
LINGO結合分子およびその医薬用使用
【課題】ラット、カニクイザルおよびヒトLINGOポリペプチドに結合することができる結合分子、かかる結合分子をコードするポリヌクレオチド;該ポリヌクレオチドを含む発現ベクター;結合分子を生成することができるポリヌクレオチドを含む発現系;前記で定義されたような発現系を含む単離された宿主細胞;特に軸索再生/可塑性を促進するための処置におけるかかる結合分子の医薬品としての使用;該結合分子を含む医薬組成物;および軸索変性および脱髄に関連する疾患の処置の方法を提供する。
【解決手段】LINGO−1の特定の領域またはエピトープに対する結合分子を提供する。本発明の結合分子は解離定数(KD)<1000nM、さらに好ましくはKD<100nM、最も好ましくはKD<10nMでラットLINGO−1、カニクイザルLINGO−1およびヒトLINGO−1の成熟外部ドメイン(残基34−550)に結合する。
【解決手段】LINGO−1の特定の領域またはエピトープに対する結合分子を提供する。本発明の結合分子は解離定数(KD)<1000nM、さらに好ましくはKD<100nM、最も好ましくはKD<10nMでラットLINGO−1、カニクイザルLINGO−1およびヒトLINGO−1の成熟外部ドメイン(残基34−550)に結合する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は例えばモノクローナル抗体またはそのFabフラグメントのようなLINGO結合分子、および中枢神経系に対する傷害を有する患者を処置するためのかかる結合分子の使用に関する。
【背景技術】
【0002】
高等脊椎動物成体の中枢神経系(CNS)に対する傷害の後の機能的回復は例外的に限定的であり、四肢の運動および感覚の喪失のような持続的な神経障害を招く。今のところ脊髄傷害(SCI)および脳皮質傷害のようなCNS傷害を有するヒトを処置するための有効な治療は欠如している。成体のCNSニューロンは一般的に軸索切断から生き延びるが、軸索再生は一過性であり、そして限局的な部分にわたってのみ生じ、それ故に機能的に関連するシナプス結合の再形成を遅延させる。さらに成体CNSの可塑性能力もまた制限され、故に傷害により切断されたものを機能的に補償するための非傷害経路の再編成を妨げる。逆説的に末梢神経系(PNS)で軸索切断された軸索は長きにわたって再生し、そして頻繁に機能的に意味のある連結を確立する高い能力を有する(Schwab, Curr Opin Neurobiol 14:118−124(2004))。この軸索再生/可塑性における制限はある程度、神経突起伸長の強力な阻害剤であることが示されているいくつかのタンパク質、すなわちNogo−A(Chen et al., Nature 403:434−439(2000);GrandPre et al., Nature 403:439−444(2000);Prinjha et al., Nature 403:383−384(2000))、ミエリン関連糖タンパク質(MAG)およびオリゴデンドロサイトミエリン糖タンパク質(OMgp)(McKerracher et al., Neuron 13:805−811(1994);Wang et al., Nature 417:941−944(2002))(図1A)の有髄化オリゴデンドロサイトにおける発現のためである。
【0003】
Nogo−Aはオリゴデンドロサイトの表面に暴露された複数の神経突起伸長阻止ドメインを含有する:二つはアミノ−末端領域(アミノ−Nogo−A)内に、そして1つはC−末端領域(Nogo−66)に位置する(Oertle et al., J Neurosci 23:5393−5406(2003))。Nogo−66はNogo−66受容体(NgR)として公知のニューロン表面上のグリコシルホスファチジルイノシトール(GPI)アンカードロイシンリッチリピート(LRR)含有受容体を介して結合およびシグナル発生する(Fournier et al., Nature 409:341−346(2001))。構造的には関連しないが、MAGおよびOMgpもまたNgRを介して結合およびシグナル発生する(Domeniconi et al., Neuron 35:283−290(2002);Liu et al., Science 297:1190−1193(2002);Wang et al., Nature 417:941−944(2002))。NgRを介するシグナル発生は小型GTPase RhoAの活性化に至り、それが今度はRho関連キナーゼ(ROCK)を活性化し、アクチン細胞骨格の硬直化および軸索伸長の阻止に至る(Niederoest et al., J Neurosci 22:10368−10376(2002);Schweigreiter et al., Mol Cell Neurosci 27:163−174(2004))。三つのリガンド全てがNgRのLRR領域内に結合し、そして部分的に重複する結合部位を有する(Fournier et al., J Neurosci 22:8876−8883(2002);Liu et al. Science 297:1190−1193(2002);Wang et al., Nature 417:941−944(2002);Barton et al., EMBO J 22:3291−3302(2003))。アミノ−Nogo−A内の阻止ドメインに関する(複数の)受容体は未知であるが、NgRとは区別されることが示されている(Schweigreiter et al., Mol Cell Neurosci 27:163−174(2004))。MAGはNgR2として公知のNgRの密接した相同体を介してシグナル発生することもまた見出されている(Pignot et al., J Neurochem 85:717−728(2003);Venkatesh et al., J Neurosci 25:808−822(2005))。
【0004】
NgRは細胞質ドメインを欠如するので、それはシグナル伝達のためにいくつかの膜貫通タンパク質、すなわち低親和性ニューロトロフィン受容体p75NTR、TROY(TAJとも称される)およびLINGO−1(LRRおよびIgドメイン含有、Nogo受容体相互作用タンパク質、LRRN6AまたはLERN1とも称される)を利用する(Wang et al. Nature 420:74−78(2002);Carim−Todd et al., Eur J Neurosci 18:3167−3182(2003);Mi et al., Nat Neurosci 7:221−228(2004);Park et al., Neuron 45:345−351(2005);Shao et al., Neuron 45:353−359(2005))。TROYおよびp75NTRはNgR受容体複合体において互いに機能的に置き換えることができるが、一方LINGO−1の存在は生じるシグナル発生のための絶対的な必要条件である。NgR受容体複合体はそれ故にリガンド結合サブユニットとしてNgR、およびp75NTRまたはTROYのいずれかと協調的に作用する一般的なシグナル伝達サブユニットとしてLINGO−1を含む三元複合体として認められる。
【0005】
LINGO−1はニューロンおよびオリゴデンドロサイト上で優先的に、CNS内で独占的に発現される単一の膜貫通タンパク質である。LINGO−1の発現は出生後早期にピークに達し、そして傷害時の成体脊髄において上方調節される。LINGO−1の外部ドメインはN−およびC−末端サブドメイン、続いて塩基性領域およびIgドメインによりフランキングされる12個のタンデムLRRを含有する(図1B)。LINGO−1外部ドメインのAP融合がNgRもしくはp75NTRまたは双方を発現するCOS−7細胞に結合し、そして同様に、LINGO−1が三つ全てのタンパク質を発現する細胞におけるNgRまたはp75NTRと共沈する場合、LINGO−1は双方と同時に相互作用することによりNgRおよびp75NTRと三元複合体を形成する可能性が最も高い。
【0006】
ニューロン上で発現されることに加えて、LINGO−1はまた成体CNSのオリゴデンドロサイトにおいても発現される(Mi et al., Nat Neurosci 8:745−751(2005))。LINGO−1−Fcでのいずれかの処理によるオリゴデンドロサイト培養物中のLINGO−1シグナル発生の阻止、RNAiでのタンパク質の下方調節、またはDN−LINGO−1の過剰発現はOPCの有髄化オリゴデンドロサイトへの分化を増強した。さらに、マウスにおけるLINGO−1の遺伝手術により成熟オリゴデンドロサイトおよび対応して脊髄における有髄軸索の数が増加する。LINGO−1シグナル発生の阻止はRhoAの活性化を低下させ、そしてFynキナーゼの作用を増大させ、その双方はオリゴデンドロサイト分化を促進させると報告されているが、LINGO−1シグナル発生の活性化に寄与する実際のリガンド/相互作用は未だに例示されていない。これにより、LINGO−1がミエリン形成の負のレギュレーターであるという結論に至っている。
【0007】
多発性硬化症(MS)は脱髄および軸索変性を特徴とし、多発性神経障害に至るCNSの慢性炎症性疾患である。軸索のミエリン再形成は疾患の早期に生じ、いくつかの時点で、ミエリン再形成は完全に失敗して軸索変性の加速および非可逆的損傷に至る。ミエリン再形成は活動性病変の辺縁部に遊走する成体オリゴデンドロサイト前駆細胞(OPC)の分化から生じる可能性が最も高い。LINGO−1はミエリン形成を負に調節するので、LINGO−1の遮断はミエリン再形成を増強し、軸索変性を減弱し、軸索再生を促進し、そして故にMSのような脱髄性疾患の進行を減弱、停止または逆転さえし得る。
【0008】
LINGO−1の遮断はまたドーパミン作動性ニューロンの生存性を改善し、そしてパーキンソン病のげっ歯類モデルにおける行動異常を低下させることも示されている(Inoue et al., Proc Natl Acad Sci USA 104:14430−14435(2007))。
【発明の概要】
【発明が解決しようとする課題】
【0009】
(発明の要旨)
驚くべきことに、LINGO−1に対する新規モノクローナルヒト抗体(本明細書後記の抗体4784、および抗体4785としても公知)がLINGO−1とNgRとの会合を有意に阻止し、そしてインビトロでnM以下の濃度でラット成体の脊髄ミエリンの神経突起伸長阻止活性を有意に減弱することが見出されている。加えて、該抗体はインビトロで一次オリゴデンドロサイトの分化を有意に増大させ、そして生存細胞における細胞表面LINGO−1を有意に下方調節することが示されている。これらの抗体での処理は軸索再生/可塑性を増大させ、そしてSCIおよび脳皮質傷害のような急性CNS傷害の後の機能的回復を改善すると予期される。さらに、オリゴデンドログリア細胞における該抗体を用いるLINGO−1シグナル発生の遮断は、MSのような脱髄性疾患における軸索のミエリン再形成を増強し、疾患進行の減弱に至る可能性がある。一致して、該抗体でのニューロンにおけるLINGO−1シグナル発生の阻止は軸索再生および神経可塑性を改善し、そして疾患の経過中の神経学的機能喪失の回復を促進すると予期できる。最後に、該抗体でのLINGO−1の遮断はパーキンソン病の病因を減弱すると予期できる。
【0010】
さらに本発明はLINGO−1の特異的エピトープに結合する結合分子を提供する。
抗体はラット、カニクイザルおよびヒトLINGO−1外部ドメインに対してnM以下のKDを有し、ラット成体脊髄ミエリンの神経突起伸長阻止活性をnM以下の濃度で有意に減弱し、そしてインビトロでオリゴデンドロサイト分化を有意に増大させた。さらに今では該抗体と同一の可変領域を有するその他のLINGO−1結合分子を構築することが可能である。
【課題を解決するための手段】
【0011】
(発明の詳細な説明)
したがって、本発明はLINGO−1の特定の領域またはエピトープに対する結合分子(以後「本発明の結合分子」または単に「結合分子」と称する)を提供する。
本発明の結合分子は解離定数(KD)<1000nM、さらに好ましくはKD<100nM、最も好ましくはKD<10nMでラットLINGO−1(配列番号1)、カニクイザルLINGO−1(配列番号2)およびヒトLINGO−1(配列番号3)の成熟外部ドメイン(残基34−550)に結合する。結合反応を例えば実施例に記載されるFACS法を含む標準的な方法(定性アッセイ)により示すことができる。加えてラット、カニクイザルおよびヒトLINGO−1に対する結合、ならびに効率もまた以下で記載されるような神経突起伸長アッセイおよびオリゴデンドロサイトアッセイにおいて示すことができる。
【0012】
故にさらに好ましい実施態様では、結合分子は(100nM、好ましくは10nM、さらに好ましくは1nM、なおさらに好ましくは0.1nMの濃度で)ラット成体脊髄ミエリンの基質で成長させたラット小脳顆粒細胞の細胞あたりの平均神経突起長を、ラット、カニクイザルおよびヒトLINGO−1外部ドメインに結合しない対照抗体で処理されたラット小脳顆粒細胞の細胞あたりの平均神経突起長と比較して、少なくとも20%、好ましくは50%、最も好ましくは60%まで増大させる。
【0013】
ペプチドマイクロアレイを使用することにより、本発明の結合分子が結合する特異的エピトープを当分野において周知の方法にしたがって決定する。結果的に別の実施態様では本発明は配列番号46−51により定義されるような少なくとも1つのLINGO−1エピトープに結合する結合分子を提供する。配列番号46:KIVILLDYMFQD、配列番号47:AIRDYSFKRLYR、配列番号48:LKVLEISHWPYL、配列番号49:NLTAVPYLAVRHLVY、配列番号50:YFTCRRARIまたは配列番号51:DVLLPNYFTCRRARI。
【0014】
別の実施態様では、本発明の結合分子は1つまたはそれより多い以下のCDR配列、例えばそこで言及された抗体4784の全てまたは抗体4785配列の全てを含む:
配列番号12
(抗体4784CDR−H1)
SSGVGVG
配列番号13
(抗体4784CDR−H2)
HIGSDDDKYYSTSLKT
配列番号14
(抗体4784CDR−H3)
NQQYGDGYPGYFDY
配列番号15
(抗体4784CDR−L1)
SGDNIGNYYVY
配列番号16
(抗体4784CDR−L2)
EDTNRPS
配列番号17
(抗体4784CDR−L3)
QSYDNLHEQV
配列番号18
(抗体4785CDR’−H1)
DNSAAWS
配列番号19
(抗体4785CDR’−H2)
LIYLRSKWDNDYAVSVKS
配列番号20
(抗体4785CDR’−H3)
TGRADEFDV
配列番号21
(抗体4785CDR’−L1)
SGSSSNIGNNYVS
配列番号22
(抗体4785CDR’−L2)
RNSKRPS
配列番号23
(抗体4785CDR’−L3)
STYDTFSIV
【0015】
さらに好ましくは、結合分子は配列番号12、13、14、15、16および/もしくは17を有する抗体4784に関する;または配列番号18、19、20、21、22および/もしくは23を有する抗体4785に関する1つまたはそれより多い前記の配列を含む。
【0016】
4784または4785を変化させることができることは当業者には理解され、それを通してそれらはいくつかの、さらに好ましくは1個またはそれより多いアミノ酸、好ましくは3個まで、例えば1または2個の前記のSDRを、特に1個もしくはそれより多い、またはそれらの全て、例えばそれらの1もしくは2個で変化させるか、または生成物の形式の代替えの翻訳後修飾を提供し、同一もしくは実質的に類似する抗Lingo−1結合挙動を実証する治療薬を導く。
別の実施態様では、本発明の結合分子は;配列番号5もしくは配列番号7に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である配列、および;
配列番号4もしくは配列番号6に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である配列、またはその直接的な均等物;からなる群から選択される少なくとも1つの抗原結合部位を含む。
【0017】
1つの実施態様では、結合分子は配列番号5または配列番号7、および;
配列番号4または配列番号6;
からなる群から選択される少なくとも1つの結合部位を含む。
本発明はさらに配列番号5に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である第1の配列、および配列番号4に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である第2の配列、またはその直接的な均等物を含む結合分子を提供する。
【0018】
本発明はさらに配列番号7に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である第1の配列、および配列番号6に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である第2の配列、またはその直接的な均等物を含む結合分子を提供する。
【0019】
1つの実施態様では、本発明は少なくとも:
a)
(i)配列番号5または配列番号7を含む可変ドメイン、および;
(ii)ヒト重鎖の定常部分またはそのフラグメント;
を含む1つの免疫グロブリン重鎖またはそのフラグメント;ならびに
b)
(i)配列番号4または配列番号6を含む可変ドメイン、および;
(ii)ヒト軽鎖の定常部分またはそのフラグメント;
を含む1つの免疫グロブリン軽鎖またはそのフラグメント;
またはその直接的な均等物;例えばa)もしくはb)の下で与えられた鎖の各々の2つもしくは3つ;
を含む上記の結合分子を提供する。
【0020】
配列は配列番号4−7に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%相同でよい。重要な因子は、各々の場合で好ましくは実施例または残りの記載にて記載されるように、かかるバリアントがLINGO−1に対する結合能力、脱抑制効果(特にラット成体脊髄ミエリンの神経突起伸長阻止活性をnM以下の濃度で減弱する能力)を保持し、そして/またはSCIの機能的回復を改善する(特にラットモデルにおいて)ことである。
【0021】
1つの実施態様では、本発明は配列番号4−7もしくは配列番号12−23による1つもしくはそれより多い配列を含む抗体またはそのフラグメント、またはその直接的な均等物である結合分子を提供する。
さらなる実施態様では、抗体としての結合分子はγ4型のヒト重鎖の定常部分またはそのフラグメントおよびλ型であるヒト軽鎖の定常部分またはそのフラグメントを有する。
【0022】
さらなる実施態様では、抗体としての結合分子はγ4型のヒト重鎖の定常部分またはそのフラグメントおよびκ型であるヒト軽鎖の定常部分またはそのフラグメントを有する。
さらなる実施態様では、結合分子はヒトまたはキメラまたはヒト化モノクローナル抗体である。
さらなる実施態様では、結合分子はhumaneered抗体である。
本発明はまた前記で定義されたような結合分子をコードするポリヌクレオチドをも提供する。
【0023】
ポリヌクレオチドを配列番号8および配列番号9からなる群から;または配列番号10および配列番号11からなる群から選択できる。
本発明はまた配列番号8−11による1つまたはそれより多いポリヌクレオチドを含む発現ベクターをも提供する。
さらに本発明は配列番号8−11によるポリヌクレオチドを含む発現ベクターを提供し、ここで該発現系またはその部分が適合する宿主細胞中に存在する場合、該発現系またはその部分は前記で示されたような結合分子を生成することができる。本発明はまたはかかる発現系を含む単離された宿主細胞をも提供する。
【0024】
本発明はまたは医薬品としての前記で示されたような結合分子の使用をも提供する。
本発明はまたはCNS傷害の処置のための医薬品の調製における前記で示されたような結合分子の使用をも提供する。
本発明はまたは前記で示されたような結合分子を少なくとも1つの薬学的に許容される担体または希釈剤と一緒に含む医薬組成物をも提供する。
【0025】
さらに本発明は有効量の前記で示されたような結合分子をかかる処置を必要とする対象に投与することを含む、軸索再生/可塑性の促進に関連する疾患の処置の方法を提供する。
本発明はまたは有効量の請求項1から10のいずれかに記載の結合分子をかかる処置を必要とする対象に投与することを含む、軸索再生/可塑性の促進に関連する疾患の処置の方法をも提供する。
抗原結合部位が第1および第2の双方のドメインを含む場合、これらは同一のポリペプチドに位置してよいか、または好ましくは各ドメインが異なる鎖に位置してよく、第1のドメインは免疫グロブリン重鎖の部分またはそのフラグメントであり、そして第2のドメインは免疫グロブリン軽鎖の部分またはそのフラグメントである。
【0026】
本発明の結合分子の実例には、ファージディスプレイにより生成されるような抗体およびヒトもしくはキメラヒト化抗体、またはさらにhumaneered抗体、またはその任意のフラグメント、例えばF(ab’)2;およびFabフラグメント、ならびに一本鎖または単一ドメイン抗体が含まれる。「抗体」なる用語はかかる結合分子を含むと意図される。
一本鎖抗体は通常10から30個のアミノ酸、好ましくは15から25個のアミノ酸からなるペプチドリンカーにより共有結合した抗体重および軽鎖の可変ドメインからなる。それ故に、かかる構造は重および軽鎖の定常部分を含まず、そして小型ペプチドスペーサーは定常部分全体よりも抗原性が低いはずであると考えられている。「キメラ抗体」とは重もしくは軽鎖または双方の定常領域がヒト起源であるが、重および軽鎖の双方の可変ドメインが非ヒト(例えばマウス)起源である抗体を意味する。「ヒト化抗体」とは超可変領域(CDR)は非ヒト(例えばマウス)起源であるが、免疫グロブリンの全てまたは実質的に全てのその他の部分、例えば定常領域および可変ドメインの高度に保存された部分、すなわちフレームワーク領域はヒト起源である抗体を意味する。しかしながらヒト化抗体は超可変領域に隣接するフレームワーク領域の部分においてネズミ配列の数個のアミノ酸を保持し得る。
【0027】
超可変領域は任意の種類のフレームワーク領域、好ましくはネズミまたはヒト起源のものに関連し得る。適当なフレームワーク領域は「Sequences of proteins of immunological interest」(Kabat E.A. et al、米国保健社会福祉省国立衛生研究所公衆衛生局、好ましくは、特にフレームワーク領域に関して出典明示により本明細書の一部とする)に記載されている。好ましくは結合分子のヒト重鎖の定常部分はサブタイプを含むγ4型のものでよく、好ましくはヒト軽鎖の定常部分はκまたはλ型のもの、さらに好ましくはλ型のものでよい。
【0028】
天然発生「抗体」はジスルフィド結合により相互連結された少なくとも2つの重(H)鎖および2つの軽(L)鎖を含む糖タンパク質である。各重鎖は重鎖可変領域(本明細書ではVHと略する)および重鎖定常領域からなる。重鎖定常領域はCH1、CH2およびCH3の3つのドメインからなる。各軽鎖は軽鎖可変領域(本明細書ではVLと略する)および軽鎖定常領域からなる。軽鎖定常領域は1つのドメイン、CLからなる。VHおよびVL領域を、フレームワーク領域(FR)と称されるさらに保存されている領域がちりばめられた相補性決定領域(CDR)と称される超可変性の領域にさらに細分することができる。各VHおよびVLは以下の順でアミノ末端からカルボキシ末端に配列した3つのCDRおよび4つのFRから構成される:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4。重および軽鎖の可変領域は抗原と相互作用する結合ドメインを含有する。抗体の定常領域は、免疫系の種々の細胞(例えばエフェクター細胞)および古典的は補体系の第1の構成要素(C1q)を含む宿主組織または因子に対する免疫グロブリンの結合を媒介し得る。
【0029】
抗体の「抗原結合部分」(または単に「抗原部分」)なる用語は、本明細書で使用される際には、抗原に特異的に結合する能力を保持する抗体の全長または1つもしくはそれより多いフラグメント(例えばLINGO−1および/またはLINGO−2)を指す。全長抗体のフラグメントにより抗体の抗原結合機能を実施できることが示されている。抗体の「抗原結合部分」なる用語に包含される結合フラグメントの実例には、Fabフラグメント、VL、VH、CLおよびCH1ドメインからなる1価フラグメント;F(ab’)2フラグメント、ヒンジ領域でジスルフィド橋により連結された2つのFabフラグメントを含む2価フラグメント;VHおよびCH1ドメインからなるFdフラグメント;抗体の単一のアームのVLおよびVHドメインからなるFvフラグメント;VHドメインからなるdAbフラグメント(Ward et al., Nature 341:544−546(1989));ならびに単離された相補性決定領域(CDR)が含まれる。
【0030】
「モノクローナル抗体」または「モノクローナル抗体組成物」なる用語は、本明細書で使用される際には、単一分子組成物の抗体分子の調製物(すなわち全て単一の親細胞のクローンである免疫細胞の1つの型により生成されるのでそれらは同一である)を指す。モノクローナル抗体組成物は(本質的に)単一の結合特異性および特定のエピトープに関する親和性を表示する。
【0031】
「ヒト抗体」なる用語は、本明細書で使用される際には、フレームワークおよびCDR領域の双方がヒト起源の配列から誘導される可変領域を有する抗体を含むと意図される。さらに抗体が定常領域を含有する場合、定常領域もまたかかるヒト配列、例えばヒト生殖細胞配列、またはヒト生殖細胞配列の変異したものから誘導される。本発明のヒト抗体はヒト配列によりコードされないアミノ酸残基(例えばインビトロで無作為もしくは部位特異的変異誘発により、またはインビボで体細胞変異により導入された変異)を含み得る。しかしながら、「ヒト抗体」なる用語は、本明細書で使用される際には、マウスのような別の哺乳動物種の生殖細胞系から誘導されたCDR配列がヒトフレームワーク配列に移植されている抗体を含むとは意図されない。
【0032】
「ヒトモノクローナル抗体」なる用語は、フレームワークおよびCDR領域の双方がヒトの配列から誘導される可変領域を有する(本質的に)単一の結合特異性を表示する抗体を指す。1つの実施態様ではヒトモノクローナル抗体は、不死化細胞に融合されたヒト重鎖導入遺伝子および軽鎖導入遺伝子を含むゲノムを有するトランスジェニック非ヒト動物、例えばトランスジェニックマウスから得られたB細胞を含むハイブリドーマにより生成される。
「組換えヒト抗体」なる用語は本明細書で使用される際には、ヒト免疫グロブリン遺伝子に関して遺伝子導入もしくは染色体導入された動物(例えばマウス)またはそこから調製されたハイブリドーマから単離された抗体、ヒト抗体を発現するように形質転換された宿主細胞、例えばトランスフェクトーマ(transfectoma)から単離された抗体、組換え体から単離された抗体、コンビナトリアルヒト抗体ライブラリーのような組換え手段により調製、発現、創成または単離された全てのヒト抗体、およびヒト免疫グロブリン遺伝子の全てまたは一部の、配列をその他のDNA配列にするスプライシングを伴う任意のその他の手段により調製、発現、創成または単離された抗体を含む。かかる組換えヒト抗体は、フレームワークおよびCDR領域がヒト生殖細胞系免疫グロブリン配列から誘導される可変領域を有する。しかしながら特定の実施態様では、かかる組換えヒト抗体をインビトロ変異誘発(またはヒトIg配列に関して遺伝子導入された動物を用いる場合、インビボ体細胞変異誘発)に供することができ、そして故に組換え抗体のVHおよびVL領域のアミノ酸配列は、ヒト生殖細胞系VHおよびVL配列から誘導され、そしてそれに関連するが、インビボでヒト抗体生殖細胞系レパートリー内に天然には存在し得ない配列である。
【0033】
本明細書で使用される際には、「アイソタイプ」とは重鎖定常領域遺伝子により提供される抗体クラス(例えばIgM、IgE、IgG1またはIgG4のようなIgG)を指す。
本明細書で使用される際には、「親和性」なる用語は単一の抗原性部位での抗体と抗原との間の相互作用の強さを指す。各抗原性部位内では、抗体「アーム」の可変領域は弱い非共有結合力を介して多くの部位で抗原と相互作用し;より相互作用すると、親和性がより強くなる。
「KD」なる用語は、本明細書で使用される際には、Kaに対するKdの比(解離率に対する会合率)(すなわちKd/Ka)から得られる解離定数を指すと意図され、そしてモル濃度(M)として表現される。当分野において十分に確立された方法を用いて抗体に関するKD値を決定することができる。抗体のKDを決定するための方法は表面プラズモン共鳴法の使用、またはBiacore(登録商標)システムのようなバイオセンサーシステムの使用による。
【0034】
本発明による結合分子は好ましくは「単離された抗体」であり、それは本明細書で使用される際には、異なる抗原特異性を有するその他の抗体を実質的に含まない抗体を指す(例えばLINGO−1、LINGO−2またはLINGO−1およびLINGO−2に特異的に結合する単離された抗体は言及されたもの以外の抗原に特異的に結合する抗体を実質的に含まない)。しかしながら特異的に結合する単離された抗体はその他の種からのLINGO−1またはLINGO−2分子のようなその他の抗原に対する交差反応性を有し得る。さらに単離された抗体は好ましくはその他の細胞材料および/または化学物質を実質的に含まない。
本発明はまたは:
a)アミノ酸配列(配列番号5)を有する重鎖の可変配列を含む第1ドメイン;
b)アミノ酸配列(配列番号4)を有する軽鎖の可変配列を含む第2ドメイン;
c)第1ドメインのN末端および第2ドメインのC末端もしくは第1ドメインのC末端および第2ドメインのN末端のいずれかに結合したペプチドリンカー;
またはその直接的な均等物;
を含む抗体4784の抗原結合部位(特に抗体4784に関して前記されたCDRを伴う)を含む一本鎖結合分子から選択できる本発明の結合分子を提供する。
本発明の結合分子を
a)アミノ酸配列(配列番号7)を有する重鎖の可変配列を含む第1ドメイン;
b)アミノ酸配列(配列番号6)を有する軽鎖の可変配列を含む第2ドメイン;
c)第1ドメインのN末端および第2ドメインのC末端もしくは第1ドメインのC末端および第2ドメインのN末端のいずれかに結合したペプチドリンカー;
またはその直接的な均等物;
を含む抗体4785の抗原結合部位(特に抗体4785に関して前記されたCDRを伴う)を含む一本鎖結合分子から選択できる。
周知のように、1つまたはいくつかのアミノ酸の欠失、付加または置換のようなアミノ酸配列におけるわずかな変化は、実質的に同一の特性を有する元来のタンパク質のアレル形態に至り得る。故に「その直接的な均等物」なる用語は:
(i)結合分子の可変領域(例えば配列番号4、5、6または7)は、配列番号4および配列番号5の各々の直接的な均等物を含む軽および重鎖または配列番号6および配列番号7の各々の直接的な均等物を含む軽および重鎖の均等な可変領域に対して少なくとも50または80%相同、好ましくは少なくとも90%相同、さらに好ましくは少なくとも95、96、97、98、99%相同であり;
(ii)ラットLINGO−1(配列番号1)、カニクイザルLINGO−1(配列番号2)およびヒトLINGO−1(配列番号3)の外部ドメイン(残基34−550)に、好ましくは解離定数(KD)<1000nM、さらに好ましくはKD<100nM、最も好ましくはKD<10nMで結合できる;
本発明のいずれかの任意の単一ドメイン結合分子(分子X)、または結合部位あたり少なくとも2つのドメインを有する本発明の任意の結合分子(分子X’)を意味する。
【0035】
故に本発明のさらなる実施態様は例えば解離定数<1000nMでラット、カニクイザルおよび/またはヒトLINGO−1の外部ドメインに結合でき、そして少なくとも1つの抗原結合部位を含み、該抗原結合部位は配列に4784の軽および重鎖(各々配列番号4および配列番号5)または4785の軽および重鎖(各々配列番号6および配列番号7)の均等な可変領域に少なくとも50%、好ましくは80、90、95、96、97、98、99%相同である可変領域を含む。
【0036】
別の実施態様では、結合分子は配列番号12−23からなる群から選択される少なくとも1つのアミノ酸配列、またはこれらの配列に対して少なくとも50%、好ましくは80、90、95、96、97、98、99%相同である配列を含む。
この解離定数を例えば実施例で記載されるFACS法を含む種々のアッセイにおいて都合よく試験できる。加えて結合分子の結合および機能的影響をバイオアッセイ、例えば以下に記載されるような神経突起伸長アッセイにおいて示すことができる。
【0037】
ヒト重鎖の定常部分はγ1;γ2;γ3;γ4;α1;α2;δまたはε型、好ましくはγ型、さらに好ましくはγ4型のものでよいが、一方ヒト軽鎖の定常部分はκまたはλ型(それはλ1;λ2;およびλ3サブタイプを含む)でよいが、好ましくはλ型のものである。これらの全ての定常部分のアミノ酸配列はKabat et al(Supra)で与えられる。
本発明の結合分子の抱合体、例えば酵素または毒素または放射性同位体抱合体もまた本発明の範囲内に含まれる。
【0038】
「ポリペプチド」には、本明細書にて特記しない場合、ペプチド結合により互いに結合したアミノ酸を含み、N末端で始まり、そしてC末端で終わるアミノ酸配列を有する任意のペプチドまたはタンパク質が含まれる。好ましくは本発明のペプチドはモノクローナル抗体であり、さらに好ましくはキメラ(Vグラフティングとも称される)またはヒト化(CDRグラフティングとも称される)モノクローナル抗体である。ヒト化(CDRグラフティング)モノクローナル抗体はアクセプター抗体のフレームワーク(FR)配列に導入されたさらなる変異を含んでも含まなくてもよい。
【0039】
ポリペプチドの機能的誘導体は本明細書で使用される際には、質的に本発明のポリペプチドに共通した生物学的活性を有する、すなわちラット、カニクイザルおよびヒトLINGO−1の外部ドメインに結合する能力を有する分子が含まれる。
機能的誘導体には本発明によるポリペプチドのフラグメントおよびペプチド類似体が含まれる。それはまた「直接的な誘導体」をも含む。
【0040】
フラグメントは本発明による、例えば特定された配列のポリペプチドの配列内の領域を含む。結合分子の、特に抗体のフラグメントは機能的なフラグメントである、すなわちそれらはLINGO−1および/またはLINGO−2に、特に配列番号46、47、48、49、50および51により与えられる少なくとも1つのエピトープに、好ましくは、前記でまたは実施例で特に好ましいとして言及された結合親和性(KD)で結合できる少なくとも1つの部分を含む。
「誘導体」なる用語は本発明による、例えば特定された配列のポリペプチドのアミノ酸配列バリアント、および共有結合性修飾を定義するために用いられる。本発明による、例えば特定された配列のポリペプチドの例えば軽および重鎖の超可変領域の機能的誘導体は好ましくは、本発明による、例えば特定された配列のポリペプチドのアミノ酸配列と少なくとも約65%、さらに好ましくは少なくとも約75%、なおさらに好ましくは少なくとも約85%、最も好ましくは少なくとも約95、96、97、98、99%の配列全体にわたる相同性を有し、そしてラット、カニクイザルおよびヒトLINGO−1(および場合によっては加えてLINGO−2)の外部ドメインに結合する能力を実質的に保持している。
【0041】
「共有結合性修飾」なる用語には、本発明による、例えば特定された配列のポリペプチド;またはそのフラグメントの有機タンパク質性または非タンパク質性誘導体化剤での修飾、異種性ポリペプチドへの融合および翻訳後修飾が含まれる。例えば特定された配列の共有結合性修飾されたポリペプチドは依然ラット、カニクイザルおよびヒトLINGO−1の外部ドメインに結合する能力を有する。共有結合性修飾は伝統的には標的化されたアミノ酸残基を、選択された側鎖または末端残基と反応できる有機誘導体化剤と反応させることにより、または選択された組換え宿主細胞において機能する翻訳後修飾のメカニズムを利用することにより導入される。特定の翻訳後修飾は発現されたポリペプチドにおける組換え宿主細胞の作用の結果である。グルタミニルおよびアスパラギニル残基は対応するグルタミルおよびアスパルチル残基に頻繁に翻訳後脱アミド化される。これに代えて、これらの残基は穏やかな酸性条件下で脱アミノ化される、その他の翻訳後修飾にはプロリンおよびリジンのヒドロキシル化、セリル、チロシンまたはスレオニル残基のヒドロキシル基のリン酸化、リジン、アルギニンおよびヒスチジン側鎖のα−アミノ基のメチル化が含まれる(例えばT. E. Creighton, Proteins:Structure and Molecular Properties, W. H. Freeman & Co., San Francisco, pp. 79−86(1983)参照)。共有結合性修飾には、例えばイムノアドヘシンのような、例えば本発明による、例えば特定された配列のポリペプチド含む融合タンパク質およびそのアミノ酸バリアントを、ならびに異種性シグナル配列に対するN末端融合が含まれる。
【0042】
自然のポリペプチドおよびその機能的誘導体に関する「相同性」(または「同一性」)は本明細書では、必要により最大相同性パーセントを達成するために配列をアラインし、そしてギャップを誘導した後、対応する自然のポリペプチドの残基と同一である、候補配列におけるアミノ酸残基のパーセンテージとして定義され、そして任意の保存置換を配列同一性の一部ではないと考える。NもしくはC末端伸長も挿入も同一性または相同性を低下させるものではないと解釈しなければならない。アラインメントのための方法およびコンピュータープログラムは周知である。
【0043】
好ましくは本明細書で使用される際には、2つのアミノ酸配列または2つのヌクレオチド配列間の相同性パーセントは2つの配列間の同一性パーセントと均等である。2つの配列間の同一性パーセントは配列により共有される同一の位置の数の関数であり(すなわち相同性%=同一の位置の数/位置の全数×100)、2つの配列の最適なアラインメントのために導入される必要のあるギャップの数および各ギャップの長さを考慮する。配列の比較および2つの配列間の同一性パーセントの決定を、以下の非限定例で記載されるような数学的アルゴリズムを用いて達成することができる:
【0044】
2つのアミノ酸配列間の同一性パーセントを、ALIGNプログラム(バージョン2.0)に組み入れられているE. MeyersおよびW. Millerのアルゴリズム(Comput. Appl. Biosci., 4:11−17(1988))を用いて、PAM120残基重み付け表(weight residue table)、ギャップ長ペナルティー12およびギャップペナルティー4を用いて決定することができる。加えて、2つのアミノ酸配列間の同一性パーセントを、GCGソフトウェアパッケージのGAPプログラム(http://www.gcg.comで利用可能)に組み入れられているNeedlemanおよびWunsch(J. Mol, Biol. 48:444−453(1970))アルゴリズムを用いて、Blossom 62マトリクスまたはPAM250マトリクスのいずれか、ならびにギャップ重み付け16、14、12、10、8、6または4および長さ重み付け1、2、3、4、5または6を用いて決定することができる。
【0045】
加えてまたはこれに代えて、本発明のタンパク質配列をさらに「クエリー配列」として用いて公開されたデータベースに対して検索を実施し、例えば関係する配列を同定することができる。Altschul, et al., J.Mol. Biol. 215:403−10(1990)のXBLASTプログラム(バージョン2.0)を用いてかかる検索を実施することができる。BLASTタンパク質検索をXBLASTプログラム、スコア=50、語長=3で実施して、本発明の抗体分子に相同なアミノ酸配列を得ることができる。比較目的のためにギャップの入ったアラインメントを得るために、Altschul et al., Nucleic Acids Res. 25(17):3389−3402(1997)に記載されるようなGapped BLASTを利用することができる。BLASTおよびGapped BLASTプログラムを利用する場合、各プログラムのデフォルトパラメーター(例えばXBLASTおよびNBLAST)を用いることができる。http:www.ncbi.nhn.nih.govを参照のこと。
「(複数の)アミノ酸」は全ての天然発生の例えばL−α−アミノ酸を指し、そしてD−アミノ酸を含む。アミノ酸は周知の一文字表記または三文字表記のいずれかにより同定される。
【0046】
「アミノ酸配列バリアント」なる用語は、本発明による、例えば特定された配列のポリペプチドと比較してそのアミノ酸配列においていくつかの差異を有する分子を指す。本発明による、例えば特定された配列のポリペプチドのアミノ酸配列バリアントは依然ラット、カニクイザルおよびヒトLINGO−1の外部ドメインに対する結合能力を有する。置換バリアントは、本発明による、例えば特定された配列のポリペプチドにおいて少なくとも1つのアミノ酸残基が除去され、そして同一位置でその場所に異なるアミノ酸が挿入されたものである。これらの置換は単一でよく、その場合分子の1個のアミノ酸のみが置換されているか、またはそれらは複数でよく、その場合2個またはそれより多い、例えば1から10個。好ましくは1から5個、さらに好ましくは1から3個のアミノ酸が同一分子内で置換されている。挿入バリアントは、1つまたはそれより多い、例えば1から10個のような1から100個のアミノ酸が、本発明による、例えば特定された配列のポリペプチドにおける特定の位置のアミノ酸に直ぐ隣接して挿入されたものである。アミノ酸に直ぐ隣接してとは、アミノ酸のα−カルボキシまたはα−アミノ官能基のいずれかに連結されていることを意味する。欠失バリアントは本発明による、例えば特定された配列のポリペプチドにおいて1個またはそれより多い、例えば1から10個または1から5個のような1から100個のアミノ酸が除去されているものである。通常欠失バリアントは分子の特定の領域において1または2個のアミノ酸が欠失されている。
【0047】
本発明の結合分子を組換えDNA技術により生成できる。これに鑑みて、結合分子をコードする1つまたはそれより多いDNA分子は構築され、適切な制御配列下に置かれ、そして発現のために適当な宿主生物に移されなければならない。
【0048】
したがって非常に一般的な様式では:
(i)本発明の単一ドメイン結合分子、本発明の一本鎖結合分子、本発明の結合分子の重もしくは軽鎖またはそのフラグメントをコードするDNA分子;および
(ii)組換え手段による本発明の結合分子の生成のための本発明のDNA分子の使用;
が提供される。
【0049】
当分野の現状では、当業者は本明細書にて提供される情報、すなわち超可変領域のアミノ酸配列およびそれをコードするDNA配列を考慮して、本発明のDNA分子を合成できる。可変ドメイン遺伝子を構築する方法は例えば欧州特許第239400号(好ましくは特に可変ドメイン遺伝子を構築するための方法に関して、出典明示により本明細書の一部とする)に記載されており、そして以下のように簡単に要約できる:どのような特異性のものでもモノクローナル抗体の可変ドメインをコードする遺伝子はクローン化される。フレームワークおよび超可変領域をコードするDNAセグメントを決定し、そして超可変領域をコードするDNAセグメントを除去し、フレームワーク領域をコードするDNAセグメントを接合部で適当な制限部位と一緒に融合する。標準的な手順によりDNA分子の変異誘発により適切な位置で制限部位を作成することができる。前記の配列にしたがってDNA合成により二本鎖合成可変領域カセットを調製する。免疫グロブリン可変ドメインをコードするDNA分子を達成するための標準的なプロトコールにより、これらのカセットに付着末端が提供され、それらを接合部でフレームワークにライゲートできるようになる。
【0050】
さらに本発明のモノクローナル抗体をコードするDNA構築物を得るために、生成ハイブリドーマ細胞系からmRNAを入手する必要はない。故にPCT出願第WO90/07861号(好ましくは特にモノクローナル抗体の生成に関して、出典明示により本明細書の一部とする)により、遺伝子のヌクレオチド配列に関して記載された情報のみを考慮して組換えDNA技術によりモノクローナル抗体を生成するための全ての指示が与えられる。
その方法は多数のオリゴヌクレオチドの合成、PCR法によるその増幅および望ましいDNA配列を与えるためのそのスプライシングを含む。
【0051】
適当なプロモーターまたは重および軽鎖定常部分をコードする遺伝子を含む発現ベクターは公的に入手可能である。故に一度本発明のDNA分子が調製されると、それを適切な発現ベクターに都合よく移すことができる。
一本鎖抗体をコードするDNA分子を標準的な方法により、例えば第WO88/1649号(好ましくは特に一本鎖抗体をコードするDNA分子に関して、出典明示により本明細書の一部とする)に記載されるように調製することもできる。
本発明の特定の実施態様では、本発明の結合分子のいくつかの生成のための組換え手段には、以下に記載されるような第1および第2のDNA構築物が含まれる:
【0052】
第1のDNA構築物は重鎖またはそのフラグメントをコードし、そして:
a)抗体4784、DNA−4784VH(配列番号8)、または抗体4785、DNA−4785VH(配列番号9)のいずれかの重鎖の可変ドメインをコードする第1の部分;この第1の部分は可変ドメインの最初のアミノ酸をコードするコドンで始まり、そして可変ドメインの最後のアミノ酸をコードするコドンで終わる;および
b)重鎖の定常部分の最初のアミノ酸をコードするコドンで始まり、そして定常部分またはそのフラグメントの最後のアミノ酸をコードするコドンで終わり、ナンセンスコドンが続く重鎖定常部分またはそのフラグメントをコードする第2の部分;
を含む。
好ましくは第2の部分はヒト重鎖の定常部分、さらに好ましくはヒトγ4鎖の定常部分をコードする。この第2の部分はゲノム起源のDNAフラグメント(イントロンを含む)またはcDNAフラグメント(イントロンを含まない)でよい。
【0053】
第2のDNA構築物は軽鎖またはそのフラグメントをコードし、そして:
a)抗体4784、DNA−4784VL(配列番号10)、または抗体4785、DNA−4785VL(配列番号11)のいずれかの軽鎖の可変ドメインをコードする第1の部分;この第1の部分は可変ドメインの最初のアミノ酸をコードするコドンで始まり、そして可変ドメインの最後のアミノ酸をコードするコドンで終わる;および
b)軽鎖の定常部分の最初のアミノ酸をコードするコドンで始まり、そして定常部分またはそのフラグメントの最後のアミノ酸をコードするコドンで終わり、ナンセンスコドンが続く軽鎖定常部分またはそのフラグメントをコードする第2の部分;
を含む。
好ましくは第2の部分はヒト軽鎖の定常部分、さらに好ましくはヒトκ鎖の定常部分をコードする。
【0054】
DNA構築物の各々を適当な制御配列の制御下に、とりわけ適当なプロモーターの制御下に置く。DNA構築物を発現のために移す宿主生物に適合するならば、任意の種類のプロモーターを使用できる。しかしながら哺乳動物細胞において発現を行うことになっている場合、免疫グロブリン遺伝子のプロモーターを使用するのがとりわけ好ましい。
【0055】
細胞培養またはトランスジェニック動物において望ましい抗体を生成できる。適当な制御配列の下に置かれた第1および第2のDNA構築物を卵にマイクロインジェクトし、そのように調製された卵を適切な偽妊娠の雌に移し、そして望ましい抗体を発現する子孫を選択することを含む標準的な方法により、適当なトランスジェニック動物を得ることができる。
抗体鎖が細胞培養中に生成されなければならない場合、DNA構築物を単一の発現ベクターまたは2つの別個であるが適合する発現ベクターのいずれかに最初に挿入すべきであり、恐らく後者が好ましい。
【0056】
したがって本発明はまた前記で記載されたDNA構築物の少なくとも1つを含む原核細胞系または真核細胞系において複製できる発現ベクターをも提供する。
次いでDNA構築物を含有する各発現ベクターを適当な宿主生物に移す。DNA構築物を2つの発現ベクターに別個に挿入する場合、それらを別個に、すなわち細胞あたり1つの型のベクターに移すか、または同時に移すことができ、恐らくこの後者が好ましい。適当な宿主生物は細菌、酵母または哺乳動物細胞系でよく、この後者が好ましい。さらに好ましくは、哺乳動物細胞系はリンパ系起源、例えば骨髄腫、ハイブリドーマまたは通常の不死化B細胞であるが、内因性抗体重または軽鎖を全く発現しない。
【0057】
宿主生物が細胞あたりに多数のベクターのコピーを含有するのも好ましい。宿主生物が哺乳動物細胞系である場合、標準的な方法にしたがってコピーの数を増幅することにより望ましい目標に到達することができる。増幅方法は通常薬物に対して高められた抵抗性に関して選択することからなり、該抵抗性は発現ベクターによりコードされる。
本発明の別の態様では、本発明の多鎖結合分子を生成するための方法が提供され、それは(i)本発明の第1および第2の構築物で形質転換された生物を培養すること;ならびに(ii)培養物から本発明の活性な結合分子を回収すること;を含む。
【0058】
これに代えて重および軽鎖を別個に回収し、そしてインビトロでリフォールディングした後、活性な結合分子に再構成することができる。再構成の方法は当分野において周知である;方法の実例はとりわけ欧州特許第120674号または欧州特許第125023号にて提供される。
それ故に方法はまた:
(i)本発明の第1のDNA構築物で形質転換された第1の生物を培養し、そして培養物から該重鎖またはそのフラグメントを回収すること;ならびに
(ii)本発明の第2のDNA構築物で形質転換された第2の生物を培養し、そして培養物から該軽鎖またはそのフラグメントを回収すること;ならびに
(iii)(i)で得られた重鎖またはそのフラグメントおよび(ii)で得られた軽鎖またはそのフラグメントから本発明の活性な結合分子をインビトロで再構成すること;
をも含む。
【0059】
類似の様式で、
(i)本発明の一本鎖または単一ドメイン結合分子を各々コードするDNA構築物で形質転換された生物を培養すること;および
(ii)培養物から該分子を回収すること;
を含む本発明の一本鎖または単一ドメイン結合分子を生成するための方法もまた提供される。
【0060】
本発明の結合分子は以下に例示されるようにLINGO−1のNgRに対する結合を有意に阻止し、ラット成体脊髄ミエリンの神経突起伸長阻止活性をnM以下の濃度で有意に減弱し、そしてオリゴデンドロサイト分化をインビトロで有意に増大させる。
【図面の簡単な説明】
【0061】
【図1】NgR:SH−SY5Y細胞に対するAP−LINGO−1結合に及ぼすFab4784および4785の影響:2μMの指示された抗LINGO−1Fabまたは抗ニワトリリゾチームFab3207の不在下または存在下で懸濁液中のNgR:SH−SY5Y細胞を1nM APまたはAP−LINGO−1のいずれかと共にインキュベートする。1−StepTM PNPPと共に30分間インキュベートした後、細胞における結合AP活性を405nmでの吸光度として測定する。AP−LINGO−1の特異的結合をAP−LINGO−1結合の全量とAP単独との結合の量との間の差異として計算する。特異的結合の平均阻止パーセンテージ(n=3、±STD)をFab3207の存在下および抗LINGO−1 Fabの存在下のAP−LINGO−1の特異的結合の量の間の差異パーセンタイルとして計算する。
【図2】抗LINGO−1 IgG4抗体4784および4785による脊髄ミエリンの脱抑制:A)脊髄ミエリンを伴わずにコーティングされたウェル(SCなし、白色棒)または脊髄ミエリンでコーティングされたウェル上で抗LINGO−1 IgG4抗体、対照抗リゾチームIgG4抗体3207(緑色棒)もしくは1μm ROCK阻害剤Y27632(黄色棒)の不在下(SC、赤色棒)または存在下、P7 CGN細胞を16時間インキュベートする。ROCKは全てではないが、たいていの、NgR受容体複合体を介してシグナル発生しないものを含むミエリン関連神経突起伸長阻害剤のシグナル発生経路における2次メッセンジャーであり、それでY27632処理が脊髄ミエリンの神経突起伸長阻止活性の減弱のための陽性対照として用いられる(図1)。プレート毎にSCを伴う、およびSCを伴わない条件で3つの96ウェルプレートで実験を実施し、そのプレート上の抗体の影響を比較し、そしてニューロンあたりの平均神経突起長(μm)を10回反復でウェルあたり500ニューロンに関して計算する。阻止パーセンテージ(白色文字)を、SCを伴って、および伴わずにコーティングされたウェル上に置かれた細胞間の平均神経突起長/ニューロンにおける差異パーセンタイルとして計算する。脱抑制パーセンテージ(黒色斜字体文字)を、抗LINGO−1抗体の存在下および不在下でSC上に置かれた細胞間の平均神経突起長における差異として、SCを伴って、および伴わずにコーティングされたウェル上に置かれた細胞間における差異パーセンタイルとして計算する。*p<0.05、**p<0.01(一元配置分散分析、抗体不在下で脊髄ミエリン上に置かれた細胞に関する平均神経突起長/ニューロンに対するHolm−Sidak比較)。B)脊髄ミエリンを伴わずに(SCなし)コーティングされたウェル上で、および脊髄ミエリンを伴って(SC)コーティングされたウェル上で、1nM 対照IgG4 3207もしくは抗LINGO−1 IgG4 4784の不在下(SC)または存在下でインキュベートされた細胞の代表的な視野の蛍光画像。4784の存在下の脊髄ミエリン上で成長させた細胞は、抗体の不在下または対照抗体3207の存在下で成長させた細胞よりも細胞あたりの神経突起が可視的により長く、そしてより多い。
【図3】抗LINGO−1 IgG4抗体IIによる脊髄ミエリンの脱抑制 A)脊髄ミエリンを伴わずにコーティングされたウェル(SCなし、白色棒)または脊髄ミエリンでコーティングされたウェル上で抗LINGO−1 IgG4抗体もしくは対照抗リゾチームIgG4抗体3207の不在下(SC、赤色棒)または存在下、P7 CGN細胞を8時間インキュベートする。プレート毎にSCを伴う、およびSCを伴わない条件で3つの96ウェルプレートで実験を実施し、そのプレート上の抗体の影響を比較し、そしてニューロンあたりの平均神経突起長(μm)を10回反復でウェルあたり500ニューロンに関して計算する。阻止パーセンテージ(白色文字)および脱抑制パーセンテージ(黒色斜字体文字)を前記のように計算する。**p<0.01(一元配置分散分析、抗体不在下で脊髄ミエリン上に置かれた細胞に関する平均神経突起長/ニューロンに対するHolm−Sidak比較)。
【図4】抗LINGO−1 IgG4抗体IIIによる脊髄ミエリンの脱抑制:A)脊髄ミエリンを伴わずにコーティングされたウェル(SCなし)または脊髄ミエリンでコーティングされたウェル上で指示された濃度の抗LINGO−1 IgG4抗体4784もしくは4785、対照抗リゾチームIgG4抗体3207もしくは1μM Y27632(ROCK)の不在下(SC)または存在下、P7 CGN細胞を8時間インキュベートする。プレート毎にSCを伴う、およびSCを伴わない条件で3つの96ウェルプレートで実験を実施し、そのプレート上の抗体の影響を比較し、そしてニューロンあたりの平均神経突起長(μm)を10回反復でウェルあたり500ニューロンに関して計算する。阻止パーセンテージ(白色文字)および脱抑制パーセンテージ(黒色斜字体文字)を前記のように計算する。*p<0.05、**p<0.01(一元配置分散分析、抗体不在下で脊髄ミエリン上に置かれた細胞に関する平均神経突起長/ニューロンに対するHolm−Sidak比較)。
【図5A】抗LINGO−1抗体は未成熟オリゴデンドロサイトの分化を有意に増大させる:A)新たに単離されたOPCをDMEM/CNTF/T3培地中100nM 4784、4785または対照IgG4 3207で3日間処理し、続いて抗O4抗体で染色して未成熟および成熟オリゴデンドロサイト(より大きければ、標識がより拡散される)を可視化し、そして核酸色素DAPI(4’,6−ジアミジン−2’−フェニル−インドール二塩酸塩)で細胞核(より小さい円形ドット)を可視化する。高度に樹状化し、そして伸長した突起およびミエリンシート状構造を担持するオリゴデンドロサイトは成熟形態を有していると考えられ、そして白色矢印で示される。抗LINGO−1抗体処理の結果、成熟形態を有するO4陽性細胞の比率の増大を招くが、一方対照IgG4 3207での処理には影響がない。
【図5B】全(左グラフ)および成熟(右グラフ)オリゴデンドロサイトの比率を3つの独立した実験(1、2、3)で定量する。左の棒グラフはO4染色に関連するDAPI染色された核のパーセンテージを表し、そして右の棒グラフは成熟形態を有するO4陽性細胞のパーセンテージを表す(3検体の平均+STD)。各棒グラフでは、最左の棒は処理なしのものであり、左から2番目の棒は対照IgGでの対照、次は4784での処理、そして最右は4785での処理を表す。抗LINGO−1抗体はオリゴデンドロサイトである細胞の比率に影響しないが、成熟形態を有するオリゴデンドロサイトの比率を有意に増大させる。*p<0.05、**p<0.01、対照IgG4 3207の存在下の成熟オリゴデンドロサイトの比率に対するHolm−Sidak比較を伴う一元配置分散分析。
【図6】抗LINGO−1抗体は細胞表面LINGO−1を下方調節する:A)トランスフェクトされていないCHO−K1またはCHO−K1−hLINGO−1細胞を100nM 4784、4785および3207と共に37℃で24時間インキュベートし、そして抗V5抗体と共に室温で30分間さらにインキュベートすることにより細胞表面でLINGO−1を検出する。細胞を4%PFAで固定し、BSAで遮断し、そして結合抗V5抗体を抗マウスIgG(Fc特異的)−POD抱合体を用いて検出し、それをその後1−Step(商標)Turbo TMB ELISAキットを用いて展開する。450nmでの吸光度は細胞表面でのLINGO−1の量の測定値と考えられる(3検体の平均+STD)。トランスフェクトされていないCHO−K1細胞に対して非常に低レベルのV5抗体結合が観察される。CHO−K1−hLINGO−1細胞を、抗LINGO−1抗体を伴うが、対照IgG4 3207を伴わないでインキュベートすることにより、結果的に細胞表面でのLINGO−1の量の有意な低下を招く。**p<0.01、対照IgG4 3207とのインキュベーションの後の吸光度に対するHolm−Sidak比較を伴う一元配置分散分析。B)トランスフェクトされていないCHO−K1またはCHO−K1−hLINGO−1細胞上の細胞表面タンパク質を4℃でビオチン化し、そして細胞を100nM 4784、4785および3207を伴って、または伴わずに37℃で指示された時間、インキュベートする。インキュベーション期間の終わりに、アガロースビーズに結合させた抗V5抗体を用いて細胞ライゼートからLINGO−1を沈殿させ、そしてビオチン化(細胞表面)LINGO−1をウェスタンブロット分析により抗ビオチン抗体を用いて検出する。トランスフェクトされていないCHO−K1細胞ではビオチン化LINGO−1に関するシグナルは検出されない。CHO−K1−hLINGO−1細胞を抗LINGO−1抗体と共にインキュベートすることにより、細胞表面のLINGO−1の分解離率が増大する。
【図7】ELSIAによる抗LINGO−1 Fabの特徴付け:ELISA分析に関する値を相対蛍光単位(RFU)の平均値として与える。これらのクローンの結合親和性はFACS飽和アッセイにより特徴付けされる。
【発明を実施するための形態】
【0062】
本発明はまた哺乳動物神経系、とりわけヒト神経系の軸索再生/可塑性の促進における本発明の結合分子の使用をも提供する。
本発明はまた有効量の本発明の結合分子をかかる処置を必要とする患者に投与することを含む、哺乳動物神経系、とりわけヒト神経系の軸索再生/可塑性を促進する方法をも提供する。
本発明はまた本発明の結合分子および薬学的に許容される担体または希釈剤を含む、哺乳動物神経系、とりわけヒト神経系の軸索再生/可塑性を促進するための医薬組成物をも提供する。
【0063】
とりわけ本発明の結合分子は、CNS傷害の軸索再生および可塑性を促進するのに有用である(傷害なる用語は本出願では、特に機械的もしくは化学的影響により引き起こされるか、または例えばニューロンの変性に至る疾患もしくは障害による傷害、特に例えばアルツハイマー病もしくはパーキンソン病のような神経学的疾患または以下に言及されるような障害もしくは疾患におけるその構造または形態を指す)。故に本発明の分子はとりわけヒト対象のために広い有用性を有する。例えば本発明の結合分子は末梢(PNS)および中枢(CNS)神経系の種々の疾患の処置において、すなわちさらにとりわけアルツハイマー病、パーキンソン病、筋萎縮性側索硬化症(ALS)、レビー様病理またはその他の一般的な認知症、頭蓋、大脳または脊髄外傷後の疾患および脳卒中のような神経変性疾患において有用である。さらにLINGO−1がミエリン形成の負のレギュレーターであることを考慮すると、本発明の結合分子は、限定するものではないが多発性硬化症、単相性脱髄、脳脊髄炎、多巣性白質脳症、全脳炎、マルキアファーヴァ・ビニャミ病、橋ミエリン崩壊、副腎白質ジストロフィー、ペリツェウス・メルツバッハー病、海綿状変性、アレキサンダー病、カナヴァン病、異染性白質ジストロフィーおよびクラッベ病を含む脱髄性疾患における軸索再生/可塑性の促進と協調してミエリン再形成を促進するのに有用である。一例では、本発明の結合分子を発現する細胞を脊髄傷害部位に移植して、傷害を受けた部位全体にわたって軸索成長を促すことができる。かかる移植細胞は傷害または外傷後の脊髄機能を回復するための手段を提供するであろう。かかる細胞は嗅神経鞘細胞および胎児神経または組織移植片の様々な系列の幹細胞を含み得る。
【0064】
加えて本発明の結合分子は一般的な虚血性網膜症、前眼部虚血性視神経症、視神経炎の全形態、加齢性黄斑変性症、糖尿病性網膜症、嚢胞様黄斑浮腫(CME)、網膜色素変性症、スタルガルト病、ベスト病の卵黄様網膜変性、レーベル先天黒内障およびその他の遺伝性網膜変性、病的近視、未熟児網膜症、およびレーベル遺伝性視神経症、角膜移植または屈折矯正角膜手術の後遺症、ならびにヘルペス角膜炎を含む網膜または角膜の変性を直接的または間接的に伴い得る眼変性傷害の処置に有用である。
さらに本発明の結合分子は精神医学的症状、とりわけ統合失調症および抑うつの処置に有用である。
【0065】
これらの適応症のための適切な投薬量はもちろん例えば用いられることになっている本発明の特定の分子、投与の様式ならびに処置される症状の性質および重篤度に依存して異なるであろう。一般的には、投薬量は好ましくは1μg/kg/日から1mg/kg/日の範囲であろう。本発明の結合分子は治療薬として病変部またはその近くに都合よくポンプにより投与されるかまたは注射され、例えばそれらをCNSに頭蓋内に、または脊椎に髄腔内に直接病変部に投与できる。しかしながら全身投与はここでは排除されない。本発明の結合分子を単独で、またはその他の薬剤との組み合わせで、または逐次的組み合わせで提供することができる。例えば本発明の結合分子を、限定するものではないが脳卒中または脊髄傷害後のさらなるニューロン損傷および軸索再生の阻止を遮断するための手段としてのコルチコステロイド、NGF、BDNFのような神経栄養因子またはExelon(商標)もしくはレボドパのような神経変性疾患のためのその他の薬物のような、抗Nogo−A抗体または抗炎症薬との組み合わせで投与することができる。脳卒中の処置のためのその他の適当な組み合わせパートナーはアルテプラーゼおよびデスモテプラーゼ(DSPA、例えば第WO90/09438号に開示される)である。1つの実施態様では本発明は、本発明の結合分子およびデスモテプラーゼを含む組み合わせをとりわけ脳卒中の処置のために、および該組み合わせを含む医薬組成物として提供する。本明細書で使用される際には、2つの薬剤が同時に投与されるか、または薬剤が同時に作用するような様式で独立して投与される場合、2つの薬剤は組み合わせで投与されると考えられる。
【0066】
コード番号、一般名または商品名により同定される活性成分の構造を標準的な事典「The Merck Index」の最新版から、またはデータベース、例えばPatents International(例えばIMS World Publications)もしくはIMS Healthにより提供されるその他のデータベースから取ることができる。その対応する内容を出典明示により本明細書の一部とする。任意の当業者が活性成分を同定することは十分に可能であり、そしてこれらの参照文献に基づいて、同様にインビトロおよびインビボの双方で標準的な試験モデルにおいて医薬的適応および特性を製造および試験することが可能である。
【0067】
都合のよい様式で本発明の医薬組成物を製造できる。例えば本発明の分子を含む本発明による組成物は好ましくは凍結乾燥形態で提供される。即時的投与のために、それを適当な水性担体、例えば注射用滅菌水または滅菌緩衝生理学的食塩水に溶解する。
適当な組成物を作り上げるのを助けるために、本発明の結合分子および場合によっては本発明の結合分子の影響を増強する第2の薬物を、混合または併用投与のための説明書と共に同一容器内に別個に包装することができる。最適な第2の薬物候補は前記で提供した。
【0068】
本発明の結合分子およびNGFのような成長因子の組み合わせの相乗効果をインビボで脊髄傷害モデルにより実証できる。
以下の実施例を参照して本発明はさらに十分に理解されよう。しかしながらそれらは本発明の範囲を限定するとは解釈されるべきではない。
実施例において注目を集めるモノクローナル抗体は抗体4784に関して軽鎖の可変部分(配列番号4)および重鎖の可変部分(配列番号5)を含有し、そして4785に関して軽鎖の可変部分(配列番号6)および重鎖の可変部分(配列番号7)を含む本発明による結合分子である。
【0069】
以下の略語を用いる:
AP ヒト胎盤アルカリ性ホスファターゼ
CDR 相補性決定領域
cDNA 相補的DNA
ELISA 酵素結合免疫吸着アッセイ
FACS 蛍光活性化セルソーティング
FBS ウシ胎仔血清
HCMV ヒトサイトメガロウイルスプロモーター
IgG 免疫グロブリンアイソタイプG
PBS リン酸塩緩衝生理食塩水
PCR ポリメラーゼ連鎖反応
PFA パラホルムアルデヒド
PNPP リン酸パラニトロフェニル
【実施例1】
【0070】
全長ラット、カニクイザルまたはヒトLINGO−1およびヒトLINGO−2を発現するCHO−K1細胞の作成
無作為およびオリゴdTプライマーを用いてユニバーサルヒト参照RNA(Stratagene)のRT−PCRによりヒトcDNAライブラリーを作成する。無作為およびオリゴdTプライマーを用いて凍結カニクイザル脳から単離されたポリA RNAのRT−PCRにより、カニクイザル脳cDNAライブラリーを作成する。Marathon−readyラット脳cDNAライブラリーをClontechから入手する。5’−XbaIおよび3’−XhoI部位によりフランキングされたヒトLINGO−1(配列番号27)、カニクイザルLINGO−1(配列番号28)およびラットLINGO−1(配列番号29)の成熟配列をコードするcDNA(残基34−614)を正方向プライマーDM14、5’−CTACGTCTAGAACGGGCTGCCCGCCCCGCT−3’(配列番号30)および逆方向プライマーDM15、5’−GGTTTCTCGAGTCATATCATCTTCATGTTGAACTTGCGG−3’(配列番号31)を用いて各々のライブラリーからPCR増幅する。PCR生成物をXbaIおよびXhoIで切断し、そしてベクターpSecTag2−V5(配列番号32)の各々の部位に挿入して、各々hLINGO−1−pSecTag2−V5、cmLINGO−1−pSecTag2−V5およびrLINGO−1−pSecTag2−V5を作成する。予測されるタンパク質生成物はN末端で2アミノ酸残基リンカーを介して14アミノ酸残基V5エピトープタグに融合されたLINGO−1の成熟配列である。5’−XbaIおよび3’−XhoI部位によりフランキングされたヒトLINGO−2(配列番号33)の成熟配列をコードするcDNA(残基26−606)を正方向プライマーDM16、5’−CTACGTCTAGAATTGGCTGCCCCGCTCGCT−3’(配列番号34)および逆方向プライマーDM17、5’−GGTTTCTCGAGTCAAATCATTTTCATGTTGAACCTCCTG−3’(配列番号35)を用いてMarathon−readyヒト脳cDNAライブラリー(Clontech)からPCR増幅する。PCR生成物をXbaIおよびXhoIで切断し、そしてベクターpSecTag2−V5の各々の部位に挿入して、hLINGO−2−pSecTag2−V5を作成する。予測されるタンパク質生成物はN末端で2アミノ酸残基リンカーを介して14アミノ酸残基V5エピトープタグに融合されたLINGO−2の成熟配列である。lipofectamine−2000(Invitrogen)を用いて製造者の説明書にしたがって、hLINGO−1−pSecTag2−V5、cmLINGO−1−pSecTag2−V5、rLINGO−1−pSecTag2−V5およびhLINGO−2−pSecTag2−V5で各々細胞をトランスフェクトすることにより、ヒトLINGO−1(CHO−K1−hLINGO−1)、カニクイザルLINGO−1(CHO−K1−cmLINGO−1)、ラットLINGO−1(CHO−K1−rLINGO−1)およびヒトLINGO−2(CHO−K1−hLINGO−2)を安定して発現するCHO−K1細胞を作成する。安定して発現するトランスフェクト体を1mg/mlゼオシン(Invivogen)で選択し、そして96ウェルプレートへの連続希釈か、またはクローンリング(clonal rings)の使用かのいずれかにより単一のクローンを単離する。細胞表面での構築物の発現を、抗V5抗体を用いる免疫蛍光分析により確認する(InvitroGen)。
【実施例2】
【0071】
ヒトLINGO−1−FcおよびヒトLINGO−1ΔLRR−Fcの作成および発現
ヒトLINGO−1をコードするMGC mRNA(クローンMGC:17422 IMAGE:4214343)をPCR増幅のための鋳型として使用する。Pwo1ポリメラーゼ(Roche Diagnostics)ならびに標的配列の5’末端でHindIII制限部位およびKozakコンセンサス配列および3’末端で標的配列の最後のコドンの直後にXhoI制限部位を加えられたプライマーで、ヒトLINGO−1の天然のシグナル配列により先行される細胞外ドメイン(ECD)(aa1−550)をPCRにより増幅する。PCR生成物をHindIIIおよびXhoIで消化し、ゲル精製し、そして以前に同一の酵素で消化されたプラスミドpRS5a−IgG(配列番号36)に挿入する。得られた発現クローン(天然リーダー(natleader)−hsLINGO−1−Fc/pRS5a、配列番号37)における挿入された配列、完全なFcおよびフランキング領域の正確さをDNAシークエンシングにより確認する。
【0072】
同一のMGCクローンをLRRドメインを欠くヒトLINGO−1(aa34−65+aa354−550)に関する発現プラスミドの遺伝子SOEによる構築のための鋳型として提供する。成熟LINGO−1に融合した異種性分泌シグナルをコードする部分配列で5’末端を伸長し、そして3’末端でC末端フラグメントの最初の7個のアミノ酸をコードする配列を加えたプライマーでヒトECD LINGO−1のN末端領域(aa34−65)をPCRにより増幅する。N末端フラグメントの最後の7個のアミノ酸をコードする配列で5’末端を伸長し、そして3’末端で標的配列の最後のコドンの直後にXhoI部位を加えたプライマーでヒトECD LINGO−1のC末端領域(aa354−550)をPCRにより増幅する。2つのPCR生成物をゲル精製し、混合し、そして5’末端でHindIII制限部位、Kozakコンセンサス配列を加え、そして異種性分泌シグナル配列を完成するプライマー、および3’末端で以前にC末端フラグメントを増幅するために用いられた外部プライマーを用いる第2のPCR増幅のための鋳型として提供する。PCR生成物をHindIIIおよびXhoIで消化し、ゲル精製し、そして以前に同一酵素で消化されたプラスミドpRS5a−IgGに挿入する。得られた発現クローン(Igリーダー−hsLINGO−1−ΔLRR−Fc/pRS5a、配列番号38)における挿入された配列、完全なFcおよびフランキング領域の正確さをDNAシークエンシングにより確認する。
【0073】
最初の発現評価として双方の構築物を小規模な実験で試験する。HEK.EBNA細胞(Invitrogen、以前のカタログ番号R620−07)をアタッチモード(attached mode)で組織培養フラスコで、25mM Hepes(Gibco/Life Technologies カタログ番号42430−025)で緩衝され、そしてさらに10%ウシ胎仔血清でさらに富化されたダルベッコ変法イーグル培地(DMEM)中培養し;培養物を加湿雰囲気下37℃および5%CO2で維持する。小規模なトランスフェクション実験のために、トランスフェクションの1日前に4×105セルをポリ−D−リジンコーティングした6ウェル(プレート)に播種する。ウェルあたりプラスミドDNA3μgおよびLipofectamine2000試薬(Invitrogenカタログ番号11668−019)6μlを用いて本質的には製造供給元により記載されるようにトランスフェクションを実施する。トランスフェクション後3日に、細胞上澄を収集し、そして細胞不含上澄をタンパク質分析、すなわちプロテインGカラムの免疫親和性HPLC分析に供する。構築物天然リーダー−hsLINGO−1−Fc/pRS5aに関して8mg/lと構築物Igリーダー−hsLINGO−1−ΔLRR−Fc/pRS5aに関して40mg/lとの間の範囲で力価を決定する。その後に双方のプラスミドに関して、HEK.EBNA懸濁培養物中で大容量規模での一過性のトランスフェクションを可能にするために大規模プラスミド調製物準備する。
規模を拡大した天然リーダー−hsLINGO−1−Fcの生成のために、1.4×106セル/mlの密度のHEK.EBNA細胞培養物2.9lをDNA:PEI溶液(DNA 1μg:PEI 2μg/ml)1.1lと混合する。4時間インキュベートした後、培養物にExCell VPRO培地(SAFC、以前はJRH、Lenexa, KS)4lを供給する。培養の6日後に細胞培養上澄を収集し、そして10kDaカットオフの使い捨てHemoflow F10HPSフィルター(Fresenius Medical Care, Germany)を用いてダイアフィルトレーションにより1lまで濃縮する。Igリーダー−hsLINGO−1−ΔLRR−Fcタンパク質を作成するために行う第2の関連するタンパク質生成を類似の様式で行う。大規模トランスフェクションに関する詳細、DNA:PEI比率、細胞密度、供給および収集は前記されたものと全く同一である。
【0074】
a)天然リーダー−hsLINGO−1−Fc
濃縮物1l(培養上澄8lから)を20mlプロテインAセファロースのクロマトグラフィーにより分析する。100mM NaPi、pH7.3のベースライン洗浄の後、結合材料を50mMクエン酸、140mM NaCl、pH2.7で溶出し、中和し、そして滅菌ろ過する。溶出された分画さらに濃縮し、そしてPBS中Superdex 75でゲルろ過し、1.2mg/mlの濃度の生成物8.2mgを生じる。
b)Igリーダー−hsLINGO−1−ΔLRR−Fc
濃縮物1l(培養上澄8lから)を20mlプロテインAセファロースのクロマトグラフィーにより分析する。100mM NaPi、pH7.3のベースライン洗浄の後、結合材料を50mMクエン酸、140mM NaCl、pH2.7で溶出し、中和し、そして滅菌ろ過して、1.5mg/mlの濃度の生成物52.5mgを生じる。
精製されたタンパク質を還元/アルキル化およびトリプシン消化の後、N末端シークエンシングおよびMALDIペプチド質量分析により詳しく特徴付けする。
【実施例3】
【0075】
AP−LINGO−1結合アッセイ
LINGO−1のNgRに対する結合の遮断は神経突起伸長の3つのミエリン関連阻害剤、すなわちNogo−66、MAGおよびOMgpのシグナル発生を防御し、そしてしたがってCNSミエリンの神経突起伸長阻止活性を減弱し、故に軸索再生/可塑性の増大および急性CNS傷害後の機能的回復の改善に至ると予期される。抗LINGO−1抗体がLINGO−1のNgRに対する結合を遮断することを実証するために、ヒト胎盤アルカリ性ホスファターゼ(AP)−タグ化ラットLINGO−1外部ドメイン(AP−LINGO−1)の、NgRを安定して発現するSH−SY5Y細胞(NgR−SH−SY5Y、Walmsley et. al., J Cell Sci 117:4591−4602(2004))への結合を測定するアッセイを用いることができる。
【0076】
5’−Xho Iおよび3’−Xba I部位によりフランキングされたラットLINGO−1外部ドメイン(残基34−532)の大部分をコードするcDNAを、正方向プライマーDM22、5’−GGTTATCTCGAGACCGGCTGCCCGCCCC−3’(配列番号24)、および逆方向プライマーDM23、5’−GGCCCTTCTAGATCACTCGCCTGGCTGGTTGGAGATG−3’(配列番号25)を用いてrLINGO−1−pSecTag2−V5からPCR増幅する。PCR生成物をXhoIおよびXbaIで切断し、そしてベクターAPtag−5−NHIS(配列番号26)の各々の部位に挿入して、APtag−5−NHIS−solrLINGO−1を作成する。予測されるタンパク質生成物はN末端で3アミノ酸残基リンカーを介してヒト胎盤アルカリ性ホスファターゼの残基23−511に融合したラットLINGO−1外部ドメインの大部分である。lipofectamine2000を用いて製造者の説明書にしたがって、HEK293T細胞をAPtag−5−NHIS−solrLINGO−1でトランスフェクトする。トランスフェクションの4時間後にトランスフェクション培地を除去し、そしてフェノールレッド(Invitrogen)を含まないOptiMEM Iで置き換える。24時間後に培地を収集し、置き換え、そしてさらに24時間後に再度収集する。13000×gで5分間遠心することにより培地を清澄化し、そしてCentriprepフィルター装置(Millipore)を用いて製造者の説明書にしたがって上澄をほぼ15倍に濃縮する。濃縮された上澄のAP活性を1−Step(商標)PNPP(Pierce)を用いて405nmでの経時的な吸光度の変化として測定し、そして以下の等式を用いて濃度に変換する(200μl PNPP/セルで96ウェル形式に適用):
【数1】

【0077】
濃縮された上澄を記載されるようにSDS−PAGEゲル電気泳動およびウェスタンブロットに供する(Walmsley et. al., J Cell Sci 117:4591−4602(2004))。ECL(商標)システム(GE Healthcare)を用いて0.1%(容量/容量)抗ペンタヒスチジン抗体(Qiagen)、続いて0.02%(容量/容量)ペルオキシダーゼ抱合抗マウスIgG抗体(Sigma)でAP−LINGO−1を検出する。AP−LINGO−1をおよそ110kDaのバンドとして可視化し、その予測される分子量、112kDaに類似する。N末端分解生成物は観察されない。
【0078】
NgR:SH−SY5Y細胞を周密度50%で酵素不含解離バッファー(Invitrogen)で収集してNgRのような細胞表面タンパク質を保存する。1nM AP、1nM AP−LINGO−1、または2μM抗LINGO−1Fabもしくはニワトリ卵白リゾチームに対する対照Fab3207の存在下の1nM AP−LINGO−1をOptiMEM(Invitrogen)中30分間プレインキュベートし、そしてその後懸濁液中NgR:SH−SY5Y細胞と共に一定の攪拌を行いながら1.5時間インキュベートする。細胞をHBH(Hanks緩衝生理食塩水中20mM HEPES pH7.4/1%ウシ血清アルブミン)で6回洗浄し、そしてPBS中4%パラホルムアルデヒド(PFA)/5%スクロース中で15分間固定する。Hanks緩衝生理食塩水中20mM HEPES pH7.4中、65℃で1時間インキュベートすることによる内因性AP活性の不活化に続いて、1−Step(商標)PNPP(Pierce)と共に製造者の説明書にしたがって30分間インキュベートした後、細胞結合性AP活性を405nmでの吸光度として定量する。
【0079】
Fabを2μMの濃度で使用してAP−LINGO−1を結合性Fabで飽和し、そしてしたがって結合を阻止するその能力に関して、その親和性の影響を最小にする。このための理由は、その結合部位の位置よりもむしろその低親和性のために結合を阻止し損ねたFabを時期尚早にさらなる実験から破棄する可能性を排除するためであるが、かかるFabの親和性が親和性成熟化およびIgG4転換により後の段階で増大し得るためである。1nM AP−LINGO−1を対照Fab3207または抗LINGO−1 Fab4784および4785のいずれかとともにプレインキュベートし、そして次に懸濁液中Fabの存在下、NgR:SH−SY5Y細胞への結合を可能にする(図1)。抗LINGO−1 Fabの存在下、特異的AP−LINGO−1結合の阻止パーセンテージをFab3207に関して正規化する。4784および4785は、細胞に対するAP−LINGO−1結合を有意に阻止する(p<0.01、一元配置分散分析、対照Fab3207の存在下、AP−LINGO−1の特異的結合に対するHolm−Sidak比較)。
【0080】
NgRに対するLINGO−1の結合の遮断は、ミエリン関連阻害剤Nogo−66、MAGおよびOMgpのシグナル発生を防御してCNSミエリンの神経突起伸長阻止活性の低下に至ると予測される。その点で、4784および4785Fabは最終的なIgG4形式(実施例8参照)に転換され、そしてラット成体脊髄ミエリン上で成長させた出生後7日のラット小脳顆粒ニューロンからの神経突起伸長の阻止を減弱するその能力に関して評価される。
【実施例4】
【0081】
神経突起伸長阻止アッセイ
インビボ軸索再生/可塑性に及ぼす抗LINGO−1抗体の影響を予測する最も関連性のあるインビトロアッセイは、CNSミエリンの神経突起伸長阻止活性を減弱するその能力である。このアッセイでは、成体ラットから抽出された全脊髄ミエリンでコーティングされたウェル中で出生後7日のラット小脳顆粒ニューロン(CGN)を成長させ、そして神経突起伸長を自動ArrayScan(登録商標)HCSリーダー(Cellomics)により定量する。
抗LINGO−1 IgG4抗体4784および4785の脱抑制活性を該神経突起伸長アッセイにおいて評価する(図2)。
【0082】
ラット成体からの新たなラット脊髄組織を3容量(重量/容量)抽出バッファー(60mM Chaps、20mM Tris pH8.0、1mM EDTA、プロテアーゼ阻害剤カクテル)中でホモジナイズし、4℃で30分間インキュベートし、そして170000×gで、4℃で30分間遠心することにより清澄化する。96ウェルプレートの各ウェルをMeOH中ニトロセルロース5μl(MeOH12ml中5cm2ニトロセルロース)でコーティングし、空気乾燥し、そして5μg/mlポリ−D−リジン100μlで、37℃で4時間インキュベートすることによりコーティングする。水で3回洗浄した後、プレートを1時間空気乾燥し、そして次に37℃で一晩インキュベートすることにより、60μg/cm2脊髄抽出物でコーティングする。以前に記載されたように(Schweigreiter et al., 2004)出生後7日のラット小脳組織のトリプシン分離物からCGN細胞を新たに精製する。LINGO−1を検出するために、2μg/ml(または13.3nM)抗LINGO−1ポリクローナル抗体(Upstate)、続いて0.02%(容量/容量)ペルオキシダーゼ抱合抗ウサギIgG抗体(Sigma)を用いてV5−タグ化ラットLINGO−1を発現するCHO−K1細胞またはP7 CGN細胞からのライゼートに関してウェスタンブロット分析を実施する。脊髄ミエリンでコーティングされていないかまたはコーティングされたウェル上で、CGN細胞(35000セル/ウェル)を37℃で30分間インキュベートした後、0−100nM抗LINGO−1 IgG4抗体または対照3207 IgG4抗体のいずれかを添加する。37℃で8−16時間インキュベートした後、細胞を4%PFAで固定し、そして核の可視化のためにHoechst 3342(Invitrogen)、および抗β−チューブリンIII抗体(R&D Systems)、続いてAlexa Fluor 546抱合抗マウスIgG抗体(Invitrogen)で染色してニューロンを特異的に可視化する。神経突起伸長のパラメーターをArrayScan(登録商標)HCSリーダー(Cellomics)を用いて決定する。ArrayScan(登録商標)IIは96ウェルマイクロタイタープレート内の細胞のフィールドを自動的に位置決定し、焦点を合わせ、そして暴露する。ArrayScan(登録商標)は高解像度光学系、対応したシングルバンド励起フィルターを伴う多帯域発光フィルター(XF100)、フレーム取り込み装置を有するCCDカメラ、および専用アプリケーションソフトウェアからなる。このアッセイではExtended Neurite Outgrowth Bioapplicationを用いる。励起フィルターホイールおよび多帯域発光フィルターを用いることにより、同一細胞における2つのフルオロフォアからの蛍光の多チャンネル撮像が可能になる。Hoechst 33342標識された核の帯域画像を獲得して別々の細胞を同定し、そして次にAlexa Fluor 488の帯域画像を獲得して抗チューブリン抗体で標識された細胞の広がりを同定する(Alexa Fluor 488に二次抱合されたものを用いる)。細胞内の不適切な物体を分析から自動的に排除して、Hoechstおよびベータ−チューブリンを重複する細胞体のみを分析する。プレートの各ウェルで5個の別々の350μm2フィールドに関して二重発光画像を獲得する。10倍対物を用いて、これにより結果的にウェルあたり400−500セルが分析される。次いでExtended Neurite Outgrowth Bioapplicationにより細胞あたりの神経突起の神経突起長数、細胞体面積、ならびに分岐および交差点を含む単一の細胞に関するニューロン形態のいくつかの定量的測定が報告される。ニューロンあたりの平均神経突起長(μm)を10回反復でウェルあたり500ニューロンに関して計算する。
【0083】
前記の神経突起伸長アッセイでは、抗LINGO−1 IgG4抗体4784および4785は1および10nMで脱抑制性であるが、一方リゾチームに対する対照IgG4抗体は双方の濃度で脱抑制を示さない(図2)。双方の濃度での4784および4785存在下で脊髄ミエリン上のニューロンあたりの神経突起の平均の長さは抗体不在下のものよりも統計的に長い。抗LINGO−1抗体4784および4785と比較して、ROCK阻害剤Y27632でより大きなレベルの脱抑制が達成されることが予期されるが、この化合物がNgR受容体複合体を介してシグナル発生するもの以外のさらなるミエリン関連神経突起伸長阻害剤のシグナル発生経路を阻止するためである。
【0084】
前記の結果を確認するために、神経突起伸長アッセイを繰り返す(図3)。再度抗LINGO−1抗体4784および4785が1nMおよび10nMで脱抑性であるが、一方リゾチームに対する対照IgG4抗体は双方の濃度で脱抑制を示さない。双方の濃度で4784および4785存在下で脊髄ミエリン上のニューロンあたりの神経突起の平均の長さは抗体不在下のものよりも統計的に長い。
【0085】
抗LINGO−1抗体4784および4785の効力をさらに確立するために、nM以下の濃度の抗体の神経突起伸長阻止に及ぼす影響を評価する(図4)。4784および4785により0.1nMほどの低濃度で脊髄ミエリンの有意な脱抑制(各々38−51%および51−57%)が示されるが、一方対照抗リゾチーム抗体は影響を及ぼさない。予期されるように、再度ROCK阻害剤Y27632により、抗LINGO−1 IgG4抗体よりも高度な脱抑制(65−74%)が示される。
【実施例5】
【0086】
一次オリゴデンドロサイト分化アッセイ
遺伝的手段による、または受容体アンタゴニストでの処理によるLINGO−1機能の遮断により、精製されたOPC培養物から生じる成熟オリゴデンドロサイトの比率が増大することが報告されている(Mi et al., Nat Neurosci 8:745−751(2005))。抗LINGO−1抗体がOPC培養物におけるLINGO−1機能を遮断し、そしてオリゴデンドロサイト成熟化を促進する能力を評価するために、新たに単離されたラットOPCをDMEM/CNTF/T3培地中4784、4785または対照IgG4 3207と共に3日間インキュベートし、続いて抗O4抗体で染色して未成熟および成熟双方のオリゴデンドロサイトを標識する(図5)。オリゴデンドロサイト成熟の程度を、成熟形態を呈するO4陽性細胞の比率として測定する。
【0087】
OPCの富化集団をOFA P3ラットから単離する。簡単には、脳を切開し、そして終脳を0.15%MgSO4含有氷冷Hankバッファー生理食塩水(HBSS、Invitrogen)中に置く。組織を1:1 HBBS/トリプシン−EDTA(Invitrogen)および100μg/ml DNAse I(Roche)と共に37℃で10分間インキュベートし、そしてFCS(Invitrogen)を最終濃度10%で添加してトリプシンを不活化する。組織懸濁液を890rpmで10分間遠心し、そしてペレットを10%ウマ血清(Invitrogen)を含むBasal Medium Eagle(BME、Invitrogen)に再懸濁する。懸濁液を、40μmセルストレーナー(BD Falcon)を通してろ過し、そして細胞をポリ−D−リジンプレコーティングされた80cm2組織培養フラスコ(BD Falcon)にフラスコあたり脳1個で置く。細胞をBME/10%ウマ血清中37℃で11日間培養する。5mM L−ロイシン−メチルエステルを添加することにより小グリア細胞を死滅させ、そしてフラスコを140rpmで2時間振盪することにより攪拌する。フラスコを200rpmで、37℃で一晩振盪することによりOPCを収集し、そして10cm細菌培養皿上で37℃で2時間予め付着させることにより、上澄に残る任意のアストロサイトをOPCからさらに分離する。非接着細胞を集め、890rpmで10分間遠心し、そしておよそ3×104セル/ウェルでポリ−D−リジンコーティングされた8ウェルチャンバースライド(BD Falcon)に置く。10ng/ml Ciliary Neurotrophic Factor(R&D Systems)および15nM トリヨードチロニン(Sigma)を含有するDMEM(Invitrogen)からなるDMEM/T3/CNTF培地中、または10μg/ml トランスフェリン(Sigma)、10μg/mlインスリン(Sigma)、100μMプトレシン(Sigma)、200nMプロゲステロン(Sigma)、520nMチロキシン(Sigma)、500pMトリヨードチロニン(Sigma)、220nM亜セレン酸ナトリウム(Sigma)、25μg/mlゲンタマイシン(Sigma)および1%HS(Invitrogen)を含有するDMEM(Invitrogen)からなるSATO培地中のいずれかで3日間培養物を維持する。オリゴデンドロサイト系列に関して培養物の純度を評価するために、抗O4抗体で染色された細胞のパーセンテージをSATO培地中7日の培養の後に定量する。典型的には細胞の80−95%が抗O4抗体で染色され、それは培養物中の細胞の大部分がオリゴデンドロサイト系列であることを実証している。
【0088】
オリゴデンドロサイト形態に基づいてオリゴデンドロサイト成熟化を評価するために、新たに単離されたOPC培養物をDMEM/T3/CNTF培地中、100nM 4784、4785または対照IgG4 3207の不在下または存在下で3日間インキュベートし、続いて抗O4抗体で染色して未成熟および成熟の双方のオリゴデンドロサイトを標識し、そしてDAPIで細胞核を標識する。明確に定義された短い突起を有するO4陽性細胞は未成熟オリゴデンドロサイトを表すが、一方ミエリンシート様構造を伴い、伸長し、そして高度に分枝した突起を担持するO4陽性細胞は成熟オリゴデンドロサイトを表すと考えられる。成熟形態を有するO4陽性細胞の比率を処理あたり3検体ずつ、ほぼ300−1300セルに関して定量し、そして対照IgG4 3207の存在下の成熟オリゴデンドロサイトの比率に対するHolm−Sidak比較を伴う一元配置分散分析を用いて有意性を決定する。培養物中の全(未成熟および成熟)オリゴデンドロサイトの比率に及ぼす抗体処理の影響を評価するために、O4染色に関連するDAPI核の比率を定量する。
【0089】
3つの独立した実験では、抗LINGO−1抗体4784および4785での処理は、広い面積にわたって伸長する高度に分枝した突起およびミエリンシート様構造を担持する細胞により表されるような成熟形態のオリゴデンドロサイトの比率を有意に増大させる(図5)。対照IgG4抗体3207での処理は培養物中の成熟オリゴデンドロサイトの比率に影響を及ぼさない。O4染色に関連するDAPI染色された核の比率は全処理に関して類似し、抗LINGO−1抗体が未成熟および成熟の双方のオリゴデンドロサイトに対応する細胞の比率に影響を及ぼさないことが実証される。
【0090】
抗LINGO−1抗体処理は全オリゴデンドロサイトの比率に影響を及ぼさないが、成熟オリゴデンドロサイトの比率の増大は、OPCの未成熟オリゴデンドロサイトへの分化の率における増大よりもむしろ未成熟オリゴデンドロサイトの成熟オリゴデンドロサイトへの分化の率の増大のために生じる可能性が最も高い。
【実施例6】
【0091】
細胞表面LINGO−1の抗LINGO−1抗体媒介の下方調節
多価抗体の細胞表面標的に対する結合は抗体:標的複合体の内部移行およびその後のエンドサイトーシス経路内での標的の分解に至り得る(Weinmann et al., Mol Cell Neurosci 32:161−173(2006))。
抗LINGO−1抗体の細胞表面LINGO−1の量に及ぼす影響を決定するために、トランスフェクトされていないCHO−K1またはCHO−K1−hLINGO−1細胞(実施例1参照)を100nM 4784、4785または3207と共に37℃で24時間インキュベートし、そしてその後細胞表面LINGO−1を抗V5抗体、続いて1−Step(商標)Turbo TMB−ELISAキット(Pierce)で展開される抗マウスIgG(Fc特異的)−POD抱合体で検出する(図6A)。
【0092】
CHO−K1−hLINGO−1細胞における細胞表面LINGO−1の量は、抗LINGO−1抗体4784および4785を伴う24時間のインキュベーションの後に有意に低下するが、一方対照IgG4 3207を伴うインキュベーションは影響を及ぼさない。加えて4785でのインキュベーションにより細胞表面LINGO−1は4784よりも大きい程度まで低下する。
【0093】
抗LINGO−1抗体の細胞表面LINGO−1の分解に及ぼす影響を評価するために、トランスフェクトされていないCHO−K1またはCHO−K1−hLINGO−1細胞上の細胞表面タンパク質を記載されるように4℃でビオチン化し(Walmsley et al., J Cell Sci 117:4591−4602(2004))、そして細胞を37℃で180分の期間にわたって種々の時間に100nM 4784、4785または3207を伴って、または伴わずにインキュベートする(図6B)。インキュベーション期間の終わりにアガロースビーズに結合させた抗V5抗体を用いて細胞ライゼートからLINGO−1を免疫沈殿し、そして沈殿物中のビオチン化LINGO−1をウェスタンブロット分析により抗ビオチン抗体(Sigma)を用いて検出する。
【0094】
ビオチン化(およびしたがって細胞表面)LINGO−1に対応するバンドの強度は、抗体を伴わずに、または対照IgG4 3207と共にインキュベートされた細胞においてよりも、抗LINGO−1抗体4784および4785と共にインキュベートされたCHO−K1−hLINGO−1細胞において、より急速に減少する。加えて4785とのインキュベーションにより細胞表面LINGO−1の分解の率は4784よりも大きい程度まで増大する。
【0095】
これらの結果により累積的に、抗LINGO−1抗体4784および4785が、最も可能性が高いのは内部移行およびタンパク質の分解を増強することにより、細胞表面でLINGO−1を有意に下方調節することが示される。この特性はLINGO−1機能の遮断におけるこれらの抗体の効率に寄与すると予期される。
【実施例7】
【0096】
酵素結合免疫吸着アッセイ(ELISA)およびFACS技術
ヒト組換えLINGO−1−Fc融合タンパク質を、直接固定されたヤギ抗ヒトIgG Fc抗体を介するFc部分の捕捉により間接的にMaxisorpプレート96または384ウェルに室温で1時間固定する(PBS中10μg/mlでコーティングされた100μlまたは20μl)。
PBS中5μg/mlの抗原20μlのコーティングの後、ウェルをPBS/0.05%Tween(PBS−T)/5%粉乳で、室温で1時間遮断する。ウェルをPBS−T BEL抽出物で洗浄した後、精製されたFabまたは対照IgGをPBSで希釈し、ウェルに加え、そして室温で1時間インキュベートする。一次抗体を検出するために、以下の二次抗体を適用する:アルカリ性ホスファターゼ(AP)抱合AffiniPureヤギF(ab’)2フラグメント抗ヒトIgGまたは抗マウスIgG(Jackson ImmunoResearch)。AP抱合体の検出のためにAttoPhos(Roche)のような蛍光発生基質を製造者の説明書にしたがって使用する。全インキュベーション工程の間に、マイクロタイタープレートのウェルをPBS−Tで5回、および二次抗体との最終インキュベーションの後に5回洗浄する。TECAN Spectrafluorプレートリーダーで蛍光を測定する。
【0097】
トランスフェクトされたCHO−K1細胞の細胞表面上に発現されたLINGO−1に結合する抗体のFACS分析
丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)中ウェルあたり2×105セルで全染色を実施する。各々の細胞系の細胞をPBS/3%FCS/0.02%NaN3(FACSバッファー)に再懸濁し、そしてa)周辺質抽出物もしくはBELライゼートからの抗体またはb)精製されたFabフラグメントまたはc)FACSバッファーで希釈された、精製されたIgGと混合し、そして4℃で30−60分間インキュベートする。次いで細胞をFACSバッファー/ウェル150μlで1回洗浄し、そしてFACSバッファー中1:200で希釈されている、フィコエリスリン標識された二次抗体(R−PE抱合されたヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに取る。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSCalibur(商標)またはFACSArray(商標)(Becton Dickinson)においてFL2蛍光強度により測定する。
【0098】
LINGO−1特異的抗体の同定のために、CHO−K1−cmLINGO−1またはCHO−K1−rLINGO−1を用いて並行して染色を行う。トランスフェクトされていないCHO−K1細胞をさらなる対照として提供する。カニクイザルおよびラットLINGO−1発現細胞は、これらの種オルソログがヒトLINGO−1タンパク質と数個のアミノ酸しか異ならないので、スクリーニングのために選択される。これらのクローンのみが、トランスフェクトされていないCHO−K1細胞において陰性であり、そしてLINGO−1発現細胞系においてバックグラウンドを超えて≧5倍であるLINGO−1特異的であると判断される。ヒトLINGO−1およびその他のオルソログ(カニクイザルLINGO−1、ラットLINGO−1)に対する、およびヒトLINGO−2パラログに対する交差反応性を逐次的に試験する。
【0099】
配列分析の後、FACS分析で細胞表面に発現されたヒトLINGO−1に対して強力な結合を示す31個の独特なクローンが同定される。12個の結合剤はELISAにおいて捕捉されたヒトLINGO−1−Fcに対する強力な結合を示し(シグナル:ノイズ比は10:1より大きい)、そして7個はELISAにおいて中程度の結合を示す(シグナル:ノイズ比は5:1より大きい)。結合剤の4個はELISAにおいて捕捉されたヒトNgR−Fc融合タンパク質(R&D Systems)に対する強力な結合を示し、そして中断される。結合剤の別の3個はLINGO−1の3つの種の全てと交差反応せず、そして中断される。ヒト/カニクイザル/ラットLINGO−1と交差反応するが、ヒトNgR−Fcとは交差反応しない残りの24個のクローンを発現させ、精製し、そしてLINGO−1のNgRに対する結合を有意に阻止する能力に関して試験し(図1参照)、そしてインビトロで脊髄ミエリンの神経突起伸長阻止活性を脱抑制し(図2−4参照)、さらなる分析のためにFab4784および4785の選択に至る。ELISAでは、4784および4785は捕捉されたヒトLINGO−1−Fcに結合するが、無関係のFc対照と比較して、ヒトLINGO−1−ΔLRR−FcまたはヒトNgR−Fcに対する結合は観察されない(表1および図7参照)。これは4784および4785がLINGO−1のLRR領域(残基66−353)内にあるエピトープを有することを示している。
【0100】
表1:ELISAによる抗LINGO−1 Fabの特徴付け
【表1】
ELISA分析に関する値は相対蛍光単位の平均値として与えられる。
【0101】
FACS飽和分析を用いる選択された抗LINGO−1 Fabの親和性決定
抗LINGO−1特異的抗体の細胞基盤の親和性をFACS飽和結合実験により決定する。染色するために試料中に存在する抗原の濃度は見かけのKD値に影響するので、FACS飽和結合実験における抗原濃度を低下させるために2×105セル/ウェルと対比して1.25×104セル/ウェルのみを用いる。その他の点は前記されたFACS染色手順と同一に染色手順を行う。
【0102】
詳細には、CHO−K1−hLINGO−1、CHO−K1−cmLINGO−1またはCHO−K1−rLINGO−1をベルセンにより培養フラスコから剥がし、FACSバッファーで洗浄し、そしてFACSバッファーに再懸濁する。精製された抗LINGO−1 FabをFACSバッファーで連続希釈し、そして丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)に広げる。各濃度に関して、2検体ずつのウェルを1.25×104セルと共に全容量100μlで、氷上で30−60分間インキュベートする。FACSバッファー150μlを適用し、そして400×gで5分間遠心することによる洗浄工程の後、細胞ペレットをフィコエリスリン標識二次抗体(R−PE抱合ヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに再懸濁し、それをFACSバッファーで1:200に希釈する。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSArray(商標)(Becton Dickinson)において細胞のFL2蛍光強度により測定する。見かけのKD値/EC50値をGraphPad Prism v3.03ソフトウェアまたはGraphPad Prism v4.03を用いて飽和結合曲線から決定し、非線形回帰曲線フィットに適用する。
【0103】
このアッセイを用いて以下の見かけのKD値を決定することができる(表2)。Fab形式では、クローン4784はヒトLINGO−1、カニクイザルLINGO−1およびラットLINGO−1に対してかなり弱い親和性を有する(各々14.07nM、27.11および24.03nM)。しかしながらクローン4784はFab形式でヒトLINGO−2に結合しない。Fab形式では、クローン4785はヒトLINGO−1、カニクイザルLINGO−1およびラットLINGO−1に対してナノモル以下の結合親和性(すなわち見かけのKD値は1×10−9M未満である)を示す。クローン4785はFab形式で低ナノモルからナノモル以下の親和性でヒトLINGO−2に対して交差反応性を示す。LINGO−2機能および分布は依然未知であるので、LINGO−2に対する交差反応性の結果は記載している時点で評価できない。しかしながら有利な影響は排除できない。
【0104】
表2:CHO−K1細胞により発現されたLINGO−1またはLINGO−2抗LINGO−1 Fabに対する見かけのKD値
【表2】
与えられた値は見かけのKD値の平均値(nM)である。nbは結合しない。
【実施例8】
【0105】
HuCAL(登録商標)のIgG4クローニング、発現および精製
IgG形式への転換
全長免疫グロブリン(Ig)を発現するために、重(VH)および軽鎖(VL)の可変ドメインフラグメントをpMORPH(登録商標)X9 MH(配列番号39)Fab発現ベクターからヒトIgG4のためのpMORPH(登録商標) h Ig(配列番号40−42)またはpMORPH(登録商標)2 h Ig(配列番号43−45)ベクターシリーズのいずれかにサブクローニングする。
【0106】
制限酵素EcoRI、MfeIおよびBlpIを用いてVHドメインフラグメントのpMORPH(登録商標) h IgG4(配列番号40)にサブクローニングする:EcoRI/BlpI消化および6400bpフラグメントの抽出によりベクター骨格を作成するが、一方MfeIおよびBlpIでの消化およびその後の精製によりVHフラグメント(350bp)を生成する。ベクターおよびインサートを、各々EcoRIおよびMfeI消化により作成された適合するオーバーハングを介して、およびBlpI部位を介してライゲートする。それによりEcoRIおよびMfeI制限部位の双方が破壊される。
【0107】
制限酵素MfeIおよびBlpIを用いて、VHドメインフラグメントをpMORPH(登録商標)2 h IgG4(配列番号43)にサブクローニングする。このIgGベクターの新たな作成では、その他の修飾時にEcoRI部位(それは適合するオーバーハングを介するサブクローニングのみを可能にする)はMfeI部位により置き換えられ、故にベクターおよびインサートの双方のMfeI/BlpI消化が可能になる。
VLドメインフラグメントのpMORPH(登録商標) h Igκ(配列番号42)およびpMORPH(登録商標)2 h Igκ(配列番号45)へのサブクローニングをEcoRVおよびBsiWI部位を介して実施するが、一方pMORPH(登録商標) h Igλ(配列番号41)およびpMORPH(登録商標)2 h Igλ2(配列番号43)へのサブクローニングをEcoRVおよびHpaIを用いて行う。
【0108】
ヒトIgGの一過性の発現および精製
HEK293細胞を等モル量のIgG重および軽鎖発現ベクターでトランスフェクトする。トランスフェクション後4または5日に、細胞培養上澄を収集する。上澄のpHを8.0に調整し、そして滅菌ろ過した後、溶液を標準的なプロテインAカラムクロマトグラフィー(Poros 20A、PE Biosystems)に供する。
【実施例9】
【0109】
FACS飽和分析を用いる選択された抗LINGO−1 IgG4の親和性決定
抗LINGO−1特異的抗体の細胞基盤の親和性をFACS飽和結合実験により決定する。見かけのKD値の決定は、抗LINGO−1Fab抗体を用いて前記された手順と同一に実施する。
詳細には、CHO−K1−hLINGO−1、CHO−K1−cmLINGO−1またはCHO−K1−rLINGO−1をベルセンにより培養フラスコから剥がし、FACSバッファーで洗浄し、そしてFACSバッファーに再懸濁する。精製された抗LINGO−1 IgG4をFACSバッファーで連続希釈し、そして丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)に広げる。各濃度に関して、2検体ずつのウェルを1.25×104セルと共に全容量100μlで、氷上で30−60分間インキュベートする。FACSバッファー150μlを適用し、そして400×gで5分間遠心することによる洗浄工程の後、細胞ペレットをフィコエリスリン標識二次抗体(R−PE抱合ヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに再懸濁し、それをFACSバッファーで1:200に希釈する。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSArray(商標)(Becton Dickinson)において細胞のFL2蛍光強度により測定する。見かけのKD値/EC50値をGraphPad Prism v3.03ソフトウェアまたはGraphPad Prism v4.03を用いて飽和結合曲線から決定し、非線形回帰曲線フィットに適用する。このアッセイを用いて以下の見かけのKD値を決定することができる(表3)。
【0110】
pMORPH(登録商標)2 h Igベクターシリーズを用いて生成された4784および4785 IgG4抗体の親和性を表3に示す。IgG4形式の4784および4785はヒト、カニクイザルおよびラットLINGO−1に対して明らかに1nMを下まわる見かけのKD値を有する。4784は4785よりもヒトLINGO−2に対してはるかに低い交差反応を有する。
【0111】
表3:CHO−K1細胞により発現されたLINGO−1またはLINGO−2に対する抗LINGO−1 IgG4の見かけのKD値
【表3】
与えられた値は見かけのKD値の平均値(nM)である。
【実施例10】
【0112】
FACS分析を用いる選択された抗LINGO−1 IgG4のヒトLINGO−1に対する結合に及ぼすヒト脳脊髄液の影響
抗LINGO−1 IgG4のヒトLINGO−1に対する結合に及ぼすヒト脳脊髄液の影響をFACS飽和結合実験により試験する。4784および4785の連続希釈物を調製する。CHO−K1−hLINGO−1に対する結合を50%ヒト脳脊髄液の存在下で試験する。前記されたFACS染色にしたがって細胞をヒトCSFの存在下、これらのIgG4抗体で染色する。
【0113】
詳細には、CHO−K1−hLINGO−1をベルセンにより培養フラスコから剥がし、FACSバッファーで洗浄し、そしてFACSバッファーに再懸濁する。精製された抗LINGO−1 IgG4をFACSバッファー+50%ヒト血清で連続希釈し、そして4℃で60分間インキュベートする。対照としてIgG4形式の候補結合剤の連続希釈物をFACSバッファー中、ヒト脳脊髄液のタンパク質含量に類似する2.6%BSAと共に4℃で60分間インキュベートする。インキュベーションの後、連続希釈物を丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)に広げる。各濃度に関して、2検体ずつのウェルを1.25×104セルと共に全容量100μlで、氷上で30−60分間インキュベートする。FACSバッファー150μlを適用し、そして400×gで5分間遠心することによる3回の洗浄工程の後、細胞ペレットをフィコエリスリン標識二次抗体(R−PE抱合ヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに再懸濁し、それをFACSバッファーで1:200に希釈する。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSArray(商標)(Becton Dickinson)において細胞のFL2蛍光強度により測定する。見かけのKD値/EC50値をGraphPad Prism v3.03ソフトウェアまたはGraphPad Prism v4.03を用いて飽和結合曲線から決定し、非線形回帰曲線フィットに適用する。
【0114】
このアッセイを用いて50%ヒト脳脊髄液の影響を対照と比較することができる(表4)。50%ヒト脳脊髄液中でのインキュベーションは差次的に影響する全ての結合剤との結合親和性の低下に至る。ヒト脳脊髄液の存在により結合親和性に及ぼす最強の影響力が4784に関して認められ、それは0.43nMから1.57nMの、73%までの親和性の低下を示す。
【0115】
表4:CHO−K1細胞により発現されたLINGO−1に対する抗LINGO−1 IgG4の見かけのKD値に及ぼすヒト脳脊髄液の影響
【表4】
与えられた値は見かけのKD値の平均値(nM)である。
【実施例11】
【0116】
FACS分析を用いる選択された抗LINGO−1 IgG4のヒトLINGO−1に対する結合に及ぼすヒト血清の影響
抗LINGO−1 IgG4のヒトLINGO−1に対する結合に及ぼすヒト血清の影響をFACS飽和結合実験により試験する。4784および4785の連続希釈物を50容量/容量%ヒト血清の存在下で調製する。細胞を60分間インキュベートした後、前記されたFACS染色にしたがってこれらのプレインキュベートされたIgG4抗体で細胞を染色する。
【0117】
詳細には、CHO−K1−hLINGO−1をベルセンにより培養フラスコから剥がし、FACSバッファーで洗浄し、そしてFACSバッファーに再懸濁する。精製された抗LINGO−1 IgG4をFACSバッファー+50%ヒト血清で連続希釈し、そして4℃で60分間インキュベートする。対照としてIgG4形式の候補結合剤の連続希釈物をFACSバッファー+2.6%BSA(ヒト血清のタンパク質含量に類似する)中でインキュベートするか、またはFACSバッファー単独中、4℃で60分間インキュベートする。インキュベーションの後、連続希釈物を丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)に広げる。各濃度に関して、2検体ずつのウェルを1.25×104セルと共に全容量100μlで、氷上で30−60分間インキュベートする。FACSバッファー150μlを適用し、そして400×gで5分間遠心することによる3回の洗浄工程の後、細胞ペレットをフィコエリスリン標識二次抗体(R−PE抱合ヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに再懸濁し、それをFACSバッファーで1:200に希釈する。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSArray(商標)(Becton Dickinson)において細胞のFL2蛍光強度により測定する。見かけのKD値/EC50値をGraphPad Prism v3.03ソフトウェアまたはGraphPad Prism v4.03を用いて飽和結合曲線から決定し、非線形回帰曲線フィットに適用する。
【0118】
このアッセイを用いて50%ヒト血清中でのプレインキュベーションの影響を対照と比較することができる(表5)。ヒト血清の存在下での1時間のインキュベーションは4784および4785のKD値に影響しない。したがってこれらの抗体はこの時間にわたって血清中で安定であり、さらにそのKDは変化しないので、血清構成要素とは交差反応しないと思われる。
【0119】
表5:CHO−K1細胞により発現されたLINGO−1に対する抗LINGO−1 IgG4の見かけのKD値に及ぼすヒト血清の影響
【表5】
与えられた値は見かけのKD値の平均値(nM)である。
【0120】
配列表と簡単な説明
配列番号1
ラット成熟LINGO−1外部ドメイン(残基34−550)
【化1】
【0121】
配列番号2
カニクイザル成熟LINGO−1外部ドメイン(残基34−550)
【化2】
【0122】
配列番号3
ヒト成熟LINGO−1外部ドメイン(残基34−550)
【化3】
【0123】
配列番号4
4784VL
【化4】
【0124】
配列番号5
4784VH
【化5】
【0125】
配列番号6
4785VL
【化6】
【0126】
配列番号7
4785VH
【化7】
【0127】
配列番号8
DNA−4784VH
【化8】
【0128】
配列番号9
DNA−4785VH
【化9】
【0129】
配列番号10
DNA−4784VL
【化10】
【0130】
配列番号11
DNA−4785VL
【化11】
【0131】
配列番号12
抗体4784CDR−H1
【化12】
【0132】
配列番号13
抗体4784CDR−H2
【化13】
【0133】
配列番号14
抗体4784CDR−H3
【化14】
【0134】
配列番号15
抗体4784CDR−L1
【化15】
【0135】
配列番号16
抗体4784CDR−L2
【化16】
【0136】
配列番号17
抗体4784CDR−L3
【化17】
【0137】
配列番号18
抗体4785CDR’−H1
【化18】
【0138】
配列番号19
抗体4785CDR’−H2
【化19】
【0139】
配列番号20
抗体4785CDR’−H3
【化20】
【0140】
配列番号21
抗体4785CDR’−L1
【化21】
【0141】
配列番号22
抗体4785CDR’−L2
【化22】
【0142】
配列番号23
抗体4785CDR’−L3
【化23】
【0143】
配列番号24
正方向プライマーDM22
【化24】
【0144】
配列番号25
逆方向プライマーDM23
【化25】
【0145】
配列番号26
APタグ−5−NHISベクター
【化26】
【化27】
【0146】
配列番号27
ヒトLINGO−1成熟DNA配列
【化28】
【0147】
配列番号28
カニクイザルLINGO−1成熟DNA配列
【化29】
【0148】
配列番号29
ラットLINGO−1成熟DNA配列
【化30】
【0149】
配列番号30
正方向プライマーDM14
【化31】
【0150】
配列番号31
逆方向プライマーDM15
【化32】
【0151】
配列番号32
pSecTag2−V5ベクター
【化33】
【化34】
【0152】
配列番号33
ヒトLINGO−2成熟DNA配列
【化35】
【0153】
配列番号34
正方向プライマーDM16
【化36】
【0154】
配列番号35
逆方向プライマーDM17
【化37】
【0155】
配列番号36
pRS5a−IgG
【化38】
【化39】
【化40】
【0156】
配列番号37
天然リーダー(natleader)−hsLINGO−1−Fc/pRS5a
【化41】
【化42】
【化43】
【化44】
【0157】
配列番号38
Igリーダー−hsLINGO−1−ΔLRR−Fc/pRS5a
【化45】
【化46】
【化47】
【化48】
【0158】
配列番号39
Fab発現ベクターpMORPH(登録商標)X9 MH
【化49】
【化50】
【化51】
【化52】
【0159】
配列番号40
IgG4発現ベクターpMORPH(登録商標)h Igγ4
【化53】
【化54】
【化55】
【化56】
【化57】
【0160】
配列番号41
IgGラムダ鎖発現ベクターpMORPH(登録商標)h Ig ラムダ
【化58】
【化59】
【化60】
【化61】
【0161】
配列番号42
IgGカッパ鎖発現ベクターpMORPH(登録商標)h Ig カッパ
【化62】
【化63】
【化64】
【化65】
【0162】
配列番号43
IgG4発現ベクターpMORPH2(登録商標) h Igγ4
【化66】
【化67】
【化68】
【化69】
【化70】
【化71】
【0163】
配列番号44
IgGラムダ鎖発現ベクターpMORPH(登録商標)2 h Ig ラムダ2
【化72】
【化73】
【化74】
【化75】
【化76】
【0164】
配列番号45
IgGカッパ鎖発現ベクターpMORPH(登録商標)2 hIg カッパ
【化77】
【化78】
【化79】
【化80】
【化81】
【技術分野】
【0001】
本発明は例えばモノクローナル抗体またはそのFabフラグメントのようなLINGO結合分子、および中枢神経系に対する傷害を有する患者を処置するためのかかる結合分子の使用に関する。
【背景技術】
【0002】
高等脊椎動物成体の中枢神経系(CNS)に対する傷害の後の機能的回復は例外的に限定的であり、四肢の運動および感覚の喪失のような持続的な神経障害を招く。今のところ脊髄傷害(SCI)および脳皮質傷害のようなCNS傷害を有するヒトを処置するための有効な治療は欠如している。成体のCNSニューロンは一般的に軸索切断から生き延びるが、軸索再生は一過性であり、そして限局的な部分にわたってのみ生じ、それ故に機能的に関連するシナプス結合の再形成を遅延させる。さらに成体CNSの可塑性能力もまた制限され、故に傷害により切断されたものを機能的に補償するための非傷害経路の再編成を妨げる。逆説的に末梢神経系(PNS)で軸索切断された軸索は長きにわたって再生し、そして頻繁に機能的に意味のある連結を確立する高い能力を有する(Schwab, Curr Opin Neurobiol 14:118−124(2004))。この軸索再生/可塑性における制限はある程度、神経突起伸長の強力な阻害剤であることが示されているいくつかのタンパク質、すなわちNogo−A(Chen et al., Nature 403:434−439(2000);GrandPre et al., Nature 403:439−444(2000);Prinjha et al., Nature 403:383−384(2000))、ミエリン関連糖タンパク質(MAG)およびオリゴデンドロサイトミエリン糖タンパク質(OMgp)(McKerracher et al., Neuron 13:805−811(1994);Wang et al., Nature 417:941−944(2002))(図1A)の有髄化オリゴデンドロサイトにおける発現のためである。
【0003】
Nogo−Aはオリゴデンドロサイトの表面に暴露された複数の神経突起伸長阻止ドメインを含有する:二つはアミノ−末端領域(アミノ−Nogo−A)内に、そして1つはC−末端領域(Nogo−66)に位置する(Oertle et al., J Neurosci 23:5393−5406(2003))。Nogo−66はNogo−66受容体(NgR)として公知のニューロン表面上のグリコシルホスファチジルイノシトール(GPI)アンカードロイシンリッチリピート(LRR)含有受容体を介して結合およびシグナル発生する(Fournier et al., Nature 409:341−346(2001))。構造的には関連しないが、MAGおよびOMgpもまたNgRを介して結合およびシグナル発生する(Domeniconi et al., Neuron 35:283−290(2002);Liu et al., Science 297:1190−1193(2002);Wang et al., Nature 417:941−944(2002))。NgRを介するシグナル発生は小型GTPase RhoAの活性化に至り、それが今度はRho関連キナーゼ(ROCK)を活性化し、アクチン細胞骨格の硬直化および軸索伸長の阻止に至る(Niederoest et al., J Neurosci 22:10368−10376(2002);Schweigreiter et al., Mol Cell Neurosci 27:163−174(2004))。三つのリガンド全てがNgRのLRR領域内に結合し、そして部分的に重複する結合部位を有する(Fournier et al., J Neurosci 22:8876−8883(2002);Liu et al. Science 297:1190−1193(2002);Wang et al., Nature 417:941−944(2002);Barton et al., EMBO J 22:3291−3302(2003))。アミノ−Nogo−A内の阻止ドメインに関する(複数の)受容体は未知であるが、NgRとは区別されることが示されている(Schweigreiter et al., Mol Cell Neurosci 27:163−174(2004))。MAGはNgR2として公知のNgRの密接した相同体を介してシグナル発生することもまた見出されている(Pignot et al., J Neurochem 85:717−728(2003);Venkatesh et al., J Neurosci 25:808−822(2005))。
【0004】
NgRは細胞質ドメインを欠如するので、それはシグナル伝達のためにいくつかの膜貫通タンパク質、すなわち低親和性ニューロトロフィン受容体p75NTR、TROY(TAJとも称される)およびLINGO−1(LRRおよびIgドメイン含有、Nogo受容体相互作用タンパク質、LRRN6AまたはLERN1とも称される)を利用する(Wang et al. Nature 420:74−78(2002);Carim−Todd et al., Eur J Neurosci 18:3167−3182(2003);Mi et al., Nat Neurosci 7:221−228(2004);Park et al., Neuron 45:345−351(2005);Shao et al., Neuron 45:353−359(2005))。TROYおよびp75NTRはNgR受容体複合体において互いに機能的に置き換えることができるが、一方LINGO−1の存在は生じるシグナル発生のための絶対的な必要条件である。NgR受容体複合体はそれ故にリガンド結合サブユニットとしてNgR、およびp75NTRまたはTROYのいずれかと協調的に作用する一般的なシグナル伝達サブユニットとしてLINGO−1を含む三元複合体として認められる。
【0005】
LINGO−1はニューロンおよびオリゴデンドロサイト上で優先的に、CNS内で独占的に発現される単一の膜貫通タンパク質である。LINGO−1の発現は出生後早期にピークに達し、そして傷害時の成体脊髄において上方調節される。LINGO−1の外部ドメインはN−およびC−末端サブドメイン、続いて塩基性領域およびIgドメインによりフランキングされる12個のタンデムLRRを含有する(図1B)。LINGO−1外部ドメインのAP融合がNgRもしくはp75NTRまたは双方を発現するCOS−7細胞に結合し、そして同様に、LINGO−1が三つ全てのタンパク質を発現する細胞におけるNgRまたはp75NTRと共沈する場合、LINGO−1は双方と同時に相互作用することによりNgRおよびp75NTRと三元複合体を形成する可能性が最も高い。
【0006】
ニューロン上で発現されることに加えて、LINGO−1はまた成体CNSのオリゴデンドロサイトにおいても発現される(Mi et al., Nat Neurosci 8:745−751(2005))。LINGO−1−Fcでのいずれかの処理によるオリゴデンドロサイト培養物中のLINGO−1シグナル発生の阻止、RNAiでのタンパク質の下方調節、またはDN−LINGO−1の過剰発現はOPCの有髄化オリゴデンドロサイトへの分化を増強した。さらに、マウスにおけるLINGO−1の遺伝手術により成熟オリゴデンドロサイトおよび対応して脊髄における有髄軸索の数が増加する。LINGO−1シグナル発生の阻止はRhoAの活性化を低下させ、そしてFynキナーゼの作用を増大させ、その双方はオリゴデンドロサイト分化を促進させると報告されているが、LINGO−1シグナル発生の活性化に寄与する実際のリガンド/相互作用は未だに例示されていない。これにより、LINGO−1がミエリン形成の負のレギュレーターであるという結論に至っている。
【0007】
多発性硬化症(MS)は脱髄および軸索変性を特徴とし、多発性神経障害に至るCNSの慢性炎症性疾患である。軸索のミエリン再形成は疾患の早期に生じ、いくつかの時点で、ミエリン再形成は完全に失敗して軸索変性の加速および非可逆的損傷に至る。ミエリン再形成は活動性病変の辺縁部に遊走する成体オリゴデンドロサイト前駆細胞(OPC)の分化から生じる可能性が最も高い。LINGO−1はミエリン形成を負に調節するので、LINGO−1の遮断はミエリン再形成を増強し、軸索変性を減弱し、軸索再生を促進し、そして故にMSのような脱髄性疾患の進行を減弱、停止または逆転さえし得る。
【0008】
LINGO−1の遮断はまたドーパミン作動性ニューロンの生存性を改善し、そしてパーキンソン病のげっ歯類モデルにおける行動異常を低下させることも示されている(Inoue et al., Proc Natl Acad Sci USA 104:14430−14435(2007))。
【発明の概要】
【発明が解決しようとする課題】
【0009】
(発明の要旨)
驚くべきことに、LINGO−1に対する新規モノクローナルヒト抗体(本明細書後記の抗体4784、および抗体4785としても公知)がLINGO−1とNgRとの会合を有意に阻止し、そしてインビトロでnM以下の濃度でラット成体の脊髄ミエリンの神経突起伸長阻止活性を有意に減弱することが見出されている。加えて、該抗体はインビトロで一次オリゴデンドロサイトの分化を有意に増大させ、そして生存細胞における細胞表面LINGO−1を有意に下方調節することが示されている。これらの抗体での処理は軸索再生/可塑性を増大させ、そしてSCIおよび脳皮質傷害のような急性CNS傷害の後の機能的回復を改善すると予期される。さらに、オリゴデンドログリア細胞における該抗体を用いるLINGO−1シグナル発生の遮断は、MSのような脱髄性疾患における軸索のミエリン再形成を増強し、疾患進行の減弱に至る可能性がある。一致して、該抗体でのニューロンにおけるLINGO−1シグナル発生の阻止は軸索再生および神経可塑性を改善し、そして疾患の経過中の神経学的機能喪失の回復を促進すると予期できる。最後に、該抗体でのLINGO−1の遮断はパーキンソン病の病因を減弱すると予期できる。
【0010】
さらに本発明はLINGO−1の特異的エピトープに結合する結合分子を提供する。
抗体はラット、カニクイザルおよびヒトLINGO−1外部ドメインに対してnM以下のKDを有し、ラット成体脊髄ミエリンの神経突起伸長阻止活性をnM以下の濃度で有意に減弱し、そしてインビトロでオリゴデンドロサイト分化を有意に増大させた。さらに今では該抗体と同一の可変領域を有するその他のLINGO−1結合分子を構築することが可能である。
【課題を解決するための手段】
【0011】
(発明の詳細な説明)
したがって、本発明はLINGO−1の特定の領域またはエピトープに対する結合分子(以後「本発明の結合分子」または単に「結合分子」と称する)を提供する。
本発明の結合分子は解離定数(KD)<1000nM、さらに好ましくはKD<100nM、最も好ましくはKD<10nMでラットLINGO−1(配列番号1)、カニクイザルLINGO−1(配列番号2)およびヒトLINGO−1(配列番号3)の成熟外部ドメイン(残基34−550)に結合する。結合反応を例えば実施例に記載されるFACS法を含む標準的な方法(定性アッセイ)により示すことができる。加えてラット、カニクイザルおよびヒトLINGO−1に対する結合、ならびに効率もまた以下で記載されるような神経突起伸長アッセイおよびオリゴデンドロサイトアッセイにおいて示すことができる。
【0012】
故にさらに好ましい実施態様では、結合分子は(100nM、好ましくは10nM、さらに好ましくは1nM、なおさらに好ましくは0.1nMの濃度で)ラット成体脊髄ミエリンの基質で成長させたラット小脳顆粒細胞の細胞あたりの平均神経突起長を、ラット、カニクイザルおよびヒトLINGO−1外部ドメインに結合しない対照抗体で処理されたラット小脳顆粒細胞の細胞あたりの平均神経突起長と比較して、少なくとも20%、好ましくは50%、最も好ましくは60%まで増大させる。
【0013】
ペプチドマイクロアレイを使用することにより、本発明の結合分子が結合する特異的エピトープを当分野において周知の方法にしたがって決定する。結果的に別の実施態様では本発明は配列番号46−51により定義されるような少なくとも1つのLINGO−1エピトープに結合する結合分子を提供する。配列番号46:KIVILLDYMFQD、配列番号47:AIRDYSFKRLYR、配列番号48:LKVLEISHWPYL、配列番号49:NLTAVPYLAVRHLVY、配列番号50:YFTCRRARIまたは配列番号51:DVLLPNYFTCRRARI。
【0014】
別の実施態様では、本発明の結合分子は1つまたはそれより多い以下のCDR配列、例えばそこで言及された抗体4784の全てまたは抗体4785配列の全てを含む:
配列番号12
(抗体4784CDR−H1)
SSGVGVG
配列番号13
(抗体4784CDR−H2)
HIGSDDDKYYSTSLKT
配列番号14
(抗体4784CDR−H3)
NQQYGDGYPGYFDY
配列番号15
(抗体4784CDR−L1)
SGDNIGNYYVY
配列番号16
(抗体4784CDR−L2)
EDTNRPS
配列番号17
(抗体4784CDR−L3)
QSYDNLHEQV
配列番号18
(抗体4785CDR’−H1)
DNSAAWS
配列番号19
(抗体4785CDR’−H2)
LIYLRSKWDNDYAVSVKS
配列番号20
(抗体4785CDR’−H3)
TGRADEFDV
配列番号21
(抗体4785CDR’−L1)
SGSSSNIGNNYVS
配列番号22
(抗体4785CDR’−L2)
RNSKRPS
配列番号23
(抗体4785CDR’−L3)
STYDTFSIV
【0015】
さらに好ましくは、結合分子は配列番号12、13、14、15、16および/もしくは17を有する抗体4784に関する;または配列番号18、19、20、21、22および/もしくは23を有する抗体4785に関する1つまたはそれより多い前記の配列を含む。
【0016】
4784または4785を変化させることができることは当業者には理解され、それを通してそれらはいくつかの、さらに好ましくは1個またはそれより多いアミノ酸、好ましくは3個まで、例えば1または2個の前記のSDRを、特に1個もしくはそれより多い、またはそれらの全て、例えばそれらの1もしくは2個で変化させるか、または生成物の形式の代替えの翻訳後修飾を提供し、同一もしくは実質的に類似する抗Lingo−1結合挙動を実証する治療薬を導く。
別の実施態様では、本発明の結合分子は;配列番号5もしくは配列番号7に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である配列、および;
配列番号4もしくは配列番号6に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である配列、またはその直接的な均等物;からなる群から選択される少なくとも1つの抗原結合部位を含む。
【0017】
1つの実施態様では、結合分子は配列番号5または配列番号7、および;
配列番号4または配列番号6;
からなる群から選択される少なくとも1つの結合部位を含む。
本発明はさらに配列番号5に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である第1の配列、および配列番号4に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である第2の配列、またはその直接的な均等物を含む結合分子を提供する。
【0018】
本発明はさらに配列番号7に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である第1の配列、および配列番号6に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%もしくは少なくとも99%相同である第2の配列、またはその直接的な均等物を含む結合分子を提供する。
【0019】
1つの実施態様では、本発明は少なくとも:
a)
(i)配列番号5または配列番号7を含む可変ドメイン、および;
(ii)ヒト重鎖の定常部分またはそのフラグメント;
を含む1つの免疫グロブリン重鎖またはそのフラグメント;ならびに
b)
(i)配列番号4または配列番号6を含む可変ドメイン、および;
(ii)ヒト軽鎖の定常部分またはそのフラグメント;
を含む1つの免疫グロブリン軽鎖またはそのフラグメント;
またはその直接的な均等物;例えばa)もしくはb)の下で与えられた鎖の各々の2つもしくは3つ;
を含む上記の結合分子を提供する。
【0020】
配列は配列番号4−7に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%相同でよい。重要な因子は、各々の場合で好ましくは実施例または残りの記載にて記載されるように、かかるバリアントがLINGO−1に対する結合能力、脱抑制効果(特にラット成体脊髄ミエリンの神経突起伸長阻止活性をnM以下の濃度で減弱する能力)を保持し、そして/またはSCIの機能的回復を改善する(特にラットモデルにおいて)ことである。
【0021】
1つの実施態様では、本発明は配列番号4−7もしくは配列番号12−23による1つもしくはそれより多い配列を含む抗体またはそのフラグメント、またはその直接的な均等物である結合分子を提供する。
さらなる実施態様では、抗体としての結合分子はγ4型のヒト重鎖の定常部分またはそのフラグメントおよびλ型であるヒト軽鎖の定常部分またはそのフラグメントを有する。
【0022】
さらなる実施態様では、抗体としての結合分子はγ4型のヒト重鎖の定常部分またはそのフラグメントおよびκ型であるヒト軽鎖の定常部分またはそのフラグメントを有する。
さらなる実施態様では、結合分子はヒトまたはキメラまたはヒト化モノクローナル抗体である。
さらなる実施態様では、結合分子はhumaneered抗体である。
本発明はまた前記で定義されたような結合分子をコードするポリヌクレオチドをも提供する。
【0023】
ポリヌクレオチドを配列番号8および配列番号9からなる群から;または配列番号10および配列番号11からなる群から選択できる。
本発明はまた配列番号8−11による1つまたはそれより多いポリヌクレオチドを含む発現ベクターをも提供する。
さらに本発明は配列番号8−11によるポリヌクレオチドを含む発現ベクターを提供し、ここで該発現系またはその部分が適合する宿主細胞中に存在する場合、該発現系またはその部分は前記で示されたような結合分子を生成することができる。本発明はまたはかかる発現系を含む単離された宿主細胞をも提供する。
【0024】
本発明はまたは医薬品としての前記で示されたような結合分子の使用をも提供する。
本発明はまたはCNS傷害の処置のための医薬品の調製における前記で示されたような結合分子の使用をも提供する。
本発明はまたは前記で示されたような結合分子を少なくとも1つの薬学的に許容される担体または希釈剤と一緒に含む医薬組成物をも提供する。
【0025】
さらに本発明は有効量の前記で示されたような結合分子をかかる処置を必要とする対象に投与することを含む、軸索再生/可塑性の促進に関連する疾患の処置の方法を提供する。
本発明はまたは有効量の請求項1から10のいずれかに記載の結合分子をかかる処置を必要とする対象に投与することを含む、軸索再生/可塑性の促進に関連する疾患の処置の方法をも提供する。
抗原結合部位が第1および第2の双方のドメインを含む場合、これらは同一のポリペプチドに位置してよいか、または好ましくは各ドメインが異なる鎖に位置してよく、第1のドメインは免疫グロブリン重鎖の部分またはそのフラグメントであり、そして第2のドメインは免疫グロブリン軽鎖の部分またはそのフラグメントである。
【0026】
本発明の結合分子の実例には、ファージディスプレイにより生成されるような抗体およびヒトもしくはキメラヒト化抗体、またはさらにhumaneered抗体、またはその任意のフラグメント、例えばF(ab’)2;およびFabフラグメント、ならびに一本鎖または単一ドメイン抗体が含まれる。「抗体」なる用語はかかる結合分子を含むと意図される。
一本鎖抗体は通常10から30個のアミノ酸、好ましくは15から25個のアミノ酸からなるペプチドリンカーにより共有結合した抗体重および軽鎖の可変ドメインからなる。それ故に、かかる構造は重および軽鎖の定常部分を含まず、そして小型ペプチドスペーサーは定常部分全体よりも抗原性が低いはずであると考えられている。「キメラ抗体」とは重もしくは軽鎖または双方の定常領域がヒト起源であるが、重および軽鎖の双方の可変ドメインが非ヒト(例えばマウス)起源である抗体を意味する。「ヒト化抗体」とは超可変領域(CDR)は非ヒト(例えばマウス)起源であるが、免疫グロブリンの全てまたは実質的に全てのその他の部分、例えば定常領域および可変ドメインの高度に保存された部分、すなわちフレームワーク領域はヒト起源である抗体を意味する。しかしながらヒト化抗体は超可変領域に隣接するフレームワーク領域の部分においてネズミ配列の数個のアミノ酸を保持し得る。
【0027】
超可変領域は任意の種類のフレームワーク領域、好ましくはネズミまたはヒト起源のものに関連し得る。適当なフレームワーク領域は「Sequences of proteins of immunological interest」(Kabat E.A. et al、米国保健社会福祉省国立衛生研究所公衆衛生局、好ましくは、特にフレームワーク領域に関して出典明示により本明細書の一部とする)に記載されている。好ましくは結合分子のヒト重鎖の定常部分はサブタイプを含むγ4型のものでよく、好ましくはヒト軽鎖の定常部分はκまたはλ型のもの、さらに好ましくはλ型のものでよい。
【0028】
天然発生「抗体」はジスルフィド結合により相互連結された少なくとも2つの重(H)鎖および2つの軽(L)鎖を含む糖タンパク質である。各重鎖は重鎖可変領域(本明細書ではVHと略する)および重鎖定常領域からなる。重鎖定常領域はCH1、CH2およびCH3の3つのドメインからなる。各軽鎖は軽鎖可変領域(本明細書ではVLと略する)および軽鎖定常領域からなる。軽鎖定常領域は1つのドメイン、CLからなる。VHおよびVL領域を、フレームワーク領域(FR)と称されるさらに保存されている領域がちりばめられた相補性決定領域(CDR)と称される超可変性の領域にさらに細分することができる。各VHおよびVLは以下の順でアミノ末端からカルボキシ末端に配列した3つのCDRおよび4つのFRから構成される:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4。重および軽鎖の可変領域は抗原と相互作用する結合ドメインを含有する。抗体の定常領域は、免疫系の種々の細胞(例えばエフェクター細胞)および古典的は補体系の第1の構成要素(C1q)を含む宿主組織または因子に対する免疫グロブリンの結合を媒介し得る。
【0029】
抗体の「抗原結合部分」(または単に「抗原部分」)なる用語は、本明細書で使用される際には、抗原に特異的に結合する能力を保持する抗体の全長または1つもしくはそれより多いフラグメント(例えばLINGO−1および/またはLINGO−2)を指す。全長抗体のフラグメントにより抗体の抗原結合機能を実施できることが示されている。抗体の「抗原結合部分」なる用語に包含される結合フラグメントの実例には、Fabフラグメント、VL、VH、CLおよびCH1ドメインからなる1価フラグメント;F(ab’)2フラグメント、ヒンジ領域でジスルフィド橋により連結された2つのFabフラグメントを含む2価フラグメント;VHおよびCH1ドメインからなるFdフラグメント;抗体の単一のアームのVLおよびVHドメインからなるFvフラグメント;VHドメインからなるdAbフラグメント(Ward et al., Nature 341:544−546(1989));ならびに単離された相補性決定領域(CDR)が含まれる。
【0030】
「モノクローナル抗体」または「モノクローナル抗体組成物」なる用語は、本明細書で使用される際には、単一分子組成物の抗体分子の調製物(すなわち全て単一の親細胞のクローンである免疫細胞の1つの型により生成されるのでそれらは同一である)を指す。モノクローナル抗体組成物は(本質的に)単一の結合特異性および特定のエピトープに関する親和性を表示する。
【0031】
「ヒト抗体」なる用語は、本明細書で使用される際には、フレームワークおよびCDR領域の双方がヒト起源の配列から誘導される可変領域を有する抗体を含むと意図される。さらに抗体が定常領域を含有する場合、定常領域もまたかかるヒト配列、例えばヒト生殖細胞配列、またはヒト生殖細胞配列の変異したものから誘導される。本発明のヒト抗体はヒト配列によりコードされないアミノ酸残基(例えばインビトロで無作為もしくは部位特異的変異誘発により、またはインビボで体細胞変異により導入された変異)を含み得る。しかしながら、「ヒト抗体」なる用語は、本明細書で使用される際には、マウスのような別の哺乳動物種の生殖細胞系から誘導されたCDR配列がヒトフレームワーク配列に移植されている抗体を含むとは意図されない。
【0032】
「ヒトモノクローナル抗体」なる用語は、フレームワークおよびCDR領域の双方がヒトの配列から誘導される可変領域を有する(本質的に)単一の結合特異性を表示する抗体を指す。1つの実施態様ではヒトモノクローナル抗体は、不死化細胞に融合されたヒト重鎖導入遺伝子および軽鎖導入遺伝子を含むゲノムを有するトランスジェニック非ヒト動物、例えばトランスジェニックマウスから得られたB細胞を含むハイブリドーマにより生成される。
「組換えヒト抗体」なる用語は本明細書で使用される際には、ヒト免疫グロブリン遺伝子に関して遺伝子導入もしくは染色体導入された動物(例えばマウス)またはそこから調製されたハイブリドーマから単離された抗体、ヒト抗体を発現するように形質転換された宿主細胞、例えばトランスフェクトーマ(transfectoma)から単離された抗体、組換え体から単離された抗体、コンビナトリアルヒト抗体ライブラリーのような組換え手段により調製、発現、創成または単離された全てのヒト抗体、およびヒト免疫グロブリン遺伝子の全てまたは一部の、配列をその他のDNA配列にするスプライシングを伴う任意のその他の手段により調製、発現、創成または単離された抗体を含む。かかる組換えヒト抗体は、フレームワークおよびCDR領域がヒト生殖細胞系免疫グロブリン配列から誘導される可変領域を有する。しかしながら特定の実施態様では、かかる組換えヒト抗体をインビトロ変異誘発(またはヒトIg配列に関して遺伝子導入された動物を用いる場合、インビボ体細胞変異誘発)に供することができ、そして故に組換え抗体のVHおよびVL領域のアミノ酸配列は、ヒト生殖細胞系VHおよびVL配列から誘導され、そしてそれに関連するが、インビボでヒト抗体生殖細胞系レパートリー内に天然には存在し得ない配列である。
【0033】
本明細書で使用される際には、「アイソタイプ」とは重鎖定常領域遺伝子により提供される抗体クラス(例えばIgM、IgE、IgG1またはIgG4のようなIgG)を指す。
本明細書で使用される際には、「親和性」なる用語は単一の抗原性部位での抗体と抗原との間の相互作用の強さを指す。各抗原性部位内では、抗体「アーム」の可変領域は弱い非共有結合力を介して多くの部位で抗原と相互作用し;より相互作用すると、親和性がより強くなる。
「KD」なる用語は、本明細書で使用される際には、Kaに対するKdの比(解離率に対する会合率)(すなわちKd/Ka)から得られる解離定数を指すと意図され、そしてモル濃度(M)として表現される。当分野において十分に確立された方法を用いて抗体に関するKD値を決定することができる。抗体のKDを決定するための方法は表面プラズモン共鳴法の使用、またはBiacore(登録商標)システムのようなバイオセンサーシステムの使用による。
【0034】
本発明による結合分子は好ましくは「単離された抗体」であり、それは本明細書で使用される際には、異なる抗原特異性を有するその他の抗体を実質的に含まない抗体を指す(例えばLINGO−1、LINGO−2またはLINGO−1およびLINGO−2に特異的に結合する単離された抗体は言及されたもの以外の抗原に特異的に結合する抗体を実質的に含まない)。しかしながら特異的に結合する単離された抗体はその他の種からのLINGO−1またはLINGO−2分子のようなその他の抗原に対する交差反応性を有し得る。さらに単離された抗体は好ましくはその他の細胞材料および/または化学物質を実質的に含まない。
本発明はまたは:
a)アミノ酸配列(配列番号5)を有する重鎖の可変配列を含む第1ドメイン;
b)アミノ酸配列(配列番号4)を有する軽鎖の可変配列を含む第2ドメイン;
c)第1ドメインのN末端および第2ドメインのC末端もしくは第1ドメインのC末端および第2ドメインのN末端のいずれかに結合したペプチドリンカー;
またはその直接的な均等物;
を含む抗体4784の抗原結合部位(特に抗体4784に関して前記されたCDRを伴う)を含む一本鎖結合分子から選択できる本発明の結合分子を提供する。
本発明の結合分子を
a)アミノ酸配列(配列番号7)を有する重鎖の可変配列を含む第1ドメイン;
b)アミノ酸配列(配列番号6)を有する軽鎖の可変配列を含む第2ドメイン;
c)第1ドメインのN末端および第2ドメインのC末端もしくは第1ドメインのC末端および第2ドメインのN末端のいずれかに結合したペプチドリンカー;
またはその直接的な均等物;
を含む抗体4785の抗原結合部位(特に抗体4785に関して前記されたCDRを伴う)を含む一本鎖結合分子から選択できる。
周知のように、1つまたはいくつかのアミノ酸の欠失、付加または置換のようなアミノ酸配列におけるわずかな変化は、実質的に同一の特性を有する元来のタンパク質のアレル形態に至り得る。故に「その直接的な均等物」なる用語は:
(i)結合分子の可変領域(例えば配列番号4、5、6または7)は、配列番号4および配列番号5の各々の直接的な均等物を含む軽および重鎖または配列番号6および配列番号7の各々の直接的な均等物を含む軽および重鎖の均等な可変領域に対して少なくとも50または80%相同、好ましくは少なくとも90%相同、さらに好ましくは少なくとも95、96、97、98、99%相同であり;
(ii)ラットLINGO−1(配列番号1)、カニクイザルLINGO−1(配列番号2)およびヒトLINGO−1(配列番号3)の外部ドメイン(残基34−550)に、好ましくは解離定数(KD)<1000nM、さらに好ましくはKD<100nM、最も好ましくはKD<10nMで結合できる;
本発明のいずれかの任意の単一ドメイン結合分子(分子X)、または結合部位あたり少なくとも2つのドメインを有する本発明の任意の結合分子(分子X’)を意味する。
【0035】
故に本発明のさらなる実施態様は例えば解離定数<1000nMでラット、カニクイザルおよび/またはヒトLINGO−1の外部ドメインに結合でき、そして少なくとも1つの抗原結合部位を含み、該抗原結合部位は配列に4784の軽および重鎖(各々配列番号4および配列番号5)または4785の軽および重鎖(各々配列番号6および配列番号7)の均等な可変領域に少なくとも50%、好ましくは80、90、95、96、97、98、99%相同である可変領域を含む。
【0036】
別の実施態様では、結合分子は配列番号12−23からなる群から選択される少なくとも1つのアミノ酸配列、またはこれらの配列に対して少なくとも50%、好ましくは80、90、95、96、97、98、99%相同である配列を含む。
この解離定数を例えば実施例で記載されるFACS法を含む種々のアッセイにおいて都合よく試験できる。加えて結合分子の結合および機能的影響をバイオアッセイ、例えば以下に記載されるような神経突起伸長アッセイにおいて示すことができる。
【0037】
ヒト重鎖の定常部分はγ1;γ2;γ3;γ4;α1;α2;δまたはε型、好ましくはγ型、さらに好ましくはγ4型のものでよいが、一方ヒト軽鎖の定常部分はκまたはλ型(それはλ1;λ2;およびλ3サブタイプを含む)でよいが、好ましくはλ型のものである。これらの全ての定常部分のアミノ酸配列はKabat et al(Supra)で与えられる。
本発明の結合分子の抱合体、例えば酵素または毒素または放射性同位体抱合体もまた本発明の範囲内に含まれる。
【0038】
「ポリペプチド」には、本明細書にて特記しない場合、ペプチド結合により互いに結合したアミノ酸を含み、N末端で始まり、そしてC末端で終わるアミノ酸配列を有する任意のペプチドまたはタンパク質が含まれる。好ましくは本発明のペプチドはモノクローナル抗体であり、さらに好ましくはキメラ(Vグラフティングとも称される)またはヒト化(CDRグラフティングとも称される)モノクローナル抗体である。ヒト化(CDRグラフティング)モノクローナル抗体はアクセプター抗体のフレームワーク(FR)配列に導入されたさらなる変異を含んでも含まなくてもよい。
【0039】
ポリペプチドの機能的誘導体は本明細書で使用される際には、質的に本発明のポリペプチドに共通した生物学的活性を有する、すなわちラット、カニクイザルおよびヒトLINGO−1の外部ドメインに結合する能力を有する分子が含まれる。
機能的誘導体には本発明によるポリペプチドのフラグメントおよびペプチド類似体が含まれる。それはまた「直接的な誘導体」をも含む。
【0040】
フラグメントは本発明による、例えば特定された配列のポリペプチドの配列内の領域を含む。結合分子の、特に抗体のフラグメントは機能的なフラグメントである、すなわちそれらはLINGO−1および/またはLINGO−2に、特に配列番号46、47、48、49、50および51により与えられる少なくとも1つのエピトープに、好ましくは、前記でまたは実施例で特に好ましいとして言及された結合親和性(KD)で結合できる少なくとも1つの部分を含む。
「誘導体」なる用語は本発明による、例えば特定された配列のポリペプチドのアミノ酸配列バリアント、および共有結合性修飾を定義するために用いられる。本発明による、例えば特定された配列のポリペプチドの例えば軽および重鎖の超可変領域の機能的誘導体は好ましくは、本発明による、例えば特定された配列のポリペプチドのアミノ酸配列と少なくとも約65%、さらに好ましくは少なくとも約75%、なおさらに好ましくは少なくとも約85%、最も好ましくは少なくとも約95、96、97、98、99%の配列全体にわたる相同性を有し、そしてラット、カニクイザルおよびヒトLINGO−1(および場合によっては加えてLINGO−2)の外部ドメインに結合する能力を実質的に保持している。
【0041】
「共有結合性修飾」なる用語には、本発明による、例えば特定された配列のポリペプチド;またはそのフラグメントの有機タンパク質性または非タンパク質性誘導体化剤での修飾、異種性ポリペプチドへの融合および翻訳後修飾が含まれる。例えば特定された配列の共有結合性修飾されたポリペプチドは依然ラット、カニクイザルおよびヒトLINGO−1の外部ドメインに結合する能力を有する。共有結合性修飾は伝統的には標的化されたアミノ酸残基を、選択された側鎖または末端残基と反応できる有機誘導体化剤と反応させることにより、または選択された組換え宿主細胞において機能する翻訳後修飾のメカニズムを利用することにより導入される。特定の翻訳後修飾は発現されたポリペプチドにおける組換え宿主細胞の作用の結果である。グルタミニルおよびアスパラギニル残基は対応するグルタミルおよびアスパルチル残基に頻繁に翻訳後脱アミド化される。これに代えて、これらの残基は穏やかな酸性条件下で脱アミノ化される、その他の翻訳後修飾にはプロリンおよびリジンのヒドロキシル化、セリル、チロシンまたはスレオニル残基のヒドロキシル基のリン酸化、リジン、アルギニンおよびヒスチジン側鎖のα−アミノ基のメチル化が含まれる(例えばT. E. Creighton, Proteins:Structure and Molecular Properties, W. H. Freeman & Co., San Francisco, pp. 79−86(1983)参照)。共有結合性修飾には、例えばイムノアドヘシンのような、例えば本発明による、例えば特定された配列のポリペプチド含む融合タンパク質およびそのアミノ酸バリアントを、ならびに異種性シグナル配列に対するN末端融合が含まれる。
【0042】
自然のポリペプチドおよびその機能的誘導体に関する「相同性」(または「同一性」)は本明細書では、必要により最大相同性パーセントを達成するために配列をアラインし、そしてギャップを誘導した後、対応する自然のポリペプチドの残基と同一である、候補配列におけるアミノ酸残基のパーセンテージとして定義され、そして任意の保存置換を配列同一性の一部ではないと考える。NもしくはC末端伸長も挿入も同一性または相同性を低下させるものではないと解釈しなければならない。アラインメントのための方法およびコンピュータープログラムは周知である。
【0043】
好ましくは本明細書で使用される際には、2つのアミノ酸配列または2つのヌクレオチド配列間の相同性パーセントは2つの配列間の同一性パーセントと均等である。2つの配列間の同一性パーセントは配列により共有される同一の位置の数の関数であり(すなわち相同性%=同一の位置の数/位置の全数×100)、2つの配列の最適なアラインメントのために導入される必要のあるギャップの数および各ギャップの長さを考慮する。配列の比較および2つの配列間の同一性パーセントの決定を、以下の非限定例で記載されるような数学的アルゴリズムを用いて達成することができる:
【0044】
2つのアミノ酸配列間の同一性パーセントを、ALIGNプログラム(バージョン2.0)に組み入れられているE. MeyersおよびW. Millerのアルゴリズム(Comput. Appl. Biosci., 4:11−17(1988))を用いて、PAM120残基重み付け表(weight residue table)、ギャップ長ペナルティー12およびギャップペナルティー4を用いて決定することができる。加えて、2つのアミノ酸配列間の同一性パーセントを、GCGソフトウェアパッケージのGAPプログラム(http://www.gcg.comで利用可能)に組み入れられているNeedlemanおよびWunsch(J. Mol, Biol. 48:444−453(1970))アルゴリズムを用いて、Blossom 62マトリクスまたはPAM250マトリクスのいずれか、ならびにギャップ重み付け16、14、12、10、8、6または4および長さ重み付け1、2、3、4、5または6を用いて決定することができる。
【0045】
加えてまたはこれに代えて、本発明のタンパク質配列をさらに「クエリー配列」として用いて公開されたデータベースに対して検索を実施し、例えば関係する配列を同定することができる。Altschul, et al., J.Mol. Biol. 215:403−10(1990)のXBLASTプログラム(バージョン2.0)を用いてかかる検索を実施することができる。BLASTタンパク質検索をXBLASTプログラム、スコア=50、語長=3で実施して、本発明の抗体分子に相同なアミノ酸配列を得ることができる。比較目的のためにギャップの入ったアラインメントを得るために、Altschul et al., Nucleic Acids Res. 25(17):3389−3402(1997)に記載されるようなGapped BLASTを利用することができる。BLASTおよびGapped BLASTプログラムを利用する場合、各プログラムのデフォルトパラメーター(例えばXBLASTおよびNBLAST)を用いることができる。http:www.ncbi.nhn.nih.govを参照のこと。
「(複数の)アミノ酸」は全ての天然発生の例えばL−α−アミノ酸を指し、そしてD−アミノ酸を含む。アミノ酸は周知の一文字表記または三文字表記のいずれかにより同定される。
【0046】
「アミノ酸配列バリアント」なる用語は、本発明による、例えば特定された配列のポリペプチドと比較してそのアミノ酸配列においていくつかの差異を有する分子を指す。本発明による、例えば特定された配列のポリペプチドのアミノ酸配列バリアントは依然ラット、カニクイザルおよびヒトLINGO−1の外部ドメインに対する結合能力を有する。置換バリアントは、本発明による、例えば特定された配列のポリペプチドにおいて少なくとも1つのアミノ酸残基が除去され、そして同一位置でその場所に異なるアミノ酸が挿入されたものである。これらの置換は単一でよく、その場合分子の1個のアミノ酸のみが置換されているか、またはそれらは複数でよく、その場合2個またはそれより多い、例えば1から10個。好ましくは1から5個、さらに好ましくは1から3個のアミノ酸が同一分子内で置換されている。挿入バリアントは、1つまたはそれより多い、例えば1から10個のような1から100個のアミノ酸が、本発明による、例えば特定された配列のポリペプチドにおける特定の位置のアミノ酸に直ぐ隣接して挿入されたものである。アミノ酸に直ぐ隣接してとは、アミノ酸のα−カルボキシまたはα−アミノ官能基のいずれかに連結されていることを意味する。欠失バリアントは本発明による、例えば特定された配列のポリペプチドにおいて1個またはそれより多い、例えば1から10個または1から5個のような1から100個のアミノ酸が除去されているものである。通常欠失バリアントは分子の特定の領域において1または2個のアミノ酸が欠失されている。
【0047】
本発明の結合分子を組換えDNA技術により生成できる。これに鑑みて、結合分子をコードする1つまたはそれより多いDNA分子は構築され、適切な制御配列下に置かれ、そして発現のために適当な宿主生物に移されなければならない。
【0048】
したがって非常に一般的な様式では:
(i)本発明の単一ドメイン結合分子、本発明の一本鎖結合分子、本発明の結合分子の重もしくは軽鎖またはそのフラグメントをコードするDNA分子;および
(ii)組換え手段による本発明の結合分子の生成のための本発明のDNA分子の使用;
が提供される。
【0049】
当分野の現状では、当業者は本明細書にて提供される情報、すなわち超可変領域のアミノ酸配列およびそれをコードするDNA配列を考慮して、本発明のDNA分子を合成できる。可変ドメイン遺伝子を構築する方法は例えば欧州特許第239400号(好ましくは特に可変ドメイン遺伝子を構築するための方法に関して、出典明示により本明細書の一部とする)に記載されており、そして以下のように簡単に要約できる:どのような特異性のものでもモノクローナル抗体の可変ドメインをコードする遺伝子はクローン化される。フレームワークおよび超可変領域をコードするDNAセグメントを決定し、そして超可変領域をコードするDNAセグメントを除去し、フレームワーク領域をコードするDNAセグメントを接合部で適当な制限部位と一緒に融合する。標準的な手順によりDNA分子の変異誘発により適切な位置で制限部位を作成することができる。前記の配列にしたがってDNA合成により二本鎖合成可変領域カセットを調製する。免疫グロブリン可変ドメインをコードするDNA分子を達成するための標準的なプロトコールにより、これらのカセットに付着末端が提供され、それらを接合部でフレームワークにライゲートできるようになる。
【0050】
さらに本発明のモノクローナル抗体をコードするDNA構築物を得るために、生成ハイブリドーマ細胞系からmRNAを入手する必要はない。故にPCT出願第WO90/07861号(好ましくは特にモノクローナル抗体の生成に関して、出典明示により本明細書の一部とする)により、遺伝子のヌクレオチド配列に関して記載された情報のみを考慮して組換えDNA技術によりモノクローナル抗体を生成するための全ての指示が与えられる。
その方法は多数のオリゴヌクレオチドの合成、PCR法によるその増幅および望ましいDNA配列を与えるためのそのスプライシングを含む。
【0051】
適当なプロモーターまたは重および軽鎖定常部分をコードする遺伝子を含む発現ベクターは公的に入手可能である。故に一度本発明のDNA分子が調製されると、それを適切な発現ベクターに都合よく移すことができる。
一本鎖抗体をコードするDNA分子を標準的な方法により、例えば第WO88/1649号(好ましくは特に一本鎖抗体をコードするDNA分子に関して、出典明示により本明細書の一部とする)に記載されるように調製することもできる。
本発明の特定の実施態様では、本発明の結合分子のいくつかの生成のための組換え手段には、以下に記載されるような第1および第2のDNA構築物が含まれる:
【0052】
第1のDNA構築物は重鎖またはそのフラグメントをコードし、そして:
a)抗体4784、DNA−4784VH(配列番号8)、または抗体4785、DNA−4785VH(配列番号9)のいずれかの重鎖の可変ドメインをコードする第1の部分;この第1の部分は可変ドメインの最初のアミノ酸をコードするコドンで始まり、そして可変ドメインの最後のアミノ酸をコードするコドンで終わる;および
b)重鎖の定常部分の最初のアミノ酸をコードするコドンで始まり、そして定常部分またはそのフラグメントの最後のアミノ酸をコードするコドンで終わり、ナンセンスコドンが続く重鎖定常部分またはそのフラグメントをコードする第2の部分;
を含む。
好ましくは第2の部分はヒト重鎖の定常部分、さらに好ましくはヒトγ4鎖の定常部分をコードする。この第2の部分はゲノム起源のDNAフラグメント(イントロンを含む)またはcDNAフラグメント(イントロンを含まない)でよい。
【0053】
第2のDNA構築物は軽鎖またはそのフラグメントをコードし、そして:
a)抗体4784、DNA−4784VL(配列番号10)、または抗体4785、DNA−4785VL(配列番号11)のいずれかの軽鎖の可変ドメインをコードする第1の部分;この第1の部分は可変ドメインの最初のアミノ酸をコードするコドンで始まり、そして可変ドメインの最後のアミノ酸をコードするコドンで終わる;および
b)軽鎖の定常部分の最初のアミノ酸をコードするコドンで始まり、そして定常部分またはそのフラグメントの最後のアミノ酸をコードするコドンで終わり、ナンセンスコドンが続く軽鎖定常部分またはそのフラグメントをコードする第2の部分;
を含む。
好ましくは第2の部分はヒト軽鎖の定常部分、さらに好ましくはヒトκ鎖の定常部分をコードする。
【0054】
DNA構築物の各々を適当な制御配列の制御下に、とりわけ適当なプロモーターの制御下に置く。DNA構築物を発現のために移す宿主生物に適合するならば、任意の種類のプロモーターを使用できる。しかしながら哺乳動物細胞において発現を行うことになっている場合、免疫グロブリン遺伝子のプロモーターを使用するのがとりわけ好ましい。
【0055】
細胞培養またはトランスジェニック動物において望ましい抗体を生成できる。適当な制御配列の下に置かれた第1および第2のDNA構築物を卵にマイクロインジェクトし、そのように調製された卵を適切な偽妊娠の雌に移し、そして望ましい抗体を発現する子孫を選択することを含む標準的な方法により、適当なトランスジェニック動物を得ることができる。
抗体鎖が細胞培養中に生成されなければならない場合、DNA構築物を単一の発現ベクターまたは2つの別個であるが適合する発現ベクターのいずれかに最初に挿入すべきであり、恐らく後者が好ましい。
【0056】
したがって本発明はまた前記で記載されたDNA構築物の少なくとも1つを含む原核細胞系または真核細胞系において複製できる発現ベクターをも提供する。
次いでDNA構築物を含有する各発現ベクターを適当な宿主生物に移す。DNA構築物を2つの発現ベクターに別個に挿入する場合、それらを別個に、すなわち細胞あたり1つの型のベクターに移すか、または同時に移すことができ、恐らくこの後者が好ましい。適当な宿主生物は細菌、酵母または哺乳動物細胞系でよく、この後者が好ましい。さらに好ましくは、哺乳動物細胞系はリンパ系起源、例えば骨髄腫、ハイブリドーマまたは通常の不死化B細胞であるが、内因性抗体重または軽鎖を全く発現しない。
【0057】
宿主生物が細胞あたりに多数のベクターのコピーを含有するのも好ましい。宿主生物が哺乳動物細胞系である場合、標準的な方法にしたがってコピーの数を増幅することにより望ましい目標に到達することができる。増幅方法は通常薬物に対して高められた抵抗性に関して選択することからなり、該抵抗性は発現ベクターによりコードされる。
本発明の別の態様では、本発明の多鎖結合分子を生成するための方法が提供され、それは(i)本発明の第1および第2の構築物で形質転換された生物を培養すること;ならびに(ii)培養物から本発明の活性な結合分子を回収すること;を含む。
【0058】
これに代えて重および軽鎖を別個に回収し、そしてインビトロでリフォールディングした後、活性な結合分子に再構成することができる。再構成の方法は当分野において周知である;方法の実例はとりわけ欧州特許第120674号または欧州特許第125023号にて提供される。
それ故に方法はまた:
(i)本発明の第1のDNA構築物で形質転換された第1の生物を培養し、そして培養物から該重鎖またはそのフラグメントを回収すること;ならびに
(ii)本発明の第2のDNA構築物で形質転換された第2の生物を培養し、そして培養物から該軽鎖またはそのフラグメントを回収すること;ならびに
(iii)(i)で得られた重鎖またはそのフラグメントおよび(ii)で得られた軽鎖またはそのフラグメントから本発明の活性な結合分子をインビトロで再構成すること;
をも含む。
【0059】
類似の様式で、
(i)本発明の一本鎖または単一ドメイン結合分子を各々コードするDNA構築物で形質転換された生物を培養すること;および
(ii)培養物から該分子を回収すること;
を含む本発明の一本鎖または単一ドメイン結合分子を生成するための方法もまた提供される。
【0060】
本発明の結合分子は以下に例示されるようにLINGO−1のNgRに対する結合を有意に阻止し、ラット成体脊髄ミエリンの神経突起伸長阻止活性をnM以下の濃度で有意に減弱し、そしてオリゴデンドロサイト分化をインビトロで有意に増大させる。
【図面の簡単な説明】
【0061】
【図1】NgR:SH−SY5Y細胞に対するAP−LINGO−1結合に及ぼすFab4784および4785の影響:2μMの指示された抗LINGO−1Fabまたは抗ニワトリリゾチームFab3207の不在下または存在下で懸濁液中のNgR:SH−SY5Y細胞を1nM APまたはAP−LINGO−1のいずれかと共にインキュベートする。1−StepTM PNPPと共に30分間インキュベートした後、細胞における結合AP活性を405nmでの吸光度として測定する。AP−LINGO−1の特異的結合をAP−LINGO−1結合の全量とAP単独との結合の量との間の差異として計算する。特異的結合の平均阻止パーセンテージ(n=3、±STD)をFab3207の存在下および抗LINGO−1 Fabの存在下のAP−LINGO−1の特異的結合の量の間の差異パーセンタイルとして計算する。
【図2】抗LINGO−1 IgG4抗体4784および4785による脊髄ミエリンの脱抑制:A)脊髄ミエリンを伴わずにコーティングされたウェル(SCなし、白色棒)または脊髄ミエリンでコーティングされたウェル上で抗LINGO−1 IgG4抗体、対照抗リゾチームIgG4抗体3207(緑色棒)もしくは1μm ROCK阻害剤Y27632(黄色棒)の不在下(SC、赤色棒)または存在下、P7 CGN細胞を16時間インキュベートする。ROCKは全てではないが、たいていの、NgR受容体複合体を介してシグナル発生しないものを含むミエリン関連神経突起伸長阻害剤のシグナル発生経路における2次メッセンジャーであり、それでY27632処理が脊髄ミエリンの神経突起伸長阻止活性の減弱のための陽性対照として用いられる(図1)。プレート毎にSCを伴う、およびSCを伴わない条件で3つの96ウェルプレートで実験を実施し、そのプレート上の抗体の影響を比較し、そしてニューロンあたりの平均神経突起長(μm)を10回反復でウェルあたり500ニューロンに関して計算する。阻止パーセンテージ(白色文字)を、SCを伴って、および伴わずにコーティングされたウェル上に置かれた細胞間の平均神経突起長/ニューロンにおける差異パーセンタイルとして計算する。脱抑制パーセンテージ(黒色斜字体文字)を、抗LINGO−1抗体の存在下および不在下でSC上に置かれた細胞間の平均神経突起長における差異として、SCを伴って、および伴わずにコーティングされたウェル上に置かれた細胞間における差異パーセンタイルとして計算する。*p<0.05、**p<0.01(一元配置分散分析、抗体不在下で脊髄ミエリン上に置かれた細胞に関する平均神経突起長/ニューロンに対するHolm−Sidak比較)。B)脊髄ミエリンを伴わずに(SCなし)コーティングされたウェル上で、および脊髄ミエリンを伴って(SC)コーティングされたウェル上で、1nM 対照IgG4 3207もしくは抗LINGO−1 IgG4 4784の不在下(SC)または存在下でインキュベートされた細胞の代表的な視野の蛍光画像。4784の存在下の脊髄ミエリン上で成長させた細胞は、抗体の不在下または対照抗体3207の存在下で成長させた細胞よりも細胞あたりの神経突起が可視的により長く、そしてより多い。
【図3】抗LINGO−1 IgG4抗体IIによる脊髄ミエリンの脱抑制 A)脊髄ミエリンを伴わずにコーティングされたウェル(SCなし、白色棒)または脊髄ミエリンでコーティングされたウェル上で抗LINGO−1 IgG4抗体もしくは対照抗リゾチームIgG4抗体3207の不在下(SC、赤色棒)または存在下、P7 CGN細胞を8時間インキュベートする。プレート毎にSCを伴う、およびSCを伴わない条件で3つの96ウェルプレートで実験を実施し、そのプレート上の抗体の影響を比較し、そしてニューロンあたりの平均神経突起長(μm)を10回反復でウェルあたり500ニューロンに関して計算する。阻止パーセンテージ(白色文字)および脱抑制パーセンテージ(黒色斜字体文字)を前記のように計算する。**p<0.01(一元配置分散分析、抗体不在下で脊髄ミエリン上に置かれた細胞に関する平均神経突起長/ニューロンに対するHolm−Sidak比較)。
【図4】抗LINGO−1 IgG4抗体IIIによる脊髄ミエリンの脱抑制:A)脊髄ミエリンを伴わずにコーティングされたウェル(SCなし)または脊髄ミエリンでコーティングされたウェル上で指示された濃度の抗LINGO−1 IgG4抗体4784もしくは4785、対照抗リゾチームIgG4抗体3207もしくは1μM Y27632(ROCK)の不在下(SC)または存在下、P7 CGN細胞を8時間インキュベートする。プレート毎にSCを伴う、およびSCを伴わない条件で3つの96ウェルプレートで実験を実施し、そのプレート上の抗体の影響を比較し、そしてニューロンあたりの平均神経突起長(μm)を10回反復でウェルあたり500ニューロンに関して計算する。阻止パーセンテージ(白色文字)および脱抑制パーセンテージ(黒色斜字体文字)を前記のように計算する。*p<0.05、**p<0.01(一元配置分散分析、抗体不在下で脊髄ミエリン上に置かれた細胞に関する平均神経突起長/ニューロンに対するHolm−Sidak比較)。
【図5A】抗LINGO−1抗体は未成熟オリゴデンドロサイトの分化を有意に増大させる:A)新たに単離されたOPCをDMEM/CNTF/T3培地中100nM 4784、4785または対照IgG4 3207で3日間処理し、続いて抗O4抗体で染色して未成熟および成熟オリゴデンドロサイト(より大きければ、標識がより拡散される)を可視化し、そして核酸色素DAPI(4’,6−ジアミジン−2’−フェニル−インドール二塩酸塩)で細胞核(より小さい円形ドット)を可視化する。高度に樹状化し、そして伸長した突起およびミエリンシート状構造を担持するオリゴデンドロサイトは成熟形態を有していると考えられ、そして白色矢印で示される。抗LINGO−1抗体処理の結果、成熟形態を有するO4陽性細胞の比率の増大を招くが、一方対照IgG4 3207での処理には影響がない。
【図5B】全(左グラフ)および成熟(右グラフ)オリゴデンドロサイトの比率を3つの独立した実験(1、2、3)で定量する。左の棒グラフはO4染色に関連するDAPI染色された核のパーセンテージを表し、そして右の棒グラフは成熟形態を有するO4陽性細胞のパーセンテージを表す(3検体の平均+STD)。各棒グラフでは、最左の棒は処理なしのものであり、左から2番目の棒は対照IgGでの対照、次は4784での処理、そして最右は4785での処理を表す。抗LINGO−1抗体はオリゴデンドロサイトである細胞の比率に影響しないが、成熟形態を有するオリゴデンドロサイトの比率を有意に増大させる。*p<0.05、**p<0.01、対照IgG4 3207の存在下の成熟オリゴデンドロサイトの比率に対するHolm−Sidak比較を伴う一元配置分散分析。
【図6】抗LINGO−1抗体は細胞表面LINGO−1を下方調節する:A)トランスフェクトされていないCHO−K1またはCHO−K1−hLINGO−1細胞を100nM 4784、4785および3207と共に37℃で24時間インキュベートし、そして抗V5抗体と共に室温で30分間さらにインキュベートすることにより細胞表面でLINGO−1を検出する。細胞を4%PFAで固定し、BSAで遮断し、そして結合抗V5抗体を抗マウスIgG(Fc特異的)−POD抱合体を用いて検出し、それをその後1−Step(商標)Turbo TMB ELISAキットを用いて展開する。450nmでの吸光度は細胞表面でのLINGO−1の量の測定値と考えられる(3検体の平均+STD)。トランスフェクトされていないCHO−K1細胞に対して非常に低レベルのV5抗体結合が観察される。CHO−K1−hLINGO−1細胞を、抗LINGO−1抗体を伴うが、対照IgG4 3207を伴わないでインキュベートすることにより、結果的に細胞表面でのLINGO−1の量の有意な低下を招く。**p<0.01、対照IgG4 3207とのインキュベーションの後の吸光度に対するHolm−Sidak比較を伴う一元配置分散分析。B)トランスフェクトされていないCHO−K1またはCHO−K1−hLINGO−1細胞上の細胞表面タンパク質を4℃でビオチン化し、そして細胞を100nM 4784、4785および3207を伴って、または伴わずに37℃で指示された時間、インキュベートする。インキュベーション期間の終わりに、アガロースビーズに結合させた抗V5抗体を用いて細胞ライゼートからLINGO−1を沈殿させ、そしてビオチン化(細胞表面)LINGO−1をウェスタンブロット分析により抗ビオチン抗体を用いて検出する。トランスフェクトされていないCHO−K1細胞ではビオチン化LINGO−1に関するシグナルは検出されない。CHO−K1−hLINGO−1細胞を抗LINGO−1抗体と共にインキュベートすることにより、細胞表面のLINGO−1の分解離率が増大する。
【図7】ELSIAによる抗LINGO−1 Fabの特徴付け:ELISA分析に関する値を相対蛍光単位(RFU)の平均値として与える。これらのクローンの結合親和性はFACS飽和アッセイにより特徴付けされる。
【発明を実施するための形態】
【0062】
本発明はまた哺乳動物神経系、とりわけヒト神経系の軸索再生/可塑性の促進における本発明の結合分子の使用をも提供する。
本発明はまた有効量の本発明の結合分子をかかる処置を必要とする患者に投与することを含む、哺乳動物神経系、とりわけヒト神経系の軸索再生/可塑性を促進する方法をも提供する。
本発明はまた本発明の結合分子および薬学的に許容される担体または希釈剤を含む、哺乳動物神経系、とりわけヒト神経系の軸索再生/可塑性を促進するための医薬組成物をも提供する。
【0063】
とりわけ本発明の結合分子は、CNS傷害の軸索再生および可塑性を促進するのに有用である(傷害なる用語は本出願では、特に機械的もしくは化学的影響により引き起こされるか、または例えばニューロンの変性に至る疾患もしくは障害による傷害、特に例えばアルツハイマー病もしくはパーキンソン病のような神経学的疾患または以下に言及されるような障害もしくは疾患におけるその構造または形態を指す)。故に本発明の分子はとりわけヒト対象のために広い有用性を有する。例えば本発明の結合分子は末梢(PNS)および中枢(CNS)神経系の種々の疾患の処置において、すなわちさらにとりわけアルツハイマー病、パーキンソン病、筋萎縮性側索硬化症(ALS)、レビー様病理またはその他の一般的な認知症、頭蓋、大脳または脊髄外傷後の疾患および脳卒中のような神経変性疾患において有用である。さらにLINGO−1がミエリン形成の負のレギュレーターであることを考慮すると、本発明の結合分子は、限定するものではないが多発性硬化症、単相性脱髄、脳脊髄炎、多巣性白質脳症、全脳炎、マルキアファーヴァ・ビニャミ病、橋ミエリン崩壊、副腎白質ジストロフィー、ペリツェウス・メルツバッハー病、海綿状変性、アレキサンダー病、カナヴァン病、異染性白質ジストロフィーおよびクラッベ病を含む脱髄性疾患における軸索再生/可塑性の促進と協調してミエリン再形成を促進するのに有用である。一例では、本発明の結合分子を発現する細胞を脊髄傷害部位に移植して、傷害を受けた部位全体にわたって軸索成長を促すことができる。かかる移植細胞は傷害または外傷後の脊髄機能を回復するための手段を提供するであろう。かかる細胞は嗅神経鞘細胞および胎児神経または組織移植片の様々な系列の幹細胞を含み得る。
【0064】
加えて本発明の結合分子は一般的な虚血性網膜症、前眼部虚血性視神経症、視神経炎の全形態、加齢性黄斑変性症、糖尿病性網膜症、嚢胞様黄斑浮腫(CME)、網膜色素変性症、スタルガルト病、ベスト病の卵黄様網膜変性、レーベル先天黒内障およびその他の遺伝性網膜変性、病的近視、未熟児網膜症、およびレーベル遺伝性視神経症、角膜移植または屈折矯正角膜手術の後遺症、ならびにヘルペス角膜炎を含む網膜または角膜の変性を直接的または間接的に伴い得る眼変性傷害の処置に有用である。
さらに本発明の結合分子は精神医学的症状、とりわけ統合失調症および抑うつの処置に有用である。
【0065】
これらの適応症のための適切な投薬量はもちろん例えば用いられることになっている本発明の特定の分子、投与の様式ならびに処置される症状の性質および重篤度に依存して異なるであろう。一般的には、投薬量は好ましくは1μg/kg/日から1mg/kg/日の範囲であろう。本発明の結合分子は治療薬として病変部またはその近くに都合よくポンプにより投与されるかまたは注射され、例えばそれらをCNSに頭蓋内に、または脊椎に髄腔内に直接病変部に投与できる。しかしながら全身投与はここでは排除されない。本発明の結合分子を単独で、またはその他の薬剤との組み合わせで、または逐次的組み合わせで提供することができる。例えば本発明の結合分子を、限定するものではないが脳卒中または脊髄傷害後のさらなるニューロン損傷および軸索再生の阻止を遮断するための手段としてのコルチコステロイド、NGF、BDNFのような神経栄養因子またはExelon(商標)もしくはレボドパのような神経変性疾患のためのその他の薬物のような、抗Nogo−A抗体または抗炎症薬との組み合わせで投与することができる。脳卒中の処置のためのその他の適当な組み合わせパートナーはアルテプラーゼおよびデスモテプラーゼ(DSPA、例えば第WO90/09438号に開示される)である。1つの実施態様では本発明は、本発明の結合分子およびデスモテプラーゼを含む組み合わせをとりわけ脳卒中の処置のために、および該組み合わせを含む医薬組成物として提供する。本明細書で使用される際には、2つの薬剤が同時に投与されるか、または薬剤が同時に作用するような様式で独立して投与される場合、2つの薬剤は組み合わせで投与されると考えられる。
【0066】
コード番号、一般名または商品名により同定される活性成分の構造を標準的な事典「The Merck Index」の最新版から、またはデータベース、例えばPatents International(例えばIMS World Publications)もしくはIMS Healthにより提供されるその他のデータベースから取ることができる。その対応する内容を出典明示により本明細書の一部とする。任意の当業者が活性成分を同定することは十分に可能であり、そしてこれらの参照文献に基づいて、同様にインビトロおよびインビボの双方で標準的な試験モデルにおいて医薬的適応および特性を製造および試験することが可能である。
【0067】
都合のよい様式で本発明の医薬組成物を製造できる。例えば本発明の分子を含む本発明による組成物は好ましくは凍結乾燥形態で提供される。即時的投与のために、それを適当な水性担体、例えば注射用滅菌水または滅菌緩衝生理学的食塩水に溶解する。
適当な組成物を作り上げるのを助けるために、本発明の結合分子および場合によっては本発明の結合分子の影響を増強する第2の薬物を、混合または併用投与のための説明書と共に同一容器内に別個に包装することができる。最適な第2の薬物候補は前記で提供した。
【0068】
本発明の結合分子およびNGFのような成長因子の組み合わせの相乗効果をインビボで脊髄傷害モデルにより実証できる。
以下の実施例を参照して本発明はさらに十分に理解されよう。しかしながらそれらは本発明の範囲を限定するとは解釈されるべきではない。
実施例において注目を集めるモノクローナル抗体は抗体4784に関して軽鎖の可変部分(配列番号4)および重鎖の可変部分(配列番号5)を含有し、そして4785に関して軽鎖の可変部分(配列番号6)および重鎖の可変部分(配列番号7)を含む本発明による結合分子である。
【0069】
以下の略語を用いる:
AP ヒト胎盤アルカリ性ホスファターゼ
CDR 相補性決定領域
cDNA 相補的DNA
ELISA 酵素結合免疫吸着アッセイ
FACS 蛍光活性化セルソーティング
FBS ウシ胎仔血清
HCMV ヒトサイトメガロウイルスプロモーター
IgG 免疫グロブリンアイソタイプG
PBS リン酸塩緩衝生理食塩水
PCR ポリメラーゼ連鎖反応
PFA パラホルムアルデヒド
PNPP リン酸パラニトロフェニル
【実施例1】
【0070】
全長ラット、カニクイザルまたはヒトLINGO−1およびヒトLINGO−2を発現するCHO−K1細胞の作成
無作為およびオリゴdTプライマーを用いてユニバーサルヒト参照RNA(Stratagene)のRT−PCRによりヒトcDNAライブラリーを作成する。無作為およびオリゴdTプライマーを用いて凍結カニクイザル脳から単離されたポリA RNAのRT−PCRにより、カニクイザル脳cDNAライブラリーを作成する。Marathon−readyラット脳cDNAライブラリーをClontechから入手する。5’−XbaIおよび3’−XhoI部位によりフランキングされたヒトLINGO−1(配列番号27)、カニクイザルLINGO−1(配列番号28)およびラットLINGO−1(配列番号29)の成熟配列をコードするcDNA(残基34−614)を正方向プライマーDM14、5’−CTACGTCTAGAACGGGCTGCCCGCCCCGCT−3’(配列番号30)および逆方向プライマーDM15、5’−GGTTTCTCGAGTCATATCATCTTCATGTTGAACTTGCGG−3’(配列番号31)を用いて各々のライブラリーからPCR増幅する。PCR生成物をXbaIおよびXhoIで切断し、そしてベクターpSecTag2−V5(配列番号32)の各々の部位に挿入して、各々hLINGO−1−pSecTag2−V5、cmLINGO−1−pSecTag2−V5およびrLINGO−1−pSecTag2−V5を作成する。予測されるタンパク質生成物はN末端で2アミノ酸残基リンカーを介して14アミノ酸残基V5エピトープタグに融合されたLINGO−1の成熟配列である。5’−XbaIおよび3’−XhoI部位によりフランキングされたヒトLINGO−2(配列番号33)の成熟配列をコードするcDNA(残基26−606)を正方向プライマーDM16、5’−CTACGTCTAGAATTGGCTGCCCCGCTCGCT−3’(配列番号34)および逆方向プライマーDM17、5’−GGTTTCTCGAGTCAAATCATTTTCATGTTGAACCTCCTG−3’(配列番号35)を用いてMarathon−readyヒト脳cDNAライブラリー(Clontech)からPCR増幅する。PCR生成物をXbaIおよびXhoIで切断し、そしてベクターpSecTag2−V5の各々の部位に挿入して、hLINGO−2−pSecTag2−V5を作成する。予測されるタンパク質生成物はN末端で2アミノ酸残基リンカーを介して14アミノ酸残基V5エピトープタグに融合されたLINGO−2の成熟配列である。lipofectamine−2000(Invitrogen)を用いて製造者の説明書にしたがって、hLINGO−1−pSecTag2−V5、cmLINGO−1−pSecTag2−V5、rLINGO−1−pSecTag2−V5およびhLINGO−2−pSecTag2−V5で各々細胞をトランスフェクトすることにより、ヒトLINGO−1(CHO−K1−hLINGO−1)、カニクイザルLINGO−1(CHO−K1−cmLINGO−1)、ラットLINGO−1(CHO−K1−rLINGO−1)およびヒトLINGO−2(CHO−K1−hLINGO−2)を安定して発現するCHO−K1細胞を作成する。安定して発現するトランスフェクト体を1mg/mlゼオシン(Invivogen)で選択し、そして96ウェルプレートへの連続希釈か、またはクローンリング(clonal rings)の使用かのいずれかにより単一のクローンを単離する。細胞表面での構築物の発現を、抗V5抗体を用いる免疫蛍光分析により確認する(InvitroGen)。
【実施例2】
【0071】
ヒトLINGO−1−FcおよびヒトLINGO−1ΔLRR−Fcの作成および発現
ヒトLINGO−1をコードするMGC mRNA(クローンMGC:17422 IMAGE:4214343)をPCR増幅のための鋳型として使用する。Pwo1ポリメラーゼ(Roche Diagnostics)ならびに標的配列の5’末端でHindIII制限部位およびKozakコンセンサス配列および3’末端で標的配列の最後のコドンの直後にXhoI制限部位を加えられたプライマーで、ヒトLINGO−1の天然のシグナル配列により先行される細胞外ドメイン(ECD)(aa1−550)をPCRにより増幅する。PCR生成物をHindIIIおよびXhoIで消化し、ゲル精製し、そして以前に同一の酵素で消化されたプラスミドpRS5a−IgG(配列番号36)に挿入する。得られた発現クローン(天然リーダー(natleader)−hsLINGO−1−Fc/pRS5a、配列番号37)における挿入された配列、完全なFcおよびフランキング領域の正確さをDNAシークエンシングにより確認する。
【0072】
同一のMGCクローンをLRRドメインを欠くヒトLINGO−1(aa34−65+aa354−550)に関する発現プラスミドの遺伝子SOEによる構築のための鋳型として提供する。成熟LINGO−1に融合した異種性分泌シグナルをコードする部分配列で5’末端を伸長し、そして3’末端でC末端フラグメントの最初の7個のアミノ酸をコードする配列を加えたプライマーでヒトECD LINGO−1のN末端領域(aa34−65)をPCRにより増幅する。N末端フラグメントの最後の7個のアミノ酸をコードする配列で5’末端を伸長し、そして3’末端で標的配列の最後のコドンの直後にXhoI部位を加えたプライマーでヒトECD LINGO−1のC末端領域(aa354−550)をPCRにより増幅する。2つのPCR生成物をゲル精製し、混合し、そして5’末端でHindIII制限部位、Kozakコンセンサス配列を加え、そして異種性分泌シグナル配列を完成するプライマー、および3’末端で以前にC末端フラグメントを増幅するために用いられた外部プライマーを用いる第2のPCR増幅のための鋳型として提供する。PCR生成物をHindIIIおよびXhoIで消化し、ゲル精製し、そして以前に同一酵素で消化されたプラスミドpRS5a−IgGに挿入する。得られた発現クローン(Igリーダー−hsLINGO−1−ΔLRR−Fc/pRS5a、配列番号38)における挿入された配列、完全なFcおよびフランキング領域の正確さをDNAシークエンシングにより確認する。
【0073】
最初の発現評価として双方の構築物を小規模な実験で試験する。HEK.EBNA細胞(Invitrogen、以前のカタログ番号R620−07)をアタッチモード(attached mode)で組織培養フラスコで、25mM Hepes(Gibco/Life Technologies カタログ番号42430−025)で緩衝され、そしてさらに10%ウシ胎仔血清でさらに富化されたダルベッコ変法イーグル培地(DMEM)中培養し;培養物を加湿雰囲気下37℃および5%CO2で維持する。小規模なトランスフェクション実験のために、トランスフェクションの1日前に4×105セルをポリ−D−リジンコーティングした6ウェル(プレート)に播種する。ウェルあたりプラスミドDNA3μgおよびLipofectamine2000試薬(Invitrogenカタログ番号11668−019)6μlを用いて本質的には製造供給元により記載されるようにトランスフェクションを実施する。トランスフェクション後3日に、細胞上澄を収集し、そして細胞不含上澄をタンパク質分析、すなわちプロテインGカラムの免疫親和性HPLC分析に供する。構築物天然リーダー−hsLINGO−1−Fc/pRS5aに関して8mg/lと構築物Igリーダー−hsLINGO−1−ΔLRR−Fc/pRS5aに関して40mg/lとの間の範囲で力価を決定する。その後に双方のプラスミドに関して、HEK.EBNA懸濁培養物中で大容量規模での一過性のトランスフェクションを可能にするために大規模プラスミド調製物準備する。
規模を拡大した天然リーダー−hsLINGO−1−Fcの生成のために、1.4×106セル/mlの密度のHEK.EBNA細胞培養物2.9lをDNA:PEI溶液(DNA 1μg:PEI 2μg/ml)1.1lと混合する。4時間インキュベートした後、培養物にExCell VPRO培地(SAFC、以前はJRH、Lenexa, KS)4lを供給する。培養の6日後に細胞培養上澄を収集し、そして10kDaカットオフの使い捨てHemoflow F10HPSフィルター(Fresenius Medical Care, Germany)を用いてダイアフィルトレーションにより1lまで濃縮する。Igリーダー−hsLINGO−1−ΔLRR−Fcタンパク質を作成するために行う第2の関連するタンパク質生成を類似の様式で行う。大規模トランスフェクションに関する詳細、DNA:PEI比率、細胞密度、供給および収集は前記されたものと全く同一である。
【0074】
a)天然リーダー−hsLINGO−1−Fc
濃縮物1l(培養上澄8lから)を20mlプロテインAセファロースのクロマトグラフィーにより分析する。100mM NaPi、pH7.3のベースライン洗浄の後、結合材料を50mMクエン酸、140mM NaCl、pH2.7で溶出し、中和し、そして滅菌ろ過する。溶出された分画さらに濃縮し、そしてPBS中Superdex 75でゲルろ過し、1.2mg/mlの濃度の生成物8.2mgを生じる。
b)Igリーダー−hsLINGO−1−ΔLRR−Fc
濃縮物1l(培養上澄8lから)を20mlプロテインAセファロースのクロマトグラフィーにより分析する。100mM NaPi、pH7.3のベースライン洗浄の後、結合材料を50mMクエン酸、140mM NaCl、pH2.7で溶出し、中和し、そして滅菌ろ過して、1.5mg/mlの濃度の生成物52.5mgを生じる。
精製されたタンパク質を還元/アルキル化およびトリプシン消化の後、N末端シークエンシングおよびMALDIペプチド質量分析により詳しく特徴付けする。
【実施例3】
【0075】
AP−LINGO−1結合アッセイ
LINGO−1のNgRに対する結合の遮断は神経突起伸長の3つのミエリン関連阻害剤、すなわちNogo−66、MAGおよびOMgpのシグナル発生を防御し、そしてしたがってCNSミエリンの神経突起伸長阻止活性を減弱し、故に軸索再生/可塑性の増大および急性CNS傷害後の機能的回復の改善に至ると予期される。抗LINGO−1抗体がLINGO−1のNgRに対する結合を遮断することを実証するために、ヒト胎盤アルカリ性ホスファターゼ(AP)−タグ化ラットLINGO−1外部ドメイン(AP−LINGO−1)の、NgRを安定して発現するSH−SY5Y細胞(NgR−SH−SY5Y、Walmsley et. al., J Cell Sci 117:4591−4602(2004))への結合を測定するアッセイを用いることができる。
【0076】
5’−Xho Iおよび3’−Xba I部位によりフランキングされたラットLINGO−1外部ドメイン(残基34−532)の大部分をコードするcDNAを、正方向プライマーDM22、5’−GGTTATCTCGAGACCGGCTGCCCGCCCC−3’(配列番号24)、および逆方向プライマーDM23、5’−GGCCCTTCTAGATCACTCGCCTGGCTGGTTGGAGATG−3’(配列番号25)を用いてrLINGO−1−pSecTag2−V5からPCR増幅する。PCR生成物をXhoIおよびXbaIで切断し、そしてベクターAPtag−5−NHIS(配列番号26)の各々の部位に挿入して、APtag−5−NHIS−solrLINGO−1を作成する。予測されるタンパク質生成物はN末端で3アミノ酸残基リンカーを介してヒト胎盤アルカリ性ホスファターゼの残基23−511に融合したラットLINGO−1外部ドメインの大部分である。lipofectamine2000を用いて製造者の説明書にしたがって、HEK293T細胞をAPtag−5−NHIS−solrLINGO−1でトランスフェクトする。トランスフェクションの4時間後にトランスフェクション培地を除去し、そしてフェノールレッド(Invitrogen)を含まないOptiMEM Iで置き換える。24時間後に培地を収集し、置き換え、そしてさらに24時間後に再度収集する。13000×gで5分間遠心することにより培地を清澄化し、そしてCentriprepフィルター装置(Millipore)を用いて製造者の説明書にしたがって上澄をほぼ15倍に濃縮する。濃縮された上澄のAP活性を1−Step(商標)PNPP(Pierce)を用いて405nmでの経時的な吸光度の変化として測定し、そして以下の等式を用いて濃度に変換する(200μl PNPP/セルで96ウェル形式に適用):
【数1】
【0077】
濃縮された上澄を記載されるようにSDS−PAGEゲル電気泳動およびウェスタンブロットに供する(Walmsley et. al., J Cell Sci 117:4591−4602(2004))。ECL(商標)システム(GE Healthcare)を用いて0.1%(容量/容量)抗ペンタヒスチジン抗体(Qiagen)、続いて0.02%(容量/容量)ペルオキシダーゼ抱合抗マウスIgG抗体(Sigma)でAP−LINGO−1を検出する。AP−LINGO−1をおよそ110kDaのバンドとして可視化し、その予測される分子量、112kDaに類似する。N末端分解生成物は観察されない。
【0078】
NgR:SH−SY5Y細胞を周密度50%で酵素不含解離バッファー(Invitrogen)で収集してNgRのような細胞表面タンパク質を保存する。1nM AP、1nM AP−LINGO−1、または2μM抗LINGO−1Fabもしくはニワトリ卵白リゾチームに対する対照Fab3207の存在下の1nM AP−LINGO−1をOptiMEM(Invitrogen)中30分間プレインキュベートし、そしてその後懸濁液中NgR:SH−SY5Y細胞と共に一定の攪拌を行いながら1.5時間インキュベートする。細胞をHBH(Hanks緩衝生理食塩水中20mM HEPES pH7.4/1%ウシ血清アルブミン)で6回洗浄し、そしてPBS中4%パラホルムアルデヒド(PFA)/5%スクロース中で15分間固定する。Hanks緩衝生理食塩水中20mM HEPES pH7.4中、65℃で1時間インキュベートすることによる内因性AP活性の不活化に続いて、1−Step(商標)PNPP(Pierce)と共に製造者の説明書にしたがって30分間インキュベートした後、細胞結合性AP活性を405nmでの吸光度として定量する。
【0079】
Fabを2μMの濃度で使用してAP−LINGO−1を結合性Fabで飽和し、そしてしたがって結合を阻止するその能力に関して、その親和性の影響を最小にする。このための理由は、その結合部位の位置よりもむしろその低親和性のために結合を阻止し損ねたFabを時期尚早にさらなる実験から破棄する可能性を排除するためであるが、かかるFabの親和性が親和性成熟化およびIgG4転換により後の段階で増大し得るためである。1nM AP−LINGO−1を対照Fab3207または抗LINGO−1 Fab4784および4785のいずれかとともにプレインキュベートし、そして次に懸濁液中Fabの存在下、NgR:SH−SY5Y細胞への結合を可能にする(図1)。抗LINGO−1 Fabの存在下、特異的AP−LINGO−1結合の阻止パーセンテージをFab3207に関して正規化する。4784および4785は、細胞に対するAP−LINGO−1結合を有意に阻止する(p<0.01、一元配置分散分析、対照Fab3207の存在下、AP−LINGO−1の特異的結合に対するHolm−Sidak比較)。
【0080】
NgRに対するLINGO−1の結合の遮断は、ミエリン関連阻害剤Nogo−66、MAGおよびOMgpのシグナル発生を防御してCNSミエリンの神経突起伸長阻止活性の低下に至ると予測される。その点で、4784および4785Fabは最終的なIgG4形式(実施例8参照)に転換され、そしてラット成体脊髄ミエリン上で成長させた出生後7日のラット小脳顆粒ニューロンからの神経突起伸長の阻止を減弱するその能力に関して評価される。
【実施例4】
【0081】
神経突起伸長阻止アッセイ
インビボ軸索再生/可塑性に及ぼす抗LINGO−1抗体の影響を予測する最も関連性のあるインビトロアッセイは、CNSミエリンの神経突起伸長阻止活性を減弱するその能力である。このアッセイでは、成体ラットから抽出された全脊髄ミエリンでコーティングされたウェル中で出生後7日のラット小脳顆粒ニューロン(CGN)を成長させ、そして神経突起伸長を自動ArrayScan(登録商標)HCSリーダー(Cellomics)により定量する。
抗LINGO−1 IgG4抗体4784および4785の脱抑制活性を該神経突起伸長アッセイにおいて評価する(図2)。
【0082】
ラット成体からの新たなラット脊髄組織を3容量(重量/容量)抽出バッファー(60mM Chaps、20mM Tris pH8.0、1mM EDTA、プロテアーゼ阻害剤カクテル)中でホモジナイズし、4℃で30分間インキュベートし、そして170000×gで、4℃で30分間遠心することにより清澄化する。96ウェルプレートの各ウェルをMeOH中ニトロセルロース5μl(MeOH12ml中5cm2ニトロセルロース)でコーティングし、空気乾燥し、そして5μg/mlポリ−D−リジン100μlで、37℃で4時間インキュベートすることによりコーティングする。水で3回洗浄した後、プレートを1時間空気乾燥し、そして次に37℃で一晩インキュベートすることにより、60μg/cm2脊髄抽出物でコーティングする。以前に記載されたように(Schweigreiter et al., 2004)出生後7日のラット小脳組織のトリプシン分離物からCGN細胞を新たに精製する。LINGO−1を検出するために、2μg/ml(または13.3nM)抗LINGO−1ポリクローナル抗体(Upstate)、続いて0.02%(容量/容量)ペルオキシダーゼ抱合抗ウサギIgG抗体(Sigma)を用いてV5−タグ化ラットLINGO−1を発現するCHO−K1細胞またはP7 CGN細胞からのライゼートに関してウェスタンブロット分析を実施する。脊髄ミエリンでコーティングされていないかまたはコーティングされたウェル上で、CGN細胞(35000セル/ウェル)を37℃で30分間インキュベートした後、0−100nM抗LINGO−1 IgG4抗体または対照3207 IgG4抗体のいずれかを添加する。37℃で8−16時間インキュベートした後、細胞を4%PFAで固定し、そして核の可視化のためにHoechst 3342(Invitrogen)、および抗β−チューブリンIII抗体(R&D Systems)、続いてAlexa Fluor 546抱合抗マウスIgG抗体(Invitrogen)で染色してニューロンを特異的に可視化する。神経突起伸長のパラメーターをArrayScan(登録商標)HCSリーダー(Cellomics)を用いて決定する。ArrayScan(登録商標)IIは96ウェルマイクロタイタープレート内の細胞のフィールドを自動的に位置決定し、焦点を合わせ、そして暴露する。ArrayScan(登録商標)は高解像度光学系、対応したシングルバンド励起フィルターを伴う多帯域発光フィルター(XF100)、フレーム取り込み装置を有するCCDカメラ、および専用アプリケーションソフトウェアからなる。このアッセイではExtended Neurite Outgrowth Bioapplicationを用いる。励起フィルターホイールおよび多帯域発光フィルターを用いることにより、同一細胞における2つのフルオロフォアからの蛍光の多チャンネル撮像が可能になる。Hoechst 33342標識された核の帯域画像を獲得して別々の細胞を同定し、そして次にAlexa Fluor 488の帯域画像を獲得して抗チューブリン抗体で標識された細胞の広がりを同定する(Alexa Fluor 488に二次抱合されたものを用いる)。細胞内の不適切な物体を分析から自動的に排除して、Hoechstおよびベータ−チューブリンを重複する細胞体のみを分析する。プレートの各ウェルで5個の別々の350μm2フィールドに関して二重発光画像を獲得する。10倍対物を用いて、これにより結果的にウェルあたり400−500セルが分析される。次いでExtended Neurite Outgrowth Bioapplicationにより細胞あたりの神経突起の神経突起長数、細胞体面積、ならびに分岐および交差点を含む単一の細胞に関するニューロン形態のいくつかの定量的測定が報告される。ニューロンあたりの平均神経突起長(μm)を10回反復でウェルあたり500ニューロンに関して計算する。
【0083】
前記の神経突起伸長アッセイでは、抗LINGO−1 IgG4抗体4784および4785は1および10nMで脱抑制性であるが、一方リゾチームに対する対照IgG4抗体は双方の濃度で脱抑制を示さない(図2)。双方の濃度での4784および4785存在下で脊髄ミエリン上のニューロンあたりの神経突起の平均の長さは抗体不在下のものよりも統計的に長い。抗LINGO−1抗体4784および4785と比較して、ROCK阻害剤Y27632でより大きなレベルの脱抑制が達成されることが予期されるが、この化合物がNgR受容体複合体を介してシグナル発生するもの以外のさらなるミエリン関連神経突起伸長阻害剤のシグナル発生経路を阻止するためである。
【0084】
前記の結果を確認するために、神経突起伸長アッセイを繰り返す(図3)。再度抗LINGO−1抗体4784および4785が1nMおよび10nMで脱抑性であるが、一方リゾチームに対する対照IgG4抗体は双方の濃度で脱抑制を示さない。双方の濃度で4784および4785存在下で脊髄ミエリン上のニューロンあたりの神経突起の平均の長さは抗体不在下のものよりも統計的に長い。
【0085】
抗LINGO−1抗体4784および4785の効力をさらに確立するために、nM以下の濃度の抗体の神経突起伸長阻止に及ぼす影響を評価する(図4)。4784および4785により0.1nMほどの低濃度で脊髄ミエリンの有意な脱抑制(各々38−51%および51−57%)が示されるが、一方対照抗リゾチーム抗体は影響を及ぼさない。予期されるように、再度ROCK阻害剤Y27632により、抗LINGO−1 IgG4抗体よりも高度な脱抑制(65−74%)が示される。
【実施例5】
【0086】
一次オリゴデンドロサイト分化アッセイ
遺伝的手段による、または受容体アンタゴニストでの処理によるLINGO−1機能の遮断により、精製されたOPC培養物から生じる成熟オリゴデンドロサイトの比率が増大することが報告されている(Mi et al., Nat Neurosci 8:745−751(2005))。抗LINGO−1抗体がOPC培養物におけるLINGO−1機能を遮断し、そしてオリゴデンドロサイト成熟化を促進する能力を評価するために、新たに単離されたラットOPCをDMEM/CNTF/T3培地中4784、4785または対照IgG4 3207と共に3日間インキュベートし、続いて抗O4抗体で染色して未成熟および成熟双方のオリゴデンドロサイトを標識する(図5)。オリゴデンドロサイト成熟の程度を、成熟形態を呈するO4陽性細胞の比率として測定する。
【0087】
OPCの富化集団をOFA P3ラットから単離する。簡単には、脳を切開し、そして終脳を0.15%MgSO4含有氷冷Hankバッファー生理食塩水(HBSS、Invitrogen)中に置く。組織を1:1 HBBS/トリプシン−EDTA(Invitrogen)および100μg/ml DNAse I(Roche)と共に37℃で10分間インキュベートし、そしてFCS(Invitrogen)を最終濃度10%で添加してトリプシンを不活化する。組織懸濁液を890rpmで10分間遠心し、そしてペレットを10%ウマ血清(Invitrogen)を含むBasal Medium Eagle(BME、Invitrogen)に再懸濁する。懸濁液を、40μmセルストレーナー(BD Falcon)を通してろ過し、そして細胞をポリ−D−リジンプレコーティングされた80cm2組織培養フラスコ(BD Falcon)にフラスコあたり脳1個で置く。細胞をBME/10%ウマ血清中37℃で11日間培養する。5mM L−ロイシン−メチルエステルを添加することにより小グリア細胞を死滅させ、そしてフラスコを140rpmで2時間振盪することにより攪拌する。フラスコを200rpmで、37℃で一晩振盪することによりOPCを収集し、そして10cm細菌培養皿上で37℃で2時間予め付着させることにより、上澄に残る任意のアストロサイトをOPCからさらに分離する。非接着細胞を集め、890rpmで10分間遠心し、そしておよそ3×104セル/ウェルでポリ−D−リジンコーティングされた8ウェルチャンバースライド(BD Falcon)に置く。10ng/ml Ciliary Neurotrophic Factor(R&D Systems)および15nM トリヨードチロニン(Sigma)を含有するDMEM(Invitrogen)からなるDMEM/T3/CNTF培地中、または10μg/ml トランスフェリン(Sigma)、10μg/mlインスリン(Sigma)、100μMプトレシン(Sigma)、200nMプロゲステロン(Sigma)、520nMチロキシン(Sigma)、500pMトリヨードチロニン(Sigma)、220nM亜セレン酸ナトリウム(Sigma)、25μg/mlゲンタマイシン(Sigma)および1%HS(Invitrogen)を含有するDMEM(Invitrogen)からなるSATO培地中のいずれかで3日間培養物を維持する。オリゴデンドロサイト系列に関して培養物の純度を評価するために、抗O4抗体で染色された細胞のパーセンテージをSATO培地中7日の培養の後に定量する。典型的には細胞の80−95%が抗O4抗体で染色され、それは培養物中の細胞の大部分がオリゴデンドロサイト系列であることを実証している。
【0088】
オリゴデンドロサイト形態に基づいてオリゴデンドロサイト成熟化を評価するために、新たに単離されたOPC培養物をDMEM/T3/CNTF培地中、100nM 4784、4785または対照IgG4 3207の不在下または存在下で3日間インキュベートし、続いて抗O4抗体で染色して未成熟および成熟の双方のオリゴデンドロサイトを標識し、そしてDAPIで細胞核を標識する。明確に定義された短い突起を有するO4陽性細胞は未成熟オリゴデンドロサイトを表すが、一方ミエリンシート様構造を伴い、伸長し、そして高度に分枝した突起を担持するO4陽性細胞は成熟オリゴデンドロサイトを表すと考えられる。成熟形態を有するO4陽性細胞の比率を処理あたり3検体ずつ、ほぼ300−1300セルに関して定量し、そして対照IgG4 3207の存在下の成熟オリゴデンドロサイトの比率に対するHolm−Sidak比較を伴う一元配置分散分析を用いて有意性を決定する。培養物中の全(未成熟および成熟)オリゴデンドロサイトの比率に及ぼす抗体処理の影響を評価するために、O4染色に関連するDAPI核の比率を定量する。
【0089】
3つの独立した実験では、抗LINGO−1抗体4784および4785での処理は、広い面積にわたって伸長する高度に分枝した突起およびミエリンシート様構造を担持する細胞により表されるような成熟形態のオリゴデンドロサイトの比率を有意に増大させる(図5)。対照IgG4抗体3207での処理は培養物中の成熟オリゴデンドロサイトの比率に影響を及ぼさない。O4染色に関連するDAPI染色された核の比率は全処理に関して類似し、抗LINGO−1抗体が未成熟および成熟の双方のオリゴデンドロサイトに対応する細胞の比率に影響を及ぼさないことが実証される。
【0090】
抗LINGO−1抗体処理は全オリゴデンドロサイトの比率に影響を及ぼさないが、成熟オリゴデンドロサイトの比率の増大は、OPCの未成熟オリゴデンドロサイトへの分化の率における増大よりもむしろ未成熟オリゴデンドロサイトの成熟オリゴデンドロサイトへの分化の率の増大のために生じる可能性が最も高い。
【実施例6】
【0091】
細胞表面LINGO−1の抗LINGO−1抗体媒介の下方調節
多価抗体の細胞表面標的に対する結合は抗体:標的複合体の内部移行およびその後のエンドサイトーシス経路内での標的の分解に至り得る(Weinmann et al., Mol Cell Neurosci 32:161−173(2006))。
抗LINGO−1抗体の細胞表面LINGO−1の量に及ぼす影響を決定するために、トランスフェクトされていないCHO−K1またはCHO−K1−hLINGO−1細胞(実施例1参照)を100nM 4784、4785または3207と共に37℃で24時間インキュベートし、そしてその後細胞表面LINGO−1を抗V5抗体、続いて1−Step(商標)Turbo TMB−ELISAキット(Pierce)で展開される抗マウスIgG(Fc特異的)−POD抱合体で検出する(図6A)。
【0092】
CHO−K1−hLINGO−1細胞における細胞表面LINGO−1の量は、抗LINGO−1抗体4784および4785を伴う24時間のインキュベーションの後に有意に低下するが、一方対照IgG4 3207を伴うインキュベーションは影響を及ぼさない。加えて4785でのインキュベーションにより細胞表面LINGO−1は4784よりも大きい程度まで低下する。
【0093】
抗LINGO−1抗体の細胞表面LINGO−1の分解に及ぼす影響を評価するために、トランスフェクトされていないCHO−K1またはCHO−K1−hLINGO−1細胞上の細胞表面タンパク質を記載されるように4℃でビオチン化し(Walmsley et al., J Cell Sci 117:4591−4602(2004))、そして細胞を37℃で180分の期間にわたって種々の時間に100nM 4784、4785または3207を伴って、または伴わずにインキュベートする(図6B)。インキュベーション期間の終わりにアガロースビーズに結合させた抗V5抗体を用いて細胞ライゼートからLINGO−1を免疫沈殿し、そして沈殿物中のビオチン化LINGO−1をウェスタンブロット分析により抗ビオチン抗体(Sigma)を用いて検出する。
【0094】
ビオチン化(およびしたがって細胞表面)LINGO−1に対応するバンドの強度は、抗体を伴わずに、または対照IgG4 3207と共にインキュベートされた細胞においてよりも、抗LINGO−1抗体4784および4785と共にインキュベートされたCHO−K1−hLINGO−1細胞において、より急速に減少する。加えて4785とのインキュベーションにより細胞表面LINGO−1の分解の率は4784よりも大きい程度まで増大する。
【0095】
これらの結果により累積的に、抗LINGO−1抗体4784および4785が、最も可能性が高いのは内部移行およびタンパク質の分解を増強することにより、細胞表面でLINGO−1を有意に下方調節することが示される。この特性はLINGO−1機能の遮断におけるこれらの抗体の効率に寄与すると予期される。
【実施例7】
【0096】
酵素結合免疫吸着アッセイ(ELISA)およびFACS技術
ヒト組換えLINGO−1−Fc融合タンパク質を、直接固定されたヤギ抗ヒトIgG Fc抗体を介するFc部分の捕捉により間接的にMaxisorpプレート96または384ウェルに室温で1時間固定する(PBS中10μg/mlでコーティングされた100μlまたは20μl)。
PBS中5μg/mlの抗原20μlのコーティングの後、ウェルをPBS/0.05%Tween(PBS−T)/5%粉乳で、室温で1時間遮断する。ウェルをPBS−T BEL抽出物で洗浄した後、精製されたFabまたは対照IgGをPBSで希釈し、ウェルに加え、そして室温で1時間インキュベートする。一次抗体を検出するために、以下の二次抗体を適用する:アルカリ性ホスファターゼ(AP)抱合AffiniPureヤギF(ab’)2フラグメント抗ヒトIgGまたは抗マウスIgG(Jackson ImmunoResearch)。AP抱合体の検出のためにAttoPhos(Roche)のような蛍光発生基質を製造者の説明書にしたがって使用する。全インキュベーション工程の間に、マイクロタイタープレートのウェルをPBS−Tで5回、および二次抗体との最終インキュベーションの後に5回洗浄する。TECAN Spectrafluorプレートリーダーで蛍光を測定する。
【0097】
トランスフェクトされたCHO−K1細胞の細胞表面上に発現されたLINGO−1に結合する抗体のFACS分析
丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)中ウェルあたり2×105セルで全染色を実施する。各々の細胞系の細胞をPBS/3%FCS/0.02%NaN3(FACSバッファー)に再懸濁し、そしてa)周辺質抽出物もしくはBELライゼートからの抗体またはb)精製されたFabフラグメントまたはc)FACSバッファーで希釈された、精製されたIgGと混合し、そして4℃で30−60分間インキュベートする。次いで細胞をFACSバッファー/ウェル150μlで1回洗浄し、そしてFACSバッファー中1:200で希釈されている、フィコエリスリン標識された二次抗体(R−PE抱合されたヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに取る。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSCalibur(商標)またはFACSArray(商標)(Becton Dickinson)においてFL2蛍光強度により測定する。
【0098】
LINGO−1特異的抗体の同定のために、CHO−K1−cmLINGO−1またはCHO−K1−rLINGO−1を用いて並行して染色を行う。トランスフェクトされていないCHO−K1細胞をさらなる対照として提供する。カニクイザルおよびラットLINGO−1発現細胞は、これらの種オルソログがヒトLINGO−1タンパク質と数個のアミノ酸しか異ならないので、スクリーニングのために選択される。これらのクローンのみが、トランスフェクトされていないCHO−K1細胞において陰性であり、そしてLINGO−1発現細胞系においてバックグラウンドを超えて≧5倍であるLINGO−1特異的であると判断される。ヒトLINGO−1およびその他のオルソログ(カニクイザルLINGO−1、ラットLINGO−1)に対する、およびヒトLINGO−2パラログに対する交差反応性を逐次的に試験する。
【0099】
配列分析の後、FACS分析で細胞表面に発現されたヒトLINGO−1に対して強力な結合を示す31個の独特なクローンが同定される。12個の結合剤はELISAにおいて捕捉されたヒトLINGO−1−Fcに対する強力な結合を示し(シグナル:ノイズ比は10:1より大きい)、そして7個はELISAにおいて中程度の結合を示す(シグナル:ノイズ比は5:1より大きい)。結合剤の4個はELISAにおいて捕捉されたヒトNgR−Fc融合タンパク質(R&D Systems)に対する強力な結合を示し、そして中断される。結合剤の別の3個はLINGO−1の3つの種の全てと交差反応せず、そして中断される。ヒト/カニクイザル/ラットLINGO−1と交差反応するが、ヒトNgR−Fcとは交差反応しない残りの24個のクローンを発現させ、精製し、そしてLINGO−1のNgRに対する結合を有意に阻止する能力に関して試験し(図1参照)、そしてインビトロで脊髄ミエリンの神経突起伸長阻止活性を脱抑制し(図2−4参照)、さらなる分析のためにFab4784および4785の選択に至る。ELISAでは、4784および4785は捕捉されたヒトLINGO−1−Fcに結合するが、無関係のFc対照と比較して、ヒトLINGO−1−ΔLRR−FcまたはヒトNgR−Fcに対する結合は観察されない(表1および図7参照)。これは4784および4785がLINGO−1のLRR領域(残基66−353)内にあるエピトープを有することを示している。
【0100】
表1:ELISAによる抗LINGO−1 Fabの特徴付け
【表1】
ELISA分析に関する値は相対蛍光単位の平均値として与えられる。
【0101】
FACS飽和分析を用いる選択された抗LINGO−1 Fabの親和性決定
抗LINGO−1特異的抗体の細胞基盤の親和性をFACS飽和結合実験により決定する。染色するために試料中に存在する抗原の濃度は見かけのKD値に影響するので、FACS飽和結合実験における抗原濃度を低下させるために2×105セル/ウェルと対比して1.25×104セル/ウェルのみを用いる。その他の点は前記されたFACS染色手順と同一に染色手順を行う。
【0102】
詳細には、CHO−K1−hLINGO−1、CHO−K1−cmLINGO−1またはCHO−K1−rLINGO−1をベルセンにより培養フラスコから剥がし、FACSバッファーで洗浄し、そしてFACSバッファーに再懸濁する。精製された抗LINGO−1 FabをFACSバッファーで連続希釈し、そして丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)に広げる。各濃度に関して、2検体ずつのウェルを1.25×104セルと共に全容量100μlで、氷上で30−60分間インキュベートする。FACSバッファー150μlを適用し、そして400×gで5分間遠心することによる洗浄工程の後、細胞ペレットをフィコエリスリン標識二次抗体(R−PE抱合ヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに再懸濁し、それをFACSバッファーで1:200に希釈する。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSArray(商標)(Becton Dickinson)において細胞のFL2蛍光強度により測定する。見かけのKD値/EC50値をGraphPad Prism v3.03ソフトウェアまたはGraphPad Prism v4.03を用いて飽和結合曲線から決定し、非線形回帰曲線フィットに適用する。
【0103】
このアッセイを用いて以下の見かけのKD値を決定することができる(表2)。Fab形式では、クローン4784はヒトLINGO−1、カニクイザルLINGO−1およびラットLINGO−1に対してかなり弱い親和性を有する(各々14.07nM、27.11および24.03nM)。しかしながらクローン4784はFab形式でヒトLINGO−2に結合しない。Fab形式では、クローン4785はヒトLINGO−1、カニクイザルLINGO−1およびラットLINGO−1に対してナノモル以下の結合親和性(すなわち見かけのKD値は1×10−9M未満である)を示す。クローン4785はFab形式で低ナノモルからナノモル以下の親和性でヒトLINGO−2に対して交差反応性を示す。LINGO−2機能および分布は依然未知であるので、LINGO−2に対する交差反応性の結果は記載している時点で評価できない。しかしながら有利な影響は排除できない。
【0104】
表2:CHO−K1細胞により発現されたLINGO−1またはLINGO−2抗LINGO−1 Fabに対する見かけのKD値
【表2】
与えられた値は見かけのKD値の平均値(nM)である。nbは結合しない。
【実施例8】
【0105】
HuCAL(登録商標)のIgG4クローニング、発現および精製
IgG形式への転換
全長免疫グロブリン(Ig)を発現するために、重(VH)および軽鎖(VL)の可変ドメインフラグメントをpMORPH(登録商標)X9 MH(配列番号39)Fab発現ベクターからヒトIgG4のためのpMORPH(登録商標) h Ig(配列番号40−42)またはpMORPH(登録商標)2 h Ig(配列番号43−45)ベクターシリーズのいずれかにサブクローニングする。
【0106】
制限酵素EcoRI、MfeIおよびBlpIを用いてVHドメインフラグメントのpMORPH(登録商標) h IgG4(配列番号40)にサブクローニングする:EcoRI/BlpI消化および6400bpフラグメントの抽出によりベクター骨格を作成するが、一方MfeIおよびBlpIでの消化およびその後の精製によりVHフラグメント(350bp)を生成する。ベクターおよびインサートを、各々EcoRIおよびMfeI消化により作成された適合するオーバーハングを介して、およびBlpI部位を介してライゲートする。それによりEcoRIおよびMfeI制限部位の双方が破壊される。
【0107】
制限酵素MfeIおよびBlpIを用いて、VHドメインフラグメントをpMORPH(登録商標)2 h IgG4(配列番号43)にサブクローニングする。このIgGベクターの新たな作成では、その他の修飾時にEcoRI部位(それは適合するオーバーハングを介するサブクローニングのみを可能にする)はMfeI部位により置き換えられ、故にベクターおよびインサートの双方のMfeI/BlpI消化が可能になる。
VLドメインフラグメントのpMORPH(登録商標) h Igκ(配列番号42)およびpMORPH(登録商標)2 h Igκ(配列番号45)へのサブクローニングをEcoRVおよびBsiWI部位を介して実施するが、一方pMORPH(登録商標) h Igλ(配列番号41)およびpMORPH(登録商標)2 h Igλ2(配列番号43)へのサブクローニングをEcoRVおよびHpaIを用いて行う。
【0108】
ヒトIgGの一過性の発現および精製
HEK293細胞を等モル量のIgG重および軽鎖発現ベクターでトランスフェクトする。トランスフェクション後4または5日に、細胞培養上澄を収集する。上澄のpHを8.0に調整し、そして滅菌ろ過した後、溶液を標準的なプロテインAカラムクロマトグラフィー(Poros 20A、PE Biosystems)に供する。
【実施例9】
【0109】
FACS飽和分析を用いる選択された抗LINGO−1 IgG4の親和性決定
抗LINGO−1特異的抗体の細胞基盤の親和性をFACS飽和結合実験により決定する。見かけのKD値の決定は、抗LINGO−1Fab抗体を用いて前記された手順と同一に実施する。
詳細には、CHO−K1−hLINGO−1、CHO−K1−cmLINGO−1またはCHO−K1−rLINGO−1をベルセンにより培養フラスコから剥がし、FACSバッファーで洗浄し、そしてFACSバッファーに再懸濁する。精製された抗LINGO−1 IgG4をFACSバッファーで連続希釈し、そして丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)に広げる。各濃度に関して、2検体ずつのウェルを1.25×104セルと共に全容量100μlで、氷上で30−60分間インキュベートする。FACSバッファー150μlを適用し、そして400×gで5分間遠心することによる洗浄工程の後、細胞ペレットをフィコエリスリン標識二次抗体(R−PE抱合ヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに再懸濁し、それをFACSバッファーで1:200に希釈する。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSArray(商標)(Becton Dickinson)において細胞のFL2蛍光強度により測定する。見かけのKD値/EC50値をGraphPad Prism v3.03ソフトウェアまたはGraphPad Prism v4.03を用いて飽和結合曲線から決定し、非線形回帰曲線フィットに適用する。このアッセイを用いて以下の見かけのKD値を決定することができる(表3)。
【0110】
pMORPH(登録商標)2 h Igベクターシリーズを用いて生成された4784および4785 IgG4抗体の親和性を表3に示す。IgG4形式の4784および4785はヒト、カニクイザルおよびラットLINGO−1に対して明らかに1nMを下まわる見かけのKD値を有する。4784は4785よりもヒトLINGO−2に対してはるかに低い交差反応を有する。
【0111】
表3:CHO−K1細胞により発現されたLINGO−1またはLINGO−2に対する抗LINGO−1 IgG4の見かけのKD値
【表3】
与えられた値は見かけのKD値の平均値(nM)である。
【実施例10】
【0112】
FACS分析を用いる選択された抗LINGO−1 IgG4のヒトLINGO−1に対する結合に及ぼすヒト脳脊髄液の影響
抗LINGO−1 IgG4のヒトLINGO−1に対する結合に及ぼすヒト脳脊髄液の影響をFACS飽和結合実験により試験する。4784および4785の連続希釈物を調製する。CHO−K1−hLINGO−1に対する結合を50%ヒト脳脊髄液の存在下で試験する。前記されたFACS染色にしたがって細胞をヒトCSFの存在下、これらのIgG4抗体で染色する。
【0113】
詳細には、CHO−K1−hLINGO−1をベルセンにより培養フラスコから剥がし、FACSバッファーで洗浄し、そしてFACSバッファーに再懸濁する。精製された抗LINGO−1 IgG4をFACSバッファー+50%ヒト血清で連続希釈し、そして4℃で60分間インキュベートする。対照としてIgG4形式の候補結合剤の連続希釈物をFACSバッファー中、ヒト脳脊髄液のタンパク質含量に類似する2.6%BSAと共に4℃で60分間インキュベートする。インキュベーションの後、連続希釈物を丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)に広げる。各濃度に関して、2検体ずつのウェルを1.25×104セルと共に全容量100μlで、氷上で30−60分間インキュベートする。FACSバッファー150μlを適用し、そして400×gで5分間遠心することによる3回の洗浄工程の後、細胞ペレットをフィコエリスリン標識二次抗体(R−PE抱合ヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに再懸濁し、それをFACSバッファーで1:200に希釈する。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSArray(商標)(Becton Dickinson)において細胞のFL2蛍光強度により測定する。見かけのKD値/EC50値をGraphPad Prism v3.03ソフトウェアまたはGraphPad Prism v4.03を用いて飽和結合曲線から決定し、非線形回帰曲線フィットに適用する。
【0114】
このアッセイを用いて50%ヒト脳脊髄液の影響を対照と比較することができる(表4)。50%ヒト脳脊髄液中でのインキュベーションは差次的に影響する全ての結合剤との結合親和性の低下に至る。ヒト脳脊髄液の存在により結合親和性に及ぼす最強の影響力が4784に関して認められ、それは0.43nMから1.57nMの、73%までの親和性の低下を示す。
【0115】
表4:CHO−K1細胞により発現されたLINGO−1に対する抗LINGO−1 IgG4の見かけのKD値に及ぼすヒト脳脊髄液の影響
【表4】
与えられた値は見かけのKD値の平均値(nM)である。
【実施例11】
【0116】
FACS分析を用いる選択された抗LINGO−1 IgG4のヒトLINGO−1に対する結合に及ぼすヒト血清の影響
抗LINGO−1 IgG4のヒトLINGO−1に対する結合に及ぼすヒト血清の影響をFACS飽和結合実験により試験する。4784および4785の連続希釈物を50容量/容量%ヒト血清の存在下で調製する。細胞を60分間インキュベートした後、前記されたFACS染色にしたがってこれらのプレインキュベートされたIgG4抗体で細胞を染色する。
【0117】
詳細には、CHO−K1−hLINGO−1をベルセンにより培養フラスコから剥がし、FACSバッファーで洗浄し、そしてFACSバッファーに再懸濁する。精製された抗LINGO−1 IgG4をFACSバッファー+50%ヒト血清で連続希釈し、そして4℃で60分間インキュベートする。対照としてIgG4形式の候補結合剤の連続希釈物をFACSバッファー+2.6%BSA(ヒト血清のタンパク質含量に類似する)中でインキュベートするか、またはFACSバッファー単独中、4℃で60分間インキュベートする。インキュベーションの後、連続希釈物を丸底96ウェルマイクロタイタープレート(NUNC(商標)、Wiesbaden, Germany)に広げる。各濃度に関して、2検体ずつのウェルを1.25×104セルと共に全容量100μlで、氷上で30−60分間インキュベートする。FACSバッファー150μlを適用し、そして400×gで5分間遠心することによる3回の洗浄工程の後、細胞ペレットをフィコエリスリン標識二次抗体(R−PE抱合ヤギ抗ヒトIgG(H+L)(Jackson ImmunoResearch)100μlに再懸濁し、それをFACSバッファーで1:200に希釈する。4℃で30−60分間インキュベートした後、細胞をFACSバッファーで1回洗浄し、FACSバッファー100μlに再懸濁し、そしてLINGO−1特異的抗体の結合をFACSArray(商標)(Becton Dickinson)において細胞のFL2蛍光強度により測定する。見かけのKD値/EC50値をGraphPad Prism v3.03ソフトウェアまたはGraphPad Prism v4.03を用いて飽和結合曲線から決定し、非線形回帰曲線フィットに適用する。
【0118】
このアッセイを用いて50%ヒト血清中でのプレインキュベーションの影響を対照と比較することができる(表5)。ヒト血清の存在下での1時間のインキュベーションは4784および4785のKD値に影響しない。したがってこれらの抗体はこの時間にわたって血清中で安定であり、さらにそのKDは変化しないので、血清構成要素とは交差反応しないと思われる。
【0119】
表5:CHO−K1細胞により発現されたLINGO−1に対する抗LINGO−1 IgG4の見かけのKD値に及ぼすヒト血清の影響
【表5】
与えられた値は見かけのKD値の平均値(nM)である。
【0120】
配列表と簡単な説明
配列番号1
ラット成熟LINGO−1外部ドメイン(残基34−550)
【化1】
【0121】
配列番号2
カニクイザル成熟LINGO−1外部ドメイン(残基34−550)
【化2】
【0122】
配列番号3
ヒト成熟LINGO−1外部ドメイン(残基34−550)
【化3】
【0123】
配列番号4
4784VL
【化4】
【0124】
配列番号5
4784VH
【化5】
【0125】
配列番号6
4785VL
【化6】
【0126】
配列番号7
4785VH
【化7】
【0127】
配列番号8
DNA−4784VH
【化8】
【0128】
配列番号9
DNA−4785VH
【化9】
【0129】
配列番号10
DNA−4784VL
【化10】
【0130】
配列番号11
DNA−4785VL
【化11】
【0131】
配列番号12
抗体4784CDR−H1
【化12】
【0132】
配列番号13
抗体4784CDR−H2
【化13】
【0133】
配列番号14
抗体4784CDR−H3
【化14】
【0134】
配列番号15
抗体4784CDR−L1
【化15】
【0135】
配列番号16
抗体4784CDR−L2
【化16】
【0136】
配列番号17
抗体4784CDR−L3
【化17】
【0137】
配列番号18
抗体4785CDR’−H1
【化18】
【0138】
配列番号19
抗体4785CDR’−H2
【化19】
【0139】
配列番号20
抗体4785CDR’−H3
【化20】
【0140】
配列番号21
抗体4785CDR’−L1
【化21】
【0141】
配列番号22
抗体4785CDR’−L2
【化22】
【0142】
配列番号23
抗体4785CDR’−L3
【化23】
【0143】
配列番号24
正方向プライマーDM22
【化24】
【0144】
配列番号25
逆方向プライマーDM23
【化25】
【0145】
配列番号26
APタグ−5−NHISベクター
【化26】
【化27】
【0146】
配列番号27
ヒトLINGO−1成熟DNA配列
【化28】
【0147】
配列番号28
カニクイザルLINGO−1成熟DNA配列
【化29】
【0148】
配列番号29
ラットLINGO−1成熟DNA配列
【化30】
【0149】
配列番号30
正方向プライマーDM14
【化31】
【0150】
配列番号31
逆方向プライマーDM15
【化32】
【0151】
配列番号32
pSecTag2−V5ベクター
【化33】
【化34】
【0152】
配列番号33
ヒトLINGO−2成熟DNA配列
【化35】
【0153】
配列番号34
正方向プライマーDM16
【化36】
【0154】
配列番号35
逆方向プライマーDM17
【化37】
【0155】
配列番号36
pRS5a−IgG
【化38】
【化39】
【化40】
【0156】
配列番号37
天然リーダー(natleader)−hsLINGO−1−Fc/pRS5a
【化41】
【化42】
【化43】
【化44】
【0157】
配列番号38
Igリーダー−hsLINGO−1−ΔLRR−Fc/pRS5a
【化45】
【化46】
【化47】
【化48】
【0158】
配列番号39
Fab発現ベクターpMORPH(登録商標)X9 MH
【化49】
【化50】
【化51】
【化52】
【0159】
配列番号40
IgG4発現ベクターpMORPH(登録商標)h Igγ4
【化53】
【化54】
【化55】
【化56】
【化57】
【0160】
配列番号41
IgGラムダ鎖発現ベクターpMORPH(登録商標)h Ig ラムダ
【化58】
【化59】
【化60】
【化61】
【0161】
配列番号42
IgGカッパ鎖発現ベクターpMORPH(登録商標)h Ig カッパ
【化62】
【化63】
【化64】
【化65】
【0162】
配列番号43
IgG4発現ベクターpMORPH2(登録商標) h Igγ4
【化66】
【化67】
【化68】
【化69】
【化70】
【化71】
【0163】
配列番号44
IgGラムダ鎖発現ベクターpMORPH(登録商標)2 h Ig ラムダ2
【化72】
【化73】
【化74】
【化75】
【化76】
【0164】
配列番号45
IgGカッパ鎖発現ベクターpMORPH(登録商標)2 hIg カッパ
【化77】
【化78】
【化79】
【化80】
【化81】
【特許請求の範囲】
【請求項1】
解離定数<1000nMで配列番号1、配列番号2または配列番号3によるタンパク質に結合することができる結合分子。
【請求項2】
配列番号46−51からなる群から選択される1つまたはそれより多い配列に結合する請求項1に記載の結合分子。
【請求項3】
20nM未満の濃度で脊髄ミエリンを脱抑制することができる請求項1または2に記載の結合分子。
【請求項4】
ラット成体脊髄ミエリンの基質上で成長させたラット小脳顆粒細胞の細胞あたりの平均神経突起長を少なくとも20%まで増加させることができる請求項1、2または3に記載の結合分子。
【請求項5】
配列番号12−17または配列番号18−23からなる群から選択される1つまたはそれより多いアミノ酸配列を含む請求項1、2、3または4のいずれかに記載の結合分子。
【請求項6】
配列番号5または配列番号7に少なくとも50%相同である配列;および
配列番号4または配列番号6に少なくとも50%相同である配列、またはその直接的な均等物;
からなる群から選択される少なくとも1つの抗原結合部位を含む請求項5に記載の結合分子。
【請求項7】
配列番号5または配列番号7に少なくとも50%相同である第1の配列および配列番号4または配列番号6に少なくとも50%相同である第2の配列またはその直接的な均等物を含む請求項6に記載の結合分子。
【請求項8】
少なくともa)
(i)配列番号5または配列番号7を含む可変ドメイン、および
(ii)ヒト重鎖の定常部分またはそのフラグメント;
を含む1つの免疫グロブリン重鎖またはそのフラグメントならびに
b)
(i)配列番号4または配列番号6を含む可変ドメイン、および
(ii)ヒト軽鎖の定常部分またはそのフラグメント;
を含む1つの免疫グロブリン軽鎖またはそのフラグメント;または
その直接的な均等物を含む請求項1−7のいずれかに記載の結合分子。
【請求項9】
抗体もしくはそのフラグメントまたはその直接的な均等物である請求項1−8のいずれかに記載の結合分子。
【請求項10】
ヒト重鎖の定常部分またはそのフラグメントがγ4型のものであり、そしてヒト軽鎖の定常部分またはそのフラグメントがκ型のものである請求項9に記載の結合分子。
【請求項11】
ヒトまたはキメラまたはヒト化モノクローナル抗体である請求項1−9のいずれかに記載の結合分子。
【請求項12】
請求項1−9のいずれかに記載の結合分子をコードするポリヌクレオチド。
【請求項13】
配列番号8および配列番号9からなる群から;または配列番号10および配列番号11からなる群から選択されるポリヌクレオチド。
【請求項14】
請求項12または13のいずれかに記載の1つまたはそれより多いポリヌクレオチドを含む発現ベクター。
【請求項15】
該発現系またはその部分が適合する宿主細胞に存在する場合、該発現系またはその部分が請求項1から11のいずれかに記載のポリペプチドを生成することができる請求項12または13のいずれかに記載のポリヌクレオチドを含む発現系。
【請求項16】
請求項15に記載の発現系を含む単離された宿主細胞。
【請求項17】
請求項1から11のいずれかに記載の結合分子の医薬品としての使用。
【請求項18】
CNS傷害の処置のための医薬品の調製における請求項1から11のいずれかに記載の結合分子の使用。
【請求項19】
CNS傷害の処置における請求項1から11のいずれかに記載の結合分子の使用。
【請求項20】
請求項1から11のいずれかに記載の結合分子を少なくとも1つの薬学的に許容される担体または希釈剤と一緒に含む医薬組成物。
【請求項21】
有効量の請求項1から11のいずれかに記載の結合分子をかかる処置を必要とする対象に投与することを含む軸索再生/可塑性の促進に関連する疾患を処置する方法。
【請求項1】
解離定数<1000nMで配列番号1、配列番号2または配列番号3によるタンパク質に結合することができる結合分子。
【請求項2】
配列番号46−51からなる群から選択される1つまたはそれより多い配列に結合する請求項1に記載の結合分子。
【請求項3】
20nM未満の濃度で脊髄ミエリンを脱抑制することができる請求項1または2に記載の結合分子。
【請求項4】
ラット成体脊髄ミエリンの基質上で成長させたラット小脳顆粒細胞の細胞あたりの平均神経突起長を少なくとも20%まで増加させることができる請求項1、2または3に記載の結合分子。
【請求項5】
配列番号12−17または配列番号18−23からなる群から選択される1つまたはそれより多いアミノ酸配列を含む請求項1、2、3または4のいずれかに記載の結合分子。
【請求項6】
配列番号5または配列番号7に少なくとも50%相同である配列;および
配列番号4または配列番号6に少なくとも50%相同である配列、またはその直接的な均等物;
からなる群から選択される少なくとも1つの抗原結合部位を含む請求項5に記載の結合分子。
【請求項7】
配列番号5または配列番号7に少なくとも50%相同である第1の配列および配列番号4または配列番号6に少なくとも50%相同である第2の配列またはその直接的な均等物を含む請求項6に記載の結合分子。
【請求項8】
少なくともa)
(i)配列番号5または配列番号7を含む可変ドメイン、および
(ii)ヒト重鎖の定常部分またはそのフラグメント;
を含む1つの免疫グロブリン重鎖またはそのフラグメントならびに
b)
(i)配列番号4または配列番号6を含む可変ドメイン、および
(ii)ヒト軽鎖の定常部分またはそのフラグメント;
を含む1つの免疫グロブリン軽鎖またはそのフラグメント;または
その直接的な均等物を含む請求項1−7のいずれかに記載の結合分子。
【請求項9】
抗体もしくはそのフラグメントまたはその直接的な均等物である請求項1−8のいずれかに記載の結合分子。
【請求項10】
ヒト重鎖の定常部分またはそのフラグメントがγ4型のものであり、そしてヒト軽鎖の定常部分またはそのフラグメントがκ型のものである請求項9に記載の結合分子。
【請求項11】
ヒトまたはキメラまたはヒト化モノクローナル抗体である請求項1−9のいずれかに記載の結合分子。
【請求項12】
請求項1−9のいずれかに記載の結合分子をコードするポリヌクレオチド。
【請求項13】
配列番号8および配列番号9からなる群から;または配列番号10および配列番号11からなる群から選択されるポリヌクレオチド。
【請求項14】
請求項12または13のいずれかに記載の1つまたはそれより多いポリヌクレオチドを含む発現ベクター。
【請求項15】
該発現系またはその部分が適合する宿主細胞に存在する場合、該発現系またはその部分が請求項1から11のいずれかに記載のポリペプチドを生成することができる請求項12または13のいずれかに記載のポリヌクレオチドを含む発現系。
【請求項16】
請求項15に記載の発現系を含む単離された宿主細胞。
【請求項17】
請求項1から11のいずれかに記載の結合分子の医薬品としての使用。
【請求項18】
CNS傷害の処置のための医薬品の調製における請求項1から11のいずれかに記載の結合分子の使用。
【請求項19】
CNS傷害の処置における請求項1から11のいずれかに記載の結合分子の使用。
【請求項20】
請求項1から11のいずれかに記載の結合分子を少なくとも1つの薬学的に許容される担体または希釈剤と一緒に含む医薬組成物。
【請求項21】
有効量の請求項1から11のいずれかに記載の結合分子をかかる処置を必要とする対象に投与することを含む軸索再生/可塑性の促進に関連する疾患を処置する方法。
【図1】

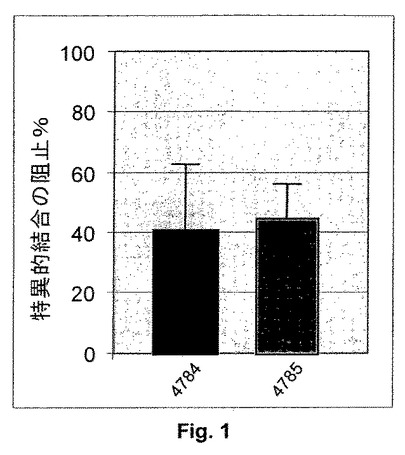
【図2】
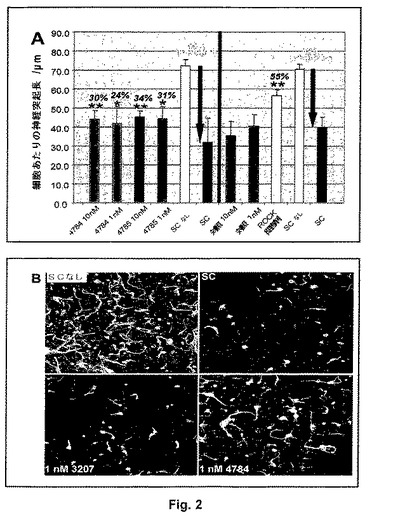
【図3】
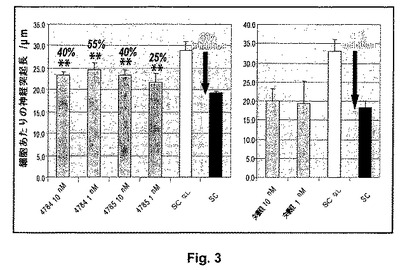
【図4】
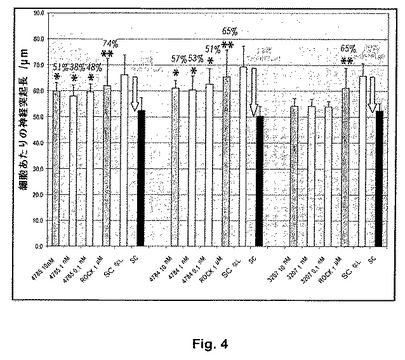
【図5A】
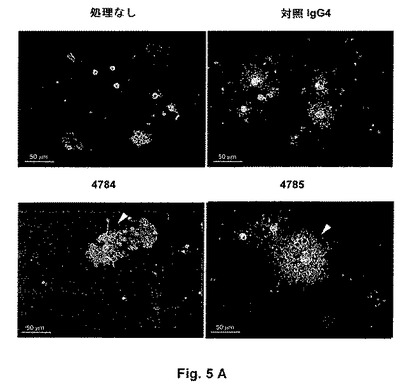
【図5B】
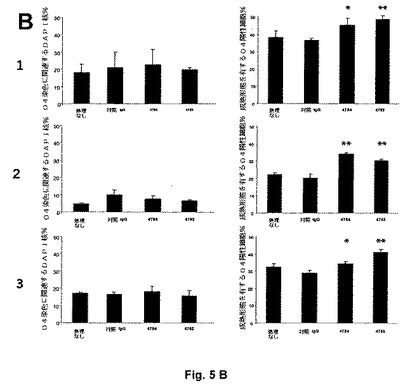
【図6】
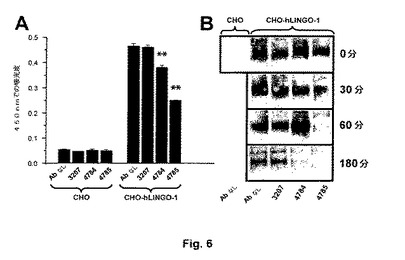
【図7】
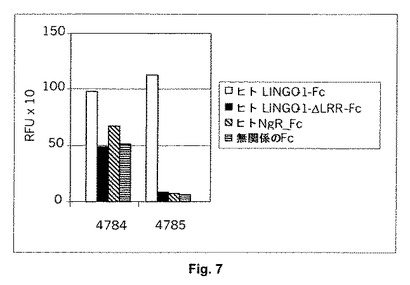
【図2】
【図3】
【図4】
【図5A】
【図5B】
【図6】
【図7】
【公開番号】特開2013−56888(P2013−56888A)
【公開日】平成25年3月28日(2013.3.28)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−222370(P2012−222370)
【出願日】平成24年10月4日(2012.10.4)
【分割の表示】特願2009−536654(P2009−536654)の分割
【原出願日】平成19年11月15日(2007.11.15)
【出願人】(504389991)ノバルティス アーゲー (806)
【Fターム(参考)】
【公開日】平成25年3月28日(2013.3.28)
【国際特許分類】
【出願番号】特願2012−222370(P2012−222370)
【出願日】平成24年10月4日(2012.10.4)
【分割の表示】特願2009−536654(P2009−536654)の分割
【原出願日】平成19年11月15日(2007.11.15)
【出願人】(504389991)ノバルティス アーゲー (806)
【Fターム(参考)】
[ Back to top ]