
LNA修飾化ヘプタマー型スモールガイド核酸
【課題】標的RNAの切断効率を許容範囲内に維持し、かつ、生体安定性に優れたヘプタマー型スモールガイド核酸を提供する。
【解決手段】標的RNAの少なくとも連続5塩基と相補的な塩基配列を含む7塩基からなり、その配列中の1から4塩基がLNA(locked nucleic acid)修飾されており、細胞内においてtRNaseZLの作用により標的RNAを切断することによってこの標的RNAを転写する遺伝子の活性を低下または消滅させるヘプタマー型スモールガイド核酸。
【解決手段】標的RNAの少なくとも連続5塩基と相補的な塩基配列を含む7塩基からなり、その配列中の1から4塩基がLNA(locked nucleic acid)修飾されており、細胞内においてtRNaseZLの作用により標的RNAを切断することによってこの標的RNAを転写する遺伝子の活性を低下または消滅させるヘプタマー型スモールガイド核酸。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、標的として切断するRNAを転写する遺伝子発現を効果的に抑制または消失させ、かつ生体内安定性に優れていることによって、その遺伝子発現等を原因とする疾患の治療または予防に有効なヘプタマー型スモールガイド核酸に関するものである。
【背景技術】
【0002】
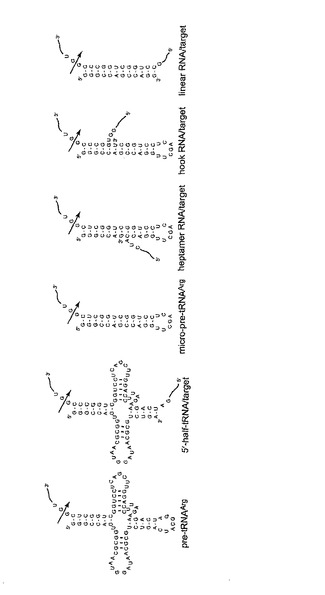
本発明の発明者らは、TRUEジーンサイレンシング(tRNase ZL-utilizing efficacious gene silencing)と名付けた新しい遺伝子発現抑制技術を開発している(特許文献1、非特許文献1−4)。tRNase ZLは、tRNAの3’をプロセシングするエンドリボヌクレアーゼ(tRNase Zまたは3’tRNase)の分子量の大きい種類の酵素であり、前駆体tRNAの3’末端部を切除する(特許文献1、非特許文献5、6)。このTRUEジーンサイレンシングは、標的RNAと合成スモールガイド核酸とによって形成されるpre-tRNA様またはマイクロpre-tRNA様複合体を認識することによって、いかなるRNAの任意箇所をも切断することができるという哺乳動物tRNase ZLのユニークな酵素的特性に基づいている(非特許文献7−13)。スモールガイド核酸(特にスモールガイドRNA)は、以下の4種:5’-half tRNA(非特許文献8)、〜12-16-nt直鎖RNA(非特許文献13)、ヘプタマー型RNA(非特許文献9)およびフック型RNA(非特許文献12)に分けられる(図1)。
【0003】
発明者らは、様々なmRNAを標的とするスモールガイド核酸(以下、sg核酸と記載することがある)を、その発現プラスミドまたは2’-O-メチル RNAsを導入することによって、様々な哺乳動物細胞におけるTRUEジーンサイレンシングの有効性を実証している(非特許文献1−4)。例えば、Jurkat細胞におけるHIV-1発現は、5’-half tRNA型sgRNAによって18日間以上抑制され(非特許文献2)、マウス肝臓におけるルシフェラーゼ発現はヘプタマー型sg核酸によって抑制された(非特許文献3)。さらに、TRUEジーンサイレンシングの効果は、RNA干渉(RNA interference: RNAi)と同等であり(非特許文献3、14)、幾つかの場合ではRNAiの効果を凌ぐことも確認されている(非特許文献4)。
【0004】
また最近、発明者らは、tRNase ZLが核内だけでなくサイトゾルにも存在すること、そしてsg核酸として使用していたのと同一の5’-half tRNAが細胞質に存在することを見出している(非特許文献15)。また、発明者らは、ヒトサイトゾルtRNase ZLが、5’-half tRNAの指揮のもとでmRNAを切断することによって遺伝子発現を調節すること、そしてPPM1F mRNAがtRNase ZLの真の標的の一つであることを見出している(非特許文献15)。さらにまた、発明者らは、miR-103を含むマイクロRNAの一部がフック型sg核酸として働き、サイトゾルtRNase ZLによるmRNA切断を介して遺伝子発現を下方調節しえることを証明している(非特許文献16)。以上のことから、TRUEジーンサイレンシングは、細胞内の小さな非コードRNAによって作動されるサイトゾルtRNase ZLの新たに解明された生理学的役割を基礎として機能していることは明らかである。
【0005】
TRUEジーンサイレンシングの最終的な目的の一つは、特定遺伝子の発現を原因とする疾患治療剤または予防剤としてのsg核酸の使用である。例えば、発明者らは既に、ガン治療の分子標的として有望視されているBcl-2およびVEGFをコードする細胞内mRNAレベルが、5’-half tRNA型sg核酸、14-nt直鎖型sg核酸またはヘプタマー型sg核酸によって発現抑制されることを見出している(非特許文献1、4、17)。
【0006】
sg核酸を薬剤成分とする場合、薬理学的な視点からは、ヘプタマー型sg核酸が最も好ましい。何故ならば、ヘプタマー(7塩基核酸)は長鎖のsg核酸よりも遙かに簡便かつ安価に合成することができる。また、ヘプタマー型sg核酸は、刺激剤(トランスフェクション試薬等)を用いることなく容易に細胞内に取込ませることができる(非特許文献18)。
【0007】
なお、ヘプタマー型sg核酸は、tRNase ZLによる標的RNAの切断誘導能や生体内での安定性の向上を目的として、様々な化学的修飾が検討されている。発明者らはかつて、2’-O-メチル RNA、ホスホロチオエートRNA、DNAおよびホスホロチオエートDNAについて調べた結果、酵素に対する各々の切断誘導能と安定性から、2’-O-メチル修飾したsg核酸がTRUEジーンサイレンシングのためには最も好ましいとの結論を得ている(非特許文献1)。
【0008】
一方、2’位の酸素原子と4’位の炭素原子がメチレンを介して架橋したロックト人工核酸(Locked Nucleic Acid:LNA)の出現が、核酸研究に広範な影響を及ぼしている(非特許文献19、20)。すなわち、LNAは生体安定性と温度安定性の両方を核酸に付与するため、アンチセンスRNAやsiRNAの化学修飾として利用されている。
【特許文献1】特許第3660718号公報
【非特許文献1】Tamura,M., Nashimoto,C., Miyake,N., Daikuhara,Y., Ochi,K. and Nashimoto,M.(2003) Intracellular mRNA cleavage by 3′ tRNase under the direction of 2′-O-methylRNA heptamers. Nucleic Acids Res., 31, 4354-4360.
【非特許文献2】Habu,Y., Miyano-Kurosaki,N., Kitano,M., Endo,Y., Yukita,M., Ohira,S., Takaku,H., Nashimoto,M. and Takaku,H. (2005) Inhibition of HIV-1 gene expression by retroviral vector-mediated small-guide RNAs that direct specific RNA cleavage by tRNase ZL. Nucleic Acids Res., 33, 235-243.
【非特許文献3】Nakashima,A., Takaku,H., Shibata,H.S., Negishi,Y., Takagi,M., Tamura,M. and Nashimoto,M. (2007) Gene-silencing by the tRNA maturase tRNase ZL under the direction of small guide RNA. Gene Therapy, 14, 78-85.
【非特許文献4】Elbarbary,R.A., Takaku,H., Tamura,M. and Nashimoto,M. (2009) Inhibition of vascular endothelial growth factor expression by TRUE gene silencing. Biochem.and Biophys. Res. Commun., 379, 924-927.
【非特許文献5】Nashimoto,M. (1997) Distribution of both lengths and 5′ terminal nucleotides of mammalian pre-tRNA 3′ trailers reflects properties of 3′ processing endoribonuclease. Nucleic Acids Res., 25, 1148-1155.
【非特許文献6】Takaku,H., Minagawa,A., Masamichi,T. and Nashimoto,M. (2003) A candidateprostate cancer susceptibility gene encodes tRNA 3′ processing endoriobonuclease. Nucleic Acids Res., 31, 2272-2278.
【非特許文献7】Nashimoto,M. (1995) Conversion of mammalian tRNA 3′ processing endoribonuclease to four-base-recognizing RNA cutters. Nucleic Acids Res., 23, 3642-3647.
【非特許文献8】Nashimoto,M. (1996) Specific cleavage of target RNAs from HIV-1 with 5′ half tRNA by mammalian tRNA 3′ processing endoribonuclease. RNA, 2, 2523-2524.
【非特許文献9】Nashimoto,M., Geary,S., Tamura,M. and Kasper,R. (1998) RNA heptamers that directs RNA cleavage by mammalian tRNA 3′ processing endoribonuclease. Nucleic Acids Res., 26, 2565-2571.
【非特許文献10】Nashimoto,M. (2000) Anomalous RNA substrates for mammalian tRNA 3′processing endoribonuclease. FEBS Letters, 472, 179-186.
【非特許文献11】Takaku,H., Minagawa,A., Takagi,M. and Nashimoto,M. (2004) The N-terminal half-domain of the long form of tRNase Z is required for the RNase 65 activity. Nucleic Acids Res., 32, 4429-4438.
【非特許文献12】Takaku,H., Minagawa,A., Takagi,M. and Nashimoto,M. (2004) A novel four-base-recognizing RNA cutter that can remove the single 3′ terminal nucleotides from RNA molecules. Nucleic Acids Res., 32, e91.
【非特許文献13】Shibata,H.S., Takaku,H., Takagi,M. and Nashimoto,M. (2005) The T loop structure is dispensable for substrate recognition by tRNase ZL. J. Biol. Chem., 280, 22326-22334.
【非特許文献14】Appasani,K. (2005) RNA interference technology: from basic science to drug development. Cambridge University Press, Cambridge.
【非特許文献15】Elbarbary,R.A., Takaku,H., Uchiumi,N., Tamiya,H., Abe,M., Takahashi,M., Nishida,H. and Nashimoto,M. (2009) Modulation of gene expression by human cytosolic tRNase ZL through 5′-half-tRNA. PLoS ONE, 4, e5908.
【非特許文献16】Elbarbary,R.A., Takaku,H., Uchiumi,N., Tamiya,H., Abe,M., Nishida,H. and Nashimoto,M. (2009) Human cytosolic tRNase ZL can downregulate gene expression through miRNA. FEBS Lett., 583, 3241-3246.
【非特許文献17】Hu,W. and Kavanagh,J.J. (2003) Anticancer therapy targeting the apoptotic pathway. Lancet Oncol., 4, 721-729.
【非特許文献18】Loke,S.L., Stein,C.A., Zhang X.H., Mori,K., Nakanishi,M., Subasinghe,C., Cohen,J.S. and Neckers,L.M. (1989) Characterization of oligonucleotide transport into living cells. Proc. Natl. Acad. Sci. USA, 86, 3474-3478.
【非特許文献19】Koch,T. and Orum,H. (2008) Locked nucleic acid. In Crooke,S.T. (ed.), Antisense Drug Technology. CRC Press, Boca Raton, pp. 519-564.
【非特許文献20】Straarup,E.M., Fisker,N., Hedtjarn,M., Lindholm,M.W., Rosenbohm,C., Aarup,V., Hansen,H.F., Orum,H., Hansen,J.B. and Koch,T. (2010) Short locked nucleic acid antisense oligonucleotides potently reduce apolipoprotein B mRNA and serum cholesterol in mice and non-human primates. Nucleic Acids Res., 38, 7100-7111.
【発明の概要】
【発明が解決しようとする課題】
【0009】
ヘプタマー型sg核酸を薬剤成分とする場合の一つの課題は、生体内に投与された場合にその主効果(tRNase ZLによる標的RNAの切断誘導)が持続的に発揮されるように、生体安定性および温度安定性を向上させることである。このような課題に対して、アンチセンスRNAやsiRNAに対する修飾として実績のあるLNA修飾は有望であると考えられる。
【0010】
しかしながら、LNA修飾によってヘプタマー型sg核酸の主効果(tRNase ZLによる標的RNAの切断誘導)がどのように変容するかは一切知られていない。仮に、LNA修飾によってヘプタマー型sg核酸の主効果が大幅に損なわれることがあれば、LNA修飾による安定性の向上は薬剤成分としてのsg核酸においては好ましいものでない。
【0011】
本願発明は、ヘプタマー型sg核酸の主効果と安定性との観点から、ヘプタマー型sg核酸が生体内でその主効果を最大限に発揮することのできるLNA修飾条件を提供することによって、ヘプタマー型sg核酸の薬剤成分としての価値をさらに向上させることを課題としている。
【課題を解決するための手段】
【0012】
本発明は、前記の課題を解決するものとして、以下の発明を提供する。
(1)標的RNAの少なくとも連続5塩基と相補的な塩基配列を含む7塩基からなり、その配列中の1から4塩基がLNA(locked nucleic acid)修飾されており、細胞内においてtTNaseZLの作用により標的RNAを切断することによってこの標的RNAを転写する遺伝子の活性を低下または消滅させるヘプタマー型スモールガイド核酸。
(2)LNA修飾された塩基以外の塩基が2’-Oメチル化修飾されている前記(1)のヘプタマー型スモールガイド核酸。
(3)標的RNAが疾患に関連する遺伝子から転写されたRNAである前記(1)または(2)のヘプタマー型スモールガイド核酸。
(4)疾患に関連する遺伝子がガンの発症に関連する遺伝子である前記(3)のヘプタマー型スモールガイド核酸。
(5)前記(3)または(4)のヘプタマー型スモールガイド核酸の有効量を含有する薬剤。
【発明の効果】
【0013】
本発明(1)によれば、標的RNAの切断効率を許容範囲内に維持し、かつ、生体内での安定性に極めて優れてたヘプタマー型スモールガイド核酸が提供される。このヘプタマー型スモールガイド核酸は、薬剤成分として生体内に投与した場合に、その優れた安定性により標的RNAの切断効果を最大限に発揮することができる。
【0014】
本発明(2)によれば、LNA修飾塩基以外の塩基がさらに2’-Oメチル化修飾されているため、標的RNAの切断効率がさらに優れたヘプタマー型スモールガイド核酸が提供される。
【0015】
本発明(3)から(5)によれば、例えばガン等の疾患治療または予防のための手段が提供される。
【図面の簡単な説明】
【0016】
【図1】4種類のスモールガイド核酸(左から2、4、5、6番目)の例。各塩基配列のうち、5’NNNN3’の領域がスモールガイド核酸。
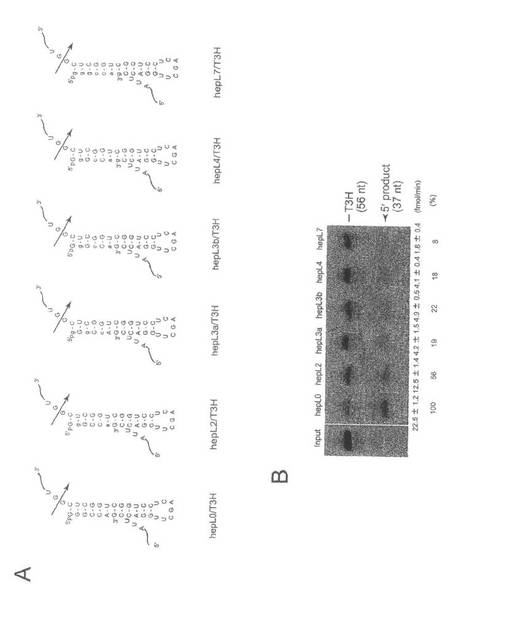
【図2】Aは実施例1で使用したヘプタマー型sgRNA/T3H RNA複合体の2次構造。各塩基配列のうち、5’NNNN3’の領域がプタマー型sgRNA。大文字は2’-Oメチル化修飾塩基、小文字はLNA修飾塩基である。矢印は標的RNAの切断箇所である。Bは変性の10%ポリアクリルアミドゲルでの標的RNA(T3H)の切断産物の電気泳動像。下に切断効率と相対的なパーセンテージを示す。
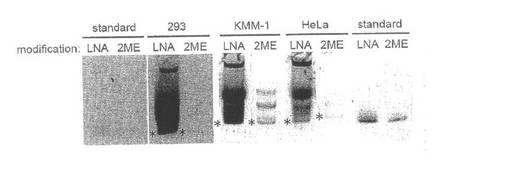
【図3】実施例2の結果である。2ME: すべて2′-O-メチル化された3′-FITC標識hepL0、LNA:2、4、6番目のヌクレオチドをLNA修飾(その他4か所は2′-O-メチル化)した3′-FITC標識hepL3b。*:細胞から分解を受けずに抽出されたヘプタマー型sgRNA。その上のバンドはヘプタマー型sgRNAと細胞内の他のRNAとの複合体であると思われる。
【発明を実施するための形態】
【0017】
本発明(1)のヘプタマー型スモールガイド核酸は、標的RNAの少なくとも連続5塩基と相補的な塩基配列を含む7塩基からなり、その配列中の1から4塩基がLNA(locked nucleic acid)修飾されている。
【0018】
LNAは、2’位の酸素原子と4’位の炭素原子がメチレンを介して架橋した人工核酸であり、例えば、2′-O-4′C-methylene-β-D-ribofuranoseを含む核酸である。
【0019】
標的RNAは、その発現を抑制または消失させる対象となる遺伝子(標的遺伝子)の転写産物RNA、特にmRNAである。標的遺伝子の発現抑制とは、その遺伝子から転写合成されるタンパク質量が、25%以下、好ましくは50%以下、さらに好ましくは75%以下、特に好ましくは95%以下に低減することである。標的遺伝子は、その発現抑制または発現消失が産業上の利用価値を有する遺伝子であり、とくにその遺伝子の発現亢進が特定の疾患の原因となるような遺伝子(以下、病原遺伝子と記載することがある)。そのような病原遺伝子の例としては、例えば、ras、erbb2、myc、apc;brca1、rb、Bcl-2、BGEF癌遺伝子、レニンなどの高血圧に関与する遺伝子、インスリンなどの糖尿病関連遺伝子、LDLレセプターなどの高脂血症関連遺伝子、レプチンなどの肥満関連遺伝子、アンギオテンシンなどの動脈硬化疾患関連遺伝子、痴呆の原因として知られているアポリポタンパク質およびプレセリニン、老化に関連する遺伝子などが例示される。これらの遺伝子およびそのmRNA配列は公知のデータベース(NCBIヌクレオチドデータベース等)において公知である。
【0020】
ヘプタマー型sg核酸は、塩基対合を介して標的RNAとtRNAアクセプターステム様二重鎖を形成することによって、tRNase ZLによる標的RNAの特異的切断を可能とするため、ヘプタマー型sg核酸の標的部位は、tRNA T-armに類似する安定したヘアピン構造領域の直下となり、ヘプタマー型sg核酸はその7塩基だけでなく、安定したヘアピン構造を形成する約12塩基の特異性によって効果的にRNAを切断する。従って、ヘプタマー型sg核酸の塩基配列は、これらの標的遺伝子のmRNA配列情報から決定することができる。
【0021】
配列を決定したヘプタマー型sg核酸は、公知の化学合成を用いる方法、あるいは酵素的転写法等にて製造することができる。公知の化学合成を用いる方法として、ホスホロアミダイト法、ホスフォロチオエート法、ホスホトリエステル法等をあげることができ、例えば、ABI3900ハイスループット核酸合成機(アプライドバイオシステムズ社製)やNTS H-6核酸合成機(日本テクノサービス社製)、Oligoilot10核酸合成機(GEヘルスケア製)により合成することができる。酵素的転写法としては、目的の塩基配列を有するプラスミドまたはDNAを鋳型として、T7、T3、SP6RNAポリメラーゼ等のRNAポリメラーゼを用いた転写をあげることができる。合成法または転写法により製造したヘプタマー型sg核酸は、次いでHPLC等にて精製する。例えばHPLC精製時には、triethylammonium acetate (TEAA)またはhexylammonium acetate(HAA)とacetonitrileの混合溶液を用いて、ヘプタマー型sg核酸をカラムから溶出する。その後、溶出体積の1000倍量の蒸留水で溶出溶液を10時間透析し、透析溶液を凍結乾燥した後、使用時まで冷凍保存する。使用時には蒸留水で最終濃度が100 μM程度になるように溶解する。
【0022】
本発明のヘプタマー型sg核酸に用いられるLNA修飾核酸以外の核酸としては、ヌクレオチドまたはそのヌクレオチドと同等の機能を有する分子が重合した分子であればいかなるものでもよい。ヌクレオチドとしては、例えばリボヌクレオチドの重合体であるRNA、デオキシリボヌクレオチドの重合体であるDNA、RNAおよびDNAが混合した重合体、ヌクレオチド類似体を含むヌクレオチド重合体が、それぞれあげられる。特にRNAが好ましい。
【0023】
ヌクレオチド類似体としては、例えばRNAまたはDNAと比較して、ヌクレアーゼ耐性の向上または安定化させるため、相補鎖核酸とのアフィニティーをあげるため、細胞透過性をあげるため、あるいは可視化させるために、リボヌクレオチド、デオキシリボヌクレオチド、RNAまたはDNAに修飾を施した分子を挙げるこことができる。例えば、糖部修飾ヌクレオチド類似体やリン酸ジエステル結合修飾ヌクレオチド類似体等があげられる。
【0024】
糖部修飾ヌクレオチド類似体とは、ヌクレオチドの糖の化学構造の一部あるいは全てに対し、任意の化学構造物質を付加あるいは置換したものであればいかなるものでもよく、例えば、2’−O−メチルリボースで置換されたヌクレオチド類似体、2’−O−プロピルリボースで置換されたヌクレオチド類似体、2’−メトキシエトキシリボースで置換されたヌクレオチド類似体、2’−O−メトキシエチルリボースで置換されたヌクレオチド類似体、2’−O−[2−(グアニジウム)エチル]リボースで置換されたヌクレオチド類似体、2’−O−フルオロリボースで置換されたヌクレオチド類似体、エチレン架橋構造型人工核酸(Ethylene bridged nucleic acid:ENA)[Nucleic Acid Research, 32, e175 (2004)]等があげられ、さらにペプチド核酸(PNA)[Acc. Chem. Res., 32, 624 (1999)]、オキシペプチド核酸(OPNA)[J. Am. Chem. Soc., 123, 4653 (2001)]、およびペプチドリボ核酸(PRNA)[J. Am. Chem. Soc., 122, 6900 (2000)]等をあげることができる。特に、2’−O−メチルリボースで置換されたヌクレオチド類似体(2’-Oメチル化修飾された核酸)が好ましい。
【0025】
リン酸ジエステル結合修飾ヌクレオチド類似体としては、ヌクレオチドのリン酸ジエステル結合の化学構造の一部あるいは全てに対し、任意の化学物質を付加あるいは置換したものであればいかなるものでもよく、例えば、ホスフォロチオエート結合に置換されたヌクレオチド類似体、N3'-P5'ホスフォアミデート結合に置換されたヌクレオチド類似体等をあげることができる[細胞工学, 16, 1463-1473 (1997)][RNAi法とアンチセンス法、講談社(2005)]。
【0026】
ヌクレオチド類似体としては、その他に、核酸の塩基部分、リボース部分、リン酸ジエステル結合部分等の原子(例えば、水素原子、酸素原子)もしくは官能基(例えば、水酸基、アミノ基)が他の原子(例えば、水素原子、硫黄原子)、官能基(例えば、アミノ基)、もしくは炭素数1〜6のアルキル基で置換されたものまたは保護基(例えばメチル基またはアシル基)で保護されたもの、核酸に、例えば脂質、リン脂質、フェナジン、フォレート、フェナントリジン、アントラキノン、アクリジン、フルオレセイン、ローダミン、クマリン、色素など、別の化学物質を付加した分子等を用いてもよい。
【0027】
核酸に別の化学物質を付加した分子としては、例えば、5’−ポリアミン付加誘導体、コレステロール付加誘導体、ステロイド付加誘導体、胆汁酸付加誘導体、ビタミン付加誘導体、Cy5付加誘導体、Cy3付加誘導体、6−FAM付加誘導体、およびビオチン付加誘導体等をあげることができる。
【0028】
本発明(5)は、前記のヘプタマー型ガイド核酸の有効量を含有する薬剤である。この薬剤は、本発明のヘプタマー型ガイド核酸それ単独で製剤化することもできるが、通常は薬理学的に許容される1つあるいはそれ以上の担体と一緒に混合し、製剤学の技術分野においてよく知られる任意の方法により製造した医薬製剤として投与するのが望ましい。
【0029】
投与経路は、治療に際し最も効果的なものを使用するのが望ましく、経口投与、または口腔内、気道内、直腸内、皮下、筋肉内および静脈内などの非経口投与をあげることができ、望ましくは静脈内投与をあげることができる。
【0030】
経口投与に適当な製剤としては、乳剤、シロップ剤、カプセル剤、錠剤、散剤、顆粒剤などがあげられる。乳剤およびシロップ剤のような液体調製物は、水、ショ糖、ソルビトール、果糖などの糖類、ポリエチレングリコール、プロピレングリコールなどのグリコール類、ごま油、オリーブ油、大豆油などの油類、p-ヒドロキシ安息香酸エステル類などの防腐剤、ストロベリーフレーバー、ペパーミントなどのフレーバー類などを添加剤として用いて製造できる。カプセル剤、錠剤、散剤、顆粒剤などは、乳糖、ブドウ糖、ショ糖、マンニトールなどの賦形剤、デンプン、アルギン酸ナトリウムなどの崩壊剤、ステアリン酸マグネシウム、タルクなどの滑沢剤、ポリビニルアルコール、ヒドロキシプロピルセルロース、ゼラチンなどの結合剤、脂肪酸エステルなどの界面活性剤、グリセリンなどの可塑剤などを添加剤として用いて製造できる。
【0031】
非経口投与に適当な製剤としては、注射剤、座剤、噴霧剤などがあげられる。注射剤は、塩溶液、ブドウ糖溶液あるいは両者の混合物からなる担体などを用いて調製される。座剤はカカオ脂、水素化脂肪またはカルボン酸などの担体を用いて調製される。また、噴霧剤は受容者の口腔および気道粘膜を刺激せず、かつ有効成分を微細な粒子として分散させ吸収を容易にさせる担体などを用いて調製される。
【0032】
担体として具体的には乳糖、グリセリンなどが例示される。本発明で用いる核酸、さらには用いる担体の性質により、エアロゾル、ドライパウダーなどの製剤が可能である。また、これらの非経口剤においても経口剤で添加剤として例示した成分を添加することもできる。
【0033】
投与量または投与回数は、目的とする治療効果、投与方法、治療期間、年齢、体重などにより異なるが、通常成人1日当たり10 μg/kg〜20 mg/kgである。
【0034】
以下、実施例を示して本発明をさらに詳細かつ具体的に説明するが、本発明は以下の例に限定されるものではない。
【実施例1】
【0035】
LNA修飾したヘプタマー型sgRNAの試験管内での切断活性と安定性試験
(1)材料と方法
(1-1)組換えヒトtRNase ZLの調製
ヒスチジン標識したヒトΔ30 tRNase ZLを、E. coli株Rosetta (DE3) pLysS(Novagen)において発現プラスミドpQE-80L(Quigen)から過剰発現させ、ニッケル−アガロースビーズを用いて精製した(非特許文献15)。
(1-2)RNA合成
標的RNA、T3H:
5’-GAUCAGAAGAUUCCAGGUUCGACUCCUGGCUGGCUCGGUGUAAGCAGGGUCGUUUU-3:配列番号1
はT7 RNAポリメラーゼ(Promega)を用い、対応する合成DNAから合成した(非特許文献8)。転写反応は製造者(Promega)が推奨する条件の下で行ない、RNAは変性ゲル電気泳動で精製した。次いで、T3HのRNA転写産物は製造者(Amersham Pharmacia Biotech)のプロトコールに従い蛍光色素で標識した(非特許文献13)。すなわち、バクテリアアルカリホスファターゼ(Takara Shuzo)でRNA転写産物の5’リン酸を除去したのち、T4ポリヌクレオチドキナーゼ(Takara Shuzo)およびATPγSを用いてRNAをリン酸化した。次いで、単一の蛍光色素部分を5’リン酸化部位に付加し、この蛍光色素標識したRNAをゲル精製した使用した。
【0036】
以下の、5’をリン酸化し、2’-Oメチル化および/またはLNA修飾したsgRNAを化学的に合成した。
hepL0:5′-pGGGCCAG-3′(配列番号2)
hepL2:5′-pGgGCCaG-3′(配列番号3)
hepL3a:5′-pgGgCcAG-3′(配列番号4)
hepL3b:5′-pGgGcCaG-3′(配列番号5)
hepL4:5′-pGgGcCag-3′(配列番号6)
hepL7:5′-pgggccag-3′(配列番号7)
上記の各sgRNAにおいて、大文字は2’-Oメチル化修飾塩基、小文字はLNA修飾塩基を示す。
(1-3)In vitro RNA切断アッセイ
10 mM Tris-HCl(pH 7.5)および15 mM MgCl2を含有する溶液(6 μl)に非標識RNAと組換えヒトΔ30 tRNase ZLを混合し、蛍光色素で標識した標的RNAのin vitro切断アッセイを行なった(非特許文献15、16)。5%または10%のポリアクリルアミド-8 M尿素ゲル上で反応産物を分解した後、Typhoon 9210 (Amersham Pharmacia Biotech)でゲルを分析した。
(1-4)Kd定量のためのゲルシフトアッセイ
ヘプタマー/標的RNA複合体の解離定数を決定するため、蛍光標識したT3H(0.1pmol)を、無標識の各sgRNA(hepL0、hepL2、hepL3a、hepL3b、hepL4、hepL7:各4-100pmol)と37℃で10分間インキュベートした。反応溶液(6 μl)は10 mM Tris-HCl(pH 7.5)および10 mM MgCl2を含む。インキュベーションの後、サンプルは等量のローディングバッファー(10 mM Tris-HCl, pH 7.5, 1 mM EDTA, 0.05% bromophenol blue, 0.05% xylene cyanol FF, 50% glycerol)と混合し、次いで、TBEバッファー(90 mM Tris base, 90 mM boric acid, 1.5 mM EDTA, pH 8.3)を用い、8%非変性ポリアクリルアミドゲル上で電気泳動した。電気泳動の後、Typhoon 9210を用いて標識化RNAを定量した。同様にして、基質/tRNase ZL複合体の乖離定数をゲルシフトアッセイにより測定した(非特許文献13)。蛍光標識したT3H (0.1 pmol)と各sgRNA(10 pmol)とのRNA複合体は、様々な量(0.5-2 μg)の組換えヒトΔ30 tRNase ZLにより、前記バッファー中で4℃、10分間インキュベートし、前記と同様の方法で分析した。
(2)結果
LNA修飾を含むヘプタマー型sgRNAがヒトtRNase ZLによる標的RNAの切断を誘導するかどうかを調べるため、2から7箇所のLNA修飾核酸を有する5個のヘプタマー型sgRNAを調製し、それぞれの切断誘導能を調べた(図2)。この目的のため、micro-pre-tRNAと類似するヘプタマー/T3H RNA複合体を試験基質として用いた(非特許文献8、13)。ヘプタマー型sgRNAのhepL2、hepL3a、hepL3bおよびhepL4は、それぞれ2-、3-、3-および4-核酸位置がLNA修飾されており、他の核酸の2’リボース炭素はO-メリル化修飾されている(図2A)。hepL7は全核酸がLNA修飾されており、コントロール(hepL0)は全核酸が2′-Oメチル化修飾されている。
【0037】
RNA切断アッセイは、組換えヒトtRNase ZLを用いて、単一ターンオーバー条件で行なった。hepL2による標的RNA T3Hの切断が観察されたが、その切断率は、hepL0の56%であった(図2B)。全体として、切断率はLNA修飾核酸の増加に伴い減少し、hepL7の切断率はhepL0のわずか8%であった(図2B)。
【0038】
次に、全反応中の各ステップがどのように切断効率に影響するかを明らかにするため、ヘプタマー型sgRNA/標的RNA複合体の解離定数、および基質/tRNase ZL複合体の解離定数を決定し、切断効率定数を調べた。ヘプタマー型sgRNA/標的RNA複合体の解離定数はゲルシフトアッセイにより決定し、これら複合体の有効濃度はそれらの推定された解離定数を用いた算出した。観察された切断効率定数は、基質の推定された有効濃度に基づき、単一ターンオーバー条件で得た。基質/tRNase ZL複合体の解離定数はまた、ヘプタマー型sgRNA/標的RNA複合体の推定された乖離定数を用いて、ゲルシフトアッセイにより決定した。
【0039】
hepL2/T3H複合体に対するKd値(0.39 μM)はhepL0/T3H複合体に対するそれ(0.44 μM)と比肩するものであった。全体として、ヘプタマー型sgRNA/標的RNA複合体のKd値はLNA修飾の数が増加するに従って増加し、hepL7/T3H複合体では3.43 μM以上であった(表1)。同様に、hepL2-T3H/tRNase ZL複合体のKd値(0.40μM)はhepL0-T3H/tRNase ZL複合体のそれ(0.32 μM)と比肩するものでった。基質/酵素複合体のKd値は、LNA修飾の数が増加するに従って増加し、hepL7-T3H/tRNase ZLでは2.89 μM以上であった(表1)。hepL0/T3H複合体のkobs値は0.40 min-1であり、hepL2/T3H複合体は0.20 min-1であるのに対して、他の複合体のkobs値は0.05 min-1以下に低下した(表1)。
【0040】
以上の詳細な分析に基づけば、各ステップがどのように切断効率に影響するかを理解することができる(図2B)。hepL2によるT3Hの切断効率が、hepL0による切断効率の56%に減少したことは、kobs値が50%に減少したことに依存していると思われる(表1)。hepL3a、hepL3bおよびhepL4によるT3H切断効率が18-22%に減少したことは、ヘプタマー/T3H複合体のKd値が32〜70%に増加した、基質/tRNase ZL複合体のKd値が3倍以上に増加したこと、そしてkobs値が10%以下に減少したことが原因であると考えられる(表1)。hepL7による切断効率が8%まで劇的に減少したことは、hepL7/T3H複合体のKd値が8倍に増加し、基質/tRNase ZL複合体のKd値が9倍に増加し、そしてKobs値が13%に減少したことが原因であると考えられる(表1)。
【0041】
以上のとおり、LNA修飾したヘプタマー型RNAは、tRNase ZL用のsgRNAとして機能することが示された。ただし、切断効率としてみたそれらの有効性は、全体として、LNA修飾数の増加とともに減少し、ヘプタマー型sgRNA/標的RNA複合体のKd値、基質/tRNase ZL複合体のKd値、およびKobs値に影響を及ぼした(図2、表1)。
【0042】
しかしながら、血清および生細胞におけるLNA修飾化ヘプタマー型RNAの高い安定性を考慮すれば、試験した2〜4 LNA修飾したヘプタマー型sgRNAの切断効率が18〜56%に減少することは、それらを薬剤成分として使用する場合に十分な許容範囲である。
【0043】
【表1】

【実施例2】
【0044】
LNA修飾したヘプタマー型sgRNAの細胞内安定性試験
(1)方法
ヒト293細胞、KMM-1細胞、またはHeLa細胞(0.8-1 X 105細胞/ウエル)を、24ウエルプレートにてheptamer型sgRNA(1 μM)を加えた培地で培養した。ヘプタマー型sgRNAは、いずれも3’末端をFITC標識した、
hepL0:5′-pGGGCCAG-3′(配列番号2)
hepL3b:5′-pGgGcCaG-3′(配列番号5)
を用いた。
【0045】
培養24時間後に細胞を集めPBSにて4回洗浄した。その後、細胞から全RNAをISOGENを用いて抽出し、20%ポリアクリルアミドー8 Mウレアゲル上で分離した。ゲルはTyphoon9210にて解析した。
(2)結果
図3に示されているように、LNA修飾されたヘプタマー型sgRNA(LNA)は、2′-O-メチル化のみのものよりも細胞内でより安定に存在できることが確認された。
【産業上の利用可能性】
【0046】
本発明によって、例えば、特定遺伝子の過剰発現を原因とする疾患の有効な治療薬剤の有効成分としの安定性に優れたヘプタマー型sg核酸が提供される。
【技術分野】
【0001】
本発明は、標的として切断するRNAを転写する遺伝子発現を効果的に抑制または消失させ、かつ生体内安定性に優れていることによって、その遺伝子発現等を原因とする疾患の治療または予防に有効なヘプタマー型スモールガイド核酸に関するものである。
【背景技術】
【0002】
本発明の発明者らは、TRUEジーンサイレンシング(tRNase ZL-utilizing efficacious gene silencing)と名付けた新しい遺伝子発現抑制技術を開発している(特許文献1、非特許文献1−4)。tRNase ZLは、tRNAの3’をプロセシングするエンドリボヌクレアーゼ(tRNase Zまたは3’tRNase)の分子量の大きい種類の酵素であり、前駆体tRNAの3’末端部を切除する(特許文献1、非特許文献5、6)。このTRUEジーンサイレンシングは、標的RNAと合成スモールガイド核酸とによって形成されるpre-tRNA様またはマイクロpre-tRNA様複合体を認識することによって、いかなるRNAの任意箇所をも切断することができるという哺乳動物tRNase ZLのユニークな酵素的特性に基づいている(非特許文献7−13)。スモールガイド核酸(特にスモールガイドRNA)は、以下の4種:5’-half tRNA(非特許文献8)、〜12-16-nt直鎖RNA(非特許文献13)、ヘプタマー型RNA(非特許文献9)およびフック型RNA(非特許文献12)に分けられる(図1)。
【0003】
発明者らは、様々なmRNAを標的とするスモールガイド核酸(以下、sg核酸と記載することがある)を、その発現プラスミドまたは2’-O-メチル RNAsを導入することによって、様々な哺乳動物細胞におけるTRUEジーンサイレンシングの有効性を実証している(非特許文献1−4)。例えば、Jurkat細胞におけるHIV-1発現は、5’-half tRNA型sgRNAによって18日間以上抑制され(非特許文献2)、マウス肝臓におけるルシフェラーゼ発現はヘプタマー型sg核酸によって抑制された(非特許文献3)。さらに、TRUEジーンサイレンシングの効果は、RNA干渉(RNA interference: RNAi)と同等であり(非特許文献3、14)、幾つかの場合ではRNAiの効果を凌ぐことも確認されている(非特許文献4)。
【0004】
また最近、発明者らは、tRNase ZLが核内だけでなくサイトゾルにも存在すること、そしてsg核酸として使用していたのと同一の5’-half tRNAが細胞質に存在することを見出している(非特許文献15)。また、発明者らは、ヒトサイトゾルtRNase ZLが、5’-half tRNAの指揮のもとでmRNAを切断することによって遺伝子発現を調節すること、そしてPPM1F mRNAがtRNase ZLの真の標的の一つであることを見出している(非特許文献15)。さらにまた、発明者らは、miR-103を含むマイクロRNAの一部がフック型sg核酸として働き、サイトゾルtRNase ZLによるmRNA切断を介して遺伝子発現を下方調節しえることを証明している(非特許文献16)。以上のことから、TRUEジーンサイレンシングは、細胞内の小さな非コードRNAによって作動されるサイトゾルtRNase ZLの新たに解明された生理学的役割を基礎として機能していることは明らかである。
【0005】
TRUEジーンサイレンシングの最終的な目的の一つは、特定遺伝子の発現を原因とする疾患治療剤または予防剤としてのsg核酸の使用である。例えば、発明者らは既に、ガン治療の分子標的として有望視されているBcl-2およびVEGFをコードする細胞内mRNAレベルが、5’-half tRNA型sg核酸、14-nt直鎖型sg核酸またはヘプタマー型sg核酸によって発現抑制されることを見出している(非特許文献1、4、17)。
【0006】
sg核酸を薬剤成分とする場合、薬理学的な視点からは、ヘプタマー型sg核酸が最も好ましい。何故ならば、ヘプタマー(7塩基核酸)は長鎖のsg核酸よりも遙かに簡便かつ安価に合成することができる。また、ヘプタマー型sg核酸は、刺激剤(トランスフェクション試薬等)を用いることなく容易に細胞内に取込ませることができる(非特許文献18)。
【0007】
なお、ヘプタマー型sg核酸は、tRNase ZLによる標的RNAの切断誘導能や生体内での安定性の向上を目的として、様々な化学的修飾が検討されている。発明者らはかつて、2’-O-メチル RNA、ホスホロチオエートRNA、DNAおよびホスホロチオエートDNAについて調べた結果、酵素に対する各々の切断誘導能と安定性から、2’-O-メチル修飾したsg核酸がTRUEジーンサイレンシングのためには最も好ましいとの結論を得ている(非特許文献1)。
【0008】
一方、2’位の酸素原子と4’位の炭素原子がメチレンを介して架橋したロックト人工核酸(Locked Nucleic Acid:LNA)の出現が、核酸研究に広範な影響を及ぼしている(非特許文献19、20)。すなわち、LNAは生体安定性と温度安定性の両方を核酸に付与するため、アンチセンスRNAやsiRNAの化学修飾として利用されている。
【特許文献1】特許第3660718号公報
【非特許文献1】Tamura,M., Nashimoto,C., Miyake,N., Daikuhara,Y., Ochi,K. and Nashimoto,M.(2003) Intracellular mRNA cleavage by 3′ tRNase under the direction of 2′-O-methylRNA heptamers. Nucleic Acids Res., 31, 4354-4360.
【非特許文献2】Habu,Y., Miyano-Kurosaki,N., Kitano,M., Endo,Y., Yukita,M., Ohira,S., Takaku,H., Nashimoto,M. and Takaku,H. (2005) Inhibition of HIV-1 gene expression by retroviral vector-mediated small-guide RNAs that direct specific RNA cleavage by tRNase ZL. Nucleic Acids Res., 33, 235-243.
【非特許文献3】Nakashima,A., Takaku,H., Shibata,H.S., Negishi,Y., Takagi,M., Tamura,M. and Nashimoto,M. (2007) Gene-silencing by the tRNA maturase tRNase ZL under the direction of small guide RNA. Gene Therapy, 14, 78-85.
【非特許文献4】Elbarbary,R.A., Takaku,H., Tamura,M. and Nashimoto,M. (2009) Inhibition of vascular endothelial growth factor expression by TRUE gene silencing. Biochem.and Biophys. Res. Commun., 379, 924-927.
【非特許文献5】Nashimoto,M. (1997) Distribution of both lengths and 5′ terminal nucleotides of mammalian pre-tRNA 3′ trailers reflects properties of 3′ processing endoribonuclease. Nucleic Acids Res., 25, 1148-1155.
【非特許文献6】Takaku,H., Minagawa,A., Masamichi,T. and Nashimoto,M. (2003) A candidateprostate cancer susceptibility gene encodes tRNA 3′ processing endoriobonuclease. Nucleic Acids Res., 31, 2272-2278.
【非特許文献7】Nashimoto,M. (1995) Conversion of mammalian tRNA 3′ processing endoribonuclease to four-base-recognizing RNA cutters. Nucleic Acids Res., 23, 3642-3647.
【非特許文献8】Nashimoto,M. (1996) Specific cleavage of target RNAs from HIV-1 with 5′ half tRNA by mammalian tRNA 3′ processing endoribonuclease. RNA, 2, 2523-2524.
【非特許文献9】Nashimoto,M., Geary,S., Tamura,M. and Kasper,R. (1998) RNA heptamers that directs RNA cleavage by mammalian tRNA 3′ processing endoribonuclease. Nucleic Acids Res., 26, 2565-2571.
【非特許文献10】Nashimoto,M. (2000) Anomalous RNA substrates for mammalian tRNA 3′processing endoribonuclease. FEBS Letters, 472, 179-186.
【非特許文献11】Takaku,H., Minagawa,A., Takagi,M. and Nashimoto,M. (2004) The N-terminal half-domain of the long form of tRNase Z is required for the RNase 65 activity. Nucleic Acids Res., 32, 4429-4438.
【非特許文献12】Takaku,H., Minagawa,A., Takagi,M. and Nashimoto,M. (2004) A novel four-base-recognizing RNA cutter that can remove the single 3′ terminal nucleotides from RNA molecules. Nucleic Acids Res., 32, e91.
【非特許文献13】Shibata,H.S., Takaku,H., Takagi,M. and Nashimoto,M. (2005) The T loop structure is dispensable for substrate recognition by tRNase ZL. J. Biol. Chem., 280, 22326-22334.
【非特許文献14】Appasani,K. (2005) RNA interference technology: from basic science to drug development. Cambridge University Press, Cambridge.
【非特許文献15】Elbarbary,R.A., Takaku,H., Uchiumi,N., Tamiya,H., Abe,M., Takahashi,M., Nishida,H. and Nashimoto,M. (2009) Modulation of gene expression by human cytosolic tRNase ZL through 5′-half-tRNA. PLoS ONE, 4, e5908.
【非特許文献16】Elbarbary,R.A., Takaku,H., Uchiumi,N., Tamiya,H., Abe,M., Nishida,H. and Nashimoto,M. (2009) Human cytosolic tRNase ZL can downregulate gene expression through miRNA. FEBS Lett., 583, 3241-3246.
【非特許文献17】Hu,W. and Kavanagh,J.J. (2003) Anticancer therapy targeting the apoptotic pathway. Lancet Oncol., 4, 721-729.
【非特許文献18】Loke,S.L., Stein,C.A., Zhang X.H., Mori,K., Nakanishi,M., Subasinghe,C., Cohen,J.S. and Neckers,L.M. (1989) Characterization of oligonucleotide transport into living cells. Proc. Natl. Acad. Sci. USA, 86, 3474-3478.
【非特許文献19】Koch,T. and Orum,H. (2008) Locked nucleic acid. In Crooke,S.T. (ed.), Antisense Drug Technology. CRC Press, Boca Raton, pp. 519-564.
【非特許文献20】Straarup,E.M., Fisker,N., Hedtjarn,M., Lindholm,M.W., Rosenbohm,C., Aarup,V., Hansen,H.F., Orum,H., Hansen,J.B. and Koch,T. (2010) Short locked nucleic acid antisense oligonucleotides potently reduce apolipoprotein B mRNA and serum cholesterol in mice and non-human primates. Nucleic Acids Res., 38, 7100-7111.
【発明の概要】
【発明が解決しようとする課題】
【0009】
ヘプタマー型sg核酸を薬剤成分とする場合の一つの課題は、生体内に投与された場合にその主効果(tRNase ZLによる標的RNAの切断誘導)が持続的に発揮されるように、生体安定性および温度安定性を向上させることである。このような課題に対して、アンチセンスRNAやsiRNAに対する修飾として実績のあるLNA修飾は有望であると考えられる。
【0010】
しかしながら、LNA修飾によってヘプタマー型sg核酸の主効果(tRNase ZLによる標的RNAの切断誘導)がどのように変容するかは一切知られていない。仮に、LNA修飾によってヘプタマー型sg核酸の主効果が大幅に損なわれることがあれば、LNA修飾による安定性の向上は薬剤成分としてのsg核酸においては好ましいものでない。
【0011】
本願発明は、ヘプタマー型sg核酸の主効果と安定性との観点から、ヘプタマー型sg核酸が生体内でその主効果を最大限に発揮することのできるLNA修飾条件を提供することによって、ヘプタマー型sg核酸の薬剤成分としての価値をさらに向上させることを課題としている。
【課題を解決するための手段】
【0012】
本発明は、前記の課題を解決するものとして、以下の発明を提供する。
(1)標的RNAの少なくとも連続5塩基と相補的な塩基配列を含む7塩基からなり、その配列中の1から4塩基がLNA(locked nucleic acid)修飾されており、細胞内においてtTNaseZLの作用により標的RNAを切断することによってこの標的RNAを転写する遺伝子の活性を低下または消滅させるヘプタマー型スモールガイド核酸。
(2)LNA修飾された塩基以外の塩基が2’-Oメチル化修飾されている前記(1)のヘプタマー型スモールガイド核酸。
(3)標的RNAが疾患に関連する遺伝子から転写されたRNAである前記(1)または(2)のヘプタマー型スモールガイド核酸。
(4)疾患に関連する遺伝子がガンの発症に関連する遺伝子である前記(3)のヘプタマー型スモールガイド核酸。
(5)前記(3)または(4)のヘプタマー型スモールガイド核酸の有効量を含有する薬剤。
【発明の効果】
【0013】
本発明(1)によれば、標的RNAの切断効率を許容範囲内に維持し、かつ、生体内での安定性に極めて優れてたヘプタマー型スモールガイド核酸が提供される。このヘプタマー型スモールガイド核酸は、薬剤成分として生体内に投与した場合に、その優れた安定性により標的RNAの切断効果を最大限に発揮することができる。
【0014】
本発明(2)によれば、LNA修飾塩基以外の塩基がさらに2’-Oメチル化修飾されているため、標的RNAの切断効率がさらに優れたヘプタマー型スモールガイド核酸が提供される。
【0015】
本発明(3)から(5)によれば、例えばガン等の疾患治療または予防のための手段が提供される。
【図面の簡単な説明】
【0016】
【図1】4種類のスモールガイド核酸(左から2、4、5、6番目)の例。各塩基配列のうち、5’NNNN3’の領域がスモールガイド核酸。
【図2】Aは実施例1で使用したヘプタマー型sgRNA/T3H RNA複合体の2次構造。各塩基配列のうち、5’NNNN3’の領域がプタマー型sgRNA。大文字は2’-Oメチル化修飾塩基、小文字はLNA修飾塩基である。矢印は標的RNAの切断箇所である。Bは変性の10%ポリアクリルアミドゲルでの標的RNA(T3H)の切断産物の電気泳動像。下に切断効率と相対的なパーセンテージを示す。
【図3】実施例2の結果である。2ME: すべて2′-O-メチル化された3′-FITC標識hepL0、LNA:2、4、6番目のヌクレオチドをLNA修飾(その他4か所は2′-O-メチル化)した3′-FITC標識hepL3b。*:細胞から分解を受けずに抽出されたヘプタマー型sgRNA。その上のバンドはヘプタマー型sgRNAと細胞内の他のRNAとの複合体であると思われる。
【発明を実施するための形態】
【0017】
本発明(1)のヘプタマー型スモールガイド核酸は、標的RNAの少なくとも連続5塩基と相補的な塩基配列を含む7塩基からなり、その配列中の1から4塩基がLNA(locked nucleic acid)修飾されている。
【0018】
LNAは、2’位の酸素原子と4’位の炭素原子がメチレンを介して架橋した人工核酸であり、例えば、2′-O-4′C-methylene-β-D-ribofuranoseを含む核酸である。
【0019】
標的RNAは、その発現を抑制または消失させる対象となる遺伝子(標的遺伝子)の転写産物RNA、特にmRNAである。標的遺伝子の発現抑制とは、その遺伝子から転写合成されるタンパク質量が、25%以下、好ましくは50%以下、さらに好ましくは75%以下、特に好ましくは95%以下に低減することである。標的遺伝子は、その発現抑制または発現消失が産業上の利用価値を有する遺伝子であり、とくにその遺伝子の発現亢進が特定の疾患の原因となるような遺伝子(以下、病原遺伝子と記載することがある)。そのような病原遺伝子の例としては、例えば、ras、erbb2、myc、apc;brca1、rb、Bcl-2、BGEF癌遺伝子、レニンなどの高血圧に関与する遺伝子、インスリンなどの糖尿病関連遺伝子、LDLレセプターなどの高脂血症関連遺伝子、レプチンなどの肥満関連遺伝子、アンギオテンシンなどの動脈硬化疾患関連遺伝子、痴呆の原因として知られているアポリポタンパク質およびプレセリニン、老化に関連する遺伝子などが例示される。これらの遺伝子およびそのmRNA配列は公知のデータベース(NCBIヌクレオチドデータベース等)において公知である。
【0020】
ヘプタマー型sg核酸は、塩基対合を介して標的RNAとtRNAアクセプターステム様二重鎖を形成することによって、tRNase ZLによる標的RNAの特異的切断を可能とするため、ヘプタマー型sg核酸の標的部位は、tRNA T-armに類似する安定したヘアピン構造領域の直下となり、ヘプタマー型sg核酸はその7塩基だけでなく、安定したヘアピン構造を形成する約12塩基の特異性によって効果的にRNAを切断する。従って、ヘプタマー型sg核酸の塩基配列は、これらの標的遺伝子のmRNA配列情報から決定することができる。
【0021】
配列を決定したヘプタマー型sg核酸は、公知の化学合成を用いる方法、あるいは酵素的転写法等にて製造することができる。公知の化学合成を用いる方法として、ホスホロアミダイト法、ホスフォロチオエート法、ホスホトリエステル法等をあげることができ、例えば、ABI3900ハイスループット核酸合成機(アプライドバイオシステムズ社製)やNTS H-6核酸合成機(日本テクノサービス社製)、Oligoilot10核酸合成機(GEヘルスケア製)により合成することができる。酵素的転写法としては、目的の塩基配列を有するプラスミドまたはDNAを鋳型として、T7、T3、SP6RNAポリメラーゼ等のRNAポリメラーゼを用いた転写をあげることができる。合成法または転写法により製造したヘプタマー型sg核酸は、次いでHPLC等にて精製する。例えばHPLC精製時には、triethylammonium acetate (TEAA)またはhexylammonium acetate(HAA)とacetonitrileの混合溶液を用いて、ヘプタマー型sg核酸をカラムから溶出する。その後、溶出体積の1000倍量の蒸留水で溶出溶液を10時間透析し、透析溶液を凍結乾燥した後、使用時まで冷凍保存する。使用時には蒸留水で最終濃度が100 μM程度になるように溶解する。
【0022】
本発明のヘプタマー型sg核酸に用いられるLNA修飾核酸以外の核酸としては、ヌクレオチドまたはそのヌクレオチドと同等の機能を有する分子が重合した分子であればいかなるものでもよい。ヌクレオチドとしては、例えばリボヌクレオチドの重合体であるRNA、デオキシリボヌクレオチドの重合体であるDNA、RNAおよびDNAが混合した重合体、ヌクレオチド類似体を含むヌクレオチド重合体が、それぞれあげられる。特にRNAが好ましい。
【0023】
ヌクレオチド類似体としては、例えばRNAまたはDNAと比較して、ヌクレアーゼ耐性の向上または安定化させるため、相補鎖核酸とのアフィニティーをあげるため、細胞透過性をあげるため、あるいは可視化させるために、リボヌクレオチド、デオキシリボヌクレオチド、RNAまたはDNAに修飾を施した分子を挙げるこことができる。例えば、糖部修飾ヌクレオチド類似体やリン酸ジエステル結合修飾ヌクレオチド類似体等があげられる。
【0024】
糖部修飾ヌクレオチド類似体とは、ヌクレオチドの糖の化学構造の一部あるいは全てに対し、任意の化学構造物質を付加あるいは置換したものであればいかなるものでもよく、例えば、2’−O−メチルリボースで置換されたヌクレオチド類似体、2’−O−プロピルリボースで置換されたヌクレオチド類似体、2’−メトキシエトキシリボースで置換されたヌクレオチド類似体、2’−O−メトキシエチルリボースで置換されたヌクレオチド類似体、2’−O−[2−(グアニジウム)エチル]リボースで置換されたヌクレオチド類似体、2’−O−フルオロリボースで置換されたヌクレオチド類似体、エチレン架橋構造型人工核酸(Ethylene bridged nucleic acid:ENA)[Nucleic Acid Research, 32, e175 (2004)]等があげられ、さらにペプチド核酸(PNA)[Acc. Chem. Res., 32, 624 (1999)]、オキシペプチド核酸(OPNA)[J. Am. Chem. Soc., 123, 4653 (2001)]、およびペプチドリボ核酸(PRNA)[J. Am. Chem. Soc., 122, 6900 (2000)]等をあげることができる。特に、2’−O−メチルリボースで置換されたヌクレオチド類似体(2’-Oメチル化修飾された核酸)が好ましい。
【0025】
リン酸ジエステル結合修飾ヌクレオチド類似体としては、ヌクレオチドのリン酸ジエステル結合の化学構造の一部あるいは全てに対し、任意の化学物質を付加あるいは置換したものであればいかなるものでもよく、例えば、ホスフォロチオエート結合に置換されたヌクレオチド類似体、N3'-P5'ホスフォアミデート結合に置換されたヌクレオチド類似体等をあげることができる[細胞工学, 16, 1463-1473 (1997)][RNAi法とアンチセンス法、講談社(2005)]。
【0026】
ヌクレオチド類似体としては、その他に、核酸の塩基部分、リボース部分、リン酸ジエステル結合部分等の原子(例えば、水素原子、酸素原子)もしくは官能基(例えば、水酸基、アミノ基)が他の原子(例えば、水素原子、硫黄原子)、官能基(例えば、アミノ基)、もしくは炭素数1〜6のアルキル基で置換されたものまたは保護基(例えばメチル基またはアシル基)で保護されたもの、核酸に、例えば脂質、リン脂質、フェナジン、フォレート、フェナントリジン、アントラキノン、アクリジン、フルオレセイン、ローダミン、クマリン、色素など、別の化学物質を付加した分子等を用いてもよい。
【0027】
核酸に別の化学物質を付加した分子としては、例えば、5’−ポリアミン付加誘導体、コレステロール付加誘導体、ステロイド付加誘導体、胆汁酸付加誘導体、ビタミン付加誘導体、Cy5付加誘導体、Cy3付加誘導体、6−FAM付加誘導体、およびビオチン付加誘導体等をあげることができる。
【0028】
本発明(5)は、前記のヘプタマー型ガイド核酸の有効量を含有する薬剤である。この薬剤は、本発明のヘプタマー型ガイド核酸それ単独で製剤化することもできるが、通常は薬理学的に許容される1つあるいはそれ以上の担体と一緒に混合し、製剤学の技術分野においてよく知られる任意の方法により製造した医薬製剤として投与するのが望ましい。
【0029】
投与経路は、治療に際し最も効果的なものを使用するのが望ましく、経口投与、または口腔内、気道内、直腸内、皮下、筋肉内および静脈内などの非経口投与をあげることができ、望ましくは静脈内投与をあげることができる。
【0030】
経口投与に適当な製剤としては、乳剤、シロップ剤、カプセル剤、錠剤、散剤、顆粒剤などがあげられる。乳剤およびシロップ剤のような液体調製物は、水、ショ糖、ソルビトール、果糖などの糖類、ポリエチレングリコール、プロピレングリコールなどのグリコール類、ごま油、オリーブ油、大豆油などの油類、p-ヒドロキシ安息香酸エステル類などの防腐剤、ストロベリーフレーバー、ペパーミントなどのフレーバー類などを添加剤として用いて製造できる。カプセル剤、錠剤、散剤、顆粒剤などは、乳糖、ブドウ糖、ショ糖、マンニトールなどの賦形剤、デンプン、アルギン酸ナトリウムなどの崩壊剤、ステアリン酸マグネシウム、タルクなどの滑沢剤、ポリビニルアルコール、ヒドロキシプロピルセルロース、ゼラチンなどの結合剤、脂肪酸エステルなどの界面活性剤、グリセリンなどの可塑剤などを添加剤として用いて製造できる。
【0031】
非経口投与に適当な製剤としては、注射剤、座剤、噴霧剤などがあげられる。注射剤は、塩溶液、ブドウ糖溶液あるいは両者の混合物からなる担体などを用いて調製される。座剤はカカオ脂、水素化脂肪またはカルボン酸などの担体を用いて調製される。また、噴霧剤は受容者の口腔および気道粘膜を刺激せず、かつ有効成分を微細な粒子として分散させ吸収を容易にさせる担体などを用いて調製される。
【0032】
担体として具体的には乳糖、グリセリンなどが例示される。本発明で用いる核酸、さらには用いる担体の性質により、エアロゾル、ドライパウダーなどの製剤が可能である。また、これらの非経口剤においても経口剤で添加剤として例示した成分を添加することもできる。
【0033】
投与量または投与回数は、目的とする治療効果、投与方法、治療期間、年齢、体重などにより異なるが、通常成人1日当たり10 μg/kg〜20 mg/kgである。
【0034】
以下、実施例を示して本発明をさらに詳細かつ具体的に説明するが、本発明は以下の例に限定されるものではない。
【実施例1】
【0035】
LNA修飾したヘプタマー型sgRNAの試験管内での切断活性と安定性試験
(1)材料と方法
(1-1)組換えヒトtRNase ZLの調製
ヒスチジン標識したヒトΔ30 tRNase ZLを、E. coli株Rosetta (DE3) pLysS(Novagen)において発現プラスミドpQE-80L(Quigen)から過剰発現させ、ニッケル−アガロースビーズを用いて精製した(非特許文献15)。
(1-2)RNA合成
標的RNA、T3H:
5’-GAUCAGAAGAUUCCAGGUUCGACUCCUGGCUGGCUCGGUGUAAGCAGGGUCGUUUU-3:配列番号1
はT7 RNAポリメラーゼ(Promega)を用い、対応する合成DNAから合成した(非特許文献8)。転写反応は製造者(Promega)が推奨する条件の下で行ない、RNAは変性ゲル電気泳動で精製した。次いで、T3HのRNA転写産物は製造者(Amersham Pharmacia Biotech)のプロトコールに従い蛍光色素で標識した(非特許文献13)。すなわち、バクテリアアルカリホスファターゼ(Takara Shuzo)でRNA転写産物の5’リン酸を除去したのち、T4ポリヌクレオチドキナーゼ(Takara Shuzo)およびATPγSを用いてRNAをリン酸化した。次いで、単一の蛍光色素部分を5’リン酸化部位に付加し、この蛍光色素標識したRNAをゲル精製した使用した。
【0036】
以下の、5’をリン酸化し、2’-Oメチル化および/またはLNA修飾したsgRNAを化学的に合成した。
hepL0:5′-pGGGCCAG-3′(配列番号2)
hepL2:5′-pGgGCCaG-3′(配列番号3)
hepL3a:5′-pgGgCcAG-3′(配列番号4)
hepL3b:5′-pGgGcCaG-3′(配列番号5)
hepL4:5′-pGgGcCag-3′(配列番号6)
hepL7:5′-pgggccag-3′(配列番号7)
上記の各sgRNAにおいて、大文字は2’-Oメチル化修飾塩基、小文字はLNA修飾塩基を示す。
(1-3)In vitro RNA切断アッセイ
10 mM Tris-HCl(pH 7.5)および15 mM MgCl2を含有する溶液(6 μl)に非標識RNAと組換えヒトΔ30 tRNase ZLを混合し、蛍光色素で標識した標的RNAのin vitro切断アッセイを行なった(非特許文献15、16)。5%または10%のポリアクリルアミド-8 M尿素ゲル上で反応産物を分解した後、Typhoon 9210 (Amersham Pharmacia Biotech)でゲルを分析した。
(1-4)Kd定量のためのゲルシフトアッセイ
ヘプタマー/標的RNA複合体の解離定数を決定するため、蛍光標識したT3H(0.1pmol)を、無標識の各sgRNA(hepL0、hepL2、hepL3a、hepL3b、hepL4、hepL7:各4-100pmol)と37℃で10分間インキュベートした。反応溶液(6 μl)は10 mM Tris-HCl(pH 7.5)および10 mM MgCl2を含む。インキュベーションの後、サンプルは等量のローディングバッファー(10 mM Tris-HCl, pH 7.5, 1 mM EDTA, 0.05% bromophenol blue, 0.05% xylene cyanol FF, 50% glycerol)と混合し、次いで、TBEバッファー(90 mM Tris base, 90 mM boric acid, 1.5 mM EDTA, pH 8.3)を用い、8%非変性ポリアクリルアミドゲル上で電気泳動した。電気泳動の後、Typhoon 9210を用いて標識化RNAを定量した。同様にして、基質/tRNase ZL複合体の乖離定数をゲルシフトアッセイにより測定した(非特許文献13)。蛍光標識したT3H (0.1 pmol)と各sgRNA(10 pmol)とのRNA複合体は、様々な量(0.5-2 μg)の組換えヒトΔ30 tRNase ZLにより、前記バッファー中で4℃、10分間インキュベートし、前記と同様の方法で分析した。
(2)結果
LNA修飾を含むヘプタマー型sgRNAがヒトtRNase ZLによる標的RNAの切断を誘導するかどうかを調べるため、2から7箇所のLNA修飾核酸を有する5個のヘプタマー型sgRNAを調製し、それぞれの切断誘導能を調べた(図2)。この目的のため、micro-pre-tRNAと類似するヘプタマー/T3H RNA複合体を試験基質として用いた(非特許文献8、13)。ヘプタマー型sgRNAのhepL2、hepL3a、hepL3bおよびhepL4は、それぞれ2-、3-、3-および4-核酸位置がLNA修飾されており、他の核酸の2’リボース炭素はO-メリル化修飾されている(図2A)。hepL7は全核酸がLNA修飾されており、コントロール(hepL0)は全核酸が2′-Oメチル化修飾されている。
【0037】
RNA切断アッセイは、組換えヒトtRNase ZLを用いて、単一ターンオーバー条件で行なった。hepL2による標的RNA T3Hの切断が観察されたが、その切断率は、hepL0の56%であった(図2B)。全体として、切断率はLNA修飾核酸の増加に伴い減少し、hepL7の切断率はhepL0のわずか8%であった(図2B)。
【0038】
次に、全反応中の各ステップがどのように切断効率に影響するかを明らかにするため、ヘプタマー型sgRNA/標的RNA複合体の解離定数、および基質/tRNase ZL複合体の解離定数を決定し、切断効率定数を調べた。ヘプタマー型sgRNA/標的RNA複合体の解離定数はゲルシフトアッセイにより決定し、これら複合体の有効濃度はそれらの推定された解離定数を用いた算出した。観察された切断効率定数は、基質の推定された有効濃度に基づき、単一ターンオーバー条件で得た。基質/tRNase ZL複合体の解離定数はまた、ヘプタマー型sgRNA/標的RNA複合体の推定された乖離定数を用いて、ゲルシフトアッセイにより決定した。
【0039】
hepL2/T3H複合体に対するKd値(0.39 μM)はhepL0/T3H複合体に対するそれ(0.44 μM)と比肩するものであった。全体として、ヘプタマー型sgRNA/標的RNA複合体のKd値はLNA修飾の数が増加するに従って増加し、hepL7/T3H複合体では3.43 μM以上であった(表1)。同様に、hepL2-T3H/tRNase ZL複合体のKd値(0.40μM)はhepL0-T3H/tRNase ZL複合体のそれ(0.32 μM)と比肩するものでった。基質/酵素複合体のKd値は、LNA修飾の数が増加するに従って増加し、hepL7-T3H/tRNase ZLでは2.89 μM以上であった(表1)。hepL0/T3H複合体のkobs値は0.40 min-1であり、hepL2/T3H複合体は0.20 min-1であるのに対して、他の複合体のkobs値は0.05 min-1以下に低下した(表1)。
【0040】
以上の詳細な分析に基づけば、各ステップがどのように切断効率に影響するかを理解することができる(図2B)。hepL2によるT3Hの切断効率が、hepL0による切断効率の56%に減少したことは、kobs値が50%に減少したことに依存していると思われる(表1)。hepL3a、hepL3bおよびhepL4によるT3H切断効率が18-22%に減少したことは、ヘプタマー/T3H複合体のKd値が32〜70%に増加した、基質/tRNase ZL複合体のKd値が3倍以上に増加したこと、そしてkobs値が10%以下に減少したことが原因であると考えられる(表1)。hepL7による切断効率が8%まで劇的に減少したことは、hepL7/T3H複合体のKd値が8倍に増加し、基質/tRNase ZL複合体のKd値が9倍に増加し、そしてKobs値が13%に減少したことが原因であると考えられる(表1)。
【0041】
以上のとおり、LNA修飾したヘプタマー型RNAは、tRNase ZL用のsgRNAとして機能することが示された。ただし、切断効率としてみたそれらの有効性は、全体として、LNA修飾数の増加とともに減少し、ヘプタマー型sgRNA/標的RNA複合体のKd値、基質/tRNase ZL複合体のKd値、およびKobs値に影響を及ぼした(図2、表1)。
【0042】
しかしながら、血清および生細胞におけるLNA修飾化ヘプタマー型RNAの高い安定性を考慮すれば、試験した2〜4 LNA修飾したヘプタマー型sgRNAの切断効率が18〜56%に減少することは、それらを薬剤成分として使用する場合に十分な許容範囲である。
【0043】
【表1】
【実施例2】
【0044】
LNA修飾したヘプタマー型sgRNAの細胞内安定性試験
(1)方法
ヒト293細胞、KMM-1細胞、またはHeLa細胞(0.8-1 X 105細胞/ウエル)を、24ウエルプレートにてheptamer型sgRNA(1 μM)を加えた培地で培養した。ヘプタマー型sgRNAは、いずれも3’末端をFITC標識した、
hepL0:5′-pGGGCCAG-3′(配列番号2)
hepL3b:5′-pGgGcCaG-3′(配列番号5)
を用いた。
【0045】
培養24時間後に細胞を集めPBSにて4回洗浄した。その後、細胞から全RNAをISOGENを用いて抽出し、20%ポリアクリルアミドー8 Mウレアゲル上で分離した。ゲルはTyphoon9210にて解析した。
(2)結果
図3に示されているように、LNA修飾されたヘプタマー型sgRNA(LNA)は、2′-O-メチル化のみのものよりも細胞内でより安定に存在できることが確認された。
【産業上の利用可能性】
【0046】
本発明によって、例えば、特定遺伝子の過剰発現を原因とする疾患の有効な治療薬剤の有効成分としの安定性に優れたヘプタマー型sg核酸が提供される。
【特許請求の範囲】
【請求項1】
標的RNAの少なくとも連続5塩基と相補的な塩基配列を含む7塩基からなり、その配列中の1から4塩基がLNA(locked nucleic acid)修飾されており、細胞内においてtRNaseZLの作用により標的RNAを切断することによってこの標的RNAを転写する遺伝子の活性を低下または消滅させるヘプタマー型スモールガイド核酸。
【請求項2】
LNA修飾された塩基以外の塩基が2’-Oメチル化修飾されている請求項1のヘプタマー型スモールガイド核酸。
【請求項3】
標的RNAが疾患に関連する遺伝子から転写されたRNAである請求項1または2のヘプタマー型スモールガイド核酸。
【請求項4】
疾患に関連する遺伝子がガンの発症に関連する遺伝子である請求項3のヘプタマー型スモールガイド核酸。
【請求項5】
請求項3または4のヘプタマー型スモールガイド核酸の有効量を含有する薬剤。
【請求項1】
標的RNAの少なくとも連続5塩基と相補的な塩基配列を含む7塩基からなり、その配列中の1から4塩基がLNA(locked nucleic acid)修飾されており、細胞内においてtRNaseZLの作用により標的RNAを切断することによってこの標的RNAを転写する遺伝子の活性を低下または消滅させるヘプタマー型スモールガイド核酸。
【請求項2】
LNA修飾された塩基以外の塩基が2’-Oメチル化修飾されている請求項1のヘプタマー型スモールガイド核酸。
【請求項3】
標的RNAが疾患に関連する遺伝子から転写されたRNAである請求項1または2のヘプタマー型スモールガイド核酸。
【請求項4】
疾患に関連する遺伝子がガンの発症に関連する遺伝子である請求項3のヘプタマー型スモールガイド核酸。
【請求項5】
請求項3または4のヘプタマー型スモールガイド核酸の有効量を含有する薬剤。
【図1】

【図2】
【図3】
【図2】
【図3】
【公開番号】特開2012−217421(P2012−217421A)
【公開日】平成24年11月12日(2012.11.12)
【国際特許分類】
【出願番号】特願2011−88479(P2011−88479)
【出願日】平成23年4月12日(2011.4.12)
【出願人】(505111982)学校法人 新潟科学技術学園 新潟薬科大学 (7)
【Fターム(参考)】
【公開日】平成24年11月12日(2012.11.12)
【国際特許分類】
【出願日】平成23年4月12日(2011.4.12)
【出願人】(505111982)学校法人 新潟科学技術学園 新潟薬科大学 (7)
【Fターム(参考)】
[ Back to top ]