
LCEによる肥満治療に有効な化合物の評価方法
肥満又は痩せの検査において、LCE遺伝子又はタンパク質の被検組織又は被検細胞における発現レベルや、当該遺伝子における多型等に基づいた検査をする。また、肥満又は痩せの治療薬のスクリーニング等をはじめとする化合物の評価において、LCE遺伝子又はタンパク質の性質を利用して当該評価をする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、LCE(long chain fatty acyl elongase)遺伝子又はタンパク質を用いた肥満の治療又は予防に有効な化合物の評価方法に関する。また、本発明は、前記遺伝子又はタンパク質を用いた肥満の検査方法に関する。
【背景技術】
【0002】
肥満は、高血圧症、糖尿病、高脂血症、虚血性心疾患等に代表される種々の成人病の危険因子である。また、これらの多くは慢性疾患であることから、将来的には医療費の高騰の原因になると考えられ、社会的にも大きな問題となっている。
【0003】
このような肥満を防止するために抗肥満薬の開発が進められており、現在では、食欲抑制剤や脂質吸収阻害剤が臨床的に利用されている。ここで、抗肥満薬研究の標的分子としては、これまでにレプチン、PPARγ、ニューロペプチドY等が知られているが、肥満の原因は非常に多様であるため、創薬標的として作用機序の異なった標的分子が待望されている。
【0004】
また、このような肥満状態あるいはその原因を適切に診断することは、その後の適切な治療にとって不可欠であるため、簡便で精度の高い肥満マーカーの出現が望まれている。また、近年、投与した薬剤の効果が被投与者の遺伝子多型等の遺伝子型に影響を受ける現象が見出されており、薬剤の開発段階における臨床試験やいわゆるオーダーメイド医療において、分子レベルでの検査や診断のマーカーが待望されている。
【0005】
一方、脂肪酸の生合成はアセチルCoAカルボキシラーゼと脂肪酸合成酵素によって行われる。LCE(Accession No.NM_024090(ヒト:配列番号1);NM_130450(マウス:配列番号2))は、脂肪酸合成酵素の一種であり、アセチルCoAを基質として合成が始まる脂肪酸合成経路において、ラウリン酸からミリスチン酸、ミリスチン酸からパルミチン酸、パルミチン酸からステアリン酸、パルミトレイン酸からワクセン酸等、主に炭素数が12以上の脂肪酸の炭素鎖の伸長を司ることが知られている(J.Biol.Chem.,276(48),45358−45366(2001);非特許文献1)。
【0006】
例えば、WO02/44320号公報(特許文献1)には、ELG5(LCE)が多不飽和脂肪酸(PUFA)を基質としてエロンゲース(elongase)としての活性を有することが記載されている。また、糖尿病モデルラットであるSTZ induced diabetic ratの肝臓においてエロンゲースの活性が亢進する旨の報告(Suneja et al.,1990,Biochem.Biophys.Acta,1042:81−85;非特許文献2)を引用して、エロンゲースが糖尿病等の疾患に関与するとの記載がある。
【0007】
また、摂餌によりマウスFACE(LCE)の発現量が変化する旨の報告もある(Matsuzaka T.et al.,J.Lipid Res.,43(6):911−20(2002);非特許文献3)。
【0008】
【特許文献1】WO02/44320号
【非特許文献1】J.Biol.Chem.,276(48),45358−45366,(2001)
【非特許文献2】Suneja et al.,Biochem.Biophys.Acta,1042:81−85(1990)
【非特許文献3】Matsuzaka T.et al.,J.Lipid Res.,43(6):911−20(2002)
【発明の開示】
【発明が解決しようとする課題】
【0009】
しかしながら、前記文献には、LCEと肥満とについて直接的に示したデータは開示されておらず、また、その際、LCEの飽和脂肪酸に対するエロンゲース活性が重要な役割を果たすことについては何ら示されていなかった。
【0010】
本発明は、上記従来技術の有する課題に鑑みてなされたものであり、LCEと肥満との直接的な関係を示すことにより、分子レベルで判断可能な肥満又は痩せの検査方法及び当該分子を用いた肥満及び痩せの検査薬等を提供することを目的とする。また、肥満又は痩せの治療薬や診断薬をスクリーニングする等、化合物の評価方法を提供することを目的とする。さらに、脂肪合成の抑制や肥満の抑制をする方法を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、上記目的を達成すべく鋭意研究を重ねた結果、体重変化とLCEの発現量との間に一定の相関関係があることを見出し、本発明を完成した。
【0012】
すなわち、本発明は、以下の肥満の治療又は予防に有効な化合物の評価方法(1)〜(4)を提供する。
(1)被検化合物を、被検動物又は被検細胞に、投与又は接触させる工程と、該被検化合物が、該被検動物又は該被検細胞中でLCE遺伝子あるいは該遺伝子と機能的に等価な遺伝子の発現レベルを調節するか否かを確認する工程と、を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
(2)被験化合物を、LCE遺伝子の発現調節領域とレポーター遺伝子との融合遺伝子を有する被検動物又は被検細胞に接触させる工程と、該レポーター遺伝子の被検動物又は被検細胞中での発現レベルを測定する工程と、を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
(3)被検化合物を、LCEタンパク質に接触させる工程と、該被検化合物が、該タンパク質の活性に影響を与えるか否かを確認する工程と、を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
(4)被検化合物を、LCEを含む複数種のエロンゲースタンパク質に接触させる工程と、該複数種のエロンゲースタンパク質の活性を測定する工程と、該複数種のエロンゲースタンパク質においてLCEの活性を阻害する被検化合物を選択する工程と、を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【0013】
ここで、前述した本発明の肥満の治療又は予防に有効な化合物の評価方法によって得られた化合物を有効成分として含有する肥満の治療又は予防剤も本発明に含まれる。
【0014】
本発明は、また、LCEの脂肪酸合成活性を抑制することを特徴とする脂肪合成抑制方法を提供する。ここで、LCEの脂肪酸合成活性を抑制する手段は特に制限されないが、RNAi(RNA interference)により抑制することが好ましい。RNAiは、配列番号13及び14に記載の核酸からなるsiRNA、配列番号15及び16に記載の核酸からなるsiRNA(small interfering RNA)、配列番号17及び18に記載の核酸からなるsiRNA、配列番号19及び20に記載の核酸からなるsiRNA、配列番号21及び22に記載の核酸からなるsiRNA、配列番号23及び24に記載の核酸からなるsiRNA、配列番号25及び26に記載の核酸からなるsiRNA、配列番号27及び28に記載の核酸からなるsiRNA、配列番号29及び30に記載の核酸からなるsiRNA、配列番号31及び32に記載の核酸からなるsiRNA、配列番号33及び34に記載の核酸からなるsiRNA、配列番号35及び36に記載の核酸からなるsiRNA、配列番号37及び38に記載の核酸からなるsiRNA、配列番号49及び50に記載の核酸からなるsiRNA、配列番号51及び51に記載の核酸からなるsiRNA及び配列番号53及び54に記載の核酸からなるsiRNAからなる群から選択される一以上のsiRNAを使用することにより達成されるが、特に、配列番号23及び24に記載の核酸からなるsiRNAを使用することが好ましい。
【0015】
本発明は、また、LCEの脂肪酸合成活性をRNAiにより抑制することを特徴とする肥満の治療又は予防方法を提供する。ここで、LCEの脂肪酸合成活性を抑制する手段は特に制限されないが、RNAiにより抑制することが好ましい。RNAiは、上述のsiRNAを使用することにより達成されるが、特に配列番号23及び24に記載の核酸からなるsiRNAを使用することが好ましい。
【0016】
本発明は、また、以下の肥満の検査方法(1)〜(4)を提供する。
(1)被検組織又は被検細胞におけるLCE遺伝子の発現レベル又は発現レベルの変化を測定することを特徴とする肥満の検査方法。
(2)被検組織又は被検細胞におけるLCEタンパク質の発現レベル又は発現レベルの変化を測定することを特徴とする肥満の検査方法。
(3)被検組織又は被検細胞におけるLCE遺伝子に存在する多型を検出することを特徴とする肥満の検査方法。
(4)LCEタンパク質と相互作用することによりLCE遺伝子の発現量に影響を及ぼすタンパク質の発現量又は活性を検出することを特徴とする肥満の検査方法。
【0017】
本発明は、さらに、配列番号23及び24に記載の核酸からなることを特徴とするsiRNAを提供し、当該siRNAを含むことを特徴とするLCE発現抑制剤、脂肪合成抑制剤及び肥満の治療又は予防剤を提供する。
【発明の効果】
【0018】
本発明の化合物の評価方法等によれば、LCEと肥満との直接的な関係が明らかとなり、分子レベルで判断可能な肥満又は痩せの検査方法及び当該分子を用いた肥満及び痩せの検査薬等を提供することが可能となる。また、肥満又は痩せの治療薬や診断薬のスクリーニング等、化合物の評価方法を提供することが可能となる。さらに、脂肪合成の抑制や肥満の抑制をする方法を提供することが可能となる。
【図面の簡単な説明】
【0019】
【図1】図1は、LCEを強制発現させた細胞における、(a)LCE mRNAの発現量、及び(b)LCE活性を表すグラフである。HEK293は非処理の細胞を、A5はLCEが中程度発現している細胞を、D8はLCEが高発現している細胞を、それぞれ表す。
【図2】図2は、LCEを強制発現させた細胞における脂肪酸組成を表すグラフである。黒色部分は炭素鎖の炭素数が18以上の脂肪酸を、白色部分は炭素鎖の炭素数が16以下の脂肪酸を表す。(a)〜(c)はトリグリセリドを構成する脂肪酸の組成を表し、(d)〜(f)はコレステロールエステルを構成する脂肪酸の組成を表し、(g)〜(i)はリン脂質を構成する脂肪酸の組成を表す。wtは非処理のHEK293細胞を表す。
【図3】図3は、LCEを強制発現させた細胞における詳細な脂肪酸組成を表すグラフである。(a)はトリグリセリドを構成する脂肪酸の組成を表し、(b)はコレステロールエステルを構成する脂肪酸の組成を表し、(c)はリン脂質を構成する脂肪酸の組成を表す。
【図4】図4は、RNAiによるLCEの発現抑制の結果を表す図である。(a)はLCE遺伝子における各siRNAの対応部分を表し、(b)は各siRNAを導入した場合のLCEmRNAの発現量を表している。
【図5】図5は、RNAiによるLCEの活性抑制の結果を表すグラフである。(a)はLCE mRNAの発現量を表し、(b)はFatty Acyl CoAの伸長活性を表している。LCERNAiは、hLCE−siRNA−6を用いてLCEのRNAiを行ったことを表す(以下、特に断らない限り同様である)。
【図6】図6は、siRNAを導入したHepG2細胞におけるLCEmRNAの発現量を表すグラフである。(a):24時間後、(b):48時間後。
【図7】図7は、siRNAを導入したHepG2細胞における(a)LCE、(b)FAS及び(c)SCDのmRNAの発現量を表すグラフである。
【図8】図8は、RNAiによるマウスLCEの発現抑制の結果を表す図である。(a)はマウスLCE遺伝子における各siRNAの対応部分を表し、(b)は各siRNAを導入した場合のマウスLCEmRNAの発現量を表し、(c)は各siRNAを導入した場合のマウスFAS mRNAの発現量を表している。
【図9】図9は、RNAiによる(a)LCE及び(b)FASの発現抑制並びに(c)脂肪酸合成抑制の結果を表す図である。
【図10】図10は、RNAiによる(a)LCEの発現抑制及び(b)アポリポタンパク質Bの分泌抑制の結果を表す図である。
【図11】図11は、siRNA投与による(a)体重、(b)血糖値及び(c)血漿インスリンの変化を表す図である。scramble RNAiは、哺乳類では効果を示さないscramblesiRNAを用いてRNAiを行ったコントロールを表す。
【図12】siRNAを導入したHepG2細胞におけるSREBP−1及びSREBP−2のmRNAの発現量を表すグラフである。
【図13】図13は、siRNAを導入したHepG2細胞における(a)脂肪酸合成能及び(b)中性脂肪合成能を表す図である。
【図14】図14は、siRNAを導入したHepG2細胞中の中性脂肪量を表すグラフである。
【図15】図15は、siRNAを導入したHepG2細胞における(a)CO2産生量、(b)ケトン体産生量及び(c)パルミチン酸の中性脂肪への取り込み量を表すグラフである。
【図16】図16は、siRNAを導入したHepG2細胞におけるCPT−1 mRNAの発現量を表すグラフである。
【図17】図17は、高スクロース食を負荷したマウスにおける(a)体重、(b)精巣上体の脂肪重量及び(c)肝臓におけるLCE mRNAの発現量を表すグラフである。CA−1は通常食を与えたマウスを表し、HSD(3days)及びHSD(10days)は、それぞれ、高スクロース食を3日間及び10日間与えたマウスを表す。
【図18】図18は、siRNAを投与したマウスの(a)体脂肪量、(b)脂肪重量/体重比及び(c)血漿レプチン濃度を表すグラフである。HSD SCR−RNAiは、高スクロース食を負荷したマウスに哺乳類では効果を示さないscramblesiRNAを投与したことを表す。HSD LCE−RNAiは高スクロース食を負荷したマウスにhLCE−siRNA−6を投与したことを表す。
【図19】図19は、siRNAを投与したマウスの(a)LCE mRNAの発現量、(b)FAS mRNAの発現量及び(c)肝臓における中性脂肪含量を表すグラフである。
【図20】図20は、siRNAを投与したマウスの肝臓におけるLCEタンパク質量の変化を表す図である。
【図21】図21は、siRNAを投与したマウスの肝臓における(a)ACC、FAS、SCD1、SREBP−1c、IRS−2及び(b)LCEのmRNAの発現量を表すグラフである。
【図22】図22は、LCE変異体のLCE活性を表すグラフである。(a)はシステインをアラニンに置換した変異体のLCE活性を表し、(b)はヒスチジンをアラニンに置換した変異体のLCE活性を表している。
【発明を実施するための最良の形態】
【0020】
以下、本発明の好適な実施形態について詳細に説明する。
【0021】
本発明における「発現レベル」とはLCE遺伝子の転写産物の絶対量又は相対量をいう。この場合、当該遺伝子は、DNA又はmRNAのいずれをも含む。また、発現の検出対象がタンパク質の場合、その「発現レベル」とは、LCE遺伝子の翻訳産物の絶対量又は相対量をいう。
【0022】
また、本発明における「被検動物」とは、化合物の評価に使用できる動物であれば、その種は特に限定されないが、具体的には、例えば、マウス、ラット、モルモット、ウサギ、イヌ、サルが挙げられる。
【0023】
また、本発明における「被検組織」とは、肥満又は痩せの検査を行う際に生体から抽出可能な組織であれば、その種類は特に限定されないが、肥満又は痩せの影響が反映されやすいとの観点から、例えば、肝臓組織、脂肪組織、筋肉組織、血液組織であることが好ましい。また、組織の単離が容易であるとの観点から、前記組織の中でも血液組織であることが好ましい。ここで、これらの組織の由来となる動物種については特に制限されないが、本発明の主たる用途がヒトの臨床的使用であることから、ヒトであることが好ましい。
【0024】
また、本発明における「被検細胞」についても、肥満又は痩せの検査を行う際に生体から抽出可能な細胞であれば、その種類は特に限定されないが、肥満又は痩せの影響が反映されやすいとの観点から、例えば、肝細胞、脂肪細胞(白色脂肪細胞、褐色脂肪細胞等)、筋肉細胞(筋芽細胞、骨格筋細胞、平滑筋細胞等)、膵細胞(膵島細胞等)、血球細胞であることが好ましい。ここで、かかる組織の由来となる動物種については特に限定されないが、本発明の主たる用途がヒトの臨床的使用であることから、ヒトであることが好ましい。
【0025】
さらに、本発明における「肥満」とは、脂肪組織が過剰に蓄積した状態と定義される一般的な肥満に加え、これに糖尿病や高血圧等の合併症又は内臓脂肪が伴う、いわゆる「肥満症」も含む。また、本発明における「肥満」は、薬物投与等による体重のコントロールを受けた場合に、もとの体重と比較して相対的に体重が増加した状態をも意味する。
【0026】
また、本発明における「検査」とは、肥満又は痩せであることを単に判断するのみならず、将来的な肥満又は痩せを「予測」する場合をも含む。
【0027】
また、本発明における「エロンゲース活性」とは、脂肪酸又はFatty acyl−CoAの炭素鎖を伸長させる活性を意味する。
【0028】
(1)肥満の治療又は予防に有効な化合物の評価方法
本発明の肥満の治療又は予防に有効な化合物の評価方法について説明する。被検化合物を被検動物や被検細胞に投与、接触させることにより変動するLCE遺伝子の発現量を測定したり、被検化合物をLCEタンパク質に接触させて当該タンパク質の活性に及ぼす影響を検討したりすることにより、当該被検化合物の評価を行うことが可能となる。
【0029】
すなわち、このような被検化合物の中には、細胞や組織に作用することにより、LCE遺伝子の発現レベルやLCEタンパク質の活性を正常化あるいはコントロールし、脂肪の蓄積や食欲のコントロール等、肥満の原因となるメカニズムの正常化を図ることができるものがあると考えられる。従って、以下に説明するような評価方法により、肥満の治療薬又は予防に有効な化合物を評価することが可能となる。
【0030】
(A)LCE遺伝子の発現レベル調節能を指標とする評価方法
被検化合物を被検動物又は被検細胞に投与又は接触させ、当該被検化合物が被検動物又は被検細胞中でLCE遺伝子あるいは当該遺伝子と機能的に等価な遺伝子の発現レベルを調節するか否かを確認することにより、肥満の治療又は予防に有効な化合物を評価することが可能となる。
【0031】
具体的には、以下の手順で被検化合物の評価を行う。
先ず、被検化合物を被検動物又は被検細胞に投与又は接触させる。ここで、被検化合物としては、肥満の治療又は予防薬の候補化合物であれば、その構造や性質は問わず、化合物種も限定されない。被検化合物を被検動物に投与する方法としては特に制限はなく、具体的には、例えば、経口投与、非経口投与(例えば、経皮投与、筋肉内注射、静脈内注射、皮下注射)が挙げられる。また、被検化合物を被検細胞に接触させる方法としても特に制限はなく、具体的には、例えば、培養液や緩衝液(リン酸緩衝液等)等の溶液中で混合し両者を接触させる方法が挙げられる。
【0032】
次に、被検化合物が被検動物又は被検細胞中でLCE遺伝子あるいは当該遺伝子と機能的に等価な遺伝子の発現レベルを調節するか否かを確認する。
【0033】
前記遺伝子の発現レベルの調節の有無の確認法としては、特に制限はなく、前述の投与又は接触の前を対照とし、当該遺伝子の発現量の変化をRT−PCRのような遺伝子増幅法、DNAマイクロアレイを用いる方法又はノーザンハイブリダイゼーション法等によって検出することにより実施することができる。また、前記遺伝子の発現調節領域とレポーター遺伝子との融合遺伝子を人為的に導入した動物又は細胞を用いてもよい。この場合、レポーター遺伝子としては、具体的には、例えば、β−ガラクトシダーゼ遺伝子、ルシフェラーゼ遺伝子又はグリーンフルオレッセンスプロテイン遺伝子が挙げられる。
【0034】
ここで、「LCE遺伝子と機能的に等価な遺伝子」とは、LCE遺伝子と塩基配列は異なるものの、比較的高い同性を示し、LCEと同じ又は類似の活性を有する遺伝子を示す。ここで、前記相同性は、遺伝子の機能が等価であれば特に制限はないが、塩基配列の相同性が70〜100%であることが好ましく、80〜100%であることがより好ましく、90〜100%であることがさらに好ましく、95〜100%であることが特に好ましい。相同性が前記下限より低い場合には、LCEと同じ又は類似の機能を示さない可能性が高い傾向にある。しかしながら、塩基配列の相同性が前記下限未満であっても、LCEに特有の機能を有するドメインと、当該ドメインに対応する塩基配列との相同性が高い場合にはLCE遺伝子と同様又は類似の機能を有する場合がある。このような遺伝子は、塩基配列の相同性が前記範囲外であっても好適に使用可能である。また、比較的相同性の高い遺伝子としては、LCE遺伝子における1又は2以上の塩基が自然若しくは人工的に置換、欠失、不可及び/又は挿入したものであってもよい。
【0035】
被検化合物を投与又は接触させない場合に比べて、被検化合物を投与又は接触させた場合のLCE遺伝子又はLCE遺伝子と機能的に等価な遺伝子の発現レベルが20%以上、好ましくは50%以上低下した場合、当該被検化合物は肥満の治療又は予防に有効な化合物と評価できる。
【0036】
(B)LCEタンパク質の活性を指標とする評価方法
被検化合物をLCEタンパク質に接触させ、被検化合物が当該タンパク質の活性に影響を与えるか否かを確認することにより、肥満の治療又は予防に有効な化合物を評価することが可能となる。
【0037】
具体的には、以下の手順で被検化合物の評価を行う。
先ず、被検化合物をLCEタンパク質に接触させる。このようなタンパク質と被検化合物を接触させる方法としては、特に制限はなく、具体的には、例えば、緩衝液(リン酸緩衝液等)等の溶液中で混合し接触させる方法が挙げられる。
【0038】
次に、被検化合物が当該タンパク質の活性に影響を与えるか否かを確認する。タンパク質の活性測定における条件は、使用するタンパク質の性質により適宜設定すればよい。このような条件としては、具体的には、例えば、LCEタンパク質の場合には、エロンゲース活性を指標とすることができ、具体的には、例えば、細胞から抽出したミクロソーム画分を、NADPH、パルミトイルCoA、14C標識マロニルCoA等を含む溶液中で混合してインキュベートし、脂肪酸を抽出した後、脂肪酸に含まれる放射性比活性を測定することによりエロンゲース活性を測定することができる。また、J.Biol.Chem.,276(48),45358−45366,(2001)を参照して行うこともできる。
【0039】
被検化合物を投与又は接触させない場合に比べて、被検化合物を投与又は接触させた場合のLCEタンパク質の活性が20%以上、好ましくは50%以上低下した場合、当該被検化合物は肥満の治療又は予防に有効な化合物と評価できる。
【0040】
以上説明したような本発明の肥満の治療又は予防に有効な化合物の評価方法により、肥満の治療薬や診断薬のスクリーニングや、これらの薬剤の有効性又は安全性の評価、さらには、オーダーメイド治療における適切な薬剤の選択が可能となる。
【0041】
(C)LCEタンパク質を阻害する化合物の評価方法
被検化合物を、LCEを含む複数種のエロンゲースタンパク質に接触させ、当該複数種のエロンゲースタンパク質の活性を測定した後、LCEの活性を阻害する被検化合物を選択することにより、エロンゲースの中でもLCEのエロンゲース活性を阻害する化合物を評価・選択することが可能となる。
【0042】
具体的には、以下の手順で評価を行う。
先ず、被検化合物を、LCEを含む複数種のエロンゲースタンパク質にそれぞれ接触させる。このようなタンパク質と被検化合物を接触させる方法としては、特に制限はなく、具体的には、例えば、緩衝液(リン酸緩衝液等)等の溶液中で混合し接触させる方法が挙げられる。ここで、複数種のエロンゲースとは、エロンゲース活性を有するものであればその種類は特に制限されないが、具体的には、例えば、FAS(Fatty Acid Synthase)、ELO−1が挙げられる。
【0043】
次に、被検化合物がそれぞれのタンパク質の活性に影響を与えるか否かを確認する。タンパク質の活性測定における条件は、使用するタンパク質の性質により適宜設定すればよい。このような条件としては、例えば、LCEタンパク質の場合には、エロンゲース活性を指標とすることができ、具体的には、J.Biol.Chem.,276(48),45358−45366,(2001)を参照して行うことができる。また、他のエロンゲースにおいては、LCEと同様にエロンゲース活性を指標にすればよく、公知の方法に従って活性測定を行えばよいが、例えば、前記のJ.Biol.Chem.,276(48),45358−45366,(2001)を参照して実施することができる。
【0044】
以上説明したような本発明の肥満の治療又は予防に有効な化合物の評価方法により、肥満の治療薬又は診断薬のスクリーニングや、これらの薬剤の有効性又は安全性の評価、さらには、オーダーメイド治療における適切な薬剤の選択が可能となる。
【0045】
(2)脂肪合成抑制方法及び肥満の治療又は予防方法
次に、本発明の脂肪合成抑制方法及び肥満の治療又は予防方法について説明する。LCEは脂肪の構成成分である脂肪酸の合成酵素であることから、かかる酵素活性を阻害することにより脂肪酸の合成を阻害することが可能となり、ひいては、脂肪の合成を阻害することが可能となる。
【0046】
具体的には、以下の手順で脂肪合成抑制を行う。
先ず、LCEの活性を阻害する物質を選択する。かかる物質としては、例えば、LCE阻害剤として機能する化合物、LCEの抗体、アンチセンスヌクレオチド及びRNAiに使用するsiRNA(small interfering RNA;センスRNA・アンチセンスRNAの二本鎖RNA)が挙げられる。
【0047】
次に、LCEが存在する個体、組織、細胞に対して前記の物質を導入する。具体的には、対象が個体であれば、導入方法としては特に制限はないが、前記化合物等を、動脈内注射、静脈内注射、皮下注射等の他、鼻腔内的、経気管支的、筋肉的又は経口的に導入する方法が挙げられる。また、対象が組織であれば、導入方法としては特に制限はないが、組織中への注入、緩衝液中での混合により導入する方法が挙げられる。さらに、対象が細胞であれば、導入方法としては特に制限はないが、緩衝液中での混合、エレクトロポレーション等が挙げられる。
【0048】
より具体的には、RNAiの場合、例えば、リポソームに封入したsiRNAを細胞培養液に添加し細胞に接触させることで細胞内にsiRNAを導入でき、RNAiを起こすことができる(Nature,411,494−498,(2001)、J.Cell Sci.,114(Pt 24),4557−4565,(2001)、Biochem.Biophys.Res.Commun.,301(3),804−809,2003)。LCEのRNAiには、以下のsiRNAを用いることができる。hLCE−siRNA−1(配列番号13及び14)、hLCE−siRNA−2(配列番号15及び16)、hLCE−siRNA−3(配列番号17及び18)、hLCE−siRNA−4(配列番号19及び20)、hLCE−siRNA−5(配列番号21及び22)、hLCE−siRNA−6(配列番号23及び24)、LCE−siRNA−2(配列番号25及び26)、hLCE−siRNA−7(配列番号27及び28)、hLCE−siRNA−8(配列番号29及び30)、hLCE−siRNA−9(配列番号31及び32)、hLCE−siRNA−10(配列番号33及び34)、hLCE−siRNA−11(配列番号35及び36)、hLCE−siRNA−12(配列番号37及び38)、hLCE−siRNA−6(配列番号49及び50)、mLCE−siRNA−7(配列番号51及び52)及びmLCE−siRNA−11(配列番号53及び54)。これらのsiRNAを複数組み合わせて使用することによりRNAiを生じさせてもよい。これらの中でも、hLCE−siRNA−6(配列番号23及び24)はLCEの発現抑制作用が特に強いため、LCEのRNAiに適している。
【0049】
このようにLCEの活性を阻害することにより、脂肪酸の炭素鎖の伸張反応が抑制され、脂肪酸の生合成が抑制される。
【0050】
また、かかる脂肪合成抑制方法を肥満の治療又は予防に応用することができる。すなわち、生体内でLCEの活性を阻害することにより脂肪酸の合成を抑制し、結果的に脂質の合成を抑制することにより肥満の治療又は予防が可能となる。
【0051】
具体的には、以下の手順で肥満の治療又は予防を行う。
先ず、LCEの活性を阻害する物質を選択する。かかる物質としては、例えば、LCE阻害剤として機能する化合物、LCEの抗体、アンチセンスヌクレオチド及びRNAiに使用するsiRNAが挙げられる。
【0052】
次に、このような物質を生体内に投与する。投与方法としては、特に制限はないが、例えば、動脈内注射、静脈内注射、皮下注射等の他、鼻腔内的、経気管支的、筋肉的又は経口的な投与方法が挙げられる。RNAiを用いた具体的な方法については、脂肪合成抑制方法で説明した通りである。
【0053】
(3)肥満又は痩せの検査方法
次に、本発明の肥満又は痩せの検査方法について説明する。
【0054】
(A)LCE遺伝子の発現レベルを測定することによる肥満または痩せの検査方法被検組織又は被検細胞におけるLCE遺伝子の発現レベルの変化を検出することにより、又は、発現レベルを測定することにより、当該被検組織又は被検細胞を抽出した生体(例えばヒト)が肥満であるか否かを検査・診断することが可能である。また、単に検査時の肥満の状態を検査するのみならず、将来的に肥満又は痩せになりうるかを予測することも可能である。
【0055】
以下に、このような検査の具体的な方法について説明する。
先ず、検査対象となる生体より被検組織又は被検細胞を抽出する。このような抽出の方法としては特に制限はなく、公知の方法により抽出することができる。
【0056】
次に、抽出された被検組織又は被検細胞から発現レベルの測定の対象となる遺伝子を調製する。LCE遺伝子の発現レベルを測定するには、先ず、被検組織又は被検細胞からLCEのRNA(total RNA又はmRNA)を調製する必要がある。このようなRNAの調製は、公知の方法によって行うことができるが、例えば、Molecular cloning A LABORATORY MANUAL 2nd EDITION(1989)(T.Maniatis著:Cold Spring Harbor Laboratory Press)7.3−7.36を参照して行うことができる。こうして調製したRNAを用いて、例えば、RT−PCRのような遺伝子増幅法、DNAマイクロアレイ(例えば、Affymetrix社製DNAチップ)を用いる方法、ノーザンハイブリダイゼーション法により、その発現量を測定することができる。また、被検組織又は被検細胞を用いたイン サイチュ ハイブリダイゼーション(in situ hybridization)等により、その発現量を測定することもできる。
【0057】
また、LCE遺伝子の発現レベルの変化を検出するには、前記の発現量の測定を当該発現量が変化すると予測される期間の前後(例えば、肥満治療薬の投与の前後)について行い、発現量の差を測定すればよい。具体的には、被検組織又は被検細胞において、前述したLCE遺伝子の発現量が変化すると予測される期間の前後で、その発現レベルが有意に上昇した場合に、体重の増加があった又は将来的に増加する可能性があると診断できる。
【0058】
(B)LCEタンパク質の発現レベルを測定することによる肥満又は痩せの検査方法被検組織又は被検細胞におけるLCEタンパク質の発現レベルの変化を検出することにより、又は、発現レベルを測定することにより、当該被検組織又は被検細胞を抽出した生体(例えばヒト)が肥満であるか否かを検査・診断することが可能である。また、単に検査時の肥満状態を検査するのみならず、将来的に肥満又は痩せになりうるかを予測することも可能である。
【0059】
以下、このような検査の具体的な方法について説明する。
タンパク質の発現レベルを測定する方法としては、生体から単離したタンパク質を定量する方法やタンパク質の血中濃度を測定する方法があり、具体的な方法としては特に限定されない。生体から単離したタンパク質を定量する方法の具体例としては、以下のとおりである。先ず、被検組織又は被検細胞からLCEタンパク質を調製する。このようなタンパク質の調製は、公知の方法によって行うことができる。こうして調製したタンパク質から、例えば、プロテインチップ(例えば、CIPHERGEN社製プロテインチップシステム)を用いる方法、免疫学的方法(例えば、ELISA、EIA法、ウェスタンブロッティング法)により、その発現量を測定することができる。また、被検組織又は被検細胞を用いた免疫染色等によって、その発現量を測定することもできる。一方、タンパク質の血中濃度を測定する方法の具体例としては、生体から採取した血液を用いて、上記免疫学的方法等により、LCEタンパク質を定量する方法が挙げられる。
【0060】
以上のようにして、LCEの遺伝子又はタンパク質の発現レベルを測定した後、その結果を解析することにより、被検体の肥満を検査できる。すなわち、本発明より、LCEタンパク質の発現レベルと体重は一定の相関関係を有することが明らかになったため、上記検査結果と対照群(健常人等)におけるLCEタンパク質の発現量とを比較することにより、肥満の程度を判断することが可能となる。また、本発明の検査方法によれば、単に検査時の肥満の状態を検査するのみならず、将来的な肥満又は痩せの可能性の予測も可能となる。
【0061】
また、LCEタンパク質の発現レベルの変化を検出するには、前記の発現量の測定を当該発現量が変化すると予測される期間の前後(例えば、肥満治療薬の投与の前後)について行い、発現量の差を測定すればよい。具体的には、被検組織又は被検細胞において、前述したLCEタンパク質の発現量が変化すると予測される期間の前後でその発現レベルが有意に上昇した場合に、体重の増加があった又は将来的に増加する可能性があると診断できる。
【0062】
(C)LCE遺伝子の遺伝的多型を検出する肥満又は痩せの検査方法
LCE遺伝子に遺伝的多型が存在する場合、その多型の有無や種類によりLCE遺伝子又はタンパク質の発現レベルが変化したり、当該タンパク質の活性に異常が生じたりする場合がある。従って、このような遺伝的多型を検出することによりLCEの発現や活性に関する知見を得、さらに、被検組織や被検細胞の由来となった被検体の肥満の検査を行うことができる。このような遺伝的多型としては、具体的には、例えば、ミニサテライト、マイクロサテライト、SNP(single nucleotide polymorphism:一塩基多型)が挙げられる。
【0063】
LCE遺伝子における多型の検出は以下のようにして行うことができる。すなわち、LCE遺伝子において、その発現量を制御する領域を検査対象となる肥満の被検体を対象として塩基配列を決定し、多型部位を検出する。検出された多型部位の対立遺伝子頻度を算出し、被検体集団において有意に増加又は減少している対立遺伝子を見出すことにより肥満と相関する多型を同定する。このようにして検出された遺伝的多型は、例えば、被検体由来のゲノムDNAについて、多型部位の塩基配列の解析、多型部位に存在する塩基の種類に依存して変化するDNAの物理化学的性質の差や制限酵素部位の相違を利用する方法、当該多型部位の検出に適当な検出用プローブを利用する方法及び質量分析法を利用した方法等によって臨床的に検出可能である。
【0064】
(D)LCEタンパク質と相互作用することによりLCE遺伝子の発現量に影響を及ぼすタンパク質の発現量又は活性を検出することによる肥満の検査方法
生体内において、多くのタンパク質は他のタンパク質と相互作用することにより、所定の生理機能を発揮する。LCEについても同様に、例えば、その発現を制御する転写因子等の作用を受けることにより発現量が制御され、所定の機能を発揮している。LCEタンパク質と、当該LCEタンパク質と相互作用することによりLCE遺伝子の発現量に影響を及ぼすタンパク質の発現量やその活性とは、一定の相関関係を有し、いずれか一方の挙動を検出することにより、他方の挙動を推測できる関係にある。
【0065】
ここで、「相互作用」とは、LCEタンパク質と別のタンパク質が直接的又は間接的に作用することをいい、例えば、LCEタンパク質が別のタンパク質と物理的に接触することによりアミノ酸の修飾等を生じるような作用や、第3のタンパク質を介して相互作用し、間接的にLCEタンパク質の発現に影響を及ぼすような作用が挙げられる。このようなタンパク質としては、例えば、LCEタンパク質を介するシグナル伝達において、LCEタンパク質の上流又は下流で生理的機能を発揮するタンパク質が挙げられる。このようなタンパク質の発現量又は活性を検出する方法としては、対象となるタンパク質の種類に応じて好適な手段を適宜選択すればよく、具体的な手段としては特に限定されない。
【0066】
以上の(A)〜(D)で説明したような本発明の肥満の検査方法によって、分子レベルで肥満の診断が可能となるばかりか、将来的に肥満になる可能性についても予測できることとなり、従来の診断方法と比較して、より的確な診断が可能となる。
【0067】
(4)肥満の治療、予防剤
LCE遺伝子は、その発現量と体重とが相関関係を示す。従って、当該遺伝子の発現レベルを正常レベルへと調節する化合物は肥満の治療又は予防に有用であるのみならず、例えば、痩せ、糖尿病、高血圧症、高脂血症、虚血性心疾患にも応用可能である。このような化合物としては、上述したような本発明の化合物の評価方法によって選択された化合物が挙げられる。これらの化合物を薬剤として使用するには、当該化合物を直接患者へ投与する以外に、公知の製剤学的方法により製剤化した医薬組成物として投与することもできる。製剤化するに際し、薬理学上許容される担体若しくは媒体としては、具体的には、例えば、滅菌水、生理食塩水、植物油、乳化剤、懸濁剤、界面活性剤、安定化剤、結合剤、滑沢剤、甘味料、香料又は着色剤が挙げられる。また、このような医薬組成物を患者に投与する方法としては、例えば、動脈内注射、静脈内注射、皮下注射等の他、鼻腔内的、経気管支的、筋肉的又は経口的な投与が挙げられる。医薬組成物の投与量は、患者の体重、年齢又は投与方法等により変動するが、当業者であれば適当な投与量を適宜選択することが可能である。
【0068】
(5)肥満の検査薬、検査キット
LCEタンパク質の発現量は、肥満に基づく体重変化と相関関係を有する。従って、当該タンパク質に対する抗体を使用して被検細胞や被検組織中のタンパク質量を検出、測定することにより、肥満の検査を簡便に行うことができる。ここで、「抗体」とは、抗原であるLCE遺伝子産物に結合しうる抗体分子全体又はその断片をいう。このような抗体は、公知の方法によって製造することができ、モノクローナル抗体又はポリクローナル抗体のいずれであってもよい。また、当該抗体を用いた免疫学的測定法としては、公知の方法を使用すればよく、具体的には、例えば、蛍光抗体法、酵素抗体法が挙げられる。
【0069】
また、このような抗体を含んだキットを製造し、本発明を実施することも可能である。キットの構成としては、当該抗体に加え、例えば、抗体を検出するために蛍光標識やラジオアイソトープで標識された2次抗体や抗原抗体反応を行う際に使用する緩衝液を備えていてもよい。
【0070】
このような肥満の検査薬を使用することにより、分子レベルで肥満の診断が可能となるばかりか、将来的に肥満になる可能性についても予測できることとなり、従来の診断方法と比較して、より的確な診断が可能となる。また、本発明の肥満の検査キットを使用することにより、前述したような的確な診断を非常に簡便に実施することが可能となる。
【0071】
(6)hLCE−siRNA−6(配列番号23及び24の核酸からなるsiRNA)及びそれを含むLCE発現抑制剤、脂肪合成抑制剤、肥満の治療又は予防剤
既に述べたように、配列番号23及び24の核酸からなるsiRNAであるhLCE−siRNA−6は、LCEの発現を強く抑制する。したがって、hLCE−siRNA−6はLCE発現抑制剤として用いることができ、また、脂肪合成抑制剤として用いることもでき、さらに、肥満の治療又は予防剤として用いることも可能である。
【実施例】
【0072】
(肥満モデル動物の作製)
製造例1:ニューロペプチドY(Neuropeptide Y:NPY)Y5アゴニスト脳質内(i.c.v.)投与マウス)
【0073】
NPY Y5アゴニストを投与することにより肥満を呈するモデルマウスを以下の要領で作製した。9〜12週齢の雄マウス(C57BL/6J:クレア社製)を、室温23±2℃、湿度55±15%の条件下、1プラスチックゲージに1匹ずつ飼育した。また、飼育時の明暗のサイクルは12時間とし、午前7時に点灯し、午後7時に消灯した。また、マウスには、飼料(CE−2(タンパク質:25.4重量%、炭水化物:50.3重量%、脂質:4.4重量%):クレア社製)と水を自由に摂取させた。
【0074】
マウスを80mg/kgペントバルビタールナトリウム(ダイナボット社製)で麻酔し、滅菌された28ゲージの脳注入カニューレ(アルゼ(Alzet)社製)を右側脳質へ定位的に移植した。カニューレを、ブレグマより後方へ0.4mm、側方へ0.8mm、深さ2mmの位置に、頭蓋骨に対し垂直に歯科用セメントで固定した。カニューレを、0.05%ウシ血清アルブミン(BSA)を含む10mMのリン酸緩衝液で満たした浸透圧ポンプ(モデルナンバー2002:アルゼ社製)にポリビニルクロライドチューブで接続した。10mM PBS(0.05%BSA含む)にD−Try34NPY(5マイクログラム/日になるように調製)を溶解した溶液をポンプに満たした後、マウスの背中の皮下に埋め込み、抗生物質(50mg/kgのセファメジン(Cefamedine):藤沢薬品製)を皮下注射した。
【0075】
これらのマウスを、平均体重を一致させた3つのグループ(溶媒のみを注入したグループ(vehicle group)、D−Try34NPY(NPY Y5アゴニスト)を注入したグループ(ad lib fed group)、D−Try34NPYを注入しペアフィードとしたグループ(pair−fed group))に分けた。
【0076】
製造例2:MCH投与マウス
MCH(melanin−concentrating hormone)を投与することにより肥満を呈するモデルマウスを以下の要領で作製した。
【0077】
13週齢の雄マウス(C57BL/6J:クレア社製)を、室温23±2℃、湿度55±15%の条件下、1プラスチックゲージに1匹ずつ飼育した。また、飼育時の明暗のサイクルは12時間とし、午前7時に点灯し、午後7時に消灯した。また、マウスには、飼料(CE−2(タンパク質:25.4重量%、炭水化物:50.3重量%、脂質:4.4重量%):クレア社製)と水は自由に摂取させた。マウスが環境に適応した頃、飼料としてMHF(タンパク質:15.0重量%、炭水化物:52.4重量%、脂質:32.6重量%、オリエンタルバイオサービス社製)を与えた。
【0078】
マウスを80mg/kgペントバルビタールナトリウム(ダイナボット社製)で麻酔し、滅菌された28ゲージの脳注入カニューレ(アルゼ(Alzet)社製)を右側脳質へ定位的に移植した。カニューレはブレグマより後方へ0.4mm、側方へ0.8mm、深さ2mmの位置に、頭蓋骨に対し垂直に歯科用セメントで固定した。カニューレを、30%プロピレングリコールで満たした浸透圧ポンプ(モデルナンバー2002:アルゼ社製)にポリビニルクロライドチューブで接続した。ポンプをマウスの背中の皮下に埋め込み、抗生物質を皮下注射した。
【0079】
これらのマウスを、平均体重を一致させた3つのグループ(溶媒のみを注入したグループ(vehicle group)、MCHを注入したグループ(ad lib fed group)、MCHを注入しペアフィードとしたグループ(pair−fed group))に分けた。続いて、エーテル麻酔下で、ポンプをMCH(3マイクログラム/日)又は溶媒(30%プロピレングリコール)に置き換えた。
【0080】
製造例3:DIO(Diet induced obesity)マウス
18週齢のマウス(C57BL/6J:クレア社製)を、室温23±2℃、湿度55±15%の条件下、1プラスチックゲージに1匹ずつ飼育した。このマウスに高カロリー食であるMHF(タンパク質:18.2重量%、炭水化物:55.6重量%、脂質:15.5重量%)を6ヶ月間に渡って与え、肥満を呈するモデルマウス(DIOマウス)を作製した。なお、実施例中、「established MFD」は、これ以上体重が増えないようになるまでMHPを与えて飼育したマウスを指す。
【0081】
また、前記のマウスにMHFよりさらに高い脂肪を含有する高カロリー食であるHFD(タンパク質:20.8重量%、炭水化物:38.59重量%、脂質:32.88重量%)を与えたDIOマウス(HPD)も作製した。
【0082】
製造例4:食事制限をしたマウス
マウス(C57BL/6N、17週齢)を1ケージに1匹ずつ個別に飼育した。また、エサは普通食(CA−1、CLEA)を与えた。摂食制限は、以下のようなスケジュールで行った。すなわち、エサ(CA−1)を1日につき3時間(10:00〜13:00)だけ与え、水は自由に摂取できるようにした。摂食時間の前後で餌の重量を測定し、その差を摂食量とした。また、摂食制限をしている期間は、体重、外見の観察等をモニターした。なお、条件付けに失敗したと思われるマウス(短期間に極度な体重減少(例えば20%程度の減少)が見られるマウス)は実験には使用しなかった。かかる条件下でマウスを7日間飼育した後、白色脂肪細胞を摘出した。
【0083】
実施例1〜5及び比較例1:白色脂肪細胞におけるLCEの発現
製造例1〜4において製造したモデルマウスを用いて、肝臓及び白色脂肪細胞(WAT)におけるLCE遺伝子の発現量を測定した。発現量の測定は、各モデルマウスの白色脂肪細胞から抽出したRNAを、mouse U74Aチップ(Affymetrix社製)を用いて処理することにより行った。
【0084】
非処理のC57BL/6Nの肝臓又はWATそれぞれにおけるLCEの発現量を1.00とした場合の、DIOマウス(DIO)、D−Try34NPY投与マウス(NPY(FF))、D−Try34NPY pair feeding投与群マウス(NPY(PF))、MCH投与マウス(MCH(FF))、MCH pair feeding投与群マウス(MCH(PF))、食事制限をしたマウス(Fasting)及びNPY Y5 antagonist投与マウス(Y5ant)におけるLCE遺伝子の発現量を表1に示す。
【0085】
表1より明らかなように、肥満モデルマウスではLCE遺伝子の発現量が増加する傾向にあり、食事制限をしたマウスではその発現量が低下していた。したがって、LCEの発現量と体重とが相関関係を有していることが明らかとなった。
【0086】
【表1】

【0087】
実施例6:HEK293細胞におけるマウスLCEmRNAの発現量測定
1.LCE発現亢進細胞の作製
マウス肝臓より抽出したRNAを用いて、RT−PCR法によりマウスLCEのcDNAを増幅した。得られたPCR産物を発現ベクターpCDNA3.1にサブクローニングした後、その塩基配列を確認した。次に、マウスLCEがサブクローニングされた発現ベクターを制限酵素ScaIで直線化し、HEK293細胞に導入した。この細胞を1mg/ml G418を含む選択培地で培養することにより、マウスLCE遺伝子の安定高発現細胞株を得た。
【0088】
以下に、RT−PCRに用いたプライマーの塩基配列を示す。
mLCE−exF:5’−
GCC ACC ATG GGC AAC ATG TCA GTG TTG ACT TTA C−3’(配列番号3)
mLCE−exR:5’−
CTA CTC AGC CTT CGT GGC TTT CTT−3’(配列番号4)
【0089】
2.HEK293細胞におけるマウスLCE mRNAの発現量測定
HEK293細胞からtotal RNAを精製し、逆転写反応によりcDNAを得た。ABI PRISM 7700 Sequence Detector SystemによるTaqMan PCR法を用いてマウスLCE mRNA及びヒトLCE mRNAの発現量を測定した。マウスLCEの標準曲線は、前記のマウスLCEがサブクローニングされた発現ベクターを制限酵素ScaIにより直鎖化したDNAを用いた発現解析により作成した。また、ヒトLCEの標準曲線は、PCRにより調製したヒトLCEのcDNA断片をpcDNA3.1にサブクローニングし、ScaIにより直鎖化したDNA断片を用いた発現解析により作成した。次に、マウスLCEの発現量をヒトLCEの発現量にて除し、導入されたマウスLCE遺伝子の発現量と内在性ヒトLCE遺伝子の発現量の比を求めた。
【0090】
以下に使用したプライマー及びプローブの塩基配列を記す。
マウスLCE用TaqMan probe
mLCE−P:5’−
CTT TCC TGT TTT CTG CGC TGT ACG CTG−3’(配列番号5)
マウスLCE用TaqMan primer
mLCE−F:5’−
GGA TGC AGG AAA ACT GGA AGA A−3’(配列番号6)
mLCE−R:5’−
TGC CGA CCA CCA AAG ATA AAG−3’(配列番号7)
ヒトLCE用TaqMan probe
hLCE−P2:5’−
ATC ACT GTG CTC CTG TAC T−3’(配列番号8)
ヒトLCE用TaqMan primer
hLCE−F2:5’−
AGC TGA TCT TCC TGC ACT GGT AT−3’(配列番号9)
hLCE−R2:5’−
GGC AAC CAT GTC TTT GTA GGA GTA−3’(配列番号10)
ヒトLCE用PCR primer
mLCE−exF:5’−
GCC ACC ATG GGC AAC ATG TCA GTG TTG ACT TTA C−3’(配列番号11)
hLCE−exR:5’−
CTA TTC AGC TTT CGT TGT TTT CCT C−3’(配列番号12)。
【0091】
3.HEK293細胞におけるLCE活性の測定
HEK293細胞を超音波破砕した後に、超遠心操作によりミクロソーム画分を調製した。得られたミクロソーム画分を用いて以下の方法によりLCE活性を測定した。反応に必要とされるNADPH、パルミトイルCoA、14C標識マロニルCoAを含むリン酸緩衝液中にミクロソーム画分を加えて、37℃で5分間インキュベートした。次に、15%水酸化カリウム−メタノール溶液を加え、75℃で45分間加熱してけん化処理を行なった。これに5N塩酸を加えた後、ヘキサンを用いて脂肪酸を抽出した。得られた脂肪酸に含まれる放射性比活性を測定し、脂肪酸延伸反応により脂肪酸に取り込まれたマロニルCoAの量を求めた。
【0092】
図1は、LCEを強制発現させた細胞における、(a)LCE mRNAの発現量、及び(b)LCE活性を表すグラフである。図1に示すように、LCEの発現が亢進した細胞株を取得できたことが確認されるとともに、これらの細胞株ではLCEの活性も亢進していることが確認できた。
【0093】
実施例7:HEK293細胞中に含まれる脂肪酸組成の測定
HEK293細胞をリン酸緩衝液中にて超音波破砕し、内部標準物質としてC17:0の中性脂肪(triglycerides)、コレステリルエステル(cholesteryl esters)及びリン脂質(phospholipids)を加えた後に、クロロホルム−メタノール(2:1)により脂質成分を抽出した。得られた脂質を窒素気流下にて乾固した後、silica gel Gを用いた薄層クロマトグラフによる分画(ヘキサン:ジエチルエーテル:酢酸=80:20:1)を行い、中性脂肪、コレステリルエステル及びリン脂質を分離した。これら3画分それぞれを5%塩酸−メタノールにより脂肪酸残基をメチル化し、ガスクロマトグラフ(GC−FID)を用いて脂肪酸組成の分析を行なった。
【0094】
炭素鎖長18以上の脂肪酸と16以下の脂肪酸を比較した結果を図2に示す。図2から明らかなように、LCEの発現が亢進した細胞株ではC18以上の脂肪酸の構成比率が増加する傾向にあり、炭素鎖の伸長反応はLCEの活性の増強に比例して進行することが確認できた。また、細胞内では、脂肪酸のほとんどがトリグリセリド、コレステリルエステル、リン脂質といったエステル体として存在する。これら全てのエステル体においてLCEの発現亢進によりC18以上の脂肪酸の構成比率が増加する傾向が認められることから、LCEの活性変化は細胞全体の脂肪酸構成比率を変更すると考えられ、LCEは細胞の脂肪酸組成を決定する重要因子であると、本発明者らは考えている。
【0095】
また、各脂肪酸の組成を比較した結果を図3に示す。図3から明らかなように、LCEの発現が亢進した細胞株ではC18以上の脂肪酸の構成比率が増加する傾向にあることが確認できた。
【0096】
実施例8:RNAiによるヒトLCEの発現抑制
1.発現抑制実験に使用するsiRNAの検討
ヒトLCE cDNAの塩基配列情報に基づいて、下記配列からなるsiRNA(small interfering RNA)を合成した。合成したsiRNAそれぞれをHepG2細胞に導入し、24時間後に細胞からtotal RNAを調製した。次に、逆転写反応によりcDNAを作製し、ABI PRISM 7700 Sequence Detector SystemによるTaqMan PCR法によりヒトLCE mRNAの発現量を測定した。図4(a)に、LCE遺伝子における各siRNAの対応部分を示した。図4(b)は、各siRNAを導入した場合のLCE mRNAの発現量を表すグラフである。図4(b)に示すように、siRNA(hLCE−siRNA−6)を用いた場合にLCEの発現抑制の効果が高いことが確認できた。
【0097】
使用したsiRNAの配列を以下に示す。
hLCE−siRNA−1
(図4における1のsiRNA)
5’−GACCGCAAGGCAUUCAUUUUU−3’(配列番号13)
3’−UUCUGGCGUUCCGUAAGUAAA−5’(配列番号14)
hLCE−siRNA−2
(図4における2のsiRNA)
5’−CACUCGAAAUCAAGCGCUUUU−3’(配列番号15)
3’−UUGUGAGCUUUAGUUCGCGAA−5’(配列番号16)
hLCE−siRNA−3
(図4における3のsiRNA)
5’−CACGUAGCGACUCCGAAGAUU−3’(配列番号17)
3’−UUGUGCAUCGCUGAGGCUUCU−5’(配列番号18)
hLCE−siRNA−4
(図4における4のsiRNA)
5’−UGAAGCCAUCCAAUGGAUGUU−3’(配列番号19)
3’−UUACUUCGGUAGGUUACCUAC−5’(配列番号20)
hLCE−siRNA−5
(図4における5のsiRNA)
5’−GCCAUUAGUGCUCUGGUCUUU−3’(配列番号21)
3’−UUCGGUAAUCACGAGACCAGA−5’(配列番号22)
hLCE−siRNA−6
(図4における6のsiRNA)
5’−AGGCCUGAAGCAGUCAGUUUU−3’(配列番号23)
3’−UUUCCGGACUUCGUCAGUCAA−5’(配列番号24)
LCE−siRNA−2
(図4における7のsiRNA)
5’−UGGACCUGUCAGCAAAUUCUU−3’(配列番号25)
3’−UUACCUGGACAGUCGUUUAAG−5’(配列番号26)
hLCE−siRNA−7
(図4における8のsiRNA)
5’−AGCACCCGAACUAGGAGAUUU−3’(配列番号27)
3’−UUUCGUGGGCUUGAUCCUCUA−5’(配列番号28)
hLCE−siRNA−8
(図4における9のsiRNA)
5’−CAUCUUCUGGUCCUCACUCUU−3’(配列番号29)
3’−UUGUAGAAGACCAGGAGUGAG−5’(配列番号30)
hLCE−siRNA−9
(図4における10のsiRNA)
5’−UCACACGUGGUGCAGCUAAUU−3’(配列番号31)
3’−UUAGUGUGCACCACGUCGAUU−5’(配列番号32)
hLCE−siRNA−10
(図4における11のsiRNA)
5’−GCACUGCUGCUGGAAGACCUU−3’(配列番号33)
3’−UUCGUGACGACGACCUUCUGG−5’(配列番号34)
hLCE−siRNA−11
(図4における12のsiRNA)
5’−ACUGUGCGAGCACAACACAUU−3’(配列番号35)
3’−UUUGACACGCUCGUGUUGUGU−5’(配列番号36)
hLCE−siRNA−12
(図4における13のsiRNA)
5’−AGGGGGUGAAUACUUCCCCUU−3’(配列番号37)
3’−UUUCCCCCACUUAUGAAGGGG−5’(配列番号38)。
【0098】
2.HepG2細胞におけるsiRNAのLCE活性低下作用
siRNA(hLCE−siRNA−6)を導入したHepG2細胞を超音波破砕した後に、超遠心操作によりミクロソーム画分を調製した。次に、得られたミクロソーム画分のLCE活性(Fatty Acyl CoAの伸長活性)を測定した。対照には哺乳類遺伝子に相同性を持たない配列であるsiRNA(scramble siRNA Duplex:Dharmacon,Inc.)を導入したHepG2細胞を用いた。図5(a)は、LCE mRNAの発現量を表すグラフであり、図5(b)は、伸長活性を表すグラフである。図5(a),(b)に示すように、LCEの発現が特異的に抑制されるとともに、LCEの活性が抑制されることが確認できた。
【0099】
また、HepG2細胞に2種類のsiRNA(発現抑制効果が強いもの:hLCE−siRNA−6、発現抑制効果が中程度のもの:LCE−siRNA−2)をそれぞれ導入した。siRNAの導入後24時間及び48時間の時点で、細胞からtotal RNAを調製し、DNAチップ(Affymetrix社製)を用いて遺伝子発現解析を行ない、siRNAの導入により発現が低下している遺伝子を選択した。図6は、siRNA導入後のLCE mRNAの発現量を表すグラフである((a):24時間後、(b):48時間後)。siRNAの導入によりLCEの発現が抑制されていることが確認された。
【0100】
siRNAの導入により発現が低下した遺伝子は、24時間で5遺伝子、48時間で64遺伝子であった。発現が低下した遺伝子の中で、LCEと同じ脂肪酸合成に関与するFAS(Fatty Acid Synthase)及びSCD(stearoyl CoA desaturase)に注目し、その発現量を検討した。具体的には、siRNA(hLCE−siRNA−6)を導入したHepG2細胞からtotal RNAを精製し、逆転写反応によりcDNAを得た。次に、ABI PRISM 7700 Sequence Detector Systemを用いたTaqMan PCR法により、ヒトFAS mRNA及びヒトSCD mRNAの発現を測定した。標準曲線は、PCRにより調製したヒトFAS cDNA断片及びヒトSCD cDNA断片をそれぞれ用いて作成した。FAS及びSCDの発現量はβ−アクチンの発現量で規格化した。図7は、siRNAを導入したHepG2細胞における(a)LCE、(b)FAS及び(c)SCDのmRNAの発現量を表すグラフである。図7に示すとおり、siRNAの導入により、LCEの発現は顕著に低下する一方、FAS及びSCDはLCEほどの発現の低下は見られないものの、40〜60%程度の発現低下が見られることが確認された。
【0101】
以下に、測定に用いたプライマー及びプローブの塩基配列を示す。
ヒトFAS用TaqMan probe
hFAS−P:5’−
ACC CGC TCG GCA TGG CTA TCTT−3’(配列番号39)
ヒトFAS用TaqMan primer
hFAS−F:5’−
GCA AAT TCG ACC TTT CTC AGA AC−3’(配列番号40)
hFAS−R:5’−
GGA CCC CGT GGA ATG TCA−3’(配列番号41)
ヒトFAS cDNA作成用PCR primer
hFAS−4823S:5’−
TAC GCC TCC CTC AAC TTC CG−3’(配列番号42)
hFAS−5604A:5’−
CAC TTG AGG GGC CGT ACC AC−3’(配列番号43)
ヒトSCD用TaqMan probe
hSCD−P:5’−
CAC ATG CTG ATC CTC ATA ATT CCC GAC G−3’(配列番号44)
ヒトSCD用TaqMan primer
hSCD−F:5’−
GCC CAC CAC AAG TTT TCA GAA−3’(配列番号45)
hSCD−R:5’−
CCA CGT GAG AGA AGA AAA AGC C−3’(配列番号46)
ヒトSCD cDNA作成用PCR primer
hSCD−600S:5’−
TGT GGA GCC ACC GCT CTT AC−3’(配列番号47)
hSCD−931A:5’−
AAG CGT GGG CAG GAT GAA GC−3’(配列番号48)。
【0102】
実施例9:RNAiによるマウスLCEの発現抑制
実施例8と同様の実験をマウスLCEについても行った。マウスLCE cDNAの塩基配列情報に基づいて、下記配列からなるsiRNAを合成した。合成したsiRNAそれぞれを3T3−L1細胞に導入し、24時間後に細胞からtotal RNAを調製した。次に、逆転写反応によりcDNAを作製し、ABI PRISM 7700 Sequence Detector SystemによるTaqMan PCR法によりマウスLCE mRNAの発現量を測定した。
【0103】
図8は、RNAiによるマウスLCEの発現抑制の結果を表す図である。(a)はマウスLCE遺伝子における各siRNAの対応部分を示す図であり、(b)は各siRNAを導入した場合のマウスLCE mRNAの発現量を表すグラフであり、(c)は各siRNAを導入した場合のマウスFAS mRNAの発現量を表すグラフである。図8に示すように、各siRNAによりLCE及びFASの発現が抑制されることが確認された。
【0104】
実験に用いたsiRNAを以下に示す。
hLCE−siRNA−6(先述したヒトLCEに対するsiRNAであるhLCE−siRNA−6と同一)
(図8における6のsiRNA)
5’−AGGCCUGAAGCAGUCAGUUUU−3’(配列番号49)
3’−UUUCCGGACUUCGUCAGUCAA−5’(配列番号50)
mLCE−siRNA−7(図8におけるm7のsiRNA)
5’−UCCCAUAUGGUGCAGCUAAUU−3’(配列番号51)
3’−UUAGGGUAUACCACGUCGAUU−5’(配列番号52)
mLCE−siRNA−11(図8におけるm11のsiRNA)
5’−GCAUCCGUUGUUCAGUUGCUU−3’(配列番号53)
3’−UUCGUAGGCAACAAGUCAACG−5’(配列番号54)。
【0105】
実施例10:3T3−L1細胞においてLCE RNAiがFAS mRNAの発現に与える影響
脂肪細胞に分化した3T3−L1細胞にsiRNA(hLCE−siRNA−6)を導入し、24時間後に細胞からtotal RNAを精製した。得られたtotal RNAより逆転写反応によりcDNAを調製し、ABI PRISM 7700 Sequence Detector Systemを用いたTaqMan PCR法によりマウスFAS mRNAの発現量を測定した。標準曲線は、PCRにより調製したマウスFAS cDNA断片を用いて作成した。FASの発現量はβ−アクチンの発現量で規格化した。
【0106】
測定に用いたプライマーおよびプローブは下記の通りである。
マウスFAS用TaqMan probe
mFAS−P2:5’−
ATG CTG GCC AAA CTA ACT ACG GCT TCG−3’(配列番号55)
マウスFAS用TaqMan primer
mFAS−F2:5’−
TGG CCT TCT CCT CTG TAA GCT G−3’(配列番号56)
mFAS−R2:5’−
CTG TTC ACA TAT ACG CTC CAT GG−3’(配列番号57)
マウスFAS cDNA作成用PCR primer
mFAS−5541S:5’−
TTC CGC TAC ATG GCT CAG GG−3’(配列番号58)
mFAS−7551A:5’−
CCC GTA CAC TCA CTC GTG GC−3’(配列番号59)
【0107】
また、脂肪細胞に分化した3T3−L1細胞に、siRNA(hLCE−siRNA−6)を導入し、24時間後に培地に14C標識酢酸ナトリウムを添加した。添加してから4時間後に細胞を0.1%SDSにて溶解し、15%水酸化カリウム−メタノール溶液を加え75℃45分間加熱によるけん化処理を行った。これに5N塩酸を加えた後、クロロホルム−メタノール(2:1)にて脂質成分を抽出した。抽出した脂質成分をSilica gel Gを用いた薄層クロマトグラフにより分画(ヘキサン:ジエチルエーテル:酢酸=80:20:1)し、脂肪酸画分に取り込まれた14C酢酸を測定することで脂肪酸合成能を測定した。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc.)を導入した3T3−L1細胞を用いた。
【0108】
図9は、RNAiによる(a)LCE及び(b)FASの発現抑制並びに(c)脂肪酸合成抑制の結果を表す図である。図9に示すように、LCEの発現を抑制することにより、脂肪酸の合成が抑制されることが確認できた。
【0109】
実施例11:HepG2細胞においてLCE RNAiがアポリポタンパク質Bの分泌に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し48時間後に培地を交換した。培地交換後48時間インキュベートし、培養上清を回収し、培地中に分泌されたアポリポタンパク質Bを定量した。アポリポタンパク質Bの定量には、マイクロプレートのEIA法(Exocell,Inc社製:APO B TEST)を用いた。アポリポタンパク質B標準液を用いて標準曲線を作成し、培養上清中のアポリポタンパク質Bの濃度を求めた。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc.社製)を導入したHepG2細胞を用いた。
【0110】
図10は、RNAiによる(a)LCEの発現抑制及び(b)アポリポタンパク質Bの分泌抑制の結果を表す図である。図10に示すように、LCEの発現を抑制することによりアポリポタンパク質Bの分泌が抑制されることが確認できた。
【0111】
以上より、LCEの活性を抑制することにより、抗肥満効果を示すことが示唆された。
【0112】
実施例12:DIOマウスにおけるLCE RNAiが与える影響
7週齢のマウス(ICR、雌)を高カロリー食であるMHFdietにて23週間飼育し、肥満を形成させた。先ず、siRNAの投与開始前にマウスの体重を測定した。siRNA(hLCE−siRNAi−6)を、HVJ−リポソームをキャリアとして用いマウスの尾静脈から投与した(40μg/mouse/injection)。投与は1日おきに5回行い、最終投与2日後にマウスの体重を測定し、siRNA投与の前後での体重変化を求めた。眼窩静脈叢より採血し血糖値を測定した。腹部大静脈より採血し血漿インスリン濃度を測定した。対照には、哺乳類細胞では効果を示さないscramble siRNAを投与したマウスを用いた。
【0113】
図11は、siRNA投与による(a)体重、(b)血糖値及び(c)血漿インシュリンレベルの変化を表す図である。図11に示すように、LCEに対するsiRNAをマウスに投与することにより体重が減少した。また、血糖値および血漿インシュリンレベルのいずれもが減少し、LCEに対するsiRNAがLCEの活性を抑制し、肥満改善効果を示すことが個体レベルでも確認できた。
【0114】
また、これらマウスの肝臓からtotal RNAを精製し、逆転写反応によりcDNAを得た。
ABI PRISM 7700 Sequence Detector SystemによるTaqMan PCR法を用いてマウスLCE mRNA、マウスacetyl−CoA carboxylase(ACC)mRNA、マウスFAS mRNA、マウスSCD−1 mRNA、マウスSREBP−1c mRNA、マウスinsulin receptor substrate(IRS)−2 mRNAの発現量を測定した。標準曲線はPCRにより調製した各遺伝子のcDNA断片をそれぞれ用いて作製した。各遺伝子の発現量はβ−アクチンの発現量で規格化した。
【0115】
測定に用いたプライマーおよびプローブの塩基配列を示す。
マウスACC1用TaqMan probe
5’−
AGCTGCAAGCCTGTCATCCTCAATATCG−3’(配列番号73)
マウスACC1 TaqMan PCR primer
forward:
5’−TTCTGAATGTGGCTATCAAGACTGA−3’(配列番号74)
reverse:
5’−TGCTGGGTGAACTCTCTGAACA−3’(配列番号75)
マウスACC1用cDNA作製用primer
forward:
5’−TAGTGTCAGCGATGTTCTGT−3’(配列番号76)
reverse:
5’−AAATCTCTGATCCACCTCAC−3’(配列番号77)
マウスSCD−1用TaqMan probe
ACTCGCCTACACCAACGGGCTCC(配列番号78)
マウスSCD−1用TaqMan primer
forward:
5’−TTTCCAAGCGCAGTTCCG−3’(配列番号79)
reverse:
5’−ATCGAGCGTGGACTTCGGT−3’(配列番号80)
マウスSCD−1 cDNA作製用PCR primer
forward:
5’−CACCCATCCCGAGAGTCAGG−3’(配列番号81)
reverse:
5’−GTGGGCCGGCATGATGATAG−3’(配列番号82)
マウスSREBP−1c用TaqMan probe
5’−
CTTCAAATGTGCAATCCATGGCTCCGT−3’(配列番号83)
マウスSREBP−1c用TaqMan primer
forward:
5’−GTAGCGTCTGCACGCCCTA−3’(配列番号84)
reverse:
5’−CTTGGTTGTTGATGAGCTGGAG−3’(配列番号85)
マウスSREBP−1c cDNA作製用PCR primer
forward:
5’−AAGCTGTCGGGGTAGCGTCT−3’(配列番号86)
reverse:
5’−AGGCTCGAGTAACCCAGCAC−3’(配列番号87)
マウスIRS−2用TaqMan probe
5’−
ACTTAGCCGCTTCAAGCCCGATGTG−3’(配列番号88)
マウスIRS−2 TaqMan PCR用primer
forward:
5’−AGAAGGTGCCCGAGTGGC−3’(配列番号89)
reverse:
5’−CCCCAGATACCTGATCCATGA−3’(配列番号90)
マウスIRS−2 cDNA作製用primer
forward:
5’−CAGTAGGCTCCATGGATGGC−3’(配列番号91)
reverse:
5’−ATGACCTTAGCACCCCGGTG−3’(配列番号92)
【0116】
図21は、siRNAを投与したマウスの肝臓における(a)ACC、FAS、SCD1、SREBP−1c、IRS−2及び(b)LCEのmRNAの発現量を表すグラフである。LCEの発現を抑制することにより、脂肪酸合成に関わる酵素であるACC、FAS、SCD1の発現が低下しており、また脂肪酸合成を調節する転写因子であるSREBP−1cの発現量も低下していることが確認された。すなわち、DIOマウスの肝臓におけるLCEの発現抑制により、肝臓での脂肪合成が抑制されることが示唆された。
【0117】
また、IRS−2(insulin receptor substrate−2)の発現がLCE RNAi投与マウスの肝臓で亢進しているのは、肝臓のLCEの発現抑制が肝臓のインスリン感受性を亢進させることを示唆する。これは、実施例12で示したLCEの発現抑制による血糖値及び血漿インスリン濃度の低下を引き起こす原因になっていると発明者らは考えている。
【0118】
実施例13:HepG2細胞においてLCE RNAiが細胞の脂肪酸組成に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し72時間後に細胞を回収した。その細胞をリン酸緩衝液中にて超音波破砕し、内部標準としてC17:0の中性脂肪、コレステリルエステル、リン脂質を加えた後にBligh−Dyer法により脂質成分を抽出した。得られた脂質を窒素気流下にて乾固した後、silica gel Gを用いた薄層クロマトグラフによる分画(ヘキサン:ジエチルエーテル:酢酸=80:20:1)を行い、中性脂肪、コレステリルエステル、リン脂質を分離した。これら3画分それぞれを5%塩酸−メタノールにより脂肪酸残基をメチル化し、ガスクロマトグラフ(GC−FID)を用いて脂肪酸組成の分析を行った。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。
【0119】
脂肪酸組成の分析結果を表2に示す。表2に示した結果から分かるように、RNAiによりLCEの発現を抑制すると、トリグリセリド、コレステリルエステル及びリン脂質のいずれにおいてもC18以上の脂肪酸の構成比率が減少することが確認できた。
【0120】
【表2】
【0121】
実施例14:HepG2細胞においてLCE RNAiが他遺伝子の発現に与える影響
脂肪酸合成に関与するSREBP−1(sterol regulatory element binding protein−1)及びコレステロール合成に関与するSREBP−2(sterol regulatory element binding protein−2)に対して、LCE発現抑制が与える影響について調べた。siRNA(hLCE−siRNA−6)を導入したHepG2細胞からtotal RNAを精製し、逆転写反応によりcDNAを得た。ABI PRISM 7700 Sequence Detector Systemを用いたTaqMan PCR法によりヒトSREBP−1 mRNA、ヒトSREBP−2 mRNAの発現を測定した。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。標準曲線はPCRにより調製したヒトSREBP−1 cDNA断片、ヒトSREBP−2 cDNA断片をそれぞれ用いて作製した。各遺伝子の発現量はβ−アクチンの発現量で規格化した。
【0122】
測定に用いたプライマーの塩基配列を示す。
ヒトSREBP−1用TaqMan primer
forward:
5’−CAACACAGCAACCAGAAACTCAAG−3’(配列番号60)
reverse:
5’−TTGCTTTTGTGGACAGCAGTG−3’(配列番号61)
ヒトSREBP−1 cDNA作製用PCR primer
forward:5’−CGGAGAAGCTGCCTATCAAC−3’(配列番号62)
reverse:5’−GGTCAGTGTGTCCTCCACCT−3’(配列番号63)
ヒトSREBP−2用TaqMan primer
forward:
5’−GATATCGCTCCTCCATCAATGAC−3’(配列番号64)
reverse:
5’−ACTTGTGCATCTTGGCGTCTG−3’(配列番号65)
ヒトSREBP−2 cDNA作製用PCR primer
forward:
5’−CATTCTGACCACAATGCCTG−3’(配列番号66)
reverse:
5’−AGTAGGGAGAGAAGCCAGCC−3’(配列番号67)
【0123】
図12は、siRNAを導入したHepG2細胞におけるSREBP−1及びSREBP−2のmRNAの発現量を表すグラフである。図12に示すとおり、siRNAの導入により、SREBP−1の発現は顕著に低下する一方、SREBP−2の発現は変化しないことが確認された。
【0124】
実施例15:HepG2細胞においてLCE RNAiが細胞の脂質合成に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し、72時間後に培地に14C標識酢酸ナトリウムを添加した。添加4時間後に細胞を0.1%SDSにて溶解した。細胞溶解液の一部に15%水酸化カリウム−メタノール溶液を加え75℃45分間加熱によるけん化処理行った。5N塩酸を加えた後に、クロロホルム−メタノール(2:1)にて脂質成分を抽出した。抽出した脂質成分をsilica gel Gを用いた薄層クロマトグラフにより分画(ヘキサン:ジエチルエーテル:酢酸=80:20:1)し、脂肪酸画分に取り込まれた14C酢酸を測定することで脂肪酸合成能を測定した。
【0125】
また、上記細胞溶解液の一部から直接、クロロホルム−メタノール(2:1)にて脂質成分を抽出した。得られた脂質成分を薄層クロマトグラフにより分画し、中性脂肪画分に取り込まれた14C酢酸を測定することで中性脂肪合成能を測定した。得られた脂肪酸合成能および中性脂肪合成能の値は、細胞溶解液中のタンパク質量により規格化した。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。
【0126】
図13は、siRNAを導入したHepG2細胞における(a)脂肪酸合成能及び(b)中性脂肪合成能を表す図である。図13に示したように、siRNAを導入してLCEの発現を抑制することで、脂肪酸合成能及び中性脂肪合成能が低下することが確認された。
【0127】
実施例16:HepG2細胞においてLCE RNAiが細胞の中性脂肪含量に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し72時間後に細胞を回収した。その細胞をリン酸緩衝液中にて超音波破砕し、その一部からBligh−Dyer法により脂質成分を抽出した。得られた脂質を窒素気流下にて乾固した後、2−プロパノールに溶解し、酵素法により中性脂肪量を測定した。得られた中性脂肪量の値は、細胞の超音波破砕液中のタンパク質量にて規格化した。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。
【0128】
図14は、siRNAを導入したHepG2細胞中の中性脂肪量を表すグラフである。図14に示したように、siRNAを導入してLCEの発現を抑制することで、細胞内の中性脂肪含量が低下することが確認された。
【0129】
実施例17:HepG2細胞においてLCE RNAiが細胞の脂肪酸酸化に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し、72時間後に培地に14Cパルミチン酸を添加した。30分間インキュベーションした後に、培地を別チューブに移し10%トリクロロ酢酸を加えた。培地から遊離したCO2を10%水酸化ナトリウム水溶液によりトラップした。トラップされたCO2の放射比活性を測定し、培地に加えたパルミチン酸からβ酸化により作られたCO2量を求めた。またCO2を遊離させた後の培地を遠心して、その上清中の酸可溶性画分の放射比活性を測定し、培地に加えたパルミチン酸からβ酸化により作られたケトン体量を求めた。インキュベーション後のHepG2細胞は培地を取り除いた後に0.1%SDSにて溶解した。細胞溶解液の一部からクロロホルム−メタノール(2:1)にて脂質成分を抽出し、薄層クロマトグラフにより分画した。そして中性脂肪画分の放射比活性を測定し、培地に加えたパルミチン酸が細胞中の中性脂肪に取り込まれた量を求めた。得られたCO2量、ケトン体量、中性脂肪への取り込み量の値は、細胞溶解液中のタンパク質量にて規格化した。対照には、哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。
【0130】
図15は、siRNAを導入したHepG2細胞における(a)CO2産生量、(b)ケトン体産生量及び(c)パルミチン酸の中性脂肪への取り込み量を表すグラフである。図15に示したように、siRNAを導入してLCEの発現を抑制すると、CO2産生量及びケトン体産生量が増加し、パルミチン酸の中性脂肪への取り込みが低下することが確認された。脂肪酸の燃焼にともなってCO2及びケトン体が産生するため、この結果は、LCEの発現抑制は、脂肪酸の燃焼を促進することを示唆している。
【0131】
実施例18:HepG2細胞においてLCE RNAiが細胞のCPT−1の発現に与える影響
脂肪酸をミトコンドリアに取り込むトランスポーターであるCPT−1(carnitine palmitoyl transferase−1)に対して、LCE発現抑制が与える影響について調べた。HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し72時間後に細胞を回収した。この細胞からtotal RNAを精製し、逆転写反応によりcDNAを得た。ABI PRISM 7700 Sequence Detector Systemを用いたTaqMan PCR法によりヒトCPT−1 mRNAの発現を測定した。対照には、哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。標準曲線はPCRにより調製したヒトCPT−1 cDNA断片を用いて作製した。得られた値はβ−アクチンの発現量で規格化した。
【0132】
測定に用いたプライマーおよびプローブの塩基配列を示す。
ヒトCPT−1 TaqMan probe
5’−
CCGGGAGGAAATCAAACCAATTCGTC−3’(配列番号68)
ヒトCPT−1用TaqMan primer
forward:
5’−TGCTTTACAGGCGCAAACTG−3’(配列番号69)
reverse:
5’−TGGAATCGTGGATCCCAAA−3’(配列番号70)
ヒトCPT−1 cDNA作製用PCR primer
forward:
5’−ATTTGAAGTTAAAATCCTGGTGGGC−3’(配列番号71)
reverse:
5’−TTCCCACGTCCAAAATAGGC−3’(配列番号72)
【0133】
図16は、siRNAを導入したHepG2細胞におけるCPT−1 mRNAの発現量を表すグラフである。図16に示したように、siRNAを導入してLCEの発現を抑制すると、CPT−1の発現量が増加することが確認された。
【0134】
実施例19:高スクロース食負荷マウスにおける肝臓でのLCE発現の変化8週齢のマウス(ICR、雄)を高スクロース食(67%スクロース)にて3日間、および7週齢のマウス(ICR、雄)を高スクロース食にて10日間飼育した。体重測定後にマウスを安楽死させ、精巣上体脂肪を摘出し重量を測定した。また、マウスから肝臓を摘出しtotal RNAを精製し、Taq Man PCR法を用いて肝臓でのLCE mRNAの発現量を測定した。LCE mRNAの発現量はβ−アクチンの発現量で規格化した。対照には通常食(CA−1)で飼育したマウスを用いた。
【0135】
図17は、高スクロース食を負荷したマウスにおける(a)体重、(b)精巣上体の脂肪重量及び(c)肝臓におけるLCE mRNAの発現量を表すグラフである。図17に示したように、高スクロース負荷することにより、マウスの体重及び脂肪重量は増加し、肝臓におけるLCE mRNAの発現量が増加することが確認された。
【0136】
実施例20:高スクロース食負荷マウスにおけるLCE RNAi投与の作用
HVJ−リポソームをキャリアとして用い、siRNA(hLCE−siRNAi−6)を、マウス(7週齢、ICR、雄)に尾静脈から投与した(40μg/mouse/injection)。投与開始直後から、マウスを高スクロース食にて飼育した。対照には、哺乳類細胞では効果を示さないscramble siRNAを投与したマウスを用いた。siRNAの投与は1日おきに5回行い、最終投与の翌日にマウスの体重を測定し、体脂肪量をNMR analyzer(Minispec;mq7.5)にて測定し、脂肪重量/体重比を求めた。腹部大静脈より採血し、血漿レプチン濃度を測定した。
【0137】
図18は、siRNAを投与したマウスの(a)体脂肪量、(b)脂肪重量/体重比及び(c)血漿レプチン濃度を表すグラフである。LCE RNAiの投与によるLCE発現抑制は、高スクロース負荷による体脂肪量の増加を抑制した。同様に脂肪重量/体重比の増加を抑制した。これはLCEの発現抑制が、高スクロース摂食による肥満形成を阻害すること(抗肥満効果)を示す。また、レプチンは脂肪組織から分泌されるので、脂肪重量が増加すると分泌されるレプチン濃度が増加する。この実験において、高スクロース食負荷によりレプチンが増加しているのは、脂肪量が増加したことを意味し、LCE RNAiの投与群でレプチン濃度上昇が抑えられているのは、LCE RNAi投与で脂肪重量の増加が抑制されたことを意味する。
【0138】
また、マウスを安楽死させた後に肝臓を摘出し、その一部からtotal RNAを精製し、TaqMan PCR法を用いて肝臓でのLCE mRNA、FAS mRNAの発現量を測定した。LCE mRNA、FAS mRNAの発現量はβ−アクチンの発現量で規格化した。また、肝臓の一部をリン酸緩衝液中においてホモジナイズし、Bligh−Dyer法により脂質成分を抽出した。得られた脂質を窒素気流下にて乾固し、2−プロパノールに溶解し、酵素法により中性脂肪量を測定した。得られた中性脂肪量の値は肝臓ホモジナイズ液中に含まれるタンパク質量で規格化した。さらに、肝臓組織からミクロソーム画分を調製し、ウェスタンブロッティング法によりLCEのタンパク質量の変化を測定した。ウェスタンブロッティングには、マウスLCEタンパク質の一次構造に含まれるアミノ酸配列”FEAYIGKVKKATKAE”に基づいて合成した合成ペプチド”CFEAYIGKVKKATKAE”をウサギ(SPF,Japanese White Rabbit)に免疫して得た抗LCEポリクローナル抗体を用いた。
【0139】
図19は、siRNAを投与したマウスの(a)LCE mRNAの発現量、(b)FAS mRNAの発現量及び(c)肝臓における中性脂肪含量を表すグラフである。図19(a)より、高スクロース食負荷により肝臓のLCEの発現が亢進すること、すなわち肥満形成の過程で肝臓のLCE発現が亢進していることが確認された。また、LCE RNAiの投与がマウスの肝臓でのLCEの発現亢進を抑制していることが確認された。
【0140】
図19(b)より、高スクロース食負荷により肝臓のFASの発現が亢進すること、すなわち肝臓での脂肪酸合成が亢進していることが示唆された。また、LCE RNAi投与によるLCE発現抑制によりFASの発現亢進が抑制され、脂肪酸合成亢進が抑制されることが示唆された。
【0141】
図19(c)より、高スクロース食負荷により肝臓への脂肪の蓄積が亢進していることが確認された。また、LCE RNAi投与によるLCE発現抑制が肥満形成の過程での肝臓での脂肪蓄積を抑制することが確認された。
【0142】
以上より、高スクロース食負荷により肝臓での脂肪合成が亢進すること(肥満形成の促進過程)と、LCEの発現抑制がこの脂肪合成亢進を阻害(肥満形成過程を抑制)することが確認された。
【0143】
図20は、siRNAを投与したマウスの肝臓におけるLCEタンパク質量の変化を表す図である。図19に示した脂肪酸合成とLCEの発現量との関係をLCEタンパク質レベルでも確認することができた。
【0144】
実施例21:ヒトLCEのアミノ酸置換及びそれによるLCE活性の変化
ヒトLCE cDNAを、BamHIサイトを付加したforward primer(hLCE−5BamHI)およびXhoIサイトを付加したreverse primer(hLCE−3XhoI)を用いたPCRにより作製した。得られたcDNA断片を、付加したBamHIサイトとXhoIサイトを用いて、プラスミドpCMV−Tag2B(Stratagene)にサブクローニングした(wild−type LCE construct)。
【0145】
用いたプライマーの配列は以下の通りである。
hLCE−5BBamHI:
GGATCCAACATGTCAGTGTTGACTT(配列番号93)
hLCE−3XhoI:
CTCGAGCTATTCAGCTTTCGTTGTT(配列番号94)
【0146】
このプラスミドを鋳型として、PCR法を用いた点突然変異導入により変異LCEをコードするcDNAを作成した。点突然変異の導入方法を以下に記す。最初にForward primerとしてhLCE−F4を、reverse primerとして各変異導入用のmutagenic primerを用いたPCRにより変異導入LCE cDNAの5’側部分を作製した。またForward primerとしてhLCE−510Sを、reverse primerとしてT7を用いたPCRによりLCE cDNAの3’側部分を作製した。得られた各変異導入LCE cDNA 5’側断片とLCE cDNA3’側断片を混合したものを鋳型として、forward primerにhLCE−F4を、reverse primerにT7を用いたPCRにより、変異を導入したLCEのコーディング領域全長を含むcDNA断片を得た。
【0147】
用いたプライマーの配列は以下の通りである。
hLCE−F4:
AACATGTCAGTGTTGACTTTAC(配列番号95)
hLCE−510S:
GTGCTCTTCGAACTGGTGCT(配列番号96)
T7:
TAATACGACTCACTATAGGG(配列番号97)
【0148】
作製した突然変異導入LCEは下記の通りである。ヒトLCEに導入した突然変異は、99番目のシステインをアラニンに置換したもの(C99A)、225番目のシステインをアラニンに置換したもの(C225A)、141番目のヒスチジンをアラニンにしたもの(H141A)、144番目のヒスチジンをアラニンにしたもの(H144A)、145番目のヒスチジンをアラニンにしたもの(H145A)、及び174番目のヒスチジンをアラニンにしたもの(H147A)である。
【0149】
これら変異導入に使用したプライマーの配列は以下の通りである。
C99A:
CCCTGGTCGGCAACTGACTGCTTC(配列番号98)
C225A:
GTGAGAGTGGGCCTGGTCATGCTG(配列番号99)
H141A:
GTGATACCAGGCCAGGAAGATC(配列番号100)
H144A:
GTGATGTGGGCATACCAGTGC(配列番号101)
H145A:
CACAGTGATGGCGTGATACCAG(配列番号102)
H174A:
CATCACGGCGGCCACGCCATAG(配列番号103)
【0150】
得られた変異LCE cDNA断片は、制限酵素EcoRI及びXhoIを利用してプラスミドpCMV−Tag2Bにサブクローニングし、変異LCE発現コンストラクトを作製した。これらのコンストラクトをLipofectamine2000(Invitrogen)を用いてHEK293細胞にトランスフェクトし、変異LCEを発現させた。トランスフェクトから2日後に細胞を回収し、ミクロソーム画分を調製し、ミクロソーム中のLCE活性を測定した。なお、LCEの活性測定は、実施例6に記載の方法と同様の方法により行った。
【0151】
図22は、LCE変異体のLCE活性を表すグラフである。(a)はシステインをアラニンに置換した変異体のLCE活性を表し、(b)はヒスチジンをアラニンに置換した変異体のLCE活性を表している。いずれの変異体も野生型と比較して半分以下にLCE活性が低下している。
【産業上の利用可能性】
【0152】
本発明によれば、長鎖脂肪鎖伸長酵素活性を抑制する活性を有する物質(例えば、siRNA、低分子化合物、タンパク質、抗体など)を用いた代謝系疾患、循環器系疾患、中枢神経系疾患などの治療・予防方法を提供することが可能となり、また、かかる物質を含有する治療・予防剤を提供することが可能となる。代謝系疾患としては、例えば、肥満症、糖尿病、ホルモン分泌異常、高脂血症、通風、脂肪肝などが挙げられる。循環器疾患としては、例えば、狭心症、急性・うっ血性心不全、心筋梗塞、冠状動脈硬化症、高血圧、腎臓病、電解質異常などが挙げられる。中枢神経系疾患としては、過食症などが挙げられる。
【技術分野】
【0001】
本発明は、LCE(long chain fatty acyl elongase)遺伝子又はタンパク質を用いた肥満の治療又は予防に有効な化合物の評価方法に関する。また、本発明は、前記遺伝子又はタンパク質を用いた肥満の検査方法に関する。
【背景技術】
【0002】
肥満は、高血圧症、糖尿病、高脂血症、虚血性心疾患等に代表される種々の成人病の危険因子である。また、これらの多くは慢性疾患であることから、将来的には医療費の高騰の原因になると考えられ、社会的にも大きな問題となっている。
【0003】
このような肥満を防止するために抗肥満薬の開発が進められており、現在では、食欲抑制剤や脂質吸収阻害剤が臨床的に利用されている。ここで、抗肥満薬研究の標的分子としては、これまでにレプチン、PPARγ、ニューロペプチドY等が知られているが、肥満の原因は非常に多様であるため、創薬標的として作用機序の異なった標的分子が待望されている。
【0004】
また、このような肥満状態あるいはその原因を適切に診断することは、その後の適切な治療にとって不可欠であるため、簡便で精度の高い肥満マーカーの出現が望まれている。また、近年、投与した薬剤の効果が被投与者の遺伝子多型等の遺伝子型に影響を受ける現象が見出されており、薬剤の開発段階における臨床試験やいわゆるオーダーメイド医療において、分子レベルでの検査や診断のマーカーが待望されている。
【0005】
一方、脂肪酸の生合成はアセチルCoAカルボキシラーゼと脂肪酸合成酵素によって行われる。LCE(Accession No.NM_024090(ヒト:配列番号1);NM_130450(マウス:配列番号2))は、脂肪酸合成酵素の一種であり、アセチルCoAを基質として合成が始まる脂肪酸合成経路において、ラウリン酸からミリスチン酸、ミリスチン酸からパルミチン酸、パルミチン酸からステアリン酸、パルミトレイン酸からワクセン酸等、主に炭素数が12以上の脂肪酸の炭素鎖の伸長を司ることが知られている(J.Biol.Chem.,276(48),45358−45366(2001);非特許文献1)。
【0006】
例えば、WO02/44320号公報(特許文献1)には、ELG5(LCE)が多不飽和脂肪酸(PUFA)を基質としてエロンゲース(elongase)としての活性を有することが記載されている。また、糖尿病モデルラットであるSTZ induced diabetic ratの肝臓においてエロンゲースの活性が亢進する旨の報告(Suneja et al.,1990,Biochem.Biophys.Acta,1042:81−85;非特許文献2)を引用して、エロンゲースが糖尿病等の疾患に関与するとの記載がある。
【0007】
また、摂餌によりマウスFACE(LCE)の発現量が変化する旨の報告もある(Matsuzaka T.et al.,J.Lipid Res.,43(6):911−20(2002);非特許文献3)。
【0008】
【特許文献1】WO02/44320号
【非特許文献1】J.Biol.Chem.,276(48),45358−45366,(2001)
【非特許文献2】Suneja et al.,Biochem.Biophys.Acta,1042:81−85(1990)
【非特許文献3】Matsuzaka T.et al.,J.Lipid Res.,43(6):911−20(2002)
【発明の開示】
【発明が解決しようとする課題】
【0009】
しかしながら、前記文献には、LCEと肥満とについて直接的に示したデータは開示されておらず、また、その際、LCEの飽和脂肪酸に対するエロンゲース活性が重要な役割を果たすことについては何ら示されていなかった。
【0010】
本発明は、上記従来技術の有する課題に鑑みてなされたものであり、LCEと肥満との直接的な関係を示すことにより、分子レベルで判断可能な肥満又は痩せの検査方法及び当該分子を用いた肥満及び痩せの検査薬等を提供することを目的とする。また、肥満又は痩せの治療薬や診断薬をスクリーニングする等、化合物の評価方法を提供することを目的とする。さらに、脂肪合成の抑制や肥満の抑制をする方法を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、上記目的を達成すべく鋭意研究を重ねた結果、体重変化とLCEの発現量との間に一定の相関関係があることを見出し、本発明を完成した。
【0012】
すなわち、本発明は、以下の肥満の治療又は予防に有効な化合物の評価方法(1)〜(4)を提供する。
(1)被検化合物を、被検動物又は被検細胞に、投与又は接触させる工程と、該被検化合物が、該被検動物又は該被検細胞中でLCE遺伝子あるいは該遺伝子と機能的に等価な遺伝子の発現レベルを調節するか否かを確認する工程と、を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
(2)被験化合物を、LCE遺伝子の発現調節領域とレポーター遺伝子との融合遺伝子を有する被検動物又は被検細胞に接触させる工程と、該レポーター遺伝子の被検動物又は被検細胞中での発現レベルを測定する工程と、を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
(3)被検化合物を、LCEタンパク質に接触させる工程と、該被検化合物が、該タンパク質の活性に影響を与えるか否かを確認する工程と、を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
(4)被検化合物を、LCEを含む複数種のエロンゲースタンパク質に接触させる工程と、該複数種のエロンゲースタンパク質の活性を測定する工程と、該複数種のエロンゲースタンパク質においてLCEの活性を阻害する被検化合物を選択する工程と、を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【0013】
ここで、前述した本発明の肥満の治療又は予防に有効な化合物の評価方法によって得られた化合物を有効成分として含有する肥満の治療又は予防剤も本発明に含まれる。
【0014】
本発明は、また、LCEの脂肪酸合成活性を抑制することを特徴とする脂肪合成抑制方法を提供する。ここで、LCEの脂肪酸合成活性を抑制する手段は特に制限されないが、RNAi(RNA interference)により抑制することが好ましい。RNAiは、配列番号13及び14に記載の核酸からなるsiRNA、配列番号15及び16に記載の核酸からなるsiRNA(small interfering RNA)、配列番号17及び18に記載の核酸からなるsiRNA、配列番号19及び20に記載の核酸からなるsiRNA、配列番号21及び22に記載の核酸からなるsiRNA、配列番号23及び24に記載の核酸からなるsiRNA、配列番号25及び26に記載の核酸からなるsiRNA、配列番号27及び28に記載の核酸からなるsiRNA、配列番号29及び30に記載の核酸からなるsiRNA、配列番号31及び32に記載の核酸からなるsiRNA、配列番号33及び34に記載の核酸からなるsiRNA、配列番号35及び36に記載の核酸からなるsiRNA、配列番号37及び38に記載の核酸からなるsiRNA、配列番号49及び50に記載の核酸からなるsiRNA、配列番号51及び51に記載の核酸からなるsiRNA及び配列番号53及び54に記載の核酸からなるsiRNAからなる群から選択される一以上のsiRNAを使用することにより達成されるが、特に、配列番号23及び24に記載の核酸からなるsiRNAを使用することが好ましい。
【0015】
本発明は、また、LCEの脂肪酸合成活性をRNAiにより抑制することを特徴とする肥満の治療又は予防方法を提供する。ここで、LCEの脂肪酸合成活性を抑制する手段は特に制限されないが、RNAiにより抑制することが好ましい。RNAiは、上述のsiRNAを使用することにより達成されるが、特に配列番号23及び24に記載の核酸からなるsiRNAを使用することが好ましい。
【0016】
本発明は、また、以下の肥満の検査方法(1)〜(4)を提供する。
(1)被検組織又は被検細胞におけるLCE遺伝子の発現レベル又は発現レベルの変化を測定することを特徴とする肥満の検査方法。
(2)被検組織又は被検細胞におけるLCEタンパク質の発現レベル又は発現レベルの変化を測定することを特徴とする肥満の検査方法。
(3)被検組織又は被検細胞におけるLCE遺伝子に存在する多型を検出することを特徴とする肥満の検査方法。
(4)LCEタンパク質と相互作用することによりLCE遺伝子の発現量に影響を及ぼすタンパク質の発現量又は活性を検出することを特徴とする肥満の検査方法。
【0017】
本発明は、さらに、配列番号23及び24に記載の核酸からなることを特徴とするsiRNAを提供し、当該siRNAを含むことを特徴とするLCE発現抑制剤、脂肪合成抑制剤及び肥満の治療又は予防剤を提供する。
【発明の効果】
【0018】
本発明の化合物の評価方法等によれば、LCEと肥満との直接的な関係が明らかとなり、分子レベルで判断可能な肥満又は痩せの検査方法及び当該分子を用いた肥満及び痩せの検査薬等を提供することが可能となる。また、肥満又は痩せの治療薬や診断薬のスクリーニング等、化合物の評価方法を提供することが可能となる。さらに、脂肪合成の抑制や肥満の抑制をする方法を提供することが可能となる。
【図面の簡単な説明】
【0019】
【図1】図1は、LCEを強制発現させた細胞における、(a)LCE mRNAの発現量、及び(b)LCE活性を表すグラフである。HEK293は非処理の細胞を、A5はLCEが中程度発現している細胞を、D8はLCEが高発現している細胞を、それぞれ表す。
【図2】図2は、LCEを強制発現させた細胞における脂肪酸組成を表すグラフである。黒色部分は炭素鎖の炭素数が18以上の脂肪酸を、白色部分は炭素鎖の炭素数が16以下の脂肪酸を表す。(a)〜(c)はトリグリセリドを構成する脂肪酸の組成を表し、(d)〜(f)はコレステロールエステルを構成する脂肪酸の組成を表し、(g)〜(i)はリン脂質を構成する脂肪酸の組成を表す。wtは非処理のHEK293細胞を表す。
【図3】図3は、LCEを強制発現させた細胞における詳細な脂肪酸組成を表すグラフである。(a)はトリグリセリドを構成する脂肪酸の組成を表し、(b)はコレステロールエステルを構成する脂肪酸の組成を表し、(c)はリン脂質を構成する脂肪酸の組成を表す。
【図4】図4は、RNAiによるLCEの発現抑制の結果を表す図である。(a)はLCE遺伝子における各siRNAの対応部分を表し、(b)は各siRNAを導入した場合のLCEmRNAの発現量を表している。
【図5】図5は、RNAiによるLCEの活性抑制の結果を表すグラフである。(a)はLCE mRNAの発現量を表し、(b)はFatty Acyl CoAの伸長活性を表している。LCERNAiは、hLCE−siRNA−6を用いてLCEのRNAiを行ったことを表す(以下、特に断らない限り同様である)。
【図6】図6は、siRNAを導入したHepG2細胞におけるLCEmRNAの発現量を表すグラフである。(a):24時間後、(b):48時間後。
【図7】図7は、siRNAを導入したHepG2細胞における(a)LCE、(b)FAS及び(c)SCDのmRNAの発現量を表すグラフである。
【図8】図8は、RNAiによるマウスLCEの発現抑制の結果を表す図である。(a)はマウスLCE遺伝子における各siRNAの対応部分を表し、(b)は各siRNAを導入した場合のマウスLCEmRNAの発現量を表し、(c)は各siRNAを導入した場合のマウスFAS mRNAの発現量を表している。
【図9】図9は、RNAiによる(a)LCE及び(b)FASの発現抑制並びに(c)脂肪酸合成抑制の結果を表す図である。
【図10】図10は、RNAiによる(a)LCEの発現抑制及び(b)アポリポタンパク質Bの分泌抑制の結果を表す図である。
【図11】図11は、siRNA投与による(a)体重、(b)血糖値及び(c)血漿インスリンの変化を表す図である。scramble RNAiは、哺乳類では効果を示さないscramblesiRNAを用いてRNAiを行ったコントロールを表す。
【図12】siRNAを導入したHepG2細胞におけるSREBP−1及びSREBP−2のmRNAの発現量を表すグラフである。
【図13】図13は、siRNAを導入したHepG2細胞における(a)脂肪酸合成能及び(b)中性脂肪合成能を表す図である。
【図14】図14は、siRNAを導入したHepG2細胞中の中性脂肪量を表すグラフである。
【図15】図15は、siRNAを導入したHepG2細胞における(a)CO2産生量、(b)ケトン体産生量及び(c)パルミチン酸の中性脂肪への取り込み量を表すグラフである。
【図16】図16は、siRNAを導入したHepG2細胞におけるCPT−1 mRNAの発現量を表すグラフである。
【図17】図17は、高スクロース食を負荷したマウスにおける(a)体重、(b)精巣上体の脂肪重量及び(c)肝臓におけるLCE mRNAの発現量を表すグラフである。CA−1は通常食を与えたマウスを表し、HSD(3days)及びHSD(10days)は、それぞれ、高スクロース食を3日間及び10日間与えたマウスを表す。
【図18】図18は、siRNAを投与したマウスの(a)体脂肪量、(b)脂肪重量/体重比及び(c)血漿レプチン濃度を表すグラフである。HSD SCR−RNAiは、高スクロース食を負荷したマウスに哺乳類では効果を示さないscramblesiRNAを投与したことを表す。HSD LCE−RNAiは高スクロース食を負荷したマウスにhLCE−siRNA−6を投与したことを表す。
【図19】図19は、siRNAを投与したマウスの(a)LCE mRNAの発現量、(b)FAS mRNAの発現量及び(c)肝臓における中性脂肪含量を表すグラフである。
【図20】図20は、siRNAを投与したマウスの肝臓におけるLCEタンパク質量の変化を表す図である。
【図21】図21は、siRNAを投与したマウスの肝臓における(a)ACC、FAS、SCD1、SREBP−1c、IRS−2及び(b)LCEのmRNAの発現量を表すグラフである。
【図22】図22は、LCE変異体のLCE活性を表すグラフである。(a)はシステインをアラニンに置換した変異体のLCE活性を表し、(b)はヒスチジンをアラニンに置換した変異体のLCE活性を表している。
【発明を実施するための最良の形態】
【0020】
以下、本発明の好適な実施形態について詳細に説明する。
【0021】
本発明における「発現レベル」とはLCE遺伝子の転写産物の絶対量又は相対量をいう。この場合、当該遺伝子は、DNA又はmRNAのいずれをも含む。また、発現の検出対象がタンパク質の場合、その「発現レベル」とは、LCE遺伝子の翻訳産物の絶対量又は相対量をいう。
【0022】
また、本発明における「被検動物」とは、化合物の評価に使用できる動物であれば、その種は特に限定されないが、具体的には、例えば、マウス、ラット、モルモット、ウサギ、イヌ、サルが挙げられる。
【0023】
また、本発明における「被検組織」とは、肥満又は痩せの検査を行う際に生体から抽出可能な組織であれば、その種類は特に限定されないが、肥満又は痩せの影響が反映されやすいとの観点から、例えば、肝臓組織、脂肪組織、筋肉組織、血液組織であることが好ましい。また、組織の単離が容易であるとの観点から、前記組織の中でも血液組織であることが好ましい。ここで、これらの組織の由来となる動物種については特に制限されないが、本発明の主たる用途がヒトの臨床的使用であることから、ヒトであることが好ましい。
【0024】
また、本発明における「被検細胞」についても、肥満又は痩せの検査を行う際に生体から抽出可能な細胞であれば、その種類は特に限定されないが、肥満又は痩せの影響が反映されやすいとの観点から、例えば、肝細胞、脂肪細胞(白色脂肪細胞、褐色脂肪細胞等)、筋肉細胞(筋芽細胞、骨格筋細胞、平滑筋細胞等)、膵細胞(膵島細胞等)、血球細胞であることが好ましい。ここで、かかる組織の由来となる動物種については特に限定されないが、本発明の主たる用途がヒトの臨床的使用であることから、ヒトであることが好ましい。
【0025】
さらに、本発明における「肥満」とは、脂肪組織が過剰に蓄積した状態と定義される一般的な肥満に加え、これに糖尿病や高血圧等の合併症又は内臓脂肪が伴う、いわゆる「肥満症」も含む。また、本発明における「肥満」は、薬物投与等による体重のコントロールを受けた場合に、もとの体重と比較して相対的に体重が増加した状態をも意味する。
【0026】
また、本発明における「検査」とは、肥満又は痩せであることを単に判断するのみならず、将来的な肥満又は痩せを「予測」する場合をも含む。
【0027】
また、本発明における「エロンゲース活性」とは、脂肪酸又はFatty acyl−CoAの炭素鎖を伸長させる活性を意味する。
【0028】
(1)肥満の治療又は予防に有効な化合物の評価方法
本発明の肥満の治療又は予防に有効な化合物の評価方法について説明する。被検化合物を被検動物や被検細胞に投与、接触させることにより変動するLCE遺伝子の発現量を測定したり、被検化合物をLCEタンパク質に接触させて当該タンパク質の活性に及ぼす影響を検討したりすることにより、当該被検化合物の評価を行うことが可能となる。
【0029】
すなわち、このような被検化合物の中には、細胞や組織に作用することにより、LCE遺伝子の発現レベルやLCEタンパク質の活性を正常化あるいはコントロールし、脂肪の蓄積や食欲のコントロール等、肥満の原因となるメカニズムの正常化を図ることができるものがあると考えられる。従って、以下に説明するような評価方法により、肥満の治療薬又は予防に有効な化合物を評価することが可能となる。
【0030】
(A)LCE遺伝子の発現レベル調節能を指標とする評価方法
被検化合物を被検動物又は被検細胞に投与又は接触させ、当該被検化合物が被検動物又は被検細胞中でLCE遺伝子あるいは当該遺伝子と機能的に等価な遺伝子の発現レベルを調節するか否かを確認することにより、肥満の治療又は予防に有効な化合物を評価することが可能となる。
【0031】
具体的には、以下の手順で被検化合物の評価を行う。
先ず、被検化合物を被検動物又は被検細胞に投与又は接触させる。ここで、被検化合物としては、肥満の治療又は予防薬の候補化合物であれば、その構造や性質は問わず、化合物種も限定されない。被検化合物を被検動物に投与する方法としては特に制限はなく、具体的には、例えば、経口投与、非経口投与(例えば、経皮投与、筋肉内注射、静脈内注射、皮下注射)が挙げられる。また、被検化合物を被検細胞に接触させる方法としても特に制限はなく、具体的には、例えば、培養液や緩衝液(リン酸緩衝液等)等の溶液中で混合し両者を接触させる方法が挙げられる。
【0032】
次に、被検化合物が被検動物又は被検細胞中でLCE遺伝子あるいは当該遺伝子と機能的に等価な遺伝子の発現レベルを調節するか否かを確認する。
【0033】
前記遺伝子の発現レベルの調節の有無の確認法としては、特に制限はなく、前述の投与又は接触の前を対照とし、当該遺伝子の発現量の変化をRT−PCRのような遺伝子増幅法、DNAマイクロアレイを用いる方法又はノーザンハイブリダイゼーション法等によって検出することにより実施することができる。また、前記遺伝子の発現調節領域とレポーター遺伝子との融合遺伝子を人為的に導入した動物又は細胞を用いてもよい。この場合、レポーター遺伝子としては、具体的には、例えば、β−ガラクトシダーゼ遺伝子、ルシフェラーゼ遺伝子又はグリーンフルオレッセンスプロテイン遺伝子が挙げられる。
【0034】
ここで、「LCE遺伝子と機能的に等価な遺伝子」とは、LCE遺伝子と塩基配列は異なるものの、比較的高い同性を示し、LCEと同じ又は類似の活性を有する遺伝子を示す。ここで、前記相同性は、遺伝子の機能が等価であれば特に制限はないが、塩基配列の相同性が70〜100%であることが好ましく、80〜100%であることがより好ましく、90〜100%であることがさらに好ましく、95〜100%であることが特に好ましい。相同性が前記下限より低い場合には、LCEと同じ又は類似の機能を示さない可能性が高い傾向にある。しかしながら、塩基配列の相同性が前記下限未満であっても、LCEに特有の機能を有するドメインと、当該ドメインに対応する塩基配列との相同性が高い場合にはLCE遺伝子と同様又は類似の機能を有する場合がある。このような遺伝子は、塩基配列の相同性が前記範囲外であっても好適に使用可能である。また、比較的相同性の高い遺伝子としては、LCE遺伝子における1又は2以上の塩基が自然若しくは人工的に置換、欠失、不可及び/又は挿入したものであってもよい。
【0035】
被検化合物を投与又は接触させない場合に比べて、被検化合物を投与又は接触させた場合のLCE遺伝子又はLCE遺伝子と機能的に等価な遺伝子の発現レベルが20%以上、好ましくは50%以上低下した場合、当該被検化合物は肥満の治療又は予防に有効な化合物と評価できる。
【0036】
(B)LCEタンパク質の活性を指標とする評価方法
被検化合物をLCEタンパク質に接触させ、被検化合物が当該タンパク質の活性に影響を与えるか否かを確認することにより、肥満の治療又は予防に有効な化合物を評価することが可能となる。
【0037】
具体的には、以下の手順で被検化合物の評価を行う。
先ず、被検化合物をLCEタンパク質に接触させる。このようなタンパク質と被検化合物を接触させる方法としては、特に制限はなく、具体的には、例えば、緩衝液(リン酸緩衝液等)等の溶液中で混合し接触させる方法が挙げられる。
【0038】
次に、被検化合物が当該タンパク質の活性に影響を与えるか否かを確認する。タンパク質の活性測定における条件は、使用するタンパク質の性質により適宜設定すればよい。このような条件としては、具体的には、例えば、LCEタンパク質の場合には、エロンゲース活性を指標とすることができ、具体的には、例えば、細胞から抽出したミクロソーム画分を、NADPH、パルミトイルCoA、14C標識マロニルCoA等を含む溶液中で混合してインキュベートし、脂肪酸を抽出した後、脂肪酸に含まれる放射性比活性を測定することによりエロンゲース活性を測定することができる。また、J.Biol.Chem.,276(48),45358−45366,(2001)を参照して行うこともできる。
【0039】
被検化合物を投与又は接触させない場合に比べて、被検化合物を投与又は接触させた場合のLCEタンパク質の活性が20%以上、好ましくは50%以上低下した場合、当該被検化合物は肥満の治療又は予防に有効な化合物と評価できる。
【0040】
以上説明したような本発明の肥満の治療又は予防に有効な化合物の評価方法により、肥満の治療薬や診断薬のスクリーニングや、これらの薬剤の有効性又は安全性の評価、さらには、オーダーメイド治療における適切な薬剤の選択が可能となる。
【0041】
(C)LCEタンパク質を阻害する化合物の評価方法
被検化合物を、LCEを含む複数種のエロンゲースタンパク質に接触させ、当該複数種のエロンゲースタンパク質の活性を測定した後、LCEの活性を阻害する被検化合物を選択することにより、エロンゲースの中でもLCEのエロンゲース活性を阻害する化合物を評価・選択することが可能となる。
【0042】
具体的には、以下の手順で評価を行う。
先ず、被検化合物を、LCEを含む複数種のエロンゲースタンパク質にそれぞれ接触させる。このようなタンパク質と被検化合物を接触させる方法としては、特に制限はなく、具体的には、例えば、緩衝液(リン酸緩衝液等)等の溶液中で混合し接触させる方法が挙げられる。ここで、複数種のエロンゲースとは、エロンゲース活性を有するものであればその種類は特に制限されないが、具体的には、例えば、FAS(Fatty Acid Synthase)、ELO−1が挙げられる。
【0043】
次に、被検化合物がそれぞれのタンパク質の活性に影響を与えるか否かを確認する。タンパク質の活性測定における条件は、使用するタンパク質の性質により適宜設定すればよい。このような条件としては、例えば、LCEタンパク質の場合には、エロンゲース活性を指標とすることができ、具体的には、J.Biol.Chem.,276(48),45358−45366,(2001)を参照して行うことができる。また、他のエロンゲースにおいては、LCEと同様にエロンゲース活性を指標にすればよく、公知の方法に従って活性測定を行えばよいが、例えば、前記のJ.Biol.Chem.,276(48),45358−45366,(2001)を参照して実施することができる。
【0044】
以上説明したような本発明の肥満の治療又は予防に有効な化合物の評価方法により、肥満の治療薬又は診断薬のスクリーニングや、これらの薬剤の有効性又は安全性の評価、さらには、オーダーメイド治療における適切な薬剤の選択が可能となる。
【0045】
(2)脂肪合成抑制方法及び肥満の治療又は予防方法
次に、本発明の脂肪合成抑制方法及び肥満の治療又は予防方法について説明する。LCEは脂肪の構成成分である脂肪酸の合成酵素であることから、かかる酵素活性を阻害することにより脂肪酸の合成を阻害することが可能となり、ひいては、脂肪の合成を阻害することが可能となる。
【0046】
具体的には、以下の手順で脂肪合成抑制を行う。
先ず、LCEの活性を阻害する物質を選択する。かかる物質としては、例えば、LCE阻害剤として機能する化合物、LCEの抗体、アンチセンスヌクレオチド及びRNAiに使用するsiRNA(small interfering RNA;センスRNA・アンチセンスRNAの二本鎖RNA)が挙げられる。
【0047】
次に、LCEが存在する個体、組織、細胞に対して前記の物質を導入する。具体的には、対象が個体であれば、導入方法としては特に制限はないが、前記化合物等を、動脈内注射、静脈内注射、皮下注射等の他、鼻腔内的、経気管支的、筋肉的又は経口的に導入する方法が挙げられる。また、対象が組織であれば、導入方法としては特に制限はないが、組織中への注入、緩衝液中での混合により導入する方法が挙げられる。さらに、対象が細胞であれば、導入方法としては特に制限はないが、緩衝液中での混合、エレクトロポレーション等が挙げられる。
【0048】
より具体的には、RNAiの場合、例えば、リポソームに封入したsiRNAを細胞培養液に添加し細胞に接触させることで細胞内にsiRNAを導入でき、RNAiを起こすことができる(Nature,411,494−498,(2001)、J.Cell Sci.,114(Pt 24),4557−4565,(2001)、Biochem.Biophys.Res.Commun.,301(3),804−809,2003)。LCEのRNAiには、以下のsiRNAを用いることができる。hLCE−siRNA−1(配列番号13及び14)、hLCE−siRNA−2(配列番号15及び16)、hLCE−siRNA−3(配列番号17及び18)、hLCE−siRNA−4(配列番号19及び20)、hLCE−siRNA−5(配列番号21及び22)、hLCE−siRNA−6(配列番号23及び24)、LCE−siRNA−2(配列番号25及び26)、hLCE−siRNA−7(配列番号27及び28)、hLCE−siRNA−8(配列番号29及び30)、hLCE−siRNA−9(配列番号31及び32)、hLCE−siRNA−10(配列番号33及び34)、hLCE−siRNA−11(配列番号35及び36)、hLCE−siRNA−12(配列番号37及び38)、hLCE−siRNA−6(配列番号49及び50)、mLCE−siRNA−7(配列番号51及び52)及びmLCE−siRNA−11(配列番号53及び54)。これらのsiRNAを複数組み合わせて使用することによりRNAiを生じさせてもよい。これらの中でも、hLCE−siRNA−6(配列番号23及び24)はLCEの発現抑制作用が特に強いため、LCEのRNAiに適している。
【0049】
このようにLCEの活性を阻害することにより、脂肪酸の炭素鎖の伸張反応が抑制され、脂肪酸の生合成が抑制される。
【0050】
また、かかる脂肪合成抑制方法を肥満の治療又は予防に応用することができる。すなわち、生体内でLCEの活性を阻害することにより脂肪酸の合成を抑制し、結果的に脂質の合成を抑制することにより肥満の治療又は予防が可能となる。
【0051】
具体的には、以下の手順で肥満の治療又は予防を行う。
先ず、LCEの活性を阻害する物質を選択する。かかる物質としては、例えば、LCE阻害剤として機能する化合物、LCEの抗体、アンチセンスヌクレオチド及びRNAiに使用するsiRNAが挙げられる。
【0052】
次に、このような物質を生体内に投与する。投与方法としては、特に制限はないが、例えば、動脈内注射、静脈内注射、皮下注射等の他、鼻腔内的、経気管支的、筋肉的又は経口的な投与方法が挙げられる。RNAiを用いた具体的な方法については、脂肪合成抑制方法で説明した通りである。
【0053】
(3)肥満又は痩せの検査方法
次に、本発明の肥満又は痩せの検査方法について説明する。
【0054】
(A)LCE遺伝子の発現レベルを測定することによる肥満または痩せの検査方法被検組織又は被検細胞におけるLCE遺伝子の発現レベルの変化を検出することにより、又は、発現レベルを測定することにより、当該被検組織又は被検細胞を抽出した生体(例えばヒト)が肥満であるか否かを検査・診断することが可能である。また、単に検査時の肥満の状態を検査するのみならず、将来的に肥満又は痩せになりうるかを予測することも可能である。
【0055】
以下に、このような検査の具体的な方法について説明する。
先ず、検査対象となる生体より被検組織又は被検細胞を抽出する。このような抽出の方法としては特に制限はなく、公知の方法により抽出することができる。
【0056】
次に、抽出された被検組織又は被検細胞から発現レベルの測定の対象となる遺伝子を調製する。LCE遺伝子の発現レベルを測定するには、先ず、被検組織又は被検細胞からLCEのRNA(total RNA又はmRNA)を調製する必要がある。このようなRNAの調製は、公知の方法によって行うことができるが、例えば、Molecular cloning A LABORATORY MANUAL 2nd EDITION(1989)(T.Maniatis著:Cold Spring Harbor Laboratory Press)7.3−7.36を参照して行うことができる。こうして調製したRNAを用いて、例えば、RT−PCRのような遺伝子増幅法、DNAマイクロアレイ(例えば、Affymetrix社製DNAチップ)を用いる方法、ノーザンハイブリダイゼーション法により、その発現量を測定することができる。また、被検組織又は被検細胞を用いたイン サイチュ ハイブリダイゼーション(in situ hybridization)等により、その発現量を測定することもできる。
【0057】
また、LCE遺伝子の発現レベルの変化を検出するには、前記の発現量の測定を当該発現量が変化すると予測される期間の前後(例えば、肥満治療薬の投与の前後)について行い、発現量の差を測定すればよい。具体的には、被検組織又は被検細胞において、前述したLCE遺伝子の発現量が変化すると予測される期間の前後で、その発現レベルが有意に上昇した場合に、体重の増加があった又は将来的に増加する可能性があると診断できる。
【0058】
(B)LCEタンパク質の発現レベルを測定することによる肥満又は痩せの検査方法被検組織又は被検細胞におけるLCEタンパク質の発現レベルの変化を検出することにより、又は、発現レベルを測定することにより、当該被検組織又は被検細胞を抽出した生体(例えばヒト)が肥満であるか否かを検査・診断することが可能である。また、単に検査時の肥満状態を検査するのみならず、将来的に肥満又は痩せになりうるかを予測することも可能である。
【0059】
以下、このような検査の具体的な方法について説明する。
タンパク質の発現レベルを測定する方法としては、生体から単離したタンパク質を定量する方法やタンパク質の血中濃度を測定する方法があり、具体的な方法としては特に限定されない。生体から単離したタンパク質を定量する方法の具体例としては、以下のとおりである。先ず、被検組織又は被検細胞からLCEタンパク質を調製する。このようなタンパク質の調製は、公知の方法によって行うことができる。こうして調製したタンパク質から、例えば、プロテインチップ(例えば、CIPHERGEN社製プロテインチップシステム)を用いる方法、免疫学的方法(例えば、ELISA、EIA法、ウェスタンブロッティング法)により、その発現量を測定することができる。また、被検組織又は被検細胞を用いた免疫染色等によって、その発現量を測定することもできる。一方、タンパク質の血中濃度を測定する方法の具体例としては、生体から採取した血液を用いて、上記免疫学的方法等により、LCEタンパク質を定量する方法が挙げられる。
【0060】
以上のようにして、LCEの遺伝子又はタンパク質の発現レベルを測定した後、その結果を解析することにより、被検体の肥満を検査できる。すなわち、本発明より、LCEタンパク質の発現レベルと体重は一定の相関関係を有することが明らかになったため、上記検査結果と対照群(健常人等)におけるLCEタンパク質の発現量とを比較することにより、肥満の程度を判断することが可能となる。また、本発明の検査方法によれば、単に検査時の肥満の状態を検査するのみならず、将来的な肥満又は痩せの可能性の予測も可能となる。
【0061】
また、LCEタンパク質の発現レベルの変化を検出するには、前記の発現量の測定を当該発現量が変化すると予測される期間の前後(例えば、肥満治療薬の投与の前後)について行い、発現量の差を測定すればよい。具体的には、被検組織又は被検細胞において、前述したLCEタンパク質の発現量が変化すると予測される期間の前後でその発現レベルが有意に上昇した場合に、体重の増加があった又は将来的に増加する可能性があると診断できる。
【0062】
(C)LCE遺伝子の遺伝的多型を検出する肥満又は痩せの検査方法
LCE遺伝子に遺伝的多型が存在する場合、その多型の有無や種類によりLCE遺伝子又はタンパク質の発現レベルが変化したり、当該タンパク質の活性に異常が生じたりする場合がある。従って、このような遺伝的多型を検出することによりLCEの発現や活性に関する知見を得、さらに、被検組織や被検細胞の由来となった被検体の肥満の検査を行うことができる。このような遺伝的多型としては、具体的には、例えば、ミニサテライト、マイクロサテライト、SNP(single nucleotide polymorphism:一塩基多型)が挙げられる。
【0063】
LCE遺伝子における多型の検出は以下のようにして行うことができる。すなわち、LCE遺伝子において、その発現量を制御する領域を検査対象となる肥満の被検体を対象として塩基配列を決定し、多型部位を検出する。検出された多型部位の対立遺伝子頻度を算出し、被検体集団において有意に増加又は減少している対立遺伝子を見出すことにより肥満と相関する多型を同定する。このようにして検出された遺伝的多型は、例えば、被検体由来のゲノムDNAについて、多型部位の塩基配列の解析、多型部位に存在する塩基の種類に依存して変化するDNAの物理化学的性質の差や制限酵素部位の相違を利用する方法、当該多型部位の検出に適当な検出用プローブを利用する方法及び質量分析法を利用した方法等によって臨床的に検出可能である。
【0064】
(D)LCEタンパク質と相互作用することによりLCE遺伝子の発現量に影響を及ぼすタンパク質の発現量又は活性を検出することによる肥満の検査方法
生体内において、多くのタンパク質は他のタンパク質と相互作用することにより、所定の生理機能を発揮する。LCEについても同様に、例えば、その発現を制御する転写因子等の作用を受けることにより発現量が制御され、所定の機能を発揮している。LCEタンパク質と、当該LCEタンパク質と相互作用することによりLCE遺伝子の発現量に影響を及ぼすタンパク質の発現量やその活性とは、一定の相関関係を有し、いずれか一方の挙動を検出することにより、他方の挙動を推測できる関係にある。
【0065】
ここで、「相互作用」とは、LCEタンパク質と別のタンパク質が直接的又は間接的に作用することをいい、例えば、LCEタンパク質が別のタンパク質と物理的に接触することによりアミノ酸の修飾等を生じるような作用や、第3のタンパク質を介して相互作用し、間接的にLCEタンパク質の発現に影響を及ぼすような作用が挙げられる。このようなタンパク質としては、例えば、LCEタンパク質を介するシグナル伝達において、LCEタンパク質の上流又は下流で生理的機能を発揮するタンパク質が挙げられる。このようなタンパク質の発現量又は活性を検出する方法としては、対象となるタンパク質の種類に応じて好適な手段を適宜選択すればよく、具体的な手段としては特に限定されない。
【0066】
以上の(A)〜(D)で説明したような本発明の肥満の検査方法によって、分子レベルで肥満の診断が可能となるばかりか、将来的に肥満になる可能性についても予測できることとなり、従来の診断方法と比較して、より的確な診断が可能となる。
【0067】
(4)肥満の治療、予防剤
LCE遺伝子は、その発現量と体重とが相関関係を示す。従って、当該遺伝子の発現レベルを正常レベルへと調節する化合物は肥満の治療又は予防に有用であるのみならず、例えば、痩せ、糖尿病、高血圧症、高脂血症、虚血性心疾患にも応用可能である。このような化合物としては、上述したような本発明の化合物の評価方法によって選択された化合物が挙げられる。これらの化合物を薬剤として使用するには、当該化合物を直接患者へ投与する以外に、公知の製剤学的方法により製剤化した医薬組成物として投与することもできる。製剤化するに際し、薬理学上許容される担体若しくは媒体としては、具体的には、例えば、滅菌水、生理食塩水、植物油、乳化剤、懸濁剤、界面活性剤、安定化剤、結合剤、滑沢剤、甘味料、香料又は着色剤が挙げられる。また、このような医薬組成物を患者に投与する方法としては、例えば、動脈内注射、静脈内注射、皮下注射等の他、鼻腔内的、経気管支的、筋肉的又は経口的な投与が挙げられる。医薬組成物の投与量は、患者の体重、年齢又は投与方法等により変動するが、当業者であれば適当な投与量を適宜選択することが可能である。
【0068】
(5)肥満の検査薬、検査キット
LCEタンパク質の発現量は、肥満に基づく体重変化と相関関係を有する。従って、当該タンパク質に対する抗体を使用して被検細胞や被検組織中のタンパク質量を検出、測定することにより、肥満の検査を簡便に行うことができる。ここで、「抗体」とは、抗原であるLCE遺伝子産物に結合しうる抗体分子全体又はその断片をいう。このような抗体は、公知の方法によって製造することができ、モノクローナル抗体又はポリクローナル抗体のいずれであってもよい。また、当該抗体を用いた免疫学的測定法としては、公知の方法を使用すればよく、具体的には、例えば、蛍光抗体法、酵素抗体法が挙げられる。
【0069】
また、このような抗体を含んだキットを製造し、本発明を実施することも可能である。キットの構成としては、当該抗体に加え、例えば、抗体を検出するために蛍光標識やラジオアイソトープで標識された2次抗体や抗原抗体反応を行う際に使用する緩衝液を備えていてもよい。
【0070】
このような肥満の検査薬を使用することにより、分子レベルで肥満の診断が可能となるばかりか、将来的に肥満になる可能性についても予測できることとなり、従来の診断方法と比較して、より的確な診断が可能となる。また、本発明の肥満の検査キットを使用することにより、前述したような的確な診断を非常に簡便に実施することが可能となる。
【0071】
(6)hLCE−siRNA−6(配列番号23及び24の核酸からなるsiRNA)及びそれを含むLCE発現抑制剤、脂肪合成抑制剤、肥満の治療又は予防剤
既に述べたように、配列番号23及び24の核酸からなるsiRNAであるhLCE−siRNA−6は、LCEの発現を強く抑制する。したがって、hLCE−siRNA−6はLCE発現抑制剤として用いることができ、また、脂肪合成抑制剤として用いることもでき、さらに、肥満の治療又は予防剤として用いることも可能である。
【実施例】
【0072】
(肥満モデル動物の作製)
製造例1:ニューロペプチドY(Neuropeptide Y:NPY)Y5アゴニスト脳質内(i.c.v.)投与マウス)
【0073】
NPY Y5アゴニストを投与することにより肥満を呈するモデルマウスを以下の要領で作製した。9〜12週齢の雄マウス(C57BL/6J:クレア社製)を、室温23±2℃、湿度55±15%の条件下、1プラスチックゲージに1匹ずつ飼育した。また、飼育時の明暗のサイクルは12時間とし、午前7時に点灯し、午後7時に消灯した。また、マウスには、飼料(CE−2(タンパク質:25.4重量%、炭水化物:50.3重量%、脂質:4.4重量%):クレア社製)と水を自由に摂取させた。
【0074】
マウスを80mg/kgペントバルビタールナトリウム(ダイナボット社製)で麻酔し、滅菌された28ゲージの脳注入カニューレ(アルゼ(Alzet)社製)を右側脳質へ定位的に移植した。カニューレを、ブレグマより後方へ0.4mm、側方へ0.8mm、深さ2mmの位置に、頭蓋骨に対し垂直に歯科用セメントで固定した。カニューレを、0.05%ウシ血清アルブミン(BSA)を含む10mMのリン酸緩衝液で満たした浸透圧ポンプ(モデルナンバー2002:アルゼ社製)にポリビニルクロライドチューブで接続した。10mM PBS(0.05%BSA含む)にD−Try34NPY(5マイクログラム/日になるように調製)を溶解した溶液をポンプに満たした後、マウスの背中の皮下に埋め込み、抗生物質(50mg/kgのセファメジン(Cefamedine):藤沢薬品製)を皮下注射した。
【0075】
これらのマウスを、平均体重を一致させた3つのグループ(溶媒のみを注入したグループ(vehicle group)、D−Try34NPY(NPY Y5アゴニスト)を注入したグループ(ad lib fed group)、D−Try34NPYを注入しペアフィードとしたグループ(pair−fed group))に分けた。
【0076】
製造例2:MCH投与マウス
MCH(melanin−concentrating hormone)を投与することにより肥満を呈するモデルマウスを以下の要領で作製した。
【0077】
13週齢の雄マウス(C57BL/6J:クレア社製)を、室温23±2℃、湿度55±15%の条件下、1プラスチックゲージに1匹ずつ飼育した。また、飼育時の明暗のサイクルは12時間とし、午前7時に点灯し、午後7時に消灯した。また、マウスには、飼料(CE−2(タンパク質:25.4重量%、炭水化物:50.3重量%、脂質:4.4重量%):クレア社製)と水は自由に摂取させた。マウスが環境に適応した頃、飼料としてMHF(タンパク質:15.0重量%、炭水化物:52.4重量%、脂質:32.6重量%、オリエンタルバイオサービス社製)を与えた。
【0078】
マウスを80mg/kgペントバルビタールナトリウム(ダイナボット社製)で麻酔し、滅菌された28ゲージの脳注入カニューレ(アルゼ(Alzet)社製)を右側脳質へ定位的に移植した。カニューレはブレグマより後方へ0.4mm、側方へ0.8mm、深さ2mmの位置に、頭蓋骨に対し垂直に歯科用セメントで固定した。カニューレを、30%プロピレングリコールで満たした浸透圧ポンプ(モデルナンバー2002:アルゼ社製)にポリビニルクロライドチューブで接続した。ポンプをマウスの背中の皮下に埋め込み、抗生物質を皮下注射した。
【0079】
これらのマウスを、平均体重を一致させた3つのグループ(溶媒のみを注入したグループ(vehicle group)、MCHを注入したグループ(ad lib fed group)、MCHを注入しペアフィードとしたグループ(pair−fed group))に分けた。続いて、エーテル麻酔下で、ポンプをMCH(3マイクログラム/日)又は溶媒(30%プロピレングリコール)に置き換えた。
【0080】
製造例3:DIO(Diet induced obesity)マウス
18週齢のマウス(C57BL/6J:クレア社製)を、室温23±2℃、湿度55±15%の条件下、1プラスチックゲージに1匹ずつ飼育した。このマウスに高カロリー食であるMHF(タンパク質:18.2重量%、炭水化物:55.6重量%、脂質:15.5重量%)を6ヶ月間に渡って与え、肥満を呈するモデルマウス(DIOマウス)を作製した。なお、実施例中、「established MFD」は、これ以上体重が増えないようになるまでMHPを与えて飼育したマウスを指す。
【0081】
また、前記のマウスにMHFよりさらに高い脂肪を含有する高カロリー食であるHFD(タンパク質:20.8重量%、炭水化物:38.59重量%、脂質:32.88重量%)を与えたDIOマウス(HPD)も作製した。
【0082】
製造例4:食事制限をしたマウス
マウス(C57BL/6N、17週齢)を1ケージに1匹ずつ個別に飼育した。また、エサは普通食(CA−1、CLEA)を与えた。摂食制限は、以下のようなスケジュールで行った。すなわち、エサ(CA−1)を1日につき3時間(10:00〜13:00)だけ与え、水は自由に摂取できるようにした。摂食時間の前後で餌の重量を測定し、その差を摂食量とした。また、摂食制限をしている期間は、体重、外見の観察等をモニターした。なお、条件付けに失敗したと思われるマウス(短期間に極度な体重減少(例えば20%程度の減少)が見られるマウス)は実験には使用しなかった。かかる条件下でマウスを7日間飼育した後、白色脂肪細胞を摘出した。
【0083】
実施例1〜5及び比較例1:白色脂肪細胞におけるLCEの発現
製造例1〜4において製造したモデルマウスを用いて、肝臓及び白色脂肪細胞(WAT)におけるLCE遺伝子の発現量を測定した。発現量の測定は、各モデルマウスの白色脂肪細胞から抽出したRNAを、mouse U74Aチップ(Affymetrix社製)を用いて処理することにより行った。
【0084】
非処理のC57BL/6Nの肝臓又はWATそれぞれにおけるLCEの発現量を1.00とした場合の、DIOマウス(DIO)、D−Try34NPY投与マウス(NPY(FF))、D−Try34NPY pair feeding投与群マウス(NPY(PF))、MCH投与マウス(MCH(FF))、MCH pair feeding投与群マウス(MCH(PF))、食事制限をしたマウス(Fasting)及びNPY Y5 antagonist投与マウス(Y5ant)におけるLCE遺伝子の発現量を表1に示す。
【0085】
表1より明らかなように、肥満モデルマウスではLCE遺伝子の発現量が増加する傾向にあり、食事制限をしたマウスではその発現量が低下していた。したがって、LCEの発現量と体重とが相関関係を有していることが明らかとなった。
【0086】
【表1】
【0087】
実施例6:HEK293細胞におけるマウスLCEmRNAの発現量測定
1.LCE発現亢進細胞の作製
マウス肝臓より抽出したRNAを用いて、RT−PCR法によりマウスLCEのcDNAを増幅した。得られたPCR産物を発現ベクターpCDNA3.1にサブクローニングした後、その塩基配列を確認した。次に、マウスLCEがサブクローニングされた発現ベクターを制限酵素ScaIで直線化し、HEK293細胞に導入した。この細胞を1mg/ml G418を含む選択培地で培養することにより、マウスLCE遺伝子の安定高発現細胞株を得た。
【0088】
以下に、RT−PCRに用いたプライマーの塩基配列を示す。
mLCE−exF:5’−
GCC ACC ATG GGC AAC ATG TCA GTG TTG ACT TTA C−3’(配列番号3)
mLCE−exR:5’−
CTA CTC AGC CTT CGT GGC TTT CTT−3’(配列番号4)
【0089】
2.HEK293細胞におけるマウスLCE mRNAの発現量測定
HEK293細胞からtotal RNAを精製し、逆転写反応によりcDNAを得た。ABI PRISM 7700 Sequence Detector SystemによるTaqMan PCR法を用いてマウスLCE mRNA及びヒトLCE mRNAの発現量を測定した。マウスLCEの標準曲線は、前記のマウスLCEがサブクローニングされた発現ベクターを制限酵素ScaIにより直鎖化したDNAを用いた発現解析により作成した。また、ヒトLCEの標準曲線は、PCRにより調製したヒトLCEのcDNA断片をpcDNA3.1にサブクローニングし、ScaIにより直鎖化したDNA断片を用いた発現解析により作成した。次に、マウスLCEの発現量をヒトLCEの発現量にて除し、導入されたマウスLCE遺伝子の発現量と内在性ヒトLCE遺伝子の発現量の比を求めた。
【0090】
以下に使用したプライマー及びプローブの塩基配列を記す。
マウスLCE用TaqMan probe
mLCE−P:5’−
CTT TCC TGT TTT CTG CGC TGT ACG CTG−3’(配列番号5)
マウスLCE用TaqMan primer
mLCE−F:5’−
GGA TGC AGG AAA ACT GGA AGA A−3’(配列番号6)
mLCE−R:5’−
TGC CGA CCA CCA AAG ATA AAG−3’(配列番号7)
ヒトLCE用TaqMan probe
hLCE−P2:5’−
ATC ACT GTG CTC CTG TAC T−3’(配列番号8)
ヒトLCE用TaqMan primer
hLCE−F2:5’−
AGC TGA TCT TCC TGC ACT GGT AT−3’(配列番号9)
hLCE−R2:5’−
GGC AAC CAT GTC TTT GTA GGA GTA−3’(配列番号10)
ヒトLCE用PCR primer
mLCE−exF:5’−
GCC ACC ATG GGC AAC ATG TCA GTG TTG ACT TTA C−3’(配列番号11)
hLCE−exR:5’−
CTA TTC AGC TTT CGT TGT TTT CCT C−3’(配列番号12)。
【0091】
3.HEK293細胞におけるLCE活性の測定
HEK293細胞を超音波破砕した後に、超遠心操作によりミクロソーム画分を調製した。得られたミクロソーム画分を用いて以下の方法によりLCE活性を測定した。反応に必要とされるNADPH、パルミトイルCoA、14C標識マロニルCoAを含むリン酸緩衝液中にミクロソーム画分を加えて、37℃で5分間インキュベートした。次に、15%水酸化カリウム−メタノール溶液を加え、75℃で45分間加熱してけん化処理を行なった。これに5N塩酸を加えた後、ヘキサンを用いて脂肪酸を抽出した。得られた脂肪酸に含まれる放射性比活性を測定し、脂肪酸延伸反応により脂肪酸に取り込まれたマロニルCoAの量を求めた。
【0092】
図1は、LCEを強制発現させた細胞における、(a)LCE mRNAの発現量、及び(b)LCE活性を表すグラフである。図1に示すように、LCEの発現が亢進した細胞株を取得できたことが確認されるとともに、これらの細胞株ではLCEの活性も亢進していることが確認できた。
【0093】
実施例7:HEK293細胞中に含まれる脂肪酸組成の測定
HEK293細胞をリン酸緩衝液中にて超音波破砕し、内部標準物質としてC17:0の中性脂肪(triglycerides)、コレステリルエステル(cholesteryl esters)及びリン脂質(phospholipids)を加えた後に、クロロホルム−メタノール(2:1)により脂質成分を抽出した。得られた脂質を窒素気流下にて乾固した後、silica gel Gを用いた薄層クロマトグラフによる分画(ヘキサン:ジエチルエーテル:酢酸=80:20:1)を行い、中性脂肪、コレステリルエステル及びリン脂質を分離した。これら3画分それぞれを5%塩酸−メタノールにより脂肪酸残基をメチル化し、ガスクロマトグラフ(GC−FID)を用いて脂肪酸組成の分析を行なった。
【0094】
炭素鎖長18以上の脂肪酸と16以下の脂肪酸を比較した結果を図2に示す。図2から明らかなように、LCEの発現が亢進した細胞株ではC18以上の脂肪酸の構成比率が増加する傾向にあり、炭素鎖の伸長反応はLCEの活性の増強に比例して進行することが確認できた。また、細胞内では、脂肪酸のほとんどがトリグリセリド、コレステリルエステル、リン脂質といったエステル体として存在する。これら全てのエステル体においてLCEの発現亢進によりC18以上の脂肪酸の構成比率が増加する傾向が認められることから、LCEの活性変化は細胞全体の脂肪酸構成比率を変更すると考えられ、LCEは細胞の脂肪酸組成を決定する重要因子であると、本発明者らは考えている。
【0095】
また、各脂肪酸の組成を比較した結果を図3に示す。図3から明らかなように、LCEの発現が亢進した細胞株ではC18以上の脂肪酸の構成比率が増加する傾向にあることが確認できた。
【0096】
実施例8:RNAiによるヒトLCEの発現抑制
1.発現抑制実験に使用するsiRNAの検討
ヒトLCE cDNAの塩基配列情報に基づいて、下記配列からなるsiRNA(small interfering RNA)を合成した。合成したsiRNAそれぞれをHepG2細胞に導入し、24時間後に細胞からtotal RNAを調製した。次に、逆転写反応によりcDNAを作製し、ABI PRISM 7700 Sequence Detector SystemによるTaqMan PCR法によりヒトLCE mRNAの発現量を測定した。図4(a)に、LCE遺伝子における各siRNAの対応部分を示した。図4(b)は、各siRNAを導入した場合のLCE mRNAの発現量を表すグラフである。図4(b)に示すように、siRNA(hLCE−siRNA−6)を用いた場合にLCEの発現抑制の効果が高いことが確認できた。
【0097】
使用したsiRNAの配列を以下に示す。
hLCE−siRNA−1
(図4における1のsiRNA)
5’−GACCGCAAGGCAUUCAUUUUU−3’(配列番号13)
3’−UUCUGGCGUUCCGUAAGUAAA−5’(配列番号14)
hLCE−siRNA−2
(図4における2のsiRNA)
5’−CACUCGAAAUCAAGCGCUUUU−3’(配列番号15)
3’−UUGUGAGCUUUAGUUCGCGAA−5’(配列番号16)
hLCE−siRNA−3
(図4における3のsiRNA)
5’−CACGUAGCGACUCCGAAGAUU−3’(配列番号17)
3’−UUGUGCAUCGCUGAGGCUUCU−5’(配列番号18)
hLCE−siRNA−4
(図4における4のsiRNA)
5’−UGAAGCCAUCCAAUGGAUGUU−3’(配列番号19)
3’−UUACUUCGGUAGGUUACCUAC−5’(配列番号20)
hLCE−siRNA−5
(図4における5のsiRNA)
5’−GCCAUUAGUGCUCUGGUCUUU−3’(配列番号21)
3’−UUCGGUAAUCACGAGACCAGA−5’(配列番号22)
hLCE−siRNA−6
(図4における6のsiRNA)
5’−AGGCCUGAAGCAGUCAGUUUU−3’(配列番号23)
3’−UUUCCGGACUUCGUCAGUCAA−5’(配列番号24)
LCE−siRNA−2
(図4における7のsiRNA)
5’−UGGACCUGUCAGCAAAUUCUU−3’(配列番号25)
3’−UUACCUGGACAGUCGUUUAAG−5’(配列番号26)
hLCE−siRNA−7
(図4における8のsiRNA)
5’−AGCACCCGAACUAGGAGAUUU−3’(配列番号27)
3’−UUUCGUGGGCUUGAUCCUCUA−5’(配列番号28)
hLCE−siRNA−8
(図4における9のsiRNA)
5’−CAUCUUCUGGUCCUCACUCUU−3’(配列番号29)
3’−UUGUAGAAGACCAGGAGUGAG−5’(配列番号30)
hLCE−siRNA−9
(図4における10のsiRNA)
5’−UCACACGUGGUGCAGCUAAUU−3’(配列番号31)
3’−UUAGUGUGCACCACGUCGAUU−5’(配列番号32)
hLCE−siRNA−10
(図4における11のsiRNA)
5’−GCACUGCUGCUGGAAGACCUU−3’(配列番号33)
3’−UUCGUGACGACGACCUUCUGG−5’(配列番号34)
hLCE−siRNA−11
(図4における12のsiRNA)
5’−ACUGUGCGAGCACAACACAUU−3’(配列番号35)
3’−UUUGACACGCUCGUGUUGUGU−5’(配列番号36)
hLCE−siRNA−12
(図4における13のsiRNA)
5’−AGGGGGUGAAUACUUCCCCUU−3’(配列番号37)
3’−UUUCCCCCACUUAUGAAGGGG−5’(配列番号38)。
【0098】
2.HepG2細胞におけるsiRNAのLCE活性低下作用
siRNA(hLCE−siRNA−6)を導入したHepG2細胞を超音波破砕した後に、超遠心操作によりミクロソーム画分を調製した。次に、得られたミクロソーム画分のLCE活性(Fatty Acyl CoAの伸長活性)を測定した。対照には哺乳類遺伝子に相同性を持たない配列であるsiRNA(scramble siRNA Duplex:Dharmacon,Inc.)を導入したHepG2細胞を用いた。図5(a)は、LCE mRNAの発現量を表すグラフであり、図5(b)は、伸長活性を表すグラフである。図5(a),(b)に示すように、LCEの発現が特異的に抑制されるとともに、LCEの活性が抑制されることが確認できた。
【0099】
また、HepG2細胞に2種類のsiRNA(発現抑制効果が強いもの:hLCE−siRNA−6、発現抑制効果が中程度のもの:LCE−siRNA−2)をそれぞれ導入した。siRNAの導入後24時間及び48時間の時点で、細胞からtotal RNAを調製し、DNAチップ(Affymetrix社製)を用いて遺伝子発現解析を行ない、siRNAの導入により発現が低下している遺伝子を選択した。図6は、siRNA導入後のLCE mRNAの発現量を表すグラフである((a):24時間後、(b):48時間後)。siRNAの導入によりLCEの発現が抑制されていることが確認された。
【0100】
siRNAの導入により発現が低下した遺伝子は、24時間で5遺伝子、48時間で64遺伝子であった。発現が低下した遺伝子の中で、LCEと同じ脂肪酸合成に関与するFAS(Fatty Acid Synthase)及びSCD(stearoyl CoA desaturase)に注目し、その発現量を検討した。具体的には、siRNA(hLCE−siRNA−6)を導入したHepG2細胞からtotal RNAを精製し、逆転写反応によりcDNAを得た。次に、ABI PRISM 7700 Sequence Detector Systemを用いたTaqMan PCR法により、ヒトFAS mRNA及びヒトSCD mRNAの発現を測定した。標準曲線は、PCRにより調製したヒトFAS cDNA断片及びヒトSCD cDNA断片をそれぞれ用いて作成した。FAS及びSCDの発現量はβ−アクチンの発現量で規格化した。図7は、siRNAを導入したHepG2細胞における(a)LCE、(b)FAS及び(c)SCDのmRNAの発現量を表すグラフである。図7に示すとおり、siRNAの導入により、LCEの発現は顕著に低下する一方、FAS及びSCDはLCEほどの発現の低下は見られないものの、40〜60%程度の発現低下が見られることが確認された。
【0101】
以下に、測定に用いたプライマー及びプローブの塩基配列を示す。
ヒトFAS用TaqMan probe
hFAS−P:5’−
ACC CGC TCG GCA TGG CTA TCTT−3’(配列番号39)
ヒトFAS用TaqMan primer
hFAS−F:5’−
GCA AAT TCG ACC TTT CTC AGA AC−3’(配列番号40)
hFAS−R:5’−
GGA CCC CGT GGA ATG TCA−3’(配列番号41)
ヒトFAS cDNA作成用PCR primer
hFAS−4823S:5’−
TAC GCC TCC CTC AAC TTC CG−3’(配列番号42)
hFAS−5604A:5’−
CAC TTG AGG GGC CGT ACC AC−3’(配列番号43)
ヒトSCD用TaqMan probe
hSCD−P:5’−
CAC ATG CTG ATC CTC ATA ATT CCC GAC G−3’(配列番号44)
ヒトSCD用TaqMan primer
hSCD−F:5’−
GCC CAC CAC AAG TTT TCA GAA−3’(配列番号45)
hSCD−R:5’−
CCA CGT GAG AGA AGA AAA AGC C−3’(配列番号46)
ヒトSCD cDNA作成用PCR primer
hSCD−600S:5’−
TGT GGA GCC ACC GCT CTT AC−3’(配列番号47)
hSCD−931A:5’−
AAG CGT GGG CAG GAT GAA GC−3’(配列番号48)。
【0102】
実施例9:RNAiによるマウスLCEの発現抑制
実施例8と同様の実験をマウスLCEについても行った。マウスLCE cDNAの塩基配列情報に基づいて、下記配列からなるsiRNAを合成した。合成したsiRNAそれぞれを3T3−L1細胞に導入し、24時間後に細胞からtotal RNAを調製した。次に、逆転写反応によりcDNAを作製し、ABI PRISM 7700 Sequence Detector SystemによるTaqMan PCR法によりマウスLCE mRNAの発現量を測定した。
【0103】
図8は、RNAiによるマウスLCEの発現抑制の結果を表す図である。(a)はマウスLCE遺伝子における各siRNAの対応部分を示す図であり、(b)は各siRNAを導入した場合のマウスLCE mRNAの発現量を表すグラフであり、(c)は各siRNAを導入した場合のマウスFAS mRNAの発現量を表すグラフである。図8に示すように、各siRNAによりLCE及びFASの発現が抑制されることが確認された。
【0104】
実験に用いたsiRNAを以下に示す。
hLCE−siRNA−6(先述したヒトLCEに対するsiRNAであるhLCE−siRNA−6と同一)
(図8における6のsiRNA)
5’−AGGCCUGAAGCAGUCAGUUUU−3’(配列番号49)
3’−UUUCCGGACUUCGUCAGUCAA−5’(配列番号50)
mLCE−siRNA−7(図8におけるm7のsiRNA)
5’−UCCCAUAUGGUGCAGCUAAUU−3’(配列番号51)
3’−UUAGGGUAUACCACGUCGAUU−5’(配列番号52)
mLCE−siRNA−11(図8におけるm11のsiRNA)
5’−GCAUCCGUUGUUCAGUUGCUU−3’(配列番号53)
3’−UUCGUAGGCAACAAGUCAACG−5’(配列番号54)。
【0105】
実施例10:3T3−L1細胞においてLCE RNAiがFAS mRNAの発現に与える影響
脂肪細胞に分化した3T3−L1細胞にsiRNA(hLCE−siRNA−6)を導入し、24時間後に細胞からtotal RNAを精製した。得られたtotal RNAより逆転写反応によりcDNAを調製し、ABI PRISM 7700 Sequence Detector Systemを用いたTaqMan PCR法によりマウスFAS mRNAの発現量を測定した。標準曲線は、PCRにより調製したマウスFAS cDNA断片を用いて作成した。FASの発現量はβ−アクチンの発現量で規格化した。
【0106】
測定に用いたプライマーおよびプローブは下記の通りである。
マウスFAS用TaqMan probe
mFAS−P2:5’−
ATG CTG GCC AAA CTA ACT ACG GCT TCG−3’(配列番号55)
マウスFAS用TaqMan primer
mFAS−F2:5’−
TGG CCT TCT CCT CTG TAA GCT G−3’(配列番号56)
mFAS−R2:5’−
CTG TTC ACA TAT ACG CTC CAT GG−3’(配列番号57)
マウスFAS cDNA作成用PCR primer
mFAS−5541S:5’−
TTC CGC TAC ATG GCT CAG GG−3’(配列番号58)
mFAS−7551A:5’−
CCC GTA CAC TCA CTC GTG GC−3’(配列番号59)
【0107】
また、脂肪細胞に分化した3T3−L1細胞に、siRNA(hLCE−siRNA−6)を導入し、24時間後に培地に14C標識酢酸ナトリウムを添加した。添加してから4時間後に細胞を0.1%SDSにて溶解し、15%水酸化カリウム−メタノール溶液を加え75℃45分間加熱によるけん化処理を行った。これに5N塩酸を加えた後、クロロホルム−メタノール(2:1)にて脂質成分を抽出した。抽出した脂質成分をSilica gel Gを用いた薄層クロマトグラフにより分画(ヘキサン:ジエチルエーテル:酢酸=80:20:1)し、脂肪酸画分に取り込まれた14C酢酸を測定することで脂肪酸合成能を測定した。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc.)を導入した3T3−L1細胞を用いた。
【0108】
図9は、RNAiによる(a)LCE及び(b)FASの発現抑制並びに(c)脂肪酸合成抑制の結果を表す図である。図9に示すように、LCEの発現を抑制することにより、脂肪酸の合成が抑制されることが確認できた。
【0109】
実施例11:HepG2細胞においてLCE RNAiがアポリポタンパク質Bの分泌に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し48時間後に培地を交換した。培地交換後48時間インキュベートし、培養上清を回収し、培地中に分泌されたアポリポタンパク質Bを定量した。アポリポタンパク質Bの定量には、マイクロプレートのEIA法(Exocell,Inc社製:APO B TEST)を用いた。アポリポタンパク質B標準液を用いて標準曲線を作成し、培養上清中のアポリポタンパク質Bの濃度を求めた。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc.社製)を導入したHepG2細胞を用いた。
【0110】
図10は、RNAiによる(a)LCEの発現抑制及び(b)アポリポタンパク質Bの分泌抑制の結果を表す図である。図10に示すように、LCEの発現を抑制することによりアポリポタンパク質Bの分泌が抑制されることが確認できた。
【0111】
以上より、LCEの活性を抑制することにより、抗肥満効果を示すことが示唆された。
【0112】
実施例12:DIOマウスにおけるLCE RNAiが与える影響
7週齢のマウス(ICR、雌)を高カロリー食であるMHFdietにて23週間飼育し、肥満を形成させた。先ず、siRNAの投与開始前にマウスの体重を測定した。siRNA(hLCE−siRNAi−6)を、HVJ−リポソームをキャリアとして用いマウスの尾静脈から投与した(40μg/mouse/injection)。投与は1日おきに5回行い、最終投与2日後にマウスの体重を測定し、siRNA投与の前後での体重変化を求めた。眼窩静脈叢より採血し血糖値を測定した。腹部大静脈より採血し血漿インスリン濃度を測定した。対照には、哺乳類細胞では効果を示さないscramble siRNAを投与したマウスを用いた。
【0113】
図11は、siRNA投与による(a)体重、(b)血糖値及び(c)血漿インシュリンレベルの変化を表す図である。図11に示すように、LCEに対するsiRNAをマウスに投与することにより体重が減少した。また、血糖値および血漿インシュリンレベルのいずれもが減少し、LCEに対するsiRNAがLCEの活性を抑制し、肥満改善効果を示すことが個体レベルでも確認できた。
【0114】
また、これらマウスの肝臓からtotal RNAを精製し、逆転写反応によりcDNAを得た。
ABI PRISM 7700 Sequence Detector SystemによるTaqMan PCR法を用いてマウスLCE mRNA、マウスacetyl−CoA carboxylase(ACC)mRNA、マウスFAS mRNA、マウスSCD−1 mRNA、マウスSREBP−1c mRNA、マウスinsulin receptor substrate(IRS)−2 mRNAの発現量を測定した。標準曲線はPCRにより調製した各遺伝子のcDNA断片をそれぞれ用いて作製した。各遺伝子の発現量はβ−アクチンの発現量で規格化した。
【0115】
測定に用いたプライマーおよびプローブの塩基配列を示す。
マウスACC1用TaqMan probe
5’−
AGCTGCAAGCCTGTCATCCTCAATATCG−3’(配列番号73)
マウスACC1 TaqMan PCR primer
forward:
5’−TTCTGAATGTGGCTATCAAGACTGA−3’(配列番号74)
reverse:
5’−TGCTGGGTGAACTCTCTGAACA−3’(配列番号75)
マウスACC1用cDNA作製用primer
forward:
5’−TAGTGTCAGCGATGTTCTGT−3’(配列番号76)
reverse:
5’−AAATCTCTGATCCACCTCAC−3’(配列番号77)
マウスSCD−1用TaqMan probe
ACTCGCCTACACCAACGGGCTCC(配列番号78)
マウスSCD−1用TaqMan primer
forward:
5’−TTTCCAAGCGCAGTTCCG−3’(配列番号79)
reverse:
5’−ATCGAGCGTGGACTTCGGT−3’(配列番号80)
マウスSCD−1 cDNA作製用PCR primer
forward:
5’−CACCCATCCCGAGAGTCAGG−3’(配列番号81)
reverse:
5’−GTGGGCCGGCATGATGATAG−3’(配列番号82)
マウスSREBP−1c用TaqMan probe
5’−
CTTCAAATGTGCAATCCATGGCTCCGT−3’(配列番号83)
マウスSREBP−1c用TaqMan primer
forward:
5’−GTAGCGTCTGCACGCCCTA−3’(配列番号84)
reverse:
5’−CTTGGTTGTTGATGAGCTGGAG−3’(配列番号85)
マウスSREBP−1c cDNA作製用PCR primer
forward:
5’−AAGCTGTCGGGGTAGCGTCT−3’(配列番号86)
reverse:
5’−AGGCTCGAGTAACCCAGCAC−3’(配列番号87)
マウスIRS−2用TaqMan probe
5’−
ACTTAGCCGCTTCAAGCCCGATGTG−3’(配列番号88)
マウスIRS−2 TaqMan PCR用primer
forward:
5’−AGAAGGTGCCCGAGTGGC−3’(配列番号89)
reverse:
5’−CCCCAGATACCTGATCCATGA−3’(配列番号90)
マウスIRS−2 cDNA作製用primer
forward:
5’−CAGTAGGCTCCATGGATGGC−3’(配列番号91)
reverse:
5’−ATGACCTTAGCACCCCGGTG−3’(配列番号92)
【0116】
図21は、siRNAを投与したマウスの肝臓における(a)ACC、FAS、SCD1、SREBP−1c、IRS−2及び(b)LCEのmRNAの発現量を表すグラフである。LCEの発現を抑制することにより、脂肪酸合成に関わる酵素であるACC、FAS、SCD1の発現が低下しており、また脂肪酸合成を調節する転写因子であるSREBP−1cの発現量も低下していることが確認された。すなわち、DIOマウスの肝臓におけるLCEの発現抑制により、肝臓での脂肪合成が抑制されることが示唆された。
【0117】
また、IRS−2(insulin receptor substrate−2)の発現がLCE RNAi投与マウスの肝臓で亢進しているのは、肝臓のLCEの発現抑制が肝臓のインスリン感受性を亢進させることを示唆する。これは、実施例12で示したLCEの発現抑制による血糖値及び血漿インスリン濃度の低下を引き起こす原因になっていると発明者らは考えている。
【0118】
実施例13:HepG2細胞においてLCE RNAiが細胞の脂肪酸組成に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し72時間後に細胞を回収した。その細胞をリン酸緩衝液中にて超音波破砕し、内部標準としてC17:0の中性脂肪、コレステリルエステル、リン脂質を加えた後にBligh−Dyer法により脂質成分を抽出した。得られた脂質を窒素気流下にて乾固した後、silica gel Gを用いた薄層クロマトグラフによる分画(ヘキサン:ジエチルエーテル:酢酸=80:20:1)を行い、中性脂肪、コレステリルエステル、リン脂質を分離した。これら3画分それぞれを5%塩酸−メタノールにより脂肪酸残基をメチル化し、ガスクロマトグラフ(GC−FID)を用いて脂肪酸組成の分析を行った。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。
【0119】
脂肪酸組成の分析結果を表2に示す。表2に示した結果から分かるように、RNAiによりLCEの発現を抑制すると、トリグリセリド、コレステリルエステル及びリン脂質のいずれにおいてもC18以上の脂肪酸の構成比率が減少することが確認できた。
【0120】
【表2】
【0121】
実施例14:HepG2細胞においてLCE RNAiが他遺伝子の発現に与える影響
脂肪酸合成に関与するSREBP−1(sterol regulatory element binding protein−1)及びコレステロール合成に関与するSREBP−2(sterol regulatory element binding protein−2)に対して、LCE発現抑制が与える影響について調べた。siRNA(hLCE−siRNA−6)を導入したHepG2細胞からtotal RNAを精製し、逆転写反応によりcDNAを得た。ABI PRISM 7700 Sequence Detector Systemを用いたTaqMan PCR法によりヒトSREBP−1 mRNA、ヒトSREBP−2 mRNAの発現を測定した。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。標準曲線はPCRにより調製したヒトSREBP−1 cDNA断片、ヒトSREBP−2 cDNA断片をそれぞれ用いて作製した。各遺伝子の発現量はβ−アクチンの発現量で規格化した。
【0122】
測定に用いたプライマーの塩基配列を示す。
ヒトSREBP−1用TaqMan primer
forward:
5’−CAACACAGCAACCAGAAACTCAAG−3’(配列番号60)
reverse:
5’−TTGCTTTTGTGGACAGCAGTG−3’(配列番号61)
ヒトSREBP−1 cDNA作製用PCR primer
forward:5’−CGGAGAAGCTGCCTATCAAC−3’(配列番号62)
reverse:5’−GGTCAGTGTGTCCTCCACCT−3’(配列番号63)
ヒトSREBP−2用TaqMan primer
forward:
5’−GATATCGCTCCTCCATCAATGAC−3’(配列番号64)
reverse:
5’−ACTTGTGCATCTTGGCGTCTG−3’(配列番号65)
ヒトSREBP−2 cDNA作製用PCR primer
forward:
5’−CATTCTGACCACAATGCCTG−3’(配列番号66)
reverse:
5’−AGTAGGGAGAGAAGCCAGCC−3’(配列番号67)
【0123】
図12は、siRNAを導入したHepG2細胞におけるSREBP−1及びSREBP−2のmRNAの発現量を表すグラフである。図12に示すとおり、siRNAの導入により、SREBP−1の発現は顕著に低下する一方、SREBP−2の発現は変化しないことが確認された。
【0124】
実施例15:HepG2細胞においてLCE RNAiが細胞の脂質合成に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し、72時間後に培地に14C標識酢酸ナトリウムを添加した。添加4時間後に細胞を0.1%SDSにて溶解した。細胞溶解液の一部に15%水酸化カリウム−メタノール溶液を加え75℃45分間加熱によるけん化処理行った。5N塩酸を加えた後に、クロロホルム−メタノール(2:1)にて脂質成分を抽出した。抽出した脂質成分をsilica gel Gを用いた薄層クロマトグラフにより分画(ヘキサン:ジエチルエーテル:酢酸=80:20:1)し、脂肪酸画分に取り込まれた14C酢酸を測定することで脂肪酸合成能を測定した。
【0125】
また、上記細胞溶解液の一部から直接、クロロホルム−メタノール(2:1)にて脂質成分を抽出した。得られた脂質成分を薄層クロマトグラフにより分画し、中性脂肪画分に取り込まれた14C酢酸を測定することで中性脂肪合成能を測定した。得られた脂肪酸合成能および中性脂肪合成能の値は、細胞溶解液中のタンパク質量により規格化した。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。
【0126】
図13は、siRNAを導入したHepG2細胞における(a)脂肪酸合成能及び(b)中性脂肪合成能を表す図である。図13に示したように、siRNAを導入してLCEの発現を抑制することで、脂肪酸合成能及び中性脂肪合成能が低下することが確認された。
【0127】
実施例16:HepG2細胞においてLCE RNAiが細胞の中性脂肪含量に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し72時間後に細胞を回収した。その細胞をリン酸緩衝液中にて超音波破砕し、その一部からBligh−Dyer法により脂質成分を抽出した。得られた脂質を窒素気流下にて乾固した後、2−プロパノールに溶解し、酵素法により中性脂肪量を測定した。得られた中性脂肪量の値は、細胞の超音波破砕液中のタンパク質量にて規格化した。対照には哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。
【0128】
図14は、siRNAを導入したHepG2細胞中の中性脂肪量を表すグラフである。図14に示したように、siRNAを導入してLCEの発現を抑制することで、細胞内の中性脂肪含量が低下することが確認された。
【0129】
実施例17:HepG2細胞においてLCE RNAiが細胞の脂肪酸酸化に与える影響
HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し、72時間後に培地に14Cパルミチン酸を添加した。30分間インキュベーションした後に、培地を別チューブに移し10%トリクロロ酢酸を加えた。培地から遊離したCO2を10%水酸化ナトリウム水溶液によりトラップした。トラップされたCO2の放射比活性を測定し、培地に加えたパルミチン酸からβ酸化により作られたCO2量を求めた。またCO2を遊離させた後の培地を遠心して、その上清中の酸可溶性画分の放射比活性を測定し、培地に加えたパルミチン酸からβ酸化により作られたケトン体量を求めた。インキュベーション後のHepG2細胞は培地を取り除いた後に0.1%SDSにて溶解した。細胞溶解液の一部からクロロホルム−メタノール(2:1)にて脂質成分を抽出し、薄層クロマトグラフにより分画した。そして中性脂肪画分の放射比活性を測定し、培地に加えたパルミチン酸が細胞中の中性脂肪に取り込まれた量を求めた。得られたCO2量、ケトン体量、中性脂肪への取り込み量の値は、細胞溶解液中のタンパク質量にて規格化した。対照には、哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。
【0130】
図15は、siRNAを導入したHepG2細胞における(a)CO2産生量、(b)ケトン体産生量及び(c)パルミチン酸の中性脂肪への取り込み量を表すグラフである。図15に示したように、siRNAを導入してLCEの発現を抑制すると、CO2産生量及びケトン体産生量が増加し、パルミチン酸の中性脂肪への取り込みが低下することが確認された。脂肪酸の燃焼にともなってCO2及びケトン体が産生するため、この結果は、LCEの発現抑制は、脂肪酸の燃焼を促進することを示唆している。
【0131】
実施例18:HepG2細胞においてLCE RNAiが細胞のCPT−1の発現に与える影響
脂肪酸をミトコンドリアに取り込むトランスポーターであるCPT−1(carnitine palmitoyl transferase−1)に対して、LCE発現抑制が与える影響について調べた。HepG2細胞にsiRNA(hLCE−siRNA−6)を導入し72時間後に細胞を回収した。この細胞からtotal RNAを精製し、逆転写反応によりcDNAを得た。ABI PRISM 7700 Sequence Detector Systemを用いたTaqMan PCR法によりヒトCPT−1 mRNAの発現を測定した。対照には、哺乳類遺伝子に相同性がない配列のsiRNA(scramble siRNA Duplex:Dharmacon,Inc)を導入したHepG2細胞を用いた。標準曲線はPCRにより調製したヒトCPT−1 cDNA断片を用いて作製した。得られた値はβ−アクチンの発現量で規格化した。
【0132】
測定に用いたプライマーおよびプローブの塩基配列を示す。
ヒトCPT−1 TaqMan probe
5’−
CCGGGAGGAAATCAAACCAATTCGTC−3’(配列番号68)
ヒトCPT−1用TaqMan primer
forward:
5’−TGCTTTACAGGCGCAAACTG−3’(配列番号69)
reverse:
5’−TGGAATCGTGGATCCCAAA−3’(配列番号70)
ヒトCPT−1 cDNA作製用PCR primer
forward:
5’−ATTTGAAGTTAAAATCCTGGTGGGC−3’(配列番号71)
reverse:
5’−TTCCCACGTCCAAAATAGGC−3’(配列番号72)
【0133】
図16は、siRNAを導入したHepG2細胞におけるCPT−1 mRNAの発現量を表すグラフである。図16に示したように、siRNAを導入してLCEの発現を抑制すると、CPT−1の発現量が増加することが確認された。
【0134】
実施例19:高スクロース食負荷マウスにおける肝臓でのLCE発現の変化8週齢のマウス(ICR、雄)を高スクロース食(67%スクロース)にて3日間、および7週齢のマウス(ICR、雄)を高スクロース食にて10日間飼育した。体重測定後にマウスを安楽死させ、精巣上体脂肪を摘出し重量を測定した。また、マウスから肝臓を摘出しtotal RNAを精製し、Taq Man PCR法を用いて肝臓でのLCE mRNAの発現量を測定した。LCE mRNAの発現量はβ−アクチンの発現量で規格化した。対照には通常食(CA−1)で飼育したマウスを用いた。
【0135】
図17は、高スクロース食を負荷したマウスにおける(a)体重、(b)精巣上体の脂肪重量及び(c)肝臓におけるLCE mRNAの発現量を表すグラフである。図17に示したように、高スクロース負荷することにより、マウスの体重及び脂肪重量は増加し、肝臓におけるLCE mRNAの発現量が増加することが確認された。
【0136】
実施例20:高スクロース食負荷マウスにおけるLCE RNAi投与の作用
HVJ−リポソームをキャリアとして用い、siRNA(hLCE−siRNAi−6)を、マウス(7週齢、ICR、雄)に尾静脈から投与した(40μg/mouse/injection)。投与開始直後から、マウスを高スクロース食にて飼育した。対照には、哺乳類細胞では効果を示さないscramble siRNAを投与したマウスを用いた。siRNAの投与は1日おきに5回行い、最終投与の翌日にマウスの体重を測定し、体脂肪量をNMR analyzer(Minispec;mq7.5)にて測定し、脂肪重量/体重比を求めた。腹部大静脈より採血し、血漿レプチン濃度を測定した。
【0137】
図18は、siRNAを投与したマウスの(a)体脂肪量、(b)脂肪重量/体重比及び(c)血漿レプチン濃度を表すグラフである。LCE RNAiの投与によるLCE発現抑制は、高スクロース負荷による体脂肪量の増加を抑制した。同様に脂肪重量/体重比の増加を抑制した。これはLCEの発現抑制が、高スクロース摂食による肥満形成を阻害すること(抗肥満効果)を示す。また、レプチンは脂肪組織から分泌されるので、脂肪重量が増加すると分泌されるレプチン濃度が増加する。この実験において、高スクロース食負荷によりレプチンが増加しているのは、脂肪量が増加したことを意味し、LCE RNAiの投与群でレプチン濃度上昇が抑えられているのは、LCE RNAi投与で脂肪重量の増加が抑制されたことを意味する。
【0138】
また、マウスを安楽死させた後に肝臓を摘出し、その一部からtotal RNAを精製し、TaqMan PCR法を用いて肝臓でのLCE mRNA、FAS mRNAの発現量を測定した。LCE mRNA、FAS mRNAの発現量はβ−アクチンの発現量で規格化した。また、肝臓の一部をリン酸緩衝液中においてホモジナイズし、Bligh−Dyer法により脂質成分を抽出した。得られた脂質を窒素気流下にて乾固し、2−プロパノールに溶解し、酵素法により中性脂肪量を測定した。得られた中性脂肪量の値は肝臓ホモジナイズ液中に含まれるタンパク質量で規格化した。さらに、肝臓組織からミクロソーム画分を調製し、ウェスタンブロッティング法によりLCEのタンパク質量の変化を測定した。ウェスタンブロッティングには、マウスLCEタンパク質の一次構造に含まれるアミノ酸配列”FEAYIGKVKKATKAE”に基づいて合成した合成ペプチド”CFEAYIGKVKKATKAE”をウサギ(SPF,Japanese White Rabbit)に免疫して得た抗LCEポリクローナル抗体を用いた。
【0139】
図19は、siRNAを投与したマウスの(a)LCE mRNAの発現量、(b)FAS mRNAの発現量及び(c)肝臓における中性脂肪含量を表すグラフである。図19(a)より、高スクロース食負荷により肝臓のLCEの発現が亢進すること、すなわち肥満形成の過程で肝臓のLCE発現が亢進していることが確認された。また、LCE RNAiの投与がマウスの肝臓でのLCEの発現亢進を抑制していることが確認された。
【0140】
図19(b)より、高スクロース食負荷により肝臓のFASの発現が亢進すること、すなわち肝臓での脂肪酸合成が亢進していることが示唆された。また、LCE RNAi投与によるLCE発現抑制によりFASの発現亢進が抑制され、脂肪酸合成亢進が抑制されることが示唆された。
【0141】
図19(c)より、高スクロース食負荷により肝臓への脂肪の蓄積が亢進していることが確認された。また、LCE RNAi投与によるLCE発現抑制が肥満形成の過程での肝臓での脂肪蓄積を抑制することが確認された。
【0142】
以上より、高スクロース食負荷により肝臓での脂肪合成が亢進すること(肥満形成の促進過程)と、LCEの発現抑制がこの脂肪合成亢進を阻害(肥満形成過程を抑制)することが確認された。
【0143】
図20は、siRNAを投与したマウスの肝臓におけるLCEタンパク質量の変化を表す図である。図19に示した脂肪酸合成とLCEの発現量との関係をLCEタンパク質レベルでも確認することができた。
【0144】
実施例21:ヒトLCEのアミノ酸置換及びそれによるLCE活性の変化
ヒトLCE cDNAを、BamHIサイトを付加したforward primer(hLCE−5BamHI)およびXhoIサイトを付加したreverse primer(hLCE−3XhoI)を用いたPCRにより作製した。得られたcDNA断片を、付加したBamHIサイトとXhoIサイトを用いて、プラスミドpCMV−Tag2B(Stratagene)にサブクローニングした(wild−type LCE construct)。
【0145】
用いたプライマーの配列は以下の通りである。
hLCE−5BBamHI:
GGATCCAACATGTCAGTGTTGACTT(配列番号93)
hLCE−3XhoI:
CTCGAGCTATTCAGCTTTCGTTGTT(配列番号94)
【0146】
このプラスミドを鋳型として、PCR法を用いた点突然変異導入により変異LCEをコードするcDNAを作成した。点突然変異の導入方法を以下に記す。最初にForward primerとしてhLCE−F4を、reverse primerとして各変異導入用のmutagenic primerを用いたPCRにより変異導入LCE cDNAの5’側部分を作製した。またForward primerとしてhLCE−510Sを、reverse primerとしてT7を用いたPCRによりLCE cDNAの3’側部分を作製した。得られた各変異導入LCE cDNA 5’側断片とLCE cDNA3’側断片を混合したものを鋳型として、forward primerにhLCE−F4を、reverse primerにT7を用いたPCRにより、変異を導入したLCEのコーディング領域全長を含むcDNA断片を得た。
【0147】
用いたプライマーの配列は以下の通りである。
hLCE−F4:
AACATGTCAGTGTTGACTTTAC(配列番号95)
hLCE−510S:
GTGCTCTTCGAACTGGTGCT(配列番号96)
T7:
TAATACGACTCACTATAGGG(配列番号97)
【0148】
作製した突然変異導入LCEは下記の通りである。ヒトLCEに導入した突然変異は、99番目のシステインをアラニンに置換したもの(C99A)、225番目のシステインをアラニンに置換したもの(C225A)、141番目のヒスチジンをアラニンにしたもの(H141A)、144番目のヒスチジンをアラニンにしたもの(H144A)、145番目のヒスチジンをアラニンにしたもの(H145A)、及び174番目のヒスチジンをアラニンにしたもの(H147A)である。
【0149】
これら変異導入に使用したプライマーの配列は以下の通りである。
C99A:
CCCTGGTCGGCAACTGACTGCTTC(配列番号98)
C225A:
GTGAGAGTGGGCCTGGTCATGCTG(配列番号99)
H141A:
GTGATACCAGGCCAGGAAGATC(配列番号100)
H144A:
GTGATGTGGGCATACCAGTGC(配列番号101)
H145A:
CACAGTGATGGCGTGATACCAG(配列番号102)
H174A:
CATCACGGCGGCCACGCCATAG(配列番号103)
【0150】
得られた変異LCE cDNA断片は、制限酵素EcoRI及びXhoIを利用してプラスミドpCMV−Tag2Bにサブクローニングし、変異LCE発現コンストラクトを作製した。これらのコンストラクトをLipofectamine2000(Invitrogen)を用いてHEK293細胞にトランスフェクトし、変異LCEを発現させた。トランスフェクトから2日後に細胞を回収し、ミクロソーム画分を調製し、ミクロソーム中のLCE活性を測定した。なお、LCEの活性測定は、実施例6に記載の方法と同様の方法により行った。
【0151】
図22は、LCE変異体のLCE活性を表すグラフである。(a)はシステインをアラニンに置換した変異体のLCE活性を表し、(b)はヒスチジンをアラニンに置換した変異体のLCE活性を表している。いずれの変異体も野生型と比較して半分以下にLCE活性が低下している。
【産業上の利用可能性】
【0152】
本発明によれば、長鎖脂肪鎖伸長酵素活性を抑制する活性を有する物質(例えば、siRNA、低分子化合物、タンパク質、抗体など)を用いた代謝系疾患、循環器系疾患、中枢神経系疾患などの治療・予防方法を提供することが可能となり、また、かかる物質を含有する治療・予防剤を提供することが可能となる。代謝系疾患としては、例えば、肥満症、糖尿病、ホルモン分泌異常、高脂血症、通風、脂肪肝などが挙げられる。循環器疾患としては、例えば、狭心症、急性・うっ血性心不全、心筋梗塞、冠状動脈硬化症、高血圧、腎臓病、電解質異常などが挙げられる。中枢神経系疾患としては、過食症などが挙げられる。
【特許請求の範囲】
【請求項1】
被検化合物を、被検動物又は被検細胞に、投与又は接触させる工程と、
該被検化合物が、該被検動物又は該被検細胞中でLCE遺伝子あるいは該遺伝子と機能的に等価な遺伝子の発現レベルを調節するか否かを確認する工程と、
を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【請求項2】
被験化合物を、LCE遺伝子の発現調節領域とレポーター遺伝子との融合遺伝子を有する被検動物又は被検細胞に接触させる工程と、
該レポーター遺伝子の被検動物又は被検細胞中での発現レベルを測定する工程と、
を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【請求項3】
被検化合物を、LCEタンパク質に接触させる工程と、
該被検化合物が、該タンパク質の活性に影響を与えるか否かを確認する工程と、
を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【請求項4】
被検化合物を、LCEを含む複数種のエロンゲースタンパク質に接触させる工程と、
該複数種のエロンゲースタンパク質の活性を測定する工程と、
該複数種のエロンゲースタンパク質においてLCEの活性を阻害する被検化合物を選択する工程と、
を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【請求項5】
請求項1〜4のいずれか一項に記載の評価方法によって得られた化合物を有効成分として含有する肥満の治療又は予防剤。
【請求項6】
LCEの脂肪酸合成活性を抑制することを特徴とする脂肪合成抑制方法。
【請求項7】
LCEの脂肪酸合成活性をRNAiにより抑制することを特徴とする脂肪合成抑制方法。
【請求項8】
前記RNAiが、配列番号13及び14に記載の核酸からなるsiRNA、配列番号15及び16に記載の核酸からなるsiRNA、配列番号17及び18に記載の核酸からなるsiRNA、配列番号19及び20に記載の核酸からなるsiRNA、配列番号21及び22に記載の核酸からなるsiRNA、配列番号23及び24に記載の核酸からなるsiRNA、配列番号25及び26に記載の核酸からなるsiRNA、配列番号27及び28に記載の核酸からなるsiRNA、配列番号29及び30に記載の核酸からなるsiRNA、配列番号31及び32に記載の核酸からなるsiRNA、配列番号33及び34に記載の核酸からなるsiRNA、配列番号35及び36に記載の核酸からなるsiRNA、配列番号37及び38に記載の核酸からなるsiRNA、配列番号49及び50に記載の核酸からなるsiRNA、配列番号51及び51に記載の核酸からなるsiRNA及び配列番号53及び54に記載の核酸からなるsiRNAからなる群から選択される一以上のsiRNAを使用したものであることを特徴とする請求項7に記載の脂肪合成抑制方法。
【請求項9】
前記RNAiが、配列番号23及び24に記載の核酸からなるsiRNAを使用したものであることを特徴とする請求項7に記載の脂肪合成抑制方法。
【請求項10】
LCEの脂肪酸合成活性を抑制する工程を含むことを特徴とする肥満の治療又は予防方法。
【請求項11】
LCEの脂肪酸合成活性をRNAiにより抑制することを特徴とする肥満の治療又は予防方法。
【請求項12】
前記RNAiが、配列番号13及び14に記載の核酸からなるsiRNA、配列番号15及び16に記載の核酸からなるsiRNA、配列番号17及び18に記載の核酸からなるsiRNA、配列番号19及び20に記載の核酸からなるsiRNA、配列番号21及び22に記載の核酸からなるsiRNA、配列番号23及び24に記載の核酸からなるsiRNA、配列番号25及び26に記載の核酸からなるsiRNA、配列番号27及び28に記載の核酸からなるsiRNA、配列番号29及び30に記載の核酸からなるsiRNA、配列番号31及び32に記載の核酸からなるsiRNA、配列番号33及び34に記載の核酸からなるsiRNA、配列番号35及び36に記載の核酸からなるsiRNA、配列番号37及び38に記載の核酸からなるsiRNA、配列番号49及び50に記載の核酸からなるsiRNA、配列番号51及び51に記載の核酸からなるsiRNA及び配列番号53及び54に記載の核酸からなるsiRNAからなる群から選択される一以上のsiRNAを使用したものであることを特徴とする請求項11に記載の肥満の治療又は予防方法。
【請求項13】
前記RNAiが、配列番号23及び24に記載の核酸からなるsiRNAを使用したものであることを特徴とする請求項11に記載の肥満の治療又は予防方法。
【請求項14】
被検組織又は被検細胞におけるLCE遺伝子の発現レベル又は発現レベルの変化を測定することを特徴とする肥満の検査方法。
【請求項15】
被検組織又は被検細胞におけるLCEタンパク質の発現レベル又は発現レベルの変化を測定することを特徴とする肥満の検査方法。
【請求項16】
被検組織又は被検細胞におけるLCE遺伝子に存在する多型を検出することを特徴とする肥満の検査方法。
【請求項17】
LCEタンパク質と相互作用することによりLCE遺伝子の発現量に影響を及ぼすタンパク質の発現量又は活性を検出することを特徴とする肥満の検査方法。
【請求項18】
配列番号23及び24に記載の核酸からなることを特徴とするsiRNA。
【請求項19】
請求項18に記載のsiRNAを含むことを特徴とするLCE発現抑制剤。
【請求項20】
請求項18に記載のsiRNAを含むことを特徴とする脂肪合成抑制剤。
【請求項21】
請求項18に記載のsiRNAを含むことを特徴とする肥満の治療又は予防剤。
【請求項1】
被検化合物を、被検動物又は被検細胞に、投与又は接触させる工程と、
該被検化合物が、該被検動物又は該被検細胞中でLCE遺伝子あるいは該遺伝子と機能的に等価な遺伝子の発現レベルを調節するか否かを確認する工程と、
を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【請求項2】
被験化合物を、LCE遺伝子の発現調節領域とレポーター遺伝子との融合遺伝子を有する被検動物又は被検細胞に接触させる工程と、
該レポーター遺伝子の被検動物又は被検細胞中での発現レベルを測定する工程と、
を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【請求項3】
被検化合物を、LCEタンパク質に接触させる工程と、
該被検化合物が、該タンパク質の活性に影響を与えるか否かを確認する工程と、
を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【請求項4】
被検化合物を、LCEを含む複数種のエロンゲースタンパク質に接触させる工程と、
該複数種のエロンゲースタンパク質の活性を測定する工程と、
該複数種のエロンゲースタンパク質においてLCEの活性を阻害する被検化合物を選択する工程と、
を含むことを特徴とする肥満の治療又は予防に有効な化合物の評価方法。
【請求項5】
請求項1〜4のいずれか一項に記載の評価方法によって得られた化合物を有効成分として含有する肥満の治療又は予防剤。
【請求項6】
LCEの脂肪酸合成活性を抑制することを特徴とする脂肪合成抑制方法。
【請求項7】
LCEの脂肪酸合成活性をRNAiにより抑制することを特徴とする脂肪合成抑制方法。
【請求項8】
前記RNAiが、配列番号13及び14に記載の核酸からなるsiRNA、配列番号15及び16に記載の核酸からなるsiRNA、配列番号17及び18に記載の核酸からなるsiRNA、配列番号19及び20に記載の核酸からなるsiRNA、配列番号21及び22に記載の核酸からなるsiRNA、配列番号23及び24に記載の核酸からなるsiRNA、配列番号25及び26に記載の核酸からなるsiRNA、配列番号27及び28に記載の核酸からなるsiRNA、配列番号29及び30に記載の核酸からなるsiRNA、配列番号31及び32に記載の核酸からなるsiRNA、配列番号33及び34に記載の核酸からなるsiRNA、配列番号35及び36に記載の核酸からなるsiRNA、配列番号37及び38に記載の核酸からなるsiRNA、配列番号49及び50に記載の核酸からなるsiRNA、配列番号51及び51に記載の核酸からなるsiRNA及び配列番号53及び54に記載の核酸からなるsiRNAからなる群から選択される一以上のsiRNAを使用したものであることを特徴とする請求項7に記載の脂肪合成抑制方法。
【請求項9】
前記RNAiが、配列番号23及び24に記載の核酸からなるsiRNAを使用したものであることを特徴とする請求項7に記載の脂肪合成抑制方法。
【請求項10】
LCEの脂肪酸合成活性を抑制する工程を含むことを特徴とする肥満の治療又は予防方法。
【請求項11】
LCEの脂肪酸合成活性をRNAiにより抑制することを特徴とする肥満の治療又は予防方法。
【請求項12】
前記RNAiが、配列番号13及び14に記載の核酸からなるsiRNA、配列番号15及び16に記載の核酸からなるsiRNA、配列番号17及び18に記載の核酸からなるsiRNA、配列番号19及び20に記載の核酸からなるsiRNA、配列番号21及び22に記載の核酸からなるsiRNA、配列番号23及び24に記載の核酸からなるsiRNA、配列番号25及び26に記載の核酸からなるsiRNA、配列番号27及び28に記載の核酸からなるsiRNA、配列番号29及び30に記載の核酸からなるsiRNA、配列番号31及び32に記載の核酸からなるsiRNA、配列番号33及び34に記載の核酸からなるsiRNA、配列番号35及び36に記載の核酸からなるsiRNA、配列番号37及び38に記載の核酸からなるsiRNA、配列番号49及び50に記載の核酸からなるsiRNA、配列番号51及び51に記載の核酸からなるsiRNA及び配列番号53及び54に記載の核酸からなるsiRNAからなる群から選択される一以上のsiRNAを使用したものであることを特徴とする請求項11に記載の肥満の治療又は予防方法。
【請求項13】
前記RNAiが、配列番号23及び24に記載の核酸からなるsiRNAを使用したものであることを特徴とする請求項11に記載の肥満の治療又は予防方法。
【請求項14】
被検組織又は被検細胞におけるLCE遺伝子の発現レベル又は発現レベルの変化を測定することを特徴とする肥満の検査方法。
【請求項15】
被検組織又は被検細胞におけるLCEタンパク質の発現レベル又は発現レベルの変化を測定することを特徴とする肥満の検査方法。
【請求項16】
被検組織又は被検細胞におけるLCE遺伝子に存在する多型を検出することを特徴とする肥満の検査方法。
【請求項17】
LCEタンパク質と相互作用することによりLCE遺伝子の発現量に影響を及ぼすタンパク質の発現量又は活性を検出することを特徴とする肥満の検査方法。
【請求項18】
配列番号23及び24に記載の核酸からなることを特徴とするsiRNA。
【請求項19】
請求項18に記載のsiRNAを含むことを特徴とするLCE発現抑制剤。
【請求項20】
請求項18に記載のsiRNAを含むことを特徴とする脂肪合成抑制剤。
【請求項21】
請求項18に記載のsiRNAを含むことを特徴とする肥満の治療又は予防剤。
【図1】


【図2】

【図3】
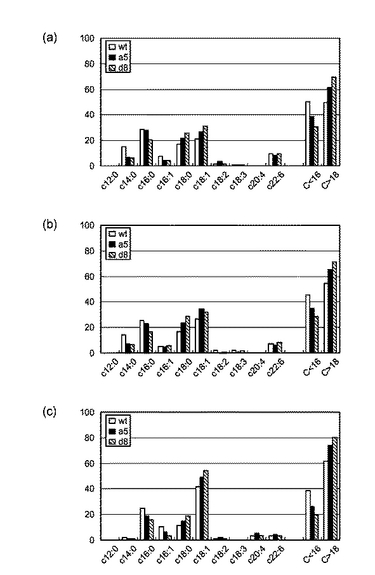
【図4】
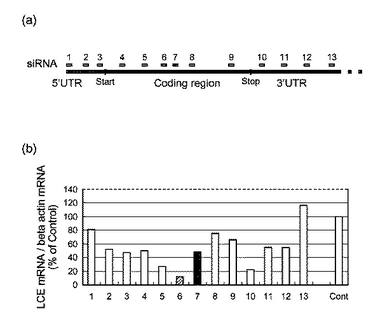
【図5】

【図6】
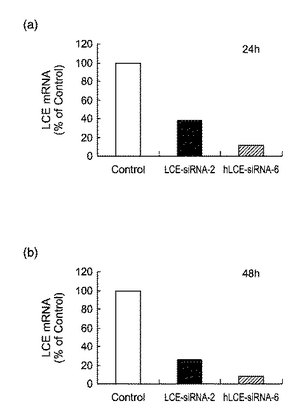
【図7】
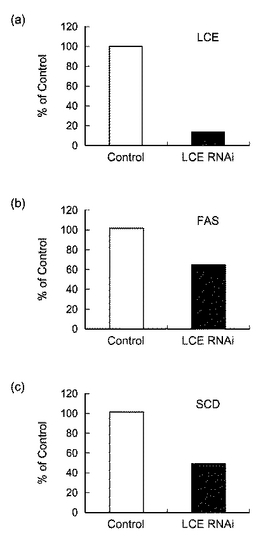
【図8】
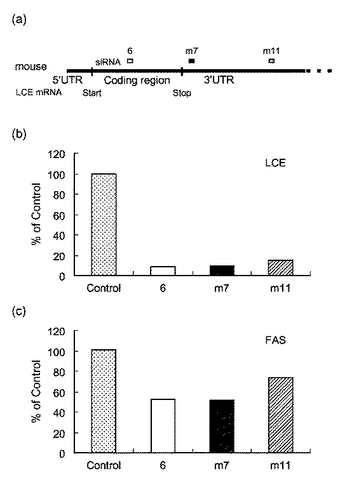
【図9】
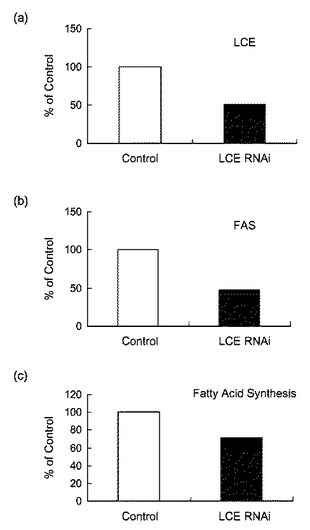
【図10】
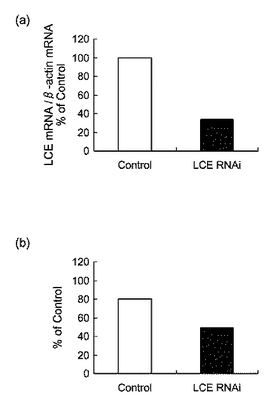
【図11】
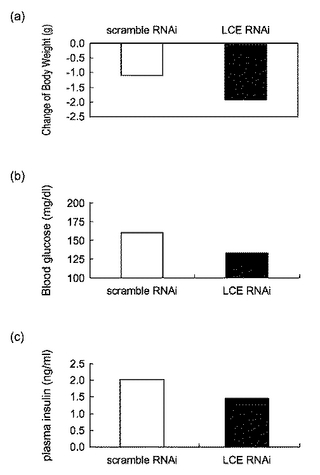
【図12】

【図13】

【図14】

【図15】
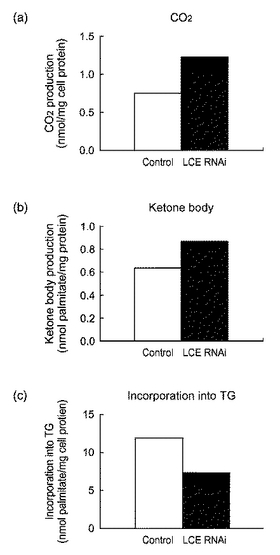
【図16】
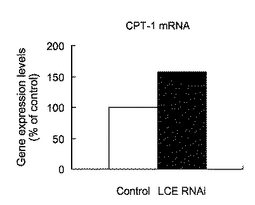
【図17】
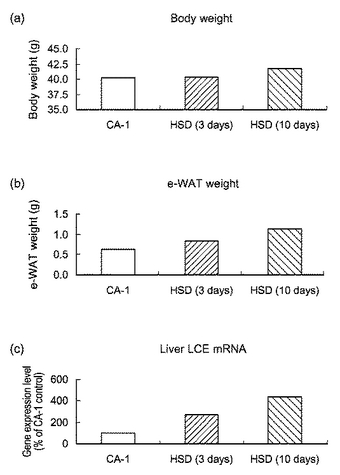
【図18】

【図19】

【図20】

【図21】
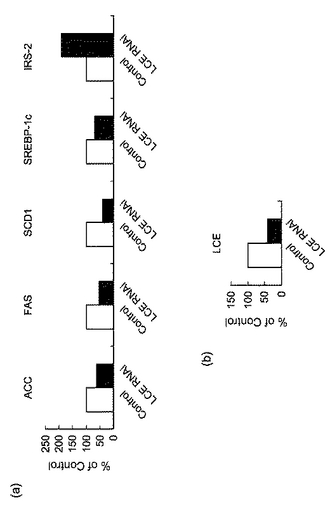
【図22】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【国際公開番号】WO2005/005665
【国際公開日】平成17年1月20日(2005.1.20)
【発行日】平成18年11月9日(2006.11.9)
【国際特許分類】
【出願番号】特願2005−511544(P2005−511544)
【国際出願番号】PCT/JP2004/009834
【国際出願日】平成16年7月9日(2004.7.9)
【出願人】(000005072)萬有製薬株式会社 (51)
【Fターム(参考)】
【国際公開日】平成17年1月20日(2005.1.20)
【発行日】平成18年11月9日(2006.11.9)
【国際特許分類】
【国際出願番号】PCT/JP2004/009834
【国際出願日】平成16年7月9日(2004.7.9)
【出願人】(000005072)萬有製薬株式会社 (51)
【Fターム(参考)】
[ Back to top ]