
M13ファージのpIXへの融合を介する代替の足場タンパク質融合ファージ提示
本発明は、M13ファージのpIXを使用して、非抗体足場タンパク質融合を産生するためのpIXファージ提示ライブラリを作成し、使用するための組成物及び方法に関する。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、2007年12月19日に出願された米国特許仮出願第61/014,778号の利益を主張し、その全体を参照として本明細書に組み込む。
【0002】
(発明の分野)
本発明は、M13ファージのpIXを使用して、非抗体足場タンパク質融合を産生するためのpIXファージ提示ライブラリを作成し、使用するための組成物及び方法に関する。
【背景技術】
【0003】
ファージ又はファージミド系における融合パートナーとしてpIII、pVII、pVIII、pIX、及びそれらの組み合わせを使用する糸状ファージ提示は、特に、デノボタンパク質及びペプチド単離並びに親和性成熟に関するタンパク質工学のための技術として使用されている(例えば、Janda et al,米国特許第7,078,166号を参照)。無作為ペプチド及びタンパク質は、関心のある対象のタンパク質標的に対するパニングを介してペプチド配列の変異を提示するファージライブラリから作成及び単離することができる。これまでに使用されたヒトタンパク質デノボライブラリは、合成的に作製されている。合成ライブラリにおいて、潜在的タンパク質をコードする推定上有用なDNA配列は、既知の配列又はモチーフに基づいて設計され、合成される。合成ペプチドライブラリに加えて、ライブラリはまた、ヒト組織に由来するタンパク質コードDNAの組み合わせクローニングによって作製することもできる。そのようなライブラリは、潜在的に有用なペプチドを提供するため、かつ連続回のパニング及び成熟又は修飾を実行して、選択された標的タンパク質の阻害生物活性等の所望の特性を有する非抗体ペプチド又はタンパク質ペプチドを見つけることを試みるために使用されている。
【0004】
ヒトペプチド、及び模倣タンパク質又は標的タンパク質に結合するか、若しくはそれを模倣する既知のタンパク質突然変異等のタンパク質は、ファージ提示pIII又はpVIIIペプチドライブラリから単離されている。特定の標的に結合するいくつかのタンパク質を単離することに成功しているが、そのようなファージ提示ライブラリ法は、ライブラリ作成のプロセスを繰り返さなければならないこと、所望の特徴を有するペプチド又はタンパク質を単離するためのパニング及び成熟の必要条件、及び他の既知の制限の問題に見舞われる。そのようなファージライブラリは、ヒトに存在するペプチド多様性の範囲、アミノ酸変異の位置及び程度、並びに異なるヒト遺伝子からの生物活性タンパク質の相対存在量を完全に包含又は模倣しないという問題にも見舞われる。天然レパートリからの合成タンパク質の偏位は、人において治療用に使用される場合、好ましくない生化学的特性及び免疫原性のリスクを増加させる可能性があり、この問題には、そのようなライブラリからの選択に対して、どのようにして配列が作成され、スクリーニングされるのかということによって潜在的に対処し得る。
【0005】
モノクローナル抗体は、標的分子に対する高親和性及び特異性が望ましい時に最も広く使用されるクラスの治療用タンパク質である。しかしながら、非抗体抗原結合ペプチド又はタンパク質は、治療法又は診断法における使用に対して高い関心のある対象であるそのような標的に結合するように操作することができる。生体分子に結合することが可能なそのようなタンパク質又はペプチドは、より小さいサイズ、ジスルフィド結合の欠如、原核宿主において発現される能力、精製の新規の方法、高い安定性、薬物/毒素への結合の容易性、及び知的財産利点等であるが、それらに限定されない、伝統的な抗体に対するいくつかの潜在的利点を有し得る。
【0006】
1つの種類の足場は、免疫グロブリン(Ig)折り畳みである。この折り畳みは、抗体の可変領域、並びに数千もの非抗体ペプチド又はタンパク質において見られる。1つのそのようなIgタンパク質、ヒトフィブロネクチンからの10番目のフィブロネクチン3型繰り返し体は、全体的なIg折り畳み構造を保持しつつ、表面に露出したループにおける多数の突然変異に耐えることができるということが示されている。そのため、アミノ酸変異体のライブラリは、これらのループ、及び多数の異なる標的に選択される特定の特定の結合剤に組み込まれている。そのような操作されたFn3ドメインは、重要な生物物理学的特性を保持しつつ、適度に高い親和性を伴う標的に結合することが見出されている。
【発明の概要】
【発明が解決しようとする課題】
【0007】
ファージライブラリのこれまでの使用は、抗体ベースのタンパク質融合ライブラリを含んできた。しかしながら、高親和性及び活性、高生産性、良い溶液物性、及び人に投与する際の低免疫応答の傾向のヒト治療抗体の決定的要素を同時に送達する、合成の代替の足場又は非抗体推定抗原結合ペプチド又はタンパク質融合ライブラリ及び方法の必要性が存在する。非抗体ペプチド又はタンパク質発見の資源コストを低下させ、及び生物的評価のための抗体の送達を加速するために、現在の方法に対して、合成ライブラリからの非抗体ペプチド又はタンパク質単離の効率性を増加させる必要性が更に存在する。本発明のライブラリ及び方法は、総合的な設計、アセンブリ技術、及びファージpIX非抗体ペプチド又はタンパク質提示を連結することによって、これらの必要性を満たす。
【課題を解決するための手段】
【0008】
先行技術の教示と対照的に、pVII及びpIXは、非抗体ペプチド又はタンパク質のための効率的かつ迅速な基本骨格、及び治療抗体の非抗体ペプチド又はタンパク質フラグメント作成及び選択を提供するために、例えば、任意にインライン成熟を伴う、突然変異作成又は他の多様性産生技術を使用して、M13ファージからのpIXを使用して高親和性非抗体ペプチド又はタンパク質ライブラリを作成するために成功裏に使用することができることが現在発見されている。本発明によると、所望の生体分子又は抗原に結合することが可能である非抗体ペプチド又はタンパク質は、任意に、代表的なヘテロ二量体モチーフにおいて、機能性非抗体ペプチド又はタンパク質を提示するために、ファージ表面上の動的相互作用において係合されるpVII及びpIXに融合される。したがって、非抗体ペプチド又はタンパク質結合剤のファージ上の提示は、メンバーが単量体又は二量体人工非抗体ペプチド又はタンパク質種として機能し、新規又は所望の生物活性の選択を可能にすることができる、組み合わせヘテロ二量体アレイの多様なライブラリの提示及び検定のための好適かつ好ましい方法である。
【0009】
本発明は、糸状ファージのpIXタンパク質、広く使用されるpIII及びpVIIIタンパク質と異なるファージ表面タンパク質に融合される非抗体ペプチド又はタンパク質デノボライブラリの設計及び提示を提供する。我々は、個々の足場ライブラリを別々に構築し、足場ライブラリのそれぞれの活性を系統的に検査し、抗原認識のための構造トポロジーを評価するためのファージ選択プロセスを開発した。
【0010】
本発明は、様々な改善された新しいpIX及びpVIIファージ提示デノボライブラリ作成方法、及び(i)pIX又はpVIIファージタンパク質に融合される、設計され、提示された非抗体ペプチド又はタンパク質デノボライブラリ、(ii)M13ファージの広く使用されるpIII及びpVIIIと異なるファージ表面タンパク質の使用、(iii)非抗体タンパク質足場に融合されるペプチドのライブラリをスクリーニングするためのpIXファージ提示システムの使用、(iv)抗原認識のための設計された配列及び構造トポロジーの効果の系統的検査を可能にする非抗体ペプチド又はタンパク質選択プロセス、(v)ライブラリ選択の一部としての流線形親和性成熟及びインライン成熟プロセス、のうちの1つ以上であるがそれらに限定されない、構成要素を提供する。ライブラリの個々又は群のライブラリ設計、選択、最適化及び成熟のそのような新しいシステムは、非抗体ペプチド又はタンパク質デノボ発見の成功のための再現可能かつ確実なシステムを提供し、また抗原相互作用に対する非抗体ペプチド又はタンパク質の構造機能相関の理解を容易にする。
【0011】
上述されるヒト非抗体ペプチド又はタンパク質デノボライブラリは、M13ファージのpIX遺伝子を介する提示によって現在の抗体ライブラリ現況技術と異なる。非抗体ペプチド又はタンパク質は、本発明によると、pIX融合タンパク質としてM13ファージの表面上に成功裏に提示することができる。両方の足場タンパク質は、それらの天然の状態において対応する足場タンパク質によって結合しない特定のタンパク質標的に結合するように操作することができる。これらの操作された足場は、ファージ上で提示されつつ、この特定の標的への結合を保持した。加えて、アミノ酸変異体のライブラリは、それぞれの足場において、本発明によって作製することができ、特定の標的に結合するそれらの能力は、pIX融合としてファージ上にライブラリメンバーを提示すること、及び標的タンパク質に対するパニングによって対処される。
【0012】
本発明において使用される人工抗体又は足場タンパク質は、非抗体ペプチド又はタンパク質分子の機能的戦略を使用するが、(1)標的抗原の配列相同性及び毒性、(2)宿主におけるか、又は非抗体ペプチド又はタンパク質を回復させるために使用されるハイブリドーマ培養における作成された非抗体ペプチド又はタンパク質の生物学的インパクト、及び(3)所望の活性に対するスクリーニング対選択、を含む、生体内拘束がない状態で作成することができる、大きい多様性のタンパク質モチーフとして本明細書において定義される。
【0013】
そのため、本発明は、多様性に富む単量体又はヘテロ二量体ポリペプチドアレイの構成のための組み合わせファージ提示形式を説明する。特に、本発明は、融合ポリペプチドをコードするゲノムを封入する糸状ファージ粒子を説明し、融合ポリペプチドは、糸状ファージpVII又はpIXタンパク質のアミノ末端に融合される非抗体足場タンパク質を含む。好ましくは、ファージ粒子は、ファージ粒子の表面上の発現された融合タンパク質を含む。
【0014】
関連する実施形態では、本発明は、融合タンパク質を発現するためのカセットを含む糸状ファージの表面上に融合タンパク質を発現するためのベクターを説明する。カセットは、挿入DNA、即ち、ポリリンカーの方向性連結反応のために適合されるヌクレオチドの配列を介して作動可能に連結される上流及び下流の翻訳可能なDNA配列を含み、上流配列は、原核分泌信号をコードし、下流配列は、pVII又はpIX糸状ファージタンパク質をコードする。翻訳可能なDNA配列は、融合ポリペプチドの部分としての翻訳可能なDNA配列の発現のための一連のDNA発現信号に作動可能に連結される。好ましい変異において、ベクターは、糸状ファージの表面上に第2の融合タンパク質を発現するための第2のカセットを更に含み、第2のカセットは、第1の融合タンパク質発現カセットがpVIIタンパク質をコードし、第2の融合タンパク質発現カセットがpIXタンパク質をコードするという条件で、第1のカセットの構造を有する。ベクターは、ファージ粒子の表面上にヘテロ二量体タンパク質複合体を発現するためにファージゲノムとして使用され、ヘテロ二量体の2つのポリペプチドは、第1のファージタンパク質及び第2のファージタンパク質、pVII及びpIXへの融合によって、ファージ粒子上でそれぞれ固定される。
【0015】
別の実施形態では、本発明は、本発明によるファージ粒子のライブラリ、即ち、組み合わせライブラリを意図し、ライブラリにおける代表的な粒子は、それぞれ、異なる融合タンパク質を提示する。粒子がヘテロ二量体タンパク質複合体を提示する場合、ライブラリは、ヘテロ二量体の組み合わせライブラリを含む。好ましいライブラリは、融合タンパク質の異なるか、又は同様の種の、少なくとも103、104、105、106、107、108、109、1010、1011、1012、1013、又はその中の任意の範囲又は値の組み合わせ多様性を有する。
【0016】
関連する実施形態は、第1のポリペプチド及び第2のポリペプチドを含む融合タンパク質を説明し、第1のポリペプチドは、非抗体足場タンパク質であり、第2のポリペプチドは、糸状ファージpVII又はpIXタンパク質であり、非抗体足場タンパク質は、糸状ファージタンパク質のアミノ末端に融合される。
【0017】
更に、本発明は、非抗体足場タンパク質をコードする遺伝子のレパートリを本発明のベクターにクローニングし、突然変異作成によって、第1の融合タンパク質及び第2の融合タンパク質ライブラリの集団の無作為的組み合わせによって、ライブラリの多様性を変化させるための標的及び親和性選択(「パニング」)等によってライブラリにおける非抗体足場タンパク質の構造を修正することによる、を含む、ファージの組み合わせライブラリを産生するための様々な方法を意図する。そのような実施形態は、本明細書において例示されるように、第2のポリペプチドに融合される代替可能な非抗体足場タンパク質を有する融合ポリペプチドを含むことができる。例えば、一実施形態では、代替可能な非抗体足場タンパク質は、TeFN3であり得、第2のポリペプチドは、pIXであり得、TeFN3のF:Gループは、突然変異作成、又はシステイン拘束されたEGFR結合ペプチド、PHPEP190等の非天然ポリペプチドによる野生型F:Gループアミノ酸残基の標的置換によって変化する。代替の実施形態では、代替可能な非抗体足場タンパク質は、配列番号2〜23、25、又は28によって特定される少なくとも1つのポリペプチドから作製されるF:Gループを有することができる。非抗体足場タンパク質が、操作された核酸ファージベクターによってコードされ、上皮成長因子受容体又は生物活性配位子等の生体分子に結合する実施形態も本明細書に開示する。
【0018】
改善された又は新規の機能を伴うタンパク質の設計は、様々な医薬、産業、環境、及び基礎研究の用途に伴う重要な目標である。組み合わせ非抗体ペプチド又はタンパク質ライブラリの開発に続き、強力な次の工程は、人工非抗体ペプチド又はタンパク質作製物、及び二量体種が野生型であるか、又は機能的であり得る他のタンパク質モチーフへの進化である。
【0019】
本発明は、pVII及びpIXが、単量体又は二量体種を形成する融合タンパク質の提示のために利用される、組み合わせ非抗体ポリペプチドアレイの構成のためのファージ提示形式を提供することによって、これらの課題に取り組む。pIX又はpVII上の発現を使用して機能的相互作用のライブラリを作成するために、1つ又は2つのタンパク質モチーフを近接して独立して提示することができるため、これが全く新しい方法論であることに留意することが重要である。
【0020】
更に、ライブラリを形成するための配列無作為化、及びハイブリッド種を形成するための鎖シャフリングプロトコルは、新規のタンパク質のサブセットにつながり得る。例えば、ホモ二量体又はヘテロ二量体型における亜鉛フィンガードメインのアレイの提示及び修正は、特定のDNA相互作用を所有する構造を産生する。加えて、全く新しい作製物が、非抗体ペプチド又はタンパク質鎖等の事前形成された足場内の所望のコードフラグメントの挿入を介して可能である。可能な挿入は、酵素特徴的配列又はリプレッサ結合タンパク質を含む。
【0021】
前述の概要及び以下の詳細な説明の両方とも、例示及び説明のためのみであり、主張される本発明を制限するものではないことが理解されるべきである。
【図面の簡単な説明】
【0022】
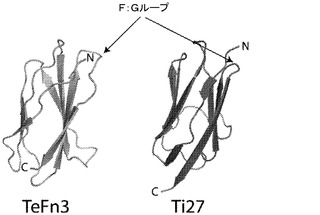
【図1】テネイシンの第3のFn3ドメイン(残基802〜893、PDB=1TEN、(Leachy,et al.,Science 258:987〜91,1992))及びタイチンのI27ドメイン(残基5253〜5341、PDB=1TIT、(Improta,et al.,Structure 4:323〜37,1996))の構造。矢印は、F:Gループ(テネイシン及びタイチンそれぞれの残基876〜881及び5258〜5259)の場所を指し、それは、EGFR結合配列を収容するように修正された。
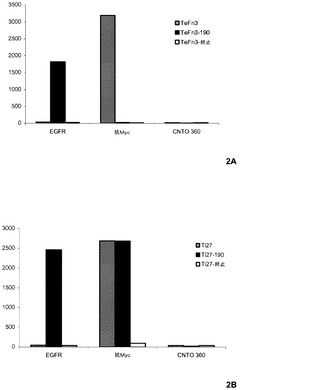
【図2】pIX上のテネイシン及びタイチン足場タンパク質の提示に関する図式的結果。
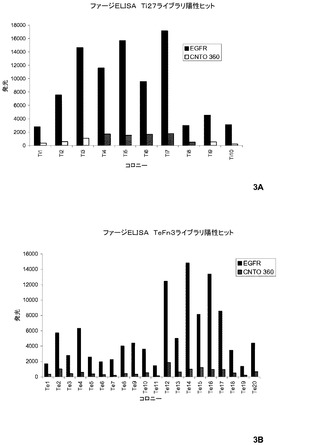
【図3】pIXに提示されるTeFn3−190及びTi27−190ライブラリからのEGFR結合作製物のためのパニングの図式的結果。
【発明を実施するための形態】
【0023】
本発明は、様々な新しいファージ提示デノボライブラリ作成方法及び構成要素を提供し、(i)pIX又は他のファージタンパク質に融合される、設計され、提示された非抗体ペプチド又はタンパク質ライブラリ、(ii)M13ファージの広く使用されるpIII及びpVIIIと異なるファージ表面タンパク質の使用、(iii)抗原認識のための設計された配列及び構造トポロジーの効果の系統的検査を可能にする非抗体ペプチド又はタンパク質選択プロセスにおける改善され、設計された組み合わせ多様性を提供するための、ライブラリ足場としてのそのようなファージ構成要素の使用、(iv)ライブラリ選択の一部としての流線形親和性成熟プロセス、等を含むが、それらに限定されない。ライブラリの個々又は群のライブラリ設計、選択、最適化及び成熟のそのような新しいシステムは、非抗体ペプチド又はタンパク質デノボ発見の成功のための再現可能かつ確実なシステムを提供し、また抗原相互作用に対する非抗体ペプチド又はタンパク質の構造機能相関の理解を容易にする。
【0024】
上述されるヒト非抗体ペプチド又はタンパク質デノボライブラリは、M13ファージのpIX又はpVII遺伝子を介する提示によって現在の抗体ライブラリ現況技術と異なる。
【0025】
用語の定義:
融合ポリペプチド:少なくとも2つのポリペプチド、及び2つのポリペプチドを1つの連続的なポリペプチドに作動可能に連結する連結配列からなるポリペプチド。融合ポリペプチドにおいて連結される2つのポリペプチドは、典型的に、2つの独立した源に由来するため、融合ポリペプチドは、通常は天然において連結して見られない2つの連結されたポリペプチドを含む。
【0026】
シストロン:アミノ酸残基配列のためにコードし、上流及び下流DNA発現制御要素を含む、DNA分子におけるヌクレオチドの配列。
【0027】
生体分子:生体によって産生される任意の有機分子、又は生体分子を産生するための生体によって使用されるプロセスによって生体外で作製される任意の有機分子であり、そのようなプロセスは、例えば、転写、翻訳、生物有機化学反応等を含む。
【0028】
非抗体足場タンパク質:少なくとも2つのベータシートを形成する表面に露出したループによって接続される6つ以上のβストランドを有する、任意の非抗体タンパク質、タンパク質断片、又はペプチド。そのような非抗体タンパク質足場の例としては、ヒトテネイシン(TeFN3)の第3のフィブロネクチンIII型ドメイン、又はヒトタイチン(Ti27)のI27ドメイン等の、フィブロネクチンIII型ドメイン又はIgドメインを含むタンパク質が挙げられる。
【0029】
糸状ファージ
本発明は、融合タンパク質(タンパク質)をコードするゲノムを封入するタンパク質のマトリックスを含む糸状ファージを意図する。融合タンパク質は、糸状ファージpVII又はpIXタンパク質のアミノ末端に融合される非抗体足場タンパク質部分を含む。
【0030】
糸状ファージは、実施例において記載されるように、ファージ粒子の表面上に提示される融合タンパク質を更に含有する。
【0031】
本発明のファージ上に存在する融合タンパク質において、非抗体足場タンパク質と糸状ファージpVII又はpIXタンパク質との間の「融合」は、典型的なアミド連鎖を含んでもよいか、又は実施例において記載されるリンカーポリペプチド(即ち、「リンカー」)を含んでもよい。典型的に一続きの長さ約5〜50のアミノ酸である様々なリンカーのいずれを使用してもよい。特に好ましいリンカーは、リンカーの点にある融合タンパク質への高度の可動性を提供する。
【0032】
ライブラリ設計:これまでの合成ライブラリは、以下のうちのいくつかを組み込んでいるが、総合的に全てを含んでいるものはない。
【0033】
発現、生化学的、及び生物物理学的特性。好ましい非抗体ペプチド又はタンパク質は、所望の生物学的及び結合活性を有するだけでなく、様々な宿主から効率的に産生され、安定であり、かつ良い溶液物性も有する。操作された組み換え核酸ファージベクターによって融合タンパク質として発現されるか、又は足場のみとして発現されるかいずれかの開示される足場タンパク質は、抗原、受容体、配位子、細胞表面タンパク質マーカ等の生体分子に結合する能力を有することが理解される。好ましい実施形態では、本明細書において記載される足場タンパク質は、上皮成長因子受容体に結合する。高頻度の生殖系列遺伝子の使用も哺乳類系における良い発現を示す。加えて、選択又はスクリーニングの細菌性ファージ提示方法によってライブラリから回復されるそのような融合タンパク質は、細菌性宿主においてよく発現されるはずである。本発明のライブラリは、よく発現され、標準的な組み換え哺乳類宿主(例えば、HEK293及びCHO細胞)及び細菌性宿主から精製されるヒト生殖細胞系列由来の鋳型に基づき、高い安定性及び良い溶液物性を有する。
【0034】
成熟。それぞれの位置において最大20の異なるアミノ酸の潜在的変異により連結される、抗原又は配位子結合の認識に影響を与えることができる代替の足場配列を模倣するV領域における位置の大きい数は、単一のライブラリに全ての変異を含む実用性を除外する。ヒト抗体は、体細胞突然変異の進行性プロセスによって高親和性及び特異性に到達する。本発明のライブラリは、それらが、ヒト抗体の特色及び特徴と同様のそれらを反映するように、それぞれの非抗体ペプチド又はタンパク質鎖の配列一体性を維持しつつ、平行の選択及び標的変異を許可するように設計され、順序付けられる。
【0035】
代替の設計。上記の設計は、天然のヒト抗体の特徴ををシミュレートする。システムのモジュラー性質は、位置の任意の集合でのアミノ酸の任意の集合の組込みに適している。
【0036】
ライブラリアセンブリ技術。好ましい非抗体ペプチド又はタンパク質ライブラリは、低いか、又は高い多様性(>1010)であり、変更に適し、組み立て易く、かつ低い望ましくない配列のバックグラウンドを有する。これらのバックグラウンド配列は、親鋳型及び低標的多様性を含む。以下の方法に従って連結することは、ライブラリアセンブリを加速し、低いバックグラウンドにつながる。(a)Kunkleによる一本鎖突然変異作成、(b)制限部位を伴うパリンドロームループ、(c)メガプライマーによるPCR。
【0037】
pIX非抗体ペプチド又はタンパク質ファージ提示。選択された非抗体ペプチド又はタンパク質鋳型とのpIXの組み合わせは、他の関連分子への変換後にそれらの選択された特性を保持する非抗体ペプチド又はタンパク質を回復するための効率的な選択システムである。
【0038】
ファージミド提示。発現された分子は、ファージpIX外被タンパク質に対して大きいため、細菌細胞において産生される全てのpIXタンパク質に連結される場合、組み換えファージ粒子のアセンブリに干渉する可能性がある。この干渉を回避する1つの方法は、本明細書において記載される当該技術分野において既知であるもの等のpIXファージミド系を使用することであり、それによって、非抗体ペプチド又はタンパク質連結pIX又はpVIIタンパク質を組み換えファージ粒子に組み込むことができる。好ましい用途において、本発明のライブラリは、ファージミド系におけるpIXによって提示される。
【0039】
提示のためのファージ外被タンパク質pIX。pIIIと同様に、pIXは、ファージ上に低いコピー数で存在し、提示された非抗体ペプチド又はタンパク質の親和性選択に適している。しかしながら、pIIIタンパク質は、感染プロセスに非常に関与し、このタンパク質上に提示されるタンパク質は、感染の効率性に干渉し得る。pIXタンパク質上に提示される本発明のライブラリは、選択及び/又はスクリーニングのために効果的に複製され、提示されると予測される。
【0040】
非抗体ペプチド又はタンパク質pIX発現。ファージライブラリから回収した非抗体ペプチド又はタンパク質をスクリーニングする1つの方法は、提示のために非抗体ペプチド又はタンパク質分子に連結されるファージ外被タンパク質を除去することである。pIXタンパク質の小さいサイズは、この工程を伴わずに直接、非抗体ペプチド又はタンパク質のスクリーニングの産生の選択肢を提供する。
【0041】
治療的使用。本明細書において記載されるように、開示される足場は、抗体の代替として使用してよい。したがって、開示される足場は、治療用途を有する。したがって、本明細書において記載される足場タンパク質は、治療用組成物を形成するために使用してよいと考えられる。1つのそのような組成物は、本明細書において記載される足場、及び製薬学的に許容できる担体を含むことができる。別の実施形態では、組成物は、そのF:Gループが非天然ポリペプチドで置換された、TeFN3等の単離されたEGFR特異的非抗体タンパク質足場、及び製薬学的に許容できる担体を含む。代替の実施形態では、組成物は、配列番号2〜23、25、又は28によって特定される少なくとも1つのポリペプチドから作製されるF:Gループを有する代替可能な非抗体足場タンパク質、及び製薬学的に許容できる担体を含む。非抗体足場タンパク質が、操作された核酸ファージベクターによってコードされ、上皮成長因子受容体等の生体分子に結合し、製薬学的に許容できる担体と組み合わされる実施形態も本明細書に開示する。
【0042】
設計ライブラリ足場。ライブラリ足場は、生殖細胞系列VH及びVL遺伝子の構造及び機能を模倣する一連のヒトタンパク質配列から作製される。
【0043】
ライブラリ足場の発現及び提示能力。ライブラリ足場非抗体ペプチド又はタンパク質の良い発現及び提示能力は、足場遺伝子の上で開発されるライブラリの質に直接関連する。ライブラリ足場非抗体ペプチド又はタンパク質発現及び提示能力は、ライブラリ構成の前に検査された。発現したが、提示しなかったか、又は不十分に提示した少数の足場非抗体ペプチド又はタンパク質は、ライブラリ構成から除外された。よく発現され、提示されたライブラリ足場は、ライブラリにおける非抗体ペプチド又はタンパク質の高い割合が機能的であり、天然源から遺伝的に増幅されるVH及びVL遺伝子の組み合わせクローニングに由来するライブラリより優れていることを確実にする。
【0044】
ライブラリ作成のための方法。修正されたKunkel突然変異作成方法を本発明に従って使用することができ、それは、数十億もの大腸菌コロニーを効率的に作成し、それぞれ、異なる非抗体ペプチド又はタンパク質配列を持ち、それは、非抗体ペプチド又はタンパク質ライブラリの精製のために使用することができる。効率的であるが、非突然変異親DNAの割合は、高度配列複合体ライブラリの作成において適合される時に増加する。加えて、長いオリゴヌクレオチドの合成の技術的制限は、遠位領域において配列多様性を含有するライブラリを作製する方法の有効性を低下させることがある。そのような制限を克服するために、オリゴヌクレオチド>350塩基(メガプライマー)を作成する、及び突然変異作成鋳型において制限酵素認識部位を含有するステムループ配列の作製の付加的な技術は、Kunkel突然変異作成方法等の技術と組み合わせて任意に使用することができる。ライブラリ作成のために他者によって使用される、制限クローニング、ファージ組み換え、及び配列特異的組み換え等の他の既知のライブラリ技術と比較して、改善されたKunkelによる方法は、1つのライブラリにつき>109配列の作成においてより効果的であり得、標的DNA上の任意の場所において配列多様性を導入する際により万能である。
【0045】
インライン親和性成熟。統合親和性成熟プロセス、又はインライン親和性成熟は、ライブラリから選択される非抗体ペプチド又はタンパク質の結合親和性の設計及び改善のために、本発明に従って使用することができる。パニング後の所望の非抗体ペプチド又はタンパク質の結合親和性を改善することは、治療用非抗体ペプチド又はタンパク質リードの特定の成功を増加させることができる。ライブラリ作成のためのKunkelの方法の使用は、単純かつ連続的なプロセスにおける配列多様化戦略の効果的な実行を改善できる。改善されたKunkel突然変異作成方法の使用の設計戦略及び技術的利点は、結果の効率性及び有効性を低下させる単調なライブラリ作成方法が使用される、他の共同成熟戦略より優れた方法を提供できる。
【0046】
平行ライブラリパニング。自動又は半自動機器を使用する平行パニングプロセスを使用して、個々に作製されたサブライブラリを処理することができる。平行パニングは、所望の特徴のペプチド又はタンパク質を有する所望のライブラリを提供するために、好適に多様な一連の非抗体ペプチド又はタンパク質のを発見する可能性を最大化できる。インライン親和性成熟における平行パニングの効果的な使用はまた、そのようなタンパク質が、改善された親和性又は生物活性等のいくつかの同時に改善された特徴を有することも可能にする。機械によるパニングシステムの開発はまた、異なるパニング条件の系統的な監視及び調製が、所望の特性を有する非抗体ペプチド又はタンパク質をより迅速にスクリーニングし、単離することも可能にする。
【0047】
親和性順位。親和性による結合検定は、更なる特徴付けのための最良の結合を選択するために、大きい、多様な、かつ高親和性の配位子又は抗原特異的結合ペプチド若しくはタンパク質に適用される。ELISA等の標準的な生化学的方法、並びに多数の試料を処理するために好適な親和性測定機器、例えば、BIAcore、Octet及びBINDを、本目的のために、単独か又は組み合わせて使用する。
【0048】
本発明は一般条件で記述されてきているが、本発明の実施形態は、特許請求の範囲を限定するように解釈されるべきではない以下の実施例で更に開示される。
【実施例】
【0049】
実施例1:非抗体タンパク質の提示:2つの非抗体ペプチド又はタンパク質は、pIX融合タンパク質としてM13ファージの表面上に成功裏に提示された。両方の足場タンパク質は、それらの天然の状態における足場タンパク質によって結合しない特定のタンパク質標的に結合するように操作した。これらの操作された足場は、ファージ上で提示されつつ、この特定の標的への結合を保持した。加えて、アミノ酸変異体のライブラリはそれぞれの足場において作製し、特定の標的に結合するそれらの能力は、pIX融合としてファージ上にライブラリメンバーを提示すること及び標的タンパク質に対するパニングによって対処した。両方のライブラリは、多数の陽性ヒットをもたらした。
【0050】
2つの非抗体ペプチド又はタンパク質、免疫グロブリン(Ig)ドメインタンパク質は、pIX融合タンパク質としてM13ファージの表面上にEGFR結合ペプチドを提示するように操作される候補として選択した。以下の基準を、候補の選択のために使用した。
【0051】
PDBデータベースの中で利用可能な高解像度原子構造
a.堆積した構造は、単一の単離されたIgドメインを表す
b.完全なヒト配列
c.配列は、ジスルフィド結合残基を含有しない
d.公開された発現及び精製条件
e.再折り畳み工程の必要性を伴わず、かつ高収率で大腸菌から成功裏に発現され、精製されることを証明した。
【0052】
PDBデータベースは、単一Igドメインを含有する全てのタンパク質及びタンパク質フラグメントを調査するために、手動で、並びに構造による整合プログラムの援助により検索した。その後、公開された発現及び精製条件を決定するために、科学文献を調査した。
【0053】
上記の基準を使用して、成功のための最高の機会を有する分子を選択したが、これらの基準に厳密に満たない他のIg分子も、ペプチド提示のための成功する足場として使用することができることが想定される。複数のIgドメイン又は単一Igドメインの繰り返し体からなる足場は、成功する足場として使用することができることも想定される。上記の分析は、更なる研究のための、2つのIg分子、ヒトテネイシンからの第3のFn3ドメイン(残基802〜893、TeFn3)及びヒトタイチンのI27ドメイン(残基5253〜5341、Ti27)の選択をもたらした。これらのタンパク質フラグメントをコードする遺伝子は、Blue Heronによって合成し、標準PCR及び制限消化方法によってプラスミドpPEP9−BbsIにサブクローンした。Ti27におけるN5331G、及びTeFn3におけるS881Gの突然変異は、ペプチドの挿入のための制限部位を導入するために含んだ(以下参照)。
【0054】
その後、両方の足場のF:Gループ(TeFn3の残基876〜881及びTi27の5258〜5259)(図1)は、標準的な制限部位クローニング方法を使用して、システイン拘束EGFR結合ペプチド、PHPEP190(配列 DPCTWEVWGRECLQ)(Wang)の一部と置換し、Ti27−190及びTeFn3−190作製物を作製した。多数の対照作製物もまた作製し、非EGFR結合F:Gループ配列(Ti27及びTeFn3と称される)を含有し、及びF:Gループに6つの終止コドン(Ti27−終止及びTeFn3−終止)を含有するものを含む。表1は、これらの作製物を要約する。
【0055】
【表1】
【0056】
それぞれの作製物は、足場タンパク質のC末端とpIXのN末端との間のEDSGGSGG(配列番号30)リンカー配列を伴うファージミドベクターからの糸状ファージの小さい外被タンパク質pIXへの融合として発現された。全ての作製物は、EGFR結合の非存在下でのタンパク質発現及び提示に対して検定するために、N末端Mycタグにより発現された。M13ファージの表面上の成功する提示、及びTeFn3−190及びTi27−190タンパク質の特定の結合活性は、ファージELISAによって検証した。0.5μgのEGFR細胞外ドメイン(sEGFR−MMB)(Wang)のFc−融合型、抗Myc非抗体ペプチド若しくはタンパク質、又は対照Fc分子(CNTO 360)を、96ウェルMaxiSorop(商標)プレート上で固定し、StartingBlock(商標)T−20により遮断した。表1に記載される作製物を提示するファージは、それぞれのプレートへの300μLのファージ含有上清の添加前に連続的に希釈し、続いて、室温で1時間培養した。それぞれのプレートは、続いてTBSTで洗浄し、蛍光プレートリーダを使用して、抗M13 HRP結合非抗体ペプチド又はタンパク質により検出されるファージに結合させた。図2は、TeFn3−190及びTi27−190が、CNTO 360への大きな結合の欠如によって証明される低いレベルのバックグラウンド結合によりプレート結合Fc−EGFRに選択的に結合したことを示す。TeFn3及びTi27はまた、抗Mycコーティングされたプレートへの結合によって示されるファージ表面上でも提示された。しかしながら、これらのタンパク質は、EGFRに結合しなかった。不明の理由で、TeFn3−190は、EGFR結合によって示されるように明らかに発現されるが、抗Mycコーティングされたプレートに結合しなかった。予想通りに、TeFn3−終止及びTi27−終止は、ファージ上に提示されなかった。
【0057】
ライブラリは、TeFn3及びTi27に関連してPHPEP190のシステインの必要性を決定し、そのようなライブラリを、pIX上のこれらの足場内で効率的に提示し、標的タンパク質に対してパニングすることができることを確認するために構築した。このライブラリは、Ti27−終止及びTeFn3−終止作製物において終止コドンを含有する全ての6つの位置を無作為化することによって構築した(表1)。Kunkelの方法(Kunkel,et al.,Methods Enzymol 154:367〜82,1987)による突然変異作成の後の100の無作為で採取したコロニーのシークエンシングは、突然変異が、Ti27−190及びTeFn3−190ライブラリからのコロニーのそれぞれ51%及び64%において見られたことを確認した。その後、これらのライブラリは、室温で1時間10μg(1回目)又は5μg(2及び3回目)のビオチン化EGFRにより100μLのファージ溶液を培養することによって、EGFR−Fc融合に対して3回パニングした。結合複合体は、ストレプトアビジンコーティングされた磁気ビーズで引き下げ、TBSTで3回洗浄した。その後、結合ファージを使用して、TG1細胞を再感染させ、表2に記載されるファージ力価をもたらした。
【0058】
【表2】
【0059】
それぞれのライブラリからの3回目のパニングからの96のコロニーは、ファージELISAによってEGFR−Fcへの結合を試験するために無作為に選択した。図3は、Ti27−190及びTeFn3−190ライブラリからの10及び20のクローンそれぞれが、EGFRへの有意な結合を示したことを示す(EGFR:CNTO 360からの総数の比率は、5より大きい)。全ての陽性ヒットからのF:Gループの配列を表3に示す。全ての配列したヒットは、本来のPHPEP190と同様に同一の位置でシステイン残基を含有し、このペプチドのシステイン拘束された性質が機能のために必要であることを示した。
【0060】
【表3】
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、2007年12月19日に出願された米国特許仮出願第61/014,778号の利益を主張し、その全体を参照として本明細書に組み込む。
【0002】
(発明の分野)
本発明は、M13ファージのpIXを使用して、非抗体足場タンパク質融合を産生するためのpIXファージ提示ライブラリを作成し、使用するための組成物及び方法に関する。
【背景技術】
【0003】
ファージ又はファージミド系における融合パートナーとしてpIII、pVII、pVIII、pIX、及びそれらの組み合わせを使用する糸状ファージ提示は、特に、デノボタンパク質及びペプチド単離並びに親和性成熟に関するタンパク質工学のための技術として使用されている(例えば、Janda et al,米国特許第7,078,166号を参照)。無作為ペプチド及びタンパク質は、関心のある対象のタンパク質標的に対するパニングを介してペプチド配列の変異を提示するファージライブラリから作成及び単離することができる。これまでに使用されたヒトタンパク質デノボライブラリは、合成的に作製されている。合成ライブラリにおいて、潜在的タンパク質をコードする推定上有用なDNA配列は、既知の配列又はモチーフに基づいて設計され、合成される。合成ペプチドライブラリに加えて、ライブラリはまた、ヒト組織に由来するタンパク質コードDNAの組み合わせクローニングによって作製することもできる。そのようなライブラリは、潜在的に有用なペプチドを提供するため、かつ連続回のパニング及び成熟又は修飾を実行して、選択された標的タンパク質の阻害生物活性等の所望の特性を有する非抗体ペプチド又はタンパク質ペプチドを見つけることを試みるために使用されている。
【0004】
ヒトペプチド、及び模倣タンパク質又は標的タンパク質に結合するか、若しくはそれを模倣する既知のタンパク質突然変異等のタンパク質は、ファージ提示pIII又はpVIIIペプチドライブラリから単離されている。特定の標的に結合するいくつかのタンパク質を単離することに成功しているが、そのようなファージ提示ライブラリ法は、ライブラリ作成のプロセスを繰り返さなければならないこと、所望の特徴を有するペプチド又はタンパク質を単離するためのパニング及び成熟の必要条件、及び他の既知の制限の問題に見舞われる。そのようなファージライブラリは、ヒトに存在するペプチド多様性の範囲、アミノ酸変異の位置及び程度、並びに異なるヒト遺伝子からの生物活性タンパク質の相対存在量を完全に包含又は模倣しないという問題にも見舞われる。天然レパートリからの合成タンパク質の偏位は、人において治療用に使用される場合、好ましくない生化学的特性及び免疫原性のリスクを増加させる可能性があり、この問題には、そのようなライブラリからの選択に対して、どのようにして配列が作成され、スクリーニングされるのかということによって潜在的に対処し得る。
【0005】
モノクローナル抗体は、標的分子に対する高親和性及び特異性が望ましい時に最も広く使用されるクラスの治療用タンパク質である。しかしながら、非抗体抗原結合ペプチド又はタンパク質は、治療法又は診断法における使用に対して高い関心のある対象であるそのような標的に結合するように操作することができる。生体分子に結合することが可能なそのようなタンパク質又はペプチドは、より小さいサイズ、ジスルフィド結合の欠如、原核宿主において発現される能力、精製の新規の方法、高い安定性、薬物/毒素への結合の容易性、及び知的財産利点等であるが、それらに限定されない、伝統的な抗体に対するいくつかの潜在的利点を有し得る。
【0006】
1つの種類の足場は、免疫グロブリン(Ig)折り畳みである。この折り畳みは、抗体の可変領域、並びに数千もの非抗体ペプチド又はタンパク質において見られる。1つのそのようなIgタンパク質、ヒトフィブロネクチンからの10番目のフィブロネクチン3型繰り返し体は、全体的なIg折り畳み構造を保持しつつ、表面に露出したループにおける多数の突然変異に耐えることができるということが示されている。そのため、アミノ酸変異体のライブラリは、これらのループ、及び多数の異なる標的に選択される特定の特定の結合剤に組み込まれている。そのような操作されたFn3ドメインは、重要な生物物理学的特性を保持しつつ、適度に高い親和性を伴う標的に結合することが見出されている。
【発明の概要】
【発明が解決しようとする課題】
【0007】
ファージライブラリのこれまでの使用は、抗体ベースのタンパク質融合ライブラリを含んできた。しかしながら、高親和性及び活性、高生産性、良い溶液物性、及び人に投与する際の低免疫応答の傾向のヒト治療抗体の決定的要素を同時に送達する、合成の代替の足場又は非抗体推定抗原結合ペプチド又はタンパク質融合ライブラリ及び方法の必要性が存在する。非抗体ペプチド又はタンパク質発見の資源コストを低下させ、及び生物的評価のための抗体の送達を加速するために、現在の方法に対して、合成ライブラリからの非抗体ペプチド又はタンパク質単離の効率性を増加させる必要性が更に存在する。本発明のライブラリ及び方法は、総合的な設計、アセンブリ技術、及びファージpIX非抗体ペプチド又はタンパク質提示を連結することによって、これらの必要性を満たす。
【課題を解決するための手段】
【0008】
先行技術の教示と対照的に、pVII及びpIXは、非抗体ペプチド又はタンパク質のための効率的かつ迅速な基本骨格、及び治療抗体の非抗体ペプチド又はタンパク質フラグメント作成及び選択を提供するために、例えば、任意にインライン成熟を伴う、突然変異作成又は他の多様性産生技術を使用して、M13ファージからのpIXを使用して高親和性非抗体ペプチド又はタンパク質ライブラリを作成するために成功裏に使用することができることが現在発見されている。本発明によると、所望の生体分子又は抗原に結合することが可能である非抗体ペプチド又はタンパク質は、任意に、代表的なヘテロ二量体モチーフにおいて、機能性非抗体ペプチド又はタンパク質を提示するために、ファージ表面上の動的相互作用において係合されるpVII及びpIXに融合される。したがって、非抗体ペプチド又はタンパク質結合剤のファージ上の提示は、メンバーが単量体又は二量体人工非抗体ペプチド又はタンパク質種として機能し、新規又は所望の生物活性の選択を可能にすることができる、組み合わせヘテロ二量体アレイの多様なライブラリの提示及び検定のための好適かつ好ましい方法である。
【0009】
本発明は、糸状ファージのpIXタンパク質、広く使用されるpIII及びpVIIIタンパク質と異なるファージ表面タンパク質に融合される非抗体ペプチド又はタンパク質デノボライブラリの設計及び提示を提供する。我々は、個々の足場ライブラリを別々に構築し、足場ライブラリのそれぞれの活性を系統的に検査し、抗原認識のための構造トポロジーを評価するためのファージ選択プロセスを開発した。
【0010】
本発明は、様々な改善された新しいpIX及びpVIIファージ提示デノボライブラリ作成方法、及び(i)pIX又はpVIIファージタンパク質に融合される、設計され、提示された非抗体ペプチド又はタンパク質デノボライブラリ、(ii)M13ファージの広く使用されるpIII及びpVIIIと異なるファージ表面タンパク質の使用、(iii)非抗体タンパク質足場に融合されるペプチドのライブラリをスクリーニングするためのpIXファージ提示システムの使用、(iv)抗原認識のための設計された配列及び構造トポロジーの効果の系統的検査を可能にする非抗体ペプチド又はタンパク質選択プロセス、(v)ライブラリ選択の一部としての流線形親和性成熟及びインライン成熟プロセス、のうちの1つ以上であるがそれらに限定されない、構成要素を提供する。ライブラリの個々又は群のライブラリ設計、選択、最適化及び成熟のそのような新しいシステムは、非抗体ペプチド又はタンパク質デノボ発見の成功のための再現可能かつ確実なシステムを提供し、また抗原相互作用に対する非抗体ペプチド又はタンパク質の構造機能相関の理解を容易にする。
【0011】
上述されるヒト非抗体ペプチド又はタンパク質デノボライブラリは、M13ファージのpIX遺伝子を介する提示によって現在の抗体ライブラリ現況技術と異なる。非抗体ペプチド又はタンパク質は、本発明によると、pIX融合タンパク質としてM13ファージの表面上に成功裏に提示することができる。両方の足場タンパク質は、それらの天然の状態において対応する足場タンパク質によって結合しない特定のタンパク質標的に結合するように操作することができる。これらの操作された足場は、ファージ上で提示されつつ、この特定の標的への結合を保持した。加えて、アミノ酸変異体のライブラリは、それぞれの足場において、本発明によって作製することができ、特定の標的に結合するそれらの能力は、pIX融合としてファージ上にライブラリメンバーを提示すること、及び標的タンパク質に対するパニングによって対処される。
【0012】
本発明において使用される人工抗体又は足場タンパク質は、非抗体ペプチド又はタンパク質分子の機能的戦略を使用するが、(1)標的抗原の配列相同性及び毒性、(2)宿主におけるか、又は非抗体ペプチド又はタンパク質を回復させるために使用されるハイブリドーマ培養における作成された非抗体ペプチド又はタンパク質の生物学的インパクト、及び(3)所望の活性に対するスクリーニング対選択、を含む、生体内拘束がない状態で作成することができる、大きい多様性のタンパク質モチーフとして本明細書において定義される。
【0013】
そのため、本発明は、多様性に富む単量体又はヘテロ二量体ポリペプチドアレイの構成のための組み合わせファージ提示形式を説明する。特に、本発明は、融合ポリペプチドをコードするゲノムを封入する糸状ファージ粒子を説明し、融合ポリペプチドは、糸状ファージpVII又はpIXタンパク質のアミノ末端に融合される非抗体足場タンパク質を含む。好ましくは、ファージ粒子は、ファージ粒子の表面上の発現された融合タンパク質を含む。
【0014】
関連する実施形態では、本発明は、融合タンパク質を発現するためのカセットを含む糸状ファージの表面上に融合タンパク質を発現するためのベクターを説明する。カセットは、挿入DNA、即ち、ポリリンカーの方向性連結反応のために適合されるヌクレオチドの配列を介して作動可能に連結される上流及び下流の翻訳可能なDNA配列を含み、上流配列は、原核分泌信号をコードし、下流配列は、pVII又はpIX糸状ファージタンパク質をコードする。翻訳可能なDNA配列は、融合ポリペプチドの部分としての翻訳可能なDNA配列の発現のための一連のDNA発現信号に作動可能に連結される。好ましい変異において、ベクターは、糸状ファージの表面上に第2の融合タンパク質を発現するための第2のカセットを更に含み、第2のカセットは、第1の融合タンパク質発現カセットがpVIIタンパク質をコードし、第2の融合タンパク質発現カセットがpIXタンパク質をコードするという条件で、第1のカセットの構造を有する。ベクターは、ファージ粒子の表面上にヘテロ二量体タンパク質複合体を発現するためにファージゲノムとして使用され、ヘテロ二量体の2つのポリペプチドは、第1のファージタンパク質及び第2のファージタンパク質、pVII及びpIXへの融合によって、ファージ粒子上でそれぞれ固定される。
【0015】
別の実施形態では、本発明は、本発明によるファージ粒子のライブラリ、即ち、組み合わせライブラリを意図し、ライブラリにおける代表的な粒子は、それぞれ、異なる融合タンパク質を提示する。粒子がヘテロ二量体タンパク質複合体を提示する場合、ライブラリは、ヘテロ二量体の組み合わせライブラリを含む。好ましいライブラリは、融合タンパク質の異なるか、又は同様の種の、少なくとも103、104、105、106、107、108、109、1010、1011、1012、1013、又はその中の任意の範囲又は値の組み合わせ多様性を有する。
【0016】
関連する実施形態は、第1のポリペプチド及び第2のポリペプチドを含む融合タンパク質を説明し、第1のポリペプチドは、非抗体足場タンパク質であり、第2のポリペプチドは、糸状ファージpVII又はpIXタンパク質であり、非抗体足場タンパク質は、糸状ファージタンパク質のアミノ末端に融合される。
【0017】
更に、本発明は、非抗体足場タンパク質をコードする遺伝子のレパートリを本発明のベクターにクローニングし、突然変異作成によって、第1の融合タンパク質及び第2の融合タンパク質ライブラリの集団の無作為的組み合わせによって、ライブラリの多様性を変化させるための標的及び親和性選択(「パニング」)等によってライブラリにおける非抗体足場タンパク質の構造を修正することによる、を含む、ファージの組み合わせライブラリを産生するための様々な方法を意図する。そのような実施形態は、本明細書において例示されるように、第2のポリペプチドに融合される代替可能な非抗体足場タンパク質を有する融合ポリペプチドを含むことができる。例えば、一実施形態では、代替可能な非抗体足場タンパク質は、TeFN3であり得、第2のポリペプチドは、pIXであり得、TeFN3のF:Gループは、突然変異作成、又はシステイン拘束されたEGFR結合ペプチド、PHPEP190等の非天然ポリペプチドによる野生型F:Gループアミノ酸残基の標的置換によって変化する。代替の実施形態では、代替可能な非抗体足場タンパク質は、配列番号2〜23、25、又は28によって特定される少なくとも1つのポリペプチドから作製されるF:Gループを有することができる。非抗体足場タンパク質が、操作された核酸ファージベクターによってコードされ、上皮成長因子受容体又は生物活性配位子等の生体分子に結合する実施形態も本明細書に開示する。
【0018】
改善された又は新規の機能を伴うタンパク質の設計は、様々な医薬、産業、環境、及び基礎研究の用途に伴う重要な目標である。組み合わせ非抗体ペプチド又はタンパク質ライブラリの開発に続き、強力な次の工程は、人工非抗体ペプチド又はタンパク質作製物、及び二量体種が野生型であるか、又は機能的であり得る他のタンパク質モチーフへの進化である。
【0019】
本発明は、pVII及びpIXが、単量体又は二量体種を形成する融合タンパク質の提示のために利用される、組み合わせ非抗体ポリペプチドアレイの構成のためのファージ提示形式を提供することによって、これらの課題に取り組む。pIX又はpVII上の発現を使用して機能的相互作用のライブラリを作成するために、1つ又は2つのタンパク質モチーフを近接して独立して提示することができるため、これが全く新しい方法論であることに留意することが重要である。
【0020】
更に、ライブラリを形成するための配列無作為化、及びハイブリッド種を形成するための鎖シャフリングプロトコルは、新規のタンパク質のサブセットにつながり得る。例えば、ホモ二量体又はヘテロ二量体型における亜鉛フィンガードメインのアレイの提示及び修正は、特定のDNA相互作用を所有する構造を産生する。加えて、全く新しい作製物が、非抗体ペプチド又はタンパク質鎖等の事前形成された足場内の所望のコードフラグメントの挿入を介して可能である。可能な挿入は、酵素特徴的配列又はリプレッサ結合タンパク質を含む。
【0021】
前述の概要及び以下の詳細な説明の両方とも、例示及び説明のためのみであり、主張される本発明を制限するものではないことが理解されるべきである。
【図面の簡単な説明】
【0022】
【図1】テネイシンの第3のFn3ドメイン(残基802〜893、PDB=1TEN、(Leachy,et al.,Science 258:987〜91,1992))及びタイチンのI27ドメイン(残基5253〜5341、PDB=1TIT、(Improta,et al.,Structure 4:323〜37,1996))の構造。矢印は、F:Gループ(テネイシン及びタイチンそれぞれの残基876〜881及び5258〜5259)の場所を指し、それは、EGFR結合配列を収容するように修正された。
【図2】pIX上のテネイシン及びタイチン足場タンパク質の提示に関する図式的結果。
【図3】pIXに提示されるTeFn3−190及びTi27−190ライブラリからのEGFR結合作製物のためのパニングの図式的結果。
【発明を実施するための形態】
【0023】
本発明は、様々な新しいファージ提示デノボライブラリ作成方法及び構成要素を提供し、(i)pIX又は他のファージタンパク質に融合される、設計され、提示された非抗体ペプチド又はタンパク質ライブラリ、(ii)M13ファージの広く使用されるpIII及びpVIIIと異なるファージ表面タンパク質の使用、(iii)抗原認識のための設計された配列及び構造トポロジーの効果の系統的検査を可能にする非抗体ペプチド又はタンパク質選択プロセスにおける改善され、設計された組み合わせ多様性を提供するための、ライブラリ足場としてのそのようなファージ構成要素の使用、(iv)ライブラリ選択の一部としての流線形親和性成熟プロセス、等を含むが、それらに限定されない。ライブラリの個々又は群のライブラリ設計、選択、最適化及び成熟のそのような新しいシステムは、非抗体ペプチド又はタンパク質デノボ発見の成功のための再現可能かつ確実なシステムを提供し、また抗原相互作用に対する非抗体ペプチド又はタンパク質の構造機能相関の理解を容易にする。
【0024】
上述されるヒト非抗体ペプチド又はタンパク質デノボライブラリは、M13ファージのpIX又はpVII遺伝子を介する提示によって現在の抗体ライブラリ現況技術と異なる。
【0025】
用語の定義:
融合ポリペプチド:少なくとも2つのポリペプチド、及び2つのポリペプチドを1つの連続的なポリペプチドに作動可能に連結する連結配列からなるポリペプチド。融合ポリペプチドにおいて連結される2つのポリペプチドは、典型的に、2つの独立した源に由来するため、融合ポリペプチドは、通常は天然において連結して見られない2つの連結されたポリペプチドを含む。
【0026】
シストロン:アミノ酸残基配列のためにコードし、上流及び下流DNA発現制御要素を含む、DNA分子におけるヌクレオチドの配列。
【0027】
生体分子:生体によって産生される任意の有機分子、又は生体分子を産生するための生体によって使用されるプロセスによって生体外で作製される任意の有機分子であり、そのようなプロセスは、例えば、転写、翻訳、生物有機化学反応等を含む。
【0028】
非抗体足場タンパク質:少なくとも2つのベータシートを形成する表面に露出したループによって接続される6つ以上のβストランドを有する、任意の非抗体タンパク質、タンパク質断片、又はペプチド。そのような非抗体タンパク質足場の例としては、ヒトテネイシン(TeFN3)の第3のフィブロネクチンIII型ドメイン、又はヒトタイチン(Ti27)のI27ドメイン等の、フィブロネクチンIII型ドメイン又はIgドメインを含むタンパク質が挙げられる。
【0029】
糸状ファージ
本発明は、融合タンパク質(タンパク質)をコードするゲノムを封入するタンパク質のマトリックスを含む糸状ファージを意図する。融合タンパク質は、糸状ファージpVII又はpIXタンパク質のアミノ末端に融合される非抗体足場タンパク質部分を含む。
【0030】
糸状ファージは、実施例において記載されるように、ファージ粒子の表面上に提示される融合タンパク質を更に含有する。
【0031】
本発明のファージ上に存在する融合タンパク質において、非抗体足場タンパク質と糸状ファージpVII又はpIXタンパク質との間の「融合」は、典型的なアミド連鎖を含んでもよいか、又は実施例において記載されるリンカーポリペプチド(即ち、「リンカー」)を含んでもよい。典型的に一続きの長さ約5〜50のアミノ酸である様々なリンカーのいずれを使用してもよい。特に好ましいリンカーは、リンカーの点にある融合タンパク質への高度の可動性を提供する。
【0032】
ライブラリ設計:これまでの合成ライブラリは、以下のうちのいくつかを組み込んでいるが、総合的に全てを含んでいるものはない。
【0033】
発現、生化学的、及び生物物理学的特性。好ましい非抗体ペプチド又はタンパク質は、所望の生物学的及び結合活性を有するだけでなく、様々な宿主から効率的に産生され、安定であり、かつ良い溶液物性も有する。操作された組み換え核酸ファージベクターによって融合タンパク質として発現されるか、又は足場のみとして発現されるかいずれかの開示される足場タンパク質は、抗原、受容体、配位子、細胞表面タンパク質マーカ等の生体分子に結合する能力を有することが理解される。好ましい実施形態では、本明細書において記載される足場タンパク質は、上皮成長因子受容体に結合する。高頻度の生殖系列遺伝子の使用も哺乳類系における良い発現を示す。加えて、選択又はスクリーニングの細菌性ファージ提示方法によってライブラリから回復されるそのような融合タンパク質は、細菌性宿主においてよく発現されるはずである。本発明のライブラリは、よく発現され、標準的な組み換え哺乳類宿主(例えば、HEK293及びCHO細胞)及び細菌性宿主から精製されるヒト生殖細胞系列由来の鋳型に基づき、高い安定性及び良い溶液物性を有する。
【0034】
成熟。それぞれの位置において最大20の異なるアミノ酸の潜在的変異により連結される、抗原又は配位子結合の認識に影響を与えることができる代替の足場配列を模倣するV領域における位置の大きい数は、単一のライブラリに全ての変異を含む実用性を除外する。ヒト抗体は、体細胞突然変異の進行性プロセスによって高親和性及び特異性に到達する。本発明のライブラリは、それらが、ヒト抗体の特色及び特徴と同様のそれらを反映するように、それぞれの非抗体ペプチド又はタンパク質鎖の配列一体性を維持しつつ、平行の選択及び標的変異を許可するように設計され、順序付けられる。
【0035】
代替の設計。上記の設計は、天然のヒト抗体の特徴ををシミュレートする。システムのモジュラー性質は、位置の任意の集合でのアミノ酸の任意の集合の組込みに適している。
【0036】
ライブラリアセンブリ技術。好ましい非抗体ペプチド又はタンパク質ライブラリは、低いか、又は高い多様性(>1010)であり、変更に適し、組み立て易く、かつ低い望ましくない配列のバックグラウンドを有する。これらのバックグラウンド配列は、親鋳型及び低標的多様性を含む。以下の方法に従って連結することは、ライブラリアセンブリを加速し、低いバックグラウンドにつながる。(a)Kunkleによる一本鎖突然変異作成、(b)制限部位を伴うパリンドロームループ、(c)メガプライマーによるPCR。
【0037】
pIX非抗体ペプチド又はタンパク質ファージ提示。選択された非抗体ペプチド又はタンパク質鋳型とのpIXの組み合わせは、他の関連分子への変換後にそれらの選択された特性を保持する非抗体ペプチド又はタンパク質を回復するための効率的な選択システムである。
【0038】
ファージミド提示。発現された分子は、ファージpIX外被タンパク質に対して大きいため、細菌細胞において産生される全てのpIXタンパク質に連結される場合、組み換えファージ粒子のアセンブリに干渉する可能性がある。この干渉を回避する1つの方法は、本明細書において記載される当該技術分野において既知であるもの等のpIXファージミド系を使用することであり、それによって、非抗体ペプチド又はタンパク質連結pIX又はpVIIタンパク質を組み換えファージ粒子に組み込むことができる。好ましい用途において、本発明のライブラリは、ファージミド系におけるpIXによって提示される。
【0039】
提示のためのファージ外被タンパク質pIX。pIIIと同様に、pIXは、ファージ上に低いコピー数で存在し、提示された非抗体ペプチド又はタンパク質の親和性選択に適している。しかしながら、pIIIタンパク質は、感染プロセスに非常に関与し、このタンパク質上に提示されるタンパク質は、感染の効率性に干渉し得る。pIXタンパク質上に提示される本発明のライブラリは、選択及び/又はスクリーニングのために効果的に複製され、提示されると予測される。
【0040】
非抗体ペプチド又はタンパク質pIX発現。ファージライブラリから回収した非抗体ペプチド又はタンパク質をスクリーニングする1つの方法は、提示のために非抗体ペプチド又はタンパク質分子に連結されるファージ外被タンパク質を除去することである。pIXタンパク質の小さいサイズは、この工程を伴わずに直接、非抗体ペプチド又はタンパク質のスクリーニングの産生の選択肢を提供する。
【0041】
治療的使用。本明細書において記載されるように、開示される足場は、抗体の代替として使用してよい。したがって、開示される足場は、治療用途を有する。したがって、本明細書において記載される足場タンパク質は、治療用組成物を形成するために使用してよいと考えられる。1つのそのような組成物は、本明細書において記載される足場、及び製薬学的に許容できる担体を含むことができる。別の実施形態では、組成物は、そのF:Gループが非天然ポリペプチドで置換された、TeFN3等の単離されたEGFR特異的非抗体タンパク質足場、及び製薬学的に許容できる担体を含む。代替の実施形態では、組成物は、配列番号2〜23、25、又は28によって特定される少なくとも1つのポリペプチドから作製されるF:Gループを有する代替可能な非抗体足場タンパク質、及び製薬学的に許容できる担体を含む。非抗体足場タンパク質が、操作された核酸ファージベクターによってコードされ、上皮成長因子受容体等の生体分子に結合し、製薬学的に許容できる担体と組み合わされる実施形態も本明細書に開示する。
【0042】
設計ライブラリ足場。ライブラリ足場は、生殖細胞系列VH及びVL遺伝子の構造及び機能を模倣する一連のヒトタンパク質配列から作製される。
【0043】
ライブラリ足場の発現及び提示能力。ライブラリ足場非抗体ペプチド又はタンパク質の良い発現及び提示能力は、足場遺伝子の上で開発されるライブラリの質に直接関連する。ライブラリ足場非抗体ペプチド又はタンパク質発現及び提示能力は、ライブラリ構成の前に検査された。発現したが、提示しなかったか、又は不十分に提示した少数の足場非抗体ペプチド又はタンパク質は、ライブラリ構成から除外された。よく発現され、提示されたライブラリ足場は、ライブラリにおける非抗体ペプチド又はタンパク質の高い割合が機能的であり、天然源から遺伝的に増幅されるVH及びVL遺伝子の組み合わせクローニングに由来するライブラリより優れていることを確実にする。
【0044】
ライブラリ作成のための方法。修正されたKunkel突然変異作成方法を本発明に従って使用することができ、それは、数十億もの大腸菌コロニーを効率的に作成し、それぞれ、異なる非抗体ペプチド又はタンパク質配列を持ち、それは、非抗体ペプチド又はタンパク質ライブラリの精製のために使用することができる。効率的であるが、非突然変異親DNAの割合は、高度配列複合体ライブラリの作成において適合される時に増加する。加えて、長いオリゴヌクレオチドの合成の技術的制限は、遠位領域において配列多様性を含有するライブラリを作製する方法の有効性を低下させることがある。そのような制限を克服するために、オリゴヌクレオチド>350塩基(メガプライマー)を作成する、及び突然変異作成鋳型において制限酵素認識部位を含有するステムループ配列の作製の付加的な技術は、Kunkel突然変異作成方法等の技術と組み合わせて任意に使用することができる。ライブラリ作成のために他者によって使用される、制限クローニング、ファージ組み換え、及び配列特異的組み換え等の他の既知のライブラリ技術と比較して、改善されたKunkelによる方法は、1つのライブラリにつき>109配列の作成においてより効果的であり得、標的DNA上の任意の場所において配列多様性を導入する際により万能である。
【0045】
インライン親和性成熟。統合親和性成熟プロセス、又はインライン親和性成熟は、ライブラリから選択される非抗体ペプチド又はタンパク質の結合親和性の設計及び改善のために、本発明に従って使用することができる。パニング後の所望の非抗体ペプチド又はタンパク質の結合親和性を改善することは、治療用非抗体ペプチド又はタンパク質リードの特定の成功を増加させることができる。ライブラリ作成のためのKunkelの方法の使用は、単純かつ連続的なプロセスにおける配列多様化戦略の効果的な実行を改善できる。改善されたKunkel突然変異作成方法の使用の設計戦略及び技術的利点は、結果の効率性及び有効性を低下させる単調なライブラリ作成方法が使用される、他の共同成熟戦略より優れた方法を提供できる。
【0046】
平行ライブラリパニング。自動又は半自動機器を使用する平行パニングプロセスを使用して、個々に作製されたサブライブラリを処理することができる。平行パニングは、所望の特徴のペプチド又はタンパク質を有する所望のライブラリを提供するために、好適に多様な一連の非抗体ペプチド又はタンパク質のを発見する可能性を最大化できる。インライン親和性成熟における平行パニングの効果的な使用はまた、そのようなタンパク質が、改善された親和性又は生物活性等のいくつかの同時に改善された特徴を有することも可能にする。機械によるパニングシステムの開発はまた、異なるパニング条件の系統的な監視及び調製が、所望の特性を有する非抗体ペプチド又はタンパク質をより迅速にスクリーニングし、単離することも可能にする。
【0047】
親和性順位。親和性による結合検定は、更なる特徴付けのための最良の結合を選択するために、大きい、多様な、かつ高親和性の配位子又は抗原特異的結合ペプチド若しくはタンパク質に適用される。ELISA等の標準的な生化学的方法、並びに多数の試料を処理するために好適な親和性測定機器、例えば、BIAcore、Octet及びBINDを、本目的のために、単独か又は組み合わせて使用する。
【0048】
本発明は一般条件で記述されてきているが、本発明の実施形態は、特許請求の範囲を限定するように解釈されるべきではない以下の実施例で更に開示される。
【実施例】
【0049】
実施例1:非抗体タンパク質の提示:2つの非抗体ペプチド又はタンパク質は、pIX融合タンパク質としてM13ファージの表面上に成功裏に提示された。両方の足場タンパク質は、それらの天然の状態における足場タンパク質によって結合しない特定のタンパク質標的に結合するように操作した。これらの操作された足場は、ファージ上で提示されつつ、この特定の標的への結合を保持した。加えて、アミノ酸変異体のライブラリはそれぞれの足場において作製し、特定の標的に結合するそれらの能力は、pIX融合としてファージ上にライブラリメンバーを提示すること及び標的タンパク質に対するパニングによって対処した。両方のライブラリは、多数の陽性ヒットをもたらした。
【0050】
2つの非抗体ペプチド又はタンパク質、免疫グロブリン(Ig)ドメインタンパク質は、pIX融合タンパク質としてM13ファージの表面上にEGFR結合ペプチドを提示するように操作される候補として選択した。以下の基準を、候補の選択のために使用した。
【0051】
PDBデータベースの中で利用可能な高解像度原子構造
a.堆積した構造は、単一の単離されたIgドメインを表す
b.完全なヒト配列
c.配列は、ジスルフィド結合残基を含有しない
d.公開された発現及び精製条件
e.再折り畳み工程の必要性を伴わず、かつ高収率で大腸菌から成功裏に発現され、精製されることを証明した。
【0052】
PDBデータベースは、単一Igドメインを含有する全てのタンパク質及びタンパク質フラグメントを調査するために、手動で、並びに構造による整合プログラムの援助により検索した。その後、公開された発現及び精製条件を決定するために、科学文献を調査した。
【0053】
上記の基準を使用して、成功のための最高の機会を有する分子を選択したが、これらの基準に厳密に満たない他のIg分子も、ペプチド提示のための成功する足場として使用することができることが想定される。複数のIgドメイン又は単一Igドメインの繰り返し体からなる足場は、成功する足場として使用することができることも想定される。上記の分析は、更なる研究のための、2つのIg分子、ヒトテネイシンからの第3のFn3ドメイン(残基802〜893、TeFn3)及びヒトタイチンのI27ドメイン(残基5253〜5341、Ti27)の選択をもたらした。これらのタンパク質フラグメントをコードする遺伝子は、Blue Heronによって合成し、標準PCR及び制限消化方法によってプラスミドpPEP9−BbsIにサブクローンした。Ti27におけるN5331G、及びTeFn3におけるS881Gの突然変異は、ペプチドの挿入のための制限部位を導入するために含んだ(以下参照)。
【0054】
その後、両方の足場のF:Gループ(TeFn3の残基876〜881及びTi27の5258〜5259)(図1)は、標準的な制限部位クローニング方法を使用して、システイン拘束EGFR結合ペプチド、PHPEP190(配列 DPCTWEVWGRECLQ)(Wang)の一部と置換し、Ti27−190及びTeFn3−190作製物を作製した。多数の対照作製物もまた作製し、非EGFR結合F:Gループ配列(Ti27及びTeFn3と称される)を含有し、及びF:Gループに6つの終止コドン(Ti27−終止及びTeFn3−終止)を含有するものを含む。表1は、これらの作製物を要約する。
【0055】
【表1】
【0056】
それぞれの作製物は、足場タンパク質のC末端とpIXのN末端との間のEDSGGSGG(配列番号30)リンカー配列を伴うファージミドベクターからの糸状ファージの小さい外被タンパク質pIXへの融合として発現された。全ての作製物は、EGFR結合の非存在下でのタンパク質発現及び提示に対して検定するために、N末端Mycタグにより発現された。M13ファージの表面上の成功する提示、及びTeFn3−190及びTi27−190タンパク質の特定の結合活性は、ファージELISAによって検証した。0.5μgのEGFR細胞外ドメイン(sEGFR−MMB)(Wang)のFc−融合型、抗Myc非抗体ペプチド若しくはタンパク質、又は対照Fc分子(CNTO 360)を、96ウェルMaxiSorop(商標)プレート上で固定し、StartingBlock(商標)T−20により遮断した。表1に記載される作製物を提示するファージは、それぞれのプレートへの300μLのファージ含有上清の添加前に連続的に希釈し、続いて、室温で1時間培養した。それぞれのプレートは、続いてTBSTで洗浄し、蛍光プレートリーダを使用して、抗M13 HRP結合非抗体ペプチド又はタンパク質により検出されるファージに結合させた。図2は、TeFn3−190及びTi27−190が、CNTO 360への大きな結合の欠如によって証明される低いレベルのバックグラウンド結合によりプレート結合Fc−EGFRに選択的に結合したことを示す。TeFn3及びTi27はまた、抗Mycコーティングされたプレートへの結合によって示されるファージ表面上でも提示された。しかしながら、これらのタンパク質は、EGFRに結合しなかった。不明の理由で、TeFn3−190は、EGFR結合によって示されるように明らかに発現されるが、抗Mycコーティングされたプレートに結合しなかった。予想通りに、TeFn3−終止及びTi27−終止は、ファージ上に提示されなかった。
【0057】
ライブラリは、TeFn3及びTi27に関連してPHPEP190のシステインの必要性を決定し、そのようなライブラリを、pIX上のこれらの足場内で効率的に提示し、標的タンパク質に対してパニングすることができることを確認するために構築した。このライブラリは、Ti27−終止及びTeFn3−終止作製物において終止コドンを含有する全ての6つの位置を無作為化することによって構築した(表1)。Kunkelの方法(Kunkel,et al.,Methods Enzymol 154:367〜82,1987)による突然変異作成の後の100の無作為で採取したコロニーのシークエンシングは、突然変異が、Ti27−190及びTeFn3−190ライブラリからのコロニーのそれぞれ51%及び64%において見られたことを確認した。その後、これらのライブラリは、室温で1時間10μg(1回目)又は5μg(2及び3回目)のビオチン化EGFRにより100μLのファージ溶液を培養することによって、EGFR−Fc融合に対して3回パニングした。結合複合体は、ストレプトアビジンコーティングされた磁気ビーズで引き下げ、TBSTで3回洗浄した。その後、結合ファージを使用して、TG1細胞を再感染させ、表2に記載されるファージ力価をもたらした。
【0058】
【表2】
【0059】
それぞれのライブラリからの3回目のパニングからの96のコロニーは、ファージELISAによってEGFR−Fcへの結合を試験するために無作為に選択した。図3は、Ti27−190及びTeFn3−190ライブラリからの10及び20のクローンそれぞれが、EGFRへの有意な結合を示したことを示す(EGFR:CNTO 360からの総数の比率は、5より大きい)。全ての陽性ヒットからのF:Gループの配列を表3に示す。全ての配列したヒットは、本来のPHPEP190と同様に同一の位置でシステイン残基を含有し、このペプチドのシステイン拘束された性質が機能のために必要であることを示した。
【0060】
【表3】
【特許請求の範囲】
【請求項1】
生体分子に選択的に結合するファージ提示融合タンパク質を発現するための操作された組み換え核酸ファージベクターであって、
a.以下に作動可能に連結する、組み換えファージリーダーコード核酸配列と、
b.以下に作動可能に連結する、組み換え制限部位と、
c.aに作動可能に連結する、ペプチドリンカーコード核酸配列と、
d.生体分子に選択的に結合する非抗体足場タンパク質をコードするヌクレオチド配列と、
を含む、操作された組み換え核酸ファージベクター。
【請求項2】
前記生体分子が、上皮成長因子受容体である、請求項1に記載の生体分子。
【請求項3】
前記ファージリーダーコード配列が、pelB配列である、請求項1に記載の操作された核酸ファージベクター。
【請求項4】
請求項1に記載の操作された核酸ファージベクターによってコードされる非抗体足場タンパク質であって、配列番号32〜53、55及び58から選択される、非抗体足場タンパク質。
【請求項5】
請求項1に記載の操作された核酸ファージベクターによってコードされる非抗体足場タンパク質であって、上皮成長因子受容体に結合する、非抗体足場タンパク質。
【請求項6】
前記ベクターが、前記非抗体足場タンパク質に融合される第2の外因性ペプチドをコードする、請求項1に記載の操作された核酸ファージベクター。
【請求項7】
前記第2の外因性ペプチドが、前記生体分子に結合する、請求項6に記載の操作された核酸ファージベクター。
【請求項8】
前記第2の外因性ペプチドが、上皮成長因子受容体に結合する、請求項7に記載の操作された核酸ファージベクター。
【請求項9】
請求項1に記載の操作された核酸ファージベクターを含む、細菌性宿主細胞。
【請求項10】
請求項9に記載の細菌性宿主細胞によって発現される、生物活性融合タンパク質。
【請求項11】
請求項10に記載の前記融合タンパク質に由来する、生物活性ペプチド又はタンパク質。
【請求項12】
請求項1に記載の複数の操作された核酸ファージベクターを含む、ファージライブラリ。
【請求項13】
前記非抗体足場タンパク質の変異型が発現される、請求項12に記載のファージライブラリ。
【請求項14】
所望の生物活性を有するペプチド又はタンパク質に対し、ファージ非抗体ペプチド又はタンパク質ライブラリをスクリーニングするための方法であって、(a)請求項13に記載のファージライブラリから前記ペプチド又はタンパク質を発現する工程と、(b)前記所望の生物活性を有する前記ペプチド又はタンパク質を発現する細菌性細胞を選択する工程と、を含む方法。
【請求項15】
請求項14に記載の方法から得られる、ペプチド又はタンパク質コード核酸。
【請求項16】
生体分子への結合を媒介する配列番号2〜23、25、又は28の少なくとも一部分を含む、単離された非抗体足場タンパク質。
【請求項17】
生体分子への結合を媒介する前記部分が、上皮成長因子受容体に結合する、請求項16に記載の単離された非抗体足場タンパク質。
【請求項18】
請求項16に記載の前記非抗体足場タンパク質、及び製薬学的に許容できる担体を含む、医薬組成物。
【請求項19】
配列番号2〜23、25及び28のうちの少なくとも1つから選択されるポリペプチドを含む、単離された非抗体足場タンパク質。
【請求項20】
前記ポリペプチドが、上皮成長因子受容体に結合する、請求項19に記載の単離された非抗体足場タンパク質。
【請求項21】
請求項19に記載の前記非抗体足場タンパク質、及び製薬学的に許容できる担体を含む、医薬組成物。
【請求項1】
生体分子に選択的に結合するファージ提示融合タンパク質を発現するための操作された組み換え核酸ファージベクターであって、
a.以下に作動可能に連結する、組み換えファージリーダーコード核酸配列と、
b.以下に作動可能に連結する、組み換え制限部位と、
c.aに作動可能に連結する、ペプチドリンカーコード核酸配列と、
d.生体分子に選択的に結合する非抗体足場タンパク質をコードするヌクレオチド配列と、
を含む、操作された組み換え核酸ファージベクター。
【請求項2】
前記生体分子が、上皮成長因子受容体である、請求項1に記載の生体分子。
【請求項3】
前記ファージリーダーコード配列が、pelB配列である、請求項1に記載の操作された核酸ファージベクター。
【請求項4】
請求項1に記載の操作された核酸ファージベクターによってコードされる非抗体足場タンパク質であって、配列番号32〜53、55及び58から選択される、非抗体足場タンパク質。
【請求項5】
請求項1に記載の操作された核酸ファージベクターによってコードされる非抗体足場タンパク質であって、上皮成長因子受容体に結合する、非抗体足場タンパク質。
【請求項6】
前記ベクターが、前記非抗体足場タンパク質に融合される第2の外因性ペプチドをコードする、請求項1に記載の操作された核酸ファージベクター。
【請求項7】
前記第2の外因性ペプチドが、前記生体分子に結合する、請求項6に記載の操作された核酸ファージベクター。
【請求項8】
前記第2の外因性ペプチドが、上皮成長因子受容体に結合する、請求項7に記載の操作された核酸ファージベクター。
【請求項9】
請求項1に記載の操作された核酸ファージベクターを含む、細菌性宿主細胞。
【請求項10】
請求項9に記載の細菌性宿主細胞によって発現される、生物活性融合タンパク質。
【請求項11】
請求項10に記載の前記融合タンパク質に由来する、生物活性ペプチド又はタンパク質。
【請求項12】
請求項1に記載の複数の操作された核酸ファージベクターを含む、ファージライブラリ。
【請求項13】
前記非抗体足場タンパク質の変異型が発現される、請求項12に記載のファージライブラリ。
【請求項14】
所望の生物活性を有するペプチド又はタンパク質に対し、ファージ非抗体ペプチド又はタンパク質ライブラリをスクリーニングするための方法であって、(a)請求項13に記載のファージライブラリから前記ペプチド又はタンパク質を発現する工程と、(b)前記所望の生物活性を有する前記ペプチド又はタンパク質を発現する細菌性細胞を選択する工程と、を含む方法。
【請求項15】
請求項14に記載の方法から得られる、ペプチド又はタンパク質コード核酸。
【請求項16】
生体分子への結合を媒介する配列番号2〜23、25、又は28の少なくとも一部分を含む、単離された非抗体足場タンパク質。
【請求項17】
生体分子への結合を媒介する前記部分が、上皮成長因子受容体に結合する、請求項16に記載の単離された非抗体足場タンパク質。
【請求項18】
請求項16に記載の前記非抗体足場タンパク質、及び製薬学的に許容できる担体を含む、医薬組成物。
【請求項19】
配列番号2〜23、25及び28のうちの少なくとも1つから選択されるポリペプチドを含む、単離された非抗体足場タンパク質。
【請求項20】
前記ポリペプチドが、上皮成長因子受容体に結合する、請求項19に記載の単離された非抗体足場タンパク質。
【請求項21】
請求項19に記載の前記非抗体足場タンパク質、及び製薬学的に許容できる担体を含む、医薬組成物。
【図1】

【図2】
【図3】
【図2】
【図3】
【公表番号】特表2011−507529(P2011−507529A)
【公表日】平成23年3月10日(2011.3.10)
【国際特許分類】
【出願番号】特願2010−539875(P2010−539875)
【出願日】平成20年12月19日(2008.12.19)
【国際出願番号】PCT/US2008/087696
【国際公開番号】WO2009/086116
【国際公開日】平成21年7月9日(2009.7.9)
【出願人】(509087759)セントコア・オーソ・バイオテツク・インコーポレーテツド (77)
【Fターム(参考)】
【公表日】平成23年3月10日(2011.3.10)
【国際特許分類】
【出願日】平成20年12月19日(2008.12.19)
【国際出願番号】PCT/US2008/087696
【国際公開番号】WO2009/086116
【国際公開日】平成21年7月9日(2009.7.9)
【出願人】(509087759)セントコア・オーソ・バイオテツク・インコーポレーテツド (77)
【Fターム(参考)】
[ Back to top ]