用する場合、好ましくはシュードモナス属、ノルカジア属、クロモバクテリウム属に属する微生物菌株で、より好ましくは、シュードモナスアウロゲノサ(Pseudomonas aurogenosa)、ノルカジアファルシニカ(NorcardiaFarcinica)、シュードモナスプチダ(Pseudomonas putida)、クロモバクテリウムビオラシウム(Chromobacterium Violaceum)に属する微生物菌株に由来するシスタチオニンγ合成酵素またはO−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼが使用される。
【0057】
第2工程において、O−アセチルホモセリンをL−メチオニン前駆体として使用する場合、好ましくはレプトスピラ属、クロモバクテリウム属、ヒポモナス属に属する微生物菌株であって、より好ましくは、レプトスピラアウロゲノサ(Leptospira aurogenosa)、ヒポモナスネプチュニウム(HyphomonasNeptunium)、クロモバクテリウムビオラシウム(Chromobacterium Violaceum)に属する微生物菌株に由来するシスタチオニンγ合成酵素またはO−スクシニルホモセリンサルフヒドリラーゼまたはO−アセチルホモセリンサルフヒドリラーゼが使用される。
【0058】
前記の反応は下記示すとおりであり、その構造式は図2に示される。
CH3SH+O−スクシニル−L−ホモセリン <=> コハク酸+メチオニン
CH3SH+O−アセチル−L−ホモセリン <=> 酢酸+メチオニン
【0059】
上記反応では図2に表記された化学式のようにメチルメルカプタンのCH3S-残基がO−スクシニルホモセリンまたはO−アセチルホモセリンのコハク酸または酢酸の残基と置換されてメチオニンを生成するようになる。反応時にメチルメルカプタン(CH3SH)は様々な形態で添加が可能である。
【0060】
前記活性を有する酵素をコードする遺伝子の配列はNCBI(米国国立バイオテクノロジー情報センター)および日本のDNAデータバンク(KEGG)のようなデータベースからも得られる。
【0061】
生物学的転換反応のために得られた遺伝子配列から遺伝子をクローニングして発現ベクターに挿入した後、組換え菌株由来の酵素を活性型で発現させる。酵素発現菌株および発現された酵素の両方を前記反応に直接用いることができる。
【0062】
上記遺伝子を発現させた酵素および酵素発現菌株はL−メチオニン前駆体を蓄積した発酵上澄液または発酵液と直接混合または部分的な混合反応を用いて反応を導く。本発明の好ましい実施態様において、発酵液に蓄積されたO−スクシニルホモセリンまたはO−アセチルホモセリンはシュードモナス属、クロモバクテリウム属、レプトスピラ属、ヒポモナス属由来のシスタチオニンγ合成酵素またはO−アセチルホモセリンサルフヒドリラーゼまたはO−スクシニルホモセリンサルフヒドリラーゼによってメチオニンに転換することができる。
【0063】
より好ましくは、発酵液に蓄積されたO−スクシニルホモセリンはシュードモナスアウロゲノサまたはシュードモナスプチダまたはクロモバクテリウムビオラシウム由来のシスタチオニンγ合成酵素およびO−アセチルホモセリンサルフヒドリラーゼおよびO−スクシニルホモセリンサルフヒドリラーゼによってメチオニンに転換することができる。
【0064】
または発酵液に蓄積されたO−アセチルホモセリンはレプトスピラメイエリ(Leptospira meyeri)、ヒポモナスネプチュニウム(Hyphomonas Neptunium)、クロモバクテリウムビオラシウム(ChromobacteriumViolaceum)由来のシスタチオニンγ合成酵素およびO−アセチルホモセリンサルフヒドリラーゼおよびO−スクシニルホモセリンサルフヒドリラーゼによってメチオニンに転換することができる。
【0065】
各遺伝子を大腸菌発現ベクターであるpCL−CJ1ベクター(韓国CJ社)で発現させ、超音波破砕を行い、細胞溶解液から酵素液を調製し、発現蛋白を得た。得られた酵素液をO−スクシニルホモセリンおよびO−アセチルホモセリンが蓄積された発酵液に添加し、これと共にメチルメルカプタン溶液を共に添加して反応を誘導した。反応はDTNB(5,5-dithiobis(2-nitro-benzoic acid, 米国sigma社)を用いて確認し、反応産物はHPLCを用いて分析した。
【0066】
本発明はO−スクシニルホモセリン、O−アセチルホモセリンとCH3SHとの反応を通じてL−メチオニンの他にもO−スクシニルホモセリンの場合はコハク酸を、O−アセチルホモセリンの場合は酢酸を別途の産生工程無しに副産物として得ることができる。
【図面の簡単な説明】
【0067】
【図1】メチオニン前駆体産生菌株の遺伝子操作図である。
【図2】第2工程でメチオニンを産生するための化学構造図である。
【図3】metA遺伝子の発現のためのベクターpMetA-CLの模式図である。
【図4】metB遺伝子の発現のためのベクターpCJ-MetB-CLの模式図である。
【図5】多様な酵素のO−スクシニルホモセリン消費に対する反応曲線である。各番号に使用された酵素液の由来は次のとおりである。酵素液21番は特定遺伝子を含まない細胞抽出液を用いた。
【表1】
【図6】多様な酵素のO−アセチルホモセリン消費に対する反応曲線である。各番号は図5と同じである。
【図7】転換反応に使用された各酵素のアミノ酸配列をDNAstarのmegalign方法を用いて整列した図である。
【図8】転換反応に使用された各酵素のアミノ酸配列をDNAstarのmegalign方法を用いて整列した図である。
【発明を実施するための形態】
【0068】
以下、本発明の現時点での現実的で好ましい実施態様を以下の実施例に示すとおり説明する。しかし、下記実施例は本発明を例示して説明するためのものであって、本発明の権利範囲が下記実施例によって限定されるものではない。
【実施例1】
【0069】
実施例1:メチオニン前駆体産生菌株の作製
【0070】
<1−1> metB遺伝子の欠損
大腸菌株においてシスタチオニンシンターゼをコードする遺伝子であるMetB遺伝子を欠損させるために、FRT-one-step PCR deletion方法を使用した(PNAS (2000)vol97:P6640-6645)。配列番号1、2番のプライマーを用いてpKD3ベクター(PNAS (2000) vol97:P6640-6645)を鋳型にしてPCR反応で欠損カセットを作製した。変性(denaturation)工程は94℃で30秒、アニーリング(annealing)工程は55℃で30秒、伸長(extension)工程は72℃で1分間施し、これを30サイクルでPCR行った。
【0071】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.2kbpサイズのバンドからDNAを精製した。回収されたDNA断片はpKD46ベクター(PNAS (2000) vol97:P6640-6645)を予め形質転換させたE.coli(K12)W3110菌株にエレクトロポレーションを行った。エレクトロポレーションの前に、pKD46で形質転換を行ったW3110菌株は100μg/Lアンピシリンと5mMの1−アラビノースが含まれたLB培地を用いて30℃でOD600が0.6になるまで培養した。その後に、培養した菌株を滅菌蒸留水で2回、10%グリセロールで1回洗浄した。エレクトロポレーションは2500Vで行った。回収した菌株は25μg/Lクロラムフェニコールを含むLBプレート培地に塗布して37℃で一夜培養した後、耐性を示す菌株を選別した。
【0072】
上記と同様の条件で同じプライマーを鋳型として選別した菌株に対してPCRを行った。1.0%アガロースゲル上で遺伝子のサイズが1.2Kbに観察されるものを確認することによってmetB遺伝子の欠損を同定した。次に、確認された菌株はpCP20ベクター(PNAS (2000) vol97:P6640-6645)で形質転換を行い、LB培地で培養した。最終的に、同じ条件でPCRを行い、1.0%アガロースゲル上で遺伝子のサイズが150bpまで小さくなったmetB遺伝子の欠損菌株を作製し、クロラムフェニコールマーカーが除去されたことを確認した。作製された菌株をW3−Bと命名した。
【0073】
<1−2> thrB遺伝子の欠損
ホモセリンキナーゼをコードするthrB遺伝子を欠損させることによって、ホモセリンからO−スクシニルホモセリンの合成量を増加させようとした。特にトレオニン産生菌株を用いる場合、ホモセリンを利用する活性が強力なため、この遺伝子の欠損は必ず必要である。上記のように作製されたW3−B菌株でthrB遺伝子を欠損させるために、上記metB遺伝子の欠損と同様にFRT-one-step PCR deletionを行った。
【0074】
thrB欠損カセットを作製するために、pKD4ベクター(PNAS (2000) vol97:P6640-6645)を鋳型にして、配列番号3および4のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で1分間施し、これを30サイクル行うPCRを行った。その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.6kbpサイズのバンドからDNAを精製した。回収したDNA断片をpKD46ベクターで予め形質転換させたW3−B菌株にエレクトロポレーションによって導入した。回収された菌株を50μg/Lカナマイシンを含むLBプレート培地に塗布して37℃で一夜培養した後、耐性を示す菌株を選別した。
【0075】
上記と同一の条件で、選別された菌株を鋳型にして配列番号3、4のプライマーを用いてPCRを行った後、1.0%アガロースゲル上で遺伝子のサイズが1.6Kbに確認される菌株を選別することによってthrB遺伝子の欠損を確認した。次に、確認された菌株をpCP20ベクターで形質転換してLB培地で培養した。最終的に、同一条件下のPCRを行い1.0%アガロースゲル上で遺伝子のサイズが150bpに小さくなったthrB遺伝子の欠損菌株を作製し、カナマイシンマーカーが除去されたことを確認した。作製された菌株はW3−BT菌株と命名した。
【0076】
<1−3> metJ遺伝子の欠損
メチオニン前駆体の合成に関与するmetA遺伝子の調節遺伝子であるmetJを欠損させるために、metB遺伝子の欠損と同様にFRT-one-step PCR deletion法を行った。
【0077】
metJ遺伝子欠損カセットを作製するために配列番号5、6のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で1分間施し、これを30サイクルでPCRを行った。
【0078】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.2kbpサイズのバンドからDNAを精製した。回収されたDNA断片をpKD46ベクターで予め形質転換させたW3−BT菌株にエレクトロポレーションによって導入した。回収された菌株はクロラムフェニコールを含むLBプレート培地に塗布して37℃で一夜培養した後、耐性を示す菌株を選別した。
【0079】
選別された菌株を鋳型にして配列番号7および8のプライマーを用いて同じ条件でPCRした後、1.0%アガロースゲル上で遺伝子のサイズが1.6Kbに変化することを確認することによってmetJ遺伝子の欠損を確認した。確認された菌株は再びpCP20ベクターで形質転換してLB培地で培養し、再び同一な条件のPCRを通じて1.0%アガロースゲル上で遺伝子のサイズが600bpに小さくなった最終metJ遺伝子欠損菌株を作製し、クロラムフェニコールマーカーが除去されたことを確認した。作製された菌株をW3−BTJと命名した。
【0080】
<1−4−1> metA遺伝子の過剰発現
より多いメチオニン前駆体の合成のために、メチオニン前駆体のO−スクシニルホモセリン合成に関与するホモセリンO−スクシニルトランスフェラーゼ酵素をコードするmetA遺伝子を過剰発現を行った。
【0081】
これのためにE.coli W3110の染色体を鋳型にして、配列番号9および10番のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを25サイクル行うPCR反応を施した。
【0082】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.2kbpサイズのバンドからDNAを精製した。回収されたDNA断片はpCL1920ベクターをSmaIで切断して得られたDNA断片と連結させた。連結されたベクターをE.coliに形質転換させて50μg/Lスペクチノマイシンが含まれたLB培地で培養した後に選別した。このように作製されたベクターをpMetA-CLと命名した。pMetA-CLベクターの模式図は図3のとおりである。上記ベクターをW3-BTJ菌株に形質転換させて作製された菌株をW3-BTJ/pMetA-CLと命名し、O−スクシニルホモセリンの増加を観察した。
【0083】
metA遺伝子の発現をさらに増加させるための他の方法として、前記のmetA遺伝子をpCL1920ベクターにCJ1プロモータ(韓国、CJ社)とEcoRV制限酵素を用いて連結させた。連結されたベクターをE.coli菌株に形質転換させ50μg/Lスペクチノマイシンが含まれたLB培地において培養した後に選別した。このように作製されたベクターをpCJ-MetA-CLと命名した。前記ベクターをW3-BTJ菌株に形質転換させて作製された菌株をW3-BTJ/pCJ-MetA-CLと命名し、O−スクシニルホモセリンの増加を観察した。
【0084】
<1−4−2> metX遺伝子の過剰発現
O−アセチルホモセリンの合成のために、メチオニン前駆体のO−アセチルホモセリン合成遺伝子であるホモセリンO−アセチルトランスフェラーゼをコードするmetX遺伝子の過剰発現を行った。
【0085】
これのためにレプトスピラメイエリの染色体を鋳型にして配列番号11および12番のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを25サイクル行うPCR反応を施した。
【0086】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.1kbpサイズのバンドからDNAを精製した。回収されたDNA断片をpCL1920ベクターにCJ1プロモータとEcoRV制限酵素を用いて連結させた。連結されたベクターをE.coliに形質転換させて50μg/Lスペクチノマイシンが含まれたLB培地で培養した後に選別した。このように作製されたベクターをpCJ1-MetXlme-CLと命名した。前記ベクターをW3-BTJ菌株に形質転換させて作製された菌株をW3-BTJ/pCJ-MetXlme-CLと命名し、O−アセチルホモセリンの増加を観察した。
【0087】
metX遺伝子の過剰発現のための他の方法として、コリネバクテリウム染色体を鋳型にして配列番号68、69番のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを25サイクル行うPCR反応を施した。
【0088】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、DNAを精製した。回収されたDNA断片をpCL1920ベクターにCJ1プロモータとEcoRV制限酵素を用いて連結させた。連結されたベクターをE.coliに形質転換させて50μg/Lスペクチノマイシンが含まれたLB培地で培養した後に選別した。このように作製されたベクターをpCJ-MetXcgl-CLと命名した。前記ベクターをW3-BTJ菌株に形質転換させて作製された菌株をW3-BTJ/pCJ-MetXcgl-CLと命名し、O−アセチルホモセリンの増加を観察した。
【0089】
<1−4−3> metA遺伝子の欠損
O−アセチルホモセリンの産生量を増加させるために、W3-BTJ菌株でホモセリンO−スクシニルトランスフェラーゼをコードする遺伝子であるmetAを欠損させた。metA遺伝子のみを導入した場合、一定量のO−スクシニルホモセリンが蓄積されることが観察されたから、metA遺伝子を欠損させる場合最も多い量のO−アセチルホモセリンを蓄積することができると推定した(表4参照)。metA遺伝子を欠損させるためにFRT one step PCR deletion方法を使用した。
【0090】
metA遺伝子欠損カセットを作製するために配列番号70および71のプライマーを用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で1分間施し、これを30サイクルでPCR行った。
【0091】
その結果得られたPCR産物を1.0%アガロースゲルで電気泳動した後、1.2kbpサイズのバンドからDNAを精製した。回収されたDNA断片をpKD46ベクターで予め形質転換させたE.coli W3-BTJ菌株にエレクトロポレーションによって導入した。回収された菌株をクロラムフェニコールを含むLBプレート培地に塗布して37℃で一夜培養した後、耐性を示す菌株を選別した。
【0092】
選別された菌株を鋳型にして配列番号70および71のプライマーを用いて同じ条件でPCRした後、1.0%アガロースゲル上で遺伝子のサイズが1.1Kbに変化されることを確認することによってmetA遺伝子の欠損を確認した。確認された菌株はpCP20ベクターで形質転換させてLB培地で培養し、同一な条件のPCRを通じて1.0%アガロースゲル上で遺伝子のサイズが100bpに小さくなった最終metA遺伝子欠損菌株を作製し、クロラムフェニコールマーカーが除去されたことを確認した。作製された菌株をW3−BTJAと命名した。W3−BTJA菌株を前記pCJ-MetXlme-CLベクターで形質転換させて作製された菌株をW3-BTJA/pCJ-MetX-CLと命名した。前記菌株を同一な方法で培養した結果、O−スクシニルホモセリンの蓄積は観察されず、O−アセチルホモセリンの産生量がW3-BTJに比べて約20%有意に増加することを確認した。
【0093】
<1−5> L−トレオニン産生菌株の転換
メチオニン要求性が解除されたL−トレオニン産生菌株の大腸菌CJM002(KCCM-10568号)を用いて前記1−1〜1−3と同一な方法でメチオニン前駆体産生菌株を作製した。作製された菌株はそれぞれCJM-BTJ、CJM-BTJ/pMetA-CLおよびCJM-BTJ/pCJ-MetA-CLと命名した。また、このCJM-BTJ菌株を用いて前記<1−4−3>の方法でmetA遺伝子を欠損させた菌株を作製し、作製された菌株をCJM-BTJAと命名した。
【実施例2】
【0094】
実施例2:L−メチオニン前駆体の産生のための発酵
【0095】
<2−1> フラスコ培養実験
実施例1から作製された菌株のメチオニン前駆体の産生量を実験するために三角フラスコ培養を施した。抗生剤スペクチノマイシンが含有されたLBプレート培地にW3-BTJとCJM-BTJおよびmetA、metX発現ベクターで形質転換させたW3-BTJ菌株を播種して31℃で一夜培養した後、単一コロニーをスペクチノマイシンが含まれた3mlのLB培地に播種した後、31℃で5時間培養し、再び25mlメチオニン前駆体の産生培地を含む250ml三角フラスコに200倍希釈して31℃200rpmで64時間培養してHPLC分析を通じてメチオニン前駆体の産生量を比較した(表3および表4)。その結果、メチオニン要求性が解除されたトレオニン産生菌株を使用して作製されたメチオニン前駆体産生菌株の場合、メチオニン前駆体の産生量が著しく増加したことがわかった。
【0096】
【表2】
【0097】
【表3】
【0098】
【表4】
【0099】
<2−2> 大型発酵槽実験
実施例1で最も優れたメチオニン前駆体の産生能を見せた菌株を用いてメチオニン前駆体を大量生産するために、5L発酵槽培養を施した。抗生剤スペクチノマイシンが含有されたLBプレート培地にCJM-BTJ/pCJ-metA-CL菌株またはCJM-BTJA/pCJ-metXlme-CL菌株を播種して31℃で一夜培養した。その後、単一コロニーをスペクチノマイシンが含まれた10mlLB培地に播種した後、31℃で5時間培養し、再び200mlメチオニン前駆体のシード培地を含む1000ml三角フラスコに100倍希釈して31℃、200rpmで3〜10時間培養した後、5L発酵槽に播種して流加式培養(Fed batch)発酵法で50〜100時間培養した。このように培養した発酵液においてメチオニン前駆体の濃度をHPLCで分析した結果は表6の通りである。
【0100】
【表5】
【0101】
【表6】
【実施例3】
【0102】
実施例3:メチオニン転換酵素の産生
【0103】
<3−1> E.coli由来のシスタチオニンγ合成酵素
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるE.coli由来のシスタチオニンγ合成酵素をコードしているmetB遺伝子をクローニングした。
【0104】
E.coliの染色体を鋳型にしてプライマー配列番号13および14を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを25サイクル行うPCR反応を通じて獲得した。
【0105】
獲得したDNA断片をNcol/HindIIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。最終作製されたベクターをpCL-MetB-CLと命名した。模式図は図4のとおりである。クローニングされたベクターをE.coli W3110細胞に形質転換させ、50μg/Lのスペクチノマイシンが含まれたLBプレート培地で培養した後、コロニーを選別した。選別されたコロニーを50μg/Lのスペクチノマイシンが含まれたLB培地3mlに播種して一夜の間37℃で培養した。培養された細胞は再び回収して0.1Mリン酸カリウム緩衝液(Potassium phosphate, pH7.5)で洗浄して再び200μlのリン酸カリウム緩衝液に懸濁し、30秒おきに5回超音波処理をして細胞を破砕した。破砕された細胞抽出液を12000rpmで10分間遠心分離した後、上澄液を取ってBio-Rad蛋白質定量液(米国BIO-Rad社)を用いて蛋白質の総量を定量した。また、SDS-PAGE法を用いて蛋白質の発現を確認した。回収された細胞抽出液の上澄液を酵素転換反応に使用した。
【0106】
<3−2> シュードモナス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるシュードモナス属(Pseudomonas Sp.)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。シュードモナスはシュードモナスアルギノサおよびシュードモナスプチダの2種を使用した。
【0107】
それぞれの菌株の染色体を鋳型にしてシュードモナスアルギノサの場合はプライマー配列番号15および16を、シュードモナスプチダの場合は配列番号17および18を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0108】
獲得したDNA断片をNdeI/PacIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0109】
<3−3> シュードモナス属由来のO−アセチルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるシュードモナス属由来のO−アセチルホモセリンサルフヒドリラーゼをコードするmetY遺伝子をクローニングした。
【0110】
シュードモナスアルギノサ菌株の染色体を鋳型にし、配列番号19および20を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0111】
獲得したDNA断片をNdeI/PacIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0112】
<3−4> コリネバクテリウムグルタミクム由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるコリネバクテリウムグルタミクム(Corynebacterium glutamicum)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0113】
コリネバクテリウムグルタミクムの染色体を鋳型にしてプライマー配列番号21および22を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0114】
獲得したDNA断片をNcoI/Hind3で切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0115】
<3−5> コリネバクテリウムグルタミクム由来のO−アセチルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるコリネバクテリウムグルタミクム由来のO−アセチルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0116】
コリネバクテリウムグルタミクムの染色体を鋳型にしてプライマー配列番号23および24を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0117】
獲得したDNA断片をNdeI/Avr2で切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0118】
<3−6> レプトスピラ属由来のO−アセチルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるレプトスピラメイエリ(Leptospira meyeri)由来のO−アセチルホモセリンサルフヒドリラーゼをコードするmetY遺伝子をクローニングした。
【0119】
レプトスピラメイエリの染色体を鋳型にしてプライマー配列番号25および26を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0120】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0121】
<3−7> サッカロミセス属由来のO−アセチルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用される出芽酵母(Saccharomyces cerevisiae)由来のO−アセチルホモセリンサルフヒドリラーゼをコードするmet25遺伝子をクローニングした。
【0122】
出芽酵母の染色体を鋳型にしてプライマー配列番号27および28を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0123】
獲得したDNA断片をNdeI/PacIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0124】
<3−8> クロモバクテリウム属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるクロモバクテリウムビオラシウム(Chromobacterium Violaceum)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0125】
クロモバクテリウムビオラシウムの染色体を鋳型にしてプライマー配列番号29および30を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0126】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)のにクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0127】
<3−9> ノルカジア属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるノルカジアファルシニカ(Norcardia Farcinica)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0128】
ノルカジアファルシニカの染色体を鋳型にしてプライマー配列番号31および32を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応通じて獲得した。
【0129】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0130】
<3−10> ブラジリゾビウム属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるブラジリゾビウムジャポニクム(Bradyrhizobium Japonicum)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0131】
ブラジリゾビウムジャポニクムの染色体を鋳型にしてプライマー配列33および34を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0132】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0133】
<3−11> ヒポモナス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるヒポモナスネプチュニウム(Hyphomonas Neptunium)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0134】
ヒポモナスネプチュニウムの染色体を鋳型にしてプライマー配列番号35および36を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0135】
獲得したDNA断片をBamHII/HindIIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0136】
<3−12> メチロコックス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるメチロコックスカプスラツス(Methylococcus Capsulatus)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0137】
メチロコックスカプスラツスの染色体を鋳型にしてプライマー配列番号37および38を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0138】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0139】
<3−13> メチロバシルス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるメチロバシルスフラゲラツス(Methylobacillus Flagellatus)由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0140】
メチロバシルスフラゲラツスの染色体を鋳型にしてプライマー配列番号39および40を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0141】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0142】
<3−14> ニトロソモナス属由来のO−スクシニルホモセリンサルフヒドリラーゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるニトロソモナスヨーロパエア由来のO−スクシニルホモセリンサルフヒドリラーゼをコードするmetZ遺伝子をクローニングした。
【0143】
ニトロソモナスヨーロパエアの染色体を鋳型にしてプライマー配列番号41および42を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0144】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0145】
<3−15> クレシエラ属由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるクレシエラニュモニアエ(Klesiella Pneumoniae)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0146】
クレシエラニュモニアエの染色体を鋳型にしてプライマー配列番号43および44を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0147】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0148】
<3−16> バシルス属由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用される枯草菌(Bacillus Subtilis)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0149】
枯草菌の染色体を鋳型にしてプライマー配列番号45および46を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0150】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0151】
<3−17> シゲラ属由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるシゲラフレキシネリ2457T(Shigella Flexneri 2457T)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0152】
シゲラフレキシネリ2457Tの染色体を鋳型にしてプライマー配列番号47および48を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0153】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0154】
<3−18> コルウェリア属由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるコルウェリアサイクレリトラエア(Colwellia Psychrerythraea)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0155】
コルウェリアサイクレリトラエアの染色体を鋳型にしてプライマー配列番号49および50を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0156】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0157】
<3−19> サルモネラ由来のシスタチオニンシンターゼ
メチオニン前駆体であるO−スクシニルホモセリンまたはO−アセチルホモセリンをメチオニンに転換させる酵素として使用されるサルモネラエンタリカ血清型パラチフス(Salmonella enterica serovar Paratyphi A)由来のシスタチオニンシンターゼをコードするmetB遺伝子をクローニングした。
【0158】
サルモネラエンタリカ血清型パラチフスの染色体を鋳型にしてプライマー配列番号51および52を用いて変性工程は94℃で30秒、アニーリング工程は55℃で30秒、伸長工程は72℃で2分間施し、これを30サイクル行うPCR反応を通じて獲得した。
【0159】
獲得したDNA断片をNdeI/AvrIIで切断して、同一に切断したpCL-CJ1ベクター(韓国、CJ社)にクローニングした。クローニングされたベクターを前記<3−1>と同じ方法を使用して細胞抽出液の上澄液を回収し、酵素転換反応に使用した。
【0160】
<3−20> OSHSを基質にした転換酵素の活性比較
前記の<3−1>から<3−19>の方法で獲得したそれぞれの酵素液を活性比較して最適のメチオニン転換酵素を選別した。
【0161】
このために、O−スクシニルホモセリン(Sigma社)を3mMの濃度で0.1Mリン酸カリウム緩衝液(pH7.5)に溶解した。また、この反応液に助酵素で使用されるピリドキサールホスフェート(Pyridoxal5'-phosphate、米国Sigma社)を最終濃度10μMになるように添加した。反応液にもう一つの基質として使用されるメチルメルカプタン(Methylmercaptan、日本東京化成工業株式会社)を最終濃度2mMで添加した。この反応液1mlを37℃に位置した後、各酵素抽出液を蛋白質濃度5mg/mlに合わせて10μl添加した。反応の進行如何は毎5〜10分おきに100μlの反応液を回収して4mg/mlのDTNB(米国Sigma社)溶液900μlに添加した後、OD415における吸光度を測定することによって確認した。
【0162】
DTNBは反応液中に残存するメチルメルカプタンのSH基と反応して黄色いの物質を合成する。従って、反応液状のメチルメルカプタンが反応によってメチオニンに転換されることで反応液の黄色が消えるものであって、反応の進行が確認できる。
【0163】
実験結果、図5のようにクロモバクテリウム由来のO−スクシニルホモセリンサルフヒドリラーゼ、ノルカジア由来のO−スクシニルホモセリンサルフヒドリラーゼ、シュードモナス属由来のO−スクシニルホモセリンサルフヒドリラーゼおよびO−アセチルホモセリンサルフヒドリラーゼが非常に高い活性を見せた。他の酵素も全てある程度の活性は見せているが、反応進行速度が相対的に遅いことが確認された。各酵素の基質に対する反応性を要約した表は表7のとおりである。また、1時間の反応が終了された後に選別された反応液からHPLC分析を通じて最終生成されたメチオニンおよびコハク酸の生成を確認した結果は表8に示した。
【0164】
<3−21> OAHSを基質にした転換酵素の活性比較。
前記<3−20>と同一の反応をO−アセチルホモセリンを用いて反応させた。O−アセチルホモセリンは発酵上澄液から精製した。O−スクシニルホモセリンと同一な反応液および同一な酵素液を用いて反応させた結果、図6のようにヒポモナス由来のO−スクシニルホモセリンサルフヒドリラーゼ、シュードモナス属由来のO−アセチルホモセリンサルフヒドリラーゼ、クロモバクテリウム属由来のO−スクシニルホモセリンサルフヒドリラーゼおよびレプトスピラ属由来のO−アセチルホモセリンサルフヒドリラーゼが高い活性を見せた。他の酵素も全てある程度の活性は見せているが、反応進行速度が相対的に遅いことが確認された。各酵素の基質に対する反応性を要約して表7に示した。また、1時間の反応が終了された後に選別された反応液からHPLC分析を通じて最終生成されたメチオニンおよび酢酸の量を確認した結果は表9に示した。
【0165】
【表7】
【0166】
【表8】
【0167】
【表9】
【0168】
<3−22> 転換酵素のフィードバック阻害確認
前記3−20および21と同一の方法を使用してメチオニンの存在有無によるフィードバックの阻害反応を確認した。前記と同一な反応液を作製し、各反応液にそれぞれ5g/Lのメチオニンを添加または非添加して同一な反応進行を行った。メチオニンを添加しない場合の反応進行速度を100%にしたとき、メチオニンの存在時の残存活性を%で表記して表10に示した。
【0169】
その結果、シュードモナス由来のO−アセチルホモセリンサルフヒドリラーゼ、ノルカジア由来のO−スクシニルホモセリンサルフヒドリラーゼおよびレプトスピラ由来のO−アセチルホモセリンサルフヒドリラーゼがメチオニンによる活性阻害を見せたことを確認した。従って、これらの酵素はメチオニンによるフィードバック阻害を有しているものと推定され、後ほど反応にはフィードバック阻害がない酵素を使用した。前記の実施例においてフィードバック阻害を受ける酵素もフィードバック阻害が解除される変異株を使用する場合、同一な反応に使用できると推定された。
【0170】
【表10】
【0171】
<3−23> 転換酵素の相同性比較
転換反応に使用された各酵素間の相同性を比較してO−スクシニルホモセリンおよびO−アセチルホモセリンに対する反応性およびフィードバック調節阻害との相関関係を確認した。
【0172】
使用された酵素の相同性を比較した結果、O−スクシニルホモセリンサルフヒドリラーゼで命名されるmetZ間の相同性およびO−アセチルホモセリンサルフヒドリラーゼで命名されるmetY間の相同性が相互間の相同性に比べて相対的に高いことがわかった。前記の実施例と連関してみてO−スクシニルホモセリンサルフヒドリラーゼで命名されたmetZの場合にはフィードバック調節阻害を見せていない場合が最も多いのに比べて、O−アセチルホモセリンサルフヒドリラーゼで命名されたmetYの場合には実施例に使用された酵素の全てがフィードバック調節阻害を見せて相対的に高いフィードバック調節阻害を有していることと確認された。O−スクシニルホモセリンおよびO−アセチルホモセリンに対する選択性について、metZ遺伝子群はO−スクシニルホモセリンに対する選択性が高く、metY遺伝子群はO−アセチルホモセリンに対する選択性は高かったが、シュードモナスプチダ由来のmetYおよびクロモバクテリウムビオラシウム由来のmetZは2つの両基質に対して特異的に高い反応性を見せた。
【0173】
実施例に使用された全ての酵素をアミノ酸配列に基づいて配列整列プログラムのクラスタルWプログラム(ClustalW program; DNAstar)で整列(align)した結果、次のような配列で明記されるドメインを共通的に有していることが確認された。
【0174】
ドメイン1:Y-(S, I, T, V)-R-X-X-(N,S)
ドメイン2:
(V,A,I)-(V,L,I)-D-N-X-(F,V,M,I)-X-(T,S)-(P,A)-X-(L,I)-(Q,C,V)-X-(P,G)-(L,F)
-X-(L,M,H)-G-(A,V)-(D,H)
ドメイン3:
(S,A,G,P)-(P,A,V)-F-(N,D)-(A,S)-(W,F,Y)-X-X-X-(K,Q,R,S)-G-(L,M,V,I,M)-(
E,K,D,R)-T-(L,M)
ドメイン5:(H,Y)-(P,A)-(A,S)-(T,S)-(T,M,Q)-(T,S)-H-
ドメイン6:(V,I,L)-R-(V,I,L,F)-(S,A)-(V,I,T)-G-(L,I)-E-
【実施例4】
【0175】
実施例4:メチオニン転換酵素によるメチオニン転換反応
【0176】
<4−1> 転換酵素の大量産生
実施例2(2−2および2−8)から作製されたメチオニン転換酵素の産生菌株を大量産生するために1L発酵槽培養を施した。抗生剤スペクチノマイシンが含有されたLBプレート培地にW3110をシュードモナス由来metZ発現ベクターで形質転換またはヒポモナス由来metZ発現ベクターで形質転換させた菌株をそれぞれ播種して30〜40℃で一夜培養した後、単一コロニーをスペクチノマイシンが含まれた40mlLB培地に播種した後、30〜40℃で5時間培養した。このように培養されたシュードモナス由来metZ発現菌株およびヒポモナス由来metZ発現菌株を1L発酵槽で600〜900rpm、30〜40℃、15〜30時間培養した。培養に使用された培地組成は表11に示した。
【0177】
培養が終わったメチオニン転換酵素発酵液を超音波電動を用いて細胞を破砕してメチオニン転換酵素液を作製した。
【0178】
【表11】
【0179】
<4−2> メチオニン転換反応
前記の実施例2(2−2)から産生されたO−スクシニルホモセリン発酵液とO−アセチルホモセリン発酵液とにそれぞれ実施例4(4−1)から作製されたシュードモナス由来O−スクシニルホモセリン転換酵素液およびヒホモナス由来のO−アセチルホモセリン転換酵素液を使用してメチオニン転換反応を行った。
【0180】
細胞を除去しないメチオニン前駆体発酵液2.0Lに破砕した酵素培養液0.1Lを添加した後、15%Na-メチルメルカプタン0.3Lを添加して反応を開始し、2時間経過後に発酵液を回収して細胞を除去した後、HPLCを通じてメチオニン生成を確認した。その結果を表12に示した。
【0181】
【表12】
【0182】
前記の結果からわかるように、既存の発明では10g/L以下の低い濃度のL−メチオニンを産生することに比べて前記の発明では30g/L以上のL−メチオニンを生成してL−メチオニンを大量に産生可能であることが確認できた。
【産業上の利用可能性】
【0183】
L−メチオニンを選択的に産生することによって飼料および食品添加剤、医薬用および医薬品の原料などの多様な分野で広く使用することができる。上述のとおり本発明の具体例をついて詳細に説明したが、本発明の範疇および技術思想範囲内で当業者にとって多様な変形および修正が可能であることは明らかであり、このような変形および修正が添付された本発明の特許請求範囲に属することも当然である。
【特許請求の範囲】
【請求項1】
O−スクシニル−L−ホモセリンの分解に関連するシスタチオニンガンマシンターゼの活性を欠損し、同時にホモセリンO−アセチルトランスフェラーゼの活性を強化させた、エシェリキア属から選択されることを特徴とするO−アセチル−L−ホモセリンを生産する菌株。
【請求項2】
前記シスタチオニンガンマシンターゼはmetB遺伝子でコードされていて、ホモセリンO−アセチルトランスフェラーゼはmetX遺伝子でコードされることを特徴とする請求項1に記載の菌株。
【請求項3】
前記菌株は、ホモセリンキナーゼ活性を弱化または欠損させたことを特徴とする請求項1に記載の菌株。
【請求項4】
前記菌株は、ホモセリンキナーゼをコードするthrB遺伝子を弱化または欠損させたことを特徴とする請求項1に記載の菌株。
【請求項5】
前記O−アセチル−L−ホモセリンの合成を強化するために、微生物菌株固有のホモセリンO−スクシニルトランスフェラーゼをコードする遺伝子を欠損または弱化させた後、新規なホモセリンO−アセチルトランスフェラーゼをコードする遺伝子を導入することを特徴とする請求項1に記載の菌株。
【請求項6】
前記菌株は、エシェリキアコリ菌株であることを特徴とする請求項1に記載の菌株。
【請求項7】
前記菌株は、メチオニン要求性が解除されたトレオニン生産菌株であるEscherichia coli MF001(受託番号:KCCM-10568)から由来したことを特徴とする請求項1に記載の菌株。
【請求項8】
前記菌株は、O−アセチルホモセリン生産能を有するEscherichia coliCJM-BTJA(pCJ-MetX-CL)(受託番号:KCCM-10873)であることを特徴とする請求項1に記載の菌株。
【請求項1】
O−スクシニル−L−ホモセリンの分解に関連するシスタチオニンガンマシンターゼの活性を欠損し、同時にホモセリンO−アセチルトランスフェラーゼの活性を強化させた、エシェリキア属から選択されることを特徴とするO−アセチル−L−ホモセリンを生産する菌株。
【請求項2】
前記シスタチオニンガンマシンターゼはmetB遺伝子でコードされていて、ホモセリンO−アセチルトランスフェラーゼはmetX遺伝子でコードされることを特徴とする請求項1に記載の菌株。
【請求項3】
前記菌株は、ホモセリンキナーゼ活性を弱化または欠損させたことを特徴とする請求項1に記載の菌株。
【請求項4】
前記菌株は、ホモセリンキナーゼをコードするthrB遺伝子を弱化または欠損させたことを特徴とする請求項1に記載の菌株。
【請求項5】
前記O−アセチル−L−ホモセリンの合成を強化するために、微生物菌株固有のホモセリンO−スクシニルトランスフェラーゼをコードする遺伝子を欠損または弱化させた後、新規なホモセリンO−アセチルトランスフェラーゼをコードする遺伝子を導入することを特徴とする請求項1に記載の菌株。
【請求項6】
前記菌株は、エシェリキアコリ菌株であることを特徴とする請求項1に記載の菌株。
【請求項7】
前記菌株は、メチオニン要求性が解除されたトレオニン生産菌株であるEscherichia coli MF001(受託番号:KCCM-10568)から由来したことを特徴とする請求項1に記載の菌株。
【請求項8】
前記菌株は、O−アセチルホモセリン生産能を有するEscherichia coliCJM-BTJA(pCJ-MetX-CL)(受託番号:KCCM-10873)であることを特徴とする請求項1に記載の菌株。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】

【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2012−213401(P2012−213401A)
【公開日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願番号】特願2012−148175(P2012−148175)
【出願日】平成24年7月2日(2012.7.2)
【分割の表示】特願2009−521706(P2009−521706)の分割
【原出願日】平成19年7月30日(2007.7.30)
【出願人】(508089369)
【Fターム(参考)】
【公開日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願日】平成24年7月2日(2012.7.2)
【分割の表示】特願2009−521706(P2009−521706)の分割
【原出願日】平成19年7月30日(2007.7.30)
【出願人】(508089369)
【Fターム(参考)】
[ Back to top ]