
LPA4受容体ノックアウト動物の解析
【課題】 本発明は、LPA4受容体を介したLPAの作用を解明し、その作用を利用した薬剤スクリーニングの方法を提供することを課題とする。
【解決手段】 本発明者らは、LPA4受容体のノックアウト動物を作出し、そのノックアウト動物を詳細に検討した結果、LPA4受容体を介したLPAからのシグナルの欠損が、抗糖尿病作用を示すことを見出し、その結果に基づいて、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法についての発明を完成した。
【解決手段】 本発明者らは、LPA4受容体のノックアウト動物を作出し、そのノックアウト動物を詳細に検討した結果、LPA4受容体を介したLPAからのシグナルの欠損が、抗糖尿病作用を示すことを見出し、その結果に基づいて、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法についての発明を完成した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、LPA4受容体を介したLPAの作用を解明し、その作用を利用した薬剤スクリーニングの方法を提供することに関する。
【背景技術】
【0002】
糖尿病は、発症機序により1型糖尿病と2型糖尿病に分類される。1型糖尿病の発症機序は、膵臓の膵島β細胞が自己免疫により破壊されて、そのインスリン産生が完全に停止することによる、インスリンの分泌低下により生じることが知られており、全病態のうち5%を占める病態である。この1型糖尿病は、外部からインスリンを投与することにより治療する。
【0003】
一方、2型糖尿病は、インスリンの分泌不足とインスリン抵抗性がベースにあり、結果としてインスリンの作用不足により生じることが知られており、全病態のうち95%を占める病態である。
【0004】
現在、2型糖尿病の治療のためには、血糖を降下させる薬剤を投与することが一般的であり、そのような血糖を降下させる薬剤としては、インスリンの分泌を促進させる薬剤;腸管からの糖質の分解・吸収を遅延させて食後高血糖を抑制する薬剤;インスリン抵抗性を改善する薬剤;等が使用されている。しかしながら、どの薬にも副作用が報告され、問題となっている。
【0005】
具体的には、インスリンの分泌を促進させる薬剤の場合には、長期に飲み続けていると飲み始めた頃に比べ、効果が薄れてきたり効かなくなってくる(すなわち、膵臓のβ細胞が疲弊する)こと、そして投与後にインスリンが分泌過剰になって低血糖に陥り、食欲亢進により体重が増加しやすい等の問題点が指摘されている。
【0006】
腸管からの糖質の分解・吸収を遅延させて食後高血糖を抑制する薬剤の場合には、食後高血糖を抑制する効果はそれほど強力ではなく、また分解が遅れて腸管に残った糖類が発酵してガスが出たり、お腹が張ったり、下痢を引き起こす等の問題点や、稀に肝障害が起こる等の問題点が指摘されている。
【0007】
インスリン抵抗性を改善する薬剤の場合には、むくみやすく、心不全の患者には投与できないこと、体重が増加しやすく、稀に肝障害や貧血が生じる等の問題点が指摘されている。
【0008】
さらに、いずれの場合にも、現在存在している血糖降下薬を適切に処方しても、2型糖尿病患者の治療の良好例は、患者の約30%程度にしか見られないという、これらの副作用以外の問題点も存在している。
【0009】
リゾホスファチジン酸(lysophosphatidic acid:LPA)は、多数の細胞種により天然に合成される生物活性を持ったグリセロリン脂質の1種であり、グリセリン骨格のひとつの水酸基に脂肪酸が結合した物質である。LPAは、グリセロリン脂質がホスホリパーゼなどの作用により加水分解されて生ずる物質であり、1-アシルグリセロール-3-リン酸、2-アシルグリセロール-3-リン酸、1-アルケニル-グリセロール-3-リン酸などがLPAの代表例として知られており、細胞増殖、細胞遊走、平滑筋収縮、細胞生存、免疫応答などを含む複数の生理学的プロセスに関与している(非特許文献1〜3など)。その多様な生物学的作用の中でも、LPAは、脂肪細胞の増殖と機能を媒介すること、そしてグルコース代謝を制御すること、が示されてきた(非特許文献4〜6)。
【0010】
LPAに対する4番目のLPA受容体(LPA4受容体、GPR23とも呼ばれる)は、それ以前に見つかっていたLPA1〜LPA3の3種類のLPA受容体との間で、わずか20〜24%のアミノ酸同一性しか有さなかった(特許文献1、非特許文献7)。
【0011】
LPA4受容体(GPR23)は、生殖組織、脳組織、脂肪組織において高レベルで発現される。LPAは、LPA4受容体(GPR23)内在化、G12/13-媒介性およびRho-媒介性の神経突起退縮およびストレスファイバー形成、Gq-媒介性Ca2+動員およびGi-媒介性Ca2+動員、Gs-媒介性cAMP増加を誘導することが示された(非特許文献8)。LPA4受容体(GPR23)は、ニューロン新生やニューロン移動等の神経発生プロセスにおいて極めて重要な役割を果たしている可能性があると考えられている(非特許文献9)。しかしながら、LPA4受容体(GPR23)の生理学的機能および生物学的機能については、非常に限定的な情報しか得られていない。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】WO2004/063224
【非特許文献】
【0013】
【非特許文献1】van Corven EJ, et al., Cell, 1989, 59, 45-54 .
【非特許文献2】Amano M, et al., Science, 1997, 275, 1308-1311 .
【非特許文献3】Moolenaar WH, Curr. Opin. Cell. Biol., 1995, 7, 203-210 .
【非特許文献4】Valet P, et al., J. Clin. Invest., 1998, 101, 1431-1438 .
【非特許文献5】Simon MF, et al., J. Biol. Chem., 2005, 280, 14656-14662 .
【非特許文献6】Yea K, et al., J. Mol. Med., 2008, 86, 211-220 .
【非特許文献7】Noguchi K, et al., J. Biol. Chem., 2003, 278, 25600-25606 .
【非特許文献8】Lee CW, et al., J. Biol. Chem., 2007, 282, 4310-4317 .
【非特許文献9】Yanagida K, et al., J. Biol. Chem., 2007, 282, 5814-5824 .
【発明の概要】
【発明が解決しようとする課題】
【0014】
本発明は、LPA4受容体を介したLPAの作用を解明し、その作用を利用した薬剤スクリーニングの方法を提供することを課題とする。
【課題を解決するための手段】
【0015】
本発明者らは、LPA4受容体のノックアウト動物を作出し、そのノックアウト動物を詳細に検討した結果、LPA4受容体を介したLPAからのシグナルの欠損が、抗糖尿病作用を示すことを見出し、その結果に基づいて、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法についての発明を完成した。
【0016】
具体的には、本発明において、高脂肪食負荷モデルの非ヒト動物に対して、LPA4受容体に結合する被検化合物を投与する工程;被検化合物を投与した高脂肪食負荷モデルの非ヒト動物において、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せを調べる工程;抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を、LPA4受容体に結合するLPA4アンタゴニストであると同定する工程;を行うことにより、LPA4受容体に結合するLPA4アンタゴニストである化合物をスクリーニングすることができることを見出した。
【発明の効果】
【0017】
本発明により、複数存在するLPA受容体のうちLPA4受容体を介して、LPAがどの様なメカニズムで生体内において機能を果たしているかを明らかにすることができた。また、そのような知見に基づいて、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を選択することができるスクリーニング方法を開発することができた。
【図面の簡単な説明】
【0018】
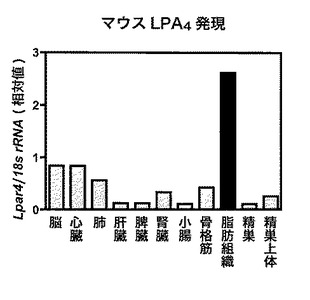
【図1】図1は、野生型マウス(WTマウス)の各臓器における、LPA4受容体の相対遺伝子発現値を示す。
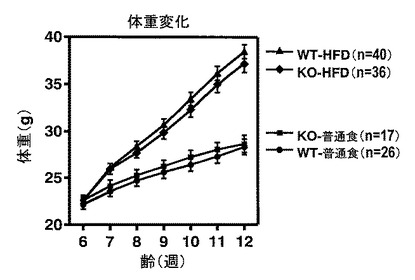
【図2】図2は、WTマウスおよびLPA4受容体欠損マウス(KOマウス)のそれぞれについて、「普通食」群および「高脂肪食(HFD)」群のそれぞれにおける体重変化を示す。
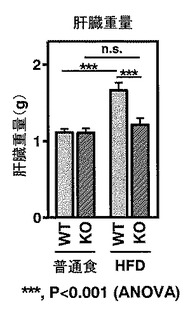
【図3】図3は、WTマウスおよびKOマウスのそれぞれについて、「普通食」群および「HFD」群のそれぞれにおける肝臓重量を示す。
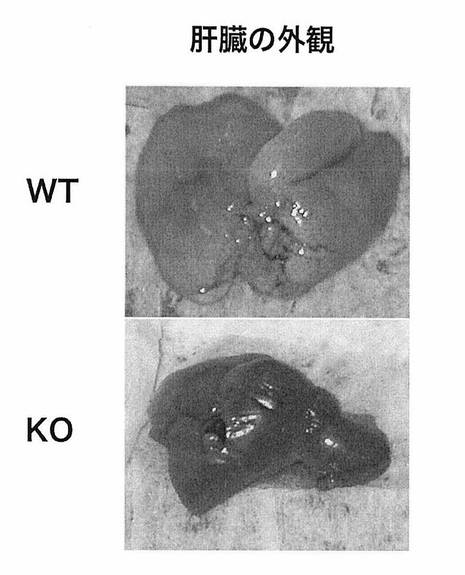
【図4】図4は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれについて、肝臓の外観を示す。
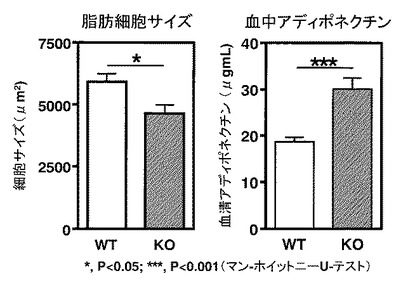
【図5】図5は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれにおける、脂肪細胞のサイズ(図5左)および血中アディポネクチン濃度(図5右)を示す。
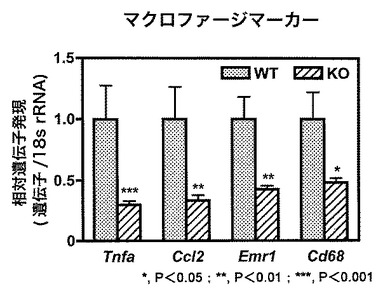
【図6】図6は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれの脂肪組織における、TNF-α(「Tnfa」)、CCL2、F4/80、CD68のそれぞれのマーカーの相対遺伝子発現値を示す。
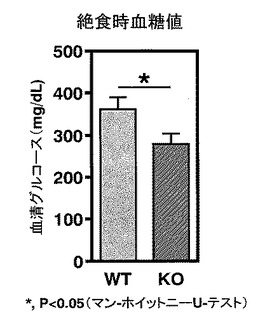
【図7】図7は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれにおける、絶食時の血糖値を示す。
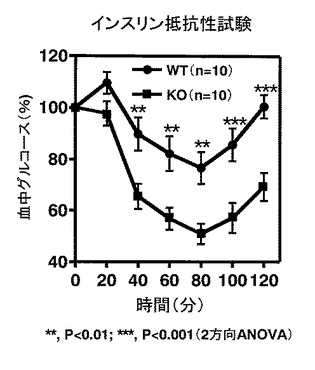
【図8】図8は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれにおける、インスリン抵抗性試験の結果を示す。
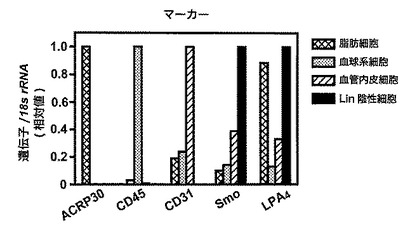
【図9】図9は、WTマウスの脂肪細胞、血球系細胞、血管内皮細胞そしてLin陰性細胞(すなわち、前駆脂肪細胞)のそれぞれにおける、各種細胞マーカー(すなわち、脂肪細胞マーカーであるACRP30(アディポネクチン)、血球系細胞マーカーであるCD45、血管内皮細胞マーカーであるCD31、前駆脂肪細胞マーカーであるSmoothened(Smo))及びLPA4受容体の遺伝子発現を示す。
【図10】図10は、WTマウス由来の胎児線維芽(MEF)細胞およびKOマウス由来のMEF細胞のそれぞれにおける、アディポネクチン(ACRP30)を指標とした脂肪細胞分化能の比較を示す。
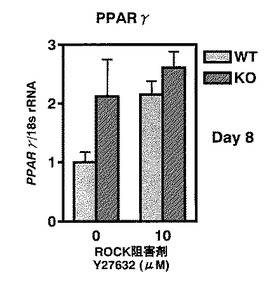
【図11】図11は、Rho結合キナーゼの活性を薬剤(Y27632;10μM)により阻害した場合、WTマウス由来のMEF細胞から脂肪細胞への分化がKOマウス由来のMEF細胞と同等までに亢進することを示す。
【発明を実施するための形態】
【0019】
リゾホスファチジン酸(lysophosphatidic acid:LPA)は、多数の細胞種により天然に合成される生物活性を持ったグリセロリン脂質の1種であり、グリセリン骨格のひとつの水酸基に脂肪酸が結合した物質であり、以下の構造式を有する化合物である。
【0020】
【化1】

【0021】
本発明者らは以前に、LPAに対する4番目のLPA受容体(LPA4受容体)の遺伝子およびアミノ酸配列を同定した(特許文献1)。しかしながら、これまでは、LPAがLPA4受容体を介してどの様な作用を発揮するのかについて、知られていなかった。本発明の発明者らは、LPA4受容体が関与するLPAからのシグナルが、どの様な作用を有しているかを明らかにするため、まずLPA4受容体のノックアウト動物を作出した。そして、このノックアウト動物に高脂肪食を負荷した場合における様々な生理学的反応および組織学的反応を検索した結果、このノックアウト動物においては、以下の様な特徴:
・LPA4受容体の欠損は、肥満そのものの進行には影響しないこと(例えば、図2);
・LPA4受容体の欠損により、脂肪肝が軽減したこと(例えば、図3および図4);
・LPA4受容体の欠損により、脂肪細胞の小型化が生じたこと(例えば、図5);
・LPA4受容体の欠損により、血中アディポネクチン濃度が上昇したこと(例えば、図5);
・LPA4受容体の欠損により、脂肪組織の炎症が軽減されたこと(例えば、図6);
・LPA4受容体の欠損により、インスリン抵抗性が改善したこと(例えば、図7および図8);
などが示されることが明らかになった。これらの特徴から、LPA4受容体を介したLPAからのシグナルの欠損が、抗糖尿病作用を示すことが明らかになった。
【0022】
これらの知見に基づいて、本発明は一態様において、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法を提供する。このスクリーニング方法においては、LPA4受容体に結合する被検化合物のうち、投与した際に、LPA4受容体の欠損の結果として生じた上述した特徴(図2〜図8に示したもの)のいずれかまたはそれらの組合せと同一の特徴を示すことができる化合物を、LPA4アンタゴニストとして選択することを特徴とする。
【0023】
具体的には、本発明は、高脂肪食負荷モデルの非ヒト動物に対して、LPA4受容体に結合する被検化合物を投与する工程;被検化合物を投与した高脂肪食負荷モデルの非ヒト動物において、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せを調べる工程;抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を、LPA4受容体に結合するLPA4アンタゴニストであると同定する工程;を含む、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法を提供する。
【0024】
上述した実施例の結果から推定されるLPA4受容体を介する生理学的メカニズムは、以下の通りである。脂肪細胞から分泌されたLPA産生酵素(オートタキシン:ATX)は細胞外でLPAを産生する。このLPAは、野生型の細胞の場合(すなわち、LPA4受容体が正常に存在する場合)には、前駆脂肪細胞に働き、脂肪細胞への分化を抑制する。マウスが脂肪負荷された場合、脂肪細胞は脂肪を蓄積する必要に迫られるが、脂肪細胞が増えないこのような状況下では一つ一つの脂肪細胞に過負荷が掛かるために細胞は徐々に肥大し、悪玉脂肪細胞へと変貌する。その結果、脂肪組織では炎症が起こり、また血中アディポネクチン量が低下する。このことが原因となり、マウスはインスリン抵抗性や脂肪肝といった生活習慣病を発症することになる。
【0025】
逆にLPA4受容体が欠損している場合には、LPA4受容体が欠損した前駆脂肪細胞は、LPAによって脂肪細胞への分化が抑制されない。そのため、マウスに脂肪が負荷された場合、脂肪細胞が容易に増えることができる。したがって、脂肪細胞一つあたりの脂肪蓄積の負担は軽く、細胞の肥大は最小限に抑えられる(善玉脂肪細胞)。このことは則ち、脂肪組織の量(重さ)は同じでも脂肪細胞の質が維持されることを意味し、生活習慣病がLPA4-KOマウスには発症しないこととなる。
【0026】
本発明のスクリーニング方法において選択することができるLPA4アンタゴニストは、まず、LPA4受容体に結合することができるという特徴を有する。これは、目的とするLPA4アンタゴニストの特徴が、LPA4受容体の欠損の結果として生じた上述した特徴(図2〜図8に示したもの)と同じものであることから、LPAがLPA4受容体と結合することを介して伝達されるシグナルが拮抗(アンタゴナイズ)されることが必要だからである。なお、本願発明において目的とする選択すべき化合物は、LPA4のインバースアゴニスト(逆作動薬)も含む。
【0027】
本発明者らの研究により、LPAがLPA4受容体に結合することにより細胞内にシグナルが伝達され、そのシグナル伝達が前駆脂肪細胞内におけるPPARγの発現を抑制していると考えられること;そしてその一方で、LPA4受容体が欠損することにより、すなわちLPA4が拮抗(アンタゴナイズ)されることにより、LPAのLPA4受容体に対する結合によって伝達されるシグナルがなくなることにより、PPARγの発現の抑制効果が失われ、PPARγの発現が亢進されること;が明らかになった。
【0028】
本発明の上述したスクリーニング方法においては、被検化合物を高脂肪食負荷モデルの非ヒト動物に対して投与した後、当該非ヒト動物体内において、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せが生じたかどうかを調べる工程を、行うことを特徴とする。
【0029】
抗糖尿病作用、抗動脈硬化作用および/または抗肥満作用を調べるため、例えば、脂肪細胞内におけるPPARγの発現亢進の検出、脂肪細胞過形成の検出、血中アディポネクチン濃度の増加の検出、またはインスリン抵抗性の改善の検出、を単独でまたはいずれかの組合せで行うことにより、LPA4受容体の欠損の結果として生じた上述した特徴(図2〜図8に示したもの)と同じ特徴を検出することができる。
【0030】
ここで、脂肪細胞内におけるPPARγの発現亢進を検出する場合、例えば、脂肪組織サンプルにおけるPPARγ mRNA発現の検出、PPARγに対する抗体を用いた免疫アッセイ、PPARγが標的とする遺伝子由来のmRNA発現の検出により行うことができる。
【0031】
また、脂肪細胞過形成を検出する場合、例えば、脂肪組織の組織学的解析、細胞数測定、DNA含有量測定により行うことができる。
【0032】
また、血中アディポネクチン濃度の増加を検出する場合、例えば、血液サンプルにおける抗アディポネクチン抗体を用いた免疫アッセイやアディポネクチン受容体への結合アッセイにより行うことができる。
【0033】
また、インスリン抵抗性の改善を検出する場合、例えば、HOMA-IRの測定、血中インスリンレベルの測定、OGTT法による評価、インスリン抵抗性試験による評価、正常血糖高インスリンクランプ法による評価により行うことができる。
【0034】
これらの方法はいずれも、当該技術分野において公知の方法であり、市販のキット、様々な書籍(例えば、糖尿病研究ストラテジー(秀潤社))、公知文献(例えば、Kubota N, et al., Mol. Cell, 1999, 4, 597-609.)等にしたがって行うことができる。
【0035】
本発明のスクリーニング方法に従って選択された被検化合物は、LPA4受容体に結合してLPAの作用を拮抗することができるとともに、脂肪肝を軽減すること、脂肪細胞の小型化を生じること、血中アディポネクチン濃度を上昇させること、脂肪組織の炎症が軽減されること、インスリン抵抗性が改善されること、等の作用を発揮することができる。
【0036】
本発明を以下の実施例により更に詳細に説明する。しかしながら、以下の実施例は、本発明の例示であって、本発明を限定することを意図するものではない。
【実施例】
【0037】
実施例1:脂肪組織におけるLPA4の高発現
本実施例においては、LPA4受容体を介したLPAからのシグナルが、どの様な作用を有するのかを明らかにすることを目的として、まずマウス各種臓器におけるLPA4受容体の遺伝子発現を調べた。
【0038】
具体的には12週齢の野生型マウス(WTマウス)から、脂肪組織を含む11臓器を採取し各臓器から全RNAを抽出し(RNeasy, Qiagen)、逆転写酵素(Superscript III, Invitrogen)によりcDNAを作製した後、これを鋳型として、ファストスタートSYBR Greenマスター(Roche)を使用して、定量PCRシステム(LightCycler, Roche)によりLPA4と18S rRNA(内部標準)の発現を定量した。
【0039】
PCRにおいて遺伝子発現を増幅するために、それぞれの遺伝子について、それぞれのヌクレオチド配列:
LPA4受容体(GenBank Accession No. NM_175271)
フォワードプライマー:gtcaacaatg cgaccaccac(SEQ ID NO: 1)
リバースプライマー:aagcaccaca gaagaacaag aaaca(SEQ ID NO: 2)
18S rRNA(GenBank Accession No. NR_003278)
フォワードプライマー:tcaacacggg aaacctcacc(SEQ ID NO: 3)
リバースプライマー:cagacaaatc gctccaccaa c(SEQ ID NO: 4)
を有するプライマー対を使用した。
【0040】
結果を図1に示す。この図において、各種臓器のLPA4の発現の相対値を示す。
その結果、LPA4は脂肪組織に強く発現することが明らかとなった。一方で、インスリン感受組織である肝臓や骨格筋においてはLPA4の発現は極めて低いことがわかった。
【0041】
実施例2:LPA4受容体欠損マウス(KOマウス)の作出
本実施例においては、LPA4受容体を介したLPAからのシグナルが、どの様な作用を有するのかを明らかにすることを目的として、LPA4受容体欠損マウスを作出することとした。
【0042】
まず、C57BL/6マウスのゲノムにおけるLPA4遺伝子のオープンリーディングフレーム(ORF)上流のEcoO65I/NcoI断片(4.0 kb)とLacZ DNA断片、ホスホグリセリン酸キナーゼプロモーターにトランスポゾンTn5ネオマイシン耐性遺伝子を連結したDNA断片(pgk/neor)、LPA4遺伝子のORF下流のXbaI/EcoO65I断片(6.3 kb)を使って、pBluescript KS M13(+)(Stratagene)をベクターとしてターゲッティングベクターを構築した。
【0043】
次に、C57BL/6マウス由来のES細胞株であるRENKA株にEco81Iで直線化した上記ターゲッティングベクターをエレクトロポレーションで導入し、形質転換株をG418(Gibco)で選択したのち、この中から相同組換えES細胞をサザンブロット解析によってスクリーニングした。
【0044】
さらに、この相同組換えES細胞をICRマウスの胚盤胞に注入してキメラマウスを作り、そのうちの雄マウスを雌のC57BL/6マウスと交配させることで雌のLPA4のヘテロザイゴートマウス(Lpar4+/-マウス)を得た。雄のC57BL/6マウスとLpar4+/-マウスを交配させることで、同腹仔である雄の野生型マウス(Lpar4+/Y; WTマウス)とLPA4受容体欠損マウス(Lpar4-/Y; KOマウス)を作出した。
【0045】
実施例3:高脂肪食負荷モデルにおける体重変化
本実施例においては、肥満とそれに伴うメタボリックシンドロームの進行におけるLPA4受容体の機能について検討すべく、野生型マウス(WTマウス)とLPA4受容体欠損マウス(KOマウス)を用いた高脂肪食負荷モデルでの体重の変化を測定した。
【0046】
具体的には、WTマウスおよびKOマウスのそれぞれについて、4週齢時から単独飼育し、6週齢時より普通食を6週間摂食させた「普通食」群、または6週齢時より高脂肪食(HFD32; 日本クレア社)を6週間摂食させた高脂肪食負荷モデル(マウスの高脂肪食負荷モデル、「HFD」群)を採用した。ここで、WTマウスの「普通食」群(WT-普通食)はn=26で実験を行い(●)、WTマウスの「HFD」群(WT-HFD)はn=40で実験を行い(▲)、KOマウスの「普通食」群(KO-普通食)はn=17で実験を行い(■)、そしてKOマウスの「HFD」群(KO-HFD)はn=36で実験を行った(◆)。結果を図2に示す。図2は、WTマウスおよびKOマウスのそれぞれについて、「普通食」群および「HFD」群のそれぞれにおける体重変化を示す。
【0047】
図2のグラフに示すように、「HFD」群は「普通食」群に比較して、著しい体重増加を認めた。しかし、その著しい体重増加について、WTマウスとKOマウスとの間に差異は認められなかった。この結果から、LPA4受容体を介したLPAシグナルの導入は、肥満そのものの進行には寄与しないことが示唆された。
【0048】
実施例4:LPA4受容体欠損マウスにおける脂肪肝の軽減
本実施例においては、実施例3において示された高脂肪食負荷モデル(「HFD」群)での著しい体重増加に関して、脂肪組織に高発現するLPA4受容体が脂肪の質的変化に対して何らかの影響を及ぼすことにより、メタボリックシンドロームの進行に関与する可能性を考え、まず肝臓に注目して脂肪肝の程度について検討した。
【0049】
内蔵脂肪型肥満は、インスリン抵抗性や脂肪肝などを引き起こし、これらが引き金となって糖尿病、高脂血症、高血圧につながり、最終的には動脈硬化の進行を加速させると考えられている。これがメタボリックシンドロームの概念である。一方、肥満つまり脂肪の「量的」増加だけではなく、その脂肪の「質的」変化がこのメタボリックシンドロームの進行に強く関わっていることが、最近の研究により明らかとなってきている。
【0050】
実施例3において記載したようにWTマウスの「普通食」群、WTマウスの「HFD」群、KOマウスの「普通食」群、そしてKOマウスの「HFD」群のマウスを飼育し、その後16時間絶食後、麻酔下で開腹・安楽死させた後、肝臓を採取しその重量および外観を比較した。統計処理をANOVAにより行った。結果を図3および図4に示す。
【0051】
WTマウスにおいて、「HFD」群は「普通食」群と比較して、肝臓重量が大幅に増加していた(***, P<0.001)(図3)。また、WTマウスの「HFD」群における肝臓の外観を調べたところ、白みを帯び、肥大化しており、脂肪肝の様相を呈していた(図4上)。
【0052】
一方、KOマウスにおいては、「HFD」群においても、重量が「普通食」群に近く、外観も脂肪肝の程度が著しく減弱していることがわかった(図3、図4下)。この結果から、LPA4受容体を介したLPAからのシグナルの導入が、脂肪肝の進行に強く関与することが明らかとなった。
【0053】
実施例5:LPA4受容体欠損マウスにおける脂肪細胞小型化と血中アディポネクチン高値
本実施例においては、LPA4受容体欠損マウスにおけるLPA4受容体の欠損が、メタボリックシンドロームに関わる他の生理学的指標に対してどの様な作用を有するかを調べることを目的として行った。
【0054】
実施例3および4において示したように、高脂肪食負荷モデル(「HFD」群)において、LPA4受容体は肥満そのものの進行には寄与しないが、脂肪肝の進行に寄与することがわかった(図2〜4)。一方で、実施例2において示したように、LPA4受容体の発現そのものは肝臓では極めて低く、脂肪組織において非常に強い(図1)。脂肪肝の進行には脂肪組織の「質的」変容が強く関与していることが、近年の研究により明らかになってきている。これらの知見を綜合して、脂肪組織におけるLPA4受容体が、脂肪組織の「質」に影響し結果としてWTマウスとKOマウスとの間での脂肪肝の程度の違いにつながっている可能性が予想された。その可能性について検討すべく、WTマウスおよびKOマウスの脂肪組織(重量や脂肪細胞サイズ)を比較した。
【0055】
6週齢時より高脂肪食(HFD32; 日本クレア社)を8週間摂食させた以外は実施例2に記載したように飼育した「HFD」群の精巣上体周囲の脂肪組織を採取し、その重量を比較した。そして、得られた脂肪組織をホルマリン固定した後、パラフィン中に包埋し、厚さ4μmの薄切切片を作製し、ヘマトキシリン・エオジン法により染色し、顕微鏡観察により脂肪細胞の大きさを測定した。この実験においては、各サンプル(WTマウスについての「HFD」群およびKOマウスについての「HFD」群)につき、200個以上の細胞を測定し、平均を算出した。統計処理をマン・ホイットニーのUテストにより行った。
【0056】
この結果、まず、WTマウスとKOマウスとの間に脂肪組織重量の差は認められなかった(グラフは示されていない)。WTマウスの「HFD」群における脂肪細胞のサイズと比較して、KOマウスの「HFD」群における脂肪細胞のサイズが、有意に小型であることが示された(*, P<0.05)(図5左)。
【0057】
一般に、小型化した脂肪細胞は、脂肪肝やインスリン抵抗性を改善させる善玉ホルモンであるアディポネクチンの産生能が高いこと、そして肥大した脂肪細胞においてはその産生が著しく減弱していることが知られている。そこで、脂肪細胞サイズの違いが循環アディポネクチンの増加をもたらしている可能性があるため、血中アディポネクチン量を検討した。統計処理をマン・ホイットニーのUテストにより行った。
【0058】
上述したWTマウス(「HFD」群)およびKOマウス(「HFD」群)から採取した血液より血清を調製し、血清中のアディポネクチン濃度をELISA(Mouse Adiponectin Elisa kit, 大塚製薬社)を用いて検討した。この結果、WTマウス(「HFD」群)と比較して、KOマウス(「HFD」群)において、有意に高いアディポネクチン濃度が検出された(***, P<0.001)(図5右)。
【0059】
実施例6:LPA4受容体欠損マウスにおける脂肪組織炎症の軽減
一般に、肥大した脂肪細胞ではアディポネクチンの産生が低下するが、代わりにMCP-1(CCL2)やTNF-αといった炎症性のサイトカインの産生が亢進することが知られている。これにより、マクロファージを中心とした炎症性細胞が脂肪組織に浸潤し、炎症が起こる。脂肪肝の背景にこの脂肪組織の炎症があることが最近の研究により明らかになってきている。
【0060】
実施例5において脂肪細胞のサイズが低下し、アディポネクチン濃度が上昇したことから、本実施例においては、KOマウスにおけるマクロファージを中心とした炎症性細胞の浸潤や炎症の程度を確認した。
【0061】
実施例5と同様に6週齢時より高脂肪食(HFD32; 日本クレア社)を8週間摂食させたWTマウスおよびKOマウスのそれぞれの「HFD」群のマウスの脂肪組織から、全RNAを抽出し(RNeasy, Qiagen)、逆転写酵素(Superscript III, Invitrogen)によりcDNAを作製した後、これを鋳型として、炎症のマーカーであるTNF-α(図6中では「Tnfa」と示す)、CCL2、マクロファージマーカーであるF4/80、CD68のそれぞれについて、蛍光試薬としてファストスタートSYBR Greenマスター(Roche)を使用して、定量PCRシステム(LightCycler, Roche)により炎症に関係する遺伝子発現を定量した。
【0062】
PCRにおいてそれぞれの遺伝子発現を増幅するために、それぞれヌクレオチド配列:
TNF-α(GenBank Accession No. NM_013693)
フォワードプライマー:ccagaccctc acactcagat cat(SEQ ID NO: 5);
リバースプライマー:tggcaccact agttggttgt ctt(SEQ ID NO: 6);
CCL2(GenBank Accession No. NM_0111333)
フォワードプライマー:ccactcacct gctgctactc att(SEQ ID NO: 7);
リバースプライマー:cagcacagac ctctctcttg agc(SEQ ID NO: 8);
F4/80(GenBank Accession No. NM_010130)
フォワードプライマー:ggcttcagtg ggatgtacag atc(SEQ ID NO: 9);
リバースプライマー:tgaaagttgg tttgtccatt gct(SEQ ID NO: 10);
CD68(GenBank Accession No. NM_009853)
フォワードプライマー:ctccctgtgt gtctgatctt gct(SEQ ID NO: 11);
リバースプライマー:ctgtgctttc tgtggctgta ggt(SEQ ID NO: 12);
を有するプライマー対を使用した。
【0063】
結果を図6に示す。この図において、それぞれのマーカーについて、WTマウス「HFD」群における発現に対するKOマウス「HFD」群における発現の相対値を示す。統計処理をマン・ホイットニーのUテストにより行った。
【0064】
その結果、KOマウス「HFD」群において、全てのマーカーの相対遺伝子発現レベルが有意に低いことが明らかになった(*, P<0.05;**, P<0.01;***, P<0.001)。このことから、実際にKOマウスの脂肪組織においてマクロファージを中心とした炎症が軽減していることがわかった(図6)。
【0065】
実施例7:LPA4受容体欠損マウスにおける糖尿病・インスリン抵抗性の改善
これまでに示した実験結果から、KOマウスにおいて、高脂肪食負荷(「HFD」群)による脂肪細胞の肥大化が軽減しており(実施例5、図5左)、これが血中アディポネクチン濃度の亢進(実施例5、図5右)、脂肪組織炎症の軽減(実施例6、図6)につながり、脂肪肝の軽減(実施例4、図3および4)をもたらしていることが予想された。
【0066】
インスリン抵抗性は肥満による糖尿病の原因であり、メタボリックシンドロームの基盤となっている病態であり、そして脂肪肝をもたらすと考えられているアディポネクチンレベルの減少、脂肪組織炎症の亢進といった現象は、ともにインスリン抵抗性を引き起こす原因と考えられている。
【0067】
そこで、前述の実施例の結果からKOマウスにおいてインスリン抵抗性が改善している可能性が高いと考えられたことから、本実施例においては、WTマウス「HFD」群およびKOマウス「HFD」群を対象として、そのインスリン抵抗性について検討した。
【0068】
まず、実施例5と同様に6週齢時より高脂肪食(HFD32; 日本クレア社)を8週間摂食させたWTマウスおよびKOマウスのそれぞれの「HFD」群のマウスを飼育し、その後それぞれのマウスを16時間絶食後、血清を調製し、血清中に含有されるグルコース濃度を、酵素法(テストワコー, Wako)により定量した(絶食時血糖値)。統計処理をマン・ホイットニーのUテストにより行った。
【0069】
この結果、WTマウス「HFD」群と比較して、KOマウス「HFD」群では、有意に絶食時血糖値が低く、糖尿病の改善が認められた(*, P<0.05)(図7)。
【0070】
次に、実施例2に記載したように6週齢時より高脂肪食(HFD32; 日本クレア社)を6週間摂食させたWTマウスおよびKOマウス(それぞれ、n=10)を対象に、4時間の絶食の後インスリン抵抗性試験を行った。インスリン抵抗性は、血液を尾静脈より採取し、血糖値を自動血糖測定装置(グルテストNeo, 三和化学研究所)で測定することにより評価した。インスリン(ノボリンR, Novo Nordisk Pharma Ltd.)の投与は腹腔内注射により行った。血糖値はインスリン投与後から20分間隔で計2時間測定した。統計処理を2方向ANOVAにより行った。
【0071】
WTマウス「HFD」群に比べて、KOマウス「HFD」群はインスリン投与による血糖値降下が良好であり、つまりKOマウス「HFD」群ではインスリン抵抗性が軽減していることがわかった(**, P<0.01;***, P<0.001)(図8)。
【0072】
実施例8:前駆脂肪細胞におけるLPA4受容体の高発現
脂肪組織は、脂肪細胞の他、その前駆細胞である前駆脂肪細胞、血管を構成する血管内皮細胞、炎症に関わるリンパ球やマクロファージなど複数の細胞から構成される。実施例2において示したように、LPA4受容体は脂肪組織に強い発現を認めるが、これらの複数の細胞種のうちどの細胞種に発現しているかは不明であった。
【0073】
そこで、本実施例において、KOマウスが上記の表現型を示す機序を解明すべく、脂肪組織を構成する細胞を分離し、LPA4受容体の発現レベルを調べた。
【0074】
まず普通食で飼育した6週齢のWTマウスから採取した脂肪組織を、撹拌しながらコラゲナーゼで2時間処理し、低速遠心により浮遊してくる脂肪細胞と沈殿する間質・血管系画分(SVF)に分離した。引き続き得られたSVFを、フィコエリスリン(PE)ラベルされた抗CD45(血球系細胞マーカー)抗体、抗Ter-119(赤血球マーカー)抗体(ともにBiolegend)と反応させ、抗PE磁気ビーズ(Miltenyi Biotec)とAutoMACS(磁気による細胞ソーティングシステム;Miltenyi Biotec)を用いることで、血球系細胞(すなわちCD45+またはTer-119+)と非血球系細胞(すなわちCD45-/Ter-119-)に分離した。これにより分離された非血球系細胞をPEラベルされた抗CD31(血管内皮細胞マーカー)抗体(Biolegend)を用いて、上記と同様の方法で、血管内皮細胞(すなわちCD45-/Ter-119-/CD31+)とLineage陰性細胞(以下Lin-細胞(すなわち、CD45-/Ter-119-/CD31-);前駆脂肪細胞から構成される)に分離した。
【0075】
次いで、脂肪細胞、血球系細胞、血管内皮細胞、Lin-細胞から全RNAを回収し、逆転写酵素によりcDNAを作製した後、これを鋳型として、各種細胞マーカー(すなわち、脂肪細胞マーカーであるACRP30(アディポネクチン)、血球系細胞マーカーであるCD45、血管内皮細胞マーカーであるCD31、前駆脂肪細胞マーカーであるSmoothened(Smo))の遺伝子発現とLPA4受容体の遺伝子発現とを、蛍光試薬としてファストスタートSYBR Greenマスター(Roche)を使用して、定量PCRにより確認した。
【0076】
PCRにおいてそれぞれの遺伝子発現を増幅するために、それぞれヌクレオチド配列:
ACRP30(アディポネクチン)(GenBank Accession No. NM_009605)
フォワードプライマー:aaaggagatg caggtcttct tgg(SEQ ID NO: 13);
リバースプライマー:tgaacgctga gcgatacaca taa(SEQ ID NO: 14);
CD45(GenBank Accession No. NM_001111316)
フォワードプライマー:gggttgttct gtgccttgtt(SEQ ID NO: 15);
リバースプライマー:tggctgtttg ttttcttgac ttg(SEQ ID NO: 16);
CD31(GenBank Accession No. NM_001032378)
フォワードプライマー:aatcaaaata aagaccccca gaaca(SEQ ID NO: 17);
リバースプライマー:cacaggactc tcgcaatcca(SEQ ID NO: 18);
Smo(GenBank Accession No. NM_176996)
フォワードプライマー:gcaggaatgg cagagacagg(SEQ ID NO: 19);
リバースプライマー:ggcacagaaa agggcaaaca(SEQ ID NO: 20);
LPA4受容体(GenBank Accession No. NM_175271)
フォワードプライマー:gtcaacaatg cgaccaccac(SEQ ID NO: 1);
リバースプライマー:aagcaccaca gaagaacaag aaaca(SEQ ID NO: 2);
を有するプライマー対を使用した。
【0077】
結果を図9に示す。この結果から、脂肪細胞マーカーであるACRP30(アディポネクチン)、血球系細胞マーカーであるCD45、マクロファージマーカーであるCD68、血管内皮細胞マーカーであるCD31、前駆脂肪細胞マーカーであるSmoothened(Smo)の遺伝子発現を検討することで、細胞分離が的確に行なわれていることが確認された。そして、LPA4受容体の遺伝子発現を調べたところ、Lin-細胞つまり前駆脂肪細胞に最も発現が高く、次いで脂肪細胞に高く発現していることがわかった。
【0078】
従って、LPA4受容体が前駆脂肪細胞から脂肪細胞への分化に影響することで、本実施例で認められたWTマウスとKOマウスとの違いをもたらしている可能性が考えられた。
【0079】
実施例9:LPA4受容体欠損マウス由来線維芽細胞における脂肪細胞分化の亢進
実施例8で示したように、LPA4受容体が前駆脂肪細胞に強く発現することがわかっため、LPA4受容体が脂肪細胞への分化に関与している可能性がさらに示唆された。
【0080】
そこで、本実施例において、脂肪細胞分化を検討する一般的な手法である、マウス胎児線維芽細胞(MEF細胞)を用いた脂肪細胞分化実験を行なった。MEF細胞は、前駆脂肪細胞と同様にin vitroで脂肪細胞へ分化させることができることが知られている。
【0081】
WTマウスおよびKOマウスの胎齢13.5日(E13.5)のマウス胎児より、MEF細胞をそれぞれ採取し、ウシ胎児血清(Gibco)を10%含有したDMEM(Nacalai tesque)により細胞培養ディッシュ(Corning)上で培養した。それぞれのMEF細胞は、コンフルエントまで培養してから2日後にインスリン(10μg/ml)、デキサメサゾン(0.25μM)、IBMX(0.5 mM)(全てSigma)の3種類の薬剤で処理することにより、脂肪細胞へと分化させた。
【0082】
上述した3種類の薬剤(インスリン、デキサメサゾン、IBMX)処理を開始した時点をday0とし、day12まで分化を進行させ、day0、day4、day8、day12の各時点において全RNAを回収し、逆転写酵素を使いcDNAを作製した。脂肪細胞への分化の程度については、脂肪細胞特異的に発現するアディポネクチン(ACRP30)の発現量を上記cDNAにより定量PCRで評価した(実施例8を参照)。
【0083】
成熟脂肪細胞のマーカーであるアディポネクチン(ACRP30)の発現レベルの時間変化を図10に示す。WTマウス由来MEF細胞およびKOマウス由来MEF細胞のそれぞれを、脂肪細胞へと分化させると、KOマウス由来のMEF細胞は、WTマウスのMEF細胞に比べて、高い脂肪細胞分化能をもつことがわかった(図10)。このことから、LPA4受容体欠損マウス由来のMEF細胞は野生型マウス由来のMEF細胞よりも早く脂肪細胞へと成熟することが明らかとなった。
【0084】
また、本発明者らは以前に、LPA4受容体が、Rhoと呼ばれる因子とその下流で働くキナーゼであるRho結合キナーゼ(ROCK)を活性化することを明らかにしており(非特許文献9)、このシグナル伝達経路が脂肪細胞分化に寄与する可能性が考えられた。そこで、本実施例においては、脂肪細胞分化実験におけるROCK阻害剤であるY27632の効果について検討した。
【0085】
上記と同様の分化実験を、10μM Y27632(Calbiochem)の存在下または非存在下の2条件で行なった。上述した3種類の薬剤(インスリン、デキサメサゾン、IBMX)処理を開始した時点をday0とし、day8まで分化を進行させ、全RNAを回収し、逆転写酵素を使いcDNAを作製した。脂肪細胞への分化の程度については、脂肪細胞特異的に発現するPPARγ(GenBank Accession No. NM_011146)の発現量を、フォワードプライマー:gaagaccact cgcattcctt tg(SEQ ID NO: 21);およびリバースプライマー:gcaggttcta ctttgatcgc actt(SEQ ID NO: 22);を有するプライマー対を得られたcDNAに対して使用して、実施例8に記載するように定量PCRにより評価した。
【0086】
結果を図11に示す。図10の結果と矛盾せず、KOマウス由来の細胞はWTマウス由来の細胞よりも、脂肪細胞マーカーであるPPARγの発現レベルが高かった。一方、LPA4受容体はROCKを活性化するシグナルを細胞内へ送るが、ROCKの活性を阻害剤(Y27632;10μM)で抑制すると、WTマウス由来の細胞のPPARγの発現レベルは、KOマウス由来の細胞と同程度まで亢進することが明らかとなった(図11)。しかもKOマウス由来の細胞は薬剤による影響を大きくは受けないことも示された。
【0087】
以上の結果から、前駆脂肪細胞では、LPA4受容体を介して伝達されるシグナルが、ROCKを活性化させることを通じてPPARγの発現を抑制しており、その結果として脂肪細胞の分化を抑制する役割を持つことが示された。
【産業上の利用可能性】
【0088】
本発明により、LPAが複数存在するLPA受容体のうち、LPA4受容体を介して、どの様なメカニズムで生体内において機能を果たしているかを明らかにすることができた。また、そのような知見に基づいて、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を選択することができるスクリーニング方法を開発することができた。
【配列表フリーテキスト】
【0089】
SEQ ID NO: 1:Forward primer for amplifying LPA4 receptor gene.
SEQ ID NO: 2:Reverse primer for amplifying LPA4 receptor gene.
SEQ ID NO: 3:Forward primer for amplifying 18S rRNA gene.
SEQ ID NO: 4:Reverse primer for amplifying 18S rRNA gene.
SEQ ID NO: 5:Forward primer for amplifying TNF-alpha gene.
SEQ ID NO: 6:Reverse primer for amplifying TNF-alpha gene.
SEQ ID NO: 7:Forward primer for amplifying CCL2 gene.
SEQ ID NO: 8:Reverse primer for amplifying CCL2 gene.
SEQ ID NO: 9:Forward primer for amplifying Emr1 gene.
SEQ ID NO: 10:Reverse primer for amplifying Emr1 gene.
SEQ ID NO: 11:Forward primer for amplifying CD68 gene.
SEQ ID NO: 12:Reverse primer for amplifying CD68 gene.
SEQ ID NO: 13:Forward primer for amplifying ACRP30 gene.
SEQ ID NO: 14:Reverse primer for amplifying ACRP30 gene.
SEQ ID NO: 15:Forward primer for amplifying CD45 gene.
SEQ ID NO: 16:Reverse primer for amplifying CD45 gene.
SEQ ID NO: 17:Forward primer for amplifying CD31 gene.
SEQ ID NO: 18:Reverse primer for amplifying CD31 gene.
SEQ ID NO: 19:Forward primer for amplifying Smo gene.
SEQ ID NO: 20:Reverse primer for amplifying Smo gene.
SEQ ID NO: 21:Forward primer for amplifying PPAR gamma gene.
SEQ ID NO: 22:Reverse primer for amplifying PPAR gamma gene.
【技術分野】
【0001】
本発明は、LPA4受容体を介したLPAの作用を解明し、その作用を利用した薬剤スクリーニングの方法を提供することに関する。
【背景技術】
【0002】
糖尿病は、発症機序により1型糖尿病と2型糖尿病に分類される。1型糖尿病の発症機序は、膵臓の膵島β細胞が自己免疫により破壊されて、そのインスリン産生が完全に停止することによる、インスリンの分泌低下により生じることが知られており、全病態のうち5%を占める病態である。この1型糖尿病は、外部からインスリンを投与することにより治療する。
【0003】
一方、2型糖尿病は、インスリンの分泌不足とインスリン抵抗性がベースにあり、結果としてインスリンの作用不足により生じることが知られており、全病態のうち95%を占める病態である。
【0004】
現在、2型糖尿病の治療のためには、血糖を降下させる薬剤を投与することが一般的であり、そのような血糖を降下させる薬剤としては、インスリンの分泌を促進させる薬剤;腸管からの糖質の分解・吸収を遅延させて食後高血糖を抑制する薬剤;インスリン抵抗性を改善する薬剤;等が使用されている。しかしながら、どの薬にも副作用が報告され、問題となっている。
【0005】
具体的には、インスリンの分泌を促進させる薬剤の場合には、長期に飲み続けていると飲み始めた頃に比べ、効果が薄れてきたり効かなくなってくる(すなわち、膵臓のβ細胞が疲弊する)こと、そして投与後にインスリンが分泌過剰になって低血糖に陥り、食欲亢進により体重が増加しやすい等の問題点が指摘されている。
【0006】
腸管からの糖質の分解・吸収を遅延させて食後高血糖を抑制する薬剤の場合には、食後高血糖を抑制する効果はそれほど強力ではなく、また分解が遅れて腸管に残った糖類が発酵してガスが出たり、お腹が張ったり、下痢を引き起こす等の問題点や、稀に肝障害が起こる等の問題点が指摘されている。
【0007】
インスリン抵抗性を改善する薬剤の場合には、むくみやすく、心不全の患者には投与できないこと、体重が増加しやすく、稀に肝障害や貧血が生じる等の問題点が指摘されている。
【0008】
さらに、いずれの場合にも、現在存在している血糖降下薬を適切に処方しても、2型糖尿病患者の治療の良好例は、患者の約30%程度にしか見られないという、これらの副作用以外の問題点も存在している。
【0009】
リゾホスファチジン酸(lysophosphatidic acid:LPA)は、多数の細胞種により天然に合成される生物活性を持ったグリセロリン脂質の1種であり、グリセリン骨格のひとつの水酸基に脂肪酸が結合した物質である。LPAは、グリセロリン脂質がホスホリパーゼなどの作用により加水分解されて生ずる物質であり、1-アシルグリセロール-3-リン酸、2-アシルグリセロール-3-リン酸、1-アルケニル-グリセロール-3-リン酸などがLPAの代表例として知られており、細胞増殖、細胞遊走、平滑筋収縮、細胞生存、免疫応答などを含む複数の生理学的プロセスに関与している(非特許文献1〜3など)。その多様な生物学的作用の中でも、LPAは、脂肪細胞の増殖と機能を媒介すること、そしてグルコース代謝を制御すること、が示されてきた(非特許文献4〜6)。
【0010】
LPAに対する4番目のLPA受容体(LPA4受容体、GPR23とも呼ばれる)は、それ以前に見つかっていたLPA1〜LPA3の3種類のLPA受容体との間で、わずか20〜24%のアミノ酸同一性しか有さなかった(特許文献1、非特許文献7)。
【0011】
LPA4受容体(GPR23)は、生殖組織、脳組織、脂肪組織において高レベルで発現される。LPAは、LPA4受容体(GPR23)内在化、G12/13-媒介性およびRho-媒介性の神経突起退縮およびストレスファイバー形成、Gq-媒介性Ca2+動員およびGi-媒介性Ca2+動員、Gs-媒介性cAMP増加を誘導することが示された(非特許文献8)。LPA4受容体(GPR23)は、ニューロン新生やニューロン移動等の神経発生プロセスにおいて極めて重要な役割を果たしている可能性があると考えられている(非特許文献9)。しかしながら、LPA4受容体(GPR23)の生理学的機能および生物学的機能については、非常に限定的な情報しか得られていない。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】WO2004/063224
【非特許文献】
【0013】
【非特許文献1】van Corven EJ, et al., Cell, 1989, 59, 45-54 .
【非特許文献2】Amano M, et al., Science, 1997, 275, 1308-1311 .
【非特許文献3】Moolenaar WH, Curr. Opin. Cell. Biol., 1995, 7, 203-210 .
【非特許文献4】Valet P, et al., J. Clin. Invest., 1998, 101, 1431-1438 .
【非特許文献5】Simon MF, et al., J. Biol. Chem., 2005, 280, 14656-14662 .
【非特許文献6】Yea K, et al., J. Mol. Med., 2008, 86, 211-220 .
【非特許文献7】Noguchi K, et al., J. Biol. Chem., 2003, 278, 25600-25606 .
【非特許文献8】Lee CW, et al., J. Biol. Chem., 2007, 282, 4310-4317 .
【非特許文献9】Yanagida K, et al., J. Biol. Chem., 2007, 282, 5814-5824 .
【発明の概要】
【発明が解決しようとする課題】
【0014】
本発明は、LPA4受容体を介したLPAの作用を解明し、その作用を利用した薬剤スクリーニングの方法を提供することを課題とする。
【課題を解決するための手段】
【0015】
本発明者らは、LPA4受容体のノックアウト動物を作出し、そのノックアウト動物を詳細に検討した結果、LPA4受容体を介したLPAからのシグナルの欠損が、抗糖尿病作用を示すことを見出し、その結果に基づいて、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法についての発明を完成した。
【0016】
具体的には、本発明において、高脂肪食負荷モデルの非ヒト動物に対して、LPA4受容体に結合する被検化合物を投与する工程;被検化合物を投与した高脂肪食負荷モデルの非ヒト動物において、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せを調べる工程;抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を、LPA4受容体に結合するLPA4アンタゴニストであると同定する工程;を行うことにより、LPA4受容体に結合するLPA4アンタゴニストである化合物をスクリーニングすることができることを見出した。
【発明の効果】
【0017】
本発明により、複数存在するLPA受容体のうちLPA4受容体を介して、LPAがどの様なメカニズムで生体内において機能を果たしているかを明らかにすることができた。また、そのような知見に基づいて、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を選択することができるスクリーニング方法を開発することができた。
【図面の簡単な説明】
【0018】
【図1】図1は、野生型マウス(WTマウス)の各臓器における、LPA4受容体の相対遺伝子発現値を示す。
【図2】図2は、WTマウスおよびLPA4受容体欠損マウス(KOマウス)のそれぞれについて、「普通食」群および「高脂肪食(HFD)」群のそれぞれにおける体重変化を示す。
【図3】図3は、WTマウスおよびKOマウスのそれぞれについて、「普通食」群および「HFD」群のそれぞれにおける肝臓重量を示す。
【図4】図4は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれについて、肝臓の外観を示す。
【図5】図5は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれにおける、脂肪細胞のサイズ(図5左)および血中アディポネクチン濃度(図5右)を示す。
【図6】図6は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれの脂肪組織における、TNF-α(「Tnfa」)、CCL2、F4/80、CD68のそれぞれのマーカーの相対遺伝子発現値を示す。
【図7】図7は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれにおける、絶食時の血糖値を示す。
【図8】図8は、WTマウス「HFD」群およびKOマウス「HFD」群のそれぞれにおける、インスリン抵抗性試験の結果を示す。
【図9】図9は、WTマウスの脂肪細胞、血球系細胞、血管内皮細胞そしてLin陰性細胞(すなわち、前駆脂肪細胞)のそれぞれにおける、各種細胞マーカー(すなわち、脂肪細胞マーカーであるACRP30(アディポネクチン)、血球系細胞マーカーであるCD45、血管内皮細胞マーカーであるCD31、前駆脂肪細胞マーカーであるSmoothened(Smo))及びLPA4受容体の遺伝子発現を示す。
【図10】図10は、WTマウス由来の胎児線維芽(MEF)細胞およびKOマウス由来のMEF細胞のそれぞれにおける、アディポネクチン(ACRP30)を指標とした脂肪細胞分化能の比較を示す。
【図11】図11は、Rho結合キナーゼの活性を薬剤(Y27632;10μM)により阻害した場合、WTマウス由来のMEF細胞から脂肪細胞への分化がKOマウス由来のMEF細胞と同等までに亢進することを示す。
【発明を実施するための形態】
【0019】
リゾホスファチジン酸(lysophosphatidic acid:LPA)は、多数の細胞種により天然に合成される生物活性を持ったグリセロリン脂質の1種であり、グリセリン骨格のひとつの水酸基に脂肪酸が結合した物質であり、以下の構造式を有する化合物である。
【0020】
【化1】
【0021】
本発明者らは以前に、LPAに対する4番目のLPA受容体(LPA4受容体)の遺伝子およびアミノ酸配列を同定した(特許文献1)。しかしながら、これまでは、LPAがLPA4受容体を介してどの様な作用を発揮するのかについて、知られていなかった。本発明の発明者らは、LPA4受容体が関与するLPAからのシグナルが、どの様な作用を有しているかを明らかにするため、まずLPA4受容体のノックアウト動物を作出した。そして、このノックアウト動物に高脂肪食を負荷した場合における様々な生理学的反応および組織学的反応を検索した結果、このノックアウト動物においては、以下の様な特徴:
・LPA4受容体の欠損は、肥満そのものの進行には影響しないこと(例えば、図2);
・LPA4受容体の欠損により、脂肪肝が軽減したこと(例えば、図3および図4);
・LPA4受容体の欠損により、脂肪細胞の小型化が生じたこと(例えば、図5);
・LPA4受容体の欠損により、血中アディポネクチン濃度が上昇したこと(例えば、図5);
・LPA4受容体の欠損により、脂肪組織の炎症が軽減されたこと(例えば、図6);
・LPA4受容体の欠損により、インスリン抵抗性が改善したこと(例えば、図7および図8);
などが示されることが明らかになった。これらの特徴から、LPA4受容体を介したLPAからのシグナルの欠損が、抗糖尿病作用を示すことが明らかになった。
【0022】
これらの知見に基づいて、本発明は一態様において、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法を提供する。このスクリーニング方法においては、LPA4受容体に結合する被検化合物のうち、投与した際に、LPA4受容体の欠損の結果として生じた上述した特徴(図2〜図8に示したもの)のいずれかまたはそれらの組合せと同一の特徴を示すことができる化合物を、LPA4アンタゴニストとして選択することを特徴とする。
【0023】
具体的には、本発明は、高脂肪食負荷モデルの非ヒト動物に対して、LPA4受容体に結合する被検化合物を投与する工程;被検化合物を投与した高脂肪食負荷モデルの非ヒト動物において、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せを調べる工程;抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を、LPA4受容体に結合するLPA4アンタゴニストであると同定する工程;を含む、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法を提供する。
【0024】
上述した実施例の結果から推定されるLPA4受容体を介する生理学的メカニズムは、以下の通りである。脂肪細胞から分泌されたLPA産生酵素(オートタキシン:ATX)は細胞外でLPAを産生する。このLPAは、野生型の細胞の場合(すなわち、LPA4受容体が正常に存在する場合)には、前駆脂肪細胞に働き、脂肪細胞への分化を抑制する。マウスが脂肪負荷された場合、脂肪細胞は脂肪を蓄積する必要に迫られるが、脂肪細胞が増えないこのような状況下では一つ一つの脂肪細胞に過負荷が掛かるために細胞は徐々に肥大し、悪玉脂肪細胞へと変貌する。その結果、脂肪組織では炎症が起こり、また血中アディポネクチン量が低下する。このことが原因となり、マウスはインスリン抵抗性や脂肪肝といった生活習慣病を発症することになる。
【0025】
逆にLPA4受容体が欠損している場合には、LPA4受容体が欠損した前駆脂肪細胞は、LPAによって脂肪細胞への分化が抑制されない。そのため、マウスに脂肪が負荷された場合、脂肪細胞が容易に増えることができる。したがって、脂肪細胞一つあたりの脂肪蓄積の負担は軽く、細胞の肥大は最小限に抑えられる(善玉脂肪細胞)。このことは則ち、脂肪組織の量(重さ)は同じでも脂肪細胞の質が維持されることを意味し、生活習慣病がLPA4-KOマウスには発症しないこととなる。
【0026】
本発明のスクリーニング方法において選択することができるLPA4アンタゴニストは、まず、LPA4受容体に結合することができるという特徴を有する。これは、目的とするLPA4アンタゴニストの特徴が、LPA4受容体の欠損の結果として生じた上述した特徴(図2〜図8に示したもの)と同じものであることから、LPAがLPA4受容体と結合することを介して伝達されるシグナルが拮抗(アンタゴナイズ)されることが必要だからである。なお、本願発明において目的とする選択すべき化合物は、LPA4のインバースアゴニスト(逆作動薬)も含む。
【0027】
本発明者らの研究により、LPAがLPA4受容体に結合することにより細胞内にシグナルが伝達され、そのシグナル伝達が前駆脂肪細胞内におけるPPARγの発現を抑制していると考えられること;そしてその一方で、LPA4受容体が欠損することにより、すなわちLPA4が拮抗(アンタゴナイズ)されることにより、LPAのLPA4受容体に対する結合によって伝達されるシグナルがなくなることにより、PPARγの発現の抑制効果が失われ、PPARγの発現が亢進されること;が明らかになった。
【0028】
本発明の上述したスクリーニング方法においては、被検化合物を高脂肪食負荷モデルの非ヒト動物に対して投与した後、当該非ヒト動物体内において、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せが生じたかどうかを調べる工程を、行うことを特徴とする。
【0029】
抗糖尿病作用、抗動脈硬化作用および/または抗肥満作用を調べるため、例えば、脂肪細胞内におけるPPARγの発現亢進の検出、脂肪細胞過形成の検出、血中アディポネクチン濃度の増加の検出、またはインスリン抵抗性の改善の検出、を単独でまたはいずれかの組合せで行うことにより、LPA4受容体の欠損の結果として生じた上述した特徴(図2〜図8に示したもの)と同じ特徴を検出することができる。
【0030】
ここで、脂肪細胞内におけるPPARγの発現亢進を検出する場合、例えば、脂肪組織サンプルにおけるPPARγ mRNA発現の検出、PPARγに対する抗体を用いた免疫アッセイ、PPARγが標的とする遺伝子由来のmRNA発現の検出により行うことができる。
【0031】
また、脂肪細胞過形成を検出する場合、例えば、脂肪組織の組織学的解析、細胞数測定、DNA含有量測定により行うことができる。
【0032】
また、血中アディポネクチン濃度の増加を検出する場合、例えば、血液サンプルにおける抗アディポネクチン抗体を用いた免疫アッセイやアディポネクチン受容体への結合アッセイにより行うことができる。
【0033】
また、インスリン抵抗性の改善を検出する場合、例えば、HOMA-IRの測定、血中インスリンレベルの測定、OGTT法による評価、インスリン抵抗性試験による評価、正常血糖高インスリンクランプ法による評価により行うことができる。
【0034】
これらの方法はいずれも、当該技術分野において公知の方法であり、市販のキット、様々な書籍(例えば、糖尿病研究ストラテジー(秀潤社))、公知文献(例えば、Kubota N, et al., Mol. Cell, 1999, 4, 597-609.)等にしたがって行うことができる。
【0035】
本発明のスクリーニング方法に従って選択された被検化合物は、LPA4受容体に結合してLPAの作用を拮抗することができるとともに、脂肪肝を軽減すること、脂肪細胞の小型化を生じること、血中アディポネクチン濃度を上昇させること、脂肪組織の炎症が軽減されること、インスリン抵抗性が改善されること、等の作用を発揮することができる。
【0036】
本発明を以下の実施例により更に詳細に説明する。しかしながら、以下の実施例は、本発明の例示であって、本発明を限定することを意図するものではない。
【実施例】
【0037】
実施例1:脂肪組織におけるLPA4の高発現
本実施例においては、LPA4受容体を介したLPAからのシグナルが、どの様な作用を有するのかを明らかにすることを目的として、まずマウス各種臓器におけるLPA4受容体の遺伝子発現を調べた。
【0038】
具体的には12週齢の野生型マウス(WTマウス)から、脂肪組織を含む11臓器を採取し各臓器から全RNAを抽出し(RNeasy, Qiagen)、逆転写酵素(Superscript III, Invitrogen)によりcDNAを作製した後、これを鋳型として、ファストスタートSYBR Greenマスター(Roche)を使用して、定量PCRシステム(LightCycler, Roche)によりLPA4と18S rRNA(内部標準)の発現を定量した。
【0039】
PCRにおいて遺伝子発現を増幅するために、それぞれの遺伝子について、それぞれのヌクレオチド配列:
LPA4受容体(GenBank Accession No. NM_175271)
フォワードプライマー:gtcaacaatg cgaccaccac(SEQ ID NO: 1)
リバースプライマー:aagcaccaca gaagaacaag aaaca(SEQ ID NO: 2)
18S rRNA(GenBank Accession No. NR_003278)
フォワードプライマー:tcaacacggg aaacctcacc(SEQ ID NO: 3)
リバースプライマー:cagacaaatc gctccaccaa c(SEQ ID NO: 4)
を有するプライマー対を使用した。
【0040】
結果を図1に示す。この図において、各種臓器のLPA4の発現の相対値を示す。
その結果、LPA4は脂肪組織に強く発現することが明らかとなった。一方で、インスリン感受組織である肝臓や骨格筋においてはLPA4の発現は極めて低いことがわかった。
【0041】
実施例2:LPA4受容体欠損マウス(KOマウス)の作出
本実施例においては、LPA4受容体を介したLPAからのシグナルが、どの様な作用を有するのかを明らかにすることを目的として、LPA4受容体欠損マウスを作出することとした。
【0042】
まず、C57BL/6マウスのゲノムにおけるLPA4遺伝子のオープンリーディングフレーム(ORF)上流のEcoO65I/NcoI断片(4.0 kb)とLacZ DNA断片、ホスホグリセリン酸キナーゼプロモーターにトランスポゾンTn5ネオマイシン耐性遺伝子を連結したDNA断片(pgk/neor)、LPA4遺伝子のORF下流のXbaI/EcoO65I断片(6.3 kb)を使って、pBluescript KS M13(+)(Stratagene)をベクターとしてターゲッティングベクターを構築した。
【0043】
次に、C57BL/6マウス由来のES細胞株であるRENKA株にEco81Iで直線化した上記ターゲッティングベクターをエレクトロポレーションで導入し、形質転換株をG418(Gibco)で選択したのち、この中から相同組換えES細胞をサザンブロット解析によってスクリーニングした。
【0044】
さらに、この相同組換えES細胞をICRマウスの胚盤胞に注入してキメラマウスを作り、そのうちの雄マウスを雌のC57BL/6マウスと交配させることで雌のLPA4のヘテロザイゴートマウス(Lpar4+/-マウス)を得た。雄のC57BL/6マウスとLpar4+/-マウスを交配させることで、同腹仔である雄の野生型マウス(Lpar4+/Y; WTマウス)とLPA4受容体欠損マウス(Lpar4-/Y; KOマウス)を作出した。
【0045】
実施例3:高脂肪食負荷モデルにおける体重変化
本実施例においては、肥満とそれに伴うメタボリックシンドロームの進行におけるLPA4受容体の機能について検討すべく、野生型マウス(WTマウス)とLPA4受容体欠損マウス(KOマウス)を用いた高脂肪食負荷モデルでの体重の変化を測定した。
【0046】
具体的には、WTマウスおよびKOマウスのそれぞれについて、4週齢時から単独飼育し、6週齢時より普通食を6週間摂食させた「普通食」群、または6週齢時より高脂肪食(HFD32; 日本クレア社)を6週間摂食させた高脂肪食負荷モデル(マウスの高脂肪食負荷モデル、「HFD」群)を採用した。ここで、WTマウスの「普通食」群(WT-普通食)はn=26で実験を行い(●)、WTマウスの「HFD」群(WT-HFD)はn=40で実験を行い(▲)、KOマウスの「普通食」群(KO-普通食)はn=17で実験を行い(■)、そしてKOマウスの「HFD」群(KO-HFD)はn=36で実験を行った(◆)。結果を図2に示す。図2は、WTマウスおよびKOマウスのそれぞれについて、「普通食」群および「HFD」群のそれぞれにおける体重変化を示す。
【0047】
図2のグラフに示すように、「HFD」群は「普通食」群に比較して、著しい体重増加を認めた。しかし、その著しい体重増加について、WTマウスとKOマウスとの間に差異は認められなかった。この結果から、LPA4受容体を介したLPAシグナルの導入は、肥満そのものの進行には寄与しないことが示唆された。
【0048】
実施例4:LPA4受容体欠損マウスにおける脂肪肝の軽減
本実施例においては、実施例3において示された高脂肪食負荷モデル(「HFD」群)での著しい体重増加に関して、脂肪組織に高発現するLPA4受容体が脂肪の質的変化に対して何らかの影響を及ぼすことにより、メタボリックシンドロームの進行に関与する可能性を考え、まず肝臓に注目して脂肪肝の程度について検討した。
【0049】
内蔵脂肪型肥満は、インスリン抵抗性や脂肪肝などを引き起こし、これらが引き金となって糖尿病、高脂血症、高血圧につながり、最終的には動脈硬化の進行を加速させると考えられている。これがメタボリックシンドロームの概念である。一方、肥満つまり脂肪の「量的」増加だけではなく、その脂肪の「質的」変化がこのメタボリックシンドロームの進行に強く関わっていることが、最近の研究により明らかとなってきている。
【0050】
実施例3において記載したようにWTマウスの「普通食」群、WTマウスの「HFD」群、KOマウスの「普通食」群、そしてKOマウスの「HFD」群のマウスを飼育し、その後16時間絶食後、麻酔下で開腹・安楽死させた後、肝臓を採取しその重量および外観を比較した。統計処理をANOVAにより行った。結果を図3および図4に示す。
【0051】
WTマウスにおいて、「HFD」群は「普通食」群と比較して、肝臓重量が大幅に増加していた(***, P<0.001)(図3)。また、WTマウスの「HFD」群における肝臓の外観を調べたところ、白みを帯び、肥大化しており、脂肪肝の様相を呈していた(図4上)。
【0052】
一方、KOマウスにおいては、「HFD」群においても、重量が「普通食」群に近く、外観も脂肪肝の程度が著しく減弱していることがわかった(図3、図4下)。この結果から、LPA4受容体を介したLPAからのシグナルの導入が、脂肪肝の進行に強く関与することが明らかとなった。
【0053】
実施例5:LPA4受容体欠損マウスにおける脂肪細胞小型化と血中アディポネクチン高値
本実施例においては、LPA4受容体欠損マウスにおけるLPA4受容体の欠損が、メタボリックシンドロームに関わる他の生理学的指標に対してどの様な作用を有するかを調べることを目的として行った。
【0054】
実施例3および4において示したように、高脂肪食負荷モデル(「HFD」群)において、LPA4受容体は肥満そのものの進行には寄与しないが、脂肪肝の進行に寄与することがわかった(図2〜4)。一方で、実施例2において示したように、LPA4受容体の発現そのものは肝臓では極めて低く、脂肪組織において非常に強い(図1)。脂肪肝の進行には脂肪組織の「質的」変容が強く関与していることが、近年の研究により明らかになってきている。これらの知見を綜合して、脂肪組織におけるLPA4受容体が、脂肪組織の「質」に影響し結果としてWTマウスとKOマウスとの間での脂肪肝の程度の違いにつながっている可能性が予想された。その可能性について検討すべく、WTマウスおよびKOマウスの脂肪組織(重量や脂肪細胞サイズ)を比較した。
【0055】
6週齢時より高脂肪食(HFD32; 日本クレア社)を8週間摂食させた以外は実施例2に記載したように飼育した「HFD」群の精巣上体周囲の脂肪組織を採取し、その重量を比較した。そして、得られた脂肪組織をホルマリン固定した後、パラフィン中に包埋し、厚さ4μmの薄切切片を作製し、ヘマトキシリン・エオジン法により染色し、顕微鏡観察により脂肪細胞の大きさを測定した。この実験においては、各サンプル(WTマウスについての「HFD」群およびKOマウスについての「HFD」群)につき、200個以上の細胞を測定し、平均を算出した。統計処理をマン・ホイットニーのUテストにより行った。
【0056】
この結果、まず、WTマウスとKOマウスとの間に脂肪組織重量の差は認められなかった(グラフは示されていない)。WTマウスの「HFD」群における脂肪細胞のサイズと比較して、KOマウスの「HFD」群における脂肪細胞のサイズが、有意に小型であることが示された(*, P<0.05)(図5左)。
【0057】
一般に、小型化した脂肪細胞は、脂肪肝やインスリン抵抗性を改善させる善玉ホルモンであるアディポネクチンの産生能が高いこと、そして肥大した脂肪細胞においてはその産生が著しく減弱していることが知られている。そこで、脂肪細胞サイズの違いが循環アディポネクチンの増加をもたらしている可能性があるため、血中アディポネクチン量を検討した。統計処理をマン・ホイットニーのUテストにより行った。
【0058】
上述したWTマウス(「HFD」群)およびKOマウス(「HFD」群)から採取した血液より血清を調製し、血清中のアディポネクチン濃度をELISA(Mouse Adiponectin Elisa kit, 大塚製薬社)を用いて検討した。この結果、WTマウス(「HFD」群)と比較して、KOマウス(「HFD」群)において、有意に高いアディポネクチン濃度が検出された(***, P<0.001)(図5右)。
【0059】
実施例6:LPA4受容体欠損マウスにおける脂肪組織炎症の軽減
一般に、肥大した脂肪細胞ではアディポネクチンの産生が低下するが、代わりにMCP-1(CCL2)やTNF-αといった炎症性のサイトカインの産生が亢進することが知られている。これにより、マクロファージを中心とした炎症性細胞が脂肪組織に浸潤し、炎症が起こる。脂肪肝の背景にこの脂肪組織の炎症があることが最近の研究により明らかになってきている。
【0060】
実施例5において脂肪細胞のサイズが低下し、アディポネクチン濃度が上昇したことから、本実施例においては、KOマウスにおけるマクロファージを中心とした炎症性細胞の浸潤や炎症の程度を確認した。
【0061】
実施例5と同様に6週齢時より高脂肪食(HFD32; 日本クレア社)を8週間摂食させたWTマウスおよびKOマウスのそれぞれの「HFD」群のマウスの脂肪組織から、全RNAを抽出し(RNeasy, Qiagen)、逆転写酵素(Superscript III, Invitrogen)によりcDNAを作製した後、これを鋳型として、炎症のマーカーであるTNF-α(図6中では「Tnfa」と示す)、CCL2、マクロファージマーカーであるF4/80、CD68のそれぞれについて、蛍光試薬としてファストスタートSYBR Greenマスター(Roche)を使用して、定量PCRシステム(LightCycler, Roche)により炎症に関係する遺伝子発現を定量した。
【0062】
PCRにおいてそれぞれの遺伝子発現を増幅するために、それぞれヌクレオチド配列:
TNF-α(GenBank Accession No. NM_013693)
フォワードプライマー:ccagaccctc acactcagat cat(SEQ ID NO: 5);
リバースプライマー:tggcaccact agttggttgt ctt(SEQ ID NO: 6);
CCL2(GenBank Accession No. NM_0111333)
フォワードプライマー:ccactcacct gctgctactc att(SEQ ID NO: 7);
リバースプライマー:cagcacagac ctctctcttg agc(SEQ ID NO: 8);
F4/80(GenBank Accession No. NM_010130)
フォワードプライマー:ggcttcagtg ggatgtacag atc(SEQ ID NO: 9);
リバースプライマー:tgaaagttgg tttgtccatt gct(SEQ ID NO: 10);
CD68(GenBank Accession No. NM_009853)
フォワードプライマー:ctccctgtgt gtctgatctt gct(SEQ ID NO: 11);
リバースプライマー:ctgtgctttc tgtggctgta ggt(SEQ ID NO: 12);
を有するプライマー対を使用した。
【0063】
結果を図6に示す。この図において、それぞれのマーカーについて、WTマウス「HFD」群における発現に対するKOマウス「HFD」群における発現の相対値を示す。統計処理をマン・ホイットニーのUテストにより行った。
【0064】
その結果、KOマウス「HFD」群において、全てのマーカーの相対遺伝子発現レベルが有意に低いことが明らかになった(*, P<0.05;**, P<0.01;***, P<0.001)。このことから、実際にKOマウスの脂肪組織においてマクロファージを中心とした炎症が軽減していることがわかった(図6)。
【0065】
実施例7:LPA4受容体欠損マウスにおける糖尿病・インスリン抵抗性の改善
これまでに示した実験結果から、KOマウスにおいて、高脂肪食負荷(「HFD」群)による脂肪細胞の肥大化が軽減しており(実施例5、図5左)、これが血中アディポネクチン濃度の亢進(実施例5、図5右)、脂肪組織炎症の軽減(実施例6、図6)につながり、脂肪肝の軽減(実施例4、図3および4)をもたらしていることが予想された。
【0066】
インスリン抵抗性は肥満による糖尿病の原因であり、メタボリックシンドロームの基盤となっている病態であり、そして脂肪肝をもたらすと考えられているアディポネクチンレベルの減少、脂肪組織炎症の亢進といった現象は、ともにインスリン抵抗性を引き起こす原因と考えられている。
【0067】
そこで、前述の実施例の結果からKOマウスにおいてインスリン抵抗性が改善している可能性が高いと考えられたことから、本実施例においては、WTマウス「HFD」群およびKOマウス「HFD」群を対象として、そのインスリン抵抗性について検討した。
【0068】
まず、実施例5と同様に6週齢時より高脂肪食(HFD32; 日本クレア社)を8週間摂食させたWTマウスおよびKOマウスのそれぞれの「HFD」群のマウスを飼育し、その後それぞれのマウスを16時間絶食後、血清を調製し、血清中に含有されるグルコース濃度を、酵素法(テストワコー, Wako)により定量した(絶食時血糖値)。統計処理をマン・ホイットニーのUテストにより行った。
【0069】
この結果、WTマウス「HFD」群と比較して、KOマウス「HFD」群では、有意に絶食時血糖値が低く、糖尿病の改善が認められた(*, P<0.05)(図7)。
【0070】
次に、実施例2に記載したように6週齢時より高脂肪食(HFD32; 日本クレア社)を6週間摂食させたWTマウスおよびKOマウス(それぞれ、n=10)を対象に、4時間の絶食の後インスリン抵抗性試験を行った。インスリン抵抗性は、血液を尾静脈より採取し、血糖値を自動血糖測定装置(グルテストNeo, 三和化学研究所)で測定することにより評価した。インスリン(ノボリンR, Novo Nordisk Pharma Ltd.)の投与は腹腔内注射により行った。血糖値はインスリン投与後から20分間隔で計2時間測定した。統計処理を2方向ANOVAにより行った。
【0071】
WTマウス「HFD」群に比べて、KOマウス「HFD」群はインスリン投与による血糖値降下が良好であり、つまりKOマウス「HFD」群ではインスリン抵抗性が軽減していることがわかった(**, P<0.01;***, P<0.001)(図8)。
【0072】
実施例8:前駆脂肪細胞におけるLPA4受容体の高発現
脂肪組織は、脂肪細胞の他、その前駆細胞である前駆脂肪細胞、血管を構成する血管内皮細胞、炎症に関わるリンパ球やマクロファージなど複数の細胞から構成される。実施例2において示したように、LPA4受容体は脂肪組織に強い発現を認めるが、これらの複数の細胞種のうちどの細胞種に発現しているかは不明であった。
【0073】
そこで、本実施例において、KOマウスが上記の表現型を示す機序を解明すべく、脂肪組織を構成する細胞を分離し、LPA4受容体の発現レベルを調べた。
【0074】
まず普通食で飼育した6週齢のWTマウスから採取した脂肪組織を、撹拌しながらコラゲナーゼで2時間処理し、低速遠心により浮遊してくる脂肪細胞と沈殿する間質・血管系画分(SVF)に分離した。引き続き得られたSVFを、フィコエリスリン(PE)ラベルされた抗CD45(血球系細胞マーカー)抗体、抗Ter-119(赤血球マーカー)抗体(ともにBiolegend)と反応させ、抗PE磁気ビーズ(Miltenyi Biotec)とAutoMACS(磁気による細胞ソーティングシステム;Miltenyi Biotec)を用いることで、血球系細胞(すなわちCD45+またはTer-119+)と非血球系細胞(すなわちCD45-/Ter-119-)に分離した。これにより分離された非血球系細胞をPEラベルされた抗CD31(血管内皮細胞マーカー)抗体(Biolegend)を用いて、上記と同様の方法で、血管内皮細胞(すなわちCD45-/Ter-119-/CD31+)とLineage陰性細胞(以下Lin-細胞(すなわち、CD45-/Ter-119-/CD31-);前駆脂肪細胞から構成される)に分離した。
【0075】
次いで、脂肪細胞、血球系細胞、血管内皮細胞、Lin-細胞から全RNAを回収し、逆転写酵素によりcDNAを作製した後、これを鋳型として、各種細胞マーカー(すなわち、脂肪細胞マーカーであるACRP30(アディポネクチン)、血球系細胞マーカーであるCD45、血管内皮細胞マーカーであるCD31、前駆脂肪細胞マーカーであるSmoothened(Smo))の遺伝子発現とLPA4受容体の遺伝子発現とを、蛍光試薬としてファストスタートSYBR Greenマスター(Roche)を使用して、定量PCRにより確認した。
【0076】
PCRにおいてそれぞれの遺伝子発現を増幅するために、それぞれヌクレオチド配列:
ACRP30(アディポネクチン)(GenBank Accession No. NM_009605)
フォワードプライマー:aaaggagatg caggtcttct tgg(SEQ ID NO: 13);
リバースプライマー:tgaacgctga gcgatacaca taa(SEQ ID NO: 14);
CD45(GenBank Accession No. NM_001111316)
フォワードプライマー:gggttgttct gtgccttgtt(SEQ ID NO: 15);
リバースプライマー:tggctgtttg ttttcttgac ttg(SEQ ID NO: 16);
CD31(GenBank Accession No. NM_001032378)
フォワードプライマー:aatcaaaata aagaccccca gaaca(SEQ ID NO: 17);
リバースプライマー:cacaggactc tcgcaatcca(SEQ ID NO: 18);
Smo(GenBank Accession No. NM_176996)
フォワードプライマー:gcaggaatgg cagagacagg(SEQ ID NO: 19);
リバースプライマー:ggcacagaaa agggcaaaca(SEQ ID NO: 20);
LPA4受容体(GenBank Accession No. NM_175271)
フォワードプライマー:gtcaacaatg cgaccaccac(SEQ ID NO: 1);
リバースプライマー:aagcaccaca gaagaacaag aaaca(SEQ ID NO: 2);
を有するプライマー対を使用した。
【0077】
結果を図9に示す。この結果から、脂肪細胞マーカーであるACRP30(アディポネクチン)、血球系細胞マーカーであるCD45、マクロファージマーカーであるCD68、血管内皮細胞マーカーであるCD31、前駆脂肪細胞マーカーであるSmoothened(Smo)の遺伝子発現を検討することで、細胞分離が的確に行なわれていることが確認された。そして、LPA4受容体の遺伝子発現を調べたところ、Lin-細胞つまり前駆脂肪細胞に最も発現が高く、次いで脂肪細胞に高く発現していることがわかった。
【0078】
従って、LPA4受容体が前駆脂肪細胞から脂肪細胞への分化に影響することで、本実施例で認められたWTマウスとKOマウスとの違いをもたらしている可能性が考えられた。
【0079】
実施例9:LPA4受容体欠損マウス由来線維芽細胞における脂肪細胞分化の亢進
実施例8で示したように、LPA4受容体が前駆脂肪細胞に強く発現することがわかっため、LPA4受容体が脂肪細胞への分化に関与している可能性がさらに示唆された。
【0080】
そこで、本実施例において、脂肪細胞分化を検討する一般的な手法である、マウス胎児線維芽細胞(MEF細胞)を用いた脂肪細胞分化実験を行なった。MEF細胞は、前駆脂肪細胞と同様にin vitroで脂肪細胞へ分化させることができることが知られている。
【0081】
WTマウスおよびKOマウスの胎齢13.5日(E13.5)のマウス胎児より、MEF細胞をそれぞれ採取し、ウシ胎児血清(Gibco)を10%含有したDMEM(Nacalai tesque)により細胞培養ディッシュ(Corning)上で培養した。それぞれのMEF細胞は、コンフルエントまで培養してから2日後にインスリン(10μg/ml)、デキサメサゾン(0.25μM)、IBMX(0.5 mM)(全てSigma)の3種類の薬剤で処理することにより、脂肪細胞へと分化させた。
【0082】
上述した3種類の薬剤(インスリン、デキサメサゾン、IBMX)処理を開始した時点をday0とし、day12まで分化を進行させ、day0、day4、day8、day12の各時点において全RNAを回収し、逆転写酵素を使いcDNAを作製した。脂肪細胞への分化の程度については、脂肪細胞特異的に発現するアディポネクチン(ACRP30)の発現量を上記cDNAにより定量PCRで評価した(実施例8を参照)。
【0083】
成熟脂肪細胞のマーカーであるアディポネクチン(ACRP30)の発現レベルの時間変化を図10に示す。WTマウス由来MEF細胞およびKOマウス由来MEF細胞のそれぞれを、脂肪細胞へと分化させると、KOマウス由来のMEF細胞は、WTマウスのMEF細胞に比べて、高い脂肪細胞分化能をもつことがわかった(図10)。このことから、LPA4受容体欠損マウス由来のMEF細胞は野生型マウス由来のMEF細胞よりも早く脂肪細胞へと成熟することが明らかとなった。
【0084】
また、本発明者らは以前に、LPA4受容体が、Rhoと呼ばれる因子とその下流で働くキナーゼであるRho結合キナーゼ(ROCK)を活性化することを明らかにしており(非特許文献9)、このシグナル伝達経路が脂肪細胞分化に寄与する可能性が考えられた。そこで、本実施例においては、脂肪細胞分化実験におけるROCK阻害剤であるY27632の効果について検討した。
【0085】
上記と同様の分化実験を、10μM Y27632(Calbiochem)の存在下または非存在下の2条件で行なった。上述した3種類の薬剤(インスリン、デキサメサゾン、IBMX)処理を開始した時点をday0とし、day8まで分化を進行させ、全RNAを回収し、逆転写酵素を使いcDNAを作製した。脂肪細胞への分化の程度については、脂肪細胞特異的に発現するPPARγ(GenBank Accession No. NM_011146)の発現量を、フォワードプライマー:gaagaccact cgcattcctt tg(SEQ ID NO: 21);およびリバースプライマー:gcaggttcta ctttgatcgc actt(SEQ ID NO: 22);を有するプライマー対を得られたcDNAに対して使用して、実施例8に記載するように定量PCRにより評価した。
【0086】
結果を図11に示す。図10の結果と矛盾せず、KOマウス由来の細胞はWTマウス由来の細胞よりも、脂肪細胞マーカーであるPPARγの発現レベルが高かった。一方、LPA4受容体はROCKを活性化するシグナルを細胞内へ送るが、ROCKの活性を阻害剤(Y27632;10μM)で抑制すると、WTマウス由来の細胞のPPARγの発現レベルは、KOマウス由来の細胞と同程度まで亢進することが明らかとなった(図11)。しかもKOマウス由来の細胞は薬剤による影響を大きくは受けないことも示された。
【0087】
以上の結果から、前駆脂肪細胞では、LPA4受容体を介して伝達されるシグナルが、ROCKを活性化させることを通じてPPARγの発現を抑制しており、その結果として脂肪細胞の分化を抑制する役割を持つことが示された。
【産業上の利用可能性】
【0088】
本発明により、LPAが複数存在するLPA受容体のうち、LPA4受容体を介して、どの様なメカニズムで生体内において機能を果たしているかを明らかにすることができた。また、そのような知見に基づいて、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を選択することができるスクリーニング方法を開発することができた。
【配列表フリーテキスト】
【0089】
SEQ ID NO: 1:Forward primer for amplifying LPA4 receptor gene.
SEQ ID NO: 2:Reverse primer for amplifying LPA4 receptor gene.
SEQ ID NO: 3:Forward primer for amplifying 18S rRNA gene.
SEQ ID NO: 4:Reverse primer for amplifying 18S rRNA gene.
SEQ ID NO: 5:Forward primer for amplifying TNF-alpha gene.
SEQ ID NO: 6:Reverse primer for amplifying TNF-alpha gene.
SEQ ID NO: 7:Forward primer for amplifying CCL2 gene.
SEQ ID NO: 8:Reverse primer for amplifying CCL2 gene.
SEQ ID NO: 9:Forward primer for amplifying Emr1 gene.
SEQ ID NO: 10:Reverse primer for amplifying Emr1 gene.
SEQ ID NO: 11:Forward primer for amplifying CD68 gene.
SEQ ID NO: 12:Reverse primer for amplifying CD68 gene.
SEQ ID NO: 13:Forward primer for amplifying ACRP30 gene.
SEQ ID NO: 14:Reverse primer for amplifying ACRP30 gene.
SEQ ID NO: 15:Forward primer for amplifying CD45 gene.
SEQ ID NO: 16:Reverse primer for amplifying CD45 gene.
SEQ ID NO: 17:Forward primer for amplifying CD31 gene.
SEQ ID NO: 18:Reverse primer for amplifying CD31 gene.
SEQ ID NO: 19:Forward primer for amplifying Smo gene.
SEQ ID NO: 20:Reverse primer for amplifying Smo gene.
SEQ ID NO: 21:Forward primer for amplifying PPAR gamma gene.
SEQ ID NO: 22:Reverse primer for amplifying PPAR gamma gene.
【特許請求の範囲】
【請求項1】
高脂肪食負荷モデルの非ヒト動物に対して、LPA4受容体に結合する被検化合物を投与する工程;
被検化合物を投与した高脂肪食負荷モデルの非ヒト動物において、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せを調べる工程;
抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を、LPA4受容体に結合するLPA4アンタゴニストであると同定する工程;
を含む、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法。
【請求項2】
抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せを調べる工程を、脂肪細胞内におけるPPARγの発現亢進の検出、脂肪細胞過形成の検出、血中アディポネクチン濃度の増加の検出、またはインスリン抵抗性の改善の検出、のいずれかにより行う、請求項1に記載のスクリーニング方法。
【請求項3】
脂肪細胞内におけるPPARγの産生亢進の検出を、脂肪組織サンプルにおけるPPARγ mRNA発現の検出、PPARγに対する抗体を用いた免疫アッセイ、PPARγが標的とする遺伝子由来のmRNA発現の検出により行う、請求項1または2に記載の方法。
【請求項4】
脂肪細胞過形成の検出を、組織学的解析、細胞数測定、DNA含有量測定により行う、請求項1または2に記載の方法。
【請求項5】
血中アディポネクチン濃度の増加の検出を、血液サンプルにおける抗アディポネクチン抗体を用いた免疫アッセイ、アディポネクチン受容体への結合アッセイにより行う、請求項1または2に記載の方法。
【請求項6】
インスリン抵抗性の改善の検出を、HOMA-IRの測定、血中インスリンレベルの測定、OGTT法による評価、インスリン抵抗性試験による評価、正常血糖高インスリンクランプ法による評価により行う、請求項1または2に記載の方法。
【請求項1】
高脂肪食負荷モデルの非ヒト動物に対して、LPA4受容体に結合する被検化合物を投与する工程;
被検化合物を投与した高脂肪食負荷モデルの非ヒト動物において、抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せを調べる工程;
抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せの作用を有する被検化合物を、LPA4受容体に結合するLPA4アンタゴニストであると同定する工程;
を含む、LPA4受容体に結合するLPA4アンタゴニストである化合物のスクリーニング方法。
【請求項2】
抗糖尿病作用、抗動脈硬化作用、抗肥満作用、またはこれらのいずれかの組合せを調べる工程を、脂肪細胞内におけるPPARγの発現亢進の検出、脂肪細胞過形成の検出、血中アディポネクチン濃度の増加の検出、またはインスリン抵抗性の改善の検出、のいずれかにより行う、請求項1に記載のスクリーニング方法。
【請求項3】
脂肪細胞内におけるPPARγの産生亢進の検出を、脂肪組織サンプルにおけるPPARγ mRNA発現の検出、PPARγに対する抗体を用いた免疫アッセイ、PPARγが標的とする遺伝子由来のmRNA発現の検出により行う、請求項1または2に記載の方法。
【請求項4】
脂肪細胞過形成の検出を、組織学的解析、細胞数測定、DNA含有量測定により行う、請求項1または2に記載の方法。
【請求項5】
血中アディポネクチン濃度の増加の検出を、血液サンプルにおける抗アディポネクチン抗体を用いた免疫アッセイ、アディポネクチン受容体への結合アッセイにより行う、請求項1または2に記載の方法。
【請求項6】
インスリン抵抗性の改善の検出を、HOMA-IRの測定、血中インスリンレベルの測定、OGTT法による評価、インスリン抵抗性試験による評価、正常血糖高インスリンクランプ法による評価により行う、請求項1または2に記載の方法。
【図1】

【図2】
【図3】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
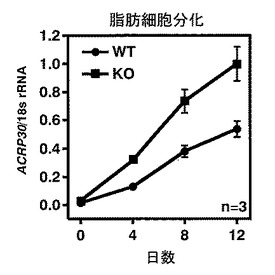
【図11】
【図4】
【図2】
【図3】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図4】
【公開番号】特開2012−239450(P2012−239450A)
【公開日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願番号】特願2011−115175(P2011−115175)
【出願日】平成23年5月23日(2011.5.23)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度、産業技術力強化法第19条の適用を受けるもの。
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【Fターム(参考)】
【公開日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願日】平成23年5月23日(2011.5.23)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度、産業技術力強化法第19条の適用を受けるもの。
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【Fターム(参考)】
[ Back to top ]