
LAMP法による急性ウイルス血症原因ウイルスの検出方法及び検出試薬キット
【課題】急性ウイルス血症原因ウイルス(WSDV)を感染初期に高感度で且つ定量的に検出できる方法及び当該検出方法に使用するための検出試薬キットを提供する。
【解決手段】wsv069遺伝子に対して設計したLAMP用プライマーを含む、LAMPにおけるWSDV検出用プライマーセット。
【解決手段】wsv069遺伝子に対して設計したLAMP用プライマーを含む、LAMPにおけるWSDV検出用プライマーセット。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、例えば環状型等温増幅反応(Loop-Mediated Isothermal Amplification:以下、「LAMP」という)法による急性ウイルス血症原因ウイルス(White spot disease virus:以下、「WSDV」という)の検出方法及び当該検出方法で使用するための検出試薬キットに関する。
【背景技術】
【0002】
WSDVは、エビ等の甲殻類に強い病原性を有するウイルスである。海外からの当該ウイルスの伝播を防ぐべく、当該ウイルスに対する高感度で、且つ定量的な検出方法の開発が望まれている。
【0003】
一方、従来より遺伝子の検出には、遺伝子増幅方法であるPCRが用いられてきた。しかしながら、PCRは感度が低いといった問題があった。
【0004】
そこで、高感度な遺伝子増幅方法として、LAMP法が開発された(特許文献1及び2)。当該LAMP法は、特定の遺伝子領域を増幅するための複数のプライマー、鋳型DNA、鎖置換型DNA合成酵素、dNTP等を混合し、一定温度(65℃付近)で一定時間反応させ、反応液の濁度により増幅を検出する方法である。
【0005】
本願発明者らは、LAMP法を用いてWSDVを検出すべく、WSDVの特定の遺伝子領域に特異的なLAMP用プライマーセットを設計し、検出を行った(特許文献3)。しかしながら、当該WSDVのLAMP用プライマーセットを使用したLAMP法は、WSDVのゲノム上の遺伝子を検出しており、不活性なウイルス(例えば、死滅したウイルスやウイルスDNAのみ)までも検出してしまう可能性があった。また、WSDVのゲノム上の遺伝子を検出するためには、ウイルス粒子の数が100個程度必要であるため、感染の極めて初期に検出することは困難であった。
【0006】
このように、WSDVを感染初期に高感度で且つ定量的に検出できる方法が望まれていた。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許第3313358号公報
【特許文献2】特開2001-242169号公報
【特許文献3】特開2009-183228号公報
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、上述した実情に鑑み、WSDVを感染初期に高感度で且つ定量的に検出できる方法及び当該検出方法に使用するための検出試薬キットを提供することを目的とする。
【課題を解決するための手段】
【0009】
上記課題を解決するため鋭意研究を行った結果、WSDV感染初期に発現する前初期タンパク遺伝子(wsv069=ie1)のmRNAを検出するようにLAMP用プライマーセットを設計し、当該LAMP用プライマーセットをLAMPに用いることで、感染し、且つ増殖している活性のあるWSDVのみを感染初期に検出できることを見出し、本発明を完成するに至った。
【0010】
本発明は、以下を包含する。
(1)配列番号1〜4に示される塩基配列を有する4つのプライマーを含む、LAMPにおけるWSDV検出用プライマーセット。
(2)配列番号5及び6に示される塩基配列を有する2つのプライマーをさらに含む、(1)記載のプライマーセット。
(3)(1)又は(2)記載のプライマーセットを含む、LAMPにおけるWSDV検出用試薬キット。
【0011】
(4)(1)又は(2)記載のプライマーセットと検体用核酸とを含有するサンプルをLAMPに供する工程と、前記LAMPによって増幅産物が産生されたか否かを検出する工程とを含む、LAMPにおけるWSDV検出方法。
(5)前記検体用核酸がmRNAである、(4)記載の方法。
(6)前記検出が定量的検出である、(4)又は(5)記載の方法。
【発明の効果】
【0012】
本発明によれば、感染し、且つ増殖している活性のあるWSDVのみを感染初期に高感度で且つ定量的に検出することができる。また、本発明によれば、国内外のエビ等の甲殻類の輸入/輸出や漁業の現場において、簡便且つ迅速にWSDVの感染の有無を検出することができる。
【図面の簡単な説明】
【0013】
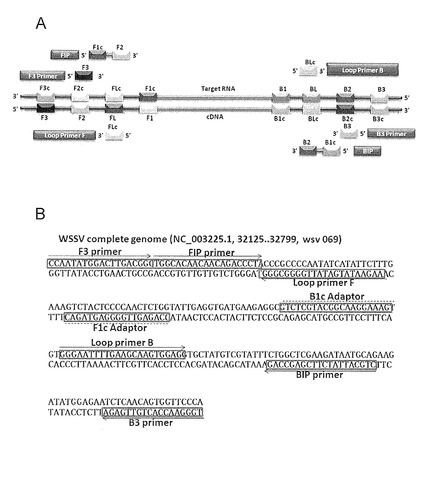
【図1】本発明に係るLAMPにおけるWSDV検出用プライマーセットに含まれる各プライマーのWSDVゲノムDNAから転写されたmRNA(Target RNA)に対する位置を示す模式図である。
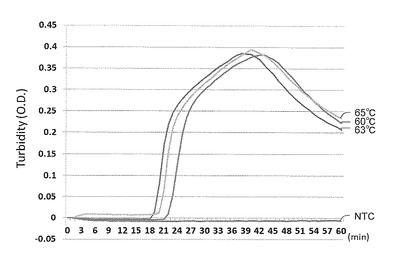
【図2】リアルタイムWSDV-LAMPにおける反応温度の検討結果を示す特性図である。
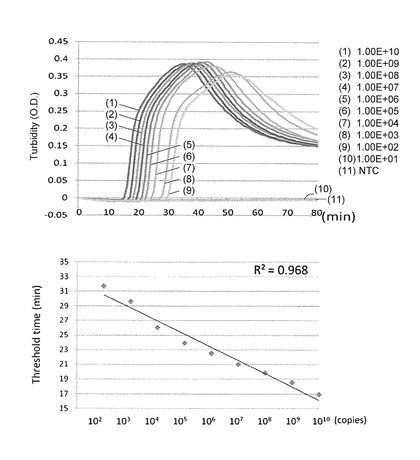
【図3】リアルタイムWSDV-LAMPにおける検量線を示す特性図である。
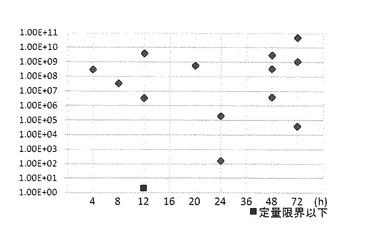
【図4】WSDVに浸漬感染させた供試エビの血リンパからのリアルタイムLAMPによるwsv069遺伝子の検出結果を示す特性図である。
【図5】WSDVに浸漬感染させた供試エビの心臓からのリアルタイムLAMPによるwsv069遺伝子の検出結果を示す特性図である。
【発明を実施するための形態】
【0014】
以下、本発明を詳細に説明する。
本発明に係るプライマーセットは、WSDVの感染初期に発現する前初期タンパク遺伝子wsv069に対して設計した複数のプライマーを含む、LAMPにおけるWSDV検出用プライマーセットである。本発明に係るLAMPにおけるWSDV検出用プライマーセット(以下、「WSDV検出用プライマーセット」と称する)は、FIP(配列番号1)、BIP(配列番号2)、F3 primer(配列番号3)及びB3 primer(配列番号4)を含む。さらに、当該WSDV検出用プライマーセットは、ループプライマー(Loop Primer)と呼ばれるLF primer(配列番号5)及びLB primer(配列番号6)を含むことで、LAMPにおいて増幅時間を大幅に短縮することができる。
【0015】
WSDVは、Nimaviridae科Whispovirus属に属するウイルスであり、二本鎖DNAウイルスである。WSDVは、またWSSV(white spot syndrome virus)とも呼ばれる。WSDVに感染すると、急性ウイルス血症を発症することとなる。
【0016】
本発明では、プライマー設計支援ソフトPrimerExplorer V4(https://primerexplorer.jp/lamp4.0.0/index.html)を用いて、WSDVゲノムDNA(GenBank accession number: NC_003225.1)上のwsv069遺伝子対応領域の塩基番号32125〜32799(GenBank accession number: AAL33073)(配列番号7)に対してLAMP用のプライマーを設計することで、WSDV検出用プライマーセットを得た。
【0017】
図1には、WSDV検出用プライマーセットに含まれる各プライマーのWSDVゲノムDNAから転写されたmRNA(Target RNA)に対するアニーリング位置を示す。パネル(A)は、FIP、BIP、F3、B3、LF及びLBプライマーのプライマー設計の概要を示す。パネル(B)は、プライマーに用いたWSDVゲノムの塩基配列を示す。プライマー設計に用いたDNA配列を囲みで示す。
【0018】
下記では、図1を用いてWSDVゲノムDNAから転写されたmRNA(Target RNA)から増幅するRT-LAMPの原理を説明する。なお、LAMPの原理は、特許文献1及び栄研化学株式会社のホームページ(http://loopamp.eiken.co.jp/lamp/index.html)に詳細に説明されている。
【0019】
ここで、「内部(内側)」、「外部(外側)」とは、WSDVゲノムDNA上の塩基番号32125〜32799の増幅標的配列の中心に対する位置を意味する。
【0020】
図1に示すように、LAMPでは、先ず検体用核酸であるWSDVゲノムDNAから転写されたmRNAに対して、FIP(配列番号1)(「内部プライマー」に相当する)がアニーリングし、そこから逆転写酵素の作用により相補的なcDNAが合成される。ここで、FIPは、Target RNA上の「F2c領域」に相補的な「F2領域」を3'末端側に有し、且つ5'末端側にTarget RNA上の「F1c領域」を有する。
【0021】
次いで、FIPの外側に位置するF3 primer(配列番号3)(「外部プライマー」に相当する)が相補的な配列(「F3c領域」)に対してアニーリングし、逆転写酵素の作用により、既に合成されたFIPから伸長したcDNAを剥がす一方で、F3 primerより新たなcDNAが合成される。F3 primerは、Target RNA上の「F3c領域」に相補的な「F3領域」を有する。剥がされたFIPからの一本鎖cDNAは、FIPが有する5'末端側の「F1c領域」と伸長部分の「F1領域」とが相補的であることから、自己アニーリングすることでループを形成することとなる。
【0022】
当該ループを形成した一本鎖cDNAに、もう一方の内部プライマーであるBIP(配列番号2)がアニーリングし、そこから鎖置換型DNAポリメラーゼの作用により伸長する。BIPは、Target RNA上の「B2領域」を3'末端側に有し、且つ5'末端側にTarget RNA上の「B1領域」に相補的な「B1c領域」を有する。ここで、鎖置換型DNAポリメラーゼとは、伸長方向に二本鎖領域が存在する場合に、当該二本鎖を解離しながら、相補鎖合成を行うことができるDNA合成酵素を意味する。
【0023】
さらに、BIPの外側に位置するB3 primer(配列番号4)が当該ループを形成した一本鎖DNAにアニーリングし、鎖置換型DNAポリメラーゼの作用により、既に合成されたBIPからの一本鎖DNAを剥がす一方で、B3 primerより伸長する。B3 primerは、Target RNA上の「B3領域」を有する。剥がされたBIPから伸長した一本鎖DNAは、両端でそれぞれ相補的な配列を有するので(すなわち、3'末端側における「F1c領域」と「F1領域」及び5'末端側における「B1領域」と「B1c領域」)、それぞれが自己アニーリングすることでループを形成することとなる。すなわち、当該剥がされたBIPから伸長した一本鎖DNAは、両端にループを形成することでダンベル型構造となる。このダンベル型構造が、LAMPにおける増幅サイクルの起点構造となる。当該ダンベル型構造を起点として、一連のLAMP増幅サイクルが行われる。
【0024】
さらに、ループプライマーLF primer(配列番号5:図1において「Loop Primer F」)及びLB primer(配列番号6:図1において「Loop Primer B」)をプライマーセットに含むことで、DNA合成の起点を増やすことができる。当該ループプライマーは、上述のダンベル型構造の5'末端側のループの一本鎖部分に相補的な配列を有するものである。LF primerは、Target RNA上の「F1c領域」と「F2c領域」との間の「FLc領域」を有する。一方、LB primerは、Target RNA上の「B1領域」と「B2領域」との間の「BL領域」に相補的な「BLc領域」を有する。
【0025】
なお、以上に説明したWSDV検出用プライマーセットの各プライマーは、例えば化学合成により作製することができる。さらに、WSDV検出用プライマーセットには、上述した各プライマーの配列番号で示される塩基配列において1又は数個(例えば、1〜10個、好ましくは1〜5個、特に好ましくは1〜3個)の塩基が欠失、置換及び/又は付加した塩基配列を有し、且つLAMPにおけるそれぞれのプライマー機能を有するプライマーを代替的に含むことができる。
【0026】
また、WSDV検出用プライマーセットは、単独で、又はLAMPで必要な試薬と共に、LAMPにおけるWSDV検出用試薬キットとして提供することができる。LAMPで必要な試薬としては、例えば鎖置換型DNA合成酵素、dNTP、緩衝液、蒸留水等が挙げられる。また、検体用核酸がmRNAである場合、逆転写酵素を試薬キットに含めることができる。特に、下記で説明するように、本発明では、感染初期におけるWSDVを検出すべく、mRNAを検体用核酸とすることが好ましい。
【0027】
一方、本発明に係るLAMPにおけるWSDV検出方法(以下、「本方法」と称する)では、WSDV検出用プライマーセットを用いてLAMPを行う。本方法では、先ず検体用核酸を準備する。検体用核酸は、例えば、上述のWSDVに感染しているか否かを検査すべき甲殻類、又は当該WSDVに感染しているが、感染の程度を確認すべき甲殻類から得られる核酸である。ここで、検体用核酸が由来するエビ類としては、上述したWSDVに感染するものであればいずれの種類であってよく、例えば、クルマエビ、ウシエビ、バナメイエビ、コウライエビ、シバエビ等が挙げられる。また、例えば、カニ類、ザリガニ類、動物性プランクトン等のエビ類以外の甲殻類及びそれらの幼生も上述したWSDVに感染するので、これら甲殻類から得た核酸も検体用核酸とすることができる。
【0028】
検体用核酸は、例えば、検査対象である甲殻類から常法により抽出されたゲノムDNA等のDNAとすることができる。あるいは、検査対象より抽出したRNAを、逆転写酵素を用いたRT-PCRに供する。このようにして得られたcDNAを検体用核酸とすることができる。
【0029】
本発明では、WSDV感染初期に発現する前初期タンパク遺伝子wsv069を検出対象として使用する。従って、wsv069遺伝子から転写されたmRNAを検体用核酸とすることで、感染初期におけるWSDVを検出することができる。従って、本発明ではmRNAを検体用核酸とすることが特に好ましい。この場合、逆転写酵素をLAMPサンプルに添加することで、逆転写酵素によるcDNAの作製と同時にLAMPを行うことができる。
【0030】
次いで、本方法では、WSDV検出用プライマーセット、検体用核酸、鎖置換型DNA合成酵素及びdNTP(並びに検体用核酸がRNAである場合には、逆転写酵素)を含有するサンプルを準備する。鎖置換型DNA合成酵素としては、例えば、BST DNA polymerase(栄研化学株式会社)、Bst DNA Polymerase, Large Fragment (New England Biolabs)、Bca BEST DNA Polymerase (TaKaRa)等が挙げられる。また、サンプル中のこれら構成成分の組成は適宜決定することができるが、例えば、サンプル25μl当たり、WSDV検出用プライマーセットに含まれる各プライマー1.0〜50pmol(好ましくは5〜40pmol)、検体用核酸10fg〜30μg(好ましくは0.1〜1μg)、鎖置換型DNA合成酵素0.5〜15U(ユニット)(好ましくは6.0〜10U)、最終濃度1.0〜10mM(好ましくは4.0〜6.0mM)のdNTPが挙げられる。また、逆転写酵素がサンプルに含まれる場合には、例えばサンプル25μl当たり逆転写酵素1.0〜10U(好ましくは4.0〜6.0U)とする。
【0031】
さらに、本方法では、準備したサンプルをLAMPに供することで、増幅産物を産生する。LAMPの条件としては、例えば、温度60℃〜65℃(好ましくは63℃)で15〜80分(好ましくは20〜60分)が挙げられる。
【0032】
LAMP後のサンプルを各検出手段に供することで、サンプル中の増幅産物を検出することできる。例えば、サンプルをアガロースゲル/エチジウムブロマイド電気泳動に供することにより、目視により増幅産物を検出することができる。また、サンプルを濁度計に供することで、濁度を指標として増幅産物を検出することができる。さらに、サンプルをLAMPに供する際にリアルタイム濁度測定装置を用いて、リアルタイムで濁度を測定することで、増幅産物を検出することができる。このように検出した増幅産物の有無により、WSDV感染の有無を判断することができる。
【0033】
また、陽性対照サンプル(例えば、増幅対象の標的配列を含むプラスミドを含む)を、様々な核酸濃度でリアルタイム濁度測定装置(例えば、栄研化学株式会社製のリアルタイム濁度測定装置(LA-200))を用いたLAMPに供する。得られた各核酸濃度サンプルの濁度を縦軸とし、当該核酸濃度を横軸とすることで、検量線を作成する。次いで、当該検量線に対して、LAMP後の検体用核酸を含むサンプルの濁度を比較することで、定量的に増幅産物(すなわち、ウイルス量)を検出することができる。
【0034】
以上に説明した本発明によれば、高感度でWSDVを検出することができる。また、本発明ではLAMPを用いることで、増幅ステップが一定温度で一定時間であるので、変性、アニーリング及び増幅の段階のサイクルを行うPCRと比較して簡便である。さらに、mRNAを検体用核酸とすることで、cDNAの作製からLAMPによる増幅(すなわち、RT-LAMP)を1本のチューブにおいて60分以内で行うことができ、従来のRT-PCR法より優れている。
【0035】
また、本発明によれば、前初期タンパク質遺伝子wsv069遺伝子から転写されたmRNAを検体用核酸として使用することで、感染し、且つ増殖している活性のあるWSDVのみを感染初期に高感度で且つ定量的に検出することができる。この場合には、サンプル内にWSDV粒子が100個以下でも検出できると考えられる。
【0036】
さらに、本発明によれば、定量的にWSDVを検出でき、感染の程度を知ることができる。このことによって、上述したWSDVの蔓延防止や急性ウイルス血症の予防に本発明は貢献することができる。従って、上述のWSDV検出用プライマーセット、WSDV検出用試薬キット及び本方法は、それぞれ急性ウイルス血症診断用プライマーセット、急性ウイルス血症診断用試薬キット、急性ウイルス血症診断方法として使用することもできる。
【0037】
(実施例)
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲はこれら実施例に限定されるものではない。
リアルタイムLAMP法によるクルマエビ類WSDVのwsv069遺伝子の検出と定量
【0038】
〔材料と方法〕
1. 供試エビ
平均体重12.0gのクルマエビにWSDVを人為感染させた個体を用いた。WSDV感染エビの心臓よりRNAiso Plus(Takara,Japan)を用い、RNA抽出を行った。
【0039】
2. Total RNAの抽出
サンプル約25 mgを1.5 mlエッペンドルフチューブに入れ、300μlのRNAiso Plusを加え、よくホモジナイズし、再び700μlのRNAiso Plusを加え1 mlシリンジ(20 G針付)で20回程度攪拌し、22℃で5分間静置した。
【0040】
次に、クロロフォルム(Wako, Japan)200μlを加え、転倒混和によって十分に混合した後、22℃で3分間静置し、さらに4℃、14,000 rpmで15分間遠心分離を行った。
【0041】
遠心分離後、上清を新しい1.5 mlエッペンドルフチューブに移し、イソプロピルアルコール(Wako, Japan)500μlを加え、4℃で30分間静置した後、4℃、14,000 rpmで30分間遠心分離を行った。
【0042】
遠心分離後、上清を除去し、80%エタノール(Wako, Japan)1 mlを加え、緩やかに混合した後、4℃、14,000 rpmで5分間遠心分離を行った。
【0043】
遠心分離後、上清を除き、5分間風乾させた後、ペレットを20μlのDiethyl Pyrocarbonate(DEPC; SIGMA, Japan)処理水に溶解して、total RNA溶液とした。
【0044】
3. cDNAの合成
抽出したtotal RNAをテンプレートとして、ReverTra Dash(TOYOBO, Japan)を用いて逆転写反応を行った。反応溶液を作製し、primerにOligo (dt) 20-P7を用い、42℃で20分間、99℃で5分間及び4℃で5分間の反応を、サーマルサイクラー(BIO RAD, Mycycler)を用いて行った。
【0045】
4. 検量線作成のためのプラスミドの作製
4-1. プラスミドの構築
プライマーF(5'-CCAATATGGACTTGACGGC-3'(配列番号8))及プライマーR(5'-TGGGAACCACTGTTGAGA-3'(配列番号9))を用い、WSDV-LAMPにおける標的遺伝子配列のPCRを行った。得られたPCR産物をライゲーションに用いた。
【0046】
4-2. ベクターへのライゲーション
Tベクター(PGEM-T Easy Vector; Promega, USA)へのライゲーション反応は、下記表1に示す試薬を混合し、4℃で16時間行った。反応溶液の組成を表1に示す。
【0047】
【表1】

【0048】
4-3. コンピテントセルへのトランスフォーメーション
トランスフォーメーションは、大腸菌DH5α(Promega,USA)を用いて行った。-80℃に保存していたコンピテントセルを氷上で融解し、ライゲーション産物2μlを加えた1.5 mlエッペンドルフチューブにコンピテントセル50μlを添加し、穏やかに混和し、氷上で20分間静置した。
【0049】
静置後、反応液を42℃のウォーターバスで50秒加熱し、2分間氷上に静置した。次いで、SOC培地(表2)950μlを加えた後、37℃のウォーターバスで1時間半振盪培養した(185 rpm)。
【0050】
振盪培養後、0.1%アンピシリン添加マッコンキー寒天培地(日水製薬)に得られた培養物100μlをプレーティングし、37℃で15時間培養した。
【0051】
培養後、形成したコロニーをレッドホワイトセレクションにより選択し、インサートの確認に用いた。
【0052】
【表2】
【0053】
4-4. コロニーPCRによるインサートの確認
マスターミックス溶液(表3)を20μlずつ0.5 mlチューブに分注し、滅菌爪楊枝を用いて生育したコロニーを少量かきとり、溶液内に懸濁させた。PCRは、T7 (5'-TAATACGACTCACTATAGGG-3'(配列番号10))及びSP6(5'-ATTTAGGTGACACTATAGAA-3'(配列番号11))プライマーを用い、94℃で30秒間の熱処理後、94℃で30秒の熱変性、55℃で30秒間のアニーリング及び72℃で30秒間の伸長反応の30サイクル、72℃、5分間で反応の停止並びに4℃で保存の反応をサーマルサイクラーMyCycler(BIO-RAD,USA)を用いて行った。
【0054】
【表3】
【0055】
反応終了後、PCR産物(10μl)を、エチジウムブロマイド(終濃度:1μg/ml)を含む1.5%アガロースゲルで電気泳動し、インサートの有無及びサイズの確認を行った。
4-5. プラスミドDNAの抽出
大腸菌を100μg/mlのアンピシリンを含む3 mlのLB培地(表4)で16時間培養し、8,000 rpmで3分間の遠心分離により集菌した。これを、QIAprep Spin Miniprep Kit(QIAGEN)を用い、マニュアルに従いプラスミド抽出を行った。250μlのBufferP1を加え、菌体ペレットを再懸濁した。250μlのBufferP2を添加した後、5、6回転倒させ完全に混和させた。350μlのBufferN3を添加した後、5、6回転倒させ完全に混和させ、13,000 rpm、4℃で10分間遠心分離した。
【0056】
遠心分離後、上清をデカントし、ピペッティングによりスピンカラムにアプライし、13,000 rpm、4℃で1分間遠心分離した後、フロースルー液を捨てた。Buffer PB 0.5 mlを添加した後、13,000 rpm、4℃で1分間遠心分離し、フロースルー液を捨てた。Buffer PE 0.75 mlを添加した後、13,000 rpm、4℃で1分間遠心分離し、フロースルー液を捨てた。残留している洗浄バッファーを除去するために、さらに13,000 rpm、4℃で1分間遠心分離した。
【0057】
次いで、カラムを新しい1.5 mlエッペンチューブにセットし、Buffer EBをカラムの中央にアプライし、1分間放置した後、13,000 rpm、4℃で1分間遠心分離して、得られた抽出液を精製プラスミドとした。
【0058】
【表4】
【0059】
プラスミドのコピー数を以下の方法を基に算出し、0.5×107〜1010(copies/μl)のプラスミド溶液を作製した。
1μl当たりのコピー数=DNAの量(ng)×アボガドロ数(6.022×1023)/テンプレートの長さ(bp)×109×一塩基対の平均重量(650)
【0060】
5. プライマーの設計
5-1 WSDV-LAMP用プライマーの設計
PrimerExplorer V4(https://primerexplorer.jp/lamp4.0.0/index.html)により、図1に示すように、WSDVのゲノムDNA(GenBank accession number: NC_003225.1)のwsv069遺伝子に対応する塩基番号32125〜32799(配列番号7)に対するLAMP用プライマーを作製した。設計したプライマーは次の表5に示す通りである。
【0061】
【表5】
【0062】
5-2. リアルタイムLAMP法
Loopampリアルタイム濁度測定装置(LA-200)を用いて、ウイルスの定量的検出を行った。
【0063】
5-3. 反応条件の検討
作製したcDNA溶液をテンプレートとし、WSDV-LAMPにおける最適反応温度の検討を行った。
【0064】
LAMP反応溶液を下記の表6に示す。LAMP法は、Loopamp DNA増幅試薬キット(栄研化学)を用いた。2×Reaction Mix(40 mM Tris-HCl,20 mM KCl,16 mM MgSO4,20 mM (NH4)2SO4,0.2%Tween 20,1.6 M Betaine ,2.8 mM dNTPs)12.5μl、FIPプライマーとBIPプライマーをそれぞれ1.0μl(40 pmol)、F3プライマーとB3プライマーをそれぞれ1.0μl(5 pmol)、LFプライマーとLBプライマーをそれぞれ1.0μl(5 pmol)及び蒸留水4.5μlの合計23μlを混合した。
【0065】
この混液を97℃で3分間加熱し、その後氷中で急冷し、Bst DNAポリメラーゼ(Eiken Chemical Co.Ltd., Japan)1.0μl(8U)を加えた。次いで、テンプレートcDNA溶液を1μl加えて、リアルタイム濁度測定装置(LA-200)を用い、60℃,63℃及び65℃で60分間反応させた。
【0066】
【表6】
【0067】
5-5 検量線の作成
定量LAMPの検量線作成のため、F3及びB3プライマーを用い、WSDV-LAMPにおける標的遺伝子配列を含むプラスミドの構築を行った。プラスミドのコピー数を以下の方法を基に算出し、0.5×101〜109(copies/μl)のプラスミド溶液を作製した。
1μl当たりのコピー数=OD260測定で得られたプラスミド濃度(ng/μl)/プラスミド分子量×6.02×1014(copies/μl)
このプラスミド溶液をテンプレートとし、上記と同様の方法で反応液を作製し、リアルタイム濁度測定装置を用いて、定量WSDV-LAMPにおける検量線の作成を行った。
【0068】
5-6. 感染エビからの検出
WSDVに浸漬感染させた供試エビを継時的にサンプリングし、血リンパ及び心臓からtotal RNAを抽出し、cDNAを作製した。
各cDNAをリアルタイム濁度測定装置(LA-200)を用いて、63℃で60分間反応させた。
【0069】
〔結果〕
1. リアルタイムLAMP法の反応条件検討
WSDV-LAMPにおける最適反応温度の検討結果を図2に示す。図2に示す結果から63℃における反応が最も早いことが明らかとなった。
【0070】
また、図3に示すように、wsv069遺伝子検出用LAMPの検量線について検討した結果、102〜1010copies/μlで、直線性の高い検量線が得られた。検出限界は102copies/μlであった。なお、図2及び3において、「NTC(non template control)」は、cDNAの代わりにD.W.を添加したサンプルを意味する。
【0071】
2. WSDVに浸漬感染させた供試エビの血リンパ及び心臓からのWSDV検出結果
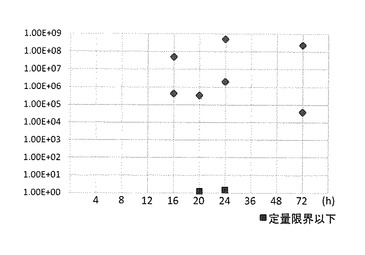
図4に示すように、WSDVに浸漬感染させた供試エビの血リンパから、wsv069プライマーにより、浸漬4時間後から検出され、検出率は51.8%であった。
【0072】
また、図5に示すように、WSDVに浸漬感染させた供試エビの心臓から、wsv069プライマーにより、浸漬16時間後から検出され、検出率は33.3%であった。
【0073】
以上の実験結果より、wsv069遺伝子に対するプライマーを用いることで、感染初期に、より高感度な検出が可能になった。
【0074】
一方、WSDVのVP-28遺伝子は、外被タンパク質をコードする遺伝子である。WSDVのVP-28遺伝子について、LAMPを用いて本実験と同じサンプルから検出を試みた結果、血リンパと心臓のいずれからもVP-28遺伝子を検出できなかった。そこで、VP-28遺伝子検出用LAMP法より感度が良いリアルタイムPCR法(qPCR)を用いてVP-28遺伝子を検出した結果、血リンパで3.7%及び心臓で14.8%と低い検出率であった。
【0075】
表7に、上記の検出率についてまとめた。本実施例で用いたwsv069遺伝子をターゲットとするLAMP法とVP-28遺伝子検出用qPCR法を比べると、明らかにwsv069-LAMP法の検出率が高い。このことから、LAMP法は、従来感度が高いとされていたqPCR法よりも感度が優れていることが明らかとなった。
【0076】
また、wsv069-LAMP法を用いた場合、血リンパでの検出率が心臓のそれよりも高い。このことから、血リンパを用いて高感度のウイルス検査が可能であることが明らかとなった。このことは、血リンパを用いることによって非破壊検査が可能となり、エビの産卵前にウイルスの存在を明らかにすることができると考えられる。
【0077】
【表7】
【技術分野】
【0001】
本発明は、例えば環状型等温増幅反応(Loop-Mediated Isothermal Amplification:以下、「LAMP」という)法による急性ウイルス血症原因ウイルス(White spot disease virus:以下、「WSDV」という)の検出方法及び当該検出方法で使用するための検出試薬キットに関する。
【背景技術】
【0002】
WSDVは、エビ等の甲殻類に強い病原性を有するウイルスである。海外からの当該ウイルスの伝播を防ぐべく、当該ウイルスに対する高感度で、且つ定量的な検出方法の開発が望まれている。
【0003】
一方、従来より遺伝子の検出には、遺伝子増幅方法であるPCRが用いられてきた。しかしながら、PCRは感度が低いといった問題があった。
【0004】
そこで、高感度な遺伝子増幅方法として、LAMP法が開発された(特許文献1及び2)。当該LAMP法は、特定の遺伝子領域を増幅するための複数のプライマー、鋳型DNA、鎖置換型DNA合成酵素、dNTP等を混合し、一定温度(65℃付近)で一定時間反応させ、反応液の濁度により増幅を検出する方法である。
【0005】
本願発明者らは、LAMP法を用いてWSDVを検出すべく、WSDVの特定の遺伝子領域に特異的なLAMP用プライマーセットを設計し、検出を行った(特許文献3)。しかしながら、当該WSDVのLAMP用プライマーセットを使用したLAMP法は、WSDVのゲノム上の遺伝子を検出しており、不活性なウイルス(例えば、死滅したウイルスやウイルスDNAのみ)までも検出してしまう可能性があった。また、WSDVのゲノム上の遺伝子を検出するためには、ウイルス粒子の数が100個程度必要であるため、感染の極めて初期に検出することは困難であった。
【0006】
このように、WSDVを感染初期に高感度で且つ定量的に検出できる方法が望まれていた。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許第3313358号公報
【特許文献2】特開2001-242169号公報
【特許文献3】特開2009-183228号公報
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、上述した実情に鑑み、WSDVを感染初期に高感度で且つ定量的に検出できる方法及び当該検出方法に使用するための検出試薬キットを提供することを目的とする。
【課題を解決するための手段】
【0009】
上記課題を解決するため鋭意研究を行った結果、WSDV感染初期に発現する前初期タンパク遺伝子(wsv069=ie1)のmRNAを検出するようにLAMP用プライマーセットを設計し、当該LAMP用プライマーセットをLAMPに用いることで、感染し、且つ増殖している活性のあるWSDVのみを感染初期に検出できることを見出し、本発明を完成するに至った。
【0010】
本発明は、以下を包含する。
(1)配列番号1〜4に示される塩基配列を有する4つのプライマーを含む、LAMPにおけるWSDV検出用プライマーセット。
(2)配列番号5及び6に示される塩基配列を有する2つのプライマーをさらに含む、(1)記載のプライマーセット。
(3)(1)又は(2)記載のプライマーセットを含む、LAMPにおけるWSDV検出用試薬キット。
【0011】
(4)(1)又は(2)記載のプライマーセットと検体用核酸とを含有するサンプルをLAMPに供する工程と、前記LAMPによって増幅産物が産生されたか否かを検出する工程とを含む、LAMPにおけるWSDV検出方法。
(5)前記検体用核酸がmRNAである、(4)記載の方法。
(6)前記検出が定量的検出である、(4)又は(5)記載の方法。
【発明の効果】
【0012】
本発明によれば、感染し、且つ増殖している活性のあるWSDVのみを感染初期に高感度で且つ定量的に検出することができる。また、本発明によれば、国内外のエビ等の甲殻類の輸入/輸出や漁業の現場において、簡便且つ迅速にWSDVの感染の有無を検出することができる。
【図面の簡単な説明】
【0013】
【図1】本発明に係るLAMPにおけるWSDV検出用プライマーセットに含まれる各プライマーのWSDVゲノムDNAから転写されたmRNA(Target RNA)に対する位置を示す模式図である。
【図2】リアルタイムWSDV-LAMPにおける反応温度の検討結果を示す特性図である。
【図3】リアルタイムWSDV-LAMPにおける検量線を示す特性図である。
【図4】WSDVに浸漬感染させた供試エビの血リンパからのリアルタイムLAMPによるwsv069遺伝子の検出結果を示す特性図である。
【図5】WSDVに浸漬感染させた供試エビの心臓からのリアルタイムLAMPによるwsv069遺伝子の検出結果を示す特性図である。
【発明を実施するための形態】
【0014】
以下、本発明を詳細に説明する。
本発明に係るプライマーセットは、WSDVの感染初期に発現する前初期タンパク遺伝子wsv069に対して設計した複数のプライマーを含む、LAMPにおけるWSDV検出用プライマーセットである。本発明に係るLAMPにおけるWSDV検出用プライマーセット(以下、「WSDV検出用プライマーセット」と称する)は、FIP(配列番号1)、BIP(配列番号2)、F3 primer(配列番号3)及びB3 primer(配列番号4)を含む。さらに、当該WSDV検出用プライマーセットは、ループプライマー(Loop Primer)と呼ばれるLF primer(配列番号5)及びLB primer(配列番号6)を含むことで、LAMPにおいて増幅時間を大幅に短縮することができる。
【0015】
WSDVは、Nimaviridae科Whispovirus属に属するウイルスであり、二本鎖DNAウイルスである。WSDVは、またWSSV(white spot syndrome virus)とも呼ばれる。WSDVに感染すると、急性ウイルス血症を発症することとなる。
【0016】
本発明では、プライマー設計支援ソフトPrimerExplorer V4(https://primerexplorer.jp/lamp4.0.0/index.html)を用いて、WSDVゲノムDNA(GenBank accession number: NC_003225.1)上のwsv069遺伝子対応領域の塩基番号32125〜32799(GenBank accession number: AAL33073)(配列番号7)に対してLAMP用のプライマーを設計することで、WSDV検出用プライマーセットを得た。
【0017】
図1には、WSDV検出用プライマーセットに含まれる各プライマーのWSDVゲノムDNAから転写されたmRNA(Target RNA)に対するアニーリング位置を示す。パネル(A)は、FIP、BIP、F3、B3、LF及びLBプライマーのプライマー設計の概要を示す。パネル(B)は、プライマーに用いたWSDVゲノムの塩基配列を示す。プライマー設計に用いたDNA配列を囲みで示す。
【0018】
下記では、図1を用いてWSDVゲノムDNAから転写されたmRNA(Target RNA)から増幅するRT-LAMPの原理を説明する。なお、LAMPの原理は、特許文献1及び栄研化学株式会社のホームページ(http://loopamp.eiken.co.jp/lamp/index.html)に詳細に説明されている。
【0019】
ここで、「内部(内側)」、「外部(外側)」とは、WSDVゲノムDNA上の塩基番号32125〜32799の増幅標的配列の中心に対する位置を意味する。
【0020】
図1に示すように、LAMPでは、先ず検体用核酸であるWSDVゲノムDNAから転写されたmRNAに対して、FIP(配列番号1)(「内部プライマー」に相当する)がアニーリングし、そこから逆転写酵素の作用により相補的なcDNAが合成される。ここで、FIPは、Target RNA上の「F2c領域」に相補的な「F2領域」を3'末端側に有し、且つ5'末端側にTarget RNA上の「F1c領域」を有する。
【0021】
次いで、FIPの外側に位置するF3 primer(配列番号3)(「外部プライマー」に相当する)が相補的な配列(「F3c領域」)に対してアニーリングし、逆転写酵素の作用により、既に合成されたFIPから伸長したcDNAを剥がす一方で、F3 primerより新たなcDNAが合成される。F3 primerは、Target RNA上の「F3c領域」に相補的な「F3領域」を有する。剥がされたFIPからの一本鎖cDNAは、FIPが有する5'末端側の「F1c領域」と伸長部分の「F1領域」とが相補的であることから、自己アニーリングすることでループを形成することとなる。
【0022】
当該ループを形成した一本鎖cDNAに、もう一方の内部プライマーであるBIP(配列番号2)がアニーリングし、そこから鎖置換型DNAポリメラーゼの作用により伸長する。BIPは、Target RNA上の「B2領域」を3'末端側に有し、且つ5'末端側にTarget RNA上の「B1領域」に相補的な「B1c領域」を有する。ここで、鎖置換型DNAポリメラーゼとは、伸長方向に二本鎖領域が存在する場合に、当該二本鎖を解離しながら、相補鎖合成を行うことができるDNA合成酵素を意味する。
【0023】
さらに、BIPの外側に位置するB3 primer(配列番号4)が当該ループを形成した一本鎖DNAにアニーリングし、鎖置換型DNAポリメラーゼの作用により、既に合成されたBIPからの一本鎖DNAを剥がす一方で、B3 primerより伸長する。B3 primerは、Target RNA上の「B3領域」を有する。剥がされたBIPから伸長した一本鎖DNAは、両端でそれぞれ相補的な配列を有するので(すなわち、3'末端側における「F1c領域」と「F1領域」及び5'末端側における「B1領域」と「B1c領域」)、それぞれが自己アニーリングすることでループを形成することとなる。すなわち、当該剥がされたBIPから伸長した一本鎖DNAは、両端にループを形成することでダンベル型構造となる。このダンベル型構造が、LAMPにおける増幅サイクルの起点構造となる。当該ダンベル型構造を起点として、一連のLAMP増幅サイクルが行われる。
【0024】
さらに、ループプライマーLF primer(配列番号5:図1において「Loop Primer F」)及びLB primer(配列番号6:図1において「Loop Primer B」)をプライマーセットに含むことで、DNA合成の起点を増やすことができる。当該ループプライマーは、上述のダンベル型構造の5'末端側のループの一本鎖部分に相補的な配列を有するものである。LF primerは、Target RNA上の「F1c領域」と「F2c領域」との間の「FLc領域」を有する。一方、LB primerは、Target RNA上の「B1領域」と「B2領域」との間の「BL領域」に相補的な「BLc領域」を有する。
【0025】
なお、以上に説明したWSDV検出用プライマーセットの各プライマーは、例えば化学合成により作製することができる。さらに、WSDV検出用プライマーセットには、上述した各プライマーの配列番号で示される塩基配列において1又は数個(例えば、1〜10個、好ましくは1〜5個、特に好ましくは1〜3個)の塩基が欠失、置換及び/又は付加した塩基配列を有し、且つLAMPにおけるそれぞれのプライマー機能を有するプライマーを代替的に含むことができる。
【0026】
また、WSDV検出用プライマーセットは、単独で、又はLAMPで必要な試薬と共に、LAMPにおけるWSDV検出用試薬キットとして提供することができる。LAMPで必要な試薬としては、例えば鎖置換型DNA合成酵素、dNTP、緩衝液、蒸留水等が挙げられる。また、検体用核酸がmRNAである場合、逆転写酵素を試薬キットに含めることができる。特に、下記で説明するように、本発明では、感染初期におけるWSDVを検出すべく、mRNAを検体用核酸とすることが好ましい。
【0027】
一方、本発明に係るLAMPにおけるWSDV検出方法(以下、「本方法」と称する)では、WSDV検出用プライマーセットを用いてLAMPを行う。本方法では、先ず検体用核酸を準備する。検体用核酸は、例えば、上述のWSDVに感染しているか否かを検査すべき甲殻類、又は当該WSDVに感染しているが、感染の程度を確認すべき甲殻類から得られる核酸である。ここで、検体用核酸が由来するエビ類としては、上述したWSDVに感染するものであればいずれの種類であってよく、例えば、クルマエビ、ウシエビ、バナメイエビ、コウライエビ、シバエビ等が挙げられる。また、例えば、カニ類、ザリガニ類、動物性プランクトン等のエビ類以外の甲殻類及びそれらの幼生も上述したWSDVに感染するので、これら甲殻類から得た核酸も検体用核酸とすることができる。
【0028】
検体用核酸は、例えば、検査対象である甲殻類から常法により抽出されたゲノムDNA等のDNAとすることができる。あるいは、検査対象より抽出したRNAを、逆転写酵素を用いたRT-PCRに供する。このようにして得られたcDNAを検体用核酸とすることができる。
【0029】
本発明では、WSDV感染初期に発現する前初期タンパク遺伝子wsv069を検出対象として使用する。従って、wsv069遺伝子から転写されたmRNAを検体用核酸とすることで、感染初期におけるWSDVを検出することができる。従って、本発明ではmRNAを検体用核酸とすることが特に好ましい。この場合、逆転写酵素をLAMPサンプルに添加することで、逆転写酵素によるcDNAの作製と同時にLAMPを行うことができる。
【0030】
次いで、本方法では、WSDV検出用プライマーセット、検体用核酸、鎖置換型DNA合成酵素及びdNTP(並びに検体用核酸がRNAである場合には、逆転写酵素)を含有するサンプルを準備する。鎖置換型DNA合成酵素としては、例えば、BST DNA polymerase(栄研化学株式会社)、Bst DNA Polymerase, Large Fragment (New England Biolabs)、Bca BEST DNA Polymerase (TaKaRa)等が挙げられる。また、サンプル中のこれら構成成分の組成は適宜決定することができるが、例えば、サンプル25μl当たり、WSDV検出用プライマーセットに含まれる各プライマー1.0〜50pmol(好ましくは5〜40pmol)、検体用核酸10fg〜30μg(好ましくは0.1〜1μg)、鎖置換型DNA合成酵素0.5〜15U(ユニット)(好ましくは6.0〜10U)、最終濃度1.0〜10mM(好ましくは4.0〜6.0mM)のdNTPが挙げられる。また、逆転写酵素がサンプルに含まれる場合には、例えばサンプル25μl当たり逆転写酵素1.0〜10U(好ましくは4.0〜6.0U)とする。
【0031】
さらに、本方法では、準備したサンプルをLAMPに供することで、増幅産物を産生する。LAMPの条件としては、例えば、温度60℃〜65℃(好ましくは63℃)で15〜80分(好ましくは20〜60分)が挙げられる。
【0032】
LAMP後のサンプルを各検出手段に供することで、サンプル中の増幅産物を検出することできる。例えば、サンプルをアガロースゲル/エチジウムブロマイド電気泳動に供することにより、目視により増幅産物を検出することができる。また、サンプルを濁度計に供することで、濁度を指標として増幅産物を検出することができる。さらに、サンプルをLAMPに供する際にリアルタイム濁度測定装置を用いて、リアルタイムで濁度を測定することで、増幅産物を検出することができる。このように検出した増幅産物の有無により、WSDV感染の有無を判断することができる。
【0033】
また、陽性対照サンプル(例えば、増幅対象の標的配列を含むプラスミドを含む)を、様々な核酸濃度でリアルタイム濁度測定装置(例えば、栄研化学株式会社製のリアルタイム濁度測定装置(LA-200))を用いたLAMPに供する。得られた各核酸濃度サンプルの濁度を縦軸とし、当該核酸濃度を横軸とすることで、検量線を作成する。次いで、当該検量線に対して、LAMP後の検体用核酸を含むサンプルの濁度を比較することで、定量的に増幅産物(すなわち、ウイルス量)を検出することができる。
【0034】
以上に説明した本発明によれば、高感度でWSDVを検出することができる。また、本発明ではLAMPを用いることで、増幅ステップが一定温度で一定時間であるので、変性、アニーリング及び増幅の段階のサイクルを行うPCRと比較して簡便である。さらに、mRNAを検体用核酸とすることで、cDNAの作製からLAMPによる増幅(すなわち、RT-LAMP)を1本のチューブにおいて60分以内で行うことができ、従来のRT-PCR法より優れている。
【0035】
また、本発明によれば、前初期タンパク質遺伝子wsv069遺伝子から転写されたmRNAを検体用核酸として使用することで、感染し、且つ増殖している活性のあるWSDVのみを感染初期に高感度で且つ定量的に検出することができる。この場合には、サンプル内にWSDV粒子が100個以下でも検出できると考えられる。
【0036】
さらに、本発明によれば、定量的にWSDVを検出でき、感染の程度を知ることができる。このことによって、上述したWSDVの蔓延防止や急性ウイルス血症の予防に本発明は貢献することができる。従って、上述のWSDV検出用プライマーセット、WSDV検出用試薬キット及び本方法は、それぞれ急性ウイルス血症診断用プライマーセット、急性ウイルス血症診断用試薬キット、急性ウイルス血症診断方法として使用することもできる。
【0037】
(実施例)
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲はこれら実施例に限定されるものではない。
リアルタイムLAMP法によるクルマエビ類WSDVのwsv069遺伝子の検出と定量
【0038】
〔材料と方法〕
1. 供試エビ
平均体重12.0gのクルマエビにWSDVを人為感染させた個体を用いた。WSDV感染エビの心臓よりRNAiso Plus(Takara,Japan)を用い、RNA抽出を行った。
【0039】
2. Total RNAの抽出
サンプル約25 mgを1.5 mlエッペンドルフチューブに入れ、300μlのRNAiso Plusを加え、よくホモジナイズし、再び700μlのRNAiso Plusを加え1 mlシリンジ(20 G針付)で20回程度攪拌し、22℃で5分間静置した。
【0040】
次に、クロロフォルム(Wako, Japan)200μlを加え、転倒混和によって十分に混合した後、22℃で3分間静置し、さらに4℃、14,000 rpmで15分間遠心分離を行った。
【0041】
遠心分離後、上清を新しい1.5 mlエッペンドルフチューブに移し、イソプロピルアルコール(Wako, Japan)500μlを加え、4℃で30分間静置した後、4℃、14,000 rpmで30分間遠心分離を行った。
【0042】
遠心分離後、上清を除去し、80%エタノール(Wako, Japan)1 mlを加え、緩やかに混合した後、4℃、14,000 rpmで5分間遠心分離を行った。
【0043】
遠心分離後、上清を除き、5分間風乾させた後、ペレットを20μlのDiethyl Pyrocarbonate(DEPC; SIGMA, Japan)処理水に溶解して、total RNA溶液とした。
【0044】
3. cDNAの合成
抽出したtotal RNAをテンプレートとして、ReverTra Dash(TOYOBO, Japan)を用いて逆転写反応を行った。反応溶液を作製し、primerにOligo (dt) 20-P7を用い、42℃で20分間、99℃で5分間及び4℃で5分間の反応を、サーマルサイクラー(BIO RAD, Mycycler)を用いて行った。
【0045】
4. 検量線作成のためのプラスミドの作製
4-1. プラスミドの構築
プライマーF(5'-CCAATATGGACTTGACGGC-3'(配列番号8))及プライマーR(5'-TGGGAACCACTGTTGAGA-3'(配列番号9))を用い、WSDV-LAMPにおける標的遺伝子配列のPCRを行った。得られたPCR産物をライゲーションに用いた。
【0046】
4-2. ベクターへのライゲーション
Tベクター(PGEM-T Easy Vector; Promega, USA)へのライゲーション反応は、下記表1に示す試薬を混合し、4℃で16時間行った。反応溶液の組成を表1に示す。
【0047】
【表1】
【0048】
4-3. コンピテントセルへのトランスフォーメーション
トランスフォーメーションは、大腸菌DH5α(Promega,USA)を用いて行った。-80℃に保存していたコンピテントセルを氷上で融解し、ライゲーション産物2μlを加えた1.5 mlエッペンドルフチューブにコンピテントセル50μlを添加し、穏やかに混和し、氷上で20分間静置した。
【0049】
静置後、反応液を42℃のウォーターバスで50秒加熱し、2分間氷上に静置した。次いで、SOC培地(表2)950μlを加えた後、37℃のウォーターバスで1時間半振盪培養した(185 rpm)。
【0050】
振盪培養後、0.1%アンピシリン添加マッコンキー寒天培地(日水製薬)に得られた培養物100μlをプレーティングし、37℃で15時間培養した。
【0051】
培養後、形成したコロニーをレッドホワイトセレクションにより選択し、インサートの確認に用いた。
【0052】
【表2】
【0053】
4-4. コロニーPCRによるインサートの確認
マスターミックス溶液(表3)を20μlずつ0.5 mlチューブに分注し、滅菌爪楊枝を用いて生育したコロニーを少量かきとり、溶液内に懸濁させた。PCRは、T7 (5'-TAATACGACTCACTATAGGG-3'(配列番号10))及びSP6(5'-ATTTAGGTGACACTATAGAA-3'(配列番号11))プライマーを用い、94℃で30秒間の熱処理後、94℃で30秒の熱変性、55℃で30秒間のアニーリング及び72℃で30秒間の伸長反応の30サイクル、72℃、5分間で反応の停止並びに4℃で保存の反応をサーマルサイクラーMyCycler(BIO-RAD,USA)を用いて行った。
【0054】
【表3】
【0055】
反応終了後、PCR産物(10μl)を、エチジウムブロマイド(終濃度:1μg/ml)を含む1.5%アガロースゲルで電気泳動し、インサートの有無及びサイズの確認を行った。
4-5. プラスミドDNAの抽出
大腸菌を100μg/mlのアンピシリンを含む3 mlのLB培地(表4)で16時間培養し、8,000 rpmで3分間の遠心分離により集菌した。これを、QIAprep Spin Miniprep Kit(QIAGEN)を用い、マニュアルに従いプラスミド抽出を行った。250μlのBufferP1を加え、菌体ペレットを再懸濁した。250μlのBufferP2を添加した後、5、6回転倒させ完全に混和させた。350μlのBufferN3を添加した後、5、6回転倒させ完全に混和させ、13,000 rpm、4℃で10分間遠心分離した。
【0056】
遠心分離後、上清をデカントし、ピペッティングによりスピンカラムにアプライし、13,000 rpm、4℃で1分間遠心分離した後、フロースルー液を捨てた。Buffer PB 0.5 mlを添加した後、13,000 rpm、4℃で1分間遠心分離し、フロースルー液を捨てた。Buffer PE 0.75 mlを添加した後、13,000 rpm、4℃で1分間遠心分離し、フロースルー液を捨てた。残留している洗浄バッファーを除去するために、さらに13,000 rpm、4℃で1分間遠心分離した。
【0057】
次いで、カラムを新しい1.5 mlエッペンチューブにセットし、Buffer EBをカラムの中央にアプライし、1分間放置した後、13,000 rpm、4℃で1分間遠心分離して、得られた抽出液を精製プラスミドとした。
【0058】
【表4】
【0059】
プラスミドのコピー数を以下の方法を基に算出し、0.5×107〜1010(copies/μl)のプラスミド溶液を作製した。
1μl当たりのコピー数=DNAの量(ng)×アボガドロ数(6.022×1023)/テンプレートの長さ(bp)×109×一塩基対の平均重量(650)
【0060】
5. プライマーの設計
5-1 WSDV-LAMP用プライマーの設計
PrimerExplorer V4(https://primerexplorer.jp/lamp4.0.0/index.html)により、図1に示すように、WSDVのゲノムDNA(GenBank accession number: NC_003225.1)のwsv069遺伝子に対応する塩基番号32125〜32799(配列番号7)に対するLAMP用プライマーを作製した。設計したプライマーは次の表5に示す通りである。
【0061】
【表5】
【0062】
5-2. リアルタイムLAMP法
Loopampリアルタイム濁度測定装置(LA-200)を用いて、ウイルスの定量的検出を行った。
【0063】
5-3. 反応条件の検討
作製したcDNA溶液をテンプレートとし、WSDV-LAMPにおける最適反応温度の検討を行った。
【0064】
LAMP反応溶液を下記の表6に示す。LAMP法は、Loopamp DNA増幅試薬キット(栄研化学)を用いた。2×Reaction Mix(40 mM Tris-HCl,20 mM KCl,16 mM MgSO4,20 mM (NH4)2SO4,0.2%Tween 20,1.6 M Betaine ,2.8 mM dNTPs)12.5μl、FIPプライマーとBIPプライマーをそれぞれ1.0μl(40 pmol)、F3プライマーとB3プライマーをそれぞれ1.0μl(5 pmol)、LFプライマーとLBプライマーをそれぞれ1.0μl(5 pmol)及び蒸留水4.5μlの合計23μlを混合した。
【0065】
この混液を97℃で3分間加熱し、その後氷中で急冷し、Bst DNAポリメラーゼ(Eiken Chemical Co.Ltd., Japan)1.0μl(8U)を加えた。次いで、テンプレートcDNA溶液を1μl加えて、リアルタイム濁度測定装置(LA-200)を用い、60℃,63℃及び65℃で60分間反応させた。
【0066】
【表6】
【0067】
5-5 検量線の作成
定量LAMPの検量線作成のため、F3及びB3プライマーを用い、WSDV-LAMPにおける標的遺伝子配列を含むプラスミドの構築を行った。プラスミドのコピー数を以下の方法を基に算出し、0.5×101〜109(copies/μl)のプラスミド溶液を作製した。
1μl当たりのコピー数=OD260測定で得られたプラスミド濃度(ng/μl)/プラスミド分子量×6.02×1014(copies/μl)
このプラスミド溶液をテンプレートとし、上記と同様の方法で反応液を作製し、リアルタイム濁度測定装置を用いて、定量WSDV-LAMPにおける検量線の作成を行った。
【0068】
5-6. 感染エビからの検出
WSDVに浸漬感染させた供試エビを継時的にサンプリングし、血リンパ及び心臓からtotal RNAを抽出し、cDNAを作製した。
各cDNAをリアルタイム濁度測定装置(LA-200)を用いて、63℃で60分間反応させた。
【0069】
〔結果〕
1. リアルタイムLAMP法の反応条件検討
WSDV-LAMPにおける最適反応温度の検討結果を図2に示す。図2に示す結果から63℃における反応が最も早いことが明らかとなった。
【0070】
また、図3に示すように、wsv069遺伝子検出用LAMPの検量線について検討した結果、102〜1010copies/μlで、直線性の高い検量線が得られた。検出限界は102copies/μlであった。なお、図2及び3において、「NTC(non template control)」は、cDNAの代わりにD.W.を添加したサンプルを意味する。
【0071】
2. WSDVに浸漬感染させた供試エビの血リンパ及び心臓からのWSDV検出結果
図4に示すように、WSDVに浸漬感染させた供試エビの血リンパから、wsv069プライマーにより、浸漬4時間後から検出され、検出率は51.8%であった。
【0072】
また、図5に示すように、WSDVに浸漬感染させた供試エビの心臓から、wsv069プライマーにより、浸漬16時間後から検出され、検出率は33.3%であった。
【0073】
以上の実験結果より、wsv069遺伝子に対するプライマーを用いることで、感染初期に、より高感度な検出が可能になった。
【0074】
一方、WSDVのVP-28遺伝子は、外被タンパク質をコードする遺伝子である。WSDVのVP-28遺伝子について、LAMPを用いて本実験と同じサンプルから検出を試みた結果、血リンパと心臓のいずれからもVP-28遺伝子を検出できなかった。そこで、VP-28遺伝子検出用LAMP法より感度が良いリアルタイムPCR法(qPCR)を用いてVP-28遺伝子を検出した結果、血リンパで3.7%及び心臓で14.8%と低い検出率であった。
【0075】
表7に、上記の検出率についてまとめた。本実施例で用いたwsv069遺伝子をターゲットとするLAMP法とVP-28遺伝子検出用qPCR法を比べると、明らかにwsv069-LAMP法の検出率が高い。このことから、LAMP法は、従来感度が高いとされていたqPCR法よりも感度が優れていることが明らかとなった。
【0076】
また、wsv069-LAMP法を用いた場合、血リンパでの検出率が心臓のそれよりも高い。このことから、血リンパを用いて高感度のウイルス検査が可能であることが明らかとなった。このことは、血リンパを用いることによって非破壊検査が可能となり、エビの産卵前にウイルスの存在を明らかにすることができると考えられる。
【0077】
【表7】
【特許請求の範囲】
【請求項1】
配列番号1〜4に示される塩基配列を有する4つのプライマーを含む、環状型等温増幅反応における急性ウイルス血症原因ウイルス検出用プライマーセット。
【請求項2】
配列番号5及び6に示される塩基配列を有する2つのプライマーをさらに含む、請求項1記載のプライマーセット。
【請求項3】
請求項1又は2記載のプライマーセットを含む、環状型等温増幅反応における急性ウイルス血症原因ウイルス検出用試薬キット。
【請求項4】
請求項1又は2記載のプライマーセットと検体用核酸とを含有するサンプルを環状型等温増幅反応に供する工程と、
前記環状型等温増幅反応によって増幅産物が産生されたか否かを検出する工程と、
を含む、環状型等温増幅反応における急性ウイルス血症原因ウイルス検出方法。
【請求項5】
前記検体用核酸がmRNAである、請求項4記載の方法。
【請求項6】
前記検出が定量的検出である、請求項4又は5記載の方法。
【請求項1】
配列番号1〜4に示される塩基配列を有する4つのプライマーを含む、環状型等温増幅反応における急性ウイルス血症原因ウイルス検出用プライマーセット。
【請求項2】
配列番号5及び6に示される塩基配列を有する2つのプライマーをさらに含む、請求項1記載のプライマーセット。
【請求項3】
請求項1又は2記載のプライマーセットを含む、環状型等温増幅反応における急性ウイルス血症原因ウイルス検出用試薬キット。
【請求項4】
請求項1又は2記載のプライマーセットと検体用核酸とを含有するサンプルを環状型等温増幅反応に供する工程と、
前記環状型等温増幅反応によって増幅産物が産生されたか否かを検出する工程と、
を含む、環状型等温増幅反応における急性ウイルス血症原因ウイルス検出方法。
【請求項5】
前記検体用核酸がmRNAである、請求項4記載の方法。
【請求項6】
前記検出が定量的検出である、請求項4又は5記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−183047(P2012−183047A)
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2011−50110(P2011−50110)
【出願日】平成23年3月8日(2011.3.8)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人農業・食品産業技術総合研究機構、生物系産業創出のための異分野融合研究支援事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(504224153)国立大学法人 宮崎大学 (239)
【Fターム(参考)】
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願日】平成23年3月8日(2011.3.8)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人農業・食品産業技術総合研究機構、生物系産業創出のための異分野融合研究支援事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(504224153)国立大学法人 宮崎大学 (239)
【Fターム(参考)】
[ Back to top ]